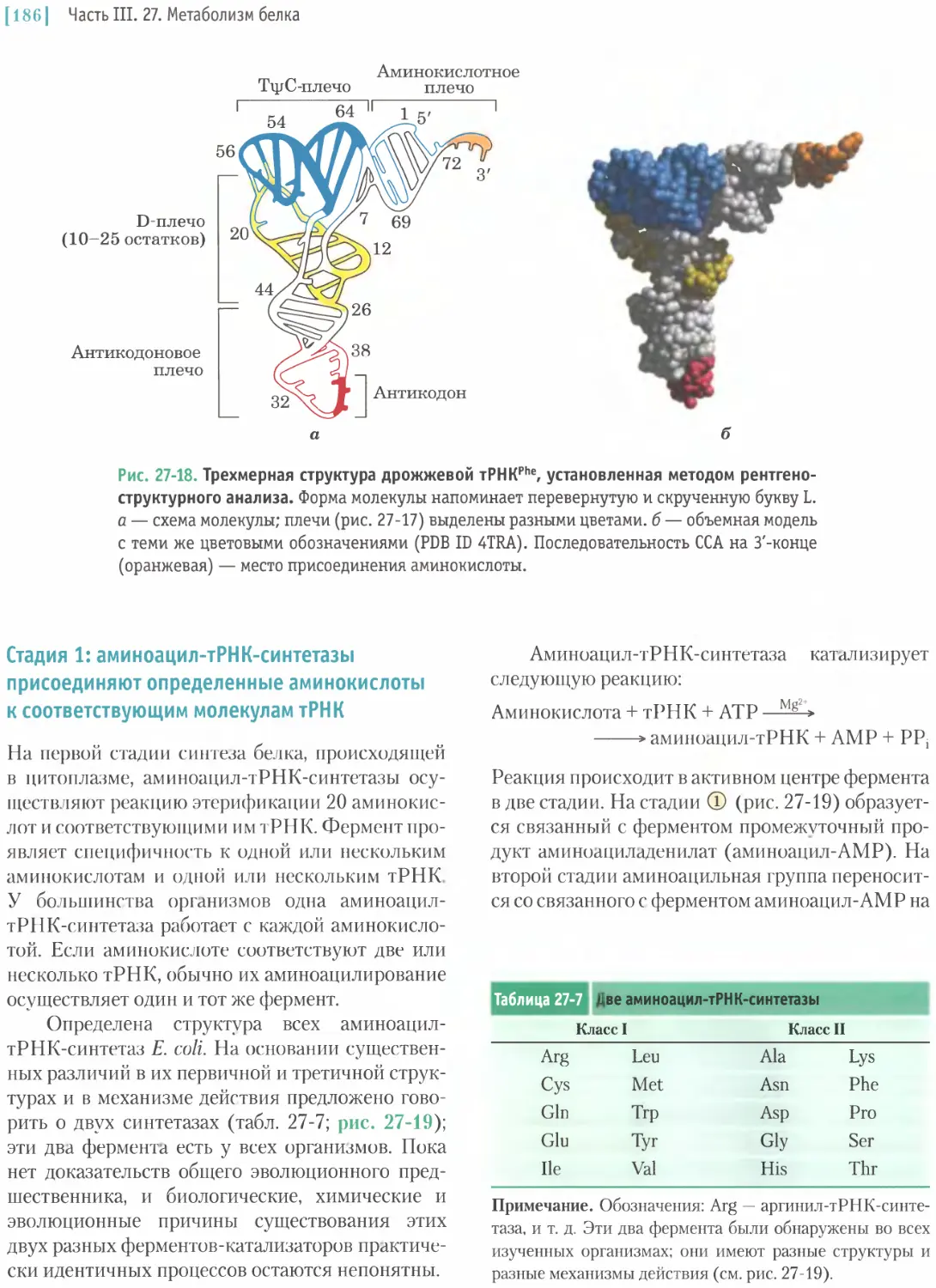
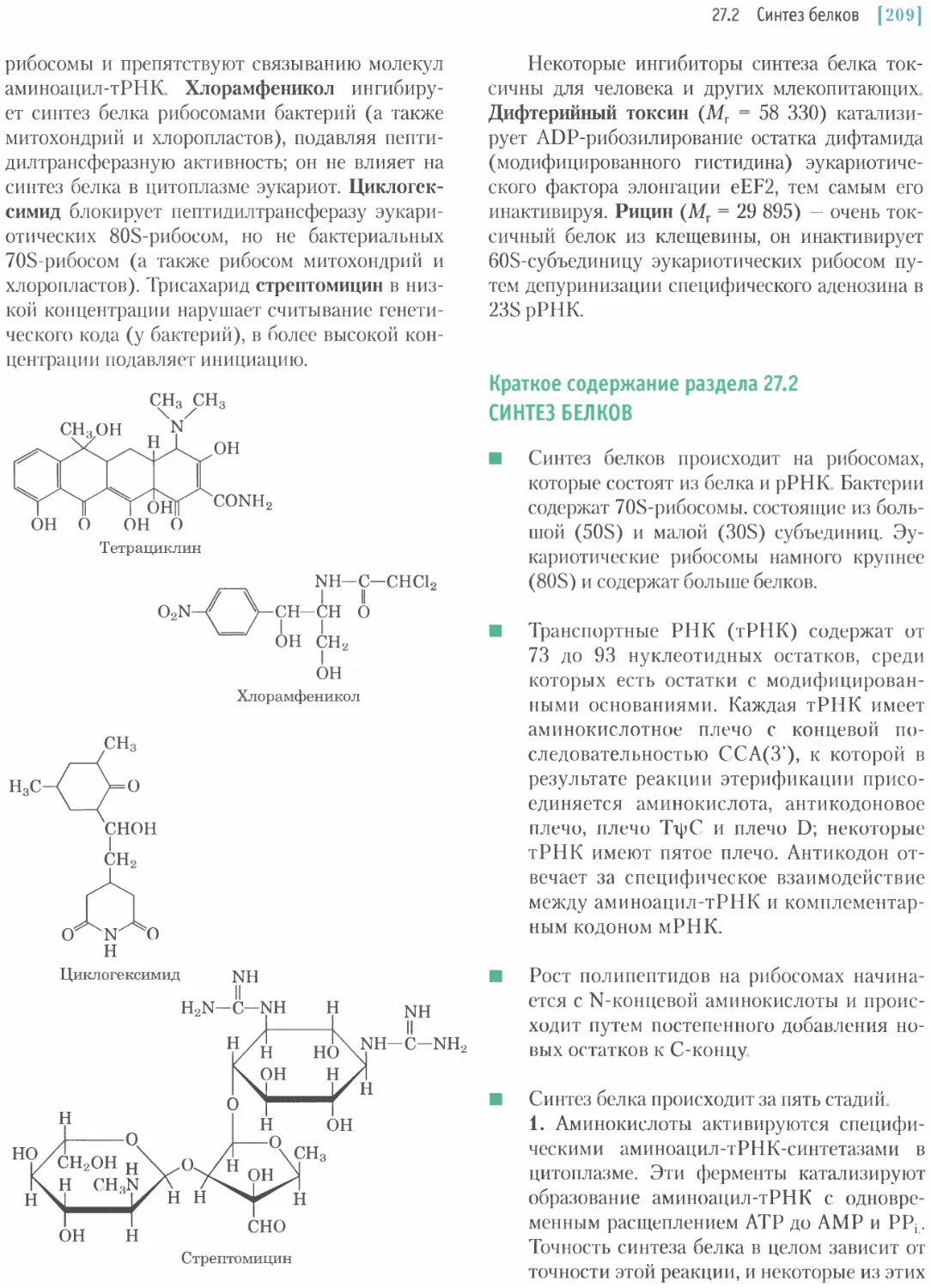
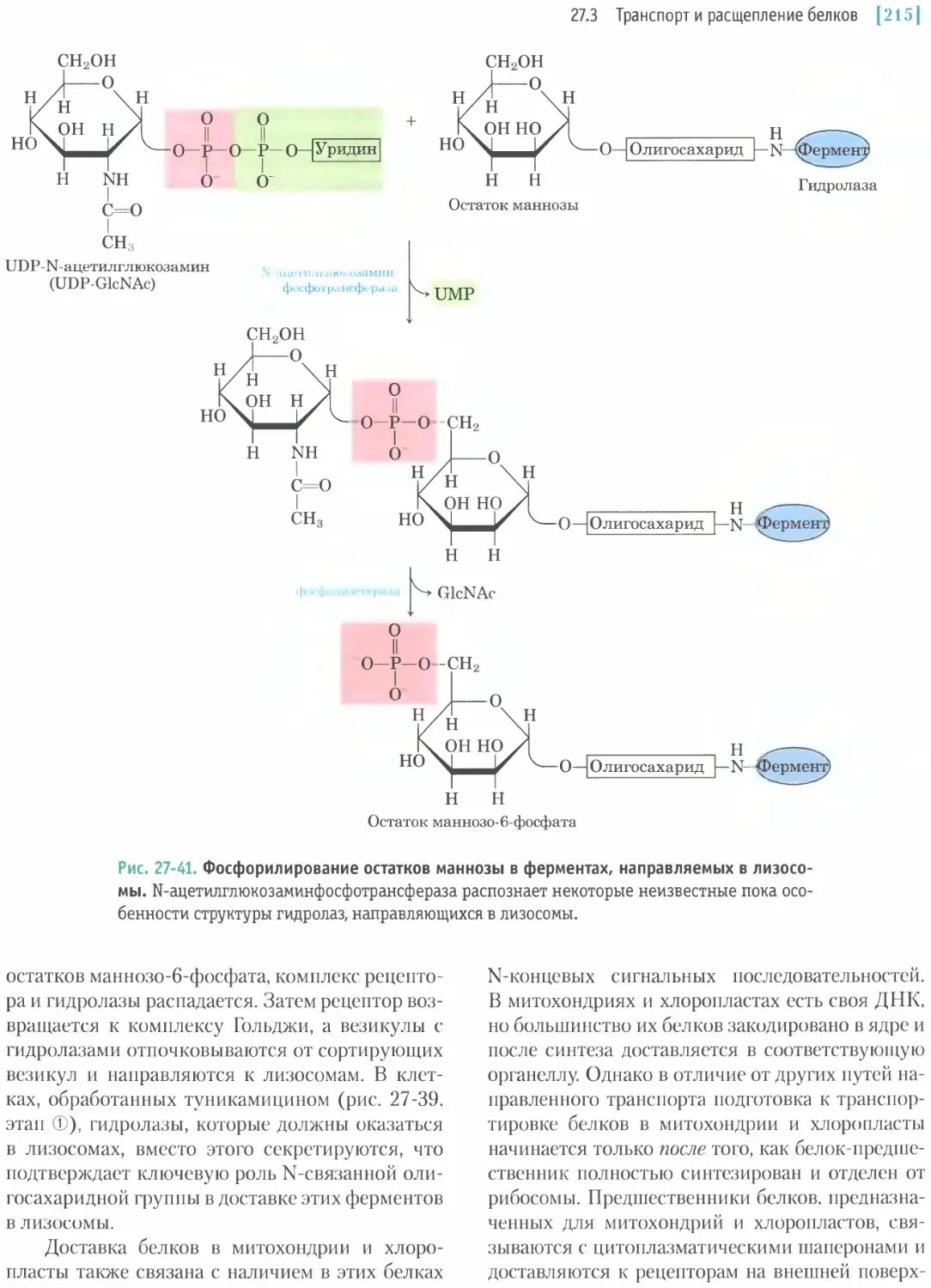
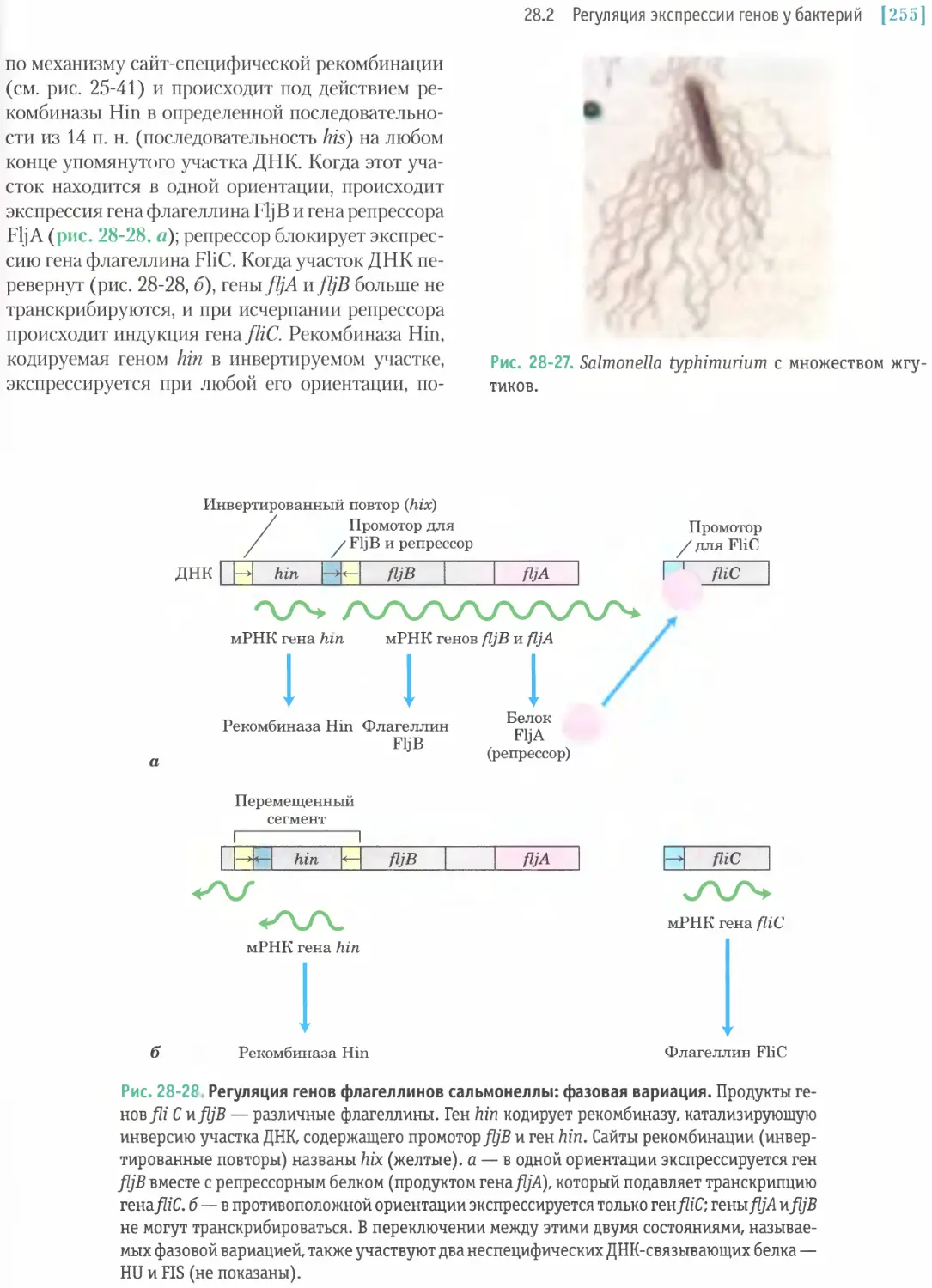
/


















































































































































































































































































































































































































































Author: Нельсон Д. Кокс М.
Tags: вирусология общая биохимия биохимия генетика молекулярная биология
ISBN: 978-5-94774-367-8
Year: 2015
Similar




Text
УДК 578.1
ББК 28.072я73
Н49
Серия основана в 2006 г.
Нельсон Д.
Н49 Основы биохимии Ленинджера : в 3 т. Т. 3 : Пути передачи
информации / Д. Нельсон, М. Кокс ; пер. с англ. —М. : БИНОМ.
Лаборатория знаний, 2015.—448 с. : ил.— (Лучший зарубежный
учебник).
ISBN 978-5-94774-367-8 (Т. 3)
ISBN 978-5-94774-364-7
В учебном издании, написанном американскими учеными, которые по-
лучили признание как талантливые преподаватели университетского уровня,
рассмотрены современные концепции биохимии в соответствии с изменив-
шейся идеологией этой науки.
В том 3 вошла часть III «Пути передачи информации», ответы на вопро-
сы, решения задач и предметно-именной указатель по материалу томов 1-3,
а также принятые сокращения и словарь терминов. Обсуждаются основ-
ная догма молекулярной биологии и ее современное понимание, процессы
передачи и хранения генетической информации как у бактерий, так и у эу-
кариот (репликация, транскрипция, трансляция, репарация и рекомбинация),
строение хромосом, механизмы ферментативных процессов, функции раз-
личных РНК в клетке, рибозимы, сплайсинг, альтернативный сплайсинг,
процессинг. Подробно описан биосинтез белка, его транспортировка к ме-
сту использования и дальнейшее разрушение, регуляция экспрессии генов.
В каждой главе (как в томах 1 и 2) приведены примеры из медицины,
молекулярной биологии и смежных областей, а также интересные задания
и вопросы.
Для студентов биологических, химических и медицинских вузов, а также
научных работников.
УДК 578.1
ББК 28.072я73
Учебное издание
Серия: «Лучший зарубежный учебник»
Нельсон Дэвид, Кокс Майкл
ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА
В трех томах
Том 3
ПУТИ ПЕРЕДАЧИ ИНФОРМАЦИИ
Ведущий редактор канд. хим. наук Т. И. Почкаева. Редактор канд. биол. наук Т. Е. Толстихина
Художник Н. А. Новак. Технический редактор Е. В. Денюкова. Компьютерная верстка: Т. Э. Внукова
Подписано в печать 23.10.14. Формат 84x108/16.
Усл. печ. л. 47,04.
Издательство «БИНОМ. Лаборатория знаний»
125167, Москва, проезд Аэропорта, д. 3
Телефон: (499)157-5272, e-mail: binom@Lbz.ru, http://www.Lbz.ru
Отпечатано в типографии «БАЛТО принт», Литва
ISBN 978-5-94774-367-8 (Т. 3)
ISBN 978-5-94774-364-7
First published in the United States
by W. H. FREEMAN AND COM PANY,
New York
Copyright © 2008 by W. H. Freeman and Company
All rights reserved.
© Перевод на русский язык,
БИНОМ. Лаборатория знаний, 2015
ЧАСТЬ III
Пути передачи
информации
24 Гены и хромосомы 7
25 Метаболизм ДНК 43
26 Метаболизм РНК 105
27 Метаболизм белка 165
28 Регуляция экспрессии генов 229
Третья, заключительная, часть книги по-
священа биохимическим механизмам,
обеспечивающим процессы передачи на-
следственной информации и эволюции живых
существ.
Какова молекулярная основа генетическо-
го материала? Как генетическая информация с
высокой точностью передается из поколения в
поколение? Как возникают редкие изменения в
генетическом материале, которые служат исход-
ным материалом для эволюции? Как генетиче-
ская информация превращается в аминокислот-
ные последовательности белковых молекул?
Современное понимание метаболических
путей передачи информации сформировалось
на стыке генетики, физики и химии — основ со-
временной биохимии. Джеймс Уотсон и Френсис
Крик в 1953 г. сформулировали гипотезу двух-
цепочечной структуры ДНК (см. рис. 8-15 в т. 1).
Генетическая теория помогла сформировать кон-
цепцию кодирования информации в генах. Вы-
дающиеся открытия в физике позволили устано-
вить молекулярную структуру гена. Вклад химии
в генетическую теорию состоял в определении
состава ДНК. Особая ценность гипотезы Уот-
сона-Крика заключалась в том, что она смогла
обобщить разнообразные наблюдения, получен-
ные в различных областях науки.
Революция в наших представлениях о
структуре ДНК неизбежно породила вопросы о
функции ДНК. Двухцепочечная структура сама
подсказывает механизм копирования ДНК, по-
зволяющий передавать из поколения в поколе-
ние закодированную информацию. Открытие
матричной и транспортной РНК, а также рас-
шифровка генетического кода позволили по-
нять, как ДНК преобразуется в функциональ-
ные белки.
Эти и другие открытия позволили сформу-
лировать основную догму молекулярной био-
логии, отражающую три главных процесса об-
работки генетической информации в клетке.
Первый процесс — репликация, суть которой
заключается в копировании ДНК и создании
на основе материнской ДНК молекулы дочер-
ней ДНК с идентичной последовательностью?
Второй процесс — транскрипция, в результате
которой часть генетической информации, зако-
дированной в ДНК, превращается в молекулы
РНК. Третий процесс — трансляция, при кото-
рой генетическая информация, закодированная
в РНК, переносится на рибосомы, где трансли-
руется в полипептид с определенной последова-
тельностью аминокислот.
Основная догма молекулярной биологии
указывает направление передачи информации в
[6] Часть III. Пути передачи информации
Репликация ДНК
Транскрипция
РНК
Трансляция
Белок
Основная концепция (догма) молекулярной биологии, ко-
торая объясняет главные метаболические пути передачи
информации — репликацию, транскрипцию и трансляцию.
Говорить, что это «догма», не совсем верно. Ведь эта концеп-
ция как догма была предложена Френсисом Криком, когда
было мало доказательств, подтверждающих выдвинутые
идеи, позже ставшие хорошо обоснованной теорией.
клетке: от репликации к транскрипции и транс-
ляции. Термин «догма» не совсем верен и сохра-
нился только по историческим причинам. Он
был введен Френсисом Криком в то время, когда
существовало мало доказательств, подтверждав-
ших выдвинутые идеи, позже ставшие хорошо
обоснованной теорией
В части II] обсуждаются эти и другие про-
цессы, связанные с передачей информации. В
главе 24 мы рассмотрим структуру, топологию и
упаковку хромосом и генов. Процессы, лежащие
в основе догмы, рассматриваются в главах 25-27.
В заключение мы рассмотрим процесс регуляции
экспрессии генетической информации (глава 28).
Важнейший вопрос, обсуждающийся во всех
этих главах, касается сложных процессов био-
синтеза информационных макромолекул. Сборка
нуклеотидов и аминокислот в определенные по-
следовательности нуклеиновых кислот и белков
служит для сохранения и точного копирования
матрицы, а ведь на этом основана сама жизнь.
Можно подумать, что образование фосфодиэфир-
ных связей в ДНК или пептидных связей в бел-
ках — тривиальная задача для клеток, обладающих
целым арсеналом ферментативных и химических
инструментов, описанных в части II. Однако, что-
бы учесть механизмы сохранения и передачи ин-
формации, нам придется значительно расширить
систему наших взглядов, сформулированную на
основе анализа метаболических путей. Химиче-
ские связи должны возникать между конкретными
субъединицами информационных биополимеров
с минимальной вероятностью появления и закре-
пления ошибок. Это требование оказывает очень
серьезное влияние на термодинамику, химию и
энзимологию процессов биосинтеза. Образование
пептидной связи требует затраты энергии, при-
мерно равной 21 кДж/моль, и может происходить
с участием относительно простых ферментов,
выступающих в качестве катализаторов. Но для
синтеза связи между двумя определенными ами-
нокислотами в конкретной точке полипептида
требуется примерно 125 кДж/моль, причем в этом
процессе задействовано более 200 ферментов, мо-
лекул РНК и специфических белков. Химический
процесс образования пептидной связи тот же са-
мый, но здесь подключаются дополнительные
процессы, гарантирующие образование этой связи
строго между определенными аминокислотами
Информация стоит дорого.
Еще одна важная тема, затрагиваемая в ча-
сти III, касается динамического взаимодействия
между нуклеиновыми кислотами и белками. За
исключением тех редких случаев, когда в роли ка-
тализаторов выступают молекулы РНК (эта тема
обсуждается в гл. 26 и 27), метаболические про-
цессы, связанные с передачей информации, ката-
лизируются и регулируются белками. Изучение
этих ферментов и других белков имеет не только
научное, но и прикладное значение, поскольку
позволяет применять их в технологиях, основан-
ных па рекомбинантных ДНК (см. гл. 9 в т. 1).
И вновь возвратимся к теме эволюции. Мно-
гие процессы, рассмотренные в части III, возникли
миллиарды лет назад, а некоторые прослеживают-
ся вплоть до последнего универсального общего
предшественника — LUCA (от англ, last universal
common ancestor). Рибосомы, практически весь
трансляционный аппарат и некоторые элементы
транскрипционного аппарата есть у всех живых
организмов на нашей планете. Генетическую ин-
формацию можно рассматривать в качестве сво-
еобразных молекулярных часов, которые позво-
ляют установить родственные отношения между
видами. Общие информационные пути связывают
человека со всеми ныне живущими на Земле ор-
ганизмами, а также со всеми прежде существо-
вавшими видами. Изучение этих путей помогает
ученым приоткрыть занавес в первом акте пьесы,
повествующей о возникновении жизни на Земле.
Суперскручивание значит для ДНК больше, чем просто силовой фактор;
оно сохраняет буйную раскидистую ДНК в узких рамках внутриклеточ-
ных условий.
— Николас Коззарелли, Лекции в Харви, 1993
24
Гены и хромосомы
24.1. Элементы хромосом 7
24.2. Сверхспирализация ДНК 15
24.3. Структура хромосом 27
Размер ДНК предлагает нам интересную био-
логическую загадку, поскольку молекулы
ДНК обычно значительно крупнее самих
клеток или вирусных частиц (рис. 24-1). Возника-
ет вопрос, как такие крупные молекулы упакованы
в клетке. Чтобы ответить на этот вопрос, следует
перейти от рассмотрения вторичной структуры
ДНК (см. гл. 8) к ее удивительной третичной
структуре, лежащей в основе строения хромо-
сом — хранилища генетической информации.
В начале главы мы рассмотрим основные элемен-
ты хромосом, а затем остановимся на обсуждении
их размера и организации. Далее мы обратимся к
топологии ДНК и обсудим варианты скручивания
и суперскручивания молекул В заключение мы
обсудим взаимодействия ДНК с белками, способ-
ствующие компактной укладке хромосом.
24.1. Элементы хромосом
Клеточная ДНК содержит гены и межгенные об-
ласти; и те и другие могут выполнять жизненно
важные функции. Более сложные геномы, напри-
мер геномы эукариот, нуждаются в более слож-
ен, о мкм
Рис. 24-1. Белковый капсид бактериофага Т2 окружа-
ет единственную линейную молекулу ДНК этого фага.
В результате лизиса частиц бактериофага в дистиллиро-
ванной воде ДНК вышла из капсида и распространилась
по поверхности воды. Частица бактериофага Т2 состоит
из головки и хвоста, с помощью которого бактериофаг
прикрепляется к внешней поверхности бактериаль-
ной клетки. Вся ДНК, показанная на этой электронной
микрофотографии, обычно содержится внутри головки
фага.
[8] Часть III. 24. Гены и хромосомы
ных уровнях организации хромосом. Мы начнем
с рассмотрения различных типов последователь-
ностей ДНК и структурных элементов хромосом.
Гены — это участки молекул ДНК,
кодирующие полипептиды и молекулы РНК
За последнее столетие наше представление о
генах существенно изменилось. Ранее геном на-
зывали участок хромосомы, кодирующий или
определяющий один признак или фенотипиче-
ское (видимое) свойство, например цвет глаз.
В 1940 г. Джордж Бидл и Эдвард Тейтем пред-
ложили молекулярное определение гена. Уче-
ные обрабатывали споры гриба Neurospora crassa
рентгеновским излучением и другими агентами,
вызывающими изменения в последовательности
ДНК (мутации), и обнаружили мутантные штам-
мы гриба, утратившие некоторые специфические
ферменты, что в некоторых случаях приводило к
нарушению целою метаболического пути. Бидл
и Тейтем пришли к выводу, что ген — это уча-
сток генетического материала, который опреде-
ляет или кодирует один фермент. Так появилась
гипотеза «один ген — один фермент». Позднее
эта концепция была расширена до определения
«один ген — один полипептид», поскольку мно-
гие гены кодируют белки, не являющиеся фер-
ментами, а полипептид может оказаться субъеди-
ницей сложного белкового комплекса.
Современное биохимическое определение
гена еще более конкретно. Генами называются все
участки ДНК, кодирующие первичную после-
довательность конечных продуктов, к которым
относятся полипептиды или РНК, обладающие
ДНК мРНК
G С
Т А
G С
G С
А Т
Т А
А Т
С G
А Т
С G
Т А
Т А
Т А
Т А
G С
С G
С G
G С
Т А
Т А
Т А
С G
I
С
G
U
G
G
А
U
А
С
А
С
и
и
и
и
G
С
С
G
U
U
U
С
и
Полипептид
|| N-конец
Arg
Gly
Туг
Thr
Phe
Ala
Vai
Ser
. С-конец
Транскрибируемая цепь
Рис. 24-2. Соответствие между кодирующими участками
ДНК, мРНК и аминокислотной последовательностью по-
липептидной цепи. Триплеты нуклеотидов в ДНК опре-
деляют аминокислотную последовательность белка при
посредничестве мРНК. Одна из цепей ДНК играет роль
матрицы для синтеза мРНК, нуклеотидные триплеты (ко-
доны) которой комплементарны триплетам ДНК. У неко-
торых бактерий и многих эукариот кодирующие после-
довательности прерываются некодирующими участками
(так называемыми интронами).
Джордж Бидл. 1903-1989
структурной или каталитической функцией. На-
ряду с генами ДНК содержит и другие последо-
вательности, выполняющие исключительно ре-
гуляторную функцию. Регуляторные последо-
вательности могут обозначать начало или конец
генов, влиять на транскрипцию или указывать
место инициации репликации или рекомбина-
ции (гл. 28). Некоторые гены могут экспрессиро-
ваться разными путями, при этом один и тот же
участок ДНК служит матрицей для образования
разных продуктов. Соответствующие механизмы
транскрипции и трансляции описаны в гл. 26-28.
24.1 Элементы хромосом [9]
Мы можем приблизительно рассчитать ми-
нимальный размер гена, кодирующего средний
белок. В гл. 27 подробно рассказано о том, что
каждая аминокислота в полипептидной цепи ко-
дируется последовательностью из трех нуклео-
тидов ( ис. 24- ); последовательности этих три-
плетов (кодонов) соответствуют цепочке ами-
нокислот в полипептиде, который кодируется
данным геном. Полипептидная цепь из 350 ами-
нокислотных остатков (цепь средней длины)
соответствует последовательности из 1050 п. н.
Однако многие гены эукариот и некоторые гены
прокариот прерываются сегментами ДНК, не
несущими информации о белке, и поэтому ока-
зываются значительно длиннее, чем показывает
простой расчет.
Сколько генов в одной хромосоме? Хро-
мосома прокариота Escherichia coli, чей геном
полностью расшифрован, представляет собой
кольцевую молекулу ДНК (на самом деле, это
не правильный круг, а скорее петля без начала
и конца), состоящую из 1 639 675 и. н. В этой
последовательности содержится примерно
4300 генов белков и еще 157 генов стабильных
молекул РНК В геноме человека примерно
3,1 млрд пар нуклеотидов, соответствующих
почти 29 000 генам, расположенным на 24 раз-
ных хромосомах.
Молекулы ДНК гораздо крупнее,
чем клеточные или вирусные структуры,
в которые они упакованы
Молекулы хромосомной ДНК обычно на мно-
го порядков длиннее, чем клетки или вирусные
частицы, в которых они содержатся (рис 24-1;
табл. 24-1). Это относится ко всем классам орга-
низмов и к вирусам.
Вирусы. Вирусы не могут жить вне другого ор-
ганизма, вне жизнеспособной клетки. Скорее их
можно назвать внутриклеточными паразитами,
использующими ресурсы клетки хозяина для
размножения. Многие вирусные частицы состоят
только из генома (обычно одной молекулы РНК
или ДНК), окруженного белковым чехлом
Геномы почти всех вирусов растений и неко-
торых вирусов бактерий и животных состоят из
РНК. Такие геномы обычно небольшого размера.
Например, геномы ретровирусов млекопитаю-
щих, таких как ВИЧ. содержат около 9000 нукле-
отидов, а бактериофаг QP — 4220 нуклеотидов.
Геномы обоих вирусов представляют собой одно-
цепочечную РНК
Геномы ДНК-содержащих вирусов намного
крупнее (табл. 24-1). Многие молекулы ДНК ви-
русов какую-то часть жизненного цикла находят-
ся в замкнутой кольцевой форме. При репликации
вируса в клетке хозяина могут появляться спец-
ифические формы вирусных ДНК, называемые
репликативными формами; например, многие ли-
нейные молекулы ДНК становятся кольцевыми, а
одноцепочечные ДНК образуют димеры. Типич-
ным ДНК-содержащим вирусом среднего размера
является бактериофаг X, инфицирующий Е. coli.
Репликативная форма ДНК фага X внутри клеток
представлена кольцевой двухцепочечной спира-
лью. Двухцепочечная ДНК содержит 48 502 п. и.,
а длина ее контура составляет 17,5 мкм. Геном
бактериофага фХ 174 тоже содержит ДНК, но его
размер намного меньше; в вирусной частице ДНК
представлена одноцепочечной кольцевой молеку-
лой, а двухцепочечная репликативная форма со-
держит 5386 и. н. Хотя вирусные геномы малень-
кие, длина их ДНК намного больше, чем размер
самих вирусных частиц, содержащих эти молеку-
лы ДНК (табл. 24-1).
[таблица 24-1 Размеры ДНК и вирусных частиц некоторых вирусов бактерий (бактериофагов)
Вирус Размер вирусной ДНК, и. н. Длина вирусной ДНК, нм Длина вирусной частицы, нм
фХ 174 5 386 1939 25
Т7 39 936 14377 78
X 48 502 17460 190
Т4 168 889 60800 210
Примечание. Размер ДНК указан для репликативной (двухцепочечной) формы Длину
ДНК оценивали, считая размер пары нуклеотидов равным 3,4 А (см. рис. 8-13 в т. 1).
[10] Часть III. 24. Гены и хромосомы
Бактерии. В одной клетке Е. coli содержится при-
мерно в 100 раз больше ДНК. чем в частице бак-
териофага X. Бактерия Е. coli имеет одну двухце-
почечную кольцевую молекулу ДНК. Она состо-
ит из 4 639 675 п. н. и достигает в длину примерно
1,7 мм, что превышает длину самой клетки Е. coli
приблизительно в 850 раз (рис. 24-3). Помимо
крупной кольцевой хромосомы в составе нуклео-
ида многие бактерии содержат одну или несколь-
ко маленьких кольцевых молекул ДНК, свобод-
но располагающихся в цитозоле. Такие внех-
ромосомные элементы называют плазмидами
ис. 24-3. Хромосома Е. coli (длиной 1,7 мм) представлена в линейной форме;
рядом приведена клетка Е. coli (2
Клетка Е. coli
ДНК Е. coli
Рис. 24-4. ДНК из лизированной клетки Е. coli. Бе-
лыми стрелками отмечены кольцевые молекулы
плазмид. Белые и черные пятна — артефакты.
(>ис. 24-4; см. также с. 439 в т. 1). Большин-
ство плазмид состоит всего из нескольких ты-
сяч пар нуклеотидов, некоторые содержат более
10 000 п. н. Они несут генетическую информацию
и реплицируются с образованием дочерних плаз-
мид, которые попадают в дочерние клетки в про-
цессе деления родительской клетки. Плазмиды
обнаружены не только в бактериях, но также в
дрожжах и других грибах.
Во многих случаях плазмиды не дают ника-
ких преимуществ клеткам-хозяевам, и их един-
ственная задача - независимое воспроизведение.
Однако некоторые плазмиды несут полезные для
хозяина гены. Например, содержащиеся в плазми-
дах гены могут придавать клеткам бактерий устой-
чивость к антибактериальным агентам. Плазмиды,
несущие ген [3-лактамазы, обеспечивают устой-
чивость к р-лактамным антибиотикам, таким как
пенициллин и амоксициллин (см. рис. 6-28 в т. 1).
Плазмиды могут переходить от клеток, устойчи-
вых к антибиотикам, к другим клеткам того же или
другого вида бактерий, в результате чего эти клет-
ки также становятся резистентными. Интенсив-
ное применение антибиотиков является мощным
селективным фактором, способствующим распро-
странению плазмид, кодирующих устойчивость к
антибиотикам (а также транспозонов, которые ко-
дируют аналогичные гены) среди болезнетворных
бактерий, и приводит к появлению бактериальных
штаммов с устойчивостью к нескольким антибио-
тикам. Врачи начинают понимать опасность ши-
рокого использования антибиотиков и назначают
их только в случае острой необходимости. По ана-
логичным причинам ограничивается широкое ис-
пользование антибиотиков для лечения сельско-
хозяйственных животных.
24.1 Элементы хромосом [111
Эукариоты. В клетке дрожжей, одних из самых
маленьких эукариот, в 2,6 раза больше ДНК,
чем в клетке Е. coll (табл. 24-2). Клетки плодо-
вой мушки Drosophila, классического объекта
генетических исследований, содержат в 35 раз
больше ДНК, а клетки человека — примерно в
700 раз больше ДНК, чем клетки Е. coli. Мно-
гие растения и амфибии содержат еще больше
ДНК. Генетический материал клеток эукариот
организован в виде хромосом. Диплоидный на-
бор хромосом (2 п) зависит от вида организма
(табл. 24-2). Например, в соматической клетке
ДНК. гены и хромосомы некоторых организмов
Общая ДНК, п. н. Число хромосом3 Примерное число генов
Escherichia coli (бактерия) 4 639 675 1 4 435
Saccharomyces cerevisiae (дрожжи) 12 080 ООО 166 5 860
Caenorhabditis elegans (нематода) 90 269 800 12в 23 000
Arahidopsis thaliana (рас гение) 119 186 200 10 33 000
Drosophila melanogaster (плодовая мушка) 120 367 260 18 20000
Oryza sativa (рис) 480 000 000 24 57 000
Mus musculus (мышь) 2 634 266 500 40 27 000
Homo sapiens (человек) 3 070 128 600 46 29 000
Примечание. Информация постоянно обновляется; для получения более свежей информации об-
ратитесь к сайтам, посвященным отдельным геномным проектам.
а Для всех эукариот, кроме дрожжей, приводится диплоидный набор хромосом
6 Гаплоидный набор. Дикие штаммы дрожжей обычно имеют восемь (октаплоидный) или больше
наборов таких хромосом.
в Для самок с двумя X хромосомами У самцов есть X хромосома, но пег Y. т. е. всего 11 хромосом.
человека 46 хромосом (рис. 24-5). Каждая хро-
мосома эукариотической клетки, как показано
на рис. 24-5, а, содержит одну очень крупную
двухспиральную молекулу ДНК. Двадцать
четыре хромосомы человека (22 парные хро-
мосомы и две половые хромосомы X и Y) раз-
личаются по длине более чем в 25 раз. Каждая
хромосома эукариот содержит определенный
набор генов.
Если соединить между собой молекулы ДНК
человеческого генома (22 хромосомы и хромосо-
мы X и Y или X и X), получится последователь-
Рис. 24-5. Хромосомы эукариот, а — пара связанных и конденсированных сестринских хроматид
из хромосомы человека. В такой форме эукариотические хромосомы пребывают после реплика-
ции и в метафазе в процессе митоза, б — полный набор хромосом из лейкоцита одного из авторов
книги. В каждой нормальной соматической клетке человека содержится 46 хромосом.
[12] Часть III. 24. Гены и хромосомы
ность длиной около одного метра. Большинство
клеток человека диплоидны, поэтому общая дли-
на ДНК таких клеток около 2 м. У взрослого чело-
века примерно 1014 клеток, таким образом, общая
длина всех молекул ДНК составляет 2 • 1011 км.
Для сравнения, окружность Земли — 4 • 101 км,
а расстояние от Земли до Солнца - 1,5 • 10й км.
Вот как удивительно компактно упакована ДНК
в наших клетках!
В клетках эукариот есть и другие орынел-
лы, содержащие ДНК, - это митохондрии и хло-
ропласты. Молекулы митохондриальной ДНК
(мтДНК) намного меньше ядерных хромосом.
Размер двухцепочечных кольцевых мтДНК в
клетках животных составляет менее 20 000 п. н.
(в митохондриях человека молекула мтДНК
состоит из 16 569 п. н.). Каждая митохондрия
обычно несет от двух до десяти копий моле-
кул мтДНК; их число может возрасти до сотен
в некоторых клетках, например в дифференци-
рующихся клетках эмбриона. У некоторых ор-
ганизмов (например, тринаносом) каждая ми-
тохондрия содержит тысячи копий мтДНК; ми-
тохондрии организованы в сложный комплекс,
называемый кинетопластом. Размеры мтДНК
растительных клеток варьируют от 200 000 до
2 500 000 п. н. Молекулы ДНК хлоропластов
(хпДНК) тоже двухцепочечные и кольцевые,
Рис. 24-6 Делящаяся митохондрия. Некоторые мито-
хондриальные белки и молекулы РНК (на фотографии
не видны) кодируются одной копией митохондриальной
ДНК. Митохондриальная ДНК (мтДНК) реплицируется
каждый раз, когда делится митохондрия, что предшеству-
ет делению клетки.
их длина колеблется от 120 000 до 160 000 п. н.
Выдвигалось множество гипотез относительно
происхождения ДНК митохондрий и хлоро-
пластов. Общепризнанная сегодня точка зре-
ния заключается в том, что они представляют
собой рудименты хромосом древних бактерий,
которые проникли в цитоплазму хозяйских кле-
ток и стали предшественниками этих органелл
(см. рис. 1-36 вт. 1). Митохондриальная ДНК
кодирует митохондриальные тРНК и рРНК, а
также несколько митохондриальных белков. Бо-
лее 95% митохондриальных белков кодируется
ядерной ДНК. Митохондрии и хлоропласты де-
лятся вместе с клетками. ДНК этих органелл ре-
плицируется перед делением клетки и в процес-
се деления, а затем дочерние молекулы мтДНК
попадают в органеллы дочерних клеток.
Гены и хромосомы эукариот
очень сложно организованы
У бактерий многих видов всего одна хромосома,
и почти во всех случаях в каждой хромосоме при-
сутствует по одной копии каждого гена. Лишь не-
многие гены, например гены рРНК, содержатся
в нескольких копиях. Гены и регуляторные по-
следовательности состав 1яют практически весь
геном прокариот. Более того, почти каждый ген
строю соответствует аминокислотной последова-
тельности (или последовательности РНК), кото-
рую он кодирует (рис. 24-2).
Структурная и функциональная организа-
ция генов эукариот гораздо сложнее. Исследо-
вание хромосом эукариот, а позднее секвени-
рование полных последовательностей геномов
эукариот принесло много сюрпризов. Многие,
если не большинство, генов эукариот обладают
интересной особенностью: их нуклеотидные по-
следовательности содержат один или несколько
участков ДНК, в которых не кодируется амино-
кислотная последовательность полипептидного
продукта. Такие нетранслируемые вставки нару-
шают прямое соответствие между нуклеотидной
последовательностью гена и аминокислотной
последовательностью кодируемого полипеп-
тида. Эти нетранслируемые сегменты в составе
генов называют интронами, или встроенными
последовательностями, а кодирующие сегмен-
ты — экзонами. У прокариот лишь немногие
гены содержат интроны.
24.1 Элементы хромосом [13]
12 3 4
Ген яичного Д
альбумина
1
90 п. н.
В С D Е F G
2
222 п. н.
Экзон
3 Интрон г.
126 п. н.
р -Субъединица7
гемоглобина
В
851 п. н.
Рис. 24-7. Интроны в двух эукариотических генах. Ген яичного альбумина содержит семь
интронов (от А до G), разделяющих кодирующие последовательности восьми экзонов (L,
от 1 до 7). Ген р-субъединицы гемоглобина несет два интрона и три экзона, включая один
интрон, содержащий более половины пар нуклеотидов данного гена.
В типичных генах высших эукариот последо-
вательности интронов обычно намного длиннее,
чем последовательности экзонов. Например, в
гене одной полипептидной цепи яичного белка
овальбумина ( >ис. 24 ) интроны намного длин-
нее экзонов: семь интронов вместе составляют
85% ДНК гена. В гене р-субъелиницы гемогло-
бина более половины ДНК содержится в един-
ственном интроне. Ген мышечного белка титина
чемпион по количеству интронов - их 178. В ге-
нах гистонов, по-видимому, интронов нет. В боль-
шинстве случаев функция интронов не установ-
лена. В целом, лишь около 1,5% ДНК человека
являются «кодирующими», т. е. несут информа-
цию о белках или РНК. Однако с учетом крупных
интронов получается, что ДНК человека на 30%
состоит из генов.
Поскольку гены составляют относитель-
но небольшую долю в геноме человека, значи-
тельная часть ДНК остается неучтенной. На
рис. 24-8 на круговой диаграмме представлены
типы последовательностей в геноме. Большая
часть некодирующей ДНК существует в форме
повторяющихся последовательностей несколь-
ких типов. Самое удивительное, что примерно
половина генома человека состоит из повторя-
ющихся последовательностей подвижных гене-
тических элементов — участков ДНК размером
от нескольких сотен до нескольких тысяч пар
оснований, которые перемещаются с места на
место внутри генома. Эти подвижные генетиче-
ские элементы (транспозоны) — пример моле-
кулярных паразитов, эффективно расселяющих-
ся внутри генома хозяина. Многие транспозоны
несут гены белков, которые катализируют про-
цесс транспозиции, описанный более подробно
в гл. 25 и 26. Некоторые транспозоны в геноме
человека активны и перемещаются с небольшой
частотой, но большинство являются неактивны-
ми реликтами, измененными в результате мута-
ций в процессе эволюции. Хотя чаще всего такие
элементы не кодирую! функциональные белки
или РНК, они сыграли важную роль в эволюции
человека, поскольку перемещение транспозонов
привело к перераспределению других геномных
последовательностей.
Еще примерно 3% генома человека составля-
ют часто повторяющиеся последовательности,
пазываемые простыми последовательностями
ДНК или повторами простых последовательно-
стей (SSR — от англ, simple sequence repeats). Эти
короткие последовательности размером обычно
менее 10 и. и. иногда повторяются в клегке мил-
лионы раз. Простые последовательности ДНК
также называют сателлитной ДНК, поскольку
при центрифугировании фрагментов клеточной
ДНК в градиенте плотности хлорида цезия из-за
необычного состава оснований они часто мигри-
руют в виде отдельных полос (как спутники, «са-
теллиты»), сопровождающих остальную ДНК.
Простые последовательности ДНК не кодируют
белки или РНК. Но в отличие от подвижных ге-
нетических элементов сателлитная ДНК может
выполнять в клетках человека определенные
функции, поскольку многие простые последова-
тельности ДНК локализованы в двух специфи-
ческих участках эукариотических хромосом -
в центромере и теломерах.
[14] Часть III. 24. Гены и хромосомы
45%
Транспозону
Рис. 24-8 Типы последовательностей в геноме челове-
ка. На диаграмме представлены три главные составляю-
щие генома — транспозоны (подвижные генетические
элементы), гены и смешанные последовательности. Из-
вестно четыре основных класса транспозонов (на диа-
грамме представлены три из них). Длинные рассеянные
повторы (LINE) размером от 6 до 8 т. п. н. (1 т. п. н. =
1000 п. н.) обычно содержат несколько генов белков, ка-
тализирующих транспозицию. В геноме насчитывается
примерно 850 000 LINE. Длина коротких рассеянных по-
второв (SINE) примерно от 100 до 300 п. н. В геноме чело-
века их содержится около 1,5 млн, из которых более 1 млн
составляют элементы Alu, названные так по той причине,
что они обычно содержат один сайт рестрикции для эн-
донуклеазы рестрикции Alul (см. рис. 9-2 в т. 1). В геноме
также обнаружено 450 000 копий транпозонов, близких
ретровирусам, длиной от 1,5 до 11 т. п. н. Хотя они «за-
хвачены» геномом и не могут перемещаться из клетки в
клетку, эволюционно они близки ретровирусам (гл. 26),
к которым относится ВИЧ. Еще один класс транспозонов
(составляющий <3%, здесь не показан) представлен раз-
личающимися по длине фрагментами транспозонов.
Примерно 30% генома составляют последовательно-
сти, содержащие гены белков, но только малая часть этой
ДНК находится в экзонах (кодирующих последователь-
ностях). К смешанным последовательностям относятся
простые короткие повторы (SSR) и повторы крупных сег-
ментов (SD); последние встречаются более чем в одной
копии в различных положениях. Как отмечается в гл. 26,
почти весь геном в результате транскрипции превраща-
ется в последовательность РНК. причем многие участ-
ки РНК еще не охарактеризованы. Кроме того, в геноме
содержатся остатки транспозонов, которые в процессе
эволюции изменились настолько сильно, что их трудно
идентифицировать.
Центромера ( >ис. 24- ) представляет собой по-
следовательность ДНК, к которой в процессе
деления клетки прикрепляются белки, связыва-
ющие хромосому с митотическим веретеном. Это
взаимодействие важно для равномерного и точ-
ного разделения наборов хромосом по дочерним
клеткам. Были выделены и изучены центромеры
Saccharomyces cerevisiae. Важнейшие для функци-
онирования центромер последовательности име-
ют длину около 130 п. н. и содержат большое ко-
личество пар А = Т Центромеры высших эукари-
от намного длиннее и, в отличие от дрожжевых,
обычно содержат простую последовательность
ДНК, состоящую из тысяч тандемных копий од-
ной или нескольких коротких последовательно-
стей из 5 10 и. н. в одинаковой ориентации. Роль
простых повторов при функционировании цен-
тромер пока точно не установлена.
Теломеры (от греч. telos — конец) - последова-
тельности на концах хромосом эукариот, стаби-
лизирующие хромосомы. Теломеры заканчива-
ются многократно повторяющимися последова-
тельностями вида
(5')(TrG^)„
(3')(AaC,)w
где х и у обычно имеют значения от 1 до 4
(табл. 24-3). Число теломерных повторов п у
большинства одноклеточных эукариот состав-
ляет от 20 до 100, а у млекопитающих, как пра-
вило, превышает 1500. Концы линейной молеку-
лы ДНК не могут полностью реплицироваться
обычным способом клеточным репликацион-
ным аппаратом (возможно, это одна из причин
кольцевой структуры ДНК у бактерий). Тело-
мерные повторы присоединяются к концам эу-
кариотических хромосом в основном с помощью
фермента теломеразы (см. рис. 26-39).
Теломера Центромера Теломера
Уникальные последовательности (гены),
рассеянные повторы, множественные
точки начала репликации
Рис. 24-9. Важные особенности строения хромосомы
дрожжей.
24.2 Сверхспирализация ДНК [15]
Теломерные последовательности
Последовательность
теломерных повторов
Таблица 24-3
Homo sapiens (человек) (TTAGGG)„
Tetrahymena thermophila (реснитчатое простейшее) (TTGGGGX
Saccharomyces cerevisiae (дрожжи) ((TG)t 3(TG)2 зк
Arabidopsis thaliana (растение) (TTTAGGGX
Для изучения функциональной роли струк-
турных элементов эукариотических хромосом
были сконструированы искусственные хромо-
сомы (гл. 9 в т. 1). Для функционирования до-
статочно стабильной линейной искусственной
хромосомы нужны лишь три компонента: цен-
тромера, теломеры на каждом конце последова-
тельности и участок, в котором происходит ини-
циация репликации. Искусственные хромосомы
дрожжей (YAC; см. рис. 9-7) были созданы как
инструмент для биотехнологических исследо-
ваний. Для лечения генетических заболеваний
с помощью соматической генной терапии были
разработаны искусственные хромосомы челове-
ка (НАС; см. доп. 9-2).
Краткое содержание раздела 24.1
ЭЛЕМЕНТЫ ХРОМОСОМ
Ген участок хромосомы, который песет ин-
формацию о функциональном полипептиде
или молекуле РНК. Наряду с генами хромо-
сомы содержат разнообразные регуляторные
последовательности, участвующие в репли-
кации, транскрипции и других процессах.
Геномная ДНК и геномная РНК вирусов
обычно на несколько порядков длиннее со-
держащих их клеток или вирусных частиц.
Многие гены эукариот (но лишь некоторые
гены бактерий и архей) прерываются неко-
дирующими последовательностями — ин-
тронами. Кодирующие участки, разделенные
интронами, называют экзонами.
Менее трети геномной ДНК человека состав-
ляют гены. В остальной ДНК много повторя-
ющихся последовательностей различных ти-
пов. Паразитирующие нуклеиновые кисло-
ты, известные как транспозоны, составляют
примерно половину генома человека.
Хромосомы эукариот содержат две важные
повторяющиеся последовательности ДНК
со специфической функцией: центромеры
(участки прикрепления митотического вере-
тена) и теломеры, расположенные на концах
хромосом.
24.2. Сверхспирализация ДНК
Клеточная ДНК. как мы видели, чрезвычайно
компактна и, следовательно, обладает высокой
степенью структурной организации Механизм
укладки служит не только для упаковывания
£ — Суперскрученная
J спираль
Рис. 24-10. Сверхспирализация. Обычный телефонный
шнур закручен в спираль, как ДНК и сам может образовы-
вать дополнительные петли сверхспирали. Эта аналогия
уместна еще и по той причине, что именно телефонный
шнур помог Джерому Винограду и его коллегам понять,
что многие свойства маленьких кольцевых молекул ДНК
могут объясняться суперскручиванием. Впервые они об-
наружили явление сверхспирализвции ДНК в 1965 г. в
образцах маленьких молекул вирусной ДНК.
[16] Часть III. 24. Гены и хромосомы
ДНК, но и для обеспечения доступа к содержа-
щейся в ней информации. Прежде чем рассмо-
треть работу этого механизма в таких клеточных
процессах, как репликация и транскрипция, мы
должны обсудить важную структурную особен-
ность ДНК сверхспирализацию.
Сверхспирализация — скручивание уже су-
ществующей спирали. Примером такой струк-
туры является скрученный телефонный шнур.
На пути между трубкой и аппаратом часто об-
разуется одна или несколько сверхспиралей
( >ис. 24-10). Молекула ДНК скручена в двой-
ную спираль, в которой обе цепи ДНК закру-
чены вокруг единой оси. Дальнейшее закручи-
вание такой оси вокруг самой себя ( ис. 24-1 )
приводит к образованию сверхспирализованной
ДНК. Как мы увидим далее, сверхспирализация
обычно является результатом структурного на-
пряжения. Если ДНК не закручена вокруг своей
оси, ее называют релаксированной.
Логично предположить, что при упаковыва-
нии ДНК возникают различные варианты сверх-
спирализации. Менее очевидно, что репликация
и транскрипция ДНК тоже влияют на сверхсии-
Двойная
спираль ДНК
Рис. 24-11 Сверхспирализация ДНК. Когда двойная спи-
раль ДНК закручивается вокруг своей оси, образуется
новая спираль (сверхспираль). Суперскрученную ДНК
обычно называют сверхспиралью.
‘ > ;• - - - ~
Рис. 24-12 При разделении нитей спиральной структуры
происходит сверхспирализация. Закрутите две резинки
в правозакрученную двойную спираль, как показано на
верхнем рисунке. Попросите кого-нибудь подержать
один конец и попробуйте разделить нить с другого конца.
Вы будете наблюдать суперскручивание.
рализацию и сами зависят от нее. Оба процесса
связаны с разделением нитей ДНК, способству-
ющим образованию дополнительных витков спи-
рали (как показано на шс. 24-1' ).
То, что плотно упакованная клеточная ДНК
закручивается вокруг самой себя, приобретая
сверхспиральную структуру, кажется логичным и
даже тривиальным, если бы не одно наблюдение:
многие клеточные кольцевые молекулы ДНК оста-
ются в высокой степени суперскрученными даже
после того, как их экстрагируют и очищают, осво-
бождая от белков и других клеточных компонен-
тов. Из этого следует, что сверхспирализация —
неотъемлемое свойство четвертичной структуры
ДНК. Оно характерно для всех молекул клеточной
ДНК и строго регулируется каждой клеткой.
Сверхспирали характеризуются нескольки-
ми параметрами, которые можно измерить; на
основе этих данных были глубже поняты струк-
туру и функции ДНК. Исследования во многом
базируются на концепциях топологии — одного
из разделов математики; топология исследует
свойства объекта при непрерывной деформации.
В случае ДНК непрерывная деформация включа-
ет изменения конформации из-за теплового дви-
жения и взаимодействия с белками или другими
молекулами; прерывистая деформация приводит
к разрыву цепей ДНК. Для кольцевых молекул
ДНК топологические свойства — это те свой-
ства, которые не меняются при деформации це-
пей ДНК до тех пор, пока не возникают разрывы.
24.2 Сверхспирализация ДНК [17]
Топологические свойства изменяются только в
результате разрыва и последующего соединения
одной или обеих цепей ДНК.
Теперь рассмотрим фундаментальные свой-
ства и физическую основу сверхспирализации.
Большинство клеточных ДНК раскручены
Чтобы понять суть сверхспирализации, сосредо-
точимся сначала на свойствах небольших коль-
цевых молекул ДНК, таких как плазмиды и не-
большие ДНК-содержащие вирусы. Если такие
молекулы ДНК не имеют разрывов ни в одной из
цепей, они называются замкнутыми кольцевыми
молекулами ДНК. Если форма замкнутой коль-
цевой ДНК близка по структуре В-форме ДНК
(структуре Уотсона- Крика; см. рис. 8-13 в т. 1)
с одним оборотом двойной спирали на 10,5 п. н..
она скорее релаксирована, чем суперскручена
( >ис. 24-1 ). Суперскручивание происходит при
некотором напряжении структуры. Очищенная
замкнутая кольцевая ДНК редко бывает релак-
сированной, независимо от ее биологического
происхождения. Более того, молекулы ДНК, по-
лученные из определенного клеточного источни-
ка, имеют характерную именно для них степень
сверхспирализации. Следовательно, ДНК напря-
жена в такой степени, чтобы в ней возникали су-
первитки, и это состояние регулируется клеткой.
Практически в любой момент времени на-
пряжения двойной спирали ДНК возника-
ют из-за ее частичного раскручивания. Ина-
че говоря, в ДНК оказывается меньше витков
спирали, чем в В-форме. Влияние частично-
го раскручивания обобщено на рис. 24-14.
Участок кольцевой ДНК размером 84 и. н. в релак-
сированной форме может содержать восемь витков
двойной спирали, или один виток на 10,5 п. н. Если
один из таких витков удален, получается 84 п. н.: 7
= 12,0 п. н. на виток, т. е. больше, чем 10,5 — вели-
чина, характерная для В-ДНК (рис. 24-14, б). Это
отклонение от наиболее стабильной формы ДНК
приводит к термодинамическому напряжению в
молекуле, т. е. к неустойчивой форме ДНК. Отча-
сти напряжение может быть снято при скручива-
нии ДНК вокруг собственной оси с образованием
сверхспирали (рис. 24-14, в; некоторая часть напря-
жения на участке из 84 п. н. может просто распреде-
ляться по раскрученной структуре более крупного
фрагмента ДНК). В принципе напряжение может
быть компенсировано при разделении двух цепей
ДНК на расстоянии около 10 п. н. (рис. 24-14, г).
В изолированной замкнутой кольцевой ДНК на-
пряжение, вызванное частичным раскручивани-
ем, обычно в большей степени компенсируется
сверхспирализацией, чем при разделении цепей,
поскольку скручивание оси ДНК, как правило,
требует меньше энергии, чем разрыв водородных
связей между парами нуклеотидов. Однако следу-
ет отметить, что после частичного раскручивания
ДНК in vivo ее нити легче разделяются, что позво-
ляет считывать содержащуюся в них информацию.
Каждая клетка активно осуществляет рас-
кручивание своей ДНК с помощью ферментатив-
ных процессов (описано ниже), и возникающее в
результате напряженное состояние служит для
Рис. 24-13 Релаксированная и сверхспирализованная плазмидная ДНК. На крайней левой
электронной микрофотографии показана релаксированная молекула; степень сверхспира-
лизации возрастает слева направо.
[18] Часть III. 24. Гены и хромосомы
а Релаксированная ДНК (8 витков)
б Растянутая ДНК (7 витков)
в Суперскрученная ДНК
г Расплетение цепей
Рис. 24-14 Результаты раскручивания ДНК. а — участок
ДНК замкнутой кольцевой молекулы длиной 84 п. н. в
релаксированной форме с восемью оборотами спирали.
б — удаление одного витка вызывает напряжение струк-
туры. в — обычно напряжение сглаживается при супер-
скручивании. г — частичное раскручивание ДНК несколь-
ко облегчает разъединение цепей. Показано, что теоре-
тически раскручивание на один оборот может облегчить
разделение цепей на участке длиной около 10 п. н. Однако
обычно разделению нитей на таком коротком участке пре-
пятствуют водородные связи между парами нуклеотидов,
и эффект становится значимым только для более длинных
участков ДНК и более сильного раскручивания.
лекуле замкнутой кольцевой ДНК число витков
спирали нельзя изменить без хотя бы временного
разрыва одной из цепей ДНК. По этой причине
число витков спирали в молекуле ДНК является
характеристикой сверхспирализации.
Степень скручивания ДНК
определяется топологическим параметром —
порядком зацепления
Топология предлагает некоторые концепции, кото-
рые помогут нам в обсуждении сверхспирализации
ДНК, в частности концепцию порядка зацепления.
Порядок зацепления — топологическое свойство
двухцепочечной ДНК, поскольку оно не изменяет-
ся при сгибании или деформации ДНК до тех пор,
пока целы обе цепи. Число зацеплений обознача-
ют как Lk — от англ, linking number ( по. 24-15).
Давайте рассмотрим процесс разделения це-
пей двухцепочечной кольцевой ДНК. Если две
цепи связаны так, как показано на рис. 24-15, а,
их прочную связь можно назвать топологиче-
ской связью. Даже если разрушаются все водо-
родные связи и стэкинговые взаимодействия
между основаниями, так что нарушается физи-
ческий контакт между цепями, они по-прежнему
a Lk = l
запасания энергии. Клетки поддерживают ДНК в
частично раскрученном состоянии для того, что-
бы облегчить ее компактную упаковку. Раскручи-
вание ДНК также важно для функционирования
ферментов, участвующих в метаболизме ДНК,
которые для выполнения своей задачи должны
разделять две нити ДНК.
В таком раскрученном состоянии ДНК может
существовать лишь в том случае, если она замкну-
та в кольцо или связана и стабилизирована бел-
ками таким образом, что цепи не могут свободно
вращаться относительно друг друга. Если одна из
цепей в изолированной, свободной от белка коль-
цевой ДНК разорвана, в этой точке спонтанно воз-
никает свободное вращение и раскрученная ДНК
переходит в релаксированное состояние. В мо-
б Lk = 6
Рис. 24-15 Порядок зацепления (Lk). Здесь, как обычно,
каждая голубая лента обозначает одну из цепей двухце-
почечной ДНК. Для молекулы на рис. a Lk = 1, для моле-
кулы на рис. б Lk = 6. Одна из нитей на рис. б изображена
незакрученной, чтобы яснее показать границу вообража-
емой поверхности (бледно-голубого цвета). Число пере-
сечений данной поверхности спиральной нитью соответ-
ствует порядку зацепления.
24.2 Сверхспирализация ДНК [19]
остаются связанными топологическим образом.
Представим себе, что одна из кольцевых це-
пей ограничивает некую поверхность (напри-
мер, такую, как поверхность мыльной пленки
внутри кольца перед тем, как из него выдувают
мыльный пузырь). Порядок зацепления можно
определить как число пересечений этой поверх-
ности второй цепью. В молекуле на рис. 24-15,
a Lk = 1; в молекуле на рис. 24-15, б Lk = 6. Для
замкнутой кольцевой ДНК порядок зацепления
всегда описывается целым числом. Принято, что
в правозакрученной спирали порядок зацепле-
ния - положительное число (+), а в левозакру-
ченной спирали - отрицательное (-). В ДНК от-
рицательный порядок зацепления не встречается.
Применим эти рассуждения к молекуле зам-
кнутой кольцевой ДНК, состоящей из 2100 и. и.
( шс. 24-16, < ). Когда молекула релаксирована,
порядок зацепления найти просто: это отноше-
нию числа всех пар оснований к числу пар осно-
ваний, приходящихся на один оборот спирали
(примерно 10,5 п.н.); следовательно, в данном
случае Lk = 200. Чтобы кольцевая молекула ДНК
могла быть характеризована определенным по-
рядком зацепления, ни одна из ее цепей не должна
содержать разрывов. Если хоть в одной цепи есть
разрыв, спирали можно полностью разделить на
две цепи. В этом случае топологические связи от-
сутствуют, и Lk нельзя определить (рис. 24-16, б).
Теперь мы можем описать скрученность
ДНК, оперируя изменением порядка зацепления.
За точку отсчета принимают порядок зацеления
в релаксированной ДНК, Lk^ Для молекулы на
рис. 24-16, a, LkQ = 200; если из молекулы удалить
два витка, Lk = 198.
bLk = Lk-Lko (24-1)
198-200 = 2
Обычно удобно выражать изменение порядка за-
цепления через параметр, не зависящий от дли-
ны молекулы ДНК. Этот параметр называют
плотностью сверхспирализации (а), или специ-
фическим изменением порядка зацепления;
плотность сверхспирализации равна отношению
изменения числа витков спирали к их числу в ре-
лаксированной ДНК:
o = bLk/Lkv (24-2)
В примере на рис. 24-16, в о = -0,01; это озна-
чает, что удален 1% (2 из 200) витков спирали
Рис. 24-16. Порядок зацепления на примере молекул
замкнутой кольцевой ДНК. Кольцевая ДНК размером
2100 п. н. показана в трех формах: а — релаксированная.
Lk = 200; б — релаксированная с одноцепочечным раз-
рывом (ником), Lk нельзя определить; в — частично, на
два витка, раскрученная ДНК, Lk = 198. Частично раскру-
ченная молекула обычно существует в суперскрученном
состоянии, но частичное раскручивание также может об-
легчить разъединения цепей ДНК.
в ДНК (в В-форме). Обычно степень раскручи-
вания клеточной ДНК составляет 5-7%; т. е. о
от - 0,05 до 0,07 Отрицательное значение по-
казывает, что изменение порядка зацепления
связано с раскручиванием ДНК. Таким образом,
сверхспирализация, вызванная частичным рас-
кручиванием, — это отрицательная сверхспира-
лизация. И, наоборот, при некоторых обстоятель-
ствах ДНК может быть перекручена, что выража-
ется положительным значением сверхспирализа-
ции. Отметим, что в случае, когда ДНК частично
расплетена (отрицательная сверхспирализация),
закрученная вокруг оси часть спирали ДНК яв-
ляется зеркальным отражением перекрученной
спирали ДНК (положительная сверхспирали-
зация) ( ис. 24-17). Сверхспирализация не слу-
чайный процесс; характер сверхспирализации по
большей части описывается деформацией круче-
ния, возникающей в ДНК при уменьшении или
увеличении порядка зацепления по сравнению с
В-формой ДНК.
[20] Часть III. 24. Гены и хромосомы
Релаксированная ДНК
Lk = 200
Поскольку сверхспирализации отрицательная,
молекула ДНК несет отрицательные сверхвитки.
Если для той же молекулы ДНК Lk = 412,
Д££=412 — 400 = 12, следовательно, а = 12/400 = 0,03.
Сверхспирализации положительная, молекула
ДНК несет положительные сверхвитки.
Рис. 24-17. Отрицательная и положительная сверх-
спирализации. Для релаксированной молекулы ДНК,
показанной на рис. 24-16, а, частичное раскручивание
или перекручивание на два оборота спирали (Lk = 198
или 202) приводит к отрицательной или положительной
сверхспирализации, соответственно. Заметьте, что ось
спирали ДНК и в том и в другом случаях закручивается
в противоположных направлениях.
Порядок зацепления может изменяться на
± 1 при разрыве одной цени ДНК, поворота од-
ного из концов на 360° вокруг второй цепи и со-
единения разорванных концов. Такое изменение
не влияет на число пар нуклеотидов или число
атомов в кольцевой молекуле ДНК. Две формы
кольцевой ДНК, отличающиеся только таким
топологическим свойством, как порядок зацепле-
ния, называются топоизомерами.
Пример 24-1 ОПРЕДЕЛЕНИЕ
ПЛОТНОСТИ СВЕРХСПИРАЛИЗАЦИИ
Какова плотность сверхспирализации о замкну-
той кольцевой молекулы ДНК длиной 4200 п. н.,
для которой порядок зацепления Lk = 374? Ка-
кова плотность сверхспирализации ДНК той же
длины при Lk = 412? Сверхвитки этих молекул
положительны или отрицательны?
Решение. Найдем LkQ. Для этого длину коль-
цевой ДНК (в п. н.) делим на 10,5 п. н./виток:
(4200 п. н.) / (10,5 п. н. / виток) = 400. Теперь по
формуле 24-1 можно найти ALk: \Lk = Lk — LkQ =
= 374 — 100 = -26 Подставим значения Д£& и LkQ
в формулу 24-2: о = \Lk/LkQ = -26/400 = -0,065.
Порядок зацепления имеет две структурные
составляющие — кручение (коэффициент закру-
ченности; Tw - от англ, twist) и райзинг (изгиб;
Wr - от англ, writhe) (рис. 24-18). Их гораздо
труднее описать, чем порядок зацепления, но
кручение можно рассматривать как меру скручи-
вания (число витков) оси спирали, а райзинг как
меру локального изгиба или пространственной
связи соседних пар оснований (число сверхвит-
ков). При изменении порядка зацепления часть
возникающего напряжения обычно компенсиру-
ется при закручивании (сверхспирализации) и
изгибе, что выражается уравнением:
Lk - Tw + Wr
Параметры Tw и Wr не обязательно целочис-
ленные. Кручение и райзинг имеют скорее гео-
метрическую, чем топологическую природу, по-
скольку изменяются при деформации замкнутой
кольцевой молекулы ДНК.
Частичное раскручивание ДНК способству-
ет сверхспирализации и в определенной мере
Прямая лента (релаксированная ДНК)
Большой виток.
незначительные изменения скручивания
Виток нулевого радиуса,
значительные изменения в скручивании
Рис. 24-18. Кручение и райзинг на модели резинового
шнура. Розовый резиновый шнур изображает ось релак-
сированной молекулы ДНК. Напряжение, возникающее
при закручивании резинки (как при частичном раскру-
чивании ДНК), проявляется в виде кручения или рай-
зинга. Топологические изменения порядка зацепления
обычно сопровождаются изменением как кручения, так
и райзинга.
24.2 Сверхспирализация ДНК [21]
Релаксированная ДНК
Топоизомеразы катализируют
изменение порядка зацепления в ДНК
Частично раскрученная ДНК
Крестообразная структура ДНК
Рис. 24-19. Облегчение образования крестообразных
структур при частичном раскручивании ДНК. В принципе,
крестообразные структуры могут формироваться на па-
линдромных последовательностях (см. рис. 8-19 в т. 1), но
они редко возникают в релаксированной ДНК, поскольку
в линейной ДНК размещается больше пар оснований, чем
в крестообразных структурах. Частичное раскручивание
ДНК облегчает разделение цепей и формирование кре-
стообразной структуры на соответствующих участках по-
следовательности.
упрощает разъединение цепей, а также облег-
чает ряд структурных изменений в молекуле.
Физиологическое значение этих изменений не
столь велико, но они помогают проиллюстри-
ровать последствия частичного раскручивания.
Напомним, что крестообразная структура (см.
рис. 8-19 в т. 1) обычно содержит несколько не-
спаренных оснований, и именно недостаточная
скрученность ДНК помогает сохранять разде-
ление нитей (рис. 24-19). Частичное раскручи-
вание правозакрученной спирали ДНК также
облегчает образование коротких отрезков лево-
закрученной Z-ДНК в тех местах, где последо-
вательность нуклеотидов позволяет реализовать
эту форму (гл. 8).
Сверхспирализация ДНК — строго регулируе-
мый процесс, который влияет на разные аспекты
метаболизма ДНК. В каждой клетке есть фермен-
ты, функция которых заключается исключитель-
но в частичном раскручивании и/или релаксации
Рис. 24-20. Обнаружение топоизомеров. В этом экспе-
рименте все молекулы ДНК содержат одинаковое число
пар оснований, но отличаются по степени сверхспира-
лизации. Поскольку молекулы сверхспирализованной
ДНК компактнее, чем релаксированные молекулы, они
движутся быстрее при электрофорезе в геле. На пред-
ставленных гелях разделены топоизомеры с близкими
значениями плотности сверхспирализации (молекулы
движутся сверху вниз). На дорожке 1 сильно сверхспи-
рализованная ДНК движется в виде единой полосы, хотя,
возможно, в ДНК присутствуют разные топоизомеры.
На дорожках 2 и 3 видны результаты обработки сверх-
спирализованной ДНК топоизомеразой типа I; ДНК на
дорожке 3 подвергалась воздействию фермента дольше,
чем ДНК на дорожке 2. Когда плотность сверхспирали-
зации снижается до значения, начиная с которого гель
способен разделять отдельные топоизомеры, становятся
видны дискретные полосы. В каждой полосе, выделен-
ной скобкой справа от дорожки 3, содержатся кольцевые
молекулы ДНК с одинаковым порядком зацепления; для
двух соседних полос порядок зацепления различается на
единицу.
[22] Часть III. 24. Гены и хромосомы
I
Закрытая
конформация
О
I
б Фермент принимает
открытую конформацию и
5 3 целая нить ДНК проходит 3 3
в разрыв первой нити.
А ® . 4
Туг в активном центре атакует и 9 | Основание |
разрушает фосфодиэфирную связь q
в одной из цепей ДНК и образует | 3 ' А.
ковалентную 5 '-фосфотирозильную у
связь белок-ДНК. \______у
О
I
Открытая 3 5 3 5
конформация
нового цикла.
Рис. 24-21. МЕХАНИЗМ РЕАКЦИИ. Бактериальные топоизомеразы типа I изменяют поря-
док зацепления. Показана последовательность реакций топоизомеразы типа I. Фермент
может находиться в закрытой и открытой конформации, а — молекула ДНК связывается с
ферментом в закрытой конформации, и одна цепь ДНК разрезается. 6 — фермент переходит
в открытую конформацию, и вторая цепь ДНК проникает через разрыв в первой цепи, в —
фермент находится в закрытой конформации, цепь ДНК восстанавливается.
24.2 Сверхспирализация ДНК [23]
ДНК. Ферменты, повышающие или понижаю-
щие степень частичного раскручивания ДНК,
называются топоизомеразами; они изменяют по-
рядок зацепления в ДНК. Особенно важную роль
эти ферменты играют в процессах репликации и
упаковки ДНК. Существует два класса топоизо-
мераз. Топоизомеразы I типа на короткое время
разрывают одну из двух цепей ДНК, затем про-
пускают целую цепь сквозь разрыв и соединяют
разорванные концы; они изменяют Lk с шагом в 1.
Топоизомеразы II типа разрывают обе нити ДНК
и изменяют Lk с шагом в 2.
Действие этих ферментов можно продемон-
стрировать с помощью электрофореза в агарозном
геле (рис. 24-20). Группа идентичных плазмид
Рис. 24-22. Возможный механизм изменения порядка за-
цепления под действием эукариотических топоизомераз
типа ПА. ф Фермент, состоящий из многих субъединиц,
связывает одну молекулу ДНК (синие полосы). Раскры-
тые полости выше и ниже связанной ДНК называются N- и
С-воротами. ® Второй сегмент той же молекулы ДНК (крас-
ные полосы) связывается в N-воротах и оказывается запер-
тым ®. Обе цепи первого сегмента ДНК расщепляются (ме-
ханизм этой реакции аналогичен механизму, изображен-
ному на рис. 24-20, б), и @ второй сегмент ДНК проходит
сквозь разрыв. 5 Разорванная ДНК восстанавливается, и
второй сегмент ДНК высвобождается через С-ворота. В ходе
данного цикла связываются и гидролизуются две молекулы
АТР; одна молекула АТР, вероятно, гидролизуется на стадии
образования комплекса на этапе @. Остальные подробности
гидролиза АТР в данной реакции еще предстоит установить.
с одинаковым порядком зацепления движется в
геле в виде дискретных полос. Метод позволяет
разделить топоизомеры со значениями Lk, разли-
чающимися всего на 1, и обнаружить изменения
порядка зацепления, вызванные топоизомеразами.
Клетки Е. coli имеют не менее четырех раз-
ных топоизомераз (от I до IV). Топоизомеразы
типа I (топоизомеразы I и III) обычно релакси-
руют ДНК, удаляя отрицательные сверхвитки
(повышают Lk). Механизм, с помощью которого
бактериальные топоизомеразы типа I изменя-
ют порядок зацепления, показан на рис. 24-21.
Бактериальный фермент типа II (топоизомераза
типа II, или ДНК-гираза) может вызвать отрица-
тельное суперскручивание (уменьшает Lk). Этот
фермент для осуществления реакции использует
энергию АТР. Для изменения порядка зацепле-
ния ДНК топоизомеразы типа II разрывают обе
цепи молекулы ДНК и пропускают в образовав-
шуюся брешь другой дуплекс. Степень сверхспи-
рализации бактериальной ДНК поддерживается
путем регуляции суммарной активности топои-
зомераз I и II.
В эукариотических клетках юже есть то-
поизомеразы типов I и II. Топоизомеразы I и
III — ферменты типа I, а единственный фермент
типа II у позвоночных присутствует в двух изо-
формах — Па и Пр. Большинство топоизомераз
типа II, включая ДНК-гиразу архей, похожи и
принадлежат к одному семейству — к типу ПА.
У архей, кроме того, есть необычные топоизоме-
разы IV, составляющие семейство типа ПВ. Эу-
кариотические топоизомеразы тина II не могут
частично раскручивать ДНК (формировать отри-
цательные сверхвитки), но могут релаксировать
как положительное, так и отрицательное супер-
скручивание (рис. 24-22).
Как мы увидим в следующих главах, топо-
изомеразы играют ключевую роль в метаболиз-
ме ДНК, поэтому они важные мишени лекарств,
предназначенных для борьбы с бактериальными
инфекциями и новообразованиями (доп. 24-1).
Для компактной упаковки ДНК
нужна особая форма сверхспирализации
У молекул сверхспирализованной ДНК есть
несколько общих признаков. В молекуле с от-
рицательной сверхспирализацией сверхвитки
закручены вправо (рис. 24-17) и легче образуют
[24] Часть III. 24. Гены и хромосомы
не компактные, а тонкие и длинные петли, часто
с множеством ответвлений ( ис. 24-2<^). При ха-
рактерной для клетки плотности сверхспирали-
зации длина суперскрученных участков с учетом
ответвлений составляет примерно 40% длины
ДНК. Такой тип сверхспирализации называется
плектонемическим (от греч. plektos — закручен-
ный и пета — нить). Этим термином можно обо-
значить любую структуру с нитями, перекручен-
ными неким простым и регулярным образом, и он
хорошо объясняет общую структуру сверхспира-
лизованной ДНК в растворе.
Плектонемическая сверхспирализация, а
именно в такой форме выделяется ДНК в ла-
бораторных условиях, недостаточно компактна
для упаковки ДНК в клетке. В другой сверх-
Дополнение 24—1
МЕДИЦИНА
Лечение заболеваний путем ингибирования топоизомераз
Топология клеточной ДНК непосредственно связана
с ее функцией. Без помощи топоизомераз клетки не
могут реплицировать и упаковывать свою ДНК и экс-
прессировать гены и поэтому погибают. Следователь-
но, ингибиторы топоизомерз могут стать важными
фармацевтическими средствами для борьбы с инфи-
цирующими агентами и злокачественными клетками.
В качестве антибиотиков используются два клас-
са ингибиторов бактериальных топоизомераз. Кума-
рины, среди которых можно назвать новобиоцин и ку-
мермицин А1, представляют собой природные продук-
ты, выделяемые из стрептомицетов. Они ингибируют
связывание АТР бактериальными топоизомеразами
типа II, ДНК-гиразой и топоизомеразой IV. Эти анти-
биотики довольно редко используются для лечения
инфекционных заболеваний человека, однако поиск
эффективных антибиотиков такого рода для клиниче-
ского применения продолжается.
Антибиотики хинолонового ряда, которые также
ингибируют бактериальную ДНК-гиразу и топоизо-
меразу IV, впервые вошли в медицинскую практику в
1962 г., когда начала применяться налидиксовая кис-
лота Это вещество обладает ограниченной эффектив-
ностью и в США уже не применяется в клинической
практике, однако активное изучение данного класса
веществ привело к появлению фторхинолонов, приме-
ром которых может служить ципрофлоксацин (ципро).
Действие хинолонов основано на ингибировании по-
следней стадии топоизомеразной реакции, заключаю-
щейся в устранении разрывов в нитях ДНК. Ципроф-
локсацин — антибиотик широкого спектра действия.
Это один из немногих препаратов, который эффективен
против возбудителей сибирской язвы и рассматривает-
ся в качестве мощного средства защиты от биологиче-
ского оружия. Хинолоны действу ют исключительно на
бактериальные топоизомеразы, а ингибирование эука-
риотических ферментов начинается при концентрации
на несколько порядков выше терапевтической дозы.
Среди наиболее мощных химиотерапевтических
препаратов для лечения онкологических больных есть
ингибиторы топоизомераз человека. В опухолевых
клетках обычно наблюдается повышенная концентра-
ция топоизомераз, поэтому препараты, ингибирующие
Топотекан
24.2 Сверхспирализация ДНК [25]
спиральной форме, так называемой соленоид-
ной форме ( нс. 24-2 ), может существовать
частично раскрученная ДНК. В отличие от вы-
тянутой правозакрученной плектонемической
формы сверхспирали для соленоидной формы
характерны плотные левозакрученные петли.
Эта структура напоминает по форме садовый
шланг, аккуратно намотанный на катушку. Хотя
плектонемическая и соленоидная формы сверх-
спирали принципиально отличаются по струк-
туре, обе эти формы отрицательной сверхспи-
рализации могут возникнуть на одном и том же
участке частично раскрученной ДНК и могут
переходить друг в друга. Плектонемическая
форма более стабильна в растворе, соленоидная
стабилизируется белками и встречается внутри
активность этих ферментов, гораздо более токсичны
для опухолей, чем для здоровых тканей. В качестве
противоопухолевых препаратов используются инги-
биторы топоизомераз типа I и II.
Камптотецин выделенный из декоративного ки-
тайского растения и впервые испытанный в клиниче-
ской практике в 1970-х гг., ингибирует эукариотиче-
скую топоизомеразу типа I. Клинические испытания
выявили ограниченную эффективность препарата, хотя
в доклинических испытаниях на мышах препарат по-
казал хорошие результаты. Однако позднее, в 1990-х гг.,
были созданы два эффективных производных этого
ОН
Доксорубицин
вещества — иринотекан (камнто) и топотекан (гикам-
тин), применяемые для лечения больных раком прямой
и толстой кишки и раком яичников соответствен но.
Возможно, в ближайшее время будут выпущены дру-
гие родственные препараты. Все лекарства этой группы
действуют за счет связывания комплекса топоизомера-
зы с ДНК, в котором ДНК находится в расщепленном
состоянии, и ингибируют лигирование.
Целый ряд противоопухолевых препаратов дей-
ствует на человеческие топоизомеразы типа II; среди
них можно назвать доксорубицин (адриамицин), это-
позид (этопофос) и эллиптицин. Доксорубицин, отно-
сящийся к классу антрациклипов, эффективен против
нескольких видов рака. Большинство препаратов этой
группы стабилизируют ковалентные связи в комплек-
се между топоизомеразой и расщепленной ДНК.
Обычно все перечисленные противоопухолевые
препараты повышают количество повреждений ДНК
в быстро растущих опухолевых тканях. Однако, к со-
жалению, могут затрагиваться и здоровые ткани, что
приводит к общему токсическому эффекту и неже-
лательному побочному действию, с которым прихо-
дится бороться в ходе курса химиотерапии. По мере
повышения эффективности противоопухолевой тера-
пии и улучшения прогноза для онкологических боль-
ных на передний план выступает проблема появления
новых независимых опухолей. По-видимому, топои-
зомеразы останутся одним из интенсивно изучаемых
объектов в поиске новых лекарственных препаратов
для борьбы с раком.
Эллиптицин
[26] Часть III. 24. Гены и хромосомы
Рис. 24-23. Плектонемическая сверхспирализация. а — электронная микрофотография
плектонемической сверхспирализованной плазмидной ДНК; б — интерпретация наблюда-
емой картины; фиолетовыми линиями обозначены оси сверхвитков; обратите внимание на
разветвления спирали, в — упрощенный вид данной структуры.
хроматина. ДНК в этой форме гораздо более
компактна (рис. 24-24. б). Соленоидная сверх-
спирализация — механизм, с помощью которого
частичное раскручивание вносит свой вклад в
компактную упаковку ДНК.
Рис. 24-24. Плектонемическая и соленоидная формы
сверхспирали, п-в плектонемической сверхспира-
ли ДНК имеет форму протяженных правозакрученных
витков. В соленоидной отрицательной сверхспирали
эта молекула по форме напоминает трубку из плотных
левозакрученных витков. Обе формы могут переходить
друг в друга, хотя соленоидная форма обычно не реали-
зуется, пока ДНК не связывается с определенными бел-
ками. б — плектонемическая (верхняя) и соленоидная
формы сверхспирали одной и той же молекулы ДНК раз-
личаются по размеру. ДНК в соленоидной форме гораздо
компактнее.
Краткое содержание раздела 24.2
СВЕРХСПИРАЛИЗАЦИЯ ДНК
Большая часть ДНК в клетке образует сверх-
спираль. При частичном раскручивании
уменьшается общее число витков спира-
ли ДНК по сравнению с релаксированной
В-формой. Для поддержания состояния ча-
стичного раскручивания ДНК должна быть
замкнута в кольцо или связана с белком. Ча-
стичное раскручивание количественно оце-
нивается с помощью такого топологического
параметра, как порядок зацепления Lk
Частичное раскручивание измеряют с помо-
щью плотности сверхспирализации о (спец-
ифического изменения зацеплений), которое
выражается как (Lk — Lk0) / Lk0. Для клеточ-
ной ДНК величина о обычно составляет от
-0,05 до -0,07, это означает, что в ДНК не
хватает от 5% до 7% витков. Частичное рас-
кручивание ДНК облегчает разделение це-
пей ферментами метаболизма ДНК.
Молекулы ДНК, отличающиеся только по-
рядком зацепления, называются топоизоме-
рами. Ферменты, обеспечивающие частичное
раскручивание или релаксацию ДНК, называ-
ются топоизомеразами; они катализируют из-
менение порядка зацепления. Топоизомеразы
типа I и II за один акт катализа изменяют Lk
на одну или две единицы соответственно
24.3 Структура хромосом [27]
24.3. Структура хромосом
Хромосомами называют молекулы нуклеиновых
кислот, которые служат хранилищем генетиче-
ской информации у вирусов, бактерий, клеток
прокариот или органелл эукариот. Этот термин
также относится к интенсивно окрашиваемым
тельцам, которые видны в световой микроскоп,
после обработки красителями в ядрах эукариоти-
ческих клеток.
Хроматин состоит из ДНК и белков
В клеточном цикле эукариот (см. рис. 12-43 в
т. 1) структура хромосом изменяется удиви-
тельным образом (рис. 24-25). В неделящих-
ся клетках эукариот (стадия GO) и на стадии
интерфазы при делении (Gl, S и G2) материал
хромосом, хроматин, аморфен и, вероятно, ха-
отически распределен по центру ядра. В фазе S
интерфазы ДНК в таком аморфном виде репли-
цируется, при этом каждая хромосома образует
две сестринские хромосомы (они называются
сестринскими хроматидами), которые остаются
связанными друг с другом после завершения ре-
пликации. Хромосомы становятся намного бо-
лее конденсированными во время профазы ми-
тоза и принимают характерную форму, которая
зависит от вида организма (рис. 24-5).
Хроматин состоит из нитей, содержащих
белок и ДНК примерно в равной пропорции
(по массе) и небольшое количество РНК. Мо-
лекула ДНК в хроматине очень плотно связана
с белками гистонами, которые упаковывают и
упорядочивают ДНК в структурные единицы —
нуклеосомы (рис. 24-26). В хроматине также
обнаружено много негистоновых белков — одни
из них помогают поддерживать структуру хро-
мосом, а другие регулируют экспрессию спец-
ифических генов (гл. 28). Начиная с нуклеосом,
ДНК эукариот образует более высокоорганизо-
ванные структуры, которые в итоге формируют
компактную хромосому, видимую в световом
микроскопе. Рассмотрим структуру хромосом
эукариот и сопоставим ее с упаковкой ДНК в
клетках бактерий.
Рис. 24-25. Изменения структуры хромосомы в
ходе клеточного цикла эукариот. Относительная
длительность фаз изображена здесь только для
удобства. Длительность каждой фазы различна в
разных типах клеток и при разных условиях роста
(для одноклеточных организмов) или состояния
метаболизма (для многоклеточных). Самой корот-
кой обычно бывает фаза митоза. Как схематично
показано на диаграмме, в интерфазе клеточная ДНК
деконденсирована. В интерфазе (см. рис. 12-43 в
т. 1) можно отдельно выделить фазу G1 (от англ.
дар — промежуток), фазу S (синтез), когда проис-
ходит репликация ДНК, и фазу G2, во время которой
реплицированные хромосомы (хроматиды) сходят-
ся друг с другом. В митозе можно выделить четыре
фазы. Конденсация ДНК происходит в профазе. Во
время метафазы конденсированные хромосомы вы-
страиваются парами в центре веретена деления.
Каждая хромосома в каждой паре хромосом связа-
на с веретеном деления, направленным к полюсу
клетки, через микротрубочки, которые расположе-
ны между веретеном и центромерой. Сестринские
хроматиды разделяются в анафазе, каждая из них
движется к полюсу, с которым она соединена вере-
теном. После завершения деления клетки хромосо-
мы деконденсируются, и цикл начинается заново.
[28] Часть III. 24. Гены и хромосомы
Гистоновое ядро Линкерная ДНК
нуклеосомы нуклеосомы
Рис. 24-26. Нуклеосомы. Разделенные равными проме-
жутками нуклеосомы состоят из гистоновых комплексов,
связанных с ДНК. а — схематичное изображение; б —
электронная микрофотография.
Гистоны — небольшие основные белки
Гистоны, с молекулярной массой от И ООО до
21 000, обнаружены во всех клетках эукариот; эти
белки отличаются высоким содержанием основ-
ных аминокислот аргинина и лизина (которые
вместе составляют четверть всех аминокислот).
Все эукариотические клетки содержат пять ос-
новных классов гистонов, различающихся моле-
кулярной массой и аминокислотным составом
(табл. 24-4). Гистоны НЗ имеют почти идентич-
ную аминокислотную последовательность в клет-
ках всех эукариот, как и гистоны Н4, что указыва-
ет на консервативность их функций. Например,
молекулы гистонов Н4 гороха и коровы различа-
ются только двумя аминокислотными остатками
из 102; молекулы гистонов H i человека и дрож-
жей различаются восемью остатками. Последо-
вательности гистонов Hl, Н2А и Н2В у разных
видов эукариот менее схожее.
Гистоны каждого класса подвергаются фер-
ментативной модификации — метилированию,
ADP-рибозилированию, фосфорилированию, гли-
козилированию или ацетилированию. Такие мо-
дификации влияют на суммарный электрический
заряд, форму и другие свойства гистонов, на струк-
турные и функциональные свойства хроматина, а
также важны для регуляции транскрипции (гл. 28).
Кроме того, некоторые гистоны эукариот су-
ществуют в виде нескольких вариантов, особенно
это касается гистонов Н2А и НЗ, которые более
подробно описаны ниже. Эти гистоны и их моди-
фикации играют в метаболизме ДНК особую роль.
Нуклеосомы — основные
структурные единицы хроматина
Эукариотические хромосомы, изображенные на
рис. 24-5, представляют собой компактную уклад-
ку молекулы ДНК длиной примерно 105 мкм в
ядре клетки, диаметр которого обычно составля-
ет от 5 до 10 мкм. Такая компактность достигается
благодаря многоуровневой организации укладки.
При частичном нарушении укладки хромосомы
обнаруживается структура, в которой ДНК проч-
но связана с глобулами (шариками) белка, ча-
сто разделенными регулярными промежутками
(рис. 24-27). Шарики в этих «бусах» — это гисто-
ны в комплексе с ДНК Шарик и нить ДНК, сое-
диняющая его со следующим шариком, образуют
нуклеосому — основную структурную единицу,
I Таблица 24-4 Классы гистонов и их свойства
Гистон Молекулярная масса Число аминокислотных остатков Доля основных аминокислот (% от общего числа)
Lys Arg
Н1* 21 130 223 29,5 11,3
Н2А* 13 960 129 10,9 19,3
Н2В* 13 774 125 16,0 16,4
НЗ 15 273 135 19,6 13,3
Н4 И 236 102 10,8 13,7
* Размеры этих гистонов слегка различаются у разных видов Приведенные данные относят-
ся к гистонам быка.
24.3 Структура хромосом [29]
на которой происходит дальнейшая укладка хро-
матина. Глобула каждой нуклеосомы содержит
восемь гистонов: по две копии Н2А, Н2В, НЗ и
Н4. Фрагменты ДНК между шариками пред-
ставляют собой повторяющиеся единицы, обыч-
но длиной 200 п. н., из которых 146 и. н. плотно
обмотаны вокруг гистонового ядра из восьми
частей, а оставшаяся часть ДНК соединяет гло-
булы между собой. Гистон Н1 взаимодействует
с соединяющей (линкерной) ДНК. Неглубокая
обработка хроматина расщепляющими ДНК
ферментами вызывает в первую очередь дегра-
дацию линкерной ДНК; в результате происходит
высвобождение гистоновых частиц, содержащих
146 п. и. связанной ДНК, которая была защищена
от расщепления. Исследователи кристаллизова-
ли полученные таким образом нуклеосомные ча-
стицы и с помощью рентгеноструктурного анали-
за установили, что ДНК обмотана вокруг восьми
молекул гистонов в виде левозакрученной соле-
ноидной сверхспирали (рис. 24-27).
Детальное изучение данной структуры объ-
яснило, почему ДНК эукариот частично раскру-
чивается даже в отсутствие ферментов в эукари-
отических клетках, осуществляющих частичное
раскручивание ДНК. Вспомним, что соленоидное
скручивание ДНК в нуклеосомах - одна из двух
форм сверхсиирали, которую может принимать
частично раскрученная (отрицательно сверхспи-
рализованная) ДНК. Плотное обматывание ДНК
вокруг гистонового ядра требует удаления из ДНК
примерно одного витка. Когда белковая сердцеви-
на нуклеосомы связывается in vitro с релаксиро-
ванной замкнутой кольцевой ДНК, происходит
отрицательная сверхспирализация. Поскольку
такой процесс связывания нс разрывает ДНК и
не изменяет порядок зацепления, формирование
Рис. 24-27. ДНК обвивается вокруг ядра нуклеосомы.
а — шаровая модель белков нуклеосомной частицы из
африканской лягушки Xenopus laevis; разными цвета-
ми показаны разные гистоны (PDB ID 1A0I). 6 и в — вид
сверху и вид сбоку на кристаллическую структуру ну-
клеосомы (контур поверхности показан серым цветом) с
обвитой связанной ДНК (синяя) длиной 146 п. н. Молеку-
ла ДНК связывается в виде левозакрученной соленоид-
ной сверхспирали, опоясывающей гистоновый комплекс
1,8 раз. Для наглядности представлено схематичное изо-
бражение нуклеосомы (в — внизу справа).
[30] Часть III. 24. Гены и хромосомы
отрицательной соленоидной сверхспирали долж-
но компенсироваться образованием положитель-
ной сверхснирали на участке несвязанной ДНК
(рис. 24-28). Как упоминалось ранее, эукариоти-
ческие топоизомеразы могут вызывать релакса-
цию положительной сверхспирали. Релаксация
несвязанной положительно сверхспирализован-
ной ДНК приводит к фиксированию отрицатель-
но свсрхспирализованной ДНК (при ее связыва-
(соленоидный) положительный
супервиток
(плектонемический)
Один (общий)
отрицательный супервиток
Рис. 24-28. Сборка хроматина, а — релаксированная
замкнутая кольцевая ДНК. б — при связывании гистоно-
вой сердцевины и формировании нуклеосомы возника-
ет один отрицательный сверхвиток. Если ни в одной из
цепей нет разрывов, где-нибудь в ДНК возникает поло-
жительный сверхвиток (LLk = 0). в — релаксация такого
положительного сверхвитка топоизомеразой оставляет
один отрицательный сверхвиток (KLk = -1).
Рис. 24-29. Расположение нуклеосомы, при котором опти-
мально используются пары нуклеотидов А=Т в участках
соприкосновения гистоновой сердцевины с малой бо-
роздкой спирали ДНК.
нии с гистоновой сердцевиной) и выражается в
общем уменьшении порядка зацепления. Было по-
казано, что топоизомеразы необходимы для сбор-
ки хроматина из очищенных гистонов и замкнутой
кольцевой ДНК in vitro.
Еще одним фактором, влияющим на связы-
вание ДНК с гистонами в нуклеосомах, является
последовательность ДНК. Гистоновая сердцевина
связывается не с любым, а с определенным участ-
ком ДНК. Принцип выбора такого участка до кон-
ца не ясен, но в некоторых случаях, вероятно, он
определяется количеством нар А=Т в том участке
спирали ДНК, где она соприкасается с гистонами
(рис. 24-29). Сужение малой бороздки спирали в
этих точках способствует плотной упаковке ДНК
вокруг гистонового ядра; сочетание двух или трех
пар нуклеотидов А=Т увеличивает вероятность
такого сужения. Особенно охотно нуклеосомы
связываются с такими последовательностями, в
которых динуклеотиды АА, АТ или ТТ встречают-
ся с интервалом 10 п. н., и именно таким свойством
обладают до 50% последовательностей, связываю-
щихся с гистонами in vivo.
Для правильного расположения некото-
рых нуклеосомных частиц на ДНК необходимы
другие белки У некоторых организмов опреде-
ленные белки связываются со специфической
последовательностью ДНК, способствуя фор-
мированию поблизости нуклеосомной частицы.
24.3 Структура хромосом [31]
В ходе репликации или в других процессах, тре-
бующих временного перемещения нуклеосом,
нуклеосомы остаются связанными с ДНК. Это
связывание осуществляется ступенчатым обра-
зом. Сначала происходит связывание тетрамера,
образованного двумя гистонами НЗ и двумя ги-
стонами Н4, вслед за которыми связываются ди-
меры Н2А Н2В. Связывание нуклеосом с хромо-
сомами после репликации хромосом происходит
при участии белкового комплекса RCAF (от англ.
replication-coupling assembly factor}. Ком11 лекс
RCAF состоит из ацетилированных гистонов НЗ
и Н4, грехсубъединичного белка, называемого
фактором сборки хроматина-1 (англ, chromatin
assembly factor 1, CAF1), и белка ASF1 (от англ.
anti-silencing factor 1). Механизм связывания ну-
клеосом изучен не полностью, хотя известно, ч го
элементы комплекса RCAF непосредственно вза-
имодействуют с элементами репликационного
аппарата. Когда после репарации ДНК или дру-
гого процесса возникает необходимость сборки
нуклеосом, RCAF заменяется другим специали-
зированным белковым комплексом, опосредую-
щим связывание. В некоторых случаях факторы
обмена гистонов позволяют осуществлять замену
основных гистонов на другие варианты. Правиль-
ное расположение этих вариантов гистонов очень
важно. Например, при отсутствии одного из ва-
риантов гистонов эмбрионы мыши погибают на
ранних стадиях развития (дон. 24-2). Точное рас-
положение нуклеосом ных частиц также может
иметь значение для экспрессии некоторых эука-
риотических генов (1Л. 28).
Нуклеосомы образуют структуры
с более сложной организацией
Закручивание вокруг нуклеосомной частицы
уменьшает длину ДНК примерно в семь раз. Но
в целом в хромосоме длина ДНК сокращается
более чем в 10 000 раз, следовательно, существу-
ют и другие уровни организации ДНК. Очень
мягкие методы выделения хромосом позволяют
обнаружить, что ядра нуклеосом организованы
в структуры, называемые 30-нм фибриллами
(рис. 24-30). Для такой упаковки требуется одна
молекула гистона Н1 на нуклеосому. Однако
участки такой укладки прерываются участками
ДНК, связанными со специфическими (негисто-
иовыми) белками. Кроме того, наличие 30-нм
фибрилл зависит от интенсивности транскрип-
ции участка ДНК. Участки активной транскрип-
ции генов находятся в менее организованном со-
стоянии и содержат очень мало гистона Н1 (или
вовсе его не содержат).
30-нм фибриллы представляют собой второй
уровень организации хроматина, обеспечиваю-
щий 100-кратное сокращение длины ДНК Более
высокие уровни организации ДНК пока не иссле-
дованы; вероятно, определенные участки ДНК
связаны с хромосомным остовом (рис. 24-31).
Ассоциированные с остовом участки разделены
петлями ДНК размером от 20 до 100 т. и. н. ДНК
в этой петле может содержать набор родственных
генов. Например, у дрозофил полный набор ге-
нов, кодирующих гистоны, сосредоточен в вило
кластеров в петлях, которые присоединены к хро-
Рис. 24-30.30-нм фибриллы — более высокий уровень организации нуклеосом, а — схе-
ма предполагаемого строения фибриллы, б — электронная микрофотография.
[32] Часть III. 24. Гены и хромосомы
Эпигенетика, структура нуклеосом и варианты гистонов
МЕДИЦИНА
Дополнение 24-2
Информация, которая передается от одного поколения
к другому (дочерним клеткам при делении клеток или
детям от родителей), но не кодируется в ДНК, назы-
вается эпигенетической. Большая часть этой инфор-
мации содержится в виде ковалентных модификаций
гистонов и/или в виде размещения вариантов гистонов
в хромосомах.
Участки хроматина, в которых происходит актив-
ная экспрессия генов (транскрипция), обычно нахо-
дятся в неконденсированной форме и называются эух-
роматином. В этих участках гистоны НЗ и Н2А часто
заменены на варианты гистонов НЗ.З и H2AZ соответ-
ственно (рис. 1). Комплексы, осуществляющие связы-
вание ДНК с нуклеосомами, содержащими варианты
гистонов, сходны с теми, что отвечают за связывание с
нуклеосомами содержащими основные гистоны. Свя-
зывание нуклеосом, содержащих гистон НЗ.З, осущест-
вляется комплексом, в котором фактор сборки хрома-
тина-1 (CAF1) заменен белком HIRA (название проис-
ходит от класса белков HIR, от англ, histone repressor).
Белки CAF1 и HIRA можно рассматривать в качестве
гистоновых шаперонов, помогающих обеспечить пра-
вильную сборку и размещение нуклеосом. Гистон НЗ.З
отличается от НЗ лишь четырьмя аминокислотными
остатками, однако эти остатки играют ключевую роль в
связывании гистона.
Подобно гистону НЗ.З, гистон H2AZ связан с опре-
деленным связывающим комплексом и обычно ассоци-
ирован с активно транскрибирующимися участками
хроматина. Включение H2AZ стабилизирует нуклео-
сомный октамер, но препятствует некоторым коопе-
ративным взаимодействиям между нуклеосомами, не-
обходимым для формирования компактной структуры
хромосомы. Это приводит к образованию хромосомы с
более открытой структурой, что облегчает экспрессию
генов в области H2AZ. Ген, кодирующий H2AZ, играет
важную роль в развитии млекопитающих. У дрозофил
отсутствие H2AZ препятствует развитию организма по-
сле стадии личинки.
Другой вариант гистона Н2А Н2АХ связан с
репарацией ДНК и генетической рекомбинацией. У мы-
шей отсутствие Н2АХ приводит к геномной нестабиль-
ности и бесплодию у самцов. Небольшие количества
Н2АХ, по-видимому, распределены по всему геному
При двухцепочечном разрыве ДНК находящиеся по-
близости молекулы Н2АХ фосфорилируются по остат-
ку Ser139 на С-конце белка. Если блокировать фосфори-
лирование экспериментальным путем, ингибируется
образование белковых комплексов, необходимых для
репарации ДНК.
Вариант гистона НЗ, известный как CENPA, свя-
зан с повторяющимися последовательностями ДНК в
центромерах. Хроматин в области центромеры содер-
жит гистоновые шапероны CAF1 и HIRA; по-видимому,
оба белка участвуют в связывании нуклеосом, содержа-
щих CENPA. Удаление гена CENPA у мышей приводит
Рис. 1. Известно несколько вариантов гистонов H3Z Н2А и
Н2В. Здесь показаны основные гистоны и несколько из-
вестных вариантов. Выделены участки метилирования
остатков Lys/Arg и фосфорилирования остатков Ser. Уча-
сток HFD (от англ, histone-fold domain) — структурный
домен, присутствующий во всех основных гистонах.
к летальному исходу.
Для изучения функций и расположения вариантов
гистонов можно применять методы геномики Одним
из полезных методов является иммунопреципитация
хроматина (ChIP, от англ, chromatin immunoprecipita-
tion) Суть метода состоит в осаждении нуклеосом, со-
держащих определенный вариант гистона, при связыва-
нии этого варианта со специфическими антителами. В
принципе нуклеосомы можно изучать отдельно от ДНК,
но чаще при исследовании связанную ДНК сохраняют,
что позволяет определить участок связывания нуклео-
сомы. Молекулу ДНК можно пометить и использовать
24.3 Структура хромосом [33]
Рис. 2. Метод иммунопреципитации хроматина на микрочипе позволяет определить после-
довательности генома, с которыми связывается тот или иной вариант гистона, а — вариант
гистона с меткой-эпитопом (белком или химической структурой, распознаваемой антителом;
см. гл. 5 и 9 в т. 1) вводят в определенный тип клеток, где он встраивается в нуклеосомы. (В
некоторых случаях в таких метках нет нужды, поскольку существуют антитела, связывающи-
еся непосредственно с модифицированным участком изучаемого гистона.) Затем из клеток
выделяют хроматин и расщепляют его микрококковой нуклеазой. Связанная с нуклеосомами
ДНК защищена от расщепления, однако линкерная ДНК расщепляется, что приводит к вы-
свобождению сегментов ДНК, связанных с одной или двумя нуклеосомами. При добавлении
антител происходит селективное осаждение нуклеосом, содержащих меченый вариант гисто-
на. Из осадка экстрагируют связанную с этими нуклеосомами ДНК, помечают ее и используют
в качестве зонда для анализа на микрочипах, содержащих геномные последовательности дан-
ного типа клеток. 6 — гибридизация на микрочипе позволяет обнаружить последовательно-
сти ДНК, связанные с нуклеосомами с определенным вариантом гистона. В данном примере
(верхняя панель) представлена гибридизация ДНК, связанной с гистоном НЗ.З в небольшом
участке генома дрозофилы. Аннотированные гены данного участка генома представлены на
нижней панели (широкие полосы). С гистоном НЗ.З связываются участки ДНК, расположен-
ные вблизи активно транскрибирующихся генов (красные полосы).
в качестве зонда в методе анализа на микрочипах (см.
рис. 9-22 в т. 1) для получения карты геномной последо-
вательности, с которой связывается каждая конкретная
нуклеосома. Этот метод часто называют иммунопреци-
питацией на чипе (ChIP on chip; рис. 2).
Варианты гистонов, а также многие ковалентные
модификации гистонов, позволяют определить функ-
ции хроматина. Они размечают хроматин, облегчая или
подавляя специфические функции, такие как расхожде-
ние хромосом, транскрипция и репарация ДНК. Моди-
фикации гистонов не исчезают при делении клеток или
при мейозе, и таким образом они становятся частью ин-
формации, передаваемой от поколения к поколению во
всех эукариотических организмах.
[34] Часть III. 24. Гены и хромосомы
Рис. 24-31. Частично расплетенная хромосома человека;
видны многочисленные петли ДНК, прикрепленные к
хромосомному остову.
Две
хроматиды
(по 10 витков
в каждой)
мосомному остову (рис. 24-32). Сам осьов также
содержит несколько белков, особенно в большом
количестве гистон Н1 (расположенный на вну-
тренней стороне волокна) и топоизомеразу II.
Присутствие топоизомеразы II еще раз указывает
на связь между частичным раскручиванием ДНК
и структурой хроматина. Топоизомераза II на-
Рис. 24-32. Петли хромосомной ДНК, прикрепленные к
хромосомному остову. Молекулы ДНК в петлях упакова-
ны в виде 30-нм фибрилл, поэтому петли — следующий
уровень организации хромосом. Петли часто содержат
группы генов с родственными функциями. Полные набо-
ры генов гистонов, как показано на данной схеме, образу-
ют кластеры в петлях такого рода. В отличие от большин-
ства других гены гистонов во многих геномах эукариот
присутствуют в большом количестве копий.
ДНК
Один виток
(30 розеток)
Одна петля
(~75 ООО п н.)
Одна розетка
(6 петель)
30-нм
фибрилла
Структура
хроматина
типа «бусины
на нитке»
Остов
хромосомы
Рис. 24-33. Компактная упаковка ДНК в хромосоме эу-
кариот. Модель демонстрирует уровни организации ДНК
эукариот, которые могут объяснить компактную упаков-
ку хромосом. В клетке уровни более высокой организа-
ции (выше уровня 30-нм фибрилл), скорее всего, не столь
единообразны, как показано на данной схеме.
24.3 Структура хромосом [35]
столько важна для поддержания структуры хро-
матина, что ингибиторы этого фермента могут
быстро уничтожить делящиеся клетки. Несколь-
ко химиотерапевтических препаратов являются
ингибиторами топоизомеразы II - фермент мо-
жет инициировать разрыв нитей, но не позволя-
ют ему зашить разрыв.
Получены доказательства еще более сложной
организации хромосом эукариот, причем каждый
уровень организации значительно повышает ком-
пактность упаковки. Одна из моделей, объясняю-
щих компактность структуры ДНК, показана на
рис. 24-33. Более высокоорганизованные струк-
туры хроматина, вероятно, различаются в разных
хромосомах, в разных участках одной хромосомы
и в разные моменты жизненного цикла клетки. Ни
одна модель не может адекватно описать все эти
структуры. Тем не менее принцип ясен: компакт-
ность ДНК эукариотических хромосом обеспечи-
вается укладкой витка на виток, витка на виток...
в Трехмерная упаковка хромосомного остова
Структура конденсированных хромосом
поддерживается SMC-белками
Наряду с гистонами и топоизомеразами суще-
ствует третий важный класс белков хроматина —
это так называемые SMC-белки (от англ, struc-
tural maintenance of chromosomes — белки, под-
держивающие структуру хромосом). Первичная
структура SMC-белков состоит из пяти разли-
чимых доменов (рис. 24-34, а). Концевые глобу-
лярные домены N и С (на N- и С-конце), каждый
из которых содержит часть активного центра, в
котором происходит гидролиз АТР, соединены с
двумя «-спиральными участками (см. рис. 4-10 в
т. 1), между которыми располагается шарнирный
домен. Белки этого класса обычно образуют ди-
мерный V-образный комплекс, соединяясь друг с
другом через шарнирный домен (рис. 24-34, б, в).
Один N- и один С-домен сближаются, образуя
функциональный центр гидролиза АТР на каж-
дом свободном конце V-образного комплекса.
Рис. 24-34. Структура SMC-белков, а — пять доменов первичной структуры SMC. Буквами
N и С обозначены N- и С-концевые последовательности соответственно, б — каждый поли-
пептид сложен таким образом, что два спиральных домена закручены относительно друг
друга, так что N- и С-домены соединяются, образуя полноценный ATP-связывающий сайт.
Два таких домена связываются шарнирной областью с образованием V-образного димера.
в — электронная микрофотография SMC-белков из Bacillus subtilis. г — когезины состоят
из пар белков SMC1 и SMC3, а конденсины — из пар SMC2-SMC4. Эти эукариотические
SMC-белки связываются с клейзином и некоторыми другими регуляторными белками (не
показаны), д - гидролиз АТР может служить для открытия и закрытия АТРазного домена
SMC-димера, который остается связанным с клейзином и другими белками (не показано).
[36] Часть III. 24. Гены и хромосомы
---G14
S-------------
Репликация
телофаза
G1
Рис. 24-35. Моделирование роли когезинов и конденсинов в клеточном цикле эукариот.
Когезины присоединяются к хромосомам в фазе G1 и удерживают вместе сестринские хромати-
ды в процессе репликации. При митозе с хроматидами связываются конденсины и удерживают
их в конденсированном состоянии. В ходе анафазы связи с когезинами удаляются под действи-
ем так называемой сепаразной активности. После разделения хроматид конденсины начинают
отделяться, и дочерние хромосомы возвращаются в неконденсированное состояние.
Белки семейства SM(' обнаружены во всех
организмах — от бактерии до человека. Эукариоты
имеют два основных типа SMC-бслков — когезины
и конденсины, причем оба типа этих белков связы-
ваются с регуляторными и вспомогательными бел-
ками (рис. 24-34, г). Когезины играют существен-
ную роль в связывании между собой сестринских
хроматид сразу после репликации и удерживании
их в таком состоянии в процессе конденсации хро-
мосом при переходе к стадии метафазы. Связыва-
ние важно для правильного расхождения хромосом
при клеточном делении. Когезины вместе с белком
третьего типа клейзином, по-видимому, образуют
вокруг реплицирующихся хромосом кольцо, кото-
рое удерживает хромосомы вместе до того момента,
когда в ходе клеточного деления начнется расхож-
дение хромосом. Это кольцо может растягиваться и
сжиматься в ответ на гидролиз АТР (рис. 24-34, д).
Конденсины важны для конденсации хромосом
при переходе клетки к митозу. В лабораторных
условиях конденсины связываются с ДНК таким
образом, что образуются положительные сверх-
витки — связывание конденсина вызывает переход
ДНК в сверхспирализованное состояние, в отличие
от состояния частичного раскручивания, вызыва-
емого связыванием нуклеосом. Пока еще не ясно,
как это помогает компактной укладке хромати-
на, однако закручивание ДНК вокруг конденсина
может играть определенную роль в конденсации
ДНК. Когезины и конденсины опосредуют многие
изменения в структуре хромосом в клеточном ци-
кле эукариот (рис. 24-35).
Бактериальная ДНК тоже сложно организована
Теперь вкратце обсудим строение бактериаль-
ных хромосом Бактериальная ДНК упакована
в структуру, называемую нуклеоидом, нуклео-
ид может занимать значительную часть объема
клетки (рис. 24-36). Установлено, что ДНК в
Рис. 24-36. Нуклеоид Е. colL Молекулы ДНК этих клеток
обработаны красителем, который флуоресцирует при
УФ-облучении. Светящиеся области представляют собой
нуклеоиды. Обратите внимание, что ДНК в некоторых
клетках реплицировалась, но клетки еще не разделились
и поэтому содержат несколько нуклеоидов.
24.3 Структура хромосом 137]
одной или нескольких точках прикрепляется к
внутренней поверхности плазматической мем-
браны. О структуре нуклеоида известно гораздо
меньше, чем о структуре хроматина эукариот,
но постепенно проясняются детали его сложно-
го строения. В клетках Е. coli существует свое-
образный каркас, позволяющий организовать
кольцевую хромосому в виде примерно 500 пет-
левых доменов, каждый в среднем по 10 000 п. н.
(рис. 24-37), как описано выше для хроматина.
Эти домены топологически ограничены; напри-
мер, если происходит разрыв ДНК в одном до-
мене, только в этом домене она и окажется ре-
лаксирована. Домены не имеют фиксированных
концов. Напротив, в ходе репликации их грани-
цы постоянно смещаются по последовательно-
сти ДНК. Бактериальная ДНК, по-видимому, не
имеет структуры, сопоставимой с нуклеосомами
эукариот. В клетках Е. coli содержится много
гистоноподобных белков (наиболее хорошо из-
учен состоящий из двух субъединиц белок HU
Мг 19 000), но эти белки связываются и диссо-
циируют в течение нескольких минут, а регуляр-
ных стабильных ДНК-гистоновых комплексов
обнаружено не было. Динамичные изменения
в структуре бактериальной хромосомы могут
быть связаны с потребностью в более быстром
Рис. 24-37. Петли в хромосоме £. colL Длина каждого та-
кого домена около 10 000 п. н. Домены не статичны, а пе-
ремещаются по последовательности ДНК по мере продви-
жения репликации. Релаксации ДНК за границами того
домена, в котором произошел разрыв нити, препятствуют
находящиеся на границах домена элементы неизвестного
состава. Эти граничные комплексы показаны в виде се-
рых овалов. Стрелки указывают перемещение ДНК через
граничные комплексы.
доступе к генетической информации. Клеточ-
ный цикл бактериальной клетки может зани-
мать всего 15 мин, в то время как типичная эука-
риотическая клетка не делится несколько часов
или даже месяцев. Кроме того, у прокариот в
кодировании РНК и/или белков задействована
гораздо большая часть ДНК. Более интенсив-
ные темпы клеточного метаболизма у бактерий
означают, что за один и тот же отрезок времени в
клетках бактерий транскрибируется или репли-
цируется значительно большая часть ДНК, чем
у большинства эукариотических клеток.
Мы рассмотрели сложное строение ДНК и
теперь готовы перейти к следующей главе, в ко-
торой обсуждается метаболизм ДНК.
Краткое содержание раздела 24.3
СТРУКТУРА ХРОМОСОМ
Основная единица организации хроматина
в клетках эукариот — нуклеосома, которая
состоит из гистонов и сегмента ДНК длиной
200 п. н. Белковое ядро нуклеосомы, содер-
жащее восемь гистонов (по две копии каж-
дого из гистонов Н2А, Н2В, НЗ и Н4), обер-
нуто сегментом ДНК (примерно 146 и. н.) в
форме левозакрученной соленоидной сверх-
спирали.
Нуклеосомы образуют 30-нм фибриллы, ко-
торые упакованы таким образом, что дости-
гается 10 000-кратная компактизация ДНК,
необходимая для размещения обычной эука-
риотической хромосомы в клеточном ядре.
Следующие уровни организации основаны
на присоединении к хромосомному остову,
состоящему из гистона Н1, топоизомеразы
II и SMC-белков. Белки SMC, главным обра-
зом когезины и конденсины, играют важную
роль в поддержании структуры хромосом на
каждой стадии клеточного цикла.
Бактериальные хромосомы в нуклеоиде
тоже находятся в компактной упаковке, но
хромосома прокариот имеет гораздо более
динамичную и нерегулярную структуру, чем
хроматин эукариот, что связано с краткостью
клеточного цикла и высокой интенсивно-
стью метаболизма бактерий.
13s ] Часть III. 24. Гены и хромосомы
Ключевые термины
Термины, выделенные жирным шрифтом, объясня-
ются в глоссарии.
30-нм фибрилла 31 Регуляторная последова-
SMC-белки 35 тельность 8
Ген 8 Релаксированная ДНК 16
Гистоны 27 Сателлитная ДНК 13
Интрон 12 Сверхспираль 16
Когезины 36 Соленоидный 25
Конденсины 36 Теломера 14
Кручение 20 Топоизомеры 20
Мутация 8 Топология 16
Нуклеоид 36 Фенотип 8
Нуклеосома 27 Хроматин 27
Плазмида 10 Хромосома 27
Плектонемический 24 Центромера 14
Плотность сверхспирали- Частичное раскручивание
зации о 19 17
Порядок зацепления 18 Экзон 12
Простая последователь- Топоизомеразы 23
ность ДНК 13 Эпигенетический 32
Райзинг 20 Эухроматин 32
Дополнительная литература
для дальнейшего изучения
Общая jii'iература
Blattner, Е R., Plunkett, G. Ill, Bloch, С. А., Рета, N. Т.,
Borland, V., Riley, М., Collado-Vides, J., Glasner, J. D.,
Rode, С. K., Mayhew, G. E, et al. (1997) The complete ge-
nome sequence of Escherichia coli K-12. Science 277,1453-1474.
Новые секреты этого обычного лабораторного орга-
низма.
Cozzarelli, N. R. & Wang, J. С. (eds). (1990) DNA Topolo-
gy and Its Biological Effects, Cold Spring Harbor Laboratory
Press, Cold Spring Harbor, NY.
Kornberg, A. & Baker, T. A. (1991) DNA Replication, 2nd
edn, W. H. Freeman & Company, New York.
Хорошая книга для углубления знаний о структуре и
функциях ДНК.
Lodish, Н., Berk, A., Kaiser, С. A., Krieger, М., Scott, М. Р,
Bretscher, A., Plough, Н., & Matsudaira, Р. (2008) Molecular
Cell Biology, 6th edn, W. H. Freeman & Company, New York.
Отличный источник информации по данной теме.
Гены и хромосомы
Bromham, L. (2002) The human zoo: endogenous retroviruses
in the human genome. Trends Ecol. Evol. 17,91-97.
Подробное описание одного из классов транспозонов,
который составляет заметную часть генома человека.
Goffeau, A., Barrell, В. G., Bussey, Н., Davis, R. W, Du-
jon, В., Feldmann, Н., Galibert, Е, Hoheisel, J. D., Jacq, C.,
Johnston, M., et al. (1996) Life with 6000 genes. Science 274,
546,563-567.
Отчет о первом полностью расшифрованном эукариоти-
ческом геноме — геноме дрожжей Saccharomyces cerevisiae.
Long, М., de Souza, S. J., & Gilbert, W. (1995) Evolution
of the intron-exon structure of eukaryotic genes. Curr. Opin.
Genet. Dev. 5,774-778.
McEachern, M. J., Krauskopf, A., & Blackbum, E. H. (2000)
Telomeres and their control. Annu. Rev. Genet. 34,331-358.
Schmid, C. W. (1996) Alu: structure, origin, evolution, signifi-
cance and function of one-tenth of human DNA. Prog. Nucleic
Acid Res. Mol. Biol. 53, 283-319.
Tyler-Smith, C. & Floridia, G. (2000) Many paths to the top
of the mountain: diverse evolutionary solutions to centromere
structure. Cell 102,5-8.
Современные данные о разнообразии центромерных
структур у разных организмов.
Vega, L.R., Mateyak, М. К., & Zakian, V. А. (2003) Getting
to the end: telomerase access in yeast and humans. Nat. Rev.
Mol. Cell Biol.4,948-959.
Сверхспирализация и топоизомеразы
Berger, J. M. (1998) Type II DNA topoisomerases. Curr. Opin.
Struct. Biol. 8,26-32.
Boles, T. C., White, J. H., & Cozzarelli, N. R. (1990) Struc-
ture of plectonemically supercoiled DNA. J. Mol. Biol. 213,
931-951.
Об основных свойствах сверхспирализованной ДНК.
Champoux, J. J. (2001) DNA topoisomerases: structure, func-
tion, and mechanism. Annu. Rev. Biochem. 70,369-413.
Прекрасный обзор классов топоизомераз.
Garcia Н. G., Grayson, P., Han, L., Inamdar, M., Kondev,
J., Nelson, P.C., Phillips, R., Widom, W, & Wiggins, P. A.
(2007) Biological consequences of tightly bent DNA: the other
life of a macromolecular celebrity. Biopolymers 85,115-130.
Понятное описание физических основ укладки ДНК.
Lebowitz, J. (1990) Through the looking glass: the discovery
of supercoiled DNA. Trends Biochem. Sci. 15, 202-207.
Краткая и интересная историческая заметка.
Pommier, Y. (2006) Topoisomerase I inhibitors: camptothc-
cins and beyond. Nat. Rev. Cancer 6, 789-802.
Wang, J. C. (2002) Cellular roles of DNA topoisomerases: a
molecular perspective. Nat. Rev. Mol. Cell Biol. 3,430-440.
Хроматин и нуклеосомы
Kornberg, R. D. (1974) Chromatin structure: a repeating unit
of histones and DNA. Science 184,868-871.
Классическая статья, в которой представлена субъеди-
ничная модель хроматина.
Вопросы и задачи [39 ]
Losada, А. & Hirano, Т. (2005) Dynamic molecular linkers of
the genome: the first decade of SMC proteins. Genes Dev. 19,
1269-1287.
Nasmyth, K. & Haering, С. H. (2005) The structure and
function of SMC and kleisin complexes. Annu. Rev. Biochem.
74,595-648.
Rando, O. J. (2007) Chromatin structure in the genomics era.
Trends Genet. 23,67-73.
Описание методов визуализации, которые применя-
лись для изучения модификации нуклеосом, определения
местоположения нуклеосом и других аспектов строения
хромосом в масштабах геномов.
Sanna, К. & Reinberg, D. (2005) Histone variants meet their
match. Nat. Rev. Mol. Cell Biol. 6, 139-149.
Segal, E., Fondufe-Mittendorf, Y., Chen, L., Thastrom, A.,
Field, Y, Moore, I. K., Wang, J. Z., & Widom, J. (2006) A
genomic code for nucleosome positioning. Nature 442,772-778.
Thanbichler, M., Wang, S. C., & Shapiro, L. (2005) The
bacterial nucleoid: a highly organized and dynamic structure.
J. Cell. Biochem. 96, 506-521.
Wyman, C. & Kanaar, R. (2002) Chromosome organization:
reaching out to embrace new models. Curr. Biol. 12, R446-R448.
Хороший краткий обзор строения и функций SMC-
белков.
