/










































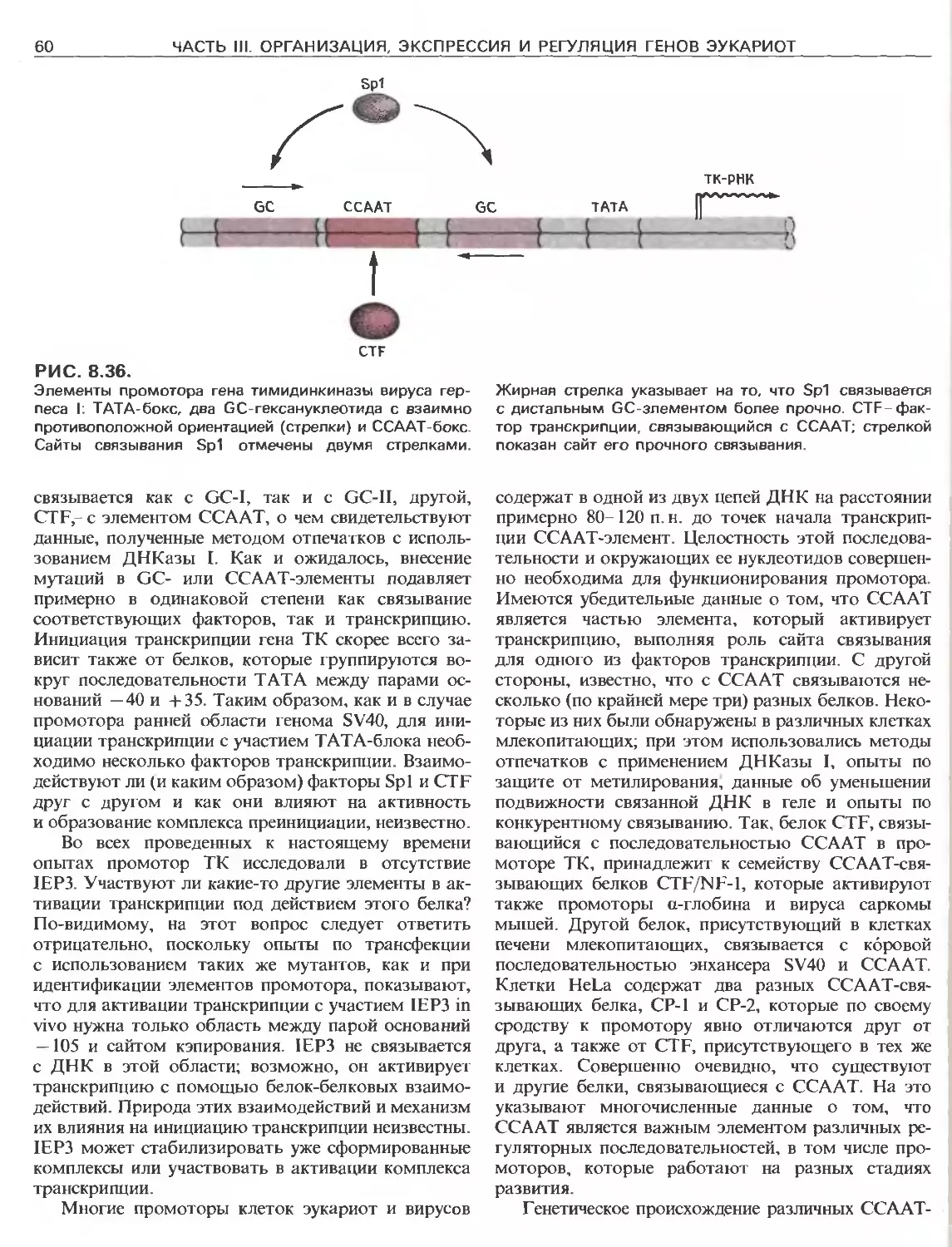

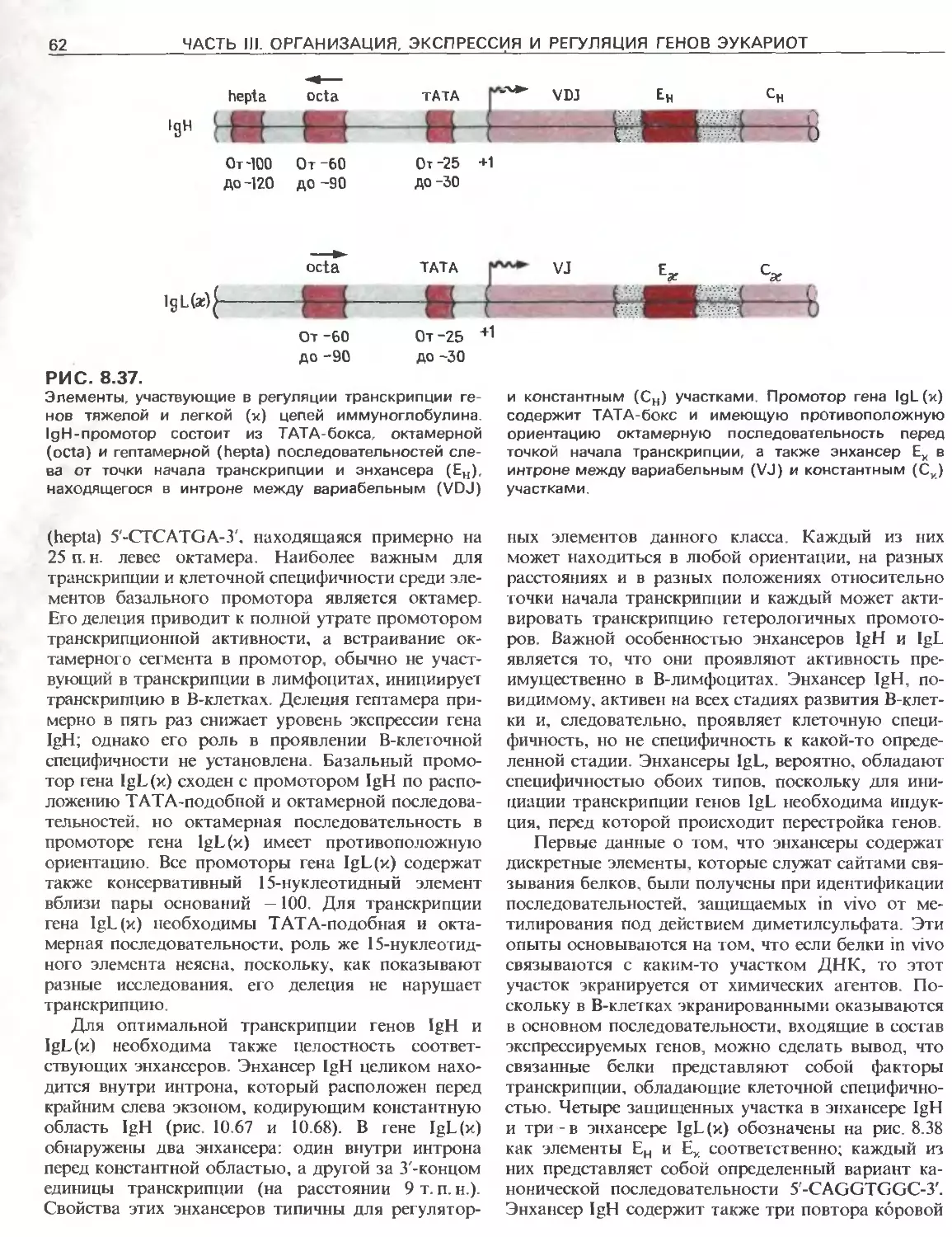



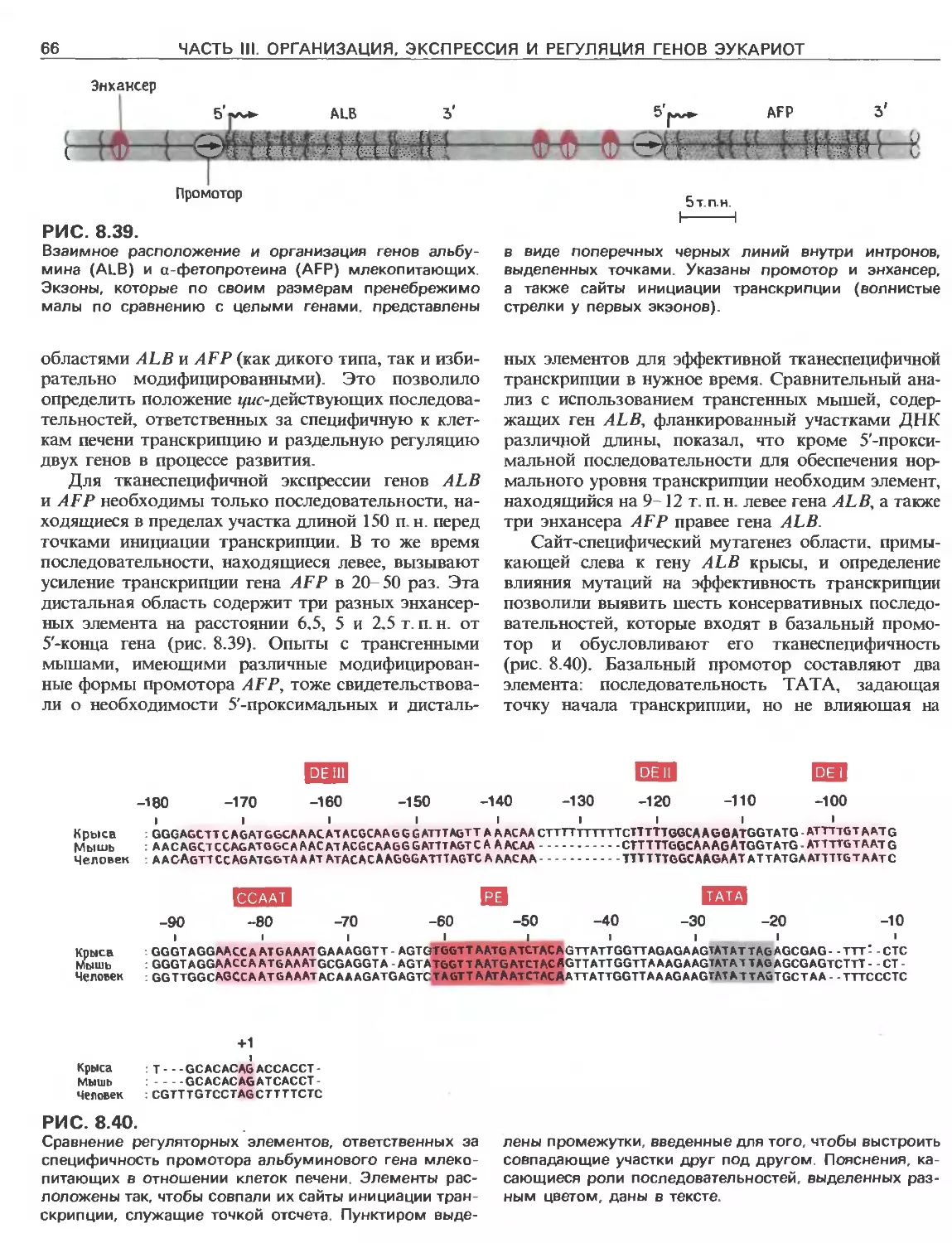






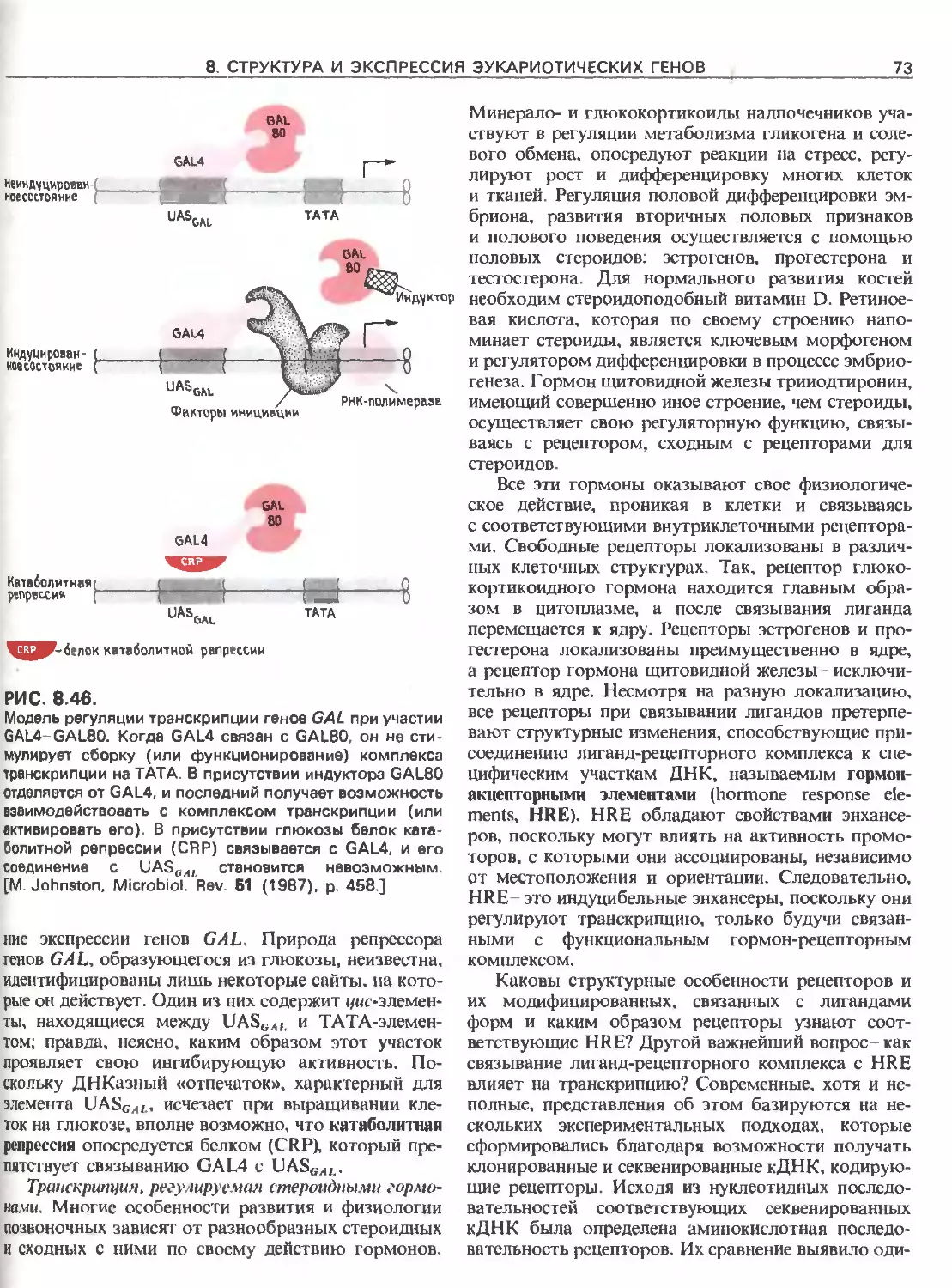










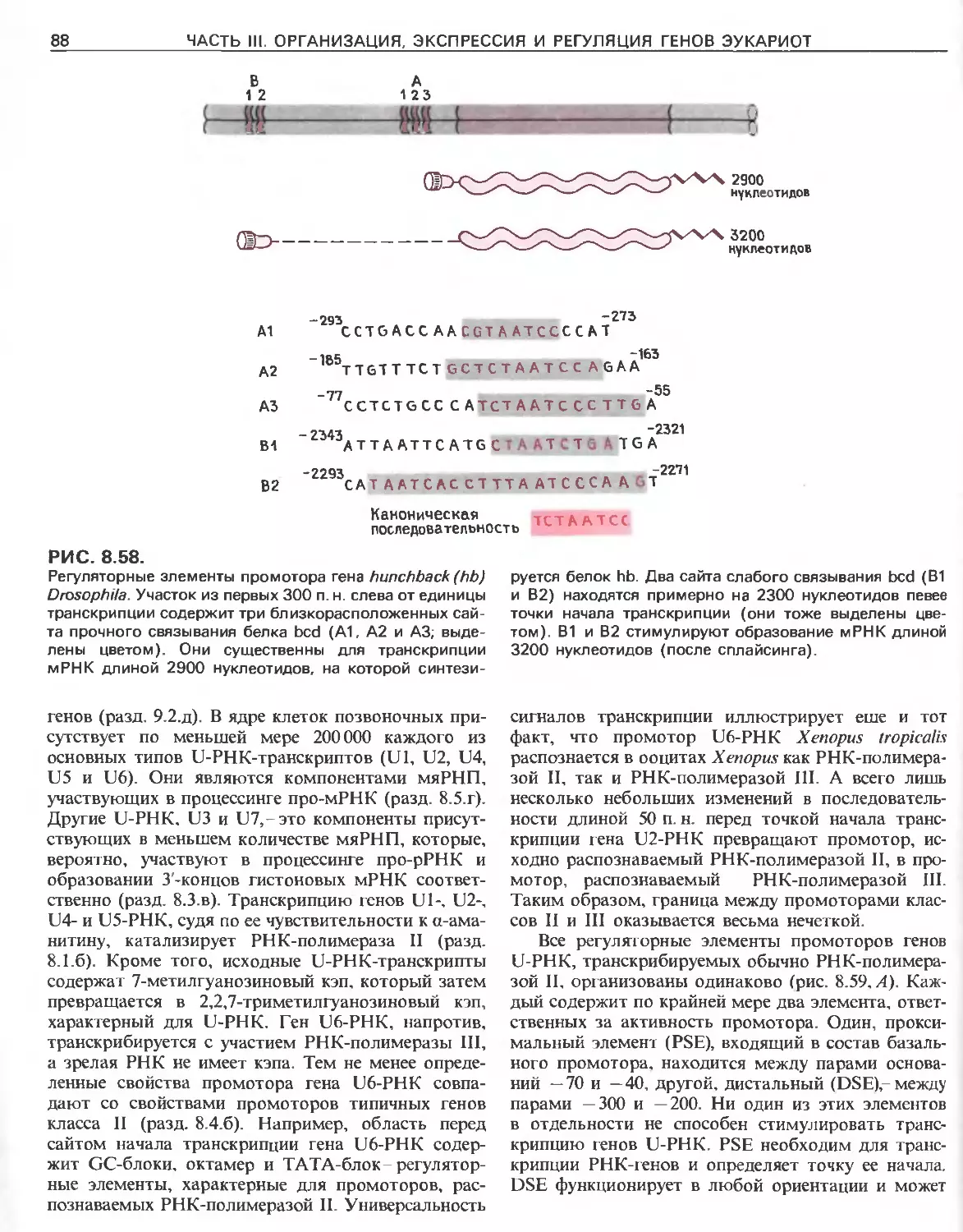
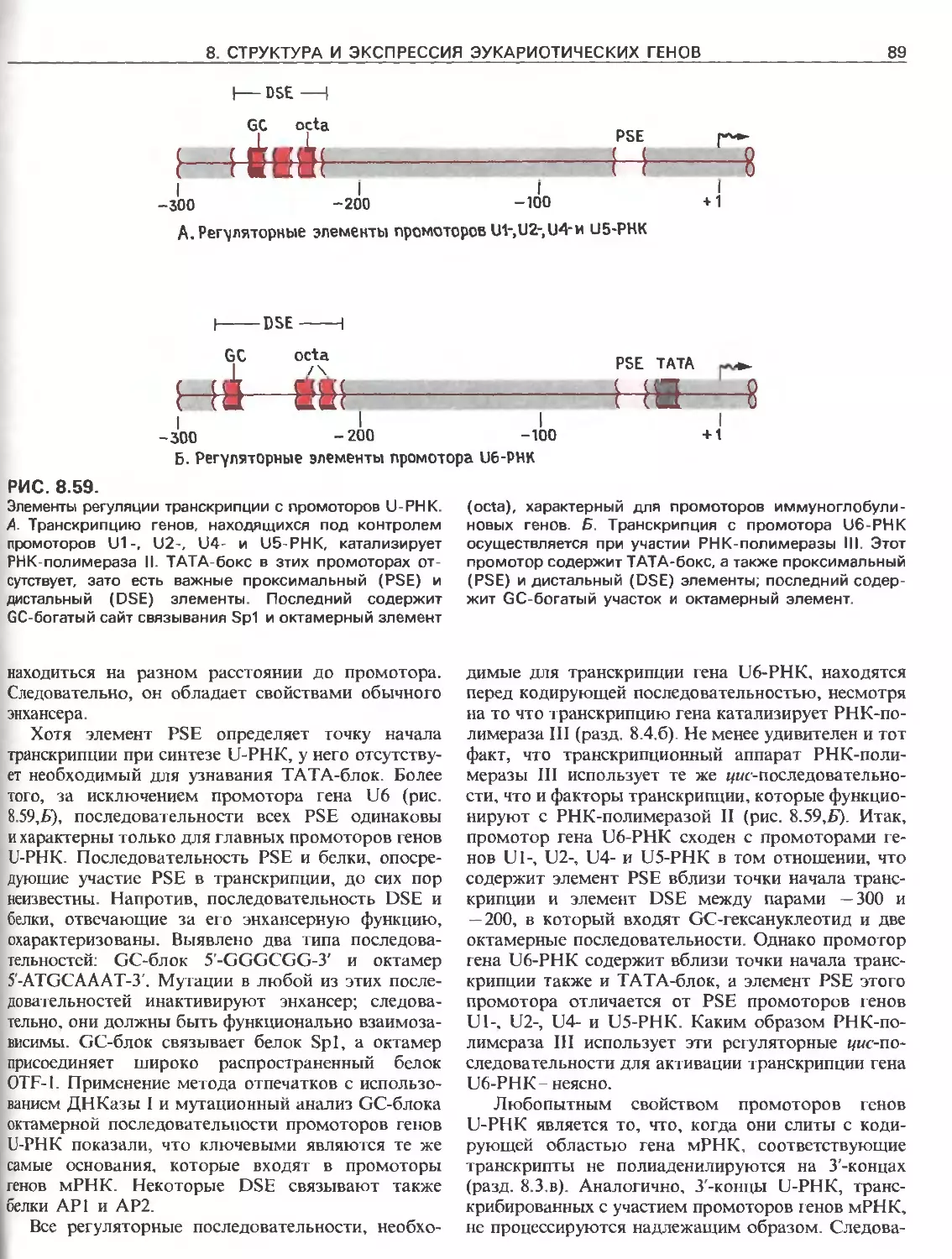



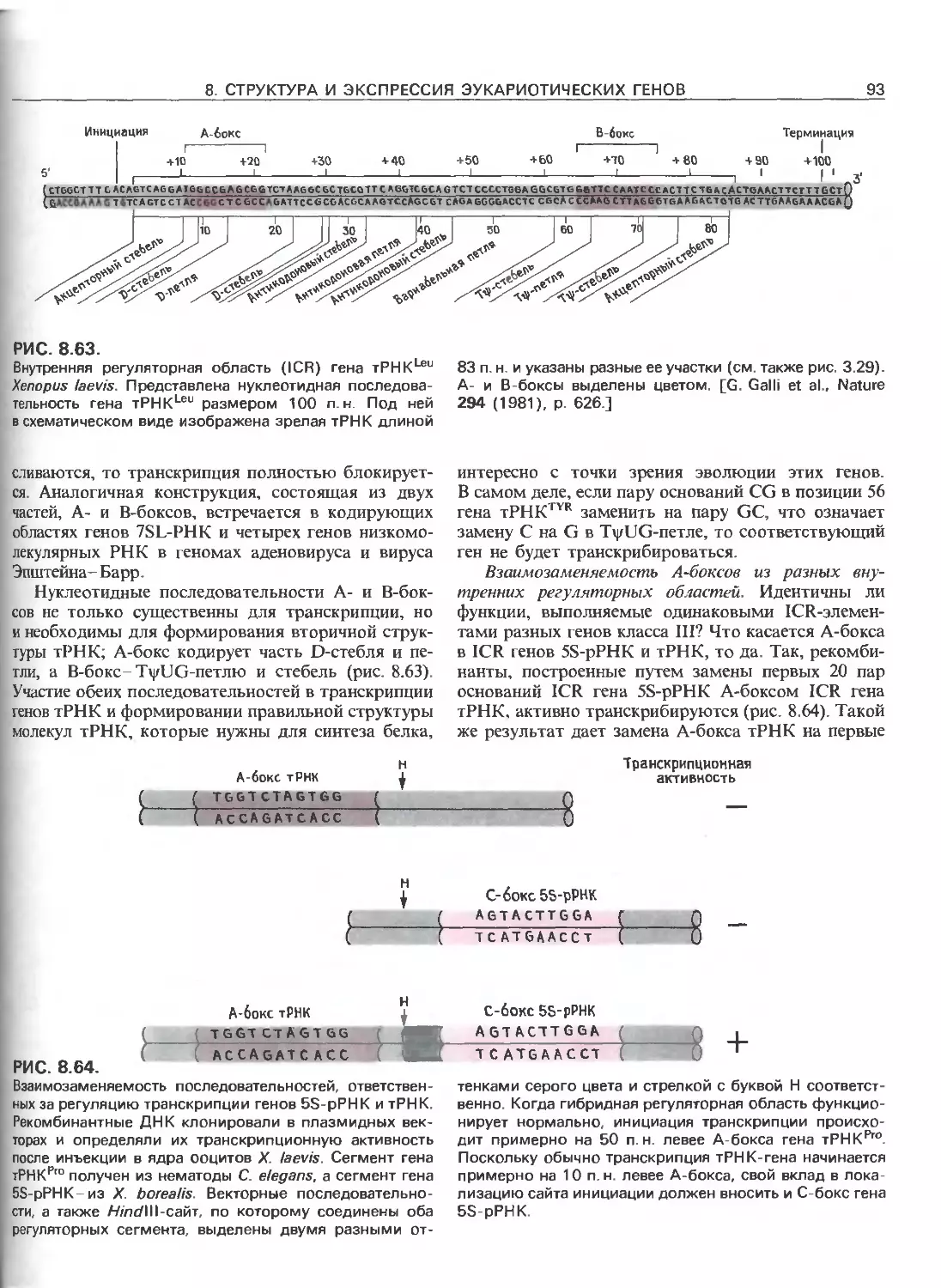


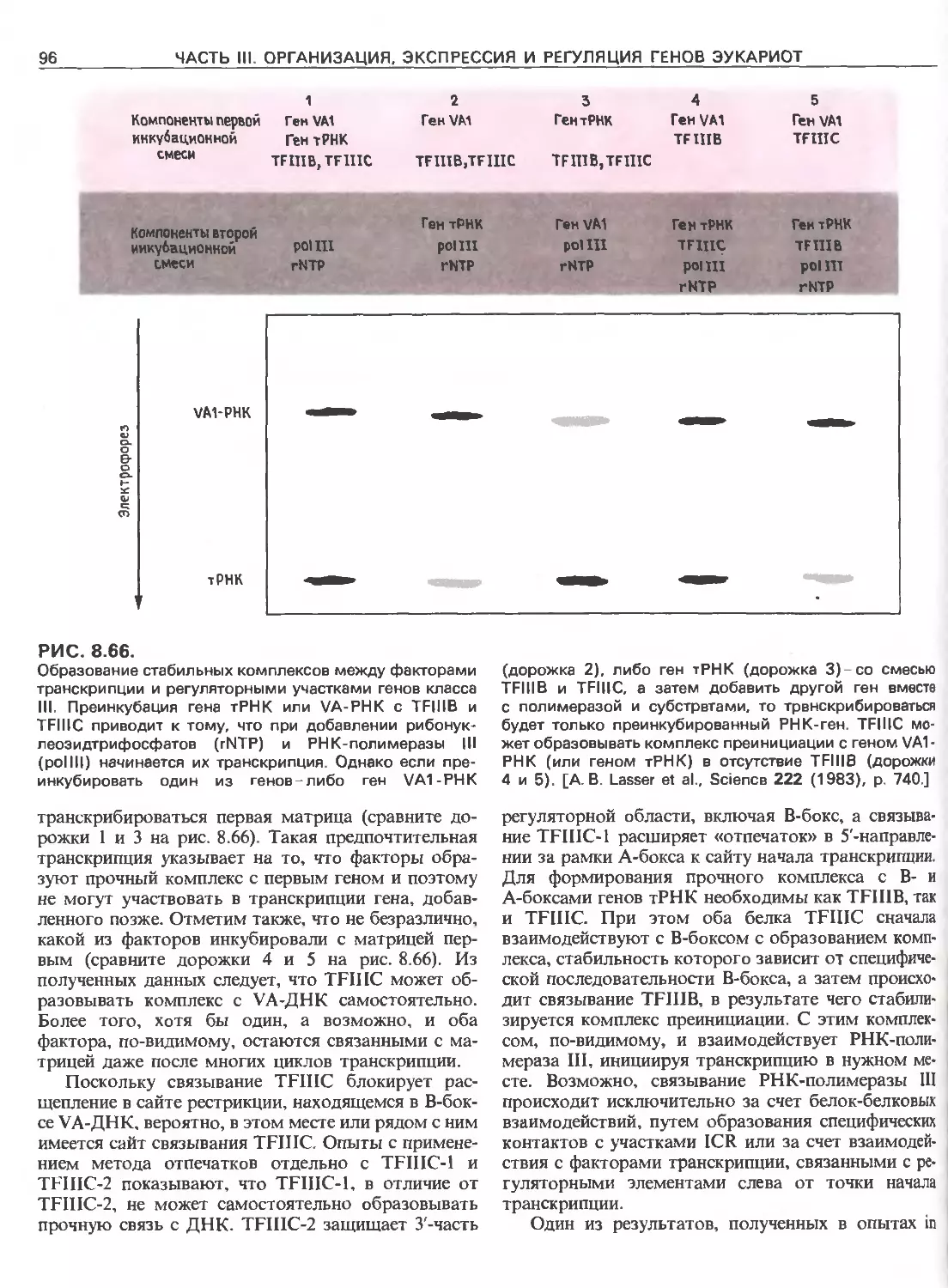

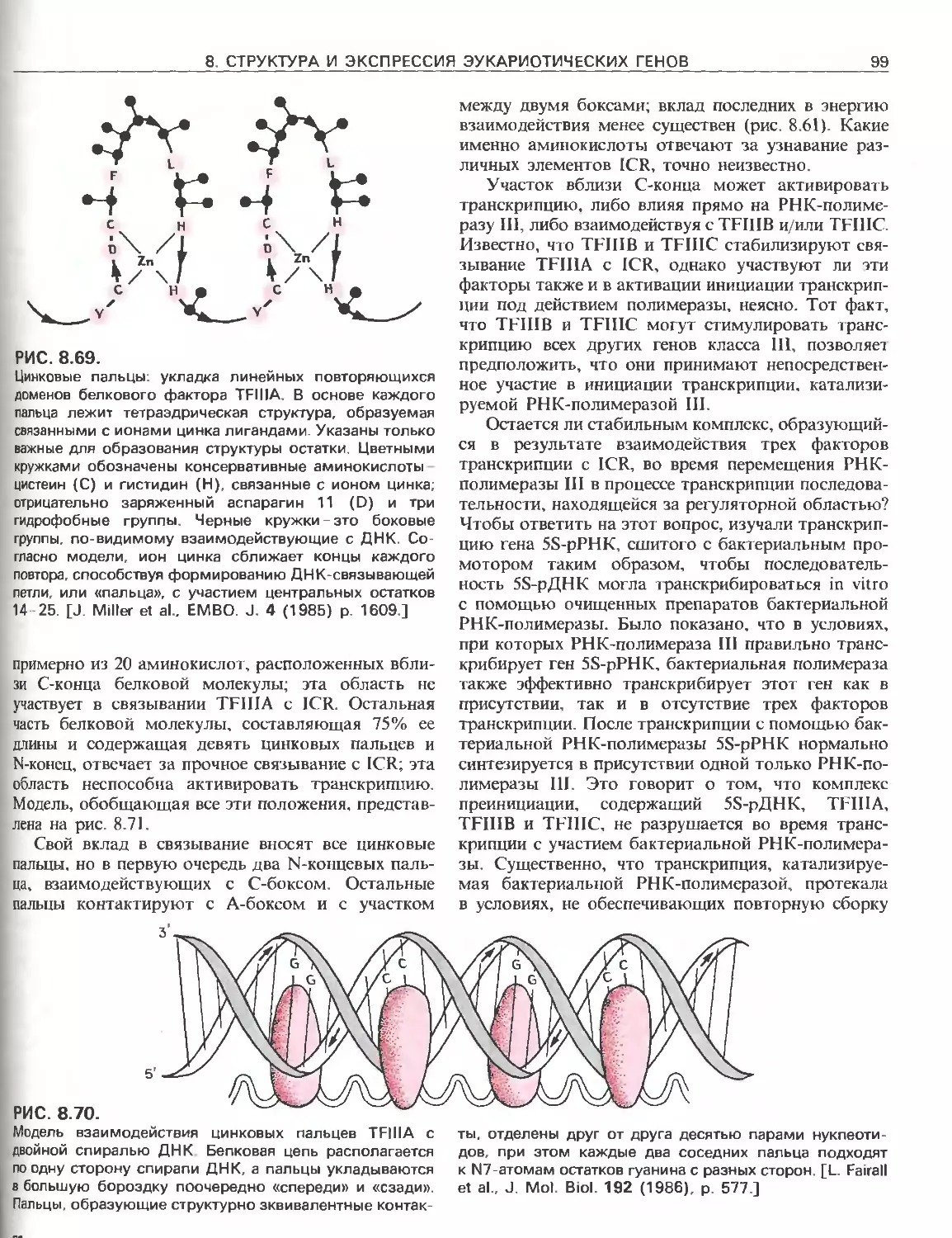
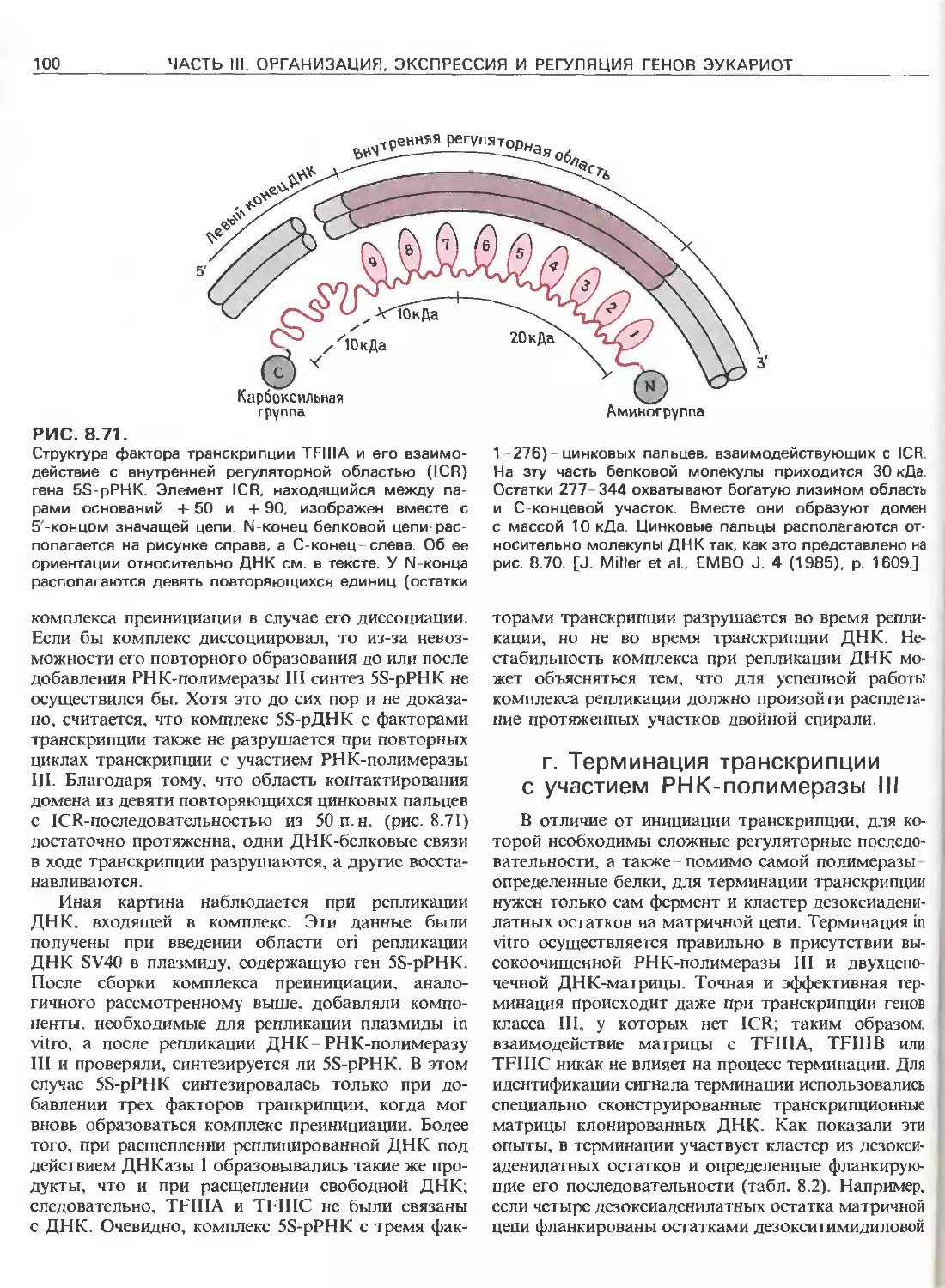






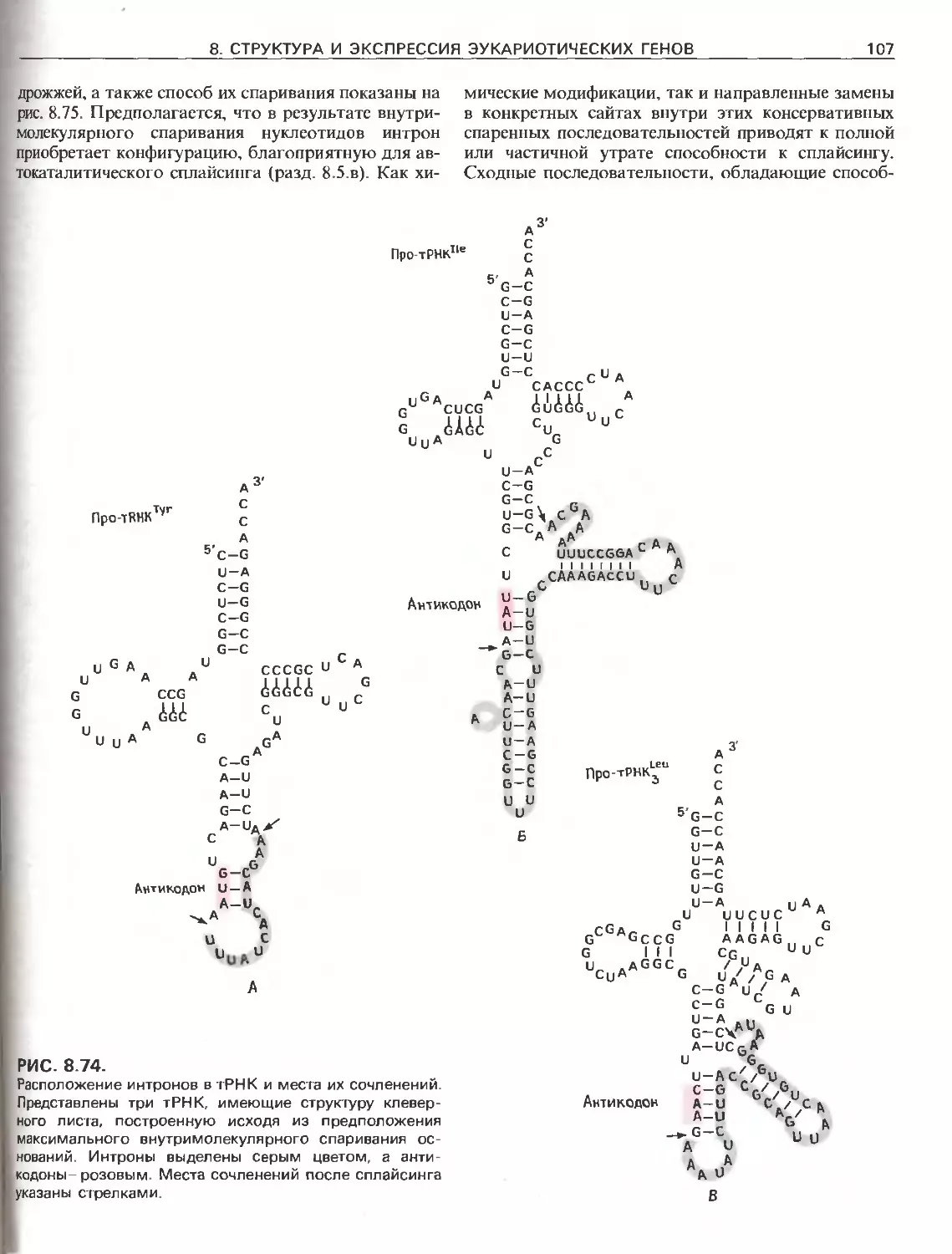


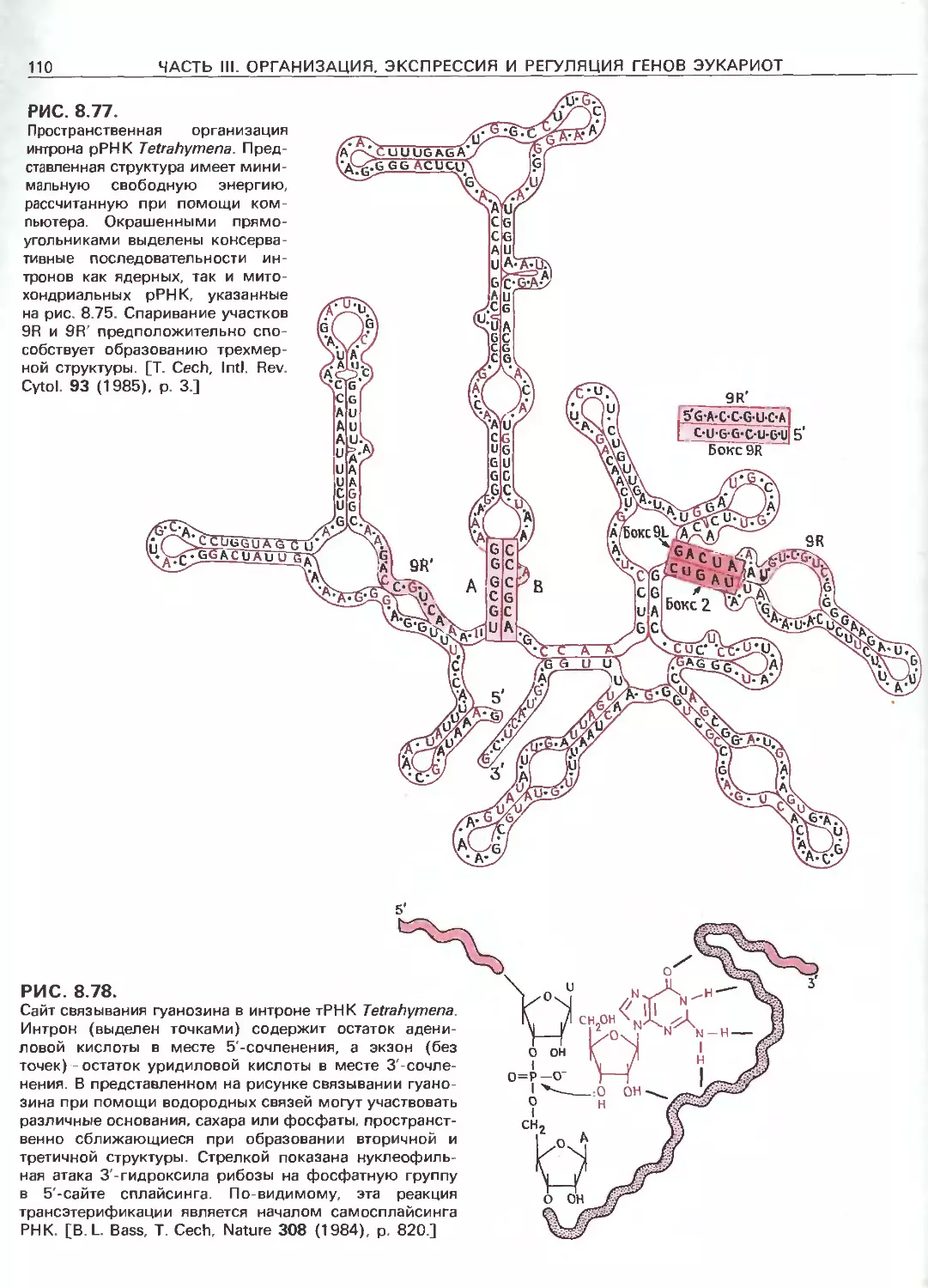
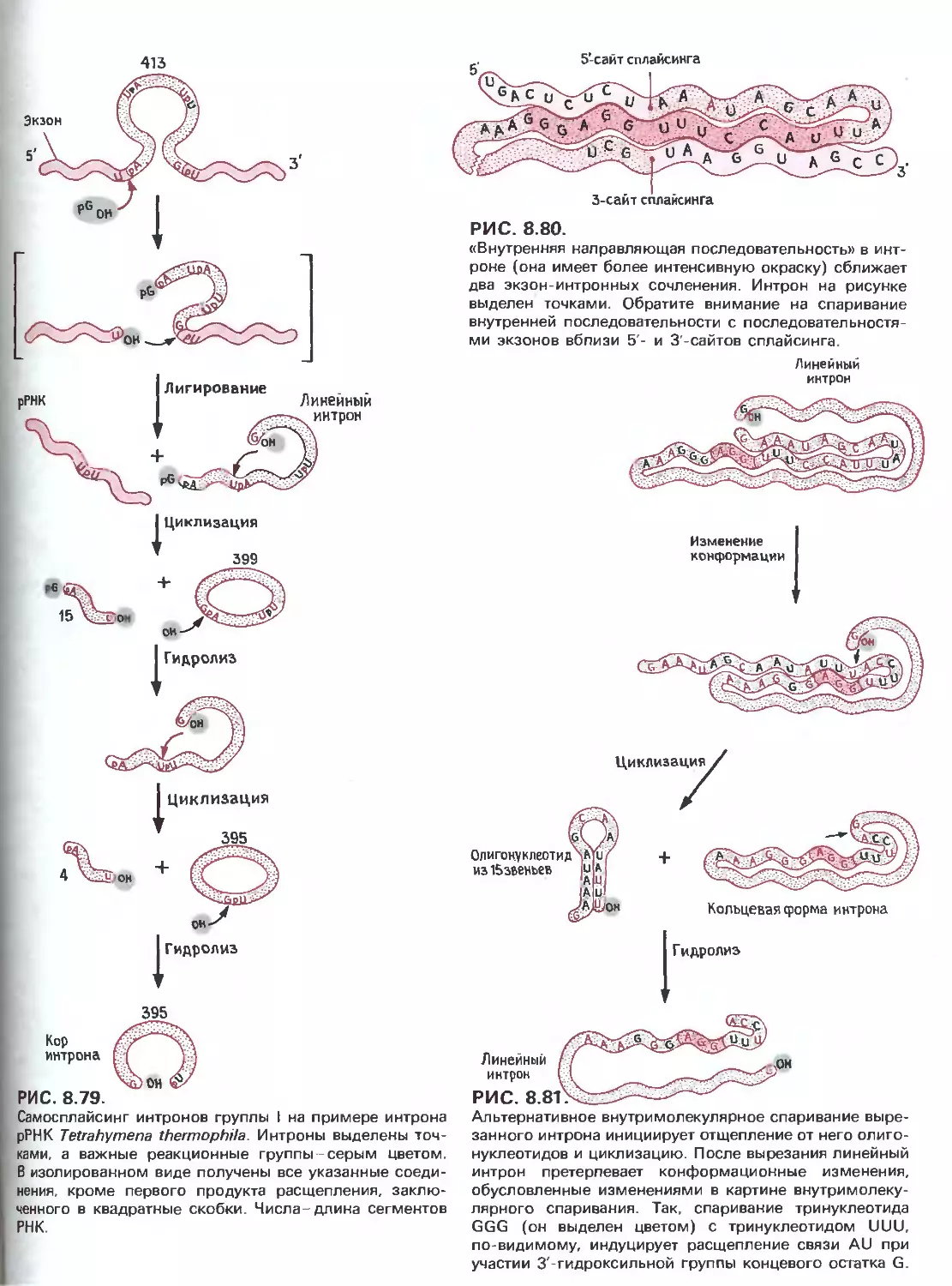


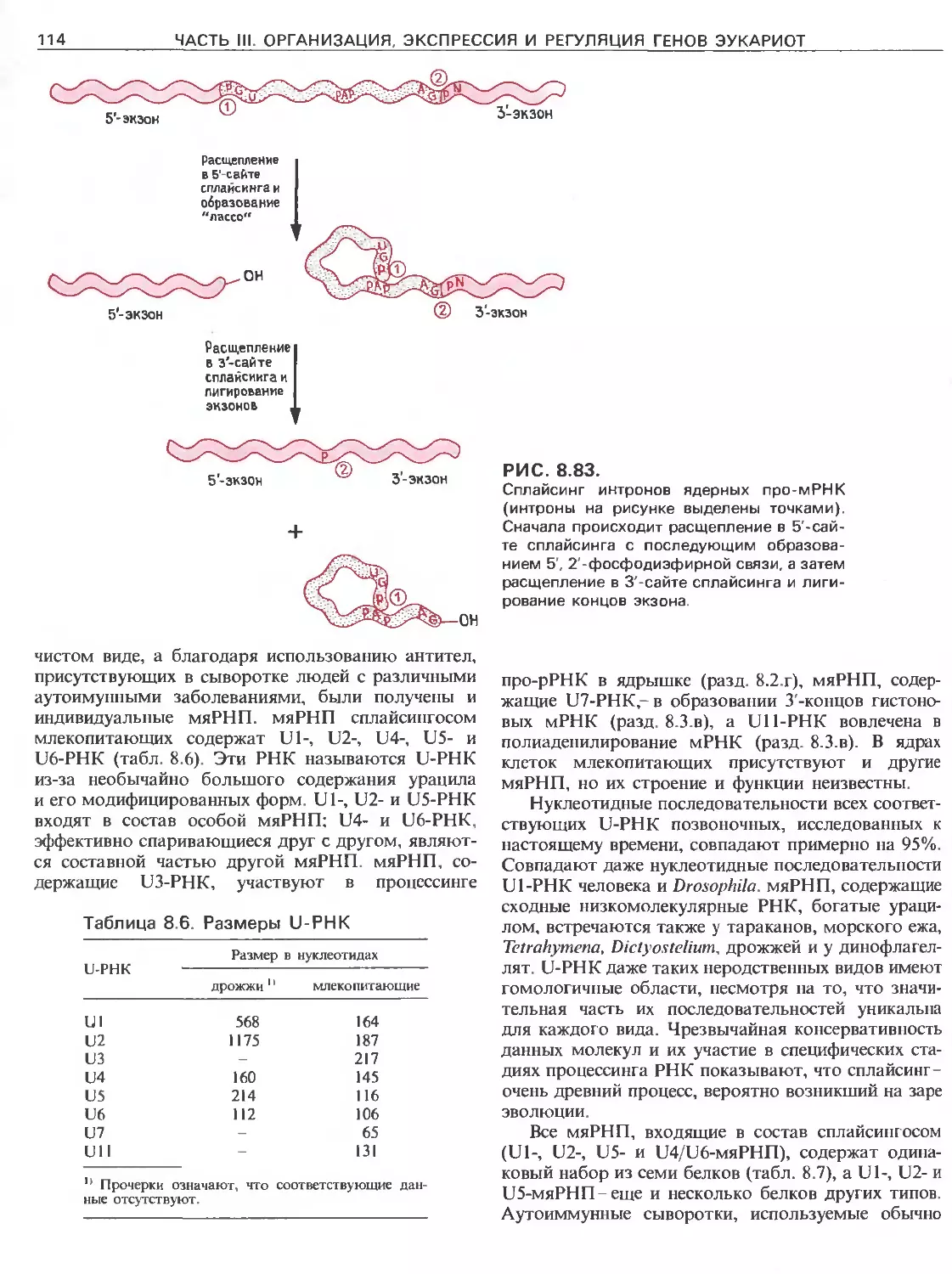



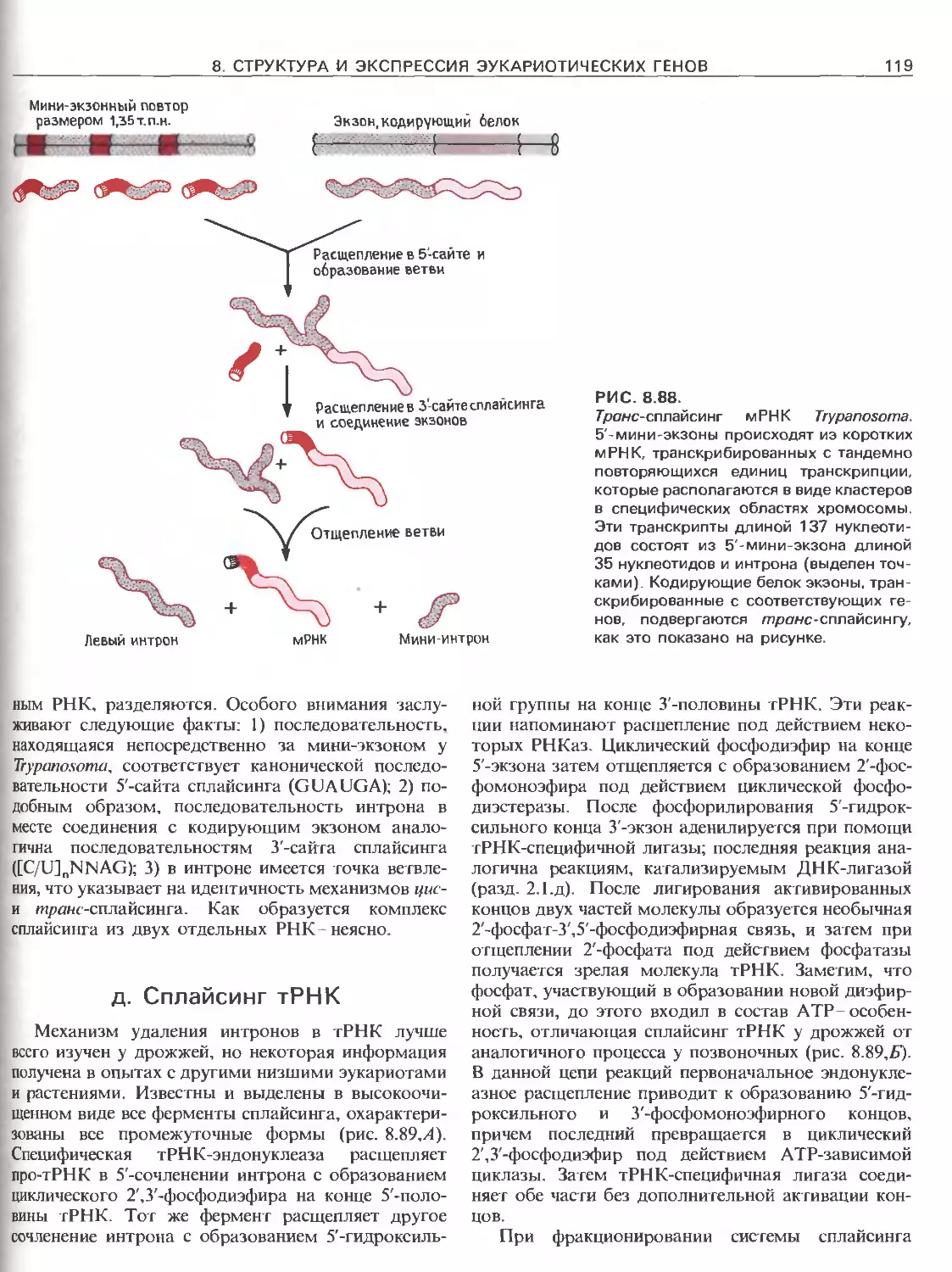
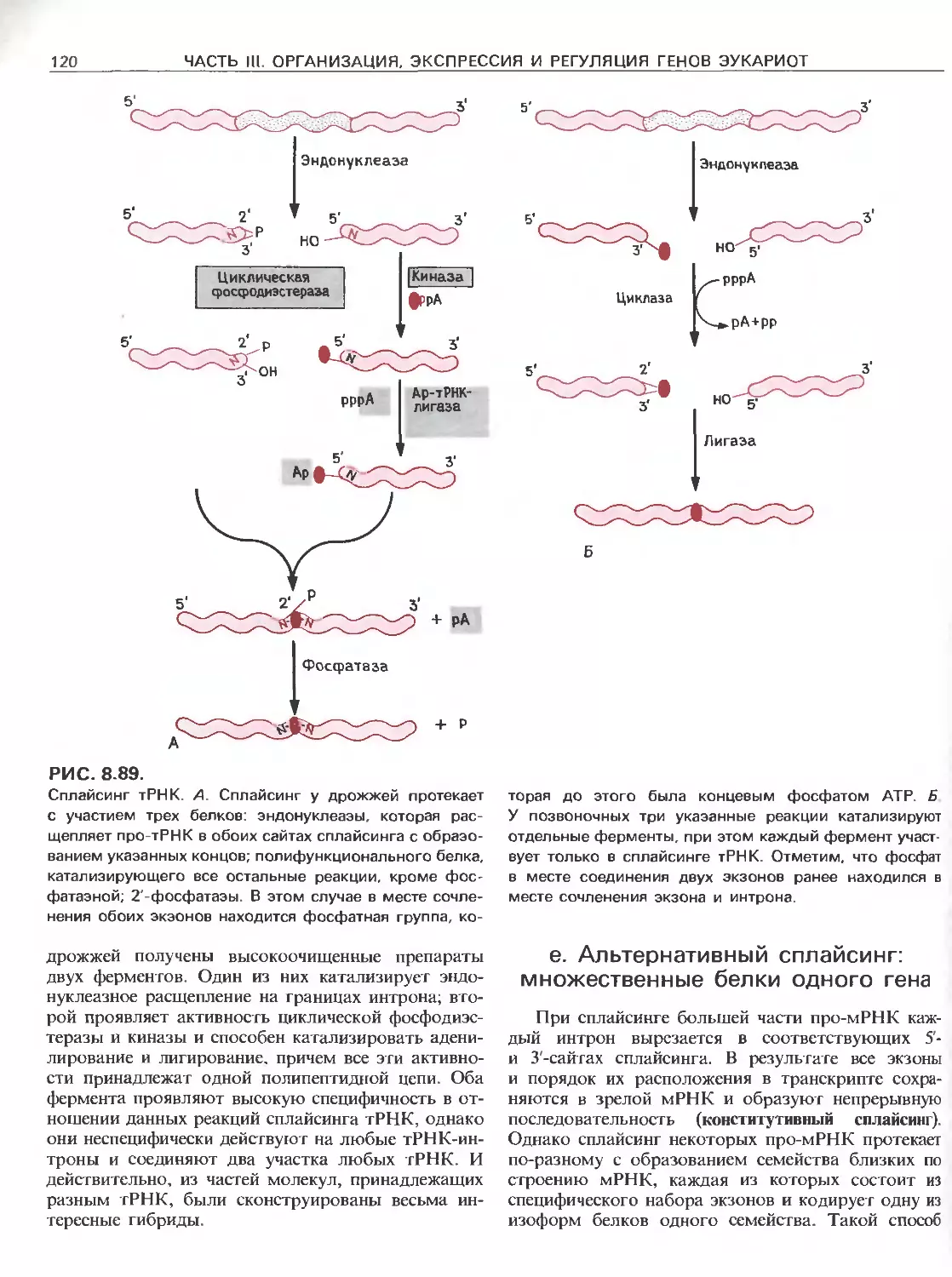

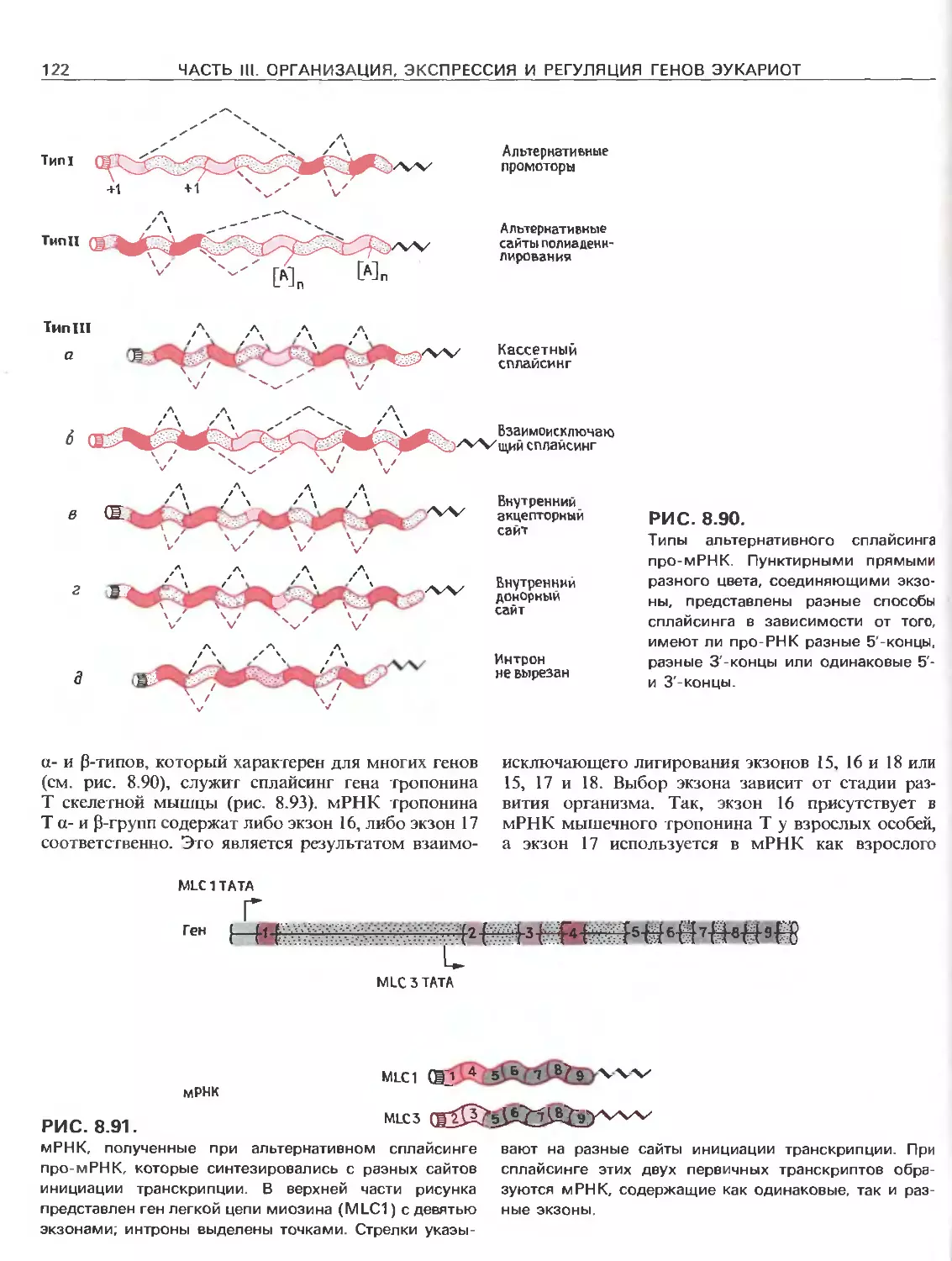
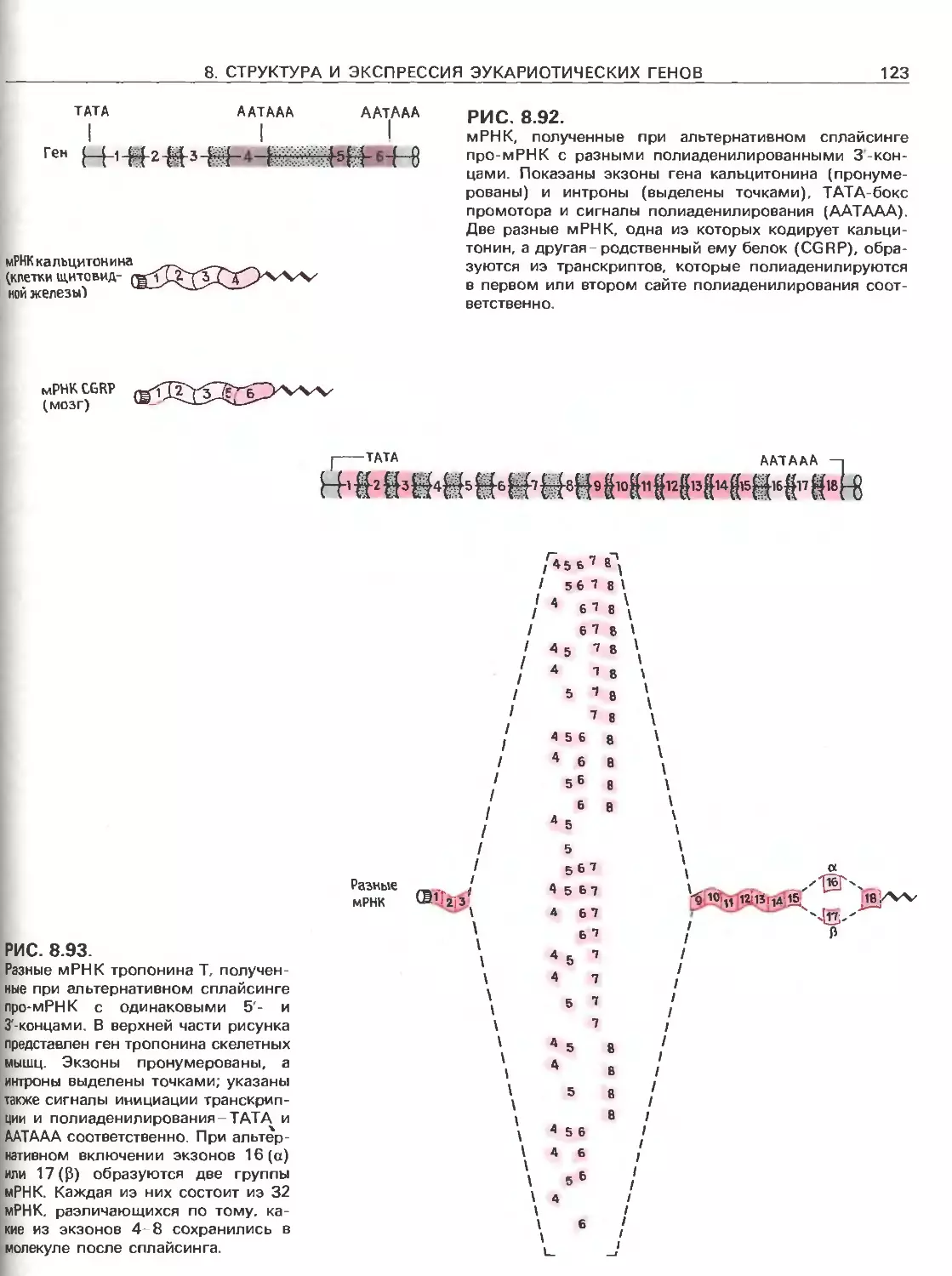
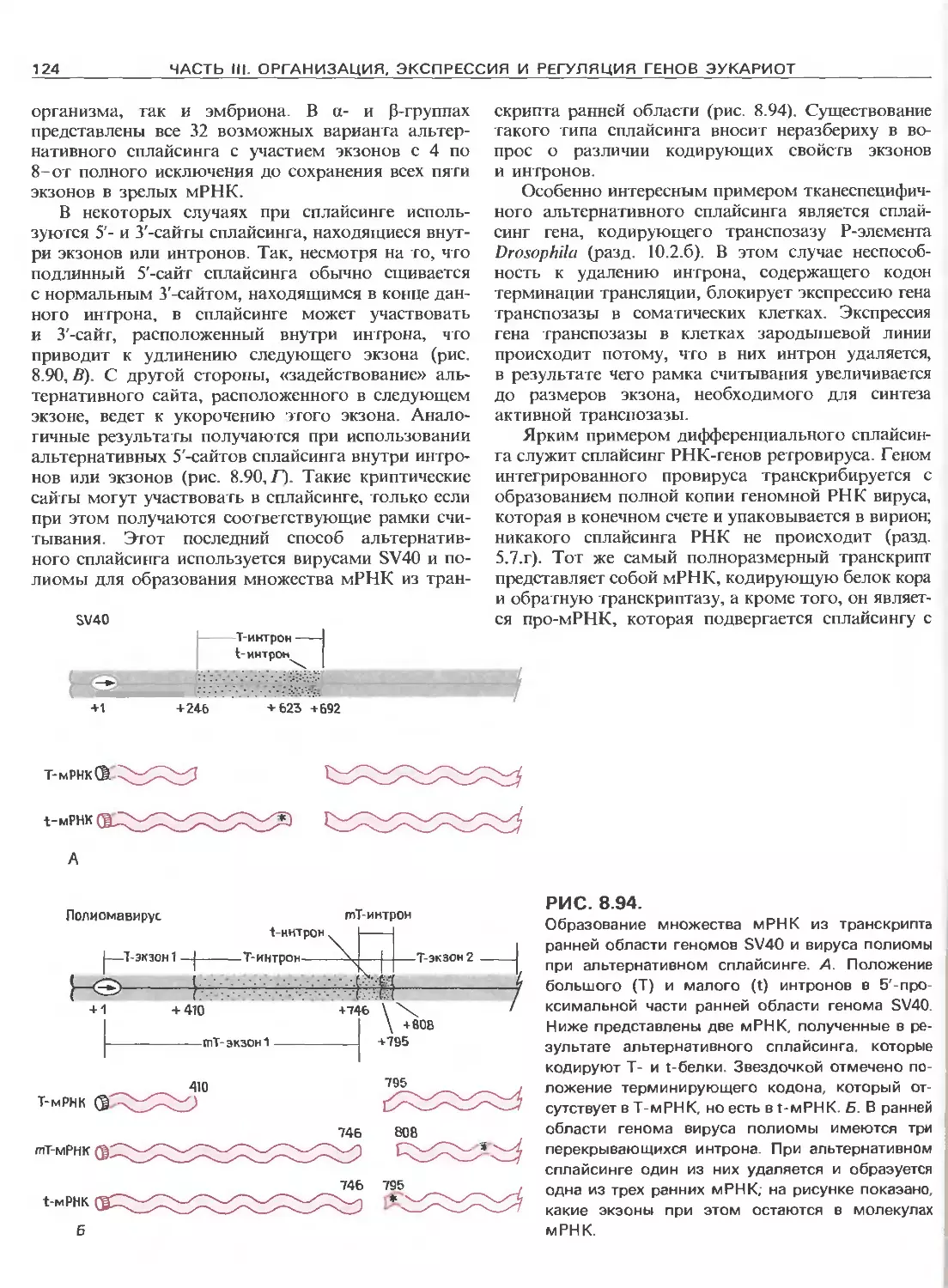

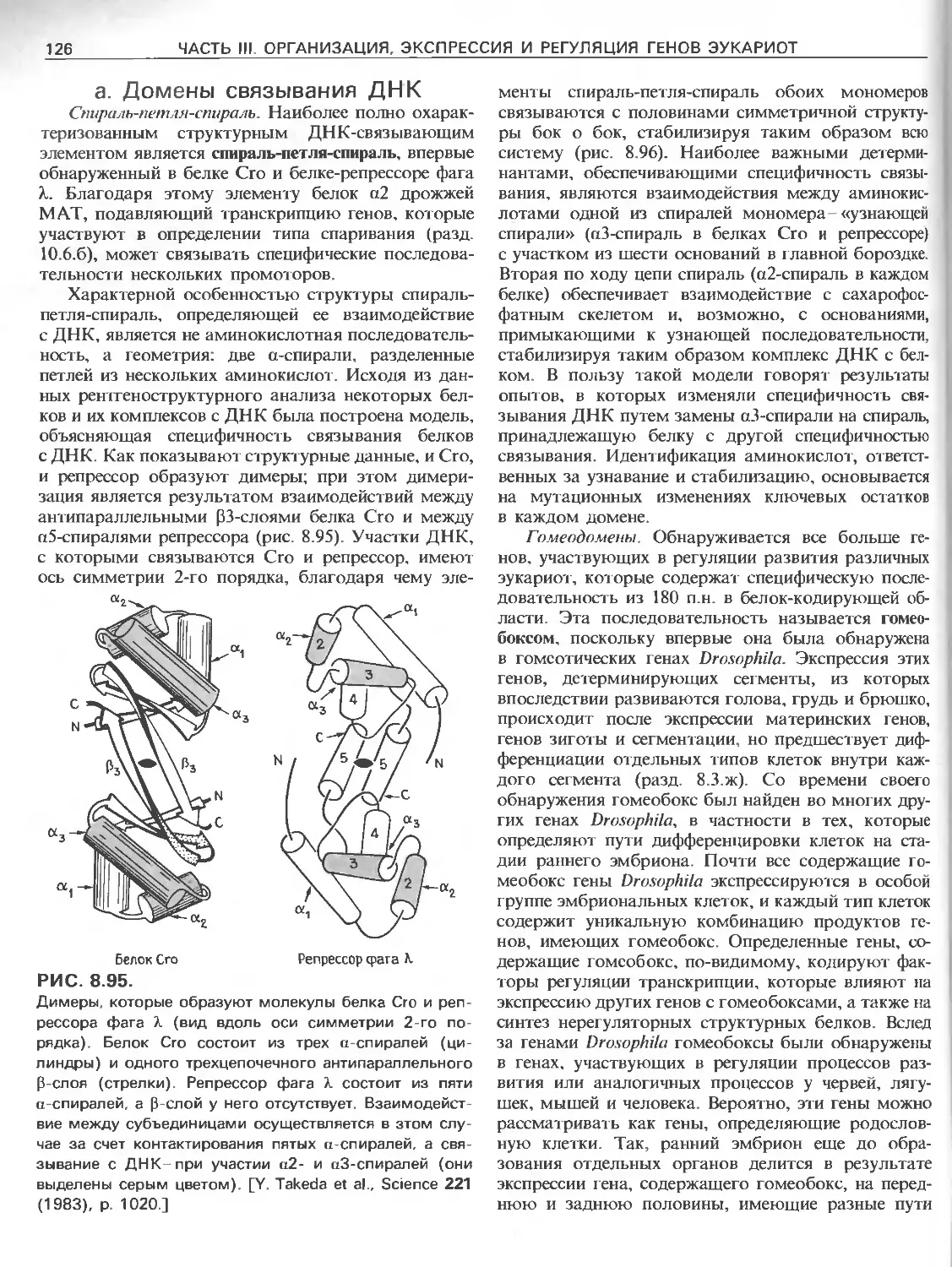
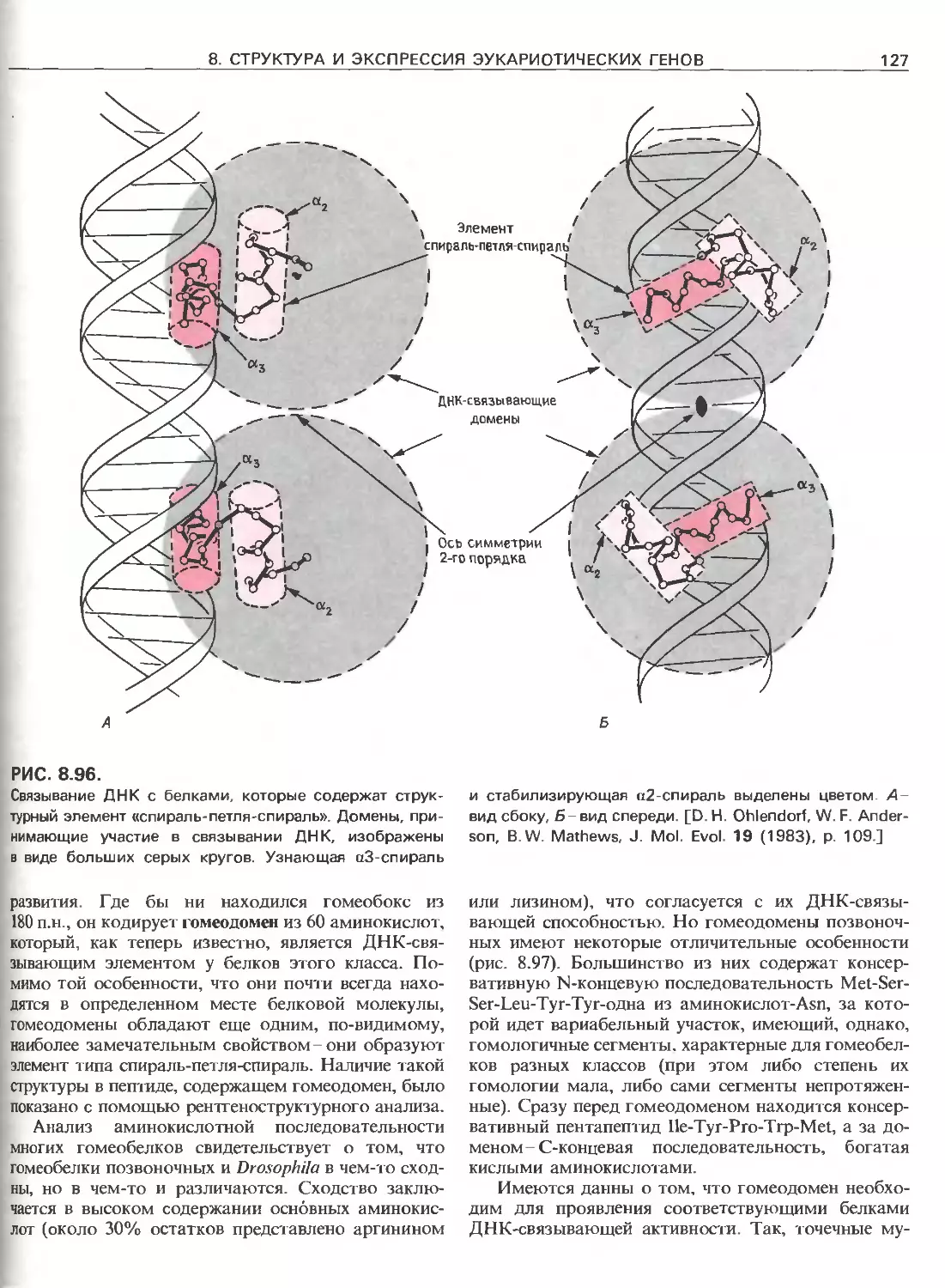

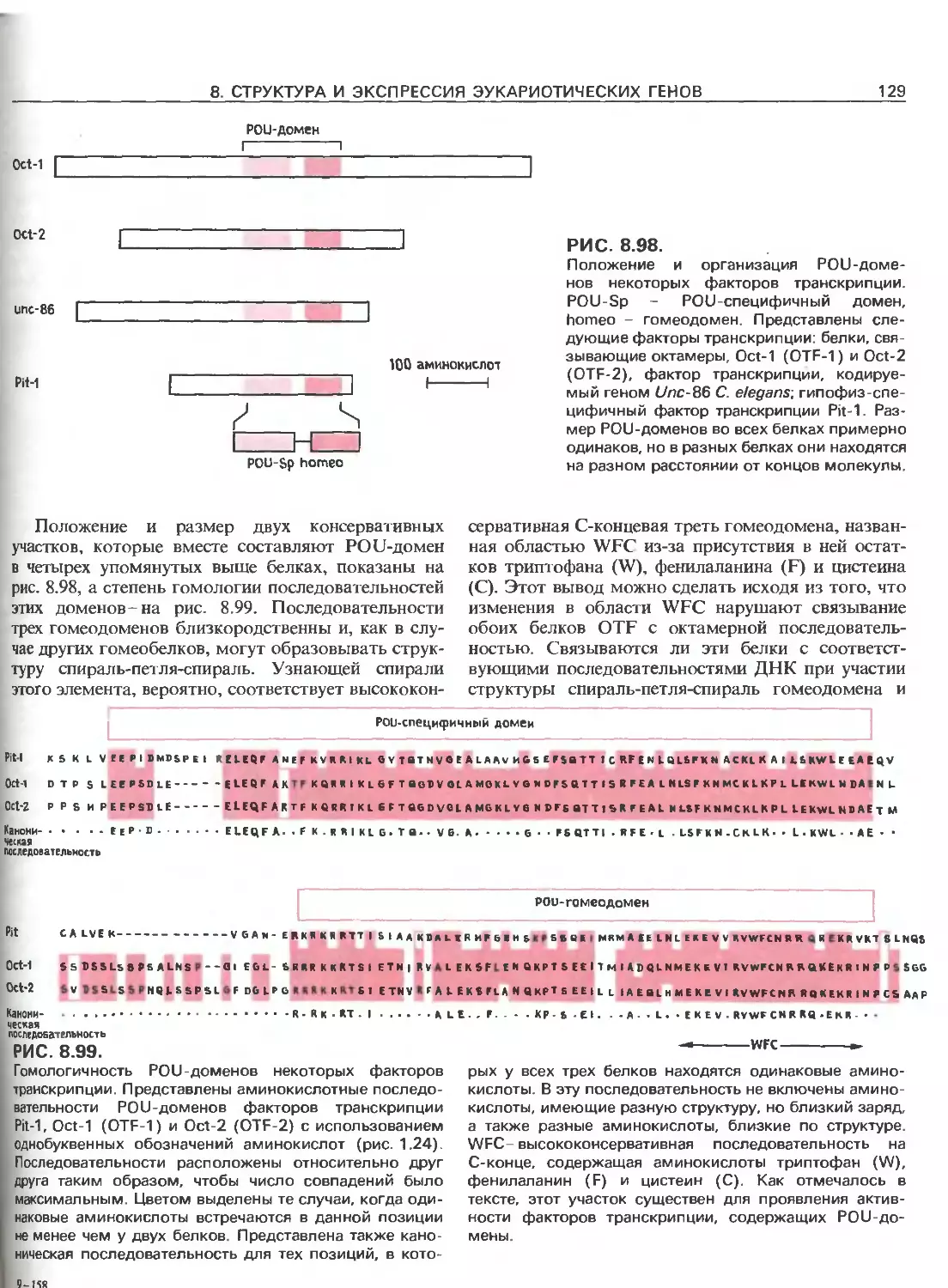











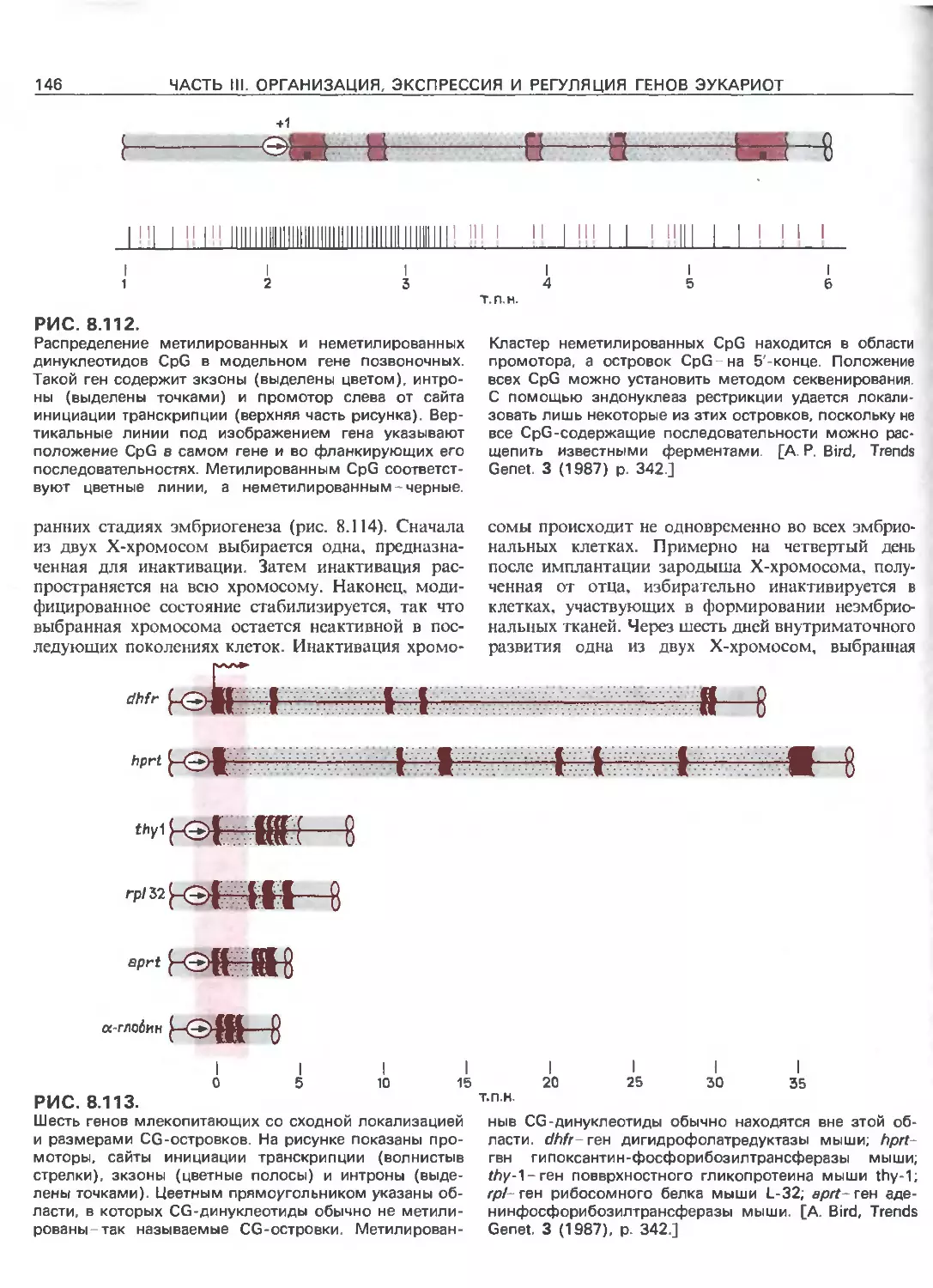




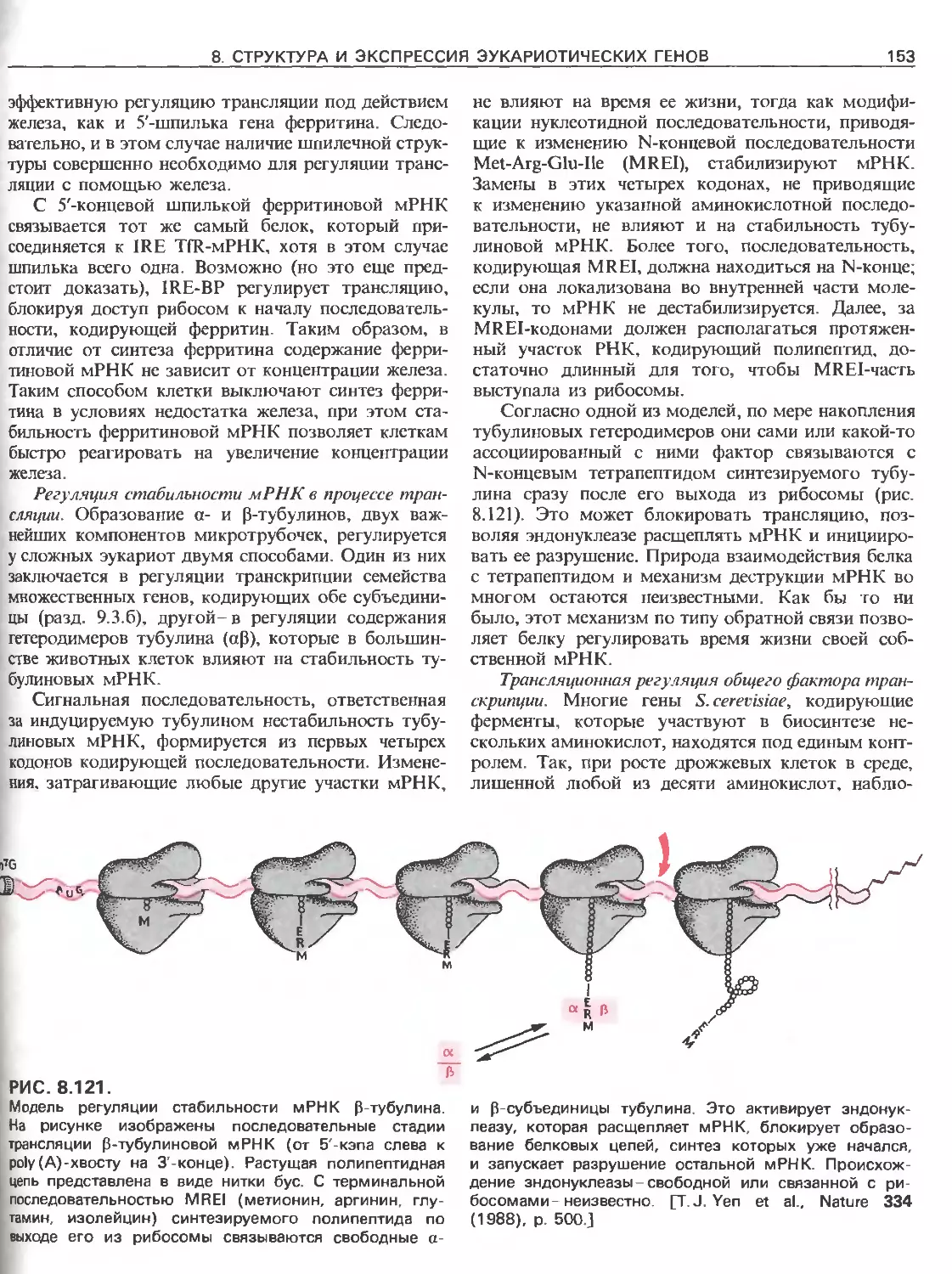









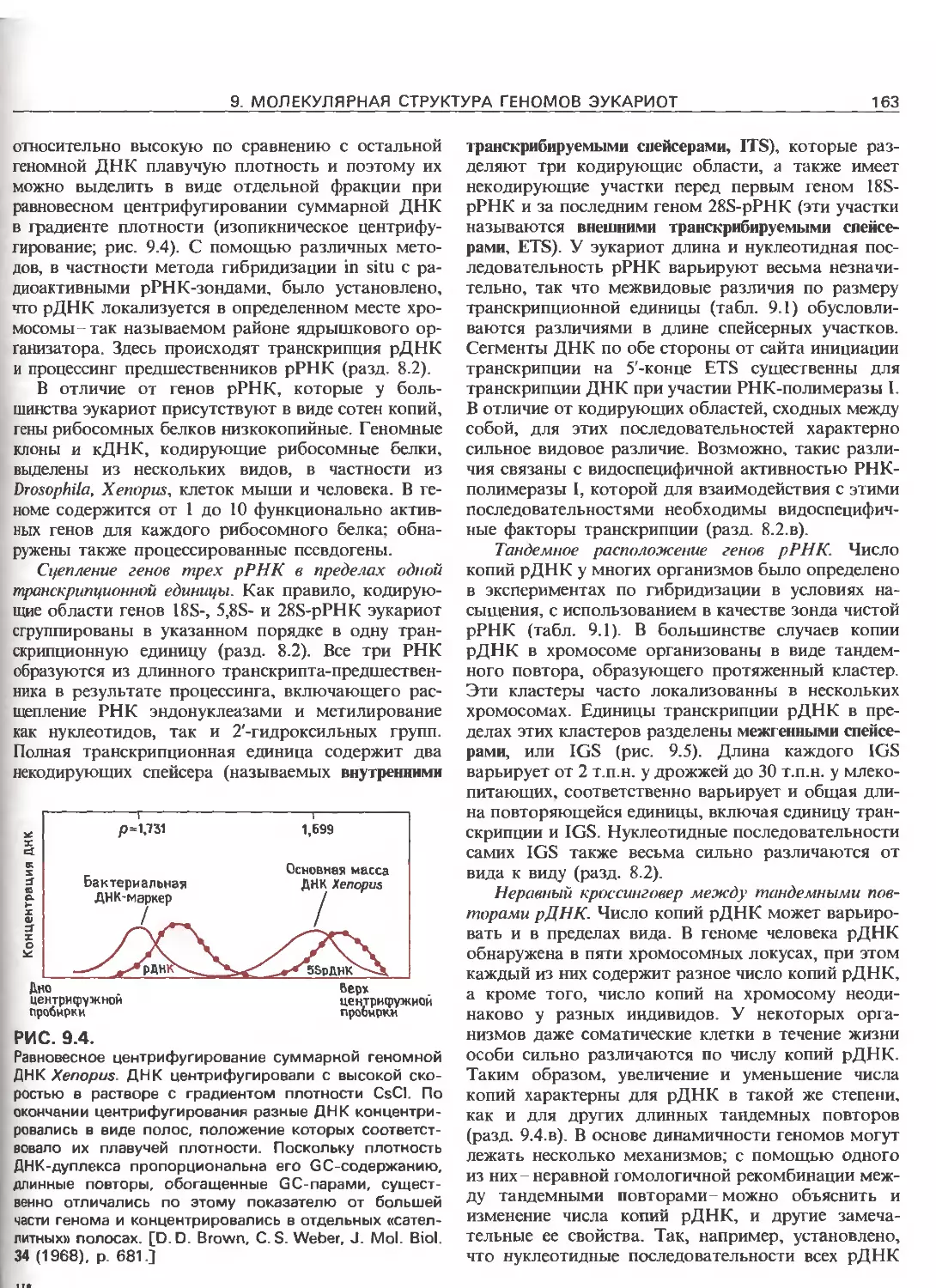

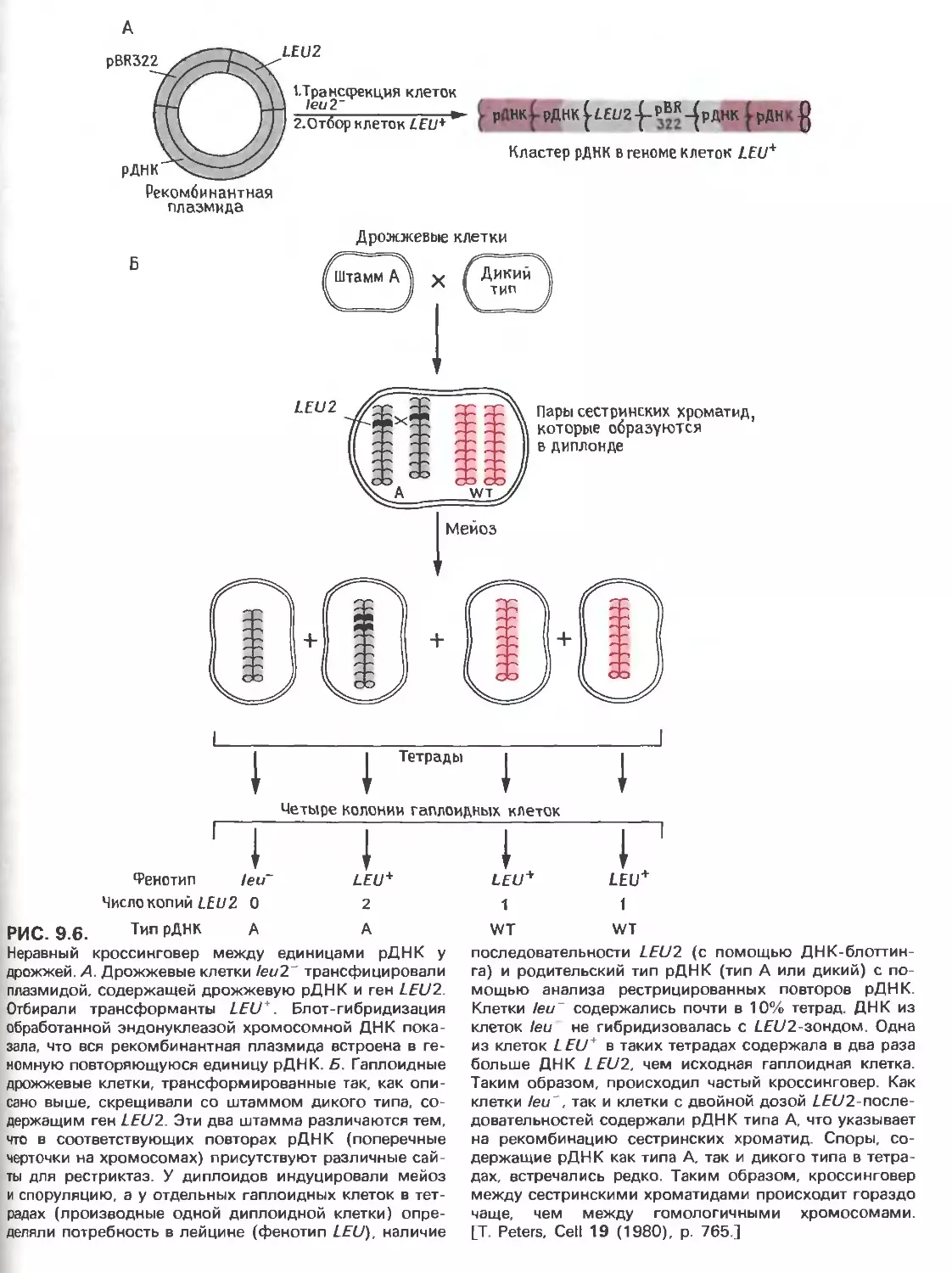





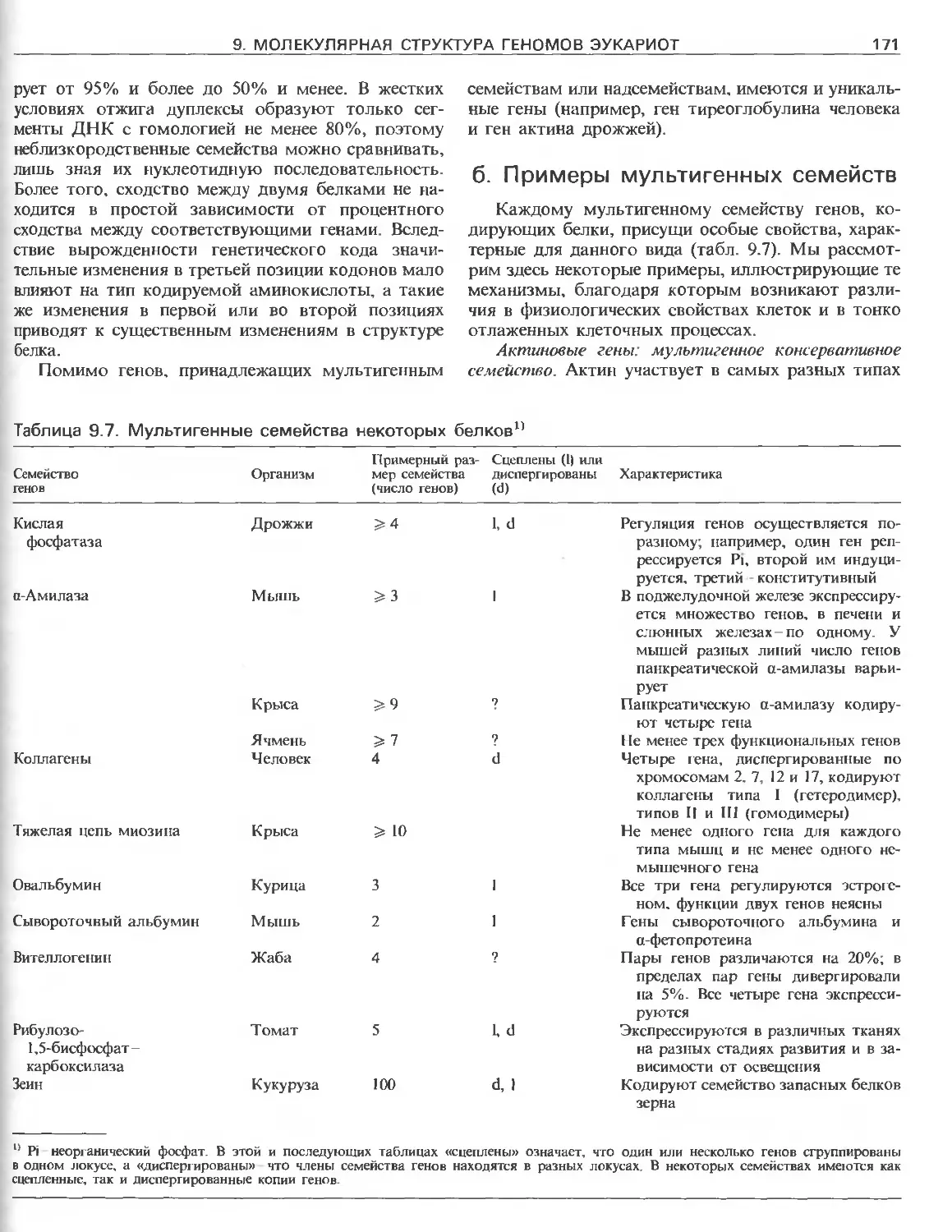



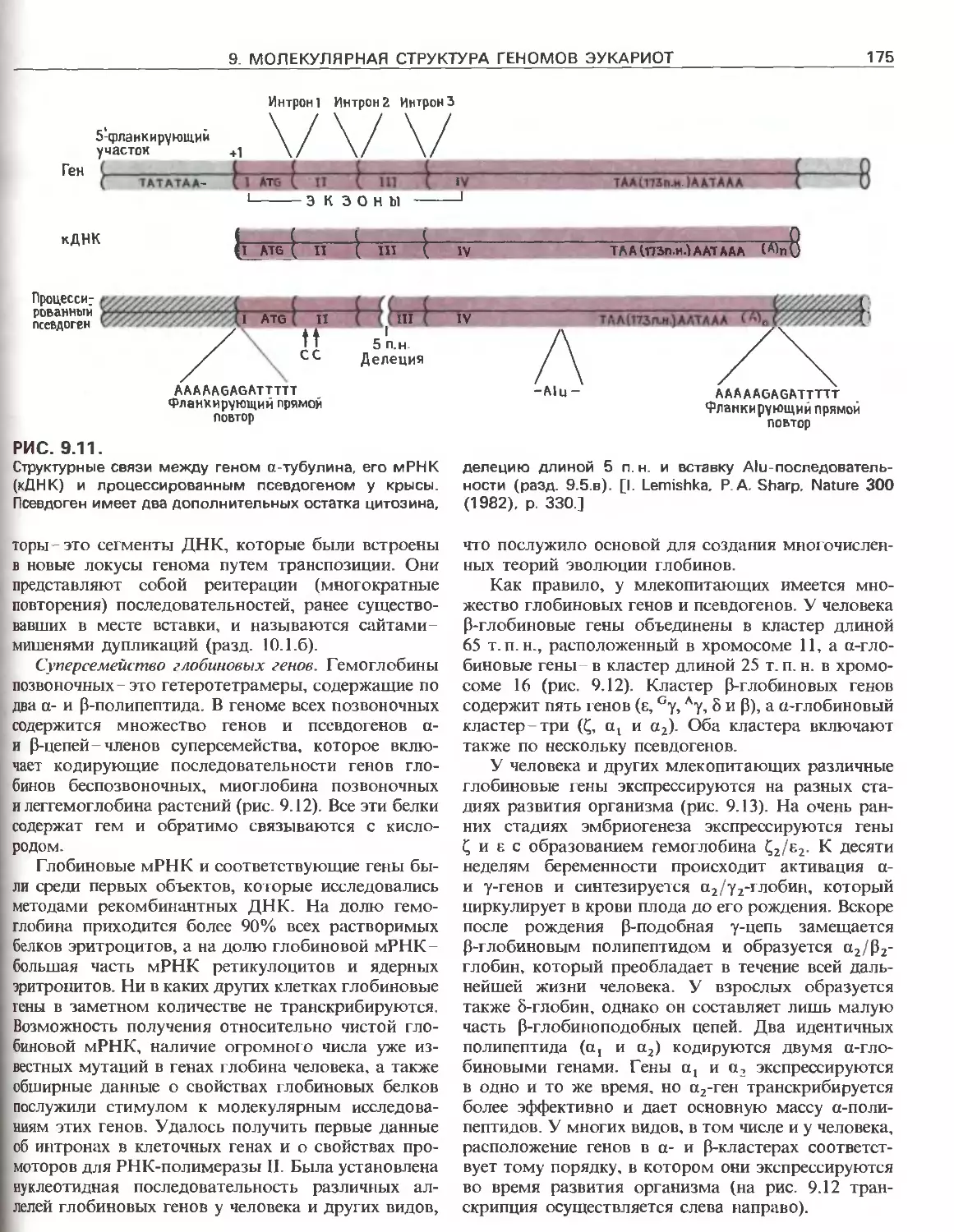
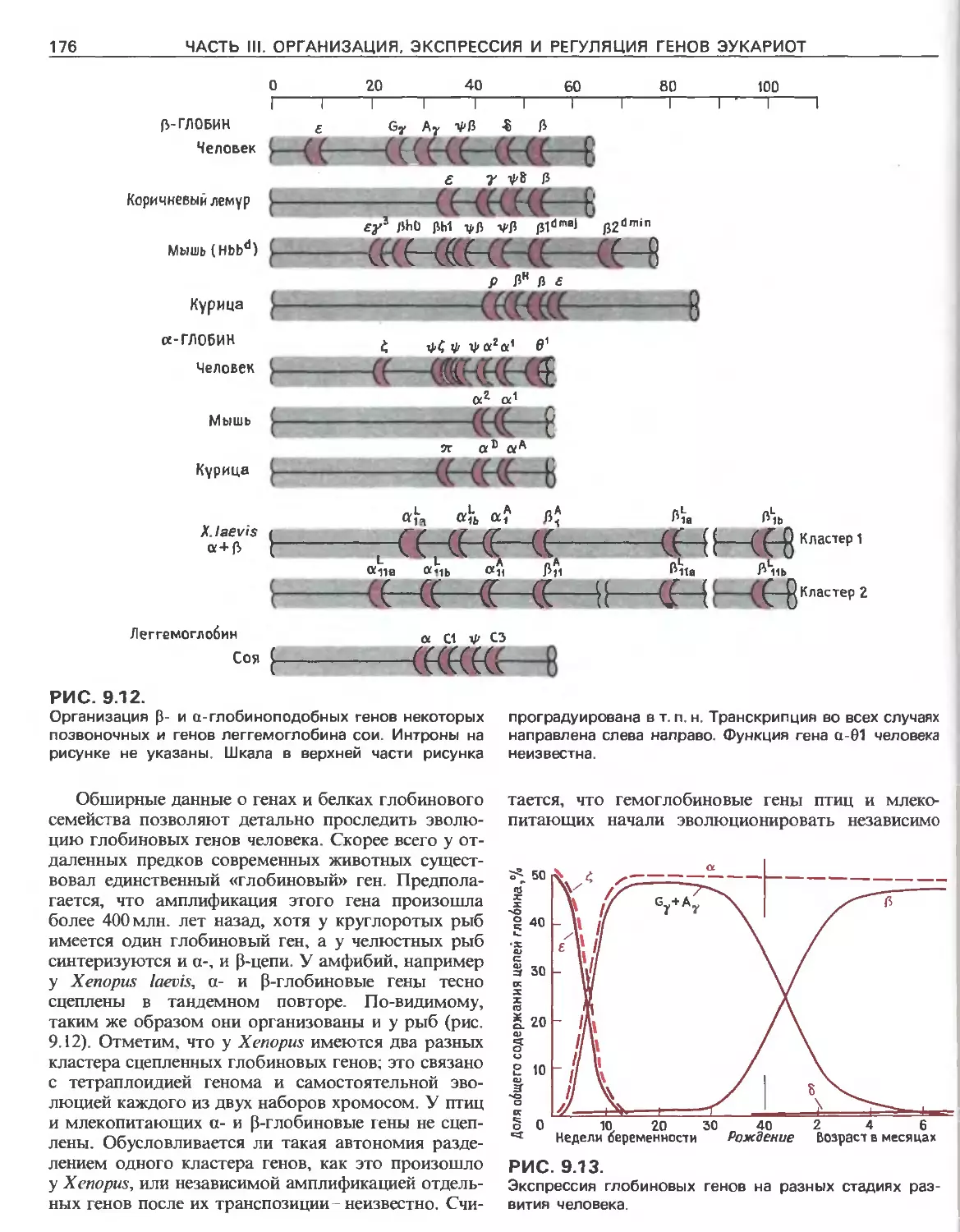
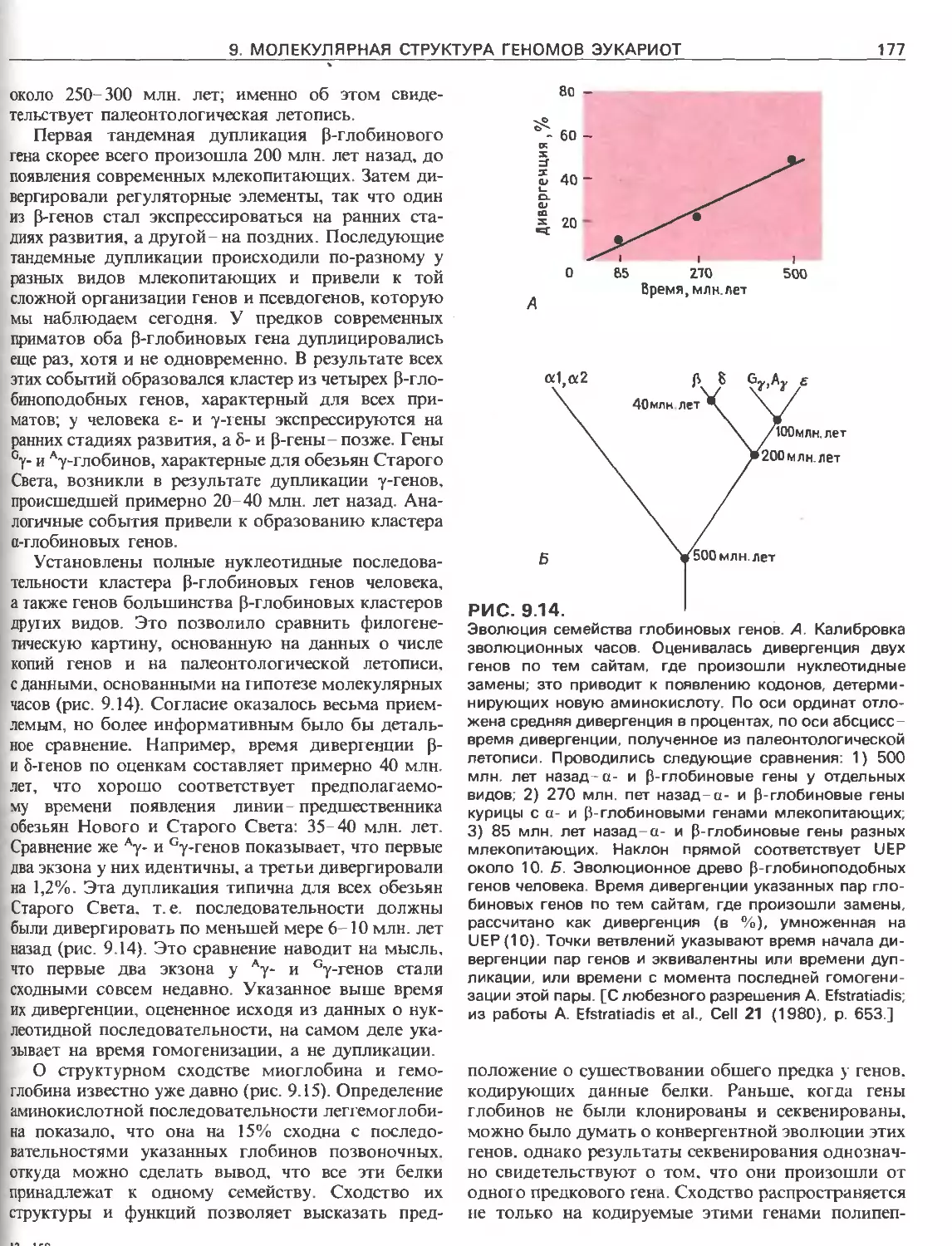

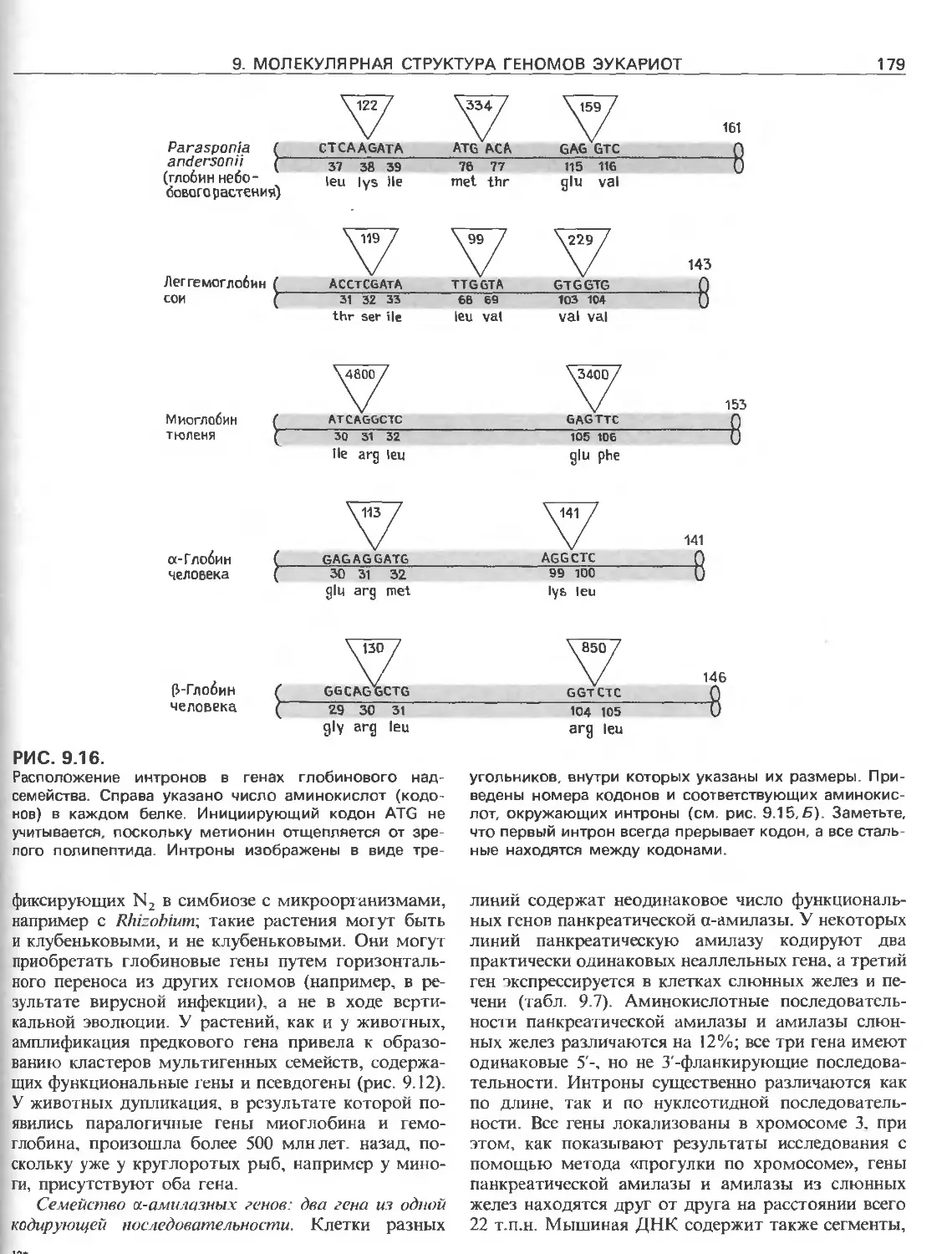





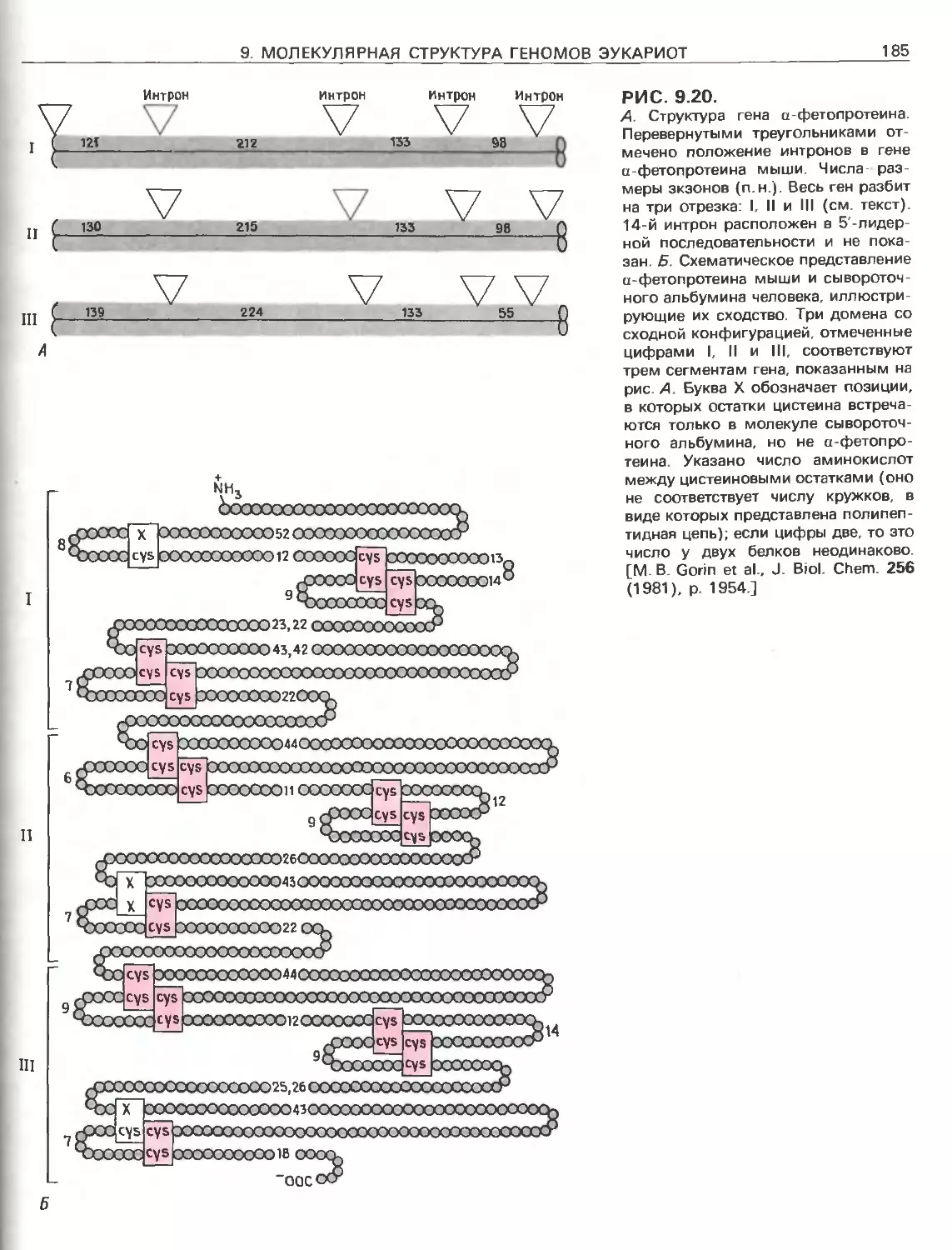

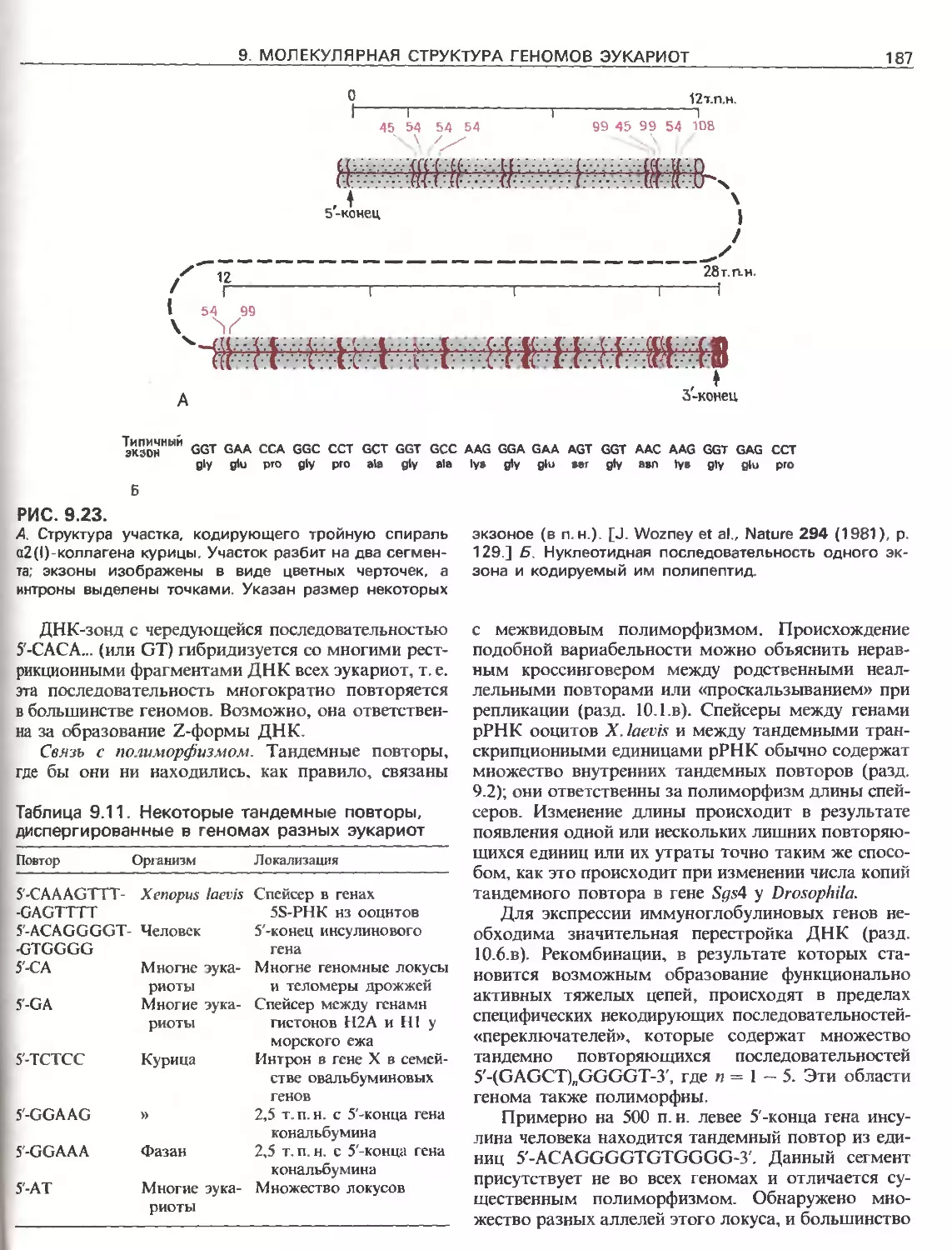

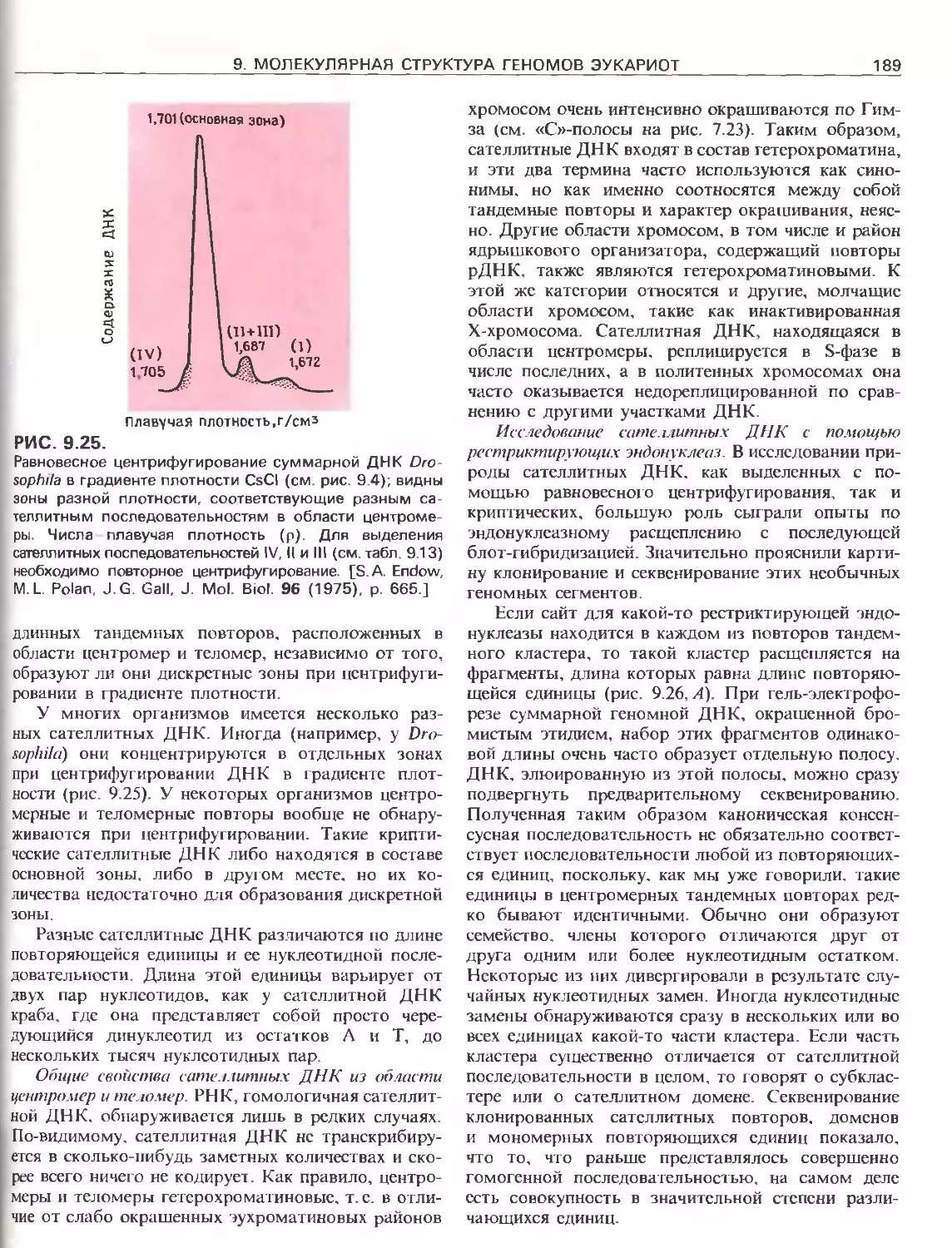






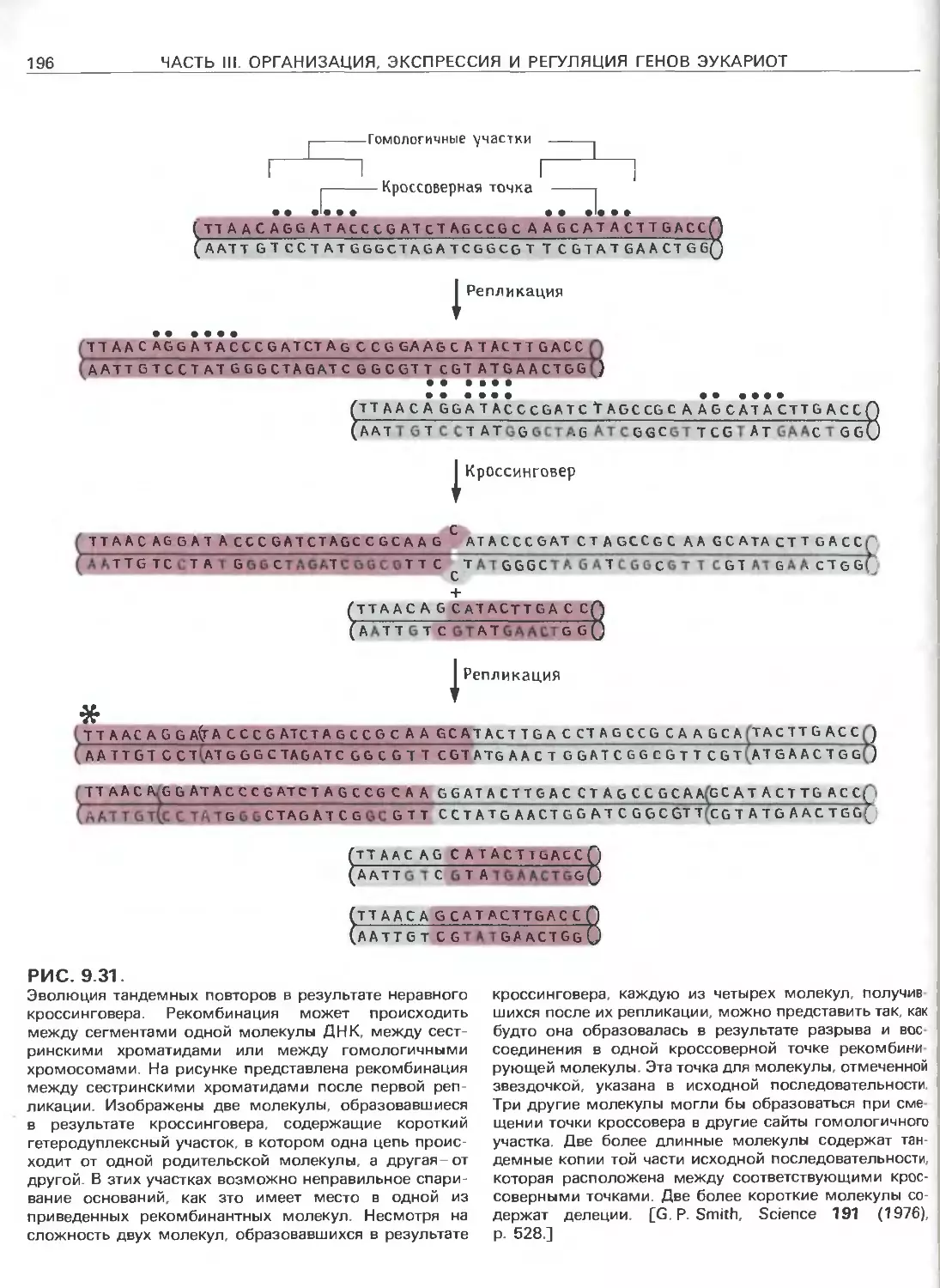
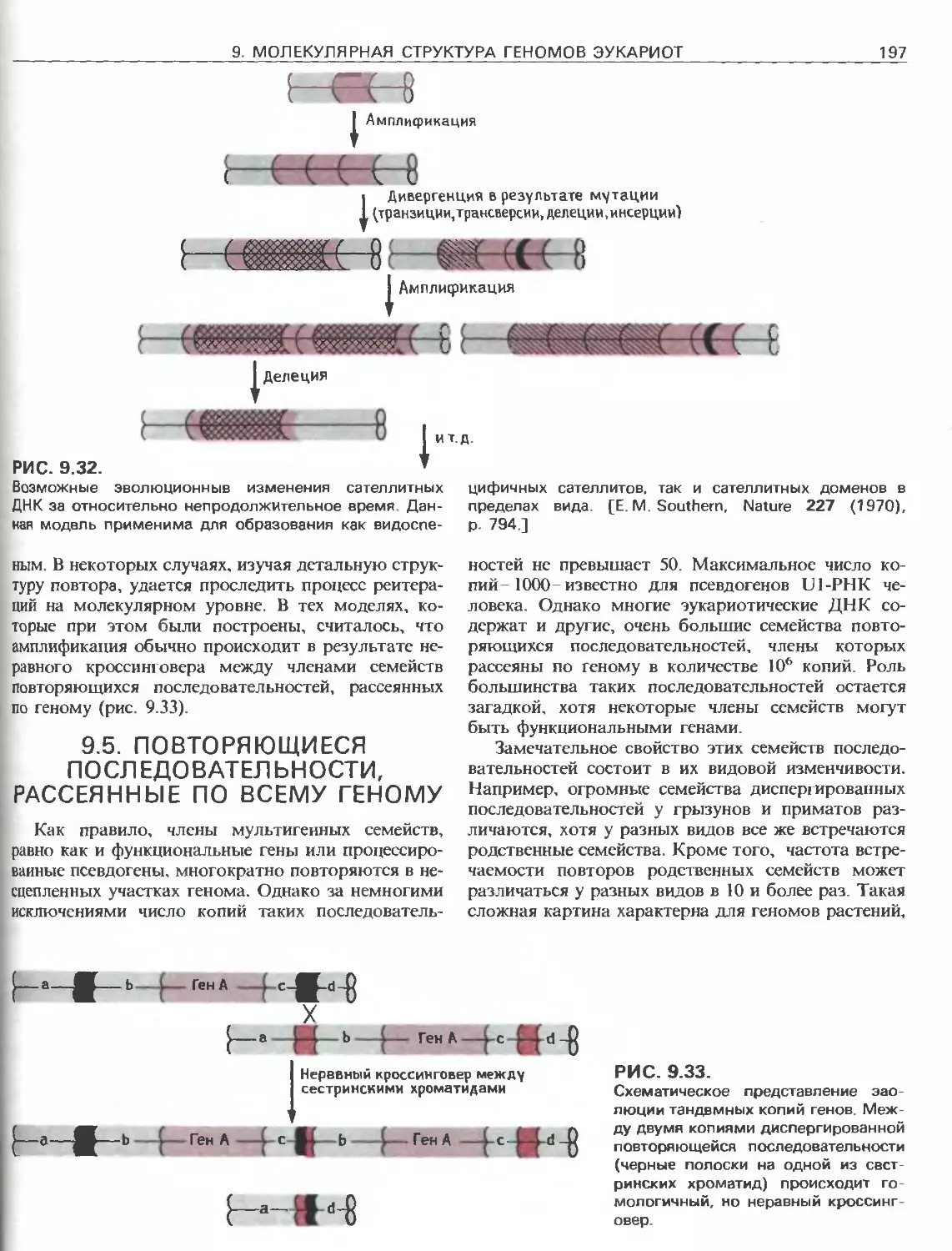

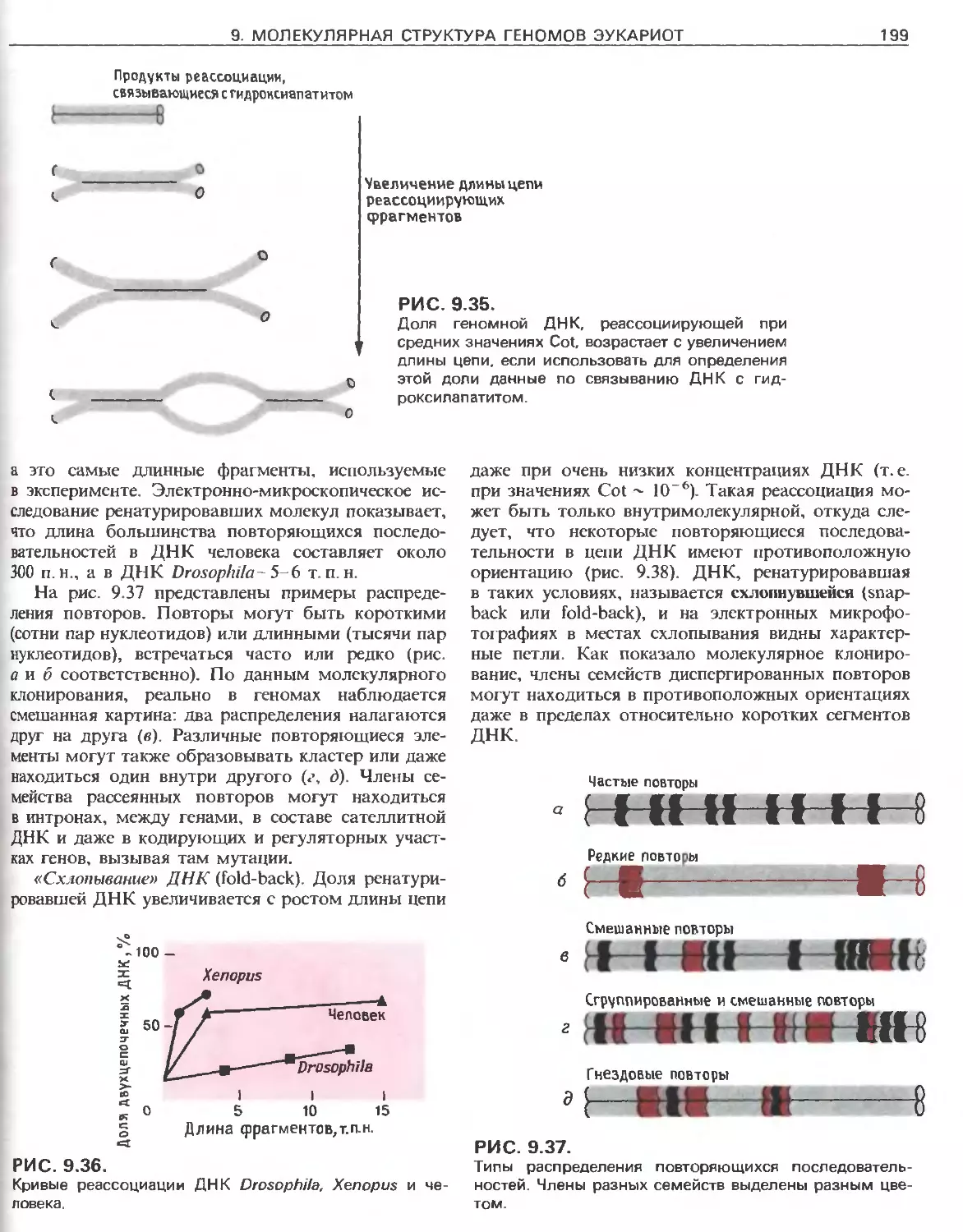








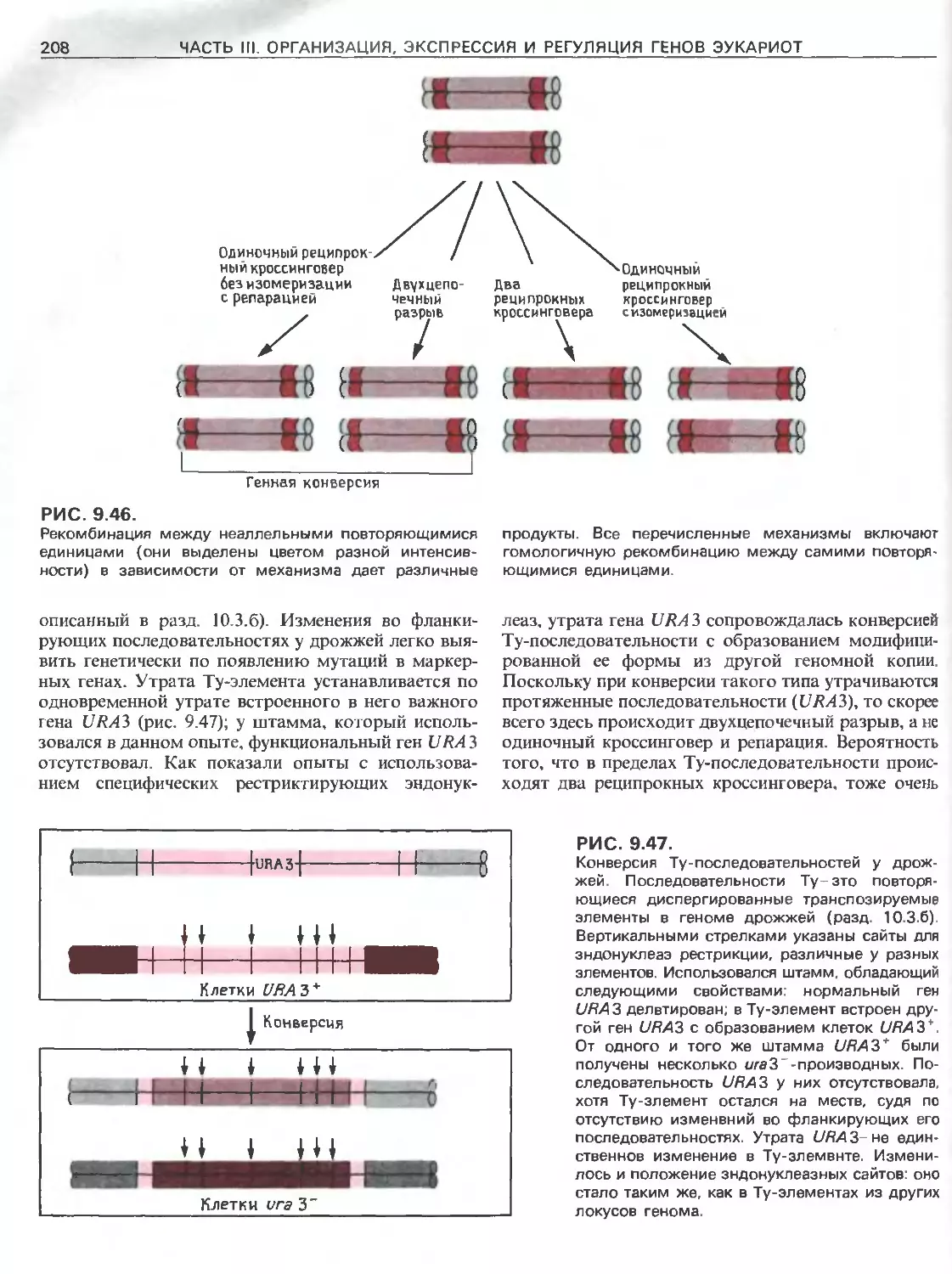


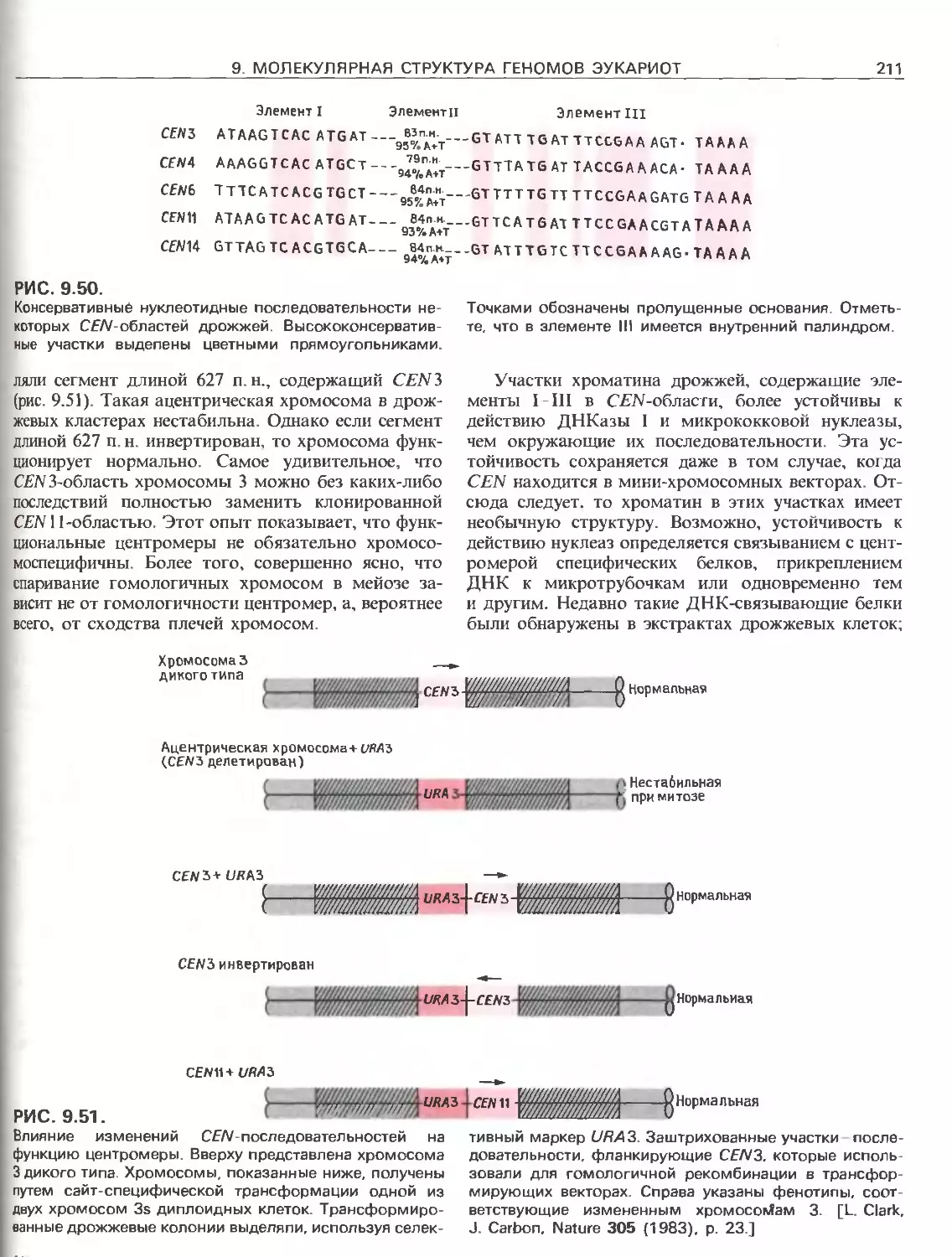










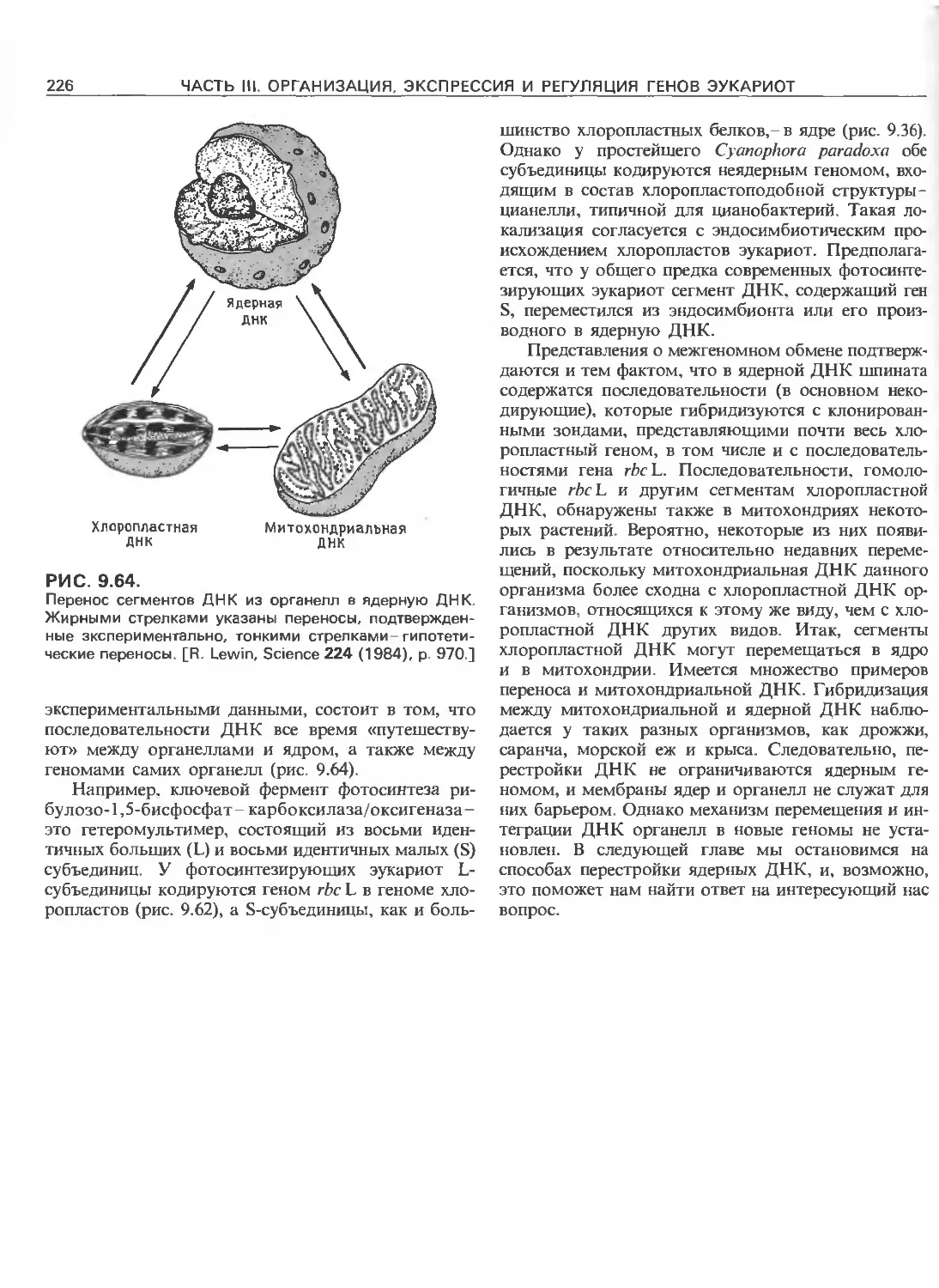






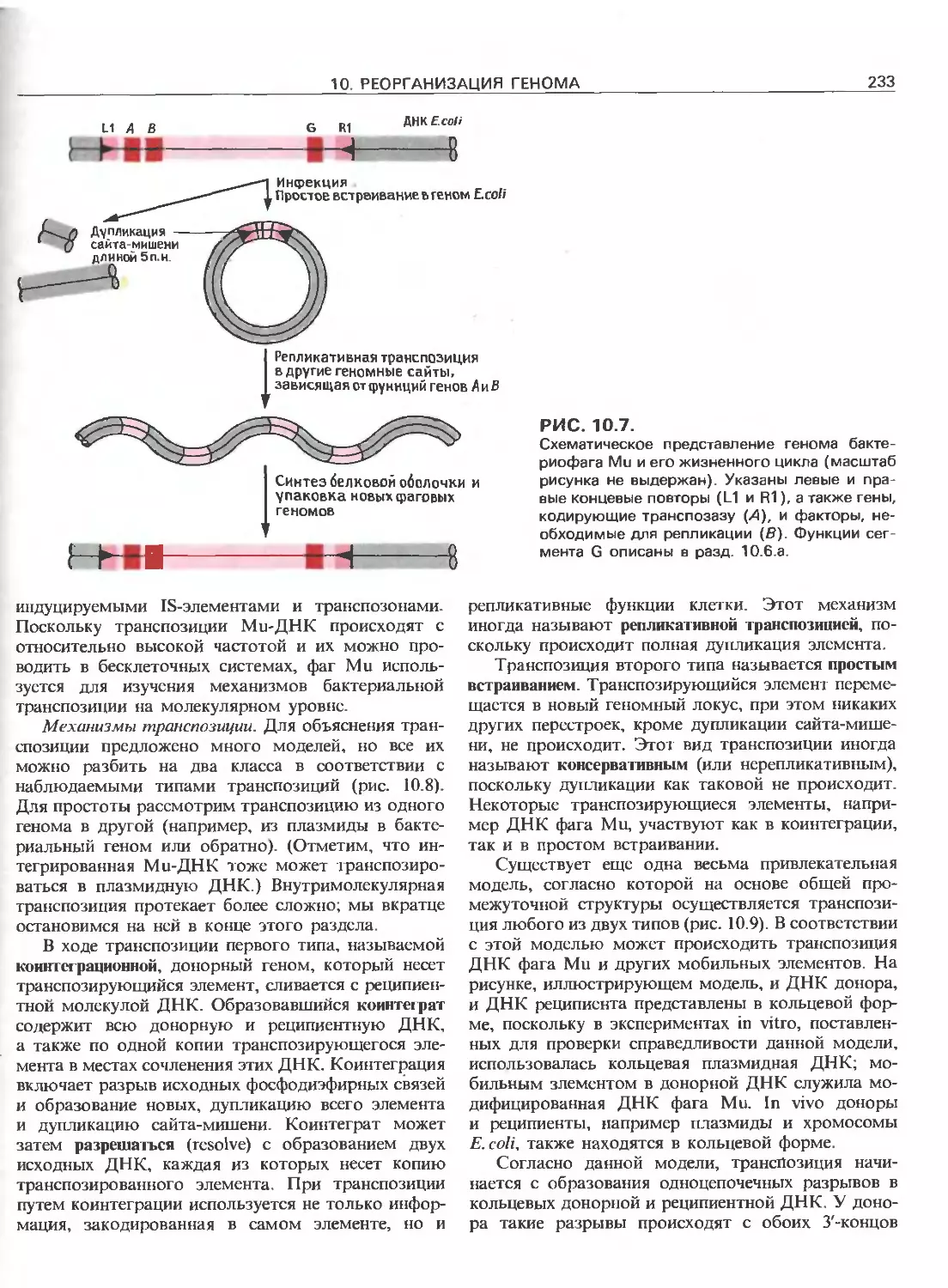

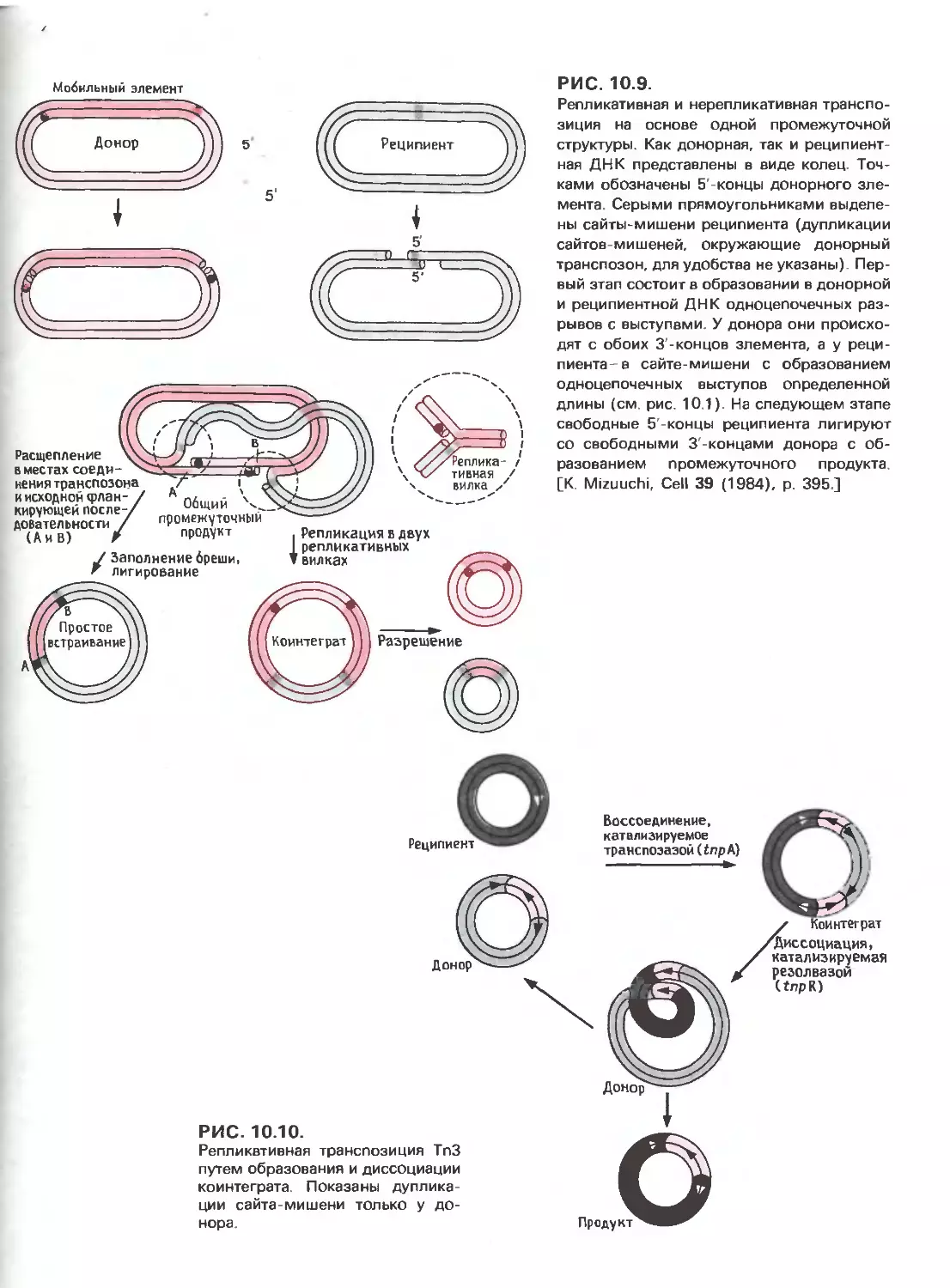
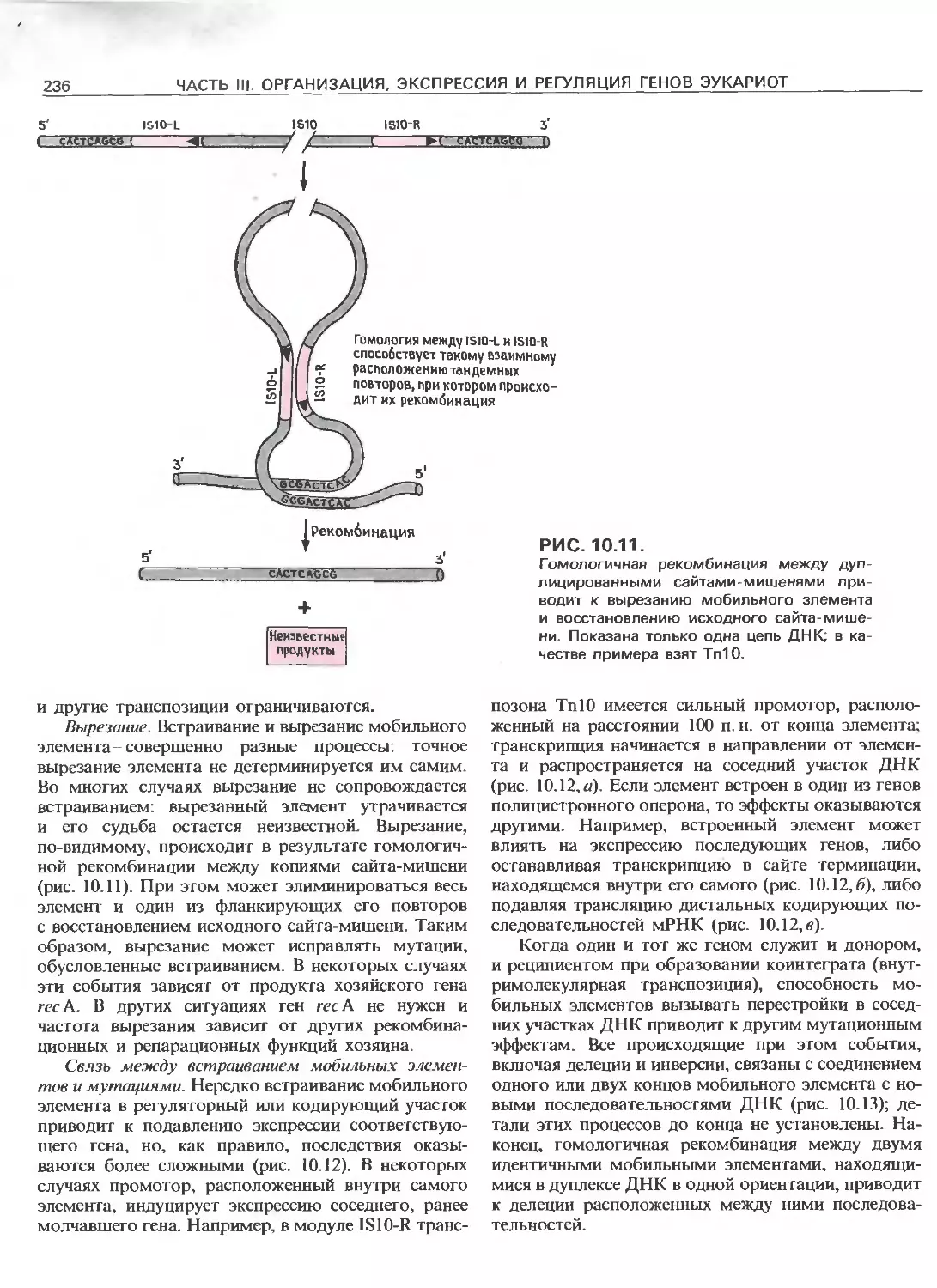


















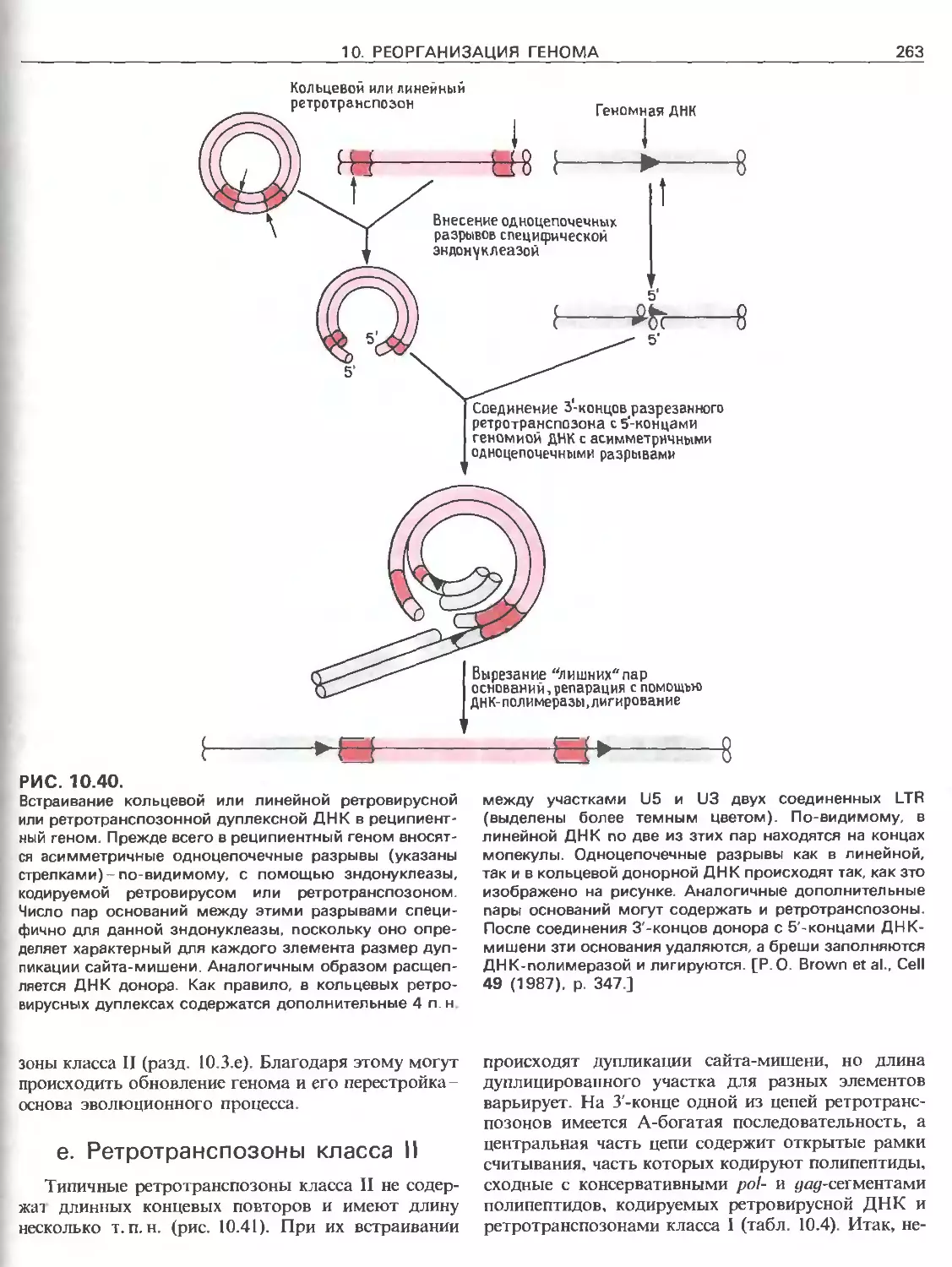
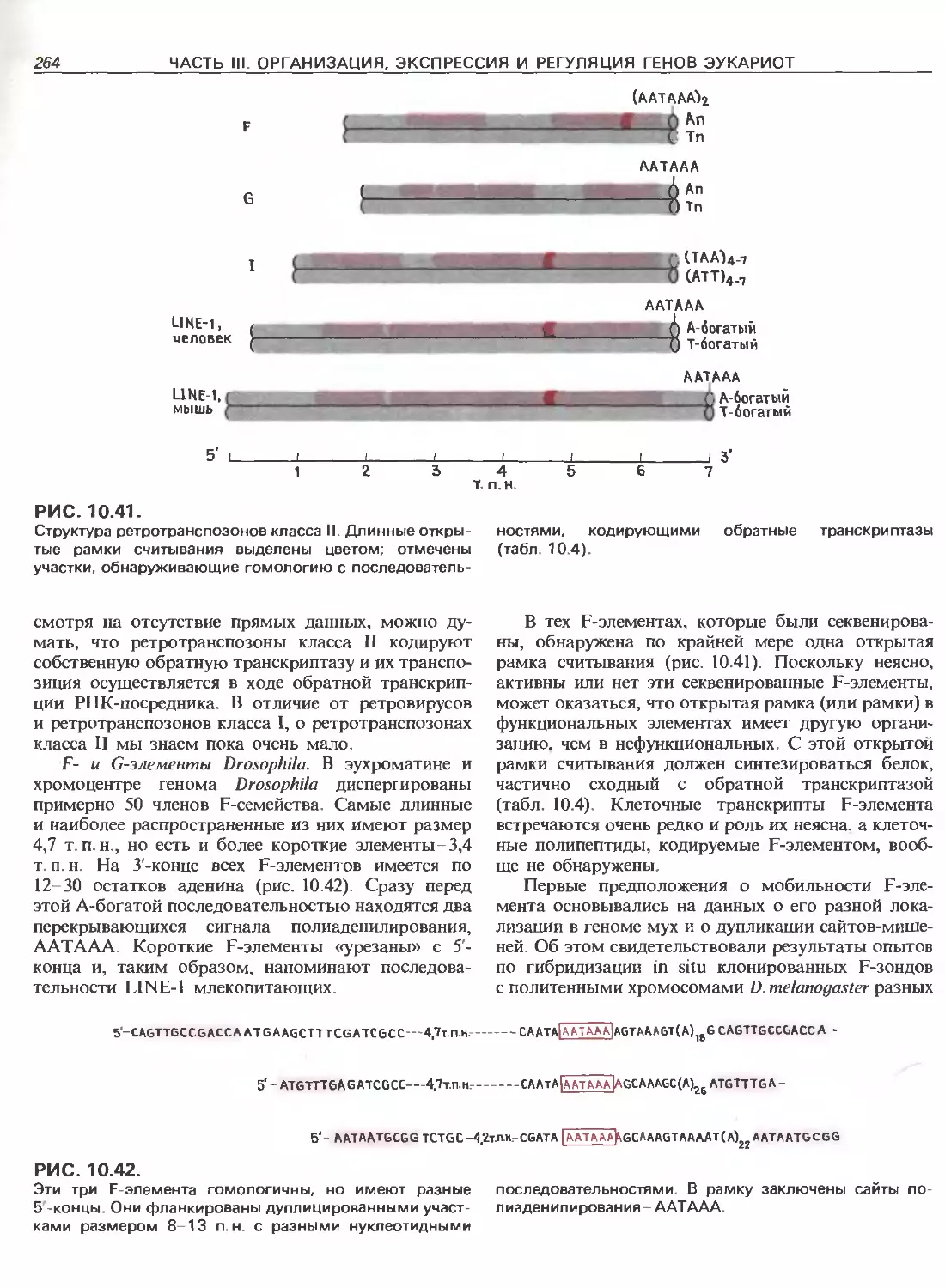



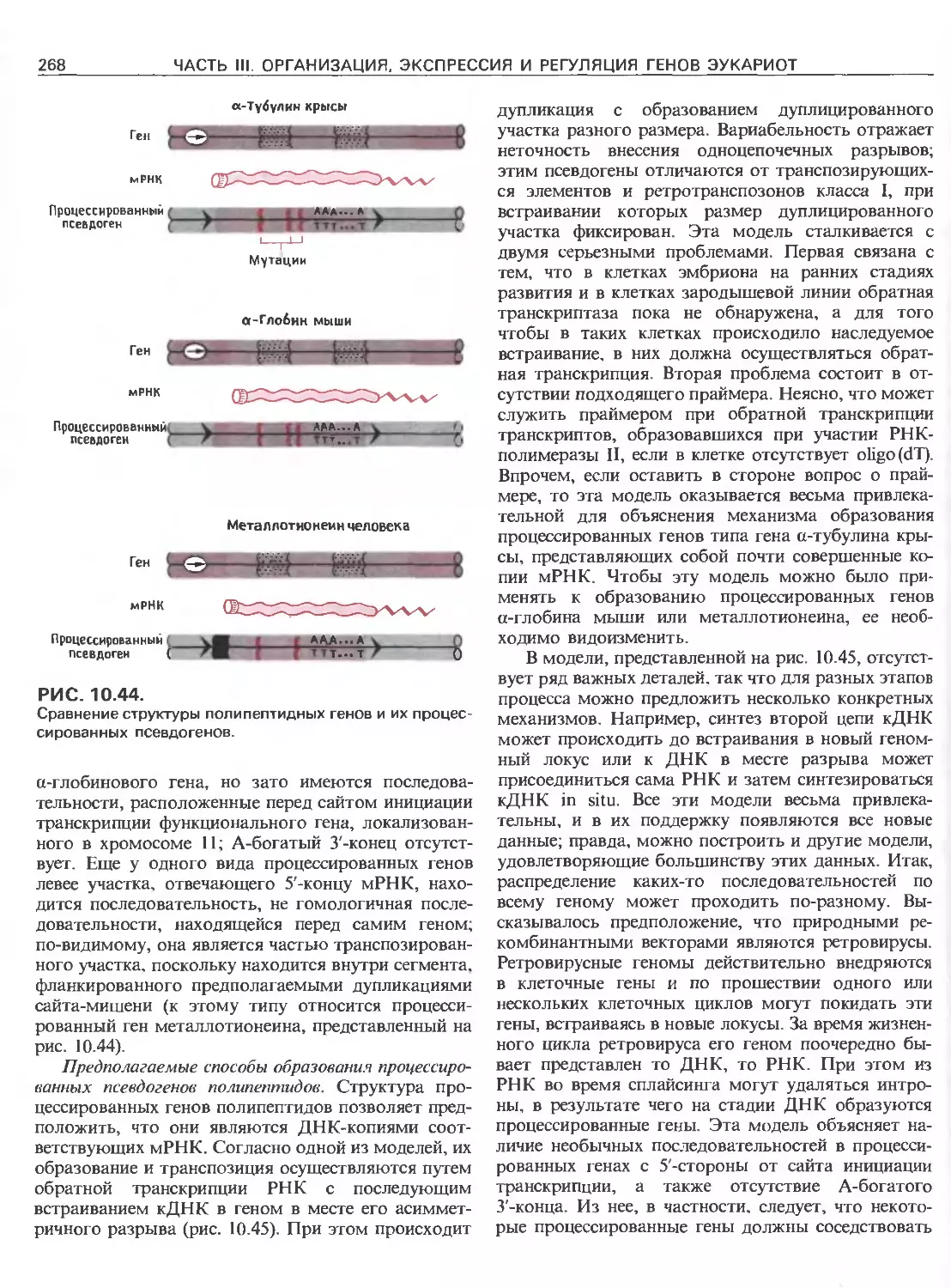
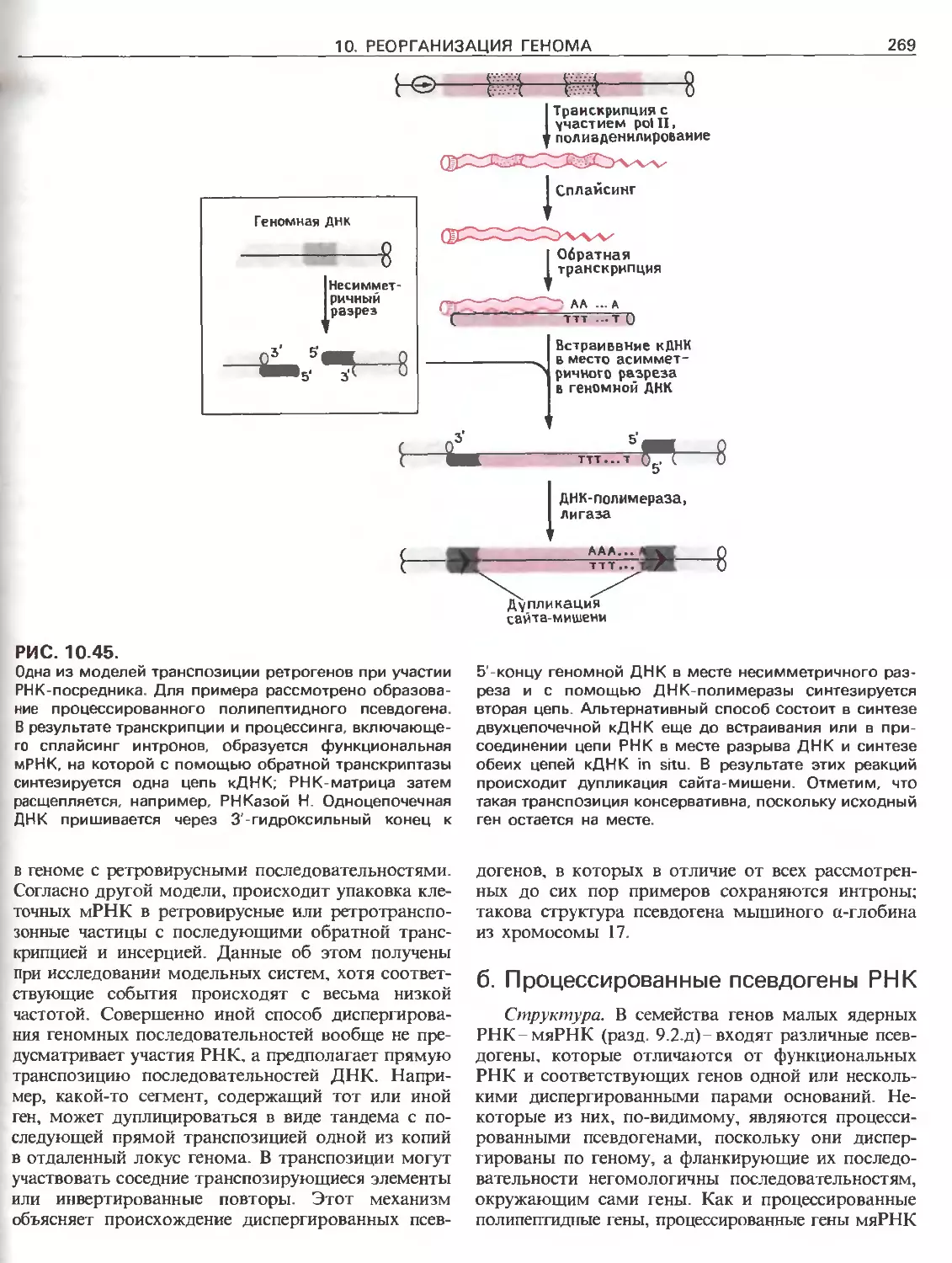
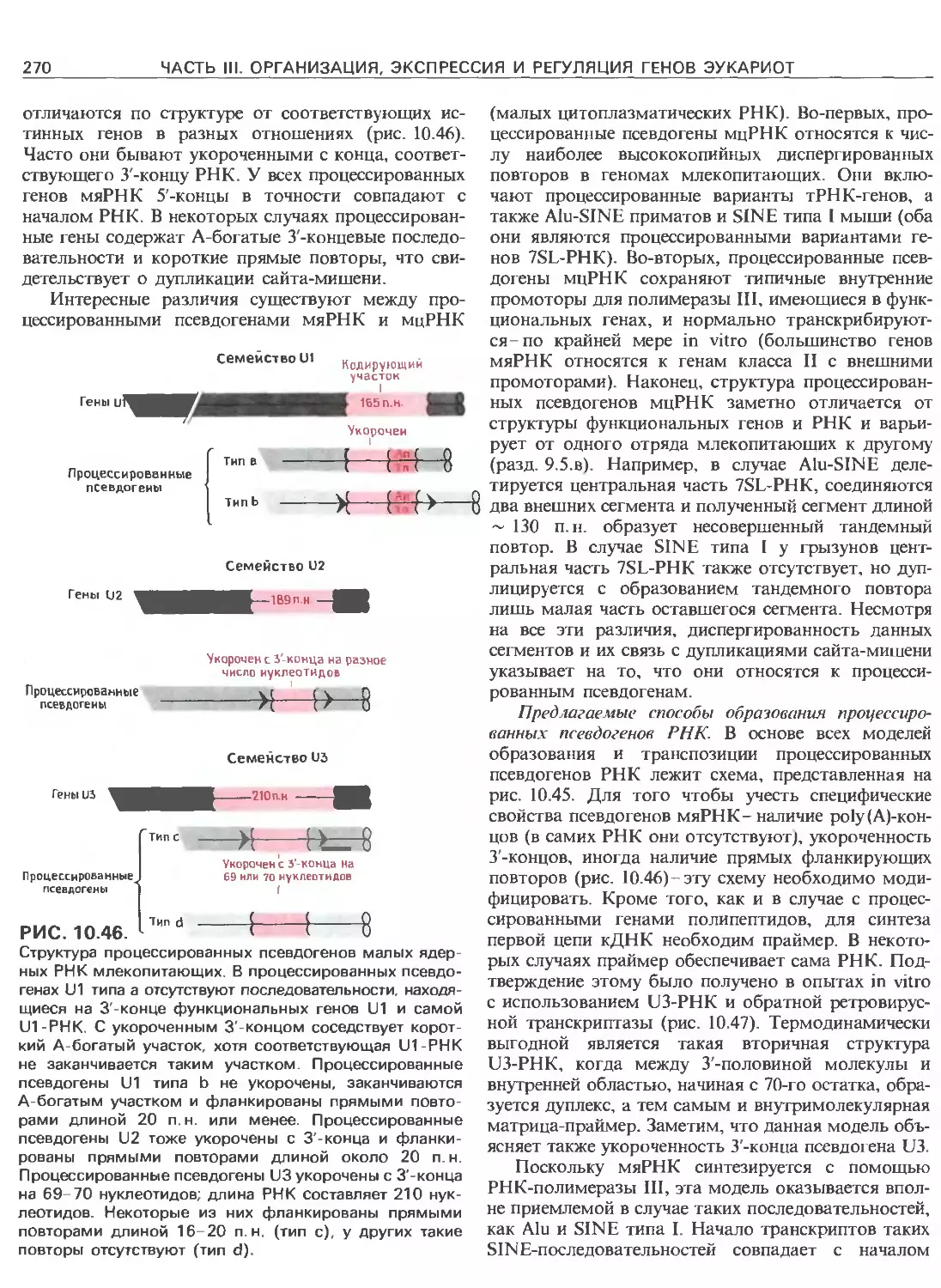


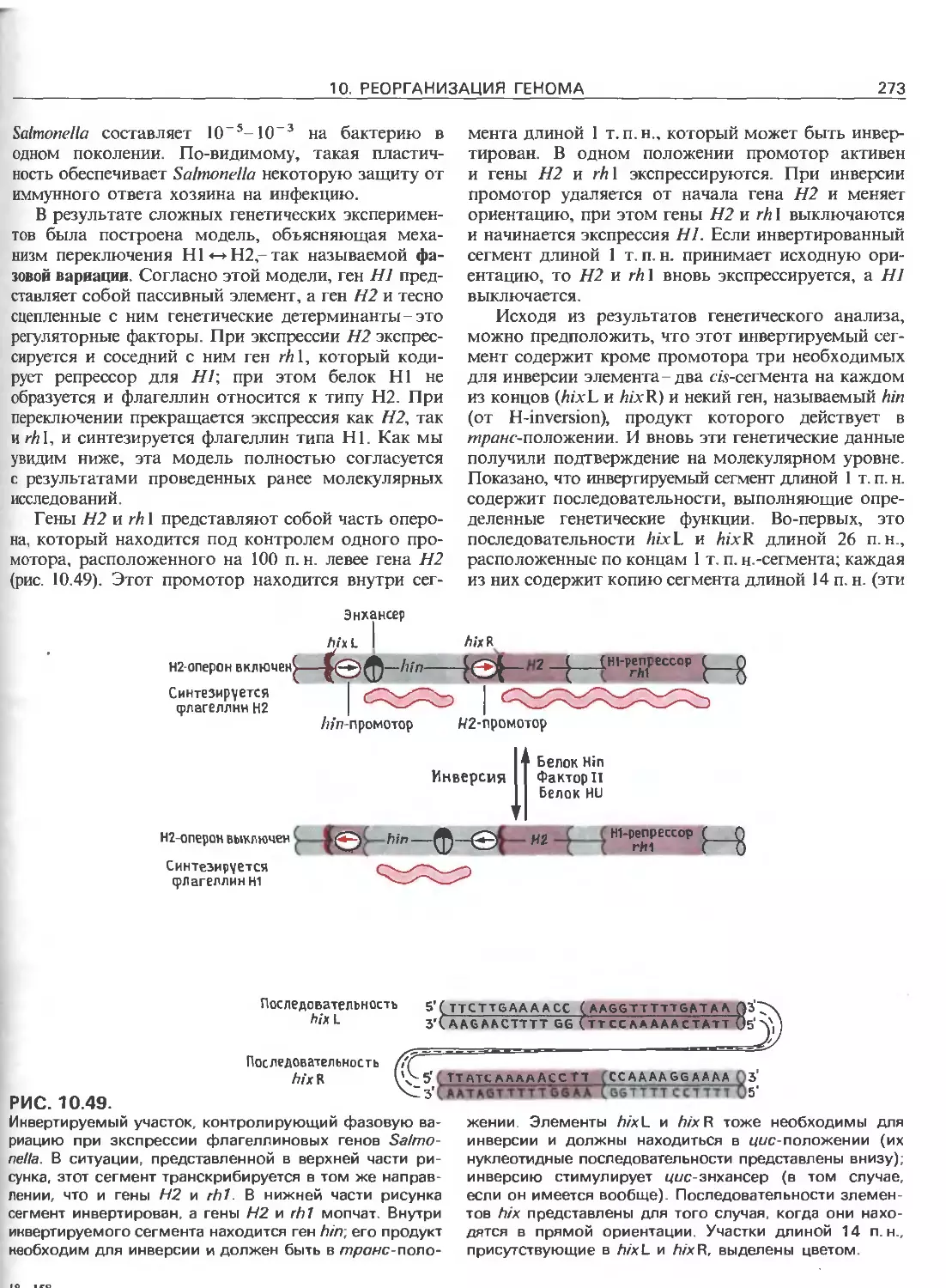



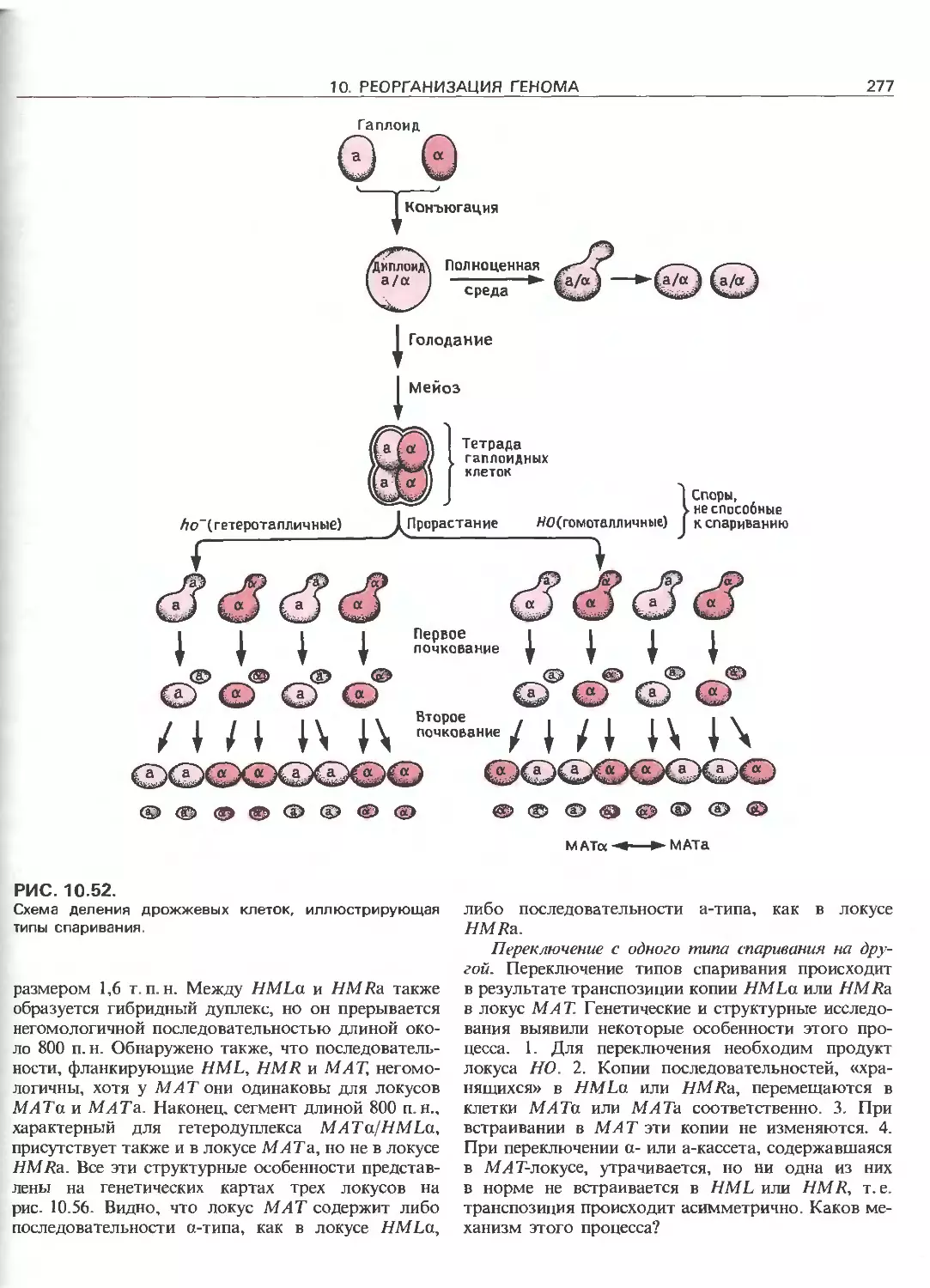
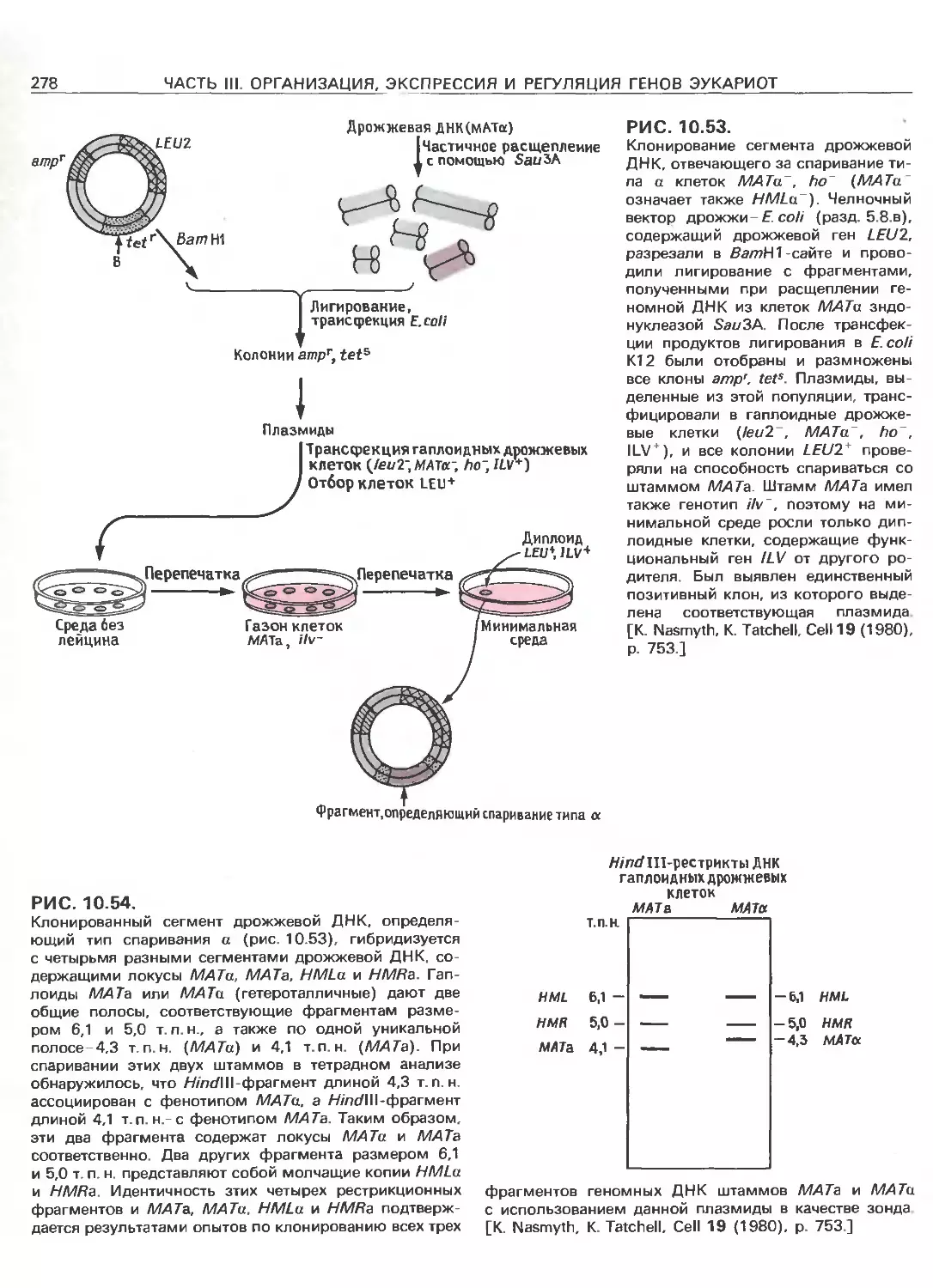
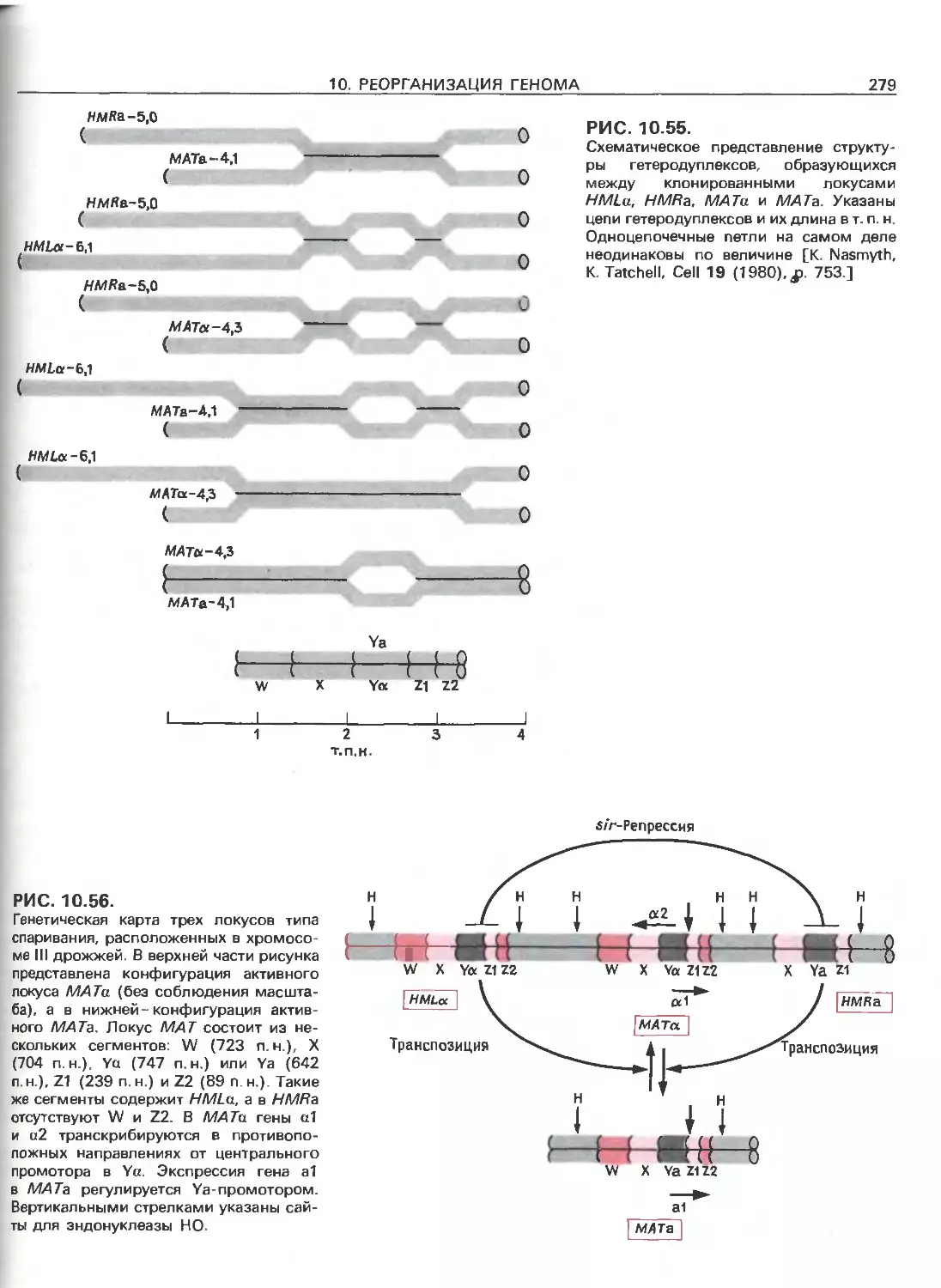

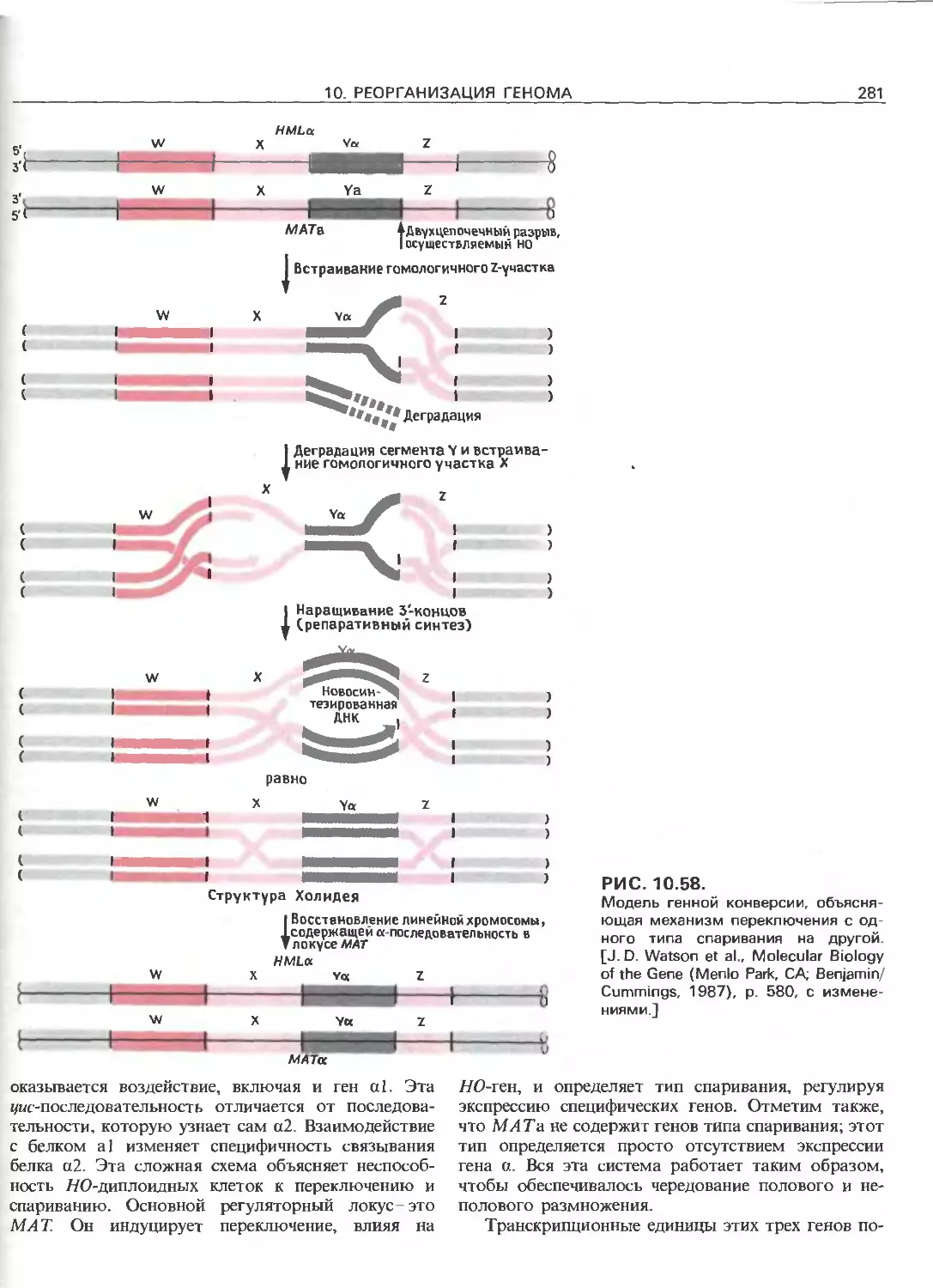
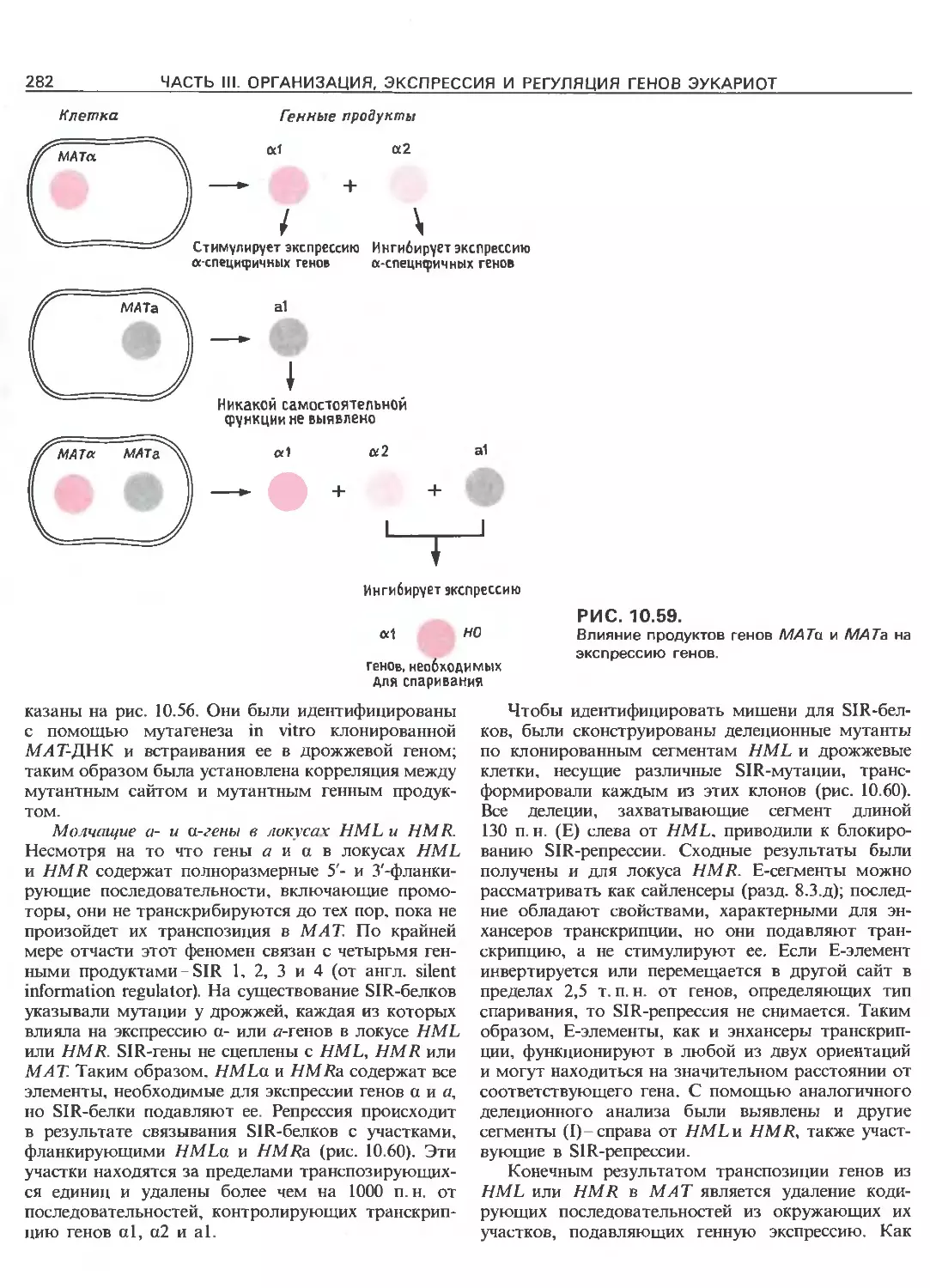

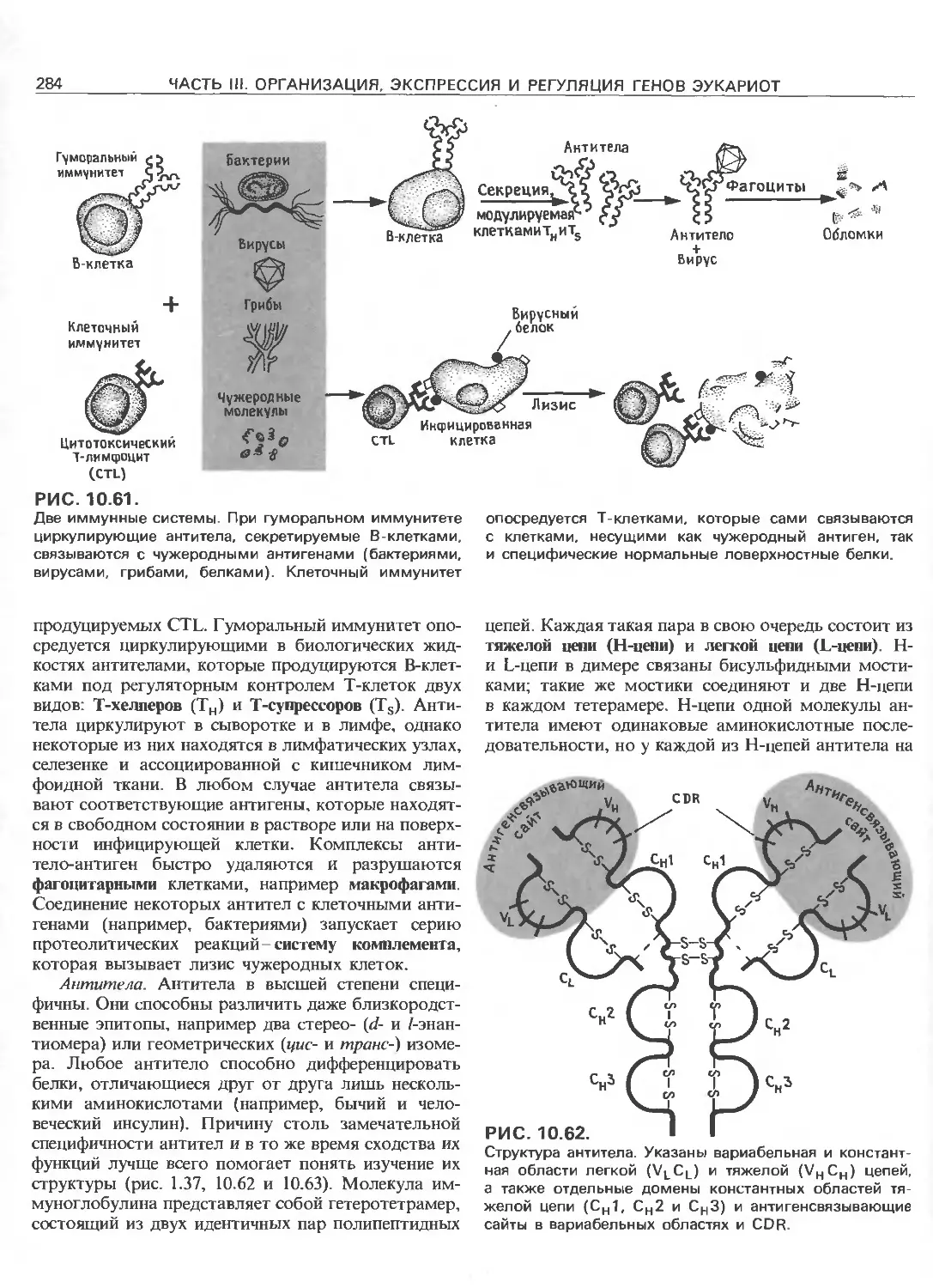

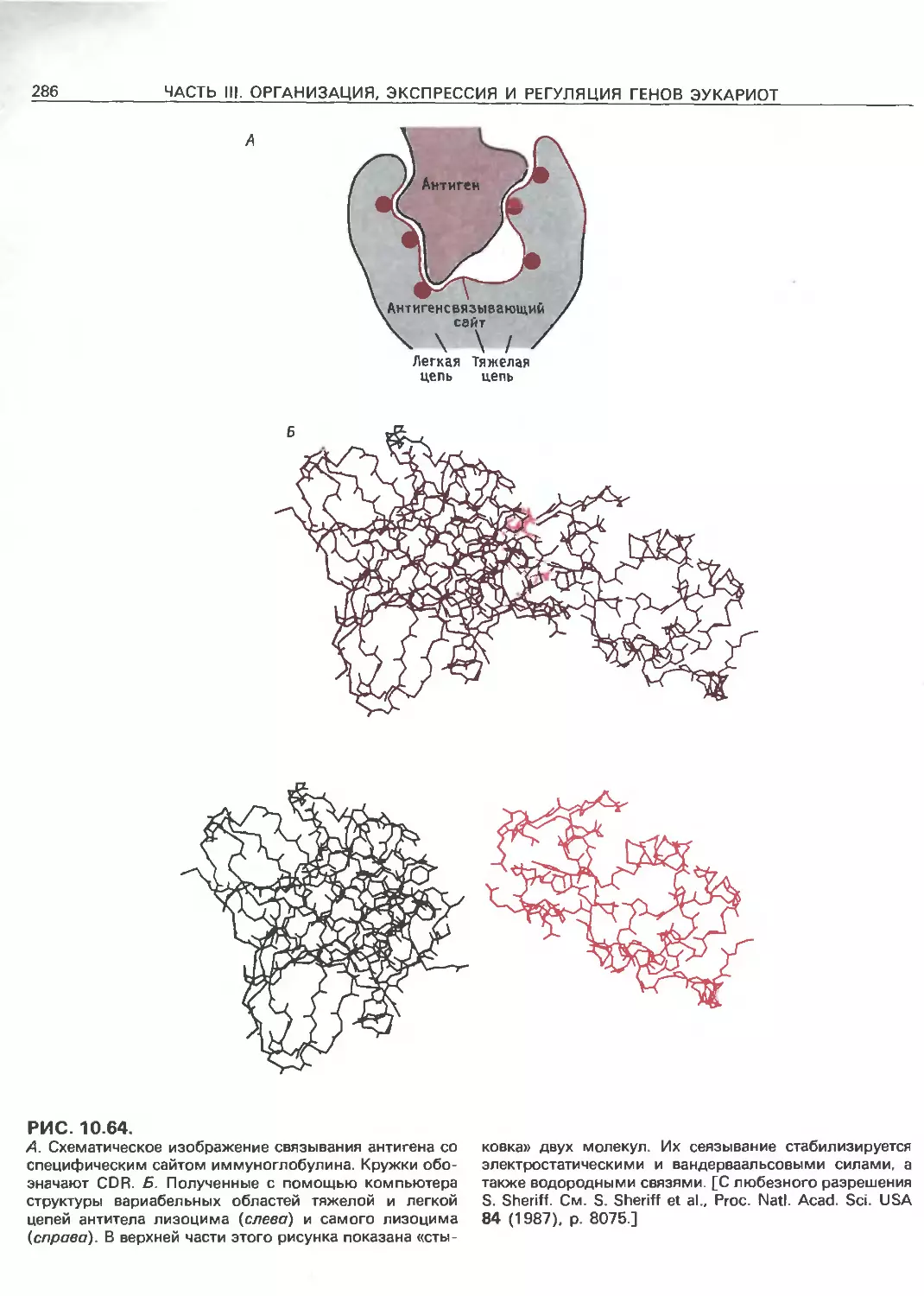

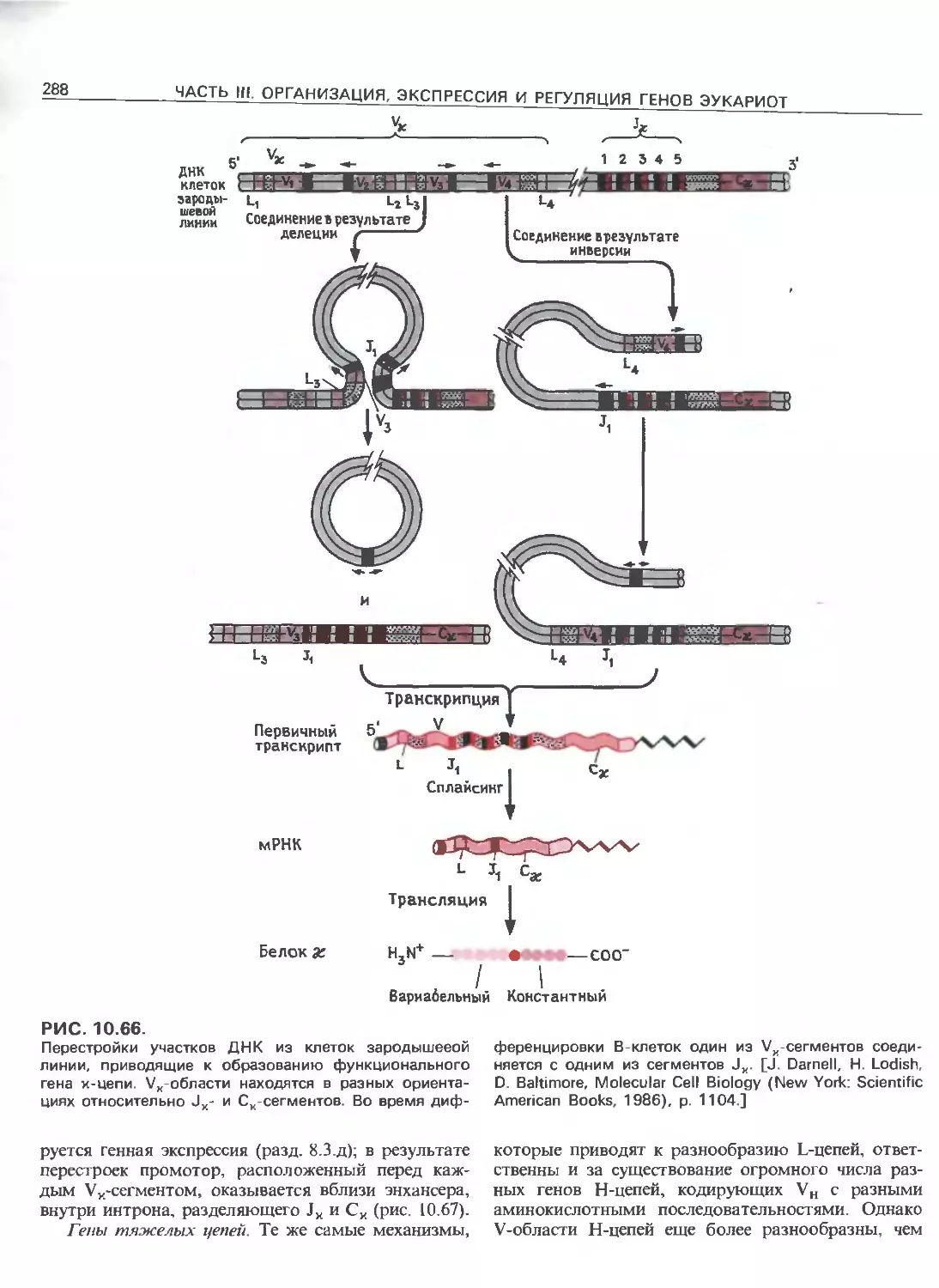

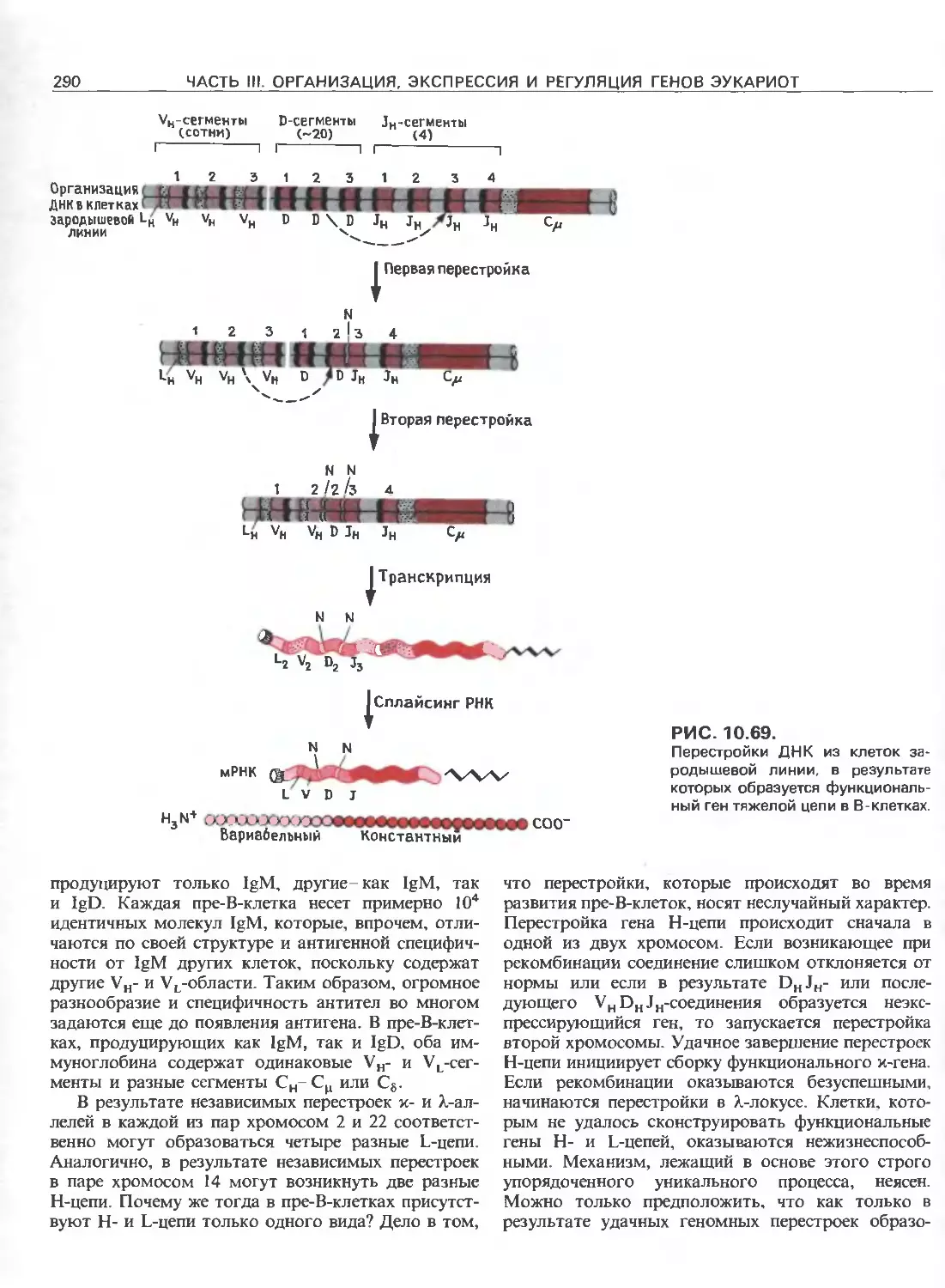
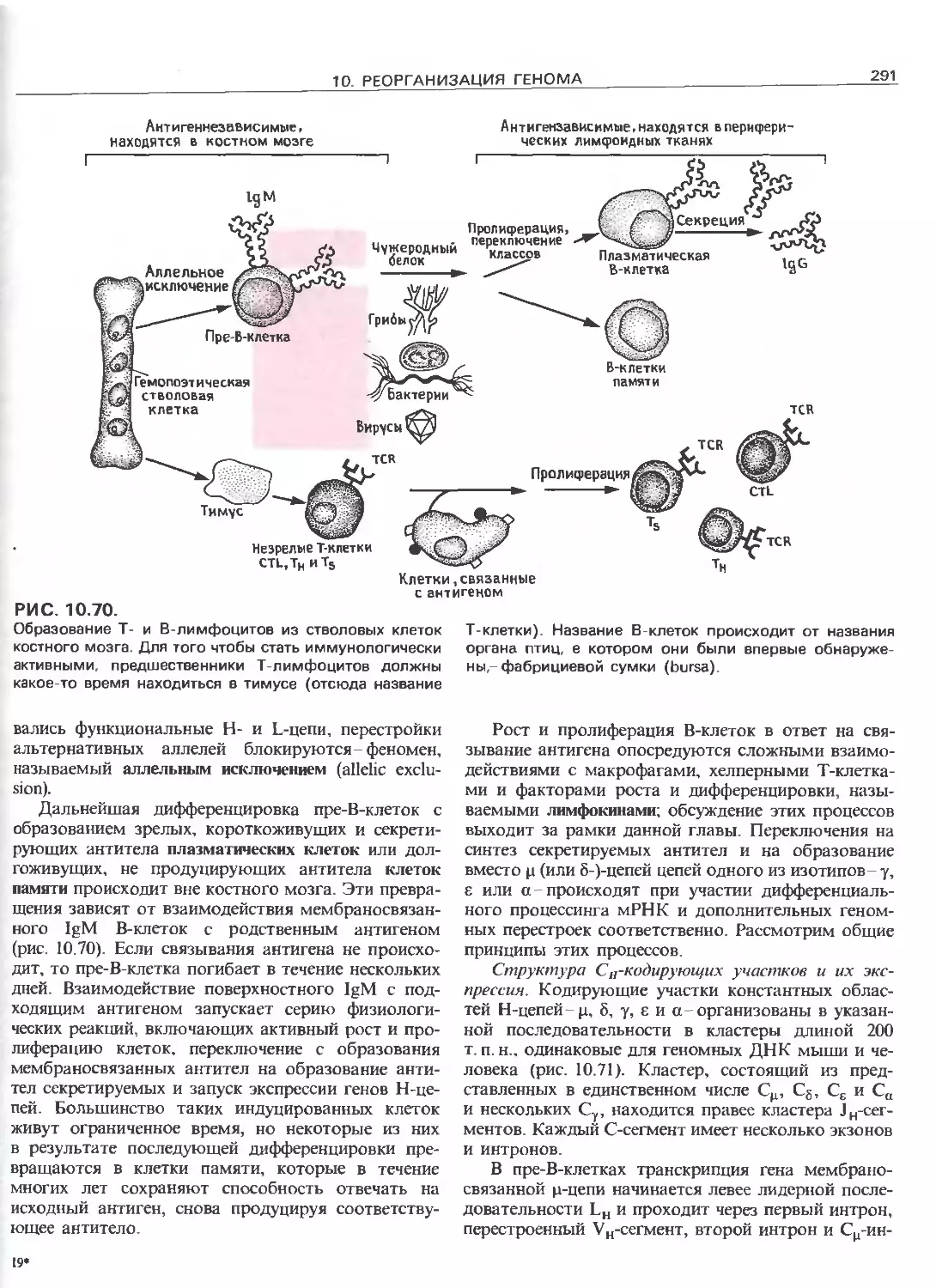








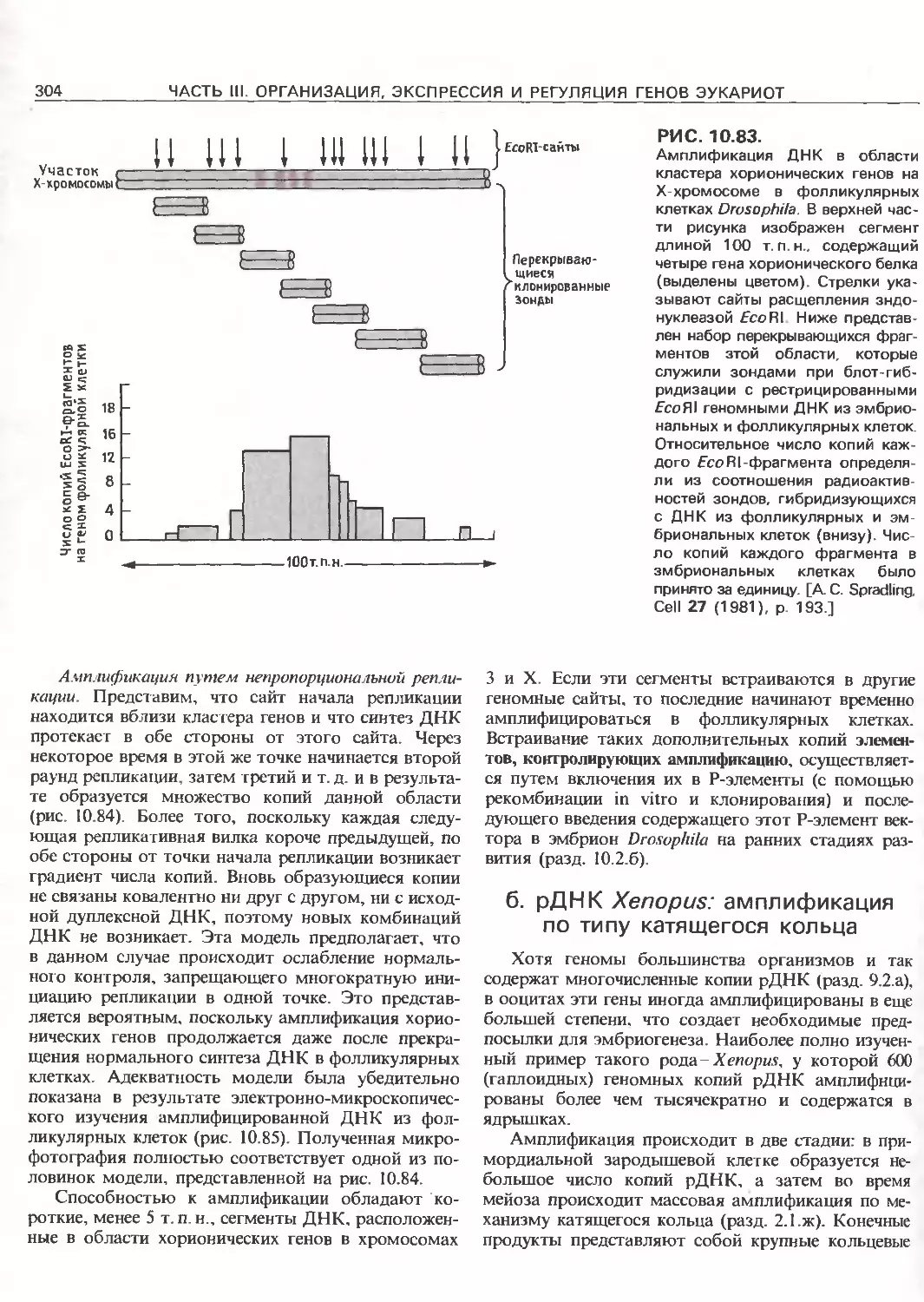
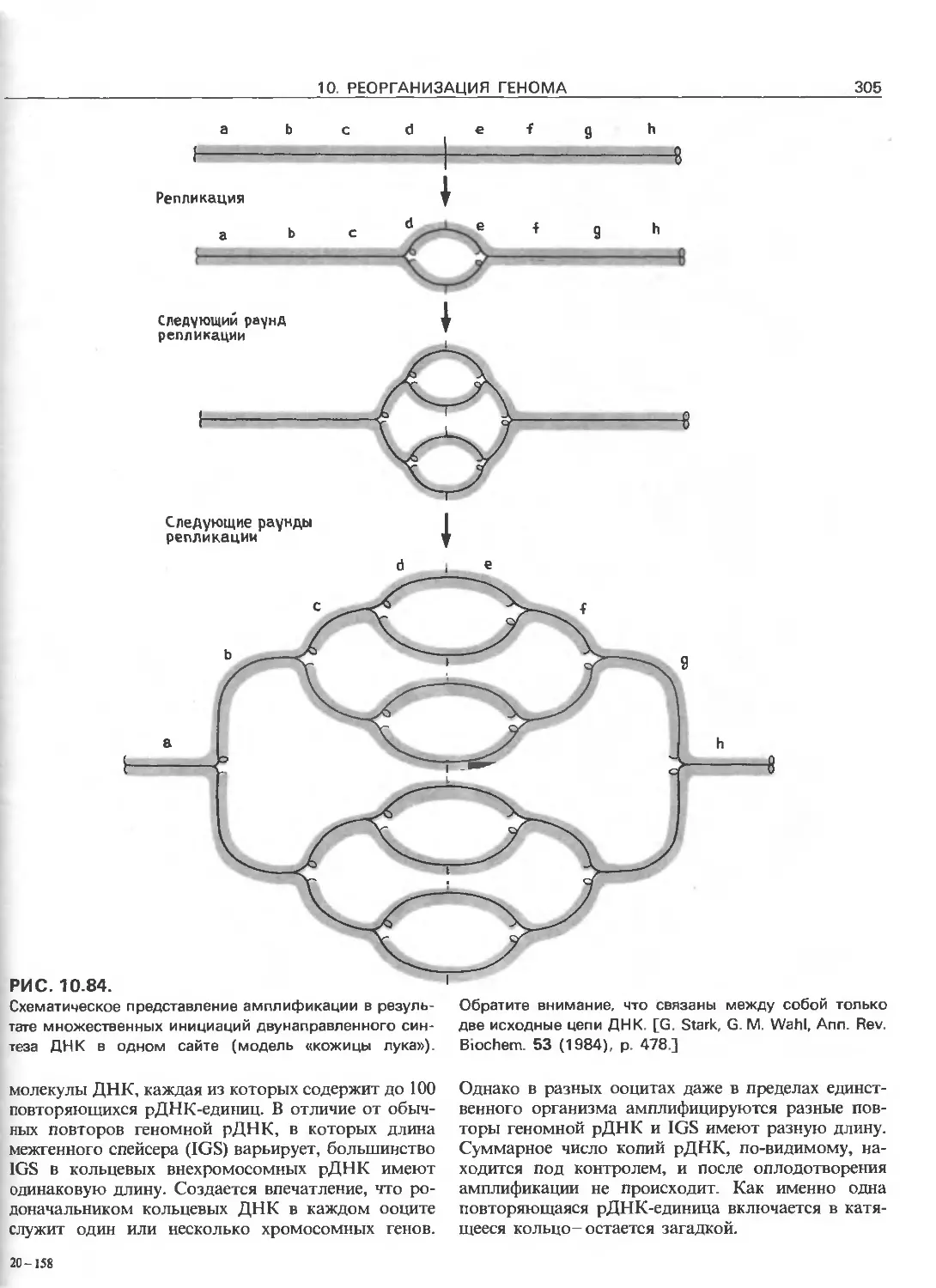
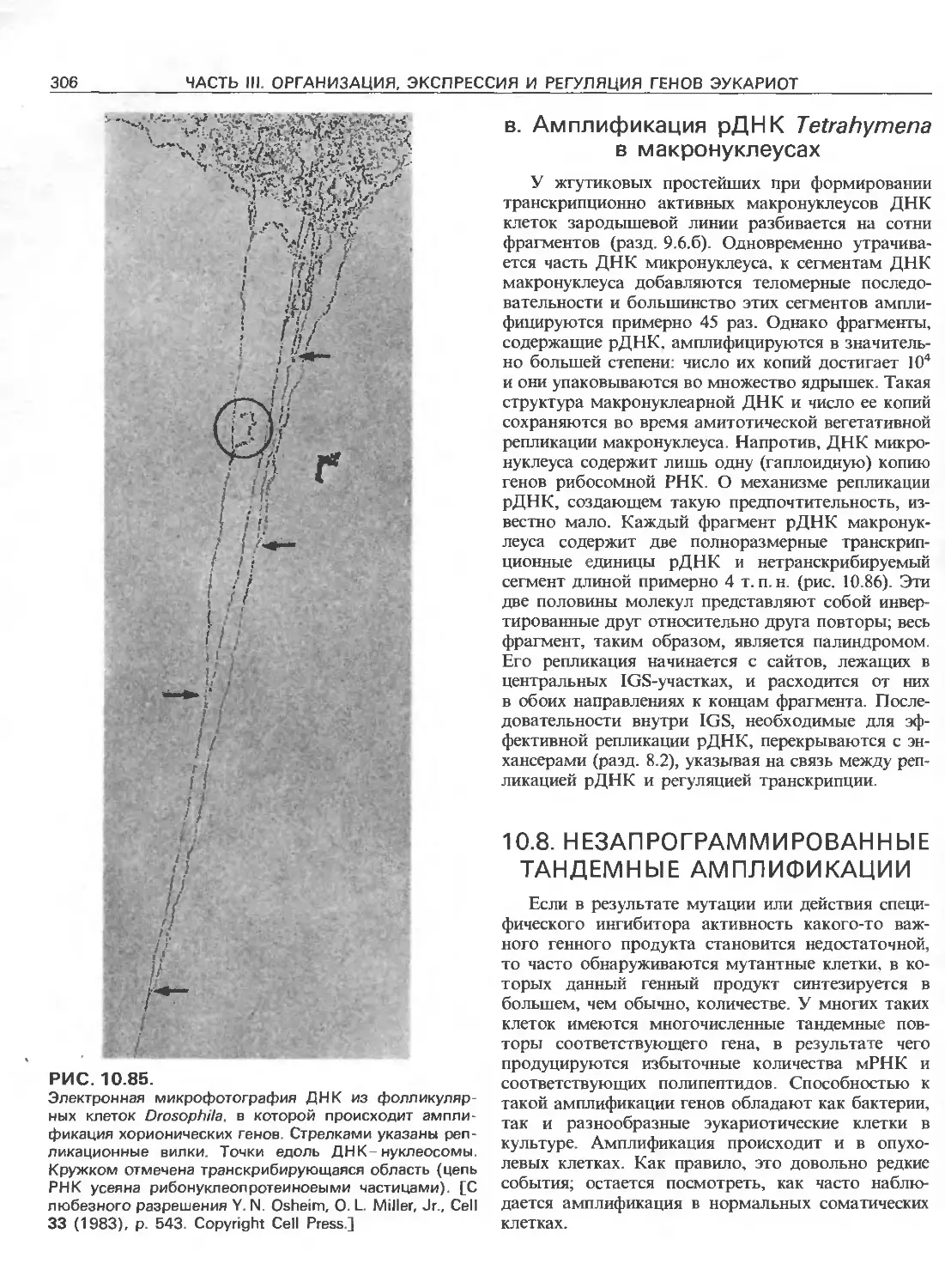
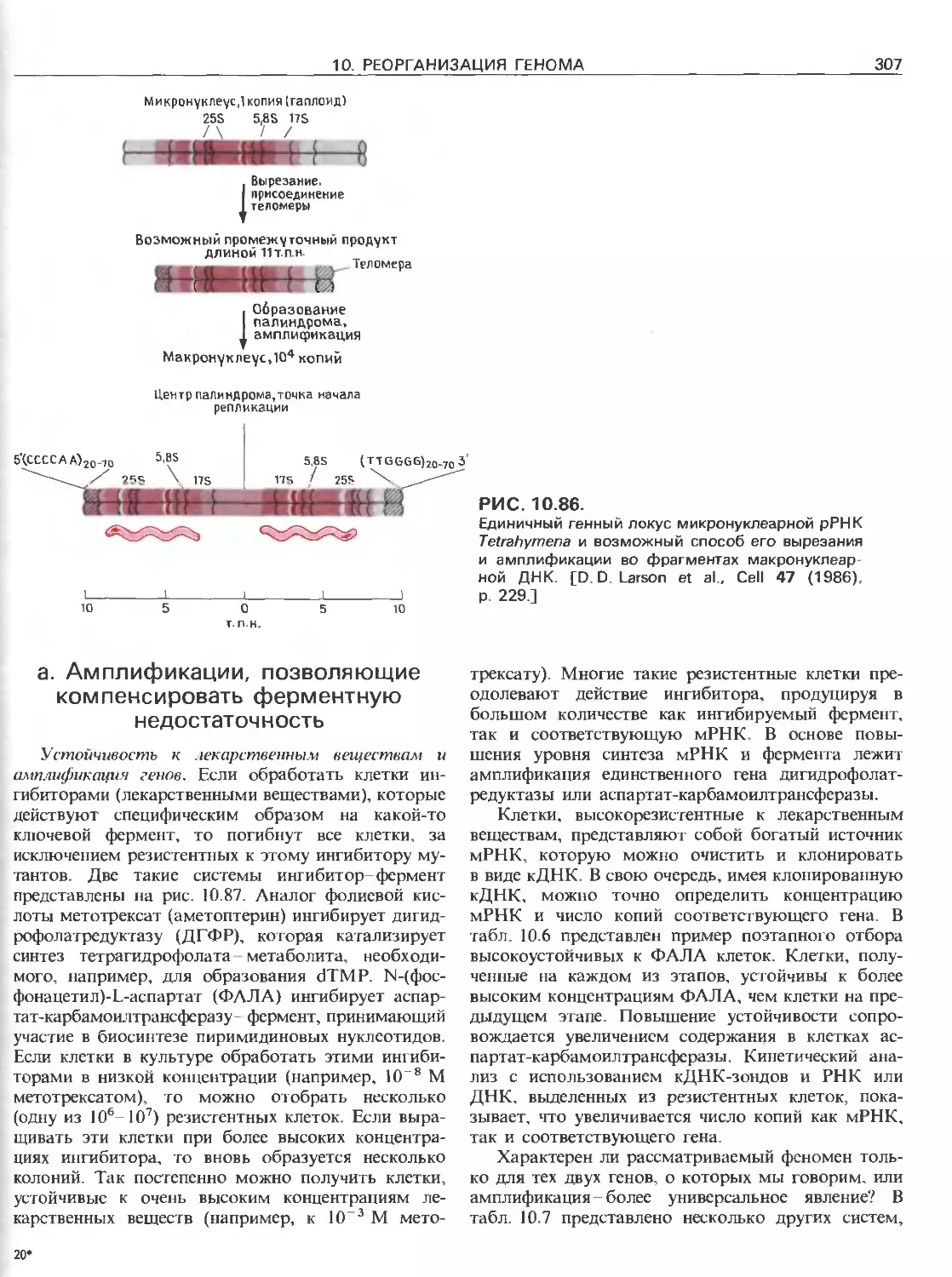



































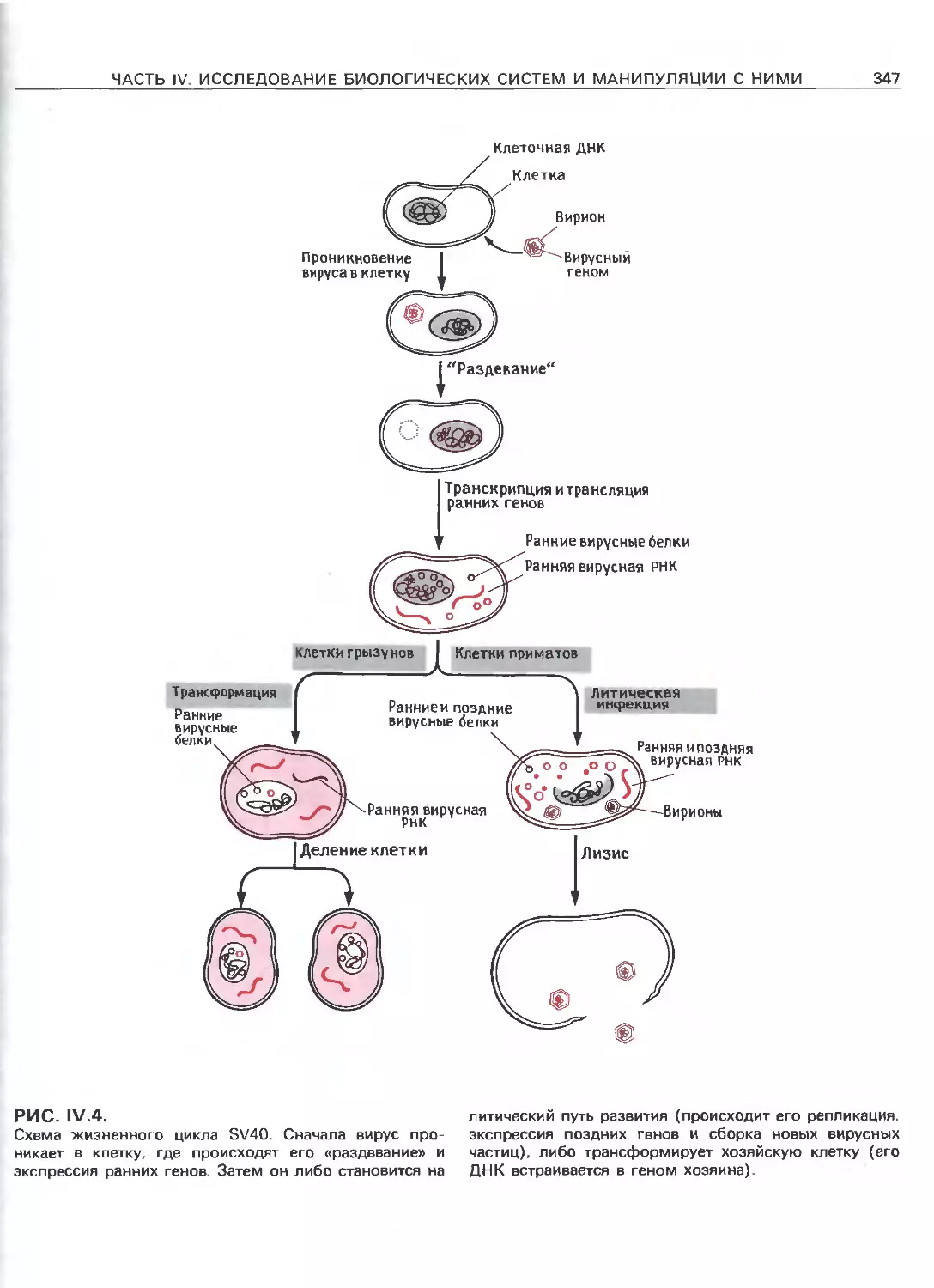
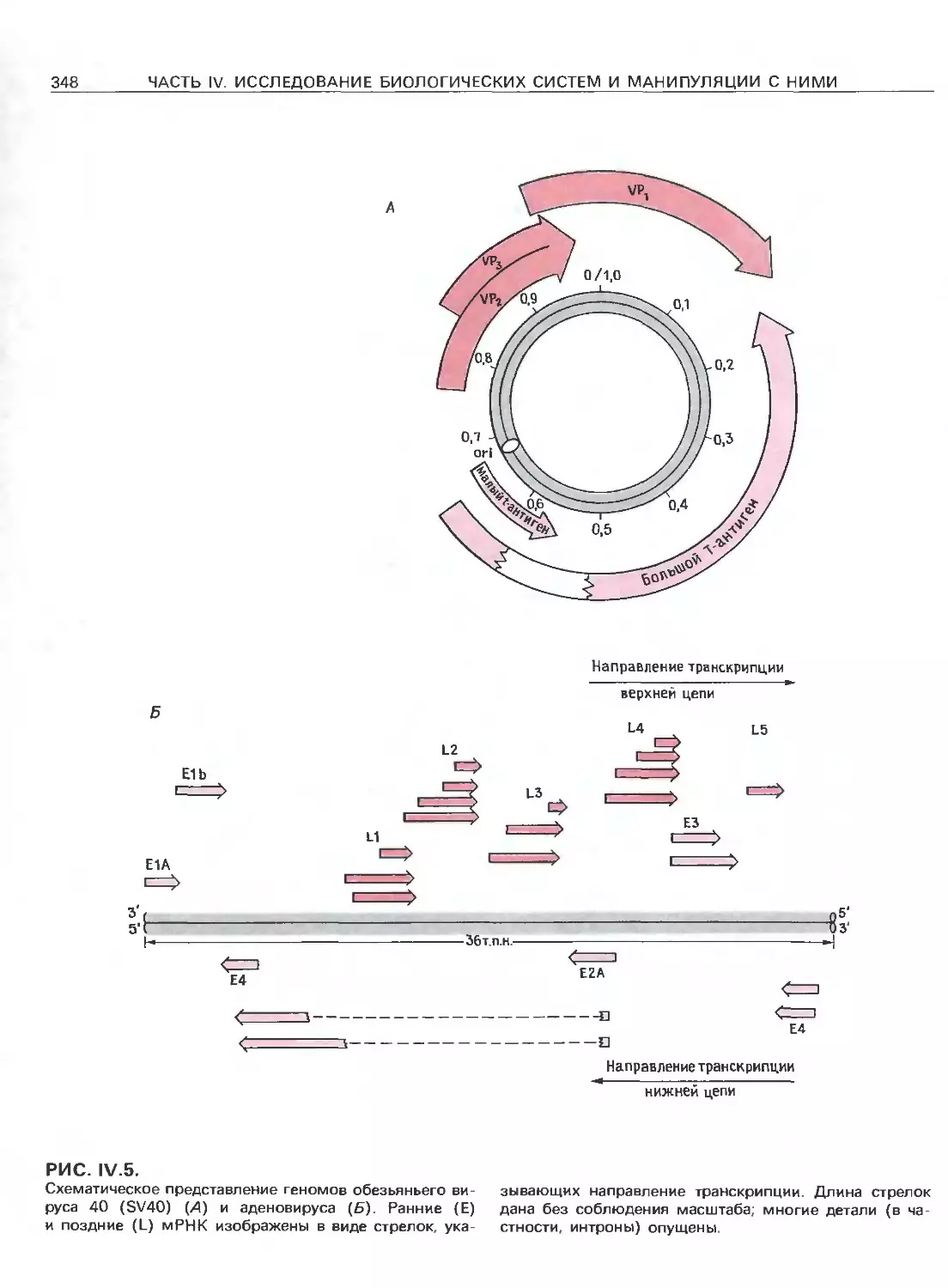
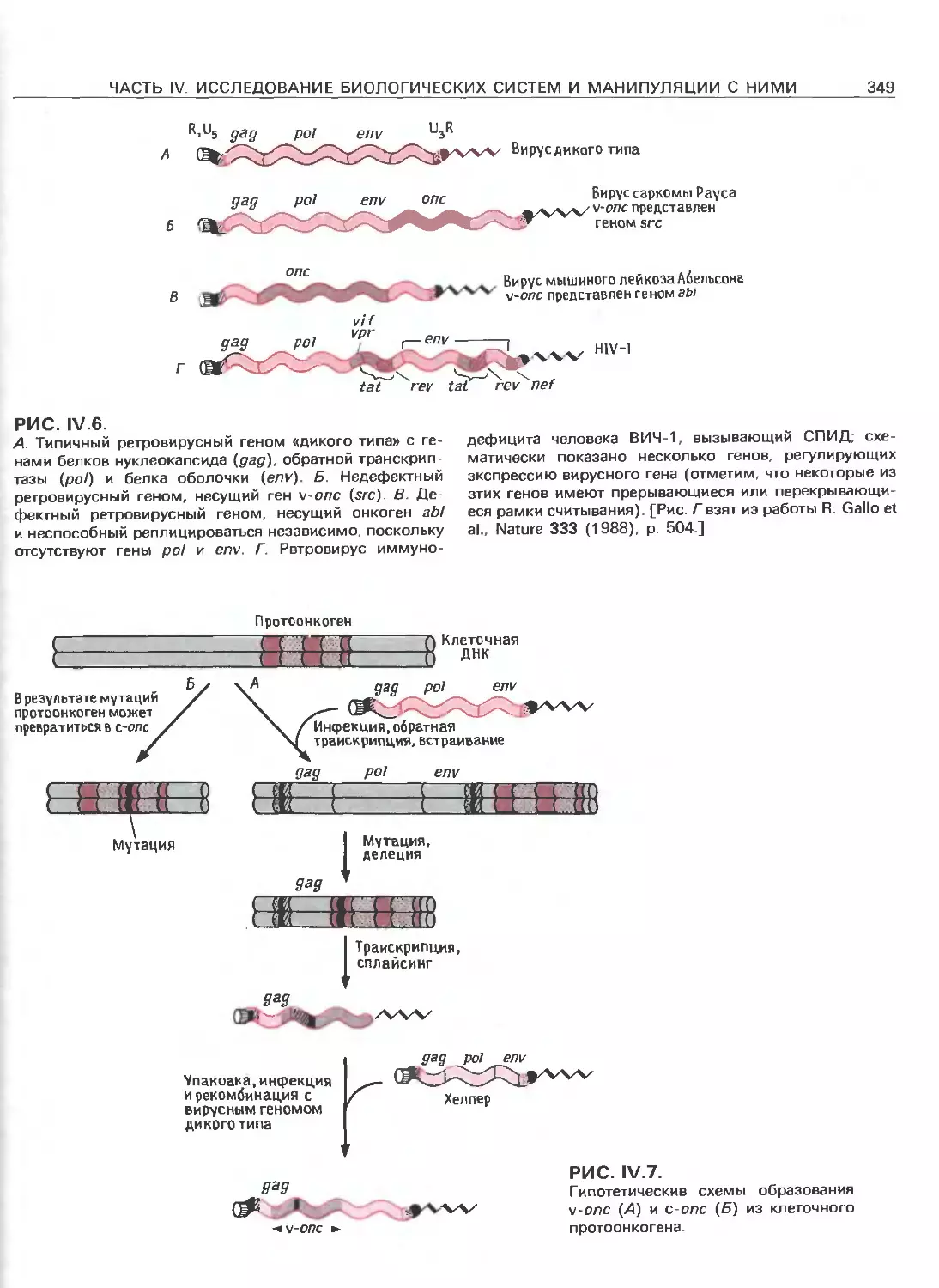


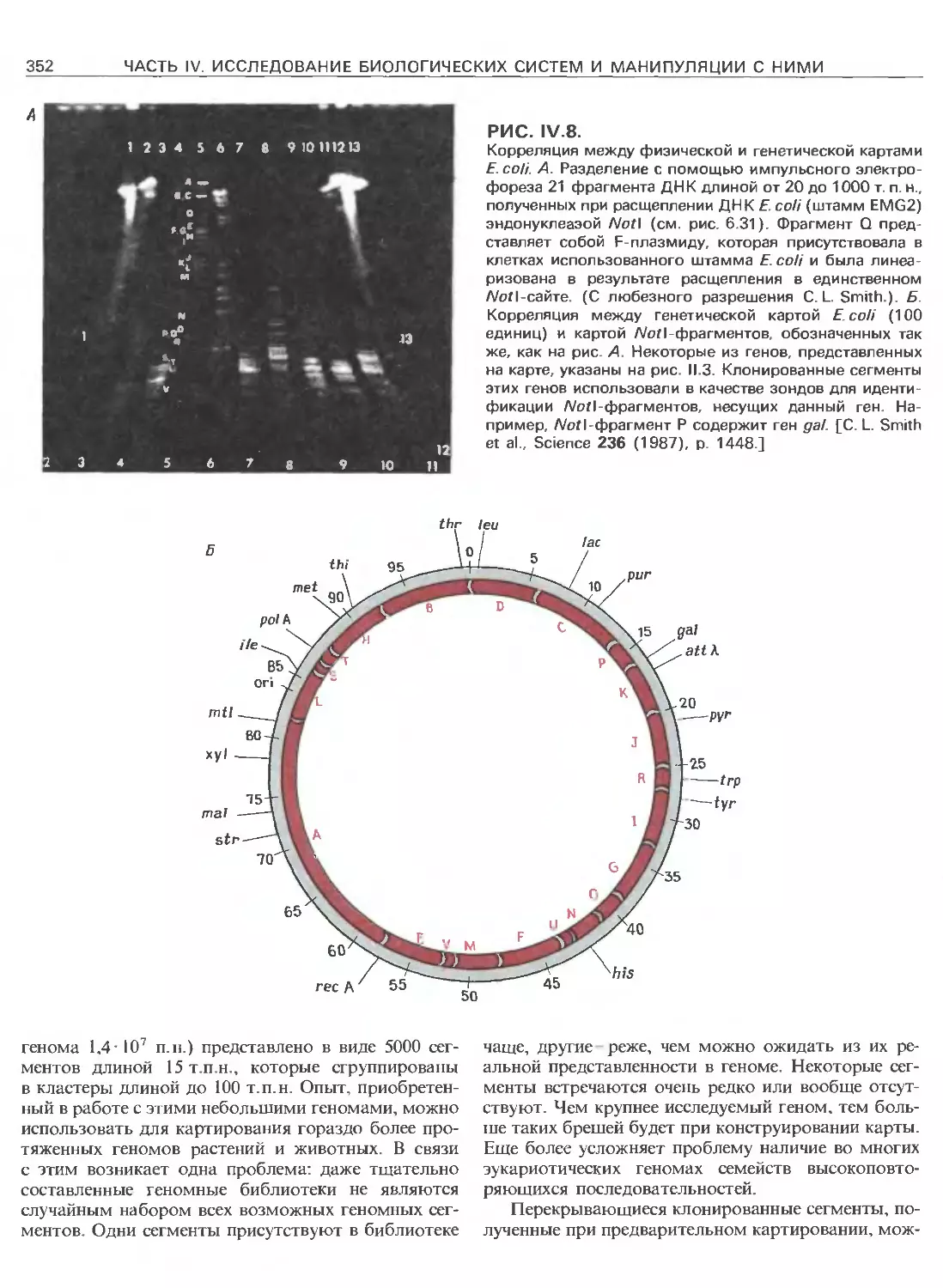







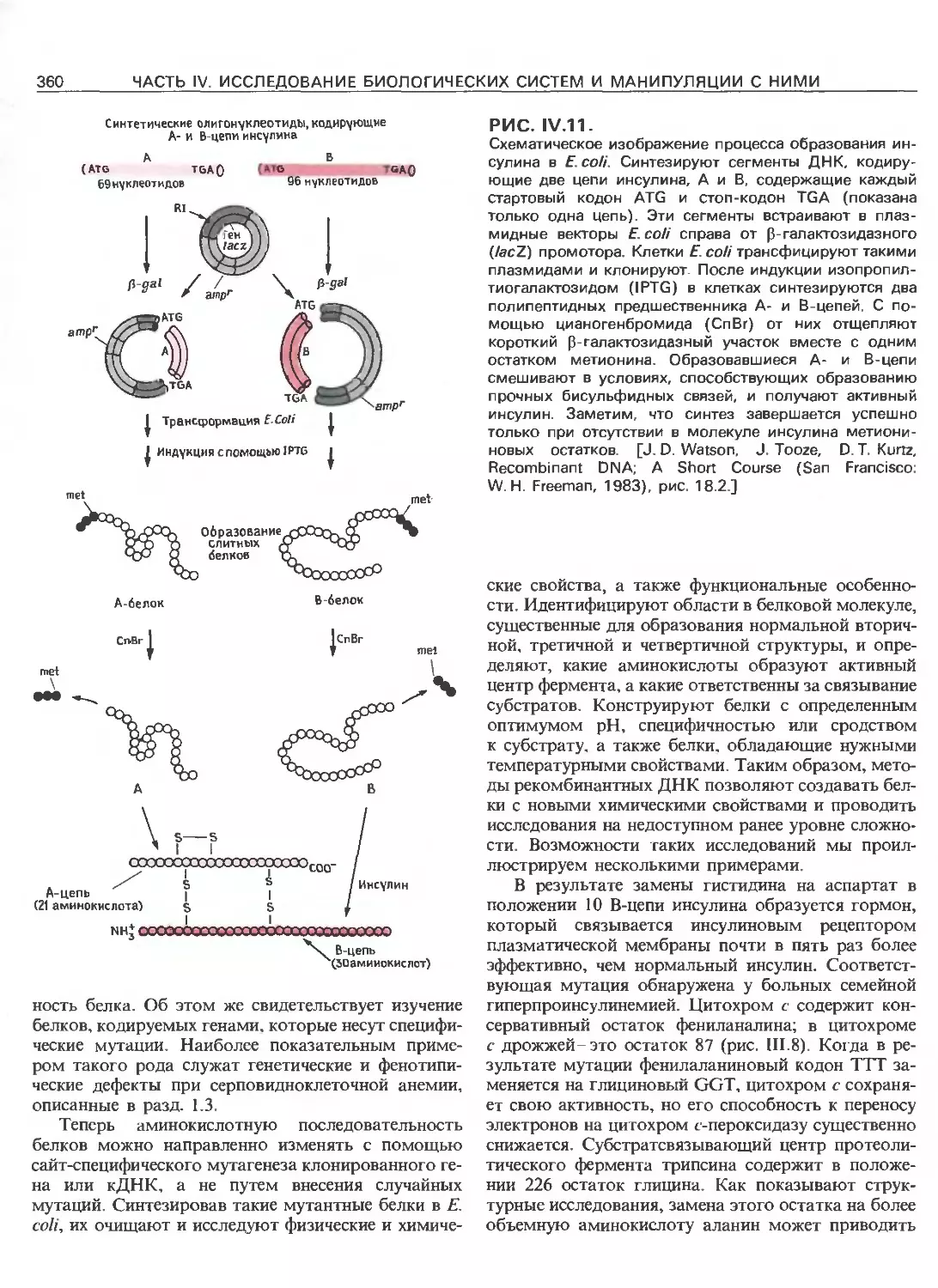




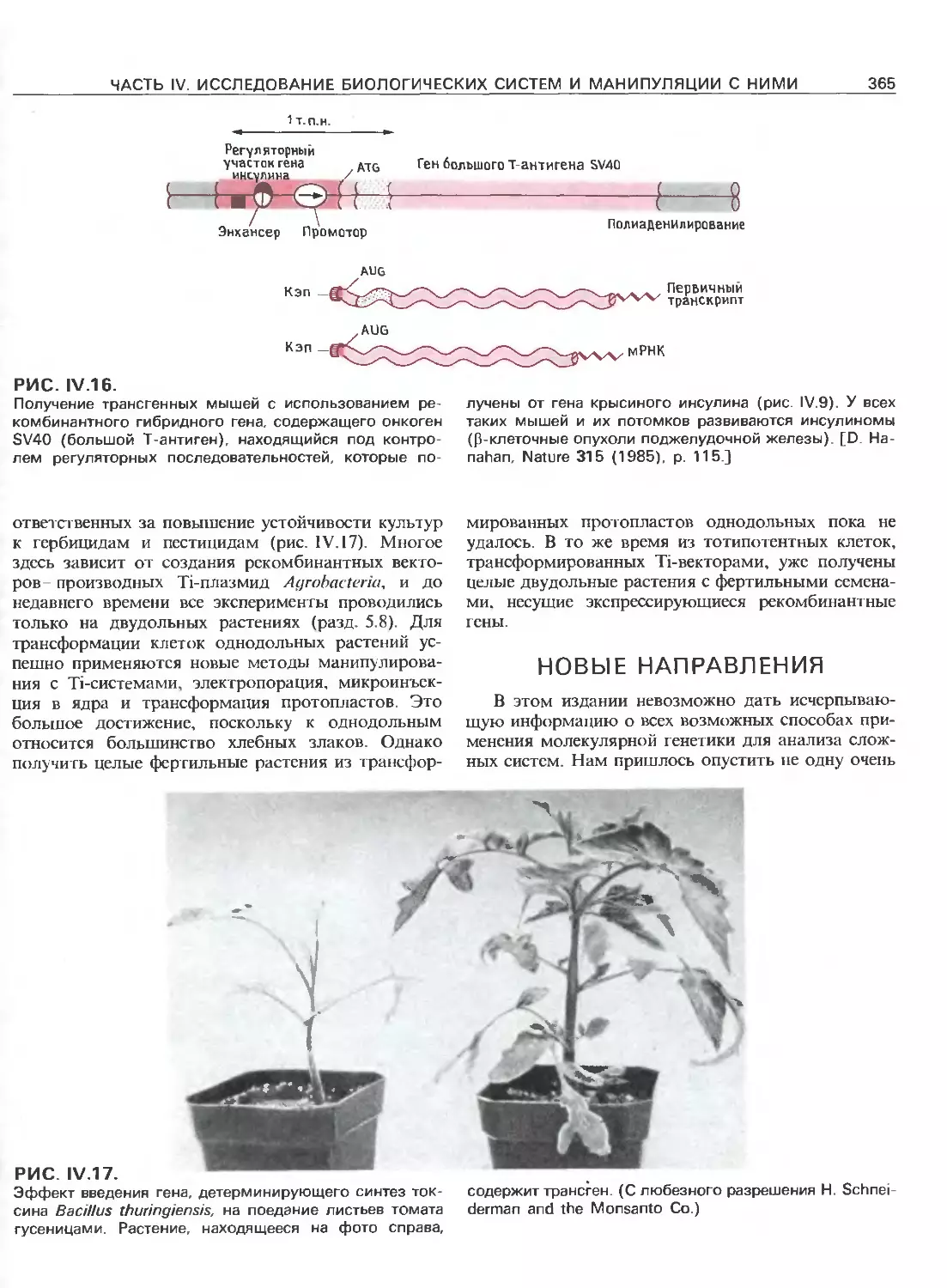
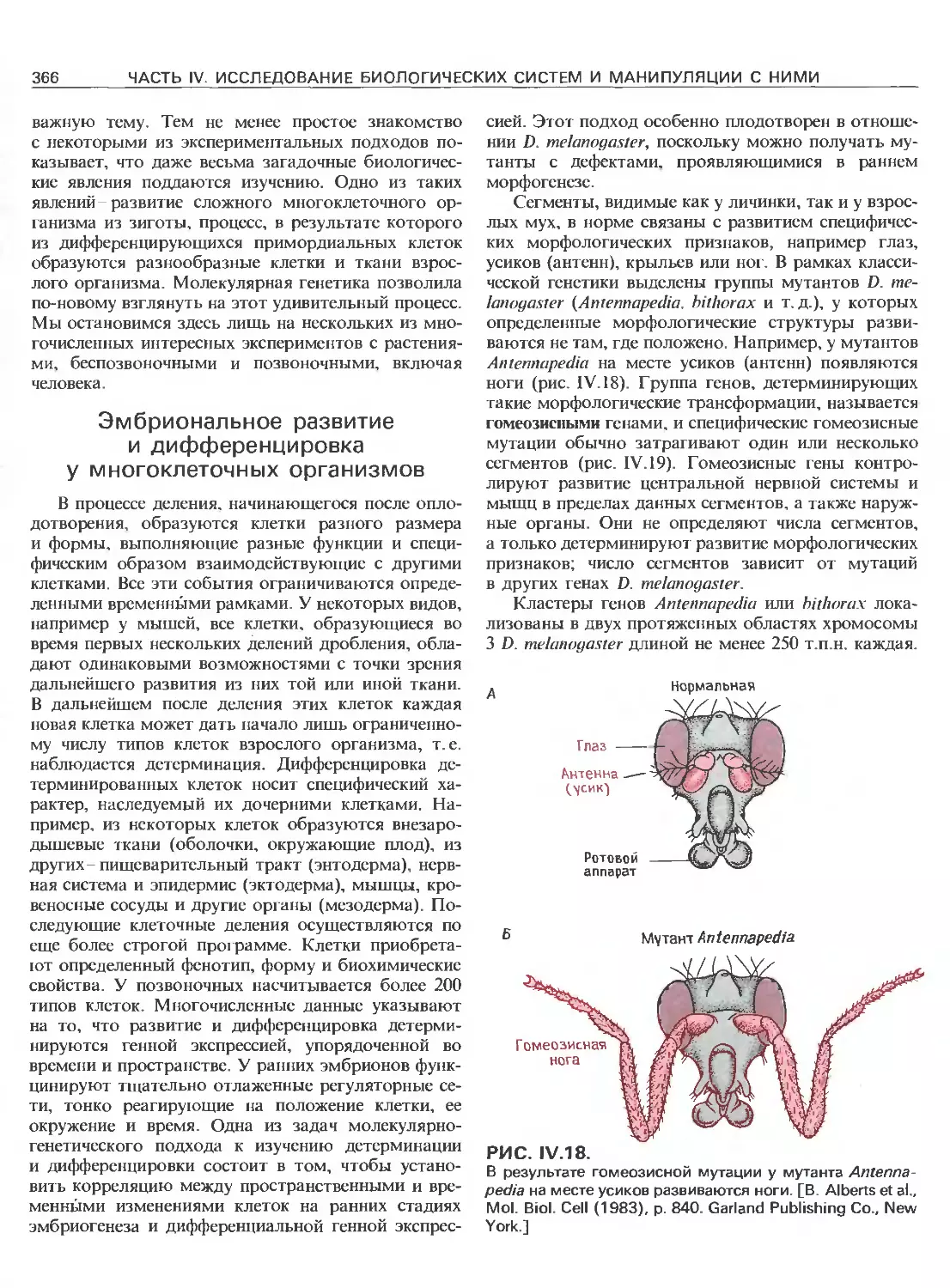



























Tags: общая генетика общая цитогенетика иммуногенетика эволюционное учение видообразование филогенез биология генетика молекулярная биология
ISBN: 5-03-002850-1
Year: 1998