Вопросы и задачи_______________________________
1. Упаковка ДНК в вирусах. Молекулярная мас-
са ДНК бактериофага Т2 составляет 120 • 104 * 6; эта
молекула ДНК упакована в головку бактерио-
фага длиной 210 нм. Подсчитайте длину ДНК
(определите молекулярную массу, зная, что в ге-
номе 650 пар нуклеотидов) и сравните ее с раз-
мером головки Т2.
2. ДНК фага М13. Нуклеотидный состав ДНК фага
М13:23% А, 36% Т, 21% G, 20% С. Что на основании
этих данных можно сказать о ДНК фага М13?
3. Геном Mycoplasma. Полный геном самой ма-
ленькой бактерии Mycoplasma genitalium состоит
из 580 070 п. н. и организован в виде кольцевой
молекулы ДНК. Определите молекулярную массу
и длину (в релаксированном состоянии) этой мо-
лекулы. Чему равно LkQ хромосомы Mycoplasma?
Если о = -0,06, чему равно Lk?
4. Размер генов эукариот. Фермент, выделенный
из печени крысы, содержит 120 аминокислотных
остатков и кодируется геном, размер которого
1440 п. н. Объясните связь между числом ами-
нокислотных остатков в ферменте и числом пар
нуклеотидов в его гене.
5. Порядок зацепления. Молекула замкнутой
кольцевой ДНК в релаксированной форме имеет
Lk = 500. Оцените приблизительно число пар ну-
клеотидов в этой ДНК. Как изменяется порядок
зацепления (увеличивается, уменьшается, не из-
меняется, нельзя определить), если: (а) белковый
комплекс присоединен к нуклеосоме; (б) одна
нить ДНК разорвана; (в) к раствору ДНК добав-
лены ДНК-гираза и АТР; (г) двойная спираль де-
натурирована нагревом?
6. Топология ДНК. В присутствии эукариоти-
ческого конденсина и топоизомеразы типа II Lk
релаксированной замкнутой кольцевой ДНК не
изменяется. Однако в ней появляется множество
узлов.
Образование узлов требует разрыва ДНК, про-
хождения участка ДНК через разрыв и лигирова-
ния под действием топоизомеразы. Учитывая, что
топоизомеразная реакция должна сопровождать-
ся изменением порядка зацепления, объясните
сохранение величины Lk.
7. Плотность суперскрученной ДНК. Бакте-
риофаг X инфицирует Е. coli, интегрируя свою
ДНК в бактериальную хромосому. Успех та-
кой рекомбинации зависит от топологии ДНК
Е. coli. Если плотность сверхвитков ДНК Е. coli
о > -0,045, вероятность интеграции менее 20%;
если о < -0,06 — вероятность превышает 70%.
Установлено, что длина плазмидной ДНК, вы-
деленной из культуры Е. coli, составляет 13 800
п. н., aLk = 1222. Рассчитайте о для этой ДНК и
предскажите вероятность того, что бактериофаг X
сможет инфицировать эти клетки.
8. Изменение порядка зацепления, (а) Чему
равно Lk кольцевой двухцепочечной молекулы
[40] Часть III. 24. Гены и хромосомы
ДНК размером 5000 п. н. с разрывом (ником) в
одной цепи? (б) Чему равно Lk молекулы из за-
дания (а) после лигирования разрыва (релакса-
ции)? (в) Как могло бы измениться Lk молекулы
из задания (б) в результате действия одной моле-
кулы топоизомеразы I Е. coli? (г) Чему равно Lk
молекулы из задания (б) после восьми актов дей-
ствия одной молекулы фермента ДНК-гиразы в
присутствии АТР? (д) Чему равно Lk молекулы
из задания (г) после четырех актов действия од-
ной молекулы бактериальной топоизомеразы I?
(е) Чему равно Lk молекулы из задания (г) после
связывания одной нуклеосомы?
9. Хроматин. Экспериментальные доказатель-
ства, которые помогли определить структуру ну-
клеосом, были получены методом электрофореза
на агарозном геле (электрофореграмма приведе-
на ниже), полосы в котором соответствуют моле-
кулам ДНК. Образцы для электрофореза были
получены путем мягкой обработки хроматина
ферментом, разрушающим ДНК, с последующим
удалением всех белков. Сбоку указаны позиции,
до которых перемещаются линейные фрагменты
ДНК соответствующего размера. Что на основа-
нии этих данных можно сказать о структуре хро-
матина? Почему полосы ДНК толстые и размы-
тые, а не узкие и концентрированные?
1,000 п. н.
800 п. н.
600 п. н.
400 п. н.
200 п. н.
10. Структура ДНК. Объясните, как частичное
раскручивание спирали В-ДНК может облегчить
или стабилизировать формирование Z-ДНК.
И. Поддержание структуры ДНК. (а) Назовите
два структурных свойства, необходимых для под-
держания отрицательной сверхспирализации мо-
лекулы ДНК. (б) Назовите три изменения струк-
туры, которые происходят легче при отрицатель-
ной сверхспирализации ДНК. (в) Какой фермент
при участии АТР может вносить в ДНК отрица-
тельные супервитки? (г) Опишите физический
механизм действия этого фермента.
12. Искусственные хромосомы дрожжей (YAC).
YAC применяются для клонирования крупных
участков ДНК в дрожжевых клетках. Какие три
типа последовательностей ДНК необходимы для
правильной репликации и воспроизведения YAC
в дрожжевой клетке?
13. Структура бактериального нуклеоида. У
бактерий транскрипция групп генов зависит от
топологии ДНК: при релаксации ДНК экспрес-
сия может усиливаться, но чаще ослабевает. При
расщеплении бактериальной хромосомы спец-
ифическим ферментом рестрикции (который
узнает и расщепляет длинные и, следовательно,
редко встречающиеся последовательности) про-
исходит усиление или ослабление экспрессии
только расположенных поблизости генов (в пре-
делах 10 000 п. н.). Транскрипция других генов
хромосомы не изменяется. Объясните этот факт.
(Подсказка. См. рис. 24-37).
14. Топоизомеры ДНК. При электрофоретиче-
ском разделении ДНК в агарозном геле короткие
фрагменты движутся быстрее длинных фрагмен-
тов. Кольцевые молекулы ДНК одинакового раз-
мера, но с разным числом сшивок также можно
разделить в агарозном геле: сильно скрученные
и, следовательно, более компактные топоизомеры
движутся быстрее (сверху вниз на гелях справа).
К гелям добавили краситель хлорохин, который
встраивается между парами оснований и стаби-
лизирует наиболее слабо скрученные молекулы
ДНК. При связывании красителя с релаксиро-
ванной формой кольцевой ДНК происходит ос-
лабление скрученности ДНК в тех участках, где
связался краситель, а в других участках возника-
ют компенсирующие положительные сверхвитки.
В представленном здесь эксперименте с помощью
топоизомераз готовили образцы одной и той же
кольцевой ДНК с разной плотностью сверхспи-
рализации (а). Полностью релаксированная ДНК
мигрировала до положения N (nicked), а сильно
скрученная ДНК (за пределом, где можно разли-
чить отдельные топоизомеры) — до положения X.
5 мкг/мл хлорохина
Движется Движется
как при +ALk как при &Lk
Гель В
а) Почему в геле А на дорожке с о = 0 (т. е. в
образце ДНК, в котором в среднем о = 0) видно
несколько полос?
б) Отрицательные или положительные
сверхвитки несет ДНК в геле В из образца с а = 0
в присутствии интеркалирующего красителя?
в) В обоих гелях на дорожке с а = -0,115 вид-
ны две полосы: одна соответствует сверхспирали-
зованной ДНК, другая — релаксированной ДНК.
Объясните присутствие релаксированной ДНК в
этом и других образцах.
г) Образец нативной ДНК (крайняя левая
дорожка в каждом геле) представляет собой ту же
кольцевую ДНК, выделенную из бактериальных
клеток, но не обработанную топоизомеразами.
Какова приблизительная плотность сверхспира-
лизации этой нативной ДНК?
Анализ экспериметнальных данных [41]
Анализ экспериментальных данных___________
15. Функциональные элементы дрожжевых
хромосом. На рис. 24-9 представлены основные
структурные элементы хромосом пивных дрож-
жей (Saccharomyces cerevisiae). Хейтер, Манн,
Снайдер и Дэвис (1985) определили свойства не-
которых из этих элементов. Их эксперимент был
основан на наблюдении, что в клетках дрожжей
при митозе плазмиды (содержащие гены и точку
начала репликации) ведут себя не так, как хромо-
сомы (содержащие те же элементы, а также цен-
тромеры и теломеры). Плазмиды не контролиру-
ются митотическим аппаратом и распределяются
между дочерними клетками случайным образом.
В отсутствие селективного маркера, заставляю-
щего хозяйскую клетку удерживать плазмиды
(см. рис. 9-4 вт. 1), они быстро теряются. Напро-
тив, хромосомы контролируются митотическим
аппаратом и даже без селективных маркеров
утрачиваются крайне редко (с частотой около
10 5 на одно деление).
Хейтер с коллегами решили выявить наиболее
важные элементы дрожжевых хромосом. Для это-
го они сконструировали плазмиды, содержащие
различные части хромосом, и наблюдали за тем,
правильно ли осуществляется расхождение этих
«синтетических хромосом» при митозе. Для опре-
деления количества аномально распределенных
хромосом потребовался быстрый метод анализа,
позволяющий подсчитывать число копий син-
тетических хромосом в различных клетках. Они
выбрали метод, основанный на том, что колонии
дрожжей дикого типа на питательной среде имеют
белую окраску, тогда как колонии некоторых нуж-
дающихся в аденине (ade-) мутантов окрашены
в красный цвет. В частности, в клетках ade2 нет
функциональной AlR-карбоксилазы (фермент,
катализирующий стадию 6а на рис. 22- 33) и в их
цитоплазме накапливается AIR (5-аминоимида-
золрибонуклеотид). Избыток AIR превращается в
красный пигмент. Кроме того, в исследовании ис-
пользовали ген SUP11, кодирующий охра-супрес-
сор (тип нонсенс-супрессора; см. доп. 27-4), пода-
вляющий проявление фенотипа некоторых ade2
мутантов. Хейтер с соавторами начали с изучения
диплоидного штамма дрожжей, гомозиготного
по мутации ade2\ у этих клеток красная окраска.
Если эти мутантные клетки несут одну копию
SUP11, метаболический дефект частично подавля-
[42] Часть III. 24. Гены и хромосомы
ется, и клетки становятся розовыми. Если в клетке
две или более копий SUP11, дефект подавляется
полностью, и клетки окрашены в белый цвет.
Исследователи вставляли копию SUP11 в син-
тетические хромосомы, содержащие различные
элементы, которые считались необходимыми для
функционирования хромосом, а затем наблюдали
за тем, насколько успешно хромосомы передаются
от одного поколения к следующему. На неселектив-
ные среды высевали розовые клетки и наблюдали
за поведением синтетических хромосом. В частно-
сти, были обнаружены колонии, в которых синте-
тические хромосомы распределялись неправильно
при первом делении после высева, давая колонии, в
которых половина генов относилась к одному типу,
а половина — к другому. Поскольку клетки дрож-
жей неподвижны, колонии были окрашены секто-
рами: половина в один цвет, половина — в другой.
а) Один из механизмов, объясняющих нерас-
хождение хромосом при митозе, состоит в том, что
хромосомы реплицируются, однако сестринские
хроматиды не могут разделиться, так что обе ко-
пии хромосомы попадают в одну и ту же дочернюю
клетку. Объясните, каким образом иерасхождение
синтетических хромосом могло бы привести к об-
разованию двухцветных красно-белых колоний.
(б ) Другой вариант нарушения митоза — по-
теря хромосомы', хромосома не входит в ядро до-
черней клетки или не реплицируется. Объясните,
каким образом потеря синтетической хромосомы
могла бы привести к образованию двухцветных
красно-розовых колоний.
Подсчет колоний разного типа позволил Хей-
теру с соавторами оценить частоту нарушений ми-
тоза для различных типов синтетических хромосом.
Сначала они попытались установить необходимый
размер центромерного фрагмента с помощью син-
тетических хромосом со вставками ДНК разной
длины, содержащими известную центромеру. Ре-
зультаты эксперимента представлены ниже.
Синтетическая хромосома Размер фрагмента, содержащего центромеру, т. и. н. хр Потеря юмосомы, Иерасхождение, % %
1 Отсутствует — >50
2 0,63 ' 1,6 1,1
3 1,6 1,9 0,4
4 3 1,7 0,35
5 6 1,6 0,35
в) На основании представленных данных
сделайте заключение о размере центромерного
фрагмента, необходимого для нормального рас-
хождения хромосом. Объясните свои рассуж-
дения.
г) Интересно, что все созданные в этих экс-
периментах синтетические хромосомы были
кольцевыми и не содержали теломер. Объясните,
каким образом они могли более или менее пра-
вильно реплицироваться.
Далее Хейтер с коллегами сконструировали
набор линейных синтетических хромосом, содер-
жащих функциональные центромерные после-
довательности и теломеры, и определяли общее
количество нарушений митоза (% потерь хромо-
сомы + % нерасхождений) как функцию размера
хромосомы. Результаты эксперимента представ-
лены ниже.
Синтетическая хромосома Размер, т. и. н. Общее число ошибок, %
6 15 И
7 55 15
8 95 0,44
9 137 0,14
д) На основании этих данных сделайте вы-
вод о размере хромосомы, необходимом для
нормального расхождения. Объясните свои
рассуждения.
с) Нормальные хромосомы дрожжей линей-
ны, их длина составляет от 250 до 2000 т. п. н., а
частота нарушений митоза не превышает 10 5 на
одно клеточное деление. Используя эти данные,
ответьте на вопрос: обеспечивали ли использо-
вавшиеся в экспериментах центромерные и тело-
мерные последовательности стабильность митоза
нормальных дрожжевых хромосом, или для это-
го процесса нужны каких-то другие элементы?
Объясните свои рассуждения (Подсказка. Рас-
смотрите график зависимости логарифма числа
ошибок от длины хромосомы.)
Литература
Heiter, Р., Mann, С., Snyder, М., & Davis,
R.W. (1985) Mitotic stability of yeast chromo-
somes: a colony color assay that measures nondis-
junction and chromosome loss. Cell 40, 381-392.
Скучных ферментов не бывает.
— Артур Корнберг. «С любовью к ферментам», 1975
Метаболизм ДНК
25.1. Репликация ДНК 45
25.2. Репарация ДНК 66
25.3. Рекомбинация ДНК 81
В качестве хранилища генетической информа-
ции ДНК занимает центральное место среди
биологических макромолекул. Нуклеотид-
ная последовательность ДНК кодирует первич-
ную структуру всех молекул клеточной РНК и
белков и через ферменты опосредованно влияет
на синтез всех других клеточных компонентов.
Переход информации от ДНК к РНК и белку
определяет размер, форму и жизнедеятельность
всех живых существ.
ДНК прекрасно обеспечивает стабильное
хранение генетической информации Однако по-
нятие «стабильное хранение» означает статичную
неизменную картину. Оно не отражает сложности
процессов, с помощью которых генетическая ин-
формация сохраняется и передается от одного
поколения клеток к следующему. Метаболизм
ДНК включает как процессы точного копирова-
ния молекул (репликация), так и процессы, кото-
рые влияют на саму структуру, несущую инфор-
мацию (репарация и рекомбинация). Именно эти
процессы мы и рассмотрим в данной главе.
Метаболизм ДНК должен происходить иде-
ально точно. Химические реакции присоедине-
ния одного нуклеотида к другому при репли-
кации ДНК элегантны и на удивление просты.
Сложности, как мы увидим далее, в фермен-
тативном аппарате, обеспечивающем точность
передачи генетической информации. Ошибки,
возникающие при синтезе ДНК, могут привести
к серьезным последствиям, причем не только по-
тому, что они изменяют или нарушают функцию
определенного гена, но и потому, что такое изме-
нение передается по наследству.
Ферменты, участвующие в синтезе ДНК, ко-
пируют молекулы ДНК, содержащие миллионы
оснований. Они проделывают это с необыкно-
венной точностью и скоростью, несмотря на то
что ДНК очень компактно упакована и связана с
другими белками. Образование фосфодиэфир-
ных связей между нуклеотидами в растущей цепи
ДНК — только часть сложного процесса, в котором
задействовано множество белков и ферментов.
Задача сохранения генетической информа-
ции лежит в основе процесса репарации ДНК.
В гл. 8 (т. 1) подробно описана чувствительность
ДНК ко многим разрушительным воздействиям.
Такие воздействия происходят нечасто, но они
играют очень важную роль, поскольку устойчи-
вость организмов к изменениям последователь-
ности ДНК очень низкая. ДНК — единственная
макромолекула, для которой предусмотрена си-
стема репарации; количество, разнообразие и
сложность механизмов репарации отражает мно-
гообразие пагубных для ДНК воздействий.
Клетки могут перестраивать хранящуюся в
них генетическую информацию в процессах под
общим названием «рекомбинация». Казалось бы,
процесс рекомбинации подрывает принцип перво-
степенного значения стабильности и целостности
генетической информации. Однако на самом деле
большинство перестроек ДНК играет конструк-
[44] Часть III. 25. Метаболизм ДНК
тивную роль в поддержании целостности генома,
специфическим образом влияя на репликацию и
репарацию ДНК и расхождение хромосом.
В этой главе особое внимание уделяется фер-
ментам метаболизма ДНК. Они заслуживают
внимательного изучения не только ввиду их боль-
шого биологического значения и чисто научного
интереса, но и в связи с их возрастающей ролью в
качестве лекарственных препаратов в медицине и
в качестве инструментов в широком круге совре-
Белок репарации ошибочно
спаренных оснований mutL
Белок, связывающий ss^
одноцепочечную ДНК
Репарация ДНК uvrA
Хеликаза dnaB
Субъединицы РНК- ГгроВЭ
полимеразы \jpoCJ
ДНК-полимераза ] polA —
ДНК-хеликаза/
репарация ошибочно —
спаренных оснований UVrD
Хеликаза 3'—>5' гер
Точка начала репликации |ortC
Инициация репликации
Рекомбинационная
репарация
Субъединица
ДНК-гиразы
Сборка праймосомы
Метилирование
Субъединицы РНК- ГгроАЭ
полимеразы \rpoDj
priA
dam
gyrB
Г dnaN~\
\dnaA J
recF
100/0
75
holC Субъединица ДНК-полимеразы III
holD Субъединица
ДНК-полимеразы III
dnaC Компонент праймосомы
polB ДНК-полимераза II
mutT Разложение В-оксо-dGTP
— polC (dnaE) Субъединица
ДНК-полимеразы III
dnaQ Субъединица
ДНК-полимеразы III
dinB ДНК-полимераза IV
recR Рекомбинационная репарация
— holA Субъединица
ДНК-полимеразы III
phr ДНК-фотолиаза
uvrB Репарация ДНК
holB Субъединица
ДНК-полимеразы III
Праймаза dnaG
Белки репарации г . ТТ
ошибочно fmut^
спаренных оснований
Рекомбинация и
рекомбинационная
репарация
Рекомбинация и
рекомбинационная гесА
репарация
Урацилгликозилаза ung
ogt 06-С-алкилтрансфераза
25 —---ГитиСЛ ДНК-полимераза V
\_umuDj
икации
ДНК-лигаза hg
uvrC Репарация ДНК
sbcB Экзонуклеаза I
АР-эндонуклеаза
------------xthA Корректирующая
holE Субъединица АР’эндонуклеаза
ДНК-полимеразы III
ruvCЭ Рекомбинация и
ruvA > рекомбинационная
ruvBj репарация
гесО Рекомбинаци-
онная репарация
Субъединица
ДНК-гиразы gyrA nfo
Рис. 254. Хромосомная карта Escherichia coli. Показано расположение генов, кодирующих
многие важные для метаболизма ДНК белки. Количество известных генов, участвующих в
метаболизме ДНК, свидетельствует о сложности этих процессов. Числа от 0 до 100 внутри
кольцевой хромосомы соответствуют генетическим единицам измерения, называемым ми-
нутами. Каждая минута соответствует отрезку молекулы ДНК длиной примерно 40 000 п. н.
Трехбуквеные названия генов обычно отражают некоторые аспекты их функции, к примеру
mut — мутагенез; dna — репликация ДНК; pol — ДНК-полимераза; гро — РНК-полимераза;
uvr — устойчивость к УФ; гес — рекомбинация; dam — метилирование аденина; hg —
ДНК-лигаза; Тег — терминация репликации; ori — точка начала репликации (у Е. coli это
oriC, как на рисунке).
25.1 Репликация ДНК [45]
менных биохимических технологий. Многие цен-
ные открытия в области метаболизма ДНК были
сделаны на клетках Escherichia coli, поэтому для
объяснения основ метаболизма обычно в качестве
примера служат хорошо известные ферменты этой
бактерии. Беглый взгляд на некоторые важные
гены на генетической карте Е. coli (рис. 25-1) по-
зволяет представить себе сложность ферментных
систем, участвующих в метаболизме ДНК.
Прежде чем перейти к детальному изучению
процесса репликации, сделаем короткое отступле-
ние и поговорим о принятых сокращениях назва-
ний бактериальных генов и белков, поскольку со
многими из них нам предстоит встретиться в этой
и последующих главах. Аналогичные договорен-
ности используются и в обозначениях эукариоти-
ческих генов, хотя конкретное сокращение может
зависеть от вида организма, и единого правила для
всех эукариотических систем не существует.
КЛЮЧЕВЫЕ ДОГОВОРЕННОСТИ. Бактериальные гены
обычно обозначают тремя строчными буквами
курсивом, и, как правило, название отражает функ-
цию этих генов. Например, dna, uvr и rec гены оз-
начают репликацию ДНК, устойчивость к поража-
ющему действию УФ-облучения и рекомбинацию
соответственно. Если несколько генов отвечают за
одну и ту же функцию, они дополнительно обо-
значаются буквами А, В, С и т. д., например dnaA,
dnaB, dnaQ, что обычно указывает на порядок их
открытия, а не порядок участия в цепи реакций.
Использование сокращений в названиях бел-
ков менее очевидно. В ходе генетических исследо-
ваний обычно выделяют и характеризуют белковые
продукты каждого гена. Многие бактериальные
гены были идентифицированы и названы до того,
как выяснилось значение их белковых продуктов.
Иногда оказывается, что продукт гена — это уже из-
вестный белок, и его приходится переименовывать.
Однако часто бывает, что продукт гена еще неизве-
стен и обладает активностью, которую нельзя опи-
сать обычным названием фермента.
КЛЮЧЕВЫЕ ДОГОВОРЕННОСТИ. Бактериальные бел-
ки часто сохраняют названия соответствующих
генов. Названия белков Е. coli пишутся прямым
шрифтом с заглавной буквы, например белковые
продукты генов dnaA и гесА называются DnaA и
RecA соответственно.
25.1. Репликация ДНК
Задолго до установления структуры ДНК ученых
удивляла способность организмов воссоздавать
самих себя и способность клеток образовывать
много идентичных копий крупных и сложных
макромолекул. Рассуждения на эту тему концен-
трировались вокруг концепции матрицы — струк-
туры, которая позволяет молекулам соединяться
в определенном порядке и образовывать макро-
молекулу с уникальной последовательностью и
функцией. В 1940-е гг. уже сложилось представ-
ление о том, что носителем генетической инфор-
мации является ДНК, но только когда Джеймс
Уотсон и Фрэнсис Крик установили ее структуру,
стало понятно, каким образом ДНК играет роль
матрицы для репликации и передает генетиче-
скую информацию: дело в том, что одна ее цепь
комплементарна другой. Основания образуют
пары по определенным правилам, и каждая цепь
является матрицей для новой цепи с предсказуе-
мой комплементарной последовательностью (см.
рис. 8-14, 8-15 в т. 1). 0 Нуклеотиды: строитель-
ные блоки нуклеиновых кислот
Было доказано, что основные свойства про-
цесса репликации ДНК и каталитические меха-
низмы этого процесса в значительной степени
идентичны у всех видов организмов. Именно
это единство механизмов мы постараемся под-
черкнуть в нашем обсуждении, продвигаясь от
общих основ процесса репликации к ферментам
репликации Е. coli и, наконец, к репликации эу-
кариот.
Основные принципы репликации ДНК
В ранних исследованиях репликации бактериаль-
ной ДНК и ее ферментов было установлено не-
сколько базовых принципов, на которых основан
синтез ДНК у всех живых существ.
Репликация ДНК полуконсервативна. Каждая
цепь ДНК служит матрицей для синтеза новой
цепи, при этом образуются две новые двухцепо-
чечные молекулы ДНК, каждая из которых со-
стоит из одной новой и одной старой цепей. По-
этому процесс называется полуконсервативной
репликацией.
Уотсон и Крик выдвинули гипотезу полу-
консервативной репликации вскоре после публи-
[46] Часть III. 25. Метаболизм ДНК
кации своей статьи о структуре ДНК с 1953 г.; в
1957 г. эта гипотеза была подтверждена Мэтью
Мезельсоном и Франклином Сталем, которые
искусно провели следующие эксперименты. Ме-
зельсон и Сталь культивировали клетки Е. coli на
протяжении многих генераций в среде, в которой
единственный источник азота (NH4C1) содержал
тяжелый изотоп азота 15N вместо наиболее рас-
пространенного легкого изотопа 14N. Плотность
выделенной из этих клеток ДНК примерно на 1 °о
больше, чем у обычной [14N] ДНК (рис. 25-2, а).
ДНК экстрагировали
и центрифугировали
в градиенте плотности Csd
Тяжелая
ДНК (15N) —
Исходная
родительская молекула
Гибридная
ДНК (15N-14N) —
первой генерации клеток
Легкая ДНК
(14N) —
Гибридная --
ДНК
второй генерации клеток
Рис. 25-2. а — несколько поколений клеток выращива-
ли в среде, содержащей только тяжелый азот 15N, поэтому
в ДНК этих клеток содержался исключительно азот 15N,
что показано в виде одной (синей) полосы, образующей-
ся при центрифугировании в градиенте плотности CsCl.
б — клетки переносили на среду, содержащую только
легкий азот 14N, и после первого деления клеток выделя-
ли ДНК, которая в градиенте хлорида цезия образовывала
более высокую (фиолетовую) полосу, в — после второго
цикла репликации ДНК разделялась на две полосы: ги-
бридную (фиолетовую) и еще более легкую (красную),
содержащую только ДНК с 14N, что подтверждало полу-
консервативный характер репликации.
Несмотря на столь небольшое различие, смесь тя-
желой [15N] ДНК и легкой [14N] ДНК можно раз-
делить центрифугированием в градиенте плотно-
сти хлорида цезия.
Клетки Е. coli, выращенные на среде с 15N,
переносили на свежую питательную среду, со-
держащую только изотоп 14N, где клетки про-
должали расти до тех пор, пока численность по-
пуляции не удваивалась. ДНК, выделенную из
этой первой генерации клеток, центрифугирова-
ли в градиенте CsCl и наблюдали единственную
полосу, положение которой свидетельствовало о
том, что двухспиральная ДНК дочерних клеток
представляет собой гибрид, содержащий одну
новую цепь с l4N и одну родительскую цепь с 15N
(рис. 25-2, б).
Представленные результаты опровергли аль-
тернативную теорию консервативной реплика-
ции, согласно которой одна дочерняя молекула
ДНК содержит две вновь синтезированные цепи,
а другая дочерняя молекула — две родительские
цепи; если бы эта теория была верна, в экспери-
менте Мсзельсона -Сталя нс было бы гибридной
ДНК. Гипотеза полуконсервативной репликации
дополнительно подтверждалась на следующем
этапе эксперимента (рис. 25-2, в). После второго
деления и удвоения клеток на среде с 14N из них
была выделена ДНК, разделившаяся в градиенте
хлорида цезия на две полосы: одна по плотности
соответствовала легкой ДНК, а другая — гибрид-
ной ДНК, полученной после первого деления
клеток.
Репликация начинается в точке начала реплика-
ции и обычно идет в двух направлениях. После
доказательства полуконсервативного механизма
репликации возникло множество вопросов. Пол-
ностью ли раскручивается родительская ДНК до
начала репликации каждой цепи? Начинается ли
репликация в случайной или в какой-то особой
точке? После инициации в какой-то точке ре-
пликация происходит в одном направлении или
в двух?
В ранних исследованиях Джона Кэрнса, про-
веденным с помощью метода авторадиографии,
удалось доказать, что репликация — высококоор-
динированный процесс, в котором родительские
цепи одновременно расплетаются и реплициру-
ются. Кэрнс получил радиоактивную ДНК Е. coli,
для чего культивировал эту бактерию на среде с
25.1 Репликация ДНК [47]
Рис. 25-3. Выявление двунаправленного характера
репликации ДНК. а — во время репликации кольцевой
хромосомы образуется структура, напоминающая гре-
ческую букву тета (0), так как обе нити реплицируются
одновременно (новая нить показана красным цветом).
б — репликация может происходить в одном или в обо-
их направлениях, что можно установить с помощью ав-
торадиографии: при внесении 3Н на короткий период
непосредственно перед прекращением реакции метка
(отмечена красным) обнаруживается в одной или в двух
репликативных вилках соответственно. Так была проде-
монстрирована двунаправленность репликации у Е. coli,
Bacillus subtilis и других бактерий. На авторадиограмме
показан репликационный «глазок» в ДНК В. subtilis. Са-
мая большая плотность зерен серебра (стрелки) наблю-
дается именно в тех двух местах, где происходит репли-
кация. Не подвергшаяся репликации часть хромосомы,
находящаяся вне глазка, не содержит метки и поэтому
невидима.
меченным тритием (3Н) тимидином. ДНК была
аккуратно выделена, покрыта слоем фотографи-
ческой эмульсии и оставлена так на несколько
недель; за это время радиоактивный тимидин
оставил в фотоэмульсии «следы» — зерна сере-
бра, создав фотоотпечаток молекулы ДНК. По
этим отпечаткам видно, что интактная хромосо-
ма Е. coli представляет собой единую гигантскую
кольцевую структуру длиной 1,7 мм. В радиоак-
тивной ДНК, выделенной из клеток во время ре-
пликации. кроме того, была обнаружена дополни-
тельная петля (рис. 25-3, «). Кэрнс сделал вывод,
что петля образуется в результате формирования
двух радиоактивных дочерних цепей, каждая из
которых комплементарна родительской цепи.
На одном или на обоих концах петли движется
репликативная вилка, в которой родительская
ДНК раскручивается, позволяя реплицировать-
ся разделившимся цепям. Согласно полученным
Кэрнсом данным, обе цепи ДНК реплицируются
одновременно, а различные варианты его экспе-
римента (рис. 25-3, б) показали, что репликация
бактериальных хромосом двунаправленная: на
обоих концах петли расположены активные ре-
пликативные вилки.
Чтобы выяснить, образуются ли реплика-
тивные вилки в каком-то определенном участке
ДНК, необходимо было установить по всей дли-
не молекулы некие реперные точки. Это было
сделано с помощью метода денатурирующего
картирования, разработанного Россом Инманом
с коллегами. С помощью хромосомы бактериофа-
га X длиной 48 502 п. н. было показано, что можно
вызвать избирательную денатурирацию ДНК в
последовательностях с необычно высоким со-
держанием пар А=Т, получая воспроизводимую
картину одноцепочечных глазков (см. рис. 8-28 в
т. 1). Таким же образом можно частично денату-
рировать выделенную ДНК, содержащую репли-
кативные петли. Это позволяет измерить и кар-
тировать положение и движение репликативных
вилок, используя денатурированные области в
качестве реперных точек. С помощью данного ме-
[48] Часть III. 25. Метаболизм ДНК
тода было показано, что в этой системе реплика-
тивные петли всегда возникают в особых точках,
которые были названы точками начала репли-
кации (ориджинами). Получил подтверждение
и выявленный ранее двунаправленный характер
репликации. У кольцевых молекул ДНК две ре-
пликативные вилки встречаются в точке, распо-
ложенной напротив точки начала репликации.
Специфические точки начала репликации с тех
пор были идентифицированы и охарактеризова-
ны у бактерий и низших эукариот.
Синтез ДНК наполовину прерывистый и прохо-
дит в направлении 5'—>3'. Новая цепь ДНК всегда
синтезируется в направлении 5'—>3', т. е. прира-
щение ДНК происходит со стороны свободной
ОН-группы на З'-конце (строение 5'- и З'-концов
цепи ДНК см. на рис. 8-7 в т. 1). Поскольку две
цепи ДНК антипараллельны, цепь, служащая в
качестве матрицы, считывается с З’-конца в на-
правлении к 5'-концу.
Но если синтез ДНК всегда происходит в на-
правлении 5'-*3', как могут обе цепи синтезиро-
ваться одновременно? Если обе цепи синтезиру-
ются непрерывно, пока движется репликативная
вилка, одна цепь должна синтезироваться в на-
правлении 3'—>5'. Эту проблему в 1960-х гг. раз-
решил Рейджи Оказаки с коллегами. Они обна-
ружили, что одна из новых цепей ДНК синтези-
руется в виде коротких отрезков, которые теперь
называются фрагментами Оказаки. В результате
их исследований стало ясно, что одна цепь син-
тезируется непрерывно, а вторая — прерывисто
(рис. 25-4). Направление синтеза непрерывной,
или лидирующей, цепи совпадает с направлени-
ем движения репликативной вилки. Направле-
ние синтеза прерывистой, или отстающей, цепи
противоположно направлению движения вилки.
Фрагменты Оказаки различаются по длине от не-
скольких сотен до нескольких тысяч нуклеотидов
в зависимости от типа клеток. Ниже мы увидим,
что синтез лидирующей и отстающей цепей четко
координируется.
ДНК разрушается нуклеазами
Чтобы объяснить энзимологию процесса репли-
кации ДНК, сначала рассмотрим ферменты, ко-
торые разрушают, а не синтезируют ДНК. Эти
ферменты называют нуклеазами или ДНКазами,
если они обладают большей специфичностью к
ДНК, чем к РНК. Каждая клетка содержит не-
сколько разных нуклеаз, принадлежащих к двум
большим классам: экзонуклеазы и эндонуклеазы.
Экзонуклеазы расщепляют нуклеиновые кисло-
ты с одного конца молекулы. Многие из них ра-
ботают только в направлении 5'-*3' или только
в направлении 3'—>5', удаляя нуклеотиды либо
с 5'-конца, либо с З'-конца одной цепи двухце-
почечной молекулы нуклеиновой кислоты или
одноцепочечной ДНК. Эндонуклеазы начинают
расщепление нуклеиновой кислоты со специфи-
ческих внутренних сайтов, расщепляя цепь на
более мелкие фрагменты. Некоторые экзонукле-
азы и эндонуклеазы расщепляют только одноце-
почечные ДНК. Существует несколько важных
классов эндонуклеаз, которые разрушают только
определенные нуклеотидные последовательно-
сти (например, эндонуклеазы рестрикции, кото-
рые имеют очень большое значение в биотехно-
логии; см. гл. 9, рис. 9-2 вт. 1). В этой и в последу-
ющих главах мы встретим много типов нуклеаз.
Рис. 25-4. Различия цепей ДНК на репликатив-
ной вилке. Новая цепь ДНК (красная) всегда
синтезируется в направлении 5Z->3Z. Матрица
считывается в противоположном направлении.
3Z->5Z. Лидирующая цепь непрерывно синтезиру-
ется по направлению движения репликативной
вилки. Другая цепь, отстающая, синтезируется
прерывисто в виде коротких отрезков (фрагмен-
тов Оказаки) в направлении, противоположном
тому, в котором движется репликативная вилка.
Фрагменты Оказаки сшиваются ДНК-лигазой.
У бактерий длина фрагментов Оказаки составляет
примерно от 1000 до 2000 нуклеотидов. В клетках
эукариот они короче: от 150 до 200 нуклеотидов.
25.1 Репликация ДНК [49]
ДНК синтезируется ДНК-полимеразами
Поиск фермента, который может синтезировать
ДНК, начался в 1955 г. Артуру Корнбергу с кол-
легами удалось очистить и охарактеризовать
ДНК-полимеразу из клеток Е. coli. Этот фермент,
состоящий из одного полипептида, теперь называ-
ют ДНК-полимеразой I (Мг =103 000; кодируется
геном polA). Гораздо позже исследователи обнару-
жили, что Е. coli содержит еще как минимум четы-
ре другие ДНК-полимеразы, описанные ниже.
При детальном изучении ДНК-полимеразы 1
были установлены общие для всех ДНК-полиме-
раз характеристики процесса синтеза ДНК. В ос-
новной реакции происходит перенос фосфориль-
ных групп. При этом З'-гидроксильная группа ну-
клеотида на З'-конце растущей цепи выступает в
роли нуклеофила, атакующего а-фосфор присо-
единяющегося дезоксинуклеозид-5'-трифосфата
(рис. 25-5). В результате высвобождается неорга-
Артур Корнберг, 1918-2007
нический пирофосфат. Основная реакция выгля-
дит следующим образом:
(dNMP)„ + dNTP------> (dNMP)„+1 + РР{ (25-1)
ДНК Удлиненная
ДНК
где dNMP и dNTP - дезоксинуклеозид-5'-моно-
фосфат и дезоксинуклеозид-5'-трифосфат соот-
ветственно. Реакция происходит с минималь-
ным изменением свободной энергии, поскольку
а
Матричная Растущая цепь
цепь ДНК
ДНК (праймер)
а " ЛЯ.3' о
{aJ о~рД>
5' И[О~> НО
С G
б
Приходящий
дезоксинуклеозид-
5 -трифосфат
О О
р О р О’
О’ О’
а-Фосфат поступающего dNTP
Рис. 25-5. МЕХАНИЗМ РЕАКЦИИ. Элонгация цепи ДНК. а — ДНК-полимераза I нуждается
в одной неспаренной цепи в качестве матрицы и в праймере, несущем на З'-конце свобод-
ную гидроксильную группу, к которой присоединяется новое нуклеотидное звено. Каждый
присоединяющийся нуклеотид выбирается по принципу комплементарности по отноше-
нию к соответствующему нуклеотиду в матричной цепи. Продукт реакции вновь содержит
свободную З'-гидроксильную группу, позволяющую присоединять следующий нуклеотид.
б — для катализа, вероятно, требуется два иона Мд2+, координированных с двумя фосфат-
ными группами присоединяющегося нуклеозидтрифосфата и с тремя остатками Asp, два из
которых консервативны у всех ДНК-полимераз. Ион Мд2+, изображенный справа, облегчает
ф
атаку З'-гидроксильной группы праймера на а-фосфат нуклеозидтрифосфата другой ион
Мд2+ способствует удалению пирофосфата. Оба иона стабилизируют структуру пятикоорди-
национного переходного состояния. По схожему механизму действует и РНК-полимераза
(см. рис. 26-1,6). □ Полимеризация нуклеотидов ДНК-полимеразой
[50] Часть III. 25. Метаболизм ДНК
одна фосфодиэфирная связь образуется из ме-
нее стабильного фосфоангидрида. Нековалент-
ные стэкинговые взаимодействия оснований и
их спаривание обеспечивают дополнительную
стабилизацию удлиняющейся ДНК по сравне-
нию со свободными нуклеотидами. Кроме того,
синтез ДНК приводит к высвобождению в клетке
19 кДж/моль энергии в результате последующего
гидролиза пирофосфата под действием фермента
пирофосфатазы (с. 38 в т. 2).
Еще в ранних исследованиях ДНК-полиме-
разы I удалось определить два основных условия,
необходимых для полимеризации ДНК. Во-первых,
все ДНК-полимеразы нуждаются в матрице. Реак-
ция полимеризации происходит на матрице ДНК
согласно правилам комплементарности оснований,
установленным Уотсоном и Криком: если в матри-
це присутствует гуанин, к новой цепи присоединя-
ется дезоксинуклеотид цитозина и т. д. Это откры-
тие имеет очень важное значение не только потому,
что оно объясняет химическую основу точной по-
луконсервативной репликации ДНК, но и потому,
что это был первый пример использования матри-
цы для проведения реакции биосинтеза.
Во-вторых, полимеразы нуждаются в прайме-
ре. Праймер — это участок цепи (комплементар-
ный матрице) со свободной 3'-гидроксильной груп-
пой, к которой может присоединиться нуклеотид.
Иначе говоря, перед началом синтеза часть новой
цепи уже должна существовать: все ДНК-полиме-
разы могут присоединять нуклеотиды только к уже
существующей цепи. Многие праймеры представ-
ляют собой олигонуклеотиды РНК, а не ДНК; спе-
циальные ферменты синтезируют праймеры там и
тогда, где и когда они требуются.
После присоединения нуклеотида к растущей
цепи ДНК ДНК-полимераза либо диссоциирует,
либо продвигается вдоль матрицы и присоединя-
ет следующий нуклеотид. Диссоциация и обрат-
ное присоединение полимеразы может ограни-
чивать среднюю скорость полимеризации — про-
цесс обычно протекает быстрее, если полимераза
присоединяет нуклеотиды, не покидая матрицы.
Среднее число нуклеотидов, присоединенных
до диссоциации полимеразы, определяет ее про-
цессивность. ДНК-полимеразы существенно
различаются по процессивности; некоторые при-
соединяют всего несколько нуклеотидов до дис-
социации, другие присоединяют многие тысячи.
• Полимеризация нуклеотидов ДНК-полимеразой
Репликация — очень точный процесс
Репликация осуществляется с исключительной
точностью. У Е. coli происходит всего одна ошиб-
ка на каждые 109-1010 присоединенных нукле-
отидов. Для хромосомы Е. coli, размер которой
составляет примерно 4,6 • 106 п. н., это означает,
что за 1000-10 000 циклов репликации случает-
ся лишь одна ошибка. Во время полимеризации
выбор между правильным и неправильным ну-
клеотидом осуществляется не только по специ-
фичности водородных связей между комплемен-
тарными основаниями в парах, но также по гео-
метрии стандартных А=Т и G=C пар оснований
(рис. 25-6). В активном центре ДНК-полимеразы
I могут разместиться только пары оснований с
правильной геометрией. Неподходящий нуклео-
тид может образовать водородные связи с нукле-
отидом в матрице, но такая пара обычно не попа-
дает в активный центр фермента. Это позволяет
отбраковать неправильное основание до того, как
сформируется фосфодиэфирная связь.
Однако, чтобы обеспечить высокую степень
точности репликации, одной только точности ре-
акции полимеризации недостаточно. Измерения
in vitro показали, что ДНК-полимераза вставляет
один неправильный нуклеотид на 104-105 пра-
вильных нуклеотидов. Иногда эти ошибки воз-
никают из-за того, что основание кратковременно
находится в нехарактерной таутомерной форме
(см. рис. 8-9 вт. 1), что позволяет ему образовать
водородные связи не с тем партнером. Число
ошибок in vivo уменьшается благодаря дополни-
тельным ферментативным механизмам.
Один механизм сокращения числа оши-
бок свойствен практически всем ДНК-
полимеразами и заключается в независимой
3'—>5'-экзонуклеазной активности, благодаря
которой каждый нуклеотид после присоедине-
ния проверяется дважды. Эта нуклеазная ак-
тивность чрезвычайно чувствительна к ано-
мальному спариванию оснований, что позволя-
ет ферменту удалять ошибочно присоединен-
ный нуклеотид (рис. 25-7). Если полимераза
присоединила неправильный нуклеотид, про-
исходит ингибирование ее перемещения к по-
зиции, в которой должен присоединяться сле-
дующий нуклеотид. Эта пауза создает возмож-
ность для исправления ошибки. С помощью
3'—>5'-экзонуклеазной активности полимераза
25.1 Репликация ДНК [51]
ДНК-полимераза I
—— Активный центр
ДНК-полимеразы
—L-j- Центр корректирующей
3'—>5'-экзонуклеазной
[§] редкая таутомерная
форма цитозина (С*),
которая образует пару
с А и встраивается
в растущую цепь
активности
Рис. 25-6. Значение геометрии пар оснований для точ-
ности репликации ДНК. а — стандартные пары основа-
ний А=Т и G=C очень схожи по геометрии, и, если актив-
ный центр фермента может вместить одну пару (синяя
рамка), то обычно он вмещает и другую, б — геометрия
аномальных пар отличается, и они не помещаются в ак-
тивном центре ДНК-полимеразы I.
Перед движением
полимеразы цитозин
претерпевает таутомерное
изменение от С* к С
Теперь новый нуклеотид
присоединен ошибочно
Ошибка связывания на
З'-конце растущей цепи
блокирует дальнейшую
элонгацию. ДНК-полимераза
скользит назад таким образом
чтобы ошибочное основание
оказалось в активным центре
3' —>5' -экзонуклеазы.
Неверно спаренный
нуклеотид удаляется.
Рис. 25-7. Пример исправления ошибки благодаря
3'->5'-экзонуклеазной активности ДНК-полимеразы I.
При изучении структуры фермента было установлено,
что при ориентации фермента по ходу движения ДНК-
центр его экзонуклеазной активности локализован
впереди центра полимеразной активности. Ошибочное
спаривание (здесь ошибка С-А) препятствует трансло-
кации ДНК-полимеразы I в следующее положение. Фер-
мент скользит назад, исправляет ошибку благодаря своей
3'-* 5'-экзо нуклеазной активности, а затем возобновляет
полимеразную активность в направлении 5'—>3'.
[52] Часть III. 25. Метаболизм ДНК
удаляет неправильный нуклеотид и продолжа-
ет удлинение цепи. Данная активность, назы-
ваемая корректирующей, не просто обращение
полимеразной реакции (уравнение 25-1), ведь
в ней не участвует пирофосфат. Полимеразную
и корректирующую активности ДНК-полиме-
разы можно измерить независимо. Корректиру-
ющая активность дополнительно увеличивает
точность реакции полимеризации в 102-103 раз.
У мономерной ДНК-полимеразы I полимераз-
ная и корректирующая активности локализова-
ны в разных активных центрах в одном и том же
полипептиде.
При тщательном подборе оснований в соче-
тании с исправлением ошибок ДНК-полимераза
совершает примерно одну ошибку на 106-108
присоединенных оснований В реальности из-
меряемая точность репликации Е. coli еще выше.
Дополнительная точность обеспечивается неза-
висимой ферментативной системой, которая за-
меняет аномальные пары оснований, оставшиеся
после репликации. Данный процесс исправления
ошибок, наряду с другими процессами репарации
ДНК, описан в разд. 25.2.
У Е. coli не менее пяти ДНК-полимераз
Более 90% ДНК-полимеразной активности в экс-
трактах Е. coli связано с функцией ДНК-поли-
меразы I. Однако вскоре после выделения этого
фермента в 1955 г. стали накапливаться сведения
о том, что он не может осуществлять репликацию
крупной хромосомы Е. coli. Во-первых, скорость,
с которой фермент присоединяет нуклеотиды
(600 нуклеотидов/мин), не соответствует ско-
рости движения репликативной вилки в бакте-
риальной клетке (скорость движения реплика-
тивной вилки, как минимум, в 100 раз выше). Во-
вторых, ДНК-полимераза I обладает относитель-
но низкой процессивностью. В-третьих, генети-
ческие исследования показали, что в репликации
участвуют многие гены и, следовательно, многие
белки: очевидно, что ДНК-полимераза I действу-
ет не в одиночку. В-четвертых, и это особенно
важно, в 1969 г. Джон Кэрнс выделил бактериаль-
ный штамм с поврежденным геном ДНК-поли-
меразы I, с которого синтезировался неактивный
фермент. Хотя этот штамм был чрезвычайно чув-
ствителен к повреждениям ДНК, тем не менее он
был жизнеспособен!
Поиск других ДНК-полимераз привел к от-
крытию в начале 1970-х гг. ДНК-полимеразы II и
ДНК-полимеразы III у Е. coli. ДНК-полимераза II
участвует в одном из механизмов репарации ДНК
(разд. 25.3). ДНК-полимераза III — главный фер-
мент репликации у Е. coli. Свойства трех ДНК-по-
лимераз Е coli сравниваются в табл. 25-1. ДНК-по-
лимеразы IV и V, идентифицированные в 1999 г.,
принимают участие в необычном варианте репара-
ции ДНК (разд. 25.2).
ДНК-полимераза I, таким образом, не глав-
ный фермент в репликации; она выполняет функ-
П Сопоставление ДНК-полимераз Е. coli
I ДНК-полимераза II III
Структурный гена polA polB polC (dnaE)
Субъединицы (количество разных типов) 1 7 а10
Относительная молекулярная масса (Мг) 103 000 88 000б 791 500
3'->5'-экзонуклеаза (коррекция ошибок) Да Да Да
5'->3'-экзонуклеаза Да Нет Нет
Скорость полимеризации (нуклеотиды/с) 16-20 10 250- 1000
Процессивность (число нуклеотидов, присоединенных до диссоциации полимеразы) 3-200 1500 ь500 000
а Для субъединичных ферментов назван ген, кодирующий субъединицу с полимеразной ак-
тивностью. Ген dnaE в настоящее время называется polC.
6 Только субъединица, отвечающая за полимеризацию. В составе ДНК-полимеразы II есть
те же субъединицы, что и у ДНК-полимеразы III: р-, у-, б-, 6-, %- и ф-субъединицы (см
табл. 25-2).
25.1 Репликация ДНК |53]
Рис. 25-8. Большой фрагмент ДНК-полимеразы I (фраг-
мент Клёнова). Эта полимераза широко распространена
у бактерий. Фрагмент Клёнова, образующийся в резуль-
тате протеолитической обработки полимеразы, сохраня-
ет полимеразную и корректирующую активности. Пред-
ставленный на рисунке фрагмент Клёнова получен из
термофильной бактерии Bacillus stearothermophilus (PDB
ID 3BDP). Активный центр полимеразы находится глубо-
ко в щели около дальнего конца связанной ДНК (синяя;
темно-синим цветом изображена матричная цепь).
цию проверки и поддержания порядка во время
репликации, рекомбинации и репарации. Особые
функции полимеразы дополняются ее 5'—>3'-экзо-
нуклеазной активностью. Эта активность, отлич-
ная от корректирующей 3'—>5'-экзонуклеазной ак-
тивности (рис. 25-7), локализована в структурном
домене, который может быть отщеплен от фер-
мента в условиях мягкой обработкой протеазами
При удалении 5'—>3'-экзонуклеазного домена
остающийся фрагмент (Мг = 68 000), называемый
большим фрагментом, или фрагментом Клёнова
(рис. 25-8), сохраняет полимеразную и корректи-
рующую активность. Интактная ДНК-полимераза
I благодаря своей 5'—>3'-экзонуклеазной активно-
сти в ходе так называемой ник-трансляции может
заменить участок ДНК (или РНК), связанный с
матричной цепью (рис. 25-9). Большинство дру-
гих ДНК-полимераз не имеют 5'—^'-экзонуклеаз-
ной активности.
ДНК-полимераза III устроена намного слож-
нее, чем ДНК-полимераза I; она состоит из субъе-
диниц десяти разных типов (табл. 25-2). Ее поли-
меразная и корректирующая активности связаны
с а- и Е-субъединицами соответственно. При объ-
единении 0-субъединицы с а- и Е-субъединицами
образуется кор-фермент, который уже может
осуществлять синтез ДНК. но с ограниченной
процессивностью. Два кор-фермента полимера-
зы могут связываться друг с другом посредством
еще одного субъединичного комплекса, называ-
емого погрузчиком зажима, или у-комплексом,
Ник
/
он® РНК или ДНК
5' “ГП—I—ГТ тп—| | | | | | | | ।—ГТ 3
3, „и, | | | | | ! | | | | | | 1„ 5,
Матричная
цепь ДНК
ДНК- V
полимераза I
он®
I I I I I I I I I I I I ГI 1111 3'
3Н 1 1 I I I I I I I I I I 1 I 1 I I 5,
Рис. 25-9. Ник-трансляция. В ходе этого процесса уча-
сток РНК или ДНК, спаренный с матричной ДНК, одновре-
менно разрушается под действием 5'->3'-экзонуклеазной
активности ДНК-полимеразы I и заменяется на новый
под действием полимеразной активности того же фер-
мента. Эта функция реализуется в процессе репарации
ДНК и при удалении РНК-праймеров в ходе репликации
(оба процесса описаны ниже). Цепь нуклеиновой кисло-
ты (ДНК или РНК), подлежащая удалению, показана зе-
леным, новая цепь — красным. Синтез ДНК начинается с
ника (разрыва фосфодиэфирной связи, освобождающего
З'-гидроксильную и 5'-фосфатную группы). Полимера-
за I удлиняет нематричную цепь ДНК и перемещает ник
вдоль ДНК. Этот процесс называется ник-трансляцией. В
месте диссоциации ДНК-полимеразы ник сохраняется и
позднее зашивается другим ферментом.
[54] Часть III. 25. Метаболизм ДНК
Таблица 25-2 Субъединицы ДНК-полимеразы III Е, coli
Число субъединиц
Субъединица на целый фермент Мт субъединицы Ген Функция субъединицы
а 2 129 900 polC (dnaE) Полимеризная активность
£ 2 27500 dnaQ(mutD) Корректирующая Кор-фермент
3'-*5'-экзонуклеаза полимеразы
0 2 8600 holE Стабилизация е-субъединицы
т 2 71 100 dnaX Стабильное связывание Погрузчик зажима
с матрицей; димеризация кор-фермента (у-комплекс), ко- торый погружает
Y 1 47 500 dnaXa Загружает зажим |3-субъединицы
6 1 38 700 holA Открывает зажим на каждый фрагмент Оказаки отстающей
6' 1 36900 holB Загружает зажим цепи
X 1 16600 holC Взаимодействие с SSB
ip 1 15200 holD Взаимодействие с у и х
Р 4 40 600 dnaN Зажим ДНК; необходим для
оптимальной процессивности
а у-Субъединицу кодирует часть гена т-субъединицы, так что 66% т-субъединицы с N-конца
совпадают по аминокислотной последовательности с у-субъединицей у-Субъединица обра-
зуется в результате смещения рамки считывания при трансляции (см. рис. 27-9), что приво-
дит к преждевременной остановке трансляции.
состоящим из пяти субъединиц четырех раз-
ных типов т2убб'. Кор-ферменты связаны через
т-субъединицы. Две дополнительные единицы х
и гр, присоединены к погрузчику зажима. Полный
набор из 13 белковых субъединиц (девяти раз-
ных типов) называется ДНК-полимеразой III*
(рис. 25-10, а}.
ДНК-полимераза III* способна полимери-
зовать ДНК, но с намного меньшей процессив-
ностью, чем нужно для организованной реплика-
ции целой хромосомы. Необходимое увеличение
процессивности обеспечивается присоединением
четырех |3-субъединиц с образованием полного
фермента ДНК-полимеразы III. |3-Субъединицы
объединяются в пары и формируют структуры в
форме бубликов, которые охватывают молекулы
ДНК, как зажимы (рис. 25-10, б). Каждый димер
связывается с кор-ферментом полимеразы III*
(один димерный зажим на кор-фермент) и во
время репликации скользит вдоль ДНК. Скольз-
ящий р-зажим предупреждает диссоциацию
ДНК-полимеразы III из комплекса с ДНК, резко
увеличивая процессивность до 500 000 нуклеоти-
дов и более (табл. 25-1).
В репликации ДНК участвует множество
ферментов и белковых факторов
Для репликации Е coli нужна не только ДНК-по-
лимераза, но также не менее 20 различных фер-
ментов и белков, каждый из которых выполняет
определенную задачу. Весь этот комплекс был
назван ДНК-репликативной системой, или ре-
плисомой. Сложность ферментативного аппара-
та репликации связана со структурой ДНК и с
требованиями высокой точности синтеза. Основ-
ные классы ферментов репликации перечислены
ниже в соответствии с теми функциями, которые
они выполняют.
Чтобы обеспечить доступ к цепям ДНК, ко-
торые служат в качестве матрицы, две родитель-
ские цепи нужно разделить. Обычно этот про-
цесс осуществляют хеликазы — ферменты, кото-
рые движутся вдоль ДНК и разделяют ее цепи,
затрачивая при этом энергию АТР. Разделение
цепей создает топологическое напряжение в спи-
ральной структуре ДНК (см. рис. 24-12), которое
снимается топоизомеразами. Разделенные цепи
стабилизируются ДНК-связывающими белка-
25.1 Репликация ДНК [55]
Вид снизу
Рис. 25-10. ДНК-полимераза III. а — строение бакте-
риальной ДНК-полимеразы III. Два центральных домена
(кор-ферменты), состоящие из субъединиц а, е и 0, свя-
заны с у-комплексом (погрузчиком зажима), состоящим
из пяти субъединиц т2удд'. Субъединицы у и т кодиру-
ются одним и тем же геном. Субъединица у представля-
ет собой укороченную субъединицу т: субъединица т
содержит домен, идентичный у, и дополнительный сег-
мент, взаимодействующий с кор-ферментом. Две другие
субъединицы х и ip (не показаны) ДНК-полимеразы III*
также связаны с у-комплексом. Два [3-зажима присоеди-
няются к двум кор-ферментам; каждый зажим — это ди-
мер [3-субъединицы. Комплекс взаимодействует с DnaB-
хеликазой через т-субъединицу. б — две (3-субъединицы
полимеразы III Е. coli образуют зажим в виде кольца,
которое окружает ДНК. Зажим скользит по молекуле
ДНК, увеличивая процессивность холофермента ДНК-по-
лимеразы III до 500 000 нуклеотидов и более, не позво-
ляя ДНК диссоциировать. Вид снизу демонстрирует две
(3-субъединицы, изображенные в виде лент голубого и се-
рого цвета, окружающих СРК-модель молекулы ДНК. Вид
сбоку демонстрирует контур поверхности (3-субъединиц
(серого цвета), которые окружают двойную спираль ДНК
(голубой и синий цвет), изображенную в виде стрежне-
вой модели (по PDB ID 2P0L).
Вид сбоку
б
ми. Как уже было сказано, прежде чем ДНК-по-
лимеразы смогут начать синтез ДНК, на матри-
це должны находиться праймеры — обычно это
короткие сегменты РНК. синтезированные фер-
ментами праймазами. Позднее РНК-праймеры
удаляются и заменяются на ДНК; у Е. coli это
действие — одна из основных функций ДНК-по-
лимеразы I. После вырезания РНК-праймера и
встраивания ДНК в последовательности ДНК
сохраняется ник в форме разорванной фосфо-
диэфирной связи. Эти ники зашивают ДНК-
лигазы. Все перечисленные процессы нуждают-
ся в координации и регуляции, которые хорошо
исследованы в системе Е. coli.
[56] Часть III. 25. Метаболизм ДНК
Репликация хромосомы Е. coli
происходит постадийно
В синтезе молекулы ДНК можно выделить три
стадии: инициация, элонгация и терминация, ко-
торые различаются как по химической сути, так
и по составу ферментов. В этой и двух последу-
ющих главах рассматриваются именно эти три
общие стадии синтеза крупных информацион-
ных полимеров — ДНК, РНК и белков, а также
специфические особенности каждого биосин-
тетического пути. Описываемые далее события
базируются на информации, первоначально по-
лученной в экспериментах in vitro на очищенных
белках Е. coli, хотя принципы репликации весьма
консервативны во всех системах.
Инициация. Точка начала репликации Е. coli
(oriC) состоит из 245 п. н.; она содержит элементы
последовательности, консервативные во многих
бактериальных точках начала репликации. Стан-
дартный набор консервативных последователь-
ностей показан на рис. 25-11. Наибольший инте-
рес представляют два типа последовательностей:
пять повторов размером по 9 п. н. (R-сайты) -
участки связывания ключевого инициаторно-
го белка DnaA, а также А=Т-богатый участок,
называемый ДНК-расплетающим элементом
DUE (от англ. DNA unwinding element). Кроме
того, существуют три дополнительных участка
связывания DnaA (1-сайты) и участки связыва-
ния белков IHF (от англ, integration host factor) и
FIS (от англ, factor for inversion stimulation). Эти
два белка необходимы для некоторых реакций
рекомбинации, описанных ниже в данной главе,
а их названия отражают их функции. Еще один
ДНК-связывающий белок HU (гистоноподоб-
ный бактериальный белок, ранее называвшийся
фактором U) также участвует в процессе, но не
имеет специфического участка связывания.
В инициации репликации участвуют по
меньшей мере 10 разных ферментов и белков (см.
табл. 25-3). Они расплетают спираль ДНК в точке
начала репликации и собирают препраймирую-
щий комплекс для последующих реакций. Клю-
чевым элементом в процессе инициации высту-
пает белок DnaA — входит в белковое семейство
ААА+ АТРаз (от англ. ATPases associated with
diverse cellular activities). Многие AAA+ АТРазы,
включая DnaA, образуют олигомерные структу-
ры и сравнительно медленно гидролизуют АТР.
Гидролиз АТР служит переключателем, опосре-
дующим переходы одной формы белка в другую.
В случае DnaA ATP-связанная форма активна, а
ADP-связанная форма неактивна.
Восемь молекул белка DnaA. каждая из ко-
торых связана с АТР, образуют спиральный
комплекс, охватывающий R- и 1-сайты в oriC
(рис. 25-12). DnaA имеет более высокое сродство
к R-сайтам, чем к 1-сайтам, и этот белок связыва-
ется с R-сайтами одинаково хорошо как в АТР-,
так и в ADP-форме. Напротив, 1-сайты, в которых
связывается только ATP-связанный DnaA, позво-
ляют различать активную и неактивную форму
DnaA. Молекула ДНК плотно закручивается в
правую сторону вокруг этого комплекса, образуя
Тандемное расположение Сайты связывания белка DnaA,
трех консенсусных пять консенсусных
последовательностей последовательностей
GATCTNTTNTTTT TT(A/T)TNCACC
из 13 нуклеотидов из 9 нуклеотидов
|---1----1 1----------------------1------------------J I 1
DUE R1 IHF R5 II 12 R2 FIS R3 13 R4
Рис. 25-11. Расположение последовательностей в точке начала репликации £. coli (оп'С).
Показаны консенсусные последовательности (см. с. 156 в т. 1) ключевых повторов. N обо-
значает любой из четырех нуклеотидов. Горизонтальные стрелки показывают ориентацию
нуклеотидных последовательностей (стрелки слева направо указывают на последователь-
ность в верхней цепи, а справа налево — в нижней). Участки FIS и IHF — сайты связывания
белков, описанных в тексте. С R-сайтами связан белок DnaA. Сайты I — дополнительные
участки связывания DnaA (у них другие последовательности), DnaA взаимодействует с
ними только в комплексе с АТР.
25.1 Репликация ДНК [57]
DnaA-ATP
DnaC-зависимое
элемента DUE DnaC + ADP + P*
Рис. 25-12. Модель инициации репликации в точке начала репликации Е. coli (priC).
Восемь связанных с АТР молекул белка DnaA присоединяются к точке начала репликации в
R- и 1-сайтах (см. рис. 25-11). ДНК закручивается вокруг этого комплекса с образованием
правозакрученной спирали. Богатый остатками А=Т участок DUE денатурирует из-за на-
пряжения, возникающего в цепи в результате связывания DnaA. Образованию спирального
комплекса с DnaA способствуют белки HU IHF и FIS (здесь не показаны, так как подробно их
структура и роль еще не определены). Гексамеры белка DnaB связываются с каждой цепью
ДНК с помощью белка DnaC. Благодаря своей хеликазной активности белок DnaB раскручи-
вает ДНК, подготавливая ее для посадки праймера и синтеза ДНК.
положительную суперспираль (см. гл. 24). Воз-
никшее в соседних участках ДНК напряжение
вызывает денатурацию А=Т-богатого участка
DUE. Кроме того, образующийся в точке начала
репликации комплекс также содержит несколько
ДНК-связываюших белков — HU, IHF и FIS. ко-
торые облегчают изгиб ДНК.
Затем другая ААА+ АТРаза, DnaC, загружа-
ет белок DnaB на разделенные цепи ДНК в дена-
турированном участке. Гексамер, состоящий из
молекул DnaC, каждая из которых связана с АТР,
образует прочный комплекс с гексамерной коль-
цевой DnaB-хеликазой. При взаимодействии
DnaC с DnaB кольцо DnaB раскрывается, чему
также способствует дальнейшее взаимодействие
между DnaB и DnaA. Два кольцевых гексамера
DnaB связываются с DUE — по одному на каж-
дую нить ДНК. Связанная с DnaC молекула АТР
гидролизуется, что приводит к высвобождению
DnaC, a DnaB остается связанным с ДНК.
Присоединение хеликазы DnaB — ключевой
момент в инициации репликации. Далее репли-
кативная хеликаза DnaB перемещается вдоль
одноцепочечной ДНК в направлении 5'->3', рас-
кручивая ДНК по мере продвижения, Таким об-
разом, связанные с двумя цепями ДНК молекулы
хеликазы DnaB перемещаются в противополож-
ных направлениях, создавая две репликативные
вилки. Все другие белки репликативной вилки
также напрямую или опосредованно связаны с
DnaB. Холофермент ДНК-полимеразы III связан
через т-субъединицы. Другие взаимодействия
DnaB описаны ниже. В начале репликации, когда
нити ДНК разделяются в области репликатив-
ной вилки, с ними связываются, стабилизируя
их, многочисленные молекулы белка, связываю-
щегося с одноцепочечной ДНК {single-stranded
DNA-binding protein, SSB), а ДНК-гираза (ДНК-
топоизомераза II) снимает топологическое на-
пряжение, возникающее впереди вилки из-за рас-
кручивания ДНК.
Инициация — единственная фаза реплика-
ции ДНК. про которую известно, что она регули-
руется, притом таким образом, что репликация
происходит только один раз за клеточный цикл.
Механизм регуляции пока еще точно не установ-
лен. но в ходе генетических и биохимических ис-
следований было выявлено несколько независи-'
мых элементов регуляции.
Как только с ДНК связывается ДНК-по-
лимераза III. а также |3-субъединипы (что сви-
детельствует о завершении фазы инициации),
с |3-субъединицами связывается белок Hda, ко-
торый взаимодействует с DnaA и стимулирует
гидролиз связанного с ним АТР. Белок Hda (на-
звание этого фермента означает «гомологичный
DnaA») — еще одна ААА+ АТРаза, родственная
DnaA. Гидролиз АТР приводит к разборке ком-
[58] Часть III. 25. Метаболизм ДНК
Белки, необходимые для инициации репликации в точке начала репликации £. coli
Белок Мг Число субъединиц Функция
Белок DnaA 52 000 1 Распознает последовательности в точке начала
Белок DnaB (хеликаза) 300 000 6* репликации; раскрывает цепи ДНК в специальных сайтах точки начала репликации Раскручивает ДНК
Белок DnaC 29 000 6* Связывает DnaB в точке начала репликации
ни 19000 2 Гистоноподобный ДНК-связывающий белок;
FIS 22 500 2* стимулирует инициацию ДНК-связывающий белок; стимулирует инициацию
IHF 22 000 2 ДНК-связывающий белок; стимулирует инициацию
Праймаза (белок DnaG) 60 000 1 Синтезирует РНК-праймеры
Белок, связывающий одноцепочечную ДНК (SSB) 75 600 4* Связывает одноцепочечную ДНК
ДНК-гираза (ДНК-топоизомераза II) 100 000 4 Ослабляет торсионное напряжение, вызываемое раскручиванием ДНК
Dam-метилаза 32 000 1 Метилирует последовательность (5')GATC в oriC
* В данном случае субъединицы одинаковые.
плекса DnaA в точке начала репликации. Мед-
ленное высвобождение ADP и связывание новых
молекул АТР замыкает цикл превращений между
неактивной (со связанным ADP) и активной (со
связанным АТР) формами белка, который длится
от 20 до 40 мин.
На ход инициации репликации влияет мети-
лирование ДНК и взаимодействия с бактериаль-
ной плазматической мембраной. ДНК в области
опС метилируется Dam-метилазой (от англ DNA
adenine methylation:, табл. 25-3), которая вводит
метильную группу по положению N6 в остатке
аденина в палиндромной последовательности
(5’)GATC. Точка начала репликации oriC у Е. coli
содержит особенно много повторов GATC — их
Ив последовательности из 245 п. н., тогда как
в среднем во всей хромосоме Е. coli этот повтор
встречается один раз на 256 п. н.
Непосредственно после репликации ДНК
оказывается наполовину метилированной: в ро-
дительских цепях последовательности oriC ме-
тилированы, а во вновь синтезированных — нет.
Эти наполовину метилированные последователь-
ности oriC изолированы за счет взаимодействия
с плазматической мембраной (механизм неизве-
стен) и связывания с белком SeqA. Через некото-
рое время последовательность oriC отделяется от
плазматической мембраны, SeqA диссоциирует,
и Dam-метилаза полностью метилирует ДНК до
того, как она вновь связывает DnaA в новом раун-
де репликации.
Элонгация. Фаза элонгации репликации включа-
ет две связанные между собой операции: синтез
лидирующей цепи и синтез отстающей цепи. Для
синтеза обеих цепей важно присутствие в репли-
катвной вилке нескольких ферментов. Сначала
исходная ДНК раскручивается ДНК-хеликазой,
а возникающее топологическое напряжение
снимается топоизомеразами. Затем белок SSB
стабилизирует каждую отдельную цепь. С этого
момента синтез лидирующей цепи и синтез от-
стающей цепи кардинально различаются.
Синтез лидирующей цепи происходит про-
ще: он начинается с синтеза праймазой (белок
DnaG) короткого (от 10 до 60 нуклеотидов) РНК-
праймера в точке начала репликации. Для осущест-
вления данной реакции DnaG взаимодействует с
хеликазой DnaB. причем праймер синтезируется
в направлении, которое противоположно направ-
лению движения хеликазы DnaB. На самом деле
хеликаза DnaB перемещается вдоль цепи, которая
становится отстающей при синтезе ДНК. Однако
первый праймер, синтезированный при первом
взаимодействии DnaG-DnaB, служит для запуска
синтеза лидирующей цепи ДНК в противополож-
25.1 Репликация ДНК [59]
ном направлении. Дезоксирибонуклеотиды до-
бавляются к этому праймеру ДНК-полимеразой
III, связанной с DnaB-хеликазой, расположенной
на противоположной цепи ДНК. Синтез лидиру-
ющей цепи происходит непрерывно со скоростью,
соответствующей скорости раскручивания ДНК в
репликативной вилке.
Как уже было отмечено, отстающая цепь син-
тезируется в виде коротких фрагментов Оказаки
(рис. 25-13, а). Сначала праймаза синтезирует
РНК-праймер. Затем, как и в процессе синтеза ли-
дирующей цепи, ДНК-полимераза III связывается
с праймером и присоединяет дезоксирибонуклео-
тиды (рис. 25-13, б). На этом этапе синтез каждого
фрагмента Оказаки кажется простым, но на самом
деле это весьма сложный процесс. Сложность за-
ключается в координации синтезов лидирующей
и отстающей цепей: обе цепи синтезируются од-
ним асимметричным димером ДНК-полимеразы
III. что достигается при образовании петли ДНК
из отстающей цепи (рис. 25-14), сближающей две
точки полимеризации.
Синтез фрагментов Оказаки на отстающей
цепи происходит с участием удивительного фер-
ментативного аппарата. DnaB-хеликаза и DnaG-
праймаза образуют отдельный функциональный
блок, праймосому, в составе репликативного
комплекса. ДНК-полимераза III использует один
б
(ДНК-полимераза IID
Рис. 25-13. Синтез фрагментов Оказаки, а — на определенном расстоянии праймаза синтези-
рует РНК-праймер для нового фрагмента Оказаки. Если рассматривать две параллельные ма-
тричные цепи, синтез отстающей цепи формально происходит в направлении, противополож-
ном направлению движения вилки, б — каждый праймер удлиняется ДНК-полимеразой III.
в — синтез ДНК продолжается до тех пор, пока полимераза не достигнет праймера пре-
дыдущего фрагмента Оказаки. Для возобновления процесса вблизи репликативной вилки
синтезируется новый праймер.
[60] Часть III. 25. Метаболизм ДНК
набор кор-ферментов (коровая полимераза) для
непрерывного синтеза лидирующей цепи, в то
время как другой набор кор-ферментов осущест-
вляет циклы синтеза фрагментов Оказаки на пет-
ле отстающей цепи. DnaB-хеликаза, находящаяся
перед ДНК-полимеразой III, раскручивает ДНК в
репликативной вилке (рис. 25-14, а), продвигаясь
вдоль матрицы отстающей цепи в направлении
5'-*3'. ДНК-праймаза время от времени связыва-
ется с DnaB-хеликазой и синтезирует короткий
Постоянный синтез
лидирующей цепи
происходит по мере
' раскручивания ДНК
_ с помощью
Лидирующая ПпаВ-хрпиюпы
UlldD AvJlMKcL)bl-
ЦеПЬ
Комплекс погрузчика
зажима с открытым
скользящим р-зажимом
Коровый домен
РНК-праймер
предыдущего
фрагмента
Оказаки
DnaB
Отстающая цепь
Рис. 25-14. Синтез ДНК на лидирующей и отстающей
цепях. События, происходящие в репликативной вилке,
координируются одним димером ДНК-полимеразы III в
комплексе с DnaB-хеликазой. Здесь показан уже идущий
процесс репликации (стадии от а до д обсуждаются в
тексте). Отстающая цепь изогнута петлей, так что синтез
ДНК происходит одновременно на лидирующей и на от-
стающей матрице. Красные стрелки указывают З'-концы
двух новых цепей и направление синтеза ДНК, толстые
черные стрелки — направление движения родительской
ДНК через комплекс. На отстающей цепи начинает син-
тезироваться фрагмент Оказаки. Цветовое обозначение
субъединиц и функции погрузчика зажима объясняются
на рис. 25-15. 3 Синтез ДНК
Синтез фрагмента
Оказаки практически
завершен. Праймер
предыдущего
фрагмен га Оказаки
подходит к коровым
субъединицам.
Ираймаза
связывается с 1)паВ
синтезирует новый
праймер, затем
диссоциирует.
25.1 Репликация ДНК [61]
Белки реплисомы Е. coli
Белок Мг Число субъединиц Функция
SSB 75 600 4 Связывание с одноцепочечной ДНК
Белок DnaB (хеликаза) 300 000 6 Раскручивание ДНК; компонент праймосомы
Праймаза (белок DnaG) 60 000 1 Синтез РНК-праймера; компонент праймосомы
ДНК-полимераза III 791 500 17 Элонгация новой цепи
ДНК-полимераза I 103 000 1 Заполнение брешей; вырезание праймеров
ДНК-лигаза 74 000 1 Лигирование
ДНК-гираза (ДНК-топоизомераза II) 400 000 4 Суперскручивание
Источник. По Kornberg, А. (1982) Supplement to DNA replication, Table SI 1-2, W. H. Freeman
and Company, New York; с изм.
РНК-праймер (рис. 25-14, б). Затем погрузчик
зажима ДНК-полимеразы III закрепляет на прай-
мере новый скользящий р-зажим (рис. 25-14. в).
Когда синтез фрагмента Оказаки заканчивается,
репликация приостанавливается, кор-фермент
ДНК-полимеразы III отсоединяется от одного за-
жима (а также от законченного фрагмента Оказа-
ки) и связывается с новым зажимом (рис. 25-14,
г, д}. Это инициирует синтез нового фрагмента
Оказаки. Как отмечалось ранее, весь комплекс,
ответственный за координированный синтез
ДНК в репликатиной вилке, называют реплисо-
мой. Белки, задействованные в репликативной
вилке, перечислены в табл. 25-4.
Погрузчик зажима ДНК-полимеразы III.
состоящий из частей двухт-субъедипиц, а также
из у, S и S'-субъединиц, функционирует, кроме
того, как ААА+ АТРаза. Этот комплекс связыва-
ется с АТР и с новым скользящим р-зажимом.
Связывание приводит к возникновению напря-
жения в димерном зажиме, в результате чего на
границе одной из субъединиц открывается коль-
цо (рис. 25-15). Отстающая цепь с только что
присоединенным праймером проскальзывает в
кольцо через образующийся разрыв Затем по-
грузчик зажима гидролизует АТР, высвобождая
скользящий р-зажим и позволяя ему сомкнуть-
ся вокруг ДНК.
Реплисома осуществляет синтез ДНК бы-
стро, присоединяя примерно по 1000 нуклео-
тидов в секунду к каждой цепи (лидирующей и
отстающей). После завершения сборки одного
фрагмента Оказаки его РНК-праймер удаляется
и замещается последовательностью ДНК с помо-
Погрузчик
Рис. 25-15. Комплекс погрузчика зажима ДНК-полимеразы III. Комплекс образован пятью
субъединицами: субъединица у, S и S', а также N-концевыми доменами обеих т-субъединиц
(см. рис. 25-10). Комплекс связывает три молекулы АТР и димерный р-зажим. Это связы-
вание приводит к открыванию р-зажима на границе одной из двух субъединиц. Гидролиз
связанного АТР позволяет р-зажиму вновь замкнуться вокруг ДНК.
[62] Часть III. 25. Метаболизм ДНК
3'-
Отстающая
цепь
----------5'
5'
3'
Ник
NMP
ДНК-полиме]
АТР (или NAD+)
АМР + РРх (или NMN) <
ДНК
Рис. 2546. Заключительные стадии синтеза фраг-
ментов отстающей цепи. ДНК-полимераза I благодаря
своей 5'->3'-экзонуклеазной активности удаляет РНК-
праймеры из отстающей цепи и замещает их на ДНК.
Оставшийся ник (разрыв) ликвидирует ДНК-лигаза. Роль
АТР и NAD+ показана на рис. 25-17.
щью ДНК-полимеразы I, а оставшийся ник лик-
видируется ДНК-лигазой (рис. 25-16).
ДНК-лигаза катализирует образование фос-
фодиэфирной связи между З’-гидроксильной
группой на конце одной цепи ДНК и 5’-фосфатом
на конце другой цепи. Фосфат должен быть пред-
варительно активирован путем аденилирования.
ДНК-лигазы, выделенные из вирусов и эукари-
от, используют для этой цели АТР. ДНК-лигазы
бактерий (рис. 25-17) отличаются тем, что в ка-
честве источника АМР они используют NAD* -
кофактор, который обычно участвует в реакциях
с переносом гидрид-иона (см. рис. 13-24). ДНК-
лигаза — еще один фермент метаболизма ДНК,
который стал важным инструментом в экспери-
ментах с рекомбинантными ДНК (см. рис. 9-1).
Терминация. Наконец, две репликативные вилки
кольцевой хромосомы Е. coli встречаются в точке
терминации, содержащей множество копий по-
следовательности из 20 п. н., называемой Тег (от
лат. terminus — конец) (рис. 25-18). Последова-
3'
5'
Фермент NH3
ДНК-лигаза
Аденилирование
ДНК-лигазы
Фермент
О
О
и
NH2 р
о
О
Р О- (Рибоза рркденин!
О
АМР из АТР (R = PPi)
или NAD (R = NMN)
PPi (из АТР)
или
NMN (из NAD )
Фермент-АМР
ОН О
Р
О’ О“
Ник в ДНК
@ Активирование
5'-фосфата
в нике
Рис. 25-17. МЕХАНИЗМ РЕАКЦИИ. Действие ДНК-лигазы.
На каждой из трех стадий одна фосфодиэфирная связь
образуется за счет другой. Стадии D и 2 приводят к акти-
вации 5'-фосфатной группы в нике. АМР сначала перено-
сится на остаток лизина в молекуле фермента, а затем на
5'-фосфат в нике. На стадии З'-гидроксильная группа
атакует этот фосфат и вытесняет АМР, образуя фосфоди-
эфирную связь и закрывая тем самым разрыв. У £. coli
АМР в реакции ДНК-лигазы происходит из NAD+. ДНК-
лигазы, выделенные из некоторых вирусных и эукарио-
тических источников, используют АТР а не NAD+, и на
стадии 1 они высвобождают пирофосфат, а не никотина-
мидмононуклеотид (NMN).
Фермент^ NHJ
L
(3) УдалениеАМР
закрывает ник
О
ДНК без разрывов
Рибоза Аденин
25.1 Репликация ДНК [63]
тельности Тег расположены на хромосоме таким
образом, что создают что-то вроде ловушки, в
которую репликативная вилка может войти, но
не может из нее выйти. Тег-последовательности
служат участками связывания белка Tus (от англ.
terminus utilization substance}. Комплекс Tus Ter
может задержать репликативную вилку движу-
щуюся только в одном направлении. В каждом
цикле репликации действует только один ком-
плекс Tus- Тег — первый из тех, с которым стол-
кнется первая вилка. Учитывая, что движущиеся
в противоположных направлениях репликатив-
ные вилки обычно останавливаются при встрече,
может показаться, что Тег-последовательности -
необязательные элементы. Однако они могут
предотвратить избыточную репликацию на од-
ной из репликативных вилок, в случае если дру-
гая задерживается или останавливается при стол-
кновении с повреждением ДНК или каким-либо
другим препятствием.
Таким образом, когда одна из репликатив-
ных вилок встречает функциональный комплекс
Tus-Ter, она останавливается; а другая вилка
останавливается, когда встречается с первой
(задержанной) вилкой. Затем реплицируются
последние несколько сотен пар оснований ДНК
между этими крупными белковыми комплекса-
ми (по неизвестному механизму), достраивая две
топологически сцепленные (катенированные)
кольцевые хромосомы (рис. 25-19). Кольцевые
ДНК, связанные таким образом, называются
Точка инициации
репликации
Репликативная вилка ' Репликативная вилка
движется против движется по часовой
часовой стрелки 4 \ стрелке
Репликативная вилка
Репликативная вилка
движется против
часовой стрелки
движется по
часовой стрелке
Катенированные
(сцепленные) хромосомы
(НК .
w.
Разделенные
хромосомы
Рис. 2549. Роль топоизомераз в терминации реплика-
ции. В результате репликации ДНК на двух противопо-
ложно направленных репликативных вилках образуют-
ся полноценные хромосомы, соединенные как катанены
(топологически соединенные кольца). Кольца не связаны
ковалентной связью, но поскольку они переплетены и
каждое из них замкнуто ковалентно, их нельзя разде-
лить без помощи топоизомераз. У Е. coli основную роль
в разделении катенированных хромосом играет топоизо-
мераза типа II (ДНК-топоизомераза IV), которая вносит
кратковременный разрыв в обе цепи ДНК в одной хромо-
соме, позволяя другой хромосоме пройти сквозь разрыв.
TerF
Ловушка для вилки,
двигающейся против
часовой стрелки
?TerD /
ТегА /
ТегВ ТегС ху
Ловушка для вилки,
двигающейся по
часовой стрелке
Рис. 2548. Терминация репликации хромосомы Е. coli.
Последовательности Тег (от ТегА до TerF) располагаются
на хромосоме в виде двух кластеров с противоположной
ориентацией.
катенанами. Функцию разделения катениро-
ванных колец у Е. coli выполняет топоизомераза
IV (топоизомераза II типа). Затем при клеточ-
ном делении разделенные хррмосомы расходят-
ся по дочерним клеткам. Заключительная фаза
репликации других кольцевых хромосом, вклю-
чая многие из ДНК-содержащих вирусов, инфи-
цирующих эукариотические клетки, происходит
по аналогичной схеме.
[64] Часть III. 25. Метаболизм ДНК
Репликация в эукариотических клетках
происходит по похожей схеме, но сложнее
Молекулы ДНК в эукариотических клетках зна-
чительно крупнее, чем в бактериях, и образуют
сложные нуклеопротеиновые структуры (хро-
матин; разд. 24.3). Основная схема репликации
ДНК у эукариот такая же, как у бактерий, и мно-
гие белковые комплексы похожи по функциям и
структуре. Однако репликация у эукариот регу-
лируется и координируется в соответствии с кле-
точным циклом, что усложняет процесс.
Участки начала репликации достаточно хо-
рошо охарактеризованы у некоторых низших эу-
кариот, но значительно хуже у высших эукариот.
У позвоночных для инициации репликации могут
использоваться разные А=Т-богатые последова-
тельности, причем участки начала репликации
могут меняться при каждом клеточном делении.
Репликация дрожжей Saccharomyces cerevisiae на-
чинается в определенной области, называемой
автономно реплицирующейся последователь-
ностью (АРП) или репликатором. Дрожжевые
репликаторы имеют длину примерно 150 п. н.
и содержат несколько важных консервативных
последовательностей. В 16 хромосомах гаплоид-
ного генома дрожжей рассредоточено около 400
репликаторов.
Механизмы регуляции обеспечивают одно-
кратную репликацию всей клеточной ДНК в каж-
дом клеточном цикле. Важную роль в этой регу-
ляции играют белки, называемые циклинами, и
циклин-зависимые киназы (CDK), с которыми
они образуют комплексы (с. 660 в т. 1). Цикли-
ны быстро разрушаются в результате убиквитин-
зависимого протеолиза в конце М-фазы (фазы
митоза), а отсутствие циклинов способствует ор-
ганизации пререпликативных комплексов (рге-
RC) в участках инициации репликации. В быстро
растущих клетках комплекс pre-RC образуется в
конце М-фазы. В медленно растущих клетках
этот комплекс не образуется до конца фазы G1.
Формирование комплексов pre-RC позволяет
клетке начать репликацию (этот этап иногда на-
зывают лицензированием).
Как и у бактерий, ключевой момент в ини-
циации репликации у всех эукариот — присо-
единение репликативной хеликазы, гетерогек-
самерного белкового комплекса МСМ (от англ.
minichromosome maintenance', субъединицы от
МСМ2 до МСМ7). Кольцевая хеликаза, состо-
ящая из субъединиц МСМ2-7, функционирует
подобно бактериальной хеликазе DnaB и связы-
вается с ДНК при помощи другого комплекса из
шести белков, называемого комплексом распоз-
навания точки инициации репликации (ORC;
от англ, origin recognition complex', рис. 25-20).
Комплекс ORC состоит из пяти ААА+ АТРазных
доменов и по своим функциям напоминает бакте-
риальный комплекс DnaA. Кроме того, для при-
соединения комплекса МСМ2-7 необходимы два
белка — CDC6 (от англ, cell division cycle) и CDT1
(от англ. CDC10-dependent transcript 1), причем
дрожжевой белок CDC6 представляет собой еще
одну ААА+ АТРазу.
Для осуществления репликации в S-фазе
происходит синтез и активация комплексов ци-
клин-CDK (таких, как комплекс циклин Е-
CDK2; см. рис. 12-45 в т. 1) и CDC7-DBF4. Оба
типа комплексов помогают активизировать ре-
пликацию путем связывания и фосфорилирова-
ния некоторых белков пререпликативных ком-
плексов. Другие циклины и CDK ингибируют
образование дополнительных pre-RC комплексов
после начала репликации. Например, CDK2 свя-
зывается с циклином А, когда уровень циклина Е
снижается в ходе S-фазы, что ингибирует актив-
ность CDK2 и предотвращает образование до-
полнительных pre-RC-комплексов.
Скорость репликативной вилки у эукариот
(около 50 нуклеотидов в секунду) примерно в
20 раз меньше, чем в Е. coli. Если бы репликация
хромосомы человека начиналась в единственной
точке инициации репликации, при такой скоро-
сти процесса репликация средней хромосомы
продолжалась бы более 500 ч. На самом деле ре-
пликация хромосом человека происходит в двух
направлениях и начинается во многих точках,
удаленных друг от друга на расстояние от 30 до
300 т. п. н. Хромосомы эукариот почти всегда на-
много длиннее, чем хромосомы бактерий, поэто-
му наличие многих точек инициации реплика-
ции, вероятно, общее свойство клеток эукариот.
Как и у бактерий, у эукариот есть несколько
типов ДНК-полимераз. Некоторые из них, воз-
можно, выполняют особые функции, например
репликацию митохондриальной ДНК. В репли-
кации ядерных хромосом участвует ДНК-по-
лимераза а в комплексе с ДНК-полимеразой 5.
Субъединичная ДНК-полимераза а имеет похо-(
25.1 Репликация ДНК [65]
жую структуру и свойства во всех эукариотиче-
ских клетках. Одна из ее субъединиц выполняет
функцию праймазы, а самая крупная субъеди-
ница (Мг ~ 180 000) обладает полимеразной ак-
тивностью. Однако данная полимераза не имеет
Рис. 25-20. Сборка пререпликативного комплекса в точке
начала репликации у эукариот. В участке инициации ре-
пликации связываются белки ORC, CDC6 и CDT1. Эти белки,
многие из которых являются ААА+ АТРазами, способству-
ют присоединению репликационной хеликазы МСМ2-7
в реакции, аналогичной реакции присоединения бакте-
риальной хеликазы DnaB с помощью белка DnaC. В резуль-
тате связывания хеликазного комплекса МСМ с ДНК обра-
зуется пререпликативный комплекс pre-RC, и именно эта
стадия оказывается ключевой для инициации репликации.
корректирующей 3'-*5'-экзонуклеазной актив-
ности и не может обеспечить высокой точности
репликации. Предполагается, что ДНК-полиме-
раза а нужна только для синтеза коротких прай-
меров (либо РНК, либо ДНК) для фрагментов
Оказаки на отстающей цепи. Удлинение этих
праймеров осуществляет мультисубъединичная
ДНК-полимераза 6. Этот фермент связывает-
ся с белком, называемым ядерным антигеном
пролиферирующих клеток (PCNA — от англ.
proliferating cell nuclear antigen] Mr = 29 000), и
стимулируется им; этот белок в большом коли-
честве содержится в ядрах делящихся клеток.
Трехмерная структура PCNA удивительным
образом напоминает структуру |3-субъединицы
ДНК-полимеразы III Е. coli (рис. 25-10, б), хотя
первичные последовательности этих белков не
имеют выраженной гомологии. Функционально
PCNA представляет собой аналог р-субъединиц;
он формирует кольцевой зажим, который су-
щественно увеличивает процессивность поли-
меразы. ДНК-полимераза 5 обладает коррек-
тирующей 3'-*5'-экзонуклеазной активностью
и, предположительно, обеспечивает синтез как
лидирующей, так и отстающей цепи в комплек-
се, напоминающем димерную бактериальную
ДНК-полимеразу III.
Еще одна полимераза, ДНК-полимераза е,
в некоторых ситуациях заменяет ДНК-полиме-
разу 5, например при репарации ДНК. ДНК-по-
лимсраза е также может действовать в реплика-
тивной вилке, возможно, аналогично бактериаль-
ной ДНК-полимеразе I, удаляя праймеры фраг-
ментов Оказаки на отстающей цепи.
В репликации ДНК у эукариот задействова-
ны еще два белковых комплекса. Белок RPA (о г
англ, replication protein А) связывает одноцепо-
чечную ДНК, и его функция аналогична функ-
ции белка SSB в клетках Е. coli. Белок RFC (от
англ, replication factor С) представляет собой по-
грузчик зажима для PCNA, который облегчает
сборку активных репликационных комплексов.
Аминокислотные последовательности субъеди-
ниц комплекса RFC имеют значительное сход-
ство с последовательностями субъединиц бакте-
риального у-комплекса.
Для терминации репликации линейных эу-
кариотических хромосом на концах каждой хро-
мосомы синтезируются специальные структуры,
называемые теломерами (см. гл. 26).
[66] Часть III. 25. Метаболизм ДНК
Вирусные ДНК-полимеразы являются
мишенями для противовирусной терапии
0 Многие ДНК-co держащие вирусы кодируют
собственные ДНК-полимеразы, и некоторые
из них становятся мишенями для лекарственных
средств. Например, ДНК-полимераза вируса про-
стого герпеса ингибируется ацикловиром, разра-
ботанным Гертрудой Элайон (см. с. 558 в т. 2).
Ацикловир состоит из гуанина, к которому при-
соединено неполное кольцо рибозы.
О
Ацикловир
Ацикловир фосфорилируется вирусной тими-
динкиназой. Сродство ацикловира к вирусному
ферменту в 200 раз выше сродства к клеточной
тимидинкиназе. В результате фосфорилирование
происходит преимущественно в клетках, инфици-
рованных вирусом. Клеточные киназы превраща-
ют образовавшийся ацикло-GMP в ацикло-GTP,
который одновременно является и ингибитором,
и субстратом ДНК-полимераз. Ацикло-GTP силь-
нее ингибирует ДНК-полимеразу вируса герпе-
са, чем клеточные ДНК-полимеразы. Поскольку
ацикло-GTP не имеет 3'-гидроксильной группы,
его включение в цепь ДНК воспринимается как
сигнал терминации. Таким образом, репликация
вируса ингибируется па нескольких стадиях.
Краткое содержание раздела 25.1
РЕПЛИКАЦИЯ ДНК
Репликация ДНК происходит с очень высо-
кой точностью и в определенной фазе кле-
точною цикла. Репликация полуконсерва-
тивна, каждая цепь выступает матрицей для
новой дочерней цепи. Репликация протекает
в три стадии: инициация, элонгация и тер-
минация. В бактериях реакция начинается в
точке инициации репликации и обычно про-
исходит в двух направлениях.
ДНК синтезируется ДНК-полимеразами в
направлении 5'—*3'. В репликативной вилке
лидирующая цепь синтезируется непрерыв-
но по ходу движения репликативной вилки;
отстающая цепь синтезируется прерывисто
в виде фрагментов Оказаки, которые затем
сшиваются
Точность репликации ДНК достигается
(1) путем выбора оснований полимеразой.
(2) с помощью корректирующей 3'->5'-эк-
зонуклеазной активности, которой обладает
большинство ДНК-полимераз, и (3) с по-
мощью специфических систем репарации,
исправляющими ошибки, оставшиеся после
репликации.
Большинство клеток имеет несколько
ДНК-полимераз. У Е. coli основной фермент
репликации — ДНК-полимераза III. ДНК-по-
лимераза I выполняет особые функции в ходе
репликации, рекомбинации и репарации.
На стадиях инициации, элонгации и терми-
нации репликации ДНК работает множество
ферментов и белковых факторов, многие из
них принадлежат к семейству ААА+ АТРаз.
Белки бактерий, участвующие в репликации,
организованы в крупные комплексы, внутри
которых матрица ДНК протягивается через
две реплисомы, связанные с плазматической
мембраной.
25.2. Репарация ДНК
Большинство клеток содержит только одинар-
ный или двойной набор геномной ДНК. Повреж-
денные белки и молекулы РНК могут быстро
заменяться новыми благодаря информации, за-
кодированной в ДНК. но сами молекулы ДНК
заменяться не могут. Поддержание целостности
информации в ДНК — важнейшая задача клетки,
осуществляемая с помощью сложного набора си-
стем репарации ДНК. ДНК может разрушиться в
результате разных процессов; некоторые из них
спонтанны, другие катализируются факторами
внешней среды (см. гл. 8 в т. 1). Даже сам процесс
репликации может иногда искажать содержащу-
юся в ДНК информацию, если в результате ошиб-
ки образуются неправильные пары оснований
(например, пара G-T).
25.2 Репарация ДНК [67]
Химия повреждений ДНК разнообразна и
сложна. Клеточный ответ на конкретное повреж-
дение связан с активностью широкого круга фер-
ментных систем, катализирующих одни из наи-
более интересных химических трансформаций
в метаболизме ДНК. Сначала мы рассмотрим
последствия изменений в последовательности
ДНК. а затем специфические системы репарации.
Онкологические заболевания
связаны с мутациями
0 Лучший способ проиллюстрировать значе-
ние репарации ДНК — рассмотреть послед-
ствия неисправленных повреждений. Наиболее
опасны такие изменения ДНК, которые в резуль-
тате репликации и передачи новым поколениям
клеток становятся постоянными. Стабильное из-
менение нуклеотидной последовательности ДНК
называется мутацией Мутации могут выражать-
ся в замене одной пары оснований на другую
(однонуклеотидные замены) или же происходит
вставка и выпадение одной или нескольких пар
оснований (инсерции и делении). Если мутация
затрагивает несущественный участок ДНК или
не имеет принципиального значения для работы
гена, ее называют молчащей мутацией. Изредка
мутации обеспечивают своему хозяину биоло-
гическое преимущество. Однако в большинстве
случаев проявляющиеся мутации являются ней-
тральными или вредоносными.
У млекопитающих наблюдается четкая корре-
ляция между накоплением мутаций и раком. В ходе
простого теста, разработанного Брюсом Эймсом,
можно выявить способность химического соеди-
нения вызывать определенные легко выявляемые
мутации в специальных бактериальных штаммах
(рис. 25-21). Согласно этому тесту, совсем немно-
гие вещества, которые мы используем повседневно,
являются мутагенами. Однако более 90% соедине-
ний, в экспериментальных условиях оказывающих
канцерогенное действие на животных, в тесте Эйм-
са проявляют мутагенную активность. Учитывая
четкую корреляцию между мутагенезом и канцеро-
генезом, тест Эймса для бактериальных мутагенов
широко применяется в качестве быстрого и дешево-
го метода выявления канцерогенов человека.
В геноме типичной клетки млекопитающего
за 24 ч аккумулируется много тысяч поврежде-
ний, Однако в результате репарации ДНК менее
одного повреждения из 1000 становится мутаци-
ей Молекула ДНК относительно стабильна, но
при отсутствии систем репарации кумулятивный
эффект многих нечастых, но разрушительных ре-
акций может привести к гибели организма.
Рис. 25-21 Тест Эймса для выявления канцерогенов, ос-
нованный на мутагенности. Штамм Salmonella typhimuri-
ит, несущий мутацию, инактивирующую фермент мета-
болического пути биосинтеза гистидина, высевают на
среду без гистидина. Растут только единичные клетки.
а — несколько мелких колоний S. typhimurium, которые,
тем не менее, вырастают на не содержащей гистидин сре-
де, несут спонтанные обратные мутации, восстанавли-
вающие биосинтез гистидина. Три чашки б, в и г с такой
средой засевают равным числом клеток. Затем в центр
каждой чашки помещают диски фильтровальной бумаги
с разной концентрацией мутагена. Мутаген значительно
увеличивает вероятность возникновения обратной мута-
ции и, соответственно, число выросших клеток в колонии.
Чистая зона вокруг фильтровальной бумаги показывает,
что концентрация мутагена здесь настолько высокая, что
оказывается летальной для клеток. По мере того как му-
таген диффундирует, удаляясь от диска фильтровальной
бумаги, его концентрация понижается до сублетальных
концентраций, которые индуцируют обратные мутации.
Мутагены можно сравнивать на основании их влияния на
частоту возникновения мутаций. Поскольку после про-
никновения в клетку многие соединения претерпевают
разнообразные химические превращения, соединения
иногда тестируют на мутагенность после их предвари-
тельной инкубации с экстрактом печени. Мутагенность
некоторых веществ проявляется только после такой об-
работки.
[68] Часть III. 25. Метаболизм ДНК
Все клетки имеют
несколько систем репарации ДНК
Количество и разнообразие систем репарации
отражает важность репарации ДНК для выжи-
вания клетки, а также многообразие способов
повреждения ДНК (табл. 25-5). Некоторые рас-
пространенные типы повреждений, например
пиримидиновые димеры (см. рис. 8-31), могут
устраняться с помощью нескольких систем ре-
парации. Многие процессы репарации ДНК
очень неэффективны энергетически в отличие
от большинства метаболических путей, в кото-
рых обычно каждая молекула АТР на счету и
используется оптимальным образом. Но когда
на карту поставлена целостность генетической
информации, энергетические затраты не имеют
значения.
Репарация ДНК возможна в значительной
степени благодаря тому, что молекула ДНК со-
стоит из двух комплементарных цепей. Повреж-
дение ДНК в одной цепи может быть удалено и
аккуратно исправлено благодаря использованию
неповрежденной комплементарной цепи в каче-
стве матрицы. Далее мы рассмотрим основные
типы систем репарации, начиная с репарации
редких неправильно спаренных нуклеотидов,
остающихся после репликации.
Репарация ошибочно спаренных оснований.
Коррекция редких ошибочно спаренных осно-
ваний, остающихся после репликации в Е. coli,
увеличивает общую точность репликации еще
на 2-3 порядка. Неправильно спаренные ос-
нования почти всегда корректируются в со-
ответствии с информацией в старой (матрич-
ной) цепи, поэтому система репарации должна
каким-то образом различать исходную и заново
синтезированную цепи. Для этого клетка метит
матричную ДНК метильными группами. Систе-
ма репарации ошибочно спаренных оснований
Е. coli включает по меньшей мере 12 белковых
компонентов (табл. 25-5), которые либо дискри-
минируют цепи, либо участвуют в самом процес-
се репарации.
Механизм дискриминации цепей не уста-
новлен для большинства бактерий и эукариот,
но хорошо изучен у Е. coli и некоторых родствен-
ных бактерий. У этих прокариот дискримина-
ция цепей основана на действии Dam-метилазы,
Таблица 25-5
Типы систем репарации ДНК у Е. coli
Ферменты/белки
Тип нарушения
Репарация ошибочного
спаривания оснований
Dam-метилаза
Белки MutH, MutL, MutS
ДНК-хеликаза II
SSB
ДНК-полимераза III
Экзонуклеаза I
Экзонуклеаза VII
RecJ-нуклеаза
Экзонуклеаза X
ДНК-лигаза
Эксцизионная репарация
оснований
Ошибочное спаривание
оснований
Д Н К-гл икози л азы
АР-эндонуклеазы
ДНК-полимераза I
ДНК-лигаза
Неправильные основания
(урацил, гипоксантин,
ксантин); алкилиро-
ванные основания; пи-
римидиновые димеры
у некоторых других
организмов)
Эксцизионная репарация
нуклеотидов
АВС-эксцизионная
нуклеаза
ДНК-полимераза I
ДНК-лигаза
Прямая репарация
ДНК-фотолиазы
О6- Метилгуанин-
ДНК-метилтрансфераза
Белок AlkB
Повреждения ДНК,
вызывающие крупные
системные изменения
(например, пиримиди-
новые димеры)
Пиримидиновые димеры
О6-Метил гуанин
1-Метилгуанин,
3-метилцитозин
которая, как вы помните, метилирует ДНК в
положении N6 остатка аденина в составе после-
довательностей (5’)GATC. Сразу после прохож-
дения репликативной вилки наступает короткий
период (несколько секунд или минут), когда ма-
тричная цепь метилируется, а вновь синтезиро-
ванная цепь нет (рис. 25-22). Временно немети-
лированные последовательности GATC во вновь
синтезированной цепи позволяют отличить но-
вую цепь от матричной. Ошибочно встроенные
основания вблизи наполовину метилированной
последовательности GATC устраняются исходя
25.2 Репарация ДНК [69]
из информации, содержащейся в метилирован-
ной родительской (матричной) цепи. Иссле-
дования in vitro показали, что, если последова-
тельности GATC метилированы в обеих цепях,
исправляется небольшое число ошибок; если ни
одна из цепей не метилирована, репарация про-
исходит, но ни одна из цепей не имеет преиму-
ществ. Клеточные системы репарации, основан-
ные на метилировании, эффективно устраняют
однонуклеотидные замены на расстоянии до
1000 п. н. от наполовину метилированной после-
довательности GATC.
Как такие относительно удаленные последо-
вательности GATC направляют коррекцию оши-
бочно спаренных оснований? Механизм показан
на рис. 25-23. Белок MutL образует комплекс
с белком MutS, и этот комплекс связывается со
всеми аномальными парами оснований (кроме
С-С). Белок MutH связывается с MutL и после-
довательностями GATC, с которыми встретился
комплекс MutL-MutS. ДНК с обеих сторон от
аномальной пары протягивается через комплекс
MutL-MutS, создавая петлю. Одновременное
продвижение обоих концов петли через комплекс
равнозначно движению комплекса вдоль ДНК
одновременно в двух направлениях. Белок MutH
обладает сайт-специфической эндонуклеазной
активностью, которая не проявляется до тех пор,
пока комплекс не встречает наполовину метили-
рованную последовательность GATC. В этом ме-
сте MutH катализирует расщепление неметили-
рованной цепи на 5’-конце остатка G в составе по-
следовательности GATC, что помечает цепь для
последующей репарации. Дальнейшие стадии
метаболического пути зависят от того, где имен-
но располагается ошибочно спаренное основание
относительно места расщепления (рис. 25-24).
Рис. 25-22. Метилирование и репарация ошибочно спа-
ренных оснований. Метилирование цепей ДНК у Е. coli
может служить для дискриминации родительских (ма-
тричных) цепей и вновь синтезированных цепей, что
имеет решающее значение для репарации однонуклео-
тидных замен; см. рис. 25-23. Метилирование происхо-
дит по положению N6 остатков аденина в последователь-
ностях 5'(GATC). Эта последовательность представляет
собой палиндром (см. рис. 8-18 в т. 1), расположенный на
двух цепях в противоположных ориентациях.
сн3
GATC
С Т A G
сн3
репликация
сн3
GATC
С Т A G
GATC
С Т A G
сн3
Вскоре после
репликации
матричная цепь
метилируется,
а новая нет
сн3
GATC
С Т A G
Полуметилированная ДНК
GATC
5' 3'
3' С t A G 5'
сн3
Через несколько минут
новая цепь метилируется,
Dam-меЪцгаза и две цРПИ становятся
неразличимы
▼
СН3
GATC
5' 3'
3' С Т A G 5'
сн3
СН3
GATC
5' 3'
3' С Т A G 5
СН3
[70] Часть III. 25. Метаболизм ДНК
Если ошибочно спаренное основание нахо-
дится со стороны 5'-конца от места расщепления
(рис. 25-24, справа), неметилированная цепь рас-
кручивается и разрушается в направлении 3'-*5'
от места расщепления, захватывая ошибочно спа-
ренное основание, и этот участок замещается но-
вой ДНК. Данный процесс требует совместного
сн3
G A T С
-гт~т"гГ..I—ГТ
, I.I 1.,
\ С Т А G /
Ошибка \ z
спаривания \ /
5'--1-------------------------4=4^--------3'
3'-------------------7---------1=1--------5'
действия ДНК-хеликазы II, SSB, экзонуклеазы I
или экзонуклеазы X (обе разрушают цепи ДНК
в направлении 3'-*5'), ДНК-полимеразы III и
ДНК-лигазы. Метаболический путь репарации
ошибочно спаренных оснований, находящихся со
стороны З'-конца от точки расщепления, отлича-
ется лишь тем, что в нем участвует экзонуклеаза
VII (которая расщепляет одноцепочечную ДНК в
направлении 5'-*3' или 3'-*5') или RecJ-нуклеаза
(которая расщепляет одноцепочечную ДНК в на-
правлении 5'—*3')-
При оценке энергетической стоимости репа-
рации однонуклеотидных замен у Е. coli выясняет-
ся, что это очень затратный процесс. Аномальная
пара оснований может находиться на расстоянии
1000 и. н. и более от GATC. Расщепление и восста-
новление сегмента такой длины требует огромно-
го количества активированных предшественников
дезоксинуклеотидов для репарации одной нуклео-
тидной замены Это еще раз подчеркивает значе-
ние целостности генома для клетки.
Все эукариотические клетки содержат не-
сколько белков, структурно и функционально ана-
логичных бактериальным белкам MutS и MutL (но
не MutH). У человека изменения в генах, кодирую-
щих белки этого типа, вызывают некоторые наибо-
лее распространенные наследуемые синдромы, свя-
занные с предрасположенностью к онкологическим
заболеваниям (доп. 25-1), что еще раз демонстри-
рует значение систем репарации для организма. У
большинства эукариот, от дрожжей до человека,
Рис. 25-23. Модель ранних стадий репарации, осно-
ванной на метилировании ДНК. Участвующие в данном
процессе белки Е, coli (см. табл. 25-5) были выделены и
очищены. Узнавание последовательности 5'(GATC) и рас-
познавание ошибочно спаренного основания — специ-
фические функции белков MutL и MutS соответственно.
Белок MutH образует комплекс с MutS в точке аномально-
го спаривания. ДНК проходит через этот комплекс таким
образом, что комплекс движется одновременно в двух
направлениях вдоль ДНК до тех пор, пока не придет в
соприкосновение с белком MutH, связанным с полумети-
лированной последовательностью GATC. Белок MutH рас-
щепляет неметилированную цепь по остатку гуанина в
этой последовательности со стороны 5'-конца. Затем ком-
плекс, включающий ДНК-хеликазу II и одну из экзону-
клеаз, расщепляет неметилированную цепь от этой точки
до ошибочно спаренного основания (см. рис. 25-24).
25.2 Репарация ДНК [71]
сн3 сн3
3-___________।______________Л_______________।__________5'
5' у 3'
V----------- --------------V
АТР-.
ADP+Pj*^
MutL-MutS
ДНК-х< mi i II
.1 зону] • । ! VII
или
р . tea <а Rt..
АГР
ADP+Pi^r
MutL-MutS
ДНК-xt mi иа II
экзонур iea.< а I
или
экзонур
сн3
___________I_______________
ДНК-полим. । , а III
SSB
ДНК-полим( । III
SSB
сн3 сн3
--------------------------I-------------------------I___
________________________________________________________+_
сн3
J___
сн3
J___
Рис. 25-24. Завершение репарации, основанной на метилировании ДНК. Действуя со-
вместно, ДНК-хеликаза II, SSB и одна из четырех экзонуклеаз удаляют участок новой цепи
между местом расщепления белком MutH и точкой, расположенной сразу за ошибочно спа-
ренным основанием. Выбор экзонуклеазы зависит от расположения места расщепления от-
носительно аномальной пары оснований. На схеме показаны альтернативные варианты.
Образующуюся брешь заполняет ДНК-полимераза III (пунктирная линия), а ник ликвиди-
рует ДНК-лигаза (не показано).
основные гомологи MutS - белки MSH2 (от MutS
homolog 2), MSH3 и MSH6. Гетеродимеры MSH2
и MSH6 обычно связываются с одиночными ано-
мальными парами оснований, но хуже связывают-
ся с чуть более длинными неправильно спаренны-
ми петлями. У многих организмов более длинные
участки аномального спаривания (от 2 до 6 п. н.)
связываются с гетеродимерами MSH2/MSH3 или
с гетеродимерами двух типов в тандеме. Гомологи
MutL, особенно гетеродимер MLH1 и PMS1 (от.
postmeiotic segregation), соединяются с комплекса-
ми MSH и стабилизируют их. Детальные исследо-
вания процесса репарации ошибочно спаренных
оснований у эукариот активно продолжаются. В
частности, пока неизвестен механизм, с помощью
которого идентифицируются вновь синтезирован-
ные цепи ДНК, хотя было установлено, что после-
довательности GATC в этом участия не принимают.
Эксцизионная репарация оснований. Каждая
клетка содержит класс ферментов, называемых
ДНК-гликозилазами; они распознают наиболее
распространенные повреждения ДНК (например,
продукты дезаминирования цитозина и аденина;
см. рис. 8-30. а) и удаляют поврежденные осно-
вания посредством расщепления N-гликозидной
связи. Такое вырезание основания приводит к
возникновению апуринового или апиримидино-
вого сайта в ДНК, который называют АР-сайтом
или сайтом, лишенным основания. Обычно каж-
дая ДНК-гликозилаза специфична к одному типу
нарушений.
[72] Часть III. 25. Метаболизм ДНК
Так, урацил-ДНК-гликозилаза, обнаружен-
ная в большинстве клеток, специфически удаля-
ет из ДНК урацил, который образуется при спон-
танном дезаминировании цитозина. В мутантных
клетках, утративших этот фермент, повышается
доля мутаций в парах G=C по сравнению с мута-
циями в парах А=Т. Эта гликозилаза не удаляет
днк-
гликозилаза
Ф
I Поврежденное
основание
ДНК молекулы NTP
полимераза I ч Дезоксирибозофосфат
+ молекулы dNMP
ДНК-
лигаза
4
остатки урацила из РНК или остатки тимина из
ДНК. Необходимость дискриминации тимина и
урацила (продукта дезаминирования цитозина)
для селективной репарации цитозина может быть
одной из причин, по которой в состав ДНК вхо-
дит тимин, а не урацил (см. с. 416 в т. 1).
У большинства бактерий есть только один
тип урацил-ДНК-гликозилазы, а у человека суще-
ствует по меньшей мере четыре типа с различной
специфичностью — это показатель важности уда-
ления урацила из ДНК. Наиболее распространен-
ная урацил-гликозилаза человека, UNG, связы-
вается с реплисомой и вырезает остатки урацила,
случайно встроенные вместо тимина в ходе репли-
кации. Дезаминирование остатков цитозина про-
исходит в 100 раз быстрее в одноцепочечной ДНК,
чем в двухцепочечной, и у человека есть специаль-
ный фермент hSMUGl, который удаляет любые
остатки урацила в одноцепочечной ДНК в ходе
репликации или транскрипции. Две другие ДНК-
гликозилазы человека, TDG и MBD4, удаляют
спаренные с гуанином остатки урацила и тимина,
образованные дезаминированием цитозина или
5-метилцитозина соответственно.
Другие ДНК-гликозилазы распознают и уда-
ляют различные поврежденные основания, вклю-
чая формамидопиримидин и 8-гидроксигуанин
(оба образуются при окислении пурина), гипок-
сантин (образуется при дезаминировании адени-
на) и алкилированные основания, такие как 3-ме-
тиладенин и 7-метилгуанин. В некоторых классах
организмов также были идентифицированы гли-
козидазы, которые распознают другие дефекты,
включая пиримидиновые димеры. Вспомните, что
АР-сайты образуются также в результате медлен-
ного спонтанного гидролиза N-гликозидных свя-
зей в ДНК (см. рис. 8-30, б в т. 1).
Образованный ДНК-гликозилазой АР-сайт
устраняют другие ферменты. Репарация проис-
Рис. 25-25. Эксцизионная репарация оснований. Ф
ДНК-гликозилаза распознает поврежденное основание
и отщепляет его от дезоксирибозы в цепи ДНК. (D АР-
эндонуклеаза расщепляет фосфодиэфирную связь около
АР-сайта. з ДНК-полимераза I инициирует репарацион-
ный синтез, начиная от свободной З'-гидроксильной груп-
пы в нике, при этом удаляется (с помощью 5'^>3'-экзону-
клазной активности) часть поврежденной цепи и проис-
ходит замена ее на новую ДНК. @ Ник, оставшийся после
диссоциации ДНК-полимеразы I, ликвидирует ДНК-лигаза.
25.2 Репарация ДНК [73]
ходит не просто путем присоединения нового ос-
нования и восстановления N-гликозидной связи.
Вместо этого удаляется оставшийся дезокси-
рибозо-5'-фосфат и встраивается новый нуклео-
тид. Этот процесс начинает одна из АР-эндо-
нуклеаз, расщепляющих цепь ДНК, в которой
содержится АР-сайт. Положение разреза отно-
сительно АР-сайта (со стороны 3'- или 5'-конца)
зависит от типа АР-эндонуклеазы. Затем участок
ДНК, включающий АР-сайт, удаляется, ДНК-по-
лимераза 1 достраивает ДНК, а ДНК-лигаза лик-
видирует оставшийся ник (рис. 25-25). У эукариот
замену нуклеотидов выполняют специализиро-
ванные полимеразы, как описано ниже.
Эксцизионная репарация нуклеотидов. Повреж-
дения ДНК, которые вызывают значительные ис-
кажения спиральной структуры, обычно восста-
навливаются системой эксцизионной репарации
нуклеотидов. Этот путь репарации принципиаль-
но важен для выживания всех свободноживущих
организмов. В этой системе репарации (рис. 25-26)
мультисубъединичный фермент гидролизует две
фосфодиэфирные связи — по одной с каждой
стороны от дефекта. У Е. coli и других бактерий
ферментная система гидролизует 5-фосфодиэ-
фирную связь с З'-конца и 8-фосфодиэфирную
связь с 5'-конца, создавая фрагмент из 12-13 ну-
клеотидов (в зависимости от того, одно или два
Повреждение ДНК
ДНК
полимера^й
ДНК шази
ДНК-лига. 1
I ...........I 1 I I I I I I I I I I 1 Г—Г
III « I I 11 I I I 1 I I 1 I I I I I I I t
Рис. 25-26. Эксцизионная репарация нуклеотидов у Е. coli и человека. Основной путь ре-
парации путем вырезания нуклеотидов почти одинаковый у всех организмов, ф Эксцизи-
онная нуклеаза связывается с ДНК в месте крупного повреждения и расщепляет повреж-
денную цепь ДНК с обеих сторон от повреждения. @ Фрагмент ДНК длиной 13 нуклеотидов
(13-мер) или 29 нуклеотидов (29-мер) удаляется с помощью хеликазы. ® Брешь заполняет-
ся ДНК-полимеразой, а @ оставшийся ник ликвидируется ДНК-лигазой.
[74] Часть III. 25. Метаболизм ДНК
основания повреждено). У человека и других
эукариот ферментная система гидролизует 6-ю
фосфодиэфирную связь с З'-конца и 22-ю фосфо-
диэфирную связь с 5'-конца, образуя фрагмент из
27-29 нуклеотидов. В результате двустороннего
разреза из дуплекса высвобождается нуклеотид с
поврежденным участком, а образующаяся брешь
заполняется у Е. coli ДНК-полимеразой I, а у че-
ловека — ДНК-полимеразой е. Ник запечатывает
ДНК-лигаза
У Е. coli ключевую роль в этом процессе ис-
полняет эксцизионная нуклеаза АВС, состоящая
из трех субъединиц: UvrA (Л/г = 104 000), UvrB
(Л/г = 78 000) и UvrC (Мг = 68 000). Определение
«эксцизионная нуклеаза» описывает уникальную
способность этого ферментного комплекса катали-
зировать двойное специфическое расщепление, что
отличает его от обычных эндонуклеаз. Комплекс
белков UvrA и UvrB (А2В) сканирует ДНК и свя-
зывается в месте повреждения. Затем димер UvrA
диссоциирует, оставляя прочный комплекс UvrB -
ДНК. После этого с UvrB связывается белок UvrC,
и UvrB производит расщепление 5-й фосфодиэ-
фирной связи со стороны З’-конца от повреждения.
Далее следует UvrC-опосредованное расщепление
8-й фосфодиэфирной связи со стороны 5'-конца.
Образующийся фрагмент из 12-13 нуклеотидов
удаляется ДНК-хеликазой. Получившаяся не-
большая брешь устраняется ДНК-полимеразой I
и ДНК-лигазой. Это основной путь репарации для
многих типов повреждений (см. рис. 25-26. слева),
включая циклобутановые пиримидиновые димеры,
6,4-фотопродукты (см. рис. 8-31 в т. 1) и некоторые
другие аддукты, включая бензо[я]ниренгуанин, ко-
торый образуется в ДНК под воздействием табач-
ного дыма. Нуклеолитическая активность эксци-
зионной нуклеазы АВС отличается от активности
других эндонуклеаз тем, что она одновременно про-
изводит два разреза в ДНК.
Механизм работы эукариотических эксцизи-
онных нуклеаз чрезвычайно схож с аналогичным
механизмом бактериального фермента, хотя для
двойной экспизии в клетках эукариот необходимы
16 полипептидов, не имеющих сходства с субъеди-
ницами эксцизионной нуклеазы Е. coli. Как описа-
но в гл. 26, эксцизионная репарация нуклеотидов и
оснований у эукариот тесно связана с транскрип-
цией. Генетические дефекты системы эксцизион-
ной репарации нуклеотидов — причина различных
серьезных заболеваний человека (см. доп. 25-1).
Прямая репарация. Некоторые типы поврежде-
ний устраняются без удаления нуклеотида или
основания. Наиболее характерный пример -
прямая фотореактивация циклобутановых пи-
римидиновых димеров, которая осуществляется
ДНК-фотолиазами. Пиримидиновые димеры
образуются в индуцируемой УФ-облучением
реакции, и фотолиазы используют энергию по-
глощенного света для устранения повреждения
(рис. 25-27). Фотолиазы обычно содержат два
кофактора, которые действуют как светопогло-
щающие агенты (хромофоры). Одним из хромо-
форов всегда является FADH . У Е. coli и дрож-
жей другой хромофор — фолат В ходе реакции
образуются свободные радикалы. В клетках пла-
центарных млекопитающих (в т.ч. у человека)
ДНК-фотолиазы отсутствуют.
Другим примером может служить репа-
рация нуклеотидов, дефект в которых вызван
алкилированием. Модифицированный нуклео-
тид О6-метилгуанин образуется в присутствии
алкилирующих агентов — это широко распро-
страненное и сильно мутагенное повреждение
(см. с. 419 в т. 1). В ходе репликации он легче
образует пару с тимином, чем с цитозином, и
в результате происходят замена пары G=C
на А=Т (рис. 25-28). Прямую репарацию О6-
метилгуанина производит О6-метилгуанин-
ДНК-метилтрансфераза, которая катализирует
перенос метильной группы на один из своих
остатков Cys. Эта метилтрансфераза не истин-
ный фермент, поскольку в результате однократ-
ного переноса метильной группы она остается
метилированной и выбывает из процесса. Рас-
ход целой белковой молекулы для исправления
одного поврежденного основания — очередная
яркая иллюстрация приоритета целостности
клеточной ДНК.
О6-Метилгуаниновый Гуаниновый
нуклеотид нуклеотид
25.2 Репарация ДНК [75]
По иному, но тоже прямому механизму идет
репарация 1-метиладенина и 3-метилцитозина.
Аминогруппы остатков А и С иногда метилируются
(обычно в одноцепочечной ДНК), и это влияет на
правильность спаривания оснований. У Е. coli окис-
лительное деметилирование подобных алкилиро-
ванных нуклеотидов опосредует белок AlkB — пред-
ставитель суперсемейства а-кетоглутарат- Fe24 -
зависимых диоксигеназ (рис. 25-29). (Описание
другого фермента из этой группы см. в доп. 4-3 в т. 1.)
свет
MTHFpoly(Glu)
MTHFpoly(Glu)
поглощает фотон
синего света
(300-500 нм).
н
HN
О
NH
'О
Циклобутановый пиримидиновый димер
*MTHFpoly(Glu)
н3с
Возбужденный
флавин (*FADH“)
предоставляет
электрон /
пиримидиновому
димеру, что
приводит к
образованию
нестабильного q —
димер-радикала i
Н3с
H
*FADH“
N. /N“ О
NH
R
Н3С
Энергия
(2) возбуждения
передается на
FADH" в
активном
центре
Hi
О
О
NH
t)
0“
NH
t)
Перегруппировки
электронной плотности
приводят к
восстановлению
мономерных
пиримидинов.
Н3С
Н3с
N. /N~ О
NH
HN
R
N' V
н о
Флавиновый
радикал
FADH-
NH
Н3с
N. /N“ О
FADH“
Электрон переносится
обратно на
флавин-радикал для
восстановления FADH~
HN
NH
'O
Мономерные пиримидины
в репарированной ДНК
Рис. 25-27 МЕХАНИЗМ РЕАКЦИИ. Устранение пиримидиновых димеров фотолиазой. Энер-
гия поглощенного света используется для ликвидации последствий фотореакции, вызвав-
шей повреждение. У Е. coli совместно работают два хромофора фотолиазы = 54 000):
№,Ы10-метенилтетрагидрофолилполиглутамат (MTHFpolyGlu) и FADH". MTHFpolyGlu выпол-
няет функцию антенны, поглощая фотоны синего света (300-500 нм). Энергия возбуждения
передается на FADH-, и возбужденный флавин (*FADH_) отдает электрон пиримидиновому
димеру, устраняя повреждение.
R
О
N' V
н о
[76] Часть III. 25. Метаболизм ДНК
Цитозин
метилирование и
репликация
метилирование
О6-Метилгуанин
Рис. 25-28. Пример образования мутаций в результа-
те повреждения ДНК. а — продукт метилирования 06-
метилгуанин легче образует пару с тимином, чем с ци-
тозином. б — при отсутствии репарации это приводит к
мутации: после репликации G=C заменяется на А=Т.
Взаимодействие репликативных вилок
с повреждением в ДНК может запустить
подверженный ошибкам
синтез ДНК через повреждение
Рассмотренные выше пути репарации обычно дей-
ствуют в случае повреждений в двухцепочечной
ДНК, при этом точность восстановления генетиче-
ской информации поврежденной цепи до первона-
чального состояния обеспечивает неповрежденная
цепь. Однако при некоторых типах повреждений,
например при двунитевых разрывах, поперечных
сшивках или повреждениях в одноцепочечной
ДНК, комплементарная цепь также оказывается
разрушенной или отсутствует. Двунитевые разры-
вы и повреждения в одноцепочечной ДНК чаще
всего возникают в тех случаях, когда репликатив-
ная вилка наталкивается на нерепарированное по-
вреждение ДНК (рис. 25-30). Такие повреждения
и поперечные сшивки ДНК, кроме того, могут
быть результатом воздействия ионизирующей ра-
диации и окислительных реакций.
При застопоривании бактериальной репли-
кативной вилки возможны два пути репарации. В
отсутствие второй цепи информация, необходи-
мая для точной репарации, должна поступить от
гомологичной хромосомы. Таким образом, систе-
ма репарации использует гомологичную генетиче-
скую рекомбинацию. Рекомбинационная репара-
ция ДНК подробно рассматривается в разд. 25.3.
При некоторых обстоятельствах реализуется вто-
рой путь репарации — подверженный ошибкам
синтез ДНК через повреждение (TLS — от англ.
translesion synthesis). При активации этого пути ре-
парация ДНК становится существенно менее точ-
ной и связана с высокой частотой мутаций. У бак-
терий подверженный ошибкам синтез ДНК через
повреждение — часть стрессового ответа клетки
на сильное повреждение ДНК, который называют
SOS-ответом, Некоторые SOS-белки, например
уже описанные UvrA и UvrB (табл. 25-6), всегда
25.2 Репарация ДНК [77]
1 -Метиладенин
О2 + соо со2 + СОО- 1
сн2 сн2
сн2 сн2
с=о соо
1 Сукцинат
соо а-Кетоглутарат
А1кГ Fe2+
Рис. 25-29. Прямая репарация алкилированных основа-
ний белком AlkB. Белок AlkB — а-кетоглутарат-Бе2+-за-
висимая диоксигеназа; катализирует окислительное
деметилирование остатков 1-метиладенина и 3-метил-
цитозина.