Text
ГЕНЫ И ГЕНОМЫ
GENES & GENOMES
A changing Perspective
Maxine Singer
President, Carnegie Institution of
Washington Scientist Emeritus,
National Institutes of Health
Paul Berg
Wilison Professor of Biochemistry
Director, Beckman Center for Molecular
and Genetic Medicine^
Stanford University School of Medicine
University Science Books
Mill Valley, California
М.СИНГЕР, П.БЕРГ
НИШ
В двух томах
Том 2
Перевод с английского
канд. биол. наук Е. А. Кабановой,
канд. биол. наук А. А. Лушниковой
под редакцией
д-ра биол. наук Н. К. Янковского
Москва «МИР» 1998
УДК 575.113/. 118
ББК 28.04
С38
Авторы: Сингер М., Берг П.
С38 Гены и геномы: В 2-х т. Т. 2. Пер. с англ.-М.: Мир,
1998.-391 с., ил.
ISBN 5-03-002850-1
Университетское руководство по молекулярной биологии, написанное
выдающимися американскими учеными, членами Национальной академии
наук (П. Берг-лауреат Нобелевской премии). Книгу отличают общебиоло-
гический подход, глубина теоретических обобщений, изящная и наглядная
форма подачи материала.
Во 2 томе рассматриваются следующие вопросы: структура и экспрес-
сия генов эукариот, молекулярная структура геномов эукариот, геномные
перестройки с участием разнообразных мобильных элементов, происходя-
щие запрограммированным и случайным образом.
Для молекулярных биологов, преподавателей и студентов универси-
тетов, специалистов.
ББК 28.04
Редакция литературы по биологии
Издание осуществлено при поддержке
Российского фонда фундаментальных исследований
по проекту № 98-04-62107
ISBN 5-03-002848-Х (русск.)
ISBN 5-03-002850-1
ISBN 0-935702-17-2 (англ.)
© 1991 by University Science Books
© перевод на русский язык издательство
«Мир», 1998
Часть III
МОЛЕКУЛЯРНАЯ ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ
И РЕГУЛЯЦИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
ВВЕДЕНИЕ
Молекулярное клонирование создало предпо-
сылки для изучения практически любой части любо-
го генома и позволило решить очень серьезные
проблемы, связанные со сложностью геномов эука-
риот и трудностью их генетического анализа. Сей-
час мы можем получать в чистом виде нужные нам
фрагменты ДНК самых разных геномов в количе-
стве, достаточном для их исчерпывающего химиче-
ского анализа, причем применение эндонуклеаз ре-
стрикции и методов определения нуклеотидных по-
следовательностей делает эту работу почти рутин-
ной. Локализация перекрывающихся участков кло-
нированных сегментов ДНК уже позволила устано-
вить нуклеотидные последовательности областей
генома млекопитающих длиной более 150 т. п. н.
И хотя геномы эукариот имеют огромные размеры
и чрезвычайно сложное строение (табл. III. 1), де-
тальное изучение их молекулярной организации уже
не представляется невозможным; теперь это в ос-
новном вопрос времени и средств. На расшифровку
последовательности генома фага X длиной 50 т. п. н.
потребовалось около пяти человеко-лет. Благодаря
использованию усовершенствованных методов сек-
венирования ДНК удалось достичь больших успе-
хов в определении нуклеотидной последовательно-
сти хромосомы Е. coll, состоящей из четырех мил-
лионов пар оснований, а сейчас проводится иссле-
дование короткой хромосомы человека, которая по
длине только в десять раз больше хромосомы Е.
coli.
Зная нуклеотидную последовательность того или
иного гена и примыкающих к нему сегментов ДНК,
мы еще ничего не можем сказать о том, как работает
этот ген, как осуществляется регуляция его экспрес-
сии при развитии и дифференцировке или при ответе
на изменения окружающей среды. Нуклеотидная
последовательность сама по себе не говорит ни
о способе координации генной экспрессии, обес-
печивающей сложное физиологическое равновесие,
характерное для клеток здорового организма, ни
о том, каким образом продукт поврежденного гена
или изменение скорости экспрессии гена нарушают
нормальное функционирование организма или при-
водят к тому или иному заболеванию. Для того
чтобы ответить на все эти вопросы, необходимы
специальные биологические исследования. И в этом
случае мощным инструментом оказывается техно-
логия рекомбинантных ДНК. Если клонированные
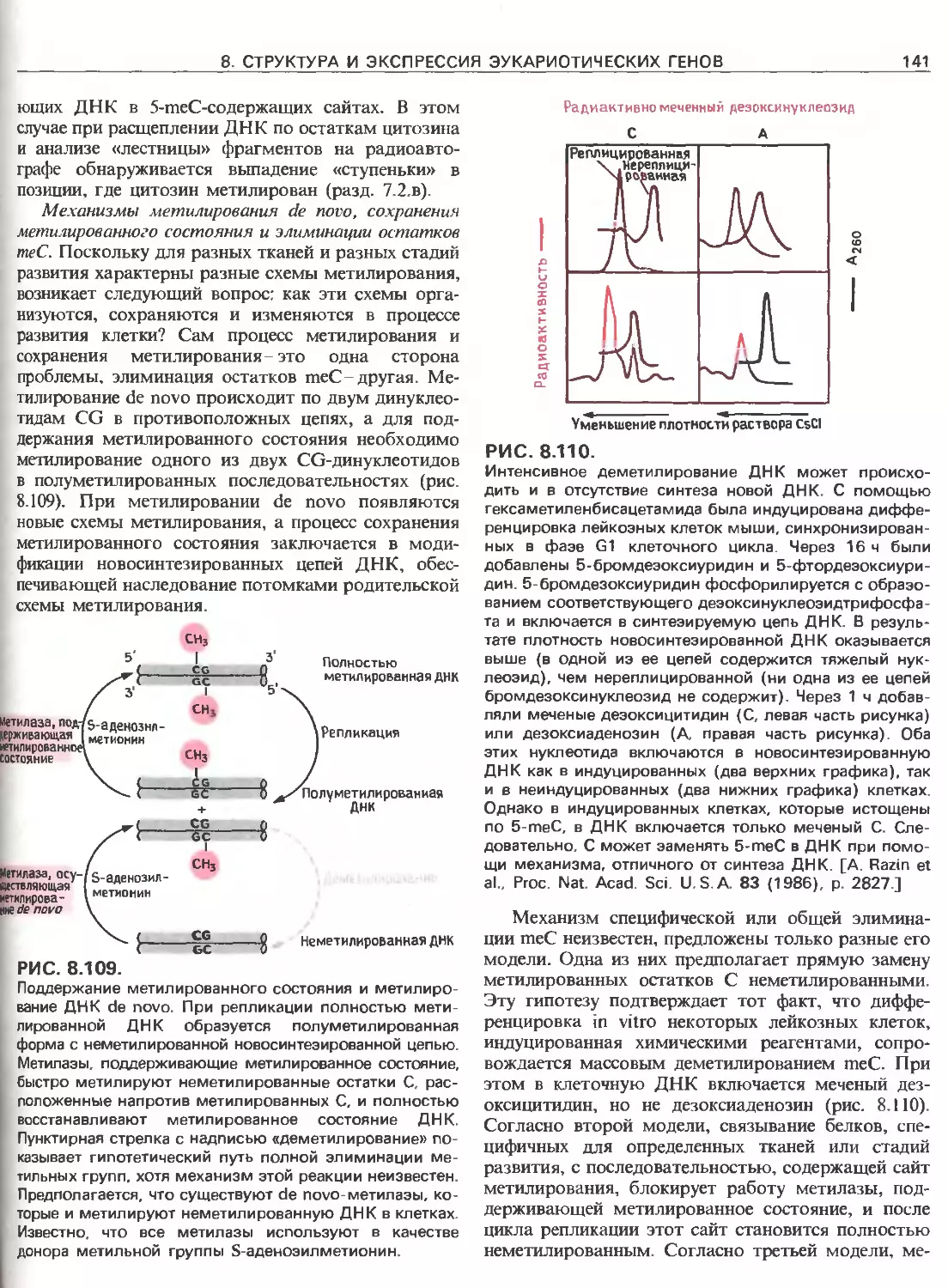
гены ввести в клетки при помощи трансфекции или
Таблица 111.1. Размер некоторых эукариотических
геномов
Организм Примерный размер Гаплоидное
гаплоидного генома, число
П.Н. хромосом
Дрожжи (Saccharomices cerevisiae) 1,35 IO7 16
Миксомицеты (Dictyoste- lium discoides) 7-107 7
Трипаносома (Trypanoso- ma brucei) 8-107 Неизвестно
Нематода (Caenorhabditis elegans) 8-107 11/12
Шелкопряд (Bombyx mori) 5 108 28
Плодовая мушка (Drosop- hila melanogaster) 1,65-IO8 4
Морской еж (Strongylocen- trotus purpuratus) 8-108 21
Шпорцевая лягушка (Xe- nopus laevis) 3 109 18
Протей (Necturus maculo- sis) 5-1O10 19
Курица (Gallus domesticus) 1,2-109 39
Мышь (Mus musculus) 3 109 20
Корова (Bovis domesticus) 3,1 109 60
Человек (Homo sapiens) 2,9-109 23
Кукуруза (Zea mays) 5 IO9 10
Лук (Allium сера) 1,5- IO10 8
Арабидопсис (Arabidopsis thaliana) 7 107 5
8
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Таблица III.2. Вставочные последовательности в некоторых генах
Продукт гена Организм Экзоны суммарная длина, п.н. Интроны
число суммарная длина, п.н.
Аденозиндезаминаза Человек 1500 11 30000
Аполипопротеин В « 14000 28 29000
Р-Глобин Мышь 432 2 762
Цитохром b Дрожжи (митохондрии) 2200 6 5100
Дигидрофо латредуктаза Мышь 568 5 31 500
Эритропоэтин Человек 582 4 1562
Фактор VIII « 9000 25 177000
Фиброин (шелк) Шелкопряд 18000 1 970
Г ипоксантин-фосфорибо- зилтрансфераза Мышь 1307 8 32000
а-Интерферон Человек 600 0 0
Рецептор липополипроте- ина низкой плотности « 5100 17 40000
Фазеолин Фасоль 1263 5 515
Тиреоглобулин Человек 8500 >40 100000
тРНКТу' Дрожжи 76 1 14
Субъединица уриказы Соя 300 7 4500
Вителлогенин Шпорцевая лягушка 6300 33 20000
Зеин Кукуруза 700 0 0
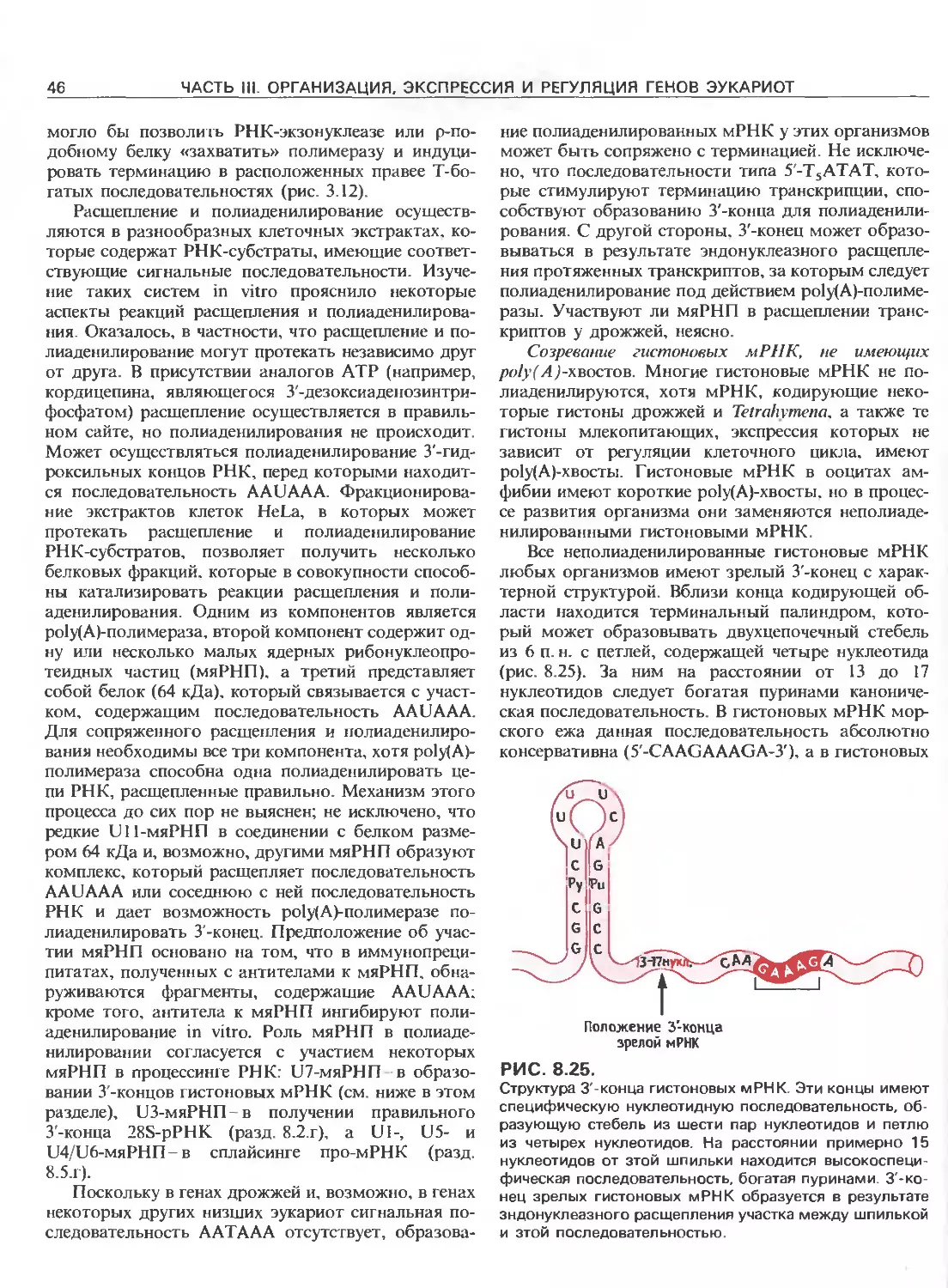
Точка
Регуляторный инициации
участок транскрипции
E.coli
poHI
-35 -10
poll
Разное
расстояние
Разное
расстояние
polIII +1
1~Т 1( —(------- ц
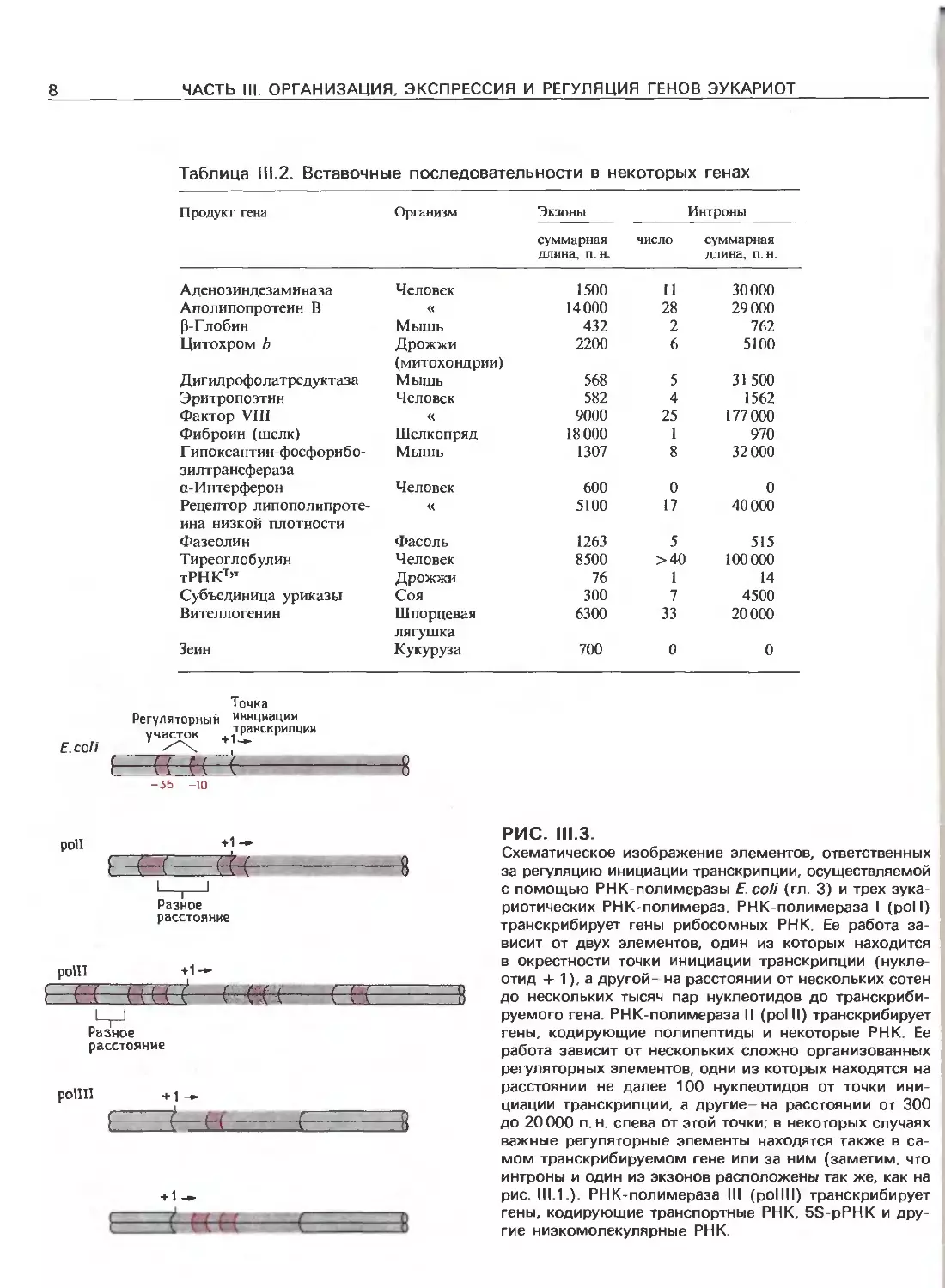
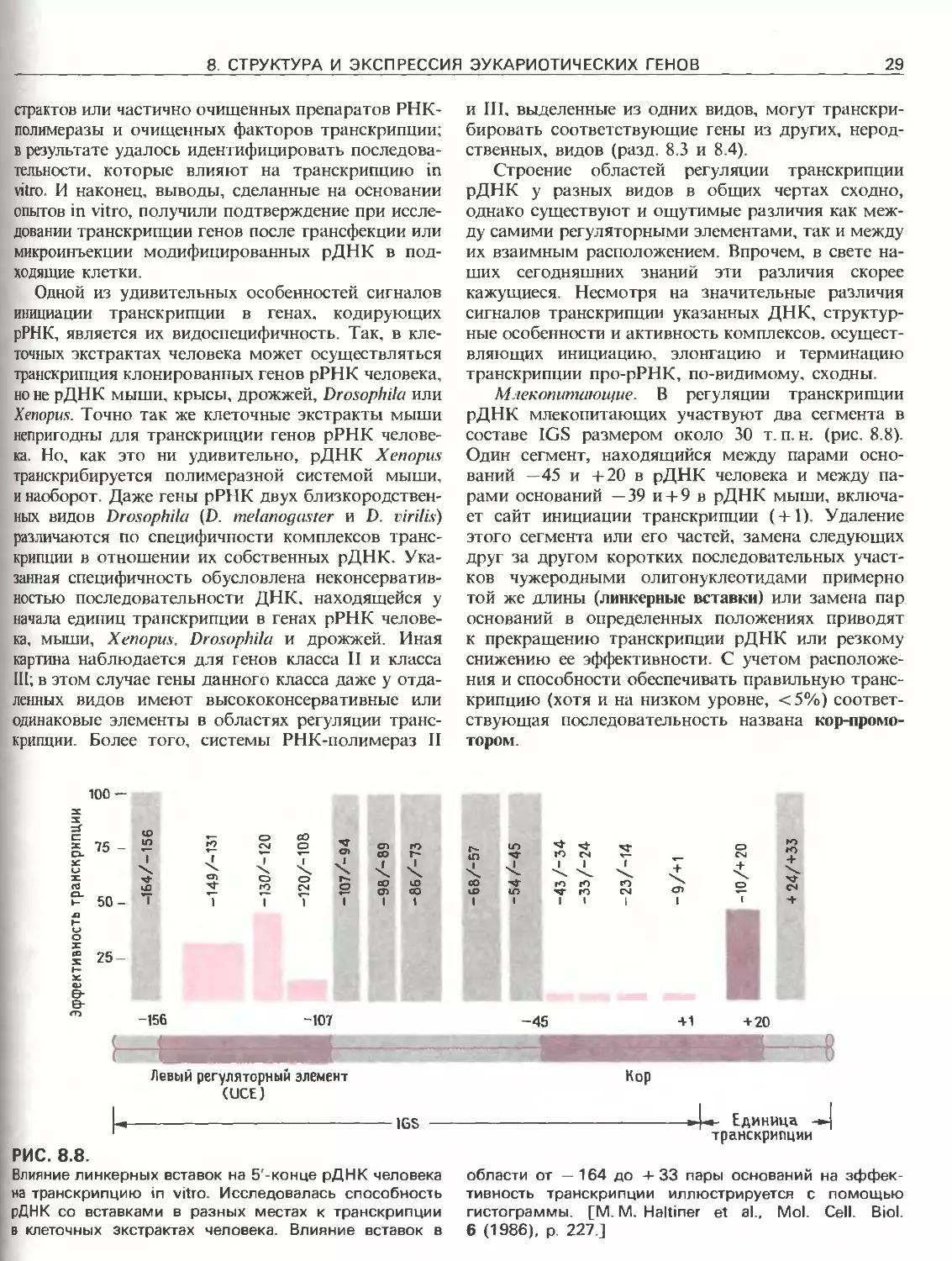
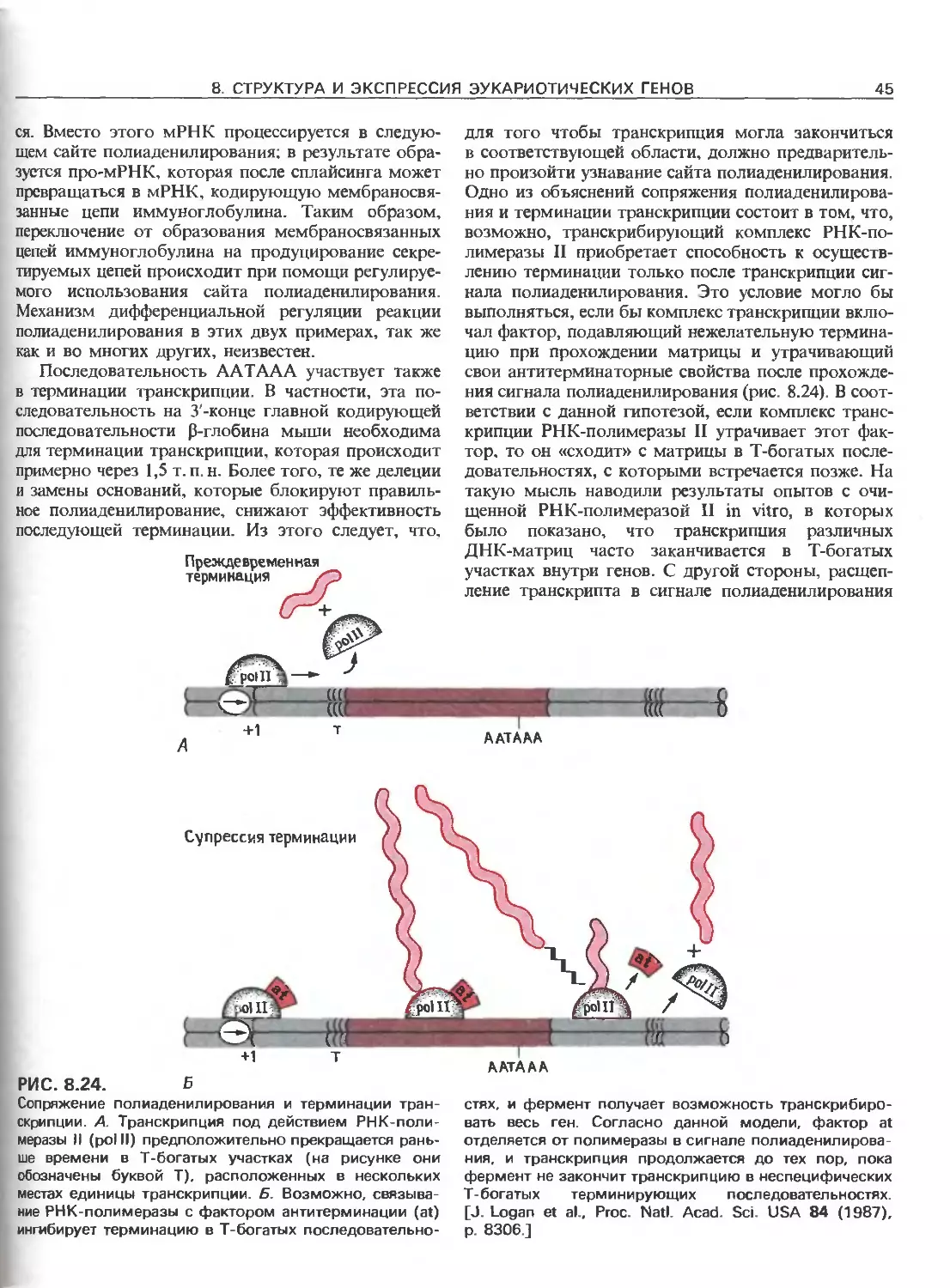
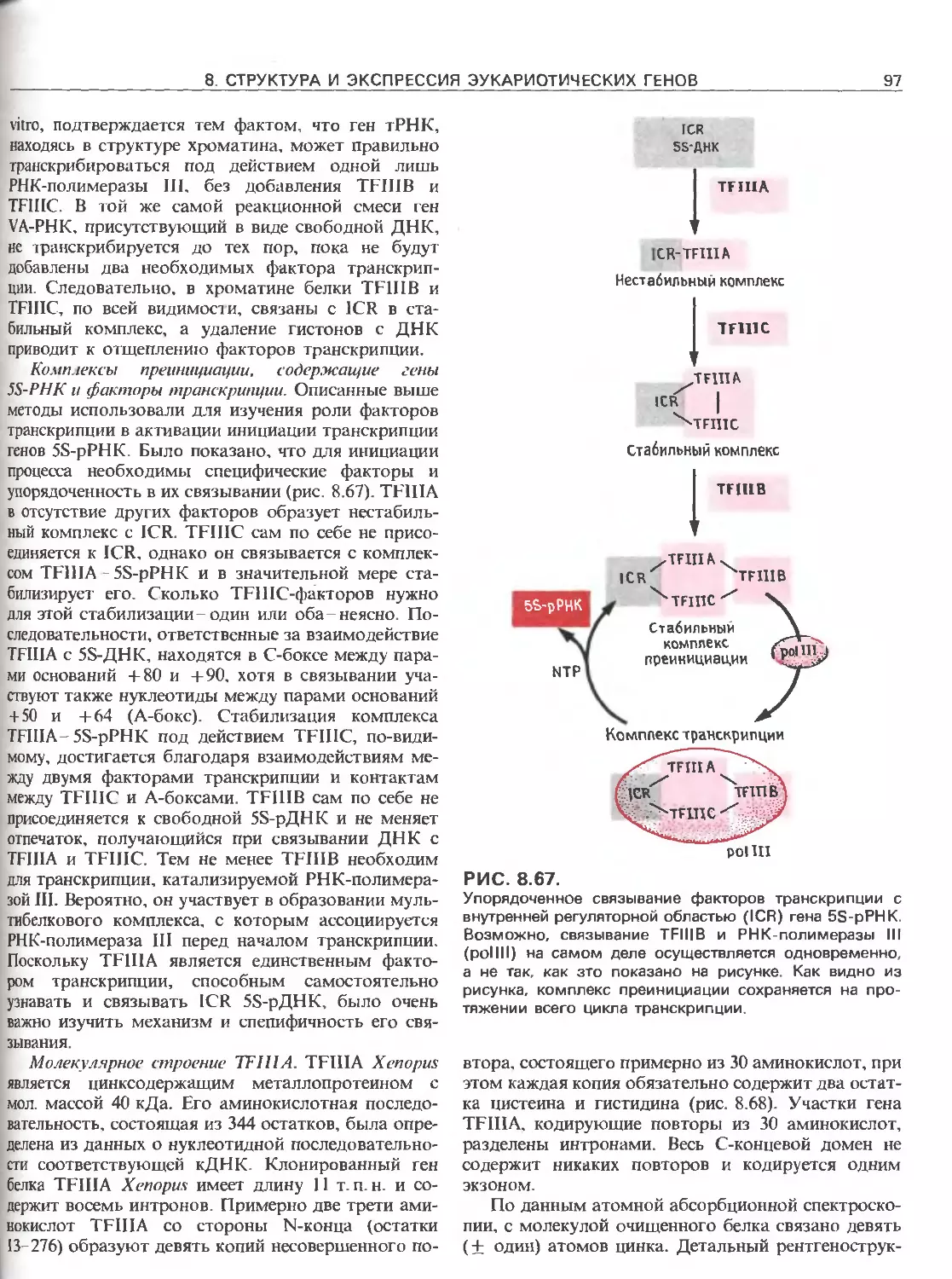
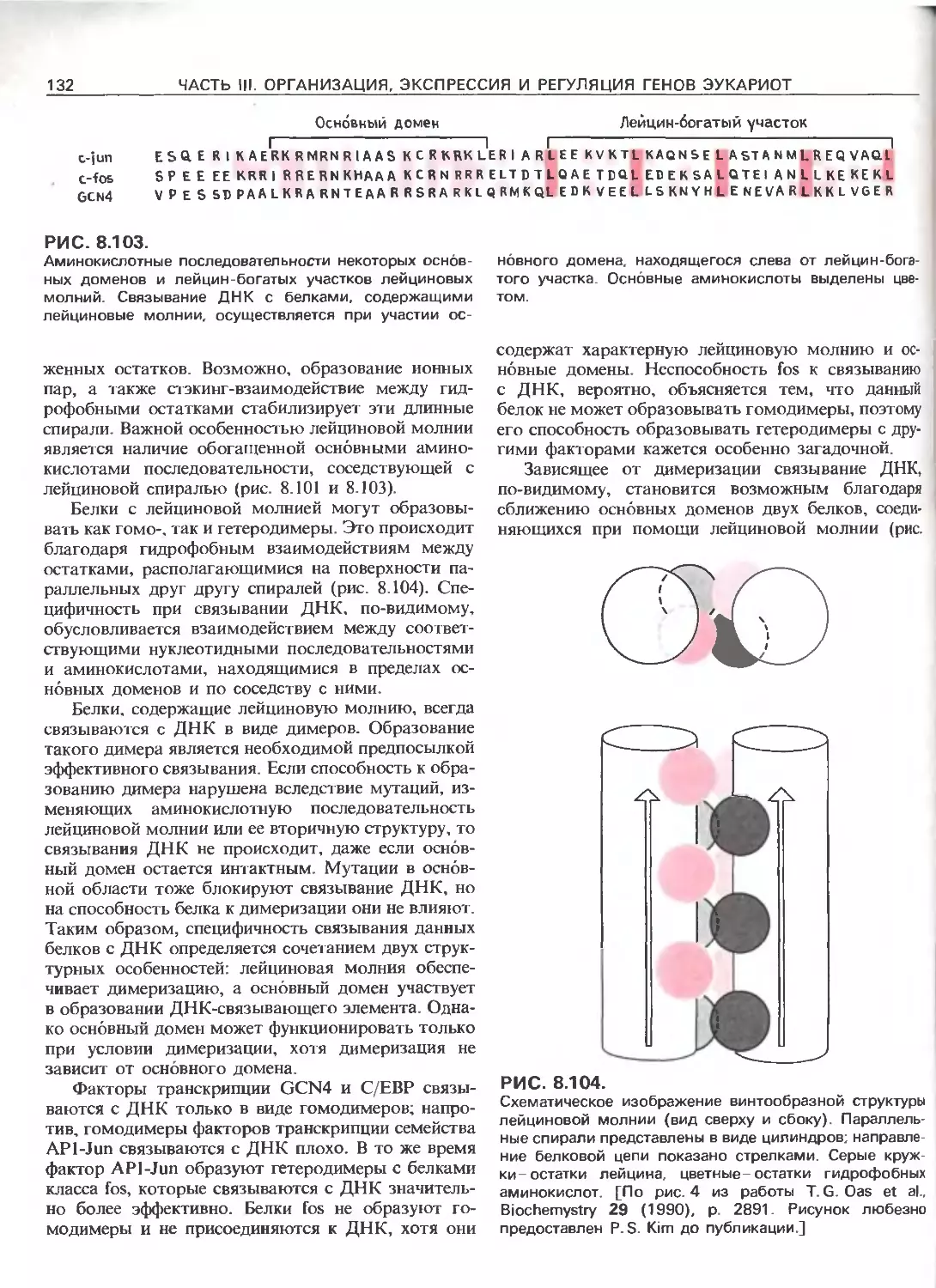
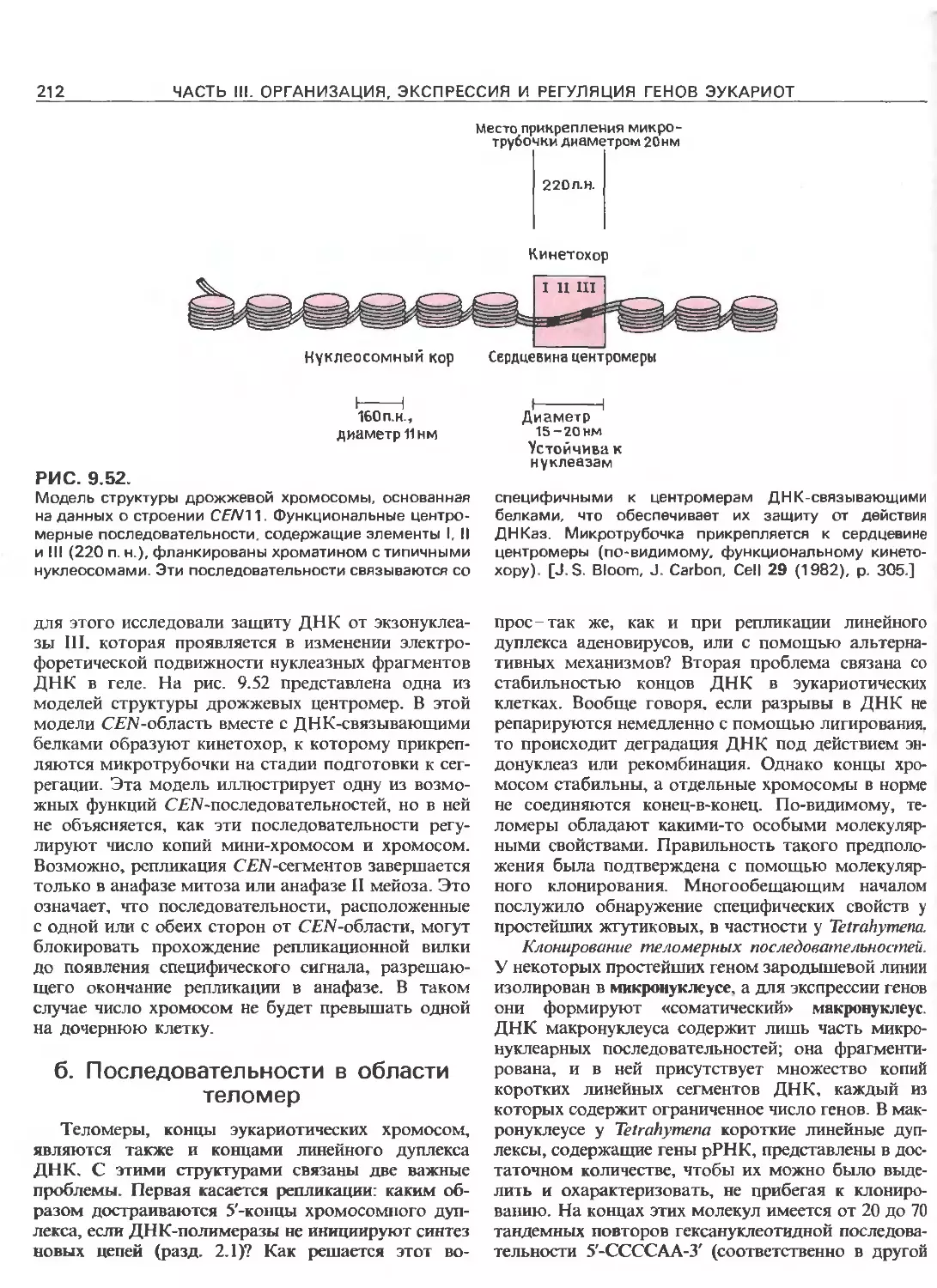
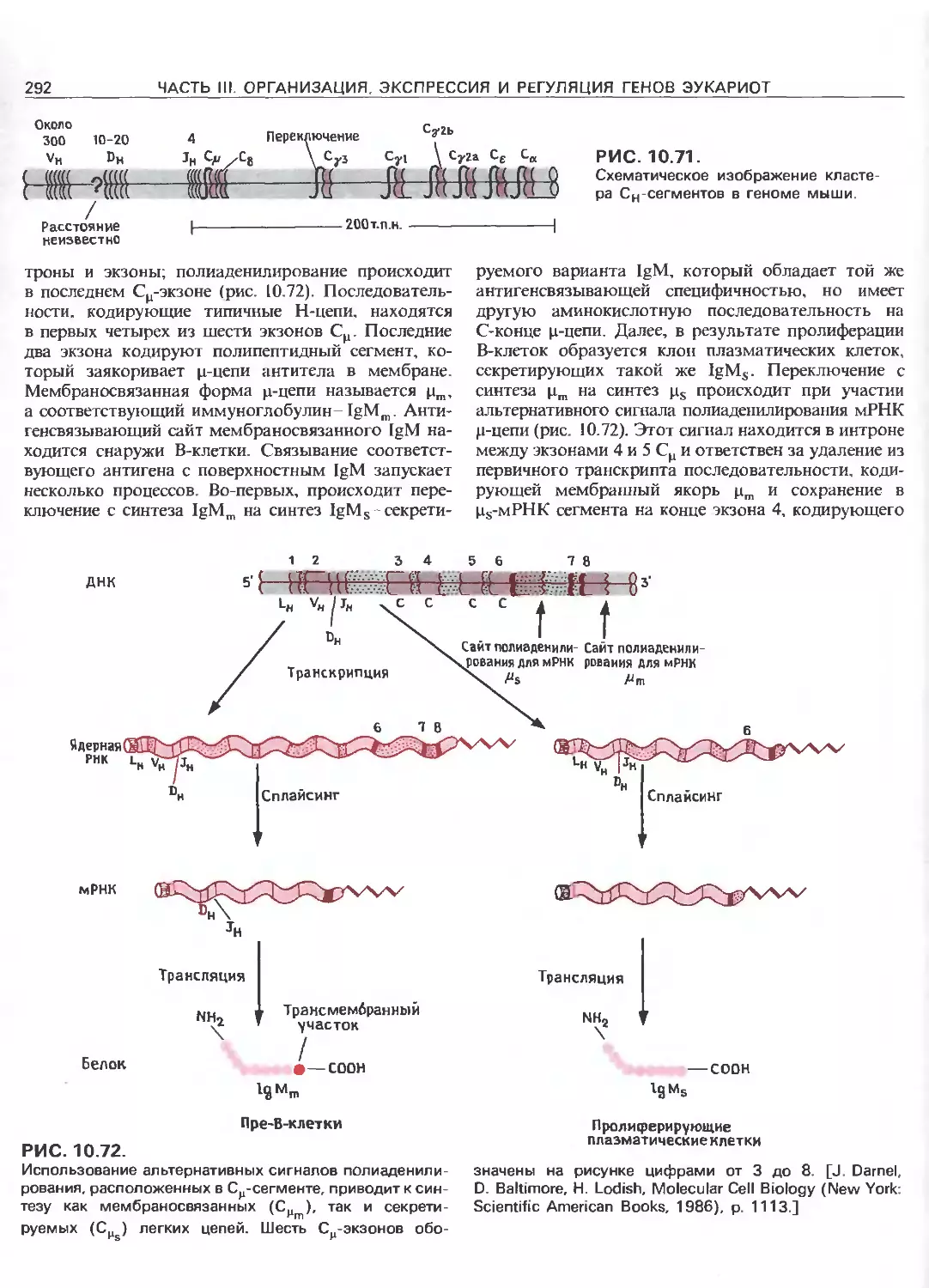
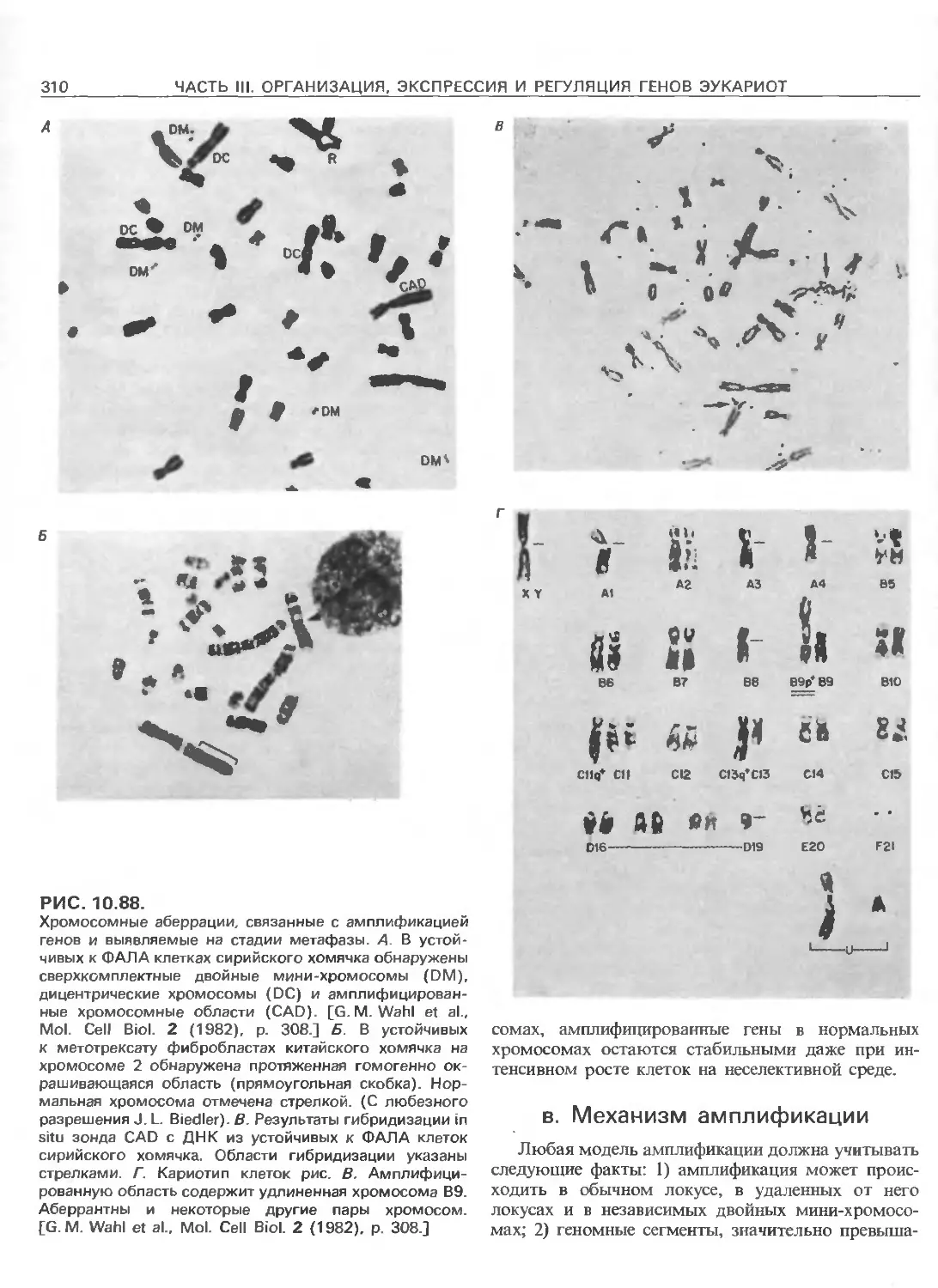
РИС. 111.3.
Схематическое изображение элементов, ответственных
за регуляцию инициации транскрипции, осуществляемой
с помощью РНК-полимеразы Е.соП (гл. 3) и трех эука-
риотических РНК-полимераз. РНК-полимераза I (poll)
транскрибирует гены рибосомных РНК. Ее работа за-
висит от двух элементов, один из которых находится
в окрестности точки инициации транскрипции (нукле-
отид + 1), а другой- на расстоянии от нескольких сотен
до нескольких тысяч пар нуклеотидов до транскриби-
руемого гена. РНК-полимераза II (polll) транскрибирует
гены, кодирующие полипептиды и некоторые РНК. Ее
работа зависит от нескольких сложно организованных
регуляторных элементов, одни из которых находятся на
расстоянии не далее 100 нуклеотидов от точки ини-
циации транскрипции, а другие-на расстоянии от 300
до 20000 п.н. слева от этой точки; в некоторых случаях
важные регуляторные элементы находятся также в са-
мом транскрибируемом гене или за ним (заметим, что
интроны и один иэ экзонов расположены так же, как на
рис. 111.1.). РНК-полимераза III (pollll) транскрибирует
гены, кодирующие транспортные РНК, 5S-pPHK и дру-
гие низкомолекулярные РНК.
ВВЕДЕНИЕ
9
ментов. К элементам первого типа относятся по-
следовательности, которые задают начало транс-
крипции; характерной особенностью промоторов Е.
coli является наличие двух участков ДНК, которые
расположены на расстоянии примерно 10 и 35 пар
оснований от точки начала транскрипции. Элемен-
ты второго типа задают конец гена или группы
генов и обусловливают терминацию транскрипции.
И наконец, существуют последовательности ДНК,
примыкающие к промотору, перекрывающиеся с
ним или располагающиеся сразу после него, кото-
рые узнаются специфическими эффекторами - ре-
прессорами, активаторами и антитерминаторами,-
регулирующими транскрипцию. Влияние всех этих
регуляторных последовательностей на экспрессию
соседствующих с ними кодирующих областей за-
висит от их взаимодействий с белками.
Последовательности ДНК, ответственные за ре-
гуляцию экспрессии эукариотических генов, нахо-
дятся в разных местах молекулы, на различных рас-
стояниях и по разные стороны от сайтов начала и кон-
ца транскрипции. Более того, три РНК-полимеразы,
I, II и III, транскрибируют три разных класса генов,
и для каждого класса характерны свои специфиче-
ские способы регуляции транскрипции и свои сигна-
лы терминации. Молекулярно-генетические и биохи-
мические исследования позволили установить меха-
низмы регуляции транскрипции и создали предпо-
сылки для выяснения природы дифференциальной
экспрессии генов. Итак, регуляторные последова-
тельности состоят из сложных наборов относитель-
но коротких последовательностей ДНК. С каждой
такой последовательностью связывается специфиче-
ский белок, фактор транскрипции. Например, гены,
которые транскрибируются только в лимфоидных
клетках, содержат набор последовательностей, узна-
ваемых факторами транскрипции, характерными
только для лимфоидных клеток. Аналогично этому
гены, экспрессируемые в строго определенное время
или при определенных условиях окружающей среды,
содержат регуляторные элементы, взаимодействую-
щие с соответствующими белками, которые присут-
ствуют в клетке или активируются именно в это
время или при данных условиях. Таким образом,
регуляция транскрипции каждого гена определяется
специфическим набором расположенных особым об-
разом элементов, наличием соответствующих фак-
торов транскрипции и белковых факторов, влияю-
щих на инициацию транскрипции. Связывание раз-
личных факторов транскрипции с регуляторными
последовательностями приводит либо к сборке
транскрипционного комплекса, включающего соот-
ветствующую РНК-полимеразу, либо к активации
такого комплекса, либо и к тому, и к другому.
Замечательной особенностью этих механизмов
является их универсальность. Удивительно, что со-
ответствующие факторы транскрипции из таких
разных источников, как дрожжи, Drosophila и мле-
копитающие, оказались взаимозаменяемыми. Так,
фактор транскрипции дрожжей и Drosophila может
взаимодействовать с регуляторными элементами
и с белками транскрипционного комплекса млеко-
питающих так же, как и соответствующий фактор
млекопитающих. Часто наблюдается и обратное.
Отсюда можно сделать вывод, что структура спе-
цифических молекул в ходе эволюции в основном
сохраняется.
Механизм терминации транскрипции уникален
для каждой из трех РНК-полимераз; каждый фер-
мент узнает определенную последовательность на
конце транскрипционной единицы или вблизи него.
Эти последовательности тоже взаимодействуют со
специфическими белками-факторами терминации.
Регуляция терминации зависит от расположения
указанных последовательностей ДНК и от образо-
вания мультибелковых комплексов, которые содей-
ствуют терминации транскрипции и модификации
конца молекулы РНК.
На экспрессию некоторых генов влияют эпигене-
тические факторы, которые изменяют способность
гена к экспрессии, не затрагивая базовой нуклеотид-
ной последовательности. Одна из установленных
форм эпигенетического изменения связана со сте-
пенью метилирования остатков цитозина в после-
довательностях 5'-CG, фланкирующих 5'-область
гена. Повышение степени метилирования в этом
участке коррелирует с уменьшением или полным
подавлением экспрессии соседнего гена, а пониже-
ние с повышением уровня экспрессии. На экспрес-
сию генов могут влиять и структурные изменения
в самом хроматине, которые до сих пор слабо
изучены; при этом высокий уровень экспрессии,
по-видимому, наблюдается только в «развернутых»
участках хроматина.
Все эти весьма неожиданные свойства эукарио-
тических генов послужили поводом к дискуссии
о том, что же такое единица наследственности-ген.
Ни одно из нескольких предложенных определений
нельзя признать полностью удовлетворительным
или пригодным для любого гена. Исходя из целей,
поставленных в этой книге, мы выбрали молекуляр-
ное определение гена (рис. III.4): эукариотический
ген- это совокупность сегментов ДНК, составляю-
щих экспрессируемую единицу, которая дает начало
одному или нескольким специфическим функцио-
нальным продуктам-молекулам РНК либо поли-
пептидам. Сегментами гена являются: 1) транскри-
бируемая область (единица транскрипции), которая
включает кодирующие последовательности, вста-
вочные последовательности, любые 5'-лидерные и
З'-трейлерные последовательности, находящиеся по
краям кодирующих последовательностей, и любые
10
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
+ Интроны
*« ?( fa Я 1?------------------------------
5-<рланки- -»----Единица транскрипции------^3-фланки-
рующий участок рующии участок
5-лидерная
последовательность
Транскрипция
3-трейлерная
последовательность
РНК-транскрипт q
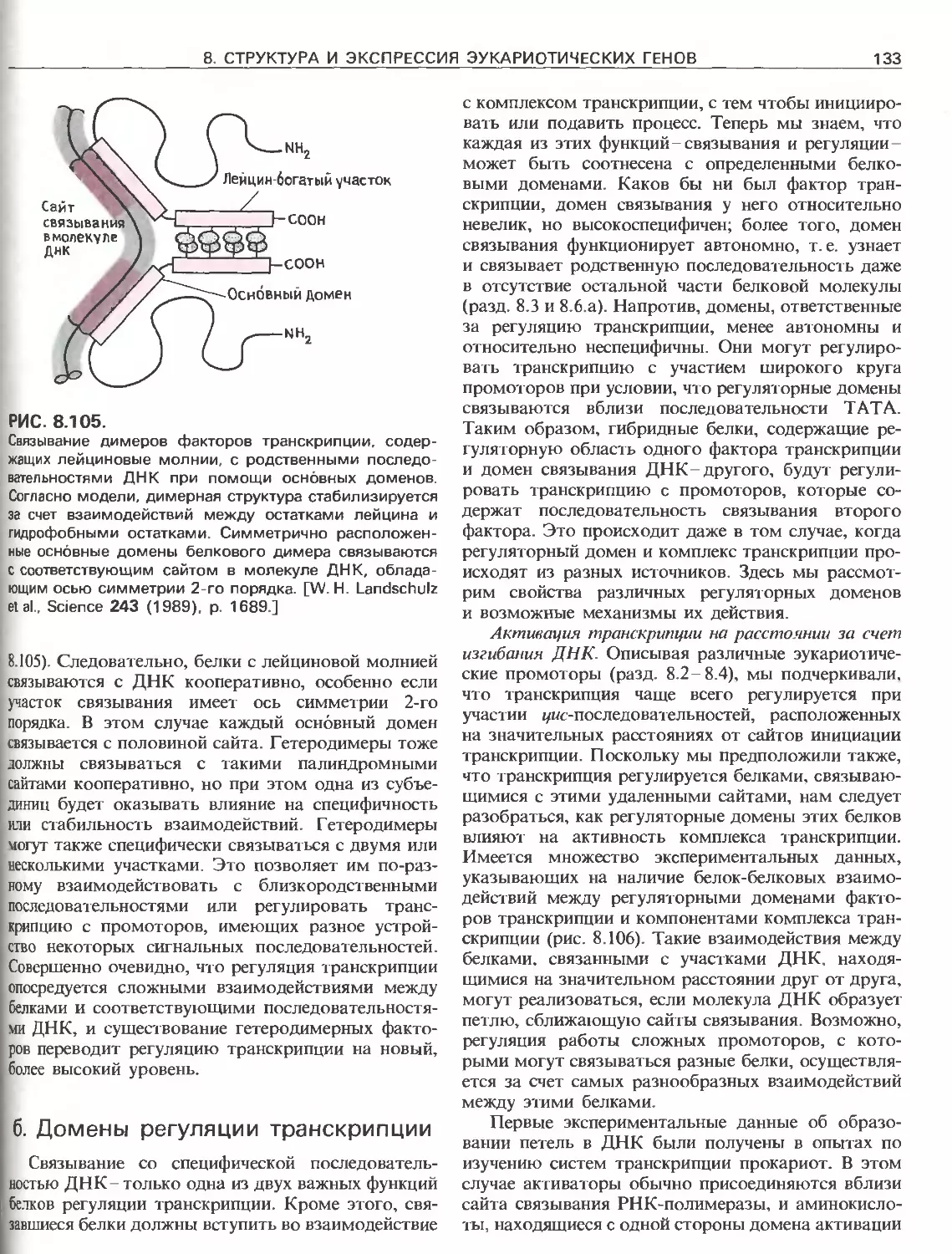
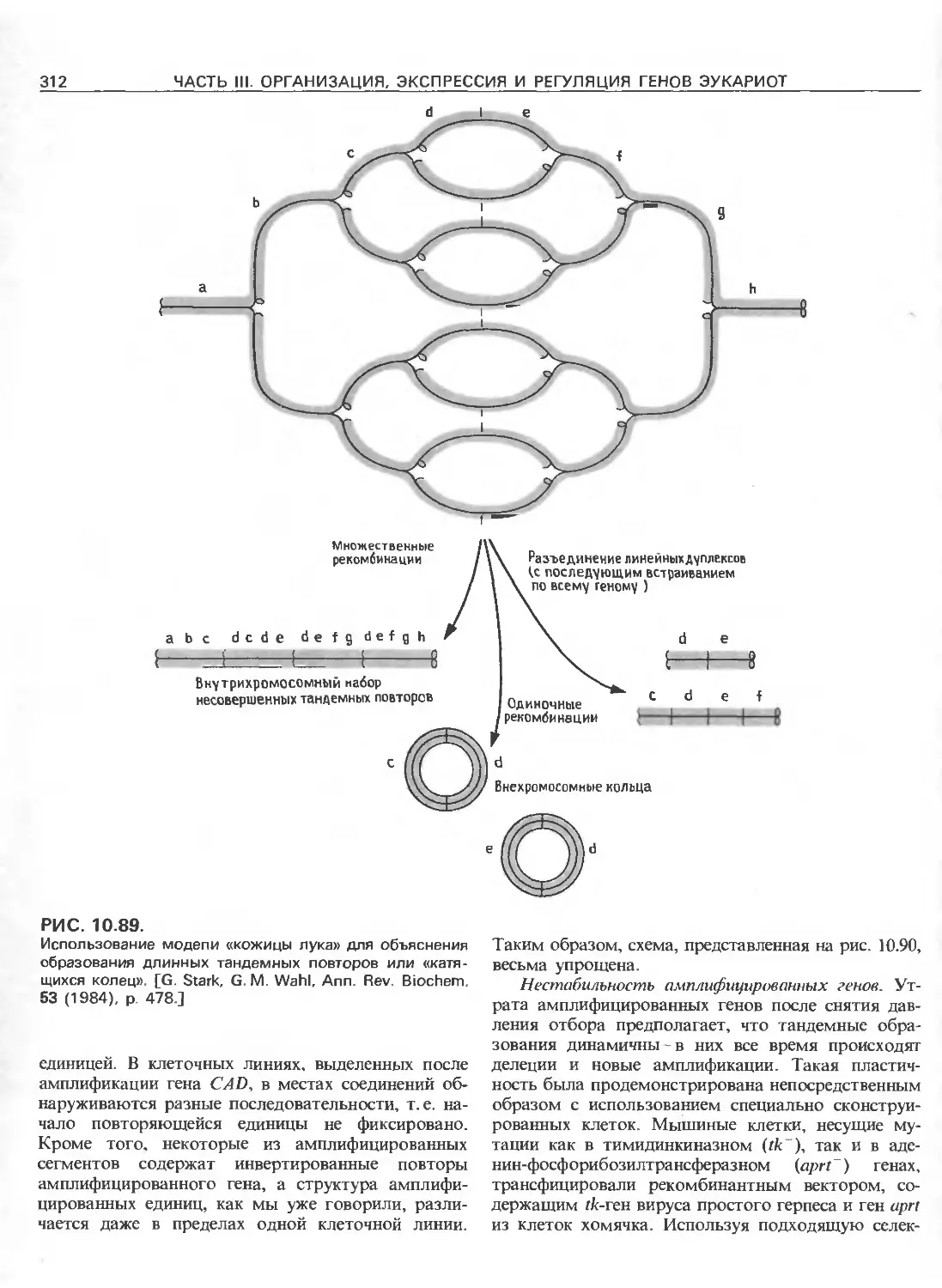
РИС. III.4.
Сегменты ДНК, составляющие ген: 1) единица тран-
скрипции, которая содержит 5'-лидерный и З'-трейлер-
ный сегменты, а также интроны; 2) регуляторные эле-
менты, которые могут находиться вне единицы тран-
скрипции или внутри нее. Гены принято изображать
слева направо в направлении транскрипции. Цепь ДНК.
имеющая ту же последовательность, что и матричная
РНК, называется значащей, и ее 5'-конец изображают
слева, а З'-конец- справа. Для удобства часто указывают
только одну эту цепь. Цепь ДНК, комплементарная
матричной РНК, называется незначащей или матричной.
Названия «5'- и З'-фланкирующие последовательности»
относятся к участкам, предшествующим транскрибиру-
емым областям или располагающимся сразу за ними
соответственно.
регуляторные элементы, содержащиеся в единице
транскрипции; 2) фланкирующие единицу транс-
крипции регуляторные последовательности, необхо-
димые для специфической экспрессии.
У прокариот основным способом регуляции экс-
прессии гена является, по-видимому, включение
выключение транскрипции, хотя в некоторых случаях
вступают в действие другие механизмы, например
аттенуация, терминация, антитерминация, контроль
трансляции и регуляция метаболизма матричных
РНК и белков. Подобно этому элементы, осуществ-
ляющие включение и выключение транскрипции,
играют решающую роль в регуляции экспрессии
генов также у эукариот. От сплайсинга, особенно от
его способа, зависит, какие именно образуются ма-
тричные РНК и соответственно кодируемые ими
белки. Кроме того, имеются данные о регуляции
при помощи аттенуации или терминации транскрип-
ции, а также указания на то, что трансляция и мета-
болизм РНК тоже подвержены регуляции в процессе
синтеза белков. Уникальные свойства эукариоти-
ческих клеток и структура их генов и геномов
обеспечивают дополнительные возможности для
контроля передачи генетической информации. На-
пример, полный транскрипт гена остается нефунк-
циональным до тех пор, пока не будут правильно
вырезаны входящие в него интроны. Далее транс-
крипты предшественники матричных РНК-
должны быть модифицированы на 5'- и З'-концах,
а сами матричные РНК должны перейти из ядра
в цитоплазму, прежде чем они смогут участвовать
в трансляции. В принципе регуляция может осу-
ществляться на каждом из этих этапов.
СТРОЕНИЕ ГЕНОМА ЭУКАРИОТ
Точно так же. как различается строение эукарио-
тических генов и генов прокариот, различается и ор-
ганизация геномов этих двух типов организмов. В
бактериальном геноме гены почти непрерывно сле-
дуют один за другим по всей длине молекулы ДНК,
а в некоторых случаях даже перекрываются. Гены,
кодирующие ферменты одного метаболического пу-
ти. или ферменты, активности которых так или
иначе связаны между собой, часто образуют одну
единицу транскрипции. Это способствует наиболее
эффективному использованию нуклеотидной после-
довательности ДНК. Но, по-видимому, для эволю-
ции эукариот принцип экономии был не столь ва-
жен, как для эволюции прокариот. Не говоря уже
о том, что большую часть ДНК занимают интроны,
эукариотические гены разделены длинными некоди-
руюшими участками. Как мы уже упоминали, боль-
шое число генов в одной единице транскрипции
встречается редко.
При обсуждении организации эукариотического
генома нельзя не отметить тот основополагающий
факт, что сама ДНК распределена по нескольким
хромосомам. Клеточные хромосомы, вероятно, все-
гда содержат линейную дуплексную ДНК, хотя
вдоль линейного остова встречаются двухцепоче-
чные петли. Каждая хромосома в интерфазе со-
держит одну двойную спираль ДНК, как и каждая
из двух сестринских хроматид метафазной хромо-
сомы. Геномы многих вирусов эукариот, а также
хромосомы хлоропластов и многих митохондрий
представлены кольцевой ДНК.
ВВЕДЕНИЕ
11
Хромосомные карты
В распределении генов по хромосомам не было
обнаружено очевидных закономерностей. Об этом
свидетельствовали еще данные по детальному кар-
тированию генов D. melanogaster генетическими и
цитогенетическими методами. Впрочем, эти карты
были получены лишь для небольшой части всей
ДНК, а генетические карты геномов млекопитаю-
щих были изучены еще меньше. К 1973 г. на ауто-
сомных хромосомах человека было локализовано
всего около 60 генов, а к 1981 г .-уже 400. В настоя-
щее время благодаря простоте получения фрагмен-
тов клонированной ДНК их число превысило 1000
(табл. III.3). Однако даже с построением весьма
протяженных карт никакой системы в расположении
генов выявлено не было. Родственные гены или
гены, представленные несколькими копиями, могут
находиться рядом, а могут быть разбросаны по
разным хромосомам. Например, гены, кодирующие
актин сердечной и скелетной мышц человека, на-
Таблица II 1.3. Локализация некоторых генов в
хромосомах человека
Гены Хромосома
Кластер генов а-глобинов 16
Кластер генов Р-глобинов 11
Гены иммуноглобулина
к (легкая цепь) 2
X (легкая цепь) 22
Тяжелая цепь 14
Псевдогены 9, 32, 15, 18
Гомологи вирусных онкогенов
c-sis 22
c-mos 8
c-mvb 6
c-IIa-ra.s-1 11
с-/ёл- 15
Ген S-аденозилмоноцистеингидролазы 20
Ген аденозиндезаминазы 20
Гены интерферонов
а-Кластср 9
Р-Кластер 9
Y 12
Кластер генов гормона роста 17
Ген пролактина 6
Гены главного комплекса гистосовмести- 6
мости (I. II, Ill)
Гены тяжелых цепей саркомерного мио- 17
зина
Ген тимидинкиназы 17
Ген галактокиназы 17
Ген инсулина 11
Ген паратиреоидного гормона 11
Ген фактора роста нервной ткани. 1
Р-субъединица
ф фа2 Фа1 а2 аЗ
[--------------------------------------8
Хромосома 16
а-Глобиновый локус
^/32 £ Gy Ay 8 0
Хромосома 11
B-Глобиновый локус
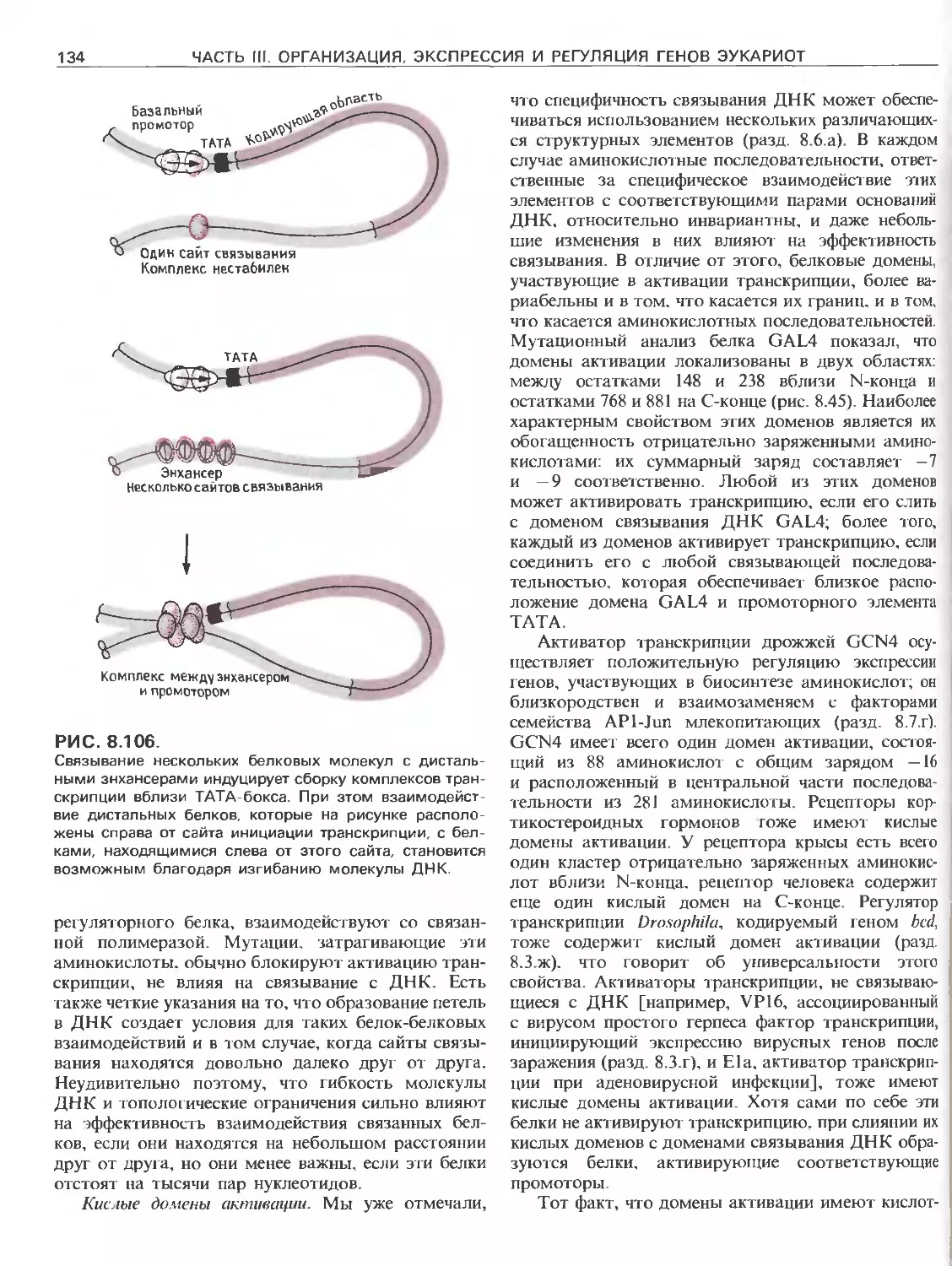
РИС. 111.5.
Кластеры генов а- и р-глобинов человека. В обоих
кластерах присутствуют псевдогены (у).
ходится в 15-й и 1-й хромосомах соответственно.
Далее, гены, кодирующие пять различных типов
Р-глобина человека, тандемно расположены в 11-й
хромосоме, а гены, кодирующие а-глобины, сгруп-
пированы в 16-й хромосоме. Все эти гены род-
ственны друг другу, и для образования глобинов,
входящих в состав гемоглобина, требуются поли-
пептиды обоих типов (рис. III.5).
Клонируемые сегменты часто содержат не толь-
ко определенный ген, но и протяженные участки
примыкающей к нему ДНК. Если использовать
в качестве зондов однокопийные фланкирующие
фрагменты ДНК, то из геномной библиотеки мож-
но выделить перекрывающиеся фрагменты. Это по-
зволяет охватить протяженные области хромосомы
и получить их детальные карты. В этих картах
содержится информация и о структуре генов, и о их
функции. Так, например, в виде перекрывающихся
фрагментов был представлен сегмент ДНК человека
размером 65 т. п. н., включающий пять генов Р-гло-
бина человека, что позволило определить его нук-
леотидную последовательность и локализовать ко-
дирующие, регуляторные и вставочные последова-
тельности.
Для анализа различий нуклеотидных последова-
тельностей аллелей чрезвычайно полезны карты
сайтов эндонуклеаз рестрикции. Например, исполь-
зуя в качестве зонда для гибридизации клонирован-
ный фрагмент из кластера Р-глобиновых генов
человека, при помощи эндонуклеазного расщепле-
ния и ДНК-блоттинга можно исследовать структу-
ру р-глобиновой области из разных источников
(рис. 111.6). Если размер фрагмента генома, который
гибридизуется с зондом, различен, это означает, что
различна и последовательность нуклеотидов в дан-
ном сайте рестрикции или между двумя сайтами
вследствие делеции либо инсерции; отсутствие гиб-
ридизующегося фрагмента означает, что делегиро-
вана целая область. Такие изменения, или поли-
12
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
ДНК с нормальным
р-глобиновым геном
D
CTGAG
GACTC
н-Зонд---—i
ДНК смутантным р-глобиновым
геном, ответственным за
серповидно-клеточную анемию
(_________CTGTG_________f
I GACAC С
i--Зонд----1
Эндонуклеаза Ddel
Электрофорез и блоттинг
Гибридизация с зондом
201 п.н.
1Т5п.н.
376 п.н.
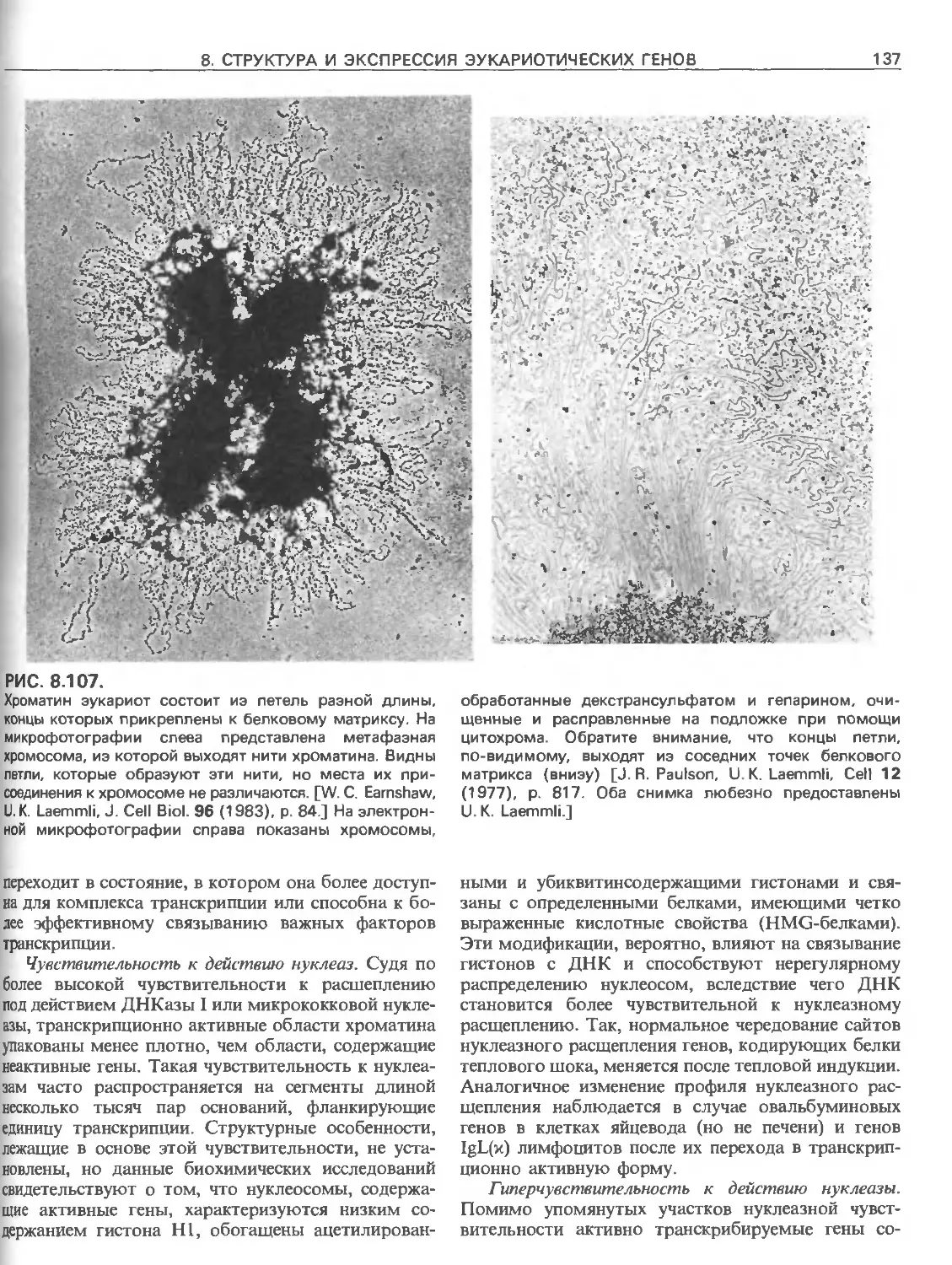
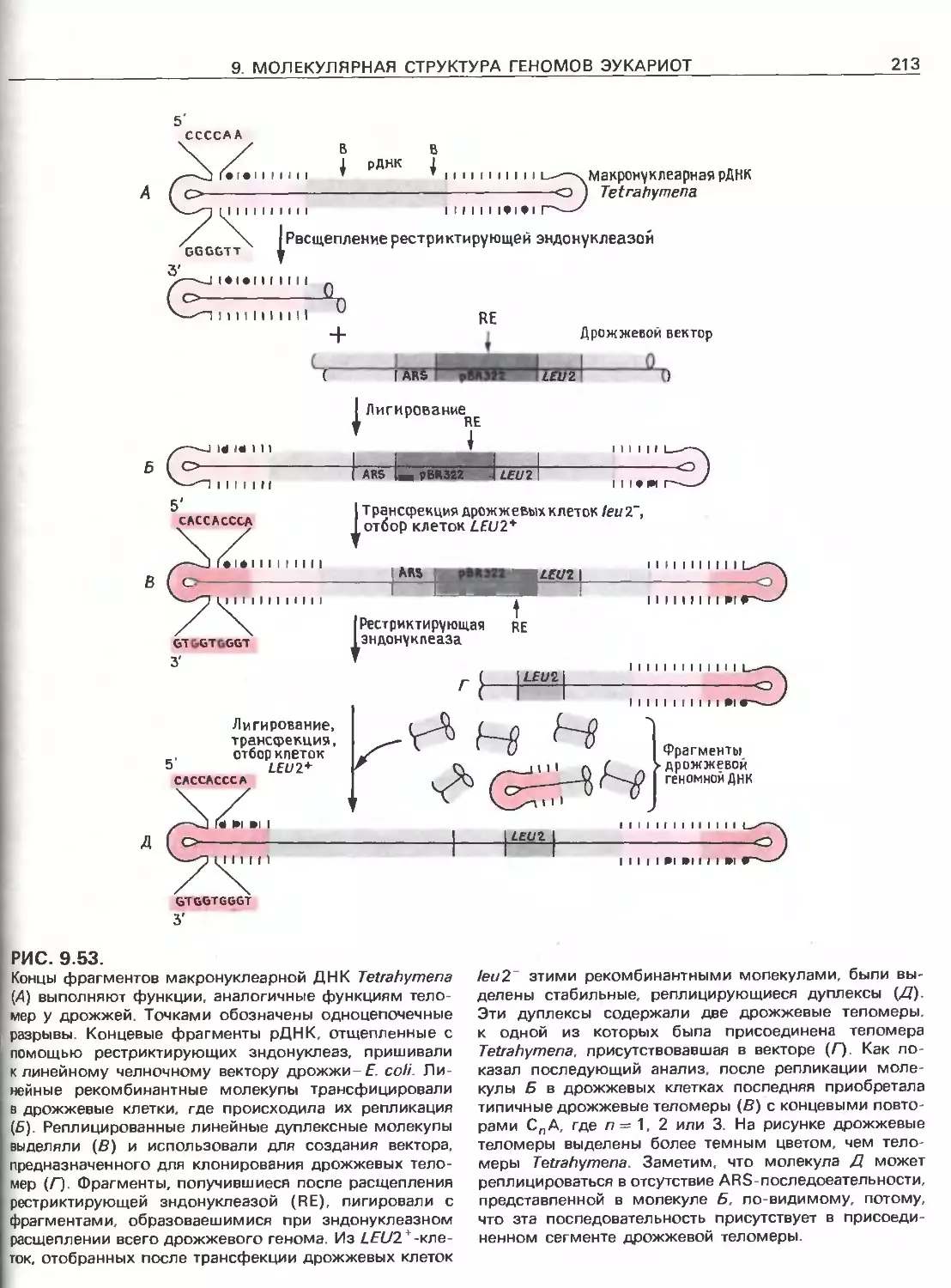
РИС. 111.6.
Идентификация мутации в р-глобиновом гене, ответст-
венной за серповидно-клеточную анемию, при помощи
зндонуклеазного расщепления Изображены нормаль-
ная и мутантная ДНК. Замена той пары оснований,
которая обусловливает данное заболевание, приводит
к разрушению сайта узнавания для рестриктазы Dde\.
После расщепления с помощью Odel, гель-злектрофо-
реза и ДНК-блоттинга с радиоактивным зондом, комп-
лементарным данному участку р-глобинового гена,
нормальная ДНК дает две полосы, а мутантная-одну.
морфизм длины рестрикционных фрагментов (ПДРФ),
даже если они не отражаются на фенотипе целого
организма, служат генетическими маркерами, не
менее полезными, чем классические маркеры, ис-
пользующиеся для генетического анализа больших
семей и популяций человека. Этот подход применял-
ся при изучении наследования гемоглобинопатий
и других генетических заболеваний. Более того,
наборы полиморфных фрагментов рестрикции из
разных частей данной хромосомы могут исполь-
зоваться в качестве маркеров для анализа обычного
генетического сцепления, что позволит конструи-
ровать генетические карты даже в отсутствие тради-
ционных фенотипических маркеров. Такой подход
применялся для установления порядка расположе-
ния нескольких генов на коротком плече 11-й хро-
мосомы человека: центромера каталаза-паратирео-
идный гормон- е-глобин- Р-глобин- инсулин пепси-
ноген.
Одно из удивительных открытий, сделанных в
ходе этих исследований,- очень высокая частота раз-
личий нуклеотидных последовательностей у разных
индивидов. Классический генетический анализ му-
таций по определению имел дело с изменениями
генома, приводящими к явным фенотипическим из-
менениям. Казалось разумным предположить, что
геномы нормальных особей (или особей «дикого
типа») очень мало отличаются друг от друга. Это
упрощенное представление часто подкреплялось
ссылками на доминантные и рецессивные аллели,
что подразумевало существование всего двух или
очень небольшого числа аллелей одного гена. В дей-
ствительности же полиморфизм длины рестрикци-
онных фрагментов- частое явление как в кодирую-
щих, так и в некодирующих областях, что указывает
на большое разнообразие геномов у различных
особей одного вида без заметных фенотипических
проявлений. Небольшие различия, обусловленные
полиморфизмом, не исключены, но большинство из
них скорее всего несущественны.
Размеры эукариотических геномов
Молекулярный анализ геномов эукариот объяс-
няет также тот кажущийся парадокс, что эукариоти-
ческие геномы содержат гораздо больше ДНК, чем
это представляется необходимым. По оценкам, сде-
ланным более 20 лет назад, если бы эукариоты
содержали 100000 генов, то при среднем размере
кодирующего участка гена 1200 п.н, геном должен
был бы содержать всего лишь ~ 12-107 п. н. Размер
генома некоторых грибов действительно не превы-
ВВЕДЕНИЕ
13
шает указанной величины, но в основном геномы
млекопитающих и растений содержат в 20-50 раз
больше ДНК (табл. III.1). Существование «избы-
точной» ДНК отчасти можно объяснить тем, что
в геноме имеются вставочные последовательности
(табл. III.2) и протяженные участки ДНК между
генами.
Большой размер генома эукариот может объяс-
няться и тем, что некоторые гены встречаются
в геноме много раз. Существование большого числа
копий отдельных генов свойственно не только эука-
риотам: так, Е. coli содержит примерно семь
генов рибосомной РНК, что можно объяснить необ-
ходимостью быстрого синтеза большого числа ри-
босом. Той же цели может служить и наличие
множественных копий одного гена у эукариот; на-
пример, у млекопитающих имеется несколько сотен
генов рибосомной ДНК. Свой вклад в «избыточ-
ную» ДНК вносят также нефункциональные копии
генов. Эти дополнительные копии называются псев-
догенами и обычно не способны к передаче полезной
информации из-за делеций или других изменений
в их последовательности. Число псевдогенов кон-
кретного гена очень сильно варьирует от особи
к особи. Таким образом, в геноме эукариот ген
может быть представлен в единственном числе или
являться частью множества повторяющихся после-
довательностей, собранных вместе или разобщен-
ных. Повторяющиеся последовательности могут со-
держать либо несколько близкородственных функ-
циональных генов, которые, однако, экспрессируют-
ся на разных стадиях развития или в разных тканях
(клетках), либо псевдогены, либо и то и другое
(рис. III.5).
Большой размер эукариотических геномов обус-
ловлен также наличием в них множества повторяю-
щихся последовательностей ДНК с неизвестными
функциями. На их долю приходится обычно 10%,
а в некоторых случаях почти 50% генома. Более
15 лет назад при помощи кинетического анализа
реассоциации денатурированной эукариотической
ДНК было показано, что значительная часть ДНК
ренатурирует гораздо быстрее, чем можно было бы
ожидать для уникальных последовательностей ДНК.
Высокая скорость реассоциации говорит о том, что
такие геномы содержат фрагменты, повторяющиеся
от сотен тысяч до миллионов раз. В настоящее
время с помощью молекулярного клонирования
и секвенирования подтверждено существование та-
ких последовательностей ДНК с большим числом
повторов, которые, подобно повторяющимся генам,
могут располагаться либо тандемно, либо изоли-
рованно друг от друга среди неродственных геном-
ных локусов.
Связь между специфическими
последовательностями ДНК и
морфологией хромосомы
Как показал молекулярный анализ, определен-
ные типы последовательностей ДНК коррелируют
с некоторыми отличительными морфологическими
признаками хромосом. Участки хромосом, назы-
ваемые ядрышковыми организаторами, несут гены
рибосомной РНК. Области теломер и центромер
большей части эукариотических хромосом содержат
тандемно повторяющиеся нуклеотидные последова-
тельности. Хотя на долю этих повторяющихся по-
следовательностей часто приходится 10 и более
процентов всей геномной ДНК, их функция до
конца не выяснена. Возможно, что такие после-
довательности несущественны для функции центро-
меры, поскольку они отсутствуют в хромосомах
дрожжей (Saccharomyces cerevisiae), где центромеры
нормально функционируют и в митозе, и в мейозе.
ПЕРЕСТРОЙКИ
ПОСЛЕДОВАТЕЛЬНОСТЕЙ ДНК
Расположение генов и большей части других
последовательностей в хромосоме остается практи-
чески постоянным, и это создает основу для по-
строения хромосомных карт. Генетическая инфор-
мация может перераспределяться между гомоло-
гичными хромосомами при реципрокном кроссин-
говере в мейозе и в митозе. Однако, несмотря на то
что при этом возникают новые комбинации алле-
лей, порядок расположения последовательностей
ДНК не меняется. И в самом деле, многие связи
между генами значительно старше, чем отдельные
виды, и гены, являющиеся синтенными (т. е. нахо-
дящимися на одной хромосоме) у одних млекопи-
тающих, часто остаются синтенными даже у млеко-
питающих разных отрядов. Тем не менее стабиль-
ность, лежащая в основе организации генома, соче-
тается со способностью к перераспределению гене-
тической информации. Перестройки могут происхо-
дить случайно, часто приводя к вредным мутациям.
Но некоторые перестройки - нормальный процесс,
изменяющий экспрессию гена упорядоченным и
запрограммированным способом.
Хромосомные аберрации, например разрыв хро-
мосомы, транслокация частей одной хромосомы
в другую, негомологичную хромосому или образо-
вание дицентрических и ацентрических хромосом,
известны давно. На молекулярном уровне все эти
морфологические изменения проявляются в изме-
нении строения ДНК. Генетическую информацию
о ферментативном аппарате, ответственном за пе-
рестройки генов, содержат все организмы (разд.
14
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
2.4). Процессы перестройки включают в себя: 1)
нереципрокный и неаллельный кроссинговер и кон-
версию генов; 2) транспозицию фрагментов ДНК из
одного локуса генома в другой; 3) образование
дополнительных копий последовательностей ДНК,
которые располагаются либо тандемно, либо в раз-
ных участках генома; этот процесс называется амп-
лификацией; 4) делецию участков ДНК; 5) негомо-
логичную рекомбинацию. Изменчивость организа-
ции генома-одно из важнейших открытий моле-
кулярной генетики, особенно ценное ввиду того, что
перестройки последовательностей ДНК относятся
к очень редким событиям. В значительной мере это
открытие стало возможным благодаря точности
методов молекулярной генетики в сочетании с вы-
сокой избирательностью методов классического ге-
нетического анализа.
Случайные перестройки генома
Непрограммируемые перестройки обычно про-
исходят в случайный момент времени и/или в слу-
чайном месте и часто приводят к вредным мута-
циям. Однако они могут также вызывать и, воз-
можно, вызывали в прошлом мутации, позволяю-
щие организму по-новому реагировать на условия
окружающей среды, способствуя таким образом
эволюционным изменениям. Конечно, для того что-
бы случайные перестройки имели эволюционное
значение, они должны происходить в линиях заро-
дышевых клеток или их предшественников; подоб-
ные процессы, протекающие в отдельной сомати-
ческой клетке, могут не иметь никаких последствий
для целого организма. Если же соматическая клетка
является стволовой, то случайные перестройки мо-
гут воспрепятствовать появлению функционально
активного дифференцированного потомства; кроме
того, изменение может быть онкогенным, т. е. при-
водить к образованию опухоли.
Особенно сильно поражает воображение один из
видов случайных геномных перестроек-перемеще-
ние фрагментов ДНК из одного места генома в дру-
гое. О «прыгающих генах» много говорилось даже
в газетах и журналах. Предположение о существо-
вании мобильных элементов генома было выска-
зано в середине 1940-х гг. исходя из результатов
опытов по генетике и цитогенетике кукурузы. Спу-
стя 20 лет ученые, занимающиеся исследованием
бактерий, получили экспериментальные данные,
подтверждающие это предположение: определенные
фрагменты ДНК действительно перемещаются. В
конце концов было установлено, что как в эука-
риотических, так и в прокариотических геномах
существует много типов мобильных элементов. С
помощью молекулярного клонирования удалось
определить структуру многих мобильных элементов
и попытаться расшифровать реальные механизмы
их перемещения из одного места генома в другое.
Мобильные элементы-это генетические «наруши-
тели спокойствия», вызывающие разнообразные
мутации у про- и эукариот. Если они внедряются
в кодирующую последовательность гена, то ген
утрачивает свою функцию. Если перемещенный
элемент окажется рядом с геном, то экспрессия
этого гена может изменяться неоднозначным обра-
зом. Многие мутации, весьма полезные для изуче-
ния генетики D. melanogaster в начале века, на самом
деле были связаны с внедрением мобильных эле-
ментов в гены.
Программируемые перестройки
Известно, что экспрессия генов может сущест-
венным образом зависеть от тех изменений, кото-
рые происходят во фланкирующих участках. В со-
I
Ч Jk С
(Транскрипция
S—S S—S
N Н2 <Х2Оо6ООООО©@0©0@@-соон
V; Jk с
РИС. III.7.
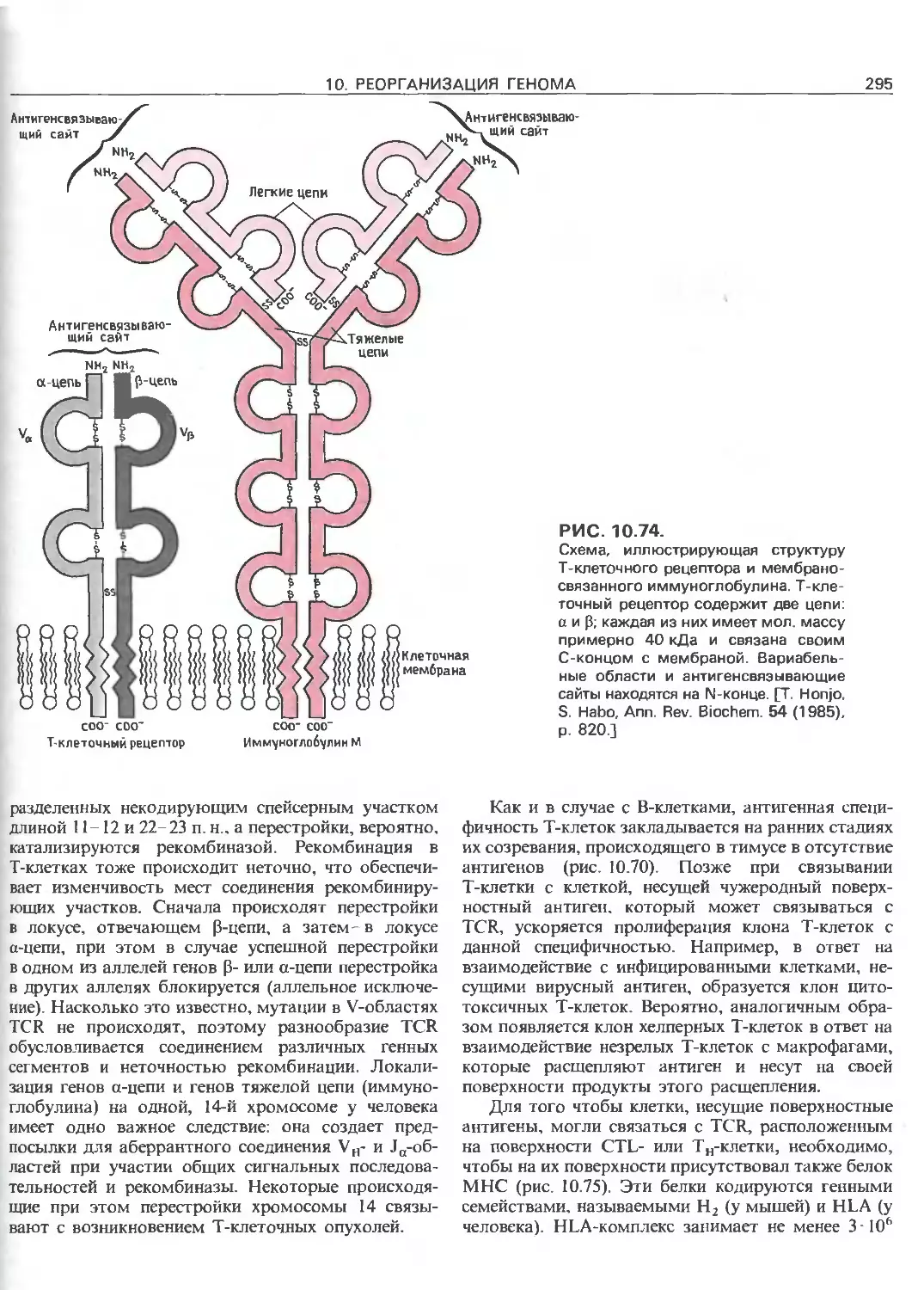
Перестройки, приводящие к образованию генов, коди-
рующих различные антитела. Эти перестройки осу-
ществляются в В-лимфоцитах путем соединения сег-
ментов ДНК, далеко отстоящих друг от друга в ДНК
клеток зародышевой линии и различных соматических
клеток. В качестве примера рассмотрено образование
гена легкой цепи иммуноглобулина из трех сегментов
ДНК (V, J и С), которые вместе кодируют данный
полипептид. Каждый из множества сегментов V и
J генома соединяется с единственным сегментом С.
Гены иммуноглобулинов отличаются друг от друга сег-
ментами V и J. В данном примере с С соединены V; и Jk.
Разнообразие белковых антител отчасти обусловливает-
ся различием аминокислот, кодируемых разными Vs и Js
(вариабельная область).
ВВЕДЕНИЕ
15
седстве с одними последовательностями ген может
«молчать», а при изменениях в них-экспрессиро-
ваться. Такие перестройки используются организ-
мами для регуляции экспрессии генов, что создает
необходимые условия для выполнения дифференци-
рованными клетками специфических функций.
Таким образом, существуют генетические програм-
мы. которые вызывают перестройки в особых сег-
ментах ДНК в определенное время и в определен-
ных клетках.
Задолго до того, как появилась возможность
исследовать гены и геномы на молекулярном уров-
не, генетики высказывали предположение, что неко-
торые загадочные биологические явления можно
объяснить специфическими перестройками генов.
Одно из таких явлений - наличие разных типов жгу-
тиков у разных особей Salmonella typhimurium. Дру-
гое, еще более замечательнее явление- необычайное
разнообразие антител, которые могут вырабаты-
ваться у млекопитающих. Как показал молекуляр-
ный анализ, в основе всех этих явлений лежат
программируемые перестройки последовательностей
ДНК (пример с иммуноглобулином схематически
представлен на рис. III.7). В обоих случаях реорга-
низация генома предпринимается для защиты орга-
низма от опасности. Меняя жгутики. Salmonella от-
ражает иммунологическую «атаку» организма хо-
зяина. Создавая новые антитела, организм защи-
щается от действия чужеродных агентов.
ЭВОЛЮЦИОННЫЕ АСПЕКТЫ
История эволюции организмов записана в их
геномах. Мутации и перестройки обусловливают
Приматы
Пшеница
Дрожжи
Приматы
Пшеница
Дрожжи
Приматы
Пшеница
Дрожжи
Приматы
Пшеница
Дрожжи
Приматы
Пшеница
Дрожжи
РИС. III.8.
Аминокислотные последовательности цитохромов с у
дрожжей, пшеницы и приматов сходны (эти последо-
вательности следует читать в каждой строке слева на-
NHj- gly asp val glu lys gly
pro pro gly asn pro asp ala gly
lys ala gly ser ala lys lys gly
glu lys gly gly lys his lys thr
asp ala gly ala gly his lys gin
glu lys gly gly pro his lys val
pro gly tyr ser tyr thr ala ala
ala gly tyr ser tyr ser ala ala
glu gly tyr ser tyr thr asp ala
tyr leu glu asn pro lys lys tyr
tyr leu leu asn pro lys lys tyr
tyr leu thr asn pro lys lys tyr
glu glu arg ala asp leu ile ala
gin asp arg ala asp leu ile ala
lys asp arg asn asp leu ile thr
NH2- ala ser phe ser glu ala
NH2- thr glu phe
cys ser gly cys his thr val
cys ala gly cys his thr val
cys leu gly cys his thr val
gly arg lys thr gly gin ala
glV ar9 0ln ser gly thr thr
gly arg his ser gly gin ala
gly glu asp thr leu met glu
glu glu asn thr leu tyr asp
asp glu asn asn met ser glu
phe val gly lie lys lys lys
phe pro gly leu lys lys pro
Phe gly gly leu lys lys glu
генетические изменения, лежащие в основе эволю-
ционного процесса. В прошлом эволюционные свя-
зи можно было изучать только по фенотипическим
признакам организма, которые были связаны с ге-
нетической информацией, записанной в его геноме.
Так, вначале изучение эволюционных связей осно-
вывалось на анатомическом исследовании ископае-
мых остатков, а также на сравнительной анатомии,
физиологии и эмбриологии современных видов. В
середине нашего столетия появился новый метод
изучения эволюционных взаимосвязей на уровне
фенотипа-сравнительная химия белков. Как ока-
залось, строение некоторых белков у многх орга-
низмов в высшей степени консервативно. Это по-
могло выявить многие недостающие эволюционные
связи, что было невозможно при изучении ископае-
мых остатков. Например, цитохром с содержится
в митохондриях и животных, и растений. Он играет
ключевую роль в процессах дыхания и поэтому
должен был появиться в числе первых среди белков.
В ходе эволюции цитохром с изменился очень мало
(рис. III.8); так, у цитохромов с дрожжей и человека
различаются только 35 из 104 аминокислотных ос-
татков (не считая дополнительных аминокислот на
N-конце молекулы цитохрома с дрожжей).
Для изучения эволюционных связей между близ-
кородственными видами цитохром с и другие высо-
коконсервативные белки не очень полезны, посколь-
ку их структура у таких видов может различаться
слишком мало. Однако некоторые белки эволю-
ционировали более быстрыми темпами. Один из
таких белков-карбоангидраза; ее строение у при-
матов различается достаточно сильно для построе-
ния филогенетического древа. Метод построения
lys lys ile phe ile met lys
ala lys ile phe lys thr lys
ala thr leu phe lys thr arg
gly pro asn leu his gly leu phe
gly pro asn leu his gly leu phe
gly pro asn leu his gly ile phe
asn lys asn lys gly ile ile trp
asn lys asn lys ala val glu trp
asn ile lys lys asn val leu trp
ile pro gly thr lys met ile
ile pro gly thr lys met val
ile pro gly thr lys met ala
tyr leu lys lys ala thr asn glu-<
tyr |eu lys lys ala thr ser ser-(
tyr leu lys lys ala cys glu-COC
право). Одинаковые аминокислоты у всех трех видов
выделены цветом. Вероятность того, что подобное
сходство является случайным, очень мала.
16
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
основан на наблюдении, что число различающихся
аминокислот у этих белков примерно пропорцио-
нально времени, прошедшему с момента диверген-
ции двух организмов от общего предшественника,
определенному исходя из палеонтологических дан-
ных.
В заключение необходимо отметить, что все
методы эволюционного анализа, основанные на изу-
чении фенотипа, по существу относятся к непрямым
методам: ведь в основе появления новых феноти-
пических признаков в ходе эволюции лежат изме-
нения именно генов и геномов. И хотя изучение
древних видов продолжает основываться на иссле-
довании морфологических особенностей ископае-
мых остатков, для установления связей между со-
временными видами чаще всего сравнивают строе-
ние ДНК. Таким образом, технология рекомби-
нантной ДНК уже внесла большой вклад в развитие
эволюционной биологии и, по-видимому, будет
применяться в этой области все более широко.
Сравнительная
молекулярная генетика
Ценность сравнения нуклеотидных последова-
тельностей ДНК для изучения эволюционных связей
стала очевидной еще до появления методов клони-
рования и секвенирования фрагментов ДНК, и были
разработаны методы оценки степени родства раз-
ных геномов. К числу этих методов относится срав-
нение стабильности дуплексной ДНК, состоящей из
цепей ДНК двух разных видов (гетеродуплекс), со
стабильностью двух цепочечной ДНК, в которой обе
цепи принадлежат одному виду (гомодуплекс). На
рис. Ш.9 представлены результаты таких опытов
с ДНК из нескольких видов Drosophila. Подобные
данные дают представление о средней дивергенции
между двумя ДНК. Они менее информативны, чем
данные сравнительной анатомии и белковой химии.
Однако прямое сравнение нуклеотидных последова-
тельностей основного объекта эволюции, ДНК, по-
зволяет гораздо глубже проникнуть в тайны эволю-
ционного процесса, а кроме того, освещает те его
стороны, которые прежде не поддавались изучению.
Более того, технически гораздо проще клонировать
и секвенировать группу гомологичных генов, чем
выделять их белковые продукты и определять их
аминокислотную последовательность.
Анализ нуклеотидных последовательностей ко-
дирующих участков ДНК более информативен, чем
определение аминокислотной последовательности
полипептидов, поскольку из-за вырожденности ге-
нетического кода изменение нуклеотидной последо-
вательности гена может не сопровождаться измене-
ниями аминокислотной последовательности соот-
ветствующего белка. И даже если известно, что
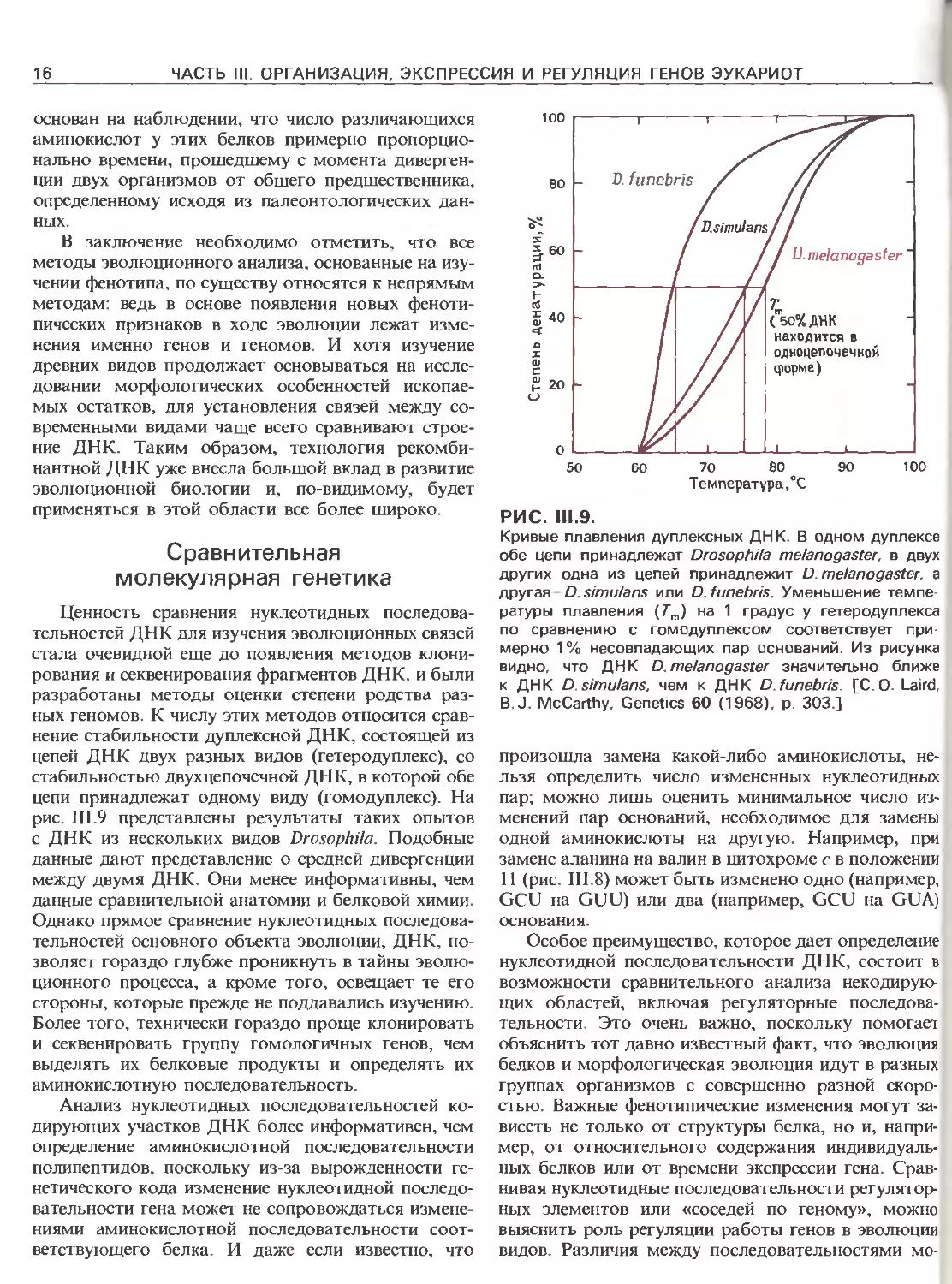
РИС. 111.9.
Кривые плавления дуплексных ДНК. В одном дуплексе
обе цепи принадлежат Drosophila melanogaster, в двух
других одна из цепей принадлежит D. melanogaster, а
другая D.simulans или D.funebris. Уменьшение темпе-
ратуры плавления (Гт) на 1 градус у гетеродуплекса
по сравнению с гомодуплексом соответствует при-
мерно 1 % несовпадающих пар оснований. Из рисунка
видно, что ДНК D. melanogaster значительно ближе
к ДНК D.simulans, чем к ДНК D.funebris. [С. О. Laird,
B.J. McCarthy, Genetics 60 (1968), р. 303.]
произошла замена какой-либо аминокислоты, не-
льзя определить число измененных нуклеотидных
пар; можно лишь оценить минимальное число из-
менений пар оснований, необходимое для замены
одной аминокислоты на другую. Например, при
замене аланина на валин в цитохроме с в положении
11 (рис. III.8) может быть изменено одно (например,
GCU на GUU) или два (например, GCU на GUA)
основания.
Особое преимущество, которое дает определение
нуклеотидной последовательности ДНК, состоит в
возможности сравнительного анализа некодирую-
щих областей, включая регуляторные последова-
тельности. Это очень важно, поскольку помогает
объяснить тот давно известный факт, что эволюция
белков и морфологическая эволюция идут в разных
группах организмов с совершенно разной скоро-
стью. Важные фенотипические изменения могут за-
висеть не только от структуры белка, но и, напри-
мер, от относительного содержания индивидуаль-
ных белков или от времени экспрессии гена. Срав-
нивая нуклеотидные последовательности регулятор-
ных элементов или «соседей по геному», можно
выяснить роль регуляции работы генов в эволюции
видов. Различия между последовательностями мо-
ВВЕДЕНИЕ
17
гут касаться одной-единственной пары оснований,
но могут быть обусловлены также геномными пе-
рестройками и транспозицией мобильных элемен-
тов.
Особенно полезным для изучения эволюционных
взаимосвязей оказалось определение нуклеотидных
последовательностей рибосомных РНК. в частности
16S- 18S-PHK малых рибосомных субъединиц. По-
скольку рибосомы в большом количестве присут-
ствуют во всех организмах, можно выделить функ-
циональные РНК и секвенировать их после синтеза
ДНК-копий с помощью обратной транскриптазы.
Хотя функции РНК малой субъединицы у всех
живых организмов одинаковы, степень консерва-
тивности разных ее сегментов неодинакова. На-
иболее консервативные участки можно использо-
вать для сравнения организмов с отдаленным род-
ством, а быстроэволюционирующие сегменты-
близкородственных видов. Таким образом, сравни-
тельный анализ всех видов можно проводить, ис-
пользуя всего один тип молекул с высококонсерва-
тивной функцией. Так были исследованы представи-
тели практически всех таксономических групп, что
позволило получить детальную картину филогене-
тических взаимоотношений и по-новому взглянуть
на процесс эволюции.
Эволюционная история генов
До сих пор, говоря об эволюции, мы имели
в виду эволюцию различных видов. Но в молекуле
ДНК записана и другая история - история проис-
хождения генов как таковых. Во многих случаях два
(или более) родственных гена (или последователь-
ности ДНК) в пределах отдельных видов образуют-
ся из одной последовательности-предшественника.
Предположим, что сегмент ДНК был амплифици-
рован (в результате случайных событий), а для
выполнения полезной функции достаточно одной
его копии. В этом случае дополнительные копии
будут свободны от тех ограничений, которые нала-
гаются на число мутаций в функциональном гене.
Мутации в дополнительных копиях могут приводить
к появлению новых функций, которые в свою оче-
редь могут сохраниться при отборе. С другой сто-
роны, эти копии могут попасть под контроль новых
или измененных регуляторных сигналов и экспрес-
сироваться на разных этапах развития или в особым
образом дифференцированных тканях. Кроме того,
дополнительные копии, в которых произошли вред-
ные мутации, могут сохраниться в качестве псевдо-
генов. Все эти три возможности иллюстрирует се-
мейство глобиновых генов (рис. III.5).
Некоторые гены образуются в результате соеди-
нения копий кодирующих последовательностей (т. е.
зкзонов). Иногда при этом происходит тандемная
амплификация одного из экзонов, что приводит
к множественным повторам соответствующих пе-
птидных фрагментов в кодируемом белке (рис.
III. 10). Гены могут иметь мозаичное строение, т. е.
состоять из перемешанных копий отдельных экзо-
нов различных генов (рис. Ш.П). Так, экзоны, про-
исходящие от общего предшественника, иногда
встречаются в составе явно неродственных генов.
Одинаковые экзоны, входящие в кодирующие по-
следовательности разных белков, по-видимому, ко-
дируют полипептидные домены, которые обуслов-
ливают одинаковое строение или одинаковые функ-
циональные свойства этих белков, например особый
тип вторичной структуры или способность связы-
ваться со специфическими лигандами (скажем, с
АТР).
Высказывалась интересная гипотеза о том, что
соединение экзонов между собой при их амплифи-
кации и перемешивании происходит вследствие ре-
комбинаций в интронах, благодаря чему кодирую-
щие участки генов остаются интактными. Эта гипо-
теза предполагает, что интроны являются древними
структурами, которые присутствовали в самых ран-
них генах и клетках и были утрачены в ходе эволю-
ции прокариот. Согласно другой гипотезе, интроны,
напротив, представляют собой вставки (инсерции)
в ранее существовавшие кодирующие области при
помощи механизма, более присущего эукариотам,
чем прокариотам. Правда, с инсерционной моделью
связаны некоторые проблемы, не возникающие в
том случае, если принимается гипотеза о древнем
происхождении интронов. В частности, встает во-
прос о том, каковы последствия мутаций, вызван-
ных широко распространенными случайными встав-
ками некодирующих сегментов ДНК, и вопрос о не-
обходимости одновременной эволюции механизма
сплайсинга. С другой стороны, если интроны при-
сутствовали в самых ранних геномах, то и сплай-
синг тоже должен быть очень древним процессом.
Данные о том, что некоторые интроны катализи-
________Экзон Интрон
| Амплификация
| Амплификация
—-в
РИС. 111.10.
Некоторые гены зволюционировапи путем тандемной
амплификации одного-единственного зкзона. В качест-
ве примера можно привести гены коллагена и а-фето-
протеина млекопитающих (разд. 9.4.а).
2-I58
Глава 8
СТРУКТУРА И РЕГУЛИРУЕМАЯ
ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
Успехи молекулярной биологии в пятидесятых-
шестидесятых годах породили у большинства уче-
ных уверенность в том, что прокариотические и
эукариотические гены имеют по существу одинако-
вое строение и одинаковый механизм экспрессии.
Было уже ясно, что оба типа организмов имеют
одинаковый генетический код и используют РНК-
полимеразу для транскрипции, а мРНК, рибосомы
и тРНК-для трансляции своих генетических сооб-
щений. Но даже скудная информация о работе
эукариотических генов, накопленная к 70-м годам,
указывала на то, что эукариоты используют более
сложные, а возможно, и совершенно иные механиз-
мы экспрессии и регуляции. Например, было пока-
зано, что эукариотические мРНК отличаются от
прокариотических необычными модификациями
5'-концов: вместо трифосфатной группы, связанной
с первым нуклеотидом на концах цепей, они содер-
жат так называемые кэп-структуры. Более того,
эукариотические мРНК имеют длинные полиаде-
нилатные последовательности на З'-концах. Кроме
данных об этих структурных особенностях имелись
указания и на коренные различия механизма биоге-
неза РНК у организмов двух типов. Например, для
синтеза различных типов РНК эукариотические
клетки используют три разных вида РНК-полиме-
разы, а прокариотические - только один. Более того,
лишь 5% РНК, синтезированной в ядре, переходит
в цитоплазму. Особенно удивительным казалось су-
ществование очень длинных молекул ядерных РНК,
содержащих те же самые нуклеотидные последова-
тельности и имеющих такие же модификации 5'-
и З'-концов, что и меньшие по размеру цитоплазма-
тические мРНК.
Предположения, возникшие в связи с этими не-
ожиданными открытиями, было трудно оценить, не
имея надежной информации о строении генов и не
располагая методами изучения их экспрессии. Но
с разработкой методов молекулярного клонирова-
ния и изучением на их основе структуры и функции
многих эукариотических генов удалось выявить но-
вые неожиданные особенности строения эукарио-
тических генов и их регуляции. В данной главе мы
расскажем об этих открытиях и их значении для
понимания сложных свойств эукариотических орга-
низмов.
Основную информацию о строении гена полу-
чают при помощи молекулярного клонирования
и секвенирования ДНК. Однако для идентификации
нуклеотидных последовательностей, ответственных
за правильную экспрессию и регуляцию генов, ис-
пользуется другой подход, так называемая обратная
генетика. В ней применяются клонированные гены
в качестве субстратов для конструирования in vitro
измененных генов, функцию которых можно изу-
чать как in vitro, так и in vivo (разд. 7.6).
К наиболее полезным для анализа модифика-
циям в структуре гена относятся: замена одного
нуклеотида или группы нуклеотидов, делеции или
вставки нескольких нуклеотидов или протяженных
участков ДНК и перестройки внутри гена. Ниже мы
обсудим, каким образом эти модификации исполь-
зуются для идентификации регуляторных последо-
вательностей, которые обеспечивают правильную
экспрессию гена и отвечают за его тканеспецифич-
ную и зависящую от времени регуляцию. Кроме
того, изучение новых генов, образующихся при
слиянии частей различных генов, очень облегчает
идентификацию последовательностей, ответствен-
ных за правильную экспрессию. Например, слияние
промотора SV40 и различных его производных с
последовательностями, кодирующими легко иденти-
фицируемые бактериальные или эукариотические
клеточные белки, позволяет выяснить, какие после-
довательности промотора обеспечивают правиль-
ную инициацию, эффективность и регуляцию транс-
крипции гена SV40. Аналогичные химерные гены,
содержащие, например, промоторы генов инсулина
или эластазы, слитые с областью, кодирующей
Т-антиген SV40, позволяют идентифицировать эле-
менты, ограничивающие экспрессию генов инсулина
или эластазы исключительно р-клетками островков
Лангерганса или ацинарными клетками соответст-
венно. Для применения методов обратной генетики
необходимо, чтобы существовал один или лучше
несколько способов определения фенотипического
проявления измененного гена. Соответствующие
бесклеточные системы, с помощью которых можно
определять эффективность транскрипции нормаль-
ных и модифицированных генов, а также изучать
процессинг или трансляцию РНК, дают прекрасную
возможность для анализа функции генов и послед-
ствий отдельных изменений в них. Трансфицируя
нормальные и модифицированные гены с помощью
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
21
подходящих векторов в культивируемые клетки,
можно установить последовательности генов, кото-
рые нужны для экспрессии и регуляции трансфици-
рованных генов в их собственных клетках или в род-
ственных клетках хозяина in vivo, например генов
иммуноглобулина в лимфоидных клетках, а генов
глобина в предшественниках эритроцитов. Вводя
гены дикого типа, а также измененные гены в сома-
тические или половые клетки либо в эмбрионы на
очень ранних стадиях развития, получают трансген-
ные растения и животные, содержащие эти гены во
всех соматических клетках и клетках зародышевой
линии. Трансгенные организмы особенно ценны для
идентификации структурных элементов, участвую-
щих в тканеспецифичной экспрессии гена в процессе
развития.
Гены, выбранные нами в качестве иллюстрации,
происходят из разных эукариотических организмов.
Большой интерес представляют гены дрожжей, в
основном Saccharomyces cerevisiae. Во-первых, они
обладают некоторыми свойствами, характерными
для генов бактерий, растений, беспозвоночных и
позвоночных. Во-вторых, связывающиеся с дрож-
жевой ДНК белки, ответственные за многие процес-
сы регуляции транскрипции у дрожжей, могут быть
заменены белками других организмов или работают
совместно с соответствующими сигналами и белка-
ми из других организмов, в том числе млекопитаю-
щих. В-третьих, глубокое изучение генетики дрож-
жей и замена нормальных генов модифицирован-
ными (разд. 5.6.в) увеличивают возможности обрат-
ной генетики. Широко представлены в данной главе
и гены вирусов млекопитающих, поскольку по сво-
им структурным и функциональным характеристи-
кам они часто коррелируют с генами своих хозяев.
Действительно, многие неизвестные ранее особен-
ности строения и регуляции эукариотических генов
были выявлены при изучении именно вирусных
геномов. В данной главе рассмотрены также неко-
торые гены растений, беспозвоночных (морского
ежа и Drosophila) и позвоночных (амфибий, птиц
и млекопитающих, включая приматов), поскольку
это помогает понять сложные процессы развития
многоклеточных организмов.
8.1. СРАВНЕНИЕ СТРУКТУРНЫХ
ОСОБЕННОСТЕЙ
ПРОКАРИОТИЧЕСКИХ И
ЭУКАРИОТИЧЕСКИХ ГЕНОВ
а. Прокариотические гены
5'-концы генов прокариот, кодирующих тот или
иной белок или стабильные РНК-продукты, имеют
характерную организацию регуляторных элементов.
К этим элементам относятся промоторные последо-
вательности, с которыми связывается РНК-поли-
мераза, и последовательности, влияющие на ско-
рость инициации транскрипции благодаря связы-
ванию со специфическими белками, репрессорами
или активаторами (рис. 8.1 и разд. 3.2.в). Все гены
прокариот транскрибируются с помощью един-
ственной РНК-полимеразы, которая при своем
функционировании связывается с сигма(о)-факто-
рами, поэтому организация разных промоторов у
прокариот одинакова (рис. 3.8). Все последователь-
ности, входящие в регуляторную область, распола-
гаются в основном на расстоянии 50-75 пар основа-
ний до точки инициации транскрипции и лишь
иногда элементы, ответственные за позитивную или
негативную регуляцию, находятся еще дальше от
этой точки либо в пределах транскрибируемого
участка или даже за его З'-концом.
Нуклеотидные последовательности, ответствен-
ные за терминацию транскрипции и высвобождение
транскрипта, располагаются на З'-конце гена. Учас-
ток ДНК между сайтами, в которых начинается
и кончается транскрипция, называется единицей
транскрипции. Большая часть белок-кодирующих ге-
нов содержит различное число нуклеотидов, пред-
шествующих белок-кодирующей последователь-
ности (5'-лидер) или следующих за ней (З'-трей-
лер). В некоторых случаях в 5'-лидерную область
входят последовательности, обеспечивающие до-
полнительный контроль транскрипции и трансляции
(в разд. 3.11.г и 3.11.е обсуждаются аттенуация и ре-
гуляция трансляции соответственно). Нуклеотидные
последовательности белок-кодирующих генов, как
правило, коллинеарны последовательностям соот-
ветствующих мРНК, а последовательности РНК-ге-
нов коллинеарны последовательностям рРНК и
тРНК, которые они кодируют; исключение состав-
ляют ген тимидилатсинтетазы фага Т4 и ген
TPHKSer у штаммов Acanthamoeba, каждый из кото-
рых содержит интрон.
Моноцистронные единицы транскрипции содер-
жат последовательность, кодирующую только один
белок или стабильную РНК (рис. 8.1, Л). Однако
единицы транскрипции прокариот, как правило, яв-
ляются полицистронными и содержат последователь-
ности. кодирующие не один, а несколько типов
белков или РНК (рис. 8.1,51 и В). Обычно все коди-
рующие сегменты в полицистронной единице транс-
крипции имеют общие 5'- и З'-регуляторные после-
довательности, что обеспечивает координированную
регуляцию и экспрессию множественных генов. По-
следовательности, кодирующие один или несколько
полипептидов, транслируются без какой бы то ни
было модификации или процессинга РНК. Напро-
тив, последовательности, кодирующие многочислен-
ные РНК, например три вида рРНК или множест-
24
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
упоминалось, гены прокариот обычно организова-
ны в опероны, и их экспрессия опосредуется полици-
стронными мРНК (разд. 3.11.6). В отличие от этого,
в эукариотических клетках образуются мРНК, ко-
дирующие только один белок. Даже если ген коди-
рует не один вид мРНК, как при альтернативных
способах удаления интронов (разд. 8.5), каждая зре-
лая мРНК имеет только одну транслируемую коди-
рующую последовательность. Несмотря на то что
гены 5S-pPHK располагаются тандемно, каждый ген
транскрибируется со своего собственного промото-
ра с образованием РНК, имеющей только одну
последовательность 5S-pPHK на молекулу. Напро-
тив, транскрипты кластеров генов 18S-, 5,8S- и
28S-pPHK, образующиеся с помощью РНК-поли-
меразы I, представляют собой длинные молекулы,
содержащие каждую из последовательностей рРНК
и промежуточные последовательности, а зрелые
молекулы рРНК меньшего размера образуются при
посттранскрипционном расщеплении.
Определение эукариотического гена. Хотя строе-
ние прокариотических и эукариотических генов в
принципе одинаково, молекулярные детали их
структуры существенно различаются. Новое опре-
деление эукариотического гена, возможно, поможет
обобщить существо этих различий. Следует при-
знать, что ни одно конкретное определение эукари-
отического гена не может полностью удовлетворить
всех и быть приемлемым во всех случаях. Определе-
ние, предложенное нами, основывается на молеку-
лярном строении генов, выполняющих разнообраз-
ные функции в разных эукариотических организмах.
Оно учитывает особенности местоположения и ти-
пов элементов последовательности ДНК, влияющих
на экспрессию гена, а также то, что изменение
фенотипических признаков обусловлено изменением
как кодирующих последовательностей, так и регу-
ляторных элементов.
Мы определяем ген как совокупность сегментов
ДНК, которые вместе составляют экспрессируемую
единицу, обусловливающую образование специфи-
ческого функционального продукта-либо молеку-
лы РНК, либо полипептида. К сегментам ДНК,
составляющим ген, относятся:
1. Единица транскрипции, которая представляет
собой протяженный участок ДНК, кодирующий по-
следовательность первичного транскрипта; в нее
входят а) последовательность, кодирующая либо
зрелую РНК, либо белковый продукт; б) интроны;
в) 5'-лидерная и З'-трейлерная последовательности,
которые присутствуют в зрелых мРНК, а также
промежуточные последовательности (спейсеры), ко-
торые удаляются в ходе процессинга первичных
транскриптов генов, кодирующих РНК.
2. Минимальные последовательности, необходи-
мые для начала правильной транскрипции (промо-
тор) и для образования правильного З'-конца зрелой
РНК.
3. Последовательности, регулирующие частоту
инициации транскрипции; к ним относятся последо-
вательности, ответственные за индуцибельность и
репрессию транскрипции, а также клеточную, ткане-
вую и временную специфичность транскрипции. Эти
области так разнообразны по строению, положению
и функциям, что для них трудно найти одно простое
и емкое название. К их числу относятся энхансеры
и сайленсеры-последовательности, которые оказы-
вают дистанционное влияние на инициацию транс-
крипции независимо от своей ориентации относи-
тельно точки начала транскрипции (разд. 8.2.в и
8.3.г).
В наше определение гена не включены ни после-
довательности ДНК, которые влияют на простран-
ственную конфигурацию гена в хроматине, ни по-
следовательности, которые регулируют его тополо-
гию. Не вошли сюда и последовательности ДНК,
которые кодируют белки или РНК, влияющие на
экспрессию отдельных единиц транскрипции.
На рис. 8.3 представлены основные структурные
особенности эукариотического гена, кодирующего
белок. Начало гена на этом рисунке находится
слева, а конец- справа в соответствии с общеприня-
тым соглашением, по которому транскрипция идет
слева направо. Это означает, что у цепи ДНК,
имеющей ту же последовательность, что и РНК-
транскрипт, т. е. у значащей цепи (верхней на рисун-
ке), левый конец 5', а правый - 3'. Служащая матри-
цей при транскрипции незначащая цепь (нижняя на
рисунке) имеет противоположное направление. Для
удобства обычно изображают только значащую
цепь. Положение первого нуклеотида транскрипта
обозначают числом +1, а нуклеотидов, расположе-
нных справа (downstream) от него (т. е. внутри еди-
ницы транскрипции), большими положительными
числами (например, + 16). Нуклеотиды, находящие-
ся слева (upstream) от +1 (нетранскрибируемые
последовательности), обозначаются отрицательны-
ми числами (например, —25).
Корреляцию между строением генов эукариот
и их экспрессией и регуляцией удобно проследить на
примере отдельных генов в связи с работой РНК-по-
лимеразы, инициирующей их экспрессию. Мы обсу-
дим по очереди особенности строения генов классов
I (разд. 8.2), II (разд. 8.3) и III (разд. 8.4); свойства
соответствующих РНК-полимераз; механизмы ре-
гуляции транскрипции указанных генов. Затем мы
рассмотрим процессинг первичных РНК-транскрип-
тов, в частности сплайсинг, с образованием зрелых
мРНК, рРНК и тРНК (разд. 8.5). В разд. 8.6 мы
остановимся на некоторых элементах последова-
тельностей, которые помогают белковым факторам
транскрипции узнавать специфические последова-
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
25
Начало РНК
(+1)
Интрон!
Интрон 2 Интрон 3 Конец мРНК
?Ь®Ив
I Левые I
регуляторные
элементы
Экзон 2 / Экзон 3
Экзон 4
5-лидерная
лоследова-
Внутренние
тельное™ регуляторные
элементы
3-трейлерная
последова-
тельность
3'
5'
Правые
регуляторные
элементы
РИС. 8.3.
Структурные особенности типичного эукариотического
гена, кодирующего белок. Единица транскрипции-это
сегмент от точки инициации транскрипции (+1) до
области ее окончания. Последняя находится за тем
сайтом, который соответствует концу последователь-
ности зрелой мРНК, но сама транскрипция редко закан-
чивается в каком-то специфическом сайте. Последова-
тельности, необходимые для инициации транскрипции.
Единица транскрипции
//--------
обычно располагаются левее сайта + 1 на участке дли-
ной до 100 п. н„ но могут находиться и за сотни и даже
тысячи пар оснований. Кроме промотора и левых регу-
ляторных элементов, у многих генов есть регуляторные
последовательности, находящиеся внутри интронов или
за З'-концом зрелой мРНК. Разные регуляторные эле-
менты на рисунке выделены по-разному, чтобы проил-
люстрировать их взаимное расположение.
тельности ДНК и взаимодействовать с ними, а так-
же друг с другом. В конце главы рассматриваются
более общие механизмы регуляции (такие, напри-
мер, которые влияют на время жизни мРНК), а
также способы влияния модификаций ДНК или
хроматина, в котором упакованы гены, на экспрес-
сию больших областей генома (разд. 8.7).
8.2. СТРОЕНИЕ ГЕНОВ КЛАССА I
И ИХ ЭКСПРЕССИЯ
Почти половина общей транскрипционной ак-
тивности в большинстве эукариотических клеток
приходится на долю генов класса I, транскрибируе-
мых с участием РНК-полимеразы I. Единственным
продуктом этого процесса является предшественник
рибосомной РНК (про-рРНК), который подвергает-
ся последовательному процессингу с образованием
зрелых 5,8S-, 18S- и 28S-pPHK. Четвертый тип
рРНК, 5S-pPHK, кодируется генами класса III
(разд. 8.4). Число генов, кодирующих рРНК, ко-
леблется от сотен до нескольких тысяч в зависимо-
сти от вида эукариот (разд. 9.2.а-в). Они находятся
в одной или нескольких хромосомах в областях
с характерной морфологией, названных ядрышко-
выми организаторами (рис. 1.8). Во время интерфазы
эти участки объединяются в ядрышки-структуры,
в которых идет активная транскрипция генов рРНК,
процессинг про-рРНК и сборка рибосом.
Гены рРНК практически всех эукариот построе-
ны в виде длинных повторов типа «голова к хвосту»
(разд. 9.2.а). На электронных микрофотографиях
фрагмента ДНК ядрышек в момент транскрипции
видны несколько кластеров генов рРНК (рис. 8.4).
а. Единицы транскрипции
У всех видов каждая единица транскрипции
рРНК кодирует 18S- 5,8S- и 28S-pPHK, перечислен-
ные в порядке их расположения, начиная с 5'-конца
(рис. 8.5). Три рРНК-кодирующие последовательно-
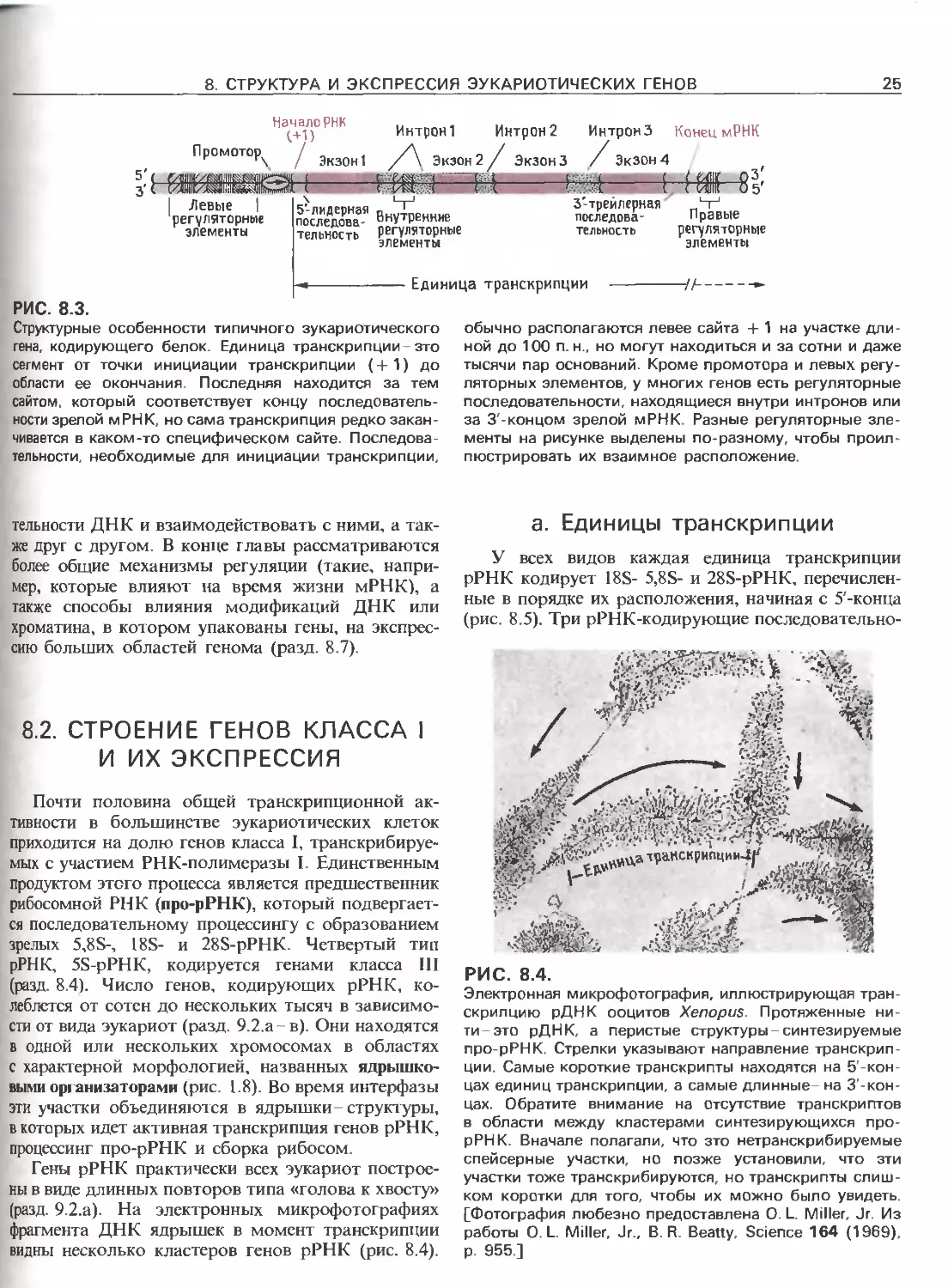
РИС. 8.4.
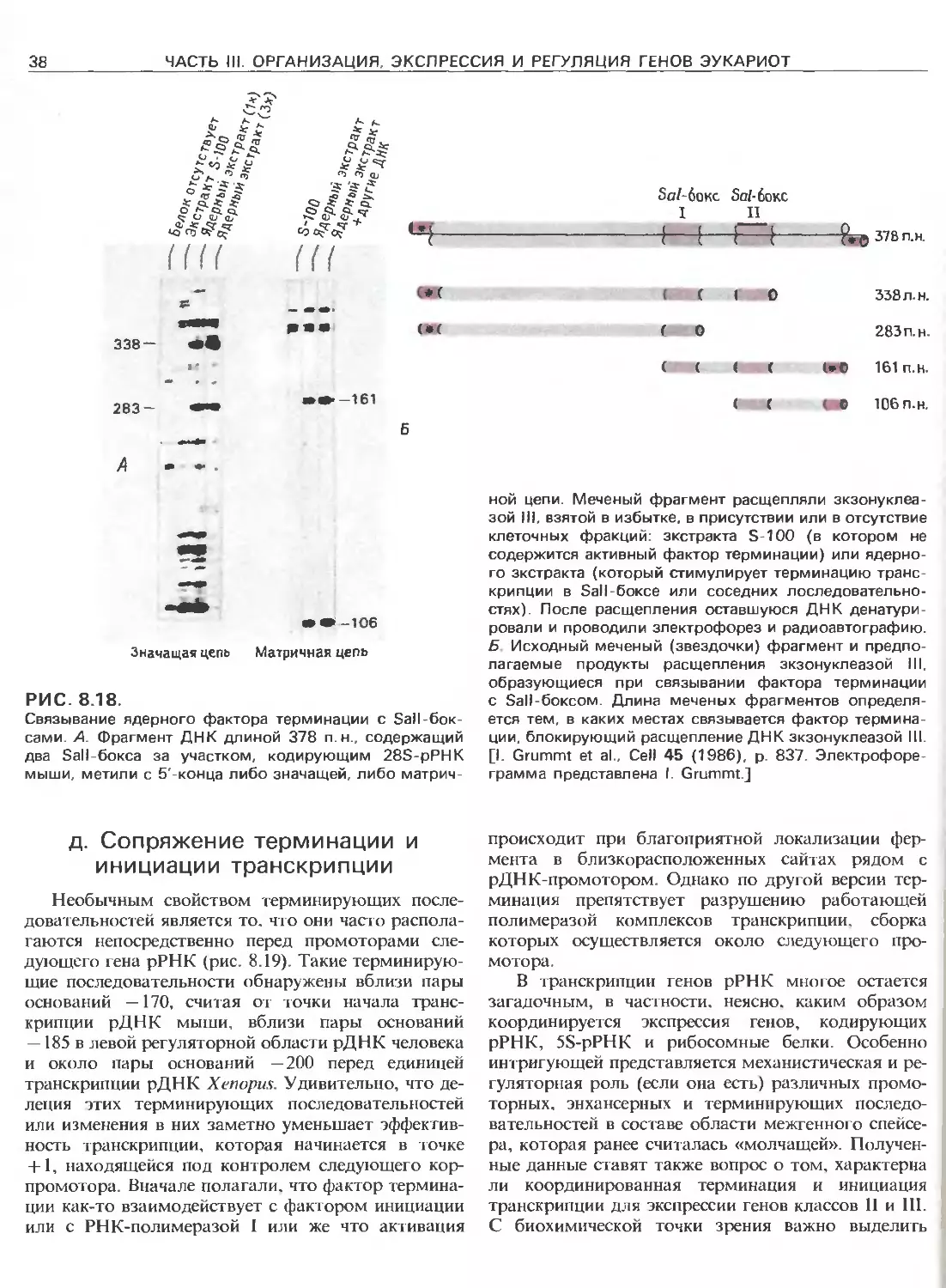
Электронная микрофотография, иллюстрирующая тран-
скрипцию рДНК ооцитов Xenopus. Протяженные ни-
ти-это рДНК, а перистые структуры-синтезируемые
про-рРНК. Стрелки указывают направление транскрип-
ции. Самые короткие транскрипты находятся на 5'-кон-
цах единиц транскрипции, а самые длинные- на З'-кон-
цах. Обратите внимание на отсутствие транскриптов
в области между кластерами синтезирующихся про-
рРНК. Вначале полагали, что это нетранскрибируемые
спейсерные участки, но позже установили, что эти
участки тоже транскрибируются, но транскрипты слиш-
ком коротки для того, чтобы их можно было увидеть.
[Фотография любезно предоставлена О. L. Miller, Jr. Из
работы О. L. Miller, Jr., В. R. Beatty, Science 164 (1969),
p. 955.]
26
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
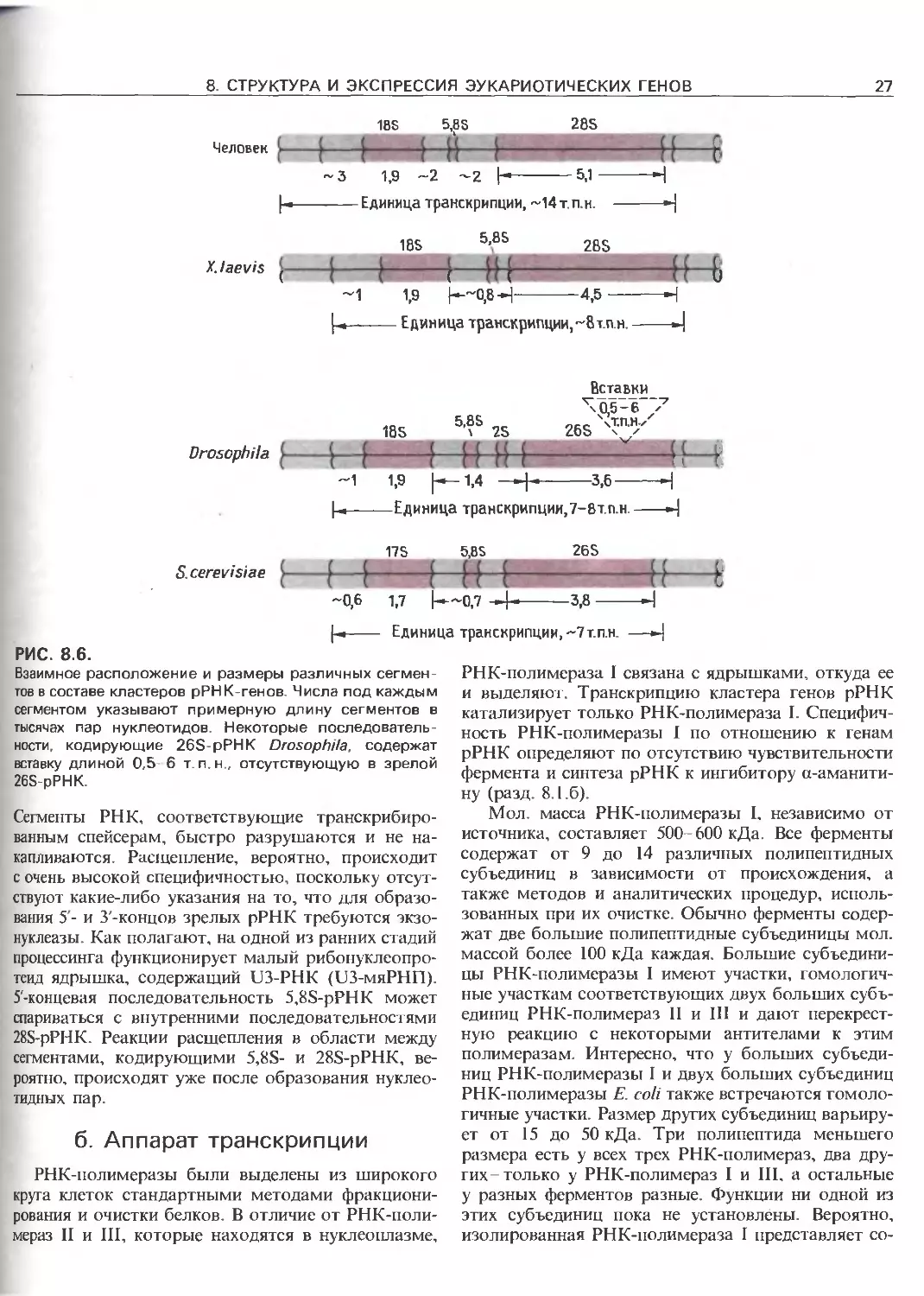
Кластер Межгенный Кластер 1Иежгенный Кластер
рРНК-генов спейсер р РНК-генов спейсер рРНК-генов
/ \
/ \
/
IGS /5 ETS 1SS 5'1TS 5,85 3’ITS 28S SETS' IGS
7-------f-----1--------1-----H-f--------------------H
2300 n.H. 156П.М. 4200 OH.
|-«-----Единица транскрипции про-рРНК --------J
РИС. 8.5.
Организация кластеров рРНК-генов эукариот. А. Каж-
дый кластер рРНК-генов длиной от 8 до 14 т.п.н. (в
зависимости от вида) отделен от других кластеров
межгенными спейсерами (IGS), длина которых у разных
видов сильно различается, но в пределах одного вида
относительно постоянна. Б. Каждый кластер рРНК-ге-
нов составляет отдельную единицу транскрипции, со-
стоящую из 5'-внешнего транскрибируемого спейсера
(5'-ETS), последовательности, кодирующей 18S-pPHK,
сти фланкируются и разграничиваются промежу-
точными транскрибируемыми сегментами. Они на-
зываются соответственно внешними (5'- и З'-ETS) и
внутренними (5'- и З'-ITS) транскрибируемыми спейсе-
рами. Область между тандемно повторяющимися еди-
ницами транскрипции протяженностью от несколь-
ких т. п. н. до почти 30 т. п. н. первоначально называ-
лась нетранскрибируемым спейсером (NTS) Посколь-
ку. как теперь стало известно, подобные спейсеры на
самом деле транскрибируются и транскрипция в
этих областях, возможно, является особенностью
регуляции экспрессии рДНК (разд. 8.2.в), примени-
тельно к ним стали использовать более подходящее
название- межгенные спейсеры (IGS).
Размеры и нуклеотидная последовательность об-
ластей, кодирующих рРНК, весьма незначительно
варьируют среди широкого круга эукариот, при
этом оценку степени гомологии нуклеотидных по-
следовательностей рРНК можно использовать при
определении их эволюционного родства. Напротив,
длина ITS изменяется в широких пределах в зависи-
мости от вида; соответственно изменяется и размер
единиц транскрипции (рис. 8.6). Так, единица транс-
крипции рДНК в ДНК человека имеет размер около
5 -внутреннего транскрибируемого спейсера (5'-ITS),
последовательности, кодирующей 5,8S-pPHK, З'-внут-
реннего транскрибируемого спейсера (З'-ITS), после-
довательности, кодирующей 28S-pPHK, и З'-ETS. Каж-
дый кластер рРНК-генов фланкируют одинаковые меж-
генные регуляторные последовательности. Единица
транскрипции про-рРНК начинается от места соеди-
нения IGS с 5'-ETS и кончается в разных сайтах IGS за
последовательностью, кодирующей 28S-pPHK.
14 т. п. н., приближаясь к самой длинной из извест-
ных, а соответствующая единица транскрипции у S.
cerevisiae- одина из самых маленьких, между 6 и 7
т. п. н. Примерно половина областей рДНК Dro-
sophila. кодирующих 28S-pPHK, имеет вставки раз-
личных размеров; они не являются интронами,
и единицы транскрипции с такими вставками не
транскрибируются. Последовательности, кодирую-
щие 28S-pPHK у Tetrahymena и у некоторых других
видов, прерываются интронами размером 400-1000
п. н., которые вырезаются из транскриптов про-
рРНК перед дальнейшим процессингом, приводя-
щим к образованию трех зрелых рРНК (разд. 8.5.6).
Во время транскрипции новообразованная
про-РНК остается постоянно связанной с соответ-
ствующими рибосомными белками и метилируется
по специфическим основаниям и рибозным остат-
кам (рис. 8.7). После завершения транскрипции
нуклеазы ядрышка расщепляют про-рРНК рибону-
клеопротеидного комплекса сначала на 5'-конце
5,8S-pPHK, а затем на 5'-конце 18S-pPHK и З'-конпе
28S-pPHK соответственно. В результате после-
довательных расщеплений под действием эндо-
нуклеазы образуются три зрелые рРНК.
^СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
27
18S 5,88 28S
Человеч [-----[------( | |(-------ЯВВ^--------------------Н-----6
~ 3 1,9 -2 -2 h--------------5,1---------Н
---------Единица транскрипции, ~14 т. п.н. --------
X. laevis
18S 5,8S 2BS
I—t—I-----------H4H---------------------4H>
~1 1,9 H-~0,8-H--------4,5--------4
- — Единица транскрипции,~8т.п.н.-
Drosophila
Вставки
~1 1,9 (—1,4 —f-------3,6
------Единица транскрипции,7-8Т.П.Н.------
17S 5,8S 26S
S.cerevisiae j-----f----(------f----(-{—[--------------------
-0,6 1,7 |—0,7 -I- ---------3,8------
(-.---- Единица транскрипции,-7T.n.H. —»-|
РИС. 8.6.
Взаимное расположение и размеры различных сегмен-
тов в составе кластеров рРНК-генов. Числа под каждым
сегментом указывают примерную длину сегментов в
тысячах пар нуклеотидов. Некоторые последователь-
ности, кодирующие 26S-pPHK Drosophila, содержат
вставку длиной 0,5- 6 т.п.н., отсутствующую в зрелой
26S-pPHK.
Сегменты РНК, соответствующие транскрибиро-
ванным спейсерам, быстро разрушаются и не на-
капливаются. Расщепление, вероятно, происходит
с очень высокой специфичностью, поскольку отсут-
ствуют какие-либо указания на то, что для образо-
вания 5'- и З'-концов зрелых рРНК требуются экзо-
нуклеазы. Как полагают, на одной из ранних стадий
процессинга функционирует малый рибонуклеопро-
теид ядрышка, содержащий U3-PHK (НЗ-мяРНП).
5'-концевая последовательность 5,8S-pPHK может
спариваться с внутренними последовательностями
28S-pPHK. Реакции расщепления в области между
сегментами, кодирующими 5,8S- и 28S-pPHK, ве-
роятно, происходят уже после образования нуклео-
тидных пар.
б. Аппарат транскрипции
РНК-полимеразы были выделены из широкого
круга клеток стандартными методами фракциони-
рования и очистки белков. В отличие от РНК-поли-
мераз II и III, которые находятся в нуклеоплазме.
РНК-полимераза I связана с ядрышками, откуда ее
и выделяют. Транскрипцию кластера генов рРНК
катализирует только РНК-полимераза I. Специфич-
ность РНК-полимеразы I по отношению к генам
рРНК определяют по отсутствию чувствительности
фермента и синтеза рРНК к ингибитору а-аманити-
ну (разд. 8.1.б).
Мол. масса РНК-полимеразы I, независимо от
источника, составляет 500-600 кДа. Все ферменты
содержат от 9 до 14 различных полипептидных
субъединиц в зависимости от происхождения, а
также методов и аналитических процедур, исполь-
зованных при их очистке. Обычно ферменты содер-
жат две большие полипептидные субъединицы мол.
массой более 100 кДа каждая. Большие субъедини-
цы РНК-полимеразы I имеют участки, гомологич-
ные участкам соответствующих двух больших субъ-
единиц РНК-полимераз II и III и дают перекрест-
ную реакцию с некоторыми антителами к этим
полимеразам. Интересно, что у больших субъеди-
ниц РНК-полимеразы I и двух больших субъединиц
РНК-полимеразы Е. coli также встречаются гомоло-
гичные участки. Размер других субъединиц варьиру-
ет от 15 до 50 кДа. Три полипептида меньшего
размера есть у всех трех РНК-полимераз, два дру-
гих-только у РНК-полимераз I и III, а остальные
у разных ферментов разные. Функции ни одной из
этих субъединиц пока не установлены. Вероятно,
изолированная РНК-полимераза I представляет со-
28
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Субъединица рибосомы
РИС. 8.7.
Схематическое представление процессинга
про-рРНК с образованием зрелых рРНК-ком-
понентов рибосом. Метилирование и связы-
вание с рибосомными белками про-рРНК
происходят непосредственно во время тран-
скрипции. Расщепление начинается на 5'-кон-
це 5,8S-pPHK и далее происходит в местах
соединений 18S- и 28S-pPHK со спейсерами.
Сайты расщепления показаны стрелками, на-
правленными вверх и пронумерованными в
соответствии с предполагаемой очередностью
расщепления. Сборка рибосомы завершается
связыванием соответствующих рибосомных
белков.
бой минимальную структуру, необходимую для
транскрипции кластера генов рРНК, поскольку для
точной и эффективной инициации и терминации
транскрипции кластера генов рРНК нужны специ-
фические вспомогательные белки (разд. 8.2.в). Дей-
ствуют ли эти белки независимо или при своем
функционировании временно связываются с поли-
меразой, неизвестно. Имеются убедительные дан-
ные о том, что РНК-полимераза I существует в ак-
тивной и неактивной формах; различие между ними
заключается либо в обратимой ковалентной моди-
фикации одной из известных субъединиц, либо во
временном присоединении и удалении субъединицы,
необходимой для эффективной транскрипции рРНК
(напомним о роли о-фактора в функционировании
РНК-полимеразы Е. coli; разд. 3.2.6). Клетки, в ко-
торых не транскрибируется кластер генов рРНК,
содержат только неактивную форму РНК-полиме-
разы I. но после реинициации синтеза рРНК в них
вновь появляется активная форма фермента, свя-
занная с ДНК. У простейшего Acanthamoeba castella-
nii, например, активный фермент обнаруживается
в растущих трофозоитах, но не в нерастущих ци-
стах.
в. Области регуляции транскрипции
РДНК
Механизм и регуляцию транскрипции рДНК изу-
чали с помощью разных экспериментальных подхо-
дов. Во-первых, сравнивали последовательности
ДНК клонированных рРНК-генов из одного и того
же генома и из геномов родственных и неродствен-
ных видов; это позволило выявить встречающиеся
на определенных расстояниях от единицы транскрип-
ции характерные нуклеотидные последовательно-
сти. Во-вторых, оценивали эффективность транс-
крипции генов рРНК дикого типа и структурно
измененных клонированных генов рРНК с исполь-
зованием зависимых от матрицы клеточных эк-
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
29
страктов или частично очищенных препаратов РНК-
полимеразы и очищенных факторов транскрипции;
в результате удалось идентифицировать последова-
тельности, которые влияют на транскрипцию in
vitro. И наконец, выводы, сделанные на основании
опытов in vitro, получили подтверждение при иссле-
довании транскрипции генов после трансфекции или
микроинъекции модифицированных рДНК в под-
ходящие клетки.
Одной из удивительных особенностей сигналов
инициации транскрипции в генах, кодирующих
рРНК, является их видоспецифичность. Так, в кле-
точных экстрактах человека может осуществляться
транскрипция клонированных генов рРНК человека,
ноне рДНК мыши, крысы, дрожжей, Drosophila или
Xenopus. Точно так же клеточные экстракты мыши
непригодны для транскрипции генов рРНК челове-
ка. Но, как это ни удивительно, рДНК Xenopus
транскрибируется полимеразной системой мыши,
и наоборот. Даже гены рРНК двух близкородствен-
ных видов Drosophila (D. melanogaster и D. virilis)
различаются по специфичности комплексов транс-
крипции в отношении их собственных рДНК. Ука-
занная специфичность обусловлена неконсерватив-
ностью последовательности ДНК, находящейся у
начала единиц транскрипции в генах рРНК челове-
ка, мыши, Xenopus. Drosophila и дрожжей. Иная
картина наблюдается для генов класса II и класса
III; в этом случае гены данного класса даже у отда-
ленных видов имеют высококонсервативные или
одинаковые элементы в областях регуляции транс-
крипции. Более того, системы РНК-полимераз II
и III, выделенные из одних видов, могут транскри-
бировать соответствующие гены из других, нерод-
ственных, видов (разд. 8.3 и 8.4).
Строение областей регуляции транскрипции
рДНК у разных видов в общих чертах сходно,
однако существуют и ощутимые различия как меж-
ду самими регуляторными элементами, так и между
их взаимным расположением. Впрочем, в свете на-
ших сегодняшних знаний эти различия скорее
кажущиеся. Несмотря на значительные различия
сигналов транскрипции указанных ДНК, структур-
ные особенности и активность комплексов, осущест-
вляющих инициацию, элонгацию и терминацию
транскрипции про-рРНК, по-видимому, сходны.
Млекопитающие. В регуляции транскрипции
РДНК млекопитающих участвуют два сегмента в
составе IGS размером около 30 т. п. н. (рис. 8.8).
Один сегмент, находящийся между парами осно-
ваний —45 и +20 в рДНК человека и между па-
рами оснований —39 и + 9 в рДНК мыши, включа-
ет сайт инициации транскрипции ( + 1). Удаление
этого сегмента или его частей, замена следующих
друг за другом коротких последовательных участ-
ков чужеродными олигонуклеотидами примерно
той же длины (линкерные вставки) или замена пар
оснований в определенных положениях приводят
к прекращению транскрипции рДНК или резкому
снижению ее эффективности. С учетом расположе-
ния и способности обеспечивать правильную транс-
крипцию (хотя и на низком уровне, <5%) соответ-
ствующая последовательность названа кор-промо-
тором.
Кор
Левый регуляторный элемент
(UCE)
--------------------------------1GS
Единица
транскрипции
РИС. 8.8.
Влияние линкерных вставок на 5'-конце рДНК человека
на транскрипцию in vitro. Исследовалась способность
рДНК со вставками в разных местах к транскрипции
в клеточных экстрактах человека. Влияние вставок в
области от — 164 до + 33 пары оснований на эффек-
тивность транскрипции иллюстрируется с помощью
гистограммы. [М. М. Haltiner et al.. Mol. Cell. Biol.
6 (1986), p. 227.]
30
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Второй сегмент, куда входят последовательно-
сти между парами оснований — 156 и — 107, значи-
тельно повышает уровень транскрипции с кор-про-
мотора как in vitro, так и in vivo. Об этом четко
свидетельствуют результаты опытов по трансфек-
ции и инъекции модифицированных рДНК в подхо-
дящие клетки. Делеции и вставки линкеров, нару-
шающие последовательность оснований в этой об-
ласти, приводят к уменьшению уровня транскрип-
ции рДНК до 100 раз. Применение более точных
методов исследования, чем те, которые использо-
вались для получения данных, представленных на
рис. 8.8, помогло установить, что упомянутая по-
следовательность простирается до основания —186.
Хотя нуклеотидная последовательность, располо-
женная между этим регуляторным элементом
(upstream control element, UCE) и кор-промото-
ром, по-видимому, не является существенной, из-
менение ее длины или взаимной ориентации ука-
занных элементов уменьшает уровень транскрип-
ции. Это наводит на мысль, что обе регуляторные
последовательности работают согласованно, обес-
печивая эффективную транскрипцию с участием
РНК-полимеразы I. UCE, вероятно, состоит из бо-
лее мелких элементов, выполняющих различные
функции.
Каким же образом эти две регуляторные области
выполняют свою функцию? Ключом к разгадке
послужило обнаружение стабильных комплексов
рДНК. Если к соответствующей системе транскрип-
ции in vitro добавить одновременно две рДНК,
кодирующие две разные рРНК, то образуются обе
рРНК (рис. 8.9). Однако если одну рДНК добавить
даже всего на несколько минут раньше, чем другую,
то транскрибируется только первая ДНК независи-
мо от того, какая именно рДНК-матрица добавлена
первой. Такого предпочтения нет, если у ранее
добавленной ДНК отсутствует область регуляции
транскрипции рДНК или же если UCE инактиви-
рован путем делеций или замен единичных пар
оснований. Отсюда следует, что рДНК взаимодей-
ствует с одним или несколькими факторами, на-
ходящимися в экстракте, вследствие чего они стано-
вятся недоступными для другой рДНК даже после
повторных циклов транскрипции. Это подтверж-
дают опыты по разделению клеточных экстрактов
на фракции, соответствующие различным белковым
факторам, которые сами по себе ни по отдельности,
ни даже все вместе не стимулируют синтез про-
рРНК, но зато помогают активной форме РНК-по-
лимеразы инициировать транскрипцию рДНК.
Поиск факторов транскрипции и их выделение из
клеток человека и мыши основывается на двух
аналитических подходах, обусловленных: 1) способ-
ностью белковых факторов восстанавливать транс-
крипционную активность экстрактов, истощенных
по факторам транскрипции; 2) способностью таких
факторов оставлять «отпечаток» (футпринт) на по-
следовательности ДНК, с которой они связываются.
Метод отпечатков (футпринтинг)-это универсаль-
Транскрипция рДНК происходит с образованием ста-
бильных комплексов. рДНК1 и рДНК2- разные, но оди-
наково эффективно трансбирующиеся матрицы, на ко-
торых in vitro синтезируются рРНК. Если эти рДНК
введены в систему транскрипции одновременно, то обе
рРНК образуются примерно в равных количествах. Од-
нако если одна из рДНК введена раньше другой то
транскрибируется преимущественно рДНК, введенная
первой. Если в первой рДНК имеются мутации в об-
ласти промотора, то транскрипция второй рДНК не
подавляется.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
31
Значащая цепь
РИС. 8.10.
Гидролиз рДНК млекопитающих, связанной с фактором
транскрипции UBF-1, под действием ДНКазы I (фут-
принтинг). Для определения характера связывания
UBF-1 со значащей и матричной цепями использовали
соответственно сегмент промотора рДНК человека
размером 297 п.н. (между парами оснований —199
и +78, меченный 5'-32Р по остатку —199) и сегмент
размером 525 п. н. (между парами оснований — 500
ный способ, позволяющий выявить присоединение
специфического белка к ДНК путем сравнения набо-
ра фрагментов, образующихся после расщепления
ДНКазой I связанной с белком и свободной ДНК
(рис. 8.10). Если белок защищает ДНК от действия
ДНКазы, то его положение на молекуле ДНК мож-
но установить по отсутствию характерных фраг-
ментов.
Достаточно полная картина процесса инициации
транскрипции рДНК была получена в результате
проведения опытов с системами транскрипции чело-
века и грызунов как in vitro, так и in vivo. В обоих
случаях в основе опыта лежит узнавание специфиче-
скими факторами транскрипции элементов, примы-
кающих к кор-промотору. Клеточные экстракты
человека содержат два белка, UBF-1 и SL-1, которые
совместно с РНК-полимеразой I катализируют
транскрипцию с промотора гена рРНК человека.
Оба белка получены в почти гомогенном состоянии,
исследован механизм их действия. UBF-1 самостоя-
тельно связывается с З'-областью UCE (последова-
тельность между парами оснований —75 и —115)
и экранирует ее (рис. 8.10 и 8.11). Связавшийся
с ДНК UBF-1 контактирует также с участком кор-
Матричная цепь
UBF
m
В Т’15
—. -96
4 5 6
и +24. меченный по остатку +24). Дорожки 1, 3,
4 и 6-продукты гидролиза, полученные в отсутствие
UBF-1, дорожки 2 и 5- продукты гидролиза, образу-
ющиеся в присутствии очищенного UBF-1 в насыща-
ющих концентрациях. Нечувствительные к действию
нуклеазы области отмечены квадратными скобками,
а сайты эффективного расщепления-звездочками.
[S. Р. Bell ei al., Science, 241 (1988), p. 1192.]
промотора в окрестности -21-й пары оснований.
SL-1 самостоятельно не связывается с промоторной
областью гена рРНК. Однако при одновременном
присутствии UBF-1 и SL-1 участок, защищенный от
ДНКазы, удлиняется до — 165-й пары оснований.
SL-1 способствует также связыванию UBF-1 с кор-
промотором, расширяя область связывания вплоть
до пары +1. Таким образом, SL-1 и UBF-1 сов-
местно образуют область контакта ДНК с белком,
охватывающую кор и UCE. Связывается ли непо-
средственно SL-1 с ДНК в присутствии UBF-1 или
же придает UBF-1 способность узнавать дополни-
тельный участок ДНК, неизвестно. Мутационные
изменения, затрагивающие UCE, блокируют связы-
вание UBF-1 даже в присутствии SL-1. Мутации
в кор-промоторе такого эффекта не дают, но нару-
шают способность факторов SL-1 и UBF-1 ак-
тивировать транскрипцию с участием РНК-поли-
меразы I.
Все эти данные говорят о том, что в результате
взаимодействия между SL-1 и UBF-1 образуется
комплекс, который связывается с двумя промотор-
ными элементами рДНК, облегчая таким образом
связывание РНК-полимеразы I (рис. 8.12). Частично
очищенный фактор транскрипции с промотора гена
32
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Матричная цепь
РИС. 8.11.
Влияние SL-1 на связывание UBF-1 с премотор-
ным участком рДНК. Тот же самый фрагмент длиной
525 п. н. с меткой на конце, который представлен на
рис. 8.10, расщепляли ДНКазой I в присутствии в реак-
ционной смеси UBF-1 и SL-1 либо по отдельности, либо
вместе. Знаки «+» и «—» означают наличие или от-
сутствие указанных факторов соответственно. Два знака
«+» указывают на то, что данный фактор присутствует
в удвоенной концентрации. Слева показано относитель-
ное положение элементов промотора. Защищенные
участки отмечены квадратными скобками, а сайты эф-
фективного расщепления-звездочками. Если в реак-
ционной смеси присутствуют оба белка, то различия
между электрофореграммами выражены сильнее, а за-
щищенная область простирается на довольно большое
расстояние в глубь UCE.
рРНК мыши (называемый TIF-1B или фактором D)
связывается с аналогичным участком кор-промото-
ра примерно между парами оснований —40 и —15.
Каковы взаимоотношения этого фактора с UBF-1
и каким образом его присоединение активирует
транскрипцию с участием РНК-полимеразы I, не-
ясно.
Последовательности, находящиеся левее, перед
-165-й парой оснований, также играют важную
роль в активации транскрипции. Поскольку функция
этих последовательностей тесно связана с термина-
цией транскрипции рДНК, мы отложим обсуждение
этого вопроса до разд. 8.2.г.
Как мы уже отмечали, транскрипция может идти
только при условии, что рДНК и комплекс транс-
крипции происходят из одного и того же или из
близкородственных видов. Так, экстракты из клеток
грызунов не катализируют транскрипцию рДНК
приматов, и наоборот. Природу этой видовой спе-
цифичности можно понять, если сконструировать
гибридные области регуляции транскрипции, в ко-
торых кор-промотор рДНК мыши соединен с UCE
человека или кор-промотор человека соединен с
UCE мыши, и исследовать транскрипцию этих ДНК
с использованием экстрактов приматов (рис. 8.13).
Поскольку промотор мыши не связывается с бел-
ками UBF-1/SL-1, его неспособность участвовать
в транскрипции легко объяснима. Гибридный про-
мотор, составленный из LJCE мыши и кор-промото-
ра человека, неактивен по той же причине. Однако
промотор, построенный из регуляторного элемента
человека и кор-элемента мыши, также не транскри-
бируется РНК-полимеразой I приматов, хотя метод
отпечатков показывает, что факторы UBF-1/SL-1
человека связываются с UCE. Следовательно, не-
смотря на то что связывание UBF-1 и SL-1 с UCE
необходимо для активации транскрипции в системе
транскрипции приматов, указанные факторы, при-
соединившись к ДНК, не могут стимулировать
транскрипцию, если UCE связан с чужеродным кор-
промотором. Для того чтобы SL-1 мог содейство-
вать связыванию РНК-полимеразы I и/или инициа-
ции транскрипции, он, вероятно, должен соединить-
ся со своей собственной видоспецифичной последо-
Предполагаемая схема взаимодействия белков UBF-1
и SL-1 с элементами промотора рДНК при активации
и транскрипции с участием РНК-полимеразы I. UBF-1
и SL-1 связываются как с кором, так и с UCE и,
по-видимому, взаимодействуют друг с другом, либо
активируя при этом РНК-полимеразу I, либо облегчая
ее связывание.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
33
100
Эффективность
транскрипции
Клетки Мо
Связывание
UBF-1.SL-I
РИС. 8.13.
Транскрипция, в отличие от связывания UBF-1 и SL-1,
осуществляется только в том случае, если UCE и коро-
вая последовательность промотора происходят от одно-
го вида. Эффективность транскрипции с промоторов
рДНК человека (Ни) и мыши (Мо) и с каждого из
гибридных промоторов исследовали, трансфицируя
плазмиду, содержащую промотор, в культуры клеток
приматов или мышей и определяя количество синтези-
руемых рРНК. Эффективность транскрипции выражали
в процентах от эффективности транскрипции при нор-
мальном расположении сегментов. Связывание ДНК
оценивали при помощи метода отпечатков, как показано
на рис. 8.10.
вательностью кор-промотора. Условие правильного
сочетания UCE и кор-промотора для связывания
факторов транскрипции, по-видимому, и определяет
видовую специфичность транскрипции.
Drosophila. Как мы уже упоминали, гены рРНК
у D. melanogaster и D. virilis не являются взаимозаме-
няемыми при использовании их в качестве матриц
с перекрестными системами транскрипции ни in
vitro, ни in vivo. Более того, последовательности,
расположенные непосредственно перед точкой ини-
циации транскрипции (т.е. в IGS), негомологичны
(или степень их гомологии невелика). Тем не менее
структура этой области у обоих видов сходна: и в
том, и в другом случаях в ней содержатся от пяти до
двенадцати повторов сегментов длиной примерно
240 п. н. оснований и они находятся непосредствен-
но перед парой +1. У разных особей данного вида
повторы имеют почти одинаковые нуклеотидные
последовательности, однако у представителей раз-
ных видов они различаются.
Исследования ядерных и цитоплазматических
транскриптов генов рРНК Drosophila обоих видов
привели к следующим выводам. Как показал анализ
генов с делециями в регуляторных элементах, ана-
логичных описанным для промотора рДНК млеко-
питающих, кор-промотор занимает участок между
парами нуклеотидов — 43 и + 20. Каждый из упомя-
нутых повторов размером 240 п.н. в IGS также
может в какой-то степени стимулировать транс-
крипцию, которая начинается через несколько пар
за его З'-концом и заканчивается примерно через
175 п.н. внутри соседнего повтора размером
240 п.н. Однако более чем в 95% случаев транс-
крипция начинается в области, находящейся сразу
же за последним из повторов (у пары + 1) и охваты-
вает всю единицу транскрипции. Поскольку корот-
кие транскрипты встречаются редко, а в цитоплаз-
матической РНК вообще отсутствуют, их функция
остается неясной. Но определенная организация и
стабильное сохранение повторов в геноме двух вы-
шеупомянутых видов Drosophila, как и наличие их
в генах рРНК Xenopus, говорит о том, что присут-
ствие кластера промоторов и терминаторов в IGS
перед «истинным» промотором имеет какое-то зна-
чение для регуляции эффективности транскрипции
РДНК.
Xenopus. Типичный порядок расположения но-
вых повторов в IGS, разделяющем единицы транс-
крипции у Xenopus laevis, показан на рис. 8.14. Для
инициации правильной транскрипции рДНК необ-
ходим кор-промотор, занимающий участок между
парами —147 и +4. В зависимости от того, какая
именно из многочисленных областей межгенных
спейсеров рассматривается, последовательность
кор-промотора размером 150 п.н. повторяется
внутри спейсера от двух до шести раз. Эти
3-158
34
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Участок 0 Участок 1
Спейсерный
промотор
Повторы из
60/81 п.н.
Спейсерный
промотор
Повторы из
60/81 п.н.
।__
-4000
Промотор
гена
5'ETS
w -8
I
-3000
I
-2000
____I__
-1000
РИС. 8.14.
Типичный межгенный спейсер рДНКХ. taevis (IGS-об-
ласть, расположенная между З'-концом последователь-
ности, кодирующей 28S-pPHK, и сайтом, в котором
начинается транскрипция про-рРНК в начале 5'-ETS).
Овалами со стрелками обозначены спейсерные про-
моторы; участки, выделенные черным цветом, гомопо-
повторы разделяются областями из чередующихся
повторов размером 60 и 81 п. н. Интересно, что
каждый из повторов имеет общую последователь-
ность длиной примерно 40 п. н. с кор-промотором
и его копиями в IGS. В крайней левой части межген-
ного спейсера расположены еще два типа повторяю-
щихся элементов (область 0 и область 1). отличаю-
щихся от последовательностей размером 60 и 81 п. н.
Спейсеры, разделяющие единицы транскрипции
рДНК, различаются в основном числом и располо-
жением различных повторов (разд. 9.2.а).
Функциональная роль разных повторов до конца
не выяснена. Некоторую ясность в этот вопрос
внесли опыты по транскрипции рДНК с участием
РНК-полимеразы I и необходимых факторов in
vitro, а также эксперименты in vivo, в которых
подходящие матрицы клонированных рДНК вводи-
ли в яйцеклетки Xenopus. Были установлены сле-
дующие факты: 1. Элемент размером 150 п.н., ко-
торый примыкает к точке начала транскрипции,
скорее всего, ответствен за инициацию транскрип-
ции рДНК, поскольку он присутствует в данной
области рДНК у всех видов. 2. Повторы длиной
150 п. н., расположенные слева, стимулируют ини-
циацию транскрипции внутри спейсера, но эта
транскрипция не распространяется на внутреннюю
область единицы транскрипции. 3. Последователь-
ности размером 40 п. н., которые характерны для
кор-промотора длиной 150 п.н. и встречаются в
повторах 60/81, вероятно, не функционируют как
промоторы. По-видимому, они повышают эффек-
тивность инициации транскрипции, при этом эффект
не зависит от ориентации данных последователь-
ностей относительно кор-промотора, а также от
расстояния между ними и кор-промотором (см.
разд. 8.3.г, в котором обсуждаются энхансеры).
4. Множественность энхансеров обусловлена повто-
гичные повторы из 60/81 п. н. Символами Т1 и Т2
отмечены неэффективный и эффективный терминаторы
транскрипции соответственно. Функции участков, обо-
значенных цифрами 0 и 1, неизвестны. [По данным
работы Р. Leibhart, R. Н. Reeder, Cell 45 (1986), р. 431.]
рами 60/81 п. н., которые оказывают разное влияние
на эффективность инициации транскрипции с уча-
стием кор-промотора в зависимости от своего
числа и расположения. Функция областей 0 и 1 не-
ясна. Итак, транскрипция рДНК Xenopus, по-види-
мому, регулируется кластером промоторов/энхан-
серов. расположенным в области межгенного спей-
сера.
Дрожжи. Гены рРНК дрожжей тоже организо-
ваны как тандемные единицы транскрипции, коди-
рующие 18S-, 5,8S- и 258-последовательности. раз-
деленные межгенными спейсерами. Своеобразие
генных кластеров дрожжей заключается в том, что
они содержат ген 5S-pPHK, относящийся к генам
класса III. который присутствует в каждом из
повторов рДНК (разд. 9.5.6); однако этот ген
транскрибируется с противоположной цепи ДНК
и не РНК-полимеразой I, а РНК-полимеразой III
(разд. 8.4).
Последовательности, влияющие на экспрессию
единицы транскрипции рДНК дрожжей, выявлены
как в опытах с системами транскрипции in vitro, так
и в опытах по трансформации дрожжевых клеток
плазмидами, содержащими ген псевдо-рРНК. Одна
из таких единиц транскрипции гена псевдо-рРНК
была сконструирована путем замещения большей
части единицы транскрипции рДНК фрагментом ДНК
бактериофага Т7. Последовательность ДНК фага
Т7 располагалась в гибридной молекуле на 130 п.н.
правее точки начала транскрипции про-рРНК и на
500 п. н. левее конца последовательности, кодирую-
щей 25S-pPHK (рис. 8.15). Этот ген псевдо-рРНК
был встроен в плазмиду, несущую ARS и центро-
меру, которая сохранялась в виде самореплицирую-
щейся плазмиды в трансформированных клетках
дрожжей (разд. 5.6.6). Транскрипты этих химерных
единиц транскрипции можно было отличить от
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
35
Активатор
транскрипции
25S
5S
«
18S-AHKtparaT7(600n.H.) 25S
—НИЯИИИв—4--------------(----4
I
+1
(-«----------1GS (2 т. п. н.)-
РИС. 8.15.
Рекомбинантный ген псевдо-рРНК, сконструированный
для идентификации сигналов транскрипции в рРНК-ге-
нах дрожжей. Рекомбинантный ген псевдо-рРНК со-
стоит из сегмента ДНК IGS длиной 2 т. п.н., 5'-коди-
рующего участка 18S-pPHK длиной 130 п.н., сегмента
ДНК фага Т7 длиной 600 п.н., 600 п. н.-сегмента, со-
держащего 500 п. н. последовательности, кодирующей
25S-pPHK, З'-фланкирующего сегмента длиной 100 п.н.
транскриптов генов эндогенной рРНК дрожжей по
наличию последовательностей ДНК фага Т7.
Благодаря использованию производных гена
псевдо-рРНК, имеющих делении и перестройки в
1GS, были выявлены три отдельные области в со-
ставе межгенного спейсера. которые влияют на ско-
рость и точность инициации транскрипции. Во-пер-
вых, последовательность между парами оснований
-149 и +15 способна самостоятельно поддержи-
вать (хотя и на низком уровне) транскрипцию псев-
до-рРНК. Особенно важен для эффективной ини-
циации транскрипции участок между парами осно-
ваний — 149 и —133. Во-вторых, область между
парами оснований —210 и —2230 отвечает за то, что
инициация транскрипции осуществляется в правиль-
ном сайте, и одновременно увеличивает ее эффек-
тивность примерно в два раза. В-третьих, добавле-
ние участка длиной 190 п.н., охватывающего после-
довательность от —2230 до —2420 на 5'-конце IGS
(рис. 8.15), повышает эффективность инициации
транскрипции в правильном сайте примерно в де-
сять раз. В то же время, как и в случае последова-
тельностей 60/81 в IGS Xenopus, этот элемент функ-
ционирует в любой ориентации, т. е. его стимули-
рующий эффект не зависит от его ориентации отно-
сительно точки начала транскрипции. Отсюда сле-
дует, что сегмент размером 190 п.н. действует как
энхансер.
Транскрипты, соответствующие межгенному
спейсеру, не обнаруживаются in vivo, и, казалось бы,
левый элемент размером 190 п. н. на 5'-конце IGS не
стимулирует инициацию транскрипции. Но опыты по
транскрипции in vitro с экстрактами дрожжей ясно
показали, что синтез новых молекул РНК иниции-
руется внутри указанного элемента. Последователь-
ность, необходимая для активации синтеза РНК
в этой области, находится в составе сегмента разме-
ром 22 п.н. (пары от —32 до —11), расположенного
Указано также попожение 55-рДНК в IGS, хотя зто
и несущественно для данного опыта. Гибридный ген,
содержащий предполагаемый активатор транскрипции
длиной 190 п.н. на 5'-конце межгенного спейсера или
не содержащий его. включали непосредственно в хро-
мосому дрожжей или вводили в клетку в составе репли-
цирующейся плазмиды. За транскрипцией гена следили
по образованию Т7-подобной рРНК.
перед эктопической (смещенной) точкой начала
транскрипции. Так, у дрожжей транскрипция класте-
ра генов рРНК может инициироваться на 2 т. п. н.
левее зрелого 5'-конца про-рРНК. Однако образова-
ние транскриптов, инициированное на 5'-конце IGS,
вероятно, завершается недалеко от начала последо-
вательности про-рРНК, а правильные 5'-концы
про-рРНК, по всей видимости, получаются при
реинициации транскрипции. Напомним, что у Droso-
phila и Xenopus транскрипция также начинается и за-
канчивается внутри областей межгенных спейсеров
генов рРНК. Как именно терминации в этой обла-
сти влияют на инициацию транскрипции, мы обсу-
дим в следующем разделе.
г. Терминация транскрипции рДНК
Поскольку про-рРНК-транскрипты заканчива-
ются последовательностью, находящейся на З'-кон-
це 28S-pPHK или рядом с ним, раньше полагали,
что эта концевая последовательность и представля-
ет собой терминатор транскрипции с участием
РНК-полимеразы I. Но последующие эксперименты
по изучению транскрипции in vitro специально скон-
струированных генов рРНК показали, что транс-
крипция проходит З'-конец зрелой 28S-pPHK, закан-
чиваясь далеко за ним. Транскрипция у Xenopus
охватывает всю область межгенного спейсера, за-
канчиваясь в сайте, находящемся примерно на
200 п. н. левее кор-промотора следующей, 35S-npo-
рРНК; зрелый З'-конец образуется в результате
эндонуклеазного расщепления РНК в сайте, нахо-
дящемся сразу же за концом последовательности,
кодирующей 28S-pPHK. У дрожжей терминация
транскрипции, инициированной с кор-промотора,
происходит в межгенном спейсере за геном 5S-
рРНК, не доходя примерно 300 п. н. до начала
следующей последовательности про-рРНК.
з»
36
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
У мышей транскрипция рДНК заканчивается
между 550 и 600 п. н. за З'-концом последовательно-
сти 28S-pPHK. Сигналом к терминации транскрип-
ции с участием РНК-полимеразы I в этой системе
служит высококонсервативная последовательность
значащей цепи длиной 18 п.н. (5'-AGGTCGACCA-
ОдуМТССО-З'. так называемый Sail-бокс), которой
обычно предшествует один или несколько пирими-
диновых кластеров. Восемь таких повторов встре-
чаются в области между 500 и 1200 п.н., считая от
З'-конца 288-рДНК (рис. 8.16), но обычно термина-
ция происходит внутри Т-участков, предшествую-
щих первому Sail-боксу. Консервативная последо-
вательность из 18 п. н. может сама вызывать терми-
нацию транскрипции, но более эффективная и точ-
ная терминация осуществляется в том случае, когда
как перед ней, так и после нее имеются Т-богатые
участки. Небольшие делеции, вставки и замены пар
оснований внутри сегмента размером 18 п.н. или
инверсия этого сегмента приводят к утрате его
способности терминировать транскрипцию. Отча-
сти похожий порядок расположения консерватив-
ных повторов (5'-GGGTTGACCA-3'), по-видимому,
служит сигналом терминации транскрипции при
синтезе про-рРНК человека. Консервативные по-
вторяющиеся последовательности (5'-GACTTGC-3',
которым предшествуют Т-повторы) встречаются в
областях межгенных спейсеров у Xenopus, при этом
терминация чаще всего осуществляется в повторе,
находящемся в З'-концевом участке спейсера сразу
перед промотором.
Вначале имелись лишь косвенные данные о том,
что терминация в указанных выше сайтах происхо-
дит при связывании неких белков со специфически-
ми последовательностями ДНК. Эти данные полу-
чили подтверждение после того, как в экстрактах
ядер были обнаружены белковые факторы, связы-
вающиеся с терминирующей последовательностью
размером 18 п.н. Связывание белков-лигандов с
терминирующей последовательностью идентифици-
ровали по защите 5'-меченого фрагмента, содержа-
щего эту сигнальную последовательность, от рас-
щепления экзонуклеазой III (рис. 8.17). Расщепление
происходило только до тех пор, пока фермент не
встречал на своем пути связанный белок. Напом-
ним, что экзонуклеаза III расщепляет фрагменты
двухцепочечной ДНК, начиная с З'-концов (табл.
4.1). В результате образуются цепи ДНК с мечены-
ми 5'-концами, причем их длина определяется тем,
в каком месте связанный белок останавливает экзо-
нуклеазное расщепление. Поскольку 5'-конец каж-
дой цепи фрагмента можно пометить независимо,
нетрудно установить границы защищенного участ-
ка. В опытах такого рода с сегментами, содержа-
CTCTGCGGGC TTTCCCGTCG CACGCCCGCT_CGCTCGCAC6 CGACCGTGTC GCCGCCCGGG CGTCACGGGG GCGGTCGCCT CGGCCCCCGC GGGGTtGCCC
GAACGACCGT GTGGTGGTTG GGGGGGGGAT CGTCU£TCC_TCCGT_CTCCC GAGGACGGTT_CGTTrCTCTT_TCCCCTrCCG_TCG£TCrcCr_TGGGTGTGGG
AGCCTCGTGC CGTCGCGACC GCGGCCTGCC GTCGCCTGCC GCCGCAGCCC CTTGCCCTCC GGCCTTGGCC AAGCCGGAGG GCGGAGGAGG GGGATCGGCG
GCGGCGGCGA CCGCGGCGCG GTGACGCACG GTGGGATCCC CATCCTCGGC GCGTCCGTCG GGGACGGCCG GTTGGAGGGGCGG6AGGGGT TTTTCCCGTG
AACGCCGCGT TCGGCGCCAG GCCTCTGGCG GCCGGGGGG CGCTCTCTCC GCCCGAGCA£CCCCACTCCC_GCCCCTCCTC_TTCGCGCGCC GCGGCGGCGA
CGTGCGTACG AGGGGAGGAT GTCGCGGTGT GGAGGCGGAG AGGGTCCGGC GCGGCGCCTC ТТССАТТГТТ TCCCCCCCAA CTTCGGAGCT CGACCAGTAC
______________ 1
TCCGGGCGAC ACTTTGTTTT ТТТТТТТТСС CCCGATGCTG GAGGTCGACC AGATGTCCGA AAGTGTCCCC CCCCCCCCC£_CCCCCCGGCG CGGAGCGGCG
2
GGGCCACTCT GGACTCTTTT__JTTTTTTTTTТТТТТТТПТТТАААТТССТ
GACCAGTACT ccgggtggta ctttgtcttt TTCTGAAAAT CCCAGAGGTC
4 --------------
TTCTTT_TTT_TTTJ_r £TTT_ TTTGGTGT_GC_CTCTTTTTGA_CTTATATACA
TCCGGGCGAC ACTUGTTTT_TTTT_rrrTTT TCCACC6AT6 ATGGAGGTCG
GCG6GCTCAC TCTGGACTC£ TTmT^TTTjnTTnTn_T_TTAAATTTC
РИС. 8.16.
Нуклеотидная последовательность терминирующей об-
ласти рДНК мыши длиной 1168 нуклеотидов, начина-
ющаяся с первой позиции справа от З'-конца гена
зрелой 28S-pPHK. З'-транскрибируемый спейсер (565
п. н.) выделен серым цветом. Цветными прямоуголь-
никами с номерами указаны элементы длиной 18 п. н.
GGAACCTTTA GGTCGACCAG TTGTCCGTCT TTTACTCCTT CATATAGGTC
3 “
GACCAGATAT CCGAAAGTCC ТСТСТТТССС ТТТАСТСТТС CCCACAGC6A
5
TGTAAATAGT GTGTACGTTT АТАТАСТ AGGAG6AGGT CGACCAGTAC
6
ACCAGATGTC CGAAAGT6TC CCGTCCCCCC CCTCCCCCCC CCGCGACGCG
7 ----------------------------
TGGAACCTTA AGGTCGAC
8
(содержащие сайты для Sa/I), находящиеся в межген-
ном спейсере сразу за сайтами терминации. Пунктиром
выделены богатые пиримидинами участки длиной не
менее десяти нуклеотидов с содержанием пиримидинов
80%. [Crummt et al.. Cell 43 (1985), p. 801.]
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
37
Расщепление /
рестрнктазой /
Hin dill /
--------------------------8
Мечение 5'-конца
при помощи киназы
и узгр.дчр;
расщепление
рестрнктазой
Есо RI
3'
5'
Расщепление
рестрнктазой
\ Есо RI
X)
5'
Мечение 5-конца
при помощи киназы
и 7эгР-АТР;
расщепление
рестрнктазой
Hi п dill
3’
Инкубирование с
ДНК-связывающими белками,
расщепление экзонуклеазой Ш,
денатурация,электрофорез и
радиоавтография
РИС. 8.17.
••Экзонуклеаза
ДНК-связываю-
щнй белок
Изучение связывания белка с помощью защиты ДНК от
расщепления под действием экзонуклеазы III. Рест-
рикционный фрагмент ДНК известной длины метили
32Р по 5'-концу. После инкубации меченых фрагментов
с клетками, ядерными экстрактами или очищенными
ДНК-связывающими белками предполагаемые образо-
вавшиеся комплексы расщепляли экзонуклеазой III
в условиях, при которых обеспечивалась стабильность
щими две предполагаемые терминирующие после-
довательности (Sail-боксы и окружающие их после-
довательности, находящиеся между парами основа-
ний 583 и 665 за З'-концом 28S-pPHK), картина
распределения экзонуклеазных фрагментов изменя-
лась после предварительной инкубации препарата
с ядерным экстрактом (рис. 8.18). Это изменение
свидетельствовало о том, что с Sail-боксом связан
какой-то белок (или белки). Изменения в пределах
одного из консервативных участков Sail-бокса или
изменение расстояния между двумя наиболее кон-
комплексов. Далее проводили денатурацию, гель-злек-
трофорез и радиоавтографию и по положению полос
определяли размер недеградированной (защищенной)
части цепи и, таким образом, устанавливали внешние
границы сайта связывания белка. В приведенном при-
мере белок связывается ближе к EcoRI-сайту, чем к
сайту Hind\\\.
сервативными участками мешают связыванию бел-
ков и нарушают терминацию. Более того, при вклю-
чении в единицу транскрипции синтетических олиго-
нуклеотидов, соответствующих консенсусной после-
довательности Sail-бокса, увеличивается эффектив-
ность терминации транскрипции с участием РНК-
полимеразы I in vitro. Белок, связывающийся с Sail-
боксом, получен в чистом виде, хотя механизм его
действия до сих пор не установлен.
38
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 8.18.
Связывание ядерного фактора терминации с Sall-бок-
сами. А. Фрагмент ДНК длиной 378 п.н., содержащий
два Sail-бокса за участком, кодирующим 28S-pPHK
мыши, метили с 5'-конца либо значащей, либо матрич-
8а/-6окс Sal- бокс
1
4-----£----1-----{---------37В п.н.
((СО 338л.н.
( О 283 п.н.
( ( ( ( НМ 161 п.н.
( ( СЯС 106 п.н.
ной цепи. Меченый фрагмент расщепляли экзонуклеа-
зой III, взятой в избытке, в присутствии или в отсутствие
клеточных фракций: экстракта S-100 (в котором не
содержится активный фактор терминации) или ядерно-
го экстракта (который стимулирует терминацию транс-
крипции в Sail-боксе или соседних последовательно-
стях). После расщепления оставшуюся ДНК денатури-
ровали и проводили электрофорез и радиоавтографию.
Б Исходный меченый (звездочки) фрагмент и предпо-
лагаемые продукты расщепления экзонуклеазой III.
образующиеся при связывании фактора терминации
с Sall-боксом. Длина меченых фрагментов определя-
ется тем, в каких местах связывается фактор термина-
ции, блокирующий расщепление ДНК экзонуклеазой III.
[I. Grummt et al.. Cell 45 (1986), p. 837. Электрофоре-
грамма представлена I. Grummt.]
д. Сопряжение терминации и
инициации транскрипции
Необычным свойством терминирующих после-
довательностей является то. что они часто распола-
гаются непосредственно перед промоторами сле-
дующего гена рРНК (рис. 8.19). Такие терминирую-
щие последовательности обнаружены вблизи пары
оснований —170, считая от точки начала транс-
крипции рДНК мыши, вблизи пары оснований
— 185 в левой регуляторной области рДНК человека
и около пары оснований —200 перед единицей
транскрипции рДНК Xenopus. Удивительно, что де-
ления этих терминирующих последовательностей
или изменения в них заметно уменьшает эффектив-
ность транскрипции, которая начинается в точке
+1, находящейся под контролем следующего кор-
промотора. Вначале полагали, что фактор термина-
ции как-то взаимодействует с фактором инициации
или с РНК-полимеразой I или же что активация
происходит при благоприятной локализации фер-
мента в близкорасположенных сайтах рядом с
рДНК-промотором. Однако по другой версии тер-
минация препятствует разрушению работающей
полимеразой комплексов транскрипции, сборка
которых осуществляется около следующего про-
мотора.
В транскрипции генов рРНК многое остается
загадочным, в частности, неясно, каким образом
координируется экспрессия генов, кодирующих
рРНК, 5S-pPHK и рибосомные белки. Особенно
интригующей представляется механистическая и ре-
гуляторная роль (если она есть) различных промо-
торных, энхансерных и терминирующих последо-
вательностей в составе области межгенного спейсе-
ра, которая ранее считалась «молчащей». Получен-
ные данные ставят также вопрос о том, характерна
ли координированная терминация и инициация
транскрипции для экспрессии генов классов 11 и III.
С биохимической точки зрения важно выделить
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
39
>30т.п.н.
285-рРНК
565 п.н.
процессинга
про-рРНК
3-конец про-рРНК
Промоторы 1GS
Т 5-ETS 185-рРНК
РИС. 8.19.
Промоторы межгенного спейсера (IGS) и терминаторы
(Т) кластера генов рРНК мыши. Транскрипция после-
довательности, кодирующей 28S-pPHK, заканчивается
внутри кластера терминаторов (Т1-Т8); получившийся
транскрипт подвергается процессингу с образованием
З'-конца зрелой 28S-pPHK. Транскрипция начинается
с промоторов IGS, а заканчивается в терминаторе, на-
ходящемся непосредственно перед промотором еди-
ницы транскрипции, расположенной справа. [I. Grummt
et al.. Cell 47 (1986), p. 901.]
и охарактеризовать взаимодействующие со специфи-
ческими последовательностями ДНК факторы транс-
крипции и терминации, ассоциированные с РНК-
полимеразой I. Это поможет понять молекулярную
основу их функционирования и взаимодействия.
8.3. СТРОЕНИЕ И ЭКСПРЕССИЯ
ГЕНОВ КЛАССА II
а. Общие положения
Свойства РНК, кодируемых генами класса II.
Гены класса II кодируют все цитоплазматические
мРНК и все, за исключением одной (U6), U-PHK,
входящие в состав мяРНП. Все транскрипты генов
класса II имеют характерные группировки (кэпы) на
5'-конце: 7-метилгуанозин (в мРНК) и 2,2,7-триметил-
гуанозин (в U-PHK). В обоих случаях метилирован-
ный гуанозин кэпа связан с первым нуклеотидом
транскрипта трифосфатным мостиком между соот-
ветствующими 5'-группами (рис. 3.53). Почти у всех
мРНК сразу за кодирующей последовательностью
находится полиаденилатный хвост разной длины,
ро!у(А); он не кодируется генами, с которых транс-
крибированы мРНК, а присоединяется в ходе отде-
льной посттранскрипционной реакции. У дрожжей
и других низших эукариот длина ро1у(А)-хвостов
варьирует от 75 до 125 остатков. Ядерные мРНК
большинства позвоночных имеют ро!у(А)-хвосты
длиной от 200 до 300 нуклеотидов, а ро1у(А)-участки
цитоплазматических мРНК меньше по размеру
и имеют разную длину у разных мРНК. Многие
гистоновые мРНК и U-PHK, напротив, не содержат
ро1у(А)-хвостов. В разд. 8.3.в мы обсудим способы
образования 5'-кэпов, З'-концов и ро1у(А)-хвостов
у всех этих РНК. Некоторые мРНК содержат
М6-метилированные остатки аденина, а все U-PHK
отличаются тем, что имеют большое количество
разным образом модифицированных остатков ура-
цила. Возможно, эти модификации осуществляются
котранскрипционно или посттранскрипционно, од-
нако их физиологическая роль неясна.
За редким исключением, мРНК и U-PHK обра-
зуются из первичных транскриптов, длина и после-
довательность которых коллинеарны ДНК, с кото-
рой они транскрибированы. Помимо кэпирования
и расщепления на З'-конце, за которыми следует
полиаденилирование, важнейшей стадией процес-
синга, необходимой для образования мРНК, явля-
ется сплайсинг. Про-U-PHK ввиду отсутствия ин-
тронов сплайсингу не подвергается, но и в этом
случае для образования зрелых З'-концов должно
произойти эндо- и экзонуклеолитическое расщеп-
ление первичных транскриптов.
Уровни регуляции экспрессии генов класса 11. Фе-
нотип дифференцированной клетки в большой сте-
пени определяется количеством и разнообразием
содержащихся в ней белков, что обычно связано
с разнообразием синтезируемых мРНК. Поэтому
основным механизмом создания разных фенотипов
является дифференциальная экспрессия генов класса
II. Эта принципиальная связь между экспрессией
гена и фенотипом была установлена при помощи
сравнительного анализа белков и мРНК на разных
стадиях развития в разных дифференцированных
тканях одного и того же организма и даже в оди-
наковых клетках в разных физиологических со-
стояниях.
В принципе (и, возможно, в действительности)
регуляция экспрессии гена может происходить на
любой стадии биогенеза мРНК или во время транс-
ляции мРНК при синтезе белка. Множество фактов
указывает на то, что количество мРНК зависит
прежде всего от частоты инициации транскрипции.
Этот процесс регулируется в основном с помощью
взаимодействий между специфическими элементами
последовательностей, которые находятся в сайте
инициации, но часто располагаются и на различных
40
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
расстояниях от него (i/uc-элементы), и набором спе-
цифических белков, многие из которых связываются
со специфическими последовательностями ДНК
(транс-действующие факторы транскрипции). Таким
образом, как временная, так и тканевая специфич-
ность регуляции транскрипции определяется нали-
чием и концентрацией различных транс-действую-
щих факторов. Второй наиболее существенный фак-
тор регуляции количества мРНК (разд. 8.5)-ско-
рость и механизм сплайсинга про-мРНК. Как пока-
зано в разд. 8.7, экспрессия по крайней мере некото-
рых генов может регулироваться также дифферен-
циальным разрушением мРНК. Регуляция транс-
порта мРНК из ядра в цитоплазму, в места транс-
ляции, вероятно, также может влиять на экспрессию
генов, кодирующих белки, но данных в пользу этой
гипотезы очень мало. Кроме того, регулируется
и процесс трансляции мРНК (убедительные данные
на этот счет представлены в разд. 8.7). Свой вклад
в регуляцию содержания белка вносит избиратель-
ная деградация белков, однако неясно, в какой
степени. Если бы время жизни белков влияло на
равновесные концен! рации ключевых компонентов
комплекса транскрипции (например, транс-дей-
ствующих факторов транскрипции), это могло бы
иметь огромное значение для регуляции биогенеза
мРНК.
б. Аппарат транскрипции
РНК-полимераза II. Гены класса II транскриби-
руются в ядре с участием РНК-полимеразы II. Этот
фермент выделен из широкого круга организмов,
включая дрожжи. Physarum, Aspergillis, Acanlhamoe-
ba, высшие растения, насекомых, Xenopus и млеко-
питающих. Субъединичное строение ферментов из
разных организмов весьма сходно и часто не раз-
личается, если судить по физическим критерям
(рис. 8.20). По-видимому, число субъединиц равно
девяти или десяти. Все ферменты имеют две боль-
шие субъединицы. Одна из них мол. массой при-
мерно 220 кДа является самым крупным из всех
полипептидов, входящих в состав РНК-полимераз
трех типов. Другая имеет мол. массу около 140 кДа.
Еще три субъединицы меньшего размера одинаковы
у всех трех РНК-полимераз; остальные четыре или
пять субъединиц характерны только для РНК-по-
лимеразы II. Структурное и химическое сходство
двух самых крупных субъединиц всех трех эукарио-
тических РНК-полимераз, на которое раньше ука-
зывали перекрестные иммунологические реакции и
наличие общих пептидов, теперь установлено путем
сравнения кодирующих последовательностей их
клонированных генов. Более того, аминокислотные
последовательности двух самых больших субъеди-
ниц в каждой из трех эукариотических РНК-полиме-
РИС. 8.20.
Белковые компоненты ядерных РНК-полимераз млеко-
питающих. Очищенные препараты РНК-полимеразы I
(а), РНК-полимеразы II (б) и РНК-полимеразы III (в)
подвергали электрофорезу в ПААГ в денатурирующих
условиях. Полосы соответствуют отдельным полипеп-
тидам. которые входят в состав соответствующих РНК-
полимераз. (Фотография предоставлена R. Young.)
раз на удивление близки к аминокислотным после-
довательностям двух больших субъединиц РНК-
полимеразы Е. coli.
Из трех РНК-полимераз эукариот наиболее чув-
ствительна к а-аманитину РНК-полимераза II. Ток-
син не мешает инициации синтеза цепей ДНК, но
блокирует их элонгацию. Участок связывания
а-аманитина, по-видимому, локализован на самой
большой субъединице. Во всяком случае, мутации,
обусловливающие устойчивость Drosophila и S. сеге-
visiae к а-аманитину, картируются в гене, кодирую-
щем субъединицу размером 220 кДа. Самая крупная
субъединица РНК-полимеразы II является также
объектом физиологического фосфорилирования-де-
фосфорилирования и имеет множество сайтов, в
которых эти реакции протекают. Высокофосфори-
лированная субъединица, обнаруженная в составе
РНК-полимеразы II, участвующей в транскрипции
in vivo, значительно более активна, чем нефосфори-
лированный фермент, использующийся в опытах in
vitro совместно с основным промотором поздней
области генома аденовируса.
Гены, кодирующие самую большую субъедини-
цу РНК-полимеразы II дрожжей. Drosophila и мле-
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
41
копитающих, кодируют С-концевые полипептидные
домены, содержащие множественные тандемные
повторы гептапептида Tyr-Ser-Pro-Thr-Ser-Pro-Ser.
Число повторов варьирует от 26 (дрожжи) до 52
(мышь). Эта повторяющаяся последовательность
уникальна и важна для активности РНК-полимера-
зы. Опыты с генами дрожжей по изменению этой
последовательности показывают, что субъединицы,
имеющие менее 12 или 13 таких повторов, не функ-
циональны in vivo. Возможно, повышенное содер-
жание серинов и треонинов в этой области создает
условия для фосфорилирования, о котором шла
речь, но точная роль повторяющейся гептамерной
последовательности неизвестна.
Факторы транскрипции. РНК-полимераза II, как
и другие РНК-полимеразы (разд. 8.2 и 8.4), нужда-
ется во вспомогательных белках для формирования
функционального комплекса транскрипции. Многие
из них являются ДНК-связывающими белками,
узнающими один или несколько элементов, вместе
составляющих промотор гена. Некоторые факторы
транскрипции, вероятно, участвуют в белок-белко-
вом взаимодействии с другими факторами и, таким
образом, изменяют сродство и специфичность их
связывания. Взаимодействуя с различными элемен-
тами последовательностей ДНК и друг с другом,
разные факторы транскрипции формируют сложные
белковые комплексы, которые регулируют способ-
ность РНК-полимеразы II инициировать транскрип-
цию. Большая часть комплексов положительно
влияет на транскрипцию, усиливая ее инициацию,
но известны и такие, которые осуществляют отри-
цательную регуляцию и, таким образом, выступают
в роли репрессоров. Существуют даже факторы,
которые стимулируют транскрипцию одного гена,
одновременно подавляя транскрипцию другого.
Многие факторы специфичны для определенных
клеток или тканей либо действуют только на опре-
деленных этапах развития, что обеспечивает диффе-
ренциальную транскрипцию генов в разных тканях
или в разное время. Образно говоря, РНК-полиме-
раза II представляет собой инструмент транскрип-
ции, а взаимодействие факторов с регуляторными
сигнальными последовательностями ДНК и друг
с другом определяет, в каком месте и с какой
скоростью этот инструмент будет работать.
Число белковых факторов, которые относят к
транс-действующим, все возрастает, хотя не все они
до конца охарактеризованы. Функционирование лю-
бого отдельного фактора зависит от типов и орга-
низации определенных участков ДНК-i/uc-элемен-
гов, входящих в состав регуляторной области гена.
Эта зависимость очень сложна. Так, некоторые
факторы транскрипции связываются не с одним
типом элементов ДНК, а отдельные последователь-
ности ДНК могут связывать факторы нескольких
видов. Более того, тандемно расположенные или
перекрывающиеся элементы часто образуют допол-
нительные сайты связывания, не представленные ни
в одном из отдельно взятых элементов, или облег-
чают связывание какого-то одного фактора по срав-
нению с другим. Реальная сложность этих взаимо-
действий наряду с отсутствием многих деталей не
позволяет всесторонне обсуждать этот важный во-
прос. Более того, для систематизации факторов
транскрипции используются разные и часто не свя-
занные друг с другом подходы. Как правило, в ос-
нову классификации положены типы тканей или
клеток, вырабатывающих факторы транскрипции,
основные характеристики связывающих их после-
довательностей или просто вкусы исследователя.
Поэтому мы ограничимся описанием характерных
i/wc-действующих элементов некоторых регулятор-
ных областей в модельных системах (разд. 8.3.г-з)
и тех факторов, взаимодействие которых с эти-
ми областями наиболее полно изучено. Ниже, в
разд. 8.6, мы более подробно обсудим особенности
строения некоторых факторов транскрипции поли-
пептидной природы, обеспечивающие их способ-
ность взаимодействовать с ДНК и активировать
транскрипцию.
в. Созревание мРНК и U-PHK
Кэпирование. Все мРНК клеток эукариот и виру-
сов, за исключением мРНК пикорнавирусов, имеют
на 5'-конце кэпы. Кэп-структуры как в ядерных, так
и в цитоплазматических транскриптах РНК-поли-
меразы II ассоциированы со специфическими кэп-
связывающими белками, однако роль кэпов и свя-
занных с ними белков в метаболизме мРНК неясна.
Тем не менее установлено, что кэп-структура специ-
фически взаимодействует с факторами инициации
трансляции при сборке трансляционного комплекса.
Кэпирование про-мРНК на 5'-конце транскрип-
тов РНК-полимеразы II осуществляется сразу же по-
сле инициации; даже самые короткие из проанали-
зированных цепочек РНК оказались кэпированны-
ми. Одним из субстратов фермента гуанилилтраис-
феразы, катализирующего кэпирование, является
GTP, другим - дифосфорилированный конец, обра-
зующийся при отщеплении крайнего фосфата от три-
фосфатной группы первого нуклеотида синтезируе-
мой РНК (рис. 8.21). В реакции кэпирования крайний
фосфат трифосфатной группы синтезируемой про-
мРНК замешается на остаток гуанилила из молеку-
лы GTP. Про-мРНК, имеющая на 5'-конце только мо-
нофосфат, не вступает в реакцию. Таким образом,
кэп помечает первый нуклеотид, с которого начина-
ется транскрипция. Присоединенный остаток гуанина
метилируется в положении 7 пуринового кольца под
42
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
О=Р-О-!
Гуанозин-7- I о- \
метилтрансфераза! SAM 4
Т про-мРНК
7-метил-гуанозиновый кэп
2-0-метил - I
трансфераза ’ SAM
2-0-метильные заместители у нуклеотидов +1
или +1 и +2 про-мРНК
действием гуанозил-7-метилтрансферазы; 2'-гидро-
ксильные группы первого и второго нуклеотидов
в синтезируемой РНК метилируются позже при
участии 2'-О-метилтрансферазы. Сам 7-метил-ОТР
не является субстратом в реакции кэпирования.
U-PHK предположительно кэпируются сходным об-
разом, но в этом случае последующее метилирова-
ние приводит к образованию 2,2,7-триметилгуано-
зиновых кэпов, а также 2'-О-метилзамещенных
групп в нескольких первых нуклеотидах транскрип-
та. Поскольку про-мРНК-транскрипты, синтезиро-
ванные в клеточных экстрактах или с участием
очищенной РНК-полимеразы II, кэпируются пра-
вильно, ответственные за это ферментативные ак-
тивности, по-видимому, связаны с самой полимера-
зой. Реакции инициации транскрипции и кэпирова-
ния под действием РНК-полимеразы II, вероятно,
строго сопряжены, так как ядерные транскрипты,
образованные при участии РНК-полимеразы III,
РИС. 8.21.
Образование 5'-кзп-структур эукариоти-
ческих мРНК. После отщепления кон-
цевого фосфата 5'-трифосфатного кон-
ца синтезированной про-мРНК гуани-
лилтрансфераза присоединяет гуаниль-
ный остаток гуанозинтрифосфата к ди-
фосфорилированному концу про-мРНК,
в результате чего образуется предшест-
венник 5'-кзпа. Специфические метил-
трансферазы, использующие S-адено-
зилметионин (SAM), метилируют гуано-
зиновый кзп в положении 7 и 2'-гидро-
ксильные группы первого и второго ос-
татков рибозы про-мРНК.
которые имеют на конце трифосфат (например,
U6-PHK, 5S-pPHK и про-тРНК), не кэпируются.
Поэтому возникает вопрос, что же определяет раз-
ные виды кэпов у про-мРНК и npo-U-PHK?
Роль кэпов в синтезе и созревании первичных
транскриптов с образованием мРНК не выяснена.
В частности, спорными являются данные об участии
кэпов в сплайсинге. Вопрос об участии кэпов в обра-
зовании ядерных рибонуклеопротеиновых комплек-
сов (яРНП), в составе которых мРНК транспортиру-
ются из ядра в цитоплазму, или в сборке цитоплаз-
матических рибонуклеопротеиновых частиц (цРНП)
остается невыясненным. Имеются указания на то,
что кэпы и связанные с ними белки защищают
мРНК от расщепления со стороны 5'-концов. Одна-
ко более вероятно, что кэпы необходимы для присо-
единения рибосом к мРНК перед инициацией транс-
ляции. Один из факторов инициации, eIF-4E, выпол-
няет специфическую функцию узнавания кэпа при
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
43
ассоциации 408-субъединицы рибосомы с 5'-концом
мРНК (разд. 3.8.6). Отсутствие структуры кэпа у
РНК, функциональных в других отношениях, делает
невозможной их трансляцию. Единственным извест-
ным исключением из правила, согласно которому
кэп необходим для трансляции, являются пикор-
навирусы (например, полиовирус), геномная РНК
которых функционирует в качестве мРНК всех ко-
дируемых вирусом белков. Более того, клетки, ин-
фицированные полиовирусом, прекращают синтез
собственных белков, поскольку кодируемый виру-
сом белок инактивирует клеточный кэп-связываю-
щий белок.
Полиаденилирование и терминация транскрипции.
Транскрипция многих генов белков эукариот за-
канчивается далеко за сайтом, в котором происхо-
дит расщепление РНК с образованием З'-конца зре-
лой РНК. Терминация осуществляется во множе-
ственных сайтах последовательности ДНК протя-
женностью в сотни, а иногда и тысячи пар нуклео-
тидов. Например, находящаяся за сайтом полиадени-
лирования область, в которой заканчивается транс-
крипция гена р-глобина позвоночных, имеет длину
более чем 1 т. п. н. Аналогичным образом, транс-
крипция первого из двух тандемно расположенных
генов а-глобина млекопитающих заканчивается в
разделяющей эти два гена промежуточной области
размером почти 2 т. п. н. Еще более ярким приме-
ром служит транскрипция поздней области генома
аденовирусов. В этом случае непрерывная транс-
крипция области, содержащей пять участков поли-
аденилирования, заканчивается только в конце ви-
русной ДНК.
Некоторые гены белков эукариот, напротив, име-
ют четко определенные сигналы терминации транс-
крипции. В гене гастрина человека, например, сиг-
нал терминации находится на 192 п.н. правее сайта
полиаденилирования мРНК гастрина. Эта последо-
вательность, которая останавливает транскрипцию
как in vivo, так и in vitro, богата остатками Т:
5'-TqA2T5AT4AT4AT5-3'. Транскрипция ДНК закан-
чивается как раз перед этой сигнальной последова-
тельностью, в каком бы месте ДНК она ни находи-
лась. Такая же последовательность- с небольшими
отличиями - найдена и в друз их генах позвоночных
как до, так и после единицы транскрипции. Этот
предполагаемый сигнал терминации, а также неко-
торые Т-богатые последовательности в генах дрож-
жей, 5'-CAATCTTG-3' и 5'-Т5АТА-3', напоминают
p-зависимые сигналы терминации у Е. coli
(разд. З.8.Г.).
Транскрипция обычно не останавливается в сай-
тах полиаденилирования. З'-полиаденилированные
концы должны образоваться позже при эндонукле-
азном расщеплении и последующем присоединении
остатков адениловой кислоты к З'-гидроксильной
группе, образовавшейся в сайте расщепления. Сайт
расщепления-полиаденилирования образуют две до-
вольно близко расположенные друг к другу после-
довательности. Одна из них-фактически неизмен-
ная последовательность ААТААА, находящаяся на
10-30 п. н. левее динуклеотида С А, локализованного
в сайте, где часто осуществляется расщепление и
полиаденилирование, или вблизи него. Замена Т на
G в последовательности ААТААА заметно пони-
жает эффективность полиаденилирования, хотя не-
большое количество образовавшихся концов поли-
аденилируется. Мутация, состоящая в замене после-
днего А на G в последовательности ААТААА на
конце гена а-глобина человека, нарушает синтез
нормальной а-глобиновой мРНК. Эти и другие
экспериментальные данные указывают на то, что
последовательность ААТААА совершенно необхо-
дима. Однако эти последовательности присутству-
ют также в кодирующих областях многих генов
в сайтах, где полиаденилирование не происходит,
поэтому для запуска реакций, ведущих к расщепле-
нию и полиаденилированию, необходима вторая
последовательность. Положение второго сигнала
определяли, исследуя влияние на полиаденилирова-
ние делеций вблизи сайта полиаденилирования.
Полученные данные показали, что точная последо-
вательность и положение нужного сигнала зависят
от конкретного гена. Наиболее вероятный сигнал(ы)
во многих генах белков позвоночных - сегмент, бо-
гатый GT или Т, 5'-YGTGTGYY-3' (Y-любой пири-
мидин), находящийся правее сайта расщепления в
интервале примерно из 25 п.н., за которым на
разном расстоянии часто встречаются короткие
Т-повторы (рис. 8.22).
В некоторых случаях даже подлинные сайты
полиаденилирования не функционируют. При про-
цессинге поздних мРНК аденовируса 2 используется
лишь какой-то один из пяти возможных сайтов
полиаденилирования первичного транскрипта
(рис. 8.23). Относительное содержание разных позд-
них мРНК аденовируса 2 отражает частоту, с кото-
рой происходит расщепление и полиаденилирование
первичного транскрипта в разных сайтах. В данном
случае соотношение количеств различных мРНК,
образованных из общего транскрипта, изменяется
в ходе инфекции, из чего следует, что выбор сайта
полиаденилирования регулируется. Вторым приме-
ром может служить переключение клеток с синтеза
мембраносвязанных цепей иммуноглобулина на об-
разование соответствующих секретируемых форм
(рис. 10.72). Различие между двумя этими формами
касается тяжелых цепей иммуноглобулина. В этих
случаях полиаденилирование по первой сигнальной
паре последовательностей в про-мРНК, которое
необходимо для образования мРНК, кодирующей
секретируемую цепь иммуноглобулина, подавляет-
Ген а-глобина человека
Ген [Ь-глобина человека
HLA-ген человека
Ген a-IFN человека
Ген y-IFN человека
Ген гормона роста
человека
Ген препроинсулина
человека
Ген металлотионеина
мыши
Ранняя область генома
SV4O
Поздняя область генома
SV40
Ad12Ela
Ad2 L2
HSV-1 ТК
ASV
A-M11LV
♦
TCTTTGAATAAAGTCTGAGTGGGCGGCAGCCTGTGTGTGCCTGGGTTCTCTCTGTCCCGGAATGTGCCAACAATGGA
CTGCCTAATAAAAAACATTTATTTTCATTGCAATGATGTATTTAAATTATTTCTGAATATTTTACTAAAAAGGGAAT
agacaaaataaatggaacacatgagaaccttcAgagtccatgtgtttcttgtgctgatttgttgcagggg
TTTTTCATTAAATTTTTACTATACAAAATTTCTTGTGTTTGTTTATTTTTTAAGATTAAATGCCAAGCCTGACTGTA
------------ ♦ --------------------------
TATATGAATAAAGTGTAAGTTCACAACTACTTATGCTGTGTTGGACTTTTTCTAAGTGAGACCTGGAGTGAAAGAAC
TGTCCTAATAAAATTAAGTTGCATCATTTTGTCTGACTAGGTGTCCTCTATAATATTATGGGGTGGAGGGGGGTGGT
AGATGGAATAAAGCCCTTGAACCAGCCCTGCTGTGCCGTCTGTGTGTCTTGGGGGCCCTGGGCCAAGCCCCACTTCC
GTTAATAATAAAAGCCTGTTTGAGTCTAACTCTGGTTTCTTGGTGTGGTTTGGCAATAAGAAACTGGGGTGACTTGA
TTCACAAATAAAOCATTTTTTTCACTGCATTCTAGTTGTGGTTTGTCCAAACTCATCAATGTATCTTATCATGTCTG
agctgcaataaacaagttaacaaAacaattgcattcattttatgtttcaggttcagggggaggtgtgggaggtttt
atgtataataaaactggtttcggttgaagtgtcttgttaatgtttgtttgggcgtggttaaacagggatataaagct
aatcaaaataaaaagtctggagtctcacgctcgcttggtcctgtaactattttgtagaatggaagacatcaactttg
gacggcaataaaaagacagaataaa/cgcacgggtgttgggtcgtttgttcataaacgcggggttcggtcccagggc
cgatacaataaacgccatttgaccattcaccacAtggtgtgtgcacctgggttcatggctggaccgtcgat ГСССТ
gtatccaataaaccctcttgc^gttgcatccgacttgtggtctcgctgttccttgggagggtctcctctgagtgcgt
РИС. 8.22.
Нуклеотидные последовательности З'-концов значащих
цепей ДНК, кодирующих разные мРНК млекопитающих
и вирусов эукариот. Подчеркнуты инвариантные олиго-
нуклеотиды ААТААА и GT-богатые участки. Стрелками
указаны сайты полиаденилирования. HLA-один из ге-
нов, кодирующих главный антиген гистосовместимости;
a-IFN и y-IFN - а- и у-интерфероны соответственно;
Ad12 Е1А и Ad2 L2-трансформирующий белок Е1А
аденовируса 12 и структурный белок аденовируса 2
соответственно; HSV-1 ТК-ген тимидинкиназы вируса
герпеса типа 1; ASV- провирусная последовательность,
кодирующая З'-конец РНК вируса саркомы птиц;
A-MuLV-провирусная последовательность, кодирующая
вирус лейкоза мышей Абельсона. [I. McLauchlan et al..
Nucleic Acids Res. 13 (1985), p. 1347.]
I---------1--------1---------1---------1---------1---------1---------I---------1--------1---------1
0 10 20 30 40 50 60 70 80 90 100
Первичный
транскрипт
Разные мРНК,
образовавшиеся
нз первичного
транскрипта
©•— •-------------
РИС. 8.23.
Разные полиаденилированные мРНК аденовируса, об-
разовавшиеся из одного первичного транскрипта. Числа
вверху-длина ДНК в процентах. Представленный на
рисунке первичный транскрипт поздней области начи-
нает синтезироваться с главного промотора поздней
области (примерно 19%), кзпируется и продолжает
синтезироваться до конца ДНК. Образовавшиеся про-
мРНК расщепляются по одному из указанных сайтов,
и к ним добавляются poly (А)-хвосты (волнистые черные
линии). Участки, которые удаляются при сплайсинге
каждой полиаденилированной формы, показаны пунк-
тирными линиями.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
45
ся. Вместо этого мРНК процессируется в следую-
щем сайте полиаденилирования; в результате обра-
зуется про-мРНК, которая после сплайсинга может
превращаться в мРНК, кодирующую мембраносвя-
занные цепи иммуноглобулина. Таким образом,
переключение от образования мембраносвязанных
цепей иммуноглобулина на продуцирование секре-
тируемых цепей происходит при помощи регулируе-
мого использования сайта полиаденилирования.
Механизм дифференциальной регуляции реакции
полиаденилирования в этих двух примерах, так же
как и во многих других, неизвестен.
Последовательность ААТААА участвует также
в терминации транскрипции. В частности, эта по-
следовательность на З'-конце главной кодирующей
последовательности Р-глобина мыши необходима
для терминации транскрипции, которая происходит
примерно через 1,5 т. п. н. Более того, те же делеции
и замены оснований, которые блокируют правиль-
ное полиаденилирование, снижают эффективность
последующей терминации. Из этого следует, что.
для того чтобы транскрипция могла закончиться
в соответствующей области, должно предваритель-
но произойти узнавание сайта полиаденилирования.
Одно из объяснений сопряжения полиаденилирова-
ния и терминации транскрипции состоит в том, что,
возможно, транскрибирующий комплекс РНК-по-
лимеразы II приобретает способность к осуществ-
лению терминации только после транскрипции сиг-
нала полиаденилирования. Это условие могло бы
выполняться, если бы комплекс транскрипции вклю-
чал фактор, подавляющий нежелательную термина-
цию при прохождении матрицы и утрачивающий
свои антитерминаторные свойства после прохожде-
ния сигнала полиаденилирования (рис. 8.24). В соот-
ветствии с данной гипотезой, если комплекс транс-
крипции РНК-полимеразы II утрачивает этот фак-
тор. то он «сходит» с матрицы в Т-богатых после-
довательностях, с которыми встречается позже. На
такую мысль наводили результаты опытов с очи-
щенной РНК-полимеразой II in vitro, в которых
было показано, что транскрипция различных
ДНК-матриц часто заканчивается в Т-богатых
участках внутри генов. С другой стороны, расщеп-
ление транскрипта в сигнале полиаденилирования
®—8
ААТААА
РИС. 8.24.
Сопряжение полиаденилирования и терминации тран-
скрипции. А. Транскрипция под действием РНК-поли-
меразы II (polll) предположительно прекращается рань-
ше времени в Т-богатых участках (на рисунке они
обозначены буквой Т). расположенных в нескольких
местах единицы транскрипции. Б. Возможно, связыва-
ние РНК-полимеразы с фактором антитерминации (at)
ингибирует терминацию в Т-богатых последовательно-
стях, и фермент получает возможность транскрибиро-
вать весь ген. Согласно данной модели, фактор at
отделяется от полимеразы в сигнале полиаденилирова-
ния, и транскрипция продолжается до тех пор, пока
фермент не закончит транскрипцию в неспецифических
Т-богатых терминирующих последовательностях.
[J. Logan et al.. Proc. Natl. Acad. Sci. USA 84 (1987),
p. 8306.]
46
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
могло бы позволить РНК-экзонуклеазе или р-по-
добному белку «захватить» полимеразу и индуци-
ровать терминацию в расположенных правее Т-бо-
гатых последовательностях (рис. 3.12).
Расщепление и полиаденилирование осуществ-
ляются в разнообразных клеточных экстрактах, ко-
торые содержат РНК-субстраты, имеющие соответ-
ствующие сигнальные последовательности. Изуче-
ние таких систем in vitro прояснило некоторые
аспекты реакций расщепления и полиаденилирова-
ния. Оказалось, в частности, что расщепление и по-
лиаденилирование могут протекать независимо друг
от друга. В присутствии аналогов АТР (например,
кордицепина, являющегося З'-дезоксиаденозинтри-
фосфатом) расщепление осуществляется в правиль-
ном сайте, но полиаденилирования не происходит.
Может осуществляться полиаденилирование З'-гид-
роксильных концов РНК, перед которыми находит-
ся последовательность AAUAAA. Фракционирова-
ние экстрактов клеток HeLa, в которых может
протекать расщепление и полиаденилирование
РНК-субстратов, позволяет получить несколько
белковых фракций, которые в совокупности способ-
ны катализировать реакции расщепления и поли-
аденилирования. Одним из компонентов является
ро1у(А)-полимераза, второй компонент содержит од-
ну или несколько малых ядерных рибонуклеопро-
теидных частиц (мяРНП), а третий представляет
собой белок (64 кДа), который связывается с участ-
ком, содержащим последовательность AAUAAA.
Для сопряженного расщепления и полиаденилиро-
вания необходимы все три компонента, хотя ро1у(А)-
полимераза способна одна полиаденилировать це-
пи РНК, расщепленные правильно. Механизм этого
процесса до сих пор не выяснен; не исключено, что
редкие Ull-мяРНП в соединении с белком разме-
ром 64 кДа и, возможно, другими мяРНП образуют
комплекс, который расщепляет последовательность
AAUAAA или соседнюю с ней последовательность
РНК и дает возможность ро1у(А)-полимеразе по-
лиаденилировать З'-конец. Предположение об учас-
тии мяРНП основано на том, что в иммунопреци-
питатах, полученных с антителами к мяРНП, обна-
руживаются фрагменты, содержащие AAUAAA;
кроме того, антитела к мяРНП ингибируют поли-
аденилирование in vitro. Роль мяРНП в полиаде-
нилировании согласуется с участием некоторых
мяРНП в процессинге РНК: Ш-мяРНП в образо-
вании З'-концов гистоновых мРНК (см. ниже в этом
разделе), иЗ-мяРНП-в получении правильного
З'-конца 28S-pPHK (разд. 8.2.г), a U1-, U5- и
и4/иб-мяРНП-в сплайсинге про-мРНК (разд.
8.5.Г).
Поскольку в генах дрожжей и, возможно, в генах
некоторых других низших эукариот сигнальная по-
следовательность ААТААА отсутствует, образова-
ние полиаденилированных мРНК у этих организмов
может быть сопряжено с терминацией. Не исключе-
но, что последовательности типа 5'-Т5АТАТ, кото-
рые стимулируют терминацию транскрипции, спо-
собствуют образованию З'-конца для полиаденили-
рования. С другой стороны, З'-конец может образо-
вываться в результате эндонуклеазного расщепле-
ния протяженных транскриптов, за которым следует
полиаденилирование под действием ро1у(А)-полиме-
разы. Участвуют ли мяРНП в расщеплении транс-
криптов у дрожжей, неясно.
Созревание гистоновых мРНК, не имеющих
poly ( А )-хвостов. Многие гистоновые мРНК не по-
лиаденилируются, хотя мРНК, кодирующие неко-
торые гистоны дрожжей и Tetrahymena, а также те
гистоны млекопитающих, экспрессия которых не
зависит от регуляции клеточного цикла, имеют
ро1у(А)-хвосты. Гистоновые мРНК в ооцитах ам-
фибии имеют короткие ро1у(А)-хвосты. но в процес-
се развития организма они заменяются неполиаде-
нилированными гистоновыми мРНК.
Все неполиаденилированные гистоновые мРНК
любых организмов имеют зрелый З'-конец с харак-
терной структурой. Вблизи конца кодирующей об-
ласти находится терминальный палиндром, кото-
рый может образовывать двухцепочечный стебель
из 6 п. н. с петлей, содержащей четыре нуклеотида
(рис. 8.25). За ним на расстоянии от 13 до 17
нуклеотидов следует богатая пуринами канониче-
ская последовательность. В гистоновых мРНК мор-
ского ежа данная последовательность абсолютно
консервативна (5'-CAAGAAAGA-3'), а в гистоновых
зрелой мРНК
РИС. 8.25.
Структура З'-конца гистоновых мРНК. Эти концы имеют
специфическую нуклеотидную последовательность, об-
разующую стебель из шести пар нуклеотидов и петлю
из четырех нуклеотидов. На расстоянии примерно 15
нуклеотидов от этой шпильки находится высокоспеци-
фическая последовательность, богатая пуринами. З'-ко-
нец зрелых гистоновых мРНК образуется в результате
эндонуклеазного расщепления участка между шпилькой
и этой последовательностью.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
47
мРНК позвоночных она менее консервативна (кано-
ническая последовательность: 5'-PuAAAGAGCUG-
3')- Нуклеотид, оказывающийся на З'-конце зрелой
гистоновой мРНК, располагается между шпилькой
и участком, богатым пуринами. Данные опытов по
изучению образования правильных З'-концов ряда
мутантных РНК свидетельствуют о том, что заме-
ны, препятствующие формированию дуплексного
стебля, блокируют нормальный процессинг, тогда
как замены пар оснований, которые не нарушают
способность РНК к образованию шпильки, не влия-
ют на образование З'-конца. Точно так же мутации
в последовательности, богатой пуринами, или даже
небольшие изменения расстояния между ней и па-
линдромом в значительной степени подавляют
З'-процессинг. Итак, для образования правильного
З'-конца необходимы две близкорасположенные об-
ласти, одна из которых находится перед сайтом
расщепления, а другая за ним. Неожиданным ока-
залось то, что процессинг на З'-концах гистоновых
мРНК с кэпированными 5'-концами осуществляется
легче, чем процессинг мРНК без кэпов.
Как показало изучение З'-эндонуклеазного рас-
щепления гистоновых мРНК морского ежа и млеко-
питающих in vitro и in vivo, для образования пра-
вильных З'-концов нужны относительно редкие
мяРНП, содержащие U7-PHK. Изменяя нуклеотид-
ную последовательность РНК 157-мяРНП, устано-
вили, какие именно нуклеотиды важны для расщеп-
ления гистоновой мРНК у З'-конца, эти нуклеотиды
находятся вблизи 5'-конца U7-PHK. Для процессин-
га, помимо 1)7-мяРНП, необходим также другой,
пока не охарактеризованный фактор. Этот фактор,
нечувствительный к действию нуклеазы (и поэтому,
по всей видимости, не относящийся к мяРНП),
связывается с петлей шпилечной структуры и спо-
собствует взаимодействию между Ш-мяРНП и бо-
гатой пуринами консервативной последовательно-
стью, находящейся за зрелым З’-концом. Сравнение
последовательностей гистоновой про-мРНК и
U7-PHK наводит на мысль, что участок, изобилую-
щий пуринами, спаривается с богатой пиримидина-
ми последовательностью длиной 10 нуклеотидов,
которая находится между 10-м и 20-м нуклеотида-
ми, считая от 5'-конца U7-PHK (рис. 8.26). Как
осуществляется расщепление, приводящее к появле-
нию З'-концов, и как оно регулируется, неизвестно.
Образование З'-концов U-PHK. Все U-PHK, син-
тезированные при участии РНК-полимеразы II, ли-
шены ро1у(А)-хвостов. Их З'-гидроксильные концы
образуются по крайней мере в две стадии. На
первой в результате терминации транскрипции или
при расщеплении длинного транскрипта образуется
предшественник U-PHK с небольшой добавочной
З'-последовательностью; затем этот предшествен-
117-МЯРНК 3' -GAUUUUCUCGACAUUGUGNNpppG
ческм™?Х'- °aaagagcug-3 Триметил
вательностъ л
3 'GAUUUUCUCGACAUUGUGNN
hH2A AUUAAAUGAGCUGUUGUG-3'
3•GAUUUUCUCGACAUUGUGNN
hH2B AAAGAAAGAGCUGGUGC-3’
3 'GAUUUUCUCGACAUUGUGNN
hH3 AACGAAAGUAGCUGUG-3’
3 'GAUUUUCUCGACAUUGUGNN
hH4 GGUGGUAACACUGGGA-3'
3 'GAUUUUCUCGACAUUGUGNN
hH4 CUGAGGAGUUGGACACUU-3'
3'GAUUUUCUCGACAUUGUGNN
mH2B AAAAUUGGAGCUUUAAU-3'
3 'GAUUUUCUCGACAUUGUGNN
mH3 AAAAAAAGAACUGUGCGC-3'
3'GAUUUUCUCGACAUUGUGNN
mH3 CAAAGAUGCUUGAAGU-3'
3'GAUUUUCUCGACAUUGUGNN
mH3 GGAAAGAGCUGUUU-3'
3 'GAUUUUCUCGACAUUGUGNN
mH4 UGAAGGUGGUAACACUGA-3'
Структурные особенности З'-концов гистоновых мРНК
и их связывание с и7-мяРНК. Спаривание нуклеотидной
последовательности вблизи 5'-конца U7-mhPHK и рас-
положенной справа от шпильки богатой пуринами по-
следовательности гистоновой мРНК происходит на
весьма протяженном участке. З'-конец зрелой гисто-
новой мРНК образуется при зндонуклеазном расщеп-
лении в указанной позиции. В правой части рисунка
показаны нуклеотиды З'-концов разных гистоновых
мРНК человека (h) и мыши (т), спаривающиеся с
5'-концом и7-мяРНК. Наверху приведена аналогич-
ная картина для канонической последовательности.
[К. L. Mowry, J.A Steiz, Trends Biochem. Sci. 13 (1988),
p. 447.]
48
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
ник укорачивается, возможно под действием экзо-
нуклеазы, и формируется зрелый З'-конец. Опре-
деляя эффективность образования правильного
З'-конца при различных структурных модифика-
циях в генах U1- и 152-мяРНК, обнаружили важ-
ную для этого процесса короткую консерватив-
ную последовательность на значащей цепи,
GTTTN0_3AAAPuNNAGA (Pu-любой пурин,
N-любое основание). Эта последовательность рас-
положена на расстоянии от 9 до 19 нуклеотидов за
зрелыми концами U1-, U2- и Ш-мяРНК, выделен-
ных из разных источников. Наличие инвертирован-
ного повтора, который может образовывать шпиль-
ку в про-мяРНК, по-видимому, необязательно. За
что именно отвечает упомянутая последователь-
ность в предшественнике мяРНК-терминацию
транскрипции или эндонуклеазное расщепление -
неизвестно.
Для образования З'-концов U-PHK несуществен-
ны ни кодирующая U-PHK последовательность, ни
триметилгуанозиновый кэп, однако - и в этом за-
ключается любопытная особенность биогенеза
U-PHK-правильные З'-концы образуются только
в том случае, если транскрипция РНК инициируется
с промоторов U-PHK. Если промотор U-PHK заме-
нить на промотор мРНК, то процесс образования
правильного З'-конца нарушается. Более того,
транскрипты, синтезированные с промоторов генов
мяРНК, не полиаденилируются даже тогда, когда
матрицы содержат подлинный сигнал полиаденили-
рования. Наоборот, транскрипты, синтезированные
с промоторов мРНК, полиаденилируются в соответ-
ствующих сайтах даже тогда, когда имеется вставка
в последовательности U-PHK, подвергающейся
процессингу. Это говорит о том, что, возможно,
специфические ферменты процессинга или факторы
терминации транскрипции связываются с комплек-
сами транскрипции в тот момент, когда они иниции-
руют транскрипцию с промоторов генов мРНК или
U-PHK. Подобным же образом в комплексы транс-
крипции генов мРНК и U-PHK, вероятно, входят
разные метилазы кэпирования.
г. Регулируемая экспрессия вирусных
генов
Многие методы и подходы, с помощью которых
сформировались основные представления о ре-
гуляции экспрессии клеточных генов, были заим-
ствованы из области изучения регуляции транскрип-
ции у вирусов. Насколько известно, клетки и
ДНК-содержащие вирусы регулируют содержание
мРНК главным образом путем изменения частоты
и места инициации транскрипции с участием
РНК-полимеразы II. Более того, регуляция транс-
крипции вирусных генов, как и клеточных генов,
зависит от специфических взаимодействий между
сложно организованными уис-действующими эле-
ментами и большим набором шрпис-действующих
факторов транскрипции. Некоторые вирусы коди-
руют также необычные факторы транскрипции, ко-
ординирующие экспрессию групп вирусных генов
или влияющие на транскрипцию отдельных клеточ-
ных генов. В действительности взаимодействие ви-
русных и клеточных шрдис-действующих факторов
может по-разному влиять на транскрипцию клеточ-
ных и вирусных генов и даже оказывать положи-
тельное влияние на одни и отрицательное-на дру-
гие гены.
В основе регуляции экспрессии генов вирусов
млекопитающих лежат те же принципы, что и в ос-
нове регуляции экспрессии клеточных генов. Как
и промоторы большей части эукариотических генов
класса И, промоторы вирусов состоят из «прокси-
мальных» регуляторных элементов, как правило,
входящих в последовательность, непосредственно
окружающую точку начала транскрипции и прости-
рающуюся до пары оснований- 100, и «дистальных»
регуляторных элементов, чаще всего расположен-
ных между парами оснований — 100 и — 300, кото-
рые обеспечивают экспрессию гена, специфичную
для данной ткани или данной стадии развития
организма (рис. 8.27). Регуляторные последователь-
ности, находящиеся внутри единицы транскрипции
или за ней, насколько известно, не входят в состав
вирусных промоторов.
Здесь мы детально рассмотрим две системы
транскрипции вирусов, которые были и остаются
наиболее информативными: 1) транскрипция ранней
области генома SV40 и 2) транскрипция гена тими-
динкиназы вируса герпеса (HSTK).
Транскрипция ранней области генома SV40. Основ-
ные особенности инфекционного цикла SV40 в раз-
ных хозяевах суммированы в разд. 5.7.6. Располо-
жение последовательностей, кодирующих мРНК
и белки, а также областей регуляции транскрипции
и репликации, показано на физической карте генома
SV40 (рис. 5.34). Ранняя область занимает около
половины кольцевого генома длиной 5243 п.н. При
транскрипции этой области вскоре после заражения
синтезируется одна-единственная про-мРНК. После-
дующий дифференциальный сплайсинг транскрипта
дает две мРНК примерно равной длины, кодирую-
щие два родственных, но разных белка: большой
и малый Т-антигены. Большой Т-антиген (Tag) не-
обходим для репликации вирусной ДНК и для
регуляции транскрипции ранней и поздней областей,
тогда как функция малого Т-антигена (tag) в жиз-
ненном цикле вируса остается невыясненной. Па-
раллельное увеличение содержания Tag и числа
дочерних молекул вирусных ДНК активирует транс-
крипцию поздней области. Дифференциальный
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
49
I —100 41 Г*
-50-40 |+10
: cl.:"—
Регуляторные элементы Базальный промотор )
(специфичны для определенных генов и (одинаковый у многих генов
типов клеток) и типов клеток)
РИС. 8.27.
Элементы, характерные для промоторов генов класса II.
Два (или несколько больше) специфических элемента,
которые находятся на участке примерно 100 п.н. левее
точки начала транскрипции (+ 1) ипи иногда заходят за
эту точку, составляют так называемый базальный про-
сплайсинг первичного транскрипта поздней области
приводит к появлению двух классов поздних мРНК,
кодирующих три структурных белка (VP1, VP2,
VP3), входящих в состав капсида вируса.
Таким образом, вирусный геном содержит две
противоположно ориентированные и регулируемые
во времени единицы транскрипции, на каждую из
которых приходится примерно половина последо-
вательности ДНК. Мы подробно рассмотрим учас-
ток длиной примерно 300 п.н., разделяющий два
сайта начала транскрипции. Внутри его рассеяны
уыс-действующие сигнальные последовательности,
которые ответственны за регуляцию транскрипции
ранней и поздней областей, а также репликации.
Они были идентифицированы при изучении влияния
разнообразных модификаций (например, делеций,
инсерций, замен пар оснований и перестроек) участ-
ка длиной 300 п. н. на транскрипцию и репликацию
как in vitro, так и in vivo. Эти опыты, в которых
исследовались взаимодействия между факторами
транскрипции, репликации и специфическими по-
следовательностями ДНК, помогли прояснить не-
которые тонкие детали процесса размножения SV40,
а также основные свойства регуляции транскрипции
у эукариот.
Организация i/ис-действующих сигналов регуля-
ции репликации и транскрипции SV40 схематически
показана на рис. 8.28 и более детально на рис. 8.29.
Транскрипция противоположно ориентированных
ранней и поздней областей инициируется в позици-
ях, указанных стрелками. Репликация, которую мы
рассмотрим в первую очередь, начинается внутри
палиндромной последовательности протяженно-
стью 27 п. н., обозначенной как ori. Делении, встав-
ки или замены пар оснований в палиндромной
последовательности блокируют инициацию репли-
кации. Для того чтобы в ori началась эффективная
репликация, необходима также целостность приле-
гающего АТ-участка из 17 пар оснований. Положи-
тельное влияние на функцию ori оказывают и по-
вторы из 21/22 п.н. вблизи АТ-участка со стороны
мотор. Между парами оснований —400 и —100 нахо-
дятся дополнительные регуляторные элементы, функ-
ционирование которых обычно зависит от типа ткани
и стадии развития.
поздней области, хотя они и не являются сущест-
венными. Как мы увидим позже, эти GC-богатые
повторы из 21/22 п.н. и соседствующая с ними
АТ-последовательность являются важными компо-
нентами сигналов транскрипции ранней области. Их
роль в усилении репликации может быть связана
с той функцией, которую они выполняют в регуля-
ции транскрипции.
Для того чтобы репликация началась, с ori дол-
жен связаться кодируемый SV40 белок Tag. Как
показали исследования, Tag связывается также с
последовательностями, примыкающими к ori. Эти
последовательности впервые были идентифициро-
ваны как области, которые не расщепляются под
действием ДНКазы I после связывания с ними Tag.
При низких начальных концентрациях Tag, вскоре
после заражения, белок преимущественно связыва-
ется с последовательностью, кодирующей 5'-конец
ранней про-мРНК (сайт Tl). С увеличением содер-
жания Tag начинается его связывание с ori (сайт Т2).
Мутации в сайте Т1, препятствующие присоедине-
нию Tag, слабо влияют на репликацию. Всевозмож-
ные изменения палиндромной последовательности,
напротив, не только предотвращают связывание
Tag с ori, но и блокируют репликацию. (Мутации
как в сайте Т1, так и в сайте Т2 также нарушают
репрессию транскрипции ранней области под дей-
ствием Tag.)
Связанный в сайте Т2 Tag работает как АТР-за-
висимая геликаза, которая в комплексе с клеточной
ДНК-праймазой и ДНК-полимеразой (а возможно,
и с другими белками) способствует инициации син-
теза новых цепей ДНК. Не исключено, что АТ-
и GC-богатые элементы влияют на репликацию
сами по себе благодаря своей близости к стартовой
точке; с другой стороны, их влияние может быть
связано с тем, что повышение частоты инициации
транскрипции в этой области содействует началу
репликации.
Каждый из сайтов связывания Tag содержит по
крайней мере две копии пентануклеотидной канони-
И-158
50
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 8.28.
Положение и организация промоторов ранней и позд-
ней областей генома SV40. В верхней части рисунка
изображена геномная ДНК SV40 с указанием последо-
вательностей, ответственных за регуляцию репликации,
а также транскрипции ранней и поздней областей.
Стрелками показаны направления транскрипции ранней
и поздней областей и сайты, в которых она начинается.
ТЗ Т2 Т1
В нижней части рисунка указаны сайт начала репли-
кации ДНК (ori), два элемента базального промотора
(АТ-бокс и повторы из 21/22 п.н.), а также тандемные
повторы длиной 72 п.н. в составе энхансера. Tt. Т2
и Т3-места связывания Т-антигена; по своему сродст-
ву к антигену они убывают в следующем порядке;
т, >т2»т3.
ческой последовательности 5'-GAGGC-3'. Очевидно,
остатки G в этой последовательности участвуют во
взаимодействии с Tag, поскольку присоединение Tag
защищает их от метилирования под действием ди-
метилсульфата. Мутационный анализ наиболее важ-
ных нуклеотидов в сайте Т1 показывает, что сигнал
связывания Tag имеет длину 17 п. н. и содержит две
пентануклеотидные последовательности, разделен-
ные АТ-участком длиной не более 6 7 пар нуклео-
тидов. Такая организация сайтов связывания, как
полагают, позволяет димеру Tag реагировать с
двумя соответствующим образом ориентирован-
ными пентануклеотидными последовательностями.
Теперь мы вернемся к рассмотрению различных
элементов последовательности, которые влияют на
инициацию транскрипции ранней области (рис. 8.28
и 8.29). Один из этих элементов, в составе базального
промотора, ассоциирован с тем же АТ-участком из
17 пар оснований, который необходим для функ-
ционирования ori. Мутационные изменения или
даже делеции всего этого участка слабо влияют на
частоту инициации транскрипции ранней области,
но нарушают точность инициации, приводя к обра-
зованию мРНК с разными кэпированными 5'-конца-
ми. Таким образом, ТАТА-подобная последова-
тельность нужна для того, чтобы инициация осуще-
ствлялась в ограниченном числе сайтов, но она
не оказывает существенного влияния на скорость
транскрипции. В отличие от этого во многих эука-
риотических генах несколько более короткая ТАТА-
подобная последовательность (ТАТААА), располо-
женная в области примерно от —25 до —35 пары
оснований, совершенно необходима для инициации
транскрипции. С другой стороны, известны эукарио-
тические промоторы, у которых отсутствует после-
довательность. сколько-нибудь напоминающая
последовательность ТАТА (например, промотор
поздней области генома SV40).
Промотор ранней области генома SV40 содер-
жит также три прямых GC-богатых повтора разме-
ром 21/22 п.н., которые дополняют функцию ori.
Они находятся между парами нуклеотидов —45 и
— 110 в области сайтов инициации транскрипции
ранних РНК (рис. 8.28 и 8.29). Каждый из повторов
длиной 21/22 п.н. содержит две копии последова-
5-GGGCGG-3'
тельности из шести пар нуклеотидов rc(CGCC5-
Деления всего участка ДНК SV40 протяженностью
65 п. н. блокирует транскрипцию как in vitro, так
и in vivo, даже если окружающие последователь-
ности сохраняются. Тем не менее даже одна ко-
пия повторяющегося элемента длиной 21/22 п.н.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
51
может поддерживать (правда, на невысоком уровне)
транскрипцию ранней области, а для обеспечения
максимальных скоростей транскрипции достаточно
двух копий. Опыты по изучению мутационных из-
менений каждого основания в повторяющемся сег-
менте из 2I пары оснований ясно показали, что
упомянутая выше гексамерная последовательность
является функционально важным элементом. Имеет
ли столь большой избыток GC-элементов в регуля-
торной области SV40 физиологическое значение,
неизвестно.
GC-богатый повтор, аналогичный присутствую-
щему в ДНК SV40, встречается во многих других
клеточных и вирусных регуляторных областях,
включая области регуляции генов HIV-1, тимидин-
киназы вируса герпеса, дигидрофолатредуктазы, ме-
таллотионеина-1л, гидроксиметилглутарил-СоА -
редуктазы и гипоксантин-гуанин фосфорибозил-
трансферазы. Этот элемент функционирует как сайт
связывания специфического фактора транскрипции
Spl, который в больших количествах содержится
в клетках многих типов и рассматривается в данном
разделе ниже.
Наличие элементов размером 21/22 п. н. является
необходимым, но не достаточным условием актива-
ции транскрипции. Кроме них нужны еще последо-
вательности, расположенные в пределах и по сосед-
ству с двумя копиями последовательности из
72 п. н., находящейся между парами оснований
— НО и —300 (рис. 8.29). Некоторые штаммы SV40
имеют только одну копию сегмента длиной 72 п. н.
и фланкирующей ее последовательности (рис. 8.29),
и этого достаточно для нормальной транскрипции.
Мутационный анализ сегмента длиной 72 п. н. и
последовательности, фланкирующей его слева, по-
казал, что эта область протяженностью примерно
90 п. н. состоит из субэлементов нескольких разных
типов (рис. 8.29). Как правило, по отдельности эти
субэлементы-слабые стимуляторы, но, действуя
вместе в разных сочетаниях, они обеспечивают вы-
сокий уровень транскрипции.
Для эффективной транскрипции ранней области
необходимы GC-богатые элементы размером
21/22 п.н. и левый сегмент длиной приблизительно
90 п. н., однако их ориентация относительно друг
друга и сайтов начала транскрипции несущественна.
Так, весь повтор из 21/22 п.н. может быть инвер-
тирован без снижения эффективности транскрипции.
Точно так же левый элемент размером 72 п. н. сти-
мулирует правильную инициацию транскрипции,
находясь в любой ориентации. В то же время,
в отличие от повтора из 21/22 п.н., который функ-
ционирует только в том случае, когда он находится
на своем обычном месте, положение сегмента из
72 п. н. не играет роли. Транскрипция ранней обла-
сти начинается в правильном месте с нормальной
эффективностью, даже если этот сегмент перемещен
в интрон ранней области или на сотни и даже
тысячи пар оснований вправо или влево от единицы
транскрипции и находится в любой ориентации.
Однако важно отметить, что сам по себе элемент из
72 п. н. не стимулирует транскрипцию, как и в том
случае, если он и элемент из 21/22 п.н. содержатся
в разных молекулах ДНК.
Несмотря на то что элементы размером 72 п. н.
и 21/22 п.н. эффективно функционируют, даже если
I-----------------------В
Регуляторные GT GT ТС ТС SphSph GC GC GC GC GC GC
элементы ЦТ] [Г] gf] [Т] {ВДЗ [Р] [б] [5] [4] [з] jzj Ш [ТатА~|
octa
РИС. 8.29.
-------72п.н
I
270 178
250
107
179
103
octa : 5-ATGCAAAG-3'
ТАТА : 5'-ТАТТТАТ-3'
GC : 5' ССбССС-З’
Р : б'-ТТАЬТСА-З'
ТС I + I1:5'-TCCCCAG-3r
GT II : 5'-GCTGTGGAATGT-3'
GT I : 5’-GGTGTGGAAAGT-3*
Sph II : 5'- AAGTATGCA-3’
Sph I :5'-AAGCATGCA-3’
Элементы базального промотора и энхансера ранней
области генома SV40. Показаны сами элементы, а также
их расположение в пределах базального промотора
и энхансера. Числа под изображением молекулы ДНК-
это расстояние в парах нуклеотидов от точки начала
транскрипции (ori). Сегменту длиной 72 п.н. соответст-
вуют два набора чисел; это означает, что данный сег-
мент тандемно повторяется. А и В домены энхансера,
которые, по-видимому, функционируют кооперативно.
52
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
они разделены сотнями и тысячами пар оснований,
в том случае, когда они находятся на близком
расстоянии друг от друга, существуют положения,
в которых их действие оптимально. Так, если эти
повторы разделены короткими сегментами ДНК,
соответствующими по длине какому-то числу пол-
ных витков спирали ДНК (на один виток приходит-
ся 10 п. н.), то активность транскрипции изменяется
незначительно. Если же разделяющие участки соот-
ветствуют по длине нечетному числу полувитков
спирали (например, 5 или 15 п. н.), то активность
транскрипции понижается (рис. 8.30). Влияние вста-
вок между повторами размером 21/22 п.н. и АТ-бо-
гатым сегментом длиной 17 п.н. зависит от их
длины; наблюдаемые эффекты весьма сложны и
различаются для разных сайтов инициации транс-
крипции. Эти данные свидетельствуют о том, что
эффективность транскрипции зависит от стереоспе-
цифических соответствий между повторяющимися
последовательностями длиной 21/22 п.н., элемента-
ми в составе сегмента размером 72 п. н. и другими
последовательностями, расположенными вблизи
сайта инициации транскрипции. Соблюдение этих
соответствий необходимо для присоединения спе-
цифических белков к различным элементам; взаимо-
действия между белками важны для инициации
транскрипции и зависят от их правильной взаимной
пространственной ориентации.
Энхансеры. Сегмент ДНК SV40 размером 72 п. н.
повышает также эффективность транскрипции дру-
гих генов. Например, ген 0-глобина млекопитаю-
щих в составе плазмиды, который обычно не транс-
крибируется после трансфекции в культивируемые
фибробласты, активируется, если к нему в любой
ориентации и на любом расстоянии присоединен
сегмент из 72 п. н. Благодаря своей способности
стимулировать транскрипцию, находящуюся под
контролем разных регуляторных элементов, неза-
висимо от расстояния и ориентации относительно
точек инициации этот элемент получил название
энхансера транскрипции. Обнаруженные впервые в
геноме SV40 энхансеры с очень близкими свойства-
ми, но с разными последовательностями являются,
как теперь установлено, важными компонентами
сигналов регуляции транскрипции многих генов
класса II всех исследованных эукариотических орга-
низмов (разд. 8.3. д-з). Энхансеры встречаются так-
же в регуляторных областях генов рРНК, транскри-
бируемых РНК-полимеразой I (разд. 8.2). Сущест-
вование энхансеров, часто находящихся на расстоя-
нии в тысячи пар оснований справа или слева от
точки инициации транскрипции, усложняет иденти-
фикацию промоторных элементов гена.
Обычно энхансеры совершенно разных генов
класса II взаимозаменяемы. В качестве примера
можно привести энхансеры регуляторных областей
вируса лейкоза мышей и SV40. Более того, энхансер
SV40 может функционировать в самых разных диф-
ференцированных тканях млекопитающих; он также
стимулирует транскрипцию у амфибий, в клетках
растений и даже у дрожжей S. pombe. Энхансер часто
обусловливает временную и клеточную специфич-
ность активации транскрипции гена. Например, как
мы увидим позже, при обсуждении регуляции транс-
крипции клеточных генов, энхансеры, ассоциирован-
ные с некоторыми генами, оказываются активными
только на определенных стадиях развития (разд.
8.3.д и ж). Кроме того, энхансер, необходимый для
транскрипции вируса полиомы, не может функцио-
нировать в недифференцированных эмбриональных
72п.н
Относительная эффективность транскрипции Размер вставки, п.н.
100 0
12 5
55 10
13 15
46 21
23 23
РИС. 8.30.
Оптимальное расстояние между регуляторными эле-
ментами промотора ранней области генома SV40. В од-
ну из позиций, указанных стрелками, встраивали сег-
менты ДНК разной длины и наблюдали за ходом
Ранние РНК
ori
Относительная
Размер Эффективность
вставки, транскрипции
П.Н. ф ©
0 100 100
-4 96 90
4 50 15
10 5 54
15 17 11
21 2 19
транскрипции, начинающейся в точках 1 и 2. Внизу
указана эффективность транскрипции при разных мо-
дификациях промотора в процентах от эффективности
транскрипции при немодифицированном промоторе.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
53
стволовых клетках, но активируется после диффе-
ренцировки, а энхансеры некоторых генов действуют
только в присутствии индукторов (разд. 8.3.е).
Можно предположить, что функция энхансеров
состоит в увеличении частоты инициации тран-
скрипции с участием РНК-полимеразы II. На это
указывают результаты опытов по включению мече-
ных предшественников в ген-специфичную РНК в
изолированных ядрах, в которых РНК продолжают
синтезировать только те молекулы полимеразы,
которые уже участвуют в транскрипции. В опытах,
о которых идет речь, трансдуцированный ген 0-гло-
бина, «сшитый» с энхансером SV40, продуцировал
больше 0-глобинспецифичной РНК, чем ген, у кото-
рого энхансер отсутствовал.
Энхансер - это наиболее сложный элемент, участ-
вующий в регуляции транскрипции SV40 (рис. 8.29).
Он состоит из нескольких различающихся коротких
элементов, расположенных в пределах повтора дли-
ной 72 п. н., и примыкающего к нему слева участка
из 25 п. н. Каждый элемент сам по себе обладает
очень низкой энхансерной активностью. Точная
идентификация элементов энхансера осложняется
тем, что их активность можно оценить только по их
влиянию на транскрипцию у определенного хозяина
и в отдельном опыте. Некоторые элементы вносят
примерно одинаковый вклад в общую активность
энхансера независимо от хозяина или системы
транскрипции, а другие функционируют только в
определенных клетках. Кроме того, мутации в ка-
ком-нибудь из этих элементов обычно понижают
активность энхансера в 5- 10 раз, в то время как при
делении всего энхансера эффективность транскрип-
ции уменьшается в сотни раз. Это говорит о том,
что энхансер включает большое число сигнальных
последовательностей и каждая из них вносит вклад
в его суммарную функциональную активность. Не
исключено, что степень и характер этой избыточно-
сти имеют реальное физиологическое значение, ко-
торое пока не удалось установить.
Три элемента энхансера, Р, Sph и октамер (octa),
образуют один домен (А), а спаренные GT- и
ТС-элементы-второй (В). Эти домены могут быть
инвертированы относительно друг друга или могут
находиться один от другого на расстоянии до
100 п. н. без заметного снижения активности энхан-
сера. Однако делеции в Sph или в октамере А-доме-
на либо GT- и ТС-элементах В-домена снижают
активность энхансера, причем степень этого сни-
жения зависит от того, в каких именно клетках
измеряется активность транскрипции. Элемент Р не-
существен для функционирования энхансера, но в
его присутствии транскрипция становится чувстви-
тельной к стимуляции форболовыми эфирами-ве-
ществами, активирующими клеточную протеинки-
назу.
Элемент Sph состоит из тандемно повторяю-
щихся последовательностей длиной 9 п. н.:
5'-AAGjATGCA-3'. Свое название этот элемент по-
лучил благодаря наличию сайта рестрикции для
Sphl в одном из повторов. При соединении двух
Sph-повторов длиной 9 п.н. образуется октамерная
последовательность (5'-ATGCAAAG-3')- Составные
Sph-октамерные элементы обеспечивают функцио-
нирование энхансера в самых разных клетках, вклю-
чая лимфоциты. При изменении участков двух
Sph-элементов, не входящих в состав октамера,
подавляется активность энхансера в фибробластах
или эпителиальных клетках, но его активность в
лимфоцитах остается на прежнем уровне. Если же
мутационные изменения затрагивают октамер, то
подавляется активность энхансера в лимфоидных
клетках, а способность энхансера активировать
транскрипцию в других клетках не меняется. Следо-
вательно, эти элементы регулируют как тканеспе-
цифичную, так и тканенеспецифичную транскрип-
цию.
Левее в энхансере находятся два ТС-повтора,
5'-TCCCCAG-3', и GT-богатая повторяющаяся по-
следовательность, 5'-GgTGTGGA£TGT-3'. Влияние
GT-I и GT-II на активность энхансера проявляется
в тех же самых клетках, что и влияние элементов
Sph. Мутации в любом из GT-элементов или в
ТС-П, а также вставки между ними уменьшают
активность энхансера; это наводит на мысль о ко-
оперативности обоих элементов. Области перекры-
вания GT- и ТС-элементов и их варианты с неболь-
шими отклонениями иногда называют кором энхан-
сера (5'-GTGGt?tG-3')- поскольку они встречаются
во многих вирусных и клеточных энхансерах. Одна-
ко сам по себе кор не может служить энхансером.
Любопытной особенностью некоторых элемен-
тов энхансера является то, что по отдельности или
даже попарно они не обладают энхансерной актив-
ностью, в то время как их мультимеры активны.
Например, тетрамеры, состоящие из спаренных
GT-элементов, почти так же активны, как весь
энхансер. Мультимеризация октамера приводит к
образованию структуры, которая может заменять
иммуноглобулиновый энхансер в лимфоидных клет-
ках, но эта структура неактивна в фибробластах.
Кор-мультимеры тоже могут функционировать в
качестве энхансера.
Каждый из указанных элементов энхансера SV40
наряду с другими i/ис-действующими регуляторны-
ми последовательностями представляет собой сайт
связывания одного или нескольких факторов транс-
крипции. По-видимому, все белки, связывающиеся
с ДНК, и вспомогательные белки, участвующие
в белок-белковых взаимодействиях, формируют
комплекс, который активирует РНК-полимеразу II
с тем, чтобы она могла инициировать транскрип-
54
ЧАСТЬ III ОРГАНИЗАЦИЯ. ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
цию. Состав и строение таких комплексов транс-
крипции, вероятно, не являются строго фиксирован-
ными, поскольку сочетания элементов, неактивные
сами по себе, становятся активными после мульти-
меризации. Кроме того, мутации, препятствующие
соединению фактора транскрипции с каким-либо
элементом, уменьшают активность энхансера, но
редко полностью блокируют его работу. Очевидно,
для того чтобы понять, как образуется комплекс
транскрипции и как он функционирует в процессе
инициации транскрипции, мы должны идентифици-
ровать и охарактеризовать различные транс-рей-
ствующие факторы транскрипции, а также изучить
их взаимодействие с различными элементами и друг
с другом.
Полипептидные факторы транскрипции. Боль-
шое число факторов, предположительно участвую-
щих в транскрипции ранней области SV40, было
выделено из клеточных экстрактов, способных осу-
ществлять транскрипцию с промотора ранней об-
ласти. Для их идентификации, очистки и изучения
применялись разные методы. Некоторые факторы
специфически связываются с конкретным элемен-
том, о чем свидетельствует их способность защи-
щать определенные основания в этом элементе от
расщепления ДНКазой I (футпринтинг; разд. 8.2.в),
а также уменьшение электрофоретической подвиж-
ности ДНК, содержащих такой элемент, при связы-
вании его с белком (смещение полосы). Имеются
данные о восстановлении транскрипционной актив-
ности в экстрактах, обедненных определенными
факторами, при добавлении последних, а также об
отсутствии такого восстановления в том случае,
когда сайты связывания разрушены. Это указывает
на то, что связывание данных факторов активирует
транскрипцию. Клонированы и секвенированы
гены, кодирующие некоторые факторы транскрип-
ции. Их экспрессия в Е. coli и способность синтези-
рованных продуктов активировать транскрипцию in
vitro позволяют идентифицировать важные домены
белков.
Кроме факторов, необходимых для функциони-
рования энхансеров и различающихся для разных
генов, в транскрипции большинства клеточных и
вирусных генов класса II участвуют дополнитель-
ные белки, которые помогают «направлять» РНК-
полимеразу II к соответствующим сайтам инициа-
ции. Вопрос о том, находятся ли эти белки среди
субъединиц, которые обычно бывают связаны с
РНК-полимеразой II, или они относятся к числу
вспомогательных факторов транскрипции, которые
присоединяются и отсоединяются во время инициа-
ции, пока не решен. Относительная эффективность
формирования комплекса инициации и его стабиль-
ность зависят от присутствия ТАТА-подобной по-
следовательности в окрестности пары оснований
— 25, а также от последовательности, в которую
встроен ТАТА-бокс, простирающейся от -40-й до
+• 10-й пары оснований. У млекопитающих широко
распространен белок, обозначаемый TF1ID и на-
зываемый «фактором связывания последовательно-
сти ТАТА». Он связывается с АТ-богатой областью
промотора SV40, о чем свидетельствует его способ-
ность защищать элемент АТ от расщепления ДНКа-
зой I (рис. 8.31). К этому исходному комплексу
присоединяется второй фактор, TFI1A, который ста-
билизирует связь TFIID с последовательностью
ТАТА. Далее к комплексу ДНК-TFIID-TFIIA по-
следовательно присоединяются фактор TFIIB и
РНК-полимераза II. Связывание еще двух вспомо-
гательных факторов, TFIIE и TFIIF, завершает
формирование комплекса преинициации. В присут-
ствии четырех рибонуклеозидтрифосфатов этот
-50 -зо -10 +ю +30
Этап 1
£ Этап 2
РИС. 8.31.
Сборка комплекса транскрипции из отдельных факто-
ров транскрипции (ТЕНА, В. D, Е, F) и РНК-полимеразы
II. Установлено, что сначала фактор TFIID связывается
с ТАТА-боксом, а затем присоединяются TFIIA и TFIIB
(в какой именно последовательности до конца неиз-
вестно). Участок связывания включает сайт инициации
транскрипции и прилегающие области (они указаны
числами в верхней части рисунка). [S. Buratowski et al..
Cell 56 (1989), p. 549.]
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
55
комплекс осуществляет инициацию и элонгацию
транскрипции. TF11A. TFIIB, TF1IE и TFIIF. а также
РНК-полимераза 11 не связываются с данной об-
ластью промотора в отсутствие TFIID; однако эти
белки, возможно, контактируют с соседними по-
следовательностями ДНК. Так, после формирова-
ния комплекса преинициации и начала транскрип-
ции область, защищенная от расщепления ДНКа-
зой, расширяется до —45- +30 пар оснований.
Факторы, подобные белкам TFIID и TFIIA мле-
копитающих, идентифицированы у дрожжей. Фак-
тор связывания ТАТА, напоминающий соответ-
ствующие факторы млекопитающих и дрожжей,
выделен также из Drosophila. Замечательно, что бел-
ки TFIID и TFIIA из дрожжей могут заменять
аналогичные белки млекопитающих при сборке
комплекса транскрипции, который обеспечивает
инициацию в правильных сайтах. Эта взаимозаме-
няемость факторов транскрипции (при незначитель-
ном изменении эффективности или специфичности
транскрипции) у совершенно неродственных орга-
низмов присуща аппаратам транскрипции всех эука-
риот. Очевидно, структура и функция белков, вхо-
дящих в состав многокомпонентных комплексов,
в процессе эволюции не изменились.
Для оптимальной транскрипции кроме белков,
составляющих описанный комплекс преинициации,
необходим фактор транскрипции Spl, который свя-
зывается с GC-богатыми элементами размером
21/22 п. н., а также ряд других факторов транскрип-
ции (рис. 8.32). Эти дополнительные факторы зави-
сят от типа клеток и условий, в которых происходит
экспрессия промотора ранней области SV40.
Фактор транскрипции Spl в виде мономера свя-
зывается в области, охватывающей три GC-богатых
повтора из 21/22 п.н. Для этого связывания, кото-
рое осуществляется в отсутствие других белков или
нуклеозидтрифосфатов, нужна только гексамерная
последовательность. При связывании Spl с промо-
тором ранней области гексамерные последователь-
ности 5-GGGCGG-3', присутствующие в каждом
повторе, оказываются защищенными от расщепле-
ния ДНКазой I, а остатки G не метилируются
диметилсульфатом (рис. 8.33). Контакты между Spl
и GC-повтором ограничиваются большим желоб-
ком молекулы ДНК и остатками G матричной цепи.
Сродство Spl к гексамерным повторам неодинако-
во, возможно из-за различий в их расположении
и последовательности (рис. 8.33). Элиминация дан-
ной области или мутационные изменения специфи-
ческих гексамерных последовательностей приводят
к изменению как связывания Spl, так и эффективно-
сти транскрипции. Таким образом, присоединение
Spl к этому элементу последовательности имеет
важное значение для транскрипции.
Если принять во внимание частоту, с которой
встречаются сайты связывания Spl в генах млекопи-
тающих, то относительная избыточность Spl не
кажется удивительной. Например, в одной клетке
HeLa (часто используемая линия клеток, полученная
из опухоли шейки матки человека) содержится от
5000 до 10000 молекул Spl. Благодаря высокой
концентрации Spl удается получать этот белок в
почти гомогенном виде, а также клонировать коди-
рующую его ДНК. Размер белковой молекулы,
оцененный исходя из нуклеотидной последователь-
ности клонированной ДНК, равен 80 кДа. хотя вы-
деленный белок Spl имеет эффективную мол. массу
90 100 кДа. Возможно, это связано с гликозилиро-
ванием Spl in vivo, хотя для ядерных белков такая
модификация не характерна. Мутационный анализ
различных участков последовательности, кодирую-
щей Spl, показал, что один из доменов этого белка
необходим для присоединения к ДНК, а другие для
активации транскрипции (рис. 8.34). Домен связы-
вания ДНК находится вблизи С-конца и имеет три
участка. Каждый участок содержит расположенные
определенным образом остатки цистеина и гисти-
дина, которые образуют хелатную связь с Zn2 + (так
называемые «цинковые пальцы»; разд. 8.7.а). Изме-
Кор тс-11,1
„О Sph Sph
АР5 NF-кВ SphSph
C/EBP otf‘1
OTF-2
РИС. 8.32.
Факторы транскрипции и сайты их связывания в про-
моторе ранней области генома SV40. Разные регуля-
торные элементы выделены разными цветами и квад-
ратными скобками. Внизу указаны белки, связыва-
ющиеся с родственными последовательностями. Комп-
Spl Spl Spl Spl Spl Sp1 TFHD-(TFnA;B,E,F)
TFHD-dFIlA.BE.F)
леке транскрипции, состоящий из указанных на ри-
сунке факторов, занимает АТ-участок, ori и область
начала транскрипции. Представлены Т-антигены, распо-
ложенные под двумя сайтами наиболее прочного свя-
зывания.
56
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Взаимодействие фактора транскрипции Spl с GC-бо-
гатыми кластерами повторов из 21 '22 п. н. Сегменты
ДНК, содержащие повторы из 21/22 п.н., инкубировали
с очищенным белком Sp1 и затем обрабатывали диме-
тилсульфатом. Остатки G в трех GC-богатых кластерах
(они окрашены наиболее интенсивно) оказываются
полностью защищенными от метилирования. Остатки
гуанина (д), содержащиеся в следующем по интенсив-
ности окраски сегменте, были защищены частично, а
остатки в бледно окрашенных сегментах не защищены
совсем. Все защищенные остатки G находятся в одной
цепи ДНК.
нения в последовательности отдельных пальцев или
в их числе и расположении уменьшают эффектив-
ность связывания и активации транскрипции. Кроме
домена связывания ДНК, для оптимальной актива-
ции транскрипции нужны два участка, богатых глу-
тамином, которые находятся в N-концевой части
полипептида, и короткие последовательности, флан-
кирующие цинковые пальцы (рис. 8.34). Роль участ-
ков, богатых серином и треонином, пока не уста-
новлена.
Элемент, находящийся у З'-концов обоих повто-
ров размером 72 п. н. (обозначенный на рис. 8.29
буквой Р), служит сайтом связывания белка, назван-
ного API (рис. 8.32). Какую роль играет связывание
API с этой последовательностью, неясно, поскольку
мутации в P-элементе, ингибирующие присоедине-
ние API, не влияют на активность энхансера; об
этом свидетельствуют результаты исследования
транскрипции in vivo. Возможно, взаимодействие
API с ДНК необходимо для транскрипции ранней
области SV40 только в определенных условиях,
например в отсутствие других факторов связывания
энхансера или при определенных внешних воздей-
ствиях.
API принадлежит к семейству близкородствен-
ных белков с мол. массой примерно 40 кДа, специ-
фически связывающихся с канонической последова-
тельностью 5'-TGANTCA-3' (N- любое основание).
У млекопитающих белки типа API кодируются
множественными генами; некоторые из них клони-
рованы. Расшифровка аминокислотных последова-
тельностей некоторых API-белков показывает, что
ДНК-связывающие домены этих белков сохрани-
лись в процессе эволюции, в то время как другие
участки претерпели изменения. Интерес к физиоло-
гической роли API усилился после того, как были
сделаны два открытия. Во-первых, у трансформи-
рующего вируса кур был обнаружен онкоген N-jun,
Домены, богатые глутамином
РИС. 8.34.
Структурные домены белка Sp1. Схема построена ис-
ходя из данных о нуклеотидной последовательности
кодирующего данный белок участка ДНК. ДНК-связы-
вающий домен включает три цинковых пальца, рас-
положенных вблизи С-конца белковой молекулы. Два
глутамин-богатых домена вблизи N-конца и два корот-
ДНК-связывающнй
домен
ких участка, фланкирующих цинковые пальцы, необхо-
димы для активации транскрипции. Домены, богатые
серином/треонином, несущественны для связывания
ДНК или для активации, но, возможно, влияют на один
или на оба этих процесса, поскольку содержат сайты, по
которым может происходить модификация белка.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
57
кодирующий API-подобный белок. Клеточная ко-
пия v-jun, c-Jun, относится к семейству генов API.
Вероятно, каждый из API-подобных белков играет
свою регуляторную роль, которая может зависеть
от того, в каком окружении находится родственная
связывающая последовательность в регуляторной
области гена. Во-вторых, было показано, что у
дрожжей содержится ген, белковый продукт которо-
го, GCN4, гомологичен белкам семейства API мле-
копитающих. Так же как белок связывания ТАТА из
дрожжей может заменять свой аналог TFIID у мле-
копитающих, GCN4 может активировать гены мле-
копитающих, которые зависят от API, а белок API
из млекопитающих может заменять GCN4 при
транскрипции дрожжевых генов, которым необхо-
дим этот фактор. Помимо гена GCN4 дрожжи
содержат также дополнительные гены, кодирующие
семейство API-подобных белков. Их характерис-
тики и способность активировать транскрипцию
в настоящее время изучаются.
Сами по себе API-белки плохо связываются
с узнаваемым ими элементом. Чтобы связывание
было прочным, API должен соединиться с другим
белком, Fos. Особенности строения белков Fos и
API и их взаимодействие друг с другом, благодаря
которому они могут влиять на эффективность
транскрипции, мы рассмотрим в разд. 8.6.6.
Повторяющиеся элементы последовательности
Spb (рис. 8.29), ответственные за функционирование
энхансера в нелимфоидных клетках (например, клет-
ках HeLa), влияют на активность энхансера, связы-
ваясь с каким-то не охарактеризованным пока бел-
ком. Мутации в элементе Spb приводят к парал-
лельному уменьшению активности энхансера и бе-
локсвязывающей способности. Интересно, что бе-
лок, присоединяющийся к элементу Sph, отсутствует
в лимфоидных клетках, в которых этот элемент не
участвует в функционировании энхансера, а тот же
самый частично очищенный белок связывается так-
же с элементом GTII, неродственным Sph и распо-
ложенным левее его. Влияет ли эта двойная спе-
цифичность (и если да, то каким образом) на актив-
ность энхансера, неясно. Таким образом, обнаружи-
вается одна странная, но характерная особенность
некоторых факторов транскрипции: способность
связывать два разных и неродственных элемента
последовательности.
Как мы уже отмечали, при перекрывании эле-
ментов Sph образуется октамерная последователь-
ность 5'-ATGCAAAG-3', целостность которой важна
для функционирования энхансера в лимфоидных
тканях. Такая последовательность-иногда с незна-
чительными изменениями-часто встречается в об-
ласти регуляции транскрипции широкого круга ге-
нов у самых разных организмов. Ее функциониро-
вание опосредуется связыванием с белками двух
классов, OTF-1 и OTF-2. Эти ДНК-связывающие
белки узнают каноническую последовательность
5'-ATGCAAAT-3' или близкородственные последо-
вательности, как правило находящиеся в окружении
других i/izc-действующих элементов. Белок OTF-1
(мол. масса примерно 90 кДа, в литературе известен
под разными названиями) распространен очень ши-
роко и участвует в активации транскрипции ряда
клеточных и вирусных промоторов. В то же время
OTF-2 (мол. масса примерно 60 кДа) присутствует
только в клетках лимфоидного происхождения, где
взаимодействует с октамерными элементами в об-
ластях энхансеров-промоторов генов иммуноглобу-
линов и других, специфичных для лимфоидной тка-
ни генов, стимулируя их транскрипцию (разд. 8.3.д).
Поскольку функционирование энхансера SV40 в не-
лимфоидных клетках опосредуется Sph-связываю-
щим белком, OTF-1 и последовательность, с кото-
рой он связывается, не обязательны. В лимфоцитах
же, у которых фактор связывания Sph отсутствует,
для транскрипции с промотора ранней области ге-
нома SV40 необходимо связывание OTF-2 с окта-
мерной последовательностью.
Несмотря на то что нуклеотидные последова-
тельности элементов ТС-I и ТС-И идентичны, белок
АР2 связывается только с ТС-П. Мутационные из-
менения в ТС-П блокируют связывание АР2 и по-
нижают активность энхансера. Соседние элементы
ТС-П и GT-I вместе образуют кор, который являет-
ся сайтом связывания фактора транскрипции АРЗ,
а в клетках печени-фактора С/ЕВР (рис. 8.32). Му-
тационные изменения в нуклеотидной последова-
тельности кора тоже блокируют связывание АРЗ
и нарушают работу энхансера. Необходимо ли для
функционирования энхансера присоединение обоих
факторов, АР2 и АРЗ, или каждый из них может
самостоятельно активировать транскрипцию в за-
висимости от типа клеток или физиологического
статуса, неясно. Частично очищенные факторы
транскрипции АР2 и АРЗ способны к связыванию
специфических последовательностей и могут акти-
вировать транскрипцию с промоторов, содержащих
родственные элементы.
В различных клеточных экстрактах обнаружены
белки, принадлежащие к семейству GT-связы-
вающих белков. Они были идентифицированы
по своей способности связываться с элементом
GT-П и с прилегающими последовательностями
(5'-CTGTGGAATGT-3'). Эти белковые факторы от-
личаются друг от друга клеточной специфичностью
и сайтами связывания, которые были выявлены при
помощи метода отпечатков с использованием
ДНКазы I и защиты от метилирования, а также при
изучении влияния мутаций в последовательности на
связывание. Один из этих факторов, АР5, присут-
ствует в клетках HeLa, но не в лимфоцитах. Воз-
58
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
можно, именно этим объясняется, почему интактная
последовательность элемента GT-П необходима для
функционирования энхансера в клетках HeLa, но не
в лимфоцитах. Второй фактор, АР4, связывается
с последовательностью 5'-CAGCTGTGG-3', которая
перекрывается с элементом GT-П. Роль АР4 в функ-
ционировании энхансера неясна. Хотя последова-
тельности GT-I и GT-П имеют 10 одинаковых пар
оснований из 12, они связываются с разными белка-
ми. АРЗ, ядерный фактор клеток HeLa, присоеди-
няется к GT-1, но не к GT-П. Мутации в GT-1,
мешающие связыванию АРЗ, приводят к пониже-
нию активности энхансера.
Природа надмолекулярного комплекса, обра-
зующегося при соединении разнообразных белко-
вых факторов с соответствующими участками, и его
роль в стимуляции ранней транскрипции у SV40
в настоящее время до конца не установлены. Тем не
менее изучение этой системы уже внесло значитель-
ный вклад в выяснение механизмов регуляции
транскрипции, поскольку она представляет собой
хорошую модель для детального исследования про-
цессов трансактивации и дифференциальной экс-
прессии генов.
Т-антиген: многоцелевой реггл.чтор транскрип-
ции. После трансляции в цитоплазме одной из двух
ранних мРНК образовавшийся Tag транспортирует-
ся в ядро, где стимулирует инициацию репликации
вирусной ДНК. а также участвует в репрессии и ак-
тивации ранней и поздней областей транскрипции
соответственно. Эту важную регуляторную роль
Tag выполняет в комплексе со специфическими по-
следовательностями ДНК и с другими белками.
Tag связывается с ДНК SV40 в трех сайтах,
обозначенных на рис. 8.28 как Tl, Т2 и ТЗ. Сайт Т1,
обладающий наибольшим сродством к Tag, нахо-
дится на участке ДНК, который кодирует 5'-не-
транслируемую последовательность ранних транс-
криптов. Т2, обладающий меньшим сродством к
Tag, охватывает палиндром длиной 27 п. н. в ori. ТЗ
обладает самым низким сродством к Tag и состоит
из GC-богатых повторов длиной 22 п. н., примы-
кающих к блоку АТ. По-видимому, главными де-
терминантами связывания белка с этими сайтами
являются средний и С-концевой домены Tag, по-
скольку N-конец для этого процесса несуществен.
Вместе с тем фосфорилирование и олигомеризация
полипептида Tag влияют на связывание, хотя меха-
низм этого влияния неясен.
В опытах in vivo и in vitro показано, что связыва-
ние Tag с сайтами Т1 и Т2 блокирует инициацию
транскрипции в большинстве сайтов инициации
транскрипции ранней области, расположенных пра-
вее. Вероятнее всего, присоединение Tag к этим
сайтам мешает сборке комплекса преинициации на
участке от —45 до +30 пары оснований (рис. 8.31).
Кроме того, возможно, что Tag влияет на экспрес-
сию, инициируемую в расположенных справа сай-
тах, связываясь с API и таким образом мешая
взаимодействию фактора транскрипции с родствен-
ной ему последовательностью энхансера. Взаимо-
действие Tag и API может влиять и на транскрип-
цию клеточных генов, для которой необходим API.
Транскрипция ранних мРНК начинается от сай-
тов, находящихся в ori, в отсутствие Tag (предранние
мРНК). Синтез задержанных ранних РНК иниции-
руется левее, в месте соединения АТ-элемента с
GC-богатым повтором длиной 22 п. н. (или рядом
с ним). Любопытно, что, когда Tag связан с сайтами
Т1 и Т2. транскрипция предранних мРНК блокиру-
ется, зато транскрипция задержанных ранних мРНК
активируется. Транскрипция с этих расположенных
левее сайтов зависит от целостности сайтов связы-
вания Spl, но. что удивительно, мало изменяется
при делециях последовательностей энхансера. Более
того, мутации в Tag или в ori, которые блокируют
репликацию, но не влияют на транскрипцию пред-
ранней мРНК, предотвращают переход транскрип-
ции к более далеким сайтам инициации. Каковы
физиологические последствия сдвига точек инициа-
ции транскрипции неясно; возможно, они связаны
со сниженной эффективностью трансляции при син-
тезе Tag на более длинной мРНК.
Что же происходит при реализации этой слож-
ной стратегической программы транскрипции? Сра-
зу после заражения Tag начинает образовываться
с большой скоростью; его накопление в клетке
приводит к усилению репликации ДНК и к образо-
ванию большого числа матриц для транскрипции.
Скорость накопления Tag регулируется связывани-
ем его с сайтами Т1 и Т2 и, возможно, взаимодей-
ствием между Tag и API. С началом репликации
ДНК общая скорость транскрипции ранней области
уменьшается; число транскриптов, образованных
при участии сайтов инициации транскрипции, рас-
положенных слева, в 5 10 раз превышает число
транскриптов, образованных при участии сайтов,
расположенных справа. Но поскольку трансляция
более длинных мРНК неэффективна, накопление
Tag снижается еще больше. Наряду с понижением
эффективности транскрипции ранней области проис-
ходит усиление транскрипции поздней области, в
результате чего транскриптов поздней области об-
разуется в 20 раз больше, чем ранней; заметим, что
мРНК поздней области необходимы для синтеза
белков капсида вириона. Таким образом, сложное
взаимодействие вирусных и клеточных факторов
транскрипции обеспечивает упорядоченное и эффек-
тивное развитие инфекционного процесса.
Транскрипция поздней области протекает в на-
правлении, противоположном направлению транс-
крипции ранней области. Инициация осуществляет-
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
59
ся в разных сайтах, главным образом в GC-богатых
повторах и в энхансере. Относительно малоизучен-
ный промо гор поздней области не имеет ТАТА-по-
добной последовательности, но содержит сайты свя-
зывания Spl и до сих пор не охарактеризованные
области в энхансере. Механизм активации транс-
крипции поздней области под действием Tag неиз-
вестен. Существует точка зрения, что активация
результат белок-белкового взаимодействия, а не
связывания с ДНК. Возможно, взаимодействие Tag
с API или с другим, пока неизвестным фактором
транскрипции обусловливает активацию транскрип-
ции поздней области.
Промотор гена тимидинкиназы вируса герпеса.
Как мы уже отмечали, ранняя область генома SV40
транскрибируется сразу после заражения, при этом
никакие вирусные белки не требуются. В то же
время для транскрипции генов предранних белков
(IEP) вируса герпеса (HSV) необходим фактор
транскрипции, кодируемый вирусом и входящий
в состав инфицирующего вириона. IEP-это регу-
ляторные белки, которые активируют транскрип-
цию нескольких задержанных ранних генов, име-
ющих отношение к репликации ДНК. Транскрип-
цию поздних генов, многие из которых кодируют
структурные белки вириона, IEP могут активиро-
вать только после начала репликации.
Для оптимальной транскрипции гена тимидин-
киназы (ТК) HSV, одного из задержанных ранних
генов, необходим один из пяти 1ЕР, а именно IEP3,
представляющий собой высокомолекулярный (при-
мерно 175 кДа) фосфорилированный белок. Об этом
свидетельствует тот факт, что уровень экспрессии
гена ТК, трансфицированного в клетки, в которых
может экспрессироваться ген IEP3, в 10 100 раз
выше, чем в клетках, где IEP3 отсутствует. Таким
образом, промотор гена ТК HSV-это еще одна
удобная модельная система для анализа взаимодей-
ствия клеточных и вирусных факторов в регуляции
транскрипции вирусных генов.
Элементы промотора ТК-гена HSV были иден-
тифицированы при помощи мутаций (делеций и
замен), систематически вносимых в участок длиной
100 п. н., расположенный непосредственно перед
точкой начала транскрипции. Способность изменен-
ных промоторов к активации транскрипции иссле-
довали в опытах по введению их в ооциты Xenopus
(разд. 7.7.6), при трансфекции клеток млекопи-
тающих в культуре, а также путем измерений эф-
фективности транскрипции в экстрактах из неинфи-
цированных клеток. Между парой оснований —105
и сайтом кэпирования идентифицировано четыре
существенных участка (рис. 8.35). Один из них, рас-
положенный вблизи точки начала транскрипции,
представляет собой вездесущую ТАТА-подобную
последовательность; три других расположены
дальше от этой точки и простираются примерно до
— 105-й пары оснований. Дистальные последова-
тельности состоят из трех элементов; два из них
представляют собой GC-богатые гексануклеотиды,
сходные с обнаруженными в 21/22 п. н.-повторах
промотора ранней области SV40. Между ними на-
ходится последовательность, содержащая элемент
5'-ССААТ-3'.
Изменение гомологии ТАТА нарушает точность
и эффективность инициации транскрипции, тогда
как модификации трех дистальных элементов при-
водят к более серьезным нарушениям процесса
транскрипции. Следует отметить, что мутации в ле-
вом GC-элементе, GC-II. подавляют транскрипцию
в большей степени, чем аналогичные мутации в
GC-I; кроме того, делеция GC-I не так заметно
отражается на активности транскрипции, как деле-
ция GC-П. При замене GC-I сегментом, содержа-
щим GC-II и ССААТ (пары оснований примерно от
— 75 до —105) в противоположной ориентации,
образуется даже более активный промотор. Воз-
можно, различие в активности двух GC-элементов
связано с тем, что GC-II находится ближе к после-
довательности ССААТ; впрочем, локальные разли-
чия в последовательностях, окружающих GC-эле-
менты, также могут играть роль.
Фракционирование белков, участвующих в син-
тезе мРНК ТК in vitro, показало, что для осу-
ществления транскрипции необходимы по крайней
мере два фактора (рис. 8.36). Один из них, Spl,
-106 -97 , । । -88 -81 I I -55 -47 I 1 -28 -21 I I +1
’ ( ACCCCGCCCA ( (TCMTGGcf fG GGOcOGCG( (CATATTAAf
, (TCGGGCGGGT ( (agtaaccg( (СССС6СС6С( (gtataatt(
GC-II ССААТ- бокс GC-I TATA- бокс
РИС. 8.35.
Элементы, существенные для транскрипционной актив-
ности промотора гена тимидинкиназы вируса герпеса I.
Кроме последовательности ТАТА, расположенной при-
мерно на 25 п.н. левее начальной точки (+1), для
функционирования промотора HSV-ТК необходим
ССААТ-бокс матричной цепи, фланкированный GC-бо-
гатыми последовательностями с взаимно противопо-
ложной ориентацией.
60
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 8.36.
Элементы промотора гена тимидинкиназы вируса гер-
песа I: ТАТА-бокс, два GC-гексануклеОтида с взаимно
противоположной ориентацией (стрелки) и ССААТ-бокс.
Сайты связывания Spl отмечены двумя стрелками.
Жирная стрелка указывает на то, что Sp1 связывается
с дистальным GC-злементом более прочно. CTF-фак-
тор транскрипции, связывающийся с ССААТ; стрелкой
показан сайт его прочного связывания.
связывается как с GC-I, так и с GC-II, другой,
CTF,- с элементом ССААТ, о чем свидетельствуют
данные, полученные методом отпечатков с исполь-
зованием ДНКазы I. Как и ожидалось, внесение
мутаций в GC- или ССААТ-элементы подавляет
примерно в одинаковой степени как связывание
соответствующих факторов, так и транскрипцию.
Инициация транскрипции гена ТК скорее всего за-
висит также от белков, которые группируются во-
круг последовательности ТАТА между парами ос-
нований — 40 и +35. Таким образом, как и в случае
промотора ранней области генома SV40, для ини-
циации транскрипции с участием ТАТА-блока необ-
ходимо несколько факторов транскрипции. Взаимо-
действуют ли (и каким образом) факторы Spl и CTF
друг с другом и как они влияют на активность
и образование комплекса преинициации, неизвестно.
Во всех проведенных к настоящему времени
опытах промотор ТК исследовали в отсутствие
IEP3. Участвуют ли какие-то другие элементы в ак-
тивации транскрипции под действием этого белка?
По-видимому, на этот вопрос следует ответить
отрицательно, поскольку опыты по трансфекции
с использованием таких же мутантов, как и при
идентификации элементов промотора, показывают,
что для активации транскрипции с участием IEP3 in
vivo нужна только область между парой оснований
—105 и сайтом кэпирования. IEP3 не связывается
с ДНК в этой области; возможно, он активирует
транскрипцию с помощью белок-белковых взаимо-
действий. Природа этих взаимодействий и механизм
их влияния на инициацию транскрипции неизвестны.
IEP3 может стабилизировать уже сформированные
комплексы или участвовать в активации комплекса
транскрипции.
Многие промоторы клеток эукариот и вирусов
содержат в одной из двух цепей ДНК на расстоянии
примерно 80-120 п.н. до точек начала транскрип-
ции ССААТ-элемент. Целостность этой последова-
тельности и окружающих ее нуклеотидов совершен-
но необходима для функционирования промотора.
Имеются убедительные данные о том, что ССААТ
является частью элемента, который активирует
транскрипцию, выполняя роль сайта связывания
для одного из факторов транскрипции. С другой
стороны, известно, что с ССААТ связываются не-
сколько (по крайней мере три) разных белков. Неко-
торые из них были обнаружены в различных клетках
млекопитающих; при этом использовались методы
отпечатков с применением ДНКазы I, опыты по
защите от метилирования, данные об уменьшении
подвижности связанной ДНК в геле и опыты по
конкурентному связыванию. Так, белок CTF, связы-
вающийся с последовательностью ССААТ в про-
моторе ТК, принадлежит к семейству ССААТ-свя-
зывающих белков CTF/NF-1, которые активируют
также промоторы а-глобина и вируса саркомы
мышей. Другой белок, присутствующий в клетках
печени млекопитающих, связывается с коровой
последовательностью энхансера SV40 и ССААТ.
Клетки HeLa содержат два разных ССААТ-свя-
зываюших белка, СР-1 и СР-2, которые по своему
сродству к промотору явно отличаются друг от
друга, а также от CTF, присутствующего в тех же
клетках. Совершенно очевидно, что существуют
и другие белки, связывающиеся с ССААТ. На это
указывают многочисленные данные о том, что
ССААТ является важным элементом различных ре-
гуляторных последовательностей, в том числе про-
моторов, которые работают на разных стадиях
развития.
Генетическое происхождение различных ССААТ-
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
61
связывающих белков неизвестно. Возможно, неко-
торые из них являются продуктами отдельных
генов, однако три формы CTF в клетках HeLa
кодируются разными мРНК, транскрибируемыми
с одного гена и образующимися в результате диф-
ференциального сплайсинга транскрипта (разд.
8.5.е). Дифференциальный сплайсинг приводит к
образованию различных форм одного фактора
транскрипции; по-видимому, эти формы связыва-
ются с одним и тем же элементом, но выполняют
разные регуляторные функции. У дрожжей тоже
есть ССААТ-элементы и родственные им белки.
Один из таких белков, димер, состоящий из двух
разных субъединиц (НАР2 и НАРЗ), активирует
транскрипцию гена, кодирующего изо-2-цитохром с.
Комплекс НАР2/НАРЗ при связывании с последова-
тельностью ССААТ в промоторе гена цитохрома
с дрожжей контактирует с ДНК таким же образом,
как и белок млекопитающих СР-1. Более того,
мутации в элементе ССААТ влияют на связывание
обоих белков одинаковым образом. Удивительно,
что компоненты СР-1 и НАР2/НАРЗ в функцио-
нальном отношении взаимозаменяемы. Так. комп-
лексы, состоящие из компонентов НАР2 или НАРЗ
и СР-1, сохраняют способность связываться с ССААТ
и активировать транскрипцию. Это еще раз показы-
вает, что белки, участвующие в регуляции транс-
крипции. не изменяются в ходе эволюции.
д. Регуляция генов, специфичная
в отношении типа клеток
и стадии развития
Многоклеточные растения и животные состоят
из самых разных клеток, имеющих характерную
морфологию и выполняющих специализированные
функции. Эта специализация возникает на разных
стадиях эмбрионального развития. Она поддержи-
вается автономно либо проявляется в ответ на
специфические межклеточные контакты или внекле-
точные стимулы. Так или иначе, фенотипическое
разнообразие клеток обусловливается дифференци-
альным характером экспрессии генов, приводящим
к накоплению и распространению различных генных
продуктов. Одна из ключевых проблем биологии
развития заключается в следующем: каков меха-
низм дифференциальной экспрессии генов в клетках,
содержащих по существу одинаковый геном? Мы
знаем, что установление и поддержание дифферен-
цированного состояния осуществляются, в частно-
сти, путем регуляции транскрипции. Так. в клетках
одного типа транскрипция определенных генов ре-
гулируется совершенно одинаковым специфическим
образом на протяжении всей жизни организма, на-
чиная с эмбрионального уровня (разд. 8.3.ж). Гены,
активно транскрибирующиеся в одних клетках (на-
пример, в клетках мозга или в лимфоцитах), совсем
не транскрибируются в других. Более того, возмож-
но, что транскрипция различных генов инициирует-
ся в клетках или тканях разных типов в ответ на
одни и те же сигналы. Не исключено также, что один
и тот же сигнал может инициировать транскрипцию
одних генов и подавлять транскрипцию других.
В этом разделе мы рассмотрим примеры дифферен-
циальной регуляции транскрипции, обусловливаю-
щей тканеспецифичную экспрессию и экспрессию,
специфичную для определенной стадии развития.
Иммуноглобулиновые гены: цис-элементы. Отли-
чительным свойством генов иммуноглобулинов (1g)
и рецепторов Т-клеток (TCR) является то, что их
i/wc-действующие регуляторные элементы далеко
отстоят друг от друга в геноме клеток зародышевой
линии и сближаются в результате рекомбинации на
определенных стадиях развития лимфоидных клеток
(разд. Ю.б.в). Такие перестройки приводят к пере-
мещению сигналов транскрипции, находящихся на
5'-концах каждого гена 1g, к элементам энхансера,
расположенным в интронах, в случае генов как
тяжелых (IgH), так и легких (IgL) цепей 1g (рис. 10.67
и 10.68). Аналогичные перестройки приводят к сбли-
жению в процессе онтогенеза Т-клеток соответ-
ствующих промоторов и энхансеров, которые регу-
лируют транскрипцию генов, кодирующих две по-
липептидные цепи TCR (рис. 10.74). Таким образом,
специфичность экспрессии генов 1g и TCR регулиру-
ется с помощью двух механизмов. Один из них
определяет, когда и в каких клетках должны про-
изойти перестройки; этот этап является важной
предпосылкой активации транскрипции. Второй ме-
ханизм зависит от взаимодействий между перестро-
енными элементами и факторами транскрипции,
доступность и активность которых дифференциаль-
но регулируются в ходе созревания В- и Т-клеток.
Здесь мы остановимся на втором способе регуляции
генов IgH и IgL(x) в В-клетках. Мы рассмотрим
природу и организацию элементов, участвующих
в регуляции транскрипции генов 1g, факторы транс-
крипции, которые связываются с этими элементами,
и те молекулярные особенности, которые обуслов-
ливают способность факторов активировать транс-
крипцию. Аналогичные элементы и факторы участ-
вуют в регуляции транскрипции генов а- и р-цепей
TCR в Т-клетках.
Несколько специфических элементов образуют
так называемый базальный промотор гена IgH
(рис. 8.37). Это ТАТА-подобная последователь-
ность, находящаяся на расстоянии 20 30 п.н. от
точки начала транскрипции, консервативная окта-
мерная последовательность (octa) 5'-ATGCAAAT-3',
расположенная еще примерно на 30- 60 п. н. левее,
и каноническая гептамерная последовательность
62
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
От'100 От-60
до -120 до -90
От -25 +1
до-30
От -60 От -25 +1
до -90 до -30
РИС. 8.37.
Элементы, участвующие в регуляции транскрипции ге-
нов тяжелой и легкой (и) цепей иммуноглобулина.
lgH-промотор состоит из ТАТА-бокса, октамерной
(octa) и гептамерной (hepta) последовательностей сле-
ва от точки начала транскрипции и энхансера (Ен),
находящегося в интроне между вариабельным (VDJ)
и константным (Сн) участками. Промотор гена IgL (и)
содержит ТАТА-бОкс и имеющую противоположную
ориентацию октамерную последовательность перед
точкой начала транскрипции, а также энхансер Еу в
интроне между вариабельным (VJ) и константным (Ск)
участками.
(hepta) 5'-CTCATGA-3', находящаяся примерно на
25 п.н. левее октамера. Наиболее важным для
транскрипции и клеточной специфичности среди эле-
ментов базального промотора является октамер.
Его делеция приводит к полной утрате промотором
транскрипционной активности, а встраивание ок-
тамерного сегмента в промотор, обычно не участ-
вующий в транскрипции в лимфоцитах, инициирует
транскрипцию в В-клетках. Делеция гептамера при-
мерно в пять раз снижает уровень экспрессии гена
IgH; однако его роль в проявлении В-клеточной
специфичности не установлена. Базальный промо-
тор гена IgL(x) сходен с промотором IgH по распо-
ложению ТАТА-подобной и октамерной последова-
тельностей. но октамерная последовательность в
промоторе гена IgL(x) имеет противоположную
ориентацию. Все промоторы гена IgL(x) содержат
также консервативный 15-нуклеотидный элемент
вблизи пары оснований —100. Для транскрипции
гена IgL(x) необходимы ТАТА-подобная и окта-
мерная последовательности, роль же 15-нуклеотид-
ного элемента неясна, поскольку, как показывают
разные исследования, его делеция не нарушает
транскрипцию.
Для оптимальной транскрипции генов IgH и
IgL(x) необходима также целостность соответ-
ствующих энхансеров. Энхансер IgH целиком нахо-
дится внутри интрона, который расположен перед
крайним слева экзоном, кодирующим константную
область IgH (рис. 10.67 и 10.68). В гене IgL(x)
обнаружены два энхансера: один внутри интрона
перед константной областью, а другой за З'-концом
единицы транскрипции (на расстоянии 9 т. п. н.).
Свойства этих энхансеров типичны для регулятор-
ных элементов данного класса. Каждый из них
может находиться в любой ориентации, на разных
расстояниях и в разных положениях относительно
точки начала транскрипции и каждый может акти-
вировать транскрипцию гетерологичных промото-
ров. Важной особенностью энхансеров IgH и IgL
является то, что они проявляют активность пре-
имущественно в В-лимфоцитах. Энхансер IgH, по-
видимому, активен на всех стадиях развития В-клет-
ки и, следовательно, проявляет клеточную специ-
фичность, но не специфичность к какой-то опреде-
ленной стадии. Энхансеры IgL, вероятно, обладают
специфичностью обоих типов, поскольку для ини-
циации транскрипции генов IgL необходима индук-
ция, перед которой происходит перестройка генов.
Первые данные о том, что энхансеры содержат
дискретные элементы, которые служат сайтами свя-
зывания белков, были получены при идентификации
последовательностей, защищаемых in vivo от ме-
тилирования под действием диметилсульфата. Эти
опыты основываются на том, что если белки in vivo
связываются с каким-то участком ДНК, то этот
участок экранируется от химических агентов. По-
скольку в В-клетках экранированными оказываются
в основном последовательности, входящие в состав
экспрессируемых генов, можно сделать вывод, что
связанные белки представляют собой факторы
транскрипции, обладающие клеточной специфично-
стью. Четыре защищенных участка в энхансере IgH
и три-в энхансере IgL(x) обозначены на рис. 8.38
как элементы Ен и Ек соответственно; каждый из
них представляет собой определенный вариант ка-
нонической последовательности 5'-CAGGTGGC-3'.
Энхансер IgH содержит также три повтора коровой
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
63
LVD1
Ен3
СН2СН3 Ен4 оста
LVJ
хВ EX1
РИС. 8.38.
Элементы, из которых состоят энхансеры генов тяжелой
и легкой цепей иммуноглобулина. Интроны выделены
точками. Элементы энхансера гена тяжелой цепи обо-
значены символами Ец 1-4 и Сн 1-3; последние пред-
ставляют собой кор-последовательности, обнаружен-
ные во многих вирусных энхансерах (разд. 8.3.г). Эле-
менты энхансера легкой цепи (х) обозначены симво-
лами иВ и Ех 1-3.
последовательности (Сн 1 - 3), которая была описана
при рассмотрении энхансера ранней области SV40
(рис. 8.29), и октамерную последовательность, иден-
тичную последовательности, обнаруженной в ба-
зальном промоторе. В области энхансера есть также
последовательности, ответственные за ингибиро-
вание транскрипции в клетках нелимфоидной ткани;
на это указывают данные о том, что некоторые
делении в области энхансера сопровождаются уси-
лением транскрипции в фибробластах, в то время
как в лимфоцитах активность энхансера прак-
тически не изменяется. В регуляции дифферен-
циальной экспрессии, вероятно, участвуют так
называемые сайленсеры. их свойства пока изучены
слабо.
Результаты исследований влияния различных
мутаций на активность энхансера IgH показывают,
что октамер-это единственный элемент, по-настоя-
щему необходимый для специфической активации
транскрипции в В-клетках. Отдельные элементы Ен
и Сн, по-видимому, не играют существенной роли
в функционировании энхансера. Впрочем, не исклю-
чено. что такой вывод не совсем справедлив, по-
скольку одновременные изменения в октамере и в
определенных элементах Ен приводят к более зна-
чительному нарушению функций энхансера, чем из-
менения в каком-то одном из этих элементов. Более
того, мультимеры, состоящие только из элементов
Ен или С„, обеспечивают избирательную актив-
ность энхансера в В-лимфоцитах. Следует помнить,
что на нынешнем этапе изучения энхансеров IgH
активность транскрипции часто определяют в опы-
тах по транзиенгной трансфекции, условия которых,
возможно, не соответствуют нормальным условиям
функционирования этих элементов. Поэтому, веро-
ятно, преждевременно говорить о том, что те или
иные участки выполняют какие-то специализиро-
ванные функции.
Энхансер IgL(x) содержит два типа элемен-
тов. Один из них, Ек(рис. 8.38), был идентифи-
цирован в опытах по защите от метилирования
in vivo. Другой, так называемый элемент В
(5'-GGAAAGTCCCC-3'), является сайтом связыва-
ния ядерного белка В-лимфоцитов (NF-xB). По-ви-
димому, активность энхансера зависит в основном
от элемента В, хотя какой-то вклад вносит и после-
довательность Ех2. Важной особенностью экспрес-
сии гена IgL(x) является то, чго она индуцируется
под действием бактериального липополисахарида,
циклогексимида и форболовых эфиров; при внесе-
нии мутаций в элемент В это свойство утрачивается.
Таким образом, последовательность В является
ключевым элементом, от которого зависит как ак-
тивность энхансера, так и индуцибельность транс-
крипции гена IgL(x). Интересно, что элемент В
встречается также в промоторах и энхансерах, функ-
ционирующих в нелимфоидных тканях, причем в
некоторых случаях эти промоторы индуцируются
теми же самыми соединениями, которые активиру-
ют экспрессию гена IgL(x) в лимфоцитах. Возмож-
но, это обусловлено свойствами NF-xB.
В транскрипции, специфичной для В-лимфоци-
тов, участвуют все проксимальные промоторы и эн-
хансеры иммуноглобулиновых генов. Наиболее ва-
жными детерминантами этой клеточной специфич-
ности как в базальном промоторе, так и в энхансере
генов IgH являются октамерные последовательно-
сти. Клетки, у которых отсутствует специфичный
для В-клеток белок, связывающийся с октамерной
последовательностью, не могут использовать ни
промотор IgH, ни его энхансер. Аналогично, В-кле-
точная специфичность энхансера IgL(x) зависит от
последовательности базального промотора, а его
активация, специфичная для поздних стадий разви-
тия В-клеток от взаимодействия элемента хВ с
белком NF-xB.
Гены иммуноглобулинов: транс-действующие
факторы. Мы уже говорили, что для сборки ком-
плекса транскрипции вблизи последовательности
ТАТА и точки инициации транскрипции необхо-
димы белковые факторы. Помимо этих факторов
для активации базальных промоторов IgH и IgL(x)
64
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
нужен лимфоцит-специфичный белок, который свя-
зывается с октамером. Напомним, что существуют
два типа таких белков: один (OTF-1) присутствует
в большинстве клеток млекопитающих, а другой
(OTF-2) только в лимфоцитах (разд. 8.3.г). Послед-
ний играет важную роль в активации тканеспеци-
фичной транскрипции с базальных промоторов IgH
и IgL(x) в лимфоцитах. Белки обоих типов связы-
ваются с октамерной последовательностью пример-
но с одинаковой эффективностью и защищают от
расщепления ДНКазой I и от метилирования ди-
метилсульфатом одни и те же остатки. Поскольку
в лимфоцитах присутствуют факторы транскрипции
обоих типов (и OTF-1, и OTF-2), неудивительно, что
в них могут экспрессироваться разнообразные гены,
промоторы которых содержат октамерный элемент.
Однако, как это ни парадоксально, промоторы IgH
и IgL(x) не функционируют в клетках, содержащих
OTF-1 и не содержащих OTF-2. Возможно, это
связано с тем, что для начала транскрипции доста-
точно присоединения к октамерному элементу 1g
фактора OTF-2 и недостаточно присоединения од-
ного только OTF-1. Вероятно, для инициации транс-
крипции под действием OTF-1 последний должен
связаться с одним или несколькими вспомогатель-
ными факторами, соединенными в свою очередь
с соседними элементами. Например, в промоторе
ранних генов SV40 и других промоторах рядом
с октамерной последовательностью находятся эле-
менты, являющиеся сайтами связывания некоторых
факторов транскрипции. Не исключено также, что
расстояние между последовательностью ТАТА и
октамером в базальных промоторах 1g отличается
от соответствующего расстояния в промоторах, ра-
ботающих в самых разных клетках.
Клонированы кДНК, кодирующие как OTF-1,
так и OTF-2. При помощи РНК-блоттинга кле-
точных мРНК с использованием кДНК в качестве
зондов было показано, что экспрессия генов OTF-1
действительно происходит в самых разных клетках,
а генов OTF-2-на всех стадиях развития лимфо-
цитов. Если фибробласты трансформировать плаз-
мидами, кодирующими белок OTF-2, то клетки
приобретают также способность к экспрессии гена
IgH, введенного совместно. В отсутствие плазмиды,
кодирующей OTF-2, ген IgH не экспрессируется. Это
указывает на то, что наличие OTF-2 совершенно
необходимо для экспрессии гена IgH в нелимфоид-
ных клетках. Кроме октамерной последовательно-
сти, OTF-2 связывается также с гептамерной после-
довательностью в базальном промоторе IgH. Воз-
можно, именно это дополнительное взаимодействие
ответственно за преимущественную экспрессию IgH
по сравнению с IgL на ранних стадиях развития
В-клеток.
Как показало определение нуклеотидных после-
довательностей клонированных кДНК, кодирую-
щих белки OTF-1 и OTF-2, аминокислотные после-
довательности их доменов, связывающихся с ДНК,
почти одинаковы в отличие от последовательностей
других участков этих белковых молекул. Возможно,
существование таких различающихся участков и
обусловливает способность этих белков взаимодей-
ствовать с факторами, специфичными в отношении
определенных генов или клеток. Более того, зависи-
мость сродства октамера к OTF-1 от фланкирую-
щих его последовательностей предполагает, что
OTF-1 обладает способностью оказывать разносто-
роннее влияние на транскрипцию многих генов.
Более подробно молекулярные особенности OTF-1
и OTF-2 и их связь с ДНК-белковыми взаимодейст-
виями и способностью к активации транскрипции
мы рассмотрим в разд. 8.6.
Три элемента Е в энхансере IgH взаимодейству-
ют с разными ядерными белками, причем измене-
ния нуклеотидной последовательности этих элемен-
тов блокируют связывание. Каков вклад указанных
взаимодействий в активацию транскрипции, специ-
фичную для клеток определенного типа, неясно,
поскольку данные белки присутствуют во многих
тканях. В целом имеющиеся данные указывают на
то, что элементы Ен не оказывают прямого влияния
на клеточную специфичность, а действуют на актив-
ность энхансера. Белки, связывающиеся с последо-
вательностью Ен, связываются также и с другими
промоторами, из чего следует, что их влияние на
транскрипцию генов 1g носит количественный, а не
качественный характер. Итак, можно сказать, что
детерминантой лимфоидной специфичности энхан-
сера IgH является октамерная последовательность.
Активация транскрипции генов IgL(x) опосреду-
ется белками, которые связываются с элементами
Е и В в энхансере. Эти белки, NF-xE2, NF-xE3
и NF-xB, были идентифицированы в ядерных эк-
страктах при помощи метода сдвига полосы (т. е. по
их способности уменьшать электрофоретическую
подвижность радиоактивно меченных коротких
фрагментов ДНК, содержащих определенную по-
следовательность; разд. 8.3.г). Возможно, к элемен-
там Еу2 и Ек3 присоединяются те же самые белки,
которые связываются с энхансером IgH и влияют на
его активность. Поскольку сайт хВ энхансера в ко-
личественном отношении является самой важной
детерминантой транскрипции IgL(x) и отвечает за
индуцибельность, а также за тканеспецифичность
экспрессии гена и специфичность ее к определенной
стадии развития, мы остановимся на свойствах бел-
ка NF-xB и его взаимодействии с сайтом хВ.
Вначале белковые комплексы, взаимодействую-
щие с элементом хВ, были обнаружены только
в ядерных экстрактах зрелых В-лимфоцитов. Мута-
ции в сайте хВ, которые блокировали экспрессию
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
65
и приводили к утрате индуцибельности генов IgL(x),
подавляли связывание NF-xB. Таким образом, мож-
но было предположить, что NF-xB скорее всего
является обычным тканеспецифичным фактором
транскрипции. Однако в этом пришлось усомнить-
ся, когда NF-xB обнаружили в экстрактах незрелых
В-лимфоцитов, в которых их собственные гены
IgL(x) не экспрессируются. Теперь мы знаем, что
NF-xB присутствует в составе неактивного комплек-
са в цитозоле самых разных клеток. Когда такие
клетки подвергаются действию соединений, индуци-
рующих экспрессию IgL(x) в В-клетках (например,
липополисахаридов, циклогексимида и форболовых
эфиров), этот комплекс разрушается и NF-xB появ-
ляется в ядре. По-видимому, NF-xB связан в цито-
золе с лабильным белковым ингибитором (1-хВ).
В самом деле, полностью активный NF-xB можно
инактировать, смешав его с I-хВ. Механизм актива-
ции NF-xB под действием индукторов in vivo до
конца не установлен. Вполне возможно, что цикло-
гексимид блокирует синтез лабильного ингибитора,
а липополисахарид и форболовые эфиры активиру-
ют ферменты, которые индуцируют модификации
в I-хВ, вызывающие диссоциацию комплекса и по-
ступление NF-xB в ядро.
NF-xB выделен в гомогенном виде из лимфоци-
тов быка и человека; по-видимому, он представляет
собой гетеродимер, состоящий из полипептидов с
мол. массой 50 и 65 кДа. За связывание с ДНК
ответственна субъединица 50 кДа, а субъединица
65 кДа необходима для инактивации NF-xB под
действием I-хВ. Независимо от типа клеток NF-xB
связывается с канонической последовательностью
5'-GGGRtTYY?C-3' (где R и Y-пурин и пиримидин
соответственно). Необычным свойством NF-xB яв-
ляется то, что его связывание с последовательно-
стью хВ стимулируется нуклеозидтрифосфатами.
Строение, функционирование и механизм образова-
ния NF-xB можно будет понять только после выде-
ления гена, кодирующего этот белок.
Элемент хВ (иногда с небольшими изменениями)
встречается во многих промоторах клеток млекопи-
тающих и вирусов. Поэтому функциональное со-
стояние NF-xB и его индуцибельность очень важны
для экспрессии соответствующих промоторов. На-
пример, взаимодействие Т-хелперных клеток с анти-
геном сопровождается активацией NF-xB, которая
инициирует образование фактора роста IL2, а также
рецептора IL2, необходимых для пролиферации
Т-клеток при нормальном иммунном ответе. Уси-
ленная транскрипция генов IL2 и рецептора IL2
опосредуется связыванием NF-xB с элементами хВ
в соответствующих промоторах. Транскрипция ге-
нов вируса иммунодефицита человека типа I (HIV-I)
зависит от связывания NF-xB с хВ-элементом в
вирусном промоторе. Экспрессия и размножение
вируса, в частности стабильность латентного со-
стояния, когда гены HIV-I не экспрессируются, ве-
роятно, зависит от состояния NF-xB в клетке и,
следовательно, от антигенной стимуляции таких
латентно инфицированных клеток. Активация
NF-xB может происходить не только в лимфоцитах,
но и во многих других клетках при поступлении
различных сигналов. Так, синтез Р-интерферона по-
сле заражения фибробластов вирусом индуцируется
путем активации и связывания NF-xB, приводящих
к инициации транскрипции.
Обратимая активация-инактивация NF-xB обес-
печивает эффективную регуляцию клеточного от-
вета на разнообразные сигналы. Известные нам
немногочисленные примеры активации NF-xB гово-
рят о том, что мы пока далеки от полного понима-
ния роли NF-xB как регулятора экспрессии раз-
личных генов.
Г?ны, экспрессирующиеся только в клетках пече-
ни. Клетки печени синтезируют многие ферменты
и секретируют белки, не образующиеся в других
клетках. Такая тканеспецифичность регулируется в
основном на уровне инициации транскрипции. Здесь
мы рассмотрим механизм этой регуляции на при-
мере двух генов, один из которых кодирует альбу-
мин (ALB), а другой-а-фетопротеин (AFP). Эти
гены образуют небольшое семейство эволюционно
родственных генов. Они дивергировали от 300 до
500 млн. лет назад, оставшись тем не менее тесно
сцепленными, и имеют одинаковую организацию
в геноме как человека, так и мыши (разд. 9.4.а).
В процессе эмбрионального развития оба гена экс-
прессируются в висцеральной эндодерме на стадии
желточного мешка, в печени и в желудочно-кишеч-
ном тракте плода. На протяжении всего развития
плода уровень экспрессии генов ALB и AFP в печени
и в кишечнике остается примерно одинаковым, но
вскоре после рождения экспрессия гена AFP в этих
органах прекращается, а содержание ЛГР-мРНК
уменьшается в 10000 раз за три-четыре недели.
С другой стороны, уровень экспрессии гена ALB
и содержание ЯТЛ-мРНК в печени остаются высо-
кими на протяжении всей жизни взрослого организ-
ма, а уменьшаются только в кишечнике. Каков
механизм тканеспецифичной транскрипции этих
двух генов и чем объясняется резкое различие в их
транскрипции после рождения?
Гены ALB и AFP мыши расположены тандемно;
расстояние между ними составляет 15 т. п. н.
(рис. 8.39). Каждый ген длиной от 20 до 22 т. п. н.
состоит в основном из интронов и содержит 15
относительно коротких экзонов. Исследовалась
экспрессия этих генов в культуре клеток с подходящи-
ми маркерными генами (например, с генами хлорам-
фениколацетилтрансферазы [CAT] и гормонов
роста) с присоединенными к ним регуляторными
66
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 8.39.
Взаимное расположение и организация генов альбу-
мина (ALB) и а-фетопротеина (AFP) млекопитающих.
Экзоны, которые по своим размерам пренебрежимо
малы по сравнению с целыми генами, представлены
в виде поперечных черных линий внутри интронов,
выделенных точками. Указаны промотор и энхансер,
а также сайты инициации транскрипции (волнистые
стрелки у первых экзонов).
областями ALB и AFP (как дикого типа, так и изби-
рательно модифицированными). Это позволило
определить положение i/нс-действующих последова-
тельностей, ответственных за специфичную к клет-
кам печени транскрипцию и раздельную регуляцию
двух генов в процессе развития.
Для тканеспецифичной экспрессии генов ALB
и AFP необходимы только последовательности, на-
ходящиеся в пределах участка длиной 150 п. н. перед
точками инициации транскрипции. В то же время
последовательности, находящиеся левее, вызывают
усиление транскрипции гена AFP в 20-50 раз. Эта
дистальная область содержит три разных энхансер-
ных элемента на расстоянии 6,5, 5 и 2,5 т. п. н. от
5'-конца гена (рис. 8.39). Опыты с трансгенными
мышами, имеющими различные модифицирован-
ные формы промотора AFP, тоже свидетельствова-
ли о необходимости 5'-проксимальных и дисталь-
ных элементов для эффективной тканеспецифичной
транскрипции в нужное время. Сравнительный ана-
лиз с использованием трансгенных мышей, содер-
жащих ген ALB, фланкированный участками ДНК
различной длины, показал, что кроме 5'-прокси-
мальной последовательности для обеспечения нор-
мального уровня транскрипции необходим элемент,
находящийся на 9-12 т. п. н. левее гена ALB, а также
три энхансера AFP правее гена ALB.
Сайт-специфический мутагенез области, примы-
кающей слева к гену ALB крысы, и определение
влияния мутаций на эффективность транскрипции
позволили выявить шесть консервативных последо-
вательностей, которые входят в базальный промо-
тор и обусловливают его тканеспецифичность
(рис. 8.40). Базальный промотор составляют два
элемента: последовательность ТАТА, задающая
точку начала транскрипции, но не влияющая на
DEII1
-180 -170 -160 -150 -140 -130 -120 -110 -100
। । । i । i i ।
Крыса : GGGAGCTT С AGATGGCAAACATACGCAAGGG ATTTAGTT А А АСАА CTTTTTTTTTTCTTTTTGGCAAGGATGGTATG - ATTTTGT AATG
Мышь : AACAGCTCCAGATGGCAAACATACGCAAGGGATTTAGTCA ААСАА............CTTTTTGGCAAAGATGGTATG-ATTTTGTAATG
Человек : A ACAgtT СС AG ATGGTA А Ат АтАСАС A AGGGATTTAGTC А ААСАА.HTTTTGGCAAGAAT ATTATGA ATTTTGT ААтС
ССААТ
-90 -80 -70 -60 -50 -40 -30 -20 -10
I I
Крыса - GGGTAGGAAC.C A ATGAAATGAAAGGTT - AGTGtpGTT AATG АТСТАСAGTTATTGGTTAGAGAAGTATATTAG AGCGAG- -ТТН -СТС
Мышь : GGGT) GGAACCAATGAAATGCGAGGTA-AGTAVjGTTAATGATCT/ejGTTATTGGTTAAAGAAGTATATTAGAGCGAGTCTTT- CT-
MenoeeK : ggttggcagccaatgaaatacaaagatgagtgBagtt aatAatctacaattattggttaaagaagtatattagtgctaa--ТТТСССТС
+1
I
крыса : Т - - - GCACACAG АССАССТ -
МЫШЬ : - - - -GCACACAGАТСАССТ-
Человек : CGTTTGTCCTAGCTTTTCTC
РИС. 8.40.
Сравнение регуляторных элементов, ответственных за
специфичность промотора альбуминового гена млеко-
питающих в отношении клеток печени. Элементы рас-
положены так, чтобы совпали их сайты инициации тран-
скрипции, служащие точкой отсчета. Пунктиром выде-
лены промежутки, введенные для того, чтобы выстроить
совпадающие участки друг под другом. Пояснения, ка-
сающиеся роли последовательностей, выделенных раз-
ным цветом, даны в тексте.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
67
частоту инициации транскрипции, и элемент
ССААТ, повреждение которого заметно уменьшает
эффективность транскрипции во всех исследованных
клетках. Специфичность в отношении клеток печени
зависит от четырех вспомогательных последова-
тельностей: элемента РЕ, расположенного между
элементами ТАТА и ССААТ, и трех дистальных
элементов, DEI, DEII и DEIIL Наиболее серьезные
изменения специфичности промотора ALB в клетках
печени происходят при модификации РЕ. Другим
важным элементом, определяющим тканеспецифич-
ность транскрипции, является DEI. Модифика-
ции DEII или DEIII приводят лишь к умерен-
ному уменьшению эффективности тканеспецифич-
ной транскрипции, тогда как удаление сразу обоих
элементов сопровождается снижением активности
промотора в 25 раз.
Каждый из унс-элементов содержит сайт связы-
вания какого-то специфического фактора транскрип-
ции (рис. 8.41). Последовательность, входящая в
PE-элемент мыши, 5'-GTTAATGATCTAC-3', род-
ственна последовательности, общей для многих ге-
нов, экспрессирующихся только в клетках печени
(табл. 8.1). Эта последовательность служит сайтом
связывания для белка HNF-1, присутствующего
искючительно в клетках печени. Очищенный HNF-1
(88 кДа) связывается и «оставляет отпечатки» в
PE-области промотора ALB, а также связывается
с гомологичной последовательностью гена AFP.
Более того, различные гены, специфичные для кле-
ток печени, и промотор вируса гепатита, которые
содержат несколько модифицированный элемент
РЕ, также связывают HNF-1. Таким образом, учас-
тие РЕ в тканеспецифичной экспрессии гена ALB
определяется присутствием в соответствующих
клетках его лиганда, HNF-1. Другие регуляторные
элементы промотора ALB, ССААТ и ТАТА, акти-
вируют транскрипцию при связывании узнающих их
белков (разд. 8.3.г); однако ни ТАТА-, ни ССААТ-
связывающие белки не являются специфичными для
клеток печени. Белки, связывающиеся с DEI и DEI1,
С/ЕВР и NF-1 соответственно, участвуют в актива-
ции различных клеточных и вирусных промоторов
и тоже присутствуют не только в клетках печени
(разд. 8.3.г).
Как объяснить участие широко распространен-
ных белков, связывающихся с ДНК, в специфичной
для клеток печени экспрессии промоторов гена
ALB! Возможно, в основе этого феномена лежат
особенности присоединения неспецифических фак-
торов, а именно - их связывание по соседству с бел-
ком HNF-1, специфичным для клеток печени. Дру-
гое возможное объяснение заключается в том, что
DE-связывающие белки являются гепатоцитспеци-
фичными факторами транскрипции, относящимися
к семействам С/ЕВР и NF-1. Напомним в связи
Стимулирующее действие
указанного элемента
15х
ATACGCAAGGGATTTAGTTAAACAACTTTTTTTTTTCTTTTTGGCAAGGATGGTATGATTTTGTAATGGGGT
I III
-160 -140 -120 -100
CTF/NFI
I------1 I---------------1 I-------1
AGGAACCAATGAAATGAAAGGTTAGTGTGGTTAATGATCTACAGTTATTGGTTAGAGAAGTATATTAGAGCGAGTTTCTCTGCACACAG
I I I I I
-80 -60 -40 -20 +1
NF-Y
HNF-I
РИС. 8.41.
Влияние разных элементов и соответствующих белков
на активность промотора альбуминового гена млеко-
питающих, определяющего специфическую транскрип-
цию в клетках печени. Представлены все элементы
и соответствующие белки, связывающиеся с наиболее
важными из них. Вклад каждого элемента в ткане-
специфичную экспрессию (он указан над элементами)
определяли при помощи делеционного анализа.
68
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Таблица 8.1. Распределение сайтов связывания HNF-1 в генах, экспрессирующихся
в клетках печени 11
Пара основа-
ний
Белок
Вид
Последовательность
а-Цепь фибриногена Крыса G G Т G ATGATTAAC —47
P-Цепь фибриногена » G Т С А ААТАТТААС -84
Альбумин » G Т Т А ATGATCTAC -53
Альбумин Человек G Т Т А АТААТСТАС -51
а-Фетопротеин Крыса G Т Т А CTAGTTAAC — 49
а-Фетопротеин » G Т Т А ATTATTGGC -115
а-Фетопротеин Человек G Т Т А CTAGTTAAC —47
а-Фетопротеин » G Т Т А ATTATTGGC -118
Транстиретин Крыса G Т Т А СТТАТТСТС -116
Транстиретин Человек G Т Т А СТТАТТСТС -116
а1-Антитрипсин Мышь G Т Т А АТ - АТТСАТ -63
а, -Антитрипсин Человек G Т Т А АТ-АТТСАС -63
Про-S 1 вируса
гепатита » G Т Т А АТСАТТАСТ -75
Каноническая последователь-
ность 2) G Т Т А ATNATTAAC
° G. Courtois et al., Proc. Nat. Acad» Sci. 85 (1988), p. 7937.
2) Приведена общая для всех представленных промоторов последовательность из 13 п.н., начинающаяся с указан-
ной пары оснований перед точкой начала транскрипции. Палиндромная последовательность высококонсервативна;
исключением является пара оснований в центре.
с этим, что существуют как широко распространен-
ные, так и тканеспецифичные формы белков OTF.
Итак, экспрессия гена ALB регулируется относи-
тельно небольшой последовательностью в пределах
участка из 150 п.н., непосредственно примыкающе-
го к сайту начала транскрипции. Эта область содер-
жит множество сайтов связывания специфических
белков; одни из них обусловливают тканеспецифич-
ность экспрессии промотора, а другие регулируют
уровень экспрессии. Как именно этот комплекс свя-
занных белков влияет на скорость транскрипции,
неизвестно. Неясно также, каким образом энхан-
серы, находящиеся примерно на 10 т. п. н. левее
и более чем на 30 т. п. н. правее сайта инициации
транскрипции, активируют базальный промотор.
При помощи аналогичных экспериментальных
подходов идентифицирован единственный элемент,
ответственный за тканеспецифичную транскрипцию
промотора AFP в клетках печени. Этот элемент
находится в пределах сайта связывания HNF-1 меж-
ду парами оснований —61 и —48. Высокий уровень
экспрессии в трансфицированных клетках печени
и у трансгенных мышей обеспечивают также три
отдельных энхансерных элемента, находящихся на
значительном расстоянии от базального промотора.
При этом необходимы все три элемента, поскольку
каждый из них вносит свой вклад в общую актив-
ность. Размер элементов варьирует от 200 до
300 п. н.; каждый из них содержит как одинаковые,
так и уникальные последовательности. Некоторые
из этих элементов (или элементы, сходные с ними)
встречаются в других клеточных и вирусных энхан-
серах. Идентификация энхансерных элементов еще
не завершена, но уже можно сказать, что одни из
них присутствуют в большом количестве, а другие
весьма немногочисленны и обеспечивают позитив-
ную и негативную регуляцию транскрипции.
Почему прекращается синтез AFP после рожде-
ния животного? Для ответа на этот вопрос прово-
дили опыты с трансгенными мышами, несущими
различные структурные варианты генов ALB и AFP.
Например, у трансгенных мышей экспрессия искус-
ственного гена, содержащего три ЛГР-энхансера,
соединенных с геном ALB и его базальным промо-
тором, не прекращалась сразу после рождения.
Трансгены, в которых ЛТВ-энхансер был присоеди-
нен слева к гену AFP, содержащему собственный
5'-пограничный участок размером всего 1 т. п. н.,
нормально экспрессировались в процессе развития
плода и переставали экспрессироваться вскоре после
рождения. Эти данные говорят о том, что информа-
ция о регуляции экспрессии AFP, специфичной в от-
ношении определенной стадии развития, содержится
либо в участке, примыкающем к базальному про-
мотору слева, либо внутри промотора, либо внутри
самой единицы транскрипции.
Согласованность действия фактора тканеспеци-
фичной транскрипции и неспецифических факторов.
Наиболее важным в регуляции экспрессии генов,
о которых шла речь выше, является взаимодействие
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
69
между i/uc-элементами последовательностей и
/иранс-действующими белками ядерной транскрип-
ции. Это взаимодействие лежит в основе механизма
транскрипции многих, а возможно и всех генов.
Можно было бы предположить, что в каждой кон-
кретной ткани и на каждой стадии развития исполь-
зуется один или несколько уникальных факторов
транскрипции, взаимодействующих с соответствую-
щим числом специфических элементов, что и обу-
словливает характерные особенности процесса
транскрипции. На самом же деле элементы, отве-
чающие за специфическую транскрипцию эукариоти-
ческих генов, состоят из относительно небольшого
числа последовательностей и набора широко рас-
пространенных белковых факторов транскрипции,
которые встречаются в самых разных тканях и
функционируют на многих стадиях развития. Ука-
занные последовательности находятся в пределах
участка размером от 100 до 400 п. н., примыкающе-
го непосредственно к точке инициации транскрип-
ции со стороны 5'-конца, в дистальных элементах
энхансера, а чаще одновременно в обоих местах.
Обычно основными для транскрипции являются
один или, возможно, два фактора транскрипции,
присутствие которых характерно только для опре-
деленной ткани или стадии развития. Но именно
сочетанное взаимодействие таких уникальных и дру-
гих, неспецифических, белков и сайтов их связыва-
ния обусловливает дифференциальную экспрессию
генов. Элементы хВ и октамерные последователь-
ности генов IgL(x) есть во многих промоторах, но
присутствие OTF-2 и индуцибельная активация
NF-xB именно в лимфоцитах обеспечивает специ-
фическую экспрессию промоторов генов 1g. Функ-
ционирование промотора гена ALB зависит от
взаимодействий между несколькими часто встре-
чающимися последовательностями и не менее ши-
роко распространенными факторами транскрипции,
однако специфичность транскрипции в клетках пече-
ни определяется связыванием фактора транскрип-
ции HNF-1, специфичного именно для этих клеток.
Следует добавить, что экспрессия многих других
генов тоже зависит от связывания некоего белка
транскрипции, который присутствует или активи-
руется в определенных клетках, на определенной
стадии развития или в определенной фазе клеточ-
ного цикла.
е. Индукция и
репрессия транскрипции
Уровень транскрипции отдельных генов часто
меняется под действием внешних сигналов. Так,
в зависимости от содержания питательных веществ
(например, источников углерода, азота и энергии)
и взаимодействия с различными гормонами, при
вирусной инфекции или воздействии металлов, све-
та, высокой температуры и радиации транскрипция
специфических генов или групп генов может либо
индуцироваться, либо подавляться. Во многих слу-
чаях повышение или понижение эффективности
транскрипции происходит в ответ на изменения
доступности необходимых факторов транскрипции
или их способности связываться со специфическими
регуляторными элементами. Рассмотрим несколько
примеров, иллюстрирующих этот механизм регуля-
ции транскрипции.
Регуляция транскрипции дрожжевых GAL-генов.
На рис. 8.42 представлены метаболические пути ути-
лизации галактозы у S. cerevisiae и гены, контроли-
рующие этот процесс. Ген GAL2 кодирует пермеазу,
необходимую для поглощения внеклеточной галак-
тозы. GALI кодирует галактокиназу, которая ката-
лизирует превращение галактозы в галактозо-1-фос-
фат, a GAL7-галактозо-1-фосфат-уридилтрансфе-
разу, катализирующую образование уридиндифос-
фогалактозы (UDPGal) из галактозо-1-фосфата и
уридиндифосфоглюкозы (UDPG). Эпимераза, спо-
собствующая регенерации UDPG из UDPGal, ко-
дируется геном G/1L10. Гены GALI, GAL7 и G/1L10
все расположены на хромосоме II, а ген СЛГ2-на
хромосоме XII (рис. 8.43). Эти четыре гена, которые
транскрибируются раздельно с индивидуальных
промоторов, не экспрессируются в клетках, расту-
щих на среде без галактозы. При выращивании
в присутствии галактозы уровень их экспрессии
GAL2
GAL1
GAL7
Пермеаза Киназа
Галактозавнеши. --------►Галактозавнутр ---------
Трансфераза
►Галактозо-1-Р - —Глюкозо-1-Р
РИС. 8.42.
Метаболические пути утилизации галактозы у S. cere-
visiae. Утилизацию галактозы, добавленной в питатель-
ную среду, контролируют четыре гена (цветные прямо-
угольники). Над (или под) обозначениями генов при-
ведены тривиальные названия кодируемых ими фер-
ментов. UDPG и UDPGal-уридиндифосфоглюкоза и
уридиндифосфогалактоза соответственно.
UDPG
Эпимераза
GAL1Q
UDPGal
------»- Гликолиз
70
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Галактоза
РИС. 8.43.
Компоненты регуляторной системы GAZ- генов. Пока-
зано положение в хромосомах относительно центромер
(О) четырех генов, кодирующих ферменты GAL, и двух
генов, кодирующих регуляторные белки GAL4 и GAL80
Внутриклеточная галактоза превращается в индуктор
(не идентифицирован), который вызывает диссоциацию
транскрипционно неактивного комплекса GAL4 GAL80
[М Johnston, Microbiol Rev 51 (1987), р. 458 ]
согласованно возрастает примерно в 1000 раз.
Индукция и репрессия ферментов GAL регули-
руется на уровне инициации транскрипции под дей-
ствием неизвестного индуктора, образующегося из
галактозы, и белков, кодируемых двумя несцеплен-
ными генами, GAL4 и G/1L80 (рис. 8.43). Мутации
в гене GA L4 блокируют в присутствии галактозы
индукцию транскрипции генов GALI, GAL2, GAU1
и GAL 10; это наводит на мысль, что GAL4 кодирует
какой-то положительный регулятор этих генов.
Многие мутации в GAL80 приводят к тому, что
транскрипция генов G/1L1, GAL2, GAL1 и GAL10
становится конститутивной (т. е. независимой от
индуктора), а в результате некоторых мутаций
транскрипция генов GAL становится неиндуцибель-
ной. Такое явление можно объяснить тем, что ин-
дуктор нейтрализует ингибирующее действие про-
дукта гена G/1L80 на активацию транскрипции,
опосредованную геном GAL4. Если это так, то
активация транскрипции генов GAL под действием
белка гена GAL4 может блокироваться при его
связывании с белком гена GAL80. Индуктор либо
вызывает диссоциацию неактивного комплекса, ли-
бо препятствует его образованию. Ключом к рас-
шифровке работы этой регуляторной системы слу-
жит изучение белка GAL4 и его взаимодействий
с промоторами генов GAL, с аппаратом транскрип-
ции и с белком GAL80.
Для САЕ4-зависимой активации транскрипции
генов GAL необходимы специфические последова-
тельности промоторов GAL\-, GAL2-, GAL2-,
GAL10- и (7ЛЕ80-генов. Подобные t/uc-регулятор-
ные элементы обозначают UAS (upstream activating
sequences); это общее обозначение, которое исполь-
зуют применительно к таким элементам в генах
дрожжей. Они напоминают энхансеры тем, что их
функция не зависит от ориентации и относительно
мало зависит от расстояния до точки начала транс-
крипции. Элемент UASC?1L был впервые обнаружен
в сегменте ДНК между генами GALI и G/1L10 по
способности обеспечивать 6Л£4-зависимую актива-
цию транскрипции с промотора GAL1. Делецион-
ный анализ этой области показал, что важными
элементами являются многочисленные копии па-
линдромной последовательности длиной 17 п. н.
(рис. 8.44). В пределах участка длиной 200 400 п. н.
перед генами GALI, GAL2, GAL7, GAL10 и GAL80
находятся от одного до четырех таких элементов,
хотя для максимальной активации достаточно двух.
Высокий уровень САЬ4-зависимой экспрессии обес-
печивает любая из последовательностей UASC?1L или
синтетический олигонуклеотид, содержащий упомя-
нутую выше каноническую последовательность,
если они помещены перед различными базальными
промоторами дрожжей.
GAL4 кодирует белок мол. массой 100 кДа, ко-
торый присутствует в очень низких концентрациях
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
71
Каноническая последовательность UASe>(L
5'-C66A^GACAGTC^TCCG-3'
3'-GCCt£cTGTCAGgAGGC-5'
РИС. 8.44.
Локализация последовательностей UAS (см. текст) в
промоторах генов GAL . Показаны ТАТА-бокс, сайты
инициации транскрипции (волнистые стрелки) и эле-
менты UAS. Внизу приведена каноническая последова-
тельность для 13 элементов UASCXL. [М. Johnston,
Microbiol. Rev. 51 (1987), р. 458.]
как в индуцированных, так и в неиндуцированных
клетках. Белок, образующийся в результате экспрес-
сии гена, клонированного в Е. colt или в дрожжевых
клетках, специфически связывается с элементом
UASgtil и защищает его палиндромную последова-
тельность из 17 п. н. как от действия ДНКазы I, так
и от метилирования диметилсульфатом. Учитывая,
что последовательность (JASgxl имеет ось симмет-
рии 2-го порядка, можно предположить, что белок
GAL4 связывается с ней в форме димера. В резуль-
тате анализа аминокислотной последовательности
GAL4 и его мутантных форм были идентифициро-
ваны белковые домены, обусловливающие связыва-
ние с UASghl, активацию транскрипции и взаимо-
действие с белком GAL80 (рис. 8.45).
За специфическое узнавание элемента UASghl
и связывание с ним ответствен участок белковой
молекулы между аминокислотными остатками 10
и 51, однако сам по себе этот участок не может
активировать транскрипцию. Расположение остат-
ков цистеина и других аминокислот на участке
между аминокислотами 10 и 32 такое же, как у цин-
ковых пальцев других ДНК-связывающих белков
(рис. 8.45, Б) (разд. 8.6.а). В данном случае цинко-
вый палец может как стабилизировать комплекс
белка и ДНК, так и обеспечивать специфичность
связывания ДНК, поскольку способность белка
узнавать элемент UASGhl определяется восемнад-
цатью аминокислотами по соседству с цинковым
пальцем. За активацию транскрипции отвечают два
участка, один из которых примыкает к домену
связывания ДНК, а другой находится в пределах
сегмента из 120 аминокислот на С-конце (рис. 8.45, А).
Хотя последний участок и необходим для активации
транскрипции, речь идет не о какой-то специфиче-
ской последовательности. Активация транскрипции
обусловлена высокой плотностью кислых амино-
кислотных остатков в этой области, что свойственно
также другим белкам-активаторам транскрипции
(например, репрессору фага X и активатору транс-
крипции дрожжей GCN4 [разд. 8.6.6]). Факторы
транскрипции с повышенным содержанием кислых
аминокислот в упомянутых участках являются наи-
более эффективными активаторами. Более того,
белки, содержащие САЬ4-участок активации транс-
крипции, связанный с чужеродными ДНК-связы-
вающими доменами, активируют транскрипцию с
промоторов, содержащих элементы, которые соот-
ветствуют ДНК-связывающему домену.
Каков механизм активации транскрипции с уча-
стием связанного белка GAL4- неизвестно. Возмож-
но, на сборку активного комплекса инициации в
сайте начала транскрипции влияют кислые домены
белка GAL4, расположенные вблизи ТАТА. Такое
влияние может быть опосредовано либо самим
ТАТА-связывающим белком, либо другими белка-
ми комплекса. Об этом свидетельствует тот факт,
что GAL4 изменяет полученный с использованием
ДНКазы I отпечаток, характерный для сайта начала
транскрипции при сборке на нем комплекса транс-
крипции дрожжей (рис. 8.46). Если UASgml встраи-
вается в регуляторный участок промотора млеко-
питающих (например, промотора вируса рака мо-
лочной железы мышей), то транскрипция с этого
промотора активируется в присутствии GAL4 in
vivo. Аналогично, включение UASghl недалеко от
ТАТА-блока (в пределах 50 п. н.) промотора адено-
вируса Е4 приводит к тому, что этот промотор
становится чувствительным к активации под дейст-
вием GAL4 in vitro. В обоих случаях активация
транскрипции под действием GAL4 синергична
с активацией, обусловленной связыванием факторов
транскрипции млекопитающих с родственными им
сайтами в этих промоторах. Видоизмененные фор-
мы белка GAL4, которые могут связываться с UASghl,
но неспособны активировать транскрипцию, не сти-