Н2С=О + Н+
Аденин
СО2
а-Кетоглутарат
О2
Сукцинат
AlkB Fe2+
Н2С=О + н
Формальдегид
Цитозин
Нерепарированное
Нерепарированный
Двух цепочечный
разрыв
I
Рекомбинационная репарация ДНК
или подверженная ошибкам репарация
Двух цепочечный
разрыв
Рис. 25-30. Повреждения ДНК и их влияние на репликацию. Если репликативная вилка
наталкивается на неисправленное повреждение или разорванную цепь, репликация обыч-
но останавливается. Слева: повреждение, оставшееся в нереплицированном одноцепо-
чечном сегменте ДНК; справа: двухцепочечный разрыв цепи. В любом случае поврежде-
ние в одной цепи не может быть устранено ранее описанными механизмами, поскольку
комплементарная цепь, необходимая для точной репарации, повреждена или отсутствует.
В таких случаях есть два возможных пути репарации: рекомбинационная репарация ДНК
(см. рис. 25-37) или при слишком обширных повреждениях подверженная ошибкам репа-
рация через повреждение. В последнем механизме задействована другая ДНК-полимераза
(ДНК-полимераза V, кодируемая генами итиС и шпиП), которая может реплицировать, хотя
и с ошибками, участки ДНК с различными типами повреждений. Такой механизм называет-
ся репарацией, «подверженной ошибкам», из-за частого возникновения мутаций.
[78] Часть III. 25. Метаболизм ДНК
Гены, индуцируемые как часть SOS-ответа у Е, coli
Название гена Кодируемый белок и/или его роль в репарации ДНК
Гены с известной функцией polB (dinA) Кодирует субъединицу ДНК-полимеразы 11 с полимеразной активностью, необходимую для возобновления рекомбинационной репарации
uvrA 1 uvrB | итиС 1 umuD] sulA Кодируют субъединицы UvrA и UvrB АВС-экзонуклеазы Кодируют ДНК-полимеразу V Кодирует белок, ингибирующий клеточное деление, возможно, высвобождая время для репарации ДНК
recA Кодирует RecA белок, необходимый для подверженной ошибкам репарации и рекомбинационной репарации
dinB himA Кодирует ДНК-полимеразу IV Кодирует субъединицу клеточного фактора интеграции (IHF), участвующего в сайт-специфической рекомбинации, репликации, транспозиции фагов, регуляции экспрессии генов
Гены метаболизма ДНК с неизвестной функцией в репарации ssb uvrD recN Гены, функция которых не установлена dinD dinF Кодирует белок, связывающийся с одноцепочечной ДНК (SSB) Кодирует ДНК-хеликазу II (раскручивает ДНК) Необходим для рекомбинационной репарации
Некоторые из этих генов и их функции обсуждаются в гл. 28.
присутствуют в клетке, но при запуске SOS-ответа
их уровень значительно повышается. В подвер-
женной ошибкам репарации участвуют и другие
SOS-белки, в том числе UmuC и UmuD («Umu» —
от англ, unmutahle — немугируемый; без функци-
онального гена ити подверженная ошибкам репа-
рация отсутствует). В SOS-регулируемом процес-
се белок UmuD превращается в более короткий
фрагмент UmuD’, который в комплексе с UmuC
образует специализированную ДНК-полимеразу
(ДНК-полимеразу V), способную восстанавли-
вать многие повреждения ДНК, блокирующие
репликацию. В дефектном участке правильное
спаривание оснований часто оказывается невоз-
можным, поэтому такая репликация через повреж-
дение сопровождается ошибками.
В этой главе мы многократно подчеркивали
особое значение сохранности генома, и поэтому
существование системы, повышающей вероят-
ность возникновения мутаций, может показаться
странным. Использование этой системы мож-
но воспринимать, как некий шаг отчаянья Гены
итиС и umuD полностью индуцируются только
на поздней стадии SOS-ответа, они не активиру-
ются для синтеза, инициируемого расщеплением
UmuD, до тех пор, пока уровень повреждений
ДНК не становится слишком высоким, и блоки-
руются все репликативные вилки. Мутации, воз-
никающие в результате такой репликации, приво-
дят к гибели одних клеток и опасны для других,
но эту биологическую цену организм платит за
то, чтобы не допустить прекращения реплика-
ции, поскольку это позволяет выжить хотя бы не-
скольким мутантным дочерним клеткам.
Наряду с ДНК-полимеразы V для репли-
кации через поврежденный участок необходим
белок RecA. Филаменты RecA, связанные с одно-
цепочечной ДНК в одном участке хромосомы, мо-
гут активировать комплексы ДНК-полимеразы V,
связанные в отдаленных участках хромосомы. Ре-
25.2 Репарация ДНК [79]
ализации этого так называемого трянс-механизма
способствует образование петель, сближающих
в пространстве удаленные друг от друга последо-
вательности хромосом. В ходе SOS-ответа инду-
цируется еще одна ДНК-полимераза — ДНК-по-
лимераза IV (продукт гена dinB). Репликация под
действием ДНК-полимеразы IV также происходит
с большим количеством ошибок. Бактериальные
ДНК-полимеразы [V и V относятся к семейству
TLS-полимераз, найденному у всех организмов.
Эти ферменты не имеют корректирующей экзону-
клеазной активности, а точность в выборе основа-
ний при репликации в 102 раз ниже, что уменьшает
точность репликации в целом примерно до одной
ошибки на 1000 нуклеотидов.
У млекопитающих есть много ДНК-поли-
мераз из семейства TLS-полимераз, отличаю-
щихся низкой точностью синтеза. Однако при-
сутствие этих ферментов не является причиной
повышенной частоты мутаций, поскольку боль-
шинство из них выполняют особые функции
при репарации. ДНК-полимераза г], например,
относится к TLS-полимеразам и обнаружена у
всех эукариот. Она осуществляет синтез через
поврежденные участки, главным образом, через
циклобутановые Т-Т-димеры. В этом случае
образуется немного мутаций, поскольку фер-
мент предпочтительно вставляет два остатка А
напротив связанных остатков Т. У эукариот не-
сколько полимераз, включая ДНК-полимеразы
|3, I и X, выполняют специальные функции с
низкой точностью при эксцизионной репарации
оснований. Каждый из этих ферментов наряду
с полимеразной активностью имеет активность
5'-дезоксирибозофосфат-лиазы. После удаления
основания гликозилазой и расщепления осто-
ва молекулы АР-эндонуклеазой эти ферменты
удаляют АР-сайт (5'-дезоксирибозофосфат) и
заполняют очень короткую брешь. Количество
мутаций, возникающих при действии ДНК-по-
лимеразы г], невелико по той причине, что она
синтезирует очень короткие участки ДНК (ча-
сто в один нуклеотид).
В результате исследования систем репара-
ции клеточной ДНК стало понятно, что метабо-
лизм ДНК поддерживает целостность генома с
помощью многочисленных и часто избыточных
систем. В геноме человека более 130 генов ко-
дируют белки, связанные с репарацией ДНК.
Во многих случаях потеря функции одного
гена приводит к нестабильности генома и повы-
шению вероятности опухолевых заболеваний
(доп. 25-1). Эти системы репарации часто рабо-
тают в комплексе с системами репликации ДНК
и дополняются системами рекомбинации, кото-
рые будут рассмотрены ниже.
Дополнение 25-1
МЕДИЦИНА
Репарация ДНК и рак
Рак у человека возникает, когда определенные гены,
регулирующие нормальное клеточное деление (онко-
гены и опухолевые супрессоры; см. гл. 12 в т. 1), пере-
стают нормально функционировать, изменяются или
активируются не вовремя. В результате рост клеток
может выйти из-под контроля — возникает опухоль.
Гены, контролирующие клеточное деление, могут быть
повреждены спонтанной мутацией или перекрываться
инвазией опухолевого вируса (1Л. 26). Неудивительно,
что изменения в генах репарации ДНК, приводящие к
усилению мутагенеза, могут значительно повысить ин-
дивидуальную чувствительность к раку. Все дефекты
генов белков, участвующих в эксцизионной репарации
оснований, репарации ошибочно спаренных основа-
ний, рекомбинационной репарации и SOS-репарации,
связаны с онкологическими заболеваниями. Итак, от
репарации ДНК может зависеть жизнь и смерть.
Эксцизионная репарация оснований у человека, в
отличие от бактерий, нуждается в большем числе бел-
ков, хотя в целом механизмы очень похожи. Генетиче-
ские дефекты, сопровождающиеся инактивацией экс-
цизионной репарации оснований, связаны с несколь-
кими генетическими заболеваниями, среди которых
лучше других исследована пигментная ксеродерма,
или ХР (от лат. xeroderma pigmentosum). Поскольку у
человека эксцизионная репарация оснований — един-
ственный способ репарации пиримидиновых димеров,
люди с ХР очень чувствительны к свету и часто заболе-
вают раком кожи, который индуцируется солнечным
светом. У большинства больных ХР также выраже-
ны неврологические расстройства, предположитель-
но из-за отсутствия у них репарации определенных
Продолжение на с. 80
[80] Часть III. 25. Метаболизм ДНК
Дополнение 25-1
гг МЕДИЦИНА
Репарация ДНК и рак (продолжение)
повреждений, вызванных высокой активностью
окислительного метаболизма в нейронах. Дефекты в
генах, кодирующих один из семи (как минимум) раз-
ных белковых компонентов системы эксцизионной
репарации оснований, могут привести к семи генети-
ческим вариантам ХР (от ХРА до XPG). Некоторые
из этих белков (особенно те, которые повреждены
при ХРВ, XPD и XPG) также участвуют в сопряжен-
ной с транскрипцией репарации окислительных по-
вреждений, как описано в гл. 26.
У большинства микроорганизмов есть дополни-
тельные пути репарации циклобутановых пирими-
диновых димеров — они могут использовать ДНК-
фотолиазную активность, а иногда и эксцизионную
репарацию оснований в качестве альтернативного меха-
низма эксцизионной репарации нуклеотидов, но у чело-
века и других плацентарных млекопитающих такой воз-
можности нет Отсутствие альтернатив эксцизионной
репарации нуклеотидов для удаления пиримидиновых
димеров вызвало предположение, что на ранней стадии
эволюции млекопитающие были мелкими животны-
ми, были покрыты мехом и вели ночной образ жизни,
не очень нуждаясь в репарации повреждений, вызван-
ных УФ-облучением. Однако у млекопитающих есть
путь синтеза, позволяющий обойти пиклобутановые
пиримидиновые димеры, и в нем участвует ДНК-по-
лимераза т]. Этот фермент предпочтительно вставляет
два остатка А напротив пиримидинового димера Т Т,
минимизируя вероятность мутации. Люди, у которых
в силу генетических нарушений не работает ДНК-по-
лимераза т], страдают заболеванием, похожим на пиг-
ментную ксеродерму (ХР-вариант V, или XPV). Клини-
ческие проявления XPV напоминают симптомы клас-
сической формы ХР, хотя уровень мутаций при XPV
бывает выше, когда клетки подвергаются воздействию
УФ-облучения. По-видимому, в нормальных клетках
система эксцизионной репарации нуклеотидов рабо-
тает совместно с ДНК-полимеразой ц, восстанавливая
и или обходя пиримидиновые димеры, что необходимо
для нормального клеточного роста и репликации ДНК.
УФ-облучение приводит к появлению множества пи-
римидиновых димеров, и для поддержания репликации
хотя бы часть из них приходится обходить путем синтеза
через повреждение Если одна из систем отсутствует, ее
частично компенсирует другая система. Отсутствие ак-
тивности полимеразы ц приводит к остановке реплика-
тивных вилок и исправлению повреждений от УФ-света
TLS-полимеразами, которые допускают гораздо больше
ошибок. И если отсутствуют другие системы репарации
ДНК, увеличение числа мутаций часто приводит к раку.
Один из распространенных наследуемых синдро-
мов предрасположенности к возникновению злокаче-
ственных опухолей наследственный неполипозный
рак толстой кишки (ННРТК). Этот синдром связан с на-
рушением репарации ошибочно спаренных оснований.
В клетках человека и других эукариот есть несколько
белковых аналогов бактериальных белков MutL и MutS
(см. рис. 25-23). Вероятность развития ННРТК могут
повысить дефекты по крайней мере в пяти разных генах
репарации ошибочно спаренных оснований. Особенно
распространены дефекты в hMLH1 (человеческий гомо-
лог 1 MutL) и hMSH2 (человеческий гомолог 2 MutS)
У человека с ННРТК рак обычно развивается в раннем
возрасте, и чаще всего это бывает рак толстой кишки.
Большинство случаев рака молочной железы у
женщин возникает без известной генетической пред-
расположенности. Однако в 10% случаев это заболе-
вание связано с наследственным дефектом двух ге-
нов — BRCA1 и BRCA2. У человека белки BRCA1 и
BRCA2 это очень большие молекулы (1834 и 3418
аминокислотных остатков соответственно), которые
взаимодействуют с многими другими белками, уча-
ствующими в транскрипции, поддержании хромосом,
репарации ДНК и контроле клеточного цикла. Кроме
того, белок BRCA2 связан с рекомбинационной репа-
рацией двухцепочечных разрывов. Однако точная мо-
лекулярная функция BRCA1 и BRCA2 в этих клеточ-
ных процессах до сих пор не установлена. Вероятность
заболевания раком груди у женщин с дефектами в гене
BRCA1 или BRCA2 превышает 80%
Краткое содержание раздела 25.2
РЕПАРАЦИЯ ДНК
Клетки имеют несколько систем репарации
ДНК. У Е. coli репарация ошибочно спарен-
ных оснований осуществляется при времен-
ном отсутствии метилирования последова-
тельностей (5')GATC во вновь синтезиро-
ванной цепи.
Системы эксцизионной репарации распоз-
нают и устраняют повреждения, вызванные
25.3 Рекомбинация ДНК [81]
факторами внешней среды (такими, как ра-
диация и алкилирующие агенты) и спонтан-
ными реакциями нуклеотидов. Некоторые
системы репарации распознают и вырезают
только поврежденные или неправильные ос-
нования, оставляя АР-сайт (лишенный осно-
вания сайт) в ДНК, который вырезается и за-
полняется новой ДНК под действием других
ферментов.
Системы эксцизионной репарации нуклеоти-
дов распознают и удаляют различные круп-
ные повреждения и пиримидиновые димеры.
Они вырезают участок цепи ДНК, содержа-
щий повреждение, и оставляют брешь, кото-
рую заделывают ДНК-полимераза и лигаза.
Некоторые повреждения ДНК ликвиди-
руются в реакциях, обратных тем, которые
вызывали дефект: пиримидиновые димеры
превращаются в мономерные пиримидины с
помощью фотодиазы, а метильная группа О6-
метилгуанина удаляется метилтрансферазой.
При очень сильных повреждениях бакте-
риальной ДНК происходит синтез новой
последовательности через повреждение с
участием TLS-полимеразы, допускающей
достаточно много ошибок. Эукариоты имеют
похожие полимеразы, выполняющие специ-
ализированные функции в репарации ДНК,
что минимизирует количество мутаций.
25.3. Рекомбинация ДНК
Перестройка заключенной в молекулах ДНК ге-
нетической информации осуществляется благо-
даря протеканию многих разнообразных процес-
сов, обобщенно называемых генетической реком-
бинацией. В настоящее время генетические пере-
группировки находят практическое применение
для модификаций все большего числа геномов
(гл. 9 вт. 1)
Выделяют три основных типа генетической
рекомбинации. Гомологичная генетическая ре-
комбинация (общая рекомбинация) представляет
собой обмен генами между двумя любыми моле-
кулами ДНК (или участками одной и той же мо-
лекулы), которые содержат протяженные отрезки
Барбара МакКлинток
1902-1992
почти идентичной последовательности. Последо-
вательности могут быть любыми, главное, чтобы
они были похожими. При сайт-специфической
рекомбинации происходят обмен только опреде-
ленных последовательностей ДНК. Транспозиция
ДНК отличается от двух других типов генетиче-
ской рекомбинации тем, что в ней обычно участву-
ет короткий участок ДНК, обладающий способно-
стью перемещаться по хромосоме с одного сайта
на другой. Такие «прыгающие» гены впервые были
обнаружены Барбарой МакКлинток в 1940-х гг. в
геноме кукурузы. Кроме того, существуют и раз-
ные другие необычные генетические перестройки,
механизм и значение которых пока не установле-
ны. В этой книге мы рассмотрим только три основ-
ных типа рекомбинации.
Функции систем генетической рекомбинации
так же разнообразны, как и их механизмы. К ним
относится участие в специализированных систе-
мах репарации ДНК, определенная активность
при репликации ДНК, регуляция экспрессии не-
которых генов, облегчение расхождения хромосом
при делении эукариотических клеток, поддержа-
ние генетического разнообразия и реализация за-
программированных генетических перестроек в
ходе эмбрионального развития. В большинстве
случаев генетическая рекомбинация тесно связа-
на с другими процессами метаболизма ДНК, и эта
связь будет рассмотрена далее.
Гомологичная генетическая рекомбинация
выполняет несколько функций
У бактерий гомологичная генетическая рекомбина-
ция служит для репарации ДНК и, как отмечалось в
разд. 25.2, называется рекомбинационной репара-
цией ДНК Обычно она направлена на восстанов-
[82] Часть III. 25. Метаболизм ДНК
ление репликативных вилок, застопорившихся на
месте повреждений в ДНК. Гомологичная генетиче-
ская рекомбинация также может происходить при
конъюгации (скрещивании), когда хромосомная
ДНК переносится из одной бактериальной клетки
(клетки донора) в другую (клетку реципиента). Ре-
комбинация в ходе конъюгации, хотя и редко про-
исходит в популяциях бактерий, вносит вклад в
генетическое разнообразие.
У эукариот гомологичная генетическая ре-
комбинация выполняет несколько функций в
процессах репликации и деления клеток, вклю-
чая репарацию застопорившихся репликативных
вилок. Чаще всего рекомбинация происходит
в ходе мейоза — процесса, с помощью которого
зародышевые клетки с двойным набором хромо-
сом делятся с образованием гаплоидных гамет
(сперматозоидов или яйцеклеток у животных
или гаплоидных спор у растений); каждая гаме-
та получает по одной хромосоме из каждой пары
(рис. 25-31). Мейоз начинается с репликации
ДНК в родительской клетке, так что в какой-то
момент каждая молекула ДНК присутствует в че-
тырех копиях. Затем клетки проходят две стадии
Рис. 25-31. Мейоз в зародышевой линии клеток живот-
ных. Хромосомы гипотетической диплоидной клетки
зародышевой линии (шесть хромосом; три пары гомоло-
гов) реплицируются и остаются соединенными центро-
мерами. Реплицированные двухцепочечные молекулы
ДНК называются хроматидами (сестринские хроматиды).
В профазе I непосредственно перед первым мейотиче-
ским делением три набора гомологичных хроматид вы-
страиваются в линию с образованием тетрад и остаются
соединенными вместе ковалентными связями в местах
гомологичных соединений (хиазм). В хиазмах происхо-
дит кроссинговер (рис. 25-32). Такие временные ассоци-
ации между гомологами обеспечивают правильность рас-
хождения хромосом на следующей стадии, когда они сме-
щаются к противоположным полюсам делящейся клетки
в первом мейотическом делении. Продуктами такого де-
ления являются две дочерние клетки, каждая из которых
содержит по три пары хроматид. Затем пары выстраива-
ются на экваторе клетки и подготавливаются к разделе-
нию хроматид (теперь они называются хромосомами).
В результате второго мейотического деления образуются
четыре гаплоидные дочерние клетки, которые могут вы-
ступать в роли гамет. Каждая из этих клеток содержит по
три хромосомы — половинный набор диплоидной клет-
ки-предшественницы. Хромосомы подверглись перерас-
пределению и рекомбинации.
клеточного деления без промежуточной стадии
репликации ДНК В результате в каждой гамете
оказывается гаплоидное количество ДНК.
После репликации ДНК в профазе перво-
го мейотического деления образовавшиеся се-
Диплоидная
клетка-
предшественница
половых клеток
репликация
25.3 Рекомбинация ДНК [83]
Г омо логичная
пара
Центромера
Гомолог
__Сестринские
хроматиды
Тетрада
Точка
перекреста
(хиазма)
Рис. 25-32. Кроссинговер. а — кроссинговер часто приводит к обмену генетическим мате-
риалом. б — гомологичные хромосомы кузнечиков показаны на стадии профазы I мейоза.
Между двумя гомологичными парами хроматид видно несколько точек контакта (хиазм).
Эти хиазмы ясно указывают на произошедшую перед этим гомологичную рекомбинацию
(кроссинговер).
стринские хроматиды остаются связанными
своими центромерами. На этой стадии каждый
набор из четырех гомологичных хромосом (те-
трада) существует в виде двух пар хроматид.
Между тесно связанными гомологичными хро-
матидами происходит обмен генетической ин-
формацией путем гомологичной генетической
рекомбинации, в ходе которой происходят раз-
рывы и воссоединение последовательностей
ДНК (рис. 25-32). Этот процесс, который иначе
называют кроссинговером, можно наблюдать в
световой микроскоп. Кроссинговер связывает
вместе две пары сестринских хроматид в точках,
называемых хиазмами.
Кроссинговер эффективно физически свя-
зывает вместе все четыре гомологичные хромати-
ды, что важно для правильного расхождения хро-
мосом в последующих мейотических клеточных
делениях. Совершенно ясно, что кроссинговер -
неслучайный процесс, его «горячие точки» были
обнаружены на многих эукариотических хромо-
сомах. Однако представление о том, что кроссин-
говер может происходить почти в любой точке
гомологичных хромосом, вполне допустимо как
приближение при генетическом картировании.
Вероятность гомологичной рекомбинации на
любом участке, разделяющем две точки на хро-
мосоме, примерно пропорциональна расстоянию
между этими точками, и это позволяет опреде-
лить относительное положение генов и расстоя-
ние между разными генами.
Итак, гомологичная рекомбинация, выпол-
няет по меньшей мере три функции: (1) она ис-
пользуется в репарации нескольких типов по-
вреждений ДНК; (2) в эукариотических клетках
она обеспечивает временную физическую взаи-
мосвязь между хроматидами, необходимую для
точного расхождения хромосом на первой стадии
мейотического деления; и (3) усиливает генети-
ческое разнообразие в популяции.
[84] Часть III. 25. Метаболизм ДНК
Рекомбинация в ходе мейоза
начинается с двухцепочечных разрывов
Возможный путь гомологичной рекомбинации в
ходе мейоза рассмотрен на рис. 25-33, а. В этой
модели отражены четыре ключевых момента
Ген А Ген Б
5' 3' 5' 3'
3' " 5Л 3 “ 5'
Двухцепочечный разрыв одного
(Т) из двух гомологов преобразуется
под действием эндонуклеаз в
двухцепочечную брешь.
Последовательности на З'-конце
разрушаются медленнее, чем
на 5'-концах, поэтому
образуются выступающие
одноцепочечные З’-концевые
участки.
Во-первых, гомологичные хромосомы выравни-
ваются. Во-вторых, экзонуклеаза увеличивает
двухцепочечный разрыв в молекуле ДНК, остав-
ляя одноцепочечные участки с З'-гидроксиль-
ной группой на конце (стадия (!•). В-третьих,
эти З'-концы захватывают интактную дуплекс-
Рис. 25-33. Рекомбинация в ходе мейоза. а — модель ре-
парации двухцепочечного разрыва в ходе гомологичной
генетической рекомбинации. Две гомологичные хромо-
сомы (одна показана голубым, другая красным цветом),
участвующие в рекомбинации, имеют похожие после-
довательности. Каждый из двух показанных генов при-
сутствует на двух хромосомах в виде разных аллелей.
Стадии процесса описаны в тексте, б — между двумя бак-
териальными плазмидами in vivo образовалась структура
Холлидея; фотография сделана с помощью электронно-
го микроскопа. Эти структуры названы в честь Робина
Холлидея, впервые предсказавшего их существование в
1964 г.
@ Выступающий 3 '-конец спаривается
с комплементарным участком целого
гомолога. Другая цепь дуплекса
смещается.
® Внедряющийся 3 '-конец удлиняется
под действием ДНК-полимеразы и
миграции ветви, в итоге образуется
молекула ДНК с двумя перекрестами,
называемая промежуточной
структурой Холлидея.
(3) При дальнейшей репликации ДНК
достраивается участок исходного
двух цепочечного разрыва.
Расщепление структуры Холлидея
*2/ специальными нуклеазами приводит
к образованию продукта рекомбинации
(1 или 2). ДНК в наборе продуктов 2.
претерпевшая репарацию с каждой
стороны этого участка, подверглась
рекомбинации.
б
а Набор продуктов 1
Набор продуктов 2
25.3 Рекомбинация ДНК [85]
ную ДНК гомолога, после чего происходит ми-
грация ветви (рис. 25-34) и/или репликация
для создания пары кроссинговерных структур
(структур Холлидея; рис. 25-33, а, стадии ®-@).
В-четвертых, расщепление двух таких структур
приводит к образованию двух конечных реком-
бинантных продуктов (стадия ®).
В данной модели репарации двухцепочеч-
ных разрывов З'-концы выполняют функцию
инициаторов генетического обмена при реком-
бинации. При их спаривании с комплементарной
цепью интактного гомолога создается область ги-
бридной ДНК, которая содержит комплементар-
ные цепи от двух разных родительских ДНК (про-
дукт стадии ® на рис. 25-33, а). Затем каждый из
З'-концов может действовать в качестве праймера
для репликации ДНК. Образующиеся при этом
структуры, называемые структурами Холлидея
(рис. 25-33, б), — признак гомологичной генетиче-
ской рекомбинации во всех организмах.
У разных видов гомологичная рекомбинация
может различаться только в некоторых деталях,
большинство же описанных выше стадий обычно
присутствует в том или ином виде Существуют
два способа расщепления, или «разрешения»,
структуры Холлидея, приводящие к тому же
порядку расположения генов после рекомбина-
ции, что и в исходных хромосомах (стадия ® на
рис. 25-33, а). При одном способе расщепления
область, фланкирующая гибридную ДНК, ока-
зывается рекомбинантной; при другом способе —
нерекомбинантной. Оба результата наблюдаются
in vivo у эукариот и прокариот.
Гомологичная рекомбинация, изображенная
на рис. 25-33, — сложный процесс, который вносит
вклад в генетическое разнообразие. Чтобы понять,
каким образом это происходит, следует вспом-
нить, что две подвергающиеся рекомбинации го-
мологичные хромосомы не обязательно идентич-
ны. Линейный порядок генов в них может быть
одинаковым, но в некоторых генах последователь-
ности оснований могут незначительно отличаться
(в разных аллелях). Например, у человека одна
хромосома может содержать аллель гемоглобина
А (обычный гемоглобин), а другая — аллель гемо-
глобина S (серповидно-клеточная мутация). Раз-
личие может быть всего в одной паре оснований
на миллион. Гомологичная рекомбинация не изме-
няет линейного порядка генов, но она может опре-
делять, какие аллели окажутся связанными друг с
другом в одной хромосоме и вместе передадутся
следующему поколению.
Рис. 25-34. Миграция ветви. Когда матричная цепь спа-
ривается с двумя разными комплементарными цепями, в
точке соединения трех цепей образуется разветвление.
Это разветвление «мигрирует» по мере того, как нару-
шается спаривание между основаниями в комплемен-
тарных цепях и возникает спаривание между одной из
комплементарных цепей и новой цепью. В отсутствие
направляющего фермента ветвь может двигаться само-
произвольно в любом направлении. Такая самопроиз-
вольная миграция ветви продолжается до тех пор, пока
в одной из комплементарных цепей не встречается не-
идентичная последовательность.
В рекомбинации участвует
множество ферментов и других белков
Ферменты, стимулирующие разные стадии го-
мологичной рекомбинации, были выделены
как у прокариот, так и у эукариот. У Е. coli гены
гесВ, гесС и recD кодируют гетеротримерный
фермент recBCD, который проявляет и хели-
казную, и нуклеазную активности. Белок RecA
стимулирует все ключевые стадии гомологич-
ной рекомбинации: спаривание двух ДНК, об-
разование промежуточных структур Холлидея
и миграцию ветви (как описано ниже). Белки
RuvA и RuvB (от англ, repair of UV damage)
образуют комплекс, который связывается с
промежуточными структурами Холлидея, вы-
тесняют белок RecA и стимулируют миграцию
ветви с большей скоростью, чем это делает бе-
лок RecA. Нуклеазы, которые специфически
разделяют структуры Холлидея, часто называ-
ют резолвазами; они были выделены из бакте-
рий и дрожжей, белок RuvC — один из двух, по
меньшей мере, таких нуклеаз у Е. coli.
[86] Часть III. 25. Метаболизм ДНК
Фермент RecBCD связывается с линейной
ДНК со стороны свободного (расщепленного)
конца и движется вдоль двойной спирали, раскру-
чивая и расщепляя ДНК в реакции, сопряженной с
гидролизом АТР (рис. 25-35). Субъединицы RecB
и RecD — это моторы геликазы: RecB движется
по одной цепи в направлении 3'—*5', a RecD — по
другой цепи в направлении 5'—*3'. Активность
фермента изменяется, когда он взаимодействует
с последовательностью (5')GCTGGTGG, называ-
емой chi-последовательностью С этого момента
расщепление цепи со свободным З'-концом суще-
ственно замедляется, а разрушение цепи со сво-
бодным 5'-концом ускоряется. В результате обра-
зуется одноцепочечная ДНК со свободным З'-кон-
цом, которая используется в последующих ста-
диях рекомбинации (рис. 25-33). В геноме Е. coli
рассредоточено 1009 chi-последовательностей,
которые в 5-10 раз повышают частоту рекомбина-
ции на расстояниях до 1000 п. н. вокруг. По мере
удаления от chi-последовательности их влияние
ослабевает. У некоторых других организмов тоже
были идентифицированы последовательности,
увеличивающие частоту рекомбинации.
Белок RecA выделяется среди других белков
метаболизма ДНК тем, что его активная форма
представляет собой упорядоченную спиральную
нить, содержащую до нескольких тысяч моно-
меров RecA, которые собираны вместе на ДНК
(рис. 25-36). Нить белка RecA в норме образуется
на одноцепочечной ДНК, например, под действи-
ем фермента RecBCD, но также может быть соби-
рана на двухцепочечной ДНК с одноцепочечной
брешью; в этом случае первые мономеры RecA
связываются с одноцепочечной ДНК в бреши, по-
сле чего образующаяся нить быстро закручивается
вокруг прилегающей дуплексной ДНК. Сборку и
разборку нитей RecA регулируют другие белки,
включая RecX, Dini, RccF, RecO и RecR.
Роль нити RecA в рекомбинации можно про-
иллюстрировать на модели замены цепи ДНК in
vitro (рис. 25-37). Сначала RecA связывает оди-
ночную цепь ДНК с образованием нуклеопроте-
иновой нити. Затем нить RecA захватывает го-
chi-узнающий
RecB сайт
RecC
АТР
3'_Нуклеазный
домен RecB
RecD
ADP+Pi
Под действием хеликазной
активности RecB и RecD расплетают
ДНК. Нуклеаза RecB разрушает
ДНК, и ДНК с 3'-конца разрезается
на более мелкие части.
chi-узнающий сайт связывается с
chi-последовательностью так, что
3'-конец цепи перестает
разрушаться. Другая цепь
продолжает разрушаться, образуя
одноцепочечный 3'-конец.
Рис. 25-35. Хеликазная и нуклеазная актив-
ности фермента RecBCD. Начиная с двухце-
почечного конца, RecBCD раскручивает и рас-
щепляет последовательность ДНК. Нуклеазный
домен субъединицы RecB располагается так,
что может разрушать либо одну цепь ДНК, либо
другую. Субъединица RecB движется в направ-
лении 3'->5' по одноцепочечной ДНК со сво-
бодным З'-концом, действуя как АТР-зависимая
хеликаза. Субъединица RecD сходным образом
движется по второй цепи в направлении 5'-*3'.
Активный центр субъединицы RecC узнает и
связывает chi-последовательности. Цепь, со-
держащая chi-последовательность, образует
петлю, и для нуклеазного домена RecB остается
доступной только цепь со свободным 5'-кон-
цом. Предполагается, что RecBCD инициирует
генетическую рекомбинацию у Е. coli. Он также
участвует в репарации двухцепочечных разры-
вов в местах разрушения репликативных вилок.
Петля оцДНК
chi связывается в узнающем сайте
ОН 3'
5'
3'
25.3 Рекомбинация ДНК [87]
в
Рис. 25-36. Белок RecA. а — нуклеопротеиновый ком-
плекс белка RecA с одноцепочечной ДНК (электронная
микрофотография). Поперечные черточки свидетель-
ствуют о том, что структура нити имеет вид правозакру-
ченной спирали, б — объемная модель нити из 24 субъе-
диниц RecA. Нить содержит шесть субъединиц на виток.
Одна субъединица окрашена в красный цвет, чтобы ее
можно было разглядеть на фоне остальных (по PDB ID
2REB). в — после лимитирующей стадии посадки ком-
плекса RecA на одноцепочечной ДНК нити удлиняются
в направлении 5'->3'. Разборка происходит также в на-
правлении 5'->3' со стороны, противоположной прира-
щению цепи, г — в сборке нитей участвуют белки RecE
RecO и RecR (общее обозначение RecFOR). Белок RecX
ингибирует удлинение нитей RecA. Белок Dini стабили-
зирует RecA, предотвращая разборку.
0DinI
88 88
мологичную двухцепочечную ДНК и соединяет
ее со связанной одиночной цепью. Для образова-
ния активной нити RecA, которая соединяет две
молекулы ДНК, необходимо связывание (но не
гидролиз) АТР. После этого две молекулы ДНК
обмениваются цепями, и в результате возникает
гибридная ДНК. Обмен происходит со скоростью
6 и. н./с в направлении 5'—*3' относительно одно-
цепочечной ДНК в составе нити RecA. В этой
реакция могут участвовать три или четыре цепи
ДНК (рис. 25-37); в последнем случае в ходе про-
цесса образуются структуры Холлидея.
Когда ДНК-дуй леке включается в состав
нити RecA и выравнивается рядом со связанной
одиночной цепью ДНК на участке протяженно-
стью в сотни пар оснований, одна из цепей ду-
плекса может поменять партнера (рис. 25-38,
стадия ®). Поскольку ДНК обладает спираль-
ной структурой, непрерывный обмен цепей тре-
бует упорядоченного вращения двух соседних
ДНК. Это осуществляется путем перемотки
(стадии 3) и ®), в результате чего точка ветвле-
ния смещается вдоль спирали. Заключительные
стадии обмена цепей ДНК, во время которых
происходит удлинение гибридной ДНК, образо-
вавшейся на первой стадии, происходят за счет
гидролиза АТР. Механизм сопряжения реакций
пока неясен.
[88] Часть III. 25. Метаболизм ДНК
Кольцевая
одноцепочечная ДНК
Кольцевая
дуплексная ДНК с
одноцепочечной брешью
Белок RecA
Гомологичная
линейная
дуплексная ДНК
Белок RecA
Белок RecA
Белок RecA связывается с одноцепочечнои ДНК
или с брешью в ДНК. Комплементарная цепь
линейной ДНК спаривается с кольцевой одинарной
цепью. Другая линейная цепь отводится в сторону
(слева) или спаривается с комплементарной цепью в
кольцевом дуплексе с образованием структуры
Холлидея (справа).
Г омо логичный
ДНК-дуплекс
АТР
Белок
RecA
АТР
ADP + Р,
ADP + Pi
Перемещение разветвления приводит к образованию
кольцевого дуплекса с ником и к образованию либо
ранее отведенной линейной цепи (слева),
либо частично одноцепочечного линейного
дуплекса (справа).
Рис. 25-37. Обмен цепей ДНК, индуцируемый белком
RecA in vitro. Замена цепи включает отделение одной
цепи дуплекса от комплементарной цепи и ее перенос на
другую комплементарную цепь для формирования ново-
го дуплекса (гетеродуплекса) ДНК. В этом процессе об-
разуется разветвленная промежуточная структура. Обра-
зование конечного продукта зависит от миграции ветви,
которой способствует RecA. В реакции могут участвовать
три цепи (слева) или при взаимообмене между двумя
гомологичными дуплексами четыре цепи (справа). В по-
следнем случае интермедиат представляет собой струк-
туру Холлидея. Белок RecA способствует миграции ветви,
используя энергию гидролиза АТР.
Рис. 25-38. Модель RecA-опосредованного обмена цепей
ДНК. Показана реакция с участием трех цепей ДНК. Раз-
мер шариков, которыми обозначены молекулы белка RecA,
непропорционально мал по сравнению с толщиной ДНК,
что позволяет продемонстрировать происходящие в ДНК
изменения. 3 Белок RecA образует спираль вокруг одно-
цепочечной ДНК. 2 В комплекс включается гомологичный
ДНК-дуплекс. з По мере того как в результате вращения
начало трехцепочечного участка смещается слева на-
право, одна из цепей дуплекса занимает место одиноч-
ной цепи, исходно связанной с нитью белка. Другая цепь
дуплекса вытесняется, а внутри нити белка формируется
новый дуплекс. По мере продолжения вращения (<£ и @)
вытесненная цепь полностью высвобождается. В данной
модели гидролиз АТР белком RecA приводит к повороту
двух молекул ДНК относительно друг друга и тем самым
продвигает обмен цепей в направлении слева направо.
25.3 Рекомбинация ДНК [89]
После образования структуры Холлидея для
завершения рекомбинации необходима целая
группа ферментов: топоизомеразы, белок мигра-
ции витка RuvAB, резолваза и другие нуклеазы,
ДНК-полимераза I или III и ДНК-лигаза. В клет-
ках Е. coli белок RuvC (Afr = 20 000) расщепляет
структуры Холлидея с образованием полнораз-
мерных неразветвленных хромосом.
Для репарации заблокированных
репликативных вилок используются
все возможности метаболизма ДНК
Как и во всех других клетках, у бактерий уровень
повреждений ДНК достаточно высокий даже в
нормальных условиях роста. Большинство по-
вреждений ДНК быстро репарируется путем экс-
цизионной репарации оснований, нуклеотидов
и другими описанными ранее способами. Тем не
менее почти каждая бактериальная репликатив-
ная вилка наталкивается на нерепарированное
повреждение ДНК или разрыв на пути от точ-
ки начала репликации до точки ее окончания
(рис. 25-30). ДНК-полимераза III не может прой-
ти мимо многих типов повреждений ДНК, что
вынуждает ее оставить эти повреждения в виде
одноцепочечной бреши. Когда полимераза встре-
чается с разрывом цепи ДНК, возникает двухце-
почечный разрыв. В обеих ситуациях необходима
рекомбинационная репарация ДНК (рис. 25-39).
В нормальных условиях роста застопорившиеся
репликативные вилки реактивируются с помо-
щью сложного пути репарации, включающего
рекомбинационную репарацию ДНК, повторный
старт репликации и репарацию всех пропущен-
ных повреждений. В этом процессе объединяют-
ся все пути метаболизма ДНК.
Остановившаяся репликативная вилка мо-
жет быть снова запущена по меньшей мере двумя
сложными способами, для каждого из которых
требуется белок RecA. Путь репарации повреж-
дений, содержащих бреши в ДНК, задействует
также белки RecF, RecO и RecR. Для репарации
двухцепочечных разрывов необходим фермент
RecBCD (рис. 25-39). После дополнительных
стадий рекомбинации следует процесс, назы-
ваемый возобновлением репликации, незави-
симым от точки инициации репликации, когда
с помощью комплекса из семи белков (PriA, В
и С, а также DnaB, С, G и Т) снова собирается
репликативная вилка. Этот комплекс, первона-
чально открытый в процессе репликации ДНК
фага фХ174 in vitro, теперь называется праймо-
сомой восстановления репликации. Для вос-
становления репликативной вилки необходима
также ДНК-полимераза II, причем ее роль пока
не установлена; активность ДНК-полимеразы II
открывает дорогу ДНК-полимеразе III для ре-
пликации протяженных участков, что необхо-
димо для завершения построения хромосомы.
Иногда происходит возобновление репликации
ниже точки повреждения еще до репарации по-
врежденного участка.
Репарация застопорившихся вилок преду-
сматривает координированный переход между
репликацией и рекомбинацией. Стадии реком-
бинации нужны для заполнения бреши в ДНК
или соединения разорванной цепи, чтобы вос-
создать разветвленную структуру ДНК в ре-
пликативной вилке. Повреждения, оставшиеся
позади дуплекса ДНК, устраняются при по-
мощи эксцизионной репарации оснований или
нуклеотидов. Таким образом, в репарации за-
стопорившихся репликативных вилок участвует
широкий круг ферментов, действующих на каж-
дой стадии метаболизма ДНК. Этот способ ре-
парации, очевидно, является главной функцией
системы гомологичной рекомбинации в каждой
клетке, и нарушения рекомбинационной репара-
ции ДНК играют важную роль в возникновении
заболеваний человека (доп. 25-1).
Сайт-специфическая рекомбинация
приводит к точным перестройкам ДНК
Гомологичная генетическая рекомбинация, ко-
торую мы обсуждали, может происходить меж-
ду любыми двумя гомологичными последова-
тельностями. Второй тип рекомбинации, сайт-
специфическая рекомбинация, принципиально
отличается тем, что ограничивается только спец-
ифическими последовательностями. Реакции
рекомбинации данного типа происходят почти в
каждой клетке, и их функции различны в разных
видах организмов. Например, таков механизм
регуляции экспрессии некоторых генов и меха-
низм активации программируемых перестроек
ДНК при эмбриональном развитии или в циклах
репликации некоторых вирусных и плазмид-
ных ДНК. Каждая система сайт-специфической
[90] Часть III. 25. Метаболизм ДНК
Повторный старт
репликации, независимо
от точки инициации
Рис. 25-39. Модели рекомбинационной репарации для восстановления застопорившихся
репликативных вилок. Репликативная вилка застопоривается при столкновении с повреж-
дением ДНК (слева) или с разрывом ДНК (справа). Для воссоздания разветвленной структу-
ры ДНК в репликативной вилке необходимы ферменты рекомбинации, облегчающие обмен
цепями ДНК. Одноцепочечные разрывы устраняются с помощью белков RecE RecO и RecR.
Двухцепочечные разрывы требуют участия фермента RecBCD. В обоих процессах задей-
ствован белок RecA. Промежуточные продукты рекомбинации процессируются благодаря
активности дополнительных ферментов (например, для разрешения структур Холлидея
нужны белки RuvA, RuvB и RuvC). Повреждения в двухцепочечной ДНК восстанавливаются
с помощью эксцизионной репарации нуклеотидов или другими способами. Репликативная
вилка перестраивается с привлечением ферментов, катализирующих повторный старт ре-
пликации, независимый от точки начала репликации, и репликация хромосомы завершает-
ся. Весь процесс требует точной координации всех путей метаболизма бактериальной ДНК.
25.3 Рекомбинация ДНК [91]
рекомбинации состоит из фермента рекомбина-
зы и короткой (от 20 до 200 и. н.) уникальной
последовательности ДНК, с которой взаимодей-
ствует рекомбиназа (сайт рекомбинации). Ход
и результат реакции регулирует один или не-
сколько вспомогательных белков.
Существуют два основных класса систем
сайт-специфической рекомбинации, различаю-
щихся наличием в активном центре ферментов
остатков Туг или Ser. При изучении многих си-
стем сайт-специфической рекомбинации тиро-
зинового класса in vitro удалось выявить основ-
ные механизмы и общие принципы процесса
(рис. 25-40, а). Некоторые из ферментов были
получены в кристаллической форме, что позво-
лило детально исследовать их структуру. Вы-
деленная рекомбиназа распознает и связывает
каждый из двух сайтов рекомбинации на двух
разных молекулах ДНК или в одной и той же
ДНК. В каждом сайте одна цепь ДНК расщепля-
ется в специфической точке, после чего реком-
Рис. 25-40. Реакция сайт-специфической рекомби-
нации. а — показанная здесь реакция характерна для
сайт-специфических рекомбиназ класса интеграз (на-
званы после открытия интегразы бактериофага к —
первой описанной рекомбиназы). Этот фермент имеет в
активном центре остатки Туг, которые играют роль ну-
клеофилов. Рекомбиназа представляет собой тетрамер,
состоящий из идентичных субъединиц, которые связы-
ваются со специфической последовательностью — сай-
том рекомбинации. 1 Одна цепь каждой молекулы ДНК
разрезается в определенных точках этой последователь-
ности. Нуклеофильным агентом является группа -ОН
остатка тирозина в активном центре, и в результате ре-
акции возникает ковалентная фосфотирозиновая связь
между белком и ДНК. ® Расщепленные цепи связывают-
ся с новыми партнерами, образуя структуру Холлидея.
Завершающие стадии @ и i напоминают две первые.
Исходная последовательность сайта рекомбинации вос-
станавливается после рекомбинации ДНК, фланкиру-
ющей сайт. Эти стадии происходят при участии мулъ-
тисубъединичного комплекса рекомбиназы, а иногда
и других, не показанных здесь белков, б — объемная
модель четырехсубъединичной Сге-рекомбиназы класса
интеграз, связанной с структурой Холлидея (изображе-
на в виде голубых и синих спиралей). Белок изображен
прозрачным, чтобы сквозь него была видна ДНК (по PDB
ID 3CRX). В активном центре рекомбиназ, относящихся к
семейству резолваз-инвертаз, роль нуклеофила испол-
няет остаток серина.
[92] Часть III. 25. Метаболизм ДНК
Инверсия
1
Рис. 25-41. Результат сайт-специфической рекомбинации. Результат сайт-специфической
рекомбинации зависит от локализации и ориентации сайтов рекомбинации (красные и
зеленые) в молекуле двухцепочечной ДНК. В данном случае под ориентацией (показана
стрелками) понимают порядок расположения нуклеотидов в сайте рекомбинации, а не на-
правление 5'“*3'. а — сайты рекомбинации с противоположной ориентацией в одной и той
же молекуле ДНК. В результате происходит инверсия, б — сайты рекомбинации с одинако-
вой ориентацией, расположенные в одной молекуле ДНК (результат — делеция) или в двух
молекулах ДНК (результат — инсерция).
инсерция деления
б
биназа ковалентно связывается с ДНК в месте
расщепления через фосфотирозиновую связь
(стадия ;Г). Временная связь белок ДНК со-
храняет фосфодиэфирную связь, которая утра-
чивается при расщеплении ДНК, поэтому на по-
следующих стадиях не требуются богатые энер-
гией кофакторы, такие как АТР. Расщепленные
цепи ДНК соединяются с новыми партнерами
с образованием структуры Холлидея, в которой
новые фосфодиэфирные связи создаются за счет
Бактериальный сайт
присоединения (attB)
Интеграция:
А-интеграза
Выщепл ение:
А-интеграза
FIS + XIS
Рис. 25-42. Интеграция и выщепление ДНК бактериофага X. в специфическом сайте хромо-
сомы. Сайт присоединения в ДНК фага X (attP) имеет лишь 15 п. н., полностью гомологичных
нуклеотидам в бактериальной ДНК (attB) в области перекреста. В результате реакции образу-
ются два новых сайта присоединения (attR и attL) с двух сторон от интегрированной ДНК фага.
Рекомбиназа фага является Х-интегразой (белок INT). Для интеграции и выщепления исполь-
зуются разные сайты присоединения и различные вспомогательные белки. В выщеплении уча-
ствуют белки XIS, кодируемые бактериофагом, и белки FIS, кодируемые бактериями. Для обеих
реакций нужен белок IHF (от англ, integration host factor — клеточный фактор интеграции),
кодируемый бактериями.
25.3 Рекомбинация ДНК [93]
связи белок-ДНК (стадия @). Для завершения
реакции процесс должен повториться во вто-
рой точке в каждом из двух сайтов рекомбина-
ции (стадии @ и ©). В системах с остатком Ser
в активном центре обе цепи в каждом сайте ре-
комбинации одновременно разрезаются и зано-
во объединяются в новые пары без образования
структур Холлидея. Но в обоих случаях обмен
всегда обоюдный и точный, и при завершении
реакции сайты рекомбинации восстанавлива-
ются. Рекомбиназа, таким образом, выступает
в качестве сайт-специфической эндонуклеазы и
лигазы одновременно.
Последовательности сайтов рекомбинации,
распознаваемые сайт-специфическими рекомби-
назами, частично асимметричны (не являются
палиндромами); в ходе реакции два сайта реком-
бинации выстраиваются в одной и той же ориен-
тации. Результат зависит от локализации и ори-
ентации сайтов рекомбинации (рис. 25-41). Если
два сайта находятся в одной и той же молекуле
ДНК, реакция либо инвертирует, либо делетиру-
ет промежуточную ДНК в зависимости от того,
имеют сайты рекомбинации противоположную
или одинаковую ориентацию соответственно.
Если сайты расположены на разных молекулах
ДНК, рекомбинация становится межмолекуляр-
ной; если одна или обе ДНК кольцевые, происхо-
дит инсерция. Некоторые рекомбиназные систе-
мы специфичны для одного из этих типов реак-
ций и действуют только на сайты в определенной
ориентации.
Первой исследованной in vitro системой
сайт-специфической рекомбинации была систе-
ма, кодируемая бактериофагом X. Когда ДНК
фага X попадает в клетку Е. coli, сложная регу-
лируемая последовательность стадий приводит
к одному из двух результатов. Либо ДНК фага
реплицируется и образует больше бактерио-
фагов (разрушающих клетку хозяина), либо
она встраивается в хромосому хозяина (в виде
профага) и пассивно реплицируется в составе
хромосомы на протяжении многих клеточных
генераций. Интеграцию осуществляет кодируе-
мая фагом рекомбиназа (Х-интеграза), которая
действует в сайтах рекомбинации в фаговой и
бактериальной ДНК - в сайтах присоединения
attP и attB соответственно (рис. 25-42). Роль
сайт-специфической рекомбинации в регуляции
экспрессии рассматривается в гл. 28.
Для полной репликации хромосомы
может потребоваться
сайт-специфическая рекомбинация
При рекомбинационной репарации ДНК коль-
цевой бактериальной хромосомы иногда об-
разуются вредные побочные продукты. При
разрешении структуры Холлидея с помощью
нуклеазы, например RuvC, и последующем за-
вершении репликации могут образоваться либо
две обычные мономерные хромосомы, либо сце-
пленная димерная хромосома (рис. 25-43) В по-
Вилка, проходящая
репарацию
терминация
репликации
переход к мономерам
с помощью системы
XerCD
Рис. 25-43. Делеция ДНК для ликвидации опасных по-
следствий рекомбинационной репарации ДНК. Разре-
шение структуры Холлидея в ходе рекомбинационной
репарации ДНК (места расщепления указаны красными
стрелками) может приводить к образованию димер-
ной хромосомы. У Е. coli специализированная сайт-
специфическая рекомбиназа XerCD превращает димер в
мономеры, способствуя расхождению хромосом и нор-
мальному клеточному делению.
[94] Часть III. 25. Метаболизм ДНК
следнем случае ковалентно связанные хромосо-
мы не могут при клеточном делении разойтись в
дочерние клетки, и деление клетки блокируется
на этой стадии цикла. У Е. coli существует специ-
альная система сайт-специфической рекомбина-
ции XerCD, которая превращает димерные хро-
мосомы в мономерные и обеспечивает нормаль-
ное клеточное деление. Эта сайт-специфическая
делеция (рис. 25-41, б) является еще одним при-
мером тесной связи между рекомбинацией ДНК
и другими процессами метаболизма ДНК.
Подвижные генетические элементы
перемещаются из одного участка ДНК в другой
Теперь рассмотрим третий важный тип рекомби-
нации, при котором происходит перенос подвиж-
ных элементов, или транспозонов. Эти участки
ДНК, обнаруженные практически во всех клет-
ках, движутся или «прыгают» из одной точки
хромосомы (донорский сайт) в другую точку на
той же или другой хромосоме (целевой сайт). Го-
мология последовательностей ДНК не обязатель-
ное условие для этого перемещения, называемого
транспозицией; новое расположение транспозона
определяется более или менее случайно. Встраи-
вание транспозона в ген может привести к гибели
клетки, поэтому транспозиция жестко регулиру-
ется и обычно происходит очень редко. Вероят-
но, транспозоны - самые простые молекулярные
паразиты, которые приспособились к пассивной
репликации в хромосомах клеток хозяина. В не-
которых случаях они несут гены, полезные для
клетки хозяина, и, таким образом, существуют в
симбиотической связи с ней.
У бактерий есть два класса транспозонов.
Инсерционные последовательности (IS-
элементы или простые транспозоны) содержат
только последовательности, необходимые для
транспозиции, и гены белков (транспозаз), ко-
торые осуществляют этот процесс. Сложные
транспозоны содержат один или несколько до-
полнительных генов помимо тех, которые не-
обходимы для транспозиции. Эти избыточные
гены могут, например, отвечать за устойчивость
к антибиотикам и тем самым повышать вероят-
ность выживания клетки хозяина. Транспози-
ция — одна из причин распространения устой-
чивости к антибиотикам в популяциях бакте-
рий, вызывающих болезни, в результате чего
применение некоторых антибиотиков становит-
ся неэффективным (с. 10).
Бактериальные транспозоны различны по
структуре, но большинство на каждом конце со-
держит короткие повторы, которые связывают
транспозазы. При транспозиции короткая после-
довательность в целевом сайте (от 5 до 10 п. н.)
удваивается с образованием дополнительного ко-
роткого повтора, который фланкирует встроен-
ный транспозон с двух сторон (рис. 25-44). Эти
повторы образуются благодаря расщеплению ну-
клеотидной последовательности и встраиванию
транспозона на новом месте.
У бактерий существуют два основных пути
транспозиции. При прямой (простой) транспо-
зиции (рис. 25-45, слева) расщепление ДНК с
двух сторон от транспозона приводит к его вы-
резанию, и транспозон перемещается на новое
место. В ДНК донора остается двухцепочечный
разрыв, который должен быть ликвидирован.
В целевом сайте производится ступенчатый раз-
Концевые
повторы
Транспозон
Транспозаза делает
ступенчатые надрезы
в ДНК.
ДНК-мишень
Транспозон встраивается
в разрез.
Бреши заполняются при репликации, при которой
последовательности, фланкирующие
транспозон, удваиваются.
Рис. 25-44. Удвоение последовательности ДНК в целе-
вом участке при встраивании транспозона. Удвоенные
после встраиваниия транспозона последовательности
показаны красным цветом. Обычно эти последователь-
ности содержат только несколько пар оснований, здесь
их размер сильно увеличен по сравнению с размером ти-
пичного транспозона.
25.3 Рекомбинация ДНК [95]
Прямая
транспозиция
! I
t 1
Репликативная
транспозиция
I
Расщепление
присоединяются
к ДНК-мишени
Рис. 25-45. Два основных способа транспозиции: пря-
мой (простой) и репликативный. CD Сначала происхо-
дит расщепление ДНК с каждой стороны от транспозона
(стрелки). (D Освободившиеся З'-гидроксильные группы
на концах транспозона действуют как нуклеофилы, на-
прямую атакуя фосфодиэфирные связи в ДНК-мишени.
Эти фосфодиэфирные связи располагаются в двух цепях
ДНК ступенчато (не непосредственно друг под другом),
з Транспозон связывается с ДНК-мишенью. При пря-
мой транспозиции (слева) репликация заполняет бре-
ши на каждом конце. При репликативной транспозиции
(справа) реплицируется весь транспозон, и образуется
коинтеграт. @ Коинтеграт часто разрушается позднее с
помощью особой системы сайт-специфической рекомби-
нации. Хозяйская ДНК, оставшаяся расщепленной после
прямой транспозиции, либо восстанавливается путем со-
единения концов ДНК, либо разрушается (не показано).
В последнем случае последствия могут быть летальными
для организма.
(слева) или реплицируется
целый транспозон (справа)
ДНК-ги'лим'лря
ДНК-лиг.
репликация
рез (как на рис. 25-44), в который встраивается
транспозон, а бреши заполняются в процессе
репликации, что приводит к удвоению последо-
вательности целевого сайта. При репликативной
транспозиции (рис. 25-45, справа) реплициру-
ется целый транспозон, оставляя в ДНК донора
одну свою копию. Промежуточная структура,
в составе которой имеется донор, ковалентно
связанный с ДНК в целевом сайте, называется
коинтегратом. В коинтеграте содержатся две
полные копии транспозона в одинаковой ориен-
тации. В случае некоторых хороню изученных
транспозонов промежуточное коинтегратное
соединение превращается в продукты путем
сайт-специфической рекомбинации, при кото-
рой делецию осуществляют специализирован-
ные рекомбиназы.
У эукариот тоже есть транспозоны, которые
по структуре напоминают бактериальные транс-
позоны, и некоторые из них используют похо-
жий механизм транспозиции. Однако в других
случаях в качестве посредников в транспозиции
участвует РНК. Эволюция таких транспозонов
имеет много общего с эволюцией определенных
классов РНК-вирусов. И те, и другие подробнее
рассматриваются в следующей главе.
Сборка генов иммуноглобулинов
происходит путем рекомбинации
Некоторые перегруппировки ДНК запрограмми-
рованы в развитии эукариотических организмов.
Важный пример — сборка полных генов имму-
ноглобулинов из отдельных сегментов в геномах
позвоночных. Человек, как и другие млекопитаю-
щие, способен производить миллионы различных
[96] Часть III. 25. Метаболизм ДНК
иммуноглобулинов (антител) с разной специ-
фичностью. несмотря на то что геном человека со-
держит лишь около 29 000 генов. Рекомбинация
позволяет организму образовывать необыкновен-
ное разнообразие антител при ограниченных воз-
можностях ДНК в кодировании информации. При
исследовании механизмов рекомбинации была
выявлена ее тесная связь с транспозицией ДНК;
возможно, эта система обеспечения разнообразия
антител возникла в глубокой древности как ре-
зультат проникновения в клетки транспозонов.
Рассмотрим механизм создания разнообраз-
ных антител на примере человеческих генов, ко-
дирующих иммуноглобулины G (IgG). Иммуно-
глобулины состоят из двух тяжелых и двух лег-
ких полинептидных цепей (см. рис. 5-21 в т. 1).
Каждая цепь содержит две области: вариабель-
ную с последовательностью, которая значительно
отличается от одного иммуноглобулина к друго-
му, и константную, фактически постоянную для
данного класса иммуноглобулинов. Существуют
также два разных семейства легких цепей — к- и
X,-. которые несколько различаются последова-
тельностями константных областей. Разнообра-
зие вариабельных областей всех трех типов по-
липептидной цепи (тяжелой цепи, легких к- и
k-цепей) возникает по схожему механизму. Гены
этих полипептидов делятся на сегменты, а геном
содержит кластеры с множеством версий каждо-
го сегмента. Соединение по одной версии каждо-
го из сегментов создает полный ген.
На рис. 25-46 показана организация ДНК,
кодирующей легкие к-цепи IgG человека; ме-
ханизм образования зрелой легкой к-цепи объ-
ДНК
линии
—TV <У П-fv Д^У-ГУ8Т~Т«Г Jj5~H С |— в-лимфоцита
транскрипция
3'| С I
Первичный
транскрипт
удаление последовательностей
между J4 и С при
сплайсинге мРНК
| У84 I J4| С —I
П роцессированная
мРНК
трансляция
Рис. 25-46. Рекомбинация V и J сегментов гена легкой
к-цепи IgG человека. Данный процесс обеспечивает
разнообразие антител. Вверху показана организация ко-
дирующих последовательностей IgG в стволовой клетке
костного мозга. При рекомбинации происходит удаление
последовательности ДНК между определенным сегмен-
том V и определенным сегментом J. После транскрипции
происходит сплайсинг РНК-транскрипта как описано в
гл. 26. В ходе трансляции образуется легкая цепь, кото-
рая может комбинироваться с любой из 5000 возможных
тяжелых цепей с образованием готового антитела.
Вариабельная
область
Полипептид
легкой цепи
укладка и сборка белка
Постоянная
область
Легкая цепь
Тяжелая цепь
Антитело
25.3 Рекомбинация ДНК [97]
ясняется именно организацией ДНК. В недиф-
ференцированных клетках информация об этой
полипептидной цепи разделена на три сегмента.
Сегмент V (вариабельный) кодирует первые 95
аминокислотных остатков вариабельной области,
сегмент J (соединяющий) кодирует оставшиеся
12 остатков вариабельной области, а сегмент С
кодирует константную область. Геном содержит
примерно 300 вариантов сегмента V, четыре ва-
рианта сегмента J и один сегмент С.
После дифференцировки стволовой клетки
костного мозга в зрелый В-лимфоцит один сег-
мент V и один сегмент J соединяются с помощью
специальной системы рекомбинации (рис. 25-46).
В ходе этой запрограммированной делеции вся
промежуточная ДНК отбрасывается. Существует
примерно 300 • 4 = 1200 возможных комбинаций
V-J. Процесс рекомбинации не так точен, как
описанная выше сайт-специфическая рекомби-
нация, поэтому место соединения сегментов V
и J может служить источником дополнительной
вариабельности последовательности. Это увели-
чивает число возможных вариантов примерно в
2,5 раза, поэтому клетки могут образовывать при-
мерно 2,5 • 1200 = 3000 различных комбинаций сег-
ментов V J. Наконец, после транскрипции процесс
сплайсинга РНК приводит к соединению области
V-J с областью С (эти процессы описаны в гл. 26).
Механизм рекомбинации при соединении
сегментов V и J показан на рис. 25-47. Непосред-
ственно после каждого сегмента V и непосред-
ственно перед каждым сегментом J находятся
сигнальные последовательности рекомбинации
(RSS; от англ, recombination signal sequences), с
которыми связаны белки RAG1 и RAG2 (от англ.
recombination activating gene). Белки RAG ката-
лизируют образование двухцепочечного разры-
ва между сигнальными последовательностями
и сегментами V (или J), которые должны соеди-
ниться. Сегменты V и J затем соединяются с по-
мощью второго комплекса белков.
Гены тяжелых цепей и легких Х-цепей образу-
ются в аналогичных процессах. Для тяжелых це-
пей существует больше генных сегментов, чем для
легких цепей, и для них возможно более 5000 ком-
бинаций. Поскольку при образовании иммуногло-
булина любая тяжелая цепь может комбиниро-
ваться с любой легкой цепью, у каждого человека
может быть по меньшей мере 3000 • 5000 = 1,5 • 107
различных IgG. Дополнительное разнообразие
Промежуточная
ДНК
репарация
двухцепочечного разрыва
путем соединения концов
V J
Рис. 25-47. Механизм перестройки гена иммуноглобу-
лина. Белки RAG1 и RAG2 связываются с сигнальными
последовательностями рекомбинации (RSS) и расще-
пляют одну цепь ДНК между RSS и сегментами V (или
J), которые будут впоследствии соединены. После этого
высвобожденная З'-гидроксильная группа действует в
качестве нуклеофила, атакует фосфодиэфирную связь в
другой цепи и создает двухцепочечный разрыв. Образую-
щиеся шпильки в области сегментов V и J расщепляются,
а их концы ковалентно связываются комплексом белков,
специализирующихся на репарации путем соединения
концов в двухцепочечных разрывах. Стадии образования
двухцепочечного разрыва, катализируемые RAG1 и RAG2,
напоминают стадии реакций транспозиции.
возникает из-за высокой частоты мутаций (меха-
низм неизвестен) генов V-сегментов в ходе диф-
ференцировки В-лимфоцитов. Каждый зрелый
В-лимфоцит образует антитела только одного
типа, но набор антител, образуемых разными клет-
ками, очень большой.
Действительно ли иммунная система отчасти
происходит от древних транспозонов? Механизм
образования двухцепочечных разрывов с помощью
RAG1 и RAG2 на самом деле идентичен нескольким
стадиям транспозиции (рис. 25-47). Кроме того, вы-
198] Часть III. 25. Метаболизм ДНК
резаемая ДНК с RSS на концах имеет структуру,
обнаруженную в большинстве транспозонов. В экс-
перименте in vitro белки RAG1 и RAG2 могут вза-
имодействовать с этой вырезанной ДНК и встраи-
вать ее, как транспозон, в другие молекулы ДНК (в
В-лимфоцитах эта реакция, вероятно, происходит
довольно редко). Мы не знаем точно, но система пе-
рестройки генов иммуноглобулинов, возможно, яв-
ляется результатом эволюционного процесса, унич-
тожившего различия между паразитом и хозяином.
Краткое содержание раздела 25.3
РЕКОМБИНАЦИЯ ДНК
Последовательности ДНК подвергаются пе-
рестройкам в реакциях рекомбинации, кото-
рые обычно четко координированы с репли-
кацией или репарацией ДНК.
Гомологичная генетическая рекомбинация мо-
жет происходить между любыми двумя моле-
кулами ДНК с гомологичными последователь-
ностями. При мейозе (у эукариот) этот тип ре-
комбинации помогает обеспечить правильное
расхождение хромосом и увеличить генетиче-
ское разнообразие. И у бактерий, и у эукари-
от рекомбинация служит для восстановления
блокированных репликативных вилок. В про-
цессе гомологичной рекомбинации формиру-
ется промежуточная структура Холлидея.
Сайт-специфическая рекомбинация проис-
ходит только на специфических последова-
тельностях и может протекать также через
образование структуры Холлидея. Реком-
биназы расщепляют ДНК в специфических
точках и связывают цепи с новыми партне-
рами. Этот тип рекомбинации обнаружен
практически во всех клетках; к его много-
численным функциям относятся в том числе
интеграция ДНК и регуляция экспрессии.
Транспозоны, существующие практически
во всех клетках, передвигаются внутри или
между хромосомами посредством рекомби-
нации. У позвоночных с помощью запро-
граммированной рекомбинации, напомина-
ющей транспозицию, происходит соедине-
ние сегментов генов иммуноглобулинов в
ходе дифференцировки В-лимфоцитов.
Ключевые термины
Термины, выделенные жирным шрифтом, объясня-
ются в глоссарии.
ААА+ АТРазы 56
АР-сайт 71
АР-эндонуклеазы 68
МСМ-комплекс 64
ORC-комплекс (комплекс
распознавания точки
инициации реплика-
ции) 64
SOS-ответ 76
Гомологичная генетиче-
ская рекомбинация
81
ДНК-гликозилазы 68. 71
ДНК-лигазы 55
ДНК-полимераза I 49
ДНК-полимераза III 52
ДНК-полимераза а 54
ДНК-полимераза 5 54
ДНК-полимераза е 54
ДНК-расплетающий эле-
мент (DUE) 56
ДНК-транспозиция 81
ДНК-фотолиазы 74
Инсерционная последова-
тельность 94
Катенаны 63
Коинтеграт 95
Коррекция 52
Лидирующая цепь 48
Лицензирование 64
Матрица 45
Мейоз 82
Миграция ветви 85
Модель репарации двуце-
почечного разрыва 84
Мутация 67
Нуклеазы 48
Отстающая цепь 48
Подверженный ошибкам
синтез ДНК через
повреждение 76
Полуконсервативная ре-
пликация 45
Праймазы 55
Праймер 50
Праймосома 59
Пререпликативный ком-
плекс (pre-RC) 64
Процессивность 50
Рекомбинационная репа-
рация ДНК 81
Репликативная вилка 47
Реплисома 54
Сайт-специфическая ре-
комбинация 81
Структура Холлидея 85
Топоизомеразы 54
Точка инициации репли-
кации (ориджин) 48
Транспозиция 81
Транспозоны 94
Фрагменты Оказаки 48
Хеликазы 54
Экзонуклеаза 48
Эксцизионная репарация
оснований 81
Эндонуклеаза 48
Дополнительная литература
для дальнейшего изучения
Общая литература
Friedberg, Е. С., Walker, G. С., Siede, W, Wood, R. D.,
Schultz, R. A., & Ellenberger, T. (2006) DNA Repair and Mu-
tagenesis, 2nd edn, American Society for Microbiology, Wash-
ington, DC.
Всестороннее исследование метаболизма ДНК и хоро-
шая отправная точка для начала экспериментирования в
этой области.
Дополнительная литература для дальнейшего изучения 199 ]
Kornberg, А. & Baker, Т. А. (1991) DNA Replication, 2nd
edn, W. Н. Freeman and Company, New York.
Прекрасный источник информации по всем аспектам
метаболизма ДНК.
Репликация ДНК
Benkovic, S. J., Valentine, А. М., & Salinas, Е (2001)
Replisome-mediated DNA replication. Annu. Rev. Biochem. 70,
181-208.
В обзоре описаны схожие механизмы и ферменты ре-
пликации ДНК в разных классах организмов.
Bloom, L. В. (2006) Dynamics of loading the Escherichia coli
DNA polymerase processivity clamp. Crit. Rev. Biochem. Mol.
BiolAt,\lV-2№>.
Frick, D. N. & Richardson, С. C. (2001) DNA primases.
Annu. Rev. Biochem. 70,39-80.
Heller, R. C. & Marians, K. J. (2006) Replisome assembly
and the direct restart of stalled replication forks. Nat. Rev. Mol.
CellBiol.l, 932-943.
Механизмы перезапуска репликативной вилки до ре-
парации повреждения в ДНК.
Hiibscher, U., Maga, G., & Spadari, S. (2002) Eukaryotic
DNA polymerases. Annu. Rev. Biochem. 71,133-163.
Обзор свойств и функций более десяти известных
ДНК-полимераз эукариот.
Indiani, С. & O’Donnell, М. (2006) The replication clamp-
loading machine at work in the three domains of life. Nat. Rev.
Mol. Cell Biol. 7,751-761.
Kamada, K., Horiuchi, T., Ohsumi, K., Shimamoto, N., &
Morikawa, K. (1996) Structure of a replication-terminator
protein complexed with DNA. Nature 383,598-603.
О структуре комплекса Tus-Ter.
Kool, E. T. (2002) Active site tightness and substrate fit in
DNA replication. Annu. Rev. Biochem. 71,191-219.
Прекрасный обзор, посвященный молекулярным осно-
вам точности ДНК-полимеразы — геометрии пар основа-
ний и образованию водородных связей.
O’Donnell, М. (2006) Replisome architecture and dynamics
in Escherichia coli. J. Biol. Chem. 281,10,653-10,656.
Очень хороший обзор реакций в репликативной вилке.
Stillman, В. (2005) Origin recognition and the chromosome
cycle. FEBS Lett. 579,877-884.
Хороший обзор по инициации репликации ДНК у эу-
кариот.
Репарация ДНК
Begley, Т. J. & Samson, L. D. (2003) AlkB mystery solved:
oxidative demethylation of N1-methyladenine and N3-meth-
ylcytosine adducts by a direct reversal mechanism. Trends Bio-
chem. Sci. 28,2-5.
Erzberger, J. P. & Berger, J. M. (2006) Evolutionary relation-
ships and structural mechanisms of AAA+ proteins. Annu Rev.
Biophys. Biomol. Struct. 35,93-114.
Friedberg, E. C., Fischhaber, P. L., & Kisker, C. (2001) Er-
ror-prone DNA polymerases: novel structures and the benefits
of infidelity. Cell 107,9-12.
Goodman, M. E (2002) Error-prone repair DNA polymer-
ases in prokaryotes and eukaryotes. Annu. Rev. Biochem. 71,
17-50.
Обзор свойств ДНК-полимераз, число которых про-
должает расти.
Kunkel, Т. А. & Erie, D. А. (2005) DNA mismatch repair.
Annu. Rev. Biochem. 74,681-710.
Lindahl, T. & Wood, R. D. (1999) Quality control by DNA
repair. Science 286,1897-1905.
Mamett, L. J. & Plastaras, J. P. (2001) Endogenous DNA
damage and mutation. Trends Genet. 17,214-221.
Sancar, A. (1996) DNA excision repair. Annu. Rev. Biochem.
65,43-81.
Sutton, M. D., Smith, В. T., Godoy, V. G., & Walker, G. C.
(2000) The SOS response: recent insights into umuDC-de-
pendent mutagenesis and DNA damage tolerance. Annu. Rev.
Genet. 34,479-497.
Wilson, D. M. Ill & Bohr, V. A. (2007) The mechanics of base
excision repair, and its relation to aging and disease. DNA Re-
pair 6,544-559.
Wood, R. D., Mitchell, M., Sgouros, J., & Lindahl, T. (2001)
Human DNA repair genes. Science 291,1284-1289.
Что расшифровка генома человека позволила узнать о
репарации ДНК.
Рекомбинация Д11К
Сох, М. М. (2001) Historical overview: searching for replica-
tion help in all of the rec places. Proc. Natl. Acad. Sci. USA 98,
8173-8180.
О том, как было показано, что рекомбинация участвует
в репарации репликативной вилки.
Сох, М. М. (2007) Regulation of bacterial RecA protein func-
tion. Crit. Rev. Biochem. Mol. Biol. 42,41-63.
Craig, N. L. (1995) Unity in transposition reactions. Science
270, 253-254.
Gellert, M. (2002) V(D)J recombination: RAG proteins,
repair factors, and regulation. Annu. Rev. Biochem. 71, 101 —
132.
Grindley, N. D. E, Whiteson, K. L., & Rice, P. A. (2006)
Mechanisms of site-specific recombination. Annu. Rev. Bio-
chem. 75,567-605.
Hallet, B. & Sherratt, D. J. (1997) Transposition and site-
specific recombination: adapting DNA cut-and-paste mecha-
11 001 Часть III. 25. Метаболизм ДНК
nisms to a variety of genetic rearrangements. FEMS Microbiol.
^.21,157-178.
Haniford, D. B. (2006) Transpososome dynamics and regu-
lation in TnlO transposition. Crit. Rev. Biochem. Mol. Biol. 41,
407-424.
Детальное описание одного из хорошо исследованных
бактериальных транспозонов.
Lusetti, S. L. & Сох, М. М. (2002) The bacterial RecA pro-
tein and the recombinational DNA repair of stalled replication
forks. Annu. Rev. Biochem. 71, 71-100.
Paques, F. & Haber, J. E. (1999) Multiple pathways of recom-
bination induced by double-strand breaks in Saccharomyces
cerevisiae. Microbiol. Mol. Biol. Rev. 63,349-404.
Singleton, M. R., Dillingham, M. S., Gaudier, M., Kowal-
czykowski, S. C., & Wigley, D. B. (2004) Crystal structure
of RecBCD enzyme reveals a machine for processing DNA
breaks. Nature 432,187-193.
Van Duyne, G. D. (2001) A structural view of Cre-loxP site-
specific recombination. Annu. Rev. Biophys. Biomol. Struct. 30,
87-104.
Удачный структурный анализ системы сайт-специ-
фичсской рекомбинации.
Вопросы и задачи________________________
1. Выводы из эксперимента Мезельсона-Сталя.
Эксперимент Мезельсона-Сталя (рис. 25-2) до-
казывает, что ДНК в клетках Е. coli претерпевает
полуконсервативную репликацию. В «дисперси-
онной» модели репликации цепи родительской
ДНК расщепляются на участки случайной дли-
ны, затем связываются с фрагментами заново
реплицированной ДНК, что приводит к образо-
ванию дуплексов. Объясните, почему результаты
эксперимента Мезельсона и Сталя исключают
такую модель.
2. Анализ репликации ДНК с помощью тяже-
лых изотопов. Культуру Е. coli, растущую на
среде с 15NH4C1, переносят на среду, содержа-
щую 14NH4C1, где клетки делятся три раза (т. е.
численность клеток возрастает в восемь раз).
Каково молярное соотношение гибридной ДНК
(15N-14N) к легкой ДНК (14N-14N) в образовав-
шейся культуре?
3. Репликация хромосомы Е. coli. Хромосома
Е. coli содержит 4 639 221 п. н.
а) Сколько витков двойной спирали рас-
кручивается при репликации хромосомы
Е. coli?
б) Исходя из сведений, представленных в
данной главе, определите, сколько времени по-
требуется для репликации хромосомы Е. coli при
37 °C, если две репликативные вилки начинают
движение в точке начала репликации? Допустим,
что скорость репликации составляет 1000 п. н./с.
В этих условиях клетки Е. coli делятся каждые
20 мин. Возможно ли это?
в) Сколько (примерно) фрагментов Оказаки
образуется при репликации хромосомы Е. coli?
Какие факторы гарантируют правильную сборку
многочисленных фрагментов Оказаки в новую
ДНК?
4. Состав оснований ДНК, образованных на
одноцепочечных матрицах. Оцените состав
оснований всей ДНК, синтезированной ДНК-
полимеразой на матрицах из эквимолярной
смеси двух комплементарных цепей бакте-
риофага фХ174 (кольцевые молекулы ДНК).
Состав оснований одной цепи: А — 24,7%; G —
24,1%; С — 18,5%; Т — 32,7%. Какое допущение
необходимо сделать, чтобы ответить на этот
вопрос?
5. Репликация ДНК. Корнберг с коллегами ин-
кубировали растворимые экстракты Е. coli со сме-
сью dATP, dTTP, dGTP и dCTP, меченных 32Р по
а-фосфатным группам. Через некоторое время
инкубационную смесь обрабатывали трихлорук-
сусной кислотой, которая осаждает ДНК, но не
осаждает нуклеотиды-предшественники. Осадок
собирали и по количеству радиоактивной метки
в осадке определяли количество предшественни-
ков, встроившихся в ДНК.
а) Если из инкубационной смеси удалить
один из четырех предшественников, будет ли
радиоактивная метка обнаружена в осадке? По-
ясните ответ.
б) Может ли 32Р включаться в состав ДНК,
если метку содержит только dTTP? Поясните
ответ.
в) Будет ли обнаружена радиоактивность
в осадке, если метку 32Р включать в [3- или
у-фосфатные, а не в а-фосфатные группы? По-
ясните.
Вопросы и задачи [ 1011
6. Химия репликации ДНК. Все ДНК-полиме-
разы синтезируют новую цепь ДНК в направ-
лении 5'-*3'. В определенном смысле реплика-
ция антипараллельных цепей ДНК-дуплекса
была бы проще при наличии второго типа по-
лимераз, способных синтезировать ДНК в на-
правлении 3'-*5'. В принципе два типа поли-
мераз могли бы координировать синтез ДНК
без тех сложных механизмов, которые требу-
ются для репликации отстающей цепи. Одна-
ко таких 3'-*5' синтезирующих ферментов не
обнаружено. Предложите два возможных ме-
ханизма синтеза ДНК в направлении 3'-*5'.
Одним из продуктов в обоих случаях был бы
пирофосфат. Осуществимы ли эти механизмы
в клетке? Почему да или почему нет? (Под-
сказка. Можете предположить существование
предшественников ДНК, которых нет в совре-
менных клетках.)
7. Лидирующая и отстающая цепи. Составьте
таблицу, в которой сравните репликацию ли-
дирующей и отстающей цепей ДНК у Е. coli.
Перечислите названия и функции предше-
ственников, ферментов и других необходимых
белков.
8. Функция ДНК-лигазы. Некоторые мутанты
Е. coli содержат дефектную ДНК-лигазу. Если
такие мутанты обработать 3Н-меченым тими-
ном и осадить полученную ДНК в градиенте
плотности щелочной сахарозы, появляются
две радиоактивные полосы. Одна соответству-
ет высокомолекулярной фракции, другая —
низкомолекулярной фракции. Объясните эти
данные.
9. Точность репликации ДНК. Какие факторы
увеличивают точность репликации во время
синтеза лидирующей цепи ДНК? Можно ли
ожидать, что отстающая цепь будет образовы-
ваться с такой же точностью. Поясните свой
ответ.
10. Важная роль ДНК-топоизомераз в репли-
кации ДНК. Раскручивание ДНК, например,
при репликации влияет на плотность сверх-
спирализации ДНК. В отсутствие топоизоме-
раз ДНК перед репликативной вилкой была
бы перекручена, поскольку позади вилки ДНК
раскручена. Бактериальная репликативная
вилка останавливается, когда плотность сверх-
спирализации (а) ДНК перед вилкой достигает
величины +0,14 (см. гл. 24).
Двунаправленная репликация плазмиды
размером 6000 п. н. в условиях in vitro иниции-
руется в точке начала репликации без участия
топоизомераз. До начала репликации а = -0,06.
Сколько пар оснований раскрутит и реплициру-
ет каждая репликативная вилка до остановки?
Считайте, что все вилки движутся с одинаковой
скоростью и имеют все необходимое для элонга-
ции, кроме топоизомеразы.
11. Тест Эймса. На тонкий слой агаризованной
питательной среды, в которой отсутствовал ги-
стидин, высеяли примерно 109 бактерий Salmo-
nella typhimurium, ауксотрофных по гистидину
(мутантные клетки, которые нуждаются в гисти-
дине для выживания), и через двое суток инку-
бации при 37 °C получили примерно 13 колоний
(рис. 25-21). Как могли появиться колонии на
среде без гистидина? Эксперимент повторили
в присутствии 0,4 мкг 2-аминоантрацена. Через
двое суток образовалось более 10 000 колоний.
Какой вывод можно сделать о свойствах 2-ами-
ноантрацена? Что можно сказать о канцероген-
ности этого вещества?
12. Механизмы репарации ДНК. Клетки по-
звоночных и растений часто метилируют ци-
тозин в ДНК с образованием 5-метилцитозина
(см. рис. 8-5, а). В тех же клетках специализи-
рованная система репарации распознает оши-
бочные пары G-Т и заменяет их парой G=C.
Какую пользу клетке приносит такая система
репарации? (Объясните, учитывая роль 5-ме-
тилцитозина в ДНК).
13. Репарация ДНК у людей с пигментной
ксеродермой. Пигментная ксеродерма (ХР)
возникает в результате мутаций по меньшей мере
в семи разных генах человека. Нарушения обыч-
но затрагивают гены, кодирующие ферменты экс-
цизионной репарации нуклеотидов. Существует
несколько типов заболевания, которые обознача-
ют буквами от А до G (ХРА, ХРВ и т. д.), и отдель-
ный тип XPV.
[102] Часть III. 25. Метаболизм ДНК
Культуры фибробластов здоровых людей и боль-
ных ХР облучали УФ-светом. ДНК выделяли и
денатурировали, а полученные одноцепочечные
молекулы ДНК анализировали путем ультрацен-
трифугирования.
а) В образцах нормальных фибробластов
после облучения существенно снижена средняя
молекулярная масса одноцепочечных молекул
ДНК, а в образцах фибробластов пациентов с
XPG такого снижения не наблюдается. Почему?
б) Если считать, что в фибробластах действу-
ет система эксцизионной репарации нуклеоти-
дов, какой этап нарушен в фибробластах пациен-
тов с XPG? Объясните.
14. Структуры Холлидея. Чем образование
структур Холлидея при гомологичной генетиче-
ской рекомбинации отличается от их образова-
ния при сайт-специфической рекомбинации?
15. Связь между репликацией и сайт-
специфической рекомбинацией. Большинство
диких штаммов Saccharomyces cerevisiae несет мно-
жество копий кольцевой плазмиды 2р (названной
так из-за того, что длина ее окружности составляет
около 2 мкм), состоящей из -6300 п. н. Плазмида
использует хозяйскую систему репликации, при-
чем реплицируется под тем же строгим контролем,
что и хозяйская хромосома, всего один раз за кле-
точный цикл. Репликация плазмиды идет в двух
направлениях, а обе репликативные вилки воз-
никают в одной и той же хорошо изученной точке
начала репликации. Однако один цикл реплика-
ции плазмиды 2р может привести к образованию
более чем двух копий плазмиды, что способству-
ет увеличению копийности плазмиды (увеличи-
вается число копий плазмиды в одной клетке),
чтобы при расхождении плазмид в ходе деления
клетки ни одна из дочерних клеток не получила
меньше копий плазмиды, чем должно быть в нор-
ме. Для амплификации требуется система сайт-
специфической рекомбинации, закодированная
в плазмиде, которая позволяет перевернуть одну
часть плазмиды по отношению к другой части.
Объясните, каким образом сайт-специфическая
инверсия может приводить к увеличению копий-
ности плазмиды. (Подсказка. Рассмотрите ситуа-
цию, когда репликативные вилки удваивают толь-
ко один из двух сайтов рекомбинации.)
Анализ экспериментальных данных___
16. Мутагенез в клетках Escherichia coli. Мно-
гие мутагенные соединения действуют путем ал-
килирования оснований ДНК. Алкилирующий
агент R7000 (7-метокси-2-нитронафто[2,1-/^фу-
ран) - очень мощный мутаген.
В условиях in vivo R7000 активируется фер-
ментом нитроредуктазой, и возникающая более
активная форма образует ковалентную связь с
ДНК, главным образом с парами G=C, но не
только. В 1996 г. Кийарде, Туати и Хофнунг из-
учали механизм мутагенного действия R7000 на
клетки Е. coli. Они сравнили генотоксическую
активность R7000 к двум штаммам Е. coli\ штам-
му дикого типа (uvr1) и мутантному штамму, не
имеющему активности uvrA (uvr ; см. табл. 25-6).
Сначала исследователи определяли степень мута-
генности. Антибиотик рифампицин ингибирует
РНК-полимеразу (см. гл. 26); в его присутствии
клетки могут расти только при условии, что в
гене РНК-полимеразы произошли определенные
мутации. Таким образом, появление устойчивых
к рифампицину колоний может являться мерой
мутагенности.
Было проанализировано действие R7000 в
разной концентрации; результаты эксперимента
представлены ниже.
Анализ экспериментальных данных [ЮЗ]
а) Почему мутации возникают даже при от-
сутствии R7000?
Кийарде с соавторами также определяли вы-
живаемость бактерий, обработанных R7000 в раз-
личных концентрациях.
б) Объясните, почему обработка R7000 уби-
вает клетки бактерий.
в) Объясните различия в степени мутаген-
ности R7000 и выживаемости для двух штаммов
бактерий — uvr' и uvr (см. данные).
Далее исследователи стали измерять количе-
ство R7000, образовавшего ковалентную связь с
ДНК в штаммах uvr+ и uvr . Они инкубировали
бактерии с [3H]R7000 в течение 10 или 70 мин,
экстрагировали ДНК и определяли в ней уровень
3Н в имп./мин в расчете на микрограмм ДНК.
3Н в ДНК, имп./мин • мкг
Время, мин uvr+ uvr
10 76 159
70 69 228
г) Объясните, почему со временем содержа-
ние 3Н уменьшается в штамме uvr4 и увеличива-
ется в штамме uvr .
Кийарде с коллегами исследовали также из-
менения последовательности ДНК, вызванные
воздействием R7000 в штаммах uvr4 и uvr. Для
этого они использовали шесть различных штам-
мов Е. coli, каждый из которых содержал спец-
ифическую точечную мутацию в гене lacZ, ко-
торый кодирует |3-галактозидазу (этот фермент
катализирует ту же реакцию, что и лактаза; см.
рис. 14-10 в т. 2). Клетки с любой из этих мутаций
имеют нефункциональную [3-галактозидазу и не
могут метаболизировать лактозу (фенотип Lac ).
Для восстановления функции гена lacZ и фено-
типа Lac ’ в каждом конкретном мутанте требова-
лась специфическая обратная мутация. Высевая
клетки на среду, содержащую лактозу в качестве
единственного источника углерода, можно про-
изводить селективный отбор таких Lac+-клеток,
претерпевших обратную мутацию. Подсчет числа
Иас+-клеток после мутагенеза каждого штамма
позволяет определить частоту мутаций каждого
типа.
Сначала исследователи проанализировали
спектр мутаций в клетках uvr . В таблице пред-
ставлены результаты для шести штаммов - от
СС101 до СС106 (в скобках указаны точечные
мутации, необходимые для образования феноти-
па Lac+ в каждом случае).
Количество клеток Lac+
(среднее ± стандартное отклонение)
R7000, мкг/мл СС101 (А=Т на C-G) СС102 (ОС на А=Т) СС103 (ОС на OG) СС104 ((ОС на Т=А) СС105 (А=Т на Т=А) СС106 (А=Т на (ОС)
0 6±3 11±9 2±1 5±3 2±1 1±1
0,075 24±19 34±3 8±4 82±23 40±14 4±2
0,15 24±4 26±2 9±5 180+71 130±50 3±2
д) При каких мутациях количество клеток
с фенотипом Lac увеличивается в значитель-
ной степени в результате обработки препаратом
R7000? Объясните, почему у одних штаммов ча-
стота возникновения такого фенотипа выше, чем
у других.
е) Могут ли все перечисленные мутации объ-
ясняться ковалентным связыванием R7000 с па-
рой GC? Поясните свой ответ.
ж) На рис. 25-28, б показано, как метилиро-
вание остатков гуанина может приводить к заме-
не GC на АТ. Используя тот же путь, покажите,
каким образом аддукт G-R7000 мог бы приво-
дить к замене GC на АТ или на ТА в приведенном
выше примере. Какое основание образует пару с
аддуктом G -R7000?
Результаты для штаммов uvr+ представлены
ниже.
110 11 Часть III. 25. Метаболизм ДНК
Количество клеток Lac+ (среднее ± стандартное отклонение)
R7000, мкг/мл СС101 (А=Т на OG) СС102 (G=C на А=Т) СС103 (G=C на OG) СС104 (G=C на Т=А) СС105 (А=Т на Т=А) СС106 (А=Т на G^C)
0 2±2 10±9 3±3 4±2 6±1 0,5±1
1 7±6 21±9 8±3 23±15 13±1 1±1
5 4±3 15±7 22±2 68±25 67±14 1±1
Литература
Quillardet, Р., Touati, Е., & Hofnung, М. (1996) In-
fluence of the uvr-dependent nucleotide excision repair
on DNA adducts formation and mutagenic spectrum of a
potent genotoxic agent: 7-methoxy-2-nitronaptho[2,l-/?]
furan (R7000). Mutat. Res. 358, 113-122.
з) Указывают ли данные результаты на то, что
все типы мутаций исправляются с одинаковой
точностью? Объясните свой ответ.
В клетке РНК содержится и в ядре, и в частицах цитоплазмы, и в качестве
«растворимой» РНК в клеточном соке; многими специалистами было по-
казано, что все эти три фракции РНК возобновляются по-разному. При
любом обсуждении роли РНК в клетке важно представлять, что РНК край-
не неоднородна с точки зрения ее метаболизма и, возможно, в связи с
этим принадлежит более чем к одному типу.
— Фрэнсис Крик, статья в материалах Симпозиума
Общества экспериментальной биологии, 1958
Метаболизм РНК
26.1. ДНК-зависимый синтез РНК 106
26.2. Процессинг РНК 122
26.3. РНК-зависимый синтез РНК и ДНК 144
Экспрессия заложенной в гене информации
обычно происходит через образование мо-
лекулы РНК, транскрибируемой с матрицы
ДНК. Последовательности РНК и ДНК могут
показаться, на первый взгляд, очень похожими,
по химическому строению они различаются толь-
ко тем, что в РНК урацил заменен на тимин, а у
альдопентозы в положении 2' находится гидрок-
сильная группа. Однако в отличие от ДНК боль-
шинство молекул РНК выполняет свои функции
в виде одноцепочечных молекул, которые склады-
ваются различными способами и по сравнению с
ДНК гораздо более разнообразны (см. гл. 8 в т. 1).
Таким образом, РНК приспособлена к выполне-
нию различных клеточных функций.
РНК — единственная известная макромоле-
кула, которая участвует как в хранении, так и в
передаче информации, а кроме того, и в катализе;
это позволило выдвинуть гипотезу о ее участии
в развитии жизни на нашей планете в качестве
самого важного молекулярного предшественни-
ка. Открытие каталитических РНК, или рибози-
мов, изменило само представление о ферментах,
функцию которых, как считалось ранее, могут
выполнять только белки. Тем не менее белки не-
обходимы для функционирования РНК. В совре-
менной клетке все нуклеиновые кислоты, вклю-
чая молекулы РНК, связаны с белками. Некото-
рые из этих комплексов очень сложно устроены,
и входящая в них РНК может совмещать струк-
турную и каталитическую функции.
Все молекулы РНК, исключая геномные
РНК некоторых вирусов, переносят информа-
цию, которая постоянно хранится в форме ДНК.
В ходе транскрипции ферментная система пре-
образует генетическую информацию на участке
двухцепочечной ДНК в цепь РНК с последова-
тельностью оснований, комплементарной одной
из цепей ДНК. Образуется три основных типа
РНК. Матричная РНК (мРНК) кодирует ами-
нокислотную последовательность одного или
нескольких полипептидов, определяемую геном
или набором генов. Транспортная РНК (тРНК)
считывает информацию, закодированную в
мРНК, и переносит соответствующую аминокис-
лоту на растущую полипептидную цепь в ходе
синтеза белка. Рибосомная РНК (рРНК) входит
в состав рибосом — сложных клеточных струк-
тур, осуществляющих синтез белков. Многие до-
полнительные специализированные молекулы
РНК осуществляют регуляторные или каталити-
ческие функции или являются предшественника-
ми РНК трех выделенных выше типов. Эти РНК
больше не рассматриваются в качестве минорных
разновидностей в списке клеточных РНК. У по-
звоночных гораздо больше типов РНК, чем про-
сто «классические» мРНК, тРНК или рРНК.
При репликации обычно копируется целая
хромосома, транскрипция же более избиратель-
на. Одномоментно транскрибируются только от-
дельные гены или группы генов, а некоторые ча-
сти генома ДНК не транскрибируются никогда.
[106] Часть III. 26. Метаболизм РНК
Клетка ограничивает экспрессию генетической
информации, благодаря чему образуются лишь те
продукты генов, которые необходимы в данный
момент. Специфические регуляторные последо-
вательности отмечают начало и окончание транс-
крибируемых участков ДНК, а также то, какая
цепь дуплексной ДНК должна использоваться в
качестве матрицы. Подробно регуляция транс-
крипции описана в гл. 28.
Всю совокупность молекул РНК, производи-
мых клеткой в определенных условиях, называют
транскриптомом клетки Учитывая, что только
сравнительно небольшая часть генома человека ко-
дирует белки, можно было бы ожидать, что транс-
крибируется лишь малая часть генома. Однако это
не так. С помощью современных методов анализа
транскрипции на микрочипах удалось установить,
что значительная часть генома человека и других
млекопитающих транскрибируется в РНК, причем
основными продуктами являются не мРНК, тРНК
или рРНК, а множество различных РНК со специ-
фическими функциями. По-видимому, многие из
них участвуют в регуляции экспрессии генов, одна-
ко роль большинства остается неизвестной.
В этой главе рассматривается процесс синтеза
РНК на матрице ДНК, последующий процессинг
и оборот молекул РНК. Обсуждаются многие
специфические функции РНК, в том числе ката-
литические. Интересно, что субстратами для ката-
литических РНК часто служат другие молекулы
РНК. Мы также рассмотрим системы, в которых
молекула РНК служит матрицей, а ДНК — про-
дуктом, а не наоборот. Информационные метабо-
лические пути, таким образом, образуют полный
цикл, демонстрируя, что синтез нуклеиновой
кислоты на матрице происходит по стандартным
правилам независимо от природы матрицы или
продукта (РНК или ДНК). Обсуждение взаимных
превращений ДНК и РНК в качестве носителей
информации подводит к вопросу об эволюцион-
ном происхождении биологической информации.
26.1. ДНК-зависимый синтез РНК
Рассмотрение синтеза РНК мы начнем с сопо-
ставления процессов транскрипции и репли-
кации ДНК (гл. 25). Транскрипция похожа на
репликацию своим основополагающим химиче-
ским механизмом, своей полярностью (направ-
лением синтеза) и наличием матрицы. Подобно
репликации, транскрипция состоит из этапов
инициации, элонгации и терминации, хотя в
литературе при описании транскрипции ини-
циацию подразделяют на два самостоятельных
этапа — связывание ДНК и инициацию синтеза
РНК. Транскрипция отличается от репликации
тем, что для нее не требуется праймер и, как пра-
~8 п. н. центр
Направление транскрипции
Рис. 26-1. МЕХАНИЗМ РЕАКЦИИ. Транскрипция у
Е. coli, осуществляемая РНК-полимеразой. ДНК
временно раскручивается для синтеза цепи РНК
комплементарной одной из двух цепей в двойной
спирали, а — в любой момент времени в раскру-
ченном состоянии находится участок длиной око-
ло 17 п. н. РНК-полимераза и транскрипционный
пузырек по мере синтеза РНК двигаются слева на-
право вдоль ДНК как показано на рисунке. Моле-
кула ДНК раскручивается впереди и закручивается
позади пузырька. Красные стрелки указывают на-
правление, в котором ДНК должна вращаться для
обеспечения этого процесса. После обратного за-
кручивания ДНК гибрид РНК-ДНК распадается, и
цепь РНК высвобождается. РНК-полимераза на-
ходится в тесном контакте с ДНК впереди транс-
крипционного пузырька, а также с разделенными
цепями ДНК и РНК внутри и сразу за пузырьком.
Через канал в белке к активному центру полимера-
зы поступают новые нуклеозидтрифосфаты (NTP).
В процессе элонгации полимераза покрывает со-
бой участок ДНК длиной около 35 п. н.
26.1 ДНК-зависимый синтез РНК [107]
в
Направление транскрипции
Рис. 264. Продолжение, б — механизм синтеза РНК с помощью РНК-полимеразы похож на
механизм действия ДНК-полимеразы (см. рис. 25-5, б). В реакции принимают участие два
иона Мд2+, координированных по фосфатным группам поступающего NTP и трем остаткам
Asp (Asp460, Asp462 и Asp464 в Р'-субъединице РНК-полимеразы £. coli), которые консерватив-
ны в РНК-полимеразах у всех видов. Один ион Мд2+ способствует присоединению З'-гидрок-
сила к сх-фосфату NTP; другой Мд2+ обеспечивает уход пирофосфата; и тот и другой Мд2+
временно стабилизируют пятикоординационное состояние, в — при транскрипции проис-
ходит изменение супер скручивания ДНК. Движение РНК-полимеразы вдоль ДНК вызывает
появление положительных сверхвитков (перекрученная ДНК) впереди транскрипционного
пузырька и отрицательных сверхвитков (частично раскрученная ДНК) позади него. Топои-
зомеразы быстро удаляют положительные сверхвитки и регулируют отрицательное супер-
скручивание (гл. 24).
[108] Часть III. 26. Метаболизм РНК
вило, в ней участвуют только отдельные участки
молекулы ДНК. Кроме того, матрицей для каж-
дой конкретной молекулы РНК при транскрип-
ции служит только одна цепь ДНК.
РНК синтезирует РНК-полимераза
Открытие ДНК-полимеразы, зависящей от матри-
цы ДНК, подстегнуло поиск фермента, синтези-
рующего молекулу РНК, комплементарную цепи
ДНК. К 1960 г. четыре группы исследователей не-
зависимо друг от друга обнаружили в клеточных
экстрактах фермент, который может формировать
полимер РНК из рибонуклеозид-5'-трифосфатов.
Последующее изучение очищенной РНК-полиме-
разы Escherichia coli помогло установить основные
принципы транскрипции (рис. 26-1). Для ДНК-
зависимой РНК-полимеразы необходимы, поми-
мо матрицы ДНК, все четыре рибонуклеозид-5'-
трифосфата (АТР, GTP, UTP и СТР) в качестве ну-
клеотидных предшественников РНК, а также ионы
Mg2+. Фермент, кроме того, связывает один катион
Zn2+. По химической сути и механизму синтез РНК
очень напоминают синтез ДНК ДНК-полимеразой
(см. рис. 25-5). РНК-полимераза удлиняет цепь
РНК, добавляя рибонуклеотидные звенья к З'-ги-
дроксильному концу и надстраивая РНК в направ-
лении 5'-*3'. Гидроксильная группа на З'-конце
действует как нуклеофил, атакуя а-фосфат посту-
пающего рибонуклеозидтрифосфата (рис. 26-1, б)
и высвобождая пирофосфат. В общем виде реакция
выглядит следующим образом:
(NMP)„ + NTP------> (NMP)„+1 + PPi
РНК Удлиненная РНК
РНК-полимераза нуждается в ДНК, и ее актив-
ность максимальна при связывании с двухцепо-
чечной ДНК. Выше отмечалось, что матрицей
служит только одна цепь ДНК. Матричная цепь
ДНК копируется в направлении 3'-*5' (противо-
положном росту новой цепи РНК), как и при ре-
пликации ДНК. Каждый нуклеотид в новой РНК
присоединяется по правилу Уотсона-Крика: оста-
ток U в РНК встраивается в пару к остатку А в ма-
трице ДНК, а остаток G — в пару к остатку С. При
выборе пары может иметь значение геометрия пар
оснований (см. рис. 25-6).
В отличие от ДНК-полимеразы РНК-поли-
мераза не нуждается в праймере для инициации
синтеза. Инициация происходит при связывании
РНК-полимеразы со специфическими последо-
вательностями ДНК — промоторами (описаны
ниже). 5'-Трифосфатная группа первого остатка в
синтезируемой молекуле РНК не расщепляется с
высвобождением РРН а сохраняется на протяжении
всего процесса транскрипции. На стадии элонга-
ции основания на растущем конце новой цепи РНК
временно образуют пары с основаниями в матрице
ДНК, что приводит к формированию короткого
участка гибридной двойной спирали РНК-ДНК
длиной около 8 п. н. (рис. 26-1, а). Вскоре после об-
разования этой структуры РНК покидает дуплекс,
и заново формируется дуплекс ДНК.
Чтобы РНК-полимераза могла синтезировать
цепь РНК, комплементарную одной из двух цепей
ДНК, ДНК-дуплекс на коротком участке распле-
тается, образуя транскрипционный «пузырек».
В клетках Е. coli РНК-полимераза обычно остав-
ляет раскрученными примерно 17 п. н., из них
8 п. н. образуют гибрид РНК-ДНК. Удлинение
транскрипта РНК-полимеразой Е. coli происходит
со скоростью от 50 до 90 нуклеотидов в секунду.
Поскольку ДНК находится в виде спирали, для
продвижения пузырька требуется поворот цепей
молекул нуклеиновой кислоты. В большинстве
молекул ДНК вращение цепи ограничивается
ДНК-связывающими белками и другими струк-
турными барьерами. В результате движущаяся
РНК-полимераза генерирует серию положитель-
ных сверхвитков впереди транскрипционного пу-
зырька и отрицательных сверхвитков позади него
(рис. 26-1, в). Этот процесс происходит как in vitro,
так и in vivo (у бактерий). В клетке топологические
проблемы, связанные с транскрипцией, разреша-
ются благодаря действию топоизомераз (гл. 24).
КЛЮЧЕВЫЕ ДОГОВОРЕННОСТИ. При транскрипции две
комплементарные цепи ДНК выполняют разные
функции. Цепь, которая служит матрицей для син-
теза РНК, называется матричной цепью. Компле-
ментарная ей нематричная, или кодирующая, цепь
идентична по нуклеотидной последовательности
транскрибируемой с данного гена РНК, с тем лишь
отличием, что РНКсодержит U вместоТ (рис. 26-2).
Для конкретного гена кодирующей цепью может
быть любая цепь данной хромосомы (как показано
на рис. 26-3 для вируса). По договоренности, регу-
ляторные последовательности, которые контроли-
руют транскрипцию (описаны ниже в этой главе),
обозначаются на кодирующей цепи.
26.1 ДНК-зависимый синтез РНК [109]
(5Z) CGCTATAGCGTTT(3')
(3') GCGATATCGCAAA(5')
(5') CGCUAUAGCGUUU(3')
Нематричная (кодирующая) цепь ДНК
Матричная цепь ДНК
РНК-транскрипт
Рис. 26-2. Матричная и нематричная (кодирующая) цепи ДНК. Две комплементарные цепи
ДНК выполняют в ходе транскрипции разные функции. Транскрипт РНК, синтезированный
на матричной цепи, идентичен последовательности нематричной, или кодирующей, цепи
(с учетом замены Т на U).
РНК-транскрипты
ДНК ......
3,6 • 104 п. н.
Рис. 26- Кодирование информации в геноме аденовируса. Генетическая информация ге-
нома аденовируса (самый простой пример) закодирована в молекуле двухцепочечной ДНК
размером 36 000 п. н., обе цепи которой кодируют белки. Информация о большинстве бел-
ков закодирована в верхней цепи по договоренности цепь ориентирована слева направо
от 5'-конца к З'-концу. Для этих транскриптов нижняя цепь играет роль матрицы. Однако
некоторые белки кодируются нижней цепью ДНК, которая транскрибируется в противопо-
ложном направлении (для них матрицей служит верхняя цепь). В реальности синтез моле-
кул мРНК аденовируса намного сложнее. Многие молекулы мРНК, показанные на верхней
цепи, сначала синтезируются в виде единого длинного транскрипта (из 25 000 нуклеоти-
дов), который затем подвергается процессингу, превращаясь в отдельные молекулы мРНК.
Аденовирусы вызывают инфекции верхних дыхательных путей у некоторых позвоночных.
ДНК-зависимая РНК-полимераза Е. coli -
большой сложный фермент, состоящий из пяти
субъединиц a2PP'w (называется кор-частицей:
Мг = 390 000) и шестой субъединицы а, имею-
щей различающиеся по размеру (по молекуляр-
ной массе) варианты. Субъединица о временно
связывается с кор-частицей и направляет фер-
мент к специфическому участку в молекуле ДНК
(показано ниже). Эти шесть субъединиц состав-
ляют холофермент РНК-полимеразы (рис. 26-4).
Холофермент РНК-полимеразы Е. coli существу-
ет, таким образом, в нескольких формах в зави-
симости от типа о-субъединицы. Обычно это
субъединица о70 (Мх = 70 000), и далее мы будем
говорить именно об этом варианте холофермента
РНК-полимеразы.
Рис. 26-4. Строение холофермента РНК-полимеразы бак-
терии Thermus aquaticus (на основании PDB ID 1IW7). В
целом строение фермента очень напоминает структуру
РНК-полимеразы Е. coli (здесь не показаны ни РНК, ни
ДНК). р-Субъединица выделена темно-серым цветом, Р'-
субъединица светло-серым; две сх-субъединицы показа-
ны разными оттенками красного; со-субъединица желтая;
о-субъединица бежевая. Изображение слева ориентиро-
вано так же, как на рис. 26-6. При повороте структуры на
180° вокруг оси у (справа) становится видна небольшая
субъединица со.
[110] Часть III. 26. Метаболизм РНК
РНК-полимеразы не имеют независимой
3'->5'-экзонуклеазной активности для исправления
ошибок (как многие ДНК-полимеразы), и частота
ошибок при транскрипции выше, чем при реплика-
ции хромосомной ДНК — примерно одна ошибка
на каждые 104-105 встроенных рибонуклеотидов.
Поскольку с одного гена обычно образуется мно-
жество копий РНК, и все молекулы РНК в конеч-
ном итоге разрушаются и удаляются, ошибка в мо-
лекуле РНК не имеет для клетки столь серьезных
последствий, как ошибки в ДНК — постоянном
источнике информации. Многие РНК-полиме-
разы, в том числе бактериальная РНК-полимераза
и эукариотическая РНК-полимераза II (обсуждает-
ся ниже), останавливаются, если в ходе транскрип-
ции добавляется неправильное основание, и могут
удалять аномальные нуклеотиды с З'-конца транс-
крипта благодаря реакции, обратной полимеразной
реакции. Но пока неизвестно, является ли эта ак-
тивность истинно корректирующей активностью и
каков ее вклад в точность транскрипции.
Синтез РНК начинается с промоторов
Инициация синтеза РНК в случайных точках мо-
лекулы ДНК была бы недопустимой расточитель-
ностью. Поэтому РНК-полимераза связывается
со специфическими последовательностями ДНК,
называемыми промоторами, которые направляют
транскрипцию прилежащих участков ДНК (генов).
Последовательности ДНК для связывания РНК-
полимераз достаточно разнообразны, идентифика-
ции этих последовательностей, определяющих функ-
цию промотора, посвящены многие исследования.
Связывание РНК-полимеразы Е. coli про-
исходит в области, начинающейся примерно за
70 п. н. до точки начала транскрипции и заканчи-
вающейся через 30 п. н. после нее. По договорен-
ности, пары оснований ДНК, соответствующие
началу молекулы РНК, имеют положительную
нумерацию, а расположенные выше точки начала
транскрипции — отрицательную. Таким образом,
область промотора расположена на участке меж-
ду позициями -70 и +30. Сравнительный анализ
бактериальных промоторов наиболее распростра-
ненного класса (распознаваемых холоферментом
РНК-полимеразы, содержащим о70) выявил ха-
рактерные последовательности около позиций
10 и 35 (рис. 26-5). Эти последовательности
важны для взаимодействия с субъединицей о70.
Хотя эти последовательности не идентичны для
всех бактериальных промоторов данного клас-
са, некоторые нуклеотиды, которые чаще всего
встречаются в конкретной позиции, образуют
1 Элемент UP | |область -35| Спенсер | Область —10| Спейсер | Начало РНК
+1
Консенсусная | NNAAA^T^TTTTNNAAAANNN последов ате л ьность |n[ TTGACA-] N17 1 ТАТААТ-! N6
ггпВ Pl I AGAAAATTATTTTAAATTTCCT |N GTGTCA N16 ТАТААТ N8 А
trp TTGACA n17 TTAACT n7 1 A
lac TTTACA n17 [ TATGTT 1 Ne A
recA TTGATA N16 1 TATAAT | n7 ГА
araBAD b JCTGACG | n18 | TACTGT 1 N6 La __
Рис. 26-5. Типичные промоторы Е. coli, распознаваемые холоферментом РНК-полимеразы,
содержащим о70. Последовательности нематричной цепи представлены обычным способом
(в направлении 5'->3'). Последовательности разных промоторов различаются, но при срав-
нении многих промоторов выявляется некоторое сходство, особенно в позициях -10 и -35.
UP-элемент, имеющийся не во всех промоторах Е. coli, обнаружен в промоторе Р1 активно
экспрессирующейся рРНК гена ттВ. UP-элементы, обычно расположенные в области от -40
до -60, эффективно стимулируют транскрипцию с содержащих эти элементы промоторов.
В промоторе rrnB Р1 UP-элемент охватывает область от -38 до -59. Консенсусная последо-
вательность для промоторов Е. coli, распознаваемая а70, показана второй сверху. В промежу-
точных участках (спейсерах) слабо различающееся число нуклеотидов (N). Показан только
первый нуклеотид последовательности, кодирующей транскрипт (позиция +1).
26.1 ДНК-зависимый синтез РНК [111]
Дополнение 26-1
ПРАКТИЧЕСКАЯ БИОХИМИЯ
РНК-пилимеразг оставляет свой след на промоторе
Метод футпринтинга основан на принципах, приме-
няемых при секвенировании ДНК, и позволяет иден-
тифицировать последовательности ДНК. связанные с
конкретным белком. Выделяют фрагмент ДНК, кото-
рый предположительно содержит последовательности,
распознаваемые ДНК-связывающим белком, и радиоак-
тивно метят один конец одной из цепей (рис. 1). Затем с
помощью химических реагентов или ферментов во фраг-
мент ДНК вносят несколько разрывов (в среднем один
разрыв на молекулу). При разделении меченых продук-
Смесь фрагментов ДНК
с радиоактивной меткой *
на одном конце одной цепи.
Обработка ДНКазой
в условиях, при которых
каждая цепь разрезается *
один раз (в среднем). *
Там, где связана
РНК-полимераза,
разрезы не
возникают.
Место
разреза
ДНКазой.
*
Выделенные меченые
фрагменты ДНК денатурируют.
На следующей стадии
анализируют только
* меченые цепи.
*
*
*
*
*
*
*
*
*
*
При электрофорезе в полиакриламидном геле
фрагменты с радиоактивной меткой проявляются
как полосы на пленке, чувствительной
к рентгеновскому излучению.
Неразрезанный
фрагмент ДНК
Отсутствие полос
свидетельствует о том,
что РНК-полимераза
была связана с ДНК.
Направление
движения ДНК
Рис. 1. Метод футпринтинга для идентификации участка
связывания полимеразы на фрагменте ДНК. Разделение
проводят в присутствии (+) и в отсутствие (-) полимеразы.
[112] Часть III. 26. Метаболизм РНК
Дополнение 26-1
ПРАКТИЧЕСКАЯ БИОХИМИЯ
РНК-полимераза оставляет свой след... (продолжение)
тов расщепления (расщепленные фрагменты разной длины) ме-
тодом электрофореза с высоким разрешением образуется «забор»
радиоактивных полос. В отдельной пробирке повторяют такую же
процедуру расщепления того же фрагмента ДНК, но в присутствии
ДНК-связывающего белка. Затем два набора продуктов подверга-
ют электрофорезу, нанося на соседние дорожки, и сравнивают полу-
ченные результаты. Пробел («футпринт») в серии радиоактивных
полос в образце ДНК с белком соответствует фрагменту, защищен-
ному ДНК-связывающим белком. Так можно определить последо-
вательности, с которыми связывается белок.
Точное положение участка связывания белка можно устано-
вить прямым секвенированием (рис. 8-34 в т. 1) того же фрагмен-
та ДНК с последующим разделением продуктов в том же геле (не
показано). На рис. 2 представлены результаты связывания РНК-
полимеразы с фрагментом ДНК. содержащим промотор. Поли-
мераза покрывает участок длиной от 60 до 80 п. н.; защищенными
оказываются, в том числе, области 10 и - 35.
Рис. 2. Определение участка связывания РНК-полимеразы с Zac-промотором (см. рис. 26-5).
В этом эксперименте 5'-конец нематричной цепи содержал радиоактивную метку. На до-
рожке К нанесен контроль, в котором меченую ДНК расщепляли химическим реагентом, что
позволило получить картину с равномерным распределением полос.
консенсусную последовательность (вспомните
последовательность опС у Е. coli\ см. рис. 25-11).
Консенсусная последовательность в области - 10:
(5')ТАТААТ(3'); консенсусная последователь-
ность в области 35: (5')TTGACA(3'). Третий,
богатый основаниями АТ фрагмент называет-
ся UP-элементом (от англ, upstream promoter —
выше промотора); он находится между позици-
ями 40 и 60 в промоторах некоторых активно
экспрессируемых генов. UP-элемент связывается
Рис. 26-6. Инициация и элонгация транскрипции РНК-полимеразой Е. coli. а — инициация
транскрипции обычно включает две основные стадии — связывание и инициацию. При
связывании инициирующее взаимодействие РНК-полимеразы с промотором приводит к за-
крытому комплексу, в котором промоторная ДНК прочно связывается, но не раскручивается.
Затем фрагмент ДНК размером 12-15 п. н. в области от -10 до +2 или +3 раскручивается, об-
разуя открытый комплекс. На стадиях, предшествующих закрытому и открытому комплек-
сам, были обнаружены дополнительные промежуточные соединения и несколько конфор-
мационных изменений белков (не показаны). При инициации транскрипции происходит
высвобождение промотора. После синтеза первых 8 или 9 нуклеотидов новой РНК субъе-
диница о высвобождается, полимераза покидает промотор и осуществляет элонгацию РНК.
б — строение кор-частицы РНК-полимеразы Е. coli на стадии элонгации. Окраска субъ-
единиц такая же, как на рис. 26-4: р- и р'-субъединицы — темно- и светло-серые;
а-субъединицы красные; со-субъединица в этом ракурсе не видна. о-Субъединица от-
сутствует, так как она отделилась после инициации. Вверху показан комплекс в целом, с
фрагментами ДНК и РНК. Активный центр транскрипции находится в щели между р- и р -
субъединицами. На среднем рисунке р-субъединица не показана, так что видны активный
центр и область гибридизации ДНК-РНК. В активном центре находится ион Мд2+ (красная
точка). На нижнем рисунке все белки удалены, чтобы показать, как ДНК и РНК протягива-
ются через комплекс.
26.1 ДНК-зависимый синтез РНК [113]
с а-субъединицей РНК-полимеразы. Эффектив-
ность, с которой РНК-полимераза связывается с
промотором и инициирует транскрипцию, в зна-
чительной степени определяется этими после-
довательностями, расстоянием между ними и их
удаленностью от точки начала транскрипции.
Функциональное значение последовательно-
стей в позициях 10 и -35 подтверждается множе-
ством независимых доказательств Мутации, вли-
яющие на работу промотора, часто затрагивают
основания из этих областей. Изменения в консен-
сусной последовательности также влияют на эф-
фективность связывания РНК-полимеразы и ини-
циацию транскрипции. Замена лишь одной пары
оснований может снизить эффективность связы-
вания на несколько порядков. Таким образом, про-
моторная последовательность задает базовый уро-
вень экспрессии, который для разных генов Е. coli
а
[114] Часть III. 26. Метаболизм РНК
может существенно различаться. Метод изучения
взаимодействия между РНК-полимеразой и про-
моторами обсуждается в доп. 26-1.
Постепенно проясняется механизм ини-
циации транскрипции и роль о-субъединицы
(рис. 26-6, а). Процесс инициации состоит из
двух основных этапов, связывания и собственно
инициации, причем каждый этап осуществляется
в несколько стадий. Сначала полимераза, направ-
ляемая фактором о. связывается с промотором,
в результате чего последовательно образуются
закрытый комплекс (в котором связанная ДНК
остается интактной) и открытый комплекс (в
котором связанная ДНК частично раскручи-
вается около позиции - 10). Затем в комплексе
начинается транскрипция, что приводит к кон-
формационному изменению — комплекс пере-
ходит в форму, характерную для элонгации, по-
Рис. 26-7. Роль п-факторов в транскрипции. РНК-поли-
мераза, направляемая связанной и-субъединицей, связы-
вается с ДНК в области промотора. Как только происхо-
дит инициация синтеза РНК, о-субъединица отсоединя-
ется и замещается белком NusA. Когда РНК-полимераза
достигает терминирующей последовательности, синтез
РНК прекращается, NusA высвобождается из комплек-
са с полимеразой, а РНК-полимераза отсоединяется от
ДНК. Свободная полимераза может связаться с любой
о-субъединицей. Тип связанной субъединицы определя-
ет, с каким промотором свяжется РНК-полимераза в сле-
дующем раунде синтеза.
инициация
◄-------------
элонгация
Терминаторная
последов ате льность
IJ 2^2
Промоторная
последовательность nY
( NusA^)
отделение
РНК-
полимеразы
Готовая мРНК
26.1 ДНК-зависимый синтез РНК [115]
Семь типов субъединиц а у Escherichia coli
Субъединица о Kd (нмоль/л) Число молекул на клетку* Доля этого холофермента ( % ) * Функция
о70 0,26 700 78 Гены домашнего хозяйства
О54 0,30 НО 8 Регуляция уровня азота в клетке
О38 4,26 <1 0 Гены стационарной фазы
О32 1,24 <10 0 Гены теплового шока
О28 0,74 370 14 Гены жгутика и хемотаксиса
а24 2,43 <10 0 Внецитоплазматические функции; участие в реакции на тепловой шок
о18 1,73 <1 0 Внецитоплазматические функции, включая транспорт цитрата железа
Источник. По Maeda, Н., Fujita, N., & Ishihama, А. (2000) Nucleic. Acids Res. 28,3500; с изм.
Замечание. о-Факторы широко распространены у бактерий; их число варьирует от одного
о-фактора у Mycoplasma genitalium до 63 у Streptomyces coelicolor.
* Приблизительное число о-субъединиц каждого типа в клетке, а также доля холофермента
РНК-полимеразы, образующего комплекс с этой о-субъединицей в экспоненциальной фазе
роста. Эти значения зависят от условий роста Доля молекул РНК-полимеразы, образую-
щих комплекс с каждой о-субъединицей, отражает как количество субъединиц этого типа,
так и их сродство к ферменту
еле чего сдвигается с промотора (высвобождение
промотора). Каждая из этих стадий зависит от
специфической последовательности промотора.
При переходе полимеразы к стадии элонгации
субъединица g спонтанно диссоциирует. Участие
РНК-полимеразы в элонгации схематично от-
ражено на рис. 26-6, б. Белок NusA (Мг = 54 430)
связывается с РНК-полимеразой, конкурируя с
о-субъединицей. После окончания транскрипции
NusA отсоединяется от фермента, РНК-полиме-
раза диссоциирует из комплекса с ДНК, а фактор
g (G70 или другой) может связываться с фермен-
том, чтобы снова инициировать транскрипцию;
этот цикл иногда называют g-циклом (рис. 26-7).
В ДНК Е. coli есть и другие классы промо-
торов, с которыми связываются холоферменты
РНК-полимеразы с другими G-субъединицами
(табл. 26-1), например, промоторы генов теплово-
го шока. Продукты этих генов образуются в более
высокой концентрации, если клетка подвергает-
ся сильному воздействию, такому как внезапное
повышение температуры. РНК-полимераза свя-
зывается с промоторами этих генов, только ког-
да субъединица G70 заменяется субъединицей G32
(Мг = 32 000), специфичной к промоторам белков
теплового шока (см. рис. 28-3). С помощью раз-
личных G-субъединиц клетка может координи-
ровать экспрессию, что позволяет ей значительно
изменять свое физиологическое состояние. Какой
именно набор генов будет экспрессироваться, за-
висит от доступности различных G-субъединиц,
что в свою очередь определяется рядом факторов:
регулируемой скоростью синтеза и деградации,
постсинтетическими модификациями, перево-
дящими отдельные G-субъединипы из активной
формы в неактивную и обратно, а также специ-
ализированным классом анти-о-белков, связы-
вающихся с определенным типом G-субъединиц
и делающих их недоступными для инициации
транскрипции.
Транскрипция регулируется
на нескольких уровнях
Потребности в продуктах любых генов изменя-
ются в зависимости от состояния клетки и стадии
развития, и транскрипция каждого гена строго
регулируется, чтобы обеспечить синтез конкрет-
ного продукта в строго определенном количестве.
Регуляция может происходить на любой стадии
транскрипции, включая элонгацию и терминацию.
Однако часто регуляции подвергаются стадии свя-
зывания полимеразы и инициации транскрипции,
изображенные на рис. 26-6, а. Один из примеров
[116] Часть III. 26. Метаболизм РНК
многоуровневого контроля — наличие промоторов
с различными последовательностями.
Связывание белков с последовательностями
вблизи и вдали от промотора также оказывает
влияние на уровень экспрессии генов. Связыва-
ние белка может активировать транскрипцию,
облегчая связывание РНК-полимеразы, или по-
давлять транскрипцию, блокируя активность по-
лимеразы. У Е. coli одним из активирующих транс-
крипцию белков является сАМР-рецепторный
белок (CRP), который при отсутствии глюкозы
в среде роста усиливает транскрипцию генов, ко-
дирующих ферменты метаболизма отличных от
глюкозы сахаров. Белки-репрессоры блокируют
синтез РНК специфических генов. Например,
Lac-penpeccop (гл. 28) блокирует транскрипцию
генов ферментов, участвующих в метаболизме
лактозы, когда лактоза недоступна.
Транскрипция — первая стадия в сложном и
энергозатратном процессе синтеза белка, поэто-
му регуляция концентрации белков как у бакте-
рий, так и у эукариот часто осуществляется на
уровне транскрипции, особенно на ее ранних
стадиях. В гл. 28 рассмотрены различные меха-
низмы, с помощью которых осуществляется та-
кая регуляция.
Специфические последовательности
подают сигнал прекращения синтеза РНК
Синтез РНК происходит с высокой пропсссивно-
стью (см. с. 50), поскольку, если РНК-полимераза
высвободит транскрипт РНК преждевременно,
она не сможет закончить синтез, и ей придется
начинать заново. Однако наличие определенных
последовательностей ДНК вызывает остановку,
а иногда и прекращение (терминацию) синтеза
РНК. Механизм терминации у эукариот еще не
до конца изучен, поэтому мы обратимся к бакте-
риям. Клетки Е. coli имеют по меньшей мере два
типа сигналов терминации: один связан с белко-
вым фактором р, другой р-независимый.
Большинство p-независимых терминаторов
отличаются двумя свойствами. Во-первых, они
содержат последовательность, транскрипт кото-
рой имеет комплементарные участки, образую-
щие структуры в виде шпильки (см. рис. 8-19, а
вт. 1), расположенные на расстоянии 15-20 нукле-
отидов от дальнего конца цепи РНК. Во-вторых,
их матричная цепь содержит консервативную по-
Изомеризация
Терминация
Отделение
Рис. 26-8. Модель p-независимой терминации транс-
крипции в клетках Е. coli. РНК-полимераза останавли-
вается в разных участках последовательности ДНК, в
том числе в терминаторах. Это может приводить к двум
результатам: либо полимераза обходит препятствие и
продолжает движение, либо комплекс претерпевает кон-
формационные изменения и изомеризуется. В последнем
случае спаривание внутримолекулярных комплементар-
ных участков в только что синтезированном транскрипте
может приводить к образованию шпильки, разрушающей
РНК-ДНК-гибрид и/или взаимодействию между РНК и
полимеразой, что вызывает изомеризацию. Нестабиль-
ным является AUU-гибридный участок на З'-конце ново-
го транскрипта, и РНК полностью выходит из комплекса,
что приводит к терминации транскрипции. Именно это
обычно происходит при встрече РНК-полимеразы с тер-
минаторами. После изомеризации комплекс может про-
должить синтез РНК в других участках.
следовательность из трех остатков А, которые при
транскрипции превращаются в остатки U вблизи
З'-конца шпильки. Когда полимераза достигает
сайта терминации, который содержит эту струк-
туру, она останавливается (рис. 26-8). При фор-
мировании шпильки в РНК разрывается несколь-
ко пар оснований A=U в гибриде РНК-ДНК и
нарушаются важные контакты между РНК и
РНК-полимеразой, что облегчает диссоциацию
транскрипта
26.1 ДНК-зависимый синтез РНК [117]
В p-зависимых терминаторах нет повто-
ряющихся остатков А в матричной цепи, но
обычно присутствует СА-богатая последова-
тельность, называемая n/г-элементом (от англ.
rho utilization — «ро»-использующая последо-
вательность). Белок р связывается со специфи-
ческим участком РНК и перемещается в на-
правлении 5'-*3' до тех пор, пока не достигнет
транскрипционного комплекса, остановившего-
ся на сайте терминации. Здесь он способствует
высвобождению РНК-транскрипта. Белок р об-
ладает ATP-зависимой РНК-ДНК-хеликазной
активностью, которая способствует перемеще-
нию белка вдоль РНК; при терминации р-белок
осуществляет гидролиз АТР. Детальный меха-
низм, посредством которого этот белок способ-
ствует высвобождению РНК-транскрипта, пока
неизвестен.
В клетках эукариот содержатся
РНК-полимеразы трех типов
Механизм транскрипции в ядрах эукариотиче-
ских клеток намного сложнее, чем у бактерий.
Эукариоты имеют три РНК-полимеразы (I, II и
III), которые различаются составом комплекса,
но имеют некоторые общие субъединицы. Каж-
дая полимераза выполняет особую функцию и
присоединяется к специфической промоторной
последовательности.