мулируют гетерологичные промоторы. Эти данные
говорят о том, что белок GAL4 может взаимодейст-
вовать с белками транскрипции млекопитающих,
72
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Связывание
АНК
Активация
транскрипции
148 196
788 6В1
856 881
Взаимодействие
с GALВО
I_______I________I________I_______I_______I________I________I_______I________I
О 100 200 300 400 500 600 700 800 900
N-конец Аминокислоты GAL4 С-коиец
А
р---------------Цистеиновый цинковый палец —Домен,ответственный за специфичность---------
10 20 30 40 50
AliCysAiplltCytArglculyslyilculyiCysSirlysGlulviProlyiCvs AlilyiCytleulyiAinAinTrpGluCyiArgTyrSerProlysThrlyiArgSirPreleuThr Arg
6
РИС. 8.45.
Функциональные домены белка GAL4. А. Домены, от-
ветственные за связывание ДНК, акти ацию транскрип-
ции и азаимодействив с GAL80 а полипептидной цепи
из 881 аминокислоты. Числа по краям прямоугольни-
ков-номера аминокислот, начиная с N-конца. Б, Ами-
повышая эффективность транскрипции, и еще раз
подтверждают универсальность механизмов акти-
вации.
Свою регуляторную функцию эелок GAL80 вы-
полняет, взаимодействуя с белком GAL4. О наличии
такого взаимодействия свидетельствует копреципи-
тация этих белков с антителами к GAL4. Кроме
того, хотя сам GAL80 не связывается с последова
тельностью UASC4t, он связывается с комплексом
GAL4-(JASCj4L и изменяет его электрофоретическую
подвижность. Для взаимодействия с белком GAL80
нужно всего 25 аминокислот на С-конце молекулы
GAL4. В самом деле, синтез пептида, содержащего
только эту последовательность, вызывает конститу-
тивную экспрессию генов GAL в клетках дрожжей,
очевидно, благодаря тому, что этот пептид связы-
вает белок GAL80 и высвобождает транскрипцион-
но активный GAL4. С другой стороны, если даже
белок GAL4, лишенный С-концевой последователь-
ности, образуется в избытке, для синтеза белков
GAL все равно необходим индуктор: избыток де-
фектного белка неспособен вызвать диссоциацию
комплекса GAL80 с интактным белком GAL4.
Вероятно, белок GAL80 блокирует активацию
нокислотные последовательности домена, связывающе-
гося с ДНК (один из цинковых пальцев, обогащенный
остатками цистеина; разд. 8.6.в), и домена, ответственного
за специфическое связывание с UASGjU . [М. Johnston,
Microbiol. Rev. 61 (1987), р. 468.]
транскрипции, опосредованную GAL4, препятствуя
необходимому взаимодействию между GAL4 и
комплексом транскрипции, а не связыванию GAL4
с UAS04L. Тот факт, что GAL4 связывается с по-
следовательностью (JASC4L в отсутствие индуктора,
объясняет, почему in vivo «отпечаток» этой после-
довательности одинаков и в присутствии, и в от-
сутствие индуктора. Предполагают, что неиденти-
фицированный пока индуктор, образующийся из
галактозы, связывается с белком GAL80 и вызывает
диссоциацию его комплекса с GAL4. Распад комп-
лекса GAL4-GJL80 приводит к обнажению домена
активации GAL4, стимулируя таким образом транс
крипцию. Этот механизм индукции отличается от
механизма, используемого для регуляции оперонов
lac и gal Е. coll, когда связывание индуктора пре-
дотвращает связывание репрессора с его операто-
ром в ДНК (разд. 3.11.в и г).
Экспрессия генов GAL регулируется также с
помощью других используемых клеткой сахаров.
Так, дрожжи S. cerevisiae, растущие на глюкозе,
в присутствии галактозы не синтезируют белки
GAL. Более того, добавление глюкозы к клеткам,
растущим на галактозе, вызывает быстрое подавле-
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
73
GAL
80
GAL4
Катаболитная (____t______и__________(__1_______О
репрессия (-------f------f { ---------)
UAS6al ТАТА
белок катаболитной репрессии
РИС. 8.46.
Модель регуляции транскрипции геное GAL при участии
GAL4-GAL80. Когда GAL4 связан с GAL80, он не сти-
мулирует сборку (или функционирование) комплекса
транскрипции на ТАТА. В присутствии индуктора GAL80
отделяется от GAL4, и последний получает возможность
взаимодействовать с комплексом транскрипции (или
активировать его). В присутствии глюкозы белок ката-
болитной репрессии (CRP) связывается с GAL4, и его
соединение с UAS(M, становится невозможным.
[М. Johnston, Microbiol. Rev. 61 (1987), р. 458.]
ние экспрессии генов GAL. Природа репрессора
генов GAL, образующегося из глюкозы, неизвестна,
идентифицированы лишь некоторые сайты, на кото-
рые он действует. Один из них содержит t/uc-элемен-
ты, находящиеся между UASGXt и ТАТА-элемен-
том; правда, неясно, каким образом этот участок
проявляет свою ингибирующую активность. По-
скольку ДНКазный «отпечаток», характерный для
элемента UASCXI., исчезает при выращивании кле-
ток на глюкозе, вполне возможно, что катаболитная
репрессия опосредуется белком (CRP), который пре-
пятствует связыванию GAL4 с UAS(M/ .
Транскрипция, регулируемая стероидными гормо-
нами. Многие особенности развития и физиологии
позвоночных зависят от разнообразных стероидных
и сходных с ними по своему действию гормонов.
Минерало- и глюкокортикоиды надпочечников уча-
ствуют в регуляции метаболизма гликогена и соле-
вого обмена, опосредуют реакции на стресс, регу-
лируют рост и дифференцировку многих клеток
и тканей. Регуляция половой дифференцировки эм-
бриона, развития вторичных половых признаков
и полового поведения осуществляется с помощью
половых стероидов: эстрогенов, прогестерона и
тестостерона. Для нормального развития костей
необходим стероидоподобный витамин D. Ретиное-
вая кислота, которая по своему строению напо-
минает стероиды, является ключевым морфогеном
и регулятором дифференцировки в процессе эмбрио-
генеза. Гормон щитовидной железы трииодтиронин,
имеющий совершенно иное строение, чем стероиды,
осуществляет свою регуляторную функцию, связы-
ваясь с рецептором, сходным с рецепторами для
стероидов.
Все эти гормоны оказывают свое физиологиче-
ское действие, проникая в клетки и связываясь
с соответствующими внутриклеточными рецептора-
ми. Свободные рецепторы локализованы в различ-
ных клеточных структурах. Так, рецептор глюко-
кортикоидного гормона находится главным обра-
зом в цитоплазме, а после связывания лиганда
перемещается к ядру. Рецепторы эстрогенов и про-
гестерона локализованы преимущественно в ядре,
а рецептор гормона щитовидной железы - исключи-
тельно в ядре. Несмотря на разную локализацию,
все рецепторы при связывании лигандов претерпе-
вают структурные изменения, способствующие при-
соединению лиганд-рецепторного комплекса к спе-
цифическим участкам ДНК, называемым гормон-
акцепторными элементами (hormone response ele-
ments, HRE). HRE обладают свойствами энхансе-
ров, поскольку могут влиять на активность промо-
торов, с которыми они ассоциированы, независимо
от местоположения и ориентации. Следовательно,
HRE- это индуцибельные энхансеры, поскольку они
регулируют транскрипцию, только будучи связан-
ными с функциональным гормон-рецепторным
комплексом.
Каковы структурные особенности рецепторов и
их модифицированных, связанных с лигандами
форм и каким образом рецепторы узнают соот-
ветствующие HRE? Другой важнейший вопрос-как
связывание лиганд-рецепторного комплекса с HRE
влияет на транскрипцию? Современные, хотя и не-
полные, представления об этом базируются на не-
скольких экспериментальных подходах, которые
сформировались благодаря возможности получать
клонированные и секвенированные кДНК, кодирую-
щие рецепторы. Исходя из нуклеотидных последо-
вательностей соответствующих секвенированных
кДНК была определена аминокислотная последо-
вательность рецепторов. Их сравнение выявило оди-
74
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
наковые и различающиеся области, что в свою
очередь помогло идентифицировать функционально
важные части рецептора. И все же наиболее значи-
мым и продуктивным подходом к изучению взаимо-
связи структуры и функции рецепторов оказался
анализ влияния мутационных изменений к ДНК ре-
цепторов на рецепторные свойства последних. Ана-
логичные подходы применялись при исследовании
гормон-акцепорных промоторов, что помогло опре-
делить и полностью охарактеризовать HRE. Были
созданы химерные гены, кодирующие гибридные
рецепторы, и это еще раз убедительно показало, что
рецепторные функции выполняют специфические
участки белковых молекул и даже определенные
аминокислоты.
Как сравнение аминокислотных последователь-
ностей, так и мутационный анализ гормон-акцеп-
торных промоторов выявили необходимые для гор-
мональной регуляции короткие палиндромные по-
следовательности, находящиеся на разном расстоя-
нии перед сайтом начала транскрипции. Гены,
транскрипция которых регулируется одними и теми
GRE
MMTV - 134 TGGTTTGGTATCAAATGT Т CTGATC TG
hGH + 87 CCTTTGGGCACAATGTGTCCTGAGGGG
hMTUA - 268 GCACCCGGTACACTGTGTCCTCCCGCT
TO - 439 CTCATATGCACAGCGAGTTCTAGTGAG
TAT -2420 TACGCAGGACTTGTTTGTTCTAGTCTT
Каноническая GGTACANNNTGTTCT
последовательность
ERE
X.vit - 338 AAAGTCAGGTCACAGTGACCTGATCA
Овальбумин - 173 T TATTCAGGTAACAATGTGT T T T CTG
Каноническая GGTCANNNTG*CC
последовательность т
TRE
rGH - 185 GTAAGATCAGGGACGTGACCGCAGG
- 108
+ 113
- 242
- 413
-2446
- 315
- 199
- 161
Каноническая
последовательность
GATCANNNNNNTGACC
РИС. 8.47.
Нуклеотидные последовательности некоторых гормон-
акцепторных элементов (HRE). Числа у начала и конца
каждой из последовательностей-это номера нуклеоти-
дов относительно сайта инициации транскрипции.
N-любой из четырех нуклеотидов; противоположно
направленные стрелки над каноническими последова-
тельностями указывают, что последние имеют ось сим-
метрии 2-го порядка (черные кружки). Представлены
глюкокортикоид-акцепторные элементы (GRE) вируса
рака молочной железы мышей (MMTV), генов гормона
роста (hGH) и металлотионеина (hMTUA) человека,
генов тирозиноксидазы (ТО) и тироэинаминотрансфе-
раэы (ТАТ). а также эстроген-акцепторные элементы
(ERE) генов вителлогенина Xenopus (X. vit.) и оваль-
бумина курицы (овальбумин). Тиреоид-акцепторный
элемент (TRE)-3to энхансер гена гормона
роста крыс (rGH).
же гормонами, имеют либо сходные, либо одинако-
вые HRE. Связывание рецепторов с генетически
идентифицированными HRE было продемонстриро-
вано также с помощью метода отпечатков в опытах
с очищенными рецепторами глюкокортикоидов,
эстрогенов, прогестерона и гормона щитовидной
железы. Несмотря на разную локализацию в про-
моторе, все HRE имеют сходные последовательно-
сти: они состоят из различного числа неспецифиче-
ских пар оснований, фланкированных пятью парами
специфических оснований палиндрома, и содержат
от 12 до 16 п.н. (рис. 8.47). Именно благодаря сход-
ству последовательностей HRE эстроген-акцептор-
ный элемент можно превратить в глюкокортикоид-
акцепторный, заменив всего две пары оснований
в палиндроме. Наличие у HRE оси симметрии 2-го
порядка предполагает, что они связываются с ре-
цепторами, находящимися в форме димеров.
Анализ аминокислотных последовательностей
рецепторов эстрогенов и глюкокортикоидов, уста-
новленных по данным о нуклеотидных последова-
тельностях соответствующих к ДНК, показал, что
вблизи центра каждого рецептора находится высо-
коконсервативный короткий участок (область С на
рис. 8.48). По данным скрининга продуктов расще-
пления геномной ДНК при помощи олигонуклео-
тидных зондов, синтезированных на основе этой
консервативной последовательности, подобная об-
ласть характерна для большого семейства генов,
очевидно кодирующих другие рецепторы. Неудиви-
тельно, что с помощью тех же самых зондов были
выделены к ДНК еще некоторых рецепторов; это
указывает на важность данного домена для функ-
ционирования рецепторов. Белки, кодируемые неко-
торыми из этих кДНК, только предположительно
можно отнести к рецепторам, поскольку их лиганды
или HRE неизвестны. Отдельные гены рецепторов
стероидов клонированы, частично секвенированы
и отнесены к соответствующим хромосомам
(рис. 8.48).
Сравнение аминокислотных последовательно-
стей разных рецепторов выявило несколько участ-
ков с той или иной степенью взаимной гомологии.
На рис. 8.48 они обозначены A/В, С, D, Е и F.
Участок A/В имеет самую низкую степень гомоло-
гии у разных рецепторов и даже у одинаковых
рецепторов из разных видов. Последовательности
участка С, напротив, во всех рецепторах высококон-
сервативны и, как показал дальнейший анализ,
представляют собой сайты связывания ДНК, т. е.
выполняют функцию, присущую всем рецепторам.
Участки D и F различаются как по длине, так и по
аминокислотной последовательности в разных ре-
цепторах и даже в одинаковых рецепторах нерод-
ственных видов. Делеционный анализ этих участков
показал, что они несущественны для функциониро-
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
75
hER
hGR
hMR
EAR
РИС. 8.48.
Функциональные домены рецепторов стероидных и ти-
реоидных гормонов. На верхней диаграмме показаны
белковые домены типичного рецептора. Обозначения:
hER - рецептор эстрогена человека; hG Я - рецептор глю-
кокортикоидного гормона человека; h MR-рецептор
минералокортикоидного гормона человека; hAR-ре-
цептор андрогена человека; hVitD3 рецептор витамина
D3 человека; hRARa-рецептор a-ретиноевой кислоты
человека; hTRa- рецептор a-тиреоидного гормона. Раз-
личные домены обозначены буквами A/В, С, D, Е и F,
начиная с N-конца белковой молекулы. Домен С лю-
бого рецептора содержит 66 или 68 аминокислот; раз-
меры других доменов неодинаковы и могут быть опре-
делены из цифр, приведенных на рисунке.
вания рецептора. Несмотря на то что участки Е у
разных рецепторов отличаются друг от друга, их
последовательности в рецепторах эстрогенов чело-
века и курицы на удивление сходны. Это позволяет
предположить, что Е относится к доменам связыва-
ния лиганда. Гипотезы относительно роли участков
Е и С вначале основывались только на результатах
сравнения аминокислотных последовательностей,
но более детальный анализ подтвердил их правиль-
ность.
Участок из 66- 68 аминокислот, присутствующий
во всех рецепторах (область С), отвечает за связы-
вание родственных HRE. Мутации, затрагивающие
эту область, не мешают связыванию с гормоном, но
препятствуют связыванию с HRE. Более того, уко-
роченные рецепторы, лишенные областей A/В, D,
Е и F, все еще связываются с родственными HRE,
хотя и неспособны регулировать транскрипцию.
Наиболее убедительно автономность действия
участка С при связывании ДНК была продемон-
стрирована в опытах, где область С из глюкокорти-
коидного рецептора заменяла соответствующую об-
ласть в другом рецепторе (например, рецепторе
эстрогена). Химерный рецептор активировал гены,
чувствительные к глюкокортикоидам, в присутствии
эстрогенов (рис. 8.49). Интересно, что интактные
рецепторы не присоединяются к своим HRE в от-
сутствие соответствующих лигандов, но могут при-
соединяться к ним, если удалить домен связывания
лиганда. Отсюда следует, что домен связывания
ДНК у рецептора неспособен к взаимодействию
с HRE, если молекула содержит незанятый домен
связывания лиганда; но присоединение лиганда или
удаление его связывающего домена каким-то обра-
зом обеспечивает взаимодействие области связы-
вания с HRE.
ER | A/В | | D | [ F |
I обмен
1 С-доменамн
GR(ER)
Активирует эстрогеи-
А/В
Евк
чувствительмые промо-
торы в присутствии глю-
кокортикоидных
гормонов
ER (GR) | А/В
Активирует глюкокор-
тикоидчувствительные
промоторы ворисутствни
эстрогена
Сбк
РИС. 8.49.
Образование химерных рецепторов стероидных гормо-
нов. Обмен сегментами, кодирующими С-домены, меж-
ду кДНК рецептора глюкокортикоидного гормона (GR)
и кДНК рецептора эстрогена (ER) приводит к образова-
нию двух химерных рецепторов; GR (ER)— рецептора
глюкокортикоидного гормона с ДНК-связывающим
доменом рецептора эстрогена и ER (GR)— рецептора
эстрогена с ДНК-связывающим доменом рецептора
глюкокортикоидного гормона. Трансфекция данных
кДНК в соответствующие клетки приводила к резуль-
татам, описанным на рисунке.
76
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Исследование областей С многих рецепторов
выявило характерное расположение остатков ци-
стеина, гидрофобных и основных аминокислот, при
котором они образуют два цинковых пальца
(рис. 8.50). В очищенном глюкокортикоидном ре-
цепторе с этой областью связаны два иона цинка.
Для связывания с ДНК важны все четыре остатка
цистеина, участвующих в образовании цинкового
пальца, следовательно, каждый ион цинка коорди-
национно связан с четырьмя остатками цистеина,
а не с остатками цистеина и гистидина, как в неко-
торых других факторах транскрипции (разд. 8.6.а).
Об автономном характере участка С свидетельству-
ет и то, что кодирующая его последовательность
отделена от последовательностей, кодирующих об-
ласти A/В и D, интронами; интересно, что цинковые
пальцы кодируются разными экзонами.
Для специфичного связывания с HRE необходи-
мы оба цинковых пальца. Однако если у разных
рецепторов произвести взаимную замену любых
цинковых пальцев, то химерные рецепторы будут
иметь специфичность по отношению к гену-мишени,
присущую цинковому пальцу I рецептора. Мутации,
затрагивающие последовательность, которая нахо-
дится перед цинковым пальцем I, или сам этот
палец не влияют на специфичность связывания ДНК
рецептором. Однако некоторые изменения последо-
вательности, примыкающей к последнему цистеи-
новому остатку цинкового пальца 1 рецептора
эстрогенов, нарушают связывание этого рецептора
с эстроген-акцепторным элементом, и он приобре-
тает способность связываться с глюкокортикоид-
акцепторным элементом. Таким образом, диффе-
ренциальное узнавание рецептором эстрогена эстро-
ген-акцепторного, но не глюкокортикоид-акцептор-
ного элемента, палиндромные последовательности
которых различаются только двумя парами основа-
ний, в значительной мере определяется тремя ами-
нокислотами цинкового пальца I рецептора эстро-
гена. Следовательно, специфические взаимодей-
ствия между рецептором и соответствующим HRE,
по-видимому, зависят от порядка расположения ос-
татков гидрофобных аминокислот в цинковом паль-
це I. Кроме того, в стабилизацию комплекса вносит
вклад взаимодействие между скелетом молекулы
ДНК и цинковым пальцем II, а также основными
аминокислотами.
Палиндромный характер HRE предполагает, что
рецепторы связываются с этим элементом в димер-
ной форме. Димеризация рецепторов происходит
при участии двух рецепторных доменов. Более силь-
ное взаимодействие, возникающее после присоеди-
нения лиганда, осуществляется при участии после-
довательностей из области Е. В более слабых взаи-
модействиях участвуют последовательности цинко-
вого пальца И. Очевидно, прочность и специфич-
ность связывания рецептора в форме димера выше,
чем в форме мономера. Механизм индуцированной
гормоном димеризации неизвестен, но не исключе-
но, что он является частью того же самого процесса,
который позволяет рецептору, уже связанному с
гормоном, присоединяться к своему HRE. И диме-
ризация, и связывание HRE, вероятно, происходят
после конформационного изменения рецептора, ин-
|-----Специфическое связывание ДНК----1 [•
Неспецифическое связывание ДНК --------------1
|--------------------- Кор
РИС. 8.50.
Цинковые пальцы ДНК-связывающего домена рецеп-
тора эстрогена человека. Аминокислотные последова-
тельности ДНК-связывающего домена (кора) и при-
легающих участков представлены с использованием
однобуквенных обозначений аминокислот (рис. 1.24).
, , Участок,ответственный,
I I— за ядерную —I
локализацию
Основные аминокислоты выделены цветом, а наиболее
высококонсервативные аминокислоты всех рецепторов
заключены в кружки. Предполагают, что область между
остатками аминокислот 252 и 262 детерминирует лока-
лизацию рецептора в ядре.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИ ЧЕСКИХГЕНОВ
77
аудированного присоединением гормона. Имеются
также указания на то, что в некоторых случаях
связывание гормона вызывает диссоциацию инги-
битора, который предотвращает спонтанную диме-
ризацию рецептора.
Связывание гормона нужно как для специфиче-
ского и эффективного присоединения рецепторов
к соответствующим HRE, так и для регуляции
транскрипции генов. Необходимым условием свя-
зывания гормона с рецептором является целост-
ность области Е, поскольку делеции, вставки или
замены оснований в гене белка Е предотвращают
связывание. Интересно, что способность рецептора
связывать лиганд сохраняется и в отсутствие обла-
стей A/В, С и D, вероятно, потому, что Е может
связывать лиганд независимо от остальной части
L молекулы белка. Об этой независимости области
Е свидетельствуют также результаты опытов по ее
[замене у разных рецепторов, аналогичных опытам
по замене доменов связывания ДНК (рис. 8.49). Так,
если область Е рецептора эстрогенов заменить на
область Е рецептора глюкокортикоидов, то химер-
ный рецептор будет активировать транскрипцию
экстроген-акцепторных генов в ответ на действие
глюкокортикоидов, а не эстрогенов.
Интактные рецепторы, не связанные с гормоном,
не присоединяются к соответствующим HRE, одна-
ко рецепторы, у которых область Е делегирована,
легко присоединяются к своим HRE в отсутствие
гормона. Отсюда следует, что структурные измене-
ния, сопровождающие связывание гормона, либо
демаскируют, либо активируют ДНК-связывающий
домен рецептора, возможно, стимулируя димериза-
цию последнего. Структурные изменения, индуци-
рованные в глюкокортикоидных рецепторах при
связывании гормона, служат также сигналом к пере-
носу комплекса лиганд-рецептор из цитоплазмы
в ядро. Кроме того, доменсвязывающий лиганд
участвует в активации транскрипции под действием
рецепторов после присоединения к соответствую-
щим HRE. Так, у дрожжей, синтезирующих химер-
ный белок, который состоит из домена связывания
гормона глюкокортикоидного рецептора и домена
связывания UASc^z. белка GAL4 дрожжей, экспрес-
сия генов GAL индуцируется при добавлении глю-
кокортикоидов. Поскольку связывание только од-
ного указанного домена белка GAL4 с UASc^t не
активирует транскрипцию, полученный результат
означает, что область Е, связанная с гормоном,
может активировать транскрипцию в дрожжах.
Действие антигормонов - соединений, которые
нарушают физиологический ответ на действие гор-
монов,- вероятно, связано с тем что они соединяют-
ся с рецептором, не вызывая при этом структурных
изменений, необходимых для димеризации, для
взаимодействия с HRE или для влияния на транс-
крипцию гормон-акцепторных генов.
В результате связывания активированных гор-
мон-рецепторных комплексов с соответствующими
HRE осуществляется регуляция транскрипции, ко-
торая в зависимости от промотора, вероятно, со-
стоит в активации или ингибировании инициации
этого процесса. Один из подходов к изучению меха-
низма активации или подавления транскрипции при
связывании рецептора заключается в идентифика-
ции областей рецептора, ответственных за влияние
на транскрипцию. В этом случае тоже применяли
подход, основанный на внесении мутаций в кДНК,
кодирующие рецепторы стероидов, гормонов щито-
видной железы, витамина D3 и ретиноида, и на
сопоставлении их с изменениями в регуляции транс-
крипции с участием соответствующих HRE. Пред-
посылкой регуляции транскрипции является связы-
вание с ДНК комплекса рецептор-гормон, поэтому
возникает вопрос: какие именно структуры рецепто-
ра опосредуют регуляторный эффект? Результаты
исследований в целом свидетельствуют о том, что
в регуляции участвуют многие области рецептора,
причем наиболее важны области A/В и Е. Однако
относительная значимость этих двух областей
варьирует у разных рецепторов и среди разных
промоторов, содержащих одинаковые HRE. Напри-
мер, если область Е рецептора эстрогена удалена, то
рецептор сохраняет способность связываться с
ДНК, но не может нормально активировать транс-
крипцию с некоторых промоторов, чувствительных
к эстрогенам. В то же время, несмотря на то, что
делеции в областях Е некоторых рецепторов препят-
ствуют связыванию с гормоном, не мешая при этом
связыванию с ДНК, их влияние на активацию транс-
крипции носит разный характер в зависимости от
промотора, а в ряде случаев и от клеток, в которых
исследуется действие рецептора. Мутации в области
A/В уменьшают эффективность активации транс-
крипции с участием связанного рецептора, но сте-
пень этого уменьшения зависит от рецептора и гена-
мишени. Так. рецептор эстрогенов, целиком утра-
тивший область A/В, все еще связывается с эстро-
ген-акцепторным элементом двух исследованных
промоторов, но в одном случае эффективность ак-
тивации транскрипции почти нормальная, а в дру-
гом активации вообще не происходит.
Влияние областей A/В и Е на активацию транс-
крипции четко продемонстрировано в опытах с ис-
пользованием химерных рецепторов, содержащих
только домены связывания регуляторного белка
GAL4, сшитые либо с доменом A/В, либо с доменом
Е некоторых стероидных рецепторов. Когда химер-
ные гены, кодирующие такие белки и содержащие
области либо А/В-СЛЕ4, либо Е-6ЛЕ4, переносятся
в клетки млекопитающих вместе с геном-маркером
(с геном Р-галактозидазы), промотор которого со-
78
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
UASGaL GAL1
0-Галактозидаза
Ген Р-галактозидазы экспрессируется только
Б присутствии галактозы
ERE GALI
i-Галактозидаза
Ген р-галактозидазыне экспрессируется вприсутствии
галактозы или эстрогена
(^-Галактозидаза
PGK Рецептор эстрогена
Ген р-галактозидазы экспрессируется вприсутствии
эстрогена, но не в его отсутствие
РИС. 8.51.
Рецептор человека может активировать тран-
скрипцию генов у S. cerevisiae. А. Ген Р-галак-
тозидазы, находящийся под контролем про-
мотора GAL 1. Трансфекция этого гена в дрож-
жевые клетки приводит к тому, что последние
приобретают способность к индуцибельной
экспрессии Р-галактозидазного гена. Б. Эле-
мент ЦА5СЛ1 GAL 1 -промотора заменен эстро-
ген-акцепторным элементом (ERE). Трансфек-
ция этого гена в дрожжевые клетки не приво-
дит к экспрессии р-галактозидазного гена ни
при добавлении эстрогена, ни при добавлении
галактозы. В. Одновременная трансфекция ге-
на, изображенного на рисунке 8.51, Б, и гена,
кодирующего рецептор эстрогена человека,
транскрипция которого находится под контро-
лем промотора гена глицерофосфаткиназы
дрожжей (GPK), приводит к зстрогензависи-
мой экспрессии гена Р-галактозидазы.
держит UASGXL вблизи ТАТА-блока, транскрипция
гена Р-галактозидазы активируется. Мутации в об-
ластях A/В или Е, подавляющие транскрипционную
активность рецептора, предотвращают и активацию
под действием химерных белков. Следовательно,
вполне вероятно, что если область A/В или Е связы-
вается вблизи промотора млекопитающих, то она
активирует транскрипцию.
Домены стероидных рецепторов, участвующие
в активации транскрипции, могут трансактивиро-
вать также промоторы дрожжей. Если промотор
GALI, включающий UASGXL, сшить с последова-
тельностью, кодирующей Р-галактозидазу, то при
добавлении галактозы в дрожжевых клетках инду-
цируется синтез Р-галактозидазы (рис. 8.51, Л).
Заменив UASC^L на эстроген-акцепторный элемент,
можно проверить, способен ли рецептор эстроге-
нов активировать комплекс транскрипции дрожжей
и является ли такая активация эстрогензависи-
мой. Оказалось, что Р-галактозидаза в этом слу-
чае не синтезируется даже в присутствии эстро-
генов, поскольку отсутствует рецептор последних
(рис. 8.51,5). Однако, когда в результате экспрес-
сии соответствующей кДНК в клетках, у которых
между промотором GALI и геном Р-галактози-
дазы находится эстроген-акцепторный элемент,
синтезируется функциональный рецептор эстроге-
нов, транскрипция целиком зависит от наличия
эстрогенов и не зависит от добавления галактозы
(рис. 8.51, В). Таким образом, совершенно очевидно,
что рецептор эстрогенов может взаимодействовать
с комплексом транскрипции дрожжей и влиять на
него.
На рис. 8.52 представлена модель, иллюстри-
рующая различные структурные особенности рецеп-
торов гормонов и предположительный механизм их
участия в активации транскрипции. Согласно этой
модели, свободный рецептор может находиться в
двух формах. В одной из них (I) домен Е свободен
и не участвует ни в каких взаимодействиях. Вторая
форма (И) представляет собой комплекс, состоящий
из рецептора и ингибитора. Предполагается, что
формы I и II не могут связываться с HRE. Присо-
единение гормона к рецептору, находящемуся в
форме I, приводит к изменению конформации до-
мена Е, димеризации рецептора (форма III) и его
кооперативному связыванию с HRE (форма IV).
Если же гормон соединяется с рецептором, находя-
щимся в форме II, то белок-ингибитор отделяется
от рецептора, последний димеризуется (форма III)
и связывается с HRE. Данная модель предполагает,
что димер образуется в результате взаимодействий
между двумя связанными с лигандами доменами
Е и двумя ДНК-связывающими доменами С.
Обсуждая вопрос о том, как именно гормоны
влияют на транскрипцию с участием гормончув-
ствительных промоторов, необходимо помнить, что
HRE-это не единственный регуляторный элемент
в данных промоторах. Большая часть таких промо-
торов содержит много разных регуляторных после-
довательностей, которые связывают разные факто-
ры транскрипции. Так, если глюкокортикоид- или
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
79
РИС. 8.52.
Модель индуцируемой гормонами димеризации сте-
роидных рецепторов и их связывания со стероид-ак-
цепторными элементами. I-мономерная форма рецеп-
тора стероидных гормонов с доменами активации
транскрипции (А/В), связывания ДНК (С) и связывания
стероидных гормонов (Е). Неактивный цитоплазмати-
ческий комплекс, форма II, предположительно обра-
зуется при связывании рецептора стероидных гормонов
с ингибитором; природа этого связывания неизвестна.
Связывание гормона (Н) с доменом Е индуцирует ди-
меризацию рецептора, происходящую при участии
сильных взаимодействий между доменами связывания
гормона и более слабых взаимодействий между ДНК-
связывающими доменами. Связывание же гормона с
неактивной формой II приводит к отделению ингиби-
тора и последующей димеризации. Форма III может
проникать в ядро и связываться с родственным гор-
мон-акцепторным элементом. Разнонаправленные
стрелки на ДНК указывают, что сайт связывания имеет
ось симметрии 2-го порядка. [S. Green, Р. Chambon,
Trends Genet. 4 (1988), р. 309.]
эстроген-акцепторный элемент включить в промо-
тор, в норме не активируемый ни одним из этих
гормонов, то промотор приобретает чувствитель-
ность к стероиду в присутствии соответствующего
рецептора. Вообще говоря, активация транскрипции
синергична, и эффективность транскрипции зависит
от характера соседних элементов и их положения
относительно HRE.
До сих пор мы говорили в основном о таких
случах, когда связывание рецепторов с соответ-
ствующими HRE активирует транскрипцию, однако
в некоторых ситуациях при таком связывании
транскрипция, напротив, подавляется. Например,
промотор гена пролактина содержит многочислен-
ные сайты связывания гормон-рецепторных комп-
лексов, но присоединение таких комплексов не ста-
билизирует, а подавляет транскрипцию. Последова-
тельности элементов, опосредующих ингибирующее
действие глюкокортикоидов, отличаются от после-
довательностей элементов, активирующих транс-
крипцию. Следовательно, ДНК-связывающий до-
мен глюкокортикоидного рецептора может соеди-
няться с глюкокортикоид-акцепторными элемента-
ми, опосредующими как ингибирующий, так и ак-
тивирующий эффекты. Анализ имеющихся данных
показывает, что связывание гормон-рецепторного
80
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
комплекса с элементом, отвечающим за негативную
регуляцию, мешает связыванию других активирую-
щих факторов транскрипции.
Регуляция генов GAL дрожжей и генов млекопи-
тающих, транскрипция которых регулируется сте-
роидами, во многом сходна. Гены GAL не транс-
крибируются до тех пор, пока к элементам UASCX,
не присоединится белок GAL4. Аналогично, для
транскрипции генов, индуцируемых гормонами, не-
обходимо, чтобы с соответствующим HRE связался
рецептор. В обоих случаях связывание активатора
транскрипции с ДНК осуществляется с помощью
индуктора; для рецептора это соответствующий
гормон, а для GAL4 некое вещество, производное
галактозы. Кроме того, у дрожжей связывание ак-
тиватора с промотором блокируется в отсутствие
индуктора путем присоединения GAL80 к GAL4,
а у млекопитающих-путем связывания ингибитора
с доменом Е рецептора. Важное различие двух
индуцибельных систем заключается в том, что ком-
понентами позитивного и негативного контроля
СЛЕ-системы являются разные полипептиды (белки
GAL4 и GAL80), а соответствующие компоненты
регуляции гормончувствительных генов принадле-
жат одному и тому же белку- рецептору. Как имен-
но активируется комплекс транскрипции в обеих
системах- неизвестно; установлено лишь, что доме-
ны активации и в том, и в другом случаях содержат
кластеры кислых аминокислот, состав и последова-
тельность которых до конца не установлены. Не
менее удивительным является тот факт, что домены
активации у дрожжей и млекопитающих взаимоза-
меняемы.
Регуляция гена ^-интерферона. Интерфероны
(IFN)-3TO секретируемые видоспецицичные поли-
пептиды, которые защищают клетки от вирусной
инфекции. IFN присутствуют в лейкоцитах (а), фи-
бробластах (Р) и в клетках иммунной системы (у)
позвоночных. В нормальных фибробластах они не
обнаруживаются, однако в течение 4-8 ч после ви-
русной инфекции или при обработке некоторых
клеток двухцепочечной РНК в них начинается син-
тез больших количеств мРНК IFN-Р; по-видимому,
такие двухцепочечные РНК образуются при раз-
множении вируса. Подобное накопление мРНК
IFN-р обусловлено в значительной мере усилением
транскрипции, индуцируемым вирусной инфекцией.
Последовательности, ответственные за регуля-
цию транскрипции гена IFN-Р, находятся между
парами оснований —204 и +1 (рис. 8.53). Вездесу-
щий ТАТА-блок локализован, как обычно, вблизи
пары —25, а элементы, определяющие индуцибель-
ность IFN-P (IRE),-между парами —200 и —37. IRE
состоит из пяти субэлементов, два из которых отве-
чают за негативную регуляцию (NRD1 и NRD2),
а три опосредуют индуцированную активацию
транскрипции (PRD I, II и III). Отметим, что NRD1
перекрывается с PRDII, a NRD2 либо граничит
с PRDIII, либо перекрывается с ним. Элементы
NRD1 и NRD2 были идентифицированы с помощью
опытов по мутационным изменениям их последова-
тельностей, в результате которых наблюдалось
уменьшение роли индукции или необходимость в
ней вообще отпадала (например, транскрипция гена
INF-Р становилась конститутивной). С другой сто-
роны, мутационные изменения в любом из трех
элементов PRD блокировали активацию транскрип-
ции под действием вируса. Таким образом, регуля-
ция экспрессии гена IFN-P может быть как позитив-
ной, так и негативной.
Опыты по защите от метилирования in vivo
показывают, что в неинфицированных клетках оба
NRD-элемента защищены, но после инфицирования
или индукции приобретают способность к метили-
рованию. Напротив, три PRD-элемента легко мети-
лируются в неинфицированных клетках, а после
заражения становятся полностью защищенными от
метилирования. Это говорит о том, что указанные
элементы по-разному связываются с ядерными фак-
торами: те, которые участвуют в репрессии транс-
крипции, связываются в неиндуцированном состоя-
нии, а медиаторы индуцированного ответа-после
индукции (рис. 8.54).
Ядерные экстракты из инфицированных или ин-
дуцированных клеток содержат разные белки, свя-
зывающиеся с каждым из элементов PRD. По всей
вероятности, в неиндуцированных экстрактах эти
белки присутствуют в неактивной форме, поскольку
для индукции транскрипции гена IFN-0 не требуется
синтез новых белков. PRDII содержит последова-
тельность, очень напоминающую сайт хВ в энхансе-
ре IgL(x), и, как и ожидалось, связывает NF-xB.
Мутации, блокирующие связывание NF-xB с
PRDII, ослабляют и индуцибельную активацию
РИС. 8.53.
Организация регуляторных элементов промотора IFN-0. PRDII и PRDIII-элементы позитивного контроля,
NRD1 и МРО2-злементы негативного контроля, PRDI, + 1-сайт инициации транскрипции.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
81
Неинауцированное
состояние
PRDIII PRDI PRDII NRD1 ТАТА
РИС. 8.54.
Сайты связывания регуляторных белков в промоторе
гена IFN-p. По-видимому, в неиндуцированном состоя-
нии с NRD1- и МЯО2-сайтами связываются регулятор-
ные белки, которые перекрывают PRDII- и PRDIII-сайты
соответственно. После индукции NRD-связывающиеся
белки отделяются, три PRD-сайта занимают соответст-
вующие белки, и происходит активация транскрипции.
промотора. Напомним, что NF-xB-это широко
распространенный белок, присутствующий главным
образом в цитоплазме в виде неактивного комплек-
са, но способный активироваться под действием
ряда факторов, в том числе под действием вирусной
инфекции. Таким образом, активация NF-xB при
заражении вирусом, вероятно, играет важную роль
в индукции транскрипции гена IFN-p. Ни индуци-
бельные факторы, которые присоединяются к PRDIII
и PRDI, ни белки, связывающиеся с элементами
NRD, пока не идентифицированы. Весьма правдо-
подобная, но пока окончательно не принятая мо-
дель регуляции гена IFN-P предполагает, что свя-
зывание факторов NRD мешает связыванию белков
PRDII и PRDIII с родственными им последователь-
ностями, поддерживая таким образом неактивное
состояние промотора (рис. 8.54). Вероятно, при ин-
дукции инактивируются факторы NRD и происхо-
дит их отделение от соответствующих сайтов свя-
зывания; не исключено также, что белки NRD вытес-
няются при связывании активированных белков PRD.
Регуляция транскрипции под действием тепла,
света и металлов. Тепловой шок и другие стрессо-
вые воздействия инициируют типичную физиологи-
ческую реакцию, названную ответом на тепловой
шок. Такой ответ может давать любой эукариотиче-
ский и прокариотический организм. Как правило,
при этом синтез нормальных белков мгновенно
прекращается, и начинают синтезироваться специ-
фические белки теплового шока (HSP). По-видимо-
му, подобная реакция необходима для выживания
организма в изменившихся условиях, поскольку му-
тации, блокирующие ее у дрожжей, детальны. Ме-
ханизм индукции генов теплового шока у Drosophila,
Xenopus, дрожжей и млекопитающих в высшей сте-
пени консервативен. Например, транскрипция гена
HSP70 Drosophila (кодирующего белок теплового
шока с мол. массой 70 кДа) при введении в клетки
Xenopus, мыши и обезьяны активируется под дей-
ствием тепла так же эффективно, как и транскрип-
ция эндогенных генов теплового шока.
Синтез HSP зависит от активации транскрипции
генов теплового шока. Индукция происходит безот-
лагательно. при этом в течение часа теплового
воздействия количество мРНК возрастает с менее
чем одной молекулы на клетку до тысяч молекул.
Некоторые гены теплового шока регулируются в
процессе развития и не нуждаются для активации
своей транскрипции в тепловом шоке. Клонирова-
ние и секвенирование генов теплового шока из
разных организмов позволило выявить основ-
ные особенности транскрипционного ответа на
это воздействие. Как правило, промоторы, осу-
ществляющие регуляцию транскрипции в ответ
на тепловой шок, состоят из нескольких повторов
относительно короткой последовательности (эле-
менты теплового шока-HSE), рассеянных внутри
области протяженностью 200 п. н. перед ТАТА. Эта
каноническая последовательность, обнаруженная
в регуляторных областях многих промоторов теп-
лового шока у разных организмов, имеет вид
5'-CNGAANTTCNG-3', где N-любое из четырех
оснований. HSE-элементы, помещенные перед
ТАТА-блоком гетерологичных промоторов (напри-
мер, промотора гена ТК вируса герпеса или про-
мотора гена Р-глобина), обусловливают чувстви-
тельность транскрипции с этих промоторов к тепло-
вой индукции.
6-158
82
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Как и в случае i/ис-элементов в других промото-
рах, HSE являются сайтами связывания регулятор-
ных белков - факторов транскрипции генов теплово-
го шока (HSTF). HSTF из разных видов сходны по
своим физическим и функциональным свойствам;
так, HSTF Drosophila и дрожжей имеют одинаковые
размеры и одинаково связываются с ДНК. Данные
многих опытов свидетельствуют о том, что HSTF
прочно связаны с HSE во время (но не до начала)
теплового шока. Следовательно, HSTF в нормаль-
ных клетках присутствуют в неактивном состоя-
нии, а тепловой шок стимулирует их присоедине-
ние к HSE всех промоторов, индуцируемых теплом.
Клонирован ген, кодирующий HSTF дрожжей,
что открывает пути к анализу его связывания с
HSE и изучению механизма активации транскрип-
ции с участием различных промоторов теплового
шока.
Высшие растения имеют множество фоторецеп-
торов, которые опосредуют их реакцию на свет.
Свет разных длин волн служит регуляторным си-
гналом сложных процессов развития и контроля за
развитием. Идея о том, что одной из функций
фоторецепторов является активация или репрессия
экспрессии генов, не нова. Экспериментальные под-
тверждения этому дали недавние исследования с ис-
пользованием молекулярно-генетических подходов,
проведенные для идентификации регуляторных эле-
ментов промоторов генов растений. Исследовалась
индуцированная светом активация ядерного гена,
кодирующего малую субъединицу рибозо-1,5-бис-
фосфат-карбоксилазы (rbcS) (рис. 6.36). Для опре-
деления активности промотора использовали уча-
сток 5'-последовательности гена размером 1 т. п. н.,
сшитый с геном хлорамфеникол-ацетилтрансфера-
зы. Этот химерный ген, а также другие гены с раз-
личными перестройками и делециями вводили в
клетки табака при помощи трансформации Ti-плаз-
мидой (разд. 5.8.в). Функционирование промотора
изучали в каллюсах табака, выдержанных на свету
и в темноте. Низкая эффективность транскрипции
с промотора rbcS связана с последовательностью,
расположенной в пределах 90 п. н. перед ТАТА-бло-
ком, а для достижения максимальной эффективно-
сти транскрипции, активируемой светом, необходим
набор энхансеров, находящихся левее, между пара-
ми оснований —975 и —90. Укорочение энхансерных
последовательностей сопровождается постепенным
снижением эффективности световой индукции. Ана-
логичная структура из ТАТА-проксимального эле-
мента световой чувствительности и левого энхан-
сера присутствует в промоторе гена, кодирующего
связанный с хлорофиллом белок, который аккуму-
лирует световую энергию и активируется под дей-
ствием света. В обоих случаях описаны специфиче-
ские элементы, ответственные за световую чувстви-
тельность, но не идентифицированы факторы транс-
крипции, с которыми они взаимодействуют. Более
того, неизвестен и механизм передачи светового
сигнала от фоторецепторов к регуляторному аппа-
рату транскрипции.
Точно так же как живые организмы реагируют
на тепло, свет и некоторые другие физические фак-
торы (например, на действие радиации) путем изме-
нения характера транскрипции определенных генов,
они реагируют и на действие металлов с помощью
гомеостатических реакций. Одни металлы токсичны
(например, кадмий и ртуть), другие являются необ-
ходимыми компонентами пищи, но ядовиты в высо-
ких концентрациях (например, медь и цинк). Глав-
ная роль в ответе на действие металлов принадле-
жит активации транскрипции генов металлотионеи-
на. Эти гены кодируют небольшие полипептиды,
богатые цистеином, которые связывают как необхо-
димые для организма, так и токсичные тяжелые
металлы.
У млекопитающих разных видов имеется не-
сколько различающихся, но близких по строению
генов металлотионеина, которые располагаются
один за другим на одной и той же хромосоме
и имеют одинаковое расположение экзонов и ин-
тронов. Те же самые тяжелые металлы, которые
связываются с белком, вызывают преходящую ин-
дукцию транскрипции гена металлотионеина, ле-
жащую в основе механизма защиты клеток от ток-
сического действия металлов. Индуцибельный ответ
на действие металлов характерен для всех клеток,
причем активируется каждый из нескольких разных
генов металлотионеина, которые могут присутство-
вать в одном организме. Гены млекопитающих,
кодирующие металлотионеины, индуцируются так-
же при стрессе и под действием некоторых ядовитых
веществ, при этом индукция опосредуется высво-
бождением глюкокортикоидов.
У S. cerevisiae имеется всего один ген металло-
тионеина. Он не является жизненно необходимым,
но защищает дрожжи от токсического действия
меди. Исследование промотора позволило иденти-
фицировать два чувствительных к меди элемента,
находящихся между парами —200 и —100, и обыч-
ный ТАТА-блок вблизи пары —30, считая от точки
начала транскрипции (рис. 8.55, Л). Эти два элемен-
та, которые активируются медью (UASCu), распо-
ложены перед промотором и представлены несовер-
шенными повторами длиной от 32 до 34 п. н. Они
являются сайтами связывания фактора медь-инду-
цируемой транскрипции (CITF). Фактор CITF, ген
которого (АСЕ-1) удалось клонировать, секвениро-
вать и экспрессировать, присоединяется к UASCu
только в комплексе с медью. Домены связывания
меди и ДНК у CITF локализованы на N-конце
молекулы белка. Зависимость связывания белка с
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
83
А.Промотор гена металлотионеина S.cerevisiae
dUASCu pUASCu
TATA
ТАТА-ЗЭВИСИМЫИ
комплекс
транскрипции
б.Промотор гена металлотионенна Пд человека
MRE MRE MRE MRF
ВЕЕ BLE 6С ТАТА
GRE
РИС. 8.55.
Организация регуляторных элементов промотора гена
металлотионенна дрожжей и млекопитающих. А. Про-
мотор гена металлотионенна S. cerevisiae’. кроме ТАТА-
бокса для индуцируемой медью транскрипции необхо-
димы проксимальный и дистальный элементы (UASCu
и dUASCu соответственно). Элемент ТАТА детермини-
рует сборку комплекса транскрипции, а два UASCu-3rie-
мента связывают фактор транскрипции CITF. Б. Про-
мотор гена металлотионеина 11А человека: фоновая ак-
тивность этого промотора, т. е. активность в отсутствие
металлов, зависит от ТАТА-бокса и комплекса тран-
скрипции (белка Sp1, связанного с GC-богатым гекса-
мерным элементом, и факторов транскрипции АР1 и
АР2, связанных с двумя энхансерами фоновой актив-
ности [BLE]). Для индуцируемой металлами транскрип-
ции необходим кластер металлозависимых элементов
(MRE), которые активируются белком MRE-BP. Ответ
на глюкокортикоиды зависит от связывания рецептора
глюкокортикоида (GR) с глюкокортикоид-акцепторным
элементом (GRE).
UASCu от меди можно объяснить тем, что форми-
рование правильного домена связывания индуци-
руется только в том случае, когда несколько ионов
меди соединяются при помощи хелатных связей
с остатками цистеина, расположенными в этой час-
ти белковой молекулы. Интересно, что С-концевая
половина CITF имеет четко выраженный кислый
характер, что присуще и другим факторам актива-
ции транскрипции (разд. 8.6.6).
Регуляторные элементы, контролирующие
транскрипцию промотора металлотионеина II чело-
века, были идентифицированы при помощи мута-
ционного анализа и исследования транскрипции с
модифицированных промоторов in vivo (рис.
8.55, Б). Перед точкой начала транскрипции в преде-
лах участка размером 175 п.н. находятся элементы,
необходимые для обеспечения фонового уровня
транскрипции (т. е. транскрипции в отсутствие ин-
дуктора); к ним относятся элементы ТАТА и GC,
а также два энхансера фонового уровня (BLE).
Помимо фактора связывания ТАТА и вспомога-
тельных факторов транскрипции, которые присо-
единяются к ТАТА-блоку и соседним последова-
тельностям, для фоновой транскрипции необходи-
мы фактор Spl, связывающийся с элементом GC,
и факторы API и АР2, связывающиеся с BLE. Для
84
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
индукции транскрипции под действием металлов
необходимы, кроме того, металл-акцепторные эле-
менты (MRE) и белок, присоединение которого к
MRE зависит от присутствия металлов. Таким
образом, фактором, ответственным за регуляцию
транскрипционной активности промоторов, являет-
ся концентрация координационно связанного с
ионами металла фактора связывания MRE, завися-
щая от концентрации свободного металла. Транс-
крипционный ответ на действие глюкокортикоидов,
не зависящий от металлов-индукторов, обусловли-
вается присоединением глюкокортикоидного рецеп-
тора в комплексе с лигандом к глюкокортикоид-ак-
цепторному элементу промотора.
ж. Транскрипционная регуляция
морфогенеза
Из всего сказанного выше можно сделать вывод,
что регуляция транскрипции опосредуется связыва-
нием варьирующего числа разнообразных факторов
транскрипции со специфическими родственными им
элементами промоторов и энхансеров. По-видимо-
му, эффективность инициации транскрипции зави-
сит от взаимодействий между этими связанными
с ДНК белками (и, возможно, другими связанными
с ними белками) и одним или более компонентами
основного аппарата транскрипции, хотя детали этой
регуляции на молекулярном уровне неизвестны. Со-
гласно этой модели, уровень экспрессии гена зави-
сит от доступности, концентрации и активности
одного или нескольких факторов, участвующих в
его транскрипции. Такой механизм в значительной
степени объясняет широкое разнообразие феноти-
пов дифференцированных клеток и тканей и их
способность реагировать на изменения окружающей
среды. Тот же самый принцип, вероятно, использу-
ется также при регуляции упорядоченного во вре-
мени и пространстве эмбрионального развития.
Следовательно, сложная временная и пространст-
венная картина экспрессии при эмбриогенезе
создается в результате координированного взаимо-
действия как повсеместно присутствующих, так и
ограниченных определенными структурами специ-
фических факторов транскрипции.
Более детальное обсуждение вопроса о роли
регуляторов транскрипции как детерминант эм-
брионального развития выходит за рамки данного
раздела. Здесь мы лишь попытаемся показать, как
с помощью факторов транскрипции создается
иерархический каскад экспрессии генов, реализую-
щийся во времени и пространстве в зависимости от
того, когда и где возникает инициация этого каскада.
Наиболее детально разработанной моделью, по-
зволяющей анализировать молекулярные процессы,
регулирующие ранние стадии эмбрионального раз-
вития, является Drosophila. Это насекомое относится
к метамерным организмам (т. е. его тело построено
из серийно повторяющихся сегментов, каждый из
которых дифференцируется в отдельные структуры
и формы согласно положению, занимаемому в орга-
низме). Образование и последовательная дифферен-
циация таких сегментов в ходе эмбриогенеза зависят
прежде всего от дробления эмбриона на повторяю-
щиеся единицы и далее от определения путей их
дифференцировки. Эмбриогенез начинается с быст-
рых делений ядра, за которыми следует миграция
ядер к наружной оболочке яйца, после чего вокруг
Время.мин
0-15
Деление ядер
15-18
90-150
Клеточная бластодерма
РИС. 8.56.
Морфологические изменения яйца Drosophila вплоть до
образования клеточной бластодермы. Свежеотложен-
ное яйцо содержит одно ядро, которое делится в тече-
ние первого часа с образованием большого числа про-
извольно распределенных ядер (ядра синцития). При-
мерно к 90 мин многие ядра мигрируют к наружной
оболочке яйца, и у заднего его полюса начинается
дифференцировка полюсных клеток. Миграция ядер
продолжается примерно еще час, после чего образуется
клеточная бластодерма.
150-195
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
85
каждого ядра образуется мембрана. В результате
формируется клеточная бластодерма (рис. 8.56).
Однако рамки для последующего воплощения плана
развития тела и участь каждой клетки устанавли-
ваются еще до образования клеточной структуры
бластодермы. Это отличает эмбриогенез Drosophila
от эмбриогенеза млекопитающих, у которых план
строения тела и дифференцированные структуры на
стадии многоклеточного бластоциста еще не опре-
делены.
Основными детерминантами формирования пе-
редне-задней конфигурации эмбриона являются
продукты генов bicoid (bed) и nanos (nos). Транскрип-
ция гена bed осуществляется в специализированных
клетках яичника, которые группируются вокруг од-
ного полюса развивающегося ооцита. мРНК гена
bed переходит в ооцит и захватывается тем его
концом, который должен стать передним полюсом
эмбриона. Продукты группы генов nos локализуют-
ся у заднего полюса яйца. Ко времени откладывания
яйца оно уже оказывается поляризованным как на
молекулярном, так и на морфологическом уровне
(рис. 8.57). Во время трансляции мРНК гена bed
образующиеся белки диффундируют от переднего
полюса и постепенно распределяются по яйцу при-
мерно до его середины. Одновременно белки-про-
дукты мРНК гена nos- перемещаются к переднему
полюсу. Итак, еще до формирования клеточной
структуры два разных продукта материнских мРНК
неравномерно распределяются вдоль будущей пе-
редне-задней оси эмбриона.
Результатом перекрывания градиентов продук-
тов генов bed и nos является локальная активация
группы генов, названных генами gap, которые
отвечают за образование семи исходных сегментов
эмбриона. Три главных члена группы генов дар,
hunchback (hb), kruppel (Кг) и knirps (kni), также
экспрессируются в определенных сегментах эмбрио-
на. Экспрессия гена hb происходит только в двух
областях: одна находится в задней четверти яйца,
а другая простирается от переднего полюса почти
до середины яйца. Ген Кг сначала экспрессируется
в одной широкой зоне в середине эмбриона, а про-
дукт гена kni появляется как спереди, так и сзади от
этой зоны. Изучение мутантов, не способных синте-
зировать продукты генов bed и nos, показывает, что
экспрессия этих трех генов регулируется продуктами
генов bed и nos и что пространственная локализация
продуктов генов дар определяется относительными
концентрациями продуктов генов bed и nos вдоль
передне-задней оси.
Белок гена bed действует как позитивный регуля-
тор экспрессии hb, вызывая накопление hb у перед-
него конца эмбриона. Продукт гена nos, по-видимо-
му, ингибирует активность белка hb. Более того,
продукты генов bed и nos блокируют экспрессию
гена Кг в передней и задней областях эмбриона
соответственно, поэтому мРНК гена Кг накаплива-
ется в центральной части эмбриона (рис. 8.57). Со
временем границы транскрипционных областей ге-
нов дар сужаются, поскольку их продукты влияют
на транскрипцию генов друг друга: так, продукт
гена Кг ингибирует транскрипцию гена hb, а про-
дукты генов kni и hb подавляют экспрессию гена Кг.
Следовательно, формирование областей стабильной
экспрессии генов дар происходит в два этапа. Сна-
чала экспрессия генов дар дифференциально регули-
руется при помощи распределения материнских де-
терминант, а позже она регулируется взаимной
репрессией.
Пространственная локализация продуктов генов
дар опосредует специфическую локальную регуля-
цию следующей группы генов в иерархическом ряду,
генов класса pair rule. Их преходящая экспрессия
происходит в семи или восьми транскрипционных
областях. Процесс периодической смены профилей
экспрессии приводит к локальной экспрессии генов
полярности сегмента; некоторые из этих генов (fushi
terazu [ftz], even skipped [ere] и engrailed [ew<y])
участвуют в дальнейшем дроблении сегментов на
отделы (рис. 8.57). Гены каждого класса явно нахо-
дятся под воздействием генов, функционирующих
на более ранних стадиях, и некоторых генов своего
класса. Так, смена стадий формирования тела Dro-
sophila в процессе эмбриогенеза определяется иерар-
хической регуляцией, при которой продукты мате-
ринских генов действуют на функционирование
впервые экспрессирующихся генов зиготы, продук-
ты которых влияют как на синтез друг друга, так
и на функционирование групп генов, экспрессирую-
щихся впоследствии взаимозависимым образом. Ре-
зультатом этой системы взаимодействий является
то, что ко времени завершения формирования кле-
точного строения эмбриона, когда все клетки ка-
жутся морфологически одинаковыми, участь клеток,
находящихся в разных частях эмбриона, оказывает-
ся уже предрешенной, поскольку в них экспрессиру-
ются разные комбинации генов класса pair rule
и генов полярности сегмента. Различные сочетания
продуктов этих генов в каждом сегменте определя-
ют экспрессию гомеотических генов ultrabithorax
(ubx) и antennapedia (antp), которые регулируют бо-
лее поздние стадии развития и дифференцировки
тканей мухи. Гомеотические гены-это гены, мута-
ции в которых трансформируют одну часть взро-
слого организма в другую, совершенно не похожую,
структуру. Например, гомеотическая мутация An-
tenna pedia превращает усик на голове мухи в ногу
нормального строения. Следовательно, один из
продуктов гена Antp, по-видимому, детерминирует
путь развития определенных клеток.
Каким образом возник и утвердился такой
86
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
gap-гены
(зигота)
Материнские
гены
Материнские гены
hunchback
Образование клеточной структуры
anieniiafjeaia штинхпогах
engrailed
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
87
иерархический каскад пространственно заданной
экспрессии генов, в особенности на стадии, когда
эмбрион содержит только неизолированные ядра?
Детальная картина этого процесса пока не установ-
лена, но уже ясно, что дифференциальная экспрессия
генов на ранних стадиях эмбрионального развития
отчасти связана с регуляцией транскрипции. Клони-
рование и секвенирование генов, которые транскри-
бируются на разных иерархических уровнях, пока-
зывают, что некоторые из них кодируют связываю-
щиеся со специфическими последовательностями
ДНК белки, которые либо активируют, либо инги-
бируют транскрипцию генов своего класса или ге-
нов, стоящих ниже в иерархическом ряду.
Как мы уже упоминали, индуцированные в зиго-
те транскрипты гена ЛЬ концентрируются на самом
краю передней части яйца-в области с наиболее
высоким содержанием белка гена bed. Поскольку
у большинства мутантов по гену bed ген hb не
экспрессируется, предполагалось, что продукт bed
нужен для активации транскрипции hb. Возмож-
ность проверить эту гипотезу появилась после того,
как был клонирован ген bed и получен его белковый
продукт, а также клонирован ген hb и идентифици-
рована область регуляции его транскрипции. Оказа-
лось, что белок bed связывается с разным сродством
со многими элементами промотора гена hb вблизи
точки начала его транскрипции (рис. 8.58). Канони-
ческая последовательность трех сайтов сильного
РИС. 8.57.
Экспрессия генов на ранних этапах развития Drosophila.
Схематически представлена временная и пространст-
венная экспрессия некоторых генов, ответственных за
организацию эмбриона Drosophila вдоль передне-зад-
ней оси в первые три часа после оплодотворения.
Формирование клеточной структуры эмбриона проис-
ходит по мере экспрессии генов antennapedia и ultra-
bithcrax. Экспрессия gap-генов hunchback, kruppel и
knirps зависит от содержания продуктов материнских
генов в яйце (например, bicoid и nanos) и происходит
в соответствии с градиентом концентрации мРНК и бел-
ка гена bicoid. Эта концентрация уменьшается в направ-
лении от переднего полюса яйца к заднему. Экспрессия
зависит также от влияния продуктов gap-генов на синтез
друг друга. К тому времени, когда устанавливается
определенная картина экспрессии генов hunchback,
kruppel и knirps. начинают экспрессироваться некоторые
гены класса pair rule-hairy и runt. Сначала зоны их
экспрессии разделяются нечетко, а затем образуются
строго чередующиеся полосы. Несколько позже экс-
прессируются другие гены класса pair rule- evenskipped
и fushi terazu, каждый в виде семи чередующихся полос.
Этот набор продуктов ранних генов индуцирует экспрес-
сию гена engrailed и синтез продуктов генов antennape-
dia и ultrabithorax, главным образом в двух зонах в пе-
редней части яйца.
связывания, которые расположены с интервалом
100 п.н. между парами оснований —293 и —55,
вероятно, имеет вид 5'-ТСТААТСС-3'. Участие бел-
ка bed в регуляции транскрипции с промотора hb
было продемонстрировано в опытах in vivo. Так,
инъекция в ранний эмбрион Drosophila химерного
гена, содержащего промотор hb, сшитый с кодирую-
щей областью хлорамфеникол-ацетилтрансферазы,
вызывает экспрессию гена в эмбрионах дикого типа,
но не в мутантных по bed самках. С помощью
последовательных делений элементов промотора hb
удалось установить, что участок между парами
оснований — 300 и +1 всецело определяет hcJ-зави-
симую и специфичную для данной области экспрес-
сию. Этот участок содержит три близко располо-
женные копии сайтов сильного связывания белка
bed. Два других, более слабых сайта связывания bed
находятся левее, вблизи пары —2300. Эти сайты
стимулируют образование hb-мРНК большего раз-
мера.
Когда при помощи трансформации с использо-
ванием Р-элемента (разд. 10.2.6) в эмбрионы были
введены химерные гены, содержащие последова-
тельности сильного связывания белка bed. сшитые
с кодирующей Р-галактозидазу последовательно-
стью, оказалось, что ген р-галактозидазы экспрес-
сируется в области, занимающей почти всю перед-
нюю половину эмбриона. В тех случаях, когда для
активации промотора маркерного гена использова-
лись последовательности слабого связывания bed,
экспрессия ограничивалась первой передней четвер-
тью эмбриона. Отсюда следует, что промотор hb
может реагировать на градиент bed двумя спосо-
бами с образованием мРНК двух видов.
ДНК-связывающий домен белка bed представля-
ет собой последовательность из 60 аминокислот
и называется гомеодоменом, поскольку кодирующая
его область была впервые обнаружена в гомеотиче-
ских генах Drosophila. Этот домен (разд. 8.6.а) встре-
чается в белках, кодируемых большой группой ре-
гуляторных генов в геномах как беспозвоночных,
так и позвоночных. Указанные гены почти всегда
присутствуют в регуляторах транскрипции. Гомео-
домены кодируются многими генами Drosophila из
уже упоминавшихся в связи с регуляторной иерар-
хией. Некоторые кодируемые ими белки содержат
цинковые пальцы и, возможно, именно поэтому
могут связываться с различными последователь-
ностями ДНК.
з. Регуляция транскрипции генов
U-PHK
Богатые урацилом малые ядерные РНК
(U-мяРНК) у всех организмов кодируются семей-
ством активно экспрессирующихся многокопийных
88
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
В
1 2
А
123
2900
нуклеотидов
3200
нуклеотидов
-293 -21Ъ
А1 CCTGACC AACGT А АТССССАТ
— 1R4 “163
А2 TTGTTTCT GCTCTA АТСС AGAA
аз ~77cctctgcc с атст а ат с се т т g А55
— 2343 “2321
В1 ATTAATTC ATGCT А АТСТй A TGA
-2293 -2221
В2 CAT А АТС АС СТ ТТ А АТ С СС A A GT
Каноническая тгтдатсс
последовательность v
РИС. 8.58.
Регуляторные элементы промотора гена hunchback (hb)
Drosophila. Участок из первых 300 п. н. слева от единицы
транскрипции содержит три близкорасположенных сай-
та прочного связывания белка bed (А1, А2 и АЗ; выде-
лены цветом). Они существенны для транскрипции
мРНК длиной 2900 нуклеотидов, на которой синтези-
руется белок hb. Два сайта слабого связывания bed (В1
и В2) находятся примерно на 2300 нуклеотидов левее
точки начала транскрипции (они тоже выделены цве-
том). В1 и В2 стимулируют образование мРНК длиной
3200 нуклеотидов (после сплайсинга).
генов (разд. 9.2.д). В ядре клеток позвоночных при-
сутствует по меньшей мере 200000 каждого из
основных типов U-PHK-транскриптов (Ul, U2, U4,
U5 и U6). Они являются компонентами мяРНП,
участвующих в процессинге про-мРНК (разд. 8.5.г).
Другие U-PHK, U3 и U7,-это компоненты присут-
ствующих в меньшем количестве мяРНП, которые,
вероятно, участвуют в процессинге про-рРНК и
образовании З'-концов гистоновых мРНК соответ-
ственно (разд. 8.3.в). Транскрипцию генов U1-, U2-,
U4- и U5-PHK, судя по ее чувствительности к а-ама-
нитину, катализирует РНК-полимераза II (разд.
8.1.6). Кроме того, исходные U-PHK-транскрипты
содержат 7-метилгуанозиновый кэп, который затем
превращается в 2,2,7-триметилгуанозиновый кэп,
характерный для U-PHK. Ген U6-PHK, напротив,
транскрибируется с участием РНК-полимеразы III,
а зрелая РНК не имеет кэпа. Тем не менее опреде-
ленные свойства промотора гена U6-PHK совпа-
дают со свойствами промоторов типичных генов
класса II (разд. 8.4.6). Например, область перед
сайтом начала транскрипции гена U6-PHK содер-
жит GC-блоки, октамер и ТАТА-блок регулятор-
ные элементы, характерные для промоторов, рас-
познаваемых РНК-полимеразой II. Универсальность
сигналов транскрипции иллюстрирует еше и тот
факт, что промотор U6-PHK Xenopus tropicalis
распознается в ооцитах Xenopus как РНК-полимера-
зой II, так и РНК-полимеразой III. А всего лишь
несколько небольших изменений в последователь-
ности длиной 50 п. н. перед точкой начала транс-
крипции гена U2-PHK превращают промотор, ис-
ходно распознаваемый РНК-полимеразой II, в про-
мотор, распознаваемый РНК-полимеразой III.
Таким образом, граница между промоторами клас-
сов II и III оказывается весьма нечеткой.
Все регуляторные элементы промоторов генов
U-PHK, транскрибируемых обычно РНК-полимера-
зой II, организованы одинаково (рис. 8.59, Л). Каж-
дый содержит по крайней мере два элемента, ответ-
ственных за активность промотора. Один, прокси-
мальный элемент (PSE), входящий в состав базаль-
ного промотора, находится между парами основа-
ний — 70 и —40, другой, дистальный (DSE),-между
парами —300 и —200. Ни один из этих элементов
в отдельности не способен стимулировать транс-
крипцию генов U-PHK. PSE необходим для транс-
крипции РНК-генов и определяет точку ее начала.
DSE функционирует в любой ориентации и может
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
89
А. Регуляторные элементы промоторов U1-,U2-И4-и U5-PHK
I----DSE-----1
-300 - 200 -100 +1
Б. Регуляторные элементы промотора U6-PHK
РИС. 8.59.
Элементы регуляции транскрипции с промоторов U-PHK.
А. Транскрипцию генов, находящихся под контролем
промоторов U1-, U2-, U4- и U5-PHK, катализирует
РНК-полимераза II. ТАТА-бокс в этих промоторах от-
сутствует, зато есть важные проксимальный (PSE) и
дистальный (DSE) элементы. Последний содержит
GC-богатый сайт связывания Sp1 и октамерный элемент
(octa), характерный для промоторов иммуноглобули-
новых генов. Б. Транскрипция с промотора U6-PHK
осуществляется при участии РНК-полимеразы III. Этот
промотор содержит ТАТА-бокс, а также проксимальный
(PSE) и дистальный (DSE) элементы; последний содер-
жит GC-богатый участок и октамерный элемент.
находиться на разном расстоянии до промотора.
Следовательно, он обладает свойствами обычного
энхансера.
Хотя элемент PSE определяет точку начала
транскрипции при синтезе U-PHK, у него отсутству-
ет необходимый для узнавания ТАТА-блок. Более
того, за исключением промотора гена U6 (рис.
8.59,Б), последовательности всех PSE одинаковы
и характерны только для главных промоторов генов
U-PHK. Последовательность PSE и белки, опосре-
дующие участие PSE в транскрипции, до сих пор
неизвестны. Напротив, последовательность DSE и
белки, отвечающие за его энхансерную функцию,
охарактеризованы. Выявлено два типа последова-
тельностей: GC-блок 5'-GGGCGG-3' и октамер
5'-ATGCAAAT-3'. Мутации в любой из этих после-
довательностей инактивируют энхансер; следова-
тельно, они должны быть функционально взаимоза-
висимы. GC-блок связывает белок Spl, а октамер
присоединяет широко распространенный белок
OTF-L Применение метода отпечатков с использо-
ванием ДНКазы I и мутационный анализ GC-блока
октамерной последовательности промоторов генов
U-PHK показали, что ключевыми являются те же
самые основания, которые входят в промоторы
генов мРНК. Некоторые DSE связывают также
белки API и АР2.
Все регуляторные последовательности, необхо-
димые для транскрипции гена U6-PHK, находятся
перед кодирующей последовательностью, несмотря
на то что транскрипцию гена катализирует РНК-по-
лимераза III (разд. 8.4.6). Не менее удивителен и тот
факт, что транскрипционный аппарат РНК-поли-
меразы III использует те же i/ис-последовательно-
сти, что и факторы транскрипции, которые функцио-
нируют с РНК-полимеразой II (рис. 8.59,Б). Итак,
промотор гена U6-PHK сходен с промоторами ге-
нов U1-, U2-, U4- и U5-PHK в том отношении, что
содержит элемент PSE вблизи точки начала транс-
крипции и элемент DSE между парами —300 и
— 200, в который входят GC-гексануклеотид и две
октамерные последовательности. Однако промотор
гена LJ6-PHK содержит вблизи точки начала транс-
крипции также и ТАТА-блок, а элемент PSE этого
промотора отличается от PSE промоторов генов
U1-. U2-, U4- и U5-PHK. Каким образом РНК-по-
лимераза III использует эти регуляторные i/ис-по-
следовательности для активации транскрипции гена
U6-PHK- неясно.
Любопытным свойством промоторов генов
U-PHK является то, что, когда они слиты с коди-
рующей областью гена мРНК, соответствующие
транскрипты не полиаденилируются на З'-концах
(разд. 8.3.в). Аналогично, З'-концы U-PHK, транс-
крибированных с участием промоторов генов мРНК,
не процессируются надлежащим образом. Следова-
90
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
тельно, можно предположить, что механизм ини-
циации транскрипции и события, которые обуслов-
ливают созревание З'-концов мРНК и U-PHK,
каким-то образом координированы. Другой пара-
докс заключается в том. что DSE промоторов
U-PHK не стимулируют транскрипцию ни с основ-
ного промотора гена ТК, ни с промотора глобино-
вого гена, хотя в случае гетерологичных промото-
ров энхансеры действуют эффективно в обоих слу-
чаях. Из этого следует, что факторы транскрипции,
связанные с DSE, не могут взаимодействовать с
ТАТА-ассоциированными комплексами или активи-
ровать их. Вероятно, комплекс транскрипции, ассо-
циированный с элементом PSE, отличается от комп-
лекса, связанного с ТАТА-блоком, и эти комплексы
не являются взаимозаменяемыми. Комплекс, нахо-
дящийся в стартовой точке транскрипции, может
влиять на процессы, происходящие на З'-конце еди-
ницы транскрипции.
8.4. СТРУКТУРА И ЭКСПРЕССИЯ
ГЕНОВ КЛАССА III
а. Основные свойства
РНК, кодируемые генами класса III. Продук-
тами генов класса III являются низкомолекуляр-
ные РНК, которые участвуют в синтезе белков
(тРНК и 5S-pPHK). их внутриклеточном транспорте
(7SL-PHK) и в посттранскрипционном процессинге
(U6-PHK). Гены класса III кодируют также некото-
рые другие РНК с неустановленными физиологиче-
скими функциями (7SK-PHK и Alu-PHK). Геном
аденовирусов содержит два гена класса III-VA1
и VA2; их РНК-продукты (длиной 157 и 140 нуклео-
тидов соответственно) переключают аппарат транс-
крипции инфицированных клеток на эффективный
синтез структурных вирусных белков. При экспрес-
сии генов класса III, входящих в состав генома
герпесвируса Эпштейна-Барр, также образуются
две низкомолекулярные РНК, EBERI и EBERII
(длиной 166 и 172 нуклеотида соответственно).
Гены класса III делятся на две группы. В первую
из них входят гены, кодирующие 5S-pPHK, 7SL-,
7SK-, U6- и VA-РНК, причем первичные транскрип-
ты имеют такую же длину, что и зрелые функцио-
нальные РНК, т.е. процессинг транскриптов в дан-
ном случае не нужен. 5'-трифосфатная концевая
группа, которая образуется при инициации транс-
крипции, становится 5'-концом, а последний из
транскрибированных остатков - З'-концом зрелой
РНК. Ко второй группе генов класса III относятся
гены, кодирующие тРНК, зрелые формы которых
образуются в результате отщепления нуклеотидов
как с 5'-, так и с З'-концов первичных транскриптов
под действием специфических эндонуклеаз. З'-кон-
цевая CCA-последовательность, необходимая для
функционирования всех тРНК, не закодирована в
соответствующем гене, а присоединяется при учас-
тии особого фермента. Поскольку некоторые гены
тРНК содержат интрон, для получения зрелых
тРНК- продуктов этих генов- необходим сплайсинг
(разд. 8.5).
Сигналы транскрипции генов класса III. Сигналы
инициации и терминации транскрипции генов класса
III высококонсервативны у таких разных эукариоти-
ческих организмов, как дрожжи, Drosophila, Xenopus
и млекопитающие. По этой причине гены тРНК,
5S-pPHK и других низкомолекулярных клеточных
и вирусных РНК из разных источников правильно
транскрибируются гетерологичными системами
транскрипции.
Отличительным свойством последовательно-
стей, ответственных за регуляцию инициации транс-
крипции многих генов класса III, является то, что
они локализованы внутри кодирующей области.
Эти внутренние регуляторные участки (ICR) немного
различаются у разных генов класса III, но в целом
они на удивление сходны между собой. У некоторых
генов класса III в пределах участка из 30 п. н. перед
точкой инициации транскрипции находятся допол-
нительные регуляторные последовательности. Дру-
гие гены класса III (например, гены U6-PHK и 7SK-
РНК млекопитающих) не содержат ICR и имеют
только 5'-регуляторные последовательности, кото-
рые целиком находятся перед точкой инициации
транскрипции. Все нуклеотидные последовательно-
сти, определяющие точку терминации транскрипции
на З'-конце генов класса III, сходны между собой;
есть лишь небольшие различия, которые влияют на
эффективность терминации.
Транскрипционный комплекс с участием РНК-по-
лимеразы III. РНК-полимераза III, которая содер-
жится в нуклеоплазме эукариотических клеток, была
выделена из самых разных организмов: дрожжей,
насекомых, амфибий, растений и некоторых млеко-
питающих. В случае дрожжей, Xenopus и млекопи-
тающих получены практически гомогенные формы
фермента. Все эти ферменты имеют большую мол.
массу, ~ 650 к Да, и содержат от 10 до 15 субъединиц
в зависимости от источника и метода выделения.
Две субъединицы имеют мол. массу в пределах от
140 до 170 кДа, а их аминокислотные последова-
тельности сходны с последовательностями соответ-
ствующих больших субъединиц РНК-полимераз I и
II, а также р- и Р'-субъединиц РНК-полимеразы Е
соН. Остальные субъединицы, по-видимому, присут-
ствуют в эквимолярных количествах, а их размер
колеблется от 10 до 90 кДа. Субъединица с мол.
массой ~ 90 кДа, вероятно, имеется только в РНК-
полимеразе III, а три субъединицы меньшего раз-
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
91
мера сходны с субъединицами соответствующих
размеров очищенных РНК-полимераз I и И, если
судить по гомологии аминокислотных последова-
тельностей и перекрестным иммунологическим ре-
акциям.
Для точной и эффективной транскрипции генов
класса III при участии препаратов очищенной РНК-
полимеразы III нужны еще некоторые белки. Кон-
кретный набор необходимых факторов транскрип-
ции варьирует среди различных генов класса III
и зависит от локализации и типа элементов, ответ-
ственных за регуляцию транскрипционной активно-
сти генов. В большинстве случаев необходимые
факторы транскрипции относятся к уникальным
факторам, характерным только для экспрессии ге-
нов класса III. Но некоторые гены класса III, зави-
сящие от 5'-регуляторных элементов (например,
гены тРНК и малых ядерных и цитоплазматических
РНК), нуждаются в дополнительных факторах
транскрипции, вероятно родственных тем белкам,
которые, как полагали ранее, специфичны в отно-
шении транскрипционных комплексов с участием
РНК-полимеразы II (разд. 8.3.з). Терминация транс-
крипции генов 5S-pPHK, по-видимому, опосредует-
ся только РНК-полимеразой III, хотя для других
генов класса III. возможно, необходимы связанные
с ней факторы транскрипции.
Активность
б. Последовательности,
регулирующие инициацию
транскрипции
Внутренние регуляторные последовательности
генов 5S-pPHK Xenopus. Клонированная ДНК Xeno-
pus, содержащая участок, кодирующий 5S-pPHK,
и фланкирующие его 5'- и З'-последовательности,
правильно транскрибируется после введения в ядра
ооцитов Xenopus или после инкубации с нефракцио-
нированными экстрактами ооцитов. При этом обра-
зуется в основном 5S-pPHK длиной 120 нуклеоти-
дов. Это ключевое открытие позволило проана-
лизировать с помощью обратной генетики, какие
структуры необходимы для точной и эффективной
транскрипции.
Определение активности транскрипции генов 5S-
рРНК с делециями разной длины на 5'-конце показа-
ло, что последовательность, фланкирующая З'-ко-
нец единицы транскрипции, и последовательность,
занимающая более первой трети кодирующей об-
ласти, несущественны для правильной инициации
транскрипции (рис. 8.60). В результате транскрип-
ции таких укороченных генов образуются РНК дли-
ной примерно 120 нуклеотидов; из-за делеций транс-
крипция в таких условиях начинается с плазмидной
последовательности, сшитой с 5'-концом оставшей-
ся части кодирующей последовательности. Анало-
гично, точная и по существу столь же эффективная
инициация транскрипции осуществляется при уда-
лении З'-фланкирующей последовательности и од-
ной трети кодирующей области со стороны З'-кон-
ца. Однако, если удалить примерно 50 п. н. и более
на 5'-конце или более 30 п. н. на З'-конце кодирую-
щей области, то транскрипции не происходит. В ре-
зультате этого эксперимента, впервые продемонст-
рировавшего наличие внутригенного сигнала транс-
11CR
+ 80 +120
РИС. 8.60. Ген 5S‘PPHK ! ------1
Влияние делеций в гене 5S-pPHK и вблизи него на I
инициацию транскрипции. +1 и + 120-сайты инициа- i
ции и терминации транскрипции соответственно. Де- [
леции, представленные прямыми линиями, начинаю- _____
щимися с 5'- или З'-конца клонированного гена, полу- __
чены путем мутагенеза in vitro и повторного клонирова-
ния. Транскрипционную активность каждого мутантного
гена определяли с использованием ядер ооцитов Xe-
nopus или бесклеточных экстрактов.
Активность
92
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
I---------------- ICR ------------------1
Промежуточный
A-бокс сегмент С-бокс
+1 +50 +64 +67
РИС. 8.61.
Структура внутренней регуляторной области (ICR) гена
SS-pPHK X. laevis. Эта область содержит A-бокс, про-
межуточный сегмент и С-бокс. Они выделены более
темным цветом, чем кодирующая часть гена 5S-pPHK.
крипции, было установлено, что внутренняя регуля-
торная область (ICR) гена 5S-pPHK Xenopus нахо-
дится примерно между парами оснований +50 и
+90. В самом деле, оказалось, что клонированный
сегмент 5S-pPHK, находящийся между парами осно-
ваний + 41 и +87, обеспечивает транскрипцию с
участием РНК-полимеразы III in vitro, при этом
инициация осуществляется в точке, расположенной
примерно на 50 п. н. левее места соединения вектора
и клонированного сегмента.
Более детально структура ICR указанного гена
была изучена путем внесения небольших делеций.
последующих вставок вместо этих делеций или за-
мен специфических пар оснований и исследования их
влияния на транскрипцию in vitro и in vivo. Эти
опыты показали, что ICR состоит из трех функцио-
нально различающихся областей (рис. 8.61). Одна из
них охватывает участок от +50-Й до +64-Й пары
оснований и по своей последовательности и функ-
ции сходна с так называемым A-блоком, который,
как теперь известно, присутствует в ICR всех генов
класса III. Вторая область простирается от +80-Й
до +90-Й пары оснований и присуща только ICR
генов 5S-pPHK (С-белок). В функционировании ICR
участвует также сегмент между парами оснований
+ 67 и +72. Изменения в этой области слабо влия-
ют на транскрипцию; более серьезные последствия
имеет изменение расстояния между А- и С-блоками.
Таким образом, для регуляции инициации транс-
крипции необходимы и достаточны последователь-
ности, находящиеся в пределах кодирующей обла-
сти генов 5S-pPHK Xenopus. Вопрос о том, как
+72 +60 +90 +120
именно эти сигналы выполняют данную функцию,
рассматривается в разд. 8.4.в.
Широкая распространенность внутренних регу-
ляторных областей генов класса III. Мутационный
анализ, аналогичный упомянутому выше, позволил
идентифицировать ICR в генах 5S-pPHK и тРНК
всех исследованных эукариот. Например, было по-
казано, что линкерные вставки в ген 5S-pPHK Dro-
sophila между парами +46 и +65, а также между
парами +76 и +98 подавляют его транскрипцию;
напротив, вставки между парами +3 и +20 лишь
незначительно уменьшают активность транскрип-
ции. ICR примерно с такими же границами обнару-
жены в генах 5S-pPHK Bombyx, Neurospora и Saccha-
romyces.
Подобные исследования с модифицированными
генами тРНК и использованием систем транскрип-
ции in vitro или ооцитов Xenopus помогли устано-
вить, что для транскрипции этих генов также нужны
ICR. Нуклеотидные последовательности на 5'- и
З'-концах кодирующей последовательности несуще-
ственны для инициации транскрипции. При помощи
делеций и линкерных вставок идентифицированы
две существенные области, положение которых
мало различается у генов разных тРНК независимо
от вида (рис. 8.62). Одна из них, A-бокс, локали-
зована в области примерно от +10 до +20 пар
оснований, а другая, В-бокс,-между парами осно-
ваний + 50 и +65. Нуклеотидные последователь-
ности А- и В-боксов варьируют, а их каноническими
последовательностями являются соответственно
5'-TGGCNNAGTGG-3’ и 5'-GGTCGANNC-3'. Ну-
клеотидная последовательность участка между А-
и В-боксами несущественна, важна лишь его длина.
Если расстояние между боксами становится меньше
25 или больше ~60 пар оснований, то активность
транскрипции снижается, а если это расстояние пре-
вышает 100 пар оснований или если А- и В-боксы
I------------------ ICR--------------------1
А-бокс
I-----(—ЯИЦ-------------
+ 1 +10 +20
РИС. 8.62.
Внутренняя регуляторная область (ICR) гена тРНК.
Показано положение А- и В-боксов, которое в разных
тРНК слегка различается. +1-сайт инициации тран-
скрипции. Такое же попарное расположение А- и В-бок-
В-бокс
от +T0
до +90
сов характерно для генов VA-PHK, EBER-PHK и
7SL-PHK. Канонические последовательности А- и В-бок-
сов имеют вид S'-TGGCNNAGTGG-З' и 5'-GGTTCGANNC-3'
соответственно (N-любой нуклеотид).
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
93
РИС. 8.63.
Внутренняя регуляторная область (ICR) гена TPHKLeu
Xenopus laevis. Представлена нуклеотидная последова-
тельность гена TPHKLeu размером 100 п.н. Под ней
в схематическом виде изображена зрелая тРНК длиной
83 п. н. и указаны разные ее участки (см. также рис. 3.29).
А- и В-боксы выделены цветом. [G. Galli et al.. Nature
294 (1981), p. 626.]
сливаются, то транскрипция полностью блокирует-
ся. Аналогичная конструкция, состоящая из двух
частей, А- и В-боксов, встречается в кодирующих
областях генов 7SL-PHK и четырех генов низкомо-
лекулярных РНК в геномах аденовируса и вируса
Эпштейна- Барр.
Нуклеотидные последовательности А- и В-бок-
сов не только существенны для транскрипции, но
и необходимы для формирования вторичной струк-
туры тРНК; A-бокс кодирует часть D-стебля и пе-
тли, а В-бокс-TyUG-петлю и стебель (рис. 8.63).
Участие обеих последовательностей в транскрипции
генов тРНК и формировании правильной структуры
молекул тРНК, которые нужны для синтеза белка,
интересно с точки зрения эволюции этих генов.
В самом деле, если пару оснований CG в позиции 56
гена tPHKtyr заменить на пару GC, что означает
замену С на G в Ту U G-петле, то соответствующий
ген не будет транскрибироваться.
Взаимозаменяемость A-боксов из разных вну-
тренних регуляторных областей. Идентичны ли
функции, выполняемые одинаковыми ICR-элемен-
тами разных генов класса III? Что касается А-бокса
в ICR генов 5S-pPHK и тРНК, то да. Так, рекомби-
нанты, построенные путем замены первых 20 пар
оснований ICR гена 5S-pPHK A-боксом ICR гена
тРНК, активно транскрибируются (рис. 8.64). Такой
же результат дает замена A-бокса тРНК на первые
РИС. 8.64.
A-бокс тРНК
TGGT CTAGTGG
ACCAGATC АСС
Взаимозаменяемость последовательностей, ответствен-
ных за регуляцию транскрипции генов 5S-pPHK и тРНК.
Рекомбинантные ДНК клонировали в плазмидных век-
торах и определяли их транскрипционную активность
после инъекции в ядра ооцитов X. laevis. Сегмент гена
тРНКРго получен из нематоды С. elegans, а сегмент гена
5S-pPHK- из X. borealis. Векторные последовательно-
сти, а также A/Zndlll-сайт, по которому соединены оба
регуляторных сегмента, выделены двумя разными от-
С-бокс 5S-pPHK
AGTACTTGGA /
ТСATGAACCT (
тенками серого цвета и стрелкой с буквой Н соответст-
венно. Когда гибридная регуляторная область функцио-
нирует нормально, инициация транскрипции происхо-
дит примерно на 50 п.н. левее A-бокса гена тРНКРго.
Поскольку обычно транскрипция тРНК-гена начинается
примерно на 10 п. н. левее A-бокса, свой вклад в лока-
лизацию сайта инициации должен вносить и С-бокс гена
SS-pPHK.
94
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
20 пар оснований ICR гена 5S-pPHK. Эти данные
свидетельствуют о том, что хотя ICR разных генов
класса III различаются, они, по всей вероятности,
представляют собой разновидности одного регуля-
торного элемента.
Влияние последовательностей, находящихся сле-
ва от тонки инициации транскрипции (левых после-
довательностей), на регуляцию транскрипции неко-
торых генов класса III. Исходя из того факта, что
присутствия ICR достаточно для инициации транс-
крипции генов 5S-pPHK ооцитов Xenopus, было
высказано предположение, что по тому же принципу
организованы все гены класса III. Однако это пред-
положение оказалось не совсем верным. Для транс-
крипции генов других 5S-pPHK (например, из Xeno-
pus [соматических], Bombyx, Drosophila, Neurospora,
Saccharomyces и некоторых видов млекопитающих)
необходима последовательность, находящаяся слева
от точки инициации транскрипции. Так, делеция или
замена последовательностей между парами —34
и — 11 в гене 5S-pPHK Bombyx mori и между парами
— 39 и —26 в гене 5S-pPHK Drosophila приводит
к блокированию их транскрипции. Для точной и
эффективной инициации транскрипции большинства
генов тРНК всех исследованных видов (за исклю-
чением Xenopus) тоже нужны левые 5'-последова-
тельности, которые, по-видимому, характерны для
данного вида, а не для гена. Так, левые 5'-участки
генов многих тРНК и генов 5S-pPHK Bombyx гомо-
логичны. К другим генам класса III, которым для
точной и эффективной транскрипции нужны одно-
временно и левые 5'-последовательности, и элемент
ICR, относятся гены вирусных VA- и EBER-PHK
и ген 7SL-PHK.
Регуляция транскрипции некоторых генов класса
III при помощи одних только левых последователь-
ностей. Как это ни удивительно, но сделанный
вначале вывод о том, что ICR являются единствен-
ными детерминантами регуляции транскрипции с
участием РНК-полимеразы III, нашел свое неожи-
данное продолжение в случае генов 7SK-, U6-PHK
и некоторых низкомолекулярных РНК (EBER) в ге-
номе вируса Эпштейна-Барр: для их транскрипции
используются только последовательности, находя-
щиеся перед кодирующими областями. Кодирую-
щая область гена U6-PHK содержит A-бокс, харак-
терный для генов тРНК и для 5'-области ICR гена
5S-pPHK, но мутации, затрагивающие этот сегмент,
или его полное удаление не влияют на транскрип-
цию, стимулированную левой 5'-последовательно-
стью. В гене 7SK-PHK аналог A-бокса полностью
отсутствует, поэтому неудивительно, что ген, полу-
ченный при слиянии сегмента из 243 п. н. левой
последовательности гена 7SK-PHK (пары оснований
от —245 до —3) с совершенно неродственной после-
довательностью, транскрибируется с участием
РНК-полимеразы III in vitro и in vivo столь же
эффективно, как и сам ген 7SK-PHK; аналогичные
опыты с левыми последовательностями гена
U6-PHK дают такой же результат. С другой сторо-
ны, для оптимальной транскрипции EBER-генов
нужны как А- и В-боксы ICR, так и некоторые
5'-элементы, характерные для промоторов-энхансе-
ров РНК-полимеразы II.
Если удалить последовательности между парами
оснований —37 и —3 левого элемента гена
7SK-PHK, то способность к транскрипции утрачива-
ется (рис. 8.65). Показано также, что левая часть
гена LJ6-PHK длиной не менее 79 п.н. необходима
для активации транскрипции соседней последова-
тельности in vitro. В обоих случаях для поддержания
нормального уровня транскрипции in vivo нужны
последовательности, находящиеся еще левее, между
парами оснований —243 и —37 в 5'-фланкирующей
последовательности гена 7SK-PHK и между парами
оснований —315 и —79 в 5'-фланкирующей последо-
вательности гена U6-PHK (разд. 8.3.ж). Интересно,
что 5'-фланкирующие последовательности генов U6-
и 7SK-PHK функционально взаимозаменяемы. Од-
нако 5'-фланкирующая последовательность гена
7SL-PHK, для транскрипции которого нужны как
внутренние, так и 5'-фланкирующие последователь-
ности, не может заменить соответствующую об-
ласть 78К-гена даже несмотря на то, что тран-
скрипцию обоих генов катализирует РНК-полиме-
раза III.
Весьма интересной особенностью левых обла-
стей генов U6- и 7SK-PHK является то, что они
содержат последовательности, характерные для
многих генов класса II (рис. 8.65). Так, у обоих генов
между парами оснований —29 и —24 находится
ТАТА-бокс. необходимый для эффективной инициа-
ции транскрипции. Более того, промоторы генов
П6- и 7SK-PHK и промоторы генов других U-PHK
содержат сходные элементы между парами —40
и — 50. Кроме того, в пределах нескольких сотен пар
оснований левее промоторов генов U6- и 7SK-PHK
встречаются элементы, характерные для промото-
ров и энхансеров генов класса II (разд. 8.3.з); в каче-
стве примера можно привести GC-бокс (5'-
GGGCGG-3') и октамер (5'-ATGCAAAT-3'). На-
сколько необходимы проксимальные и дистальные
элементы промоторов и какова их физиологическая
роль-неизвестно. Непонятно также, имеет ли ка-
кое-либо значение удивительное сходство между
левыми регуляторными последовательностями про-
моторов генов U6- и 7SK-PHK. Вызывает интерес
и высокая гомологичность между регуляторными
последовательностями, находящимися левее гена
U6-PHK, который относится к классу III, и регуля-
торными последовательностями генов, кодирующих
РНК от U1 до U5 и относящихся к генам класса II
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
95
CTGCAGTАТТТAGCATGCCCCACCCATCTGCAAGGCATTCTGGATAGTGTCAAAACAGCCGGAAATCAA
I
-243
(100%)
GTCCGTTTATCTCAAACTTTAGCATTTTGGGAATAAATGATATTTGCTATGCTGGTTAAATTAGATTTT
I
-154
(63%)
AGTTAAATTTCCTGCTGAAGCTCTAGTACGATAAGCAACTTGACCTAAGTGTAAAGTTGAGACTTCCTT
I I I
-59 -45 -37
(25,3%) (24%) (21%)
CAGGTTTATatagcttgtgcgccgcttgggtacctcg
I I I I I
-26 -15 -8 -3 *1
(<1%) (<1%) (< 1%) (<1%)
РИС. 8.65.
Нуклеотидная последовательность участка, фланкиру-
ющего ген 7SK-PHK с 5'-конца и начинающегося от
пары оснований — 243. Делеции в этом участке полу-
чали при помощи эндонуклеазы Ва/ЗЛ. Положение кон-
цевых нуклеотидов делегированных участков (считая с
(разд. 8.3.з). Возможно, существуют факторы транс-
крипции, взаимодействующие с одинаковыми для
генов классов II и III регуляторными последова-
тельностями и осуществляющие совместную регу-
ляцию разных наборов генов.
Еще одним указанием на физиологическую связь
между сигналами транскрипции, осуществляемой
с помощью РНК-полимераз II и III, служит тот
факт, что в ДНК, кодирующей нетранслируемую
лидерную последовательность гена класса II (гена
с-тус), находится промотор для РНК-полимеразы
III. Более того, сигнальные последовательности
промоторов для обеих РНК-полимераз (II и III)
инициируют транскрипцию, начинающуюся в од-
ном и том же сайте. Однако транскрипция с участи-
ем РНК-полимеразы III прекращается в конце пер-
вого экзона, в то время как транскрипция, катали-
зируемая РНК-полимеразой II, идет до конца. Пер-
вый экзон гена с-тус содержит A-бокс, но не содер-
жит В-бокса, а кроме того, у него есть левые
5-последовательности, напоминающие те, которые
необходимы для транскрипции генов 5S-pPHK и
ItPHK Bombyx mori. Контролируется ли транскрип-
ция с-тус в клеточном цикле транскрипционными
сигналами от обеих РНК-полимераз (II и III), пока
неясно; неизвестна также и роль преждевременной
терминации транскрипции, катализируемой РНК-
полимеразой III.
в. Влияние факторов транскрипции
на инициацию транскрипции
с участием РНК-полимеразы III
Для инициации транскрипции при участии
РНК-полимеразы III нужны дополнительные белки,
5'-конца) отмечено отрицательными числами. Так, му-
тант — 243 содержит 243 п. н. левее сайта инициации
транскрипции (+1), а мутант — 154- только 154 п.н.
Числа в скобках-относительная эффективность тран-
скрипции. [S. Murphy et aL, Cell 51 (1987), p. 81.]
три из которых уже идентифицированы. Для транс-
крипции всех генов класса III, зависящей от ICR,
требуются факторы TFIIIB и TFII1C (последний,
как теперь известно, представляет собой смесь
TFIIIC-1 и TFIIIC-2). TFIIIA нужен исключительно
для транскрипции генов 5S-pPHK. Данные факторы
транскрипции встречаются у всех эукариот; ни один
из них не проявляет сколько-нибудь заметной видо-
вой специфичности. Так, гены 5S-pPHK человека
после введения в ядра ооцитов Xenopus транскриби-
руются столь же эффективно и точно, как и гены
Xenopus', подобно этому, в клеточных экстрактах
человека, содержащих РНК-полимеразу III и необ-
ходимые факторы, одинаково хорошо транскриби-
руются обе 5S-pPHK. Вначале к регуляторным
участкам присоединяется не сама РНК-полимераза
III, а вспомогательные факторы, которые форми-
руют стабильные комплексы преинициации. О су-
ществовании таких комплексов свидетельствуют ре-
зультаты, полученные в опытах по конкуренции
матриц (см. разд. 8.2.в) и по связыванию с нитро-
целлюлозными фильтрами, а также данные, полу-
ченные с помощью метода отпечатков и других
методов.
Взаимодействие TFIIIB и TFIIIC с А- и В-бокса-
ми. Опыты по конкуренции матриц помогли выя-
вить некоторые особенности формирования комп-
лекса между TFIIIB и TFIIIC, с одной стороны, и А-
и В-боксами генов класса III-с другой. При сов-
местной инкубации генов тРНК и VAI-РНК в си-
стеме транскрипции in vitro образуются оба
РНК-продукта (рис. 8.66). Однако если перед добав-
лением РНК-полимеразы III одну из ДНК проинку-
бировать с TFIIIB и TFIIIC, то при последующем
добавлении второго гена будет преимущественно
96
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Компоненты первой инкубационной смеси 1 Ген VA1 Ген тРНК TFIIIB, TFIIIC 2 Ген VA1 TFIIIB,TFIIIC 3 ГентРНК TFIIIB,TFIIIC 4 Ген VA1 TFIIIB 5 Ген VA1 TFIIIC
Компоненты второй инкубационной Ген тРНК Ген VAI Ген тРНК Ген тРНК
pollll pel HI pollll TFIIIC TFIIIB
смеси rNTP rNTP rNTP pollll rNTP pollll rNTP
РИС. 8.66.
Образование стабильных комплексов между факторами
транскрипции и регуляторными участками генов класса
III. Преинкубация гена тРНК или VA-РНК с TFIIIB и
TFIIIC приводит к тому, что при добавлении рибонук-
леозидтрифосфатов (rNTP) и РНК-полимеразы III
(pollll) начинается их транскрипция. Однако если пре-
инкубировать один из генов-либо ген VA1-PHK
транскрибироваться первая матрица (сравните до-
рожки 1 и 3 на рис. 8.66). Такая предпочтительная
транскрипция указывает на то, что факторы обра-
зуют прочный комплекс с первым геном и поэтому
не могут участвовать в транскрипции гена, добав-
ленного позже. Отметим также, что не безразлично,
какой из факторов инкубировали с матрицей пер-
вым (сравните дорожки 4 и 5 на рис. 8.66). Из
полученных данных следует, что TFIIIC может об-
разовывать комплекс с VA-ДНК самостоятельно.
Более того, хотя бы один, а возможно, и оба
фактора, по-видимому, остаются связанными с ма-
трицей даже после многих циклов транскрипции.
Поскольку связывание TFIIIC блокирует рас-
щепление в сайте рестрикции, находящемся в В-бок-
се VA-ДНК, вероятно, в этом месте или рядом с ним
имеется сайт связывания TFIIIC. Опыты с примене-
нием метода отпечатков отдельно с TFIIIC-1 и
TFIIIC-2 показывают, что TFIIIC-1, в отличие от
TFIIIC-2, не может самостоятельно образовывать
прочную связь с ДНК. TFIIIC-2 защищает З'-часть
(дорожка 2), либо ген тРНК (дорожка 3)-со смесью
TFIIIB и TFIIIC, а затем добавить другой ген вместе
с полимеразой и субстратами, то транскрибироваться
будет только проинкубированный РНК-ген. TFIIIC мо-
жет образовывать комплекс преинициации с геном VA1-
РНК (или геном тРНК) в отсутствие TFIIIB (дорожки
4 и 5). [А. В. Lasser et al.. Science 222 (1983), p. 740.]
регуляторной области, включая В-бокс, а связыва-
ние TFIIIC-1 расширяет «отпечаток» в 5'-направле-
нии за рамки A-бокса к сайту начала транскрипции.
Для формирования прочного комплекса с В- и
A-боксами генов тРНК необходимы как TFIIIB, так
и TFIIIC. При этом оба белка TFIIIC сначала
взаимодействуют с В-боксом с образованием комп-
лекса, стабильность которого зависит от специфиче-
ской последовательности В-бокса, а затем происхо-
дит связывание TFIIIB, в результате чего стабили-
зируется комплекс преинициации. С этим комплек-
сом, по-видимому, и взаимодействует РНК-поли-
мераза III, инициируя транскрипцию в нужном ме-
сте. Возможно, связывание РНК-полимеразы III
происходит исключительно за счет белок-белковых
взаимодействий, путем образования специфических
контактов с участками ICR или за счет взаимодей-
ствия с факторами транскрипции, связанными с ре-
гуляторными элементами слева от точки начала
транскрипции.
Один из результатов, полученных в опытах in
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
97
vitro, подтверждается тем фактом, что ген тРНК,
находясь в структуре хроматина, может правильно
транскрибироваться под действием одной лишь
РНК-полимеразы III, без добавления TFIIIB и
TFIIIC. В той же самой реакционной смеси ген
VA-РНК, присутствующий в виде свободной ДНК,
не транскрибируется до тех пор, пока не будут
добавлены два необходимых фактора транскрип-
ции. Следовательно, в хроматине белки TFIIIB и
TFIIIC, по всей видимости, связаны с 1CR в ста-
бильный комплекс, а удаление гистонов с ДНК
приводит к отщеплению факторов транскрипции.
Комплексы преинициации, содержащие гены
5S-PHK и факторы транскрипции. Описанные выше
методы использовали для изучения роли факторов
транскрипции в активации инициации транскрипции
генов 5S-pPHK. Было показано, что для инициации
процесса необходимы специфические факторы и
упорядоченность в их связывании (рис. 8.67). TFIIIA
в отсутствие других факторов образует нестабиль-
ный комплекс с ICR. TFIIIC сам по себе не присо-
единяется к ICR, однако он связывается с комплек-
сом TFIIIA -5S-pPHK и в значительной мере ста-
билизирует его. Сколько TF1 ПС-факторов нужно
для этой стабилизации-один или оба-неясно. По-
следовательности, ответственные за взаимодействие
TFIIIA с 58-ДНК, находятся в С-боксе между пара-
ми оснований +80 и +90, хотя в связывании уча-
ствуют также нуклеотиды между парами оснований
+ 50 и +64 (A-бокс). Стабилизация комплекса
TFIIIA- 5S-pPHK под действием TFIIIC, по-види-
мому, достигается благодаря взаимодействиям ме-
жду двумя факторами транскрипции и контактам
между TFIIIC и A-боксами. TFIIIB сам по себе не
присоединяется к свободной SS-рДНК и не меняет
отпечаток, получающийся при связывании ДНК с
TFIIIA и TFIIIC. Тем не менее TFIIIB необходим
для транскрипции, катализируемой РНК-полимера-
зой III. Вероятно, он участвует в образовании муль-
тибелкового комплекса, с которым ассоциируется
РНК-полимераза III перед началом транскрипции.
Поскольку TFIIIA является единственным факто-
ром транскрипции, способным самостоятельно
узнавать и связывать ICR 58-рДНК, было очень
важно изучить механизм и специфичность его свя-
зывания.
Молекулярное строение TFIIIA. TFIIIA Xenopus
является цинксодержащим металлопротеином с
мол. массой 40 кДа. Его аминокислотная последо-
вательность, состоящая из 344 остатков, была опре-
делена из данных о нуклеотидной последовательно-
сти соответствующей кДНК. Клонированный ген
белка TFIIIA Xenopus имеет длину 11 т. п. н. и со-
держит восемь интронов. Примерно две трети ами-
нокислот TFIIIA со стороны N-конца (остатки
13-276) образуют девять копий несовершенного no-
ICR
5S-AHK
TFIIIA
ICR-ТЕША
Нестабильный комплекс
TFIIIC
TFIIIA
ICR |
^TFIIIC
Стабильный комплекс
TFIIIB
РИС. 8.67.
Упорядоченное связывание факторов транскрипции с
внутренней регуляторной областью (ICR) гена 5S-pPHK.
Возможно, связывание TFIIIB и РНК-полимеразы III
(pollll) на самом деле осуществляется одновременно,
а не так, как зто показано на рисунке. Как видно из
рисунка, комплекс преинициации сохраняется на про-
тяжении всего цикла транскрипции.
втора, состоящего примерно из 30 аминокислот, при
этом каждая копия обязательно содержит два остат-
ка цистеина и гистидина (рис. 8.68). Участки гена
TFIIIA, кодирующие повторы из 30 аминокислот,
разделены интронами. Весь С-концевой домен не
содержит никаких повторов и кодируется одним
экзоном.
По данным атомной абсорбционной спектроско-
пии, с молекулой очищенного белка связано девять
(+ один) атомов цинка. Детальный рентгенострук-
98
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Каноническая
последовательност» т G Е к
т>
277
13
23
26
30
SCDKR F Т
1 MG
Е КА
13
38
68
99
130
160
189
215
247
312
Р V V Y К Rj
F А
GTE
12
DC
GC
DG С
ENC
GC
D S
VC
GC
NR
VG
F R
LAS
NWKL
ANMK
N О L
TNGS
37
67
98
VH
R Н
L КН
129
159
188
214
246
276
311
344
8
I С
S
2
3
5
6
8
9
D Р Е
G Е
G Е
N
К
К
N F
Р С
Т С
К
Е
Е
G А
Е К
G
F
N
D
S
D L
R
к к
к
R Н
Н
К
S
К
Н Н L
О
А Н
L С
R Н
К Н
S
F
N
I К
Q Q
A G -
D -
Q
Е
Е
К
S
К Е
Е О
R К
V S
С
L А
V С
Е С
Р С
V С
R Т
L С
R
Р
К
Н
Р
К
Р
F
н
К
R
Е
О
О
О
V С
Е К С
К Т D
Е
Р
S
Н A G С
R Р К
L V К
R
G К
D К
А
R
С S
о
G
N
R
К
К
S
К
F
К
К
Н
S
К
н
Р
S R L
К
К
Q
Е
К
S Y T
С F А
Р S G
К Н
Т Н
R F Н
S Н
V Н
Т W Т L
D Y L
N L
К
R
V
А
Е
С н
А
М
К
F
R
К
S L
Е
R L
Т G
Р
D Н Q К
S Н I о
• R
Н S V
Р К
V L
S
Т Н
F Н
V
Н
S К
D К
Е
К
N
А
Q
РИС. 8.68.
Аминокислотная последовательность фактора TFIIIA из
X. laevis (использовано однобуквенное обозначение
аминокислот; см. рис. 1.24). Последовательность рас-
положена на рисунке таким образом, чтобы выделить
повторяющиеся элементы. Молекула состоит из сле-
дующих сегментов: N-концевого участка (остатки 1-12),
девяти повторов (остатки 13 276), богатого лизи-
ном участка вблизи С-конца и короткого концевого
участка. Вверху представлена каноническая последова-
тельность-пронумерованные от 1 до 30 обозначения,
заключенные в кружки,- которая иллюстрирует харак-
терные особенности типичного повтора. Конечная точка
турный анализ с высоким разрешением показал, что
каждый такой атом образует хелатные связи с па-
рами цистеиновых и гистидиновых остатков, нахо-
дящимися в одном из повторов (рис. 8.69). Наи-
большее предпочтение отдается структурной моде-
ли TFIIIA, в соответствии с которой белок имеет
девять цинковых пальцев, в каждом из которых
атом цинка координационно связан с четырьмя
аминокислотными остатками (два остатка цистеина
и два-гистидина) (рис. 8.69). Участки между этими
спаренными остатками цистеина и гистидина всегда
содержат фенилаланин (или тирозин) и лейцин (или
метионин) и разное число основных аминокислот
(рис. 8.68 и 8.69). Перед каждым цинковым пальцем
обязательно находится остаток тирозина или фе-
нилаланина, а между двумя цистеинами обычно
расположен кислый остаток. Основные и другие
остатки, входящие в состав пальцев, образуют
ДНК-связывающий участок TFIIIA.
По-видимому, TFIIIA связывается с последова-
тельностью длиной примерно 50 п. н., входящей в
ICR гена 5S-pPHK, при этом каждый из девяти
цинковых пальцев контактирует с участком длиной
~ 5 п. н. Как показали опыты по определению чув-
ствительности ДНК в составе ДНК-белкового
комплекса к действию диметилсульфата и микро-
каждой единицы выбрана произвольно после НЗО.
Наиболее высококонсервативные остатки выделены
цветом. Предполагается, что С8, С13. Н26 и НЗО связы-
вают ион Zn2+. Гидрофобные остатки Y6, F7 и L23,
вероятно, образуют внутренний каркас предполагаемой
мультидоменной структуры. Звездочками (*) отмечены
позиции, в которых иногда в норме присутствуют встав-
ки, а черточками в повторах-пропуски аминокислот на
рисунке. Точками отмечены вариабельные участки по-
следовательности. [J. Miller et al., EMBO J. 4 (1985),
p. 1609.]
кокковой нуклеазы, контакты между TFIIIA и ДНК
осуществляются через каждые 5 п. н., что равно
половине витка спирали. Однако, за исключением
возможной периодичности в расположении остат-
ков гуанина, в ICR нет никаких явных повторов,
которые могли бы формировать одинаковые сайты
связывания для каждого из девяти пальцев. В на-
стоящее время принята модель связывания, предпо-
лагающая, что TFIIIA взаимодействует с кластера-
ми гуанина, располагаясь вдоль одной стороны
двойной спирали (рис. 8.70). Существование таких
множественных контактов позволяет объяснить ста-
бильность комплекса TFIIIA-TF1IIC-ICR и сохране-
ние его целостности при многократных циклах
транскрипции, во время которых РНК-полимераза
перемещается вдоль ДНК, отсоединяя последова-
тельно один или несколько пальцев. Некоторые
факторы транскрипции, осуществляемой с помо-
щью РНК-полимеразы II, для связывания с род-
ственными регуляторными последовательностями
тоже используют цинковые пальцы (разд. 8.3.д и е;
8.6.а).
Был проведен неполный протеолиз белка TFIIIA
и определена активность его укороченных форм
разной длины. Оказалось, что область, отвечающая
за активацию транскрипции генов 5S-pPHK, состоит
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
99
РИС. 8.69.
Цинковые пальцы: укладка линейных повторяющихся
доменов белкового фактора TFIIIA. В основе каждого
пальца лежит тетраэдрическая структура, образуемая
связанными с ионами цинка лигандами. Указаны только
важные для образования структуры остатки. Цветными
кружками обозначены консервативные аминокислоты
цистеин (С) и гистидин (Н), связанные с ионом цинка;
отрицательно заряженный аспарагин 11 (D) и три
гидрофобные группы. Черные кружки-это боковые
группы, по-видимому взаимодействующие с ДНК. Со-
гласно модели, ион цинка сближает концы каждого
повтора, способствуя формированию ДНК-связывающей
петли, или «пальца», с участием центральных остатков
14-25. [J. Miller et al.. EMBO. J. 4 (1985) p. 1609.]
примерно из 20 аминокислот, расположенных вбли-
зи С-конца белковой молекулы; эта область не
участвует в связывании TFIIIA с ICR. Остальная
часть белковой молекулы, составляющая 75% ее
длины и содержащая девять цинковых пальцев и
N-конец, отвечает за прочное связывание с ICR; эта
область неспособна активировать транскрипцию.
Модель, обобщающая все эти положения, представ-
лена на рис. 8.71.
Свой вклад в связывание вносят все цинковые
пальцы, но в первую очередь два N-концевых паль-
ца, взаимодействующих с С-боксом. Остальные
пальцы контактируют с A-боксом и с участком
между двумя боксами; вклад последних в энергию
взаимодействия менее существен (рис. 8.61). Какие
именно аминокислоты отвечают за узнавание раз-
личных элементов 1CR, точно неизвестно.
Участок вблизи С-конца может активировать
транскрипцию, либо влияя прямо на РНК-полиме-
разу III, либо взаимодействуя с TFIIIB и/или TFIIIC.
Известно, что TFIIIB и TFIIIC стабилизируют свя-
зывание TFIIIA с ICR, однако участвуют ли эти
факторы также и в активации инициации транскрип-
ции под действием полимеразы, неясно. Тот факт,
что TFIIIB и TFIIIC могут стимулировать транс-
крипцию всех других генов класса III, позволяет
предположить, что они принимают непосредствен-
ное участие в инициации транскрипции, катализи-
руемой РНК-полимеразой III.
Остается ли стабильным комплекс, образующий-
ся в результате взаимодействия трех факторов
транскрипции с ICR, во время перемещения РНК-
полимеразы III в процессе транскрипции последова-
тельности, находящейся за регуляторной областью?
Чтобы ответить на этот вопрос, изучали транскрип-
цию гена 5S-pPHK, сшитого с бактериальным про-
мотором таким образом, чтобы последователь-
ность 58-рДНК могла транскрибироваться in vitro
с помощью очищенных препаратов бактериальной
РНК-полимеразы. Было показано, что в условиях,
при которых РНК-полимераза III правильно транс-
крибирует ген 5S-pPHK, бактериальная полимераза
также эффективно транскрибирует этот ген как в
присутствии, так и в отсутствие трех факторов
транскрипции. После транскрипции с помощью бак-
териальной РНК-полимеразы 5S-pPHK нормально
синтезируется в присутствии одной только РНК-по-
лимеразы III. Это говорит о том, что комплекс
преинициации, содержащий 58-рДНК, TFIIIA,
TFIIIB и TFIIIC, не разрушается во время транс-
крипции с участием бактериальной РНК-полимера-
зы. Существенно, что транскрипция, катализируе-
мая бактериальной РНК-полимеразой, протекала
в условиях, не обеспечивающих повторную сборку
Модель взаимодействия
цинковых пальцев TFIIIA с
ты, отделены друг от друга десятью парами нукпеоти-
двойной спиралью ДНК
Белковая цепь располагается
дов, при этом каждые два соседних пальца подходят
по одну сторону спирали ДНК, а пальцы укладываются
в большую бороздку поочередно «спереди» и «сзади».
Пальцы, образующие структурно эквивалентные контак-
к Ю-атомам остатков гуанина с разных сторон. [L. Fairall
et al., J. Mol. Biol. 192 (1986), p. 577.]
100
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 8.71.
Структура фактора транскрипции TFIIIA и его взаимо-
действие с внутренней регуляторной областью (ICR)
гена 5S-pPHK. Элемент ICR, находящийся между па-
рами оснований + 50 и + 90, изображен вместе с
5'-концом значащей цепи. N-конец белковой цепи-рас-
полагается на рисунке справа, а С-конец слева. Об ее
ориентации относительно ДНК см. в тексте. У N-конца
располагаются девять повторяющихся единиц (остатки
1 -276)-цинковых пальцев, взаимодействующих с ICR.
На эту часть белковой молекулы приходится 30 кДа.
Остатки 277-344 охватывают богатую лизином область
и С-концевой участок. Вместе они образуют домен
с массой 10 кДа. Цинковые пальцы располагаются от-
носительно молекулы ДНК так, как это представлено на
рис. 8.70. [J. Miller et al., EMBO J. 4 (1985), p. 1609]
комплекса преинициации в случае его диссоциации.
Если бы комплекс диссоциировал, то из-за невоз-
можности его повторного образования до или после
добавления РНК-полимеразы III синтез 5S-pPHK не
осуществился бы. Хотя это до сих пор и не доказа-
но, считается, что комплекс 58-рДНК с факторами
транскрипции также не разрушается при повторных
циклах транскрипции с участием РНК-полимеразы
III. Благодаря тому, что область контактирования
домена из девяти повторяющихся цинковых пальцев
с ICR-последовательностью из 50 п.н. (рис. 8.71)
достаточно протяженна, одни ДНК-белковые связи
в ходе транскрипции разрушаются, а другие восста-
навливаются.
Иная картина наблюдается при репликации
ДНК. входящей в комплекс. Эти данные были
получены при введении области ori репликации
ДНК SV40 в плазмиду, содержащую ген 5S-pPHK.
После сборки комплекса преинициации, анало-
гичного рассмотренному выше, добавляли компо-
ненты, необходимые для репликации плазмиды in
vitro, а после репликации ДНК-РНК-полимеразу
III и проверяли, синтезируется ли 5S-pPHK. В этом
случае 5S-pPHK синтезировалась только при до-
бавлении трех факторов транкрипции, когда мог
вновь образоваться комплекс преинициации. Более
того, при расщеплении реплицированной ДНК под
действием ДНКазы 1 образовывались такие же про-
дукты, что и при расщеплении свободной ДНК;
следовательно, TFIIIA и TFIIIC не были связаны
с ДНК. Очевидно, комплекс 5S-pPHK с тремя фак-
торами транскрипции разрушается во время репли-
кации, но не во время транскрипции ДНК. Не-
стабильность комплекса при репликации ДНК мо-
жет объясняться тем, что для успешной работы
комплекса репликации должно произойти расплета-
ние протяженных участков двойной спирали.
г. Терминация транскрипции
с участием РНК-полимеразы III
В отличие от инициации транскрипции, для ко-
торой необходимы сложные регуляторные последо-
вательности, а также- помимо самой полимеразы
определенные белки, для терминации транскрипции
нужен только сам фермент и кластер дезоксиадени-
латных остатков на матричной цепи. Терминация in
vitro осуществляется правильно в присутствии вы-
сокоочищенной РНК-полимеразы III и двухцепо-
чечной ДНК-матрицы. Точная и эффективная тер-
минация происходит даже при транскрипции генов
класса III, у которых нет ICR; таким образом,
взаимодействие матрицы с TFIIIA, TFIIIB или
TFIIIC никак не влияет на процесс терминации. Для
идентификации сигнала терминации использовались
специально сконструированные транскрипционные
матрицы клонированных ДНК. Как показали эти
опыты, в терминации участвует кластер из дезокси-
аденилатных остатков и определенные фланкирую-
щие его последовательности (табл. 8.2). Например,
если четыре дезоксиаденилатных остатка матричной
цепи фланкированы остатками дезокситимидиловой
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
101
Таблица 8.2. Сайты терминации транскрипции при
участии РНК-полимеразы III °
Сайты эффективной терми-
нации
Xenopus borealis (гены со-
матических клеток)
Xenopus borealis (гены
ооцитов, тип I)
Xenopus borealis (гены
ооцитов, тип 3)
Аденовирус VAI
tPHKa1j Bombyx mori
Сайты относительно неэф-
фективной терминации
Xenopus laevis (гены
ооцитов, тип I)
Сайты неэффективной тер-
минации
Xenopus laevis (гены
ооцитов, гип 2)
Xenopus laevis (псевлоген
ооцитов)
5-TAGGCTTTTGCACT-3'
- TAGGCTTTTAGACT
- TAGTCTTTTTCCAG
GCTCCTTTTGGCTT
- ACGATTTTGTTAT
- TAGGCTTTTCAAAG
- TAGGCTTTCCAAAG
- AGAAGTTTTCAAAG
" Сайты терминации для РНК-полимеразы III находятся на
З'-конце последовательности, кодирующей 5S-pPHK, за З'-конЦом
последовательности, кодирующей тРНК. Как правило, терминация
происходит в пределах кластера Т (подчеркнут), а не у какого-то
определенного остатка Т. Сайты относительно неэффективной
терминации содержат короткие A-повторы, расположенные перед
кластерами Т или за ними. Транскрипт 5S-pPHK не подвергается
процессии! у на З'-конце, но у транскриптов тРНК-генов нуклеоти-
ды с З'-конца должны быть удалены, чтобы произошло присоеди-
нение З'-САА-копца и образовались зрелые тРНК. [D. F. Bogen-
bagen. D.D. Brown, Cell 24. (1981), p. 261].
кислоты, то терминация происходит неэффективно,
в отличие от того случая, когда с одной стороны
дезоксиаденилатного кластера находится динуклео-
тид CG, а с другой GC (5'-GCAAAAGC-3'). По-
следнее имеет место в случае гена 5S-pPHK сомати-
ческих клеток Xenopus borealis. Мутации, приводя-
щие к изменению числа аденилатных остатков в
кластере, уменьшают эффективность терминации.
Синтез РНК прекращается при спаривании остатка
уридиловой кислоты с одним из дезоксиаденилат-
ных остатков. Ни отдаленные последовательности,
ни вторичная структура РНК, по-видимому, на тер-
минацию не влияют. Если в клонированном гене
делетирован З'-конец гена 5S-pPHK, то транскрип-
ция продолжается до тех пор, пока РНК-полимераза
III не дойдет до соответствующего случайного си-
гнала во фланкирующей вектор последовательно-
сти. А если аденилатный кластер ввести в произ-
вольное место матричной цепи какого-либо гена
класса III. то терминация часто осуществляется
в этом модифицированном сайте.
д. Регуляция экспрессии генов
5S-pPHK
в процессе развития организма
Два вида генов 5S-pPHK Xenopus. Существенные
изменения в экспрессии генов 5S-pPHK наблюдают-
ся в процессе дифференцировки и развития Xenopus
(табл. 8.3). На ранних этапах оогенеза происходят
быстрый рост ооцитов и синтез большого количе-
ства 5S-pPHK, необходимой для удовлетворения
потребности в рибосомах в ходе раннего эмбриоге-
неза. В зрелом яйце, напротив, никакие РНК не
синтезируются. Синтез всех РНК, в том числе и
5S-pPHK, возобновляется на стадии бластулы (4000
клеток) или чуть раньше и продолжается затем при
жизни взрослого организма.
Два разных набора генов 5S-pPHK экспрессиру-
ются на нескольких стадиях развития. Одни экспрес-
сируются почти исключительно на стадии ооцита
(гены 5S-pPHK ооцита), вторые- как в ооцитах, так
и в соматических клетках развивающегося эмбрио-
на, а также в тканях взрослого организма (гены
соматической 5S-pPHK). Таким образом, несмотря
на примерно одинаковое число транскриптов ооци-
тарной и соматической 5S-pPHK в бластуле, на
стадии головастика и последующих стадиях более
98% 5S-pPHK относятся к соматическому типу. Эта
Таблица 8.3. Изменение соотношения между
ооцитарной и соматической 5S-pPHK и содер-
жание TFIIIA в клетках в процессе развития
Xenopus* III 11
Стадия развития Преобладающий Число
тип синтезируемой молекул
5S-pPHK TFIIIA на ген
5S-pPHK
Незрелый ооцит
1 рост
Зрелый ооцит
[мейоз I
Неоплодотворенное
яйцо
мейоз II
оплодотворение
J- деление клетки
Эмбрион на стадии
бластулы
(4000 клеток)
деление клетки
дифференцировка
Эмбрион на стадии
гаструлы
деление клетки
дифференцировка
Плавающий головастик
Ооцитарная 5 I О7
» 5I06
РНК не синтези- 4 10s
руется
Поровну 10
Соматическая 2
» 0,4
11 W. М. Wormington et al.. Cold Spring Harbor Symposium 47 (1982),
p. 879.
102
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
разница в экспрессии становится особенно впечат-
ляющей, если учесть, что в геноме Xenopus содер-
жится около 20000 генов 5S-pPHK ооцита и только
около 400 генов соматической 5S-pPHK (разд. 9.2.6).
Чем же объясняется преимущественная экспрессия
генов 5S-pPHK ооцита в ооцитах и их репрессия
в соматических клетках? Каким образом гены сома-
тической 5S-pPHK «одерживают верх» и становятся
почти единственной матрицей для синтеза 5S-pPHK
в соматических клетках? Теперь мы знаем, что
дифференциальное функционирование генов опосре-
дуется регуляцией транскрипции, причем эта регу-
ляция, по-видимому, зависит от нескольких пара-
метров. К ним относятся: меняющееся в процессе
развития содержание факторов транскрипции (осо-
бенно TFIIIA, но, возможно, и других), разная
стабильность комплексов ооцитарных и соматиче-
ских генов с этими факторами, структура хрома-
тина, в который упакованы 58-рДНК обоих типов,
время репликации 58-рДНК каждого типа в течение
S-фазы.
Дифференциальная экспрессия генов in vitro. Не-
смотря на упомянутые выше количественные разли-
чия, в структурном отношении гены ооцитарной
и соматической 5S-pPHK различаются мало. Эти
различия затрагивают главным образом от трех до
шести пар оснований (в зависимости от вида) в по-
следовательности, находящейся вблизи 5'-конца ICR
(рис. 8.72). Транскрипция клонированных генов со-
матической 5S-pPHK примерно в пять раз более
эффективна, чем транскрипция клонированных ге-
нов ооцитарной 5S-pPHK при использовании и тех
и других в качестве матриц либо с экстрактами
соматических клеток или ооцитов, либо с очищен-
ной РНК-полимеразой III в присутствии TFIIIA,
TFIIIB и TFIIIC. Это не очень большое различие
Ген 5’
клеток ] зо
соматических GCCTACGGCCACACCACCCTGAAAGTGCCC
ооцитов Т
60
соматических GATCTCGTCTGATCTCGGA AGCC A A GCAGG
ооцитов A G ТА
90
соматических GTT С GGGCCTGGTT AGT А СТ TGG ATGGGAG
ооцитов С
120
coMaTHMecKHXACCGCCTGGGAATACCAGGTGTCGTAGGCTT
ооцитов G
РИС. 8.72.
Нуклеотидные последовательности соматических и
ооцитарных генов 5S-pPHK X. laevis. Приведены только
первые 120 нуклеотидов значащих цепей с указанием
оснований, разных в генах двух типов. Закрашенные
прямоугольники внутренняя регуляторная область.
[P.J. Ford, R. D. Brown, Cell 8 (1976), р. 485.]
в эффективности транскрипции, вероятно, отражает
несколько более высокое сродство TFIIIA к ICR
соматических генов. Но, поскольку в клетках число
генов ооцитарной 5S-pPHK в 50 раз превышает
число генов соматической 5S-pPHK, кажется мало-
вероятным, что несколько более высокое сродство
TFIIIA к ICR генов соматических клеток может
объяснить дифференциальную экспрессию, наблю-
даемую in vivo.
После того как было показано, что картина
дифференциальной экспрессии генов может быть
воспроизведена с использованием очищенного хро-
матина в качестве матрицы для РНК-полимеразы
III. появилась возможность исследовать регулятор
экспрессии in vitro. Неоплодотворенные яйца или
клетки на ранних стадиях развития бластулы во-
обще не синтезируют РНК, а хроматин, выделенный
из таких клеток, не поддерживает синтез 5S-pPHK
при инкубации с РНК-полимеразой III. Напротив,
при транскрипции хроматина, выделенного из ооци-
тов, образуются 5S-pPHK в основном ооцитарного
типа, тогда как при транскрипции в тех же условиях
хроматина из соматических клеток преимущест-
венно соматическая 5S-pPHK. Для синтеза ооцитар-
ной или соматической 5S-pPHK при использовании
соответственно хроматина ооцитов или хроматина
соматических клеток в качестве матриц необходимо
добавить лишь РНК-полимеразу III. Таким обра-
зом, гены в хроматине, вероятно, ассоциированы
с тремя факторами транскрипции, образуя комп-
лекс преинициации. Но даже при добавлении этих
факторов вместе с полимеразой ооцитарные гены не
транскрибируются в хроматине соматических кле-
ток. Однако если хроматин соматических клеток
вначале экстрагировать 0,6 М NaCl, а затем доба-
вить три фактора транскрипции вместе с полиме-
разой, то может произойти транскрипция генов
ооцитарной 5S-pPHK в составе хроматина сомати-
ческих клеток. Добавление соли не влияет на транс-
крипцию генов соматической 5S-PHK, для которой
по-прежнему требуется только РНК-полимераза III.
Этот феномен можно объяснить, в частности, тем,
что гены ооцитов в репрессированном хроматине
не содержат требуемых факторов транскрипции и
невосприимчивы к факторам, добавленным извне,
или тем, что в репрессированном хроматине имеет-
ся потенциально активный комплекс, но он недо-
ступен для полимеразы. Возможно, разрушение хро-
матиновой структуры при солевой экстракции «рас-
крывает» хроматин или удаляет какие-то белки,
и в результате гены ооцитов ассоциируют с добав-
ленными факторами и РНК-полимеразой III и вос-
станавливают свою транскрипционную активность.
Гистоны нуклеосом Н2А. Н2В. НЗ и Н4 не
отщепляются при солевой экстракции, однако такая
обработка приводит к эффективному удалению ги-
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
103
стона Н1 из хроматина. Возможно, именно гистон
Н1 поддерживает гены 5S-pPHK ооцитов в репрес-
сированном состоянии в хроматине соматических
клеток и при его удалении гены ооцитарной
5S-pPHK становятся способными к транскрипции.
Об этом свидетельствует тот факт, что добавление
гистона Н1 к хроматину соматических клеток, ис-
тощенному по гистону Н1, блокирует транскрипцию
генов 5S-pPHK ооцитов. Следовательно, дифферен-
циальная экспрессия генов 5S-pPHK в соматических
клетках зависит от опосредованного гистоном Н1
уплотнения хроматина, содержащего гены 5S-pPHK
ооцитов.
Ответ на вопрос о том, каким образом гистон Н1
ограничивает связывание факторов транскрипции
с ICR, дают результаты анализа конкурентного
связывания нуклеосом (октамеров гистонов) и
TFIIIA. Эксперименты с использованием метода
отпечатков показывают, что кор нуклеосомы связан
с 5'-концом гена 5S-pPHK. включая две трети длины
ICR (рис. 8.73). Тем не менее TFIIIA связывается со
свободной частью ICR и, возможно, вытесняет ну-
клеосому из комплекса с 5'-концом регуляторной
области. Этого, вероятно, достаточно для того,
чтобы TFIIIB, TFIIIC и РНК-полимераза III связа-
лись с геном, и в надлежащем сайте началась
транскрипция. Поскольку гистон Н1 связан с ну-
I Н2А Н2В К4 НЗ | НЗ Н4 К2В Н2А | Нукпеосомныи
-3-6-5-4-3-2-1 0 1 2 3 4 5 6 1
РИС. 8.73.
Конкуренция TFIIIA с нуклеосомами за связывание
с ICR 5'-рДНК. А. Ген 5S-pPHK X. laevis (выделен
цветом) и фланкирующие его последовательности; ука-
зано положение ICR. Вверху представлены восемь
гистонов, связывающихся с геном и фланкирующей его
последовательностью. Б. Схематическое представление
накручивания ДНК на нуклеосомный кор (не показан).
Поскольку гистон Н1 связывается с ДНК на входе ее
в нуклеосому и на выходе из нее, можно ожидать, что
его связывание препятствует присоединению TFIIIA к
ICR. [D. Rhodes, EMBO J. 4 (1985), р. 3473.)
клеосомами на входе ДНК в октамер и на выходе из
него (рис. 1.17), его присутствие в хроматине, веро-
ятно, препятствует связыванию TFIIIA и, таким
образом, блокирует доступ TFIIIC, TFIIIB и
РНК-полимеразы III к матрице.
Переключение экспрессии генов 5S-pPHK в процес-
се развития Xenopus. Если присоединение гистона
Н1 к ооцитарным генам обусловливает дифферен-
циальную экспрессию в дифференцированных сома-
тических клетках, возникает вопрос: когда и каким
образом в процессе развития осуществляется пере-
ход от экспрессии преимущественно ооцитарных
генов к экспрессии почти исключительно соматиче-
ских генов? В созревающих ооцитах параллельно
с активацией генов ооцитарной 5S-pPHK синтезиру-
ется большое количество TFIIIA (табл. 8.3). Это не
удивительно, потому что TFIIIA необходим для
транскрипции. TFIIIA не только связывается с ге-
нами 5S-pPHK, но и образует 78-рибонуклеопро-
теидную частицу с новосинтезированной 5S-pPHK.
По-видимому, эти частицы являются формой хра-
нения 5S-pPHK до ее последующего включения
в рибосомы в оогенезе. При образовании таких
частиц 5S-pPHK конкурирует со своими собствен-
ными генами за связывание TFIIIA, участвуя таким
образом в регуляции синтеза РНК по типу обратной
связи.
После начального всплеска синтеза TFIIIA ско-
рость его образования снижается и продолжает
уменьшаться после оплодотворения и начала эм-
бриогенеза. Постепенное подавление экспрессии ге-
на 5S-pPHK ооцитов после стадии средней бластулы
коррелирует с дальнейшим понижением внутрикле-
точной концентрации TFIIIA (табл. 8.3). Итак, не-
зрелые ооциты содержат большой избыток TFIIIA
по сравнению с общим числом генов 5S-pPHK
и синтезируют большое количество 5S-pPHK ооци-
тарного типа; в соматических клетках на каждые
2-3 гена 5S-pPHK приходится всего одна молекула
TFIIIA. На конкуренцию между генами 5S-pPHK
ооцитарного и соматического типов за связывание
TFI11A сильно влияет более высокая нестабильность
комплексов транскрипции генов 5S-pPHK ооцитов
в оплодотворенных яйцах; подобный феномен на-
блюдается также в экстрактах ранних бластул. В
результате комплексы транскрипции генов ооцитар-
ной 5S-pPHK разрушаются, в то время как аналогич-
ные комплексы генов соматической 5S-pPHK преи-
мущественно остаются. Снижение скорости синтеза
TFIIIA по мере развития эмбриона и связывание его
с 5S-pPHK ограничивают участие TFIIIA в образо-
вании комплекса транскрипции. Большая лабиль-
ность комплекса транскрипции в случае генов ооци-
тарной 5S-pPHK наряду с предпочтительным свя-
зыванием TFIIIA и TFIIIC с генами соматической
5S-pPHK приводит к постоянному оттоку факторов
104
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
транскрипции от генов ооцитарной 5S-pPHK к ге-
нам соматической 5S-pPHK. Поскольку гены ооци-
тарной 5S-pPHK теряют факторы транскрипции,
данная область, вероятно, «выключается» в резуль-
тате уплотнения хроматина с участием гистона Н1.
Но каким образом это состояние преимущест-
венной активации генов соматической 5S-pPHK под-
держивается во время репликации ДНК при после-
довательных делениях клетки? При репликации
комплексы транскрипции разрушаются, и для во-
зобновления транскрипции они должны образовать-
ся вновь. Интересно, что в соматических клетках
Xenopus гены соматической 5S-pPHK реплицируют-
ся во время первой четверти S-фазы, а гены ооци-
тарной 5S-pPHK на более поздних стадиях в S-фа-
зе. Благодаря такой поэтапной репликации хромо-
сомы гены соматической 5S-pPHK могут повторно
образовывать функциональные комплексы транс-
крипции. не конкурируя с ооцитарными генами.
Наши представления о регуляции генов 5S-pPHK
Xenopus в процессе развития весьма неполны. Тем
не менее можно выделить некоторые характерные
моменты: 1) гиперпродуцирование TFIIIA в течение
оогенеза, позволяющее генам 5S-pPHK обоих типов
экспрессироваться пропорционально их количеству;
2) прекращение образования TFIIIA в процессе со-
зревания яйца и на последующих стадиях эмбриоге-
неза и весьма низкое его содержание в соматических
клетках; 3) влияние содержания TFIIIA на взаимо-
действие TFIIIA с 5S-pPHK в процессе развития;
4) преимущественное разрушение комплексов транс-
крипции генов ооцитарной 5S-pPHK и преимущест-
венное накопление-за счет конкуренции - комплек-
сов транскрипции генов соматической 5S-pPHK;
5) преимущественная конденсация хроматина, со-
держащего «пустые» гены ооцитарной 5S-pPHK.
при помощи гистона Н1; 6) более ранняя реплика-
ция генов соматической 5S-pPHK во время S-фазы,
что позволяет повторно образовываться в первую
очередь комплексам транскрипции генов соматиче-
ской 5S-pPHK. Дополнительная регуляция может
состоять: 1) в дифференциальной регуляции экспрес-
сии гена TFIIIA на ранних стадиях оогенеза и на
поздних стадиях эмбриогенеза и в соматических
клетках; 2) в наличии функционально активных
TFIIIC и TFIIIB; 3) во влиянии факторов транскрип-
ции, которые реагируют с регуляторными последо-
вательностями, расположенными перед генами.
8.5. ИНТРОНЫ
Кодирующие последовательности эукариотиче-
ских генов (экзоны) часто перемежаются некодирую-
щими (интронами). Это поразительное открытие по-
ставило перед учеными несколько вопросов. 1. Где,
когда и как появились интроны в генах, какую
роль они сыграли в биологической эволюции?
2. Каким образом первичные транскрипты таких
мозаичных генов превращаются в матричные РНК,
у которых интроны отсутствуют? 3. Какое значение
имеет сплайсинг (а также его механизм) для регуля-
ции экспрессии генов? 4. Какую функцию (если она
существует) выполняют интроны в мириадах опера-
ций, совершаемых в современных геномах? Напри-
мер, являются ли интроны рудиментом истории
эволюции генов или они играют важную роль в жиз-
ни сегодняшних организмов? Некоторые аспекты
первого из перечисленных вопросов обсуждаются во
введении к части III и в гл. 9. Мы же здесь ограни-
чимся рассмотрением того, что мы знаем о строе-
нии интронов, механизме их удаления в процессе
биогенеза зрелых функциональных РНК и о роли
сплайсинга в экспрессии эукариотических генов.
а. Частота встречаемости интронов
Частота, с которой встречаются прерывистые
гены, заметно варьирует у разных видов эукариот.
Наиболее обильно они представлены в геномах
растений и животных, а также в геномах инфици-
рующих их вирусов. У беспозвоночных (например,
у Drosophila, нематоды и морского ежа) интроны
встречаются реже. Однако некоторые гены, ответ-
ственные за регуляцию морфогенеза у Drosophila
(например, группы ultrahithorax и antennapedia),
имеют сложную картину чередования экзонов и
интронов. В большей части ядерных генов S. cerevi-
siae (за исключением генов актина и некоторых
генов тРНК) интроны отсутствуют, но зато преры-
вистые гены в изобилии имеются у их дальнего
родственника S. pombe. Интроны, редко встречаю-
щиеся в ядерных генах S. cerevisiae. широко пред-
ставлены в митохондриальных генах данного орга-
низма. Сразу после обнаружения интронов полага-
ли, что они присутствуют только в эукариотических
генах. Однако позже интрон был выявлен в прока-
риотическом гене тимидилатсинтетазы бактериофа-
га Т4, а затем- в гене tPHKs" и в гене высокомоле-
кулярной рРНК Archaebacteria. Поэтому мы не уди-
вимся, если при более полном исследовании гено-
мов прокариот обнаружатся новые гены с интрона-
ми.
Интроны присутствуют в ядерных генах, коди-
рующих мРНК. рРНК (правда, пока они обнаруже-
ны только в генах высокомолекулярных рРНК низ-
ших эукариот) и отдельные типы тРНК. Преры-
вистые гены часто встречаются в генах хлороплас-
тов и митохондрий, которые кодируют информа-
ционные, рибосомные и транспортные РНК этих
органелл. В то же время среди генов млекопи-
тающих, которые почти всегда содержат интроны,
встречаются исключения. Так, в большинстве генов.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
105
кодирующих пять гистонов, интроны отсутствуют;
это же относится и к генам двух семейств, коди-
рующим а- и Р-интерфероны. Напротив, единствен-
ный ген у-интерферона содержит множество интро-
нов (рис. 8.2). Не обнаружены интроны в генах 5,8S-
и 5S-pPHK, а также ни в одном из генов U-PHK,
7SL-PHK и 7SK-PHK.
б. Различные виды интронов
Все интроны транскрибируются в составе РНК-
предшественника и впоследствии удаляются в про-
цессе разрыва-воссоединения, который называется
сплайсингом. Особенности строения интронов и со-
ответствующие механизмы сплайсинга лежат в ос-
нове классификации этих элементов (табл. 8.4).
Интроны генов ядерных мРНК. Первыми были
обнаружены интроны в ядерных генах, кодирующих
белки. Их размер варьирует от 100 п. н. до 10 т. п. н.
и более. Интроны соответствующих генов позво-
ночных одного вида могут отличаться друг от друга
по величине и нуклеотидной последовательности
в такой же степени, как два интрона из неродствен-
ных генов. Размеры экзонов в основном группиру-
ются вблизи величин 52, 140, 223 и 299 п. н„ причем
преобладают экзоны последней группы. В то же
время известны экзоны длиной всего от 15 до
30 п. н. и, напротив, в несколько сотен или тысяч пар
оснований. Наиболее характерной отличительной
чертой всех интронов является наличие специфиче-
ских последовательностей вблизи их 5'- (левой, или
донорной) и 3'- (правой, или акцепторной) концов
(т.е. на стыках интронов и экзонов, или в сайтах
сплайсинга).
Нуклеотидные последовательности в местах со-
единения экзонов и интронов весьма консервативны
и практически одинаковы во всех генах ядерных
мРНК почти у всех изученных видов (табл. 8.4).
5'-сайт сплайсинга чаще всего фланкирует последо-
вательность CRG (где R пурин), а З'-сайт всего
один остаток G. Тем не менее последовательности,
фланкирующие интроны извне, могут значительно
варьировать, а мутации в них никогда не предот-
вращают сплайсинг, хотя и могут влиять на его
скорость. Нуклеотидные последовательности, флан-
кирующие интроны изнутри, отличаются большим
постоянством. Первыми двумя нуклеотидами на
5'-конце интрона в РНК почти всегда являются GU
(исключение, GC, встречается всего в двух случаях);
следующие четыре нуклеотида могут немного ва-
рьировать, но, по-видимому, канонической является
последовательность gAGU. Замена остатка G или
U в месте сочленения обычно блокирует сплайсинг,
а замены соседних оснований влияют на сплайсинг
по-разному. Указанные шесть нуклеотидов на 5'-
конце интрона и определяют специфическую функ-
цию 5'-сайта сплайсинга. Даже криптические (скры-
тые) сайты сплайсинга (разд. 8.5.е), которые исполь-
зуются только иногда или в тех случаях, когда
основные сайты повреждены или пропущены, содер-
жат элемент GU на 5'-конце вырезаемой последова-
тельности. На З'-конце интрона всегда находится
пара AG. перед которой в интронах млекопитаю-
щих чаще всего находится богатый пиримидином
участок (Y„NYAG). Мутации, приводящие к замене
константных А и G на другие основания, также
блокируют сплайсинг в этом сайте.
Остаток А вблизи З'-конца интрона играет важ-
Таблица 8.4. Канонические последовательности вблизи сайтов сплай-
синга разного типа 11
Тип интрона 5'-сайт сплайсинга Вблизи З'-сайта сплайсинга З'-сайт сплайсинга
Ядерная про-мРНК CRG1GUAAGU А Y.AG1N
(суммарная) Ядерная про-мРНК 1GUAUGU UACUAAC Y„AG 1N
(дрожжи) тРНК Группа I (ядерная рРНК, митохондриальная мРНК и рРНК) Группа II (митохондриальная мРНК) мРНК хлоропластов (Euglena) 11 Представлены последовательности NjN У! 1GUGCG IGUGf.G РНК. Y пиримидин, п N|N G1 Y„AU 1 Y„Acl число пиримидинов, R пурин.
Инвариантные основания подчеркнуты; стрелками показаны сайты сплайсинга. [Т. Cech, Cell 44
(1986), р. 207.]
106
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
ную роль в сплайсинге ядерных про-мРНК
(разд. 8.5.г). В интронах млекопитающих этот оста-
ток не находится в фиксированном положении или
в какой-либо определенной последовательности, по-
скольку его роль, вероятно, может играть любой из
нескольких остатков А, расположенных на участке
от 18 до 37 нуклеотида перед З'-сайтом сплайсинга.
Однако мутации, которые затрагивают соседствую-
щие с указанным остатком аденина последователь-
ности, приводят к существенному уменьшению эф-
фективности сплайсинга in vitro; следовательно, хо-
тя этот остаток и не принадлежит какой-то опреде-
ленной последовательности, его окружение влияет
на сплайсинг. Что касается ядерных про-РНК дрож-
жей, то, напротив, в этом случае активный остаток
А входит в состав специфической гептануклеотид-
ной последовательности 5'-UACUAAC-3', располо-
женной между 6 и 69 нуклеотидами, считая от
З'-конца сайта сплайсинга.
Наличие канонической последовательности в
сайте сплайсинга не всегда означает, что данный
интрон может быть удален. Иногда одна или обе
указанные последовательности сайтов сплайсинга
находятся в интронах и экзонах в положениях, по
которым сплайсинг не происходит. Однако при
некоторых условиях такие криптические сайты все-
таки функционируют (например, когда подлинные
сайты изменены или отсутствуют).
Иногда интроны имеют несколько 5'- или З'-сай-
тов сплайсинга, и тогда возможен альтернативный
сплайсинг. Например, в ранней области генома
SV40 содержатся два интрона, имеющих один и тот
же 3'-, но разные 5'-сайты сплайсинга, в результате
чего из одного и того же первичного транскрипта
образуются две разные мРНК. В некоторых случаях
выбор сайта сплайсинга зависит от стадии развития
или типа ткани (разд. 8.5.е). Имеющиеся сайты мо-
гут и не использоваться. Например, геномы ретро-
вирусов образуются из транскриптов провирусной
ДНК без участия сплайсинга, однако для образова-
ния мРНК, кодирующих определенные вирусные
белки, сплайсинг необходим. В этом случае, вероят-
но, одни и те же транскрипты либо подвергаются
сплайсингу, либо остаются интактными.
Как мы уже говорили, в интронах могут содер-
жаться разные генетические элементы, например
энхансеры, другие гены, возможно, сигналы репли-
кации и упаковки хромосомы или последовательно-
сти, необходимые для упаковки про-мРНК в рибо-
нуклеопротеидные частицы.
Интроны в генах тРНК. Размер интронов в ге-
нах тРНК колеблется от 14 до примерно 60 нуклео-
тидов, но они локализуются всегда в одном и том
же месте: через один нуклеотид от З'-конца анти-
кодона (рис. 8.74). Как правило, если ген данной
тРНК имеет интрон, то все другие гены в пределах
вида, кодирующие эту тРНК, тоже содержат такой
интрон. Однако у генов, кодирующих разные тРНК,
внутренние и фланговые участки интронов заметно
различаются. Принимая во внимание ту роль, кото-
рую играют внутригенные участки в регуляции
транскрипции генов тРНК с участием РНК-поли-
меразы III (разд. 8.4.6), можно задать вопрос: не
влияют ли интроны в генах некоторых тРНК на их
транскрипцию? Установлено, что удаление интрона
гена супрессора тРНКТ),г при помощи направленно-
го мутагенеза не влияет на способность к экспрессии
при введении в клетки. И все же, несмотря на то что
эта тРНК транскрибируется и процессируется нор-
мально, остаток U в антикодоне не модифицирует-
ся, как обычно, с образованием у. Является ли это
указанием на роль интронов в посттранскрипцион-
ной модификации тРНК или мы имеем дело с уни-
кальным свойством тРНКТуг неясно.
Особые типы интронов: группа I. Гены ядерных
рРНК некоторых низших эукариот (например, Tetra-
hymena thermophila и Physarum polycephalum) содер-
жат особые интроны и имеют уникальный механизм
сплайсинга. Эти интроны, названные интронами
группы I, встречаются также в генах рРНК мито-
хондрий, хлоропластов, дрожжей и грибов, в неко-
торых митохондриальных генах мРНК дрожжей
и грибов и тРНК хлоропластов высших растений.
К этой же группе относится и необычный интров
в гене тимидилатсинтетазы фага Т4. Подобные
интроны обнаружены во многих генах, но ни один
из них не был выявлен в генах позвоночных.
Хотя интроны группы I отличаются друг от
друга по размеру-интрон про-рРНК Tetrahymena
состоит примерно из 400 остатков, а длина неко-
торых интронов митохондриальных мРНК превы-
шает тысячу нуклеотидов,- они имеют ряд общих
свойств. 1. Они сами катализируют свой сплайсинг,
который может протекать in vitro в отсутствие
каких бы то ни было белков. 2. Информация, необ-
ходимая для сплайсинга, содержится во множестве
относительно коротких внутренних последователь-
ностей внутри интрона, которые обеспечивают
укладку молекулы с образованием характерной про-
странственной структуры. 3. Сплайсинг иницииру-
ется свободным гуанозином или любым из его
5'-фосфорилированных производных. 4. Конечными
продуктами сплайсинга являются рРНК и линейная
РНК, размер которых несколько меньше, чем раз-
мер интрона.
Основные особенности строения всех интронов
класса I определяются четырьмя нуклеотидными
последовательностями длиной примерно 10 нуклео-
тидов каждая (табл. 8.5). Относительное располо-
жение этих элементов в интроне рРНК Tetrahymena
и в интроне гена цитохрома b митохондрий из
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
107
дрожжей, а также способ их спаривания показаны на
рис. 8.75. Предполагается, что в результате внутри-
молекулярного спаривания нуклеотидов интрон
приобретает конфигурацию, благоприятную для ав-
токаталитического сплайсинга (разд. 8.5.в). Как хи-
мические модификации, так и направленные замены
в конкретных сайтах внутри этих консервативных
спаренных последовательностей приводят к полной
или частичной утрате способности к сплайсингу.
Сходные последовательности, обладающие способ-
3'
Про-TRHK
Туг
Про-тРНк1|е
guGacucg'
g aw
Uu А
с
C-G
U-A
C-G
G-C
U-U
и
А
и
U
U
G
G
U
U
G
А
А
и
А
А3'
С
С
А
5’c-g
U-A
C-G
U-G
C-G
G-C
G-C
Антикодон
G-C _
I САССС
ч
C-G
G-C _
U-G\ CGA
G-C.A А
Л аа
с UUUCCGGA
I I I I I I I I
cAAAgaccu
С U и
U
и
А
А
С
С А
U
А
А
CCG
W6
G
С—G
A-U
A-U
G-C
CCCGC
WWi
с
и
A=“
и
и
С А
G
ис
А
С А
.1 А
G — С
Антикодон и —А
A—U
СА
U С
%AU
U
U-g'
A-U
U-G
A-U
"G-C
C U
A-U
A-U
C-G
U-A
U —A
C-G
G-C
G-C
U U
U
6
А
А
С
Leu
Про-тРНК5
А
и
GcGaGccG
G III
и aaGGC
С (J A G
А3'
С
С
А
5'g-c
G-C
U-A
U-A
G-C
U-G
U-A и
UUCUC
РИС. 8.74.
Расположение интронов в тРНК и места их сочленений.
Представлены три тРНК, имеющие структуру клевер-
ного листа, построенную исходя из предположения
максимального внутримолекулярного спаривания ос-
нований. Интроны выделены серым цветом, а анти-
кодоны-розовым. Места сочленений после сплайсинга
. C-G
Антикодон а-и
А-и
С
А
А
указаны стрелками.
А А
G
ис
A AG AG
САиА
и/ / 6
С-G Аи/
C-G %
U“A AU
g-cxava
A-UCgA
U °G
U-A с'/Би
6гЛиг
СА</С*
Ь А
U U
U
A
A U
В
и
А
А
U
108
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Таблица 8.5. Консервативные элементы, присут-
ствующие во многих интронах группы 111
Элемент Синоним Каноническая последовательность Комнлемен- гарность
9R' E He консервативен Eokc9R
A P AUGCUGGAAA В
В Q AAUCAGCAGG А
Eokc9L R UCAGAGACUACA Бокс2
Bokc9R E' He консервативен 9R'
Бокс2 S AAGAUAUAGUCC Eokc9L
” Подчеркнута та часть канонической последовательности, ко-
торая предположительно участвует в образовании нуклеотидных пар. [Т Cech, Int. Rev. of Cytology 93 (1985), p. 3J
ностью к такому же внутримолекулярному спари-
ванию, обнаружены в интроне группы I гена цито-
хромоксидазы дрожжевых митохондрий. И в этом
случае мутации, затрагивающие нуклеотиды, кото-
рые участвуют в образовании вторичной структуры,
тоже препятствуют сплайсингу и экспрессии окси-
дазного гена. Дальнейшие пояснения, касающиеся
структурных особенностей интронов группы I, бу-
дут даны при обсуждении механизма сплайсинга
(разд. 8.5.в).
Особые типы интронов: группа II. Интроны груп-
пы II распространены менее широко, чем интроны
группы I. Они обнаружены в двух митохондриаль-
ных генах дрожжей, кодирующих одну из субъеди-
ниц цитохромоксидазы и цитохром Ь; интересно,
что в этих генах присутствуют также интроны груп-
пы I. Интроны группы II не содержат канонических
А. Интрон гена рРНК Tetrahymena
9R'A
последовательностей, характерных для интронов
группы I, но они тоже имеют специфическую вто-
ричную структуру, образующуюся благодаря вну-
тримолекулярному спариванию оснований. Интро-
ны группы II также подвергаются самосплайсингу in
vitro, но в этом случае реакция инициируется не
экзогенным гуанозином, а остатком, входящим в
состав самого интрона. Третье ключевое отличие
интронов группы II от интронов группы I заключа-
ется в структуре вырезанного интрона. Если при
сплайсинге в случае интронов группы I образуются
промежуточные кольцевые формы и линейные ко-
нечные продукты, то интроны группы II. высвобож-
денные в результате сплайсинга, представляют со-
бой лассоподобные структуры, в которых 5'-кон-
цевой фосфат РНК интрона соединен фосфодиэфир-
ной связью с 2'-гидроксильной группой внутреннего
нуклеотида (рис. 8.76).
в. Интроны, вырезаемые при помощи
автокаталитического сплайсинга
Самосплайсиш - это поразительный феномен,
но, как показали дальнейшие исследования, не менее
поразителен его механизм. Изучение этого меха-
низма помогло понять, почему для сплайсинга ядер-
ных про-мРНК нужны вспомогательные белки и
особые рибонуклеопротеиды (мяРНП). Самосплай-
синг - это упорядоченный процесс, состоящий из
последовательных автокаталитических реакций фос-
фодиэфирного обмена (трансэтерификации); РНК
функционирует в нем как некий фермент рибозим,
создавая необходимые внутримолекулярные усло-
БоксЭ
В L R Бокс2
РИС. 8.75.
Организация консервативных последовательностей ми-
тохондриальных и ядерных интронов класса I. Интроны
выделены точками, а на участках, отвечающих экзонам,
точек нет. Одинаково окрашенные участки внутри каж-
дого интрона взаимно комплементарны и, как полагают,
могут образовывать нуклеотидные пары (рис. 8.77). А.
Области взаимодействия, выявленные путем прямого
биохимического анализа и компьютерной обработки
данных о последовательности вырезанной РНК. Б.
Области взаимодействия, предполагаемые исходя из
филогенетических и генетических данных. [Т. Cech, Inti.
Rev. Cytol. 93 (1985), p. 3.]
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
109
РИС. 8.76.
Образование 2'.5'-фосфодиэфирных связей между
нуклеотидами, расположенными на значительном рас-
стоянии друг от друга вдоль цепи. При связывании
5'-фосфатной группы с 2'-гидроксильной группой нук-
леотида, находящегося внутри цепи, образуется струк-
тура типа лассо.
вия и поставляя реакционные группы для целой
серии упорядоченных специфических реакций транс-
этерификации.
Самоспмйсинг интронов группы I. Сплайсинг
интрона про-рРНК Tetrahymena, прототипа интрона
группы 1, осуществляется при участии последова-
тельных реакций трансэтерификации, в которых ак-
ты фосфодиэфирного обмена не сопровождаются
гидролизом. Во все акты трансэтерификации, за
исключением инициации с участием свободного гуа-
нозина или его нуклеотида, вовлечены реакционные
группы содержащиеся в самом интроне. Более того,
специфичность этих актов определяется простран-
ственной организацией интрона, которая зависит от
внутримолекулярного спаривания удаленных друг
от друга консервативных последовательностей ин-
трона (рис. 8.75 и 8.77).
Первой стадией в каскаде реакций сплайсинга
является присоединение к интрону гуанозина
(рис. 8.78). Далее неподеленная пара электронов
З'-гидроксильной группы связанного гуанозина ата-
кует фосфатную группу в месте 5'-сочленения ин-
трона с экзоном ( — UpA —). что приводит к нуклео-
фильному расщеплению молекулы РНК (рис. 8.79).
Образовавшаяся в сайте расщепления (конец 5'-эк-
зона) З'-гидроксильная группа реагирует с фосфат-
ной группой в месте З'-сочленений интрона и экзона,
далее происходит лигирование двух участков экзона
с высвобождением линейного интрона длиной 413
нуклеотидов. Затем этот линейный интрон претер-
певает две последовательные реакции внутримоле-
кулярной трансэтерификации, которые сопровожда-
ются гидролизом, приводящим к отщеплению сна-
чала 15, а затем четырех нуклеотидов с образовани-
ем процессированного интрона. Следует отметить,
что вся реакция идет почти без затрат энергии;
разрушение одной диэфирной связи компенсируется
одновременным образованием другой такой же
связи.
Гуанозин или одно из его 5'-фосфорилированных
производных инициирует реакцию сплайсинга спе-
цифическим образом. Если происходит модифика-
ция 2'- или З'-гидроксильных групп или нарушается
способность остатка гуанозина к связыванию с
остатками цитозина, то эффективность инициации
сплайсинга снижается. Очевидно, что спаривание
гуанозина с комплементарным остатком интрона
необходимо для правильной ориентации З'-гидро-
ксильной группы гуанозина относительно фосфат-
ной группы в 5'-сочленении.
Для того чтобы могло произойти лигирование
двух экзонов после расщепления цепи при участии
гуанозина, два экзон-интронных сочленения должны
сблизиться. Такому сближению способствует изги-
бание интрона, при котором места двух сочленений
оказываются расположенными поблизости один от
другого, по-видимому, около сайта связывания гуа-
нозина. Можно предположить, что интрон содержит
последовательность, которая способна спариваться
с последовательностями экзона в местах двух со-
членений (рис. 8.80). Наличие такой «внутренней
направляющей последовательности» могло бы со-
здать условия для реакции З'-гидроксильной груп-
пы, образовавшейся при расщеплении 5'-экзон-ин-
тронного сочленения, с фосфатной группой, нахо-
дящейся в З'-сайте сплайсинга. Этот сайт образуют
остаток G, которым заканчивается интрон, и сосед-
ние нуклеотиды. В высвободившемся линейном ин-
троне происходят конформационные изменения, ко-
торые, как полагают, облегчают циклизацию этого
элемента, сопровождающуюся отщеплением корот-
ких олигонуклеотидов (рис. 8.81).
Возможно, такой механизм самосплайсинга, ко-
торый реализуется в случае интрона гена рРНК
Tetrahymena, характерен и для других интронов
группы 1. Все они содержат четыре одинаковые
консервативные последовательности, обозначаемые
А, В, 9L и 2, а также две неконсервативные, но
комплементарные последовательности, обозначае-
мые 9R и 9R' (табл. 8.5 и рис. 8.75). Эти шесть
элементов всегда расположены в интроне в одном
и том же порядке: 5'-9R'-A-B-9L-9R-2-3'. Кроме того,
большинство интронов группы I содержат последо-
вательность, которая функционирует как некий вну-
тренний «гид». Как известно, мутации, блокирую-
щие сплайсинг, встречаются в ряде генов мРНК
и рРНК митохондрий. Такие мутации, как правило,
мешают спариванию оснований. Например, способ-
ность к сплайсингу, утраченную в результате изме-
нений в одной из двух последовательностей, 9R или
9R', можно восстановить, индуцировав компенси-
рующие изменения в другой последовательности.
Каталитические или ферментоподобные свойства
110
ЧАСТЬ III. ОРГАНИЗАЦИЯ. ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 8.77.
Пространственная организация
интрона рРНК Tetrahymena. Пред-
ставленная структура имеет мини-
мальную свободную энергию,
рассчитанную при помощи ком-
пьютера. Окрашенными прямо-
угольниками выделены консерва-
тивные последовательности ин-
тронов как ядерных, так и мито-
хондриальных рРНК, указанные
на рис. 8.75. Спаривание участков
9R и 9R' предположительно спо-
собствует образованию трехмер-
ной структуры. [Т. Cech, Inti. Rev.
Cytol. 93 (1985), p. 3.]
A Z~<C UUUGAG A 7
'A^-£-G GG ACUCtlS
~\g\
c
c
A
U
G
A
]G<
C I
Jc
/ЛС'
7Т?V
G'
7и/
G
G
U
U
6
А
С
G
6
9R'
С
и
G
G
G
IG
i U
iC
А
А
А
G
G
5’
U
5’
5'G-A-C-C-G-UC-A|
C-U-GG-C-U-&U| 5‘
Бокс 9R
и
и
с
и
G
G
G
С
G
U
с
с
G
С
А
c a ~a
6 и u
РИС. 8.78.
Сайт связывания гуанозина в интроне тРНК Tetrahymena.
Интрон (выделен точками) содержит остаток адени-
ловой кислоты в месте 5'-сочленения, а экзон (без
точек) - остаток уридиловой кислоты в месте З'-сочле-
нения. В представленном на рисунке связывании гуано-
зина при помощи водородных связей могут участвовать
различные основания, сахара или фосфаты, пространст-
венно сближающиеся при образовании вторичной и
третичной структуры. Стрелкой показана нуклеофиль-
ная атака З'-гидроксила рибозы на фосфатную группу
в 5'-сайте сплайсинга. По-видимому, эта реакция
трансэтерификации является началом самосплайсинга
РНК. [В. L. Bass, Т. Cech, Nature 308 (1984), р. 820.]
Самосплайсинг интронов группы I на примере интрона
рРНК Tetrahymena thermophila. Интроны выделены точ-
ками, а важные реакционные группы-серым цветом.
В изолированном виде получены все указанные соеди-
нения, кроме первого продукта расщепления, заклю-
ченного в квадратные скобки. Числа-длина сегментов
РНК.
РИС. 8.80.
«Внутренняя направляющая последовательность» в инт-
роне (она имеет более интенсивную окраску) сближает
два экзон-интронных сочленения. Интрон на рисунке
выделен точками. Обратите внимание на спаривание
внутренней последовательности с последовательностя-
ми экзонов вблизи 5'- и З'-сайтов сплайсинга.
Линейный
интрон
Изменение
конформации
ЛинеЙНЫЙ Н пн
интрон II
РИС.
Альтернативное внутримолекулярное спаривание выре-
занного интрона инициирует отщепление от него олиго-
нуклеотидов и циклизацию. После вырезания линейный
интрон претерпевает конформационные изменения,
обусловленные изменениями в картине внутримолеку-
лярного спаривания. Так, спаривание тринуклеотида
GGG (он выделен цветом) с тринуклеотидом UUU,
по-видимому, индуцирует расщепление связи AU при
участии З'-гидроксильной группы концевого остатка G.
112
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
интронов, выщепляющихся путем самосплайсинга,
рассматриваются более подробно в разд. 8.5.г.
Сплайсинг in vivo в случае некоторых интронов
группы I генов про-мРНК митохондрий дрожжей
зависит от консервативных последовательностей,
которые участвуют в сплайсинге про-рРНК Tetra-
hymena:, аналогичны и продукты сплайсинга. Однако
in vitro эти про-мРНК к самосплайсингу неспособ-
ны. Выщепление интронов в этом случае зависит от
транс-действующих белков, которые кодируются
внутренними последовательностями тех же самых
или родственных генов и, как правило, имеют об-
щую рамку считывания для экзонов и интронов
(разд. 9.7.а). Такие белки, названные матуразами,
относятся к короткоживущим и, вероятно, помога-
ют укладке интрона про-мРНК с образованием
структуры, облегчающей сплайсинг под действием
РНК. Это объясняет, почему нонсенс-мутации или
мутации со сдвигом рамки в интроне, кодирующем
матуразу, предотвращают сплайсинг в случае этого
интрона, а также в случае интронов группы I в дру-
гих митохондриальных генах. Данная модель дает
также рациональный ответ на вопрос, почему су-
прессорные мутации или мутации, компенсирующие
сдвиг рамки, которые приводят к восстановлению
синтеза матуразы, восстанавливают также правиль-
ный сплайсинг. Таким образом, правильная укладка
некоторых интронов группы I, вероятно, происхо-
дит спонтанно с образованием структуры, стабиль-
ной in vitro; в то же время образование и/или
стабилизация каталитически активной структуры
других интронов могут происходить только при
участии белков. Играет ли какую-нибудь роль в
укладке интрона белок-белковые взаимодействия -
неизвестно.
Еи/е один тип самосплайсинга. Интроны группы
И про-мРНК митохондрий дрожжей также имеют
консервативные последовательности, а возможно,
и специфическую вторичную структуру, но она от-
личается от структуры интронов группы I. Интроны
этого типа также способны к самовырезанию, но об
особенностях их вторичной и третичной структуры
и соответственно о молекулярных деталях сплай-
синга известно меньше. Очевидны две особенности
механизма сплайсинга: 1) в отличие от самосплай-
синга интронов группы I для сплайсинга интронов
группы II не нужен нуклеозид-инициатор; 2) в ре-
зультате сплайсинга образуется структура типа лас-
со (рис. 8.76).
Каким образом интрон группы II вырезает сам
себя и чем обусловлено образование нового РНК-
продукта, имеющего форму лассо? Как и в случае
интронов группы I, этот процесс, по-видимому,
протекает при участии трансэтерификации; на пер-
вой стадии происходит расщепление 5'-сайта сплай-
синга, а на второй-расщепление З'-сайта сплайсин-
га и лигирование двух экзонов. Однако природа
атакующего нуклеофила совершенно иная: это не
гуанозин, как в случае интронов группы I, а 2'-ги-
дроксильная группа нуклеотида, находящегося где-
то внутри самого интрона (рис. 8.82). Лассоподоб-
ная структура возникает при образовании новой
2', 5'-фосфодиэфирной связи в средней части после-
довательности РНК.
Трехмерная структура интрона, формирующаяся
либо спонтанно, либо при участии белков, должна
быть такой, чтобы обеспечивалось сближение
2'-гидроксильной группы и 5'-сайта сплайсинга, не-
обходимое для активации трансэтерификации. Вы-
бор подлежащих расщеплению межнуклеотидных
связей, вероятно, определяется последовательно-
З’ОН
РИС. 8.82.
Сплайсинг интронов группы II. 2'-гидроксильная группа
остатка А осуществляет нуклеофильную атаку на оста-
ток G и инициирует расщепление в 5'-сайте сплайсинга.
Далее З'-гидроксильная группа 5'-экзона вступает в
реакцию с фосфатной группой З'-экзона, завершая про-
цесс сплайсинга с высвобождением молекулы РНК,
имеющей форму лассо.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
113
стью GUGCG на 5'-конце интрона, специфичной
для интронов группы II, и последовательностью
YAU на З'-конце интрона (табл. 8.4). Как показано
в разд. 8.5.г, этот механизм сплайсинга очень напо-
минает таковой в случае ядерных про-мРНК. Обя-
зательное участие в процессе шрпнс-активных рибо-
нуклеопротеидных частиц может быть связано с не-
обходимостью более сложной и специфической
укладки про-мРНК разных видов, обеспечивающих
сплайсинг в нужном месте.
г. Сплайсинг ядерной про-мРНК
Основную информацию о механизме сплайсинга
ядерной про-мРНК удалось получить благодаря
тому, что в распоряжении исследователей имелись
клонированные прерывистые гены. Особенно полез-
ным оказалось изучение модифицированных форм
этих генов. При установлении структурных особен-
ностей, необходимых для успешного сплайсинга,
используют также специально сконструированные
элементы, состоящие из соединенных специфических
последовательностей экзонов и интронов. Но са-
мым информативным оказался биохимический под-
ход с использованием клеточных экстрактов, обес-
печивающих сплайсинг, и последующей очисткой
компонентов аппарата сплайсинга. Особенно важ-
ную роль сыграло применение кэпированных РНК-
субстратов, полученных в результате транскрипции
специальным образом сконструированных ДНК-
матриц, встроенных справа от промотора фага SP6,
при помощи специфической РНК-полимеразы SP6
(рис. 6.23).
Общие свойства. Сплайсинг ядерных про-мРНК
осуществляется в ядре, возможно, одновременно
с транскрипцией для одних генов и лишь после
завершения транскрипции для других. Есть свиде-
тельства в пользу того, что на этот процесс влияет
кэпирование 5'-конца транскрипта, хотя неясно, по-
чему: в полиаденилировании РНК явно нет необхо-
димости. Главная проблема, и концептуальная, и
практическая, состоит в том, каким образом проис-
ходит сплайсинг про-мРНК с множественными ин-
тронами, в результате которого соседние экзоны
соединяются друг с другом. Поскольку известно,
что 5'-сайт сплайсинга одного интрона может при-
соединяться к З'-сайту сплайсинга другого, важно
понять, каким образом это предотвращается в про-
цессе нормального вырезания интронов и как стиму-
лируется при альтернативных способах сплайсинга
(разд. 8.5.е).
Последовательность этапов сплайсинга. Сначала
мы рассмотрим реакции расщепления и лигирова-
ния. Первым этапом сплайсинга является сборка
комплекса сплайсинга. Самые ранние продукты,
обнаруживаемые в процессе сплайсинга in vitro.
образуются в результате точного расщепления в
5'-сайте сплайсинга; один из них содержит 5'-экзон,
а другой-интрон и З'-экзон (рис. 8.83). Поскольку
никакие молекулы РНК с изменениями в З'-сайте
сплайсинга, кроме конечного продукта сплайсинга,
не выявлены, расщепление в 5'-сайте должно пред-
шествовать расщеплению в З'-сайте. В ходе реакции
накапливаются два продукта: правильно лигиро-
ванные экзоны и свободный целый интрон. Как
продукт начального расщепления, так и вырезанный
интрон содержат структуры типа лассо, аналогич-
ные описанным выше структурам митохондриаль-
ных интронов самосплайсинга группы II (рис. 8.83).
Сначала вывод о лассообразности структуры ин-
трона был сделан на основании данных об аномаль-
ной электрофоретической подвижности вырезанного
интрона, о блокировании обратной транскрипции
последовательности за точкой, находящейся у
З'-конца интрона, и об устойчивости интрона к пол-
ному расщеплению под действием некоторых рибо-
нуклеаз. Позже существование этой структуры было
подтверждено при выделении разветвленных ну-
клеотидных структур. Точка ветвления всегда со-
держала 5'-фосфат концевого гуанозина интрона,
присоединенного диэфирной связью к 2'-гидрок-
сильной группе остатка аденозина, независимо от
последовательности вырезанного интрона. Тем са-
мым наконец-то получил объяснение удивительный
факт существования ядерных РНК с разветвления-
ми, отходящими от 2'-гидроксильных групп остат-
ков адениловой кислоты.
Сплайсинг про-мРНК в сплайсингосомах. В общих
чертах сплайсинг ядерных про-мРНК и сплайсинг
РНК, содержащих интроны группы II, приводят
к одинаковому результату: вырезанию интрона в
форме лассо и лигированию двух экзонов. Отличие,
однако, состоит в том, что для сплайсинга ядерных
про-мРНК требуется множество ядерных факто-
ров-белков и рибонуклеопротеидных комплексов
(мяРНП). Комплекс, состоящий из множества субъ-
единиц, который катализирует сплайсинг, называют
сплайсингосомой. Сплайсингосома, упорядоченная
сборка которой осуществляется на интроне перед
расщеплением в 5'-сайте сплайсинга, состоит из
интрона, связанного по меньшей мере с пятью
разными мяРНП и некоторыми вспомогательными
белками, обычно не связанными с этими мяРНП.
Сплайсингосомы образуются путем спаривания мо-
лекул РНК, присоединения белков к РНК и связы-
вания этих белков друг с другом.
мя-РНП. Установлению роли мяРНП-частиц в
ядерном сплайсинге предшествовали их идентифи-
кация и изучение свойств. После усовершенствова-
ния методов фракционирования благодаря получе-
нию антител к уникальному триметилгуанозино-
вому кэпу мяРНК они были выделены в достаточно
8-158
114
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Расщепление
в з'-сайте
сплайсинга и
лигирование
экзонов ,
РИС. 8.83.
Сплайсинг интронов ядерных про-мРНК
(интроны на рисунке выделены точками).
Сначала происходит расщепление в 5'-сай-
те сплайсинга с последующим образова-
нием 5', 2'-фосфодиэфирной связи, а затем
расщепление в З'-сайте сплайсинга и лиги-
рование концов экзона.
чистом виде, а благодаря использованию антител,
присутствующих в сыворотке людей с различными
аутоимунными заболеваниями, были получены и
индивидуальные мяРНП. мяРНП сплайсингосом
млекопитающих содержат U1-, U2-, U4-, U5- и
U6-PHK (табл. 8.6). Эти РНК называются U-PHK
из-за необычайно большого содержания урацила
и его модифицированных форм. U1-, U2- и U5-PHK
входят в состав особой мяРНП; U4- и U6-PHK,
эффективно спаривающиеся друг с другом, являют-
ся составной частью другой мяРНП. мяРНП, со-
держащие U3-PHK, участвуют в процессинге
Таблица 8.6. Размеры U-PHK
U-PHK Размер в нуклеотидах
дрожжи 11 млекопитающие
ы 568 164
U2 1175 187
из — 217
U4 160 145
U5 214 116
U6 112 106
U7 — 65
ин - 131
” Прочерки означают, что соответствующие дан-
ные отсутствуют.
про-рРНК в ядрышке (разд. 8.2.г), мяРНП, содер-
жащие U7-PHK-B образовании З'-концов гистоно-
вых мРНК (разд. 8.3.в), а U11-PHK вовлечена в
полиаденилирование мРНК (разд. 8.3.в). В ядрах
клеток млекопитающих присутствуют и другие
мяРНП, но их строение и функции неизвестны.
Нуклеотидные последовательности всех соответ-
ствующих U-РНК позвоночных, исследованных к
настоящему времени, совпадают примерно на 95%.
Совпадают даже нуклеотидные последовательности
U1-PHK человека и Drosophila. мяРНП, содержащие
сходные низкомолекулярные РНК, богатые ураци-
лом, встречаются также у тараканов, морского ежа,
Tetrahymena, Diclyostelium, дрожжей и у динофлагел-
лят. U-РНК даже таких неродственных видов имеют
гомологичные области, несмотря на то, что значи-
тельная часть их последовательностей уникальна
для каждого вида. Чрезвычайная консервативность
данных молекул и их участие в специфических ста-
диях процессинга РНК показывают, что сплайсинг-
очень древний процесс, вероятно возникший на заре
эволюции.
Все мяРНП, входящие в состав сплайсингосом
(U1-, U2-, U5- и и4/иб-мяРНП), содержат одина-
ковый набор из семи белков (табл. 8.7), a U1-, U2- и
П5-мяРНП-еще и несколько белков других типов.
Аутоиммунные сыворотки, используемые обычно
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
115
Таблица 8.7. Белки, входящие в состав мяРНП
млекопитающих 11
Ветки мяРНП. содержащие Мол. масса. кДа
указанные L1-PHK
U1 U2 U3 U5 1)471)6
В, В, D, D, Е. T T — 4-
F, G
70К, А. С +
А', В" +
25К, IBP +
(34К, 74К. 59К +
ЗОК, 13К, 125К) +
+ 28; 29; 16; 15.5: 12;
II; 9
70; 34; 22
33; 28
25; 100 или 70
34; 74; 59
30; 13; 125
11 Белковые компоненты мяРНП от U7 до U11 не идентифициро-
ваны из-за их низкого содержания. Белки обозначены буквами или
числами, отвечающими их кажущейся мол. массе. Мол. масса
белков, входящих в состав U1-, L)2-, U5- и и4/иб-мяРНП, рассчи-
тана исходя из их аминокисло гного состава, который был опреде-
лен на основании данных о нуклеотидных последовательностях
клонированных кДНК.
для выделения и классификации мяРНП, часто бы-
вают специфичны к одному или нескольким белкам
частицы. иЗ-мяРНП, которая участвует в процес-
синге рРНК. содержит по крайней мере шесть бел-
ков, не представленных в мяРНП, участвующих
в сплайсинге. Белки более редких U7- и Ull-РНП
изучены хуже.
Взаимодействие м.чРНП с интроном. Первые
соображения о способе участия мяРНП в сплайсинге
можно было высказать после того, как обнаружи-
лось, что нуклеотидная последовательность на
5'-конце U1-PHK комплементарна канонической по-
следовательности в 5'-сайтах сплайсинга всех интро-
нов (рис. 8.84). Кроме того, Ul-мяРНП специфиче-
ски связывается с 5'-сочленением in vitro, защищая
РИС. 8.84.
Связывание U1-PHK с последовательностями в местах
экзон-интронных сочленений в мРНК. Показано спа-
ривание последовательности в месте сочленений интро-
на (выделен точками) с экзоном в двух мРНК адено-
вируса Е1 с 5'-концом U1-PHK.
при этом последовательность интрона примерно из
17 нуклеотидов в 5'-сайте сплайсинга от растепле-
ния РНКазой; РНК при этом связывании не моди-
фицируется и не разрушается. 02-мяРНП связыва-
ется с областью из 40 нуклеотидов, которая содер-
жит нуклеотид А, участвующий в образовании раз-
ветвленной лассообразной структуры, и защищает
ее. Молекулярный механизм связывания 1)2-мяРНП
с этой областью интронов млекопитающих не впол-
не ясен, поскольку последовательность не является
высококонсервативной. Однако у дрожжей, где по-
следовательность, содержащая остаток А, высоко-
консервативна, комплементарность между 5'-кон-
цом U2-PHK и последовательностью интрона уста-
новлена при помощи генетических экспериментов со
всей очевидностью. Участок примерно из 15 нуклео-
тидов на З'-конце интрона связывается с и5-мяРНП.
Однако, возможно, в физиологических условиях
этот сайт связывается и активируется комплексом
US-мяРНП и С14/иб-мяРНП. Для связывания как
U2-, так и 05-мяРНП с соответствующими после-
довательностями-мишенями нужны вспомогатель-
ные белки. Один из них (AF) соединяется с З'-сочле-
нением интрона (участок, богатый AG и пиримиди-
нами) и способствует связыванию и2-мяРНП по
соседству. Для связывания комплекса U4/U6- и
С5-мяРНП также требуется белок, который стаби-
лизирует его прикрепление к З'-сочленению интрона.
В образовании этих комплексов, по-видимому,
играют роль белок-белковые и РНК-белковые вза-
имодействия.
Окончательный вывод об участии ЕЛ- и U2-
мяРНП в сплайсинге был сделан после того, как
было показано, что иммунопреципитация U1- и
О2-мяРПН из экстрактов, катализирующих сплай-
синг про-мРНК, при взаимодействии с соответ-
ствующими антителами ингибирует сплайсинг. Бо-
лее того, способность фракционированных опреде-
ленным образом экстрактов катализировать сплай-
синг зависит от добавления U1- и Ш-мяРНП. Ана-
логично, при проведении сплайсинга в реакционную
смесь необходимо добавлять эндогенные мяРНП,
если собственные мяРНП экстрактов разрушены
направленным расщеплением под действием эндо-
нуклеазы (РНКазы Н) с использованием коротких
олигодезоксинуклеотидов, комплементарных каж-
дой из U-PHK.
Сборка сп.гайсингосом. Сплайсинг осуществля-
ется в сплайсингосомах. Рассмотрим поэтому их
структуру, сборку и механизм действия. Сплайсин-
госомы, выделенные из реакционной смеси в усло-
виях, когда стадия расщепления заблокирована,
представляют собой частицы в форме эллипса раз-
мером примерно 25 х 50 мкм (рис. 8.85). В этих
частицах интрон связан с U1-, U2-, U4/U6- и
05-мяРНП одновременно. Функционально актив-
116
ЧАСТЬ 111. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 8.85.
Электронная микрофотография сплайсингосом. [Я. Reed
et al.. Cell 53 (1988), p. 949, с любезного разрешения
J. Griffith.]
ные сплайсингосомы содержат и другие белки, в
частности те, которые участвуют во взаимодействии
мяРНП с сайтами-мишенями или обычно связаны
с ядерной про-мРНК.
Для присоединения мяРНП к интрону, по-види-
мому, необходима лишь очень небольшая часть
экзона. Как видно из рис. 8.86, на котором пред-
ставлена сборка сплайсингосомы, с 5'-сайтом сплай-
синга сначала связывается U1-мяРНП независимо
от присутствия функциональных сайтов связывания
U2- и П5-мяРНП; но этого недостаточно для после-
дующего расщепления в 5'-сайте сплайсинга. Затем
присоединяются U2-, U5- и и4/1)6-мяРНП, для чего
требуется АТР. Вероятно, присоединение П2-мяРНП
к соответствующему сайту в интроне осуществля-
ется путем образования нуклеотидных пар; однако
включение в сплайсингосому комплекса, содержа-
щего U4/U6- и П5-мяРНП, зависит от взаимодейст-
вий между мяРНП. По сходной схеме происходит
сборка сплайсингосомы при сплайсинге про-мРНК
дрожжей. Судьба сплайсингосомы после лигирова-
ния экзонов неясна, но отдельные мяРНП или их
комплексы, вероятно, повторно используются для
сплайсинга других интронов, в то время как РНК
интрона разрушается. Описанная последователь-
ность реакций представляет лишь один из возмож-
ных вариантов сборки сплайсингосомы. Для того
чтобы однозначно установить эту последователь-
ность, необходимо исследовать взаимоотношения
между предшественником и продуктом. В данной
модели не объясняется также, каким образом рас-
щепляется З'-сайт сплайсинга, как происходит лиги-
рование и от чего зависит правильное соединение
экзона.
Самосплайсинг и сплайсинг с участием сплайсин-
госом. Конечный результат сплайсинга в случае
интронов группы II и в случае интронов про-мРНК
одинаков: интрон вырезается, а два фланкирующих
его экзона соединяются. Более того, одинаков и хи-
мизм этих двух процессов. В обоих случаях 2'-ги-
дроксильная группа в составе интрона выступает
в роли нуклеофила, способствуя расщеплению
5'-сайта сплайсинга, а З'-гидроксильная группа лево-
го экзона является нуклеофилом, расщепляющим
З'-сайт сплайсинга с образованием связи между
экзонами. Ключевое различие состоит в том, что
в случае по крайней мере некоторых интронов груп-
пы II может происходить самосплайсинг in vitro,
а для сплайсинга ядерных про-мРНК необходим
сложный комплекс нескольких мяРНП и вспомога-
тельных белков. Даже в случае интронов группы II
для сплайсинга in vitro необходимы матуразы, хотя
вполне вероятно, что и нуклеотиды, и каталитиче-
ские центры, участвующие в автокаталитическом
сплайсинге и в сплайсинге, опосредованном матура-
зами, одинаковы. Если для автокаталитического
сплайсинга нужна особая укладка ДНК. то матура-
зы, вероятно, необходимы для складывания или
стабилизации структуры, которая может подвер-
гаться правильным последовательным актам транс-
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
117
АДР
I та U2.AF
I * Ц2-МЯРНП
РИС. 8.86.
Упорядоченная сборка сплайсинго-
сомы в процессе сплайсинга про-
мРНК. Что происходит со сплайсин-
госомой после лигирования двух эк-
зонов и высвобождения интрона в
форме лассо (петли), на рисунке не
показано.
этерификации. Таким образом, единственное, чем,
возможно, отличается сплайсинг с участием матураз
от автокаталитического сплайсинга,- это способ об-
разования каталитически активной конформации
интрона.
Вероятно, то же самое справедливо для сплай-
синга с участием сплайсингосом. Возможно, мяРНП
и ассоциированные с ними факторы создают что-то
вроде «каркаса», на котором интрон «крепится»
таким образом, что два его конца и нуклеотидный
остаток из точки разветвления оказываются рядом
друг с другом и могут активироваться. В этом
смысле роль сплайсингосомы в случае ядерных
про-мРНК аналогична роли матураз при сплайсинге
некоторых интронов группы II. Возникает интерес-
ный вопрос: почему для укладки интронов ядерных
про-мРНК необходим такой сложный комплекс
мяРНП, если для сплайсинга некоторых интронов
группы II достаточно одного белка-матуразы?
По-видимому, это связано со сложностью феномена
сплайсинга. Все митохондриальные интроны груп-
пы II похожи друг на друга; каждый из них содер-
жит несколько высококонсервативных участков, и,
возможно, для образования и поддержания актив-
ной конформации интрона в этом случае нужен
всего один или два белка. Но размеры и последова-
тельности интронов ядерных про-мРНК необычай-
но разнообразны, очень велико и число разных
интронов. Это создает дополнительные проблемы.
Никакой отдельный белок, по-видимому, не спосо-
бен придать всем интронам каталитически актив-
ную форму. Однако не исключено, что высококон-
сервативные мяРНП. которые могут спариваться
всего с двумя или тремя консервативными последо-
вательностями во всех интронах, а также связывать-
ся друг с другом, способны придать любому интро-
ну конформацию, благоприятную для протекания
двух реакций трансэтерификации, катализируемых
РНК.
Транс-сплайсинг. До сих пор, говоря о сплайсин-
ге, мы рассматривали внутримолекулярные, или
t/wc-реакции. А существует ли межмолекулярный,
или тдаис-сплайсинг? Иными словами, может ли
происходить лигирование двух экзонов, находящих-
ся в разных молекулах РНК, с одновременным
удалением фланкирующих их интронов? Принци-
118
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
пиальная возможность межмолекулярного сплай-
синга действительно продемонстрирована в опытах
in vitro со специально сконструированными РНК-
субстратами. Более того, как оказалось, транс-
сплайсинг является важным этапом внутриклеточ-
ного образования всех мРНК у Trypanosoma и трех
из четырех мРНК активных генов у С. elegans.
В каждом случае сайты сочленений относятся к ка-
ноническому типу: GU в 5'-сочленении и AG в
Экзон 1 Интрон Экзон 2
5'.сайт сплайсинга З'-сайт сплайсинга
РИС. 8.87.
Тронс-сплайсинг мРНК in vitro. А. Сплайсинг с участием
экзонов, находящихся в разных молекулах РНК, иссле-
довали в экстракте, катализирующем сплайсинг про-
мРНК. Одна РНК состоит из экзона и интрона, соеди-
ненных в 5'-сайте сплайсинга, а другая из экзона и
интрона, соединенных в З'-сайте сплайсинга. Компле-
ментарные участки этих РНК спарены. Вырезанные
интроны имеют структуру типа лассо. Б. Две разные
мРНК, интроны которых комплементарны почти по всей
своей длине, использовались в качестве субстратов для
сплайсинга in vitro. Ad это 5'- и З'-экэоны в мРНК
аденовируса 2, GI-5'- и З'-экзоны в глобиновой мРНК.
Интроны выделены точками. Внизу приведены четыре
возможных продукта сплайсинга; два из них образова-
лись в результате транс-сплайсинга, а два-в резуль-
тате цас-сплайсинга. В рамки заключены основные
продукты сплайсинга.
З'-сочленении. Более того, в качестве продукта об-
разуется разветвленная структура.
Для демонстрации транс-сплайсинга in vitro бы-
ли разработаны два экспериментальных подхода.
В первом случае 5'-экзон и сегмент фланкирующего
его интрона находятся на одной РНК, а З'-экзон
и связанный с ним интрон- на второй (рис. 8.87,Л).
Эти РНК удерживаются вместе благодаря компле-
ментарному спариванию двух сегментов интронов.
Эффективность правильного соединения экзонов и
удаления сегментов интронов в клеточных экстрак-
тах составляет примерно 15% от эффективности,
наблюдаемой в том случае, когда интроны находят-
ся в одной и той же молекуле РНК. Итак, ни
наличие вторичной структуры, ни ковалентные раз-
рывы в интроне не препятствуют сплайсингу. Вто-
рой эксперимент также основан на комплементар-
ности последовательностей в двух интронах, что
позволяет соединить соответствующие РНК
(рис. 8.87,£). В этом случае могут образоваться
четыре продукта сплайсинга: два в результате
транс-сплайсинга и два в результате г/ис-сплайсинга.
При сплайсинге in vitro такого субстрата получают-
ся 5'-адено-адено-3'- (транс-сплайсинг) и 5'-глобин-
адено-3'- (г/нс-сплайсинг)-продукты. Продукты цис-
сплайсинга 5'-адено-глобин-3' и транс-сплайсинга
5'-глобин-глобин-3' тоже образуются, но в меньших
количествах, вероятно, из-за близости стебля из
спаренных оснований к точке разветвления интрона.
С помощью транс-сплайсинга образуются функ-
циональная актиновая мРНК и некоторые другие
мРНК в клетках С. elegans. В случае актина кэп
и первые 22 нуклеотида на 5'-коннах актиновых
мРНК, специфичных для мышечной ткани, обра-
зуются из 5'-конца РНК длиной 100 нуклеотидов,
транскрибированной с повтора 58-рДНК, локализо-
ванного на другой хромосоме. Однако лучше всего
изучен транс-сплайсинг при образовании мРНК
Trypanosoma (разд. Ю.б.г). У всех мРНК Trypano-
soma перед непрерывной кодирующей последова-
тельностью находится одинаковая нетранслируемая
5'-последовательность из 35 нуклеотидов. Эти 5'-ми-
ни-экзоны образуются из 5'-концевого участка РНК
длиной 137 нуклеотидов, который транскрибируется
с сотен тандемных копий соответствующего сегмен-
та длиной 137 п. н. геномной ДНК (рис. 8.88). В ходе
транс-сплайсинга 5'-мини-экзон длиной 35 нуклео-
тидов от одной РНК соединяется с кодирующим
белок экзоном другой РНК. Сначала при соедине-
нии двух экзонов образуется разветвленная молеку-
ла РНК. На следующей стадии З'-сайт сплайсинга
расщепляется, и два экзона соединяются. Затем
разветвленный сегмент, содержащий «интронные»
последовательности от двух отдельных РНК, рас-
щепляется под действием особой дебранчинг-гидро-
лазы, и интроны, которые принадлежали двум раз-
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
119
Мини-экзонный повтор
размером 1,35 т. п.н.
Экзон, кодирующий белок
РИС. 8.88.
Транс-сплайсинг мРНК Trypanosoma.
5'-мини-экзоны происходят иэ коротких
мРНК, транскрибированных с тандемно
повторяющихся единиц транскрипции,
которые располагаются в виде кластеров
в специфических областях хромосомы.
Эти транскрипты длиной 137 нуклеоти-
дов состоят из 5'-мини-экзона длиной
35 нуклеотидов и интрона (выделен точ-
ками). Кодирующие белок экзоны, тран-
скрибированные с соответствующих ге-
нов, подвергаются транс-сплайсингу,
как это показано на рисунке.
ным РНК, разделяются. Особого внимания заслу-
живают следующие факты: 1) последовательность,
находящаяся непосредственно за мини-экзоном у
Trypanosoma, соответствует канонической последо-
вательности 5'-сайта сплайсинга (GUAUGA); 2) по-
добным образом, последовательность интрона в
месте соединения с кодирующим экзоном анало-
гична последовательностям З'-сайта сплайсинга
([C/U]nNNAG); 3) в интроне имеется точка ветвле-
ния, что указывает на идентичность механизмов цис-
и транс-сплайсинга. Как образуется комплекс
сплайсинга из двух отдельных РНК-неясно.
д. Сплайсинг тРНК
Механизм удаления интронов в тРНК лучше
всего изучен у дрожжей, но некоторая информация
получена в опытах с другими низшими эукариотами
и растениями. Известны и выделены в высокоочи-
щенном виде все ферменты сплайсинга, охарактери-
зованы все промежуточные формы (рис. 8.89,Л).
Специфическая тРНК-эндонуклеаза расщепляет
про-тРНК в 5'-сочленении интрона с образованием
циклического 2',3'-фосфодиэфира на конце 5'-поло-
вины тРНК. Тот же фермент расщепляет другое
сочленение интрона с образованием З'-гидроксиль-
ной группы на конце З'-половины тРНК. Эти реак-
ции напоминают расщепление под действием неко-
торых РНКаз. Циклический фосфодиэфир на конце
5'-экзона затем отщепляется с образованием 2'-фос-
фомоноэфира под действием циклической фосфо-
диэстеразы. После фосфорилирования 5'-гидрок-
сильного конца З'-экзон аденилируется при помощи
тРНК-специфичной лигазы; последняя реакция ана-
логична реакциям, катализируемым ДНК-лигазой
(разд. 2.1.д). После лигирования активированных
концов двух частей молекулы образуется необычная
2'-фосфат-3',5'-фосфодиэфирная связь, и затем при
отщеплении 2'-фосфата под действием фосфатазы
получается зрелая молекула тРНК. Заметим, что
фосфат, участвующий в образовании новой диэфир-
ной связи, до этого входил в состав АТР-особен-
ность, отличающая сплайсинг тРНК у дрожжей от
аналогичного процесса у позвоночных (рис. 8.89,5).
В данной цепи реакций первоначальное эндонукле-
азное расщепление приводит к образованию 5'-гид-
роксильного и З'-фосфомоноэфирного концов,
причем последний превращается в циклический
2',3'-фосфодиэфир под действием АТР-зависимой
циклазы. Затем тРНК-специфичная лигаза соеди-
няет обе части без дополнительной активации кон-
цов.
При фракционировании системы сплайсинга
120
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 8.89.
Сплайсинг тРНК. А. Сплайсинг у дрожжей протекает
с участием трех белков: эндонуклеазы, которая рас-
щепляет про-тРНК в обоих сайтах сплайсинга с образо-
ванием указанных концов; полифункционального белка,
катализирующего все остальные реакции, кроме фос-
фатаэной; 2'-фосфатаэы. В этом случае в месте сочле-
нения обоих экэонов находится фосфатная группа, ко-
дрожжей получены высокоочищенные препараты
двух ферментов. Один из них катализирует эндо-
нуклеазное расщепление на границах интрона; вто-
рой проявляет активность циклической фосфодиэс-
теразы и киназы и способен катализировать адени-
лирование и лигирование, причем все эти активно-
сти принадлежат одной полипептидной цепи. Оба
фермента проявляют высокую специфичность в от-
ношении данных реакций сплайсинга тРНК, однако
они неспецифически действуют на любые тРНК-ин-
троны и соединяют два участка любых тРНК. И
действительно, из частей молекул, принадлежащих
разным тРНК, были сконструированы весьма ин-
тересные гибриды.
торая до этого была концевым фосфатом АТР. Б
У позвоночных три указанные реакции катализируют
отдельные ферменты, при этом каждый фермент участ-
вует только в сплайсинге тРНК. Отметим, что фосфат
в месте соединения двух экзонов ранее находился в
месте сочленения экзона и интрона.
е. Альтернативный сплайсинг:
множественные белки одного гена
При сплайсинге большей части про-мРНК каж-
дый интрон вырезается в соответствующих 5'-
и З'-сайтах сплайсинга. В результате все экзоны
и порядок их расположения в транскрипте сохра-
няются в зрелой мРНК и образуют непрерывную
последовательность (конститутивный сплайсинг).
Однако сплайсинг некоторых про-мРНК протекает
по-разному с образованием семейства близких по
строению мРНК, каждая из которых состоит из
специфического набора экзонов и кодирует одну из
изоформ белков одного семейства. Такой способ
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
121
процессинга РНК называется альтернативным
сплайсингом. Растет число генов из разных орга-
низмов, от Drosophila до человека и их вирусов,
о которых известно, что при созревании их про-
мРНК используется альтернативный сплайсинг. Эти
гены кодируют многие белки, в том числе некото-
рые белки, участвующие в формировании цитоске-
лета, мышечном сокращении, сборке мембранных
рецепторов, пептидных гормонов, в промежуточном
метаболизме и транспозиции ДНК.
Альтернативный сплайсинг позволяет получать
разные продукты от одного-единственного гена без
изменения его геномной организации. Он представ-
ляет собой эффективный способ продуцирования
мРНК, кодирующих структурно родственные белки,
в которых одинаковые аминокислотные последо-
вательности определяются общими экзонами, а ин-
дивидуальные последовательности - экзонами, участ-
вующими в альтернативном сплайсинге. В неко-
торых случаях разные мРНК при альтернативном
сплайсинге образуются одновременно, при этом
отдельные изоформы белка могут выполнять как
одинаковые, так и разные функции. Все четыре
изоформы основного белка миелина являются ком-
понентами миелиновых оболочек нейронов цент-
ральной нервной системы; иммуноглобулины пред-
ставлены двумя изоформами - мембраносвязанной
и секретируемой (разд. 10.6.в). Сплайсинг некото-
рых транскриптов генов в разных тканях протекает
по-разному; например, при экспрессии единствен-
ного гена кальцитонина в щитовидной железе син-
тезируется кальцитонин, а в мозгу его изоформа-
белок, родственный кальцитонину; при этом каж-
дый белок синтезируется на своей собственной
мРНК, образующейся в результате дифференциаль-
ного сплайсинга. В некоторых системах генов (в
качестве примера можно привести ген тропонина
Т млекопитающих и комплекс ultrabithorax у Dro-
sophila) относительное содержание и типы мРНК,
образующихся в результате альтернативного сплай-
синга, регулируются в процессе развития. Альтер-
нативный сплайсинг играет важную роль в ста-
новлении пола во время эмбриогенеза Drosophila.
Так, разная активность регуляторных генов у сам-
цов и самок обусловлена в большей степени специ-
фическими половыми различиями в сплайсинге
РНК, которые ведут к образованию функционально
разных транскриптов у разных полов. Более того,
отдельные гены в этой регуляторной иерархии не
только самостоятельно контролируют свою актив-
ность на уровне сплайсинга, но и определяют способ
сплайсинга транскриптов, функционирующих позд-
нее, на следующих этапах.
Типы альтернативного сплайсинга. Различают
три основных типа альтернативного сплайсинга
(рис. 8.90). Сплайсинг типа I основывается на
использовании разных промоторов для образования
родственных про-мРНК с неодинаковыми 5'-прокси-
мальными областями разной длины. Представите-
лями генов, сплайсинг которых осуществляется по
этому типу, являются ген а-амилазы мыши (разд.
9.3.6) и ген легкой цепи миозина (MLC) позвоноч-
ных. Ген MLC транскрибируется с двух промо-
торов, находящихся на расстоянии около 10 т.п.н.
друг от друга (рис. 8.91). Более длинная про-мРНК
содержит все экзоны, а более короткая лишена
первого экзона. При альтернативном сплайсинге
образуются две мРНК примерно одинакового раз-
мера; обе они имеют пять З'-проксимальных консти-
тутивных экзонов, но различаются своими 5'-экзо-
нами. При сплайсинге более длинной про-мРНК
экзон 1 присоединяется к экзону 4, «игнорируя»
З'-сайты сплайсинга экзонов 2 и 3. Напротив, при
сплайсинге более короткой про-мРНК соединяются
экзоны 2, 3 и 5, а экзоны 3 и 4- нет. Анализ
нуклеотидных последовательностей показал, что
З'-сайт сплайсинга, фланкирующий экзон 2, повреж-
ден, чем и объясняется невозможность соединения
экзонов 1 и 2. Невозможность соединения экзонов
1 и 3, а также экзонов 3 и 4 объясняется иными
причинами, которые обсуждаются ниже в этом раз-
деле.
В альтернативном сплайсинге типа II участвуют
про-мРНК, имеющие неодинаковые З'-проксималь-
ные последовательности различной длины, что, как
правило, является результатом полиаденилирова-
ния транскрипта в разных местах (рис. 8.90). При
альтернативном сплайсинге, основанном на исполь-
зовании пяти разных сайтов полиаденилирования,
образуются пять разных групп поздних мРНК
аденовируса 2. Таким же способом образуются два
вида мРНК тяжелой цепи иммуноглобулина (разд.
Ю.б.в). Один и тот же ген кальцитонина млеко-
питающих кодирует две мРНК (рис. 8.92). Одна
получается из более короткой про-мРНК, а другая
кодирует пептид, родственный кальцитонину. Все
четыре экзона в более короткой про-мРНК каль-
цитонина сохраняются. Более длинная про-мРНК
содержит два добавочных экзона, 5 и 6, которые
остаются в гене кальцитонина после сплайсинга
соответствующей мРНК, а экзон 4 утрачивается.
З'-сайт сплайсинга интрона 4 лучше всего подходит
для соединения с 5'-сайтом сплайсинга третьего
интрона. В данном случае характер сплайсинга
определяют разные тканеспецифичные сигналы по-
лиаденилирования.
При альтернативном сплайсинге типа III, наибо-
лее разнообразном и загадочном, из одинаковых
про-мРНК образуются разные функциональные
мРНК (рис. 8.90). В про-мРНК присутствуют все
потенциальные экзоны, и при сплайсинге между
ними делается выбор. Ярким примером сплайсинга
122
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Альтернативные
промоторы
Альтернативные
сайты полиаденн-
лирования
Тип III
а
Кассетный
сплайсинг
Интрон
не вырезан
РИС. 8.90.
Типы альтернативного сплайсинга
про-мРНК. Пунктирными прямыми
разного цвета, соединяющими экзо-
ны, представлены разные способы
сплайсинга в зависимости от того,
имеют ли про-PH К разные 5'-концы,
разные З'-концы или одинаковые 5'-
и З'-концы.
а- и P-типов, который характерен для многих генов
(см. рис. 8.90), служит сплайсинг гена тропонина
Т скелетной мышцы (рис. 8.93). мРНК тропонина
Т а- и p-групп содержат либо экзон 16, либо экзон 17
соответственно. Это является результатом взаимо-
исключающего лигирования экзонов 15, 16 и 18 или
15, 17 и 18. Выбор экзона зависит от стадии раз-
вития организма. Так, экзон 16 присутствует в
мРНК мышечного тропонина Т у взрослых особей,
а экзон 17 используется в мРНК как взрослого
MLC1 ТАТА
MLC 3 ТАТА
мРНК
РИС. 8.91.
MLC1 0'1
MLC3
мРНК, полученные при альтернативном сплайсинге
про-мРНК, которые синтезировались с разных сайтов
инициации транскрипции. В верхней части рисунка
представлен ген легкой цепи миозина (MLC1) с девятью
экзонами; интроны выделены точками. Стрелки указы-
вают на разные сайты инициации транскрипции. При
сплайсинге этих двух первичных транскриптов обра-
зуются мРНК, содержащие как одинаковые, так и раз-
ные экзоны.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
123
ТАТА
ААТААА
ААТААА
мРНКкалъцитонина
(клеткищитовид-
ной железы)
РИС. 8.92.
мРНК, полученные при альтернативном сплайсинге
про-мРНК с разными полиаденилированными 3-кон-
цами. Показаны экзоны гена кальцитонина (пронуме-
рованы) и интроны (выделены точками), ТАТА-бокс
промотора и сигналы полиаденилирования (ААТААА).
Две разные мРНК, одна иэ которых кодирует кальци-
тонин, а другая-родственный ему белок (CGRP), обра-
зуются иэ транскриптов, которые полиаденилируются
в первом или втором сайте полиаденилирования соот-
ветственно.
мРНК CGRP
(мозг)
I--ТАТА ААТААА —i
H,f2i3&l5l6l7M8f9fi0ll1f12f,3&14t,5l,6f17f8{-8
45 6 7 8
56 1
6 1
6 7
iel/w
РИС. 8.93.
Разные мРНК тропонина Т, получен-
ные при альтернативном сплайсинге
про-мРНК с одинаковыми 5'- и
З'-концами. В верхней части рисунка
представлен ген тропонина скелетных
мышц. Экзоны пронумерованы, а
интроны выделены точками; указаны
также сигналы инициации транскрип-
ции и полиаденилирования-ТАТА и
ААТААА соответственно. При альтер-
нативном включении экзонов 16(a)
или 17 (Р) образуются две группы
мРНК. Каждая иэ них состоит иэ 32
мРНК. различающихся по тому, ка-
кие из экзонов 4-8 сохранились в
молекуле после сплайсинга.
5
5
Разные ,
мрнк СВ’Т2]з'
5 6
5 е
6
5 6
6
5
5
56Т
5 67
Б 7
Б 1
124
ЧАСТЬ III. ОРГАНИЗАЦИЯ. ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
организма, гак и эмбриона. В а- и Р-группах
представлены все 32 возможных варианта альтер-
нативного сплайсинга с участием экзонов с 4 по
8-от полного исключения до сохранения всех пяти
экзонов в зрелых мРНК.
В некоторых случаях при сплайсинге исполь-
зуются 5'- и З'-сайты сплайсинга, находящиеся внут-
ри экзонов или интронов. Так, несмотря на то, что
подлинный 5'-сайт сплайсинга обычно сшивается
с нормальным З'-сайтом, находящимся в конце дан-
ного интрона, в сплайсинге может участвовать
и З'-сайт, расположенный внутри интрона, что
приводит к удлинению следующего экзона (рис.
8.90, В). С другой стороны, «задействование» аль-
тернативного сайта, расположенного в следующем
экзоне, ведет к укорочению этого экзона. Анало-
гичные результаты получаются при использовании
альтернативных 5'-сайтов сплайсинга внутри интро-
нов или экзонов (рис. 8.90, Г}. Такие криптические
сайты могут участвовать в сплайсинге, только если
при этом получаются соответствующие рамки счи-
тывания. Этот последний способ альтернатив-
ного сплайсинга используется вирусами SV40 и по-
лиомы для образования множества мРНК из тран-
SV40
скрипта ранней области (рис. 8.94). Существование
такого типа сплайсинга вносит неразбериху в во-
прос о различии кодирующих свойств экзонов
и интронов.
Особенно интересным примером тканеспецифич-
ного альтернативного сплайсинга является сплай-
синг гена, кодирующего транспозазу Р-элемента
Drosophila (разд. 10.2.6). В этом случае неспособ-
ность к удалению интрона, содержащего кодон
терминации трансляции, блокирует экспрессию гена
транспозазы в соматических клетках. Экспрессия
гена транспозазы в клетках зародышевой линии
происходит потому, что в них интрон удаляется,
в результате чего рамка считывания увеличивается
до размеров экзона, необходимого для синтеза
активной транспозазы.
Ярким примером дифференциального сплайсин-
га служит сплайсинг РНК-генов ретровируса. Геном
интегрированного провируса транскрибируется с
образованием полной копии геномной РНК вируса,
которая в конечном счете и упаковывается в вирион;
никакого сплайсинга РНК не происходит (разд.
5.7.г). Тот же самый полноразмерный транскрипт
представляет собой мРНК, кодирующую белок кора
и обратную транскриптазу, а кроме того, он являет-
ся про-мРНК, которая подвергается сплайсингу с
А
РИС. 8.94.
Образование множества мРНК из транскрипта
ранней области геномов SV40 и вируса полиомы
при альтернативном сплайсинге. А. Положение
большого (Т) и малого (t) интронов в 5'-про-
ксимальной части ранней области генома SV40.
Ниже представлены две мРНК, полученные в ре-
зультате альтернативного сплайсинга, которые
кодируют Т- и t-белки. Звездочкой отмечено по-
ложение терминирующего кодона, который от-
сутствует в T-мРНК, но есть в t-мРНК. Б. В ранней
области генома вируса полиомы имеются три
перекрывающихся интрона. При альтернативном
сплайсинге один из них удаляется и образуется
одна из трех ранних мРНК; на рисунке показано,
какие экзоны при этом остаются в молекулах
мРНК.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
125
образованием мРНК, кодирующей белок оболочки
вируса. В случае вирусов Т-клеточного лейкоза
и иммунодефицита человека благодаря разным
способам сплайсинга первичного транскрипта обра-
зуется целый набор мРНК, каждая из которых
кодирует особый белок, необходимый для размно-
жения вируса и развития инфекционного процесса
(рис. IV.6).
Выбор сайта сп.шйсинга. Самой большой загад-
кой как конститутивного, так и альтернативного
сплайсинга про-мРНК является механизм выбора
пар соединяемых сегментов. Все объяснения можно
разбить на две группы. Одни считают, что основная
роль принадлежит структурным элементам самой
про-мРНК или промежуточных продуктов, обра-
зующихся в процессе сплайсинга; они названы
yuc-факторами. Другие - что основную роль играют
троис-факторы, белки или другие РНК, которые
влияют на выбор интронов, подлежащих удалению
при сплайсинге. Использование сайтов сплайсинга
почти наверняка носит иерархический характер,
и эта иерархия, возможно, отражает относительное
конкурентное преимущество наиболее предпочти-
тельных 5'- и З'-сайтов сплайсинга в процессе
образования сплайсингосом. При сплайсинге мРНК
антигенов SV40 выбор между двумя путями альтер-
нативного сплайсинга определяется выбором точки
разветвления в интроне. Имеется несколько убеди-
тельных примеров, когда мутации в одном сайте
сплайсинга приводили к тому, что вместо него
начинал функционировать другой, обычно не ис-
пользующийся сайт. Есть все основания полагать,
что щр«нс-факторы также влияют на характер
сплайсинга, особенно в тех случаях, когда пред-
положительно одинаковые про-мРНК подвергаются
альтернативному сплайсингу, зависящему от физио-
логических условий или от типа ткани. Возможно,
на характер сплайсинга влияют особые мяРНП или
подобные матуразам белки, синтез или функциони-
рование которых зависит от типа ткани или стадии
развития организма. Ответ на некоторые из этих
вопросов мы сможем получить в ближайшие не-
сколько лет.
8.6. НОВЫЕ СТРУКТУРНЫЕ
ЭЛЕМЕНТЫ В ФАКТОРАХ
ТРАНСКРИПЦИИ
Факторы транскрипции должны выполнять по
крайней мере две важные функции: связываться со
специфическими последовательностями ДНК и
влиять на инициацию транскрипции. Опыты с раз-
нообразными белками, связывающимися с ДНК,
показали, что специфичность связывания опреде-
ляется кооперативными аддитивными взаимодейст-
виями между какими-то областями ДНК и участ-
ками белковой молекулы с характерной структурой.
Один из таких участков благодаря особой амино-
кислотной последовательности обеспечивает взаимо-
действие с ДНК, зависящее не от нуклеотидной
последовательности этой молекулы, а от ее струк-
турных особенностей (например, связывание может
происходить с сахарофосфатным остовом В-спира-
ли). Специфичность связывания обычно обусловли-
вается неким структурным элементом, обеспечи-
вающим взаимодействие между боковыми группа-
ми определенных аминокислот и парами оснований
в главном желобке сайта связывания ДНК с по-
мощью водородных связей и вандерваальсовых сил.
Установлены два вида участков и аминокислоты,
ответственные в некоторых случаях за сильное
специфическое взаимодействие: при связывании
белка-репрессора и белка Сго фага А, и при свя-
зывании белка-репрессора trp и белка CAP E.coli.
Вряд ли можно предположить, что стерические фак-
торы определяют специфические ДНК-белковые
взаимодействия подобно тому, как это происходит
при образовании пурин-пиримидиновых пар в нук-
леиновых кислотах. В самом деле, изучение упомя-
нутых ранее ДНК-белковых взаимодействии свиде-
тельствует о том, что отдельная аминокислота мо-
жет образовывать связи с разными основаниями, а
отдельное основание-с разными аминокислотами.
Но одного только связывания факторов тран-
скрипции с ДНК еще недостаточно для регуляции
транскрипции, хотя оно и необходимо. Белок дол-
жен иметь также регуляторный домен. Связываю-
щий и регуляторный домены могут быть частью
одного и того же белка или принадлежать разным
белкам. Эти два белка, взаимодействуя друг с дру-
гом, обеспечивают образование функционального,
промоторспецифичного фактора регуляции тран-
скрипции.
Изучение факторов транскрипции эукариот еще
только начинается. Тем не менее уже определены
некоторые структурные домены, создающие прост-
ранственный каркас, который обеспечивает специ-
фические взаимодействия аминокислот с парами
оснований. В каждом белке имеются два домена,
один из которых отвечает за связывание ДНК,
а второй-за регуляцию транскрипции. Домены не
являются взаимозависимыми; можно соединить
домен связывания ДНК и регуляторный домен из
разных белков и образовать факторы транскрипции
со свойствами, характерными для каждого домена.
Более того, если связывающаяся с белком после-
довательность располагается вблизи ТАТА-блока,
то такие химерные белки могут присоединяться
к ДНК и регулировать транскрипцию у целого ряда
эукариот (например, дрожжей, Drosophila, лягушек
и млекопитающих).
126
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
а. Домены связывания ДНК
Спиралъ-петля-спираль. Наиболее полно охарак-
теризованным структурным ДНК-связывающим
элементом является спираль-петля-спираль, впервые
обнаруженный в белке Сто и белке-репрессоре фага
А. Благодаря этому элементу белок а2 дрожжей
МАТ, подавляющий транскрипцию генов, которые
участвуют в определении типа спаривания (разд.
10.6.6), может связывать специфические последова-
тельности нескольких промоторов.
Характерной особенностью структуры спираль-
петля-спираль, определяющей ее взаимодействие
с ДНК, является не аминокислотная последователь-
ность, а геометрия: две а-спирали, разделенные
петлей из нескольких аминокислот. Исходя из дан-
ных рентгеноструктурного анализа некоторых бел-
ков и их комплексов с ДНК была построена модель,
объясняющая специфичность связывания белков
с ДНК. Как показывают структурные данные, и Сго,
и репрессор образуют димеры; при этом димери-
зация является результатом взаимодействий между
антипараллельными Р3-слоями белка Сго и между
а5-спиралями репрессора (рис. 8.95). Участки ДНК,
с которыми связываются Сго и репрессор, имеют
ось симметрии 2-го порядка, благодаря чему эле-
белок Сго Репрессор фага К
РИС. 8.95.
Димеры, которые образуют молекулы белка Сго и реп-
рессора фага X (вид вдоль оси симметрии 2-го по-
рядка). Белок Сго состоит из трех а-спиралей (ци-
линдры) и одного трехцепочечного антипараллельного
P-слоя (стрелки). Репрессор фага X состоит из пяти
а-спиралей, а p-слой у него отсутствует. Взаимодейст-
вие между субъединицами осуществляется в этом слу-
чае за счет контактирования пятых а-спиралей, а свя-
зывание с ДНК-при участии а2- и аЗ-спиралей (они
выделены серым цветом). [Y. Takeda et al.. Science 221
(1983), p. 1020.]
менты спираль-петля-спираль обоих мономеров
связываются с половинами симметричной структу-
ры бок о бок, стабилизируя таким образом всю
систему (рис. 8.96). Наиболее важными детерми-
нантами, обеспечивающими специфичность связы-
вания, являются взаимодействия между аминокис-
лотами одной из спиралей мономера-«узнающей
спирали» (аЗ-спираль в белках Сго и репрессоре)
с участком из шести оснований в главной бороздке.
Вторая по ходу цепи спираль (а2-спираль в каждом
белке) обеспечивает взаимодействие с сахарофос-
фатным скелетом и, возможно, с основаниями,
примыкающими к узнающей последовательности,
стабилизируя таким образом комплекс ДНК с бел-
ком. В пользу такой модели говорят результаты
опытов, в которых изменяли специфичность свя-
зывания ДНК путем замены аЗ-спирали на спираль,
принадлежащую белку с другой специфичностью
связывания. Идентификация аминокислот, ответст-
венных за узнавание и стабилизацию, основывается
на мутационных изменениях ключевых остатков
в каждом домене.
Гомеодомены. Обнаруживается все больше ге-
нов, участвующих в регуляции развития различных
эукариот, которые содержат специфическую после-
довательность из 180 п.н. в белок-кодирующей об-
ласти. Эта последовательность называется гомео-
боксом, поскольку впервые она была обнаружена
в гомеотических генах Drosophila. Экспрессия этих
генов, детерминирующих сегменты, из которых
впоследствии развиваются голова, грудь и брюшко,
происходит после экспрессии материнских генов,
генов зиготы и сегментации, но предшествует диф-
ференциации отдельных типов клеток внутри каж-
дого сегмента (разд. 8.3.ж). Со времени своего
обнаружения гомеобокс был найден во многих дру-
гих генах Drosophila, в частности в тех, которые
определяют пути дифференцировки клеток на ста-
дии раннего эмбриона. Почти все содержащие го-
меобокс гены Drosophila экспрессируются в особой
группе эмбриональных клеток, и каждый тип клеток
содержит уникальную комбинацию продуктов ге-
нов, имеющих гомеобокс. Определенные гены, со-
держащие гомеобокс, по-видимому, кодируют фак-
торы регуляции транскрипции, которые влияют на
экспрессию других генов с гомеобоксами, а также на
синтез нерегуляторных структурных белков. Вслед
за генами Drosophila гомеобоксы были обнаружены
в генах, участвующих в регуляции процессов раз-
вития или аналогичных процессов у червей, лягу-
шек, мышей и человека. Вероятно, эти гены можно
рассматривать как гены, определяющие родослов-
ную клетки. Так, ранний эмбрион еще до обра-
зования отдельных органов делится в результате
экспрессии гена, содержащего гомеобокс, на перед-
нюю и заднюю половины, имеющие разные пути
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
127
и стабилизирующая а2-спираль выделены цветом. А-
вид сбоку, 6-вид спереди. [D. Н. Ohlendorf, W. F. Ander-
son, B.W. Mathews, J. Mol. Evol. 19 (1983), p. 109.]
РИС. 8.96.
Связывание ДНК с белками, которые содержат струк-
турный элемент «спираль-петля-спираль». Домены, при-
нимающие участие в связывании ДНК, изображены
в виде больших серых кругов. Узнающая аЗ-спираль
развития. Где бы ни находился гомеобокс из
180 п.н., он кодирует гомеодомен из 60 аминокислот,
который, как теперь известно, является ДНК-свя-
зывающим элементом у белков этого класса. По-
мимо той особенности, что они почти всегда нахо-
дятся в определенном месте белковой молекулы,
гомеодомены обладают еще одним, по-видимому,
наиболее замечательным свойством-они образуют
элемент типа спираль-петля-спираль. Наличие такой
структуры в пептиде, содержащем гомеодомен, было
показано с помощью рентгеноструктурного анализа.
Анализ аминокислотной последовательности
многих гомеобелков свидетельствует о том, что
гомеобелки позвоночных и Drosophila в чем-то сход-
ны, но в чем-то и различаются. Сходство заклю-
чается в высоком содержании основных аминокис-
лот (около 30% остатков представлено аргинином
или лизином), что согласуется с их ДНК-связы-
вающей способностью. Но гомеодомены позвоноч-
ных имеют некоторые отличительные особенности
(рис. 8.97). Большинство из них содержат консер-
вативную N-концевую последовательность Met-Ser-
Ser-Leu-Tyr-Tyr-одна из аминокислот-Asn, за кото-
рой идет вариабельный участок, имеющий, однако,
гомологичные сегменты, характерные для гомеобел-
ков разных классов (при этом либо степень их
гомологии мала, либо сами сегменты непротяжен-
ные). Сразу перед гомеодоменом находится консер-
вативный пентапептид Ile-Tyr-Pro-Trp-Met, а за до-
меном- С-концевая последовательность, богатая
кислыми аминокислотами.
Имеются данны о том, что гомеодомен необхо-
дим для проявления соответствующими белками
ДНК-связываюгцей активности. Так, точечные му-
128
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Консервативный
N-коиец
Интрон
[MSSLYY-N [Вариабельный участок
Гомеодомен
1YPWM
гомео пептид
1 2 3
а-Спирали
Кислый
конец
Arg Lys Arg Gly Arg Gin Thr Tyr Thr Arg Tyr Gin Thr Leu Glu Leu Glu Lys Gly Phe
21 40
His Phe Asn Arg Tyr Leu Thr Arg Arg Arg Arg He Glu He Ala His Ala Leu Cys Leu
Thr Glu Arg G|n He Lys He Trp Phe Gin Asn Arg Arg Met Lys Trp Lys Lys Glu Asn
РИС. 8.97.
Особенности строения белков, содержащих гомеодо-
мены. В верхней части рисунка представлены основные
структурные элементы таких белков. Многие из них
имеют характерную N-концевую последовательность,
содержащую метионин (М), серин (S), лейцин (L),
тирозин (Y), какую-нибудь одну из аминокислот ( )
и аспарагин (N). Далее находится участок, длина и
аминокислотная последовательность которого варьи-
руют, а за ним короткий инвариантный участок из пяти
аминокислот: изолейцина (I), тирозина (Y), пролина
(Р), триптофана (W) и метионина (М). Гомеодомен
тации и мутации со сдвигом рамки в гомеодомене
полностью подавляют ДНК-связываюгцую актив-
ность белков, в то время как мутации в других
участках белка дают менее выраженные эффекты.
При замене гомеодоменов связывающая способ-
ность нового белка соответствует связывающей
способности белка, откуда произошел этот гомео-
домен. Участки ДНК, с которыми связываются
гомеобелки, обычно не имеют оси симметрии 2-го
порядка, поэтому можно предположить, что свя-
зываются только мономеры. Важным свойством
некоторых гомеобелков является их способность
связывать неодинаковые, но родственные последо-
вательности ДНК, хотя и с разной эффективностью.
Возможно, такая специфичность связывания поз-
воляет регулировать транскрипцию нескольких ви-
дов генов с помощью одного гомеобелка.
Детали специфического связывания с ДНК бел-
ков, имеющих гомеодомены, неизвестны. Широко
распространено мнение, что специфичность опре-
деляется организацией аминокислот в пределах
самого элемента спираль-петля-спираль и вокруг
него. Вероятно, на связывание влияют и другие
консервативные особенности белков, но это влия-
ние, по-видимому, невелико. Не исключено, что
способность данных белков связывать ДНК и/или
с ярко выраженными основными свойствами, состоящим
из 60 аминокислот, расположен вблизи кислого (бога-
того остатками глутаминовой кислоты) С-конца моле-
кулы. Он может образовывать три а-спирали, из кото-
рых формируется ДНК-связывающий элемент спираль-
петля-спираль. В нижней части рисунка приведена
аминокислотная последовательность гомеодомена бел-
ка, кодируемого геном antennapedia Drosophila. Основ-
ные аминокислоты выделены цветом. [C.V. Е. Wright et
al.. Trends Biochem. Sci. 14 (1989), p. 52; W. J. Gehring,
Sci. American 253 (October 1985), p. 153.]
регулировать транскрипцию опосредуется действи-
ем небольших молекул. Еще более вероятно, что
у этих белков есть участки, которые позволяют им
взаимодействовать с другими белками с образо-
ванием функционально активного белкового комп-
лекса. Связанные с ДНК гомеобелки влияют на
транскрипцию со своих промоторов-мишеней неиз-
вестным пока способом. Возможно, в этом прини-
мают участие кислые С-концы и N-концевые ва-
риабельные области, богатые пролином и глута-
мином (разд. 8.6.6).
P0U-домены. Некоторые факторы транскрип-
ции, принимающие участие в специфичной для дан-
ной ткани или стадии развития регуляции, имеют
сложную регуляторную область примерно из 160
аминокислот, куда входит характерный гомеодо-
мен. Эта область высококонсервативна в факторе
транскрипции (Pit-1), необходимом для активации
транскрипции генов пролактина и гормона роста
у крыс, в двух белках, связывающих октамерную
последовательность, как широко распространенном,
так и лимфоидспецифичном: OTF-1 и OTF-2, а так-
же в транскрипционном регуляторе развития нейро-
нов у C.elegans (Unc-86). Сокращение POU относится
к белкам, в которых эта регуляторная область была
впервые найдена.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
129
Oct-1
POU-домен
I------------1
unc-86 | |
100 аминокислот
Pit-1 | “] I--------1
POU-Sp homeo
РИС. 8.98.
Положение и организация POU-доме-
нов некоторых факторов транскрипции.
POU-Sp - POU-специфичный домен,
homeo - гомеодомен. Представлены сле-
дующие факторы транскрипции: белки, свя-
зывающие октамеры, Oct-1 (OTF-1) и Oct-2
(OTF-2), фактор транскрипции, кодируе-
мый геном Unc-86 С. elegans; гипофиз-спе-
цифичный фактор транскрипции Pit-1. Раз-
мер POU-доменов во всех белках примерно
одинаков, но в разных белках они находятся
на разном расстоянии от концов молекулы.
Положение и размер двух консервативных
участков, которые вместе составляют POU-домен
в четырех упомянутых выше белках, показаны на
рис. 8.98, а степень гомологии последовательностей
этих доменов-на рис. 8.99. Последовательности
трех гомеодоменов близкородственны и, как в слу-
чае других гомеобелков, могут образовывать струк-
туру спираль-петля-спираль. Узнающей спирали
этого элемента, вероятно, соответствует высококон-
сервативная С-концевая треть гомеодомена, назван-
ная областью WFC из-за присутствия в ней остат-
ков триптофана (W), фенилаланина (F) и цистеина
(С). Этот вывод можно сделать исходя из того, что
изменения в области WFC нарушают связывание
обоих белков OTF с октамерной последователь-
ностью. Связываются ли эти белки с соответст-
вующими последовательностями ДНК при участии
структуры спираль-петля-спираль гомеодомена и
Рои-специфичный домен
’it-l к 5 к lveepiimdspei Rt LEQF А н е г к v r ri kl о v таг н v о е A l aav hgs е fsh тт i с RF Е n l qlsfk h ackl к a i lskwl e EA E qv
Oct-т D TP s LEEPSDLE-------------------------------------------------------------------------------------------------------ELEQF АКТЕ К OR R I К L 6 F TBOtVOl AMOKLVS H DFSftTT IS It F EA L К LS F KNMCKLKP L LIKWLHDAl N L
1-2 ₽ PS И PEEP St) LE-ELEQF AR t F К QRR 1 К L 6 F T 06 D VOL A M6KL V в H DFS ЯТ T 1 SR F E Al И LSF KNMCKLK P L LEKWLNIAEt M
«ОНИ............ttP-I........................................................................................................ELEQF A- . F К . R R I KL 6. Ta.. VS. A.6 • Ft IlTTI . RFE . L . LSFKH .CALK. L.KWl . . AE •
РОи-гомеодомен
V 6 A H - ERKRKRRTT I S I A A KIA II ER И F 6 С И SI R S al I MRM AIELNLEKEVV RVWFCN R R Я RE KR VKT S LHQS
Oct-1 SSSSSLs8PSALNSF--ai EGL- SRRR К К RTS I ETH | KVA L EK SFl ER QKPT S EE IT M IADQLNMEKE VI RVWFCN R R CLKE К R 1 H P PS SGG
Oct-2 S V V S SIS S F HQ L S S P S L 6 F DG L F С IR А К К AT 8 I E TN V I F A L E К S FL A H Q KPT s E E I L L IA E fl L н м E К E V I RV WEEN R RQKEKRINPCSAAP
Каноми- R.RK-RT.I...........A L В . , r. - - - Kp - 6 - E I . ..A-'L«'EKEV. RVWF CNRRQ-EKR - • -
ческая
последовательность
РИС. 8.99. -------WFC---------*
Гомологичность POU-доменов некоторых факторов
транскрипции. Представлены аминокислотные последо-
вательности POU-доменов факторов транскрипции
Pit-1, Oct-1 (OTF-1) и Oct-2 (OTF-2) с использованием
однобуквенных обозначений аминокислот (рис. 1.24).
Последовательности расположены относительно друг
друга таким образом, чтобы число совпадений было
максимальным. Цветом выделены те случаи, когда оди-
наковые аминокислоты встречаются в данной позиции
не менее чем у двух белков. Представлена также кано-
ническая последовательность для тех позиций, в кото-
рых у всех трех белков находятся одинаковые амино-
кислоты. В эту последовательность не включены амино-
кислоты, имеющие разную структуру, но близкий заряд,
а также разные аминокислоты, близкие по структуре.
WFC- высококонсервативная последовательность на
С-конце, содержащая аминокислоты триптофан (W),
фенилаланин (F) и цистеин (С). Как отмечалось в
тексте, этот участок существен для проявления актив-
ности факторов транскрипции, содержащих POU-до-
мены.
Q-KS
130
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
какова роль POU-специфичного домена в этом свя-
зывании и/или регуляции транскрипции неясно.
Полагают, что POU-специфичная область обеспе-
чивает взаимодействие этих белков с другими бел-
ками, при котором изменяется их регуляторный
эффект.
Цинковые пальцы. Исследование аминокислотной
последовательности TFIIIA, одного из факторов
транскрипции 58-рРНК-генов, показало, что в доме-
не связывания ДНК присутствуют так называемые
цинковые пальцы (разд. 8.4.в). С тех пор было
обнаружено множество белков, содержащих анало-
гичные последовательности, которые могут образо-
вывать хелатные связи с ионом цинка. Различное
число цинковых пальцев содержится в белке GAL4
дрожжей (разд. 8.3.е), в факторе Spl млекопитаю-
щих (разд. 8.3.г) и в рецепторах стероидных гормо-
нов (разд. 8.3.е). У Drosophila белок, который содер-
жит несколько таких пальцев, кодируется (разд.
8.3.ж) дар-геном hunchback. Белок млекопитающих,
индуцируемый стеролом, который регулирует ра-
боту генов, участвующих в биосинтезе стерола,
и гена, кодирующего рецептор липопротеина низкой
плотности, содержит шесть цинковых пальцев, ко-
торые связываются со стерол-акцепторными эле-
ментами в промоторах этих генов. Корреляция
между способностью к регуляции транскрипции
и наличием цинковых пальцев становится настолько
очевидной, что обнаружение таких структурных
элементов в белке, кодируемом неким геном, может
служить основанием для того, чтобы говорить
о регуляторной роли продукта этого гена в тран-
скрипции.
Известны по крайней мере три типа цинковых
пальцев. Они различаются числом и локализацией
остатков цистеина и гистидина, координационно
связанных с атомом цинка. Основное структурное
свойство одного из типов цинковых пальцев (тип
TFIIIA) наличие петли, образующейся при коорди-
национном связывании атома цинка с парами ос-
татков цистеина и гистидина, разделенных 12 ами-
нокислотами (рис. 8.69). Цинковые пальцы второго
типа характеризуются тем, что в координационном
связывании участвует разное число остатков цис-
теина. Например, цинковый палец GAL4 содержит
шесть цистеиновых остатков, из которых только
четыре связаны с атомом цинка; цинковые пальцы
ДНК-связывающих доменов в рецепторах стероид-
ных гормонов содержат четыре цистеина. У цинко-
вых пальцев третьего типа в связывании с атомом
цинка участвуют три остатка цистеина и один - гис-
тидина. Но не исключено, что для образования
цинкового пальца подходит любая комбинация ос-
татков цистеина и гистидина. Вероятно, с этими
остатками могут связываться и атомы других ме-
таллов, а когда имеется более четырех потенциаль-
ных сайтов координационного связывания, металлы
образуют координационные связи с альтернатив-
ными парами сайтов.
Для изучения структуры цинковых пальцев ис-
пользовали разные физические методы, в частности
метод кругового дихроизма и ЯМР; исследовали
также трехмерную структуру пептида, содержащего
один цинковый палец. Было показано, что атом
цинка погружен в глубь молекулы и находится
в окружении а-спиралей и других спиральных струк-
тур, связанных друг с другом при помощи Р-слоев
(рис. 8.100).
Фактор TFIIIA может связываться с внутренней
регуляторной областью гена 5S-pPHK только при
наличии интактных цинковых пальцев, а для свя-
зывания Spl с соответствующей последователь-
ностью ДНК необходимо, чтобы в белковой моле-
куле присутствовал цинк. Далее, мутации в облас-
тях, прилегающих к цинковому пальцу в белке
GAL4 или к пальцу в рецепторе кортикостероидного
гормона, блокируют их связывание с соответст-
вующими последовательностями-мишенями. Нали-
чие у цинковых пальцев ДНК-связывающей спо-
собности подтверждается двумя фактами: 1) уко-
роченные белки TFIIIA или Spl, содержащие только
сами пальцы, сохраняют способность специфически
связываться с ДНК; 2) при замене цинковых пальцев
РИС. 8.100.
Модель цинкового пальца, построенная с помощью
компьютерной графики по данным ЯМР. Углеродный
скелет представлен в виде трубки. Атом цинка (шар)
соединен при помощи координационных связей с двумя
остатками цистеина и двумя остатками гистидина. По-
казаны также остатки фенилаланина и лейцина. [М. Lee
et al.. Science 245 (1989), р. 635. Фотография предостав-
лена М. Е. Pique, Р. Е. Wright.]
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
131
, Участок,
Основный богатый
участок лейцином
c-jun
CZZ
Основный богатый Связывание ДНК
участок лейцином
c-fos
IFZ
РИС. 8.101.
Положение и организация элементов, получив-
ших название «лейциновая молния», в факторах
транскрипции млекопитающих и дрожжей. Осно-
вой элемента является а-спираль, в которой каж-
дый седьмой остаток представлен лейцином
(рис. 8.103). Еще один компонент-участок с ярко
выраженными основными свойствами, ответствен-
ный за связывание белка с ДНК.
6СН4
СвязываниеДНК „ , . участок.
Основный богатый
участок лейцином
Связывание ДНК
лейцина в пределах относительно протяженного
а-спирального участка (рис. 8.102). Если мы по-
смотрим на а-спираль с торца, то она будет иметь
эстрогена на соответствующие цинковые пальцы
рецептора кортикостероидного гормона образуется
гибридный рецептор, который соединяется с эле-
ментом, чувствительным к кортикоиду, а не с сай-
том связывания эстрогенового рецептора.
Механизм связывания цинковых пальцев с ДНК
до конца не выяснен. Известно, что связывание
происходит по главной бороздке ДНК, но остатки,
которые определяют специфичность и прочность
связывания, пока не установлены. В некоторых
случаях за специфичность ответственны аминокис-
лотные остатки пальца, образующие петлю. В ре-
цепторе экстрогена определяющую роль в специ-
фичности связывания ДНК играет один из двух
пальцев; более того, некоторые остатки, влияющие
на специфичность связывания, находятся у осно-
вания пальца (разд. 8.3.е). Так, единственный цин-
ковый палец в белке GAL4 можно заменить на
палец, кодируемый чужеродным геном, и модифи-
цированный белок все равно будет связываться
с UASgal. Итак, цинковые пальцы образуют струк-
туру типа петли, что обеспечивает специфичность
взаимодействия отдельных остатков с соответст-
вующими сайтами связывания ДНК. Возможно,
пальцы участвуют и в неспецифических взаимодей-
ствиях, стабилизирующих ДНК-белковый комплекс,
т.е. играют роль, аналогичную роли второй спи-
рали в элементе спираль-виток-спираль.
«Лейциновые молнии». Некоторые факторы
транскрипции, в частности С/ЕВР, ДНК-связываю-
щие регуляторы транскрипции млекопитающих
семейства APl-Jun и родственные GCN4 факто-
ры транскрипции дрожжей, содержат одинаковый
структурный элемент, состоящий из двух компо-
нентов и названный «лейциновой молнией» (рис.
8.101). Такой структурный элемент несут и неко-
торые факторы транскрипции млекопитающих, в
основном кодируемые генами с-тус и c-fos, хотя
сами эти факторы не связываются с ДНК.
Характерной особенностью лейциновой молнии
вид кольца, от которого отходят боковые группы
аминокислот, при этом остатки лейцина окажутся
с каждым новым витком на одной и той же стороне
спирали. На противоположной же стороне спирали
обнаружится необычно много отрицательно заря-
РИС, 8.102.
Представление а-спиральных участков в лейциновых
молниях в виде колец. Рисунок иллюстрирует поло-
жение остатков лейцина (L) в лейциновых молниях
следующих белков: белка, связывающего энхансер
(С/ЕВР); белка тус (с-тус) мыши; белка, кодируемого
вирусным онкогеном v-jun; белка GCN4 дрожжей. При-
ведены также другие аминокислотные остатки, входя-
щие в состав а-спирального участка лейциновой мол-
нии. Номера аминокислот возрастают по мере их уда-
является периодичность в расположении остатков
ления от читателя.
132
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Основный домен Лейцин-богатый участок
I------------------1 I--------------------------------1
c-jun ESQ Е RIKAERK RMRN RIAAS KCRKRKLERIARLEEKVKTL KAQNSE L A ST A N М L R EQ VAQ.I
C-fos SP Е Е ЕЕ KRR I RRERNKHAAA KCRNRRRELTDTLQAETDQLEDEKSALQTEIANLLKEKEKL
GCN4 VPESSBPAALKRARNTEAARRSRARKLQRMKQLEDKVEELLSKNYHLENEVARLKKLV6ER
РИС. 8.103.
Аминокислотные последовательности некоторых основ-
ных доменов и лейцин-богатых участков лейциновых
молний. Связывание ДНК с белками, содержащими
лейциновые молнии, осуществляется при участии ос-
женных остатков. Возможно, образование ионных
пар, а также стэкинг-взаимодействие между гид-
рофобными остатками стабилизирует эти длинные
спирали. Важной особенностью лейциновой молнии
является наличие обогащенной основными амино-
кислотами последовательности, соседствующей с
лейциновой спиралью (рис. 8.Ю1 и 8.103).
Белки с лейциновой молнией могут образовы-
вать как гомо-, так и гетеродимеры. Это происходит
благодаря гидрофобным взаимодействиям между
остатками, располагающимися на поверхности па-
раллельных друг другу спиралей (рис. 8.104). Спе-
цифичность при связывании ДНК, по-видимому,
обусловливается взаимодействием между соответ-
ствующими нуклеотидными последовательностями
и аминокислотами, находящимися в пределах ос-
новных доменов и по соседству с ними.
Белки, содержащие лейциновую молнию, всегда
связываются с ДНК в виде димеров. Образование
такого димера является необходимой предпосылкой
эффективного связывания. Если способность к обра-
зованию димера нарушена вследствие мутаций, из-
меняющих аминокислотную последовательность
лейциновой молнии или ее вторичную структуру, то
связывания ДНК не происходит, даже если основ-
ный домен остается интактным. Мутации в основ-
ной области тоже блокируют связывание ДНК, но
на способность белка к димеризации они не влияют.
Таким образом, специфичность связывания данных
белков с ДНК определяется сочетанием двух струк-
турных особенностей: лейциновая молния обеспе-
чивает димеризацию, а основный домен участвует
в образовании ДНК-связывающего элемента. Одна-
ко основный домен может функционировать только
при условии димеризации, хотя димеризация не
зависит от основного домена.
Факторы транскрипции GCN4 и С/ЕВР связы-
ваются с ДНК только в виде гомодимеров; напро-
тив, гомодимеры факторов транскрипции семейства
API-Jun связываются с ДНК плохо. В то же время
фактор APl-Jun образуют гетеродимеры с белками
класса fos, которые связываются с ДНК значитель-
но более эффективно. Белки fos не образуют го-
модимеры и не присоединяются к ДНК, хотя они
новного домена, находящегося слева от лейцин-бога-
того участка. Основные аминокислоты выделены цве-
том.
содержат характерную лейциновую молнию и ос-
новные домены. Неспособность fos к связыванию
с ДНК, вероятно, объясняется тем, что данный
белок не может образовывать гомодимеры, поэтому
его способность образовывать гетеродимеры с дру-
гими факторами кажется особенно загадочной.
Зависящее от димеризации связывание ДНК,
по-видимому, становится возможным благодаря
сближению основных доменов двух белков, соеди-
няющихся при помощи лейциновой молнии (рис.
РИС. 8.104.
Схематическое изображение винтообразной структуры
лейциновой молнии (вид сверху и сбоку). Параллель-
ные спирали представлены в виде цилиндров; направле-
ние белковой цепи показано стрелками. Серые круж-
ки-остатки лейцина, цветные-остатки гидрофобных
аминокислот. [По рис. 4 из работы Т. G. Oas et al.,
Biochemystry 29 (1990), p. 2891. Рисунок любезно
предоставлен Р. S. Kim до публикации.]
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
133
РИС. 8.105.
Связывание димеров факторов транскрипции, содер-
жащих лейциновые молнии, с родственными последо-
вательностями ДНК при помощи основных доменов.
Согласно модели, димерная структура стабилизируется
за счет взаимодействий между остатками лейцина и
гидрофобными остатками. Симметрично расположен-
ные основные домены белкового димера связываются
с соответствующим сайтом в молекуле ДНК, облада-
ющим осью симметрии 2-го порядка. [W. Н. Landschulz
et al.. Science 243 (1989), p. 1689.]
8.105) . Следовательно, белки с лейциновой молнией
связываются с ДНК кооперативно, особенно если
участок связывания имеет ось симметрии 2-го
порядка. В этом случае каждый основный домен
связывается с половиной сайта. Гетеродимеры тоже
должны связываться с такими палиндромными
сайтами кооперативно, но при этом одна из субъе-
диниц будет оказывать влияние на специфичность
изи стабильность взаимодействий. Гетеродимеры
могут также специфически связываться с двумя или
несколькими участками. Это позволяет им по-раз-
ному взаимодействовать с близкородственными
последовательностями или регулировать транс-
крипцию с промоторов, имеющих разное устрой-
ство некоторых сигнальных последовательностей.
Совершенно очевидно, что регуляция транскрипции
опосредуется сложными взаимодействиями между
белками и соответствующими последовательностя-
ми ДНК, и существование гетеродимерных факто-
ров переводит регуляцию транскрипции на новый,
более высокий уровень.
б. Домены регуляции транскрипции
Связывание со специфической последователь-
ностью ДНК - только одна из двух важных функций
белков регуляции транскрипции. Кроме этого, свя-
завшиеся белки должны вступить во взаимодействие
с комплексом транскрипции, с тем чтобы иницииро-
вать или подавить процесс. Теперь мы знаем, что
каждая из этих функций-связывания и регуляции-
может быть соотнесена с определенными белко-
выми доменами. Каков бы ни был фактор тран-
скрипции, домен связывания у него относительно
невелик, но высокоспецифичен; более того, домен
связывания функционирует автономно, т. е. узнает
и связывает родственную последовательность даже
в отсутствие остальной части белковой молекулы
(разд. 8.3 и 8.6.а). Напротив, домены, ответственные
за регуляцию транскрипции, менее автономны и
относительно неспецифичны. Они могут регулиро-
вать транскрипцию с участием широкого круга
промоторов при условии, что регуляторные домены
связываются вблизи последовательности ТАТА.
Таким образом, гибридные белки, содержащие ре-
гуляторную область одного фактора транскрипции
и домен связывания ДНК-другого, будут регули-
ровать транскрипцию с промоторов, которые со-
держат последовательность связывания второго
фактора. Это происходит даже в том случае, когда
регуляторный домен и комплекс транскрипции про-
исходят из разных источников. Здесь мы рассмот-
рим свойства различных регуляторных доменов
и возможные механизмы их действия.
Активация транскрипции на расстоянии за счет
изгибания ДНК. Описывая различные эукариотиче-
ские промоторы (разд. 8.2-8.4), мы подчеркивали,
что транскрипция чаще всего регулируется при
участии z/wc-последовательностей, расположенных
на значительных расстояниях от сайтов инициации
транскрипции. Поскольку мы предположили также,
что транскрипция регулируется белками, связываю-
щимися с этими удаленными сайтами, нам следует
разобраться, как регуляторные домены этих белков
влияют на активность комплекса транскрипции.
Имеется множество экспериментальных данных,
указывающих на наличие белок-белковых взаимо-
действий между регуляторными доменами факто-
ров транскрипции и компонентами комплекса тран-
скрипции (рис. 8.106). Такие взаимодействия между
белками, связанными с участками ДНК, находя-
щимися на значительном расстоянии друг от друга,
могут реализоваться, если молекула ДНК образует
петлю, сближающую сайты связывания. Возможно,
регуляция работы сложных промоторов, с кото-
рыми могут связываться разные белки, осуществля-
ется за счет самых разнообразных взаимодействий
между этими белками.
Первые экспериментальные данные об образо-
вании петель в ДНК были получены в опытах по
изучению систем транскрипции прокариот. В этом
случае активаторы обычно присоединяются вблизи
сайта связывания РНК-полимеразы, и аминокисло-
ты, находящиеся с одной стороны домена активации
134
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 8.106.
Связывание нескольких белковых молекул с дисталь-
ными энхансерами индуцирует сборку комплексов тран-
скрипции вблизи ТАТА-бокса. При этом взаимодейст-
вие дистальных белков, которые на рисунке располо-
жены справа от сайта инициации транскрипции, с бел-
ками, находящимися слева от этого сайта, становится
возможным благодаря изгибанию молекулы ДНК.
регуляторного белка, взаимодействуют со связан-
ной полимеразой. Мутации, затрагивающие эти
аминокислоты, обычно блокируют активацию тран-
скрипции, не влияя на связывание с ДНК. Есть
также четкие указания на то, что образование петель
в ДНК создает условия для таких белок-белковых
взаимодействий и в том случае, когда сайты связы-
вания находятся довольно далеко друг от друга.
Неудивительно поэтому, что гибкость молекулы
ДНК и топологические ограничения сильно влияют
на эффективность взаимодействия связанных бел-
ков, если они находятся на небольшом расстоянии
друг от друга, но они менее важны, если эти белки
отстоят на тысячи пар нуклеотидов.
Кислые домены активации. Мы уже отмечали,
что специфичность связывания ДНК может обеспе-
чиваться использованием нескольких различающих-
ся структурных элементов (разд. 8.6.а). В каждом
случае аминокислотные последовательности, ответ-
ственные за специфическое взаимодействие этих
элементов с соответствующими парами оснований
ДНК, относительно инвариантны, и даже неболь-
шие изменения в них влияют на эффективность
связывания. В отличие от этого, белковые домены,
участвующие в активации транскрипции, более ва-
риабельны и в том. что касается их границ, и в том,
что касается аминокислотных последовательностей.
Мутационный анализ белка GAL4 показал, что
домены активации локализованы в двух областях:
между остатками 148 и 238 вблизи N-конца и
остатками 768 и 881 на С-конце (рис. 8.45). Наиболее
характерным свойством этих доменов является их
обогащенность отрицательно заряженными амино-
кислотами: их суммарный заряд составляет —7
и —9 соответственно. Любой из этих доменов
может активировать транскрипцию, если его слить
с доменом связывания ДНК GAL4; более того,
каждый из доменов активирует транскрипцию, если
соединить его с любой связывающей последова-
тельностью, которая обеспечивает близкое распо-
ложение домена GAL4 и промоторного элемента
ТАТА.
Активатор транскрипции дрожжей GCN4 осу-
ществляет положительную регуляцию экспрессии
генов, участвующих в биосинтезе аминокислот; он
близкородствен и взаимозаменяем с факторами
семейства APl-Jun млекопитающих (разд. 8.7.г).
GCN4 имеет всего один домен активации, состоя-
щий из 88 аминокислот с общим зарядом —16
и расположенный в центральной части последова-
тельности из 281 аминокислоты. Рецепторы кор-
тикостероидных гормонов гоже имеют кислые
домены активации. У рецептора крысы есть всего
один кластер отрицательно заряженных аминокис-
лот вблизи N-конца. рецептор человека содержит
еще один кислый домен на С-конце. Регулятор
транскрипции Drosophila, кодируемый геном bed,
тоже содержит кислый домен активации (разд.
8.3.ж). что говорит об универсальности этого
свойства. Активаторы транскрипции, не связываю-
щиеся с ДНК [например, VP16, ассоциированный
с вирусом простого герпеса фактор транскрипции,
инициирующий экспрессию вирусных генов после
заражения (разд. 8.3.г), и Е1а, активатор транскрип-
ции при аденовирусной инфекции], тоже имеют
кислые домены активации. Хотя сами по себе эти
белки не активируют транскрипцию, при слиянии их
кислых доменов с доменами связывания ДНК обра-
зуются белки, активирующие соответствующие
промоторы.
Тот факт, что домены активации имеют кислот-
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
135
ные свойства, становится понятным, если учесть,
что при повышении суммарного отрицательного за-
ряда одного из таких доменов наблюдается пропор-
циональное усиление активации. В самом деле, тран-
скрипцию генов GAL могут активировать даже
сегменты белков E.coli, просто обогащенные кис-
лыми аминокислотами, если соединить их с доме-
ном связывания ДНК GAL4. Например, белок
GAL80 можно превратить из ингибитора зависящей
от GAL4 транскрипции в активатор, если соединить
его с каким-либо кислым сегментом белка E.coli.
В этом случае активацию обеспечивает сегмент
E.coli, а домен активации GAL4 блокируется в ре-
зультате ассоциации его с GAL80.
Хотя активация транскрипции не зависит от
наличия конкретной аминокислотной последова-
тельности, по-видимому, для нее более предпочти-
тельны последовательности, которые могут обра-
зовывать алифатические а-спирали. В таких спи-
ралях вдоль одной их стороны располагаются отри-
цательно заряженные остатки, а вдоль другой-
гидрофобные, как в лейциновых молниях (разд.
8.6.а). Например, синтетический пептид из 15 ами-
нокислот, способный принять спиральную конфи-
гурацию с описанными свойствами, активирует
транскрипцию при слиянии его с соответствующим
доменом связывания ДНК в отличие от пептида,
состоящего из тех же самых аминокислот, соеди-
ненных в произвольном порядке.
Пока остается нерешенным главный вопрос:
каков механизм влияния кислых доменов активации
на транскрипцию? Поскольку уже упоминавшиеся
домены активации функционируют в самых разных
гетерологичных системах (например, в клетках
Drosophila, растений, дрожжей и млекопитающих),
они, по-видимому, взаимодействуют с каким-то
одним компонентом комплекса транскрипции. Так,
активация могла бы происходить в результате
связывания кислых доменов с основными гистонами
в нуклеосомах, которое влияет на поведение кислых
доменов при их взаимодействии с промоторами. Не
исключено также, что домены активации контак-
тируют с основным доменом РНК-полимеразы II
или одного из белков комплекса транскрипции
и стимулируют сборку этого комплекса или по-
вышают его активность. Полученный с помощью
ДНКазы I «отпечаток» транскрипционного комп-
лекса в области ТАТА-блока изменяется, когда
по соседству связываются факторы транскрипции
млекопитающих. Аналогичные изменения в «отпе-
чатке» наблюдаются в том случае, когда рядом
с ТАТА-блоком находится последовательность
UASgal, а ДНК-связывающий домен GAL4 при-
соединяется к встроенному в ДНК синтетическому
сегменту.
Независимо от того, на какой компонент комп-
лекса транскрипции влияет локальное связывание
белков с кислыми доменами, можно сказать, что
в основе этого взаимодействия, по-видимому, лежат
электростатические эффекты, а не стереоспецифи-
ческие или комплементарные взаимодействия. По-
скольку факторы транскрипции из дрожжей или
млекопитающих работают так же хорошо с чу-
жеродными комплексами транскрипции, не исклю-
чено, что такая относительная нетребовательность
к аминокислотной последовательности выработа-
лась в процессе эволюции. Эффективность тран-
скрипции прямо пропорциональна суммарному от-
рицательному заряду, и, изменяя этот заряд, можно
модулировать регуляторный ответ. Суммарный от-
рицательный заряд домена активации изменяется,
например, при амидировании остатков глутамино-
вой или аспарагиновой кислоты.
Другие домены активации. Функциональные до-
мены активации некоторых факторов не содержат
четко выраженных кластеров кислых аминокислот.
Например, у Spl они обогащены глутамином,
а также серином и треонином. Каким образом эти
домены активируют транскрипцию-неясно. Воз-
можно, какую-то роль играет фосфорилирование
и дефосфорилирование гидроксильных групп сери-
на, в результате которого изменяется содержание
кислых остатков. Кроме того, увеличивать или
уменьшать отрицательный заряд может обратимое
амидирование и дезамидирование остатков глута-
мина. Недавно было показано, что Spl и другие
факторы транскрипции могут гликозилироваться;
это приводит к изменению их заряда и тем са-
мым-к изменению эффективности транскрипции.
8.7. ГЛОБАЛЬНЫЕ МЕХАНИЗМЫ
РЕГУЛЯЦИИ
ЭКСПРЕССИИ ГЕНОВ
До сих пор, рассматривая экспрессию генов, мы
ограничивались вопросами регуляции транскрип-
ции отдельных генов и процессинга транскриптов
с образованием зрелых молекул РНК. Скорость,
время и место протекания этих процессов регу-
лируются белками, которые узнают относительно
короткие специфические участки молекулы ДНК
и/или характерные элементы других белков, свя-
зываются с ними и опосредуют инициацию и тер-
минацию транскрипции, а также процессинг. От
присутствия и функционального состояния этих бел-
ков часто и зависит экспрессия генов. Однако
имеются четкие данные о том, что на экспрессию
генов, в частности на их транскрипцию, могут
влиять обратимые изменения в структуре ДНК. Это
могут быть, например, изменения степени сверх-
спиральности ДНК, т. е. ее топологии, а следова-
136
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
тельно, изменения конформации (переход от В- к Z-
или другим формам) (разд. 1.1.в). Регуляция может
осуществляться и путем изменения состояния хро-
матина, в частности изменения упаковки нуклеосом
(разд. 1.1.ж). Это создает еще один уровень ре-
гуляции экспрессии. Как правило, активация тран-
скрипции коррелирует с переходом высококонден-
сированных форм хроматина в менее плотно упа-
кованные «открытые» формы. Активность транск-
рипции зависит также от степени метилирования ци-
.. 5'-CG-3'
тозина в динуклеотидных парах основании
Подавление транскрипции часто бывает связано
с гиперметилированием ДНК, а активация-с ги-
пометилированием.
Здесь мы остановимся на некоторых из упомя-
нутых модификаций и рассмотрим их влияние на
экспрессию генов. Поскольку данные регуляторные
механизмы не являются ген-специфичными, а влия-
ют на экспрессию многих генов, мы назовем их
глобальными. Для иллюстрацйи участия нетран-
скрипционных механизмов в регуляции содержания
мРНК мы приведем два примера. В одном случае
речь идет о посттранскрипционном избирательном
разрушении мРНК, в другом- о новом способе
регуляции синтеза дрожжевого фактора транскрип-
ции на уровне трансляции.
а. Упаковка ДНК
Основной структурной единицей хроматина
эукариот является нуклеосома-сверхспиральный
сегмент ДНК длиной 145 п. н., намотанный с обра-
зованием двух левых витков на гистоновый окта-
мер. Последний состоит из четырех белков, Н2А,
Н2В, НЗ и Н4, каждый из которых представлен
в октамере дважды (разд. 1.1.ж). Нуклеосомы обра-
зуют фибриллы диаметром 10 нм при участии
гистона Н1, который связывается с ДНК на входе
в нуклеосому и на выходе из нее (рис. 1.17). Далее
эти фибриллы толщиной 10 нм скручиваются в спи-
раль из шести нуклеосом на виток с образованием
соленоида диаметром 30 нм (рис. 1.18). При кон-
денсации спиралей диаметром 30 нм образуются
еще более сложные структуры, однако детали их
строения неизвестны.
Электронно-микроскопические данные свиде-
тельствуют о том, что хроматин эукариот состоит
из отдельных участков-петель, или доменов, дли-
ной примерно 30-300 т. п. н., прикрепленных к белко-
вому матриксу (рис. 8.107). Каждая петля, по-ви-
димому, имеет одну точку начала репликации и
функционирует как отдельная система. Петли яв-
ляются автономными сверхспиральными единица-
ми, их топология не зависит от состояния соседних
петель. Возможно, это определяется тем, что концы
всех петель жестко закреплены в матриксе. Каждая
петля содержит множество единиц транскрипции, но
функционирование всех их, по-видимому, коорди-
нируется, т. е. вся петля находится либо в активном,
либо в неактивном состоянии.
Репрессированный хроматин. Большая часть ге-
нов многоклеточных организмов эукариот экспрес-
сируется только на определенных стадиях развития
и/или только в определенных тканях. Это означает,
что клетки должны располагать эффективными
средствами подавления транскрипции всех генов,
кроме тех, которые функционируют в клетках дан-
ного вида. Репрессия больших групп генов в прин-
ципе может осуществляться путем ограничения дос-
тупности необходимых факторов транскрипции, но,
по-видимому, такой механизм маловероятен. Об
этом свидетельствует тот факт, что гены, обычно не
экспрессирующиеся в клетках определенного типа,
могут экспрессироваться при введении их в клетки
путем трансфекции. Например, гены глобина после
трансфекции в фибробласты экспрессируются в 1000
раз эффективнее, чем эндогенные глобиновые гены
в составе хромосом; более того, это различие сохра-
няется при последовательных клеточных делениях
даже несмотря на то, что трансфицированные гены
встраиваются в хромосомы. Как правило (хотя и не
всегда), активность транскрипции генов, передан-
ных путем трансфекции, бывает на порядок выше,
чем у соответствующих эндогенных генов. Этот
феномен объясняют тем, что репрессия происходит
в результате изоляции молчащих генов в хрома-
тиновых структурах более высокого порядка, из-за
чего они становятся недоступными для комплексов
транскрипции.
Поскольку мы не располагаем детальной ин-
формацией о таких высокоупорядоченных хрома-
тиновых структурах, мы не можем описать процесс
перехода репрессированного хроматина в экспрес-
сирующийся. Один из подходов к решению этой
проблемы мог бы состоять в установлении корре-
ляции между периодом клеточного цикла, когда
происходит репликация конкретного гена, и его
способностью участвовать в транскрипции. Везде-
сущие гены «домашнего хозяйства», экспрессирую-
щиеся во всех тканях (например, гены дигидрофо-
латредуктазы, цитоплазматического актина и глю-
козо-6-фосфат-дегидрогеназы) реплицируются в
первой половине S-фазы, а неэкспрессируемые гены,
как правило, реплицируются позднее. Более того,
гены имеют тенденцию к ранней репликации в тех
тканях, где они экспрессируются, и к поздней - в тех
тканях, где они молчат. Аналогичным образом,
у млекопитающих неактивная Х-хромосома репли-
цируется позже, чем транскрипционно активная.
Такая закономерность говорит о том, что при ран-
ней репликации во время S-фазы ДНК, вероятно,
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
137
РИС. 8.107.
Хроматин эукариот состоит иэ петель разной длины,
концы которых прикреплены к белковому матриксу. На
микрофотографии слееа представлена метафаэная
хромосома, иэ которой выходят нити хроматина. Видны
петли, которые образуют эти нити, но места их при-
соедин ния к хромосоме не различаются. [W. С. Earnshaw,
U.K. Laemmli, J. Cell Biol. 96 (1983), p. 84.] На электрон-
ной микрофотографии справа показаны хромосомы.
обработанные декстрансульфатом и гепарином, очи-
щенные и расправленные на подложке при помощи
цитохрома. Обратите внимание, что концы петли,
по-видимому, выходят иэ соседних точек белкового
матрикса (внизу) [J. Я. Paulson, U.K. Laemmli, Cell 12
(1977), p. 817. Оба снимка любезно предоставлены
U. К. Laemmli.]
переходит в состояние, в котором она более доступ-
на для комплекса транскрипции или способна к бо-
лее эффективному связыванию важных факторов
транскрипции.
Чувствительность к действию нуклеаз. Судя по
более высокой чувствительности к растеплению
под действием ДНКазы I или микрококковой нукле-
азы, транскрипционно активные области хроматина
упакованы менее плотно, чем области, содержащие
неактивные гены. Такая чувствительность к нуклеа-
зам часто распространяется на сегменты длиной
несколько тысяч пар оснований, фланкирующие
единицу транскрипции. Структурные особенности,
лежащие в основе этой чувствительности, не уста-
новлены, но данные биохимических исследований
свидетельствуют о том, что нуклеосомы, содержа-
щие активные гены, характеризуются низким со-
держанием гистона Н1, обогащены ацетилирован-
ными и убиквитинсодержащими гистонами и свя-
заны с определенными белками, имеющими четко
выраженные кислотные свойства (HMG-белками).
Эти модификации, вероятно, влияют на связывание
гистонов с ДНК и способствуют нерегулярному
распределению нуклеосом, вследствие чего ДНК
становится более чувствительной к нуклеазному
расщеплению. Так, нормальное чередование сайтов
нуклеазного расщепления генов, кодирующих белки
теплового шока, меняется после тепловой индукции.
Аналогичное изменение профиля нуклеазного рас-
щепления наблюдается в случае овальбуминовых
генов в клетках яйцевода (но не печени) и генов
IgL(x) лимфоцитов после их перехода в транскрип-
ционно активную форму.
Гиперчувствительность к действию нуклеазы.
Помимо упомянутых участков нуклеазной чувст-
вительности активно транскрибируемые гены со-
138
ЧАСТЬ III, ОРГАНИЗАЦИЯ. ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
держат гиперчувствительные сайты, которые нахо-
дятся в пределах единиц транскрипции и вблизи них.
В очищенной ДНК такие сайты гиперчувствитель-
ности к ДНКазе I отсутствуют. Сайты гиперчувст-
вительности обычно располагаются непосредствен-
но перед точкой начала транскрипции гена, но
иногда они встречаются внутри единицы транскрип-
ции или на ее З'-конце; напомним, что в этих местах
могут находиться энхансер и терминатор транскрип-
ции. Более того, в некоторых генах сайты гипер-
чувствительности к ДНКазе I находятся на зна-
чительных расстояниях слева от сайта кэпирования
(например, на несколько тысяч пар оснований левее
кластера генов овальбумина птиц и более чем на
20 т. п. н. левее гена Р-глобина человека). В боль-
шинстве случаев сайты гиперчувствительности к
действию ДНКазы I присутствуют только в ДНК
тканей или клеток, в которых либо происходит,
либо запрограммирована экспрессия определенного
гена. Например, сайт гиперчувствительности на
5'-конце гена препроинсулина обнаружен в панкреа-
тических Р-островковых клетках поджелудочной
железы и отсутствует в клетках печени. Аналогично,
сайты гиперчувствительности имеются в перестро-
енных иммуноглобулиновых генах В-клеток, но от-
сутствуют в генах клеток некроветворных тканей.
Области, разделяющие нуклеосомы. Сайты ги-
перчувствительности к нуклеазам, вероятно, нахо-
дятся в областях, разделяющих нуклеосомы. Впер-
вые на это обратили внимание, когда обнаружилась
зависимость между положениями сайтов гиперчувст-
вительности и не содержащей нуклеосом областью
в геноме SV40. Такая область видна на электрон-
но-микроскопических фотографиях очищенных ми-
ни-хромосом SV40 (рис. 8.108). Она охватывает
участок между точкой начала транскрипции ранних
генов и 5'-концом энхансера. Если этот сегмент
длиной 400 п. н. транслоцировать в другие области
генома SV40 или в чужеродные плазмидные ДНК,
то в последних появляются области, не содержа-
щие нуклеосом, и сайты гиперчувствительности
к ДНКазе I. Отсюда можно сделать вывод, что оба
эти свойства, по-видимому, обусловливаются осо-
бенностями нуклеотидной последовательности дан-
ного участка ДНК. В то же время на гиперчувстви-
тельность межнуклеосомной области к действию
нуклеаз, вероятно, влияют негистоновые белки,
связанные с этой областью,- возможно, белки,
взаимодействующие с Z-ДНК (разд. 8.7.6), и фак-
торы транскрипции, ассоциированные с энхансером
(разд. 8.3.г).
Такие же сайты гиперчувствительности и сво-
бодные от нуклеосом области имеются в энхансерах
генов IgH и IgL(x) (разд. 8.3.д), а также в UAS-noc-
ледовательностях, ассоциированных с 5'-концом
генов GAL и металлотионеина (разд. 8.3.е). Сайты
РИС. 8.108.
Мини-хромосомы SV40, выделенные из ядер заражен-
ных клеток, окрашенные водным раствором уранилаце-
тата и сфотографированные при помощи электронного
микроскопа в темном поле. Четко видно, что в мини-
хромосомах имеется участок ДНК, свободный от нукле-
осом. [S. Saragosti, G. Moyne, М. Yaniv, Cell 20 (1980),
р. 65. Фотографии любезно предоставлены М. Yaniv.]
гиперчувствительности к нуклеазному расщеплению
появляются в глюкокортикоид-акцепторном элемен-
те (разд. 8.3.е) через несколько минут после введения
в клетки глюкокортикоида и исчезают вскоре после
удаления гормона. Возможно, сайты гиперчувстви-
тельности к ДНКазе I связаны также с активацией
транскрипции генов рРНК, поскольку такие сайты
встречаются как перед точками инициации тран-
скрипции, так и по краям спейсеров рРНК-генов.
Гиперчувствительность к нуклеазе стала своего
рода маркером областей регуляции транскрипции.
Поскольку положение областей регуляции тран-
скрипции согласуется с положением как сайтов ги-