РНК-полимераза I (Pol I) отвечает за синтез
только одного типа РНК прерибосомной РНК
(пре-рРНК), которая содержит предшественни-
ков 18S, 5,8S и 28S рРНК (см. рис. 26-25). После-
довательности промоторов Pol I существенно раз-
личаются у разных видов Важная функция РНК-
полимеразы II (Pol П) — синтез молекул мРНК
и некоторых специализированных молекул РНК.
Этот фермент распознает тысячи промоторов с
сильно различающимися последовательностями.
Многие промоторы Pol II обладают несколькими
общими свойствами, в том числе содержат ТАТА-
бокс (эукариотическая консенсусная последова-
тельность ТАТААА) вблизи позиции -30 и Inr-
последовательность (инициатор) вблизи точки
начала синтеза РНК в позиции +1 (рис. 26-9).
РНК-полимераза III (Pol III) образует мо-
лекулы тРНК, 5S рРНК и некоторые другие не-
большие специализированные молекулы РНК.
Промоторы, распознаваемые Pol III. хорошо
охарактеризованы. Интересно, что некоторые
последовательности, необходимые полимеразе
III для осуществления регулируемой инициа-
ции транскрипции, расположены в самом гене,
а другие локализованы более традиционно
выше точки начала транскрипции (гл. 28).
-30
ТАТААА
Различные
регуляторные
последов ате льности
ТАТА-бокс
+ 1
yyanJyy
Inr
Рис. 26-9. Распространенные последовательности промоторов, распознаваемые эукари-
отической РНК-полимеразой II. Точкой сборки белков преинициаторных комплексов Pol П
является ТАТА-бокс. ДНК в инициаторной последовательности (Inr) расплетается, а точка на-
чала транскрипции обычно находится в этой последовательности или в непосредственной
близости от нее. В представленной здесь консенсусной последовательности Inr N обозначает
любой нуклеотид, a Y — пиримидиновый нуклеотид. Многие дополнительные последователь-
ности служат участками связывания для большого числа белков, влияющих на активность
Pol П. Эти последовательности важны для регуляции промоторов Pol II и значительно различа-
ются по типу и количеству; как правило, промоторы эукариот намного сложнее, чем показано
здесь (см. рис. 15-23 в т. 2). Многие последовательности расположены на расстоянии несколь-
ких сотен пар оснований от ТАТА-бокса с 5'-стороны, другие могут быть удалены на тысячи пар
оснований. Представленные здесь консенсусные последовательности эукариотических промо-
торов Pol II различаются намного сильнее, чем в промоторах Е. coli (см. рис. 26-5). Во многих
промоторах Pol II отсутствует либо ТАТА-бокс или элемент Inr, либо обе последовательности.
Один или несколько транскрипционных факторов распознают дополнительные последователь-
ности около ТАТА-бокса или ниже по последовательности (справа в данном случае).
[118] Часть III. 26. Метаболизм РНК
Для проявления активности
РНК-полимеразы II требуются
другие белковые факторы
РНК-полимераза II играет ведущую роль в экс-
прессии эукариотических генов и поэтому ак-
тивно изучалась. Хотя эта полимераза значи-
тельно сложнее своего бактериального аналога,
за сложностью скрывается удивительная кон-
сервативность структуры, функции и механиз-
ма действия. Фермент Pol II из дрожжей имеет
гигантский размер и состоит из 12 субъединиц.
Самая крупная субъединица RBP1 обладает зна-
чительной гомологией с |3'-субъединицей бакте-
риальной РНК-полимеразы. Другая субъединица
RBP2 по структуре похожа на бактериальную
р-субъединицу, еще две субъединицы RBP3 и
RP11 имеют некоторую структурную гомологию
с двумя бактериальными а-субъединицами. По-
лимеразе Pol II приходится действовать в гораздо
более сложных геномах и взаимодействовать с
гораздо более сложно упакованными молекула-
ми ДНК. чем у бактерий. Дополнительная слож-
ность полимеразы эукариот объясняется необхо-
димостью ориентироваться в лабиринте из мно-
жества белковых факторов и вступать с ними в
белок-белковые взаимодействия.
Самая крупная субъединица Pol II облада-
ет необычным свойством: на С-конце она имеет
длинную аминокислотную последовательность,
состоящую из многократно повторяющейся кон-
сенсусной последовательности семи аминокис-
лот -YSPTSPS-. В ферменте дрожжей содер-
жится 27 таких повторов (18 из них в точности
соответствуют консенсусной последовательно-
сти), а в ферментах мыши и человека — по 52 по-
втора (21 точный). Этот С-концевой домен CTD
(от англ, carboxyl-teminal domain} отделен от
основного тела фермента неструктурированной
линкерной последовательностью. Как показано
Белки, необходимые для инициации транскрипции с промоторов РНК-полимеразы II (Pol II) у эукариот
Белок Количество субъединиц Мт субъединиц Функции
Инициация Pol II 12 10 000-220 000 Катализирует синтез РНК
ТВР (ТАТА- 1 38 000 Распознает ТАТА-бокс
связывающий белок) TFIIA 3 12 000.19 000,35 000 Стабилизирует связывание TFIIB и ТВР с промотором
TFIIB 1 35 000 Связывается с ТВР; собирает комплекс Pol II-TFIIF
TFIIE 2 34 000,57 000 Собирает TFIIH; обладает АТРазной и хеликазной
TFIIF 2 30 000,74 000 активностями Прочно связывается с Pol II; связывается с TFIIB
TFIIH 12 35 000-89 000 и препятствует неспецифическому связыванию Pol II с ДНК Раскручивает ДНК у промотора (хеликазная
Элонгация3 ELL6 1 80 000 активность); фосфорилирует Pol II (в CTD); собирает белки эксцизионной репарации
pTEFb 2 43 000,124 000 Фосфорилирует Pol 11 (в CTD)
SII (TFIIS) 1 38 000
Элонгин (SIII) 3 15 000,18 000,110 000
а Функция всех факторов элонгации заключается в преодолении остановки или прекраще-
ния транскрипции комплексом Pol II-TFIIF.
6 Сокращение от англ, eleven-nineteen lysine-rich leukemia. Ген ELL часто подвергается реком-
бинации при остром миелоидном лейкозе.
26.1 ДНК-зависимый синтез РНК [119]
со ел
-30 +1
| ТАТА | | Inr | ДНК
ТВР (и/или TFIIA)
TFIIB
TFIIF-Pol П
I TFIIE
I //
TFIIH
TFIIF Закрытый комплекс
TFIIB Ро111
mpp TFIIE
б
Pol II и
дефосфорилирование
TFIIH
TFIIA
ДНК
раскручивается
с образованием
открытого
комплекса
Рис. 26-10. Транскрипция с промоторов РНК-полимеразы II. а — последовательное при-
соединение ТВР (часто вместе с TFIIA), TFIIB, TFIIF, затем Pol II, TFIIE и TFIIH приводит к
формированию закрытого комплекса. Внутри комплекса происходит раскручивание ДНК в
области Inr за счет активности TFIIH и, возможно, TFIIE, что приводит к открытию комплек-
са. С-концевой домен самой крупной субъединицы Pol II фосфорилируется под действи-
ем TFIIH, затем полимераза оставляет промотор и начинает транскрипцию. Элонгация со-
провождается высвобождением многих факторов транскрипции и усиливается факторами
элонгации (см. табл. 26-2). После терминации Pol II высвобождается, дефосфорилируется
и может участвовать в новом синтезе. 6 — ТВР человека (серый цвет) связан с ДНК (синий
и голубой цвета) (PDB ID 1TGH). в — схема элонгации транскрипции, катализируемой кор-
ферментом полимеразы II.
[120] Часть III. 26. Метаболизм РНК
ниже, CTD необходим для выполнения множе-
ства функций Pol II.
Для формирования активного транскрип-
ционного комплекса РНК-полимераза II нуж-
дается в ряде других белков, называемых фак-
торами транскрипции. Основные факторы
транскрипции для каждого промотора Pol II
(обычно обозначаются TFII с дополнительной
идентификацией) достаточно консервативны
у всех эукариот (табл. 26-2). В процессе транс-
крипции под действием фермента Pol II можно
выделить несколько стадий: сборка, инициация,
элонгация, терминация; на каждой стадии уча-
ствуют определенные белки (рис. 26-10). Шаг
за шагом описанный ниже процесс приводит к
активной транскрипции in vitro. В клетке многие
белки могут присутствовать в виде более круп-
ных предварительно организованных комплек-
сов, облегчая сборку комплексов на промоторах.
Факторы, участвующие в трансляции, перечис-
лены на рис. 26-10 и в табл. 26-2.
Сборка РНК-полимеразы и факторов транс-
крипции на промоторе. Образование закрытого
комплекса начинается с взаимодействия ТАТА-
связывающего белка (ТВР) с ТАТА-боксом
(рис. 26-10, б). В свою очередь ТВР связывается
с фактором транскрипции TFIIB, который тоже
связывается с ДНК с обеих сторон от ТВР. Свя-
зывание TFIIA не всегда существенно, но может
стабилизировать комплекс TFIIB-TBP на ДНК,
что может иметь большое значение на неконсен-
сусных промоторах, где связывание ДНК с ТВР
сравнительно слабое. Затем комплекс TFIIB-TBP
соединяется с другим комплексом, состоящим из
TFIIF и Pol II. Фактор TFIIF помогает точной
стыковке Pol II с промотором, как путем взаимо-
действия с TFIIB, так и путем ослабления связы-
вания полимеразы с неспецифическим участками
ДНК. Наконец, присоединяются TFIIE и TFIIH, и
образуется закрытый комплекс. Фактор TFIIH об-
ладает ДНК-хеликазной активностью и начинает
раскручивание ДНК вблизи точки начала транс-
крипции РНК (процесс нуждается в гидролизе
АТР), создавая открытый комплекс. С учетом всех
субъединиц разных факторов (не считая TFIIA),
этот минимальный активный комплекс состоит
из 30 или более полипептидов. Структурные ис-
следования, выполненные Роджером Корнбергом
и его сотрудниками, позволили более детально
изучить строение кор-фермента РНК полимера-
зы II при элонгации (рис. 26-10, в).
Инициация цепи РНК и высвобождение про-
мотора. На стадии инициации TFIIH выполняет
дополнительную функцию. Киназная активность
одной из его субъединиц фосфорилирует после-
довательность CTD в Pol II во многих местах
(рис. 26-10, а). Несколько других протеинкиназ,
включая CDK9 (циклин-зависимая киназа 9), ко-
торая является частью комплекса pTEFb (от англ.
positive transcription elongation factor b — положи-
тельный фактор транскрипции и элонгации Ь), так-
же фосфорилируют CTD, особенно остатки серина.
Это приводит к конформационным изменениям
всего комплекса и инициирует транскрипцию.
Фосфорилирование CTD также имеет большое
значение для следующей стадии элонгации, причем
степень фосфорилирования CTD изменяется по
мере продвижения транскрипции. Эти изменения
влияют на взаимодействия между транскрипцион-
ным комплексом и другими ферментами, так что
при инициации транскрипции в комплексе связа-
ны другие белки, нежели на более поздних стадиях.
Некоторые из этих белков участвуют в процессинге
транскрипта (как описано ниже).
По мере синтеза первых 60-70 нуклеотидов
сначала из комплекса выходит TFIIE, потом TFIIH,
и Pol II начинает стадию элонгации.
Элонгация, терминация и высвобождение. На
стадии элонгации TFIIF остается связанным с
Pol II. На этой стадии активность полимеразы
значительно усиливается белками, называемы-
ми факторами элонгации (табл. 26-2). Факторы
элонгации, некоторые из них связаны с фосфо-
рилированным CTD, препятствуют остановке
транскрипции, а также координируют взаимо-
действия между белковыми комплексами, вовле-
ченными в посттранскрипционный процессинг
молекул мРНК. Как только синтез транскрипта
РНК завершен, транскрипция прекращается.
Полимераза II дефосфорилируется, после чего
она готова инициировать транскрипцию другого
участка (рис. 26-10, а).
Регуляция активности РНК-полимеразы II. Ре-
гуляция транскрипции под действием Pol II до-
статочно сложна. Она включает взаимодействие
широкого круга белков с прединициаторным ком-
26.1 ДНК-зависимый синтез РНК [121]
плексом. Некоторые из этих регуляторных бел-
ков взаимодействуют с факторами транскрипции,
другие — с самой Pol II. Регуляция транскрипции
более подробно рассматривается в гл. 28.
Различные функции TFIIH. У эукариот репа-
рация поврежденной ДНК (см. табл. 25-5) более
эффективна в активно транскрибируемых генах,
чем в других поврежденных участках ДНК, и
матричная цепь репарируется эффективнее не-
матричной цепи. Эти удивительные наблюдения
объясняются тем, что субъединицы TFIIH выпол-
няют и другие функции. Фактор TFIIH не только
принимает участие в формировании закрытого
комплекса в ходе сборки транскрипционного
комплекса (как описано выше), но некоторые из
его субъединиц также выступают важными ком-
понентами независимого эксцизионного репара-
ционного комплекса (см. рис. 25-26).
В Когда транскрипция под действием полиме-
разы II приостанавливается из-за поврежде-
ния ДНК, TFIIH может связаться с поврежденным
участком и собрать вокруг себя целый эксцизион-
ный репарационный комплекс. У человека генети-
ческий дефект некоторых субъединиц TFIIH мо-
жет привести к развитию заболеваний, например к
пигментной ксеролерме (см. доп. 25-1) и синдрому
Коккейна, которые характеризуются прекращени-
ем роста, повышенной светочувствительностью и
неврологическими расстройствами.
Возможно селективное ингибирование
ДНК-зависимой РНК-полимеразы
Элонгация цепей РНК под действием РНК-по-
лимеразы и у бактерий, и у эукариот ингибирует-
ся антибиотиком актиномицином D (рис. 26-11).
Плоская часть этой молекулы встраивается (ин-
Sar
L-Pro L-meVal
Sar
L-Pro L-meVal
D-Val О
"b-Thr
D-Val
^L-Thr
Акридин
a
Рис. 26-11. Ингибиторы транскрипции ДНК актиномицин D и акридин, а — обведенная
рамкой часть молекулы актиномицина D плоская и встраивается между двумя последо-
вательно расположенными парами оснований G^C в дуплексной ДНК. Две циклические
пептидные структуры актиномицина D связываются с малой бороздкой двойной спирали.
Саркозин (Sar) — это N-метилглицин; meVal — метилвалин. Акридин также встраивается в
ДНК. б — комплекс актиномицина D с ДНК (PDB ID 1DSC). Остов ДНК показан синим цветом,
основания — серым, встроившаяся часть актиномицина (обведена рамкой на рис. а) —
оранжевым цветом, оставшаяся часть актиномицина — красным. ДНК изогнута в результа-
те связывания актиномицина.
[122] Часть III. 26. Метаболизм РНК
теркалирует) в двойную спираль ДНК между
расположенными друг за другом парами основа-
ний G=C, вызывая деформацию ДНК. Это пре-
пятствует движению полимеразы вдоль матрицы.
Поскольку актиномицин D ингибирует элонга-
цию РНК в интактных клетках, а также в клеточ-
ных экстрактах, его используют для идентифика-
ции клеточных процессов, связанных с синтезом
РНК. Акридин ингибирует синтез РНК сходным
образом (рис. 26-11).
Рифампицин подавляет синтез бактериальной
РНК, связываясь с р-субъединицей бактериаль-
ных РНК-полимераз, препятствуя высвобождению
промотора при транкрипции (рис. 26-6). Иногда его
используют в качестве антибиотика.
Бледная поганка {Amanita phalloides) обла-
дает очень эффективным механизмом защиты от
животных. Она синтезирует а-аманитин, кото-
рый прерывает образование мРНК в клетках жи-
вотных, блокируя Pol II, а в высоких концентра-
циях и Pol III. Ни Pol I, ни бактериальная РНК-
полимераза нечувствительны к а-аманитину, но
главное, к нему нечувствительна РНК-полиме-
раза II самого гриба A. phalloidesl
Краткое содержание раздела 26.1
ДНК-ЗАВИСИМЫЙ СИНТЕЗ РНК
Транскрипцию катализируют ДНК-зави-
симые РНК-полимеразы, которые использу-
ют рибонуклеозид-5'-трифосфаты для синте-
за молекул РНК, комплементарных матрич-
ной цепи ДНК-дуплекса. Транскрипция осу-
ществляется в несколько этапов: связывание
РНК-полимеразы с промоторным участком
ДНК, инициация синтеза транскрипта, элон-
гация и терминация.
Для распознавания промотора бактериаль-
ная РНК-полимераза нуждается в специ-
альной субъединице. Связывание РНК-по-
лимеразы с промотором и инициация транс-
крипции тесно взаимосвязаны и составляют
первый этап транскрипции. Транскрипция
прекращается на последовательностях ДНК,
называемых терминаторами.
В эукариотических клетках есть три типа
РНК-полимераз. Для связывания РНК-по-
лимеразы II с ее промоторами необходимы
белковые факторы транскрипции. Факто-
ры элонгации участвуют в фазе элонгации.
Длинный С-концевой домен самой крупной
субъединицы Pol II фосфорилирован на ста-
диях инициации и элонгации.
26.2. Процессинг РНК
Многие молекулы РНК бактерий и практически
все молекулы РНК эукариот после синтеза под-
вергаются в той или иной степени процессингу. В
метаболизме РНК интересные превращения про-
исходят именно в ходе постсинтетического про-
цессинга. Удивительно, что некоторые ферменты,
катализирующие эти реакции, состоят из РНК, а
не из белка. Открытие таких каталитических РНК,
называемых рибозимами, произвело революцию в
наших представлениях о функции РНК и о проис-
хождении жизни.
Вновь синтезированная молекула РНК на-
зывается первичным транскриптом. Вероятно,
наиболее интенсивному процессингу подверга-
ются первичные транскрипты мРНК эукариот и
тРНК бактерий и эукариот. Молекулы РНК со
специфическими функциями также подвергают-
ся процессингу.
Первичные транскрипты эукариотических
мРНК обычно содержат один ген, но последова-
тельности, кодирующие полипептид, могут быть
разделены другими фрагментами. Эти некодиру-
ющие участки, которые прерывают кодирующие
последовательности, называют интронами, а ко-
дирующие участки — экзонами (об интронах и эк-
зонах в ДНК см. в гл. 24). В процессе сплайсинга
интроны удаляются из первичного транскрипта,
а экзоны соединяются с образованием непрерыв-
ной последовательности, которая соответствует
функциональному полипептиду. Кроме того, мо-
дифицируются концы эукариотической мРНК.
К 5'-концу присоединяется модифицированный
остаток, называемый 5'-кэпом. З'-Конец отщепля-
ется, а взамен присоединяется последователь-
ность из 80-250 остатков А, образующая поли(А)-
«хвост». Иногда сложные белковые комплексы,
которые принимают участие в каждой из этих
трех реакций процессинга мРНК, действуют не
самостоятельно, а связываются друг с другом и с
фосфорилированной CTD-последовательностью
Pol II; каждый комплекс влияет на функции
26.2 Процессинг РНК [123]
ДНК
। Экзон Интрон
5'-Кэп
Транскрипция и
5 -кэпирование
Pol II
завершение сборки
первичного
„ транскрипта
Первичный Некодирующая
транскрипт концевая последовательность
Рис. 2642. Образование первичного транскрипта и его процессинг в ходе созревания
мРНК в эукариотических клетках. Перед завершением синтеза первичного транскрипта
на 5'-конце появляется кэп (показан красным цветом). Некодирующая последовательность
(интрон), расположенная за последним экзоном, выделена оранжевым цветом. Сплайсинг
может происходить либо до, либо после стадий расщепления и полиаденилирования. Все
изображенные здесь процессы происходят в ядре.
других. Другие белки, участвующие в транспорте
мРНК в цитоплазму, связаны с мРНК и в ядре, и
процессинг транскрипта связан с его транспортом.
В сущности, синтезированная эукариотическая
мРНК находится в сложном многокомпонентном
комплексе, состоящем из десятков белков. Состав
комплекса изменяется по ходу процессинга пер-
вичного транскрипта, транспорта в цитоплазму и
высвобождения мРНК на рибосоме для трансля-
ции. Эти белковые комплексы модулируют актив-
ность и перемещение мРНК. Данные процессы от-
ражены на рис. 26-12 и подробнее описаны ниже.
Первичные транскрипты прокариотиче-
ских и эукариотических тРНК в результате про-
цессинга теряют концевые последовательности
(отщепление), а иногда и интроны (сплайсинг).
Многие основания и сахара в тРНК модифици-
руются; зрелая тРНК содержит много необыч-
ных оснований, которые не были обнаружены в
других нуклеиновых кислотах (см. рис. 26-23).
Многие РНК со специальной функцией также
подвергаются сложному процессингу, часто с
удалением фрагментов с одного или обоих кон-
цов последовательности.
Итак, все молекулы РНК подвергаются пол-
ному регулируемому расщеплению. Скорость
оборота молекул РНК определяет их стационар-
ное содержание в клегке и скорость, с которой
клетки могут остановить экспрессию гена, в про-
дукте которого больше не нуждаются. Например,
в ходе развития многоклеточных организмов
определенные белки экспрессируются только на
одной стадии развития, и молекулы мРНК, коди-
рующие эти белки, образуются и разрушаются в
строго определенное время.
К 5'-концу эукариотической мРНК
присоединяется кэп
У эукариот в большинстве молекул мРНК
имеется 5'-кэп — остаток 7-метилгуанозина, свя-
занный с 5'-концевым остатком мРНК через не-
обычную 5',5'-трифосфатную связь (рис. 26-13).
Функция 5'-кэпа — защита мРНК от рибону-
клеаз. Кроме того, кэп соединяется со специфи-
ческим кэп-связывающим комплексом белков и
участвует во взаимодействии мРНК с рибосомой
для инициации трансляции (гл. 27).
[124] Часть III. 26. Метаболизм РНК
5'-конец РНК с
трифосфатной группой
pppNp ——————
})осфо1 идрс
ppNp
РР,
GpppNp
Gppp GTP
O-Mei ил i рансфера.
. 1НИН-
ii гил трансфер ua
adoMet
adoHcy
m7GpppNp
adoMet
adoHcy
m7GpppmNp
5'-Конец РНК с кэпом
Рис. 26-13. 5'-Кэп на мРНК. а — почти во всех молекулах эукариотической мРНК к 5'-кон-
цу необычной 5',5'-трифосфатной связью присоединен остаток 7-метилгуанозина (m7G).
Часто в положении 2' первого и второго нуклеотидов также обнаруживаются метильные
группы (показаны розовым цветом). Молекулы РНК в клетках дрожжей не имеют 2'-ме-
тильных групп. Во втором нуклеотиде 2'-метильная группа обычно встречается только
в РНК у позвоночных, б — формирование 5'-кэпа происходит за 4-5 отдельных стадий
(adoHcy — S-аденозилгомоцистеин). в — синтез кэпа осуществляют ферменты, связан-
ные с С-концевой последовательностью (CTD) полимеразы II. Кэп связан с CTD через кэп-
связывающий комплекс (СВС).
5'-Кэп образуется путем конденсации моле-
кулы GTP с трифосфатом на 5'-конце транскрип-
та. Затем гуанин метилируется по положению
N-7, кроме того, метилированию часто подвер-
гаются 2'-гидроксильные группы двух соседних
с кэпом нуклеотидов (рис. 26-13, а). Метильные
группы предоставляет S-аденозилметионин. Эти
реакции происходят в самом начале транскрип-
ции после присоединения первых 20-30 нукле-
отидов транскрипта. Пока кэп не синтезирован,
все три осуществляющих кэпирование фермента
и через них 5'-конец самого транскрипта ассо-
циированы с С-концевым доменом РНК-поли-
меразы II. Затем копированный 5'-конец осво-
бождается от ферментов и соединяется с кэп-
связывающим комплексом (рис. 26-13, в).
26.2 Процессинг РНК [125]
Из ДНК в РНК транскрибируются
и интроны, и экзоны
У бактерий полипептидная цепь обычно колли-
неарна кодирующей ее последовательности ДНК,
которая составляет непрерывный отрезок матри-
цы ДНК, необходимый для точного воспроизведе-
ния полипептида. Однако идея о том, что все гены
непрерывны, была опровергнута еще в 1977 г., ког-
да Филипп Шарп и Ричард Робертс независимо
друг от друга установили, что многие гены эукари-
от имеют прерывистое строение и содержат неко-
дирующие последовательности (интроны).
Подавляющее большинство генов позвоноч-
ных содержат интроны; среди немногих исключе-
ний — гены гистонов. В других эукариотах ситуа-
ция может быть различной. Во многих генах дрож-
жей Saccharomyces cerevisiae интронов нет, но в дру-
гих видах дрожжей интроны встречаются довольно
часто. Интроны также обнаружены в нескольких
генах бактерий и архей. Интроны в ДНК транскри-
бируются РНК-полимеразами вместе с остальны-
ми частями генов. Затем интроны вырезаются из
первичного транскрипта в процессе сплайсинга, а
экзоны соединяются с образованием зрелой функ-
циональной РНК. В молекулах мРНК эукариот
длина большинства экзонов не превышает 1000 ну-
клеотидов. размер многих составляет всего лишь
от 100 до 200 нуклеотидов; эти экзоны кодируют
фрагменты из 30 60 аминокислот в составе более
длинного полипептида. Интроны имеют размер от
50 до 20 000 нуклеотидов. Гены высших эукариот,
включая человека, обычно содержат гораздо боль-
ше ДНК в составе интронов, чем в составе экзонов.
Многие гены содержат интроны, в некоторых генах
они исчисляются десятками.
РНК катализирует сплайсинг интронов
Существует четыре группы интронов. Интроны,
относящиеся к группам 1 и II. различаются де-
талями механизма сплайсинга, но имеют одно
общее удивительное свойство: они осуществляют
аутосплайсинг, т. е. сплайсинг без участия фер-
ментов белковой природы. Интроны группы I
присутствуют в некоторых ядерных и митохон-
дриальных генах и в генах хлоропластов, кото-
рые кодируют рРНК, мРНК и тРНК. Интроны
группы II обычно обнаруживаются в первичных
транскриптах мРНК митохондрий и хлоропла-
стов у грибов, водорослей и растений. Интро-
ны групп I и II также обнаружены у некоторых
бактерий. Таким интронам для сплайсинга не
требую гея высокоэнергетические кофакторы
(такие как АТР). Механизм сплайсинга в груп-
пах I и II включает две стадии трансэтерифи-
кации (рис. 26-14), в процессе которых 2'- или
З'-гидроксильные группы рибозы осуществляют
Рис. 26-14. Реакция трансэтерификации. Первая стадия сплайсинга интронов группы I.
В данном примере З'-ОН-группа молекулы гуанозина выступает в роли нуклеофила и атаку-
ет фосфодиэфирную связь между остатками U и А на стыке экзон-интрон в молекуле мРНК
(см. рис. 26-15).
[126] Часть III. 26. Метаболизм РНК
нуклеофильную атаку на атом фосфора, и вместо
старой фосфодиэфирной связи образуется новая,
поддерживая энергетический баланс. Эти реак-
ции очень напоминают реакции разрыва и вос-
соединения ДНК, катализируемые топоизоме-
разами (см. рис. 24-21) и сайт-специфическими
рекомбиназами (см. рис. 25-40).
Реакция сплайсинга интронов группы I про-
текает с участием гуанидинового нуклеозида или
нуклеотидного кофактора, но кофактор исполь-
зуется не в качестве источника энергии: З'-гид-
роксильная группа гуанозина выступает в роли
нуклеофила на первом этапе сплайсинга. 3'-Гид-
роксильная группа гуанозина образует обычную
3',5'-фосфодиэфирную связь с 5'-концом интрона
(рис. 26-15). Затем З'-гидроксильная группа экзо-
на, которая на этой стадии вытесняется, выступает
в роли нуклеофила в похожей реакции на З'-конце
интрона. В результате происходит точное выреза-
ние интрона и соединение концов экзонов.
Сплайсинг интронов группы II происходит
по тому же принципу, только в качестве нуклео-
фила на первой стадии выступает 2'-гидроксиль-
ная группа остатка А внутри интрона (рис. 26-16).
В качестве интермедиата образуется разветвлен-
ная структура в форме лассо (лариат).
Аутосплайсинг интронов впервые был обна-
ружен Томасом Чеком с сотрудниками в 1982 г.
при изучении механизма сплайсинга интрона
группы I в рРНК ресничного простейшего Tet-
rahymena thermophila. Исследователи транскри-
Томас Чек
Первичный
транскрипт
Рис. 26-15. Механизм сплайсинга интронов группы 1. Нуклео-
филом на первой стадии может быть гуанозин, GMP, GDP или
GTP. Вырезанный интрон полностью распадается.
5-
5'-Экзон
иРд
тт 3'-Экзон
GpU------------------з
pG lOH
З'-ОН гуанозина
действует как нуклеофил,
атакуя фосфат по 5 '-сайту
сплайсинга.
pGp.\
Интермедиат
U ОН GpU-----------------3'
В 5'-экзоне З'-ОН
становится нуклеофилом,
завершающим реакцию.
Сплайсированная РНК
5'------------UpU--------------3'
26.2 Процессинг РНК [127]
Интрон
2'-ОН в специфическом
аденозине. В интроне эта группа
действует как нуклеофил,
атакуя 5'-сайт сплайсинга,
чтобы образовалась
структура в форме лассо.
3'
Интермедиат
5’ .........
У аденозина в лассо-
подобной структуре три
фосфодиэфирные связи
В 5'-экзоне З'-ОН действует
как нуклеофил, завершая
реакцию.
Сплайсированная РНК
UpU ------------------------ 3 ОН(З')
Рис. 26-16. Механизм сплайсинга интронов группы II. Химические принципы механизма в
данном случае такие же, как и при сплайсинге интронов группы I. Отличие заключается в
нуклеофиле на первой стадии и в образовании лассоподобного интермедиата, одна из вет-
вей которого представляет собой 2',5'-фосфодиэфирную связь.
бировали содержащую интрон ДНК Tetrahymena
in vitro, используя очищенную бактериальную
РНК-нолимеразу. Сплайсинг РНК происходил
с высокой точностью без участия ферментов Tet-
rahymena. Открытие молекул РНК с каталитиче-
скими функциями стало важным этапом в фор-
мировании нашего понимания устройства биоло-
гических систем.
Большинство интронов не подвергаются ау-
тосплайсингу и не обозначаются групповыми но-
мерами. Третья и самая обширная группа интро-
нов обнаружена в первичных транскриптах ядер-
ной мРНК. Они называются сплайсосомными
интронами, поскольку их удаление происходит
внутри крупного белкового комплекса сплайсо-
сомы и катализируется ею. Внутри сплайсосомы
интроны вырезаются по тому же механизму, что и
интроны группы II (через образование лариата).
Сплайсосомы состоят из специализированных
РНК-белковых комплексов — малых ядерных ри-
бонуклеопротеинов (мяРНП). Каждый мяРНП
содержит одну молекулу эукариотической РНК
длиной 100-200 нуклеотидов, называемых малы-
ми ядерными РНК (мяРНК). В реакциях сплай-
[128] Часть III. 26. Метаболизм РНК
синга участвуют пять мяРНК (Ul, U2, U4, U5 и
U6), которые обычно в большом количестве об-
наруживаются в ядрах эукариот. Белки и РНК в
мяРНП высококонсервативны у всех эукариот
(от дрожжей до человека), в Сплайсинг мРНК
Сплайсосомные интроны обычно имеют GU-
последовательность на 5'-конце и AG-последо-
вательность на З'-конце. Эти последовательно-
сти указывают место сплайсинга. Молекула U1-
мяРНК содержит последовательность, компле-
ментарную последовательностям вблизи 5'-кон-
цевого сайта сплайсинга интронов ядерной мРНК
(рис. 26-17, а), и Ul-мяРНП связывается с этой
областью в первичном транскрипте. Присоеди-
нение U2, U4, U5 и U6 приводит к формирова-
нию сплайсосомы (рис. 26-17, б). За счет мяРНП
сплайсосома получает пять молекул РНК и при-
мерно 50 белков, надмолекулярная структура
сплайсосомы почти такая же сложная, как у рибо-
сомы (см. гл. 27). Еще примерно 50 белков связы-
ваются со сплайсосомой на разных стадиях сплай-
синга, причем многие из этих белков участвуют
в нескольких процессах, таких как сплайсинг,
перенос мРНК в цитоплазму, трансляция и рас-
щепление мРНК. Для сборки сплайсосомы требу-
ется АТР, но реакции расщепления-лигировапия
РНК, по-видимому, в АТР не нуждаются. Неко-
торые интроны в мРНК удаляются с помощью
менее распространенной сплайсосомы, в которой
U1 и U2 заменены на U11 и U12. Сплайсосомы,
содержащие U1 и U2, удаляют интроны с (5')GU-
и АС(3')-концевыми последовательностями (см.
рис. 26-16), а сплайсосомы, содержащие U11 и
U12, вырезают редкие интроны группы с (5')AU- и
АС(3')-концевыми последовательностями. Сплай-
сосомы, участвующие в сплайсинге ядерной РНК,
могли эволюционировать из более древних интро-
нов II группы, а мяРНП заменили каталитические
домены их самосплайсирующихся предков.
Некоторые элементы аппарата сплайсинга,
вероятно, связаны с CTD РНК-полимеразы II,
и это очень интересный механизм реакции
(рис. 26-17, в). Когда синтезируется первый
участок сплайсинга, он соединяется с CTD-
«хвостом» сплайсосомы. Затем этим комплек-
сом захватывается второй синтезированный
участок сплайсинга, что облегчает сближение
концов интрона и его вырезание. После сплай-
синга интрон остается в ядре и в конце концов
разрушается.
Интроны IV группы, найденные в некото-
рых тРНК, отличаются от интронов I и II групп
тем, что для их сплайсинга требуются АТР и эн-
донуклеаза. Сплайсирующая эндонуклеаза рас-
щепляет фосфодиэфирные связи на двух концах
интрона, и два экзона соединяются по механиз-
му, напоминающему реакцию ДНК-лигазы (см.
рис. 25-17).
Сплайсосомные интроны встречаются, по-
видимому, только у эукариот, а интроны других
групп повсеместно. Гены с интронами I и II групп
в настоящее время обнаружены и у бактерий, и
у бактериальных вирусов. Бактериофаг Т4, на-
пример, содержит несколько генов с интронами
I группы. Возможно, интроны чаще встречаются
у архей, чем у бактерий.
Рис. 26-17. Механизм сплайсинга в первичных транскриптах мРНК. а — спаривание участ-
ков РНК при формировании сплайсосомных комплексов. Вблизи 5'-конца Ul-мяРНК есть
последовательность, комплементарная сайту сплайсинга на 5'-конце интрона. Спаривание
оснований U1 с этим участком первичного транскрипта помогает выявить 5'-сайт сплай-
синга при сборке сплайсосомы (гр — псевдоуридин; см. рис. 26-24). Затем U2 связывается
с участком интрона, содержащим остаток А (розовый), который осуществляет нуклеофиль-
ную атаку в реакции сплайсинга. При связывании П2-мяРНК создается петля, которая пере-
мещается и помогает активировать аденилат, 2'-0Н-группа которого может формировать
лассоподобную структуру через 2',5'-фосфодиэфирную связь, б — сборка сплайсосомы.
Сначала соединяются U1- и П2-мяРНП, затем с ними связываются остальные мяРНП (ком-
плекс U4/U6 и U5) с образованием неактивной сплайсосомы. После внутренних перестроек
этот комплекс преобразуется в активную сплайсосому, из которой исключаются U1 и U4,
a U6 соединяется одновременно с 5'-сайтом сплайсинга и с U2. Затем следуют каталити-
ческие стадии, аналогичные каталитическим стадиям сплайсинга интронов II группы (см.
рис. 26-15). в — координация сплайсинга с транскрипцией обеспечивает механизм сбли-
жения двух сайтов сплайсинга. Подробности см. в тексте.
26.2 Процессинг РНК [129]
< U1 мяРНП
АТР
ADP + Р^
U2 мяРНП
Неактивная
и4-и . сплайсосома
1GU А
WR U5^
U1 <yU2
АТР
ADP + Pi
Ul, U4
б
[130] Часть III. 26. Метаболизм РНК
На З'-конце молекулы мРНК
имеются характерные структуры
У эукариот в большинстве молекул мРНК иа
З'-конце располагается последовательность из
80-250 остатков аденина, образующих так назы-
ваемый поли(А)-«хвост» (последовательность
полиаденилирования). Этот «хвост» служит для
связывания одного или нескольких специфиче-
ских белков. Поли(А)-«хвост» и ассоциированные
с ним белки, вероятно, помогают защитить мРНК
от ферментативного расщепления. Многие мо-
лекулы мРНК прокариот также имеют поли(А)-
«хвосты», но эти «хвосты» скорее стимулируют
распад мРНК, чем защищают ее от расщепления.
Присоединение поли(А)-«хвоста» — много-
стадийный процесс. Транскрипт синтезирует-
ся дальше границы участка, к которому должен
быть добавлен «хвост», а потом расщепляется
Матрица
ДНК
5'- \
Кэп '
РНК
I AAUAAA-]
Ферментный
комплекс
5 —..— 1 'чГААЦААА |
5' I AAUAAA | -ОН(З')
5'------------ГААЦААА | — ААА(А)П—ОН(3 )
по этому положению с помощью эндонуклеаз-
ной активности крупного ферментного ком-
плекса, связанного с CTD РНК-полимеразы П
(рис. 26-18). Участок мРНК, по которому проис-
ходит расщепление, содержит два характерных
элемента: консервативную последовательность
(5')AAUAAA(3'), расположенную на 10 -30 ну-
клеотидов выше точки расщепления (ближе к
5'-концу), и менее охарактеризованную после-
довательность, содержащую много остатков G и
U, на расстоянии 20 40 нуклеотидов ниже точки
расщепления. При расщеплении образуется сво-
бодная З'-гидроксильная группа на конце мРНК.
к которому полиаденилатполимераза немедлен-
но присоединяет остатки А по реакции
РНК + п АТР РНК-(АМР)„ +п PPj
где п = 80 4- 250. Этот фермент не нуждается в ма-
трице, но в качестве праймера использует расще-
пленную мРНК.
Весь комплекс реакций процессинга типичной
эукариотической мРНК представлен на рис. 26-19.
В некоторых случаях кодирующая область мРНК
тоже подвергается «редактированию» (подробнее
см. разд. 27.1). Это могут быть реакции внесения
или удаления оснований в кодирующих областях
первичных транскриптов или изменения последо-
вательности (например, ферментативное дезами-
нирование остатка С. приводящее к образованию
остатка U). Один из удивительных примеров
процессинг у паразитических простейших три-
паносом: длинные последовательности их мРНК
синтезируются вообще без уридилата, а остатки U
встраиваются позднее в процессе редактирования.
Рис. 26-18. Добавление поли(А)-«хвоста» к первично-
му транскрипту РНК у эукариот. Pol II синтезирует РНК,
продвигаясь дальше сигнальных последовательностей,
указывающих участок расщепления, включая консерва-
тивную последовательность (5')AAUAAA, расположен-
ную выше точки расщепления. £ Сигнальная последо-
вательность связывается с ферментным комплексом, в
состав которого входит эндонуклеаза, полиаденилатпо-
лимераза и несколько других мультисубъединичных бел-
ков, участвующих в распознавании последовательности,
облегчении расщепления и регуляции длины поли(А)-
«хвоста». 2 РНК ниже (в направлении к З'-концу) после-
довательности AAUAAA расщепляется эндонуклеазой на
10-30 нуклеотидов, з Полиаденилатполимераза синте-
зирует поли(А)-«хвост» длиной 80-250 нуклеотидов, на-
чиная с места расщепления транскрипта.
26.2 Процессинг РНК [131]
транскрипция и
5'-копирование
Первичный
транскрипт
Избыточная РНК
Кэп
Зрелая
мРНК
L1 23 4 5 6 7
ниш и ААА(А),
|<---- 1872 ------>|
нуклеотида
Рис. 26-19. Схема процессинга эукариотической мРНК. Ген яичного альбумина содержит
интроны от А до G и экзоны от 1 до 7, а также последовательность L (L кодирует сигналь-
ный пептид, направляющий белок за пределы клетки; см. рис. 27-34). В ходе процессинга
удаляется примерно три четверти РНК. До остановки транскрипции Pol II успевает син-
тезировать первичный транскрипт значительно дальше участка расщепления и сайта по-
лиаденилирования (избыточная РНК). Сигналы терминации для Pol II пока не обнаружены.
Альтернативный процессинг РНК
приводит к образованию
нескольких продуктов одного гена
Один из парадоксов современной геномики за-
ключается в том, что сложность организмов ни-
как не коррелирует с количеством генов, кодиру-
ющих белки, или даже с количеством геномной
ДНК (с. 160 вт. 1). Однако при традиционном
подходе, когда основное внимание уделяется
генам белков, не учитывается сложность транс-
криптома организма. По мере расширения наших
знаний о функциях РНК в строении генома вы-
являются новые сложные элементы.
Из некоторых транскриптов эукариотичес-
ких мРНК образуется только одна зрелая мРНК и
один соответствующий полипептид, но процессинг
других может происходить несколькими способа-
ми и приводить к образованию различных мРНК
и различных полипептидов. Первичный транск-
рипт содержит молекулярные сигналы для всех
альтернативных путей процессинга, а предпочти-
тельный для данной клетки путь определяется фак-
торами процессинга — РНК-связывающими белка-
ми, которые запускают один конкретный путь.
Сложные транскрипты могут иметь либо бо-
лее одного сайта расщепления и полиаденилиро-
вания, либо альтернативные сайты сплайсинга,
либо и то, и другое. При наличии двух или не-
скольких сайтов расщепления и полиаденилиро-
вания, чем ближе к 5'-концу выбирается сайт, тем
большая часть последовательности первичного
транскрипта удаляется (рис. 26-20, а). Такой
механизм, называемый выбором сайта поли(А),
создает разнообразие вариабельных доменов тя-
желых цепей иммуноглобулинов (см. рис. 25-46).
Путем альтернативного сплайсинга на разных
стадиях развития плодовой мушки (рис. 26-20, б)
из общего первичного транскрипта образуются
три разные формы тяжелой цепи миозина. Аль-
тернативные варианты процессинга РНК могут
использоваться одновременно для синтеза двух
разных гормонов: регулирующего кальциевый
обмен гормона кальцитонина в щитовидной же-
лезе крысы и пептида, связанного с геном каль-
цитонина (CGRP) в мозге крысы (рис. 26-21).
Существует множество других вариантов альтер-
нативного сплайсинга (рис. 26-22). Многие гены
в геноме млекопитающих (возможно, большин-
ство) подвергаются альтернативному сплайсингу,
[1321 Часть III. 26. Метаболизм РНК
Сайты поли(А)
разрезание разрезание
и полиаденили- и полиаденили-
рование по А] рование по А2
-ААА(А)П
5' сайт 3' сайт
11111111«»?о°оУааа(а)п 1ПШШШ-ААА(А),
Зрелая мРНК
Зрелая мРНК
ААА(А)„
Рис. 26-2С Два механизма альтернативного процессинга сложных транскриптов у эукариот.
а — альтернативные варианты расщепления и полиаденилирования. Показаны два сайта по-
лиаденилирования — А1 и А2. 6 — альтернативные способы сплайсинга. Показаны два раз-
ных З'-концевых сайта расщепления. Оба механизма позволяют получить разные зрелые мРНК
с одного и того же транскрипта.
Первичный
транскрипт
Сайт поли(А) Сайт поли(А)
разрезание и
.полиаденилирование
/разрезание и
полиаденилирование
Щитовидная
железа
1 2 3 4 1
Ш в]ААА(А)п
12 3 4
Зрелая мРНК 1 ААА(А)П
I действие
I протеазы
V
Кальцитонин
Г“
1 2 3 5 6
Зрелая мРНК ААА(А)П
I действие
I протеазы
CGRP
Рис. 26-21 Альтернативный процессинг транскрипта гена кальцитонина у крыс. Первич-
ный транскрипт содержит два сайта полиаденилирования; один чаще используется в клет-
ках мозга, другой — в щитовидной железе. В мозге при сплайсинге элиминируется экзон
кальцитонина (экзон 4); в щитовидной железе этот экзон остается. Образующиеся пептиды
процессируются далее с образованием конечных гормонов: связанного с геном кальцито-
нина пептида (CGRP) в мозге крысы и кальцитонина в щитовидной железе.
26.2 Процессинг РНК [133]
Альтернативные экзоны
Альтернативные 5 '-сайты
сплайсинга
Альтернативные 3 '-сайты
сплайсинга
Отсутствие удаления интрона
Взаимоисключающие экзоны
Альтернативный промотор
и первый экзон
Альтернативный сайт полиаде-
нилирования и концевой экзон
Рис. 26-22. Общая схема альтернативного сплайсинга.
Экзоны изображены в виде светло- и темно-зеленых пря-
моугольников, а интроны и нетранслируемые области - в
виде желтых линий. Звездочками отмечены позиции, по
которым должен присоединяться полиаденозин. Черными
линиями в левой части схемы обозначены возможные ва-
рианты соединения экзонов, приводящие к соответствую-
щим мРНК (в правой части схемы). Варианты соединения,
показанные выше и ниже транскрипта, образуют верхний
или нижний продукт соответственно. Красными и оранже-
выми прямоугольниками обозначены 5'-кэпы и З'-нетранс-
лируемые области соответственно.
что значительно увеличивает количество коди-
руемых генами белков. Этот процесс у низших
эукариот играет гораздо менее заметную роль, к
примеру у дрожжей лишь несколько генов под-
вергаются альтернативному сплайсингу.
Молекулы рРНК и тРНК
также подвергаются процессингу
Посттранскрипционному процессингу подверга-
ется не только мРНК. Рибосомные РНК в клет-
ках прокариот, архей и эукариот образуются из
более длинных предшественников, называемых
4-Тиоуридин (S4U)
1-Метилгуанозин (mTG)
| Рибоза |
ZCH3
nh-ch2-ch=c
СНз
I 1>
ЧЬГ N
| Рибоза |
Ы6-Изопентиладенозин (i6A)
Риботимидин (Т)
Дигидроуридин (щ)
Дигидроуридин (D)
Рис. 26-23. Некоторые модифицированные основания в рРНК и тРНК, возникающие
в ходе посттранскрипционных реакций. В скобках указаны сокращенные названия
оснований. Обратите внимание на необычное место присоединения рибозы в псевдо-
уридине. Здесь представлено лишь несколько примеров из 96 известных модификаций
нуклеотидов, обнаруженных в различных РНК (на данный момент известен 81 тип мо-
дификаций в тРНК и 30 типов в рРНК). Полный список модифицированных оснований
можно найти в базе данных по модификациям РНК (http://mods.rna.albany.edu/mods/).
[134] Часть III. 26. Метаболизм РНК
Рис. 26-24. Процессинг транскриптов пре-РНК у бактерий, i Перед расщеплением пред-
шественник 30S РНК метилируется по определенным основаниям (красные черточки), а
некоторые остатки уридина превращаются в псевдоуридин (синие черточки) или дигид-
роуридин (черные черточки). Метилированию иногда подвергаются основания или же
2'-гидроксильные группы. @ При расщеплении высвобождаются предшественники рРНК и
тРНК. Расщепление в точках 1,2 и 3 осуществляют ферменты РНКаза III, РНКаза Р и РНКаза Е
соответственно. Как объясняется далее, РНКаза Р представляет собой рибозим. 3? Конеч-
ные продукты 16S, 23S и 5S рРНК образуются в результате действия разных специфических
нуклеаз. Семь копий гена пре-рРНК в хромосоме Е. coh различаются числом, расположе-
нием и типом входящих в первичный транскрипт тРНК. Некоторые копии гена несут до-
полнительные участки гена тРНК между сегментами 16S и 23S рРНК, а также на З'-конце
первичного транскрипта.
Зрелые РНК
5S рРНК
5S
прерибосомными РНК или пре-рРНК. Транс-
портные РНК тоже происходят из более длинных
предшественников. Эти молекулы РНК также
могут содержать различные модифицированные
нуклеозиды; некоторые примеры представлены
на рис. 26-23.
Рибосомные РНК. У бактерий 16S, 23S и 5S
рРНК (а также некоторые тРНК, хотя большин-
ство тРНК кодируются в другом месте) образуют-
ся из единого предшественника — 30S РНК дли-
ной около 6500 нуклеотидов. При процессинге от
30S РНК удаляются концевые фрагменты, а так-
же участки между последовательностями рРНК
(рис. 26-24). В 16S и 23S рРНК содержатся моди-
фицированные нуклеозиды. У Е. coli обнаружено
11 модификаций в последовательности 16S рРНК:
один псевдоуридин и 10 нуклеозидов, метилиро-
ванных по основанию и/или но 2'-гидроксильной
группе. В 23S рРНК содержится 10 псевдоуриди-
нов, один дигидроуридип и 12 метилированных
нуклеозидов. У бактерий каждую модификацию
обычно катализирует особый фермент. В реакции
метилирования в качестве кофактора использует-
ся S-аденозинметионин. Для образования псевдо-
уридина кофакторов не требуется.
В геноме Е. coli закодировано семь молекул
пре-рРНК. Все эти гены имеют похожие рРНК-
кодирующие области, но участки, расположен-
ные между ними, различаются. На участке между
генами 16S и 23S рРНК обычно закодирована
одна или две молекулы тРНК. причем из раз-
ных пре-рРНК транскриптов получаются разные
тРНК. Последовательности, кодирующие моле-
26.2 Процессинг РНК [135]
кулы тРНК, также были обнаружены на З'-конце
5S рРНК в некоторых первичных транскриптах.
У эукариот ситуация сложнее. РНК-поли-
мераза I синтезирует транскрипт 45S пре-рРНК,
который в ядрышке расщепляется на 18S, 28S и
5,8S рРНК, характерные для эукариотических
рибосом (рис. 26-25). Как и у бактерий, процес-
синг включает в себя реакции расщепления под
действием эндо- или экзонуклеаз, а также реак-
ции модификации нуклеозидов. В некоторых
пре-рРНК есть интроны, которые нужно выре-
зать. Процесс в целом инициируется в ядрыш-
ке, в больших комплексах, собирающихся на
предшественнике рРНК по мере его синтеза под
действием Pol I. Существует тесная связь меж-
ду транскрипцией рРНК, созреванием рРНК и
сборкой рибосом в ядрышке. В каждом комплек-
се содержатся рибонуклеазы, расщепляющие
предшественников рРНК, ферменты, модифи-
цирующие определенные основания, большое
количество малой ядрышковой РНК (мякРНК),
направляющей модификацию нуклеозидов и
некоторые реакции расщепления, а также рибо-
сомные белки У дрожжей в процессе участву-
ют пре-рРНК, более 170 нерибосомных белков,
мякРНК для модификации каждого нуклеозида
(всего -70, поскольку некоторые опосредуют два
типа модификаций), а также 78 рибосомных бел-
ков. У человека обнаружено еще больше моди-
фицированных нуклеозидов — около 200, и, сле-
довательно, больше соответствующих мякРНК.
В ходе сборки рибосом состав комплексов может
изменяться, и многие промежуточные комплек-
сы не уступают по сложности самой рибосоме и
мяРНК. У большинства эукариот 5S рРНК обра-
зуется другой полимеразой (Pol III) в виде неза-
висимого транскрипта.
При модификации нуклеозидов в эукари-
отической рРНК чаще всего происходят пре-
вращения уридина в псевдоуридин и adoMet-
зависимое метилирование (как правило, по 2'-ги-
дроксильной группе). Эти реакции опосредованы
комплексами мякРНП — комплексами белков
с мякРНК, состоящими из мякРНК и четырех
или пяти белков, в том числе и осуществляю-
щего модификацию фермента. Существует два
класса таких комплексов, различающихся клю-
чевыми консервативными элементами после-
довательности (боксами). Комплексы с боксом
Н/АСА участвуют в образовании псевдоуридина,
Рис. 26-25. Процессинг транскриптов пре-РНК у позво-
ночных. В ходе транскрипции первичный транскрипт
45S включается в ядрышковый прерибосомный ком-
плекс 90S, в котором происходит сопряжение процес-
синга рРНК и сборки рибосом. & Предшественник 45S
метилируется более чем по 100 нуклеотидам из 14 000
(по основаниям или по 2'-0Н-группам), несколько остат-
ков уридина превращаются в псевдоуридин, а также
происходят некоторые другие модификации. Серия
ферментативных расщеплений 45S приводит к образо-
ванию 18S, 5,8S и 28S рРНК, и постепенно при присо-
единении рибосомных белков начинают формироваться
субъединицы рибосом. В реакциях расщепления и всех
реакциях модификации участвует малая ядрышковая
РНК (мякРНК), входящая в состав белковых комплексов
(мякРНП) в ядрышке, напоминающих сплайсосомы. Мо-
лекулы 5S рРНК образуются независимо.
[136] Часть III. 26. Метаболизм РНК
а комплексы с боксом С/ D — в метилировании по
2'-ОН-группе. В отличие от бактерий у эукариот
один и тот же фермент может участвовать в моди-
фикациях различных участков под руководством
соответствующих мякРНК.
Эти мякРНК состоят из 60-300 нуклеоти-
дов. Многие закодированы в интронах других
генов и транскрибируются совместно с этими ге-
нами. Каждая мякРНК содержит последователь-
ность из 10-21 нуклеотидов, которая полностью
комплементарна определенному участку рРНК.
Консервативные последовательности в осталь-
ной части мякРНК образуют структуры, кото-
рые связываются с соответствующими белками в
мякРНП (рис. 26-26).
Транспортные РНК. В большинстве клеток син-
тезируется от 40 до 50 различных тРНК, причем
эукариотические клетки содержат множество ко-
пий многих генов тРНК. Молекулы транспорт-
ной РНК образуются из более длинных пред-
шественников путем ферментативного удаления
нуклеотидов с 5'- и З'-концов (рис. 26-27). У эу-
кариот в транскриптах нескольких тРНК присут-
ствуют интроны, которые должны быть удалены.
Если в одном первичном транскрипте содержатся
две или несколько разных тРНК, они разделяют-
ся путем ферментативного расщепления. Эндо-
нуклеаза РНКаза Р, обнаруженная у всех орга-
низмов, удаляет РНК с 5'-конца молекул тРНК.
Этот фермент содержит и белок, и РНК РНК-
компонент необходим для проявления фермен-
тативной активности, и в бактериальных клетках
фермент может осуществлять процессинг даже
без участия белкового компонента. Таким об-
разом, РНКаза Р — еще один пример каталити-
ческих РНК, действие которых более подробно
рассматривается ниже. З'-Конец молекул тРНК
модифицируется одной или несколькими нукле-
азами, в том числе экзонуклеазой РНКазой D.
....ОН(3)
Рис. 26-26. Роль мякРНК в модификации рРНК. а — для осуществления реакций метилирова-
ния РНК спаривается с мякРНК с боксом С/D. Участки метилирования в рРНК-мишени (темно-
зеленые) расположены в области спаривания с C/D-боксом мякРНК. Высококонсервативные
последовательности боксов С и D (а также С' и D') являются участками связывания белков,
составляющих наиболее крупный комплекс мякРНП. б — РНК спаривается с мякРНК с боксом
Н/АСА для превращения уридина в псевдоуридин. Участки превращения уридина в псевдо-
уридин в рРНК-мишени (зеленые) также находятся в областях, спаривающихся с мякРНК, а
консервативные последовательности Н/АСА-боксов являются участками связывания белков.
26.2 Процессинг РНК [137]
5
Первичный
транскрипт
3'
OH
Интермедиат
3
ОН
Зрелая тРНКТуг
pGUUAUCAGUUAAUUG Л .
и Разрез РНКазой D
A^
G
5
с
с
Разрез РНКазой Р с
G
G
UUGa ди
G aACCG а
G AAGGCr
V ии с
G
G
G
С
С
u G
С C CGC
G
А
С
и
G
U
GGGC G
/и
GAGA
и
и
с
и
Uu
G Г
С модификация
оснований,
разрезание с 5'-конца,
разрезание с З'-конца,
присоединение ССА
D D G
mG
G
D
рС
и
с
и
с
G
G
и
AC CGmG
G
G
G
G
С
С
С CCGC
G
с
do
aaggc
I mG
C
G
с
и
G
Ф
А
U
U
U A
и
с
А
С
с
и
и
GGGC G
mC
. D
GaGA
U
U
C
Ф
G
и
с
U A
с
с
5 рС
и
с
и
с
G
С л n G
mA D D G . U
G mG AACCGmG
* ° сплайсинг Gd DAAGGCmG
U C
A
A
G
A
C
3'
OH
I
A
c
c
A
G
A
G
G
G
C
С у C
С c CGC
GGGC G
mC Ф
. D
gaga
и
и
с
ф
А
Рис. 26-27 Процессинг тРНК у бактерий и эукариот. Проиллюстрируем важные этапы про-
цессинга на примере дрожжевой тРНКтуг (тРНК, специфически связывающая тирозин; см.
гл. 27). Нуклеотидная последовательность, показанная желтым цветом, удаляется из пер-
вичного транскрипта. Сначала происходит процессинг концов: в первую очередь 5'-конца,
затем З'-конца. Потом к З'-концу присоединяется последовательность ССА; эта стадия необ-
ходима в процессинге эукариотической тРНК и тех бактериальных тРНК, которые не имеют
этой последовательности в первичном транскрипте. Одновременно с процессингом кон-
цов происходит модификация специфических оснований в оставшейся части транскрип-
та (рис. 26-23). Для показанной здесь эукариотической тРНК заключительная стадия —
сплайсинг интрона из 14 нуклеотидов. Интроны встречаются в некоторых эукариотических
тРНК, но не обнаружены в бактериальной тРНК.
Предшественники тРНК могут подвергать-
ся дальнейшему посттранскрипционному про-
цессингу. З'-Концевой тринуклеотид ССА (3'), к
которому в ходе синтеза белка присоединяется
аминокислота (гл. 27), отсутствует у некоторых
бактериальных и у всех эукариотических предше-
ственников тРНК и поэтому добавляется в ходе
процессинга (рис. 26-27). Эту функцию выполня-
ет тРНК-нуклеотидилтрансфераза — необычный
фермент, который связывает три рибонуклеозид-
трифосфата в отдельных активных центрах и ка-
тализирует формирование фосфодиэфирных свя-
зей с образованием последовательности ССА (3').
С интез этой последовательности, таким образом,
не нуждается в матрице ДНК или РНК — матри-
цей служит сайт связывания фермента.
Последний тип процессинга тРНК заклю-
чается в модификации некоторых оснований в
реакциях метилирования, дезаминирования или
восстановления (рис. 26-23). В случае псевдоури-
дина основание (урацил) удаляется и вновь при-
соединяется к остатку сахара по положению С-5.
Некоторые из модифицированных оснований на-
ходятся в специфических позициях во всех моле-
кулах тРНК (рис. 26-27).
РНК со специализированными функциями
подвергаются различным вариантам процессинга
Число идентифицированных классов специали-
зированных РНК быстро растет, и параллельно с
этим расширяются наши знания об их функциях.
Многие из этих молекул РНК подвергаются про-
цессингу.
Малая ядерная и малая ядрышковая РНК
не только облегчают процессинг РНК, но и сами
синтезируются в виде более крупных предше-
ственников, а затем подвергаются процессингу.
Многие мякРНК кодируются в интронах дру-
гих генов. По мере вырезания интронов из пре-
мРНК белки комплекса мякРНП связываются с
последовательностями мякРНК. а рибонуклеазы
удаляют лишние участки РНК с 5'- и З'-концов.
Направляющиеся в сплайсосомы мяРНК синте-
[138] Часть III. 26. Метаболизм РНК
Рис. 26-28. Синтез и процессинг микроРНК. Первичный
транскрипт микроРНК представляет собой более длинную
последовательность РНК вариабельного размера, назы-
ваемую при-микроРНК. В ее процессинге активную роль
принимают две эндорибонуклеазы из семейства РНКаз III
Drosha и Dicer. В ядре при-микроРНК сначала сокращает-
ся до предшественника микроРНК из 70-80 нуклеотидов
(пре-микроРНК) в результате действия белкового ком-
плекса, состоящего из Drosha и белка DGCR8. Далее пре-
микроРНК переносится в цитоплазму, где на нее действует
белок Dicer, в результате чего образуется практически зре-
лая микроРНК спаренная с коротким фрагментом РНК. Эта
комплементарная последовательность РНК удаляется РНК-
хеликазой, и зрелая микроРНК включается в белковые
комплексы, такие как RISC (от англ. RNA-induced silencing
complex), которые затем связываются с мРНК-мишенью.
При достаточной комплементарности микроРНК и ее ми-
шени происходит расщепление мРНК-мишени, а если ком-
плементарностъ лишь частичная, образовавшийся ком-
плекс блокирует трансляцию мРНК-мишени.
тарны определенным участкам мРНК. Они регу-
лируют функции мРНК путем ее расщепления
или подавления ее трансляции. МикроРНК обна-
ружены у многих многоклеточных эукариот — от
червей и плодовых мушек до растений и млекопи-
тающих. Возможно, до 1% генома человека коди-
рует микроРНК, причем микроРНК могут взаи-
модействовать с третью человеческих мРНК. Их
функция в регуляции генов обсуждается в гл. 28.
МикроРНК синтезируются из гораздо более
крупных предшественников за несколько ста-
дий (рис. 26-28). Первичные транскрипты для
микроРНК (при-микроРНК) значительно раз-
личаются по размеру; некоторые закодированы
в интронах других генов и экспрессируются со-
вместно с этими генами. Их роль в регуляции ге-
нов также подробно обсуждается в гл. 28.
зируются в виде пре-мяРНК с помощью РНК-
полимеразы II, а рибонуклеазы удаляют лишние
фрагменты РНК с обоих концов. Определенные
нуклеозиды в мяРНК могут быть подвергнуты 11
разным модификациям, причем чаще всего про-
исходит 2'-О-метилирование и превращение ури-
дина в псевдоуридин.
МикроРНК это специализированный
класс РНК, участвующих в регуляции экспрессии
генов. Некодирующие последовательности этих
РНК длиной около 22 нуклеотидов комплемен-
Каталитические РНК осуществляют
некоторые реакции метаболизма РНК
Исследование посттранскрипционного про-
цессинга молекул РНК привело к одному из са-
мых захватывающих открытий в современной
биохимии — открытию каталитических РНК
(рибозимов). Лучше других охарактеризованы
такие рибозимы, как аутосплайсирующиеся ин-
троны группы 1, РНКаза Р и молотоголовый ри-
бозим (см. ниже). Активность этих рибозимов
26.2 Процессинг РНК [139]
U А
G А
C-G
Рис. 26-29. Молотоголовый рибозим. Вирусоподобные
элементы, называемые вирусоидами, имеют маленький
РНК-геном и обычно нуждаются в другом вирусе, кото-
рый помогает им реплицироваться и/или упаковывать-
ся. Некоторые молекулы вирусоидных РНК содержат
небольшие участки, которые запускают реакции сайт-
специфического расщепления РНК, связанные с репли-
кацией. Эти участки называются молотоголовыми рибо-
зимами, поскольку их вторичная структура напоминает
головку молотка, а — минимальная последовательность
рибозима, обладающего каталитической активностью.
Обведенные рамкой консервативные нуклеотиды не-
обходимы для проявления каталитической активности.
Стрелка указывает сайт саморасщепления. 6 — трех-
мерная структура (PDB ID1MME, на рис. 8-25, б в т. 1
представлена в виде СРК-модели). Цепи окрашены так
же, как на рис. а. Молотоголовые рибозимы относятся
к металлоферментам; для проявления активности им
необходимы ионы Мд2+. Фосфодиэфирная связь в сайте
саморасщепления показана стрелкой, в Молотоголо-
вый рибозим
связана с двумя основными реакциями — реакци-
ей трансэтерификации (рис. 26-14) и гидролизом
фосфодиэфирных связей. Часто субстратом ри-
бозимов является молекула РНК, которая может
быть даже частью самого рибозима. Между РНК-
субстратом и РНК-катализагором может проис-
ходить спаривание оснований, наилучшим обра-
зом подготавливающее субстрат к реакции.
Рибозимы значительно различаются по раз-
мерам. Аутосплайсирующиеся интроны группы I
могут содержать более 400 нуклеотидов. Моло-
тоголовый рибозим состоит из двух цепей РНК и
содержит всего 41 нуклеотид (рис. 26-29). Как и в
случае белковых ферментов, для функций рибози-
мов важное значение имеет их трехмерная струк-
тура. Рибозимы инактивируются при нагревании
выше температуры плавления, в присутствии де-
натурующих агентов или комплементарных оли-
гонуклеотидов, которые нарушают нормальное
спаривание оснований. Рибозимы также могут
инактивироваться при модификации некоторых
важных нуклеотидов в их составе. Вторичная
структура аутосплайсирующихся интронов груп-
пы 1 из предшественника 26S рРНК у Tetrahymena
изображена на рис. 26-30.
Ферментативные свойства интронов группы I
Кроме способности повышать скорость реак-
ции, аутосплайсирующиеся интроны группы I
имеют и другие общие с ферментами свойства,
в том числе кинетическое поведение и специ-
фичность. Связывание гуанозинового кофак-
тора (рис. 26-14) с интроном I группы из рРНК
Tetrahymena (рис. 26-26) достигает насыщения
(Км « 30 мкмоль/л) и подвержено конкурентному
ингибированию З'-дезоксигуанозином. Интрон
очень точно осуществляет реакцию расщепле-
ния, в основном благодаря так называемой внут-
ренней адаптерной последовательности, кото-
рая может спариваться с последовательностями
экзона вблизи 5'-сайта сплайсинга (рис. 26-30).
Такое спаривание способствует правильному рас-
положению специфических связей, которые под-
вергаются расщеплению и воссоединению.
Поскольку интрон сам химически изменя-
ется во время реакции сплайсинга (его концы
расщепляются), казалось бы, он лишен важней-
шей характеристики фермента — способности
многократно осуществлять катализ. Детальное
[140] Часть III. 26. Метаболизм РНК
uGC
А
G (
'А
А
G
G
Р5аЯ
G
°
с
180-^
Р5с GUAg
AaAGGG u
-с
GUUCC G G
с
A GACA
йи-
юо-Е с
G • U-120
nrU • G
Р5с G
~ G
G
А
А
А
G
G
G
С
G
U
с
и
А
А
260
СА
U
А-140 А
U-G
иси
А
G
340-U
ag
Р9.1 с
G
А
Р9 . Е
augggag
А •
UUCCUC
А
-А А
А У
G £
и • G
X иР2
8
G
G
А
(К
с 360
G
С
X. U
У • G ( Р9 2 380
gAACUAAUUUGUAUGCGa
UUGAUUAGuUAUAUGAA
С А
augc
Рис. 26-30. Вторичная структура аутосплайси-
рующегося интрона рРНК из Tetrahymena. По-
следовательности интрона отмечены желтым
цветом, экзона — зеленым. Желтыми линиями
обозначены связи между соседними нуклеоти-
дами в последовательности (это вынужденная
мера, позволяющая представить объемную мо-
лекулу в двух измерениях; длинная голубая
линия между остатками С и G — нормальное
спаривание оснований); изображены все ну-
клеотиды последовательности. Каталитическое
ядро, обладающее аутосплайсирующей актив-
ностью, выделена серым прямоугольником. Не-
которые участки со спаренными основаниями
пронумерованы (Р1, РЗ, Р2.1, Р5а и т. п.) в со-
ответствии с принятыми договоренностями для
данной молекулы РНК. В области Р1, которая
содержит внутреннюю адаптерную последо-
вательность (в рамке), находится 5'-конец ин-
трона (красная стрелка). Часть этой последова-
тельности спаривается с З'-концом экзона, что
приводит к непосредственному контакту 3'- и
5'-сайтов сплайсинга (красная и голубая стрел-
ки). Трехмерная структура крупного фрагмента
этого интрона показана на рис. 8-25, в в т. 1.
G
U
А
и
исследование показало, что интрон из 414 нукле-
отидов, вырезанный из рРНК Tetrahymena, может
(in vitro) действовать как истинный фермент (in
vivo он быстро разрушается). Серия внутримоле-
кулярных реакций циклизации и расщепления
в вырезанном интроне приводит к удалению 19
нуклеотидов с его 5'-конца. Линейная РНК из
оставшихся 395 нуклеотидов (названная L-19
IVS от англ, intervening sequence lacking 19 nucleo-
tides) способствует реакциям переноса нуклео-
тидов, в которых некоторые олигонуклеотиды
удлиняются за счет других (рис. 26-31). Самыми
хорошими субстратами этой РНК являются та-
кие олигонуклеотиды, как синтетический олиго-
мер (С)5, который может спариваться с такой же
богатой гуанином внутренней адаптерной после-
довательностью, которая ориентировала 5'-конец
экзона для аутосплайсинга.
Ферментативная активность рибозима L-19
IVS проявляется в цикле реакций трансэтерифи-
кации, по механизму напоминающих аутосплай-
синг. Каждая молекула рибозима может обрабо-
тать примерно 100 молекул субстрата в час и не из-
меняется в ходе реакции; таким образом, интрон
действует как катализатор. Его действие соот-
ветствует кинетике Михаэлиса-Ментен, специ-
фично по отношению к олигонуклеотидным
РНК в качестве субстратов и может быть пол-
26.2 Процессинг РНК [141]
ностью блокировано ингибитором. Отношение
Kcat/KM (константа специфичности) составляет
103 м 1 • с \ что меньше, чем у многих ферментов,
но по сравнению с некатализируемой реакцией
рибозим повышает скорость гидролиза примерно
в 1010 раз. Такое ускорение достигается благодаря
ориентации субстрата, ковалентного катализа и
катализа ионом металла — так действуют и бел-
ковые ферменты.
Характеристика других рибозимов. РНКаза Р
Е coli состоит из РНК (Ml РНК из 377 нукле-
отидов) и белка (Мг = 17 500). В 1983 г. Сидней
Олтман и Норман Пейс с коллегами обнаружили,
что при определенных условиях М1 РНК способ-
на сама осуществлять катализ, расщепляя пред-
шественников тРНК в совершенно определенном
месте. Белковый компонент, вероятно, стабили-
зирует РНК или облегчает ее функционирование
(5') G A A A U A G С A A U A U и|и А С С U U U G GaTg G Gl А
G ОН(З')
Сплайсированный интрон
рРНК
19 нуклеотидов с 5'-конца
L-19 IVS (5') , U U G G A G G G | А ==*....? - G ОН (3 )
G
(З')НО '<Ц (С)5
\ но с с с с с
(5 ) U U G G A G G G А V_______>
сг
Рис. 26-31. Каталитическая активность L-19 IVS in vitro, а — L-19 IVS образуется путем
аутокаталитического удаления 19 нуклеотидов с 5'-конца вырезаемого интрона у Tetrahy-
тепа. Сайт расщепления во внутренней адаптерной последовательности (в рамке) показан
стрелкой. Остаток G (на розовом фоне), присоединенный на первой стадии реакции сплай-
синга (см. рис. 26-15), входит в удаляемую последовательность. Часть внутренней адаптер-
ной последовательности остается на 5'-конце L-19 IVS. 6 — L-19 IVS удлиняет некоторые
олигонуклеотиды РНК за счет других в цикле реакций трансэтерификации (стадии D-@).
Ключевую роль в этом цикле играет группа З'-ОН в остатке G на З'-конце L-19 IVS (следует
подчеркнуть, что это не остаток G, добавленный в процессе сплайсинга). Одним из лучших
субстратов рибозима оказывается (С)5, поскольку он может связываться с адаптерной по-
следовательностью, сохраняющейся в интроне. Хотя такая каталитическая активность не
реализуется в клетке, она имеет большое значение для формирования современной гипо-
тезы эволюции, которая обсуждается в конце данной главы.
114 21 Часть III. 26. Метаболизм РНК
in vivo. Рибозим РНКаза P распознает трехмер-
ную форму субстрата (пре-тРНК), а также по-
следовательность ССА, и таким образом может
отщепить 5'-концевые последовательности от
разных молекул тРНК (рис. 26-27).
Список известных рибозимов продолжает
расти. Некоторые вирусоиды, мелкие молекулы
РНК, ассоциированные с РНК-содержащими
вирусами растений, содержат структуру, кото-
рая запускает реакцию саморасщепления. К это-
му классу каталитических молекул относится
изображенный на рис. 26-29 молотоголовый ри-
бозим, катализирующий гидролиз внутренней
фосфодиэфирной связи. Реакция сплайсинга в
сплайсосоме, по-видимому, осуществляется в ка-
талитическом центре, сформированном U2, U5 и
U6 мяРНК (рис. 26-17). И, вероятно, самое глав-
ное — PH К-компонент рибосом катализирует
синтез белка (см. гл. 27).
Обнаружение каталитических молекул РНК
расширило наши представления о каталитиче-
ской функции в целом и сыграло важную роль
в формировании наших представлений о проис-
хождении и эволюции жизни на нашей планете
(см. разд. 26.3).
мРНК в клетке разрушаются
с разной скоростью
Экспрессия генов регулируется на многих уров-
нях. Решающий фактор, управляющий экспрес-
сией генов, — концентрация в клетке соответству-
ющей мРНК. Концентрация любой молекулы
зависит от двух факторов: скорости синтеза и
скорости распада. Когда скорости синтеза и рас-
пада мРНК становятся равными, мРНК нахо-
дится в стационарной концентрации. Изменение
скорости синтеза или скорости распада приводит
к накоплению или исчерпанию мРНК. Метаболи-
ческие пути расщепления предотвращают нако-
пление мРНК в клетке и синтез ненужных белков.
Скорости расщепления мРНК разных эука-
риотических генов в значительной степени разли-
чаются. Время полужизни мРНК тех продуктов,
которые нужны недолго, может составлять не-
сколько минут или даже секунд. Напротив, мРНК
тех генов, чьи продукты требуются постоянно, мо-
гут быть стабильны на протяжении многих клеточ-
ных генераций. Среднее время полужизни мРНК
в клетках позвоночных составляет примерно 3 часа,
а пул каждого типа мРНК на протяжении одной
клеточной генерации оборачивается примерно
10 раз. Время полужизни бактериальной мРНК
намного меньше — лишь около 1,5 мин; возможно,
это обусловлено регуляцией метаболизма.
Матричная РНК разрушается рибонуклеа-
зами, присутствующими во всех клетках. В Е. coli
процесс начинается с одного или нескольких раз-
рывов, которые вносит эндорибонуклеаза, с после-
дующим 3'-*5'-расщеплением экзорибонуклеаза-
ми. У низших эукариот основной метаболический
путь начинается с укорочения поли(А)-«хвоста» с
последующим «декапированием» 5'-конца и рас-
щеплением мРНК в направлении 5'-*3'. Путь рас-
щепления в направлении 3'—*5' также существует
и, возможно, является основным путем у высших
эукариот. У всех эукариот существует комплекс,
называемый экзосомой, содержащий до 10 кон-
сервативных 3'—*5'-экзорибонуклеаз, которые уча-
ствуют в процессинге З'-конца рРНК и тРНК, а
также в расщеплении мРНК.
Напоминающая шпильку структура бакте-
риальной мРНК с р-независимым терминатором
(рис. 26-8) обеспечивает защиту от расщепления.
Схожие шпилечные структуры стабилизируют
некоторые части первичного транскрипта, что яв-
ляется причиной неравномерного расщепления
транскриптов. В клетках эукариот важное зна-
чение для стабильности многих молекул мРНК
имеют 3'-поли(А)-«хвост» и 5'-кэп. в Жизненный
цикл мРНК
Полинуклеотидфосфорилаза создает
случайные РНК-подобные полимеры
В 1955 г. Марианна Грюнберг-Манаго и Северо
Очоа открыли бактериальный фермент поли-
нуклеотидфосфорилазу, которая in vitro катали-
зирует реакцию:
(NMP)„ + NDP — (NMP)„+1 + Pj
Удлиненный
полинуклеотид
Полинуклеотидфосфорилаза была обнаружена
первой среди всех ферментов, синтезирующих
нуклеиновые кислоты (вскоре после этого Артур
Корнберг открыл ДНК-полимеразу). Реакция,
катализируемая полинуклеотидфосфорилазой,
принципиально отличается от всех упомянутых
выше полимеразных реакций тем, что не зависит
26.2 Процессинг РНК [143]
Марианна Грюнберг-
Манаго, 1921-2013
Севере Очоа,
1905-1993
от матрицы. Этот фермент использует в качестве
субстратов 5'-дифосфаты рибонуклеозидов и не
действует на гомологичные 5'-трифосфаты или на
дезоксирибонуклеозид-5'-дифосфаты. Полимер
РНК, синтезированный полинуклеотидфосфори-
лазой, содержит обычные 3',5'-фосфодиэфирные
связи, которые расщепляются рибонуклеазой. Эта
реакция легко обратима, и равновесие смещается
в сторону расщепления полирибонуклеотида при
повышении концентрации фосфата. Предполагае-
мая роль фермента в клетке заключается в разру-
шении молекул мРНК до иуклеозиддифосфатов.
Поскольку реакция фосфорилирования по-
линуклеотидов не нуждается в матрице, созда-
ваемый полимер нс содержит специфических
последовательностей оснований. Реакция про-
текает одинаково хорошо с любым из четырех
или со всеми четырьмя нуклеотиддифосфатами,
а состав образующегося полимера зависит лишь
от относительной концентрации 5'-дифосфатов
(субстратов) в среде.
Полинуклеотидфосфорилаза может при-
меняться в лаборатории для синтеза полимеров
РНК с самыми разными последовательностями
оснований. Синтетические полимеры РНК тако-
го рода сыграли важнейшую роль при выяснении
генетического кода аминокислот (гл. 27).
Краткое содержание раздела 26.2
ПРОЦЕССИНГ РНК
Эукариотические молекулы мРНК моди-
фицируются путем присоединения остатка
7-метилгуанозииа на 5'-конце, а также рас-
щепления и полиаденилирования З'-конца с
формированием длинного поли(А)-«хвоста».
Многие первичные транскрипты мРНК со-
держат интроны (некодирующие области),
которые удаляются сплайсингом. Выреза-
ние интронов группы I, обнаруженных в
некоторых молекулах рРНК, нуждается в
гуанозиновом кофакторе. Некоторые интро-
ны групп [ и II способны к аутосплайсингу
без участия белковых ферментов. Ядерные
предшественники мРНК содержат интроны
III группы (самой многочисленной), кото-
рые вырезаются с помощью РНК-белковых
комплексов, называемых мяРНП, организо-
ванных в сплайсосомы. Интроны IV группы
осуществляют сплайсинг белковых фермен-
тов; эти интроны обнаружены в некоторых
молекулах тРНК.
Функционирование многих эукариотиче-
ских мРНК регулируется комплементарной
микроРНК. Молекулы микроРНК образу-
ются из более длинных предшественников в
результате серии реакций процессинга.
Молекулы рибосомной и транспортной РНК
образуются из более длинных молекул пред-
шественников, расщепляемых нуклеазами.
В ходе созревания некоторые основания
модифицируются ферментами. Некоторые
модификации нуклеозидов регулируются
мякРНК в белковых комплексах мякРНП.
Аутосплайсирующиеся интроны и РНК-
компонент РНКазы Р (отщепляет 5'-консц
предшественников тРНК) — примеры ри-
бозимов. Эти биологические катализаторы
обладаю! свойствами истинных ферментов.
Обычно они катализируют гидролитиче-
ское расщепление и трансэтерификацию,
используя в качестве субстрата РНК. Со-
четанию таких реакций способствует выре-
занный интрон группы I из рРНК Teirahy-
тепа, образующийся в ходе реакции РНК-
полимеризации.
Полинуклеотидфосфорилаза в обратимой
реакции синтезирует РНК-подобные по-
лимеры из рибонуклеозид-5'-дифосфатов,
добавляя или удаляя рибонуклеотиды на
З'-гидроксильном конце полимера. Фермент
разрушает РНК in vivo.
[144] Часть III. 26. Метаболизм РНК
26.3. РНК-зависимый синтез РНК и ДНК
До сих пор при обсуждении синтеза ДНК и РНК
мы рассматривали в роли матрицы исключитель-
но молекулы ДНК. Однако некоторые ферменты
для синтеза нуклеиновых кислот используют ма-
трицу РНК. За очень важным исключением, каса-
ющимся вирусов с геномом из РНК, эти ферменты
играют в информационных путях лишь скромную
роль Именно РНК-содержащие вирусы являются
источником большинства известных к настояще-
му времени РНК-зависимых полимераз.
Существование репликации РНК требует
пересмотра основной догмы молекулярной био-
логии (рис. 26-32; сравните с диаграммой на с. 5).
Ферменты, участвующие в репликации РНК, по-
могают пролить свет на природу самореплициру-
ющихся молекул, которые могли существовать в
пребиотические времена.
Обратная транскриптаза
синтезирует ДНК с матрицы вирусной РНК
Некоторые РНК-содержащие вирусы, инфициру-
ющие клетки животных, содержат в вирусных ча-
стицах РНК-зависимую ДНК-полимеразу, назы-
ваемую обратной транскриптазой. При инфици-
ровании фермент попадает в клетку хозяина вме-
сте с одноцепочечным вирусным РНК-геномом
(~10 ООО нуклеотидов). Обратная транскриптаза
сначала катализирует синтез цени ДНК, компле-
ментарной вирусной РНК (рис. 26-33), затем раз-
рушает цепь РНК в гибриде РНК-ДНК и заме-
няет ее на ДНК. Образовавшаяся двухцепочечная
ДНК часто встраивается в геном клетки хозяина.
Такие интегрированные (и молчащие) вирусные
Репликация
ДНК /
Х^ДНК
Транскрипция Обратная
транскрипция
РНК
Репликация
РНК
Трансляция
Белок
Рис. 26-32. Расширение основной догмы молекулярной
биологии с учетом РНК-зависимого синтеза РНК и ДНК.
Рис. 26-33. Ретровирусная инфекция в клетке млекопи-
тающего и интеграция ретровируса в хромосому хозяина.
Вирусные частицы, проникающие в клетку хозяина, несут
вирусную обратную транскриптазу и клеточную тРНК (за-
хваченную из предыдущей клетки-хозяина) уже в паре с
вирусной РНК. Молекулы тРНК опосредуют немедленное
превращение вирусной РНК в двухцепочечную ДНК при
участии обратной транскриптазы, как описано в тексте.
Двухцепочечная ДНК проникает в ядро и встраивается в
геном хозяина. Интеграцию катализирует вирусная интег-
раза. Механизм интеграции ДНК вируса в ДНК хозяина на-
поминает механизм встраивания транспозонов в бактери-
альные хромосомы (см. рис. 25-45). Например, на участке
интеграции несколько пар оснований ДНК хозяина удва-
иваются, формируя короткие повторы из 4-6 п. н. с обеих
сторон от вставки ретровирусной ДНК (не показано).
гены могут активироваться и транскрибироваться,
а продукты этих генов вирусные белки, а также
сам РНК-геном вируса могут упаковываться в но-
вые вирусные частицы. РНК-вирусы, содержащие
обратные транскриптазы, называют ретровируса-
ми (лат. приставка retro означает «обратно»).
Существование обратных транскриптаз РНК-
сод ержащих вирусов в 1962 г. предсказал Говард
Темин, и в 1970 г. эти ферменты были обнаружены
Теминым и независимо Дейвидом Балтимором.
Открытие привлекло большое внимание, посколь-
ку пошатнуло основную догму и показало, что
генетическая информация может переходить «на-
зад» от РНК к ДНК.
Ретровирусы обычно несут три гена: gag (от
исторически сложившегося англ, названия group
associated antigen — групповые ассоциированные
26.3 РНК-зависимый синтез РНК и ДНК [145]
Говард Темин, 1934-1994
Дейвид Балтимор
антигены), pol и ет (рис. 26-34). Транскрипт, со-
держащий gag и pol, транслируется в длинный
«полипротеин» — полипептид, который расще-
пляется на шесть белков с различными функ-
циями. Белки, кодируемые геном gag, образуют
сердцевину вирусной частицы. Ген pol кодирует
протеазу, расщепляющую длинный полипептид,
интегразу, которая встраивает вирусную ДНК в
хромосому хозяина, и обратную транскриптазу.
Многие обратные транскриптазы состоят из двух
субъединиц а и р. Генpol кодирует р-субъединицу
(Мг = 90 000), а а-субъединица (Л/г = 65 000) —
это протеолитический фрагмент р-субъединицы.
Ген ет кодирует белки вирусной оболочки. На
каждом конце линейного РНК-генома находятся
длинные концевые повторы (LTR), состоящие из
нескольких сотен нуклеотидов. Транскрибиро-
ванные в двухцепочечную ДНК, эти последова-
тельности облегчают интеграцию вирусной хро-
мосомы в ДНК хозяина и содержат промоторы
для экспрессии генов вируса.
Обратные транскриптазы катализируют три
разные реакции: (1) РНК-зависимый синтез ДНК,
(2) расщепление РНК и (3) ДНК-зависимый син-
тез ДНК. Подобно многим ДНК- и РНК-полимера-
зам, обратные транскриптазы содержат Zn2+. Любая
транскриптаза наиболее активна с РНК собствен-
ного вируса, но все они могут применяться в экспе-
рименте для получения молекул ДНК, комплемен-
тарных разнообразным молекулам РНК. В синтезе
ДНК и РНК и в расщеплении РНК задействованы
разные активные центры белка. Для начала синте-
за ДНК обратной транскриптазе нужен праймер,
клеточная тРНК, захваченная у предыдущего хо-
зяина и переносимая в составе вирусной частицы.
З'-Конец этой тРНК спарен с комплементарной
последовательностью вирусной РНК. Новая цепь
ДНК синтезируется в направлении 5'—*3', как во
Рис. 26-34. Структура и продукты интегри-
рованного ретровирусного генома. Длин-
ные концевые повторы (LTR) содержат
последовательности, необходимые для ре-
гуляции и инициации транскрипции. После-
довательность гр нужна для упаковки моле-
кул ретровирусной РНК в зрелые вирусные
частицы. Транскрипция ретровирусной ДНК
приводит к образованию первичного транс-
крипта, включающего гены дад, pol и env.
В результате трансляции (гл. 27) образует-
ся длинный полипротеин — полипептид,
кодируемый генами дад и pol, который рас-
щепляется на шесть белков с различными
функциями. После сплайсинга первичного
транскрипта, синтезируемого на основе гена
env, образуется мРНК, которая также транс-
лируется в полипротеин, расщепляющийся
затем с образованием белков вирусной обо-
лочки.
J протеолитическое
4 протеолитическое
расщепление
о Белки
%°5оо° ВИРУСНОИ
* оболочки
Структурные
белки вируса
Обратная
' ' • транскриптаза
[146] Часть III. 26. Метаболизм РНК
всех реакциях с участием РНК- и ДНК-полимераз.
Обратные транскриптазы, как и РНК-полимеразы,
не имеют корректирующей экзонуклеазной актив-
ности в направлении 3'-*5'. Частота ошибок, допу-
скаемых этими ферментами, составляет примерно
1 на 20 000 присоединенных нуклеотидов. Такая
высокая частота ошибок совершенно необычна
для репликации ДНК и, вероятно, свойственна
большинству ферментов, реплицирующих геномы
РНК-содержащих вирусов. Результатом является
большая частота мутаций и высокий теми эволю-
ции вирусов, что приводит к частому появлению
новых болезнетворных штаммов ретровирусов.
Обратные транскриптазы стали важными
инструментами в изучении взаимодействий меж-
ду ДНК и РНК, а также в методах клонирования
ДНК. Они позволяют синтезировать ДНК, ком-
плементарную матрице мРНК; приготовленная
таким способом синтетическая ДНК называется
комплементарной ДНК (кДНК) и может ис-
пользоваться для клонирования клеточных генов
(см. рис. 9-14 в т. 1).
Некоторые ретровирусы вызывают рак и СПИД
Изучение ретровирусов позволило сильно
продвинуться в понимании молекулярной
природы рака. Большинство ретровирусов не
убивают хозяйские клетки, а остаются встроен-
ными в клеточную ДНК и реплицируются при
делении клетки. Некоторые ретровирусы, отно-
сящиеся к РНК-содержащим опухолевым виру-
сам, содержат онкоген, который может вызвать
неконтролируемый рост клеток. Первым из-
ученным вирусом этого типа был вирус саркомы
Рауса (другое название - вирус саркомы птиц;
рис. 26-35), названный в честь Ф. Пейтона Рауса,
который изучал опухоли кур, вызываемые этим
вирусом. После открытия Харолдом Вармусом
и Майклом Бишопом онкогенов в ретровирусах
были обнаружены десятки таких генов.
Вирус иммунодефицита человека (ВИЧ), вы-
зывающий синдром приобретенного иммунного
дефицита (СПИД), тоже относится к ретровиру-
сам. Вирус иммунодефицита человека, идентифи-
цированный в 1983 г., имеет РНК-геном со стан-
дартным набором ретровирусных генов наряду с
некоторыми необычными генам (рис. 26-36). В от-
личие от многих других ретровирусов ВИЧ чаще
убивает инфицированные им клетки (главным
образом Т-лимфоциты), чем вызывает формиро-
вание опухолей. Это постепенно приводит к по-
давлению иммунной системы хозяина. Обратная
транскриптаза ВИЧ допускает еще больше оши-
бок, чем другие известные обратные транскрипта-
зы, — в 10 или более раз, что приводит к высокой
частоте мутаций вируса. При каждой репликации
генома вируса ВИЧ обычно появляется одна или
несколько ошибок, поэтому любые две молекулы
РНК этого вируса могут различаться между собой.
Многие современные вакцины, эффектив-
ные в отношении вирусных инфекций, состоят
из одного или нескольких белков оболочки ви-
руса; их получают способами, описанными в гл. 9
(в т. 1). Сами эти белки не вызывают зараже-
ния, но они стимулируют иммунную систему и
помогают ей распознать вирус при возможной
инфекции и справиться с ним (гл. 5 в т. 1). Из-
за высокой частоты ошибок в реакции обрат-
ной транскриптазы ген env ВИЧ (и весь геном)
очень быстро мутирует, что затрудняет создание
эффективной вакцины. Однако для распростра-
нения ВИЧ-инфекции необходимы повторные
циклы заражения клеток и репликация, поэтому
самый эффективный современный способ лече-
ния основан на ингибировании вирусных фер-
ментов. ВИЧ-протеаза является мишенью для
лекарственных препаратов, называемых ингиби-
торами протеазы (см. доп. 6-3 в т. 1). Обратная
транскриптаза является мишенью для некоторых
других лекарств, широко применяемых для лече-
ния ВИЧ-инфицированных (см. доп. 26-2).
LTR
LTR
gag
pot env src
Рис. 26-35. Геном вируса саркомы Рауса. Ген src кодирует тирозинкиназу, относящуюся к
классу ферментов, влияющих на клеточное деление, межклеточные взаимодействия и пере-
дачу внутриклеточных сигналов (гл. 12 в т. 1). Этот же ген обнаружен в ДНК кур (обычный
хозяин данного вируса) и в геномах многих эукариот, включая человека. Под воздействием
вируса саркомы Рауса этот онкоген начинает активно экспрессироваться, способствуя не-
контролируемому клеточному делению и возникновению рака.
26.3 РНК-зависимый синтез РНК и ДНК [117]
Дополнение 26-2
rev
| vif | | tat \vpu\
| gag
| nef |
LTR
pol
LTR
Рис. 26-36. Геном ВИЧ. Помимо типичных генов ретровирусов ВИЧ содержит несколько
небольших генов с различными функциями (не все они идентифицированы и здесь не рас-
сматриваются). Некоторые из этих генов перекрываются (см. с. 175). В результате альтер-
нативного сплайсинга из этого маленького генома (9,7 • 103 нуклеотидов) образуется много
разных белков.
МЕДИЦИНА
Борьба со СПИДом с помощью ингибиторов обратной транскриптазы
Жизненный цикл и структура вируса иммунодефи-
цита человека (ВИЧ) были установлены в результате
изучения химии зависимого от матрицы биосинтеза
нуклеиновых кислот современными методами молеку-
лярной биологии. Через несколько лет после выделе-
ния ВИЧ эти исследования позволили создать лекар-
ства, продлевающие жизнь людей, инфицированных
этим вирусом.
Первый препарат, допущенный к клиническим
испытаниям — азидотимидин (AZT) — структурный
аналог дезокситимидина. Впервые AZT был синте-
зирован в 1964 г. Джеромом Хорвицем. В 1985 г. вы-
яснилось, что это неудачное противоопухолевое сред-
ство (препарат был создан для борьбы с раком) может
оказаться полезным для лечения больных СПИДом.
Наиболее чувствительные к ВИЧ-инфекции клетки
иммунной системы Т-лимфоциты захватывают AZT
и преобразуют его в AZT-трифосфат. (Непосред-
ственное введение AZT-трифосфата неэффективно,
3 ’ - Азидо- 2', 3' - дидезокси-
тимидин (AZT)
2',3'-Дидезоксиинозин (DDI)
поскольку это соединение не может проходить через
плазматическую мембрану.) Обратная транскрип-
таза ВИЧ обладает повышенным сродством к AZT-
трифосфату по сравнению с dTTP. и связывание AZT-
трифосфата с ферментом конкурентно ингибирует
связывание dTTP. Когда к З'-концу растущей цепи
ДНК присоединяется AZT, отсутствие З'-гидроксиль-
ной группы приводит к тому, что синтез вирусной
ДНК прерывается.
AZT-трифосфат нетоксичен для самих Т-лим-
фоцитов, поскольку клеточная ДНК-полимераза
имеет пониженное сродство к этому соединению по
сравнению с dTTP. В концентрации 1 -5 мкмоль AZT
воздействует на обратную транскрипцию ВИЧ, но
не влияет на репликацию клеточной ДНК. К сожале-
нию. AZT оказался токсичным для клеток костного
мозга - предшественников эритроцитов, поэтому у
многих пациентов, принимающих AZT, развивается
анемия. Применение AZT может увеличить время
жизни людей с хронической формой ВИЧ-инфекции
примерно на год и заметно отодвигает проявления
симптомов СПИДа у пациентов на ранней стадии
инфицирования ВИЧ. Некоторые другие средства
против ВИЧ-инфекции, например дидезоксиинозин
(DDI), имеют схожий механизм действия Более со-
временные лекарственные средства инактивируют
ВИЧ-протеазу. Из-за высокой степени ошибок об-
ратной транскриптазы и связанной с этим быстрой
эволюцией вируса большинство эффективных спо-
собов лечения основано на комбинации лекарствен-
ных средств, подавляющих и протеазу, и обратную
транскриптазу.
[148] Часть III. 26. Метаболизм РНК
Многие транспозоны, ретровирусы
и интроны могут иметь
общее эволюционное происхождение
Структура некоторые хорошо изученных транс-
позонов из столь разных источников, как дрожжи
и дрозофила, очень напоминает структуру ретро-
вирусов; иногда их называют ретротранспозо-
нами (рис. 26-37). Ретротранспозоны кодируют
фермент, гомологичный ретровирусной обратной
транскриптазе, и их кодирующие области флан-
кированы последовательностями LTR. Они пере-
носятся внутри клеточного генома из одного по-
ложения в другое с помощью РНК-посредника,
используя обратную транскриптазу для создания
ДНК-копии РНК с последующей интеграцией
ДНК в новый сайт. Большинство транспозонов
эукариот для транспозиции использует этот ме-
ханизм, что отличает их от транспозонов бакте-
рий. которые перемещаются непосредственно в
виде фрагментов ДНК из одного участка хромо-
сомы в другой (см. рис. 25-45).
Ретротранспозоны утратили ген env и не мо-
гут образовывать вирусные частицы. Поэтому их
следует рассматривать как дефектные вирусы,
оказавшиеся в клетке, как в ловушке. Сравнение
ретровирусов с эукариотическими транспозона-
ми показывает, что обратная транскриптаза
очень древний фермент, появившийся до возник-
новения многоклеточных организмов.
Интересно, что многие интроны групп ] и II
тоже являются подвижными генетическими эле-
ментами. Они не только способны осуществлять
аутосплайсинг, но и кодируют эндонуклеазы
ДНК, способствующие их перемещению. В про-
Элемент Ту
(Saccharomyces)
(LTR) (gag)_____(pol)____(LTR)
| gj TYA || TYB | g |
LTR LTR
Элемент Сорт । | рд дур др | 1
(Drosophila) I--1 - -------------------1---1
Рис. 26-37. Транспозоны эукариот. Ту-элемент дрожжей
Saccharomyces и copia-элемент плодовой мухи Droso-
phila — примеры транспозонов эукариот со структурой,
напоминающей ретровирусы, но утратившие ген env.
Последовательности 8 элемента Ту — функциональный
эквивалент ретровирусных последовательностей LTR.
В copia-элементе последовательности INT и RT гомоло-
гичны участкам интегразы и обратной транскриптазы в
составе гена pol.
цессе обмена генами между клетками одного вида
или при внедрении в клетку ДНК под действием
паразитов или иным способом эти эндонуклеазы
способствуют встраиванию интрона в идентич-
ный участок в другой копии ДНК гомологичного
гена, который не содержит этого интрона. Данный
процесс называется хомингом (рис. 26-38). Хо-
минг интронов группы I осуществляется на основе
ДНК, тогда как хоминг интронов группы I] — че-
рез РНК-посредника. Эндонуклеазы интронов
группы II обладают также активностью обратной
транскриптазы. Эти белки могут образовывать
комплексы с РНК интронов после того, как ин-
троны вырезаны из первичных транскриптов. По-
скольку процесс хоминга включает встраивание
РНК интрона в ДНК и обратную транскрипцию
интрона, перемещение этих интронов называют
ретрохомингом В популяции каждая копия кон-
кретного гена со временем может приобрести ин-
трон. Гораздо реже интрон самостоятельно встра-
ивается в новое место в неродственном гене. Если
при этом хозяйская клетка не погибает, это может
быть началом эволюционных изменений и при-
вести к распространению интрона в новом месте.
Структуры подвижных интронов и механизмы их
распространения подтверждают предположение,
что по крайней мере некоторые из них возникли
как молекулярные паразиты, чья эволюция восхо-
дит к ретровирусам и транспозонам.
Теломераза — специализированная
обратная транскриптаза
Теломеры — структуры на концах линейных хро-
мосом эукариот (см. рис. 24-9); обычно они со-
стоят из многих тандемных повторов короткой
олигонуклеотидной последовательности. Эта по-
следовательность, как правило, имеет вид TXG^
в одной цепи и СуАх в комплементарной цепи,
где значения х и у колеблются от 1 до 4 (с. 13).
Теломеры различаются по длине от нескольких
десятков пар оснований у некоторых ресничных
простейших до десятков тысяч пар оснований у
млекопитающих. Цепь TG длиннее, чем компле-
ментарная ей цепь, поэтому на З'-конце имеется
участок одноцепочечной ДНК длиной до не-
скольких сотен нуклеотидов.
Репликация концов линейной хромосомы
клеточными ДНК-полимеразами — процесс до-
статочно сложный Для репликации ДНК нуж-
26.3 РНК-зависимый синтез РНК и ДНК [149]
а Образование хоминг-эндонуклеазы
Интрон I группы
б Хоминг
транскрипция
сплайсинг
трансляция
Продукт гена X
ДНК гена X,
аллель а
Первичный
транскрипт
Сплайсированный
интрон I группы
М Хоминг-
эндонуклеаза
Рис. 26-38. Подвижные интроны: хоминг и ретрохо-
минг. Некоторые интроны содержат ген (красный) фер-
мента, который активирует хоминг (некоторые интроны
I группы) или ретрохоминг (некоторые интроны II груп-
пы). а — ген внутри вырезаемого интрона связывается
рибосомой и транслируется. Хоминг-интроны I группы
кодируют сайт-специфическую эндонуклеазу, называ-
емую хоминг-эндонуклеазой. Ретрохоминг-интроны II
группы кодируют белок с активностью эндонуклеазы и
обратной транскриптазы одновременно, б — хоминг. Ал-
лель а генаХ, содержащего хоминг-интрон I группы, при-
сутствует в клетке, содержащей аллель b того же гена, но
без интрона. Хоминг-эндонуклеаза, образованная а, рас-
щепляет Ъ в положении, соответствующем интрону в а, а
затем в результате репарации двухцепочечного разрыва
(рекомбинация с аллелем а; см. рис. 25-31, а) возникает
новая копия интрона в Ь. в — ретрохоминг. Аллель а гена
У содержит ретрохоминг-интрон II группы; аллель b не
имеет интрона. Вырезанный интрон встраивается в коди-
рующую цепь b в реакции, обратной реакции сплайсин-
га, когда интрон вырезается из первичного транскрипта
(см. рис. 26-15); однако здесь происходит встраивание в
ДНК, а не в РНК. Некодирующая цепь ДНК в аллеле b рас-
щепляется закодированной в интроне эндонуклеазой/
обратной транскриптазой. Этот же фермент использует
встроенную РНК в качестве матрицы для синтеза компле-
ментарной цепи ДНК. Затем РНК разрушается клеточны-
ми рибонуклеазами и заменяется на ДНК.
в
Ген X, аллель а с интроном
ДНК гена X,
аллель Ь,
без интрона
хоминг-эндонуклеаза
репарация
двухцепочечного разрыва
Ретрохоминг
Интрон II группы
Сплайсированный
интрон
обратный
сплайсинг
транскрипция
сплайсинг
44 шдонукл
а с интроном
b с интроном
ДНК гена У,
аллель а, донор
(Эндонуклеаза/
обратная транскриптаза
!м ......
ДНК гена У,
аллель Ь,
реципиент
РНК замещается ДНК лигирование
Ь с интроном
ны матрица и праймер, а за пределами линейной
молекулы ДНК нет матрицы для посадки РНК-
праймера. Без специального механизма реплика-
ции концов хромосомы становились бы короче и
короче с каждым клеточным делением. Фермент
теломераза решает эту проблему добавляя к кон-
цам хромосомы теломеры.
Существование такого фермента, возможно,
не вызывает удивления, но механизм его действия
весьма необычен. Теломераза, подобно некоторым
другим ферментам, описанным в этой главе, со-
стоит из РНК и белка Длина РНК-компонента
составляет примерно 150 нуклеотидов, и в нем
содержится около 1.5 копий теломерного повто-
[1501 Часть III. 26. Метаболизм РНК
а
ДНК 5'
3'
Теломераза
РНК-матрица
TTTTGGGGTTTJG ОН(З') /
^ССААААССССААААГ
AG 7
и
с
А
А
А
ДНК-связывающие
белки теломерного
уплекса
Цепь-
СА
®| полимеризация
I и гибридизация
5'
3'
TTTTGGGGTTTTGGGGTTTTr ч
। । G ОН(3)
ССААААСС С CAAAA(L
U А
С А
TRF1и TRF2
zjs I транслокация и
I повторная гибридизация
5' TTTTGGGGTTTTGGGGTTTTG ОН(З')
3' сСААААССССАААд
AG G.
Дальнейшая полимеризация
Рис. 26-39. Цепь TG и Т-петля в теломерах, а — собствен-
ная РНК-матрица теломеразы связывается и образует пары
оснований с TG-праймером ДНК. Сь Теломераза присоеди-
няет к TG-праймеру дополнительные остатки Т и G, затем
2х перемещает собственную матрицу РНК, чтобы з про-
должить присоединение оснований Т и G. Комплементар-
ная цепь синтезируется клеточными ДНК-полимеразами
(не показано). 6 — предполагаемая структура Т-петель в
теломерах. Одноцепочечный «хвост», синтезированный
теломеразой, загибается назад и спаривается с компле-
ментарной последовательностью двухцепочечного участ-
ка. Теломера связывается с несколькими белками, включая
TRF1 и TRF2 (от англ, telomere repeat binding factors — фак-
торы связывания теломерных повторов), в — электронная
микрофотография Т-петли на конце хромосомы, выделен-
ной из гепатоцитов мыши. Длина черного отрезка внизу
фотографии соответствует 5000 п. н.
pa C^AV. Эта область РНК действует как матрица
для синтеза цепи TtG^ в теломере. Теломераза, та-
ким образом, выступает в клетке в роли обратной
транскриптазы, в активном центре которой проис-
ходит РНК-зависимый синтез ДНК. В отличие от
обратной транскриптазы ретровирусов теломера-
за копирует только маленький участок РНК, ко-
торый она содержит внутри самой себя. Для син-
теза теломеры в качестве праймера используется
З'-конец хромосомы; синтез протекает в обычном
направлении 5'-*3'. Синтезировав одну копию по-
втора, фермент перемещается, чтобы продолжить
удлинение теломеры (рис. 26-39, а).
После продления теломеразой цепи TXG^ ком-
плементарная ей цепь СуАх синтезируется клеточ-
ными ДНК-полимеразами, начиная с праймера
РНК (см. рис. 25-13). У многих низших эукариот,
особенно тех видов, теломеры которых содержат
не больше нескольких сотен пар оснований, одно-
цепочечную область защищают специальные свя-
зывающие белки. У высших эукариот (включая
млекопитающих) с теломерами длиной в несколь-
ко тысяч пар оснований одноцепочечный конец
образует особую структуру, называемую Т-петлей
(рис. 26-39, б). Одноцепочечный конец загибается
назад и спаривается с комплементарным участ-
26.3 РНК-зависимый синтез РНК и ДНК [151]
ком в двухцепочечной части теломеры. При об-
разовании Т-петли З'-конец одноцепочечной те-
ломеры внедряется в ДНК-дуплекс, возможно, по
механизму, аналогичному механизму инициации
гомологичной генетической рекомбинации (см.
рис. 25-33). У млекопитающих петля ДНК связа-
на с двумя белками TRF1 и TRF2, причем послед-
ний белок участвует в формировании Т-петли.
Т-петли защищают З'-концы хромосом, делая их
недоступными для нуклеаз и ферментов репара-
ции двухцепочечных разрывов.
У простейших (таких как Tetrahymena) по-
теря теломеразной активности приводит к посте-
пенному укорачиванию теломер при каждом кле-
точном делении и неизбежно заканчивается гибе-
лью клеточной линии. Похожая связь между дли-
ной теломер и старением клеток (прекращением
клеточного деления) наблюдается и у человека. В
линиях зародышевых клеток, обладающих тело-
меразной активностью, теломеры имеют постоян-
ную длину; в соматических клетках, утративших
теломеразу, теломеры укорачиваются. Существу-
ет обратная линейная зависимость между длиной
теломер в культивируемых фибробластах и воз-
растом людей, у которых эти фибробласты были
взяты: чем старше человек, тем короче теломеры
в его соматических клетках. Если в условиях in
vitro в соматические клетки человека ввести тело-
меразную обратную транскриптазу, теломеразная
активность восстанавливается, а продолжитель-
ность жизни клеток значительно увеличивается.
Является ли постепенное укорочение тело-
мер ключом к пониманию процесса старения?
Определяет ли длина теломер при рождении про-
должительность нашей жизни? Дальнейшие ис-
следования в этой области могут привести к уди-
вительным открытиям.
Некоторые вирусные РНК реплицируются
РНК-зависимой РНК-полимеразой
Некоторые бактериофаги Е. coli, включая f2, MS2,
R17 и QP, а также некоторые вирусы эукариот (в
том числе вирус гриппа и вирус Синдбис, вызы-
вающий одну из форм энцефалита) имеют РНК-
геномы. Хромосомы этих вирусов представляют
собой одноцепочечные молекулы РНК и функци-
онируют также в качестве мРНК для синтеза ви-
русных белков; они реплицируются в клетках хо-
зяина с помощью РНК-зависимой РНК-полиме-
разы (РНК-репликазы). Все РНК-содержащие
вирусы, за исключением ретровирусов, должны
кодировать белок с активностью РНК-зависимой
РНК-полимеразы, поскольку клетки хозяина не
образуют этот фермент.
РНК-репликаза большинства РНК-содер-
жащих бактериофагов состоит из четырех субъ-
единиц и имеет молекулярную массу около
210 000. Одна субъединица (Мг = 65 000) — про-
дукт гена репликазы, содержащегося в вирусной
РНК; она содержит активный центр для репли-
кации. Другие три субъединицы — это белки хо-
зяина, обычно участвующие в синтезе клеточных
белков: факторы элонгации Е. coli Ти (Мг = 30 000)
и Ts (Мг = 45 000) (которые доставляют молекулы
аминоацил-тРНК к рибосомам) и белок S1 (часть
рибосомной субъединицы 30S). Эти три белка
f хозяина помогают РНК-репликазе связаться с
З'-концами молекул вирусных РНК.
РНК-репликаза из клеток Е. coli, инфициро-
ванных QP, катализирует образование РНК, ком-
плементарной вирусной РНК, по реакции, анало-
гичной реакции ДНК-зависимых РНК-полимераз.
Синтез новых цепей РНК происходит в направле-
нии 5'-*3'; механизм идентичен механизму всех
других реакций синтеза нуклеиновых кислот на
матрице. РНК-репликаза использует в качестве
матрицы РНК и не работает с ДНК. Она не име-
ет независимой корректирующей эндонуклеаз-
ной активности и допускает примерно столько же
ошибок, сколько и РНК-полимераза. В отличие от
ДНК- и РНК-полимераз РНК-репликазы специ-
фичны к РНК из собственного вируса и обычно
не реплицируют РНК клетки-хозяина. Вот поче-
му в хозяйских клетках, содержащих много других
молекул РНК, предпочтительно реплицируются
именно РНК-содержащие вирусы.
Синтез РНК открывает важный подход
к изучению биохимической эволюции
Сложность строения и высокая организация —
вот, что отличает живые организмы от неживых
систем, и это же главное в проявлении жизнен-
ных процессов. Поддержание жизни требует,
чтобы определенные химические превращения
происходили очень быстро — особенно те, кото-
рые используют внешние источники энергии для
синтеза сложных и специализированных клеточ-
ных макромолекул. Жизнь зависит от мощных и
[152] Часть III. 26. Метаболизм РНК
Карл Бёзе, 1928-2012
Френсис Крик, 1916-2004
Лесли Оргел, 1927-2007
избирательных катализаторов (ферментов) и от
информационных систем, способных надежно
сохранять копии этих ферментов и точно их вос-
производить для передачи из поколения в поко-
ление. Хромосомы кодируют не модель клетки,
а модели ферментов, которые создают и поддер-
живают клетку Одновременная потребность в
информации и в катализе — прямо-таки класси-
ческая головоломка на тему: что было сначала
информация, для которой нужна специфическая
структура, или ферменты, необходимые для под-
держания и передачи информации?
Разгадка структурной и функциональной
сложности РНК привела в 1960-х гг. Карла Бёзе,
Френсиса Крика и Лесли Оргела к заключению,
что макромолекулы могут быть и носителями ин-
формации, и катализаторами. Открытие катали-
тических молекул РНК перевело это утверждение
из разряда догадок в разряд гипотез и послужило
причиной широкого распространения теории о
том, что «мир РНК» мог иметь большое значение
для перехода от пребиотической химии к жизни
(см. рис. 1-34 вт. 1). Предком всех живых существ
на нашей планете, который мог воспроизводить
себя через поколения от самого зарождения жизни
до настоящего времени, могла быть самореплици-
рующаяся РНК или же другой полимер с эквива-
лентными химическими характеристиками.
Как такой самореплицирующийся полимер
мог возникнуть? Как он мог сохраниться во внеш-
ней среде, в которой не было в изобилии предше-
ственников для его синтеза? Как, начав с такого
полимера, эволюция могла создать современный
ДНК-белковый мир? Для ответа на такие сложные
вопросы нужны очень тщательные эксперименты,
которые, возможно, позволят понять, как на плане-
те Земля зародилась и эволюционировала жизнь.
Предполагаемое происхождение пуриновых
и пиримидиновых оснований подтверждается
экспериментами, разработанными для проверки
гипотез о химическом составе пребиотического
мира (с. 55-56 в т. 1). Начиная с простых моле-
кул, которые могли присутствовать в древней
атмосфере нашей планеты (СН4, NH3, Н2О, Н2),
под действием электрических разрядов (напри-
мер, молнии) возникали более реакционноспо-
собные молекулы, такие как HCN и альдегиды,
а затем совокупность аминокислот и органиче-
ских кислот (см. рис. 1-33 вт. 1). По-видимому,
когда появилось много таких молекул, как HCN,
начали синтезироваться пурины и пиримидины.
Ведь уже через несколько суток в концентриро-
ванном растворе цианида аммония, нагреваемо-
го в сосуде с обратным холодильником, образу-
ется до 0,5% аденина (рис. 26-40). Аденин мог
быть первым и преобладающим нуклеотидным
компонентом, появившимся на Земле. Интерес-
но, что большинство кофакторов ферментов со-
держат в своем составе аденозин, хотя он напря-
мую не участвует в их функционировании (см.
рис. 8-38 вт. 1). Возможно, этот простой синтез
аденина из цианида подтверждает глубокие эво-
люционные связи.
холодильником
ТТ/ТК! ____________:
нагревание
(NH4CN) с обратным
Рис. 26-40. Возможная реакция синтеза аденина из ци-
анида аммония в пребиотической атмосфере. Аденин
образуется из пяти молекул цианида (выделены розовым
цветом).
26.3 РНК-зависимый синтез РНК и ДНК [153]
G
\ AGCa,
5-UGACUCUCUAAAU а
I 111 • 111 S
--IGGGAGGUUUX т?
3 Pl
Внутренняя
Рибозим гидовая
5'
'G
X ttagcaa
5'-UGACUCUCU ААи ~
I•I I I • Il V
^IGGGAGGUUUX^ t?
3' Pi
Расщепленный рибозим
последовательность
б
Матричная
РНК
Комплементарные
цепи олиго-РНК
\ / G
GGAGUACCAC AGUAGCAC
I I I I I I I I I I I I I I I I I I
CCUCAUGGUGCCUCAUCGUG
GUACCACGGAGUAGCAC
I I I I I I I I I I I I I I I II
CAUGGUGCCUCAUCGUG
Рис. 26-41. РНК-зависимый синтез полимера РНК из олигонуклеотидных предшествен-
ников. а — на первой стадии удаления аутосплайсирующегося интрона группы I из пред-
шественника рРНК у Tetrahymena происходит обратимая атака остатка гуанозина на 5'-сайт
сплайсинга. Детально изображена только область Р1 включающая в себя внутреннюю
адаптерную (гидовую или направляющую) последовательность (в рамке) и 5'-сайт сплай-
синга; остальная часть рибозима изображена в виде серо-зеленой глобулы. Полная вторич-
ная структура рибозима представлена на рис. 26-30. б — если область Р1 удаляется (что
показано в виде темно-зеленой полости), рибозим сохраняет как свою трехмерную структу-
ру, так и каталитическую способность. Новая молекула РНК, внесенная in vitro, может свя-
зываться с рибозимом таким же образом, как внутренняя адаптерная последовательность
Р1 на рис. а. Это обеспечивает матрицу для дальнейших реакций полимеризации РНК, в
которых происходит спаривание внесенной РНК и комплементарных ей олигонуклеотидов.
Рибозим может связывать эти олигонуклеотиды в реакции, обратной той, что показана на
рис. а. На рис. б показана только одна такая реакция, но повторяющиеся акты связывания и
катализа могут привести к РНК-зависимому синтезу длинных полимеров РНК.
Гипотеза «мира РНК» предполагает, что ну-
клеотидный полимер способен сам себя воспро-
изводить. Может ли рибозим осуществлять свой
собственный синтез на матрице? Аутосплайсиру-
ющийся интрон из рРНК Tetrahymena (рис. 26-30)
катализирует обратимую атаку остатка гуанозина
на фосфодиэфирную связь в 5'-сайте сплайсинга
(рис. 26-41). Если 5'-сайт сплайсинга и внутрен-
няя адаптерная последовательность удаляются из
интрона, остаток интрона может связывать цепи
[154] Часть III. 26. Метаболизм РНК
Дополнение 26-3
ПРАКТИЧЕСКАЯ БИОХИМИЯ
Метод SELEX для получения РНК
с заданными свойствами
Метод SELEX (от англ, systematic evolution of ligands
by exponential enrichment — систематическая эволюция
лигандов путем экспоненциального обогащения) ис-
пользуется для отбора аптамеров - олигинуклеотид-
ных последовательностей, которые прочно связывают-
ся со специфической молекулярной мишенью. С помо-
щью этой технологии при работе в автоматическом ре-
жиме удается идентифицировать один или несколько
антамеров с желаемой специфичностью связывания
На рис. 1 показано, как по технологии SELEX
произвести отбор РНК-последовательностей, кото-
рые связываются с АТР. На стадии Ф случайную смесь
РНК-последовательностей подвергают «искусствен-
ному отбору», пропуская через смолу со связанным на
ней АТР На практике работают со смесью, содержа-
щей примерно 10^ разных последовательностей, что
соответствует полному набору последовательностей
из 25 нуклеотидов (425 « 1015). В случае более длин-
ных молекул РНК используемый пул РНК не всегда
содержит все возможные РНК-последовагельности.
® РНК, которые не связываются на колонке, отбра-
сывают; © молекулы, которые связываются с АТР,
смывают с колонки солевым раствором и собирают.
@ Отобранные РНК амплифинируют с помощью об-
ратной транскриптазы, получая большое количество
комплементарных молекул ДНК, а затем с помощью
РНК-полимеразы синтезируют РНК, комплементар-
ные полученным молекулам ДНК. © Новый пул РНК
подвергают той же процедуре отбора, и весь цикл опе-
1015 вариантов
последовательностей
РНК
РНК, которые
не связались с АТР
Рис. 1. Метод SELEX.
РНК, спаренные с короткими олигонуклеотидами.
Часть оставшегося интактного интрона эффектив-
но действует в качестве матрицы для выравнива-
ния и легирования коротких олигонуклеотидов.
По сути, это реакция, обратная атаке гуанозина на
5'-сайт сплайсинга, но ее результат — синтез ко-
ротких РНК-последовательностей по матрице.
Самореплецирующиеся макромолекулы
быстро расходуют предшественников, которые в
условиях пребиотической атмосферы поставля-
лись медленными процессами. Таким образом,
для эффективного синтеза предшественников
уже на ранних стадиях эволюции требовались
метаболические пути, где катализаторами, воз-
можно, были рибозимы. Известные сейчас ри-
бозимы имеют ограниченные функции, и от тех
«древних» рибозимов, существование которых
можно только предполагать, сейчас не осталось
и следа. Чтобы глубже исследовать гипотезу
«мира РНК», мы должны понять, способна ли
РНК катализировать реакции, которые проис-
ходили в примитивных метаболических путях.
Обнаружению молекул РНК с новыми ката-
литическими функциями помог метод быстрого
скрининга, позволяющий находить и выделять в
смеси олигонуклеотидов последовательности, ко-
торые обладают определенными активностями.
Этот метод известен как SELEX-технология -
ускоренная эволюция в пробирке (см. доп. 26-3).
Метод использовали для получения молекул РНК,
26.3 РНК-зависимый синтез РНК и ДНК [155]
Рис. 2. Схема строения аптамера РНК, обладающего срод-
ством к АТР. Нуклеотиды, обозначенные буквами, необхо-
димы для связывания РНК с АТР.
отбора и амплификации специфических олигонуклео-
тидных последовательностей из очень сложного пула
дает возможность получения новых эффективных
терапевтических средств. Например, можно отобрать
РНК, которые прочно связываются с рецептором бел-
ка, выступающего над поверхностью клеточных мем-
бран, в частности над клетками опухолей. Блокируя
активность рецептора или направляя в клетку опухо-
ли токсин, который связывается с аптамером, можно
уничтожить клетки опухоли. Метод SELEX был ис-
пользован для отбора аптамеров ДНК при обнаруже-
нии спор сибирской язвы. Изучаются и другие интерес-
ные применения этой технологии.
раций повторяется 10 и более раз. В конце остается
лишь несколько аптамеров (РНК-последовательности
с выраженным сродством к АТР).
Структурные элементы, которые необходимы для
связывания с АТР, показаны на рис 2. Такие молекулы
связываются с АТР (и с другими аденозиновыми нукле-
отидами), Xd < 50 мкмоль. На рис. 3 приведена трехмер-
ная структура комплекса аптамера из 36 нуклеотидов с
АМР, полученного методом SELEX. Схема на рис. 2 как
раз и соответствует этому аптамеру РНК
Помимо исследований функций РНК метод
SELEX имеет важное практическое значение при
идентификации коротких молекул РНК фармацев-
тического назначения. Разумеется, нельзя получить
аптамеры, специфически связывающиеся со всеми по-
тенциальными мишенями. Но эта технология быстрого
Рис. 3. (Взято из PDB ID 1RAW). Комплекс РНК-аптамера
с АМР. В консервативных нуклеотидных последователь-
ностях (формирующих полость, где и связывается АМР)
основания показаны серым цветом, красный — связан-
ный АМР.
которые связываются с аминокислотами, органи-
ческими красителями, нуклеотидами, цианокоба-
ламииом и другими молекулами. Исследователи
выделили рибозимы, катализирующие образова-
ние сложноэфириой и амидной связей, протекание
SN2-peaKnnfi, а также металлирование порфиринов
(введение в молекулы ионов металла) и образова-
ние углерод-углеродных связей. Благодаря эво-
люции кофакторов ферментов с нуклеотидной
«ручкой», что облегчает связывание с рибозимом,
спектр доступных для изучения химических про-
цессов, протекание которых предполагается в
примитивных системах, расширился.
Как мы увидим в следующей главе, син-
тез пептидных связей катализируют некото-
рые природные молекулы РНК; поэтому, по-
видимому, вполне справедливо утверждение
о том. что мир РНК изменился благодаря вы-
сокому каталитическому потенциалу белков.
Синтез белков мог стать прорывом в эволюции
мира РНК, но мог и ускорить его гибель. Роль
носителя информации могла перейти от РНК к
ДНК, поскольку ДНК химически более стабиль-
на. РНК-репликаза и обратная транскриптаза,
возможно, это сегодняшние версии ферментов,
которые когда-то играли важную роль в осу-
ществлении перехода к современной биосисте-
ме, основанной на ДНК.
Молекулярные паразиты тоже могли прийти
из мира РНК С появлением первых неэффек-
[156] Часть III. 26. Метаболизм РНК
Дополнение 26-4 Расширяющийся мир РНК, или транскрипты с неизвестной функцией
В книге многократно приводились современные оце-
ночные данные относительно количества генов в гено-
ме человека и многих других организмов. Эти оценки
предполагают, что ученые знают о существовании гена,
если они его «видят» на основании современных пред-
ставлений о ДНК, РНК и белках. Но насколько спра-
ведлив такой подход?
Как отмечалось в гл. 9, по-видимому, менее 2%
генома человека кодирует белки. Даже с учетом ин-
тронов получается, что лишь небольшая часть генома
транскрибируется в РНК, главным образом мРНК, ко-
дирующую эти белки. Остальную часть генома иногда
называют мусорной ДНК. Однако определение «му-
сорная» указывает лишь на недостаточность наших
знаний, поскольку в настоящее время мы постепенно
начинаем понимать, что большая часть генома полно-
стью функциональна.
В попытках точнее определить размеры челове-
ческого транскриптома ученые изобрели новые ин-
струменты, позволяющие с высокой точностью иден-
тифицировать последовательности генома, которые
транскрибируются в РНК. Результаты исследований
оказались неожиданными. Выяснилось, что транс-
крибируется значительно большая часть нашего гено-
ма, чем считали ранее. Большая часть образующейся
РНК, по-видимому, не кодирует белков. Многие виды
РНК нс имеют базовых структурных элементов (на-
пример, З'-поли(А)-последовательности), характери-
зующих мРНК. Зачем же нужна эта РНК?
Большую часть методов, применяемых для ре-
шения таких задач, можно отнести к одной из двух
крупных категорий: клонирование кДНК и анализ
на микрочипах. Создание библиотек кДНК для из-
учения транскрибируемых генов какого-либо эука-
риотического генома описано в гл. 9 (см. рис. 9-14 в
т. 1). Однако классические методы получения кДНК
часто приводят к клонированию только части после-
довательности определенного транскрипта. Посколь-
ку обратная транскриптаза может останавливаться
в участках с определенной вторичной структурой
мРНК или просто диссоциировать, обычно полно-
размерные ДНК составляют не более 20% клонов в
библиотеке кДНК. Это затрудняет использование
таких библиотек для построения карт участков на-
чала транскрипции (TSS — от англ, transcription start
sites) и для изучения тех частей генов, которые ко-
дируют N-концевые последовательности белков. На
рис. 1 проиллюстрирован один из многочисленных
методов, разработанных для преодоления этих огра-
ничений. Усовершенствованные методы позволяют
создавать такие библиотеки кДНК, в которых полно-
размерные клоны составляют более 95%. что значи-
тельно расширяет возможности получения информа-
ции о составе клеточной РНК Однако кДНК обычно
создаются на основе транскриптов РНК, содержащих
поли( А)-последовательности. Применение мето-
дов анализа на микрочипах в сочетании с методами
приготовления кДНК, не связанными с наличием
поли(А)-последовательностей (рис. 2), показало, что
значительная часть РНК в клетках эукариот не имеет
характерных концевых структур.
Полная картина еще не ясна, однако уже можно
сделать некоторые выводы. Если исключить повто-
ряющиеся последовательности (например, транспо-
зоны), составляющие до половины генома млекопи-
тающих, не менее 40% (а, возможно, и подавляющая
часть) оставшейся геномной ДНК транскрибируется
в РНК. По-видимому, существует больше видов РНК
без поли(А)-«хвоста», чем видов РНК, имеющих эту
последовательность. Большая часть таких РНК не
тивных самореплицирующихся молекул транс-
позиция оказалась потенциально возможной аль-
тернативой репликации в качестве стратегии для
успешного воспроизводства и выживания. Древ-
ние молекулы паразитической РНК могли про-
сто встроиться в самореплицирующуюся моле-
кулу путем каталитической трансэтерификации,
а затем пассивно подвергаться репликации. Есте-
ственный отбор мог привести к тому, что транс-
позиция стала сайт-специфической и затрагивала
последовательности, которые не влияли на ката-
литическую активность РНК хозяина. Реплика-
торы и РНК-транспозоны могли существовать в
примитивной симбиотической взаимосвязи, при-
чем одни способствовали эволюции других. Со-
временные интроны, ретровирусы и транспозоны
могут быть рудиментами этого «путешествия на
чужом горбу» древних паразитических молекул
РНК. И сейчас эти элементы продолжают влиять
на эволюцию своих хозяев.
26.3 РНК-зависимый синтез РНК и ДНК [157]
В
В
В
в
Ткань
выделение РНК
Различные РНК-
' связывающие белки
удаление белков
иммунопреципитация
удаление белков
синтез кДНК
(праймеры и
ревертаза)
ковалентное
присоединение
биотина (В) к кэпу
расщепление РНК,
не связанной с кДНК
РНК-азой I
В--------
Б--------
В--- В
выделение
полноразмерной кДНК,
связанной с биотином
транспортируется в цитоплазму, а концентрируется
исключительно в ядре. Многие участки генома транс-
крибируются с обеих цепей: один транскрипт компле-
ментарен другому (такие последовательности называ-
ют антисмысловыми). Часто антисмысловые РНК бы-
вают задействованы в регуляции тех РНК, с которыми
они спариваются. Многие виды РНК образуются лишь
в одной или нескольких тканях, причем при анализе
новых тканей каждый раз выявляются новые транс-
крипты. Таким образом, на настоящий момент еще не
описан полный транскриптом ни одного организма.
Более того, часто новые виды РНК транскрибируются
с таких последовательностей генома (как те, что про-
иллюстрированы на рис. 9-20), которые расположены
одинаково у нескольких организмов (так называемая
синтения — связь генов с определенными хромосома-
ми). Такая эволюционная консервативность указывает
на важную функцию этих РНК.
Среди новых видов РНК, о существовании кото-
рых стало известно лишь в последние 20 лет, можно
назвать мякРНК, мяРНК и микроРНК. Были откры-
ты новые TSS. Появляются новые классы молекул
РНК. Обнаружены новые варианты альтернативного
сплайсинга. И все эти открытия изменяют наши пред-
ставления о генах. В геномах человека и мыши даже
количество кодирующих белки транскриптов мРНК
может оказаться гораздо больше, чем считалось до
сих пор, и число идентифицированных генов белков
тоже может возрасти. Однако функции большинства
недавно обнаруженных транскриптов остаются неиз-
вестными, их называют просто транскриптами с не-
известной функцией (TUF — от англ, transcripts of
unknown function). Открытие целого мира этих новых
РНК позволяет надеяться, что мы сможем лучше по-
нять механизмы работы эукариотической клетки и,
возможно, лучше представить себе мир РНК, суще-
ствовавший в далеком прошлом.
в
в
Рис. 1. Стратегия клонирования полноразмерных кДНК. Из образца ткани изолиру-
ют пул мРНК. В некоторых случаях мРНК, связанную с определенным белком, можно
получить путем иммунопреципитации этого белка с последующим выделением свя-
занной мРНК. Например, биотин ковалентно связывается с 5'-концом молекул мРНК,
используя особые свойства 5'-кэпа. Чтобы запустить обратную транскрипцию молекул
мРНК, применяют праймер поли(сГГ). РНКаза I расщепляет РНК, не входящую в состав
гибридов ДНК-РНК, таким образом, уничтожаются неполные пары кДНК-РНК. Пол-
норазмерные гибриды кДНК-РНК выделяют с помощью стрептавидиновых шариков
(которые связываются с биотином), превращают в двухцепочечные ДНК и клонируют.
[158] Часть III. 26. Метаболизм РНК
Дополнение 26-4 Расширяющийся мир РНК, или транскрипты с неизвестной функцией (продолжение)
а Геномная последовательность
AAGCTTATGGTCCCTAATCATAACGTAGCTTCCCTAATCTGA
Олигонуклеотиды, синтезированные для микрочипа:
GCTTATGGTCCCTAATCATAACGTA
TGGTCCCTAATCATAACGTAGCTTC
-----CCTAATCATAACGTAGCTTCCCTAA
TCATAACGTAGCTTCCCTAATCTGA
ДНК-
микрочип
Ткань
Ядерная РНК
ро!у(А)+
ро1у(А)~
Цитоплазматическая РНК
ро!у(А)+
ро!у(А)~
Синтез кДНК
Синтез кДНК
Синтез кДНК
Синтез кДНК
(poly(dT)-праймеры) (случайные праймеры) (poly(dT)-праймеры) (случайные праймеры)
Рис. 2. Анализ транскриптома на микрочипах, а — на основе неповторяющихся участков
генома синтезированы геномные микрочипы. Соседние олигонуклеотиды на микрочипе
перекрываются по своим последовательностям, так что любой из нуклеотидов (например,
Т, выделенный синим цветом) может быть представлен многократно. 6 — образец ткани
фракционировали для разделения ядер и цитоплазмы, и из каждой фракции выделяли РНК.
Образцы РНК, содержащие поли(А)-«хвост», отделяли от образцов, не имеющих этой последо-
вательности. пропуская РНК через колонку со связанным поли(бТ). РНК с поли(А)-«хвостом»
превращали в кДНК (рис. 1). Используя случайные праймеры, РНК без поли(А)-«хвоста»
также превращали в кДНК. Не все образующиеся фрагменты ДНК по длине соответствуют
тем РНК, на основе которых они были получены, однако общий набор ДНК содержит боль-
шую часть последовательностей исходных РНК. Затем кДНК метили и использовали в ка-
честве зондов в микрочипах. Флуоресцентные метки на микрочипе позволяют определить
транскрибированные РНК-последовательности.
Дополнительная литература для дальнейшего изучения [ 1 59]
«Мир РНК» — все еще гипотеза, в которой
многое требует объяснения, однако эксперимен-
тальные доказательства подтверждают растущий
список ключевых положений этой теории. По-
следующие эксперименты расширят наше по-
нимание мироустройства. Важные недостающие
фрагменты этой мозаики будут получены в об-
ласти фундаментальной химии, в исследованиях
живых клеток, возможно, даже на других плане-
тах. Тем временем мир РНК продолжает расши-
ряться (доп. 26-4).
Краткое содержание раздела 26.3
РНК-ЗАВИСИМЫЙ СИНТЕЗ РНК И ДНК
РНК-зависимая ДНК-полимераза (обратная
транскриптаза) была впервые обнаружена
у ретровирусов, которые в процессе своего
жизненного цикла должны превращать свой
РНК-геном в двухцепочечную ДНК. Эти
ферменты транскрибируют вирусную РНК в
ДНК; данный процесс применяется для по-
лучения комплементарной ДНК.
Многие эукариотические транспозоны род-
ственны ретровирусам, и их транспозиция
проходит через стадию образования РНК.
Теломераза — фермент, который синтезиру-
ет теломерные концы линейных хромосом, —
специализированная обратная транскрипта-
за, содержащая внутреннюю матрицу РНК.
РНК-зависимые РНК-полимеразы, такие
как репликазы РНК бактериофагов, спец-
ифичны по отношению к вирусной РНК, ко-
торую используют в качестве матрицы.
Открытие молекул каталитической РНК и
метаболических путей взаимного превраще-
ния РНК и ДНК позволило предположить,
что важным этапом эволюции было появление
РНК (или равнозначного полимера), способ-
ной катализировать собственную реплика-
цию. Биохимический потенциал молекул РНК
можно изучать методом SELEX, позволяющим
быстро отбирать последовательности РНК со
специфическим сродством или определенны-
ми каталитическими свойствами.
Ключевые термины
Термины, выделенные жирным шрифтом, объясни-
ются в глоссарии.
5'-кэп 123
сАМР-рецепторный бе-
лок (CRP) 116
аптамер 154
ДНК-зависимая РНК-по-
лимераза 108
комплементарная ДНК
(кДНК) 146
консенсусная последова-
тельность 112
матричная РНК
(мРНК) 105
микроРНК 138
мякРНК 135
мяРНК 127
обратная транскриптаза
144
первичный транскрипт
122
поли(А)-«хвост» 130
промотор 110
репрессор 116
ретровирус 144
рибозим 122
рибосомная РНК
(рРНК) 105,134
РНК-зависимая РНК-
полимераза (РНК-
репликаза) 151
сплайсинг РНК 122
сплайсосома 127
теломераза 149
транскриптом 106
транскрипция 105
транспортная РНК
(тРНК) 105,136
факторы транскрипции
120
футпринтинг 111
хоминг 148
Дополнительная литература
для дальнейшего изучения
Общая литература
Jacob, Е & Monod, J. (1961) Genetic regulatory mechanisms
in the synthesis of proteins./ Mol. Biol. 3, 318-356.
Классическая статья, в которой озвучено много важ-
ных идей.
Lodish, Н., Berk, A., Kaiser, С. A., Krieger, М., Scott,
М. Р., Bretscher, A., Ploegh, Н., & Matsudaira, Р. (2008)
Molecular Cell Biology, 6th edn, W. H. Freeman & Company,
New York.
ДНК-зависимый синтез РНК
Boeger, H., Bushnell, D., Davis, R., Griesenbeck, J., Lorch,
Y., Strattan, J., Westover, K., & Kornberg, R. (2005) Struc-
tural basis of eukaryotic gene transcription. FEBS Lett. 579,
899-903.
Bourbon, H. M., Aguilera, A., Ansari, A. Z., Asturias,
E J., Berk, A. J., Bjorklund, S., Blackwell, T. K., Borg-
grefe, T., Carey, M., Carlson, M., et al. (2004) A unified
nomenclature for protein subunits of Mediator complexes
linking transcriptional regulators to RNA polymerase II.
Mol. Cell 14, 553-557.
DeHaseth, P. L., Zupancic, M. L., & Record, M. T., Jr.
(1998) RNA polymerase-promoter interactions: the comings
and goings of RNA polymerase./ Bacteriol. 180, 3019-3025.
11601 Часть III. 26. Метаболизм РНК
Gruber, T. M. & Gross, С. А. (2003) Multiple sigma subunits
and the partitioning of bacterial transcription space. Annu. Rev.
Microbiol. 57,441-466.
Laine, J. P. & Egly, J. M. (2006) When transcription and re-
pair meet: a complex system. Trends Genet. 22,430-436.
Mooney, R. A., Darst, S. A., & Landick, R. (2005) Sigma
and RNA polymerase: an on-again, off-again relationship? Mol.
Cell 20,335-345.
Murakami, K. S. & Darst, S. A. (2003) Bacterial RNA poly-
merases: the wholo story. Curr. Opin. Struct. Biol. 13, 31-39.
Svejstrup, J. Q., Conaway, R. C., & Conaway, J. W. (2006)
RNA polymerase II: a “Nobel” enzyme demystified. Mol. Cell
24, 637-642.
Thomas, M. C. & Chiang, С. M. (2006) The general tran-
scription machinery and general cofactors. Crit. Rev. Biochem.
Mol. Biol. 41,105-178.
Процессинг РНК
Blencowe, В. J. (2006) Alternative splicing: new insights
from global analyses. Cell 126, 37-47.
Butcher, S. E. & Brow, D. A. (2005) Towards understanding
the catalytic core structure of the spliceosome. Biochem. Soc.
Trans. 33,447-449.
Decatur, W. A. & Fournier, M. J. (2003) RNA-guided nucleo-
tide modification of ribosomal and other RNAs./ Biol. Chem.
278,695-698.
Gogarten, J. P. & Hilario, E. (2006) Inteins, introns, and
homing endonucleases: recent revelations about the life cycle
of parasitic genetic elements. BMC Evol. Biol. 6,94.
Granneman, S. & Baserga, S. J. (2005) Crosstalk in gene ex-
pression: coupling and co-regulation of rDNA transcription,
pre-ribosome assembly, and pre-rRNA processing. Curr. Opin.
Cell Biol. 17,281-286.
Huang, Y. Q. & Steitz, J. A. (2005) SRprises along a messen-
ger's journey. Mol. Cell 17,613-615.
Kaberdin, V. R. & Blasi, U. (2006) Translation initiation
and the fate of bacterial mRNAs. FEMS Microbiol. Rev. 30,
967-979.
Parker, R. & Song, H. W. (2004) The enzymes and control of
eukaryotic mRNA turnover. Nat. Struct. Mol. Biol. 11,121-127.
Rodriguez-Trelles, E, Tarrio, R., & Ayala, F. J. (2006) Ori-
gins and evolution of spliceosomal introns. Annu. Rev. Genet.
40,47-76.
Schneider, D. A., Michel, A., Sikes, M. L., Vu, L., Dodd,
J. A., Salgia, S., Osheim, Y. S., Beyer, A. L., & Nomura,
M. (2007) Transcription elongation by RNA polymerase I is
linked to efficient rRNA processing and ribosome assembly.
Mol. Cell26, 217-229.
Wienholds, E. & Plasterk, R. H. A. (2005) MicroRNA func-
tion in animal development. FEBS Lett. 579, 5911-5922.
РНК-зависимый синтез РНК или ДНК
Blackbum, Е. Н., Greider, С. W, & Szostak, J. W. (2006)
Telomeres and telomerase: the path from maize, Tetrahymena
and yeast to human cancer and aging. Nat. Med. 12,1133-1138.
Boeke, J. D. & Devine, S. E. (1998) Yeast retrotransposons:
finding a nice, quiet neighborhood. Cell 93,1087-1089.
Cech, T. R. (2004) Beginning to understand the end of the
chromosome. Cell 116,273-279.
Frankel, A. D. & Young, J. A. T. (1998) HIV-1: fifteen pro-
teins and an RNA. Annu. Rev. Biochem. 67,1-25.
Griffith, J. D., Comeau, L., Rosenfield, S., Stansel, R. M.,
Bianchi, A., Moss, H., & de Lange, T. (1999) Mammalian
telomeres end in a large duplex loop. Cell 97,503-514.
Temin, H. M. (1976) The DNA provirus hypothesis: the es-
tablishment and implications of RNA-directed DNA synthesis.
Science 192,1075-1080.
Обсуждение первоначального значения обратной
транскрипции у ретровирусов.
Рибозимы и эволюция
Caminci, Р. (2007) Constructing the landscape of the mam-
malian genome./ Exp. Biol. 210,1497-1506.
Хороший обзор работ, показывающий, что транскрип-
томы млекопитающих гораздо более обширны, чем счита-
лось раньше.
DeRose, V. J. (2002) Two decades of RNA catalysis. Chem.
Biol. 9,961-969.
Doudna, J. A. & Lorsch, J. R. (2005) Ribozyme catalysis:
not different, just worse. Nat. Struct. Mol. Biol. 12, 395—
402.
Green, R. & Doudna, J. A. (2006) RNAs regulate biology.
ACS Chem. Biol. 1, 335-338.
Huttenhofer, A. & Vogel, J. (2006) Experimental ap-
proaches to identify non-coding RNAs. Nucleic Acids Res.
34, 635-646.
Joyce, G. E (2002) The antiquity of RNA-based evolution.
Nature 418, 214-221.
Kazantsev, A. V. & Pace, N. R. (2006) Bacterial RNase
P: a new view of an ancient enzyme. Nat. Rev. Microbiol. 4,
729-740.
Miiller, U. E (2006) Re-creating an RNA world. Cell. Mol. Life
Sci. 63,1278-1293.
Narlikar, G. J. & Herschlag, D. (1997) Mechanistic aspects
of enzymatic catalysis: lessons from comparison of RNA and
protein enzymes. Annu. Rev. Biochem. 66,19-59.
Willingham, A. T. & Gingeras, T. R. (2006) TUF love for
“junk” DNA. CeZZ 125,1215-1220.
Yams, M. (2002) Primordial genetics: phenotype of the ribo-
cyte. Annu. Rev. Genet. 36,125-151.
Подробное обсуждение того, какой могла бы быть
жизнь на основе РНК, и хороший обзор эксперименталь-
ных результатов, лежащих в основе этой гипотезы.
Вопросы и задачи 11611
Вопросы и задачи________________________
1. РНК-полимераза. (а) Сколько времени по-
требуется РНК-полимеразе Е. coli для синте-
за первичного транскрипта генов Е. coli, ко-
дирующих ферменты метаболизма лактозы
(/б/с-оперон размером 5300 и. н., рассмотрен
в гл. 28)? (б) На какое расстояние вдоль цепи
ДНК может продвинуться транскрипционный
«пузырек», образованный РНК-полимеразой,
за 10 секунд?
2. Коррекция ошибок РНК-полимеразой. ДНК-
полимеразы могут корректировать ошибки, а
корректирующая способность РНК-полимеразы
весьма ограниченна. Учитывая, что замена един-
ственного основания при репликации или транс-
крипции может привести к ошибке в синтезе бел-
ка, приведите возможное биологическое объясне-
ние такого поразительного различия.
3. Посттранскрипционный процессинг РНК.
Предскажите вероятные последствия мутации в
последовательности (5')-AAUAAA в транскрипте
эукариотической мРНК.
4. Кодирующая и матричная последователь-
ности. РНК-геном фага Q(3 представляет собой
нематричную, или кодирующую, цепь, и при по-
падании в клетку он функционирует в качестве
мРНК. Предположим, что РНК-репликаза фага
QP синтезирует главным образом матричную
цепь РНК и включает именно ее, а не кодирую-
щую цепь, в состав вирусных частиц. Что будет с
матричными цепями, когда они попадут в новую
клетку? Какой фермент нужно включить в состав
вирусных частиц для успешного инфицирования
клетки-хозяина?
5. Химия биосинтеза нуклеиновых кислот.
Опишите три общих свойства реакций, катализи-
руемых ДНК-полимеразой, РНК-полимеразой,
обратной транскриптазой и РНК-репликазой.
Чем фермент полинуклеотидфосфорилаза похож
на эти три фермента и чем отличается от них?
6. Сплайсинг РНК. Каково минимальное число
реакций трансэтерификации, необходимое для
сплайсинга интрона из транскрипта мРНК? По-
ясните ответ.
7. Процессинг РНК. Если в клетках позвоноч-
ных блокируется сплайсинг мРНК, блокируются
и реакции модификации рРНК. Объясните это
наблюдение.
8. РНК-геномы. У РНК-содержащих вирусов от-
носительно маленькие геномы. Например, одно-
цепочечные молекулы РНК ретровирусов состо-
ят примерно из 10 000 нуклеотидов, а РНК фага
QP содержит лишь 4220 нуклеотидов. Зная свой-
ства обратной транскриптазы и РНК-репликазы,
описанные в этой главе, объясните, почему у это-
го вируса такой маленький геном.
9. Скрининг молекул РНК методом SELEX.
Максимальное число разных последователь-
ностей РНК, которые можно подвергнуть
скринингу методом SELEX, составляет 1015.
(а) Предположим, вы работаете с олигонукле-
отидами длиной 32 нуклеотида. Сколько вари-
антов этих молекул может быть в их случайном
наборе, содержащем все возможные последова-
тельности? (б) Какую часть из них (в %) можно
проанализировать методом SELEX? (в) Пред-
положим, вы хотите выбрать молекулу РНК,
которая катализирует гидролиз определенного
эфира. На основании того, что вам известно о
катализе (гл. 6 в т. 1), предложите стратегию,
которая позволит вам выбрать подходящий ка-
тализатор.
10. Отравление бледной поганкой. Бледная по-
ганка {Amanita phalloides) содержит несколько
опасных соединений, включая смертельный яд
а-аманитин. Этот токсин блокирует элонгацию
РНК путем связывания с эукариотической РНК-
полимеразой II с очень высоким сродством; он
смертельно опасен уже в такой низкой концен-
трации, как 10-8 М. Сначала у человека, съевше-
го этот гриб, проявляется желудочно-кишечное
расстройство (вызываемое некоторыми другими
токсинами). Потом эти симптомы исчезают, но
через 48 ч человек умирает, обычно от поражения
печени. Объясните, почему а-аманитину нужно
столько времени, чтобы погубить человека?
11. Выявление штаммов возбудителя ту-
беркулеза, устойчивых к рифампицину. Ри-
фампицин — важный антибиотик, применяемый
для борьбы с туберкулезом и другими заболева-
11621 Часть III. 26. Метаболизм РНК
ниями, вызываемыми микобактериями. Некото-
рые штаммы Mycobacterium tuberculosis (возбуди-
тель туберкулеза) устойчивы к рифампицину Эти
штаммы приобрели устойчивость в результате
мутации гена гроВ, кодирующего р-субъединицу
РНК-полимеразы. Рифампицин не может свя-
заться с мутантной РНК-полимеразой и поэтому
не блокирует инициацию транскрипции. Было об-
наружено, что последовательности ДНК из боль-
шого числа рифампицин-устойчивых штаммов
М. tuberculosis несут мутации в специфической
области из 69 пар оснований гена гроВ. В одном
хорошо изученном штамме с устойчивостью к ри-
фампицину заменена одна пара оснований в гене
гроВ, и это привело к единичной аминокислот-
ной замене в кодирующий р-субъединице: вместо
остатка His появился остаток Asp.
а) Основываясь на знаниях химии белков
(гл. 3 и 4 в т. 1), предложите способ, который позво-
ляет выявить рифампицин-устойчивый штамм,
содержащий этот конкретный мутантный белок.
б) Основываясь на знаниях химии нуклеино-
вых кислот (гл. 8 в т. 1), предложите способ иден-
тификации мутантного гена гроВ.
Биохимия в Интернете
12. Ген рибонуклеазы. Рибонуклеаза поджелу-
дочной железы человека состоит из 128 амино-
кислотных остатков.
а) Какое минимальное число пар нуклеоти-
дов требуется для кодирования данного белка?
б) мРНК из клеток поджелудочной желе-
зы копировали обратной транскриптазой для
создания библиотеки ДНК человека. После-
довательность мРНК, кодирующую панкреа-
тическую рибонуклеазу человека, определяли
путем секвенирования комплементарной ДНК
(кДНК) из этой библиотеки, которая включает
открытую рамку считывания для этого белка.
Используйте базу данных Entrez database sys-
tem (www.ncbi.nlm.nih.gov/Entrez), чтобы най-
ти опубликованную последовательность этой
мРНК (изучите базу данных CoreNucleotide
для учетного номера D26129). Какова длина
этой мРНК?
в) Как объяснить несоответствие между раз-
мером, который определили вы (пункт а), и ре-
альным размером мРНК?
Анализ экспериментальных данных__________
13. Пример редактирования РНК. Рецептор
АМРА (а-амино-3-гидрокси-5-метил-4-изокса-
золпропионовой кислоты) — важный компонент
нервной системы человека. Он присутствует
в нескольких формах в разных нейронах, при-
чем отчасти это разнообразие объясняется пост-
транскрипционными модификациями. Данная
задача посвящена изучению механизмов такого
редактирования РНК.
В 1991 г. Соммер с соавторами анализиро-
вали последовательность, кодирующую важ-
ный остаток Arg в молекуле рецептора АМРА.
В последовательности кДНК (см. рис. 9-14 в
т. 1) рецептора АМРА остатку аргинина соот-
ветствовал кодон CGG (см. рис. 27-7). Однако
в геномной ДНК в этой позиции стоял кодон
CAG (Gin).
а) Объясните, как такой результат согла-
суется с наличием посттраискрипционной мо-
дификации мРНК рецептора АМРА. Рютер с
коллегами (1995) подробно исследовали дан-
ный вопрос. Сначала они разработали метод,
позволяющий различать модифицированные
и немодифицированные транскрипты с по-
мощью секвенирования ДНК по Сенгеру (см.
рис. 8-33 вт. 1). Они модифицировали метод,
чтобы определить, было ли это основание А
(как в CAG) или нет. Они синтезировали два
ДНК-праймера на основе последовательности
геномной ДНК в этом участке гена рецептора
АМРА. Ниже представлены праймеры, а также
последовательность геномной ДНК нематрич-
ной цепи для соответствующего участка гена
рецептора АМРА; модифицированное основа-
ние А выделено красным цветом.
Чтобы определить, сохранилось ли осно-
вание А или было заменено на какое-то другое,
Рютер с соавторами использовали описанную
ниже процедуру.
1. Готовили кДНК, комплементарную мРНК, ис-
пользуя праймер 1, обратную транскриптазу,
dATP, dGTP, dCTP и dTTP.
2. Удаляли мРНК.
3. Отжигали праймер 2, меченный 32Р, с кДНК
и проводили реакцию с ДНК-полимеразой,
Анализ экспериментальных данных 1163]
dGTP, dCTP, dTTP и ddATP (дидезокси-АТР;
см. рис. 8-33).
4. Денатурировали образующиеся дуплексы и
разделяли их с помощью электрофореза в по-
лиакриламидном геле (с. 136 в т. 1).
5. Детектировали меченную 32Р ДНК методом ав-
торадиографии.
Ученые обнаружили, что из модифицирован-
ной мРНК образовалась [32Р]ДНК длиной
22 нуклеотида, тогда как из немодифициро-
ванной мРНК образовалась [32Р]ДНК длиной
19 нуклеотидов.
б) Используя представленные ниже последо-
вательности, объясните, почему модифицирован-
ная и немодифицированная мРНК давали разные
продукты.
Используя тот же подход для измерения
доли транскриптов, подвергающихся редакти-
рованию при различных условиях, исследова-
тели обнаружили, что в экстрактах культуры
эпителиальных клеток (так называемых клеток
HeLa) происходит активное редактирование
мРНК. Для изучения механизмов этого процес-
са ученые приготовили активный экстракт кле-
ток HeLa, как описано в таблице, и определяли
его способность модифицировать мРНК рецеп-
тора АМРА. Протеиназа К расщепляет толь-
ко белки, а микрококковая нуклеаза — только
ДНК.
Образец Предобработка Доля редактированной мРНК, %
1 Нет 18
2 Протеиназа К 5
3 Нагревание до 65 °C 3
4 Нагревание до 85 °C 3
5 Микрококковая нуклеаза 17
в) Используйте эти данные для доказательства
того, что в процессах редактирования мРНК при-
нимают участие белки. В чем заключается наибо-
лее слабое место этого доказательства?
Для точной идентификации редактирован-
ного основания Рютер с коллегами использовали
описанную ниже процедуру.
1. Синтезировали мРНК, добавляя в реакцион-
ную смесь [а-32Р]АТР.
2. Модифицировали меченую мРНК путем инку-
бации с экстрактом клеток HeLa.
3. Гидролизовали модифицированную мРНК до
отдельных нуклеотидмонофосфатов с помо-
щью нуклеазы Р1.
4. Разделяли нуклеотидмонофосфаты методом
тонкослойной хроматографии (см. рис. 10-24 в
т. 1).
5. Идентифицировали образующиеся 32Р-мече-
ные нуклеотидмонофосфаты методом автора-
диографии.
В неизмененной мРНК исследователи обна-
ружили только [32Р] АМР, а в модифицированной
мРНК содержался главным образом [32Р]АМР,
но также некоторое количество [32Р]1МР (ино-
зинмонофосфат, см. рис. 22-34 в т. 2).
г) Почему в этом эксперименте использова-
ли [а-32Р]АТР, а не [0-32Р]АТР или [у-32Р]АТР?
д) Почему использовали [а-32Р]АТР, а не
[a-32P]GTP, [a-32P]CTP или [a-32P]UTP?
е) Как полученные результаты исключают
возможность удаления всего нуклеотида А (са-
хар, основание и фосфат) и его заменены на ну-
клеотид I?
Далее исследователи модифицировали
мРНК, меченную [2,8-3Н]АТР, и повторили опи-
санную выше процедуру. Метка 3Н содержалась
лишь в мононуклеотидах АМР и IMP.
(5 ) ...GTCTCTGGTTTTCCTTGGGTGCCTTTATGCAGCAAGGATGCGATATTTCGCCAAG...
Праймер 1: С GTT С СТ AC GCT AT AAAGC GGTT С (5')
Праймер 2: (5Z) CCTTGGGTGCCTTTA
[НИ ] Часть III. 26. Метаболизм РНК
ж) Как этот результат исключает возмож-
ность удаления основания А (при сохранении
сахарофосфатного остова) при замене на основа-
ние I? Каков в этом случае наиболее вероятный
механизм редактирования?
з) Как замена остатка А на I в мРНК объяс-
няет замену Gin на Arg в белковой последователь-
ности двух форм рецептора АМРА? (Подсказка.
См. рис. 27-8).
Литература
Rueter, S. М., Bums, С. М., Coode, S. A., Mookheijee, Р.,
& Emesont, R. В. (1995) Glutamate receptor RNA editing in
vitro by enzymatic conversion of adenosine to inosine. Science
267,1491-1494.
Sommer, B., Kohler, M., Sprengel, R., & Seeburg, P. H.
(1991) RNA editing in brain controls a determinant of ion flow
in glutamate-gated channels. Cell 67,11-19.
Очевидно, что открытия Гарри [Ноллера] не объясняют, как началась
жизнь, и не дают ответа на вопрос, что же было до РНК. Но накапливается
все больше косвенных доказательств, что на нашей планете была жизнь и
до нас, вот от нее мы и произошли — это факт!
— Джеральд Джойс, из комментария в Science, 1992
Метаболизм белка
27.1. Генетический код 166
27.2. Синтез белков 178
27.3. Транспорт и расщепление белков 210
Белки — конечный продукт большинства ин-
формационных метаболических путей. В
каждый момент времени клетке требуются
тысячи разных белков. Они должны синтезиро-
ваться в соответствии с потребностями клетки,
доставляться к месту локализации и разрушать-
ся, если в них больше нет нужды.
Установление механизма биосинтеза бел-
ка, очень сложного жизненно важного процес-
са, — один из самых замечательных успехов
биохимии. У эукариот в синтезе белка участву-
ет более 70 различных рибосомных белков, 20
или более ферментов для активации аминокис-
лот, 10 или более вспомогательных ферментов
и других белковых факторов для инициации,
элонгации и терминации синтеза полипепти-
дов, около 100 дополнительных ферментов для
заключительного процессинга белков и 40 или
более типов транспортных и рибосомных РНК.
В целом в синтезе полипептидов задействовано
почти 300 разных макромолекул. Многие из этих
макромолекул включены в сложные трехмерные
структуры рибосом.
Значение синтеза белка для клетки можно по-
нять, если оценить клеточные ресурсы, направлен-
ные на реализацию этого процесса. На синтез белка
может расходоваться до 90% химической энергии,
затрачиваемой клеткой на все реакции биосинтеза.
В каждой клетке бактерий, архей или эукариот со-
держится от нескольких единиц до нескольких ты-
сяч копий многих белков и молекул РНК. В типич-
ной бактериальной клетке 15 000 рибосом, 100 000
белковых факторов и ферментов, связанных с син-
тезом белка, и 200 000 молекул тРНК, которые со-
ставляют более 35% сухой массы клетки.
Несмотря на то что синтез белка — очень
сложный процесс, он происходит исключитель-
но быстро. Полипептид из 100 аминокислотных
остатков синтезируется в клетке Escherichia coli
(при 37 °C) примерно за 5 секунд. Синтез тысяч
различных белков в клетке регулируется таким
образом, что их количество точно соответствует
текущему метаболическому состоянию. Для под-
держания необходимого состава и концентрации
белков скорости процессов транспорта и расще-
пления должны соответствовать скорости синте-
за. Благодаря многочисленным исследованиям
мы постепенно приоткрываем занавес, скрыва-
ющий слаженный молекулярный танец, в ходе
которого каждый белок занимает свое место в
клетке и избирательно разрушается, если он уже
не нужен.
В процессе изучения синтеза белка мы от-
крыли мир каталитических молекул РНК, ко-
торый мог существовать до того, как зародилась
жизнь в ее современном виде. Исследователи
установили строение бактериальных рибосом,
что позволило понять процессы синтеза белка в
клетке во всех изумительных подробностях. Что
же они обнаружили? Белки синтезируются ги-
гантскими РНК-ферментами!
[166] Часть III. 27. Метаболизм белка
27.1. Генетический код
Современные представления о биосинтезе белка
основаны на трех больших научных открытиях.
В начале 1950-х гг. Пол Замечник с сотрудниками
провели серию экспериментов, чтобы выяснить,
где в клетке происходит синтез белков. Они вво-
дили крысам путем инъекции радиоактивно ме-
ченные аминокислоты, а через разные интервалы
времени после инъекции у мертвого животного
Пол Замечник, 1912-2009
брали печень, гомогенизировали ее, фракциони-
ровали гомогенат центрифугированием и иссле-
довали субклеточные фракции для обнаружения
радиоактивного белка. Через несколько часов
и суток после инъекции меченых аминокислот
все субклеточные фракции содержали меченые
белки. Однако если после инъекции проходили
минуты, меченые белки обнаруживались только
во фракции, содержащей маленькие рибонукле-
опротеиновые частицы. Эти частицы, видимые
в тканях животных с помощью электронного
микроскопа, затем были идентифицированы как
место синтеза белка из аминокислот; позднее их
назвали рибосомами (рис. 27-1)
Малой Хогланд и Замечник сделали и второе
ключевое открытие, обнаружив, что аминокис-
лоты «активировались» при инкубации с АТР и
цитозольной фракцией клеток печени Амино-
кислоты оказывались присоединенными к тер-
мостабильной растворимой РНК определенного
типа, которая была открыта и описана Робертом
Холли и позднее названа транспортной РНК
(тРНК); в результате образовывались молекулы
аминоацил-тРНК. Ферменты, которые катали-
зируют этот процесс, называются аминоацил-
тРНК-синтетазами.
Цитоплазма Просвет ЭР
Рис. 27-1. Рибосомы и эндоплазматический ретикулум.
Электронная микрофотография и схематическое изо-
бражение фрагмента клетки поджелудочной железы, на
которых показано присоединение рибосом к внешней
(обращенной к цитоплазме) поверхности эндоплазмати-
ческого ретикулума (ЭР). Рибосомы выглядят как много-
численные маленькие точки по краям параллельно рас-
положенных слоев мембран.
мРНК
Нуклеотидный триплет,
кодирующий аминокислоту
Рис. 27-2. Адаптерная гипотеза Крика. Теперь мы знаем,
что аминокислота ковалентно связывается с З'-концом
молекулы тРНК, а специфический нуклеотидный три-
плет в молекуле тРНК взаимодействует с определенным
триплетным кодоном в мРНК, образуя водородные связи
между комплементарными основаниями.
27.1 Генетический код [167]
Третье важное открытие появилось в резуль-
тате исследований Френсиса Крика. Он поставил
важный вопрос: каким образом генетическая ин-
формация, закодированная в четырехбуквенном
языке нуклеиновых кислот, может переводиться
на 20-буквенный язык белков? Роль адаптера мог-
ли бы выполнять маленькие молекулы нуклеино-
вой кислоты (возможно, РНК) — одна часть адап-
терной молекулы связывается со специфической
аминокислотой, другая часть распознает нукле-
отидную последовательность, кодирующую дан-
ную аминокислоту в составе мРНК (рис. 27-2).
Вскоре удалось проверить эту гипотезу. Адаптер-
ная молекула тРНК переводила (транслировала)
нуклеотидную последовательность мРНК в ами-
нокислотную последовательность полипептида.
Весь процесс направляемого мРНК синтеза бел-
ка часто называют просто трансляцией.
Вскоре после этих открытий были выявлены
основные стадии синтеза белка и расшифрован
генетический код, который определяет каждую
аминокислоту.
Генетический код был расшифрован
с помощью искусственных мРНК
К 1960-м гг. стало очевидно, что для кодирования
каждой аминокислоты необходимы по меньшей
мере три нуклеотидных остатка в ДНК. Четыре
буквы кода ДНК (А, Т, G, С), сгруппированные по
две, могут дать только 42 = 16 различных комби-
наций, что недостаточно для кодирования 20 ами-
нокислот. При группировке буквенного кода ДНК
по 3 возникают 64 комбинации (43 = 64).
Несколько ключевых принципов кодирова-
ния было установлено уже в ранних генетических
исследованиях (рис. 27-3, 27-4). Кодон — триплет
нуклеотидов, кодирующий определенную амино-
кислоту. Трансляция осуществляется таким обра-
зом, что эти нуклеотидные триплеты считываются
в виде непрерывной последовательности. Первый
кодон в последовательности определяет рамку
считывания, в пределах которой каждый новый
кодон начинается через три нуклеотидных остатка.
Знаков пунктуации между кодонами нет. Амино-
кислотная последовательность белка определяется
линейной последовательностью соседних трипле-
тов. В принципе каждая одноцепочечная последо-
вательность ДНК или мРНК может иметь три рам-
ки считывания. Каждая рамка считывания содер-
Неперекрещива- ,А U ANC G ANG U С,,_____
ющийся код 12 3
Перекрещива- AUACGAGUC
ющийся код 1
2
3
Рис. 27-3. Перекрывающийся и неперекрывающийся
генетический код. В неперекрывающемся коде кодоны
(пронумерованы последовательно) расположены подряд,
т. е. не разделены нуклеотидами. В перекрывающемся коде
некоторые нуклеотиды в мРНК относятся к нескольким
разным кодонам. При триплетном коде с максимальным
перекрыванием многие нуклеотиды (например, третий
слева (А)) относятся к трем кодонам. Обратите внимание,
что в перекрывающемся коде триплетная последователь-
ность первого кодона ограничивает возможные варианты
последовательностей для второго кодона. Неперекрываю-
щийся код обеспечивает значительно большую вариабель-
ность нуклеотидов в соседних кодонах и, соответственно,
возможных аминокислотных последовательностей, обо-
значаемых этим кодом. У всех известных в настоящее вре-
мя организмов генетический код неперекрывающийся.
жит свою последовательность кодонов (рис. 27-5),
но только одна из них соответствует конкретному
белку. Остается вопрос: какие трехбуквенные кодо-
ны каким аминокислотам соответствуют?
В 1961 г. Маршалл Ниренберг и Генрих Мат-
теи опубликовали работу, ставшую первым про-
рывом в расшифровке генетического кода. Они
инкубировали синтетический полиуридилат
(полиН) с экстрактом Е. coli, GTP, АТР и смесью
20 аминокислот в 20 разных пробирках — каждая
пробирка содержала одну радиоактивно меченную
аминокислоту. Поскольку мРНК полиН состоит
из большого количества триплетов UUU, на ней,
Маршалл Ниренберг,
1927-2010
[168] Часть III. 27. Метаболизм белка
мРНК 5-- GUAGC С U AC GGA U----------------------3'
(+)
Инсерция -----^(3 U A [G С t?][U С А1] С G G|^A U----
(-)
Делеция — G U А^С~ С U||A С G|1G А Ц|------------
(+) (-)
Инсерция __„д и а][А_~(Г C][C^~_Cj[G G А| U--------
и делеция ।___>
Рамка считывания
восстанавливается
Рис. 27-4. Неперекрывающийся триплетный код. Принципы организации генетическо-
го кода были установлены с помощью многочисленных и разнообразных экспериментов, в
том числе генетических экспериментов по изучению мутаций типа инсерций или делеций.
Инсерция или делеция одной пары оснований (показано на примере транскрипции мРНК)
изменяет последовательность триплетов неперекрывающегося кода; это изменение отража-
ется на всех последующих аминокислотах, кодируемых мРНК. Сочетание инсерции и делеции
изменяет отдельные аминокислоты, но в результате может восстановить исходную аминокис-
лотную последовательность. Встраивание или удаление трех нуклеотидов (не показано) не
меняет состава триплетов, и это доказывает, что кодон состоит из трех, а не из четырех или
пяти нуклеотидов. Серым цветом отмечены кодоны, транскрибируемые с исходного гена; го-
лубым цветом показаны новые кодоны, возникшие в результате инсерции или делеции.
как на матрице, может синтезироваться только по-
липептид, состоящий из одной аминокислоты, ко-
дируемой триплетом ULT U. В самом деле, радио-
активный полипептид образовался только в одной
пробирке, содержащей радиоактивный фенил-
аланин. Ниренберг и Маттеи сделали вывод, что
триплет UUU кодирует фенилаланин. С помощью
аналогичного подхода было доказано, что полици-
тидилат (полиС) кодирует полипептид, состоя-
щий исключительно из пролина (полипролин), а
полиаденилат (полиА) кодирует полилизин. По-
лигуанилат не образует каких-либо пептидов в та-
ком экстракте, поскольку он спонтанно формиру-
ет четырехцепочечные структуры (см. рис 8-20),
которые не могут связываться рибосомами.
Синтетические полинуклеотиды для этих
экспериментов были получены с помощью по-
линуклеотидфосфорилазы (с. 26.2), которая
катализирует образование РНК-полимеров, на-
чиная с ADP, UDP, CDP и GDP. Этот фермент,
открытый Северо Очоа, не нуждается в матрице
и создает макромолекулы, состав которых зави-
сит только от относительного содержания в среде
нуклеозид-5'-дифосфатов. В присутствии одного
UDP фермент синтезирует только полиП. Если
же его добавить к смеси из пяти частей ADP и од-
ной части CDP, он образует полимер, в котором
пять шестых составляют аденилаты, а одну ше-
стую — цитидилаты. Это нерегулярный полимер,
содержащий много триплетов ААА, меньше три-
Рамка считывания 1 5'-- UUC U С GGA С C~U GG A G A~U U с”А С" A G U1--3'
Рамка считывания 2 ----U||U С U][C~ G G|[A С C~||U G G][A G~AnPu U C||A C~T[G~U~--
Рамка считывания 3 ----U U| |C U C| |G G Ai IС C U] |G G A |G A U| [U C~A |C~A~G | [ЁГ--
Рис. 27-5. Рамки считывания и генетический код. В неперекрывающемся триплетном коде
все молекулы мРНК могут иметь до трех рамок считывания, которые здесь изображены раз-
ными цветами. Триплеты и, соответственно, кодируемые ими аминокислоты разные в каж-
дой рамке считывания.
27.1 Генетический код [169]
Включение аминокислот в состав полипептидов в присутствии нерегулярных полимеров — РНК
Аминокислота Наблюдаемая частота Предполагаемый включения (Lys=100) нуклеотидный состав* * соответствующего кодона Ожидаемая частота включения, основанная на условной частоте для Lys = 100
Аспарагин 24 А2С Глутамин 24 А2С Гистидин 6 АС2 Лизин 100 ААА 20 20 4 100
Пролин 7 АС2, ССС Треонин 26 А2С, АС2 4,8 24
Примечание. Представленные данные получены в одном из первых экспериментов по рас-
шифровке генетического кода. Полипептид синтезировали на матрице синтетической РНК,
содержащей только остатки А и С в соотношении 5:1, после чего определяли количествен-
ный и качественный состав аминокислот в составе полипептида. Далее, исходя из относи-
тельной частоты встречаемости остатков А и С в составе синтетической РНК и условно при-
нимая частоту встречаемости кодона ААА (самый распространенный кодон) за 100, было
подсчитано, что должно быть три варианта кодонов состава А2С, каждый с частотой встре-
чаемости 20; три варианта кодонов состава АС2, каждый с частотой встречаемости 4, а от-
носительная частота встречаемости кодона ССС должна составлять 0,8. Смысл кодона ССС
установлен в предыдущих экспериментах с использованием последовательности поли(С)
* Эти данные ничего не говорят о последовательности нуклеотидов в составе кодона (конеч-
но, за исключением кодонов ААА и ССС).
плетов ААС, АСА и САА, относительно немного
ACC, ССА и С АС и совсем мало триплетов ССС
(табл. 27-1). С помощью разнообразных искус-
ственных молекул мРНК, созданных с помощью
полинуклеотидфосфорилазы из разных старто-
вых смесей ADP, GDP, UDP и CDP, исследовате-
ли вскоре идентифицировали состав триплетов,
кодирующих почти все аминокислоты. Хотя эти
эксперименты позволили выявить состав основа-
ний в триплетах, последовательность оснований
этим способом установить не удалось.
КЛЮЧЕВЫЕ ДОГОВОРЕННОСТИ. Далее в нашем из-
ложении мы многократно будем встречаться с
тРНК. Специфичность каждой тРНК к амино-
кислоте обозначается в виде верхнего индек-
са, например тРНКА1а, а аминоацилированная
тРНК пишется через дефис: аланил-тРНКА1а или
А1а-тРНКА1а.
В 1964 г. Ниренберг и Филипп Ледер сде-
лали другое важное открытие. Изолированные
рибосомы Е. coli могут связывать специфиче-
скую аминоацил-тРНК в присутствии соответ-
ствующей синтетической полинуклеотидной ма-
трицы. Например, рибосомы, инкубированные с
поли(Н) и фенилаланил-тРНКРЬе(РЬе-тРНКр11с),
связывают молекулы РНК обоих типов, но,
если рибосомы инкубируют с поли(П) и неко-
торыми другими аминоацил-тРНК, молекулы
аминоацил-тРНК не связываются, поскольку
не распознают триплеты в поли(И) (табл. 27-2).
Даже последовательности из трех нуклеотидов
могут специфически связываться с соответству-
Таблица 27-2 Тринуклеотиды, способные индуцировать специфическое связывание молекул аминоацил-тРНК с рибосомами
Относительное увеличение
количества меченой аминоацил-тРНК,
связанной с рибосомой*
Тринуклеотиды РЬе-тРНКРЬе Lys-TPHKLys Рго-тРНКРго
иии 4.6 0 0
ААА 0 7,7 0
ССС 0 0 3,1
Исгочник. По Nirenberg, М. & Leder. Р. (1964) RNA code
words and protein synthesis. Science 145.1399. с изм.
* Числа указывают, во сколько раз возросло количество
связанного 14С в присутствии соответствующего тринук-
леотида по сравнению с контролем (без тринуклеотида).
[170] Часть III. Z1. Метаболизм белка
ющими молекулами тРНК, поэтому такие экс-
перименты можно проводить даже с короткими
синтетическими молекулами олигонуклеотидов.
С помощью данного метода удалось определить,
какие аминоацил-тРНК связываются пример-
но с 50 из 64 возможных триплетных кодонов.
Некоторым кодонам не соответствуют никакие
молекулы аминоацил-тРНК или соответствует
несколько. Для завершения расшифровки и под-
тверждения генетического кода требовался дру-
гой метод.
Примерно в то же время дополнительный
метод анализа предложил X. Гобинд Корана,
который придумал химические способы синте-
за полирибонуклеотидов с определенными по-
вторяющимися последовательностями от двух
X. Гобинд Корана,
1922-2011
до четырех оснований. Полипептиды, состоя-
щие из этих молекул мРНК, содержат повто-
ряющиеся последовательности из небольшого
числа аминокислот. Эти последовательности в
сочетании с информацией о случайных поли-
мерах. синтезированных Ниренбергом с колле-
гами, позволяли точно определить соответствие
кодонов и аминокислот. Например, в сополи-
мере (АС)Л повторяются кодоны АСА и С АС:
АСАСАСАСАСАСАСА. Полипептид, синтезиро-
ванный на такой матрице, содержит равные ко-
личества треонина и гистидина. Поскольку уста-
новлено, что кодон гистидина содержит один А
и два С (табл. 27-1), С АС должен кодировать
гистидин, а АСА — треонин.
Результаты многочисленных экспериментов
позволили установить природу 61 из 64 возмож-
ных кодонов. Оставшиеся три кодона оказались
стоп-кодонами — они прерывали синтез бел-
ка на основе синтетической полимерной РНК-
матрицы (рис. 27-6). Назначение каждого три-
плетного кодона (рис. 27-7) было установлено
к 1966 г. и подтверждено различными методами.
Расшифровка генетического кода оценивается
как одно из самых значимых открытий XX в.
Кодоны играют ключевую роль в трансляции
генетической информации и определяют синтез
специфических белков. Рамка считывания уста-
навливается на первом этапе трансляции мРНК
и сохраняется на всем протяжении процесса при
переходе от одного триплета к следующему. Если
исходная рамка считывания теряет одно или два
основания, или если трансляция каким-то об-
разом перескакивает через нуклеотид в мРНК,
нарушается порядок считывания всех последу-
ющих кодонов; в результате обычно образуется
«бессмысленный» белок с искаженной аминокис-
лотной последовательностью.
Несколько кодонов выполняют особые
функции (рис. 27-7). Инициаторный кодон
AUG — самый распространенный сигнал начала
синтеза полипептида во всех клетках, а внутри
последовательности он же кодирует метионин.
Стоп-кодоны (UAA, UAG и UGA), иначе называ-
емые терминирующими кодонами или нонсенс-
кодонами, дают сигнал окончания синтеза по-
липептида и не кодируют никаких аминокислот
Некоторые исключения из этого правила обсуж-
даются в доп. 27-1.
Рамка считывания 1
Рамка считывания 2
Рамка считывания 3
5'-- GITA A G U] А~А G^U А~А U Ai|A G U А А --3’
----G [U A A ||G U А||А~G цЦа A G U A A Jg lTa]A-----------
----G U |А A G„U A A„G u7a^|A G U||A A G ;Гц~Д~~аГ|------
Рис. 27-6. Роль стоп-кодона в повторяющемся тетра нуклеотиде. Стоп-кодоны (розовый
цвет) располагаются после каждого четвертого кодона в трех разных рамках считывания
(показаны разными цветами). В зависимости от первоначального места связывания рибо-
сомы происходит синтез дипептидов или трипептидов.
27.1 Генетический код [171]
Первая буква кодона (5'-конец)
Вторая
буква кодона
У U_________________С А______________G
иии Phe UCU Ser UAU Tyr UGU Cys
иис Phe UCC Ser UAC Tyr UGC Cys
UUA Leu UCA Ser UAA Crop UGA Crop
UUG Leu UCG Ser UAG Crop UGG Trp
сии Leu ecu Pro CAU His CGU Arg
сис Leu CCC Pro CAC His CGC Arg
CUA Leu CCA Pro CAA Gin CGA Arg
CUG Leu CCG Pro CAG Gin CGG Arg
AUU He ACU Thr AAU Asn AGU Ser
AUC He ACC Thr AAC Asn AGC Ser
AUA He АСА Thr AAA Lys AGA Arg
AUG Met ACG Thr AAG Lys AGG Arg
GUU Vai GCU Ala GAU Asp GGU Gly
GUC Vai GCC Ala GAC Asp GGC Gly
GUA Vai GCA Ala GAA Glu GGA Gly
GUG Vai GCG Ala GAG Glu GGG Gly
Рис. 27-7. «Словарь» аминокислотного кода в молекулах
мРНК. Кодоны записывают в направлении 5Z-*3Z. Третье
основание каждого кодона (выделено жирным шрифтом)
играет менее важную роль в спецификации аминокис-
лоты, чем первые два. Три стоп-кодона изображены на
розовом фоне, инициаторный кодон AUG — на зеленом.
Всем аминокислотам, за исключением метионина и трип-
тофана, соответствует более одного кодона. Как правило,
кодоны, определяющие одну и ту же аминокислоту, от-
личаются только третьим основанием.
Как описано в разд. 27.2, инициация синтеза
белка в клетке — сложный процесс, зависящий
от инициаторных кодонов и других сигналов в
мРНК. Теперь понятно, что эксперименты Ни-
ренберга, Кораны и их коллег по выявлению
функции кодонов не были бы успешными без
инициаторных кодонов. К счастью, условия экс-
периментов были не очень строгими. В истории
биохимии достаточно часто революционные про-
рывы в исследованиях происходили благодаря
старанию людей и счастливому случаю.
В случайной последовательности нуклеоти-
дов в среднем один из 20 кодонов в каждой рам-
ке считывания оказывается концевым кодоном.
Рамку считывания из 50 и более кодонов, не пре-
рывающуюся стоп-кодоном, называют открытой
рамкой считывания (ОРС) Длинные открытые
рамки считывания обычно соответствуют генам,
кодирующим белки. Для анализа баз данных ну-
клеотидных последовательностей с целью поиска
открытых рамок считывания среди гигантского
объема некодирующей информации существуют
сложные компьютерные программы. Для считы-
вания непрерывного гена, кодирующего типичный
белок с молекулярной массой 60 000, нужна от-
крытая рамка из 500 или большего числа кодонов.
Удивительное свойство генетического кода за-
ключается в том. что одной аминокислоте может со-
ответствовать более одного кодона, поэтому генети-
ческий код называют вырожденным. Это не озна-
чает, что код некорректен: хотя одна аминокислота
может записываться с помощью двух или несколь-
ких кодонов, каждый кодон обозначает только одну
аминокислоту. Вырожденность кодона неоднород-
на. Метионин и триптофан имеют по одному кодо-
ну, три аминокислоты (Leu, Ser, Arg) кодируются
шестью кодонами каждая, у пяти аминокислот по
четыре кодона, изолейцин имеет три кодона и де-
вять аминокислот - два кодона (табл. 27-3).
Генетический код почти универсален. За ис-
ключением некоторых минорных вариаций в
митохондриях, некоторых бактериях и некото-
рых одноклеточных эукариотах (доп. 27-1), кодо-
ны аминокислот идентичны у всех известных на
сегодняшний день видов организмов. Человек,
кишечная палочка, табак, земноводные и вирусы
используют один и тот же генетический код. Это
позволяет предположить, что все живые существа
имеют общего предка, чей генетический код сохра-
нился на протяжении биологической эволюции.
Даже исключения подтверждают общее правило.
Таблица 27-3 Вырожденность генетического кода
Аминокислот Число а кодонов Аминокислота Число кодонов
Met 1 Туг 2
Trp 1 Не 3
Asn 2 Ala 4
Asp 2 Gly 4
Cys 2 Pro 4
Gin 2 Thr 4
Glu 2 Vai 4
His 2 Arg 6
Lys 2 Leu 6
Phe 2 Ser 6
[172] Часть III. 27. Метаболизм белка
Дополнение 27-1 Исключение, подтверждающее правило: природные вариации генетического кода
В биохимии, как и в других дисциплинах, исключения из
общих правил затрудняют преподавание и расстраивают
студентов. В то же время исключения из правил учат нас,
что жизнь сложна, и вдохновляют на удивительные от-
крытия. Кроме того, понимание сути исключений может
неожиданным образом подтвердить правило.
Казалось бы, в генетическом коде немного возмож-
ностей для вариаций. Даже замена одной аминокислоты
может оказать разрушительное действие на структуру
белка. Тем не менее вариации кода действительно наблю-
даются у некоторых организмов, и эти вариации интерес-
ны и поучительны. Типы вариаций и их исключитель-
ность являются убедительным подтверждением общего
эволюционного происхождения всех живых существ.
Для изменения кода модификации должны затро-
нуть гены одной или нескольких тРНК, причем таким
образом, чтобы изменился антикодон. В результате та-
кого изменения специфическая аминокислота встраива-
ется в аминокислотную последовательность всякий раз,
когда в последовательности мРНК встречается данный
кодон, который в норме (см. рис. 27-7) не кодирует эту
аминокислоту. Генетический код определяется двумя
элементами: (1) антикодонами тРНК, которые указыва-
ют, где в растущем полипептиде размещается конкретная
аминокислота, и (2) специфичностью аминоапил-тРНК-
синтстаз, подбирающих аминокислоты для каждой моле-
кулы тРНК.
Большинство внезапных изменений кода может
иметь катастрофические последствия для клеточных
белков, поэтому модификации наиболее вероятны там,
где их эффект минимален, например, в маленьких гено-
мах, кодирующих лишь несколько белков. Кроме того,
биологические последствия изменения кода сравнитель-
но невелики, если они происходят в трех стоп-кодонах,
которые обычно не встречаются внутри генов (см. доп.
27-4 по поводу исключений из этого правила). Именно
такая картина и наблюдается в действительности.
Среди немногих известных вариаций генетическо-
го кода большинство наблюдается в митохондриальной
ДНК (мтДНК), в которой закодировано всего от 10 до
20 белков. Митохондрии содержат собственные моле-
кулы тРНК, поэтому вариации их генетического кода
не оказывают существенного влияния на геном клетки.
Большинство известных изменений в митохондриях (и
единственные изменения кода, обнаруженные в клеточ-
ных геномах) затрагивает стоп-кодоны. Эти изменения
влияют на синтез продуктов только некоторых генов, а
иногда их действие сведено к минимуму из-за наличия в
генах большого (избыточного) количества стоп-кодонов.
У позвоночных мтДНК содержат гены 13 белков,
двух рРНК и 22 тРНК (см. рис. 19-38 в т. 2). Редкие
случаи изменения кода и необычный набор качающих-
ся оснований в кодонах позволяют считывать митохон-
дриальные гены с помощью лишь 22 тРНК, в отличие
от нормального кода, для которого требуется 32 тРНК.
Эти изменения в митохондриях можно воспринимать
как оптимизацию генома, поскольку уменьшение раз-
мера генома дает органелле преимущества в репликации.
Для четырех семейств кодонов, в которых аминокислота
полностью определяется первыми двумя нуклеотидами,
достаточно по одной тРНК с остатком U в первом (кача-
ющемся) положении в антикодоне: либо U спаривается
каким-то образом с любым из четырех возможных ос-
нований в третьем положении кодона, либо реализуется
механизм «два из трех», когда спаривание в третьем по-
ложении не требуется Другая группа тРНК распознает
кодоны либо с А, либо с G в третьем положении, а третья
группа распознает U или С, таким образом, практически
все тРНК распознают или два, или четыре кодона.
В нормальном коде только двум аминокислотам
соответствует по одному кодону: метионину и трип-
тофану (см. табл. 27-3). Если все митохондриальные
тРНК распознают два кодона, следует предположить,
что в митохондриях для Met и Тгр существуют допол-
нительные кодоны. И действительно, одна наиболее
распространенная вариация кода касается изменения
смысла обычного стоп-кодона UGA, который в данном
случае кодирует триптофан. Молекула тРНКТгр встраи-
«Качание» позволяет некоторым молекулам
тРНК распознавать более одного кодона
Если несколько разных кодонов определяют одну
аминокислоту, они обычно различаются только
третьим основанием (расположенным на З'-кон-
це). Например, аланин кодируется триплетами
GCU, GCC, GCA и GCG. Кодоны для большин-
ства аминокислот можно обозначить условной
формулой XYag или XY1 с. Именно две первые
буквы в каждом кодоне определяют аминокис-
лоту, и эта особенность имеет ряд интересных по-
следствий
27.1 Генетический код [173]
I Таблица 1 Известные варианты кодонов в митохондриях
Кодоны*
UGA AUA AGA AGG CUN CGG
Нормальное назначение кодона Стоп Не Arg Leu Arg
Животные
Позвоночные Тгр Met Стоп + +
Drosophila Тгр Met Ser + +
Дрожжи
Saccharomyces cerevisiae Тгр Met + Thr +
Torulopsis glabrata Тгр Met + Thr ?
Schizosaccharomyces pomhe Тгр + + + +
Мицелиальные грибы Тгр + + + +
Трипаносомы Тгр + + + +
Высшие растения + + + + Trp
Clamydomonas reinhardtii ? + + + ?
* N обозначает любой нуклеотид; + - кодон имеет т о же назначение, чт о и в нормальном коде;
? кодон не встречается в данном митохондриальном геноме.
васт остаток Тгр как на месте UGA, так и на месте стан-
дартного кодона Тгр, а именно, UGG. Второе наиболее
распространенное изменение касается кодона AUA,
который в данном случае означает не Не, a Met; нор-
мальный кодон метионина AUG тоже действует, и одна
тРНК распознает оба варианта Известные вариации
кода в митохондриях обобщены в табл. 1.
Если обратить внимание на наиболее редкие из-
менения кода клеточных геномов (отличающиеся от
митохондриальных), мы обнаружим, что у бактерий
единственная известная вариация опять заключается в
использовании UGA для кодирования остатков Тгр. Это
изменение встречается в самых простых свободноживу-
щих клетках — в бактерии Mycoplasma capricolum. Среди
эукариот вариации кода вне митохондрий наблюдаются
у немногочисленных видов ресничных простейших, у
которых оба стоп-кодона UAA и UAG могут кодировать
глутамин. Известны также редкие, но интересные слу-
чаи, когда стоп-кодоны используются для кодирования
аминокислот, не входящих в число 20 стандартных ами-
нокислот (см. доп. 27-3).
Изменения не обязательно касаются всех специфи-
ческих кодонов в данном геноме: кодон не всегда коди-
рует одну и ту же аминокислоту. Например, у большин-
ства бактерий, включая Е. coli, кодон GUG (Vai) иногда
используется в качестве инициаторного кодона, означа-
ющего метионин. Это происходит только в тех генах, в
которых колон GUG расположен надлежащим образом
по отношению к инициирующим трансляцию последова-
тельностям мРНК (разд. 27.2).
Эти вариации свидетельствуют о том, что код не
столь универсален, как считалось ранее, но его вариа-
бельность строго ограничивается. Вариации являются
очевидными производными обычного кода, и примеров
совсем другого кода пока не обнаружено. Ограниченное
число вариантов кода подтверждает идею, что вся жизнь
на нашей планете эволюционировала из единого (мало
вариабельного) генетического кода.
Спаривание транспортных РНК с кодонами
мРНК осуществляется через последовательность
трех оснований тРНК, называемую антикодоном
Первое основание кодона в мРНК (в направлении
5'->3') образует пару с третьим основанием анти-
кодона ( не. 27- ). Если триплет антикодона
тРНК распознает только один кодон в мРНК,
который по всем трем позициям удовлетворяет
уотсон-криковскому принципу соответствия ос-
нований, клетки должны иметь разные молекулы
тРНК для каждого кодона. Однако это не так, по-
скольку в некоторых тРНК антикодоны содержат
[174] Часть III. 27. Метаболизм белка
мРНК 5
|1 2 3|
Кодон
6Г
3 2 1
Антикодон (3') G-C-1
Кодон (5') C-G-A
12 3
3 2 1
G-C-I
C-G-U
1 2 3
3 2 1
G-C-I (5 )
C-G-C (3')
12 3
Рис. 27-8. Связывание кодона с антикодоном, а — моле-
кулы РНК имеют противоположную направленность. Мо-
лекула тРНК изображена традиционным образом в конфи-
гурации клеверного листа, б — три варианта спаривания
кодона с антикодоном при наличии инозината в составе
антикодона в тРНК.
нуклеотид инозинат (I), имеющий необычное ос-
нование гипоксантин (см. рис. 8-5, б в т. 1). Ино-
зинат может образовывать водородные связи с
тремя нуклеотидами (U, С и А; рис. 27-8, б), хотя
связи в таких парах намного слабее, чем водо-
родные связи в уотсон-криковских парах (G=C
и A=U). У дрожжей одна молекула T₽HKArg име-
ет антикодон (5')-ICG, который распознает три
кодона аргинина: (5')-CGA, (5')-CGU, (5')-CGC.
Первые два основания в этих кодонах идентичны
(CG) и образуют пары с соответствующими осно-
ваниями антикодона строго по правилу Уотсона-
Крика, но третье основание (A, U или С) образует
слабые водородные связи с остатком I в первой
позиции антикодона.
Изучение спаривания этих и других кодонов с
антикодонами привело Крика к выводу, что третье
основание в большинстве кодонов часто достаточ-
но слабо связано с соответствующим основанием
антикодона; иными словами, третье основание та-
ких кодонов и первое основание соответствующих
антикодонов проявляют нестрогое соответствие,
или «качаются». Крик сформулировал четыре по-
ложения, названных гипотезой качания:
1. Первые два основания кодона мРНК всегда
формируют канонические пары (пары Уот-
сона-Крика) с основаниями антикодона в
тРНК и в основном определяют соответству-
ющую аминокислоту
2. Первое основание антикодона (при считы-
вании в направлении 5'-*3'; это основание
спаривается с третьим основанием кодона)
определяет число кодонов, распознаваемых
молекулой тРНК. Если первое основание
Таблица 27-4
Как качающиеся основания антикодона
определяют число кодонов.
которые может распознать тРНК
1. Один распознаваемый кодон:
Антикодон (3') X - Y - С (У) (3') X - Y - А (5')
Кодон (5') X - Y - G (3') (5') X - Y - U (3')
2. Два распознаваемых кодона:
Антикодон (З')Х- Y-U(5') (3')X- Y- G(5')
Кодон (5')X-Y £(3') (5')X- Y- у (3')
3. Три распознаваемых кодона
Антикодон (3') X - Y -1 (5')
Кодон (5') X - Y - и (5')
Примечание. X и Y основания, комплементарные осно-
ваниям X' и Y' соответственно; образуют с ними уотсон-
криковские пары. Качающиеся пары оснований в З'-поло-
жении кодонов и в 5 -положении антикодонов выделены
розовым цветом.
антикодона С или А, спаривание оснований
специфично, и такая тРНК распознает толь-
ко один кодон. Если первое основание U или
G, связывание менее специфичное, и тРНК
может узнавать два разных кодона Если пер-
вым (качающимся) нуклеотидом антикодона
является инозин (I), тРНК может распознать
три разных кодона — максимальное число
для любой тРНК. Эти принципы обобщены в
табл. 27-4.
3 Если аминокислота определяется нескольки-
ми кодонами и если эти кодоны различаются
первым или вторым основанием, то для каж-
дого кодона нужна своя тРНК.
4. Для трансляции всех кодонов (их 61) нужно
как минимум 32 тРНК (31 для кодирования
аминокислот и 1 для инициации).
Качающееся (третье) основание кодона
вносит определенный вклад в специфичность
связывания, но, поскольку связывается с соот-
ветствующим основанием антикодона достаточ-
но слабо, это позволяет тРНК быстро диссоции-
ровать из комплекса с кодоном в ходе синтеза
белка. Если бы все три основания кодона обра-
зовывали с тремя основаниями антикодона пра-
вильные уотсон-криковские пары, тРНК слиш-
ком медленно покидала бы комплекс и это могло
значительно ограничить скорость синтеза белка.
При взаимодействии кодона и антикодона со-
блюдается баланс между точностью и скоростью
синтеза.
Генетический код позволяет понять, каким
образом информация о последовательности бел-
ка сохраняется в нуклеиновых кислотах, и дает
ключ к разгадке того, как эта информация транс-
лируется в белок. Теперь обратимся к молекуляр-
ным механизмам процесса трансляции.
27.1 Генетический код [175]
Считывание последовательности зависит от
сдвига рамки и редактирования РНК
После того как в процессе синтеза белка уста-
навливается рамка считывания, кодоны начи-
нают транслироваться без перекрывания и про-
пусков до момента, пока рибосомный комплекс
не встречает стоп-кодон. Две другие возможные
рамки считывания обычно не содержат полез-
ной генетической информации, однако некото-
рые гены, их немного, устроены таким образом,
что на определенной стадии трансляции мРНК
рибосомы ошибаются, что приводит к сдвигу
рамки считывания, начиная с этой точки. Воз-
можно, этот механизм позволяет синтезировать
с одного транскрипта несколько родственных,
но не полностью идентичных белков или регу-
лировать синтез белка.
Один из наиболее хорошо изученных слу-
чаев сдвига рамки считывания — трансляция
мРНК перекрывающихся генов gag и pol вируса
саркомы Рауса (см. рис. 26-35). Рамка считыва-
ния гена pol отстоит на одну пару оснований вле-
во по сравнению с рамкой считывания гена gag
(рис. 27-9).
Продукт гена pol (обратная транскрипта-
за) образуется на основе той же мРНК, что ис-
пользуется для синтеза продукта гена gag (см.
рис. 26-34). Образующийся длинный полипро-
теин, или белок gag-pol, затем подвергается
протеолитическому расщеплению, укорачива-
ется, превращаясь в зрелый белок — обратную
транскриптазу. Для образования полипротеина
необходим сдвиг рамки считывания в области
перекрывания генов, поскольку это позволяет
рибосоме миновать стоп-кодон UAG на конце
гена gag (выделен розовым цветом на рис. 27-9).
Сдвиг рамки происходит приблизительно в
5% случаев трансляции на этой мРНК, так что по-
липротеин gag-pol (в итоге — обратная транскрип-
Рамка считывания gag
--Leu — Gly — Leu — Arg — Leu — Thr — Asn — Leu Стоп
5'—|c u a||g~g g||c u c]|c g c|[u u g||a c a||a a u|[u u a||u a g|g g a g G GC C A-3
--CUAGGGCUCCGCUUGACAAAUUU |A U A|[G~G G||A G G]|G C C| A----
Рамка считывания pol lie — Gly — Arg — Ala -
Рис. 27-9. Сдвиг рамки считывания в транскриптах у ретровирусов. Представлен участок
перекрывания генов дад-pol в РНК вируса саркомы Рауса.
[176] Часть III. 27. Метаболизм белка
таза) синтезируется в 20 раз реже продукта гена
gag, но этого вполне достаточно для эффективной
репродукции вируса. У некоторых ретровирусов
сдвиг рамки считывания позволяет транслировать
еще более длинный полипротеин, в состав которо-
го входит продукт гена env, слитый с продуктами
генов gag и pol (см. рис. 26-34). Аналогичный ме-
ханизм позволяет синтезировать субъединицы т
и у ДНК-полимеразы III Е. coli на основе одного
транскрипта гена dnaX (см. табл. 25-2).
Иногда перед трансляцией происходит ре-
дактирование мРНК. Редактирование РНК
может заключаться в добавлении, удалении или
замене нуклеотидов, в результате чего изменяет-
ся смысл транскрипта. Удаление или добавление
нуклеотидов чаще всего наблюдается в РНК из
геномов митохондрий и хлоропластов эукари-
от. В этом механизме задействован особый тип
молекул РНК. кодируемых в тех же органел-
лах. Последовательности этих РНК комплемен-
тарны последовательностям модифицируемых
мРНК. Эти руководящие (гидовые) молекулы
РНК (рис. 27-10) служат матрицей при редак-
тировании.
Примером редактирования путем введения
дополнительных нуклеотидов могут служить
первичные транскрипты генов, кодирующих
субъединицу II цитохромоксидазы в митохон-
дриях некоторых протестов. Эти транскрипты
не соответствуют полностью той последователь-
ности, которая должна быть на С-конпе белково-
го продукта. В ходе посттранскрипционной мо-
дификации в последовательность встраиваются
четыре остатка урацила, что сдвигает рамку счи-
тывания. На рис. 27-10 изображены встроенные
остатки U в небольшом фрагменте транскрипта,
подвергающемся редактированию. Заметьте, что
при спаривании оснований первичного транс-
крипта и руководящей РНК образуется несколь-
ко пар G=U (синие точки), которые часто встре-
чаются в молекулах РНК.
Редактирование РНК путем замены нуклео-
тидов чаще всего состоит в ферментативном деза-
минировании остатков аденозина или цитидина,
в результате чего образуются инозин или уридин
соответственно (рис. 27-11), хотя встречаются
замены и других оснований. В ходе трансляции
инозин воспринимается как G. Реакции дезами-
нирования аденозина катализируют ферменты
семейства ADAR (от англ, adenosine deaminases
that act on RNA). Дезаминирование цитидина
происходит под действием семейства пептидов
АРОВЕС (ароВ mRNA editing catalytic peptide), к
которому относятся также родственные дезами-
назы AID {activation-induced deaminase). Эти две
группы дезаминирующих ферментов содержат
гомологичные каталитические домены, коорди-
нирующие атом цинка.
Хорошо изученным примером такого рода
реакции является дезаминирование гена аполи-
попротеина В, входящего в состав липопротеина
низкой плотности у позвоночных. Одна форма
Кодирующая
цепь ДНК 5' —-|А A "a||G Т A|[G A G|[A~A С||С Т G||G Т А|------3
--- Lys — Vai Glu Asn Leu Vai ----------------------
Отредактированная
мРНК
---|A A a|[g~u a||g a u]|u g u||a u a||c c u||g g u] —
---- Lys — Vai — Asp — Cys
1--->
a
He — Pro — Gly ------------
мРНК 5-----A
Гидовая U
PHK
3
A AGUAGAU U
U AUAUCUAA
G U A U А С C
• 1 1 i i
U A U A U G G
UGG U---3
A U A U
5
6
Рис. 27-10. Редактирование транскрипта гена субъединицы II цитохромоксидазы из мито-
хондрий Trypanosoma brucei. а — встраивание четырех остатков урацила (розовые) при-
водит к изменению рамки считывания. 6 — направляющие (гидовые или адаптерные) РНК.
комплементарные модифицированному продукту, служат матрицей в ходе редактирова-
ния. Обратите внимание на присутствие не уотсон-криковских пар G=U (синие).
27.1 Генетический код [177]
\ тт
\ Инозин
Рис. 27-11. Реакции дезаминирования в ре-
дактировании РНК. а — превращение адено-
зиннуклеотидов в инозиннуклеотиды ката-
лизируют ферменты ADAR, б — превращение
цитидина в уридин катализируют ферменты
семейства АРОВЕС.
О ОН
' Цитидин
О ОН
' Уридин
аполипопротеина В, ароВ-100 (М, = 513 000),
синтезируется в печени, вторая форма, ароВ-48
(Мг = 250 000), — в кишечнике; мРНК обеих форм
синтезируется на основе гена ароВ-100. Цитидин-
дезаминаза АРОВЕС, обнаруженная только в ки-
шечнике, связывается с мРНК на уровне кодона
для аминокислотного остатка 2153 (CAA=Gln)
и превращает С в U, в результате чего образует-
ся стоп-кодон UAA. Итак, в кишечнике на основе
этой модифицированной мРНК образуется белок
ароВ-48 — укороченная (N-концевая) форма бел-
ка ароВ-100 (рис. 27-12). Эта реакция — пример
тканеспецифичного синтеза двух разных белков
с одного гена.
Замена основания А на I при участии белков
ADAR чаще всего встречается в транскриптах
генов приматов, причем свыше 90% замен про-
исходит в коротких диспергированных повторах
(short interspersed elements, SINE), называемых Alu-
элементами (см. рис. 24-8). В ДНК человека насчи-
тывается свыше миллиона Alu-элементов длиной
300 п. н., что составляет около 10% всего генома.
Они концентрируются вблизи генов белков, на-
пример, в интронах и нетранслируемых участках
Номер аминокислотного остатка
2146 2148 2150 2152 2154 2156
Печень __________ ________ _________
человека 5'- CAACUG |С A G][A~C aTu a"u’ A U G A U A |C A A| UUUGAUCAGUAU- -3
(apoB-100) - Gin - Leu — Gin - Thr - Tyr — Met — He - Gin - Phe — Asp - Gin — Tyr -
Кишечник
человека
(ароВ-48)
CAACUG |C A G~H~A C A| UAUAUGAUA |U A A|!U U"u||G AUCAGUAU-
— Gin — Leu — Gin — Thr — Tyr — Met — lie Стоп
Рис. 27-12. Редактирование транскрипта гена ароВ-100 — компонента Л ПНП. Дезаминиро-
вание, которое происходит только в кишечнике, приводит к превращению специфического
цитидина в уридин, что влечет за собой замену кодона Gin на стоп-кодон и синтез укоро-
ченного белка.
[ 1 / (S ] Часть III. 27. Метаболизм белка
на 3'- и 5'-концах транскриптов. В только что син-
тезированной (до процессинга) мРНК человека
в среднем содержится от 10 до 20 Alu-элементов.
Ферменты ADAR связываются только с двух-
цепочечной РНК и осуществляют замену А на I.
Наличие множества Alu-элементов предоставляет
большие возможности для внутримолекулярного
спаривания оснований в транскриптах, обеспе-
чивая необходимые для действия ADAR дуплек-
сы. Иногда модификации изменяют кодирующие
последовательности генов. Нарушения функции
ADAR бывают связаны с различными невроло-
гическими состояниями, включая латеральный
амиотрофический склероз (болезнь Шарко), эпи-
лепсию и клиническую депрессию.
Геномы позвоночных содержат множество
SINE-элементов, причем у большинства организ-
мов много разных типов SINE. Alu-элементы пре-
обладают только у приматов. Детальный анализ
генов и транскриптов показывает, что у людей
замена А на I происходит в 30-40 раз чаще, чем
у мышей, в значительной степени благодаря при-
сутствию многочисленных Alu-элементов. Срав-
нительно частые замены А на I, а также распро-
страненность альтернативного сплайсинга (см.
рис. 26-22) — две особенности, отличающие гено-
мы приматов от геномов других млекопитающих.
Пока неясно, случайно ли произошли эти изме-
нения или они играли важную роль в эволюции
приматов и, в итоге, в появлении человека.
Краткое содержание раздела 27.1
ГЕНЕТИЧЕСКИЙ КОД
Специфическая аминокислотная последова-
тельность белка создается путем трансляции
информации, закодированной в мРНК. Этот
процесс происходит на рибосомах.
Аминокислоты определяются кодонами
мРНК, которые представляют собой нукле-
отидные триплеты. Для трансляции нужны
адаптерные молекулы тРНК, которые рас-
познают кодоны и встраивают аминокисло-
ты в соответствующем порядке в синтезиру-
емый полипептид.
Последовательности оснований в кодонах
были установлены с помощью синтетиче-
ских мРНК с известным составом и последо-
вательностью.
Кодон AUG указывает начало трансляции,
кодоны UAA, UAG и UGA являются сигнала-
ми окончания синтеза белка.
и Генетический код вырожден: почти все ами-
нокислоты кодируются несколькими кодо-
нами.
Стандартный генетический код одинаков у
всех организмов, за исключением некоторых
минорных вариаций в митохондриях и в не-
которых одноклеточных организмах.
Третье положение в каждом кодоне гораздо
менее специфичное, чем первое и второе; ну-
клеотиды в этом положении называют кача-
ющимися нуклеотидами.
Сдвиг рамки считывания и редактирование
РНК влияют на считывание генетического
кода при трансляции.
27.2. Синтез белков
Ранее, описывая синтез ДНК и РНК (гл. 25 и
26), мы показали, что в синтезе полимерных био-
молекул можно выделить стадии инициации,
элонгации и терминации. Обычно эти основные
процессы дополняются еще двумя стадиями: ак-
тивация предшественников перед синтезом и
постсинтетический процессинг собранных по-
лимеров. Синтез белка следует тем же путем.
Активация аминокислот перед их включением
в полипептиды и посттрансляционный процес-
синг собранного полипептида играют очень важ-
ную роль, гарантируя как точность синтеза, так
и нормальное функционирование готового про-
дукта. Клеточные компоненты, участвующие в
этих пяти стадиях синтеза белка у Е. coli и других
бактерий, перечислены в табл. 27-5; потребности
эукариотических клеток примерно такие же, хотя
иногда эукариотам требуются дополнительные
компоненты. Сначала рассмотрим стадии синтеза
белка в целом, что поможет нам в последующей
дискуссии.
Z12 Синтез белков [179]
Клеточные компоненты, участвующие в пяти основных стадиях синтеза белка у Е. coli
Стадия Основные компоненты
1. Активация аминокислот 20 аминокислот 20 аминоацил-тРНК-синтетаз 32 или более типов тРНК АТР Mg24
2. Инициация мРНК А-формилметионил-тРНК^01 Инициаторный кодон в мРНК (AUG) 305-субъединица рибосомы 505-субъединица рибосомы Факторы инициации (IF-1, IF-2, IF-3) GTP Mg2+
3. Элонгация Функциональная 705-рибосома (инициаторный комплекс) Специфические аминоацил-тРНК Факторы элонгации (EF-Tu, F-Ts, EF-G) GTP Mg24
4. Терминация и высвобождение Стоп-кодон в мРНК Факторы терминации (RF-1, RF-2, RF-3) EF-G IF-3
5. Укладка и посттрансляционный процессинг Специфические ферменты, кофакторы и другие компоненты для удаления инициаторного остатка и сигнальных последовательностей, дополнительный протеолитический процессинг, модификация концевых остатков и присоединение ацетильной, фосфорильной, метильной, карбоксильной, углеводной или простетической группы.
Синтез белка происходит в пять стадий
Стадия 1: активация аминокислот. Для синтеза
полипептида со специфической последователь-
ностью необходимо соблюдение двух условий:
(1) карбоксильная группа каждой аминокисло-
ты должна быть активирована для облегчения
формирования пептидной связи; (2) необхо-
димо однозначное соответствие между каждой
новой аминокислотой и кодирующей ее мРНК.
Оба эти требования удовлетворяются путем
присоединения аминокислоты к тРНК на пер-
вой стадии синтеза белка. Принципиально важ-
но соединить правильную аминокислоту с пра-
вильной тРНК. Эта реакция происходит в цито-
плазме, а не на рибосоме. Каждая из 20 амино-
кислот ковалентно связывается со специфиче-
ской тРНК с затратой энергии АТР с помощью
активирующих Mg24 -зависимых ферментов,
называемых аминоацил-тРНК-синтетазами.
Связанную с соответствующей аминокислотой
(аминоацетилированную) тРНК называют «на-
груженной».
Стадия 2: инициация. Кодирующая полипептид
мРНК связывается с меньшей из двух субъединиц
рибосомы и с инициаторной аминоацил-тРНК.
Затем к ним присоединяется большая субъеди-
ница рибосомы, и формируется инициаторный
комплекс. Инициаторная аминоацил-тРНК спа-
ривается с кодоном AUG в мРНК, который обо-
значает начало полипептида. Этот процесс требу-
ет затраты GTP и запускается белками цитоплаз-
мы, называемыми факторами инициации.
[180] Часть III. 27. Метаболизм белка
Стадия 3: элонгация. Полипептид удлиняется
путем последовательного ковалентного связы-
вания аминокислот, которые доставляются к ри-
босоме и точно размещаются специфическими
молекулами тРНК путем спаривания с основани-
ями соответствующих кодонов в мРНК. Элонга-
ция требует участия цитоплазматических белков,
называемых факторами элонгации. Связыванию
каждой поступающей молекулы аминоацил-
тРНК и продвижению рибосом вдоль мРНК спо-
собствует гидролиз GTP.
Стадия 4: терминация и высвобождение. Сиг-
налом окончания синтеза полипептидной цепи
является стоп-кодон в мРНК. Новый полипеп-
тид высвобождается из комплекса с рибосомой с
помощью факторов высвобождения, а рибосома
готовится для синтеза следующего белка.
Стадия 5: укладка и посттрансляционный про-
цессинг. Для перехода в активную форму новый
полипептид должен принять свою специфиче-
скую трехмерную конформацию. До или после
соответствующей упаковки полипептид может
подвергаться ферментативным превращениям,
таким как удаление одной или нескольких ами-
нокислот (обычно с N-конца), присоединение
ацетильной, фосфорильной, метильной, карбок-
сильной или других групп к определенным ами-
нокислотным остаткам, протеолитическое рас-
щепление и или присоединение олигосахаридов
или просте гических групп.
Прежде чем перейти к детальному обсуждению
этих пяти стадий, следует обратить внимание на
два ключевых элемента в биосинтезе белка: рибо-
сомы и молекулы тРНК.
Рибосома — сложная
надмолекулярная машина
Каждая клетка Е. coli имеет 15 000 или более ри-
босом, на них приходится до четверти сухой мас-
сы клетки. Бактериальные рибосомы содержат
около 65% рРНК и 35% белка: их диаметр при-
мерно 18 нм, они состоят из двух неравных субъ-
единиц с коэффициентами седиментации 30S и
50S (общий коэффициент седиментации 70S).
Обе субъединицы содержат десятки рибосомных
белков и, как минимум, одну крупную молекулу
рРНК (табл. 27-6).
Рибосомы стали активно изучать после
установления Замечником их роли в синтезе
белка и после расшифровки генетического кода.
В конце 1960-х гг. Масаясу Номура с коллегами
показали, что обе субъединицы рибосомы можно
разъединить на составляющие их РНК и белок и
вновь собрать in vitro. При определенных усло-
виях эксперимента происходит самопроизволь-
ная сборка РНК и белка с образованием 30S- и
505-субъединиц, практически идентичных по
Масаясу Номура,
1927-2011
I Таблица 27-6 РНК и белковые компоненты рибосомы £. coli
Субъединица Число различных белков Общее 1 число белков Обозначение белков Число и тип молекул рРНК
30S 21 21 S1-S21 1 (16SpPHK)
50S 33 36 L1-L36* 2 (5S и 23S рРНК)
* Обозначения белков от L1 до L36 не соответствуют 36 разным белкам. Белок, первона-
чально обозначенный L7. на самом деле является модифицированной формой L12, a L8 -
это комплекс трех других белков. Установлено также, что L26 и S20 один и тот же белок
и он не входит в состав 508-субъединицы. В большой субъединице 33 разных белка. Белок
L7 L12 содержится в четырех копиях, и вместе с тремя дополнительными копиями общее
количество белков равно 36.
27.2 Синтез белков [181]
50S 30S
Рис. 27-13. Бактериальная рибосома. Мы сильно продвинулись в понимании строения ри-
босом благодаря множеству изображений выского разрешения бактериальной рибосомы
и ее субъединиц получены несколькими группами исследователей. Некоторые из этих
структур приведены здесь, а — субъединицы 50S и 30S бактериальной рибосомы разъеди-
нены, чтобы показать поверхности которые взаимодействуют в активной рибосоме. Слева
показано строение 505-субъединицы (по PDB ID 20W8, 1VSA и 1GIX) с молекулами тРНК
(зеленые), связанными с сайтами Е, Р и А (описаны далее в тексте); антикодоны тРНК выде-
лены красным цветом. Белки изображены в виде синих спиралей; рРНК изображена серым
цветом, видна структура ее поверхности. Справа представлено строение 305-субъединицы
(по PDB ID 20W8). Белки изображены в виде коричневых спиралей; рРНК бежевая, виден
рельеф ее поверхности. Часть мРНК, взаимодействующая с антикодонами тРНК, окрашена
в красный цвет. Остальная мРНК (не показано) протягивается через бороздки или каналы
на поверхности 305-субъединицы. 6 — активная бактериальная рибосома; вид снизу на бо-
роздку, разделяющую субъединицы (по PDB ID 20W8,1VSA и 1GIX). Все элементы окрашены,
как и на рис. а. в — рибосома показана в том же ракурсе, как на рис. б, но все составляющие
показаны в объеме. Справа (рис. в) отсутствуют молекулы тРНК, так что видна полость, где
и происходит синтез белка, г — 505-субъединица бактериальной рибосомы (PDB ID 1Q7Y).
Она показана со стороны, которая присоединяется к 305-субъединице,. Активный центр,
в котором происходит образование пептидной связи (в результате активности пептидил-
трансферазы), располагается глубоко в бороздке и далеко от всех белков; с активным цен-
тром связан ингибитор пуромицин (красный).
[182] Часть III. 27. Метаболизм белка
Дополнение 27-2
Из мира РНК в мир белка
Известные в настоящее время рибозимы обычно за-
пускают реакцию одного из двух типов: гидролити-
ческое расщепление фосфодиэфирных связей или
трансфосфорилирование (гл. 26). И в том и другом
случаях субстраты этих реакций тоже молекулы РНК
Рибосомные РНК значительно расширяют диапазон
каталитических возможностей рибозимов. По мере
продвижения лабораторных исследований каталити-
ческих функций РНК (см. доп. 26-3) идея мира РНК
как предшественника современных форм жизни ста-
новится все более популярной.
Жизнеспособный мир РНК должен иметь воз-
можность независимо реплицироваться, обладать
примитивным метаболизмом для синтеза необходи-
мых рибонуклеотидных предшественников и иметь
клеточные границы, чтобы накапливать субстраты и
изолировать их от окружающей среды. Для осущест-
вления реакций между разнообразными метаболи-
тами и макромолекулами могли возникнуть более
крупные и более сложные РНК-катализаторы. Боль-
шое количество отрицательно заряженных фосфо-
рильных групп в каркасе молекулы РНК понижает ее
стабильность и тем самым ограничивает размер моле-
кул РНК. В мире РНК для поддержания стабильно-
сти крупных молекул в их состав могли включаться
двухвалентные катионы или другие положительно
заряженные группы.
Способствовать стабилизации крупных моле-
кул РНК могут некоторые пептиды Например, мно-
гие рибосомные белки в клетках современных эука-
риот — это длинные молекулы, не формирующиие вто-
ричной структуры; вытягиваясь вдоль молекул рРНК,
они вызывают их стабилизацию (рис. 1). Весьма ве-
роятно, что появление катализируемого рибозимами
синтеза пептидов исходно имело своей целью стаби-
лизацию структуры больших молекул РНК. Возмож-
но, синтез пептидов помогал стабилизовать крупные
рибозимы, но это усовершенствование стало началом
конца мира РНК Как только стал возможен синтез
пептидов, более высокий каталитический потенциал
белков обеспечил необратимый переход к метаболиз-
му, основанному на функциях белков.
В конце концов в ведение белков перешло боль-
шинство каталитических функций, но не все. У всех
современных организмов важная задача синтеза бел-
ков решена с помощью рибозимов. По-видимому в
активном центре рибозима существует единствен-
ная оптимальная комбинация нуклеотидов (или
этих комбинаций совсем не много), которая может
участвовать в катализе синтеза пептидов. У всех
видов организмов остатки рРНК больших субъе-
диниц, которые, как предполагается, вовлечены в
пептидилтрансферазную активность рибосом, вы-
сококонсервативны. Используя метод «эволюции in
vitro» (SELEX; см. доп. 26-3), исследователи выде-
лили искусственные рибозимы для осуществления
синтеза пептидов. Интересно, что в большинстве
этих биополимеров имеется высококонсервативная
последовательность из восьми рубонуклеотидов
(5')AUAACAGG(3'), которая также обнаружена в
пептидилтрансферазном центре рибосом всех кле-
ток. По-видимому, есть только одно оптимальное
решение общей задачи химического синтеза белков
со специфической последовательностью с помощью
рибозимов. Эволюция однажды нашла это решение,
и ни в каких формах жизни оно не было улучшено.
Рис. 1. 505-субъединица бактериальной рибосомы (PDB
ID 1NKW). Синие нити — молекулы белка; структурные
элементы рРНК — прозрачные. Неструктурированные
протяженные участки многих рибосомных белков впле-
таются в структуры рРНК, помогая их стабилизировать.
212 Синтез белков [183]
структуре и активности нативным субъедини-
цам. Это открытие открыло дорогу исследова-
ниям в области функции и структуры рибосом-
ных РНК и белков, которые продолжаются уже
несколько десятилетий. Одновременно были
достигнуты успехи в развитии структурных ме-
тодов исследования, благодаря чему строение
рибосом было изучено в деталях.
В начале нового тысячелетия структура
субъединиц бактериальных рибосом была ис-
следована с очень высоким разрешением, и это
принесло много «сюрпризов» (рис. 27-13). Во-
первых, изменилось традиционное мнение о
ведущей роли белковых компонентов рибосом.
Субъединицы рибосом представлены очень
большими (гигантскими) молекулами РНК.
Сердцевину 508-субъединицы формируют 5S- и
23S-pPHK. В этом комплексе белки как бы вто-
ричны — они покрывают его поверхность. Во-
вторых, и это самое важное, на расстоянии 18 А
от активного центра отсутствует белок, который
мог бы сформировать пептидную связь. Струк-
турные исследования с высоким разрешением
подтвердили то, о чем высказывались предпо-
ложения более 10 лет назад: рибосома это и есть
рибозим. Кроме того, понимание механизма
синтеза белка (описанного ниже) и определение
детального строения рибосомы и ее субъединиц
породили новые взгляды на эволюцию жизни
(доп. 27-2).
Бактериальная рибосома имеет слож-
ное строение; ее общая молекулярная масса
-2,7 млн Д. Две рибосомные субъединицы не-
правильной формы объединены таким образом,
что между ними образуется щель, через кото-
рую при трансляции проходит молекула мРНК
(рис. 27-13, б). В бактериальных рибосомах 57
белков, которые очень сильно различаются по
размеру и структуре; молекулярная масса этих
белков от 6000 до 75 000 Большинство белков
имеет глобулярные домены на поверхности ри-
босомы. У некоторых белков есть также протя-
женные неструктурированные хвосты, которые
проникают в состоящую из рРНК сердцевину
рибосомы, стабилизируя ее структуру. Назна-
чение некоторых из этих белков до настоящего
времени неясно, но многие, судя по всему, вы-
полняют структурную функцию.
У многих организмов последовательности
молекул рРНК установлены. Каждая из трех
5S рРНК
Рис. 27-14. Бактериальные рРНК. Вторичная структура
16S- и 5S-pPHK Е. coli. Отмечены (и в 16S рРНК оцифро-
ваны — цифры в скобках) первый (на 5'-конце) и послед-
ний (на З'-конце) остатки рибонуклеотидов.
одноцепочечных рРНК Е. coli имеет специфи-
ческую трехмерную конформацию с протяжен-
ными участками спаренных оснований внутри
цепи. Предсказанная вторичная структура моле-
кул рРНК (рис. 27-14) в значительной степени
была подтверждена на моделях, но она не может
объяснить сложные взаимодействия в третич-
ной структуре.
Рибосомы эукариотических клеток (кото-
рые отличаются от рибосом митохондрий и хло-
ропластов) крупнее и сложнее бактериальных
рибосом (рис. 27-15), их диаметр -23 нм, а коэф-
фициент седиментации -80S. Они также состо-
ят из двух субъединиц, которые отличаются по
размеру у разных видов, но в среднем 60S и 40S.
[184] Часть III. 27. Метаболизм белка
В сумме рибосомы эукариот содержат более 80
различных белков. Рибосомы митохондрий и
хлоропластов меньше и проще бактериальных
рибосом. Однако у всех организмов и во всех
органеллах структура и функции рибосом по-
разительно похожи.
Бактериальная
рибосома
70S
50S
Мг 0,9 х 106
16S рРНК
(1540 нуклеотидов)
21 белок
Мг 1,8 х 106
5S рРНК (120
нуклеотидов)
23S рРНК (3200
нуклеотидов)
36 белков
Эукариотическая
рибосома
80S
18S рРНК
(1900 нуклеотидов)
~ 33 белка
Рис. 27-15. Масса и состав рибосом бактерий и эукариот.
Субъединицы рибосом характеризуются коэффициента-
ми седиментации в единицах Сведберга S, которые от-
ражают скорость их осаждения при центрифугировании.
Значения S для рибосом не всегда аддитивны, поскольку
скорость осаждения определяется не только их массой,
но и формой.
Транспортные РНК
имеют специфическую структуру
Для того чтобы понять, каким образом молеку-
лы тРНК могут переводить информацию с язы-
ка нуклеиновых кислот на язык белков, нужно
подробнее проанализировать их структуру Мо-
лекулы транспортной РНК относительно малы
и состоят из одной цепи РНК со строго опреде-
ленной трехмерной структурой (см. рис. 8-25, а).
тРНК бактерий и в цитозоле эукариот содержат
от 73 до 93 нуклеотидных остатков; общая мо-
лекулярная масса от 24 000 до 31 000. тРНК ми-
тохондрий и хлоропластов меньше по размеру. В
клетке для каждой аминокислоты присутствует
хотя бы один тип тРНК. Для распознавания ко-
донов всех аминокислот требуются различные
РНК не менее 32 типов (некоторые распознают
более одного кодона), но нередко в клетке задей-
ствовано более 32 типов тРНК.
Дрожжевая тРНК для аланина (тРНКА1а;
первая нуклеиновая кислота, последователь-
ность которой была полностью секвенирована;
рис. 27-16) состоит из 76 нуклеотидных остатков,
из которых 10 имеют модифицированные осно-
вания. Сопоставление тРНК разных видов вы-
явило много общих структурных особенностей
(рис. 27-17). Не менее восьми нуклеотидных
остатков содержат модифицированные основа-
ния и сахара, многие нуклеотиды представлены
метилированными производными обычных ну-
клеотидов. Большинство молекул тРНК несет
на 5'-конце остаток гуанилата (pG), а на З'-кон-
це все РНК имеют последовательность ССА(З').
На плоскости структура всех гРНК с водород-
ными связями изображается в виде клеверного
листа с четырьмя плечами, более длинные мо-
лекулы тРНК имеют короткое дополнительное
пятое плечо (рис. 27-17). В трехмерном изобра-
жении тРНК имеет форму скрученной буквы L
(рис. 27-18).
Для выполнения адаптерной функции прин-
ципиальное значение имеют два плеча тРНК.
Аминокислотное плечо может нести специфиче-
скую аминокислоту, карбоксильная группа кото-
рой соединена эфирной связью с 2'- или З'-гидрок-
сильной группой остатка А на З'-конце тРНК. Ан-
тикодоновое плечо содержит антикодон. Другие
крупные плечи — это плечо D, которое содержит
необычный нуклеотид дигидроуридин D, и плечо
212 Синтез белков [185]
А Сайт
С присоединения
С аминокислоты
5' А
pG С
G С
G U
С G
G С
U U
G с U U а
D G U A G G С С А
С A U G С G тЧ} g
U С С G G р
G AGCGC С V
G D m2G D
С - iG А G G
U А
С G
С G
С G
U
и щ1!
ТфС, содержащие риботимидин Т, которого в мо-
лекулах РНК, как правило, нет, и псевдоуридин гр
с необычной углерод-углеродной связью между
основанием и рибозой (см. рис. 26-23). Плечи D и
Tip С обеспечивают важные взаимодействия при
укладке молекул тРНК, а плечо ТгрС взаимодей-
ствует с более тяжелой субъединицей рРНК.
Ознакомившись со структурой рибосом и
тРНК, начнем подробное обсуждение пяти ста-
дий синтеза белка.
зх
А
С Аминокислотное
С плечо (САА-«хвост»)
Триплет
антикодона
Рис. 27-16. Нуклеотидная последовательность дрож-
жевой тРНКА1а. Эта структура была определена в 1965 г.
Робертом Холли с коллегами; молекула изображена в
конформации клеверного листа, при которой достига-
ется максимальное спаривание оснований. В модифи-
цированных нуклеотидах (на розовом фоне) исполь-
зованы следующие обозначения: гр — псевдоуридин;
I — инозин; Т — риботимидин; D — 5,6-дигидроуридин;
тЧ — 1-метилинозин; nfG — 1-метилгуанозин; m2G —
N2-диметил гуано зин (см. рис. 26-23). Голубые линии меж-
ду параллельными фрагментами обозначают уотсон-кри-
ковские пары оснований. В молекулах РНК гуанозин часто
спаривается с уридином, хотя пара G=U не так стабильна,
как пара G^C (гл. 8 в т. 1). Антикодон может распознать
три кодона аланина (GCA, GCU и GCC). Другие структурные
элементы тРНК показаны на рис. 27-17 и 27-18.
5х Ри
Содержит два
или три остатка
D в разных
положениях
ТфС-плечо
Дополнительное
плечо
Варьирует в размере,
встречается
не во всех тРНК
Антикодоновое
плечо
Положение
нестрогого
соответствия
Антикодон
Роберт Холли,
1922-1993
Рис. 27-17. Схема вторичной структуры тРНК в форме
клеверного листа. Точками внутри последовательности
обозначены нуклеотидные остатки; голубые линии обо-
значают пары оснований. Нуклеотиды на розовом фоне
характерны и/или инвариантны во всех тРНК. Транс-
портные РНК имеют длину от 73 до 93 нуклеотидов. До-
полнительные нуклеотиды образуют D-плечо. В конце
антикодонового плеча формируется петля, в которой
всегда содержится семь неспаренных нуклеотидов.
Плечо D в разных тРНК содержит два или три остатка
D (5,6-дигидроуридин). В некоторых тРНК только три
пары оснований плеча D связаны водородными связями.
Основные символы расшифрованы на рис. 27-16; Ри —
пуриновый нуклеотид; Ру — пиримидиновый нуклеотид;
G* — гуанилат или 2'-0-метилгуанилат.
[1861 Часть III. 27. Метаболизм белка
Рис. 27-18. Трехмерная структура дрожжевой тРНКРИе, установленная методом рентгено-
структурного анализа. Форма молекулы напоминает перевернутую и скрученную букву L.
а — схема молекулы; плечи (рис. 27-17) выделены разными цветами, б — объемная модель
с теми же цветовыми обозначениями (PDB ID 4TRA). Последовательность ССА на З'-конце
(оранжевая) — место присоединения аминокислоты.
Стадия 1: аминоацил-тРНК-синтетазы
присоединяют определенные аминокислоты
к соответствующим молекулам тРНК
На первой стадии синтеза белка, происходящей
в цитоплазме, аминоацил-тРНК-синтетазы осу-
ществляют реакцию этерификации 20 аминокис-
лот и соответствующими им тРНК. Фермент про-
являет специфичность к одной или нескольким
аминокислотам и одной или нескольким тРНК.
У большинства организмов одна аминоацил-
тРНК-синтетаза работает с каждой аминокисло-
той. Если аминокислоте соответствуют две или
несколько тРНК, обычно их аминоацилирование
осуществляет один и тот же фермент.
Определена структура всех аминоацил-
тРНК-синтетаз Е. coli. На основании существен-
ных различий в их первичной и третичной струк-
турах и в механизме действия предложено гово-
рить о двух синтетазах (табл. 27-7; рис. 27-19);
эти два фермента есть у всех организмов. Пока
нет доказательств общего эволюционного пред-
шественника, и биологические, химические и
эволюционные причины существования этих
двух разных ферментов-катализаторов практиче-
ски идентичных процессов остаются непонятны.
Аминоацил-тРНК-синтетаза катализирует
следующую реакцию:
Аминокислота + тРНК + АТР —
----> аминоацил-тРНК + АМР + РР,
Реакция происходит в активном центре фермента
в две стадии. На стадии ф (рис. 27-19) образует-
ся связанный с ферментом промежуточный про-
дукт аминоациладенилат (аминоацил-АМР). На
второй стадии аминоацильная группа переносит-
ся со связанного с ферментом аминоацил-АМР на
I Таблица 27-7 Две аминоацил-тРНК-синтетазы
Класс I Класс II
Arg Leu Ala Lys
Cys Met Asn Phe
Gin Trp Asp Pro
Glu Tyr Gly Ser
He Vai His Thr
Примечание. Обозначения: Arg — аргинил-тРНК-синте-
таза, и т. д. Эти два фермента были обнаружены во всех
изученных организмах; они имеют разные структуры и
разные механизмы действия (см. рис. 27-19).
27.2 Синтез белков [187]
R-C---С—О
О—Р—О—Р—О—Р—О—сц
NH3 О
О
I «и I
0“ о
Аминокислота
АТР
а-Карбоксил
аминокислоты атакует
а-фосфат АТР, образуя
5 '-аминоацил аденил ат
о
Н ||
R—С С—О—Р—О—СН2
NH3 О
о
PPi
ОН он
12
ОН ОН
| Аденин~|
| Аденин~|
Рис. 27-19. МЕХАНИЗМ РЕАКЦИИ. Амино-
ацилирование тРНК аминоацил-тРНК-
синтетазами. На стадии ф образуется
аминоациладенилат, который сохраняет
связь с активным центром. На второй ста-
дии аминоацилъная группа переносится
на тРНК. Механизм этой реакции несколь-
ко отличается для двух аминоацил-тРНК-
синтетаз (см. табл. 27-7). Аминоацилсин-
тетаза I @ сначала переносит аминоа-
цильную группу на 2'-гидроксил аденина
на З'-конце РНК, затем ® на З'-гидроксил
того же остатка (трансэтерификация).
Амино ацил синтетаза II @ переносит ами-
ноацильную группу прямо на З'-гидрок-
сильную группу концевого аденилата.
5 '-Аминоациладенилат
(аминоацил-АМР)
. 1ИНС ци
[188] Часть III. 27. Метаболизм белка
соответствующую специфическую тРНК. Проте-
кание второй стадии зависит от синтетазы I или II
(реакции @ и @ на рис. 27-19). Гидролиз образо-
вавшейся эфирной связи между аминокислотой
и тРНК (рис. 27-20) характеризуется большим
отрицательным значением изменения стандарт-
ной свободной энергии (AG'° = - 29 кДж/моль).
Получившийся в результате реакции активации
пирофосфат гидролизуется до фосфата под дей-
ствием неорганической пирофосфатазы. Таким
образом, на активацию каждой аминокислоты
расходуются две высокоэнергетические фосфат-
ные связи, и суммарная реакция активации ами-
нокислоты необратима:
Аминокислота + тРНК + АТР —— »
----> аминоацил-тРНК + АМР + 2 Р,
AG'°« -29 кДж/моль
Корректирующая функция аминоацил-тРНК-
синтетаз. Аминоацилирование тРНК приводит к
двум результатам: (1) происходит активация ами-
нокислоты для формирования пептидной связи и
(2) аминокислота присоединяется к адаптерной
тРНК, что обеспечивает правильное положение
аминокислоты в растущем полипептиде. На рибо-
Рис. 27-20. Общая структура молекул аминоацил-тРНК.
Аминоацилъная группа присоединяется по З'-положению
концевого остатка А. Сложноэфирная связь, которая ак-
тивирует аминокислоту и связывает ее с тРНК отмечена
розовым цветом.
соме присоединяемая аминокислота не идентифи-
цируется, поэтому для синтеза определенной по-
следовательности важно контролировать связыва-
ние тРНК с правильной аминокислотой.
В гл. 6 (т. 1) мы говорили о том, что специ-
фичность фермента определяется энергией свя-
зывания фермента с субстратом. Способность раз-
личать две похожие аминокислоты была подробно
изучена на примере Пе-тРНК-синтетазы, которая
узнает валин и изолейцин, различие между кото-
рыми — всего одна метиленовая группа ( СН2-).
соо соо
+ I .1
H3N- С-Н H3N-С-Н
Н—С—СН3 Н—С—СН3
сн3 сн2
сн3
Валин Изолейцин
По-видимому, метиленовая группа (в Не) спо-
собна усилить связывание субстрата (с образова-
нием Пе-АМР), поэтому изолейцин связывается
в активном центре фермента, давая Пе-тРНК-
синтетазу в 200 раз чаще, чем валин. Однако в дей-
ствительности валин по ошибке встраивается на
место изолейцина не чаще одного раза из 3000. Что
повыщает точность работы фермента более чем в
10 раз? Дело в том, что подобно некоторым другим
аминоацил-тРНК-синтетазам Пе-тРНК-синтетаза
проявляет корректирующую активность.
Вспомним общий принцип, который мы
обсуждали при оценке корректирующей актив-
ности ДНК-полимераз (табл. 25-1): если взаимо-
действия фермента с двумя разными субстрата-
ми не приводят к тому, что фермент достоверно
дифференцирует эти субстраты, необходимой
специфичности можно достичь путем двухсту-
пенчатого взаимодействия. Усиление эффекта от
использования двух последовательных фильтров
оказывается мультипликативным. В случае Пе-
тРНК-синтетазы первым таким фильтром служит
исходное связывание аминокислоты с фермен-
том и ее активация до аминоацил-АМР Второй
фильтр - это связывание любых неправильных
активированных аминокислот с другим активным
центром фермента; субстрат, связывающийся со
вторым активным центром, подвергается гидро-
лизу. У валина размер R-группы немного меньше,
чем у изолейцина, поэтому Vai-АМР помещается
в гидролитическом (корректирующем) центре
27.2 Синтез белков [189]
Пе-тРНК-синтетазы, а Пе-АМР нет. Благодаря
этому Val-AMP в корректирующем центре расще-
пляется на валин и АМР, а связанная с ферментом
тРНК не подвергается аминоацилированию не-
правильной аминокислотой.
Кроме корректирующей активности боль-
шинство аминоацил-тРНК-синтетаз после об-
разования аминоацил-АМР могут расщеплять
сложноэфирную связь между аминокислотой и
тРНК в аминоацил-тРНК. Такой гидролиз осо-
бенно быстро происходит в случае неправильно
нагруженных молекул тРНК. что обеспечивает
третий фильтр для повышения точности всего
процесса. Немногочисленные аминоацил-тРНК-
синтетазы (например, Cys-тРНК-синтетаза), ко-
торые активируют аминокислоты, не имеющие
близких структурных аналогов, практически не
имеют корректирующей активности; в таких слу-
чаях активный центр аминоацилирования доста-
точно точно различает истинный субстрат и лю-
бую неправильную аминокислоту.
Общее число ошибок при синтезе белка
(~1 ошибка на 104 встроенных аминокислот) зна-
чительно больше, чем при репликации ДНК. Это
объясняется тем, что повреждения исчезают при
расщеплении белка и не передаются следующим
поколениям. Точность синтеза белка достаточна
для того, чтобы обеспечить правильность сборки
большинства белков, и потерь большого количе-
ства энергии, как это требуется при синтезе бел-
ка, обычно не происходит. Одна дефектная копия
во множестве нормальных копий белка обычно
не играет никакой роли.
Взаимодействие аминоацил-тРНКсинтетазы
и тРНК: «второй генетический код». Каждая
аминоацил-тРНК-синтетаза специфична не толь-
ко к определенной аминокислоте, но также к опре-
деленным тРНК. Найти правильную тРНК среди
десятков других так же важно для точности син-
теза белка, как и найти свою аминокислоту. Взаи-
модействия между аминоацил-тРНК-синтетазами
и тРНК называют «вторым генетическим кодом»,
подчеркивая их важную роль в обеспечении точ-
ности синтеза белка. Правила этого «второго»
кода сложнее правил «первого» кода.
На рис. 27-21 собрана информация о нукле-
отидах, важных для распознавания некоторыми
аминоацил-тРНК-синтетазами. Некоторые ну-
клеотиды консервативны во всех тРНК и, one-
s'
*
• Аминокислотное
• плечо
5'
м—В
Т1ДС- плечо
Антикодоновое
плечо
Антикодон
Рис. 27-21. Положения нуклеотидов в молекулах тРНК,
которые распознаются аминоацил-тРНК-синтетазами.
Некоторые положения (голубые точки) консерватив-
ны во всех молекулах тРНК и поэтому не являются их
отличительными признаками. Другие положения рас-
познаются одной (желтые) или несколькими (зеленые)
аминоацил-тРНК-синтетазами. Для некоторых синтетаз
при распознавании важна не последовательность тРНК.
а другие особенности строения.
довательно, не участвуют в выборе правильных
последовательностей. Проанализировав вариа-
ции нуклеотидов в тРНК, которые определяют
субстратную специфичность аминоацил-тРНК-
синтетаз, исследователи выявили те позиции, ко-
торые отвечают за дискриминацию субстратов.
Предполагается, что эти позиции сосредоточе-
ны в аминокислотном и антикодоновом плечах,
включая последовательность самого антикодо-
на, но важные позиции есть и в других частях
молекулы тРНК. Расшифровка кристалличе-
ских структур аминоацил-тРНК-синтетаз — не-
оценимый вклад в наше понимание взаимодей-
ствий при образовании косплекса тРНК и АТР
(рис. 27-22).
В распознавании тРНК ее аминоацил-тРНК-
синтетазой задействовано 10 или более спец-
ифических нуклеотидов. Но в некоторых случаях
механизм распознавания очень простой. У всех
[190] Часть III. ZJ. Метаболизм белка
Рис. 27-22. Аминоацил-тРНК-синтетазы. Для обеих
синтетаз приведены комплексы с соответствующи-
ми тРНК (зеленые стержневые структуры). Связан-
ный АТР (красный) находится в активном центре
в аминоацилъном плече, а — Gln-тРНК-синтетаза
организмов от бактерий до человека в распознава-
нии тРНК А1а-тРНК-синтетазами главную роль
играет одна-единственная пара оснований G=U
в аминокислотном плече тРНКА1а (рис. 27-23, а).
Короткая РНК всего из 7 и. н., образующих про-
из Е. coli, типичный пример мономерной синте-
тазы I (PDB ID 1QRT). 6 — Asp-тРНК-синтетаза из
дрожжей, типичный пример димерной синтетазы
II (PDB ID 1ASZ).
Взаимодействие аминоацил-тРНК-синтетаз
с соответствующими тРНК играет важную
стую миниснираль в виде шпильки, успешно ами-
ноацил ируется А1а-тРНК-синтетазой при нали-
чии в РНК ключевой пары G=U (рис. 27-23, б).
Такая простая система распознавания аланина,
возможно, сохранилась с того времени, когда оли-
гонуклеотиды-нредшественники РНК, подвер-
гались аминоацилированию для синтеза белка в
примитивной системе.
роль в правильном считывании генетическо-
го кода. Любое расширение кода с включением
новой аминокислоты неизбежно потребовало
бы образования новой пары аминоацил-тРНК-
синтетаза/тРНК. В природе расширение гене-
тического кода наблюдается редко, чаще этот
случай может быть реализован в лабораторных
условиях (доп. 27-3).
Рис. 27-23. Структурные элементы тРНКА1а, необ-
ходимые для распознавания А1а-тРНК-син-
тетазой. а — структурные элементы тРНКА1а,
распознаваемые Ala-тРНК-синтетазой. Пара осно-
ваний G=U (выделена розовым цветом) — един-
ственный элемент, необходимый для специфи-
ческого связывания и аминоацилирования, б —
3' короткая миниспиралъ РНК, содержащая ключе-
• 76 вую пару оснований G=U, но без большой части
• структуры тРНК. Эта спираль специфически взаи-
• модействует с Ala-тРНК-синтетазой почти так же
е эффективно, как целая молекула тРНКА1а.
•
И 70
• 66 А Удаленные
• 13 ^нуклеотиды
27.2 Синтез белков [191]
Дополнение 27-3 Естественное и искусственное расширение генетического кода
Как мы видели, 20 аминокислот, которые обычно
встречаются в белках, обладают лишь ограниченной
химической функциональностью. Живые системы
обычно преодолевают эти ограничения, используя
кофакторы ферментов или модифицируя отдельные
аминокислоты после их включения в белки. Расшире-
ние генетического кода для включения в белки новых
аминокислот предлагает принципиально другой путь
изменения функциональности, однако реализовать
этот путь достаточно сложно. Подобное изменение мо-
жет инактивировать тысячи клеточных белков.
Расширение генетического кода для включения
новой аминокислоты требует ряда изменений в клет-
ке В частности, необходима новая аминоацил-тРНК-
синтетаза, а также соответствующая тРНК. Оба эти
компонента должны быть высокоспецифичными и
взаимодействовать исключительно между собой и с
новой аминокислотой. В клетке должно присутство-
вать достаточное количество новой аминокислоты, что
может привести к появлению новых метаболических
путей Как подчеркивав гея в доп. 27-1, антикодон на
тРНК с большей вероятностью спаривался бы с ко-
доном, который в норме вызывает терминацию транс-
ляции. Выполнение всех этих условий в живой клетке
кажется маловероятным, однако такие ситуации все
же встречаются в природе и могут быть созданы искус-
ственным путем в лабораторной практике.
В настоящее время известно уже не 20, а 22 ами-
нокислоты, которым соответствует определенный
генетический код. Две дополнительные аминокисло-
ты селеноцистеин и пирролизин; обе встречаются
лишь в нескольких белках, но позволяют узнать кое-
что новое1 об эволюции генетического кода
СОО
+ I
H3N—СН
сн2
SeH
Селеноцистеин
СОО
. I
HjN—СН
сн2
сн2
сн2
Пирролизин
Веэ всех известных клетках лишь немногим белкам
(таким как формиатдегидрогеназа бактерий и глутати-
онпероксидаза млекопитающих) для проявления ката-
литической (ферментативной) активности требуется се-
леноцистеин. В клетках Е. coli селеноцистеин встраива-
ется в формиатдегидрогеназу в процессе трансляции на
место находящегося в рамке считывания кодона UGA.
Особая Ser-тРНК, количество которой в клетке меньше,
чем других Ser-тРНК, распознает исключительно ко-
дон UGA. Эта тРНК нагружается серином при помощи
обычной сериновой аминоацил-тРНК-синтетазы, а под
действием другого фермента серин превращается в се-
леноцистеин до синтеза пептидной связи на рибосоме.
Нагруженная тРНК узнает далеко не все кодоны UGA;
какие-то еще не до конца изученные сигналы в мРНК
отвечают за то, что эта тРНК лишь в некоторых генах
распознает некоторые кодоны VGA, которые восприни-
маются как селеноцистеин. На самом деле UGA играет
роль одновременно стоп-кодона и (очень редко) кодона
селеноцистеина. Такое расширение генетического кода
потребовало появления соответствующей тРНК, но не
специфической аминоацил-тРНК-синтетазы. Итак, об-
разуется селеноцистеин, теперь можно предположить,
что весь этот процесс является промежуточным этапом
образования совершенно нового кодона.
Пирролизин содержится в группе анаэробных
архей, называемых метаногенами (см. доп. 22-1). Про-
дуктом метаболизма этих микроорганизмов являет-
ся меган, а некоторые из них, относящиеся к группе
метаносарцин, в качестве субстрата для метаногенеза
используют метиламины. Для образования метана из
монометиламина необходима специфическая метил-
трансфераза. В рамке считывания гена этого фермента
содержится стоп-кодон UAG. Структура данной метил-
трансферазы была установлена в 2002 г., при этом вы-
яснилось, что в ней содержится новая аминокислота
пирролизин, соответствующая этому кодону UAG. По-
следующие эксперименты показали, что в отличие от
селеноцистеина пирролизин прямо присоединяется к
соответствующей тРНК с помощью пирролизил-тРНК-
синтетазы. Метаболический путь синтеза пиррол из ина
в этих клетках еще предстоит исследовать. В данном
случае имеются все признаки, необходимые для присво-
ения кодону UAG новой функции, однако кодон UAG
транслируется в пирролизин лишь в одном конкретном
Продолжение на с. 192
[192] Часть III. 27. Метаболизм белка
Дополнение 27-3 Естественное и искусственное расширение генетического кода (продолжение)
гене Как и в случае селеноцистеина, возможно, суще-
ствуют определенные сигналы, направляющие данную
тРНК к правильному кодону UAG.
Могут ли ученые воссоздать этот эволюционный
путь искусственным путем? Модификации белков с
различными функциональными группами могут зна-
чительно расширить наши представления об их актив-
ности и структуре. Однако модификация белков часто
представляет собой достаточно сложную проблему На-
пример, если требуется присоединить новую группу к
определенному остатку цистеина, нужно каким-то об-
разом защитить все остальные остатки Cys в этом белке.
Но если бы удалось адаптировать генетический код та-
ким образом, чтобы заставить клетку вводить модифи-
цированное основание в конкретное положение в белке,
работать с такой системой стало бы гораздо удобнее.
Питер Шульц с коллегами действовали именно так.
Чтобы создать кодон для новой аминокислоты,
требуется новая аминоацил-тРНК-синтетаза и соот-
ветствующая новая тРНК, которые предназначены для
работы исключительно с новой аминокислотой. Ученые
пытались создать такой «искусственный код» в клет-
ках Е. coli. Для кодирования новой аминокислоты был
выбран кодон UAG. Этот кодон в качестве стоп-кодона
используется реже двух других, и штаммы с тРНК к
этому кодону (см. доп. 27-4) нормально растут. Для
создания новой тРНК и новой гРНК-синтетазы из ар-
хей Methanococcus jannaschii были взяты гены тирозил-
тРНК и соответствующей тирозил-тРНК-синтетазы
(Л//тРНКТуг и ЛТ/Туг-тРНК-синтетаза). ЛТ/Туг-тРНК-
синтетаза не связывается с антикодоновой петлей
Л7/тРНКТуг. что позволило изменить антикодон на CUA
(комплементарен UAG), не нарушая взаимодействия.
Системы трансляции архей и бактерий ортологичны,
поэтому модифицированные компоненты системы ар-
хей можно перенести в клетки Е. coli без нарушения соб-
ственной системы трансляции бактериальных клеток.
Во-первых, нужно было модифицировать ген
Л7/тРНКТуг таким образом, чтобы создать идеаль-
ную тРНК. которую не распознавали бы эндогенные
аминоацил-тРНК-синтетазы Е. coli, но которая подвер-
галась бы аминоацилированию под действием М/Туг-
тРНК-синтетазы. Для поиска такого варианта была
предпринята серия циклов положительного и отрица-
тельного отбора, что позволило провести эффективный
скрининг вариантов генов тРНК (рис. 1). Ген А7/тРНКТуг
был подвергнут случайному мутагенезу, что позволило
создать библиотеку клонов, экспрессирующих различ-
ные версии тРНК. Кроме того, был сконструирован ген
барназы (токсичной для Е coli рибонуклеазы), мРНК
которого содержал несколько кодонов UAG, и этот ген
также был введен в клетки бактерии на плазмиде. Если
Продолжение на с. 194
Рис. 1. Отбор вариантов ЯутРНК1^, которые взаимодействуют только со специфической
тирозил-тРНК-синтетазой (M/Tyr-тРНК-синтетазой). В последовательность гена М/тРНК1^
встроенного в плазмиду, внесено 11 точечных мутаций в тех местах, которые не влияют на вза-
имодействие с/$Туг-тРНК-синтетазой (красные точки). Плазмиды с мутантным геном вводили
в клетки Е. coli для создания библиотеки, содержащей миллионы вариантов ДО/тРНК1^, которые
здесь представлены шестью клетками. Для отрицательного отбора на другой плазмиде в клетки
был введен токсичный для клеток ген барназы. В этот ген была встроена последовательность
TAG, так что транскрипт гена приобрел кодон UAG. Если ген барназы экспрессируется, клетки
погибают. Ген мог экспрессироваться лишь в том случае, если вариант М/тРНК1^, экспресси-
руемый определенной клеткой, подвергался аминоацилированию под действием эндогенных
(т. е. принадлежащих Е. coli) аминоацил-тРНК-синтетаз, что приводило к встраиванию амино-
кислоты, а не к остановке трансляции. Кроме того, в клетки был введен еще на одной плазми-
де модифицированный ген р-лактамазы, также с последовательностью TAG, образующей при
транскрипции стоп-кодон UAG. На той же плазмиде содержался ген ТИ/Туг-тРНК-синтетазы. Эта
система использовалась для положительного отбора среди оставшихся вариантов ТИ/тРНК1^. Те
варианты, которые подвергались аминоацилированию под действием М/Туг-тРНК-синтетазы
экспрессировали ген р-лактамазы, что позволяло клеткам расти на среде с ампициллином. Че-
рез несколько циклов положительного и отрицательного отбора были выделены лучшие ва-
рианты ]И/’тРНКТуг, которые подвергались аминоацилированию исключительно под действием
TfjTyr-тРНК-синтетазы и эффективно использовались для трансляции.
212 Синтез белков [193]
Библиотека
Клетки, содержащие
Ген барназы
Введение точечных мутаций в
последовательности 7И/тРНКтуг по
11 позициям, трансформация
клеток для создания библиотеки
Добавление плазмиды,
несущей ген барназы.
О' О О ~ 0’010(00?
аминоацилированные эндогенными
тРНК-синтетазами последовательности
Л//тРНКтуг, погибают. В выживших
клетках последовательность Л/утРНКТу1-
не аминоацилирована.
Ген A/yTyrRS
Удаление
плазмиды
с барназой.
Добавление
плазмиды,
несущей
A/yTyrRS и
ген
/3-лактамазы.
Рост на среде с Положительный
ампициллином. отбор
Выживают только клетки, содержащие
А/утРНКтУг, аминоацилированную
A/yTyrRS.
[194] Часть III. 27. Метаболизм белка
Дополнение 27-3 Естественное и искусственное расширение генетического кода (продолжение)
вариант Л//тРНКТуг, экспрессирующийся в определен-
ных клетках из библиотеки, подвергался аминоацили-
рованию эндогенной тРНК-синтетазой, он также экс-
прессировал ген барназы, и такие клетки погибали (от-
рицательный отбор). Выжившие клетки должны были
содержать варианты тРНК, которые не подвергались
аминоацилированию эндогенной тРНК-синтетазой,
но, в принципе, могли подвергаться аминоацилиро-
ванию Л7/Туг-тРНК-синтетазой. Для положительного
отбора (рис. 1) был использован генно-инженерный
вариант гена р-лактамазы (сообщает клеткам устой-
чивость к антибиотику ампициллину), транскрипт
которого содержал несколько кодонов UAG. Этот
ген был введен в клетки вместе с геном Л//Туг-тРНК-
синтетазы. Те варианты М/тРПК1уг, которые могли
подвергаться аминоацилированию иод действием
Л^Туг-тРНК-синтетазы, росли на среде с ампицилли-
ном только в том случае, если в них также экспресси-
ровалась Л//Туг-тРНК-синтетаза. Несколько циклов
отрицательного и положительного отбора позволили
идентифицировать новый вариант Л7/тРПКТуг, на ко-
торый не действовали эндогенные ферменты, но кото-
рый подвергался аминоацилированию под действием
Л^/Туг-тРНК-синтетазы и нормально функционировал
при трансляции.
Во-вторых, нужно было модифицировать Л//Туг-
тРПК-синтетазу, чтобы она распознавала новую
аминокислоту. Для создания большой библиотеки
вариантов ген Л//Туг-тРНК-синтетазы подвергли му-
тагенезу. Те варианты, которые аминоацилировали
новый вариант Л//тРНКТуг с помощью эндогенных
аминокислот, были исключены методом отбора с ис-
пользованием гена барназы. Был проведен второй ва-
риант положительного отбора (аналогичный отбору
с помощью ампициллина), в результате клетки мог-
ли выживать лишь при условии, что новый вариант
Л//тРНКТугподвергался аминоацилированию только в
присутствии неприродной аминокислоты. После не-
скольких циклов отрицательного и положительного
отбора была найдена пара тРНК-сиптстаза/тРПК,
которая узнавала только неприродную аминокислоту.
Эти молекулы были переименованы, чтобы указать
на неприродное происхождение аминокислоты, кото-
рая была использована для отбора.
С помощью данного подхода было сконструиро-
вано много штаммов Е. coli, каждый из которых вое
принимал кодон UAG как сигнал к встраиванию толь-
ко одной ненриродной аминокислоты. Тот же подход
применяли для искусственного расширения генети-
ческого кода дрожжей и даже клеток млекопитаю-
щих. В определенные положения в последователь-
ности клонированных белков удалось ввести свыше
30 различных аминокислот (рис. 2). Полученные
результаты предоставили очень полезный и гибкий
инструмент для углубленного изучения структуры и
функций белка.
СОО
СОО
СОО
+ I
H3N—СН
СОО
+ I
H3N— СН
СН2
СН2
сн2
сн2
SH
ЙРфимеры неприродных аминокислот, кодоны для которых добавлены в генетиче-
ежмй чадгВ^этих неприродных аминокислотах содержатся не совсем обычные химические
г^’/г->Л50Рмв«етогруппа; б — азидная группа; в — функциональная группа, способная об-
ра . < /бшютздг^крестные сшивки с соседними группами под действием света; г — амино-
« л. ^СТ1ДЛфЛ9«ф*гцентными свойствами; д — аминокислота с тяжелым атомом Вг для кри-
- _ : исследований; е — длинноцепочечный аналог цистеина способный
дисульфидные мостики.
212 Синтез белков [195]
Стадия 2: синтез белка инициирует
определенная аминокислота
Синтез белка начинается с N-конца и происходит
путем последовательного присоединения амино-
кислот к С-концу растущего полипептида, что
было установлено Говардом Динцисом в 1961 г.