перчувствительности, так и областей, свободных от
нуклеосом, можно предположить, что все эти эле-
менты взаимосвязаны. Возможно, присоединение
факторов транскрипции к соответствующим после-
довательностям приводит к вытеснению нуклеосом
или изменяет их упаковку, при этом образуются
свободные от нуклеосом участки, которые можно
визуализировать, или даже менее заметные нару-
шения, обеспечивающие нуклеазное расщепление.
Если, однако, плотноупакованные нуклеосомы ста-
билизированы. например, гистоном Н1 или путем
образования структур более высокого порядка, то
доступ белков транскрипции к сигнальным после-
довательностям затрудняется, и транскрипция бло-
кируется.
Структурные особенности активного и репресси-
рованного хроматина и механизм регуляции его
перехода из одной формы в другую во многом
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
139
остаются невыясненными. Нам известно только, что
участки хроматина могут находиться в конденси-
рованном, не экспрессируемом состоянии в течение
многих раундов репликации и переходить в менее
конденсированное, способное к экспрессии состоя-
ние под действием специфических экзогенных и эн-
догенных стимулов. Как происходит активация
участков хроматина- при помощи белков позитив-
ного контроля или в результате удаления белков-
репрессоров, а возможно, при участии и того
и другого еще предстоит определить.
б. Топология и конформация ДНК
Значение сверхспиральности для регуляции тран-
скрипции. Имеются достоверные данные о том, что
экспрессия прокариотических генов зависит от со-
стояния сверхспиральности ДНК. Факторы, влия-
ющие на этот параметр, мутации или ингибиторы
топоизомераз - активируют транскрипцию одних
генов и одновременно подавляют транскрипцию
других. Поскольку связывание белков со сверхспи-
ральной ДНК с отрицательными сверхвитками энер-
гетически выгодно, по-видимому, изменения состоя-
ния сверхспиральности ДНК сказываются на спо-
собности РНК-полимеразы или белковых актива-
торов и репрессоров связываться с регуляторными
последовательностями. Изменения в состоянии или
степени сверхспиральности могут отразиться и на
способности ДНК образовывать петли и, таким
образом, на возможности факторов транскрипции,
связанных с отдаленными друг от друга участками
ДНК, взаимодействовать между собой.
Как указывалось в разд. 8.7.а, у эукариот хро-
матин представляет собой систему петель разной
длины, прикрепленных к белковому матриксу (рис.
8.107). Возможно, такая конфигурация обеспечивает
независимость плотности сверхвитков в каждой
петле от торсионного состояния соседних петель.
Интересно, что ДНК-топоизомераза II, фермент,
ответственный за релаксацию сверхспиральной
ДНК (разд. 2.1.е), -это один из основных белков
хромосомного остова, к которому и прикреплены
петли. Более того, сайты гиперчуветви гельности
к ДНКазе I, сходные с участками регуляции тран-
скрипции, часто проявляют чувствительность к
нуклеазам, специфически расщепляющим одноцепо-
чечные ДНК; это наводит на мысль, что данные
области находятся в доменах с отрицательной
сверхспирализацией. Далее, топоизомераза I. по-ви-
димому, связывается с сайтами гиперчувствитель-
ности к нуклеазам. Влияние на транскрипцию (как
положительное, так и отрицательное) соединений,
интеркалирующих в ДНК или ингибирующих ак-
тивность топоизомераз и в обоих случаях изме-
няющих степень сверхспиральности ДНК, тоже го-
ворит о том, что топология ДНК важный фактор
регуляции транскрипции. Тем не менее у мутантов
дрожжей, не имеющих ДНК-топоизомеразы I или
II, транскрипция не нарушена. Это означает, что для
транскрипции достаточно одной из этих двух эука-
риотических ДНК-топоизомераз.
Большинство данных, указывающих на влияние
топоизомераз на топологию ДНК в процессе ре-
гуляции транскрипции у эукариот, носят косвенный
характер и неполны. Прямые доказательства того,
что изменения в плотности сверхвигков в хромо-
сомных петлях влияют на активность транскрипции
целой области или генов, расположенных в этой
области, фактически отсутствуют.
В- и Z-ДНК и их транскрипция. ДНК может
иметь как правоспиральную (В), так и левоспи-
ральную (Z) конформацию (разд. 1.1.в и рис. 1.8).
Соотношение между этими двумя формами зависит
от нуклеотидной последовательности ДНК и сте-
пени ее сверхспиральности, а также от ионной силы
раствора, температуры и содержания ионов метал-
лов. При определенных условиях право- и лево-
спиральные участки могут чередоваться, а после-
довательности в местах сочленений или рядом
с этими местами обладать необычными свойства-
ми субстратов для различных ферментов.
Образование Z-ДНК in vitro было подтверждено
при помощи нескольких физических методов, но
существование такой структуры in vivo до недавнего
времени подвергалось сомнению. Теперь эти формы
обнаружены как в про-, так и в эукариотических
генах. Как и ожидалось, содержание Z-ДНК in vivo
сильно зависит от степени сверхспиральности ДНК.
Ее образование облегчается также в тех участках,
где пурины и пиримидины чередуются. Таковы,
например, повторы d(CA)n • d(GT)n в середине мо-
лекул. Образованию Z-формы способствует и ме-
тилирование остатков С в местах чередования
С и G (т. е. 5'-CGCGCG-3'). Имеются также данные
о том, что последовательности, способные прини-
мать Z-конформацию, соответствуют сайтам гипер-
чувствительности к нуклеазе, находящимся в об-
ласти регуляции транскрипции некоторых генов.
Идентифицированы и очищены белки, специфич-
ные для Z-ДНК. Например, некоторые из этих
белков связываются с энхансером промотора ран-
ней области генома SV40. И все же, несмотря на
существование Z-ДНК-сегментов в области регу-
ляции транскрипции или по соседству с ней и бел-
ков, узнающих Z-ДНК, вопрос об участии этой
формы ДНК в регуляции транскрипции пока ос-
тается открытым. Известно об одном случае учас-
тия белка, связывающего Z-ДНК, в рекомбинации
ДНК, но и здесь конкретный механизм влияния
ДНК-связывающего белка и конформации на ре-
комбинацию остается неясным.
140
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
в. Метилирование ДНК
Установлено, что экспрессия генов позвоночных
коррелирует со степенью метилирования специфи-
ческих остатков цитозина внутри регуляторных об-
ластей генов и вокруг них. За исключением всего
нескольких важных случаев, при наличии метили-
рованных групп (гиперметилированне) эффектив-
ность генной экспрессии уменьшается, а часто
экспрессии вообще не происходит. И наоборот,
активное состояние генов обычно сочетается с
отсутствием метилирования или понижением сте-
пени метилирования (гнпометилирование) в тех же
самых сайтах.
Одним из загадочных аспектов метилирования
является то, что это состояние сохраняется в ряду
многих поколений клеток. Так, гены, экспрессирую-
щиеся тканеспецифичным образом, остаются гипер-
метилированными на протяжении всего периода
эмбрионального развития во всех соматических
тканях, за исключением одной, в которой данный
ген экспрессируется. И наоборот, области, содер-
жащие гены «домашнего хозяйства», которые экс-
прессируются в процессе развития во всех тканях,
остаются гипометилированными. Более того, пе-
реданные путем трансфекции неметилированные
гены обычно экспрессируются нерегулярно даже
после интеграции в хромосомы и последовательных
клеточных делений. Переданные путем трансфекции
метилированные ДНК, напротив, неспособны к
преходящей экспрессии или к экспрессии после
интеграции в хромосому. Замечательно, что интег-
рированные неметилированные ДНК остаются не-
метилированными и экспрессируются во многих
поколениях клеток даже тогда, когда соответст-
вующие клеточные гены метилированы и не экс-
прессируются.
Еще одно указание на связь между метилирова-
нием и экспрессией получено в опытах с ретро-
вирусом мышей. Этот вирус не может размножать-
ся и экспрессировать свои гены в эмбриональных
клетках мыши. Однако в том случае, когда эмбрио-
нальные клетки инфицируются после индукции их
дифференцировки, происходят и размножение ви-
руса, и экспрессия его генов. Одно из существенных
различий между этими клеточными системами
заключается в том, что интегрированная ДНК про-
вируса в недифференцированных клетках подвер-
гается метилированию и в дальнейшем сохраняет
это состояние, тогда как в индуцированных клетках
ДНК вируса не метилируется. Существование об-
ратной связи между активностью генов и состоя-
нием их метилирования и сохранение их статуса
в определенных тканях и на определенных стадиях
развития в значительной степени стимулировало
исследование механизма генной регуляции.
Метилирование специфических остатков цито-
зина. Как правило, метилирование происходит по
положению 5 остатков цитозина (5-шеС) в ди-
нуклеотиде 5'-CG-3'. (У высших растений иногда
метилируется крайний 5'-цитозин в тринуклеотиде
5'-CNG-3', где N-любой нуклеотид.) Напомним,
что у позвоночных динуклеотид CG встречается
с частотой примерно в пять раз меньшей, чем это
следует из нуклеотидного состава ДНК (разд. 1.1).
Так, при случайном распределении оснований и
GC-содержании ДНК млекопитающих 40% частота,
с которой встречаются CG-динуклеотиды, должна
быть равна примерно 0,04-такой же величине, как
и для динуклеотидов GC. Однако в действитель-
ности она составляет 0,008.
По-видимому, пониженное содержание динук-
леотидов CG и высокая частота их метилирования
взаимосвязаны. Возможно, такая корреляция явля-
ется следствием спонтанного дезаминирования ос-
татков цитозина в ДНК. Это дезаминирование лег-
ко обращается при помощи механизма репарации,
узнающего остатки урацила (разд. 2.3.а). Однако
при дезаминировании 5-теС образуется Т, который
нельзя превратить в С, поэтому несущественные
CG-динуклеотиды, подвергшиеся метилированию,
могут элиминироваться. Тот факт, что последова-
тельности, содержащие метилированные динуклео-
тиды CG в регуляторных областях, сохраняются,
вероятно, объясняется их особой ролью в экспрес-
сии генов.
У позвоночных метилированы от 50 до 90%
остатков С в динуклеотидах CG. Однако этот
процент неодинаков для разных тканей, разных
стадий развития и разных участков ДНК. Общую
степень метилирования можно оценить, определив
соотношение между 5-теС и общим содержанием
остатков цитозина. Однако большую ценность
представляют данные о метилировании специфи-
ческих участков ДНК. Их можно получить, исполь-
зуя две рестриктазы, которые различаются по своей
способности расщеплять ДНК в зависимости от
того, какой остаток содержит сайт расщепления:
С или 5-теС (разд. 4.2.д). Например, последова-
тельность 5'-CCGG-3' расщепляется обеими эндо-
нуклеазами, Hpall и MspI, но 5'-CmeCGG-3'- только
эндонуклеазой MspI. Наоборот, Hpall расщепляет
ДНК в сайте 5'-meCCGG-3', а MspI-нел. Сравнивая
рестриктные карты различных участков ДНК, по-
лученные с помошью Hpall и MspI, можно иден-
тифицировать сайты метилирования. Содержащие
5-теС CG-динуклеотиды, которые не входят в со-
став сайта рестрикции или которые не удается
выявить при помощи имеющихся эндонуклеазных
пар-изошизомеров, можно идентифицировать пу-
тем прямого секвенирования геномной ДНК с ис-
пользованием химических реагентов, не расщепля-
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
141
ющих ДНК в 5-теС-содержащих сайтах. В этом
случае при расщеплении ДНК по остаткам цитозина
и анализе «лестницы» фрагментов на радиоавто-
графе обнаруживается выпадение «ступеньки» в
позиции, где цитозин метилирован (разд. 7.2.в).
Механизмы метилирования de novo, сохранения
метилированного состояния и элиминации остатков
теС. Поскольку для разных тканей и разных стадий
развития характерны разные схемы метилирования,
возникает следующий вопрос: как эти схемы орга-
низуются, сохраняются и изменяются в процессе
развития клетки? Сам процесс метилирования и
сохранения метилирования-это одна сторона
проблемы, элиминация остатков теС- другая. Ме-
тилирование de novo происходит по двум динуклео-
тидам CG в противоположных цепях, а для под-
держания метилированного состояния необходимо
метилирование одного из двух CG-динуклеотидов
в полуметилированных последовательностях (рис.
8.109). При метилировании de novo появляются
новые схемы метилирования, а процесс сохранения
метилированного состояния заключается в моди-
фикации новосинтезированных цепей ДНК, обес-
печивающей наследование потомками родительской
схемы метилирования.
Поддержание метилированного состояния и метилиро-
вание ДНК de novo. При репликации полностью мети-
лированной ДНК образуется полуметилированная
форма с неметилированной новосинтеэированной цепью.
Метилаэы, поддерживающие метилированное состояние,
быстро метилируют неметилированные остатки С, рас-
положенные напротив метилированных С, и полностью
восстанавливают метилированное состояние ДНК.
Пунктирная стрелка с надписью «деметилирование» по-
казывает гипотетический путь полной элиминации ме-
тильных групп, хотя механизм этой реакции неизвестен.
Предполагается, что существуют de novo-метилаэы, ко-
торые и метилируют неметилированную ДНК в клетках.
Известно, что все метилаэы используют в качестве
донора метильной группы S-аденоэилметионин.
Радиактивно меченный дезоксинуклеозид
Уменьшение плотности раствора CsCI
РИС. 8.110.
Интенсивное деметилирование ДНК может происхо-
дить и в отсутствие синтеза новой ДНК. С помощью
гексаметиленбисацетамида была индуцирована диффе-
ренцировка лейкозных клеток мыши, синхронизирован-
ных в фазе G1 клеточного цикла. Через 16 ч были
добавлены 5-бромдеэоксиуридин и 5-фтордеэоксиури-
дин. 5-бромдезоксиуридин фосфорилируется с образо-
ванием соответствующего деэоксинуклеоэидтрифосфа-
та и включается в синтезируемую цепь ДНК. В резуль-
тате плотность новосинтеэированной ДНК оказывается
выше (в одной из ее цепей содержится тяжелый нук-
леозид), чем нереплицированной (ни одна из ее цепей
бром дезокси нуклеозид не содержит). Через 1 ч добав-
ляли меченые деэоксицитидин (С, левая часть рисунка)
или дезоксиаденозин (А, правая часть рисунка). Оба
этих нуклеотида включаются в новосинтеэированную
ДНК как в индуцированных (два верхних графика), так
и в неиндуцированных (два нижних графика) клетках.
Однако в индуцированных клетках, которые истощены
по 5-теС, в ДНК включается только меченый С. Сле-
довательно. С может заменять 5-теС в ДНК при помо-
щи механизма, отличного от синтеза ДНК. [A. Razin et
al.. Proc. Nat. Acad. Sci. U.S.A. 83 (1986), p. 2827.]
Механизм специфической или общей элимина-
ции теС неизвестен, предложены только разные его
модели. Одна из них предполагает прямую замену
метилированных остатков С неметилированными.
Эту гипотезу подтверждает тот факт, что диффе-
ренцировка in vitro некоторых лейкозных клеток,
индуцированная химическими реагентами, сопро-
вождается массовым деметилированием теС. При
этом в клеточную ДНК включается меченый дез-
оксицитидин, но не дезоксиаденозин (рис. 8.110).
Согласно второй модели, связывание белков, спе-
цифичных для определенных тканей или стадий
развития, с последовательностью, содержащей сайт
метилирования, блокирует работу метилаэы, под-
держивающей метилированное состояние, и после
цикла репликации этот сайт становится полностью
неметилированным. Согласно третьей модели, ме-
142
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
тилирование происходит в определенных CG-сайтах
в высококонденсированном хроматине, стабилизи-
руя таким образом транскрнпционно неактивную
форму ДНК, и элиминация теС происходит после
релаксации ДНК с переходом ее в активное состоя-
ние.
Изучение ферментов, ответственных за метили-
рование ДНК, только начинается. Очищены неко-
торые метилтрансферазы ДНК млекопитающих,
клонирован и секвенирован ген. кодирующий одну
из них. Мол. масса этих ферментов лежит в диапа-
зоне от 135 до 175 кДа. Роль донора метильной
группы выполняет S-аденозилметионин. Выделен-
ные ферменты метилируют полуметилированную
ДНК с гораздо большей эффективностью, чем
неметилированную. Следовательно, они относятся
к метилазам, поддерживающим метилированное
состояние. В опытах с использованием в качестве
субстрата части промотора гена р-глобина мыши
показано, что очищенная ДНК-метилтрансфераза
млекопитающих имеет сложную субстратную спе-
цифичность. Указанная часть промотора содержит
десять CG-динуклеотидов на участке длиной
368 п. н. Из них метилируются лишь пять или шесть
динуклеотидов, которые группируются на участке
из 29 п.н., находящемся примерно на 100 п.н. левее
точки начала транскрипции, причем они метили-
руются в разной степени и с разной скоростью.
По-видимому, на характер метилирования влияют
последовательности, окружающие динуклеотиды
CG, a in vivo-еще и состояние хроматина. Никакие
другие факторы, особенно регулирующие метили-
рование и деметилирование, пока не обнаружены.
Метилирование и экспрессия гена. Многочислен-
ные опыты с вирусными геномами, а также с гено-
мами эукариот невирусной природы тоже свиде-
тельствуют о том, что гиперметилирование корре-
лирует с ингибированием или инактивацией про-
моторов, тогда как промоторы, активно участвую-
щие в транскрипции, часто бывают гипометилиро-
ваны или не метилированы вообще. Эту корреля-
цию иллюстрируют опыты с клетками, трансфор-
мированными аденовирусом 12. ДНК аденовируса
12 не метилирована; после проникновения в клетки
хомяка она встраивается в клеточные хромосомы,
и одни ее гены экспрессируются, а другие-нет. Как
правило (хотя и не всегда), неэкспрессируемые гены
находятся в метилированном состоянии, а экспрес-
сируемые остаются неметилированными. Детальное
исследование области Е2А аденовируса показало,
что в линии НЕ1 трансформированных клеток она
нормально транскрибируется и транслируется, при
этом ни одна из 14 последовательностей 5'-CCGG-3'
области Е2А не метилирована (табл. 8.8). Однако
в линиях трансформированных клеток НЕ2 и НЕЗ
ген Е2А не экспрессируется, и все 14 CG-динуклео-
Таблица 8.8. Экспрессия и метилирование адено-
вирусного гена Е2А в трансформированных клет-
ках
Линия Присутствие Экспрес- Метили- Метилирова-
клеток интактного сия гена рование нис всех 13 CG
промотора Е2А 14 сайтов между парами
Е2А » 5'-CCGG-3'2’ нуклеотидов + 24 и -1603)
НЕ1 + + -
НЕ2 + - + +
НЕЗ + - + +
” Образование Е2А-мРНК и белка.
21 5-теС определяли при помощи дифференциального расщепле-
ния эндонуклеазами Hpall и Mspl.
31 5-теС во всех CG-динуклеотидах определяли при помощи сек-
венирования ДНК.
тидов метилированы. Используя уже упоминавший-
ся метод секвенирования геномной ДНК, можно
«просмотреть» все CG-динуклеотиды промотора
Е2А независимо от того, входят ли они в Hpall-
сайты. В линии клеток НЕ1, где промотор Е2А
активен, 13 CG-динуклеотидов между парами ос-
нований + 24 и — 160 находятся в неметилирован-
ном состоянии. В линиях клеток НЕ2 и НЕЗ тот же
промотор неактивен, а те же самые CG-последова-
тельности обеих цепей метилированы. Таким обра-
зом, для участка промотора Е2А длиной пример-
но 180 п.н. имеет место упомянутая выше корре-
ляция.
Метилирование гена Е2А in vitro метилазой
Hpali почти всегда приводит к ингибированию
транскрипционной активности при транзиентной
трансфекции или стабильной трансформации. На-
против, неметилированный промотор функциональ-
но активен в обоих случаях (рис. 8.111). Более того,
у интегрированной ДНК сохраняется характер ме-
тилирования, свойственный ДНК, передаваемой пу-
тем трансфекции. Аналогично, метилирование in
vitro при участии разных CG-специфичных метилаз
приводит к репрессии промоторов генов аденови-
руса Е1А, а-глобина, тимидинкиназы, аденинфос-
форибозилтрансферазы и IgL(x). Сайты метилиро-
вания, сопряженные с инактивацией, локализованы
в основном в области промоторов, но происходит
и избирательное метилирование более далеких ре-
гуляторных последовательностей. Активность по-
луметилированной ДНК промотора Е2А, получен-
ной путем гибридизации неметилированных оди-
ночных цепей с олигонуклеотидами, содержащими
метилированные последовательности 5'-CCGG-3',
заметно снижается, но полностью не утрачивается.
Нередко корреляция между метилированием
и репрессией оказывается более сложной, чем та,
о которой мы говорили выше. В качестве примера
рассмотрим метилирование участка ДНК человека
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
143
Промотор
Е2А CAT
н-еч------------в
Клеточные клоны
1 2 3
I I I
I I I
Экспрессия + + +
CAT
Метилирование_
промотора
СТ3
Мети паза t
Нрап * I
снз | сн3
Сн3
' >
Трансформация
клеток хомяка Клеточные клоны
12 3
I I I
Клонирование III
клеток ’
- - +
РИС. 8.111.
Промоторы, метилированные in vitro, остаются мети-
лированными и неактивными in vivo. ДНК, содержащую
промотор аденовируса Е2А, к которому пришита после-
довательность, кодирующая хлорамфеникол-ацетил-
трансферазу (CAT), метилировали in vitro при помощи
очищенной метилаэы Hpall. После введения метили-
рованных и неметилированных ДНК в клетки хомяка
исследовали экспрессию гена CAT и состояние метили-
рования промотора в независимых клонах. Знаки « + »
или «—» означают соответственно присутствие или от-
сутствие как метильных групп, так и экспрессии CAT.
размером 30 т.п.н., содержащего гены у-, 5- и
Р-глобина (рис. III.5), в клетках, в которых экспрес-
сировался тот или другой из указанных генов. Этот
участок содержит 17 сайтов узнавания для эндо-
нуклеаз рестрикции, активность которых зависит от
состояния метилирования динуклеотида CG: ///)«! I,
Hhal, Xhol и Sall (разд. 4.2.в и д). Продукты
расщепления геномной ДНК из разных клеток ана-
лизировали при помощи ДНК-блоттинга с исполь-
зованием соответствующих зондов. Полученные
данные свидетельствуют о том. что область генов
у-глобина практически не метилирована в линии
лейкозных клеток, в которых экспрессируются гены
у-глобина, и гипометилирована в предшественниках
эритроцитов из средней части печени плода, где
происходит наиболее активная экспрессия генов
у-глобина плода человека. Сайты, соседствующие
с другими глобиновыми генами, метилированы
в большей степени. Напротив, весь кластер глоби-
новых генов печени, лимфоцитов, спермы взрослого
человека и клеток мозга плода гиперметилирован.
Еще более сложная картина возникает при анализе
различий между индивидуальными сайтами расщеп-
ления. Так, некоторые сайты остаются неметили-
рованными даже в печени взрослого человека, тогда
как другие метилированы по крайней мере в неко-
торых предшественниках эритроцитов печени пло-
да. В плаценте, где глобиновые гены не экспресси-
руются, весь кластер этих генов гипометилирован.
Эти результаты говорят о том, что гипометили-
рование коррелирует с экспрессией генов и, воз-
можно, даже совершенно необходимо для нее, но
само по себе не является решающим фактором.
Корреляция между ингибированием транскрип-
ции и метилированием соответствующих регулятор-
ных областей выявлена для многих генов, но все же
не для всех. Ингибирование транскрипции может
происходить в результате нарушения связывания
факторов транскрипции при наличии остатков
5-теС. Например, метилирование CG-динуклеоти-
дов в промоторе ингибирует связывание главного
фактора поздней транскрипции аденовируса и тран-
скрипцию in vitro. Метилирование другого динук-
леотида CG расположенного на несколько осно-
ваний правее, на транскрипцию не влияет. Связы-
вание соответствующих факторов транскрипции
с промоторами генов аденовируса Е2 и тирозин-
аминотрансферазы млекопитающих также блоки-
руется при метилировании CG-динуклеотидов в
сайтах связывания факторов транскрипции. При-
сутствующий во многих индуцибельных и ткане-
специфичных генах сАМР-чувствительный элемент
(CRE) содержит CG-динуклеотид (5'-TGACGTCA-
3'). Метилирование промоторов, транскрипционная
активность которых зависит от одного или пары
CRE, всегда блокирует связывание фактора тран-
скрипции CRE и предотвращает транскрипцию со-
ответствующих генов как in vitro, так и in vivo. Все
эти наблюдения говорят о том, что метилирование
CG-динуклеотидов в участках связывания факторов
144
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
транскрипции может блокировать их связывание
и подавлять функцию промотора.
Исключением из этого правила является мети-
лирование динуклеотида CG в сайте связывания
Spl, 5'-GGGCGG-3' (разд. 8.3.г). Ни связывание Spl,
ни Spl-зависимая транскрипция как in vitro, так и in
vivo при метилировании не подавляются. Эта осо-
бенность Spl-сайтов, вероятно, определяется харак-
тером связывания белка и ДНК. Не исключено, что
она сформировалась в процессе отбора, поскольку
Spl участвует в транскрипции многих важных и пов-
семестно экспрессирующихся генов.
Еще одним указанием на участие метилирования
в регуляции экспрессии специфических генов служат
результаты опытов с 5-азацитидином (5-azaC). Этот
аналог цитидина, содержащий атом азота вместо
атома углерода в положении 5 пиримидинового
кольца, может включаться в ДНК во время репли-
кации, но не может метилироваться. Инкубация
клеток с 5-azaC сопровождается массовой элими-
нацией остатков 5-теС. Механизм этого процесса
пока неясен, но, судя по некоторым данным, остатки
5-azaC в ДНК ингибируют метилтрансферазу ДНК.
Массовая элиминация теС часто бывает временной,
но в некоторых типах клеток отдельные гены все
время остаются гипометилированными.
Наиболее замечательным свойством 5-azaC яв-
ляется его способность влиять на фенотип опре-
деленных клеток; в частности, он может изменять
состояние дифференцировки и влиять на индукцию
экспрессии специфических генов. В тех случаях,
когда репрессия эндогенных или переданных путем
трансфекции генов обусловливается их гипермети-
лированием, обработка клеток 5-azaC может при-
водить или к временной, или к стабильной де-
репрессии. По-видимому, транскрипционно актив-
ными становятся именно те гены, которые пере-
ходят в стабильное гипометилированное состояние.
Тем не менее некоторые гены, деметилирующиеся
в присутствии 5-azaC, не активируются или акти-
вируются только после дополнительной обработки.
Таким образом, несмотря на то что 5-azaC вы-
зывает временную массовую элиминацию остатков
теС генома, неясно, оказывает ли это изменение
прямое влияние на процесс активации. Не исклю-
чено, что элиминация остатков теС может проис-
ходить после событий, непосредственно участвую-
щих в активации экспрессии определенных генов.
Представлению о том, что метилированные
CG-динуклеотиды играют решающую роль в ре-
гуляции транскрипции, противоречат данные об
отсутствии теС в некоторых эукариотических ге-
номах. Так, теС не обнаружены в геномах дрожжей
и других грибов, а также у беспозвоночных, таких
как Drosophila и нематода C.elegans. При этом у
данных организмов имеются сложно организован-
ные хромосомы, и экспрессия их генов зависит от
стадии развития организма и от внешних факторов.
Таким образом, вопрос о роли метилирования ДНК
у позвоночных остается нерешенным.
Неметилированные кластеры CG. Большинство
CG-динуклеотидов в ДНК позвоночных находится
в метилированном состоянии, однако в геноме
встречаются и кластеры неметилированных CG.
Так, примерно 1% фрагментов ДНК позвоночных,
образующихся после расщепления ее эндонуклеазой
Hpall-ферментом, который разрезает неметилиро-
ванные последовательности 5'-CCGG-3',- состав-
ляют низкомолекулярные фрагменты средней дли-
ны 120 п.н., называемые мелкими ТУрдИ-фрагмен-
тами (HTF-от англ. Hpall tiny fragments).
Блот-гибридизация продуктов расщепления ге-
номной ДНК с HTF-зондом показала, что боль-
шинство гибридизовавшихся фрагментов находятся
в «островках» с неметилированными CG-динуклео-
тидами длиной 1-2 т.п.н. Всего, по-видимому,
имеется около 30000 таких островков, расположен-
ных в среднем на расстоянии 100 т.п.н. друг от
друга. Такие кластеры неметилированных CG на-
зываются CG- или HTF-островками. Их общими
свойствами являются: 1) более высокое, чем в ос-
тальной ДНК, среднее GC-содержание; 2) частота
встречаемости CG-динуклеотидов, соответсвуюгцая
GC-содержанию в островке; 3) отсутствие метиль-
ных групп в большинстве CG-динуклеотидов.
Около 70% CG-островков связаны с генами
«домашнего хозяйства», остальные встречаются
в тканеспецифичных генах (табл. 8.9). Примеры
генов, не содержащих CG-островков, приведены
в табл. 8.10. CG-островки, как правило, находятся
в тех областях, где начинается транскрипция (рис.
8.112). Так, почти у всех (а возможно, и у всех генов
домашнего хозяйства, транскрибируемых РНК-по-
лимеразой И) на 5'-концах имеются CG-островки.
Длина CG-островков примерно одинакова во всех
генах, независимо от их размеров (рис. 8.113). Ост-
ровки могут частично перекрываться с транскри-
бируемым участком, обычно с первым экзоном или
интроном.
Ни происхождение CG-островков, ни их функции
пока неизвестны, хотя уже высказано множество
интересных гипотез. Например, можно предполо-
жить, что изобилие динуклеотидов CG в местах
начала транскрипции многих генов есть результат
их защищенности от метилирования благодаря свя-
зыванию с факторами транскрипции. CG-динуклео-
тиды в других частях генома не защищены и могут
утрачиваться при метилировании и дезаминирова-
нии с превращением в TG, что объясняет их повы-
шенную концентрацию на 5'-концах генов. Согласно
этой гипотезе, островки не обязательно выполняют
какую-то специальную функцию: их метилирование
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
145
Таблица 8.9. Гены, ассоциированные с CG-остров-
ками п
Ген Вид
Гены ферментов метаболических путей
Гипоксантин-гуанозин - фосфорибозил-
трансфераза Мышь, человек
Аденин-фосфоробозилтрансфераза Мышь, хомяк
Дигидрофолатредуктаза Мышь, человек
Глюкозо-6-фосфат- дегидрогеназа Человек
Г идроксиметилглутарил-СоА-
редуктаза »
Глицеральдегид-З-фосфат-
дегидрогеназа Курица
Триозофосфатизомераза Человек
Аденозиндезаминаза »
Фосфоглицераткиназа »
Индуцибельные клеточные гены
Металлотионеин I Мышь
Супероксиддисмутаза Человек
Тепловой шок »
Клеточные копии вирусных онкогенов
с-1пН »
с-тус Человек, мышь
c-src Курица
c-fos Мышь
c-Ha-ras Человек
Гены структурных клеточных белков
Белок рибосом L-30 Мышь
Белок рибосом L-32 »
Р-Актин Крыса
Р-Тубулин Крыса, человек
Белок гистосовместимости
класса I Человек, мышь
Гены тканеспецифичных белков
а2(1у-коллаген Курица
Ретинолсвязывающий белок Человек
а-Глобин Человек, коза
Thy-I Человек, мышь
Белок гистосовместимости класса II Человек
11 Этот неполный перечень включает гены, секвепированные с их
непосредственным окружением, в которых обнаружены HTF-no-
добные области. Гены гипоксантин-гуанозин фосфорибозилтранс-
феразы и глюкозо-6-фосфат дегидрогеназы сцеплены с Х-хромо-
сомой, при этом в неактивной Х-хромосоме островки метилиро-
ваны. 5'-конец гена глюкозо-6-фосфат дегидрогеназы не иденти-
фицирован. но в гене есть HFT-подобные последовательности
вблизи З'-конца. [A. Bird, Nature 321 (1986), р. 209.]
предотвращается только потому, что они связаны
с одним или несколькими белками. По другой ги-
потезе, кластеры неметилированных CG образуют-
ся благодаря особенностям строения хроматина,
обеспечивающим возможность взаимодействия
CG-богатых участков с элементами комплекса тран-
скрипции. Это означает, что метилирование исклю-
чает связывание белковых факторов с неактивными
островками, а связывание белковых факторов ис-
ключает метилирование активных островков. Обе
гипотезы предполагают, что CG-островки играют
Таблица 8.10. Гены, в которых не обнаружены
CG-островки п
Ген Вид
Семейство р-глобиновых Человек, мышь,
генов кролик, коза
Ген плацентарного лактогена Человек
Ген фактора роста нервной ткани »
Гены а-амилазы I и II Мышь
Гены гормона роста »
Ген инсулина »
Ген а-интерферона »
Ген фибриногена Крыса
Ген 8-кристаллина Мышь
Ген миоглобина Тюлень
п В этот неполный перечень входят секвенированные гены с низ-
ким содержанием островков CG как в самом гене, так и во
фланкирующих его последовательностях. [A. Bird, Nature 321 (1986), р. 209.]
важную роль в экспрессии соответствующих генов.
Скрупулезной проверки этих гипотез не проводи-
лось, однако имеются некоторые факты в их под-
держку. При трансфекции в клетки генов, содер-
жащих CG-островки, те CG-нуклеотиды, которые
находятся в островках, остаются неметилирован-
ными, в отличие от динуклеотидов CG, находя-
щихся во фланкирующих ген участках. Более того,
если CG-динуклеотиды островков предварительно
метилировать in vitro, а затем ввести в клетки путем
трансфекции, то экспрессия генов нарушится, а ост-
ровки останутся метилированными и в дальнейшем.
Как мы уже говорили, CG-островки обнаружены
в генах, экспрессия которых происходит тканеспе-
цифичным образом (табл. 8.9). Более того, эти
области ДНК остаются неметилированными как
в тех клетках, где ген экспрессируется, так и в тех,
где он молчит. Это подкрепляет предположение, что
одного только присутствия неметилированных клас-
теров CG в промоторной области гена недостаточ-
но для его транскрипции в соответствующих клет-
ках. Для дифференциальной экспрессии генов в раз-
личных тканях необходимы тканеспецифичные фак-
торы позитивного контроля или факторы негатив-
ного контроля, связывающиеся с 5-теС.
Метилирование и процессы развития. Связь ме-
тилирования с важнейшими процессами развития
у млекопитающих иллюстрируют два поразитель-
ных феномена: инактивация Х-хромосомы у самок
и дифференциальный импринтинг аутосом, получен-
ных от отца и матери.
У плацентарных млекопитающих инактивация
одной из двух хромосом в клетках самок обеспечи-
вает эквивалентное распределение информации, со-
держащейся в Х-хромосоме, у ХХ-самок. Инакти-
вация Х-хромосомы осуществляется в три этапа на
146
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
йш----а—г —выы
I "I I UI'HIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIH1 'Н 1 11 I I I 1 “III II!111
2
I
6
РИС. 8.112.
Распределение метилированных и неметилированных
динуклеотидов CpG в модельном гене позвоночных.
Такой ген содержит экзоны (выделены цветом), интро-
ны (выделены точками) и промотор слева от сайта
инициации транскрипции (верхняя часть рисунка). Вер-
тикальные линии под изображением гена указывают
положение CpG в самом гене и во фланкирующих его
последовательностях. Метилированным CpG соответст-
вуют цветные линии, а неметилированным-черные.
ранних стадиях эмбриогенеза (рис. 8.114). Сначала
из двух Х-хромосом выбирается одна, предназна-
ченная для инактивации. Затем инактивация рас-
пространяется на всю хромосому. Наконец, моди-
фицированное состояние стабилизируется, так что
выбранная хромосома остается неактивной в пос-
ледующих поколениях клеток. Инактивация хромо-
Кластер неметилированных CpG находится в области
промотора, а островок CpG-на Б'-конце. Положение
всех CpG можно установить методом секвенирования.
С помощью эндонуклеаз рестрикции удается локали-
зовать лишь некоторые из этих островков, поскольку не
все CpG-содержащие последовательности можно рас-
щепить известными ферментами. [А. Р. Bird, Trends
Genet. 3 (1987) р. 342.]
сомы происходит не одновременно во всех эмбрио-
нальных клетках. Примерно на четвертый день
после имплантации зародыша Х-хромосома, полу-
ченная от отца, избирательно инактивируется в
клетках, участвующих в формировании неэмбрио-
нальных тканей. Через шесть дней внутриматочного
развития одна из двух Х-хромосом, выбранная
Шесть генов млекопитающих со сходной локализацией
и размерами CG-островков. На рисунке показаны про-
моторы, сайты инициации транскрипции (волнистые
стрелки), экзоны (цветные полосы) и интроны (выде-
лены точками). Цветным прямоугольником указаны об-
ласти, в которых CG-динуклеотиды обычно не метили-
рованы-так называемые CG-островки. Метилирован-
ные CG-динуклеотиды обычно находятся вне этой об-
ласти. dhfr-ген дигидрофолатредуктазы мыши; hprt-
гвн гипоксантин-фосфорибозилтрансферазы мыши;
гЛу-1-ген поверхностного гликопротеина мыши thy-1;
rpl- ген рибосомного белка мыши L-32; aprt- ген аде-
нинфосфорибозилтрансферазы мыши. [A. Bird, Trends
Genet. 3 (1987), р. 342.]
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
147
Пара Х-хромосом в
эмбрионе женского пола
до имплантации
Случайный выбор одной
хромосомы для первичного
акта инактивации
Ранний постимплан-
тационный эмбрион
I
Молчащие
гены
Экспрессирую-
щиеся гены
Экспрессирую-
щиеся гены
РИС. 8.114.
Инактивация одной из двух Х-хромосом у млекопи-
тающих. В тканях эмбриона до его имплантации обе
хромосомы активны и не метилированы. После имплан-
тации экспрессия генов одной из двух хромосом пре-
кращается, в то время как другая хромосома остается
транскрипционно активной. По-видимому, инактивация
предшествует интенсивному метилированию, которое
происходит несколько позже. Обработка клеток, содер-
жащих активную и неактивную хромосомы, азацитиди-
ном приводит к интенсивной элиминации остатков
5-теС из неактивной хромосомы и к активации экспрес-
сии ее генов.
случайно, инактивируется во всех клетках, пред-
назначенных для плода. Таким образом, плод
женского пола развивается как мозаичный орга-
низм, в клетках которого экспрессируются либо
отцовские, либо материнские Х-хромосомы. У
анеуплоидных особей, имеющих более двух Х-хро-
мосом, инактивируются все Х-хромосомы, кроме
одной.
Инактивированная Х-хромосома в интерфазных
ядрах имеет вид высококонденсированной гетеро-
хроматиновой структуры (тельце Барра). В отличие
от активной гомологичной хромосомы, неактивная
Х-хромосома реплицируется в конце S-фазы. Почти
все гены, кодируемые неактивной Х-хромосомой,
теряют способность к экспрессии. Неактивное со-
стояние очень стабильно, и только в редких случаях
происходит реактивация отдельных областей. При
нормальном развитии реактивация осуществляется
только в зародышевых клетках непосредственно
перед мейозом. Однако активацию генов Х-хро-
мосомы можно индуцировать, обработав клетки
5-azaC (рис. 8.114). Это означает, что инактивация
Х-хромосомы может обусловливаться метилирова-
нием ДНК. Об этом свидетельствуют некоторые
косвенные данные, однако прямые доказательства
того, что метилирование ДНК является причиной,
а не следствием инактивации, отсутствуют.
Одно из косвенных доказательств основано на
том, что трансформировать клетки мыши с фе-
нотипом hprt~ к фенотипу hprt+ может только
ДНК. hprt-ren которой находится в активной
Х-хромосоме (аХ). Однако после обработки 5-azaC
ДНК из прежде неактивной Х-хромосомы (iX)
приобретает способность трансформировать клетки
hprt~ в hprt+. Еще одна группа данных в пользу
упомянутой гипотезы была получена при исследо-
вании сайтов метилирования гена hprt в аХ- и
iX-хромосомах. В целом профиль метилирования
CG-содержащих последовательностей в гене hprt
аХ-хромосомы сходен во всех тканях и характерен
для других генов «домашнего хозяйства» в аХ-хро-
мосоме и в аутосомах. Так, сайты, которые нахо-
дятся в области, окружающей первый экзон гена
hprt, в аХ-хромосоме полностью неметилированы.
Напротив, те же сайты в iX-хромосоме почти цели-
ком метилированы. После обработки клеток 5-azaC
профиль метилирования iX-хромосомы становится
похожим на профиль метилирования аХ-хромосо-
мы. Однако исследование кинетики инактивации
Х-хромосомы и изменения профиля метилирования
гена hprt на определенных стадиях постимпланта-
ционного развития показало, что эти два процесса
не коррелируют. Специфические сайты гена hprt не
метилируются в большинстве эмбриональных кле-
ток в течение нескольких дней после инактивации
Х-хромосомы. Следовательно, метилирование, по-
148
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
видимому, не определяет выбор хромосомы для
инактивации и не является причиной прогрессирую-
щей инактивации хромосомы на ранних стадиях
(рис. 8.114), а служит вторичным механизмом
поддержания неактивного состояния.
Геномный импринтинг означает дифференциаль-
ную модификацию материнского и отцовского ге-
нетического материала, входящего в зиготу, кото-
рая опосредует дифференциальную экспрессию ро-
дительских аллелей в процессе развития и у взрос-
лых особей. У мышей характерная модификация
материнского и отцовского геномов, а также, ве-
роятно, специфических генов необходима для нор-
мального развития эмбриона и плода.
Природа изменений импринтинга, а также ме-
ханизмы их возникновения, сохранения и исчезно-
вения неизвестны. Интересно, однако, что содержа-
ние 5-теС в ДНК сперматозоида и ДНК яйцеклетки
различно. Это наводит на мысль, что половые
различия в степени метилирования ДНК, проявля-
ющиеся при гаметогенезе, могут вносить вклад
в процесс импринтинга. С этим согласуются данные
о том, что экспрессия и метилирование трансгенов
зависят от пола родителя-донора. Так, трансген,
находящийся под контролем вирусного промотора,
экспрессируется, если он получен от отца, но только
в тканях сердца; если же этот ген получен от матери,
он остается молчащим во всех тканях (табл. 8.11).
Определение относительной чувствительности транс-
генов к расщеплению эндонуклеазами Hpall и MspI
в сайтах 5'-CCGG-3' показало, что между экспрес-
сией трансгена и степенью его метилирования су-
ществует четкая связь. Если трансген унаследован
от отца, то он гипометилирован и экспрессируется,
а если от матери, то гиперметилирован и молчит.
Импринтинг устанавливается во время гамето-
генеза или до него и стабильно сохраняется при
делении соматической клетки во многих поколе-
ниях. Он исчезает в линии зародышевых клеток,
Таблица 8.11. Зависимость активности трансгена
и степени его метилирования от источника транс-
гена ”
Родитель. от которого получен трансген Число мышей с указанными свойствами
ген экспресси- руется ген не экспресси- руется метилирование
гипо гипер
Мать 0 42 0 59
Отец 20 0 50 0
11 Ход эксперимента описан в тексте. Число животных, использо-
ванных при определении экспрессии и степени метилирования
ДНК, различалось. [J. L. Swain et al,, Cell 50 (1987), p. 719.]
чтобы снова дифференциальным образом восста-
новиться в сперматозоиде и яйцеклетке (рис. 8.115).
Геном яйцеклетки в значительной степени гипо-
метилирован, а геном сперматозоида относительно
гиперметилирован. Во время предимплантацион-
ного состояния зародыша оба этих набора хромо-
сом частично деметилируются, затем, в процессе
развития плода, родительские хромосомы интен-
сивно, но дифференцированно метилируются. О ме-
ханизмах. последствиях и регуляции процесса мети-
лирования-деметилирования у двух полов ничего не
известно. Возможно, в этом случае метилирование
тоже представляет собой некий вторичный процесс
по отношению к более ранней стадии импринтинга
и служит только способом стабилизации конкрет-
ного состояния.
г. Регулируемая утилизация мРНК
О механизмах, регулирующих стабильность
мРНК и ее способность к трансляции, известно
сравнительно мало, хотя оба они существенным
образом влияют на экспрессию генов. Важность
процессов, при помощи которых осуществляется это
влияние, иллюстрируется несколькими хорошо изу-
ченными примерами.
Влияние концевых последовательностей на ста-
бильность мРНК. Стабильность эукариотических
мРНК в значительной степени зависит от наличия
особых последовательностей в нетранслируемых
участках мРНК. Так, poly(A) в комплексе с белком
увеличивает стабильность мРНК, защищая ее 3'-
концы от экзонуклеазного расщепления. Мутации
в сигнальной последовательности, блокирующие
полиаденилирование (разд. 8.3.в). приводят к обра-
зованию очень нестабильных транскриптов. Кроме
того, удаление ро1у(А)-хвоста у зрелых мРНК со-
провождается их быстрой деградацией после вве-
дения в ооциты Xenopus или в клетки млекопи-
тающих, а восстановление этих хвостов восстанав-
ливает и стабильность, характерную для мРНК.
Имеются данные о том, что последовательные цик-
лы трансляции мРНК сопровождаются постепен-
ным укорочением ро1у(А)-хвостов, так что на ка-
ком-то этапе З'-концы мРНК становятся чувстви-
тельными к экзонуклеазному расщеплению. Таким
образом, способность мРНК к трансляции, вероят-
но. коррелирует с ее стабильностью. Стабильность
гистоновых мРНК, у которых ро1у(А)-хвосты, как
правило, отсутствуют, обусловливается образова-
нием на их З'-концах структур типа шпильки (разд.
8.3.В).
Недавно в З'-нетранслируемых участках не-
которых эукариотических мРНК была обнаруже-
на специфическая последовательность, способст-
вующая их деградации. Эта последовательность,
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
149
Спермаюзоид
Яйцеклетка
Отцовским
'отпечаток
Материнским
отпечаток"
Два уровня дифференциаль-
ного импринтинга?
Отец мать
Оплодотворение
Гаметогенез
Стирание следов импринтинга
'Преимлантацня
тДеметилиро-
I вание
Бластоциста
Линия \
зародышевых \
клеток X.
Метилирование
tie novo не происходит
/ Метилирование\
X tie novo /ТХ
Оплодотворенное яйцо на Гр мт
стадии, предшествующей
гаструле неэмбриональные ткани
Низкая степень метилирования
Взрослая особь
РИС. 8.115.
Модель дифференциального метилирования
хромосом в гаметах и соматических клетках в
процессе развития млекопитающих. Степень ме-
тилирования ДНК на разных стадиях развития
изображена при помощи разных цветовых оттен-
ков: чем интенсивнее цвет, тем она выше. Р и
М- мужское и женское начало соответственно
Однородная окраска сегментов Р и М не означает,
что каждый из них одинаково гипер- или гипо-
метилирован. ДНК клеток, которым суждено
стать клетками зародышевой линии, в основном
не метилирована, но в ходе гаметогенеза ядерная
ДНК сперматозоидов и яйцеклеток метилируется,
хотя и по-разному. В результате оплодотворен-
ное яйцо содержит ДНК двух типов: у одной ДНК
степень метилирования характерна для сперма-
тозоидов, у другой-для яйцеклеток. На этапах
предимплантации и имплантации метильные
группы в ДНК как отцовского, так и материнского
происхождения утрачиваются, а при дальнейшем
развитии оплодотворенного яйца до стадий плода
и взрослой особи происходит специфическое ме-
тилирование хромосом, унаследованных от отца
и матери. [М. Monk, Genes and Devel. 2 (1988),
p. 921.]
J
5-AUUUA-3', встречается многократно перед сиг-
налом полиаденилирования некоторых коротко-
живущих мРНК (рис. 8.116). К последним относится
ряд мРНК, кодирующих клеточные белки, которые
регулируют рост и дифференцировку клеток, а так-
же мРНК, транскрибируемые с нормальных вариан-
тов некоторых клеточных и вирусных онкогенов.
Для всех этих мРНК характерна относительная
метаболическая нестабильность, вследствие чего
кодируемые ими белки оказывают свое физиологи-
ческое действие сравнительно недолго.
Делеция AU-богатого участка нормального кле-
точного гена c-fos повышает равновесную концент-
рацию соответствующей мРНК и превращает его
в онкоген. И наоборот, вирусный онкоген v-fos
можно превратить в неонкогенный, заменив его
UAAUAUUUAUAUAUUUAUAUUUUUAAAAUAUUUAUUUAUUUAUUUAUUUAA
UAUUUAUUUAUUUAA
UUUUGAAAUUUUUAUUAAAUUAUGAGUUAUUUUUAUUUAUUUAAAUUUUAUUUUGGAAAA
UAUUUAUUAAUAUUUAACAUUAUUUAUAU
UUAUUUUUUAAUUAUUAUUUAUAUAUGUAUUUAUAAAUAUAUUUAAGAUAAUUAUAAUAU
UAUUUAUUUAAAUAUUUAAAUUUUAUAUUUAU
GUUUUUAAUUUAUUUAUUAAGAUGGAUUCUCAGAUAUUUAUAUUUUUAUUUUAUUUUUUU
UAAUUUUUUUUAUyUAAGUACAUUUUGCUUUUUAAAGUUGAUUUUUUUCUAUUGUUUUUA
GM-CSF
IFNa
IFN0
if Nt
IL1
IL2
foS
myc
РИС. 8.116.
В З'-нетранслируемом участке некоторых мРНК лим-
фокинов и ядерных факторов транскрипции человека
часто встречаются AU-богатые последовательности,
способствующие деградации этих мРНК. В некоторых
мРНК последовательности AUUUA (подчеркнуты) пе-
рекрываются (черта сверху). Представленные мРНК че-
ловека кодируют следующие белки: GM-CSF один из
колонийстимулирующих факторов; IFNa, IFNp и IFNy-
различные классы интерферонов; IL1 и IL2-интерлей-
кины; fos и тус-белки c-fos и с-тус человека соот-
ветственно. [G. Shaw, R. Kamen, Cell 46 (1986), р. 6Б9.]
150
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
З'-конец соответствующим участком неоикогенного
гена c-fos. Вероятно, способность индуцировать
образование опухолей обусловлена повышением
содержания продукта трансляции долгоживущих
мРНК. Еще одно подтверждение дестабилизирую-
щего влияния AU-богатого участка получено в опы-
тах, в которых стабильные мРНК, например Р-гло-
биновая мРНК, становятся нестабильными после
включения в их З'-концевые участки одного или
нескольких AU-богатых повторов (рис. 8.117).
Каким образом AU-богатые последовательности
дестабилизируют мРНК-неизвестно. Возможно,
с этими последовательностями связывается какой-
то белок, в результате чего они становятся чувстви-
тельными к экзонуклеазному расщеплению. Есть,
однако, указания на то, что AU-богатые последо-
вательности оказывают свое действие, увеличивая
скорость укорочения ро1у(А)-хвостов.
Данные о влиянии 5'-нетранслируемых последо-
вательностей на стабильность мРНК весьма фраг-
ментарны. Равновесное содержание гистоновых
мРНК и синтез большинства гистоновых белков
тесно связаны с синтезом ДНК во время S-фазы
клеточного цикла. В конце S-фазы содержание гис-
тоновых мРНК уменьшается. Эта избирательная де-
стабилизация, следующая за прекращением синтеза
ДНК, является посттранскрипционной и в случае ги-
стона НЗ опосредуется участком 5'-нетранслируе-
мой области мРНК. Механизм дифференциальной де-
стабилизации гистоновой мРНК неизвестен, но, воз-
можно, какую-то роль здесь играет белок, который
образуется только после окончания синтеза ДНК.
Регуляция стабильности мРНК рецептора транс-
феррина, опосредуемая железом. Ярким примером
регулируемой деградации мРНК является опосре-
дованная железом регуляция содержания мРНК ре-
цептора трансферрина (TfR). В пролиферирующих
клетках млекопитающих железо влияет на синтез по
крайней мере двух белков, играюших ключевую
роль в его метаболизме. Поглощение железа клет-
кой зависит в первую очередь от белка TfR. Синтез
этого белка подавляется при избытке железа и сти-
мулируется при его недостатке (табл. 8.12). Ферри-
тин-это белок, участвующий в запасании железа
внутри клетки. При поступлении железа в клетку
в достаточном количестве он образуется с более
высокой скоростью, чем при недостаточном. Обра-
зование обоих упомянутых белков регулируется
с помощью железа, однако механизмы регуляции
различаются. В первом случае регуляция состоит
в изменении скорости разрушения TfR-мРНК, а во
втором-в изменении скорости трансляции ферри-
тиновой мРНК. Несмотря на различие этих двух
процессов регуляции, в них участвуют общие ре-
гуляторные элементы. Мы рассмотрим вначале
опосредованную железом регуляцию содержания
TfR-мРНК, а затем регуляцию трансляции ферри-
тиновой мРНК.
После трансфекции в клетки млекопитающих
клонированной кДНК TfR, сшитой с промотором
SV40, содержание TfR-мРНК регулируется железом,
хотя транскрипция находится под контролем про-
Кодиру кидая
Промотор последовательность
Нестабильная
мРИК
3-UTR
GQQQZQ_____L Ген h-IFN-p
ЖШАШдД С
*са°А6 {_______8^ гло^мна
Стабильная
МРНК
р-глобина
(. ГТу ( ( ( ( ( НЖЙН Q Гибридный
( ГТ ( ( QXSSSBSО р-глобина
Нестабильная^
р-глобина
РИС. 8.117.
AUUUA-последовательности в З'-нетранслиру-
емых участках мРНК способствуют их быстрой
деградации. мРНК, содержащие одну или не-
сколько таких последовательностей, быстро раз-
рушаются. Об этом свидетельствуют результаты
опыта, в котором последовательность AUUUA из
очень нестабильной мРНК Р-интерферона чело-
века (h-IFN-P) встраивали в З'-концевую область
стабильной мРНК [3-глобина. Полученная гиб-
ридная мРНК р-глобина оказывалась такой же
нестабильной, как и мРНК IFN-p. Бледно окра-
шенные участки-это экзоны, а участки, выделен-
ные точками,- интроны. Дестабилизирующая по-
следовательность РНК и кодирующий ее сегмент
гена выделены одинаковым цветом. Пунктирные
линии в мРНК-вырезанные интроны.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
1Б1
Таблица 8.12. Регуляция синтеза белков-рецеп-
тора трансферрина и ферритина- при участии
железа
Условия Рецептор трансферрина Ферритин
белок мРНК бело к мРНК
Дефицит железа 1 1 1 Не меняется
Достаточное количество железа 1 1 т Не меняется
мотора SV40, который на железо не реагирует.
Деления нетранслируемого участка на З'-конце
кДНК приводит к тому, что содержание мРНК
перестает зависеть от железа. Более того, если
нетранслируемый участок З'-конца гена TfR ввести
в нетранслируемые области на З'-концах разных
кДНК, то содержание их мРНК будет изменяться
под влиянием железа. Таким образом, наличие не-
транслируемого участка на З'-конце TfR-мРНК яв-
ляется необходимым и достаточным условием для
регуляции стабильности этой мРНК под действием
железа.
Чувствительный к железу элемент TfR-мРНК
(IRE) занимает примерно треть (680 нуклеотидов)
З'-нетранслируемого участка. Он имеет специфичес-
кую нуклеотидную последовательность, образую-
щую пять сходных по структуре шпилек (рис. 8.118).
Все шпильки содержат следующие элементы: I) пет-
лю CAGA/VGN; 2) верхний стебель из пяти пар
оснований; 3) неспаренный остаток С на расстоянии
пяти пар оснований от 5'-конца петли; 4) нижний
стебель разной длины. Этот характерный структур-
ный домен встречается в нетранслируемой З'-обла-
сти TfR-мРНК всех видов, имеющих данный ген.
Ни одна из петель по отдельности, ни первые две
или последние три петли вместе не обеспечивают
опосредованную железом регуляцию содержания
TfR-мРНК. Для этого необходимы кооперативные
взаимодействия нескольких таких структур.
Экстракты цитозоля клеток, выращенных при
дефиците железа, содержат белок, специфически
связывающийся с IRE (IRE-BP). При избытке же-
леза содержание функционально активного IRE-BP
уменьшается. Согласно одной из гипотетических
моделей регуляции IRE-зависимой деградации
TfR-мРНК под действием железа, связывание
1RE-BP (или его комплекса с другими белками)
с кластером шпилек защищает IRE или примы-
кающие к нему участки от действия эндонуклеазы
(рис. 8.119). Полагают, что первоначальное эндо-
нуклеазное расщепление приводит к быстрому эк-
зонуклеазному расщеплению остальной части мо-
РИС. 8.118.
Шпильки с петлями в мРНК Н-цепи ферритина и ре-
цептора трансферрина. Левая шпилька находится в
5'-нетранслируемой области (Б'-UTR) мРНК Н-цепи
ферритина человека, а остальные пять (А- Е) располо-
жены одна за другой в З'-нетранслируемой области
(З'-UTR) мРНК рецептора трансферрина человека (TfR).
Области, необходимые для проявления регуляторной
активности,- пять остатков, образующих петлю, и не-
спаренный остаток цитозина в стебле,- выделены более
темным цветом
152
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 8.119.
Атомы железа регулируют образование рецептора
трансферрина (TfR), влияя на стабильность мРНК TfR.
Эта стабильность определяется тем, связывается ли
специфический белок, чувствительный к железу (IRE-BP),
с пятью шпильками в З'-нетранслируемой области
(З'-UTR). Доступность или активность IRE-BP зависит
от концентрации трехвалентного железа. При избытке
железа способность IRE-BP связываться с IRE подав-
ляется и З'-UTR расщепляется под действием РНК-эн-
донуклеазы. При недостатке же железа IRE-BP связы-
вается с IRE и защищает З'-UTR от эндонуклеазного
расщепления.
лекулы мРНК. Регуляция по этой схеме поддер-
живает синтез TfR в том случае, когда клетки
вынуждены ограничиваться тем небольшим коли-
чеством железа, которое имеется в среде.
Изучение очищенного IRE-BP показывает, что
для его связывания с TfR-мРНК необходима сво-
бодная сульфгидрильная группа. Это наводит на
мысль, что при высоких концентрациях железа об-
легчается образование дисульфидной или неактив-
ной формы IRE-BP, а при низких-восстановление
дисульфидных групп белка до свободных сульфгид-
рильных групп и переход белка в активное состоя-
ние (рис. 8.120). Таким образом, скорость дегра-
дации TfR-мРНК должна зависеть от количества
IRE-BP, связанного с IRE, которое в свою очередь
определяется регулируемой железом степенью окис-
ления остатков цистеина IRE-BP. Внутриклеточная
концентрация железа играет роль своеобразного
«сульфгидрильного выключателя», который опре-
деляет, будет ли IRE-BP связываться с TfR-мРНК
и защищать ее от деградации.
Регуляция трансляции ферритиновой мРНК с
помощью железа. Синтез ферритина в клетках мле-
копитающих регулируется с помощью железа пост-
транскрипционно. Транскрипция гена ферритина,
как и стабильность соответствующей мРНК, не
зависит от концентрации железа; в этом случае
объектом регуляции является трансляция мРНК
(табл. 8.12). С помощью делеционного анализа бы-
ла идентифицирована последовательность длиной
43 п.н. внутри 5'-нетранслируемой лидерной области
ферритиновой мРНК, которая и участвовала в ре-
гуляции синтеза ферритина с помощью железа.
Компьютерная модель вторичной структуры этой
области содержит одну шпилечную структуру, очень
похожую на каноническую шпильку в IRE TfP-
мРНК (рис. 8.118). И в самом деле, оказалось, что
гены, экспрессия которых в норме не регулируется
железом, приобретают способность к такой ре-
гуляции при введении в их 5'-лидерные области
синтетического олигонуклеотида, способного обра-
зовать аналогичную структуру, или при введении
любой из шпилек, принадлежащих IRE TfR. Каждая
из этих шпилечных структур обеспечивает столь же
Неактивен
Регуляторная активность IRE-BP определяется степенью
окисления сульфгидрильных групп белка. Свободные
сульфгидрильные группы (SH) в IRE-BP-состояние при
недостатке железа- необходимы для связывания IRE-BP
со шпильками. При избытке железа сульфгидрильные
группы находятся в окисленной форме (S—S), IRE-BP
не может связаться со шпильками, и мРНК TfP разру-
шается.
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
1БЗ
эффективную регуляцию трансляции под действием
железа, как и 5'-шпилька гена ферритина. Следо-
вательно, и в этом случае наличие шпилечной струк-
туры совершенно необходимо для регуляции транс-
ляции с помощью железа.
С 5'-концевой шпилькой ферритиновой мРНК
связывается тот же самый белок, который при-
соединяется к IRE TfR-мРНК, хотя в этом случае
шпилька всего одна. Возможно (но это еще пред-
стоит доказать), IRE-BP регулирует трансляцию,
блокируя доступ рибосом к началу последователь-
ности, кодирующей ферритин. Таким образом, в
отличие от синтеза ферритина содержание ферри-
тиновой мРНК не зависит от концентрации железа.
Таким способом клетки выключают синтез ферри-
тина в условиях недостатка железа, при этом ста-
бильность ферритиновой мРНК позволяет клеткам
быстро реагировать на увеличение концентрации
железа.
Регуляция стабильности мРНК в процессе тран-
сляции. Образование а- и р-тубулинов, двух важ-
нейших компонентов микротрубочек, регулируется
у сложных эукариот двумя способами. Один из них
заключается в регуляции транскрипции семейства
множественных генов, кодирующих обе субъедини-
цы (разд. 9.3.6), другой- в регуляции содержания
гетеродимеров тубулина (оф), которые в большин-
стве животных клеток влияют на стабильность ту-
булиновых мРНК.
Сигнальная последовательность, ответственная
за индуцируемую тубулином нестабильность тубу-
линовых мРНК, формируется из первых четырех
кодонов кодирующей последовательности. Измене-
ния, затрагивающие любые другие участки мРНК,
не влияют на время ее жизни, тогда как модифи-
кации нуклеотидной последовательности, приводя-
щие к изменению N-концевой последовательности
Met-Arg-Glu-Ile (MREI), стабилизируют мРНК.
Замены в этих четырех кодонах, не приводящие
к изменению указанной аминокислотной последо-
вательности, не влияют и на стабильность тубу-
линовой мРНК. Более того, последовательность,
кодирующая MREI, должна находиться на N-конце;
если она локализована во внутренней части моле-
кулы, то мРНК не дестабилизируется. Далее, за
MREI-кодонами должен располагаться протяжен-
ный участок РНК, кодирующий полипептид, до-
статочно длинный для того, чтобы MREI-часть
выступала из рибосомы.
Согласно одной из моделей, по мере накопления
тубулиновых гетеродимеров они сами или какой-то
ассоциированный с ними фактор связываются с
N-концевым тетрапептидом синтезируемого тубу-
лина сразу после его выхода из рибосомы (рис.
8.121). Это может блокировать трансляцию, поз-
воляя эндонуклеазе расщеплять мРНК и иницииро-
вать ее разрушение. Природа взаимодействия белка
с тетрапептидом и механизм деструкции мРНК во
многом остаются неизвестными. Как бы то ни
было, этот механизм по типу обратной связи позво-
ляет белку регулировать время жизни своей соб-
ственной мРНК.
Трансляционная регуляция общего фактора тран-
скрипции. Многие гены S. cerevisiae, кодирующие
ферменты, которые участвуют в биосинтезе не-
скольких аминокислот, находятся под единым конт-
ролем. Так, при росте дрожжевых клеток в среде,
лишенной любой из десяти аминокислот, наблю-
“R Р
РИС. 8.121.
Модель регуляции стабильности мРНК Р-тубулина.
На рисунке изображены последовательные стадии
трансляции Р-тубулиновой мРНК (от Б'-кэпа слева к
poly (А)-хвосту на З’-конце). Растущая полипептидная
цепь представлена в виде нитки бус. С терминальной
последовательностью MREI (метионин, аргинин, глу-
тамин, изолейцин) синтезируемого полипептида по
выходе его из рибосомы связываются свободные а-
и р-субъединицы тубулина. Это активирует эндонук-
леазу, которая расщепляет мРНК, блокирует образо-
вание белковых цепей, синтез которых уже начался,
и запускает разрушение остальной мРНК. Происхож-
дение эндонуклеазы-свободной или связанной с ри-
босомами-неизвестно. [Т. J. Yen et al.. Nature 334
(1988), p. 500.J
154
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 8.122.
Открытые рамки считывания (ORF)
в гене дсп4 S. cerevisiae + 1 - сайт
инициации транскрипции, +1534
сайт терминации. Белок GCN4 коди-
руется последовательностью, начи-
нающейся в точке ATG (+ 591) и за-
канчивающейся в точке TGA. Четыре
открытые рамки считывания в 5'-не
транслируемой области мРНК GCN4
выделены тем же цветом, что и об-
ласть. кодирующая GCN4, ниже при-
ведены их нуклеотидные последова-
тельности.
дается стимуляция транскрипции вплоть до 30 ге-
нов, участвующих в биосинтезе соответствующих
аминокислот. Клонирование и секвенирование мно-
гих генов, работа которых зависит от содержания
в среде аминокислот, показывает, что эти гены
регулируются при участии короткой нуклеотидной
последовательности. Часто эта последовательность
повторяется несколько раз в любой ориентации
в премоторной области совместно регулируемых
генов. По своей локализации и функциям эти эле-
менты напоминают элементы UASG?1L, о которых
шла речь в разд. 8.3.е. Их каноническая последо-
вательность имеет вид 5'-RRTGACTCATTT-3', где
R любой из двух пуринов. Инвариантный сегмент,
TGACTC, является сайтом связывания фактора по-
зитивной регуляции GCN4. Напомним, что GCN4
высокогомологичен фактору транскрипции млеко-
питающих API и его онкогенному двойнику Jun
и что оба они связывают одну и ту же гексамерную
последовательность (разд. 8.3.г и 8.6.а). Неспеци-
фическая регуляция зависит от содержания GCN4,
его связывания с TGACTC-сайтом и активации
комплекса транскрипции. Два последних свойства
уже обсуждались в разд. 8.3 и 8.6. Здесь мы рас-
смотрим регуляцию образования GCN4.
Принципиальным моментом в синтезе GCN4
является новый способ его трансляционного конт-
роля, который основывается на особенностях строе-
ния мРНК GCN4. Кодирующая GCN4 последо-
вательность начинается примерно на 600 нуклео-
тидов правее 5'-конца мРНК. Необычно длинная
5'-лидерная последовательность отличается от пос-
ледовательностей большинства дрожжевых мРНК
тем, что содержит четыре короткие открытые рамки
считывания (ORF), каждая из которых имеет ини-
циирующий и терминирующий кодоны (рис. 8.122)
Такие левые AUG-кодоны обычно блокируют ини-
циацию трансляции более удаленных инициирую-
щих кодонов (разд. 3.8). Необычная особенность
синтеза GCN4 заключается в том, что его бло-
кирование может зависеть от аминокислотного
дефицита.
Делеционный анализ этой области показал, что
аминокислоты, кодируемые четырьмя левыми ORF,
несущественны ни для проявления функции GCN4 in
vivo, ни для трансляции последовательности, ко-
дирующей GCN4, как in vitro, так и in vivo. Однако
элиминация четырех левых AUG или их модифи-
кация с образованием других кодонов приводит
к тому, что происходящего в норме усиления син-
теза GCN4 в ответ на аминокислотное голодание не
происходит. Интересно, что четыре ORF неэкви-
валентны, поскольку их последовательная элими-
нация, начиная с ORF4, приводит к постепенной
утрате способности репрессировать трансляцию
GCN4 при достаточном содержании аминокислот.
Опосредованная аминокислотами регуляция син-
теза GCN4 зависит от сложных взаимодействий
между продуктами по крайней мере трех генов дсп
и десяти или более генов gcd. Для усиления синтеза
GCN4 при аминокислотном дефиците нужны белки
Ген<?сп4
GCN4-MPHK
«/«/-Гены —► GCD-mphk -------►белкиGCD
БелкиGCN •
Белки GCN4
Пути биосинтеза
аминокислот
РИС. 8.123.
Синтез белка GCN4 регулируется продуктами генов gcd
и дсп. Перемещению рибосомы и прохождению каждой
открытой рамки считывания в GCN4-MPHK препятст-
вуют белки GCD. Их блокирующий эффект снимается
белками GCN1, GCN2 и GCN3, которые ингибируют
трансляцию GCD-mPHK
8. СТРУКТУРА И ЭКСПРЕССИЯ ЭУКАРИОТИЧЕСКИХ ГЕНОВ
1ББ
GCN1, GCN2 и GCN3. Образование GCN4 при
достаточном количестве аминокислот блокируется
белками GCD. Разные последствия мутаций в генах
дсс/ и осп показывают, что белки GCD усиливают
влияние AUG-кодонов в лидерных последователь-
ностях как негативных регуляторов трансляции
GCN4, а белки GCN участвуют в позитивной
регуляции синтеза GCN4 в противовес негатив-
ному влиянию на эти AUG-кодоны белков GCD
(рис. 8.123).
Каким именно образом белки GCD и GCN
осуществляют совместную регуляцию снабжения
аминокислотами, преобразуют сигнал об их нали-
чии и либо игнорируют, либо стимулируют комп-
лекс трансляции, неизвестно. Скорее всего эти регу-
ляторные сигналы влияют на способность 408-субъ-
единицы рибосомы проходить лидерные инициирую-
щие кодоны. Рибосомы эукариот в отличие от рибо-
сом прокариот обычно начинают трансляцию в пер-
вом AUG правее 5'-кэпа (разд. 3.8). Когда они
доходят до терминирующего кодона, трансляция
прекращается, а реинициация в следующем AUG-
кодоне происходит неэффективно. Множественные,
последовательно расположенные инициирующие
и терминирующие кодоны типа тех, которые встре-
чаются в лидерной последовательности мРНК
GCN4, являются серьезным препятствием для
трансляции последовательности. кодирующей
GCN4; четыре последовательных сайта инициации-
терминации на относительно небольшом участке
позволяют осуществлять эффективную репрессию
трансляции GCN4. Возможно, белки GCD дейст-
вуют на определенные компоненты комплекса
трансляции, еще сильнее уменьшая вероятность
реинициации. Белки GCN, вероятно, являются анта-
гонистами белков GCD и вызывают противополож-
ные изменения, которые повышают способность
рибосом реинициировать трансляцию после терми-
нации. Как белки GCN «узнают» о содержании
аминокислот, неизвестно.
Глава 9
МОЛЕКУЛЯРНАЯ СТРУКТУРА
ГЕНОМОВ ЭУКАРИОТ
9.1. АРХИТЕКТУРНЫЕ ЭЛЕМЕНТЫ
Исследуя молекулярную структуру геномов, мы
ставим перед собой цель ответить на два вопроса:
в какой мере организация носителя генетической
информации влияет на его экспрессию и в конечном
счете на основные биологические феномены-раз-
витие организма, дифференцировку и генетические
аномалии? Что может сказать эта организация об
эволюции геномов и целых организмов?
Говоря об организации, мы имеем в виду три ее
уровня. Первый уровень-это отдельные гены и со-
ответствующие регуляторные элементы; об этом
шла речь в гл. 8. Здесь в результате изучения
структуры и функций генов удалось установить
наиболее важные факты и некоторые общие прин-
ципы. Второй уровень - предмет обсуждения данной
главы-это число генов и других сегментов ДНК
и их взаимное расположение. Здесь получено до-
вольно много данных, но общие принципы орга-
низации геномов окончательно не выяснены. Впро-
чем, установлен один интересный факт: протяжен-
ные GC- и АТ-богатые последовательности (изо-
хоры), обнаруженные у теплокровных животных,
находятся в R- и G-областях хромосом соответст-
венно. Лишь в очень немногих случаях относитель-
ное расположение генов подчиняется определенной
закономерности. В качестве примера можно при-
вести полицистронные опероны, характерные для
прокариот и нетипичные для эукариот. Имеющиеся
данные позволяют высказать предположение о
значительной свободе в организации генетической
информации, хотя реальная картина может ока-
заться несколько иной. Третий уровень относится
к структуре хроматина и к ее влиянию на линейную
организацию и экспрессию генов. Связь между
экспрессией генов и структурой хроматина оче-
видна. но о ее механизме известно очень мало
(разд. 8.7.а).
В этой главе, остановившись вначале на неко-
торых терминах и эволюционных вопросах, мы
рассмотрим организацию различных генов и других
сегментов ДНК в хромосомах. Вопрос одновремен-
но и сложный, и слишком упрощенный. Каждый
сегмент ДНК независимо от того, является он геном
или нет, обладает интересными специфическими
свойствами. Кроме того, геном каждого вида уни-
кален, хотя и имеет общие черты с геномами других
видов. Мы остановимся на таких отличительных
особенностях и приведем соответствующие приме-
ры. Конечно, эти примеры дают лишь слабое пред-
ставление о существующем в природе разнообра-
зии, поскольку сейчас исследована небольшая часть
сегментов ДНК.
а. Классификация сегментов ДНК
Гены. Мы рассматриваем ген как молекулярную
структуру. Ген эукариот-это совокупность сегмен-
тов ДНК, образующих экспрессирующуюся едини-
цу. В результате экспрессии образуются один
или несколько функциональных генных продуктов-
РНК или полипептидов. Каждый ген содержит один
или несколько сегментов ДНК, ответственных за
регуляцию транскрипции и, следовательно, за ре-
гуляцию экспрессии гена. Кодирующие области-
это сегменты ДНК, которые кодируют какой-либо
полипептид или функциональную РНК либо их
составляющие. Те сегменты ДНК, с которых не
транскрибируется никакой генный продукт, назы-
ваются некодирующими. Одни некодирующие об-
ласти (например, регуляторные сигналы, фланки-
рующие кодоны, или вставочные последовательно-
сти, прерывающие ген) являются составными час-
тями генов. Другие участки, имеющие отношение
к репликации ДНК или выполняющие какие-то пока
неизвестные функции, находятся между генами.
Псевдогены. Псевдогены-это участки генома,
близкие в структурном отношении специфическим
функционально активным генам, но не являющиеся
их аллельными формами и не кодирующие никаких
функциональных генных продуктов. У некоторых
псевдогенов в регуляторной или кодирующей об-
ласти или в них обоих имеются мутации. Другие
псевдогены имеют дефектные стоп-кодоны или со-
держат лишь часть кодирующей и регуляторной
областей. Таким образом, псевдогены могут быть
молчащими или с них могут транскрибироваться
и даже транслироваться дефектные полипептиды.
Как правило, псевдогены бывают тесно сцеплены
с соответствующими функциональными генами и
фланкированы последовательностями, гомологич-
ными последовательностям, которые фланкируют
функциональный ген. Обычно такие псевдогены со-
держат интроны и, по-видимому, образовались в
результате многократных тандемных повторений
(реитераций) сегментов ДНК.
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
157
Прямые повторы
5'-AAGAGNNNNNAAGAGNNNNNNNNAAGAGNNNNNNNAAGAG-3'
З'-ТТС TCNNNNN Т TCTCNNNNNNNN Т TCTCNNNNNNNT TCTC-5'
Тандемные прямые повторы
5‘-AAGAG AAGAGAAG AG AAGAG-3'
з-ттстсттстсттстсттстс-5’
РИС. 9.1.
Некоторые типы нуклеотидных последователь-
ностей (N-любой нуклеотид). Для образования
петли на конце шпильки необходимо не менее
трех-четырех нуклеотидов.
Обратные повторы
N N
G
NNCTGGAC-3'
NNGACCTG-5'
5-GTCCAGN
З'-CAGGTCN.
с
G
3'
Шпилькаспетлей
Шпилька
G
G
Палиндром
5-GAATTC-3'
3-CTTAAG-5’
Истинный (симметричный) палиндром
5-GTCAATGAAGTAACTG-3'
3'-CAGTTACTTCATTGAC-5'
Процессированные псевдогены - это подкласс
псевдогенов с определенной локализацией и струк-
турой. Как правило, они не сцеплены с соответст-
вующим функционально активным геном, а рас-
сеяны по всему геному, в том числе и по разным
хромосомам. По своей структуре они больше похо-
жи на ДНК-копии мРНК, чем на гены. Например,
процессированные псевдогены обычно не содержат
вставочных последовательностей, которые имеются
в функционально активных генах, а последователь-
ности, фланкирующие процессированный ген с 5'-
и З'-концов, как правило, отличаются от после-
довательностей, фланкирующих активные гены,
и представлены ро!у(А)-хвостом на З'-конце. Вероят-
но, процессированные псевдогены образовались в
результате обратной транскрипции и транспозиции
(разд. 10.4).
Некоторые особые типы последовательностей.
В предыдущих главах мы ввели термины, необхо-
димые для описания некоторых особых нуклеотид-
ных последовательностей. Рассмотрим их еще раз,
поскольку они понадобятся нам в дальнейшем при
обсуждении структуры эукариотических геномов
(рис. 9.1).
Повторяющиеся последовательности, ориенти-
рованные в одинаковом направлении в молекуле
ДНК, называются прямыми повторами или после-
довательностями, расположенными «голова-к-хвос-
ту». Такие прямые повторы могут входить в состав
непрерывных (тандемных) повторов или быть раз-
деленными какими-то другими последовательнос-
тями. Размер повторяющейся единицы варьирует от
двух до тысяч пар оснований.
Повторяющиеся последовательности, ориенти-
рованные в противоположных направлениях, назы-
ваются обратными (инвертированными) повторами
(или повторами с осью симметрии 2-го порядка или
повторами, расположенными «голова-к-голове» ли-
бо «хвост-к-хвосту»). Если инвертированный повтор
присутствует в одной из цепей двухцепочечной
ДНК, то такой же повтор, но имеющий противопо-
ложное направление, есть и в комплементарной
цепи. Одноцепочечная ДНК или РНК, содержащие
обратные повторы, разделенные тремя-четырьмя
основаниями, могут образовать внутримолекуляр-
ную двух цепочечную структуру. Если расстояние
между обратными повторами достаточно велико, то
при образовании дуплекса (ствола) может сформи-
роваться одноцепочечная петля; такая структура
называется «петля на стебле» (stem-and-loop). Если
же повторы разделены небольшим числом нуклеоти-
дов, то одиночная цепь ДНК образует двухцепочеч-
ную структуру, называемую шпилькой (hairpin).
Иногда обратные повторы находятся в пределах
одной структурной единицы (это могут быть, на-
пример. сайты узнавания для рестриктирующих эн-
донуклеаз, представленные на рис. 4.7); тогда соот-
ветствующая последовательность называется па-
линдромом. Такие палиндромы отличаются от ис-
тинных (симметричных) палиндромов, у которых
последовательность нуклеотидов читается одинако-
во в направлениях 5' -» 3' и 3' -» 5'. В двойной спи-
рали ДНК каждому истинному палиндрому в одной
цепи всегда соответствует комплементарный истин-
ный палиндром в другой.
б. Повторяемость
последовательностей ДНК
Значительную часть большинства эукариотиче-
ских геномов составляют последовательности, пов-
торяющиеся в неизменной или несколько модифи-
цированной форме. Число таких копий варьирует от
158
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
двух до нескольких миллионов. Ген и родственные
ему псевдогены образуют семейство повторяющих-
ся последовательностей. В данном случае семейство
содержит как кодирующие, так и некодирующие
элементы. Другие семейства, например повторяю-
щиеся гистоновые гены, состоят преимущественно
из кодирующих элементов. Такие семейства, как
сателлитные ДНК, содержат исключительно неко-
дирующие единицы. Наличие повторяющихся пос-
ледовательностей в геноме характерно не только
для эукариот. У таких прокариот, как Е. coli и
S. typhimurium, имеются короткие (от 20 до 40 п.н.)
палиндромные последовательности, повторяющиеся
в межгенных промежутках 500 и более раз, причем
на их долю приходится 0,5% всего генома. Роль
этих повторов до конца не установлена; возможно,
они служат сайтами узнавания для ДНК-гиразы,
которая обладает высоким сродством к ним. Кроме
того, ДНК прокариот содержит повторяющиеся
единицы длиной несколько тысяч пар оснований;
одни из них являются генами рибосомной РНК,
а другие-мобильными элементами (разд. 10.2.а).
Неравный кроссинговер между этими повторами,
а также между случайными очень короткими дупли-
цированными участками способствует образованию
тандемных повторов и делеций. В отличие от про-
кариот, у многих эукариот на долю повторов при-
ходится 50 и даже более процентов генома, и не
исключено, что на самом деле в геноме эукариот
содержится очень мало уникальных сегментов.
Важно различать уникальный сегмент (или после-
довательность) ДНК и уникальный ген. Специфи-
ческий функционально активный ген. представлен-
ный в гаплоидном геноме в единственном числе
и потому уникальный, может входить в семейство
повторяющихся последовательностей, которое со-
держит родственные ему гены и псевдогены.
Одни повторяющиеся последовательности пред-
ставляют собой непрерывные блоки тандемно пов-
торенных единиц (рис. 9.1). Члены других семейств
образуют кластеры прямых или обратных повто-
ряющихся единиц, разделенных другими последо-
вательностями. Члены некоторых семейств рассея-
ны по геному и разным хромосомам, иногда дис-
пергированы и сами тандемные повторы.
Вариабельность членов семейств повторяющихся
последовательностей. Нуклеотидные последова-
тельности членов данного семейства повторов мо-
гут быть как идентичными, так и различаться по
одной и более парам оснований. В последнем случае
говорят о полиморфных последовательностях. В не-
которых семействах такая дивергенция последова-
тельностей весьма значительна. Применительно
к более или менее сходным между собой членам
семейств используют термины «подсемейства» и
«надсемейства». Степень дивергенции членов разных
семейств можно определить с помощью четырех
методов, расположенных в порядке увеличения точ-
ности оценки: 1) сравнение карт рестрикции;
2) определение способности членов разных семейств
к гибридизации; 3) измерение температуры плав-
ления гибридных дуплексов, которые образуют
между собой члены семейств; 4) сравнение нуклео-
тидных последовательностей. В жестких условиях
отжига (относительно высокая температура около
65 °C - и низкая ионная сила - примерно 0.45М NaCl)
дуплексы обычно образуются только между сег-
ментами ДНК, комплементарными не менее чем на
85%. При понижении температуры и увеличении
ионной силы отжигаются сегменты с меньшей
гомологией. Стабильность образующихся гетеро-
дуплексов оценивается путем измерения их тем-
пературы плавления (разд. 1.1.е). Как правило,
понижение температуры плавления на 1 С соот-
ветствует 1% неспарившихся оснований.
Если последовательности в дуплексе идентичны,
то он называется гомологичным. Однако термин
«гомологичные» часто используется применительно
к сходным, но не идентичным сегментам ДНК.
Часто для указания степени идентичности после-
довательностей применяют качественный показа-
тель (например, высокая гомология означает вы-
сокую степень сходства). Количественная оценка
гомологии применима в тех случаях, когда известны
нуклеотидные последовательности. Так. говорят, что
два сегмента ДНК гомологичны на 70% (имеют
70%-ное сходство), если 70% пар оснований (или
нуклеотидных остатков) у них идентичны и кол-
линеарны. Такой подход вытекает из определения
гомологичности хромосом, однако он отличается от
принятого в биологической литературе по эволю-
ции. Там гомология, как правило, означает, что
морфологические структуры или последовательно-
сти имеют общего предка, и процентная гомология
теряет смысл. Хотя последовательности ДНК, го-
мологичные на 70 и даже на 30%, скорее всего
происходят от общей предковой последователь-
ности ДНК, они могут быть и неродственными.
Таким образом, термин «процентная гомология»,
используемый в молекулярно-генетической литера-
туре и в данной книге, двусмыслен. Термин «про-
цент сходства», которым мы также пользуемся,
однозначен, но встречается в литературе довольно
редко.
Размер семейств повторяющихся последователь-
ностей. Число членов семейств у разных эукариот
колеблется от двух по миллионов; неодинаково
и число копий в геномах разных видов. Число
копий-это весьма информативный параметр, од-
нако многие семейства описаны лишь как семейства
с большим, средним или малым числом повторов.
Исторически такая классификация связана с ис-
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
159
пользованием для этих целей кинетики реассоциа-
ции и показателя Cot (разд. 7.5.в). Например, для
ДНК млекопитающих с большим числом повторов
(105 копий и более) Cot равно 0,1, для ДНК со
средним числом повторов (102 105 копий) Cot ле-
жит в диапазоне от 0,1 до 100, а для ДНК с малым
числом повторов (менее 100 копий) Cot превышает
100.
Консенсусные (канонические) последовательно-
сти. Если семейство повторяющихся последова-
тельностей состоит из многих членов, каждый из
которых отличается от других лишь несколькими
нуклеотидами, то удобно построить некую усред-
ненную консенсусную (каноническую) последова-
тельность. В такой последовательности представ-
лены те основания, которые чаще всего встречаются
в данной позиции во всех членах семейства, и она не
обязательно идентична последовательности каждо-
го индивидуального члена. Консенсусные последо-
вательности получают двумя способами. Во-пер-
вых. иногда удается выделить и проанализировать
большую популяцию повторяющихся последова-
тельностей, не содержащую ни одной неродствен-
ной последовательности. Поскольку методы секве-
нирования не позволяют выявить очень малый
процент различий в каждой позиции, даже если
каждая молекула ДНК в популяции отличается от
всех других несколькими нуклеотидами, в итоге
получается уникальная последовательность (разд.
7.2). Во-вторых, можно сравнить последователь-
ности нескольких клонированных членов семейства
и определить, какие нуклеотиды встречаются в каж-
дом сайте наиболее часто.
Если результаты секвенирования отсутствуют,
то в семейство включают все последовательности,
которые гибридизуются в жестких условиях друг
с другом или с одним из клонированных членов
семейства. Вообще говоря, это означает, что иден-
тифицируются только последовательности, которые
отличаются от консенсусной не более чем на 15-
20%, а остальные членами данного семейства не
считаются. При идентификации семейств повторя-
ющихся последовательностей с помощью кинетики
ренагурации не учитываются также слишком корот-
кие для ренатурации последовательности. Напри-
мер, таким способом не выявляются последователь-
ности 5'-АСТ(А или Т)„ТА, находящиеся перед ге-
нами тРНК (разд. 9.2). Нередко последовательности,
идентичные по результатам секвенирования на 30 -
50%, не гибридизуются друг с другом и не попа-
дают в одно семейство или надсемейство.
в. Эволюционный аспект
повторяемости последовательностей
Роль амплификации последовательностей ДНК
в эволюции генома. Сходные последовательности
в пределах одного генома в принципе могут возни-
кать как независимо, так и при копировании исход-
ной уникальной последовательности ДНК-после-
довательности-«родоначальника». Вероятность то-
го, что две сходные последовательности возникли
независимо, тем меньше, чем больше их сходство
и длина. Нет сомнений, что именно увеличение
числа предковых последовательностей привело к
появлению семейств сходных последовательностей,
которые составляют значительную часть современ-
ных геномов. Увеличение числа копий сегментов
ДНК в ходе эволюции или в процессе эксперимента
называется амплификацией. За амплификацию пос-
ледовательностей в составе кластеров или после-
довательностей, рассеянных по новым геномным
локусам, отвечают разные механизмы (гл. 10). Если
основная последовательность удовлетворяет физио-
логические потребности организма, то образование
дополнительных ее копий в геноме не приводит
к особым преимуществам - подразумевается, что все
эти копии, кроме одной, не содержат мутаций,
включая нуклеотидные замены, делеции и вставки.
Одна измененная копия может быть нефункцио-
нальной или выполнять какие-то новые функции
либо служить регуляторным элементом. Если такие
измененные последовательности окажутся полез-
ными, то они сохранятся под давлением отбора.
В противном случае эти последовательности следует
отнести к псевдогенам. Таким образом, амплифи-
кация ДНК создает основу для эволюции.
Интересно сравнить отдельные гены и мульти-
генные семейства для разных видов, а также для
разных индивидов одного вида. Анализ различий
в нуклеотидных последовательностях, числе копий,
вставочных последовательностях и организации
генов может стать важным дополнением к тра-
диционным эволюционным исследованиям. В ка-
честве примера рассмотрим мультигенное семейст-
во у позвоночных, которое содержит гены гормона
роста и пролактина. Это надсемейство-хорошая
иллюстрация связи между структурой генов и эво-
люционными взаимоотношениями в пределах вида
и между видами.
Надсемейство генов гормона роста у позвоноч-
ных. Гормон роста и пролактин (которые у млеко-
питающих выполняют иные, чем у других позво-
ночных, функции) различаются приблизительно по
65% позициям, однако их молекулы имеют при-
мерно одинаковый размер (190 199 аминокислот
в зависимости от вида), а в некоторых положениях
у них находятся одинаковые аминокислотные остат-
160
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
ки. В генах этих гормонов в одинаковых сайтах
присутствуют по четыре интрона, которые, однако,
существенно различаются по длине и по фланки-
рующим их последовательностям. Каждый из генов
экспрессируется в клетках гипофиза определенного
типа, причем процесс находится под контролем
разных регуляторных элементов. В гаплоидном ге-
номе большинства позвоночных имеется только по
одному гену пролактина и гормона роста. У пла-
центарных же млекопитающих присутствует по
крайней мере еще один член данного семейства - ген
плацентарного лактогена, а возможно, и другие,
менее сходные гены. В генах гормона роста и пла-
центарного лактогена интроны находятся в оди-
наковых областях, а кодирующие последователь-
ности сходны примерно на 92% (по данным об
аминокислотных последовательностях сходство со-
ставляет 85%). Интроны высокогомологичны; 5'-
фланкирующие участки гомологичны по меньшей
мере по 500 п.н., а З'-фланкирующие последова-
тельности различаются больше. Это весьма инте-
ресно, поскольку гормон роста и плацентарный
лактоген синтезируются в разных тканях: в передней
доле гипофиза и в плаценте соответственно, и их
экспрессия контролируется независимо.
Поскольку отдаленно родственные гены плацен-
тарного лактогена и гормона роста есть у всех
позвоночных, не исключено, что их общий предок
дуплицировался еще до появления амфибий и рыб,
которые дивергировали 400 млн. лет назад (рис. 9.2),
и далее два гена эволюционировали у каждого вида
независимо. Гены, эволюционирующие таким спо-
собом, называются паралогичными. Как именно
произошла дупликация-с образованием сцеплен-
ных генов или рассеянных по всему геному копий-
неизвестно. У современных приматов гены гормона
роста и плацентарного лактогена диспергированы.
Например, у человека ген плацентарного лактогена
локализован в хромосоме 6, а ген гормона роста-в
хромосоме 17. Помимо паралогичных существуют
ортологичные гены, т. е. структурно и функциональ-
но гомологичные гены у разных видов. Например,
гены гормона роста у всех позвоночных ортоло-
гичны; кодирующие участки этих генов у курицы,
человека, быка и крысы гомологичны на 75%.
Ген плацентарного лактогена есть у всех мле-
копитающих, но у других позвоночных он отсутст-
вует, поэтому можно предположить, что дуплика-
ция гена гормона роста произошла до появления
большинства видов млекопитающих, т. е. 85-
100 млн. лет назад. Эти паралогичные гены эво-
люционировали вместе, оставаясь сцепленными.
Кроме того, у приматов имеется множество сцеп-
ленных генов как гормона роста, так и плацентар-
ного лактогена: например, у человека в хромосоме
17 локализованы два гена гормона роста и три гена
плацентарного лактогена. Это свидетельствует о
дополнительной амплификации генов, которая про-
Уникальный предковый ген
с четырьмя интронами
«н
I Дупликация >400 млн. лет назад
Позвоночные
Гормон роста Пролактин
I Дупликация >85 млн. лет назад
Плацентарные млекопитающие
Гормонроста Плацентарный лактоген Гормонроста Плацентарный
лактоген
РИС. 9.2.
Схематическое представление эволю-
ции мультигвнного семейства генов
гормона роста человека. Вариабель-
ность размера интронов не учиты-
валась. [G.S. Barsh, Р. Н. Seeburg,
R. Е. Gelinas, Nucleic Acid Res. 11
(1983), p. 3939.)
-«----------------------Примерно 50т.п.н.
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
161
РИС. 9.3.
Схематическое представление эво-
люции гена гормона роста и родст-
венных генов, которая включает тан-
демное повторение предкового экзо-
на, перестановку экзонов и утрату
интрона. [A. Barta et al.. Proc. Natl.
Acad. Sci. USA 78 (1981), p. 4367.]
изошла до обособления приматов, около 60 млн.
лет назад. Можно, конечно, предположить, что
близкие по своей организации гены возникли неза-
висимо, но это крайне маловероятно, если учесть,
насколько сходны между собой гены в пределах
надсемейства.
Роль повторов и перераспределений экзонов в эво-
люции гена гормона роста. Структура гена гормона
роста (и других генов этого надсемейства) свиде-
тельствует об амплификации экзонов. При клони-
ровании и секвенировании гена гормона роста кры-
сы было обнаружено пять экзонов (рис. 9.3). Первый
и часть второго экзона кодируют сигнальную поли-
пептидную последовательность (разд. 3.10.6). Сход-
ство второго, четвертого и пятого экзонов свиде-
тельствует о том, что все они образовались в ре-
зультате четырехкратной тандемной амплификации
предкового экзона (экзон 5 содержит две копии
гомологичных последовательностей). Экзон 3 от-
личается от других и, вероятно, появился при пе-
рераспределении экзонов - вставки копии одного из
экзонов другого гена. Действительно, пептид, ко-
дируемый экзоном 3, не стимулирует рост, хотя
и обладает противоинсулиновой активностью, в то
время как гормон роста обладает обеими этими
активностями.
Молекулярные часы. Детальное сравнение струк-
туры генов в надсемействе генов гормона роста
освещает еще один аспект молекулярной эволюции,
в которой предполагается, что мутации накапли-
ваются с более или менее постоянной скоростью.
Исходя из этого, сходство современных родствен-
ных кодирующих последовательностей, выраженное
в процентах, характеризует: 1) время с момента
дивергенции генов от единого предкового гена; 2)
скорость накопления мутаций в генах; 3) скорость
фиксации мутаций в популяции. Эти соображения
применимы как к ортологичным, так и к пара-
логичным генам. С такой общей гипотезой согла-
суются данные по многим белковым последова-
тельностям. Однако эволюционные «часы» идут
для различных белков с разной скоростью, и свя-
зано это в основном с различиями в скорости
фиксации мутаций. За единицу скорости эволюции
принимается время (исчисляемое миллионами лет),
за которое две кодирующие последовательности
дивергируют на 1% (эта единица обозначается
ЕЕР, от англ, unit evolutionary period). UEP равно 15
для умеренно консервативного цитохрома с и
400-для высококонсервативного гистона Н4. UEP
определяют, оценивая дивергенцию (в %) ортоло-
гичных кодирующих последовательностей у двух
видов в разные моменты времени от начала дивер-
генции этих видов, определяемого по палеонтоло-
гической летописи.
UEP для гормона роста и плацентарного лакто-
гена у человека и крысы составляет примерно 4,5.
Сравнивая нуклеотидные последовательности генов
гормона роста и плацентарного лактогена человека
и используя это значение UEP, можно проверить
гипотезу часов. Дивергенция указанных генов со-
ставляет 10%, откуда следует, что для амплифи-
кации и появления гена плацентарного лактогена
потребовалось примерно 45 млн. лет (10-4,5). Это
весьма неожиданный результат, поскольку согласно
палеонтологическим данным дивергенция млекопи-
тающих произошла 85-100 млн. лет назад. Можно
было бы объяснить такое несоответствие тем, что
дупликация у каждого вида млекопитающих про-
текала независимо, но это крайне маловероятно.
11 — ISR
162
ЧАСТЬ 111. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Можно также предположить, что UEP для генов
плацентарного лактогена и гормона роста человека
различаются. Но есть еще одно объяснение-неадек-
ватность простой «модели часов»: в ходе эволюции
происходят не только накопление и фиксация точ-
ковых мутаций, но и другие события, а независимая
эволюция двух генов ограничивается еще какими-то
механизмами. Здесь необходимо учесть два прин-
ципиальных факта. Во-первых, гены гормона роста
и плацентарного лактогена человека гораздо ближе
по своей организации ортологичным генам крысы
и быка. Во-вторых, интроны генов гормона роста
и плацентарного лактогена человека близки друг
другу почти в такой же степени, как кодирующие
последовательности этих генов, в то время как
обычно аналогичные интроны в паралогичных генах
существенно различаются по длине и нуклеотидным
последовательностям. Очевидно, что эволюция па-
ралогичных генов не зависит от так называемой
согласованной эволюции, или гомогенизации. Это
отмечается и в других мультигенных семействах
(например, семействах глобиновых генов; разд.
9.3.6) и в диспергированных повторяющихся после-
довательностях (разд. 9.5.д).
Какие процессы могут отвечать за гомогени-
зацию структуры пары родственных, но различаю-
щихся генов? Здесь может быть несколько альтер-
натив. Во-первых, неравный кроссинговер, который
обеспечил гомогенизацию генов рРНК по крайней
мере в нескольких геномах (разд. 9.2.а). Во-вторых,
новые делеции в генах, происходящие вслед
за амплификациями. В-третьих, генная конверсия
(разд. 2.4). Что касается отдельных однородных
семейств генов, то пока трудно отдать предпочтение
какой-то из этих альтернатив. Тем не менее ясно,
что в молекулярную эволюцию вносят вклад не-
сколько разных процессов. Несомненно, на ход
эволюционных часов влияют как рекомбинацион-
ные события, так и мутации.
Эволюция интронов. Являются ли интроны столь
же древними, как первичные кодирующие участки,
или же они появились в ходе последующей эво-
люции? В семействе генов гормона роста и в других
мультигенных семействах положение и число интро-
нов одинаково как для пара-, так и для орто-
логичных генов, хотя их длина и нуклеотидная
последовательность варьируют. Это согласуется
с предположением, что участки, прерывающие гены,
имеют столь же древнее происхождение, как и
большинство самих предковых генов, но они срав-
нительно независимы от давления отбора. Об этом
свидетельствуют, например, данные о структуре
генов инсулина млекопитающих. Как правило,
в гаплоидном геноме позвоночных присутствует
единственный ген инсулина с двумя интронами.
Однако у некоторых грызунов имеется второй
функционально активный ген инсулина с единствен-
ным интроном. Проще предположить, что второй
ген появился в результате случайной дупликации
исходного гена и потери интрона, чем считать, что
в предковом гене у всех позвоночных имелся только
один интрон.
Такие же доводы можно привести и в случае
семейства глобиновых генов, которое описано ниже.
У растений в генах леггемоглобина имеются три
интрона, два из которых расположены так же, как
интроны в генах гемоглобина позвоночных. Поло-
жение интронов в глобиновых генах иллюстрирует
еще одну общую закономерность: экзоны часто
кодируют определенные структурные и функцио-
нальные домены белков. Все эти факты свидетель-
ствуют о том, что интроны присутствовали уже
в самых первых генах. Однако наличие интронов на
ранних этапах эволюции не означает, что они не
могли внедряться в уже существующие кодирующие
области. Возможно, именно таков механизм появ-
ления некоторых интронов в генах семейства сери-
новых протеаз (например, тромбина, трипсина и
химотрипсина). Недавние эксперименты позволили
построить модели встраивания интронов. Так, ин-
троны I и II групп внедряются в сайты-мишени
с помощью генной конверсии или обратного са-
мосплайсинга. Конверсия генов I группы зависит от
белков, которые кодируются интроном.
9.2. ГЕНЫ, КОДИРУЮЩИЕ РНК
В трансляции мРНК и процессинге РНК-пред-
шественников участвует широкий спектр молекул
РНК, и многие из них требуются в относительно
больших количествах. В отличие от генов, коди-
рующих белки, где каждый транскрипт в результате
многократной трансляции дает множество генных
продуктов, транскрипт РНК-гена дает только одну
копию конечного РНК-продукта. Спрос на РНК
часто удовлетворяется за счет огромного избытка
генов.
а. Гены 18S-, 5,8S- и 28S-pPHK
Многие данные о генах рибосомных РНК
(рДНК) были получены еще до появления методов
рекомбинантных ДНК, поскольку рРНК, выделяе-
мые из очищенных рибосом, не содержат примесей
и могут использоваться в качестве зондов для
гибридизации без предварительного клонирования.
Кроме того, среднее GC-содержание рДНК обычно
выше, чем генома в целом, и в большинстве
эукариотических геномов рибосомные гены присут-
ствуют в виде протяженных тандемных повторов.
Это значит, что высокомолекулярные ДНК, содер-
жащие большое число копий рРНК-генов, имеют
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
163
относительно высокую по сравнению с остальной
геномной ДНК плавучую плотность и поэтому их
можно выделить в виде отдельной фракции при
равновесном центрифугировании суммарной ДНК
в градиенте плотности (изопикническое центрифу-
гирование; рис. 9.4). С помощью различных мето-
дов, в частности метода гибридизации in situ с ра-
диоактивными рРНК-зондами, было установлено,
что рДНК локализуется в определенном месте хро-
мосомы-так называемом районе ядрышкового ор-
ганизатора. Здесь происходят транскрипция рДНК
и процессинг предшественников рРНК (разд. 8.2).
В отличие от генов рРНК, которые у боль-
шинства эукариот присутствуют в виде сотен копий,
гены рибосомных белков низкокопийные. Геномные
клоны и кДНК, кодирующие рибосомные белки,
выделены из нескольких видов, в частности из
Drosophila, Xenopus, клеток мыши и человека. В ге-
номе содержится от 1 до 10 функционально актив-
ных генов для каждого рибосомного белка; обна-
ружены также процессированные псевдогены.
Сцепление генов трех рРНК в пределах одной
транскрипционной единицы. Как правило, кодирую-
щие области генов 18S-, 5,8S- и 28S-pPHK эукариот
сгруппированы в указанном порядке в одну тран-
скрипционную единицу (разд. 8.2). Все три РНК
образуются из длинного транскрипта-предшествен-
ника в результате процессинга, включающего рас-
щепление РНК эндонуклеазами и метилирование
как нуклеотидов, так и 2'-гидроксильных групп.
Полная транскрипционная единица содержит два
некодирующих спейсера (называемых внутренними
РИС. 9.4.
Равновесное центрифугирование суммарной геномной
ДНК Xenopus. ДНК центрифугировали с высокой ско-
ростью в растворе с градиентом плотности CsCI. По
окончании центрифугирования разные ДНК концентри-
ровались в виде полос, положение которых соответст-
вовало их плавучей плотности. Поскольку плотность
ДНК-дуплекса пропорциональна его GC-содержанию,
длинные повторы, обогащенные GC-парами, сущест-
венно отличались по этому показателю от большей
части генома и концентрировались в отдельных «сател-
литных» полосах. [D. D. Brown, С. S. Weber, J. Mol. Biol.
34 (1968), р. 681.]
транскрибируемыми спенсерами, ITS), которые раз-
деляют три кодирующие области, а также имеет
некодирующие участки перед первым геном 18S-
рРНК и за последним геном 28S-pPHK (эти участки
называются внешними транскрибируемыми спенсе-
рами, ETS). У эукариот длина и нуклеотидная пос-
ледовательность рРНК варьируют весьма незначи-
тельно, так что межвидовые различия по размеру
транскрипционной единицы (табл. 9.1) обусловли-
ваются различиями в длине спейсерных участков.
Сегменты ДНК по обе стороны от сайта инициации
транскрипции на 5'-конце ETS существенны для
транскрипции ДНК при участии РНК-полимеразы I.
В отличие от кодирующих областей, сходных между
собой, для этих последовательностей характерно
сильное видовое различие. Возможно, такие разли-
чия связаны с видоспецифичной активностью РНК-
полимеразы I, которой для взаимодействия с этими
последовательностями необходимы видоспецифич-
ные факторы транскрипции (разд. 8.2.в).
Тандемное расположение генов рРНК. Число
копий рДНК у многих организмов было определено
в экспериментах по гибридизации в условиях на-
сыщения, с использованием в качестве зонда чистой
рРНК (табл. 9.1). В большинстве случаев копии
рДНК в хромосоме организованы в виде тандем-
ного повтора, образующего протяженный кластер.
Эти кластеры часто локализованны в нескольких
хромосомах. Единицы транскрипции рДНК в пре-
делах этих кластеров разделены межгенными спенсе-
рами, или IGS (рис. 9.5). Длина каждого IGS
варьирует от 2 т.п.н. у дрожжей до 30 т.п.н. у млеко-
питающих, соответственно варьирует и общая дли-
на повторяющейся единицы, включая единицу тран-
скрипции и IGS. Нуклеотидные последовательности
самих IGS также весьма сильно различаются от
вида к виду (разд. 8.2).
Неравный кроссинговер между тандемными пов-
торами рДНК. Число копий рДНК может варьиро-
вать и в пределах вида. В геноме человека рДНК
обнаружена в пяти хромосомных локусах, при этом
каждый из них содержит разное число копий рДНК,
а кроме того, число копий на хромосому неоди-
наково у разных индивидов. У некоторых орга-
низмов даже соматические клетки в течение жизни
особи сильно различаются по числу копий рДНК.
Таким образом, увеличение и уменьшение числа
копий характерны для рДНК в такой же степени,
как и для других длинных тандемных повторов
(разд. 9.4.в). В основе динамичности геномов могут
лежать несколько механизмов; с помощью одного
из них - неравной гомологичной рекомбинации меж-
ду тандемными повторами-можно объяснить и
изменение числа копий рДНК, и другие замеча-
тельные ее свойства. Так, например, установлено,
что нуклеотидные последовательности всех рДНК
164
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Таблица 9.1. Локализация, число копий и размеры генов рРНК
Вид Номер хромосомы, в которой локализован геи Примерное число копий (на гаплоидный геном) Примерный размер единицы транскрип- ции, т.п. и.
С. elegans 1 55 6
D. discoides 42
S. cerevisiae 12 100 7
T. pyriformis 1
D. melanogaster X, Y 200 8
X. laevis (В одной из хромосом) 600 8
G. domesticus 200 10,5
M musculus ** 15, 18. 19 100 13
12, 16, 18
H. sapiens 13. 14, 15, 21. 22 150-200 14
Z. mays 6 3000-9000
11 В двух разных иибредных линиях.
человека на удивление сходны, несмотря на то что
кластеры тандемных повторов находятся в разных
хромосомах. Если бы рДНК в каждой хромосоме
были никак не связаны друг с другом, то в ре-
зультате накопления случайных мутаций они со
временем дивергировали бы, причем особенно силь-
но в области IGS. Единообразие повторов рДНК
человека говорит о том, что они как-то связаны
между собой. К образованию общего пула всех
рД НК-копий приводят неравные гомологичные
рекомбинации между повторами рДНК из разных
хромосом, а также между неаллельными повторами
сестринских хроматид. Но эти же процессы при-
водят и к изменению числа копий рДНК в клас-
терах.
Кроссинговер между рДНК проще изучать у
дрожжей, чем у млекопитающих. Если рекомби-
нантную плазмидную ДНК, содержащую маркер-
ные гены дрожжей, ввести путем трансфекции
в дрожжевые клетки, то некоторые из них включат
маркерные гены в хромосомную ДНК (разд. 5.6.в).
Трансформация происходит в результате рекомби-
нации между маркерными генами и гомологичными
геномными сегментами. Плазмида, несущая и ген
рДНК, и ген LEU2, рекомбинирует в основном
с геномной рДНК благодаря присутствию послед-
ней в большом числе копий (рис. 9.6). Такие тран-
сформанты содержат кластер тандемных повторов
рДНК, прерываемых в одном месте геном LEU2, по
которому их можно, отбирать. При спаривании
трансформантов с клетками дикого типа образуют-
ся диплоидные дрожжевые клетки, у которых можно
индуцировать мейоз и споруляцию. Продукты мейо-
за представлены в спорах в виде четырех отдельных
гаплоидных клеток, или тетрад. Эти клетки можно
разделить, вырастить и проанализировать. Резуль-
таты показывают, что неравный кроссинговер про-
исходит между цепями ДНК сестринских хроматид
во время мейоза. Из рис. 9.6 видно, что число копий
рДНК, образующихся в результате кроссинговера,
в пределах одного рД НК-кластера варьирует
(рис. 9.6).
Дивергенция в области IGS. Структура IGS весь-
ма сложна и неодинакова у разных видов как по
нуклеотидной последовательности, так и по органи-
зации (разд. 8.2.в). Этим IGS существенно отличает-
ся от высококонсервативных кодирующих областей
рДНК. Общей особенностью всех IGS является то,
что они обычно содержат прямые тандемные повто-
ры, правда различающиеся по нуклеотидным после-
довательностям у разных организмов. За исключе-
нием млекопитающих, эти последовательности
обычно сходны с теми, которые окружают сайты
начала транскрипции, т. е. сигнальные последова-
тельности для РНК-полимеразы I. Некоторые из
них служат «минорными» сайтами инициации тран-
скрипции и ответственны за присутствие небольших
количеств РНК, гомологичных IGS. Иногда такие
РИС. 9.5.
18S 5,8S 28S
18S 58S 28S
18S 5.8S 288
IGS ETS ITS ITS ETS IGS ETS ITS ITS ETS IGS ETS ITS ITS ETS
Организация рибосомных ДНК. Единицы транскрипции
рДНК обычно образуют кластеры из длинных тандем-
ных повторов. Представлены кодирующие области для
18S-, 5.8S- и 28S-pPHK. а также три типа спенсеров:
внешний транскрибируемый спейсер (ETS), внутренний
транскрибируемый спейсер (ITS) и межгенный спейсер
(IGS).
pBR322
LEU2
РДНК
1.Трансфекц,мя клеток
leu 2________________
2.0тбор клеток LEU* *
рДНКРДНК ^Е£/2|-Р^-|рДНК £рДНК$
Кластер рДНК в геноме клеток LEU*
Б
Рекомбинантная
плазмида
Дрожжевые клетки
LEU2
Пары сестринских хроматид,
которые образуются
в диплонде
Мейоз
Тетрады
Четыре колонии гаплоидных клеток
Фенотип
Число копий LEU2 0 2 1
РИС. 9.6. Тип РАНК А А
Неравный кроссинговер между единицами рДНК у
дрожжей. А. Дрожжевые клетки 1еи2~ трансфицировали
плазмидой, содержащей дрожжевую рДНК и ген LEU2.
Отбирали трансформанты LEU*. Блот-гибридизация
обработанной эндонуклеазой хромосомной ДНК пока-
зала, что вся рекомбинантная плазмида встроена в ге-
номную повторяющуюся единицу рДНК. Б. Гаплоидные
дрожжевые клетки, трансформированные так, как опи-
сано выше, скрещивали со штаммом дикого типа, со-
держащим ген LEU2. Эти два штамма различаются тем,
что в соответствующих повторах рДНК (поперечные
черточки на хромосомах) присутствуют различные сай-
ты для рестриктаз. У диплоидов индуцировали мейоз
и споруляцию, а у отдельных гаплоидных клеток в тет-
радах (производные одной диплоидной клетки) опре-
деляли потребность в лейцине (фенотип LEU), наличие
WT WT
последовательности LEU2 (с помощью ДНК-блоттин-
га) и родительский тип рДНК (тип А или дикий) с по-
мощью анализа рестрицированных повторов рДНК.
Клетки 1еи~ содержались почти в 10% тетрад. ДНК из
клеток leu не гибридизовалась с LEU2-3Onp,OM. Одна
из клеток LEU* в таких тетрадах содержала в два раза
больше ДНК LEU2, чем исходная гаплоидная клетка.
Таким образом, происходил частый кроссинговер. Как
клетки 1еи~, так и клетки с двойной дозой ££С/2-после-
довательностей содержали рДНК типа А, что указывает
на рекомбинацию сестринских хроматид. Споры, со-
держащие рДНК как типа А, так и дикого типа в тетра-
дах, встречались редко. Таким образом, кроссинговер
между сестринскими хроматидами происходит гораздо
чаще, чем между гомологичными хромосомами.
[Т. Peters, Cell 19 (1980), р. 765.]
166
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
последовательности содержат энхансеры экспрессии
рДНК. Повсеместное присутствие в IGS тандемных
повторов наводит на мысль, что они дают какие-то
преимущества при транскрипции рДНК.
Разное число актов кроссинговера, происходя-
щих между тандемно повторяющимися единицами
в IGS, приводит к различиям в числе самих этих
единиц, а также в числе полноразмерных рДНК.
Таким образом, IGS эволюционирует как согла-
сованно с рДНК-единицей, в которую он входит,
так и независимо от нее. Кроме того, в результате
кроссинговера общая длина повторов рДНК варьи-
рует от одного близкородственного вида к другому
и даже в пределах одного вида.
б. Гены 5S-pPHK
Гены 5S-pPHK, как правило, сцеплены с генами
18S-, 5,8S- и 28S-pPHK; исключения обнаружены
только среди грибов и простейших. Поскольку гены
5S-pPHK транскрибирует РНК-полимераза III,
часть их внутренних кодирующих последователь-
ностей функционирует как регуляторный элемент
(разд. 8.4.6). Гены 5S-pPHK, длина которой со-
ставляет 120 нуклеотидов, не прерываются ника-
кими интронами.
Тандемное повторение генов 5S-pPHK. Как и
кластеры генов других рРНК, гены 5S-pPHK у мно-
гих эукариот многократно повторяются и образуют
семейства длинных тандемных повторов (табл. 9.2).
Так, при изопикническом центрифугировании ДНК
Xenopus SS-рДНК концентрируется в отдельной
полосе, хотя на ее долю приходится менее I %
геномной ДНК. На электронных микрофотогра-
фиях частично денатурированных тандемных по-
второв SS-рДНК обнаруживаются легкоплавкие
АТ-богатые участки, чередующиеся с GC-богатыми
сегментами. Кодирующий участок длиной 120 п. н.-
это и есть GC-богатый сегмент, на долю которого
приходится менее 20% всего повтора; остальная его
часть представлена АТ-богатым нетранскрибируе-
мым спейсером. Вывод о вариабельности генов
5S-pPHK можно сделать исходя из данных о ге-
терогенности SS-рДНК, очищенной с помощью
центрифугирования в градиенте плотности, а также
из того, что их кривая плавления состоит из
нескольких компонент. Число копий генов 5S-pPHK
в различных геномах определяли методом гибри-
дизации с очищенной 5S-pPHK в условиях насы-
щения.
У D.melanogaster примерно 160 генов 5S-pPHK
собраны в тандемный повтор в хромосоме 2. Каж-
дая повторяющаяся единица содержит упомянутый
выше кодирующий участок длиной 120 п.н., тран-
скрибируемый спейсер длиной примерно 15 п.н. на
З'-конце транскрипционной единицы и нетранскри-
бируемый спейсер длиной 250 п.н. (рис. 9.7). По-
видимому, все 160 повторяющихся единиц образуют
единый, ничем не прерываемый кластер. Свыше 20
тандемных повторов было клонировано в виде од-
ного сегмента длиной более 8 т. п. н. АТ-богатый
участок в спейсере содержит внутренний повтор
и имеет разную длину (примерно + 5 п. н.).
Организация генов 5S-pPHK Xenopus. В отличие
от D. melanogaster и других организмов, для которых
характерна относительно простая организация ге-
нов 5S-pPHK, каждый из видов Xenopus (а также
других амфибий и рыб) имеет два семейства генов
5S-pPHK (разд. 8.4.д; табл. 9.2; рис. 9.7). Одно из
семейств содержит тысячи генных копий, которые
экспрессируются только в оогенезе. Другое семейст-
во состоит из гораздо меньшего числа генов, экс-
прессирующихся как в ооцитах, так и в сомати-
ческих клетках на разных стадиях развития. Каждое
семейство имеет уникальную кодирующую после-
довательность. но в пределах семейства кодирую-
щие последовательности гомогенны. Оба семейства
представляют собой тандемные повторы. Повто-
ряющаяся единица генов 5S-pPHK соматических
клеток устроена довольно просто: она содержит
Таблица 9.2. 5S-pPHK Локализация, число копий и размер повторяющейся единицы генов
Вид Локализация на хромосоме Примерное число колий (на гаплоидный геном) Примерный размер повторяющейся единицы, включая 1GS, п.н.
S. cerevisiae Хромосома 12 150
D. melanogaster Полоса 56EF хромосомы 2R 160 375 ± 5
X. laevis Теломерные участки многих 400-в соматических клетках 850-900
(или всех) хромосом 20000-в ооцитах 600- 1000
X. borealis 700- в соматических клетках 9000-в ооцитах 850-900 1000-2000
H. sapiens Хромосома 1 2000
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
167
Drosophila
Кодирующий
участок
Транскрибируемый спейсер
L-Нетранскрибируемый
спейсер
Xenopus borealis'.семейство из соматических клеток
GC-богатый спейсер, фиксированная длина,
нет внутренних повторов
Xenopus boreal is: семейство из ооцитов Псевдоген
шяяшшяшя-----------------------в
1-АТ-6огать1Й спейсер, варьирующая
длина,внутренние повторы
100 п.и.
РИС. 9.7.
Структура некоторых повторяющихся единиц 58-рДНК.
В ооцитах X. borealis эти единицы содержат несколько
генов и псевдогенов (их число варьирует), разделенных
спенсерами длиной около 80 п. н. Сами единицы тоже
разделены спейсерами разной длины, обогащенными
АТ-ларами. Такой спейсер содержит множественные
тандемные повторы единицы длиной 21 п. н. (5'-
CGTCGCGTCGI I I I lGTCGCG-З'), а его длина зависит
от числа копий этой единицы. Расщепление 58-рДНК
эндонуклеазой Hha\, узнающей сайт GCGC, облегчает
анализ структуры, поскольку при этом образуются от-
дельные фрагменты длиной 21 п. н. В клетке обнару-
жено несколько транскриптов псевдогена, который от-
личается от функционального гена по 15 позициям.
длинный спейсер и короткую транскрипционную
единицу. Однако у ооцитов эта единица имеет более
сложную структуру, неодинаковую даже у родст-
венных видов.
в. Сцепление всех четырех генов
рРНК у дрожжей
Отличительная особенность генов рРНК неко-
торых простейших и грибов, в частности дрожжей
5. cerevisiae, состоит в том, что все рибосомные гены,
в том числе и гены 5S-pPHK, сцеплены и находятся
в одной повторяющейся единице (рис. 9.8). Однако
гены 5S-pPHK транскрибируются с цепи ДНК,
комплементарной той, на которой синтезируется
длинный предшественник 18S-, 5,8S- и 28S-pPHK.
При этом используются две разные РНК-полиме-
разы: длинный полицистронный транскрипт обра-
зуется при участии РНК-полимеразы I, a 5S-pPHK-
РНК-полимеразы III. У другого вида дрожжей,
Shizosaccharomyces cerevisiae, и у грибов Neurospora
crassa гены 5S-pPHK не входят в состав единого
тандемного повтора, а рассеяны по всему геному.
Как показывают результаты генетического карти-
рования, большинство, если не все, повторяющиеся
единицы рДНК S. cerevisiae находятся в одном тан-
демном кластере в хромосоме 12. Известна карта
этой области: -U RA4-pPHK-PEP3-GAL2-ASP5-WHT-
ромера. (Ориентация единиц рДНК относительно
центромеры показана на рис. 9.8.)
г. Гены, кодирующие тРНК
Структура генов тРНК. Гены тРНК, как и гены
5S-pPHK, транскрибируются с помощью РНК-по-
лимеразы III, при этом промотор тоже распола-
гается внутри кодирующей последовательности
(разд. 8.4.6). Одни гены тРНК содержат интроны,
25S 5S
18S 5,8S 25S
poi Ш
poll
I I I I I I I I I I
0 1 2345678 9'
т.п.н.
К центромере
РИС. 9.8.
Повторяющаяся единица в дрожжевой рДНК. Стрелки
показывают направление транскрипции.
168
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
другие нет. И прерывистые, и непрерывные гены
могут быть даже у одного и того же вида, например
у D.melanogaster и Xenopus. Как правило, интроны
генов тРНК короткие, не более 45 п.н., и всегда
находятся с З'-стороны от антикодоновой после-
довательности.
Повторение и разные способы организации генов
тРНК. Гены тРНК в эукариотических геномах, как
и гены рРНК, обычно присутствуют в большом
числе копий (табл. 9.3). Но в отличие от рДНК с их
строго упорядоченной организацией гены тРНК
кажутся расположенными беспорядочно. Они могут
как соседствовать друг с другом, так и нет. Мно-
гокопийные гены одной тРНК могут находиться
в одном кластере или в разных. Например, при
гибридизации in situ тРНК с хромосомами D.tnela-
nogaster обнаруживается около 50 различных ло-
кусов, в которых группируются гены. Данный локус
может содержать гены разных тРНК (рис. 9.9). При
том, что гены той или иной тРНК могут находиться
в разных кластерах, некоторые гены, кодирующие
одинаковые тРНК, локализуются в одном тандем-
ном повторе. Иными словами, организация генов
тРНК весьма разнообразна.
Специфические повторяющиеся последовательно-
сти ДНК, расположенные перед генами тРНК.
Функциональные гены тРНК фланкируются самыми
разными последовательностями, но среди них часто
встречаются и специфические повторы. Функцио-
нальная роль самих повторяющихся единиц пока
неясна. У Drosophila, дрожжей и других организмов
в предшествующем генам тРНК участке длиной
100 п. н. часто обнаруживается последовательность
5'-АСТ(А или Т)„ТА (п = 0 — 5). В геноме дрожжей
примерно 30 раз встречается специфическая после-
довательность длиной 340 п. н., называемая сигма-
Таблица 9.3. Число копий и распределение генов
тРНК
Вид Образуют кластеры (с) или диспер- гированы (d) Примерное общее число копий (на гаплоидный геном)
S. cerevisiae d 360
Tetrahytnena pyroformis 800
D. tnelanogaster c,d 800 (~ 12 каждой тРНК) *»
X. laevis c,d 7000 (~200 каждой тРНК)
Rattus norvegicus c,d 6500 (~ 100 каждой тРНК)
H. sapiens c,d 1300 (10-20 каждой тРНК)
11 Известно, что некоторые из этих генов являются псевдогенами
РИС. 9.9.
Примерная организация кластера тРНК-генов в хро-
мосомной области 42А Drosophila. Стрелки указывают
направление транскрипции. [Р. Н. Yen, N. Davidson, Cell
22 (1980), р. 137.]
последовательностью. Она всегда располагается на
расстоянии не более 18 п.н. от начала тРНК-гена
и может находиться в любой из двух возможных
ориентаций относительно направления транскрип-
ции. Однако большинство генов тРНК у дрожжей не
сцеплены с сигма-последовательностью, а аллели
гена тирозиновой тРНК, соседствующие с этой
последовательностью, по своим фенотипическим
проявлениям не отличаются от аллелей, не свя-
занных с ней. Сигма-последовательность представ-
ляет собой копию длинного концевого повтора
(LTR), обнаруженного на концах дрожжевого рет-
ротранспозона ТуЗ (разд. 10.3). Такие «соло»-ЬТЯ
являются остатками ретротранспозонов, а локали-
зация сигма-последовательности именно перед ге-
нами тРНК свидетельствует о сайт-специфичности
вставок ТуЗ.
д. Гены малых ядерных
и цитоплазматических РНК
Все эукариотические клетки содержат множество
коротких стабильных молекул РНК, большинство
которых в составе нуклеопротеидных частиц (ну-
клеосом) присутствуют в ядре или в цитоплазме.
Некоторые из малых ядерных РНК (мяРНК) игра-
ют ключевую роль в процессинге первичных тран-
скриптов с образованием зрелых мРНК и рРНК
(разд. 8.5). Функции малых цитоплазматических
РНК (мцРНК), за исключением 7SL-PHK сигнал-
распознающих частиц, неизвестны (разд. 3.10.6).
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
169
Таблица 9.4. Гены некоторых мяРНК у многоклеточных
На- звание 11 Организм Длина РНК в нуклеоти- дах Кодирующие гены
примерное число копни функциональш генов диспергированы (d) >ix или собраны в кластеры (с) полиме- раза
U1 Все многоклеточные 164 30 (человек) 5 (курица) 10 (мышь) 500 (Xenopus) 1-2 (Phaseolus vulgaris) 3-4 (Drosophila) с с с (голова-к-голове) с (тандем) d п
U2 То же 188-189 20-40 (человек) 10 (мышь) 500 (Xenopus) (морской еж) 5 (Drosophila) с (тандем) с (тандем) с (тандем) c,d II
из Все многоклеточные (ядрышки) 216 6 (мышь) c,d II
U4 Все многоклеточные 142-146 4 (Drosophila) 2 (курица) d с II
U5 То же 116-118 Xenopus Тандем II
U6 » 107-108 200 (мышь) 3 (Drosophila) d с III III
U7 Морской еж 58 5 с II
4,5S, Грызуны 98-99 104 (крыса) d III
11 У Xenopus имеется не менее двух классов генов U1: около 500 генов класса I повторены тандемно и экспресси-
руются на ранних этапах эмбриогенеза, а малочисленные гены класса II диспергированы и экспрессируются
в ооцитах. У дрожжей последовательности, гомологичные U4 и U6, содержатся в одном гене.
Множественные гены и псевдогены мяРНК. В ге-
номе многоклеточных организмов присутствует
множество сегментов ДНК, которые гибридизуются
с U1-, U2-, U3-, U4-, U5-, U6- и 117-мяРНК или
с соответствующими кДНК (табл. 9.4). Одни такие
сегменты диспергированы, а другие сгруппированы
(обычно в тандемные повторы). Как и в случае генов
5S-pPHK, соответствующий кодирующий участок
может входить в состав длинного сегмента ДНК,
который образует повторяющуюся единицу. Как
правило (хотя и не всегда), функциональные гены
организованы в кластеры, а псевдогены дисперги-
рованы. Кодирующие последовательности функцио-
нальных генов комплементарны последовательнос-
тям соответствующих мяРНК. Функциональные ге-
ны данной мяРНК обычно имеют сходные после-
довательности, фланкирующие кодирующий учас-
ток и содержащие регуляторы транскрипции (разд.
8.3). У псевдогенов, которые образовались из
мяРНК в результате крупных делений, вставок или
точковых мутаций, фланкирующие последователь-
ности в значительной степени различаются. Харак-
тер расположения и число генов и псевдогенов
варьируют от одной мяРНК к другой и от вида
к виду. Гены не столь многочисленных мяРНК,
например Ull-мяРНК, пока не исследованы. За-
метим, что у дрожжей имеется только по одной
копии гена каждой из таких мяРНК и ни одного
псевдогена. Как у дрожжей, так и у многоклеточных
эукариот мяРНК участвуют в процессинге первич-
ных транскриптов и в образовании функциональных
РНК (разд. 8.5).
В качестве примера, иллюстрирующего сложную
организацию семейств рассматриваемых последо-
вательностей, можно привести U1 -семейство ДНК
человека. Оно содержит примерно 30 копий мяРНК-
генов, организованных в тандемные повторы, и
находится на коротком плече хромосомы 1. Каждый
функциональный ген в повторе входит в состав
повторяющейся единицы длиной не менее 44 т. п. н.
Псевдогены располагаются в различных локусах
генома и встречаются в 10 раз чаше, чем соот-
ветствующие гены.
Гены мцРНК. В переносе новосинтезированных
секретируемых и связанных с мембранами поли-
пептидов через липидный бислой эндоплазматиче-
ского ретикулума участвуют сигналраспознающие
частицы, основным компонентом которых являет-
ся 7SL-PHK. Нуклеотидные последовательности
7SL-PHK у грызунов и приматов практически оди-
наковы, а у дрозофилы эта РНК на 64% гомологич-
на соответствующей РНК человека. В геноме чело-
170
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Таблица 9.5. Гены некоторых мцРНК
РНК Организм Пример ная длина РНК в нуклео- тидах Число копий функциональных генов (на га- плоидный геном) РНК-по- лнмераза
7SL Позвоночные, 300 3-4 (человек) III
беспозвоноч- 2 (Drosophila)
ные 254 1 (S. pombe)
7SK Позвоночные 330 < 10 (человек) III
4,5S Грызуны 90 94 850 (мышь) 690 (крыса) III
века обнаружено три гена 7SL-PHK и несколько
сотен псевдогенов (табл. 9.5), а кроме того, пример-
но 10б рассеянных по геному последовательностей,
почти наполовину гомологичных последовательно-
сти 7SL-PHK. Половина этих последовательностей
представляет собой комбинацию 5'- и З'-концов
с делегированной центральной частью и называется
Alu-последовательиостью (разд. 9.5.в). Сходная кар-
тина наблюдается у грызунов.
Функции других мцРНК не установлены; извест-
но, что большинство этих РНК ассоциированы
с крупными семействами последовательностей, со-
держащими как гены, так и псевдогены.
9.3. ГЕНЫ, КОДИРУЮЩИЕ БЕЛКИ
а. Некоторые общие замечания
Гены, кодирующие белки, как правило, содержат
кодирующие области (экзоны), прерываемые одним
или более интронами. Однако некоторые гены (на-
пример, гистоновые или кодирующие интерфероны)
не имеют вставочных последовательностей. Число
таких последовательностей сильно варьирует от ге-
на к гену, а количество ДНК, приходящееся на долю
интронов, во много раз превышает количество ДНК
кодирующих областей (табл. III.2)-у некоторых
организмов в десятки раз. Как правило, нуклеотид-
ные последовательности аналогичных экзонов, от-
носящихся к паралогичным генам в данном геноме
или к ортологичным генам в геноме разных видов,
более консервативны, чем нуклеотидные последо-
вательности соответствующих интронов. Положе-
ние интронов, как правило, фиксировано, а по длине
и составу они варьируют.
Все известные гены, кодирующие белки, тран-
скрибируются РНК-полимеразой II и поэтому часто
имеют сходные промоторы и сигналы полиадени-
лирования. Но многие такие гены связаны с бо-
лее специфичными регуляторными последователь-
ностями, которые опосредуют действие гормональ-
ных, средовых или онтогенетических факторов
(разд. 8.3).
В отличие от генов, кодирующих РНК, поли-
пептидные гены представлены в геноме в единствен-
ном числе, однако при этом геном часто содержит
сегменты, гомологичные данному специфическому
гену. Таким образом, однокопийный ген может
входить в состав семейства близкородственных пос-
ледовательностей (например, в семейство генов
гормона роста). Члены такого семейства могут
кодировать незначительно различающиеся белки
(например, изозимы). Однако они могут иметь
разные регуляторные сигналы, ответственные за
экспрессию генов в разных тканях или на разных
стадиях развития (как в случае генов гормона роста
или плацентарного лактогена). (Напомним, что
согласно определению гена, принятому в данной
книге, ген включает регуляторные элементы, не
входящие в единицу транскрипции.) Равным обра-
зом членами семейства могут быть и псевдогены.
Условились, что два неаллельных гена считаются
идентичными, если они кодируют фактически оди-
наковые белки и находятся под общим контролем.
Близкородственные гены считаются членами одного
мультигенного семейства, даже если они неиден-
тичны (табл. 9.6).
Однокопийные гены могут также принадлежать
большому семейству отдаленно родственных пос-
ледовательностей. Несмотря на структурное сход-
ство, гены такого надсемейства кодируют совер-
шенно разные белки (как, например, уже упомя-
нутые гены пролактина и гормона роста). Распре-
деление отдельных генов по надсемействам не всег-
да удается провести вполне четко. Часто сходство
нуклеотидных последовательностей членов варьи-
Таблица 9.6. Некоторые семейства родственных
генов и белков В
В высшей степени гомологичные по структуре и функциям
р-Глобины
а-Глобины
Актины
Тубулины
Зеины
Гормон роста / Плацентарный лактоген
Рецепторы стероидных гормонов
О псины
Родственные, но различающиеся по структуре и функциям
а-Глобины/Р-Глобины
Пролактин/Гормон роста
Сывороточный альбумин / а-Фетопротеин
а-Интерфероны / Р-Интерфероны
РНК-полимеразы
Отдаленно родственные
Гаптоглобин 'Сывороточные протеазы (химотрипсин)
Антитромбин III/Овальбумин/Ангиотензиноген
Церулоплазмин/Фактор V/Фактор VIII
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
171
рует от 95% и более до 50% и менее. В жестких
условиях отжига дуплексы образуют только сег-
менты ДНК с гомологией не менее 80%, поэтому
неблизкородственные семейства можно сравнивать,
лишь зная их нуклеотидную последовательность.
Более того, сходство между двумя белками не на-
ходится в простой зависимости от процентного
сходства между соответствующими генами. Вслед-
ствие вырожденности генетического кода значи-
тельные изменения в третьей позиции кодонов мало
влияют на тип кодируемой аминокислоты, а такие
же изменения в первой или во второй позициях
приводят к существенным изменениям в структуре
белка.
Помимо генов, принадлежащих мультигенным
семействам или надсемействам, имеются и уникаль-
ные гены (например, ген тиреоглобулина человека
и ген актина дрожжей).
б. Примеры мультигенных семейств
Каждому мультигенному семейству генов, ко-
дирующих белки, присущи особые свойства, харак-
терные для данного вида (табл. 9.7). Мы рассмот-
рим здесь некоторые примеры, иллюстрирующие те
механизмы, благодаря которым возникают разли-
чия в физиологических свойствах клеток и в тонко
отлаженных клеточных процессах.
Актиновые гены: мультигенное консервативное
семейство. Актин участвует в самых разных типах
Таблица 9.7. Мультигенные семейства некоторых белков1’
Семейство генов Организм Примерный раз- мер семейства (число генов) Сцеплены (1) или диспергированы (d) Характеристика
Кислая фосфатаза Дрожжи 5- 4 1, d Регуляция генов осуществляется по- разному; например, один ген реп- рессируется Pi, второй им индуци- руется, третий конститутивный
а-Амилаза Мышь >3 1 В поджелудочной железе экспрессиру- ется множество генов, в печени и слюнных железах-по одному. У мышей разных линий число генов панкреатической а-амилазы варьи- рует
Крыса >9 ? Панкреатическую а-амилазу кодиру- ют четыре гена
Ячмень > 7 ? Не менее трех функциональных генов
Коллагены Человек 4 d Четыре гена, диспергированные по хромосомам 2. 7, 12 и 17, кодируют коллагены типа I (гетеродимер), типов II и II1 (гомодимеры)
Тяжелая цепь миозина Крыса > 10 Не менее одного гена для каждого типа мышц и не менее одного не- мышечного гена
Овальбумин Курица 3 1 Все три гена регулируются эстроге- ном. функции двух генов неясны
Сывороточный альбумин Мышь 2 1 Гены сывороточного альбумина и а-фетопротеина
Вителлогенин Жаба 4 ? Пары генов различаются на 20%; в пределах пар гены дивергировали на 5%. Все четыре гена экспресси- руются
Рибулозо- 1,5-бисфосфат- карбоксилаза Томат 5 1, d Экспрессируются в различных тканях на разных стадиях развития и в за- висимости от освещения
Зеин Кукуруза 100 d, 1 Кодируют семейство запасных белков зерна
11 Pi неорганический фосфат. В этой и последующих таблицах «сцеплены» означает, что один или несколько генов сгруппированы
в одном локусе, а «диспергированы» что члены семейства генов находятся в разных локусах. В некоторых семействах имеются как
сцепленные, так и диспергированные копии генов.
172
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
клеточного движения, в частности он обеспечивает
клеточную подвижность и мышечные сокращения.
Соответственно у эукариот имеются несколько раз-
ных типов актина и кодирующих их генов. Одни из
этих генов организованы в кластеры, другие дис-
пергированы (табл. 9.8). Единственный известный
организм, имеющий один актиновый ген,-дрожжи.
Цитоплазматические актины, обеспечивающие
клеточную подвижность, сходны по структуре у всех
эукариот; к этому же классу относятся и все актины
беспозвоночных и растений. У Drosophila и боль-
шинства других беспозвоночных основная функция
цитоплазматических актинов состоит в обеспечении
мышечных сокращений; у позвоночных эту функцию
выполняет особая группа а-актинов. У птиц и мле-
копитающих обнаружено шесть актиновых белков.
Два а-актина скелетных и сердечной мышц участ-
вуют в мышечном сокращении. Актины а и у при-
сутствуют в гладких мышцах. В цитоплазме прак-
тически всех клеток млекопитающих и птиц со-
держатся Р- и у-актины.
Несмотря на структурные различия, все актины
сходны по своей аминокислотной последователь-
ности, за исключением Ь1Н2-концевых участков. 11
Таблица 9.8. Семейства актиновых генов
Число из- Число ге- Сцеплены Тип актн-
вестных нов, вклю- (1) или на: мы-
Организм белков чая псев- дисперги- шечный
догены1’ рованы (m) или
(d) цитоплаз-
матиче-
ский (с)
S. cerevisiae 1 1 с
Dictyostelium > 2 17 I, d с
С. elegans 4 1, d с
Oxytricha > 2 3 с
Соя >2 с
Drosophila 3 6 d с
S. purpuratus 8 1, d т, с
Курица 6 8-10 d т, с
Крыса 6 >8 d т, с
Человек 6 20-30 d т, с
11 Число функциональных генов для актинов разного типа можно
оценить с помощью ДНК-зондов из З'-некодирующей области (см.
текст). Сцепление устанавливают по наличию одного или несколь-
ких генов в клонированном геномном сегменте. Нуклеотидные
последовательности различных генов Diciyosfelium различаются на
10%, не менее восьми из них экспрессируются; некоторые из этих
генов могут кодировать одинаковые белки. Все четыре актиновых
гена у нематоды С. elegans функциональные, три из них участвуют
в мышечном сокращении. У Drosophila экспрессируются все шесть
генов; один белок, несмотря на свое сходство с цитоплазмати-
ческим актином млекопитающих, специфичен для мышечной тка-
ни. У морского ежа 5. purpuratus не менее шести генов представ-
ляют собой различающиеся функциональные гены Гибридизация
соответствующих зондов с ДНК из гибридных соматических кле-
ток человек мышь показывает, что три из генов это уникальные
функциональные гены для а-актина, у-актина и а-актинов из
сердечной и скелетных мышц, локализованные в хромосомах 7, 17,
15 и 1 соответственно.
Так, Р- и у-актины человека различаются всего по
четырем аминокислотным остаткам. У большинст-
ва организмов гены, кодирующие различные акти-
ны, образуют мультигенные семейства, а кодирую-
щие области этих генов гомологичны у разных i
видов. Например, аминокислотная последователь-
ность а-актина скелетных мышц одинакова у ку-
рицы и млекопитающих, а актиновая мРНК, вы-
деленная из вегетативных клеток Dictyostelium, слу-
жит также хорошим зондом при отборе клони-
рованных актиновых генов дрожжей, кукурузы, сои
и млекопитающих.
5'- и З'-фланкирующие последовательности ак-
тиновых генов дивергировали сильнее, чем коди-
рующие области. Однако гены одинаковых актинов,
например гены Р-актинов млекопитающих, имеют
у разных видов сходные фланкирующие последо-
вательности, заметно отличающиеся от последова-
тельностей, фланкирующих у этих же видов а-акти-
новые гены. Фланкирующие последовательности
некоторых актиновых генов часто оказываются
настолько специфичными, что могут использовать-
ся в качестве гибридизационных зондов для иденти-
фикации соответствующих генов и их транскриптов.
Например, клонированные З'-концевые последова-
тельности каждого из шести различающихся функ-
циональных генов морского ежа можно использо-
вать для исследования дифференциальной транск-
рипции соответствующих генов в процессе развития
и дифференцировки. Таким образом, несмотря на
близкое родство всех кодирующих областей акти-
новых генов и самих актинов, ген каждого из типов
актина, по-видимому, связан со специфическими
некодирующими последовательностями, вероятно
выполняющими регуляторные функции. Таким об-
разом, выявляются два важных в эволюционном
отношении момента. Первый касается функцио-
нальных свойств генного продукта, второй-регу-
ляции генной экспрессии в дифференцированных
клетках.
Еще одна особенность мультигенного семейства
актиновых генов состоит в вариабельности числа
и положения интронов (рис. 9.10), обнаруживаю-
щейся при сравнении как паралогичных генов (на-
пример, генов р- и а-актинов скелетных мышц
крысы), так и ортологичных актиновых генов рас-
тений и беспозвоночных. Это существенно отли-
чается от обычной ситуации, характерной для
большинства ортологичных генов и мультигенных
семейств, где интроны располагаются одинаково.
Представляется маловероятным, что в ходе эво-
люции актиновых генов в них встраивались новые
интроны. Каждое такое событие приводило бы
к смещению соседних последовательностей и к сдви-
гу рамки считывания. Можно предположить обрат-
ное: большинство актиновых генов содержали инт-
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
173
Число кодонов
I 100 1 1 200 300
Дрожжи X—
Соевые бобы (2) —1
Кукуруза —1
Oxytrlcha ----
Dictyostelium(,Ke}----
Drosophila (1) —1
Drosophila (2)---
Морской еж (2)---
Мышцы морского еже (1)____
ос-Актин курицы ------
ос-Актин скелетных
мышц крысы
Человек, р-актин крысы ---
Человек,а-актин
сердечной мышцы ------
Человек, гладкие мышцы----
РИС. 9.10.
Число и локализация интронов в кодирующих областях
некоторых актиновых генов. Шкала в верхней части
рисунка проградуирована в числе кодонов. В круглых
скобках указано число разных генов, имеющих такую
роны во всех тех положениях, где они находятся
и в современных генах, но при эволюции разных
таксонов утрачивались различные интроны. Соглас-
но этой гипотезе, Dictyostelium и Oxytrlcha активно
избавлялись от интронов, поскольку ни одного из
них не осталось в кодирующих областях.
Актиновые гены в основном рассеяны по хро-
мосомам. У мыши, например, гены а-актина ске-
летных мышц и сердечной мышцы локализованы
в хромосомах 3 и 17 соответственно, а гены ци-
топлазматического Р-актина-в хромосоме 5.
У большинства изученных организмов число ак-
тиновых генов, по-видимому, превышает число из-
вестных актиновых белков. Хотя не исключено, что
какие-то из этих «экстрал-последовательностей ко-
дируют не обнаруженные пока формы актина, боль-
шинство из них. вероятно, являются псевдогенами,
в том числе процессированными. У человека по
меньшей мере две из двадцати актиноподобных
последовательностей представляют собой процес-
сированные псевдогены р-актина; они не содержат
интронов и несут в кодирующих областях различ-
ные мутации. Все шесть актиновых генов Drosophila
и шесть генов морского ежа функциональны и экс-
прессируются в соответствии со специфической про-
граммой развития данных организмов. Так, у
Drosophila два гена цитоплазматических актинов
экспрессируются на всех стадиях развития, два ге-
на-в основном в мышцах груди и конечностей. Как
организацию. В нетранспируемой 5'-пидерной после-
довательности (не представлена) у некоторых актино-
вых генов имеется дополнительный интрон.
правило, дифференциальная экспрессия генов на-
ходится под контролем регуляторных последова-
тельностей, находящихся во фланкирующих участ-
ках некоторых актиновых генов.
Тубулиновые гены: мулътигенное семейство, ко-
торое включает гены двух разных субъединиц гете-
родимерного белка. Микротрубочки участвуют во
многих процессах, протекающих во всех эукарио-
тических клетках: мейозе, митозе, клеточном дви-
жении и секреции. Поэтому не удивительно, что
структура тубулина-белка, из которого состоят
микротрубочки,-равно как и нуклеотидная после-
довательность гена, кодирующего тубулин, одина-
ковы у всех эукариот. кДНК, синтезированные на
тубулиновой мРНК курицы, гибридизуются с ту-
булиновыми генами таких отдаленных организмов,
как дрожжи и млекопитающие.
Тубулин-это гетеродимер, состоящий из двух
полипептидов: а и р. а- и Р-субъединицы содержат
450-451 и 445 аминокислотных остатка соответст-
венно и гомологичны примерно на 40%. Гены обеих
субъединиц относятся к одному мультигенному су-
персемейству, хотя довольно сильно различаются
и не гибридизуются.
Как правило, семейства тубулиновых генов
у каждого вида эукариот кодируют разные изотипы
а- и Р-субъединиц (табл. 9.9). Аминокислотные
последовательности различных а- или Р-субъеди-
ниц, кодируемых этими генами, обычно различают-
174
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Таблица 9.9. Семейства тубулиновых генов
Организм Число известных функциональных генов Пример- ное число Сцеплены (1)
псевдоге- нов для групп а или р или диспер- гированы (d)
S. cerevisiae 2а, 1р 0 d
Chlamidomonas 2а, 2Р 0 d
Trypanosoma Около 15а, 15Р 0 1 (тандем)
Drosophila 4а, 4р 0 d
Морской еж 2а, ЗР 10 1, d
Курица 2а, 7Р < 5 d
Млекопитающие Около 5а, 5Р 15 d
Arabidopsis 1а (возможно, 4)
thaliana
ся лишь незначительно (менее чем на 10%), и раз-
личия касаются в основном карбоксильных концов
молекул. По-видимому, вариации в структуре ту-
булинов связаны с тем, что немного различаются те
микротрубочки, которые они образуют. Последние
в свою очередь специфичны в отношении клеточных
процессов, типов клеток и стадий развития. Таким
образом, разные а- и р-изотипы могут соответство-
вать разным типам микротрубочек, выполняюших
определенные функции, хотя такая специализация
и не абсолютна. Дальнейшие изменения в структуре
тубулина происходят уже после транскрипции и
тоже способствуют функциональной специализации
микротрубочек. О такой специализации свидетель-
ствует высокая консервативность специфических
изотипов у позвоночных. Например, у различных
позвоночных преобладающий в нервной ткани
Р-тубулин имеет на карбоксильном конце совершен-
но одинаковые последовательности.
В отличие от кодирующих последовательностей,
5'- и З'-фланкирующие участки тубулиновых генов а-
и Р-мультигенных семейств значительно различают-
ся. По-видимому, это связано с независимой ре-
гуляцией экспрессии. Например, один из генов а-ту-
булина мышей экспрессируется только в семенни-
ках. У Drosophila концентрация мРНК, транскри-
бируемых с различных тубулиновых генов, неоди-
накова в разных тканях и на разных стадиях разви-
тия. Один из а-тубулиновых генов экспрессируется
только у взрослых самцов дрозофилы, другой
только в яичниках, а два экспрессируются постоян-
но. У дрожжей два гена а-тубулина, по-видимому,
кодируют одинаковые в функциональном отноше-
нии полипептиды, хотя один из генов экспрессирует-
ся эффективнее другого. Единственный ген Р-ту-
булина дрожжей существен для их роста. Оказалось,
что дрожжевые клетки, у которых блокировано
деление, несут мутацию в гене Р-тубулина (мутация
cdc).
У большинства видов тубулиновые гены рассея-
ны по геному. Например, у Drosophila все они хотя
и локализованы в хромосоме 3, но расположены
далеко друг от друга. Исключением из этого пра-
вила являются некоторые простейшие. У Tetrahy-
mena brucei примерно десять копий генов объеди-
нены в один тандемный повтор, в котором гены а-
и Р-тубулинов чередуются между собой; у Leishmar
nia enrietti гены а- и Р-тубулинов с таким же числом
копий сгруппированы в два кластера.
Некоторые последовательности, гомологичные
тубулиновым генам, представляют собой псевдоге-
ны; большинство из них процессировано. В качестве
типичного примера можно привести процессирован-
ный псевдоген а-тубулина крысы (разд. 9.1.а;
рис. 9.11). Установлена нуклеотидная последова-
тельность функционального гена, в том числе его 5'-
и З'-фланкирующих участков. Как любой типичный
тубулиновый ген позвоночных, этот ген содержит
три интрона (первый расположен непосредственно
за инициирующим кодоном ATG), а инициирующий
кодон и ТАТА-блок разделены участком длиной
примерно 120 н.п. Нуклеотидная последователь-
ность тубулиновой мРНК была установлена по
клонированной кДНК; она начинается на расстоя-
нии примерно 20 п. н. левее ТАТА-блока и занимает
следующие 100 п.н. до инициирующего кодона
ATG. Таким образом, эта последовательность иден-
тична последовательности соответствующих экзо-
нов после сплайсинга интронов. кДНК заканчивает-
ся ро1у(А)-участком, который начинается на рас-
стоянии 16 п.н. за сигналом полиаденилирования.
Процессированный ген, в отличие от функциональ-
ного гена, более сходен с кДНК. На протяжении
99 п. н. с 5'-стороны от инициирующего кодона ATG
(т. е. в 5'-лидерной последовательности) процесси-
рованный ген идентичен как функциональному гену,
так и кДНК, а далее его гомология функциональ-
ному гену становится менее полной. З'-конец про-
цессированного гена идентичен кДНК функциональ-
ного гена, синтезированного от poly(A), и таким
образом отличается от З'-фланкирующего участка
функционального гена. Более того, в процессиро-
ванном гене отсутствуют все интроны. Процесси-
рованный ген мог бы являться полной копией
кДНК, если бы не некоторые модификации в ко-
дирующей области, исключающие образование ин-
тактного а-тубулина. Эти модификации приводят
к сдвигу рамок считывания и состоят во вставке
двух остатков цитозина в экзон 2, делеции пяти
нуклеотидных пар в экзоне 3 и вставке неродствен-
ного сегмента из 110 п.н. (на рис. 9.11 обозна-
ченного Alu) в экзон 4 (Alu-последовательности
рассматриваются в разд. 9.5.в). Процессированный
ген а-тубулина фланкирован короткими прямыми
повторами-это еще одна характерная черта всех
процессированных псевдогенов. Такие прямые пов-
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
175
Интрон 1 Интрон 2 Интрон 3
5-фланкирующий
участок
111
кДНК
TAA(1Un.« 1ААТААА
ТАТАТАА- *т<5 (. П
1-------------------ЭКЗОНЫ
Процесси;
рованным
псевдоген
РИС. 9.11.
Структурные связи между геном а-тубупина, его мРНК
(кДНК) и процессированным псевдогеном у крысы.
Псевдоген имеет два дополнительных остатка цитозина.
делецию длиной 5 п. н. и вставку Alu-последователь-
ности (разд. 9.5.в). [I. Lemishka. Р. A. Sharp. Nature 300
(1982), р. 330.]
торы-это сегменты ДНК, которые были встроены
в новые локусы генома путем транспозиции. Они
представляют собой реитерации (многократные
повторения) последовательностей, ранее существо-
вавших в месте вставки, и называются сайтами-
мишенями дупликаций (разд. 10.1.6).
Суперсемейство глобиновых генов. Гемоглобины
позвоночных - это гетеротетрамеры, содержащие по
два а- и Р-полипептида. В геноме всех позвоночных
содержится множество генов и псевдогенов а-
и P-цепей-членов суперсемейства, которое вклю-
чает кодирующие последовательности генов гло-
бинов беспозвоночных, миоглобина позвоночных
и леггемоглобина растений (рис. 9.12). Все эти белки
содержат гем и обратимо связываются с кисло-
родом.
Глобиновые мРНК и соответствующие гены бы-
ли среди первых объектов, которые исследовались
методами рекомбинантных ДНК. На долю гемо-
глобина приходится более 90% всех растворимых
белков эритроцитов, а на долю глобиновой мРНК-
большая часть мРНК ретикулоцитов и ядерных
эритроцитов. Ни в каких других клетках глобиновые
гены в заметном количестве не транскрибируются.
Возможность получения относительно чистой гло-
биновой мРНК, наличие огромного числа уже из-
вестных мутаций в генах глобина человека, а также
обширные данные о свойствах глобиновых белков
послужили стимулом к молекулярным исследова-
ниям этих генов. Удалось получить первые данные
об интронах в клеточных генах и о свойствах про-
моторов для РНК-полимеразы П. Была установлена
нуклеотидная последовательность различных ал-
лелей глобиновых генов у человека и других видов,
что послужило основой для создания многочислен-
ных теорий эволюции глобинов.
Как правило, у млекопитающих имеется мно-
жество глобиновых генов и псевдогенов. У человека
Р-глобиновые гены объединены в кластер длиной
65 т.п.н., расположенный в хромосоме 11, а а-гло-
биновые гены в кластер длиной 25 т. п. н. в хромо-
соме 16 (рис. 9.12). Кластер Р-глобиновых генов
содержит пять генов (е, °у, Ау, 8 и Р), а а-глобиновый
кластер-три (£, at и а2). Оба кластера включают
также по нескольку псевдогенов.
У человека и других млекопитающих различные
глобиновые гены экспрессируются на разных ста-
диях развития организма (рис. 9.13). На очень ран-
них стадиях эмбриогенеза экспрессируются гены
£ и е с образованием гемоглобина £2/е2. К десяти
неделям беременности происходит активация а-
и у-генов и синтезируется а2/у2-глобин, который
циркулирует в крови плода до его рождения. Вскоре
после рождения P-подобная у-цепь замещается
Р-глобиновым полипептидом и образуется а2/Р2-
глобин, который преобладает в течение всей даль-
нейшей жизни человека. У взрослых образуется
также 8-глобин, однако он составляет лишь малую
часть Р-глобиноподобных цепей. Два идентичных
полипептида (а, и а2) кодируются двумя а-гло-
биновыми генами. Гены и а2 экспрессируются
в одно и то же время, но а2-ген транскрибируется
более эффективно и дает основную массу а-поли-
пептидов. У многих видов, в том числе и у человека,
расположение генов в а- и P-кластерах соответст-
вует тому порядку, в котором они экспрессируются
во время развития организма (на рис. 9.12 тран-
скрипция осуществляется слева направо).
176
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 9.12.
Организация 0- и а-глобиноподобных генов некоторых
позвоночных и генов леггемоглобина сои. Интроны на
рисунке не указаны. Шкала в верхней части рисунка
проградуирована в т. п. н. Транскрипция во всех случаях
направлена слева направо. Функция гена а-61 человека
неизвестна.
Обширные данные о генах и белках глобинового
семейства позволяют детально проследить эволю-
цию глобиновых генов человека. Скорее всего у от-
даленных предков современных животных сущест-
вовал единственный «глобиновый» ген. Предпола-
гается, что амплификация этого гена произошла
более 400 млн. лет назад, хотя у круглоротых рыб
имеется один глобиновый ген, а у челюстных рыб
синтеризуются и а-, и P-цепи. У амфибий, например
у Xenopus laevis, а- и Р-глобиновые гены тесно
сцеплены в тандемном повторе. По-видимому,
таким же образом они организованы и у рыб (рис.
9.12). Отметим, что у Xenopus имеются два разных
кластера сцепленных глобиновых генов; это связано
с тетраплоидией генома и самостоятельной эво-
люцией каждого из двух наборов хромосом. У птиц
и млекопитающих а- и р-глобиновые гены не сцеп-
лены. Обусловливается ли такая автономия разде-
лением одного кластера генов, как это произошло
у Xenopus, или независимой амплификацией отдель-
ных генов после их транспозиции-неизвестно. Счи-
тается, что гемоглобиновые гены птиц и млеко-
питающих начали эволюционировать независимо
РИС. 9.13.
Экспрессия глобиновых генов на разных стадиях раз-
вития человека.
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
177
около 250-300 млн. лет; именно об этом свиде-
тельствует палеонтологическая летопись.
Первая тандемная дупликация Р-глобинового
гена скорее всего произошла 200 млн. лет назад, до
появления современных млекопитающих. Затем ди-
вергировали регуляторные элементы, так что один
из p-генов стал экспрессироваться на ранних ста-
диях развития, а другой-на поздних. Последующие
тандемные дупликации происходили по-разному у
разных видов млекопитающих и привели к той
сложной организации генов и псевдогенов, которую
мы наблюдаем сегодня. У предков современных
приматов оба р-глобиновых гена дуплицировались
еще раз, хотя и не одновременно. В результате всех
этих событий образовался кластер из четырех Р-гло-
биноподобных генов, характерный для всех при-
матов; у человека в- и у-гены экспрессируются на
ранних стадиях развития, а 8- и p-гены - позже. Гены
°у- и Ау-глобинов, характерные для обезьян Старого
Света, возникли в результате дупликации у-генов,
происшедшей примерно 20-40 млн. лет назад. Ана-
логичные события привели к образованию кластера
а-глобиновых генов.
Установлены полные нуклеотидные последова-
тельности кластера Р-глобиновых генов человека,
а также генов большинства р-глобиновых кластеров
других видов. Это позволило сравнить филогене-
тическую картину, основанную на данных о числе
копий генов и на палеонтологической летописи,
с данными, основанными на гипотезе молекулярных
часов (рис. 9.14). Согласие оказалось весьма прием-
лемым, но более информативным было бы деталь-
ное сравнение. Например, время дивергенции р-
и 5-генов по оценкам составляет примерно 40 млн.
лет, что хорошо соответствует предполагаемо-
му времени появления линии-предшественника
обезьян Нового и Старого Света: 35-40 млн. лет.
Сравнение же Ау- и су-генов показывает, что первые
два экзона у них идентичны, а третьи дивергировали
на 1,2%. Эта дупликация типична для всех обезьян
Старого Света, т. е. последовательности должны
были дивергировать по меньшей мере 6-10 млн. лет
назад (рис. 9.14). Это сравнение наводит на мысль,
что первые два экзона у Ау- и су-генов стали
сходными совсем недавно. Указанное выше время
их дивергенции, оцененное исходя из данных о нук-
леотидной последовательности, на самом деле ука-
зывает на время гомогенизации, а не дупликации.
О структурном сходстве миоглобина и гемо-
глобина известно уже давно (рис. 9.15). Определение
аминокислотной последовательности леггемоглоби-
на показало, что она на 15% сходна с последо-
вательностями указанных глобинов позвоночных,
откуда можно сделать вывод, что все эти белки
принадлежат к одному семейству. Сходство их
структуры и функций позволяет высказать пред-
Эволюция семейства глобиновых генов. А. Калибровка
эволюционных часов. Оценивалась дивергенция двух
генов по тем сайтам, где произошли нуклеотидные
замены; это приводит к появлению кодонов, детерми-
нирующих новую аминокислоту. По оси ординат отло-
жена средняя дивергенция в процентах, по оси абсцисс-
время дивергенции, полученное из палеонтологической
летописи. Проводились следующие сравнения: 1) 500
млн. лет назад-а- и ^-глобиновые гены у отдельных
видов; 2) 270 млн. пет назад-а- и Р-глобиновые гены
курицы с а- и Р-глобиновыми генами млекопитающих;
3) 85 млн. лет назад-а- и Р-глобиновые гены разных
млекопитающих. Наклон прямой соответствует UEP
около 10. Б. Эволюционное древо Р-глобиноподобных
генов человека. Время дивергенции указанных пар гло-
биновых генов по тем сайтам, где произошли замены,
рассчитано как дивергенция (в %), умноженная на
UEP(10). Точки ветвлений указывают время начала ди-
вергенции пар генов и эквивалентны или времени дуп-
ликации, или времени с момента последней гомогени-
зации этой пары. [С любезного разрешения A. Efstratiadis;
из работы A. Efstratiadis et al.. Cell 21 (1980), p. 653.]
положение о существовании общего предка у генов,
кодирующих данные белки. Раньше, когда гены
глобинов не были клонированы и секвенированы,
можно было думать о конвергентной эволюции этих
генов, однако результаты секвенирования однознач-
но свидетельствуют о том. что они произошли от
одного предкового гена. Сходство распространяется
не только на кодируемые этими генами полипеп-
178
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 9.15.
Структура глобинов. А. Модели трехмерной структуры
леггемоглобина (слева), Р-субъединицы гемоглобина
(справа вверху) и миоглобина (справа внизу), по-
строенные с помощью компьютера. (С любезного раз-
решения Richard Feldman.) Кольцо гема почти перпен-
дикулярно плоскости рисунка. Б, Аминокислотные по-
следовательности миоглобина (сперма кашалота), а-
и р-глобинов (человек) и леггемоглобина (соя). Ис-
пользованы однобуквенные обозначения аминокислот
(рис. 1.24). Прочерк (—) означает, что аминокислота
в данном сайте отсутствует. Аминокислоты, одинаковые
для всех четырех полипептидов, заключены в рамки.
Пунктирными рамками показаны неидентичные, но
сходные аминокислоты (консервативные замены), на-
пример серин/треонин или валин/лейцин/изолейцин.
Аминокислоты, одинаковые у а- и р-глобинов, выде-
лены цветом. В конце указано общее число амино-
кислот в каждой последовательности.
А
Миоглобин
«-Глобин
В-Глобии
Леггемоглобин-а
I^Wsl
V-L'S;
vh lit;
V A F |T|
EGEWQL
PAD К T N
PEEKSA
EKQDAL
V
V
V
V
L H VWAKVEADVAGHGQDI L I RLFKSHPETLEKF
KAAWGKVGAHAGEYGAEALERMFLSF
T A L WG KV-NVDEVGGEALGRLLVVY
SSSFEAFKANIPQYSVVFYTSILEKA
P T T К T Y
PWTQRF
F
F
PAAKD L
F
DRFKHLKTEA
PHF-DL S H-
ESFGDLSTPD
S F L A NG V DP —
Миоглобин
а-Глобин
fl-Глобин
Леггемоглобин-а
EMK AS E D|LlK К
----GS aqIvIkg
A VMG N P K'VIK A
----т n p k!l:t g
H
H
H
H
GVTVL — T]A|LGA I LKKKGH —HEAELKP
G К К V A —D|AjL TNAVAHVDD-MPNALSA
G К К V L - G|A|F SDGL A H L DN-L KG T F AT
A E К L F A L[yjR DSAGQLKASGTVVADAA
L
L
L
L
AQS
SDL
SEL
GSV
H
H
H
H
ATKHK I P I KYLE
AH KL R VDPVNF К
CDKLHVDPENFR
AQKAVTDP Q—F V
Миоглобин F
«-Глобин L
Л-Глобин L
Леггемоглобин-а v
к
i tS E a; I bHVLHSRHPGNF GADIABGAMNK A L E L F RKDTAA
LjSHC|L Г"-..........-----------------
L|G N VIL . .. ~ - . . . ..... . „ ..
VlK E a!l|ljK T I KAAVGDKWSD E[LjS R AWE V A Y D E L A A AiIjK-|k
i; n v i- no n n run г и м n л i_ c l г n lz, i . м м r\
L|V T L A A H L P A E F TP AIVlH A SLDKFLASVST VlLIT s k
ViC V L A H H F G К E F T P P'vIq A A Y Q К V V AG VA N а1Ца H К
. ' T I IZ A A 1 f. fl . , , . f , , A I * . » I I 1.Z .z
Y К E L G Y QG-153
Y R-141
YH-146
A-144
Б
тиды и длину кодирующих участков, но, что наибо-
лее замечательно, на расположение интронов.
Все глобиновые и миоглобиновые гены позво-
ночных содержат по два интрона в одних и тех же
позициях (рис. 9.16). Первый интрон находится
вблизи ЗО-го кодона от 1ЧН2-конца и всегда пре-
рывает какой-либо кодон. Второй располагается
между двумя кодонами, недалеко от 100-го кодона.
Два из трех интронов гена леггемоглобина растений
находятся примерно в тех же самых позициях, что
интроны в глобиновых генах позвоночных, а тре-
тий-почти посередине кодирующей области.
Результаты анализа распределения интронов
в глобиновых генах убедительно показывают, что
разные экзоны кодируют уникальные структурно-
функциональные полипептидные домены. В глоби-
новых субъединицах позвоночных обнаруживается
несколько компактных областей, находящихся до-
вольно далеко друг от друга. Две из них соот-
ветствуют экзонам 1 и 3 (на рис. 9.15 это остатки
1-30 и 105-146 Р-глобиновой цепи), а другая пара
(остатки 40-60 и 70-100) не соответствует никаким
двум экзонам. Если бы экзоны действительно от-
вечали структурным доменам полипептида, то меж-
ду генами этих компактных областей должны были
бы располагаться интроны. Вскоре после того, как
эта гипотеза была сформулирована, удалось опре-
делить структуру леггемоглобинового гена сои,
и в предсказанной позиции действительно был об-
наружен дополнительный интрон. Общий предок
всех современных глобиновых генов, по всей ве-
роятности, содержал не менее трех интронов. На-
личие соответствия между доменами и экзонами
вполне понятно, если вспомнить о функции гло-
бинов. Например, участок между аминокислотами
65 и 96 связывает гем, а также содержит остатки,
участвующие в формировании ар-глобинового ди-
мера. В то же время в экзоне 3 сгруппированы
остатки, необходимые для образования тетрамера
а2р2-
Ответить на вопрос о существовании предкового
гена до появления царств растений и животных
довольно трудно. Гены леггемоглобина имеются
лишь у ограниченного числа двудольных растений,
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
179
Parasponia
andersonii
(глобин небо-
бового растения)
\l59/ 161
( CTCAAGATA ATG АСА GAG GTC Q
( 37 38 39 76 77 115 116 0
met thr
glu val
leu lys lie
Лег гемоглобин
сои
Миоглобин
тюленя
ATCAGGCTC
30 31 32
lie arg leu
gagttc
105 106
glu phe
153
а-Глобин
человека
GAGAGGATG__________________AGGCTC
30 31 32 99 100
glu arg met lys leu
(J-Глобин
человека
GGTCTC
146
29 30 31
gly arg leu
104 105
arg leu
РИС. 9.16.
Расположение интронов в генах глобинового над-
семейства. Справа указано число аминокислот (кодо-
нов) в каждом белке. Инициирующий кодон ATG не
учитывается, поскольку метионин отщепляется от зре-
лого полипептида. Интроны изображены в виде тре-
угольников, внутри которых указаны их размеры. При-
ведены номера кодонов и соответствующих аминокис-
лот, окружающих интроны (см. рис. 9.15, Б). Заметьте,
что первый интрон всегда прерывает кодон, а все сталь-
ные находятся между кодонами.
фиксирующих N2 в симбиозе с микроорганизмами,
например с Rhtzobiunr, такие растения могут быть
и клубеньковыми, и не клубеньковыми. Они могут
приобретать глобиновые гены путем горизонталь-
ного переноса из других геномов (например, в ре-
зультате вирусной инфекции), а не в ходе верти-
кальной эволюции. У растений, как и у животных,
амплификация предкового гена привела к образо-
ванию кластеров мультигенных семейств, содержа-
щих функциональные гены и псевдогены (рис. 9.12).
У животных дупликация, в результате которой по-
явились паралогичные гены миоглобина и гемо-
глобина, произошла более 500 млн лет. назад, по-
скольку уже у круглоротых рыб, например у мино-
ги, присутствуют оба гена.
Семейство а-амилазных генов: два гена из одной
кодирующей последовательности. Клетки разных
линий содержат неодинаковое число функциональ-
ных генов панкреатической а-амилазы. У некоторых
линий панкреатическую амилазу кодируют два
практически одинаковых неаллельных гена, а третий
ген экспрессируется в клетках слюнных желез и пе-
чени (табл. 9.7). Аминокислотные последователь-
ности панкреатической амилазы и амилазы слюн-
ных желез различаются на 12%; все три гена имеют
одинаковые 5'-, но не З'-фланкирующие последова-
тельности. Интроны существенно различаются как
по длине, так и по нуклеотидной последователь-
ности. Все гены локализованы в хромосоме 3, при
этом, как показывают результаты исследования с
помощью метода «прогулки по хромосоме», гены
панкреатической амилазы и амилазы из слюнных
желез находятся друг от друга на расстоянии всего
22 т.п.н. Мышиная ДНК содержит также сегменты,
180
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
гомологичные кодирующим областям а-амилазных
генов; по крайней мере один из них представляет
собой псевдоген, сцепленный с генным кластером
в хромосоме 3.
Экспрессия уникального а-амилазного кодирую-
щего участка в клетках околоушных слюнных желез
и печени находится под контролем двух элементов,
разделенных сегментом длиной примерно 3 т.п.н.
(рис. 9.17). Уровень экспрессии этого участка, если
последняя в обоих типах клеток находится под
контролем ближайшего регуляторного элемента,
одинаков и относительно низок. Но в околоушных
железах у взрослых мышей дальний регуляторный
элемент может активироваться, в результате чего
транскриптов образуется почти в 30 раз больше.
Именно этим объясняется высокое содержание
а-амилазы в клетках околоушных желез. Каждый
регуляторный элемент сцеплен со своим ТАТА-бло-
ком. Экзоны, находящиеся на 5'-конце первично-
го транскрипта, различаются; при последующем
сплайсинге они унифицируются и оба содержат
инициирующий кодон AUG. Таким образом, 5'-ли-
мРНК
Транскрипция в клетках
околоушной железы
Ген а-амила-
зы (Ату-1)
ТАТА А А
ТАТААА
Транскрипция в клетках
I печени и околоушной железы
AUG
мРНК
РИС. 9.17.
Экспрессия кодирующего участка а-амилазного гена
мышей (Zmy-1) регулируется с помощью двух про-
моторов. При участии одного из них с относительно
малой эффективностью образуются транскрипты в
клетках печени и околоушных желез, другой специфи-
чески и с высокой эффективностью индуцирует синтез
мРНК в клетках околоушных желез. [U. Schibler et al..
Cell 33 (1983). p. 501.]
дерные последовательности двух образующихся
мРНК различаются. Экзон, который содержит сиг-
нал инициации относительно малоэффективной
транскрипции, входит в состав большого интрона,
который при созревании специфичной для околоуш-
ных желез мРНК вырезается.
У крыс гены а-амилазы в околоушных и под-
желудочной железах, а также в печени различаются,
при этом панкреатическую а-амилазу могут коди-
ровать не менее четырех разных генов. Кроме того,
имеются псевдогены и процессированные гены. Как
и ген Ату-1 у мышей, крысиный ген, экспрессирую-
щийся в слюнных железах, имеет два независимых
промотора.
Интерфероновые гены. В ответ на разнообразные
внешние воздействия клетки многих позвоночных
секретируют полипептиды, называемые интерферо-
нами (IFN; табл. 9.10). Например, в результате
вирусной инфекции и попадания в клетку двухце-
почечной РНК в лейкоцитах индуцируется синтез
интерферонов группы а, а в фибробластах интер-
феронов группы р. Каждая группа содержит раз-
личные структурные родственные белки. Совершен-
но другой белок, уникальный IFN-y, синтезируется
в лимфоцитах при репликации ДНК, индуцирован-
ной митогенами. Все эти интерфероны в свою оче-
редь вызывают различные клеточные ответы-по-
Таблица 9.10. Интерфероновые гены человека
IFN-a IFN-P IFN-y
Название Лейкоци- тарный Фибро- бластный Иммунный
Число нсаллель- ных генов 3= 15 => 2 1
Псевдо- (или про- цессированные) гены > 3 ? ?
Хромосомная локализация 9р 9р; 2; другие 12(q24.1)
Интроны Нет Нез 3
Гомология на уровне нуклео- тидной после- довательности генов INF-a (%) 85-95 50 Мата
Гомология на уровне амино- кислотной пос- ледователь- ности IFN-a (%) 80 30 < 10
Длина молекулы11 (число амино- кислот) 155-166 166 146
11 Имеется в вид) длина молекул самих интерферонов. Соответст-
вующие гены кодируют также сигнальные пептиды длиной 20 23
аминокислоты, которые удаляются во время трансляции.
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
181
давление вирусной инфекции, иммунные реакции,
противоопухолевую активность.
Потенциальная значимость интерферонов как
терапевтических средств для лечения вирусных ин-
фекций и раковых заболеваний стимулировала их
исследование. Сначала успехи были весьма скром-
ными, поскольку активированные клетки продуци-
ровали лишь небольшие количества белка. Пробле-
му удалось решить благодаря применению методов
рекомбинантных ДНК. Был получен хороший вы-
ход продукта в клетках E.coli с использованием
клонированной кДНК. Однако само клонирование
это сложная процедура, поскольку мРНК, как
и полипептиды, не содержатся в клетках в дос-
таточно большом количестве. Например, на долю
мРНК у-интерферона в стимулированных лимфо-
цитах приходится всего 0,04% от полиаденилиро-
ванных РНК. Для клонирования таких редких
кДНК необходимы специальные методики. Сум-
марную РНК сначала фракционируют по размерам,
а затем проверяют фракции на способность служить
матрицами для синтеза IFN-y при введении в ооци-
ты Xenopus laevis. Затем функционально активную
фракцию используют для синтеза двухцепочечной
кДНК с целью клонирования в плазмидном векто-
ре. Образовавшиеся колонии подвергают дифферен-
циальному скринингу (разд. 6.5.6). IFN-у-кодирую-
щие последовательности были обнаружены в коло-
ниях, ДНК которых гибридизовалась с кДНК-зон-
дами из стимулированных лимфоцитов и не гибри-
дизовалась с ДНК нормальных лимфоцитов. Кло-
нированную кДНК использовали затем в качестве
зонда для выделения генов IFN-y. Таким способом
можно получить полноразмерные кДНК различных
интерферонов и определить аминокислотную после-
довательность данных белков, а также составить
детальную картину организации этого интересного
мультигенного семейства.
Среди клонированных сегментов ДНК человека,
содержащих IFN-a-гены, некоторые несли более од-
ного IFN-a-гсна. Эти данные, а также обнаружение
методом гибридизации in situ единственного IFN-a-
локуса в геноме человека говорят о том, что гены,
кодирующие интерфероны группы а, образуют еди-
ничный кластер. Генетический анализ показывает,
что ближайшими соседями генов IFN-a на хромо-
соме 9 являются гены IFN-p. Единственный ген
IFN-y расположен на другой хромосоме. Соседние
гены сходны по своей структуре. Например, гены
IFN-a и IFN-P, локализованные на хромосоме 9,
имеют аналогичные кодирующие и З'-фланкирую-
щие последовательности, а кроме того, в них обоих
отсутствуют интроны. В то же время ген IFN-y,
расположенный на хромосоме 12, содержит три
интрона и имеет весьма отдаленное сходство с ге-
нами IFN-a и IFN-P; различаются и соответствую-
щие полипептиды. Генные семейства интерферонов
других млекопитающих обладают аналогичными
свойствами.
в. Гистоновые гены:
консервативность кодирующих
последовательностей
и различия в организации
Общие свойства гистоновых генов. Первичная
структура гистонов у самых разных эукариот вы-
сококонсервативна. Это неудивительно, поскольку
гистоны играют ключевую роль в поддержании
структуры хроматина (разд. 1.1.ж). И все же ка-
кие-то различия между гистонами существуют. Это
касается прежде всего гистонов Н1 и в наименьшей
степени НЗ и Н4. На разных этапах развития орга-
низма, на разных стадиях клеточного цикла или
в разных тканях у представителей одного вида
могут синтезироваться немного различающиеся гис-
тоны. Например, большинство гистоновых генов
синтезируется в S-фазе клеточного цикла и, следо-
вательно, параллельно репликации ДНК. Экспрес-
сия других генов происходит с малой эффектив-
ностью в течение всего клеточного цикла. В отличие
от аминокислотных последовательностей гистонов,
достаточно консервативных у разных видов, число
копий и организация гистоновых генов варьируют
(рис. 9.18). Как и в случае тРНК-генов, это свиде-
тельствует о том. что консервативность кодирую-
щих последовательностей необязательно означает
такую же консервативность числа копий генов или
геномной организации.
Гены разных гистонов обычно сцеплены. Но,
в отличие от генов рРНК, которые объединены по
три гена в одной транскрипционной единице, все
гистоновые гены транскрибируются по отдельнос-
ти, часто с противоположных цепей ДНК. Кроме
того, опять-таки в отличие от генов рРНК, гисто-
новые гены у разных видов и даже у разных инди-
видов одного вида распределены неодинаково.
У одних видов они образуют тандемные повторы,
а у птиц и млекопитающих рассеяны по всему
геному. Большинство реплицирующихся гистоно-
вых генов лишено интронов, а у мРНК отсутствуют
3'-ро1у(А)-концы.
Еще до появления методов рекомбинантных
ДНК некоторую информацию об организации гис-
тоновых генов удалось получить с помощью двух
экспериментальных подходов. Первый из них по-
зволил установить число копий и тандемную орга-
низацию гистоновых генов у морского ежа путем
разделения сателлитных фракций гистоновых генов
после центрифугирования суммарной ДНК в гра-
диенте плотности. С помощью второго подхода
182
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Вид Группа гистоновых генов Примерная длина клас- тера, т.п. и. Примерное число копий
Дрожжи Н2АН2В 2
S.cerevisiae нз-н7 6 и 13 2
Drosophila НЗ-Н4-Н2А-Н2В-ТН 5 100
Морской еж нГ- Н4-Н2В-НЗ- Н2А 6-7 500(ранние)
5,purpuratus (рассеянные, некоторые образуют пары) Ю(лоздние)
Тритон N.viridescens Н4-Н2А Н2В НЗ Н1 Э 700
Xenopus НЗ-Н4-Н2А-Н2В-Н1В-НЗ-Н4 16 25
Н$-Й7-Н2А-Н2В 6
Курица НЗ-Н2А-Н4-Н1 •'Н2А-Н2Б 14 10
Н2А-Н4-Н2АН2В >10
НЗ- Н4 >10
Н5 (экспрессируется в эритро- цитах вместо гема Н1) 1
Человек НЗ-Н4-Н4-НЗ-Н2А-Н2В 20 10-20
Н4-НЗ-Н1-Н2В-Н2А 15 5
Н2А-Н2В-Н4 6 Неизвестно
РИС. 9.18.
Типичные примеры групп сцепленных гистоновых генов
у различных видов. Отметим, что все группы в пределах
вида одинаковы. У человека и, возможно, у некоторых
других видов часть копий представлена псевдогенами.
Направление транскрипции (там, где это известно) ука-
зано стрелкой над геном. [С. С. Hentschel, М. L. Birnstiel,
Cell 25 (1981). р. 301.]
ранними и поздними гистоновыми генами. Их орга-
низация различается у разных видов; мы рассмо-
трим в качестве примера гены Strongylocentrotus
purpuratus.
Геномные сегменты, содержащие ранние гисто-
новые гены, были клонированы из библиотеки
£'cwR [-фрагментов с использованием в качестве зон-
да суммарной гистоновой мРНК (рис. 9.19). Были
получены два клонированных фрагмента, не гибри-
дизующихся между собой. Как показало геномное
картирование, они сцеплены и находятся друг от
друга на расстоянии не более 7 т.п.н. Расположение
всех пяти гистоновых генов в этих клонах опреде-
ляли с помощью их гибридизации с выделенными
методом электрофореза мРНК и последующего
анализа образовавшихся гетеродуплексов под элект-
ронным микроскопом. Кроме того, индивидуаль-
ные мРНК отжигали с отдельными цепями клони-
рованных фрагментов. Все РНК гибридизовались
только с одной из двух цепей; это означает, что
транскрипция соответствующих генов происходила
в одном и том же направлении. Все эти данные
получили подтверждение после определения нуклео-
тидных последовательностей; были установлены
и точные размеры кодирующих участков и спейсе-
ров. Несколько сотен сцепленных ранних гистоно-
удалось получить частично очищенную гистоновую
мРНК из обогащенного препарата.
Клонирование гистоновых генов. В течение при-
мерно первых десяти часов после оплодотворения
яйцеклетка морского ежа претерпевает от 2-109 до
2-1O10 делений, которые сопровождаются быстрой
репликацией ДНК и синтезом гистонов. Около 30%
синтезированных в это время белков представляют
собой ядерные белки и в большом количестве при-
сутствуют гистоновые мРНК. Все эти мРНК имеют
одинаковый размер и коэффициент седиментации
9S. мРНК разных гистонов можно разделить с по-
мощью электрофореза в полиакриламидном геле
и затем идентифицировать по образованию соот-
ветствующих полипептидов в результате трансля-
ции in vitro. Соответствующие мРНК морского ежа
можно также использовать в качестве зондов для
клонирования гистоновых генов растений, грибов
и животных.
Гистоновые гены морского ежа. На самых ран-
них стадиях эмбриогенеза у морского ежа функцио-
нируют специфические гены, ответственные за быст-
рый синтез гистонов. После образования бластулы
начинается экспрессия других гистоновых генов,
ответственных за массовый синтез гистонов, необ-
ходимых для дальнейшего развития зародыша и
взрослого ежа. Соответствующие гены называются
I
1
НЕ Е Е Н
HI Н4 Н2В НЗ Н2А
РИС. 9.19. —— — — —
Группа пяти сцепленных ранних гистоновых генов
морского ежа (S. purpuratus). Из ЕсоR1 -геномной
библиотеки были выделены два клонированных фраг-
мента (а) и (б), которые не гибридизуются с суммарной
гистоновой мРНК, но гибридизуются друг с другом. Как
фрагмент а, так и фрагмент б гибридизуются с уни-
кальным фрагментом размером 7 т. п. н„ полученным
при расщеплении геномной ДНК рестрнктазой HindXXX
(е). После обработки геномной ДНК двумя рестрикта-
зами, EcoRI и HindXXX, фрагмент длиной 7 т.п.н., гибри-
дизующийся с фрагментом а или б, не выявлялся.
Вместо этого со смесью фрагментов а и б гибридизо-
вались два фрагмента, соответствующих по длине этим
двум. Таким образом, два фрагмента, а и б, сцеплены
между собой в пределах сегмента геномной ДНК дли-
ной 7 т.п.н.
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
183
вых генов сгруппированы в длинные тандемные
повторы.
В отличие от ранних генов, которые повторяют-
ся до 500 раз, поздние гистоновые гены повторяют-
ся всего около десяти раз. Поздние гены отличаются
от ранних и по некоторым другим признакам.
Во-первых, они никогда не бывают сцеплены в груп-
пы по пять генов и не образуют тандемных повто-
ров. Некоторые гены, например НЗ и Н4, действи-
тельно соседствуют, располагаясь на расстоянии
1 т.п.н. друг от друга, но «одиночные» гены нахо-
дятся на расстоянии не менее 6 т.п.н. от других
гистоновых генов. Ни по своему расположению, ни
по способности транскрибироваться в обоих на-
правлениях поздние гены не имеют ничего общего
с ранними. Кроме того, организация поздних генов
варьирует от одного рода морских ежей к другому.
Клонированные родственные гены ранних и
поздних гистонов не гибридизуются между собой
в жестких условиях, хотя их генные продукты имеют
сходную (или даже идентичную) структуру. Разли-
чия в кодирующих последовательностях говорят
о том, что для одних и тех же аминокислот исполь-
зуются альтернативные кодоны. Еще больше разли-
чаются последовательности, фланкирующие родст-
венные ранние и поздние гены; исключение состав-
ляют олигонуклеотиды, важные для транскрипции
с участием РНК-полимеразы II.
Гистоновые гены у других видов. С помощью
гибридизации in situ было показано, что все гисто-
новые гены Drosophila локализованы в одной об-
ласти политенной хромосомы 2. Пять разных генов
сцеплены в пределах одной единицы, повторяю-
щейся примерно 100 раз; расположение генов и на-
правление транскрипции существенно отличаются
от таковых у морского ежа (рис. 9.18). Переклю-
чение синтеза гистонов на путь их образования
в массовом количестве в ходе эмбриогенеза у Dro-
sophila, как и у морского ежа, по-видимому, связано
с экспрессией альтернативных генов, а не с мо-
дуляцией экспрессии одного и того же набора генов.
В ДНК дрожжей (S. cerevisiae) имеются две копии
сегмента, содержащего по одному гену гистонов
Н2А и Н2В, и две копии сегмента, содержащего по
одному гену НЗ и Н4 (рис. 9.18). Гомологичные
гены в каждой паре этих кластеров могут коди-
ровать как слегка различающиеся (например, Н2А),
так и одинаковые (например, НЗ) аминокислотные
последовательности. Имеет ли такая вариабель-
ность какое-либо функциональное значение-неясно.
У дрожжей гистон (или ген) Н1 не был иденти-
фицирован.
Организация гистоновых генов у позвоночных,
как и у беспозвоночных, весьма разнообразна (рис.
9.18). Например, у Xenopus имеется несколько групп
из пяти сцепленных гистоновых генов с разным их
расположением. По меньшей мере две такие группы
отличаются тем, что у них имеются особые гены Н1.
Эти гены гибридизуются со специфической мРНК
ооцитов, с каждой из которых in vitro транслируется
специфический белок Н1. Кроме того, в геноме
Xenopus есть несколько тандемных повторов групп
гистоновых генов, но в отличие от ранних гисто-
новых генов морского ежа соседние группы внутри
тандемного повтора при эндонуклеазном расщеп-
лении обнаруживают полиморфизм по сайтам рест-
рикции.
У тритона Notophthalmus viridescens имеются
примерно 700 гистоновых генов, локализация кото-
рых, по крайней мере в некоторых аспектах, не
соотносится с генной экспрессией. Гены пяти гисто-
нов объединены в сегментах ДНК длиной 9 т.п.н.,
каждый из которых входит в состав длинных тан-
демных повторов неродственных последователь-
ностей в одном из плечей хромосомы. Как правило,
группы гистоновых генов разделены участками из
тандемных повторов длиной от 50 до 100 т.п.н.
Аналогичные тандемные повторы встречаются и в
области центромер (разд. 9.4.в).
У теплокровных животных имеется относитель-
но небольшое число гистоновых генов. Они не
сгруппированы каким-то определенным образом, но
часто соседствуют друг с другом.
9.4. ТАНДЕМНЫЕ ПОВТОРЫ:
ХАРАКТЕРНАЯ ОСОБЕННОСТЬ
ВСЕХ ЭУКАРИОТИЧЕСКИХ
ГЕНОМОВ
Один из самых важных выводов, который можно
сделать из результатов изучения геномной органи-
зации, состоит в том, что родственные последова-
тельности ДНК часто образуют тандемные повто-
ры. Иногда, например в случае генов рРНК или
гистоновых генов, тандемно повторяются группы
одинаковых генов. В других случаях в тандемы
организуются такие родственные, но все же разли-
чающиеся гены, как члены мультигенных семейств
глобиновых, овальбуминовых генов и генов гор-
мона роста человека. Однако тандемные повторы
встречаются не только в целых генах, они присут-
ствуют также в их кодирующих и некодирующих
участках.
а. Гены, состоящие из тандемных
повторов сегментов ДНК
Внутренние области некоторых кодирующих
участков, по-видимому, состоят из тандемных, хотя
и несколько различающихся повторов. Это наводит
на мысль, что в эволюции генов участвовала амп-
184
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
лификация. Рассмотрим четыре примера, свидетель-
ствующие о широкой распространенности повторов
во внутренних областях кодирующих последова-
тельностей.
Гены сывороточного альбумина и а-фетопротеи-
на. У зародышей млекопитающих среди белков
сыворотки крови преобладает а-фетопротеин, а
после рождения основным белком становится сы-
вороточный альбумин (разд. 8.3.д). Оба белка син-
тезируются в печени и каждый из них кодируется
уникальным геном. У мышей эти два гена сцеплены
с образованием тандема длиной 15 т. п. н.; тандем
локализован в хромосоме 5, причем альбуминовый
ген находится перед геном а-фетопротеина. Хотя
эти гены значительно дивергировали и не гибриди-
зуются, по своей структуре они достаточно сходны
и можно предположить, что они образовались в ре-
зультате тандемных реитераций и последующей ди-
вергенции. Например, положение четырнадцати
вставочных последовательностей и длины пятнад-
цати экзонов в обоих генах очень близки (структу-
ра гена а-фетопротеина показана на рис. 9.20,Л).
У человека два упомянутых гена, сцепленных с
третьим членом семейства [геном витамин D-свя-
зывающего плазматического белка-группоспеци-
фичного компонента (Gc)], локализованы в длинном
плече хромосомы 4.
Сывороточный альбумин и а-фетопротеин дают
перекрестную иммунологическую реакцию. Они со-
стоят примерно из 580 аминокислот и сходны по
35% из них. Оба они содержат по три домена (рис.
9.20,5), на удивление сходных по структуре, поло-
жению цистеиновых остатков и числу аминокислот
между соседними цистеинами. Эти домены, по-ви-
димому, представляют собой родственные полипеп-
тиды. О родстве свидетельствует анализ гена а-фе-
топротеина мышей. На рис. 9.20,Л сегменты ДНК,
соответствующие трем доменам в белке, располо-
жены один под другим. Видно, что число и поло-
жение вставочных последовательностей во всех до-
менах примерно одинаковы. Кроме того, кодирую-
щие последовательности домена I на 39 и 35%
гомологичны кодирующим последовательностям
доменов II и III соответственно. Итак, ген а-фе-
топротеина содержит три тандемные копии сегмен-
та ДНК. В ходе эволюции эти копии дивергировали,
но и сейчас видно, что в их основе лежит одна
предковая последовательность. Аналогичную струк-
туру имеют и альбуминовые гены X.laevis, что
подтверждает гипотезу существования общего пред-
шественника у генов амфибий и млекопитающих.
Однако у Xenopus а-фетопротеин отсутствует. В
этом случае независимные реитерации привели
к появлению дополнительных альбуминовых ге-
нов.
Гены, кодирующие рецептор липопротеина низкой
плотности. В крови позвоночных холестерол цирку-
лирует в основном в виде липопротеина низкой
плотности (ЛПНП), который представляет собой
замкнутую мембранную структуру с интегральным
белком апопротеином В и содержащимися внутри
структуры молекулами эфира холестерола. ЛПНП
связывается со специфическим рецептором, распо-
ложенным на цитоплазматической мембране разно-
образных клеток, после чего происходит эндоцитоз.
Далее рецептор отсоединяется от ЛПНП и возвра-
щается в мембрану, а ЛПНП направляется к лизо-
сомам, в которых апопротеин В деградирует. Вы-
свободившийся холестерол вновь включается в мем-
брану. Кроме того, внутриклеточный холестерол
регулирует скорость собственного биосинтеза, по-
давляя транскрипцию гена, кодирующего 3-гидро-
кси-З-метил-глутарил-СоА-редуктазы, первого уни-
кального фермента в этой биосинтетической цепи.
Таким образом, рецептор ЛПНП выполняет две
функции: он участвует в регуляции скорости обра-
зования холестерола и скорости его поступления из
плазмы. Нарушения функционирования рецептора
холестерола лежат в основе заболевания семейной
гиперхолестеролемии (разд. 9.5.г).
Рецептор ЛПНП-сложный белок, состоящий из
839 аминокислот, которые образуют несколько до-
менов (рис. 9.21). Это домен, ответственный за
связывание ЛПНП и выступающий из мембраны;
еще один наружный домен с неизвестной функцией;
небольшой высокогликолизированный домен, ло-
кализованный на поверхности мембраны; малый
гидрофобный домен, пронизывающий мембрану;
домен, выступающий в цитоплазму и необходимый
для эндоцитоза.
У человека единственный ген рецептора ЛПНП
имеет 18 экзонов, соответствующих нескольким бел-
ковым доменам (рис. 9.21). Экзон 1 кодирует сиг-
нальную последовательность, которая удаляется в
просвете эндоплазматического ретикулума. Экзоны
2-6 кодируют ЛПНП-связывающий домен и содер-
жат семь тандемных повторов сегмента, кодирую-
щего обогащенный цистеином белок длиной 40 ами-
нокислот. Эти повторы различаются, но несомненно
происходят от общего предка. Кроме того, эти семь
родственных полипептидных последовательностей
гомологичны также участку из 40 аминокислот,
который повторяется несколько раз в одном из
белков системы комплемента-факторе С9. Таким
образом, в образовании ЛПНП-связывающего до-
мена принимали участие как перестановки экзонов,
так и амплификация. Эти два процесса играли
важную роль и в образовании кодирующих после-
довательностей домена с неизвестной функцией
в центре белка (экзоны 7-14). Каждый из экзонов 7,
8 и 14 кодирует полипептидные участки длиной 40
аминокислотных остатков, сходные по аминокис-
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
185
Интрон
"ООСОСг
РИС. 9.20.
А. Структура гена а-фетопротеина.
Перевернутыми треугольниками от-
мечено положение интронов в гене
а-фетопротеина мыши. Числа- раз-
меры экзонов (п.н.). Весь ген разбит
на три отрезка: I, II и III (см. текст).
14-й интрон расположен в 5'-лидер-
ной последовательности и не пока-
зан. Б. Схематическое представление
а-фетопротеина мыши и сывороточ-
ного альбумина человека, иллюстри-
рующие их сходство. Три домена со
сходной конфигурацией, отмеченные
цифрами I, II и III, соответствуют
трем сегментам гена, показанным на
рис. А. Буква X обозначает позиции,
в которых остатки цистеина встреча-
ются только в молекуле сывороточ-
ного альбумина, но не а-фетопро-
теина. Указано число аминокислот
между цистеиновыми остатками (оно
не соответствует числу кружков, в
виде которых представлена полипеп-
тидная цепь); если цифры две, то это
число у двух белков неодинаково.
[М. В. Gorin et al., J. Biol. Chem. 256
(1981), p. 1954.]
186
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Домен с неизвестной
функцией
Сигнальный Домен Связывания ЛПНП Гликозклиро- Домен,пронизываю- цитоплазмати-
пептидНЫИ Гомологичен С 9 фактора роста ванный домен щии мембрану ческии домен
Номера
экзонов
7 9
11 12 14
11
РИС. 9.21.
Схематическое представление гена рецептора ЛПНП
человека; показаны 18 экзонов и перечислены коди-
руемые ими функциональные домены. С9- один из бел-
ков системы комплемента, EGF-эпидермальный фак-
тор роста. [М. Brown, J. Goldstein, Science 232 (1986),
Р- 34.J
лотной последовательности с повторами в другом
белке-эпидермальном факторе роста.
Гены «приклеивающих» белков дрозофилы. На ли-
чиночной стадии слюнные железы Drosophila экскре-
тируют белки, с помощью которых куколка при-
клеивается к различным поверхностям. Используя
мРНК одного из этих белков, sgs4, клонировали
соответствующую кДНК и использовали ее как
зонд для выделения гена Sgs4 из геномной библио-
теки Drosophila. Вблизи 5'-конца кодирующей об-
ласти гена Sgs$ находятся 19 прямых тандемных
повторов последовательности длиной 21 п.н., ко-
торая кодирует семь аминокислот (рис. 9.22). Все
вместе эти повторы составляют почти половину
кодирующей области. Повторы несовершенны, т. е.
гептапептидная единица также дивергировала. У не-
скольких линий Drosophila ген Sgs$ имеет другую
длину вследствие изменения числа копий 21 п. н.-
повтора. У одной такой линии число повторов
равно 19, у других-22 и 27. Этот феномен может
быть связан с неравным кроссинговером.
Коллагеновые гены. Коллаген - основной внекле-
точный структурный белок позвоночных. Несмотря
на разнообразие существующих форм коллагена,
все они представляют собой тройные спирали, об-
разованные тремя не обязательно одинаковыми по-
липептидными цепями (рис. 1.30). Уникальной осо-
бенностью коллагеновых пептидов является то, что
у них каждой третьей аминокислотой является гли-
цин (gly-X-Y); кроме того, они обогащены пролино-
выми и лизиновыми остатками, многие из которых
гидроксилированы (посттрансляционная модифика-
ция).
Гены, кодирующие а2(1)-коллагеновые полипеп-
тиды у курицы, мыши и человека, сходны между
собой (рис. 9.23, Л). Как показывает электронно-
микроскопическое исследование коллагеновой
мРНК курицы, гибридизованной с клонированными
участками этого гена длиной 38 т. п. и., коллагено-
вый ген имеет весьма необычную организацию. Он
содержит не менее 52 экзонов, длина интронов
варьирует от 80 п. н. до более чем 2 т. п. н. Однако
длина всех экзонов кратна девяти нуклеотидам
и составляет 54 или 108 п.н. (рис. 9.23,Б). Таким
образом, каждый ген состоит из амплифицирован-
ных единиц длиной 9 п. н„ чем и объясняется повто-
рение в полипептидной цепи трипептида gly-X-Y.
б. Тандемные повторы
вне кодирующих областей
Тандемные повторы часто обнаруживаются меж-
ду генами и в интронах и содержат от двух нуклео-
тидов и выше. Некоторые примеры такого рода
приведены в табл. 9.11. Такие вездесущие повторы
могут быть представлены различными последова-
тельностями, например (CCG)„, (АТ)„ и (ТААССС)„,
но мы сейчас не можем приписать большинству из
них какие-либо функции. Среди наиболее простых
повторяющихся единиц особенно интересна одна.
б Каноническая
последовательность
АСА TGCАA A ACTGA6 CCA CCC
thr cys lys thr glu pro pro
g Отдельные повторы
TGCTGTAAAACTGAACCACCG
cys cys lys thr glu pro pro
AC A TGC A A A A CT G AG CCA CCC
thr cys lys thr glu pro pro
ACATGCAGAACTGAGCCACCC
thr cys arg thr glu pro pro
РИС. 9.22.
А. Структура гена Sgs4 Drosophila. Ген содержит
19 тандемных повторов единицы длиной 21 п.н.
Б. Каноническая последовательность единицы
длиной 21 п. н. В. Канонические последователь-
ности нескольких повторов. [М.А. Т. Muskavetch,
D.S. Hogness, Cell 29 (1982), р. 1041.]
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
187
12т.п.н.
-------------------1
99 45 99 54 108
28т.п.н.
З'-конец
GGT GAA CCA GGC ССТ GCT GGT GCC
gly glu pro gly pro ala gly ala
AAG GGA GAA AGT GGT AAC AAG GGT GAG CCT
lys gly glu ser gly asn lys gly glu pro
Типичный
экзон
Б
РИС. 9.23.
А. Структура участка, кодирующего тройную спираль
а2(1)-коллагена курицы. Участок разбит на два сегмен-
та; экзоны изображены в виде цветных черточек, а
интроны выделены точками. Указан размер некоторых
ДНК-зонд с чередующейся последовательностью
5'-САСА... (или GT) гибридизуется со многими рест-
рикционными фрагментами ДНК всех эукариот, т. е.
эта последовательность многократно повторяется
в большинстве геномов. Возможно, она ответствен-
на за образование Z-формы ДНК.
Связь с полиморфизмом. Тандемные повторы,
где бы они ни находились, как правило, связаны
Таблица 9.11. Некоторые тандемные повторы,
диспергированные в геномах разных эукариот
Повтор Организм Локализация
5-CAAAGTTT- -GAGTTTT Xenopus laevis Спейсер в генах 5S-PHK нз ооцитов
5-ACAGGGGT -GTGGGG Человек 5'-конец инсулинового гена
5'-СА Многие риоты эука- Многие геномные локусы и теломеры дрожжей
5'-GA Многие риоты эука- Спейсер между генами гистонов Н2А и HI у морского ежа
5-ТСТСС Курица Интрон в гене X в семей- стве овальбуминовых генов
5'-GGAAG » 2,5 т.п.н. с 5'-конца гена кональбумина
5'-GGAAA Фазан 2,5 т. п. н. с 5'-конца гена кональбумина
5'-АТ Многие риоты эука- Множество локусов
экзоное (в п. н.). [J. Wozney et al., Nature 294 (1981), p.
129.] Б. Нуклеотидная последовательность одного эк-
зона и кодируемый им полипептид.
с межвидовым полиморфизмом. Происхождение
подобной вариабельности можно объяснить нерав-
ным кроссинговером между родственными неал-
лельными повторами или «проскальзыванием» при
репликации (разд. 10.1.в). Спейсеры между генами
рРНК ооцитов X. laevis и между тандемными тран-
скрипционными единицами рРНК обычно содержат
множество внутренних тандемных повторов (разд.
9.2); они ответственны за полиморфизм длины спей-
серов. Изменение длины происходит в результате
появления одной или нескольких лишних повторяю-
щихся единиц или их утраты точно таким же спосо-
бом, как это происходит при изменении числа копий
тандемного повтора в гене Sgs4 у Drosophila.
Для экспрессии иммуноглобулиновых генов не-
обходима значительная перестройка ДНК (разд.
Ю.б.в). Рекомбинации, в результате которых ста-
новится возможным образование функционально
активных тяжелых цепей, происходят в пределах
специфических некодирующих последовательностей-
«переключателей», которые содержат множество
тандемно повторяющихся последовательностей
5'-(GAGCT)„GGGGT-3', где и = 1 - 5. Эти области
генома также полиморфны.
Примерно на 500 п.н. левее 5'-конца гена инсу-
лина человека находится тандемный повтор из еди-
ниц 5'-ACAGGGGTGTGGGG-3'. Данный сегмент
присутствует не во всех геномах и отличается су-
щественным полиморфизмом. Обнаружено мно-
жество разных аллелей этого локуса, и большинство
188
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
особей гетерозиготно по нему. Впервые такой по-
лиморфизм проявился в изменчивости длины рест-
рикционных фрагментов, которые гибридизовались
с клонированным ДНК-зондом из этой области. Он
обусловливается различиями в числе повторяющих-
ся единиц-от 26 до ~200, а также случайными
изменениями в самих этих единицах. Какую функ-
цию выполняет эта последовательность-неизвест-
но; вероятно, она не связана с локализацией ее
вблизи инсулинового гена, поскольку по соседству
с геном крысиного инсулина ни одной ее копии не
обнаружено.
Мини-сателлиты. Предрасположенность тан-
демных повторов к полиморфизму наиболее ярко
проявляется на примере так называемых мини-са-
а (AGGGCTGGAGGAGGGCTGGAGGAGGGCTGGAGG) )8
6 (AGAGGTGGGCAGGTGG) Jg
в (GGGAGG&GGGCAGGAGG) («
Индивидуумы: 8 9 10 11 12 13
г
РИС. 9.24.
Если использовать в качестве зондов клонированные
мини-сателлитные последовательности ДНК человека,
то обнаружится полиморфизм рестрикционных фраг-
ментов по длине. В верхней части рисунка приведены
нуклеотидные последовательности трех родственных
мини-сателлитов (а, б. е). Ниже приведены результаты
блот-гибридизации клонированного зонда б с ДНК,
обработанной эндонуклеазой Hinf \ (г). Четыре образца
ДНК получены от членов одной семьи: отца (квадратик),
матери (кружок) и их детей-однояйцовых близнецов
(деа квадратика). Две последние дорожки соответст-
вуют ДНК, взятой от неродственных индивидуумов
мужского пола. Стрелками отмечены «родительские»
сателлитные полосы на электрофореграммах ДНК
близнецов. [A. Jeffreys et al.. Nature 316 (1985), p. 76;
фотография любезно предоставлена A. Jeffreys.]
теллитных последовательностей. В геномах млеко-
питающих в разных местах расположено несколько
таких относительно простых тандемных повторов
разного типа (рис. 9.24). При блот-гибридизации
зондов, содержащих сегменты мини-сателлитной
ДНК, с рестрицированной геномной ДНК на элект-
рофореграмме появляется несколько полос. Их рас-
пределение у разных особей данного вида разли-
чается, но для ДНК из разных тканей одного инди-
вида остается одинаковым. Распределение полос
наследуется по Менделю, причем можно соотнести
их с тем или иным родителем. Количества ДНК,
содержащегося в нескольких каплях крови, вполне
достаточно для проведения блот-гибридизации, так
что метод, основанный на использовании мини-са-
теллитной ДНК, можно применять в судебно-меди-
цинской практике.
в. Тандемные повторы в области
центромер и теломер
Наибольшая концентрация тандемных повторов
обычно наблюдается в области центромер и тело-
мер в хромосомах животных и растений. Иногда
в одном таком участке обнаруживаются сотни и да-
же тысячи копий какой-то одной единицы, и на их
долю часто приходится значительная часть сум-
марной геномной ДНК (табл. 9.12). При денатура-
ции суммарной ДНК с последующей реассоциацией
эти последовательности составляют фракцию наибо-
лее быстро ренатурирующей ДНК (т. е. для них
характерны очень малые значения Cot). Как и дру-
гие длинные тандемные повторы, при равновесном
центрифугировании в градиенте плотности эти эле-
менты часто образуют отдельные «сателлитные зо-
ны» (рис. 9.25). Все эти ДНК, включая и ДНК из
зон, содержащих рДНК или повторяющиеся гисто-
новые гены, были названы сателлитными. Этот тер-
мин используется сейчас в основном в отношении
Таблица 9.12. Отношение центромерной сател-
литной ДНК к суммарной геномной ДНК (%)
Вид” %
Geeareinus lateralis Drosophila melanogaster Mus musculus Rattus norvegicus Bovis domesticus Cercopithecus aethiops (африканская зеленая мартышка) Homo sapiens 21 16 8 1-3 >23 13-20 Вероятно. около 5
Приведены точные видовые названия, поскольку содержание сателлитных последовательностей варьирует от вида к виду даже в пределах одного рода.
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
189
Плавучая плотность,г/смз
РИС. 9.25.
Равновесное центрифугирование суммарной ДНК Dro-
sophila в градиенте плотности CsCI (см. рис. 9.4); видны
зоны разной плотности, соответствующие разным са-
теллитным последовательностям в области центроме-
ры. Числа плавучая плотность (р). Для выделения
сателлитных последовательностей IV, II и III (см. табл. 9.13)
необходимо повторное центрифугирование. [S.A. Endow,
M.L. Polan, J.G. Gall, J. Mol. Biol. 96 (1975), p. 665.]
длинных тандемных повторов, расположенных в
области центромер и теломер, независимо от того,
образуют ли они дискретные зоны при центрифуги-
ровании в градиенте плотности.
У многих организмов имеется несколько раз-
ных сателлитных ДНК. Иногда (например, у Dro-
sophila) они концентрируются в отдельных зонах
при центрифугировании ДНК в градиенте плот-
ности (рис. 9.25). У некоторых организмов центро-
мерные и теломерные повторы вообще не обнару-
живаются при центрифугировании. Такие крити-
ческие сателлитные ДНК либо находятся в составе
основной зоны, либо в другом месте, но их ко-
личества недостаточно для образования дискретной
зоны.
Разные сателлитные ДН К различаются по длине
повторяющейся единицы и ее нуклеотидной после-
довательности. Длина этой единицы варьирует от
двух пар нуклеотидов, как у сателлитной ДНК
краба, где она представляет собой просто чере-
дующийся динуклеотид из остатков А и Т, до
нескольких тысяч нуклеотидных пар.
Общие свойства сателлитных ДНК из области
центромер и теломер. РНК, гомологичная сателлит-
ной ДНК, обнаруживается лишь в редких случаях.
По-видимому, сателлитная ДНК не транскрибиру-
ется в сколько-нибудь заметных количествах и ско-
рее всего ничего не кодирует. Как правило, центро-
меры и теломеры гетерохроматиновые, т. е. в отли-
чие от слабо окрашенных эухроматиновых районов
хромосом очень интенсивно окрашиваются по Гим-
за (см. «С»-полосы на рис. 7.23). Таким образом,
сателлитные ДНК входят в состав гетерохроматина,
и эти два термина часто используются как сино-
нимы, но как именно соотносятся между собой
тандемные повторы и характер окрашивания, неяс-
но. Другие области хромосом, в том числе и район
ядрышкового организатора, содержащий повторы
рДНК, также являются гетерохроматиновыми. К
этой же категории относятся и другие, молчащие
области хромосом, такие как инактивированная
Х-хромосома. Сателлитная ДНК, находящаяся в
области центромеры, реплицируется в S-фазе в
числе последних, а в политенных хромосомах она
часто оказывается недореплицированной по срав-
нению с другими участками ДНК.
Исследование сателлитных ДНК с помощью
рестриктирующих эндонуклеаз. В исследовании при-
роды сателлитных ДНК, как выделенных с по-
мощью равновесного центрифугирования, так и
криптических, большую роль сыграли опыты по
эндонуклеазному расщеплению с последующей
блот-гибридизацией. Значительно прояснили карти-
ну клонирование и секвенирование этих необычных
геномных сегментов.
Если сайт для какой-то рестриктирующей эндо-
нуклеазы находится в каждом из повторов тандем-
ного кластера, то такой кластер расщепляется на
фрагменты, длина которых равна длине повторяю-
щейся единицы (рис. 9.26, А). При гель-электрофо-
резе суммарной геномной ДНК, окрашенной бро-
мистым этидием, набор этих фрагментов одинако-
вой длины очень часто образует отдельную полосу.
ДНК, элюированную из этой полосы, можно сразу
подвергнуть предварительному секвенированию.
Полученная таким образом каноническая консен-
сусная последовательность не обязательно соответ-
ствует последовательности любой из повторяющих-
ся единиц, поскольку, как мы уже говорили, такие
единицы в центромерных тандемных повторах ред-
ко бывают идентичными. Обычно они образуют
семейство, члены которого отличаются друг от
друга одним или более нуклеотидным остатком.
Некоторые из них дивергировали в результате слу-
чайных нуклеотидных замен. Иногда нуклеотидные
замены обнаруживаются сразу в нескольких или во
всех единицах какой-то части кластера. Если часть
кластера существенно отличается от сателлитной
последовательности в целом, то говорят о субклас-
тере или о сателлитном домене. Секвенирование
клонированных сателлитных повторов, доменов
и мономерных повторяющихся единиц показало,
что то, что раньше представлялось совершенно
гомогенной последовательностью, на самом деле
есть совокупность в значительной степени разли-
чающихся единиц.
190
ЧАСТЬ III, ОРГАНИЗАЦИЯ. ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
| Hinfl
I Электрофорез
I НМ\
I Электрофорез
359 п.н.
1077 п.н.
718 п.н.
359 п.н.
б
РИС. 9.26.
Эндонуклеазное расщепление сателлитной последова-
тельности III из области центромеры Drosophila. При
электрофорезе рестриктов различных доменов (о, б, в,
г) обнаруживаются разные фрагменты. А. Основная
повторяющаяся единица имеет длину 359 п. н. и содер-
жит один сайт для эндонуклеазы Hinf\. При расщеп-
лении с помощью Hinf\ большая часть сателлитных
последовательностей превращается в набор фрагментов
длиной 359 п. н. в количестве, достаточном для обра-
зования отдельной полосы на электрофореграмме, чет-
ко различимой на фоне других геномных фрагментов
после окрашивания бромистым этидием. Б. В некото-
рых доменах сателлитной последовательности III у
части повторяющихся единиц сайт для Hinf\ отсутствует.
При расщеплении с помощью Hinf\ образуется «лестни-
ца» фрагментов ДНК длиной 359 п.н., 2 x 359 п.н.
и т.д. В. В некоторых доменах сателлитной последо-
вательности III повторяющиеся единицы содержат сайты
для эндонуклеазы НаеIII. При расщеплении с помощью
A/aelll образуется лестница фрагментов, длина которых
кратна 359 п. н„ вплоть до 359 х 15 п. н. (не показаны).
Г. В каждой повторяющейся единице сателлитной по-
следовательности III содержатся сайты как для Hinf, так
и для Л/аеШ.
Отметим, что если те или иные модификации
затрагивают эндонуклеазный сайт в повторяющейся
единице, то продукты рестрикции будут представле-
ны не только мономерной повторяющейся едини-
цей, но и фрагментами, кратными двум, трем и г. д.
«Лестница сегментов», которую получают при
электрофорезе, как раз и свидетельствует о наличии
сателлитной ДНК (рис. 9.26, В).
Иногда повторяющаяся единица сателлитной
последовательности содержит сайты для нескольких
рестриктаз. При расщеплении сателлитной ДНК
любым из этих ферментов образуются повторяю-
щиеся фрагменты одинакового размера. Однако при
последовательном расщеплении разными фермен-
тами сателлитов из одного и того же препарата
ДНК могут получаться фрагменты разной длины.
Таким образом, используя набор разных рестрик-
тирующих ферментов, можно провести предвари-
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
191
тельный анализ сателлитных компонентов генома.
Более детальные количественные данные можно
получить, используя клонированные или даже не-
клонированные сателлитные сегменты в качестве
ДНК-зондов. Можно использовать также суммар-
ную геномную ДНК или фракции, полученные при
ее центрифугировании в градиенте плотности. Про-
водя гибридизацию этих зондов с хромосомами in
situ, удается локализовать сателлитную центромер-
ную и теломерную ДНК. Такой анализ довольно
часто позволяет выявить множественные центро-
мерные сателлиты, специфичные для определенного
вида. Эта специфичность проявляется в длине
повторов и их нуклеотидной последовательности.
Видоспецифичные сателлитные ДНК. Каждый
вид имеет характерный набор центромерных сател-
литных последовательностей, отличный от соответ-
ствующего набора даже у близкородственного вида
данного рода. Этот факт, замечательный с точки
зрения высокой консервативности последователь-
ностей и геномной синтении, создает серьезные
трудности при объяснении функций сателлитов.
Чтобы проиллюстрировать возникающие здесь
проблемы, рассмотрим сателлитные последователь-
ности некоторых организмов.
Центромерные сателлитные последовательности
у D.melanogaster. В ДНК D.melanogaster имеются
четыре разные сателлитные последовательности
(рис. 9.25 и табл. 9.13). Все вместе они составляют
около 16% геномной ДНК. Сателлитная последо-
вательность I содержит два простых близкородст-
венных домена: один с пентамерной и другой-с
гептамерной повторяющейся единицей. Почти 80%
сателлитной последовательности II составляют тан-
демные декамерные повторы. Сателлитная после-
довательность IV, как и последовательность I, со-
держит пентамерный и гептамерный домены и, оче-
видно, родственна сателлитной последовательности
1. Сателлитная последовательность III имеет уни-
кальную структуру. Она содержит повторяющуюся
единицу длиной 359 п.н., и в большинстве таких
единиц находится сайт для эндонуклеазы Hinfl (рис.
9.26,Л). Примерно 16000 повторов в составе сател-
литной последовательности III полиморфны. Мно-
гие из них отличаются друг от друга и от ка-
нонической последовательности всего несколькими
парами оснований. Об этом свидетельствуют ре-
зультаты экспериментов по расщеплению сателлит-
ной последовательности III или ее длинных клони-
рованных сегментов рестриктазой НаеШ. После
разделения образовавшихся фрагментов с помощью
гель-электрофореза получается «лестница» (рис.
9.26,В). Это значит, что многие единицы в тандем-
ном повторе содержат сайт для НаеШ, но есть
единицы, в которых он отсутствует. Клонирование
длинных тандемных повторов сателлитной после-
довательности III показало, что они содержат от-
дельные домены, по-видимому представляющие
собой однородную сателлитную последователь-
ность. Некоторые домены содержат в каждой пов-
торяющейся единице сайты как для НпеШ. так и для
Hinfl (рис. 9.26, Г). В других доменах в каждой
единице присутствует Н infl-cam, а сайты для НаеШ
вообще отсутствуют (рис. 9.26,Я). Секвенирование
клонированных повторяющихся единиц показывает,
что члены семейства отличаются и по другим приз-
накам. Например, один домен сателлитной после-
довательности III построен из тандемных повторов
единицы, состоящей из 254 п.н. вместо обычных
359 п. н. Итак, одни центромерные сателлитные
последовательности в ДНК Drosophila устроены до-
вольно просто, а другие, например сателлитная
последовательность III, имеют сложную структуру.
При отжиге in situ радиоактивных зондов сател-
литных последовательностей 1 и IV с политенными
хромосомами Drosophila гибридизация происходит
со всеми центромерами (рис. 9.27). Однако ее эф-
фективность у разных хромосом значительно раз-
личается, что свидетельствует о вариабельности
числа повторяющихся единиц. Кроме того, каждая
сателлитная последовательность гибридизуется с
определенной областью центромеры, что говорит
о специфичности ее локализации. Сателлитная пос-
ледовательность III гибридизуется только с X- и
Y-хромосомами. Таким образом, центромерные са-
теллитные последовательности локализуются каж-
дая в определенной области. Более того, центро-
мерные сателлитные последовательности Drosophila
видоспецифичны. Об этом свидетельствует сравне-
ние сателлитных последовательностей D. virilis и
Таблица 9.13. Содержание центромерных сател- литных ДНК у D. me/anogaster и D. virilis
Сателлит D. meianogaster
% геном- ной ДНК повторяющаяся единица
1 2 5'-[ А АТ АТ] А АТ АТ А АТ АТ и т.д.
1 5'-[ААТАТАТ] ААТАТАТААТ и т.д.
II 3 5'-[AATAACATAG] ААТААС и т.д.
III 5 Повторяющиеся единицы длиной
254 и 359 п. н.
IV 4 5'-[AAGAG] AAGAGAA и т. д.
0.5 5'-[AAGAGAG]AAGAGAG и т.д.
Сателлит D. virilis
% геном- ной ДНК повторяющаяся единица
I 25 5'-[АСАААСТ] AC АА ACT АС и т.д.
II 8 5'-[АТАААСТ] АТАААСТАТАА и т.д.
III 8 5'-[АСАААТТ] АСАААТТАС и т.д.
IV 0.1 5'-[AATATAG]AATATAGAA и т.д.
192
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 9.27.
Гибридизация in situ радиоактивных зондов центромер-
ной сателлитной последовательности I (р = 1,672) Dro-
sophila с митотическими хромосомами 3 (А) и Y (Б).
Внизу (рис. В) показано положение сателлитных по-
следовательностей I и IV (р = 1,705) на хромосоме 3.
Относительное положение двух сателлитов на границе
между центромерным гетерохроматином и эухромати-
ном плеча 3R определяли с помощью электронной
микроскопии отожженных препаратов. [D. М. Steffenson,
R. Appels, W. J. Peacock, Chromosoma (Berl.), 82 (1979),
p. 525 ]
D. melanogaster (табл. 9.13). Хотя эти мухи и отно-
сятся к одному роду, их центромерные сателлитные
последовательности существенно различаются: с
повторами D. melanogaster сходен лишь повтор 1с
D.virilis. Заметим также, что три из сателлитных
последовательностей D. virilis настолько близки, что
при гибридизации in situ их трудно различить.
Сателлитные последовательности, которые не уда-
ется разделить по плавучей плотности, считаются
доменами одного сателлита.
Сателлитные последовательности у быка и мы-
ши. У быка известны восемь разных сателлитных
последовательностей (рис. 9.28). Все вместе они
составляют более 23% геномной ДНК и, судя по
кинетике реассоциации, занимают промежуточное
положение между высоко- и умеренноповторяющи-
мися последовательностями (см. разд. 9.1.6).
Повторяющиеся единицы представлены набором
взаимосвязанных последовательностей. Секвениро-
вание показало, что по меньшей мере пять из
восьми сателлитов содержат одну или более оди-
наковых последовательностей, хотя и дивергиро-
вавших настолько, что не гибридизуются между
собой, а рестрикционный анализ не выявляет их
родства. Длина повторяющейся единицы у боль-
шинства сателлитов быка превышает 1 т. п. н. Внут-
ри этих длинных повторяющихся единиц имеются
повторы, встречающиеся более часто.
Такие сложные структуры характерны не только
для бычьей сателлитной ДНК. В сателлитах у дру-
гих организмов, несмотря на их видоспецифичность,
также имеются последовательности, встречающиеся
относительно редко и более часто. Главный цент-
ромерный сателлит Mus musculus, например, имеет
в своей основе повторяющуюся единицу длиной
234 п. н. В каждой из таких единиц имеется по
В
одному сайту для эндонуклеаз 5ow96I, EcoRll и
Avail. Секвенирование показывает, что повтор раз-
мером 234 п.н. в свою очередь содержит четыре
(несовершенных) внутренних тандемных повтора
длиной 58, 60, 58 и 58 п.н. Каждый из них можно
разбить на два родственных, но различающихся
сегмента длиной 28 и 30 п.н. соответственно. Эти
сегменты также родственны и, возможно, образо-
вались из одной нуклеотидной последовательности.
а-Сателлиты приматов. У приматов имеются
сателлитные последовательности как с длинными,
так и с короткими повторяющимися единицами. По
крайней мере один класс центромерных сателлитов
у каждого вида из числа исследованных содержит
видоспецифичную повторяющуюся последователь-
ность длиной примерно 170 п.н., называемую
a-последовательностью (рис. 9.29). а-Сателлиты
различных организмов не всегда гибридизуются
между собой, но тем не менее они близкородствен-
ны, о чем свидетельствует их первичная структура.
Например, повторяющаяся единица длиной 340 п. н.
у бабуина и некоторые а-сателлиты человека пред-
ставляют собой димеры повторяющейся единицы
длиной 172 п. н., присутствующей в ДНК афри-
канской зеленой мартышки. Эти два мономера,
составляющие сателлитный димер у человека, отли-
чаются друг от друга и от соответствующей после-
довательности у мартышки на 30%. У человека
и бабуина эти последовательности также различа-
ются. В а-сателлите бабуина один раз на каждые
343 п.н. встречается сайт для эндонуклеазы В«/иН1,
а в домене а-сателлита человека один раз на каждые
340 п.н.-сайт для эндонуклеазы EcoRl. Наблю-
дается внутривидовая дивергенция повторяющихся
единиц. Нуклеотидная последовательность клони-
рованной повторяющейся единицы длиной 172 п.н.
ц-сателлита мартышки отличается от канонической
последовательности в среднем на 3%. Отмечаются
изменения двух типов. Первые, весьма редкие, за-
трагивают отдельные нуклеотидные пары и могут
обусловливаться точковыми мутациями. Вторые
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
193
ДЛИНА
ПОВТОРА, п.н.
2350 ’
3800
1413
2600
1,720а 0,1
Неизвестна
1399
Неизвестна
РИС. 9.28.
Сателлитные последовательности, выделенные при
центрифугировании геномной ДНК быка в градиенте
плотности. Гомологичные участки разных сателлитных
последовательностей выделены одинаковым цветом.
Сателлит с р = 1,711 (Ь) сходен с сателлитом с р =
= 1,715, за исключением того, что ~ 60% повторя-
ющихся единиц длиной 1399 п.н. прерываются участ-
ком длиной 1200 пар, частично гомологичных участку
сателлита с р = 1,711 (а). Последовательности с р =
= 1,723 и р = 1,709 нв родственны никаким другим.
Гомологичные участки сателлитов с р = 1,715 и р =
= 1,711 (Ь) содержат несколько внутренних тандемных
повторяющихся единиц длиной 31 п. н. Аналогично,
гомологичные сегменты в длинных повторяющихся еди-
ницах с р = 1,706 и р = 1,711(a) содержат множест-
венные внутренние тандемные повторы тех или иных
вариантов одной последовательности длиной 23 п. н„
а повторяющаяся единица длиной 46 п. н. в последова-
тельности с р = 1,720 (Ь) есть не что иное, как димер
того жв самого сегмента длиной 23 п. н.
касаются многих повторяющихся единиц; по-види-
мому, такие изменения тоже исходно затрагивали
отдельные пары оснований, но затем дивергиро-
вавшие повторяющиеся единицы амплифицирова-
лись. Например, многие мономерные единицы име-
ют уникальный сайт для EcoRI, а другие-специ-
фические сайты для Нас III; в канонической после-
довательности оба они отсутствуют. Те повторяю-
щиеся единицы, которые содержат сайты для Есо R1
или для //«till, обычно встречаются в разных тан-
демных повторах, называемых сателлитными до-
менами.
В некоторых а-сателлитных доменах человека
длина повторов значительно превышает длину ди-
меров, состоящих из двух единиц в 170 п.н. Оказа-
лось, что эти домены являются пентамерами или
даже додекамерами. Мономерные единицы в таких
длинных повторах часто отличаются от канони-
ческой последовательности на 15% и более. Клони-
рованные зонды, представляющие собой такие до-
мены, не гибридизуются с другими доменами в
жестких условиях. Используя эти специфические
зонды и линии гибридных соматических клеток или
гибридизацию in situ, удалось показать, что данные
домены находятся в центромерах специфических
хромосом. Например, домен, содержащий додека-
мерный повтор, имеет длину 12 х 171, или при-
мерно 2,1 т.п.н. (рис. 9.30). На каждые 2,1 т.п.н.
приходится по одному сайту для ВатН1 и Pst I. Как
показывают результаы опытов по гибридизации,
имеется примерно 5000 додекамерных повторов
и локализованы они в Х-хромосоме. В одной хро-
мосоме могут находиться несколько разных доме-
нов, но для каждой хромосомы характерны а-сател-
литные домены определенной длины. Такая хро-
мосомная специфичность встречается и у других
приматов; по-видимому, это общее свойство са-
теллитных ДНК.
Бабуин
(10%)
РИС. 9.29.
Африканская
зеленая
мартышка
(13-207»)
Человек
(>1%)
а-Сателлиты приматов и их содержание в геноме. Са-
теллиты африканской зеленой мартышки и бабуина-
типичные представители видоспецифичных а-сателли-
тов приматов. Приведенный здесь сателлит человека
представляет собой один домен полного а-сателлита
человека. На долю а-сателлитов может приходиться
более 5% геномной ДНК человека, но этот домен
составляет от геномной ДНК менее 1%.
194
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
171 п.н.
РИС. 9.30.
Один из доменов а-сателлита, локализованный в
Х-хромосоме человека. Каждая повторяющаяся едини-
ца длиной 2,1 т. п. н. содержит по 12 копий а-сатвллит-
ного мономера длиной 170 п. н. Эти 12 повторов диввр-
гировали между собой на 10-20%, однако в пределах
повторов более высокого порядка соответствующие
г. Возможные функции тандемных
повторов
Функции многих тандемных повторов, встреча-
ющихся в эукариотических геномах, пока не уста-
новлены. Конечно, здесь есть исключения-напри-
мер, тандемные повторы некоторых генов, тандем-
ные повторы внутри самих генов, специфические
последовательности типа «переключателей», связан-
ные с генами тяжелых цепей иммуноглобулина.
Однако функции многих диспергированных тандем-
ных повторов и огромных кластеров в районе цент-
ромер и теломер остаются загадкой. Возможно, мы
просто не знаем, каковы эти функции, но не исклю-
чено, что некоторые последовательности ДНК во-
обще не выполняют никаких определенных функ-
ций. Тандемные повторы могут образовываться
и элиминироваться благодаря таким процессам, как
репликация и рекомбинация ДНК. Если появление
и распространение этих повторов не нарушают рабо-
ту важных кодирующих и регуляторных элементов,
то повторы будут накапливаться и исчезать без
всякого влияния со стороны естественного отбора.
Полиморфизм некоторых тандемных повторов как
раз и свидетельствует об этом. Тем не менее это
положение представляется спорным, поскольку
эволюция большинства геномов происходит под
давлением естественного отбора. Геномы могут
эволюционировать и в результате нейтральных из-
менений в структуре ДНК-мутаций, никак не влия-
ющих на фенотип. Однако концепция нейтральной
молекулярной эволюции применима в основном
к изменениям на уровне единичных пар оснований
или к небольшим делециям и реитерациям, а также
к коротким диспергированным повторам с неясной
пока функцией. Что касается сателлитных после-
довательностей в области центромер и теломер, на
долю которых часто приходится весьма значитель-
ная часть суммарной геномной ДНК, то к ним
применить теорию нейтральной эволюции труднее.
2,1 т.п.н.
повторы (т.в. 1, 2, 3 и т.д.) весьма консервативны.
Сайты для SflmHI и Pst\ в пределах каждого 12-мера
располагаются в одной и той жв мономерной единице.
[J.S. Waye, Н. F. Willard, Nucleic Acid Res. 13 (1985),
p. 2731.]
Широкая распространенность сателлитных по-
следовательностей, их локализация и большой
объем содержащейся в них ДНК наводят на мысль,
что они выполняют какие-то важные функции. Для
выяснения функций этих областей хромосом долгое
время использовали цитогенетический анализ гете-
рохроматина, который обычно считают синонимом
сателлитной ДНК. Рассматривались самые разные
клеточные функции, прежде всего процессы, проис-
ходящие при митозе и в клетках зародышевой ли-
нии, в частности хромосомные перестройки, спа-
ривание и расхождение хромосом и рекомбинация.
Было высказано предположение, что сателлитные
последовательности столь же важны для функцио-
нирования генома, как и остальная его часть. Одна-
ко, несмотря на огромное количество эксперимен-
тальных работ, окончательный вывод так и не был
сделан. В настоящее время сателлитным последо-
вательностям не приписывается никаких фенотипи-
ческих эффектов и, следовательно, никаких функций.
Об отсутствии у них таких функций свидетельст-
вуют следующие факты. Функционально активные
центромеры S.cerevisiae не содержат центромерных
сателлитов (разд. 9.6). Мутанты Drosophila, у ко-
торых отсутствует большая часть или весь центро-
мерный гетерохроматин, функционируют нормаль-
но. Значительная вариабельность нуклеотидной
последовательности сателлитной ДНК даже у род-
ственных видов наводит на мысль о ненужности
этой ДНК вообще. Кроме того, недавно полученные
данные показывают, что количество сателлитной
ДНК в области центромеры и ее организация су-
щественно различаются даже у особей одного вида.
То же самое относится и к а-сателлитным после-
довательностям человека. Изменчивость минорных
центромерных сателлитов характерна также для
африканской зеленой мартышки. Эти данные позво-
ляют предположить, что для функционирования
центромер не важны ни число, ни точная нуклео-
тидная последовательность тандемных повторов.
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
195
Единственный факт, заслуживающий внимания
с точки зрения роли центромерной сателлитной
ДНК,-это ее широкая распространенность. Даже
у размножающихся путем деления дрожжей Schizo-
saccharomyces poinbe, в отличие от почкующихся
дрожжей Saccharomyces cerevisiae, имеются длинные
тандемные повторы в области центромер. Как мы
увидим ниже (разд. 9.4.Д), те же самые процессы,
которые приводят к изменению сателлитных после-
довательностей и к увеличению их числа в геноме,
могут приводить и к их элиминации, однако этого
не происходит. Возможно, тандемные повторы все
же играют какую-то роль в функционировании цент-
ромер, которая не зависит от их конкретной нуклео-
тидной последовательности. Интересное и, по-ви-
димому, имеющее отношение к обсуждаемому воп-
росу наблюдение сделано при исследовании хро-
мосом кукурузы. В 23 нецентромерных областях
этих хромосом были обнаружены гетерохромати-
новые узелки, которые обладают одним замеча-
тельным свойством: в мейозе они могут образо-
вывать дополнительные функционирующие центро-
меры. Узелковый гетерохроматин кукурузы, как
и центромерный гетерохроматин, реплицируется
в поздней S-фазе; узелки содержат длинные тан-
демные повторы последовательностей ДНК, кото-
рые отличаются от центромерных сателлитных пос-
ледовательностей. Таким образом, для функциони-
рования центромеры, по-видимому, важны тандем-
ные повторы как таковые, а не их нуклеотидная
последовательность.
д. Механизмы образования
и эволюции тандемных повторов
Реитерация последовательностей может проис-
ходить в результате общей рекомбинации между
неаллельными гомологичными участками ДНК
(разд. 2.4). Последовательные раунды такого нерав-
ного кроссинговера могут приводить к удлинению
или укорочению длинных тандемных повторов.
Вначале амплификация может захватывать доволь-
но короткие гомологичные участки (рис. 9.31)-та-
кие, как нонануклеотиды, встречающиеся с большой
частотой в сателлитной ДНК мыши. Известная
пластичность тандемных повторов в значительной
степени обусловливается неравной гомологичной
рекомбинацией, хотя нельзя исключить и такие
механизмы, как генная конверсия или «проскаль-
зывание» ДНК во время репликации.
Эволюция сателлитных ДНК. Сателлиты боль-
шинства классов ограничены по своей распростра-
ненности одним видом или группой близкородст-
венных видов; это наводит на мысль, что они
появились относительно недавно или изменялись
в ходе эволюции значительно быстрее, чем все
остальные области генома. Эти изменения, при-
водящие к образованию видоспецифичных сателли-
тов, по-видимому, представляли собой последова-
тельно сменяющие друг друга амплификации (и
делеции) при помощи неравного кроссинговера
и модификации нуклеотидных последовательностей
(нуклеотидные замены, делении и вставки) (рис.
9.32). Эта модель объясняет межвидовую изменчи-
вость, наличие специфических внутривидовых до-
менов и полиморфизм в пределах современных
видов. Наблюдаемые вариации будут зависеть от
соотношения между частотой рекомбинационных
событий и частотой изменений, фиксируемых в ходе
естественного отбора или в результате генетиче-
ского дрейфа (т. е. от вероятности фиксации). Эта
модель объясняет также образование более длинных
тандемных повторов из более коротких, однако она
не отвечает на вопрос, почему при образовании
сателлитов происходит амплификация лишь опре-
деленных последовательностей. Согласно одной
широко обсуждавшейся гипотезе, у родственных
видов имеется одинаковая специфическая библио-
тека последовательностей, способных амплифици-
роваться с образованием сателлитной ДНК. Однако
ни природа этой гипотетической библиотеки, ни
типы хранящихся в ней последовательностей не
описаны. Короче говоря, на вопрос о том, имеют ли
разные сателлитные последовательности какие-либо
общие свойства, мы не можем ответить более опре-
деленно, чем на вопрос о функции сателлитов.
Амплифицироваться и даже сохранять стабильность
в амплифицированных повторах при определенных
условиях могут (разд. 10.8) самые разные геномные
сегменты, в том числе и уникальные гены, но обыч-
но амплифицированные последовательности быва-
ют нестабильны. Сателлитные последовательности
также нестабильны в том смысле, что для них
характерна высокая изменчивость. Тем не менее
локализация, распределение и количество тандем-
ных повторов в геноме сохраняются. Какими осо-
быми свойствами обладают центромеры и тело-
меры, благодаря которым в них образуются и со-
храняются тандемные повторы с разной нуклеотид-
ной последовательностью?
Эволюция тандемных повторов, входящих в ко-
дирующие последовательности. В этой главе мы уже
приводили несколько примеров существования тан-
демных повторов внутри генов. В одних из них,
таких как рДНК, разные кодирующие сегменты
идентичны. В других, например в глобиновых генах
и в кластере генов гормона роста человека/плацен-
тарного лактогена, некоторые кодирующие сегмен-
ты относятся к разным генам. В общих чертах
механизм образования тандемных повторов путем
амплификации сегментов с последующей мутацией
одной копии гена и отбором считается установлен-
196
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Гомологичные участки
Кроссоверная точка
ТТ А А С A G G А Т А с С С G АТ С Т AG С С G С A A G С АТ А С Т Т G А С СI
(AATT GTCCTATGGGCTAGATCGGCGT TCGTATGAACTGGi
Репликация
i'TTAACAGGATACCCGATCTAgCCGGAAGCATACTTGACC
AATT GTCCTATGGGCTAGATC G GCGT Т СGТ ATGAACTGG
ttaacaggatacccgatcTagccgcaagcatacttgacc
A AT I 0 T C CT AT G i’ I .6 A T C GGCGT T CG AT • •• C GG
Кроссинговер
( ttaacaggat acccgatctagccgcaag atacccgat ctagccgc aa gcatacttgacc
(aattgtcctatgggctagatcggcgttc^tatgggctagatcggcgttcgiatgaActggi
+
TTAACAGCATACTTGACC
aattgtcgtatgaactgg
Репликация
(TTAACAGGAfrACCCG АТСТ a G CCGCAA GCATACTTGACCTAGCCG CAAGCAfTACTTGACCQ
( A A TT GT C CT(AT gggctagatcggcgtt cgtatgaact ggatcggcgtt c gt(at g a AC t ggQ
( TT AA C A^G G AT А С С C G АТС T A G С C G C A A G G АТ A С T T G А С С T A G С C G C A A(GC at A CT TG ACC(~
f А т т g t(c c tatg g g ctag at c g gc g t t cct at g aact g g AT c GGCGT t(cg t A T 6 A AC tgg(
TTAACAG CATACTTGACCQ
ААТТй 1CGTA1 G A ACTGG0
TTAACAGCATACTTGACC
AATTGTCGTATGAACTGG
РИС. 9.31.
Эволюция тандемных повторов в результате неравного
кроссинговера. Рекомбинация может происходить
между сегментами одной молекулы ДНК, между сест-
ринскими хроматидами или между гомологичными
хромосомами. На рисунке представлена рекомбинация
между сестринскими хроматидами после первой реп-
ликации. Изображены две молекулы, образовавшиеся
в результате кроссинговера, содержащие короткий
гетеродуплексный участок, в котором одна цепь проис-
ходит от одной родительской молекулы, а другая-от
другой. В этих участках возможно неправильное спари-
вание оснований, как это имеет место в одной из
приведенных рекомбинантных молекул. Несмотря на
сложность двух молекул, образовавшихся в результате
кроссинговера, каждую из четырех молекул, получив
шихся после их репликации, можно представить так, как
будто она образовалась в результате разрыва и вое
соединения в одной кроссоверной точке рекомбини
рующей молекулы. Эта точка для молекулы, отмеченной
звездочкой, указана в исходной последовательности.
Три другие молекулы могли бы образоваться при сме
щении точки кроссовера в другие сайты гомологичного
участка. Две более длинные молекулы содержат тан-
демные копии той части исходной последовательности,
которая расположена между соответствующими крос-
соверными точками. Две более короткие молекулы со-
держат делеции. [G. Р. Smith, Science 191 (1976),
р. 528.]
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
197
| Амплификация
< с с с е-8
। Дивергенция в результате мутации
1 (транзиции,трансверсии, делеции.инсерции)
! («Ж(-8г-юн
| Амплификация
РИС. 9.32.
Возможные эволюционные изменения сателлитных
ДНК за относительно непродолжительное время. Дан-
ная модель применима для образования как видоспе-
цифичных сателлитов, так и сателлитных доменов в
пределах вида. [Е. М. Southern, Nature 227 (1970),
р. 794.]
ным. В некоторых случаях, изучая детальную струк-
туру повтора, удается проследить процесс реитера-
ций на молекулярном уровне. В тех моделях, ко-
торые при этом были построены, считалось, что
амплификация обычно происходит в результате не-
равного кроссинговера между членами семейств
повторяющихся последовательностей, рассеянных
по геному (рис. 9.33).
9.5. ПОВТОРЯЮЩИЕСЯ
ПОСЛЕДОВАТЕЛЬНОСТИ,
РАССЕЯННЫЕ ПО ВСЕМУ ГЕНОМУ
Как правило, члены мультигенных семейств,
равно как и функциональные гены или процессиро-
ванные псевдогены, многократно повторяются в не-
сцепленных участках генома. Однако за немногими
исключениями число копий таких последователь-
ностей не превышает 50. Максимальное число ко-
пий-1000-известно для псевдогенов U1-PHK че-
ловека. Однако многие эукариотические ДНК со-
держат и другие, очень большие семейства повто-
ряющихся последовательностей, члены которых
рассеяны по геному в количестве 106 копий. Роль
большинства таких последовательностей остается
загадкой, хотя некоторые члены семейств могут
быть функциональными генами.
Замечательное свойство этих семейств последо-
вательностей состоит в их видовой изменчивости.
Например, огромные семейства диспергированных
последовательностей у грызунов и приматов раз-
личаются, хотя у разных видов все же встречаются
родственные семейства. Кроме того, частота встре-
чаемости повторов родственных семейств может
различаться у разных видов в 10 и более раз. Такая
сложная картина характерна для геномов растений,
Неравный кроссинговер между
сестринскими хроматидами
РИС. 9.33.
Схематическое представление эво-
люции тандвмных копий генов. Меж-
ду двумя копиями диспергированной
повторяющейся последовательности
(черные полоски на одной из сест-
ринских хроматид) происходит го-
мологичный, но неравный кроссинг-
овер.
198
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
многих беспозвоночных и позвоночных. Но плас-
тичность диспергированных повторов не ограничи-
вается только первичной структурой последователь-
ностей и числом их копий. Многие рассеянные
повторы представляют собой мобильные генети-
ческие элементы, локализация которых даже у близ-
кородственных видов может не быть одинаковой.
Более того, аллельные различия могут определяться
наличием или отсутствием какого-либо повтора.
Механизмы амплификации диспергированных пов-
торов и их транспозиции в новые локусы генома
обсуждаются в гл. 10. В этой главе мы остановимся
на их общей организации, поскольку детальная
картина вряд ли может быть получена.
а. Характер распределения повторов
Впервые диспергированные повторяющиеся по-
следовательности были обнаружены при изучении
кинетики реассоциации. Эти исследования показали,
что высокоповторяющиеся последовательности час-
то бывают встроенными в другие последователь-
ности ДНК.
Анализ кинетики ассоциации. На кривых реассо-
циации геномной ДНК повторяющимся последо-
вательностям соответствуют участки с низким и
средним значениями Cot, а уникальным- участок
с высоким Cot (гл. 7.5.6). Если для определения
доли реассоциированной ДНК используется гидро-
ксилапатит, то двухцепочечными считаются как
частично, так и полностью ренатурировавшие мо-
лекулы (рис. 9.34). В результате мы получаем ка-
жущееся увеличение доли геномной ДНК, реассо-
циирующей при низких и средних значениях Cot,
при увеличении длины фрагментов ДНК. Это про-
исходит потому, что повторяющиеся последова-
тельности, которые реассоциируют при относитель-
но низких Cot, рассеяны по геному среди после-
довательностей с низким числом копий (рис. 9.35).
В опытах, о которых идет речь, определяется за-
висимость доли ДНК, считающейся двухцепочечной
по данным связывания с гидроксилапатитом, от
длины фрагментов ДНК. Чтобы получить препара-
ты молекул ДНК разной длины, суммарную ДНК
фрагментируют случайным образом и затем раз-
деляют по размерам центрифугированием в гра-
диенте плотности. Фрагменты денатурируют и от-
жигают при фиксированных значениях Cot, слиш-
ком низких для реассоциации уникальных одно-
копийных последовательностей.
Разные типы распределения повторов. Некото-
рые типичные кривые реассоциации показаны на
рис. 9.36. Во всех трех случаях доля ДНК, связы-
вающейся с гидроксилапатитом, возрастает с уве-
личением длины цепи, однако кривые существенно
Одноцепочечные молекулы
проходят через колонку
Частично и полностью
двухцепочечные молекулы
остаются в колонке
Колонкас гидроксиапатитом,
уравновешенная 0,12 М
фоыратиым буфером
о
° Количество ДНК,связанной
Степень с гидроксиапатитом
реассоциации = Суммарное количество ДНК
РИС. 9.34.
Определение степени реассоциации денатурированной
ДНК по связыванию с гидроксилапатитом. Частично или
полностью двухцепочечные молекулы связываются с
гидроксилапатитом в 0,12 М фосфатном буфере, а од-
ноцепочечные проходят свободно.
различаются по форме. Если длина фрагментов
ДНК человека достигает ~ 2,2 т. п. н., то с гидро-
ксилапатитом связывается в целом 60% геномной
ДНК. Поскольку в геноме человека на долю по-
следовательностей с умеренным и высоким числом
повторов приходится лишь 20% (точка пересечения
кривых с осью ординат на рис. 9.36), низкокопийные
последовательности, как правило, должны флан-
кировать повторяющиеся сегменты. Излом кривой
в точке ~2,2 т. п. н. действительно говорит о том.
что в 60% ДНК человека повторяющиеся последо-
вательности разделены относительно низкокопий-
ными последовательностями со средней длиной
2,2 т. п. н. В отличие от этого соответствующая
кривая для ДНК Drosophila изломов не имеет. Сред-
нее расстояние между повторяющимися последова-
тельностями у нее составляет не менее 12 т. п. н.,
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
199
С
Продукты реассоциации,
связывающиеся с гидроксиапатитом
Увеличение длины цепи
реассоциирующих
Фрагментов
РИС. 9.35.
Доля геномной ДНК, реассоциирующей при
1 г средних значениях Cot, возрастает с увеличением
длины цепи, если использовать для определения
этой доли данные по связыванию ДНК с гид-
роксилап атитом.
а это самые длинные фрагменты, используемые
в эксперименте. Электронно-микроскопическое ис-
следование ренатурировавших молекул показывает,
что длина большинства повторяющихся последо-
вательностей в ДНК человека составляет около
300 п.н., а в ДНК Drosophila- 5-6 т.п.н.
На рис. 9.37 представлены примеры распреде-
ления повторов. Повторы могут быть короткими
(сотни пар нуклеотидов) или длинными (тысячи пар
нуклеотидов), встречаться часто или редко (рис.
а и б соответственно). По данным молекулярного
клонирования, реально в геномах наблюдается
смешанная картина: два распределения налагаются
друг на друга (в). Различные повторяющиеся эле-
менты могут также образовывать кластер или даже
находиться один внутри другого (?, д). Члены се-
мейства рассеянных повторов могут находиться
в интронах, между генами, в составе сателлитной
ДНК и даже в кодирующих и регуляторных участ-
ках генов, вызывая там мутации.
«Схлопывание» ДНК (fold-back). Доля ренатури-
ровавшей ДНК увеличивается с ростом длины цепи
даже при очень низких концентрациях ДНК (т.е.
при значениях Cot ~ 10~6). Такая реассоциация мо-
жет быть только внутримолекулярной, откуда сле-
дует, что некоторые повторяющиеся последова-
тельности в цепи ДНК имеют противоположную
ориентацию (рис. 9.38). ДНК, ренатурировавшая
в таких условиях, называется схлопнувшейся (snap-
back или fold-back), и на электронных микрофо-
тографиях в местах схлопывания видны характер-
ные петли. Как показало молекулярное клониро-
вание, члены семейств диспергированных повторов
могут находиться в противоположных ориентациях
даже в пределах относительно коротких сегментов
ДНК.
Частые повторы
Редкие повторы
РИС. 9.36.
Кривые реассоциации ДНК Drosophila, Xenopus и че-
ловека.
Смешанные повторы
Сгруппированные и смешанные повторы
Гнездовые повторы
РИС. 9.37.
Типы распределения повторяющихся последователь-
ностей. Члены разных семейств выделены разным цве-
том.
д
200
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Одиночные цепи
( о
( f-C (-( о
( (*( (♦( о
( Г-( (-( (♦( о
Отжиг Cot=10"6
( (-( о
( (-( (*( о
РИС. 9.38.
«Схлопывание» ДНК.
б. Диспергированные повторы
у беспозвоночных
Разнообразие в распределении повторов и в их
количестве можно наглядно проиллюстрировать на
примере различных беспозвоночных. Рассмотрим
два из twx- Drosophila и морского ежа.
Drosophila. На долю диспергированных повторов
в геноме Drosophila приходится около 9%. тем не
менее типичные высокоповторяющиеся последова-
тельности пока не идентифицированы. Известно
лишь, что имеются от 50 до 100 семейств, каждое из
которых содержит от 10 до нескольких сотен чле-
нов. Многие из них представлены мобильными эле-
ментами длиной несколько т.п.н. (разд. 10.2 и 10.3).
Некоторые из областей генома Drosophila содержат
кластеры из членов нескольких разных семейств.
Один такой кластер длиной 15 т.п.н. может содер-
жать несколько копий членов семнадцати разных
семейств. Составляющие кластера, по-видимому,
перемешаны между собой. Такие беспорядочные
кластеры обнаружены также в геноме у курицы
и в некоторых участках генома морского ежа.
Морской еж. В геноме морского ежа имеется
почти 1000 разных семейств диспергированных по-
вторяющихся последовательностей, большинство из
которых имеют длину 300 п.н. Размеры семейств
колеблются от 100 до 104 членов; всего в геноме
насчитывается не менее 5 • 105 таких сегментов. Не-
которые семейства состоят из весьма сходных по
своей нуклеотидной последовательности членов,
члены других семейств сильно различаются. Любое
данное семейство можно встретить сразу у несколь-
ких разных видов морских ежей, однако его содер-
жание даже у близкородственных видов может раз-
личаться в 20 раз. При сравнении видов, принадле-
жащих разным отрядам, относительное число копий
в различных семействах различается еще больше,
хотя суммарная доля таких последовательностей
в геноме остается на удивление постоянной. К тому
же, хотя то или иное семейство можно иденти-
фицировать у некоторых видов с помощью пере-
крестной гибридизации, часто такие семейства име-
ют видоспецифичную каноническую последователь-
ность. Например, какая-то особенность последова-
тельности ДНК, выявленная при расщеплении ДНК
определенными эндонуклеазами, может быть харак-
терна для всех членов семейства у одного вида, но
нехарактерна для родственных ему видов.
Многие из повторяющихся последовательностей
в геноме морского ежа транскрибируются в составе
неродственных им протяженных транскрипционных
единиц. Подобные транскрипты распространены
в ядрах и содержат как уникальные последователь-
ности, так и повторы. Таким образом, диспергиро-
ванность распространяется и на ядерную РНК.
В транскриптах, хотя обычно не в одной и той же
молекуле РНК, бывают представлены обе цепи пов-
торяющихся элементов ДНК.
Функции повторяющихся сегментов ДНК, а
также их РНК-копий неизвестны. За исключением
ооцитов морского ежа, транскрипты повторяющих-
ся последовательностей обнаруживаются в основ-
ном в ядре, а в плазматических полиаденилирован-
ных мРНК отсутствуют. Какие-либо данные о том,
что повторяющиеся последовательности ассоцииро-
ваны с функциональными мРНК, даже если это
цитоплазматические РНК ооцитов, отсутствуют.
Представляется маловероятным, что уже исследо-
ванные транскрипты кодируют какие-либо белки,
поскольку во всех трех рамках считывания они
прерываются терминирующими кодонами.
в. Многочисленные семейства
диспергированных повторов
в геномах млекопитающих
Классический анализ кинетики реассоциации по-
казал, что для геномов человека и других мле-
копитающих характерно присутствие коротких, час-
то встречающихся повторов. Об этом же свиде-
тельствуют те данные на молекулярном уровне,
которые были получены с помощью рестрикцией-
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
201
Крысы
(10® копий на геном")
GGGGCTGGGG ATTTAGCTCA G-TGGTAGAG
1111 1111III I III Hill
GGCG GCGTAGCTCA GATGGTAGAG
CGCTTGCCTA GGAAGCGCAA GGCCCTGGGT
Illi II II I I III III Illi
CGCTCGCTTA GCATGCGAGA GGTACCGGGA
TCGGTCCCCA GCTCCGAAAA AAAAAAAAAA
III I III II II I
TCGATACCCG GCGCCTCCA
Аланиновая тРНК
Мыши
(8x104 копий на геном)
GGGGCTGGAG AGATGGCTCA GTGGTTAAGA GCACCTGACT GCTCTTCCGA AGGT- ---CCTGAGT ТСААТТСССА GCAACCACAT GGTGGCTCAC
I I II Illi I lllllllll |l пи HI III I I II II II I Hill I I I
GACG AGGTGGCCGA GTGG7TAAG- GCGATGGACT GCTAA1CCAT TGTGCTCTGC ACGCGTGGGT TCGAATCCCA TCCTCGTCG
Сериновая тРНК
РИС. 9.39.
Сходство нуклеотидных последовательностей членов
семейства SINE и тРНК. [G. R. Daniels, Р. L. Deininger,
Nature 317 (1985), р. 819.]
него анализа, клонирования и секвенирования. Из
всех разнообразных семейств и подсемейств дис-
пергированных повторов лишь немногие высоко-
копийны и являются в геноме преобладающими. То
или иное доминирующее семейство может состав-
лять более 5% суммарной ДНК. а все вместе - более
20%. Члены некоторых из этих семейств имеют
длину 100-500 п.н. и называются SINE-короткие
диспергированные повторы (short interspersed re-
peats). Другие семейства представлены последова-
тельностями длиной 6 т.п.н. и более, называемыми
LINE-длинные диспергированные повторы (/ong
interspersed repeats). Диспергированные повторы,
напоминающие SINE, обнаружены у немлекопита-
ющих, в том числе слизевиков, саранчи, иглокожих,
рыб и амфибий. Они встречаются также у растений
и беспозвоночных (разд. 10.3.е).
SINE. Сначала к SINE относили все короткие
высокоповторяющиеся диспергированные' последо-
вательности млекопитающих, основываясь просто
на их длине и распространенности. Позже в основу
классификации было положено более существенное
их свойство. По-видимому, последовательности
SINE представляют собой процессированные псев-
догены, происходящие от генов класса III, которые
кодируют малые плазматические РНК, в частности
тРНК (рис. 9.39) и 7SL-PHK (рис. 9.40), т. е. после-
довательности SINE гомологичны последователь-
ностям таких РНК (и соответствующих генов). Как
правило, SINE окружены прямыми повторами -
сайтами-мишенями дупликаций (разд. 10.1.6) и со-
держат A-богатую З'-концевую последовательность.
Каждый вид (или отряд) имеет свой набор
SINE-семейств. Для иллюстрации разнообразия
и сложности таких последовательностей достаточно
привести несколько примеров. Одно из наиболее
изученных семейств SINE-это семейство А1и-по-
следовательностей у приматов Старого Света, наз-
ванное так потому, что все они содержат сайт для
эндонуклеазы Alul. Alu-элементы фланкируют гены,
они обнаружены в интронах, в сателлитной ДНК
и в составе кластеров вместе с другими вставоч-
ными последовательностями. В пределах любого
данного вида различия между Alu-последователь-
ностями достигают 15%. Alu-последовательности
5’
SINE,грызуны
7SL-PHK
3’
и, человек
300 п.н.
РИС. 9.40.
Сравнение 7SL-PHK и канонических после-
довательностей SINE типа I у грызунов и
Alu у приматов; сходные области обозна-
чены одинаковым образом. Ап-А-богатая
последовательность на З'-конце одной из
цепей каждой мономерной единицы (Тп-
комплементарная последовательность).
Гены 7SL-PHK человека имеют такую же
структуру, как представленная здесь РНК.
Центральная часть 7SL-PHK неродственна
Alu- или SINE-последовательностям. [Е. Ullu,
С. Tschudi, Nature 312 (1984), р. 171.]
202
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
разных видов приматов Старого Света различаются
примерно в такой же степени, как и Alu-после-
довательности у разных индивидов одного вида.
Типичная Alu-последовательность приматов
представляет собой димер, состоящий из двух по-
второв длиной около 130 п.н., соединенных голова-
к-хвосту (рис. 9.40). Мономерные единицы в составе
димера неодинаковы: наиболее существенное раз-
личие между ними состоит в том, что во втором из
них имеется дополнительный сегмент длиной 31 п.н.
Каждый из мономеров (З'-конец верхней цепи ДНК
на рис. 9.40) заканчивается ро!у(А)-хвостом разной
длины. Alu-мономеры содержат последовательно-
сти, гомологичные 5'- и З'-концам 7SL-PHK; цент-
ральная часть 7SL-PHK длиной 155 п.н. в А1и-по-
следовательности делегирована.
Впервые Alu-последовательности были выделе-
ны и клонированы (случайным образом) из ДНК
человека (рис. 9.41), и исходя из этих данных была
построена каноническая последовательность. Затем
клонированные члены Alu-семейства использовали
в качестве зондов для определения числа копий
методом кинетики реассоциации. Было обнаружено
9-105 копий Alu-последовательностей на гаплоид-
ный геном (или около 9% геномной ДНК человека).
В среднем на каждые 5 т.п.н. генома человека
и других приматов Старого Света приходится одна
Alu-последовательность. С Alu-зондами гибридизу-
ется ДНК более 90% рекомбинантных фагов, содер-
жащих хромосомную ДНК человека, а большинство
из клонированных сегментов длиной 15-20 т.п.н.
содержит более одного члена Alu-семейства. Сосед-
ние Alu-последовательности могут располагаться
друг относительно друга в любой ориентации, поэ-
тому некоторые из них входят во фракцию «схлоп-
нувшейся» геномной ДНК. Например, из восьми
Alu-последовательностей в пределах сегмента ДНК
длиной 65 т.п.н, которые входят в состав мультиген-
ного семейства генов 0-глобина, пять ориентированы
в одном направлении, а три - в другом (рис. 9.42).
Распространенность Alu-последовательностей
кажется удивительной, если учесть, что в ДНК
человека имеются только три функциональных гена
7SL-PHK. Более того, все Alu-последовательности
приматов должны были возникнуть уже после появ-
ления в процессе эволюции большинства отрядов
млекопитающих, поскольку SINE-семейство грызу-
нов, гомологичное 7SL-PHK. существенно отлича-
ется от Alu-семейств приматов (рис. 9.40). При-
мерно 10s членов семейства типа I у грызунов
имеют длину 130 п.н., что практически равно од-
ному из двух соединенных голова-к-хвосту моно-
меров, типичных для Alu-последовательностей при-
матов. Другие представители SINE грызунов-се-
мейства типа II и ID-гомологичны различным
тРНК-генам (рис. 9.39).
Денатурация
Отжиг Cot-68
I1--------fl
(Фракционирование по размерам
с помощью электрофореза
Количество
ДНК
РИС. 9.41.
| Ват HI-линкеры
< »G
СCCTAG
.клонирование
BE.coli
Выделение Alu-последовательностей человека. Сум-
марную ДНК человека фрагментировали до средней
длины фрагментов 2 т. п. н„ денатурировали и прово-
дили ренатурацию; Cot было равно 68, при этом после-
довательности с низким числом копий оставались од-
ноцепочечными, а повторяющиеся последовательности
со средним и высоким их числом образовывали дуп-
лексы. При обработке дуплексов эндонуклеазой, спе-
цифичной к одноцепочечным участкам, оставались пол-
ностью двухцепочечные фрагменты. После разделения
последних с помощью гель-электрофореза обнаружи-
лось, что большую их часть составляют фрагменты
длиной 300 п. н., при расщеплении которых эндонук-
леазой А!и\ образуются в основном фрагменты длиной
170 и 120 п.н. Репрезентативные члены Alu-семейства
из фракции в 300 п. н. были клонированы с помощью
Ват HI-линкеров и плазмиды pBR322. [Р. L. Deininger et
al., J. Mol. Biol. 151 (1981), p. 17.]
Члены SINE-семейства и родительские гены
класса III имеют гомологичные внутренние про-
моторы, А- и В-боксы, а SINE-единицы, которые
содержатся в клонированных сегментах ДНК, часто
транскрибируются in vitro при участии РНК-поли-
меразы III (рис. 9.43). Транскрипция начинается
строго на 5'-конце единицы и заканчивается на
первом участке из четырех или более остатков
аденина в матричной цепи ДНК. Однако, несмотря
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
203
Alu-семейство
Q Семейство LINE-I
РИС. 9.42.
Мультигенное семейство генов Р-глобина человека,
состоящее из диспергированных повторов, которые
принадлежат к разным семействам.
на то что многие SlNE-единицы эффективно тран-
скрибируются in vitro, большинство из них не слу-
жат матрицей для РНК-полимеразы III in vivo.
И все же в наиболее интересных моделях, рассмат-
ривающих амплификацию SINE-последовательно-
стей и встраивание их в ДНК, предполагается, что
происходит обратная транскрипция с участием
РНК-полимеразы III (разд. 10.4).
SIN Е-последовательности транскрибируются
также с помощью РНК-полимеразы II. Это проис-
ходит в том случае, когда они находятся в каком-
либо интроне, в 5'-лидерной или З'-трейлерной пос-
ледовательности неродственной транскрипционной
единицы класса II. Поэтому гетерогенные ядерные
РНК содержат в большом количестве диспергиро-
ванные транскрипты SINE-последовательностей.
В ходе процессинга первичных транскриптов с обра-
зованием мРНК SIN Е-последовательности обычно
удаляются, в результате чего плазматическая РНК
содержит их почти в десять раз меньше, чем
ядерная.
LINE. Кроме многочисленных SINE-семейств,
все геномы млекопитающих содержат одно крупное
семейство LINE-последовательностей, а именно се-
мейство LINE-1, члены которого могут достигать
длины 6-7 т.п.н. (рис. 9.44). В некоторых геномах
встречаются и другие семейства LINE, но их распро-
страненность гораздо ниже. У разных млекопитаю-
щих все LINE-семейства родственны, но немного
различаются: чем ближе виды, тем более они сходны.
А-бокс
GGCGTGGTGG
Фланкирующая
последовательность
Многие члены семейства LINE-1 имеют длину мень-
ше 6- 7 т. п. н. В таких укороченных сегментах обыч-
но отсутствуют 5'-концевые последовательности
(рис. 9.44), а иногда внутренние участки. Кроме того,
не все члены семейства коллинеарны. Некоторые из
них инвертированы или в них произошли какие-то
перестройки. Как в геноме мыши, так и в геномах
приматов укороченных членов семейства намного
больше, чем полноразмерных. В результате число
копий различных частей длинных LINE-1-последо-
вательностей варьирует и увеличивается от левого
конца к правому. Например, у человека содержится
около 3,5-103 копий левого конца последователь-
ностей и почти 105 копий правого конца.
Члены семейства LINE-1 фланкируют гены, они
встречаются в интронах и сателлитной ДНК. Ис-
следование членов LINE-1-семейства, содержащихся
в клонированных геномных сегментах, а также гиб-
ридизация in situ LINE-1-зондов с интактными хро-
мосомами показали, что они распределены по раз-
ным хромосомам. Носит ли это распределение слу-
чайный характер, неизвестно. Иногда LINE-1-по-
следовательности образуют кластеры. Например,
необычно много членов этого семейства содержится
в сегменте ДНК человека размером 65 т. п. н., кото-
рый содержит р-глобиновые гены (рис. 9.42).
LINE-1, как и SINE-последовательности, часто
бывают окружены прямыми повторами, которые,
вероятно, служат сайтами-мишенями дупликаций
(разд. 10.1.6), а на З'-конце у них имеется А-богатый
участок. Однако, в отличие от SINE, LINE-1-еди-
ницы, по-видимому, не являются псевдогенами
обычных генов. Скорее всего некоторые наиболее
длинные единицы LINE-1 предсталяют собой мо-
Фланкнруюцая
последовательность
В-бокс
GTTCAAGAC
ШШ !;
РИС. 9.43.