(рис. 27-24) Инициаторный кодон AUG соот-
ветствует N-концевому остатку метионина. Хотя
для метионина существует лишь один кодон -
(5')AUG, все организмы имеют для метионина
две тРНК. Одна используется только для иници-
аторного кодона (5')AUG, другая — для встраи-
вания остатка Met во внутреннее положение по-
липептидной цепи.
Направление роста цепи
Число остатков
Рис. 27-24. Доказательство того, что рост полипептид-
ной цепи происходит путем присоединения остатков
аминокислот к С-концу: эксперимент Динциса. Рети-
кулоциты (незрелые эритроциты) — клетки, в которых
происходит синтез гемоглобина, инкубировали с радио-
активно меченным лейцином (эта аминокислота была
выбрана по той причине, что она часто встречается как
в а-, так и р-глобине). Через определенные промежутки
времени из препарата ретикулоцитов отбирали сфор-
мированные сс-цепи и определяли в них распределе-
ние радиоактивности. Темно-красные зоны показывают
долю полностью сформированных сс-глобиновых цепей,
содержащих остатки Leu с радиоактивной меткой. На
четвертой минуте метка содержится лишь в нескольких
остатках со стороны С-конца цепи сс-глобина, ведь через
4 мин могли сформироваться сс-глобины только из тех
последовательностей, синтез которых был закончен на
момент добавления метки. При увеличении времени ин-
кубации появились более длинные полипептиды, содер-
жавшие меченые остатки, всегда на участке со стороны
С-конца цепи. Так было показано, что немеченый конец
полипептида (N-конец) — это инициаторный конец, и
полипептидная цепь растет путем последовательного
присоединения аминокислот к С-концу.
Различить инициаторный и внутренний ко-
доны AUG нетрудно. У бактерий есть два типа
тРНК, специфичных для метионина, которые
обозначаются как тРНКМе< и TPHKfMet Иници-
аторному кодону (5')AUG соответствует ами-
нокислота N-формилметионин (fMet). Она по-
ступает на рибосому в виде N-формилметионил-
TPHKtMet — формируется в ходе двух последо-
вательных реакций. Сначала метионин присо-
единяется к TPHKfMet ферментом Met-тРНК-
синтетазой (в клетках Е. coli этот фермент осу-
ществляет аминоацилирование и TPHKMet , и
TPHKfMet):
Метионин + TPHKfMet + АТР----->
-----> Met-TPHKfMet+ АМР + РР1
Затем трансформилаза переносит формильную
группу от Ы10-формилтетрагидрофолата на ами-
ногруппу остатка Met:
Ы10-формилтетрагидрофолат + Met-TPHKfMet->
---> тетрагидрофолат + fMet-тРНКfMet
Трансформилаза более избирательна, чем Met-
тРНК-синтетаза; этот фермент специфичен
для остатков метионина, присоединенных к
TPHKfMel, по-видимому, он распознает особенно-
сти строения этой тРНК Met-TPHKMet же встав-
ляет метионин внутри последовательностей по-
липептидов.
Н СОО
I I
Н—С—N—С—Н
II I
О сн2
СН2
S
I
СН3
N-Формилметионин
Присоединение N-формильной группы к
аминогруппе метионина под действием транс-
формилазы предотвращает встраивание fMet во
внутренние положения в молекулах полипепти-
дов, однако fMet-TPH KfMet может связываться со
специфическим инициаторным сайтом рибосо-
мы, который не присоединяет ни Met-TPHKMet,
ни какие-либо другие аминоацил-тРНК.
В клетках эукариот все полипептиды, синте-
зируемые цитоплазматическими рибосомами, на-
чинаются с остатка Met (а не с fMet), но и в этом
[196] Часть III. 27. Метаболизм белка
случае клетки используют специализированную
инициаторную тРНК, которая отличается от
TPHKMet для кодонов AUG во внутренних поло-
жениях мРНК. Полипептиды, синтезируемые на
рибосомах митохондрий и хлоропластов, начина-
ются с N-формилметионина. Это весьма убеди-
тельно подтверждает теорию, что митохондрии и
хлоропласты произошли от бактериальных пред-
ков, которые на ранней стадии эволюции были
включены в клетки-предшественницы современ-
ных эукариотических клеток (см. рис. 1-36 в т. 1).
Каким образом единственный кодон
(5')AUG может определять, встроится в данной
позиции стартовый N-формилметионин (или
метионин у эукариот) или внутренний остаток
метионина? Ответ кроется в механизме процес-
са инициации.
Три стадии инициации. Для инициации синтеза
полипептида у бактерий требуется (1) ЗОБ-субъе-
диница рибосомы, (2) мРНК синтезируемого
пептида. (3) инициаторная fMet-T₽HKfMet. (4) на-
бор из трех белков, называемых факторами
инициации (IF-1, IF-2 и IF-3), (5) GTP, (6)
505-субъединица рибосомы и (7) ионы Mg2+
Формирование инициаторного комплекса проис-
ходит в три стадии (рис. 27-25).
На стадии Ф ЗОБ-субъединица рибосомы
связывает два фактора инициации. IF-1 и IF-
2. Фактор IF-3 предупреждает преждевремен-
ное соединение субъединиц 30S и 50S. Затем с
ЗОБ-субъединицей связывается мРНК. Иници-
аторный кодон (5')AUG попадает в правильную
позицию благодаря последовательности Шай-
на-Дальгарно (в честь идентифицировавших ее
австралийских исследователей Джона Шайна и
Линн Далыарно) в мРНК. Эта консенсусная по-
следовательность из 4- 9 пуриновых оснований
расположена на расстоянии 8 13 п. н. в сторону
Субъединица
50S
Рис. 27-25. Образование инициаторного комплекса
у бактерий. Комплекс формируется в три стадии (описа-
ны в тексте) за счет энергии гидролиза GTP с образовани-
ем GDP и Р? Факторы IF-1, IF-2 и IF-3 — факторы инициа-
ции. Р — пептидильный сайт, А — аминоацильный сайт,
Е — сайт выхода. Антикодон тРНК ориентирован в на-
правлении 3'-*5' (слева направо), как на рис. 27-8, но в
противоположном направлении по сравнению с ориен-
тацией на рис. 27-21 и 27-23.
кодон
212 Синтез белков [197]
5'-конца от инициаторного кодона (рис. 27-26,
а) Эта последовательность спаривается с ком-
плементарной ей обогащенной пиримидина-
ми последовательностью, находящейся вблизи
З'-конца 16S рРНК ЗОБ-субъединицы рибосомы
(рис. 27-26, б). Взаимодействие между мРНК и
рРНК позволяет расположить инициаторную
(5')АиС-последовательность мРНК в строго
определенном участке ЗОБ-субъединицы. От дру-
гих кодонов метионина последовательность
(5')AUG, с которой должна связаться fMet-
T₽HKtMet, отличается своей близостью к последо-
вательности Шайна-Дальгарно в мРНК.
Бактериальные рибосомы имеют три сайта
связывания аминоацил-тРНК — аминоациль-
ный (А) и пептидильный (Р) , а также сайт
выхода (Е). С сайтами А и Р связываются мо-
лекулы аминоацил-тРНК, тогда как с сайтом
Е связывается исключительно ненагруженная
тРНК. Сайты А и Р образованы обеими субъе-
диницами рибосомы (30S и 50S), а сайт Е лока-
лизован в основном в 505-субъединице. Иници-
аторный кодон (5')AUG располагается в пепти-
дильном участке — только здесь может связать-
ся fMet-TPHKtMet (рис. 27-25). Единственная
аминоацил-тРНК, которая сначала связывается
с Р-сайтом, — это fMet-TPHKfMet. В процессе
элонгации все другие поступающие молекулы
аминоацил-тРНК (включая Met-TPHKMet, ко-
торая связывается только с внутренними кодо-
нами AUG) сначала связываются с сайтом А и
только затем с сайтами Р и Е. Из сайта Е «раз-
груженные» тРНК высвобождаются на стадии
элонгации. Фактор IF-1 связывается с сайтом А
и предотвращает связывание тРНК с этим сай-
том на стадии инициации.
К комплексу, состоящему из 308-субъе-
диницы рибосомы, IF-3 и мРНК, на стадии ини-
циации 2 присоединяются GTP и инициаторная
fMet-TPHKfMet (рис. 27-25). Теперь антикодон
этой тРНК спаривается с инициаторным кодо-
ном мРНК
Ген trpA Е. coli (5') A G С А
Ген araB Е. coli U U U G
CGAGGGGAAAUCU
GAUGGAGUGAAAC
Ген lacl Е. coli
Ген белка А
фага 0X174
Ген его фага X
GAU GGAACGCUAC(3)
GAUG GCGAUUGCA
UGUG AAACCAGUA
U A U G GUUCGUUCU
U A U G GAACAACGC
CAAUUCAGGGUGGUGAA
AAUCUUGGAGGCUUUUU
AUGUACUAAGGAGGUUG
Ш айн а-Д а л ьгарно;
спаривается с 16S рРНК
Инициаторный кодон;
спаривается с Ше1-тРНК£МеШе1
а
Бактериальная мРНК
с консенсусной
последовательностью 3' I
Шайна-Дальгарно ОН G
А
A U
3'-конец U с
16SpPHK U С С U С С А
(5') G A U U С С
UAGGAGGUUUGAC CUAUGCGAGCUUUUAGU (3')
б
Рис. 27-26. Последовательности мРНК, которые служат сигналами инициации синтеза
белка у бактерий, а — правильное расположение инициаторного кодона AUG (на зеленом
фоне) в 305-субъединице рибосомы отчасти зависит от расположенной выше последова-
тельности Шайна-Дальгарно (на розовом фоне). Показаны фрагменты транскриптов мРНК
пяти бактериальных генов. Обратите внимание на белок Lacl Е. coli, который начинается с
кодона GUG (Vai) (см. доп. 27-1). б — последовательность Шайна-Дальгарно в мРНК спари-
вается с последовательностью вблизи З'-конца 16S рРНК.
[198] Часть III. 27. Метаболизм белка
На стадии з этот крупный комплекс объеди-
няется с SOS-субъединицей рибосомы; в это же
время GTP, связанный с IF-2, гидролизируется до
GDP и Рь которые высвобождаются из комплек-
са. В этот момент все три фактора инициации от-
деляются от рибосомы.
Завершение стадий (рис. 27-25) приводит к
образованию функциональной 705-рибосомы,
называемой инициаторным комплексом, ко-
торая содержит мРНК и инициаторную fMet-
TPHKfMet. Правильное связывание fMet-TPHKfMet
на P-сайте в инициаторном 705-комплексе обе-
спечивается, как минимум, тремя точками рас-
познавания: взаимодействием кодона и антико-
дона, включающим фиксирование инициатор-
ного кодона AUG на P-сайте; взаимодействием
между последовательностью Шайна-Дальгарно
в мРНК и 16S рРНК; и связыванием P-сайта ри-
босомы и fMet-TPHKtMet. Теперь инициаторный
комплекс готов к элонгации.
Инициация в эукариотических клетках. Транс-
ляция в эукариотических и бактериальных
клетках происходит похожим образом; боль-
шинство различий касаются механизма инициа-
ции. Эукариотические мРНК взаимодействуют
с рибосомой в виде комплекса с несколькими
специфическими связывающими белками Не-
которые из них подтягивают друг к другу 5'- и
40S Субъединица рибосомы
Рис. 27-27. Роль белков в формировании инициаторного
комплекса у эукариот. 3'- и 5'-концы эукариотической
мРНК связаны с комплексом белков, состоящим из не-
скольких факторов инициации и поли(А)-связывающего
белка (РАВ). Факторы eIF4E и eIF4G входят в состав более
крупного комплекса eIF4F. Этот комплекс связывается с
408-субъединицей рибосомы.
З'-концы матрицы РНК. С З'-конца мРНК свя-
зывается поли(А)-связывающим белком (РАВ от
англ, poly (A) binding protein). Эукариотические
клетки содержат, как минимум, девять факторов
инициации. Комплекс eIF4F, который включает
в себя белки eIF4E, eIF4G и eIF4A, связывает-
Таблица 27-8
Белковые факторы, необходимые для инициации трансляции в бактериальных и эукариотических клетках
Фактор
Функция
Бактерии
IF1 Препятствует преждевременному связыванию молекул тРНК с сайтом А
IF2 Облегчает связывание £МеЁ-тРНКШе1 с 305-субъединицей рибосомы
IF3 Связывается с 305-субъединицей рибосомы; предупреждает преждевременное связывание
505-субъединицы; усиливает специфичность P-сайта к fMet-TPHKfMet
Эукариоты
eIF2 Облегчает связывание инициаторной Met-TPHKMet с 405-субъединицей рибосомы
eIF2B, eIF3 Первыми связываются с 405-субъединицей рибосомы; облегчают последующие стадии
eIF4A РНК-хеликазная активность изменяет вторичную структуру мРНК, позволяя
связаться с 405-субъединицей рибосомы; часть комплекса elFlF
eIF4B Связывается с мРНК; облегчает сканирование мРНК для поиска первого кодона AUG
eIF4E Связывается с 5'-кэпом мРНК; часть комплекса eIF4F
eIF4G Связывается с eIF4E и поли(А)-связывающим белком (РАВ); часть комплекса eIF4F
eIF5 Способствует диссоциации нескольких факторов инициации из комплекса с 405-субъединицей
перед присоединением 605-субъединипы для формирования 805-инипиаторного комплекса
eIF6 Облегчает диссоциацию неактивной 805-рибосомы на субъединицы 40S и 60S
27.2 Синтез белков [199]
ся с 5'-кэпом (см. рис. 26-13) через eIF4E. Белок
eIF4G взаимодействует и с eIF4E, и с РАВ, эффек-
тивно соединяя их между собой (рис. 27-27). Бе-
лок eIF4A обладает активностью РНК-хеликазы.
Комплекс eIF4F соединяется с другим фактором,
eIF3, и с dOS-субединицей рибосомы. Эффек-
тивность трансляции зависит от многих свойств
мРНК и белков данного комплекса, включая дли-
ну 3'-поли(А)-хвоста (в большинстве случаев чем
он длиннее, тем лучше). Сближение концов эука-
риотической мРНК облегчает регуляцию транс-
ляции экспрессии генов (см. гл. 28).
Инициаторный кодон (5')AUG в мРНК опре-
деляется не по его близости к последовательности
Шайна Дальгарно, а путем сканирования мРНК,
начиная от 5'-конца до первого встреченного кодо-
на AUG, который указывает начало рамки считыва-
ния. Возможно, в этом процессе участвует комплекс
elFiF, который использует РНК-хеликазную ак-
тивность elF4 для удаления вторичной структуры
в 5'-нетранслируемой области мРНК. Белок eIF4B
тоже облегчает сканирование.
Функции различных бактериальных и эу-
кариотических факторов инициации отражены
в табл. 27-8. Механизм действия этих белков ак-
тивно изучается.
Стадия 3: пептидные связи
образуются на стадии элонгации
Третья стадия синтеза белка элонгация. Для
начала мы опять сосредоточим внимание на бак-
териальных клетках. Для элонгации требуется
(1) описанный выше инициаторный комплекс,
(2) аминоацил-тРНК, (3) три растворимых ци-
топлазматических белка, называемых факторами
элонгации (EF-Tu, EF-Ts и EF-G у бактерий) и
(4) GTP. В клетке присоединение каждой амино-
Рис. 27-28. Первая стадия элонгации у бактерий: связы-
вание второй аминоацил-тРНК. Вторая аминоацил-тРНК
связывается на A-сайте рибосомы, присоединяясь к EF-Tu
(здесь просто Ти), который связан с GTP. Связывание
второй аминоацил-тРНК на A-сайте сопровождается ги-
дролизом GTP до GDP и Рк а также отделением комплекса
EF-Tu-GDP от рибосомы. Связанный GDP высвобождается,
когда комплекс EF-Tu-GDP присоединяется к EF-Ts, a EF-Ts
высвобождается, когда другая молекула GTP связывается
с EF-Tu. В результате фактор EF-Tu готов к очередному ци-
клу элонгации.
кислоты происходит в три стадии, и эти стадии
повторяются столько раз, сколько аминокислот-
ных остатков нужно присоединить.
Первая стадия элонгации: связывание амино-
ацил-тРНК. На первой стадии цикла элонгации
(рис. 27-28) соответствующая аминоацил-тРНК
[200] Часть III. 27. Метаболизм белка
взаимодействует с фактором EF-Tu, связанным
с GTP Комплекс аминоацил-тРНК-EF-Tu-GTP
связывается на сайте А 708-инициаторного ком-
плекса. Молекула GTP гидролизуется, и комплекс
EF-Tu-GDP высвобождается из 703-рибосомы.
Далее при участии фактора EF-Ts и GTP комплекс
EF-Tu-GTP регенерирует.
Вторая стадия элонгации: формирование пеп-
тидных связей. На этой стадии формируется
пептидная связь между двумя аминокислота-
ми, связанными через соответствующие тРНК
с А- и P-сайтами рибосомы. Инициаторная
N-формилметионильная группа переносится
от своей тРНК на аминогруппу второй амино-
кислоты, находящейся в сайте А (рис. 27-29).
При этом а-аминогруппа аминокислоты в сай-
те А выступает в роли нуклеофила, вытесняя
тРНК, находящуюся в сайте Р, и образуя пеп-
тидную связь. В результате на сайте А образу-
ется дипептидил-тРНК, а «разгруженная» (де-
ацилированная) тРНКШс1 остается связанной с
сайтом Р. Затем молекулы тРНК сдвигаются, за-
нимая промежуточное положение, при котором
каждая частично находится в двух сайтах рибосо-
мы, как показано на рис. 27-29.
Исторически сложилось так, что фермента-
тивную активность, ответственную за образование
пептидной связи, назвали пептидилтрансфераз-
ной и приписывали одному или нескольким бел-
кам в большой субъединице рибосомы. Теперь мы
знаем, что эту реакцию катализирует 23S рРНК
(рис. 27-13, г) — еще один рибозим. Это открытие
интересно тем, что позволяет проследить эволю-
ционные связи (см. доп. 27-2).
Третья стадия элонгации: транслокация. На за-
ключительной стадии цикла элонгации, называ-
емом транслокацией, рибосома передвигается
на один кодон по направлению к З'-концу мРНК
(рис. 27-30, а). При этом антикодон дипептидил-
тРНК, который все еще присоединен ко второму
кодону мРНК, сдвигается из сайта А в сайт Р, а
деацилированная тРНК смещается из сайта Р в
сайт Е, откуда высвобождается в цитозоль. Те-
перь в сайте А расположен третий кодон мРНК,
а в сайте Р — второй кодон. Для передвижения
рибосомы вдоль мРНК необходим фактор EF-G
(транслоказа) и энергия, которая выделяется
в результате гидролиза следующей молекулы
Сайт Е Сайт Р Сайт А
образование
пептидной
связи
Рис. 27-29. Вторая стадия элонгации у бактерий: фор-
мирование первой пептидной связи. Пептидилтрансфе-
разная активность, ответственная за осуществление этой
реакции, принадлежит рибозиму 23S рРНК. В ходе реак-
ции N-формилметионильная группа переносится на ами-
ногруппу второй аминоацил-тРНК, образуя дипептидил-
тРНК. При этом обе тРНК, оставаясь связанными с ри-
босомой, сдвигаются и занимают промежуточное поло-
жение на 508-субъединице рибосомы. «Разгруженная»
тРНК сдвигается таким образом, что ее 3'- и 5'-концы
находятся на сайте Е. Аналогичным образом, 3'- и 5'-кон-
цы пептидил-тРНК перемещаются в сайт Р. Антикодоны
остаются в сайтах А и Р.
27.2 Синтез белков [201]
Сайт Е Сайт Р Сайт А
транслокация
F-G GTP
EF-d + GDP +
движения рибосомы
GTP. Изменение трехмерной конформации ри-
босомы приводит к ее перемещению вдоль мо-
лекулы мРНК. Поскольку структура EF-G на-
поминает структуру комплекса EF-Tu-tPHK
(рис. 27-30, б), возможно, что EF-G связывается
на сайте А и смещает пептидил-тРНК.
После транслокации рибосома с присоеди-
ненными к ней дипептидил-тРНК и мРНК го-
Рис. 27-30. Третья стадия элонгации у бактерий: транс-
локация. а — рибосома сдвигается на один кодон по на-
правлению к З'-концу мРНК, используя энергию гидро-
лиза GTP, связанного с EF-G (транслоказой). Дипептидил-
тРНК полностью смещается на сайт Р, а сайт А освобожда-
ется для следующей (третьей) аминоацил-тРНК. «Разгру-
женная» тРНК отщепляется от сайта Е, и цикл элонгации
начинается снова, б — структура EF-G подобна структуре
EF-Tu в комплексе с тРНК. Слева: EF-Tu, связанный с тРНК
(зеленая) (PDF ID 1В23); справа: EF-G, связанный с GDP
(красный) (PDF ID 1DAR). С-конец EF-G (темно-серый) по
форме и по распределению заряда напоминает антикодо-
новое плечо тРНК.
това к следующему циклу элонгации и связы-
ванию третьего аминокислотного остатка. Этот
процесс осуществляется таким же образом, как
и присоединение второго остатка (рис. 27-28,
27-29 и 27-30), при продвижении рибосомы от
кодона к кодону вдоль мРНК по направлению к
З'-концу Присоединение каждого аминокислот-
ного остатка к растущей полипептидной цепи
сопровождается гидролизом двух молекул GTP
до GDP и Рр
Полипептид остается связанным с тРНК
своей последней (присоединенной) аминокис-
лотой. Таким образом сохраняется взаимосвязь
между информацией в мРНК и синтезом поли-
пептида. Сложноэфирная связь между тРНК и
[202] Часть III. 27. Метаболизм белка
С-концом растущего полипептида готова к тому,
что ее карбоксильная группа будет нуклеофиль-
но атакована следующей аминокислотой, что
приведет к образованию новой пептидной связи
(рис. 27-29). При образовании новой пептидной
связи сложноэфирная связь между полипепти-
дом и тРНК разрывается, но взаимосвязь между
полипептидом и информацией в мРНК сохраня-
ется, так как каждая новая аминокислота присо-
единяется к своей тРНК.
Цикл элонгации у эукариот напоминает цикл
элонгации у бактерий. По функциям три фактора
элонгации эукариот eEFla, eEFl|3y и eEF2 ана-
логичны факторам элонгации бактерий EF-Tu,
EF-Ts и EF-G. В рибосомах эукариот нет сайта Е;
разгруженные молекулы тРНК отщепляются от
сайта Р.
Коррекция ошибок на рибосоме. На первой
стадии элонгации в бактериальных клетках
(рис. 27-28) СТРазная активность EF-Tu во
многом определяет скорость и точность всего
процесса биосинтеза. Оба комплекса EF-Tu-GTP
и EF-Tu-GDP существуют всего несколько
миллисекунд, а потом распадаются; за это вре-
мя дважды осуществляется проверка правиль-
ности взаимодействия кодона с антикодоном.
Обычно в первый раз неправильная аминоацил-
тРНК отщепляется от сайта А. Если вместо
GTP используется его аналог гуанозин-5'-О-(3-
тиотрифосфат) (GTPyS), гидролиз замедляется,
точность синтеза повышается (благодаря более
длительной проверке), но снижается скорость
синтеза.
В ходе эволюции синтез белка (спаривание
кодона и антикодона) достиг оптимального соот-
ношения между скоростью и точностью (напри-
мер, правильность аминокислотной последова-
тельности или спаривания оснований). Точность
может замедлять скорость, а скорость может ухуд-
шить точность. Вспомните также, что механизм
коррекции ошибок на рибосоме проверяет только
правильность спаривания кодона с антикодоном,
но не выбор присоединяемой к тРНК аминокис-
лоты. Если тРНК связала неправильную амино-
кислоту (что можно сделать экспериментально),
эта аминокислота успешно встроится в белковую
последовательность в той позиции, где находится
распознаваемый этой тРНК кодон.
Стадия 4: для прекращения синтеза
полипептида нужен специальный сигнал
Элонгация продолжается до тех пор, пока рибо-
сома не добавит последнюю аминокислоту, ко-
дируемую мРНК. О переходе к заключительной
стадии биосинтеза белка, терминации, сигнали-
зирует один из трех стоп-кодонов (UAA, UAG,
UGA). который располагается сразу после ко-
дона последней аминокислоты. Мутации в анти-
кодоне тРНК, которые позволяют встраивать
аминокислоту в ответ на стоп-кодон, обычно гу-
бительны для клетки (доп. 27-4).
У бактерий, после того как стоп-кодон зани-
мает A-сайт рибосомы, подключаются три фак-
тора терминации, или фактора высвобожде-
ния, — белки RF-1. RF-2 и RF-3. Они участвуют
(1) в гидролизе концевой связи пептидил-тРНК;
(2) в высвобождении свободного полипептида
и последней разгруженной молекулы тРНК из
P-сайта: и (3) в диссоциации 705-рибосомы на
субъединицы 30S и 50S, которые готовы к началу
нового цикла синтеза полипептида ( ис. 27-31).
Фактор RF-1 распознает стоп-кодоны UAG и
UAA, a RF-2 - UGA и UAA. Либо RF-1. либо
RF-2 (в зависимости от стоп-кодона) связывает-
ся со стоп-кодоном; затем под действием пепти-
дилтрансферазы растущий полипептид связы-
вается с молекулой воды, а не с другой амино-
кислотой. По-видимому, факторы терминации
содержат домены, имитирующие структуру
тРНК, как показано на рис. 27-30, б для фактора
элонгации EF-G. Специфическая функция RF-3
пока не установлена, но предполагается, что он
участвует в высвобождении субъединицы рибо-
сомы. У эукариот один фактор терминации eRF
распознает все три стоп-кодона.
Высвобождение и распад рибосом на субъ-
единицы приводит к диссоциации компонентов
трансляционного комплекса Факторы термина-
212 Синтез белков [203]
Дополнение 27-4 Индуцированные вариации генетического кода: нонсенс-супрессия
Если в результате мутации внутри гена образуется
стоп-кодон, трансляция преждевременно прекращает-
ся, а незавершенный полипептид обычно оказывается
неактивным. Такие мутации называются нонсенс-му-
тациями (бессмысленными мутациями). Нормальная
функция гена может быть восстановлена в результате
второй мутации, которая (1) превращает ошибочный
стоп-кодон в кодон какой-нибудь аминокислоты, или
(2) подавляет действие стоп-кодона. Такие восстано-
вительные мутации называются нонсенс-супрессора-
ми; они обычно происходят в генах тРНК и приводят
к образованию измененных (супрессорных) молекул
тРНК, способных распознавать стоп-кодон и встраи-
вать на это место аминокислоту. В антикодонах боль-
шинства известных супрессорных молекул тРНК про-
изошла замена одного основания.
Супрессорные тРНК создают экспериментально
индуцированные вариации генетического кода, позво-
ляя производить считывание со стоп-кодонов, по ана-
логии с природными вариациями кода, описанными в
доп. 27-1. Нонсенс-супрессии не полностью блокиру-
ют нормальный процесс передачи информации в клет-
ке, поскольку обычно в клетке содержится несколько
копий гена каждой тРНК; некоторые из этих копий
слабо экспрессируются и составляют незначительную
часть клеточного пула конкретной тРНК. Супрессор-
ные мутации обычно затрагивают эти «минорные» ко-
пии тРНК, а основная тРНК продолжает нормально
считывать соответствующий кодон.
Например, Е. coli имеет три идентичных гена для
тРНКТуг, с каждого из которых синтезируется тРНК
с антикодоном (5’)GUA Один из этих генов экспрес-
сируется относительно интенсивно, и его продукт
представляет основной тип тРНКТуг; два других гена
транскрибируются очень слабо. В антикодоне одного
из этих двух продуктов происходит замена (5’)GUA на
(5’)СUA, приводящая к синтезу минорной тРНКТуг, ко-
торая на месте стоп-кодонов UAG встраивает тирозин.
Встраивание тирозина на месте кодона UAG происхо-
дит с низкой эффективностью, но это позволяет син-
тезировать достаточное количество полноразмерного
белка с гена, несущего нонсенс-мутацию, что позволя-
ет клетке выжить. Основная тРНКТуг продолжает нор-
мально транслировать генетический код для синтеза
большинства белков.
Мутация, приводящая к возникновению супрес-
сорной тРНК, не всегда затрагивает антикодон. Су-
прессия нонсенс-кодонов UGA обычно происходит в
тРНКТуг, которая в норме распознает HGG Измене-
ние, которое позволяет считывать UGA (и встраивать
в это положение остатки Тгр), заключается в замене G
на А в позиции 24 (в отдаленном от антикодона плече
тРНК); теперь эта тРНК может распознавать оба кодо-
на — и UGG, и VGA. Похожее изменение обнаружено
в тРНК, участвующих в наиболее распространенном
природном изменении генетического кода (VGA=Trp:
см. доп. 27-1).
Супрессия может привести к образованию боль-
шого количества аномально длинных белков, но это
происходит не всегда. Пока нам известны лишь некото-
рые подробности молекулярных событий, происходя-
щих при терминации трансляции и нонсенс-супрессии.
ции уходят из комплекса (с разгруженной тРНК в
P-сайте) и заменяются на EF-G и так называемый
фактор рециклизании рибосом (RRF; Мг 20 300).
Гидролиз GTP под действием EF-G приводит к
уходу SOS-субъединицы из комплекса 305-тРНК-
мРНК. Факторы EF-G и RRF заменяются на IF-3,
который способствует высвобождению тРНК. За-
тем отделяется мРНК. После этого комплекс IF-3
и 305-субъединицы готов инициировать новый
раунд синтеза белка (рис. 27-25).
Энергетическая стоимость точности (правиль-
ности) синтеза белка. Синтез белка на основе
закодированной в мРНК информации требует
энергетических затрат. На формирование каждой
аминоацил-тРНК затрачиваются две высокоэнер-
гетические фосфатные группы. Каждый раз. когда
ошибочно активированная аминокислота гидро-
лизируется под действием деацилирующей спо-
собности аминоацил-тРНК-синтетазы, расходу-
ется еше одна молекула АТР Одна молекула GTP
расщепляется на GDP и во время первой стадии
элонгации, и еще одна — при транслокации. Та-
ким образом, для образования каждой пептидной
связи в полипептиде требуется энергия гидролиза,
как минимум, четырех молекул NTP до NDP.
Это обеспечивает более чем достаточный
термодинамический стимул для процесса син-
[204] Часть III. 27. Метаболизм белка
Связывание
теза: по меньшей мере энергия четырех фосфо-
диэфирных связей идет на образование одной
пептидной связи, стандартная свободная энер-
гия гидролиза которой составляет лишь около
-21 кДж/моль: 4 30.5 кДж/моль =122 кДж/
моль. Таким образом, суммарное изменение сво-
бодной энергии в реакции синтеза пептидной
связи составляет -101 кДж/моль. Белки — это
Рис. 27-31. Терминация синтеза белка у бактерий. Тер-
минация происходит в ответ на стоп-кодон в сайте А.
Сначала фактор терминации RF (RF-1 или RF-2, в зави-
симости от стоп-кодона) связывается с сайтом А. Это
приводит к гидролизу эфирной связи между полипеп-
тидом и тРНК в сайте Р и высвобождению завершенного
полипептида. В конце мРНК, деацилированная тРНК, и
фактор терминации покидают рибосому, которая дис-
социирует на 30S- и 5OS-субъединицы при участии
фактора рециклизации рибосом (RRF), фактора инициа-
ции IF-3 и за счет энергии, которая выделяется при гид-
ролизе GTP, опосредованном фактором EF-G. Комплекс
305-субъединицы с IF-3 готов к началу следующего цик-
ла трансляции (см. рис. 27-25).
информационные биополимеры Биохимическая
цель реакций пептидного синтеза заключается не
просто в образовании любой пептидной связи, а
связи между двумя определенными аминокисло-
тами. Каждое из богатых энергией фосфатных
соединений, затраченных в этом процессе, играет
важную роль в обеспечении правильного соот-
ветствия между каждым новым кодоном в мРНК
и соответствующей аминокислотой в растущем
полипептиде. Затраченная энергия обеспечивает
очень высокую точность трансляции генетиче-
ской информации из мРНК в аминокислотную
последовательность белка.
Быстрая трансляция одной матрицы полисо-
мами. Из бактериальных и эукариотических
клеток можно выделить крупные кластеры из
10 100 рибосом, проявляющие высокую актив-
ность в синтезе белка. На электронных микро-
фотографиях видно, что рибосомы образуют
кластеры — полисомы, в которых соседние ри-
босомы соединены между собой тонкими во-
локнами (рис. 27-32). Эти волокна представля-
ют собой молекулы мРНК. с которых происхо-
дит трансляция белка одновременно многими
близко расположенными рибосомами, что зна-
чительно повышает эффективность процесса.
У бактерий процессы транскрипции и транс-
ляции тесно связаны между собой. Молекулы
мРНК синтезируются и транслируются в одном
и том же направлении (5'-*3'). Рибосомы начи-
нают трансляцию 5'-конпа мРНК еще до завер-
шения транскрипции (рис. 27-33). Совершенно
иная ситуация наблюдается в клетках эукариот,
где вновь синтезированные молекулы мРНК сна-
27.2 Синтез белков [205]
а
Рис. 27-32. Полисома, а — четыре рибосомы, одновременно транслирующие одну эукарио-
тическую молекулу мРНК, движутся от 5'- к З'-концу и синтезируют полипептид с N-конца
к С-концу. 6 — электронная микрофотография; схема поясняет строение полисомы из шел-
ковой железы личинки шелковичного червя. Трансляцию мРНК одновременно осуществляет
множество рибосом. Растущий полипептид удлиняется по мере передвижения рибосом в на-
правлении к З'-концу мРНК. Конечный продукт этого процесса — шелковое волокно.
Направление трансляции
Рис. 27-33. Сопряжение транскрипции и трансляции у
бактерий. Рибосомы уже начинают трансляцию мРНК,
которая еще не закончила транскрибироваться РНК-
полимеразой с матрицы ДНК. Это возможно по той при-
чине, что для встречи с рибосомами бактериальная мРНК
не должна транспортироваться из ядра в цитоплазму. На
схеме рибосомы изображены более мелкими, чем РНК-
полимераза. На самом деле рибосомы (Мг = 2,7 • 10б) на
порядок крупнее РНК-полимеразы (Мх = 3,9 • 105).
чала покидают ядро и лишь затем используются
для синтеза белка.
Молекулы бактериальной мРНК обычно су-
ществуют всего несколько минут (разд. 26.2), а
потом разрушаются нуклеазами. Для поддержа-
ния высоких темпов синтеза белка каждая мРНК
должна синтезироваться постоянно и трансли-
роваться с максимальной эффективностью. Кра-
ткость существования мРНК у бактерий позво-
ляет быстро прекратить синтез, если белок боль-
ше не нужен.
Стадия 5: вновь синтезированные полипептиды
сворачиваются и процессируются
На заключительной стадии синтеза белка об-
разующаяся цепь полипептида сворачивает-
ся в биологически активную форму. Во время
синтеза или после него полипептид постепенно
приобретает свою нативную конформацию, при
этом образуются водородные связи, проявляют-
ся ван-дер-ваальсовы, ионные и гидрофобные
взаимодействия. В результате линейная, т. е.
[206] Часть III. 27. Метаболизм белка
одномерная, генетическая информация мРНК
преобразуется в трехмерную структуру белка.
Некоторые вновь синтезированные белки бак-
терий, архей и эукариот не принимают оконча-
тельной биологически активной конформации
до тех нор, пока не подвергнутся некоторым ре-
акциям процессинга, называемым посттрансля-
ционными модификациями.
Модификации N-конца и С-конца. Первый ами-
нокислотный остаток во всех полипептидах -
N-формилметионин (у бактерий) или метионин
(у эукариот). Однако в процессе образования
функционального белка формильная группа,
N-концевой остаток метионина и несколько дру-
гих N-концевых остатков (а иногда и С-концсвых
остатков) могут удаляться ферментативным пу-
тем. Примерно у 50% эукариотических белков
аминогруппа N-концевого остатка после трансля-
ции подвергается N-ацилированию. С-концсвые
остатки тоже иногда модифицируются.
Удаление сигнальной последовательности. Как
мы увидим в разд. 27.3, у некоторых белков N-
концевая последовательность из 15 30 остатков
нужна для доставки белка к месту его назначения
в клетке. Такие сигнальные последовательности
удаляются специфическими пептидазами.
Модификация некоторых аминокислот. Гидрок-
сильные группы некоторых остатков Ser, Thr и
Туг в некоторых белках подвергаются фермента-
тивному фосфорилированию под действием АТР
(рис. 27-34, я); фосфатные группы сообщают
полипептидам отрицательный заряд. Функци-
ональное значение этих модификаций в разных
белках различно. Например, молочный белок ка-
зеин содержит много фосфатных групп, которые
связывают ионы Са2\ Кальций, фосфат и ами-
нокислоты — три важных компонента питания
новорожденных. И, как мы видели на многочис-
ленных примерах, циклы фосфорилирования -
дефосфорилирования регулируют активность
многих ферментов и регуляторных белков.
СОО соо
H3N—С—Н О 1 II СН2-О—Р—О H3N—С—Н
сн2
О
Фосфосерин о
СОО
4- | о
H3N—С—Н О
1 II О=Р О
Н—С -О- Р о
1 1 о
СН3 О Фосфотреонин Фосфотирозин
соо
+ I
H3N—С—Н
СН2
сн
оос хсоо
у - Карбоксиглутамат
б
Рис. 27-34. Некоторые модифицированные аминокис-
лотные остатки, а — фосфорилированные аминокисло-
ты. б — карбоксилированная аминокислота, в — некото-
рые метилированные аминокислоты.
соо СОО
H3N—С—Н H3N— С—Н
1 сн2 1 сн2
сн2 сн2
сн2 сн2
1 СН2 СН2
+nh2 +NH
сн3 сн3 СНз
Метиллизин Диметиллизин
СОО СОО
H3N—С—Н H3N— С—Н
сн2 । сн2
сн2 сн2
сн2 с=о
сн2 О
^N. СНз
Триметиллизин Метилглутамат
в
27.2 Синтез белков [207]
К остаткам Glu некоторых белков могут при-
соединяться дополнительные карбоксильные
группы. Например, белок системы свертывания
крови протромбин в N-концевой области содер-
жит несколько остатков у-карбоксиглутамата
(рис. 27-34, б), присоединенных витамин-К-
зависимым ферментом. Эти карбоксильные груп-
пы связывают ионы Са24, необходимые для ини-
циации механизма свертывания.
В некоторых мышечных белках и в цито-
хроме с присутствуют остатки монометил- и ди-
метиллизина (рис. 27-34, я). У большинства ви-
дов организмов белок кальмодулин содержит
один остаток триметиллизина в определенной
позиции полинептидной цепи. В других белках
карбоксильные группы некоторых остатков Glu
подвергаются метилированию, что нейтрализует
отрицательный заряд белков.
Присоединение углеводных боковых цепей. Угле-
водные боковые цепи гликопротеинов присоединя-
ются ковалентной связью во время или после син-
теза полипептида. В некоторых гликопротеинах
углеводные боковые цепи присоединяются фер-
ментативным путем к остаткам Asn (N-связанные
олигосахариды), у других - к остаткам Ser или Thr
(О-связанпые олигосахариды) (см. рис. 7-29 в т. 1).
Многие внеклеточные белки, а также протеоглика-
ны мембран клеток слизистых оболочек, содержат
олигосахаридные боковые цепи (см. рис. 7-27).
Присоединение изопренильных групп. Неко-
торые эукариотические белки модифицируют-
ся путем присоединения изопренильных групп
(производные изопрена). Между изопренильной
группой и остатком Cys в белке образуется тио-
эфирная связь (см. рис. 11-14 в г. 1). Изопрениль-
ные группы образуются из нирофосфорилиро-
ванных интермедиатов биосинтеза холестерина
(см. рис. 21-35 в т. 2), таких как фарнезилпиро-
фосфат (рис. 27-35). К таким белкам относятся
Ras-белки - продукты онкогенов и протоонкоге-
нов гах, и G-белки (и те, и другие обсуждаются в
гл. 12 в т. 1), а также ламины - белки хромосом-
ного остова. Изопренильная группа помогает за-
якорить белок на мембране. Трансформирующая
(канцерогенная) активность онкогена ras исчезает
при дефекте изопренилирования белка Ras; это
открытие стимулировало поиск ингибиторов для
применения в противоопухолевой химиотерапии.
Белок Ras
Ras
Фарнезилированный белок Ras
Рис. 27-35. Фарнезилирование остатка Cys. Тиоэфирная
связь выделена красным цветом. Белок Ras — продукт
онкогена ras.
Присоединение простетических групп. Актив-
ность многих белков бактерий и эукариот зави-
сит от наличия присоединенных ковалентной
связью простетических групп. Два характерных
примера - молекула биотина в ацетил-СоА-кар-
боксилазе и гем в гемоглобине или цитохроме с.
Протеолитический процессинг. Многие белки
синтезируются в виде крупных неактивных пред-
шественников, которые в результате протеолиза
превращаются в более короткие активные формы.
К таким белкам относятся проинсулин, некоторые
вирусные белки и протеазы, включая химотрипси-
ноген и трипсиноген (см. рис. 6-38 в т. 1).
Образование дисульфидных мостиков. В не-
которых белках, находящихся в нативной кон-
формации, образую гея внутримолекулярные
или межмолекулярные дисульфидные мостики
между остатками Cys. У эукариот дисульфидные
связи часто встречаются в белках, выступающих
за пределы клетки. Сульфидные мостики защи-
щают нативную конформацию белка от денатура-
ции во внеклеточной среде, которая может суще-
ственно отличаться от внутриклеточной среды и
обычно является окислительной.
Многие антибиотики и токсины
ингибируют синтез белка
Синтез белка — самый важный клеточный процесс
и мишень для действия многих природных анти-
биотиков и токсинов. За небольшими исключени-
ями, антибиотики ингибируют синтез белков у бак-
[208] Часть III. 27. Метаболизм белка
терий. Несмотря на совсем незначительные разли-
чия путей белкового синтеза у бактерий и эукариот,
только для клеток эукариот обсуждаемые ниже
соединения неопасны, т. е. не оказывают токсиче-
ского эффекта. Удивительно то, что синтезируемые
одними микроорганизмами соединения оказыва-
ются токсичными для других микроорганизмов.
Биосинтез белка на любой стадии может специфи-
чески подавляться каким-то антибиотиком, поэто-
му антибиотики могут предоставить нам ценный
инструмент для изучения биосинтеза белка.
Пуромицин, продуцируемый актиномице-
том Streptomyces alboniger, — один из самых из-
ученных антибиотиков-ингибиторов. Его струк-
тура очень напоминает З'-конец аминоацил-
тРНК, и поэтому он способен связываться на
сайте А рибосомы и участвовать в образовании
пептидной связи, превращаясь в пептидилпуро-
мицин (рис. 27-36). Но так как пуромицин напо-
минает только З'-конец тРНК, он не может уча-
ствовать в транслокации и выходит из рибосомы
вскоре после связывания с С-концом пептида.
Это приводит к преждевременному прекраще-
нию синтеза белка.
Тетрациклины ингибируют синтез белка в
клетках бактерий, поскольку блокируют А-сайт
Рис. 27-36. Нарушение образования пептидной связи в присутствии
пуромицина. а — структура антибиотика пуромицина напомина-
Сайт Р
пептидил-тРНК
Сайт А
пуромицин
ет структуру аминоацилъного конца нагруженной тРНК, и поэтому
он может связываться с A-сайтом рибосомы и принимать участие в
образовании пептидной связи (см. рис. 27-13, г). Но продукт этой
реакции не перемещается к сайту Р, а покидает рибосому, вызывая
преждевременный обрыв цепи. 6 — пептидилпуромицин.
а
NH
R-C-H CH2^QhOCH3
О=С-N-C-H
н С=О
NHOH
СН СН
б
27.2 Синтез белков [209]
рибосомы и препятствуют связыванию молекул
аминоацил-тРНК. Хлорамфеникол ингибиру-
ет синтез белка рибосомами бактерий (а также
митохондрий и хлоропластов), подавляя пепти-
дилтрансферазную активность; он не влияет на
синтез белка в цитоплазме эукариот. Циклогек-
симид блокирует пептидилтрансферазу эукари-
отических 805-рибосом, но не бактериальных
705-рибосом (а также рибосом митохондрий и
хлоропластов). Трисахарид стрептомицин в низ-
кой концентрации нарушает считывание генети-
ческого кода (у бактерий), в более высокой кон-
центрации подавляет инициацию.
Тетрациклин
NH—С—СНС12
I II
СН—СН о
I I
ОН СН
I
он
Хлорамфеникол
Некоторые ингибиторы синтеза белка ток-
сичны для человека и других млекопитающих
Дифтерийный токсин (Мт = 58 330) катализи-
рует ADP-рибозилирование остатка дифтамида
(модифицированного гистидина) эукариотиче-
ского фактора элонгации eEF2, тем самым его
инактивируя. Рицин (Мт = 29 895) — очень ток-
сичный белок из клещевины, он инактивирует
605-субъединицу эукариотических рибосом пу-
тем депуринизации специфического аденозина в
23S рРНК.
Краткое содержание раздела 27.2
СИНТЕЗ БЕЛКОВ
Синтез белков происходит на рибосомах,
которые состоят из белка и рРНК Бактерии
содержат 705-рибосомы, состоящие из боль-
шой (50S) и малой (30S) субъединиц. Эу-
кариотические рибосомы намного крупнее
(80S) и содержат больше белков.
Транспортные РНК (тРНК) содержат от
73 до 93 нуклеотидных остатков, среди
которых есть остатки с модифицирован-
ными основаниями. Каждая тРНК имеет
аминокислотное плечо с концевой по-
следовательностью ССА(З’), к которой в
результате реакции этерификации присо-
единяется аминокислота, антикодоновое
плечо, плечо Tip С и плечо D; некоторые
тРНК имеют пятое плечо. Антикодон от-
вечает за специфическое взаимодействие
между аминоацил-тРНК и комплементар-
ным кодоном мРНК.
Рост полипептидов на рибосомах начина-
ется с N-концевой аминокислоты и проис-
ходит путем постепенного добавления но-
вых остатков к С-концу
Синтез белка происходит за пять стадий.
1. Аминокислоты активируются специфи-
ческими аминоацил-тРНК-синтетазами в
цитоплазме. Эти ферменты катализируют
образование аминоацил-тРНК с одновре-
менным расщеплением АТР до АМР и PPL.
Точность синтеза белка в целом зависит от
точности этой реакции, и некоторые из этих
[210] Часть III. 27. Метаболизм белка
ферментов обладают независимыми центра-
ми корректирующей активности.
2. Во всех белках бактерий инициаторная
аминоацил-тРНК — N-формилметионил-
TPHKfMet. Инициация синтеза белка заклю-
чается в формировании комплекса между
ЗОБ-субъединицей рибосомы, мРНК, GTP,
fMet-TPHKfMet, тремя факторами инициации
и 505-субъединицей; GTP расщепляется до
GDP и Pj
3. В процессе элонгации для связывания сле-
дующей аминоацил-тРНК на сайте А в рибо-
соме требуются GTP и факторы элонгации.
В первой пептидилтрансферазной реакции
остаток fMel переносится на аминогруппу
следующей аминоацил-тРНК. Далее при пе-
ремещении рибосомы вдоль молекулы мРНК
происходит перенос дипептидил-тРНК с сай-
та А на сайт Р, что сопровождается гидроли-
зом молекулы GTP. Деацилированные на сай-
те Е молекулы тРНК покидают рибосому.
4. После многократного повторения циклов
элонгации синтез полипептида прекраща-
ется при участии факторов высвобождения.
Для образования каждой пептидной связи
требуется по меныпей мере четыре высоко-
энергетических фосфатных эквивалента (в
виде АТР и GTP); такие энергетические за-
траты необходимы для обеспечения высокой
точности трансляции.
5. Синтезированные полипептиды переходят
в активную трехмерную конформацию. Мпо-
1ие белки после синтеза подвергаются пост-
трансляционным модификациям.
Многие хороню изученные антибиотики и
токсины ингибируют некоторые стадии син-
теза белка.
27.3. Транспорт и расщепление белков
Эукариотическая клетка состоит из отделов и
органелл, которые выполняют специфические
функции, требующие разных наборов белков и
ферментов. Эти белки (за исключением тех, ко-
торые образуются в митохондриях и пластидах)
синтезируются на рибосомах в цитоплазме, от-
куда каким-то образом они должны попадать к
местам своего целевого назначения.
Только теперь мы начинаем понимать этот
сложный и удивительный процесс. Первые
этапы метаболизма белков, предназначенных
для секреции, интеграции в плазматическую
мембрану или заключения в лизосомы, обычно
происходят в эндоплазматическом ретикулуме.
Белки митохондрий, хлоропластов и ядер ис-
пользуют три разных транспортных пути, а ци-
тозольные белки просто остаются там, где были
синтезированы.
Очень важную роль при доставке белков к
месту их постоянной локализации играет ко-
роткая последовательность аминокислот, на-
зываемая сигнальной последовательностью.
Ее функция была впервые определена Гюнте-
ром Блобелом с коллегами в 1970 г. Сигналь-
ная последовательность направляет белок к
месту его локализации в клетке, и у многих
белков она удаляется во время транспортиров-
ки или после того, как белок достигает места
назначения. В белках, предназначенных для
транспортировки в митохондрии, хлоропла-
сты или эндоплазматическую сеть, сигналь-
ная последовательность находится на N-копце
только что синтезированного полипептида.
Во многих случаях способность конкретной
сигнальной последовательности доставлять
белок была подтверждена экспериментально
путем слияния сигнальной последовательно-
сти одного белка с другим белком. В результа-
те сигнальная последовательность направляла
второй белок к месту обычной локализации
первого белка. Избирательное расщепление
белков, которые больше не нужны клетке, тоже
направляется набором молекулярных сигна-
лов, заложенных в структуре каждого белка.
Гюнтер Блобел
27.3 Транспорт и расщепление белков [211]
В этом заключительном разделе мы рассмо-
трим процессы транспорта и расщепления бел-
ков, уделяя особое внимание молекулярным сиг-
налам и механизмам регуляции, которые играют
столь важную роль в метаболизме клетки (если
не оговаривается особо, речь пойдет об эукарио-
тических клетках)
Посттрансляционная модификация
многих эукариотических белков начинается
в эндоплазматическом ретикулуме
Вероятно, лучше всего изучены системы транс-
порта белков, которые начинаются в эндоплаз-
матическом ретикулуме (ЭР). Большинство
лизосомных, мембранных или секретируемых
белков имеют N-концевую сигнальную после-
довательность (рис. 27-37), которая маркирует
их для переноса в просвет ЭР; таких последо-
вательностей обнаружены сотни. На С-конце
сигнальной последовательности находится сайт
расщепления протеазой, которая отделяет сиг-
нальную последовательность от остального бел-
ка после того, как белок попадает в ЭР. Сигналь-
ные последовательности составляют в длину
от 13 до 36 аминокислотных остатков и имеют
общие свойства: (1) они содержат 10-15 гидро-
фобных аминокислотных остатков; (2) один или
несколько положительно заряженных остатков,
обычно между N-концом и гидрофобной после-
довательностью; и (3) короткую сравнитель-
но полярную последовательность на С-конце,
обычно содержащую аминокислотные остатки
Джордж Паладе.
1912-2008
с короткими боковыми цепями (особенно Ala),
вблизи сайта расщепления.
Джордж Паладе впервые показал, что белки
с такими сигнальными последовательностями
синтезируются на рибосомах, связанных с ЭР.
Сигнальная последовательность помогает напра-
вить рибосому к ЭР (стадий (1 -® на рис. 27-38).
® Подготовка к доставке к месту назначения на-
чинается на стадии инициации синтеза белка на
свободных рибосомах. ® Сигнальная последова-
тельность образуется в начале синтеза, поскольку
она расположена на N-конце, который синтези-
руется первым. @ Когда сигнальная последова-
тельность выходит из рибосомы, она связана с
ней крупной сигнал-распознающей частицей
(SRP от англ, signal recognition particle)} затем
SRP связывается с GTP и останавливает элон-
гацию полипептида, когда его длина достигает
Вирус гриппа А
человека
место
расщепления
Met Lys Ala Lys Leu Leu Vai Leu Leu Tyr Ala Phe Vai Ala Gly Asp Gin
Препроинсулин Met Ala Leu Trp Met Arg Leu Leu Pro Leu Leu Ala Leu Leu Ala Leu Trp Gly Pro Asp Pro Ala Ala Ala Phe Vai
человека
Met Met Ala Ala Gly Pro Arg Thr Ser Leu Leu Leu Ala Phe Ala Leu Leu Cys Leu Pro Trp Thr Gin Vai Vai Gly Ala Phe
роста
Промеллитин пчелы
Met Lys Phe Leu Vai Asn Vai Ala Leu Vai Phe Met Vai Vai Туг Пе Ser Tyr He Tyr Ala Ala Pro
Белок слюны
Drosophila
Met Lys Leu Leu Vai Vai Ala Vai He Ala Cys Met Leu Пе Gly Phe Ala Asp Pro Ala Ser Gly Cys Lys
Рис. 27-37. N-концевые сигнальные последовательности некоторых эукариотических
белков, направляющие их в ЭР. Гидрофобному участку (выделен желтым цветом) предше-
ствует один или несколько остатков основных аминокислот (синие). Обратите внимание на
наличие полярных остатков с короткими боковыми цепями, которые предшествуют сайтам
расщепления (слева от сайтов расщепления, показанных красными стрелками).
[212] Часть III. 27. Метаболизм белка
Эндоплазматический
ретикулум (ЭР)
Рис. 27-38. Доставка эукариотических белков с сигнальными последовательностями в эн-
доплазматический ретикулум. Этот процесс включает цикл SRP транслокацию и расщепле-
ние вновь образованного полипептида (см. текст). Комплекс SRP имеет форму палочки; он
содержит РНК из 300 нуклеотидов (7 SL-PHK) и шесть разных белков (общая масса 325 000).
Одна белковая субъединица SRP связывается непосредственно с сигнальной последователь-
ностью и останавливает элонгацию, блокируя доступ молекул аминоацил-тРНК и ингибируя
пептидилтрансферазу. Другая белковая субъединица связывает и гидролизирует GTP. Гете-
родимерный рецептор SRP состоит из субъединиц а (М, = 69 000) и р (Мх = 30 000), каждая
связывает и гидролизирует большое количество молекул GTP.
примерно 70 аминокислот, а сигнальная после-
довательность уже полностью вышла за пределы
рибосомы. 3 Теперь GTP-связанный SRP на-
правляет рибосому (все еще связанную с мРНК)
и незаконченный полипептид к рецепторам GTP-
связанного SRP на цитоплазматической поверх-
ности ЭР; образованный полипептид доставляет-
ся к комплексу транслокации белка в ЭР, кото-
рый может непосредственно взаимодействовать с
рибосомой. ® SRP отделяется от рибосомы, что
сопровождается гидролизом GTP как в SRP, так
и в рецепторе SRP. ® Элонгация полипептида
возобновляется при помощи АТР-зависимого
комплекса транслокации, направляющего расту-
щий полипептид в просвет ЭР до тех пор, пока не
синтезируется весь белок. Ъ Сигнальная после-
довательность удаляется сигнальной пептидазой
в просвете ЭР; ® рибосома диссоциирует и снова
готова к работе.
Гликозилирование играет ключевую роль
в транспорте белка
В просвете ЭР вновь синтезированные белки мо-
дифицируются различным образом. После удале-
ния сигнальных последовательностей происхо-
27.3 Транспорт и расщепление белков [213]
дит укладка полипептидных цепей с возможным
образованием дисульфидных связей, а многие
белки гликозилируются с образованием глико-
протеинов. Во многих гликопротеинах связь с со-
ответствующими олигосахаридами осуществля-
ется через остатки Asn. Такие N-связанные олиго-
сахариды достаточно разнообразны (гл. 7 в т. 1).
но первая стадия гликозилирования белка всегда
одинаковая. Сначала синтезируется олигосаха-
ридное ядро из 14 остатков, которое затем перено-
сится от донорной молекулы долихолфосфата на
определенные остатки Asn в белке (рис. 27-39).
Трансфераза находится на люминальной поверх-
ности ЭР и поэтому не может катализировать
гликозилирование белков в цитоплазме. После
переноса через мембрану олигосахариды в раз-
ных белках укорачиваются и модифицируются
по-разному, но пентасахаридное ядро от исходно-
го олигосахарида из 14 остатков сохраняется во
всех N-олигосахаридах. Некоторые антибиотики
блокируют одну или несколько стадий этого про-
цесса, что позволило исследовать стадии глико-
зилирования белков. Лучше других изучен анти-
биотик туникамицин, который имитирует струк-
Н N-ацетил гл юкозамин (GlcNAc)
< Манноза(Мап)
ф Глюкоза(О1с)
туникамицин
Рис. 27-39. Синтез олигосахаридного фрагмента гликопротеинов. Олигосахарид синтези-
руется путем последовательного присоединения моносахаридных единиц. (L, ® Первые
стадии происходят на цитоплазматической поверхности ЭР. ® В результате транслока-
ции незавершенный олигосахарид переносится через мембрану (механизм не показан), и
4 окончание синтеза происходит в просвете ЭР. Дополнительные остатки маннозы и глю-
козы для растущего олигосахарида поставляют производные долихолфосфата. На первой
стадии образования N-гликопротеина (В), 6 олигосахарид переносится с долихолфосфата
на остаток Asn белка в просвете ЭР. В дальнейшем олигосахаридная часть в зависимости
от конкретного белка по-разному модифицируется в ЭР или в комплексе Гольджи. Однако
пять сахарных остатков, изображенные на бежевом фоне (после стадии ®), сохраняются
в структуре всех N-связанных олигосахаридов. ® Высвободившийся долихолпирофосфат
вновь переносится через мембрану, так что пирофосфат оказывается на цитоплазматиче-
ской поверхности ЭР, где ® гидролизуется с регенерацией долихолфосфата.
[214] Часть III. 27. Метаболизм белка
ТУРУ UDP-N-ацетилглюкозамина и блокирует
первую стадию (рис. 27-39, стадия СР). Некоторые
белки в ЭР подвергаются О-гликозилированию.
однако чаще О-гликозилирование осуществля-
ется в комплексе Гольджи или в цитоплазме (для
белков, которые не попадают в ЭР).
Далее модифицированные белки могут
транспортироваться в различные клеточные
отделы. Из ЭР в комплекс Гольджи белки по-
ступают в транспортных пузырьках (везику-
лах) (рис. 27-40). В комплексе Гольджи олиго-
сахариды присоединяю 1ся к некоторым белкам
О-гликозидной связью, а N-связанные олигоса-
хариды подвергаются дальнейшим модифика-
циям. С помощью механизма, суть которого еше
полностью не установлена, аппарат 1ольджи со-
ртирует белки и направляет их к месту назначе-
ния. Сортировка белков, предназначенных для
секреции, и белков для плазматической мембра-
ны или лизосом основана на структурных осо-
бенностях, которые не связаны с сигнальными
последовательностями, поскольку последние
удаляются в просвете ЭР.
Лучше всего изучен процесс сортировки
гидролаз, направляющихся в лизосомы. При по-
падании гидролазы в комплекс Гольджи не-
кий еще не до конца исследованный элемент ее
трехмерной структуры (иногда называемый сиг-
нальным бугорком) распознается фосфотранс-
феразой, которая фосфорилирует определенные
остатки маннозы в олигосахаридной части гли-
копротеина (рис. 27-41) Наличие одного или
нескольких остатков маннозо-6-фосфата в оли-
госахаридной части гидролазы является сигна-
лом для ее отправки в лизосому. Рецепторный
белок в мембране комплекса Гольджи распозна-
ет маннозо-6-фосфат и связывает помеченную
таким образом гидролазу. Везикулы, содержа-
щие эти комплексы рецептора и гидролазы, от-
почковываются от транс-стороны Гольджи и
направляются к сортирующим везикулам. Здесь
в условиях низкого значения pH и под действи-
ем фосфатазы, удаляющей фосфатные группы с
Сортировка
Транс-сторона
комплекс
Гольджи
*-Секреторные гранулы /
Транспортные /.
пузырьки
Цитозоль
У
Лизосома
" Плазматическая мембрана
Рис. 27-40. Транспорт белков в лизосомы, плазматиче-
скую мембрану или во внеклеточное пространство. Из
ЭР белки в транспортных везикулах направляются к цис-
стороне комплекса Гольджи. Сортировка преимуществен-
но происходит на транс-стороне комплекса Гольджи.
27.3 Транспорт и расщепление белков [2151
О—j Олигосахарид"}—N-Ч^рмен^
Гидролаза
СНз
UDP-N-ацетилглюкозамин
(UDP-GlcNAc)
те г ил1 люымамин
фосфо 1 рннсферила
Остаток маннозо-6-фосфата
Рис. 27-41. Фосфорилирование остатков маннозы в ферментах, направляемых в лизосо-
мы. N-ацетилглюкозаминфосфотрансфераза распознает некоторые неизвестные пока осо-
бенности структуры гидролаз, направляющихся в лизосомы.
остатков маннозо-6-фосфата, комплекс рецепто-
ра и гидролазы распадается. Затем рецептор воз-
вращается к комплексу Гольджи, а везикулы с
гидролазами отпочковываются от сортирующих
везикул и направляются к лизосомам. В клет-
ках, обработанных туникамицином (рис. 27-39.
этап Ф), гидролазы, которые должны оказаться
в лизосомах, вместо этого секретируются, что
подтверждает ключевую роль N-связанной оли-
госахаридной группы в доставке этих ферментов
в лизосомы.
Доставка белков в митохондрии и хлоро-
пласты также связана с наличием в этих белках
N-концевых сигнальных последовательностей.
В митохондриях и хлоропластах есть своя ДНК,
но большинство их белков закодировано в ядре и
после синтеза доставляется в соответствующую
органеллу. Однако в отличие от других путей на-
правленного транспорта подготовка к транспор-
тировке белков в митохондрии и хлоропласты
начинается только после того, как белок-предше-
ственник полностью синтезирован и отделен от
рибосомы. Предшественники белков, предназна-
ченных для митохондрий и хлоропластов, свя-
зываются с цитоплазматическими шаперонами и
доставляются к рецепторам на внешней поверх-
[216] Часть III. 27. Метаболизм белка
ности органеллы. Затем с помощью специализи-
рованных систем транслокации белок транспор-
тируется к своему месту в органелле, после чего
сигнальная последовательность удаляется.
Сигнальные последовательности
ядерных белков не отщепляются
Между ядром и цитоплазмой макромолекулы
должны проходить через ядерные поры. Моле-
кулы РНК, синтезированные в ядре, переносятся
в цитоплазму. Рибосомные белки, синтезирован-
ные на рибосомах в цитоплазме, переносятся в
ядро и собираются в 60S- и 405-субъединицы ри-
босом в ядрышке; затем собранные субъединицы
экспортируются назад в цитоплазму. Разнообраз-
ные ядерные белки (РНК- и ДНК-полимеразы,
гистоны, топоизомеразы, белки-регуляторы экс-
прессии генов и т. п.) синтезируются в цитоплаз-
ме и переносятся в ядро. Этот путь управляется
сложной системой молекулярных сигналов и
транспортных белков, действие которых посте-
пенно стало более понятно.
У большинства многоклеточных эукариот
ядерная оболочка разрывается при каждом кле-
точном делении, а по окончании деления восста-
а
Цитоплазма
Ядерный
белок
NLS
GDP
Нуклеоплазма
Рис. 27-42. Транспорт ядерных белков, а — d Белок с сигналом ядерной локализации (NLS)
связывается комплексом импортинов аир.® Образующийся комплекс связывается с ядер-
ной порой и переносится в ядро. ® В ядре отщеплению импортина р способствует связы-
вание Ran-GTP. & Импортин а связывается с Ran-GTP и CAS (cellular apoptosis susceptibility
protein), высвобождая ядерный белок. 5 Импортины а и р и CAS выводятся из ядра и готовы
к новому циклу переноса. Выход в цитоплазму сопровождается гидролизом связанного с
Ran GTP. ® Ran-GDP связывается с NTF2 и возвращается в ядро. ® В ядре RanGEF облегчает
обмен GDP на GTE и Ran-GTP готов к переносу следующего комплекса импортинов с NLS-
содержащим белком, б — микрофотография поверхности ядерной оболочки, полученная
методом сканирующей электронной микроскопии; видны многочисленные ядерные поры.
Ядерная
оболочка
Ядерная
пора
Кап
GTP
0,2 мкм
27.3 Транспорт и расщепление белков [217]
навливается, при этом рассеянные в цитоплазме
ядерные белки снова импортируются в ядро.
Ввиду необходимости возвращения в ядро сиг-
нальная последовательность, которая направля-
ет белок в ядро, сигнал ядерной локализации
(NLS от англ, nuclear localization sequence), не
удаляется после того, как белок достигает своего
места назначения. В отличие от других сигналь-
ных последовательностей, NLS может находить-
ся почти в любом месте первичной последова-
тельности белка. Последовательности NLS мо-
гут существенно различаться, но многие состоят
из четырех-восьми аминокислотных остатков
и содержат последовательность из нескольких
остатков основных аминокислот (Arg или Lys).
Транспорт в ядро опосредован несколькими
белками, курсирующими между цитоплазмой и
ядром (рис. 27-42), включая импортины а и р и
небольшую СТРазу Ran (Ras-related nuclear pro-
tein). Гетеродимер импортинов аир действует в
качестве растворимого рецептора транспортиру-
емых в ядро белков, причем а-субъединица слу-
жит для связывания NLS-содсржащих белков в
цитоплазме. Комплекс NLS-содержащего белка
и импортина подходит к ядерной поре и прохо-
дит через нее, причем это сопровождается затра-
той энергии. В ядре импортин [3 связывается с
СТРазой Ran и высвобождает доставленный бе-
лок. Импортин а связывается с Ran и С AS {cellular
apoptosis susceptibility protein) и отщепляется от
NLS-со-держащего белка. Затем импортины а и
|3 в комплексах с Ran и CAS экспортируются из
ядра. В цитоплазме белок Ran гидролизует GTP,
высвобождая импортины, которые могут начи-
нать новый цикл переноса. Сам Ran также перено-
сится обратно в ядро за счет связывания Ran-GDP
с ядерным транспортным фактором 2 (NTF2). В
ядре связанный с Ran GDP под действием факто-
ра обмена гуанозиновых нуклеотидов RanGEF за-
мещается па GTP (см. доп. 12-2 в т. 1).
Бактерии тоже используют сигнальные
последовательности для транспорта белков
У бактерий белки направляются во внутренние
или внешние мембраны, в периплазматическое
пространство или во внеклеточную среду. Бакте-
риальные белки имеют N-концевьте сигнальные
последовательности (рис. 27-43), во многом на-
поминающие аналогичные последовательности
эукариотических белков, направляющихся в ЭР,
митохондрии и хлоропласты.
Белки внутренней мембраны
Основной белок вирусной
оболочки фага fd Met ЬУ8 LYS Ser Leu Val Leu LYS Ala
Минорный белок вирусной
оболочки фага fd
Белки периплазматического пространства
Щелочная фосфатаза
Лейцин-специфичный
связывающий белок
/3 -Лактамаза
плазмиды pBR322 Met
Белки внешней мембраны
Липопротеин
LamB
OmpA Met Met He
Met Lys Lys Leu
Met Lys Gin Ser Thr He Ala
Met Lys Ala Asn Ala Lys Thr He He
Ser lie Gin His Phe Arg Val Ala
Met Lys Ala Thr Lys Leu
Leu Arg Lys Leu Pro Leu Ala
Thr Met Lys Lys Thr Ala He Ala
Ser Val Ala Val Ala Thr Leu Val
Leu Phe Ala lie Pro Leu Val Val
Leu Ala Leu Leu Pro Leu Leu Phe
Ala Gly Met He Ala Leu Ala lie
Leu lie Pro Phe Phe Ala Ala Phe
Val Leu Gly Ala Val He Leu Gly
Val Ala Val Ala Ala Gly Val Met
He Ala Val Ala Leu Ala Gly Phe
место
расщепления
Pro Met Leu Ser Phe Ala* Ala Glu
Pro Phe Tyr Ser His Ser* Ala Glu
Thr Pro Val Thr Lys Ala* Arg Thr
Ser His Thr Ala Met Ala* Asp Asp
Cys Leu Pro Val Phe Ala* His Pro
Ser Thr Leu Leu Ala Gly* Cys Ser
Ser Ala Gin Ala Met Ala* Val Asp
Ala Thr Val Ala Gin Ala* Ala Pro -
Рис. 27-43. Сигнальные последовательности, направляющие бактериальные белки в
разные места локализации. Голубым цветом выделены основные аминокислоты вблизи
N-конца, желтым цветом обозначены гидрофобные аминокислоты в центральной части
сигнальных пептидов. Сайты расщепления сигнальных последовательностей показаны
красными стрелками. Обратите внимание, что белки оболочки и ДНК фага fd собираются
в фаговые частицы на внутренней мембране бактериальной клетки (см. рис. 1-6 в т. 1).
OmpA — белок внешней мембраны; LamB — рецепторный белок на поверхности клетки
бактериофага X.
[218] Часть III. 27. Метаболизм белка
Периплазматическое
пространство
Рис. 27-44. Модель экспорта белка у бактерий. 1 После завершения трансляции поли-
пептид связывается в цитоплазме с шапероном SecB, который ® доставляет его к белку
SecA, связанному с комплексом транслокации (SecYEG) в мембране бактериальной клетки,
(з SecB высвобождается, SecA встраивается в мембрану, проталкивая через комплекс
транслокации примерно 20 аминокислотных остатков белка. @ Гидролиз АТР белком
SecA обеспечивает энергию для конформационных изменений, которые позволяют SecA
высвободиться из мембраны, оставив полипептид в периплазматическом пространстве.
5 SecA связывает другую молекулу АТР и проталкивает через комплекс транслокации
следующий фрагмент белка из 20 аминокислотных остатков. Стадии и @ повторяются
до тех пор, пока ® весь белок не пройдет через мембрану и не высвободится в периплаз-
му. Электрохимический потенциал на мембране (обозначенный знаками «+» и «-») явля-
ется дополнительной движущей силой, способствующей транслокации белка.
В большинстве случаев экспорт белков из
клеток Е. coli осуществляется по механизму,
изображенному на рис. 27-44. После трансля-
ции предназначенный для экспорта белок сво-
рачивается очень медленно, поскольку этому
препятствует N-концевая сигнальная последо-
вательность. Растворимый шаперон SecB свя-
зывается с сигнальной последовательностью
или с другими участками частично свернутого
белка. В таком виде белок доставляется к белку
SecA внутренней мембраны. Белок SecA слу-
жит одновременно и рецептором, и трансло-
цирующей АТРазой. Освободившись от SecB,
связанный с SecA белок включается в комплекс
транслокации в мембране, состоящий из SecY.
SecE и SecG, и проходит мембрану через ком-
плекс SecYEG, длина которого составляет при-
мерно 20 аминокислотных остатков На каждой
стадии происходит гидролиз АТР, катализиру-
емый SecA.
Таким образом, экспорт белка через мембра-
ну опосредует белок SecA, локализованный на
внутренней поверхности мембраны, а не белок
периплазматического пространства. Этот ме-
ханизм может быть обусловлен просто тем. что
транслоцирующая АТРаза находится там, где
есть АТР. Трансмембранный электрохимический
потенциал также может способствовать трансло-
кации белка (в этом вопросе еше много неясного).
Большинство бактериальных белков поки-
дает клетку именно этим путем, но известны и
такие белки, которые экспортируются по друго-
му механизму, где принимают участие сигналь-
ные последовательности и рецепторные белки,
гомологичные компонентам белка SRP и его ре-
цепторам у эукариот (рис. 27-38).
27.3 Транспорт и расщепление белков [219]
Белки проникают в клетки путем
опосредованного рецепторами эндоцитоза
Некоторые белки поступают в клетки из окру-
жающей среды; у эукариот это липопротеины
низкой плотности (ЛПНП), переносчик железа
трансферрин, пептидные гормоны и циркулиру-
ющие белки, направляющиеся на расщепление.
Существует несколько механизмов проникно-
вения белков в клетку (рис. 27-45). В одном из
них белки связываются с рецепторами в углу-
блениях мембраны, называемых окаймленны-
ми пузырьками, где содержится значительно
больше рецепторов эндоцитоза, чем других бел-
ков клеточной поверхности. Эти углубления со
стороны цитоплазмы покрыты сеткой из белка
клатрина, который формирует замкнутые мно-
гогранные структуры (рис. 27-46). Клатриновая
сеть расширяется по мере увеличения числа ре-
цепторов, связанных с белками-мишенями, пока
полностью окруженная мембраной везикула не
отпочковывается от плазматической мембраны
с помощью крупной СТРазы динамина и вы-
ходит в цитоплазму. Клатрин быстро удаляется
ферментативным путем, и везикула сливается с
эндосомой. АТРазная активность в мембранах
эндосом понижает уровень pH, что облегчает
диссоциацию комплексов рецепторов с белка-
ми-мишенями. В похожем процессе кавеолин
вызывает инвагинацию участков мембраны, со-
держащих липидные рафты, связанные с опре-
деленными типами рецепторов (см. рис. 11-21 в
т. 1). Затем образующиеся везикулы сливаются
с содержащими кавеолин внутренними струк-
турами — кавеосомами, в которых поглощенные
вещества сортируются и перенаправляются к
другим участкам клетки, а кавеолины готовятся
к возвращению к поверхности мембраны. Кроме
того, существуют пути, не связанные с действи-
ем клатрина или кавеолина; в каких-то из них
участвует динамин, а в каких-то нет.
Далее импортируемые белки и их рецеп-
торы претерпевают различные превращения, и
их дальнейшая судьба зависит от типа клеток
и типа белков. Трансферрин и его рецептор в
конечном итоге используются вновь. Некото-
рые гормоны, факторы роста и иммунные ком-
плексы после индукции соответствующего кле-
точного ответа разрушаются вместе со своими
рецепторами Липопротеины низкой плотности
разрушаются после того, как связанный холе-
стерин доставляется к месту назначения, но
рецепторы ЛПНП используются многократно
(см. рис. 21-42 в т. 2).
Клатрин-
зависимый
эндоцитоз
Динамин
Клатрин
Сброс
клатриновой
оболочки Э
Кавеолин-
зависимый
эндоцитоз
Пути эндоцитоза без участия
кларина и кавеолина
Рис. 27-45. Пути эндоцитоза в эукариотических клетках. В путях, зависимых от клатри-
на или кавеолина, везикулы отделяются от плазматической мембраны с помощью СТРазы
динамина. В других путях не участвуют ни кавеолин, ни клатрин, а динамин может быть
задействован или нет.
[220] Часть III. 27. Метаболизм белка
Рис. 27-46. Клатрин. а — три легкие (L) цепи (Мг = 35 000) и три тяжелые (Н) цепи
(Мг = 180 000) образуют единицу клатрина (HL)3, имеющую трехлучевую структуру, на-
зываемую трискелионом. б — трискелионы собраны в многогранную сетчатую структуру.
в — электронная микрофотография окаймленного пузырька на цитоплазматической по-
верхности плазматической мембраны фибробласта.
8 ' 0,1 мкм
Расщепление белков
во всех клетках осуществляется
специализированными системами
Расщепление белков препятствует накоплению
аномальных или ненужных белков и позволяет
реутилизировать аминокислоты. Время полу-
жизни эукариотических белков составляет от
30 секунд до многих суток. Оборот большинства
белков происходит достаточно быстро по срав-
нению с продолжительностью жизни клетки,
хотя некоторые белки (например, гемоглобин)
могут сохраняться на протяжении всего времени
существования клетки (например, эритроциты
живут около 110 суток). Быстро разрушаются
дефектные белки, которые были неправильно
собраны или получили повреждения в процессе
функционирования, а также регуляторные фер-
менты, катализирующие ключевые реакции ме-
таболических путей.
Дефектные белки и белки с наиболее корот-
ким временем полужизни обычно как в бактери-
альных, так и в эукариотических клетках разру-
шаются с помощью селективных АТР-зависимых
цитоплазматических систем. Другая система,
которая у позвоночных действует в лизосомах,
высвобождает аминокислоты из мембранных
белков, внеклеточных белков и белков с большим
временем полужизни
У Е. coli многие белки расщепляются АТР-
зависимой протеазой Lon (название происходит
от «long form» - «длинных форм» белков, ко-
торые появляются в клетке только в отсутствие
данной протеазы). Протеаза Lon активируется
в присутствии дефектных белков или белков с
коротким временем полужизни; на расщепление
каждой пептидной связи расходуются две мо-
лекулы АТР. Детали этого процесса пока уста-
новить не удалось. Когда белок расщеплен на
мелкие* неактивные пептиды, завершают процесс
другие ATP-независимые протеазы.
ATP-зависимый путь деградации белков в
эукариотических клетках несколько отличается и
зависит от действия белка убиквитина. Убиквитин
(76 аминокислотных остатков) является одним из
самых высококонсервативных белков, он практи-
чески идентичен у столь разных организмов, как
дрожжи и человек. Убиквитин ковалентно связы-
вается с белками, предназначенными для расще-
пления по ATP-зависимому пути с участием трех
разных ферментов (El, Е2 и ЕЗ на рис. 27-47).
Связанные с убиквитином белки расщепля-
ются крупным белковым комплексом, называе-
мым 268-протеасомой (Мг = 2,5 • 106) (рис. 27-48).
27.3 Транспорт и расщепление белков [221]
Протеасома эукариот содержит как минимум 32
разные субъединицы, представленные в двух ко-
пиях, причем большинство этих субъединиц высо-
коконсервативно у многих видов, от дрожжей до
человека Протеасома состоит из комплексов двух
Повторение цикла
приводит к присоединению
новой молекулы убиквитина
Рис. 27-47. Трехстадийный процесс присоединения
убиквитина к белку. Процесс происходит через об-
разование двух разных промежуточных комплексов
убиквитина с ферментами. В конечном итоге свободная
карбоксильная группа С-концевого остатка Gly убикви-
тина через амидную (изопептидную) связь соединяется
с е-аминогруппой остатка Lys белка-мишени. При повто-
рении цикла образуется полиубиквитин — ковалентный
полимер из субъединиц убиквитина, который направляет
связанный эукариотический белок к месту расщепления.
типов: центральной частицы в форме бочонка и ре-
гуляторных частиц на обоих ее концах. Централь-
ная 208-частица состоит из четырех колец; внеш-
ние кольца сформированы из семи а-субъединиц,
а внутренние кольца — из семи |3-субъединиц. Три
из семи субъединиц каждого p-кольца обладают
протеазной активностью, причем все они разли-
чаются по субстратной специфичности. Кольца
центральной частицы формируют структуру в
форме бочонка, в котором происходит расщепле-
ние белков-мишеней. Регуляторные 198-частицы
на концах бочонка содержат по 18 субъединиц,
включая те, которые распознают и связывают
убиквитинированные белки Шесть субъединиц
являются ААА+ АТРазами (см. гл. 25), которые,
вероятно, раскручивают убиквитинированные
белки и направляют развернутые полипептиды
в центральную частицу для разрушения. Кроме
того, 198-частица отщепляет от разрушаемых бел-
ков убиквитин. В большинстве клеток есть допол-
нительные регуляторные комплексы, способные
заменить 198-частицу. Эти альтернативные регу-
ляторы не гидролизуют АТР и не связывают убик-
витин, однако они необходимы для расщепления
некоторых клеточных белков. В зависимости от
внутриклеточных условий 268-протеасоме могут
помогать различные регуляторные комплексы.
Мы пока не понимаем всех сигналов, инду-
цирующих присоединение убиквитина, но один
простой сигнал был обнаружен. Первая ами-
нокислота, которая остается после удаления с
N-конца остатка Met или после другого посттран-
сляционного протеолитического процессинга
N-конца, определяет время жизни многих белков
(табл. 27-9). Эти N-концевые сигналы не измени-
лись за миллиарды лет эволюции — они одинако-
вые в бактериальных системах расщепления бел-
ков и в системе убиквитинирования у человека.
Были обнаружены и более сложные сигналы (см.
рис. 12-46, т. 1).
Убиквитин-зависимый протеолиз одинако-
во важен как для регуляции клеточных процес-
сов, так и для уничтожения дефектных белков.
Многие белки, которые нужны только на одной
стадии клеточного цикла эукариотических кле-
ток, после выполнения своей функции быстро
разрушаются по убиквитин-зависимому пути.
В регуляции клеточного цикла очень важную
роль играет убиквитин-зависимое разрушение
циклина (рис. 12-46 в т. 1). Компоненты Е2 и ЕЗ,
[222] Часть III. 27. Метаболизм белка
Субстратный
белок
а Центральная частица 20S
Полиубиквитин,
присоединенный
к белку, взаимодействует
с протеасомой
Регуляторная
частица 19S
б Полностью собранная протеасома
Рис. 27-48. Трехмерная структура эукариотической протеасомы. У всех эукариот
265-протеасома высококонсервативна. Она состоит из центральной 205-частицы и двух ре-
гуляторных 195-частиц. а — Центральная частица (PDB ID 1IRU) состоит из четырех колец,
образующих структуру в форме бочонка. Два внутренних кольца содержат по семь разных
[3-субъединиц (голубые), три из которых обладают протеазной активностью (темно-синие).
Два внешних кольца состоят из семи разных а-субъединиц (серые), б — Центральный бо-
чонок с двух сторон закрыт регуляторными частицами. Центральная часть изображена
теми же цветами, что и на рис. а. Основание и вершина каждой регуляторной частицы изо-
бражены разными оттенками розового цвета. Регуляторная частица раскручивает убикви-
тинированные белки (синяя спираль) и проталкивает их в бочонок.
участвующие в реакциях убиквитинирования
(рис. 27-47), представляют два крупных семей-
ства белков. Ферменты Е2 и ЕЗ проявляют раз-
ное сродство к белкам-мишеням и регулируют
таким образом разные клеточные процессы.
Некоторые ферменты Е2 и ЕЗ локализованы в
строго определенных клеточных компартмен-
тах, что отражает их специализацию.
Неудивительно, что нарушения механизма
убиквитинирования приводят к различным
заболеваниям. Неспособность разрушать опре-
деленные белки (продукты онкогенов), которые
активируют деление клеток, может привести к
развитию опухолей, но и слишком быстрое раз-
рушение белков, которые действуют в качестве
опухолевых супрессоров, может вызвать тот же
эффект. Предполагается, что неэффективное или
очень быстрое разрушение клеточных белков
обусловливает развитие болезней почек, астму,
нейродегенеративные расстройства, такие как
паркинсонизм и болезнь Альцгеймера (при этих
болезнях обнаружены характерные белковые
Таблица 27-9 Связь между временем жизни белка и его N-концевым остатком
N-концевой остаток Время полужизни*
Стабилизирующий
Met, Gly, Ala, Ser. Thr, Vai >20ч
Дестабилизирующий
He, Gin ~ 30 мин
Tyr. Glu ~ 10 мин
Pro ~ 7 мин
Leu, Phe, Asp, Lys ~ 3 мин
Arg ~ 2 мин
Источник. По Bachmair A., Finley D., Varshavsky А. (1986)
In vivo half-life of a protein is a function of its amino-terminal
residue. Science 234, 179- 186, с изм.
* Время полужизни определяли для дрожжевой р-галак-
тозидазы, модифицированной таким образом, что в каж-
дом эксперименте фермент имел другой N-концевой
остаток. Время полужизни разных белков и в разных
организмах может быть различным, но общая тенденция,
вероятно, прослеживается повсеместно
Ключевые термины [223]
структуры в нейронах), кистозный фиброз (воз-
никает в результате слишком быстрого разруше-
ния и нарушения функции хлорных каналов; см.
доп. 11-3 в т. 1), синдрома Лиддла (натриевые
каналы в почках не разрушаются, что приводит к
избыточному всасыванию Na+ и раннему разви-
тию гипертонии) и многие другие заболевания.
При лечении некоторых из этих болезней можно
использовать ингибирующую активность проте-
асом. При изменении метаболизма разрушение
белков играет такую же важную роль для выжи-
вания клетки, как и их синтез. Еще многое пред-
стоит узнать об этих метаболических путях.
Краткое содержание раздела 27.3
ТРАНСПОРТ И РАСЩЕПЛЕНИЕ БЕЛКОВ
После синтеза многие белки направляются к
месту своей локализации в клетке. Один из
механизмов направленной доставки белка
основан на узнавании сигнального пепти-
да, чаще всего расположенного на N-конце
вновь синтезированного белка.
В клетках эукариот один класс сигнальных
последовательностей распознается сигнал-
распознающей частицей (SRP), которая свя-
зывает сигнальную последовательность, как
только она появляется на рибосоме, и пере-
носит целую рибосому и незаконченный по-
липептид в ЭР. Полипептиды с такими сиг-
нальными последовательностями по мере
синтеза перемещаются в просвет ЭР, где они
модифицируются и откуда переносятся в
комплекс Гольджи. Далее белки сортируют-
ся и направляются в лизосомы, плазматиче-
скую мембрану или транспортные везикулы.
Эукариотические белки, доставляемые в ми-
тохондрии и хлоропласты, и секретируемые
белки бактерий также имеют N-концевую
сигнальную последовательность.
Белки, предназначенные для транспорта
в ядро, содержат внутреннюю сигнальную
последовательность, которая в отличие от
других сигнальных последовательностей не
отщепляется после того, как белок доставля-
ется к месту назначения.
Некоторые клетки эукариот импортируют
белки путем опосредованного рецепторами
эндоцитоза.
Все клетки со временем разрушают белки,
используя специализированные системы
протеолиза. Дефектные белки и белки с ко-
ротким временем полужизни обычно разру-
шаются с помощью ATP-зависимой системы.
В клетках эукариот белки сначала связыва-
ются с высококонсервативным белком убик-
витином. Убиквитин-зависимый протеолиз
осуществляется протеасомами, которые тоже
высококонсервативны. Этот механизм игра-
ет очень важную роль в регуляции многих
клеточных процессов.
Ключевые термины
Термины, выделенные жирным шрифтом, объясня-
ются в глоссарии.
аминоацил-тРНК 166
аминоацил-тРНК-
синтетазы 166
аминоацильный (А) сайт
196
антикодон 173
динамин 219
дифтерийный токсин
209
инициаторный кодон
170
инициаторный комплекс
198
инициация 179, 196
кдатрин 219
кодон 167
комплекс транслокации
212
нонсенс-супрессия 203
окаймленные пузырьки
219
открытая рамка считыва-
ния (ОРС) 171
пептидилтрансфераза
200
пептидильный (Р) сайт
196
полисома 204
последовательность Шай-
на-Дальгарно 196
посттрансляционная мо-
дификация 206
протеасома 220
пуромицин 208
рамка считывания 167
редактирование РНК
176
рицин 209
сайт выхода (Е) 196
сдвиг рамки считывания
175
сигнал ядерной локализа-
ции (NLS) 217
сигнал-раснознающая ча-
стица (SRP) 211
сигнальная последова-
тельность 206 210
стоп-кодоны 170
стрептомицин 209
терминация 180, 202
тетрациклины 209
транслокация 200
трансляция 167
туникамицин 213
убиквитин 220
факторы терминации 202
факторы элонгации 199
хлорамфеникол 209
циклогексимид 209
элонгация 180, 199
[221] Часть III. 27. Метаболизм белка
Дополнительная литература
для дальнейшего изучения
Генс 1 ическиli код
Ambrogelly, A., Palioura, S., & Soil, D. (2007) Natural ex-
pansion of the genetic code. Nat. Chem. Biol. 3,29-35.
Blanc, V. & Davidson, N. O. (2003) C-to-U RNA editing:
mechanisms leading to genetic diversity. J. Biol. Chem. 278,
1395-1398.
Crick, F. H. C. (1966) The genetic code: III. Sci. Am. 215 (Oc-
tober), 55-62.
Обзор о структуре генетического кода, написанный в
то время, когда генетический код еще только начал рас-
шифровываться.
Hohn, М. J., Park, Н. S., O’Donoghue, Р, Schnitzbauer,
М., & Soli, D. (2006) Emergence of the universal genetic code
imprinted in an RNA record. Proc. Natl. Acad. Sci. USA 103,
18 095-18 100.
Klobutcher, L. A. & Farabaugh, P. J. (2002) Shifty ciliates:
frequent programmed translational frameshifting in Euplotids.
Cell 111, 763-766.
Levanon, K., Eisenberg E., Rechavi G., & Levanon, E. Y.
(2005) Letter from the editor: adenosine-to-inosine RNA
editing in Alu repeats in the human genome. EMBO Rep. 6,
831-835.
Maas, S., Rich, A., & Nishikura, K. (2003) A-to-I RNA ed-
iting: recent news and residual mysteries. J. Biol. Chem. 278,
1391-1394.
Neeman, Y., Dahary, D., & Nishikura, K. (2006) Editor
meets silencer: crosstalk between RNA editing and RNA inter-
ference. Nat. Rev. Mol. Cell Biol. 7,919-931.
Nirenberg, M. (2004) Historical review: deciphering the
genetic code — a personal account. Trends Biochem. Sci. 29,
46-54.
Schimmel, P. & Beebe, K. (2004) Molecular biology — ge-
netic code seizes pyrrolysine. Nature 431, 257-258.
Stadtman, T. C. (1996) Selenocysteine. Annu. Rev. Biochem.
65,83-100.
Vetsigian, K., Woese, C., & Goldenfeld, N. (2006) Collec-
tive evolution and the genetic code. Proc. Natl. Acad. Sci. USA
103,10 696-10 701.
Xie, J. M. & Schultz, P. G. (2006) Innovation: a chemical
toolkit for proteins — an expanded genetic code. Nat. Rev. Mol.
Cell Biol. 7,775-782.
Yanofsky, C. (2007) Establishing the triplet nature of the ge-
netic code. Cell 128,815-818.
Yanis, M., Caporaso, J. G., & Knight, R. (2005) Origins of
the genetic code: the escaped triplet theory. Annu. Rev. Bio-
chem. 74,179-198.
Синтез белка
Ban, N., Nissen, P, Hansen, J., Moore, P. B., & Steitz, T. A.
(2000) The complete atomic structure of the large ribosomal
subunit at 2.4 angstrom resolution. Science 289,905-920.
Первое описание структуры с высокой степенью разре-
шения большей субъединицы рибосомы.
Bjork, G. R., Ericson, J. U., Gustafsson, С. E. D., Hager-
vall, T. G., Jonsson, Y. H., & Wikstrom, P. M. (1987) Trans-
fer RNA modification. Annu. Rev. Biochem. 56,263-288.
Chapeville, E, Lipmann, E, von Ehrenstein, G., Weisblum,
B., Ray, W. J., Jr., & Benzer, S. (1962) On the role of soluble
ribonucleic acid in coding for amino acids. Proc. Natl. Acad. Sci.
USAM,\№£-Mm.
Классические эксперименты, подтверждающие право-
ту выдвинутой Криком адаптерной гипотезы и показыва-
ющие, что аминокислоты не проверяются после того, как
они связываются с молекулами тРНК.
Dintzis, Н. М. (1961) Assembly of the peptide chains of hemo-
globin. Proc. Natl. Acad. Sci. USA 47, 247-261.
Классический эксперимент, подтверждающий, что
синтез белка начинается с N-конца.
Giege, R., Sissler, М., & Florentz, С. (1998) Universal rules
and idiosyncratic features in tRNA identity. Nucleic Acid Res.
26,5017-5035.
Gray, N. K. & Wickens, M. (1998) Control of translation ini-
tiation in animals. Annu. Rev. Cell Dev. Biol. 14,399-458.
Ibba, M. & Soli, D. (2000) Aminoacyl-tRNA synthesis. Annu.
Rev. Biochem. 69,617-650.
Kapp, L. D. & Lorsch, J. R. (2004) The molecular me-
chanics of eukaryotic translation. Annu. Rev. Biochem. 73,
657-704.
Korostelev, A., Trakhanov, S., Laurberg, M., & Noller, H. E
(2006) Crystal structure of a 70S ribosome-tRNA complex
reveals functional interactions and rearrangements. Cell 126,
1065-1077.
Moore, P. B. & Steitz, T. A. (2003) The structural basis of
large ribosomal subunit function. Annu. Rev. Biochem. 72,813—
850.
Peske, E, Rodnina, M. V., & Wintermeyer, W. (2005) Se-
quence of steps in ribosome recycling as defined by kinetic
analysis. Mol. Cell 18,403-412.
Poehlsgaard, J. & Douthwaite, S. (2005) The bacterial ribo-
some as a target for antibiotics. Nat. Rev. Microbiol. 3,870-881.
Rodnina, M. V. & Wintermeyer, W. (2001) Fidelity of ami-
noacyl-tRNA selection on the ribosome: kinetic and structural
mechanisms. Annu. Rev. Biochem. 70,415-435.
Woese, C. R., Olsen, G. J., Ibba, M., & Soil, D. (2000) Ami-
noacyl-tRNA synthetases, the genetic code, and the evolutio-
nary process. Microbiol. Mol. Biol. Rev. 64,202-236.
Вопросы и задачи [225]
Транспорт и секреция белка
DeMartino, G. N. & Gillette, Т. G. (2007) Proteasomes: ma-
chines for all reasons. Cell 129,659-662.
Hartmann-Petersen, R., Seeger, M., & Gordon C. (2003)
Transferring substrates to the 26S proteasome. Trends Biochem.
Sci. 28,26-31.
Higgins, M. K. & McMahon, H. T. (2002) Snap-shots of clath-
rin-mediated endocytosis. Trends Biochem. Sci. 27,257-263.
Liu, C. W., Li, X. H., Thompson, D., Wooding, K., Chang,
T., Tang, Z., Yu, H., Thomas, P. J., & DeMartino, G. N.
(2006) ATP binding and ATP hydrolysis play distinct roles in
the function of 26S proteasome. Mol. Cell 24,39-50.
Luzio, J. P., Pryor, P. R., & Bright, N. A. (2007) Lysosomes:
fusion and function. Nat. Rev. Mol. Cell Biol. 8,622-632.
Mayor, S. & Pagano, R. E. (2007) Pathways of clathrin-inde-
pendent endocytosis. Nat. Rev. Mol. Cell Biol. 8, 603-612.
Neupert, W. (1997) Protein import into mitochondria. Annu.
Rev. Biochem. 66,863-917.
Pickart, С. M. & Cohen, R. E. (2004) Proteasomes and their kin:
proteases in the machine age. Nat. Rev. Mol. Cell Biol. 5,177-187.
Royle, S. J. (2006) The cellular functions of clathrin. Cell. Mol.
Life Sci. 63,1823-1832.
Schatz, G. & Dobberstein, B. (1996) Common principles of
protein translocation across membranes. Science 271,1519-1525.
Schekman, R. (2007) How sterols regulate protein sorting
and traffic. Proc. Natl. Acad. Sci. USA 104,6496-6497.
Smalle, J. & Vierstra, R. D. (2004) The ubiquitin 26S protca-
some proteolytic pathway. Annu. Rev. Plant Biol. 55,555-590.
Stewart, M. (2007) Molecular mechanism of the nuclear pro-
tein import cycle. Nat. Rev. Mol. Cell Biol. 8,195-208.
Вопросы и задачи___________________________
1. Трансляция матричной РНК. Определите
аминокислотные последовательности пептидов,
синтезируемых рибосомами на основе последо-
вательностей мРНК, при условии, что рамка счи-
тывания начинается с первых трех оснований в
каждой последовательности.
a) GGUCAGUCGCUCCUGAUU
б) UUGGAUGCGCCAUAAUUUGCU
в) CAUGAUGCCUGUUGCUAC
г) AUGGACGAA
2. Скольким разным последовательностям
мРНК может соответствовать одна аминокис-
лотная последовательность? Напишите все
возможные последовательности мРНК, кодиру-
ющие трипептид Leu-Met-Tyr. На основании
вашего ответа вы сможете оценить, сколько вари-
антов мРНК могут кодировать один полипептид.
3. Можно ли предсказать последовательность
мРНК исходя из аминокислотной последователь-
ности ее полипептидного продукта? Определен-
ная последовательность оснований мРНК будет
кодировать одну и только одну последовательность
аминокислот в полипептиде, если определена рам-
ка считывания. Можно ли, исходя из определенной
последовательности остатков аминокислот в белке,
например цитохроме с, предсказать последователь-
ность оснований в единственной мРНК, кодирую-
щей этот белок? Ответ обоснуйте.
4. Кодирование полипептида ДНК-дуплексом.
Матричная цепь участка ДНК-дуплекса содер-
жит последовательность
(5')CTTAACACCCCTGACTTCGCGCCGTCG(3')
а) Какая последовательность оснований
мРНК может транскрибироваться с этой цепи?
б) Какая аминокислотная последовательность
может кодироваться молекулой мРНК, опреде-
ленной вами в пункте (а) (начиная с 5'-конца)?
в) Если комплементарная (нематричная)
цепь этой ДНК транскрибируется и транслиру-
ется, будет ли образуемая аминокислотная по-
следовательность такой же, как в (б)? Объясните
биологическое значение своего ответа.
5. Метионину соответствует только один ко-
дон. Метионин — одна из двух аминокислот, ко-
торым соответствует только по одному кодону.
Как единственный кодон для метионина может
кодировать и первый аминокислотный остаток, и
внутренние остатки метионина в пептидах, син-
тезируемых клетками Е. coli?
6. Синтетические молекулы мРНК. Генетический
код был расшифрован благодаря полирибонукле-
отидам, синтезированным в лаборатории фермен-
тативным или химическим путем. Учитывая со-
временные знания о генетическом коде, как бы вы
синтезировали полирибонуклеотид, который мог
бы служить в качестве мРНК, кодирующий большое
количество остатков Phe и немного остатков Leu и
Ser? Какие еще аминокислоты кодировал бы этот
полирибонуклеотид, но в еще меньшем количестве?
7. Энергетическая стоимость биосинтеза белка.
Определите минимальную энергетическую сто-
имость (в эквивалентах АТР) биосинтеза р-цепи
[226] Часть III. 27. Метаболизм белка
гемоглобина (146 остатков) при наличии всех не-
обходимых аминокислот, АТР и GTP. Сравните ре-
зультат с энергетическими затратами на биосинтез
линейной цепи гликогена из 146 остатков глюкозы,
связанных al -^4-связью, при наличии глюкозы,
UTP и АТР (см. гл. 15 в т. 2). Исходя из полученного
результата оцените дополнительные энергетические
затраты на образование пептида, в котором все остат-
ки выстраиваются в определенной последовательно-
сти, по сравнению с затратой энергии на синтез по-
лисахарида, содержащего то же количество остатков,
но не несущего информационного содержания?
Помимо прямой затраты энергии на синтез
белка, есть и косвенные затраты — на производ-
ство ферментов, необходимых для синтеза белка.
Сравните уровень косвенных затрат эукариоти-
ческой клетки на биосинтез линейных (al ^4)
цепей гликогена и на биосинтез полипептидов с
учетом количества участвующих ферментов.
8. Зная кодоны, определите антикодоны. Боль-
шинство аминокислот кодируется несколькими
кодонами и связывается с несколькими тРНК —
каждая со своим антикодоном. Напишите все
возможные антикодоны для четырех кодонов
глицина: (5’)GGU, GGC, GGA и GGG.
а) На основании своего ответа определите,
какие положения в антикодонах наиболее важ-
ны для определения соответствующих кодонов в
случае глицина?
б) Какие основания кодона и антикодона об-
разуют «качающиеся» пары?
в) В каких парах кодон/антикодон все три
позиции связаны уотсон-криковскими водород-
ными связями?
9. Влияние однонуклеотидной замены на амино-
кислотную последовательность белка. Многие
важные подтверждения генетического кода были
получены путем анализа изменений аминокис-
лотной последовательности мутантных белков
после замены единственного основания в генах
этих белков. Какие из следующих аминокислот-
ных замен могут быть вызваны заменой одного
основания? Какие изменения не могут быть вы-
званы заменой одного основания? Почему?
а) Phe^Leu д) Ile^Leu
б) Lys^Ala е) His^Glu
в) Ala^Thr ж) Pro^Ser
г) Phe^Lys
10. Причина серповидноклеточной мутации. Ге-
моглобин серповидных эритроцитов содержит
остаток Vai в положении 6 |3-цепи вместо остатка
Glu, содержащегося в нормальном гемоглобине
А. Какое изменение произошло в кодоне для глу-
тамата, в результате которого остаток Glu был за-
менен на Vai?
И. Корректирующая функция аминоацил-
тРНК -синтетаз. Изолейцил-тРНК-синтетаза
имеет корректирующую активность, которая
обеспечивает точность аминоацилирования, а
гистидил-тРНК-синтетаза такой активности не
имеет. Объясните этот факт.
12. Значение «второго генетического кода».
Некоторые аминоацил-тРНК-синтетазы не рас-
познают и не связывают антикодон соответству-
ющих тРНК, и специфичность их связывания
основана на распознавании других элементов
молекул тРНК. По-видимому, к этой категории
относятся тРНК для аланина.
а) Какие элементы тРНКл1а распознаются
А1а-тРНК-синтетазой?
б) Опишите последствия замены C^G в тре-
тьей позиции антикодона тРНКЛ1а.
в) Какие другие типы мутаций могут оказы-
вать аналогичное действие?
г) Мутации такого типа не были обнару-
жены в естественных популяциях организмов.
Почему? Подсказка. Подумайте, что может про-
изойти с отдельными белками и с организмом
в целом.
13. Точность синтеза белка. Химические меха-
низмы, обеспечивающие точность синтеза бел-
ка, отличаются от механизмов, обеспечивающих
точность репликации ДНК. ДНК-полимеразы
имеют корректирующую 3'^>5'-экзонуклеазную
активность, позволяющую удалить аномально
встроенные основания из растущей цепи ДНК.
У рибосом нет аналогичной корректирующей
функции, и присоединяющиеся с помощью
тРНК аминокислоты никогда не идентифици-
руются. Гидролиз пептидной связи уже после
того, как неправильная аминокислота встро-
илась в растущий полипептид (по аналогии с
этапом коррекции последовательности ДНК-
полимеразами), оказался бы невыгодным. Поче-
му? Подсказка. Вспомните, как в процессе элон-
Анализ экспериментальных данных [227]
гации поддерживается связь между растущим
полипептидом и мРНК. См. рис. 27-20 и 27-30.
14. Предсказание клеточной локализации белка.
Ген эукариотического полипептида из 300 амино-
кислотных остатков изменен таким образом, что
распознаваемая SRP сигнальная последователь-
ность находится на N-конце полипептида, а сигнал
ядерной локализации (NLS) находится внутри,
начиная с позиции 150. В каком отделе клетки мо-
жет оказаться этот белок?
15. Что требуется для переноса белка через мем-
брану? У секретируемого бактериального белка
ОшрА есть предшественник, РгоОшрА, который
содержит необходимую для секреции N-концевую
сигнальную последовательность. Если денату-
рировать очищенный РгоОшрА с помощью 8 М
мочевины и затем мочевину удалить (например,
быстро пропустить раствор белка через колонку
для гель-фильтрации), белок переносится через
выделенные внутренние бактериальные мембра-
ны ш vitro. Однако транслокация не происходит,
если РгоОшрА предварительно инкубировать не-
сколько часов в отсутствие мочевины. Кроме того,
способность к транслокации сохраняется дольше,
если РгоОшрА предварительно инкубировать в
присутствии другого бактериального белка, назы-
ваемого триггерным фактором. Опишите возмож-
ную функцию этого фактора.
16. Способность вирусной ДНК кодировать
белок. Геном бактериофага фХ174, состоящий
из 5386 п. н., содержит 10 белков, обозначае-
мых от А до К, их размеры приведены ниже в
таблице. Сколько ДНК нужно для кодирования
этих 10 белков? Как соотносится размер генома
фХ174 с объемом его кодирующих последова-
тельностей?
Белок Количество остатков аминокислот Белок Количество остатков аминокислот
А 455 F 427
В 120 G 175
С 86 Н 328
D 152 J 38
Е 91 К 56
Анализ экспериментальных данных___
17. Конструирование белков на основании слу-
чайных последовательностей генов. Изучение
аминокислотных последовательностей и соот-
ветствующих трехмерных структур природных и
мутантных белков позволило многое узнать о тех
закономерностях, которые лежат в основе свора-
чивания белков. Для тестирования полученной
информации полезно было сконструировать бе-
лок на основании этих закономерностей и прове-
рить, соответствует ли его трехмерная структура
предсказанной структуре.
Камтекар с коллегами (1993) использовали
знание генетического кода для конструирова-
ния случайных белковых последовательностей
с определенным расположением гидрофильных
и гидрофобных остатков. Их подход к изучению
факторов, определяющих трехмерную структуру
белка, был основан на знании свойств аминокис-
лот и генетического кода.
Исследователи сконструировали ряд бел-
ков с простой структурой, состоящей из четырех
а-спиралей (см, ниже, справа; а-спирали изобра-
жены в виде цилиндров), соединенных участками
с неупорядоченной спиральной структурой (ро-
зовые). Каждая а-спираль обладала амфифиль-
ными свойствами: на одной стороне спирали
R-группы были только гидрофобными (желтые),
а на другой стороне — только гидрофильными
(голубые). Предполагалось, что белок, состоя-
щий из четырех таких спиралей, соединенных
нерегулярными спиральными последовательно-
стями, образует трехмерную структуру, в которой
гидрофильные поверхности спиралей обращены
в сторону раствора.
Пучок из четырех спиралей
Амфифильная спираль
1228 ] Часть III. 27. Метаболизм белка
а) Какие силы или взаимодействия удержи-
вают вместе четыре а-спирали в данном пучке?
На рис. 4-4, а (т. 1) представлен участок
а-спирали, состоящий из 10 аминокислотных
остатков.
б) Пронумеруйте R-группы на рис. 4-4, а
сверху (N-конец; 1) вниз (С-конец; 10). R-группы
с какими номерами выступают слева, а какие
справа?
в) Предположим, вам нужно сконструиро-
вать участок амфифильной спирали из 10 ами-
нокислотных остатков, в котором слева были бы
расположены гидрофильные остатки, а справа —
гидрофобные. Предложите последовательность
из 10 аминокислот, которые могли бы образовать
такую спираль. Учтите, что правильных ответов
на этот вопрос может быть несколько.
г) Приведите пример последовательности
двухцепочечной ДНК, которая могла бы коди-
ровать аминокислотную последовательность,
предложенную вами в ответе на вопрос (в). (Это
внутренняя последовательность, так что нет не-
обходимости вводить в нее стартовый кодон и
стоп-кодон.)
Камтекар с коллегами синтезировали бел-
ки не с определенной последовательностью, а с
почти случайной последовательностью, в кото-
рой в определенных позициях находились ги-
дрофобные и гидрофильные аминокислотные
остатки. Для этой цели ученые воспользовались
интересными особенностями генетического
кода, что позволило им создать библиотеку ис-
кусственных молекул ДНК с почти случайными
последовательностями, выстроенными опреде-
ленным образом.
Для создания последовательности ДНК, ко-
торая кодировала бы случайную последователь-
ность гидрофобных аминокислотных остатков,
исследователи начали с вырожденного кодона
NTN, в котором N может означать A, G, С или Т.
Для синтеза молекул ДНК с различными нукле-
отидами в позиции N они использовали эквимо-
лярную смесь оснований A, G, С и Т (см. рис. 8-35
вт. 1). Аналогичным образом, для создания слу-
чайной последовательности полярных аминокис-
лот они начали с вырожденного кодона NAN и ис-
пользовали для синтеза эквимолярную смесь А,
G и С (в этом случае без Т).
д) Какие аминокислоты может кодировать
триплет NTN? Все ли аминокислоты из этого на-
бора гидрофобные? Входят ли в этот набор все
гидрофобные аминокислоты?
е) Какие аминокислоты может кодировать
триплет NAN? Все ли они полярные? Входят ли в
этот набор все полярные аминокислоты?
ж) Почему при синтезе кодонов NAN реакци-
онная смесь не должна содержать основание Т?
Камтекар с коллегами клонировали библи-
отеку случайных последовательностей ДНК на
плазмидах, отобрали 48 вариантов с правильным
расположением гидрофильных и гидрофобных
аминокислотных остатков и экспрессировали их в
клетках Е. coli. Далее предстояло определить, соот-
ветствует ли укладка белков предсказанной струк-
туре. Экспрессировать и кристаллизовать каждый
белок, а затем определять его трехмерную струк-
туру было бы слишком трудоемким делом. Вместо
этого исследователи использовали процессинг
в клетках Е. coli для отсева наиболее дефектных
вариантов белков. После этого первичного скри-
нинга они сохранили лишь те клоны, в которых по
данным SDS-электрофореза в полиакриламидном
геле была белковая полоса с ожидаемой молеку-
лярной массой (см. рис. 3-18 в т. 1).
з) Почему белки с очень сильными наруше-
ниями структуры не образуют при электрофорезе
полосу с ожидаемой молекулярной массой?
Этот первичный тест прошли несколько бел-
ков, и дальнейшие исследования показали, что
они имели предсказанную структуру из четырех
спиралей.
и) Почему не все белки со случайной после-
довательностью, прошедшие первичный скри-
нинг, могли образовывать структуру из четырех
спиралей?
Литература
Kamtekar, S., Schiffer, J. М., Xiong, Н., Babik, J. М., &
Hecht, М. Н. (1993) Protein design by binary patterning
of polar and nonpolar amino acids. Science 262, 1680-1685.
Самая важная задача химической физиологии и эмбриологии — понять,
почему клетки ткани не экспрессируют постоянно все заложенные в их
геномах возможности.
— Франсуа Жакоб и Жак Моно,
из статьи в Journal of Molecular Biology, 1961
28
Регуляция экспрессии генов
28.1. Принципы регуляции генов 230
28.2. Регуляция экспрессии генов
у бактерий 243
28.3. Регуляция экспрессии генов
у эукариот 257
В клетке одновременно экспрессируется толь-
ко часть из 1000 генов типичного бакте-
риального генома или же из 20 000 генов
генома человека. Продукты некоторых генов
находятся в клетке в очень большом количестве:
например, необходимые для синтеза белка фак-
торы элонгации — очень распространенные бак-
териальные белки, а рибулозо-1,5-бифосфат-
карбоксилаза/оксигеназа (рубиско) растений и
фотосинтезирующих бактерий, по-видимому, —
самый распространенный фермент в биосфере.
Продукты других генов содержатся в гораздо
меньших количествах, например в клетке может
присутствовать несколько молекул ферментов,
исправляющих редкие повреждения ДНК. Со
временем потребность в продуктах генов может
меняться. Так, в разных метаболических путях
потребности в ферментах могут увеличиваться
и уменьшаться при изменении источников пи-
тания или по мере их истощения. На различных
стадиях развития многоклеточного организма
белки, влияющие на дифференцировку клеток,
присутствуют только в некоторых клетках и
совсем краткое время. Специализация клеточ-
ных функций влияет на потребность в продук-
тах различных генов; например, в эритроцитах
только один белок (гемоглобин) присутствует
в очень высокой концентрации. С учетом вы-
сокой энергетической стоимости синтеза белка
регуляция экспрессии генов весьма существен-
на для оптимального использования энергии.
Концентрация белка в клетке определяется
сложным равновесием, как минимум, семи про-
цессов, причем каждый процесс имеет несколько
активных точек регуляции.
1. Синтез первичных транскриптов РНК
(транскрипция).
2. Посттранскрипционная модификация
мРНК.
3. Расщепление мРНК.
4. Синтез белка (трансляция).
5. Посттрансляционная модификация белка.
6. Компартментализация и транспорт белка.
7. Расщепление белка.
Эти процессы обобщены на рис. 28-1. Некоторые
механизмы мы рассмотрели в предыдущих гла-
вах. Посттранскрипционная модификация мРНК
посредством таких процессов, как альтернатив-
ный сплайсинг (см. рис. 26-22) или редактирова-
ние РНК (см. рис. 27-10, 27-12), может влиять на
то, какие белки синтезируются на транскриптах
мРНК и в каком количестве. Скорость деграда-
ции мРНК зависит от ее нуклеотидной последо-
вательности (разд. 26.1). Скорость трансляции
мРНК и скорость посттрансляционной модифи-
кации белка, его транспорта и расщепления так-
же зависят от множества факторов (гл. 27).
[230] Часть III. 28. Регуляция экспрессии генов
Из процессов на рис. 28-1 лучше всего ис-
следованы те, которые действуют на уровне
инициации транскрипции; они в основном и
рассматриваются в данной главе. Открытия
сложных, иногда даже удивительных механиз-
мов регуляции подтверждают важность регу-
ляции на уровне посттранскрипционной мо-
дификации и трансляции, особенно в клетках
эукариот. Регуляция экспрессии многих генов
происходит по сложным и даже избыточным
Ген
ДНК--------
Определение локализации
белка и его транспорт
Рис. 284. Семь процессов, влияющих на стационарную
концентрацию белка. Каждый процесс имеет несколько
точек регуляции.
механизмам и поэтому она сопряжена с боль-
шими затратами химической энергии.
Контроль инициации транскрипции син-
хронно регулирует многочисленные гены, ко-
дирующие продукты с взаимозависимыми
функциями. Например, при сильном повреж-
дении бактериальной ДНК клеткам требуется
координированная активизация синтеза многих
ферментов репарации ДНК. Вероятно, наиболее
сложная координация реализуется в регулятор-
ных схемах, которые управляют развитием мно-
гоклеточных эукариот и используют множество
регуляторных механизмов.
Мы начнем с рассмотрения взаимодей-
ствий между белками и ДНК, которые играют
ключевую роль в регуляции транскрипции За-
тем обсудим специфические белки, влияющие
на экспрессию генов у бактерий и у эукариот.
Кроме того, там, где это необходимо, мы рас-
смотрим элементы регуляции на уровне пост-
транкрипционной модификации и трансляции,
что позволит полнее отразить сложность меха-
низмов регуляции.
28.1. Принципы регуляции генов
Гены тех продуктов, которые необходимы клет-
ке постоянно, такие как гены ферментов основ-
ных метаболических путей, экспрессируются
на более или менее постоянном уровне в каж-
дой клетке организма. Такие гены часто называ-
ют генами домашнего хозяйства. Постоянная
экспрессия гена называется конститутивной
экспрессией.
Концентрация продуктов других генов уве-
личивается и уменьшается в ответ на молекуляр-
ные сигналы; это регулируемая экспрессия гена.
Гены, концентрация продуктов которых увеличи-
вается при особых обстоятельствах, относятся к
индуцибельным; увеличение экспрессии называ-
ется индукцией. Например, экспрессия многих
генов, кодирующих ферменты репарации ДНК,
индуцируется при сильных повреждениях ДНК.
Наоборот, гены, концентрация продуктов кото-
рых уменьшается в ответ на молекулярный сиг-
нал, относят к репрессибельным, а этот процесс
называется репрессией. Например, у бактерий
избыток триптофана приводит к репрессии генов
28.1 Принципы регуляции генов [231]
тех ферментов, которые катализируют биосинтез
триптофана.
Транскрипция опосредуется и регулирует-
ся взаимодействиями белка и ДНК, в частно-
сти, с участием белковых компонентов РНК-
полимеразы (гл. 26). Сначала мы обсудим ре-
гуляцию активности РНК-полимеразы, а затем
перейдем к общему описанию белков, участву-
ющих в этом процессе. После этого мы иссле-
дуем молекулярные основы распознавания раз-
личных последовательностей ДНК специфиче-
скими белками.
РНК-полимераза связывается с ДНК
в области промоторов
РНК-полимеразы связываются с ДНК в обла-
сти промотора и инициируют транскрипцию
(см. рис. 26-5); эти участки обычно расположены
вблизи точки, в которой начинается синтез РНК
на матрице ДНК. Регуляция инициации транс-
крипции часто действует на уровне взаимодей-
ствия РНК-полимеразы с промотором.
Нуклеотидные последовательности промо-
торов значительно различаются, что определя-
ет сродство РНК-полимеразы и, следовательно,
частоту инициации транскрипции. Некоторые
гены Escherichia coli транскрибируются один раз
в секунду, другие — менее одного раза за клеточ-
ный цикл. Во многом эти различия определяются
спецификой последовательностей промоторов.
Даже в отсутствие регуляторных белков разли-
чия в последовательностях промоторов могут
объяснять более чем 1000-кратные различия в
частоте инициации транскрипции. Большинство
промоторов Е. coli имеет последовательность,
близкую к консенсусной (рис. 28-2). Мутации,
изменяющие консенсусную последовательность,
обычно снижают эффективность промотора; и,
наоборот, мутации, приближающие к консен-
сусной последовательности, обычно усиливают
промотор.
Хотя гены домашнего хозяйства экспресси-
руются конститутивно, концентрация кодиру-
емых ими продуктов в клетке изменяется в ши-
роком диапазоне. Взаимодействие РНК-полиме-
разы с промотором сильно влияет на вероятность
инициации транскрипции этих генов; различия
в последовательностях промоторов позволяют
клетке синтезировать нужное количество про-
дукта каждого конститутивного гена.
Скорость инициации транскрипции на про-
моторах неконститутивных генов также опреде-
ляется последовательностью промотора, но да-
лее их экспрессия модулируется регуляторными
белками. Многие из этих белков действуют путем
усиления или ослабления взаимодействия между
РНК-полимеразой и промотором.
Последовательности эукариотических про-
моторов более разнообразны, чем у бактерий (см.
рис. 26-9). Чтобы связаться с промотором, трем
эукариотическим РНК-полимеразам обычно ну-
жен набор основных факторов транскрипции.
Тем не менее, как и при экспрессии бактериаль-
ных генов, уровень транскрипции определяется
влиянием промоторных последовательностей на
работу РНК-полимеразы и связанных с ней фак-
торов транскрипции.
Сайт начала транскрипции
область -35
область -10
ДНК 5
UP-элемент
TTGACA
N17
ТАТААТ
N5-9
мРНК
Рис. 28-2. Консенсусные последовательности промоторов Е. coli. Большинство замен ос-
нований на участках -10 и -35 отрицательно сказывается на эффективности промотора.
Некоторые промоторы также включают в себя UP-элемент (см. рис. 26-5). По договорен-
ности. последовательности ДНК изображают в виде нематричной цепи с 5'-концом слева.
Нумерация нуклеотидов начинается в точке начала транскрипции; положительные номера
находятся справа от нее (в направлении транскрипции), а отрицательные номера — слева.
N — любой нуклеотид.
[232] Часть III. 28. Регуляция экспрессии генов
Инициация транскрипции регулируется
белками, которые связываются
с промоторами или недалеко от них
Инициацию транскрипции РНК-полимеразой
регулируют, как минимум, три типа белков: фак-
торы специфичности изменяют специфичность
РНК-полимеразы по отношению к данному про-
мотору или набору промоторов: репрессоры бло-
кируют присоединение РНК-полимеразы к про-
мотору; активаторы усиливают взаимодействие
РНК-полимеразы с промотором.
Мы уже познакомились с бактериальными
факторами специфичности в гл. 26, хотя там мы их
так не называли. Фактор специфичности, который
опосредует распознавание промотора и связыва-
ние с ним, — это о-субъединица холофермента
РНК-полимеразы Е. coli. Большинство промото-
ров Е. coli распознаются одной о-субъединицей о70
(Мг = 70 000). При определенных обстоятельствах
некоторые о7,,-субъединицы замещаются одним
из шести других факторов специфичности. В
частности, такая ситуация возникает, когда бак-
терии подвергаются тепловому шоку, что приво-
дит к замене о7() на о32 (Л/г = 32 000). Связанная
с о32 РНК-полимераза направляется к специали-
зированному набору промоторов с другими кон-
сенсусными последовательностями (рис. 28-3).
Эти промоторы контролируют экспрессию не-
скольких генов, кодирующих белки ответа на те-
пловой шок, в том числе некоторые белки-шапе-
роны (с. 212). Таким образом, путем изменения
сродства полимеразы, которое обеспечивает свя-
зывание с другими промоторами, достигается ко-
ординированная регуляция генов, участвующих
в взаимосвязанных процессах. В эукариотиче-
ских клетках в качестве факторов специфичности
можно рассматривать некоторые общие факторы
транскрипции, такие как ТАТА-связывающий бе-
лок (ТВР, см. рис. 26-9).
С определенными участками ДНК связыва-
ются белки-репрессоры. В бактериях их участки
связывания, называемые операторами, обычно
находятся вблизи промотора. Репрессор блоки-
рует связывание РНК-полимеразы или ее про-
движение вдоль ДНК. Регуляция с участием
репрессора, подавляющего транскрипцию, назы-
вается отрицательной регуляцией. Связывание
репрессора с ДНК регулируется сигнальной мо-
лекулой, или эффектором. Обычно эффектор -
это небольшая молекула или белок, который
присоединяется к репрессору и изменяет его кон-
формацию. Взаимодействие между репрессором
и эффектором либо усиливает, либо ослабляет
транскрипцию. В некоторых случаях конформа-
ционные изменения приводят к отсоединению
репрессора от оператора (рис. 28-4, а). После это-
го инициация транскрипции может продолжить-
ся В других случаях взаимодействие между не-
активным репрессором и эффектором заставляет
репрессор связаться с оператором (рис. 28-4, б).
В эукариотических клетках участок связывания
репрессора может находиться на некотором рас-
стоянии от промотора; связывание приводит к
тому же эффекту, что и в бактериальных клет-
ках, - ингибированию сборки или активности
транскрипционного комплекса на промоторе.
Активаторы оказывают противоположное
действие; они связываются с ДНК и увеличивают
активность РНК-полимеразы на промоторе. Это
пример положительной регуляции. Участки свя-
зывания активатора часто примыкают к тем про-
моторам, с которыми РНК-полимераза сама по
себе (т. е. без активаторов) не связывается совсем
или связывается очень слабо, так что без активато-
ра транскрипция осуществляется с очень низкой
Сайт начала транскрипции
ДНК 5'
TNTCNCCCTTGAA
N13-15
ССССАТТТА
N7
мРНК
Рис. 28-3. Консенсусные последовательности промоторов, которые регулируют экс-
прессию генов теплового шока в клетках Е. coli. Эта система реагирует на повышение
температуры и на некоторые другие внешние стрессы, вызывая индукцию определенных
белков. Связывание РНК-полимеразы с промоторами теплового шока опосредуется спе-
циализированной о-субъединицей полимеразы о32, которая заменяет а70 в инициаторном
комплексе РНК-полимеразы.
28.1 Принципы регуляции генов [233]
Отрицательная регуляция
(связанный репрессор подавляет транскрипцию)
а
Молекулярный сигнал
Сигнальная
Промотор
вызывает отсоединение
регуляторного белка
от ДНК
молекула
мРНК
Молекулярный сигнал
вызывает связывание
регуляторного белка
с ДНК
3'
мРНК
Положительная регуляция
(связанный активатор облегчает транскрипцию)
б
Рис. 28-4. Общие схемы регуляции инициации транскрипции. Показаны два типа отрица-
тельной регуляции, а — репрессор (розовый) связывается с оператором в отсутствие сиг-
нальной молекулы; внешний сигнал вызывает диссоциацию репрессора и тем самым разре-
шает транскрипцию, б — репрессор связывается в присутствии сигнальной молекулы; при
прекращении сигнала репрессор отделяется, и начинается транскрипция. Положительную
регуляцию опосредуют активаторы. Показаны два типа положительной регуляции, в —
активатор (зеленый) связывается с ДНК в отсутствие сигнальной молекулы, и происходит
транскрипция; когда приходит сигнал, активатор отделяется и транскрипция подавляет-
ся. г — активатор связывается с ДНК под действием сигнала и отделяется только при пре-
кращении сигнала. Обратите внимание, что термины «положительная» и «отрицательная»
регуляция относятся к регуляторному белку, участвующему в этих процессах: связанный
белок либо способствует, либо препятствует транскрипции. В любом случае влияние сиг-
нальной молекулы может усиливать или ослаблять транскрипцию в зависимости от дей-
ствия этого сигнала на регуляторный белок.
эффективностью. Некоторые эукариотические
активаторы усиливают транскрипцию, связыва-
ясь с участками ДНК. называемыми энхансерами,
которые расположены далеко от промотора (на
расстоянии до нескольких тысяч пар оснований).
Некоторые активаторы постоянно связаны с ДНК
и усиливают транскрипцию, пока связывание сиг-
нальной молекулы не заставляет их отделиться от
ДНК (рис. 28-4. в). В других случаях активатор
связывается с ДНК только после взаимодействия
с сигнальной молекулой (рис. 28-4, г). Сигнальные
молекулы, следовательно, могут усиливать или
[234] Часть III. 28. Регуляция экспрессии генов
ослаблять транскрипцию в зависимости от харак-
тера их воздействия на активатор. Как мы увидим
далее, в эукариотах чаще реализуется положитель-
ная регуляция.
Многие бактериальные гены собраны
в кластеры и регулируются в виде оперонов
У бактерий есть общий простой механизм ко-
ординации регуляции генов, продукты которых
участвуют в серии взаимосвязанных процессов:
эти гены образуют кластеры на хромосоме и
транскрибируются вместе. Многие бактериаль-
ные мРНК полицистронные (в одном транскрип-
те содержится несколько генов), и единственный
промотор, инициирующий транскрипцию всего
кластера, служит участком регуляции экспрессии
всех генов кластера. Кластер генов и промотор, а
также дополнительные последовательности, ко-
торые тоже участвуют в регуляции, называются
опероном (рис. 28-5). Обычно опероны содержат
от двух до шести генов, транскрибируемых как
единое целое; некоторые опероны объединяют 20
или более генов.
Франсуа Жакоб,
1920-2013
Сайт связывания Сайт связывания
активатора репрессора (оператор)
ДНК I I [Промотор Г/М | А I В I С I
Регуляторные
последовательности
Гены, транскрибируемые
как единое целое
Рис. 28-5. Типичный бактериальный оперон. Гены А,
В и С транскрибируются в виде одной полицистронной
мРНК. К типичным регуляторным последовательностям
относятся участки связывания белков, которые либо ак-
тивируют, либо подавляют транскрипцию с промотора.