Транскрипция in vitro SINE-последовательности с по-
мощью РНК-полимеразы III (Pol III). В качестве при-
мера рассмотрен типичный клонированный сегмент
Alu-последовательности приматов. Представлен со-
стоящий из двух частей регуляторный участок (А- и
В-боксы; разд. 8.3), а также сайт инициации транскрип-
ции, соответствующий началу повторяющейся единицы
204
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Полноразмерный член семейства UNE-1(~4-1osKonHH на гаплоидный геном человека)
К К К К Стоп-кодон ААТААА
5’ ♦ Г////// л ♦ ♦ 1 1 3' ]А-6огатая f)
шш одинаковые у всех млекопитающих ]Т-богатая Q
(у|А-богатая()
(' 'Jl-богатаяО
РИС. 9.44.
Полноразмерный член семейства LINE-1 человека
(вверху) аналогичен членам, обнаруженным у всех мле-
копитающих. Указано несколько характерных для при-
матов рестрикционных сайтов. Например, ACpnl-фраг-
менты длиной 1,5 и 1.2 т.п.н.. образующиеся после
эндонуклеазного расщепления геномной ДНК, присут-
ствуют в гидролизате в достаточно большом количест-
ве, чтобы их можно было обнаружить после электро-
фореза и окрашивания геля бромистым этидием. Учас-
ток. выделенный светло-серым цветом, аналогичен у
всех млекопитающих и содержит протяженные участки
с открытыми рамками считывания (от 5'- к З'-концу,
слева направо, верхняя цепь). 5'- и З’-концевые участки
у разных отрядов млекопитающих существенно раз-
личаются по длине и нуклеотидной последовательности
и не содержат открытых рамок считывания. Большинст-
во членов семейства укорочены с 5'-конца (нижняя
часть рисунка).
бильные элементы, которые амплифицировались
и встроились в новые геномные сайты в результате
обратной транскрипции соответствующих РНК
(разд. Ю.З.е). Обратную транскриптазу может ко-
дировать участок LINE-1-единицы размером
4 т.п.н.; у млекопитающих этот участок весьма
консервативен и имеет в одной из цепей протяжен-
ную открытую рамку считывания (рис. 9.44). Уко-
роченные единицы LINE-1 или единицы с пере-
стройками могли произойти от неполных или абер-
рантных транскриптов, от обратных транскриптов
или же появиться в результате рекомбинаций, со-
пряженных с инсерцией.
Последовательности LINE-1 транскрибируются
РНК-полимеразой II вместе с геном, в некодирую-
щий участок которого они встроены. В результате
среди гетерогенных ядерных РНК встречается до-
вольно много РНК, гомологичных обеим цепям
последовательностей LINE-1. В полиаденилирован-
ной плазматической РНК дискретных транскриптов
не обнаружено, как не обнаружено и соответствую-
щих белков. Исключение составляют клетки тера-
токарциномы человека и мышиных лимфоцитов,
которые содержат полноразмерные транскрипты
цепи с открытой рамкой считывания. Однако бел-
ков. кодируемых этими РНК, не обнаружено.
г. Функции диспергированных
повторов
Наличие огромного количества семейств повто-
ряющихся последовательностей, разбросанных по
десяткам и сотням тысяч локусов эукариотического
генома, кажется необъяснимым. Например, на до-
лю SINE- и LINE-последовательностей приходит-
ся не менее 10% геномной ДНК млекопитающих.
Какую роль могут играть столь многочисленные
повторы помимо той, которую играют члены, со-
держащие функциональные гены? На этот счет есть
две гипотезы. Первая предполагает, что повто-
ряющиеся последовательности, их транскрипты
или те и другие вместе участвуют в регуляции
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
205
генной экспрессии. Согласно второй гипотезе,
большинство таких последовательностей вообще не
выполняет никаких функций.
Семейства повторяющихся последовательностей
как регуляторы экспрессии генов. Напомним, что
в отличие от прокариот, у которых родственные
гены, имеющие отношение к определенным мета-
болическим реакциям, обычно сцеплены и регули-
руются с помощью одного оперона (разд. 3.11),
родственные эукариотические гены обычно диспер-
гированы. Например, гены а- и Р-глобинов распо-
ложены на разных хромосомах, но должны экспрес-
сироваться согласованно. Другими словами, весь
набор диспергированных генов в конкретной ткани
или на определенной стадии развития организма
должен экспрессироваться одновременно. Такая
координация возможна, если у всех членов генного
кластера имеется общий регуляторный элемент.
В таком случае включать или выключать весь набор
генов может одна эффекторная молекула, взаимо-
действующая с одинаковыми диспергированными
регуляторными элементами. Контроль за экспрес-
сией генов может осуществляться на уровне ДНК
путем регуляции транскрипции или же на уровне
РНК. Например, в гетерогенной ядерной РНК,
по-видимому, сохраняется картина распределения
повторов, характерная для геномной ДНК, и тогда
координация регуляции может осуществляться с
помощью контроля созревания первичных транск-
риптов с образованием мРНК. В то же время копии
повторяющихся последовательностей могут содер-
жаться и в зрелых мРНК, и тогда регуляция генной
экспрессии будет происходить на уровне трансля-
ции.
Существуют ли какие-либо доказательства этой
гипотезы и ее альтернативных вариантов? Известно,
что некоторые семейства повторяющихся последо-
вательностей участвуют в регуляции специфических
групп генов. Как правило, члены этого семейства
имеют длину не более 10 п.н., поэтому они исклю-
чаются из числа диспергированных последователь-
ностей, идентифицируемых методом кинетики ре-
натурации или легко выявляемых при клонирова-
нии. Например, некоторые наиболее распространен-
ные из таких последовательностей входят в состав
регуляторных участков, сцепленных с генами, ко-
торые транскрибируются с участием одной из РНК-
полимераз; формально эти последовательности от-
носятся к повторяющимся и диспергированным.
На недостаток аминокислот дрожжи отвечают
экспрессией генов, детерминирующих их биосинтез.
По крайней мере некоторые из этих генов содержат
на 5'-конце последовательность 5'-TGACTC-3'. Она
встречается один или несколько раз на участке
33-330 п.н. перед точкой инициации транскрипции
в составе негомологичных во всем остальном пос-
ледовательностей. Для экспрессии гена в ответ на
дефицит аминокислоты необходимо и достаточно
одной копии 5'-TGACTC-3', если только остальная
часть контролирующих транскрипцию элементов
интактна. Транскрипция стимулируется ДНК-свя-
зывающим белком GCN4, который распознает ука-
занный гексануклеотид (разд. 8.3.е). На 5'-фланки-
рующем участке генов Drosophila, которые активи-
руются при поышении температуры, находится дру-
гая короткая повторяющаяся последовательность,
5'-CTNGAANNTTCNAG-3'. По-видимому, она от-
стоит от точки начала транскрипции этих генов
теплового шока на 150 п.н. и необходима для
инициации ответа на повышение температуры как
у Drosophila, так и у клеток млекопитающих в куль-
туре. Кроме того, имеются некие сцепленные с инт-
ронами последовательности, существенные для пра-
вильного и эффективного сплайсинга. Эти после-
довательности содержат динуклеотиды 5'-GT-3' и
5'-AG-3'-два первых и два последних остатка в инт-
ронах эукариот (разд. 8.5.в). У дрожжей во всех
известных интронах между 20-м и 55-м остатками
слева от AG на З'-конце сайта сплайсинга при-
сутствует последовательность 5'-ТАСТААС-3' (разд.
8.5.в). Например, в единственном интроне актино-
вого гена дрожжей она находится от З'-сайта сплай-
синга на расстоянии 47 нуклеотидов (рис. 9.10). Если
эта последовательность у генов, клонированных
в дрожжевом векторе, делегирована, то мРНК в
дрожжевых клетках не образуется, поскольку соот-
ветствующие транскрипты не сплайсируются.
Известно ли что-либо о весьма многочисленных
семействах, члены которых имеют длину, значи-
тельно превышающую 10 п. н., в том числе и о се-
мействах SINE и LINE? Есть ли данные об их
регуляторных функциях? У морского ежа и у Dic-
tyostelium, например, в разных наборах транскрип-
тов, образующихся в тканях зародыша и взрослой
особи, встречаются разные повторяющиеся после-
довательности. Более того, семейства таких после-
довательностей, представленные в ядерных тран-
скриптах, сильно различаются у дифференцирован-
ных клеток разного типа. Могут ли эти РНК участ-
вовать в регуляции функциональных генов, экспрес-
сирующихся во времени одинаковым образом?
В транскриптах, выделенных из специализирован-
ных клеток млекопитающих, обнаружены SINE-
последовательности. Семейство В2 этих последова-
тельностей у мышей часто встречается в мРНК,
присутствующих в повышенной концентрации в ра-
ковых клетках и в клетках мышиных эмбрионов на
определенных стадиях развития. Такие последова-
тельности обнаружены также в З'-трейлерных участ-
ках мРНК некоторых генов главного комплекса
гистосовместимости мыши. Эти данные наводят на
мысль, что повторяющиеся элементы выполняют
206
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
какую-то специфическую функцию, но, прежде чем
утверждать это, нужно получить дополнительную
информацию.
Семейства поторяющихся последовательностей
как нефункциональные структуры. В разд. 9.4.г мы
уже говорили о том, что некоторые геномные сег-
менты не выполняют никаких генетических функций.
По-видимому, именно таковы псевдогены и процес-
сированные гены. Если учесть, что многие дисперги-
рованные семейства содержат большое число таких
генов, то можно сделать вывод о нефункционально-
сти целых семейств. Такая ДНК получила название
эгоистичной, поскольку вся ее деятельность направ-
лена на собственные амплификацию и распростра-
нение в геноме. Возможные механизмы амплифика-
ции и распространения описаны в гл. 10. Здесь мы
лишь отметим, что такие события отнюдь не безо-
бидны; распространение повторов в геноме может
приводить к инсерционным мутациям в регулятор-
ных или кодирующих последовательностях. Кроме
того, множественные диспергированные повторы
могут благоприятствовать делециям функциональ-
ных сегментов в результате гомологичной рекомби-
нации. Таким образом, ничем не органиченная эго-
истичность может иметь катастрофические послед-
ствия для функциональной части генома.
Alu-последовательности благоприятствуют де-
лециям. Способность диспергированных повторов
облегчать делегирование можно проиллюстрировать
на примере гена рецептора ЛПНП человека (разд.
9.4.а). У гомозигот по мутантным генам рецептора
ЛПНП в плазме накапливается холестерол, что
приводит к раннему атеросклерозу и даже к инфарк-
ту у детей. По оценкам, один мутантный аллель
встречается у одного из 500 человек. У гетерозигот
развивается более легкая форма семейной гиперхо-
лестеролемии. Клонирование и сравнительный струк-
турный анализ генов нескольких индивидуумов,
страдающих этим заболеванием, показывает, что
некоторые дефектные рецепторы кодируются гена-
ми с протяженными делециями. На участке ДНК
длиной 45 т. п. н., содержащем ген рецептора
ЛПНП, находится много Alu-последовательностей.
Некоторые из них расположены в З'-половине гена,
в частности три- в З'-нетранслируемой части по-
следнего экзона. Два охарактеризованных и неза-
висимых делеционных мутанта, по-видимому, обра-
зовались в результате гомологичной рекомбинации
между такими Alu-последовательностями (рис. 9.45).
Как следствие экзоны, кодирующие часть рецепто-
ра, пронизывающую мебрану, а также его цито-
плазматический участок, делегированы. Мутантные
белки не встраиваются в мембрану, а выводятся из
клетки.
д. Замечательное единообразие
диспергированных повторяющихся
последовательностей
Чем объясняется внутривидовая однородность
членов семейств диспергированных последователь-
ностей? Тандемные повторы, например кластеры
рДНК в разных хромосомах, также имеют сходные
последовательности. Один из возможных механиз-
мов, обеспечивающих такую гомогенность,- рецип-
рокный гомологичный кроссинговер между неал-
РИС. 9.45.
А
Ген рецептора ЛПНП
5' Экзон 13 14 15
/А TI 4
Alu
i—i 16 17
А. Делеционный мутант по гену рецептора ЛПНП, полу-
ченный в результате рекомбинации между Alu-nocne-
доватепьностями. З'-участок нормального гена с экзо-
нами изображен так же, как на рис. 9.21. Представлены
Alu-последоватепьности в одном интроне и в З'-кон-
цевом экзоне (экзон 18). Б. Рестриктная карта этого же
участка нормального гена. В. Рестриктная карта кло-
нированного мутантного гена. По отсутствию некоторых
рестриктных сайтов можно сделать вывод, что концы
делегированного участка находятся в Alu-поспедова-
тельностях. Об этом же говорят результаты сравнения
нуклеотидных последовательностей нормального и му-
тантного клонированных генов. [М.А. Lehrman et al..
Science 227 (1985), p. 140.]
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
207
дельными членами семейств; именно такой кроссин-
говер происходит между единицами рДНК у дрож-
жей (разд. 9.2.а). Аналогичным образом можно
объяснить и внутривидовую гомогенность других
тандемных повторов, например центромерных са-
теллитных ДНК. Однако те способы гомогениза-
ции, которые характерны для тандемных повторов,
не могут реализовываться в случае диспергирован-
ных повторов без угрозы целостности уникальных
последовательностей, окружающих эти повторы.
Возможные механизмы гомогенизации дисперги-
рованных повторов. Разумно предположить, что все
члены гомологичных SINE- и LlNE-семейств у раз-
ных организмов произошли от общей предковой
последовательности. В каждой из эволюционных
ветвей появлялись определенные варианты этих
последовательностей, ставшие весьма многочислен-
ными. Например, все Alu-последовательности при-
матов сходны между собой. У мыши нет таких
димерных Alu-элементов, зато имеется много копий
мономерных последовательностей, родственных
Alu,- SINE типа I. И все же оба этих семейства,
вероятно, образовались в процессе перестройки,
амплификации и распространения по всему геному
последовательностей, происшедших от предковых
генов 7SL-PHK.
Представляется маловероятным, что внутриви-
довая гомогенизация этих семейств могла произой-
ти в результате естественного отбора, действую-
щего на каждого члена семейства по отдельности.
Если множественные копии выполняют какие-то
функции, то их изобилие могло бы минимизировать
значимость любой единичной мутантной копии.
Большинство диспергированных повторов отлича-
ется от видоспецифичной консенсусной последова-
тельности по 5-15% позиций. Однако если боль-
шинство копий нефункциональны, то еше менее
вероятно, что гомогенизация с образованием ка-
кой-либо видоспецифичной последовательности бу-
дет происходить под давлением отбора. Скорее
всего каждая внутривидовая копия эволюциониро-
вала согласованно с другими копиями, так что все
они оказались сходными.
Рассмотрим для примера SlNE-семейство. В ка-
кой-то момент времени, до появления позвоночных,
произошли амплификация и распространение одно-
го из модифицированных генов 7SL-PHK. Поэтому
соответствующее SINE-семейство имеется у Xeno-
pus, но не у Drosophila, хотя гены 7SL-PHK есть
у обоих видов. Семейство повторяющихся после-
довательностей образовалось у каждого вида позво-
ночных, но члены предкового семейства замещались
одной видоспецифичной последовательностью, выс-
тупающей в роли новой последовательности-осно-
вателя. Все или большинство предковых копий у но-
вых видов утратились, а семейство образовалось in
toto из этой последовательности-основателя. Если
бы все это случилось одномоментно, то последствия
были бы катастрофичными для гена. Однако вна-
чале происходило длительное постепенное замеще-
ние существующих SINE-последовательностей пос-
ледовательностью-основателем. Если допустить су-
ществование нейтрального отбора, то за 70 млн.
лет, прошедших со времени дивергенции приматов
и грызунов, при скорости замещения 0.01 за год
могла бы осуществиться гомогенизация такого
большого семейства, как Alu-семейство приматов.
Замещение могло происходить, например, в ре-
зультате рекомбинации между неаллельными чле-
нами семейства повторяющихся последовательнос-
тей; при этом рекомбинировали бы только сами
повторяющиеся элементы, но не фланкирующие их
последовательности, что исключило бы слишком
обширную перестройку. Кроссинговер не приведет
к изменению фланкирующих последовательностей
и будет нереципрокным, если разрыв цепи в про-
межуточном продукте и последующее воссоедине-
ние произойдут без изомеризации и если в результа-
те репарации участка гетеродуплекса восстановится
одна из кроссоверных копий (рис. 2.61 и 9.46). Такой
же результат дает генная конверсия, происходящая
в соответствии с моделью двухцепочечного разрыва
(рис. 2.60 и 9.46). С помощью этих двух процессов
отдельные участки гомологичных последователь-
ностей или все они целиком могут стать идентич-
ными (т. е. гомогенизироваться). Если в результате
конверсии один из членов семейства приобретает
какие-то преимущества над остальными членами, то
он и становится последовательностью-основателем.
Постепенно в результате конверсии отдельных ге-
нов происходит гомогенизация всего семейства. В
рамках такой модели предполагается один источник
направленных изменений. Элементами-основателя-
ми могут быть те локусы, которые наиболее актив-
но транскрибируются в клетках зародышевой ли-
нии. Отметим, что ни два реципрокных кроссинго-
вера в пределах повтора, ни одиночный кроссовер,
образовавшийся после изомеризации, не приводит
к конверсии (рис. 9.46).
Генная конверсия у дрожжей. У дрожжей, где
генную конверсию можно изучать генетически, на-
блюдается направленность изменений. Одни локусы
более эффективны как матрицы, чем другие. При-
чина этих различий неясна, но частота конверсий
достаточно высока, так что даже при незначитель-
ном преимуществе с помощью одного такого локу-
са может произойти гомогенизация весьма мно-
гочисленного семейства. Прямые данные о превра-
щении одного члена семейства в последователь-
ность, сходную с другим членом, получены при
изучении диспергированных повторов у дрожжей,
называемых Ту-элементами (Ту-мобильный элемент,
208
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Двухцепо-
чечный
разрыв
Два
реципрокных
кроссинговера
Одиночный реципрок-
ный кроссинговер
без изомеризации
с репарацией
Одиночный
реципрокный
кроссинговер
сизомеризацией
РИС. 9.46.
Рекомбинация между неаллельными повторяющимися
единицами (они выделены цветом разной интенсив-
ности) в зависимости от механизма дает различные
№ ff—к
ft
продукты. Все перечисленные механизмы включают
гомологичную рекомбинацию между самими повторя-
ющимися единицами.
1---№
описанный в разд. 10.3.6). Изменения во фланки-
рующих последовательностях у дрожжей легко выя-
вить генетически по появлению мутаций в маркер-
ных генах. Утрата Ту-элемента устанавливается по
одновременной утрате встроенного в него важного
гена URA3 (рис. 9.47); у штамма, который исполь-
зовался в данном опыте, функциональный ген URA3
отсутствовал. Как показали опыты с использова-
нием специфических рестриктирующих эндонук-
леаз, утрата гена URA3 сопровождалась конверсией
Ту-последовательности с образованием модифици-
рованной ее формы из другой геномной копии.
Поскольку при конверсии такого типа утрачиваются
протяженные последовательности (URA3), то скорее
всего здесь происходит двухцепочечный разрыв, а не
одиночный кроссинговер и репарация. Вероятность
того, что в пределах Ту-последовательности проис-
ходят два реципрокных кроссинговера, тоже очень
РИС. 9.47.
Конверсия Ту-последовательностей у дрож-
жей. Последовательности Ту-это повторя-
ющиеся диспергированные транспозируемые
элементы в геноме дрожжей (разд. 10.3.6).
Вертикальными стрелками указаны сайты для
эндонуклеаз рестрикции, различные у разных
элементов. Использовался штамм, обладающий
следующими свойствами: нормальный ген
URA3 делвтирован; в Ту-элемент встроен дру-
гой ген URA3 с образованием клеток URA3\
От одного и того же штамма URA3* были
получены несколько игаЗ'-производных. По-
следовательность URA3 у них отсутствовала,
хотя Ту-злемент остался на меств, судя по
отсутствию изменений во фланкирующих его
последовательностях. Утрата (УНА 3-не един-
ственное изменение в Ту-злемвнте. Измени-
лось и положение зндонуклеазных сайтов: оно
стало таким же, как в Ту-элементах из других
локусов генома.
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
209
мала, поскольку наблюдаемая частота замен в де-
сять раз выше частоты даже одной реципрокной
рекомбинации в отдельном локусе.
Помимо замен, обусловленных генной конвер-
сией, гомогенизация, по-видимому, происходит так-
же в результате делеций последовательностей-пред-
шественников типа SINE и вставки новых копий.
В противном случае копийность семейств дисперги-
рованных последовательностей у разных видов
должна была бы оставаться относительно постоян-
ной, а это не так. Например, у человека и шимпанзе
число Alu-последовательностей равно соответствен-
но 9-105 и 3-105. Четкие указания на наличие
инсерций (и/или делеций) были получены после
выявления аллелей отдельных генов, которые раз-
личаются по наличию последовательности SINE
в одном из фланкирующих участков или в интроне.
Возможные механизмы встраивания или делеций
описаны в разд. 10.4.
9.6. ПОСЛЕДОВАТЕЛЬНОСТИ
В ОБЛАСТИ ЦЕНТРОМЕР
И ТЕЛОМЕР
Центромеры и теломеры-наиболее четко выра-
женные морфологические структуры хромосом.
Долгое время считалось, что их строение и функции
связаны с какими-то особыми последовательнос-
тями ДНК. Однако удалось выявить лишь одну
такую особенность на молекулярном уровне: при-
сутствие в области центромер и теломер сателлит-
ной ДНК; впрочем, о функциях этой ДНК ничего не
известно. Изучение взаимосвязи структуры и функ-
ций стало возможным с появлением методов мо-
лекулярного клонирования, которые и помогут нам
получить четкие данные на этот счет.
а. Последовательности в области
центромер
Строение центромер. У млекопитающих цент-
ромеры имеют сложную дискообразную структуру,
называемую кииетохором. С каждой стороны хро-
мосомы располагается по одному кинетохорному
диску. Во время митоза микротрубочки фибрилл
веретена прикрепляются непосредственно к плот-
ному наружному слою кинетохора, связанному с
петлями хроматина (рис. 9.48). Кинетохоры всех
млекопитающих, по-видимому, сходны по своей
структуре, поскольку все они образуют комплексы
со специфическими антителами из сыворотки боль-
ных, страдающих редким аутоиммунным заболе-
ванием-системной склеродермой. С антителами
взаимодействуют только кинетохоры митотических
клеток, однако соответствующие комплексы обра-
зуются и в специфических участках интерфазных
хромосом. Антитела не связываются с микротру-
бочками или с другими прикрепленными к ним
белками.
Хромосомы дрожжей (5. cerevisiae) малы, и к
А
РИС. 9.48.
Прикрепление микротрубочек к центромерной области митотических хро-
мосом. А. В клетках китайского хомячка микротрубочки присоединяются
непосредственно к центромерным хроматиновым фибриллам (кинето-
хору) (увеличение 40000). [Н. Ris, Р. L. Witt, Chromosoma 82 (1981), р.
153.) Б. У дрожжей S. cerevisiae одиночные микротрубочки соединяют
хромосомы с фибриллами веретена. Никакой напоминающей кинетохор
структуры в месте прикрепления микротрубочек диаметром 20 нм к хро-
матиновым фибриллам (стрелки) не обнаружено (увеличение 48000).
[J.B. Peterson, Н. Ris, J. Cell Science 22 (1976), р. 219.)
Микро-
трубочки
Хроматин
14-158
210
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
СЕЛ/11 IMET14
5,2т.п.н.
5 S3 S3 S
1,6 т.п.н.
858 п.н.
РИС. 9.49.
Получение из клонированного фрагмента дрожжевой
ДНК CEN11 -сегмента. Сконструированный плазмидный
вектор, содержащий дрожжевой ген TRPA и АЯЗ-по-
следовательность, использовали для создания геном-
ной библиотеки. После трансфекции дрожжевых клеток
mef14“ и Ггр1 ' были отобраны колонии, растущие на
среде без метионина и триптофана. Клетки из одной
такой колонии содержали плазмиду с геном МЕГ 14
в составе фрагмента дрожжевой ДНК длиной 5,2 т.п.н.
(А), в котором присутствовала также функциональная
центромера. Последующее субклонирование в векторе
такого же типа (Б, В, Г) позволило получить фрагмент
ДНК размером 858 п.н., который все еще сохранял
центромерную функцию, но не содержал гена МЕ714.
Векторные последовательности выделены серым цве-
том.
каждой митотической центромере прикрепляется
одна-единственная микротрубочка (у позвоночных
это целые пучки микротрубочек). Среди известных
эукариот только у S.cerevisiae не обнаружено цент-
ромерной сателлитной ДНК. Однако короткие сег-
менты ДНК (CEN) из центромер дрожжей все-таки
выполняют функции как митотических, так и мейо-
тических центромер в рекомбинантных векторах
(разд. 5.6.6).
Клонирование дрожжевых СEN-областей. С по-
мощью детального генетического картирования
хромосом S. cerevisiae выявлены некоторые гены,
расположенные очень близко к центромерам. На-
пример, ген МЕТЫ, необходимый для биосинтеза
метионина, тесно сцеплен с центромерой хромо-
сомы И (CEN11). Такое сцепление позволило про-
вести клонирование самой центромерной последо-
вательности. С помощью дрожжевого синтетиче-
ского плазмидного вектора была создана библио-
тека геномных последовательностей дрожжей, а за-
тем выделена колония, содержащая плазмидный
вектор с геном МЕТЫ (рис. 9.49). По данным
генетического анализа, эта плазмида, по-видимому,
содержит также и функциональную центромеру, по-
скольку она стабильно сохраняется в дрожжевых
клетках, число ее копий ограничивается одной плаз-
мидой на клетку и она сегрегирует при митозе
и мейозе как истинная «мини-хромосома». Путем
субклонирования выделен фрагмент ДНК длиной
858 п. н., выполняющий функции центромеры. Кло-
нированы центромерные сегменты и других дрож-
жевых хромосом; нуклеотидные последовательнос-
ти некоторых из них представлены на рис. 9.50.
Минимальная длина сегмента, полностью сохра-
няющего центромерную функцию, составляет при-
мерно 150 п. н.
Последовательности в области дрожжевых цент-
ромер. На рис. 9.50 суммированы общие и отличи-
тельные структурные особенности некоторых CEN-
областей. Все фланкирующие последовательности
центромер различаются. Несмотря на то что CEN-
области не гибридизуются друг с другом в жестких
условиях, они все же имеют два коротких гомоло-
гичных участка (элементы I и III на рис. 9.50).
Элементы I и III разделены АТ-богатым элементом
II. Делеция сегмента протяженностью от элемента
I по элемент III включительно приводит к утрате
плазмидой функций митотической и мейотической
центромеры. Как показывают опыты по внесению
специфических мутаций и делеций в эти сегменты,
функционирование митотической центромеры зави-
сит от целостности элемента III и в меньшей степе-
ни-от целостности элементов I и II, в то время как
для функционирования мейотической центромеры
необходимы все три элемента. Были проведены
также эксперименты, в которых из хромосомы уда-
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
211
Элемент! Элемент!! Элемент III
CEN3 ATAAGTCACATGAT —95В’ХТ—GT АТТ Тб AT TTCCGAA AGT- ТАААА
CENA AAAGGTCAC ATGCT — -9’®ПА"Т— GTTTA TG AT TACCG А А АСА • ТАААА
CENb TTTCATCACGTGCT— 956«™т—6ТТТТTGТТTTCCGAAGATGТА А А А
CEN11 ATAAGTCACATGAT-— ^^---GT ТС А Т 6 АТ Т ТСС GA A CGT А Т А А А А
CfA/14 6TTAGTCACGTGCA—- 84п.к gt ATTTGTCТТСCGAA AAGТА АА А
РИС. 9.50.
Консервативные нуклеотидные последовательности не-
которых С£А/-областей дрожжей. Высококонсерватив-
ные участки выделены цветными прямоугольниками.
ляли сегмент длиной 627 п.н., содержащий CEN3
(рис. 9.51). Такая ацентрическая хромосома в дрож-
жевых кластерах нестабильна. Однако если сегмент
длиной 627 п. н. инвертирован, то хромосома функ-
ционирует нормально. Самое удивительное, что
CEN 3-область хромосомы 3 можно без каких-либо
последствий полностью заменить клонированной
CEN 11 -областью. Этот опыт показывает, что функ-
циональные центромеры не обязательно хромосо-
моспецифичны. Более того, совершенно ясно, что
спаривание гомологичных хромосом в мейозе за-
висит не от гомологичности центромер, а, вероятнее
всего, от сходства плечей хромосом.
Хромосомаз _
Точками обозначены пропущенные основания. Отметь-
те, что в элементе III имеется внутренний палиндром.
Участки хроматина дрожжей, содержащие эле-
менты I—III в CEN-области, более устойчивы к
действию ДНКазы I и микрококковой нуклеазы,
чем окружающие их последовательности. Эта ус-
тойчивость сохраняется даже в том случае, когда
CEN находится в мини-хромосомных векторах. От-
сюда следует, то хроматин в этих участках имеет
необычную структуру. Возможно, устойчивость к
действию нуклеаз определяется связыванием с цент-
ромерой специфических белков, прикреплением
ДНК к микротрубочкам или одновременно тем
и другим. Недавно такие ДНК-связывающие белки
были обнаружены в экстрактах дрожжевых клеток;
— -
Ацентрическая хромосома* WM3
(CEN3 делетирован)
к Нестабильная
I при митозе
CEN3 + UKA3
CENi инвертирован
Нормальная
CEW11 + LIRAS
РИС. 9.51.
ШШШ
Сел 11
ШШП
Нормальная
Влияние изменений CEN-последовательностей на
функцию центромеры. Вверху представлена хромосома
3 дикого типа. Хромосомы, показанные ниже, получены
путем сайт-специфической трансформации одной из
двух хромосом 3s диплоидных клеток. Трансформиро-
ванные дрожжевые колонии выделяли, используя селек-
тивный маркер URA3. Заштрихованные участки после-
довательности, фланкирующие C£7V3. которые исполь-
зовали для гомологичной рекомбинации в трансфор-
мирующих векторах. Справа указаны фенотипы, соот-
ветствующие измененным хромосомам 3. [L. Clark,
J. Carbon, Nature 305 (1983), р. 23.]
212
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Место прикрепления микро-
трубочки диаметром 20нм
220л. н.
Диаметр
15-20 нм
Устойчива к
нуклеазам
160 п.н.,
диаметр 11 нм
РИС. 9.52.
Модель структуры дрожжевой хромосомы, основанная
на данных о строении CENA 1. Функциональные центро-
мерные последовательности, содержащие элементы I, II
и III (220 п. н.), фланкированы хроматином с типичными
нуклеосомами. Эти последовательности связываются со
специфичными к центромерам ДНК-связывающими
белками, что обеспечивает их защиту от действия
ДНКаз. Микротрубочка прикрепляется к сердцевине
центромеры (по-видимому, функциональному кинето-
хору). [J.S. Bloom, J. Carbon, Cell 29 (1982), р. 305.]
для этого исследовали защиту ДНК от экзонуклеа-
зы III. которая проявляется в изменении электро-
форетической подвижности нуклеазных фрагментов
ДНК в геле. На рис. 9.52 представлена одна из
моделей структуры дрожжевых центромер. В этой
модели CETV-область вместе с ДНК-связывающими
белками образуют кинетохор, к которому прикреп-
ляются микротрубочки на стадии подготовки к сег-
регации. Эта модель иллюстрирует одну из возмо-
жных функций CETV-последовательностей, но в ней
не объясняется, как эти последовательности регу-
лируют число копий мини-хромосом и хромосом.
Возможно, репликация CETV-сегментов завершается
только в анафазе митоза или анафазе II мейоза. Это
означает, что последовательности, расположенные
с одной или с обеих сторон от CE/V-области, могут
блокировать прохождение репликационной вилки
до появления специфического сигнала, разрешаю-
щего окончание репликации в анафазе. В таком
случае число хромосом не будет превышать одной
на дочернюю клетку.
б. Последовательности в области
теломер
Теломеры, концы эукариотических хромосом,
являются также и концами линейного дуплекса
ДНК. С этими структурами связаны две важные
проблемы. Первая касается репликации: каким об-
разом достраиваются 5'-концы хромосомного дуп-
лекса, если ДНК-полимеразы не инициируют синтез
новых цепей (разд. 2.1)? Как решается этот во-
прос-так же, как и при репликации линейного
дуплекса аденовирусов, или с помощью альтерна-
тивных механизмов? Вторая проблема связана со
стабильностью концов ДНК в эукариотических
клетках. Вообще говоря, если разрывы в ДНК не
репарируются немедленно с помощью лигирования,
то происходит деградация ДНК под действием эн-
донуклеаз или рекомбинация. Однако концы хро-
мосом стабильны, а отдельные хромосомы в норме
не соединяются конец-в-конец. По-видимому, те-
ломеры обладают какими-то особыми молекуляр-
ными свойствами. Правильность такого предполо-
жения была подтверждена с помощью молекуляр-
ного клонирования. Многообещающим началом
послужило обнаружение специфических свойств у
простейших жгутиковых, в частности у Tetrahymena.
Клонирование теломерных последовательностей.
У некоторых простейших геном зародышевой линии
изолирован в микронуклеусе, а для экспрессии генов
они формируют «соматический» макронуклеус.
ДНК макронуклеуса содержит лишь часть микро-
нуклеарных последовательностей; она фрагменти-
рована, и в ней присутствует множество копий
коротких линейных сегментов ДНК, каждый из
которых содержит ограниченное число генов. В мак-
ронуклеусе у Tetrahymena короткие линейные дуп-
лексы, содержащие гены рРНК, представлены в дос-
таточном количестве, чтобы их можно было выде-
лить и охарактеризовать, не прибегая к клониро-
ванию. На концах этих молекул имеется от 20 до 70
тандемных повторов гексануклеотидной последова-
тельности 5'-ССССАА-3' (соответственно в другой
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
213
в
5'
CCCCAА
GGGGTT
В
| РДНК |
Макронуклеарная рДНК
I Tetrahymena
II I I I I I I I I I 1111111*1*1 Г''—s
^Рвсщеплениерестриктирующей эндонуклеазой
Дрожжевой вектор
Лигирование,
трансфекция,
отбор кпеток
LEU2+
GTGGTGG6T
3'
Трансфекция дрожжевых клеток /ей2",
отбор клеток LEU2*
I Лигирование
САССАСССА
CTGGTGGGT
3'
5’
САССАСССА
Рестриктирующая
эндонуклеаза
Фрагменты
> дрожжевой
геномной ДНК
РИС. 9.53.
Концы фрагментов макронуклеарной ДНК Tetrahymena
(А) выполняют функции, аналогичные функциям тело-
мер у дрожжей. Точками обозначены одноцепочечные
разрывы. Концевые фрагменты рДНК отщепленные с
помощью рестриктирующих эндонуклеаз, пришивали
к линейному челночному вектору дрожжи-Е. coli. Ли-
нейные рекомбинантные молекулы трансфицировали
в дрожжевые клетки, где происходила их репликация
(Б). Реплицированные линейные дуплексные молекулы
выделяли (В) и использовали для создания вектора,
предназначенного для клонирования дрожжевых тело-
мер (Г). Фрагменты, получившиеся после расщепления
рестриктирующей эндонуклеазой (RE), лигировали с
фрагментами, образовавшимися при зндонуклеазном
расщеплении всего дрожжевого генома. Из LEU2'-кле-
ток, отобранных после трансфекции дрожжевых клеток
!еи2 этими рекомбинантными молекулами, были вы-
делены стабильные реплицирующиеся дуплексы (Д)
Эти дуплексы содержали две дрожжевые теломеры,
к одной из которых была присоединена теломера
Tetrahymena, присутствовавшая в векторе (Г). Как по
казал последующий анализ, после репликации моле-
кулы Б в дрожжевых клетках последняя приобретала
типичные дрожжевые теломеры (В) с концевыми повто-
рами С„А, где п= 1, 2 или 3. На рисунке дрожжевые
теломеры выделены более темным цветом, чем тело
меры Tetrahymena Заметим, что молекула Д может
реплицироваться в отсутствие ARS последовательности,
представленной в молекуле Б, по-видимому, потому,
что эта последовательность присутствует в присоеди-
ненном сегменте дрожжевой теломеры.
214
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
цепи-5'-TTGGGG-3') (рис. 9.53, Л). В определенных
местах С-богатых сегментов встречаются одноце-
почечные разрывы, а на концах молекул имеются
шпильки с петлей. Таким образом, «концы» рДНК
отличаются от нормальных концов ДНК. Эти
структуры как раз и представляют собой функцио-
нальные теломеры и проявляют себя в этом ка-
честве при введении в дрожжевые клетки в составе
линейных векторов (рис. 9.53, Б). Итак, линейные
фрагменты ДНК, содержащие дрожжевые ARS (ав-
тономно реплицирующиеся последовательности) и
концевые участки, происходящие от рДНК Tetra-
hymena, стабильно реплицируются в дрожжевых
клетках. В отсутствие таких концов линейные фраг-
менты, как правило, оказываются крайне неста-
бильными.
Выделенные из дрожжей реплицированные ли-
нейные рекомбинантные молекулы (рис. 9.53, В) ис-
пользовались в качестве векторов для клонирования
самих теломер дрожжей. Один из необычных
концов сначала удалили, расщепив дуплекс соответ-
ствующей рестриктирующей эндонуклеазой (рис.
9.53, Г). К отрезанному концу пришили рестрици-
рованную дрожжевую ДНК. После трансфекции
были отобраны клетки, содержащие молекулы, ко-
торые реплицировались в дрожжевых клетках как
стабильные линейные ДНК (рис. 9.53, Д)- Функцио-
нальные дрожжевые теломеры, выделенные путем
субклонирования, использовали для определения
числа копий этих последовательностей в геноме
дрожжей. Оказалось, что на гаплоидный геном при-
ходится 30 40 таких копий - именно столько, сколь-
ко и должно быть при условии, что гомологичные
сегменты находятся на каждом из концов всех 16
хромосом дрожжей.
Структура дрожжевой теломеры весьма сходна
со структурой теломеры у Tetrahymena. Одна из
цепей содержит тандемный повтор 5'-С„А-3' (п = 1,
2 или 3), в котором в определенном месте имеется
одпоцепочечный разрыв. Как показали дополни-
тельные эксперименты, похожие тандемные повто-
ры содержатся в теломерах многих примитивных
эукариот (например, у трипаносом имеется повтор
5'-СССТАА-3'). На самых концах хромосом, как
правило, находятся очень короткие С- и А-богатые
последовательности. У некоторых видов (например,
S. cerevisiae) очень близко к концам располагаются
другие тандемные повторы с более длинными
повторяющимися единицами. Они могут участво-
вать в различных теломерных взаимодействиях,
в частности в связывании теломер между собой
и в присоединении теломеры к ядерной мембране.
У дрожжей и жгутиковых найдены белки, которые
связываются с теломерными повторами. Вероятно,
эти белки стабилизируют концы хромосом, защи-
щая ДНК от деградации и лигирования.
Элонгация теломер во время репликации. При
репликации линейных ДНК, содержащих теломеры
из Tetrahymena, в дрожжевых клетках концы ново-
синтезированных молекул модифицируются (ср.
рис. 9.53, Б и В). После репликации они наращи-
ваются приблизительно на 200 п. н. путем добавле-
ния тандемных повторов 5'-С„А-3', характерных для
теломер дрожжей, но не для Tetrahymena. Кроме
того, одноцепочечные разрывы в этих повторах
располагаются так же, как в теломерах дрожжей,
а не Tetrahymena. Аналогичные данные были полу-
чены и при исследовании структуры теломер неко-
торых других видов с использованием гетероло-
гичных хозяйских клеток. Элонгация концов хро-
мосом, по-видимому, является частью нормальной
репликации. Так, у трипаносом клонированные те-
ломерные зонды гибридизовались с рестрикцион-
ными фрагментами ДНК, выделенными на разных
стадиях синхронизованной многократной реплика-
ции, начиная с одной клетки. В течение нескольких
сотен генераций фрагменты, которые гибридизо-
вались с зондом, увеличивались в длину на 7-10 п. н.
за каждую генерацию (и соответственно утрачи-
вали большую часть из добавленных пар осно-
ваний). Сходные данные были получены и для
Tetrahymena.
Проблемы, связанные с загадочными свойства-
ми теломер дрожжей и простейших жгутиковых,
получили частичное разрешение с открытием фер-
мента теломеразы-терминальной дезоксинуклеоти-
дилтрансферазы. Этот фермент, выделенный из
Tetrahymena, присоединяет повтор 5'-TTGGGG-3',
последовательно по одному нуклеотиду, к З'-кон-
цам специфических олигонуклеотидных праймеров
(TTGGGG)„ (Tetrahymena) и (TGTGTGGG),, (дрож-
жи). Таким образом, теломераза может строить
теломеры, при этом родительская ДНК не исполь-
зуется в качестве матрицы (рис. 9.54). Теломера-
за-это крупный рибонуклеопротеиновый комплекс;
для проявления ее ферментативной активности не-
обходимы как РНК, так и белки. Теломеразная РНК
содержит последовательность 5'-СААССССАА-3',
которая, по-видимому, и служит матрицей для син-
теза олигонуклеотида TTGGGG. Эти недавно полу-
ченные данные не проясняют механизма синтеза
С-богатой теломерной цепи. Возможно, матрицей
и затравкой здесь служат G-богатые повторы.
Шпильки с петлями содержат G-богатый участок
и стабилизируются благодаря необычным взаимо-
действиям между остатками гуанина.
Эти механизмы, открытые при изучении одно-
клеточных, вероятно, свойственны большинству
эукариот. Клонирование и секвенирование теломер-
ных последовательностей растения Arabiilopsis и
человека показывают, что они состоят из олиго-
нуклеотидных повторов, очень близких повторам
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
215
[ДАСССС)п
(С « «
(TTGGeG)n
Распрямление
З'-гидроксильного
конца цепи,содержащей
S'-(TTGGG)n-3'
Терминальная
теломерная
нуклеотидил-транссрераза,
dGIP.TTP
| (TTGGGG)nO.
Праймаза, ДНК-полимераза,
неполное лигирование,
повторное "схлопывание"
РИС. 9.54.
Гипотетическая схема образования теломеры Tetrahy-
mena. В верхней части рисунка представлено образо-
вание петли на конце цепи, содержащей последователь-
ность 5'-(TTGGGG)n-3', и одноцепочечных разрывов
в противоположной цепи, содержащей последователь-
ность 5'-(ССССАА)п-3'. К З'-концу нижней цепи с по-
мощью теломерной нуклеотидилтрансферазы (тело-
изомеразы) присоединяются последовательно, по од-
ному нуклеотиду, единицы 5'-TTGGGG-3'. Праймаза
и ДНК-полимераза копируют 5'-(TTGGGG)n-3'-uenb
с образованием новых 5-(ССССАА)п-3'-единиц.
В результате неполного лигирования в С-богатой це-
пи остаются одноцепочечные разрывы. На З'-конце
5'-(TTGGGG)n-3'-uenn вновь образуется петля, стаби-
лизируемая взаимодействиями между остатками гуано-
зина. [C.W. Greider, Е. Н. Blackburn, Cell 43 (1985).
р. 405.]
жгутиковых и дрожжей. Теломераза обнаружена
и в клетках человека.
Образование множественных теломер в макро-
нуклеусах простейших. Рассмотрим еще раз струк-
туру рДНК Tetrahymena. В диплоидной ДНК мик-
ронуклеуса Tetrahymena имеются всего два гена
рДНК. В результате их амплификации в макро-
нуклеусе образуются сотни линейных молекул, каж-
дая из которых содержит рДНК и теломерную
структуру. Однако в микронуклеусе рДНК не явля-
ется теломерной. Кроме того, большинство линей-
ных дуплексных ДНК длиной от 2 до 100 т.п.н.,
типичных для макронуклеуса Tetrahymena, имеют
одинаковые концевые последовательности. Эти мо-
лекулы содержат кроме рДНК много других генов
и образуются согласно общей программе фрагмен-
тации (и элиминации) ДНК, которая приводит к
развитию соматического макронуклеуса. Подобная
картина наблюдается у другого простейшего жгути-
кового. Oxytricha, за исключением того, что конце-
вой теломерный повтор представляет собой после-
довательность 5'-ССССАААА-3', характерную для
теломеры Oxytricha из хромосомы микронуклеуса.
Например, ген макронуклеарного актина Oxytricha,
содержащийся во фрагменте длиной 1,6 т.п.н.,
фланкирован четырьмя копиями последовательно-
сти 5'-ССССАААА-3' (или 5'-TTTTGGGG-3')- Как
образовались такие концевые сегменты? У Tetrahy-
mena в местах расщепления микронуклеарной ДНК,
происходящего при формировании макронуклеар-
ных фрагментов ДНК, находится консервативная
последовательность длиной 15 п.н. (рис. 9.55). кото-
рая, по-видимому, содержит сайт для специфиче-
ской эндонуклеазы. После (или в процессе) расщеп-
ления некоторые из окружающих этот сайт после-
довательностей ДНК удаляются, и, вероятно, с по-
мощью терминальной нуклеотидилтрансферазы
достраиваются 5'-GGGGTT-3'-KOHnbi- так, как это
описано в предыдущем абзаце. Отметим, что мик-
ронуклеарные предшественники фрагментов ДНК
макронуклеуса как у Tetrahymena, так и у Oxytricha
совсем не обязательно должны содержать флан-
кирующие 5'-СССС(А)„Т-3'-повторы. Итак, при об-
разовании макронуклеарной ДНК происходят вы-
резание сегментов из ДНК микронуклеуса, утрата
части некодирующей фланкирующей ДНК, присое-
динение олигонуклеотидных повторов, образование
шпилек и, наконец, формирование функциональных
теломер. Шпильки, вероятно, стабилизируют фраг-
менты ДНК, предотвращая их деградацию и непра-
вильное сшивание с другими дуплексами.
в. Искусственные хромосомы
дрожжей
В настоящее время в распоряжении исследователей
имеются все сегменты ДНК. необходимые для кон-
струирования функциональных дрожжевых хромо-
сом. Клонированные фрагменты, содержащие ARS-
последовательность для репликации, CEN-элемент
для формирования центромеры и теломерные сег-
менты, можно сшить между собой in vitro в нужном
порядке. Чтобы включить в хромосому один или
несколько селективных маркерных генов, например
LEU2 или HIS3, в линейный дуплекс можно
встроить соответствующие сегменты ДНК (рис.
9.56). Если суммарная длина линейной ДНК состав-
ляет менее 20 т.п.н., то синтетические хромосомы
216
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
5'„
(Тконец
3. ( рДНК
GAAATTA ТТААА
СТТТААТ ААТТТ
Микронуклеус
’ТТТТААСТТА С ( ТАААССААССТ С ТТ Т I GTTTA Л
AAAATTGAAT (k(ATTTGGTTGGAGAА^, ( CAAAT Q
/ '-------------------*----------'
11 П.Н-
5'
з'
Макронуклеус
Г ТGGGGTTGGGGIТ GGG1
А А ССССААС С Ci CCCi
ТТ< GGG
120-420 п.н., присое--------<
диняющиеся к свободному концу дуплекса
РИС. 9.55.
З'-концевыв последовательности в микро- и макронук-
леарной рДНК Tetrahymena. Фигурной скобкой обозна-
чена последовательность длиной 15 п.н., в которой
происходит расщепление. В микронуклеарной ДНК, в
отличив от рДНК макронуклеуса, присутствует только
одна копия последовательности 5'-ССССАА. [В. О. King,
М.-С. Yeo, Cell 31 (1982), р. 177; М.-С. Yao, К. Zheng,
С.-Н. Yao, Cell 48 (1987), р. 779.]
оказываются нестабильными во время митоза, и
центромера не контролирует число копий. В резуль-
тате в каждой клетке накапливается множество
молекул. Если же длина ДНК превышает 50 т. п. н.,
то поддерживается стабильность хромосом, осуще-
ствляется контроль за числом их копий и происхо-
дит правильное расхождение хромосом. Молекулы
длиной 150 т.п.н. ведут себя более или менее нор-
мально как в митозе, так и в мейозе. Очевидно, для
того чтобы хромосомы могли перемещаться при
клеточном делении, их длина должна быть не мень-
ше определенной величины. Такие искусственные
хромосомы дрожжей используются в качестве век-
торов при клонировании очень длинных сегментов
ДНК с целью картирования сложных эукариоти-
ческих геномов.
9.7. ГЕНОМЫ ОРГАНЕЛЛ
ЭУКАРИОТ: ДНК МИТОХОНДРИЙ
И ХЛОРОПЛАСТОВ
Отличительная особенность клеток эукариот
состоит в том, что часть генетической информации
у них заключена в молекулах, находящихся вне
хромосом, локализованных в ядре. Существуют два
типа таких цитоплазматических ДНК: одни нахо-
дятся в митохондриях эукариот, другие-в хлоро-
пластах зеленых растений и водорослей. Как и все
цитоплазматические элементы, они наследуются по
материнской линии, а не по законам Менделя.
Большинство белков, из которых построены
функциональные и структурные компоненты мито-
хондрий и хлоропластов, кодируются хромосомной
ДНК, синтезируются на рибосомах в цитоплазме
и транспортируются в соответствующие органеллы
(разд. З.Ю.в). Однако несколько белков кодируются
неядерной ДНК и синтезируются на особых ри-
босомах органелл. Таким образом, органеллы-это
результат объединенных усилий двух геномов и
двух трансляционных аппаратов. РНК-компоненты
рибосом органелл, а также тРНК, использующиеся
при трансляции, кодируются геномами митохонд-
рий и хлоропластов.
Очевидно, что взаимодействие между продукта-
ми хромосомных, митохондриальных и хлоропласт-
ных генов обусловлено генетически. Например, му-
тации в митохондриальной ДНК у дрожжей и куку-
рузы могут супрессироваться хромосомными мута-
циями. Определенные виды мужской стерильности
(образование стерильной пыльцы), широко распро-
страненные у растений, вызваны мутациями в мито-
хондриальной ДНК и наследуются не в соответствии
Н/53, дрожжи
Теломера
PBR322
ARS, дрожжи
77?Р1,дрожжи
CEN3t дрожжи
РИС. 9.56.
Функциональная искусственная хро-
мосома дрожжей. [A. W. Murray,
J.W. Szostak, Nature 305 (1983),
р. 189.]
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
217
с законами Менделя, а по материнской линии. Фер-
тильность таких растений может восстановиться
в результате мутаций в хромосомных генах; разные
виды мужской стерильности цитоплазматической
природы отличаются друг от друга, поскольку фер-
тильность восстанавливается в результате мутаций
в разных хромосомных генах.
В табл. 9.14 перечислены некоторые компоненты
митохондрий, кодируемые ядерными генами. Даже
отдельные ферменты могут состоять из полипепти-
дов, кодируемых двумя разными геномами. Более
того, экспрессия некоторых митохондриальных ге-
нов зависит от активности продуктов ядерных генов
(например, процессинг транскриптов дрожжевой
митохондриальной ДНК с образованием функцио-
нальных мРНК). В табл. 9.14 приведены лишь об-
щие данные; разные организмы имеют свои осо-
бенности. Ген, находящийся в митохондриальной
ДНК у одного организма, может быть хромосом-
ным у другого; иногда гены, кодирующие данный
полипептид, содержатся в обоих геномах. Анало-
гичная картина наблюдается и в хлоропластах.
а. Геномы митохондрий
Многие митохондриальные геномы представля-
ют собой замкнутые кольцевые сверхспиральные
дуплексные ДНК. Большинство из них кодируют
сходные функции. Но сделать еще какие-то обобще-
ния относительно митохондриальных ДНК доволь-
но трудно. Иногда даже близкородственные виды
Таблица 9.14. Геномный источник некоторых
компонентов митохондрий1’
Хромосомы (ядро) Митохондрии
Ферменты + факторы реп- ликации РНК-полимераза + фак- торы транскрипции Ферменты сплайсинга РНК (дрожжи) Рибосомные белки Аминоацил-тРНК- синте- тазы Факторы трансляции Субъединицы цитохром- оксидазы Субъединицы АТРазы Субъединицы NADH-де- гидрогеназы Матуразы, участвующие в сплайсинге (дрожжи) Один рибосомный белок (дрожжи) тРНК рРНК Субъединицы цитохром- оксидазы Субъединицы АТРазы Субъединицы NADH-де- гидрогеназы
11 Распределение компонентов немного различается у разных орга-
низмов. Фермент дыхательной цепи NADH-дегидрогеназа известен
также под названием комплекса I или NADH-убихинон оксидо-
редуктазы.
имеют совершенно разные рестриктные карты и
лишь частично совпадающие нуклеотидные после-
довательности. Расположение гомологичных генов
в кольцевой ДНК также различается. Иногда у от-
дельного растения присутствуют митохондриаль-
ные геномы разных размеров, при этом они пред-
ставлены как линейными, так и кольцевыми моле-
кулами. Митохондрии других организмов, включая
некоторые грибы и простейшие, имеют линейные
геномы.
Геномы митохондрий существенно различают-
ся по размеру. У животных они относительно
малы и обычно не превышают 20 т.п.н. Однако
дрожжевые митохондриальные ДНК состоят из
примерно 80 т.п.н., а длина митохондриальных
ДНК растений варьирует от нескольких сотен до
нескольких тысяч т. п. н. Все митохондриальные
ДНК обладают весьма интересным свойством - они
не метилированы. В очень крупных митохондриаль-
ных геномах подавляющую долю избыточной ДНК
составляют некодирующие последовательности
с неизвестной функцией. Напротив, митохондриаль-
ные ДНК млекопитающих организованы весьма
рационально, и между генами почти нет проме-
жутков.
Митохондриальная ДНК эволюционирует гораз-
до быстрее, чем ядерная, и мутации в ней происхо-
дят почти в десять раз чаще. В результате возникает
широкий внутривидовой полиморфизм, и ощути-
мые изменения выявляются даже в пределах не-
скольких поколений. Благодаря высокой скорости
мутирования полиморфизм эндонуклеазных сайтов
становится очень удобным параметром при попу-
ляционно-генетических исследованиях растений и
животных, в том числе и человека.
Митохондриальная ДНК млекопитающих. Нук-
леотидная последовательность митохондриальной
ДНК некоторых млекопитающих полностью уста-
новлена (рис. 9.57). Организация всех этих геномов
близка к организации геномов Drosophila и Xenopus.
Тем не менее одни локусы могут быть сходны на
20%, а другие-на 90%; значительные различия
наблюдаются даже в пределах одной кодирующей
области.
Нуклеотид, обозначенный на рис. 9.57 цифрой
1, указывает точку начала репликации митохонд-
риальной ДНК и находится в области, существен-
ной для регуляции экспрессии митохондриальных
генов. Если двигаться от этого нуклеотида по
часовой стрелке вдоль митохондриальной ДНК че-
ловека, то первым будет ген тРНКр*с; далее идет ген
РНК малой субъединицы митохондриальных рибо-
сом-ген 12S-pPHK и т.д. Большинство генов,
кодирующих РНК или белки, разделены одним или
более тРНК-генами. Промежутки между генами,
как правило, не превышают 25 п. н. и в большинстве
218
ЧАСТЬ III. ОРГАНИЗАЦИЯ. ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
16SrRNA
ND5
tRNAhls
ND4
ND4L
tRNAar9
tRNAleu
tRNAser_
lRNA,eu
tRNApr°
tRNA
cytb
tRNA9lu
ND1
16 569 п.н.
ВО 00
/tRNA'1*
-tRNABln
tRNAp-met
ND2
tRNAtr₽
-tRNAa,a
tRNAesn
ORI(L)
tRNAcVs
tRNA*Vr
C01
tRNAser
СОП tRNAasp
tRNA'V5
tRNA₽he
12SrRNA
tRNAVBl
ND3
tRNAS1*
coin
ATPase6
ATPase8
РИС. 9.57.
Митохондриальная ДНК человека. Гены, кодирующие
белки, выделены темно-серым цветом; они кодируют
три большие субъединицы цитохром с-оксидазы (COI,
СОИ. СОШ), цитохром b (cytjb), субъединицы 6 и
8 АТРазного комплекса и субъединицы NADH-дегидро-
геназного комплекса (ND). Гены, кодирующие различ-
ные рРНК, выделены светло-серым цветом, а гены
тРНК- окрашенными полосками. Каждый ген имеет
свое систематическое название, но для простоты здесь
и на рис. 9.58 указаны только сокращенные обозначения
соответствующих генных продуктов. Точками отмечена
примерная локализация сайтов инициации транскрип-
случаев составляют менее 3 п. и. Иногда (например,
в случае генов тРНК изолейцина и тРНК глутамина,
а также генов АТРазных субъединиц 6 и 8) гены
перекрываются.
Транскрипция митохондриальной ДНК млекопи-
тающих. В отличие от большинства эукариотичес-
ких генов, каждый из которых представляет собой
независимую транскрипционную единицу, обе цепи
всего митохондриального генома транскрибируют-
ся с образованием одной молекулы РНК, начи-
нающейся от сайта вблизи точки начала репликации
тяжелой цепи ДНК-ORI-H (цепь называется тяже-
лой, поскольку при равновесном центрифугирова-
нии денатурированной ДНК в градиенте плотности
CsCl она образует полосу с более высокой плот-
ностью, чем другая, легкая цепь). Кроме того,
ции двух цепей. Отметим, что с Н-цепи как матрицы
транскрибируются две РНК (по часовой стрелке). Из
короткого РНК-транскрипта образуется рРНК, а из
длинного-мРНК и большинство тРНК. На L-цепи об-
разуется только один полноразмерный транскрипт. Ген
7S-PHK транскрибируется против часовой стрелки в
непосредственной близости к сайту инициации. Воз-
можно, эта РНК играет роль регулятора транскрипции
или созревания. Участки, выделенные точками,- сайты
инициации репликации ДНК. Маленькими стрелками
обозначено 5’-» З'-направление смысловой цепи в раз-
личных генах.
образуются более короткие транскрипты Н-цепи.
заканчивающиеся на З'-конце гена 16S-pPHK. Этих
коротких транскриптов в десять раз больше, чем
длинных: после процессинга из них образуются
12S-pPHK, 16S-pPHK, а также тРНКРКе и rPHKVal.
Изобилие этих транскриптов обеспечивает быстрое
накопление рРНК. Из более длинных транскриптов
Н-цепи и транскриптов L-цепи в результате процес-
синга образуются мРНК и другие тРНК. Функцио-
нальные РНК транскрибируются с обеих цепей, хотя
одна из них, L-цепь, является основной матрицей
и гибридизуется с большинством зрелых молекул
РНК (рис. 9.57).
Цепи митохондриальной ДНК транскрибируют-
ся специфической митохондриальной РНК-полиме-
разой, которая кодируется ядерной ДНК. Ни ме-
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
219
ханизм инициации транскрипции, ни механизм пре-
вращения очень длинных первичных транскриптов
в отдельные молекулы РНК до конца не ясны.
Установлено, что промоторы содержатся в сегмен-
тах ДНК длиной 40 п.н. или менее, обрамляющих
сайты начала транскрипции. Они неодинаковы у
разных видов млекопитающих; видоспецифичны
и митохондриальные РНК-полимеразы. В современ-
ных моделях уникальная роль в созревании длинных
молекул РНК отводится тРНК-кодирующим сег-
ментам, которые фланкируют почти все гены. Дело
в том, что молекулы тРНК в длинных транскрип-
тах, по-видимому, образуют внутримолекулярные
шпилечные структуры. Эти структуры распознают-
ся специфическими рибонуклеазами, вероятно сход-
ными с РНКазой Р E.coli', фермент разрезает их
с обеих сторон, в результате чего образуются мо-
лекулы тРНК и рРНК или мРНК. И действительно,
в митохондриях млекопитающих была выявлена
РНКаза P-подобная активность. Процессированные
транскрипты претерпевают затем обычные превра-
щения (созревают): мРНК полиаденилируются,
тРНК приобретают каноническую З'-концевую пос-
ледовательность и 5'-ССА. Однако 5'-концы мРНК
не кэпируются, что необычно для эукариот. Сплай-
синга не происходит, поскольку ни один из генов не
содержит вставочных последовательностей.
Транскрипты L-цепи существенны также для
инициации синтеза ДНК в точке ORI-Н. По-види-
мому, некоторые из этих транскриптов разрезаются
в пределах участка длиной примерно 200 нуклеоти-
дов, начиная от сайта инициации транскрипции. Эти
короткие молекулы РНК служат затем затравкой
при репликации ДНК.
Необычный трансляционный аппарат митохонд-
рий м ккопитающих. Структура митохондриальных
мРНК обладает особенностями, ставящими ряд
проблем с точки зрения трансляции. В них отсутст-
вуют не только типичные кэп-структуры. но и про-
тяженные 5'-лидерные последовательности слева от
инициирующих кодонов, так что рибосомы мито-
хондрий должны узнавать и связывать мРНК ка-
ким-то другим способом. Еще одна особенность
трансляции у митохондрий состоит в использовании
необычных кодонов: инициирующими кодонами
могут быть, помимо AUG, также и метиониновые
кодоны AUA, AUU и AUC (табл. 9.15). Универ-
сальные стоп-кодоны UAA и UAG используются
и в митохондриях млекопитающих, однако еще
один обычный стоп-кодон, UGA, кодирует трип-
тофан. Универсальные аргининовые кодоны AGA
и AGG служат стоп-кодонами и не кодируют ни-
каких аминокислот.
У некоторых митохондриальных генов на конце
кодирующей последовательности вообще нет стоп-
кодона. Примером может служить ген субъединицы 6
АТРазы. В этих случаях при процессинге транск-
рипта на конце остается или U, или UA. В результа-
те последующего полиаденилирования на З'-конце
мРНК образуется стоп-кодон UAA.
Таблица 9.15. Митохондриальные кодоны и антикодоны у че-
ловека11
Кодон Антикодон Кодон Антикодон Кодон Антикодон Кодон Антикодон
Р- AAG UCU Sa ser AGb UGG UAC ‘УГ AUG UGU АГГ UGC CVS ACG
UUG leU AAU UAA Стоп. UAG UGA АГТТ UGG trP ACU
сии Sa leu GAb cue ecu ccc CCA РГ° GGU CCG GAG his GUG LAC CAA - CAG 91" CUU CGU CGA аГ3 GGU CGG
AUU , П*Г AUC "e UAG ACU ACC thr UGU АСА ACG AAG asn UUG AAC AGU . ir'r' ACC SCrUCG
AUU AUA metUAC AUG AAA . п и т AAC '*S AGA Стоп AGG
GUU GUA Va‘ GUG ecu GCC । r't-t t GCA ala CGU GCG GAU Г-ЫГ- GAC a5₽ CUG gag 8iu cbb GGU GGC i г-г-и GCA 8l> CCb GGG
° Основания в кодонах перечислены в направлении 5'-*3\ а антикодонов 3'-»5'.
У некоторых млекопитающих кодоны AUA, AUU и AUC могут быть инициирующими
и узнаются N-формилметиониновой тРНК с антикодоном 3'-UAC-5'.
220
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Необычному генетическому коду отвечает спе-
цифическое семейство тРНК. На генетической карте
рис. 9.57 представлены все 22 митохондриальные
тРНК человека, а в табл. 9.15-соответствующие
антикодоны. В отличие от универсального кода,
в котором для каждого из синонимичных кодонов
используется своя тРНК, в митохондриях для двух
или четырех таких кодонов используется одна
тРНК. Каждая тРНК, связывающаяся с двумя си-
нонимичными кодонами, использует для этого
в одном из случаев образование пары G • U. У каж-
дой тРНК, связывающейся с четырехчленным набо-
ром кодонов, на 5'-конце антикодонового участка
находится урацил. При этом либо U спаривается
с любым из четырех оснований, либо во взаимо-
действии кодон-актикодон участвуют только две
пары оснований.
Дрожжевая митохондриальная ДНК. Несмотря
на то что эта ДНК содержит примерно столько же
генов, сколько митохондриальная ДНК человека, по
своей длине она почти в пять раз больше - около 80
т.п.н. (рис. 9.58). Большая часть этой избыточной
ДНК приходится на долю АТ-богатых участков
с неизвестной функцией, а также последователь-
ностей, которые прерывают кодирующие области
и отсутствуют в митохондриальных ДНК млеко-
питающих. Различия в структуре митохондриаль-
ной ДНК у разных штаммов определяются в основ-
ном изменчивостью некодирующих участков. Одна-
ко неодинаковы и кодирующие участки, при том что
функции соответствующих генных продуктов оста-
ются практически одинаковыми.
Транскрипция и трансляция дрожжевой митохон-
дриальной ДНК. Те ограничения, которые налагают-
ся на размер митохондриального генома млекопи-
тающих (а также плоских червей, морского ежа,
насекомых и т.д.), у дрожжей отсутствуют. Эта
свобода связана с меньшей специфичностью его
структуры и функций. Гены тРНК не служат знака-
ми пунктуации, разделяющими другие гены. Тран-
скрипция начинается не с одного, а по меньшей мере
с 19 разных промоторов; ее катализируют специфи-
ческие митохондриальные РНК-полимеразы, коди-
руемые хромосомной ДНК. На 5'-конце каждой
транскрипционной единицы находится короткая про-
моторная последовательность (5'-A/TTATAAGTA-3'),
простирающаяся от -9-го до + 1-го нуклеотида
относительно точки инициации транскрипции (+1).
В результате процессинга длинных транскриптов
при участии эндонуклеазы образуются функцио-
нальные РНК. Генетический код дрожжевых мито-
хондрий тоже отличается от универсального, но эти
отличия все же меньше, чем у млекопитающих.
Длина молекул рРНК также ближе к обычной: 3200
и 1660 нуклеотидов по сравнению с 1559 и 954
нуклеотидами у человека. Хотя кодирующие после-
довательности дрожжевой митохондриальной ДНК
высокогомологичны соответствующим последова-
тельностям у человека, дрожжевые мРНК имеют
5'-лидерную и З'-трейлерную последовательности,
как и большинство мРНК (ср. гены цитохрома
b у человека и дрожжей; рис. 9.59).
Тем не менее дрожжевой ДНК присущи неко-
торые особенности. Это единственная известная
ДНК, у которой гены рРНК разделены неродствен-
ными генами. Далее, эта ДНК кодирует один ри-
РИС. 9.58.
Некоторые особенности митохондриальной
ДНК дрожжей (S. cerevisiae). Гены, коди-
рующие белки, окрашены более интенсив-
но, а гены, кодирующие РНК,- светлее.
Вставочные последовательности выделены
точками (некоторые из них содержат от-
крытые рамки считывания; часть их ука-
зана). Серым цветом выделены неохарак-
теризованные области; некоторые из них
содержат сайты начала репликации. Тран-
скрипция в основном идет по часовой
стрелке; единственное известное исключе-
ние-транскрипция генов тРНК, начина-
ющаяся в районе двух часов (отмечена
стрелкой).
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
221
ДРОЖЖИ
Ген cyt Ь
ЧЕЛОВЕК
Ген cvtb
РИС. 9.59.
Гены цитохрома b и соответствующие мРНК у дрожжей
и человека. Экзоны выделены цветом; интроны в дрож-
жевой ДНК выделены точками. [Р. Borst, L. A. Grivell,
Nature 290 (1981). р. 443.]
босомный белок (локус шг-1), а все другие рибо-
сомные белки кодируются ядерной ДНК; у млеко-
питающих все рибосомные белки митохондрий ко-
дируются ядерной ДНК. И наконец, в митохонд-
риальной ДНК дрожжей впервые обнаружены ко-
дирующие участки во вставочных последователь-
ностях.
Гены, расположенные внутри интронов дрожже-
вой митохондриальной ДНК. В митохондриальной
ДНК S. cerevisiae гены цитохрома b (cytb), 21S-pPHK
и субъединицы I цитохромоксидазы (CD-I) содержат
интроны. Некоторые из этих интронов присутст-
вуют не во всех дрожжевых штаммах. Другие су-
щественны для жизнедеятельности дрожжевых кле-
ток, поскольку в них картируются мутации, на-
рушающие функции дыхательной цепи. По данным
теза цитохрома Ь. Кроме того, некоторые мутации
в интронах плейотропны: при их наличии не только
ингибируется синтез цитохрома Ь, но и нарушается
экспрессия СО-I. Это наводит на мысль, что данный
интрон кодирует какой-то транс-белок; подтвержде-
нием этому служит обнаружение открытых рамок
считывания во многих митохондриальных интронах.
Некоторые мутации в интронах влияют на образо-
вание специфической вторичной структуры, необ-
ходимой для сплайсинга интронов (разд. 8.5.г).
Как показали результаты клонирования и секве-
нирования, у гена cytb имеются два интрона, 12 и 14,
которые содержат протяженные открытые рамки
считывания и могут кодировать белки (рис. 9.60).
Созревание первичного транскрипта начинается с
удаления II и соединения экзонов 1 и 2 (сплайсинг
таких интронов группы II описан в разд. 8.5.в).
Образовавшаяся РНК служит матрицей для синтеза
матуразы 12- белка, участвующего в дальнейшем
процессинге РНК. Последовательность, кодирую-
щая матуразу 12, начинается в первом экзоне гена
цитохрома b и распространяется на кодирующий
участок интрона 12 с открытой рамкой. Матураза 12
участвует затем в вырезании области 12, разрушая
таким образом свою собственную мРНК - уникаль-
ный регуляторный механизм! Генетические и моле-
6 7 8
J____I___I
генетического анализа, вставочные последователь-
ности, например, в гене cytb необходимы для син-
Е1 Е2 ЕЗ Е4 Е5 Е6
Я I 8г-
II I II
И 12 13 14 15
Продукт
неизвестен'*
2-й сплайсинг
1 Матураза 12
РИС. 9.60.
Ген цитохрома b дрожжей. Экзоны выделены
цветом, интроны-точками и серым цветом.
Участки интронов, выделенные редкими точками,
содержат последовательности, кодирующие ма-
туразы 12 и 14. Детали сплайсинга неизвестны.
Дополнительный сплайсинг,
матураза 14
мРНК для цитохрома Ь
222
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
кулярные исследования показали также, что про-
тяженная открытая рамка считывания интрона 14
кодирует матуразу, отличную от участвующей в
сплайсинге 14. Матураза 14 ответственна за плейо-
тропное действие упомянутых выше мутаций. Му-
тации в 14 cyib блокируют сплайсинг интрона в
транскриптах гена СО-1. В свою очередь, первый
интрон гена СО-l сам кодирует матуразу.
Интрон 215-рДНК не абсолютно необходим.
Для сплайсинга не нужны ни этот сегмент длиной
1,1 т.п.н., ни другие митохондриальные интроны.
Тем не менее данный интрон был тщательно иссле-
дован, поскольку он обладает необычными свойст-
вами. При спаривании дрожжевых штаммов, име-
ющих интрон в 215-рДНК (со+), со штаммами,
у которых он отсутствует (со-), почти все дочерние
митохондрии, включая и произошедшие от роди-
тельских митохондрий со-, оказываются со+. Это
связано с тем, что копия интрона длиной 1,1 т.п.н.
встраивается в со -215-рДНК. Встраивание проис-
ходит в строго определенном месте 215-рДНК
в последовательности 5'-GATAACAG-3'. Если в этой
последовательности-мишени возникают мутации,
то встраивания не происходит. К такому же ре-
зультату приводят и мутации в интроне 1,1 т.п.н.,
если при этом прерывается открытая рамка счи-
тывания длиной 235 кодонов. Этот интрон кодирует
сайт-специфическую эндонуклеазу, необходимую
для его встраивания в со--218-рДНК. Таким обра-
зом, он представляет собой некое подобие мобиль-
ного элемента (гл. 10). Изучая данную систему,
можно составить представление о том, как «про-
никают» интроны в уже существующие гены.
Репликация митохондриальной ДНК. Клетки
млекопитающих в культуре содержат тысячи мо-
лекул митохондриальной ДНК. Точное их число
различается у разных организмов, и даже для кле-
ток определенного типа, по-видимому, нельзя наз-
вать их точной цифры. Хотя репликация митохон-
дриальной ДНК не связана с клеточным циклом,
обычно образуется достаточное количество новых
молекул, чтобы снабдить ими все дочерние клетки.
Лучше всего изучен механизм репликации ми-
тохондриальной ДНК мышей (рис. 2.5). Напомним,
что каждая из цепей этой дуплексной молекулы
имеет свою точку начала репликации: Ori-H и Ori-L.
В мышиных клетках (но не у всех особей) большин-
ство митохондриальных ДНК содержат короткий
сегмент реплицированной Н-цепи (550-650 т.п.н.)
и так называемую D-петлю (от англ, displacement)
в области Ori-H. Другая необычная особенность
зрелых геномов состоит в наличии рассеянных по
геному рибонуклеотидов вместо дезоксирибонук-
леотидов. Возможно, эти нуклеотиды- остатки
РНК-праймеров, необходимых для репликации
ДНК, и рибонуклеотидных вставок, участвующих
в наращивании цепи. Но почему синтез митохонд-
риальной ДНК столь некорректен? В дрожжевой
митохондриальной ДНК, в отличие от ДНК мле-
копитающих, имеются семь или даже больше мест
начала репликации, рассеянных по всему геному
(рис. 9.58); каждое из таких мест содержит три
коротких GC-богатых сегмента, разделенных двумя
длинными участками, почти целиком состоящими
из АТ-пар.
За одним исключением, о ферментах или бел-
ковых факторах, участвующих в репликации ми-
тохондриальных ДНК, ничего не известно. Уста-
новлено лишь, что из трех типичных эукариоти-
ческих ДНК-полимераз в синтезе митохондриаль-
ных ДНК участвует у-полимераза.
б. Необычная митохондриальная
ДНК трипаносом
Отличительная особенность простейших из от-
ряда так называемых кинетопластид, который
включает и трипаносом, состоит в том, что в их
единственной митохондрии имеется необычная дис-
ковидная структура. Диски называются кинетоплас-
тами и содержат тысячи кольцевых молекул ДНК.
У Trypanosoma brucei 45 кольцевых молекул имеют
длину примерно 21 т.п.н. (макси-кольца); длина
остальных 5500 молекул-примерно 1 т.п.н. (ми-
ни-кольца). Кольцевые молекулы ДНК, соединяясь
в так называемые катенаны, образуют переплетен-
ную сеть (рис. 9.61). Это уникальное явление: ника-
ких других примеров образования таких сетей неиз-
вестно. Изолированные сети кинетопластов ДНК
составляют около 10 мкм в диаметре, но внутри
клетки они конденсируются с образованием дисков
в десять раз меньшего размера. Такие диски всегда
находятся в основании жгутика трипаносомы.
Мини-кольца одной сети часто различаются по
РИС. 9.61.
Кинетопласты трипаносомы {Crithidia fasciculate). На
электронной микрофотографии видны катенаны из
сцепленных мини- и макси-колец ДНК. Каждая петля-
это мини-кольцо размером 2,5 т. п. н. [С любезного
разрешения Р.Т. Englund; см. Р.Т. Englund, Ann. Rev.
Biochem. 52 (1982), p. 701.]
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
223
своим нуклеотидным последовательностям, за иск-
лючением консервативного сегмента длиной не-
сколько сотен пар оснований, но даже и этот сег-
мент не всегда идентичен у разных видов. Однако
все известные мини-кольца непременно содержат
в пределах упомянутого консервативного участка
последовательность 5'-GGGGTTGGTGTAA-3', в
которой, по-видимому, начинается репликация.
Кроме того, вблизи консервативного участка мо-
лекула ДНК имеет искривленную форму (разд.
1.1.д). Функции мини-колец неизвестны. Был иден-
тифицирован один непротяженный транскрипт дли-
ной 240 нуклеотидов, но он содержит только корот-
кие открытые рамки считывания неизвестного на-
значения, как и вся мини-кольцевая ДНК. Макси-
кольца выполняют функции митохондриального ге-
нома, аналогичные уже описанным выше. В отличие
от мини-колец, все макси-кольца в сети сходны. Как
и дрожжевая митохондриальная ДНК, они сильно
(на 80%) обогащены АТ-парами.
Репликация сложной сети кинетопластной ДНК
происходит один раз за клеточное деление. Она
начинается с расщепления катенанов и высвобожде-
ния мини-колец, вероятно, с помощью топоизоме-
разы II. Далее мини-кольца реплицируются с обра-
зованием свободных кольцевых дуплексов с про-
белами в одной цепи, по которым происходит их
присоединение к матриксу. Затем пробелы запол-
няются, и формируются замкнутые дуплексы. На-
конец, матрикс делится пополам и две сети ДНК
распределяются по дочерним клеткам. Макси-коль-
ца остаются присоединенными к сети и, по-види-
мому, реплицируются по механизму катящегося
кольца. Новосинтезированные линейные молекулы
отсоединяются от комплекса репликации и вклю-
чаются в сеть в кольцевой форме.
Редактирование на уровне РНК. У кинетопластид
обнаруживаются удивительные отклонения от уни-
версального механизма передачи генетической ин-
формации. Некоторые макси-кольца кодируют по-
следовательности, существенно отличающиеся от
последовательностей соответствующих функцио-
нальных мРНК. Например, смысловая цепь гена
СО-П T.hrucei содержит сегмент 5'-AAAGTAGAG-
AACCTGGTAGGTGTAATGAAATAA-3', однако со-
ответствующая последовательность СО-П-мРНК
(полученная исходя из данных о последовательно-
сти кДНК) имеет вид 5'-AAAGUAGAUUGUAUA-
CCUGGUAGGUGUAAUGAAAUAA-3' (в обоих
случаях приведена лишь часть рамки считывания,
начинающаяся с кодона 5'-ААА, отвечающего ли-
зину). Выделенные жирным шрифтом остатки U
были встроены в первичный транскрипт; связанный
с этим сдвиг рамки считывания ( + 4) обеспечивает
полную трансляцию РНК с образованием поли-
пептида СО-П. Наличие такого редактирования
РНК означает, что ДНК не всегда служит единст-
венным источником генетической информации. Уже
сейчас известны три вида организмов, у которых
правильная экспрессия пяти разных кинетопластид-
ных митохондриальных генов связана с редакти-
рованием РНК. Кроме сдвига рамки считывания,
некоторые вставки U приводят к появлению стар-
товых (AUG) или стоп-кодонов (UAA). В одном
случае (ген СО-Ш T.hrucei) более 50% остатков
мРНК представлены посттранскрипционно встроен-
ными остатками урацила. У других кинетопластид
в митохондриальных ДНК обнаружены последова-
тельности, гомологичные кДНК CO-111 T.hrucei, а не
самому гену СО-Ш, обедненному остатками U.
Помимо вставок при редактировании РНК мо-
жет происходить и вырезание остатков U. Возмож-
но, в некоторых случаях функциональная РНК об-
разуется в несколько этапов, с сайт-специфической
инсерцией множества остатков U и последующим
их вырезанием, за исключением остатков, необхо-
димых для функционирования мРНК. Многие де-
тали этого процесса остаются неясными. Например,
неизвестно, где содержится информация о после-
довательности конечного продукта-мРНК и как
узнается транскрипт, который подлежит редакти-
рованию. У одних видов мРНК, кодируемая дан-
ным геном, обязательно редактируется, у других
остается интактной. Вероятно, процесс редактиро-
вания РНК каким-то образом регулируется. Так,
у T.hrucei некоторые РНК (например, СО-П-РНК)
подвергаются редактированию, если данный пара-
зит обитает в организме насекомого-хозяина, и ос-
таются интактными у паразитов, обнаруженных
в крови млекопитающих. По-видимому, это связано
с тем, что простейшие, живущие в организме насе-
комых, нуждаются для образования АТР в окисли-
тельном фосфорилировании, а в крови млекопи-
тающих АТР образуется путем гликолиза.
Насколько универсален процесс редактирования
РНК? Происходит ли он только в кинетопластидах
и митохондриях? Результаты, полученные в двух
проведенных недавно исследованиях, показывают,
что редактирование РНК происходит у многих ор-
ганизмов и в ядерных транскриптах. Так, ген апо-
липопротеина у млекопитающих кодирует две
мРНК, при этом в одной из них вместо кодируемого
геномной ДНК остатка С появляется остаток U и
соответственно возникает стоп-кодон. Далее, уста-
новлено, что две мРНК, кодируемые одним геном
SV40, различаются тем, что в одной из них при-
сутствуют два «лишних» остатка гуанина. Редакти-
рование РНК не ограничивается вставкой или вы-
резанием U. Сходны ли механизмы, используемые
у млекопитающих, с механизмами, действующими
у кинетопластид, пока неясно.
224
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
в. ДНК хлоропластов
Геном хлоропластов в чем-то сходен с геномом
митохондрий. ДНК представляет собой кольцевой
дуплекс, содержащий какое-то количество (но да-
леко не все) генов, существенных для функциони-
рования хлоропластов и поддержания их структу-
ры, и имеет достаточно большую длину: как пра-
вило, 120-180 т.п.н. При этом каждый хлоропласт
может содержать десятки копий хлоропластного
генома. Полностью установлены последовательнос-
ти ДНК хлоропластов у печеночников (моховид-
ные), табака (двудольные) и риса (однодольные).
Структура хлоропластной ДНК. По данным
рестрикционного картирования, структура ДНК
хлоропластов, в отличие от митохондриальной
ДНК, в пределах вида остается довольно консерва-
тивной. Так, хлоропластная ДНК шпината (рис.
9.62) имеет структуру, типичную для соответствую-
щей ДНК всех покрытосеменных, как одно-, так
и двудольных. В пределах сегмента длиной при-
мерно 25 т.п.н. находятся четыре гена рРНК (16S,
23S, 5S и 4,5S), и этот сегмент повторен в обратной
ориентации еще раз через 15 т.п.н. Локализованы
и другие гены, в том числе ген большой субъеди-
ницы рибулозо-1,5-бисфосфат-карбоксилазы, rbcL
(ген малой субъединицы находится в хромосоме),
ген компонента тилакоидной мембраны массой
32 кДа (фотоген 32, psb А, который экспрессируется
только на свету) и гены нескольких субъединиц
хлоропластной РНК-полимеразы (гро). Всего хло-
ропластная ДНК кодирует примерно 100 полипеп-
тидов и 35 РНК (рРНК и тРНК). Продукты многих
генов с открытой рамкой считывания уже иденти-
фицированы. Единственное известное исключение из
правил организации хлоропластной ДНК покры-
тосеменных-ДНК некоторых бобовых (например,
гороха), у которых утрачена одна копия инверти-
рованного повтора. Такой повтор присутствует
даже в хлоропластной ДНК зеленой водоросли
Chlatnidomonas reinhardii, но не у Euglena. Последняя
отличается тем, что у нее имеются три тандемных
повторяющихся участка длиной 5,6 т. п. н., которые
содержат гены рРНК. В отличие от ДНК митохонд-
рий, в ДНК хлоропластов, по-видимому, исполь-
зуется универсальный генетический код. В ней со-
держится много генов тРНК, и в тех полипептидных
генах, которые были исследованы, присутствует
большинство универсальных кодонов.
Четкие данные о репликации хлоропластной
ДНК или о локализации мест начала репликации
пока отсутствуют.
Структура хлоропластных генов. Структура
хлоропластных белков и соответствующих генов
РИС. 9.62.
Диаграмма хлоропластной ДНК шпината
с указанием локализации некоторых генов.
Двунаправленные наружные стрелки отме-
чают инвертированные повторы, которые
содержат гены рибосомных РНК. Малень-
кими стрелками указано направление тран-
скрипции. Обозначения: trn- ген тРНК (со-
ответствующая аминокислота обозначается
одной буквой согласно общепринятому
правилу), psa-ген компонента фотосисте-
мы I; psb-ген компонента фотосистемы II;
pef-гены полипептидов цитохромного
комплекса; rbcL- ген большой субъедини-
цы рибулозо-1,5-бисфосфат- карбоксила-
зы/оксигеназы; afp-гены субъединиц Н + -
АТРазного комплекса; гро-гены субъеди-
ниц РНК-полимеразы. Гены рибосомных
белков не показаны. [С любезного разре-
шения W. Gruissem; из работ W. Gruissem,
G. Zuwarski, EMBO J. 4 (1985), p. 1637;
G. Sijben-MOIIer et al.. Nucleic Acids Res. 14
(1986), p. 1029.]
9. МОЛЕКУЛЯРНАЯ СТРУКТУРА ГЕНОМОВ ЭУКАРИОТ
225
у самых разных видов высококонсервативна. Ско-
рость эволюции кодирующих последовательностей
хлоропластных генов значительно ниже скорости
эволюции ядерных генов.
В генах тРНК, рРНК и полипептидов разных
видов содержатся интроны, однако картина их рас-
пределения неодинакова. Например, у Euglena в ге-
нах хлоропластных тРНК интроны отсутствуют,
в отличие от соответствующих генов у многих
организмов. У той же Euglena в полипептидных
генах интроны имеются; в то же время многие из
гомологичных мРНК цианобактерий и высших рас-
тений не нуждаются в сплайсинге. Даже среди выс-
ших растений одни полипептидные гены содержат
интроны, а другие нет. В отличие от всех других
исследованных организмов, у Chlamidomonas rein-
hardii ген 23S-pPHK прерывистый. Пока никаких
закономерностей в наличии или отсутствии интро-
нов в генах хлоропластов ДНК не обнаружено.
Хлоропластные интроны, по-видимому, анало-
гичны или интронам группы I, или интронам
группы II (разд. 8.5. в). Можно предположить, что
при сплайсинге используются сходные механизмы.
В большинстве хлоропластных интронов протяжен-
ные открытые рамки считывания отсутствуют.
Впрочем, такая рамка длиной 509 кодонов обна-
ружена в интроне гена лизиновой тРНК в хлоро-
пластах табака и в интроне гена 23S-pPHK у
C.reinhardii. В соответствующих белковых продук-
тах имеются участки, гомологичные участкам ма-
тураз, кодируемым митохондриальными интрона-
ми дрожжей. Таким образом, интроны как хло-
ропластных, так и митохондриальных ДНК могут
содержать функциональные сегменты.
Некоторые мРНК у водорослей и высших расте-
ний образуются в результате шронс-сплайсинга
(разд. 8.5.г). Например, 5'-экзон гена, кодирующего
рибосомный белок хлоропластов slz, транскриби-
руется с одной цепи ДНК, а два других экзона-с
другой цепи, на расстоянии 30 т.п.н. от этого
локуса. При шрпнс-сплайсинге эти транскрипты сое-
диняются в одну функциональную мРНК.
г. Происхождение ДНК органелл
Широко распространено мнение, что митохонд-
рии и хлоропласты произошли от прокариотических
эндосимбионтов, которые обитали в цитоплазме
предшественников эукариот. Такие функции прока-
риот, как аэробный метаболизм и фотосинтез, при-
обрели важное значение для их хозяев. Гены, от-
ветственные за эти функции, и соответствующие им
структуры сохранились и у современных организ-
мов, а гены, кодирующие другие, несущественные
для хозяев функции и структуры эндосимбионтов,
в ходе эволюции были утрачены. Эту гипотезу
подтверждают различные свойства экспрессии хло-
ропластных генов. Так, промоторы экспрессирую-
щихся хлоропластных генов напоминают типичные
-35- и -10-последовательности прокариотических
промоторов (рис. 9.63). Кроме того, рибосомы хло-
ропластов очень сходны с рибосомами прокариот.
Близки по размеру и последовательностям моле-
кулы соответствующих рРНК, а некоторые рибо-
сомные белки хлоропластов иммунологически род-
ственны рибосомным специфическим белкам Et coli.
Трансляция на рибосомах хлоропластов подавляет-
ся хлорамфениколом, который оказывает такое же
действие на трансляцию у прокариот. Сходство
генной экспрессии у митохондрий и прокариот не
столь очевидно. Трансляционный аппарат мито-
хондрий, особенно у животных, имеет лишь отда-
ленное сходство с аппаратом современных прока-
риот. Тем не менее тщательный анализ митохон-
дриальной рРНК у растений наводит на мысль
о существовании у нее общего предка с рРНК
пурпурных бактерий a-типа-группы, включающей
таких известных симбионтов растений, как Agro-
bacteria и Rhizobacteria. Возможно, рибосомы и ге-
нетический код митохондрий произошли от како-
го-то вымершего (или неизвестного) прокариота.
Гены митохондрий и хлоропластов, участвую-
щие в поддержании их структуры и функций, рас-
пределены между ядерной ДНК и ДНК органелл.
Более того, у разных организмов одни и те же гены
могут быть в одном случае цитоплазматическими,
а в другом - ядерными. Например, ген, кодирующий
субъединицу 9 митохондриального АТРазного ком-
плекса, у S.cerevisiae находится в митохондриях,
а у грибов и у человека-в ядре. Если следовать
гипотезе эндосимбиоза буквально, то все важные
в структурном и функциональном отношении гены
митохондрий и хлоропластов должны оставаться
в геноме органелл- потомке исходной прокариоти-
ческой ДНК. Почему у современных организмов
многие из этих генов обнаруживаются в ядерных
хромосомах? Одно из объяснений, подкрепленное
РИС. 9.63.
Промоторы, обеспечивающие транскрипцию
хлоропластных генов шпината, и промоторы
E.coli. [W. Gruissem, G. Zuwarski, EMBO J.
4 (1985), p. 1637.]
-35
Консенсусная после-
довательность E.coli
Ген тРНК™1 хлоро-
пластов шпината
rbcL
psbA
-последовательность -10-последовательность
TCTTGАСАТ
TATTGCTTA
GGTTGCGCC
66ТТ G ACAC
ТАТААТ
ТАТААТ
TATACA
TATACT
15-158
226
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 9.64.
Перенос сегментов ДНК из органелл в ядерную ДНК.
Жирными стрелками указаны переносы, подтвержден-
ные экспериментально, тонкими стрелками-гипотети-
ческие переносы. [R. Lewin, Science 224 (1984), р. 970.]
экспериментальными данными, состоит в том, что
последовательности ДНК все время «путешеству-
ют» между органеллами и ядром, а также между
геномами самих органелл (рис. 9.64).
Например, ключевой фермент фотосинтеза ри-
булозо-1,5-бисфосфат- карбоксилаза/оксигеназа-
это гетеромультимер, состоящий из восьми иден-
тичных больших (L) и восьми идентичных малых (S)
субъединиц. У фотосинтезирующих эукариот L-
субъединицы кодируются геном rbc L в геноме хло-
ропластов (рис. 9.62), а S-субъединицы, как и боль-
шинство хлоропластных белков,-в ядре (рис. 9.36).
Однако у простейшего Cyanophora paradoxa обе
субъединицы кодируются неядерным геномом, вхо-
дящим в состав хлоропластоподобной структуры-
цианелли, типичной для цианобактерий. Такая ло-
кализация согласуется с эндосимбиотическим про-
исхождением хлоропластов эукариот. Предполага-
ется, что у общего предка современных фотосинте-
зирующих эукариот сегмент ДНК. содержащий ген
S, переместился из эндосимбионта или его произ-
водного в ядерную ДНК.
Представления о межгеномном обмене подтверж-
даются и тем фактом, что в ядерной ДНК шпината
содержатся последовательности (в основном неко-
дирующие), которые гибридизуются с клонирован-
ными зондами, представляющими почти весь хло-
ропластный геном, в том числе и с последователь-
ностями гена rbcL. Последовательности, гомоло-
гичные rbcL и другим сегментам хлоропластной
ДНК, обнаружены также в митохондриях некото-
рых растений. Вероятно, некоторые из них появи-
лись в результате относительно недавних переме-
щений, поскольку митохондриальная ДНК данного
организма более сходна с хлоропластной ДНК ор-
ганизмов, относящихся к этому же виду, чем с хло-
ропластной ДНК других видов. Итак, сегменты
хлоропластной ДНК могут перемещаться в ядро
и в митохондрии. Имеется множество примеров
переноса и митохондриальной ДНК. Гибридизация
между митохондриальной и ядерной ДНК наблю-
дается у таких разных организмов, как дрожжи,
саранча, морской еж и крыса. Следовательно, пе-
рестройки ДНК не ограничиваются ядерным ге-
номом, и мембраны ядер и органелл не служат для
них барьером. Однако механизм перемещения и ин-
теграции ДНК органелл в новые геномы не уста-
новлен. В следующей главе мы остановимся на
способах перестройки ядерных ДНК, и, возможно,
это поможет нам найти ответ на интересующий нас
вопрос.
Глава 10
РЕОРГАНИЗАЦИЯ ГЕНОМА
В предыдущей главе мы рассмотрели организа-
цию современных геномов. Это рассмотрение по-
казывает, насколько велика эволюционная роль пе-
рестроек и дупликаций последовательностей ДНК.
Сравнивая разные геномы, можно сделать вывод,
что для их реорганизации использовались несколько
стратегий. Одна из них состояла в перемещении
сегментов из исходного локуса в различные сайты.
Другая-в дупликации сегментов в определенном
локусе с последующими тандемными реитерациями
(многократными амплификациями). Третья страте-
гия состояла в дупликации сегментов ДНК и пере-
мещении их в отдаленные сайты в геноме с по-
мощью процессов, в которых участвуют как сама
ДНК, так и РНК-продукты и обратная транскрип-
таза. И наконец, копии каких-то значимых сегмен-
тов могли мутировать и адаптироваться к новым
функциям. Результатами всех этих процессов яв-
ляются увеличение размера генома и фенотипиче-
ское разнообразие, а в сочетании с естественным
отбором эволюционная гибкость вида и его раз-
витие.
В этой главе мы рассмотрим экспериментальные
системы, как генетические, так и биохимические,
которые помогают установить механизмы геном-
ных перестроек. Выводы, следующие из этих экспе-
риментальных наблюдений, совпадают с теми пред-
ставлениями, которые можно получить, исследуя
структуру ныне существующих геномов. Описанные
здесь механизмы в меру их адекватности позволяют
строить модели процессов, протекавших в ходе
эволюции. Особого внимания заслуживает один из
этих механизмов-обратная транскрипция, посколь-
ку он играет важную роль в перестройке всех эука-
риотических геномов. Включение в геномы ДНК-
копий молекул РНК создает основу замечательной
пластичности геномов. Недавно получены данные
о том, что обратная транскрипция, возможно, осу-
ществляется и у прокариот. В полной мере значение
этого процесса пока остается неясным.
Во всех случаях, о которых шла речь, изменяется
положение какого-то сегмента ДНК относительно
окружающих его последовательностей. Подобные
события происходят повсеместно в разных вариан-
тах, однако большинство из них наблюдается от-
носительно редко. Многие геномные перестройки
происходят в результате гомологичной рекомбина-
ции между аллельными последовательностями и не
затрагивают соседних сегментов ДНК. Вообще же
говоря, структура генома вполне стабильна. Это
и не может быть иначе, поскольку необходимо
поддерживать жизнеспособность особей и видов.
Многие геномные перестройки не запрограмми-
рованы, они не связаны с каким-то специфическим
влиянием на экспрессию генов и в них есть элемент
случайности. Случайными могут быть частота та-
ких событий, сами сегменты ДНК или то и другое.
Примерами таких довольно редких событий служит
транспозиция последовательностей ДНК из одного
геномного локуса в другой или дупликация и после-
дующая амплификация сегментов ДНК. Однако
сходные транспозиции и амплификации могут быть
сопряжены также с неслучайными, запрограммиро-
ванными изменениями. Такие запрограммирован-
ные события играют ключевую роль в регуляции
экспрессии некоторых генов во время дифференци-
ровки и развития определенных типов клеток.
10.1. ОБЩИЕ СВОЙСТВА
НЕЗАПРОГРАММИРОВАННЫХ
ТРАНСПОЗИЦИЙ
Некоторые сегменты ДНК обладают способ-
ностью перемещаться в другие геномные локусы,
иногда существенно изменяя экспрессию соседних
генов. Такой эффект наблюдается, например, при
встраивании мобильного сегмента в новый геном-
ный локус, сопровождающемся изменением коди-
рующего участка или важного регуляторного эле-
мента. В простейшем случае ген перестает экспрес-
сироваться. Но нередко сами мобильные элементы
содержат регуляторные элементы. В таких случаях
экспрессия генов, соседствующих со вставкой, мо-
жет претерпевать сложные изменения, в том числе
с переходом на новые способы регуляции.
Различают несколько типов мобильных ДНК-
элементов. Лучше всего группировать эти элементы
в соответствии с механизмами их транспозиции.
Однако механизм транспозиции многих эукариоти-
ческих и прокариотических элементов пока до конца
не установлен. Поэтому мы используем классифи-
кацию, основанную прежде всего на структурных
особенностях элементов. Резонно предположить,
что общие морфологические особенности опреде-
ляют и общие механизмы транспозиции, однако так
ли это на самом деле-пока неясно.
15’
228
ЧАСТЬ 111. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
а. Различные типы мобильных
элементов
Транспозирующиесн элементы. Первые мобиль-
ные элементы, исследованные на молекулярном
уровне, происходили из геномов прокариот, а также
их фагов и плазмид. Сейчас известно несколько
разных типов таких элементов, но все они обладают
двумя общими свойствами: во-первых, несут ген
(или несколько генов), необходимый для транспо-
зиции; во-вторых, на концах содержат специфиче-
ские взаимно инвертированные повторяющиеся
последовательности, также необходимые для тран-
спозиции. Сами транспозирующиеся элементы не
кодируют никаких существенных для организма
функций, однако часто содержат специфические ге-
ны, например ген устойчивости к антибиотикам.
Транспозиция этих элементов, как правило, сопро-
вождается сильными мутагенными эффектами.
Мобильные элементы многих эукариот сходны
со своими прокариотическими «родственниками»
в том отношении, что для их транспозиции необхо-
димы гены, содержащиеся в самих элементах, и спе-
цифические концевые последовательности. Если
транспозиция протекает без участия обратной тран-
скрипции, то мы будем относить их к транспози-
рующимся элементам. К этому классу принадлежат
контролирующие элементы кукурузы первые из об-
наруженных мобильных единиц, а также некоторые
элементы Drosophila. Сходные элементы имеются
у других беспозвоночных и у некоторых цветковых
растений, где они отвечают за пеструю раскраску
лепестков.
Ретротранспозоны. Эукариотические мобильные
элементы, транспозиция которых происходит при
транскрипции или обратной транскрипции, назы-
ваются ретротранспозонами. Они содержат цент-
ральный сегмент, кодирующий среди других белков
обратную транскриптазу. У некоторых ретротранс-
позонов, называемых здесь транспозонами класса I,
этот центральный сегмент окружен длинными кон-
цевыми повторами (LTR). У ретротранспозонов
класса I на одном из концов имеются также корот-
кие инвертированные повторы. По своей структуре,
особенностям транскрипции и механизму транспо-
зиции они напоминают ретровирусные провирусы.
Отличие состоит в отсутствии жизнеспособных вне-
клеточных форм. Семейства ретротранспозонов об-
наружены у разных беспозвоночных, в частности
у дрожжей и Drosophila, а также у растений и неко-
торых млекопитающих.
Ретротранспозоны класса И не имеют концевых
повторов, а один из концов часто бывает представ-
лен АТ-богатой последовательностью. В этом от-
ношении они не похожи на ретровирусные про-
вирусы. Элементы класса II менее изучены, чем
элементы класса I. хотя они тоже широко распро-
странены среди эукариот. Удивительно, что ни один
из классов ретротранспозонов не имеет прокарио-
тических партнеров.
Ретрогены. По геному перемешаются также раз-
нообразные сегменты ДНК, не обладающие спе-
цифическими структурными и кодирующими свой-
ствами транспозонов или ретротранспозонов. В от-
личие от мобильных элементов других классов, они
весьма гетерогенны по размеру и структуре. У них
нет концевых повторов, а на одном из концов часто
присутствует АТ-богатая последовательность. К
элементам этого класса, называемым ретрогенами,
относятся процессированные псевдогены и SINE-no-
следовательности; они обнаружены у различных
эукариот, но особенно обильно представлены у мле-
копитающих. Транспозиция ретрогенов, по-видимо-
му, происходит через образование РНК с после-
дующей обратной транскрипцией. Однако этот про-
цесс скорее всего пассивный в том смысле, что
данные элементы не кодируют необходимых для
транспозиции активностей (т. е. обратную тран-
скриптазу).
б. Дупликация сайтов-мишеней
Независимо от типа мобильного элемента его
встраивание в новый генетический локус обычно
сопровождается дупликацией короткого участка
ДНК в месте встраивания (в сайте-мишени). Эти
дуплицированные сегменты фланкируют встроив-
шийся элемент (рис, 10.1). Почти обязательная дуп-
ликация сайтов-мишеней указывает на то, что при
различиях в механизме большинство вставок проис-
ходит с образованием в потенциальных сайтах-ми-
шенях смещенных одноцепочечных разрывов, как
это показано на рис. 10.1. Сайт-мишень в геноме,
содержащий мобильный элемент, условно называ-
ют «заполненным», а не содержащий такового-
«свободным».
Размер дуплицированных последовательностей
для разных мобильных элементов различается. Для
транспозонов и ретротранспозонов класса I, как
правило, характерны дупликации сайтов-мишеней
определенного размера. Например, встраивание
транспозонов ТпЗ E.coli в новые локусы всегда
сопровождается дупликацией участка размером
5 п. н.; встраивание так называемого Лс-элемента
(активатора) у кукурузы приводит к дупликации
участка размером 8 п.н., а ретротранспозона Dro-
sophila, называемого gypsy,-4 п.н. В то же время
при встраивании ретротранспозонов класса II и рет-
рогенов происходят дупликации участков разного
размера. Например, встраивание F-элемента Dro-
sophila- ретротранспозона класса II-приводит к
дупликации участка длиной от 8 до 13 п.н. Такая
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
229
5(ACGAT
3'( TGCTA
Сайт-мишень
GTCGCAGAG
XAfiCGTCTC
TATGC Q3'
ATACG Об'
Ступенчатые разрывы
Одноцепочечным X
разрыв ’
5'(ACGAT0(«W Cf*a(TATGC 03
3’(TGCTA (CAGCGT СТС O(ATACG 05'
Одноцепочечный разрыв
Сшивание 3-концов мобильного
элемента с 5-концами цепей
5'(ACGat р_________
j'(TGCTA(CAGCGTCTC
iSWSgaftAO(TATGC Q3’
5 iatacgOs'
Застраивание бреши ДНК-поли-
F меразой
Лигирование
6 (ACGAT( 8tcpc>3ag(__________(GTCGCAGAG tTATGC О 3
3'(TGCTA Ls5s£Jo.TCTC ( (taecj ttXblATACG Об'
Мобильный элемент
РИС. 10.1
Дупликация сайта-мишени после встраивания мобиль-
ного элемента. Вначале обе цепи ДНК расщепляются
эндонуклеазой. Места расщеплений отстоят друг от
друга на несколько пар оснований (в данном случае
девять), в результате чего образуются ступенчатые раз-
резы. В рассматриваемом примере концы выступают
в 5'-направлении. З'-концы мобильного элемента сши-
ваются с 5'-концами цепей в месте разрывов. Образо-
вавшиеся бреши застраиваются, в результате чего появ-
ляются две копии сайта-мишени-по одной с каждой
стороны вставки. Длина дуплицированного участка оп-
ределяется расстоянием между одноцепочечными раз-
рывами. Аналогичные процессы протекают и в том
случае, когда после одноцепочечных разрывов обра-
зуются З'-выступы.
вариабельность или, напротив, неизменность раз-
меров, вероятно, обусловливается специфичностью
ферментов, катализирующих образование смещен-
ных одноцепочечных разрывов в сайтах-мишенях.
Неизменность размеров дуплицированных участков,
характерная для транспозонов и ретротранспозонов
класса 1, по-видимому, связана с тем, что ДНК
разрезается специфическими эндонуклеазами, коди-
руемыми каждым из элементов.
в. Мобильные элементы
как диспергированные
повторяющиеся последовательности
Как правило, каждый тип мобильных элементов
встречается в геноме многократно, т. е. элементы
образуют семейства диспергированных повторяю-
щихся последовательностей. Такие семейства бла-
гоприятствуют дальнейшим геномным перестрой-
кам, создавая сайты для неаллельных гомологич-
ных рекомбинаций, что приводит к делециям, дуп-
ликациям, инверсиям и транслокациям. Способ-
ность мобильных элементов облегчать перестройки
подтверждается на примере ретротранспозона Ту
класса I у дрожжей (разд. 10.3). Если такие пере-
стройки происходят в единичных соматических
клетках многоклеточного организма, то вероят-
ность их влияния на функционирование соответст-
вующего органа очень мала. Но в клетках заро-
дышевой линии они могут вызывать серьезные му-
тационные изменения и хромосомные аберрации.
Если такие перестройки фиксируются в популяции
в результате естественного отбора или генетическо-
го дрейфа, то это имеет заметные эволюционные
последствия.
Гомологичные рекомбинации между неаллель-
ными диспергированными повторами в отдельных
семействах часто привлекают для объяснения по-
вторения сегментов ДНК (разд. 2.4.а). А некоторые
различия между гомологичными локусами у разных
млекопитающих (например, между локусами Р-гло-
биноподобных генов) можно связать с делениями
в прямых повторах. К геномным перестройкам
приводят также неравный гомологичный кроссин-
говер между диспергированными повторами в сест-
ринских хроматидах или гомологичных хромосо-
мах, внутрицепочечная рекомбинация и «проскаль-
зывание с неправильным спариванием» (“slipped
misparing”) (рис. 10.2). Молекулярно-генетический
анализ, проведенный на Drosophila, показывает, что
высокая частота неравного кроссинговера между
определенными парами аллелей-это результат при-
сутствия в каждом из членов пары мобильных
элементов, находящихся в разных позициях. Рецип-
рокная гомологичная рекомбинация между диспер-
гированными повторяющимися последовательнос-
тями из разных хромосом может приводить и к
крупным хромосомным перестройкам типа тех, что
наблюдаются при цитогенетическом исследовании
клеток млекопитающих.
Принимая во внимание высокую частоту диспер-
гированных повторяющихся последовательностей
в большинстве эукариотических геномов, можно
только удивляться стабильности хромосом. Воз-
можно, рекомбинация между этими сегментами эф-
фективно подавляется с помощью какого-то неиз-
вестного механизма. У дрожжей, где частоту таких
рекомбинаций легко измерить генетическими ме-
тодами, транслокация и другие перестройки отме-
чаются удивительно редко. Например, в клетках,
содержащих две копии гена HI S3, расположенные
на негомологичных хромосомах, реципрокный крос-
синговер обеспечивает только примерно 10% ре-
комбинационных событий в данном гене, остальные
приходятся на генную конверсию.
230
ЧАСТЬ 111, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕН О В ЭУ КАРИ ОТ
РИС. 10.2.
Делеции и амплификации в результате
проскальзывания с неправильным спарива-
нием (“slipped misparing”) могут происхо-
дить в том случае, если неправильно спари-
ваются два прямых повтора (прямоуголь-
ники) в дуплексной ДНК вблизи реплика-
тивной вилки. А. «Проскальзывание» мат-
ричной цепи и вырезание петли или репли-
кация, минуя петлю, с последующей репа-
рацией приводят к делеции. Б. Проскаль-
зывание синтезируемой цепи и последу-
ющая репарация могут привести к ампли-
фикации.
10.2. ТРАНСПОЗИРУЮЩИЕСЯ
ЭЛЕМЕНТЫ
а. Транспозирующиеся элементы
прокариот
О транспозирующихся элементах мы упоминали
в гл. 5. когда рассматривали плазмидные векторы.
R-плазмида, представленная на рис. 5.1, еще раз
более детально, со всеми транспозирующимися эле-
ментами, изображена на рис. 10.3. Первые из них,
обозначенные IS (от znsertion .sequence-инсерционная
последовательность), располагаются или изолиро-
ванно (как IS2), или по концам более длинных
единиц сложных транспозонов (например, IS 10, ок-
ружающие ТпЮ). Другие, транспозоны (например,
ТпЗ), не связаны с IS-элементами. Все эти элементы
содержат на обоих концах инвертированные повто-
ры. Последние, а также один или более генов,
содержащихся в самих мобильных элементах, необ-
ходимы для транспозиции. Кроме того, инсерция
любого из данных транспозирующихся элементов
обычно бывает связана с дупликациями сайтов-ми-
шеней, имеющих специфическую для определенного
элемента длину.
Инсерционные последовательности (IS). Такие
IS-элементы, как IS1, IS2 и IS 10, имеют определен-
ную длину и уникальную нуклеотидную последо-
вательность (табл. 10.1). Их перемещения в новые
геномные локусы часто приводят к мутациям,
заключающимся в прерывании регуляторных и ко-
дирующих участков. Частота транспозиций у раз-
ных элементов неодинакова и составляет 10-5-10-7
на поколение. При транспозиции IS в новое поло-
жение исходный IS-элемент остается на прежнем
месте; таким образом, инсерция сопровождается
точным синтезом второй копии и зависит от репли-
кативных функций хозяина. Она не зависит от ре-
комбинационного аппарата Е. coli; не требуется так-
же никакой гомологичности между IS и геномным
сайтом-мишенью.
Длина и нуклеотидная последовательность раз-
ных IS-элементов E.coli неодинаковы (табл. 10.1)
Однако на обоих концах каждого из них всегда
имеются инвертированные повторы (иногда несо-
вершенные) (рис. 10.4), которые необходимы для
транспозиции. Нуклеотидные последовательности
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
231
РИС. 10.3.
R-плазмида со всеми инсерционными последователь-
ностями (IS) и транспозонами (Тп). Гены устойчивости
к антибиотикам указаны в табл. 5.3.
этих повторов различны для разных IS и варьируют
по длине от 10 до 40 п. н. Еще одна характерная
особенность любого IS-размер дуплицированного
участка в сайте-мишени (табл. 10.1). Большая часть
ДНК в любом IS представлена одним или несколь-
кими структурными генами и соответствующими
регуляторными последовательностями. Их генный
продукт- транспозаза- необходима для транспози-
ции. В отличие от транспозонов, IS обычно не
содержат каких-либо генов, кроме нужных для
транспозиции.
IS-элементы не только ответственны за инсер-
ционную инактивацию кодирующих и регуляторных
последовательностей в сайте-мишени, но и связаны
с разнообразными мутационными событиями. На-
пример, один из IS может в 100-1000 раз повышать
частоту делеций и инверсий в соседних участках
генома. Более того, премоторные элементы внутри
самих IS тоже могут влиять на экспрессию близ-
лежащих генов.
Сложные транспозоны. Часто IS-элементы дли-
ной примерно 1 т. п. н. входят в состав более слож-
ных транспозонов (рис. 10.3 и 10.5; табл. 10.2). Их
длина может достигать тысяч пар оснований, а
центральные области могут содержать разнообраз-
ные гены, например гены устойчивости к антибио-
Таблица 10.1. Типичные IS-элементы Е.соН
IS1 768 23 9
IS2 1327 41 5
IS4 1426 18 11
IS10 1329 23 9
Обычное место-
нахождение
Хромосома Е.соН, 4-19
копий; плазмида R6,
две копии; Shigella
dysenteria, > 40 копий
Хромосома Е.соН, 0-12
копий; R6, одна копия;
F-плазмида, одна копия
Хромосома Е.соН, 1-2
копии
R-плазмиды
тикам. С обеих сторон эта центральная область
фланкируется одинаковыми IS-элементами (от IS1
до IS 10)- IS-L и IS-R с левой и правой сторон
соответственно. В различных сложных транспозо-
нах взаимная ориентация IS-L и IS-R может быть
как одинаковой, так и разной. Поскольку сами
IS-элементы несут инвертированные концевые по-
вторы, их содержат и транспозоны независимо от
того, в прямой или обратной ориентации находятся
фланкирующие IS.
Вся информация, необходимая для перемещения
сложного транспозона, содержится в его IS-части;
это та самая информация, которая используется
IS-элементом как таковым,- инвертированные кон-
цевые повторы и один или несколько генов, коди-
рующих транспозазу. Требование обязательного
наличия в транспозоне концевых повторов означает,
что наружные части IS-L и TS-R должны быть
идентичными, однако, поскольку для транспозиции
достаточно одной копии структурных генов, эта
идентичность может не распространяться на всю
длину IS-L и IS-R. Один из элементов может быть
поврежден в результате мутации, приводящей к час-
тичной или полной утрате его функции. Так, у слож-
ного транспозона Тп10 (рис. 10.5) транспозаза мо-
жет кодироваться или IS10-L, или IS10-R, но IS10-R
гораздо более эффективен. Кроме того, IS10-R мо-
жет выступать в роли независимого IS-злемента,
а ISlO-L-нет. По-видимому, за различия в актив-
ности соответствующих генных продуктов отвечают
^"Левый"конец 722 п.н. "Правынлконец
°(GbTG ft TGCMSCAACTTAC ЬБАТ_________АТС A A ftAAGTTGCA ATT ACCQ3'
tCCACT ACG ACGGTTGAATG ACTA TAGT Т АТТСААССТ С AGTAAT GG()
з' 5
РИС. 10.4. дающие пары оснований выделены цветными прямо-
Концевые инвертированные повторы у IS1. Несовпа- угольниками.
232
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Таблица 10.2. Типичные транспозоны Е.соН
Транспозон Размер, п.н. IS-концы Ориентация IS Длина концевых инвертированных повторов, п.н. Длина дуплициро- ванной мишени, п.и. Содержащиеся гены
ТпЗ 4957 Нет — 38 5 атр*
Тп9 2638 IS1 Прямая 23 9 cam*
ТпЮ 9300 IS10 Обратная 1329 9 tetR
весьма незначительные различия в последователь-
ностях этих практически одинаковых ISlO-элемен-
тов. В некоторых сложных транспозонах один из
IS-элементов полностью нефункционален (за иск-
лючением инвертированных концевых повторов).
Простые транспозоны. Некоторые транспозоны
не связаны ни с какими IS-элементами (табл. 10.2):
информация, необходимая для транспозиции, за-
кодирована в них самих. В качестве примера можно
привести транспозон ТпЗ (рис. 10.6), который со-
держит инвертированные концевые повторы длиной
38 п. н., IS-L и IS-R, необходимые для транспозиции.
Между этими повторами находятся три гена: два из
них кодируют транспозиционные белки (tnp А и
tnp R), третий- р-лактамазу (ген атр). Кроме того,
имеется некодирующий участок длиной 170 п.н.,
содержащий промоторы генов tnp А и tnpR, а также
специфический участок, называемый res и тоже не-
обходимый для транспозиции. Важное различие
между IS-элементами и простыми транспозонами
состоит в том, что последние содержат гены типа
атр, не связанные с транспозицией.
Сайты-мишени. До сих пор, говоря о транспо-
зиции у прокариот, мы рассматривали сами транс-
позирующиеся элементы. Что же представляют
собой те сегменты ДНК, в которые они встраивают-
ся,-сайты-мишени? Одни транспозирующиеся эле-
менты довольно разборчивы и охотнее встраивают-
ся в короткие геномные сегменты, гомологичные
концам самого элемента. Другие менее капризны
и не отдают явного предпочтения никаким сайтам-
мишеням, хотя наблюдается некоторая тенденция
к встраиванию их в АТ-богатые участки. Некоторые
элементы высокоспецифичны в отношении опреде-
ленных палиндромных сайтов-мишеней (например,
Концевые
инвертированные
повторы
РИС. 10.5.
Сложный транспозон Тп10 имеет две копии IS10, флан-
кирующие центральный сегмент, который несет ген
устойчивости к тетрациклину.
РИС. 10.6.
Простой транспозон ТпЗ. Стрелки указывают направле-
ние трвнскрипции трех генов.
IS5 предпочитает последовательность 5'-CTAG, а
TnlO- 5'-GCTNAGC). Поскольку для каждого транс-
позирующегося элемента характерен свой размер
дуплицированного участка, расстояние между одно-
цепочечными разрывами в сайтах-мишенях, по-ви-
димому, полностью или частично детерминируется
самими элементами.
Бактериофаг Ми. Умеренный бактериофаг Е.соН,
называемый Ми, по своему поведению сходен с
транспозонами или IS-элементами. Геном Ми пред-
ставляет собой дуплексную ДНК длиной около
39 т. п. н. с концевыми инвертированными (несовер-
шенными) повторами длиной примерно 20 п.н.,
которые фланкированы сегментами ДНК Е.соН
(рис. 10.7). После инфицирования Ми-ДНК встраи-
вается в хромосому Е. coli', при этом в месте встраи-
вания происходит дупликация участка длиной 5 п. н.
и образуется стабильный лизоген. В результате
индукции таких Mu-лизогенов может начаться реп-
ликация Mu-ДНК. Однако в отличие от событий,
происходящих при индукции лизогенов фага X, реп-
лицирующаяся ДНК бактериофага Ми остается
встроенной в хромосому, а новые фаговые геномы
образуются при репликации фаговой ДНК in situ.
Эта новосинтезированная Ми-ДНК, не связанная со
встроенной копией, внедряется в новые хромосом-
ные локусы. В конце концов хозяйский геном раз-
рушается, и Ми-ДНК, фланкированная с обоих
концов фрагментами ДНК Е. coli, упаковывается
в фаговые частицы.
Присутствие ДНК профага Ми в ДНК Е.соН
порождает разнообразные перестройки, связанные
с ее транспозицией. Они сходны с перестройками,
10, РЕОРГАНИЗАЦИЯ ГЕНОМА
233
и и в
ДНК Е.соН
Репликативная транспозиция
в другие геномные сайты,
зависящая от функций генов Л и В
Синтез белковой оболочки и
упаковка новых фаговых
геномов
РИС. 10.7.
Схематическое представление генома бакте-
риофага Ми и его жизненного цикла (масштаб
рисунка не выдержан). Указаны левые и пра-
вые концевые повторы (L1 и R1), а также гены,
кодирующие транспозазу (<4), и факторы, не-
обходимые для репликации (В). Функции сег-
мента G описаны в разд. Ю.б.а.
индуцируемыми IS-элементами и транспозонами.
Поскольку транспозиции Mu-ДНК происходят с
относительно высокой частотой и их можно про-
водить в бесклеточных системах, фаг Ми исполь-
зуется для изучения механизмов бактериальной
транспозиции на молекулярном уровне.
Механизмы транспозиции. Для объяснения тран-
спозиции предложено много моделей, но все их
можно разбить на два класса в соответствии с
наблюдаемыми типами транспозиций (рис. 10.8).
Для простоты рассмотрим транспозицию из одного
генома в другой (например, из плазмиды в бакте-
риальный геном или обратно). (Отметим, что ин-
тегрированная Mu-ДНК тоже может транспозиро-
ваться в плазмидную ДНК ) Внутримолекулярная
транспозиция протекает более сложно; мы вкратце
остановимся на ней в конце этого раздела.
В ходе транспозиции первого типа, называемой
коинтеграционной, донорный геном, который несет
транспозирующийся элемент, сливается с реципиен-
тной молекулой ДНК. Образовавшийся коинтеграт
содержит всю донорную и реципиентную ДНК,
а также по одной копии транспозирующегося эле-
мента в местах сочленения этих ДНК. Коинтеграция
включает разрыв исходных фосфодиэфирных связей
и образование новых, дупликацию всего элемента
и дупликацию сайта-мишени. Коинтеграт может
затем разрешазъея (resolve) с образованием двух
исходных ДНК, каждая из которых несет копию
транспозированного элемента. При транспозиции
путем коинтеграции используется не только инфор-
мация, закодированная в самом элементе, но и
репликативные функции клетки. Этот механизм
иногда называют репликативной транспозицией, по-
скольку происходит полная дупликация элемента.
Транспозиция второго типа называется простым
встраиванием. Транспозирующийся элемент переме-
щается в новый геномный локус, при этом никаких
других перестроек, кроме дупликации сайта-мише-
ни, не происходит. Этот вид транспозиции иногда
называют консервативным (или нерепликативным),
поскольку дупликации как таковой не происходит.
Некоторые транспозирующиеся элементы, напри-
мер ДНК фага Ми, участвуют как в коинтеграции,
так и в простом встраивании.
Существует еще одна весьма привлекательная
модель, согласно которой на основе общей про-
межуточной структуры осуществляется транспози-
ция любого из двух типов (рис. 10.9). В соответствии
с этой моделью может происходить транспозиция
ДНК фага Ми и других мобильных элементов. На
рисунке, иллюстрирующем модель, и ДНК донора,
и ДНК реципиента представлены в кольцевой фор-
ме, поскольку в экспериментах in vitro, поставлен-
ных для проверки справедливости данной модели,
использовалась кольцевая плазмидная ДНК; мо-
бильным элементом в донорной ДНК служила мо-
дифицированная ДНК фага Mu. In vivo доноры
и реципиенты, например плазмиды и хромосомы
Е. coli, также находятся в кольцевой форме.
Согласно данной модели, транспозиция начи-
нается с образования одноцепочечных разрывов в
кольцевых донорной и реципиентной ДНК. У доно-
ра такие разрывы происходят с обоих З'-концов
234
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 10.8.
Два возможных способа встраивания мобильного эле-
мента в новый локус. Донор мобильного элемента
и реципиент изображены в виде двух отдельных колец,
хотя оба они могут находиться в одной и той же
молекуле ДНК. Сам элемент (выделен цветом) изобра-
жен вместе с концевыми инвертированными повторами
и дупликациями сайта-мишени (жирные концы стре-
лок). При первом способе образуется промежуточный
коинтеграт с дупликацией мобильного элемента; по-
следний затем диссоциирует на два продукта, каждый
из которых содержит копию мобильного элемента. При
втором способе-простом встраивании-мобильный
элемент перемещается от донора к реципиенту без
репликации. Кольцевая донорная ДНК утрачивает эле-
мент, и его дальнейшая судьба неизвестна. Сайт-ми-
шень реципиента дуплицируется независимо от способа
встраивания.
элемента, у реципиента-в сайте-мишени с образо-
ванием 5'-выступов. Затем разрезанные концы мо-
бильного элемента соединяются с концами реци-
пиента, и образуется общий промежуточный про-
дукт. Если в цепях, противоположных по отноше-
нию к разрезанным первоначально, никаких допол-
нительных разрывов не происходит, то имеет место
простое встраивание В результате заполнения бре-
ши и лигирования происходит дупликация сайта-
мишени. Таким способом может осуществляться
нерепликативная транспозиция TnlO. Эту реакцию
катализирует транспозаза TnlO, которая ускоряет
разрезание на концах TnlO, а также, по-видимому,
воссоединение разорванных концов. При реплика-
тивной транспозиции общий промежуточный про-
дукт претерпевает совсем другие превращения. За-
метим, что интермедиат по своей структуре анало-
гичен кольцевой ДНК с двумя репликативными
вилками. Если репликация начинается в каждой
вилке, то конечный продукт представляет собой
коинтеграт-кольцо, содержащее и донорную, и ре-
ципиентную ДНК, а также копии транспозирующе-
гося элемента, реплицированные полуконсерватив-
ным путем. При разрешении коинтеграта в конце
концов осуществляется транспозиция новой копии
мобильного элемента в другой сайт-мишень. Раз-
решение может происходить путем гомологичной
рекомбинации между двумя копиями транспози-
рующегося элемента с участием рекомбинационного
аппарата E.coli. (рис. 10.10). Однако более эффек-
тивное разрешение наблюдается в том случае, если
оно детерминируется самими транспозирующими
элементами.
Транспозиция ТпЗ и родственных ему простых
транспозонов происходит репликативным путем
с образованием коинтеграта (рис. 10.10). Этот коин-
теграт содержит прямые повторы ТпЗ и два сочле-
нения между донорной и реципиентной ДНК. Для
его образования нужна транспозаза продукт гена
tnpk, а также элементы IR-L и IR-R; необходим
также репликативный аппарат клетки-хозяина. Раз-
решение включает внутримолекулярную сайт-спе-
цифическую рекомбинацию между двумя res-сак-
тами двух ТпЗ-элементов (рис. 10.6) и протекает при
участии продукта гена tnpR- белка резолвазы. Если
ген tnpR отсутствует, то разрешение может проис-
ходить при помощи рекомбинационного аппарата
Е. coli, но со значительно меньшей эффективностью.
Механизм действия ТпЗ-транспозазы неизвестен.
Однако реакции, катализируемые очищенной резол-
вазой. можно изучать в опытах in vitro. При этом
субстрат-дуплексная ДНК-должен представлять
собой кольцевую молекулу с отрицательными
сверхвитками и содержать два res-сайта, ориенти-
рованных в коинтеграте одинаковым образом.
Сайт-специфическая резолваза связывается с обо-
ими res-сайтами и катализирует топоизомеразопо-
добный обмен цепей ДНК с последующим кроссин-
говером и рекомбинацией. В реакции принимают
участие четыре цепи ДНК, при этом разрываются
и вновь образуются четыре фосфодиэфирные связи.
Репликации ДНК не происходит. Отметим, что
промоторы обоих генов- tnp А и tnp R- находятся
в сайте res (рис. 10.6). При связывании резолвазы
с res-сайтами экспрессия обоих генов подавляется
Мобильный элемент
3KJ
Расщепление
в местах соеди-
нения транспозона
и исходной флан-/ " о6 й \
ZXZ7 промежуточный
(Айв) / n₽°«V^
Репликация в двух
Разрешение
Коинтеграт
Простое
встраивание
Реплика- /
тивная /
вилка /
РИС. 10.9.
Репликативная и нерепликативная транспо-
зиция на основе одной промежуточной
структуры. Как донорная, так и реципиент-
ная ДНК представлены в виде колец. Точ-
ками обозначены 5'-концы донорного эле-
мента. Серыми прямоугольниками выделе-
ны сайты-мишени реципиента (дупликации
сайтов-мишеней, окружающие донорный
транспозон, для удобства не указаны). Пер-
вый этап состоит в образовании в донорной
и реципиентной ДНК одноцепочечных раз-
рывов с выступами. У донора они происхо-
дят с обоих З’-концов элемента, а у реци-
пиента-в сайте-мишени с образованием
одноцепочечных выступов определенной
длины (см. рис. 10.1). На следующем этапе
свободные 5'-концы реципиента лигируют
со свободными З'-концами донора с об-
разованием промежуточного продукта.
[К. Mizuuchi, Cell 39 (1984), р. 395.]
Воссоединение,
катализируемое
транспозазой (tnpA)
оинтеграт
Диссоциация,
катализируемая
резолвазой
(tnpR)
РИС. 10.10.
Репликативная транспозиция ТпЗ
путем образования и диссоциации
коинтеграта. Показаны дуплика-
ции сайта-мишени только у до-
нора.
Продукт
Донор
236
ЧАСТЬ III. ОРГАНИЗАЦИЯ. ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
5' IS1O-L
( CACTCAGC6 (
IS1O-R З'
( ►(" CA’tTtASee —р
5'
С
Гомология между IS10-L и 1S10-R
способствует такому взаимному
расположению тандемных
повторов, при котором происхо-
дит их рекомбинация
5'
CACTCAGC6
Неизвестные
продукты
РИС. 10.11.
Гомологичная рекомбинация между дуп-
лицированными сайтами-мишенями при-
водит к вырезанию мобильного элемента
и восстановлению исходного сайта-мише-
ни. Показана только одна цепь ДНК; в ка-
честве примера взят ТпЮ.
и другие транспозиции ограничиваются.
Вырезание. Встраивание и вырезание мобильного
элемента-совершенно разные процессы: точное
вырезание элемента не детерминируется им самим.
Во многих случаях вырезание не сопровождается
встраиванием: вырезанный элемент утрачивается
и его судьба остается неизвестной. Вырезание,
по-видимому, происходит в результате гомологич-
ной рекомбинации между копиями сайта-мишени
(рис. 10.11). При этом может элиминироваться весь
элемент и один из фланкирующих его повторов
с восстановлением исходного сайта-мишени. Таким
образом, вырезание может исправлять мутации,
обусловленные встраиванием. В некоторых случаях
эти события зависят от продукта хозяйского гена
гее А. В других ситуациях ген гее А не нужен и
частота вырезания зависит от других рекомбина-
ционных и репарационных функций хозяина.
Связь между встраиванием мобильных элемен-
тов и мутациями. Нередко встраивание мобильного
элемента в регуляторный или кодирующий участок
приводит к подавлению экспрессии соответствую-
щего гена, но, как правило, последствия оказы-
ваются более сложными (рис. 10.12). В некоторых
случаях промотор, расположенный внутри самого
элемента, индуцирует экспрессию соседнего, ранее
молчавшего гена. Например, в модуле IS10-R транс-
позона ТпЮ имеется сильный промотор, располо-
женный на расстоянии 100 п.н. от конца элемента:
транскрипция начинается в направлении от элемен-
та и распространяется на соседний участок ДНК
(рис. 10.12, а). Если элемент встроен в один из генов
полицистронного оперона, то эффекты оказываются
другими. Например, встроенный элемент может
влиять на экспрессию последующих генов, либо
останавливая транскрипцию в сайте терминации,
находящемся внутри его самого (рис. 10.12,6), либо
подавляя трансляцию дистальных кодирующих по-
следовательностей мРНК (рис. 10.12, в).
Когда один и тот же геном служит и донором,
и реципиентом при образовании коинтеграта (внут-
римолекулярная транспозиция), способность мо-
бильных элементов вызывать перестройки в сосед-
них участках ДНК приводит к другим мутационным
эффектам. Все происходящие при этом события,
включая делеции и инверсии, связаны с соединением
одного или двух концов мобильного элемента с но-
выми последовательностями ДНК (рис. 10.13); де-
тали этих процессов до конца не установлены. На-
конец, гомологичная рекомбинация между двумя
идентичными мобильными элементами, находящи-
мися в дуплексе ДНК в одной ориентации, приводит
к делеции расположенных между ними последова-
тельностей.
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
237
а. Включение экспрессии соседних генов
Слабый промотор
Незанятый
сайт
Экспрессия происходит
с низкой Эффективностью
или вообще отсутствует
Занятый
сайт
Экспрессия
6. Полярные транскрипционные эффекты в полицистронных оперонах
Незанятый
сайт
Включение
полицистронного
оперона
6. Полярные транскрипционные эффекты в полицистронном опероне
Незанятый
сайт
Рибосомы
Белок
Обычно рибосомы
отделяются,ие достигнув
кодона AUG в гене 2, и
трансляция прерывается.
I рансляция, сопряженная
с транскрипцией, также
может прерываться
ных элементов.
238
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Мобильный
элемент
I Инверсия с
I дупликацией
Перестройки в молекуле ДНК, связанные с наличием
в ней мобильных элементов. [N. Kleckner, Алл. Rev. Gen.
15 (1981), р. 34.]
б. Р-элементы дрозофилы
Генетическая особенность некоторых линий
D.melanogaster состоит в том, что для них характе-
рен гибридный дисгенез. Как мы увидим далее,
свойства систем с дисгенетическими признаками
коррелируют с присутствием в них мобильных эле-
ментов. У дрозофилы известно несколько таких
систем; одна из них-P-система-особенно хорошо
охарактеризована как на генетическом, так и на
молекулярном уровнях.
Транспозиция P-элементов. В геноме некоторых
линий D.melanogaster диспергировано примерно 40
членов Р-семейства (в расчете на гаплоидный ге-
ном). В то же время у дрозофил других линий
вообще нет P-элементов. Последовательность на-
иболее длинных P-элементов размером 2,9 т.п.н.
высококонсервативна; длина других элементов
варьирует от 0,5 до 2,9 т. п. н. и по сравнению
с полноразмерным элементом длиной 2,9 т. п. н. в
них имеются значительные делении. Во всех Р-эле-
ментах, независимо от размера, есть концевые ин-
вертированные повторы длиной 31 п.н.
Способность P-элементов к транспозициям про-
является несколькими способами. Во-первых, число
копий P-элемента варьирует от одной линии мух
к другой. Во-вторых, локализация P-элементов в ге-
номе у разных линий неодинакова. Так, при расщеп-
лении ДНК разных линий специфическими рестрик-
тазами и блот-гибридизации с использованием в ка-
честве зонда P-элементов получаются разные про-
фили. И наконец, четко показано, что встраивание
и вырезание P-элементов в специфические локусы
коррелируют соответственно с мутациями и ревер-
сиями в определенных генах.
В геноме D. melanogaster имеется локус white,
участвующий в детерминации цвета глаз насеко-
мого. Как правило, мутанты по этому локусу имеют
белые, а не красные глаза, как у обычных мух.
ДНК-блоттинг с использованием в качестве зондов
сегментов ДНК из области мутантного локуса white
и локуса дикого типа показал, что одна группа
мутаций while обусловлена встраиванием Р-элемен-
та в этот локус (рис. 10.14). Данные о том, что
P-элемент действительно ответствен за упомянутую
мутацию, были получены при исследовании ДНК из
мух-ревертантов с красными глазами. Между му-
тантной и ревертантной ДНК не было обнаружено
никаких поддающихся идентификации различий,
кроме того, что у ревертантов P-элемент был утра-
чен, но осталась единственная копия сайта-мишени
(у мутантов эти копии находятся по обоим концам
Р-элемента).
P-элементы встраиваются преимущественно в
определенные сайты. Сравнение копий сайтов-ми-
шеней в окрестности нескольких P-элементов позво-
лило получить консенсусную последовательность
такого сайта: 5'-GGCCAGAC. При встраивании
P-элемента этот сайт-мишень длиной 8 п. н. дупли-
цируется.
Мобильные элементы других видов в D.melano-
gaster также варьируют по числу копий и тоже
вызывают инсерционные мутации (разд. 10.3). Од-
нако их структура и механизм транспозиции сущес-
твенно отличаются от таковых для Р-элемента.
Свойства P-элементов, в частности наличие кон-
цевых инвертированных повторов и генов, кодиру-
ющих один или несколько необходимых для транс-
позиции факторов, наводит на мысль об их сходстве
с мобильными элементами бактерий. Более того,
P-элемент, как и ТпЗ, может сам подавлять детер-
минируемые им процессы, в частности транспози-
цию, хотя это сложный процесс, который зависит
и от других генетических факторов.
Гибридный дисгенез в линиях, содержащих Р-эле-
менты. Экспрессия генов, кодируемых Р-элемента-
ми, оказывает существенное влияние на клетки за-
родышевой линии мух-потомков от скрещивания
самцов из линий, содержащих P-элемент (Р-линий),
и самок из линий без Р-элемента (М-линий). Сово-
купность наблюдаемых эффектов и называют гиб-
ридным дисгенезом. Дисгенные мухи часто оказы-
ваются стерильными, а если они фертильны, то у их
потомков обнаруживается множество мутаций и
хромосомных аберраций, и эти признаки передают-
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
239
1тпц Направление транскрипции
, ____________________________и примерное положение
кодирующего участка
Теломера 4_
SSc X
44 х
В Xh Xh Sm Н
4- 4- 4- 4- X
в —>Центромера
РИС. 10.14.
Дикий тип,
красные глаза
Мутант,1
белые глаза
Сайт-мишень
Мутация! тРеверсия
Цс₽-эяемеит * В xh^|m J
------>--------»---------------------
I I
Б
X
Мутация «белые глаза» у Drosophila обусловливается
встраиванием P-элемента в локус white Х-хромосомы.
Для построения геномной карты данной области ис
пользовали перекрывающиеся клонированные сегмен
ты, охватывающие участок длиной более 25 т. п. н., кото-
рый содержит локус white дикого типа. Используя эти
сегменты в качестве зондов, клонировали фрагменты,
включающие гомологичные участки из рекомбинантной
библиотеки, сконструированной из ДНК мутанта white,
и построили карту мутантного локуса. Этот локус со-
держит лишние 1,4 т.п.н по сравнению с локусом
дикого типа, которые гибридизуются с ДНК клониро-
ванного P-элемента. Аналогичным образом была по
строена карта мутантного локуса для ревертанта с крас
ными глазами. У этого ревертанта P-элемент утрачен;
осталась только одна копия сайта-мишени размером
8 п. н., которая в дуплицированном виде находилась на
концах P-элемента. Таким образом, структура локуса
white у ревертанта и дикого типа одинакова.
ся следующим поколениям Кроме того, некоторые
из мутаций у дисгенных мух довольно нестабильны
и с высокой частотой ревертируют к дикому типу
или к измененным аллелям. Другая характерная
особенность систем с дисгенными свойствами со-
стоит в том, что в отличие от нормальных самцов
Drosophila у дисгенных самцов наблюдается реком-
бинация.
В основе гибридного дисгенеза лежит внедрение
отцовских хромосом, несущих P-элементы, в яйцо
М-линии. При скрещивании Р-самцов с Р-самками
или М-самцов с Р-самками гибридного дисгенеза не
наблюдается (рис. 10.15). Для его проявления необ-
ходимы как P-элементы, так и не содержащие этих
элементов клетки М-линии Согласно одной из мо-
делей, присутствие P-элемента в материнских хро-
мосомах приводит к подавлению транспозиции
P-элемента и тем самым блокированию гибридного
дисгенеза.
Высокая частота мутаций и реверсий у дисген-
ных мух в значительной степени обусловливается
встраиванием и вырезанием P-элементов. Встраива-
ние, по-видимому, происходит только в клетках
зародышевой линии с частотой примерно одно
встраивание на хромосому в одном поколении мух
(эти оценки получены с помощью гибридизации in
situ с политенными хромосомами) Как показывают
результаты реверсии мутации в локусе white, вы-
резание происходит очень точно (рис. 10.14). Час-
тоту этих событий можно оценить генетически ис-
ходя из частоты реверсий; они наблюдаются у
каждых четырех из тысячи мух, т. е. эта частота
намного выше, чем у транспозонов бактерий. У
недисгенных мух (например, в поколении от скре-
щивания Р-самцов и Р-самок) частота как инсер-
ционных мутаций, так и реверсий значительно ниже.
Для гибридного дисгенеза нужен полноразмер-
ный P-элемент. В укороченных P-элементах по-
следовательности, необходимые для транспозиции
и вырезания, делегированы. Однако P-функции ко-
ротких элементов могут восполнять в одних и тех
же клетках полноразмерные транс-элементы. Рас-
смотрим, например, мутацию sn"' в локусе singed,
вызванную встраиванием короткого Р-элемента.
Мутантные мухи имеют необычные щетинки. С по-
мощью генетических манипуляций можно получить
мух, несущих мутацию sn"’, но не содержащих дру-
гих P-элементов. При скрещивании таких самцов
с самками из М-линии гибридного дисгенеза не
наблюдается, и мутация sn" оказывается стабиль-
ной. Однако при введении в геном полноразмерного
P-элемента эта мутация становится нестабильной.
Таким образом, целый P-элемент обеспечивает вы-
резание, находясь в шранс-положении по отноше-
нию к дефектному элементу (рис. 10.16).
Функции, кодируемые P-элементом. Фенотип
особей с дисгенетическими признаками указывает,
что P-элементы кодируют несколько факторов
Один или несколько из них ответственны за тран-
спозицию (например, транспозаза) и, следователь-
но, за повышение частоты встраивания (1), за выре-
зание (2) и стерильность (3), обусловленную в ос-
новном высокой скоростью мутирования. Транс-
позазная активность, по-видимому, ограничивается
240
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Нормальные мухи
М-линия Р-линия
Гибридный
дисгенез:
стерильность,
М-линия
высокая частота
реверсий,мутаций,
хромосомных
аберраций
РИС. 10.15.
Гибридный дисгенез у Drosophila. Точки на хромосо-
мах- Р-элементы.
клетками зародышевой линии. Кроме того, супрес-
сия транспозиции и, следовательно, других фено-
типических проявлений у Р-самок позволяет пред-
полагать, что P-элементы кодируют и репрессию
функций P-элементов. Поскольку в цитоплазме
М-самок репрессор отсутствует, экспрессия генов
P-элемента и гибридный дисгенез не блокируются.
Полноразмерный P-элемент содержит четыре
открытые рамки считывания (экзона) в одной цепи
ДНК (рис. 10.17). В соматических клетках дисгенных
зародышей обнаруживаются два больших полиаде-
нилированных транскрипта P-элемента, содержа-
щих нуклеотидную последовательность указанной
выше цепи. Синтез обеих РНК начинается вблизи
5'-конца смысловой цепи P-элемента. Синтез более
короткой РНК длиной 2,5 т. п. н. заканчивается
сразу за ро1у(А)-концом, на остатке 2700, а синтез
более длинной РНК размером 3 т. п. н,-вблизи
конца P-элемента, возможно во фланкирующих ге-
номных последовательностях. Сплайсинг этих РНК
проходит одинаково. Первые два интрона удаляют-
ся с образованием одной длинной открытой рамки
считывания (ORF); четвертая ORF остается прер-
ванной и, по-видимому, не транслируется. Этот
факт весьма удивителен, поскольку мутационный
анализ полноразмерных P-элементов показал, что
транспозазе необходимы все четыре ORF. Одно из
объяснений может состоять в том, что сплайсинг
этого последнего интрона с образованием мРНК,
на которой синтезируется транспозаза, происхо-
дит только в клетках зародышевой линии. Такая
«регуляторная» модель объясняет, почему транс-
позиции P-элемента происходят только в этих
клетках.
Получить достаточное количество РНК из кле-
ток зародышевой линии D. melanogaster для прямой
проверки этой гипотезы невозможно. Поэтому были
проведены опыты по схеме, представленной на рис.
10.16 (обратная генетика). В P-элементы были вне-
сены сайт-специфические мутации в места 5'- и
З'-сочленений третьего и четвертого экзонов. В дру-
гом варианте был удален интрон между этими
двумя экзонами. Мутантные элементы инъециро-
вали в клетки мух, несущих мутацию sn"' и не
содержащих других P-элементов, и определяли ста-
бильность мутации ли"1 у потомков этих мух. Если
мутация оказывалась нестабильной, то это озна-
чало, что в клетках присутствует функциональный
P-элемент. В соответствии с данной моделью Р-эле-
менты, у которых мутация затрагивает одну пару
оснований в любом из сочленений экзонов, не де-
стабилизируют snw. Однако инъекция P-элемента, не
содержащего интрона, дестабилизирует мутацию
sn"' точно так же, как дестабилизирует ее нормаль-
ный P-элемент. Более того, не содержащий интрона
P-элемент даже облегчает транспозицию и выреза-
ние в соматических клетках, что тоже свидетель-
ствует об уникальной способности клеток зароды-
шевой линии к специфическому сплайсингу. В соот-
ветствии с этим Р-РНК, содержащая интрон 3,
обнаруживается во всех соматических клетках и на
протяжении всего развития организма. Таким об-
разом, регуляция транскрипции не отвечает за то,
что транспозазная активность оказывается прису-
щей только клеткам зародышевой линии. Вместо
этого экспрессия соответствующих генов регули-
руется на уровне процессинга первичного транс-
крипта.
Соматические клетки D. melanogaster, которые
содержат измененный P-элемент, лишенный третье-
го интрона, в отличие от клеток с нормальным
P-элементом, синтезируют белок мол. массой
87 кДа. Судя по размеру, он кодируется полностью
сплайсированной Р-РНК и, вероятно, является
транспозазой. Другой, более короткий полипептид
транслируется с Р-РНК, в которых интрон 3 сохра-
нен, т.е. с РНК длиной 2,5 и 3 т.п.н. из сома-
тических клеток. Соответствующий Р-кодируемый
полипептид мол. массой 66 кДа, по-видимому, от-
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
241
Drosophila
Зародыш дрозофилы snw
Предшествен-
ники клеток
зародышевой
линии
ДНК
snw, дефектный Р-элемент
—Н-----------8
pBR322
Взрослые мухи.зи'*
Скрещивание
с мухой из
М-линии
Потомство с высокой
частотой генотипа зл+
РИС. 10.16.
Все факторы, необходимые для транспозиции, кодирует
один P-элемент размером 2,9 т.п.н. Слева представ-
лена схема эксперимента, справа-два участка геномной
ДНК, на которых разворачиваются все события. Полно-
размерный P-элемент длиной 2,9 т. п. н. был клонирован
в плазмиде pBR322 и инъецирован в эмбрион snw на
очень ранних стадиях. Эмбрион не содержал никаких
других P-элементов, кроме короткого дефектного эле-
мента Р, детерминирующего мутацию snw. Развившиеся
из этих зародышей взрослые мухи имели стабильный
фенотип snw. Однако у потомков некоторых иэ них
мутация snw оказалась нестабильной. Кроме того, ге-
номная ДНК взрослых мух и их потомков содержала
новый полноразмерный P-элемент. Таким образом, не-
которые из инъецированных P-элементов встроились
в ДНК клеток зародышевой линии, успешно реплици-
ровались во время дифференцировки и развития и
экспрессировались (в зародышевых клетках). После-
довательности pBR322, которые были ковалентно свя-
заны с P-элементом в инъецированной ДНК, в клетках
потомков не обнаружены. Таким образом, встраивается
P-элемент не путем неспецифической рекомбинации,
а целиком переносится из вектора в геном Drosophila
с помощью собственного аппарата транспозиции
[А. С. Spradling, G. М. Rubin, Science 218 (1982), р. 341.]
ветствен за репрессию транспозиции, наблюдаемую
в соматических клетках и Р-яйцах.
Механизм встраивания и вырезания Р-элементов
неизвестен. Помимо транспозазы, в этих процессах,
по-видимому, участвует полипептид мол. массой
примерно 65 кДа, который связывается с концевыми
повторами длиной 31 п.н. После вырезания Р-эле-
мента остаются разные сайты-мишени. В одних
случаях происходит полное вырезание Р-элемента
и одной из копий сайта-мишени (рис. 10.14), в дру-
гих эта вторая копия целиком или полностью оста-
ется, иногда остается даже часть Р-элемента.
2,9 т.п.н.
।----------------------------------------------1
Р-элемент
31 п.н.
31 п.н.
Интрон! Интрон 2 Интрон 3
А I Транскрипция,полмадени-
1 лирование, сплайсинг
РИС. 10.17.
А. Структура полноразмерного Р-элемента,
содержащего концевые инвертированные пов-
торы длиной 31 п. н„ четыре экзона и три
интрона. Б. Две полиаденилированные Р-РНК,
обнаруженные в соматических клетках, и ко-
дируемый ими белок мол. массой 66 кДа.
Транскрипция начинается с 87-й пары основа-
ний. В. Предполагаемая структура полиадени-
лированной РНК, которая содержится только
в клетках зародышевой линии и кодирует
транспоэаэу мол. массой 87 кДа; все три
интрона в этой РНК удалены при сплайсинге.
[К. O'Hare, G. М. Rubin, Cell 34 (1983), р. 25;
R. Е. Karess, G. М. Rubin, Cell 38 (1984), р. 135.]
poly (А+)-РНК и белок в соматических клетках
ООООЭОСХХХХЮООООЭСЮ
Белок мол. массой ббкДа
poly (А+)-РНК и белок в клетках зародышевой линии
сооостэоооооооооооосхэо
Белок мол. массой В7 кДа
16-158
242
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Выводы. Описанные на рис. 10.16 эксперименты
показывают, что P-элементы можно использовать
в качестве векторов для введения в D. melanogaster
практически любого гена (часть IV). Кроме того,
P-система интересна и с эволюционной точки зре-
ния. Появление в геноме P-элемента может привести
к репродуктивной изоляции популяций мух, зани-
мающих одну экологическую нишу. Таким образом,
P-элементы могут служить фактором видообразо-
вания, хотя, по-видимому, и не дают никаких селек-
тивных преимуществ, поскольку мухи М-линий в
отсутствие P-элемента более жизнеспособны. Р-се-
мейство-лишь одно из нескольких семейств мо-
бильных элементов, обусловливающих гибридный
дисгенез у Drosophila, так что эволюционный по-
тенциал транспозирующихся элементов у этого ор-
ганизма весьма высок. В связи с этим важно отме-
тить, что не все мобильные элементы обладают
свойствами транспозирующихся. По меньшей мере
один из них 1-элемент-относится к ретротранспо-
зонам класса II (разд. Ю.З.е). У Drosophila имеются
также транспозирующиеся элементы, близкие Р, но
не связанные с гибридным дисгенезом (например,
элемент, называемый hobo).
в. Контролирующие элементы у кукурузы
Мобильные генетические элементы впервые бы-
ли обнаружены при генетических исследованиях ку-
курузы и получили название контролирующих эле-
ментов. Идентифицировано по меньшей мере три
семейства контролирующих элементов, а последние
данные позволяют предположить, что на самом
деле их более двенадцати. Аналогичные элементы
встречаются и у других видов растений. Члены
каждого семейства могут быть подразделены на два
класса: автономные элементы наследуются неста-
бильно и способны к независимому вырезанию и
транспозиции; неавтономные элементы сами по себе
стабильны и способны к транспозиции только в том
случае, когда в какой-либо области генома при-
сутствует автономный член того же семейства. По-
скольку автономные элементы кукурузы детерми-
нируют активности, необходимые для транспози-
ции, и обладают характерными структурными осо-
бенностями, мы объединим их в одну группу с
транспозирующимися элементами прокариот и Р-
элементом Drosophila. На самом деле функциональ-
ные различия автономных и неавтономных контро-
лирующих элементов носят такой же характер, что
и различия между полноразмерными и укорочен-
ными P-элементами. Тем не менее между этими
двумя системами есть важные различия. Например,
P-элементы функционируют только в клетках за-
родышевой линии, что проявляется при гибридном
дисгенезе, а контролирующие элементы опосредуют
мутации как в соматических, так и в половых
клетках. У кукурузы транспозиции происходят на
определенных стадиях развития с определенной час-
тотой; таким образом, их влияние на генную экс-
прессию носит регулярный, заданный во времени
характер. И хотя эти эффекты накладываются на
наследственную программу развития, определяемую
геномом, нетрудно представить, как в ходе эво-
люции они стали играть существенную роль в раз-
витии организма.
Генетическая система. Не зная об особенностях
развития кукурузы, очень трудно разобраться, как
с помощью одних только классических генетических
и цитогенетических методов были открыты контро-
лирующие элементы. Каждое зерно в початке куку-
рузы содержит зародыш и многоклеточный эндо-
сперм, в котором имеется большое количество пи-
тательных веществ, используемых во время про-
растания (рис. 10.18). Зародыш и эндосперм обра-
зуются в результате отдельных слияний гаплоидных
ядер, но обычно содержат одинаковую генетиче-
скую информацию. Эти пары мужских и женских
ядер являются сестринскими и образуются в резуль-
тате митотических делений гаплоидных предшест-
венников. Следовательно, мутации в клетках за-
родышевой линии, которые приводят к наблюда-
емым фенотипическим изменениям в эндосперме,
к счастью, сохраняются в зародыше зерна, что
позволяет проводить дальнейшие исследования.
Каждое из нескольких сотен зерен в початке пред-
ставляет собой результат отдельного скрещивания,
поэтому удается выявлять редкие мутации в клетках
зародышевой линии.
Изучение мутации в соматических клетках также
не вызывает затруднений. Из одиночных клеток-
РИС. 10.18.
Зерно кукурузы. Перикарпий (выделен черным цветом)
образуется из материнских клеток, остальная часть
имеет генетическую конституцию зародыша.
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
243
Таблица 10.3. Некоторые мутации у кукурузы, использующиеся в генетических экспериментах
Локус Мутантный фенотип Фенотип дикого типа Хромо- сома Кодируемая функция
С Бесцветный алейрон Пигментированный алейрон 9 Регуляция синтеза антоцианина
Bz (bronze) Бледный алейрон Пигментированный алейрон 9 Гликозилирование антоцианина с по- мощью UDP-глюкоза; флавонол-3- О-гликозил-трансферазы
Их (waxy) Отсутствие амилозы в эн- досперме; после обработ- ки раствором 12-К1 зерна становятся бледно- коричневыми Наличие амилозы; после об- работки 12-КТ зерна ста- новятся иссиня-черными 9 Глюкозилтрансфераза, катализиру- ющая синтез амилозы
Sh (shrunken) Морщинистые зерна, обра- зующиеся вследствие дефицита крахмала Вздутые зерна 9 Сахарозосинтетаза, катализирующая синтез крахмала
Adh\ Отсутствие алкогольде- гидрогеназы 1 Одна субъединица алкогольдегидро- геназы
предшественников в процессе дифференцировки эн-
досперма образуются локальные клоны клеток, и
мутация в единичной клетке на определенной стадии
развития проявляется затем в группе клеток с му-
тантным фенотипом, расположенной в определен-
ной области эндосперма. Многие фенотипические
варианты с большей или меньшей жизнеспособ-
ностью. наблюдаемые визуально в эндосперме.
очень удобны для экспериментальных исследований
(табл. 10.3). Рассмотрим, например, локус С. У рас-
тений, гомозиготных по рецессивной мутации в
этом локусе, нарушен синтез пигмента антоцианина,
поэтому зерна почти бесцветны (рис. 10.19, Л).
Свойства контролирующих элементов, выявля-
емые с помощью генетических методов. У растений,
гомозиготных по нестабильному рецессивному ал-
Б
РИС. 10.19.
Фенотипические признаки, определяемые С-
локусом. А. Початок с зернами дикого типа
(пурпурного цвета) и мутантными зернами
(бесцветными). Последние несут стабильную
мутацию в гене С, детерминирующем синтез
антоцианина. Б. Одни зерна бесцветны, другие
имеют мозаичную окраску, обусловленную
нестабильной мутацией в гене С, которая воз-
никает при встраивании транспозирующегося
элемента. Клоны клеток, из которых удален
этот элемент, содержат функциональный ген
и синтезируют антоцианин. Показаны зерна,
в которых транспозирующийся элемент удален
из С-локуса на ранних (В), средних (Г), и
поздних (с малой частотой) стадиях развития
зерен (Д). (С любезного разрешения Nina
Fedoroff.)
16*
244
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
лелю локуса С, зерна имеют бледную окраску и
покрыты сильно пигментированными пятнами
(рис. 10.19, Б-Д). Каждое такое пятно-это клон
клеток, произошедший из клетки, которая ревер-
тировала к дикому типу с возобновлением синтеза
антоцианина. Нестабильные рецессивные мутации
обусловлены вставкой контролирующего элемента,
а реверсия произошла в результате последующих
перестроек-таких, как вырезание элемента. Кроме
этого, контролирующий элемент способствует де-
лениям, дупликациям, инверсиям, транслокациям,
хромосомным разрывам и воссоединениям, а также
перестройкам в соседних хромосомных локусах. Все
это напоминает перестройки в соседних последова-
тельностях, связанные с образованием межмолеку-
лярных коинтегратов бактериальными транспозо-
нами. Однако сходны ли соответствующие меха-
низмы у растений и бактерий-неясно.
Автономные контролирующие элементы вызы-
вают нестабильные инсерционные мутации (рис. 10.20),
неавтономные же, как правило, связаны со ста-
бильными мутациями, которые, однако, становятся
нестабильными, когда в какой-либо области генома
присутствует автономный элемент того же семейст-
ва. Здесь имеет смысл вернуться к свойствам
P-элементов D.melanogaster. полноразмерные Р-эле-
менты можно считать автономными, поскольку они
сами детерминируют собственную транспозицию
и вырезание, а укороченные, дефектные Р-элемен-
ты-неавтономными, поскольку их способность к
транспозиции реализуется только при наличии пол-
норазмерных Р-элементов.
В одном из семейств контролирующих элемен-
тов, так называемом семействе Ac-Ds, автономным
элементом подобного рода является Ас (от англ,
activator-активатор), который обеспечивает тран-
спозицию и вырезание не только самого себя, но
и родственных неавтономных элементов, называ-
емых Ds (от англ, dissociator-диссоциатор). ^-эле-
менты нестабильны только при наличии в геноме
одного или более Лс-элсмснтов. Кроме того, они
образуются в результате каких-либо повреждений
Ас- в соответствии с предположением об аналогии
с P-элементами. Например, показано, что незави-
симые мутантные линии с Ds-элсмснтом в локусе Bz
произошли от линий, несущих в этом же локусе
элемент Ас (рис. 10.21). Они сохраняют мутантный
фенотип Bz, но остаются стабильными до тех пор,
пока в результате скрещивания в геном не будет
встроен еще один Лс-элемент (точно так же остается
стабильной мутация snv у D. melanogaster в отсутст-
вие в геноме полноразмерного Р-элемента).
При встраивании Ас- или Ds-элсмснтов рядом
с тем или иным геном наблюдается множество
разных фенотипов. При этом может изменяться
эффективность транскрипции, нарушаться регуля-
РИС. 10.20.
Семейства контролирующих элементов кукурузы со-
держат как автономные, так и неавтономные элементы.
А. Встраивание неавтономных элементов в геном, не
содержащий автономных элементов, приводит к ста-
бильной мутации. При встраивании автономных эле-
ментов возникают нестабильные мутации. Б. Инсер-
ционные мутации, обусловленные неавтономными эле-
ментами, нестабильны, если в геноме находится авто-
номный элемент этого же семейства.
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
245
Делеция в Лс
Фенотип
/^нестабильный
Встраивание Ас
стабильный
РИС. 10.21.
Ds-элементы появляются в результате мутаций
Дс-элемента. Ds-элемент активируется, если в
геном встраивается автономный Дс-элемент.
ция генной экспрессии, специфичной в отношении
определенной стадии развития или типа ткани, син-
тезироваться мутантный белок. Определенные фе-
нотипические особенности связаны также с неста-
бильностью участков, соседствующих с Ас- или
DS-элементами в присутствии еще одного Лс-эле-
мента в другом месте генома. Наиболее существен-
ные из них-разрывы хромосом, приводящие к рас-
щеплению и утрате ацентрических хромосомных
фрагментов вместе со всеми кодируемыми ими
функциями (рис. 10.22), а также появление допол-
нительных мутаций вблизи встроенного D.s-элемсн-
та. Любопытно, что эти события гораздо чаще
происходят в окружении Ds-, чем Л с-элементов. Не
все Ds-элсмснты, даже в присутствии Ас, дают весь
спектр эффектов. Одни из них сопряжены с раз-
рывами хромосом, другие вызывают такие разрывы
значительно реже. Некоторые элементы утратили
способность к транспозиции, но ревертируют и вы-
зывают мутации в соседних локусах. Разнообразие
свойств Ds-элсмснтов связано с различиями в по-
вреждениях /1 (-элементов.
Изучение Ac-Ds-мутантов на молекулярном
уровне. Выделены клонированные сегменты ДНК,
содержащие или Ас-, или Ds-элемент. Общий подход
к изучению /k-Ds-мутантов можно проиллюстри-
ровать на примере локуса waxy (табл. 10.3). Ген
waxy дикого типа кодирует белок мол. массой
65 кДа, а производный белок мол. массой 58 кДа
присутствует в гранулах крахмала. Из эндосперма
была выделена poly (А)+-мРНК, которую исполь-
зовали для получения плазмидных клонов, содер-
жащих соответствующие кДНК. Эти кДНК были
подвергнуты скринингу с помощью гибридизацион-
ной селекции (рис. 6.26) для обнаружения тех из них,
которые кодируют мРНК для белка мол. массой
65 кДа. Позитивные кДНК-клоны в свою очередь
использовали для отбора гомологичных клонов из
геномной библиотеки кукурузы. Пять таких кло-
нированных геномных сегментов представлены на
рис. 10.23. Первый произошел из локуса waxy дико-
го типа, второй получен из Лс-мутанта, третий
(Ds-1)-H3 D.v-мутанта-производного мутанта Ас,
и четвертый из ревертанта D.v-1-мутанта с диким
фенотипом. Пятый сегмент получен из Ds-2-мутанта
независимого происхождения. Лс-индуцированная
мутация возникает в результате встраивания сег-
мента длиной 4,5 т. п. н. вблизи З'-конца транскрип-
ционной единицы гена waxy; этот сегмент представ-
ляет собой сам Л (-элемент. У мутанта Ds-1, полу-
ченного из Лс-мутанта, имеется вставка точно в
таком же месте, но с пропуском нескольких сотен'
пар оснований. ДНК ревертанта дикого типа не
содержит вставки и аналогична исходной ДНК ди-
кого типа. Как показывает опыт изучения бакте-
риальных транспозирующихся элементов и Р-эле-
мента Drosophila, все эти свойства характерны для
транспозирующейся единицы. Ds-2 также содержит
вставку (2 т. п. н.) в транскрипционной единице
и’н.уу, но в другом месте. Электронно-микроскопи-
ческое исследование гетеродуплекса, образующегося
из элементов Ds-2 и Ас, показывает, что Ds-2 содер-
жит примерно по 1 т. п. н., соответствующих каж-
дому из концов Ас, но не гибридизуется с его
внутренним участком длиной 2,3 т.п.н. Некоторые
Ds-сдиницы еще короче. Например, в 5'-лидерной
последовательности мутантного локуса Adh 1 име-
ется Ds-вставка размером 405 п.н. (рис. 10.24). Та-
ким образом, внутренняя часть D.s-элемснта, по-
видимому, в значительной степени варьирует.
Установлена нуклеотидная последовательность
Лс-элемента (рис. 10.25). На концах сегмента дли-
246
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Фенотип
РИС. 10.22.
Генотип
Воссоединение
Профаза
| Разрыв
Wx Bz Bz Wx . „
»____( ( Cf (________- О A н a э a
"7 I t *—D Телосраза
Wx Bz Bz
Wx Bz Bz
I
Непрерывные циклы
разрывов-воссоединений
С, Bz, Wx
С, bz, wx
'Репликация,
воссоединение
Профаза
Wx
Wx
I
Непрерывные циклы
разрывов-воссоединений
Б
Хромосомный разрыв и перестройка, происходящие
вблизи элементов семейства Ac-Ds. А. Один из гомо-
логов хромосомы 9 содержит вставку Ds, расположен-
ную ближе к центромере, чем локус waxy. Разрыв
хромосомы в Os-сайте приводит к тому, что дочерние
клетки утрачивают ацентрический фрагмент, несущий
доминантные локусы дикого типа С, Bz и Wx. Б. При
встраивании Ds-злемента в сайт, расположенный от
центромеры дальше, чем локус waxy, сестринские хро-
матиды после разрыва в Os-сайте воссоединяются с
образованием U-образного ацентрического фрагмента
и U-образного дицентрического фрагмента, который
может разворачиваться и разрываться при последу-
ющем митозе. Образовавшиеся концы могут вновь вос-
соединиться, и так далее до бесконечности, поддержи-
вая нестабильное состояние. В области теломер вос-
соединения хромосом не происходит (разд. 9.6.6).
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
247
Фенотип Генотип
Обеспечивает функцию4с
в /77/7<7нс-положении
РИС. 10.23.
Геномные клоны кукурузы, содержащие Ас-
и Ds-мутации в локусв waxy.
Сайт-мишень (8п.н.н.)
\ adh\
Дикий тип
Концевые
Ds инвертированные
405л.н. / повторы
adh\
£ Ps-мутан
РИС. 10.24.
Геномные клоны кукурузы, содержащие ген Adh\. Вол-
нистая стрелка показывает направление транскрипции.
adh\
'i ревертанта
8 дупликация
сайта-мишени
сохраняется
2 т.п.н.
248
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
0,5 т.п.н.
I-----------1
CAGGGATGAAA
ТТТСАТСССТА
нов, кодирующих одну протяженную мРНК, и сама эта
мРНК длиной 3,5 т.п.н.
РИС. 10.25.
Структура транспозирующегося Ас-злемента кукурузы
размером 4,6 т. п. н. Показаны несовершенные концевые
повторы длиной 11 п. н. Ниже представлены пять зкзо-
ной 4,6 т. п. н. находятся концевые инвертированные
повторы длиной 11 п. н. Вблизи обоих концов име-
ются еще несколько групп инвертированных пов-
торов. Концевые повторы сохраняются и у Ds-эле-
ментов. Ас-элемент содержит пять открытых рамок
считывания, которые могут кодировать полипептид.
Делеции в нескольких экзонах приводят к тому, что
элементы становятся неспособными к транспози-
ции. В тканях растений, кодирующих активный
Ас-элемент, обнаружен Ас-транскрипт длиной 3,5
т. п. н., в котором отсутствуют четыре интрона.
Вероятно, этот транскрипт представляет собой
мРНК для Ас-транспозазы, а возможно, и других
специфических факторов.
Встраивание Ds- и Ас-элементов сопровождается
дупликацией сайта-мишени размером 8 п.н. При
утрате элемента или реверсии дуплицированный
участок обычно сохраняется. Например, у описан-
ного ранее ревертанта мутанта Adhl дополнитель-
ная копия дуплицированного участка размером
8 п. н. остается как след предыдущей вставки
(рис. 10.24). Нередко одна или обе оставшиеся ко-
пии оказываются слегка модифицированными в ре-
зультате точковых мутаций или делеций пар осно-
ваний. Таким образом, вырезание происходит не
совсем точно и не всегда приводит к реверсии на
уровне фенотипа.
Механизмы встраивания и вырезания Ас- и
Ds-элементов неизвестны. Согласно молекулярно-
генетическим данным, возможно, происходит не-
репликативное встраивание с вырезанием Ас- (или
Ds-) элемента из одного сайта и встраиванием этого
же сегмента ДНК в новое положение. Сохранение
копий сайтов-мишений на месте вырезанной вставки
говорит о сходстве механизма вырезания Ас- и
Ds-элементов, с одной стороны, и Р-элемента-с
другой.
Повторение последовательностей Ac-Ds. Субкло-
нированные сегменты Ас- и Ds-элементов гибриди-
зуются со многими фрагментами ДНК кукурузы,
полученными при ее рестрикции. Всего в геномах
некоторых линий встречается до 35 копий элемен-
тов. Однако профиль гибридизации весьма сложен
и говорит о том, что существует множество разных
типов Ds-элементов и, возможно, несколько видов
Ас-элементов. Об этом разнообразии свидетельст-
вует уже простое рассмотрение Ds-элементов; как
мы говорили, они бывают длиной 4,1; 2,0 и 0,405
т.п.н. Кроме того, в результате инсерционной му-
тации, обусловленной встраиванием Ds-элемента в
один из интронов гена сахарозосинтазы (локус
shrunken, Sh, табл. 10.3), образуется сложный эле-
мент длиной примерно 30 т. п. н., который содержит
несколько копий Ds-элемента, а также другие, не-
родственные последовательности (рис. 10.26). На
концах этого сложного элемента находятся несо-
вершенные инвертированные Ds-повторы, каждый
Последовательности
локуса Sh
Фенотип
Sh~
1 Э0Т.П.Н.
I
Ds
FE=4 И 8
РИС. 10.26.
Сложные Ds-вставки в гене сахарозосинтазы (Sh) ку-
курузы. Структуру мутантного (Sh~) и ревертантного
(Sh+) локусов определяли с помощью клонированных
сегментов геномной ДНК. У мутанта Sh~ данный гвн
прерывается инвертированными дупликациями Ds-эле-
мента, которые окружают какой-то неохарактеризован-
ный сегмент ДНК, отличный от Ds. Участок длиной
более 20 т. п. н. на 5'-концв гена дуплицирован; он
состоит из части гена и части сложной вставки. У
Sh+ -ревертанта вставка утрачена, но дупликация на
5'-конце сохраняется. [U. Courage-ТвЬЬе, Cell 34 (1983),
р. 383.]
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
249
длиной более 3 т. п. н. Это не единственная особен-
ность области мутантного локуса shrunken на хро-
мосоме 9; на 5'-конце гена присутствует дуплици-
рованная часть мутантного локуса. Этот дуплици-
рованный участок содержит часть самого гена и
часть последовательности Ds. Такая структура еще
раз подтверждает справедливость выводов, которые
можно сделать из результатов генетического ана-
лиза: присутствие Лс-йл-элементов создает предпо-
сылки для последующих перестроек в соседних
участках. Подобным же образом подтверждаются
результаты генетических исследований, проведен-
ных 30 лет назад, при молекулярном анализе ре-
вертанта данного мутанта: у таких Sh+-ревертантов
Ds-элемент сохраняется, поскольку частота хромо-
сомных разрывов в локусе остается высокой. Таким
образом, несмотря на то что клонированный ген
ревертанта Sh+ утратил всю встроенную последо-
вательность длиной 30 т.п.н. и по своей структуре
очень близок к гену дикого типа, в нем частично
сохранились дуплицированные участки самого гена
и Ds-элемента. Уже первые результаты генетических
исследований кукурузы содержали четкие указания
на существование мобильных элементов и сложных
молекулярных структур, связанных с контролиру-
ющими Лс-йл-элементами.
10.3. РЕТРОТРАНСПОЗОНЫ
а. Общие свойства ретротранспозонов
класса I
Типичные ретротранспозоны класса I состоят из
центрального сегмента длиной несколько т.п.н. и
фланкирующих его LTR длиной от сотен до тысяч
пар оснований (рис. 10.27). Несколько пар осно-
ваний в начале и в конце каждого LTR также
представляют собой инвертированные друг отно-
сительно друга повторы. Встраивание ретротранс-
позона в новый локус ДНК сопровождается дупли-
кацией сайта-мишени, причем размер дуплициро-
ванного участка (но не его последовательность) для
данного элемента всегда одинаков. Эти ретротранс-
позоны, как и все мобильные элементы, образуют
в геномах мультисемейства.
Как правило, ретротранспозоны класса I актив-
но транскрибируются и транслируются, хотя в ходе
развития и дифференцировки транскрипция обычно
регулируется. Одни транскрипты намного короче
полноразмерных элементов, другие начинаются в
одном LTR и заканчиваются в другом, копируя весь
элемент. Эти длинные транскрипты содержат на
концах прямые повторы и являются промежуточ-
ными продуктами при транспозиции. По-видимому,
они подвергаются обратной транскрипции анало-
гично транскрипции РНК ретровирусов, в резуль-
Дупликация
сайта-мишени
ин l участок LTR *
1т.п.н.
РИС. 10.27.
Типичный ретротранспозон класса I и его транскрипт.
тате которой образуется провирусная ДНК (разд. 2.2.а).
Процесс обратной транскрипции ретротранспозо-
нов в деталях не изучен. Ретротранспозоны коди-
руют несколько белков. Один из них-обратная
транскриптаза, а другие вместе со специфическими
транскриптами образуют внутриклеточные рибо-
нуклеопротеидные частицы.
б. Ту-элементы дрожжей
Структура Ту-элементов. Ту-элементы имеют
структуру, типичную для ретротранспозонов класса
I (рис. 10.28). Их длина составляет примерно 5,9
т. п. н„ а центральная область, называемая £, флан-
кирована длинными концевыми повторами разме-
ром 330 п. н., называемыми 8. На каждом из концов
8 в свою очередь находятся инвертированные пов-
торы длиной всего 2 п. н. Другая структурная осо-
бенность- наличие прямых повторов размером 5 п. н.
в местах соединения £ с 8. Встраивание Ту-элемента
всегда сопровождается дупликациями сайтов-мише-
ней размером 5 п.н.
Гаплоидный геном лабораторных штаммов
S. cerevisiae содержит примерно 35 Ту-элементов, что
составляет ~ 1% всего генома. У других штаммов
дрожжей число копий Ту меньше. Помимо полно-
размерных Ту-элементов, у S. cerevisiae имеется
примерно 100 одиночных 8-сегментов, не имеющих
отношения к Ту-элементам. Гибридизация клони-
рованных Е-зондов с рестрицированной геномной
ДНК из разных штаммов дрожжей говорит о том,
что гомологичные сегменты различаются по числу
копий и по размерам; это согласуется с представ-
лением о нефиксированной локализации Ту-элемен-
тов.
Нуклеотидные последовательности копий Ту-эле-
мента в одном геноме неодинаковы, хотя и доста-
точно близки, чтобы могла происходить их гибри-
дизация друг с другом. Полиморфизм выявляется,
например, при сравнении рестрикционных карт от-
дельных элементов. Однако два 8-сегмента, свя-
занные с одним Ту-элементом, обычно оказываются
250
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 10.28.
Ретротранспозон Ту у дрожжей.
330 П.Н.
330 п.н.
идентичными; этим они отличаются от IS-L и IS-R
у некоторых сложных бактериальных транспозонов.
Перемещения Ту-элементов. Способность Ту-
элементов перемещаться из одного геномного ло-
куса в другой была продемонстрирована неодно-
кратно. Во-первых, разные штаммы существенно
различаются по размерам и числу копий рестрик-
ционных фрагментов, которые гибридизуются с
Ту-последовательностями. Во-вторых, по данным
молекулярного клонирования, аллели одного и того
же локуса у различных штаммов дрожжей отли-
чаются друг от друга наличием или отсутствием
Ту-элементов. Фактически Ту-элемент был открыт
благодаря тому, что супрессор тирозиновой тРНК
дрожжей гибридизовался с разными по размеру
ЕсоRI-фрагментами ДНК из двух разных штаммов.
Клонирование этих фрагментов показало, что те их
участки, которые непосредственно примыкают
к гену тРНК, идентичны, чего нельзя сказать о бо-
лее отдаленной области, расположенной левее
(рис. 10.29). Эти различия обусловлены тем, что
в штамме 2 содержится Ту-элемент. Еще одна
особенность штамма 2- дупликация сайта-мишени:
с обеих сторон Ту-элемента находится пентанук-
леотид 5'-GAAAC. В обоих штаммах имеются не-
сколько одиночных 6-сегментов, локализованных
в одинаковых сайтах.
В-третьих, были получены прямые данные о
мобильности Ту-элементов. Опыты состояли в сле-
дуюшем. Одну колонию дрожжей разделили на пять
частей и культивировали в течение месяца, ежеднев-
но перенося небольшие инокуляты на свежую среду.
Через месяц провели блот-гибридизацию ДНК из
РИС. 10.29. 5'-gaaac
— t~ т т т Г~
Одни и те же локусы у двух штаммов
дрожжей различаются тем, что в од-
ном из них (штамм 2) присутствует
Ту-злемент. [J. R. Cameron, Е. Y. Loh,
R.W. Davis, Cell 16 (1979), р. 739;
J. Gafner, P. Philippsen, Nature 286
(1980), p. 414.]
каждой культуры с клонированной ДНК Ту (а). В
двух из пяти образцов на электрофореграмме обна-
ружилась новая полоса, отсутствующая в случае
исходной ДНК. В свою очередь новые полосы,
присутствующие у этих двух ДНК, тоже разли-
чались. Таким образом было продемонстрировано
перемещение Ту-элемента.
И наконец, выявление мутаций, обусловленных
транспозицией Ту-элемента, окончательно показа-
ло, что транспозиция действительно происходит.
Такие события наблюдались неоднократно, хотя
Ту-элемент перемещается в какой-то определенный
локус лишь в одной из 10® клеток. Некоторые
инсерционные Ту-мутации прерывают кодирующие
области, но в большинстве случаев они обусловли-
ваются встраиванием Ту-элемента в почему-то
предпочитаемый им 5'-регуляторный участок со-
ответствующей транскрипционной единицы. Эти
мутации приводят к разным фенотипическим эф-
фектам. В некоторых случаях прекращается тран-
скрипция генов, соседствующих со вставкой, при
этом Ту-элемент может находиться в любой ориен-
тации (рис. 10.30). В других случаях, напротив,
наблюдается 50-кратное увеличение синтеза мРНК
и продукта гена, примыкающего к вставке. В
«сверхпродуцентах» Ту-элемент всегда оказывается
встроенным так, что транскрибируется в направ-
лении, противоположном направлению транскрип-
ции соседнего гена (рис. 10.30). Сверхпродукция за-
Место встраивания
I--------------1
2 т.п.н.
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
251
1 т.п.н.
РИС. 10.30.
Встраивание Ту-злемента в промотор
гена HIS4 приводит к появлению му-
танта his4. В результате вырезания
Ту-злемента с сохранением части одно-
го из 6-сегментов происходит частичная
инверсия к HIS4'. [Р. J. Farabough.
G. R. Fink, Nature 286 (1980), р. 352.]
висит от транскрипции Ту и наблюдается лишь
в гаплоидных клетках, поскольку в диплоидных
дрожжевых клетках Ту транскрибируется неэффек-
тивно (разд. 17.5.6). Более того, влияние, которое
оказывает встраивание Ту на соседние гены, неоди-
наково для разных Ту-элементов и в некоторых
случаях вообще отсутствует.
Все эти различия зависят - по крайней мере от-
части- от присутствия на 5'-конце Ту сегмента ДНК
размером ~ 600 п. н. Замена одной пары оснований
в этом сегменте-CG на ТА-заметно уменьшает
активацию соседнего гена. По-видимому, эта акти-
вация опосредуется каким-то шранс-действующим
белковым фактором, который кодируется клеткой-
хозяином и связывается с регуляторным участком.
Кроме того, влияние Ту на экспрессию соседних
генов супрессируется не менее чем тремя другими
несцепленными между собой мутациями, назван-
ными spt (от англ, suppressor of Ту-супрессор Ту).
Продукты генов SPT необходимы для транскрип-
ции Ту.
Транскрипция Ту-элемента. В гаплоидных дрож-
жевых клетках обычно содержится огромное коли-
чество транскриптов Ту; на их долю приходится от
5 до 10% полиаденилированной РНК. Основная их
масса-это молекулы размером 5,7 т.н., почти
равные по длине целому элементу. Нуклеотидные
последовательности индивидуальных клонирован-
ных Ty-кДНК различаются очень мало, т. е., по-
видимому, большинство Ту-элементов транскриби-
руются одинаково.
5'-конец транскрипта длиной 5,7 т.н. нахо-
дится на расстоянии 245 п. н. от начала левого
5-сегмента, и транскрипт охватывает остальные его
95 нуклеотидов, весь Е-сегмент и второй 5, закан-
чиваясь на расстоянии примерно 40 п.н. от конца
правого 8-сегмента (рис. 10.31). Завершается тран-
скрипт poly (А)-хвостом, не кодируемым Ту-элемен-
том. Все эти структурные особенности наводят на
ряд интересных идей, которые можно применить
к другим ретротранспозонам класса I и к ретро-
вирусам.
Во-первых, несмотря на то что Ту-транскрипты
не содержат участков, соответствующих началу и
концу Ту-ДНК, в них представлена нуклеотидная
последовательность всего элемента. Объясняется
это тем, что пропущенные участки правого 8-сег-
мента транскрибируются с левого 8, и наоборот.
Отметим также, что участок длиной примерно 50
нуклеотидов в транскрипте дуплицирован. Первые
50 нуклеотидов на 5'-конце РНК идентичны послед-
ним 50 нуклеотидам на правом конце, перед poly (А)-
хвостом. Таким образом, на обоих концах тран-
скрипта имеются прямые повторы длиной 50 п.н.
Если предположить, что промотор, ответственный
за регуляцию транскрипции Ту-элемента, находится
внутри самого элемента (это согласуется с тем
фактом, что все Ту-элементы транскрибируются
одинаково, независимо от окружения), то можно
ожидать, что инициаторами транскрипции могут
быть оба 5-сегмента ввиду их идентичности. Однако
никаких транскриптов, начинающихся с дистально-
го 8-сегмента, не обнаружено. Далее, если дисталь-
ный 5-сегмент ответствен за терминацию транс-
крипции, полиаденилирование или оба этих про-
цесса, то почему транскрипция не заканчивается
в гомологичных последовательностях, находящихся
на 50 п.н. правее сайта инициации? Эти загадки
напоминают нам о том, что регуляция транскрип-
ции осуществляется на многих уровнях. О неко-
торых из них мы что-то знаем, о других даже не
догадываемся. В одном из способов для регуляции
252
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
OR г A- leu asn asn lys his asp leu his leu arg pro glu thr tyr stop
UUG A AC A AU A AGCACG ACCUU C AUCUUAGGCCAGAA ACUUACUGAA
ORF B- thr Ile ser thr thr phe ile leu gly gin lys leu thr glu
РИС. 10.31.
Транскрипция и трансляция Ту-злемента. Среди тран-
скриптов Ту-злемента в дрожжевых клетках в наиболь-
шем количестве присутствует РНК длиной 5,7 т.н. Этот
транскрипт содержит две перекрывающиеся открытые
рамки считывания в одной цели: ORF А и ORF В.
транскрипции Ту-элементов используется локус ти-
па спаривания (разд. 10.6.6): для диплоидов, гетеро-
зиготных по а и а, характерен низкий по сравнению
с гаплоидами уровень транскрипции Ту-элементов.
Для транскрипции Ту необходимы также продукты
генов SPT.
Трансляция Ту. Установлена нуклеотидная по-
следовательность нескольких Ту-элементов. Та цепь
ДНК, которая отвечает транскрипту длиной 5,7
т.н., имеет две длинные открытые рамки счи-
тывания, при этом З'-конец первой рамки (ORF А)
перекрывается с 5'-концом второй (ORF В) на участ-
ке длиной примерно 40 нуклеотидов (это число
варьирует у разных Ту), а считывание триплетов
в ORF А и ORF В происходит с использованием
разных рамок (рис. 10.31). Несмотря на обилие
Ту-транскриптов, их трансляция в дрожжевых клет-
ках протекает с очень низкой эффективностью. Од-
нако если в клетку вводится плазмидный вектор,
тоже содержащий Ту и обеспечивающий образо-
вание большого числа его копий, то Ту-полипеп-
тиды синтезируются в большом количестве. Рамка
ORF А длиной 1,3 т.н. транслируется с образо-
ванием полипептида мол. массой 55 кДа. Удиви-
тельно, что наряду с этим продуктом на уровне 5%
синтезируется также полипептид мол. массой при-
мерно 190 кДа, транслирующийся с обеих открытых
рамок. Для завершения синтеза этого длинного
полипептида во время трансляции происходит сдвиг
Представлены сами эти рамки и область их перекрыва-
ния. В 5'- и З'-направлениях от этой области ORF
А и ORF В соответственно не прерываются. [R.T. Elder et
al.. Proc. Natl. Acad. Sci. USA 80 (1983), p. 2432; G. Mellor
et al.. Nature 313 (1985), p. 243.)
рамки считывания-как мы уже говорили, рамки не
совпадают. Этот сдвиг осуществляется в том месте,
где рамки перекрываются. Длинный полипептид
подвергается протеолизу с образованием несколь-
ких белков. Один из них, по-видимому, соответст-
вует продукту ORFA. С-концевая часть второго
белка-продукта трансляции ORFB является об-
ратной транкриптазой. Отметим, что альтернатив-
ный механизм образования мРНК, способной
транслироваться в полипептид ORFB, мог бы реа-
лизовываться через сплайсинг первой рамки. Уди-
вительно, что Ту не использует этот широко рас-
пространенный механизм, а прибегает к необыч-
ному способу-сдвигу рамки считывания.
Продукт ORF В Ту-элемента отчасти гомоло-
гичен полипептидам, кодируемым ретровирусными
генами pol (табл. 10.4). У ретровирусов эти гены
кодируют несколько ферментов, в том числе про-
теазу, эндонуклеазу, называемую также интегразой,
и обратную транскриптазу (RT). Если в дрожжевых
клетках Ту-элемент эффективно транскрибируется
и транслируется, то в клеточных экстрактах обна-
руживается активная обратная транскриптаза, а в
частично очищенных фракциях таких экстрактов
при добавлении дезоксинуклеозидтрифосфатов и
Mg2+ синтезируется Ту-ДНК. Внесение мутации
в ORF В приводит к утрате ферментативной актив-
ности, показывая еще раз, что обратная транскрип-
таза кодируется Ту-элементом. Клетки, в которых
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
253
Таблица 10.4. Сходные участки в некоторых ретровирусных и ретротранспозонных полипептидах1’
Ретровирусы Вирус саркомы Рауса Вирус лейкоза мышей Молони V R LPQGM L Р Q G F 24 25 CM L I L L HYMDDLLLAAT
Q Y V D D LLLAAT
Ретротранспозоны класса I
Ту (ORF В) R PPPHL 52 TIC L F V D D M V L F S К
Copia R L Р QG I 58 Y V L L Y V D D V V I A T G
IAP V LPQGM 25 I L L L Y MD D I L L С H К
Ретротранспозоны класса II
F G I Р Q G S 20 TVS T F A D D T A I L S R
1 G V Р Q G S 24 К F N A Y A D D F F L I IN
LINE-1 (человек) G Т R QG С 34 К L S L F A D D M I V Y L E
4 Показаны только два из нескольких сходных участков каждого элемента. Известно, что у ретровирусов они входят в состав генов роК
кодирующих обратную транскриптазу; по-видимому, это относится и к ретротранспозонам. Аминокислоты, выделенные жирным
шрифтом, в большинстве случаев идентичны или очень сходны. Замены сходных в химическом отношении аминокислот (например,
фенилаланина на тирозин или лейцина на изолейцин) называются консервативными Указано число аминокислот, разделяющих два
полипептидных участка.
обнаруживается обратная транскриптаза, содержат
также белок мол. массой 90 кДа, связывающийся
с антителами к синтезированному химическим пу-
тем нонапептиду, структура которого получена ис-
ходя из данных о нуклеотидной последовательности
ORFB. При очистке этот белок выделяется вместе
с обратной транскриптазой. Заметим, что помимо
белка мол. массой 90 кДа в результате посттран-
сляционного протеолиза другого, более высокомо-
лекулярного белка мол. массой 190 кДа, синтези-
руемого со сдвигом рамки считывания, образуются
и несколько других ORFB-белков. Один из них-
протеаза, отвечающая за посттрансляционный про-
цессинг, а другой, вероятно, интеграза, необходимая
для транспозиции.
Появление активной обратной транскриптазы
Ту-элемента в дрожжевых клетках сопровождается
образованием цитоплазматических частиц. Эти ви-
русоподобные частицы диаметром 60 нм содержат
фермент Ту-РНК, а некоторые из них-полнораз-
мерный линейный дуплексный Ту-элемент. Основ-
ной структурный белок этих частиц представлен
полипептидом мол. массой 48 к Да, образующимся
в результате протеолиза продукта ORFA. Таким
образом, рамка ORFA аналогична гену дад ретро-
вирусов. Однако, в отличие от ретровирусов, Ту-
элементы не детерминируют образование инфек-
ционных частиц.
В нормальных дрожжевых клетках ни обратная
транскриптаза, ни Ту-частицы не обнаруживаются,
несмотря на присутствие примерно 35 Ту-элементов
и большого количества полиаденилированной Ту-
РНК. Эксперименты, описанные в предыдущих па-
раграфах, удалось провести благодаря тому, что
были сконструированы специальные плазмиды,
обеспечивающие эффективную транскрипцию и
трансляцию встроенных в них Ту-элементов (такая
плазмида описывается в следующем разделе). Боль-
шинство хромосомных Ту-элементов. по-видимому,
дефектны или репрессированы. Они не обеспечи-
вают образование измеримых количеств обратной
транскриптазы или цитоплазматических частиц и,
в отличие от плазмидных Ту, почти не способны
к транспозиции. Однако их транспозиционная ак-
тивность повышается в тех случаях, когда происхо-
дит эффективная экспрессия функционального Ту-
при наличии мультикопийных плазмид с Ту-эле-
ментами или Ту-элементов, транскрибирующихся
с индуцибельного гетерологичного промотора (см.
ниже).
РНК как промежуточное звено при транспозиции
Ту. Перенос Ту-элемента происходит с помощью
обратной транскрипции РНК-копии Ту по существу
таким же способом, каким синтезируется ретро-
вирусная ДНК на ретровирусной геномной РНК.
Подтверждение этому было получено с помощью
следующих прямых опытов (рис. 10.32).
В дрожжевые клетки his3 были введены сразу
254
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
два плазмидных вектора. Один из них содержал
Ту-элемент и служил донором ретротранспозона,
другой являлся реципиентом и содержал ген ЛмЗ
с делетированным промотором. Многие ревертанты
HIS +, выделенные из такой популяции дрожжей,
содержали Ту-элемент, транспозированный в 5'-
фланкирующий участок мутантного гена ЛйЗ в
плазмиде-мишени.
Донор Ту-элемента для этих экспериментов был
модифицирован: первые 243 п.н. с 5'-конца 8 уда-
Чужеродный
промотор
Укороченный 6
Донорная V
плазмида u
2 экзонаи 1 интрон
из другого дрож-
жевого гена
лены, а Ту-промотор заменен промотором, акти-
вирующимся в присутствии галактозы (промотор
дрожжей GAL1; разд. 8.3.е). В этих условиях тран-
скрипция Ту-элемента начинается с правильного
сайта (пара оснований 245; рис. 10.31) с образо-
ванием полноразмерной Ту-РНК, но скорость про-
цесса необычайно высока. Транспозированные в
плазмиды-мишени Ту-элементы содержат два пол-
норазмерных фланкирующих 8-повтора, хотя у до-
нора Ту отсутствовал 5'-конец 8-сегмента. Из всех
этих наблюдений можно сделать два вывода. Во-
первых, в отличие от ситуации с транспозирующи-
мися элементами, для транспозиции не нужен пол-
норазмерный 5'-8. Во-вторых, укороченный Ту-эле-
мент восстанавливается с образованием полнораз-
мерного Ту, т. е. 3'-8 должен служить матрицей для
реконструкции утраченных частей 5'-8. При этом
Плазмида-
мишень
Промотор
отсутствует
в качестве посредника может использоваться соот-
ветствующая РНК, поскольку в доноре имеется по
меньшей мере одна копия необходимых последо-
вательностей. Этот механизм фактически идентичен
механизму обратной транскрипции ретровирусной
РНК (рис. 2.41).
Участие РНК-посредника в транспозиции Ту-
элемента получило еще более убедительное под-
тверждение в опытах с использованием повторно
модифицированного донора Ту. В его центральный
участок е был встроен интрон из неродственного
дрожжевого гена вместе с короткими участками
фланкирующих его экзонов. Обычно Ту-элементы
не содержат интронов. Не содержал интрона и
Трансфекция
клеток Ызъ~
РИС. 10.32.
Опыты, иллюстрирующие участие РНК в транспозиции
Ту-злемента. В дрожжевые клетки his3' были введены
две плазмиды. Донорная плазмида содержала Ту-зле-
мент, в котором отсутствовали первые 243 пары осно-
ваний с 5'-конца 6-сегмента, а Ту-промотор замещен
Отбор клеток Н15Ъ*,
выделение плазмиды
неродственным промотором, и при этом синтезировал-
ся нормальный Ту-транскрипт (рис. 10.31). Кроме того.
в центральную часть е-сегмента Ту-злемента был
встроен дрожжевой интрон (выделен точками) и при-
мыкающие к нему экзонные последовательности (выде-
лены черным). Плазмида-мишень содержала ген HI S3
без промотора, центромерную область хромосомы
4 (для обеспечения стабильности) и ARS (для репли-
Плазмида-мишень
с встроенным Ту
кации). Из клеточной популяции были выделены коло-
нии /-//S3*, а из них-плазмида-мишень. Ту-злемент
был транспозирован из донора в 5'-фланкирующий
Полноразмерный 6
участок гена his3 и восполнил функции отсутствующего
промотора (ориентация встроенного Ту неизвестна).
Транспозированный Ту-злемент содержит два полных
6-сегмента, интрон удален при сплайсинге. [J. D. Воске
et al., Cell 40 (1985), р. 491.]
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
255
Ту-элемент, транспозированный в плазмиду. Он
был вырезан при сплайсинге, а в плазмиде остались
фланкирующие его экзоны, чужеродные для Ту-эле-
мента. Поскольку такой сплайсинг происходит
только на уровне РНК, стала ясна роль РНК и об-
ратной транскрипции в транспозиции Ту. Наличие
чужеродных экзонов с убедительностью показывает,
что Ту это элемент, транспозированный именно из
донорной плазмиды, а не какой-нибудь другой
Ту-элемент из геномной ДНК. Участие Ту-тран-
скриптов в транспозиции подтверждается также
тем, что мутанты .spz3 не могут транспозировать
хромосомные Ту-элементы, но транспозируют Ту-
элементы плазмидного происхождения, находящиеся
под транскрипционным контролем не-Ту-индуци-
бельного промотора. Недавно опыты по транспо-
зиции были проведены с использованием бесклеточ-
ной системы, которая содержала очищенные Ту-час-
тицы и соответствующую ДНК-мишень. По-види-
мому, линейная дуплексная Ту-ДНК. синтезируемая
с помощью обратной транскриптазы и обнаружен-
ная внутри частиц, служит прямым предшествен-
ником транспозированного элемента. Данные о
том, что и другие ретротранспозоны класса 1 пере-
мещаются подобным образом, будут приведены
в следующем разделе.
Вырезания и другие перестройки, связанные с
Ту-элементом. Как и транспозирующиеся элементы
бактерий. Ту вырезается случайным образом: это
событие происходит примерно в одной клетке из
10s-106 в одном поколении. У мутантов his4 выре-
зание сопровождается по крайней мере частичной
реверсией к фенотипу HIS4+ (рис. 10.30). При этом
в молекуле остается «инсерционный след» в виде
одиночного 8-сегмента, фланкированного такими
же повторами размером 5 п. н., которые выявляют-
ся у мутанта. Скорее всего такое вырезание проис-
ходит при гомологичной реципрокной рекомбина-
ции прямых повторов 8-сегментов, фланкирующих
Ту-элемент (рис. 10.33, Л). Можно также предполо-
жить, что многие одиночные 8-сегменты появились
в результате подобного вырезания полноразмерных
Ту-элементов. Второй продукт вырезания кольце-
вой Ту-элемент с одним 8-сегментом - скорее всего
утрачивается. Подобные реципрокные рекомбина-
ции в принципе могут происходить и между дуп-
лицированными сайтами-мишенями длиной 5 п.н.,
однако в действительности никаких событий, вос-
станавливающих сайты-мишени, не наблюдалось.
Геномные перестройки другого рода возникают
в результате рекомбинаций между диспергирован-
ными Ту-элементами или одиночными 8-сегмента-
ми. В одних случаях мы видим лишь генетические
последствия таких перестроек, другие описаны на
молекулярном уровне. Перестройки с участием двух
любых Ту-элементов встречаются в 1% клеток,
подвергающихся мейозу; в делящихся гаплоидных
клетках рекомбинация между этими же Ту-элемен-
тами происходит лишь с частотой 10 ~ \ Делении
возникают в результате реципрокных рекомбинаций
между Ту-элементами и одиночными 8-сегментами,
находящимися в одинаковой ориентации на одной
и той же хромосоме (рис. 10.33,5); в локусе дикого
типа, представленном на рис. 10.29, они происходят
с частотой 10“В результате реципрокных реком-
бинаций между противоположно ориентированными
Ту-элементами, находящимися на одной и той же
хромосоме (рис. 10.33, В), возникают инверсии; если
же Ту-элементы локализованы в разных хромосо-
мах, то происходят хромосомные транслокации
(рис. 10.33, Г). Если между Ту-элементами, распо-
ложенными в разных локусах гомологичных хро-
мосом диплоидного организма, происходит нерав-
ный кроссинговер, то в одной из хромосом по-
является деления, а в другой происходит дупли-
кация (рис. 10.33, Д). Отметим, что если центромеры
расположены по разные стороны от точки крос-
совера, то в результате перестроек, показанных на
рис. 10.33, Г и Д, образуются дицентрические и
ацентрические хромосомы. Наконец, генная конвер-
сия между двумя Ту-элементами может приводить
к изменениям нуклеотидной последовательности
самого элемента, не затрагивающим фланкирующие
его участки (рис. 10.33. Е).
в. Copia-подобные элементы
дрозофилы
Вся совокупность семейств ретротранспозонов
класса I D.melanogaster была объединена в группу
copia-подобных элементов. Copia-подобные элемен-
ты, ретротранспозоны класса II и транспозиру-
ющиеся элементы типа Р вместе создают харак-
терную для генома D.melanogaster картину чере-
дования повторов (разд. 9.5.а).
Группа copia. Название элементу-прототипу бы-
ло дано исходя из свойств многокопийной РНК,
гибрид и зующейся с неким семейством диспергиро-
ванных повторяющихся последовательностей ДНК.
У D.melanogaster можно обнаружить 20 отдельных
copia-подобных семейств, что составляет 10% всего
генома (по 5-100 членов в каждом семействе). Не-
которые наиболее полно описанные семейства пред-
ставлены на рис. 10.34. Как и Ту-элемент, члены
этих семейств состоят из нескольких тысяч пар
оснований и содержат прямые концевые повторы
длиной несколько сотен пар оснований. У всех
членов одного семейства LTR идентичны, а для
разных семейств они различаются. Концы каждого
LTR представлены короткими инвертированными
друг относительно друга повторами. В отличие от
Ту-элементов, отдельные копии LTR, аналогичные
256
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Б. Реципрокная рекомбинация между двумя
Ту-элементами, находящимися в одной хромо-
соме и имеющими одинаковую ориентацию
A BCD Е
I м® ж в
В. Реципрокная рекомбинация между двумя
Ту-элементами, находящимися в одной хромо-
соме и имеющими противоположную ориентацию
Г. Реципрокная рекомбинация между
Ту-элемеитами, находящимися в
негомологичных хромосомах
BCD
Е
W—я
А ВС
I Ш х И
D ЕF
D "вс
— II III II
/{.Неравный кроссинговер между двумя неаллельными
Ту-элементами, находящимися в гомологичных
хромосомах
£. Генная конверсия
А В В С
t ШЧЖ -1Ш Ш № № Я
Г III 1 I —
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
257
РИС. 10.33.
В результате рекомбинаций между Ту-злементами и
последовательностями S происходят различные хромо-
сомные перестройки. А, В, С, D, Е и F- генетические
маркеры геномной ДНК дрожжей. А. Делеция Ту с со-
хранением одного 5 в результате гомологичной ре-
ципрокной рекомбинации между фланкирующими S. Б.
Делеция в результате гомологичной реципрокной ре-
комбинации между двумя Ту-злементами, расположен-
ными в одной хромосоме и имеющими одинаковую
ориентацию. В. Инверсия в результате гомологичной
реципрокной рекомбинации между двумя Ту-злемен-
тами, находящимися в одной хромосоме и имеющими
противоположную ориентацию. Г. Реципрокная реком-
бинация между двумя Ту-злементами, находящимися
в негомологичных хромосомах. Д. Неравный кроссин-
говер в результате рекомбинации между двумя неал-
лельными Ту-злементвми, находящимися в гомологич-
ных хромосомах. Е. Гомогенизация Ту-злементов. рас-
положенных на негомологичных хромосомах, в резуль-
тате генной конверсии. (См. также рис. 9.47.)
одиночным 8, встречаются редко. Встраивание
copia-элемента в новый геномный локус сопровож-
дается дупликацией сайта-мишени, при этом нук-
леотидная последовательность дуплицированного
участка варьирует. Члены разных copia-подобных
семейств не гибридизуются между собой и не обна-
руживают заметного сходства. Что касается членов
одного семейства, то они все-таки гибридизуются,
но часто характеризуются полиморфизмом, обус-
ловленным дивергенцией последовательностей и де-
лециями. У других представителей рода Drosophila,
например у D.simulans, имеются другие copia-no-
добные семейства. Одни из них обнаруживают ог-
раниченную гомологию с членами соответствующих
семейств D. melanogaster, другие, по-видимому, ви-
доспецифичны.
Транспозиция. Положение ретротранспозонов
одного семейства в геноме у разных линий D. mela-
nogaster неодинаково, о чем убедительно свидетель-
ствуют результаты опытов по гибридизации in situ.
Рис. 10.35 иллюстрирует гибридизацию одного из
членов семейства 412 с гомологичными хромосо-
мами мухи-потомка от скрещивания двух линий
D. melanogaster. В том месте, где политенные хро-
мосомы разделены, четко видны члены семейства
412, имеющие разную локализацию (темные по-
лосы). Об изменчивости положения членов семейст-
ва у разных линий свидетельствуют и результаты
рестрикционного анализа, аналогичного описанно-
му для Ту-элементов. Изменения как локализации,
так и числа копий элементов наблюдаются при
росте клеток Drosophila в культуре: число копий
возрастает до пяти раз по сравнению с таковым
в ДНК самих мух. Частота перемещений отдельных
copia-подобных элементов в культуре клеток может
достигать 10“3-10“4 на одно поколение; это зна-
чительно меньше, чем в ДНК клеток самих мух. На
самом деле постоянство общего числа элементов
в геноме организмов поражает. Не исключено, что
в поддержании этого постоянства участвуют какие-
то активные процессы.
Copia-подобные элементы при встраивании в
гены тоже вызывают мутации. Многие из них из-
учены на молекулярном уровне. Например, пока-
зано, что встраивание copia в локус white приводит
к изменению цвета глаз у мух-они становятся
абрикосовыми (и,а) или белыми. Как мы уже отме-
чали, говоря о встраивании Ту-элементов. полу-
чающиеся мутантные фенотипы могут различаться.
Более того, иногда на инсерционный эффект нала-
гается эффект несцепленных генов Drosophila', воз-
17-158
258
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 10.35.
Гибридизация in situ одного из ретротранспозонов се-
мейства 412 с политенной хромосомой Drosophila- по-
томка от скрещивания мух двух разных линий. Четко
видна разная локализация членов семейства 412 в двух
хроматидах в асинаптической области хромосомы 2
(черные зерна серебра). [С любезного разрешения
G. Rubin, Cold Spring Harbor Symp. Quant Biol. 45 (1980),
p. 620.]
можно, это связано с тем, что последние участвуют
в регуляции функций, кодируемых самим элемен-
том. Мутации, вызываемые copia-подобными эле-
ментами, могут ревертировать при вырезании эле-
мента в процессе гомологичной рекомбинации меж-
ду двумя LTR, причем один LTR и дупликация
сайта-мишени размером 5 п.н. остаются. Однако
в отличие от Ту такие события, по-видимому,
происходят редко, поскольку в геноме обнаружи-
вается очень мало одиночных LTR.
Транскрипция элементов copia. На долю тран-
скриптов, гомологичных некоторым членам семей-
ства copia, в клетках Drosophila приходится 0,5 -3%
суммарной РНК. Соответствующие РНК обнару-
жены как в ядре, так и в цитоплазме; некоторые из
них полиаденилированы. Содержание их зависит от
стадии развития организма и типа транскрибиру-
емого элемента. Например, транскрипты В104 не
выявляются в культуре клеток, но обнаруживаются
в большом количестве во время эмбриогенеза.
Транскрипты copia присутствуют в культивируемых
клетках и на всех стадиях развития, но у личинок
и взрослых особей их больше, чем у зародышей.
Транскрипты 412 появляются у зародышей, а у
взрослых мух их содержание уменьшается.
Транскрипты copia-элементов, количество кото-
рых наиболее велико, сходны с Ту-РНК дрожжей.
Обычно по длине они почти равны целому элементу
(примерно 5 т. п. н.), начинаясь и заканчиваясь в
прямых концевых повторах, и комплементарны
лишь одной цепи ДНК. Транскрипция, по-видимо-
му, идет в одном направлении. В культуре клеток
обнаруживаются также транскрипты меньших раз-
меров (например, copia-PHK длиной 2 т. н.), но вряд
ли их роль велика, поскольку в клетках самих мух
этих РНК содержится очень мало.
Трансляция элементов copia. Большинство copia-
PHK, выделенной из культуры клеток, кэпировано
и транслируется in vitro с образованием нескольких
полипептидов мол. массой 18-51 кДа. Скорее всего
эти полипептиды кодируются мРНК, транскриби-
рованными с левой половины copia, поскольку сег-
менты, клонированные из левой половины, блоки-
руют трансляцию (разд. 6.4.6).
Почти весь участок между LTR у copia-элемента
и у copia-PHK длиной 5 т. н. составляет одну про-
тяженную открытую рамку считывания. У неко-
торых copia-подобных элементов аналогичный
участок включает несколько длинных открытых ра-
мок (такая же картина наблюдается у Ту-элемен-
тов). Генные продукты содержат участки, гомоло-
гичные генным продуктам Ту-элемента и ретро-
вирусов. Это, в частности, участки, обладающие
активностью протеазы, интегразы и обратной тран-
скриптазы (табл. 10.4). По-видимому, copia-подоб-
ные элементы кодируют один или более полипеп-
тидов, которые расщепляются в результате про-
теолиза с образованием типичных белков.
В клетках Drosophila присутствуют рибонуклео-
протеиновые частицы, содержащие длинные тран-
скрипты copia-подобных элементов. Частицы, со-
держащие транскрипты copia длиной 5 т. п. н.,
включают до шести полипептидов и четыре допол-
нительных РНК длиной менее 300 нуклеотидов,
а также обратную транскриптазу. И полипептиды,
синтезированные на copia-PHK in vitro, и полипеп-
тиды самих частиц осаждаются антителами к этим
частицам. Таким образом, copia-PHK кодируют
белки, с которыми эти антитела ассоциируют. Кро-
ме того, copia-PHK, присутствующая в частицах,
как и Ту-РНК, по-видимому, участвует в реплика-
ции и транспозиции copia, происходящей при обрат-
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
259
ной транскрипции. Посредниками в этом процессе
могут служить гибриды РНК-ДНК copia-подобных
ретротранспозонов, которые уже обнаружены.
г. IАР-последовательности у мышей
У различных позвоночных обнаружены элемен-
ты, очень сильно напоминающие ретротранспозоны
класса I. Однако охарактеризовано всего одно такое
семейство- мышиные 1АР-элементы.
А-частицы, содержащиеся в цистернах. Цито-
плазматические рибонуклеопротеиновые комплексы
из клеток мыши (А-частицы), содержащиеся в цис-
тернах и обозначаемые ТАР (intracisternal A-parti-
cles), весьма необычны. Они появляются на очень
ранних стадиях развития зародыша и в различных
раковых клетках, кроме клеток тимуса (и только
в определенных линиях мышей); в нормальных тка-
нях они почти не встречаются. И по своему составу,
и по внешнему виду эти частицы напоминают не-
которые ретровирусы, но они не инфекционны. Эти
ореховидные частицы диаметром около 80 нм от-
почковываются от мембраны эндоплазматического
ретикулума и остаются внутри цистерн.
Очищенные IAP-частицы содержат основной
структурный белок мол. массой 73 кДа, обратную
транскриптазу и несколько специфических РНК,
большинство которых гомологичны друг другу и
имеют длину 7,2 т.н. (35 S). В некоторых клетках
миеломы мышей на долю IAP-PHK приходится
примерно 8% суммарной цитоплазматической по-
лиаденилированной РНК. В бесклеточной системе
эта РНК транслируется с образованием основного
структурного белка мол. массой 73 кДа.
РНК IAP негомологична РНК большинства
мышиных ретровирусов, за исключением РНК двух
видов азиатских мышей, у которых она частично
сходна с геномом эндогенных ретровирусов.
Гены, кодирующие А-частицы. Последователь-
ности, гибридизующиеся с РНК IAP, встречаются во
всех хромосомах Mus musculus в ~ 1000 дисперги-
рованных сайтах; у других видов мышей в хромо-
сомах присутствуют всего 20-25 копий, однако
у сирийских хомячков- около 900 копий. Повто-
ряющиеся единицы имеют длину 7,1 т. п. н., хотя
встречаются и более короткие члены семейства. Как
и все диспергированные повторяющиеся элементы,
отдельные члены семейства отличаются друг от
друга. У них имеется центральный участок, флан-
кированный LTR длиной 338 п.н., каждый из ко-
торых в свою очередь имеет короткие концевые
инвертированные повторы длиной 4 п.н. Таким
образом, ДНК IAP- это типичный ретротранспозон
класса I (рис. 10.27). При его встраивании происхо-
дит дупликация сайта-мишени длиной 6 п. н. 5'-кон-
цы длинных молекул РНК, обнаруженных в IAP,
соответствуют последовательностям левых LTR
(LTR-L), т. е. транскрипция протекает аналогично
транскрипции Ту-элемента и copia.
Транспозиция IAP и их мутагенное действие.
IAP-элементы, перемещаясь в новые геномные сай-
ты, вызывают мутации как в клетках зародышевой
линии, так и в соматических клетках. Например, из
клеток-сверхпродуцентов легких цепей иммуно-
глобулинов были получены мутантные клеточные
линии, вообще не продуцирующие этих цепей, и в
некоторых из них в интроны гена легкой цепи были
встроены IAP-элементы. При этом IAP-элемент мог
находиться в любой ориентации относительно на-
правления транскрипции иммуноглобулинового ге-
на. Известен и еще один пример. Встраивание
IAP-элемента приводит к повышению уровня экс-
прессии соседнего с ним гена-явление, типичное
для многих транспозирующихся элементов. Так,
IAP-элемент, встроенный в 5'-половину кодиру-
ющей области мышиного онкогена c-mos, вызывает
ускорение транскрипции последовательностей c-mos,
расположенных в З'-направлении от вставки, при
этом, как и Ту-элемент, IAP встраивается таким
образом, что транскрибируется в противоположном
направлении от последовательностей c-mos. Анализ
аберрантных транскриптов с использованием нук-
леазы S1 наводит на мысль, что LTR-L содержит
дополнительный контролирующий элемент, кото-
рый ускоряет транскрипцию в направлении от
IAP-элемента. Наличие двунаправленных промото-
ров и энхансера в LTR-L было прямо показано
в опытах с использованием специально сконструи-
рованных векторов.
д. Сравнение с ретровирусами
Структурное сходство. Структурное сходство
ретротранспозонов класса I и интегрированной
двухцепочечной формы ретровирусного генома
(провируса) не вызывает сомнений (рис. 10.36). Оба
они имеют одинаковую структуру: длинные кон-
цевые прямые повторы (LTR), окружающие более
протяженный центральный участок. Несмотря на то
что нуклеотидные последовательности LTR у раз-
ных семейств различаются, все они начинаются
и заканчиваются короткими инвертированными
повторами, так что этими повторами начинаются
и заканчиваются и сами единицы. Более того,
первые два нуклеотида инвертированных повторов
в разных единицах одинаковы: это 5'-TG (рис. 10.37).
В некоторых случаях сходными оказываются и дру-
гие пары оснований. Кроме того, отмечается сход-
ство между U3-, R- и US-областями провирусных
LTR и LTR ретротранспозонов. Сигнальные по-
следовательности, участвующие в регуляции тран-
скрипции с помощью РНК-полимеразы II, в том
17*
260
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
LTR-R
Типичный
встроенный
ретровирус-
ный геном
(провирус)
Геномная РНК
--------pro RT endo__-------
ORF А —______________ ORF С
ORFB
Анк СВЯ1ЫВ110-
щий участок
Ш fry
ORF
pro endo
ORFB
RT
ДНК-свимва»
щийучасток-
РИС. 10.36.
Структура ретровирусного генома,
встроенного в ДНК хозяйской клетки
(провируса), в сравнении со струк-
турой Ту-элемента, copia и IAP. Про-
вирусный участок ро/ кодирует про-
теазу (pro), обратную транскриптазу
(RT) и эндонуклеазу (endo). ORF
ретротранспозонов кодируют поли-
пептиды, которые содержат участки,
гомологичные соответствующим
ретровирусным белкам. Разные ORF
перекрываются.
числе промоторная последовательность ТАТААА
и сигнал полиаденилирования ААТААА, находятся
в указанных LTR в аналогичных положениях. Более
того, многие ретротранспозоны имеют последова-
тельность, сходную с ретровирусной (Р-последова-
тельность; рис. 2.39), которая гибридизуется с
тРНК-праймером (затравкой) во время инициации
обратной транскрипции (рис. 10.37). Как и в ретро-
вирусных геномах, эта P-последовательность нахо-
дится на границе между LTR-L и центральным
сегментом ДНК. Далее, область центрального сег-
мента ретротранспозонов, примыкающая к LTR-R,
часто бывает обогащена пуринами, как и соответст-
вующая область ретровирусного генома (рис. 10.37).
Сходство процессов транскрипции и кодирования.
Основные транскрипты элементов Ту, copia и IAP
также весьма сходны с транскриптами провирусов.
Все те РНК, которые были охарактеризованы, начи-
наются в LTR-L и заканчиваются в LTR-R, так что
транскрипт, во-первых, содержит не менее одной
копии полной последовательности элемента и, во-
вторых, имеет прямые концевые повторы (рис. 10.31).
Как и у ретровирусов, у ретротранспозонов класса
I почти вся центральная часть занята открытыми
рамками считывания. Ближайший к LTR-L коди-
рующий участок детерминирует синтез белка, ко-
торый связывается с нуклеиновой кислотой; этот
участок аналогичен гену gag ретровирусов. Следу-
ющий кодирующий участок, pol, детерминирует
синтез обратной транскриптазы, а также, вероятно,
белки, гомологичные протеазе (pro), интегразе (int)
и РНКазе Н ретровирусов. Ближе к З'-концу геном
ретровируса содержит третий кодирующий участок
(env), который кодирует белок оболочки вириона.
У большинства ретротранспозонов функциональ-
ный участок env, по-видимому, отсутствует; воз-
можно, именно с этим связана их неспособность
продуцировать инфекционные частицы. ДНК IAP,
ю. РЕОРГАНИЗАЦИЯ ГЕНОМА
261
TGGTAGCG... // ...
GGTTATGG... // . ..
TGGCGACC... // .. .
TGGTGCCG... // ...
TGTTG ... ТАТАААА...
TGTTG ТАТААДТ...
TGTA--- ТАТАТТА...
TGT... ТАТААС ...
* СТСА...
* ААСА...
ААТААА ...ТАСА...
ААТААА... ААСА...
GGGTGGTA И Ту
AGGGGGCG // Copia
GAGGGAGA // 412
AGGAGAGA // ТАР
TGTA... ТАТТТАА... А АТА А А... ТТ С А... TGGTGACC... II ... GAGGGGGA // RSV
TGAAAG ... ААТАААА... А АТ А А А.. .ТТС A... TGGGGGCTC... Ц . .. AAGGGGGG // MoMLV
РИС. 10.37.
Общие структурные особенности Ту-злемента, copia,
членов семейства 412, IAP и интегрированных форм
ретровирусов (провирусов). Звездочками отмечены
в отличие от других ретротранспозонов, содержит
участок, достаточно длинный для того, чтобы ока-
заться геном eiw. Однако этот участок не может
быть функциональной кодирующей последователь-
ностью, поскольку содержит множество стоп-кодо-
нов.
У всех исследованных ретровирусов и ретро-
транспозонов класса I дад или г/од-подобные рамки
считывания (ORF А) транслируются нормально, но
затем часто происходит протеолиз с образова-
нием разных продуктов. Copia содержит только
одну рамку считывания, и все белки (например,
дяд-подобный белок и обратная транскриптаза),
по-видимому, представляют собой продукты пост-
трансляционного процессинга. У других ретро-
транспозонов и многих ретровирусов гдиу-подобные
и ро/-рамки считывания перекрываются (рис. 10.36),
но при этом используются разные наборы трипле-
тов, как в случае Ту-элементов. Иногда трансляция,
начинаясь с дяд-подобного участка, продолжается
со сдвигом рамки считывания с образованием слит-
ного белка, содержащего как дя<?-подобную, так и
/^/-последовательности. Из этого белка в резуль-
тате процессинга образуются отдельные белки.
Локализация транскриптов в рибонуклеопротеи-
новых частицах. Аналогия между ретровирусами
и ретротранспозонами распространяется и на упа-
ковку ретротранспозонных транскриптов в рибо-
нуклеопротеиновые частицы. Частицы с РНК copia-
подобных элементов и РНК Ту-элементов обнару-
жены у Drosophila и дрожжей соответственно. Эти
частицы содержат белковые компоненты, кодиру-
емые дяд-подобными и ро/-областями РНК.
Транспозиция ретротранспозонов. Имея в виду
участки, где эквивалентных последовательностей не об
наружено.
то сходство, которое существует между ретровирус-
ным геномом и ретротранспозонами класса I. мож-
но сделать вывод, что последние перемещаются
в геноме с помощью механизма, аналогичного то-
му, который используют ретровирусы при интегра-
ции в клеточную ДНК. Детали этого процесса для
каждого данного семейства ретротранспозонов
окончательно не установлены, но можно предло-
жить следующую общую модель: транскрипт рет-
ротранспозона с помощью обратной транскрипта-
зы, кодируемой самим мобильным элементом,
превращается в дуплексную ДНК, которая затем
встраивается в новый геномный локус. Обратная
транскрипция ретровирусной РНК подробно опи-
сана в разд. 2.2.а и 5.7.г.
В рамках данной модели оба LTR нового элемента
должны образовываться из одной и той же последо-
вательности, поскольку сама РНК содержит лишь
один полный набор LTR-последовательностей. Об-
щее происхождение двух LTR дает ответ на вопрос,
почему повторы, окружающие один ретротранспо-
зон, практически одинаковы, в то время как LTR
у членов разных семейств различаются. Отметим,
что многие, а возможно подавляющее большинство
членов семейства ретротранспозонов, неспособны
к независимой транспозиции. Для них характерны
нуклеотидные замены и другие изменения, затраги-
вающие как регуляторные, так и кодирующие
участки генома, которые могут прерывать тран-
скрипцию и трансляцию и блокировать образование
функциональных генных продуктов. Некоторые
элементы могут приобретать способность к транс-
позиции при наличии /нрянс-действующих обратной
транскриптазы и других активностей.
262
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 10.38.
Две кольцевые формы copia, различающиеся тем, что
первая содержит один LTR, а вторая два.
В результате обратной транскрипции ретрови-
русной РНК образуются три разные дуплексные
ДНК: одна линейная и две кольцевые, различа-
ющиеся тем, что содержат один или два LTR
(рис. 2.40). Немногочисленные кольцевые формы
Ту, IAP и copia-подобных элементов были выделены
и амплифицированы путем клонирования. Напри-
мер. были обнаружены две кольцевые ДНК copia:
длиной 5,0 и 4,7 т.п.н. (рис. 10.38), различающиеся
тем, что содержат одну или две копии LTR; этим же
различаются и кольцевые ретровирусные ДНК. Од-
нако кольцевые Ту-ДНК содержат лишь один LTR.
Возможно, кольцевые формы образуются в резуль-
тате обратной транскрипции РНК таким же спо-
собом, каким образуются ретровирусные кольцевые
молекулы. Но они могут возникать и в результате
обычных процессов рекомбинации с участием толь-
ко ДНК (рис. 10.39). В обоих этих случаях неинте-
грированная кольцевая ретротранспозонная ДНК
может утрачиваться. Как показывают эксперименты
in vitro, линейные дуплексные ДНК и в случае
Ту-элемента, и в случае ретровирусов являются
промежуточными формами при транспозиции.
Последний этап транспозиции встраивание
дуплексной ДНК в геном реципиента, которое у
ретротранспозонов и ретровирусов протекает сход-
ным образом. Согласно моделям, представленным
на рис. 10.40, в транспозиции действительно участ-
вует линейная дуплексная ДНК, причем этапы этого
процесса аналогичны описанным для простого
встраивания транспозирующихся элементов прока-
риот (рис. 10.9). Встраивание начинается с разрыва
четырех фосфодиэфирных связей: двух -у донора
и двух (несимметричных)-у реципиента. Затем
между З'-концами донора и 5'-концами реципиента
образуются новые фосфодиэфирные связи.
Общность свойств ретротранспозонов класса 1 и
ретровирусов наводит на мысль о наличии у них
общего предка. По-видимому, в ходе эволюции
ретровирусы развились в жизнеспособные внекле-
точные формы, а ретротранспозоны-в исключи-
тельно внутриклеточные. Важны оба типа элемен-
тов, поскольку оба участвуют в передаче информа-
ции от РНК к геному, как это делают ретротранспо-
Сайт- Copia в Сайт-
мишень ।------------- геноме --------------1 мишень
Гомологичная
рекомбинация
между дупли-
цированными
сайтами-мише-
нями
Гомологичная
рекомбинация
между концевыми
прямыми повторами
РИС. 10.39.
Схема образования кольцевых copia-
ДНК при вырезании.
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
263
Кольцевой или линейный
геномной ДНК с асимметричными
одноцепочечными разрывами
РИС. 10.40.
Встраивание кольцевой или линейной ретровирусной
или ретротранспозонной дуплексной ДНК в реципиент -
ный геном. Прежде всего в реципиентный геном вносят-
ся асимметричные одноцепочечные разрывы (указаны
стрелками)-по-видимому, с помощью эндонуклеазы,
кодируемой ретровирусом или ретротранспозоном.
Число пар оснований между этими разрывами специ-
фично для данной эндонуклеазы, поскольку оно опре-
деляет характерный для каждого элемента размер дуп-
ликации сайта-мишени. Аналогичным образом расщеп-
ляется ДНК донора. Как правило, в кольцевых ретро-
вирусных дуплексах содержатся дополнительные 4 п.н
между участками U5 и U3 двух соединенных LTR
(выделены более темным цветом). По-видимому, в
линейной ДНК по две из этих пар находятся на концах
молекулы. Одноцепочечные разрывы как в линейной,
так и в кольцевой донорной ДНК происходят так, как это
изображено на рисунке. Аналогичные дополнительные
пары оснований могут содержать и ретротранспозоны.
После соединения З'-концов донора с 5'-концами ДНК-
мишени эти основания удаляются, а бреши заполняются
ДНК-полимеразой и лигируются. [P.O. Brown et al., Cell
49 (1987), p. 347.]
зоны класса II (разд. Ю.З.е). Благодаря этому могут
происходить обновление генома и его перестройка -
основа эволюционного процесса.
е. Ретротранспозоны класса II
Типичные ретротранспозоны класса II не содер-
жат длинных концевых повторов и имеют длину
несколько т.п.н. (рис. 10.41). При их встраивании
происходят дупликации сайта-мишени, но длина
дуплицированного участка для разных элементов
варьирует. На З'-конце одной из цепей ретротранс-
позонов имеется A-богатая последовательность, а
центральная часть цепи содержит открытые рамки
считывания, часть которых кодируют полипептиды,
сходные с консервативными pol- и уод-сегментами
полипептидов, кодируемых ретровирусной ДНК и
ретротранспозонами класса I (табл. 10.4). Итак, не-
264
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
F
(аатааа")2
'О
Тп
G
ААТААА
Ап
Т) Тп
I (ТАА)4-,
J (АТТ)4.7
LINE-1,
человек
ААТААА
А-богатый
Т-богатый
ААТААА
А-богатый
Т-богатый
L1NE-1,
мышь
5
2
6
1
3 4
Т. П.Н.
5' L
J 3'
7
РИС. 10.41.
Структура ретротранспозонов класса II. Длинные откры-
тые рамки считывания выделены цветом; отмечены
участки, обнаруживающие гомологию с последователь-
ностями, кодирующими обратные транскриптазы
(табл. 10.4).
смотря на отсутствие прямых данных, можно ду-
мать, что ретротранспозоны класса II кодируют
собственную обратную транскриптазу и их транспо-
зиция осуществляется в ходе обратной транскрип-
ции РНК-посредника. В отличие от ретровирусов
и ретротранспозонов класса I, о ретротранспозонах
класса II мы знаем пока очень мало.
F- и G-элементы Drosophila. В эухроматине и
хромоцентре генома Drosophila диспергированы
примерно 50 членов F-семейства. Самые длинные
и наиболее распространенные из них имеют размер
4,7 т.п.н., но есть и более короткие элементы-3,4
т. п. н. На З'-конце всех F-элементов имеется по
12-30 остатков аденина (рис. 10.42). Сразу перед
этой A-богатой последовательностью находятся два
перекрывающихся сигнала полиаденилирования,
ААТААА. Короткие F-элементы «урезаны» с 5'-
конца и, таким образом, напоминают последова-
тельности LINE-1 млекопитающих.
В тех F-элементах, которые были секвенирова-
ны, обнаружена по крайней мере одна открытая
рамка считывания (рис. 10.41). Поскольку неясно,
активны или нет эти секвенированные F-элементы,
может оказаться, что открытая рамка (или рамки) в
функциональных элементах имеет другую органи-
зацию. чем в нефункциональных. С этой открытой
рамки считывания должен синтезироваться белок,
частично сходный с обратной транскриптазой
(табл. 10.4). Клеточные транскрипты F-элемента
встречаются очень редко и роль их неясна, а клеточ-
ные полипептиды, кодируемые F-элементом, вооб-
ще не обнаружены.
Первые предположения о мобильности F-эле-
мента основывались на данных о его разной лока-
лизации в геноме мух и о дупликации сайтов-мише-
ней. Об этом свидетельствовали результаты опытов
по гибридизации in situ клонированных F-зондов
с политенными хромосомами D. melanogaster разных
5'-CAGTT6CCGACCA ATGAAGCTTTCGATCGCC—4,7т.п.и.-----СААТА|АATAAA|AGTAAAGT( А)1Д G CAGTTGCCGACC А
5‘ - ATGTTTGAG ATCGCC—41т.п.н---------CAAtA|.AATAAa]aGCAAAGC(A)2e ATGTTTGА -
5‘- AATAATGCGG TCTGC-4,2t.ii.h.-CGAtA |AATAAA|AGCAAAGTAAAAT(A).,,,AATAATGCGG
РИС. 10.42.
Эти три F-элемента гомологичны, но имеют разные
5 -концы. Они фланкированы дуплицированными участ-
ками размером 8-13 п.н. с разными нуклеотидными
последовательностями. В рамку заключены сайты по
лиаденилирования- ААТААА.
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
265
линий. Локализация F-элемента оказывается неоди-
наковой даже у мух в пределах одной линии. Раз-
личия между линиями использовали при клониро-
вании и сравнении свободных и занятых сайтов-
мишеней точно так же, как в случае Р-элементов
(разд. 10.2.6). Эти эксперименты позволили устано-
вить границы полноразмерного F-элемента и пока-
зали, что размер дуплицированного участка для
разных копий F-элемента неодинаков и составляет
от 8 до 13 п.н. Мобильность F-элемента была
прямо продемонстрирована на локусе white Dro-
sophila. White-ivory (глаза цвета слоновой кости) - это
мутация, обусловленная дупликацией сегмента дли-
ной 3 т.п.н. в локусе white (рис. 10.43). У одного из
ревертантов white-ivory этот дуплицированный учас-
ток делетирован, и вместо него встроен Р-элемент.
G-элементы очень напоминают по своей струк-
туре P-элементы (рис. 10.41), но организованы в
геноме D. melanogaster совершенно по-другому. G-
элементы встречаются в тандемных повторах,
встроенных в межгенные спейсеры (IGS) в кластерах
рибосомных генов. По-видимому, эти рДНК лока-
лизованы в хромоцентрах политенных хромосом,
поскольку клонированные G-элементы гибридизу-
ются исключительно с этой областью хромосомы.
В тех G-элементах, которые были секвенирова-
ны, имеется несколько открытых рамок считывания,
на одной из которых синтезируется полипептид,
сходный с обратной транскриптазой. Генные тран-
скрипты или белковые продукты G-элемента, равно
как и F-элемента, не идентифицированы.
1-элемент Drosophila. Вторая система гибридного
дисгенеза, отличная от дисгенеза, обусловленного
P-элементами, называется I-R и обнаружена в опре-
деленных линиях Drosophila. При скрещивании меж-
ду так называемыми самцами-индукторами (I, от
inducer) и реактивными самками (R, от reactive)
получается дисгенное потомство. Как и в случае
с P-элементом, при других типах скрещивания (на-
пример, при скрещивании самцов R с самками I)
потомство оказывается нормальным (на рис. 10.15
показаны соответствующие скрещивания в Р-М-сис-
теме). Фенотипические проявления I-R-дисгенеза
сходны с таковыми при Р-М-дисгенезе.
Механизм I-R-дисгенеза аналогичен механизму
Р-М-дисгенеза. Когда транспозирующийся 1-эле-
мент попадает в цитоплазму яйца мухи R-линии,
происходит его активация. Если 1-элемент попадает
в яйцо мухи 1-линии, то активация по каким-то
причинам репрессируется. В соответствии с этим
некоторые независимые мутации, встречающиеся
в локусе white у I-R-дисгенных мух, оказываются
обусловленными встраиванием в него сегмента дли-
ной 5,4 т. п. н., т. е. 1-элемента. Каждый элемент
фланкирован копией сайта-мишени длиной от 10 до
14 п.н.
По своей структуре 1-элемент сходен с F- и
G-элементами (рис. 10.41); он не содержит ни пря-
мых, ни инвертированных концевых повторов. А-
богатый конец представлен четырьмя-семью тан-
демными повторами последовательности ТАА, но
сигнал полиаденилирования отсутствует. Цепь ДНК
с A-богатым З'-концом содержит две протяженные
открытые рамки считывания (ORF), разделенные
участком длиной 471 п.н.; никаких потенциальных
сайтов сплайсинга не выявлено. ORF, ближайшая
к З'-концу 1-элемента, кодирует полипептид, сход-
ный с обратной транскриптазой (табл. 10.4). Со-
гласно генетическим данным, 1-элементы кодируют
активности, необходимые для транспозиции и ее
Локус white (tv+)
тов к дикому типу (w+) эта дупликация утрачивается,
и вместо нее встраивается F-злемент, окруженный
дуплицированными сайтами-мишенями. [R. Е. Karess,
G. М. Rubin, Cell 30 (1982), р. 63.)
РИС. 10.43.
Данные, свидетельствующие о транспозиции F-злемен-
та. У Drosophila мутация white ivory (w') (глаза цвета
слоновой кости) обусловливается дупликацией участка
длиной 3 т. п. н. в локусе white дикого типа. У ревертан-
266
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
регуляции; возможно, первая из них является об-
ратной транскриптазой. Идентифицированы тран-
скрипты 1-элемента; встречаются они только в яич-
никах Droxophila.
1-элемент, представленный на рис. 10.41, про-
изошел от элемента, который, согласно результатам
генетических экспериментов, способен к транспо-
зиции. Кроме того, 5'-конец этого элемента иден-
тичен 5'-концам некоторых других, уже секвени-
рованных [-элементов. Эти данные позволяют
предположить, что секвенированный [-элемент-са-
модостаточная единица, содержащая все необходи-
мое для транспозиции. Из предположения, что
транспозиция происходит при участии РНК-посред-
ника, который соответствует полноразмерной еди-
нице, следует, что элементы, регулирующие тран-
скрипцию, должны находиться внутри самого I-
элемента. По-видимому, это же справедлио и для
других ретротранспозонов класса II. Действитель-
но, недавние исследования ретротранспозона класса
II Drosophila, названного Jockey, показали, что его
сегмент, расположенный перед первой (5') ORF,
обладает промоторной активностью. Если 13 п.н,
на 5'-конце делетированы, то транскрипция не
происходит. Таким образом, по-видимому, эти эле-
менты используют необычные, внутренние регуля-
торные последовательности для РНК-полимеразы
II.
Семейство LINE-1 млекопитающих. Наиболее
многочисленное семейство длинных диспергирован-
ных повторов млекопитающих, LINE-1 (разд. 9.5.в),
представляет собой семейство ретротранспозонов
класса II, сходных с Jockey и 1-элементами Dro-
sophila (рис. 10.41). О мобильности LINE-1 свиде-
тельствовал тот факт, что их встраивание в геном
сопровождалось появлением мутаций. Например,
у больных гемофилией мальчиков в гене фактора
VIII (белка, участвующего в свертывании крови),
локализованном на Х-хромосоме. имелась вставка
LINE-1, в то время как у матерей эта вставка
отсутствовала.
У многих из 104-105 копий LINE-1 в геномах
млекопитающих укорочен 5'-конец, однако несколь-
ко тысяч копий полноразмерные и составляют в
длину 6-7 т.п.н. в зависимости от вида. Дуплика-
ция сайта-мишени происходит не всегда. Полно-
размерные элементы не содержат ни прямых, ни
инвертированных концевых повторов. Их А-бога-
тые З'-концы могут быть представлены непрерыв-
ной ро!у(А)-последовательностью либо тандемны-
ми повторами ТААА или сходных единиц, как
у 1-элементов. Сравнение LINE-1-элементов млеко-
питающих, относящихся к разным отрядам, пока-
зывает, что все они имеют консервативную цент-
ральную область длиной 5 т. п. н. В одной из цепей
ДНК эта область перекрывается с протяженными
открытыми рамками считывания. В большинстве
клонированных и секвенированных LINE-1 ORF
прерываются стоп-кодонами; наблюдается также
сдвиг рамок, обусловленный небольшими деления-
ми и вставками. Однако, исходя из структуры кон-
сенсусной последовательности, можно сделать вы-
вод, что все LINE-1 произошли в результате дивер-
генции от одного исходного элемента с двумя ORF.
Итак, хотя большинство LINE-1 не содержат функ-
циональных генов, один или более полноразмерных
элементов у каждого вида могут кодировать один
или несколько полипептидов. Более короткая 5'-
ORF (ORF I) менее консервативна, чем длинная
З'-ORF (ORFII); последняя кодирует полипептид,
сходный с обратной транскриптазой (табл. 10.4 и
рис. 10.41). Сегменты, фланкирующие эти две рамки
с 5'- и З'-концов, неодинаковы по длине и нуклео-
тидной последовательности у разных видов млеко-
питающих.
В разных клетках млекопитающих ядерная РНК
представлена большим количеством гетерогенных
по длине молекул, которые гибридизуются с
LINE-1-зондами, причем с обеими цепями. Вероят-
но, многие из этих РНК являются первичными
транскриптами неродственных генов, содержащих
вставки LINE-1 (например, в интронах). В отличие
от этого полиаденилированная РНК, содержащая
полноразмерный элемент LINI-1, была обнаружена
в клетках только тератокарциномы человека. В
большинстве клеток Drosophila присутствуют РНК,
сходные с Jockey. Все эти РНК в принципе могут
быть мРНК или посредниками при транспозиции.
Они негомогенны: кДНК, синтезированные на РНК
человека длиной 6,5 т.н., различаются примерно
по 2% нуклеотидных остатков. Кроме того, многие
из них содержат нуклеотидные замены, прерыва-
ющие две рамки считывания, так что лишь отдель-
ные РНК могут быть функциональными мРНК.
Однако разрыв между двумя ORF, показанный на
рис. 10.41, сохраняется в кДНК человека (а следо-
вательно, и в соответствующих РНК), а также в
геномных LINE-1 у грызунов и человека.
Общая характеристика ретротранспозонов клас-
са II. Если оставить в стороне вопрос об отсутствии
у ретротранспозонов класса II длинных концевых
повторов, то можно сделать вывод, что эти рет-
ротранспозоны во многом сходны с ретровирусами
и ретротранспозонами класса I. Во-первых, это
способность кодировать обратную транскриптазу;
во-вторых, наличие во многих случаях двух ORF
в одной цепи. Аналогичное сходство наблюдается
между дад- (и (/ог-подобными) генами и роАоблас-
тями ретровирусов и ретротранспозонов класса I.
Однако, несмотря на то что ORF11 у всех элементов
класса II сходны в том отношении, что они кодиру-
ют обратную транскриптазу, их ORF I различаются,
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
267
за исключением ORF I у близкородственных видов.
Как и у элементов, содержащих LTR, участки, раз-
деляющие две ORF в разных семействах класса II,
варьируют (рис. 10.41). В Р-, G- и 1-элементах эти
рамки разделены несколькими сотнями пар осно-
ваний, а в L1NE-1 мыши они перекрываются-так
же как в некоторых ретровирусах и в Ту-элементе.
Для трансляции ORFII в ретротранспозонах класса
II либо должен произойти сплайсинг РНК, либо
каким-то образом устраняться или «проскакивать-
ся» разрыв между рамками в процессе трансляции.
Поскольку элементы класса II лишены LTR, их
обратная транскрипция и транспозиция должны
осуществляться с помощью механизмов, отличных
от тех, которые используются ретровирусами и рет-
ротранспозонами класса I. Как это происходит-
пока неизвестно. Возможно, посредниками при
транспозиции служат недавно обнаруженные коль-
цевые F- и LINE-1-ДНК.
10.4. РЕТРОГЕНЫ
Ретрогенами (используется также термин ретро-
позоны) мы называем разнородную группу элемен-
тов, которые, по-видимому, транспозируются с по-
мощью РНК-посредника, хотя механизм транспо-
зиции четко не установлен. К ретрогенам относятся
самые разнообразные последовательности, кото-
рые, однако, обладают тем общим свойством, что
сами не кодируют обратную транскриптазу. Их
общие структурные особенности состоят в следу-
ющем: 1) у них отсутствуют концевые повторы-как
прямые, так и инвертированные; 2) на З'-конце
одной из цепей ДНК имеется A-богатый участок;
3) длина у дуплицированного сайта-мишени варьиру-
ет. Нуклеотидной последовательностью ретрогена
условились считать последовательность цепи с
A-богатым З'-концом. Данные о мобильности рет-
рогенов носят в основном косвенный характер; это
диспергированность членов семейства по различ-
ным геномным локусам, дупликация сайтов-мише-
ней, а также наличие аллелей, различающихся при-
сутствием или отсутствием в них ретрогена.
Транспозиция ретро генов, по-видимому, осу-
ществляется пассивным образом; РНК может под-
вергаться обратной транскрипции с помощью фер-
ментов, поставляемых ретротранспозонами или
ретровирусами. Таким образом, процессированные
псевдогены-это и есть ретрогены, которые могут
быть копиями мРНК (т.е. процессированными
псевдогенами полипептидов) или небольших РНК
типа тРНК или 7S-PHK. На долю всех ретрогенов
приходится примерно 10% общей длины некоторых
геномов, т. е. они вносят существенный вклад в по-
ток информации от РНК к ДНК. Число копий
Таблица 10.5. Примерное число копий некоторых
процессированных псевдогенов
Ген Организм Примерное число копий
всех генов функцио- нальных генов
Ген актина Человек 20-30 6
Ген а-тубулина Крыса 4 1
Ген Р-тубулина Человек 15-20 3
Ген IFN-a » 15 10
Ген U1-PHK » 1100 100
Ген глицеральдегид- Курица 1 1
3-фосфатдегидро- Человек 10-30 1
геназы Мышь > 200 1
отдельных процессированных псевдогенов полипеп-
тидов относительно невелико, а число процессиро-
ванных форм любого специфического гена может
значительно варьировать от вида к виду (табл. 10.5).
Отметим, однако, что процессированные псевдоге-
ны 7SL-PHK, например семейства SINE у приматов
и мышей, встречаются с необычайно высокой часто-
той (так, в ДНК человека присутствует 5-105 копий
Alu-SINE-повторов).
а. Процессированные псевдогены
полипептидов
Структура. Процессированные псевдогены по-
липептидов образуют отдельный класс псевдогенов
(разд. 9.1 .а). Несмотря на имеющиеся различия, все
они больше похожи на ДНК-копии мРНК, чем на
соответствующие им гены (рис. 10.44). Более того,
они располагаются в геномных сайтах, далеко от-
стоящих от локусов соответствующих функцио-
нальных генов, часто даже в других хромосомах.
Любопытно, что процессированные псевдогены по-
липептидов в наибольшем количестве присутствуют
в геноме млекопитающих, реже встречаются у дру-
гих позвоночных и у растений, а у беспозвоночных
они вообще не обнаружены.
Типичный процессированный ген ц-тубулина
крысы начинается там же. где и соответствующая
мРНК, не содержит вставочных последовательнос-
тей, фланкирован последовательностями, очень
сильно отличающимися от последовательностей,
окружающих сам ген, и таким образом лишен сиг-
налов регуляции транскрипции, несет несколько му-
таций и содержит З'-концевую ро!у(А)-последова-
тельность (рис. 9. И и 10.44). Другие процессирован-
ные гены полипептидов обладают лишь некото-
рыми из этих свойств. Например, в процессирован-
ном гене а-глобина мыши, расположенном в хромо-
соме 15, отсутствуют два интрона, характерных для
268
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Сравнение структуры полипептидных генов и их процес-
сированных псевдогенов.
а-глобинового гена, но зато имеются последова-
тельности, расположенные перед сайтом инициации
транскрипции функционального гена, локализован-
ного в хромосоме 11; A-богатый З'-конец отсутст-
вует. Еще у одного вида процессированных генов
левее участка, отвечающего 5'-концу мРНК, нахо-
дится последовательность, не гомологичная после-
довательности, находящейся перед самим геном;
по-видимому, она является частью транспозирован-
ного участка, поскольку находится внутри сегмента,
фланкированного предполагаемыми дупликациями
сайта-мишени (к этому типу относится процесси-
рованный ген металлотионеина, представленный на
рис. 10.44).
Предполагаемые способы образования процессиро-
ванных псевдогенов полипептидов. Структура про-
цессированных генов полипептидов позволяет пред-
положить, что они являются ДНК-копиями соот-
ветствующих мРНК. Согласно одной из моделей, их
образование и транспозиция осуществляются путем
обратной транскрипции РНК с последующим
встраиванием кДНК в геном в месте его асиммет-
ричного разрыва (рис. 10.45). При этом происходит
дупликация с образованием дуплицированного
участка разного размера. Вариабельность отражает
неточность внесения одноцепочечных разрывов;
этим псевдогены отличаются от транспозирующих-
ся элементов и ретротранспозонов класса I, при
встраивании которых размер дуплицированного
участка фиксирован. Эта модель сталкивается с
двумя серьезными проблемами. Первая связана с
тем, что в клетках эмбриона на ранних стадиях
развития и в клетках зародышевой линии обратная
транскриптаза пока не обнаружена, а для того
чтобы в таких клетках происходило наследуемое
встраивание, в них должна осуществляться обрат-
ная транскрипция. Вторая проблема состоит в от-
сутствии подходящего праймера. Неясно, что может
служить праймером при обратной транскрипции
транскриптов, образовавшихся при участии РНК-
полимеразы II, если в клетке отсутствует oligo (dT).
Впрочем, если оставить в стороне вопрос о прай-
мере, то эта модель оказывается весьма привлека-
тельной для объяснения механизма образования
процессированных генов типа гена а-тубулина кры-
сы, представляющих собой почти совершенные ко-
пии мРНК. Чтобы эту модель можно было при-
менять к образованию процессированных генов
а-глобина мыши или металлотионеина, ее необ-
ходимо видоизменить.
В модели, представленной на рис. 10.45, отсутст-
вует ряд важных деталей, так что для разных этапов
процесса можно предложить несколько конкретных
механизмов. Например, синтез второй цепи кДНК
может происходить до встраивания в новый геном-
ный локус или к ДНК в месте разрыва может
присоединиться сама РНК и затем синтезироваться
кДНК in situ. Все эти модели весьма привлека-
тельны, и в их поддержку появляются все новые
данные; правда, можно построить и другие модели,
удовлетворяющие большинству этих данных. Итак,
распределение каких-то последовательностей по
всему геному может проходить по-разному. Вы-
сказывалось предположение, что природными ре-
комбинантными векторами являются ретровирусы.
Ретровирусные геномы действительно внедряются
в клеточные гены и по прошествии одного или
нескольких клеточных циклов могут покидать эти
гены, встраиваясь в новые локусы. За время жизнен-
ного цикла ретровируса его геном поочередно бы-
вает представлен то ДНК, то РНК. При этом из
РНК во время сплайсинга могут удаляться интро-
ны, в результате чего на стадии ДНК образуются
процессированные гены. Эта модель объясняет на-
личие необычных последовательностей в процесси-
рованных генах с 5'-стороны от сайта инициации
транскрипции, а также отсутствие А-богатого
З'-конца. Из нее, в частности, следует, что некото-
рые процессированные гены должны соседствовать
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
269
ДНК-полимераза,
лигаза
Дупликация
сайча-мишени
РИС. 10.45.
Одна из моделей транспозиции ретрогенов при участии
РНК-посредника. Для примера рассмотрено образова-
ние процессированного полипептидного псевдогена.
В результате транскрипции и процессинга, включающе-
го сплайсинг интронов, образуется функциональная
мРНК, на которой с помощью обратной транскриптазы
синтезируется одна цепь кДНК; РНК-матрица затем
расщепляется, например, РНКазой Н. Одноцепочечная
ДНК пришивается через З'-гидрОксильный конец к
5'-концу геномной ДНК в месте несимметричного раз-
реза и с помощью ДНК-полимеразы синтезируется
вторая цепь. Альтернативный способ состоит в синтезе
двухцепочечной кДНК еще до встраивания или в при-
соединении цепи РНК в месте разрыва ДНК и синтезе
обеих цепей кДНК in situ. В результате этих реакций
происходит дупликация сайта-мишени. Отметим, что
такая транспозиция консервативна, поскольку исходный
ген остается на месте.
в геноме с ретровирусными последовательностями.
Согласно другой модели, происходит упаковка кле-
точных мРНК в ретровирусные или ретротранспо-
зонные частицы с последующими обратной транс-
крипцией и инсерцией. Данные об этом получены
при исследовании модельных систем, хотя соответ-
ствующие события происходят с весьма низкой
частотой. Совершенно иной способ диспергирова-
ния геномных последовательностей вообще не пре-
дусматривает участия РНК, а предполагает прямую
транспозицию последовательностей ДНК. Напри-
мер, какой-то сегмент, содержащий тот или иной
ген, может дуплицироваться в виде тандема с по-
следующей прямой транспозицией одной из копий
в отдаленный локус генома. В транспозиции могут
участвовать соседние транспозирующиеся элементы
или инвертированные повторы. Этот механизм
объясняет происхождение диспергированных псев-
догенов, в которых в отличие от всех рассмотрен-
ных до сих пор примеров сохраняются интроны;
такова структура псевдогена мышиного а-глобина
из хромосомы 17.
б. Процессированные псевдогены РНК
Структура. В семейства генов малых ядерных
РНК-мяРНК (разд. 9.2.д)-входят различные псев-
догены. которые отличаются от функциональных
РНК и соответствующих генов одной или несколь-
кими диспергированными парами оснований. Не-
которые из них, по-видимому, являются процесси-
рованными псевдогенами, поскольку они диспер-
гированы по геному, а фланкирующие их последо-
вательности негомологичны последовательностям,
окружающим сами гены. Как и процессированные
полипептидные гены, процессированные гены мяРНК
270
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
отличаются по структуре от соответствующих ис-
тинных генов в разных отношениях (рис. 10.46).
Часто они бывают укороченными с конца, соответ-
ствующего З'-концу РНК. У всех процессированных
генов мяРНК 5'-концы в точности совпадают с
началом РНК. В некоторых случаях процессирован-
ные гены содержат A-богатые З'-концевые последо-
вательности и короткие прямые повторы, что сви-
детельствует о дупликации сайта-мишени.
Интересные различия существуют между про-
цессированными псевдогенами мяРНК и мцРНК
Семейство U1 Кодиру(Ощий
участок
Гены и
Укорочен
I
Процессированные
псевдогеиы
™па ---f—НЧ—8
Семейство U2
Укорочен с 3-конца на разное
число нуклеотидов
Процессированные ________________уг______f у И
леев доге и ы (л Q
Семейство из
Гены U3
Тип с
Процессированные
псевдогены
Укорочен с З'-конца на
69 или 70 нуклеотидов
f
РИС. 10.46. $ £ 8
Структура процессированных псевдогенов малых ядер-
ных РНК млекопитающих. В процессированных псевдо-
генах U1 типа а отсутствуют последовательности, находя-
щиеся на З'-конце функциональных генов U1 и самой
U1-PHK. С укороченным З'-концом соседствует корот-
кий A-богатый участок, хотя соответствующая U1-PHK
не заканчивается таким участком. Процессированные
псевдогены U1 типа b не укорочены, заканчиваются
A-богатым участком и фланкированы прямыми повто-
рами длиной 20 п. н. или менее. Процессированные
псевдогены U2 тоже укорочены с З'-конца и фланки-
рованы прямыми повторами длиной около 20 п. н.
Процессированные псевдогены U3 укорочены с З'-конца
на 69-70 нуклеотидов; длина РНК составляет 210 нук-
леотидов. Некоторые из них фланкированы прямыми
повторами длиной 16-20 п.н. (тип с), у других такие
повторы отсутствуют (тип d).
(малых цитоплазматических РНК). Во-первых, про-
цессированные псевдо гены мцРНК относятся к чис-
лу наиболее высококопийных диспергированных
повторов в геномах млекопитающих. Они вклю-
чают процессированные варианты тРНК-генов, а
также Alu-SINE приматов и SINE типа [ мыши (оба
они являются процессированными вариантами ге-
нов 7SL-PHK). Во-вторых, процессированные псев-
догены мцРНК сохраняют типичные внутренние
промоторы для полимеразы III, имеющиеся в функ-
циональных генах, и нормально транскрибируют-
ся-по крайней мере in vitro (большинство генов
мяРНК относятся к генам класса II с внешними
промоторами). Наконец, структура процессирован-
ных псевдогенов мцРНК заметно отличается от
структуры функциональных генов и РНК и варьи-
рует от одного отряда млекопитающих к другому
(разд. 9.5.в). Например, в случае Alu-SINE деле-
тируется центральная часть 7SL-PHK, соединяются
два внешних сегмента и полученный сегмент длиной
~ 130 п.н. образует несовершенный тандемный
повтор. В случае SINE типа I у грызунов цент-
ральная часть 7SL-PHK также отсутствует, но дуп-
лицируется с образованием тандемного повтора
лишь малая часть оставшегося сегмента. Несмотря
на все эти различия, диспергированность данных
сегментов и их связь с дупликациями сайта-мишени
указывает на то, что они относятся к процесси-
рованным псевдогенам.
Предлагаемые способы образования процессиро-
ванных псевдогенов РНК. В основе всех моделей
образования и транспозиции процессированных
псевдогенов РНК лежит схема, представленная на
рис. 10.45. Для того чтобы учесть специфические
свойства псевдогенов мяРНК-наличие ро1у(А)-кон-
цов (в самих РНК они отсутствуют), укороченность
З'-концов, иногда наличие прямых фланкирующих
повторов (рис. 10.46)-эту схему необходимо моди-
фицировать. Кроме того, как и в случае с процес-
сированными генами полипептидов, для синтеза
первой цепи кДНК необходим праймер. В некото-
рых случаях праймер обеспечивает сама РНК. Под-
тверждение этому было получено в опытах in vitro
с использованием U3-PHK и обратной ретровирус-
ной транскриптазы (рис. 10.47). Термодинамически
выгодной является такая вторичная структура
U3-PHK, когда между З'-половиной молекулы и
внутренней областью, начиная с 70-го остатка, обра-
зуется дуплекс, а тем самым и внутримолекулярная
матрица-праймер. Заметим, что данная модель объ-
ясняет также укороченность З'-конца псевдогена U3.
Поскольку мяРНК синтезируется с помощью
РНК-полимеразы III, эта модель оказывается впол-
не приемлемой в случае таких последовательностей,
как Alu и SINE типа I. Начало транскриптов таких
SINE-последовательностей совпадает с началом
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
271
РИС. 10.47.
U3-PHK сама образует праймер, необходимый для
функционирования обратной транскриптазы. З'-конец
молекулы РНК гибридизуется с комплементарной ему
внутренней частью последовательности. Синтез первой
цепи кДНК осуществляется путем присоединения dNTP
Гибрид U3 - мя РНК - кДНК
к З'-концевой гидроксильной группе (остаток 213).
кДНК комплементарна остаткам с 1 по 74-участку,
соответствующему процессированным псевдогенам U3
типов end (рис. 10.46). [L. В. Bernstein, S. М. Mount,
А. М. Werner, Cell 32 (1983), р. 461.]
элемента. Заканчиваются они в первом Т-богатом
участке, который узнает полимераза, обычно на
некотором расстоянии за концом SINE, так что
транскрипты содержат дополнительный по сравне-
нию с самим элементом участок, расположенный
между A-богатой последовательностью и U-бога-
тым концом транскрипта. Этот U-богатый З'-конец
может изгибаться и спариваться с A-богатым участ-
ком с образованием праймера, необходимого для
обратной транскрипции. Транспозиция завершается
гидролизом РНК-матрицы и встраиванием кДНК
в геном.
Новый элемент содержит последовательность,
отвечающую всей РНК, включая часть исходного
A-богатого сегмента, и не несет никаких дополни-
тельных последовательностей. Поскольку ново-
транспозированный ретроген содержит также внут-
ренние промоторы родительского гена, он может
служить родоначальником нового семейства. Этот
«автокаталитический» эффект можно объяснить вы-
сокой распространенностью AIu-элементов. Заме-
тим, что все эти рассуждения касаются лишь меха-
низма амплификации и транспозиции SINE-после-
довательностей, но не происхождения самих этих
элементов. Возможно, одна или несколько из мно-
жества геномных SINE возникли непосредственно
из генов 7SL-PHK или тРНК с помощью обратной
транскрипции (рис. 10.45). Но в таком случае их
структура должна была бы претерпеть существен-
ные изменения, в частности присоединение А-бога-
того участка с З'-конца, отсутствующего в РНК,
а в случае 7SL-PHK- делению центральной области,
а также тандемные дупликации (рис. 9.40).
10.5. ДРУГИЕ НЕОБЫЧНЫЕ
МОБИЛЬНЫЕ ЭЛЕМЕНТЫ
Большинство мобильных элементов эукариот
можно в первом приближении разбить на транспо-
зирующиеся элементы, ретротранспозоны и ретро-
гены. Однако есть и другие мобильные элементы,
обладающие уникальной структурой и необычными
свойствами. Большинство таких элементов, описан-
ных к настоящему времени, обнаружены у беспоз-
воночных, но, по-видимому, в будущем они будут
выявлены у всех эукариот. В наши задачи не входит
детальное описание каждого из этих элементов. Мы
лишь кратко остановимся на некоторых из них,
чтобы проиллюстрировать их разнообразие.
а. Fold-back-элементы дрозофилы
FB-элементы (от fold-back) Drosophila, а также
подобные им элементы Xenopus имеют длинные
концевые повторы, находящиеся друг относительно
друга в противоположной ориентации (рис. 10.48).
Для FB-элементов характерны еще две отличитель-
ные особенности. Во-первых, центральный участок
у разных FB-элементов имеет неодинаковый размер
и нуклеотидную последовательность. Во-вторых,
длина инвертированных концевых повторов также
варьирует, а прямые тандемные повторы имеют
сложную структуру. Для последних типичны деле-
ции (или вставки), возникающие в результате нерав-
ного кроссинговера. Два концевых повтора у дан-
ного члена одного семейства более сходны по длине
и нуклеотидной последовательности, чем повторы
у членов разных семейств, но все же не идентичны.
Однако концы инвертированных повторов высоко-
консервативны даже для разных FB-элементов.
Встраивание FB-элементов Drosophila вызывает
мутации, а их вырезание может сопровождаться
реверсией к дикому типу. FB-элементы перемеща-
ются не только как независимые единицы, но и
вместе с длинными сегментами генома Drosophila.
Члены одного из классов протяженных транспози-
рующихся элементов, называемых ТЕ, представля-
ют собой два FB-элемента, окружающих длинную
272
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
IR С 1R
Юп.н. I—Q 5'-CGTTTGCCCA
и Консенсусная
20П.Н. J5’-CGTTTGCCCACCCTTTAAAA последовательность
31п,н ЯВ 5-CGTTTGCCCACCCTTTAAAATTAAATTATTT
РИС. 10.48.
Схематическое представление мобильного
FB-злемента Drosophila. Оба концевых ин-
вертированных повтора (IR) и центральная
область (С) варьируют подлине: IR состав-
ляет 400-3400 п. н.. С-<100-4000 п.н.
Различаются и нуклеотидные последова-
тельности С. Диаграмма внизу иллюстри-
рует сложную структуру тандемных повто-
ров, типичных для IR.
неродственную последовательность. FB-элементы
были обнаружены на концах транспозированного
сегмента ДНК длиной несколько сотен т. п. н., кото-
рый переместился с Х-хромосомы в хромосому 2,
а оттуда-во множество отдаленных сайтов. Эта
транспозиция сопровождалась разнообразными му-
тациями и перестройками как внутри ТЕ, так и в
соседних с ними генах. По всем этим свойствам ТЕ
напоминают сложные транспозоны прокариот.
б. Вставки в рДНК Drosophila
У некоторых видов Drosophila очень большая
доля сегментов, кодирующих 28S-pPHK, прерывает-
ся последовательностями, отличными от интронов.
Эти прерывающиеся рДНК, по-видимому, нефунк-
циональны и транскрибируются с очень низкой
эффективностью, а функция самих вставок неиз-
вестна. Один из типов таких инсерционных после-
довательностей, тип I, встречается в виде тандем-
ных повторов в хромоцентре, а также в нескольких
диспергированных эухроматиновых областях и в
кодирующей части гена 28S-pPHK. Из нескольких
сотен членов семейства инсерционных последова-
тельностей типа I Drosophila почти половина пред-
ставлена вставками в специфические сайты 288-рДНК
в Х-хромосоме. Длина этих вставок составляет в
основном 5 т. п. н., однако есть и вставки длиной
0,55-1 т. п. н., а те, которые локализованы в хромо-
центре, даже более гетерогенны. Данные о мобиль-
ности единиц типа I не носят систематического
характера: не обнаружено ни типичных структурных
свойств, присущих мобильным элементам, ни экс-
периментальных доказательств их транспозиции.
Нуклеотидные последовательности, непосредствен-
но окружающие элементы типа I, как правило,
сходны, даже если эти элементы находятся в разных
геномных локусах. Таким образом, их встраивание,
по-видимому, происходит в какие-то предпочти-
тельные участки и сопровождается сайт-специфи-
ческой рекомбинацией. Этим можно объяснить,
почему вставки инсерционных последовательностей
типа I всегда обнаруживаются в одном специфи-
ческом сайте рДНК.
10.6. ЗАПРОГРАММИРОВАННЫЕ
ПЕРЕСТРОЙКИ И МОДУЛЯЦИЯ
ЭКСПРЕССИИ ГЕНОВ
Запрограммированные перестройки генов, влия-
ющие на их экспрессию, выявлены у прокариот,
дрожжей, тетрахимены, трипаносом и млекопита-
ющих. Такое разнообразие позволяет предполо-
жить, что гены, экспрессия которых изменяется
в результате аналогичных перестроек, имеются у
многих организмов. Здесь мы остановимся на не-
которых сайт-специфических перестройках у про-
кариот, опосредующих регуляцию экспрессии генов
при помощи инвертирования сегментов ДНК. Это
позволит нам ввести некоторые общие понятия,
необходимые для обсуждения соответствующих
процессов в эукариотических системах.
а. Прокариотические модели-
транслокация путем
флип-флоп-инверсий
Изменение экспрессии двух генов, кодирующих
флагеллин у Salmonella. Флагеллин - основной белок
жгутиков Salmonella- представлен двумя иммуноло-
гически различающимися формами, Н1 и Н2; они
кодируются двумя неаллельными генами, родствен-
ными, но не сцепленными друг с другом в хромосо-
ме Salmonella. В любой момент времени экспресси-
руется только один из этих генов; частота переклю-
чений с Н1 на Н2 и обратно у разных штаммов
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
273
Salmonella составляет 10 ~5-10“3 на бактерию в
одном поколении. По-видимому, такая пластич-
ность обеспечивает Salmonella некоторую защиту от
иммунного ответа хозяина на инфекцию.
В результате сложных генетических эксперимен-
тов была построена модель, объясняющая меха-
низм переключения Н1 «->Н2,-так называемой фа-
зовой вариации. Согласно этой модели, ген Н1 пред-
ставляет собой пассивный элемент, а ген Н2 и тесно
сцепленные с ним генетические детерминанты-это
регуляторные факторы. При экспрессии Н2 экспрес-
сируется и соседний с ним ген гЛ1, который коди-
рует репрессор для Н1; при этом белок Н1 не
образуется и флагеллин относится к типу Н2. При
переключении прекращается экспрессия как Н2, так
и гЛ1, и синтезируется флагеллин типа Н1. Как мы
увидим ниже, эта модель полностью согласуется
с результатами проведенных ранее молекулярных
исследований.
Гены Н2 и rh\ представляют собой часть оперо-
на, который находится под контролем одного про-
мотора, расположенного на 100 п.н. левее гена Н2
(рис. 10.49). Этот промотор находится внутри сег-
мента длиной 1 т. п. н„ который может быть инвер-
тирован. В одном положении промотор активен
и гены Н2 и rh\ экспрессируются. При инверсии
промотор удаляется от начала гена Н2 и меняет
ориентацию, при этом гены Н2 и rh I выключаются
и начинается экспрессия HI. Если инвертированный
сегмент длиной 1 т. п. н. принимает исходную ори-
ентацию, то Н2 и rhl вновь экспрессируется, a HJ
выключается.
Исходя из результатов генетического анализа,
можно предположить, что этот инвертируемый сег-
мент содержит кроме промотора три необходимых
для инверсии элемента-два сй-сегмента на каждом
из концов (AixL и hixR) и некий ген, называемый hin
(от H-inversion), продукт которого действует в
транс-положении. И вновь эти генетические данные
получили подтверждение на молекулярном уровне.
Показано, что инвертируемый сегмент длиной 1 т. п. н.
содержит последовательности, выполняющие опре-
деленные генетические функции. Во-первых, это
последовательности hixL и hixR длиной 26 п.н.,
расположенные по концам 1 т. п. н.-сегмента; каждая
из них содержит копию сегмента длиной 14 п. н. (эти
Н2-оперон включен
Синтезируется
флагеллин Н2
Н2-оперон выключен
Синтезируется
флагеллин Н1
Инверсия
Белок Hin
Фактор II
Белок ни
Н1-репрессор
ГЙ1
Последовательность
hixR
РИС. 10.49.
Инвертируемый участок, контролирующий фазовую ва-
риацию при экспрессии флагеллиновых генов Salmo-
nella. В ситуации, представленной в верхней части ри-
сунка, этот сегмент транскрибируется в том же направ-
лении, что и гены Н2 и rh1. В нижней части рисунка
сегмент инвертирован, а гены Н2 и rh1 молчат. Внутри
инвертируемого сегмента находится ген hin; его продукт
необходим для инверсии и должен быть в транс-попо-
Последовательность 5'
hix L v
CCAAAAGGAAAA Q3'
GGTTTTСС11ТТ Q5’
жении. Элементы Лй!_ и hixR тоже необходимы для
инверсии и должны находиться в щ/с-положении (их
нуклеотидные последовательности представлены внизу);
инверсию стимулирует щ/с-знхансер (в том случае,
если он имеется вообще). Последовательности элемен-
тов hix представлены для того случая, когда они нахо-
дятся в прямой ориентации. Участки длиной 14 п.н.,
присутствующие в hixL и hixR, выделены цветом.
274
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
копии находятся во взаимно противоположной
ориентации). Сами последовательности длиной 26
п. н. представляют собой несовершенные палиндро-
мы. Во-вторых, это ген hin, кодирующий полипеп-
тид из 190 аминокислотных остатков. Эксперимен-
ты in vitro с использованием очищенного белка Hin
и рекомбинантных плазмид, содержащих два hix-
сайта (но не остальную часть 1 т. п. н.-сегмента),
показывают, что белок Hin специфически связы-
вается с /n'x-сайтами. И в-третьих, для эффективной
инверсии необходимо, чтобы последовательность
внутри кодирующего участка гена hin находилась
в «/нс-положении. Этот элемент остается активным,
даже если его переместить за пределы инверти-
руемого сегмента ДНК или инвертировать. Таким
образом, он обладает свойствами энхансера транс-
крипции. Как показали эксперименты in vitro, для
инверсии необходимы еще два белка. Один из них-
неболыпой основный белок, названный фактором II
(или Fis),- связывается с энхансером. Другой пред-
ставляет собой прокариотический гистоноподобный
белок HU. Фактор И, НО и белок Hin стабили-
зируют ДНК в конфигурации, способствующей пе-
рестройкам.
Установлены нуклеотидные последовательности
самого инвервертируемого элемента длиной 1 т. п. н.
и фланкирующих его последовательностей. При
этом последовательности на концах элемента были
определены независимо для обеих ориентаций. По-
ложение промотора для гена hin точно не установ-
лено, но скорее всего ген начинается в пределах
концевого повтора; стартовый кодон ATG гена hin
находится в пределах 100 п.н. от начала повтора.
Отметим, что благодаря сходству двух концов эле-
мента промотор hin при инверсии не разрушается.
Таким образом, ген hin может экспрессироваться
в любой ориентации. С помощью секвенирования
установлено положение промотора оперона Н2:
внутри сегмента длиной 1 т. п. н., ближе к одному из
его концов. Сама кодирующая часть Н2 начинается
на расстоянии 16 п.н. от инвертируемого сегмен-
та.
Контроль специфичности по отношению к хозяину
с помощью инверсии в геноме фага Ми. Фаг Ми тоже
способен изменять экспрессию генов путем инверсии
с помощью механизма, сходного с таковым у Salmo-
nella. В геноме фага Ми размером 37 т. п. н. имеется
так называемый сегмент G длиной 3 т. п. н., способ-
ный к инверсии (рис. 10.7 и 10.50). Его фланкируют
инвертированные повторы длиной 34 п.н., , причем
26 п. н. из этих 34, аналогично повторам, флан-
кирующим соответствующий сегмент у Salmonella,
необходимы для «флип-флопа». Однако нужный для
инверсии функциональный ген gin лежит за рамками
инвертируемого сегмента, а не внутри его, как ген
hin у Salmonella. В пределах кодирующей части gin
находится рекомбинационный энхансер, аналогич-
ный энхансеру у Salmonella, а инверсия in vitro
стимулируется дополнительными клеточными бел-
ками. Инверсия сегмента G происходит с частотой
примерно 1 на 10б фаговых частиц за один лити-
ческий цикл и сопровождается изменением специ-
фичности фага Ми по отношению к хозяину. При
ориентации инвертируемого сегмента G (+) фаг ин-
фицирует Е.соН К12. Если G находится в противо-
положной ориентации, G( —), то Ми инфицирует
штамм Е. coli С, а также некоторые другие бактерии.
Сам сегмент G кодирует белки, входящие в состав
фибрилл хвостового отростка фага, и в зависимости
от его ориентации синтезируются разные белки.
Таким образом, смена круга хозяев, по-видимому,
определяется изменением способности фага при-
крепляться к реципиентным клеткам.
Сегмент G содержит кодирующие последова-
тельности для двух альтернативных наборов бел-
ков, входящих в состав фибрилл; эти последова-
тельности находятся в противоположных ориента-
циях- S и S' (рис. 10.50). В положении G(+) экспрес-
сируется S-оперон. а в положении G( — )-оперон S'.
Промотор для этих двух оперонов находится за
пределами G, в области, называемой Sc; за преде-
лами G находятся и последовательности, кодиру-
ющие 5'-концы мРНК и МН2-концевые последова-
тельности первого белка, кодируемого каждым из
оперонов. Этот ЫН2-конец одинаков у обоих белков
S и S' и кодируется Sc-областью; С-концевые части
белков различаются. Поскольку промотор S нахо-
дится за пределами G, ориентация последнего не
влияет на регуляцию экспрессии S, но зато влияет на
тип синтезируемого белка.
Сходство Hin и Gin. На удивление сходные ин-
версионные системы Salmonella и Ми, вероятнее
всего, представляют собой две адаптации одной
предковой генетической программы, поскольку белки
Hin и Gin по своей структуре очень близки и могут
замещать друг друга. Клетки Salmonella hin~, лизо-
генные по фагу Ми, подвергаются фазовой вариа-
ции, а функциональный ген hin опосредует инверсию
G-сегмента в фаге gin~. Поскольку гены Hin и Gin
действуют в тесной связи с инвертированными пов-
торами, фланкирующими инвертируемый сегмент,
неудивительно, что соседствующие с G-сегментом
фага Ми повторы частично гомологичны инверти-
рованным повторам Salmonella. В данное семейство
белков входят и другие члены. Например, в геноме
некоторых штаммов Е. coli содержится инвертиру-
емый сегмент, ассоциированный с фланкирующими
повторами и с белком, называемым Pin, который
восполняет недостающие функции фага Mu gin~.
Аминокислотные последовательности белков Hin,
Gin и Pin совпадают более чем на 60%. Кроме того,
по своей структуре они сходны с резолвазой ТпЗ.
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
275
G-сегмент
IB IR
34п.н. 34п.н.
G« -------------------------------
Sc S S gin
H-► I-------► -----1 |---
Транскрипция
Инверсия
t—^<4^™
s gin
РИС. 10.50.
Карта G-сегмента в геноме фага Mu и
фланкирующих его участков в положениях
G( + ) и G(-).
Механизм сайт-специфических флип-флоп-инвер-
сий. Механизм флип-флоп-инверсий удалось деталь-
но исследовать благодаря тому, что 1) с использо-
ванием очищенных компонентов можно осуществ-
лять рекомбинацию in vitro; 2) метод рекомбинант-
ных ДНК позволяет получать необходимые суб-
страты, в частности содержащие сайт-специфиче-
ские мутации. Сегменты ДНК, фланкированные
двумя инвертированными повторами (Л/х-элемен-
тами или гомологичными инвертированными пов-
торами G-сегмента фага Ми) и входящие в состав
сверхспиральной плазмиды, инвертируются в при-
сутствии очищенных белков Hin и Gin. Расстояние
между повторами варьирует от нескольких сотен до
тысяч пар оснований. Hin и Gin представляют собой
рекомбиназы и одновременно ДНК-связывающие
белки, которые узнают инвертированные повторы,
фланкирующие инвертируемый сегмент. Повторы
как раз и являются теми сайтами, в которых проис-
ходят перестройки ДНК, приводящие к инверсии.
Этот процесс стимулируется фактором II и белком
HU, а также энхансером в t/wc-положении, с кото-
рым специфически связывается фактор II.
По-видимому, при инверсии образуется сложный
комплекс, состоящий из инвертируемого сегмента
ДНК и рекомбиназы (Hin или Gin), расположенных
друг относительно друга в нужном положении.
Формирование комплекса облегчает фактор II, свя-
зывающийся с энхансером, а также белок HU; есть
данные о том, что последний обеспечивает изгиба-
ние ДНК, необходимое для сближения двух реком-
бинационных сайтов. Для инверсии нужно, чтобы
в одной молекуле ДНК присутствовали два реком-
бинационных сайта, находящихся в противополож-
ной ориентации. В отличие от этого резолваза ТпЗ
предпочитает, чтобы эти сайты имели одинаковую
ориентацию. Однако в обоих случаях в рекомбина-
ции участвуют четыре цепи ДНК, происходит раз-
рыв четырех фосфодиэфирных связей и четыре но-
вых соединения (рис. 10.51).
б. Типы спаривания
у дрожжей - кассетный механизм
Скрещивание гаплоидных клеток с разным типом
спаривания. Почкующиеся клетки S. cerevisiae могут
быть как гаплоидными, так и диплоидными. Ди-
плоиды гетерозиготны по локусу, называемому
локусом, определяющим тип спаривания (МЛ Г-от
англ, mating type locus); два кодоминантных аллеля
дикого типа называются МАТа и МАТа. Таким
образом, гаплоидные клетки являются либо МА Та,
либо МАТа. Для успешной конъюгации двух га-
плоидных клеток и образования диплоида необ-
ходимо, чтобы эти клетки имели противоположные
типы спаривания: одна МАТа, а другая МЛ Та.
Диплоидные клетки МАТа/МАТа неспособны спа-
риваться путем конъюгации, но конъюгацию можно
индуцировать голоданием, в результате чего клетки
подвергаются мейозу и споруляции с образованием
четырех дочерних гаплоидных клеток, составля-
ющих тетраду (рис. 10.52 и разд. 9.2.а).
Одни штаммы S. cerevisiae, называемые гетеро-
талличными, продуцируют гаплоиды со стабильным
типом спаривания, другие попадают в группу гомо-
таллнчных штаммов, гаплоиды которых нестабиль-
ны по локусу МАТ. В гомоталличных гаплоидах
аллели МАТ легко переключаются с МАТа на
МЛ Та Клетки МЛ Та дают потомство МАТа с
частотой более 80% в каждом делении (почкова-
нии); переключение с МАТа на МАТа происходит
только в тех клетках, которые почковались нс менее
одного раза. В результате нескольких клеточных
делений одна гомоталличная клетка образует попу-
ляцию клеток, способных к конъюгации и образова-
нию диплоидов (скрещивающиеся таллотипы гомо-
талличные). Однако клон гетероталличных дрожже-
вых клеток должен вступить в контакт с популяцией
клеток противоположного типа спаривания, иначе
клетки не смогут конъюгировать.
18*
276
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
5'
hixL
TTGAAAACC AAGGTTTTTGATAА CCAATGTTA
TTGAAAACC AAGGTTTTTGATA A A
AACTTTTGG TTCCAAAAACTATT T
Н2 включен
Белок
Шп
Т А AC ATTGGT Т АТС А А АА АССТТ СС AAA AGGAQ
ATTGT A ACC A AT AGTTTTTGGA AGGTTTT CCTQ
5'
hixR
РИС. 10.51.
Сайт-специфическая рекомбинация между Л/xL и Л/xR
приводит к инверсии расположенного между ними сег-
AGGATTGCT ТТАТСАААААССТТССААAAGGA
TCCTAACGA ААТ AGTTTTTGGA AGGTTTT С СТ
Н2 выключен
мента. [J. R. Zieg, М. Simon, Proc. Natl. Acad. Sci. USA 77
(1980), p. 4196.]
Какими будут дрожжевые клетки -гетеротал-
личными или гомоталличньгми,- определяется един-
ственным геном НО, продукт которого необходим
для гомоталличного фенотипа. Гаплоидные клетки,
несущие мутантный аллель НО (ho~), гетеротал-
личны и неспособны к переключению между МАТа
и МАТа. Помимо несцепленных между собой МАТ
и НО, локализованных на хромосомах 3 и 4 соот-
ветственно, для переключения требуются еще два
генетических локуса: НМ Lu. и HMRa. Генетические
эксперименты показали, что 1) локус НМ Lu. необхо-
дим для превращения МАТа в МАТа, а локус
HMRa-для обратного процесса; 2) НМ La и HMRa
локализованы на левом и правом плечах хромосо-
мы 3 соответственно, а МА Т где-то на правом
плече, левее HMRa.
Кассетная модель. На основании этих экспери-
ментов была построена модель, описывающая мо-
лекулярный механизм переключения с МАТа на
МАТа и обратно. Согласно этой модели локусы
HMLa и HMRa содержат молчащие копии генов,
ассоциированные с типами спаривания а и а соот-
ветственно. Реплики гена а или а транслоцируются
из HMLa или HMRa в МАТ, где и экспрессируются.
Одновременно происходит замещение исходной по-
следовательности в локусе МАТна альтернативную.
Согласно этой модели, переключение с одного типа
спаривания на другой аналогично смене кассеты в
кассетном магнитофоне (МАТ), за исключением
того, что копии HMLa и HMRa остаются на месте,
а их реплики перемещаются в МАТ. Изучение
структуры HMLa, HMRa и МАТ, а также моле-
кулярное клонирование полностью подтвердили
справедливость кассетной модели и добавили много
новых деталей в этот механизм клеточной диф-
ференцировки.
Структура HMLa. HMRa и МАТ. Изучение
структуры этих локусов началось с клонирования
фрагмента дрожжевой ДНК, который отвечал за
спаривание типа а у гаплоидных дрожжевых клеток
ho~ с дефектным локусом МАТа (рис. 10.53). Кло-
нированный фрагмент использовали в качестве зон-
да для скрининга различных рестрицированных
дрожжевых ДНК с помощью блот-гибридизации
(рис. 10.54). Было выявлено четыре разных гибриди-
зующихся /7ги</Ш-фрагмента, которые соответство-
вали сегментам, содержащим локусы МАТа, МАТа,
HMLa и HMRa. Каждый из этих фрагментов был
клонирован. Затем все их возможные пары денату-
рировали и гибридизовали друг с другом, а полу-
ченные гетеродуплексы исследовали с помощью
электронного микроскопа (рис. 10.55). В соответст-
вии с кассетной моделью при гибридизации МАТа
с HMLa или МА Та с HMRa образуются непрерыв-
ные гетеродуплексы, причем дуплекс МАТа/НМ La
почти на 800 п.н. длиннее дуплекса МАТ а/HMRa
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
277
Гаплоид
ho (гетероталличные)
J Конъюгация
^Прорастание ЯО(гомоталлич н ые)
Споры,
.не способные
к спариванию
Первое
почкование
©
Второе
почкование
® © ® (g) <g> © ® ®
МАТа
—►МАТа
РИС. 10.52.
Схема деления дрожжевых клеток, иллюстрирующая
типы спаривания.
размером 1,6 т.п.н. Между HMLa и HMRa также
образуется гибридный дуплекс, но он прерывается
негомологичной последовательностью длиной око-
ло 800 п.н. Обнаружено также, что последователь-
ности, фланкирующие HML, HMR и МАТ, негомо-
логичны, хотя у МАТ они одинаковы для локусов
МАТа и МАТа. Наконец, сегмент длиной 800 п.н.,
характерный для гетеродуплекса МАТ а/HMLa,
присутствует также и в локусе МАТ а, но не в локусе
HMRa. Все эти структурные особенности представ-
лены на генетических картах трех локусов на
рис. 10.56. Видно, что локус МАТ содержит либо
последовательности a-типа, как в локусе HMLa,
либо последовательности а-типа, как в локусе
HMRa.
Переключение с одного типа спаривания на дру-
гой. Переключение типов спаривания происходит
в результате транспозиции копии HMLa или HMRa
в локус МАТ. Генетические и структурные исследо-
вания выявили некоторые особенности этого про-
цесса. 1. Для переключения необходим продукт
локуса НО. 2. Копии последовательностей, «хра-
нящихся» в HMLa или HMRa, перемещаются в
клетки МАТа или МАТа соответственно. 3. При
встраивании в МАТ эти копии не изменяются. 4.
При переключении а- или а-кассета, содержавшаяся
в МЛГ-локусе, утрачивается, но ни одна из них
в норме не встраивается в HML или HMR, т. е.
транспозиция происходит асимметрично. Каков ме-
ханизм этого процесса?
278
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Дрожжевая ДНК(мАТа)
[Лигирование,
трансфекция Е.соН
Колонии amprt tets
Плазмиды
РИС. 10.53.
Клонирование сегмента дрожжевой
ДНК, отвечающего за спаривание ти-
па а клеток МАTa~, ho~ (МАТа~
означает также HMLa~). Челночный
вектор дрожжи - Е. coli (разд. 5.8.в),
содержащий дрожжевой ген LEU2,
разрезали в ва/пН1 -сайте и прово-
дили лигирование с фрагментами,
полученными при расщеплении ге-
номной ДНК из клеток МАТа эндо-
нуклеазой Sai/ЗА. После трансфек-
ции продуктов лигирования в Е. coli
К12 были отобраны и размножены
все клоны amp'. tets. Плазмиды, вы-
деленные из этой популяции, транс-
фицировали в гаплоидные дрожже-
вые клетки (/ес/2-, МАТа, ho~,
ILV + ), и все колонии LEU2+ прове-
ряли на способность спариваться со
штаммом МАТа. Штамм МАТа имел
также генотип ilv~, поэтому на ми-
нимальной среде росли только дип-
лоидные клетки, содержащие функ-
циональный ген ILV от другого ро-
дителя. Был выявлен единственный
позитивный клон, из которого выде-
лена соответствующая плазмида
[К. Nasmyth, К. Tatchell, Cell 19 (1980),
р. 753.]
РИС. 10.54.
Клонированный сегмент дрожжевой ДНК, определя-
ющий тип спаривания а (рис. 10.53), гибридизуется
с четырьмя разными сегментами дрожжевой ДНК. со-
держащими локусы МАТа, МАТа, HMLa и HMRa. Гап-
лоиды МА Та или МАТа (гетероталличные) дают две
общие полосы, соответствующие фрагментам разме-
ром 6,1 и 5,0 т. п. н„ а также по одной уникальной
полосе-4.3 т.п.н. (МАТа) и 4,1 т.п.н. (МАТа). При
спаривании этих двух штаммов в тетрадном анализе
обнаружилось, что Hind\II-фрагмент длиной 4,3 т.п.н.
ассоциирован с фенотипом МАТа. a Hind\II-фрагмент
длиной 4,1 т.п.н,-с фенотипом МАТа. Таким образом,
эти два фрагмента содержат локусы МАТа и МАТа
соответственно. Два других фрагмента размером 6,1
и 5,0 т. п. н. представляют собой молчащие копии HMLa
и HMRa. Идентичность этих четырех рестрикционных
фрагментов и МАТа, МАТа, HMLa и HMRa подтверж-
дается результатами опытов по клонированию всех трех
HML
HMR
МАТа
H/WlII-рестрикты ДНК
гаплоидных дрожжевых
клеток
фрагментов геномных ДНК штаммов МАТа и МАТа
с использованием данной плазмиды в качестве зонда
[К. Nasmyth, К. Tatchell, Cell 19 (1980), р. 753.]
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
279
HMRa-5,0
( 0
МАТа-4,1 -----------------
( О
HMRa-5,0
( О
HMLa-6,1
( О
НМ/?а-5,0
( С
МАТа-4,3 ----- -----------
( О
HMLa -6,1
( О
МАТа-4,1 -------------- -------------
( О
HMLa-6,1
( О
МАТа-4,3 ----------------------------
( О
МАта-4,3
(-----------:--------- -------------8
МАТа-4,1
РИС. 10.55.
Схематическое представление структу-
ры гетеродуплексов, образующихся
между клонированными локусами
HMLa, HMRa, МАТа и МАТа. Указаны
цепи гетеродуплексов и их длина в т. п. н.
Одноцепочечные петли на самом деле
неодинаковы по величине [К. Nasmyth,
К. Tatchell, Cell 19 (7 980), jp. 753.]
J
4
РИС. 10.56.
Генетическая карта трех локусов типа
спаривания, расположенных в хромосо-
ме III дрожжей. В верхней части рисунка
представлена конфигурация активного
локуса МАТа (без соблюдения масшта-
ба), а в нижней-конфигурация актив-
ного МАТа. Локус МАТ состоит из не-
скольких сегментов: W (723 п. н.), X
(704 п. н.). Ya (747 п.н.) или Ya (642
п.н.), Z1 (239 п. н.) и Z2 (89 п. н.). Такие
же сегменты содержит HMLa, а в HMRa
отсутствуют W и Z2. В МАТа гены al
и а2 транскрибируются в противопо-
ложных направлениях от центрального
промотора в Ya. Экспрессия гена а1
в МАТа регулируется Ya-промотором.
Вертикальными стрелками указаны сай-
ты для эндонуклеазы НО.
s/г-Репрессия
| МАТа |
280
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
---Y 21--► Расщепление эндонуклеазой но
5‘-GCTTT CCGC AACAjpGTATA-
3‘— C G A A A G G C Gpjr Т G Т С А Т Ат - МАТа
5' — ACTTCGCGC AACAjpGTATA-
3'~TGAAGC G CGP|TTGT САТАТ- МАТа
РИС. 10.57.
Эндонуклеаза НО разрезает двухцепочечную ДНК в
специфическом сайте вблизи соединения Y и Z1.
Локус НО кодирует эндонуклеазу, которая уз-
нает специфический сайт в локусе МАТ (стрелки на
рис. 10.56 и 10.57) и делает разрез. В гетероталлич-
ных дрожжевых клетках (Ло-) или диплоидных (а/а)
гомоталличных клетках эндонуклеаза НО отсутст-
вует и переключения в локусе МАТ не происходит.
У значительной части клеток, в которых происходит
переключение, 7/О-сайты расщеплены. Однако соот-
ветствующие сайты в локусах НМ Lu. и HMRa оста-
ются интактными, т. е. они каким-то образом за-
щищены от расщепления. Учитывая, что расщеп-
ление-это один из ранних очень важных этапов
переключения, можно предположить, что асиммет-
рия транспозиции по крайней мере отчасти связана
с защитой HMLa и HMRa от расщепления.
Сам ген НО был отобран из библиотеки геном-
ной дрожжевой ДНК, полученной с помощью плаз-
мидного вектора, благодаря своей способности
превращать гетероталличные клетки в гомоталлич-
ные. Отсутствие эндонуклеазы НО в гетероталлич-
ных клетках, по-видимому, обусловливается ка-
кой-то мутацией. Используя этот клонированный
ген в качестве зонда, определяли содержание
НО-мРНК при разных условиях в клетках, мутант-
ных по специфическим фазам клеточного цикла.
В диплоидных клетках a/а НО-мРНК не образуется,
и именно поэтому в них не происходит переключе-
ния. В гомоталличных гаплоидных клетках как
НО-мРНК, так и эндонуклеаза НО синтезируются
на поздних стадиях фазы G1, но ко времени начала
митоза ни мРНК, ни соответствующий фермент
в клетке не обнаруживаются. В дочерних клетках во
время Gl-фазы НО-мРНК и эндонуклеаза НО не
выявлены, что согласуется с данными о том, что,
для того чтобы приобрести способность к переклю-
чению, клетки должны пройти хотя бы одно почко-
вание (рис. 10.52). Способность клетки к синтезу
НО-мРНК приобретается ею во время прохождения
клеточного цикла. Для регуляции транскрипции
весьма сложного процесса-необходимы последова-
тельности, диспергированные в 5'-фланкирующем
участке гена НО длиной более 1400 п.н., а также
некоторые белки.
Чтобы выяснить все детали механизма транспо-
зиции, нам нужны дополнительные данные. В наи-
более интересных моделях предполагается, что
транспозиция сопряжена с нереципрокным гомоло-
гичным кроссинговером (конверсией генов)-про-
цессом, аналогичным представленному на рис. 2.60.
Кроссинговер происходит между двумя гомологич-
ными последовательностями в хромосоме 3: HMLa
и МАТа или HMRa и МАТа (рис. 10.58). Однако
ввиду утраты кассеты, локализованной в локусе
МАТ, кроссинговер должен включать какие-то до-
полнительные этапы. Отметим, что двухцепочечный
разрыв, который осуществляет в локусе МАТ эндо-
нуклеаза НО, вполне вписывается в данную модель.
Кроме того, для переключения необходимы также
продукты других генов, участвующих в репарации
и рекомбинации ДНК.
Экспрессия генов типа спаривания. Гены типа
спаривания экспрессируются только в том случае,
когда они находятся в локусе МАТ, но не в HMLa
или HMRa. До сих пор мы касались только одного
фенотипического различия между a (МАТа)- и
а (ЛЛ4 То.(-дрожжевыми клетками, а именно способ-
ности клеток типа а спариваться с клетками типа а,
но не клеток а с клетками а или клеток а с клетками
а. Как контролируется этот процесс и каковы другие
фенотипические особенности клеток МАТа и МАТа?
Известно, что а~сегмент содержит только один ген,
al, а-сегмент-два гена, al и а2. Продукты этих
генов представляют собой регуляторные белки, ко-
торые влияют на экспрессию других генов. Данные
о действии этих регуляторных элементов суммиро-
ваны на рис. 10.59. В клетках МАТа продукт гена al
действует как позитивный регулятор, запуская тран-
скрипцию группы молчащих генов, также необхо-
димых для проявления a-фенотипа. Эта группа со-
держит ген MF al, кодирующий a-фактор-секрети-
руемый из клетки олигопептид феромон. Продукт
гена а2 представляет собой негативный регулятор,
выключающий экспрессию генов в клетках МА Та.
Как al, так и а2 влияют на транскрипцию соответ-
ствующих генов-мишеней, связываясь со специфи-
ческими последовательностями вблизи 5'-концов
этих генов. Активность а2 зависит также от его
кооперативного взаимодействия с другим вездесу-
щим белком дрожжевых клеток. Кроме этого, у
диплоидов МАТа/МАТа, в которых экспрессиру-
ются оба локуса МАТ, а2, действуя совместно с а1,
1) ингибирует экспрессию al и таким образом всех
позитивно регулирующихся им генов; 2) ингибирует
экспрессию множества других генов, необходимых
для спаривания, например НО и Ту (разд. 10.3.б); 3)
индуцируют мейоз и споруляцию клеток при соб-
людении других условий. Такая координированная
репрессия нескольких генов при совместном дейст-
вии белков а2 и а 1 определяется специфической
последовательностью длиной 20 п.н., находящейся
в 5'-фланкирующей области всех генов, на которые
10, РЕОРГАНИЗАЦИЯ ГЕНОМА
281
Z
Ya
Z
МАТа
HMLa
равно
РИС. 10.58.
Модель генной конверсии, объясня-
ющая механизм переключения с од-
ного типа спаривания на другой.
[J. D. Watson et al.. Molecular Biology
of the Gene (Menlo Park, CA; Benjamin/
Cummings, 1987), p. 580, с измене-
ниями.]
t Двухцепочечный разрыв,
осуществляемый но
| Встраивание гомологичного Z-участка
Деградация
Деградация сегмента Y и встраива-
ние гомологичного участка X
I Наращивание З'-концов
1 (репаративный синтез)
Новосин-^
тезированная
ДНК
Структура Холидея
I Восстановление линейной хромосомы,
I содержащей «-последовательность в
у локусе МАТ
HMLa
оказывается воздействие, включая и ген al. Эта
i/wc-последовательность отличается от последова-
тельности, которую узнает сам а2. Взаимодействие
с белком al изменяет специфичность связывания
белка а2. Эта сложная схема объясняет неспособ-
ность //О-диплоидных клеток к переключению и
спариванию. Основной регуляторный локус-это
МА Т. Он индуцирует переключение, влияя на
//О-ген, и определяет тип спаривания, регулируя
экспрессию специфических генов. Отметим также,
что МАТа не содержит генов типа спаривания; этот
тип определяется просто отсутствием экспрессии
гена а. Вся эта система работает таким образом,
чтобы обеспечивалось чередование полового и не-
полового размножения.
Транскрипционные единицы этих трех генов по-
282
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Клетка Генные продукты
«специфичных генов «-специфичных генов
Никакой самостоятельной
Функции не выявлено
а!
Ингибирует экспрессию
<х1 но
генов, необходимых
для спаривания
РИС. 10.59.
Влияние продуктов генов МАТа и МАТа на
экспрессию генов.
казаны на рис. 10.56. Они были идентифицированы
с помощью мутагенеза in vitro клонированной
МЛ Т-ДНК и встраивания ее в дрожжевой геном;
таким образом была установлена корреляция между
мутантным сайтом и мутантным генным продук-
том.
Молчащие а- и d-гены в локусах HML и HMR.
Несмотря на то что гены а и а в локусах HML
и HMR содержат полноразмерные 5'- и З'-фланки-
рующие последовательности, включающие промо-
торы, они не транскрибируются до тех пор, пока не
произойдет их транспозиция в МАТ. По крайней
мере отчасти этот феномен связан с четырьмя ген-
ными продуктами-SIR 1, 2, 3 и 4 (от англ, silent
information regulator). На существование SIR-белков
указывали мутации у дрожжей, каждая из которых
влияла на экспрессию а- или «-генов в локусе HML
или HMR. SIR-гены не сцеплены с HML, HMR или
МАТ. Таким образом. HMLa и HMRa содержат все
элементы, необходимые для экспрессии генов а и fl,
но SIR-белки подавляют ее. Репрессия происходит
в результате связывания SIR-белков с участками,
фланкирующими HMLa и HMRa (рис. 10.60). Эти
участки находятся за пределами транспозирующих-
ся единиц и удалены более чем на 1000 п. н. от
последовательностей, контролирующих транскрип-
цию генов al, a2 и al.
Чтобы идентифицировать мишени для SIR-бел-
ков, были сконструированы делеционные мутанты
по клонированным сегментам HML и дрожжевые
клетки, несущие различные SIR-мутации, транс-
формировали каждым из этих клонов (рис. 10.60).
Все делении, захватывающие сегмент длиной
130 п.н. (Е) слева от HML, приводили к блокиро-
ванию SIR-репрессии. Сходные результаты были
получены и для локуса HMR. Е-сегменты можно
рассматривать как сайленсеры (разд. 8.3.д); послед-
ние обладают свойствами, характерными для эн-
хансеров транскрипции, но они подавляют тран-
скрипцию, а не стимулируют ее. Если Е-элемент
инвертируется или перемещается в другой сайт в
пределах 2,5 т.п.н. от генов, определяющих тип
спаривания, то SIR-репрессия не снимается. Таким
образом, Е-элементы, как и энхансеры транскрип-
ции, функционируют в любой из двух ориентаций
и могут находиться на значительном расстоянии от
соответствующего гена. С помощью аналогичного
делеционного анализа были выявлены и другие
сегменты (I)-справа от HMLa HMR, также участ-
вующие в SIR-репрессии.
Конечным результатом транспозиции генов из
HML или HMR в МАТ является удаление коди-
рующих последовательностей из окружающих их
участков, подавляющих генную экспрессию. Как
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
283
HML*
Да —
Нет Г —
РИС. 10.60.
Мутагенез in vitro, позволяющий идентифицировать
сайт-мишень для SIR-белков в локусе HML. Горизон-
тальные отрезки указывают размеры делеций, получен-
ных в результате расщепления клонированного сегмен-
та HML в сайте Xho\ и последующего гидролиза с
помощью 5AZ.31 (разд. 7.6.6). Каждый из этих деле-
ционных мутантов использовали для трансформации
дрожжевых клеток, несущих различные SIR-мутации.
Способность различных делеций репрессироваться
SIR-белками представлена в виде диаграммы внизу
слева (да или нет). К действию SIR-белков чувствителен
участок Е, выявленный в этом и других экспериментах.
именно взаимодействие SIR-белков с последова-
тельностями, так далеко отстоящими от промото-
ров, ингибирует транскрипцию, пока неясно. Воз-
можно, это связано с наличием в сайтах взаимо-
действия с SIR-белками Л RS-элементов (разд. 5.6.6),
поскольку дерепрессированные клетки должны
пройти S-фазу, чтобы в них происходила репрессия.
Определенную роль может играть и изменение
структуры хроматина. Если эти изменения обуслов-
ливают устойчивость HMLa или HMRa к расщеп-
лению с помощью НО, то становится понятным,
почему при переключении МАТ служит именно
реципиентом, а не донором. В соответствии с этой
гипотезой как HML, так и HMR являются реци-
пиентами транспозиции в штаммах sir~.
в. Гены, кодирующие иммунные белки
позвоночных
В иммунном ответе позвоночных участвует
множество специфических белков-секретируемых
антител и рецепторов, расположенных на плазмати-
ческой мембране лимфоцитов. Их роль заключается
в защите организма от инфекции вирусами, бакте-
риями, паразитами и, возможно, от пролиферации
опухолевых клеток. В отличие от генов других
клеточных белков гены, кодирующие антитела и
рецепторы, в зиготе отсутствуют. Вместо этого
в ДНК клеток зародышевой линии содержатся их
предшественники в виде отдельных сегментов, а
функциональные гены образуются в результате спе-
цифических перестроек, которые происходят только
во время развития В- и Т-лимфоцитов (называемых
также В- и Т-клетками).
По-видимому, в иммунном ответе участвуют
соединения с самой разной химической структурой.
Антигенными свойствами обладают практически все
типы встречающихся в природе веществ-белки, уг-
леводы, липиды, нуклеиновые кислоты. Иммунный
ответ могут вызывать даже химические вещества,
полученные искусственным путем. Взаимодействие
антигена с различными антителами определяется
структурными особенностями его эпитопа. Даже
один эпитоп способен стимулировать синтез разных
(но сходных по структуре) антител.
Разнообразие и специфичность иммунных систем
определяются двумя их замечательными свойства-
ми. Во-первых, иммунная система способна разли-
чать свое (химические структуры, свойственные
данному организму) и чужое. Именно этим обус-
ловливается отторжение трансплантированных кле-
ток и тканей. Аутоиммунитет (т. е. нарушение толе-
рантности к собственным антигенам) встречается
редко и обычно бывает связан с определенными
патологическими состояниями. Во-вторых, иммун-
ная система обладает памятью, т. е. организм выра-
батывает антитела более эффективно и быстро, если
встречается с антигенами во второй раз даже спустя
много лет.
Клеточный и гуморальный иммунитет. Сущест-
вуют две разные, но связанные между собой иммун-
ные системы, каждая из которых ассоциирована с
лимфоцитами определенного типа (рис. 10.61). Кле-
точный иммунитет обеспечивается Т-клетками, а гу-
моральный-как В-, так и Т-лимфоцитами. Клеточ-
ный иммунитет опосредуется специфическими Т-
клетками - цитотоксичными Т-лимфоцитами, или
«киллерами» (CTL), и направлен против клеток,
несущих на своей поверхности чужеродные анти-
гены (например, против опухолевых клеток, в кото-
рых синтезируются новые белки клеточной поверх-
ности, или вирусных частиц на поверхности инфи-
цированных клеток). Для воздействия киллера на
клетку-мишень между ними должен возникнуть
контакт, во время которого и осуществляется дейст-
вие одного или нескольких специфических белков,
284
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Клеточный
иммунитет
Бактерии
Секреция
модулируемая
В-кдетка клеткамитниТ5
Антитела
Антитело
Вирусы
Грибы
Фагоциты
Обломки
Цитотоксический
Т-лимсроцит
(CTL)
Вирус
Вирусный
белок
Чужеродные
молекулы
0* -8
Лизис
Инфицированная
клетка
РИС. 10.61.
Две иммунные системы. При гуморальном иммунитете
циркулирующие антитела, секретируемые В-клетками,
связываются с чужеродными антигенами (бактериями,
вирусами, грибами, белками). Клеточный иммунитет
опосредуется Т-клетками, которые сами связываются
с клетками, несущими как чужеродный антиген, так
и специфические нормальные поверхностные белки.
продуцируемых CTL. Гуморальный иммунитет опо-
средуется циркулирующими в биологических жид-
костях антителами, которые продуцируются В-клет-
ками под регуляторным контролем Т-клеток двух
видов: Т-хелперов (Тн) и Т-супрессоров (Ts). Анти-
тела циркулируют в сыворотке и в лимфе, однако
некоторые из них находятся в лимфатических узлах,
селезенке и ассоциированной с кишечником лим-
фоидной ткани. В любом случае антитела связы-
вают соответствующие антигены, которые находят-
ся в свободном состоянии в растворе или на поверх-
ности инфицирующей клетки. Комплексы анти-
тело-антиген быстро удаляются и разрушаются
фагоцитарными клетками, например макрофагами.
Соединение некоторых антител с клеточными анти-
генами (например, бактериями) запускает серию
протеолитических реакций - систему комплемента,
которая вызывает лизис чужеродных клеток.
Антитела. Антитела в высшей степени специ-
фичны. Они способны различить даже близкородст-
венные эпитопы, например два стерео- (d- и /-энан-
тиомера) или геометрических (цис- и транс-) изоме-
ра. Любое антитело способно дифференцировать
белки, отличающиеся друг от друга лишь несколь-
кими аминокислотами (например, бычий и чело-
веческий инсулин). Причину столь замечательной
специфичности антител и в то же время сходства их
функций лучше всего помогает понять изучение их
структуры (рис. 1.37, 10.62 и 10.63). Молекула им-
муноглобулина представляет собой гетеротетрамер,
состоящий из двух идентичных пар полипептидных
цепей. Каждая такая пара в свою очередь состоит из
тяжелой цепи (Н-цепи) и легкой цепи (L-цепи). Н-
и L-цепи в димере связаны бисульфидными мости-
ками; такие же мостики соединяют и две Н-цепи
в каждом тетерамере. Н-цепи одной молекулы ан-
титела имеют одинаковые аминокислотные после-
довательности, но у каждой из Н-цепей антитела на
Структура антитела. Указаны вариабельная и констант-
ная области легкой (VLCL) и тяжелой (Vh^h) Цепей,
а также отдельные домены константных областей тя-
желой цепи (Сн1, Сн2 и Сн3) и антигенсвязывающие
сайты в вариабельных областях и CDR.
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
285
Легкая цепь (23кДа)
CDR1 CDR2 CBR3
nh сххсжхжххжхмммвмевмв
Вариабельный участок ^Константный участок^
Тяжелая цепь (53кДа)
Вариабельный участок । Константный участок
РИС. 10.63.
Схематическое изображение
легкой и тяжелой цепей им-
муноглобулина.
N-конце имеется вариабельный участок (VH) длиной
примерно 115 аминокислотных остатков. Соответст-
венно и у L-цепей есть N-концевой вариабельный
участок (VL) длиной примерно ПО аминокислот.
С-концевая половина L-цепей (~ 110 аминокислот)
называется константной областью (CL), поскольку
независимо от последовательности VL она пред-
ставлена последовательностью одного из двух ти-
пов-и или X. Н-цепи также содержат N-концевые
константные области, Сн, состоящие из ~ 330 ами-
нокислотных остатков. Они бывают нескольких ти-
пов и определяют класс и эффекторные функции
соответствующих антител.
Известны лишь два типа легких цепей-каппа (х)
и лямбда (X). Тип цепи определяется аминокислот-
ной последовательностью Ск-сегмента и включает
L-цепи со множеством разных VL-последователь-
ностей. Тяжелые цепи более разнообразны; известно
пять типов Сн: ц, 5, у, е и а. Им соответствуют пять
классов иммуноглобулинов-так называемые изоти-
пы IgM, IgD, IgG, IgE и IgA. Каждое из этих
антител играет специфическую роль в гуморальном
иммунитете. Например, иммуноглобулины G-это
обычные циркулирующие антитела, IgE участвуют в
аллергических реакциях, a IgA, которые концентри-
руются в жидкостях, окружающих эпителиальные
клетки, по-видимому, защищают организм от мик-
робов. Каждая из тяжелых цепей связывается либо
с н-L-, либо с X-L-цепью.
Структура, образующаяся в результате взаимо-
действия VH и VL, представляет собой уникальный
сайт связывания антигенов и, таким образом, опре-
деляет специфичность антитела. Каждое антитело
бивалентно, поскольку содержит два идентичных
сайта связывания антигена. Специфичность связы-
вания обусловливается комплементарностью анти-
гена и антитела (рис. 10.64) и определяется тремя
участками VL и VH, которые называются гипер-
вариабельными и обозначаются CDR1, CDR2 и
CDR3 (от англ, complementary-determining regions).
Именно в этих участках наиболее сильно разли-
чаются области VL и VH у разных антител.
Аминокислотные последовательности самых
разных иммуноглобулинов были известны задолго
до того, как стало возможным прямое изучение
эукариотических генов. Было показано существо-
вание у этих полипептидов вариабельных и кон-
стантных областей. Для объяснения поразительного
разнообразия V-участков и существования практи-
чески неограниченных возможностей для обеспече-
ния разных антигенных специфичностей было выд-
винуто много разных гипотез. Основная проблема
состоит в том. как объяснить возникновение прак-
тически неограниченного числа генов, кодирующих
разные V-, но одинаковые С-участки. Как оказалось,
ни одна из этих гипотез сама по себе не объясняет
данного феномена. Разнообразие V-областей обе-
спечивается разными механизмами.
Гены легкой цепи. Первые данные о том, что гены
полноразмерной L-цепи образуются в результате
перестроек ДНК во время дифференцировки В-кле-
ток, были получены в результате сравнения рест-
рикционных карт последовательностей, кодиру-
ющих L-цепи в клетках, которые продуцируют ан-
титела (например, в клетках миеломы), и в клетках,
которые антител не продуцируют (например, в
клетках печени или спермиях) (рис. 10.65). Оказа-
лось, что в ДНК из клеток миеломы V- и С-области
экспрессирующегося гена L-цепи находятся в одном
рестрикционном фрагменте, в отличие от ДНК из
клеток зародышевой линии или нелимфоидных кле-
ток. С тех пор были клонированы и секвенированы
кодирующие последовательности многих генов раз-
личных х- и Х-цепей. Был не только подтвержден
сам факт перестроек, но и в какой-то мере проясне-
ны механизмы, участвующие в этих процессах.
Функциональные гены X- и х-цепей образуются
практически одинаковым способом у большинства
позвоночных. Рис. 10.66 иллюстрирует образование
функционального х-гена. При этом используются три
сегмента ДНК: VK, JK и Ск. Все они обнаружены на
хромосоме 2 человека, при этом присутствуют ~ 300
тандемных копий Уи-сегментов, 5 тандемных копий
Зу-сегментов и один Си-сегмент. В клетках зароды-
шевой линии человека и в большинстве соматических
клеток Уи-кластер удален от Jy-кластера на 23 т. п. н.,
но во время дифференцировки каждой В-клетки один
из Уи-сегментов соединяется в результате рекомби-
нации с каким-либо Jy-сегментом. При этом участок
ДНК между Уи- и Ли-сегментами утрачивается или
остается в зависимости от взаимной ориентации VK
и JK в клетках зародышевой линии.
286
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕН ОВЭУКАРИОТ
РИС. 10.64.
А. Схематическое изображение связывания антигена со
специфическим сайтом иммуноглобулина. Кружки обо-
значают CDR. Б. Полученные с помощью компьютера
структуры вариабельных областей тяжелой и легкой
цепей антитела лизоцима (слева) и самого лизоцима
(справа). В верхней части этого рисунка показана «сты-
ковка» двух молекул. Их связывание стабилизируется
электростатическими и вандерваальсовыми силами, а
также водородными связями. [С любезного разрешения
S. Sheriff. См. S. Sheriff et al.. Proc. Natl. Acad. Sci. USA
84 (1987), p. 8075.]
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
287
к ДНК-зонд;
Полноразмер-
ная КДНК
З'-конец
Образец Эмбрион Эмбрион
ДНК: Миелома Миелома
Направление
электрофореза
РИС. 10.65.
Перестройка кодирующих последовательностей гвна
легкой цвпи х в процессе созревания В-клеток. Геном-
ная ДНК была выделена иэ клеток мышиных эмбрионов
и клеток миеломы мыши. Миеломы-это В-клеточные
опухоли, представленные одним клеточным клоном и
секретирующие единственное гомогенное антитело.
ДНК расщепляли эндонуклеазой BamHI и проводили
электрофорез и блот-гибридизацию с кДНК-зондами,
отвечающими полноразмерной глобулиновой мРНК иэ
клеток миеломы или З'-концевой (Ск) части этой мРНК.
Оба зонда гибридизуются с одним рестрикционным
фрагментом ДНК из клеток миеломы, но с разными
фрагментами ДНК эмбриональных клеток (или ДНК из
других, нежели В-клетки, зрелых соматических клеток).
[N. Hozumi, S. Tonegawa, Proc. Natl. Acad. Sci. USA 73
(1976), p. 3628.]
Каждый Ух-сегмент содержит три функциональ-
ные части: промотор и два экзона, Lz и Vz (рис. 10.67).
Экзон Lz кодирует сигнальную последовательность,
которая направляет образующийся полипептид в
эндоплазматический ретикулум, a Vz кодирует учас-
ток длиной примерно 80 аминокислот V-области
х-цепи. Остальные ~ 30 аминокислот V-области
кодируются присоединенным Зх-сегментом. Пере-
строенный функциональный ген и-цепи содержит
промотор, три экзона и два интрона. Один интрон
разделяет экзоны Lz и Vz-Jz, а второй находится
между экзонами Vz-Jz и Cz. Один из CDR-сегментов
L-цепей - CDR3- кодируется последовательностью,
образующейся при соединении Vz и Jz.
Помимо альтернативного использования X- или
х-цепей за разнообразие L-цепей в молекулах анти-
тел отвечают три механизма. Во-первых, любой из
сотен Уи-сегментов может соединиться с любым
1х-сегментом, ассоциированным с Cz. Таким обра-
зом, специфичность L-цепи отчасти определяется
тем, какие именно из сегментов Vz и Jz соединяются.
Аналогично, из разных V;-, J; - и С^-кодируюших
участков образуются легкие цепи X, обладающие
разной специфичностью. Во-вторых, соединение VL-
и Зк-сегментов происходит неточно, и в результате
образуются несколько альтернативных кодирующих
последовательностей. В-третьих, после образования
функционального гена L-цепи Уг-сегмент подвер-
гается в ходе последующей пролиферации В-клеток
необычно частым мутационным изменениям. Такие
мутации еще более увеличивают разнообразие анти-
генсвязывающих областей L-цепей.
Сайт-специфические рекомбинации, в результате
которых Уь-сегмент соединяется с сегментом JL,
протекают при участии особой рекомбиназной сис-
темы. Рекомбинация зависит от двучленных олиго-
нуклеотидных сигналов узнавания, которые нахо-
дятся на З'-конце VL и на 5'-конце JL (рис. 10.68).
Сигнал состоит из двух участков длиной 7 и 9 п. н„
разделенных 11 п.н. (примерно один виток спирали)
в Vj -области и 23 п. н. (примерно два витка спирали)
в .^-области. Рекомбинация зависит от целостности
как гептамерного и нонамерного участков, так и
разделяющего их сегмента, но не от нуклеотидной
последовательности последнего. Хотя специфич-
ность этих сигналов узнавания весьма высока, на
самом деле соединение VL- и .^-сегментов происхо-
дит не очень точно: небольшое количество осно-
ваний при этом либо утрачивается, либо, напротив,
добавляется. Вследствие такой неточности иногда
рамки считывания соединяемых VL и JL как бы
оказываются в фазе, т. е. между ними возникает
транслируемый участок, однако такие случаи весьма
редки. Более того, сами «совпадающие по фазе»
соединения отличаются одно от другого, что при-
водит к изменчивости кодирующих последователь-
ностей домена CDR3 У|-области и еще более уве-
личивает разнообразие последней.
Механизм сборки гена L-цепи эволюционировал
в направлении увеличения разнообразия цепей ан-
тител даже ценой появления огромного числа де-
фектных генов. Теоретически соединение любого из
почти 300 Ух-сегментов с любым из пяти сегментов
Jz приводит к образованию примерно 1500 разных
генов Lz-uenefi. Неточность соединения может уве-
личить это число за счет образования 10 совпа-
дающих по фазе сочленений примерно до 1,5 104.
Вместе с аналогичным разнообразием Х-цепей это
приводит к огромному увеличению числа возмож-
ных легких цепей. У человека имеется несколько
разных константных Х-областей, поэтому число
разнообразных L-цепей поистине огромно.
При сборке гена легкой цепи не только происхо-
дит увеличение разнообразия цепей, но и активи-
288
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
мРНК
Н,Ы+— —СОО”
к I \
Вариабельный Константный
Белок X
РИС. 10.66.
Перестройки участков ДНК из клеток зародышевой
линии, приводящие к образованию функционального
гена и-цепи. \/к-области находятся в разных ориента-
циях относительно JK- и Ск-сегментов. Во время диф-
ференцировки В-клеток один из \/к-сегментов соеди-
няется с одним из сегментов JK. [J. Darnell, Н. Lodish,
D. Baltimore, Molecular Cell Biology (New York: Scientific
American Books, 1986), p. 1104.]
руется генная экспрессия (разд. 8.3.д); в результате
перестроек промотор, расположенный перед каж-
дым Уу-сегментом, оказывается вблизи энхансера,
внутри интрона, разделяющего JK и Ск (рис. 10.67).
Гены тяжелых цепей. Те же самые механизмы,
которые приводят к разнообразию L-цепей, ответ-
ственны и за существование огромного числа раз-
ных генов Н-цепей, кодирующих VH с разными
аминокислотными последовательностями. Однако
V-области Н-цепей еще более разнообразны, чем
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
289
РИС. 10.67.
Функциональные элементы гена Ьк-цепи, образовавше-
гося в результате перестроек.
соответствующие области L-цепей, поскольку ко-
дирующие их участки строятся не из двух, а из трех
сегментов: VH, DH и JH (рис. 10.69). У человека все
эти сегменты находятся на хромосоме 14, однако
VH-, DH- и .1,,-кластеры располагаются на большем
расстоянии друг от друга. Случайные комбинации
одного из нескольких сотен VH с одним из 20 DH
и одним из 5 JH дают 104—105 различных цепей.
Сигналы узнавания, по которым происходит ре-
комбинация, состоят из таких же олигонуклеотидов
длиной 7 и 9 п. н., которые опосредуют соединение
VL- и .^-областей; они находятся на З'-конце VH, на
5'- и З'-концах DH и на 5'-конце JH. Гептамеры
и нонамеры каждого рекомбинационного сегмента
тоже разделяются последовательностями длиной 11
и 23 п.н. Происходят два последовательных акта
соединения. Сначала один из Он-элементов соеди-
няется с одним из .(ц-элементов, а затем к уже
перестроенному сегменту DHJH пришивается сег-
мент VH. Соединения VHDH и DHJH происходят
неточно и часто сопровождаются делениями и ин-
серциями новых нуклеотидов. Встроенные, так на-
зываемые N-участки, по-видимому, появляются в
результате действия терминальной дезоксинуклео-
тидилтрансферазы-фермента, который был извес-
тен за 25 лет до того, как удалось установить его
физиологическую роль (разд. 4.8). Эти участки мо-
гут кодировать до десяти новых аминокислот
Н-цепи. В результате делений и вставок с любой из
сторон DH вариабельность Ун-области увеличивает-
ся в несколько сотен раз. Соединение различных
VH-, DH- и ,1н-сегментов дает ~ 105 комбинаций;
вариабельность N-участка увеличивает число раз-
личных Н-цепей до 107-108, а сочетание разных Н-
и L-цепей дает 10п-1012 сайтов связывания имму-
ноглобулинов.
Константные области Н-цепей (Сн) также раз-
личаются, но эти различия не влияют на CDR. Итак,
полученный в результате перестроек Ун-сегмент
может соединиться с каким-нибудь Сн-сегментом
ц, 5, у, е или а. В зависимости от этого возникают
антитела изотипов IgM, IgD, IgG, IgE и IgA соот-
ветственно. Прежде чем пытаться понять, как обра-
зуются Н-цепи с разными Сн-областями, рассмот-
рим дифференцировку В-клеток. Кроме всего про-
чего, это поможет нам понять, почему в ответ на
один антиген иммунной системой продуцируется
огромное количество специфических антител.
Дифференцировка В-клеток. Стволовые клетки-
предшественники В-клеток и всех других клеток,
участвующих в гемопоэзе,-появляются вначале в
печени и селезенке плода, а после рождения -и
в красном костном мозге. На этих первых этапах,
когда образуются циркулирующие, но непроли-
феративные пре-В-клетки, антиген отсутствует
(рис. 10.70). Тем не менее пре-В-клетки уже содер-
жат перестроенные гены функциональных Н- и
L-цепей и образуют антитела, относящиеся к мемб-
раносвязанным IgM и IgD. Некоторые такие клетки
Кодирует Уж
®( CC ТСС |cACAGTG—П —АСАААААСС
{ GG AGg|gtGTcAC-11-ТйТТТГ TSG
pro
КодируетЗх
GGTTTTTGT—23 —CACTGTG IgTGG Q
CCAAAAACA-23-GTGACAC |САСС Q
trp
Рекомбинация
РИС. 10.68.
pro trp
I---к----1
[CCT TGG 0
[6tA ACC Q
pro trp
Рамка
;cctc TGG
[GGAG ACC
сохраняется
Рамка
Соединение VK c JK происходит в результате реком-
бинации специфических гомологичных сигналов узна-
вания. Каждый такой сигнал состоит из двух олиго-
нуклеотидов длиной 7 и 9 п. н., которые в VK и JK
разделены участками размером 11 и 23 п. н. соответст-
венно. Для рекомбинации должен произойти диффе-
ренциальный сплайсинг между гептамерной и нона-
мерной единицами. Соединение происходит неточно.
нарушается
pro arg
(cctc ggq
(GGAG CcQ
Рамка
сохраняется
так что кодирующие части VK и JK утрачивают несколько
пар оснований или приобретают лишние. В результате
в некоторых случаях происходит сдвиг рамки считы-
вания, и пегкая цепь вообще не синтезируется. В других
случаях соединение VK и JK происходит «в фазе», рамка
считывания сохраняется, но само место сочленения
варьирует, что обусловливает изменчивость CDR3-O6-
ласти.
19-158
290
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Ун-сегменты D-сегменты К-сегменты
(сотни) (—20) (4)
Вторая перестройка
N N
^Сплайсинг РНК
мРНК
H3N+ .<>jrx.xXX)O——>ЧЧ||Ц1>В> соо-
Вариабельный Константный
РИС. 10.69.
Перестройки ДНК из клеток за-
родышевой линии, в результате
которых образуется функциональ-
ный ген тяжелой цепи в В-клетках.
продуцируют только IgM, другие-как IgM, так
и IgD. Каждая пре-В-клетка несет примерно 104
идентичных молекул IgM, которые, впрочем, отли-
чаются по своей структуре и антигенной специфич-
ности от IgM других клеток, поскольку содержат
другие VH- и Уь-области. Таким образом, огромное
разнообразие и специфичность антител во многом
задаются еще до появления антигена. В пре-В-клет-
ках, продуцирующих как IgM, так и IgD, оба им-
муноглобина содержат одинаковые VH- и VL-cer-
менты и разные сегменты Сн-Сц или С5.
В результате независимых перестроек х- и Х-ал-
лелей в каждой из пар хромосом 2 и 22 соответст-
венно могут образоваться четыре разные L-цепи.
Аналогично, в результате независимых перестроек
в паре хромосом 14 могут возникнуть две разные
Н-цепи. Почему же тогда в пре-В-клетках присутст-
вуют Н- и L-цепи только одного вида? Дело в том,
что перестройки, которые происходят во время
развития пре-В-клеток, носят неслучайный характер.
Перестройка гена Н-цепи происходит сначала в
одной из двух хромосом. Если возникающее при
рекомбинации соединение слишком отклоняется от
нормы или если в результате DHJH- или после-
дующего VH DHJ „-соединения образуется неэкс-
прессирующийся ген, то запускается перестройка
второй хромосомы. Удачное завершение перестроек
Н-цепи инициирует сборку функционального х-гена.
Если рекомбинации оказываются безуспешными,
начинаются перестройки в Х-локусе. Клетки, кото-
рым не удалось сконструировать функциональные
гены Н- и L-цепей, оказываются нежизнеспособ-
ными. Механизм, лежащий в основе этого строго
упорядоченного уникального процесса, неясен.
Можно только предположить, что как только в
результате удачных геномных перестроек образо-
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
291
Антигеннезависимые,
находятся в костном мозге
Антигензависимые, находятся в перифери-
ческих лимфоидных тканях
Т-клетки). Название В-клеток происходит от названия
органа птиц, е котором они были впервые обнаруже-
ны,- фабрициевой сумки (bursa).
РИС. 10.70.
Образование Т- и В-лимфоцитов из стволовых клеток
костного мозга. Для того чтобы стать иммунологически
активными, предшественники Т-лимфоцитов должны
какое-то время находиться в тимусе (отсюда название
вались функциональные Н- и L-цепи, перестройки
альтернативных аллелей блокируются-феномен,
называемый аллельным исключением (allelic exclu-
sion).
Дальнейшая дифференцировка пре-В-клеток с
образованием зрелых, короткоживущих и секрети-
рующих антитела плазматических клеток или дол-
гоживущих, не продуцирующих антитела клеток
памяти происходит вне костного мозга. Эти превра-
щения зависят от взаимодействия мембраносвязан-
ного IgM В-клеток с родственным антигеном
(рис. 10.70). Если связывания антигена не происхо-
дит, то пре-В-клетка погибает в течение нескольких
дней. Взаимодействие поверхностного IgM с под-
ходящим антигеном запускает серию физиологи-
ческих реакций, включающих активный рост и про-
лиферацию клеток, переключение с образования
мембраносвязанных антител на образование анти-
тел секретируемых и запуск экспрессии генов Н-це-
пей. Большинство таких индуцированных клеток
живут ограниченное время, но некоторые из них
в результате последующей дифференцировки пре-
вращаются в клетки памяти, которые в течение
многих лет сохраняют способность отвечать на
исходный антиген, снова продуцируя соответству-
ющее антитело.
Рост и пролиферация В-клеток в ответ на свя-
зывание антигена опосредуются сложными взаимо-
действиями с макрофагами, хелперными Т-клетка-
ми и факторами роста и дифференцировки, назы-
ваемыми лимфокинами; обсуждение этих процессов
выходит за рамки данной главы. Переключения на
синтез секретируемых антител и на образование
вместо ц (или 5-)-цепей цепей одного из изотипов-у,
е или а-происходят при участии дифференциаль-
ного процессинга мРНК и дополнительных геном-
ных перестроек соответственно. Рассмотрим общие
принципы этих процессов.
Структура С „-кодирующих участков и их экс-
прессия. Кодирующие участки константных облас-
тей Н-цепей р, 5, у, е и а-организованы в указан-
ной последовательности в кластеры длиной 200
т. п. н., одинаковые для геномных ДНК мыши и че-
ловека (рис. 10.71). Кластер, состоящий из пред-
ставленных в единственном числе Сц, Cg, СЕ и Са
и нескольких Су, находится правее кластера ^-сег-
ментов. Каждый С-сегмент имеет несколько экзонов
и интронов.
В пре-В-клетках транскрипция гена мембрано-
связанной ц-цепи начинается левее лидерной после-
довательности LH и проходит через первый интрон,
перестроенный Ун-сегмент, второй интрон и Сц-ин-
19*
292
ЧАСТЬ III. ОРГАНИЗАЦИЯ. ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 10.71.
Схематическое изображение класте-
ра Сн-сегментов в геноме мыши.
троны и экзоны; полиаденилирование происходит
в последнем Сц-экзоне (рис. 10.72). Последователь-
ности. кодирующие типичные Н-цепи, находятся
в первых четырех из шести экзонов См. Последние
два экзона кодируют полипептидный сегмент, ко-
торый заякоривает p-цепи антитела в мембране.
Мембраносвязанная форма ц-цепи называется цт,
а соответствующий иммуноглобулин-IgMm. Анти-
генсвязывающий сайт мембраносвязанного IgM на-
ходится снаружи В-клетки. Связывание соответст-
вующего антигена с поверхностным IgM запускает
несколько процессов. Во-первых, происходит пере-
ключение с синтеза IgMm на синтез IgM s- секрети-
руемого варианта IgM, который обладает той же
антигенсвязывающей специфичностью, но имеет
другую аминокислотную последовательность на
С-конце p-цепи. Далее, в результате пролиферации
В-клеток образуется клон плазматических клеток,
секретирующих такой же IgMs. Переключение с
синтеза рт на синтез ps происходит при участии
альтернативного сигнала полиаденилирования мРНК
p-цепи (рис. 10.72). Этот сигнал находится в интроне
между экзонами 4 и 5 См и ответствен за удаление из
первичного транскрипта последовательности, коди-
рующей мембранный якорь рт и сохранение в
Ps-mPHK сегмента на конце экзона 4, кодирующего
мРНК
Белок
Трансмембранный
участок
®—СООН
19 мш
Пре-В-клетки
РИС. 10.72.
Использование альтернативных сигналов полиаденили-
рования. расположенных в Ср-сегменте, приводит к син-
тезу как мембраносвязанных (СМт), так и секрети-
руемых (CMs) легких цепей. Шесть Ср-экзонов обо-
Трансляция
NH, f
\
—соон
19MS
Пролиферирующие
плазматические клетки
значены на рисунке цифрами от 3 до 8. [J. Darnel,
D. Baltimore, Н. Lodish, Molecular Cell Biology (New York:
Scientific American Books, 1986), p. 1113.]
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
293
С-конец ps. Эта дополнительная последователь-
ность удаляется из рт-мРНК при сплайсинге.
При синтезе IgD также используются альтерна-
тивные сигналы полиаденилирования и сплайсинга.
Если первичный транскрипт тяжелой цепи продол-
жается, захватывая Cg-второй сегмент Сн-облас-
ти,-и полиаденилирование происходит за Cg. то
при дифференциальном сплайсинге из перестроен-
ного VDJ первичного транскрипта может элими-
нироваться Сц и в некоторых пре-В-клетках синте-
зируется иммуноглобулин IgDm (рис. 10.71). Пере-
ключение с синтеза IgDm на синтез lgDs происходит
в результате дифференциального использования
сайтов полиаденилирования аналогично тому, как
это имеет место в случае IgM.
Индуцированная антигеном дифференцировка
пре-В-клеток с образованием плазматических В-кле-
ток и В-клеток памяти сопровождается переключе-
нием изотипа (например, с IgM на IgG). Вначале,
по-видимому, происходит альтернативный сплай-
синг очень длинных пре-мРНК, синтез которых
начинается в обычном сайте инициации транскрип-
ции на 5'-конце лидерной последовательности V-об-
ласти и продолжается до сигналов полиаденилиро-
вания на конце кодирующих областей Су, СЕ или Са
(рис. 10.71). В результате сплайсинга З'-конца сег-
мента VHDHJH с интроном, непосредственно при-
мыкающим к одному из С-сегментов, образуются
мРНК, которые кодируют полипептиды с одина-
ковыми VH, но разными Сн. Однако этот механизм
переключения изотипов заменяется основанным на
рекомбинационном соединении VH DHJ „-сегмента с
одним из Сн-сегментов, расположенных за Сц и Cg,
с утратой разделяющего их участка ДНК (рис. 10.73).
Например, в результате делеции Сц, Cg, Суз и Су1
синтезируется тяжелая цепь с константной областью
Су2Ь, т.е. образуется IgG2b, а делеция См, Cg, Су3,
Су1, Су2Ь и СЕ приводит к синтезу IgA. При этом
легкие цепи и V „-области, а также антигенная спе-
цифичность остаются прежними.
Переключение тяжелых цепей происходит в ре-
зультате гомологичной рекомбинации между по-
следовательностями ДНК в интронах, расположен-
ными на расстоянии 2-3 т.п.н. перед каждым из
Сн-сегментов, за исключением Cg. Эти последо-
вательности обозначаются буквой S (от switch-пе-
реключение). У мыши несколько копий S-последо-
вательностей образуют перед каждым С„-сегмен-
том тандемные повторы длиной обычно 40- 80 нук-
леотидов; повторы, расположенные перед SM-cer-
ментом, гораздо короче, длиной 4-6 нуклеотидов,
при этом у мыши это последовательности GAGCTG
и TGGG.
Клональный отбор. Для объяснения огромного
многообразия антител и их уникальной специфич-
ности при связывании антигенов было высказано
|--------------------200-т.п.н.--------------------1
Переключение Су2ь
\ Сг3 СУ| \ Сгга С£ С«
________________>
Рекомбинационное
переключение
Су2Ь
VDJ Cyl Су2а С£ С«
Щ л лялж
Транскрипция,
полиаденилирование,
сплайсинг
IgG^MPHK
РИС. 10.73.
Перестройки, происходящие при рекомбинационном
переключении, в результате которых вместо иммуно-
глобулинов класса IgM начинают синтезироваться им-
муноглобины класса lgGt.
много гипотез. По-видимому, наиболее адекватной
является одна из них-гипотеза клонального отбора,
в которой разнообразие антител объясняется пере-
стройками ДНК. Вкратце идею клонального отбора
можно представить следующим образом. На ранних
этапах созревания антиген-независимых В-клеток
образуется много самых разных пре-В-клеток
(рис. 10.70). Каждая из них несет специфическое
антитело в виде поверхностного IgM. Специфич-
ность определяется уникальностью аминокислот-
ных последовательностей VL- и V „-областей, из
которых образуется антигенсвязывающий сайт
(CDR). При случайном взаимодействии пре-В-клет-
ки с родственным антигеном происходит стимуля-
ция клеточной пролиферации и секреции иммуно-
глобулина. Таким образом, после связывания анти-
гена пре-В-клетка дает начало клону циркулиру-
ющих плазматических клеток, которые секретируют
иммуноглобулины с той же антигенной специфич-
ностью, что и у исходно стимулированной В-клетки.
При таком поверхностном описании клональ-
ного отбора мы опускаем многие интересные дета-
ли, в частности не упоминаем о ключевой роли
вспомогательных клеток и факторов роста, которые
облегчают трансформацию пре-В-клеток. Еще одна
очень важная особенность состоит в том, что не все
потомки активированной пре-В-клетки представля-
ют собой секретирующие антитела плазматические
клетки; некоторые из них - это «бессмертные»
В-клетки памяти, которые «помнят» обо всех ветре-
294
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
чах организма с антигенами. Эта «запись» хранится
в циркулирующих В-клетках, несущих такие мемб-
раносвязанные антитела (IgM), VL- и VH-o6ласти
которых были отобраны по признаку наилучшего
соответствия чужеродному антигену. При повтор-
ной встрече с этим же антигеном В-клетки памяти
стимулируются к пролиферации и секреции таких же
антител, какие продуцировались пре-В-клетками-
предшественниками.
Сама по себе гипотеза клонального отбора не
объясняет в полной мере ни природы разнообразия
антител, ни того факта, что качество подгонки
антитела и антигена изменяется при повторных
встречах. В основе этих свойств иммунной системы
могут лежать какие-то другие факторы, тоже обе-
спечивающие разнообразие VH и VL. В покое
В-клетки памяти накапливают мутации, возника-
ющие как в самих Ун-областях, так и в ближайшем
их окружении, причем частота этих мутаций в мил-
лион раз превышает частоту спонтанных мутаций
в других генах. В результате происходит изменение
CDR, т.е. изменение антигенной специфичности ан-
титела с уже перестроенной V „-областью. Иногда
сайт связывания изменяется настолько, что анти-
тело вообще теряет свою активность. Но мутацион-
ные изменения могут и улучшать соответствие меж-
ду антителом и антигеном. Тогда при повторном
контакте с антигеном активируются преимущест-
венно те В-клетки, которые способны продуциро-
вать наиболее прочно связывающиеся антитела.
Поскольку эта система создает клетки, «наученные»
распознавать угрозу повторной инфекции более эф-
фективно, безопасность организма существенно по-
вышается.
Роль Т-клеток. Как мы уже отмечали, Т-лимфо-
циты обеспечивают как клеточный, так и гумораль-
ный иммунитет. Эти клетки образуются в костном
мозге, но созревают в тимусе (рис. 10.70). Основные
«гаранты» клеточного иммунитета-цитотоксичные
Т-клетки (CTL) (или Т-киллеры), а гуморального-
Т-хелперы и Т-супрессоры. Все физиологические
функции, характерные для Т-клеток, реализуются
в результате взаимодействия поверхностного Т-кле-
точного рецептора (TCR) с соответствующим анти-
геном. При этом CTL убивают клетки, несущие
антиген, а Тн-клетки ускоряют пролиферацию
пре-В-клеток. Взаимодействие TCR Тн-клеток и ан-
тигена, связанного В-клеткой. приводит к тому, что
Т-клетки начинают высвобождать специфические
белки роста и дифференцировки В-клеток (лимфо-
кины), которые стимулируют пролиферацию В-кле-
ток и секрецию ими антител. Роль Ts-клеток до
конца не установлена; возможно, они «приглуша-
ют» ответную реакцию В-клеток на собственные
антигены организма.
По своей структуре и способности распознавать
антиген TCR напоминают мембраносвязанные ан-
титела (рис. 10.74), однако есть и ряд существенных
отличий. TCR остаются связанными с клеточной
плазматической мембраной и никогда не секрети-
руются. Кроме того, если поверхностные В-клеточ-
ные антитела связывают как растворимые, так и
связанные с клеткой антигены, то TCR взаимо-
действуют только с антителами, ассоциированными
с клеткой. Кроме того, TCR обычно узнают и
чужеродные антигены, но только в том случае,
когда они находятся в комплексе со специфическими
белками, локализованными на поверхности клетки,
представляющей антиген. Эти белки являются про-
дуктами генов так называемого главного комплекса
гистосовместимости (МНС). Каким образом TCR
узнают и связывают чужеродные антиген и белки
МНС и чем объясняется такой дуализм? Эти интри-
гующие вопросы еще ждут ответа. Здесь мы скон-
центрируем свое внимание на структуре TCR и тех
геномных перестройках, которые приводят к экс-
прессии генов, кодирующих составляющие TCR по-
липептиды.
Все TCR представляют собой мембраносвязан-
ные гетеродимеры (рис. 10.74). Большинство из них
состоит из двух цепей, а и ₽, но небольшая часть
TCR содержит две другие цепи-у и 5. Мол. масса
а-. 0- и 5-цепей составляет примерно 45 кДа. а
у-цепи-35 кДа. Разные типы Т-клеток несут либо
оф-, либо уб-TCR. Цепи удерживаются в паре с
помощью дисульфидных связей. Аналогично Н-
и L-цепям антител, каждая цепь TCR содержит
вариабельный N-конец, почти целиком выступа-
ющий из мембраны, и константный С-конец, от-
части заякоренный в мембране. Антигенная спе-
цифичность TCR определяется трехмерной струк-
турой. которая образуется в результате взаимо-
действия расположенных соответствующим обра-
зом Va- и Vp- или Vy- и Vg-областей.
Сходство между TCR и антителами распростра-
няется и на то. как обеспечивается разнообразие,
например, V-областей а- и 0-цепей на генетическом
уровне. Информация о V-области 0-цепи распреде-
лена между тремя отдельными участками ДНК
в хромосоме 6 мыши и хромосоме 7 человека:
с каждым из двух Ср-сегментов соединяются при-
мерно 20 Vp-, 2 Dp- и 6 Jp-сегментов. Генные
сегменты a-цепи- примерно 50 Va, 50 Ja и один
Са- у мыши и человека локализованы на хромосоме
14. Кодирующие сегменты для у-цепей также раз-
делены, но образуют четыре кластера, каждый из
которых содержит несколько V-, J- и С-сегментов;
в результате их соединения образуется ген функцио-
нальной у-цепи. Механизм перестройки до конца не
изучен, но, по-видимому, сходен с таковым для
генов антител. Здесь тоже сигналом узнавания слу-
жит комплекс из гептамерной и нонамерной единиц,
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
295
Легкие цепи
Антигенсвязываю-
щий сайт
а-цепь
Клеточная
мембрана
NH2
Антигенсвязываю-
щий сайт
Антйгенсвязываю-
щий сайт
NH2
NH, NH
6-цепь
COO- coo-
Т-клеточный рецептор
Тяжелые
цепи
COO' coo-
Иммуноглобулин M
РИС. 10.74.
Схема, иллюстрирующая структуру
Т-клеточного рецептора и мембрано-
связанного иммуноглобулина. Т-кле-
точный рецептор содержит две цепи:
а и Р; каждая из них имеет мол. массу
примерно 40 кДа и связана своим
С-концом с мембраной. Вариабель-
ные области и антигенсвяэывающие
сайты находятся на N-конце. [Т. Honjo.
S. Habo, Ann. Rev. Biochem. 54 (1985).
p. 820.]
разделенных некодирующим спейсерным участком
длиной 11-12 и 22-23 п. н.. а перестройки, вероятно,
катализируются рекомбиназой. Рекомбинация в
Т-клетках тоже происходит неточно, что обеспечи-
вает изменчивость мест соединения рекомбиниру-
ющих участков. Сначала происходят перестройки
в локусе, отвечающем P-цепи, а затем-в локусе
a-цепи, при этом в случае успешной перестройки
в одном из аллелей генов Р- или a-цепи перестройка
в других аллелях блокируется (аллельное исключе-
ние). Насколько это известно, мутации в V-областях
TCR не происходят, поэтому разнообразие TCR
обусловливается соединением различных генных
сегментов и неточностью рекомбинации. Локали-
зация генов a-цепи и генов тяжелой цепи (иммуно-
глобулина) на одной, 14-й хромосоме у человека
имеет одно важное следствие: она создает пред-
посылки для аберрантного соединения VH- и J „-об-
ластей при участии общих сигнальных последова-
тельностей и рекомбиназы. Некоторые происходя-
щие при этом перестройки хромосомы 14 связы-
вают с возникновением Т-клеточных опухолей.
Как и в случае с В-клетками, антигенная специ-
фичность Т-клеток закладывается на ранних стадиях
их созревания, происходящего в тимусе в отсутствие
антигенов (рис. 10.70). Позже при связывании
Т-клетки с клеткой, несущей чужеродный поверх-
ностный антиген, который может связываться с
TCR, ускоряется пролиферация клона Т-клеток с
данной специфичностью. Например, в ответ на
взаимодействие с инфицированными клетками, не-
сущими вирусный антиген, образуется клон цито-
токсичных Т-клеток. Вероятно, аналогичным обра-
зом появляется клон хелперных Т-клеток в ответ на
взаимодействие незрелых Т-клеток с макрофагами,
которые расщепляют антиген и несут на своей
поверхности продукты этого расщепления.
Для того чтобы клетки, несущие поверхностные
антигены, могли связаться с TCR, расположенным
на поверхности CTL- или Тн-клетки, необходимо,
чтобы на их поверхности присутствовал также белок
МНС (рис. 10.75). Эти белки кодируются генными
семействами, называемыми Н2 (у мышей) и HLA (у
человека). HLA-комплекс занимает не менее 3-106
296
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 10.75.
Т-клеточный рецептор, связанный как с антигеном, так
и с поверхностным белком МНС. (С любезного раз-
решения Н. McDevitt.)
п. н. на хромосоме 6. Для ответа Т-клетке, по-види-
мому, необходим тройной комплекс, состоящий из
МНС, антигена и Т-клеточного рецептора. Одна
группа белков МНС, называемая МНС класса 1.
присутствует на поверхности большинства сомати-
ческих клеток. Эти белки необходимы для узнавания
и связывания цитотоксичной Т-клеткой чужеродных
антигенов. Для связывания чужеродного антигена
Тн-клетками антиген должен находиться в комп-
лексе с белком МНС класса II, который присутству-
ет на поверхности как макрофага, так и В-клеток.
Гены МНС у разных особей одного вида существен-
но различаются, поэтому полиморфны и соответст-
вующие белки классов I и II. Различия особей по
белкам класса I лежат в основе того, что клетки
и ткани, взятые от какого-то индивида, оказывают-
ся неспособны расти в окружении клеток другого
индивида - явление, называемое отторжением тка-
ней. Эти различия влияют и на аутоиммунный ответ
антител на собственные антигены: нарушение то-
лерантности. Различия по белкам класса II могут
обусловливать некоторую изменчивость антигенной
специфичности среди особей одного вида.
г. Вариабельные поверхностные
антигены трипаносом
Трипаносомии. В некоторых регионах Африки
среди людей и домашних животных довольно ши-
роко распространена так называемая сонная бо-
лезнь. Ее возбудителем является простейшее Trypa-
nosoma brucei, а переносчиком-муха цеце. Паразит
размножается путем деления, достигая высокого
титра в соединительных тканях и внеклеточных
жидкостях, но более 99,9% клеток разрушаются
с помощью иммунной системы хозяина. У неко-
торых организмов, устойчивых к основным анти-
телам, через 7-10 дней наблюдается повторная
вспышка численности популяции паразита. Этот
процесс повторяется с 7- 10-дневными интервалами
в течение месяцев (рис. 10.76), причем каждый раз
происходит размножение трипаносом, несущих оп-
ределенный, отличный от предыдущих случаев ва-
риант антигена (VAT). Дикие животные могут в
конце концов побороть инфекцию, но у человека
и домашних животных, если их не лечить, исход
обычно оказывается летальным. Смерть наступает
в результате целого ряда нарушений работы им-
мунологической системы и инфекций ЦНС.
Уникальная особенность трипаносом, обеспечи-
вающая им успех, состоит в их удивительной спо-
собности избегать иммунологического надзора. У
нескольких организмов из всей многочисленной по-
пуляции (с частотой ~ 10“6 за одно клеточное де-
ление) происходит переключение на синтез нового
типа антигена, и клетки вновь начинают размно-
жаться.
Варианты типов антигенов (VAT). Единствен-
ный антигенный детерминант, представляемый ор-
ганизму-хозяину,- это поверхностный гликопро-
теин. На плазматической мембране плотно упако-
ваны примерно 107 молекул этого вещества. Каж-
дый VAT имеет уникальный поверхностный антиген
(он называется вариантом поверхностного гликопро-
теина, VSG), и потомки одной клонированной три-
паносомы продуцируют 1000 и даже более разных
VSG (рис. 10.76). Если особи данного вида инфи-
цированы одним штаммом трипаносом, то в них
обнаруживаются VAT одного типа. Трипаносомы,
размножающиеся в слюнных железах мух цеце,
которые инфицированы данным штаммом, проду-
цируют набор тех же антигенов. В отличие от
млекопитающих, у которых в данный момент про-
дуцируются VAT преимущественно одного вида,
у мух цеце трипаносомы образуют гетерогенную
в отношении антигенов популяцию. Таким образом,
геном трипаносом содержит целый набор генов,
кодирующих разные поверхностные антигены.
Переход на синтез нового VAT происходит в ре-
зультате выключения одного гена и включения дру-
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
297
РИС. 10.76.
Циклический характер инфекций у кроликов, заражен-
ных T.brucei. VAT-вариант антигена, VSG-соответст-
вующий вариант поверхностного гликопротеина. В пер-
вом цикле значительная часть паразитов несет на своей
поверхности VSG, отличные от VSG, соответствующего
VAT-2
исчезает
Юдней
VAT-N. Каждый VAT может стабильно поддерживаться
путем переноса от кроликов к мышам или крысам,
в которых зта циклическая инфекция несколько зату-
хает. В 1 мл крови крысы насчитывается 109 трипано-
сом.
того; при этом у разных клеток мухи обнаружи-
ваются разные поверхностные антигены. Разные
штаммы T.brucei имеют свои наборы поверхност-
ных антигенов. Набор, продуцируемый одним кло-
ном трипаносом, называется серодемом.
Каждый штамм Т. brucei не только экспрессирует
определенный серодем, но и синтезирует разные
антигены-члены этого серо дема-в определенном
порядке. Одни VAT появляются на ранних стадиях
инфекции, другие позже. Кроме того, когда пара-
зиты проникают в уже инфицированный организм,
этот порядок сохраняется. Как правило, самая пер-
вая многочисленная популяция паразита во вновь
инфицированном хозяине продуцирует разнообраз-
ные VAT, а последующие популяции становятся
более гомогенными (рис. 10.76).
Варианты поверхностных гликопротеинов (VSG).
Несмотря на индивидуальные особенности, все VSG
имеют общую структурную основу: это гомоди-
меры гликозилированных полипептидов длиной
примерно 500 аминокислот (рис. 10.77). Все VSG
гликозилированы по определенным позициям; к не-
которым аминокислотам в других позициях при-
соединены углеводы. У самих полипептидов можно
выделить вариабельную и относительно констант-
ную области. Вариабельная область находится на
N-конце и содержит примерно 400 аминокислот.
Константные области содержат 100 аминокислот-
ных остатков; они находятся вблизи С-конца и
сходны у всех VSG, но не идентичны. В зависимости
от степени гомологии С-концов выделяют два под-
класса VSG Т. brucei. Такая гомологичность присуща
мРНК
AUG
Мини-экзон,
35 нуклеотидов трансляция
Первичный
продукт
трансляции
Сигнальный пептид
"Хвост"
Созревание
полипептида
Зрелый VSG
сно сно
I I I
I сно сно
сно
Вариабельная
область
Константная
область
РИС. 10.77.
Типичный поверхностный гликопротеин T.brucei
(VSG), соответствующая мРНК (ее структура
была определена исходя из данных о структуре
клонированной кДНК) и первичный продукт
трансляции.
298
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
даже VSG, относящимся к разным серодемам. По
этой причине и в особенности потому, что сайты
гликозилирования находятся на С-конце VSG, на-
блюдается перекрестная реакция между антителами
к разным очищенным VSG, а не к самим трипа-
носомам. Основным антигенным детерминантом
трипаносомы является МН2-конец VSG. поскольку
именно он выступает из мембраны.
Зрелые VSG, выделенные из трипаносом, обра-
зуются из полипептидных предшественников, ко-
торые простираются как за NH2-, так и за СООН-
концы. Об этом говорят результаты сравнения
структуры VSG с соответствующей мРНК и пер-
вичным продуктом трансляции (рис. 10.77). Струк-
тура VSG-мРНК была установлена на основании
данных о структуре кДНК; на долю VSG при-
ходится примерно 10% суммарных белков трипа-
носомы, и наличие соответствующих мРНК в
больших количествах облегчает клонирование кДНК.
Идентичность клонов кДНК была доказана мето-
дом HART (см. рис. 6.25) с использованием спе-
цифических антител. Типичная кДНК содержит
5'-нетранслируемую лидерную последовательность
длиной примерно 100 нуклеотидов, за которой сле-
дует участок из 99 нуклеотидов, кодирующий ти-
пичный сигнальный пептид; последний состоит из 33
аминокислот и не обнаружен в VSG. Далее идут
последовательности, кодирующие зрелый VSG мол.
массой 50 кДа, а затем участок из 60-70 нуклеоти-
дов, кодирующий С-концевой сегмент, который, как
и сигнальный пептид, в зрелом VSG отсутствует.
Этот С-концевой сегмент обладает ярко выражен-
ными гидрофобными свойствами, что облегчает
заякоривание VSG в мембране трипаносомы, и при
соответствующих условиях может быть удален.
VSG-мультигенное семейство. Каждый из кон-
кретных серодемов, продуцируемых разными кло-
нами трипаносом, кодируется неким набором генов.
Независимо от того, какой именно VSG синтези-
руется, в каждой трипаносоме присутствуют гены
всех VSG. Есть данные, что по крайней мере часть
этих генов объединена в геноме трипаносомы в
кластеры. Примерно 103 VSG-генов могут быть
сходны или даже идентичны, но другие гены су-
щественно различаются. Наименее вариабельны
З'-концы генов. Об этом свидетельствуют резуль-
таты блот-гибридизации субклонов из различных
областей кДНК с рестрицированной геномной ДНК.
5'-концевые участки кДНК при гибридизации дают
одну или несколько полос, фрагменты, взятые из
середины тех же кодирующих последовательнос-
тей,-обычно несколько большее их число, а З'-кон-
цевые зонды-до 50 полос. Лишь небольшая часть
этих зондов представляет собой совершенные по-
вторы, поскольку с ужесточением условий гибри-
дизации многие полосы исчезают. Интересно, что
зонды, происходящие из З'-нетранслируемой об-
ласти мРНК, гибридизуются с самым большим
числом геномных фрагментов.
До сих пор мы говорили о мультигенных се-
мействах отдельных клонов T.brucei. Как соотно-
сятся между собой генные семейства, кодирующие
разные серодемы? Некоторые VSG-гены и белки
присутствуют во многих (но не во всех) серодемах,
другие характерны только для какого-то одного
серодема. Это означает, что в отличие от боль-
шинства генов и генных семейств по крайней ме-
ре некоторые VSG-гены эволюционируют очень
быстро.
Переключение экспрессии с одного гена VSG на
другой. Как только трипаносомный клон переходит
с образования одного VAT на синтез другого,
выключается один VSG-ген и включается другой.
Каждому VAT соответствует только один вид
VSG-мРНК. Ранее в этой главе мы обсуждали три
механизма переключения для быстрой регуляции
транскрипции. При синтезе флагеллина Salmonella
оперон Н2 включается и выключается в результате
обратимых инверсий промотора. При инверсии
G-сегмента в геноме фага Ми промотор остается на
месте, но определенные гены перемещаются в по-
ложение правее промотора. У дрожжей при пере-
ключении типа спаривания сегменты, содержащие
соответствующие гены и промоторы, перемещаются
в активный участок генома. Именно этот последний
механизм используют трипаносомы для переклю-
чения генов VSG, но не в чистом виде, а с некото-
рыми модификациями, еще не до конца установлен-
ными. Ясно одно: для трипаносом и других прос-
тейших характерны уникальность, с одной стороны,
и изменчивость-с другой.
Связь экспрессирующихся генов VSG с теломер-
ными областями хромосом трипаносомы. Экспрес-
сирующиеся гены VSG были идентифицированы
с помощью блот-гибридизации в жестких условиях
зондов кДНК с геномной ДНК, а также благодаря
повышенной чувствительности этих генов в составе
хроматина к ДНКазе 1 (разд. 8.7.а). Весьма свое-
образны карты этих сегментов, особенно последо-
вательностей, фланкирующих экспрессирующиеся
гены VSG (рис. 10.78).
К 5'-концу экспрессирующегося гена часто при-
мыкает участок, не содержащий эндонуклеазных
сайтов (рис. 10.78, Б). По мере удаления от 5'-конца
карты приобретают более обычный вид. Расстояние
между кодирующей частью VSG и эндонуклеазными
сайтами перед 5'-концом гена может различаться
даже для ДНК из двух разных клонов трипаносом,
кодирующих одинаковые VSG (ср. рис. 10.78, Б и В).
Эта ситуация сохраняется, несмотря на то что орга-
низация эндонуклеазных сайтов в 5'-дистальном
кластере может быть одинаковой, свидетельствуя об
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
299
В.
А. Тан демный повтор ВС в крупных хромосомах
76п.н.-повторы
б. Одна из ELC-копим vsb-гена 2
В Bg ESPvh
И НИ
>-------—
Е
♦
Экспрессия
Выкл.
Вкл.
Вторая ELC-копия VSG-гена 2
В Bq ES Pv
И *11
Вкл.
Г ВС-копия геназ, расположенная в теломерной области
В Н Xm Xh В
т Выкл.или вкл.
А - Ген 3 может переместиться в другой сайт экспрессии
Вкл.
Примерно 25 т.п.н.
РИС. 10.78.
Связь между основной (ВС) и экспрессируемой (ELC)
копиями VSG-генов (без соблюдения масштаба). Ко-
дирующие участки выделены цветом. Транспозирован-
ные единицы включают последовательности, располо-
женные к 5'-концу от кодирующего участка. Вертикаль-
ные линии - повторы длиной 76 п. н. (их число не со-
ответствует указанному на рисунке). Обычно с ELC
связано больше повторов, чем с ВС. А. Тандемный
повтор ВС (1 и 2), локализованный во внутренней
области крупной хромосомы. Б, Структура одной из
ELC-копий гена 2 после транспозиции. В. Структура
другой ELC-копии гена 2. Расстояние между геном
и кластером сайтов рестрикции на дистальном конце
теломеры (Т) различно для случаев Б и В. Находятся ли
эти две ELC на разных теломерах или на одной и той же,
неясно. Г. Структура теломерной ВС гена 3. Ген в этом
сайте может быть как молчащим, так и экспрессиру-
емым. Д. ELC гена 3 после перемещения в другую
теломеру.
одинаковом расположении VSG-генов в обоих кло-
нах. Вариабельность длины участка, примыкающего
к 5'-концу гена, связана с различием в числе тандем-
ных повторов длиной 76 п. н.
Участок, примыкающий к З'-концу экспрессиру-
ющихся генов, также содержит очень немного эндо-
нуклеазных сайтов (рис. 10.78, Б, В, Д'). Более того,
карты нельзя продолжить на расстояние более 5-10
т.п.н. от З'-конца гена VSG. Независимо от того,
какая именно эндонуклеаза используется, в резуль-
тате рестрикции получается фрагмент, один конец
которого находится в этой «конечной» позиции.
Кроме того, если перед расщеплением рестрнктазой
препараты ДНК обработать Вд/31 (разд. 4.1.в), то
получатся более короткие фрагменты. Все это
объясняется тем, что «конец» представляет собой
реальный предшествовавший конец дуплексной ДНК.
Экспрессирующиеся гены VSG всегда расположены
вблизи теломеры, и их З'-конец ориентирован в
сторону теломеры.
T.hrucei имеет примерно 100 хромосом. Многие
из них очень малы: менее 106 п.н. Экспрессиру-
ющиеся VSG-гены обнаружены по крайней мере
в трех разных теломерных областях, но точное
число потенциально активных теломерных локусов
неизвестно. Однако ясно, что локализация гена в
области теломеры - необходимое, но не достаточное
условие экспрессии. Помимо одного, экспрессиру-
ющегося в данный момент времени гена, в области
теломеры могут находиться и другие, молчащие
VSG-гены (рис. 10.78, Г). Они могут включаться без
300
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
изменения своей локализации. Существуют и другие
молчащие VSG-гены, расположенные не в области
теломеры, а во внутренних хромосомных локусах
(рис. 10.78, Л). Прежде чем экспрессироваться, ко-
пии этих генов перемещаются в теломерные сайты.
Сходным образом некоторые теломерные молча-
щие VSG-гены перед экспрессией подвергаются дуп-
ликативной транспозиции в другую теломеру
(рис. 10.78, Г, Д). Таким образом, для экспрессии
одного из VSG-генов необходимо, чтобы 1) этот ген
находился в нужной области теломеры; 2) эта об-
ласть была активирована; 3) VSG-гены в других
потенциально активных областях теломеры были
молчащими. Несмотря на то что некоторые спосо-
бы встраивания VSG-генов в потенциально экспрес-
сируемые области теломер установлены, механизм
активации и инактивации экспрессии генов в этих
областях остается неизвестным. Результаты неко-
торых недавно проведенных экспериментов наводят
на мысль, что отсутствие генной экспрессии обус-
ловливается какими-то локальными модификация-
ми остатков цитозина. В «молчащих» областях те-
ломеры, в отличие от транскрибируемых областей,
Prwll- и Pst 1-сайты не подвергаются расщеплению.
Встраивание VSG-генов в потенциально активные
теломерные области. Установлены некоторые спо-
собы встраивания VSG-генов в потенциально ак-
тивные теломерные области. Одна из копий мол-
чащего гена, так называемая основная копия (basic
сору, ВС), с помощью дупликативной транспозиции
может встроиться в потенциально активную тело-
мерную область. При этом молчащий ген может
находиться как во внутренней области хромосомы,
так и в неактивной теломере (рис. 10.79, Л или Б).
Наиболее вероятный механизм включения VSG-
гена-дупликативная транспозиция по типу генной
конверсии, которая происходит у дрожжей при пе-
реключении типа спаривания. Следовательно, штамм,
синтезирующий определенный VSG. часто имеет на
одну копию соответствующего гена больше, чем
несинтезирующий штамм. При следующем пере-
ключении эта дополнительная копия обычно, но не
всегда, утрачивается, и ее место может занять дру-
гой VSG-ген. Помимо дупликативной транспозиции
за перемещение VSG-гена из молчащей теломеры в
потенциально активную может быть ответственна
реципрокная рекомбинация (рис. 10.79, В). В этом
случае активный ген не утрачивается и может реак-
тивироваться в результате последующего переноса.
Все эти типы переключений выявляются с помошью
импульсного гель-электрофореза и ДНК-блоттинга
(рис. 10.80).
На рис. 10.78 (А. Б и В) представлена структура
основной копии VSG-гена, расположенной во внут-
ренней области хромосомы, и двух транспозирован-
ных экспрессирующихся копий (ELC; от англ,
expression-linked copies) гена из двух разных клонов
трипаносомы. Структура ВС была получена с ис-
пользованием фаговых и космидных клонов, ото-
бранных из геномных библиотек с помощью гиб-
ридизации с кДНК-зондами. Клонирование ELC из
транспозируемых VSG-генов в Е. coli затруднено
нестабильностью их последовательностей. Поэтому
данные о структуре ELC были получены отчасти
путем блот-геномной гибридизации ДНК с субкло-
нированными ВС-зондами, а отчасти с помощью
секвенирования коротких субклонированных участ-
ков ELC. Размер транспозируемых ELC-единиц
варьирует от I до 40 т. п. н., но чаще всего состав-
ляет примерно 3 т. п. н. Они содержат последова-
тельности, присутствующие в зрелой мРНК, а также
5'-фланкирующие участки разной длины, которых
нет в мРНК. З'-конец транспозируемой единицы
близок к З'-концу гена. Области ELC и ВС за
пределами самой транспозируемой единицы не го-
мологичны между собой. Отметим, что участок
вблизи 5'-конца ВС содержит несколько тандемно
повторяющихся элементов длиной 76 п. н., которые
Реципрокная
Дупликативная транспозиция транслокация
А Б В
I 8 I---------------+’ I-----------»+’
I
Молчащий
ген
I----Н-+т !----------Н-Н
| Переключение |
f----в-+т !----------м—1-т
РИС. 10.79.
Три способа встраивания генов VSG в экспрессируемые
теломерные области. Дупликативные транспозиции из
внутреннего хромосомного сайта (А) или из другой
теломеры (Б), по-видимому, сопровождаются генной
конверсией. При каждом новом встраивании ранее ак-
тивный ген или утрачивается (дупликативная транспо-
зиция), или перемещается в новый сайт (реципрокная
транслокация; В).
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
301
в гораздо большем количестве присутствуют вблизи
5'-конца ELC из теломер. Таким образом, транспо-
зиция в сайт экспрессии может сопровождаться
спариванием этих повторов, причем в рекомбина-
ционных событиях могут участвовать разные копии.
В результате даже при транспозиции в один и тот же
сайт экспрессии расстояния между VSG-геном и рас-
положенными левее эндонуклеазными сайтами мо-
гут быть разными.
З'-конец транспозируемой единицы обладает не
менее интересными особенностями, чем 5'-конец.
Напомним, например, о необычайно консервативных
З'-концевых кодирующих последовательностях у
сильно различающихся в остальном отношении
VSG-генов. Еще более консервативен З'-нетрансли-
руемый участок различных ВС, ELC и кДНК. На-
против, у множества других мультигенных семейств
гомологичные кодирующие участки ассоциированы
с высокодивергированными З'-нетранслируемыми
последовательностями. В чем смысл необычной
консервативности З'-нетранслируемых участков?
Частично ответ на этот вопрос дает детальное
сравнение З'-нетранслируемого участка мРНК (и
ELC) с соответствующими ВС. В основном кДНК
и ВС сходны, но все же не идентичны: различия
состоят в единичных нуклеотидных заменах и не-
больших делециях. Впрочем, иногда кДНК оказы-
ваются полностью идентичными соответствующим
ВС. Эти факты можно объяснить тем, что при
дупликативной транспозиции одной из копий ВС
происходит не только спаривание тандемных пов-
торов длиной 76 п. н. на 5'-конце, но и гомологичная
рекомбинация на З'-конце транспозируемой едини-
цы и З'-конце VSG-гена. Вследствие миграции ветви
точка кроссинговера может находиться в любом
месте гомологичного участка, который начинается
внутри З'-кодирующей области и продолжается до
конца нетранслируемого участка (разд. 2.4.6). Та-
ким образом, ELC могут представлять собой мо-
заику последовательностей, часть которых происхо-
дит от старого гена, а часть-от встроенного. За-
метим, что в таком случае могли бы существовать
два фактора, дающие вклад в давление отбора,
направленное на обеспечение консервативности
З'-конца: 1) сохранение структуры С-концов VSG-
генов и 2) сохранение последовательности, которая
обеспечивает гомологичность рекомбинации.
Транскрипция генов VSG. Для экспрессии генов
VSG необходимо, чтобы они находились в опреде-
ленных областях теломеры. Но, как мы уже гово-
рили, это лишь одно из условий экспрессии, по-
скольку некоторые VSG-гены остаются молчащими
даже в этом случае (рис. 10.80). Каков же механизм
переключения гена из одного состояния в другое?
Ответа на этот вопрос мы не знаем, хотя некоторые
данные на этот счет имеются. Например, экспрес-
Переключение
VAT-A VAT-B VAT-A'
VSG-A VSG-B VSG-A'
Н Зонд-VSG-A-ген
И Зонд-VSG-В-ген И
Зонд-"анонимная" ДНК
РИС. 10.80.
Геномные перестройки, приводящие к переключению
синтеза с VSG-А на VSG-В и обратно (VSG-А') в одном
из трипаносомных клонов. В каждой колонке суммиро-
ваны результаты блот-гибридизации нерасщепленных
хромосом трипаносомы, разделенных с помощью им-
пульсного гель-злектрофореза (разд. 6.5). Каждая по-
лоса отвечает сегменту ДНК, гибридизующемуся со
специфическим зондом (см. разъяснения в нижней
части рисунка). Гены VSG изображены в виде прямо-
угольников, которые выделены цветом, если ген экс-
прессируется в исследуемых VAT-клетках. В VAT-A-
клетках экспрессируется ген VSG-А, который находится
в теломерной области хромосомы размером примерно
2000 т.п.н. Вторая, молчащая копия гена VSG-А лока-
лизуется в крупной хромосоме, которая остается вверху
пластины электрофорезного геля. VAT-A содержит так-
же молчащую копию гена VSG-В, локализованную в
теломере. После переключения в фазу VAT-В тело-
мерный ген VSG-А становится молчащим, но остается
в ранее активной теломере. Копия теломерного VSG-B
встраивается в активную теломеру хромосомы длиной
340 т.п.н.; при этом, по-видимому, происходит дуп-
ликативная транспозиция участка размером 90 т. п. н.,
поскольку длина реципиентной хромосомы увеличива-
ется до 430 т. п. н. (эту хромосому выявляют на радио-
автографе по гибридизации с анонимной пробой, спе-
цифичной для данной хромосомы), и вновь начинается
экспрессия VSG-A (VAT-A'). Исходно активный ген
VSG-В перемещается с помощью рекомбинации из
экспрессируемого сайта в крупную хромосому (вверху
геля) и становится молчащим, а от ранее активной
хромосомы остается сегмент длиной 140 т.п.н. Ген
VSG-А реактивируется без изменения своей локали-
зации. [Из работы С. Shea et al., J. Biol. Chem. 261
(1986), p. 6056, с изменениями.]
302
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
сирующиеся VSG-гены находятся на большем рас-
стоянии от конца хромосомы, чем не экспресси-
рующиеся. Есть и другой, более интересный факт:
необычные свойства 5'-концов соответствующих
мРНК и длина транскрипционных единиц.
5'-фланкирующий участок VSG-мРНК сущест-
венно отличается от соответствующего участка
ELC, с которого он транскрибируется (рис. 10.81).
Во-первых, он начинается с нетранслируемого
участка длиной 35 пар оснований, ни в чем не
напоминающего область, примыкающую к ELC, и,
во-вторых, он содержит делеции. Аналогичные
участки длиной 35 п.н. присутствуют перед всеми
мРНК VSG, а также перед большинством (а воз-
можно, перед всеми) мРНК трипаносом. Они вхо-
дят в состав сегмента длиной 1.4 т.п.н., повто-
ряющегося в геноме трипаносомы примерно 200
раз. Большинство таких единиц-это прямые тан-
демные повторы, но некоторые из них дисперги-
рованы. Повторы транскрибируются in vivo с обра-
зованием кэпированных РНК длиной 140 нуклео-
тидов (medPHK, от англ, mini exon derived), несущих
на 5'-конце сегмент длиной 35 п.н. Транскрипция,
по-видимому, осуществляется с помощью РНК-по-
лимеразы II. Синтез мРНК трипаносом-будь то
мРНК VSG или других белков- сопряжен с перено-
сом кэпированного 5'-сегмента длиной 35 п.н. с
medPHK на транскрипт кодирующего участка. По-
скольку концентрация medPHK у трипаносом
обычно бывает высокой, вряд ли они участвуют
в транскрипционной регуляции экспрессии генов
VSG. По-видимому, регуляция осуществляется на
уровне транскрипции в самом сайте экспрессии.
Способные к экспрессии теломерные локусы, где
находятся VSG-гены, могут быть как транскрип-
ционно активными, так и неактивными и лишь
в очень редких случаях (во время самого переклю-
чения) в одной клетке одновременно функциониру-
РИС. 10.81.
5-конец VSG-мРНК (и других трипаносомных РНК)
является кзпированным 35-нуклеотидным мини-зкзо-
ном, который не представлен в ELC (Т-это теломера).
Помимо концевого гуанозина (m7G) модифицированы
еще четыре остатка. Два из них, А и С, 2'-0-метилиро-
ваны, а структура двух других, А и U (они отмечены
звездочками), неизвестна. [К. L. Perry, К. Р. Watkins,
N. Agabian, Proc. Natl. Acad. Sci. USA 84 (1987), p. 8190.)
ют несколько таких локусов. Транскрипция может
начаться на расстоянии до 60 т. п. н. перед VSG-геном.
Помимо VSG-области, этот длинный первичный
транскрипт содержит несколько неродственных ко-
дирующих участков. Для образования функцио-
нальной VSG-мРНК к области первичного тран-
скрипта, содержащей VSG-кодирующую последо-
вательность, должна присоединиться medPHK
(рис. 10.82). Эти две РНК соединяются с помощью
транс-сплайсинга (разд. 8.5.г), в результате прямого
сочленения экзона длиной 35 п. н. с VSG-кодиру-
ющим участком (рис. 10.82). З'-конец medPHK дли-
ной 100 нуклеотидов отделяется от полиаденили-
рованной части трипаносомной РНК с помощью
дебранчинг-фермента из клеток человека. Это пред-
определяется Y-подобной структурой побочного
продукта транс-сплайсинга (разд. 8.5.г). Аналогич-
ным образом могут образоваться функциональные
мРНК кодирующих последовательностей, располо-
женных перед VSG-геном.
Несмотря на то что образование VSG-mPHK
изучено во всех деталях, механизм включения и
выключения самого сайта экспрессии неясен. Тран-
скрипция VSG-генов не блокируется даже при очень
высоких концентрациях а-аманитина, а в области
промотора обнаружены последовательности, сход-
ные с промоторами трипаносомной рДНК. Таким
образом, возможно, в транскрипции участвует РНК-
полимераза I. Высказывалось интересное предпо-
ложение, состоящее в том, что активная транскрип-
ция осуществляется в ядрышке, где, как известно,
находится большая часть (а возможно, и вся)
РНК-полимераза I. В таком случае в качестве регу-
лятора транскрипции может выступать процесс пе-
рехода сайта экспрессии в ядрышко.
10.7. ЗАПРОГРАММИРОВАННЫЕ
АМПЛИФИКАЦИЯ И МОДУЛЯЦИЯ
ЭКСПРЕССИИ ГЕНОВ
В регуляции генной экспрессии в процессе раз-
вития иногда участвует запрограммированная амп-
лификация специфических сегментов ДНК. Это чет-
ко показано для некоторых простейших, беспозво-
ночных и позвоночных. В каждом случае в резуль-
тате амплификации в специфических клетках в опре-
деленный момент времени образуется огромное
число генных копий, в результате чего синтези-
руется большое количество важного генного про-
дукта. Однако молекулярные стратегии, использу-
емые для амплификации в конкретных случаях,
существенно различаются.
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
303
Ген VSG
Тандемные повторы medflHK
РИС. 10.82.
Образование VSG-мРНК. Матрицей для синтеза medPHK
служат тандемные повторы е medflHK. Кодирующая
область VSG-гена представлена как часть значительно
более длинного первичного транскрипта. medPHK и
длинный VSG-транскрипт соединяются друг с другом
в результате /лрамс-сплайсинга с образованием побоч-
ного продукта Y-PHK. [L.H.T. Van der Ploeg, Cell 47
(1986), p. 479.]
а. Непропорциональная репликация:
хорионические гены дрозофилы
Высокие требования к белкам оболочки яйца. За
непродолжительное время оогенеза у насекомых
фолликулярные клетки яичника, окружающие со-
зревающий ооцит, должны секретировать огромное
количество различных белков, которые формируют
хорион (оболочку яйца). У некоторых насекомых
эта краткосрочная проблема решается путем быст-
рой и эффективной транскрипции высокоповторя-
ющихся хорионических генов, которые составляют
часть генома соматических клеток и клеток заро-
дышевой линии. У Drosophila в нормальном га-
плоидном геноме имеются лишь единичные копии
генов различных белков хориона, но примерно за
18 ч до начала синтеза этих белков в фоллику-
лярных клетках начинается амплификация соответ-
ствующих генов, которые затем в течение 5 ч ин-
тенсивно экспрессируются.
Некоторые из 20-50 различных хорионических
генов Drosophila образуют два кластера; один клас-
тер из четырех тандемно расположенных генов на-
ходится на хромосоме 3, а другой из шести генов-
на Х-хромосоме (рис. 10.83). Амплификация этих
геномных областей в фолликулярных клетках была
прямо показана путем оценки изменений числа ко-
пий определенных рестрикционных фрагментов, по-
лученных из клонированных генов. Было показано,
например, что в Х-хромосоме область, содержащая
сами гены, амплифицирована почти 15 раз. Фланки-
рующие эту область последовательности амплифи-
цированы в меньшей степени, причем число копий
тем меньше, чем дальше они находятся от генного
кластера. Участки, удаленные от кластера на 40
т. п. н., амплифицированы только два раза. Другая
характерная особенность амплификации состоит в
том, что в результате нее не образуется никаких
новых рестрикционных фрагментов, которые гиб-
ридизовались бы с зондами из амплифицированной
области. Таким образом, никаких новых комби-
наций из сегментов ДНК не появляется. Наличие
градиента амплификации и отсутствие рекомбина-
ции позволяют предложить некий механизм увели-
чения числа копий хорионических генов.
304
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Перекрываю-
щиеся
^клонированные
зонды
РИС. 10.83.
Амплификация ДНК в области
кластера хорионических генов на
X хромосоме в фолликулярных
клетках Drosophila. В верхней час-
ти рисунка изображен сегмент
длиной 100 т.п.н.. содержащий
четыре гена хорионического белка
(выделены цветом). Стрелки ука-
зывают сайты расщепления эндо-
нуклеазой fcoRI Ниже представ
лен набор перекрывающихся фраг-
ментов этой области, которые
служили зондами при блот-гиб-
ридизации с рестрицированными
£соЯ1 геномными ДНК из эмбрио-
нальных и фолликулярных клеток.
Относительное число копий каж-
дого EcoRI-фрагмента определя
ли из соотношения радиоактив-
ностей зондов, гибридизующихся
с ДНК из фолликулярных и эм-
бриональных клеток (внизу). Чис-
ло копий каждого фрагмента в
эмбриональных клетках было
принято за единицу. [А. С. Spradling,
Cell 27 (1981), р. 193 ]
Амплификация путем непропорциональной репли-
кации. Представим, что сайт начала репликации
находится вблизи кластера генов и что синтез ДНК
протекает в обе стороны от этого сайта. Через
некоторое время в этой же точке начинается второй
раунд репликации, затем третий и т. д. и в результа-
те образуется множество копий данной области
(рис 10.84). Более того, поскольку каждая следу-
ющая репликативная вилка короче предыдущей, по
обе стороны от точки начала репликации возникает
градиент числа копий. Вновь образующиеся копии
не связаны ковалентно ни друг с другом, ни с исход-
ной дуплексной ДНК, поэтому новых комбинаций
ДНК не возникает. Эта модель предполагает, что
в данном случае происходит ослабление нормаль-
ного контроля, запрещающего многократную ини-
циацию репликации в одной точке. Это представ-
ляется вероятным, поскольку амплификация хорио-
нических генов продолжается даже после прекра-
щения нормального синтеза ДНК в фолликулярных
клетках. Адекватность модели была убедительно
показана в результате электронно-микроскопичес-
кого изучения амплифицированной ДНК из фол-
ликулярных клеток (рис. 10.85). Полученная микро-
фотография полностью соответствует одной из по-
ловинок модели, представленной на рис 10.84.
Способностью к амплификации обладают ко-
роткие, менее 5 т. п. н„ сегменты ДНК, расположен-
ные в области хорионических генов в хромосомах
3 и X Если эти сегменты встраиваются в другие
геномные сайты, то последние начинают временно
амплифицироваться в фолликулярных клетках.
Встраивание таких дополнительных копий элемен-
тов, контролирующих амплификацию, осуществляет-
ся путем включения их в P-элементы (с помощью
рекомбинации in vitro и клонирования) и после-
дующего введения содержащего этот P-элемент век-
тора в эмбрион Drosophila на ранних стадиях раз-
вития (разд. 10.2.6).
б. рДНК Xenopus: амплификация
по типу катящегося кольца
Хотя геномы большинства организмов и так
содержат многочисленные копии рДНК (разд. 9 2.а),
в ооцитах эти гены иногда амплифицированы в еще
большей степени, что создает необходимые пред-
посылки для эмбриогенеза. Наиболее полно изучен-
ный пример такого рода-Xenopus, у которой 600
(гаплоидных) геномных копий рДНК амплифнци-
рованы более чем тысячекратно и содержатся в
ядрышках.
Амплификация происходит в две стадии: в при-
мордиальной зародышевой клетке образуется не-
большое число копий рДНК, а затем во время
мейоза происходит массовая амплификация по ме-
ханизму катящегося кольца (разд. 2.1.ж). Конечные
продукты представляют собой крупные кольцевые
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
305
Следующие раунды
репликации
Обратите внимание, что связаны между собой только
две исходные цепи ДНК. [G. Stark, G. М. Wahl, Ann. Rev.
Biochem. 53 (1984), p. 478.]
РИС. 10.84.
Схематическое представление амплификации в резуль-
тате множественных инициаций двунаправленного син-
теза ДНК в одном сайте (модель «кожицы лука»).
молекулы ДНК, каждая из которых содержит до 100
повторяющихся рДНК-единиц. В отличие от обыч-
ных повторов геномной рДНК, в которых длина
межгенного спейсера (IGS) варьирует, большинство
1GS в кольцевых внехромосомных рДНК имеют
одинаковую длину. Создается впечатление, что ро-
доначальником кольцевых ДНК в каждом ооците
служит один или несколько хромосомных генов.
Однако в разных ооцитах даже в пределах единст-
венного организма амплифицируются разные пов-
торы геномной рДНК и IGS имеют разную длину.
Суммарное число копий рДНК, по-видимому, на-
ходится под контролем, и после оплодотворения
амплификации не происходит. Как именно одна
повторяющаяся рДНК-единица включается в катя-
щееся кольцо-остается загадкой.
20-158
306
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 10.85.
Электронная микрофотография ДНК из фолликуляр-
ных клеток Drosophila, в которой происходит ампли-
фикация хорионических генов. Стрелками указаны реп-
ликационные вилки. Точки едоль ДНК-нуклеосомы.
Кружком отмечена транскрибирующаяся область (цепь
РНК усеяна рибонуклеопротеиноеыми частицами). [С
любезного разрешения Y. N. Osheim, О. L. Miller, Jr., Cell
33 (1983), р. 543. Copyright Cell Press.]
в. Амплификация рДНК Tetrahymena
в макронуклеусах
У жгутиковых простейших при формировании
транскрипционно активных макронуклеусов ДНК
клеток зародышевой линии разбивается на сотни
фрагментов (разд. 9.6.6). Одновременно утрачива-
ется часть ДНК микронуклеуса, к сегментам ДНК
макронуклеуса добавляются теломерные последо-
вательности и большинство этих сегментов ампли-
фицируются примерно 45 раз. Однако фрагменты,
содержащие рДНК. амплифицируются в значитель-
но большей степени: число их копий достигает 104
и они упаковываются во множество ядрышек. Такая
структура макронуклеарной ДНК и число ее копий
сохраняются во время амитотической вегетативной
репликации макронуклеуса. Напротив, ДНК микро-
нуклеуса содержит лишь одну (гаплоидную) копию
генов рибосомной РНК. О механизме репликации
рДНК, создающем такую предпочтительность, из-
вестно мало. Каждый фрагмент рДНК макронук-
леуса содержит две полноразмерные транскрип-
ционные единицы рДНК и нетранскрибируемый
сегмент длиной примерно 4 т.п.н. (рис. 10.86). Эти
две половины молекул представляют собой инвер-
тированные друг относительно друга повторы; весь
фрагмент, таким образом, является палиндромом.
Его репликация начинается с сайтов, лежащих в
центральных IGS-участках, и расходится от них
в обоих направлениях к концам фрагмента. После-
довательности внутри IGS, необходимые для эф-
фективной репликации рДНК, перекрываются с эн-
хансерами (разд. 8.2), указывая на связь между реп-
ликацией рДНК и регуляцией транскрипции.
10.8. НЕЗАПРОГРАММИРОВАННЫЕ
ТАНДЕМНЫЕ АМПЛИФИКАЦИИ
Если в результате мутации или действия специ-
фического ингибитора активность какого-то важ-
ного генного продукта становится недостаточной,
то часто обнаруживаются мутантные клетки, в ко-
торых данный генный продукт синтезируется в
большем, чем обычно, количестве. У многих таких
клеток имеются многочисленные тандемные пов-
торы соответствующего гена, в результате чего
продуцируются избыточные количества мРНК и
соответствующих полипептидов. Способностью к
такой амплификации генов обладают как бактерии,
так и разнообразные эукариотические клетки в
культуре. Амплификация происходит и в опухо-
левых клетках. Как правило, это довольно редкие
события; остается посмотреть, как часто наблю-
дается амплификация в нормальных соматических
клетках.
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
307
Микронуклеус,! копия (гаплоид)
25S 5,8S 17S
Вырезание,
присоединение
теломеры
Возможный промежуточный продукт
длиной 11 т.п.н.
_ Теломера
Образование
палиндрома,
1 амплификация
Макронуклеус,104 копий
Центр палиндрома,точка начала
репликации
I-------------1______________I______________I_____________I
10 5 0 5 10
т.п.н.
РИС. 10.86.
Единичный генный локус микронуклеарной рРНК
Tetrahymena и возможный способ его вырезания
и амплификации во фрагментах макронуклеар-
ной ДНК. [D.D. Larson et al.. Cell 47 (1986),
P. 229 ]
а. Амплификации, позволяющие
компенсировать ферментную
недостаточность
Устойчивость к лекарственным веществам и
амплификация генов. Если обработать клетки ин-
гибиторами (лекарственными веществами), которые
действуют специфическим образом на какой-то
ключевой фермент, то погибнут все клетки, за
исключением резистентных к этому ингибитору му-
тантов. Две такие системы ингибитор фермент
представлены на рис. 10.87. Аналог фолиевой кис-
лоты метотрексат (аметоптерин) ингибирует дигид-
рофолатредуктазу (ДГФР), которая катализирует
синтез тетрагидрофолата метаболита, необходи-
мого, например, для образования dTMP. Ь1-(фос-
фонацетил)-Е-аспартат (ФАЛА) ингибирует аспар-
тат-карбамоилтрансферазу- фермент, принимающий
участие в биосинтезе пиримидиновых нуклеотидов.
Если клетки в культуре обработать этими ингиби-
торами в низкой концентрации (например, 10“8 М
метотрексатом), то можно отобрать несколько
(одну из 10б— 107) резистентных клеток. Если выра-
щивать эти клетки при более высоких концентра-
циях ингибитора, то вновь образуется несколько
колоний. Так постепенно можно получить клетки,
устойчивые к очень высоким концентрациям ле-
карственных веществ (например, к 10-3 М мето-
трексату). Многие такие резистентные клетки пре-
одолевают действие ингибитора, продуцируя в
большом количестве как ингибируемый фермент,
так и соответствующую мРНК. В основе повы-
шения уровня синтеза мРНК и фермента лежит
амплификация единственного гена дигидрофолат-
редуктазы или аспартат-карбамоилтрансферазы.
Клетки, высокорезистентные к лекарственным
веществам, представляют собой богатый источник
мРНК, которую можно очистить и клонировать
в виде кДНК. В свою очередь, имея клонированную
кДНК, можно точно определить концентрацию
мРНК и число копий соответствующего гена. В
табл. 10.6 представлен пример поэтапного отбора
высокоустойчивых к ФАЛА клеток. Клетки, полу-
ченные на каждом из этапов, устойчивы к более
высоким концентрациям ФАЛА, чем клетки на пре-
дыдущем этапе. Повышение устойчивости сопро-
вождается увеличением содержания в клетках ас-
партат-карбамоилтрансферазы. Кинетический ана-
лиз с использованием к ДНК-зондов и РНК или
ДНК, выделенных из резистентных клеток, пока-
зывает, что увеличивается число копий как мРНК,
так и соответствующего гена.
Характерен ли рассматриваемый феномен толь-
ко для тех двух генов, о которых мы говорим, или
амплификация более универсальное явление? В
табл. 10.7 представлено несколько других систем,
20*
308
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Аспартат
Карбамоилфосфат
МЧфосфонацетил'М.-аспартат
/(ФАЛА)
N-карбамиласпартат
Асп артат-карбамоилтрансфераза
РИС. 10.87.
Ингибирование дигидрофолатредуктазы метотрексатом
(А) и аспартат-карбамоилтрансферазы 1\1-(фосфонаце-
тил)-Ь-аспартатом (ФАЛА) (Б).
в которых были отобраны клетки с амплифици-
рованными генами. Заметим, что в одном из слу-
чаев отбор клеток с амплифицированными генами
не зависел от применяемого лекарственного вещест-
ва. Результаты всех этих опытов наводят на мысль,
что амплификация многих сегментов генома в куль-
тивируемых клетках происходит спонтанно, хотя
и с низкой частотой (10-4 10~7 в зависимости от
системы). Эта спонтанность означает, что давление
отбора-просто механизм, позволяющий выявить
эти редкие события.
Реверсия мутантного фенотипа, происходящая
в результате генной амплификации. Дополнитель-
ные указания на возможность спонтанных ампли-
фикаций ДНК в эукариотических клетках дает ана-
лиз вторичных мутаций, изменяющих мутантный
фенотип. Фермент гипоксантин-гуанин -фосфорибо-
зилтрансфераза (ГГФРТаза) ответствен за утилиза-
цию пуринов в животных клетках (рис. 5.32). Клет-
ки, мутантные по гену hprt, синтезируют малоак-
тивную форму ГГФРТазы и не растут на среде,
содержащей гипоксантин, аминоптерин и тимидин
(среда HAT), поскольку аминоптерин - близкий род-
ственник метотрексата-блокирует синтез de novo
пуриновых нуклеотидов (рис. 10.87), что препятст-
вует эффективному использованию гипоксантина.
В клетках-ревертантах, хорошо растущих на среде
HAT, ген hprt часто бывает амплифицирован, в
одном случае до 50 раз. Эти клетки синтезируют
большое количество мРНК мутантной ГГФРТазы,
и содержание данного белка в них почти в 50 раз
превышает таковое в клетках дикого типа (табл. 10.7).
На то, что этот избыточный белок представляет
собой именно мутантную форму, указывают его
необычные кинетические свойства и нестабильность.
Итак, клонированные клетки, содержащие единст-
венный мутантный ген hprt, кодирующий неста-
бильный фермент, дают ревертантов с амплифи-
цированным мутантным геном, в которых синте-
зируется огромное количество дефектного фермен-
та. Недостаток качественного фермента компенси-
руется количеством мутантной формы.
б. Структура амплифицированных
генов
Изменения нуклеотидной последовательности.
Амплифицированная единица содержит помимо са-
мого гена еще и другие последовательности. Обыч-
но амплифицированные сегменты содержат регу-
ляторные последовательности, которые фланкиру-
ют транскрипционную единицу, кодирующий учас-
ток, интроны, а также другие протяженные после-
довательности. Повторяющиеся единицы образуют
тандем, а некоторые из них содержат инвертиро-
Таблица 10.6. Отбор клеток сирийского хомячка, устойчивых к ФАЛА
Линия клеток Концентрация ФАЛА Относительное содержание - аспартат-карбамоил транс- феразы Относитель- ное число копий гена
условия, в которых про- изводится отбор условия. При которых рост подавляется на 50%
мРНК фермент
Дикий тип 0 мМ 6 мкМ 1 1 2980 1
А 0,1 мМ 1,6 мМ 6,6 6,4 225 13
В 10 мМ 17 мМ 42 25 54 55
С 25 мМ 34 мМ 68 73 25 190
” Указаны значения Cot0 5 для ДНК сирийского хомячка, полученные при отжиге с аспартат-карба-
моилтрансферазной кДНК (разд. 7.5.6). [G. М. Wahl, R. A. Padgett. G. R. Stark J. Biol. Chem. 2S4 (1978),
p. 8679.]
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
309
Таблица 10.7. Амплификация генов, компенси-
рующая ферментную недостаточность (отдель-
ные примеры)
Фермент, кодиру- Организм
емый данным геном
Признак, по которому
проводился отбор
Дигидрофолатре-
дуктаза
Аспартат-карба-
моилтрансфе-
раза
Металлотионеин
Аспарагин-
синтетаза
Г ипоксантин-гуа-
нин- фосфорибо-
зилтрансфераза
Адениндеза-
миназа
ADH411
Мышь, китай-
ский хомячок
Сирийский
хомячок
Мышь,
обезьяна
Китайский
хомячок
Мышь, китай-
ский хомячок
Крыса
Дрожжи
Устойчивость к мето-
трексату (МТХ)
Устойчивость к N-
(фосфонацетил)-Ь-
аспартату (ФАЛА)
Устойчивость к Cd2 +
Устойчивость к Р-ас-
партилгидроксамату
Реверсия клеток от
hrpt~ к hrpt+
Устойчивость к дезок-
сикоформицину
Устойчивость к анти-
мицину А
11 Ген ADH4 дрожжей кодирует либо изоформу алкогольдегидро-
геназы, либо белок, регулирующий экспрессию гена алкоголь-
дегидрогеназы.
ванные дупликации гена. Рассмотрим сначала при-
мер с аспартат-карбамоилтрансферазой. У живот-
ных этот фермент является частью единого поли-
пептида, обладающего еще двумя активностями:
карбомоил-фосфатсинтазной и дигидрооротазной.
Они вступают в действие соответственно перед ас-
партат-карбамоилтрансферазой и после нее в цепи
реакций биосинтеза пиримидиновых нуклеотидов.
Этот «трехглавый» фермент, называемый CAD, ко-
дируется одним геном. У сирийского хомячка этот
ген содержит 25 т. п. н., большая часть которых
представлена по меньшей мере 37 вставочными
последовательностями; мРНК составляет в длину
лишь 7,9 т.н. и содержит примерно 6300 коди-
рующих оснований. Однако соответствующая амп-
лифицированная единица может содержать более
500 т.п.н. Аналогично, дигидрофолатредуктазный
ген мыши имеет длину 31 т. п. н., содержит не менее
пяти интронов и кодирует белок из 186 амино-
кислот; длина соответствующей мРНК примерно
1,6 т.н. Длина амплифицированной единицы у раз-
ных резистентных клеток различается, но может
превышать 100 т.п.н.
Самое простое объяснение столь большого раз-
личия в длине повторяющихся единиц и самих генов
состоит в том, что эти длинные геномные сегменты
содержат амплифицированные гены. Однако струк-
тура повторяющихся единиц такова, что возникают
предположения о наличии более сложных процес-
сов. В некоторых случаях амплифицированные еди-
ницы включают последовательности, происходящие
из несцепленных геномных локусов, которые в нор-
ме не соседствуют с данным геном. Кроме того,
отдельные повторяющиеся единицы оказываются не
всегда идентичными даже в клонированных кле-
точных линиях. Более того, в период интенсивного
роста клеток в условиях, при которых отбор благо-
приятствует амплификации, перестройки внутри
повторяющихся единиц могут происходить непре-
рывно.
Изменение структуры хромосом. Амплификация
генов часто (но не всегда) сопровождается сущест-
венными морфологическими изменениями хромосом.
Эти изменения могут быть нескольких типов, и
какой именно из них реализуется, зависит от вида
клеток и самого гена. Иногда накапливаются очень
маленькие дополнительные хромосомы, называемые
двойными мини-хромосомами (рис. 10.88, Л), которые,
по-видимому, содержат амплифицированные после-
довательности. Двойные мини-хромосомы ацент-
ричны и при делении клетки распределяются нерав-
номерно. Этот факт, а также то, что клетки, содер-
жащие много двойных мини-хромосом, растут хуже,
чем нормальные клетки, означает, что в неселектив-
ной (не содержащей лекарственного вещества) среде
резистентность клеток постепенно утрачивается.
Морфологические изменения второго типа об-
наруживаются в нормальных во всем остальном
хромосомах и проявляются в атипичном характере
исчерченности определенных областей при диффе-
ренциальном окрашивании (рис. 7.23); эти облас-
ти называют гомогеиио окрашивающимися (HSR).
Как правило, они весьма протяженны, и хромосо-
ма, содержащая HSR, длиннее своего гомолога
(рис. 10.88, Б). В клетках, устойчивых к метотрекса-
ту, после гибридизации in situ с радиоактивным
зондом, представляющим собой клонированный
дигидрофолатредуктазный ген, и обработки фото-
чувствительной эмульсией на HSR обнаруживаются
зерна серебра. Таким образом, амплифицированные
последовательности находятся внутри HSR. В хро-
мосомах клеток сирийского хомячка, содержащих
амплифицированные гены аспартат-карбамоил-
трансферазы, также обнаруживаются «лишние»
протяженные участки. Поскольку эти участки окра-
шиваются негомогенно, они не являются HSR, но
тем не менее содержат амплифицированные сегмен-
ты ДНК (рис. 10.88, В). В некоторых случаях амп-
лификация гена CAD происходит и в нормальном
локусе CAD сирийского хомячка (в хромосоме В9р),
хотя другие хромосомы тоже могут содержать мно-
го копий этого гена и иметь разнообразные ано-
малии (рис. 10.88, Г). Могут встречаться и двойные
мини-хромосомы. Во всех исследованных случаях
амплификация касается только одной пары гомо-
логичных хромосом. В отличие от нестабильных
амплифицированных генов в двойных мини-хромо-
310
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 10.88.
Хромосомные аберрации, связанные с амплификацией
генов и выявляемые на стадии метафазы. А. В устой-
чивых к ФАЛА клетках сирийского хомячка обнаружены
сверхкомплектные двойные мини-хромосомы (DM),
дицентрические хромосомы (DC) и амплифицирован-
ные хромосомные области (CAD). [G. М. Wahl et al..
Mol. Cell Biol. 2 (1982), p. 308.] Б. В устойчивых
к метотрексату фибробластах китайского хомячка на
хромосоме 2 обнаружена протяженная гомогенно ок-
рашивающаяся область (прямоугольная скобка). Нор-
мальная хромосома отмечена стрелкой. (С любезного
разрешения J. L. Biedler). В. Результаты гибридизации in
situ зонда CAD с ДНК из устойчивых к ФАЛА клеток
сирийского хомячка. Области гибридизации указаны
стрелками. Г. Кариотип клеток рис. В. Амплифици-
рованную область содержит удлиненная хромосома В9.
Аберрантны и некоторые другие пары хромосом.
[G.M. Wahl et al.. Mol. Cell Biol. 2 (1982), p. 308.]
сомах, амплифицированные гены в нормальных
хромосомах остаются стабильными даже при ин-
тенсивном росте клеток на неселективной среде.
в. Механизм амплификации
Любая модель амплификации должна учитывать
следующие факты: 1) амплификация может проис-
ходить в обычном локусе, в удаленных от него
локусах и в независимых двойных мини-хромосо-
мах; 2) геномные сегменты, значительно превыша-
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
311
ющие по своей длине сам ген, амплифицированы; 3)
для амплифицированных участков характерны пере-
стройки исходных геномных последовательностей;
4) амплифицированные участки могут иметь разную
структуру. Эти модели должны объяснять неста-
бильность некоторых амплифицированных сегмен-
тов в неселективной среде, хотя не обязательно все
должно сводиться к простой реверсии амплифи-
кации. Кроме того, необходимо рассмотреть воп-
рос, действует ли отбор на сам процесс амплифи-
кации или его роль состоит в благоприятствовании
редким геномным событиям, а значит, возможности
их проявления.
Цитологические аспекты. В результате длитель-
ного роста устойчивых к метотрексату клеток в
присутствии данного лекарственного вещества в
популяции с нестабильным фенотипом и двойными
мини-хромосомами появляются клетки со стабиль-
ным резистентным фенотипом и HSR. Эти наблю-
дения наводят на мысль, что в некоторых случаях
вначале образуются двойные мини-хромосомы, а
затем внехромосомная ДНК с помощью рекомби-
нации вновь встраивается в хромосому. Далее мо-
жет происходить дополнительная амплификация.
В пользу этой гипотезы говорят данные о том, что
трансформация клеток чужеродными клонирован-
ными генами (например, при трансформации мы-
шиных клеток дигидрофолатредуктазным геном ки-
тайского хомячка или клеток китайского хомячка
дигидрофолатредуктазным геном мыши) с после-
дующим поэтапным отбором сопровождается
амплификацией чужеродного гена. Амплифициро-
ванная единица содержит чужеродный ген и со-
седствующие с ним векторные последовательности
(например, pBR322), а также сегменты хозяйской
ДНК, которые могут фланкировать встроенные
сегменты. Такие эксперименты показывают, что
амплификация исходного гена может сопровож-
даться целой серией событий, в том числе ампли-
фикацией in situ в исходном генном локусе, по-
следующим вырезанием, образованием внехромо-
сомного промежуточного продукта-двойных мини-
хромосом, повторным встраиванием в геном и до-
полнительной амплификацией. Это могло бы объ-
яснить тот факт, что амплификация происходит
в локусах, отличных от того, где в норме находится
данный ген; кроме того, это обусловливает сцеп-
ление данного сайта в результате рекомбинации
с удаленной областью генома. То или другое из этих
событий может чаще происходить в каких-то оп-
ределенных генах или клетках, приводя к разным
эффектам амплификации в различных системах. Все
эти сложные процессы, проявляющиеся на цито-
логическом уровне, свидетельствуют о разнообра-
зии и сложности явлений на молекулярном уровне.
Молекулярные механизмы. При запрограммиро-
ванной амплификации могут использоваться разные
молекулярные механизмы, и любой из них или все
сразу могут участвовать в случайной амплификации
генов. Кроме того, на разных стадиях поэтапной
амплификации могут действовать разные механиз-
мы. Длинные тандемные повторы образуются как
в результате непропорциональной репликации
(разд. 10.7.а), так и амплификации по типу катя-
щегося кольца (разд. 10.7.6). Непропорциональная
репликация в рамках модели, описанной на рис. 10.84,
сама по себе не приводит к появлению сцепленных
тандемных повторов. Однако можно представить,
что противоположные концы нескольких копий сег-
ментов ДНК в «кожице лука» оказываются в не-
посредственной близости друг от друга и соеди-
няются в результате рекомбинации с образованием
тандемных повторов, в том числе повторов, со-
держащих инвертированные дуплицированные гены
(рис. 10.89). В таком сцеплении могут участвовать,
например, члены семейства диспергированных пов-
торов, находящиеся на противоположных концах
повторяющейся единицы. Кроме того, противопо-
ложные концы новой копии могут соединиться с
образованием внехромосомной кольцевой структу-
ры, а затем амплифицироваться с помощью меха-
низма катящегося кольца. Все эти модели предпо-
лагают, что амплифицированная единица содержит
точку начала репликации. Об адекватности модели
этого типа свидетельствует тот факт, что в одном из
клонов устойчивых к ФАЛА клеток хомячка были
обнаружены внехромосомные замкнутые кольцевые
ДНК длиной около 250 т.п.н.. Эти молекулы,
слишком мелкие, чтобы их можно было отнести
к двойным мини-хромосомам, содержат ген CAD
и реплицируются автономно. Возможно, они яв-
ляются посредниками между исходными хромосом-
ными генами и двойными мини-хромосомами.
Длинные тандемные повторы образуются также
в результате последовательных актов неравного
кроссинговера между повторяющимися последова-
тельностями в сестринских хроматидах или гомо-
логичных хромосомах (разд. 2.4.а и 9.4.д). При этом
должно произойти несколько клеточных делений.
Однако эта модель не может объяснить амплифи-
кацию в хромосомных локусах, удаленных от ис-
ходного гена.
Из всех этих моделей следует, в частности, что
в месте соединения тандемно повторяющихся еди-
ниц появляется новый сегмент. Это не согласуется
с амплификацией хорионического гена Drosophila,
о которой шла речь в разд. 10.7.а; в этом случае
дополнительные копии ДНК не связаны ковалентно
с остальным геномом и никаких новых сегментов не
образуется. Новые последовательности в месте со-
единений обнаружены в С/4 D-системе (рис. 10.90).
Отметим, что они амплифицируются вместе с самой
312
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
РИС. 10.89.
Использование модели «кожицы лука» для объяснения
образования длинных тандемных повторов или «катя-
щихся колец». [G. Stark, G. М. Wahl, Ann. Rev. Biochem.
53 (1984), р. 478.]
единицей. В клеточных линиях, выделенных после
амплификации гена CAD, в местах соединений об-
наруживаются разные последовательности, т. е. на-
чало повторяющейся единицы не фиксировано.
Кроме того, некоторые из амплифицированных
сегментов содержат инвертированные повторы
амплифицированного гена, а структура амплифи-
цированных единиц, как мы уже говорили, разли-
чается даже в пределах одной клеточной линии.
Таким образом, схема, представленная на рис. 10.90,
весьма упрощена.
Нестабильность амплифицированных генов. Ут-
рата амплифицированных генов после снятия дав-
ления отбора предполагает, что тандемные обра-
зования динамичны-в них все время происходят
делеции и новые амплификации. Такая пластич-
ность была продемонстрирована непосредственным
образом с использованием специально сконструи-
рованных клеток. Мышиные клетки, несущие му-
тации как в тимидинкиназном (z/c~), так и в аде-
нин-фосфорибозилтрансферазном {aprt~) генах,
трансфицировали рекомбинантным вектором, со-
держащим z/c-ген вируса простого герпеса и ген aprt
из клеток хомячка. Используя подходящую селек-
10. РЕОРГАНИЗАЦИЯ ГЕНОМА
313
Нормальные Резистентные
клетки клетки
т.п.н.
8,5
5,6
-Начало
РИС. 10.90.
Возникновение и выявление тандемных повторов в ДНК
на примере амплификации гена CAD. А. Схема ампли-
фикации и образования нового рестрикционного фраг-
мента на месте соединения генов (Е1 ЕЗ-ЕсоЯ1-сайты).
Б. Блот-гибридизация по Саузерну EcoRI-фрагментов
ДНК из нормальных и С-клеток (табл. 10.6). В качестве
зонда использовали фрагмент гена CAD левее сайта Е2.
В ДНК из нормальных клеток с зондом гибридизуется
один сегмент (его длина 8,5 т. п. н.), присутствующий
тивную среду, получили клеточные линии, явля-
ющиеся одновременно и tk+, и aprt+, которые со-
держали тандемные амплификации сегмента ДНК,
включающего оба гена (рис. 10.91). Если поддер-
живали давление отбора в отношении амплифи-
цированного гена tk, но растили клетки на среде,
селективной для фенотипа aprt~, то мутанты aprt~
появлялись с той же частотой, с какой они появ-
ляются в нормальных клеточных популяциях с ге-
ном aprt дикого типа. Этот факт весьма удивителен,
поскольку он означает, что одновременно мути-
ровали все 20 содержащихся в данных клетках
в виде единственной копии на гаплоидный геном. В та-
ком же числе копий этот сегмент находится и в ДНК
С-клеток. Однако в этой ДНК присутствует еще один
сегмент, 5,6 т.п.н.. гибридизующийся с указанным зон-
дом. Он находится в месте соединения CAD-генов,
и число его копий, оцениваемое по интенсивности по-
лосы на радиоавтографе, соответствует 190-кратной
амплификации CAD.
амплифицированных копий aprt+. Однако именно
это и случилось! В некоторых клонированных aprt~,
tk+-клетках все гены aprt были делегированы, что
дало фенотип aprt~. В других клонах все 20 копий
гена aprt+ несли одну и ту же мутацию. Конкретный
тип мутации варьировал от одной клеточной линии
к другой, но общая картина была одинакова: все 20
генов aprt изменялись одним и тем же образом.
В тех случаях, когда фенотип aprt обусловливался
делециями, последние во всех копиях тандемного
повтора также находились в одинаковых позициях.
Поскольку вероятность возникновения одинаковых
314
ЧАСТЬ III ОРГАНИЗАЦИЯ. ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
-► -► -► -► -►
! *- И 1 g * g ><<-><-«
| Отбор aprt~
Деления гена Мутация в гене
aprt aprt
’’
♦ Направлениетранскрипции
Ген aprt (хомячок)
Ген4к (герпесвирус)
□ Другая последовательность
(S Мутантный ген aprt
РИС. 10.91.
Изменения в наборе тандемных повторов одинаковы
в каждом из его элементов.
случайных мутаций во всех копиях мала, напра-
шивается вывод, что изменение произошло в одной
из копий, а затем все другие копии были заменены
ею. Идентичность копий в тандемном повторе
должна обеспечиваться с помощью какого-то меха-
низма. Однако точно так же, как мы не можем
сейчас предложить какой-то конкретный механизм
спонтанной амплификации, мы не можем остано-
виться и ни на одном из механизмов гомогенизации
членов тандемных повторов. Помимо неравного
кроссинговера, благодаря которому может умень-
шаться и увеличиваться число копий в тандемных
повторах, здесь могут играть роль последователь-
ные делеции и амплификации, непропорциональная
репликация и генная конверсия.
Химическая стимуляция амплификации. Имеют-
ся данные о том, что многие сегменты генома
изредка подвергаются спонтанной случайной амп-
лификации. Частота такой амплификации в куль-
туре клеток (в отсутствие давления отбора) состав-
ляет не более IO-3~ 10-4 на каждую амплифици-
руемую единицу в одном клеточном поколении.
Если учесть, что средняя длина амплифицируемой
единицы составляет ~ 105 п.н. и, таким образом,
в геноме размером 109 п.н. содержится 104 таких
единиц и каждая из них может амплифицироваться,
то большинство вновь образующихся клеток будут
содержать одну новую амплификацию. Кроме того,
если создать определенные условия, благоприятст-
вующие амплификации (например, постепенно по-
вышать концентрацию метотрексата, в результате
чего будут амплифицироваться гены дигидрофолат-
редуктазы), то частота амплификаций будет изме-
няться. Частоту амплификаций случайных геном-
ных сегментов увеличивают некоторые соединения,
в частности ингибиторы репликации ДНК и многие
канцерогены. В клетках опухолей, либо индуциро-
ванных канцерогенами, либо возникших спонтанно,
часто обнаруживаются гомогенно окрашенные об-
ласти хромосом и двойные мини-хромосомы. Эти
и другие хромосомные аберрации связаны с ампли-
фикацией последовательностей ДНК, в частности
кодирующих известные онкогены.
Может оказаться, что разнообразные агенты,
нарушающие нормальную репликацию ДНК, спо-
собствуют незапрограммированной репликации
случайных областей генома. При подходящих се-
лективных условиях клетки с избыточными копиями
какого-либо важного гена будут иметь преимущест-
во перед обычными клетками, и после нескольких
циклов клеточных делений, особенно в условиях
усиливающегося жесткого давления отбора, может
произойти массированная амплификация соответ-
ствующего гена.
Литература
Введение
Ниже приведены ссылки на статьи, дающие представ-
ление об основных затрагиваемых здесь темах. Все работы
сгруппированы в соответствии с разделами, выделенными
в данном введении, и даны в хронологическом порядке.
Структура эукариотических генов и
их экспрессия
Р. Chambon. 1981. Split Genes., Sei. American, 244 (5), 60-71.
W. Gilbert. 1981. DNA Sequencing and Gene Structure. Nobel
Lecture, December 8, 1980. Science, 214, 1305-1312.
A. B. Lassar, P.L. Martin. R.G. Roeder. 1983. Transcription of
Class III Genes: Formation of Preinitiation Complexes.
Science, 222, 740-748.
E.J. Milner-White. 1984. Isozymes, Isoproteins and Intors.
Trends Biochem Sci., 9, 517-519.
E Setfling, M. Jasin. W. Schaffner. 1985. Enhancers and Euka-
ryotic Gene Transcription. Trends Genet., 1, 224-230.
W. Gilbert, M. Marchionni, G. Me Knight. 1986. On hhe Anti-
quity of Introns. Cell. 46, 151-154.
G. Dreyfuss, M.S. Swanson, S. Pinol-Roma. 1988. Heterogeneous
Nuclear Ribonucleoprotein Particles and the Pathway of
mRNA Formation. Trends Biochem Sci., 13, 86-89.
E. P. Geiduschek, G. P. Tocchini-Valentini. 1988. Transcription
by RNA Polymerase III. Annu. Rev. Biochem., 57,
873-914.
T. W Traut. 1988. Do Exons Code for Structural or Functional
Units in Proteins? Proc. Natl. Acad. Sci. U.S.A., 85,
2944-2948.
B. Wasylyk. 1988. Transcription Elements and Factors of RNA
Polymerase В Promoters of Higher Eukaryotes. Crit.
Rev. Biochem., 23, 77-120.
J. A. Witkowski. 1988. The Discovery of Split Genes. Trends
Biochem. Sci., 13, 110-113.
M. Belfort. 1989. Bacteriophage Introns: Parasities Within
Parasites. Trends Genet., 5, 209-213.
P.N. Benfey, N. Chua. 1989. Regulated Genes in Transgenic
Plants. Science, 244, 174-181.
R. G. Brennan, B. W. Matthews. 1989. Structural Basis of DNA-
Protein Recognition. Trends Biochem Sci., 14, 286-290.
ITS. Dynan. 1989. Modularity in Promoters and Enhancers.
Cell, 58, 1-4.
R.B. Goldberg, S.J. Barker, L. Perez-Grau. 1989. Regulation of
Gene Expression During Plant Embryogenesis. Cell, 56,
149-160.
N.D. Hastie, R.C. Allshire. 1989. Human Telomeres: Fusion
and Interstitial Sites. Trends Genet., 5, 326-331.
S. Murphy, B. Moorefield, T. Pieler. 1989. Common Mechanisms
of Promoter Recognition by RNA Polymerases II and III.
Trends Genet., 5, 123-126.
M. Ptashne. 1989. How Gene Activators Work. Sci. American,
260 (1), 40-47.
J.H. Rogers. 1989. How Were Introns Inserted into Nuclear
Genes? Trends Genet., 5, 213-216.
Строение генома эукариот
E О. Long. I. В. Dawid. 1980. Repeated Genes in Eukaryotes.
Annu. Rev. Biochem., 49, 727-764.
J.G. Gall. 1981. Chromosome Structure and the C-Value
Paradox. J. Cell Biol., 91, 3s-14s.
A.C. Spradling, G.M. Rubin. 1981. Drosophila Genome Orga-
nization: Concerved and Dynamic Aspects. Annu. Rev.
Genet., 15, 219-264.
D. Baltimore. 1985. Retroviruses and Retrotransposons: The
Role of Reverse Transcriptase in Shaping the Eukaryotic
Genome. Cell, 40, 481-482.
E. F. Vanin. 1985. Processed Pseudogenes: Characteristics and
Evolution. Annu. Rev. Genet., 19, 253-272.
J. H. Nadeau. 1989. Maps of Linkage and Synteny Homologies
Between Mouse and Man. Trends Genet., 5. 82-86.
Перестройки последовательностей ДНК
В. McClintock. 1951. Chromosome Organization and Gene
Expression. Cold Spring Harbor Symp. Quant. Biol., 16,
13-57.
J. Lederberg, T. lino. 1956. Phase Variation in Salmonella.
Genetics, 41, 743-757.
M.P. Calos, J.H. Miller. 1980. Transposable Elements. Cell,
20, 579-595.
R. T. Schimke. 1980. Gene Amplification and Drug Resistance.
Sci. American, 243 (5), 60-69.
P. Leder. 1982. The Genetics of Antibody Diversity. Sci.
American, 246 (5), 102- 115.
N. V. Federoff. 1984. Transposable Genetic Elements in Maize
Sci. American, 250 (6), 84-98.
D. Baltimore. 1985. Retroviruses and Retrotransponosons: The
Role of Reverse Transcriptase in Shaping the Eukaryotic
Genome. Cell, 40, 481-482.
S. Tonegawa. 1985. The Molecules ot the Immune System. Sci.
American, 253 (4), 122-131.
P. Borst, D. R. Greaves. 1987. Programmed Gene Rearrange-
ments Altering Gene Expression. Science, 235, 658-667.
H. Varmus. 1987. Reverse Transcription. Sci. American, 257 (3),
56-64.
D. J. Finnegan. 1989. Eukaryotic Transposable Elements and
Genome Evolution. Trends Genet., 5, 103-107.
A. M. Lambowitz. 1989. Infectious Introns. Cell, 56, 323-326.
H. E. Varmus. 1989. Reverse Transcription in Bacteria. Cell, 56,
721-724.
Эволюционные аспекты
F. Avala. 1978. The Mechanisms of Evolution. Sci. American,
239, (3), 56-69.
J. E. Darnell. Jr. 1978. Implications of RNA-RNA Splicing in
Evolution of Eukaryotic Cells. Science, 202, 1257-1260.
W. Gilbert. 1978. Why Genet in Pieces? Nature, 271, 501.
M. Kimura. 1979. The Neutral Theory of Molecular Evolution.
Sci. American, 241 (5), 98-126.
D. Reannev. 1984. Genetic Noise in Evolution? Nature, 307,
318-319.
J. E. Darnell, Jr. 1985. RNA. Sci. American, 253 (4), 68-78.
R. F. Doolittle. 1985. The Geneology of Some Recently Evolved
Vertebrate Proteins. Trends Biochem. Sci., 10, 233-237.
R. F. Doolittle. 1985. Proteins. Sci. American, 253 (4). 88-99.
W. Gilbert. 1985. Genes in Pieces Revisited. Science, 228,
823-824.
A. C. Wilson. 1985. The Molecular Basis of Evolution. Sci.
American, 253 (4), 164-173.
T.R.Cech. 1986. Biological Catalysis by RNA. Annu. Rev.
Biochem., 55, 599-629.
316
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
J.E. Darnell, W. F. Doolittle. 1986. Speculations on the Early
Course of Evolution. Proc. Natl. Acad. Sci. U.S.A., 83,
1271-1275.
G. A. Dover. 1986. Molecular Drive in Multigene Families: How
Biological Novelties Arise, Spread, and Are Assimilated.
Trends Genet., 2, 159-165.
N.R.Pace, G. J. Olsen, C.R.Woese. 1986. Ribosomal RNA
Phylogeny and the Primary Lines of Evolutionary Descent.
Cell, 45, 325-326.
W. Gilbert. 1987. The RNA World. Cold Spring Harbor Symp.
Quant. Biol., 52, 901-905.
A.M. Weiner, N. Maizels. 1987. tRNA-like Structures Tag the
Ends of Genomic RNA Molecules for Replication: Impli-
cations for the Origin of Protein Synthesis. Proc. Natl.
Acad. Sci. U.S.A., 84, 7383-7387.
U. Wintersberger, E. Wintersberger. 1987. RNA Makes DNA:
A Speculative View of the Evolution of DNA Replication
Mechanisms. Trends Genet., 3, 198-202.
L. L. Cavalli-Sforza, A. Piazza, P. Menozzi, J. Mountain. 1988.
Reconstruction of Human Evolution: Bringing Together
Genetic, Archeological, and Linguistic Data. Proc. Natl.
Acad. Sci. U.S.A., 85, 6002-6006.
C. R. Woese. 1987. Bacterial Evolution. Microbiol. Rev., 51,
221-271.
K. G. Field, G. J. Olsen, D. J. Lane, S. J. Gioannoni, M. T. Ghi-
selin, E.C. Raff, N. R. Pace, R.A. Raff. 1988. Molecular
Phylogeny of the Animal Kingdom. Science, 239, 748-
753.
R. F. Doolittle. 1989. Similar Amino Acids Revisited. Trends
Biochem. Sci., 14, 244-245.
G. F. Joyce. 1989. RNA Evolution and the Origins of Life.
Nature, 338, 217-224.
S. Pdabo, R.G. Higuchi, A.C. Wilson. 1989. Ancient DNA and
the Polymerase Chain Reaction. The Emerging Field of
Molecular Archaeology. J. Biol. Chem., 264, 9709-9712.
Статьи обзорного характера
J. Darnell. H. Lodish, D. Baltimore. 1986. Molecular Cell Bio-
logy. Scientific American Books. New York. The Molecu-
lar Biology of Homo sapiens. Cold Spring Harbor Quant.
Biol. 51, whole issue.
D. Freifelder. 1987. Molecular Biology, 2nd, ed. Jones and
Bartlett, Boston.
J.D. Watson, N.H. Hopkins. J. W. Roberts, J.A.Steitz,
A. M. Weiner. 1987. Molecular Biology of the Gene, 4th ed.
Benjamin/Cummings, Menlo Park, Ca.
B. John, G. Miklos. 1988. The Eukaryote Genome in Develop-
ment and Evolution. Allen and Unwin, London.
L. Stryer. 1988. Biochemistry, 3rd ed. W. H. Freeman. San
Francisco.
Evolution of Catalysis. 1988. Cold Spring Harbor Symp. Quant.
Biol., 52, whole issue.
B. Alberts, D. Bray, J. Lewis. M. Raff, K. Roberts, J.D. Wat-
son. 1989. Molecular Biology of the Cell, 2nd ed. Garland
Publishing, New York and London.
D. E. Berg, M.M. Howe (eds.). 1989. Mobile DNA. American
Society for Microbiology, Washington, D. C.
B. Lewin. 1990. Genes, 4th ed. Wiley, New York.
J.D. Rawn. 1988. Biochemistry. Carolina Biological Supply
Co., Burlington, North Carolina.
Статьи к соответствующим разделам глав
8.2.а
R. Н. Reeder. 1985. Mechanisms of Nucleolar Dominance in
Animals and Plants. J. Cell Biol., 101, 2013-2016.
B. Sollner-Webb. J. Tower. 1986. Transcription of Cloned
Eukaryotic Ribosomal RNA Genes. Annu. Rev. Biochem.,
55, 801-830.
R. G. Worton, J. Sutherland, J. E. Sylvester, H. F. Wllard,
S. Bodrug, 1. Dube, C. Duff. V. Kean, P. N. Ray, R. D. Schmi-
ckel. 1988. Human Ribosomal RNA Genes: Orientation of
the Tandem Array and Conservation ot the 5' End.
Science, 239. 64-67.
8.2.6
P. Chambon. 1974. Eukaryotic RNA Polymerases. In:
P. D. Boyer (ed.), The Enzymes, pp. 261-331. Academic
Press, New York.
L. A. Allison, M. Moyle. M. Shales, C.J. Ingles. 1985. Extensi-
ve Homology Among the Largest Subunits of Eukaryotic
and Prokaryotic RNA Polymerases. Cell, 42, 599-610.
J. Biggs, L.L. Searles, A.L. Greenleat. 1985. Structure of the
Eukaryotic Transcription Apparatus: Features of the Gene
for the Largest Subunit of Drosophila RNA Polymerase II.
Cell, 42, 611-621.
A. Sentenac. 1985. Eukaryotic RNA Polymerases. Crit. Rev.
Biochem., 18, 31-90.
E. Bateman, M.R. Paule. 1986. Regulation of Eukaryotic
Ribosomal RNA Transcription by RNA Polymerase Mo-
dification. Cell, 47, 445-450.
P. Bucher, E.N. Trifonov. 1986. Comrilation and Analysis of
Eukaryotic POL II Promoter Sequences. Nucleic Acids
Res., 14, 10009-10026.
J. L. Buhler, M. Riva, C. Mann, P. Thuriaux, S. Menet, J. Y. Mi-
covin, I. Threich. S. Mariotte, A. Sentenac. 1987. Eukaryo-
tic RNA Polymerases, Subunits and Genes. In: W. S. Rez-
nikoff et all. (eds.), RNA Polymerase and the Regulation of
Transcription, pp. 25-36. Elsevier, Amsterdam.
D.L. Cadena, M.E. Dahmus. 1987. Messenger RNA Synthesis
in Mammalian Cells in Catalyzed by the Phosphorylated
Form of RNA Polymerase II. J. Biol. Chem., 262, 12468-
12474.
M. Nonet, D. Sweetser, R.A. Young. 1987. Functional Redun-
dancy and Structural Polymorphism in the Large Subunit
of RNA Polymerase II. Cell, 50, 909- 915.
J. Tower, B. Sollner-Webb. 1987. Transcription of Mouse rDNA
in Regulated by ano Activated Subform of RNA Polyme-
rase 1. Cell, 50, 873-883.
8.2.B
E. A. Elion, J. R. Warner. 1984. The Major Promoter Element
of rRNA Transcription in Yeast Lies 2 kb Upstream. Cell,
39, 663-673.
R. Miesfeld, N. Arnheim. 1984. Species-Specific rDNA Trans-
cription Is Due to Promoter-Specific Binding Factors.
Mol. Cell. Biol., 4, 221-227.
R. Miesfeld, B. Sollner-Webb, C. Croce, N. Arnheim. 1984. The
Absence of a Human-Specific Ribosomal DNA Transcrip-
tion Factor Leads to Nucleolar Dominance in Mouse >
> Human Hybrid Cells. Mol. Cell. Biol., 4, 1306-1312.
R.H. Reeder. 1984. Enhancers and Ribosomal Gene Spacers.
Cell, 38, 349-351.
I. Grummt, H. Sorbaz, A. Hofmann, E. Roth. 1985. Spacer Se-
quences Downstream of the 28S RNA Coding Region Are
Part of the Mouse rDNA Transcription Unit. Nucleic
Acids Res., 13, 2293-2304.
R. M. Learned, S. Cordes, R. Tjian. 1985. Purification and
Characterization of a Transcription Factor That Confers
Promoter Specificity to Human RNA Polymerase I. Mol
Cell. Biol., 5, 1358-1369.
K. G. Miller. J. Tower, B. Sollner-Webb. 1985. A Complex Cont-
rol Region of the Mouse rRNA Gene Directs Accurate
Initiation by RNA Polymerase 1. Mol. Cell. Biol., 5,
554-562.
V. L. Murtif P. M. M. Rae. 1985. In vivo Transcription of rDNA
Spacers in Drosophila. Nucleic Acids Res., 13, 3221-3224.
M.E. Swanson, M. Yip, M.J. Holland. 1985. Characterization
of an RNA Polymerase I-Dependent Promoter Within the
ЛИТЕРАТУРА
317
Spacer Region of Yeast Ribosomal Cistrons. J. Biol.
Chem., 260, 9905-9915.
R.F.J. DeWinter, T. Moss. 1986. Spacer Promoters Are Essen-
tial for Efficient Enhancement of X. laevis Ribosomal
Transcription. Cell, 44, 313-318.
P. Labhart, R.H. Reeder. 1986. Characterization of Three Sites
of RNA 3' End Formation in the Xenopus Ribosomal
Gene Spacer. Cell, 45, 431-443.
R.M. Learned, T.K. Learned. M.M. Haltiner, R.T. T/ian. 1986.
Human rDNA Transcription is Modulated by the Coordi-
nate Binding of Two Factors to an Upstream Control
Element. Cell, 45, 847-857.
J. Windle, B. Sollner-Webb. 1986. Upstream Domains of the
Xenopus laevis rDNA Promoter Are Revealed in Micro-
injected Oocytes. Mol. Cell. Biol., 6, 1228-1234.
P. Kownin, E. Bateman, M.R.Paule. 1987. Eukaryotic RNA
Polymerase I Promoter Binding Is Directed by Protein
Contacts with Transcription Initiation Factor and Is DNA
Sequence-Independent. Cell., 50, 693-699.
E. Bateman. M. R. Paule. 1988. Promoter Occlusi on During
Ribosomal RNA Transcription. Cell, 54, 985-992.
S.P. Bell, R.M. Learned, H.M. Jantzen, R. T/ian. 1988. Func-
tional Cooperativity Between Transcription Factors UBF1
and SL1 Mediates Human Ribosomal RNA Synthesis.
Science, 241, 1192-1197.
M.H. Jones. R.M. Learned, R. Tjian. 1988. Analysis of Cluste-
red Point Mutations in the Human Ribosomal RNA Gene
Promoter by Transient Expression in Vivo. Proc. Natl.
Acad. Sci. U.S.A., 85, 669-673.
8.2.r
I. Grummt. U. Maier. A. Ohrlein, N. Hassouna, J. P. Bachelle-
rie. 1985. Transcription of Mouse rDNA Terminates
Downstream of the 3' End of 28S RNA and Involves
Interaction of Factors with Repeated Sequences in the 3'
Spacer. Cell, 43. 801-810.
S. M. Baker, T. Platt. 1986. Pol I Transcription: Which Comes
First, the End or the Begining? Cell, 47, 839 840.
I. Grummt. H. Rosenbauer, I. N iedermeyer, U. Maier, A. Ohr-
lein. 1986. A Repeated 18 bp Sequence Motif in the
Mouse rDNA Spacer Mediates Binding of a Nuclear
Factor and Transcription Termination. Cell, 45, 837-846.
P. Labhart, R.H. Reeder. 1986. Characterization of Three Sites
of RNA 3' End Formation in the Xenopus Ribosomal
Gene Spacer. Cell, 45, 431-443.
I. Bartsch. C. Schoneberg, I. Grummt. 1987. Evolutionary Chan-
ges of Sequences and Factors That Direct Transcription
Termination of Human and Mouse Ribosomal Genes.
Mol. Cell. Biol., 7, 2521-2529.
S.L.Henderson, K. Ryan. B. Sollner-Webb. 1989. The Promoter-
Proximal rDNA Terminator Augments Initiation by Pre-
venting Disruption of the Stable Transcription Complex
Caused by Polymerase Readin. Genes and Devel., 3,
212-223
A. Kuhn. I. Grummt. 1989. З'-End Formation of Mouse Pre-
rRNA Involves Both Transcription Terminatoin and a
Specific Processing Reaction. Genes and Devel., 3,
224-231.
8.2.Д
/. Grummt. A. Kuhn, I. Bartsch. H. Rosenbauer. 1986. A Trans-
cription Terminator Located Upstream of the Mouse
rDNA Initiation Site Affects rRNA Synthesis. Cell, 47,
901-911.
S. Henderson, B. Sollner-Webb. 1986. A Transcriptional Termi-
nator is a Novel Element of the Promoter of the Mouse
Ribosomal RNA Gene. Cell, 47, 891-900.
B. McStay, R. H. Reeder. 1986. A Termination Site for Xenopus
RNA Polymerase 1 Also Acts as an Element of a Adijacent
Promoter. Cell, 47, 913-920.
A. Kuhn, A. Normann, I. Bartsch, I. Grummt. 1988. The Mouse
Ribosomal Gene Terminator Consists of Three Functio-
nally Separable Sequence Elements. EMBO J., 7, 1497-
1502.
8.3.B
K.S. Zaret, F. Sherman. 1982. DNA Sequence Required for
Efficient Transcription Termination in Yeast. Cell, 28.
563-573.
C. Montell. E.F. Fisher. M. H. Caruthers, A. J. Berk. 1983.
Inhibition of RNA Cleavage but Not Polyadenylation by
a Point Mutation in mRNA 3' Consensus Sequence
AAUAAA. Nature, 305, 600 605.
A. Gil., N.J. Proudfoot. 1984. A Sequence Downstream of
AAUAAA is Required for Rabbiy Р-Globin mRNA 3'-End
Formation. Nature, 312, 473-474.
S. Henikoff, E. H. Cohen. 1984. Sequences Responsible for Trans-
cription Termination on a Gene Segment in Saccharomy-
ces cerevisiea. Mol. Cell. BioL, 4, 1515-1520.
I. Pettersson. M. Hinterberger, T. Mimori. E. Gottlieb, J. A. Steitz.
1984. The Structure of Mammalian Small Nuclear Ribo-
nucleoproteins. J. Biol. Chem., 259, 5907-5914.
B.M. Bhat. W.S.M. Wold. 1985. ААТААА as Well as Down-
stream Sequences Are Required for RNA З'-End Forma-
tion in the E3 Complex Transcription Unit of Adenovirus.
Mol. Cell. Biol., 5, 3183-3193.
M. L. Birnstiel, M. Busslinger, K. Strub. 1985. Transcription
Termination and 3' Processing: The End is in Site! Cell, 41,
349-359.
L. Conway, M. Wickens. 1985. A Sequence Downstream of
A-A-U-A-A-A is Required for Formation of Simian
Virus 40 Late mRNA 3' Termini in Frog Oocytes. Proc.
Natl. Acad. Sci. U.S.A., 82, 3949-3953.
R. P. Hart, M.A. McDevitt, H. Ali, J.R. Nevins. 1985. Defini-
tion of Essential Sequences and Functional Equivalence of
Elements Downstream of the Adenovirus E2A and the
Early Simian Virus 40 Polyadenylation Sites. Mol. Cell.
Biol., 5, 2975-2983.
J. L. Manley, H. Yu. L. Ryner. 1985. RNA Sequence Containing
Hexanucleotide AAUAAA Directs Efficient mRNA Poly-
adenylation in Vitro. Mol. Cell. Biol., 5, 373-379.
J. McLauchlan. D. Gaffney. J. L. Whitton, J.B. Clements. 1985.
The Consensus Sequence YGTGTTYY Located Down-
stream from the ААТААА Signal Is Required for Efficient
Formation of mRNA 3' Termini. Nucleic Acids Res., 13,
1347-1368.
C. L. Moore. P.A. Sharp. 1985. Accurate Cleavage and Poly-
adenylation of Exogenous RNA Substrate. Cell, 41.
845-855.
K.H. Baek. K. Sato, R. Ito. K. Agarwal. 1986. RNA Poly-
merase Il Transcription Terminates at a Specific DNA
Sequence in a HeLa Cell-Free Reaction. Proc. Natl. Acad.
Sci. U.S.A., 83, 7623-7627.
G. Ciliberto, N. Dathan, R. Frank, L. Philipson, I. W. Maitaj.
1986. Formation of the 3' End on U snRNAs Requires at
Least Three Sequence Elements. EMBO J., 5, 2931-2937.
C. Hashimoto, J. A. Steitz. 1986. A Small Nuclear Ribonucleo-
protein Associates with the AAUAAA Polyadenylation
Signal in Vitro. Cell, 45, 581-591.
N. Hernandez, A.M. Weiner. 1986. Formation of the 3' End of
U1 snRNA Requires Compatible snRNA Promoter Ele-
ments. Cell, 47, 249-258.
M.A. McDevitt, R.P. Hart, W.W. Wong, J.R. Nevins. 1986.
Sequneces Capable of Restoring Poly(A) Site Function
Define Two Distinct Downstream Elements. EMBO J., 5,
2907-2913.
T. Platt. 1986. Transcription Termination and the Regulation
of Gene Expression. Annu. Rev. Biochem., 55, 339-372.
K. Sato, R. Ito, K.H. Baek. K. Agarwal. 1986. A Specific DNA
Sequence Controls Termination of Transcription in the
318
ЧАСТЬ III, ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Gastrin Gene. Mol. Cell. Biol., 6, 1032-1043.
H.E.N. de Vegvar, E. Lund. J. E. Dahlberg. 1986. 3' End
Formation of U1 snRNA Precursors in Coupled to
Transcription from snRNA Promoters. Cell, 47, 259-266.
E. Whitelaw, N. Proudfoot. 1986. a-Thalassaemia Caused by
a Poly(A) Site Mutation Reveals Thas Transcriptional
Termination in Linked to 3' End Processing in the Human
a2 Globin Gene. EM BO J., 5, 2915-2922.
R.A. Ach. A.M. Weiner. 1987. The Highly Conserved U Small
Nuclear RNA З'-End Formation Signal is Quite Tolerant
to Mutation. Mol. Cell. Biol., 7, 2070-2079.
R.L. Dedrick, C.M.Kane, M. J. Chamberlin. 1987. Purified
RNA Polymerase II Recognized Specific Termination Sites
During Transcription in Vitro. J. Biol. Chem., 262,
9098-9108.
A. Gil, N.J. Proudfoot 1987. Position-Dependent Sequence
Elements Downstream of AAUAAA Are Required for
Efficient Rabbit Р-Globin m RNA 3' End Formation. Cell,
49, 399-406.
J. Logan. E. Falck-Pedersen, J.E. Darnell. Jr., T. Shenk. 1987.
A Poly(A) Addition Site and a Downstream Termination
Region Are Required for Efficient Cessation of Transcrip-
tion by RNA Polymerase II in the Mouse Р-Globin Gene.
Proc. Natl. Acad. Sci. U.S.A., 84, 8306-8310.
K.L. Mowry, J.A.Steitz. 1987. Both Conserved Signals on
Mammalian Histone Pre-mRNAs Associate with Small
Nuclear Ribonucleoproteins During 3' End Formation in
Vitro. Mol. Cell. Biol., 7, 1663-1672.
K.L. Mowry. J. A. Steiz. 1987. Identification of the Human U7
snRNP as One of Several Factors Involved in the 3' End
Maturation of Histone Premessenger RNA’s. Science, 38,
1682-1687.
D. Reines, D. Wells, M.J. Chamberlin, С. M. Kane. 1987. Identi-
fication of Intrinsic Termination Sites in Vitro for RNA
Polymerase II Within Eukaryotic Gene Sequences. J. Mol.
Biol., 196, 299-312.
J. S. Butler, T. Platt. 1988. RNA Processing Generates the
Mature 3' End of Yeast CYC1 Messenger RNA in Vitro.
Science, 242, 1270-1274.
G. Chistofori, W. Keller. 1988. 3' Cleavage and Polyadenylation
of mRNA Precursors in Vitro Requires a Poly(A) Polyme-
rase, a Cleavage Factor, and a snRNP. Cell, 54, 875-889.
G. M. Gilmartin, F. Schaufele. G. Schaffner, M. L. Birnstiel.
1988. Functional Analysis of the Sea Urchin U7 Small
Nuclear RNA. Mol. Cell. Biol., 8, 1076-1084.
K.L. Mowry, J. A. Steitz. 1988. snRNP Mediators of 3' End
Processing: Functional Fossils? Trends Biochem. Sci., 13,
447-451.
Y Takagaki, L.C. Ryner. J.L. Manley. 1988. Separation and
Characterization of a Poly(A) Polymerase and a Cleavage/
Specificity Factor Required for Pre-mRNA Polyadenyla-
tion. Cell, 52, 731-742.
J. Wllusz, D. I. Feig, T. Shenk. 1988. The C Proteins of Hetero-
geneous Ribonucleoprotein Complexes interact with RNA
Sequences Downstream of Polyadenylation Cleavage Sites.
Mol. Cell. Biol., 8, 4477-4483.
J. Wllusz. T. Shenk. 1988. A 64 kd Nuclear Protein Binds to
RNA Segments That Include the AAUAAA Polyadenyla-
tion Motif. Cell, 52, 221-228.
N. Levitt, D. Briggs, A. Gil, N.J. Proudfoot. 1989. Definition of
an Efficient Synthetic Poly(A) Site. Genes and Devel., 3,
1019-1025.
N.J. Proudfoot. 1989. How RNA Polymerase II Terminates
Transcription in Higher Eukaryotes. Trends Biochem. Sci.,
14, 105-110.
M.D. Sheets. M. Wlckens. 1989. Two Phases in the Addition of
a Poly(A) Tail. Genes and Devel., 3, 1401-1412.
8.3.r
ITS. Dynan, R. Tjian. 1983. The Promoter-Specific Transcrip-
tion Factor Sp I Binds to Upstream Sequences in the SV40
Early Promoter. Cell, 35, 79- 87.
M. Fromm, P. Berg. 1983. Simian Virus 40 Early- and Late-
Region Promoter Functions Are Enhanced by the 72-Ba-
se-Pair Repeat Inserted at Distant Locations and Inverted
Orientations. Mol. Cell. Biol., 3, 991-999.
M. Fromm, P. Berg. 1983. Transcription in Vivo from SV40
Early Promoter Deletion Mutants Without Repression by
Large T Antigen. J. Mol. Appl. Genet., 2, 127-135.
J. Brady, M. Radonovich, M. Thoren, G. Das. N.P. Salzman.
1984. Simian Virus 40 Major Late Promoter: An Upstre-
am DNA Sequences Required for Efficient in Vitro Trans-
cription. Mol. Cell. Biol., 4, 133-141.
A. R. Buchman, P. Berg. 1984. Unusual Regulation of Simian
Virus 40 Early-Region Transcription in Genomes Contai-
ning Two Origins of DNA Replication. Mol. Cell. Biol., 4,
1915-1928.
A. R. Buchman, M. Fromm, P. Berg. 1984. Complex Regulation
of Simian Virus 40 Early-Region Transcription from
Differen: Overlapping Promoters. Mol. Cell. Biol., 4.
1900-1914.
D. Gidoni, IKS. Dynan, R. Tjian. 1984. Multiple Specific Con-
tacts Between a Mammalian Transcription Factor and Its
Cognate Promoters. Nature, 312, 409-413.
S.L. McKnight, R.C. Kingsbury, A. Spence, M. Smith. 1984.
The Distal Transcription Signals of the Herpesvirus tk
Gene Share a Common Hexanucleotide Control Sequence.
Cell, 37, 253-262.
F. Weber, J. de Villiers, W. Schaffner. 1984. An SV40 “Enhancer
Trap” Incorporates Exogenous Enhancers from its Own
Sequences. Cell, 36, 983-992.
IKS. Dynan, R. Tjian. 1985. Control of Eukaryotic Messenger
RNA Synthesis by Sequence-Specific DNA-Binding Pro-
teins. Nature, 316, 774-778.
D. Gidoni, J. T. Kadonaga. H. Barrera-Saldana, K. Takahashi.
P. Chambon. R. Tjian. 1985. Bidirectional SV 40 Transcrip-
tion Mediated by Tandem SP1 Binding Interactions.
Science, 230, 511-517.
K. A. Jones, K. R. Yamamoto, R. Tjian. 1985. Two Distinct
Transcription Factors Bind to the HSV Thymidine Kinase
Promoter in Vitro. Cell, 42, 559-572.
J. M. Keller, J.C. Alwine. 1985. Analysis of an Activatable
Promoter: Sequences in the Simian Virus 40 Late Promo-
ter Required for T-Antigen-Mediated trans Activation.
Mol. Cell. Biol., 5, 1859-1869.
K. Ryder, E. Vakalopoulou, R. Mertz, I. Mastrangelo, P. Hough,
P. Tegtmeyer, E. Fanning. 1985. Seventeen Base Pairs of
Region I Encode a Novel Tripartite Binding Signal for
SV 40 T Antigen. Cell, 42, 539-548.
E. Serfling. M. Jasin, W. Schaffner. 1985. Enhancers and Euka-
ryotic Gene Transcription. Trends Genet., 1, 224-230.
R. Treisman. T. Maniatis. 1985. Simian Virus 40 Enhancer
Increases Number of RNA Polymerase II Molecules on
Linked DNA. Nature, 315, 72-77.
M.R. Briggs. J.T Kadonaga, S.P. Bell, R. Tjian. 1986. Puri-
fication and Biochemical Characterization of the Pro-
moter-Specific Transcription Factor, Sp 1. Science, 234,
47-52.
W. Herr, J. Clarke. 1986. The SV40 Enhancer Is Composed of
Multiple Functional Elements That Can Compensate for
One Another. Cell, 45, 461 470.
J.T. Kadonaga, K. A. Jones, R. Tjian. 1986. Promoter-Specific
Activation of RNA Polymerase 11 Transcription by Spl.
Trends Biochem. Sci., 11, 20-23.
J. T. Kadonaga, R. Tjian. 1986. Affinity Purification of Sequen-
ce-Specific DNA Binding Proteins. Proc. Natl. Acad. Sci.,
U.S.A., 83, 5889-5893.
ЛИТЕРАТУРА
319
S. McKnight, R. Tjian. 1986. Transcriptional Selectivity of Viral
Genes in Mammalian Cell. Cell, 46, 795-805.
S.E. Pion, J. C. Wang. 1986. Transcription of the Human
Р-Globin Gene Is Stimulated by an SV40 Enhancer to
Which It Is Physically Linked but Topologically Uncoup-
led. Cell, 45, 575-580.
P. Sassone-Corsi, E. Borrelli. 1986. Transcriptional Regulation
by trans-Acting Factors. Trends Genet., 2, 215-219.
K. Takahashi, M. Vigneron, H. Matthes, A. Wildeman, M. Zen-
ke, P. Chambon. 1986. Requirement of Stereospecific Align-
ments for Initiation from the Simian Virus 40 Early
Promoter. Nature, 319, 121-126.
M. Zenke, T. Grundstrom, H. Matthes, M. Wintzerith, C. Schatz,
A. Wildeman, P. Chambon. 1986. Multiple Sequence Motifs
Are Involved in SV 40 Enchancer Function. EMBO J., 5,
387-397.
A. Dorn. J. Bollekens, A. Staub, C. Benoist, D. Mathis. 1987.
A Multiciplicity of ССААТ Box-Binding Proteins. Cell, 50,
863-872.
R.D. Everett. 1987. The Regulation of Transcription of Viral
and Cellular Genes by Herpesvirus Immediate-Early Gene
Products. Anticancer Res., 7, 589-604.
L. Guarente. 1987. Regulatory Proteins in Yeast. Annu. Rev.
Genet., 21, 425-452.
W. Lee, P. Mitchell, R. Tjian. 1987. Purified Transcription
Factor AP-1 Interacts with TPA-Inducible Enhancer Ele-
ments. Cell, 49, 741-752.
E. May, F. Omilli, M. Ernoult-Lange, M. Zenke. P. Chambon.
1987. The Sequence Motifs That Are Involved in SV 40
Enhancer Function Also Control SV 40 Late Promoter
Activity. Nucleic Acids Res., 15, 2445-2462.
P. J. Mitchell, C. Wang, R. Tjian. 1987. Positive and Negative
Regulation of Transcription in Vitro: Enhancer-Binding
Protein AP-2 Is Inhibited by SV 40 T Antigen. Cell. 50.
847-861.
R. Rosales, M. Vigneron, M. Macchi, I. Davidson, J. H. Xiao,
P. Chambon. 1987. In Vitro Binding og Cell-Specific and
Ubiquitous Nuclear Protein to the Octamer Motif of the
SV 40 Enhancer and Related Motifs Present in Other
Promoters and Enhancers. EMBO J., 6, 3015-3025.
S. Schirm, J. Jiricny. W. Schaffner. 1987. The SV 40 Enchancer
Can Be Dissected into Multiple Segments, Each with
a Different Cell Type Specificity. Genes and Devel., 1,
65-74.
J. H. Xiao. I. Davidson, D. Ferrandon. R. Rosales, M. Vigneron,
M. Macchi, F. Ruffenach, P. Chambon. 1987. One Cell-
Specific and Three Ubiquitous Nuclear Proteins Bind in
Vitro to Overlapping Motifs in the Domain Bl of the SV
40 Enhancer. EMBO J., 6, 3005-3013.
S.M. Admayr, J.L. Workman. R.G. Roeder. 1987. The Pseudo-
rabies Immediate Early Protein Stimulates in Vitro Trans-
cription by Facilitating TFIID: Promoter Interactions.
Genes and Devel.. 2, 542-553.
T.J. Bos, D. Bohmann, H. Tsuchie, R. Tjian, P.K. Vogt. 1988.
v-jun Encodes a Nuclear Protein with Enhancer Binding
Properties of AP-1. Cell, 52, 705-712.
S. Buratowski, S. Hahn, P.A. Sharp, L. Guarente. 1988. Function
of a Yeast TATA Elemnet-Binding Protein in a Mammalian
Transcription System. Nature, 334, 37-42.
B. Cavallini, J. Huet, J-L. Plassat, A. Sentenac, J-M. Egly,
P. Chambon. 1988. A Yeast Activity Can Substitute for the
HeLa Cell TATA Box Factor. Nature, 334, 77-80.
L.A. Chodosh, A.S. Baldwin. R. W. Carthew, P.A. Sharp. 1988.
Human ССААТ-Binding Proteins Have Heterologous
Subunits. Cell, 33, 11 - 24.
L.A. Chodosh, J. Olesen, S. Hahn, A.S. Baldwin, L. Guarente,
P. A. Sharp. 1988. A Yeast and a Human CCAAT-Binding
Protein Have Heterologous Subunits That Are Functio-
nally Interchangeable. Cell, 53, 25-35.
L. Clark, R.M. Pollock, R.T Hay. 1988. Identification and
Purification of EBP1: A HeLa Cell Protein That Binds to
a Region Overlapping the “Core” of the SV 40 Enhancer.
Genes and Devel., 2, 991-1002.
A. J. Courey, R. Tjian. 1988. Analysis of Sp-1 in Vivo Reveals
Multiple Transcriptional Domains, Including a Novel
Glutamine-Rich Activation Motif. Cell, 55, 887-898.
C. Fromental. M. Kanno, H. Nomiyama, P. Chambon. 1988.
Cooperativity and Hierarchical Levels of Functional
Organization in the SV 40 Enhancer. Cell, 54, 943-953.
S. Hahn, L. Guarente. 1988. Yeast HAP2 and HAP3: Trans-
criptional Activators in a Heteromeric Complex. Science,
240, 317-321.
K.D. Harshman, IKS. Moye-Rowley. C.S. Parker. 1988. Trans-
criptional Activation by the SV 40 AP-1 Recognition
Element in Yeast Is Mediated by a Factor Similar to AP-1
That Is Distinct from GCN4. Cell, 53, 321-330.
N.C. Jones. P.W.J. Rigby, E.B. Ziff. 1988. Tram-Acting Pro-
teins Factors and the Regulation of Eukaryotic Trans-
cription: Lessons from Studies on DNA Tumor Viruses.
Genes and Devel., 2, 267-281.
J. T. Kadonaga, A. J. Courey, J. Ladika. R. Tjian. 1988. Distinct
Regions of SP1 Modulate DNA Binding and Transcrip-
tional Activation. Science, 242, 1566-1569.
S.N. Maity, P.T Golumbek, G. Karsenty, B. de Crombrugghe.
1988. Selective Activation of Transcription by a Novel
ССААТ Binding Factor. Science, 241, 582-584.
M. Raymondjean, S. Cereghini, M. Yaniv. 1988. Several Distinct
“ССААТ” Box Binding Proteins Coexist in Eukaryotic
Cells. Proc. Natl. Acad. Sci. U.S.A., 85, 757-761.
B. Wasylyk. 1988. Enhancers and Transcription Factors in the
Control of Gene Expression. Biochimica et Biophysica
Acta, 951, 17-35.
S. Buratowski, S. Hahn, L. Guarente, P.A. Sharp. 1989. Five
Intermediate Complexes in Transcription Initiation by
RNA Polymerase II. Cell, 56, 549-561.
P. J. Mitchell, R. Tjian. 1989. Transcriptional Regulation in
Mammalian Cells by Sequence-Specific DNA Binding
Proteins. Science, 245, 371-378.
WS. Moye-Rowley, K.D. Harshman, C.S. Parker. 1989. Yeast
YAP\ Encodes a Novel Form of the Jun Family of
Transcriptional Activator Proteins. Genes and Devel., 3,
283-292.
A. G. Saltzman, R. Weinmann. 1989. Promoter Specificity and
Modulation of RNA Polymerase II Transcription. FASEB
J., 3, 1723-1733.
8.3.Д
M. Mercola, X. Wang, J. Olsen, K. Calarne. 1983. Transcrip-
tional Enhancer Activity in the Immunoglobulin Heavy
Chain Locus. Science, 221, 663-665.
F. G. Falkner, H.G. Zachau. 1984. Correct Transcription of an
Immunoglobulin x Gene Requires an Upstream Fragment
Containing Conserved Sequence Elements. Nature, 310,
71-74.
D. Picard, W. Schaffner. 1984. A Lymphocyte-Specific Enhancer
in the Mouse Immunoglobulin x Gene. Nature, 307,
80-82.
F. W. Alt. TK. Blackwell, G.D. Yancopoulos. 1985. Immuno-
globulin Genes in Transgenic Mice. Trends Genet., 1,
231-236.
K. Calarne. 1985. Mechanisms Which Regulate Immunoglobu-
lin Gene Expression. Annu. Rev. Immunol., 3, 159-195.
R. Grosschedl, D. Baltimore. 1985. Cell-Type Specificity of
Immunoglobulin Gene Expression is Regulated by at
Least Three DNA Sequence Elements. Cell, 41, 885-897.
R. Krumlauf, R. E. Hammer, S. M. Tilghman, R. L. Brinster.
1985. Developmental Regulation of a-Fetoprotein Genes
in Transgenic Mice. Mol. Cell. Biol., 5, 1639-1648.
M. Mercola, J. Goverman, C. Mirell, K. Calarne. 1985. Immu-
noglobulin Heavy-Chain Enhancer Requires One or More
320
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Tissue-Specific Factors. Science, 227, 266-270.
D. Picard, W. Schaffner. 1985. Cell-Type Preference of Immuno-
globulin and Gene Promoters. EMBO J., 4, 2831-2838.
R. Godbout, R. Ingram, S.M. Tilghman. 1986. Multiple Regula-
tory Elements in the Intergenic Region Between the
a-Fetoprotein and Albumin Genes. Mol. Cell. Biol., 6,
477-487.
C. L. Peterson, K. Orth, K.L. Calame. 1985. Binding in Vitro of
Multiple Cellular Proteins to Immunoglobulin Heavy-
Chain Enhancer DNA. Mol. Cell. Biol., 6, 4168-4178.
R. Sen, D. Baltimore. 1986. Inducibility of x Immunoglobulin
Enhancer-Binding Protein NF-xB by a Posttranslational
Mechanism. Cell, 47, 921-928.
R. Sen, D. Baltimore. 1986. Multiple Nuclear Factors Interact
with the Immunoglobulin Enhancer Sequences. Cell, 46,
705-716.
H. Singh, R. Sen, D. Baltimore, P. A. Sharp. 1986. A Nuclear
Factor That Binds to a Conserved Sequence Motif in
Transcriptional Control Elements of Immunoglobulin
Genes. Nature, 319, 154-158.
L.M. Staudt, H. Singh, R. Sen, T. Wirth, P.A. Sharp, D. Balti-
more. 1986. A Lymphoid-Specific Protein Binding to the
Octamer Motif og Immunoglobulin Genes. Nature, 323,
640-643.
M.L. Atchison, R.P. Perry. 1987. The Role of the x Enhancer
and Its Binding Factor NF-xB in the Development
Regulation of x Gene Transcription. Cell, 48, 121-128.
L.E. Babiss, R.S. Herbst, A.L. Bennett, J.E. Darnell, Jr. 1987.
Factors That Interact with the Rat Albumin Promoter Are
Present Both in Hepatocytes and Other Cell Types. Genes
and Devel., 1, 256-267.
S. Cereghini, M. Raymondjean, A.G. Carranca, P. Herbomel.
M. Yaniv. 1987. Factors Involved in Control of Tissue-
Specific Expression of Albumin Gene. Cell, 50, 627-638.
S. Eaton, K. Calame. 1987. Multiple Elements Are Necessary
for the Function of an Immunoglobulin Heavy Chain
Promoter. Proc. Natl. Acad. Sci. U.S.A., 84, 7634-7638.
J.M. Heard, P. Herbomel, M.O. Ott, A. Mottura-Rollier,
M. Weiss, M. Yaniv. 1987. Determinants of Rat Albumin
Promoter Tissue Specificity Analyzed by an Improved
Transient Expression System. Mol. Cell. Biol., 7, 2425-
2434.
J. L. Imler, C. Lemaire. C. Wasylyk, B. Wasylyk. 1987. Negative
Regulation Contributes to Tissue Specificity of the Immu-
noglobulin Heavy-Chain Enhancer. Mol. Cell. Biol., 7,
2558-2567.
M. Lenar do. J. W. Pierce, D. Baltimore. 1987. Protein-Binding
Sites in 1g Gene Enhancers Determine Transcriptional
Activity and Inducibility. Science, 236, 1573- 1577.
S. Lichsteiner, J. Wuarin, V. Schibler. 1987. The Interplay of
DNA-Binding Proteins on the Promoter of the Mouse
Albumin Gene. Cell. 51, 963-973.
C. Peterson, K. Calame. 1987. Complex Protein-DNA Inter-
actions on the Mouse Immunoglobulin Heavy Chain
Enhancer. Mol. Cell. Biol., 7, 4194-4203.
C. Scheidereit, A. Heguy, R.G. Roeder. 1987. Identification and
Purification of a Human Lymphoid-Specific Octamer-
Binding Protein (OTF-2) That Activates Transcription of
an Immunoglobulin Promoter in Vitro. Cell, 51, 783-793.
K. Araki, H. Maeda, J. Wang. D. Kitamura, T. Watanabe. 1988.
Purification of a Nuclear Trans-Acting Factor Involved in
the Regulated Transcription of a Human Immunoglobulin
Heavy Chain Gene. Cell, 53, 723-730.
A.S. Baldwin, Jr., P. Sharp. 1988. Two Transcription Factors,
NF-xB and H2TF1, Interact with a Single Regulatory
Sequence in the Class I Major Histocompatibility Complex
Promoter. Proc. Natl. Acad. Sci. U.S.A., 85, 723-727.
K. Calame, S. Eaton. 1988. Transcriptional Controlling Ele-
ments in the Immunoglobulin and T Cell Receptor Loci.
Adv. Immunol. 43, 235-275.
S. Cereghini, M. Blumenfeld, M. Yaniv. 1988. A Liver-Specific
Factor Essential for Aibumin Transcription Differs Between
Differentiated and Dedifferentiated Rat Hepatoma Cells.
Genes and Devel., 2, 957-974.
R.H. Costa, E. Lai. D. R. Grayson, J.E. Darnell, Jr. 1988. The
Cell-Specific Enhancer of the Mouse Transthyretin (Pre-
albumin) Gene Binds a Common Factor at One Site and
a Liver-Specific Factor(s) at Two Other Sites. Mol. Cell.
Biol., 8, 81-90.
G. Courtois, S. Baumhueter, G. R. Crabtree. 1988. Purified
Hepatocyte Nuclear Factor 1 Interacts with a Family of
Hepatocyte-Specific Promoters. Proc. Natl. Acad. Sci.
U.S.A., 85. 7937-7941.
R. Godbout, R.S. Ingram, S.M. Tilghman. 1988. Fine-Structure
Mapping of the Three Mouse a-Fetoprotein Gene Enhan-
cers. Mol. Cell. Biol., 8, 1169-1178.
R. Godbout, S. M. Tilghman. 1988. Configuration of the a-Feto-
protein Regulatory Domain During Development. Genes
and Devel., 2, 949-956.
D. R. Grayson, R. H. Costa, K. G. Xanthopoulos, J. E. Darnell.
Jr. 1988. A Cell-Specific Enhancer of the Mouse al-Anti-
trypsin Gene Has Multiple Functional Regions and Cor-
responding Protein-Binding Sites. Mol. Cell, Biol., 8,
1055-1066.
D. R. Grayson, R. H. Costa, K. G. Xanthopoulos, J. E. Darnell.
1988. One Factor Recognizes the Liver-Specific Enhancers
in al-Antitrypsin and Transthyretin Genes. Science, 239,
786-788.
R. Grosschedl, M. Marx. 1988. Stable Propagation of the
Active Transcriptional State of an Immunoglobulin Gene
Requires Continuous Enhancer Function. Cell, 55,
645-654.
K. Kawakami, C. Scheidereit, R. G. Roeder. 1988. Identification
and Purification of a Human Immunoglobulin-Enhancer-
Binding Protein (NF-xB) That Activates Transcription
from a Human Immunodeficiency Virus Type 1 Promoter
in Vitro. Proc. Natl. Acad. Sci. U.S.A., 85, 4700-4704.
H. Ko, P. Fast, W. McBride, L. M. Staudt. 1988. A Human
Protein Specific for the Immunoglobulin Octamer DNA
Motif Contains a Functional Homeobox Domain. Cell,
55, 135-144.
J. H. LeBowitz, T. Kobayashi, L. Staudt, D. Baltimore, P. A. Sharp.
1988. Octamer-Binding Proteins from В or HeLa Cells
Stimulate Transcription of the Immunoglobulin Heavy-
Chain Promoter in Vitro. Genes and Devel., 2, 1227—
1237.
M.M. Muller, S. Ruppert. W. Schaffner, P. Matthias. 1988. A
Cloned Octamer Transcription Factor Stimulates Trans-
cription from Lymphoid-Specific Promoters in Non-B
Cells. Nature, 336, 544-551.
J. Perez-Mutul, M. Macchi, B. Wasylyk. 1988. Mutational
Analysis of the Contribution of Sequence Motifs Within
the IgH Enhancer to Tissue Specific Transcriptional
Activation. Nucleis Acids Res., 16, 6085-6096.
C. Scheidereit. J. A. Cromlish, T. Gerster, K. Kawakami, C. Bal-
maceda, R.A. Currie, R.G. Roeder. 1988. A Human Lymp-
hoid-Specific Transcription Factor That Activates Immu-
noglobulin Genes Is a Homeobox Protein. Nature, 336,
551-557.
H. Singh, J.H. LeBowitz, A.S. Baldwin, Jr., P.A. Sharp. 1988.
Molecular Cloning of an Enhancer Binding Protein:
Isolation by Screening of an Expression Library with
a Recognition Site DNA. Cell, 52, 415-423.
L. M. Staudt. R. G. Clerc, H. Singh, J. H. LeBowitz, P. A. Sharp,
D. Baltimore. 1988. Cloning of a Lymphoid-Specific cDNA
Encoding a Protein Binding the Regulatory Octamer
DNA Motif. Science, 241, 577-579.
B. Wasylyk. 1988. Transcription Elements and Factors of RNA
Polymerase В Promoters of Higher Eukaryotes. Crit. Rev.
Biochem., 23, 77-120.
ЛИТЕРАТУРА
321
K.L. Calame. 1989. Immunoglobulin Gene Transcription:
Molecular Mechanisms. Trends Genet., 5, 395-399.
S.A. Camper, S.M. Tighman. 1989. Postnatal Repression of the
a-Fetoprotein Gene Is Enhancer Independent. Genes and
Devel., 3, 537-546.
A. D. Friedman. W.H. Landschulz, S.L. McKnight. 1989.
ССААТ/Enhancer Binding Protein Activates the Promo-
ter of the Serum Albumin Gene in Cultured Hepatoma
Cells. Genes and Devel., 3, 1314-1322.
M. A. Garcia-Blanco, R.G. Clerc, P. A. Sharp. 1989. The
DNA-Binding Homeo Domain of the Oct-2 Protein.
Genes and Devel., 3, 739-745.
P. Herbomeb, A. Mottura, F. Tranche, M.O. Ott, M. Yaniv,
M. Weiss. 1989. The Rat Aibumin Promoter Is Composed
of Six Distinct Positive Elements Within 130 Nucleotides.
Mol. Cell. Biol., 9, 4750-4758.
I. Kemler, E. Schreiber, M.M. Muller, P. Matthias, W. Schaf-
fner. 1989. Octamer Transcription Factors Bind to Two
Different Sequence Motifs of the Immunoglobulin Heavy
Chain Promoter. EM BO J., 8, 2001-2008.
M.J. Lenardo, D. Baltimore. 1989. NF-xB: A Pleiotropic Medi-
tor of Inducible and Tissue Specific Gene Control. Cell, 58,
227-229.
К. B. Meyer, M.S. Neuberger. 1989. The Immunoglobulin x
Locus Contains a Second, Stronger B-Cell-Specific Enhan-
cer Which Is Located Downstream of the Constant
Region. EM BO J., 8, 1959-1964.
C. Murre, P.S. McCaw, D. Baltimore. 1989. A New DNA
Binding and Dimerization Motif in Immunoglobulin En-
hancer Binding, Daughterless, MyoD, and myc Proteins.
Cell, 56, 777-783.
R. H. Scheuermann, V. Chen. 1989. A Developmental-Specific
Factor Binds to Suppressor Sites Flanking the Immuno-
globulin Heavy-Chain Enhancer. Genes and Devel., 3,
1255-1266.
8.3.e
R. I. Richards, A. Heguy, M. Karin. 1984. Structural and Func-
tional Analysis of the Human Metalloothionein-1A Gene:
Differential Induction by Metal Ions and Glucocorticoids.
Cell, 37, 263-272.
C. Wu. 1984. Activating Protein Factor Binds in Vitro to
Upstream Control Sequeces in Heat Shock Gene Chro-
matin. Nature, 311, 81-84.
M. Bienz. 1985. Transient and Development Activation of
Heat-Shock Genes. Trends Biochem. Sci., 10, 157-161.
J. Simpson, M. P. Timko, A. R. Cashmore, J. Schell, M. Van-
Montagu, L. Herrera-Estrella. 1985. Light-Inducible and
Tissue-Specific Expression of a Chimaeric Gene Under
Control of the 5'-Flanking Sequence of a Pea Chlorophyll
ab-Binding Protein Gene. EMBO J., 4, 2723-2729.
M.P. Timko, A.P. Kausch. C. Castresana, J. Fussier, L. Herre-
ra-Estrella, G. Van den Broeck, M. Van Montagu, J. Schell,
A.R. Cashmore. 1985. Light Regulation of Plant Gene
Expression by an Upstream Enhancer-Like Element.
Nature, 318, 579-582.
M. Bienz, H.R.B. Pelham. 1986. Heat Shock Regulatory Ele-
ments Function as an Inducible Enhancer in the Xenopus
hs70 Gene and When Linked to a Heterologous Promoter.
Cell, 45 753-760.
S. Grenn, P. Walter, V. Kumar, A. Krust, J. Bornert, P. Argos,
P. Chambon. 1986. Human Oestrogen Receptor cDNA:
Sequence, Expression and Homology to v-erb-A. Nature,
320, 134-139.
D.H. Hamer. 1986. Metallothionein. Annu. Rev. Biochem., 55,
913-951.
R. Klemenz, W.J. Gehring. 1986. Sequence Requirement for
Expression of the Drosophila melanogaster Heat Chock
Protein hs22 Gene During Heat Shock and Normal
Development. Mol. Cell. Biol., 6, 2011-2019.
R.D. Andersen, S.J. Taplitz, S. Worg, G. Bristol, B. Larkin,
H.R. Herschman. 1987. Metal-Dependent Binding of a
Factor in Viro to the Metal-Responsive Elements of the
Metallothionein-1 Gene Promoter. Mol. Cell. Biol., 7,
3574-3581.
V. Giguere, E.S. Ong, P. Segui, F.M. Evans. 1987. Identification
of a Receptor for the Morphogen Retinoic Acid. Nature,
330, 624-629.
G. Gill, M. Ptashne. 1987. Mutants of GAL4 Protein Altered in
an Activation Function. Cell, 51, 121-126.
S. Green. P. Chambon. 1987. Oestradiol Induction of a Gluco-
corticoid-Responsive Gene by a Chimaeric Receptor. Na-
ture, 325, 75-78.
M. Johnston. 1987. A Model Fungal Gene Regulatory Mecha-
nism: The GAL Genes of Saccharomyces cerevisiae. Micro-
biol. Rev., 51, 458-476.
M. Karin, A. Haslinger, A. Heguy, T. Dietlin, T. Cooke. 1987.
Metal-Responsive Elements Act as Positive Modulators of
Human Metallothionein-IIA Enhancer Activity. Mol. Cell.
Biol., 7, 606-613.
R. E. Kingston, T. J. Schuetz, Z. Lavin. 1987. Heat-Inducible
Human Factor That Binds to a Human hspld Promoter.
Mol. Cell. Biol., 7, 1530-1534.
D. Kuhl, J. de la Fuente, M. Chaturveci, S. Parimoo, J. Ryals,
F. Meyer, C. Weissmann. 1987. Reversible Silencing of
Enhancers by Sequences Derived from the Human IFN-a
Promoter. Cell, 50, 1057- 1069.
C. Kuhlemeier, P.J. Green, N. Chua. 1987. Regulation of Gene
Expression in Higher Plants. Annu. Rev. Plant Physiol.,
38, 221-257.
V. Kumar, S. Green. G. Stack, M. Berry, J. Jin, P. Chambon.
1987. Functional Domains of the Human Estrogen Recep-
tor. Cell, 51, 941-951.
W. Lee, A. Haslinger, M. Karin, R. Tjian. 1987. Activation of
Transcription by Two Factors That Bind Promoter and
Enhancer Sequences of the Human Metallothionein Gene
and SV 40. Nature, 325, 368-372.
N.F. Lue, D.I. Chasman, A.R. Buchman, R.D. Kornberg. 1987.
Interaction of GAL4 and GAL80 Gene Regulatory Pro-
teins in Vitro. Mol. Cell. Biol., 7, 3446-3451.
J. Ma, M. Ptaschne. 1987. Deletion Analysis of GAL4 Defines
Two Transcriptional Activating Segments. Cell, 48, 847-
853.
M. Petkovich, N.J. Brand, A. Krust, P. Chambon. 1987. A
Human Retinoic Acid Receptor Which Belongs to the
Family of Nuclear Receptors. Nature, 330, 444-450.
G. Wiederrecht, D. J. Shuey, W.A.Kibbe, C.S. Parker. 1987.
The Saccharomyces and Drosophila Heat Shock Trans-
cription Factors Are Identical in Size and DNA Binding
Properties. Cell, 48, 507-515.
V. Zimarino, C. Wu. 1987. Induction of Sequence-Specific Bin-
ding of Drosophila Heat Shock Activator Protein Without
Protein Synthesis. Nature, 327, 727-730.
I. E. Akerblom, E.P. Slater, M. Geato, J.D. Baxter, P.L. Mel-
lon. 1988. Negative Regulation by Glucocorticoids Through
Interference with a cAMP Responsive Enhancer. Science,
241, 350-353.
N. Brand, M. Petkovich, A. Krust, P. Chambon, H. de The,
A. Marchio, P. Tiollais, A. Dejean. 1988. Identification of
a Second Human Retinoic Acid Receptor. Nature, 332,
850 853.
R. M. Evans. 1988. The Steroid and Thyroid Hormone Recep-
tor Superfamily. Science, 240, 889-895.
P. Furst, S. Hu, R. Hackett, D. Hamer. 1988. Copper Activates
Metallothionein Gene Transcription by Altering the Con-
formation of a Specific DNA Binding Protein. Cell, 55,
705-717.
V. Giguere, N. Yang, P. Segui, R. M. Evans. 1988. Identification
of a New Class of Steroid Hormone Receptors. Nature,
331, 91-94.
21-158
322
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
S. Green, Р. Chambon. 1988. Receptors Enhance Our Under-
standing of Transcription Regulation. Trends. Genet., 4.
309- 314.
S. Green, V. Kumar, /. Theulaz, W. Wahli, P. Chambon. 1988.
The N-Terminal DNA-Binding “Zinc Finger" of the
Oestrogen and Glucocorticoid Receptors Determines
Target Gene Specifity. EMBO J., 7, 3037-3044.
S.M. Hollenberg, R.M. Evans. 1988. Multiple and Cooperative
Trans-Activation Domains of the Human Glucocorticoid
Receptor. Cell, 55, 899-906.
M. Horikoshi, M.F. Carey. H. Kakidani. R.G. Roeder. 1988.
Mechanism of Action of a Yeast Activator: Direct Effect
of GAL4 Derivatives on Mammalian TFIID-Promoter
Interactions. Cell, 54, 665-669.
H. Kakidani, M. Ptaschne. 1988. GAL4 Activates Gene Expres-
sion in Mammalian Cells. Cell, 52, 161-167.
V. Kumar, P. Chambon. 1988. The Estrogen Receptor Binds
Tightly to Its Responsive Element as a Ligand-Induced
Homodimer. Cell, 55, 145-156.
Y. Lin. M.F. Carey, M. Ptaschne, M.R. Green. 1988. GAL4
Derivatives Function Alone and Synergistically with
Mammalian Activators in Vitro. Cell, 54, 659-664.
T. Maniatis. 1988. Mechanisms of Human p-Interferon Gene
Regulation. Harvey Lectures, Series 82, 71-104.
D. Metzger, J. H. White. P. Chambon. 1988. The Human Oest-
rogen Receptor Functions in Yeast. Nature, 334, 31-36.
A.E. Oro, S.M. Hollenberg, R.M. Evans. 1988. Transcriptional
Inhibition by a Glucocorticoid Receptor-p-Galactosidase
Fusion Protein. Cell, 55, 1109-1114.
A.E. Oro. E.S.Ong, J. S. Margolis, J.W. Posakony, M. Mc-
Keown, R.M. Evans. 1988. The Drosophila Gene Knirps-
Related Is a Member of the Steroid-Receptor Gene
Superfamily. Nature, 336, 493-496.
M. Ponglikimongkol. S. Green, P. Chambon. 1988. Genomic
Organization of the Human Oestrogen Receptor Gene.
EMBO J., 7, 3385-3388.
D.D. Sakai, S. Helms, J. Carlstedt-Duke, J. Gustafsson,
F.M. Rottman.K.R. Yamamoto. 1988. Hormone-Mediated
Repression: A Negative Glucocorticoid Response Element
from the Bovine Prolactin Gene. Genes and Devel., 2,
1144-1154.
M. Schena. K.R. Yamamoto. 1988. Mammalian Glucocorticoid
Receptor Derivatives Enhance Transcription in Yeast.
Science, 241, 965-967.
R. Schule, M. Muller, C. Kaltschmidt, R. Renkawitz. 1988.
Many Transcription Factors Interact Synergistically with
Steroid Receptors. Science, 242, 1418-1420.
L. Тога, H. Gronemeyer, B. Turcotte, M. Gaub, P. Chambon.
1988. The N-Terminal Region of the Chicken Progeste-
rone Receptor Specifies Target Gene Activation. Nature,
333, 185-188.
S. Y. Tsai, J. Carlstedt-Duke, N. L. Weigel, K. Dahlman, J. Gus-
tafsson, M. Tsai, B. W. O'Malley. 1988. Molecular Inter-
actions of Steroid Hormone Receptor with its Enhancer
Element: Evidence for Receptor Dimer Formation. Cell,
55, 361-369.
K. Umesono. V. Giguere, C.K. Glass, M.G. Rosenfeld,
R.M. Evans. 1988. Retinoic Acid and Thyroid Hormone
Induce Gene Expression Through a Common Responsive
Element. Nature, 336, 262-265.
M. L. Waterman, S. Adler, C. Nelson. G. L. Greene. R. M. Evans,
M.G. Rosenfeld. 1988. A Single Domain of the Estrogen
Receptor Confers Deoxyribonucleic Acid Binding and
Transcriptional Activation of the Rat Prolactin Gene.
Mol. Endocrin., 2, 14-21.
N. Webster, J.R. Jin. S. Green, M. Hollis, P. Chambon. 1988.
The Yeast UASC Isa Transcriptional Enhancer in Human
HeLa Cells in the Presence of the GALA Trans-Activator.
Cell, 52, 169-178.
G. Wiederrecht, D. Seto, C.S. Parker. 1988. Isolation of the
Gene Encoding the S. cerevisiae Heat Shock Transcription
Factor. Cell, 54, 841-853.
P.N. Benfey. N. Chua. 1989. Regulated Genes in Transgenic
Plants. Science, 244, 174-181.
E. A. Craig. 1989. Essential Roles of 70kDa Heat Inducible
Proteins. BioEssays, 11, 48-52.
K. Damm, С. C. Thompson, R. M. Evans. 1989. Protein Encoded
by v-erbA Functions as a Thyroid-Hormone Receptor
Antagonist. Nature, 339, 593-596.
R. B. Goldberg, S.J. Barker, L. Perez-Grau. 1989. Regulation of
Gene Expression During Plant Embryogenesis. Cell, 56,
149-160.
M.J. Lenardo. C. Fan, T. Maniatis, D. Baltimore. 1989. The
Involvement of NF-xB in P-lnterferon Gene Regulation
Reveals Its Role as a Widely Inducible Second Messenger.
Cell, 57, 287-294.
S. Mader, V. Kumar, H. de Verneuil, P. Chambon. 1989. Three
Amino Acids of the Oestrogen Receptor Are Essential to
Its Ability to Distinguish an Oestrogen from a Gluco-
corticoid-Responsive Element. Nature, 338, 271-274.
M. Meyer, H. Gronemeyer, B. Turcotte, M. Bocquel, D. Tasset,
P. Chambon. 1989. Steroid Hormone Receptors Compete
for Factors That Mediate Enhancer Function. Cell, 57,
433-442.
S. Y. Tsai, M. Tsai, B. W. O'Malley. 1989. Cooperative Binding
of Steroid Hormone Receptors Contributes to Transcrip-
tional Synergism at Target Enhancer Elements. Cell, 57,
443-448.
8.3.Ж
W.J. Gehring. 1985. The Molecular Basis of Development. Sci.
American, 253, 153-162.
W.J. Gehring. 1987. Homeo Boxes in the Study of Develop-
ment. Science, 236, 1245-1252.
M.P. Scott, S.B. Carroll. 1987. The Segmentation and Homeo-
tic Gene Network in Early Drosophila Development. Cell,
51, 689-698.
W. Driever. C. Nusslein-Volhard. 1988. The Bicoid Protein
Determines Position in the Drosophila Embryo in a Con-
centration-Dependent Manner. Cell, 54, 95- 104.
W. Driever, C. Nusselein- Volhard. 1988. A Gradient of Bicoid
Protein in Drosophila Embryos. Cell, 54, 83-93.
P. W. Ingham. 1988. The Molecular Genetics of Embryonic
Pattern Formation in Drosophila. Nature, 335, 25-34.
P. A. Lawrence. 1988. Background to Bicoid. Cell, 54, 1-2.
M.D. Biggin, R. Tjian. 1989. Transcription Factors and the
Control of Drosophila Development. Trends Genet., 5,
377-383.
W. Driever, C. Nusslein-Volhard. 1989. The Bicoid Protein Is
a Positive Regulator of Hunchback Transcription in the
Early Drosophila Embryo. Nature, 337, 138-143.
W. Driever. G. Thoma, C. Nusslein-Volhard. 1989. Determina-
tion of Spatial Domains of Zygotic Gene Expression in the
Drosophila Embryo by the Affinity of Binding Sites for the
Bicoid Morphogen. Nature, 340, 363-367.
M.J. Pankratz, M. Hoch, E. Seifert, H. Jackie. 1989. Kruppel
Requirement for Knirps Enhancement Reflects Overlap-
ping Gap Gene Activities in the Drosophila Embrvo.
Nature, 341, 337-340.
D. Stanojevic, T. Hoey, M. Levine. 1989. Sequence-Specific
DNA-Binding Activities of the Gap Proteins Encoded by
Hunchback and Kruppel in Drosophila. Nature, 341,
331-335.
J. Treisman, C. Desplan. 1989. The Products of the Drosophila
Gap Genes Hunchback and Kruppel Bind to the Hunch-
back Promoters. Nature, 341, 335-337.
A.S. Wilkins. 1989. Organizing the Drosophila Posterior Pat-
tern: Why Has the Fruit Fly Made Life So Complicated
for Itself? BioEssays, 11, 67.
ЛИТЕРАТУРА
323
С. V. Е. Wright. 1989. Vertebrate Homeodomain Proteins:
Families of Region-Specific Transcription Factors. Trends
Biochem. Sci., 14, 52-56.
8.3.3
M. Ares, Jr., J. Chung, L. Giglio, A.M. Weiner. 1987. Distinct
Factors with Spl and NF-A Specificities Bind to Adjancent
Functional Elements of the Human U2 snRNA Gene
Enhancer. Genes and Devel., 1, 808-817.
P. Carbon, S. Murgo, J. Ebel, A. Krol, G. Tebb, I. W. Mattaj.
1987. A Common Octamer Motif Binding Protein Is
Involved in the Transcription of U6 snRNA by RNA
Polymerase III and U2 snRNA by RNA Polymerase II.
Cell, 51, 71-79.
J. T. Murphy, J. T. Skuzeski, E. Lund, T. H. Steinberg, R. R. Bur-
gess, J. E. Dahlberg. 1987. Functional Elements of the
Human U1 RNA Promoter. J. Biol. Chem., 262, 1795—
1803.
G. Das. D. Henning. D. Wright, R. Reddy. 1988. Upstream Re-
gulatory Elements Are Necessary and Sufficient for Trans-
cription of a U6 RNA Gene by RNA Polymerase 111.
EMBO J., 7, 503-512.
G. R. Kunkel, T. Pederson. 1988. Upstream Elements Required
for Efficient Transcription of a Human U6 RNA Gene
Resemble Those of U1 and U2 Genes Even Though
a Different Polymerase Is Used. Genes and Devel., 2,
196-204.
I. W. Mattaj, N.A. Dathan, H.D. Parry, P. Carbon, A. Krol.
1988. Changing the RNA Polymerase Specificity of U
snRNA Gene Promoters. Cell, 55, 435-442.
M. Tanaka, U. Grossniklaus. W. Herr, N. Hernandez. 1988.
Activation of the U2 snRNA Promoter by the Octamer
Motif Defines a New Class of RNA Polymerase II
Enhancer Elements. Genes and Devel., 1, 1764-1778.
P. Weller, C. Bark. L. Janson, U. Pettersson. 1988. Transcrip-
tion Analysis of a Human U4C Gene: Involvement of
Transcription Factors Novel to snRNA Gene Expression.
Genes and Devel., 2, 1389-1399.
S. Murphy, B. Moorefield, T. Pieler. 1989. Common Mechanisms
of Promoter Recognition by RNA Polymerases II and HI.
Trends Genet., 5, 123-126.
H.D. Parry. D. Scherly, I. W. Mattaj. 1989. “Snurpogenesis”:
The Transcriptionand Assembly of U snRNA Compo-
nents. Trends Biochem. Sci., 14, 15-19.
8.4,a
G. Ciliberto, L. Castagnoli, R. Cortese. 1983. Transcription by
RNA Polymerase Ill. Genome Function, Cell Interactions,
and Differentiation. Current Topics in Dev. Biol., 18,
59-88.
A.L. Bak, A.L. Jorgensen. 1984. RNA Polymerase 111 Control
Regions in Retrovirus LTR, Alu-Type Repetitive DNA,
and Papovavirus. J. Theor. Biol., 108, 339-348.
E.P. Geiduschek, G. P. Tocchini-Valentini. 1988. Transcription
by RNA Polymerase 111. Annu. Rev. Biochem., 57,
873-914.
8.4.6
D.F. Bogenhagen, S. Sakonju, D.D. Brown. 1980. A Control
Region in the Center of the 5S RNA Gene Directs Specific
Initiation of Transcription; 11. The 3' Border of the Region.
Cell, 19, 27-35.
S. Sakonju. D. F. Bogenhagen, D.D. Brown. 1980. A Control
Region in the Center of the 5S RNA Gene Directs Specific
Initiation of Transcription: I. The 5' Border of the Region.
Cell, 19, 13-25.
G. Galli. H. Hofstetter. M.L. Birnstiel. 1981. Two Conserved
Sequence Blocks Within Eukaryotic tRNA Genes Are
Major Promoter Elements. Nature, 294, 626 631.
D. Larson, J. Bradford-Wilcox, L.S. Young, K.U. Sprague.
21*
1983. A Short 5' Flanking Region Containing Conserved
Sequences Is Required for Silkworm Alanine tRNA Gene
Activity. Proc. Natl. Acad. Sci. U.S.A., 80, 3416-3420.
D.F. Bogenhagen. 1985. The Intragenic Control Region of the
Xenopus 5S RNA Gene Contains Two Factor A Binding
Domains That Must Be Aligned Property for Efficient
Transcription Initiation. J. Biol. Chem., 260, 6466-6471.
K.C. Raymond. G. J. Raymond, J. D. Johnson. 1985. In Vivo
Modulation of Yeast tRNA Gene Expression by 5'-Flank-
ing Sequences. EMBO J., 4, 2649-2656.
S.J. Sharp, J. Schaack, I. Cooley, D.J. Burke, D. Soli. 1985.
Structure and Transcription of Eukaryotic tRNA Genes.
Crit. Rev. Biochem., 19, 107- 144.
E. Ullu. A. M. Weiner. 1985. Upstream Sequences Modulate the
Internal Promoter of the Human 7SL RNA Gene. Nature,
318, 371-375.
G.R. Kunkel, R.L. Maser. J.P. Calvet, T. Pederson. 1986. U6
Small Nuclear RNA Is Transcribed by RNA Polymerase
III. Proc. Natl. Acad. Sci. U.S.A., 83, 8575-8579.
M.J. Morry, J.D. Harding. 1986. Modulation of Transcrip-
tional Activity and Stable Complex Formation by
5'-Flanking Regions of Mouse tRNA His Genes. Mol. Cell.
Biol., 6, 105-115.
J. Chung, D.J. Sussman, R. Zeller. P. Leder. 1987. The c-mys
Gene Encodes Superimposed RNA Polymerase II and III
Promoters. Cell, 51, 1001-1008.
A.D. Garcia, A.M. O’Connell, S.J. Sharp. 1987. Formation of
an Active Transcription Complex in the Drosophila mela-
nogaster 5S RNA Gene Is Dependent on an Upstream
Region. Mol. Cell. Biol., 7, 2046-2051.
S. Murphy, C. diLiegro, M. Meili. 1987. The in Vitro Trans-
cription of the 7SK RNA Gene by RNA Polymerase Ill
Is Dependent Only on the Presence of an Upstream
Promoter. Cell, 51, 81-87.
T. Pieler, J. Hamm, R.G. Roeder. 1987. The 5S Gene Internal
Control Region Is Composed of Three Distinct Sequence
Elements, Organized at Two Functional Domains with
Variable Spacing. Cell, 48, 91 -100.
S.J. Sharp, A.D. Garcia. 1988. Transcription of the Drosophila
melanogaster 5S RNA Gene Requires an Upstream Pro-
moter and Four Intragenic Sequence Elements. Mol. Cell.
Biol., 8, 1266- 1274.
E. T. Wilson. D. P. Condliffe, K.U. Sprague. 1988. Transcrip-
tional Properties of BmX, a Moderately Repetitive Silk-
worm Gene That Is an RNA Polymerase III Template.
Mol. Cell. Biol., 8. 624-631.
8.4.B
A.B. Lassar, P.L. Martin, R.G. Roeder. 1983. Transcription of
Class III Genes: Formation of Preinitiation Complexes.
Science. 222, 740-748.
R.E. Baker, B.D. Hall. 1984. Structural Features of Yeast
tRNA Genes Which Affect Transcription Factor Binding.
EMBO J., 3, 2793-2800.
D.D. Brown. 1984. The Role of Stable Complexes That Repress
and Activate Eukaryotic Genes. Cell, 37. 359-365.
D. R. Smith, I. J. Jackson, D.D. Brown. 1984. Domains of the
Positive Transcription Factor Specific for the Xenopus 5S
RNA Gene. Cell, 37, 645-652.
T. Enver. 1985. A Pulling Out of Fingers. Nature, 317, 385-386.
W.K. Hoeffler, R.G. Roeder. 1985. Enhancement of RNA Poly-
merase Ill Transcription by the El A Gene Product of
Adenovirus. Cell, 41, 955 963.
J. Miller, A.D. McLachlan. A. Klug. 1985. Repetitive Zins-Bin-
ding Domains in the Protein Transcription Factor 111A
from Xenopus Oocytes. EMBO J., 4. 1609-1614.
D.R. Setzer, D.D. Brown. 1985. Formation and Stability of the
5S RNA Transcription Complex. J. Biol. Chem., 260,
2483-2492.
M.J. Taylor, J. Segall. 1985. Characterization of Factors and
324
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
DNA Sequences Required for Accurate Transcription of
the Saccharomyces cerevisiae 5S RNA Gene. J. Biol. Chem.,
260, 4531-4540.
L. Fairall. D. Rhodes, A. Klug. 1986. Mapping of the Sites of
Protection on a 5S RNA Gene by the Xenopus Transcrip-
tion Factor III A. J. Mol. Biol., 192, 577-591.
S.C. Harrison. 1986. Fingers and DNA Half-Turns. Nature,
322, 597-598.
D. Rhodes, A. Klug. 1986. An Underlying Repeat in Some
Transcriptional Control Sequences Corresponding to Half
a Double Helical Turn of DNA. Cell, 46, 123-132.
A.P. Wolffe, D.D. Brown. 1986. DNA Replication in Vitro
Erases a Xenopus 5S RNA Gene Transcription Complex.
Cell, 47, 217-227.
A. Klug, D. Rhodes. 1987. “Zinc Fingers”: A Novel Protein
Motif for Nucleic Acid Recognition. Trends Biochem. Sci.,
12, 464-469.
G. A. McConkey, D. F. Bogenhagen. 1987. Transition Mutations
Within the Xenopus borealis Somatic 5S RNA Gene Can
Have Independent Effects on Transcription and DFIIIA
Binding. Mol. Cell. Biol., 7, 486-494.
W. R. Folk. 1988. Changing Directions in Pol III Transcription.
Genes and Devel., 2, 373-375.
WK. Hoeffler, R. Kovelman, R.G. Roeder. 1988. Activation of
Transcription Factor IIIC by the Adenovirus E1A Protein.
Cell, 53, 907- 920.
B. Sollner-Webb. 1988. Surprises in Polymerase III Transcrip-
tion. Cell, 52, 153-154.
K.E. Vrana, M.E.A. Churchill, T.D. Tullius, D.D. Brown. 1988.
Mapping Functional Regions of Transcription Factor
TFIIIA. Mol. Cell. BioL, 8, 1684-1696.
8.4.r
N. R. Cozzarelli. S. P. Gerrard, M. Schlissel, D. D. Brown,
D. F. Bogenhagen. 1983. Purified RNA Polymarase III
Accurately and Efficiently Terminates Transcription of 5S
RNA Genes. Cell, 34, 829-835.
J. Hess, C. Perez-Stable, G.J. Wu. B. Weir. I. Tinoco, Jr.,
C.K.J. Shen. 1985. End-to-End Transcription of an Alu
Family Repeat. J. Mol. Biol., 184, 7-21.
8.4.Д
W. M. Wormington, M. Schlissel, D. D. Brown. 1982. Develop-
mental Regulation of Xenopus 5S RNA Genes. Cold
Spring. Harbor Symp. Quant. Biol., 47, 879 884.
W. M. Wormington. D.D. Brown. 1983. Onset of 5S RNA Gene
Regulation During Xenopus Embryogenesis. Devel. Biol.,
99, 248-257.
M.S. Schlissel, D.D. Brown. 1984. The Transcriptional Regula-
tion of Xenopus 5S RNA Genes in Chromatin: The Roles
of Active Stable Transcription Complexes and Histone Hl.
Cell, 37, 903-913.
D.D. Brown, M.S. Schlissel. 1985. A Positive Transcription
Factor Controls the Differential Expression of Two 5S
RNA Genes. Cell, 42, 759-767.
M. T. Andrews. D.D. Brown. 1987. Transient Activation of
Oocyte 5S RNA Genes in Xenopus Embryos by Raising
the Level of the Trans-Acting Factor TFIIIA. Cell, 51,
445-453.
L. J. Korn, D.R.Guinta, J.Y. Tso. 1987. TFIIIA Mediated
Control of 5S RNA Gene Expression in Xenopus. In:
R. A. Firtel and E. H. Davidson (eds.). Molecular Approaches
to Developmental Biology. Proceedings of the Duppon-
Genetech UCLA Symposium, Keystone, Colorado, March
30-April 6, 1986 (UCLA Symposia on Molecular and
Cellular Biology, Vol. 51), pp. 25-37. Alan R. Liss, New
York.
L.J. Peck, L. MUlstein, P. Eversole-Cire, J.M. Gottesfeld,
A. Varshavsky. 1987. Transcriptionally Inactive Oocyte-
Type 5S RNA Genes of Xenopus laevis Are Complexed
with TFIIIA in Vitro. Mol. Cell. Biol., 7, 3503-3510.
A.P. Wolffe, D.D. Brown. 1987. Differential 5S RNA Gene
Expression in Vitro. Cell, 51, 733-740.
G. A. McConkey, D.F. Bogenhagen. 1988. TFIIIA Binds with
Equal Affinity to Somatic and Major Oocyte 5S RNA
Genes. Genes and Devel., 2, 205-214.
A.P. Wolffe, D.D. Brown. 1988. Developmental Regulation of
Two 5S Ribosomal RNA Genes. Science. 241. 1626-1632
8.5.6 и в
T.R. Cech. 1983. RNA Splicing: Three Themes with Variations.
Cell, 34, 713-716.
F. K. Chu, G.F. Maley, F. Maley. M. Belfort. 1984. Intervening
Sequence in the Thymidylate Synthase Gene of Bacterio-
phage T4. Proc. Natl. Acad. Sci. U.S.A., 81, 3049-3053.
G. Garriga, A.M. Lambowitz. 1984. RNA Splocing in Neuros-
pora Mitochondria: Self-Splicing of a Mitochondrial In-
tron in Vitro. Cell, 38, 631-641.
H.F. Tabak, G. Van der Horst, K.A.Osinga, A.C. Arnberg.
1984. Splicing og Large Ribosomal Precursor RNA and
Processing of Intron RNA in Yeast Mitochondria. Cell, 39,
623-629.
M. Belfort, J. Pedersen-Lane. D. West, K. Ehrenman. G. Maley,
F. Chu, F. Maley. 1985. Processing of the Intron-Contai-
ning Thymidylate Synthase (td) Gene og Phage T4 Is at
the RNA Level. Cell, 41, 375-382.
T. R. Cech. 1985. Self-Splicing RNA: Implications for Evolution.
Inti. Rev. Cytol., 93, 3-22.
J. V. Price, T.R. Cech. 1985. Coupling of Tetrehymena Riboso-
mal RNA Splicing to p-Galactosidase Expression in Esche-
richia coli. Science, 228, 719-722.
F. J. Schmidt. 1985. RNA Splicing in Prokaryotes: Bacteriopha-
ge T4 Leads the Way. Cell, 41, 339-340.
G. van der Horst. H.F. Tabak. 1985. Self-Splicing of Yeast
Mitochondrial Ribosomal and Messenger RNA Precur-
sors. Cell, 40, 759-766.
A.C. Arnberg, G. Van der Horst. H.F. Tabak. 1986. Formation
of Lariats and Cicles in Self-Splicing of the Precursor to
the Large Ribosomal RNA of Yeast Mitochondria. Cell,
44, 235-242.
T.R. Cech, 1986. The Generality of Self-Splicing RNA: Rela-
tionship to Nuclear mRNA Splicing. Cell, 44, 207-210.
T. R. Cech. 1986. RNA as an Enzyme. Sci American, 255,
64-75.
F.K. Chu. G.F. Maley, D.K. West, M. Belfort. F. Maley. 1986.
Characterization of the Intron in the Phage T4 Thymidy-
late Synthase Gene and Evidence for Its Self-Excision from
the Primary Transcript. Cell, 45, 157-166.
G. Garriga, A. M. Lambowitz. 1986. Protein-Dependent Spli-
cing of a Group I Intron in Ribonucleoprotein Particles
and Soluble Fractions. Cell. 46, 669 680.
J.M. Gott, D.A. Shub, M. Belfort. 1986. Multiple Self-Splicing
Introns in Bacteriophage T4: Evidence from Autocatalytic
GTP Labeling of RNA in Vitro. Cell, 47, 81-87.
C.L. Peebles. P.S. Perlman, K.L. Mecklenburg, M.L. Petrillo,
J. H. Tabor, K. A. Jarrell, H.-L. Cheng. 1986. A Self-Spli-
cing RNA Excises an Intron Lariat. Cell, 44, 213 223.
C. Schmelzer. R.J. Schweyen. 1986. Self-Splicing of Group II
Introns in Vitro: Mapping of the Branch Point and
Mutational Unhibition of Lariat Formation. Cell, 46,
557-565.
H.F. Tabak, L.A. Grivell. 1986. RNA Catalysis in the Excision
of Yeast Mitochondrial Introns. Trends Genet., 2, 51-55.
R. van der Veen, A.C. Arnberg, G. van der Horst, L. Bonen,
H. F. Tabak, L. A. Grivell. 1986. Excised Group II Intron in
Yeast Mitochondria Are Lariats and Can Be Formed by
Self-Spricing in Vitro. Cell, 44, 225-234.
R. B. Waring, P. Towner, S.J. Minter, R. W. Davies. 1986. Spli-
ce-Site Selection by a Self-Splicing RNA of Tetrahymena.
Nature, 321, 133-139.
ЛИТЕРАТУРА
325
R.A. Akins. A. M. Lambowitz. 1987. A Protein Required for
Splicing Group I Introns in Neurospora Mitochondria Is
Mitochondrial Tyrosyl-tRNA Synthetase or a Derivative
Thereof. Cell, 50, 331-345.
T.R. Cech. 1987. The Chemistry of Self-Splicing RNA and
RNA Enzymes. Science, 236, 1532-1539.
D. H. Hall. С. M. Povinelli, K. Ehrenman, J. Pedersen-Lane,
F. Chu, M. Belfort. 1987. Two Domains for Splicing in the
Intron of the Phage T4 Thymidylate Synthase (td) Gene
Established by Nondirected Mutagenesis. Cell, 48, 63- 71.
J. Kjems, R. A. Garrett. 1988. Novel Splicing Mechanism for
the Ribosomal RNA Intron in the Archaebacterium Desul-
furococcus mobilis. Cell, 54, 693-703.
J. V. Price, T.R. Cech. 1988. Determinants of the 3' Splice Site
for Self-Splicing of the Tetrahymena Pre-rRNA. Genes and
Devel., 2, 1439-1447.
S.A. Woodson, T.R. Cech. 1989. Reverse Self-Splicing of the
Tetrahymena Group I Intron: Implication for the Directio-
nally of Splicing and for Intron Transposition. Cell, 57,
335-345.
S. Augustin, M.W' Muller, R.J. Schweyen. 1990. Reverse Self-
Splising of Group II Intron RNAs in Vitro. Nature, 343,
383-386.
M. Morl, C. Schmelzer. 1990. Integration of Group II Intron
bll into a Foreign RNA by Reversal of the Self-Splicing
Reaction in Vitro. Cell, 60, 629- 636.
8.5.Г
H. Busch, R. Reddy, L. Rothblum, Y.C. Choi. 1982. SnRNAs,
SnRNPs, and RNA Processing. Annu. Rev. Biochem., 51,
617-654.
H. Domdey, B. Apostol, R.J. Lin, A. Newman, E. Brody,
J. Abelson. 1984. Lariat Structures Are in Vivo Interme-
diates in Yeast Pre-mRNA Splicing. Cell, 39, 611-621.
C. Hashimoto, J. A. Steitz. 1984. U4 and U6 RNAs Coexist in
a Single Small Nuclear Ribonucleoprotein Particle. Nuc-
leic Acids Res., 12, 3283-3293.
W. Keller. 1984. The RNA Lariat: A New Ring to the Splicing
of mRNA Precursors. Cell, 39, 423-425.
J. R. Rodriguez, C. W. Pikielny, M. Rosbash. 1984. In Vivo Cha-
racterization of Yeast mRNA Processing Intermediates.
Cell, 39, 603-610.
D. L. Black, B. Chabot, J. A. Steitz. 1985. U2 as Well as U1
Small Nuclear Ribonucleoproteins Are Involved in Pre-
messenger RNA Splicing. Cell, 42, 737-750.
M.M. Konarska, R.A. Padgett, P.A. Sharp. 1985. Trans Spli-
cing of mRNA Precursors in Vitro, Cell, 42, 165-171.
A.J. Newman, R.J. Lin, S.C. Cheng, J. Abelson. 1985. Mole-
cular Consequences of Specific Intron Mutations on Yeast
mRNA Splicing in Vivo and in Vitro. Cell, 42, 335-344.
P.A. Sharp. 1985. On the Origin of RNA Splicing and Introns.
Cell, 42, 397-400.
D. Solnick. 1985. Trans Splicing of mRNA Precursors. Cell, 42,
157-164.
M. Ares, Jr. 1986. U2 RNA from Yeast Is Unexpectedly Large
and Contains Homology to Vertebrate U4, U5 and U6
Small Nuclear RNAs. Cell, 47, 49-59.
T.R. Cech. 1986. The Generally of Self-Splicing RNA: Rela-
tionship to Nuclear mRNA Splicing. Cell, 44, 207-210.
V. Gerke, J. A. Steitz. 1986. A Protein Associated with Small
Nuclear Ribonucleoprotein Particles Recognizes the 3'
Splice of Premessenger RNA. Cell, 47, 973-984.
M.R. Green. 1986. Pre-mRNA Splicing. Annu. Rev. Genet., 20,
671-708.
C. Guthrie. 1986. Finding Functions for Small Nuclear RNAs
in Yeast. Trends Biochem. Sci., 11, 430-434.
W.J. Murphy. K.P. Watkins, N. Agabian. 1986. Identification of
a Novel Y Branch Structure as an Intermediate in
Trypanasome mRNA Processing: Evidence for Trans Spli-
cing. Cell, 47, 517-525.
R. A. Padgett, P. J. Grabowski, M. M. Konarska, S. Seiler,
P.A. Sharp. 1986. Splicing of Messenger RNA Precursors.
Annu. Rev. Biochem., 55, 1119-1150.
C. W. Pikielny, M. Robash. 1986. Specific Small Nuclear RNAs
Are Associated with Yeast Spliceosomes. Cell, 45, 869-
877.
R. E. Sutton, J.C. Boothroyd. 1986. Evidence for Trans Splicing
in Trypanosomes. Cell, 47, 527-535.
J. Tazi, C. Alibert, J. Temsamani. I. Reveillaund, G. Cathala,
C. Brunel, P. Jeanteur. 1986. A Protein That Specifically
Recognizes the 3' Splice Site of Mammalian Pre-mRNA
Introns Is Associated with a Small Nuclear Ribonucleo-
protein. Cell, 47, 755-766.
L. H. T. Van der Ploeg. 1986. Discontinuous Transcription and
Splicing in Trypanasomes. Cell, 47, 479-480.
Y Zhuang, A.M. Weiner. 1986. A Compensatory Base Change
in U1 snRNA Suppresses a 5' Splice Site Mutation. Cell,
46, 827-835.
A. Bindereif, M.R. Green. 1987. An Ordered Pathway of
snRNP Binding During Mammalian Pre-mRNA Splicing
Complex Assembly. EMBO J., 6, 2415 2424.
S.C. Cheng, J. Abelson. 1987. Spliceosome Assembly in Yeast.
Genes and Devel., 1, 1014-1027.
J. R. Couto, J. Tamm, R. Parker, C. Guthrie. 1987. A Trans-
acting Suppressor Restores Splicing of a Yeast Intron with
a Branch Point Mutation. Genes and DeveL, 1, 445-455.
M.M. Konarska, P.A. Sharp. 1987. Interactions Between Small
Nuclear Ribonucleoprotein Particles in Formation of
Spliceosomes. Cell, 49, 763-774.
M. Krause, D. Hirsh. 1987. A Trans-Splised Leader Sequence
on Actin mRNA in C. elegans. Cell, 49, 753-761.
M. Lossky. G.J. Anderson. S.P. Jackson, J. Beggs. 1987. Identi-
fication of a Yeast snRNP Protein and Detection of
snRNP—snRNP Interactions. Cell, 51, 1019-1026.
R. Luhrmann. 1987. snRNP Proteins. In: M. L. Bimstiel (ed.).
Structure and Function of Major and Minor Small
Nuclear Ribonucleoprotein Particles, pp. 71-99. Springer
Verlag, Heidelberg.
T. Maniatis, R. Reed. 1987. The Role of Small Nuclear Ribo-
nucleoprotein Particles in Pre-mRNA Splicing. Nature,
325, 673-678.
R. Parker, P.G. Siliciano, C. Guthrie. 1987. Recognition of the
TACTAAC Box During mRNA Splicing in Yeast Involves
Base Pairing to the U2-like snRNA. Cell, 49, 229-239.
P.A. Sharp. 1987. Trans Splicing: Variation on a Familiar
Theme? Cell, 50, 147-148.
S. Bektesh, K. Van Doren, D. Hirsh. 1988. Presence of the
Caenorhabditis elegans Spliced Leader on Different
mRNAs and in Different Genera of Nematodes. Genes and
Devel., 2, 1277-1283.
C. Guthrie, B. Patterson. 1988. Spliceosomal snRNAs. Annu.
Rev. Genet., 22, 387-418.
D.H. Kedes, J. A. Steitz. 1988. Correct in Vivo Splicing of the
Mouse Immunuglobulin К Light-Chain Pre-mRNA Is
Dependent on 5' Splice-Site Position Even in the Absence
of Transcription. Genes and Devel., 2, 1448-1459.
A. Kramer. 1988. Presplicing Complex Formation Requires
Two Proteins and U2 snRNP. Genes and Devel., 2,
1155-1167.
R. Reed, J. Griffith, T. Maniatis. 1988. Purification and Visuali-
zation of Native Spliceosomes. Cell, 53, 949-961.
R. Reed, T. Maniatis. 1988. The Role of the Mammalian
Branchpoint Sequence in Pre-mRNA Splicing. Genes and
Devel., 2, 1268-1276.
B. Ruskin, P. D. Zamore. M. R. Green. 1988. A Factor, U2AF, Is
Required for U2 snRNP Binding and Splicing Complex
Assembly. Cell, 52, 207-219.
P. G. Silicano, C. Guthrie. 1988. 5' Splice Site Selection in Yeast:
Genetic Alterations in Base-Pairing with U1 Reveal Addi-
tional Requirements. Genes and Devel., 2, 1258-1267.
326
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
J. A. Steitz. 1988. “Snurps”. Sci. American, 258 (6), 56-63.
8.5.Д
C.L. Peebles, R.C. Ogden, G. Knapp, J. Abelson. 1979. Splicing
of Yeast tRNA Precursors: A Two-Stage Reaction. Cell,
18, 27-35.
E.M. De Robertis, P. Black, K. Nishikura. 1981. Intranuclear
Location of the tRNA Splicing Enzymes. Cell. 23, 89-93.
C.L. Greer, C.L. Peebles, P. Gegenheimer, J. Abelson. 1983.
Mechanism of Action of a Yeast RNA Ligase in tRNA
Splicing. Cell, 32, 537-546.
C.L. Peebles, D. Gegenheimer, J. Abelson. 1983. Precise Exci-
sion of Intervening Sequences from Precursor tRNAs by
a Membrane-Associated Yeast Endonuclease. Cell, 32,
525-536.
M.l. Baldi, E. Mattoccia, S. Ciafre, D.G. Attardi, G. P. Toc-
chini-Valentini. 1986. Binding and Cleavage of pre-tRNA
by the Xenopus Splicing Endonuclease: Two Separatable
Steps of the Intron Excision Reaction. Cell, 47, 965-971.
C.L. Greer. 1986. Assembly of a tRNA Splicing Complex:
Evidence for Concerted Excision and Joining Steps in
Splicing in Vitro. MoL Cell. Biol., 6, 635-644.
E.M. Phizicky, R.C. Schwartz, J. Abelson. 1986. S. cerevisiae
tRNA Ligase: Purification of the Protein and Isolation of
the Structural Gene. J. Biol. Chem., 261, 2978-2986.
C.L. Greer, D. Soil, I. Willis. 1987. Substrate Recognition and
Identification of Splice Sites by the tRNA-Splicing Endo-
nuclease and Ligase from Saccharomyces cerevisiae. Mol.
Cell. BioL, 7, 76-84.
V.M. Reyes, J. Abelson. 1988. Substrate Recognition and Splice
Site Determination in Yeast tRNA Splicing. Cell, 55,
719-730.
N. Stange, H.J. Gross. H. Beier. 1988. Wheat Germ Splicing
Endonuclease Is Highly Specific for Plant Pre-tRNAs.
EMBO J., 7, 3823-3828.
M. Winey, I. Edelman, M. R. Culbertson. 1989. A Synthetic
Intron in a Naturally Intronless Yeast Pre-tRNA Is
Spliced Efficiently in Vivo. Mol. Cell. Biol., 9, 329-331.
8.5.e
M. G. Rosenfeld. C. R. Lin, S. G. Amara, L. Stolarsky, B. A. Ro-
os, E.S. Ong, R.M. Evans. 1982. Calcitonin mRNA Poly-
morphism: Peptide Switching Associated with Alternative
RNA Splicing Events. Proc. Natl. Acad. Sci. U.S.A., 79,
1717-1721.
M. Kress, D. Glaros. G. Khoury, G. Jay. 1983. Alternative RNA
Splicing in Expression of the H-2K Gene. Nature, 306,
602-604.
D. W. Chung, E. W. Davie. 1984. у and y' Chains of Human
Fibrinogen Are Produced by Alternative mRNA Proces-
sing. Biochemistry, 23, 4232-4236.
F. de Ferra, H. Engh, L. Hudson, J. Kamholz, C. Puckett,
S. Molineaux, R.A. Lazzarini. 1985. Alternative Splicing
Accounts for the Four Forms of Myelin Basic Protein.
Cell, 43, 721-727.
A.R. Kornblith, K. Vmezawa, K. Vibe-Pedersen, F.E. Baralle.
1985. Primary Structure of Human Fibronectin: Differen-
tial Splicing May Generate at Least 10 Polypeptides from
a Single Gene. EMBO J., 4., 1755-1759.
D. Solnick. 1985. Alternative Splicing Caused by RNA Secon-
dary Structure. Cell, 43, 667-676.
R. Zamoyska, A. C. Vollmer, К. C. Sizer, C. W. Liaw, J. R. Par-
nes. 1985. Two Lyt-2 Polypeptides Arise from a Single
Gene by Alternatuve Splicing Patterns of mRNA. Cell, 43,
153-163.
У. Ben-Neriah, A. Bernards, M. Paskind, G.Q. Daley, D. Balti-
more. 1986. Alternative 5' Exons in c-abl mRNA. Cell, 44,
577-586.
S.E.Leff, M.G. Rosenfeld. 1986. Complex Transcriptional
Units: Diversity in Gene Expression by Alternative RNA
Processing. Annu. Rev. Biochem., 55, 1091-1117.
J.L Paul, J.E. Schwarzbauer, J. W. Tamkun, R.O. Hynes. 1986.
Cell-Type-Specific Fibronectin Subunits Generated by
Alternative Splicing. J. Biol. Chem., 261, 12258-12265.
R. Reed, T. Maniatis. 1986. A Role for Exon Sequences and
Splice-Site Proximity in Splice-Site Selection. Cell, 46,
681-690.
A. Andreadis, M. E. Gallego, B. Nadal-Ginard. 1987. Genera-
tion of Protein Isoform Diversity by Alternative Splicing:
Mechanistic and Biological Implications. Annu. Rev. Cell
Biol., 3, 207-242.
R. E. Breitbart, A. Andreadis. B. Nadal-Ginard. 1987. Alterna-
tive Splicing: A Ubiquitous Mechanism for the Generation
of Multiple Protein Isoforms from Single Genes. Annu.
Rev. Biochem., 56, 467-495.
R. E. Breitbart, B. Nadal-Ginard. 1987. Developmentally Indu-
ced, Muscle-Specific Trans Factors Control the Differential
Splicing of Alternative and Constitutive Troponin T
Exons. Cell, 49, 793-803.
J.C.S. Noble, C. Prives. J.L. Manley. 1988. Alternative Spli-
cing of SV 40 Early Pre-mRNA Is Determined by Branch
Site Selection. Genes and Devel., 2, 1460-1475.
J.C.S. Noble, Z.Q. Pan, C. Prives, J.L. Manley. 1987. Splicing
of SV 40 Early Pre-mRNA to Large T and Small t mRNAs
Utilizes Different Patterns of Lariat Branch Sites. Cell, 50,
227-236.
B. S. Baker. 1989. Sex in Flies: The Splice of Life. Nature, 340,
521-524.
8.6.a
С. O. Pabo, R. T. Sauer. 1984. Protein-DNA Recognition. Annu.
Rev. Biochem.. 53, 293-321.
J.E. Anderson, M. Plashne, S.C. Harrison. 1985. A Phage
Repressor-Operator Complex at 7 A Resolution. Nature,
316, 596-605.
W.J. Gehring. 1985. The Homeo Box: A Key to the Under-
standing of Development? Cell, 40, 3-5.
J.M. Berg. 1986. More Metal-Binding Fingers. Nature, 319,
264-265.
J.M. Berg. 1986. Potential Metal-Binding Domains in Nucleic
Acid Binding Proteins. Science, 232, 485-487.
U. B. Rosenberg, C. Schroder, A. Preiss, A. Kienlin, S. Cote,
I. Riede, H. Jackie. 1986. Structural Homology of the
Product of the Drosophila kruppel Gene with Xenopus
Transcription Factor I1IA. Nature, 319, 336-339.
A. Klug, D. Rhodes. 1987. “Zinc Fingers”: A Novel Protein
Motif for Nucleic Acid Recognition. Trends Biochem. Sci.,
12, 464-469.
K. Struhl. 1987. The DNA-Binding Domains of the Jun Oncop-
rotein and the Yeast CN4 Transcriptional Activator
Protein Are Functionally Homologous. Cell, 50, 841-
846.
J.M. Berg. 1988. Proposed Structure for the Zinc-Binding
Domains from Transcription Factor 1IIA and Related
Proteins. Proc. Natl. Acad. Sci. U.S.A., 85, 99-103.
M. Bodner, J. Castrillo. L. E. Theill, T. Deerinck. M. Ellisman.
M. Karin. 1988. The Pituitary-Specific Transcription
Factor GHF-1 Is a Homeobox-Containing Protein. Cell,
55, 505-518.
R. Chiu, W.J. Boyle, J. Meek, T. Smeal, T. Hunter, M. Karin.
1988. The c-Fos Protein Interacts with c-Jun/AP-1 to
Stimulate Transcription of AP-1 Responsive Genes. Cell,
54, 541-552.
R.G. Clerc, L.M. Corcoran. H.H. LeBowitz, D. Baltimore.
1988. The В Cell Specific Oct-2 Protein Contains POU-
Box and Homeo Box Type Domains. Genes and Devel., 2,
1570-1582.
T. Curran, B. R. Franza, Jr. 1988. Fos and Jun: The AP-1
Connection. Cell, 55, 395-397.
C. Desplan, J. Theis, P. H. O’Farrell. 1988. The Sequence Speci-
ЛИТЕРАТУРА
327
ficity of Homeodomain-DNA Interaction. Cell, 54, 1081 —
1090.
R.M. Evans, S.M. Hollenberg. 1988. Zinc Fingers: Gilt by
Association. Cell, 52, 1-3.
T.D. Halazonetis, K. Georgopoulos, M.E. Greenberg, P. Leder.
1988. c-Jun Dimerizes with Itself and with c-Fos, Forming
Complexes of Different DNA Binding Affinities. Cell, 55,
917-924.
W. Herr. R. A. Sturm, R. G. Clerc, L. M. Corcoran, D. Baltimore,
P. A. Sharp, H. A. Ingraham, M.G. Rosenfeld, M. Finney,
G. Ruvkun, H.R. Horvitz. 1988. The POU Domain: A
Large Conserved Region in the Mammalian Pit-1, Oct-1,
Oct-2, and Caenorhabditis elegans Unc-86 Gene Products.
Genes and Devel., 2, 1513-1516.
H. A. Ingraham, R. Chen. H. J. Mangalam, H. P. Elsholtz,
S. E. Flynn. C. R. Lin. D. M. Simmons, L. Swanson.
M. G. Rosenfeld. 1988. A Tissue-Specific Transcription
Factor Containing a Homeodomain Specifies a Pituitary
Phenotype. Cell, 55, 519-529.
T. Kouzarides, E. Ziff. 1988. The Role of the Leucine Zipper in
the Fos-Jun Interaction. Nature, 336, 646-651.
W. H. Landschulz. P. F. Johnson. S. L. McKnight. 1988. The
Leucine Zipper: A Hypothetical Structure Common to
a New Class of DNA Binding Proteins. Science, 240,
1759-1764.
M. Levine, T. Hoey. 1988. Homeobox Proteins as Sequence-
Specific Transcription Factors. Cell, 55, 537-540.
Y. Nakabeppu, K. Ryder, D. Nathans. 1988. DNA Binding Acti-
vities of Three Murine Jun Proteins: Stimulation by Fos.
Cell. 55, 907-915.
F.J. Rouscher III, P.J. Voulalas. B.R. Franza, Jr., T. Curran.
1988. Fos and Jun Bind Cooperatively to the AP-1 Site:
Reconstitution in Vitro. Genes and Devel., 2, 1687-1699.
M. Robertson. 1988. Homeo Boxes, POU Proteins and the
Limits to Promiscuity. Nature, 336, 522-524.
R. T. Sauer, D.L. Smith, A.D. Johnson. 1988. Flexibility of the
Yeast a2 Repressor Enables It to Occupy the Ends of Its
Operator, Leaving the Center Free. Genes and Devel., 2,
807-816.
R. A. Sturm. G. Das. W. Herr. 1988. The Ubiquitous Octamer-
Binding Protein Oct-1 Contains a POU Domain with
a Homeo Box Subdomain. Genes and Devel., 2, 1582—
1599.
R.G. Brennan, B. W. Matthews. 1989. Structural Basis of DNA-
Protein Recognition. Trends Biochem Sci., 14, 286-290.
W.H. Landschulz, P.F. Johnson, S.L. McKnight. 1989. The
DNA Binding Domain of the Rat Liver Nuclear Protein
C/EBP is Bipartite. Science, 243, 1681-1687.
M.S. Lee, G.P. Gippert, К. V. Soman, D.A. Case. P.E. Wright.
1989. Three-Dimensional Solution Structure of a Single
Zinc Finger DNA-Binding Domain. Science, 245, 635-637.
A. R. Oliphant, C. J. Branch, K. Struhl. 1989. Definning the
Sequence Specificity of DNA-Binding Proteins by Selec-
ting Binding Sites from Random-Sequence Oligonucleo-
tides: Analysis of Yeast GCN4 Protein. Mol. Cell. Biol., 9,
2944-2949.
E.K. O’Shea, R. Rutkowski. W.F. Stafford III, P.S. Kim. 1989.
Preferential Heterodimer Formation by Isolated Leucine
Zippers from Fos and Jun. Science, 245, 646-648.
T. B. Rajavashisth, A. K. Taylor, A. Andalibi, K. L. Svenson,
A. J. Lusis. 1989. Identification of a Zinc Finger Protein
That Binds to the Sterol Regulatiry Element. Science, 245,
640-643.
M.P. Scott, J. W. Tamkun, G. W. Hartzell HI. 1989. The Struc-
ture and Function of the Homeodomain. Biochim.
Biophys. Acta. Ser. Rev. Cancer, 989 (1), 25-48.
W. Schaffner. 1989. How Do Different Transcription Factors
Binding the Same DNA Sequence Sort Out Their Jobs?
Trends Genet., 5, 37-38.
K. Struhl. 1989. Helix-Tum-Helix, Zinc-Finger, and Leucine-
Zipper Motifs for Eukaryotic Transcriptional Regulatory
Proteins. Trends Biochem. Sci., 14, 127-140.
С. V. E. Wright, K. W. Y. Cho, G. Oliver, E. M. DeRobertis. 1989.
Vertebrate Homeodomain Proteins: Families of Region-
Specific Transcription Factors. Trends Biochem. Sci., 14,
52-56.
8.6.6
I. A. Hope, K. Struhl. 1986. Functional Dissection of a Euka-
ryotic Transcriptional Activator Protein, GCN4 of Yeast.
Cell, 46, 885-894.
L. Keegan. G. Gill, M. Ptashne. 1986. Separation of DNA
Binding from the Transcription-Activating Function of
Eukaryotic Regulatory Protein. Science, 231, 699-704.
J. Ma, M. Ptashne. 1987. A New Class of Yeast Transcriptional
Activators. Cell, 51, 113-119.
K. Struhl. 1987. Promoters, Activator Proteinns, and the Me-
chanism of Transcriptional Initiation in Yeast. Cell, 49,
295-297.
L. Guarente. 1988. UASa and Enhancers: Common Mechanism
of Transcriptional Activation in Yeast and Mammals. Cell,
52, 303-305.
I. A. Hope, S. Mahadevan, K. Struhl. 1988. Structural and
Functional Characterization of the Short Acidic Trans-
criptional Activation Region of Yeast GCN4 Protein.
Nature, 333, 635-640.
S. P. Jackson. R. Tjian. 1988. О-Glycosylation of Eukaryotic
Transcription Factors: Implications for Mechanisms of
Transcriptional Regulation. Cell, 55, 125-133.
M. Ptashne. 1988. How Eukaryotic Transcriptional Activators
Work. Nature, 335, 683-689.
K. Struhl. 1988. JUN Oncoprotein, a Vertebrate Transcription
Factor, Activates Transcription in Yeast. Nature, 332,
649-650.
J. C. Wang, G. N. Giaever. 1988. Action at a Distance Along
a DNA. Science, 240, 300-304.
J. W. Lillie. M. R. Green. 1989. Transcription Activation by the
Adenovirus Ela Protein. Nature, 338, 39-44.
C. V.E. Wright, K.W.YCho, G. Oliver, E.M. De Roberts. 1989.
Vertebrate Homeo Domain Proteins: Families of Region-
Specific Transcription Factors. Trends Biochem. Sci., 14,
52-56.
8.7.a
J. R. Paulson, J. K. Laemmli. 1977. Structure of Histone-Dep-
leted Metaphase Chromosomes. Cell, 12, 817-826.
M.P. F. Marsden, U.K. Laemmli. 1979. Metaphase Chromo-
some Structure: Evidence for a Radial Loop Model. Cell,
17, 849 858.
C. Wu, P. M. Bingham, K. J. Livak, R. Holmgren, S. C. R. Elgin.
1979. The Chromatin Structure of Specific Genes: 1.
Evidence for Higher Order Domains of Defined DNA
Sequence. Cell, 16, 797-806.
C. Wu. Y.-C. Wong, S.C.R. Elgin. 1979. The Chromatin Struc-
ture of Specific Genes: II. Disruption of Chromatin
Structure During Gene Activity. Cell, 16, 807-814.
S. Saragosti, G. Moyne, M. Yaniv. 1980. Absence of Nucleo-
somes in a Fraction of SV 40 Chromatin Between the
Origin of Replication and the Region Coding for the Late
leader RNA. Cell, 20, 65-73.
H. Weintraub. 1985. Assembly and Propagation of Repressed
and Derepressed Chromosomal States. Cell, 42, 705-711.
M. Yaniv, S. Cereghini. 1986. Structure of Transcriptionally
Active Chromatin. Crit. Rev. Biochem., 31, 1-26.
Y. Lorch. J. W. LaPointe, R.D. Kornberg. 1987. Nucleosomes
Inhibit the Initiation of Transcription but Allow Chain
Elongation with the Displacement of Histones. Cell, 49,
203- 210.
M. A. Goldman. 1988. The Chromatin Domain as a Unit of
Gene Regulation. BioEssays, 9, 51-55.
328
ЧАСТЬ 111. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Y. Lorch, J. fk LaPointe, R. D. Kornberg. 1988. On the Dis-
placement of Histones from DNA by Transcription. Cell,
55. 743-744.
8.7.6
M. Gellert. 1981. DNA Topoisomerases. Annu. Rev. Biochem.,
50, 879-910.
A. Rich, A. Mondheim, A. H.-J. Wang. 1984. The Chemistry
and Biology of Left-Handed Z-DNA. Annu. Rev. Bio-
chem., 53, 791-846.
J. C. Wang. 1985. DNA Toposiomerases. Annu. Rev. Biochem.,
54. 665-697.
A. Jaworski, W.-T. Hsieh, J. A. Blaho, J.E. Larson. R.D. Wells.
1987. Left-Handed DNA in Vivo. Science, 238, 773-777.
F. Lancillotti, M.C. Lopez, P. Arias, C. Alonso. 1987. Z-DNA
in Transcriptionally Active Chromosomes. Proc. Natl.
Acad. Sci. U.S.A., 84, 1560-1564.
R.D. Wells. 1988. Unusual DNA Structures. J. Biol. Chem., 263,
1095-1098.
W. Zacharias. A. Jaworski, J.E. Larson, R.D. Wells. 1988. The
В-to Z-DNA Equilibrium in Vivo Is Perturbed by Biolo-
gical Processes. Proc. Natl. Acad. Sci. U.S.A., 85, 7069-
7073.
H. Zhang, J.C. Wang, L.F. Liu. 1988. Involvement of DNA
Topoisomerase I in Transcription of Human Ribosomal
RNA Genes. Proc. Natl. Acad. Sci. U.S.A., 85, 1060-1064.
A.R. Rahmouni, R.D. Wells. 1989. Localized Supercoiling Sta-
bilized Short (CG) Sequences in the Z-Structure in Vivo.
Science, 246, 358-363.
B. Wittig, T. Dorbic, A. Rich. 1989. The Level of Z-DNA in
Metabolically Active, Permeabilized Mammalian Cell
Nuclei Is Regulated by Torsional Strain. J. Cell. Biol., 108,
755-764.
8.7.B
L.H. T. van der Ploeg, R.A. Flavell. 1980. DNA Methylation in
the Human ySP-Globin Locus in Erythroid and Noneryth-
roid Tissues. Cell, 19, 947-958.
M. Busslinger, J. Hurst, R.A. Flavell. 1983. DNA Methylation
and the Regulation of Globin Gene Expression. Cell, 34,
197-206.
W. Doerfler. 1983. DNA Methylation and Gene Activity. Annu.
Rev. Biochem., 52, 93-124.
A. Bolden, C. Ward, J. A. Siedlecki, A. Weissbach. 1984. DNA
Methylation. Inhibition of de Novo and Maintenance
Methylation in Vitro and Synthetic Polynucleotides.
J. Biol. Chem., 259, 12437-12443.
A. Bird, M. Taggart, M. Frommer, O.J. Miller, D. Macleod.
1985. A Fraction of the Mouse Genome That Is Derived
from Islands of Nonmethylated, CpG-Rich DNA. Cell, 40,
91-99.
G. L. Cantoni, A. Razin (eds.). 1985. Biochemistry and Biology
of DNA Methylation, vol. 198, Progress in Clinical and
Biological Research. Alan R. Liss, New York.
P. A. Jones. 1985. Altering Gene Expression with 5-Azacytidine.
Cell, 40, 485-486.
A. P. Bird. 1986. CpG-Rich Islands and the Function of DNA
Methylation. Nature, 321, 209-213.
M. Monk. 1986. Methylation and the X Chromosome. Bio-
Essays, 4, 204-208.
A. Razin, M. Szyf, T. Kafri, M. Roll, H. Giloh, S. Scarpa,
D. Carotti, G. L. Cantoni. 1986. Replacement of 3-Methyl-
cytosine by Cytosine: A Possible Mechanism for Transient
DNA Demethylation During Differentiation. Proc. Natl.
Acad. Sci. U.S.A., 83, 2827-2831.
A. P. Bird. 1987. CpG Islands as Gene Markers in the Verte-
brate Nucleus. Trends Genet., 3, 342-347.
R. Holliday. 1987. The Inheritance of Epigenetic Defects.
Science, 238, 163-170.
L.F. Lock, N. Takagi, G.R. Martin. 1987. Martin. 1987. Met-
hylation of the Hprt Gene on the Inactive X Occurs After
Chromosome Inactivation. Cell, 48, 38-46.
M. Monk. 1987. Genomic Imprinting: Memories of Mother and
Father. Nature, 328, 203-204.
J. P. Sanford, H.J. Clark, V. M. Chapmen, J. Rossant. 1987. Dif-
ferences in DNA Methylation During Oogenesis and
Spermatogenesis and Their Persistence During Early
Embryogenesis in the Mouse. Genes and Devel., 1, 1039-
1046.
J. L. Swain, T. A. Stewart, P. Leder. 1987. Parental Legacy
Determines Methylation and Expression of an Autosomal
Transgene: A Molecular Mechanism for Parental Imprin-
ting. Cell, 50, 719-727.
G. Theiss, R. Schleicher, G. Schimpff-Weiland, H. F oilman. 1987.
DNA Methylation in Wheat. Purification and Properties
of DNA Methyltransferase. J. Eur. Biochem., 167, 89-96.
H. Cedar. 1988. DNA Methylation and Gene Activity. Cell, 53,
3-4.
M. Holler, G. Westin, J. Jiricny, W. Schaffner. 1988. Spl Trans-
cription Factor Binds DNA and Activates Transcription
Even When the Binding Site Is CpG Methylated. Genes
and Devel., 2, 1127-1135.
R. Khan, X,- Y. Zhang, P. C. Supakar, К. C. Ehrlich, M. Ehrlich.
1988. Human Methylated DNA-Binding Protein. J. Biol.
Chem., 263, 14374-14383.
M. Monk. 1988. Genomic Imprinting. Genes and Devel., 2,
921-925.
W Doerfler. 1989. Complexities in Gene Regulation by Promo-
ter Methylation. Nucleic Acids and Mol. Biol., 3, 92- 119.
R. Hollidav. 1989. A Different Kind of Inheritance. Sci. Ameri-
can, 260 (6), 60-70.
S. M. M. Iguchi-Ariga, W. Schaffner. 1989. CpG Methylation
of the cAMP-Response Enhancer/Promoter Sequence
TGACGTCA Abolishes Specific Factor Binding as Well as
Transcriptional Activation. Genes and Devel., 3, 612-619.
8.7.Г
G.R. Fink. 1986. Translational Control of Transcription in
Eukaryotes. Cell, 45, 155-156.
T. Morris. F. Marashi, L. Weber, E. Hickey, D. Greenspan,
J. Bonner, J. Stein, G. Stein. 1986. Involvement of the
5'-Leader Sequence in Coupling the Stability of a Human
H3 Histone mRNA with DNA Replication. Proc. Natl.
Acad. Sci. U.S.A., 83, 981-985.
P.P. Mueller, A.G. Hinnebusch. 1986. Multiple Upstream AUG
Codons Mediate Translational Control of GCN4. Cell, 45,
201-207.
G. Shaw, R. Kamen. 1986. A Conserved AU Sequence from the
3' Untranslated Region of GM-CSF mRNA Mediates
Selective mRNA Degradation. Cell, 46, 659-667.
G. Brawerman. 1987. Determinants of Messenger RNA Stabi-
lity. Cell, 48, 5-6.
M. W. Hentze, S. W. Caughman, T. A. Roualt, J. G. Barriocanal,
A. Dancis, J.B. Harford, R.D. Klausner. 1987. Identifica-
tion of the Iron-Responsive Element for the Translational
Regulation of Human Ferritin mRNA. Science, 238, 1570-
1573.
M. W. Hentze, T. A. Rouault, S. W. Caughman, A. Dancis.
J.B. Harford, R.D. Klausner. 1987. A cis-Acting Element
Is Necessary and Sufficient for Translational Regulation of
Human Ferritin Expression in Response to Iron. Proc.
Natl. Acad. Sci. U.S.A., 84, 6730-6734.
J. S. Pachter, T. J. Yen, D. W. Cleveland. 1987. Autoregulation of
Tubulin Expression Is Achieved Through Specific Degra-
dation of Polysomal Tubulin mRNAs. Cell, 51, 283-292.
T. A. Rouault, M.W. Hentze, A. Dancis, W. Caughman,
J.B. Harford, R.D. Klausner. 1987. Influence of Altered
Transcription on the Translational Control of Human
Ferritin Expression. Proc. Natl. Acad. Sci. U.S.A., 84,
6335-6339.
ЛИТЕРАТУРА
329
J.L. Cassey. В. Di Jeso, К. Rao. R.D. Klausner, J.B. Harford.
1988. Two Genetic Loci Participate in the Regulation by
Iron of the Gene for the Human Transferrin Receptor.
Proc. Natl. Acad. Sci. U.S.A., 85, 1787-1792.
J. L. Casey. M. W. Hentze, D. M. Koeller, S. W. Caughman,
TA. Rouault. R.D. Klausner, J.B. Harford. 1988. Iron-
Responsive Elements: Regulatory RNA Sequences That
Control mRNA Levels and Translation. Science. 240,
924-927.
5. W. Caughman, M.W. Hentze, T. A. Rouault, J.B. Harford,
R.D. Klausner. 1988. The Iron-Responsive Element Is the
Single Element Responsible for Iron-Dependent Transla-
tional Regulation of Ferritin Biosynthesis. J. Biol. Chem.,
263, 19048-19052.
M.W. Hentze, S.W. Caughman, J.L. Casey, D.M. Koeller,
T. A. Rouault, J. B. Harford, R. D. Klausner. 1988. A Model
for the Structure and Functions of Iron-Responsive Ele-
ments. Gene, 72, 201-208.
A.G. Hinnebusch. 1988. Mechanisms of Gene Regulation in the
General Control of Amino Acid Biosynthesis in Saccharo-
myces cerevisiae. Microbiol. Rev., 52, 248-273.
R.D. Klausner. 1988. From Receptors to Genes- Insights from
Molecular Iron Metabolism. Clin. Res., 36, 494-500.
T A. Rouault, M. W. Hentze, S. W. Caughman. J. B. Harford.
R. D. Klausner. 1988. Binding of a Cytosolic Protein to the
Iron-Responsive Element of Human Ferritin Messenger
RNA. Science, 241, 1207-1210.
T. Wilson, R. Treisman. 1988. Removal of Poly (A) and Conse-
quent Degradation of c-fos mRNA Facilitated by 3'
AU-Rich Sequences. Nature, 336, 396-399.
T.J. Yen. D.A. Gay, J.S. Pachter, D. W. Cleveland. 1988. Auto-
regulated Changes in Stability of Polyribosome-Bound
Р-Tubulin mRNAs Are Specified by the First 13 Trans-
lated Nucleotides. Mol. Cell. Biol., 8, 1224-1235.
T.J. Yen, P.S. Machlin. D.W. Cleveland. 1988. Autoregulated
Instability of Р-Tubulin mRNAs by Recognition of the
Nascent Amino Terminus of Р-Tubulin. Nature, 334.
580-585.
M.W. Hentze, TA. Rouault, J.B. Harford, R.D. Klausner.
1989. Oxidation-Reduction and the Molecular Molecular
Mechanism of a Regulatory RNA-Protein Interaction.
Science, 244, 357-359.
C.J. Paddon, E.M. Hannig, A.G. Hinnebusch. 1989. Amino
Acid Sequence Similarity Between GCN3 and GCD2,
Positive and Negative Translational Regulators of GCN4:
Evidence for Antagonism by Competition. Genetics, 122,
551-559.
D. Tzamarias, I. Roussou. G. Thireos. 1989. Coupling of GCN4
mRNA Translational Activation with Decreased Rates of
Polypeptide Chain Initiation. Cell, 57, 947-954.
R.C. Wek, B.M. Jackson. A.G. Hinnebusch. 1989. Juxtaposi-
tion of Domains Homologous to Protein Kinases and
Histidyl-tRNA Synhetases in GCN2 Protein Suggests
a Mechanism for Coupling GCN4 Expression to Amino
Acid Availability. Proc. Natl. Acad. Sci. U.S.A., 86,
4579-4583.
9.1.a
J. Battey. E.E. Max, WO. McBride, D. Swan, P. Leder. 1982.
A Processed Human Immunoglobulin e Gene Has Moved
to Chromosome 9. Proc. Natl. Acad. Scad. Sci. U.S.A., 79.
5956- 5959.
E.F. Vanin. 1985. Processed Pseudogenes: Characteristics and
Evolution. Annu. Rev. Genet., 19, 253-272.
G. Bernardi. 1989. The Isochore Organization of the Human
Genome.Annu. Rev. Genet., 23, 637-641.
9.1.6
E O. Long, L B. Dawid. 1980. Repeated Genes in Eukaryotes.
Annu. Rev. Biochem., 49, 727-764.
E. Gilson, J.M. Clement, D. Perrin, M. Hofnung. 1987. Palind-
romic Units: A Case of Highly Repetitive DNA Sequences
in Bacteria. Trends Genet., 3, 226-230.
Y. Yang, G. F.-L. Ames. 1988. DNA Gyrase Binds to the Family
of Prokaryotic Repetitive Extragenic Palindromic Sequen-
ces. Proc. Natl. Acad. Sci. U.S.A., 85, 8850-8854.
9. Lb
A.C. Wilson, S.S. Carlson, T.J. White. 1977. Biochemical Evo-
lution. Annu. Rev. Biochem., 46, 573-639.
D. Baltimore, 1981. Gene Conversion: Some Implications for
Immunoglobulin Genes. Cell, 24, 592-594.
F. A. Perler. A. Efstratiadis, P. Lomedico. W. Gilbert, R. Kolod-
ner, J. Dodgson. 1981. The Evolution of Genes: The
Chicken Preproinsulin Gene. Cell, 20, 555-566.
V. J. Kidd, G. F. Saunders. 1982. Linkage Arrangement of
Human Placental Lactogen and Growth Hormone Genes.
J. Biol. Chem., 157, 10673-10680.
G.S. Barsch, P.H.Seeburg, R.E. Gelinas. 1983. The Human
Growth Hormone Gene Family: Structure and Evolution
of the Chromosomal Locus. Nucleic Acids Res., 11,
3939-3858.
G. A. Dover. R.B. Flavell. 1984. Molecular Coevolution: DNA
Divergence and the Maintenance of Function. Cell, 38,
622-623.
G. R. Fink, T. D. Petes. 1984. Gene Conversion in the Absebce
of Reciprocal Recombination. Nature, 310, 728-729.
C. C.F. Blake. 1985. Exons and the Evolution of Proteins. Inti.
Rev. CytoL, 93, 149-185.
D.LH.Linzer. D. Nathans. 1985. A New Member of the
Prolactin-Growth Hormone Gene Family Expressed in
Mouse Placenta. EMBO J., 4, 1419-1423.
L Strauss, W. Gilbert. 1985. Genetic Engineering in the Precam-
brian. Structure of the Chicken Triosephosphate Isome-
rase Gene. Mol. Cell. Biol., 5, 3497 3506.
G. A. Dover. 1986. Molecular Drive in Multigene Families: How
Biological Novelties Arise, Spread and Are Assimilated.
Trends Genet., 2, 159- 165.
N. Maeda, O. Smithies. 1986. The Evolution of Multigene
Families: Human Haptoglobulin Genes. Annu. Rev.
Genet., 20, 81-108.
M. Marchioni. W. Gilbert. 1986. The Triosephosphate Isome-
rase Gene from Maize: Introns Antedate the Plant —
Animal Divergence. Cell, 46, 133-141.
G.L. McKnight. P.J. O’Hara. M.L. Parker. 1986. Nucleotide
Sequence of the Triosephosphate Isomerase Gene from
Aspergillus nidulans: Implications for a Differential Loss of
Introns. Cell, 46, 143-147.
Molecular Evolutionary Clock. 1987. J. Mol. Evol. 26 (1 and 2)
special issue.
S.J. O’Brien, H.N.Seuanez, J.E. Womack. 1988. Mammalian
Genome Organization: An Evolutionary View. Annu. Rev.
Genet., 22, 323-351.
A.C. Wdson, H. Ochman, E.M. Prager. 1987. Molecular Time
Scale for Evolution. Trends Genet., 3, 241-247.
J. Felsenstein. 1988. Phylogenies from Molecular Sequences:
Interference and Reliability. Annu. Rev. Genet., 22,
521-565.
M.R. Green. 1988. Mobile RNA Catalysis. Nature, 336,
716-718.
E. Y. Chen, Y.-C. Liao, D.H. Smith, H. A. Barrera-Saldana,
R.E. Gelinas, P.H.Seeburg. 1989. The Human Growth
Hormone Locus: Nucleotide Sequence, Biology, and
Evolution. Genomics, 4, 479-497.
N.J. Dibb, A. J. Newman. 1989. Evidence That Introns Arose at
Proto-Splice. EMBO J., 8, 2015-2021.
P.S. Perlman, R.S. Butow. 1989. Mobile Introns and Intron-
Encoded Proteins. Science, 246, 1106-1109.
J. Rogers. 1989. How Were Introns Inserted into Nuclear
Genes? Trends Genet., 5, 213-216.
330
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
9.2.а
l.B. Dawid, D.D. Brown, R.H. Reeder. 1970. Composition and
Structure of Chromosomal and Amplified Ribosomal
DNAs of Xenopus laevis. J. Mol. Biol., 51, 341-360.
N. V. Federoff. 1979. On Spacers. Cell, 16, 697-710.
J. R. Bedbrook, W. L. Gerlach. 1980. Cloning of Repeated Sequ-
ence DNA from Cereal Plants. In: J.K. Setlow and
A. Hollaender (eds.). Genetic Engineering, vol. 2, pp. 1-19.
Plenum Press, New York.
E.O. Long, l.B. Dawid 1980. Repeated Genes in Eukaryotes.
Annu. Rev. Biochem., 49, 727-764.
P. D’Eustachio, O. Meyuhas, F. Ruddle, R. Perry. 1981. Chro-
mosomal Distribution of Ribosomal Protein Genes in the
Mouse. Cell, 24, 307-312.
D. M. Glover. 1981. The rDNA of Drosophila melanogaster. Cell,
26, 297-298.
M. Krystal. P. D’Eustachio, F.H. Ruddle, N. Arnheim. 1981.
Human Nucleolus Organizers on Non-Homologous Chro-
mosomes Can Share the Same Ribosomal RNA Gene
Variants. Proc. Natl. Acad. Sci. U.S.A., 78, 5744-5748.
O.L. Miller. Jr. 1981. The Nucleolus, Chromosomes, and
Visualization of Genetic Activity. J. Cell Biol., 91, 15s- 27s.
J. Cortadas, M. C. Pavon. 1982. The Organization of Riboso-
mal Genes in Vertebrates. EMBO J., 1, 1075- 1080.
S.A Gerbi. C. Jeppesen. B. Stebbins-Boaz. M. Ares, Jr. 1987.
Evolution of Eukaryotic rRNA: Constraints Imposed by
RNA Interactions. Cold Spring Harbor Symp. Quant.
Biol., 52, 709-719.
R.S. Hawley, C.H. Marcus. 1989. Recombinational Controls of
rDNA Redundancy in Drosophila. Annu. Rev. Genet., 23,
87-120.
9.2.6
D.D. Brown, K. Sugimoto. 1973. 5S DNAs of Xenopus laevis
and Xenopus mulleri: Evolution of a Gene Family. J. Mol.
Biol., 78, 397-415.
RC. Peterson, J.L. Doering, D.D. Brown. 1980. Characteriza-
tion of Two Xenopus Somatic 5S DNAs and One Minor
Oocyte-Specific 5S DNA. Cell, 20, 131 141.
J. Mao, B. Appel, J. Schaak, S. Sharp, H. Yamada, D. Soil. 1982.
The 5S RNA Genes of Schizosaccharomyces ponibe. Nuc-
leic Acids Res., 10, 487-500.
9.2.B
G. I. Bell, L. J. DeGennaro. D. H. Gelfand. R. J. Bishop, P. Va-
lenzuela. W.J. Rutter. 1977. Ribosomal RNA Genes of
Saccharomyces cerevisiae. 1. Physical Map of the Repeating
Unit and Location of the Regions Coding for 5S, 5.8S, 18S,
and 25S Ribosomal RNA. J. Biol. Chem., 252, 8118-8125.
T.D. Petes. 1979. Yeast Ribosomal Genes Are Located on
Chromosome XII. Proc. Natl. Acad. Sci. U.S.A., 76,
410-414.
T.D. Petes. 1980. Unequal Meiotic Recombination Within
Tandem Arrays of Yeast Ribosomal DNA Genes. Cell, 19,
765-774.
J.W. Szostak. R. Wu. 1980. Unequal Crossing Over in the
Ribosomal DNA of Saccharomyces cerevisiae. Nature, 284,
426-430.
T. J. Zamb, T.D. Petes. 1982. Analysis of the Junction Between
Ribosomal RNA Genes and Single-Copy Chromosomal
Sequences in the Yeast Saccharomyces cerevisiae. Cell, 28,
355-364.
9.2.r
P.H. Yen, N. Davidson. 1980. The Gross Anatomy of a tRNA
Gene Cluster at Region 42A of the D. melanogaster
Chromosome. Cell, 22, 137-148.
E. Kubli. 1982. The Genetics of Transfer RNA in Drosophila.
Adv. Genet., 21, 123-172.
S.G. Clarkson. 1983. Transfer RNA Genes. In: N. MacLean,
S. P. Gregory, and R. A. Flavell (eds.). Eukaryotic Genes:
Their Structure, Activity and Regulation, pp. 239-261.
Butterworth, London.
9.2.Д
H. Busch, R. Reddy, L. Rothblum. C. Y. Choi. 1982. SnRNAs,
SnRNPs, and RNA Processing. Annu. Rev. Biochem., 51,
617-654.
H.P. Saluz, T. Schmidt, R. Dudler. M. Altwegg, E. Stumm-Zol-
linger, E. Kubli, P.S. Chen. 1983. The Genes Coding for
4 snRNAs of Drosophila melanogaster: Localization and
Determination of Gene Numbers. Nucleic Acids Res., 11.
77-90.
J. A. Wise, D. Tollervey. D. Maloney, H. Swerdlow, E.J. Dunn,
C. Guthrie. 1983. Yeast Contains Small Nuclear RNAs
Encoded by Single Copy Genes. Cell, 35, 743-751.
E. Lund, J. E. Dahlberg. 1984. True Genes for Human U1 Small
Nuclear RNA: Cory Number, Polymorphism, and Methy-
lation. J. Biol. Chem., 259, 2013-2021.
S. W. Van Arsdell. A.M. Weiner. 1984. Human Genes for U2
Small Nuclear RNA Are Tandemly Repeated. Mol. Cell.
BioL, 4, 492-499.
R. Zeller. M.-T. Carri, I. W. Mattaj, E. M. DeRobertis. 1984.
Xenopus laevis U1 snRNA Genes: Characterization of
Transcriptionally Active Genes Reveads Major and Minor
Repeated Families. EMBO J., 3, 1075-1081.
L.B. Bernstein, T. Manser, A.M. Weiner. 1985. Human U1
Small Nuclear RNA Genes: Extensive Conservation of
Flanking Sequences Suggests Cycles of Gene Amplification
and Transposition. Mol. Cell. Biol., 5, 2159-2171.
C. Guthrie. 1986. Finding Functions for Small Nuclear RNAs
in Yeast. Trends Biochem. Sci., Il, 430-434.
K. A. Montzka, J. A. Steitz. 1988. Additional Low-Abundance
Human Small Nuclear Ribonucleopriteins: UH, U12, etc.
Proc. Natl. Acad. Sci. U.S.A., 85, 8885-8889.
9.3.6
A. Efstratiadis, J. W. Posakony, T Maniatis, R. M. Lawn,
C. O'Connell. R. A. Spritz. J.K. DeRiel, B.G. Forget,
S. M. Weissman. J. L. Slightom. A. E. Blechl. F. E. Baralle,
C.C. Shoulders, N.J. Proudfoot. 1980. The Structure and
Evolution of the Human p-Globin Gene Family. Cell, 21,
653-668.
E. F. Fritsch. C.K. Shen. R.M. Lawn, T. Maniatis. 1980. Mole-
cular Cloning and Characterization of the Human p-like
Globin Gene Cluster. Cell, 19, 959-973.
I. Lemischka. P. A. Sharp. 1982. The Sequences of an Expressed
Rat u-Tubulin Gene and a Pseudogene with an Inserted
Repetitive Element. Nature, 300, 330-335.
C.D. Wilde. L.TChow. F.C. Wefalc. N.J. Cowan. 1982. Struc-
ture of Two Human ct-Tubulin Genes. Proc. Natl. Acad.
Sci. U.S.A., 79, 96 100.
M.B. Buckingham. A. J. Minty. 1983. Contractile Protein Ge-
nes. In: N. MacLean, S. P. Gregory, and R. A. Flavell
(eds.). Eukaryotic Genes: Their Structure, Activity, and
Regulation, vol. 2, pp. 365-395. Butterworth, London.
H. Czosnek, U. Nudel, Y. Mayer, P. E. Barker, D. D. Pravtcheva,
F.H. Ruddle, D. Yaffe. 1983. The Genes Coding for the
Cardiac Muscle Actin, and Skeletal Muscle Actin, and the
Cytoplasmic Р-Actin Are Located on Three Different
Mouse Chromosomes. EMBO J., 2, 1977-1979.
S. Pestka. 1983. Interferon Genes. Arch. Biochem. Biophys.,
221, 1-37.
F. S. Collins, S.M. Weissman. 1984. The Molecular Genetics of
Human Hemoglobin. Prog. Nucleic Acid Res, and Mol.
Biol., 31, 317-462.
R. Garcia. B. Paz-Aliaga, S. G. Ernst. W. R. Crain, Jr.. 1984. Sea
Urchin Actin Genes Show Different Patterns of Expres-
sion: Muscle Specific, Embryo Specific, and Constitutive.
Mol. Cell. Biol., 4, 840-845.
ЛИТЕРАТУРА
331
A. D. Sagar, Р. В. Sehgal, L. Т. Мау, М. Inouye, D. L. Slate,
L. Shulman, F.H. Ruddle. 1984. Interferon-(3-Related DNA
Is Dispersed in the Human Genome. Science, 223, 1312-
1315.
R. J. Shott. J. J. Lee. R. J. Britten, E. H. Davidson. 1984. Diffe-
rential Expression of the Actin Gene Family of Strongy-
locentrotus purpuratus. Devel. Biol., 101, 295-306.
D.W. Cleveland, K. F. Sullivan. 1985. Molecular Biology and
Genetics of Tubulin. Annu. Rev. Biochem., 54, 331-365.
S. Karlsson, A. W. Nienhuis. 1985.. Developmental Regulation of
Human Globin Genes. Annu. Rev. Biochem., 54, 1071—
1108.
S.-Y Ng, P. Gunning, R. Eddy, P. Ponte, J. Leavitt, T. Shows.
L. Kedes. 1985. Evolution of the Functional Human
Р-Actin Gene and Its Multi-Pseudogene Family: Conser-
vation of Noncoding Regions and Chromosomal Disper-
sion of Pseudogenes. Mol. Cell. Biol., 5, 2720-2732.
M. Ohlsson, J. Feder. L.L. Cavalli-Sforza. A. von Gabain. 1985.
Close Linkage of a and (3 Interferons and Infrequent
Duplication of P Interferon in Humans. Proc. Natl. Acad.
Sci. U.S.A., 82, 4473-4476.
A.-C. Pittet, U. Schibler. 1985. Mouse u-Amylase Locus:
Amy-la and Amy-2a Are Closely Linked, j. Mol. BioL,
182, 359-365.
P. Shaw, B. Sordat, V. Schibler. 1985. The Two Promoters of
the Mouse u-Amylase Gene Amy-la Are Differentially
Activated During Parotid Gland Differentiation. Cell, 40,
907-912.
K. Wiebauer, D. L. Guntucio. J. M. Jones. R. M. Caldwell.
H. T. Hartle, M.H. Meisler. 1985. A 78-K.ilobase Region of
Mouse Chromosome 3 Contains Salivary and Pancreatic
Amylase Genes and a Rseudogene. Proc. Natl. Acad. Sci.
U.S.A., 82, 5446-5449.
S. Pestka, J. A. Langer, K.C. Zoon. С. E. Samuels. 1987. Inter-
ferons and Their Actions. Annu. Rev. Biochem., 56,
Т2.1-ТП.
H.P. Erba. R. Eddy, T. Shows, L. Kedes, P. Gunning. 1988.
Structure, Chromosomal Location, and Expression of the
Human y-Actin Gene: Differential Evolution, Location,
and Expression of the Cytoskeletal p- and y-Actin Genes.
Mol. Cell. BioL, 8, 1775-1789.
S. Esteal. 1988. Rate Constancy of Globin Gene Evolution in
Placental Mammals. Proc. Natl. Acad. Sci. U.S.A., 85,
622-7626.
G. Romeo. G. Fiorucci, G.B. Rossi. 1989. Interferons in Cell
Growth and Development. Trends Genet., 5, 19-24.
9.3.B
R.H. Cohn, J.C. Lowry, L.H. Kedes. 1976. Histone Genes of
the Sea Urchin (S. purpuratus) Cloned in E. coli: Order,
Polarity, and Strandedness of the Five Histone-Coding
and Spacer Regions. Cell, 9, 147-161.
С. C. Hentschel, M.L. Birnsteil. 1981. The Organization and
Expression of Histone Gene Families. Cell, 25. 301-313.
A. Ruiz-Carrillo, M. Affolrer, J. Renaud. 1983. Genomic
Organization of the Genes Coding for the Six Main
Histones of the Chicken: Complete Sequence of the H5
Gene. J. Mol. Biol., 170, 843-859.
M.M. Smith. K. Murray. 1983. Yeast H3 and H4 Histone
Messenger RNAs Are Transcribed from Two Non-Allelic
Gene Sets. J. Mol. Biol., 169, 641 661.
P. C. Turner. H. R. Woodland. 1983. Histone Gene Number and
Organization in Xenopus: Xenopus borealis Has a Homo-
geneous Major Cluster. Nucleic Acids Res., 11, 971-986.
L.P. Woudt. A. Pastink. A.E. Kempers-Veenstra, A.E.M. Jan-
sen, W. H. Mager. R. J. Planta. 1983. The Genes Coding for
Histone H3 and H4 in Neurospora crassa Are Unique and
Contain Intervening Sequences. Nucleic Acids. Res., 11,
5347-5360.
R. W. Old. H. R. Woodland. 1984. Histone Genes: Not So Simple
After All. Cell, 38, 624-626.
P. Tripputi. B. S. Emanuel, С. M. Croce, L. G. Green. G. S. Stern,
J. L. Stern. 1986. Human Histone Genes Map to Multiple
Chromosomes. Proc. Natl. Acad. Sci. U.S.A., 83, 3185—
3188.
D. Schumperli. 1988. Multilevel Regulation of Replication-
Dependent Histone Genes. Trends Genet., 4, 187-191.
9.4.a
F. A. Eiferman, P. R. Young, R. W. Scott, S.M. Tilghman. 1981.
Intragenic Amplification and Divergence in the Mouse
u-Fetoprotein Gene. Nature, 294, 713-718.
S. Ohno. 1981. Original Domain for the Serum Albumin Family
Arose from Repeated Sequences. Proc. Natl. Acad. Sci.
U.S.A., 78, 7657-7661.
A. Dugaiczk, S. W Law. О. E. Dennison. 1982. Nucleotide
Sequence and the Encoded Amino Acids of Human Serum
Albumin mRNA. Proc. Natl. Acad. Sci. U.S.A., 79, 71-75.
M. A. T. Muskavitch, D.S. Hogness. 1982. An Expandable Gene
that Encodes a Drosophila Glue Protein Is Not Expressed
in Variants Lacking Remote Upstrem Sequences. Cell, 29.
1041 1051.
F. Alexander, P. R. Young, S.M. Tighman. 1984. Evolution of
the Albumin: u-Fetoprotein Ancestral Gene from Amplifi-
cation of a 27 Nucleotide Sequence. J. Mol. Biol., 173,
159-176.
M.S. Brown. J.L. Goldstein. 1986. A Receptor-Mediated Path-
way for Cholesterpl Homeostasis. Science, 232, 34-47.
9.4.6
P. Gill, A. J. Jeffreys, D.J. Werrett. 1985. Forensic Application
of DNA Fingerprints. Nature, 318, 577-579.
A. J. Jeffreys. V. Wdson, S.L. Thein. 1985. Individual-Specific
“Fingerprints” of Human DNA. Nature, 316, 76-79.
A. P. Jarman, R. A. Wells. 1989. Hypervariable Minisatellites:
Recombinators or Innocent Bystanders? Trends. Genet., 5,
367 372.
9.4.B
W.J. Peacock. D. Brutlag, E. Goldring, R. Appels. C. W. Hinton,
D.L. Lindsey. 1973. The Organization of Highly Repeated
DNA Sequences inn Drosophila melanogaster Chromo-
somes. Cold Spring Harbor Symp. Quant. Biol., 38,
405-416.
W. J. Peacock. A. R. Lohe, W. L. Gerlach. P. Dunsmuir, E. S. Den-
nis, R. Appels. 1977. Fine Structure and Evolution of DNA
in Heterochromatin. Cold Spring Harbor Symp. Quant.
Biol., 42, 1121-1135.
D. Brutlag. 1980. Molecular Arrangement and Evolution of
Heterochromatic DNA. Annu. Rev. Genet., 14, 121-144.
A. J. Hilliker, R. Appels, A. Schalet. 1980. The Genetic Analysis
of D. melanogaster Heterochromatin. Cell, 21, 607-619.
M.F. Singer. 1982. Highly Repeated Sequences in Mammalian
Genomes. Inti. Rev. Cytol., 76, 67-112.
M.J.M. Pages, G.P. Roizes. 1984. Nature and organization of
the Sequence Variations in the Long-Range Periodicity
Calf Satellite DNA I. J, Mol. BioL, 173, 143- 157.
H. F. Willard, J. S. Wave. 1987. Hierarchical Order in Chromo-
somc-Spccific Human Alpha Satellite DNA. Trends
Genet., 3, 192-198.
R. S. Verma (ed.). 1988. Heterochromatin. Cambridge Univer-
sity Press, Cambridge.
S.J. Durfy, H.F. Willard. 1989. Patterns of Intra- and Interar-
ray Sequence Variation in Alpha Satellite from the Human
X Chromosome: Evidence for Short-Range Homogeniza-
tion of Tandemly Repeated DNA Sequences. Genomics, 5,
810-821.
R. Wevrick. H.F. Willard. 1989. Long Range Organization of
Tandem Arrays of a-Satellite DNA at the Centromeres of
Human Chromosomes: High Frequency Array-Lenght
332
ЧАСТЬ 111. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Polymoprhism and Meiotic Stability. Proc. Natl. Acad.
Sci. U.S.A., 86, 9394-9398.
9.4.Г
A. Pryor, K. Faulkner, M.M. Rhoades, W.J. Peacock. 1980.
Asynchronous Replication of Heterochromatin in Maize.
Proc. Natl. Acad. Sci. U.S.A., 77, 6705-6709.
9.4.Д
E.M. Southern. 1970. Base Sequence and Evolution of Guinea
Pig a-Satellite DNA. Nature, 227, 794-798.
G. P. Smith. 1976. Evolution of Repeated DNA Sequences by
Unequal Crossover. Science, 191, 528 535.
9.5.a
R.J. Britten, D.E. Kohne. 1968. Repeated Sequences in DNA.
Science, 161, 529-540.
J. E. Manning, C. W. Schmid, N. Davidson. 1975. Interspersion
of Repetitive and Nonrepetitive DNA Sequences in the
Drosophila melanogaster Genome. Cell, 4, 141-155.
C. W. Schmid, P. L. Deininger. 1975. Sequence Organization of
the Human Genome. Cell, 6, 345-358.
A. C. Spradling, G. M. Rubin. 1981. Drosophila Genome Organi-
zation: Conserved and Dynamic Aspects. Annu. Rev.
Genet., 15, 219-264.
9.5.6
D. E. Graham, B.R. Neufeld, E.H. Davidson, R.J. Britten. 1974.
Interspersion of Repetitive and Nonrepetitive DNA
Sequences in the Sea Urchin Genome. Cell, 1, 127-137.
W.H. Klein, T.L. Thomas, C. Lai, R.H. Scheller, R.J. Britten,
E.H. Davidson. 1978. Characteristics of Individual Repeti-
tive Sequence Families in the Sea Urchin Genome Studied
with Cloned Repeats. Cell, 14, 889-900.
J. W. Posakony, R. H. Scheller, D. M. Anderson, R. J. Britten,
E. H. Davidson. 1981. Repetitive Sequences of the Sea
Urchin Genome. Nucleotide Sequences of Cloned Repeat
Elements. J. Mol. Biol., 149, 41- 67.
J. W. Posakony, C. N. Flytzanis, R. J. Britten. E. H. Davidson.
1983. Interspersed Sequence Organization and Develop-
mental Representation of Cloned Poly (A) RMAs from Sea
Urchin Eggs. J. Mol. Biol, 167, 361 389.
9.5.B
P. L. Deininger, D. J. Jolly, С. M. Rubin, T. Friedmann,
C. W. Schmid. 1981. Base Sequence Studies of 300 Nucleo-
tide Renatured Repeated Human DNA Clones. J. Mol.
Biol., 151, 17-33.
P. Jagadeeswaran, B. Forget, S. M. Weissman. 1981. Short
Interspersed Repetitive DNA Elements in Eucaryotes:
Transposable DNA Elements Generated by Reverse
Transcription of RNA Pol III Transcripts? Cell, 26,
141-142.
W.R. Jelinek, C.W. Schmid. 1982. Repetitive Sequences in
Eukaryotic DNA and Their Expression. Annu. Rev.
Biochem.. 51, 813-844.
E. Ullu, C. Tschudi. 1984. Alu Sequences Are Processed 7SL
RNA Genes. Nature, 312, 171-172.
G. R. Daniels, P.L. Deininger. 1985. Several Major Mammalian
SINE Families Are Derived from tRNA Genes. Nature,
317, 819 822.
H. R. Hwu, J. W. Roberts. E. H. Davidson. R. J. Britten. 1986.
Insertion and/or Deletion of Many Repeated DNA
Sequences in Human and Higher Ape Evolution. Proc.
Natl. Acad. Sci. U.S.A., 83, 3875-3879.
K. Matsumoto, K. Murakami, N. Okada. 1986. Gene for Lysyl
tRNAl May Be a Progenitor of the Highly Repetitive and
Transcribable Sequences Present in the Salmon Genome.
Proc. Natl. Acad. Sci. U.S.A., 83, 3156-3160.
R.J. Britten, W.F. Baron, D.B. Stout, E. H. Davidson. 1988.
Sources and Evolution of Human Alu Repeated Sequen-
ces. Proc. Natl. Acad. Sci. U.S.A., 85, 4770-4774.
R. Dornburg, H.M. Temin. 1988. Retroviral Vector System for
the Study of cDNA Gene Formation. Mol. Cell. Biol., 8,
2328-2334.
TG. Fanning, M.F. Singer. 1988. LINE-1: A Mammalian
Transposable Element. Biochem. Biophys. Acta, 910,
203 212.
J.R. Korenberg, M.C. Rykowski. 1988. Human Genome Orga-
nization: Alu, LINES, and the Molecular Structure of
Metaphase Chromosome Bands. Cell, 53, 391-400.
T.L. Chen, L. Manuelidis. 1989. SINEs and LINEs Cluster in
Distinct DNA Fragments of Giemsa Band Size. Chromo-
soma (Berl), 98, 309-316.
P.L. Deininger. 1989. SINEs: Short Interspersed Repeated
DNA Elements in Higher Eucaryotes. In: D. E. Berg and
M.M. Howe (eds.), Mobile DNA, pp. 619-636. American
Society of Microbiology, Washington, D. C.
C. A. Hutchison III, S. C. Hardies, D. D. Loeb, W. R. Shehee,
M.H. Edgell. 1989. LINES and Realted Retroposons:
Long Interspersed Repeated Sequences in the Eucaryotic
Genome. In: D. E. Berg and M. M. Howe (eds.), Mobile
DNA, pp. 593-618. American Society of Microbiology,
Washington, D. C.
R. K. Moyzis, D. C. Torney, J. Meyne. J. M. Buckingham,
J.R. Wu. C. Burks, K.M. Sirotkin, W.B. Goad. 1989. The
Distribution of Interspersed Repetitive DNA Sequences in
the Human Genome. Genomics, 4. 273-289.
9.5.Г
R.J. Britten, E.H. Davidson. 1969. Gene Regulation in Higher
Cells: A Theory. Science, 165, 349-358.
F. del Rey, T. F. Donahue, G. R. Fink. 1983. The Histidine tRNA
Genes of Yeast. J. Biol. Chem., 258, 8175-8182.
G. S. Roeder. 1983. Unequal Crossing-Over Between Yeast
Transposable Elements. Mol. Gen. Genet., 190, 117-121.
M. Kress. Y. Barra. J.G. Seidman. G. Khoury. G.Jay. 1984.
Functional Insertion of an Alu Type 2 (B2 Sine) Repetitive
Sequence in Murine Class I Genes. Science, 226, 974-977.
A. Hinnebusch, G. Lucchini, G. R. Fink. 1985. A Synthetic HIS4
Regulatory Element Confers General Amino Acid Control
on the Cytochrome c Gene (YC1) of Yeast. Proc. Natl.
Acad. Sci. U.S.A, 82, 498-502.
M. A. Lehman, W. J. Schneider, T. C. Sudhof M. S. Brown,
J. L. Goldstein, D. W Russell. 1985. Mutation in LDL
Receptor. Alu - Alu Recombination Deletes Exons Enco-
ding Transmembrane and Cytoplasmic Domains. Science,
227, 140-146.
J.H. Rogers. 1985. The Origin and Evolution of Retroposons.
Inti. Rev. CytoL, 93, 187-279.
D. Tautz. M. Trick, G. A. Dover. 1986. Cryptic Simplicity in
DNA Is a Major Source of Genetic Variation. Nature, 322,
652-656.
9.5.Д
T. Nagylaki, T.D. Petes. 1982. Intrachromosomal Gene Con-
version and the Maintenance of Sequence Homogeneity
Among Repeated Genes. Genetics, 100, 315-337.
A. M. Weiner, P. L. Deininger, A. Efstratiadis. 1986. Nonviral
Retroposons: Genes, Pseudogenes and Transposable
Elements Generated by the Reverse Flow of Genetic
Information. Annu. Rev. Biochem, 55, 631-661.
9.6.a
H. Ris, P.L. Witt. 1981. Structure of the Mammalian Kineto-
chore. Chromosoma (Berl.), 82, 153- 170.
M.C. Ya, C.-H. Yao. 1981. Repeated Hexanucleotide
С— С- C—C—A—A Is Present Near Free Ends of
Macronuclear DNA of Tetrahymena. Proc. Natl. Acad. Sci.
U.S.A, 78, 7436-7439.
ЛИТЕРАТУРА
333
D.T. Stinchcomb. C. Mann. R.W. Davis. 1982. Centromeric
DNA from Saccharomyces cerevisiae. J. Mol. Biol., 158,
157-179.
L. Clarke, J. Carbon. 1983. Genomic Substitutions of Cento-
meres in Saccharomyces cerevisiae. Nature, 305, 23-28.
J. Carbon. 1984. Yeast Centromeres: Structure and Function.
Cell, 37, 351 353.
L. Clarke, J. Carbon. 1985. Structure and Function of Yeast
Centomeres. Annu. Rev. Genet., 19, 29-56.
P. Hieter, C. Mann. M. Snyder, R. W. Davis. 1985. Mitotic
Stability of Yeast Chromosomes: A Colony Color Assay
that Measures Nondisjunction and Chromosome Loss.
Cell, 40, 393-403.
P. Hieter, D. Pridmore, J. H. Hegemann, M. Thomas, R. W. Da-
vis, P. Philippsen. 1985. Functional Selection and Analysis
of Yeast Centromeric DNA. Cell, 42, 913-921.
D. Koshland, J.C. Kent. L.H. Hartwell. 1985. Genetic Analysis
of the Mitotic Transmission of Minichromosomes. Cell, 40,
381-392.
J. H. Hegemann, J. H. Shero, G. Cottarei, P. Philippsen,
P. Hieter. 1988. Mutational Analysis of Centromere DNA
from Chromosome VI of Saccharomyces cerevisiae. Mol.
Cell. Biol., 8, 2523-2535.
R. Ng, J. Carbon. 1988. Mutational and in Vitro Protein
Binding Studies on Centromere DNA from Saccharomyces
cerevisiae. Mol. Cell. Biol., 7, 4522-4534.
M. Saunders, M. Fitzgerald-Hayes, K. Bloom. 1988. Chromatin
Structure of Altered Yeast Centomeres. Proc. Natl. Acad.
Sci. U.S.A., 85, 175-179.
R.E. Baker, M. Fitzgeral-Hayes. T.C. O’Brien. 1989. Purifica-
tion of the Yeast Centromere Binding Protein CPI and
a Mutational Analysis of Its Binding Site. J. Biol. Chem.,
264, 10843 10850.
M. Cai. R. W. Davis. 1989. Purification of a Yeast Centromere-