Принципы экспрессии бактериальных генов
впервые были выявлены при изучении метабо-
лизма лактозы в клетках Е. coli, которая может
использовать этот сахар как единственный ис-
точник углерода. В 1960 г. Франсуа Жакоб и Жак
Моно опубликовали в Трудах Французской ака-
демии наук короткую статью, в которой описали,
как два соседних гена, участвующих в метабо-
Галактоза
Глюкоза
Рис. 28-6. Метаболизм лактозы в клетках Е. coli. Для погло-
щения лактозы и ее метаболизма в клетках Е. coli требуются
галактозидпермеаза и р-галактозидаза. В незначительной
степени р-галактозидаза также катализирует превращение
лактозы в аллолактозу путем трансгликозилирования.
28.1 Принципы регуляции генов [235]
лизме лактозы, подвергаются координированной
регуляции под действием генетического элемен-
та, расположенного на одном конце этого класте-
ра генов. Речь шла о гене |3-галактозидазы, кото-
рая расщепляет лактозу на галактозу и глюкозу, и
гене галактозидпермеазы (лактозопермеаза, с. 567
вт. 1), которая переносит лактозу внутрь клет-
ки (рис. 28-6). В этой статье впервые появились
термины «оперон» и «оператор». Представленная
модель оперона впервые позволила рассматри-
вать регуляцию генов на молекулярном уровне.
Отрицательная регуляция лактозного оперона
Лактозный оперон (lac) (рис. 28-7, а) содержит
гены р-галактозидазы (Z), галактозидпермеазы (У)
и тиогалактозидтрансанетилазы (Л). Фермент тио-
галактозидтрансацетилаза, по-видимому, модифи-
цирует токсичные галактозиды, чтобы облегчить
их удаление из клетки. Каждому из этих трех генов
предшествует участок связывания рибосомы (на
рис. 28-7 не показан), который направляет транс-
ляцию этого гена независимо от остальных (гл. 27).
Lac-penpeccop
Днк |р,| I
б Оператор
Рис. 28-7. Лактозный оперон. а — Zac-оперон в репрессированном состоянии. Ген I ко-
дирует Lac-penpeccop. Гены lac Z,YiaA кодируют р-галактозидазу, галактозидпермеазу и
тиогалактозидтрансацетилазу соответственно. Р — промотор Zac-генов, Р: — промотор
гена I. 0: — главный оператор Zac-оперона; 02 и 03 — дополнительные операторы с мень-
шим сродством к Lac-penpeccopy. б — Lac-penpeccop связывается с главным оператором и
с дополнительными операторами 02 или 03, что, вероятно, приводит к образованию петли
ДНК, которая обвивается вокруг репрессора, в — Lac-penpeccop, связанный с ДНК (по PDB
ID 1LBG). Видно, что белок (серый) связан с короткими участками ДНК (синие), г — изме-
нение конформации Lac-penpeccopa, вызванное его связыванием с искусственным индук-
тором изопропилтиогалактозидом (ИПТГ; по PDB ID 1LBH и 1LBG). Структура тетрамерно-
го репрессора показана без связанного ИПТГ (бледное изображение на заднем фоне) и со
связанным ИПТГ (яркое изображение; ИПТГ не показан). В отсутствие ИПТГ связанная ДНК
не показана. Когда ИПТГ связан, а ДНК не связана, ДНК-связывающие домены репрессора
сильно дезорганизованы, и их нельзя выделить в кристаллическом виде.
[236] Часть III. 28. Регуляция экспрессии генов
Регуляция /яс-оперона /яс-репрессором (Lac) про-
исходит по схеме, представленной на рис. 28-4, а.
При изучении клеток с мутациями 1ас-
оперона были установлены некоторые под-
робности его регуляции. В отсутствие лактозы
транскрипция генов /яс-оперона подавлена.
Мутации в области оператора или другого гена,
гена 7, приводят к конститутивному синтезу про-
дуктов. Если ген 1 поврежден, репрессию можно
восстановить, введя в клетку нормально функ-
ционирующий ген 1 на другой молекуле ДНК.
Это доказывает, что ген 1 кодирует способную
диффундировать молекулу, которая и подавляет
транскрипцию. Оказалось, что эта молекула —
тетрамерный белок, теперь называемый Lac-
репрессором, состоящий из идентичных субъе-
диниц. Оператор Оь с которым Lac-penpeccop
связывается прочнее всего, граничит с точкой
начала транскрипции (рис. 28-7, а). Ген 1 транс-
крибируется со своего собственного промотора
(Pt) независимо от других генов /яс-оперона.
В /яс-опероне есть два дополнительных участка
связывания Lac-penpeccopa. Центр одного из них
(О2) располагается в позиции 4 410 внутри гена,
кодирующего |3-галактозидазу (Z); центр второ-
го (О3) — в позиции -90 внутри гена I. Чтобы
подавить экспрессию оперона, Lac-penpeccop,
по-видимому, связывается одновременно с глав-
ным оператором и с одним из двух дополнитель-
ных участков, так что расположенная между
ними последовательность ДНК образует петлю
(рис. 28-7, б, в). При любом варианте связыва-
ния инициация транскрипции блокируется.
Несмотря на образование этого сложного ком-
плекса, транскрипция подавляется не полностью.
Связывание Lac-penpeccopa уменьшает вероят-
ность инициации транскрипции в 103 раз. Если
сайты О2 и О3 повреждены в результате делеции
или мутации, связывание Lac-penpeccopa с одним
только оператором Ot уменьшает транскрипцию в
102 раз. Даже при связывании репрессора в каждой
клетке есть несколько молекул |3-галактозидазы и
галактозидпермеазы, по-видимому, синтезиро-
ванных во время тех редких эпизодов, когда /яс-
репрессор на краткое время отсоединяется от опе-
раторов. Этот минимальный уровень транскрип-
ции необходим для регуляции оперона.
Когда в клетках появляется лактоза, про-
исходит индукция /яс-оперона. Молекула ин-
дуктора связывается с особым участком Lac-
penpeccopa, изменяя его конформацию (рис. 28-7,
г) и приводя к отделению репрессора от опера-
тора. Индуктором в системе /яс-оперона слу-
жит не сама лактоза, а ее изомер — аллолактоза
(рис. 28-6). Транспорт лактозы в клетки Е. coli
осуществляет присутствующий там фермент пер-
меаза; далее под действием [3-галактозидазной
активности лактоза превращается в аллолактозу
Отделение репрессора от оператора после свя-
зывания репрессора с аллолактозой позволяет
начать транскрипцию генов /яс-оперона и приво-
дит к тысячекратному увеличению концентрации
Р-галактозидазы.
Некоторые |3-галактозиды, похожие по стро-
ению на аллолактозу, индуцируют /яс-оперон,
но не являются субстратами р-галактозидазы;
другие, наоборот, являются субстратами р-галак-
тозидазы, но не индукторами. В экспериментах
часто используется очень эффективный (и при
этом нерасщепляемый) индуктор /яс-оперона
изопропилтиогалактозид (IPTG):
н он
Изопропил 7?-D-тиога л актозид
(ИПТГ)
Нерасщепляемый индуктор позволяет изучать
физиологическую роль лактозы как источника
углерода для роста клеток независимо от ее уча-
стия в регуляции экспрессии генов.
В клетках бактерий обнаружено множество
оперонов; несколько полицистронных опе-
ронов найдено и в клетках низших эукариот.
Однако в клетках высших эукариот почти все
гены, кодирующие белки, транскрибируются по
отдельности.
Механизмы регуляции оперонов могут зна-
чительно отличаться от простой модели, пред-
ставленной на рис. 28-7. Даже /яс-оперои на са-
мом деле сложнее, чем показано здесь; как мы
увидим в разд. 28.2, в системе действует еще и
активатор. Прежде чем перейти к рассмотрению
уровней регуляции экспрессии, мы исследуем
ключевые молекулярные взаимодействия между
ДНК-связывающими белками (такими как ре-
прессоры и активаторы) и последовательностями
ДНК, с которыми они связываются.
28.1 Принципы регуляции генов [237]
Регуляторные белки содержат
специальные ДНК-связывающие домены
Регуляторные белки обычно связываются со
специфическими последовательностями ДНК.
Их сродство к этим последовательностям в
Ю4-106 раз выше, чем к любым другим участ-
кам ДНК. Большинство регуляторных белков
имеет особые ДНК-связывающие домены, ко-
торые специфично и прочно связываются с
ДНК. Эти связывающие домены почти всегда
содержат одну или несколько небольших функ-
циональных групп с характерными структур-
ными мотивами.
Для специфического связывания с ДНК
регуляторные белки должны распознавать ха-
рактерные структурные элементы ДНК. Боль-
шинство функциональных групп, определя-
ющих различия между четырьмя азотистыми
основаниями, выступают в большую бороздку
ДНК, где участвуют в образовании водородных
связей (рис. 28-8). Почти все специфические
контакты между ДНК и белком обеспечива-
ются водородными связями. Примечательно,
что при С-5 пиримидинового цикла тимина (в
отличие от цитозина) имеется неполярная ме-
тильная группа, которая не может участвовать
в образовании водородных связей. Белки могут
контактировать с ДНК и в малой бороздке, но
здесь они, как правило, не различают пары ос-
нований друг от друга.
Регуляторные белки чаще всего образуют
водородные связи между основаниями ДНК и
боковыми цепями остатков Asn, Gin, Glu, Lys и
Arg. Есть ли простой код распознавания, в соот-
ветствии с которым определенная аминокислота
всегда спаривается с определенным основани-
ем? Две водородные связи, которые образуют
Asn и Gin с N6 и N-7 аденина, не могут возник-
нуть между другими основаниями. Остаток
Arg может образовать две водородные связи с
N-7 и О6 гуанина (рис. 28-9). Однако изучение
структур многих ДНК-связывающих белков по-
казало, что белок может узнавать каждую пару
оснований более чем одним способом; из этого
следует вывод, что простого кода «аминокис-
лота-основание» не существует. У некоторых
белков взаимодействие Gln-аденин может точ-
но идентифицировать пару А=Т, а у других бел-
ков карман для метильной группы тимина также
позволяет узнавать пару оснований А=Т. Даже
зная структуру ДНК-связывающего белка, пока
нельзя на этой основе предложить последова-
тельность ДНК, с которой он связывается.
Для взаимодействия с основаниями в боль-
шой бороздке ДНК белок должен содержать
относительно небольшой структурный фраг-
мент, выступающий над поверхностью белка.
ДНК-связывающие домены регуляторных бел-
ков обычно небольшого размера (60-90 амино-
кислотных остатков), а структурные фрагменты
внутри этих доменов, которые непосредственно
Рис. 28-8. Фнукциональные группы ДНК, участвующие в связывании белка. Показаны все
четыре варианта пар оснований и их функциональные группы, которые располагаются в
большой и малой бороздках ДНК. Группы, которые могут быть использованы белками для
распознавания пар оснований, выделены красным цветом.
[2381 Часть III. 28. Регуляция экспрессии генов
Тимин = Аденин
н О
Цитозин = Гуанин
Рис. 28-9. Два примера специфического взаимодей-
ствия при связывании ДНК и белка.
контактируют с ДНК, еще меньше. Многие ма-
ленькие белковые молекулы нестабильны из-за
своих ограниченных возможностей сформиро-
вать складки для прикрытия гидрофобных групп
(с. 173 вт. 1). ДНК-связывающие мотивы либо
обладают очень компактной стабильной струк-
турой, либо обеспечивают способ, позволяющий
сегменту белка выступать над его поверхностью.
В ДНК участки связывания с регуляторны-
ми белками часто представлены инвертирован-
ными повторами коротких последовательностей
(палиндромами), с которыми происходит коопе-
ративное связывание нескольких (обычно двух)
субъединиц регуляторного белка. В этом смысле
Lac-penpeccop необычен тем, что он функциони-
рует 13 виде тетрамера из двух димеров, сцеплен-
ных вместе концами, удаленными от участков
связывания ДНК (рис. 28-7, б). Клетка Е. coli
содержит около 20 тетрамеров Lac-penpeccopa.
Каждый из сцепленных димеров в отдельности
связывается с палиндромной последовательно-
стью оператора и находится в контакте с 17 из
22 и. н. /«с-оперона (рис. 28-10). Каждый из сце-
пленных димеров может независимо связывать-
ся с операторной последовательностью, причем
один обычно связывается с Оь а другой — с О2
или О3 (как на рис. 28-7, б). Последовательность
оператора Ot двух парных субъединиц Lac-
penpeccopa имеет ось симметрии второго поряд-
ка. Тетрамерный Lac-penpeccop in vivo связыва-
ется со своей операторной последовательностью;
константная диссоциации ~10 10 М. Репрессор
связывается с оператором примерно в 10ь чаще,
чем с другими последовательностями, таким об-
разом. связывание именно с этими несколькими
парами оснований из 4,6 млн п. н. в хромосоме
Е. coli высокоспецифично.
Описано несколько ДНК-связывающих мо-
тивов, но здесь мы сосредоточим внимание на
двух, которые более всего важны при связывании
Промотор Сайт начала
(связывается РНК-полимеразой) транскрипции
। i ! ।
ДНК TAGGCACCCCAGGCTTTACACTTTATGCTTCCGGCTCGTATGTTGTGTGGAATTGTGAGCGGATAACAATTTCAC
I_______________________________________________________________________I___________I
область —35 область —10
Оператор (Ор
(связывается Ьас-репрессором)
мрнк ХуХ/Х/Х/ХУХ/ХУ4*
Рис. 28-10. Связь между /ос-операторной последовательностью 0! и /ас-промотором.
На бежевом фоне выделены основания (образуют палиндром), симметричные относитель-
но оси, обозначенной вертикальной пунктирной линией.
28.1 Принципы регуляции генов [239]
Рис. 28-11 Спирал ь-поворот-спираль. а — ДНК-связывающий
домен Lac-penpeccopa (PDB ID 1LCC). Мотив спиралъ-пово-
рот-спиралъ изображен красным и оранжевым цветами; ДНК-
распознающая спираль красная, б — Lac-penpeccop целиком
(по PDB ID 1LBC). ДНК-связывающие домены — серые, уча-
ствующие в образовании тетрамера а-спирали — красные.
Оставшаяся часть белка (оттенки зеленого) содержит участки
связывания с аллолактозой, соединенные с ДНК-связывающим
доменом линкерными спиралями (желтые), в — поверхность
ДНК-связывающего домена Lac-penpeccopa (серый), связанно-
го с ДНК (синяя), г — та же ДНК-связывающая область, что и
на рис. в, но отделенная от ДНК; видны поверхности белка и
ДНК, взаимодействующие при связывании. Некоторые функци-
ональные группы белка и ДНК, образующие водородные связи,
выделены красным цветом; другие группы, которые связаны
гидрофобными взаимодействиями, оранжевые. Эта модель
позволяет увидеть только некоторые группы, участвующие в
узнавании последовательности ДНК. Комплементарность двух
поверхностей весьма заметна.
ДНК с регуляторными белками; это мотивы спи-
раль-поворот-спираль и цинковый палец. Мы
также рассмотрим ДНК-связывающий домен дру-
гого типа (гомеодомен), обнаруженный в некото-
рых эукариотических белках.
Спираль-поворот-спираль. Этот ДНК-связыва-
ющий мотив играет очень важную роль при
взаимодействиях с ДНК многих регуляторных
белков у бактерий; похожие мотивы также об-
наружены в некоторых эукариотических регу-
ляторных белках. Мотив спираль-поворот-спи-
раль состоит примерно из 20 аминокислот, об-
разующих два коротких а-спиральных сегмента,
каждый длиной 7-9 аминокислотных остатков,
разделенных p-поворотом (рис. 28-11). Сама эта
[240] Часть III. 28. Регуляция экспрессии генов
структура нестабильна; она входит в более круп-
ный ДНК-связывающий домен. Один из двух
а-спиральных сегментов называется распознаю-
щей спиралью, так как здесь обычно содержатся
аминокислоты, взаимодействующие с ДНК спец-
ифическим образом. Эта сс-спираль лежит поверх
других сегментов белковой структуры и высту-
пает над поверхностью белка. При связывании с
ДНК распознающая спираль располагается вну-
три или почти внутри большой бороздки. У Lac-
репрессора именно такой ДНК-связывающий
структурный фрагмент (рис 28-11).
Цинковый палец. В мотиве цинкового пальца
около 30 аминокислотных остатков образуют вы-
тянутую петлю, основание которой удерживается
одним ионом Zn2+, координированного с четырь-
мя аминокислотными остатками (четыре остатка
Cys или два остатка Cys и два остатка His). Ион
цинка не взаимодействует с ДНК, а служит для
стабилизации этого маленького структурного
мотива. Поддержанию стабильности также спо-
собствует наличие нескольких гидрофобных бо-
ковых цепей внутри структуры. На рис. 28-12
показано взаимодействие между ДНК и тремя
цинковыми пальцами из одного полипептида мы-
шиного регуляторного белка Zif 268.
Многие ДНК-связывающие белки эукариот
содержат мотив цинкового пальца. Взаимодей-
ствие одного цинкового пальца с ДНК обычно
слабое, и многие ДНК-связываюшие белки, как и
Zif 268, содержат множество цинковых пальцев,
что существенно усиливает связывание. Один
из ДНК-связываюгцих белков лягушки Xenopus
имеет 37 цинковых пальцев. В бактериальных
белках этот мотив встречается очень редко.
В разных белках с цинковыми пальцами
связывание с ДНК осуществляется по-разному.
Некоторые цинковые пальцы содержат амино-
кислотные остатки, важные для распознавания
последовательности ДНК, тогда как другие, по-
видимому, связываются с ДНК неспецифическим
образом (в последнем случае аминокислоты, не-
обходимые для специфического связывания, рас-
положены в других участках белка). Цинковые
пальцы могут также участвовать в связывании
РНК, например, в некоторых белках, связываю-
щихся с эукариотической мРНК и действующих
в качестве репрессоров трансляции. Мы обсудим
их функцию позднее (разд. 28.3).
Рис. 28-12. Цинковый палец. Три цинковых пальца (се-
рые) регуляторного белка Zif 268 в комплексе с ДНК (си-
ние и светло-серые) (PDB ID 1A1L). Каждый ион Zn2+ (ко-
ричневые) координирован с двумя остатками His и двумя
остатками Cys (не показано).
Рис. 28-13. Гомеодомен. Показан гомеодомен, связанный
с ДНК; можно видеть, что одна из а-спиралей (красная),
наложенная на две другие, укладывается внутри боль-
шой бороздки ДНК (PDB ID 1В81). Это только небольшая
часть гораздо более крупного белка Ultrabithorax (Ubx),
участвующего в регуляции развития плодовой мушки
(см. разд. 28.3).
28.1 Принципы регуляции генов [241]
Гомеодомен. ДНК-связывающий домен другого
типа был идентифицирован в ряде белков, уча-
ствующих в регуляции транскрипции, особен-
но в процессе развития эукариот. Этот участок
из 60 аминокислот называется гомеодоменом,
поскольку он был открыт в гомеозисных генах
(генах, регулирующих развитие тела). Этот вы-
сококонсервативный домен обнаружен в белках
широкого круга организмов, включая человека
(рис. 28-13). ДНК-связывающий участок этого
домена родственный мотиву спираль-поворот-
спираль. Последовательность ДНК, которая ко-
дирует этот домен, называется гомеобоксом.
Регуляторные белки содержат
также домены, ответственные
за взаимодействия белка с белком
Регуляторные белки содержат не только доме-
ны для связывания с ДНК. но и домены для вза-
имодействия с белками — с РНК-полимеразой,
другими регуляторными белками или с другими
субъединицами того же регуляторного белка. Это
характерно, например, для многих эукариотиче-
ских факторов транскрипции, функционирующих
в качестве активаторов генов. Они часто связы-
ваются с ДНК в виде димеров, используя ДНК-
связывающие домены с цинковыми пальцами.
Некоторые структурные домены предназначены
для взаимодействий, обеспечивающих образова-
ние димеров, если это необходимо для связывания
с ДНК. Как и в случае ДНК-связывающих моти-
вов, среди структурных мотивов, обеспечивающих
взаимодействия белка с белком, можно выделить
мотивы несколько типов. Среди них следует упо-
мянуть лейциновую молнию и основную спи-
раль-петлю-спираль. Наличие этих структурных
мотивов позволяет подразделить регуляторные
белки на различные структурные семейства.
Лейциновая молния. Этот мотив представлен
амфифильной а-спиралью, на одной стороне
которой сгруппированы гидрофобные амино-
кислотные остатки (рис. 28-14): гидрофобная
поверхность образует зону контакта между дву-
мя полипептидами димера. Особенность этих
Регуляторный
белок
Аминокислотная последовательность
Источник
Связующее звено
ДНК-связывающая область (6 аминокислот) Лейциновая молния
।-------------------------к-----о--------------------------------1
Млеко-ГС/ЕВР DKNSNEYRVRRERNNIAVRKSRDKAKQRNVETQQKVLELTSDNDRLRKRVEQLSRELDTLRG-
ПИТа’ Jun S QER I KAERKRMRNR I ААSKCRKRKLER I ARLEEKVKT LKAQNS ЕLA S Т ANMLT EQVAQLKQ-
ющие
lFos EERRRIRRIRRERNKMAAAKCRNRRRELTDTLQAETDQLEDKKSALQTEI ANLLKEKEKLЕF-
Дрожжи GCN4 реSSDРAALKRARNTЕAARRSRARKLQRMKQLЕDKVЕЕLLSKNYHLENEVARLKKLVGER
RR R
--L------L-------L-------L-------L---
Консенсусная
молекула
R RR
- R-------
К КК
Рис. 28-14. Лейциновая молния, а — сравнение
аминокислотных последовательностей нескольких
белков, содержащих лейциновые молнии. Обратите
внимание на остатки Leu (L) в каждой седьмой по-
зиции в области молнии и на ряд остатков Lys (К)
и Arg (R) в ДНК-связывающем участке, б — лейци-
новая молния из дрожжевого белка GCN4 (PDB ID
1YSA). Показаны только «застегнутые» а-спирали
(серая и голубая), относящиеся к разным субъеди-
ницам димерного белка. Две спирали оборачива-
ются одна вокруг другой с образованием суперспи-
рали. Взаимодействующие остатки Leu выделены
красным цветом.
[242] Часть III. 28. Регуляция экспрессии генов
а-спиралей заключается в наличии в каждой
седьмой позиции остатков Leu, которые образуют
прямую линию вдоль гидрофобной поверхности.
Ранее предполагали, что остатки Leu смыкают-
ся, как застежка-молния, но теперь известно, что
они выстраиваются в линию благодаря тому, что
взаимодействующие сх-спирали обвивают друг
друга, образуя суперспираль (рис. 28-14, б). В
регуляторных белках с лейциновыми молниями
часто имеется отдельный ДНК-связывающий
домен с высоким содержанием положительно за-
ряженных остатков аминокислот (Lys или Arg),
которые могут взаимодействовать с несущими
отрицательный заряд фосфатными группами ске-
лета ДНК. Лейциновые молнии обнаружены во
многих эукариотических белках и в нескольких
белках бактерий.
Основная спираль-петля-спираль. Этот струк-
турный мотив встречается в некоторых эукари-
отических регуляторных белках, участвующих в
контроле экспрессии генов при развитии много-
клеточных организмов. Эти белки содержат кон-
сервативную область длиной около 50 аминокис-
лотных остатков, которая в равной степени важна
как для связывания с ДНК, так и для димеризации
белка. Этот участок образует две короткие амфи-
фильные а-спирали, соединенные петлей различ-
ной длины. Мотив спираль-петля спираль следу-
ет отличать от мотива спираль- поворот-спираль,
который участвует в связывании ДНК. Мотивы
спираль петля спираль двух полипептидов взаи-
модействуют с образованием димеров (рис. 28-15).
Связывание этих белков с ДНК опосредуют ко-
роткие смежные аминокислотные последователь-
ности, богатые остатками основных аминокислот;
эти короткие последовательности напоминают
отдельные ДНК-связывающие участки в белках с
лейциновыми молниями.
Комбинации субъединиц в регуляторных белках
эукариот. На основании близкого структурного
сходства было выделено несколько семейств эу-
кариотических факторов транскрипции. Члены
каждого семейства могут образовывать димеры из
двух одинаковых белков (гомодимер) или из двух
разных белков, принадлежащих к одному семей-
ству (гетеродимер). Гипотетическое семейство из
четырех различных белков с лейциновыми мол-
ниями может, таким образом, образовать 10 разно-
Рис. 28-15. Спираль-петля-спираль. Человеческий транс-
крипционный фактор Мах, связанный с ДНК (PDB 1D
1HL0). Белок представляет собой димер (окрашена лишь
одна субъединица). ДНК-связывающий сегмент (розовый)
стыкуется с первой спиралью мотива спиралъ-петля-спи-
ралъ (красный). Вторая спираль стыкуется с С-концом этой
субъединицы (фиолетовая). Взаимодействие С-концевых
спиралей двух субъединиц приводит к образованию сверх-
спиральной структуры, очень напоминающей структуру
лейциновой молнии (см. рис. 28-14, б), но в данном при-
мере лишь с одной парой взаимодействующих остатков
Leu (красные боковые цепи вверху). Всю эту структуру в
целом иногда называют мотивом спираль-петля-спираль/
лейциновая молния.
видностей димеров. Во многих случаях различные
комбинации, по-видимому, имеют разные регуля-
торные и функциональные особенности.
Наряду со структурными доменами для свя-
зывания с ДНК и для димеризации (или олиго-
меризации) многие регуляторные белки должны
также содержать участки для взаимодействия
с РНК-полимеразой и/или с неродственными
регуляторными белками. Охарактеризованы до-
полнительные домены, как минимум, трех раз-
личных типов, они ответственны за межбелковые
взаимодействия (в первую очередь у эукариот):
домен, богатый глутамином, богатый пролином
и кислый домены; названия отражают природу
аминокислотных остатков, которые преобладают
в этих доменах.
28.2 Регуляция экспрессии генов у бактерий [243]
Связывание ДНК с белком — основа слож-
ных регуляторных схем функционирования ге-
нов. Теперь мы переходим к более подробному
изучению этих схем сначала в бактериальных, а
затем в эукариотических системах.
Краткое содержание раздела 28.1
ПРИНЦИПЫ РЕГУЛЯЦИИ ГЕНОВ
Экспрессию генов регулируют процессы,
влияющие на скорость синтеза и разрушения
продуктов генов. В значительной степени
регуляция осуществляется на уровне иници-
ации транскрипции посредством регулятор-
ных белков, которые либо подавляют транс-
крипцию (отрицательная регуляция), либо
активируют ее (положительная регуляция)
на специфических промоторах.
Гены бактерий, кодирующие продукты с вза-
имосвязанными функциями, часто органи-
зованы в кластеры — опероны (отдельные
транскрипционные единицы). Транскрипция
генов обычно блокируется при связывании
специфического белка-репрессора с участ-
ком ДНК, называемым оператором. Неболь-
шие молекулы (индукторы) могут вызывать
отсоединение репрессора от оператора. Эти
принципы впервые были установлены при
изучении лактозного (lac) оперона. Lac-
репрессор отделяется от fac-оператора, когда
репрессор связывается со своим индуктором
аллолактозой.
Регуляторные белки — ДНК-связывающие
белки, распознающие специфические по-
следовательности ДНК; в большинстве ре-
гулируемых белков имеется особый ДНК-
связывающий домен. Для этих доменов харак-
терны типичные структурные мотивы, ответ-
ственные за связывание с ДНК: спираль-пово-
рот-спираль, цинковый палец и гомеодомен.
Регуляторные белки также содержат домены
для межбелковых взаимодействий; это лей-
циновая молния и спираль-петля-спираль,
необходимые для димеризации, а также дру-
гие мотивы, участвующие в активации транс-
крипции.
28.2. Регуляция экспрессии генов
у бактерий
Как и во многих других биохимических исследо-
ваниях, ранее регуляция экспрессии генов изуча-
лась у бактерий, а потом уже у других организмов.
Представленные ниже примеры регуляции бак-
териальных генов выбраны из множества хорошо
изученных систем отчасти из-за их исторического
значения, но прежде всего потому, что они позво-
ляют оценить весь спектр регуляторных механиз-
мов у бактерий. Принципы регуляции бактериаль-
ных генов помогают в понимании механизмов экс-
прессии генов в эукариотических клетках.
Мы начнем с изучения лактозного и трип-
тофанового оперонов; в каждой из этих систем
участвуют регуляторные белки, но общие меха-
низмы регуляции очень различаются. Далее мы в
общих чертах рассмотрим механизм SOS-ответа
в клетках Е. coli и увидим, каким образом осу-
ществляется координированная регуляция генов,
рассеянных по всему геному. Затем мы исследуем
две совершенно разные бактериальные системы
с тем, чтобы показать разнообразие механизмов
регуляции, а именно рассмотрим, во-первых, ре-
гуляцию синтеза рибосомного белка на уровне
трансляции, в процессе которой многие регуля-
торные белки связываются с РНК, а не с ДНК; во-
вторых, регуляцию так называемой фазовой ва-
риации у сальмонеллы, которая возникает вслед-
ствие генетической рекомбинации. Наконец, мы
разберем некоторые дополнительные примеры
посттранскрипционной регуляции, когда РНК
сама модулирует свою функцию.
Положительная регуляция
лактозного оперона
Взаимодействия оператора, репрессора и индук-
тора, описанные здесь на примере fac-оперона
(рис. 28-7), представляют собой упрощенную
модель регуляции экспрессии генов по механиз-
му включения/выключения. В действительности
регуляция оперона редко бывает настолько про-
стой. Среда обитания бактерий слишком слож-
ная, чтобы их гены можно было контролировать
одним-единственным сигналом. Наряду с лак-
тозой на экспрессию fac-генов влияют и другие
факторы, например глюкоза. Для Е. coli глюко-
за, расщепляемая в процессе гликолиза, предпо-
[244] Часть III. 28. Регуляция экспрессии генов
Рис. 28-16. Гомодимер CRP (PDB ID 1RUN). Связанная
молекула сАМР показана красным цветом. Обратите
внимание, как ДНК изгибается вокруг белка. Область,
которая взаимодействует с РНК-полимеразой (желтая).
чтительна как источник энергии. Другие сахара
тоже могут служить главным или единственным
питательным субстратом, но для их включения
в гликолиз требуются дополнительные реакции,
для которых нужны специфические ферменты.
Очевидно, что экспрессия генов белков, расще-
пляющих такие сахара, как лактоза или арабино-
за, совершенно не оправдана при наличии доста-
точного количества глюкозы
А как происходит экспрессия /бгс-оперона,
если в среде одновременно присутствуют и глю-
коза, и лактоза? В присутствии глюкозы экс-
прессию генов, необходимых для катаболизма
лактозы, арабинозы и других вторичных саха-
ров, ограничивает регуляторный механизм, на-
зываемый катаболитной репрессией. Влияние
глюкозы опосредует с АМР, выступающий в ка-
честве коактиватора, и активаторный белок, на-
зываемый сАМР-рецепторным белком (CRP,
или САР — от англ, catabolite gene activator
protein белок-активатор катаболитных ге-
нов). Белок CRP представляет собой гомодимер
(масса субъединицы Мг = 22 000), содержащий
участки для связывания ДНК и с АМР. Связыва-
ние осуществляется через мотив спираль-пово-
рот спираль в ДНК-связывающем домене белка
(рис. 28-16). В отсутствие глюкозы комплекс
CRP-cAMP связывается с ДНК вблизи 1ас-
нромотора (рис. 28-17, а) и в 50 раз усиливает
транскрипцию РНК. Следовательно, комплекс
CRP-cAMP — положительный регуляторный
элемент, реагирующий на концентрацию глю-
козы, a Lac-penpeccop - отрицательный регуля-
торный элемент, реагирующий на лактозу. Оба
элемента действуют согласованно. Когда Lac-
penpeccop блокирует транскрипцию, комплекс
СайтСИР Связывается РНК-полимеразой 5 мРНК
: I I
ДНК 5-ATTAATGTGAGTTAGCTCACTCATTAGGCACCCCAGGCTTTACACTTTATGCTTCCGGCTCGTATGTTGTGTGGAATTGTGAGCGGATAACAATTTCACAC
: I______________________I
область-35 область 10
а
/ас-промотор
Консенсусная
последов ате л ьность
промотора
ТТТАСА TATGTT
область -35 область—10
TTGACA ТАТААТ
б
Рис. 28-17 Активация транскрипции Zac-оперона под действием CRP. а — участок связы-
вания комплекса CRP-cAMP находится вблизи промотора. Как и последовательность 1ас-
оператора, этот участок двухцепочечной ДНК симметричен относительно оси, обозначен-
ной штриховой линией (комплементарные основания бежевые), б — последовательность
Zac-промотора по сравнению с консенсусной последовательностью промотора. Различия
с консенсусной последовательностью обусловливает относительно слабое связывание
РНК-полимеразы с Zac-промотором до тех пор, пока ее не активирует комплекс CRP-cAMP.
28.2 Регуляция экспрессии генов у бактерий [245]
Высокий
уровень
глюкозы (низкий
уровень сАМР)
б
Рис. 28-18 Влияние глюкозы и лактозы на экспрессию /ос-оперона. а — высокий уровень
транскрипции достигается только при достаточно низкой концентрации глюкозы (когда
концентрация сАМР высокая, и комплекс CRP-cAMP связан с ДНК) и высокой концентрации
лактозы (когда Lac-penpeccop не связан с ДНК). 6 — без связанного активаторного ком-
плекса CRP-cAMP транскрипция с Zac-промотора слабая, даже когда концентрация лактозы
высокая, и Lac-penpeccop не связан с ДНК.
CRP- сАМР оказывает незначительное влия-
ние на /б?с-оперон, и диссоциация репрессора от
/яс-оператора мало изменяет транскрипцию 1ас-
оперона, если не присутствует комплекс CRP-
cAMP, облегчающий транскрипцию. Без связан-
ного CRP /яс-промотор дикого типа является
довольно слабым промотором (рис. 28-17. б). В
отсутствие комплекса CRP-cAMP открытый
комплекс РНК-полимеразы и промотора (см.
рис. 26-6) образуется с трудом. Белок CRP взаи-
модействует непосредственно с а-субъединицей
РНК-полимеразы (рис. 28-16).
Действие глюкозы на CRP опосредовано
сАМР (рис. 28-18). Наиболее активно CRP связы-
вается с ДНК при высокой концентрации сАМР.
В присутствии глюкозы синтез сАМР подавля-
ется, и стимулируется его выход из клетки. По
мере снижения концентрации сАМР ослабевает
связывание CRP с ДНК, что снижает экспрессию
/б?с-оперона. Поэтому для сильной индукции 1ас-
оперона необходимо присутствие и лактозы (для
инактивации fac-penpeccopa), и глюкозы в низкой
концентрации (для повышения концентрации
сАМР и усиления связывания сАМР с CRP).
CRP и сАМР участвуют в координирован-
ной регуляции многих оперонов, прежде всего
тех, которые кодируют ферменты метаболизма
сахаров, таких как лактоза и арабиноза. Не-
сколько оперонов с общим регулятором назы-
вается регул оном. Эта структура из сотен генов
участвует в координации и изменении клеточ-
ных функций, а также играет центральную роль
в регуляции экспрессии генов, рассеянных по ге-
ному эукариот. Приведем примеры бактериаль-
ных регулонов: система генов теплового шока,
которая реагирует на изменения температуры
(разд. 26.1), и система генов, индуцируемых в
клетках Е. coli в рамках SOS-ответа на повреж-
дения ДНК; эти регулоны описаны ниже.
Многие гены ферментов биосинтеза
аминокислот регулируются
путем аттенюации транскрипции
Для синтеза белка требуются 20 незаменимых
аминокислот в больших количествах. Е. coli мо-
жет синтезировать их все. Гены ферментов, не-
обходимых для синтеза каждой аминокислоты,
обычно образуют оперон и экспрессируются по
мере необходимости. Если аминокислоты доста-
точно, ферменты биосинтеза не нужны и экспрес-
сия оперона подавляется
[246] Часть III. 28. Регуляция экспрессии генов
Триптофановый оперон (trp) Е. coli содержит
пять генов ферментов, необходимых для превра-
щения хоризмата в триптофан (рис. 28-19). Два
из этих ферментов катализируют более одного
этапа биохимического пути. Время полужизни
мРНК /тр-оперона составляет лишь около трех
минут, поэтому-то клетка может быстро реагиро-
вать на изменяющиеся потребности в этой ами-
нокислоте. Тгр-репрессор представляет собой
гомодимер, каждая субъединица которого содер-
жит 107 аминокислотных остатков (рис. 28-20).
Когда триптофана много, он связывается с Тгр-
репрессором и изменяет его конформацию, что
позволяет репрессору связаться с trp-оператором
и блокировать экспрессию /тр-оперона. Участок
trp-оператора перекрывается с промотором, по-
этому связывание репрессора блокирует связы-
вание РНК-полимеразы.
Опять-таки, такая простая схема опосредо-
ванного репрессором механизма включения/вы-
ключения экспрессии не описывает всю систему
в полной мере. При колебании концентрации
триптофана в клетке скорость синтеза ферментов
этого пути биосинтеза меняется более чем в 700
раз. Когда действие репрессора ослабевает и на-
чинается транскрипция, скорость транскрипции
тонко регулируется с помощью второго процес-
са. называемого аттенюацией транскрипции, при
котором транскрипция начинается в обычном
порядке, но внезапно останавливается, не доходя
до генов оперона. Аттенюация транскрипции ре-
гулируется наличием триптофана и основана на
Рис. 28-19. Триптофановый оперон. Этот оперон регулируется двумя механиз-
мами: (1) при высокой концентрации триптофана репрессор (вверху слева)
связывается со своим оператором, и (2) происходит аттенюация транскрипции
Рр-мРНК (см. рис. 28-21). Внизу представлена схема биосинтеза триптофана
ферментами, закодированными в frp-опероне (также см. рис. 22-17).
Н----Регуляторная область
Регулируемые гены
Аттенюированная
мРНК (высокий
уровень триптофана)
trp-мРНК
(низкий уровень
триптофана)
(3-субъединица ^-субъединица
фосфат
пируват
Триптофан-
синтаза,
Антранилат-
синтаза,
компонент I
Антранилат-
синтаза,
компонент II
Триптофан-
синтаза,
Индол-3- <
глицерофосфат
L-трипто-
фан
Глицеральдегид- Ь-серин
3-фосфат
Триптофансинтаза
(«2Д2)
Антранилат-
синтаза.
(12> п2)
Ы-(б'-фосфорибозил)-
антранилатизомераза;
индол- 3 - гл ицеро -
фосфатсинтаза
28.2 Регуляция экспрессии генову бактерий [247]
Рис. 28-20. Триптофановый репрессор. Репрессор пред-
ставляет собой димер, обе субъединицы которого (серая
и голубая) связываются с ДНК через мотивы спиралъ-
поворот-спиралъ (1PDB ID 1TR0). Связанные молекулы
триптофана показаны красным цветом.
непосредственной связи транскрипции и транс-
ляции в клетках бактерий.
Механизм аттенюации trp-оперона исполь-
зует сигналы, закодированные в четырех по-
следовательностях внутри 162-нуклеотидного
лидерного участка на 5'-конце мРНК. предше-
ствующего инициаторному кодону первого гена
(рис. 28-21, а). Внутри лидерного участка нахо-
дится так называемый аттенюатор, состоящий
из последовательностей 3 и 4. Основания этих
последовательностей спариваются с образова-
нием С=С-богатой шпильки (стебель и петля),
сразу за которой следует серия остатков U. Ат-
тенюатор выступает в роли терминатора транс-
крипции (рис. 28-21, б). Последовательность 2
также может спариваться с последовательно-
стью 3 (рис. 28-21, в). Тогда аттенюатор не обра-
зуется. и гены ферментов биосинтеза триптофа-
на нормально транскрибируются; петля, образу-
ющаяся при спаривании последовательностей 3
и 2, не мешает транскрипции.
Регуляторная последовательность 1 играет
главную роль в механизме чувствительности
к триптофану; она определяет, спаривается ли
последовательность 3 с последовательностью 2
(что позволит продолжать транскрипцию) или
с последовательностью 4 (что приведет к атте-
нюации транскрипции). Формирование шпиль-
ки зависит от событий, которые происходят при
трансляции регуляторной последовательно-
сти 1; она кодирует лидерный пептид (назван-
ный так, потому что он кодируется лидерной
последовательностью мРНК), состоящий из 14
аминокислот, среди которых два остатка трип-
тофана. Лидерный пептид не выполняет ника-
ких других функций, его синтез нужен только
для регуляции экспрессии оперона. Этот пеп-
тид транслируется сразу же после транскрип-
ции на рибосоме, которая по мере продвиже-
ния транскрипции следует непосредственно за
РНК-полимеразой.
Когда концентрация триптофана высокая,
в клетке содержится много нагруженной трип-
тофаном тРНК (Тгр-тРНКТгР). В таких усло-
виях происходит быстрая трансляция двух ко-
донов Тгр в последовательности 1 и переход к
трансляции последовательности 2 до того, как
РНК-полимераза синтезирует последователь-
ность 3. При этом последовательность 2 закры-
та рибосомой и недоступна для спаривания с
синтезируемой последовательностью 3. В ре-
зультате при спаривании последовательностей
3 и 4 образуется аттенюатор, и транскрипция
останавливается (рис. 28-21, б, вверху}. Однако
при низкой концентрации триптофана рибосо-
ма задерживается на двух кодонах Тгр в после-
довательности 1 из-за недостатка Тгр-тРНКТгр.
В этом случае во время синтеза последователь-
ности 3 последовательность 2 остается сво-
бодной, что приводит к их спариванию и про-
должению транскрипции (рис. 28-21, б, внизу}.
Таким образом, по мере уменьшения концен-
трации триптофана уменьшается и вероятность
аттенюации транскрипции.
Во многих других оперонах биосинтеза
аминокислот также используется механизм
аттенюации, позволяющий корректировать
уровень синтеза ферментов в соответствии с
текущими потребностями клетки. Лидерный
пептид, образуемый ^/zc-опероном, состоит из
15 аминокислот и содержит семь остатков Phe.
Лидерный пептид /ew-оперона содержит четыре
расположенных подряд остатка Leu. Лидерный
пептид /zzs-оперона содержит семь расположен-
ных подряд остатков His. Заметим, что меха-
низм аттенюации в /zzs-опероне и ряде других
оперонов настолько чувствителен, что является
единственным регуляторным механизмом соот-
ветствующего пути биосинтеза.
[248] Часть III. 28. Регуляция экспрессии генов
Лидерный пептид
Met—Lys—Ala—Не —Phe —Val — »
мРНК pppAAGUUCACGUAAAAAGGGUAUCGACAAUGAAAGCAAUUUUCGUAC^
AAAUGCGUACCACUUAUGUGACGGGCAAAGUCCUUCACGCGGUGGUtJ '
2 (стер)—Ser — Thr — Arg — Trp — Trp "
139 162
Q. I I
C/ACCCAGCCCGCCUAAUGAGCGGGCUUUUUUUUGAACAAAAUUAGAGAAUAACAAUGCAAACA-*
3
1
Сайт
аттенюации
тр анскрипции
(Met - Gin - Thr —►
Полипептид TrpE
Конец
лидерного
участка (trpL)
а
Когда уровень триптофана высокий, рибосома быстро транслирует
последовательность 1 (открытая рамка считывания, кодирующая
лидерный пептид) и блокирует последовательность 2 до того.
пока не транскрибируется последовательность 3. Продолжающаяся
транскрипция приводит к аттенюации на подобной терминатору
структуре аттенюатора, образованной последовательностями 3 и 4.
AGAUAc/ UUUUUUU
I
110
Спаривание 3 : 4
(аттенюатор)
Когда уровень триптофана низкий, рибосома приостанавливается
на кодонах Тгр в последовательности 1. Спаривание
последовательностей 2 и 3 предотвращает
аттенюацию, потому что теперь последовательность 3
недоступна для образования аттенюаторной структуры
с последовательностью 4. Структура 2 : 3, в отличие от аттенюатора
из пары 3 : 4, не предотвращает транскрипцию.
А
U А
G А
C-G- 100
G-C
U-A
А
90-С
С
А
C-G
U-A
A-U
U-A-110
G-C
U
80-G
А
C-G
G-C
G-C
G-C
C-G
А С
А С
U
U
А
С
С
А
Спаривание
2:3
б
в
28.2 Регуляция экспрессии генов у бактерий [249]
Рис. 28-21. Аттенюация транскрипции в trp-опероне. Транскрипция инициируется в начале
162-нуклеотидой лидерной последовательности мРНК, которая закодирована в так называемом
frpl-участке ДНК (см. рис. 28-19). Существует механизм регуляции, который определяет, про-
изойдет ли аттенюация транскрипции в конце лидерной последовательности или считывание
продолжится в область структурных генов, а — лидерный участок frp-мРНК (trpL). В механиз-
ме аттенюации в frp-опероне задействованы последовательности 1-4 (выделены цветом), б —
последовательность 1 кодирует небольшой лидерный пептид, содержащий два остатка Trp (W);
он транслируется сразу же после начала транскрипции. Последовательности 2 и 3 комплемен-
тарны друг другу, как и последовательности 3 и 4. Аттенюатор формируется при спаривании
последовательностей 3 и 4 (вверху). По структуре и функции он напоминает терминатор транс-
крипции (см. рис. 26-8). Спаривание последовательностей 2 и 3 (внизу) предотвращает форми-
рование аттенюатора. Обратите внимание, что лидерный пептид не выполняет других функций
в клетке. Трансляция его открытой рамки считывания играет исключительно регуляторную
роль, определяя, какие комплементарные последовательности (2 и 3 или 3 и 4) образуют пары.
в — спаривание комплементарных участков лидерной последовательности frp-мРНК.
При индукции SOS-ответа происходит
разрушение репрессорных белков
Значительные повреждения ДНК в бактериаль-
ной хромосоме запускают индукцию многих ге-
нов, расположенных далеко друг от друга. Этот
процесс, называемый SOS-ответом (разд. 25.2),
является еще одним хорошим примером коорди-
нированной регуляции генов. Многие из индуци-
рованных генов участвуют в репарации ДНК (см.
табл. 25-6). Ключевые роли в этом процессе игра-
ют белок RecA и репрессор LexA.
Репрессор LexA (Мг = 22 700) подавляет
транскрипцию всех SOS-генов (рис. 28-22), по-
Рис. 28-22. SOS-ответ в клетках Е. coli.
В табл. 25-6 перечислены функции многих
генов SOS-ответа. Белок LexA выполня-
ет функцию репрессора; его операторные
участки (красные) расположены у каждого
гена. Поскольку ген гесА не полностью по-
давляется репрессором LexA нормальная
клетка содержит около 1000 мономеров
RecA. (D При значительных поврежде-
ниях ДНК (например, под действием УФ-
облучения), репликация ДНК останавлива-
ется, и число одноцепочечных разрывов в
ДНК увеличивается, d Белок RecA связыва-
ется с поврежденной одноцепочечной ДНК,
что активирует его протеазную активность.
3 В связанном с ДНК состоянии белок RecA
облегчает расщепление и инактивацию
репрессора LexA. Инактивация репрессо-
ра приводит к индукции генов SOS-ответа,
включая ген гесА; концентрация RecA уве-
личивается в 50-100 раз.
[250] Часть III. 28. Регуляция экспрессии генов
этому индукция SOS-ответа требует удаления
LexA. В данном случае происходит не просто
отсоединение репрессора от ДНК в ответ на
связывание небольшой молекулы, как в 1ас-
опероне. Репрессор LexA инактивируется, ка-
тализируя собственное специфическое расще-
пление по пептидной связи Ala-Gly с образо-
ванием двух приблизительно равных по длине
белковых фрагментов. При физиологическом
значении pH для этой автокаталитической ре-
акции расщепления необходим белок RecA.
Этот белок не протеаза в классическом смысле,
но его взаимодействие с LexA облегчает реак-
цию расщепления репрессора. Эту активность
RecA иногда называют сопутствующей проте-
азной активностью.
Белок RecA обеспечивает функциональную
связь между биологическим сигналом (повреж-
дением ДНК) и индукцией SOS-генов. Значи-
тельное повреждение ДНК приводит к много-
численным одноцепочечным разрывам в ДНК,
и только RecA, который связывается с одноце-
почечной ДНК, может облегчить расщепление
репрессора LexA (рис. 28-22, внизу). Связывание
RecA в области разрыва, в конечном итоге, ак-
тивирует его протеазную активность, что приво-
дит к расщеплению репрессора LexA и индукции
SOS-ответа.
При индукции SOS-ответа в сильно повреж-
денных клетках RecA также расщепляет и тем
самым инактивирует репрессоры, которые под-
держивают в спящем состоянии некоторые ли-
зогенные вирусы, находящиеся в бактериальной
клетке. Это замечательный пример эволюцион-
ной адаптации. Эти репрессоры, как и LexA, тоже
расщепляют сами себя по специфической пеп-
тидной связи Ala-Gly, поэтому индукция SOS-
ответа приводит к репликации вирусов и лизису
клетки с высвобождением новых вирусных ча-
стиц. Так бактериофагу удается «сбежать» из по-
врежденной клетки.
Синтез рибосомных белков происходит
координированно с синтезом рРНК
Когда бактериальной клетке требуется больше
белка, в ней не повышается активность каждой
рибосомы, а увеличивается их общее число.
С увеличением скорости роста клеток увели-
чивается и численность рибосом. При высокой
скорости роста клеток рибосомы составляют
приблизительно 45% сухой клеточной массы.
Ресурсы, предназначенные для производства
рибосом, так велики, а функция рибосом так
важна, что клеткам приходится координировать
синтез компонентов рибосом — рибосомных
белков и РНК (рРНК). Механизмы регуляции
этого процесса отличаются от описанных выше
механизмов, поскольку здесь регуляция осу-
ществляется в основном на уровне трансляции.
Рибосомные белки кодируются 52 генами,
которые расположены, как минимум, в 20 оперо-
нах по 1-11 генов в каждом. Некоторые из этих
оперонов также содержат гены субъединиц ДНК-
праймазы (см. рис. 25-13), РНК-полимеразы
(см. рис. 26-4) и факторов элонгации синтеза бел-
ка (см. рис. 27-28), что обеспечивает тесное со-
пряжение репликации, транскрипции и синтеза
белка в процессе роста клетки.
Опероны рибосомных белков регулируют-
ся главным образом на уровне трансляции по
механизму обратной связи. Один из рибосом-
ных белков, кодируемых каждым опероном,
функционирует как репрессор трансляции: он
связывается с мРНК, транскрибируемой с дан-
ного оперона, и блокирует трансляцию всех ге-
нов, закодированных этой мРНК (рис. 28-23).
Обычно рибосомный белок, который играет
роль репрессора, сам связывается с рРНК. Каж-
дый белок-репрессор трансляции связывается с
соответствующей рРНК с более высоким срод-
ством, чем со своей мРНК, поэтому связывание
мРНК и подавление трансляции происходит
только тогда, когда концентрация рибосомного
белка превышает концентрацию рРНК. В ре-
зультате трансляция мРНК рибосомных белков
подавляется только тогда, когда уровень синте-
за этих рибосомных белков превышает запросы
для сборки функциональных рибосом. Таким
образом, скорость синтеза рибосомных белков
координирована с наличием рРНК.
Участок связывания репрессора трансляции
с мРНК находится вблизи точки начала транс-
ляции одного из генов оперона, обычно первого
гена (рис. 28-23). В других оперонах это повли-
яло бы только на трансляцию этого конкретного
гена, потому что в полицистронных мРНК бак-
терий большинство генов имеет независимые
сигналы трансляции. Однако в оперонах рибо-
сомных белков трансляция каждого гена зави-
28.2 Регуляция экспрессии генову бактерий [251]
/?-оперон 5'|— | L10 \Ъ7/Ь1Д Z? 1 Z?' I I 3'
S7 11
sZr-оперон 5'1 | S12 1 S7 | EF-G | EF-7u |з'
Рис. 28-23. Принцип обратной связи в регуляции
трансляции оперонов некоторых рибосомных бел-
ков. В виде розовых кружков изображены рибосом-
ные белки, выполняющие функцию репрессоров
трансляции. Каждый репрессор трансляции блокиру-
ет трансляцию всех генов оперона, связываясь с ука-
занным участком мРНК. Гены субъединиц РНК-поли-
меразы изображены желтым цветом; гены факторов
элонгации — голубым. L1-L34 — белки большой
субъединицы (50S) рибосомы, S1-S21 — белки малой
(30S) субъединицы.
_____________ S4
t
а-оперон 5'| | S73 | 5П | S4 | а | L17 | ~|з
L4
I t
SlO-оперон 5'| | S20 | 13 | L4 | L23 | L2 | (L2ZS19) | S3 | £7g | L29 | S77 | |з
S8
t
spc-оперон 5' | | L14 | L24 | L5 | S14 | S<9 | | L18 | S5 | L30 | L15 | 13
сит от трансляции всех остальных. Подробности
механизма такого трансляционного сопряжения
пока не ясны. В некоторых случаях трансляция
многочисленных генов, по-видимому, блокиру-
ется из-за формирования сложной трехмерной
структуры мРНК, которая стабилизируется
при спаривании оснований внутри цепи (как на
рис. 8-23) и связывании белка-репрессора транс-
ляции. Когда репрессор трансляции отсутствует,
связывание рибосомы и трансляция одного или
нескольких генов разрушают трехмерную струк-
туру мРНК, что позволяет транслироваться
всем остальным генам.
Поскольку синтез рибосомных белков ко-
ординирован с уровнем рРНК, регуляция об-
разования рибосом соотносится с регуляцией
синтеза рРНК. В клетках Е. coli синтез рРНК с
семи оперонов коррелирует со скоростью кле-
точного роста и наличием основных питатель-
ных веществ, особенно аминокислот Регуля-
цию, зависящую от концентрации аминокис-
лот, называют строгим ответом (рис. 28-24).
При низкой концентрации аминокислот синтез
рРНК останавливается. Аминокислотное голо-
дание приводит к связыванию ненагруженных
тРНК с A-сайтом рибосомы; это запускает цепь
событий, которая начинается со связывания
с рибосомой фермента, называемого строгим
фактором (белком RelA). При связывании с
рибосомой строгий фактор катализирует об-
разование необычного нуклеотида (гуанозин-
тетрафосфата ppGpp, см. рис. 8-39) — пирофос-
фат присоединяется по З'-положению в GTP:
GTP + АТР = pppGpp + АМР
и фосфогидролаза отщепляет один фосфат с об-
разованием ppGpp. Резкое повышение уровня
ppGpp при недостатке аминокислот приводит
[252] Часть III. 28. Регуляция экспрессии генов
к значительному снижению синтеза рРНК, от-
части вызванному связыванием ppGpp с РНК-
полимеразой.
Как и сАМР, нуклеотид ppGpp относится к
модифицированным нуклеотидам, которые дей-
ствуют в клетке как вторичные мессенджеры
(с. 427 вт. 1). В клетках Е. coli эти два нуклеоти-
да служат сигналами голодания; они вызывают
значительные изменения клеточного метабо-
лизма путем усиления или ослабления транс-
крипции сотен генов. В эукариотах нуклеотид-
ные вторичные мессенджеры тоже выполняют
многочисленные регуляторные функции. Коор-
динация роста клетки и ее метаболизма весьма
сложна, и, несомненно, со временем нам станут
известны и другие механизмы регуляции этих
процессов
Рис. 28-24. Строгий ответ в клетках Е. coli. Ответ на ами-
нокислотное голодание запускается связыванием нена-
груженной тРНК с A-сайтом рибосомы. Белок, называе-
мый строгим фактором, связывается с рибосомой и ката-
лизирует синтез pppGpp, который под действием фосфо-
гидролазы превращается в ppGpp. Сигнал ppGpp ослабля-
ет транскрипцию одних генов и усиливает транскрипцию
других, отчасти за счет связывания с р-субъединицей
РНК-полимеразы и изменения специфичности промото-
ра этого фермента. При увеличении концентрации ppGpp
синтез рРНК ослабевает.
Функция некоторых мРНК регулируется
малыми РНК по цис- или троне-механизму
Как неоднократно отмечалось в данной главе,
белки играют важную и вполне понятную роль в
регуляции экспрессии генов. Однако существен-
ную роль играют и РНК, что становится все бо-
лее очевидным по мере обнаружения новых при-
меров регуляторных РНК. Функцию только что
синтезированной мРНК могут контролировать
либо РНК-связывающие белки, как мы видели в
случае оперонов рибосомных белков, либо другие
типы РНК. С молекулой мРНК может связаться
другая молекула РНК и регулировать ее актив-
ность; это так называемая транс-регуляция. Кро-
ме того, сама молекула мРНК может регулиро-
вать свою функцию. Регуляция функций одной
части молекулы под действием другой части той
же молекулы называется z^c-регуляцией.
Хорошо изученным примером транс-
регуляции РНК является регуляция мРНК гена
rpoS (RNA polymerase sigma factor), кодирующе-
го as — один из семи сигма-факторов Е. coli (см.
табл. 26-1). Клетки используют этот фактор в
определенных стрессовых ситуациях, например
в стационарной фазе (состояние, при котором
клетка не растет из-за недостатка питательных
веществ); as необходим для транскрипции мно-
гих генов, участвующих в ответе на стресс. Почти
в любых условиях мРНК фактора as в небольшом
количестве присутствует в клетке, но не транс-
лируется, поскольку расположенная выше коди-
рующей области крупная шпилька препятствует
связыванию с рибосомой (рис. 28-25). В некото-
рых стрессовых условиях происходит индукция
одной или обеих специализированных малых
РНК — DsrA (downstream region А) и RprA (Rpos
regulator RNA А). Обе могут спариваться с одной
цепью шпильки мРНК as, разрушая ее структуру
и тем самым позволяя осуществлять трансляцию
rpoS. Еще одна малая РНК, OxyS (oxidative stress
gene S), индуцируется в условиях окислительного
стресса и ингибирует трансляцию rpoS. вероятно,
путем спаривания с участком мРНК, ответствен-
ным за связывание с рибосомой. OxyS экспрес-
сируется при клеточном ответе на другой тип
стресса (окислительное повреждение), а не тот.
за который отвечает rpoS, и задача этой РНК со-
стоит в предотвращении ненужных в данный мо-
мент путей репарации. DsrA, RprA и OxyS пред-
28.2 Регуляция экспрессии генов у бактерий [253]
Сайт связывания
рибосомы
3'
Сайт связывания
рибосомы
Рис. 28-25. трояс-Механизм регуляции функций бакте-
риальной мРНК под действием малых РНК. Некоторые
малые РНК — DsrA, RprA и OxyS — участвуют в регу-
ляции гена rpoS. Для проявления активности им нужен
белок Hfq — РНК-шаперон, облегчающий образование
РНК-дуплекса. Белок Hfq имеет геометрию тороида с по-
рой в центре, а — DsrA стимулирует трансляцию путем
спаривания с одной цепью шпильки, что блокирует уча-
сток связывания рибосомы. Аналогичным образом дей-
ствует RprA. 6 — OxyS блокирует трансляцию, спариваясь
с участком связывания рибосомы.
ставляют собой сравнительно небольшие (менее
300 нуклеотидов) молекулы бактериальной РНК
(в эукариотах существуют другие малые РНК,
которым соответствуют другие обозначения).
Всем РНК для осуществления их функций тре-
буется белок Hfq, т. е. РНК-шаперон, облегчаю-
щий образование пар РНК-РНК. Бактериаль-
ных генов, регулируемых таким образом, извест-
но немного — всего несколько десятков. Однако
эти примеры служат прекрасными модельными
системами для изучения более сложных и более
многочисленных случаев опосредованной РНК
регуляции в эукариотах.
В регуляции по цг/с-механизму задейство-
ваны РНК, относящиеся к классу так называе-
мых «рибопереключателей». Как говорилось в
доп. 26-3. аптамерами называют синтезируемые
in vitro молекулы РНК, которые способны спец-
ифическим образом связываться с определен-
ными лигандами. Как можно ожидать, такие
лиганд-связывающие домены в молекулах РНК
существуют и в природе (в рибопереключателях)
во многих бактериальных мРНК (и даже в неко-
торых мРНК эукариот). Эти природные аптамеры
представляют собой структурные домены, распо-
ложенные в нетранслируемых участках на 5'-кон-
це некоторых бактериальных мРНК. Связыва-
ние такого домена со специфическим лигандом
приводит к изменениям конформации молекулы
мРНК, в результате чего происходит либо оста-
новка транскрипции, обусловленная стабилиза-
цией структуры, вызывающей преждевременную
остановку транскрипции, либо остановка транс-
ляции (по ф/с-механизму) в результате блокиро-
вания участка связывания рибосомы (рис. 28-26).
В большинстве случаев рибопереключатели дей-
ствуют по механизму обратной связи. Многие ре-
гулируемые таким образом гены участвуют в син-
тезе или транспорте лигандов, связывающихся с
рибопереключателями. Таким образом, при вы-
сокой концентрации лиганда рибопереключатели
ингибируют экспрессию генов, необходимых для
пополнения содержания этого лиганда.
Каждый рибопереключатель связывается
лишь с одним лигандом. Обнаружено более 10
видов рибопереключателей, которые реагируют
на такие лиганды, как тиаминпирофосфат (ТРР,
витамин Bt), кобаламин (витамин В12), флавин-
мононуклеотид. лизин, S-аденозинметионин
(adoMet), пурины, N-ацетилглюкозамин-б-фос-
фат и глицин. Вполне вероятно, что в будущем
будут найдены и другие рибопереключатели.
По-видимому, наиболее распространен рибопе-
реключагель, реагирующий на ТРР. Он обнару-
жен во многих бактериях, грибах и в некоторых
растениях. У некоторых бактерий этот участок
ингибирует трансляцию, а в других индуциру-
ет преждевременную остановку транскрипции
(рис. 28-26). В эукариотах ТРР-связывающий
рибопереключатель обнаружен в интронах не-
[254] Часть III. 28. Регуляция экспрессии генов
которых генов; его функция состоит в модуля-
ции альтернативного сплайсинга этих генов (см.
рис. 26-22). Пока неизвестно, насколько распро-
странены такие структуры. По некоторым оцен-
кам, рибопереключатели регулируют экспрессию
более 4% генов бактерии Bacillus subtilis.
По мере расширения знаний о рибопере-
ключателях ученые стали находить им ме-
дицинское применение. Например, большин-
ство описанных на настоящий момент рибопе-
реключателей, включая тот, что реагирует на
сигнал adoMet, обнаружено только в бактери-
ях. Лекарственный препарат, связывающийся с
этим переключателем и активирующий его, мог
терминирующей
транскрипцию
структуры обрывает
транскрипцию.
---------------3'
Блокирование сайта
связывания рибосомы
препятствует
транскрипции.
---------------3'
Рис. 28-26. цпс-Механизм регуляции функций бакте-
риальной мРНК при участии рибопереключателей.
Несколько примеров различных рибопереключателей,
действие которых основано на связывании распростра-
ненного природного аптамера с тиаминпирофосфатом.
Связывание тиаминпирофосфата (ТРР) с аптамером при-
водит к изменению конформации с различными послед-
ствиями, как показано на рис. (а-в).
бы отключать гены ферментов, синтезирующих
и транспортирующих adoMet, лишая бактери-
альные клетки этого важного кофактора. Лекар-
ства подобного типа, возможно, станут антибио-
тиками нового поколения.
Открытия новых функциональных типов
РНК продолжаются, что подтверждает гипотезу
о важной роли РНК в эволюции жизни (гл. 26).
Малые РНК и рибопереключатели, подобно ри-
бозимам и рибосомам, возможно, представляют
собой рудименты, оставшиеся из далекого про-
шлого мира РНК, которые до сих пор функцио-
нируют в современной биосфере Лабораторный
скрининг аптамеров и рибозимов с новыми ли-
ганд-связывающими и каталитическими функ-
циями (см. доп. 26-3) показывает, что РНК мо-
жет обладать активностями, необходимыми для
существования жизнеспособного мира РНК. От-
крытие многих типов РНК с одинаковыми функ-
циями в различных живых организмах говорит
о том, что должны существовать ключевые эле-
менты метаболизма на основе РНК. Например,
природные аптамеры (рибопереключатели) мо-
гут происходить из молекул РНК, которые мил-
лиарды лет назад связывались с кофакторами,
участвовавшими в каталитических процессах
метаболизма в мире РНК.
Некоторые гены регулируются
путем генетической рекомбинации
Теперь мы обратимся к рассмотрению другого типа
регуляции бактериальных генов, происходящей на
уровне перестройки ДНК, — рекомбинации. Бак-
терия Salmonella typhimurium, которая может по-
селяться в кишечнике млекопитающих, передви-
гается благодаря вращению находящихся на по-
верхности клетки жгутиков (рис. 28-27). Много-
численные копии белка флагеллина (Мг = 53 000),
из которых образованы жгутики, являются мише-
нью для атаки иммунной системы млекопитаю-
щих. Но клетки Salmonella выработали механизм,
предохраняющий их от уничтожения иммунной
системой: примерно через каждую тысячу поколе-
ний они переключаются с синтеза одного варианта
флагеллина на другой (FljB и FliC) при помощи
процесса, называемого фазовой вариацией.
Переключение достигается путем периоди-
ческой инверсии участка ДНК, содержащего про-
мотор гена флагеллина. Инверсия осуществляется
28.2 Регуляция экспрессии генов у бактерий [255]
по механизму сайт-специфической рекомбинации
(см. рис. 25-41) и происходит под действием ре-
комбиназы Hin в определенной последовательно-
сти из 14 п. н. (последовательность his) на любом
конце упомянутого участка ДНК. Когда этот уча-
сток находится в одной ориентации, происходит
экспрессия гена флагеллина Flj В и гена репрессора
Flj А (рис. 28-28. «); репрессор блокирует экспрес-
сию гена флагеллина FliC. Когда участок ДНК пе-
ревернут (рис. 28-28, б), гены///Л иfljB больше не
транскрибируются, и при исчерпании репрессора
происходит индукция гена fliC. Рекомбиназа Hin,
кодируемая геном hin в инвертируемом участке,
экспрессируется при любой его ориентации, по-
Рис. 28-27. Salmonella typhimurium с множеством жгу-
тиков.
а
Инвертированный повтор (hix)
Промотор для
ДНК I Н fan Н~| /ZjB I I Д/А |
Промотор
/ для FliC
I ' /вс |
Перемещенный
сегмент
I ' I________________________
| |->р| Мп |«-| /7/В | | ДМ |
Н I
мРНК гена fliC
б Рекомбиназа Hin
Флагеллин FliC
Рис. 28-28. Регуляция генов флагеллинов сальмонеллы: фазовая вариация. Продукты ге-
нов fli С и fljB — различные флагеллины. Ген hin кодирует рекомбиназу, катализирующую
инверсию участка ДНК, содержащего промотор fljB и ген hin. Сайты рекомбинации (инвер-
тированные повторы) названы hix (желтые), а — в одной ориентации экспрессируется ген
fljB вместе с репрессорным белком (продуктом гена fljA), который подавляет транскрипцию
гена/Zz’C. б — в противоположной ориентации экспрессируется только ген fliC; гены fljA vifljB
не могут транскрибироваться. В переключении между этими двумя состояниями, называе-
мых фазовой вариацией, также участвуют два неспецифических ДНК-связывающих белка —
HU и FIS (не показаны).
[256] Часть III. 28. Регуляция экспрессии генов
Примеры регуляции генов путем рекомбинации
Система Рекомбиназа/ сайт рекомбинации Тип рекомбинации Функция
Фазовая вариация (Salmonella) Hin/fex Сайт-специфическая Альтернативная экспрессия двух генов флагеллина позволяет избежать иммунного ответа хозяина
Круг хозяев (бактериофаг р) Gin/gix Сайт-специфическая Альтернативная экспрессия двух наборов генов хвостового отростка позволяет менять круг хозяев
Переключение типов Эндонуклеаза НО, спаривания (дрожжи) белок RAD52, другие белки/МАТ Нереципрокная4 В результате альтернативной экспрессии группы генов образуются клетки дрожжей с разными типами спаривания а и а, которые могут спариваться и подвергаться мейозу
Антигенная вариация (трипаносомы)6 Разные Нереципрокнаял Последовательная экспрессия различных генов, кодирующих вариабельные поверхностные гликопротеины (VSG), позволяет избежать иммунного ответа хозяина
а При нереципрокной рекомбинации (этот тип рекомбинации не обсуждался в гл. 25) гене-
тическая информация переметается из одной части генома (где она «молчит») в другую
(где она экспрессируется). Этот процесс напоминает репликативную транспозицию (см.
рис. 25-45).
6 Трипаносомы вызывают африканскую сонную болезнь и другие заболевания (см. доп. 22-3
в т. 2). Внешняя поверхность трипаносомы состоит из многочисленных копий одного только
VSG, главного поверхностного антигена. Поверхностные антигены могут принимать до 100
различных форм, избегая атаки иммунной системы хозяина.
этому клегка всегда может переключиться из од-
ного состояния в другое.
Преимущество этого типа регуляции за-
ключается в сто однозначности: экспрессия гена
невозможна, когда ген физически отделен от
своего промотора (обратите внимание на поло-
жение промотора fljB на рис. 28-28, б). Абсолют-
ное включенис/выключение экспрессии важно
по той причине, что жгутик, содержащий хотя
бы одну копию «неправильного» флагеллина,
может оказаться уязвимым для антител хозяина
против этого белка. Такая система существует
отнюдь не юлько в клетках сальмонеллы. По-
добные схемы регуляции есть у ряда других
бактерий и некоторых бактериофагов; системы
рекомбинации с похожими функциями найде-
ны и в эукариотах (табл. 28-1). Регуляция генов
путем перестановок в ДНК, в результате кото-
рых происходят перемещения генов и/или про-
моторов, особенно распространена у патогенов,
которые получают возможность менять хозяев
или менять состав своих поверхностных белков,
опережая иммунную систему хозяина.
Краткое содержание раздела 28.2
РЕГУЛЯЦИЯ ЭКСПРЕССИИ ГЕНОВ У БАКТЕРИЙ
Глс-опсрон Е. coli не только подавляется Lac-
рспрессором, но и подвергается положитель-
ной регуляции сАМР-рецепторным белком
(CRP). Когда концентрация глюкозы низкая,
концентрация с АМР высокая, и комплекс
CRP-cAMP связывается со специфическим
участком ДНК, стимулируя транскрипцию
/яс-оперона и образование ферментов мета-
болизма лактозы. При наличии глюкозы кон-
центрация с АМР снижается, что ослабляет
экспрессию lac и других генов, участвующих
в метаболизме вторичных сахаров. Группа
координированно регулируемых оперонов
называется регулоном.
Опероны, в которых закодированы фермен-
ты биосинтеза аминокислот, подчиняются
рщуляторному механизму, называемому ат-
тенюацией; в этом механизме задействован
участок терминации транскрипции (атте-
28.3 Регуляция экспрессии генов у эукариот [25/]
нюатор) в мРНК. Образование аттенюатора
модулируется путем сопряжения транскрип-
ции и трансляции и зависит от небольших
изменений концентрации аминокислот.
Действие SOS-системы заключается в одно-
временной индукции множества несвязан-
ных генов, подавляемых одним репрессором;
повреждение ДНК при посредничестве бел-
ка RecA запускает автокаталитический про-
теолиз репрессора.
При синтезе рибосомных белков один белок
в каждом опероне рибосомных белков дей-
ствует как репрессор трансляции. Он свя-
зывается с мРНК и блокирует трансляцию,
только когда находится в избытке к рРНК.
Посттранскрипционная регуляция некото-
рых мРНК опосредована малыми РНК, ко-
торые действуют по ттгрбшс-механизму, или
частями самой мРНК (рибопереключателя-
ми), которые действуют по ф/с-механизму.
Некоторые гены регулируются с помощью ге-
нетической рекомбинации, в результате кото-
рой промоторы перемещаются относительно
регулируемых генов. Кроме того, регуляция
может происходить на уровне трансляции.
28.3. Регуляция экспрессии генов
у эукариот
Инициация транскрипции — ключевая точка в
регуляции экспрессии генов в любых организ-
мах. Несмотря на то что у эукариотов и бактерий
некоторые механизмы регуляции одинаковые,
регуляция транскрипции у них принципиально
различается.
Можно определить базовый уровень транс-
крипции как активность промоторов и всего ме-
ханизма транскрипции in vivo в отсутствие регу-
ляторных последовательностей. Бактериальная
РНК-полимераза обычно имеет доступ к любому
промотору и может связываться с ним и иници-
ировать транскрипцию с некоторой эффектив-
ностью без помощи активаторов и репрессоров;
следовательно, транскрипция носит пермиссив-
ный характер. Однако обычно у эукариот силь-
ные промоторы in vivo неактивны в отсутствие
регуляторных белков, т. е. это непермиссивная
транскрипция. Это принципиальное различие яв-
ляется причиной, как минимум, четырех важных
особенностей регуляции экспрессии эукариоти-
ческих генов.
Во-первых, у эукариот доступ к промоторам
ограничен строением хроматина, и активация
транскрипции связана со многими изменениями
структуры хроматина в транскрибируемом участ-
ке. Во-вторых, хотя у эукариотических клеток
есть как положительные, так и отрицательные
регуляторные механизмы, положительные меха-
низмы преобладают во всех охарактеризованных
до сих пор системах. Таким образом, с учетом не-
пермиссивнаого характера транскрипции, чтобы
прошла транскрипция любого эукариотического
гена, нужна активация. В-третьих, по сравнению
с бактериями эукариоты имеют более крупные
и сложно устроенные регуляторные белки, как
правило, состоящие из нескольких субъединиц.
Наконец, в-четвертых, транскрипция в ядрах эу-
кариот отделена от трансляции в цитоплазме как
во времени, так и в пространстве.
Как мы увидим далее, в эукариотических
клетках схемы регуляции очень сложны. Мы за-
вершим этот раздел описанием одной из наибо-
лее подробно исследованных схем регуляции —
схемы регуляторного каскада, который контро-
лирует развитие дрозофилы.
Транскрипционно активный хроматин
по структуре отличается
от неактивного хроматина
Влияние структуры хромосом на регуляцию ге-
нов эукариот не имеет очевидной аналогии в
клетках бактерий. В клеточном цикле эукариот
интерфазные хромосомы на первый взгляд ка-
жутся дисперсными и аморфными (см. рис. 12-43
в т. 1, рис. 24-25). Тем не менее в таких хромосомах
можно обнаружить несколько форм хроматина.
В типичной эукариотической клетке около 10%
хроматина находится в более конденсированной
форме, чем остальной хроматин. Это транскрип-
ционно неактивный гетерохроматин. Гетерох-
роматин обычно связан со специальными струк-
турами хромосомы, например с центромерами.
Остальной менее конденсированный хроматин
называется эухроматином.
[258] Часть III. 28. Регуляция экспрессии генов
Транскрипция эукариотических генов вну-
три гетерохроматина подавлена. Напротив, не-
которая часть эухроматина (но не весь) активно
транскрибируется. Транскрипционно активные
области хромосом характеризуются не только
более свободной структурой хроматина, но так-
же наличием нуклеосом особого состава и вида.
В транскрипционно активном хроматине обычно
содержится меньше гистона Н1, который связы-
вается с линкерной ДНК между нуклеосомами,
но больше вариантов НЗ.З и H2AZ (см. доп. 24-2).
Гистоны из транскрипционно активного хро-
матина и гетерохроматина также отличаются по
характеру ковалентной модификации. Гистоны из
ядра нуклеосомной частицы (Н2А, Н2В, НЗ, Н4;
см. рис. 24-27) модифицируются путем метилиро-
вания остатков Lys, фосфорилирования остатков
Ser или Thr, ацетилирования (см. ниже), убикви-
тинирования (см. рис. 27-47) или сумоилирова-
ния. Все гистоны нуклеосомного ядра имеют два
разных структурных домена. Центральный домен
участвует во взаимодействии между гистонами и
в оборачивании ДНК вокруг нуклеосомы. Второй,
богатый лизином N-концевой домен обычно рас-
полагается ближе к внешней стороне собранной
нуклеосомной частицы; ковалентным модифика-
циям подвергаются специфические аминокислот-
ные остатки именно этого домена. Характер моди-
фикаций позволяет предполагать существование
гистонового кода, с помощью которого эти моди-
фикации узнаются ферментами, изменяющими
структуру хроматина. Модификации, связанные с
активацией транскрипции (главным образом, ме-
тилирование и ацетилирование), могли бы узна-
ваться ферментами, которые делают хроматин бо-
лее доступным для транскрипции. И в самом деле
некоторые из этих модификаций необходимы для
взаимодействия с белками, выполняющими важ-
ные функции при транскрипции.
Для эукариотической ДНК характерно ме-
тилирование остатков цитозина в последователь-
ностях CpG по пятому положению (с. 419 в т. 1),
но ДНК из транскрипционно активного хромати-
на часто слабо метилирована. Кроме того, сайты
CpG в определенных генах слабее метилированы
в клетках тех тканей, где эти гены экспрессиру-
ются, чем там, где они не экспрессируются. Таким
образом, подготовка хроматина к транскрипции
происходит путем удаления возможных струк-
турных преград.
Хроматин ремоделируется путем
ацетилирования и перемещения нуклеосом
Связанные с транскрипцией изменения в струк-
туре хроматина происходят в процессе ремо-
делирования хроматина. В ремоделировании
участвуют ферменты, которые катализируют
процессы модификации. Некоторые ферменты
ковалентно модифицируют гистоны нуклеосомы.
Другие ферменты используют химическую энер-
гию АТР для перестановки нуклеосом на ДНК
(табл. 28-2); третьи изменяют состав гистонов в
нуклеосомах.
Ацетилирование и деацетилирование ги-
стонов играет очень важную роль в процессах
активации хроматина. Как отмечалось выше,
N-концевые домены гистонов в ядре нуклеосомы
обычно обогащены остатками Lys и Arg. В ходе
транскрипции гистон НЗ метилируется (под дей-
ствием специфических гистонметилаз) по остатку
Lys4 в нуклеосоме вблизи 5’-конца кодирующей
области и по остатку Lys36 внутри кодирующей
области. Метилирование облегчает связывание
гистонацетилтрансфераз (HAT — от англ, histone
acetyltransferases), которые ацетилируют опреде-
ленные остатки лизина. Цитоплазматические HAT
(тип В) ацетилируют вновь синтезированные ги-
стоны до того, как они поступают из цитоплазмы
в ядро. Последующее встраивание гистонов в хро-
матин после репликации облегчается шаперонами
гистонов: CAF1 для НЗ и Н4 и NAP1 для Н2А и
Н2В (см. табл. 24-2).
В тех участках, где хроматин активирует-
ся для транскрипции, гистоны нуклеосом далее
ацетилируются под действием ядерных HAT
(тип А). Ацетилирование многочисленных
остатков Lys в N-концевых доменах гистонов
НЗ и Н4 может уменьшать сродство к ДНК всей
нуклеосомы. Ацетилирование специфических
остатков Lys играет ключевую роль во взаимо-
действиях нуклеосом с другими белками. Ког-
да транскрипция гена больше не нужна, в ходе
общего процесса сайленсинга (отключения) ге-
нов, который возвращает хроматин в транскрип-
ционно неактивное состояние, под действием
гистондеацетил азной (HDAC — от англ, histone
deacetylases) активности ацетилирование нукле-
осом в этом участке ослабевает. Наряду с удале-
нием определенных ацетильных групп происхо-
дит новая ковалентная модификация гистонов,
28.3 Регуляция экспрессии генов у эукариот [259]
Некоторые ферментные комплексы, катализирующие связанные с транскрипцией структурные изменения хроматина
Ферментный комплекс3 Олигомерная структура (количество полипептидов) Источник Функции
Модификация гистонов GCN5-ADA2-ADA3 3 Дрожжи GCN5 имеет активность HAT (тип А)
SAGA PCAF >20 Эукариоты Включает GCN5-ADA2-ADA3;
NuA4 не менее 12 Эукариоты ацетилирует остатки в НЗ и Н2В Компонент Esal имеет активность HAT;
Перемещение и замена гистонов (с затратой АТР) SWI SNF 6; общая Мг = 2 • 106 Эукариоты ацетилирует Н4, Н2А и H2AZ Ремоделирование нуклеосом;
Семейство ISWI Различная Эукариоты активация транскрипции Ремоделирование нуклеосом;
Семейство SWR1 -12 Эукариоты подавление транскрипции; в некоторых случаях активация транскрипции (NURF) Связывание H2AZ
Шапероны гистонов, которым не требуется АТР HIRA 1 Эукариоты Связывание НЗ.З в ходе транскрипции
d Смысл сокращенных названий эукариотических генов и белков гораздо менее очевиден,
чем генов бактерий. Комплекс белков GCN5 (от англ, general conrol nonderepressible) и
ADA (alteration/deficiency acivatiori) был открыт при изучении генов метаболизма азота в
дрожжах. Эти белки могут быть частью более крупного дрожжевого комплекса SAGA (SPF;
ADA2,3; GCN5; acetyltransferase). Человеческий эквивалент SAGA — PCAF (рЗОО/СВР-
associated factor). NuA4 (nucleosome acetyltransferase H4) нуклеосомная ацетилтрансфе-
раза H4, ESAI (essential SAS2-related acetyltransferase) ацетилтрансфераза, родственная
SAS2. Белковый фактор SWI (switching) необходим для экспрессии определенных генов,
участвующих в переключении типа спаривания у дрожжей, a SNF (sucrose nonfermenting) -
регулятор экспрессии дрожжевого гена сахаразы. Впоследствии были обнаружены много-
численные белки SWI и SNF, действующие в комплексе. Комплекс SWI/SNF играет роль в
экспрессии широкого круга генов; он обнаружен у многих эукариот, включая человека. ISWI
(imitation SWI) имитация SWI, NURF (nuclear remodeling^ictor) ядерный ремоделиру-
ющий фактор, SWR1 (Swi2/Snf2-relatedATPase 1) связанная c Swi2 Snf2 АТРаза 1; HIRA
(histone regulator А) регулятор гистонов A.
которая маркирует хроматин как транскрипци-
онно неактивный. Например, часто в гетерохро-
матине остаток Lys9 гистона НЗ метилирован.
Известно пять семейств ферментных ком-
плексов, которые активным образом перемеща-
ют нуклеосомы при гидролизе АТР; три из них
играют важную роль в активации транскрипции
(табл. 28-2). Комплекс SWI/SNF, обнаруженный
во всех эукариотических клетках, содержит не ме-
нее шести центральных полипептидов, которые
вместе перестраивают хроматин таким образом.,
что нуклеосомы располагаются менее регулярно,
и стимулируют связывание факторов транскрип-
ции. В состав комплекса вблизи С-конца активной
АТРазной субъединицы входит специализирован-
ный белковый домен, называемый бромодоменом,
который взаимодействует с ацетилированными
гистоновыми «хвостами». Комплекс SWI/SNF
требуется для транскрипции не всех генов. Ком-
плекс NURF из семейства ISW1 также перестраи-
вает хроматин, дополняя и перекрывая активность
SWI/SNF. Эти два ферментных комплекса играют
[260] Часть III. 28. Регуляция экспрессии генов
ключевые роли в подготовке участка хроматина к
активной транскрипции. Некоторые представи-
тели третьего семейства комплексов, SWR1, уча-
ствуют в связывании варианта гистона H2AZ с
транскрипционно активным хроматином.
Некоторые представители других семейств
преобразующих хроматин ферментов осущест-
вляют перегруппировку нуклеосом в хроматине
в процессе сайленсинга (отключения) генов. Об-
щий результат перестройки хроматина — повы-
шение доступности участка хромосомы и его «ме-
чение» (химическая модификация) для облегче-
ния связывания и действия транскрипционных
факторов, регулирующих экспрессию гена (или
генов) в этой области.
Многие эукариотические промоторы
подвергаются положительной регуляции
Как уже отмечалось, эукариотические РНК-по-
лимеразы имеют небольшое сродство к своим
промоторам (или не имеют его совсем), поэтому
инициация транскрипции почти всегда зависит
от действия многочисленных активаторных бел-
ков. Одна важная причина явного доминирова-
ния положительной регуляции кажется очевид-
ной — сохранение ДНК внутри хроматина обе-
спечивает недоступность большинства промото-
ров, поэтому в отсутствие других регуляторных
механизмов гены не экспрессируются. Структура
хроматина делает одни промоторы более доступ-
ными, чем другие, и репрессоры, которые связы-
ваются с ДНК и предотвращают доступ РНК-по-
лимеразе (отрицательная регуляция) просто не
нужны. Использование положительной регуля-
ции объясняется и другими факторами, главным
образом, двумя: гигантским размером эукариоти-
ческого генома и более высокой эффективностью
положительной регуляции по сравнению с отри-
цательной регуляцией.
В крупных геномах высших эукариот не-
специфическое связывание регуляторных бел-
ков с ДНК становится серьезной проблемой.
Кроме того, с увеличением размера генома по-
вышается вероятность того, что отдельные спец-
ифические последовательности случайно могут
оказаться в ненадлежащем месте. Специфич-
ность активации транскрипции может быть по-
вышена, если каждый белок-регулятор сначала
связывает специфическую последовательность
ДНК, а затем уже образует активационный ком-
плекс с другими белками. В многоклеточном ор-
ганизме среднее число регуляторных участков
для каждого гена, по-видимому, не менее пяти.
Необходимость связывания нескольких регуля-
торных белков со специфическими последова-
тельностями ДНК значительно снижает вероят-
ность случайного функционального совпадения
всех задействованных участков связывания.
В принципе подобная стратегия может реализо-
ваться и для многочисленных элементов отри-
цательной регуляции, но тут играет роль вторая
причина преимущественного использования
положительной регуляции. Эта вторая причи-
на — более высокая эффективность положитель-
ной регуляции. Для отрицательной регуляции
-29 000 генов человеческого генома каждая клет-
ка должна была бы постоянно синтезировать та-
кое же количество различных репрессоров (или
намного больше, если каждый промотор регули-
руется несколькими элементами) в достаточно
высокой концентрации, чтобы обеспечить спец-
ифичное связывание каждого «ненужного» гена.
При положительной регуляции большинство ге-
нов обычно находится в неактивном состоянии
(т. е. РНК-полимеразы не связываются с промо-
торами), и клетка синтезирует только белки-ак-
тиваторы, необходимые для транскрипции груп-
пы генов, необходимых в данное время. Несмотря
на эти аргументы, в эукариотах (от дрожжей до
человека) все-таки встречаются примеры отрица-
тельной регуляции, как мы увидим в дальнейшем.
ДНК-связывающие активаторы
и ко-активаторы способствуют сборке
основных факторов транскрипции
Продолжая наше исследование регуляции экс-
прессии генов эукариот, рассмотрим взаимо-
действия между промоторами и РНК-поли-
меразой II (Pol II), ответственной за синтез
эукариотических мРНК. Хотя многие (но не
все) промоторы Pol II содержат ТАТА-бокс и
Inr (инициатор) в стандартных позициях (см.
рис. 26-9), они сильно различаются и по числу,
и по расположению дополнительных после-
довательностей, необходимых для регуляции
транскрипции. В высших эукариотах эти до-
полнительные регуляторные последователь-
ности называют энхансерами, а в дрожжах —
28.3 Регуляция экспрессии генов у эукариот [261]
активирующими последовательностями UAS
(от англ, upstream activator sequences). Типич-
ный энхансер может находиться на сотни или
даже тысячи пар оснований выше точки нача-
ла транскрипции, а иногда и ниже, т. е. в самом
гене. Энхансер, связанный с соответствующими
регуляторными белками, усиливает транскрип-
цию с соседних промоторов независимо от его
ориентации в ДНК. Последовательности UAS
в дрожжах действуют аналогичным образом,
хотя обычно они располагаются на несколько
сотен пар оснований выше точки начала транс-
крипции. Промотор Pol II может находиться под
влиянием до полудюжины (в среднем) регуля-
торных последовательностей такого рода: часто
встречаются и еще более сложные промоторы
(см., например, рис. 15-23).
Успешное связывание активного холофер-
мента РНК-полимеразы II на одном из его про-
моторов обычно обеспечивают дополнительные
белки (рис. 28-29) четырех типов: (1) активато-
ры транскрипции, которые связываются с энхан-
серами или UAS и способствуют транскрипции:
(2) описанные выше белки модификации и ре-
моделирования хроматина; (3) ко-активаторы;
и (4) основные факторы транскрипции (см.
рис. 26-10. табл. 26-2). Ко-активаторы действуют
не напрямую (т. е. они не связываются с ДНК) и
требуются для координации связи между актива-
торами и комплексом из Pol II и основных фак-
торов транскрипции. Кроме того, во взаимодей-
ствие между РНК-полимеразой и активаторами
могут вмешиваться разнообразные белки-репрес-
соры, что приводит к подавлению транскрипции
(рис. 28-29. б). Далее мы подробнее рассмотрим
белковые комплексы, изображенные на рис. 28-29,
и их взаимодействия в процессе активации транс-
крипции.
Активаторы транскрипции. Разные промоторы
сильно различаются по своим потребностям в
активаторах. Известно несколько активаторов,
которые способствуют транскрипции с сотен
промоторов, но большинство специфично лишь
к некоторым промоторам. Многие активаторы
зависят от связывания сигнальных молекул, что
позволяет активировать или подавить транс-
крипцию при изменении условий. Некоторые
связывающиеся с активаторами энхансеры на-
ходятся далеко от ТАТА-бокса промотора. Как
а
Транскрипция
Inr
Белки HMG
UAS
ТАТА
[тар
Ферменты
модификации и
ремоделирования Медиатор
Энхансеры
Активаторы
транскрипции
CTD Преинициа-
торный
комплекс
ДНК
Я
б
ТАТА
Энхансеры
Рис. 28-29. Эукариотические промоторы и регулятор-
ные белки. РНК-полимераза II и ассоциированные с ней
основные факторы транскрипции образуют преиници-
аторный комплекс на последовательностях ТАТА-бок-
са и Inr в соответствующем промоторе; этому процессу
способствуют активаторы транскрипции, действующие
через медиатор, а — промотор с элементами типичных
последовательностей и белковыми комплексами, су-
ществующими как в дрожжах, так и в высших эукарио-
тах. С-концевой домен (CTD) Pol II (см. рис. 26-10) —
важный участок взаимодействия с медиатором и други-
ми белковыми комплексами. Ферменты модификации
гистонов катализируют метилирование и ацетилиро-
вание; ферменты ремоделирования изменяют состав и
расположение нуклеосом. ДНК-связывающие домены
активаторов транскрипции изображены зеленым цве-
том, активирующие домены — розовым. Взаимодействия,
обозначенные голубыми стрелками, обсуждаются в тек-
сте. 6 — репрессоры транскрипции эукариот действуют
посредством нескольких механизмов. Некоторые на-
прямую связываются с ДНК, вытесняя необходимый для
активации белковый комплекс; другие предотвращают
активацию, взаимодействуя с различными частями транс-
крипционных или активаторных комплексов. Возможные
участки взаимодействия показаны красными стрелками.
[262] Часть III. 28. Регуляция экспрессии генов
действуют такие активаторы? В большинстве
случаев, по-видимому, расположенная между
этими последовательностями ДНК образует
петлю, в результате чего различные белковые
комплексы приходят в непосредственный кон-
такт. Образованию петли способствуют неко-
торые негистоновые белки, которые в большом
количестве содержатся в хроматине и неспец-
ифическим образом связываются с ДНК. Эта
группа белков с высокой электрофоретиче-
ской подвижностью в полиакриламидном геле
(HMG — от англ, high mobility group; рис. 28-29)
играет важную роль в ремоделировании хрома-
тина и активации транскрипции.
Ко-активаторные белковые комплексы. В боль-
шинстве случаев для протекания транскрипции
требуются дополнительные белковые комплек-
сы. Были проведены биохимические и генетиче-
ские исследования некоторых важных комплек-
сов регуляторных белков, которые взаимодей-
ствуют с Pol II. Эти ко-активаторные комплексы
действуют как посредники между активаторами
и комплексом Pol II.
Эукариотический ко-активатор состоит из
20-30 или более полипептидов, собранных в
белковый комплекс, называемый медиатором
(рис. 28-29). Многие из 20 полипептидов серд-
цевины этого комплекса высококонсервативны
у разных видов организмов от грибов до челове-
ка. Дополнительный комплекс из четырех субъ-
единиц может взаимодействовать с медиатором
и подавлять инициацию транскрипции. Меди-
атор прочно связывается с С-концевым доме-
ном (CTD) самой крупной субъединицы Pol II.
Медиаторный комплекс необходим как для ба-
зовой, так и для регулируемой транскрипции с
промоторов полимеразы II; он также стимули-
рует фосфорилирование CTD под действием
основного транскрипционного фактора TFIIH.
Активаторы транскрипции взаимодействуют с
одним или несколькими компонентами медиа-
торного комплекса, причем в строго определен-
ных местах, различных для разных активаторов.
Ко-активаторные комплексы функционируют в
области ТАТА-бокса или поблизости.
Описаны и другие ко-активаторы, способ-
ствующие транскрипции одного или нескольких
генов. Некоторые функционируют в паре с меди-
атором, а другие в системах без медиаторов.
ТАТА-связывающий белок. При сборке пре-
инициаторного комплекса (PIC — от англ.
preinitiation complex) в области ТАТА-бокса ти-
пичного промотора Pol II первым присоединяет-
ся ТАТА-связывающий белок (ТВР — от англ.
TATA-bindingprotein). В полный комплекс входят
основные факторы транскрипции TFIIB, TFIIE,
TFIIF, TFIIH, Pol II и, возможно, TFIIA. Однако
этого минимального преинициаторного комплек-
са часто недостаточно для инициации транскрип-
ции, если же промотор скрыт внутри хроматина,
преинициаторный комплекс обычно вообще не
образуется. Положительную регуляцию, приво-
дящую к началу транскрипции, оказывают акти-
ваторы и ко-активаторы.
Согласованный механизм активации транскрип-
ции. Теперь мы можем воссоздать всю последова-
тельность событий активации транскрипции на
типичном промоторе Pol II (рис. 28-30). Порядок
связывания некоторых компонентов может раз-
личаться; представленная на рис. 28-30 модель
отражает общие принципы активации на приме-
ре одного типичного варианта сборки. Многие ак-
тиваторы транскрипции обладают значительным
сродством к своим участкам связывания, даже
если те находятся в области конденсированно-
го хроматина. Связывание активаторов обычно
приводит к активации промотора. Связывание
одного активатора может облегчить связывание
других, что постепенно приводит к вытеснению
нескольких нуклеосом.
Далее происходит последовательное ремо-
делирование хроматина, чему способствует вза-
имодействие активаторов с HAT или с фермент-
ными комплексами, такими как SWI/SNF (или и
с теми, и с другими). Таким образом, связанный
активатор может вовлекать другие компоненты,
необходимые при дальнейшем ремоделировании
хроматина для транскрипции специфических
генов. Связанные активаторы взаимодействуют
с большим медиаторным комплексом. В свою
очередь медиатор обеспечивает поверхность для
связывания сначала ТВР (TFIID), затем TFIIB
и других компонентов преинициаторного ком-
плекса, включая РНК-полимеразу II. Медиатор
стабилизирует связывание Pol II и соответству-
ющих транскрипционных факторов и значитель-
но облегчает формирование преинициаторного
комплекса. Сложность таких регуляторных схем
28.3 Регуляция экспрессии генов у эукариот [263]
ТАТА Inr
Активатор
ДНК
Энхансер
Медиатор
Ферменты модификации
и ремоделирования
ТАТА Inr
Ферменты
модификации и
ремоделирования
Медиатор
ТВР и TFIIB
TFIIB
Inr
ТВР
TFIIA
TFIIB
TFIIF-Pol II
TFIIE
TFIIH
TFIIF
Pol II
TFIIB
TFIIA
TBP
TFIIH
Рис. 28-30. Элементы, необходимые для активации
транскрипции. Активаторы сначала могут связываться с
ДНК. Затем они вовлекают в процесс медиатор и комплек-
сы модификации гистонов/перестройки нуклеосом. Ме-
диатор облегчает связывание ТВР и TFIIB, а также других
основных факторов транскрипции, после чего связывается
Pol II. Фосфорилирование С-концевого домена (CTD) Pol II
приводит к инициации транскрипции (не показано).
TFIIE
с многочисленными активаторами транскрипции
скорее правило, чем исключение.
Детали процесса могут различаться для раз-
ных промоторов, но большинству промоторов для
начала транскрипции требуется строго упорядо-
ченная сборка всех компонентов. Процесс сборки
не всегда проходит быстро. Для некоторых генов
он может длиться минуты, а для некоторых генов
высших эукариот — даже сутки.
Обратимость активации транскрипции. Хотя и
редко, некоторые эукариотические регуляторные
белки, которые связывают промоторы Pol II, мо-
гут действовать как репрессоры, ингибируя об-
разование активных преинициаторных комплек-
сов (рис. 28-29, б). Некоторые активаторы могут
находиться в различных конформациях, что по-
зволяет им быть либо активаторами, либо ре-
прессорами транскрипции. Например, рецепторы
некоторых стероидных гормонов (см. ниже) дей-
ствуют в ядре как активаторы, стимулирующие
транскрипцию определенных генов в присут-
ствии определенного стероидного гормона. Когда
же гормон отсутствует, рецепторные белки воз-
вращаются в конформацию репрессора, предот-
вращая образование преинициаторных комплек-
сов. В некоторых случаях в репрессии участву-
ют гистопдеацетилазы и другие белки, которые
помогают вернуть хроматин в трапскрипционно
неактивное состояние. Если медиатор содержит
субъединицы с ингибиторной функцией, он тоже
способен блокировать инициацию транскрипции.
Возможно, этот механизм обеспечивает упоря-
доченную сборку преинициаторного комплекса
(благодаря отсрочке активации транскрипции до
тех пор, пока не соберутся все необходимые фак-
торы) или помогает инактивировать промоторы,
когда транскрипция не требуется.
Гены метаболизма галактозы в дрожжах
подвергаются и положительной,
и отрицательной регуляции
Некоторые из описанных выше общих принци-
пов можно проиллюстрировать на примере одной
хорошо изученной схемы регуляции в клетках эу-
кариот (рис. 28-31). Ферменты дрожжей, необхо-
димые для доставки и метаболизма галактозы, ко-
дируются генами, разбросанными по нескольким
хромосомам (табл. 28-3). Каждый из генов GAL
[264] Часть III. 28. Регуляция экспрессии генов
Рис. 28-31. Регуляция транскрипции генов метаболизма галак-
тозы в дрожжах. Галактоза доставляется в клетку и превраща-
ется в галактозо-6-фосфат при участии шести ферментов, гены
которых расположены на трех хромосомах (см. табл. 28-3).
Транскрипция этих генов регулируется совместным действием
белков Gal4p, Gal80p и Gal3p, причем Gal4p играет центральную
роль активатора транскрипции. Комплекс Gal4p-Gal80p неак-
тивен. Связывание галактозы с Gal3p и взаимодействие Gal3p
с Gal80p вызывает изменение конформации Gal80p. Затем ком-
плекс Gal3p-Gal80p выходит из ядра, что позволяет Gal4p связать
медиатор и SAGA и участвовать в активации транскрипции.
Преинициаторный
комплекс
Gal3p +
галактоза
транскрибируется отдельно, поскольку в клетках
дрожжей, в отличие от бактерий, нет оперонов.
Однако промоторы всех генов GAL похожи и коор-
динированно регулируются общим набором бел-
ков. Промоторы генов GAL состоят из ТАТА-бокса
и последовательности Inr, а также активирующей
последовательности UASG, распознаваемой акти-
ватором транскрипции Gal4p. Регуляция экспрес-
сии генов галактозой происходит через взаимо-
действие между Gal4p и двумя другими белками
Gal80p и Gal3p (рис. 28-31). Белок Gal80p образу-
ет комплекс с Gal4p, что не позволяет Gal4p дей-
ствовать в качестве активатора промоторов GAL.
Если имеется галактоза, она связывается с белком
Gal3p, который затем взаимодействует с Gal80p, а
Gal4p может действовать в качестве активатора на
различных промоторах GAL.
^GalSp
Gal80p
экспортируется
в цитозоль
Медиатор
|SAGA
ТАТА Inr
ТВР
SAGA
Медиатор
UASg
В активации транскрипции генов GAL уча-
ствуют и другие белковые комплексы. Это ком-
плекс SAGA для ацетилирования гистонов, ком-
плекс SWI/SNF для ремоделирования нуклеосом
и медиаторный комплекс. Когда Gal4p не связан с
Таблица 28-3 Гены метаболизма галактозы в дрожжах
Размер белка Хромосома (число остатков) Относительный уровень экспрессии на разных источниках углерода
Ген Функция белка Глюкоза Глицерин Галактоза
Регулируемые гены
GAL1 Галактокиназа 11 528 — - +++
GAL2 Галактозопермеаза XII 574 - - +++
PGM2 Фосфоглюкомутаза XIII 569 + +
GAL7 Галактозо-1 -фосфат- уридилилтрансфераза 11 365 — — +++
GAL10 иВР-глюкозо-4-эпимераза II 699 - - +++
MEL1 «-Галактозидаза II 453 - + ++
Регуляторные гены GAL3 Индуктор IV 520 — + ++
GAL4 Активатор транскрипции XVI 881 +/- + +
GAL80 Ингибитор транскрипции XIII 435 + +
Источник. По Reece R. & Platt А. (1977) Signaling activation and repression of RNA polymerase II
transcription in yeast. Bioessays 19,1001-1010 (с сокр.).
28.3 Регуляция экспрессии генов у эукариот [265]
Gal880p, он привлекает эти дополнительные фак-
торы, необходимые для инициации транскрипции.
Оценить сложность белковых взаимодействий в
процессе активации транскрипции в клетках эука-
риот можно, если обратиться к рис. 28-32.
Дрожжи, как и бактерии, предпочитают ис-
пользовать в качестве источника углерода глюко-
зу. В присутствии глюкозы происходит репрессия
большинства генов GAL, независимо от наличия
галактозы. Описанная выше система регуляции
GAL эффективно подавляется сложной системой
катаболитной репрессии, в которой участвует не-
сколько белков (не показаны на рис. 28-32).
ТАТА
| TFIIA
|ТВР
Медиатор
I SWI/SNF, SAGA
TFIIA ▼
ТАТА
Комплекс РНК-
полимеразы II
Рис. 28-32. Белковые комплексы, участвующие в актива-
ции транскрипции группы родственных эукариотических
генов. Система GAL иллюстрирует сложность этого про-
цесса, причем нам известны еще не все беловые комплек-
сы, которые влияют на транскрипцию генов GAL. Обратите
внимание, что многие комплексы (SWI/SNF, GCN5-ADA2-
ADA3 и медиатор) влияют на транскрипцию множества
генов. Сборка комплексов происходит последовательно, в
несколько стадий, как показано на рис. 28-30.
Активаторы транскрипции
имеют модульное строение
Типичные активаторы транскрипции содержат
хорошо выраженный структурный домен для
специфического связывания ДНК и один или не-
сколько дополнительных доменов для активации
транскрипции или для взаимодействия с другими
регуляторными белками. Взаимодействие двух ре-
гуляторных белков часто опосредовано доменами,
содержащими лейциновые молнии (рис. 28-14)
или мотивы спираль петля спираль (рис. 28-15).
Мы рассмотрим три примера активаторов транс-
крипции, содержащих разные активирующие до-
мены (рис. 28-33, а)\ Gal4p, Spl и CTF1.
В ДНК-связывающем домене белка Gal4p
вблизи N-конца содержится структура типа
цинкового пальца; здесь находятся шесть остат-
ков Cys, координирующие два иона Zn2+. Белок
функционирует в виде гомодимера (димеризация
обусловлена взаимодействием двух суперспира-
лей) и связывается с UASG - палиндромной по-
следовательностью ДНК длиной около 17 п. н.
Белок Gal4p имеет отдельный активирующий
домен, содержащий множество остатков кислых
аминокислот. Вариации последовательности в
этом кислом активирующем домене Gal4p в экс-
периментальных условиях позволяют предполо-
жить, что кислая природа домена принципиально
важна для его работы, но сама последователь-
ность может значительно изменяться.
Белок Spl (А/, = 80 000) активирует транс-
крипцию большого числа генов высших эука-
риот. Его ДНК-связывающий GC-бокс (консен-
сусная последовательность GGGCGG) обычно
располагается в непосредственной близости от
ТАТА-бокса. ДНК-связывающий домен белка
Spl находится рядом с С-концом и содержит три
цинковых пальца. Два других домена Spl уча-
ствуют в активации и замечательны тем, что на
25% состоят из глутамина. Такой богатый глута-
мином домен есть и во многих других белках-ак-
тиваторах.
С С ААТ-связывающий фактор транскрип-
ции 1 (CTF1) относится к семейству активаторов
транскрипции, которые связываются с последова-
тельностью, называемой ССААТ-участком (кон-
сенсусная последовательность TGGN6GCCAA,
где N — любой нуклеотид). ДНК-связывающий
домен CTF1 содержит множество остатков ос-
[266] Часть III. 28. Регуляция экспрессии генов
новных аминокислот, а связывающий участок,
по-видимому, представляет собой а-спираль.
Этот белок не содержит мотивов спираль- пет-
ля спираль или цинковых пальцев, и механизм
его связывания с ДНК пока не ясен. Фактор
CTF1 содержит богатый пролином активирую-
щий домен, в котором доля пролиновых остатков
превышает 20% общего числа аминокислот.
ДНК
р р р
CTFI
Spl
GC
ДНК
б
Рис. 28-33. Активаторы транскрипции, а — типичные
активаторы, такие как CTFI, Gal4p и Spl, содержат ДНК-
связывающий и активирующий домены. Природа активи-
рующего домена показана символами: «----» — кислый;
«QQQ» — богатый глутамином; «РРР» — богатый проли-
ном. Эти белки обычно активируют транскрипцию при
помощи ко-активаторных комплексов, таких как медиа-
тор. Обратите внимание, что изображенные здесь участки
связывания не всегда расположены вместе у одного гена.
6 — химерный белок, содержащий ДНК-связывающий до-
мен Spl и активирующий домен CTF1; белок активирует
транскрипцию при наличии GC-бокса в ДНК.
Домены активации и связывания ДНК в регу-
ляторных белках часто действуют независимо, что
было продемонстрировано в экспериментах по за-
мене доменов. С помощью генно-инженерных ме-
тодов (гл. 9 в т. 1) богатый пролином домен CTF1
был соединен с ДНК-связывающим доменом Spl,
в результате чего получился белок, который по-
добно нормальному Spl связывался с GC-боксом
в ДНК и активировал транскрипцию с соседнего
промотора (как на рис. 28-33, б). Аналогичным об-
разом ДНК-связываюший домен Gal4p был заме-
нен ДНК-связывающим доменом репрессора LexA
из Е. coli (системы SOS-ответа; рис. 28-22). Этот
химерный белок не связывался с последователь-
ностью UASg и не активировал гены GAL дрожжей
(как сделал бы нормальный белок Gal4p). если
только последовательность UASG в ДНК не заме-
няли участком узнавания LexA.
Экспрессия эукариотических генов
может регулироваться внеклеточными
и внутриклеточными сигналами
Действие стероидных гормонов (а также тирео-
идных и ретиноидных гормонов, действующих
сходным образом) — это еще один хорошо из-
ученный пример модуляции действия эукарио-
тических регуляторных белков путем прямого
взаимодействия с сигнальными молекулами (см.
рис. 12-29 вт. 1). В отличие от гормонов других
типов стероидные гормоны не связываются с ре-
цепторами цитоплазматической мембраны. Они
могут взаимодействовать с внутриклеточными
рецепторами, которые сами являются активато-
рами транскрипции. Стероидные гормоны (на-
пример, эстроген, прогестерон и кортизол) слиш-
ком гидрофобные и плохо растворяются в крови,
поэтому их доставку от места синтеза к тканям-
мишеням осуществляют специфические белки-
переносчики. В тканях гормоны проходят через
цитоплазматическую мембрану путем простой
диффузии и связываются со специфическими ре-
цепторными белками в ядре. Комплекс гормона с
рецептором с высокой специфичностью связыва-
ется с последовательностями ДНК, называемыми
гормон-чувствительными элементами (HRE —
от англ, hormone response elements), влияя на экс-
прессию генов. Связывание гормонов вызывает
конформационные изменения рецепторных бел-
ков, так что они приобретают способность взаи-
28.3 Регуляция экспрессии генов у эукариот [267]
Гормон-чувствительные элементы (HRE),
с которыми связываются рецепторы стероидных гормонов
Рецептор гормона Связываемая консенсусная последовательность41
Андроген Глюкокортикоид Ретиноевая кислота Витамин D Тиреоидный гормон RX6 GG(A T)ACAN2TGTTCT GGTACAN3GTTCT AGGTCAN5AGGTCA AGGTCAN3AGGTCA AGGTCAN3AGGTCA AGGTCANAGGTCANAGGTCANAGGTCA
а N - любой нуклеотид.
6 Образует димер с рецептором ретиноевой кислоты или рецептором витамина D
модействовать с дополнительными факторами
транскрипции. Связанный гормон-рецепторный
комплекс может либо усиливать, либо подавлять
экспрессию соседних генов
Последовательности HRE. с которыми свя-
зываются комплексы разных стероидных гормо-
нов с рецепторами, похожи по длине и структуре,
но имеют разный нуклеотидный состав Каждому
рецептору соответствует консенсусная после-
довательность HRE (табл. 28-4), с которой гор-
мон-рецепторный комплекс связывается более
эффективно. Каждая такая последовательность
состоит из двух шестинуклеотидных фрагментов,
которые либо соприкасаются, либо разделены
тремя нуклеотидами и расположены в виде тан-
дема или палиндрома. Рецепторы содержат вы-
сококонсервативный ДНК-связывающий домен
с двумя цинковыми пальцами (рис. 28-34). Гор-
D R
I К
KAFFKRSIQGHNDYM
RLRKCYEVGMMKGGIRKDRRGG
MKETRY
□ соо
Активация
транскрипции
(различаются
аминокислотные
последовательности
и их длина)
Связывание
ДНК
(66-68 остатков;
высоко-
консервативная
последов ате л ьность)
Связывание гормона
(различаются аминокислотные
последовательности
и их длина)
N
К R
Рис. 28-34. Типичные рецепторы стероидных гормонов. Эти рецепторные белки содержат
участок связывания с гормоном, ДНК-связывающий домен и участок, активирующий транс-
крипцию регулируемого гена. Высококонсервативный ДНК-связывающий домен содержит
два цинковых пальца. Здесь показана последовательность рецептора эстрогена, но остатки,
выделенные жирным шрифтом, являются общими для всех рецепторов стероидных гормонов.
[268] Часть III. 28. Регуляция экспрессии генов
мон-рецепторный комплекс связывается с ДНК
в виде димера, при этом домены с цинковыми
пальцами каждого мономера узнают по одному
шестинуклеотидному фрагменту. Способность
конкретного гормона изменять экспрессию спец-
ифического гена, действуя через гормон-рецеп-
торный комплекс, зависит от нуклеотидной по-
следовательности HRE, ее расположения отно-
сительно гена и количества последовательностей
HRE, связанных с этим геном.
В отличие от ДНК-связывающего домена ли-
ганд-связывающий участок рецепторного белка
(всегда расположенный на С-конце белка) весь-
ма специфичен к конкретному рецептору Ли-
ганд-связываюший участок глюкокортикоидного
рецептора имеет лишь 30% сходства с рецептором
эстрогена и 17% — с рецептором тиреоидного гор-
мона. Размеры лиганд-связывающих участков ре-
цепторов сильно варьируют; в рецепторе витами-
на D это всего 25 аминокислотных остатков, тог-
да как в рецепторе минералокортикоидов — 630
остатков. Мутации, в результате которых про-
исходит замена только одного аминокислотного
остатка в этом участке, могут привести к исчезно-
вению реакции на специфический гормон. Мута-
ции такого типа встречаются у людей, нечувстви-
тельных к кортизолу тестостерону витамину D
или тироксину.
Некоторые рецепторы гормонов, включая
человеческий рецептор прогестерона, активиру-
ют транскрипцию при помощи необычного ко-
активатора — РНК-активатора стероидного ре-
цептора {steroid receptor RNA activator, SRA). Эта
последовательность РНК длиной -700 нуклеоти-
дов действует как часть рибонуклеопротеинового
комплекса, но для ко-активации транскрипции
нужен именно его РНК-компонент. Подробности
взаимодействия SRA с другими компонентами
систем регуляции данных генов еще предстоит
выяснить.
Регуляция может осуществляться
путем фосфорилирования
ядерных факторов транскрипции
В гл. 12 (т. 1) мы рассказывали о том, что влияние
инсулина на экспрессию генов происходит через
каскад реакций, в конечном итоге приводящих к
активации протеинкиназы в ядре, которая фос-
форилирует специфические ДНК-связывающие
белки и тем самым изменяет их способность
действовать в качестве факторов транскрипции
(см. рис. 12-6). По такому механизму действу-
ют многие нестероидные гормоны. Например,
Р-адренергический путь, ведущий к повыше-
нию уровня цитоплазматического сАМР, вы-
полняющего функцию вторичного мессенджера
и у эукариот, и у бактерий (см. рис. 12-4, в т. 1 и
рис. 28-18), также влияет на транскрипцию не-
которых генов, расположенных рядом со спец-
ифической последовательностью ДНК. называе-
мой сАМР-чувствительным элементом (CRE —
от англ. сАМР response element). Каталитическая
субъединица протеинкиназы А, которая высво-
бождается при повышении концентрации сАМР
(см рис. 12-6), проникает в ядро и фосфорили-
рует ядерный белок CREB (CRE-связывающий
белок). Фосфорилированный CREB связывает-
ся с элементами CRE рядом с определенными
генами и действует как фактор транскрипции,
запуская экспрессию этих генов.
Трансляция многих
эукариотических мРНК подавляется
В клетках эукариот регуляция экспрессии на
уровне трансляции играет гораздо более важ-
ную роль, чем в клетках бактерий, и происходит
в целом ряде ситуаций. В отличие от бактерий, в
которых транскрипция и трансляция непосред-
ственно связаны, транскрипты, образующиеся в
эукариотических ядрах, до начала трансляции
должны пройти процессинг и попасть в цито-
плазму. Все это может значительно затормозить
синтез белка. Когда клетке требуется быстро
увеличить продукцию белка, находящаяся в ци-
топлазме мРНК, трансляция которой подавлена,
может быть быстро активирована для трансля-
ции Регуляция на уровне трансляции особен-
но важна для экспрессии некоторых очень про-
тяженных эукариотических генов (некоторые
гены состоят из миллионов пар оснований),
транскрипция и процессинг мРНК которых мо-
гут длиться много часов. Некоторые гены ре-
гулируются как на стадии транскрипции, так и
на стадии трансляции, причем последний меха-
низм служит для тонкой настройки концентра-
ции клеточных белков. В некоторых безъядер-
ных клетках, таких как ретикулоциты (незрелые
эритроциты), контроль на уровне транскрипции
28.3 Регуляция экспрессии генов у эукариот [269]
невозможен, поэтому решающую роль игра-
ет контроль трансляции уже синтезированной
мРНК. Как будет рассказано далее, контроль
трансляции играет важную роль в процессе раз-
вития многоклеточного организма, когда путем
регуляции трансляции ранее синтезированной
мРНК создается локальный градиент белка.
У эукариот реализуется, как минимум, четы-
ре основных механизма регуляции трансляции.
1. Факторы инициации трансляции подверга-
ются фосфорилированию протеинкиназами.
Фосфорилированные формы обычно ме-
нее активны и вызывают общее замедление
трансляционных процессов в клетке.
2. Некоторые белки связываются непосред-
ственно с мРНК, многие — со специфически-
ми участками в З'-нетранслируемой области
(3'UTR) и действуют как репрессоры транс-
ляции. Связанные в этом участке белки вза-
имодействуют с другими факторами иници-
ации трансляции, связанными с мРНК или
с 405-субъединицей рибосомы, и предотвра-
щают инициацию трансляции (рис. 28-35,
ср. с рис. 27-27).
408-субъединица рибосомы
Нетранслируемая
3'-область (З'-UTR)
Рис. 28-35. Регуляция трансляции эукариотической мРНК.
Один из самых важных механизмов регуляции трансляции
в клетках эукариот заключается в связывании репрессо-
ров трансляции (РНК-связывающих белков) со специфи-
ческими участками в З'-нетранслируемой области (З'-UTR)
мРНК. Эти белки взаимодействуют с эукариотическими
факторами инициации или с рибосомами (см. рис. 27-27),
что предотвращает или замедляет трансляцию.
3. Некоторые белки, имеющиеся у всех эукари-
от — от дрожжей до млекопитающих, наруша-
ют взаимодействия между eIF4E и eIF4G (см
рис. 27-27). Соответствующие белки млекопи-
тающих называют 4Е-ВР (белки, связываю-
щие eIF4E). Когда клетка растет медленно, эти
белки ограничивают трансляцию, связываясь
с участком на eIF-4E, который в норме взаимо-
действует с eIF4G. Когда клеточный рост воз-
обновляется или ускоряется в ответ на сигнал
факторов роста или на другие стимулы, связы-
вающие белки инактивируются фосфорили-
рованием под действием протеинкиназ.
4. Регуляция экспрессии генов, опосредованная
РНК, которую мы обсудим чуть позже, обыч-
но заключается в подавлении трансляции.
Разнообразие механизмов регуляции трансляции
обеспечивает гибкость при осуществлении по-
давления трансляции — замедление трансляции
может касаться лишь нескольких мРНК или же
охватить глобальную регуляцию всего процесса
трансляции в клетке.
Хорошо изучен процесс регуляции трансля-
ции в ретикулоцитах. В этих клетках в регуляции
трансляции участвует фактор инициации eIF2. ко-
торый связывается с инициаторной тРНК и под-
водит ее к рибосоме. Когда Met-тРНК связывает-
ся в P-сайте, с eIF2 связывается фактор eIF2B и
высвобождает его за счет связывания и гидролиза
GTP. В ходе созревания ретикулоцитов ядро кле-
ток разрушается, и все пространство внутри плаз-
матической мембраны оказывается заполненным
гемоглобином. Матричная РНК. накопленная в
цитоплазме до исчезновения ядра, позволяет под-
держивать запасы гемоглобина. Когда ретику-
лоциты испытывают дефицит железа или гема,
происходит остановка трансляции мРНК глоби-
нов. Активируется протеинкиназа HCR (от англ.
hemin-conrolled repressor — репрессор, контроли-
руемый гемином), которая фосфорилирует eIF2.
Фосфорилированный eIF2 образует стабильный
комплекс с eIF2B, что делает невозможным уча-
стие eIF2 в трансляции. Так ретикулоциты коор-
динируют синтез глобина с наличием гема.
При изучении развития многоклеточных ор-
ганизмов было выявлено множество других при-
меров регуляции трансляции, что подробнее об-
суждается ниже.
[270] Часть III. 28. Регуляция экспрессии генов
Посттранскрипционный сайленсинг гена
опосредован РНК
В клетках высших эукариот, в том числе нематод,
плодовых мушек, растений и млекопитающих, вы-
ключение (сайленсинг) многих генов опосредуют
малые молекулы РНК, названные микро-РНК.
Эти РНК функционируют путем взаимодействия
с молекулами мРНК (часто в области 3’UTR),
приводя либо к деградации мРНК, либо к пода-
влению трансляции. В любом случае мРНК, а зна-
чит и ген, с которого она образована, отключается
(«молчит»). Этот механизм регуляции генов кон-
тролирует порядок событий в ходе развития неко-
торых организмов. Такая регуляция также исполь-
зуется для защиты от заражения РНК-вирусами
(что особенно важно для растений, у которых нет
иммунной системы) и для контроля активности
транспозонов. Кроме того, малые молекулы РНК
могут играть важную (но пока еще неопределен-
ную) роль в образовании гетерохроматина.
Многие микро-РНК существуют только на
определенных этапах развития, и иногда их отно-
сят к малым временным РНК (мвРНК). В высших
эукариотах идентифицированы тысячи различных
микро-РНК, которые могут влить на регуляцию
трети всех генов млекопитающих. Они транскри-
бируются в виде РНК-предшественников длиной
около 70 нуклеотидов с внутренними комплемен-
тарными последовательностями, которые образу-
ют структуры типа шпильки (см. рис. 26-28). Эти
предшественники расщепляются эндонуклеазами,
такими как Drosha и Dicer, с образованием корот-
ких дуплексов длиной 20-25 нуклеотидов. Одна
цепь процессированной микро-РНК связывается
с мРНК-мишепыо (или с РНК вируса или транс-
позона), что приводит к подавлению трансляции
или к деградации РНК ( ис. 28-34 ). Некоторые
микро-РНК связываются с мРНК одного вида,
управляя экспрессией единственного гена. Дру-
гие взаимодействуют со многими мРНК, являясь
ключевым элементом регулона и координируя
экспрессию множества генов.
Этот механизм регуляции генов имеет ин-
тересную и очень полезную практическую сто-
рону. Если ввести в организм РНК-дуплекс, по-
следовательность которого соответствует фак-
тически любой мРНК, эндонуклеаза Dicer рас-
щепляет эти дуплексы на короткие фрагменты,
называемые малыми интерферирующими РНК
(миРНК). Они связываются с мРНК и вызыва-
ют их сайленсинг (рис. 28-36, б). Этот процесс
называют интерференцией РНК. В растениях
фактически любой ген может быть эффективно
выключен таким способом. Нематоды способны
поглощать функциональные РНК целиком, и
поэтому простое добавление дуплексной РНК в
их корм позволяет очень эффективно отключать
гены-мишени. Такой метод быстро стал важным
инструментом в изучении функций генов, так как
он позволяет отключить ген без создания мутант-
ного организма. Этот подход может оказаться
полезным и при лечении человека. Искусствен-
но синтезированные миРНК позволяют блоки-
ровать распространение ВИЧ и полиовирусной
инфекции в культурах клеток человека примерно
в течение недели Быстрый прогресс в этой об-
ласти позволяет надеяться на внедрение методов
интерференции РНК в медицину в будущем.
Рис. 28-36. Сайленсинг генов в результате интерферен-
ции РНК. а — малые временные РНК (мвРНК) образуют-
ся под действием эндонуклеазы Dicer из более длинных
предшественников, которые содержат дуплексные участ-
ки. Затем эти мвРНК связываются с мРНК, приводя к раз-
рушению мРНК или подавлению трансляции. 6 — двух-
цепочечные РНК можно искусственно сконструировать и
ввести в клетку. Dicer превращает дуплексные РНК в ма-
лые интерферирующие РНК (миРНК), которые взаимодей-
ствуют с мРНК-мишенью. В этом случае происходит либо
расщепление мРНК, либо ингибирование ее трансляции.
28.3 Регуляция экспрессии генов у эукариот [ 2 /1 ]
У эукариот реализуется несколько
вариантов РНК-опосредованной регуляции
экспрессии генов
У эукариот к РНК со специфическими функция-
ми относят описанные выше микроРНК, мяРНК,
участвующие в сплайсинге РНК (см. рис. 26-16),
а также мяРНК, участвующие в модификации
рРНК (см. рис. 26-26). Все не кодирующие бе-
лок РНК, включая рРНК и тРНК, имеют общее
название — некодирующие РНК (нкРНК). Ве-
роятно, в геномах млекопитающих содержится
больше некодирующих, чем кодирующих РНК
(см. доп. 26-4). Неудивительно, что до сих пор
появляются новые функциональные классы не-
кодирующих РНК.
Многие из недавно обнаруженных некоди-
рующих РНК взаимодействуют не с РНК, а с
белками, и изменяют свойства связанных белков.
Один из примеров — белок SRA, который высту-
пает в роли ко-активатора генов, реагирующих
на действие стероидных гормонов; он влияет на
активацию транскрипции. Еще один пример —
белки теплового шока. Фактор теплового шока-1
(HSF-1) — это белок-активатор, который в обыч-
ных условиях существует в виде мономера, свя-
занного с шапероном Hsp90. При стрессе HSF-1
высвобождается из комплекса с Hsp90 и триме-
ризуется. Тример HSF-1 связывается с ДНК и
активирует транскрипцию генов, продукты кото-
рых необходимы для реакции на стресс. Некоди-
рующая РНК фактора HSR1 (от англ, heat shock
RNA 1; - 600 нуклеотидов) стимулирует образо-
вание тримеров HSF-1 и связывание ДНК. Фак-
тор HSR1 действует не в одиночку, а в комплексе
с фактором элонгации трансляции eEFlA.
Вспомогательные РНК влияют на ход транс-
крипции многими способами. Наряду с участием
в сплайсинге (см. рис. 26-17) мяРНК Ш прямо
связывается с фактором транскрипции TFIIH.
Функция мяРНК U1 в этом комплексе не совсем
понятна; возможно, она регулирует активность
TFIIH и/или влияет на сопряжение процессов
транскрипции и сплайсинга. Еще одна РНК с не-
определенной функцией, названная 7SK, состоит
из 331 нуклеотида; она содержится в большом ко-
личестве в клетках млекопитающих, связывается
с фактором элонгации транскрипции p-TEFb (см.
табл. 26-2) и подавляет удлинение транскрипта.
Другая нкРНК, В2 (-178 нуклеотидов) напря-
мую связывается с Pol II при тепловом шоке и
подавляет транскрипцию. Связанная с В2 поли-
мераза II образует стабильный преинициаторный
комплекс, но транскрипция при этом блокирова-
на. Таким способом при тепловом шоке РНК В2
подавляет транскрипцию многих генов, однако
существует неизвестный пока механизм, позво-
ляющий чувствительным к HSF-1 генам экспрес-
сироваться в присутствии В2.
Представления о роли нкРНК в экспрессии
генов и во многих других клеточных процессах
быстро развиваются. Одновременно с этим ин-
тересы биохимиков, изучающих регуляцию ге-
нов, все больше смещаются от белков в сторону
РНК.
Развитие контролируется
каскадами регуляторных белков
По сложности схем регуляции генов процесс
превращения зиготы в многоклеточное живот-
ное или растение не имеет себе равных. В ходе
развития происходят изменения морфологии и
белкового состава, которые зависят от точной
координации изменений в экспрессии генома.