Binding Protein That Is Able to Distinguish Single Base-
Pair Mutations in its Recognition Site. Mol. Cell. Biol., 9,
2544-2550.
G. Cottarei, J.H. Shero, P. Hieter, J.H. Hegemann. 1989. A
125-Base-Pair CEN6 DNA Fragment Is Sufficient for
Complete Meiotic and Mitotic Centromere Functions in
Saccharomyces cerevisiae. Mol. Cell. Biol., 9, 3342-3349.
9.6.6
J. G. Gall, K. Karrier, M.-C. Yao. 1977. The Ribosomal DNA of
Tetrahymena. In: P. Ts’o (ed.), The Molecular Biology of
the Mammalian Genetic Apparatus, pp. 80-85. Elsevier-
North Holland Biomedical Press, Amsterdam.
E.H. Blackburn. 1984. Telomeres: Do the Ends Justify the
Means? Cell, 37, 7-8.
E. H. Blackburn, J. W. Szostak. 1984. The Molecular Structure
of Centromeres and Telomeres. Annu. Rev. Biochem., 53,
163 194.
C. W. Grieder, E. H. Blackburn. 1987. The Telomere Transferase
of Tetrahymena Is a Ribonucleoprotein Enzyme with Two
Kinds of Primer Specificity. Cell, 51, 887-898.
E. H. Henderson, С. C. Hardin. S. K. Walk, 1. Tinoco, Jr..
E.H. Blackburn. 1987. Telomeric DNA Oligonucleotides
Form Novel Intramolecular Structures Containing Guani-
ner-Guanine Base Pairs. Cell, 51, 899-908.
M-C. Yao, K. Zheng, C-H. Yao. 1987. A Conserved Nucleotide
Sequence at the Sites of Developmentally Regulated Chro-
mosomal Breakage in Tetrahymena. Cell, 48, 779-788.
R. C. Allshire. J. R. Gosden, S. H. Cross, G. Granston, D. Rout,
N. Sugawara, J. W. Szostak. P. A. Fantes. N.D. Hastie.
1988. Telomeric Repeat from Tetrahymena Cross-Hybri-
dizes with Human Telomeres. Nature, 332, 656-659.
T. R. Cech. 1988. G.-Strings at Chromosome Ends. Nature, 332,
777-778.
R.M. McCarroll, W.L. Fangman. 1988. Time of Replication of
Yeast Centromeres and Telomeres. Cell, 54, 505-513.
R.K. Moyzis, J. M. Buckingham, L.S. Cram, M. Dani.
L. L. Deaven, M. D. Jones, J. Meyne, R. L. Ratliff, J-R. Wu.
1988. A Highly Conserved Repetitive DNA Sequence,
(TTAGGG)n, Present at the Telomeres of Human Chro-
mosomes. Proc. Natl. Acad. Sci. U.S.A., 85, 6662-6626.
E. J. Richards. F. Ausubel. 1988. Isolation of a Higher Eukaryo-
tic Telomere from Arabidopsis thaliani. Cell, 53, 127-136.
J. Shampay, E.H. Blackburn. 1988. Generation of Telomere-
Lenght Heterogeneity in Saccharomyces cerevisiae. Proc.
Natl. Acad. Sci. U.S.A., 85, 534-538.
A. M. Weiner. 1988. Eukaryotic Nuclear Telomeres: Molecular
Fossils of the RNP World? Cell, 52, 155-157.
J. Meyne, R. L. Ratliff, R. K. Moyzis. 1989. Conservation of the
Human Telomere Sequence (TTAGGG)n Among Ver-
tebrates. Proc. Natl. Acad. Sci. U.S.A., 86, 7049-7053.
M.K. Raghuraman, T.R. Cech. 1989. Assembly and Self-
Assotiation of Оxv tricha Telomeric Nucleoprotein Com-
plexes. Cell, 59, 719-728.
J. W. Szostak. 1989. The Beginning of the Ends. Nature, 337,
303-304.
V. A. Zakian. 1989. Structure and Function of Telomeres. Annu.
Rev. Genet., 23, 579 604.
V. Lundblad, E.H. Blackburn. 1990. RNA-Dependent Polyme-
rase Motifs in ESTI: Tentative Identification of a Protein
Component of an Essential Yeast Telomerase. Cell, 60,
529-530.
9.6.B
A. W. Murray. 1985. Chromosome Structure and Behavior.
Trends Biochem. Sci., 10, 112-115.
D.T. Burke, GF. Carle, M.V. Olson. 1987. Cloning of Large
Segments of Exogenous DNA into Yeast by Means of
Artifical Chromosome Vectors. Science, 236, 806-812.
A. W. Murray, J. W. Szostak. 1987. Artifical Chromosomes. Sci.
American, 257 (5), 62-68.
9.7.3
M.L. Claisse, P.P. Slonimski, J. Johnson, H.R. Mahler. 1980.
Mutations Within an Intron and Its Flanking Sites:
Patterns of Novel Polypeptides Generated by Mutants in
One Segment of the cob-box Region of Yeast Mitochon-
drial DNA. Mol. Gen. Genet., 177, 375-387.
H. Bechmann, A. Haid, R.J. Schweyen, S. Mathews, F. Kau-
dewitz. 1981. Expression of the Split Gene COB in Yeast
mtDNA: Translation of Intervening Sequences in Mutant
Strains. J. Biol. Chem., 256, 3525-3531.
B. Weiss-Brummer, G. Rodel. R.J. Schweyen, F. Kaudewitz.
1982. Expression of the Split Gene cob in Yeast: Evidence
for a Precursor of a “Maturase” Protein Translated from
Intron 4 and Preceding Exons. Cell, 29, 527-536.
C. Attardi. 1985. Animal Mitochondrial DNA: An Extreme
Example of Genetic Economy. Inti. Rev. CytoL, 93,
93 145.
T. K. Biswas. J.C. Edwards, M. Rabinowitz. G.S. Getz. 1985.
Characterization of a Yeast Mitochondrial Promoter by
Deletion Mutagenesis. Proc. Natl. Acad. Sci. U.S.A., 82,
1954-1958.
M. Douglas, M. Takeda. 1985. Nuclear Genes Encoding Mito-
chondrial Proteins in Yeast. Trends Biochem. Sci., 10.
192- 194.
J. E. Hixson, D. A. Clayton. 1985. Initiation of Transcription
from Each of the Two Human Mitochondrial Promoters
Requires Unique Nucleotides at the Transcriptional Start
Sites. Proc. Natl. Acad. Sci. U.S.A., 82. 2660-2664.
B. Jacguier, B. Dujon. 1985. An Intron-Encoded Protein Is
Active in a Gene Conversion Process that Spreads an
Intron into a Mitochondrial Gene. Cell, 41, 383-394.
I.G. Macreadie, R.M. Scott. A.R. Zinn, R. A. Butow. 1985.
Transposition of an Intron in Yeast Mitochondria Requi-
res a Protein Encoded by that Intron. Cell, 41, 395-402.
334
ЧАСТЬ 111. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Т. W. Wong, D. A. Clayton. 1985. In Vitro Replication of Human
Mitochondrial DNA: Accurate Initiation at the Origin of
Light-Strand Synthesis. Cell, 42, 951-958.
M. de Zamaroczy, G. Bernardi. 1985. Sequence Organization of
the Mitochondrial Genome of Yeast - A Review. Gene, 37,
1-17.
G. Attardi. 1986. The Elucidation of the Human Mitochondrial
Genome: A Historical Perspective. BioEssays, 5, 34- 39.
R.L. Cann, M. Stoneking, A. C. Wilson. 1986. Mitochondrial
DNA and Human Evolution. Nature, 325, 31-36.
T.D. Fox. 1986. Nuclear Products Required for Translation of
Specific Mitochondrially Coded mRNAs in Yeast. Trends
Genet., 2, 97-100.
A.L. Greenleaf, J.L. Kelly, R. Lehman. 1986. Yeast RPO41
Gene Product Is Required for Transcription and Main-
tenance of the Mitochondrial Genome. Proc. Natl. Acad.
Sci. U.S.A., 83, 3391-3394.
R. M. Mulligan, V. Walbot. 1986. Gene Expression and Recom-
bination in Plant Mitochondrial Genomes. Trends Genet.,
2, 263-266.
K. O'Hare. 1986. Genes Within Genes. Trends Genet., 2, 33.
A. Tzagaloff, A. M. Myers. 1986. Genetics of Mitochondrial
Biogenesis. Annu. Rev. Biochem., 55, 249-285.
R. Bordonne, G. Dirheimcr, R. P. Martin. 1987. Transcription
Initiation and RNA Processing of a Yeast Mitochondrial
tRNA Gene Cluster. Nucleic Acids Res., 15, 7381
7394.
K. J. Netwon. 1988. Plant Mitochondrial Genomes: Organiza-
tion, Expression and Variation. Annu. Rev. Plant Physiol,
and Mol. BioL, 39, 503- 532.
L.A.Grivell, R. J. Schweyen. 1989. RNA Splicing in Yeast
Mitochondria: Taking Out the Twists. Trends Genet., 5,
39-41.
A. Delahodde. V. Goguel. A. M. Becam. F. Creusot. J. Perea.
J. Banrogues, C. Jacg. 1989. Site-Specific DNA Endonu-
clease and RNA Maturase Activities of Two Homologous
Intron-Encoded Proteins from Yeast Mitochondria. Cell,
56, 431-441.
C.S. Levings III, G.G. Brown. 1989. Molecular Biology of
Plant Mitochondria. Cell, 56, 171-179.
J.M. Wenzlau. R.J. Saidanha. R.A. Butow. P.S. Perlman. 1989.
A Latent Intron-Encoded Maturase Is also an Endonuc-
lease Needed for Intron Mobility. Cell, 56, 421-430.
9.7.6
P. Englund et al. 1982. Kinetoplasts. Annu. Rev. Biochem., 51,
695-726.
S.L. Hajduk, V. A. Klein. P.T. Englund. 1984. Replication of
Kinctoplast DNA Maxicircles. Cell, 36, 483-492.
J. Griffith, M. Bleyman. C. A. Rauch. P. A. Kitchin, P. T. Eng-
lund. 1986. Visualization of the Bent Helix in Kinetiplast
DNA by Electron Microscopy. Cell, 46. 717-724.
H. Eisen. 1988. RNA Editing: Who’s on First? Cell. 53.
331 332.
A. I. Lamond. 1988. RNA Editing and the Mysterious Under-
cover Genes of Trypanosomatid Mitochondria. Trends
Biochem. Sci., 13, 283-284.
N. Maizels, A. Weiner. 1988. In Search of a Template. Nature.
334, 469 470.
K. A. Ryan. Т.А. Shapiro, C.A. Rauch. J.D. Griffith, P.T. Eng-
lund. 1988. A Knotted Free Minicircle in Kinetoplast
DNA. Proc. Natl. Acad. Sci. U.S.A., 85, 5844-5848.
P.S. Covello. N.W. Gray. 1989. RNA Editing in Plant Mito-
chondria. Nature, 341 662-666.
J. M. Gualberto, L. Lamattina, G. Bonnard. J-H. Weil,
J-M. Grienenherger. 1989. RNA Editing in Wheat Mito-
chondria Results in the Conservation of Protein Sequen-
ces. Nature, 341, 660-662.
K. A. Ryan. P.T. Englund. 1989. Replication of Kinetoplast
DNA in Trypanosoma eguiperdum. Minicircle H Strand
Fragments Which Map at Specific Locations. J. Biol.
Chem., 264, 823-830.
L. Simpson, J. Shaw. 1989. RNA Editing and the Mitochon-
drial Cryptogenes of Kinetoplastid Protozoa. Cell, 57,
355-366.
9.7.B
L. Bogorad. 1981. Chloroplasts. J. Cell. Biol., 91, 256s-270s.
J. C. Gingrich, R. B. Hallick. 1985. The Euglena gracilis Chloro-
plast Ribulose-1,5-Bisphosphate Carboxylase Gene. J.
Biol. Chem., 260, 16162-16168.
S.D. Kung, CM. Lin. 1985. Chloroplast Promoters from
Higher Plants. Nucleic Acids Res., 13, 7543-7549.
J.D. Palmer. 1985. Comparative Organization of Chloroplast
Genomes. Annu. Rev. Genet., 19, 325-354.
G. Van den Broeck, M.P. Timko. A.P. Kausch. A.R. Cashmore,
M. Van Montagu, L. Herrera-Estrella. 1985. Targeting of
a Foreign Protein to Chloplasts by Fusion to the Transit
Peptide from the Small Subunit of Rebulose 1,5-Bisphoap-
hate Carboxylase. Nature, 313, 358-363.
J Gray. 1986. Wonders of Chloroplast DNA. Nature, 322,
501-502.
G. W. Schmidt, M. L. Mishkind. 1986. The Transport of Proteins
into Chloroplasts. Annu. Rev. Biochem., 55, 879-912.
K. Shinozaki, M. Ohme, M. Tanaka. T. Wakasugi. N. Hayashida.
T. Matsubayashi, N. Zaita, J. Chunwongse. J. Obokata,
K. Yamaguchi-Shinozaki, C. Ohto, K. Torazawa, B. Y Meng,
M. Sugita, H. Deno, T. Kamogashira. K. Yamada. J. Ku-
suda. F. Takaiwa. A. Kato, N. Tohdoh, H. Shimada,
M. Sugiura. 1986. The Complete Nucleotide Sequence of
the Tobacco Chloroplast Genome: Its Gene Organization
and Expression. EMBO J., 5, 2043-2049.
M. Wu, J.K. Lou, D. Y. Chang, C.H. Chang. Z.Q. Hie. 1986.
Structure and Function of a Chloroplast DNA Replication
Origin of Chlamvdomonas reinhardtii. Proc. Natl. Acad.
Sci. U.S.A., 83, 6761-6765.
K. Vmesono, H. Ozeki. 1987. Chloroplast Gene Organization in
Plants. Trends Genet., 3, 281-287.
J. E. Mullet. 1988. Chloroplast Development and Gene Expres-
sion. Annu. Rev. Plant Physiol, and Plant Mol. Biol., 39,
475-502.
K. Ohyarna T. Kohchi, T. Sana, Y. Yamada. 1988. Newly Identi-
fied Groups of Genes in Chloroplasts. Trends Biochem.
Sci.. 13, 19-22.
W Gruissem. 1989. Chloroplast Gene Expression: How Plants
Turn Their Plastids On. Cell, 56, 161 170.
9.7.Г
L. Margulis. 1970. Origin of Eukaryotic Cells. Yale University
Press, New Haven, Conn.
R. Lewin. 1983. Promiscuous DNA Leaps All Barriers. Science,
219, 478-479.
D. Yang, Y. Oyaizu. G.J. Olsen. C.R. Woese. 1985. Mitochond-
rial Origins. Proc. Natl. Acad. Sci. U.S.A., 82, 4443-4447.
M-C. Shih, G. Lazar. H.M. Goodman. 1986. Evidence in Favor
of the Symbiotic Origin of Chloroplasts: Primary Structure
and Evolution of Tobacco Glyceraldehyde-3-Phosphate
Dehydrogenase. Cell, 47, 73-80.
M-C. Shih. P. Heinrich, H. M. Goodman. 1988. Intron Existence
Predated the Divergence of Eukaryotes and Prokaryotes.
Science, 242, 1164-1166.
M. W. Gray. 1989. The Evolutionary Origins of Organelles.
Trends Genet., 5, 294-299.
lO.l.a
D. J. Finnegan, D.H. Fawcett. 1986. Transposable Elements in
D. melanogaster. Oxford Surv. Eukar. Genes, 3, 1-62.
A.J. Kingsman, K.F.Chater. S.M. Kingsman feds.). 1988.
Transposition. Society for General Microbiology Sym-
posium 43, Cambridge University Press, Cambridge.
ЛИТЕРАТУРА
335
D.E. Berg, M.M. Howe. 1989. Mobile DNA. Amer. Soc.
Microbiol., Washington, D. C.
10.1. в
M.L. Goldberg. J-Y. Sheen, W.J. Gehring, M.M. Green. 1983.
Unequal Crossing-Over Associated with Asymmetrical
Synapsis Between Nomadic Elements in the Drosophila
melanogaster Genome. Proc. Natl. Acad. Sci. U.S.A., 80,
5017-5021.
G. Levinson, G. A. Gutman. 1987. Slipped-Strand Mispairing:
A Major Mechanism for DNA Sequence Evolution.
A Review. Mol. Biol. Evol., 4, 203-221.
10.2.a
N. Kleckner. 1981. Transposable Elements in Prokaryotes.
Annu. Rev. Genet., 15, 341-404.
N. Grindley. 1983. Transposition of Tn3 and Releated Trans-
posons. Cell, 32, 3-5.
S. lida. J. Meyer, W. Arber. 1983. Prokaryotic IS Elements. In:
J. A. Shapiro (ed.), Mobile Genetic Elements, pp. 159-221.
Academic Press, New York.
A. Toussaint, A. Resibois. 1983. Phage Mu: Transposition as
a Life Style. In: J. A. Shapiro (ed.), Mobile Genetic
Elements, pp. 105-158. Academic Press, New York.
R. Craigie, K. Mizuuchi. 1985. Mechanism of Transposition of
Bacteriophage Mu: Structure of a Transposition Inter-
mediate. Cell, 41, 867-876.
К. M. Derbyshire, N. D. F. Grindley. 1986. Replicative and
Conservative Transposition in Bacteria. Cell, 47, 325-327.
K. Mizuuchi. R. Craigie. 1986. Mechanism of Bacteriophage
Mu Transposition. Annu. Rev. Genet., 20, 385-429.
D. Brunier, B. Michel, S.D. Ehrlich. 1988. Copy Choice Illegi-
timate DNA Recombination. Cell, 52, 883-892.
D.J. Galas, M. Chandler. 1989. Bacterial Insertion Sequences.
In: D. E. Berg and M. M. Howe (eds.), Mobile DNA.
pp. 109 162. American Society for Microbiology, Was-
hington, D. C.
N. Kleckner. 1989. Transposon TnlO. In: D. E. Berg and
M.M. Howe (eds.), Mobile DNA, pp. 163-184. American
Society for Microbiology, Washington, D. C.
M.L. Pato. 1989. Bacteriophage Mu. In: D. E. Berg and
M.M. Howe (eds.). Mobile DNA, pp. 23-52. American
Society for Microbiology, Washington, D. C.
D. Sheratt. 1989. Tn3 and Related Transposable Elements: Site
Specific Recombination and Transposition. In: D. E. Berg
and M. M. Howe (eds.), Mobile DNA, pp. 163-184.
American Society for Microbiology, Washington, D. C.
W.M. Stark. M.R. Boocock. D.J. Sherratt. 1989. Site-Specific
Recombination by Tn3 Resolvasc. Trends Genet., 5, 304
309. 10.2.b.
P.M. Bingham, M.G. Kidwell, G.M. Rubin. 1982. The Molecu-
lar Basis of P—M Hybrid Dysgenesis: The Role of the
P Element, a P-Strain-Specific Transposon Family. Cell,
29, 995-1004.
G.M. Rubin, A.C. Spradling. 1982. Genetic Transformation of
Drosophila with Transposable Element Vectors. Science,
218, 348-353.
A.C. Spradling, G.M. Rubin. 1982. Transposition of Clined
P Elements into Drosophila Germ Line Chromosomes.
Science, 218, 314 347.
R.E. Karess, G.M. Rubin. 1984. Analysis of P Transposable
Element Functions in Drosophila. Cell, 28, 135 146.
K. O'Hare. 1985. The Mechanism and Control of P Element
Transposition in Drosophila melanogaster. Trends Genet..
1, 250-254.
D. C. Rio. G.M. Rubin. 1988. Identification and Purification of
a Drosophila Protein that Binds to the Terminal 31-Base-
Pair Inverted Repeats of the P Transposable Element.
Proc. Natl. Acad. Sci. U.S.A., 85, 82-833.
M. Snyder, WF. Doolittle. 1988. P Elemnets in Drosophila:
Selection at Many Levels. Trends Genet., 4, 147-149.
W.R. Engels. 1989. P Elements in Drosophila melanogaster. In:
D.E. Berg and M.M. Howe (eds.), Mobile DNA, pp.
437-484. American Society for Microbiology, Washing-
ton, D. C.
10.2.B
B. McClintock. 1951. Chromosome Organization and Gene
Expression. Cold Spring Harbor Symp. Quant. Biol., 16,
13-47.
B. Burr. F.A. Burr. 1982. Ds Controlling Elements of Maize at
the Shrunken Locus Are Large and Dissimilar Insertions.
Cell, 29, 977-986.
H.-P. Daring, P. Starlinger. 1984. Barbara McClintock’s Cont-
rolling Elements: Now at the DNA Level. Cell, 39,
253-259.
M. Muller-Neumann, J. I. Yoder, P. Starlinger. 1984. The DNA
Sequence of the Transposable Element AC of Zea mays L.
Mol. Gen. Genet., 198, 19-24.
WD. Sutton, W.L. Gerlach, D. Schwartz, W.J. Peacock. 1984.
Molecular Analysis of Ds Controlling Element Mutations
at the Adh Locus of Maize. Science, 223, 1265- 1268.
J. W. Schiefelbein. V. Rabox, N. V. Fedoroff. О. E. Nelson, Jr.
1985. Deletions Within a Defective Supressor-Mutator
Element in Maize Affect the Frequency and Develop-
mental Timing of Its Excision from the Bronze Locus.
Proc. Natl. Acad. Sci. U.S.A., 82, 4783-4787.
B. Baker, J. Schell, H. Lorz, N. Fedoroff. 1986. Transposition of
the Maize Controlling Element “Activator" in Tobacco.
Proc. Natl. Acad. Sci. U.S.A., 83, 4844-4848.
H. Dooner, J. English, E. Ralston, E. Week. 1986, A Single
Genetic Unit Specifies Two Transposition Functions in the
Maize Element Activator. Science, 234, 210-211.
H-P. Doring, P. Starlinger. 1986. Molecular Genetics of Trans-
posable Elements in Plants. Annu. Rev. Genet., 20, 175—
200.
S. R. Wesslcr. 1988. Phenotypic Diversity Mediated by the
Maize Transposable Elements Ac and Spin. Science, 242,
399 405.
G. Coupland, C. Plum. S. Chatterjee, A. Post, P. Starlinger.
1989. Sequences Near the Termini Are Required for
Transposition of the Maize Transposon Ac in Transgenic
Tobacco Plants. Proc. Natl. Acad. Sci. U.S.A., 86, 9385-
9388.
N. V. Fedoroff. 1989. Maize Transposable Elements. In:
D. E. Berg and M. M. Howe (eds), Mobile DNA, pp.
375-412. American Society for Microbiology, Washing-
ton, D.C.
10.3.6
R. T. Elder, E. Y. Loh, R. W. Davis. 1983. RNA from the Yeast
Transposable Element Tyl Has Both Ends in the Direct
Repeats, a Structure Similar to Retrovirus RNA. Proc.
Natl. Acad. Sci. U.S.A., 80. 2432-2436.
С. E. Paquin, V.M. Williamson. 1984. Temperature Effects on
the Rate of Ту Transposition. Science, 226, 53-55.
G. S. Roeder, M. Smith, E.J. Lambie. 1984. Intrachromosomal
Movement of Genetically Marked S. cerevisiae Trans-
posons by Gene Conversion. Mol. Cell. BioL, 4, 703-711.
S.J. Silverman, G.R. Fink. 1984. Effects of Ту Insertions on
H1S4 Transcription in Saccharomyces cerevisiae. Mol. Cell.
BioL, 4, 1246-1251.
J.D. Boeke. D.J.Garfinkel, C. A. Styles, G.R. Fink. 1985. Ту
Elements Transpose Through an RNA Intermediate. Cell.
40, 491-500.
J. Clare, P. Farabaugh. 1985. Nucleotide Sequence of a Yeast
Ту Element. Evidence for an Unusual Mechanism of Gene
Expression. Proc. Natl. Acad. Sci. U.S.A., 82, 2829 2833.
D.J.Garfinkel, J.D. Boeke, G.R. Fink. 1985. Ty-Element
336
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Transposition: Reverse Transcriptase and Virus-Like Par-
ticles. Cell, 42, 507-517.
J.D. Boeke, C.A. Styles. G.R. Fink. 1986. Saccharomyces cere-
visiae SPT3 Gene Is Required for Transposition and
Transpositional Recombination of Chromosomal Ту Ele-
ments. Mol. Cell. Biol., 6, 3575-3581.
G.R. Fink, J.D. Boeke, D.J. Garfinkel. 1986. The Mechanism
and Consequences of Retrotransposition. Trends Genet., 2,
118-123.
S. E. Adams. J. Mellor. K. Gull, S. B. Sim, M. F. Tuite,
S.M. Kingsman, A. J. Kingsman. 1987. The Functions and
Relationships of Ty-VLP Proteins in Yeast Reflect Those
of Mammalian Retroviral Proteins. Cell, 49, 111-119.
L. R. Coney, G.S. Roeder. 1988. Control of Yeast Gene Expres-
sion by Transposable Elements: Maximum Expression
Requires a Functional Ту Activator Sequence and a Defec-
tive Ту Promoter. Mol. Cell. Biol., 8, 4009-4017.
D.J. Eichinger. J.D. Boeke. 1988. The DNA Intermediate in
Yeast Tyl Element Transposition Copurifies with Virus-
Like Particles: Cell-Free Tyl Transposition. Cell, 54,
955-966.
A. Gael, R.E. Pearlman. 1988. Transposable Element- Media-
ted Enhancement of Gene Expression in Saccharomyces
cerevisiae Involves Sequence-Specific Binding of a Trans-
Acting Factor. Mol. Cell. Biol., 8, 2572-2580.
A. J. Kingsman, S. M. Kingsman. 1988. Ту: A Retroelement
Moving Forward. Cell, 53, 333-335.
J.D. Boeke. 1989. Transposable Elements in Saccharomyces
cerevisiae. In: D. E. Berg and M. M. Howe (eds.), Mobile
DNA, pp. 335-374. American Society for Microbiology,
Washington, D. C.
J.D. Boeke, EG. Corces. 1989. Transcription and Reverse
Transcription of Retrotransposons. Annu. Rev. Microbiol.,
43, 403- 434.
K. Yu. R. T. Elder. 1989. A Region Internal to the Coding
Sequences Is Essential for Transcription of the Yeast
Ty-D15 Element. Mol. Cell. Biol., 9, 3667-3678.
10.3.B
A. J. Flavell. S. W. Ruby. J. J. O’Toole. В. E. Roberts. G.M. Ru-
bin. 1980. Translation and Developmental Regulation of
RNA Encoded by the Eukaryotic Transposable Element
Copia. Proc. Natl. Acad. Sci. U.S.A, 77, 7107-7111.
N.A. Tchurikv, Y.V. Ilyin, K.G. Skryabin, E.V. Ananiev,
A. A. Bayer. Jr., A.S. Krayev, E.S. Zelentsova, EE Kul-
guskin, N. V. Lyubomirskaya, G.P. Georgiev. 1981. General
Properties of Mobile Dispersed Genetic Elements in
Drosophila melanogaster. Col Spring Harbor Symp. Quant.
Biol., 45, 655 665.
A. J. Flavell, D. Ish-Horowicz. 1983. The Origin of Extrachro-
mosomal Circular Copia Elements. Cell, 34, 415-419.
T. Shiha. K. Saigo. 1983. Retrovirus-Like Particles Containing
RNA Homologous to the Transposable Element Copia in
Drosophila melanogaster. Nature, 302, 119-124.
S.M. Mount, G.M. Rubin. 1985. Complete Nucleotide Sequen-
ce of the Drosophila Transposable Element Copia: Homo-
logy Between Copia and Retroviral Proteins. Mol. Cell.
Biol, 5, 1630 1638.
A. Flavell. 1986. Transposon Tricks Revealed. Nature, 320, 397.
T. Hazelrigg. 1987. The Drosophila White Gene: A Molecular
Update. Trends Genet, 3, 43-47.
P.M. Bingham, Z. Zachar. 1989. Retrotransposons and the FB
Transposon from Drosophila melanogaster. In: D. E. Berg
and M. M. Howe (eds.), Mobile DNA, pp. 485-502. Ame-
risan Society for Microbiology, Washington, D. C.
Ю.З.г
S. H. Wilson, E. L. Kuff. 1972. A Novel DNA Polymerase Acti-
vity Found in Association with Intracistemal A-Tupe
Particles. Proc. Natl. Acad. Sci. U.S.A, 69, 1531-1536.
E.L.Kuff, L. A. Smith, K.K. Lueders. 1981. Intracistemal A
Particle Genes in Mus musculus: A Conserved Family of
Retrovirus-Like Elements. Mol. Cell. Biol, I, 216-277.
M. Horowitz, S. Luria, G. Rechavi, D. Givol. 1984. Mechanism
of Activation of the Mouse c-mos Oncogene by the LTR of
an Intracistemal А-Particle Gene. EMBO J, 3, 2937-2941.
L. Piko, M.D. Hammons. K.D. Taylor. 1984. Amounts, Synt-
hesis, and Some Properties of Intracistemal A Particle
Related RNA in Early Mouse Embryos. Proc. Natl. Acad.
Sci. U.S.A, 81, 488-492.
E.LKuff. K.K. Lueders. 1988. The Intracistemal A-Particle
Gene Family: Structure and Functional Aspects. Adv.
Cancer Res, 51, 183-276.
10.3.Д
T. Fujiwara, K. Mizuuchi. 1988. Retroviral DNA Integration:
Structure of an Integration Intermediate. Cell, 54, 497-
504.
R.F. Doolittle, D-F. Feng, M.S. Johnson, M.A. McClure. 1989.
Origins and Evolutionary Relationships of Retroviruses.
Quart. Rev. Biol, 64, 1-30.
D. P. Grandgenett, S. R. Mumm. 1990. Unraveling Retrovirus
Integration. Cell, 60, 3-4.
l0.3.e
P. P. DiNocera, M.E. Digan, LB. Dawid. 1983. A Family of
Oligo-Adenylate-Terminated Transposable Sequences in
Drosophila melanogaster. J. Mol. Biol, 168, 715-727.
A. Bucheton, R. Paro, H.M. Sang, A. Pelisson, D.J. Finnegan.
1984. The Molecular Basis of I R Hybrid Dysgenesis in
Drosophila melanogaster. Identification, Cloning, and Pro-
perties of the I Factor. Cell, 38, 153-163.
H.M. Sang, A. Pelisson, A. Bucheton, D.J. Finnegan. 1984.
Molecular Lesions Associated with White Gene Mutations
Induced by I — R Hybrid Dysgenesis in Drosophila melano-
gaster. EMBO J, 3, 3079-3085.
D.H. Fawcett, C.K. Lister, E. Kellett, D.J. Finnegan. 1986.
Transposable Elements Controlling I—R Hybrid Dysge-
nesis in D. melanogaster Are Similar to Mammalian Lines.
Cell, 47, 1007-1015.
S. Antonarakis, H. H. Kazazian, Jr. 1988. The Molecular Basis
of Hemophilia A in Man. Trends Genet, 4, 233-237.
P. P. DiNocera. 1988. Close Relationship Between Non-Viral
Retroposons in Dtosophila melanogaster. Nucleic Acids
Res, 16, 4041 4052.
L. J. Mizrokhi, S.G. Georgieva. У V. Ilyin. 1988. Jockey, a Mo-
bile Drosophila Element Similar to Mammalian LINEs, Is
Transcribed from the Internal Promoter by RNA Polyme-
rase II. Cell, 54, 685-691.
J. Skowronski. TG. Fanning, M.F. Singer. 1988. Unit-Length
Line-1 Transcripts in Human Teratocarcinoma Cells. Mol.
Cell. Biol, 8, 1385-1397.
D.J. Finnegan. 1989. F and Related Elements in Drosophila
melanogaster. In: D. E. Berg and M. M. Howe (eds.),Mobi-
le DNA, pp. 519-522. American Society for Microbiology,
Washington, D. C.
D.J. Finnegan. 1989. The I Factor and 1 —R Hybrid Dysgene-
sis in Drosophila melanogaster. In: D. E. Berg and
M. M. Howe (eds.). Mobile DNA, pp. 503-518. American
Society for Microbiology, Washington, D. C.
C.A. Hutchison 111, S.C. Hardies. D.D. Loeb, W. R. Shehee.
M. H. Edgell. 1989. LINES and Related Retroposons:
Long Interspersed Repeated Sequences in the Eukaryotic
Genome. In: D. E. Berg and M. M. Howe (eds.). Mobile
DNA, pp. 593-618. American Society for Microbiology,
Washington, D. C.
A. Bucheton. 1990. I Transposable Elements and I—R Hybrid
Dysgenesis in Drosophila. Trends Genet, 6, 16-21.
ЛИТЕРАТУРА
337
10.4.а
J. Battey, Е.Е. Max, W.O. McBrido, D. Swan, P. Leder. 1982.
A Processed Human Immunoglobulin e Gene Has Moved
to Chromosome 9. Proc. Natl. Acad. Sci. U.S.A., 79,
5956-5960.
I. Lemischka, P. A. Sharp. 1982. The Sequences of an Expressed
Rat Alpha-Tubulin Gene and a Pseudogene with an
Inserted Repetitive Element. Nature, 300, 330-335.
P. A. Sharp. 1983. Conversion of RNA to DNA in Mammals:
Alu-Like Elements and Pseudogenes. Nature, 301, 471-
472.
A.M. Weiner, P.L. Deininger, A. Efstratiadis. 1986. Nonviral
Retroposons: Genes, Pseudogenes and Transposable Ele-
ments Generated by the Reverse Flow of Genetic Informa-
tion. Annu. Rev. Biochem., 55, 631 661.
G. Drouin, G. Dover. 1987. A Plant Processed Pseudogene,
Nature, 328, 557-558.
R. Dornburg, H. M. Temin. 1988. Retroviral Vector System for
the Study of cDNA Gene Formation. Mol. Cell. Biol., 8,
2328-2334.
10.4.6
P. Jagadeeswaran, B.G. Forget, S. M. Weissman. 1981. Short
Interspersed Repetitive DNA Elements in Eukaryotes:
Transposable DNA Elements Generated by Reverse
Transcription of RNA Pol III Transcripts? Cell, 26,
131-142.
L.B. Bernstein. S.M. Mount, A.M. Weiner. 1983. Pseudogenes
for Human Small Nuclear RNA U3 Appear to Arise by
Integration of Self-Primed Reverse Transcripts of the RNA
into New Chromosomal Sites. Cell, 32, 461-472.
C. Bark. K. Hammarstrom. G. Westin. U. Rettersen. 1985. Non-
random Integration of Human U4 RNA Pseudogenes.
Mol. Cell. Biol., 5, 943-948.
V. Lindgren, L. B. Bernstein, A.M. Weiner, U. Francke. 1985.
Human U1 Small Nuclear RNA Pseudogenes Do Not
Map to the Site of the U1 Genes in lp36 But Are Clustered
in Iql2-q22. Mol. Cell. Biol., 5, 2172-2180.
10.5.a
S. Potter, M. Truett, M. Philipps, A. Maher. 1980. Eukaryotic
Transposable Genetic Elements with Inverted Terminal
Repeats. Cell, 20, 639 647.
M.A. Truett, R.S. Jones. S.S. Potter. 1981. Unusual Structure
of the FB Family of Transposable Elements in Drosophila.
Cell, 24, 753-763.
M. Collins, G. M. Rubin. 1983. High-Frequency Precise Exci-
sion of the Drosophila Foldback Transposable Element.
Nature, 303, 259-260.
R. Paro, M.L. Goldberg, W.J. Gehring. 1983. Molecular Ana-
lysis of Large Transposable Elements Carrying the White
Locus of Drosophila melanogaster. EMBO J., 2, 853-860.
P. M. Bingham. Z. Zachar. 1989. Retrotransposons and the FB
Transposon from Drosophila melanogaster. In: D. E. Berg
and M.M. Howe (eds.). Mobile DNA, pp. 485-502. Ame-
rican Society for Microbiology, Washington, D. C.
B. Hoffman-Liebermann. D. Liebermann, S.N. Cohen. 1989. TY
Elements and Puppy Sequences. In: D. E. Berg and
M.M. Howe (eds.). Mobile DNA, pp. 575-592. American
Society for Microbiology, Washington, D. C.
10.5.6
D.M. Glover. D.S. Hogness. 1977. A Novel Arrangement of the
18S and 28S Sequeces in a Repeating Unit of Drosophila
melanogaster rDNA. Cell, 10, 167-176.
P. K. Wellauer, I. B. Dawid. 1977. The Structural Organization
of Ribosomal DNA in Drosophila melanogaster. Cell, 10,
193-212.
R. L. White, D. S. Hogness. 1977. R Looping of the 18S Sequen-
ces in the Short Repeating Units of Drosophila melano-
gaster rDNA. Cell, 10, 177-192.
LB. Dawid. E.O. Long, P. P. DiNocera, M.L. Pardue. 1981.
Ribosomal Insertion-Like Elements in Drosophila melano-
gaster Are Interspersed with Mobile Sequences. Cell, 25,
399-408.
H. Roiha. J.R. Miller, L.C. Woods. D.M. Glover. 1981. Arran-
gements and Rearragements of Sequences Flanking the
Two Types of rDNA Insertion in D. melanogaster. Nature,
290, 749-753.
M.J. Browne, D.A. Read, H. Roiha, D.M. Glover. 1984. Site
Specific Insertion of a Type I rDNA Element into a Uni-
que Sequence in the Drosophila melanogaster Genome.
Nucleic Acids. Res., 12, 9111-9122.
Y. Xiong, TH. Eickbush. 1988. The Site-Specific Ribosomal
Insertion Element RIBm Belongs to a Class of Non-Long-
Terminal Repeat Retrotransposons. Mol. Cell. Biol., 8,
144 153.
10.6.a
J. Lederberg, T. lino. 1956. Phase Variation in Salmonella.
Genetics, 41, 743 757.
M. Silverman. M. Simon. 1983. Phase Variation and Related
Systems. In: J. A. Shapiro (ed.), Mobile Genetic Elements,
pp. 537-557. Academic Press, New York.
R.H.A. Plasterk, R. Kanaar. P. van de Putte. 1984. A Genetic
Switch in Vitro. DNA Inversion by Gin Protein of Phage
Mu. Proc. Natl. Acad. Sci. U.S.A., 81, 2689-2692.
M. Watson. 1984. Prokaryote Invertible DNA Systems Are
Highly Conserved. Trends Biochem. Sci., 9, 82-83.
N.L. Craig. 1985. Site-Specific Inversions: Enhancers, Recom-
bination Proteins, and Mechanism. Cell, 41, 649-650.
R.C. Johnson, M.F. Bruist, M.L Simon. 1986. Host Protein
Requirements for in Vitro Site-Specific DNA Inversion.
Cell, 46, 531-539.
R.C. Johnson, A.C. Glasgow, M.l. Simon. 1987. Spatial Rela-
tionship of the Fis Binding Sites for Hin Recombinational
Enhancer Activity. Nature, 329, 462-465.
R. Kanaar, P. van de Putte, N. R. Cozzarelli. 1988. Gin-Media-
ted DNA Inversion: Product Structure and the Mecha-
nism of Strand Exhange. Proc. Natl. Acad. Sci. U.S.A., 85,
752-756.
A.C. Glasgow, K.T. Hughes, M.l. Simon. 1989. Bacterial DNA
Inversion Systems. In: D. E. Berg and M. M. Howe (eds.),
Mobile DNA, pp. 637 - 660. American Society for Micro-
biology, Washington. D. C.
M. Simon. J. Zieg, M. Silverman, G. Mandel, R. Doolittle. 1989.
Phase Variation: Evolution of a Controlling Element.
Science, 209, 1370-1374.
10.6.6
J. Strathem, J. Hicks, I. Herskowilz. 1981. Control of Cell Type
in Yeast by the Mating Type Locus: The al-a2 Hypo-
thesis. J. Mol. Biol., 147, 357-372.
K.A. Nasmyth. 1982. Molecular Genetics of Yeast Matting
Type. Annu. Rev. Genet., 16, 439-500.
R. Jensen, G.F. Sprague, 1. Herskowilz. 1983. Regulation of
Yeast Mating Type Interconversion: Feedback Control of
HO Gene Expression by the Mating Type Locus. Proc.
Natl. Acad. Sci. U.S.A., 80, 3035-3039.
R. Kostriken, J.N. Strathem, A.J.S. Klar, J.B. Hicks, F. Heff-
ron. 1983. A Site-Specific Endonuclease Essential for
Mating-Туре Switching in Saccharomvces cerevisiae. Cell,
35, 167-174.
J. Abraham. K.A. Nasmyth. J. N. Strathem, A.J.S. Klar,
J.B. Hicks. 1984. Regulation of Mating-Туре Information
in Yeast: Negative Control Requiring Sequences Both 5'
and 3' to the Regulated Region. J. Mol. Biol., 176,
307-331.
J.B. Feldman. J.B. Hicks. J.R. Broach. 1984. Identification of
Sites Required for Repression of a Silent Mating Type
22 -158
338
ЧАСТЬ III. ОРГАНИЗАЦИЯ, ЭКСПРЕССИЯ И РЕГУЛЯЦИЯ ГЕНОВ ЭУКАРИОТ
Locus in Yeast. J. Mol. Biol., 178, 815-834.
M. Stem. R. Jensen, I. Herskowitz. 1984. Five SW1 Genes Are
Required for Expression of the HO Gene in Yeast. J. Mol.
BioL, 178, 853-868.
K. Nasmyth. 1985. At least 1400 Base Pairs of 5'-Flanking
DNA Is Required for the Correct Expression of the HO
Gene in Yeast. Cell, 42, 213-223.
A. J. S. Klar. 1987. Determination of the Yeast Cell Lineage.
Cell, 49, 433-435.
A. R. Buchman, W.J. Kimmerly. J. Rine, R.D. Kornberg. 1988.
Two DNA-Binding Factors Recognize Specific Sequences
at Silencers, Upstream Activating Sequences, Autono-
mously Replicating Sequences, and Telomeres in Saccharo-
myces cerevisiae. Mol. Cell. Biol., 8, 210-225.
C. Goutte, A. D. Johnson. 1988. al Protein Alters the DNA
Binding Specificity of a2 Repressor. Cell, 52, 875-882.
C. A. Keleher, C. Goutte, A.D. Johnson. 1988. The Yeast Cell-
Tupe-Specific Repressor a2 Acts Cooperatively with a
Non-Cell-Type-Specific Protein. Cell, 53, 927-936.
A. J.S. Klar. 1989. The Interconversion of Yeast Mating Type:
Saccharomyces cerevisiae and Schizosaccharomyces pombe.
In: D.E. Berg and M.M. Howe (eds.), Mobile DNA, pp.
671-692. American Society for Microbiology D. C.
N. Hozumi, S. Tonegawa. 1976. Evidence for Somatic Rearran-
gement of Immunoglobulin Genes Coding for Variable
and Constant Regions. Proc. Natl. Acad. Sci. U.S.A., 73,
3628-3632.
D. Baltimore. 1981. Somatic Mutation Gains its Place Among
the Generators of Diversity. Cell, 26, 295-296.
T. Nikaido, S. Nakai, T Honjo. 1981. The Switch Region of the
Immunoglobulin Cg Gene Is Composed of Simple Tandem
Repetitive Sequences. Nature, 292, 845-848.
10.6.B
S. Tonegawa. 1983. Somatic Generation of Antibody Diversity.
Nature, 302, 575-581.
L. Hood, 1. Weissman, W. Wood, J. Wilson. 1984. Immunology,
2nd ed. Benjamin, Menlo Park, California.
G. L. Ada, G. Nossal. 1987. The Clonal Selection Theory. Sci.
American, 257 (2), 62-69.
F. W. Alt, T.K. Blackwell, G.D. Yancopoulos. 1987. Develop-
ment of the Primary Antibody Repertoire. Science, 238,
1079-1087.
E. S. Golub. 1987. Somatic Mutation: Diversity and Regulation
of the Immune Repertoire. Cell, 48, 723-724.
P. Marrack, J. Kappler. 1987. The T Cell Receptor. Science,
238, 1073-1079.
D. R. Davies, S. Sheriff, E.A. Padlan. 1988. Antibody—Antigen
Complexes. J. Biol. Chem., 263, 10541-10544.
M.M. Davis, P.J. Bjorkman. 1988. T-Cell Antigen Receptor
Genes and T-Cell Recognition. Nature, 334, 395-402.
D. Kappes, J. L. Strominger. 1988. Human Class II Major
Histocompatibility Complex Genes and Proteins. Annu.
Rev. Biochem., 57, 991-1028.
T.K. Blackwell, F.W. Alt. 1989. Mechanism and Developmen-
tal Program of Immunoglobulin Gene Rearrangement in
Mammals. Annu. Rev. Genet., 23, 605-636.
T.K. Blackwell, F. W. Alt. 1989. Molecular Characterization of
the Lymphoid V(D)J Recombination Activity. J. Biol.
Chem., 264, 10327-10330.
S. Lewis, M. Gellert. 1989. The Mechanism of Antigen Recep-
tor Gene Assembly. Cell, 59, 585-588.
S.G. Lutzker, F.W. Alt. 1989. Immunoglobulin Heavy-Chian
Class Switching. In: D. E. Berg and M. M. Howe (eds.).
Mobile DNA, pp. 693-714. American Society for Micro-
biology, Washington, D. C.
H. G. Zachau. 1989. Immunoglobulin Light-Chain Genes of the
x Type in Man and Mouse. In: T. Honjo, F.W. Alt, and
T. Rabbitts (eds.), The Immunoglobulin Genes, pp. 91-
109. Academic Press, London.
10.6.Г
P. Borst, G.A.M. Cross. 1982. Molecular Basis for Trypano-
some Antigenic Variation. Cell, 29, 291-303.
P.T. Englund, S.L. Hajduk, J.C. Marini. 1982. The Molecular
Biology of Trypanosomes. Annu. Rev. Biochem., 51,
695-726.
R. G. Nelson. M. Parsons. P. J. Barr. K. Stuart, M. Selkirk.
N. Agabian. 1983. Sequences Homologous to the Variant
Antigen mRNA Spliced Leader Are Located in Tandem
Repeats and Variable Orphons in Trypanosoma brucei.
Cell, 34, 901-909.
E. Pays, S. Van Assel, M. Laurent, M. Darville, T. Vervoort,
N. Van Meirvenne, M. Steinert. 1983. Gene Conversion as
a Mechanism for Antigenic Variation in Trypanosomes.
Cell, 34, 371-381.
J. R. Young, J.S. Shah, G. Matthyssens, R.O. Williams. 1983.
Relationships Between Multiple Copies of a T. brucei
Variable Surface Glycoprotein Gene Whose Expression Is
Not Controlled by Duplication. Cell, 32, 1149-1159.
D. A. Campbell, M.P. van Bree, J.C. Boothroyd. 1984. The
5'-Limit of Transposition and Upstream Barren Region of
a Trypanosome VSG Gene: Tandem 76 Base-Pair Repeats
Flanking (TAA)90. Nucleic Acids Res., 12, 2759-2773.
D.M. Dorfman, J. E. Donelson. 1984. Characterization of the
1.35 Kilobase DNA Repeat Unit Containing the Conser-
ved 35 Nucleotides at the 5'-Termini of Variable Surface
Glycoprotein mRNAs in Trypanosoma brucei. Nucleic
Acids Res., 12, 4907-4920.
P. A. M. Michels, L. H. T. Van der Ploeg, A. Y C. Lium. P. Borst.
1984. The Inactivation and Reactivation of an Expression-
Linked Gene Copy for a Variant Surface Glycoprotein in
Trypanosoma brucei. EMBO J., 3, 1345-1351.
M. Milhausen, R.G. Nelson, S. Sather, M. Selkirk, N. Agabian.
1984. Identification of a Small RNA Containing the
Trypanosome Spliced Leader: A Donor of Shared 5'
Sequences of Trypanosomatid mRNAs? Cell, 38, 721-729.
L. H. T. Van der Ploeg, D. C. Schwartz, C. Cantor, P. Borst.
1984. Antigenic Variation in Trypanosoma brucei Analyzed
by Electrophoretic Separation of Chromosome-Sized
DNA Molecules. Cell, 37, 77-84.
F.E.G. Cox. 1985. Chromosomes of Malaria Parasities and
Trypanosomes. Nature, 315, 280-281.
J. E. Donelson. A.C. Rice-Ficht. 1985. Molecular Biology of
Trypanosome Antigenic Variation. Microbiol. Rev., 49,
107-125.
R.M. Klug. 1985. The Role of RNA Priming in Viral and
Trypanosoma! mRNA Synthesis. Cell, 41, 651-652.
P. Borst. 1986. Discontinuous Transcription and Antigenic
Variation in Trypanosomes. Annu. Rev. Biochem., 55,
701-732.
E. Pays. 1986. Variability of Antigen Genes in African Trypa-
nosomes. Trends Genet., 2, 21-26.
L. Van der Ploeg. 1986. Discontinuous Transcription and
Splicing in Trypanosomes. Cell, 47, 479-480.
L.H.T. Van der Ploeg. 1987. Control of Variant Surface Anti-
gen Switching in Trypanosomes. Cell, 51, 159-161.
E. Pays, M. Steinert. 1988. Control of Antigen Gene Expression
in African Trypanosomes. Annu. Rev. Genet., 22, 107-126.
R.E. Sutton, J.C. Boothroyd. 1988. The Cap of Both Mini-
Exon-Derived RNA and mRNA of Trypanosomes Is
7-Methylguanosine. Mol. Cell. Biol., 8, 494-496.
J.E. Donelson. 1989. DNA Rearregements and Antigenic Va-
riation in African Trypanosomes. In: D. E. Berg and
M.M. Howe (eds.), Mobile DNA, pp. 763-782. American
Society for Microbiology, Washington, D. C.
E. Pays, P. Tebabi, A. Pays, H. Coquelet, P. Revelard, D. Sal-
mon, M. Steinert. 1989. The Genes and Transcripts of an
Antigen Gene Expression Site from T. brucei. Cell, 57,
835-845.
C. Roth, F. Bringaud, R.E. Layden, T. Baltz, H. Eisen. 1989.
ЛИТЕРАТУРА
339
Active Late-Appearing Variable Surface Antigen Genes in
Trypanosoma equiperdum Are Constructed Entirely from
Pseudogenes. Proc. Natl. Acad. Sci. U.S.A., 86, 9375- 9379.
10.7.a
A.C. Spradling, A.P. Mahowald. 1981. A Chromosome Inver-
sion Alters the Pattern of Specific DNA Replication in
Drosophila Follicle Cells. Cell, 27, 203-209.
TH. Eickbush. F.C. Kqfatos. 1982. A Walk in the Chorion
Locus of Bombyx mori. Cell, 29, 633-643.
D. V. de Cicco, A. C. Spradling. 1984. Localization of a Cis-
Acting Element Responsible for the Developmentally
Regulated Amplification of Drosophila Chorion Gene. Cell,
38, 45-54.
F.C. Kafatos. W. Orr, C. Delidakis. 1985. Developmentally
Regulated Gene Amplification. Trends Genet., 1, 301 306.
B. Wakimoto, L. J. Kalfayan, A. C. Spradling. 1986. Develop-
mentally Regulated Expression of Drosophila Chorion
Genes Introduced at Diverse Chromosomal Positions. J.
Mol. Biol., 187, 33-45.
A. Spradling, T Orr-Weaver. 1987. Regulation of DNA Replica-
tion During Drosophila Development. Annu. Rev. Genet.,
21, 373-403.
10.7.6
l.B. Dawid, D.D. Brown, R.H. Reeder. 1970. Composition and
Structure of Chromosomal and Amplified Ribosomal
DNAs Xenopus laevis. J. Mol. Biol., 51, 341-360.
S. A. Endow, K.C. Atwood. 1988. Magnification: Gene Amplifi-
cation by an Inducible System of Sister Chromatid Ex-
change. Trends Genet., 4, 348-351.
10.7в
M.A. Truett. J.C. Call. 1911. The Replication of Ribosomal
DNA in the Macronucleus of Tetrahymena. Chromosoma
(Berk), 64, 295-303.
M-C. Yao, J.G. Gall. 1977. A Single Integrated Gene for
Ribosomal RNA in a Eucaryote, Tetrahymena pyriformis.
Cell, 12, 121-132.
E. H. Blackburn, J.G. Gall. 1978. A Tandemly Repeated Se-
quence at the Termini of the Extrachromosomal Riboso-
mal RNA Genes in Tetrahymena. J. Mol. Biol., 120, 33-53.
E. H. Blaekburn, К. M. Karrer. 1986. Genomic Reorganization
in Ciliated Protozoans. Annu. Rev. Genet., 20, 501-521.
D. D. Larson, E. H. Blackburn, P. C. Yaeger. E. Orias. 1986.
Control of rDNA Replication in Tetrahymena Involves
a Cis-Acting Upstream Repeat of a Promoter Element.
Cell, 47, 229-240.
M-C. Yao, K. Zheng, C-H. Yao. 1987. A Conserved Nucleotide
Sequence at the Sites of Developmentally Regulated
Chromosomal Breakage in Tetrahymena. Cell, 48, 779
788
E. K. Robinson. P.D. Cohen, E. H. Blaekburn. 1989. A Novel
DNA Deletion-Ligation Reaction Catalyzed in Vitro by
a Developmentally Controlled Activity from Tetrahymena
Cells. Cell, 58. 887-900.
M-C. Yao. 1989. Site-Specific Chromosome Breakage and DNA
Deletion in Ciliates. In: D. E. Berg and M. M. Howe (eds.),
Mobile DNA, pp. 715-734. American Society for Micro-
biology, Washington, D. C.
10.8. a
T.D. Kempe, E. A. Swyryd, M. Brulet, G.R. Stark. 1976. Stable
Mutants of Mammalian Cells That Overproduce the First
Three Enzymes of Pyrimidine Nucleotide Biosynthesis.
Cell, 9, 541-550.
G.M. Wahl, R.A. Padgett. G.R. Stark. 1979. Gene Amplifica-
tion Causes Overproduction of First Three Enzymes of
UMP Synthesis in N-(Phosphonacetyl)-!-Aspartate-Resi-
stant Hamster Cells. J. Biol. Chem., 254, 8679 8689.
J. L. Hamlin, J. D. Milbrandt, N. H. Heintz, J. C. Azizkhan.
1984. DNA Sequence Amplification in Mammalian Cells.
Inti. Rev. Cytol., 90, 31-82.
R.T. Schimke. 1984. Gene Ampflication in Cultured Animal
Cells. Cell, 37, 705-713.
G. Stark, G.M. Wahl. 1984. Gene Amplification. Annu. Rev.
Biochem., 53, 447-491.
K. Alitalo. 1985. Amplification of Cellular Oncogenes in Can-
cer Cells. Trends Biochem. Sci., 10, 194-197.
R. T. Schimke. 1988. Gene Amplification in Cultured Cells. J.
Biol. Chem., 263, 5989-5992.
10.8.6
R.J. Kaufman, P.C. Brown. R.T. Schimke. 1979. Amplified
Dihydrofolate Reductase Genes in Unstably Methotre-
xate-Resistant Cells Are Associated with Double Minute
Chromosomes. Proc. Natl. Acad. Sci. U.S.A., 76. 5669
5673.
J. K. Cowell. 1982. Double Minutes and Homogeneously
Staining Regions: Gene Amplification in Mammalian
Cells. Annu. Rev. Genet., 16, 21-59.
M. Montoya-Zavala. J.L. Hamlin. 1985. Similar 150 Kilobase
DNA Sequences Are Amplified in Independently Derived
Methotrexate Resistant Chinese Hamster Cells. Mol. Cell.
BioL, 5, 619-627.
M. Ford, M. Fried. 1986. Large Inverted Duplications Are
Associated with Gene Amplification. Cell, 45, 425 430.
E. Giulotto, I. Saito, G. R. Stark. 1986. Structure of DNA For-
med in the First Step of CAD Gene Amplification. EMBO
J., 5, 2115-2121.
I. Saito, R. Groves, E. Giulotto, M. Rolfe, G.R. Stark. 1989.
Evolution and Stability of Chromosomal DNA Coampli-
fied with the CAD Gene. Mol. Cell. Biol., 9, 2445-2452.
10.8.B
M. Botchan, W. Topp, J. Sambrook. 1978. Studies on Simian
Virus 40 Excision from Cellular Chromosomes. Cold
Spring Harbor Symp. Quant. Biol., 43, 709-719.
W. Wigler, M. Perucho. D. Kurtz. S. Dana. A. Pellicer. R. Axel.
S. Silverstein. 1980. Transformation of Mammalian Cells
with an Amplifiable Dominant-Acting Gene. Proc. Natl.
Acad. Sci. U.S.A., 77, 3567-3570.
J.M. Roberts, R. Axel. 1982. Gene Amplification and Gene
Correction in Somatic Cells. Cell, 29, 109-119.
J. M. Roberts, L. B. Buck. R. Axel. 1983. A Structure for Ampli-
fied DNA. Cell, 33, 53-63.
K. Alitalo. 1985. Amplification of Cellular Oncogenes in Can-
cer Cells. Trends Biochem. Sci., 10, 194-197.
S. M. Carroll. M. L. DeRose. P. Gaudray. С. M. Moore.
D. R. Needham-Vandevanter. D.D. Von Hoff, G.M. Wahl.
1988. Double Minute Chromosomes Can Be Produced
from Precursors Derived from a Chromosomal Deletion.
Mol. Cell. Biol., 8, 1525-1533.
D.D. Von Hoff, D. R. Needham-VanDevanter, J. Yucel.
B E. Windle. G.M. Wahl. 1988. Amplified Human MYC
Oncogenes Localized to Replicating Submicroscopic Cir-
cular DNA Molecules. Proc. Natl. Acad. Sci. U.S.A., 85,
4804-4808.
J. C. Ruiz, K. Choi. D. D. Von Hoff, I. B. Roninson. G. M. Wahl.
1989. Autonomously Replicating Episomes Contain mdrl
Genes in a Multidrug-Resistant Human Cell Line. Mol.
Cell. Biol., 9, 109 115.
G. R. Stark. M. Debatisse, E. Giulotto, G. M. Wahl. 1989. Recent
Progress in Understanding Mechanisms of Mammalian
DNA Amplification. Cell, 57, 901-908.
22*
Часть IV
ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ
И МАНИПУЛЯЦИИ С НИМИ
В предыдущих главах, используя в качестве при-
меров хорошо изученные системы, мы пытались
показать, как с помощью современных эксперимен-
тальных методик можно получить новые данные
о структуре и функциях генов на молекулярном
уровне. В части III, где всесторонне рассмотрены
структура, организация и экспрессия эукариотиче-
ских генов и геномов, мы обращали особое внима-
ние на сходство генетических процессов у разных
организмов. При этом как бы замаскировывалось
огромное разнообразие биологических механизмов,
используемых для «превращения» генотипов в пре-
успевающие организмы. На самом деле число стра-
тегий, которые используют живые организмы, по-
видимому, безгранично. С помощью исследований,
проводимых на молекулярном уровне, мы лишь
приступаем к выявлению генетических механизмов,
которые обусловливают разнообразие живых форм,
их образа жизни, среды обитания, поведения и функ-
ций. Здесь мы познакомимся с подходами к анализу
таких сложных систем, одновременно резюмируя
то, что уже было сказано и как бы вводя читателя
в курс нашей следующей книги.
В этой книге мы подробно рассмотрим вклад
достижений молекулярной генетики эукариот в на-
ше понимание устройства и функционирования не-
которых сложных, взаимодействующих друг с дру-
гом генетических систем. Мы остановимся также на
новых концепциях и методах, используемых для
изменения фенотипа и отдельных генных продуктов
клеток и всего организма. Эти новые эксперимен-
тальные подходы резко расширяют наши возмож-
ности в изучении механизма генетической изменчи-
вости растений и животных. Такая изменчивость
вовсе не является чем-то новым. Люди давно и ус-
пешно занимаются селекцией генетически изменен-
ных организмов. Все важные в сельскохозяйствен-
ном отношении растения и животные-результат
тысячелетних скрещиваний. Одни из них фактически
созданы человеком, другие представляют собой гиб-
риды самостоятельных видов. На основе ослаблен-
ных вирусов с особыми фенотипическими свойства-
ми созданы эффективные вакцины против полио-
миелита и других вирусных заболеваний. Налаже-
но широкомасштабное культивирование различных
мутантных микроорганизмов - продуцентов антиби-
отиков. Современные методики позволяют созда-
вать и отбирать организмы с измененным феноти-
пом гораздо более эффективно. Кроме того, они
уменьшают вероятность нежелательных и непред-
сказуемых изменений, которые создают серьезные
помехи при межвидовых скрещиваниях.
ИССЛЕДОВАНИЕ СЛОЖНЫХ
БИОЛОГИЧЕСКИХ СИСТЕМ
Биохимия и генетика иллюстрируют нам так
называемый редукционистский подход к биологии:
берется один биологический объект (белок, фермент
или ген) и изучается во всех деталях с помощью
разных экспериментальных методов. Этот подход
позволяет прояснить многие фенотипические свой-
ства сложных систем, в том числе и целых орга-
низмов, если их фенотип прямо зависит от функции
единственного гена. Таков, например, феномен крас-
но-зеленой цветовой слепоты (дальтонизм) у муж-
чин.
Семейство генов зрительных
пигментов
Колбочки сетчатки - это цветочувствительные
клетки глаза позвоночных. Другие, более многочис-
ленные клетки, палочки, чувствительны к свету ма-
лой интенсивности, но не обеспечивают хроматиче-
ского восприятия. За чувствительность этих клеток
к слабому свету ответствен мембраносвязанный
зрительный пигмент родопсин - белок, который со-
держит хромофор 11-1/нс-ретиналь, производное ви-
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
341
тамина А. Когда свет падает на сетчатку, в ней
возникает электрический сигнал. Чистый родопсин
можно получить в большом количестве, что позво-
ляет детально изучить его биохимические свойства.
Более чем 40-летние исследования в этой области
позволили воссоздать процессы, протекающие в па-
лочке: поглощение света, генерация сигнала, переда-
ча возбуждения через каскад ферментативных реак-
ций нейронам. В этих реакциях участвуют специфи-
ческие белки и GTP. Был клонирован и охаракте-
ризован ген опсина белкового компонента родо-
псина- из глаза быка. Используя его в качестве
зонда, удалось выделить олеиновый ген человека
(рис. 6.38).
Наши сведения о цветочувствительных компо-
нентах колбочек не столь полны. Основные идеи
о роли пигментов в восприятии цвета еще в конце
XVIII в. высказал Джон Дальтон, сам страдавший
дальтонизмом. Однако лишь в 1911 г. было показа-
но, что ген, ответственный за красно-зеленую слепо-
ту, находится в Х-хромосоме, поскольку такого
рода дальтонизм встречается в основном у мужчин.
Колбочки содержатся в сетчатке лишь в незначи-
тельном количестве, что затрудняет выделение из
них зрительных пигментов и исследование молеку-
лярных основ цветового зрения. Недавно, используя
частичную гомологию между олеиновыми генами
и генами, кодирующими белковые компоненты пиг-
ментов, ответственных за восприятие красного, зе-
леного и синего цветов у человека, удалось клони-
ровать гены этих пигментов (рис. 6,38).
Гены всех четырех белков образуют мультиген-
ное семейство, которое, вероятно, произошло от
одного предкового гена в результате амплификаций
и мутаций как кодирующей, так и регуляторной
последовательностей (рис. IV.1). Имея в распоряже-
нии эти гены, мы можем теперь исследовать меха-
низм. с помощью которого в соседних клетках
синтезируется только один из пигментов: родопсин
в палочках, а чувствительные к красному, зеленому
или синему цвету пигменты-в различных колбоч-
ках. Клонирование этих генов дает ответ на вопрос
и о причине высокой частоты цветовой слепоты
у человека. У мужчин, не страдающих дальтониз-
мом, имеются один ген чувствительного к красному
цвету пигмента и разное число (один или три) генов
пигмента, чувствительного к зеленому цвету, кото-
рые образуют тандем на длинном плече Х-хромосо-
мы. Результаты блот-гибридизации с использовани-
ем геномной ДНК мужчин, страдающих разными
типами красно-зеленой цветовой слепоты, показы-
вают, что аберрантное цветовосприятие часто быва-
ет связано с мутациями, возникающими при реком-
бинациях, сопряженных с неравным кроссинговером
Семейство генов,кодирующих зрительные пигменты человека
Родопсин палочек Хромосома 3
Синий пигмент колбочек Хромосома 1
Красный и зеленый пигменты колбочек Х-хромосома
Разная организация-генов к раскосо и зеленого пигментов
Дальтонизм отсутствует
Фенотип
НЕН—(Srf—8
R+G +
R+G+
n u РИС. IV.1.
Организация мультигенного семейства, кодиру-
ющего зрительные пигменты человека. Все гены
r+G~ родственны друг другу. Гены, кодирующие крас-
ный и зеленый пигменты, образуют тандем на
Х-хромосоме. Благодаря их высокой гомологии
между ними часто происходит рекомбинация.
R~G+ Аминокислотная последовательность красного и
зеленого пигментов совпадает на 96%, а другие
пары пигментов (например, зеленый и синий) - на
40-45%. [J. Nathans, Annu. Rev. Neurosci. 10
R'G" (1987), p. 163.]
342
ЧАСТЬ IV ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
РИС. IV.2.
Электронные микрофотографии различных вирусных
частиц (см. также рис. 1.14). А. Вирус гепатита В
(170000 х). Б. Вирус простого герпеса типа 1 (114000 х).
В. Вирус простого герпеса типа 1, лишенный оболочки;
четко видны капсиды (127 000 х). Г. Вирус везикуляр-
ного стоматита (107 000 х). Д Вирус папилломы че
ловека (94000 х). Е. Вирус гриппа (128000 х). Ж
Почкующийся мышиный ретровирус типа С (100000 х).
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
343
РИС. IV.2 (продолжение)
3. Вирус иммунодефицита человека ВИЧ-1 (AIDS).
Фотография получена сканированием поверхности лим-
фоцитов Т4 человека (74 000 х ). И. Компьютерное изо-
бражение капсида вируса полиомы, диаметр около 500
А. Фото А Д и 3 любезно предоставлены F. A. Murphy;
фото Ж-J. Е. Dahlberg, фото И- D. М. Salunke. [Из
работы D. М. Salunke et al.. Cell. 46 (1986), p. 895).] И
или генной конверсией. Высокая гомологичность
генов цветового зрения, а также их близость друг
к другу стимулируют эти рекомбинационные собы-
тия, что приводит к высокой частоте мутаций.
Согласованные взаимодействия
между продуктами разных генов
В отличие от цветовой слепоты подавляющее
большинство других фенотипических признаков у
организмов обусловливается сложным взаимодей-
ствием продуктов многих генов. Чтобы охарактери-
зовать эти фенотипы, нужно исследовать свойства
физиологических систем (например, нервной систе-
мы), органелл, тканей, целых организмов, популя-
ций организмов, влияние на них окружающей сре-
ды. Альтернативный редукционистскому холистиче-
ский подход предусматривает, что для полного по-
нимания поведения и функций живых систем необ-
ходимо исследовать их фенотипы в контексте всего
организма и его окружения. Уже первые попытки
применения такого подхода наводят на мысль, что
для анализа столь сложных мультигенных систем
можно использовать широкие возможности моле-
кулярной генетики. Таким образом, фенотипы, де-
терминируемые многими генами, как и «моноген-
ные», можно описать в молекулярных терминах.
Среди примеров такого рода, рассмотренных в этой
книге, можно отметить регуляцию содержания хо-
лестерола в сыворотке с помощью липопротеинов
низкой плотности (ЛПНП), функционирование им-
мунной системы позвоночных и контроль типа спа-
ривания у дрожжей.
Еще один пример-образование и сборка рибо-
сом. Этот процесс детерминируется самыми раз-
ными генами, кодирующими множество РНК и бел-
ков. Более того, экспрессия этих многочисленных
генов, а также упорядоченные взаимодействия меж-
ду ними регулируются независимо. Так, вскоре по-
сле завершения митоза инициируется транскрипция
344
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
генов рибосомной РНК, которая запускает процесс
образования ядрышка и синтез локализованных в
нем примерно 80 рибосомных белков, нескольких
ферментов, малых ядерных РНК, необходимых для
созревания рибосомной РНК, а также других бел-
ков, составляющих сложную структуру ядрышка.
Транскрипция генов рибосомных белков осуществ-
ляется с помощью РНК-полимеразы II, но для
образования рибосомных РНК необходимы РНК-
полимеразы I и III. В отличие от генов рибосомной
РНК, гены рибосомных белков не относятся к высо-
кокопийным. Образование продуктов этих разных
транскрипционных систем должно быть скоордини-
ровано между собой и с трансляцией рибосомных
и других белков. Например, известно, что у дрож-
жей созревание рибосомной РНК, возможно, сопря-
жено с уровнем рибосомных белков. Наконец, сбор-
ка рибосомных частиц из соответствующих компо-
нентов осуществляется в определенном порядке.
Умение разобраться в сложных изолированных
генетических элементах-лишь первый шаг в реше-
нии проблем более высокого порядка, касающихся
живых систем. По мере воссоздания полной молеку-
лярной картины этих сложных систем острые редук-
ционистско-холистические дебаты постепенно затих-
нут. Мы будем все глубже и глубже проникать
в суть биологических феноменов, начиная с исследо-
вания фундаментальных свойств генов и геномов
и кончая анализом сложных иерархически организо-
ванных взаимоотношений живых существ. Стоящие
перед нами задачи неординарны; нам придется рас-
сматривать самые разные организмы и системы.
Например, мы остановимся на том, как ведут себя
вирусы вне рамок своего специализированного жиз-
ненного цикла; проследим за координированной
регуляцией генов во время развития и дифференци-
ровки; проанализируем механизмы достижения го-
меостаза и стабильности организмов.
Жизненный цикл вирусов
Наиболее полно связь между целым геномом
и определяемым им фенотипом изучена для неко-
торых вирусов. Вирион (вирусная частица) состоит
из молекулы нуклеиновой кислоты, заключенной
в белковую оболочку (капсид). У некоторых более
сложных вирионов капсид окружен липидным би-
слоем с погруженными в него белками (рис. IV.2).
После того как вирион проникает в клетку, он
утрачивает белковую оболочку, и начинается на-
правляемый вирусным геномом процесс образова-
ния новых вирусных частиц. По сравнению с гено-
мами других организмов вирусный геном относи-
тельно мал и кодирует лишь ограниченное число
белков, в основном белки капсида и один или не-
сколько белков, участвующих в репликации и экс-
прессии вирусного генома. Необходимые метаболи-
ты и энергия поставляются хозяйскими клетками.
Кроме того, в той или иной мере вирус пользуется
аппаратом клетки-хозяина для транскрипции, транс-
ляции и репликации. В настоящее время, используя
метод рекомбинантных ДНК и усовершенствован-
ные методы культур клеток, стало возможным про-
водить опыты с вирусами in vitro, что позволило
обойти трудности, связанные с инфицированием
животных.
Вирусный геном может быть представлен моле-
кулами ДНК или РНК, линейными либо кольцевы-
ми (рис. IV.3). Одни вирусы, реплицируясь в хо-
зяйских клетках, приводят к их гибели. Другие
встраиваются в ДНК клетки-хозяина, трансформи-
руя клетку, и постоянно присутствуют в таком виде
в инфицированной клетке и в ее потомках; во
многих случаях интегрированные вирусные геномы
сохраняют способность детерминировать образова-
ние новых вирусных частиц. Такое разнообразие
форм и функций иллюстрирует то общее положение,
что в ходе эволюции апробируются самые разные
генетические механизмы, но принимаются только
те, которые способны работать. В этом отношении
вирусы не отличаются от более сложных организ-
мов, они лишь мельче и более зависимы.
Некоторые вирусы, в частности паповавирусы,
аденовирусы и герпесвирусы, используют для реп-
ликации ДНК-полимеразы. У других, например у
вируса гепатита В и вируса мозаики цветной капус-
ты, сначала с помощью клеточной РНК-полимера-
зы II синтезируется РНК, а затем на ней как на
матрице путем обратной транскрипции образуется
новая геномная ДНК; при этом обратная тран-
скриптаза кодируется вирусным геномом. Ретрови-
русы также образуют обратную транскриптазу, ко-
торая копирует одноцепочечную геномную РНК
с образованием дуплексной ДНК, которая затем
встраивается в клеточный геном и находится там
в виде провируса; новые вирусные геномы обра-
зуются с помощью клеточной РНК-полимеразы
(разд. 2.2.а и 5.7.г). Другие РНК-содержащие виру-
сы, например полиовирусы и вирус ящура, реплици-
руются при непосредственном копировании РНК
с помощью РНК-полимераз, кодируемых вирусом
(разд. 2.5.).
Методы рекомбинантных ДНК особенно широ-
ко используются в случае вируса гипатита В, пато-
генного агента, распространенного по всему земно-
му шару. Все другие подходы использовать не
удавалось, поскольку до недавнего времени отсут-
ствовали клеточные системы, в которых можно
было бы осуществлять репликацию этого вируса in
vitro. Кроме того, из крови инфицированных живот-
ных можно было выделить лишь очень небольшое
количество кольцевой геномной ДНК. Однако этой
ЧАСТЬ IV ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
345
Вирус
(примерный размер
геномав т.п.н.ипи т.н.)
Структура генома
Вирус гепатита В(3)
Паповавирусы (5)
Частично одноцепочечная
кольцевая ДНК
Сверхспиральная замкнутая
кольцевая Днк
Аденовирус (36)
Герпесвирус(200)
Линейный дуплекс
Вирус осповакцины (200)
Линейный дуплекс с
ковалентно сшитыми
концами
Парвовирус (2)
Одноцепочечная
линейная ДНК
Один из вирусов
кукурузы (3)
Одноцепочечное
кольцо
Ретровирус (10)
Вирус гриппа (16)
РНК-содержащие
вирусы
Одноцепочечный
диплоидный геном
В разных одноцепо-
чечных молекул
Полиовирус (7)
Одноцепочечная
молекула
Реовирус(30)
10 разных двухцепо-
чечных молекул
РИС. IV.3.
Вирусные геномы могут быть представлены
ДНК или РНК, последние могут быть одно-
и двухцепочечными, кольцевыми и линей-
ными.
ДНК хватало для клонирования в системе Е. coli
и получения препаратов вирусной ДНК, пригодных
для определения химических свойств ДНК и для
получения вакцин иммуногенетическими методами.
Первой вакциной, произведенной с помощью ме-
тодов рекомбинантных ДНК и получившей в США
лицензию, позволяющую использовать эту вакцину
для введения человеку, был белок оболочки вируса
гепатита В, синтезированный в клетках дрожжей.
Была клонирована также ДНК, кодирующая белок
оболочки агента, ответственного за гепатит С. Все
это открывает пути к созданию эффективных вакцин
против такого типа инфекций.
Первой ступенькой, ведущей к исследованию
генной структуры и природы регуляторных сигна-
лов транскрипции эукариотических геномов, послу-
жили геномы обезьяньего паповавируса 40 (SV40)
и аденовируса. С их помощью была выявлена роль
многих основных регуляторных сигналов, управля-
ющих транскрипцией с участием РНК-полимеразы
II, в частности роль ТАТА-блоков, GC-богатых
единиц, энхансеров и сигналов полиаденилирования
346
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
ААТААА (разд. 8.3). В ходе работы с этими вируса-
ми были открыты интроны и сплайсинг (разд. 8.5),
а также белки, регулирующие транскрипцию, напри-
мер Spl (разд. 8.3). И до сих пор эти вирусы остают-
ся весьма ценной модельной системой при изучении
новых аспектов молекулярной генетики эукариот
и используются для конструирования эукариотиче-
ских векторов. Например, исследуются механизмы
репликации вирусных геномов; созданы системы,
в которых такая репликация осуществляется in vitro.
Эти, а также другие ДНК-co держащие вирусы могут
служить моделями для изучения дифференцировки
и развития сложных организмов, поскольку их жиз-
ненный цикл состоит из упорядоченных во времени
событий, а кроме того, для них характерны альтер-
нативные способы поведения. Например, вирус
SV40 размножается в определенных клетках при-
матов, но в клетках грызунов он не реплицирует-
ся. Вместо этого его геном встраивается в клеточ-
ную ДНК, вызывая злокачественное перерождение
(рис. IV.4).
В отличие от клеточных геномов эукариот, в ге-
номе SV40 и аденовирусов расстояние между генами
очень мало (рис. IV.5.). Иногда кодирующие и ре-
гуляторные последовательности перекрываются, а
у аденовирусов важная генетическая информация
в некоторых областях генома может быть зако-
дирована даже в обеих цепях. Аденовирусная ДНК
в семь раз длиннее ДНК паповавирусов. Такое
увеличение кодирующей емкости связано с услож-
нением жизненного цикла и структуры вириона,
хотя последняя все-таки остается достаточно про-
стой: это геном, заключенный в белковый капсид.
В отличие от этого вирион герпесвирусов окружен
липидным бислоем, который покрыт гликопротеи-
нами, кодируемыми вирусом. Информация о слож-
ном жизненном цикле герпесвирусов закодирована
в линейной дуплексной молекуле ДНК длиной 250
т.п.н.; вирион содержит примерно 30 разных ви-
русных белков (у SV40 их всего три, а у аденови-
русов-примерно 10). Семейство герпесвирусов
включает несколько вирусов, являющихся возбуди-
телями серьезных заболеваний у человека и живот-
ных. Например, вирусы простого герпеса типов
1 и 2 вызывают рецидивирующие новообразования
на лице и гениталиях, цитомегаловирусная инфек-
ция часто является причиной гибели плода, вирус
Эпштейна Барр вызывает лимфому Беркитта и
инфекционный мононуклеоз, а вирус болезни Ма-
река-опухоли у птиц.
Молекулярное клонирование широко применя-
лось и при исследовании РНК-co держащих вирусов,
поскольку нуклеотидную последовательность длин-
ных молекул вирусных РНК практически невозмож-
но определить секвенированием. Кроме того, ви-
русные гены, клонированные в виде ДНК, при
встраивании их в составе подходящих векторов
в геном бактерий или животных клеток используют
для получения вирусных белков, которые можно
затем исследовать биохимическими методами или
получать антитела. Например, член семейства пи-
корнавирусов (от "pico”-маленький и “та”-РНК) -
вирус ящура-чрезвычайно сложный для исследова-
ния объект. Его геном представлен одноцепочечной
РНК длиной 8 т.н. Этот вирус вызывает серьезное
заболевание у молодых парнокопытных животных,
и работы с ним в США строго ограничены. К сча-
стью, синтезирована и клонирована кДНК, коди-
рующая белки вирусной оболочки, не представля-
ющая никакой опасности при работе в лаборатории.
В результате ее экспрессии в Е. соН образуются
полипептиды, на основе которых получают эффек-
тивные вакцины. Кроме того, поскольку благодаря
клонированию удается определить аминокислотную
последовательность белка оболочки, можно полу-
чить синтетические полипептиды, способные вызы-
вать иммунный ответ.
В 1911 г. Пейтон Раус (Peyton Rous) показал, что
некий вирус (названный позже в его честь вирусом
саркомы Рауса, RSV) вызывает саркому у кур.
Изучению подобных онкогенных РНК-вирусов (ре-
тровирусов) позвоночных уделялось огромное вни-
мание, поскольку была надежда, что это поможет
установить механизм возникновения рака у чело-
века, хотя к началу этих работ никаких онкогенных
РНК-содержащих вирусов человека известно не бы-
ло. Сейчас мы знаем, что геном многих онкогенных
ретровирусов содержит сегменты, отличные от
обычных вирусных генов. Эти сегменты представ-
ляют собой модифицированные клеточные гены
(рис. IV.6), которые и обусловливают способность
многих ретровирусов индуцировать образование
опухолей. Они называются вирусными онкогенами
(ч-опс). Каждый из ч-опс кодирует белки, которые
в том случае, когда они синтезируются в клетке,
обусловливают ее опухолевый фенотип. Нормаль-
ные клеточные гены, от которых произошли ч-опс,
называются протоонкогенами. Обычно протоонко-
гены не приводят к образованию раковых клеток.
Другое дело, если при включении в вирусный геном
происходит их модификация.
Таким образом, ретровирусы, вызывающие опу-
холи, относятся к трансдуцирующим, подобно бак-
териофагу X. Когда-то произошла их рекомбинация
с клеточной ДНК, и теперь продукт рекомбинации
может встраиваться в геном инфицированной клет-
ки (рис. IV.7). Обычно ч-опс имеет другую струк-
туру, чем его клеточный аналог. Кроме того, тран-
скрипция провирусного ч-опс находится под кон-
тролем провирусного регуляторного элемента, а не
клеточного. Эта аномальная регуляция и/или ано-
мальный продукт экспрессии ч-опс сложным и не
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
347
Транскрипция и трансляция
ранних генов
РИС. IV.4.
Схема жизненного цикла SV40. Сначала вирус про-
никает в клетку, где происходят его «раздевание» и
экспрессия ранних генов. Затем он либо становится на
литический путь развития (происходит его репликация,
экспрессия поздних генов и сборка новых вирусных
частиц), либо трансформирует хозяйскую клетку (его
ДНК встраивается в геном хозяина).
348
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
Направление транскрипции
верхней цепи
Направление транскрипции
нижней цепи
Б
РИС. IV.5.
Схематическое представление геномов обезьяньего ви-
руса 40 (SV40) (А) и аденовируса (Б). Ранние (Е)
и поздние (L) мРНК изображены в виде стрелок, ука-
зывающих направление транскрипции. Длина стрелок
дана без соблюдения масштаба; многие детали (в ча-
стности, интроны) опущены.
ЧАСТЬ IV ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
349
Вирус саркомы Рауса
у-опс представлен
геном src
Вирус мышиного лейкоза Абельсона
у-опс представлен геном аы
РИС. IV.6.
А. Типичный ретровирусный геном «дикого типа» с ге-
нами белков нуклеокапсида (дад), обратной транскрип-
тазы (ро!) и белка оболочки (ели) Б Недефектный
ретровирусный геном, несущий ген \j onc (src) В. Де
фектный ретровирусный геном, несущий онкоген аЫ
и неспособный реплицироваться независимо, поскольку
отсутствуют гены ро! и ели. Г. Рвтровирус иммуно-
дефицита человека ВИЧ-1, вызывающий СПИД; схе-
матически показано несколько генов, регулирующих
экспрессию вирусного гена (отметим, что некоторые из
этих генов имеют прерывающиеся или перекрывающи-
еся рамки считывания). [Рис. Г взят из работы R. Gallo et
al., Nature 333 (1988), p. 504.]
Транскрипция,
сплайсинг
Упакоака, инфекция
и рекомбинация с
вирусным геномом
дикого типа
gag pol env
Хелпер
РИС. IV.7.
Гипотетические схемы образования
v-олс (Л) и с-опс (Б) из клеточного
протоонкогена.
350
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
вполне понятным образом дают раковый фенотип.
И точно так же, как рак-это не какое-то од-
но конкретное заболевание, а целый набор рас-
стройств, различные онкогены кодируют разнооб-
разные функции, а их туморогенность обеспечивает-
ся разными механизмами (табл. IV. 1).
Идентификация белков, которые кодируются не-
которыми из примерно 50 известных протоонкоге-
нов, показывает, что некоторые из них участвуют
в нормальной регуляции клеточного роста. Напри-
мер, одни протоонкогены кодируют факторы роста,
другие- мембранные рецепторы для факторов рос-
та. Гены, гомологичные некоторым онкогенам, об-
наружены у дрожжей и D. melanogaster, здесь они
выполняют важные для клетки функции. Например,
у дрожжей обнаружены два гена, родственные про-
тоонкогену H-ras. Если их удалить, то дрожжевые
клетки перестают расти, однако способность к росту
восстанавливается, если в дрожжевую ДНК встро-
ить ген H-ras человека. Белки, кодируемые вирус-
ными и клеточными rav-генами, а также ras-подоб-
ными генами дрожжей, локализуются на плазмати-
ческой мембране и прочно, с высокой специфич-
ностью связывают GTP и CDP. У дрожжей ras-no-
Таблица IV.1. Свойства некоторых онкогенов
Ген Местонахожде- ние ч-опс Местонахожде- ние протоонко- гена Характеристика ген- ного продукта
src Вирус саркомы Рауса (птиц) Позвоночные Протеинкиназа плазматической мембраны, фос- форилирующая остатки тирозина
аЫ Вирус лейкоза мышей Адельсона Позвоночные, Drosophila То же самое, что и продукт гена src
егЬВ Эритроблас- тозные клет- ки птиц Позвоночные, Drosophila Протеинкиназа, фосфорилиру- ющая остатки тирозина; нор- мальный ген ко- дирует рецептор фактора роста эпидермиса, рас- положенный на плазматической мембране
H-ras Вирус саркомы мышей Харвея Позвоночные, Drosophila, дрожжи GDP/GTP-связы- вающий белок плазматической мембраны; нор- мальный белок- ОТРаза
тус Вирус МС29 миелоцито- матоза птиц Позвоночные Находится в ядер- ном матриксе
добные белки сопряжены с системой, регулирующей
уровень циклического АМР второго посредника-
который опосредует ответ всех эукариотических
клеток на внешние воздействия, например гормоны.
Протоонкогены могут превратиться в онкогены
и без участия ретровируса; такие аномальные кле-
точные гены называются с-опс (рис. IV.7.K). В этом
случае за превращение могут быть ответственны
различные мутации, перестройки и амплификации.
Например, протоонкоген может превратиться в
с-опс в результате мутации, при которой изменяется
одна аминокислота в нормальном продукте гена
H-ra5 человека. Измененные H-rav-гены обнаружены
в некоторых опухолях человека (например, в карци-
номе мочевого пузыря). Итак, эксперименты, нача-
ло которым было положено исследованиями ретро-
вирусов позвоночных, привели к открытию прото-
онкогенов, аномальный вариант которых -с-опс-
ответствен за образование опухолей у человека.
Еще один результат этих исследований-выявле-
ние антионкогенов. Белки, кодируемые антионко-
генами, способны предотвращать трансформацию
нормальных клеток в раковые. В качестве примера
можно привести гены, ассоциированные с опухолью
Вильмса и ретинобластомой. У индивидуума, уна-
следовавшего ген ретинобластомы лишь в одной из
двух хромосом (гемизигота), образуется продукт
нормального аллеля, и опухоль не развивается.
Однако если в этом функциональном аллеле в со-
матических клетках возникает мутация, то может
начать образовываться опухоль. Природа этого фе-
номена стала понятна благодаря исследованию ге-
нетически обусловленной опухоли глаз у детей -
ретинобластомы. Клонирован и охарактеризован
ген ретинобластомы. Он кодирует ядерный ДНК-
связывающий белок. Трансфекция этого гена в опу-
холевые клетки в культуре приводит к реверсии
опухолевого фенотипа. Таким образом, продукт
антионкогена каким-то неизвестным способом по-
давляет рост опухоли-в противоположность мно-
гочисленным онкогенным продуктам, стимулирую-
щим клеточную пролиферацию.
Фундаментальные исследования РНК-содержа-
щих вирусов, вызывающих опухоли у птиц и млеко-
питающих, указали путь к решению насущных кли-
нических проблем, связанных с раковыми заболе-
ваниями человека и синдромом приобретенного
иммунного дефицита-СПИДом. СПИД вызывает
один из ретровирусов HIV (ранее он назывался
HTLV-III), который инфицирует макрофаги и один
из типов Т-лимфоцитов, выводя из строя иммунную
систему больного. Другие лимфотропные вирусы
человека- HTLV-I и HTLV-II -также инфицируют
Т-клетки, но вызывают лейкозы. HTLV-I был пер-
вым идентифицированным ретровирусом человека.
Благодаря проведенным ранее интенсивным иссле-
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
351
дованиям ретровирусов позвоночных и применению
методов рекомбинантных ДНК структуру и биоло-
гические свойства HTLV и HIV (рис. IV.6) удалось
установить довольно быстро. Это позволило кло-
нировать вирусные гены, экспрессировать их в Е. coli
и разработать диагностические тесты для выявления
инфицированных лиц по наличию у них антител,
образование которых индуцирует вирус. Используя
эти тесты, сейчас проводят скрининг всех препара-
тов крови, использующихся при трансфузии; это
позволяет избежать распространения инфекции.
Детальное изучение свойств вируса HIV позволит
найти экспериментальные подходы к терапии
СПИДа.
Исследование ретровирусов имеет большое зна-
чение для понимания природы онкогеннности неко-
торых ДНК-содержащих вирусов. Например, SV40
является онкогенным для новорожденных грызунов,
а некоторые папилломавирусы, аденовирусы и гер-
песвирусы могут быть онкогенными для человека.
Онкогенность этих вирусов тоже связана с вирусны-
ми генами, хотя, насколько известно, у \-onc ДНК-
содержащих вирусов нет нормальных клеточных
гомологов. Последние данные позволяют предпола-
гать, что по меньшей мере некоторые из \-onc таких
вирусов могут влиять на рост клеток косвенным
путем, взаимодействуя с продуктами антионкоге-
нов, аналогично тому, как это имеет место в случае
гена ретинобластомы. Возможно, при таком взаи-
модействии подавляются нормальные ингибирую-
щие рост функции антионкогенного продукта, что
приводит к клеточной пролиферации и развитию
опухоли.
Еще один важный результат, который был полу-
чен при исследовании ретровирусов,- создание на их
основе рекомбинантных векторов (разд. 5.7. г). Эти
векторы оказались особенно ценными для введения
новых генов в эмбрионы млекопитающих на ранних
стадиях развития, а значит, практически во все
клетки организма, включая клетки зародышевой
линии; получающиеся при этом животные называ-
ются трансгенными. Роль трансгенных животных
в исследовании тканеспецифичной экспрессии генов
и действия онкогенов описана в гл. 8.
КАРТИРОВАНИЕ ГЕНОМОВ
В основе плодотворного анализа вирусных гене-
тических систем лежат сравнение физической и гене-
тической карт, получение подробной информации
о структуре генома и сопоставление структурных,
генетических и биохимических данных. Все это
очень важно для создания предпосылок к изучению
более сложных генетических систем - от бактерий до
человека.
Физическая и генетическая карты
Первый шаг в построении полной физической
карты состоит в представлении генома или отдель-
ных хромосом в виде упорядоченного набора рест-
рикционных фрагментов ДНК. Полную картину
может дать лишь определение нуклеотидной после-
довательности генома. Уже построена карта рест-
рикционных фрагментов хромосомы Е. coli длиной
4,5 103 т.п.н. (рис. IV.8). Она состоит из 21 фраг-
мента, полученного при расщеплении ДНК эндо-
нуклеазой Notl, которая узнает и расщепляет спе-
цифическую последовательность из 8 п.н. Положе-
ние этих фрагментов в кольцевом геноме опреде-
ляют несколькими методами. Это блот-гибриди-
зация с использованием в качестве зондов предва-
рительно клонированных и охарактеризованных ге-
нов Е. coli', частичное расщепление с помощью Notl;
сравнение фрагментов, получающихся при рестрик-
ции ДНК дикого типа и ДНК из штаммов Е. coli,
содержащих вставки ДНК профага X в заранее
известных локусах или крупные делеции и инверсии.
На хромосоме Е. соН было локализовано свыше
1000 генов, поэтому уже проведена корреляция
между генетической и физической картами.
Аналогичные подходы можно использовать и
для построения карт более крупных и сложных
эукариотических геномов. В этих случаях создают
библиотеки клонированных фрагментов целого ге-
нома, поскольку даже такие ферменты, как Notl,
дают слишком много рестрикционных фрагментов
для разрешения их современными методами. Кло-
нированные фрагменты должны быть достаточно
крупными, чтобы облегчить процедуру их упорядо-
чивания.
По-видимому, наиболее подходящие для этого
векторы-космиды или даже искусственные хромо-
сомы дрожжей (YAC) (разд. 9.6. в). В компьютерном
банке данных уже имеются физические карты про-
тяженных участков двух небольших эукариотичес-
ких геномов: Caenorhabditis elegans и S. cerevisiae.
Для построения таких карт из геномных библиотек,
созданных с помощью фага X или космидных век-
торов, случайным образом отбирают большое чис-
ло длинных клонированных фрагментов. Размеры
фрагментов, которые получаются из каждого кло-
нированного сегмента при его расщеплении огра-
ниченным набором рестриктаз, систематизируют.
Созданы специальные компьютерные программы,
которые позволяют сравнить разные наборы фраг-
ментов и выявить, какие из клонов, скорее всего,
содержат перекрывающиеся геномные области. В
настоящее время уже 95% генома нематоды (размер
гаплоидного генома 8-Ю7 п.н.) представлено пере-
крывающимися клонированными сегментами. У
дрожжей более 95% генома (размер гаплоидного
352
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
РИС. IV.8.
Корреляция между физической и генетической картами
Е. coli. А. Разделение с помощью импульсного электро-
фореза 21 фрагмента ДНК длиной от 20 до 10ОО т. п. н.,
полученных при расщеплении ДНК Е. coli (штамм EMG2)
эндонуклеазой Not] (см. рис. 6.31). Фрагмент О пред-
ставляет собой F-плазмиду, которая присутствовала в
клетках использованного штамма Е. coli и была линеа-
ризована в результате расщепления в единственном
/Vofl-сайте. (С любезного разрешения С. L. Smith.). Б.
Корреляция между генетической картой Е.соН (100
единиц) и картой Not]-фрагментов, обозначенных так
же, как на рис. А. Некоторые из генов, представленных
на карте, указаны на рис. II.3. Клонированные сегменты
этих генов использовали в качестве зондов для иденти-
фикации A/ofl-фрагментов, несущих данный ген. На-
пример, Л/otl-фрагмент Р содержит ген gal. [С. L. Smith
et al.. Science 236 (1987), p. 1448.]
генома 1,4- IO7 п.н.) представлено в виде 5000 сег-
ментов длиной 15 т.п.н., которые сгруппированы
в кластеры длиной до 100 т.п.н. Опыт, приобретен-
ный в работе с этими небольшими геномами, можно
использовать для картирования гораздо более про-
тяженных геномов растений и животных. В связи
с этим возникает одна проблема: даже тщательно
составленные геномные библиотеки не являются
случайным набором всех возможных геномных сег-
ментов. Одни сегменты присутствуют в библиотеке
чаще, другие реже, чем можно ожидать из их ре-
альной представленности в геноме. Некоторые сег-
менты встречаются очень редко или вообще отсут-
ствуют. Чем крупнее исследуемый геном, тем боль-
ше таких брешей будет при конструировании карты.
Еще более усложняет проблему наличие во многих
эукариотических геномах семейств высокоповто-
ряющихся последовательностей.
Перекрывающиеся клонированные сегменты, по-
лученные при предварительном картировании, мож-
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ 353
но использовать для определения полной нуклеоти-
дной последовательности геномов. Между тем появ-
ляется все больше геномных сегментов, которые
были клонированы для особых целей: всего хранит-
ся более 2 • 107 таких последовательностей. К лету
1990 г. в Центральном компьютерном банке данных
имелись сведения о почти 7 106н.п. из разных
частей генома человека. Собраны также обширные
данные по геномам 5. cerevisiae, С. elegans, D. melano-
gaster и мыши. Планируется расширение банка дан-
ных путем использования более совершенных мето-
дов секвенирования, с тем чтобы получить полную
нуклеотидную последовательность геномов некото-
рых из этих видов.
Используя клонированные гены, уже картиро-
ванные с помощью рекомбинантных методов, мож-
но сопоставлять физическую и генетическую карты,
как в случае с Е. coli. Относительно подробные
генетические карты построены для таких организ-
мов, как дрожжи, нематода и D. melanogaster, и здесь
такое сравнение может оказаться весьма продуктив-
ным. У D. melanogaster корреляция упрощается бла-
годаря наличию обширных цитогенетических дан-
ных. Комбинированные молекулярно-генетические
карты позволяют клонировать гены, связанные с оп-
ределенными фенотипом и локусом, но продукт
которых неизвестен (например, так был клонирован
ген per D. melanogaster; рис. 6.39).
Картирование больших геномов
Распространение методов картирования на очень
большие, сложные геномы растений и позвоночных,
включая человека, сталкивается с серьезными проб-
лемами. Это связано с огромными размерами гено-
мов и малочисленностью маркеров, а также трудно-
стями экспериментального скрещивания (у человека
такие скрещивания вообще невозможны). Ранее при
построении генетических карт человека ученые мог-
ли использовать только редкие данные по большим
семьям, члены которых обладали специфическим
мутантным фенотипом. Число идентифицирован-
ных локусов в геноме человека было очень невелико.
Все это затрудняло не только фундаментальный
генетический анализ, но и раннюю диагностику
наследственных болезней. До недавнего времени
пренатальная диагностика была возможна лишь для
ограниченного числа болезней-тех, которые обу-
словливались значительными хромосомными пере-
стройками (например, синдром Дауна, связанный
с трисомией по хромосоме 21) или сопровождались
специфическими фенотипическими проявлениями,
которые легко обнаруживались при развитии плода
(например, болезнь Тея-Сакса, причиной которой
служит отсутствие гексозаминидазы А-гидролазы
лизосом). Ситуация стала меняться с появлением
все большего числа клонированных сегментов ДНК
человека и картированием новых специфических ло-
кусов. В разд. 7.4. б описаны два метода, благодаря
которым, используя клонированные зонды ДНК,
можно практически рутинным способом картиро-
вать на хромосомах человека уникальные одноко-
пийные последовательности. Это метод гибридиза-
ции in situ и ДНК-блоттинг с использованием гиб-
ридных соматических клеток человек/грызун. В этих
методах не используется мутационный анализ, дан-
ные по сцеплению или родословные семей.
Несмотря на то что число идентифицированных
локусов быстро увеличивалось, генетическая карта
человека до самого последнего времени почти
сплошь состояла из белых пятен. Рассмотрим такой
пример. 1000 генов, каждый из которых имеет в
среднем размер 10 т.п.н. (экзоны плюс интроны),
составляют лишь 104 т.п.н. из 3-106 т.п.н. гаплоид-
ного генома человека. Эти гены могут быть разде-
лены миллионами пар оснований, что затрудняет
применение метода «прогулки по хромосоме» или
рекомбинационного анализа, поскольку число ро-
дословных, позволяюших проводить такой анализ,
мало. Что же касается диагностики, то использо-
вание этих методов ограничивается отсутствием
информации о мутантных генах и дефектных генных
продуктах, ответственных за многие генетические
заболевания. К счастью, теперь ситуация здесь в
корне изменилась благодаря появлению нового
подхода, на котором мы остановимся ниже. Этот
подход позволяет проследить за судьбой генов в
нескольких поколениях; он пригоден для целей пре-
натальной диагностики, анализа распределения гена
в популяции, анализа сцепления и картирования.
Его можно использовать и для других организмов.
Например, таким способом картируют хромосомы
кукурузы, что имеет большое научное значение и
может найти применение в сельском хозяйстве.
Полиморфизм длины
рестрикционных фрагментов
В основе этого подхода лежит тот факт, что
разные аллели одного локуса различаются по ну-
клеотидной последовательности. Это могут быть
одиночные нуклеотидные замены, делеции, вставки
или другие перестройки, в результате которых из-
меняется сайт узнавания для той или иной рестри-
ктирующей эндонуклеазы или расстояние между
этими сайтами. Как следствие может изменяться
размер фрагмента(ов) ДНК гомологичных хромо-
сом, которые гибридизуются с данным зондом,
и наблюдается неодинаковый профиль гибридиза-
ции у разных особей одного вида [в качестве при-
мера можно привести аллели овальбуминового гена
23-158
354
ЧАСТЬ IV, ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
(разд. 7.4.а) и полиморфные мини-сателлитные об-
ласти (разд. 9.4)]. Таким образом, полиморфизм
длины рестрикционных фрагментов (ПДРП) пред-
ставляет собой маркер для аллелей определенного
локуса. Для такого анализа достаточно очень не-
больших количеств ДНК, которую можно выделить
из лимфоцитов взрослого человека или из культуры
клеток зародыша, полученных из околоплодной
жидкости или ворсинок хориона.
В простейшем случае ПДРФ обусловливается
тем же изменением нуклеотидной последовательно-
сти, которое детерминирует определенный мутант-
ный фенотип. Именно такова ситуация с мутантным
Р-глобиновым геном, сопряженным с серповидно-
клеточной анемией (рис. III.6). ПДРФ такого рода
встречается редко, однако анализ рестриктов ДНК
человека и других млекопитающих с помощью раз-
личных случайным образом отобранных клониро-
ванных зондов показал, что даже у нормальных
особей аллели данного локуса у гомологичных хро-
мосом часто обнаруживают полиморфизм. Число
таких локусов гораздо больше, чем это следует из
генетических и структурных данных. По оценкам,
полиморфной может быть каждая сотая пара осно-
ваний из 65 т.п.н., составляющих ген Р-глобина
человека.
Как правило, ПДРФ наследуется по законам
Менделя, т. е. представляет собой генетический мар-
кер, фактически аналогичный классическому фено-
типическому маркеру. Маркерами для картирова-
ния наряду с сегментами, функция которых извест-
на, могут служить даже случайные однокопийныё
сегменты ДНК, выполняющие неизвестную функ-
цию (так называемые анонимные ДНК-зонды) или
вообще не являющиеся генами. Таким образом,
благодаря ПДРФ мы имеем огромное число гене-
тических маркеров человека. Их можно использо-
вать как таковые или вместе с фенотипическими
маркерами или хромосомными аберрациями для
анализа частоты рекомбинаций и выявления сце-
пления. Они уже помогли локализовать определен-
ные гены и создали предпосылки для диагностики
болезней, обусловленных неизвестными молекуляр-
ными дефектами (мышечной дистрофии Дюшенна,
хореи Гентингтона и муковисцидоза). В случае мы-
шечной дистрофии Дюшенна молекулярный дефект
удалось выявить. После локализации гена, обуслов-
ливающего данную патологию, на коротком плече
Х-хромосомы были клонированы этот ген и кДНК,
а затем в клетках Е. coli синтезирован продукт
данного гена и получены соответствующие антите-
ла. С их помощью был выявлен белок скелетных
мышц у здоровых людей, дефектный у пациентов
с болезнью Дюшенна.
Усовершенствование этого общего подхода к кар-
тированию позволило исследовать полиморфизм
известных последовательностей, не связанный с
ПДРФ. Короткие 16-членные олигонуклеотиды спе-
цифическим образом отжигаются с комплементар-
ными геномными сегментами. Поскольку в форми-
ровании дуплекса участвует небольшое число осно-
ваний, образование даже одной неправильной пары
приведет к значительному увеличению температуры
плавления дуплекса. Гибридизуя олигомерный зонд
с геномной ДНК при строго определенных услови-
ях, можно по температуре плавления отлцчить один
аллель от другого.
ДИФФЕРЕНЦИАЛЬНАЯ
ЭКСПРЕССИЯ ГЕНОВ
В СПЕЦИФИЧЕСКИХ
КЛЕТКАХ И ТКАНЯХ
Из оплодотворенного яйца в результате последу-
ющих клеточных делений образуются тысячи типов
клеток и разные ткани. Разнообразие формы и функ-
ции дифференцированных клеток-т.е. разнообра-
зие их фенотипов-обусловливается избирательной
экспрессией специфических генов в определенных
клетках или тканях или на строго определенном
этапе развития организма. Однако многие гены,
по-видимому, экспрессируются в большинстве, а
возможно и во всех типах клеток. Эти так называе-
мые гены «домашнего хозяйства» кодируют присут-
ствующие во всех клетках продукты, например ри-
босомные белки, транспортные РНК, актин и фер-
менты, необходимые для биосинтеза нуклеотидов
и получения энергии.
Временная и пространственная регуляция экс-
прессии генов осуществляется в основном на уровне
инициации транскрипции, при сборке транскрипци-
онных комплексов вблизи сайта инициации транск-
рипции. Предполагается, что сам процесс сборки
либо транскрипционная активность собранного
комплекса зависит от взаимодействия между спе-
цифическими ДНК-связывающими белками (факто-
рами транскрипции) и короткими сегментами ДНК
с определенной последовательностью. Некоторые
факторы транскрипции осуществляют свое действие
через белок-белковые взаимодействия, а не при
связывании с ДНК. Различные комбинации относи-
тельно небольшого числа таких специфических по-
следовательностей ДНК, каждая из которых связы-
вается с уникальным набором белков, и создают
предпосылки для функционирования аппарата транс-
крипции практически неограниченным числом спо-
собов. ДНК-связывающие и активирующие транс-
крипцию свойства некоторых факторов транскрип-
ции часто зависят в свою очередь от их связывания
с небольшими молекулами (например, стероидами
или металлами). Ковалентная модификация специ-
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
355
фических аминокислот фактора транскрипции мо-
жет даже служить механизмом тонкой настройки их
способности влиять на транскрипцию. В результате
всего этого мы и наблюдаем огромное разнообра-
зие регуляторных ответов, специфичных в отноше-
нии клеток, тканей и стадий развития организма.
Имеются и другие регуляторные механизмы, напри-
мер перестройки геномной ДНК, с помощью кото-
рых происходят образование и активация экспрес-
сии иммуноглобулиновых генов и генов Т-клеточ-
ных рецепторов у млекопитающих. Как показывают
экспериментальные данные, экспрессия не регулиру-
ется по принципу «все или ничего», однако ее
уровень может изменяться на несколько порядков.
Синтез белков регулируется не только на уровне
инициации транскрипции, но и на уровне процессин-
га мРНК, ее трансляции, посттрансляционной моди-
фикации, переноса и разрушения полипептидов.
Дифференциальная генная экспрессия в специфи-
ческих клетках и тканях распространяется не только
на единичные гены, но и на наборы генов, экспрес-
сия которых включается и выключается координи-
рованно (табл. IV.2). Кроме того, отдельные члены
некоторых мультигенных семейств кодируют сход-
ные или одинаковые белки, синтезирующиеся в раз-
личных тканях или на разных стадиях развития.
Например, гены а-фетопротеина и сывороточного
альбумина мыши, расположенные один за другим
на хромосоме 5, оба экспрессируются у зародышей.
Вскоре после рождения уровень экспрессии а-фето-
протеинового гена уменьшается по крайней мере
в 104 раз под действием транс-действующего про-
дукта другого мышиного гена (разд. 8.3.д и 9.4.а).
Один из подходов к изучению механизма диффе-
ренциальной экспрессии генов на разных этапах
развития состоит в исследовании развивающихся
организмов, в частности ранних эмбрионов. Позже
в этой части мы вкратце суммируем некоторые
последние работы в этой области. Другой подход
основывается на исследовании уже дифференциро-
ванных систем. В частности, таким способом была
продемонстрирована экспрессия на разных стадиях
развития различных а- и 0-глобиновых генов в
предшественниках эритроцитов позвоночных (разд.
9.3.6). Для изучения онтогенеза лимфоцитов иденти-
фицируют перестройки в иммуноглобулиновом гене
и гене Т-клеточного рецептора на разных стадиях
развития В- и Т-клеток соответственно. Особенно
ценны в этом отношении опухолевые клетки.
После того как идентифицированы гены, экс-
прессирующиеся в клетках определенного типа,
можно приступать к выявлению белок-связываю-
щих сегментов ДНК и транс-действующих элемен-
тов, участвующих в регуляции транскрипции, а так-
же выделять и исследовать другие регуляторные
элементы, например сайты сплайсинга или пост-
Таблица IV.2. Группы генов, функционирование
которых регулируется согласованным образом
в специфических клетках или на определенных
стадиях развития организма
Белки, кодируемые данными генами Локализация
Химотрипсин, трипсин, панкреа-
тическая амилаза: одновремен-
ный синтез
Экзокринные клетки
поджелудочной же-
лезы
а-Фетопротеин и сывороточный Желточный мешок
альбумин; синтезируются в и клетки печени
желточном мешке и клетках пе- (мышь)
чени зародыша. После рожде-
ния сывороточный альбумин
синтезируется в печени
а-Актин скелетных мышц, а-ак- Скелетные и сердечная
тин сердечной мышцы, а-актин мышцы, гладкие
гладкомышечной ткани сосудов, мышцы сосудов и
а-актин гладкомышечной ткани кишечника
кишечника
Коллаген типов I и II
а- и Р-глобины: одновременный
синтез. В тканях зародыша и
взрослого организма экспресси-
руются разные типы а- и Р-
глобинов
Ферменты, катализирующие био-
синтез различных аминокис-
лот; синтезируются в условиях
голодания
Белки теплового шока; синтези-
руются при повышенных тем-
пературах
Различные типы миозина, кото-
рые синтезируются в постоян-
ной пропорции в стенке тела,
мышечной ткани и в мышцах
Внеклеточный мат-
рикс; сухожилия,
кости, кожа, гладкие
мышцы, хрящи
Эритроциты
Дрожжи (S. cerevisiae)
Многие организмы
С. elegans
глотки
трансляционных изменений. Важную роль играет
также мутационный анализ. Например, наследст-
венное заболевание человека талассемия обуслов-
ливается недостаточностью глобиновых полипеп-
тидов. При некоторых формах талассемии не син-
тезируется функциональная глобиновая мРНК; это
связано с мутациями в местах сочленений цепей
после сплайсинга. Другие талассемии обусловлены
делениями одного или нескольких глобиновых ге-
нов. В некоторых случаях при делеции Р-глобино-
вого гена, который экспрессируется у взрослых, во
взрослом состоянии продолжают экспрессироваться
зародышевые гены, что указывает на наличие вза-
имосвязанных регуляторных систем.
Использование методов обратной генетики осво-
бождает ученого от необходимости полагаться
лишь на счастливый случай. Теперь можно вносить
в нужные гены определенные мутации и вводить эти
23*
356
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
гены в клетки или в ранние эмбрионы исследуемых
организмов. Здесь мы приводим лишь несколько
примеров из широкого круга современных исследо-
ваний по регуляции генной экспрессии в дифферен-
цированных клетках.
Специфическая экспрессия
генов в поджелудочной железе
В поджелудочной железе происходят синтез и
экскреция целой группы высокоспециализированных
белков. Синтез инсулина осуществляется в измери-
мых количествах только в эндокринных (3-клетках
островков поджелудочной железы; химотрипсин,
трипсин и панкреатическая амилаза синтезируются
в экзокринных клетках. Тем не менее соответст-
вующие гены присутствуют и в клетках всех других
тканей. Чем определяется выборочная, селективная
экспрессия генов? Каким образом в процессе раз-
вития вырабатывается способность к синтезу строго
определенных продуктов? 5'-фланкирующий участок
транскрипционной единицы гена инсулина содержит
сложный энхансер, работающий только в инсулин-
продуцирующих клетках, но не, скажем, в фибро-
бластах (рис. IV.9). Две основные детерминанты
энхансерной активности находятся на расстоянии
около 100 п.н. друг от друга и формируют общую
кор-последовательность длиной 8 п.н. Каждый из
этих коровых элементов связывает специфичный
для (3-клеток транс-действующий белок. В клетках,
не синтезирующих инсулин, по-видимому, имеется
транс-действующий фактор, подавляющий энхансер.
Аналогичным образом i/мс-элементы и соответст-
вующие транс-белки регулируют синтез ферментов,
специфичных для экзокринных клеток. Итак, в про-
цессе дифференцировки способность экзокринных
и эндокринных клеток к синтезу определенных фер-
ментов реализуется путем переключения набора ре-
гуляторных элементов. Чтобы выяснить, каким
образом происходят такие переключения на разных
этапах развития организма, необходимы дальней-
шие исследования.
Другой способ регуляции синтеза инсулина ис-
пользуется во время посттрансляционного процес-
Специфический
энхансер в-клеток
Кэп
Область повышен-®^
ной чувствитель-
ности к днказе I
TGA ААТААА
\ /
100 п.н.
Транскрипция
| Сплайсинг
К эпz
Сигнальный пептид | Трансляция
NH* ^ООООООСОЭОООСХХООООО
Пре-про-инсулин
Удаление сигнального пептида
I и завершение трансляции
NH*
Про-инсулин
соо
s—s
оооооооооо +
С (связывающая) цепь
I Протеолиз
s—s
NH3 СЮОООООООООО
A-цепь |
I I
S S
+ I I
NH3 ОООООООООООООО
В-цепь
РИС. IV.9.
Типичный инсулиновый ген млекопитающих.
Указаны интрон, кодирующие последова-
тельности, специфичный для 0-клеток под-
желудочной железы энхансер и ТАТА-бокс
(промотор). Представлены также генные
транскрипты, первичный продукт трансля-
ции и функционально активная гетероди-
мерная молекула инсулина (см. также
рис. 1.27).
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
357
синга, когда из длинного предшественника образу-
ется активный гормон. Протеолитический процес-
синг полипептидных предшественников играет важ-
ную роль в регуляции концентрации многих гормо-
нов и других межклеточных полипептидных посред-
ников.
Посттрансляционные изменения как
регуляторы направленного потока
генных продуктов
Тот факт, что для превращения полипептидного
предшественника в активный продукт необходима
модификация этого предшественника, создает воз-
можности для посттрансляционной регуляции пото-
ка генных продуктов. Подобные модификации осо-
бенно широко распространены среди множества
полипептидов, которые служат межклеточными
«медиаторами» в многоклеточных организмах. Та-
кие полипептидные гормоны, как инсулин, цирку-
лируют в крови и осуществляют координацию ра-
боты отдаленных клеток. Другие пептиды «корот-
кодействующие», они влияют на активность клеток,
расположенных вблизи секретирующей клетки. На-
пример, пептидные нейромедиаторы передают ин-
формацию от одной нервной клетки другой. Все
пептидные медиаторы работают сходным образом
независимо от того, распространяется ли их дейст-
вие на большие расстояния (инсулин) или они дейст-
вуют локально (нейромедиатор энкефалин). В лю-
бом случае медиатор вначале связывается с высо-
коспецифичным рецептором, расположенным на по-
верхности определенной клетки-мишени, запуская
те или иные процессы в зависимости от свойств
клеточного рецептора. Это может быть процесс
роста, секреция другого полипептида, экспрессия
определенного гена, возбуждение нейрона, специ-
фические поведенческие реакции и т.д.
Каждая клетка получает множество сигналов, но
отвечает только на те из них, которые воспринима-
ются ее специфическими рецепторами. При этом
данный пептидный медиатор может индуцировать
разные ответы, связываясь с соответствующими
рецепторами на различных типах клеток. Хороший
тому пример - пептидный медиатор холецистоки-
нин; его секретируют определенные клетки кишеч-
ника, и с кровотоком он распространяется по всему
организму. Многие клетки на него вообще не реаги-
руют, но в желчном пузыре и головном мозге
находятся клетки, несущие соответствующие рецеп-
торы. При связывании холецистокинина клетками
желчного пузыря увеличивается поток желчи в ки-
шечник, а в отношении клеток мозга холецисто-
кинин действует как нейромедиатор. Число уже
известных многочисленных пептидных медиаторов
постоянно увеличивается. Вместе с другими, непеп-
Таблица IV.3. Энкефалины-пептидные медиаторы
tyr-gly-gly-phe-met met-Энкефалин
tyr-gly-gly-phe-leu leu-Энкефалин
тидными медиаторами (ацетилхолином, адренали-
ном, норадреналином и жирорастворимыми про-
стагландинами) они координируют работу огром-
ного числа физиологических систем.
Гены различных полипептидных межклеточных
медиаторов обычно кодируют белки длиной не-
сколько сотен аминокислот, а пептидные медиаторы
образуются из них в результате посттрансляцион-
ного процессинга. Они могут состоять всего из пяти
аминокислот, как, например, два энкефалина, кото-
рые действуют на клетки мозга аналогично природ-
ным опиатам и регулируют перистальтику кишеч-
ника (табл. IV.3). Обычно продукт трансляции генов
всех белков, предназначенных для секреции, содер-
жит N-концевой сигнальный пептид, который в
процессе прохождения полипептида в эндоплазма-
тический ретикулум отщепляется (рис. IV. 10). Как
правило, остающаяся часть полипептида гликозили-
рована. Затем в результате протеолитического рас-
щепления в специфических сайтах из длинного пред-
шественника образуется один или несколько актив-
ных пептидных медиаторов. Например, белок из 263
аминокислот, называемый пре-про-энкефалин А.
кодируется у млекопитающих единственным геном,
и из него образуются шесть молекул met-энкефа-
лина и одна-1еи-энкефалина.
Сигнальный пептид
осхэсосхххххюсххххххххххюоозо
Пре-про-полипротеин
1 Протеолитическое
расщепление
(XXXXXX^CXXXXWOCDCXDOOCDO
Про-полипротеин
Активные пептидные посредники
РИС. IV.10.
Гены пептидных медиаторов кодируют длинный пре-
про-полипротеин. В эндоплазматическом ретикулуме
сигнальный полипептид отщепляется, а оставшийся
про-полипротеин гликозилируется. В результате про-
теолитического процессинга иэ этого полипротеина
образуются один или несколько пептидных медиаторов.
358
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
Характерные особенности генов, кодирующих
межклеточные медиаторы млекопитающих, сфор-
мировались на ранних этапах эволюции. Аналогич-
ная стратегия используется для образования не-
больших полипептидов, называемых феромонами,
участвующих в спаривании дрожжевых клеток. Ана-
логичным способом образуются небольшие пепти-
ды, которые координируют поведенческие реакции у
беспозвоночных, например у морского червя Aply-
sia. В этом случае, как и у млекопитающих, целый
каскад пептидных медиаторов связывает между со-
бой клетки разного типа, запуская работу много-
численных элементов, координирующих разные ак-
тивности.
Расщепление предшественников с образованием
активных пептидных медиаторов катализируют спе-
цифические протеазы. Возможно, изменение их кон-
центрации в клеточных компартментах (например,
в аппарате Гольджи и секреторных гранулах), со-
держащих медиаторы, служит способом контроля
скорости образования пептидных медиаторов. К
сожалению, идентифицировать и охарактеризовать
эти протеазы весьма непросто. В частности, возни-
кают проблемы с отделением специфических про-
теаз от неспецифических протеолитических фермен-
тов, находящихся в лизосомах. Наиболее успешные
эксперименты были проделаны на дрожжах: у них
с помощью мутационного анализа был выявлен ген,
кодирующий протеазу, которая катализирует рас-
щепление предшественника феромона- а-фактора.
Такая же протеаза ответственна за созревание дру-
гих пре-белков, а ее аналог, по-видимому, присут-
ствует в клетках млекопитающих.
Регуляция генной экспрессии путем
перестроек последовательности ДНК
В природе широко распространена дифференци-
альная экспрессия генов, зависящая от геномных
перестроек, хотя у любого данного организма этот
механизм, по-видимому, охватывает лишь неболь-
шое число генов. Несколько примеров таких запро-
граммированных реорганизаций генома мы рассмо-
трели в гл. 10. Один из них-образование генов
иммунопротеинов (разд. Ю.б.в). В процессе участву-
ют t/подействующие сигналы рекомбинации и бел-
ки, которые кодируются геномной ДНК и образу-
ются исключительно в Т- и В-клетках. В результате
перестроек 1) образуются гены, состоящие из сег-
ментов ДНК, не соседствующих в геноме клеток
зародышевой линии (например, соединение V-, J-
и С-областей с образованием гена легкой цепи
иммуноглобулина); 2) сближаются промотор и эн-
хансер, и в результате инициируется эффективная
транскрипция; 3) образуются разнообразные сег-
менты, кодирующие вариабельные области иммуно-
протеинов, что обеспечивает огромное разнообра-
зие ответов на антигены.
К другим запланированным перестройкам отно-
сятся процессы, с помощью которых прокариоты
отвечают на изменение окружающей среды, дрож-
жевые клетки переключают тип спаривания, а три-
паносомы уклоняются от иммунного ответа хозяи-
на. В некоторых системах (гены рибосомных РНК
Xenopus и гены, кодирующие белки хориона у
D. melanogaster) для удовлетворения потребности
в генных продуктах происходит массовая амплифи-
кация специфических генов. Известны случаи, когда,
напротив, наблюдается массовая утрата ДНК. У
некоторых простейших, например у Tetrahymena,
геном зародышевой линии заключен в микрону-
клеус, а гены экспрессируются в «соматическом»
макронуклеусе. При переходе в макронуклеус может
утрачиваться 90% генома, поскольку из ДНК ис-
ключаются почти все повторяющиеся последова-
тельности. У множества многоклеточных беспоз-
воночных, в том числе у некоторых нематод, насе-
комых и ракообразных, большая часть высокопо-
вторяющихся последовательностей в соматических
стволовых клетках утрачивается, но в клетках заро-
дышевой линии сохраняется. Этот феномен впервые
наблюдали под микроскопом в 1887 г. как димину-
цию хромосом во время развития нематод. Таким
образом, утверждение, что каждая клетка целого
организма имеет ту же ДНК, что и оплодотворен-
ное яйцо, из которого она возникла, не совсем
верно. Тем не менее вклад специфических перестроек
ДНК в процесс дифференцировки соматических кле-
ток, по-видимому, невелик; подавляющее большин-
ство уже клонированных генов имеют одинаковую
структуру и в клетках зародышевой линии, и в со-
матических клетках.
ИЗМЕНЕНИЕ БИОЛОГИЧЕСКИХ
СИСТЕМ
Те принципиальные возможности, которыми об-
ладает молекулярная генетика, позволяют нам зна-
чительно лучше понять функционирование биологи-
ческих систем. Среди этих возможностей -новые
способы изменения генотипа, а также продуктов
отдельных генов, фенотипа клеток и целых орга-
низмов. По своим масштабам влияния на природу
эти воздействия далеко превосходят то, что было
доступно ученым раньше, и помимо чисто научных
проблем неизбежно затрагивают проблемы полити-
ческие, социальные и культурные. Конечно, человек
манипулирует генотипами растений и животных уже
тысячелетия, с тех пор как он начал заниматься
сельским хозяйством. Влияние на другие виды при-
суще не только человеку. Все виды оказывают воз-
действие друг на друга. Это воздействие может
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
359
быть более или менее плавным, как при отборе,
направленном на определенные признаки (напри-
мер, при отборе на способность использовать дру-
гие источники питательных веществ) или резко вы-
раженным, когда уничтожается большая часть по-
пуляции, не приспособленной к новым условиям.
Бесспорно, однако, что новейшая биология дает
человеку более мощные и специфические средства
воздействия как на него самого, так и на другие
виды. Поэтому грандиозные планы биологов долж-
ны уравновешиваться осознанием того, что в реше-
нии вопроса о генетических изменениях ныне живу-
щих организмов должны участвовать и простые
граждане.
Как именно будут использоваться концепции
и методы молекулярной генетики для изменения
биологических молекул и систем в будущем-мы
пока не знаем. Сейчас эти методы позволяют прово-
дить манипуляции начиная от синтеза измененных
белков и кончая введением новых генов в клетки
растений и животных. Далее в этой части мы вкрат-
це рассмотрим современные экспериментальные
системы.
Синтез нормальных
и модифицированных
эукариотических белков
Исходно цель опытов с использованием реком-
бинантных ДНК состояла в получении важных с ме-
дицинской и экономической точек зрения белков,
например вакцин и межклеточных пептидных по-
средников (инсулина, гормона роста и окситоцина).
Идея заключалась в клонировании гена, кодирую-
щего данный полипептид, встраивании его в плаз-
миду, которая реплицируется в Е. coli таким обра-
зом, чтобы промотор £. coli регулировал транскрип-
цию, а затем в синтезе на рибосомах Е. coli больших
количеств нужного белка. Почему эта довольно пря-
молинейная схема оказалась сложнее, чем вначале
предполагалось (разд. 7.8)? Во-первых, в большин-
стве эукариотических генов имеются интроны, а в
генах Е. coli их нет; у бактерий отсутствует меха-
низм сплайсинга, и поэтому невозможно получить
соответствующую данному эукариотическому гену
мРНК. Во-вторых, из первичных продуктов транс-
ляции многих эукариотических генов, в частности из
предшественников полипептидных гормонов, может
образоваться активный генный продукт лишь в ре-
зультате специфического посттрансляционного про-
цессинга, который в клетках Е. coli не осуществляет-
ся. Наконец, успешному получению больших коли-
честв многих эукариотических белков мешает их
токсичность для бактериальных клеток, деградация
бактериальными протеазами и нерастворимость в
цитоплазме бактериальной клетки.
Для решения первой проблемы используют кло-
нированную кДНК, а не сам ген, поскольку в кДНК
нет интронов. Вторую проблему иногда удается
решить, используя не природную кДНК, а кДНК,
кодирующую конечный процессированный продукт.
Например, инсулин представляет собой гетероди-
мерный белок, но две его полипептидные цепи
входят в состав длинного предшественника, кото-
рый расщепляется с образованием двух полипепти-
дов (рис. IV. 10). Используя две разные рекомби-
нантные плазмиды, каждая из которых содержит
кДНК, соответствующую одной из цепей молекулы
активного инсулина, в разных популяциях Е. coli
можно синтезировать разные полипептиды (рис.
IV. 11). Поскольку такие кДНК нельзя получить
с использованием в качестве матрицы природной
информационной РНК, их синтезировали химиче-
ски, основываясь на данных о нуклеотидной после-
довательности инсулина. Третья проблема была
решена с помощью индуцибельного промотора Е.
coli, который контролировал синтез чужеродного
белка: клетки выращивали в отсутствие индуктора
до высокой плотности, а затем индуцировали транс-
крипцию и трансляцию желаемого гена. Выход про-
дукта можно было увеличить, используя штаммы Е.
coli с мутантным протеазным геном. Иногда реком-
бинантный ген экспрессировался более эффективно
в дрожжевых или животных клетках, а не в Е. coli.
В эукариотических клетках могут осуществляться
соответствующие посттрансляционные модифика-
ции (например, гликозилирование) полипептида, и
можно даже сконструировать такой вектор, чтобы
происходила нормальная секреция полипептида.
Методом рекомбинантных ДНК уже получены мно-
гие ценные белковые продукты, хотя на это потребо-
валось больше времени, чем представлялось в сере-
дине 70-х годов. Мы уже упоминали вакцину против
вируса гепатита В. Проводятся клинические испыта-
ния вакцины против малярии-болезни, которой
ежегодно заболевают более 100 млн. человек. Полу-
чены человеческие инсулин и гормон роста, осуще-
ствляется тестирование лимфокинов, противорако-
вого терапевтического средства, выделен тканевой
плазминогенный активатор - сильнодействующее
средство при заболеваниях сердца.
Синтез в больших количествах специфических
полипептидов позволил не только получить важные
для клиники белки, но и исследовать их структуру
и функции. Трехмерная структура и, следовательно,
биологическая активность каждого белка зависят от
его аминокислотной последовательности. Как пока-
зали химические исследования, модификация боко-
вых групп отдельных аминокислот существенно
влияет на способность белковой молекулы к обра-
зованию специфической вторичной, третичной или
четвертичной структуры, а следовательно, на актив-
360
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
Синтетические олигонуклеотиды, кодирующие
А- и В-цепи инсулина
А в
(АТС TSAO (ATG TCAQ
69нуклеотидов 96 нуклеотидов
ность белка. Об этом же свидетельствует изучение
белков, кодируемых генами, которые несут специфи-
ческие мутации. Наиболее показательным приме-
ром такого рода служат генетические и фенотипи-
ческие дефекты при серповидноклеточной анемии,
описанные в разд. 1.3.
Теперь аминокислотную последовательность
белков можно направленно изменять с помощью
сайт-специфического мутагенеза клонированного ге-
на или кДНК, а не путем внесения случайных
мутаций. Синтезировав такие мутантные белки в Е.
coli, их очищают и исследуют физические и химиче-
РИС. IV.11.
Схематическое изображение процесса образования ин-
сулина в Е.соП. Синтезируют сегменты ДНК, кодиру-
ющие две цепи инсулина, А и В, содержащие каждый
стартовый кодон ATG и стоп-кодон TGA (показана
только одна цепь). Эти сегменты встраивают в плаз-
мидные векторы Е. coli справа от р-галактозидазного
(/acZ) промотора. Клетки Е. coli трансфицируют такими
плазмидами и клонируют. После индукции изопропил-
тиогалактозидом (IPTG) в клетках синтезируются два
полипептидных предшественника А- и В-цепей. С по-
мощью цианогенбромида (СпВг) от них отщепляют
короткий р-галактозидазный участок вместе с одним
остатком метионина. Образовавшиеся А- и В-цепи
смешивают в условиях, способствующих образованию
прочных бисульфидных связей, и получают активный
инсулин. Заметим, что синтез завершается успешно
только при отсутствии в молекуле инсулина метиони-
новых остатков. [J. D. Watson, J. Tooze, D. Т. Kurtz,
Recombinant DNA; A Short Course (San Francisco:
W. H. Freeman, 1983), рис. 18.2.]
ские свойства, а также функциональные особенно-
сти. Идентифицируют области в белковой молекуле,
существенные для образования нормальной вторич-
ной, третичной и четвертичной структуры, и опре-
деляют, какие аминокислоты образуют активный
центр фермента, а какие ответственны за связывание
субстратов. Конструируют белки с определенным
оптимумом pH, специфичностью или сродством
к субстрату, а также белки, обладающие нужными
температурными свойствами. Таким образом, мето-
ды рекомбинантных ДНК позволяют создавать бел-
ки с новыми химическими свойствами и проводить
исследования на недоступном ранее уровне сложно-
сти. Возможности таких исследований мы проил-
люстрируем несколькими примерами.
В результате замены гистидина на аспартат в
положении 10 В-цепи инсулина образуется гормон,
который связывается инсулиновым рецептором
плазматической мембраны почти в пять раз более
эффективно, чем нормальный инсулин. Соответст-
вующая мутация обнаружена у больных семейной
гиперпроинсулинемией. Цитохром с содержит кон-
сервативный остаток фениланалина; в цитохроме
с дрожжей-это остаток 87 (рис. III.8). Когда в ре-
зультате мутации фенилаланиновый кодон ТТТ за-
меняется на глициновый GGT, цитохром с сохраня-
ет свою активность, но его способность к переносу
электронов на цитохром с-пероксидазу существенно
снижается. Субстратсвязывающий центр протеоли-
тического фермента трипсина содержит в положе-
нии 226 остаток глицина. Как показывают струк-
турные исследования, замена этого остатка на более
объемную аминокислоту аланин может приводить
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
361
к изменению сродства фермента к лизиновому и
аргининовому остаткам в составе пептидных суб-
стратов; по-видимому, более длинная боковая цепь
аргинина в норме глубже проникает в карман, чем
лизин, и лучше заполняет его пространство, а при
замене глицина на аланин в кармане не хватает для
него места. Это предположение подтверждается
экспериментальными данными (рис. IV. 12).
Генетическая инженерия
С помощью направленного мутагенеза in vitro
получают гены белков, обладающих нужными свой-
Клонированный ген трипсиногена
Вс Е
11
Клонированная кДНК трипсиногена
Делеция всп-сегмента
из интрона
соединение
5'половиныгена
С 3-половиной
КДНК
Кодоны
сигнальной
последова-
тельности
делецияоставшейся части
интрона с помощью
олигонуклеотид-направ-
ленного мутаганеза
Вс Е
Клонирование в мп
Амидазная активность
Дикий тип Мутант
Субстрат
val-leu-lys-F
val-leu-arg F
Км
144
14
kcat
1308
1444
км
3665
482
kcat
172
13
Т4-полимераза
Т4-лигаза
Получение
активного
трипсина
выделение
мутантного
трипсиногена
f Трансфекция
cos-клеток
Трансформация f. coll
Скрининг М13, несущего
мутантный ген, с помощью
32₽-GTACACCGCG6G6TT
Мутантный
ген трипси-
ногена
РИС. IV.12.
Изменение центра связывания («кармана»)
субстрата в молекуле трипсина. По-видимому,
при наличии в последней в положении 226
остатка глицина карман имеет размеры, доста-
точные для размещения в нем объемных
аминокислот. Чтобы заменить глицин 226,
объединили 5 -участок клонированного гена
трипсиногена с З'-участком клонированной
кДНК. В результате был получен полнораз-
мерный ген трипсиногена, не содержащий
интронов, так что кодирующий участок приоб-
рел 1ЧН2-концевую сигнальную последова-
тельность, отсутствующую в кДНК. После
встраивания этого реконструированного гена
в вектор М13 и олигонуклеотиднаправленного
мутагенеза (рис. 7.36) изменили кодон 226
с GGT (глицин) на GCG (аланин) Полученный
мутантный ген ввели в челночный вектор
SV40-pBR322, так чтобы в cos клетках мог
транскрибироваться ген трипсиногена под
контролем раннего промотора (разд. 5.7.6).
Очищенный трипсиноген подвергали протео-
лизу и получали активный трипсин. Относи-
тельную способность мутантного и нормаль-
ного ферментов гидролизовать аргинин- и
лизинсодержащие пептиды определяли по
флуоресценции амидогруппы. Мутантный бе-
лок проявил более высокое сродство к лизин-
содержащему субстрату, если судить об этом
по изменению (измеряется в мкМ) и Af:at
(измеряется в мин-1). [С. S. Craik et al.. Science
228 (1985), p. 291.)
24-158
362
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
ствами. Большой интерес представляет изучение
влияния структуры белков, их регуляторных участ-
ков или геномной архитектуры на фенотип целых
клеток и организмов. Для этого в клетки или целые
организмы вводят измененные специфическим об-
разом гены. Здесь мощь метода рекомбинантных
ДНК проявляется во всей полноте: биология от
своего традиционного занятия-исследования ус-
тройства и функционирования живых систем-пе-
реходит в сферу манипуляций, позволяющих со-
здавать в них наследуемые изменения. Термин гене-
тическая инженерия, которого мы до сих пор избега-
ли, здесь вполне уместен. Он означает, помимо
всего прочего, что свойственный молекулярной ге-
нетике редукционистский подход позволяет исследо-
вать целые клетки и организмы, а также плейотроп-
ный эффект отдельных генов в отношении физиоло-
гических и морфологических признаков систем и их
развития.
Один из основных методов молекулярной гене-
тики-это трансформация эукариотических клеток
в культуре с помощью рекомбинантных векторов,
содержащих гены, кДНК и специфические мутант-
ные копии природных сегментов ДНК. Этот метод
лежит в основе многих экспериментов, описанных
в данной книге. Если исследуются бактериальные
или дрожжевые клетки, то манипуляции проводятся
на целых организмах, пусть и одноклеточных. Кор-
ректируя мутации, мы проводим своего рода тера-
пию на уровне генов (генотерапию). Распростране-
ние методов обратной генетики и генотерапии на
диплодные организмы, размножающиеся половым
путем, требует решения сложных методологических
и экспериментальных проблем.
Изменение соматических клеток
Один из подходов к генетическим модификациям
многоклеточных организмов заключается в измене-
нии генотипа только одного вида дифференциро-
ванных клеток: соматических. Для этого выделяют
нужные клетки, помещают их в культуральную
среду, обеспечивающую их жизнедеятельность и де-
ление, трансформируют клетки рекомбинантным
вектором, содержащим интересующий ген или соот-
ветствующую кДНК, и вновь вводят клетки в орга-
низм, из которого они были извлечены. Этот подход
лежит в основе разрабатываемых ныне методов
коррекции некоторых наследственных нарушений
у человека. В опытах используют красный костный
мозг млекопитающих, который содержит стволовые
клетки-предшественники циркулирующих клеток
крови (рис. IV. 13). Трансформацию проводят раз-
ными способами, в основном с помощью ретрови-
русных векторов, несущих специфические функцио-
нальные гены млекопитающих. В настоящее время
Животное-донор
костного
мозга
Культура
клеток
в чашке
Трансфекция
с помощью
клонирован-
ного гена
Отбор и рост
трансформи-
рованных
клеток
РИС. IV.13.
Изменение генома клеток костного мозга мыши и по-
следующее введение их в другое животное. Для более
успешного включения инъецированных клеток в кост-
ный мозг животное подвергают рентгеновскому облу-
чению.
этот общий подход апробируют на эксперименталь-
ных животных. Если будут получены обнадеживаю-
щие результаты, можно будет пытаться использо-
вать его для лечения наследственных болезней чело-
века. Некоторые наследственные дефекты частично
компенсируют введением в клетки крови соответст-
вующего функционального гена. В качестве примера
можно привести больных, гомозиготных по мутант-
ному гену аденозиндезаминазы, у которых наблю-
даются серьезные нарушения иммунитета.
Изменение клеток зародышевой
линии
Модификация соматических клеток не приводит
к наследственным изменениям многоклеточного ор-
ганизма. Для этого необходимо модифицировать
клетки зародышевой линии, и такой подход сейчас
разрабатывается на организмах трех типов: D. mela-
nogaster, экспериментальных животных и растениях.
Клетки зародышевой линии D. melanogaster можно
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
363
модифицировать с помощью транспозирующегося
P-элемента (разд. 10.2.6). Опыт состоит в следую-
щем. Рекомбинантный вектор, несущий нужный ген,
встроенный в функциональный P-элемент, инъеци-
руют в очень ранние эмбрионы D. melanogaster,
и P-элемент вместе с «лишним» геном транспозиру-
ется из вектора в геномную ДНК (рис. IV. 14). ДНК
клеток зародышевой линии взрослых мух, которые
развиваются из таких эмбрионов, часто несут Р-эле-
мент и экспрессируют новый ген. Этот метод откры-
вает широкие возможности для изучения регуляции
генной экспрессии в процессе развития и дифферен-
цировки и может дать ответ на следующие вопросы.
1. Экспрессируется ли данный ген во всех клетках
или только в тех из них, где экспрессировался бы
нормальный ген? 2. Какие регуляторные элементы
должны быть ассоциированы с введенным геном,
чтобы его экспрессия полностью имитировала экс-
прессию нормального гена? 3. Существенна ли хро-
мосомная локализация введенного гена для его
правильной регуляции в процессе развития и диф-
ференцировки?
Эти же вопросы возникают и при введении чуже-
родной ДНК в клетки зародышевой линии экспери-
ментальных млекопитающих. До настоящего време-
ни наиболее предпочтительным экспериментальным
объектом для создания таких трансгенных живот-
ных служили мыши. Проводились опыты и на ов-
цах, кроликах и свиньях, прежде всего с целью
улучшения пород этих домашних животных. Транс-
генных животных можно конструировать несколь-
кими способами, но наиболее эффективен один из
них. Клонированные гены инъецируют в пронуклеус
оплодотворенной яйцеклетки, которую импланти-
руют в матку приемной матери (рис. IV. 15). Чуже-
родная ДНК встраивается в геном реципиента на
самых ранних этапах развития, трансформируя как
клетки зародышевой линии, так и соматические
клетки, и наследуется по законам Менделя. Как
у самих трансгенных животных, так и у их потомков
можно выявлять различные аномалии и определять
время экспрессии гена и место, где она осуществля-
ется. Как правило, клонированный сегмент (транс-
ген) содержит различные промоторные элементы
и последовательности, существенные для тканеспе-
цифичной экспрессии гена. Для удобства в таких
опытах часто присоединяют подходящие регулятор-
ные элементы к последовательности, кодирующей
дикого типа
РИС. IV.14.
Изменение клеток зародышевой линии Dro-
sophila путем «трансдукции» с помощью
Р-злемента. Локус rosy кодирует ксантин-де-
гидрогеназу и детерминирует цвет глаз.
[G.M. Rubin, A. Spradling, Science 218 (1982),
р. 348.)
24*
364
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
9
d
Имплантация
в яйцевод
ВыделениеДнк
из клеток
кусочка хвоста
Эндонуклеазное расщепле-
ние, электрофорез,
ДНК-блоттинг
Идентификация инъеци-
рованной ДНКс помощью
специфического зонда
РИС. IV.15.
Получение трансгенных животных путем инъекции кло-
нированных генов в пронуклеус оплодотворенной яйце-
клетки. Мышь 3-трансгенное животное; она передаст
свой новый ген потомкам.
легко идентифицируемый продукт. Иногда больший
интерес представляет изучение действия специфиче-
ского генного продукта на трансгенное животное,
чем определение роли регуляторных сигналов. В та-
ком случае для обеспечения экспрессии гена соответ-
ствующую кодирующую последовательность присо-
единяют к гетерологичному промотору.
Возможность использования трансгенных мы-
шей для решения разнообразных биологических
проблем можно проиллюстрировать на примере
опытов с инсулиновыми генами. Ген инсулина экс-
прессируется только в 0-клетках поджелудочной
железы и регулируется цис-действующими элемента-
ми, расположенными в его 5'-фланкирующей обла-
сти (рис. IV.9). Если эту 5'-фланкирующую область
инсулинового гена крысы присоединить к кодирую-
щей области онкогена SV40 (большого Т-антигена)
и полученные рекомбинантные молекулы ввести
в ранние эмбрионы мыши, то геном потомков будет
содержать рекомбинантную ДНК (рис. IV. 16).
Большой Т-антиген синтезируется исключительно
в 0-клетках поджелудочной железы мыши, и его
онкогенность проявляется в образовании 0-клеточ-
ных опухолей-инсулином. Этот опыт подтвержда-
ют результаты экспериментов in vitro, демонстри-
руя роль регуляторных сигналов инсулиновых генов
в их тканеспецифичной экспрессии. В другом случае
данный онкоген был сцеплен с областью, регули-
рующей транскрипцию гена эластазы, который в
норме экспрессируется в экзокринных ацинарных
клетках поджелудочной железы; в этих опытах на-
блюдалось развитие аденом поджелудочной желе-
зы, а не инсулином. Эти модельные системы предо-
ставляют уникальную возможность для исследова-
ния онкогенеза и создания тонких инструментов для
генной терапии.
Основная цель экспериментального изменения
генотипов растений, как и цель соответствующих
экспериментов на животных, состоит в установле-
нии механизмов фундаментальных биологических
процессов. Что касается растений, то здесь это
дифференцировка тканей и световая индукция спе-
цифической генной экспрессии. Стимулом к таким
исследованиям служит также необходимость улуч-
шения важных сельскохозяйственных культур. Сей-
час большие усилия направлены на изучение генов,
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
365
Регуляторный
участок гена
инсулина /
Энхансер Промотор
Ген большого т-антигена SV40
Полиаденилирование
Первичный
транскрипт
Кэп _
РИС. IV.16.
Получение трансгенных мышей с использованием ре-
комбинантного гибридного гена, содержащего онкоген
SV40 (большой Т-антиген), находящийся под контро-
лем регуляторных последовательностей, которые по-
лучены от гена крысиного инсулина (рис. IV.9). У всех
таких мышей и их потомков развиваются инсулиномы
(Р-клеточные опухоли поджелудочной железы). [D. Ha-
nahan, Nature 315 (1985), р. 115.]
ответственных за повышение устойчивости культур
к гербицидам и пестицидам (рис. IV. 17). Многое
здесь зависит от создания рекомбинантных векто-
ров- производных Ti-плазмид Agrobacteria, и до
недавнего времени все эксперименты проводились
только на двудольных растениях (разд. 5.8). Для
трансформации клеток однодольных растений ус-
пешно применяются новые методы манипулирова-
ния с Ti-системами, электропорация, микроинъек-
ция в ядра и трансформация протопластов. Это
большое достижение, поскольку к однодольным
относится большинство хлебных злаков. Однако
получить целые фертильные растения из трансфор-
мированных протопластов однодольных пока не
удалось. В то же время из тотипотентных клеток,
трансформированных Ti-векторами, уже получены
целые двудольные растения с фертильными семена-
ми. несущие экспрессирующиеся рекомбинантные
гены.
НОВЫЕ НАПРАВЛЕНИЯ
В этом издании невозможно дать исчерпываю-
щую информацию о всех возможных способах при-
менения молекулярной генетики для анализа слож-
ных систем. Нам пришлось опустить не одну очень
содержит трансген. (С любезного разрешения Н. Schnei-
derman and the Monsanto Co.)
РИС. IV.17.
Эффект введения гена, детерминирующего синтез ток-
сина Bacillus thuringiensis, на поедание листьев томата
гусеницами. Растение, находящееся на фото справа.
366
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
важную тему. Тем не менее простое знакомство
с некоторыми из экспериментальных подходов по-
казывает, что даже весьма загадочные биологичес-
кие явления поддаются изучению. Одно из таких
явлений развитие сложного многоклеточного ор-
ганизма из зиготы, процесс, в результате которого
из дифференцирующихся примордиальных клеток
образуются разнообразные клетки и ткани взрос-
лого организма. Молекулярная генетика позволила
по-новому взглянуть на этот удивительный процесс.
Мы остановимся здесь лишь на нескольких из мно-
гочисленных интересных экспериментов с растения-
ми, беспозвоночными и позвоночными, включая
человека.
Эмбриональное развитие
и дифференцировка
у многоклеточных организмов
В процессе деления, начинающегося после опло-
дотворения, образуются клетки разного размера
и формы, выполняющие разные функции и специ-
фическим образом взаимодействующие с другими
клетками. Все эти события ограничиваются опреде-
ленными временными рамками. У некоторых видов,
например у мышей, все клетки, образующиеся во
время первых нескольких делений дробления, обла-
дают одинаковыми возможностями с точки зрения
дальнейшего развития из них той или иной ткани.
В дальнейшем после деления этих клеток каждая
новая клетка может дать начало лишь ограниченно-
му числу типов клеток взрослого организма, т. е.
наблюдается детерминация. Дифференцировка де-
терминированных клеток носит специфический ха-
рактер, наследуемый их дочерними клетками. На-
пример, из некоторых клеток образуются внезаро-
дышевые ткани (оболочки, окружающие плод), из
других-пищеварительный тракт (энтодерма), нерв-
ная система и эпидермис (эктодерма), мышцы, кро-
веносные сосуды и другие органы (мезодерма). По-
следующие клеточные деления осуществляются по
еще более строгой программе. Клетки приобрета-
ют определенный фенотип, форму и биохимические
свойства. У позвоночных насчитывается более 200
типов клеток. Многочисленные данные указывают
на то, что развитие и дифференцировка детерми-
нируются генной экспрессией, упорядоченной во
времени и пространстве. У ранних эмбрионов функ-
цинируют тщательно отлаженные регуляторные се-
ти, тонко реагирующие на положение клетки, ее
окружение и время. Одна из задач молекулярно-
генетического подхода к изучению детерминации
и дифференцировки состоит в том, чтобы устано-
вить корреляцию между пространственными и вре-
менными изменениями клеток на ранних стадиях
эмбриогенеза и дифференциальной генной экспрес-
сией. Этот подход особенно плодотворен в отноше-
нии D. melanogaster, поскольку можно получать му-
танты с дефектами, проявляющимися в раннем
морфогенезе.
Сегменты, видимые как у личинки, так и у взрос-
лых мух, в норме связаны с развитием специфичес-
ких морфологических признаков, например глаз,
усиков (антенн), крыльев или ног. В рамках класси-
ческой генетики выделены группы мутантов D. me-
lanogaster (Antennapedia. bithorax и т. д.), у которых
определенные морфологические структуры разви-
ваются не там, где положено. Например, у мутантов
Antennapedia на месте усиков (антенн) появляются
ноги (рис. IV. 18). Группа генов, детерминирующих
такие морфологические трансформации, называется
гомеозисными генами, и специфические гомеозисные
мутации обычно затрагивают один или несколько
сегментов (рис. IV. 19). Гомеозисные гены контро-
лируют развитие центральной нервной системы и
мышц в пределах данных сегментов, а также наруж-
ные органы. Они не определяют числа сегментов,
а только детерминируют развитие морфологических
признаков; число сегментов зависит от мутаций
в других генах D. melanogaster.
Кластеры генов Antennapedia или bithorax лока-
лизованы в двух протяженных областях хромосомы
3 D. melanogaster длиной не менее 250 т.п.н. каждая.
В результате гомеозисной мутации у мутанта Antenna-
pedia на месте усиков развиваются ноги. [В. Alberts et al..
Mol. Biol. Cell (1983), p. 840. Garland Publishing Co., New
York.]
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
367
Комплекс
Bithorax,
250 т.п.н.
ubx abd-h abd-v>
Dfd Scrffz Antp
РИС. IV.19.
Схематическое представление корреляции между го-
меозисными генами и сегментами Drosophila. Личинки
Drosophila состоят из отдельных сегментов, каждый из
которых дает начало определенной части тела взрослой
мухи. Комплексы Antennapedia и bithorax представляют
собой огромные кластеры гомеозисных генов на хро-
мосоме 3 (ubx, abd, Amp), определяющих морфогенез
передних (голова и грудь) и задних (грудь и брюшко)
сегментов соответственно. Показано соответствие меж-
ду торакальными (Т) и абдоминальными (А) сегмен-
тами личинки, с одной стороны, и частями тела взрос-
лой мухи-с другой. [К. Harding et al.. Science 229
(1985), p. 1236; W. J. Gehring, Science 236 (1987),
p. 1245.]
Комплекс
Antennapedia,
3 А1
2 3 4 5 6 7 8
В кластерах имеются множественные кодирующие
участки, ассоциированные с формированием специ-
фических сегментов. Некоторые из этих участков,
по-видимому, контролируют функционирование
всех генов кластера; в качестве примера можно
привести участок ubx кластера bithorax. Функциони-
рование тех или иных гомеозисных генов в специфи-
ческих сегментах регулируется-по крайней мере
частично-на уровне их тканеспецифичной транс-
крипции. Так, в клетках сегмента, модифицирован-
ного в результате мутации определенного гомеозис-
ного гена, транскрипты этого гена образуются с
большим избытком. Продукты гомеозисных генов-
зто ДНК-связывающие белки, специфичные для ре-
гуляторных элементов других генов. Все эти экспе-
рименты показывают, что морфогенез, как и биохи-
мическая специфичность, регулируется с помощью
дифференциальной генной экспрессии.
Гены, экспрессирующиеся на определенных ста-
диях развития, идентифицированы у таких разных
организмов, как кукуруза, нематоды, морские ежи
и мыши. Для получения клонов мРНК, экспресси-
рующейся на данной, но не на других стадиях
развития, созданы библиотеки кДНК, с помощью
которых можно получать клонированную мРНК,
синтезирующуюся на этих стадиях. Один из при-
меров такого рода представлен в разд. 6.5.6. Речь
идет о выделении кДНК, транскрибирующейся в
гаструле Xenopus, но не в яйцах. Однако вряд ли
можно ожидать быстрого прогресса в этой области
в отношении всех указанных выше организмов,
прежде всего потому, что в распоряжении ученых
имеется далеко не полная генетическая информация.
Исключение составляет нематода Caenorhabditis ele-
gans. В этом случае не только проведены обширные
генетические исследования, но и установлена пред-
история каждой из 2000 клеток взрослого червя,
начиная с зиготы. Удалось определить, какие гены
С. elegans экспрессируются и когда в процессе де-
терминации и дифференцировки обособляется та
или иная клеточная линия. Успешное введение кло-
нированных генов в клетки зародышевой линии С.
elegans (и обратные мутации) позволяет проводить
еще более тонкие эксперименты. Среди млекопи-
тающих в подобных опытах чаще всего используют-
ся мыши. В отношении их имеется обширная генети-
ческая информация и известно множество мутантов.
Кроме того, есть возможность получать мышиные
клетки на самых ранних стадиях развития зароды-
ша. Для мышей и некоторых других млекопитаю-
щих получены также ценные линии клеток ранних
эмбрионов и тератокарцином- опухолей, состоящих
из аномально быстропролиферирующих эмбрио-
нальных клеток. Однако самыми многообещающи-
ми являются два совершенно новых подхода к ис-
следованию процесса развития у мышей. Первый из
368
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
них берет начало с анализа гомеозисных генов у
D. melanogaster, второй-с изучения трансгенных
мышей.
Все кодирующие области гомеозисных генов со-
держат гомологичные последовательности длиной
примерно 180 п.н., известные под названием гомео-
боксов (рис. IV. 19). Они детерминируют синтез од-
нотипных полипептидных доменов в продуктах раз-
личных гомеозисных генов, а сами эти домены
отвечают за ДНК-связывающие свойства соответ-
ствующих белков. Существенно, что типичный го-
меобоксный домен высококонсервативен. Последо-
вательности, гомологичные гомеобоксам D. melano-
gaster, встречаются у дрожжей, беспозвоночных и
позвоночных, включая млекопитающих. Во всех
исследованных видах гомеобоксные продукты пред-
ставляют собой ДНК-связывающие белки, которые
взаимодействуют со специфическими нуклеотид-
ными последовательностями и с другими белками,
участвуя в сложных регуляторных процессах. Го-
меобоксные последовательности начинают транск-
рибироваться в эмбриональных клетках мышей
примерно на десятый день беременности. Этот факт
интересен с точки зрения обсуждения вопроса о
возможном использовании одинаковых стратегий
развития столь отдаленными друг от друга орга-
низмами, как D. melanogaster и млекопитающие.
Есть надежда, что идентификация генных продуктов
гомеобоксов послужит значительному прогрессу в
понимании морфогенеза млекопитающих.
Инсерционный мутагенез
Второй новый подход к изучению детерминации
и дифференцировки у млекопитающих основан на
использовании трансгенных мышей. Этот подход
был разработан для определения временных и про-
странственных аспектов экспрессии генов и для по-
становки экспериментов по генной терапии (рис.
IV. 15). Суть его состоит во встраивании инъециро-
ванных генов в неспецифические, случайные сайты
генома клетки-хозяина. Такое отсутствие специфич-
ности может затруднить исследование генной экс-
прессии и генотерапии, однако даст преимущества
при изучении морфогенеза. Случайное встраивание
инъецированной ДНК в кодирующие или важные
в регуляторном отношении участки может привести
к изменению экспрессии гена-мишени. В результате
может синтезироваться мутантный генный продукт
или вообще блокироваться экспрессия данного гена.
Этот метод внесения мутаций аналогичен так назы-
ваемому инсернионному мутагенезу, широко исполь-
зующемуся в генетике прокариот, дрожжей, D. mela-
nogaster и кукурузы. В последнем случае в качестве
инсерционного мутагенного фактора используют
транспозирующиеся элементы. У мышей функцию
транспозирующихся элементов при инъекции в зиго-
ту могут выполнять как рекомбинантные ретрови-
русные векторы, так и сегменты ДНК. В результате
иногда у трансгенных мышей и их потомков наблю-
даются совершенно случайные сильно выраженные
наследственные изменения фенотипа ранних эмб-
рионов и взрослых особей. Первое поколение транс-
генных мышей гетерозитно по инсерционной мута-
ции. В результате направленного инбридинга можно
получить как гетерозиготное, так и гомозиготное
потомство, и наблюдать фенотипические последст-
вия рецессивных мутаций. Поскольку встроенный
сегмент обычно содержит и чужеродную для мыши-
ного генома последовательность, например вектор,
его можно легко идентифицировать на ДНК-блотах
и в геномных библиотеках, а затем клонировать
вместе с фланкирующими последовательностями ге-
на-мишени. Таким способом была идентифицирова-
на в коллагеновом гене летальная инсерционная
мутация, прерывающая развитие зародыша на 11-й
день беременности. Другая инсерционная мутация
у мышей приводит к различным дефектам в разви-
тии костей конечностей. Ген-мишень в этом случае
представляет собой один из аллелей мутантного
гена, который был случайно обнаружен более 20 лет
назад (рис. IV.20). Клонирование этого гена и иден-
тификация генного продукта во многом должны
способствовать пониманию механизма морфогенеза
у млекопитающих.
Специфическая и случайная
модификация генома
млекопитающих
Практически все описанные выше методы осно-
ваны на способности эукариотических клеток интег-
рировать инъецированную ДНК в случайные сайты
генома. Однако нередко оказывается более удоб-
ным, а иногда и необходимым введение новой
генетической информации в определенные хромо-
сомные сайты. До недавнего времени это удавалось
осуществить лишь у дрожжей (разд. 5.6.в). Теперь
благодаря конструированию специфических векто-
ров, которые рекомбинируют с гомологичными по-
следовательностями хромосом млекопитающих,
здесь был достигнут значительный прогресс. Ис-
пользуя эти методы в сочетании с технологиями,
позволяющими изолировать нужные клетки и вво-
дить их в ранние мышиные эмбрионы, можно по-
лучать животных, несущих специфические мутации
в определенных генах, или элиминировать мутации
из дефектных генов.
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
369
I 2 3 4 5 6 7 8 9 10 II 12 13 14 15 16
wt wt wt wt wt id wt wt wt wt wt
WT WT Id Id
17 18 19 20 21 22 23 24
в
WT WT WT Id WT WT WT
РИС. IV.20.
Инсерционный мутагенез у трансгенных мышей, вызы-
вающий наследственную деформацию конечностей.
Трансгенные мыши (рис. IV.15) были получены путем
инъекции в зародыши сегмента ДНК, несущего ретро-
вирусный LTR в качестве промотора и мышиный прото-
онкоген тус в составе вектора pBR322, и последующего
инбридинга потомков. Все мыши данной линии несли
рецессивную аутосомную мутацию, как видно из фото-
графии четырехдневных гомозиготных мутантных мы-
шей (ZI) и их скелетов (£>). У гомозигот конечности
деформированы. При скрещиваниях зта аномалия ко-
нечностей (Id) сегрегирует вместе с участками ДНК,
которые гибридизуются с последовательностями LTR-
тус, как зто видно из родословной. Кружками обозна-
чены самки, квадратиками-самцы. Черным цветом
обозначены животные, гомозиготные по вставке LTR-
тус (определялось с помощью ДНК-блоттинга). Знач-
ки, зачерненные наполовину, соответствуют особям,
гетерозиготным по вставкам, а полностью белые знач-
ки-животным, не имеющим вставки LTR-myc. Буквами
внизу обозначен фенотип: дикий тип (WT) или Id.
Перечеркнутый символ означает, что животное погибло
до получения данных. С помощью гибридных сома-
тических клеток мышь/китайский хомячок было пока-
зано, что вставка LTR-myc локализована на хромосоме
2 мыши. [R. Р. Woychik et al., Nature 318 (1985), p. 36.J
Перспективы
Молекулярная генетика находится лишь в начале
своего пути. Уже накоплена обширная информация
о многих биологических системах, но этого недоста-
точно для полного или даже более или менее глубо-
кого понимания их функционирования. Мы знаем
пока гораздо меньше, чем нам предстоит узнать.
Все, что создано руками человека, не может срав-
ниться по своему разнообразию и сложности с жи-
выми существами. Никакая искусственная информа-
ционная система не приближается по объему содер-
жащейся в пей информации и по сложности регуля-
ции к биологическим системам, осуществляющим
экспрессию генов при помощи столь хитроумно
устроенных информационных сетей. Нам еще пред-
стоит открыть такие механизмы и принципы переда-
чи информации, которые трудно даже вообразить.
Пока же мы можем только очертить-да и то не
всегда - те важные области, о которых нам мало что
известно. Это могут быть такие давние проблемы,
как развитие и дифференцировка или молекулярные
370
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
основы мышления, либо проблемы относительно
новые, осознанные благодаря достижениям самой
молекулярной генетики. И конечно, мы понимаем,
что есть вещи, которые кажутся нам сегодня доста-
точно ясными, но которые в будущем могут ока-
заться гораздо более сложными. А о некоторых из
них мы вообще можем иметь ошибочное представ-
ление.
Другая очень важная область-это все, что свя-
зано с онкогенезом: v-onc, с-опс и протоонкогены.
Достигнут значительный прогресс в изучении нор-
мальных биохимических функций протоонкогенов,
однако важнейший вопрос о том, чем обусловли-
вается различие между функционированием про-
тоонкогена и его онкогенной копии с-опс, иссле-
довался лишь в самых общих чертах. У нас нет
четкого представления о том, каким образом с-опс
или v-onc превращает нормальную клетку в опу-
холевую. Возможно, в некоторых случаях в основе
этого феномена лежит активация фосфорилирова-
ния белков и постоянное пребывание переключа-
телей транскрипции во включенном состоянии.
Сколько клеточных аберраций, отличных от тех,
которые связаны с экспрессией v-onc и с-опс, должно
произойти, чтобы клетка стала опухолевой? И, что
наиболее важно, можно ли блокировать онкогенез
с помощью каких-либо прецизионных молекуляр-
ных инструментов?
Феномен генетической рекомбинации впервые
был описан в начале XX в. В то время под реком-
бинацией понимали гомологичный мейотический
кроссинговер. Теперь мы знаем, что существует
множество видов рекомбинации, осуществляющей-
ся как на стадии мейоза, так и в митотических
клетках: гомологичный кроссинговер, генная кон-
версия, транспозиция мобильных элементов, геном-
ные перестройки и т. д. Каковы их молекулярные
механизмы? Какой тип рекомбинации преобладает
у разных организмов, когда (на каком этапе разви-
тия) и где (в каких тканях)? Можно ли как-то влиять
на рекомбинацию? Эти вопросы сейчас интенсивно
исследуются.
Будущие заботы
Быстрые темпы развития современной биологии
обусловливаются запросами технологии, здравоох-
ранения, сельского хозяйства, промышленности и,
конечно же, любознательностью исследователей.
Поскольку все мы и наши дети - продукты функцио-
нальных генетических систем, чисто академический
интерес подогревается еще и личной заинтересован-
ностью. Это сочетание научного интереса и лично-
стного аспекта приводит к огромному обществен-
ному интересу к технологии рекомбинантных ДНК
и ее детищу-генетической инженерии. Достижения
биологии XX в. относятся к событиям историческо-
го значения. Это была настоящая революция, кото-
рая пока приносила положительные плоды. Мы все
глубже познаем самих себя и другие живые сущест-
ва. Получены многие важные биологические продук-
ты-гормоны, вакцины и ферменты, использующие-
ся как в исследовательских целях, так и в медицине
и промышленности. Люди научились направленно
изменять вредные микроорганизмы, превращая их
в полезных агентов окружающей среды. Однако
у этой революции есть и тревожные моменты. Необ-
ходимо помнить, что измененные «в лучшую сторо-
ну» микроорганизмы могут обладать другими, сов-
сем не полезными свойствами. Чтобы исключить
возможность использования генотерапии соматиче-
ских клеток или изощренных диагностических мето-
дов с применением новых технологий не по назначе-
нию, необходимо тщательно проанализировать по-
следствия. Ответственность здесь очень велика, по-
скольку эти последствия могут оказаться непредска-
зуемыми.
Почти ничего из написанного в этой книге не
было известно в ту пору, когда мы закончили свое
формальное образование более 30 лет назад. Очень
жаль, что сегодня мы не можем дать прогноз на
следующие 30 лет. Ясно лишь одно: будущие ис-
следования приведут к появлению новых фундамен-
тальных концепций, столь же революционных, каки-
ми были для нас интроны и мобильные элементы.
Литература
Ниже приведены ссылки на работы обзорного харак-
тера, касающиеся рассмотренных в этой части биологиче-
ских систем. В каждом разделе ссылки даны в хронологи-
ческом порядке
Исследование сложных
биологических систем
M.S. Brown. J.L. Goldstein. 1986. A Receptor-Mediated Path-
way for Cholesterol Homeostasis. Science, 232, 34- 47.
A. W. Murray. M. W. Kirschner. 1989. Dominoes and Clocks:
The Union of Two Views of the Cell Cysle. Science, 246,
614 621.
Семейство генов зрительных пигментов
J. Nathans. 1987. Molecular Biology of Visual Pigments.
Annu. Rev. Neurosci., 10, 163-194.
T. Piantanida. 1988. The Molecular Genetics of Color Vision
and Color Bindness. Trends Genet., 4, 319-323.
D. Vollrath, J. Nathans. R. W. Davis. 1988. Tandem Array of
Human Visual Pigment Genes at Xq28. Science, 240,
1669 1672.
J. Nathans. 1989. The Genes for Color Vision. Sci. American,
260 (2). 42-49.
Согласованные взаимодействия между
продуктами разных генов
J. Sommerville. 1985. Organizing the Nucleolus. Nature, 318,
410-411.
G. Jordan. 1987. At the Heart of the Nucleolus. Nature, 329,
489-490.
R.J. Planta, H.A. Raue. 1988. Control of Ribosome Biogenesis
in Yeast. Trends Genet., 4, 64-68.
S. Rose. 1988. Reflections on Reductionism. Trends Biochem.
Sci., 13, 160-162.
F. Amaldi, 1. Bozzoni, E. Beccari, P. Pierandrei-Amaldi. 1989.
Expression of Ribosomal Protein Genes and Regulation of
Ribosome Biosynthesis in Xenopus Development. Trends
Biochem. Sci., 14. 175-178.
B-Z. Shillo. 1987. Proto-Oncogenes in Drosophila melanogaster.
Trends Genet., 3, 69-72.
A.S. Fauci. 1988. The Human Immunodeficiency Virus: Infecti-
vity and Mechanisms of Pathogenesis. Science, 239, 617—
622.
D.l.H. Linzer. 1988. The Marriage of Oncogenes and Anti-
Oncogenes. Trends Genet.. 4, 245-247.
P C. Nowell. C.M. Croce. 1988. Chromosomal Approaches to
Oncogenes and Oncogenesis. FASEB J., 2, 3054-3060.
R. A. Weinberg. 1988. Finding the Anti-Oncogene. Sci. Ameri-
can, 259 (3), 4 51.
Картирование геномов
Физическая и генетическая карты
A. Coulson, J. Sulston, S. Brenner, J. Karn. 1986. Toward a
Physical Map of the Genome of the Nematode, Caenor-
habdilis elegans. Proc. Natl. Acad. Sci. U.S.A., 83, 7821 —
7825.
M. V. Olson, J. E. Dutchik. M. Y. Graham. G. M. Brodeur.
C. Helms. M. Frank, M. MacColin, R. Scheinman.
T. Frank. 1986. Random-Clone Strategy for Genomic
Restriction Mapping in Yeas. Proc. Natl. Acad. Sci.
U.S.A., 83, 7826-7830.
H. Cooke. 1987. Cloning in Yeast: An Appropriate Scale for
Mammalian Genomes. Trends Genet., 3, 173-175.
Y. Kohara. K. Akiyama. K. Isono. 1987. The Physical Map of
the Whole Ё. coli Chromosome: Application of a New
Strategy for Rapid Analysis and Sorting of a Large
Genomic Library. Cell, 50, 495-508.
C. L. Smith. C.R. Cantor. 1987. Preparation and Manipulation
of Large DNA Molecules. Trends Biochem. Sci., 12,
284-287.
C. L. Smith. J.G. Econome, A. Schutt. S. Klco, C. R. Cantor.
1987. A Physical Map of the Escherichia coli Genome.
236 11|'1 8_1 53
D. Vollrath, R. W. Davis. C. Connely, P. Hieter. 1988. Physical
Mapping of Large DNA by Chromosome Fragmentation.
Proc. Natl. Acad. Sci. U.S.A., 85, 6027-6031.
V. Knott. D.J. Blake. G.G. Brownlee. 1989. Completion of the
Detailed Restriction Map of the E. coli Genome by the
Isolation of Overlapping Cosmid Clones. Nucleic Acids
Res., 17, 5901-5912.
Жизненный цикл вирусов
J.M. Bishop. 1982. Oncogenes. Sci. American, 246 (3), 80-92.
J.M. Bishop. 1983. Cellular Oncogenes and Retroviruses.
Annu. Rev. Biochem., 52, 301 354.
R. A. Weinberg. 1983. A Molecular Basis of Cancer. Sci. Ameri-
can, 249 (5), 126-143.
H.E. Varmus. 1984. Molecular Genetics of Cellular Oncogenes.
Annu. Rev. Genet., 18, 553-612.
R. DiMarchi, G. Brooke. C. Gale, V. Cracknell. T. Doel, N. Mo-
wat. 1986. Protection of Cattle Against Foot-and-Mounth
Disease by a Synthetic Peptide. Science, 232, 639-641.
J. M. Bishop. 1987. The Molecular Genetics of Cancer. Science,
235, 305-311.
Картирование больших геномов
V.A. McKusick. 1986. The Gene Map of Homo sapiens'. Status
and Prospects. Cold Spring Harbor Symp. Quant BioL, 51.
15-27.
D.T. Burke. G.F. Carle. M.V. Olson. 1987. Cloning of Large
Segments of DNA into Yeast by Means of Artifical
Chromosome Vectors. Science, 236, 806-812.
H. Donis-Keller, P. Green, C. Helms, S. Cartinhour, B. Weijfen-
bach, K. Stephens, T. P. Keith, D. W. Bowden. D. R. Smith.
E.S. Lander, D. Botstein. G. Akots, K.S. Rediker, T. Gra-
vitts, V. A. Brown, M. B. Rising, C. Parker, J. A. Powers,
D. E. Watt, E. R. Kauffman, A. Bricker. P. Phipps, H. Mul-
ler-Kahle, T. R. Fulton, S. Ng, J. W. Schumm, J. C. Braman,
372
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
R. Knowlton. D. F. Barker. S. М. Crooks. S. E. Lincoln.
M.J. Daly, J. Abarhamson. 1987. A Genetic Linkage Map
of the Human Genome. Cell, 51, 319-327.
T. Helentjaris. 1987. A Genetic Linkage Map for Maize Based
on RFLPs. Trends Genet., 3, 217-221.
Human Gene Mapping 9. 1987. Cytogenet. Cell Genet., 46, Nos.
1-4.
V. A. McKusick. F. H. Ruddle. 1987. Editorial: Toward a Comp-
lete Map of the Human Genome. Genomics, 1, 103 106.
Y. Nakamura. M. Lathrop, P. O'Connell. M. Leppert. D. Barker.
E. Wright, M. Skolnick, S. Kondoleon. M. Litt, J-M. La-
louel. R. White. 1988. A Mapped Set of DNA Markers for
Human Chromosome 17. Genomics, 2, 302-309.
C.M. Rick. J. I. Yoder. 1988. Classical and Molecular Genetics
of Tomato: Highlights and Perspectives. Annu. Rev.
Genet., 22, 281-300.
J. Schmidtke, D. N. Copper. 1989. A Comprehensive List of
Cloned Human DNA Sequences. Nucleic Acids Res.. 17
(suppl.), r!73-r28l.
R. White, J-M. Lalouel. 1988. Linked Sets of Genetic Markers
for Human Chromosomes. Annu. Rev. Genet., 22. 259-
279.
Полиморфизм длины рестрикционных
фрагментов
A. J. Jeffreys. 1979. DNA Sequence Variants in the Gy-, Ay-, 5-
and Р-Globin Genes of Man. Cell, 18, 1-10.
S. H. Orkin. H.H. Kazazian. Jr. 1984. The Mutation and Poly-
morphism of the Human Beta-Globin Gene and Its
Surrounding DNA. Annu. Rev. Genet., 18, 131 171.
J. F. Gusclla. 1986. DNA Polymorphism and Human Disease.
Annu. Rev. Biochem.. 55, 831-854.
С. T. Caskey. 1987. Disease Diagnosis by Recombinant DNA
Methods. Science, 236, 1223-1228.
A. P. Monaco, L.M. Kunkel. 1987. A Giant Locus for the
Duchenne and Becker Muscular Dystrophy Gene. Trends
Genet., 3. 33-37.
M. Namanura, M. Leppert. P. O’Connell, R. Wolff. T. Holm.
M. Culver, C. Martin, E. Fujimoto, M. Hoff. E. Kumlin,
R. White. 1987. Variable Number of Tandem Repeat
(VNTR) Markers for Human Gene Mapping. Science. 235,
1616 1622.
M. Dean. 1988. Molecular and Genetic Analysis of Cystic
Fibrosis. Genomics, 3, 93-99.
J. A. Witkowski. 1988. The Molecular Genetics of Duchenne
Muscular Dystrophy: The Beginning of the End? Trends
Genet., 4, 27-30.
R.G. Woron. M.W. Thompson. 1988. Genetics of Duchenne
Muscular Dystrophy. Annu. Rev. Genet., 22, 601-629.
B-S. Karem, J.M. Rommens, J. A. Buchanan, D. Markiewicz.
T. K. Cox. A. Chakravarti, M. Buchwald, L-C. Tsui. 1989.
Identification of the Cystic Fibrosis Gene: Genetic Analy-
sis. Science, 245, 1073-1080.
J. L. Mandel. 1989. Dystrophin: The Gene and Its Product.
Nature, 339. 584-586.
B. Martin. J. Niehuis, G. King. A. Schaefer. 1989. Restriction
Fragment Length Polymorphysms Associated with Water
Use in Tomato. Science, 243, 1725 1728.
J. R. Riordan. J. M. Rommens, B-S. Karem, N. Alon, R. Roz-
mahel, Z. Grzelzak, J. Zielenski. S. Lok. N. Plavsic,
J-L. Chou. M.L. Drumm. M.C. lannuzzi. F.S. Collins,
L-C. Tsui. 1989. Identification of the Cystic Fibrosis Gene:
Cloning and Characterization of Complementary DNA.
Science, 245. 1066-1073.
J. M. Rommens, M. C. lannuzzi, B-S. Karem, M. L. Drumm,
G. Kelmer, M. Dean. R. Rozmahel. J.L. Cole. D. Kenne-
dy, N. Hidaka. M Zsiga. M. Buchwald, J. R. Riordan,
L-C. Tsui, F.S. Collins. 1989. Identification of the Cystic
Fibrosis Gene: Chromosome Walking and Jumping. Scien-
ce, 245, 1059- 1065.
Дифференциальная экспрессия
генов в специфических клетках
и тканях
Специфическая экспрессия генов
в поджелудочной железе
Т. Edlund, M.D. Walker. P.J. Barr, W.J. Rutter. 1985. Cell-
Specific Expression of the Rat Insulin Gene: Evidence for
Role of Two Distinct 5' Flanking Elements. Science, 230,
912 916.
O. Karlsson. T. Edlund. J. B. Moss, W. J. Rutter, M. D. Walker.
1987. A Mutational Analysis of the Insulin Gene Trans-
cription Control Region: Expression in Beta Cells Is
Dependent on Two Related Scqueces Wthin the Enhancer.
Proc. Natl. Acad. Sci. U.S.A., 84, 8819-8823.
L. G. Moss. J.B. Moss, W.J. Rutter. 1988. Systematic Binding
Analysis of the Insulin Gene Transcription Control
Region: Insulin and Immunoglobulin Enhancers Utilize
Similar Transactivators. Mol. Cell. BioL, 8, 2620 2627.
H. Ohlsson. O. Karlsson. T. Edlund. 1988. A Beta-Ccll-Specific
Protein Binds to the Two Major Regulatory Sequences of
the Insulin Gene Enhancer. Proc. Natl. Acad. Sci. U.S.A.,
85, 4228-4231.
Посттрансляционные изменения как
регуляторы направленного потока генных
продуктов
J. Douglas. О. Cirelli. Е. Herbert. 1984. Polyprotein Gene
Expression: Generation of Diversity of Neuroendocrine
Peptides. Annu. Rev. Biochem., 53, 665-715.
S. H. Snyder. 1985. The Molecular Basis of Communication
Between Cells. Sci. American, 253 (4), 132-141.
TT. Puck. F-T. Kao. 1982. Somatic Cell Genetics and Its
Application to Medicine. Annu. Rev. Genet., 16, 225-271.
J. Stanbury, J. B. Wyngaarden. D. S. Fredrickson. J. L. Gold-
stein. M.S. Brown. 1983. The Metabolic Basis of Inherited
Disease, 5th ed. McGraw-Hill, New York.
K.E. Davies (ed.). 1986. Human Genetic Diseases: A Practical
Approach. 1RL Press, Oxford and Washington, D. C.
Изменение биологических систем
Синтез нормальных и модифицированных
эукариотических белков
J. F. Young, W. Т. Hockmeyer, М. Gross, W. R. Ballou,
R. A. Wirtz. J. H. Trosper. R. L. Beaudoin, M. R. Holling-
dale, L. H. Trosper. R. L. Beaudoin. M. R. Hollingdale.
L.H. Miller. CL. Diggs, M. Rosenberg. 1985. Expression
of Plasmodium falciparum Circumsporozoite Proteins in E.
coli for Potential Use in a Human Malaria Vaccine.
Science, 228, 958-962.
A. J. Clark, P. Simons, 1. Wilmut, R. Lathe. 1987. Pharmaceuti-
cals from Transgenic Livestock. Trends Biotech., 5, 20-24.
C. S. Craik, S. Roczniak. S. Sprang, R. Fletlcrick, W. Rutter.
1987. Redesigning Trypsin Via Genetic Engineering. J.
Cell. Biochem., 33, 199-211.
J. R. Knowles. 1987. Tinkering witz Enzymes: What Are We
Learning? Science, 236, 1252- 1258.
ЛИТЕРАТУРА
373
М. Liang, G.L. Pielak, A.G. Mauk, M. Smith, B.M. Hoffman.
1987. Yeast Cytochrome c with Phenylalanine or Tyrosine
at Position 87 Transfers Electrons to (Zinc Cytochrome
c Peroxidase)* at a Rate Ten Thousand Times That of the
Serine-87 or Glycinc-87 Variants. Proc. Natl. Acad. Sci.
U.S.A.. 84, 1249 1252.
D. Shortlc. 1989. Probing the Determinants of Protein Folding
and Stability with Amino Acid Substitutions. J. Biol.
Chem., 264, 5315-5318.
Изменение соматических клеток
T. Friedman. 1983. Gene Therapy: Fact and Fiction. Cold
Spring Harbor Laboratory, Cold Spring Harbor, New
York.
WF. Anderson. 1984. Prospects for Human Gene Therapy.
Science, 226, 401- 409.
J. Ellis, A. Bernstein. 1988. Gene Targeting with Retroviral
Vectors. Cold Spring Harbor Laboratory, Cold Spring
Harbor, New York.
Y. Gluzman. S.H. Hughes. 1988. Viral Vectors. Gold Spring
Harbor Laboratory, Cold Spring Harbor, New York.
T. Friedmann. 1989. Progress Toward Human Gene Therapy.
Science, 244. 1275 1281.
Изменение клеток зародышевой линии
G.M. Rubin, A.C. Spradling. 1982. Genetic Transformation of
Drosophila with Transposable Element Vectors. Science,
218, 348-353.
A.C. Spradling, G.M. Rubin. 1982. Transposition of Cloned
P Elements into Drosophila Germ Line Chromosomes.
Science, 218, 341 347.
R. L. Chisholm. 1983. Gene Therapy in Drosophila. Trends
Biochem. Sci., 8, 191 193.
R. Juenisch. K. Harbers. A. Schnieke. J. Lohler. I. Chumakov.
D. Jahner, D. Grotkopp, E. Hoffmann. 1983. Germline
Integration of Moloney Murine Leukemia Virus at the
Mot 13 Locus Leads to Recessive Lethal Mutation and
Early Embryonic Death. Cell, 32, 209-216.
TA. Stewart. P.K. Puttengale, P. Leder. 1984. Spontaneous
Mammary Adeno-Carcinomas in Transgenic Mice that
Carry and Express MTV/mrc Fusion Genes. Cell, 38,
627-637.
J. W. Gordon, F.H. Ruddle. 1985. DNA-Mediated Genetic
Transformation of Mouse Embryos and Bone Marrow-
A. Review. Gene, 33, 121 136.
D. Hanahan. 1985. Heritable Formation of Pancreatic Beta-Cell
Tumours in Transgenic Mice Expressing Recombinant
Insulin/Simian Virus 40 Oncogenes. Nature. 315, 115-123.
R. L. Briuster. R.D. Palmiter. 1986. Introduction of Genes into
the Germ Line of Animals. In: The Harvey Lectures, pp.
1-38, series 80. Alan R. Liss, New York.
D. Hanahan. 1986. Oncogenesis in Transgenic Mice. In: T. Graf
and P. Kahn (eds.). Oncogenes and Growth Control, pp.
349-363. Spring-Vcrlag, Heidelberg.
R.D. Palmiter. R. L. Brinster. 1986. Germ-Line Transformation
of Mice. Annu. Rev. Genet., 20, 465 499.
R. B. Church. 1987. Embryo Manipulation and Gene Transfer
in Domesttic Animals. Trends Biotech., 5. 13-19.
E. C. Cocking. M. R. Davey. 1987. Gene Transfer in Cereals.
Science, 236, 1259- 1262.
R.M. Goodman. H. Hauptili, A. Crossway, КС. Knauf. 1987.
Gene Transfer in Crop Improvement. Science, 236 48-54.
D. Hanahan. 1988. Dissecting Multistep Tumorigenesis in
Transgenic Mice. Annu. Rev. Genet., 22, 479-519.
C. A. Rhodes, D. A. Pierce, I. J. Mettler, D. Mascarenhas,
J. J. Detmer. 1988. Genetically Transformed Maize Plants
from Protoplasts. Science, 240, 204 207.
L. Wilbnitzer. 1988. The Use of Transgenic Plants to Study
Plant Gene Expression. Trends Genet., 4, 13-18.
M. R. Capacchi. 1989. Altering the Genome by Homologous
Recombination. Science, 224, 1288-1292.
Cell. 56 (2). 1989. Весь выпуск посвящен растительным
системам.
C.S. Gasser. R. Т. Fraley. 1989. Genetically Engineering Plants
for Crop Improvement. Science, 244, 1293-1299.
D. Hanahan. 1989. Transgenic Mice as Probes into Complex
Systems. Science 246. 1265-1275.
S. Thompson, A. R. Clarke, A. M. Pow, M. L. Hooper,
D. fK Melton. 1989. Germ Line Transmission and Expres-
sion of a Corrected HPRT Gene Produced by Gene
Targeting in Embryonic Stem Cells. Cell, 56, 313-321.
Новые направления
Эмбриональное развитие и дифференци-
ровка многоклеточных организмов
P.W. Sternberg, H.R. Horvitz. 1984. The Genetic Control of
Cell Lineage During Nematode Development. Annu. Rev.
Genet., 18, 489-524.
W. Bender. 1985. Homeotic Gene Products as Growth Factors.
Cell, 43, 559-560.
W.J. Gehring. 1985. The Molecular Basis of Development. Sci.
American, 253 (4), 152B- 162.
E. H. Dandson. 1986. Gene Activity in Early Development, 3rd
ed. Academic Press, Orlando.
W.J. Gehring, Y. Hiromi. 1986. Homeotic Genes and the
Homeobox. Genetics, 20, 147-173.
I. Duncan. 1987. The Bithorax Complex. Annu. Rev. Genet., 21,
285-319.
W.J. Gehring. 1987. Homeo Boxes in the Study of Develop-
ment. Science, 236, 1245-1252.
M. Robertson. 1987. Towards a Biochemistry of Morphogene-
sis. Nature, 330, 420-421.
P. W. Ingham. 1988. The Molecular Genetics of Embryonic
Pattern Formation in Drosophila. Nature, 335, 25-34.
M. Akam. 1989. Hox and Hom: Homologous Gene Clusters in
Insects and Vertebrates. Cell, 57, 347-349.
G. R. Dressier. 1989. An Update on the Vertebrate Homeobox.
Trends Genet., 5, 129-131.
Инсерционный мутагенез
R. P. Woychik, TA. Stewart. L.G. Davis, P.D. Eustachio, P. Le-
der. 1985. An Inherited Limb Deformity Created by
Insertional Mutagenesis in a Transgenic Mouse. Nature,
318, 36-40.
7. Gridley. P. Soriano, R. Jaenisch. 1987. Insertional Mutagene-
sis in Mice. Trends Genet., 3, 162-166.
L. Cooley. R. Kelley, A. Spradling. 1988. Insertional Mutagene-
sis of the Drosophila Genome with Single P Elements.
Science. 239, 1121-1128.
Перспективы
Office of Technology Assessment, U.S. Congress, 1981. Impacts
of Applied Genetics: Microorganisms, Plants, and Ani-
mals. U.S. Government Printing Office, Washington, D.C.
President's Commission for the Study of Ethical Problems in
Medicine and Biomedical and Behavioral Research. 1982.
Splicing Life: The Social and Ethical Issues of Genetic
Engineering with Human Beings. U.S. Government Prin-
ting Office, Washington, D.C.
374 ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ
D. J. Weatherall. The New Genetics and Clinical Practice. 1982.
Nuffield Provincial Hospitals Trust, London.
J.G. Perpich (ed.). 1986. Biotechnology in Society: Private
Initiatives and Public Oversight. Pergamon Press, New
York.
Introduction of Recombinant DN А-Engineering Organisms into
the Environment: Key issues. 1987. National Academy
Press, Washington, D.C.
Commission on Life Sciences, National Research Council. 1988.
Mapping and Sequencing the Human Genome. National
Academy Press, Washington, D.C.
E. K. Nichols. 1988. Human Gene Therapy. Institute of Medi-
cine, National Academy of Sciences. Harvard University
Press, Cambridge, Mass.
Office of Technology Assessment, U. S. Congress. 1988. Field-
Testing Engineering Organisms: Genetic and Ecological
Issues. U.S. Government Printing Office, Washington,
D.C.
Office of Technology Assessment. U.S. Congress. 1988. Mapping
Our Genes. Genome Projects: How Big? How Fast?
U. S. Government Printing Office, Washington, D.C.
L. Walters. 1989. Report to the Biomedical Ethics Advisory
Commitee of the Biomedical Ethics Board. Washington,
D.C. Available through National Reference Center for
Bioethics Literature, Kennedy Institute of Ethics, Geor-
getown University, Washington, D.C.
Предметный указатель
Абрин 1: 162
Автономно реплицирующиеся после-
довательности (ARS) I: 253-254
Arap 1: 229
- бесцветные колонии I: 236, 241, 244
- голубые колонии I: 229, 236, 241, 244
Адапторная гипотеза I: 132
Аденилатциклаза I: 177
Адениндезаминаза II: 309
Аденовирусы I: 87-88, 91
- в качестве векторов I: 259
- ген Е2А II: 142-143
- геном II: 90, 345, 346, 348
- ДНК II: 344-346, 351
- метилирование промотора Е2А II:
143
- поздняя мРНК II: 121
- репликация ДНК I: 80, 87
- транскрипция II: 43-44
S-аденозилметионин II: 42, 141-142
Аденозинтрифосфат I: 118-119
- участие в образовании аминоацил-
тРНК I: 140
Аденозин-5'-фосфат I: 53
5-азацитидин II: 144
Активаторы I: 172-173; II: 9
Активирующие последовательности
левые (UAS) II: 70-71
Актин, псевдогены II: 173
- семейства генов II: 172-173
Актиновые гены, интроны II:
172-173
-- кодирующие области II: 172-173
— фланкирующие последовательности
II: 172
Актиномицин D I: 160 161
Аланин I: 56
Алкилирующие агенты I: 99
Алкогольдегидрогеназа I: 354
Аллели I: 25
Аллельное исключение II: 291
Альбуминовые гены, организация II:
65-66
— промоторы II: 68-69
-- экспрессия II: 355
---тканеспецифичная II: 65 -69
а-Аманитин I; 161: II: 22, 40, 88
а-Амилазные гены, семейство II: 171,
179
Аминоацил-тРНК I: 116, 138 141,
160
Аминоацил-тРНК-синтетазы I: 116,
132-133, 138, 140-142
- узнавание аминокислот и родствен-
ных тРНК I: 138 142
Аминокислоты, аминогруппы I: 56-57
- боковые группы I: 56-57, 59-61
- в пептидной цепи I: 59-62
- включение в белки I: 132- 134
- и соответствующие кодоны I: 131
132
- карбоксильные группы I: 56
-кодоны I: 116, 136-137
- связывание с соответствующими
тРНК I: 138-142
- структура I: 57-59
- трехбуквенные обозначения I:
58
- С-концевые I: 57
- N-концевые I: 57, 116
Аминоптерин I: 258
Ампициллин I: 207, 230
Амплификация II: 14, 159, 195, 230
- ДНК запрограмированная, влияние
на экспрессию геиов II: 302-306
- и устойчивость к лекарственным
веществам II: 307-308
— эволюция генома II: 17-18
изменение морфологии хромосом
II: 309-310
- коррекция ферментной недостаточ-
ности II: 306-308
- механизм II: 310-314
- незапрограмированные повторы II:
306 314
- при непропорциональной реплика-
ции II: 304-306
- рДНК по типу катящегося кольца
II: 304
- реверсия мутантного фенотипа II:
308
- стимуляция химическими агентами
II: 314
- цитологические аспекты II: 311
- элементы, ее контролирующие II:
304
Амплифицированные гены, неста-
бильность II: 312-314
— структура II: 308-310
Анафаза I: 14-18
Анизомицин I: 162
«Анонимный» ДНК-зонд II: 354
Антибиотики, устойчивость к ним I:
204-207, 230-231; II: 232
-----детерминируемая плазмидами
I: 230
Т-антиген II: 20, 48-50, 58
Антигены II: 283
Антикодоны I: 37, 138-139
- митохондрий II: 219
- модификация оснований I: 156
- мутации I: 157
Антионкогены II: 350
Антитела II: 283-286
- гены II: 14-15
- специфичность II: 284
- структура I: 65; II: 284-285
Антитерминаторы I: 127; II: 9
Антитерминация I: 181
Аполипопротеин В, ген II: 223
Аппарат Гольджи I: 164, 168-169
Арабинозный оперон I: 177
Аргинин I: 56
Аспарагин I: 56
Аспарагинсинтетаза I: 309
Аспартат I: 56
Аспартат-карбамоилтрансфераза
II: 307-309
АТРаза ДНК-зависимая I: 214
Аттенуация I: 178-181
Ауксин I: 275
Аутоиммунитет II: 283
Африканская зеленая мартышка, клет-
ки I: 260
Ацетилирование I: 355
Ацетил-СоА-карбоксилаза I: 163
Бактериальные колонии, скрининг I:
293
Бактерии I: II, 13
- введение в клетки новой информа-
ции I: 196-202
- идентификация геномов I: 363
- лизогенные I: 198
- содержащие ДНК 1: 40
- штаммы, устойчивые к лекарствен-
ным веществам I: 204-206; II:
307-308
- экспрессия генов I: 37
Бактериофаг вирулентный I: 198
- умеренный I: 198
- fd I: 71
- Ff I: 229
- G4 I: 71, 86
- M13 I: 46, 86, 89-90, 229, 243-246
- жизненный цикл I: 244
— клонирование в векторе, сконст-
руированном на его основе I: 326,
329, 331
- - репликация в Е. coli I: 242-244
- MS2 I: 185
— геном I: 148-149, 185
- Мн, геном и жизненный цикл II:
233
- - инверсия в геноме II: 274-275
- - G-сегмент II: 275
- ТЗ I: 123
- Т4 I: 28, 44. 119, 340
— ДНК-лигаза I: 80-81, 222, 287-288,
290
- - ДНК-полимераза I: 80
— интроны II: 19, 104
— тимидилатсинтетаза II: 21
- Т7 I: 80-81, 123
- - встраивание ДНК в ген псевдо-
рРНК II: 34
— ДНК-лигаза I: 80-81
- фХ174 I: 45-46, 71, 86, 89-90,
118
- XI: 28,40, 43,44,46,90-91, 181-183,
228, 238-242
— векторы I: 239-243
— геном I: 238-241
— геномные библиотеки I: 304
- - лизогенный путь I: 183-185
— литический путь I: 182-185
— нестабильность при клонировании
I: 338
— операторы I: 182-184
— промоторы 1: 182-184
— репликация ДНК I: 68
— репрессор cl I: 184
- - упаковка Х-векторных молекул I:
241-243
- - формы генома I: 182, 238-241
— электрофорез фрагментов ДНК I:
218
376
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Бактериофаги 1: 27. 40. 43. 45
- инфекция 1: 238
- рестрикция, контролируемая хозяи-
ном 1: 207
- трансфекция 1: 238
Белки 1 11. 33-34. 115-116. 131- 132
- активаторные 1: 37; 11: 21
- биологическая активность 1: 59, 62-
64
- время жизни II: 40
- гликозилирование 1: 169, 170
- глобулярные 1: 58-59
- денатурация 1: 62
- доставка к клеточным мембранам I:
164-170
информационные взаимоотношения
с ДНК и РНК 1: 31 32; 11: 132-
133
- конформация I: 59 66
- негистоновые 1: 49
- олигомерная структура 1: 62
- первичная структура 1: 59
- первичные продукты трансляции 1:
163-172
- размер и форма I: 58
- репрессорные 1: 37, 172-174; 11: 9, 21
- рибосомные 1: 186-188
— трансляция 1: 186-187
- связывающие ССААТ II: 60 61
- связывающиеся с олноцепочечными
участками ДНК 1: 82, 87, И 1
- секретируемые 1: 170
-синтез 1: 116-117, 142- 145, 152-155
- содержание модифицированных
аминокислот 1: 58
— Р-слоев 1: 59, 61-62
— а-спиралей 1: 59-64
- структура 1: 56-66; 11: 359-360
-- вторичная 1: 34, 59 66
— третичная 1: 34, 59-66
— четвертичная 1: 59-63
- теплового шока 11: 137, 205
- транспорт в клеточные органеллы 1:
170-172
- а-углеродный атом 1: 59-63
- элонгация и терминация полипеп-
тидной цепи 1: 145
Белковый блоттинг 1: 298
Белок-активатор катаболизма (БАК)
1; 175-176
- cl 1: 183, 228
- сП 1: 183-185
- сП! 1: 183-185
- САР 11; 125
- СР-1 II; 60-61
- Сго I: 182-185; 11: 125
— димеризация 11: 126
- eEF-1 1: 160
- eEF-2 1; 160
- eIF-2 1; 160
- eRF 1: 145
- Fos 11: 57
- HF I: 111
- HU 1: 52
- Lys 1: 185
- N 1: 182-184
- NF-xB 11: 64 65, 69, 80
- NF-XE2 II: 64
- NF-xE3 11: 64
- OTF-1 11; 57, 63, 89, 128-129
- OTF-2 11: 57, 63, 69, 128-129
Q 1; 183
- recA 1; 108, 110
- SL-1 11: 31-33
- Spl 11: 55-56, 60, 83, 89, 130. 135, 144,
346
- UBF-1 11: 31 33
- 11 1: 51-52
-pl: 126
N-бензоилдезоксиаденозин I: 281
N-бензоилдезоксицитидин 1: 281
Библиотеки кДНК 1: 304-3056, 11: 367
- рекомбинантных молекул 1: 302-
311, 338
Блоттинг 1: 278-279
A-боксы, их взаимозаменяемость II:
92-93
В-боксы 11: 92-93
С-боксы 11: 92
ССААТ-боксы 11; 59-61, 67
ТАТА-боксы II: 62, 71, 80-83, 89-90,
345, 356
Болезнь Тея-Сакса 11: 353
Валин 1: 56
Варианты антигенов (VAT) II: 296-297
- поверхностного гликопротеина (VSG)
II: 296-298
--гены II: 298-302
--мРНК 11: 298-299, 301-303
Векторы 1: 203, 205, 227, 231-238
- плазмидные 1: 228-229, 231. 233-
238. 249-250. 292
- челночные 1: 251, 263, 355
- экспрессирующие 1: 354-360
Вестерн-блоттинг 1: 278. 299
Вирион II: 342
Вирус болезни Марека II: 346
- везикулярного стоматита II: 343
- гепатита В человека 1; 97, 354; II:
342-345. 359
- герпеса, промотор гена тимидин-
киназы II: 59-60
- гриппа II: 343, 345
- иммунодефицита человека (ВИЧ) 1:
363; II: 65, 344. 349-351
- МС29 миелоцитоматоза птиц 11: 350
- мозаики цветной капусты 11: 97, 273
- осповакцины II: 347
- оспы крупного рогатого скота 1:
259-260
- папилломы человека I: 265; 11: 343
— Шоупа 1: 40, 43
простого герпеса 11: 59-61, 346
--типа I 11: 343
- саркомы мышей Харвея 11: 350
— Рауса 11: 346, 350
- Эпштейна-Барр I: 257, 259; II: 90,
94, 346
- РНК (EBERI и EBERII) 11: 90
- ящура 11: 344, 346
- SV40 1: 40, 259-265, 292; II: 345,
351, 364-365
— альтернативный сплайсинг 11: 124
— белки 1: 260-263; П; 48-49
— геном 1: 259-262, 332; II: 48-50.
138, 346. 348
-- ДНК 1: 40, 45, 68, 340; 11: 346-347
— секвенирование 1: 329-332
--транскрипция ранней области ге-
нома II: 48-59
-- жизненный цикл И: 346-347
-- конструирование экспрессирующих
векторов I: 360
— литический путь развития 11: 346
347
— мини-хромосомы 11: 138
— первичная структура ДНК I: 329
— поздние гены I: 261; II: 58-59
— промотор II: 20, 49-52, 55
-- ранние белки I: 260; 11: 48-50,
57-58
-- репликация I: 260-262; II: 48-50
-- сборка комплекса транскрипции 11:
54
— цис-действующие элементы 11: 49-51
- - энхансеры II: 51-54
Вирус-помощник I: 262, 292
Вирусы в качестве векторов I: 257,
259-260
— обратной генетике II: 21
- геномы 1: 45-46, 52, 68, 200-202; 11:
345
- животных I: 80; II: 7
- жизненный цикл 11: 342-351
- идентификация 1: 363
- оболочка 11: 342
- онкогенные II: 346
- растений I: 46, 273
- репликация РНК 1: 111
- РНК-содержащие 1: 38, 111-114
-ДНК 1: 38, 40, 44, 97; 11: 342,
345-346, 351
- трансдуцирующие 11: 346
- ( + )- и (-)-цепи РНК 1: 111-114
- человека лимфотропные II: 350
Витамин D 11: 73
Вителлогенин 11: 171
Внешние транскрибируемые спейсеры
П: 26, 163-164
Внутренние регуляторные участки 11:
90-92
- транскрибируемые спейсеры II: 26,
163-164
Внутрицепочечная рекомбинация 11:
229
Водородные связи в молекуле ДНК 1:
42-43
Временная экспрессия I: 257
Вставки ДНК 1: 203, 277-290
- источники для клонирования 1: 277
- клонированные 1: 317-319
- лигирование вектора I: 286-290
- определение положения данного
сегмента I; 227, 318-319
- - размера 1: 317-318
- синтетические I: 280-285
Второй посредник (сАМР) II: 350
Вырезание вирусного генома 1: 198-
200
Вырожденность генетического кода 1:
116
Р-Галактозидаза 1: 58, 174-176. 229,
241, 356
Р-Галактозидпермеаза 1: 175-176,
229
Р-Галактозидтрансацетилаза I: 175—
176
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
377
Галактозный оперон 1: 174, 177
Г алактозо-1 -фосфат-уридилтрансфе-
раза 11: 69
UDP-галактозоэпимераза I: 173
Галактокиназа II: 69
Гаметы I: 17-18; П: 148-149
Гексокиназа I: 59
Геликазы I: 79, 87-88, 101, 109, 121
Гель-электрофорез 1: 317, 320-321
Гемоглобин 1: 58-60, 62, 64, 163; II:
177
- гены II: 175
— экспрессия II: 175
Ген гастрина человека II: 43
- индуцибельный I: 175
- инсулина II; 162, 187, 356, 364
- кальцитонина II: 121, 123
- легкой цепи миозина (MLC) II: 121
- металлотионеина 11: 268, 309
- ответственный за красно-зеленую
слепоту II: 341
- плацентарного лактогена II: 159-162
- пролактина II: 159-160
- резистентности к гигромицину I: 259
- CAD II: 309-313
- c-fos II: 149-150
- c-nivc II: 95
- env I: 267-268; II: 260, 349
- gag I: 267-268; II. 260-261, 349
- HIS3 I: 252
- int II: 260
- lac\ I: 229
- lacl I: 236
- pol 1: 267-268; 11: 260, 349
- pro 11: 260
- rrpE 1: 237
Генетическая инженерия II: 361-362
- информация, перенос в клетке 1:
31-32
Генетические системы, происхожде-
ние 11: 18-19
Генетический дрейф II: 195, 229
- код I: 33-35. 115-116, 130-137
-- вырожденность 1: 116
— избыточность I: 136
— расшифровка 1: 116, 132-137
— универсальность I: 137
Генная конверсия I: 104, 108, 254;
11: 162, 207-209
- экспрессия 1: 257
— в клетках I: 299-302
----хлоропластах II: 225-226
— дифференциальная II: 61-62
— и метилирование II: 140- 148
— модуляция II: 272-302
- - перестройки и модуляция II: 272-
302
----последовательности ДНК II: 358
— повторяющиеся последовательности
как регуляторы II: 204-206
— регуляция I: 172-176; II: 9-10,
61-69
----на уровне транскрипции I: 186
Геном(ы) I: 27
- архитектурные элементы II:
156-162
- картирование II: 351-353
- локализация клонированных сег-
ментов 1: 333-339
- митохондрий 11; 217-222
25-158
- нуклеотидные последователь-
ности II: 5
- программируемые перестройки II:
14
- случайные перестройки II: 14
- физические и генетические карты II:
351
- эукариотические I: 339; II: 5, 10-13,
227
Геномные библиотеки 1: 304-305
Геномный импринтинг II: 145-148
Генотерапия II: 362, 368
Генотип I: 27
Гены I: 11, 23, 33
- глобиновые II: 11, 202-203, 354
- - анализ с помощью эндонуклеаз
рестрикции II: 143
- гормона роста П: 159-162
- «домашнего хозяйства» II: 136, 140,
144, 354
- и белки 1: 26-27
— ДНК I: 27
- иммуноглобулиновые, промоторы
II: 89
— транс-действующие факторы II:
63-65
- - энхансеры 11: 138
- класса 1: П: 23, 25-39
---аппарат транскрипции II: 27-28
---единицы транскрипции II: 25-27
---регуляция транскрипции рДНК
И: 28 35
---сопряжение терминации и транс-
крипции II: 38
---терминация транскрипции рДНК
II: 35-37
— II II: 23, 39-90
---аппарат транскрипции 11: 40
---промоторы 11: 48-49
---РНК II: 39
---созревание мРНК II: 41-48
---U-PHK II: 41-48
---уровни регуляции экспресии II:
39
---факторы транскрипции II: 41
---энхансеры II: 52-54
— Ill II: 23
---инициация транскрипции II: 91-
95
---РНК, кодируемые ими 11: 90
---сигналы транскрипции II: 90
---структура II: 90-104
---экспрессия II: 90 104
— pair rule II: 85-87
- клонирование I: 306-315
- коллагеновые II: 186-187
- маркерные I: 228. 233-234
- металлотионеина II: 82-83, 138
- митохондриальные I: 137
- молчащие II: 136
- определение II: 9
- ортологичные II: 160
- паралогичные II: 160
- полярности сегмента II: 85-87
- прерывистые II: 6-7, 104
- «прыгающие» 11: 14
- регуляторные последовательности
II: 25
- рРНК II: 162-165, 342
- тРНК 1: 128- 130; II: 92-93. 105-107,
167-168
- синтенные II: 13
- состоящие из тандемных повторов
II: 183-186
- структура II: 10
- химерные II: 20
- хлоропластные II: 224-225
- эволюционная история II: 17-18
- экспрессия I: 38
— координированная II: 355
- эукариотические 11: 5-7, 22-25, 69-
84
- - сигналы транскрипции II: 22-23
- gap II: 85-87
Гептапептид I: 57
Герпесвирусы II: 344-346, 351
Гетеродуплексы 1: 54, 56, 107-108
Гетерозиготные организмы 1: 25
Гетерохроматин, цитогенетический
анализ II: 194
Гибридизационная селекция 1: 297
Гибридизация I: 335-336; II: 163, 353
- в условиях насыщения 1: 341-342
- олигомерного зонда с геномной
ДНК II: 354
Гибридные спирали ДНК-РНК 1: 54,
56
Гибридный дисгенез II: 238-242, 265
- полипептид I: 356-357
Гигромицин I: 259
М6-гидроксидезоксицитидин-5'-трифос-
фат 1: 346-347
Гидроксилапатит I: 341-342; 11: 198—
199
Гидроксилизин I: 57-58
З-гидрокси-З-метилглутарил-СоА-ре-
дуктаза II: 184
5-гидроксиметилурацил I: 39-41
5-гидроксиметилцитозин 1: 39, 41
Гидроксипролин I: 57-58
№-гидроксицитидин 1: 347
Гипервариабельные участки II: 285,
289
Гиперметилирование II: 140
Гиперхолестеролемия семейная II:
184, 206
Гиперчувствительность к действию
нуклеаз II: 137-138
Гипоксантин I: 259; II: 308-309
Гипоксантин-ТЧ-гликозилаза I: 100
Гипометилирование II: 140
Гираза 1: 88, ПО
Гистидин I: 56
Гистидиновый оперон I: 180
Гистоновые гены II: 181-183
— клонирование II: 182-183
- морского ежа II: 182
- - свойства II: 181
- - сцепление II: 181-182
- - тандемные повторы II: 183
Гистоны I: 49-52
Главный комплекс гистосовмести-
мости (МНС) II: 294-296
N-гликозилазы I: 100- 102
Гликозилгидролазы I: 170
Гликозилирование I: 169, 170, 355
Гликопротеины I: 57
Глицин I: 56
Глобиновые гены II: 11, 17, 43, 162,
175-181, 355
378
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
— интроны II: 162, 178-179
— организация II: 175-178
-- предковый ген II: 178
— суперсемейство 11: 175-179
— экспрессия II: 175-177
Глобины II: 175-178, 267-269
Глутамат 1: 56
Глутамин I: 56
Глюкозил-НМС I: 41
Глюкокортикоид-акцепторные элемен-
ты II: 74, 83-84, 138
Гомеобоксы II: 126, 368
Гомеодомены 11: 87, 126-129
Гомеозисные мутации II: 366-367
Гомогенно окрашивающиеся области
II: 309-310, 314
Гомозиготные организмы I: 25
Гомологичная рекомбинация 1: 339
Гомология последовательностей ДНК
Г. 330, 332
Гормон роста 1: 354; II: 359
Гормон-акцепторные элементы II: 74,
78-80
Грамотрицательные организмы I: 249
Грамположительные организмы I:
249
Гуанилилтрансфераза II: 41-42
Гуаниловая кислота I: 259-260
Гуанозил-7-метилтрансфераза II: 41
Гуанозинтрифосфат I: 118-119, 150-
151, 160
Гуанозин-5'-фосфат I: 53
Гуморальный иммунитет II; 283-284
Двойные мини-хромосомы II: 309-311,
314
Дезоксиаденозинмонофосфат I: 39
Дезоксиаденозии-5'-фосфат I: 39
Дезоксирибозные остатки 1: 39
Дезоксирибонуклеотиды I: 31
Дезокситимидинмонофосфат I: 39,
258
Дезоксицитидинмонофосфат I: 39
Делеции I: 23-24; II: 229-230
Делеционные мутанты I: 343-345
Дефосфорилирование 1: 288-289
Диаминопимелиновая кислота I: 229
Дивергентная эволюция II: 19
5,6-дигидроуридин 1: 54
Дигидрофолатредуктаза II: 307-309
Дидезоксинуклеотиды, применение
для секвенирования ДНК I: 325-
328
Дидезоксирибоиуклеотиды, химиче-
ский синтез 1: 283
Диметокситритильная группа I: 280-
281
Дипептид 1: 57
Диспергированные повторы, едино-
образие II: 206-209
— механизмы гомогенизации II: 207
— мобильные элементы II: 229
Дистальный элемент II: 88-89
Дисульфидные мостики 1: 57
Дифтерийный токсин 1: 162
Дифференцировка клеток II: 366
Дицентрические хромосомы II: 310
Длинные диспергированные повторы
II: 201. 203-206
Длинный концевой повтор (LTR) I:
95-97; II; 228, 259
--в дрожжевом геноме 11: 168
----ретровирусном геноме I: 267-
268
ДНК I: 11, 20, 27-30, 38-52; П: 18
- амплификация II: 14
- в комплексе с белками I: 45, 49
- введение метки в З'-концы I: 323-
325
----5'-концы I: 322-324
- гетеродуплекс II: 16
- гомодуплекс II: 16
- гомологичные последовательности
I: 330, 332; II: 158-159
- двойная спираль I: 30-32, 41-
47, 82-83; II: 10
- двухцепочечная I: 45-47
- дезаминирование I: 97-98
- деметилирование II: 141-142
- денатурация 1: 47-48, 54
- депуринизация I: 97-98
- длина молекулы I: 44-45
- значащая цепь II; 10.24
- и белки, информационная связь меж-
ду ними I: 115
- - гены I: 27-30
- изопикническое центрифугирование
II: 163, 166
- инициация образования новых це-
пей I: 86-89
- искривление спирали I: 45
- кольцевая форма I: 45-46, 84-86, 92;
II: 10, 233
- компоненты молекулы I: 39-41
- криптические сателлитные II: 189
- лидирующая цепь 1: 73, 75
- линейная I: 45-46, 92-93
- липкие концы I: 224
- макси-кольца II: 222-223
- матричная функция I: 67-68, 73-74,
115, 118-120
- - цепь II: 10
- метилирование I: 218-220, 328; II:
140-148
- мечение цепей а-32Р-дезоксинук-
леозидтрифосфатом I: 326
- мини-кольца II: 222-223
- митохондриальная П: 217-223
- молекулярная масса I: 44-45
- набор фрагментов (фингерпринт) 1:
215, 218
- незначащая цепь II: 10, 24
- нуклеотидные последовательности
II: 157-159, 209-215, 225
--гомология II; 158-159
--отжиг II: 158
--полиморфные II: 158
- нуклеотидный состав I: 42
- одноцепочечная I: 46-47, 86, 326
- отстающая цепь I; 73-75, 77-79, 88
- перестройки I: 38, 67; II: 6, 13-15, 20,
226, 358
- пиримидиновые димеры I: 99-100
- повреждения под действием иони-
зирующей радиации I: 97
— при повышении температуры 1: 97
- повторяемость последовательнос-
тей II: 157-159
- последовательность-основатель II:
159, 207
- разделение цепей I; 326
- размер молекул 1: 44
- разнообразие форм 1: 44-47
- раскручивание спирали I: 82-86, 124
- реассоциация (ренатурация) I: 47-
48; II: 13
- рекомбинантные I: 195-210. 290-
292, 354
- ренатурация I: 47
- репарация повреждений I: 33, 38-39,
67, 77-78, 97-103, 108
- репликация I: 31-32, 38-39, 67-93,
118-121
- - образование комплекса белок-
dCMP I: 87-88
— по типу катящегося кольца I: 89-
90
- ретровирусная, интегрированная в
клеточный геном Г. 97
- рибосомная, организация II: 164
- РНК и белки, информационные взаи-
моотношения I: 31-32
-сателлитная II: 158, 189-193, 209
— видоспецифичная П: 191
-- исследование с помощью рестрик-
тирующих эндонуклеаз II: 189-191
— быка II: 192-193
— мыши II: 192
— эволюция II: 195-197
- сверхспиральная 1: 45-46, 49, 82,
343-344
- сверхспиральность и регуляция транс-
крипции II: 139
- связь с низкомолекулярными анио-
нами I: 49
- сегменты, картирование I: 218-219
- секвенирование I: 320- 328, 331 - 332
- синтез I: 280
- структура I: 27-31, 39-47
- схлопнувшаяся II: 199-200
- сшивание цепей I: 77
- типы повреждений I: 98
- топология и конформация II: 135—
136, 139
- упаковка II: 136-139
— в хромосомах I: 49-52
- УФ-индуцированные повреждения
и их репарация I: 102, 108
- ферментативная амплификация сег-
ментов I: 360-363
- ферментативное секвенирование I:
320, 324-328
- ферментативный гидролиз I: 214
- A-форма I: 42-44, 54
- В-форма I: 42, 44; II: 139
- Z-форма I: 44; II: 139, 187
- фрагменты с тупыми двухцепочеч-
ными концами I: 217
- функции I: 32
- химическое секвенирование I: 320-
325, 327
- хлоропластов II: 216-226
- хроматиновая, расплетание I: 82
- центромерная сателлитная II: 188-
194, 210
- частота встречаемости линуклеоти-
дов I: 39-40
- человека I: 40; II: 199
- число витков спирали I: 45
- шпилькообразные структуры I; 93;
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
379
II: 157
- эгоистичная II: 206
- ядерная 1: 49
кДНК, клонирование 1: 306-315; II:
359
— полноразмерных молекул I: 285-
287
- создание библиотеки 1: 304- 306; II:
367
рДНК II: 35-37, 215-216
- транскрипция II: 28-33
- человека, единообразие повторов II:
164
- 5S II: 100, 166-167
Т-ДНК I: 274-276
ДНКаза 1 : 343-344; 11: 138
ДНК-алкилтрансфераза I: 99
ДНК-блоттинг II: 353
ДНК-гираза II: 158
ДНК-зависимая РНК-полимераза I:
115, 121-123
ДНК-копия 1: 223-224, 286-287, 294
ДНК-лигаза 1: 58, 80-82, 88, 91-92,
101, ПО, 203-205, 222, 225, 287-
290, 354
- млекопитающих I: 81
ДНК-метилтрансфераза II: 142
ДНК-полимераза 1: 58-59, 361
- в вирусной репликации II: 344
- - полимеразной цепной реакции 1:
360
— эукариотических клетках I: 80
- синтез митохондриальных ДНК 11:
222
- I I: 64, 78-80, 222-225, 290, 345-347,
354
- - в химическом синтезе полидезок-
синуклеотидов 1: 282
- - превращение ДНК-фрагментов с
липкими концами в ДНК-фраг-
менты с тупыми концами 1: 223
— удлинение З'-концов I: 323-325
- II I: 78
- Ill 1: 78, 88-89, 91
ДНК-полимеразы I: 32, 77-82, 92, 94,
108, 121
- млекопитающих I: 80
ДНК-праймаза 1: 175
ДНК-сегменты, амплификация
внутри отдельных клеток I: 363
- клонированные 1: 333-334, 343-349
- определение первичной структуры I:
319
- транскрипция и трансляция в эука-
риотических системах 1: 353-354
ДНК-топоизомераза типа I 1: 82-85,
ПО
— II 1: 84-85, 92
Дот-блоттинг I: 350
Дрожжевые колонии, скрининг 1: 293
Дрожжи I: 252-253
- активатор транскрипции II: 134
- белок GAL4 II: 71-72
- ген ADH4 11: 309
— GCN4 II: 57, 205
- генная конверсия II: 207
- гены 11: 350
— GALU: 69-73, 80
- гетероталличные штаммы II: 275-
276
- гистоновые гены II: 183
- гомоталличные штаммы II: 275-276
- рДНК, кроссинговер II: 164-165
— повторяющаяся единица II: 167
- интроны II: 23, 104
- искусственные хромосомы 11: 215-
216
- локус, определяющий тип спарива-
ния (MAT) II: 275-279
— HMLa II: 276-280
— HMRa II: 276-280
— НО II: 277, 281
- микротрубочки II: 209-210
- митохондриальная ДНК II: 220-222
- молчащие а- и a-гены в локусах
HMLm HMR 11: 282-283
- CEN-области II: 210-211
- обратная генетика II: 21
- переключение с одного типа спари-
вания на другой II: 277-280
- 2 мкм-плазмидная ДНК I: 252-253,
359
- псевдо-рРНК II: 34-35
- размер генома II: 5
- сайт-специфическая трансформация
I: 254-256
-система хозяин-вектор I: 252-255
- спаривание между гаплоидными клет-
ками с разным типом спаривания
II: 275-276
- сплайсинг тРНК II: 119- 120
- стабильная трансформация I: 254-
255
- теломеры II: 214
- терминация транскрипции и поли-
аденилирование II: 46
- типы скрещивания II: 126, 275-283,
342
- транскрипция рДНК II: 34- 35
- тубулиновые гены II: 174
- фактор транскрипции II: 9
- центромеры II: 210-212
- экспрессирующие векторы I: 359-
360
- экспрессия генов типа спаривания II:
280-282
- Ту-элементы II: 249-252
Единица транскрипции II: 9-10, 21-27
Железо II: 150-153
Животные клетки, введение в них
ДНК I: 257-258
— системы «хозяин-вектор» 1: 257
— трансформация 1: 256-257
--нестабильная и стабильная I:
256-257
— трансформированные, фенотипи-
ческий отбор I: 258-259
— экспрессирующие векторы I: 360
Зеин II: 171
Зигота I: 18
Зонды при гибридизации 1: 278, 280
Зрительные пигменты II: 340-341
N-изобутирилдезоксигуанозин 1: 281
Изозимы II: 170
Изолейцин I: 56
Изопропил-Р-В-тиогалактозид I: 356
Изотипы 11: 173-174
Изоформы белка II: 120
Изошизомеры II: 216
Изохоры II: 156
Иммунитет гуморальный 11: 283-284
- клеточный II: 283-284
Иммунная система II: 283, 342
Иммунные белки II: 283-296, 358
Иммуноглобулин, гены II: 14-15
- D(lgD) II: 293
- M(IgM) II: 292-293
Иммуноглобулиновые гены, транс-
крипция II: 61-63
— i/uc-элементы II: 61-63
— экспрессия II: 61-63, 187, 355
Иммуноглобулины I: 62, 64; II: 121,
138. 284-289, 358
- гены тяжелых цепей II: 288-289
- изоформы II: 121
- легкие цепи 1: 65; II: 284-285
- мембраносвязанные II: 295
- регуляция синтеза мРНК II: 43, 45
- типы II: 284-285
- тяжелые цепи 1: 65; II: 284-285, 291 —
293
Иммунологический скрининг 1: 300,
354
Импринтинг аутосом II: 145- 148
Импульсный электрофорез I: 303-304,
317
Инактивация Х-хромосомы II: 145-147
Инверсии флип-флоп II: 272-276
Индуцибельный ген I: 175
Инициаторный кодон I: 116, 146-148,
159-160
Инициация репликации I: 68-72, 86
Инозин I: 54
Инсерционная трансформация I: 257,
275
Инсерционные последовательности 1:
231; II: 230-232
Инсерционный мутагенез I: 344; II:
368
Инсулин I: 58, 163-164, 354; II: 359
- ген II: 162, 187, 356, 364
- модификации II: 360-361
- синтез в клетках £. coli П: 359-360
Интеграза II: 252
Интерфаза I: 14-16, 21
Интерферон I: 162, 354
- гены II: 180-181
- мРНК, последовательность AUUUA
II: 149-150
Р II: 80-81
Интроны I: 319; 11: 7-8, 10. 17-18.
23-24, 104-125, 346
- автокаталитический сплайсинг II:
108-113
- альтернативное спаривание после
вырезания II: 109, 111
- в генах, кодирующих белки II: 170
--тРНК II: 105-107, 167-168
— дрожжевой митохондриальной
ДНК П: 221-222
- - прокариотическом геноме II: 19,
21, 23
— растительных генах II: 23
- - хлоропластных генах II: 225
- виды II: 105-108
- внутренняя направляющая последо-
25*
380
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
вательность II: 109, 111
- генов хлоропластной мРНК II: 105
— ядерной про-мРНК II: 105- 106
- группы I II: 105-112, 162
— И II: 105, 108, 112-113, 162
- модели встраивания II: 162
- сплайсинг тРНК II: 119
- частота встречаемости II: 104
- эволюция II: 162
- эукариот II: 205
- ядерной про-мРНК II: 113, 116-117
Инфекционный мононуклеоз II: 346
Инфекция I: 290-292
Инфузории, изменения в словаре ко-
донов 1: 137
Искусственные хромосомы дрожжей
(YAC) 11: 351
Казугамицин I: 162
Кальцитонин II: 121, 123
Канамицин 1: 162, 230, 249
Канонические последовательности II:
47, 70, 159
Капсид II: 342-344
Карбоангидраза II: 15
у-Карбоксиглутамат I: 57-58
Кариотип I: 21, 22
Картирование геномов II: 351-353
- сегментов ДНК I: 218-219
- с помощью эндонукдеазы SI (S1-
картирование) I: 352
- хромосом с использованием гиб-
ридных соматических клеток 1:
352
Кассетный мутагенез I: 347, 349
Катаболитная репрессия II: 73
Катенаны II: 222-223
Кератин I: 59
Киназы 1: 221
Кинетика реассоциации, анализ II: 198
— ДНК, исследование S1-нуклеазным
методом I: 341-342
Кинетопласты II: 222-223
Кинетохор II: 209
Кирромицин I: 162
Кислая фосфатаза, семейство генов II:
171
Кислота хоризмовая I: 178
Клетки I: 14, 16, 18
- африканской зеленой мартышки I:
260
- гаплоидные I: 14
- детерминированные II: 366
- диплоидные I: 14
- животные I: 12-13
- зародышевой линии II: 229, 240, 358,
362-365
- опухолей II: 314
- памяти II: 291, 293
- плазматические 11: 291, 293
- растительные I: 12, 273
- соматические 1: 18; II: 148-149, 358,
362
— гибридные I: 336-338
- стволовые I: 172; II: 289, 291
- ТК грызунов I: 258-261
- тотипотентные II: 365
- фагоцитарные II: 284
- COS I: 262-264
Клеточная теория I: 14
Клеточное деление 1: 16
Клеточные линии I: 257
Клеточный иммунитет II: 283-284
- цикл 1: 16, 18, 21
Т-клеточный рецептор (TCR) I: 310; II:
294-296
Клон, идентификация I: 292
Клональный отбор II: 293-294
Клонирование I: 202-204, 290-292
- вирусов I; 202-203
- клеток 1: 202-203
- условия I: 227
Клонированные ДНК-сегменты, по-
лучение мутантов I: 343-349
Коактиваторы 1: 173
Кодирующие области II: 156
Кодон-антикодоновые взаимодействия
I: 153-157
Кодон AUG I: 116-117, 136, 143, Кб-
147, 150, 159-160
Кодоны I: 116, 131-132
- инициирующие I: 137; II: 219
- митохондриальные II: 219
- мутации I: 157
- терминирующие 1: 117, 134, 137, 151
Колифаг QP I: 114
Колицины I: 230
Коллаген 1: 59-61, 163; II: 171
- а2 (I) II: 187
Коллагеновые гены II: 186-187
Комбинирование независимое Г. 25
Комплекс преинициации II: 99- 100
Комплемент 11: 284
Комплементарность антигена и анти-
тела II: 285-286
Конкатемеры I: 239-240
Контролирующие элементы автоном-
ные II: 242, 244-245
— дрозофилы II: 244-245
- - кукурузы II: 228, 242-249
---неавтономные II: 242, 244
— свойства II: 242-244
- - семейства II: 242
— Ac II: 244-249
— Ac-Ds II: 244-246
Конъюгация I: 196-198
- перенос F-фактора между бакте-
риями I: 206-207
Корепрессоры I: 173
Корова, размер генома II: 5
- сателлитная ДНК II: 192
Корончатые галлы I: 273, 275
Короткие диспергированные повторы
II: 201-206, 228, 270-271
Кор-промотор II: 29-31, 33
Космиды I: 246-247, 304, 333-334; II:
351
Котрансдукция I: 198-199
Котранскрипты рРНК-тРНК I: 128
Котрансляционный транспорт 1: 165—
166, 172
Котрансформация I: 258
Кроссинговер 1: 104; II: 13
- неравный I: 104; II: 158, 162, 195,
229
Крыса, а-амилазные гены II: 180
Ксантин-гуанин - фосфорибозил-
трансфераза 1: 259
Кукуруза, генетическая система II: 242
- контролирующие элементы II: 242-
249
- локус Adhl II: 245, 247-248
— СП: 243
- мутации II: 242-245
— в клетках зародышевой линии II:
242
— локус shrunken II: 243, 248-249
--ivaxr II: 243, 246-247
- размер генома II: 5
- семейство контролирующих элемен-
тов Ac-Ds II: 244-248
- элемент Ac II: 228
Культура клеток I: 257; II: 342
Курица, размер генома II; 5
Кэпирование мРНК эукариот II: 39,
41-42
Кэп-структуры 11: 20
P-Лактамаза I: 206, 354
Лактозный оперон, индукция I: 175-
176
— оператор 1: 175-176
Лассо-подобные структуры II:
108-109, 112
Левый регуляторный элемент (UCE)
II: 30-31
Леггемоглобин II: 175-179
Лейцин 1: 56
«Лейциновые молнии» II: 131-133
Лигирование вектора со вставкой I:
286-290
5'-лидерная последовательность II: 10,
21-22, 24
Лидерная ггр-мРНК I: 179-181
Лидерный пептид 1: 179
Лизин I: 56
Лизогения 1: 198, 200
Лизосомы I: 164
Лизоцим II: 286
Лимфокины II: 291, 359
Лимфома Беркитта I: 24; II: 346
Лимфотропные вирусы человека П:
350
Лимфоциты I: 13
В-лимфоциты II: 283-284, 289-292
Т-лимфоциты, антигенная специфич-
ность II: 283-284, 294-295
- образование из стволовых клеток II:
291
- созревание II: 294-296
- функция II: 294-296
- цитотоксичные (киллеры) II: 283-284
Линии клеток II: 367
Линкеры I: 280, 290-291; II: 29
Линкомицин I: 162
Липкие концы I: 215, 290
— присоединение 1: 289-290
— синтез I: 224-226
Липопротеин низкой плотности (ЛПНП)
II: 184-186, 206, 342
Лук, размер генома II: 5
Макаки-резусы I: 260
Макрофаги I: 13; 11: 284, 291
Макси-кольца ДНК II: 222-223
Малый рибонуклеопротеид ядрышка
II: 27, 39, 46-47, 87, 108, 113-116
Малярия 1: 354; II: 359
Маркер G-418 1: 259
Маркерные гены I: 228, 233-234
Матрица I: 31
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
381
Матуразы I: 185; II: 112, 221
Межгенные спейсеры 11: 26, 38-39,
163- 164
--дивергенция II: 164-166
Мейоз 1: 16-18, 103
Мелкие Hpal 1-фрагменты (HTF) II: 144
Металлозависимые элементы, кластер
II: 83
Металлотионеин II: 268, 309
Металлы, влияние на регуляцию
транскрипции II: 82-84
Метамерные организмы II: 84
Метафаза 1: 14-17
М6-метиладениловая кислота I: 54
№-метиладенин 1: 39-41
Метилазы 1: 214, 218-220
М7-метилгуаниловая кислота 1: 54
ЬР-метилгуанозин 1: 54
Метиленомицин А 1: 250-252
Метилирование ДНК I: 40-41, 218
Метилтрансфсразы II: 42
5-метилуридинриботимидин 1: 54
5-метилцитидин 1: 54
5-метилцитозин 1: 39-41
Метионил-тРНК- трансформилаза 1:
144
Метионин 1: 56
Метод отпечатков 1: 236, 238
- «прогулка по хромосоме» 1:334-335;
11: 179
Метотрексат II: 307-309
Миграция ветви 1: 106
Миелин II: 121
Микофеноловая кислота 1: 259, 271
Микротрубочки 11: 173, 209- 211
Миксомицеты, размер генома 11: 5
Мини-кольца ДНК II: 222-223
Мини-сателлитные последовательности
11: 188
Мини-экзоны II: 118
Миоглобин 1: 59; II: 175, 177
Миозин 1: 59
- тяжелая цепь II: 171
Миссенс-мутации 1: 157-158
Митоз 1: 14 16
Митохондрии 1: 164, 170-171; II: 216-
226
- генная экспрессия II: 225-226
ДНК 11: 217-223
- полиморфизм эндонуклеазных сай-
тов 11: 217-218
Млекопитающие, ДНК митохондриаль-
ная II: 217-220
- распределение повторов в ДНК II:
200-204
семейство LINE-1 II: 266-267
- транскрипция рДНК 11: 29 32
Мобильные элементы II: 14, 158, 222,
227-249
- вырезание 11: 236, 255
-- простое встраивание 11: 233-234
Модель «кожицы лука» II: 304-306.
311-312
Со/-модуль I: 230
Мозаичные единицы транскрипции II:
23
Молекулярная генетика 1: 30-37; 11:
16, 365, 369
- эволюция II: 161 162
Молекулярное клонирование 1: 203-205,
210; II: 20, 346
Молекулярные часы II: 161-162
Молчащие гены 11: 136
Монослой клеток I: 257
Моноцистронная транскрипция II: 21,
23-24
Морской еж II: 5, 173, 182, 200
Морфогенез, транскрипционная регу-
ляция II: 84-87
Мультигенные семейства II: 171-181,
355
Мутагенез 1: 343
- кассетный 1: 347, 349
- химический I: 344-345
Мутагенное копирование I: 345
Мутанты I: 343-349
инсерционные 1: 344; II: 368
Мутации 1: 33, 363; 11: 17
- затрагивающие различные компо-
ненты трансляционного аппарата
Г. 157
- и встраивание мобильных элемен-
тов II: 236-238
— функционирование белков Г. 63-66
- индукция бисульфитом натрия I:
345-346
- плейотропные 11: 221
- сайт-специфические I: 343, 346-348;
II: 360
- сдвиг рамки считывания 1: 131
- супрессорные 1: 157
точечные 1: 344-349
Мышечная дистрофия Дюшенна II.
354
Мыши трансгенные 11: 368-369
Мышиный ретровирус типа С II: 343
Мышь, генетические эксперименты II:
367-368
ДНК митохондриальная II: 222
- сателлитная 11: 192
- IAP-последоватсльности II: 259
- размер генома II: 5. 291-292
РНК в лимфоцитах II: 204
- семейство а-амилазных генов II:
179-180
транскрипция рДНК II: 36
тубулиновые гены II: 174
Надсемейства генов II: 170
нуклеотидных последовательностей
11: 158
Некодирующие области II: 156
Нематода II: 5, 358
Неоднозначного соответствия правило
1: 156
Неомицин I: 162, 230
Неспсцифические эндонуклеазы 1: 343
И и котинамидадсниндинуклеотид (NAD)
1: 81
Ник-трансляция 1: 78-79, 88, 222
Нитрозомочевина 1: 97, 99
Нитроцеллюлозный фильтр 1: 293 294
Нозерн-блоттинг 1: 278
Нопалин I: 274
Нуклеаза Bal31 1: 212-213
из Neurospora crassa 1: 212
- Phaseolus aureus 1: 212
- recBCD I: 109
- SI 1: 212
Нуклеазы 1: 211-213, 224; 11: 211-212
- гиперчувствительность к ним II:
137-138
- чувствительность к ним П: 137
Нуклеиновые кислоты I: 27- 33, 38- 56
- - гибридизация 1: 56
- - гомологичные последовательности
I: 54, 56
Нуклеосома I: 49-51; II: 136
Нуклеотид Г. 39
Нуклеотидные последовательности II:
5
- триплеты, соответствие с аминокис-
лотами 1: 116
Обратная генетика I: 343; II: 6, 20-21,
355
- транскриптаза 1: 94, 214, 223-224,
267: II: 227, 249, 261, 344
- - введение метки в З'-концы I: 324-
325
- - применение в химическом синтезе
полидезоксинуклеотидов 1: 282
- транскрипция 1: 31-32, 38, 351, 353;
11: 227
-- в ДНК-содержащих вирусах 1: 97
- эукариотических клетках 1: 93-97
Обратные повторы 1: 330-331; II: 157
Овальбумин I: 318-320
- гены 1: 333 334; 11: 137, 171
Одноцепочечные «хвосты» 1: 286, 289
G-окрашивание I: 336
Октамер ATGCAAAT II: 89
Октопин I: 274
Олигодезоксинуклеотиды синтетиче-
ские 1: 347-348
Онкогенез II: 364, 370
Онкогенные вирусы П: 346
Онкогены 11: 346, 350, 364, 370
c-jun 11: 57
- c-mos 11: 259
- с-опс II: 346, 349-350, 370
- v-fos 11: 149
- v-jim II: 56
- v-onc 11: 346, 349, 370
Операторы I: 174
Оперон галактозный 1: 173- 174, 177
-триптофановый I: 177-181. 201
Lnc-оперон I: 174-177, 356
- репрессия 1: 175-176
- экспрессия 1: 174-177
Опероны 1: 173-174
Опины 1: 274
Оплодотворение 1: 18
Опсины 1: 312; II: 341
Опухоль Вильмса I: 24; II: 350
Организмы гетерозиготные 1: 25
гомозиготные 1: 25
грамотрицатсльныс 1: 249
грамположительные I: 249
Ортологичные гены II: 160
Основания, комплементарное спари-
вание 1: 42, 75, 97, 115 116, 118,
120, 132, 150
- метилирование 1: 40 41
- образование пар 1: 32. 41 42, 55, 67
ошибочное спаривание 1: 74. 77,
107- 108
- спаривание в гетеродуплсксах 1: 54
- по правилам Уотсона - Крика 1: 54
- химическая модификация 1: 321-323
382
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
CG-островки II: 144-146
- гены, ассоциированные с ними II:
145
- размер II: 144- 146
-функция II: 144-145
Отжиг с комплементарным полинук-
леотидом I: 292-299
Отпечатков метод I: 236, 238
Отторжение тканей II: 296
Пактамицин I: 162
Палиндром I: 215; II: 157-158
Панкреатическая амилаза П: 356
- ДНКаза 1 I: 212
- РНКаза 1: 211-212
Папилломавирусы I: 257, 265-266,
360; II: 351
- в качестве векторов I: 259, 266, 292
Паповавирусы I: 80, 105; II: 344-346
Паралогичные гены II: 160
Парвовирусы I: 46; II: 345
Пептидилтрансфераза I: 150-151
Пептидилтрансферазная реакция I:
146, 150
Пептидные связи I: 57
Пептидный посредник II: 357
Первичная структура белков I; 33
— ДНК I: 328, 330
Первичные транскрипты I: 118, 127
Переключение изотипа II: 293
Периплазматическое пространство I:
172
Пермеаза II: 69-70
Пероксисомы I: 164
D-петля II: 222
«Петля-на-стебле» II: 157
Пикорнавирусы II: 43, 346
F-пили I: 243-244
Пиримидиновые нуклеотиды I: 39-42,
52-54
Пируватдегидрогеназный комплекс I:
58
Пируваткарбоксилаза 1: 163
Плазматические клетки II: 291, 293
Плазмидиые векторы I: 228-229, 231.
233-238, 249-250, 292
Плазмиды I: 204-207, 246-248; II:
230-231
- конъюгативные I: 230
- мобилизуемые I: 230
- модульная организация 1: 229-232
- неконъюгативные I: 230
- репликационные модули I: 230
- с двумя селективными маркерами I:
233-234, 236
— одним селективным маркером I:
232
- содержание в клетках, резистентных
к антибиотикам I: 204-206, 230
- Ар 201 I: 230
- ColEl I: 87, 230, 234
- ColV I: 230
- DF41 I: 235, 237
- Flac I: 229
-GEM 1: 294-295
- MB9 I: 235, 237
- pBR322 1: 235-237, 266, 268-269, 272
- r I: 231
- R I: 230. 249
- R100 1: 230
- RK2 I: 236-237
- RK2501 I: 235-237
- RTF I: 231
- SC101 1: 235
- SV2 1: 265
- SV3 I: 265
- Ti I: 273-276; II: 365
- UC7 I: 236
Плацентарный лактоген II: 159-162
Плейотропные мутации II: 221
Повторы, богатые GC II: 50-51, 59,
89, 156, 345
- диспергированные, пластичность II:
198
- обратные 1: 330-331; II: 157
- прямые 1: 330-331; II: 157
- семейство LINE II: 266- 267
-тандемные II: 157, 183-197
- характер распределения II: 198-200
Повторяемость последовательностей,
эволюционный аспект II: 159-162
Поджелудочная железа, дифферен-
циальная экспрессия генов II: 356
Подсемейства нуклеотидных после-
довательностей II: 158
Полиаденилирование 11: 45
Полидезоксииуклеотиды, химический
синтез 1: 280
Полидезоксирибонуклеотиды, синтез,
ферментативный метод I: 282,
285-286
-- фосфиттриэфирный метод I: 281 —
285
— химический метод I: 281-282, 284
Полилинкер I: 244-245, 344
Ро1у(А)-полимераза I: 226; II: 46
Полимеразная цепная реакция (ПЦР)
I: 360-363
---олигонуклеотидные праймеры I:
360, 363
Полимеризация без матрицы I: 224
Полиморфный маркер I: 335; II: 11-
12, 354
Полинуклеотидкиназа 1; 221, 323
Полинуклеотидные цепи I: 39-40
Полиовирус I: 112-113; II: 43, 344-345
Полиомавирусы I: 40; II: 124, 344
Полипептид гибридный I: 356-357
Полипептидные цепи, посттрансля-
ционная модификация I: 163
— сборка I: 146
— терминация синтеза у эукариот I:
160
— элонгация I: 150-151, 160
Полинуклеотидфосфорилаза 1: 282,
285
Полипептиды I: 57-58
- аминокислотная последовательность
I: 56-57, 62-66
- синтез в присутствии клонированных
сегментов эукариотической ДНК
I: 354-360
— in vitro I: 294-298
Полисомы I: 152. 153. 160
Полицистронные единицы транскрип-
ции II: 21
- опероны II: 156
Полицистронный транскрипт I: 173
Половые клетки (клетки зародышевой
линии) 1: 18; II: 24, 358, 362-365
- стероиды IE 73
- факторы I: 230
Последовательности, богатые AT IE
156
— GC IE 50-51, 59, 89, 156. 345
- ДНК II: 157-159, 209-215, 225
- IAP II: 259-262
- инсерционные (IS) I: 231; IE 230-232
- мини-сателлитные II: 188
- «переключатели» II: 187
S-последовательности IE 293
Последовательность Шайна-Дальгарно
I: 147-148, 159
- ААТААА II: 43-46, 346, 356
- AAUAAA IF 46
- Alu IE 170, 174-175, 201-202, 206-207,
270-271
- и мутационные делении IE 206
- у человека, выделение II: 201-202
- Sail-бокс П: 36-38
- Ту у дрожжей II: 208
Посттранскрипционные модификации
I: 127; II: 39
Посттрансляционные изменения 1:
169. 354; IE 357-358
Посттрансляционный перенос про-
кариотических белков 1: 172
Праймазы I: 79, 86, 88, 121
Праймер I: 73-75
- удлинение I: 353
Праймосома I: 86-88
Предранние мРНК I: 182-183
Преинициация II: 99-100
Пре-В-клетки IE 289-293
Прерывистые гены II: 6-7, 104
Прибнов-бокс I: 123
Приматы, повторяющиеся последо-
вательности II: 199
- а-сателлиты II: 192-194
Провирусы I: 38, 267; II: 259-260
Прогенота II: 19
Прогестерон II: 73
Прокариотические гены II: 7, 9, 21-22
Прокариоты I: 13, 23
- белковый синтез I: 162
- виды тРНКМе| 1: 143-145
- генетический анализ I: 195
- происхождение II: 19
- процессинг РНК I: 127-130
- транскрипция I: 1.15
— и регуляция экспрессии генов 1:
117-118
--трансляция I: 152, 154, 177
- трансляционная регуляция генной
экспрессии I: 186- 188
- трансляция мРНК I: 145-151
- транспорт белков в клетке I: 171-172
- упаковка ДНК I: 51-52
Проксимальный элемент II: 88-90
Пролактин, ген II: 159-160
Пролин I: 56
Прометафаза I: 14
Промотор I: 355-356
- кор II: 29-31, 33
- lac I: 356-357
- lac UV5 I: 356
- PL I: 356-357, 359
tac I: 356
- trp I: 356-357
Промоторы I: 121-123, 125, 174; II: 21,
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
383
24-25
- гормон-чувствительные II: 78-79
- экспрессирующихся хлоропластных
генов II: 225
Про-рРНК II: 25-26, 28
«Проскальзывание» с неправильным
спариванием II: 229-230
Протеазы II: 358
Протей, размер генома II: 5
Протеолиз 1: 355; II: 356-358
Протоонкогены II: 346, 350, 370
Протопласты I: 249-250
Профаг I: 181
Профаза I: 14-17
- II мейотическая I: 17
Процессинг РНК у прокариот I: 127-130
Прямые повторы I: 330; II: 157
Псевдогены II: 11, 156-157, 159, 167,
170, 206
- мяРНК II: 169
-процессированные II: 156-157, 163,
197, 228
— полипептидов II: 267-269
- РНК II: 269-271
Псевдоуридилат I: 130
Псевдоуридин 1: 54
Пуриновые основания I: 39-42, 52-54
Пуромицин I: 163-164
Радиоавтография I: 278-279
Растения, ДНК хлоропластов II: 216,
224
- промоторы генов II: 82
- системы хозяин-вектор I: 273-276
- хлоропластная ДНК II: 226
- экспериментальное изменение гено-
типов II: 364-365
Регуляторный белок N 1: 182-184
Редактирование на уровне РНК II: 223
Редукционистский подход II: 340
Резолваза II: 234, 275
Резушка Таля, размер генома II: 5
Рекомбиназная система II: 287
Рекомбинантные ДНК I: 354
— идентификация I: 297
— меры безопасности I: 227-228
— технология I: 11, 196, 204, 211,
227; II: 5-6, 342, 345
- клоны, отбор I: 227, 277
- молекулы, библиотеки I: 302-311
Рекомбинация I: 25, 33, 67, 77, 103-111;
II: 370
- внутримолекулярная I: 105
- внутрицепочечная II: 229
- гомологичная I: 339
- между фагом и бактериальными
хромосомами I: 27-29
- негомологичная (случайная) 1: 104-
105
- нереципрокная (генная конверсия) I:
105
- общая, или гомологичная (кроссин-
говер) I: 104-105, 108, 109; II:
195-196
- сайт-специфическая I: 104-106, 111
- типы I: 104- 108
- у бактерий и эукариот I: 103- 104
- - фагов I: 104-105
- in vitro I: 227
Реовирус II: 345
Репликазы РНК-зависимые I: 111-114
Репликативная вилка I: 68, 82, 87,
89
Репликация 1: 83, 227, 230, 234-235
- ДНК I: 38-39
- - инициация I: 68-71, 74, 76
— перенос дезоксинуклеотидов I: 74
— по типу катящегося кольца I: 89-
90
— полуконсервативная 1: 68, 72-74
— полунепрерывный механизм I: 73,
87
— поочередная цепей ДНК аденови-
руса 1: 91
— РНК-полимераза I I: 87-88, 91,
101, 108-109
— терминация I: 92-93
— топологические проблемы I: 82
- РНК I: 93-95, 111-114
- ферменты I: 78-82
Реплисома I: 88-89
Репрессированный хроматин II: 136-
139
Репрессия катаболитная II: 73
Репрессор температурочувствительный
1: 248
- cl 1: 183-185, 228; II: 125-126
- Lac I: 58, 175-176
Рестриктирующие эндонуклеазы I:
207-221, 226, 290
- - действие на специфически метили-
рованные участки ДНК II: 140—
141
— локализация метилцитозина I:
219-220
— несимметричное разрезание ДНК
I: 209-210
— расщепление геномной ДНК I: Т11
— сайты для них I: 216-217, 290-291,
318
---содержащие 5-meC II: 140-141
— специфичность I: 210
— типа 1 I: 214-215
---II I: 214-218
— Ill I: 214-215
— BamHl I: 217
— Bell I: 217
— Bgll I: 217
— BsrNl I: 220
— Clal I: 217
- Eco RI I: 215-216, 219
— £coRII I: 219
— Bae II I: 216
— //Will I: 216, 287
— Hinfl I: 216
— Hpal I: 287
— Hpall I: 217, 219-220
— Kpnl I: 287
— Mbol I: 217
— MspI I: 217, 219-220
— Notl I: 217
— Pstl I: 217
— Tag I I: 217
Рестрикция, контролируемая хозяи-
ном I: 207-208
Ретинобластома II: 350
Ретиноевая кислота II: 73
Ретровирусы I: 38, 52, 93-97, 260,
268-272, 292; II: 344-346
- геном I: 94
- дифференциальный сплайсинг II:
124-125
- жизненный цикл I: 267-268
- онкогенные 11: 346
- репликация РНК I: 111
- синтез ДНК I: 94-97
- создание на их основе рекомбинант-
ных векторов II: 351
- сравнение с ретротранспозонами II:
259-263
Ретрогены II: 228, 267-271
Ретротранспозон(ы) II: 228-229, 249-
267
- класса 1 II: 249
— II II: 263-267
- кодируемые ими полипептиды II:
252-253
- локализация транскриптов в РНП-
частицах II: 261
- транспозиция II: 261-263
- ТуЗ II: 168
Рецептор(ы) II: 283
- андрогена человека II: 74-75
- витамина D3 человека II: 74-75
- глюкокортикоидного гормона че-
ловека II: 74-75, 83-84
- минералокортикоидного гормона
человека II: 74-75
- а-ретиноевой кислоты человека II:
74-75
- стероидных гормонов II: 74-75
- а-тиреоидного гормона II: 74-75
- эстрогена человека II: 75-76
Рибозим II: 108
Рибосомные белки, гены II: 163, 342
- ДНК, организация II: 164
Рибосомы I: 36-37, 117, 138, 160, 164
- диссоциация на 50S- и 308-субъеди-
ницы I: 153
- прокариот I: 142-143
- сборка II: 26-28, 342
- синтез белка I: 142-145
- трансклокация I: 150-151
- A-участок I: 143, 145, 150
- P-участок I: 142, 145, 150
- эксперименты по реконструированию
I: 142
- эукариот I: 142-143
- Е. coli I: 144
Рибулозо-1,5-бисфосфат- карбокси-
лаза I: 309; II: 171
Рибулозо-1,5-бисфосфат - карбоксила-
за/оксигеназа II: 226
Рифамицин I: 160
Рифампицин I: 160
Рицин I: 162
РНК 1: 11, 30, 38-39, 52-56; II: 18
- гены II: 162- 166
- гетерогенная ядерная (гяРНК) I: 52;
II: 7, 204
- двухцепочечная I: 52-54
- денатурация I: 54
- информационная (матричная, мРНК)
I: 36-38, 52, 115, 132, 195-196; II:
18
— взаимоотношения с ДНК и бел-
ками 1: 31-32
- как матрица I: 111-114
- кодирующие ее гены II: 162-170
- компоненты молекулы I: 52
384
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
- копирование с образованием ДНК I:
285-286
- малая цитоплазматическая (мцРНК)
I: 52; II; 22, 270
— ядерная (мяРНК) 1: 52; II: 22, 168-
170, 269-270
- матричная (мРНК) 1: 52
— альтернативный сплайсинг II; 120
— бактериальная I: 152, 154, 155
— вирусная 1; 111-114
-- вторичная структура 1; 148-149
— гистоновая II: 46-47
— задержанная ранняя I: 183
— зонды I: 294-295
- кэпирование 1: 159, 196
- З'-нетранслируемые участки II:
148-150
— полиаденилирование II: 20
-- последовательности З'-концов II:
44
- посттранскрипционные модифика-
ции 1: 196
-- предранняя I: 182-183
- регуляция транскрипции II: 39
- утилизации II: 148- 155
— рецептора трансферрина, регуля-
ция стабильности II: 150-153
-- связывание с Ul-PHK II: 115
-- созревание II: 41
- стабильность 1: 137; 11: 148-150,
153
- считывание кодирующей последо-
вательности 1: 136
-- транскрипция 1: 173
— участие в трансляции 1: 143-151;
11: 153
- ро!у(А)-хвосты I: 196; II: 39, 148,
150
- эукариот I: 159, 195 196; II: 7,
20, 22
--копирование I: 285
- низкомолекулярные II: 90, 94
образование петель 1: 53, 55
- одноцепочечная 1: 52- 54
- полиадснилирование 1: 350-351; II:
23
- посттранскрипционная модифика-
ция I: 185
- прокариот, регуляция содержания в
процессе биосинтеза 1: 172-174
- ренатурация 1: 54
- рибосомная (рРНК) 1: 52, 127- 128.
142, 186, 196; 11: 17 18
- виды II: 25
генная экспрессия 11: 101 104
гены 11: 95-97, 162 167, 342
гомология нуклеотидных последо-
вательностей II: 26
кластеры ichob II: 25-28
у мыши II: 39
-- спейсеры II: 138
транскрипционная единица II: 163
- транскрипция II: 342
5S 1: 52, 127-128; II: 22, 24
- дифференциальная экспрессия
генов II: 102- 103
соматическая, гены II: 101-103
5,8S I: 52
16S 1: 52, 127-128
— 18S I: 52
- 23S I: 52, 127-128
— 28S 1: 52
- синтез на ДНК-матрице 1: 118-121
- стабильность 1: 173
- структура I: 52-54
- транспортная (тРНК) I: 37, 52-53,
128-130, 137, 196; 11: 18
— антикодон I: 138-140
— варьирующие последовательности
I: 138
-- вторичная структура I: 138, 140
- гены 1: 128- 130; II: 92-93, 105-107,
167-168
— изоакцепторная 1: 138, 140
— митохондриальная II: 218-220
-- модель «клеверного листа» I: 130.
140
— посттранскрипционная модифика-
ция 1: 173
-- содержание модифицированных
нуклеотидов 1: 129-130
— созревание 1: 128, 130; II: 90
— сплайсинг II: 119 120
-- структура 1: 53, 55, 138-142
-- транскрипция 11: 22-23
— трансляция более чем одного ко-
дона I: 153, 156
- - ферменты, участвующие в ее мо-
дификациях 1: 129-130
— четвертичная структура 1; 138-139
-этерификация I: 138 141
- - эукариот I: 139
- укладка цепей 1: 53
- химические связи, соединяющие ком-
поненты молекулы I: 52
цитоплазматическая 11: 168-170, 203
- эндонуклеазная активность I: 129
- ядерная 1: 196
- 7SK II: 94 95
-7SL 1: 165; 11: 169-170. 201 202.
207. 267
- U1, псевдогены 11: 197
- U6 11: 94
тРНК-гены, участие в процессинге РНК
у прокариот 1: 127-129
тРНК-метилтрансфераза 1: 175
гРНК-нуклеотидилтрансфераза I: 129
тРНК-спсцифическая лигаза II: 119
тРНК-эндонуклеаза II: 119-120
тРНКМс‘ I: 143- 145
U-PHK 11: 39, 41-48, 87-90, 114-115
РНКаза D 1: 129
Н 1: 87, 95, 212, 214, 224
- Р 1: 129
III I: 128
РНК-блоттинг I: 349-350
РНК-завпсимые репликазы 1: 111 114
РНК-затравка (праймер) 1: 74, 76, 86.
92
РНК-полимераза(ы) 1: 36. 58, 86. 115.
118 122. 182- 184; II: 20 21
активные и неактивные формы II: 28
бактерий I: 121
ДНК-зависимая 1: 115
ингибирование I: 160
кодируемая геномом Е. coli 1: 175
- кодируемые вирусами II: 344
- митохондриальная II: 218-219
- остановка по завершении процесса
транскрипции I: 126
- связывание с промотором I: 123—
124, 175-177
- сигналы транскрипции II: 95
РНК-праймер I: 86
- структура, ее модификации I: 172—
173
- субъединицы I: 121- 122; II: 27-28,
40
- эукариот 1: 118
- 1 I: 159; 11: 8-9, 27-28, 167, 342
-- субъединицы II: 27
— транскрипция 11: 22-23, 25, 31-33,
35-36, 163
- II 1: 159, 269, 308; II: 8-9, 40-41,
54, 94- 100, 342, 345
— транскрипция II: 22-23. 40 41, 53,
88, 170
--SlNE-последовательностей II: 203
-- участие в репликации вирусов 1:
344
- Ill I: 159; II: 8-9, 22-23, 42, 88-89.
166-167, 202-203, 342
-- дрожжевая II: 34
— сайты терминации 11: 101
-- субъединицы II: 90
-- терминация транскрипции II: 100—
101
— транскрипционный комплекс II:
90-91
-- факторы транскрипции 11: 91
РНК-репликазы 1: 86
РНК-содержащие вирусы 1: 286; 11:
342, 344-346
Родопсин I: 312 313; 11: 341
Сайленссры II: 24
Сайт ап В I: ПО 111
- an L 1: 111
- апР I; 110-111
- attK 1: ПО 111
Сайты для рестриктирующих эндо-
нуклеаз, локализация I: 330
- полиаденилирования И: 43-46
- сплайсинга II: 105
cos-Сайты I: 91, 239-241, 246
Сайты-мпшсни 11: 228, 232
- дупликаций II: 175, 201. 203, 228-229.
231, 234
Самосплайсинг 11: 108-112
а-Сатслтиты приматов II: 192-224
Саузерн-блоттинг 1: 278
Сегмент, богатый GT II: 43-44
- CEN 1: 254
Сегменты ДНК, классификация II:
156-157
- соответствующие им транскрипты
I: 349 354
функции 1: 348 354
- РЙК. ферментативная амплификация
1: 360 363
Сегрегация независимая I: 25
Секвенирование ДНК II: 20
Секреторные клетки 1: 21
- пузырьки (везикулы) I: 164, 170
Серин I; 56
Ссродем 11: 297
Серповидноклеточная анемия I: 63,
66; 11: 354
Сестринские хроматиды 1: 14, 17-18
Сигма-последовательность 11: 168
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
385
Сигма-фактор II: 21. 28
Сигнал-распознающая частица (СРЧ)
1: 165-167
Сигналы транскрипции в эукариоти-
ческих генах II: 22-23
- узнавания 11; 287-289
Сигнальная пептидаза 1: 167
- последовательность I: 164, 167-169,
172
Сигнальные протеазы I: 172
Сигнальный полипептид 11: 357
Синапсис 1: 16
Синдром Блума I: 103
- Дауна 1: 23-24; 11: 353
- приобретенного иммунодефицита
(СПИД) 11: 350
Синтетические вставки 1: 280- 285
- олигодезоксинуклеотиды 1: 347-348
Системы экспрессии 1: 354-355
Скрининг SV40-6j1hllick 1: 293-294
- иммунологический I: 300. 354
- клонированных популяций реком-
бинантных молекул I: 292
- рекомбинантных клонов 1: 227. 236,
277, 292-302
Согласованная эволюция II: 162
Соматостатин, ген 1: 280
Спаривание оснований комплемен-
тарное 1: 74
- - неоднозначное соответствие I: 156
- при трансляции 1: 37
Спарсомицин 1: 162
Спеисерные сегменты 1: 127
Спейсеры I): 22, 24. 26
- межгенные 11: 26, 163 164
- рРНК-генов 11: 138
Сперматозоиды I: 18, 363
Спираль-петля-спираль 11: 126- 127
Сплайсинг 1: 300-301; II: 7, 10, 17- 18,
39 40. 42, 105, 120 125. 225. 346,
355
- сайты 11: 105
- - выбор II: 125
Сплайсингосомы 11: 113, 116-117
Среда G-418 1: 263. 271
- HAT 1: 259; 11: 308
Стафилококковая нуклеаза 1: 212
Стволовые клетки I: 172; 11: 289, 291
Стероидных гормонов рецепторы II:
74-77
- димеризация 11: 76- 79
---связывание гормона II: 76- 77
Стоп-кодоны 1: 136, 145, 151, 160;
11: 219
Стреи голилигин 1: 160
Стрептомицин 1: 161-162, 230
Структуры Холлидеч 1: 106 109
Субклонирование I: 318
Супрессор Ту II: 251
Т-с}прессоры II: 284. 294
Сферопласты I: 252
Сывороточный альбумин, гены II:
171, 184 185
Талассемия II: 355
Тандемные повторы II: 157, 183- 197,
313 314
- в геномах эукариот II: 187
- гистоновых генах 11: 183
— кодирующих последовательностях
II: 195-197
---межгенных спейсерах II: 164-166
----области центромер и теломер II:
188-189
- вне кодирующих областей 11: 186—
188
-- механизм образования И: 195-197
— удлинение и укорочение 11: 195- 196
— функция II: 193-195
— эволюция П: 195-197
Теломераза 11: 214-215
Теломеры I: 19, 93, 254
- множественные в макронуклеусах
простейших 11: 215
- последовательности ДНК II: 209-215
- тандемные повторы II: 188-189
элонгация во время репликации 11:
214
Телофаза 1 мейотическая 1: 17
- II мейотическая I: 17
Тельце Барра II: 147
Температурочувствительный репрессор
1: 248
Теория «один ген один фермент» I:
27, 115
Тепловой шок II: 81
Тератокарцинома человека II: 204, 367
Терминаза 1: 241
Терминальная дезоксинуклеотидил-
трансфераза I: 224
Терминирующие белки RF-I I: 145,
151
- RF-2 1: 145, 151
- RF-3 1: 145, 151
- кодоны 1: 157- 158
Тестостерон II; 73
Тетрагидрофолат 11: 307-308
Тетрады I: 16- 17; 11: 164-165
Тетрациклин 1: 162, 230, 232, 251
Тимидинкнназа I: 258-259
Тиминовые димеры 1: 108
4-тиоуридин 1: 54
Тирозил-тРНК синтетаза 1: 354
Тирозин 1: 56
Тиострептон 1: 162
Тканевой плазминогенный активатор
11: 359
Тотипотентные клетки I: 276; II: 365
Точечные мутации 1: 344 349
Трансгенные животные I: 257
- организмы 11: 21. 351. 363-364. 368
Трансгены 11: 148. 363
Г/д»/с-действующие факторы II: 40-
41, 48, 125, 355
Трансдукция I: 196. 198 201
Трансдуцирующис фаговые частицы
I: 198-202
Транскрипты Ту 11: 251
Транскрипция I: 35-36. 38
активаторпые белки 1: 117
активация на расстоянии за счет из-
гибания ДНК 11: 133-134
вирусов, регуляция II: 48
ДПК в РНК 1: 115. 117 127
ингибиторы 1: 160 163
раскручивание спирали 1: 118. 120
121
инициация I: 37, 86- 87, 117, 121,
123 124. 174; II: 24
обратная II: 249
- регуляция 11: 24, 76-78, 81-84
--стероидами II: 73-80
- терминация I: 120, 125, 127, 179;
II: 9, 38, 43-46
- у эукариот 1: 196
Транслокации фрагментов хромосом
1; 21, 67; 11: 272-275
Трансляция I: 31, 36-37, 131, 145,
157-160
- аутогенная, регуляция 1: 186
- бактериальных мРНК I: 152, 154—
155
- ингибиторы I: 160 163
-инициация I: 116, 132, 134, 136,
145-151
- ошибочная I: 157
- «рамки считывания» I: 116
--преинициаторный комплекс 1:
160
-терминация 1: 145, 151, 157-160
- у прокариот 1: 145-151
— эукариот I: 157-160
- in vitro I: 294-298
Транспозазы II: 231, 234, 239-240
Транспозирующиеся элементы 11:
228-249
Транспозиции 11: 227-229, 233-236
Транспозоны 1: 231; II: 228, 230-235
- обратная транскрипция 11: 249
- сложные II: 230-232
Транспортные везикулы окаймленные
1: 169-170
Tprwc-сплайсинг II: 117-119, 225
Трансфекция 1: 228, 232, 241. 245, 258,-
290-292
Трансферрин 1; 314-315
Трансформация бактерий 1: 196 197,
235
- инсерггионная 1: 257, 275
- клеток животных 1: 256-261
--стабильная I: 254-257
- сайт-специфическая I: 252. 255-256
Трансэтерификация II: 108
З'-трейлерная последовательность 11:
10. 21-22. 24
Треонин 1: 56
1-(2,4,6)-триизопропилбензолсульфо-
нилхлорид (ТПС) 1: 283
Трииодтиронин 11: 73
Трипаносомии II: 296-297
Трипсин 11: 356, 360-361
- ингибитор 1: 34
Триптофан 1: 56
Триптофановый оперон 1: 177 181.
201
Триптофансинтаза 1: 58
Тропоколлаген I: 60
Тропонин Т II: 122 123
u-Тубулин II: 268
Тубулиновые гены, мультигенное се-
мейство II: 173 174
Тубулины II: 153, 173- 175
«Узнающая спираль» II: 126
Упаковка ДНК 11: 136 139
Уранил-М-гликозилаза 1: 100
Урацилтрифосфат!: 118-119
Уридилтрансфераза 1: 173
Урилиндифосфогалактоза 11: 69-70
Уридиндифосфоглюкоза 11: 69
386
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Уридин-5'-фосфат I: 53
N-участки II: 289
Фабрициева сумка II: 291
Фагоцитарные клетки II: 284
Фазовая вариация II: 273
Фактор антитерминации II: 45
- медьиндуцируемой транскрипции
II: 82
- связывания последовательности
ТАТА II: 54-55
- транскрипции API II: 56-57, 83, 89
— АР2 II: 57, 83, 89
— АРЗ II: 58
— АР4 II: 58
— АР5 II: 57
— TFIIIA II: 95-100, 102-110, 130
- D II: 32
Факторы инициации I: 145
— IF-1 I: 145-146
— IF-2 I: 151
— IF-3 I: 153, 155
- резистентности (R-факторы) I: 205-
206
- терминации II: 9
- тканеспецифичиой транскрипции II:
68
- транскрипции II: 8-9, 22, 30-31, 55
— гомеодомены II: 126-129
— домены регуляции транскрипции
II: 133-135
---связывания ДНК II: 126-133
— регуляторный домен II: 125
— связывание с ДНК II: 125
— структурные домены II: 125
— трансляционная регуляция II: 153-
155
— APl-Jun II: 132, 134, 154
— CTF II: 60-61
— GCN4 II: 132, 134, 154-155
— HNF-1 II: 69
— Pitl II: 129
— POU-доменов II: 128-130
— TFIIIB II: 95-100
— TFIIIC II: 95-100
— TIF-1B II: 32
- фертильности (F-факторы) I: 206-207
- элонгации I: 145
— EF-G I: 145, 150-151, 175
— EF-Ts I: 175
-- EF-Tu I: 150, 175
1д/с-Факторы II: 125
Фенилаланин I: 56
Фенотип I: 27, 31, 33; II: 15-16, 342
Фенотипический отбор клеток I: 258-
259, 299-300, 354
Ферментная недостаточность II: 307-
308
Феромоны II: 358
Ферритин II: 150-153
а-Фетопротеин II: 65-68, 184-185, 355
Фибриноген I: 59-60
Флагеллин II: 272-273
З'-фланкирующий участок II: 10
5'-фланкирующий участок II: 10
Флип-флоп-инверсии II: 272-276
Фокусы трансформантов I: 266, 300
2'-фосфатаза II: 119- 120
Фосфодиэстераза змеиного яда I: 212
- селезенки I: 212
Фосфодиэфирные мостики I: 30
- связи I: 118
Фосфомоноэстеразы I: 221
К-(фосфонацетил)-Е-аспартат II: 307-
309
Фосфорилирование I: 355
Фотолиазы I: 99
Фрагмент Кленова I: 78
Фрагменты ДНК I: 209-210, 304-306
— электрофорез I: 218, 277-278
- Оказаки I: 73, 77-78, 80, 86, 88
Фузидиевая кислота I: 162
Футпринтинг II: 30-31
Т-хелперы II: 284, 291, 294
Хиазма I: 26
Химерные белки II: 125
- гены II: 20
Химический мутагенез I: 344-345
Химотрипсин I: 60; II: 355
Хлорамфеникол I: 162, 207, 230
Хлоропласты I: 164
- гены II: 224-225
и-Хлорфенилфосфордихлоридат I: 281-
282
Холестерол II: 184
Холецистокинин II: 357
Хоризмовая кислота I: 178
Хроматиды I: 14, 19. 21
- сестринские, обмен I: 103-104
Хроматин I: 49-52, 118; II: 136
- активный и репрессированный II:
138-139
- сборка I: 82
- структура I: 51; II: 136-137, 156
- транскрипционно активные области
II: 137
Хромосомные карты эукариотическо-
го генома II: 11-12
Хромосомы I: 11-27, 49-52; II: 10
- аберрация 1: 23-24; II: 13
- акроцентрические I: 20
- вырезание сегментов I: 279
- гетерохроматиновые области I: 19-
20
- гомологичные пары I: 16, 18
- дицентрические II: 310
- дупликация I: 14, 70
- идиограммы I: 22, 24
- изменение структуры при амплифи-
кации II: 309-310
- интерфаза I: 21, 23
- кроссинговер при мейозе I: 25-26
- мейотические I: 21
- метафазные I: 18-21
- метацентрические I: 19
- методы окрашивания I: 19-21, 336
- морфология II: 13
- С-окрашивание I: 20, 336
- G-окрашивание I: 20
- Q-окрашивание I: 20, 336
- R-окрашивание I: 20, 336; II: 156
- плечи I: 18
- политенные I: 21, 23
- половые I: 18
- прокариот I: 13, 51-52
- прометафаза I: 21
- репликация в мейозе I: 16-18
- строение I: 18-25, 93
- трисомия I: 23-24; II: 353
- упаковка ДНК I: 49-52
- фаговые I: 28- 29
- человека I: 16
— локализация генов II: 11
— фракционирование I: 303
- эукариотические I: 13, 49
- эухроматиновые области I: 19-20
- XI: 18; II: 193-194
- Y I: 18
Хромоцентр I: 23
Центромера I: 14, 18-23; II: 13
- последовательности ДНК II: 209-
212
- строение II: 209
- тандемные повторы II: 188-193
Цианобактерии I: 52
Циклическая фосфодиэстераза II: 119
Циклический AMP I: 175-177
Циклобутановые димеры I: 99-102
Циклогексимид I: 162
«Цинковые пальцы» II: 55-56, 71-72,
76, 87, 99, 130-131
//«(-действующие элементы I: 230; II:
39-41, 48
Цистеин I: 56
Цитидин-5'-фосфат I: 53
Цитогенетика I: 23
Цитозинтрифосфат I: 118-119
Цитозоль I: 164
Цитокинез I: 16
Цитокинин I: 275
Цитомегаловирус II: 346
Цитоплазматическая РНК II: 168-170,
203
Цитоплазматические рибонуклеопро-
теиновые частицы (цРНП) II: 42
Цитохром Ь, гены II: 220-221
- с I: 354; II: 15, 360
Часы молекулярные II: 161-162
Челночный вектор в системе Т-ДНК-
Е. coli I: 275-276
-----дрожжи- Е. coli I: 251-253
— дрожжевая мини-хромосома I: 254
— на основе ретровируса I: 268
Человек, размер генома II: 5, 353
Число копий, оценка с помощью гиб-
ридизации в условиях насыщения
I: 339, 341-342
- повторов II: 157-158
Шайна-Дальгарно последовательность
I: 147-148, 159, 355
Шелкопряд, размер генома II: 5
Шовдомицин I: 162
Шпинат, хлоропластная ДНК II: 224-
225
Шпорцевая лягушка, размер генома
П: 5
Щелочная фосфатаза I: 58
Эволюция, единица скорости II: 161
- нейтральная молекулярная II: 194
- согласованная II: 162
Экзонуклеаза I I: 212
- HI I: 212, 214; II: 36-37
- VII I: 212-213
X-Экзонуклеаза I: 212
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
387
Экзонуклеазы I: 78-80, 101-102, 107,
211-213, 222
Экзоны II: 7-8, 17-18, 23, 104-105,
170
- перераспределение в ходе эволюции
гена 11: 161
Экспрессирующие векторы I: 354-360
Экспрессия вирусных генов, регуля-
ция II: 48-61
- временная I: 257
-генов I: 38, 45, 52-53, 115-117; II;
156
— глобальные механизмы регуляции
11: 135-155
- - дифференциальная I: 306; II: 354-
358, 366
— координированная II: 355
- - регуляция I: 37, 117, 172-188, 195-
196; 11: 20, 39, 358
Эластаза II: 364
Электронная микроскопия I: 319
Электропорация I: 258
Электрофорез импульсный I: 303-304,
317
Элемент(ы) мобильные II: 158, 222,
228-249
- сАМР-чувствительный II: 143
- copia, кольцевые ДНК II: 262
— сравнение с ретровирусами II: 259-
263
- - транскрипция II: 258
— трансляция II: 258
— транспозиция II: 257-258
- fold-bach II: 271-272
- 1 II: 242, 265-266
- Р, встраивание в определенные сай-
ты II: 238
— гибридный дисгенез II: 238-241
- - как векторы II: 242
— свойства II: 238
— структура II: 239-241
— транспозиция II: 238-239
— функции, ими кодируемые II: 239-
242
- sph II: 53, 57
- ТАТА II: 67
- Tfr-мРНК, чувствительный к железу
II: 151-153
- Ту, вырезание II: 255
— класса I II: 229, 249-255, 259-262
- - обратная транскриптаза II: 252
— перемещение II: 250-251
— РНК как посредник при транспо-
зиции Ту II: 253-255
— транскрипты II: 251
— транскрипция II: 251-252
— трансляция II: 252-253
- -у дрожжей II: 249-252
— хромосомные перестройки II: 255-
256
Эмбриогенез II: 84-87, 366-368
Эмбрионы животных, введение в них
ДНК I: 257
Эндонуклеаза(ы) I: 211-213
- неспецифические I: 343
- рестриктирующие I: 207-221, 226
— сайты I: 318; II: 11
- Есо RI 1: 215
- uvrABC I: 101
Эндоплазматический ретикулум I:
164-165
— гладкий I: 164
- - шероховатый I: 164
Энкефалины II: 357
Энхансеры II: 24, 54, 345
- фоновой активности II: 83
Эпигенетические факторы II: 9
Эпимераза II: 69
Эпитоп II: 283
Эритробластозные клетки птиц II: 350
Эритромицин I; 162
Эстроген-акцепторные элементы II: 74
Эстрогены II: 73
Эукариотические белки II: 359-361
- геномы 1: 339; II: 5, 10-13, 227
- гены II: 5-7, 22-25, 69-84
- клетки I: 49-51, 70, 164; II: 212
- системы хозяин- вектор I: 227, 252-
276
Эукариотический геном, повторяю-
щиеся последовательности I: 339
Эукариоты I: 13, 23, 25, 40, 43
- виды тРНКМм I: 143-144
- генетический анализ I: 195-196
- генная экспрессия I: 172, 300-301; II:
6-10, 20
- происхождение II: 19
- синтез белка I: 145, 162
- содержание ДНК-зависимых РНК-
полимераз I: 115
- транскрипция и регуляция генов I:
117-118
- трансляция мРНК I: 157-160
Ядерные рибонуклеопротеиновые ком-
плексы (яРНП) II: 42
Ядро I: 11-13, 21
Ядрышки I: 21; II: 25, 342
Ядрышковый организатор I: 19-20;
II: 25, 163
Указатель латинских названий
Acantamoeba, ген тРНК&г II: 21
- РНК-полимераза I II: 27-28
Agrobacteria II: 225, 365
Aplysia, пептидные посредники II: 358
Arabidopsis, теломеры II: 214
Archaebacteria, интроны II: 19, 104
Bacillus suhrilis I: 40, 43, 122
- ДНК-лигазы I: 81
-- система хозяин - вектор I: 227, 249,
250
Caenorhabditis elegans II: 351-353
-- генетические исследования II: 367
- карты сайтов для рестриктаз II:
351
— транскрипционный регулятор раз-
вития нейронов II: 128-129
-- гирпис-сплайсинг II: 118
Chlamydomonas 1: 40; II: 225
Crithidia fascicidata II: 222
С у anaphora paradox а II: 236
Dictyostelium, актиновые мРНК II:
'172-173
Drosophila melanogaster II: 353, 366-367
— актиновые гены II: 173
- - белки теплового шока II: 205
- - ген antennapedia II: 85-87, 104. 128
---bicoid (bed) II: 85
---engrailed (eng) II: 85-87
---even skipped (ere) II: 85-87
fushi terazu (fiz) II: 85-87
---hairy II: 86
- - hunchback (hb) II: 85-87
---knirsp (kni) II: 85-87
— kruppel (Кг) II: 85 87
---nanos (nos) II: 85
per II: 353
---runt II: 86
- ultrabithorax II: 85-86, 104
- генетическое картирование I: 334
335
гены I: 25, 312, 313; II: II, 350,
358
приклеивающих белков II: 186
гистоновые II; 183
гомеотические II: 87, 126
- гомеозисные мутации II: 366-367
- дисгснез 1-R II: 265
ДНК I: 45, 73, 340-341
- рДНК II: 26-27, 32- 33, 272
- изменения клегок зародышевой
линии II: 362 363
- кластеры тРНК-гснов II: 168
клонирование гена, регулирующе-
го ритмичность поведения I: 311
- - локус white II: 238
- морфогенез II: 84-87
- мутация Sn" II: 239 241
- непропорциональная репликация
II: 303-304
неравный кроссинговср II: 229
- повторяющиеся последовательности
II: 198 199
— copia-подобные элементы 11:255-259
— политенные хромосомы I: 21. 23,
279
— половые хромосомы I: 18
— размер генома II: 5
- - распределение повторов II: 198-
199
— рекомбинации ДНК I: 25
- - ретротранспозон gypsy II: 228
— 7SL-PHK II: 169-170
— сателлитная ДНК II: 189, 192
- - секреторные клетки I: 21
- - скорость репликации 1: 70
— сравнительная молекулярная гене-
тика II: 16
— тандемные повторы генов 5S-pPHK
II: 166 167
— транспозаза II: 124
-- транспозирующиеся элементы II:
241-242
- - тубулиновые гены II: 174
- - фактор транскрипции II; 9
- - хорионические гены II: 303-304,
311
- - хромосомы I: 16, 27
-- экспрессия ранних генов в эмбрио-
генезе II: 84-87
— элементы F II: 228, 264-265
- FB II: 271-272
--G II: 264-265
Escherishia coli II: 359
активация транскрипции гена GAL4
II: 134 135
белок-репрессор trp II: 125
- встраивание и выщепление ДНК
фага к I: 110-111
- Р-галактозидазный ген I: 200
-галактозный оперон I: 173-174.
177
-- ген ага С I: 174
gpt I: 259, 265. 271
- neo I: 259, 263, 266. 271
tip R I: 178
генетическая карта I: 197-198
генетический анализ I: 27
- геном I: 44. 175
ДНК I: 40. 43, 45. 49
- кинетика гибридизации второго
порядка I: 340
ДНК-гираза I: 85
ДНК-лигазы I: 80-82. 222. 225
- ДНК-полимеразы I: 78-80
ДНК-праймаза I; 175
-- ДНК-связывающие белки I: 51
-- инвертированные повторы опера-
торных последовательностей I:
176
- инсерционные последовательности
II: 230-231
- клонирование ретровирусных ге-
номов I: 268 273
-конъюгация I: 197-198
— метилазы I: 218 220
- нестабильность при клонировании
I: 338
— образование зрелых тРНК I: 128-130
— оперон ага 1: 174, 177
--lac 1: 174-177
- - плазмиды I: 251-252
- - полуконсервативный механизм
репликации ДНК I: 87-88
— промоторы I: 123; II: 9
- - регуляция генной экспрессии I: 172
— репликация и репарация хромо-
сомной ДНК I: 78-79
-- рестрикционные эндонуклеазные
фрагменты II: 351-352
— рибосомы I: 142
— 16S-pPHK I: 55
- Р-РНКаза II: 219
-- РНК-полимераза 1: 121; II: 8
- РНК-праймеры I; 86
-- родственные гены I: 173-174
— сайты расщепления для РНКазы
Ill в предшественниках рРНК I:
128
- системы экспрессии I: 354-355
-- скорость репликации I: 68-72
-- транскрипционные единицы рРНК
I: 127
-- транспозоны II: 232
— триптофановый оперон I: 178-181
-- экспрессирующие векторы I: 355—
360
- экспрессия эукариотических генов
I: 300-301
- эндонуклеаза ucr ABC I: 101
- К12, ген dap 1: 229
- eiidK I: 229
- hsdR I; 229
------rec К I: 229
- как хозяин в экспериментах с ре-
комбинантными ДНК I: 227- 228
- система хозяин - вектор I: 227-229
--трансфекция I: 232
--штаммы I: 228-229
Escherichia со/г-системы. плазмидно-
фаговые векторы I: 246-248
Euglena II: 225
Hemophilus influenzae I: 45, 249
Klebsiella pneumoniae I: 249
Leishnwnia enrietti. тубулиновые гены
II: 174
Micrococcus lysodeikticus I: 40
Neurospora crassa I: 212; II: 167
Notophtalmtis viridcscens II: 183
Oxytricha II: 215
Phaseohis aureus, нуклеаза I: 212
Physarum polycephalum. интроны груп-
пы I II: 106
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
389
Plasmodium falciparum I: 354
Rhizobacteria IT. 225
Saccharomvces cerevisiae I: 40, 43, 45,
252; II: 353
— активация транскрипции II: 78, 80
- - ген дсп 4 II: 154
- - гены RAD I: 102
- - механизм репарации УФ-индуци-
рованных повреждений в ДНК I:
102
- - размножение I: 252
— сателлитная ДНК II: 195
Salmonella II: 272-274
Schizosaccharomyces pombe II: 23, 104,
167, 195
Streptomyces I: 249-252
- coelicolor I: 250
- Hvidans I: 250
Strongylocentrotus purpuratus, гистоно-
вые гены II: 182
Tetrahymena, амплификация ДНК
в макронуклеусах II: 306-307
- ДНК макронуклеарная II: 212-213
- интрон гена рРНК II: 109-111
- кластеры рРНК II: 26
- макронуклеус II: 212, 306-307, 358
- микронуклеус II: 212, 358
- сплайсинг интрона про-рРНК II: 109
- тубулиновые гены II: 174
- интроны группы I II: 106
- теломеры II: 212-215
Тhermits aquaticus. ДН К-полимераза I:
361
Trypanosoma II: 5, 118-119, 222-223,
296-302
- brucei II: 222-223. 296
Xenopus, альбуминовые гены II; 184
- гены рРНК II: 90-92, 101-103, 166-
167, 358
— гистоновые II: 182-183
— глобиновые II: 176
- рДНК, амплификация по типу ка-
тящегося кольца II: 304
— изопикническое центрифугирование
II: 166
- межгенный спейсер II: 33-34
- ооциты 1: 257, 354; II: 25
- повторяющиеся последовательности
II: 199
- 7SL-PHK II: 207
-рРНК 5S, гены II: 101-103
--регуляция генов II: 104
- тРНК II: 93
-транскрипция рДНК II: 33-35
- фактор транскрипции TFIIIA II: 97-
100
- экспрессия генов I: 308
- промотор U6-PHK II: 88
Оглавление
Часть III. Молекулярная органи-
зация, экспрессия и
регуляция эукариоти-
ческих генов
Введение 5
Структура эукариотических генов и их экспрес-
сия ......................................... 6
Прерывистые гены................... 6
Регуляция экспрессии генов у эукариот 7
Строение генома эукариот . 10
Хромосомные карты............... 11
Размеры эукариотических геномов ... 12
Связь между специфическими последова-
тельностями ДНК и морфологией хромо-
сомы ..... 13
Перестройки последовательностей ДНК . 13
Случайные перестройки генома............. 14
Программируемые перестройки . . 14
Эволюционные аспекты................. 15
Сравнительная молекулярная генетика 16
Эволюционная история генов............... 17
Происхождение генетических систем . . 18
Заключение 19
Глава 8. Структура и регулируемая
экспрессия эукариотических
генов.......................................... 20
8.1. Сравнение структурных особенностей про-
кариотических и эукариотических генов 21
а. Прокариотические гены................. 21
б. Эукариотические гены................. 22
8.2. Строение генов класса I и их экспрессия 25
а. Единицы транскрипции .... 25
б. Аппарат транскрипции................. 27
в. Области регуляции транскрипции рДНК 28
г. Терминация транскрипции рДНК ... 35
д. Сопряжение терминации и инициации
транскрипции.................. ... 38
8.3. Строение и экспрессия генов класса II 39
а. Общие положения ... 39
б. Аппарат транскрипции . . 40
в. Созревание мРНК и U-PHK............... 41
г. Регулируемая экспрессия вирусных генов 48
д. Регуляция генов, специфичная в отно-
шении типа клеток и стадии развития 61
е. Индукция и репрессия транскрипции 69
ж. Транскрипционная регуляция морфоге-
неза ................................ 84
з. Регуляция транскрипции генов U-PHK 87
8.4. Структура и экспрессия генов класса III 90
а. Основные свойства ..... 90
б. Последовательности, регулирующие ини-
циацию транскрипции.................. 91
в. Влияние факторов транскрипции на ини-
циацию транскрипции с участием РНК-
полимеразы III......................... 95
г. Терминация транскрипции с участием
РНК-полимеразы III.................... 100
д. Регуляция экспрессии генов 5S-pPHK в
процессе развития организма . 101
8.5. Интроны 104
а. Частота встречаемости интронов . 104
б. Различные виды интронов............105
в. Интроны, вырезаемые при помощи ав-
токаталитического сплайсинга . 108
г. Сплайсинг ядерной про-мРНК ... 113
д. Сплайсинг тРНК ......... 119
е. Альтернативный сплайсинг: множествен-
ные белки одного гена...............120
8.6. Новые структурные элементы в факторах
транскрипции . . 125
а. Домены связывания ДНК............. 126
б. Домены регуляции транскрипции ... 133
8.7. Глобальные механизмы регуляции экспрес-
сии генов .... 135
а. Упаковка ДНК ..................... 136
б. Топология и конформация ДНК . 139
в. Метилирование ДНК................. 140
г. Регулируемая утилизация мРНК 148
Глава 9. Молекулярная структура гено-
мов эукариот . 156
9.1. Архитектурные элементы............ 156
а. Классификация сегментов ДНК ... 156
б. Повторяемость последовательностей
ДНК.................................157
в. Эволюционный аспект повторяемости
последовательностей .............. 159
9.2. Гены, кодирующие РНК .... 162
а. Гены 18S-, 5, 8S- и 28S-pPHK..... 162
б. Гены 5S-pPHK ....... 166
в. Сцепление всех четырех генов рРНК у
дрожжей............................ 167
г. Гены, кодирующие тРНК............ 167
д. Гены малых ядерных и цитоплазматиче-
ских РНК ..... 168
9.3. Гены, кодирующие белки............ 170
а. Некоторые общие замечания ... 170
б. Примеры мультигенных семейств . . 171
в. Гистоновые гены: консервативность ко-
дирующих последовательностей и раз-
личия в организации................ 181
9.4. Тандемные повторы: характерная особен-
но сть всех эукариотических геномов . . 183
а. Гены, состоящие из тандемных повторов
сегментов ДНК...................... 183
б. Тандемные повторы вне кодирующих
областей............................186
в. Тандемные повторы в области центро-
мер и теломер ......... 188
г. Возможные функции тандемных повто-
ров ................................194
д. Механизмы образования и эволюции
тандемных повторов..................195
ЧАСТЬ IV. ИССЛЕДОВАНИЕ БИОЛОГИЧЕСКИХ СИСТЕМ И МАНИПУЛЯЦИИ С НИМИ 391
9.5. Повторяющиеся последовательности, рас-
сеянные по всему геному . . 197
а. Характер распределения повторов 198
б. Диспергированные повторы у беспозво-
ночных ..................................200
в. Многочисленные семейства диспергиро-
ванных повторов в геномах млекопи-
тающих ................................200
г. Функции диспергированных повторов 204
д. Замечательное единообразие дисперги-
рованных повторяющихся последова-
тельностей ....... 206
9.6. Последовательности в области центромер
и теломер..................................209
а. Последовательности в области центро-
мер ...................................209
б. Последовательности в области теломер 212
в. Искусственные хромосомы дрожжей 215
9.7. Геномы органелл эукариот: ДНК мито-
хондрий и хлоропластов.....................216
а. Геномы митохондрий . .................217
б. Необычная митохондриальная ДНК
трипаносом.............................222
в. ДНК хлоропластов..................... 224
г. Происхождение ДНК органелл . 225
Глава 10. Реорганизация генома . . 227
10.1. Общие свойства незапрограммированных
транспозиций 227
а. Различные типы мобильных элементов 228
б. Дупликация сайтов-мишеней .... 228
в. Мобильные элементы как диспергиро-
ванные повторяющиеся последователь-
ности ...............................229
10.2. Транспозирующиеся элементы . 230
а. Транспозирующиеся элементы прока-
риот ...............................230
б. P-элементы дрозофилы................238
в. Контролирующие элементы у кукурузы 242
10.3. Ретротранспозоны . . 249
а. Общие свойства ретротранспозонов
класса I......................... 249
б. Ту-элементы дрожжей..........249
в. Copia-подобные элементы дрозофилы 255
г. IAP-последовательности у мышей 259
д. Сравнение с ретровирусами . . 259
е. Ретротранспозоны класса II . 263
10.4. Ретрогены .... ............ 267
а. Процессированные псевдогены поли-
пептидов ....................."... 267
б. Процессированные псевдогены РНК 269
10.5. Другие необычные мобильные элементы 271
а. Fold-back-элементы дрозофилы ... 271
б. Вставки в рДНК Drosophila . . 272
10.6. Запрограммированные перестройки и мо-
. дуляция экспрессии генов.................272
а. Прокариотические модели-транслока-
ция путем флип-флоп-инверсий . . . 272
б. Типы спаривания у дрожжей-кассет-
ный механизм.........................275
в. Гены, кодирующие иммунные белки
позвоночных..........................283
г. Вариабельные поверхностные антигены
трипаносомы...........................296
10.7. Запрограммированные амплификация и
мо дуляция экспрессии генов .... 302
а. Непропорциональная репликация: хо-
рионические гены дрозофилы ... 303
б. рДНК Xenopus: амплификация по типу
катящегося кольца.....................304
в. Амплификация рДНК Tetrahymena в
макронуклеусах........................306
10.8. Незапрограммированные тандемные ам-
плификации ............................ . 306
а. Амплификации, позволяющие компен-
сировать ферментную недостаточность 307
б. Структура амплифицированных генов 308
в. Механизм амплификации..............310
Литература 315
Часть IV. Исследование биоло-
гических систем и ма-
нипуляции с ними
Исследование сложных биологических систем 340
Семейство генов зрительных пигментов 340
Согласованные взаимодействия между
продуктами разных генов................342
Жизненный цикл вирусов 342
Картирование геномов.......................351
Физическая и генетическая карты . . . . 351
Картирование больших геномов .... 353
Полиморфизм длины рестрикционных
фрагментов.............................353
Дифференциальная экспрессия генов в специфи-
ческих клетках и тканях....................354
Специфическая экспрессия генов в подже-
лудочной железе........................356
Посттрансляционные изменения как регу-
ляторы направленного потока генных про-
дуктов ................................357
Регуляция генной экспрессии путем перест-
роек последовательности ДНК............358
Изменение биологических систем.............358
Синтез нормальных и модифицированных
эукариотических белков.................359
Генетическая инженерия.................361
Изменение соматических клеток .... 362
Изменение клеток зародышевой линии 362
Новые направления................... . 365
Эмбриональное развитие и дифференци-
ровка у многоклеточных организмов . 366
Инсерционный мутагенез.................368
Специфическая и случайная модификация
генома млекопитающих...................368
Перспективы . . 369
Будущие заботы . 370
Литература 371
Предметный указатель.......................375
Указатель латинских названий...............388
Максин Сингер, Пол Берг
ГЕНЫ И ГЕНОМЫ
В 2-х томах
Том 2
Зав. редакцией канд. биол. наук М.Д. Гроздова
Ведущий редактор Н. Н. Шафрановская
Редактор В. И. Николаева
Художник В. П. Медников
Технический редактор Л.П. Бирюкова
Корректор В. И. Николаева
Лицензия ЛР № 010174 от 20.05.97 г.
Сдано в набор 26.02.96 г. Подписано к печати 22.09.98 г.
Формат 84 х 108716 Бумага офсетная. Печать офсетная.
Гарнитура тайме. Объем 12,25 бум. л. Усл.печ.л. 41,16.
Уч.-изд.л. 49,43. Изд. № 4/9021. Тираж 5000 экз. Зак. 158.
С 025.
Издательство «Мир» Комитета Российской Федерации по
печати. 129820, ГСП, Москва, И-110, 1-й Рижский пер., 2.
ОАО «Можайский полиграфический комбинат» Комитета
Российской Федерации по печати. 143200, г. Можайск,
ул. Мира, 93.
9 785030 028507
-
Максин Сингер родилась в Нью-Иорке.
Окончила Йельский университет. В 1957 г.
получила степень доктора философии в
области биохимии. С 1956 по 1975 г.
работала в Национальном институте артрит„
и нарушений обмена веществ. В 1975 г.
перешла в Национальный институт рака, гдп
в 1979 г. возглавила лабораторию биохимии.
В 1988 г. д-р Сингер стала Президентом
института Карнеги, сохраняя за собой
руководство лабораторией. Как член
Национальной академии наук она
возглавляла редколлегию журнала
Proc edings of the National Academy of
Sciences. В 1988 г. получила Президентскую
премию за достижения в области науки,
что является высочайшей наградой,
присуждаемой гражданским лицам.
В настоящее время д-р Сингер занимается
изучением мобильных элементов генома
человека.
поп Берг родился в Нью-Йорке. Окончил
Университет шт. Пенсильвания. В 1952 г.
получил степень доктора философии
в области биохимии Продолжил свою
деятельность в Институте клеточной
физиологии в Копенгагене и в Университете
Вашингтона в Сент-Луисе. Затем перешел
в Стэнфордский университет, где
и продолжает работать в качестве
профессора биохимии и директора
Бекмзновс» ) ц ,-нтра по молекулярно
генетическим исследованиям в медицине.
В 1980 г. Берг получил Ласкеровскую
премию в области медицины и Нобелевскую
премию по химии за свои исследования
н/клсиноаых кислот, в частности
рекомбинантный ДНК. Занимается анализом
экспрессии генов эукариот.