В ранний период развития экспрессируется
больше генов, чем в любой другой период жиз-
ненного цикла. Например, ооцит морского ежа
содержит около 18 500 различных мРНК, а ти-
пичная дифференцированная ткань — около
6000. Матричные РНК ооцита дают начало ка-
скаду событий, которые регулируют экспрессию
многих генов во времени и в пространстве.
Некоторых животных особенно удобно ис-
пользовать в качестве модельных систем для
изучения развития, поскольку их легко содер-
жать в лабораторных условиях, и они достаточно
быстро размножаются. К таким животным отно-
сятся нематоды, плодовые мушки, рыбки-данио,
мыши и растения Arabidopsis. В этом разделе мы
сосредоточим внимание на развитии плодовых
мушек. Мы достаточно далеко продвинулись в
понимании молекулярных событий, происходя-
щих в ходе развития Drosophila melanogaster, и
имеющуюся информацию можно использовать
для иллюстрации общих схем и принципов.
Развитие дрозофилы идет с полным мета-
морфозом на пути от эмбриона к взрослой осо-
би (рис. 28-37). Самые важные характеристики
эмбриона — это полярность (передний и задний
[272] Часть III. 28. Регуляция экспрессии генов
Эмбрион на
ранней стадии —
нет сегментов
развитие
эмбриона 3
0-е сутки
Яйцо
оплодотворение
Ооцит
Эмбрион на поздней стадии сегментирован
Голова Грудь Брюшко
1-е сутки
вылупление
три личиночные стадии,
разделенные линьками
Личинка
5-е сутки
окукливание
1 мм
Рис. 28-37. Жизненный цикл плодовой мушки Drosophila melanogaster. Развитие дрозо-
филы протекает с полным метаморфозом, т. е. взрослое насекомое кардинальным образом
отличается от промежуточных форм; такая трансформация требует множества изменений в
ходе развития. К поздней эмбриональной стадии уже сформированы сегменты, содержащие
специализированные структуры, из которых впоследствии развиваются различные придат-
ки и другие структуры взрослой мухи.
Куколка
метаморфоз
конец животного легко различимы, также как
спинная и брюшная стороны) и метамерия (тело
эмбриона составлено из наборов повторяющих-
ся сегментов с характерными признаками) В
процессе развития эти сегменты превращаются
в голову, грудь и брюшко. Каждый грудной сег-
мент взрослого насекомого содержит разный на-
бор придатков. Реализация этого сложного плана
развития находится под генетическим контролем.
Уже известно множество генов, регулирующих
этот процесс и удивительным образом влияющих
на формирование тела животного.
Яйцеклетка дрозофилы и 15 клеток-ня-
нек окружены слоем фолликулярных клеток
(рис. 28-38). По мере развития яйцеклетки (до
оплодотворения) мРНК и белки, происходя-
щие из клеток-нянек и фолликулярных клеток,
откладываются в яйцеклетке, и некоторые из
них играют определяющую роль в развитии. В
определенное время самка откладывает опло-
дотворенное яйцо, которое делится, и дочерние
клетки продолжают синхронно делиться каж-
дые 6-10 минут. Вокруг образующихся ядер нет
плазматической мембраны, они распределяются
внутри цитоплазмы яйца, формируя синцитий.
Между восьмым и одиннадцатым делением ядра
мигрируют к периферии яйца, образуя моно-
слой, окружающий общую насыщенную желт-
ком цитоплазму; это синцитиальная бластодер-
ма. После еще нескольких дополнительных де-
лений мембрана образует выпячивания, которые
окружают ядра, в результате чего возникает слой
клеток, формирующих клеточную бластодерму.
На этой стадии митотические циклы в различ-
ных клетках перестают синхронизироваться.
Дальнейшее развитие каждой клетки определя-
ется мРНК и белками, запасенными клетками-
няньками и фолликулярными клетками на са-
мой ранней стадии.
Белки, которые путем изменения локальной
концентрации или активности заставляют окружа-
ющую ткань принимать определенную форму или
структуру, иногда называют морфогенами; они
являются продуктами генов, регулирующих стро-
28.3 Регуляция экспрессии генов у эукариот [273]
Кристиана Нюсляйн-Фольхард
ение тела. Как установили Кристиана Нюсляйн-
Фольхард, Эдвард Б. Льюис и Эрик Ф. Вишаус,
три главных класса генов, регулирующих строение
тела, — материнские гены, гены сегментации и го-
меотические гены — действуют на последователь-
ных стадиях развития, точно определяя основные
структуры тела эмбриона дрозофилы. Материн-
ские гены экспрессируются в неоплодотворенном
яйце, и образующиеся материнские мРНК оста-
ются спящими до оплодотворения. Они кодируют
большинство белков, необходимых на самых ран-
них стадиях развития — до образования клеточной
бластодермы. Некоторые белки, кодируемые ма-
теринскими мРНК, управляют пространственной
организацией развивающегося зародыша на ран-
них стадиях, устанавливая его полярность. Гены
сегментации, транскрибируемые после оплодот-
ворения, задают точное число сегментов тела. На
последовательных стадиях действуют, как мини-
мум, три подкласса генов сегментации: гены про-
бела (gap-гены) делят развивающийся зародыш
на несколько обширных областей; гены парного
Рис. 28-38. Ранние стадии развития дрозофилы. В процес-
се развития яйца материнские мРНК (включая транскрип-
ты генов bicoid и nanos, которые обсуждаются в тексте) и
белки синтезируются клетками-няньками и фолликуляр-
ными клетками и откладываются в развивающемся ооците
(неоплодотворенной яйцеклетке). После оплодотворения
два ядра оплодотворенного яйца синхронно делятся вну-
три общей цитоплазмы (синцитий), а затем мигрируют к
периферии. Складки мембраны окружают ядра, формируя
монослой клеток на периферии (стадия клеточной бласто-
дермы). Во время первых делений ядер несколько ядер на
заднем конце становятся полярными клетками, которые
позднее станут клетками зародышевой линии.
[274] Часть III. 28. Регуляция экспрессии генов
правила (pair-rule) вместе с генами полярности
сегментов определяют 14 полос, которые станут
14 сегментами тела нормального эмбриона. Гоме-
отические гены экспрессируются еще позже; они
определяют, какие органы и придатки разовьются
в каждом сегменте тела.
Множество регуляторных генов из этих трех
классов отвечает за появление взрослой мушки с
головой, грудью и брюшком, с надлежащим чис-
лом сегментов и с нужными придатками в каж-
дом сегменте. Хотя полный эмбриогенез занима-
ет около суток, все эти гены активируются в тече-
ние первых четырех часов развития. Некоторые
мРНК и белки появляются лишь на несколько
минут в определенных фазах этого периода раз-
вития. Некоторые гены кодируют транскрипци-
онные факторы, которые влияют на экспрессию
других генов, — возникает каскад генетических
событий. Регуляция осуществляется и на уровне
трансляции: многие регуляторные гены кодиру-
ют репрессоры трансляции, большинство из ко-
торых связываются с нетранслируемой областью
на З’-конце мРНК (рис. 28-35). Так как многие
мРНК накапливаются в яйце задолго до того, как
потребуется их трансляция, репрессия трансля-
ции оказывается очень важным механизмом ре-
гуляции развития
Материнские гены. Некоторые материнские
гены экспрессируются внутри клеток-нянек и
фолликулярных клеток, другие — в самом яйце
В неоплодотворенном яйце дрозофилы продукты
материнских генов формируют две оси тела —
передне-заднюю и спинно-брюшную; тем самым
определяется, какие области радиально симме-
тричного яйца образуют голову и брюшко, а так-
же в верх и низ тела взрослой мухи. Ключевым
событием на самых ранних стадиях развития яв-
ляется установление градиентов мРНК и белков
вдоль осей тела. Некоторые материнские мРНК
кодируют белковые продукты, которые диф-
фундируют через цитоплазму и распределяются
в яйце несимметричным образом. В результате
клетки клеточной бластодермы наследуют раз-
ные количества этих белков, что направляет клет-
ки по разным путям развития. Среди продуктов
материнских мРНК есть активаторы и репрес-
соры транскрипции, а также репрессоры транс-
Рис. 28-39. Распределение продукта мате-
ринского гена в яйце дрозофилы, а — мик-
рофотография яйца, окрашенного с помощью
антител, демонстрирует распределение про-
дукта гена bicoid (bed). Графики отражают
интенсивность окраски. Такое распреде-
ление необходимо для нормального раз-
вития структур передней части животного.
б — если у матери ген bed не экспрессируется
(мутант bcd"/bcd_), мРНК bicoid не накапли-
вается в яйце, и в результате формируется
нежизнеспособный эмбрион с двумя задними
частями.
Яйцо bed /bed
Нормальное яйцо
Расстояние от переднего
конца (% от длины яйца)
100'
g m
К й
§
Мутант bed /bed
0 50 100
Нормальная личинка
а
Расстояние от переднего
конца (% от длины яйца)
Личинка с двумя
задними концами
б
28.3 Регуляция экспрессии генов у эукариот [275]
ляции; все они регулируют экспрессию других
генов, регулирующих строение тела. Возникаю-
щие специфические схемы и последовательности
экспрессии генов различаются в разных линиях
клеток, что в конечном итоге определяет разви-
тие каждой структуры взрослой особи.
Формирование передне-задней оси зароды-
ша дрозофилы в значительной степени опреде-
ляется продуктами генов bicoid и nanos. Продукт
гена bicoid - главный морфоген переднего конца
тела, а продукт гена nanos — главный морфоген
заднего конца. Матричная РНК гена bicoid син-
тезируется клетками-няньками и откладывает-
ся в неоплодотворенном яйце вблизи переднего
конца. Нюсляйн-Фольхард обнаружила, что эта
мРНК транслируется вскоре после оплодотво-
рения, и белок Bicoid распределяется в клетке
создавая к седьмому клеточному делению гради-
ент концентрации, расходящийся радиально от
переднего полюса (рис. 28-39, а). Белок Bicoid -
фактор транскрипции, активирующий экспрес-
сию ряда генов сегментации; этот белок содержит
гомеодомен (рис. 28-13). Кроме того. Bicoid яв-
ляется репрессором трансляции и инактивирует
определенные мРНК. Количество белка Bicoid в
различных частях зародыша влияет на последу-
ющую экспрессию других генов, причем транс-
крипция генов активируется или их трансляция
подавляется, только если концентрация белка
Bicoid превышает определенный порог. Форма
градиента концентрации белка Bicoid оказывает
очень сильное влияние на строение тела мухи.
Отсутствие белка Bicoid приводит к развитию
эмбриона с двумя брюшками, но без головы и
грудного отдела (рис. 28-39. б). Однако не имею-
щий белка Bicoid эмбрион развивается нормаль-
но, если в соответствующий конец яйца ввести
достаточное количество мРНК bicoid. Ген nanos
играет аналогичную роль, но его мРНК сосредо-
точена в заднем конце яйца и максимум передне-
заднего градиента белка достигается на заднем
полюсе. Белок Nanos подавляет трансляцию.
Если взглянуть на действие материнских
генов в целом, можно обнаружить общую схему
развития. Кроме асимметрично расположен-
ных мРНК bicoid и nanos, в цитоплазме равно-
мерным образом распределены другие материн-
ские мРНК. Три из этих мРНК кодируют бел-
ки Pumilio, Hunchback и Caudal, находящиеся
под влиянием nanos и bicoid (рис. 28-40). Белки
Локализованная
мРНК фна
bicoid
Локализованная
мРНК гена
трансляция мРНК и диффузия
Белок
Bicoid
продуктов создает
градиенты концентраций
Белок
Nanos
Подавление/активация трансляции
равномерно распределенных мРНК
отражает градиент регулятора
сб
у
У
ф
К
о
й
И
S
а
ф
с
мРНК гена caudal
✓Ч/Ч ХЧ/Ч ХЧХЧ ХЧХЧ
Белок Caudal
мРНК гена hunchback
Белок Hunchback
мРНК гена pumilio
Белок Pumilio
Цитоплазма яйца
Рис. 28-40. Схема формирования передне-задней оси в
яйце дрозофилы. Матричные РНК bicoid и nanos откла-
дываются вблизи переднего и заднего концов яйца соот-
ветственно; мРНК caudal, hunchback и pumilio равномер-
но распределены в цитоплазме. Градиенты белков Bicoid
(Bed) и Nanos приводят к накоплению белка Hunchback
на переднем конце, а белка Caudal на заднем конце яйца.
Белок Pumilio функционирует только на заднем конце
яйца поскольку для его активности в качестве репрессора
трансляции гена hunchback требуется белок Nanos.
Caudal и Pumilio участвуют в развитии заднего
конца мушки: Caudal активатор транскрипции
с гомеодоменом, Pumilio — репрессор трансля-
ции. Белок Hunchback играет важную роль в раз-
витии переднего конца и регулирует транскрип-
цию многих генов: в некоторых случаях — поло-
жительно, в других - - отрицательно. Белок Bicoid
подавляет трансляцию гена caudal на переднем
конце и активирует транскрипцию гена hunchback
в клеточной бластодерме. Поскольку экспрессия
hunchback происходит и с материнских мРНК.
и с генов развивающегося яйца, его можно рас-
[276] Часть III. 28. Регуляция экспрессии генов
сматривать и как материнский ген, и как ген сег-
ментации. В результате активности Bicoid кон-
центрация Hunchback на переднем конце яйца
увеличивается. Белки Nanos и Pumilio являются
репрессорами трансляции гена hunchback, пода-
вляя синтез этого белка на заднем конце яйца.
Белок Pumilio не функционирует в отсутствие
белка Nanos, и градиент экспрессии nanos ограни-
чивает активность обоих белков на заднем конце.
Репрессия трансляции гена hunchback приводит
к деградации мРНК hunchback в задней части.
Однако отсутствие на заднем конце белка Bicoid
приводит к экспрессии гена caudal. В результате
белки Hunchback и Caudal распределены в яйце
асимметричным образом.
Рис. 28-41. Распределение продукта гена fushi tarazu
(ftz) в раннем эмбрионе дрозофилы, а — в нормальном
эмбрионе продукт гена можно обнаружить в семи поло-
сах, охватывающих тело эмбриона (показаны схематич-
но); б — на авторадиограмме продольного среза эмбри-
она эти полосы выглядят как темные пятна (радиоактив-
ная метка); в — полосы определяют переднюю границу
сегментов в позднем эмбрионе (обозначены красным
цветом).
Гены сегментации. На последовательных стади-
ях эмбрионального развития дрозофилы активи-
руются три подкласса генов сегментации: гены
пробела (gap), парного правила (pair-rule) и по-
лярности сегмента. Обычно экспрессия генов gap
регулируется продуктами одного или нескольких
материнских генов. Некоторые гены gap кодиру-
ют транскрипционные факторы, которые влия-
ют на экспрессию других генов сегментации или
(позднее) гомеотических генов.
Примером хорошо охарактеризованного
гена сегментации является ген fushi tarazu (ftz)
из подкласса генов pair-rule. При делении гена
ftz у эмбриона появляется только семь сегмен-
тов вместо положенных 14, причем каждый
сегмент вдвое шире нормы. Белок Fushi tarazu
(Ftz) — активатор транскрипции, обладающий
гомеодоменом. Матричная РНК и белки, син-
тезированные на основе нормального гена ftz.
образуют удивительный рисунок в виде семи
полос, опоясывающих эмбрион в задних двух
третях его тела (рис. 28-41). Полосы указывают
расположение формирующихся позже сегмен-
тов; если функция ftz нарушена, сегменты не об-
разуются. Белок Ftz и еще несколько подобных
регуляторных белков прямо или опосредованно
регулируют экспрессию множества генов в ка-
скаде развития.
Гомеотические гены. Группа из 8-11 гомеоти-
ческих генов связан с развитием конкретных
структур в определенных отделах тела. Эти
гены теперь чаще называют Нох-генами; на-
звание происходит от слова «homeobox», обо-
значающего консервативную последователь-
ность гена, кодирующую гомеодомен и присут-
ствующую во всех этих генах. Однако следует
учесть, что гомеодомен имеют не только белки,
связанные с развитием структур организма (к
примеру, описанный выше продукт гена bicoid
тоже содержит гомеодомен); «Нох» подразуме-
вает скорее функциональную, чем структурную
классификацию. Гены Нох организованы в кла-
стеры. У дрозофил есть один такой кластер, у
млекопитающих их четыре (рис. 28-42). Гены в
этих кластерах — от нематод до человека — уди-
вительно похожи. У дрозофил каждый из генов
Нох экспрессируется в определенном отделе за-
родыша и контролирует развитие соответству-
ющей части тела. Терминология, использую-
28.3 Регуляция экспрессии генов у эукариот [277]
а
Рис. 28-42. Гены кластера Нох и их роль в развитии
организма, а — каждый ген Нох дрозофилы отвечает
за развитие структур определенной части тела и экс-
прессируется в определенных участках зародыша, что
показано с помощью разных цветов, б — дрозофила
имеет один кластер генов Нох, в геноме человека таких
кластеров четыре. У многоклеточных животных многие
из этих генов обладают высоким уровнем консерватив-
ности. Эволюционные связи между генами кластера Нох
дрозофилы и млекопитающих показаны пунктирными
линиями. При выравнивании последовательностей ана-
логичные связи прослеживаются между четырьмя набо-
рами генов Нох млекопитающих. <
Drosophila
Нох-В
ноХ-с ч 1 I I I I I I I I I I I I I.........I...I.ГП-
С4 С5 С6 С8 С9 СЮ С11 С12 С13
4 1 Г Illi I I I I I I I I...I.I.ТУТ-
DI D3 D4 D8 D9 DIO Dll D12 D13
щаяся для обозначения генов Нох, достаточно
запутанная. Названия генов дрозофилы сложи-
лись исторически (например, ultrabithorax), в
то время как для обозначения генов млекопи-
тающих применяют систему букв (А, В, С, D) и
цифр (1,2, 3,4).
Утрата гомеотических генов в результате
мутации или делении приводит к развитию нор-
мальных придатков или структур в неправиль-
ных местах. Важным примером является ген
ultrabithorax (ubx). При потере функции Ubx
первый брюшной сегмент развивается непра-
вильно и приобретает структуру третьего груд-
ного сегмента. Другие мутации гомеотических
генов вызывают формирование дополнитель-
ной пары крыльев или двух ног вместо антенн
на голове (рис. 28-43). Многие гены Нох весьма
протяженные. Например, длина гена ubx, со-
ставляет 77 000 п. н. Более 73 000 п. н. в составе
гена приходится на интроны, один из которых
имеет длину 50 000 п. н. Транскрипция гена ubx
занимает около часа. Полагают, что задержка
[278] Часть III. 28. Регуляция экспрессии генов
Рис. 28-43. Результаты мутаций в гомеотических генах дрозофилы, а — нормальная го-
лова. 6 — гомеотический мутант (antennapedia) с ногами вместо антенн, в — нормальное
строение тела, г — гомеотический мутант (bithorax) у которого появился еще один грудной
сегмент с дополнительной парой крыльев.
Дополнение 28-1 0 плавниках, крыльях и клювах
В Южной Америке обитает несколько видов вьюр-
ков. Около 3 млн лет назад небольшая группа вьюр-
ков одного вида вылетела с континентального берега
Тихого Океана. Возможно, из-за шторма птицы поте-
ряли ориентацию и пролетели около 1000 км. Такие
мелкие птицы могли погибнуть в подобном путеше-
ствии, однако этим вьюркам повезло, и они долетели
до недавно образовавшихся вулканических островов,
которые теперь мы называем Галапагосским архи-
пелагом. Здесь было обилие растений и насекомых,
и птицы выжили. Со временем образовались новые
острова, которые были заселены новыми растениями
и насекомыми, а также вьюрками. Птицы использо-
вали биоресурсы островов, и постепенно выделялись
отдельные i руппы вьюрков и превращались в новые
виды. В 1835 г. на этих островах побывал Чарльз Дар-
вин и обнаружил множество видов вьюрков*, питав-
шихся семенами, фруктами, насекомыми, пыльцой и
даже кровью.
* Галапагосские, или дарвиновские, вьюрки подсемей-
ство семейства вьюрковых, к которому относятся 14 ви-
дов птиц, 13 из которых населяют Галапагосские острова,
один вид обитает на о. Кокос. Прим, перев.
Разнообразие живых существ поражало челове-
ка задолго до того, как ученые начали исследовать его
причины. Самая важная роль в этом исследовании
принадлежит Дарвину, в частности, его наблюдени-
ям за птицами Галапагосских островов. На основе
теории Дарвина можно объяснить само существова-
ние организмов разного вида, но возникает вопрос о
механизмах эволюционного процесса. Решение этой
проблемы ученые стали находить во второй половине
XX в. при изучении геномов и метаболизма нуклеи-
новых кислот и совсем недавно в связи с появлением
нового раздела биологии эволюционной биологии
развития (сокращенно evo-devo), которая сочетает из-
учение процессов эволюции и развития.
В современном виде теория эволюции строится
на двух основных положениях: мутации в популяции
создают генетическое разнообразие; затем эта попу-
ляция подвергается естественному отбору, который
благоприятствует распространению организмов с бо-
лее полезным генетическим инструментарием и пода-
вляет распространение остальных. В геноме каждого
организма, в каждой клетке постоянно происходят
мутации (см. гл. 8 в т. 1 и гл. 25). Мутации в одно-
клеточных организмах или в зародышевых клетках
28.3 Регуляция экспрессии генов у эукариот [279]
экспрессии гена ubx, возникающая из-за дли-
тельной транскрипции, нужна для синхрони-
зации процессов в общей временной регуляции
последующих стадий развития. Многие гены
Нох далее регулируются малыми интерфериру-
ющими РНК, закодированными между после-
довательностями генов внутри кластеров Нох.
Все продукты генов Нох являются факторами
транскрипции и регулируют экспрессию ряда
генов на следующих стадиях развития. Эта це-
почка взаимодействий все еще изучается.
Многие из рассмотренных выше принципов
развития справедливы и для других эукариот, от
нематоды до человека. Во многих видах функци-
онируют одни и те же белки. Например, продук-
ты гена Н0ХА7 мыши и гена antennapedia дрозо-
филы различаются лишь одним аминокислотным
остатком. Конечно, несмотря на сходство молеку-
лярных механизмов регуляции, пути развития
механизмов различны (у человека нет ни кры-
льев, ни антенн), что определяется генами-мише-
нями. контролируемыми генами Нох Открытие
структурных детерминант со специфическими
молекулярными функциями — первый шаг к по-
ниманию молекулярных событий, лежащих в ос-
нове развития. По мере идентификации новых
генов и их белковых продуктов все больше прояс-
няются биохимические аспекты этой гигантской
головоломки.
При обсуждении регуляции развития мы
сделали круг и вернулись к биохимическим на-
чалам — и фигурально, и буквально. Самые пер-
вые и самые последние рассуждения в данной
книге относятся к теме эволюции. Если эволю-
ция состоит в генерации изменений, приводящих
к появлению различных видов организмов, она
непременно должна затрагивать программу раз-
вития организма. Процессы эволюции и разви-
тия тесно связаны между собой (доп. 28-1). Био-
химия как наука призвана помочь человечеству
одновременно развиваться в будущем и понять
собственное происхождение в прошлом.
многоклеточных организмов могут наследоваться,
причем вероятность наследования (т. е. передача му-
тации значительной части потомства) увеличивается,
если такая мутация дает определенное преимуще-
ство. Это очевидная схема. Однако многие ученые
сомневались в том, может ли такая схема объяснить,
скажем, разнообразие формы клюва галапагосских
вьюрков или разнообразие форм и размеров млеко-
питающих. До последних десятилетий существовало
несколько широко распространенных гипотез эво-
люционного процесса. Считалось, что для появления
новой физической структуры необходимо множество
мутаций и новые гены, сложные организмы должны
иметь более крупный геном и у сильно различающих-
ся видов мало общих генов. Все эти предположения
оказались ошибочными.
Современные исследования показали, что в ге-
номе человека содержится меньше генов, чем предпо-
лагалось, — немного больше, чем в геноме дрозофи-
лы, и меньше, чем в геноме некоторых земноводных.
Геномы всех млекопитающих — от мыши до чело-
века — удивительно похожи по числу, типу и хромо-
сомному расположению генов. Однако эволюционная
биология развития объясняет, как на таком генетиче-
ском базисе могут эволюционировать сложные и очень
разнообразные создания.
В конце XIX в. английский биолог Уильям Бейт-
сон изучал различных мутантных животных, вроде
тех, что изображены на рис. 28-43. На основании сво-
их наблюдений он оспаривал утверждение Дарвина о
том, что эволюционные изменения должны быть по-
степенными. Современные исследования генов, кон-
тролирующих развитие организма, подтвердили идеи
Бейтсона. Незначительные изменения механизмов
регуляции в ходе развития, возникающие в результа-
те лишь одной или нескольких мутаций, могут при-
вести к физическим изменениям и удивительно бы-
строй эволюции.
Галапагосские вьюрки — прекрасный пример,
демонстрирующий связь между эволюцией и раз-
витием. Существует 14 (некоторые специалисты на-
зывают 15) видов галапагосских вьюрков, основные
различия между которыми заключаются в строении
клювов. У земляных вьюрков, например, широкий
тяжелый клюв, предназначенный для раскалывания
крупных и прочных зерен. У кактусовых вьюрков клюв
Продолжение на с. 280
[280] Часть III. 28. Регуляция экспрессии генов
Дополнение 28-1 0 плавниках, крыльях и клювах (продолжение)
более длинный и тонкий, он идеально подходит для
проникновения в плоды и цветки кактусов (рис. 1).
Клиффорд Табин с коллегами тщательно проанализи-
ровали набор генов, экспрессирующихся в ходе разви-
тия головы птиц. Они идентифицировали единствен-
ный ген, Втпр4, уровень экспрессии которого коррели-
рует с образованием более мощного клюва у земляных
вьюрков. В экспериментальных условиях более высо-
кий уровень экспрессии гена Втпр4 в соответствующих
тканях также вызывает образование более крепких
клювов у зародышей кур, что подтверждает значение
гена Втр4. В аналогичном исследовании было показа-
но, что формирование более длинного и тонкого клю-
ва коррелирует с экспрессией гена кальмодулина (см.
рис. 12-11 в т. 1) в определенных тканях на определен-
ной стадии развития Таким образом, значительные из-
менения формы и функций клюва могут быть вызваны
небольшими изменениями экспрессии всего двух ге-
нов, участвующих в регуляции развития. Нужно лишь
несколько мутаций, но они должны затрагивать меха-
низмы регуляции. Новые гены не нужны.
У всех позвоночных система регуляторных ге-
нов, ответственных за развитие, очень консервативна.
Повышение уровня экспрессии Втр4 в нужной ткани
и в нужное время приводит к формированию более
мощной челюсти у полосатых данио. Тот же ген игра-
ет ключевую роль в развитии зубов млекопитающих.
Развитие глаз дрозофил и млекопитающих запускает-
ся экспрессией одного и того же гена. Рахб. Мышиный
ген Рахб запускает развитие нормальных глаз мушки
у дрозофилы, а ген Рахб дрозофилы запускает разви-
тие мышиных глаз у мыши. В каждом организме эти
гены являются частью сложного регуляторного каска-
да, который в конечном итоге приводит к образованию
правильных структур в правильном месте в каждом
организме. Это очень древний каскад. Например, опи-
санные гены Нох участвовали в развитии многокле-
точных эукариот на протяжении более 500 млн лет.
Незначительные изменения регулярного каскада ока-
зывают сильное влияние на развитие и, следовательно,
на внешний вид организма. Эти же незначительные из-
менения могут стать толчком для необычайно быстрой
эволюции. Например, от 400 до 500 известных видов
цихлпд (окуневых рыб), обитающих в озерах Малави и
Виктория на Африканском континенте, происходят от
одной или от небольшого числа популяций, населяв-
ших эти озера 100- 200 тыс. лет назад. Птицы Галапа-
госских островов прошли свой заключительный путь
эволюции, который у других живых существ занял
миллиарды лет.
Смешанное питание зернами и насекомыми
Цветки/плоды кактуса
Предок
Высокая концентрация СаМ — удлиненный клюв
Низкая концентрация Вшр4 - узкий и сплющенный клюв
Крепкие крупные семена
Кактусовый
вьюрок
Большой
земляной
вьюрок
Высокая концентрация СаМ —
удлиненный, узкий
и сплющенный клюв
Низкая концентрация СаМ — короткий клюв
Высокая концентрация Вшр4 —
широкий и высокий клюв
Рис. 1. Эволюция строения клюва для приспособления к новым источникам пищи. Различие
в строении клювов у двух видов галапагосских вьюрков (кактусового вьюрка и крупного
земляного вьюрка), питающихся по-разному, в значительной степени объясняется неболь-
шим числом мутаций, которые изменили время и уровень экспрессии всего двух генов —
гена кальмодулина и гена Втр4.
Ключевые термины [281 ]
Краткое содержание раздела 28.3
РЕГУЛЯЦИЯ ЭКСПРЕССИИ ГЕНОВ У ЭУКАРИОТ
Для эукариотических генов более характерна
положительная регуляция, чем отрицательная,
и транскрипция сопровождается большими
изменениями структуры хроматина.
Типичные промоторы Pol II содержат ТАТА-
бокс и последовательность Inr, а также много-
численные участки связывания активаторов
транскрипции. Последние иногда располо-
жены за сотни или тысячи пар оснований от
ТАТА-бокса; в дрожжах они называются акти-
вирующими последовательностями (UAS), а в
высших эукариотах — энхансерами.
Для регуляции транскрипции обычно требу-
ются крупные белковые комплексы. Влияние
активаторов транскрипции на Pol II опосре-
довано ко-активаторными белковыми ком-
плексами, такими как медиатор. Активаторы
содержат отдельные домены, отвечающие
за активацию и за связывание ДНК. Другие
белковые комплексы, включая гистонаце-
тилтрансферазы и ATP-зависимые комплек-
сы, такие как SWI/SNF и NURF, обратимо
перестраивают и модифицируют структуру
хроматина.
Гормоны регулируют экспрессию генов од-
ним из двух способов. Стероидные гормоны
напрямую взаимодействуют с внутриклеточ-
ными рецепторами, которые являются ДНК-
связывающими регуляторными белками;
связывание гормона либо усиливает, либо
ослабляет транскрипцию генов-мишеней.
Нестероидные гормоны связываются с ре-
цепторами на клеточной поверхности, запу-
ская сигнальный путь, который может при-
вести к фосфорилированию регуляторного
белка и изменить его активность.
Регуляция экспрессии генов при участии мо-
лекул РНК играет важную роль в эукариоти-
ческих клетках, причем число известных ме-
ханизмов такой регуляции продолжает расти.
Развитие многоклеточного организма регу-
лируется очень сложно. Судьба клеток в ран-
нем эмбрионе определяется путем формиро-
вания передне-заднего и спинно-брюшного
градиентов белков, которые действуют как
активаторы транскрипции или репрессоры
трансляции, регулируя экспрессию генов,
необходимых для развития определенных
структур в определенных частях тела. Груп-
пы регуляторных генов последовательно
действуют во времени и пространстве, пре-
вращая отдельные зоны яйцеклетки в струк-
туры взрослого организма.
Ключевые термины
Термины, выделенные жирным шрифтом, объясня-
ются в глоссарии.
сАМР-рецепторный белок
(CRP) 244
SWI/SNF 259
ТАТА-связывающий белок
(ТВР) 262
активатор 232
активаторы транскрипции
261
активирующая последова-
тельность (UAS) 261
аттенюация транскрипции
246
белки с высокой электро-
форетической под-
вижностью (HMG)
262
гены домашнего хозяйства
230
гены парного правила
(pair-rule) 273
гены полярности сегмен-
тов 274
гены пробела (gap) 273
гены сегментации 273
гистонацетилтрансферазы
(HAT) 258
гомеобокс 241
гомеодомен 241
гомеотические гены 276
гормон-чувствительные
элементы (HRE) 266
индукция 230
интерференция РНК 270
катаболитная репрессия
244
ко-активаторы 261
лейциновая молния 241
материнские гены 273
материнские мРНК 273
медиатор 262
метамерия 272
микро-РНК 270
морфогены 272
нкРНК271
оператор 232
оперон 234
основная спираль-петля-
спираль 241
основные факторы транс-
крипции 261
огр и I щтел ь и ая ре гу л я ц и я
232
положительная регуляция
232
полярность 271
преинициаторный ком-
плекс (PIC)262
регулон 245
ремоделирование хромати-
на 258
репрессия 231
репрессор 232
репрессор трансляции 250
рибопереключатель 253
спираль-поворот-спираль
239
строгий ответ 251
фазовая вариация 254
факторы специфичности
232
цинковый палец 239
энхансеры 233
12821 Часть III. 28. Регуляция экспрессии генов
Дополнительная литература
для дальнейшего изучения
Общая .in юра гура
Carroll, S. В. (2005) Endless Forms Most Beautiful: The New
Science of Evo Devo and the Making of the Animal Kingdom, W.
W. Norton & Company, New York.
Интересное описание того, как биология развития до-
полняет эволюционную биологию.
Muller-Hill, В. (1996) The lac Operon: A Short History of a
Genetic Paradigm, Walter de Gruyter, New York.
Прекрасный подробный обзор исследований этой важ-
ной системы.
Neidhardt, Е С. (ed.). (1996) Escherichia coli and Salmonella
typhimurium, 2nd edn, Vol. 1: Cellular and Molecular Biology
(Curtiss, R., Ingraham, J. L., Lin, E. С. C., Magasanik, B., Low,
К. B., Reznikoff, W. S., Riley, M., Schaechter, M., & Umbarger,
H. E., vol. eds), American Society for Microbiology, Washing-
ton, DC.
Прекрасный обзор многих бактериальных оперонов.
Сетевая версия, EcoSal, регулярно обновляется.
Sarai, А. & Kono, Н. (2005) Protein-DNA recognition pat-
terns and predictions. Annu. Rev. Biophys. Biomol. Struct. 34,
379-398.
Описание современных методов, включая геномику, в
приложении к анализу механизмов регуляции генов.
von Hippel, Р. Н. (2007) From “simple” DNA-protein inte-
ractions to the macromolecular machines of gene expression.
Annu. Rev. Biophys. Biomol. Struct. 36,79-105.
Подробное описание биофизических закономерностей
ДНК-белковых взаимодействий и их роли в регуляции
генов.
Регуляция экспрессии iciiob бактерий
Barrick, J. Е. & Breaker, R. R. (2007) The power of riboswit-
ches: discovering relics from a lost world run by RNA mole-
cules may lead to modern tools for fighting disease. Sci. Am.
(January), 50-57.
Coppins, R. L., Hall, К. B., & Groisman, E. A. (2007) The in-
tricate world of riboswitches. Curr. Opin. Microbiol. 10,176-181.
Gourse, R. L., Gaal, T., Bartlett, M. S., Appleman, J. A.,
& Ross, W. (1996) rRNA transcription and growth rate-de-
pendent regulation of ribosome synthesis in Escherichia coli.
Annu. Rev. Microbiol. 50, 645-677.
Jacob, E & Monod, J. (1961) Genetic regulatory mechanisms
in the synthesis of proteins./ Mol. Biol. 3,318-356.
В этой исторической статье представлены модель оперо-
на и концепция матричной РНК, впервые изложенные эти-
ми авторами в Трудах Французской академии наук в 1960 г.
Johnson, R. С. (1991) Mechanism of site-specific DNA inver-
sion in bacteria. Curr. Opin. Genet. Dev. 1,404-411.
Kolb, A., Busby, S., Buc, H., Garges, S., & Adhya, S. (1993)
Transcriptional regulation by cAMP and its receptor protein.
Annu. Rev. Biochem. 62,749-795.
Romby, P. & Springer, M. (2003) Bacterial translational
control at atomic resolution. Trends Genet. 19, 155-161.
Storz, G., Altuvia, S., & Wassarman, К. M. (2005) An abun-
dance of RNA regulators. Annu. Rev. Biochem. 74,199-217.
В обзоре рассмотрены и бактерии, и эукариоты.
Winkler, W. С. & Breaker, R. R. (2005) Regulation of bacte-
rial gene expression by riboswitches. Annu. Rev. Microbiol. 59,
487-517.
Yanofsky, C., Konan, К. V., & Sarsero, J. P. (1996) Some
novel transcription attenuation mechanisms used by bacteria.
BiochimielS, 1017-1024.
Pei уляция экспрессии i снов эукарио!
Agami, R. (2002) RNAi and related mechanisms and their
potential use for therapy. Curr. Opin. Chem. Biol. 6,829-834.
Bashirullah, A., Cooperstock, R. L., & Lipshitz, H. D.
(1998) RNA localization in development. Annu. Rev. Biochem.
67,335-394.
Bjorklund, S. & Gustafsson, С. M. (2005) The yeast me-
diator complex and its regulation. Trends Biochem. Sci. 30,
240-244.
Cerutti, H. (2003) RNA interference: traveling in the cell and
gaining functions? Trends Genet. 19, 9-46.
Conaway, R. C., Brower, C. S., & Conaway, J. W. (2002)
Gene expression — emerging roles of ubiquitin in transcription
regulation. Science'll, 1254-1258.
Conaway, R. C., Sato, S., Tomomori-Sato, C., Yao, T., &
Conaway, J. W. (2005) The mammalian mediator complex
and its role in transcriptional regulation. Trends Biochem. Sci.
30, 250-255.
Gingras, A.-C., Raught, B., & Sonenberg, N. (1999) eIF4
initiation factors: effectors of mRNA recruitment to ribo-
somes and regulators of translation. Annu. Rev. Biochem. 68,
913-963.
Goodrich, J. A. & Kugel, J. E (2006) Non-coding RNA regu-
lators of RNA polymerase II transcription. Nat. Rev. Mol. Cell
Biol. 7,12-616.
Gray, N. K. & Wickens, M. (1998) Control of translation ini-
tiation in animals. Annu. Rev. Cell Dev. Biol. 14,399-458.
Hannon, G. J. (2002) RNA interference. Nature MS, 244-251.
Kapranov, P, Willingham, A. T., & Gingeras, T. R. (2007)
Genome-wide transcription and the implications for genomic
organization. Nat. Rev. Genet. 8,413-423.
О значении недавно обнаруженных уровней транс-
крипции большей части геномной ДНК млекопитающих.
Keene, J. D. (2007) RNA regulons: coordination of post-
transcriptional events. Nat. Rev. Genet. 8,533-543.
Вопросы и задачи 12831
Kim, Y.-J. & Lis, J. Т. (2005) Interactions between subunits
of Drosophila mediator and activator proteins. Trends Biochem.
Sci. 30,245-249.
Klose, R. J. & Zhang, Y. (2007) Regulation of histone methy-
lation by demethylimination and demethylation. Nat. Rev. Mol.
Cell Biol. 8, 307-318.
Kornberg, R. D. (2005) Mediator and the mechanism of
transcriptional activation. Trends Biochem. Sci. 30, 236-239.
Luger, K. (2003) Structure and dynamic behavior of nucleo-
somes. Curr. Opin. Genet. Dev. 13,127-135.
Prud’homme, B., Gompel, N., & Carroll, S. B. (2007) Emer-
ging principles of regulatory evolution. Proc. Natl. Acad. Sci.
[/5X104,8605-8612.
Reese, J. C. (2003) Basal transcription factors. Curr. Opin.
Genet. Dev. 13, 114-118.
Rivera-Pomar, R. & Jackie, H. (1996) From gradients to
stripes in Drosophila embryogenesis: filling in the gaps. Trends
Genet. 12,478-483.
Saha, A., Wittmeyer, J., & Cairns, B. R. (2006) Chromatin
remodeling: the industrial revolution of DNA around histones.
Nat. Rev. Mol. Cell. Biol. 7,437-447.
Saunders, A., Core, L. J., & Lis, J. T. (2006) Breaking bar-
riers to transcription elonga<on. Nat. Rev. Mol. Cell Biol. 7,
557-567.
Shahbazian, M. D. & Grunstein, M. (2007) Functions of site-
specific histone acetylation and deacetylation. Annu. Rev. Bio-
chem. 76,75-100.
Struhl, K. (1999) Fundamentally different logic of gene regu-
lation in eukaryotes and prokaryotes. Cell 98,1-4.
Thomas, M. C. & Chiang, C.-M. (2006) The general trans-
cription machinery and general cofactors. Crit. Rev. Biochem.
Mol.BiolAC 105-178.
Waterhouse, P. M. & Helliwell, C. A. (2003) Exploring plant
genomes by RNA-induced gene silencing. Nat. Rev. Genet. 4,
29-38.
Вопросы и задачи____________________________________
1. Влияние стабильности мРНК и белка на ре-
гуляцию. Предположим, клетки Е. coli растут в
среде, где единственным источником углерода
служит глюкоза. Затем в среду добавляют трип-
тофан. Клетки продолжают расти и делиться каж-
дые 30 мин. Опишите качественные изменения во
времени уровня активности триптофансинтазы в
клетках при следующих условиях:
а) мРНК триптофана стабильна (медленно
разрушается на протяжении многих часов).
б) мРНК триптофана быстро расщепляется,
но триптофансинтаза стабильна.
в) мРНК триптофана и триптофансинтаза
быстро разрушаются.
2. Отрицательная регуляция. Опишите воз-
можное влияние на регуляцию генов /яс-оперона
следующих мутаций: (а) в /яс-операторе делеги-
рована большая часть О^ (б) мутация гена lad,
инактивирующая репрессор; (в) в промоторе из-
менена область вокруг позиции -10.
3. Специфичность связывания ДНК с регуля-
торными белками. Типичный бактериальный
репрессорный белок в 104-106 раз эффективнее
связывается со специфическим участком ДНК
(оператором), чем с неспецифической последо-
вательностью ДНК. Для обеспечения эффектив-
ной репрессии достаточно 10 молекул репрессора
на клетку. Предположим, в клетке человека есть
очень похожий репрессор с такой же специфич-
ностью к своему участку связывания. Сколько
копий репрессора нужно, чтобы достичь такой же
эффективности репрессии, как в бактериальной
клетке? Подсказка. Геном Е. coli содержит около
4,6 млн п. н.; гаплоидный геном человека — около
3,2 млрд п. н.
4. Концентрация репрессора в Е. coli. Констан-
та диссоциации комплекса репрессор-оператор
очень низкая (около 10-13 М). Клетка Е. coli (объем
2 • 1012мл) содержит 10 копий репрессора. Вычис-
лите клеточную концентрацию белка-репрессора.
Сравните эту величину с константой диссоциации
комплекса репрессор-оператор. Каков смысл это-
го результата?
5. Катаболитная репрессия. Клетки Е. coli растут
в среде с лактозой, но без глюкозы. Укажите, по-
влияют ли перечисленные ниже изменения или
условия на экспрессию /яс-оперона (усилят, ос-
лабят или не вызовут изменений). Возможно, по-
лезно изобразить модель, описывающую каждую
ситуацию.
а) Добавление глюкозы в высокой концен-
трации.
б) Мутация, предотвращающая отсоедине-
ние Lac-penpeccopa от оператора.
в) Мутация, полностью инактивирующая
|3-галактозидазу
г) Мутация, полностью инактивирующая га-
лактозидпермеазу.
12841 Часть III. 28. Регуляция экспрессии генов
д) Мутация, препятствующая взаимодей-
ствию CRP с участком связывания вблизи про-
мотора.
6. Аттенюация транскрипции. Как изменится
транскрипция £ф-оперонаЕ. coli при следующих из-
менениях в лидерном участке мРНК триптофана?
а) Увеличение расстояния (числа основа-
ний) между геном лидерного пептида и последо-
вательностью 2.
б) Увеличение расстояния между последова-
тельностями 2 и 3.
в) Удаление последовательности 4. /
г) Замена двух кодонов Тгр в гене лидерного
пептида на кодоны His.
д) Удаление участка связывания рибосомы в
гене лидерного пептида.
е) Изменение нескольких нуклеотидов в по-
следовательности 3, в результате чего она может
спариваться с последовательностью 4, но не с по-
следовательностью 2.
7. Репрессоры и репрессия. Как может повлиять
на SOS-ответ в Е. coli мутация гена lex А, которая
предотвращает автокаталитическое расщепление
белка LexA?
8. Регуляция путем рекомбинации. Что про-
изойдет с клетками, если рекомбиназа Hin в си-
стеме фазовой вариации у Salmonella станет более
активной и будет осуществлять рекомбинацию
(инверсию ДНК) несколько раз в каждой клеточ-
ной генерации?
9. Инициация транскрипции в клетках эукариот.
В грубом экстракте из экзотического гриба была
обнаружена новая активность РНК-полимеразы.
Эта РНК-полимераза инициирует транскрипцию
с единственного специализированного промото-
ра. При очистке полимеразы ее активность посте-
пенно снижалась, а чистый фермент был полно-
стью неактивным, если только в реакционную
смесь не добавляли грубый экстракт. Объясните
эти наблюдения.
10. Функциональные домены в регуля-
торных белках. Биохимик заменил ДНК-
связывающий домен дрожжевого белка Gal4
ДНК-связывающим доменом Lac-penpeccopa и
обнаружил, что сконструированный белок не ре-
гулирует транскрипцию генов GAL в дрожжах.
Изобразите схему функциональных доменов
белка Gal4 и химерного белка. Почему химерный
белок не регулирует транскрипцию генов GAL?
Что нужно изменить в последовательности ДНК,
с которой связывается химерный белок, чтобы он
мог активировать транскрипцию генов GAL?
И. Модификация нуклеосом при активации
транскрипции. При подготовке участков генома
к транскрипции происходит ацетилирование и
метилирование специфических аминокислотных
остатков гистонов в соответствующих нуклеосо-
мах. Когда транскрипция завершена, необходимо
вернуть модифицированные остатки в прежнее
состояние. У млекопитающих метилирование
остатков Arg в гистонах снимается под действием
пептидиларгининдеиминаз. Однако в результате
этой реакции образуется не неметилированный
аргинин, а цитруллин. Каков второй продукт этой
реакции? Предложите возможный механизм.
12. Механизмы наследования в развитии. Яйца
дрозофилы с генотипом bcd /bcd могут разви-
ваться нормально, но взрослые насекомые с та-
ким генотипом не производят жизнеспособного
потомства. Объясните этот факт.
Биохимия в интернете
13. ТАТА-связывающий белок и ТАТА-бокс.
Чтобы исследовать взаимодействие между фак-
торами транскрипции и ДНК, войдите в базу дан-
ных FirstGlance в Jmol на сайте http://firstglance.
jmol.org и загрузите файл PDB ID 1TGH. Этот
файл моделирует взаимодействия между ТАТА-
связывающим белком человека и участком двух-
цепочечной ДНК. Когда структура загрузится,
выберете опцию “Spin”, чтобы молекула переста-
ла вращаться. Далее нажмите ссылку “Contacts”.
При выборе функции “Chains” кликните на лю-
бую часть белка (цепь А голубого цвета), чтобы
выбрать соответствующую цепь. Кликните “Show
Atoms Contacting Target” и в появившемся спи-
ске найдите функцию “Show putatively hydrogen-
bonded non-water” (показать возможные водород-
ные связи не с водой), чтобы увидеть водородные
связи между этим белком и ТАТА-боксом ДНК.
Затем кликните на самую правую кнопку для по-
лучения изображения (Maximum detail: Target
Анализ экспериментальных данных 1285 ]
& Contacts Balls and Sticks, Colored by Element).
Теперь вы сможете приближать и поворачивать
структуру. Ответьте на следующие вопросы.
а) Какие пары оснований в ДНК образуют
водородные связи с белком? Какие из них вносят
вклад в специфическое узнавание ТАТА-бокса
этим белком? (Длина водородной связи между
донором и акцептором водорода находится в диа-
пазоне 2,5-3,3 А).
б) Остатки каких аминокислот в белке взаи-
модействуют с этими парами оснований?
в) Какая последовательность ДНК представ-
лена в этой модели и какие части этой последова-
тельности узнаются ТАТА-связывающим белком?
г) Укажите зоны гидрофобных взаимодей-
ствий в этом комплексе. Они единичные или
многочисленные? Чтобы ответить на этот во-
прос, кликните на “Return to contacts” и восполь-
зуйтесь опцией “Show hydrophobic (apolar van der
Waals) interactions”.
Анализ экспериментальных данных___________
14. Конструирование генетического переключа-
теля в клетках Escherichia coli. Регуляцию экс-
прессии генов часто описывают в терминах «вклю-
чен/выключен», подразумевая при этом, что ген
либо экспрессируется, либо не экспрессируется.
На самом деле в подавлении и активации экс-
прессии играют роль лиганды, поэтому при про-
межуточном содержании регуляторных молекул
может наблюдаться промежуточный уровень экс-
прессии. Например, рассмотрим уравнение свя-
зывания Lac-penpeccopa, оператора и индуктора
для /яс-оперона Е. coli (см. рис. 28-7). Это сложный
кооперативный процесс, однако в определенном
приближении его можно описать с помощью сле-
дующего уравнения (R — репрессор, ИПТГ — ин-
дуктор, — изопропил-|3-В-тиогалактозид):
К,=ю <м
R + ИПТГ R•ИПТГ
Свободный репрессор R связывается с операто-
ром и предотвращает транскрипцию /яс-оперона;
комплекс R-ИПТГ не связывается с оператором
и, следовательно, транскрипция /яс-оперона про-
исходит.
а) С помощью уравнения (5-8) можно
определить относительный уровень экспрессии
белков /яс-оперона как функцию концентра-
ции индуктора [ИПТГ]. Используйте эти рас-
четы для определения диапазона концентрации
ИПТГ, когда уровень экспрессии изменяется от
10% до 90%.
б) На качественном уровне оцените содержа-
ние белков /яс-оперона в клетках Е. coli до, во вре-
мя и после действия ИПТГ. Не нужно указывать
точное количество белка в точно определенные
моменты времени, просто укажите тенденцию.
Гарднер, Кантор и Коллинс (2000) попыта-
лись создать «генетический тумблер» — систе-
му регуляции генов с двумя ключевыми поло-
жениями, как у электрического выключателя.
(А) В таком выключателе есть только два по-
ложения: включен — выключен; он не может под-
держивать какую-то пониженную освещенность.
В биохимических терминах это означает, что
ген-мишень или группа генов (оперон) либо экс-
прессируется, либо не экспрессируется совсем,
а промежуточный уровень экспрессии невозмо-
жен. (Б) Оба состояния устойчивы: вы можете
изменять положение выключателя пальцем, но
когда вы отпускаете палец, выключатель остается
в заданном положении. В биохимических терми-
нах это означает, что под воздействием индуктора
или другого сигнала экспрессия гена или оперона
включается или выключается и остается в этом
состоянии и после прекращения сигнала.
в) Объясните, почему /яс-оперон не обладает
характеристиками А и Б.
Для создания «генетического тумблера» Гар-
днер с сотрудниками сконструировали плазмиду,
состоящую из следующих элементов:
ОР|ас область оператора и промотора 1ас-
оперона Е. coli
ОРХ область оператора и промотора фага к
lad ген /лс-репрессора, Lad. В отсутствие
ИПТГ этот белок полностью подавляет
ОР)ас; в присутствии ИПТГ он позволяет
осуществлять нормальную экспрессию с
ОР,ас.
rep[s ген мутантного термочувствительного
белка-репрессора фага к, reptts. При 37 °C
этот белок полностью подавляет ОРХ;
при 42 °C он позволяет осуществлять
нормальную экспрессию с ОРХ.
GFP ген зелеЯЪго флуоресцентного белка
(GFP) — интенсивно флуоресцирующего
маркерного белка (см. рис. 9-15 в т. 1)
Т терминатор транскрипции
[286] Часть III. 28. Регуляция экспрессии генов
Эти элементы (см. рисунок) были размеще-
ны таким образом, что два промотора взаимно
подавляли друг друга: ОР1ас контролировал экс-
прессию гер[\ а ОРХ контролировал экспрессию
lad. Состояние системы оценивали по уровню
экспрессии GFP, который также находился под
контролем ОР|ас.
г) Сконструированная система могла нахо-
диться в двух состояниях: «GFP включен» (вы-
сокий уровень экспрессии) и «GFP выключен»
(низкий уровень экспрессии). Опишите, какие
белки присутствуют и какие промоторы экспрес-
сируются в каждом из двух состояний.
д) Можно было ожидать, что обработка
IPTG переведет систему из одного состояния в
другое. Из какого в какое? Объясните свои рас-
суждения.
е) Можно было ожидать, что повышение
температуры до 42 °C также вызовет переход си-
стемы из одного состояния в другое. Из какого в
какое? Объясните свои рассуждения.
ж) Почему сконструированная плазмида
должна была иметь описанные выше характери-
стики А и Б?
Для подтверждения того, что конструкция
действительно обладала указанными характери-
стиками, Гарднер с соавторами сначала показали,
что после включения выключения экспрессия
GFP (высокая низкая) остается на том же уров-
не на протяжении длительного времени (свой-
ство Б). Затем они измерили уровень GFP при
разных концентрациях ИПТГ и получили следу-
ющие результаты.
Оказалось, что средний уровень экспрессии
GFP имел некое промежуточное значение при
концентрации ИПТГ, равной X. Однако при из-
мерении уровня экспрессии GFP в индивидуаль-
ных клетках при |ИПТГ| = X выяснилось, что
уровень GFP был либо низким, либо высоким, но
ни в одной клетке не было промежуточного уров-
ня экспрессии этого белка.
з) Объясните, как это наблюдение подтверж-
дает тот факт, что система обладает свойством А?
Почему при |ИПТГ] = X в одних клетках наблю-
дается высокий, а в других низкий уровень экс-
прессии маркерного белка?
Литература
Gardner, Т. S., Cantor, С. R., & Collins, J. J. (2000)
Construction of a genetic toggle switch in Escherichia coli.
Nature 403,339-342.
Приложение А
Принятые в биохимии сокращения и аббревиатуры
А — ангстрем
А — аденин, аденозн или аденилат
А — абсорбция
Ab (АТ) — антитело
АВС — ATP-связывающая кассета
АСАТ — ацил-СоА-холестерин-ацилтрансфераза
АСС — ацетил-СоА-карбоксилаза
ACh (АХ) — ацетилхолин
АСР — ацилпереносящий белок
ACT (АКТГ) — адренокортикотропный гормон
acyl-CoA — ацил-производные кофермента А (или
ацил-S-CoA)
ADH — алкогольдегидрогеназа
adoHcy — S-аденозилгомоцистеин
adoMet — S-аденозилметионин (SAM)
AFM (ACM) — атомно-силовая микроскопия
Ag (АГ) — антиген
AGE — конечный продукт гликирования
AIDS (СПИД) — синдром приобретенного иммуноде-
фицита
АКАР — белок, заякоривающий киназу А
Ala — аланин
[а][)25 С -удельное (оптическое) вращение
AMP, ADP, АТР — аденозин-5'-моно-, -ди-, -трифос-
фат
АМРК — AMP-активируемая протеинкиназа
ANF — атриальный натрийуретический фактор
AQP — аквапорин
Arg — аргинин
ARS — автономно реплицирующаяся последователь-
ность
Asn — аспарагин
Asp — аспарагиновая кислота (аспартат)
ATCase — аспартаттранскарбамоилаза
atm (атм) — атмосфера
ATPase (АТРаза) — аденозинтрифосфатаза
В12 — кофермент В12, кобаламин
ВАС — искуственная бактериальная хромосома
ВАТ — бурая жировая ткань
BMR — базовый уровень метаболизма
Ьр (п. н.) — пары нуклеотидов
1,3-BPG (1,3-БФГ) — 1,3-бисфосфоглицерат
С — цитозин, цитидин или цитидилат
САМ — метаболизм органических кислот по типу
толстянковых
СаМ — кальмодулин
сАМР — циклический 3',5'-АМР
CD (КД) — круговой дихроизм
CDK — циклинзависимая протеинкиназа
cDNA (кДНК) — комплементарная ДНК
CFTR -белок-регулятор мембранной проводимости
хлорных каналов
cGMP — циклический 3',5'-GMP
Chi (Хлф) — хлорофилл
ChREBP — белок, связывающий углевод-чувстви-
тельный элемент (фактор транскрипции)
СКП — казеинкиназа II
CL — кардиолипин
CMP, CDP, СТР — цитидин-5'-моно-, -ди-, -трифосфат
СоА — кофермент A (CoASH)
СОНЬ — карбоксигемоглобин
CoQ — кофермент Q (убихинон, UQ)
СОХ — циклооксигеназа; цитохромоксидаза
CREB — белок, связывающий сАМР-чувствительный
элемент (фактор транскрипции)
CRP — сАМР-рецепторный белок
Cys — цистеин
Cyt — цитохром
D — дигидроуридин
D — коэффициент диффузии
d (р) — плотность
dADP, dGDP и т. д. — дезоксиаденозин-5'-дифосфат,
дезоксигуанозин-5'-дифосфат и т. д.
dAMP, dGMP и т. д. — дезоксиаденозин-5'-моно-
фосфат, дезоксигуанозин-5'-монофосфат и т. д.
dATP, dGTP и т. д. — дезоксиадснозин-5'-трифосфат,
дезоксигуанозин-5'-трифосфат и т. д.
DEAE — диэтиламиноэтил
DFP — диизопропилфлуорофосфат (DIFP)
DHAP — дигидроксиаценонфосфат
DHF — дигидрофолат (Н2-фолат)
DMS — диметилсульфат
DNA — дезоксирибонуклеиновая кислота
Dnase (ДНКаза) — дезоксирибонуклеаза
DNP (ДНФ) — 2,4-динитрофенол
Doi — долихол
dopa (дофа) — дигидроксифенилаланин
Е — электрическй потенциал
Е.С. — Комиссия по ферментама (после номера фер-
мента по его формальной классификации)
ЕСМ — внеклеточный матрикс
EDTA (ЭДТА) — этилендиаминтетраацетат
EF — фактор элонгации
EGF — эпидермальный фактор роста
ELISA (ИФА) — иммунофлуоресцентный анализ
ЕМ (ЭМ) — электронная микроскопия
emf (эдс) — электродвижущая сила
ЕРО — эритропоэтин
е — молярный коэффициент абсорбции
ER (ЭР) — эндоплазматический ретикулум
[288] Приложение A
EST — метка экспрессируемой последовательности
Г| — вязкость
Е(Ф) - фарад
f — коэффициент трения
FA (ЖК) — жирная кислота
G6PD — глюкозо-6-фосфатдегидрогеназа
FAD (ФАД), FADH‘2 (ФАДН2) — флавинадениндину-
клеотид и его восстановленная форма
FAS — синтеза жирных кислот
FBPase-1 — фру ктозо-1,6-бисфосфатаза
FBPasc-2 — фруктозо-2,6-бисфосфатаза
Fd (Фд) — ферредоксин
FDNB (DNFB) — 1-фтор-2,4-динитробензол
FOF1 — митохондриальная АТР-синтаза
FFA (СЖК) — свободная (неэтерифицированная)
жирная кислота
FH (СГ) — семейная гиперхолестеринемия
fMet — N-формилметионин
FMN, FMNH2 (ФМН, ФМН Н2) — флавинмонону-
клеотид и его восстановленная форма
FOXO1 — фактор транскрипции; от англ. forkhead box
01
FP — флавопротеин
F1P — фруктозо-1-фосфат
F6P — фруктозо-6-фосфат
FRAP — восстановление флуоресценции после фото-
выцветания
FRET — резонансный перенос энергии флуоресцен-
ции
Fru — D-фруктоза
AG — изменение свободной энергии
AG° — изменение стандартной свободной энергии
AG'° — изменение приведенной стандартной сво-
бодной энергии в биоимических стандартных
состояниях
AG* — энергия активации
AGB — энергия связывания
AGp — изменение свободной энергии гидролиза АТР
(потенциал фосфорилирования) во внутрикле-
точных условиях
G — гуанин, гуанозин или гуанилат
g — ускорение свободного падения
GABA (ГАМК) — у-аминомасляная кислота
Gal — D-галактоза
GalN — D-галактозам ин
GalNAc — М-ацетил-И-галактозамин
GAP — глицеральдегид-3-фосфат; белок-активатор
СТРазы
GDH — глутаматдегидрогеназа
GEF — фактор обмена гуаниновых нуклеотидов
GFP — зеленый флуоресцентный белок
GH (ГР) — гормон роста
GHB — гликированный гемоглобиин
GLC (ГЖХ) — газожидкостная хроматография
Glc — D-глюкоза
GlcA — D-глюконовая кислота
GlcN — D-глюкозамин
GlcNAc — М-ацетил-И-глюкозамин (NAG)
GlcUA — D-глюкуроновая кислота
Gin — глутамин
Glu — глутаминовая кислота, глутамат
GLUT — транспортер глюкозы
Gly — глицин
GMP, GDP, GTP — гуанозин-5'-моно, -ди-, трифосфат
GPCR — рецептор, сопряженный с G-белком
GPI — гликозилированное производное фосфатиди-
линозита
G1P — глюкозо-1-фосфат
G6P — глюкозо-6-фосфат
GRK — киназа, сопряженная с G-белком
GSH, GSSG — глутатион и его окисленная форма
GSK3 — киназа 3-гликогенсинтазы
АН — изменение энтальпии
НА — белок гемагглютинин
НЬ, НЬО2 — гемоглобин, оксигемоглобин, окислен-
ный гемоглобин
HDL (ЛПВП) — липопротеины высокой плотности
Н2 folate (Н2-фолат) — дигидрофолат (DHF)
Н4 folate (Н4-фолат) — тетрагидрофолат (THF)
HIF — индуцируемый гипоксией транскрипционный
фактор
His — гистидин
HIV (ВИЧ) — вирус иммунодефицита человека
HMG-CoA — р-гидрокси-р-метилглутарил-СоА
hnRNA (гяРНК) — гетерогенная ядерная РНК
HPLC (ВЭЖХ) — высокоэффективная жидкостная
хроматография
HRE — элемент гормонального ответа (гормон-чув-
ствительный элемент)
Hsp — белок теплового шока
НТН — спираль-поворот-спираль
Нур — гидроксипролин
I — инозин
IF — фактор инициации
Ig (ИГ) — иммуноглобулин (антитело)
IgG — иммуноглобулин G
Не — изолейцин
IMP, IDP, ITP — инозин-5'-моно-, -ди-, -трифосфат
1Р3 — инозит-1,4,5-трифосфат
IR (ИК) — инфракрасный
IRP — железо-регуляторный белок
IRS-1, IRS-2 — субстрат-1 и субстрат-2 инсулинового
рецептора
IS (IS-элемент)- инсерционная последовательность
J (Дж) - джоуль
JAK — Янус-киназа
Ка — константа диссоциации кислоты
Кд — константа диссоциации
Принятые в биохимии сокращения и аббревиатуры [2S9]
Кп — константа диффузии
Kcq — константа равновесия
7Ceq — константа равновесия в стандартных условиях
К} — константа ингибирования
Км — константа Михаэлиса
k — константа скорости
&cat — число оборотов
kb — тысячи [пар] нуклеотидов, килобазы
kbp (т. п. н.) — тысячи пар нуклеотидов
a-KG — а-кетоглутарат
lac — лактоза
X — длина волны
LDH (ЛДГ) — лактатдегидрогеназа
LDL (ЛПНП) — липопротеин низкой плотности
Leu — лейцин
LH (ЛГ) — лютеинизирующий гормон
LHC — светопоглощающий комплекс
Lk — порядок зацепления
In — логарифм по основанию е
1g — логарифм по основанию 10
LT — лейкотриен
LTR — длинный концевой повтор
Lys — лизин
Мг — относительная молекулярная масса
Man — D-манноза
МАРК — митоген-активируемые протеинкиназы
Mb, MbO2 — миоглобин, оксимиоглобин, окисленный
миоглобин
MCAD — ацил-СоА-дегидрогеназа средних цепей
МСМ — белки поддержания минтхромосом
mDNA (мДНК) — митохондриальная ДНК
Met — метионин
MFP — мультифункциональный белок (в окислении
жирных кислот)
miRNA — микроРНК
MODY — диабет взрослого типа у молодых
mRNA (мРНК) — матричная РНК
MS (МС) — масс-спектроскопия
MSH (МСГ) — меланоцит-стимулирующий гормон
р — электрофоретическая подвижность
Mur — мурамовая кислота
Mur2Ac — N-ацетилмурамовая кислота (NAM)
NAD+, NADH — никотинамидадениндинуклеотид и
его восстановленная форма
NADP+, NADPH — никотинамидадениндинуклеотид-
фосфат и его восстановленная форлй
NAG — N-ацетилглюкозамин (GlcNAc)
NAM — N-ацетилмурамовая кислота (Mur2Ac)
ncRNA (нкРНК) — некодирующая РНК
Neu5Ac — N-ацетилнейраминовая кислота (сиаловая
кислота)
NMN+, NMNH — никотинамидмононуклеотид и его
восстановленная форма
NMP, NDP, NTP — нуклеозидмоно-, -ди-, -трифосфат
NMR (ЯМР) — ядерный магнитный резонанс
NO — оксид азота
ОАА — оксалоацетат
ORC — комплекс распознавания точки инициации
репликации
ORF — открытая рамка считывания
Р — давление
Pi — неорганический фосфат
РАВ или РАВА (ПАВК) — и-аминобензоат (ияря-ами-
нобензойная кислота)
PAGE — электрофорез в полиакриламидном геле
РС — пластоцианин; фосфатидилхолин
PCR (ПЦР) — полимеразная цепная реакция
PDB — Protein Data Bank (база данных белков)
PDE — фосфодиэстераза циклических нуклеотидов
PDH (ПД) — пируватдегидрогеназа
РЕ — фосфатидилэтаноламин
РЕР (ФЕП) — фосфоенолпируват
РЕРСК — фосфоенолпируваткарбоксикиназа
РЕТ (ПЭТ) — позитронная эмисионная томография
PFK (ФФК) — фосфофруктокиназа
PG (ПГ) — простагландин
2PGA (2ФГ) — 2-фосфоглицерат
3PGA (ЗФГ) — 3-фосфоглицерат
pH - lg(l/[H+])
Phe — фенилаланин
PI — фосфатидилинозит
pl — изоэлектрическая точка
Р1Р2 — фосфатидилинозит-4,5-бисфосфат
РК (ПК) — протеинкиназа; пируваткиназа
pK-lg(l/K)
РКА — сАМР-зависимая протеинкиназа (протеинки-
наза А)
РКВ — протеинкиназа В (Akt)
РКС — Са2+/кальмодулин-зависимая протеинкиназа
(протеинкиназа С)
PKG — cGMP-зависимая протеинкиназа (протеинки-
наза G)
PKU (ФКУ)- фенилкетонурия
PLC — фосфолипаза С
PLP — пиридоксаль-5-фосфат
рО2 — парциальное давление кислорода
Pol — полимераза (ДНК или РНК)
PPi — неорганический пирофосфат
РР1 (ФПФаза 1) — фосфопротеинфосфатаза 1
PPAR — рецептор, активируемый пролифератором
пероксисомы
PQ (ПХ) — пластохинон
pRb — белок ретинобластомы
Pro — пролин
PRPP — 5-фосфорибозил-1-пирофосфат
PS (ФС) — фосфатидилсерин; фотосистема
АФ — трансмембранный электрический потенциал
РТВ — фосфотирозиновое связывание
1290] Приложение A
PUFA (ПНЖК) — полиненасыщенная жирная
кислота
Q — отношение действующих масс
Q, QH2 — убихинон, восстановленный убихинон
Rb — ген ретинобластомы (ген опухолевого
супрессора)
RC — фотореакционный центр
RER (ШЭР) — шероховатый эндоплазматический
ретикулум
RF — релизинг-фактор; репликативная форма
RFLP — полиморфизмы длины фрагментов
рестрикции
RGS — регулятор сигнала G-белка
RIA (РИА) — радиоиммунный анализ
Rib — D-рибоза
RLK — рецептороподобная протеинкиназа
RNA (РНК) — рибонуклеиновая кислота
RNAi (РНКи) — интерференция РНК
RNase (РНКаза) — рибонуклеаза
ROS — активные формы кислорода (АФК); наружный
сегмент палочки (сетчатка)
rRNA (рРНК) — рибосомная РНК
RSV — вирус саркомы Рауса
RTK — рецептор тирозинкиназы (рецептор Туг-
киназы)
А5 — изменение энтропии
SAM — S-аденозилметионин (adoMet)
SDS — додецил сульфат натрия
SELEX — систематическая эволюция лигандов путем
экспоненциального обогащения
SER (ГЭР) — гладкий эндоплазматический ретику-
лум
Ser — серин
SERCA — кальциевая АТРаза саркоплазматического
и эндоплазматического ретикулумов
SMC — белки, поддерживающие структуру хромосом
SNARE — растворимый рецептор связывания
N-этилмалеимид-чувствительного фактора
snoRNA (мякРНК) — малая ядрышковая РНК
STP — однонуклеотидный полиморфизм
snRNA (мяРНК) — малая ядерная РНК
snRNP (мяРНП) — малый ядерный рибонуклеопро-
теин
SREBP — белки, взаимодействующие со стерол-регу-
лируемым элементом
sRNA (маРНК) - малая РНК
SRP — сигнал-распознающая частица
STAT — преобразователь сигнала и активатор транс-
крипции
STP — стандартные температура и давление
STR — короткий тандемный повтор
stRNA (мвРНК) — малая временная РНК
STS — последовательно-меченные сайты
SUR — рецептор сульфонилмочевины
Т — тимин, тимидин или тимидилат
Т — абсолютная температура
T2DM (ИНЗСД) — сахарный диабет 2-го типа (инсу-
линнезависимый)
ТВР — ТАТА-связывающий белок
ТСА (ТКК) — трикарбоновая кислота
TFP — трифункциональный белок
THF (ТГФ) — тетрагидрофолат (Н4-фолат)
Thr — треонин
TLC (ТСХ) — тонкослойная хроматография
ТМР, TDP, ТТР — тимидин-5'-моно, -ди-, -трифосфат
TMV (ВТМ) — вирус табачной мозаики
TPI — триозофосфатизомераза
ТРР — тиаминпирофосфат
tRNA (тРНК) — транспортная РНК
Тгр — триптофан
ТХ — тромбоксан
Туг — тирозин
U — урацил, уридин или уридилат
UAS — активирующая последовательность
UCP — разобщающий белок
UDP-Gal — уридиндифосфатгалактоза (UDP-
галактоза)
UDP-Glc — уридиндифосфатглюкоза (UDP-
глюкоза)
UMP, UDP, UTP — уридин-5'-моно-, -ди-, -трифосфат
UQ — убихинон (кофермент Q; CoQ)
UV (УФ) — ультрафиолет
Vin — трансмембранный электрический потенциал
(мембранный потенциал)
Umax — максимальная скорость
Vq — начальная скорость
Vai — валин
VDR — рецептор витамина D
VEGF — фактор роста эндотелия сосудов
VLDL (ЛПОНП) — липопротеин очень низкой
плотности
WAT — белая жировая ткань
YAC — искуственная дрожжевая хромосома
Z — суммарный заряд
Приложение Б
Краткие решения задач и ответы на вопросы
Решения алгебраических задач представлены в чис-
ленном виде, причем число значащих цифр ответа обо-
сновано
д
Карбоксил
Глава 1
1. а) Диаметр увеличенной клетки составляет 500 мм;
б) 2,74012 молекул актина; в) 36 000 митохондрий;
г) 3,94010 молекул глюкозы; д) 50 молекул глюкозы
на молекулу гексокиназы.
2. а) 140 12 г = 1 пг; б) 10%; в) 5%.
3. а) 1,6 мм; в 800 раз длиннее клетки: ДНК очень
плотно упакована; б) 4000 белков.
4. а) Скорость метаболизма ограничена диффузией,
которая в свою очередь определяется площадью по-
верхности; б) 12 мкм 1 для бактерии, 0,04 мкм 1 для
амебы; у бактерии отношение площади поверхности
к объему в 300 раз выше.
5. 2406 с (около 23 суток).
6. Молекулы витамина из двух источников идентич-
ны, и организм не может отличить природную мо-
лекулу от синтетической. От источника фермента
зависит только наличие в нем тех или иных при-
Метил
сн2
сн2
NH
| Амидогруппа
с=о
Н—С—ОН Гидроксил
Н3С'—С- СН3 Метил
СН20Н Гидроксил
\ Альдегид
С
I
Н—С—NH3 Аминогруппа
Гидроксил НО—С—Н
Н—С—ОН
Н—С- ОН
Гидроксилы
СН2ОН
месей.
7. а НН
+ 1 1
H3N—С—С—ОН 1 1
1 1 н н
Амино- Гидроксил
группа
б Н
8. Два энантиомера по-разному взаимодействуют с
хиральпым биологическим «рецептором» (белком)
н—С- он
н—с- он
Гидроксилы
н—с- он
о
II .
НО—Р—О Фосфорил
I
н\ ?
/С=С—соо
Н Карбоксил
г СОО Карбоксил
b I
Амино- HoN*_C—Н
3 I v
группа тт '
Н—С—ОН Гидроксил
СН8
Метил
9. а) Только в аминокислотах есть аминогруппы, по-
этому разделение может быть основано на заряде
или реакционной способности этих групп. Жирные
кислоты хуже растворяются в воде, чем аминокис-
лоты. Кроме того, эти два типа молекул различа-
ются по размеру и по форме. Разделение может
быть основано на различиях этих параметров,
б) Молекула глюкозы по размеру меньше молеку-
лы нуклеотида, так что разделение может быть ос-
новано на разнице размеров молекул. Кроме того,
азотистое основание и фосфатная группа обеспечи-
вают нуклеотидам такие свойства (растворимость,
заряд), которые можно использовать для их отде-
ления от глюкозы.
10. Вряд ли кремний мог бы стать основой жизни, осо-
бенно в кислородной атмосфере, окружающей нашу
Землю. Длинные цепи, состоящие из атомов крем-
ния, синтезировать достаточно сложно, так что вряд
[292] Приложение Б
ли удалось бы получить полимерные макромолеку-
лы, необходимые для реализации сложных функций.
Кислород разрушает связи между атомами кремния,
а образующиеся связи между кремнием и кислоро-
дом чрезвычайно прочные, что затрудняет разрыв и
образование химических связей, что все время про-
исходит при процессах жизнедеятельности.
И. Физиологической активностью обладает лишь
один энантиомер. Препарат декседрин состоит
именно из этого энантиомера, а бензедрин пред-
ставляет собой рацемическую смесь.
12. а) Три фосфорильные группы, a-D-рибоза, гуанин;
б) холин, фосфорная кислота, глицерин, олеиновая
кислота, пальмитиновая кислота; в) тирозин, два
глицина, фенилаланин, метионин.
13. а) СН2О, С3Н6О3.
б)
он zH
н-с-с;
j’j с—он
нох
V /он
н-с-с;
j’j с-он
нох
ун он
н-с—с;
j’j с- он
н
продуктов реакции и более сильного связывания
и, следовательно, более сладкого вкуса и большего
значения относительной сладости, б) Определение
сладости на животных занимает слишком много
времени; компьютерные программы для предска-
зания сладости веществ, возможно, не всегда отли-
чаются высокой точностью, но позволяют химикам
гораздо быстрее сконструировать эффективные
подсластители. Затем такие молекулы-кандида-
ты должны быть опробованы на животных или на
людях, в) В отрезке длиной от 0,25 до 0,4 нм укла-
дывается от 1,5 до 2,5 длин «типичных» одинарных
связей. Используем изображенный ниже шаблон в
качестве линейки: любой атом, попадающий в ро-
зовый прямоугольник, находится на расстоянии от
0,25 до 0,4 нм от начала линейки.
В данных молекулах содержится множество возмож-
ных групп АН-В; ниже выделены некоторые из них.
н он
н он 'н
Дезоксисахароза
он он н н 1 он 1
но—с—с. но—с—с; н-с- -с— с\
н /С-н н pj \-он н н 1** н он
4 5 6
НО Н п 1 1 Z н—С—С—С НО он п । । Z н—с—с—с н 1 н -с- ?н Z -с—с
н н Хон 1 1 \т НН н 1 н он н
7 8 9
НО н п 1 1 н—с—с—с 1 1 \ НО О он 1 II 1 н—с—с—с- н 1 1 н 1 н-с- 1 О II -С- он 1 -с- н 1
но н н 1 1 н н 1 н он
в) X содержит хиральный центр, так что можно исклю-
чить все структуры, кроме 6 и 8; i) X содержит кислую
функциональную группу, и это позволяет исключить
структуру 8; структура 6 соответствует всем условиям;
Д) структура 6; различить два возможных энантиомера
мы не можем
14. а) Большее отрицательное значение AG° соответ-
ствует большей величине Keq для реакции связы-
вания, так что равновесие сдвигается в сторону
Алитам
Краткие решения задач и ответы на вопросы [293]
Неотам
Тетрабромсахароза
г) Во-первых, в каждой из приведенных молекул
есть несколько групп АН-В, так что трудно опреде-
лить, какие из них играют определяющую роль. Во-
вторых, группа АН-В очень простая и часто встре-
чается во многих несладких молекулах, д) Сахароза
и дезоксисахароза. Дезоксисахароза не имеет одной
из групп АН-В, имеющихся в молекуле сахарозы,
и обладает чуть более низким значением отно-
сительной сладости, как и следует ожидать, если
эти группы АН-В определяют сладость вещества,
е) Таких примеров много, вот некоторые из них:
(1) D-триптофан и 6-хлор-В-триптофан имеют одну
и ту же группу АН-В, но значения относительной
сладости для этих молекул очень сильно различа-
ются. (2) Аспартам и неотам также имеют одинако-
вые группы АН-В, но сильно различаются по отно-
сительной сладости. (3) Неотам имеет две группы
АН-В, а алитам - три, но неотам более чем в 5 раз
слаще. (4) Бром обладает менее выраженной элек-
троотрицательностью, чем кислород, и, следователь-
но, должен бы ослаблять активность групп АН-В,
однако тетрабромсахароза гораздо слаще сахарозы,
ж) При достаточной «подгонке» параметров моде-
ли к выборке данных любая модель сможет более
или менее адекватно описывать эту выборку. По-
скольку исследователи хотели создать модель, спо-
собную предсказать АС° для молекул, которые не
были протестированы in vivo, они должны были по-
казать, что модель хорошо описывает молекулы, на
которых ее не «тренировали». Степень отклонения
результатов моделирования от истинных значений
для тестируемых молекул показывает, насколько
хороша предложенная модель, з) Относительная
сладость связана с Keq, которая в свою очередь экс-
поненциальным образом зависит от AG0. Поэтому
увеличение значения АС° на постоянную соответ-
ствует умножению относительной сладости на по-
стоянную. На основании представленных значений
можно сказать, что изменение AG° на 1,3 ккал/моль
соответствует 10-кратному изменению относитель-
ной сладости.
Глава 2
1. Этиловый спирт полярная молекула, а молекула
этан нет. Гидроксильная группа этилового спирта
может участвовать в образовании водородных свя-
зей с молекулами воды.
2. а) 4,76; б) 9,19; в) 4,0; г) 4,82.
3. а) 1,51 • 10 4 М; б) 3,01 • 10 7 М: в) 7,76 • 10 12 М.
4. 1,1.
5. а) НС1 Н/ б) 3,3; в) NaOH Na+ + ОН ; г) 9,8
6. 1,1.
7. 1,7 • 10 9 моль ацетилхолина.
8. 0,1 МНС1.
9. 3,3 мл.
10. a) RCOO ; б) RNH2; в) Н2РО4; г) НСО3.
11. а) 5,06; б) 4,28; в) 5,46: г) 4,76; д) 3,76.
12. а) 0,1 М НС1; б) 0,1 М NaOH; в) 0,1 М NaOH.
13. Вариант (г). Бикарбонат слабое основание и ти-
трует -ОН-группы, превращая их в О , в резуль-
тате чего соединение становится более полярным и
лучше растворяется в воде.
14. Из желудка. Нейтральная форма аспирина, при-
сутствующая при более низких значениях pH, яв-
ляется менее полярной и легче проникает через
мембраны.
15.9 .
16. 7,4.
17. а) при pH 8,6 10.6; б) 4 5: в) 10 мл: г) pH = рКа - 2.
18. 1,4.
19. NaH2PO4 H2O - 5,8 г/л. Na2HPO4 8.2 г/л.
20. [А ]/|НА] = 0,10.
21. Смешать 150 мл 0,10 М раствора ацетата натрия и
850 мл 0,10 М уксусной кислоты.
22. Раствор уксусной кислоты; рК, уксусной кислоты
ближе всего к заданному pH.
23. а) 4,6; б) 0,1 единицы pH; в) 4 единицы pH
24. 4,3.
[294] Приложение Б
25. 0,13 М ацетат и 0,07 М уксусная кислота.
26. 1,7.
27. 7.
28’а) соон
+ I
H3N—С- Н
СН3
Полностью
протонированная
форма
ООО
I
h2n—с- н
сн3
Полностью
депротонированная
форма
б) Полностью протонирован; в) цвиттерион;
г) цвиттер-ион; д) полностью депротонирован
29. а) pH крови контролируется буферной системой
диоксид углерод/бикарбонат: СО2 + Н2О Н4 +
НСО3. При гиповентиляции легких [СО2] артери-
альной крови повышается, что приводит к повыше-
нию | И4| и снижению pH. б) При гипервентиляции
легких [СО2] артериальной крови понижается, что
приводит к снижению [Н4| и повышению pH 2* 7,4
(выше нормы), в) Лактат — кислота средней силы;
при физиологических условиях он полностью дис
социирует. снижая pH крови и жидкостей мышеч-
ной ткани. Гипервентиляция удаляет Н4, что со-
провождается повышением pH крови и тканей, т. е.
противодействует эффекту лактата.
30. 7,4.
31. Растворение дополнительного количества СО2
приводит к повышению содержания Н в крови и
внеклеточных жидкостях, что снижает pH:
CO2(d) + Н2О Н2СО3 Н+ + НСО3
32. а) Сначала использовать вещество в форме ПАВ,
что позволит перевести разлитое масло в эмуль-
сию, собрать эмульсию, а затем переключить ве-
щество в форму «пе-ПАВ». б) Равновесие реакции
сильно сдвинуто вправо. Более сильная кислота
Н2СО3 (более низкое значение рКа) отлает протон
сопряженному основанию более слабой кислоты —
амидина (более высокое значение рКа). в) Актив-
ность ПАВ зависит от гидрофильности функцио-
нальной группы в головке: чем сильнее выражены
гидрофильные свойства головки, тем более актив-
ные свойства ПАВ. Амидиний значительно более
гидрофильное соединение, чем амидин, поэтому
он является более мощным ПАВ. г) Точка А\ ами-
диний; СО2 имел достаточно времени для взаимо-
действия с амидином, чтобы перевести его в форму
амидиния. Точка В: амидин; Аг удалил СО2 из рас-
твора, оставив ПАВ в форме амидина, д) Проводи-
мость возрастает по мере того, как незаряженный
амидин взаимодействует с СО2 и превращается в
заряженный амидиний. е) Проводимость падает по
мере того, как Аг удаляет из системы СО2, сдвигая
равновесие в сторону образования незаряженного
амидина ж) Следовало бы обработать переключае-
мый ПАВ СО2 для его перевода в форму амидиния
и использовать для эмульгирования масла. Затем
обработать эмульсию Аг, чтобы удалить СО2 и вер-
нуться к неактивному амидину. Масло отделится
от воды, и его можно будет собрать.
Глава 3
1. L; следует определить абсолютную конфигурацию
заместителей у сх-атома углерода и сравнить с моле-
кулами D- и L-глицеральдегида
2. а) I; б) II; в) IV; г) II; д) IV; е) II и IV; ж) III; з) III;
и) V; к) III; л) V; м) II; н) III; о) V; п) 1, III и V.
3. а) р/ > рКа для сх-карбоксильной группы и р/ < рКа
для а-аминогруппы, так что обе группы заряже-
ны (ионизованы), б) Одна на 2,19 • 107; р/ аланина
составляет 6.01. Из таблицы 3-1 и уравнения Хен-
дерсона Хассельбаха следует, что незаряженными
остается одна карбоксильная группа из 4680 и одна
аминогруппа из 4680. Доля молекул аланина, в кото-
рых не заряжены обе группы, равна обратной вели-
чине произведения этих двух чисел.
4. а)-в)
СООН
рК\ = 1,82
—V
Н4
2
pKR = 6,0
pH Структура Суммарный заряд Движется к
1 1 +2 катоду
4 2 +1 катоду
8 3 0 не движется
12 4 - 1 аноду
Краткие решения задач и ответы на вопросы [295]
5. a) Asp; б) Met; в) Glu; г) Gly; д) Ser.
6. а) 2: б) 4: в)
СОО соо
4 i 4 i
H3N—С-Н 3 1 H3N— С— Н 3 ।
н- с—сн3 f н3с—с— н
сн2 сн2
СН3 СН3
соо СОО
Н-С- NH3 Н- С- NH3 । 3
н-с-сн3 Н3С— С— н
сн2 сн2
сн3 сн3
7. а) Структура при pH 7:
О 4 II H3N— СН— С О II -N—СН—С । । О II —N— СН—с—О
СН3 рХ2 = 8,03 н СНо 1 Н СН3 рКг = 3,39
б) Электростатическое взаимодействие между
карбоксилатным анионом и протонированной
аминогруппой цвиттер-иона аланина способству-
ет ионизации карбоксильной группы Это элек-
тростатическое взаимодействие ослабевает по
мере удлинения последовательности поли(А1а),
что выражается в увеличении рХь в) Иониза-
ция протонированной аминогруппы разрушает
электростатические взаимодействия, описанные
в пункте (б). При увеличении расстояния между
заряженными группами облегчается удаление
протона от аминогруппы в молекуле поли(А1а),
и рК2 уменьшается. Внутримолекулярное влия-
ние амидных (пептидных) связей проявляется в
уменьшении рКа по сравнению с рКа алкилзаме-
щенных аминов.
8. 75 000.
9. а) 32 000. При образовании пептидной связи выде-
ляется молекула воды, так что молекулярные мас-
сы остатка триптофана и свободного триптофана
не равны; б) 2.
10. Белок состоит из четырех субъединиц с молеку-
лярными массами 160, 90, 90 и 60 кДа Субъеди-
ницы с молекулярной массой 90 кДа (возможно,
они идентичные) связаны одной или несколькими
дисульфидными связями.
11. а) При pH 3 — +2, при pH 8 — 0, при pH И-1;
б) р/=7,8.
12. р7 ~ 1; карбоксилатные группы; Asp и Glu.
13. Lys, His и Arg; отрицательно заряженные фосфат-
ные группы молекул ДНК взаимодействуют с по-
ложительно заряженными боковыми группами
гистонов.
14. a) (Glu)20; б) (Lys-Ala)3; в) (Asn-Ser-His)5;
г) (Asn-Ser-His)5.
15. а) Удельная активность фермента: 200 ед.7 мг после
стадии 1, 2 600 ед./мг после стадии 2, 3 250 ед./мг
после стадии 3, 4 000 ед.,мг после стадии 4,
15 000 ед./мг после стадии 5, 15 000 ед./мг
после стадии 6.; б) стадия 4; в) стадия 3; г) Да. По-
скольку удельная активность на стадии 6 не увели-
чилась; провести электрофорез в полиакриламид-
ном геле в присутствии SDS.
16. a) [NaCl] = 0,5 мМ: б) [NaCl] = 0.05 мМ.
17. Первым сходит пептид С, вторым пептид В, а тре-
тьим пептид А.
18. Tyr-Gly-Gly-Phe-Leu.
19.
Orn Phe
/ \
Vai Pro
t I
Pro Vai
X /
Phe Orn
Стрелки указывают пептидную связь СО -> NH
20. 88%, 97%. Процент (х) правильных аминокислот-
ных остатков, высвобожденных в цикле п, состав-
ляет хп/х. Все остатки, высвобожденные в первом
цикле, правильные, даже если эффективность рас-
щепления неидеальная.
21. a) Y (1), F (7) и R (9); б) положения 4 и 9; К (Lys)
чаще встречается в положении 1, R (Arg) всегда
присутствует в положении 9; в) положения 5 и
10; чаще всего в обеих положениях встречается Е
(Glu); г) положение 2; S (Ser)
22. а) Белок, который предстоит выделить (цитратсин-
таза, ЦС), составляет довольно малую фракцию
всего клеточного белка. Проведение выделения на
холоду тормозит процесс деградации белка, а саха-
роза обеспечивает изотоническую среду, в которой
при гомогенизации лучше сохраняется целост-
ность органелл, б) На этой стадии клеточные орга-
неллы разделяются в зависимости от их размера,
в) Первое добавление сульфата аммония позво-
ляет удалить из гомогената некоторые балласт-
ные белки. При добавлении дополнительного
количества сульфата аммония ЦС тоже выпадает
в осадок г) Для перевода ЦС в раствор необхо-
[296] Приложение Б
димо удалить сульфат аммония, но сделать это
нужно при таких значениях pH и ионной силы,
чтобы сохранить нативную конформацию белка,
д) Молекулы ЦС больше размера пор хроматогра-
фического геля. Белок можно идентифицировать
по поглощению света с длиной волны 280 нм, что
характерно для остатков Туг и Тгр. е) ЦС несет
на себе положительный заряд и поэтому связыва-
ется с отрицательно заряженными группами ка-
тионообменника. Сначала с колонки смываются
нейтральные и отрицательно заряженные белки, а
затем ЦС смывается с помощью раствора с более
высоким значением pH, который изменяет заряд
на молекуле ЦС. ж) разные белки могут иметь
одно и то же значение р/. Но SDS-электрофорез
подтверждает, что в данной фракции содержится
единственный белок. Полностью удалить SDS из
белкового препарата трудно, и в его присутствии
нарушаются кислотно-основные свойства белка,
включая значение р/.
23. а) В любой линейной полипептидной цепи есть
лишь два типа свободных аминогрупп: единствен-
ная а-аминогруппа на N-конце цепи и с-ами-
ногруппы в боковой цепи остатков лизина. Эти
аминогруппы реагируют с FDNB с образованием
DNP-производных аминокислот. Инсулин обра-
зует два разных а-амино-DNP-производных, что
говорит о существовании двух N-концов и, следо-
вательно, двух полипептидных цепей, одна из кото-
рых имеет на N-конце остаток Gly, а другая — оста-
ток Phe. Поскольку лизин образует e-DNP-лизин,
он не является N-концевым остатком, б) Да. Цепь
А имеет на N-конце остаток Gly; цепь В имеет на
N-конце остаток Phe и неконцевой (в позиции 29)
остаток Lys. в) Phe-Val-Asp-Glu-. Пептид В1 име-
ет на N-конце остаток Phe. В пептиде В2 также
содержится остаток Vai, но поскольку DNP-Val
не образуется, Vai не является N-концевым остат-
ком, а должен находиться со стороны С-конца от
Phe. Следовательно, В2 имеет последовательность
DNP-Phe-Val. Аналогичным образом, ВЗ имеет по-
следовательность DNP-Phe-Val-Asp, и цепь В на-
чинается последовательностью Phe-Val-Asp-Glu-.
г) Нет. Известно, что В-цепь инсулина начинается
с последовательности Phe-Val-Asn-Gln-. При опре-
делении аминокислотной последовательности по
методу Сенгера Asn и Glu превращаются в Asp и
Glu соответственно по той причине, что на стадии
гидролиза (стадия 7) происходит гидролиз амид-
ных связей в Asn и Gin (как и пептидных связей),
в результате чего образуются Asp и Glu. На этой
стадии анализа Сенгер с соавторами не могли от-
личить Asp от Asn, a Glu от Gin. д) Она в точности
совпадает с последовательностью, представленной
на рис. 3-24. Для каждого представленного в табли-
це пептида приведена информация, позволяющая
определить, в каком случае Asx является аспара-
гином, а в каком аспарагиновой кислотой, a Glx —
глутамином или глутаминовой кислотой.
Ас 1: остатки 20-21. Это единственная последова-
тельность Cys-Asx в цепи А; в этом пептиде содер-
жится примерно одна амидная группа, так что это
должна быть последовательность Cys-Asn:
A-Gly-Ile-Val-Glx-Glx-Cys-Cys-Ala-Ser-Val-Cys-
1 5 10
Ser-Leu-Tyr-Glx-Leu-Glx-Asx-Tyr-Cys-Asn-C
15 20
Ар15: остатки 14-15-16. Это единственный участок
с последовательностью Туг-Glx-Leu в цепи А. В нем
содержится примерно одна амидная группа, так что
это должна быть последовательность Tyr-Gln-Leu:
V-Gly-Ile-Val-Glx-Glx-Cys-Cys-Ala-Ser- Val-Cys-
1 5 10
Ser-Leu-Tyr-Gln-Leu-Glx-Asx-Tyr-Cys-Asn-C
15 20
Ap14: остатки 14-15-16-17. Здесь содержится
примерно одна амидная группа, и нам уже извест-
но, что в положении 15 находится Gin, так что в по-
ложении 17 — Glu:
V-Gly-Ile-Val-Glx-Glx-Cys-Cys-Ala-Ser- Val-Cys-
1 5 10
Ser-Leu-Tyr-Gln-Leu-Glu-Asx-Tyr-Cys-Asn-C
15 20
АрЗ'. остатки 18-19-20-21. Здесь содержится при-
мерно две амидные группы. Мы знаем, что в положе-
нии 21 находится Asn, так что в остаток 18 также Asn:
V-Gly-Ile-Val-Glx-Glx-Cys-Cys-Ala-Ser-Val-Cys-
1 5 10
Ser-Leu-Tyr-Gln-Leu-Glu-Asn-Tyr-Cys-Asn-C
15 20
Ар1\ остатки 17-18-19-20-21. В положениях 18 и
21 находится Asn.
Ар5ра1: остатки 1-2-3-4. Здесь нет амидных групп,
значит, в положении 4 находится остаток Glu:
Leu-Tyr-Gln-Leu-Glu-Asn-Tyr-Cys-Asn-C
15 20
Leu-Tyr-Gln-Leu-Glu-Asn-Tyr-Cys-Asn-C
15 20
Ар5: остатки от 1 до 13. Здесь содержится пример-
но одна амидная группа. Нам уже известно, что в
положении 4 находится Glu, так что в положении 5
должен стоять Gin:
V-Gly-Ile-Val-Glu-Gln-Cys-Cys-Ala-Ser-Val-Cys-
1 5 10
Ser-Leu-Tyr-Gln-Leu-Glu-Asn-Tyr-Cys-Asn-C
15 20
Краткие решения задач и ответы на вопросы 129 / ]
Глава 4
1. а) Более короткие связи имеют большую кратность
(по сравнению с одинарными связями) и прочность.
Пептидная вязь С-N прочнее одинарной связи и за-
нимает промежуточное положение между простой
и двойной связью, б) Вращение вокруг пептидной
связи при физиологических условиях затруднено,
поскольку отчасти она имеет характер двойной связи.
2. а) Основной структурной единицей шерстяного
волокна (а-кератина) является виток а-спирали,
повторяющийся через каждые 5,4 А; длина витка
спирали составляет около 5,2 А. Нагревание и рас-
тягивание волокна приводит к образованию растя-
нутой p-конформации, в которой расстояние между
соседними R-группами составляет -7,0 А. Когда
полимер возвращается в исходную а-спиральную
конформацию, волокно вновь укорачивается,
б) Под действием влаги и тепла шерстяное волок-
но возвращается из p-конформации в нативную
а-спиральиую конформацию и происходит опять
укорачивание. Структура белка шелка — p-лист, где
небольшие боковые цепи аминокислотных остатков
упакованы достаточно плотно, и такая структура
оказывается прочнее структуры шерсти.
3. - 42 пептидные связи в секунду
4. При pH > 6 карбоксильные группы поли(С1и) де-
протонированы. Взаимное отталкивание между от-
рицательно заряженными карбоксильными груп-
пами приводит к разворачиванию белковой цепи.
Аналогичным образом, при pH 7,0 протонированы
аминогруппы поли(Еуз). Взаимное отталкивание
между этими положительно заряженными группа-
ми тоже приводит к разворачиванию цепи.
5. а) Дисульфидные мостики образованы ковалент-
ными связями, и поэтому они гораздо прочнее не-
ковалентных взаимодействий, стабилизирующих
большинство белковых структур. Дисульфидные
мостики образуют перекрестные сшивки между по-
липептидными цепями, повышая их жесткость и ме-
ханическую прочность, б) Наличие дисульфидных
связей между остатками цистеина предотвращает
полное разворачивание белка.
6. а) Изгибы с наибольшей вероятностью возника-
ют у остатков 7 и 19; наличие остатков Pro в ре-
конфигурации способствует возникновению пово-
ротов. б) Остатки Cys в положениях 13 и 24 могут
образовывать дисульфидные связи, в) На внешней
поверхности находятся полярные и заряженные
остатки (Asp, Gin, Lys), внутри глобулы — неполяр-
ные и алифатические остатки (Ala, Не). Хотя Thr и
является полярной аминокислотой, его индекс ги-
дропатии близок к нулю, так что он может находить-
ся как на поверхности глобулы, так и внутри.
7. 30 аминокислотных остатков; 0,87.
8. Миоглобин — это и то, и другое, и третье. Мотив
«глобиновой укладки» встречается во всех гло-
бинах. Полипептидная цепь образует один-един-
ственный домен в данном белке.
10. а) Число молей DNP-валина, образующееся из
1 моль белка, равно числу N-концевых остатков и,
следовательно, числу полипептидных цепей, б) 4;
в) При проведении SDS-электрофореза различные
полипептидные цепи, возможно, давали бы отдель-
ные полосы.
И. Пептид (а), поскольку в нем больше аминокис-
лотных остатков, способствующих образованию
а-спиральной структуры (см. Таблицу 4-1).
12. а) По-видимому, остатки ароматических ами-
нокислот играют важную роль в стабилизации
амилоидных волокон. Поэтому молекулы с арома-
тическими группами могли бы ингибировать обра-
зование амилоида, препятствуя взаимодействию
ароматических боковых цепей аминокислотных
остатков, б) При диабете II типа в поджелудочной
железе образуются амилоидные волокна, как и в
головном мозге при болезни Альцгеймера. Хотя
амилоидные волокна при этих двух заболевани-
ях состоят из разных белков, основная структура
амилоида по сути одна и та же и стабилизирована
одинаковым образом. Поэтому лекарства, способ-
ные разрушать эту структуру, возможно, эффек-
тивны в обоих случаях.
13. а) Транскрипционный фактор NFkB, также на-
зываемый трансформирующим фактором RelA.
б) Нет. Результаты похожи, но списки родствен-
ных белков разные, в) Белок состоит из двух
субъединиц. Существует множество вариантов
этих субъединиц, но лучше других охарактеризо-
ваны варианты с молекулярной массой 50, 52 или
65 кДа. Они образуют разнообразные гомо- или
гетеродимеры. Структуру некоторых из них мож-
но найти в базе данных PDB. г) Транскрипцион-
ный фактор NFkB представляет собой димерный
белок, который связывается со специфической
последовательностью ДНК, усиливая транс-
крипцию соседних генов. Среди этих генов, в
частности, ген легкой к-цепи иммуноглобулина,
в связи с чем транскрипционный фактор и полу-
чил свое название.
14. a) Aba является подходящей заменой, поскольку
Aba и Cys имеют боковые цепи примерно одинако-
вого размера и одинаковую гидрофобность. Однако
Aba не может образовывать дисульфидные связи и
поэтому не будет адекватной заменой в том случае,
когда такие связи требуются, б) Между синтезиро-
ванным белком и ВИЧ-протеазой, образующейся в
[298] Приложение Б
человеческих клетках, существует несколько важ-
ных различий — любое из них может объяснить
отсутствие активности у синтетического белка.
(1) Хотя Cys и Aba похожи по размеру и гидрофоб-
ности, этого может быть недостаточно для форми-
рования правильной укладки белка. (2) В фолдин-
ге ВИЧ-протеазы определенную роль могут играть
дисульфидные связи. (3) Фолдинг многих белков
происходит еще в процессе синтеза на рибосомах,
по данный белок складывается только после за-
вершения синтеза. (4) Белки, синтезируемые на
рибосомах, могут взаимодействовать с рибосо-
мами в процессе фолдинга, но для данного белка
это невозможно. (5) Состав цитозоля сложнее со-
става буфера, который использовался в данном
исследовании, а для фолдинга некоторых белков
могут требоваться специфические и неизвестные
пока белки. (6) Для правильного сворачивания
некоторым белкам требуются шапероны, которых
в буфере нет. (7) В клетках ВИЧ-протеаза синте-
зируется как часть более длинной цепи, которая
затем подвергается протеолитическому процес-
сингу, а белок в данном исследовании был синте-
зирован как единая молекула, в) Поскольку фер-
мент оказался функциональным даже при замене
Cys на Aba, видимо, дисульфидные связи не игра-
ют важной роли в формировании ВИЧ-протеазы.
г) Модель У: укладка, как в L-протеазе. Аргумент
«за»: ковалентная структура белков одинаковая
(за исключением хиральности), поэтому и уклад-
ка будет такой же. Аргумент «против»', хираль-
ность — важное свойство, трехмерная структура
играет ключевую роль в биологической активно-
сти молекул. Поэтому синтетический фермент
не может укладываться таким же образом, как
L-протеаза. Модель 2: образуется зеркальное отра-
жение L-протеазы. Аргумент «за»: поскольку от-
дельные элементы синтетического и природного
белков являются зеркальными отражениями друг
друга, целая молекула синтетического белка тоже
примет структуру зеркального отражения при-
родного белка. Аргумент «против»: фолдинг бел-
ка обеспечивают очень сложные взаимодействия,
так что синтетический белок, вероятно, примет
иную структуру Модель 3: иная укладка. Аргумент
«за»: фолдинг белка обеспечивают очень сложные
взаимодействия, так что синтетический белок, ве-
роятно, примет какую-то иную структуру. Аргу-
мент «против»: поскольку отдельные структуры
синтетического и природного белков являются
зеркальными отражениями друг друга, целая мо-
лекула синтетического белка тоже примет струк-
туру зеркального отражения природного белка,
д) Модель 1: Фермент оказался активным по отно-
шению к энантиомеру природного субстрата и ин-
гибируется энантиомером природного ингибитора.
Это означает, что D-протеаза является зеркаль-
ным отражением L-протеазы. е) Краситель Эван-
са — ахиральная молекула и связывается с обеими
формами фермента, ж) Нет. Химотрипсин не рас-
щепляет D-протеазу, поскольку протеазы содер-
жат только L-аминокислоты и распознают только
L-пептиды. з) Не всегда. В зависимости от особен-
ностей фермента при его синтезе могут возникать
различные проблемы, перечисленные в пункте (б),
в результате чего фермент может оказаться неак-
тивным.
Глава 5
1. Белок В обладает более высоким сродством к ли-
ганду X: для него полунасыщепие достигается при
значительно меньшей концентрации X, чем для
белка А. Для белка А К.А = 10G М \ для белка В
К.Л=1№М].
2. Во всех трех случаях < 1. При связывании ли-
ганда кажущуюся отрицательную кооператив-
ность можно объяснить наличием в белке центров
связывания двух или нескольких типов с разным
сродством к лиганду или присутствием несколь-
ких белков в растворе. Кажущаяся отрицательная
кооперативность часто наблюдается в гетероген-
ных белковых образцах. Примеров истинной от-
рицательной кооперативности известно совсем
немного.
3. а) Снижается; б) повышается; в) снижается; г) сни-
жается.
4. = 8,9 • 10 5 с
5. Кооперативность связывания кислорода гемо-
глобином обусловлена взаимодействиями между
субъединицами этого белка.
6. а) Тот факт, что гемоглобин А (НЬА, гемоглобин ма-
тери) при рО2 = 4 кПа насыщен примерно на 60%,
тогда как гемоглобин F (HbF, гемоглобин плода) в
тех же условиях насыщен более чем на 90%, указы-
вает на более высокое сродство HbF к О2 по срав-
нению с НЬА. б) Более высокое сродство HbF к
кислороду обеспечивает перемещение кислорода
из материнской крови в кровь плода через плацен-
ту Кровь плода близка к насыщению кислородом
за счет сравнительно низкого сродства НЬА к кис-
лороду в) Тот факт, что кривая насыщения НЬА
кислородом при связывании БФГ сдвигается силь-
нее, чем кривая насыщения HbF, говорит о том, что
НЬА связывает БФГ прочнее, чем HbF. Различие в
связывании БФГ двумя типами гемоглобина может
определять их различное сродство к О2.
Краткие решения задач и ответы на вопросы [299]
7. a) Hb Memphis; б) HbS, Hb Milwaukee, Hb Provi-
dence, возможно Hb Cowtown; в) Hb Providence.
8. Сильнее. Неспособность образовывать тетрамеры
ограничивает кооперативность связывания в слу-
чае этих вариантов, и кривая связывания принима-
ет более выраженный гиперболический характер.
Также повреждается центр связывания БФГ. Кис-
лород, скорее всего, связывается сильнее, посколь-
ку в отсутствие связанного БФГ белок находится в
высокоаффинном состоянии R
9. а) 1 • 10 8 М; б) 5 • 10 8 М; в) 8 • 108 М; г) 2 • 107 М.
Замечание', уравнение 5-8 можно привести к виду
[L] = 0Kd/(l - 0).
10. Скорее всего, эпитопом является некая структу-
ра. которая оказывается скрытой при связывании
G-актина с F-актином.
11. Во многих патогенах, включая ВИЧ, возникли ме-
ханизмы, с помощью которых они могут постоянно
изменять структуру поверхностных белков, с ко-
торыми изначально связывались компоненты им-
мунной системы. В результате хозяйский организм
каждый раз встречается с новыми антигенами, для
уничтожения которых иммунной системе требует-
ся время. Как только иммунная система реагирует
на один вариант, сразу же появляется другой.
12. Связывание миозина с АТР вызывает диссоциа-
цию миозина из комплекса с тонкими активновы-
ми филаментами Когда АТР отсутствует, актин и
миозин прочно связаны друг с другом.
14. a) L - легкая цепь, а Н - тяжелая цепь Fab-фраг-
мента данного антитела. Цепь Y - это лизоцим
б) В вариабельной и константной областях фрагмен-
та преобладают [3-структуры, в) В тяжелой цепи 218
остатков, в легкой цепи 214 остатков, в лизоциме 129
остатков. В контакте с Fab-фрагментом находится
менее 15% молекулы лизоцима, г) Из последователь-
ности Н-цепи в контакте с лизоцимом могут нахо-
диться Gly31, Туг32, Arg", Asp11’1’ и Туг11’1. Из последо-
вательности L-цепи - Туг32, Туг49, Туг51’ и Тгр92. В зоне
контакта антиген-антитело находятся следующие
остатки лизоцима: Asn14 * * * * 19 * *, Gly22, Туг23, Ser24, Lys116,
Gly117, Thr118, Asp119, Gin121 и Arg125. He все эти остат-
ки расположены рядом друг с другом в первичной
структуре. Центр связывания антигена образуется
из различных участков белковой последовательно-
сти при их сближении в процессе укладки.
15. а) Наибольшим сродством к лиганду L обладает
белок с Kd = 5 мкМ. В случае Kd = 10 мкМ удво-
ение [L] от 0,2 до 0,4 мкМ (что намного ниже Kd)
приводит почти к двукратному увеличению 0 (на
самом деле значение возрастает в 1,96 раза). Это
свойство гиперболической зависимости; при низ-
ких концентрациях лиганда 0 почти линейным об-
разом зависит от [L]. Напротив, удвоение [L] от 40
до 80 мкМ (это выше значения Kd; в этой области
кривая связывания приближается к своему асим-
птотическому пределу) приводит к росту 0 всего
в 1.1 раза. Идентичное увеличение фактора 0 на-
блюдается для кривых, построенных на основании
уравнения 5-11. б) 0 = 0,998. в) Результаты зависят
от значений вводимых параметров.
16. а)
Рисунок выполнен нс в масштабе. В реальности
на поверхности любой клетки связано гораздо
больше молекул миозина, б) АТР является источ-
ником энергии, приводящей систему в движение
(см. гл. 13). в) Если бы исследователи исполь-
зовали антитело, которое связывается не с «хво-
стом» миозина, а с «головкой», где расположен
участок связывания актина, это не позволило бы
актину связаться с миозином и, следовательно,
предотвратило бы перемещение. Антитело, кото-
рое связывается с актином, также мешало бы свя-
зыванию и перемещению, г) Есть два возможных
объяснения. (1) Трипсин расщепляет пептидную
связь только у остатков Lys и Arg (см. табл. 3-7),
так что в белке существует лишь небольшое чис-
ло возможных участков расщепления трипсином.
(2) Не все остатки Arg и Lys одинаково доступны
для фермента; первым расщепляется наиболее
доступный сайт, д) Модель S1. «Шарнирная мо-
дель» предсказывает, что двигаться могут только
комплексы шарик/антитела/НММ (с шарниром),
но не комплексы шарик/антитела/SHMM (без
шарнира). Модель S1 предсказывает, что дви-
гаться могут оба типа комплексов, поскольку оба
содержат «головку» S1. Тот факт, что система ша-
[300] Приложение Б
рик/SHMM (без шарнира) движется, подтверж-
дает справедливость модели S1. е) При низкой
плотности связывания молекул миозина шарики
время от времени были бы отсоединены от акти-
на. Шарики передвигались бы медленно, посколь-
ку после отсоединения одной молекулы миозина
требуется время для присоединения следующей.
При высокой плотности связывания миозина по-
сле отсоединения одной молекулы сразу же свя-
зывается вторая, и движение происходит быстрее,
ж) Выше определенной плотности молекул ми-
озина скорость перемещения шариков начинает
определяться скоростью, с которой молекулы ми-
озина способны перемещать шарики. Поскольку
молекулы миозина движутся с максимальной ско-
ростью, добавление новых молекул не приводит
к увеличению скорости, з) Поскольку движущая
сила формируется в «головке» S1, повреждение
«головки» вполне могло инактивировать моле-
кулу, и SHMM мог потерять способность пере-
мещать шарики, и) Скорее всего, формирование
«головки» S1 обеспечивают нековалентные взаи-
модействия и этого вполне достаточно для сохра-
нения функциональности этого фрагмента.
Глава 6
1. При нагревании теряется активность фермента, пре-
вращающего сахар в крахмал.
2. 2,4 • 10 6 М.
3.9,5- 108 лет.
4. Фермент-субстратный комплекс устойчивее свобод-
ного фермента.
5. а) 190 А; б) при фолдинге белка эти аминокислот-
ные остатки сближаются.
6. Скорость реакции можно определить по умень-
шению поглощения NADH при 340 нм в процес-
се реакции. Сначала определяем Км. При кон-
центрации субстрата » Км измеряем начальную
скорость реакции (скорость расходования NADH,
определяется с помощью спектрофотометра) при
нескольких концентрациях фермента и строим
график зависимости начальной скорости от кон-
центрации фермента. График должен быть линей-
ным, а угол его наклона является мерой активно-
сти ЛДГ.
7. (б), (д), (ж).
8. а) 1,7-10’3 М; б) 0,33; 0,67; 0,91; в) верхняя кривая
соответствует ферменту В (для этого фермента
[X] > Км), а нижняя кривая — ферменту А.
9. а) 400 с1; б) 10 мкМ; в) а = 2, а' = 3; г) смешанный
тип ингибирования.
10. а) 24 нМ; б) 4 мкМ (vQ ~ у2 К™, так что [А] = Км},
в) 40 мкМ (г>0 = Уг Vmax, так что [А] = 10 Км в при-
сутствии ингибитора); г) не достигает; Аса1/Км =
= 0,33/(4 • 10-6 М 1 • с ') = 8,25 • 104 М1 - с1, что
значительно ниже диффузионно контролируемого
предела.
11. Vmax~ 140 мкМ/мин, Км ~ 110 5 М.
12. a) Vinax = 51,5 мМ/мин, Км = 0,59 мМ; б) конкурент-
ное ингибирование.
13. Км = 2,2 мМ, Vniax = 0,50 мкМ/мин.
14. Кривая А.
15. £са1 = 2,0 • 107 мин L
16. Сохраняются те же приближения, что и при выводе
уравнения Михаэлиса-Ментен. Реакция находит-
ся в стационарном состоянии, поэтому скорость
реакции описывается уравнением г’о = &2[ES]. За-
пишем все известные нам соотношения для опре-
деления [ ES]:
[Et] = [Е] + |ES| + [EI] и [EI] = [£][!]/*,
[Е] можно определить, пользуясь уравнением 6-19.
Далее следует идти тем же путем, что и при выводе
уравнения Михаэлиса-Ментен.
17. Минимальное значение Мт = 29 000.
18. Активность фермента простаты определяется как
общая активность фосфатазы в крови за вычетом
активности фосфатазы в присутствии достаточно-
го количества тартрата, специфически и полностью
ингибирующего эту форму фермента.
19. Смешанный тип ингибирования. Поскольку значе-
ние Км практически не изменяется, это может быть
примером особого случая смешанного ингибирова-
ния, называемого неконкурентным ингибированием.
20. Значение [S], при котором v0 = Vniax/2a', можно
найти из правой части уравнения 6-30:
[S]/(aKM + a'[S])=%a'.
21. Оптимум ферментативной активности достигается,
когда остаток Glu35 протонирован, а остаток Asp52 де-
протонирован.
22. а) Увеличится в 1,96 раз; г>0 = 50 мкМ • с1; увели-
чится в 1,048 раз; б) Когда a = 2,0, кривая сдви-
гается вправо, поскольку Км увеличивается в
2 раза. Когда а' = 3,0, асимптота кривой (зна-
чение Vmax) снижается в 3 раза. Когда a = 2,0,
а а' = 3,0, кривая круто поднимается по отношению
к кривой, для которой a = а' = 1,0, из-за умень-
шения Км. Однако асимптота кривой низкая,
поскольку Vmax уменьшается в 3 раза, в) Когда
a = 2,0, точка пересечения кривой с осью х сдви-
гается вправо. Когда a = 2,0, а а'= 3,0, точка пере-
сечения с осью х смещается влево.
Краткие решения задач и ответы на вопросы [301]
23. а) В центре связывания фермента дикого типа суб-
страт удерживается с помощью водородных связей
и ион-дипольных взаимодействий между заряжен-
ной боковой цепью остатка Arg109 и полярной кар-
бонильной группой пирувата. В процессе катализа
заряженная боковая группа Arg109 стабилизирует
поляризованное карбонильное переходное состо-
яние. В случае мутантного фермента связывание
гораздо слабее, поскольку осуществляется исклю-
чительно водородными связями, и стабилизации
переходного состояния не происходит, что снижа-
ет каталитическую активность, б) Остатки Lys и
Arg имеют приблизительно одинаковый размер и
одинаковый положительный заряд, возможно, они
обладают похожими свойствами. Кроме того, по-
скольку пируват, по-видимому, связан с остатком
Arg171 ионными силами, можно предположить, что
замена Arg на Lys не оказывает серьезного влия-
ния на связывание субстрата в) В конфигурации
«вилки» две положительно заряженные группы
Arg могут взаимодействовать с двумя отрицатель-
но заряженными атомами кислорода в молекуле
пирувата, что способствует образованию двух
связей сметанного типа (водородных связей и
ион-дипольных взаимодействий). Если место Arg
занимает Lys, возникает лишь одна такая связь,
что ослабляет взаимодействие, и ориентирование
субстрата происходит менее точно, г) Гидрофоб-
ный остаток Не250 взаимодействует с гидрофоб-
ным циклом NADH. С гидрофильной боковой
цепью Gin такое взаимодействие невозможно,
д) Расположение аминокислотных остатков в ак-
тивном центре мутантной ЛДГ представлено ниже.
Arg102
\н
I
Arg109
Z-\
HN^NH
н н
HiS195
NH 0-0
н3(
HbT^NH 1н
н н у112
О=С Оксало
/7 NH | ацетат
\ VI С
Н 0^0
о НН
Thr246
I
!—С—ОН
Н
H3C-C-CH2
Це250
HN^+^NH
NH
A^171
Aspi68
e) Мутантный фермент выталкивает пируват, по-
скольку гидрофобная метильная группа пирува-
та не может взаимодействовать с гидрофильной
гуанидиновой группой Arg102. Связывание окса-
лоацетата обеспечивают сильные ионные взаимо-
действия между боковой цепью Arg102 и карбок-
сильной группой оксалоацетата. ж) Белок должен
быть достаточно гибким, чтобы разместить объем-
ную боковую цепь нового аминокислотного остатка
и более крупный субстрат
Глава 7
1. В результате восстановления кислорода карбонила
до гидроксила атомы С1 и СЗ стали химически экви-
валентными: молекула глицерина не имеет хираль-
ных центров.
2. Эпимеры различаются конфигурацией заместителей
лишь у одного атома углерода, а) D-альтроза (С2),
D-глюкоза (СЗ), D-гулоза (С4); б) D-идоза (С2),
D-галактоза (СЗ), D-аллоза (С4); в) D-арабиноза
(С2), D-ксилоза (СЗ).
3. При образовании озазона изменяется конфигура-
ция заместителей у атома С2 в альдозах, поэтому
альдозы, различающиеся лишь конфигурацией за-
местителей у атома С2, превращаются в одинаковые
соединения с одинаковой температурой плавления.
4. а)
a-D-Глюкоза
/З-и-Глюкоза
б) Свежеприготовленный раствор a-D-глюкозы или
p-D-глюкозы подвергается мутаротации и постепен-
но образует равновесную смесь а- и p-форм в) 36%
сх-формы, 64% р-формы.
5. Для превращения a-D-глюкозы в p-D-глюкозу нуж-
но разорвать связь между атомом С1 и гидроксиль-
ной группой у С5 (как на рис. 7-6). Для превраще-
ния D-глюкозы в D-маннозу нужно разорвать связь
атома С2 либо с - Н, либо с ОН. Превращения
конформации «кресло» не требуют разрыва связей,
в этом и заключается главное различие между кон-
фигурацией и конформацией.
6. Нет; глюкоза и галактоза различаются расположе-
нием заместителей у атома С4.
7. а) Оба полимера состоят из звеньев D-глюкозы, но
в целлюлозе эти звенья соединены (р1-*4)-связью.
а в гликогене (а!-*4)-связью. б) Оба соединения —
[302] Приложение Б
гексозы, но глюкоза — это альдогексоза, а фрук-
тоза — кетогексоза, в) Оба соединения - дисахари-
ды. но в мальтозе два остатка D-глюкозы соединены
(а!-*4)-связью. а в сахарозе остатки D-глюкозы и
D-фруктозы соединены (а2«->2Р)-связью.
9. Полуацеталь образуется при конденсации альдозы
или кетозы со спиртом; гликозид — при конденса-
ции полуацеталя со спиртом (см. рис. 7-5, т. 1, с. 343).
10. При замыкании цикла фруктоза образует либо пи-
ранозу, либо фуранозу. При повышении темпера-
туры равновесие сдвигается в сторону фуранозы, а
она менее сладкая.
11. Мальтоза; в сахарозе нет восстанавливающей
(окисляемой) группы, поскольку аномерные ато-
мы углерода обоих моносахаридных звеньев уча-
ствуют в образовании гликозидной связи.
12. Скорость мутаротации достаточно высокая, так что
по мере использования ферментом [3-D-глюкозы
все больше a-D-глюкозы превращается в р-форму,
и постепенно окисляется вся глюкоза. Глюкозоок-
сидаза специфична по отношению к глюкозе и не
взаимодействует с другими восстанавливающими
сахарами (такими как галактоза), которые реагиру-
ют с реактивом Фелинга.
13. а) Определить скорость изменения оптического
вращения; б) оптическое вращение смеси измеря-
ется относительно раствора сахарозы; в) -2,0°.
14. Смешать сахарозу с водой (начинка), добавить не-
большое количество сахаразы (инвертазы) и не-
медленно покрыть шоколадом
15. В сахарозе нет свободного аномерного атома угле-
рода, который мог бы подвергаться мутаротации.
17. Ы-Ацетил-р-О-глюкозамин — восстанавливающий
сахар, его атом С1 может окисляться (см. рис. 7-10,
т. 1, с. 347). D-Глюконат — невосстанавливаюший
сахар, его атом С1 уже находится в окисленном со-
стоянии GlcN(al«->la)Glc — невосстанавливаю-
щий сахар, поскольку аномерные атомы углерода
обоих звеньев участвуют в образовании гликозид-
ной связи.
18. В желудочно-кишечном тракте человека нет цел-
люлаз. и поэтому люди не могут расщеплять цел-
люлозу.
19. Нативная целлюлоза состоит из мономеров глю-
козы, соединенных (pi—*4)-гликозидными связя-
ми, которые придают цепи вытянутую конформа-
цию. Параллельные цепи образуют между собой
водородные связи и собираются в длинные, жест-
кие и нерастворимые волокна. Гликоген состоит
из мономеров глюкозы, соединенных (al—*4)-
гликозидными связями, из-за которых полимер-
ные цепи изгибаются и не образуют длинных во-
локон. Кроме того, гликоген сильно разветвлен и
многие из его гидроксилов находятся в контакте с
водой, поэтому он легко гидратируется и диспер-
гируется в воде.
Целлюлоза образует структурный каркас
растений, состоящий из протяженных нераство-
римых волокон. Гликоген служит запасным ве-
ществом в организме животных. Гидратирован-
ные гранулы гликогена, содержащие множество
невосстанавливающих концов, быстро гидроли-
зуются гликогенфосфорилазой с образованием
глюкозо-1 -фосфата.
20. Целлюлоза в несколько раз длиннее; она находится
в вытянутой конформации, тогда как амилоза име-
ет спиральную структуру.
21. 6000 остатков/с.
22. Ис.
23. Шаро-стержневая модель дисахарида на рис. 7-19
(т. 1) не отражает пространственных взаимодей-
ствий в молекуле, а вот СРК-модель, в которой
относительные размеры атомов соответствуют ре-
альным, может показать стерическое напряжение в
конформере 170°, -170°, которого нет в конфор-
мере 30°, - 4(Г.
24. Отрицательно заряженные группы в молекуле хон-
дроитинсульфата отталкиваются, что заставляет
молекулу принять вытянутую конформацию. По-
лярные молекулы притягивают множество моле-
кул воды, что увеличивает общий объем молекулы.
В сухом виде отрицательные заряды уравновеши-
ваются положительными противоионами, и моле-
кула укорачивается.
Краткие решения задач и ответы на вопросы [303]
25. Положительно заряженные аминокислотные
остатки связываются с отрицательно заряженны-
ми группами гепарина. Известно, что с гепарином
взаимодействует остаток Lys антитромбина III.
26. Восемь возможных последовательностей, 144 свя-
зи и 64 стереоизомера - всего 73 728 перестановок!
27.
Цепи
олигосахаридов:
О
— 1
— 2
— 3
28. Олигосахарид; мономерные звенья сахаров могут
комбинироваться большим числом способов, чем
звенья олигопептидов. Каждая гидроксильная
группа может участвовать в образовании глико-
зидной связи, и сама эта связь может иметь а- или
[3-конфигурацию. Полимер может быть как линей-
ным, так и разветвленным.
29. а) 2,3 -ди-О-метилглюкоза образуется из остат-
ков, расположенных в точках ветвления, все
остальные остатки превращаются в 2,3,6-три-О-
метилглюкозу; б) 3,75%.
30. Остатки D-глюкозы соединены (1—>6)-связями;
боковые разветвления (примерно через каждые
20 остатков) - (1->3)-связями.
31. а) Нужно растворить небольшое количество об-
разца в разных растворителях, а затем исследовать
растворы и нерастворенные осадки, чтобы устано-
вить, различаются ли они по химическому составу,
б) Все молекулы чистого вещества идентичны, по-
этому химический состав этого вещества в растворе
и осадке одинаков. В грязном веществе содержится
более одного компонента. В каком-то из раство-
рителей один из компонентов может растворяться
лучше другого. Поэтому составы раствора и осадка
различны, в) Данные количественного анализа по-
зволяют утверждать, что при деградации фермен-
тативная активность не теряется. Для определения
структуры молекулы очень важно, чтобы в образце
находились только интактные (неповрежденные)
молекулы. Если же образец подвергся частичному
разрушению, результаты структурного анализа бу-
дут неточными и, возможно, неинтерпретируемы-
ми. Количественный анализ позволяет определить
очень незначительную ферментативную актив-
ность даже при больших разрушениях образца, г)
Результаты 1 и 2. Результат 1 согласуется с извест-
ной структурой: антиген В содержит три молекулы
галактозы, а антигены А и 0 только по две. Ре-
зультат 2: антиген А содержит два аминосахара —
N-ацетилгалактозамин и N-ацетилглюкозамин, а
антигены В и 0 только один N-ацетилглюкозамин.
Результат 3 не согласуется с известными фактами:
в антигене А соотношение глюкозамина и галак-
тозамина составляет 1 : 1, а в антигене В 1:0.
д) Возможно, образцы содержали примесь или
были частично разрушены. Первые два ре-
зультата оказались верными, поскольку метод
анализа был лишь полуколичественным и не-
достаточно чувствительным, чтобы зафикси-
ровать присутствие примесей. Третий резуль-
тат количественный и, следовательно, больше
отклоняется от предсказанных значений из-за
присутствия примеси или разрушения образца,
е) Экзогликозидаза. Эндогликозидаза расщеп-
ляла бы антиген 0 на галактозу, N-ацетил-
галактозамин или N-ацетилглюкозамин, и хотя бы
один из этих сахаров должен был ингибировать рас-
щепление. Поскольку ни один из этих сахаров не
ингибировал активность фермента, это была экзо-
гликозидаза, отщеплявшая от цепи лишь концевую
группу. Концевым сахаром в молекуле антигена 0
является фукоза, так что фукоза - единственный
сахар, который мог бы ингибировать расщепление
антигена 0. ж) Эта экзогликозидаза отщепляет
N-ацетилгалактозамин от антигена А и галактозу
от антигена В. Поскольку в результате этих реак-
ций фукоза не образуется, она не ингибирует рас-
щепление, и образующиеся продукты деградации
уже не обладают активностью антигенов А и 0.
Однако продукты реакции обладают активностью
антигена 0, поскольку расщепление останавлива-
ется на фукозе, з) Все результаты согласуются с
тем, что изображено на рис. 10-15 (т. 1). На конце
антигена 0 расположена фукоза, и лишь фукоза
могла бы ингибировать его расщепление. На кон-
це антигена А находится N-ацетилгалактозамин, и
лишь этот сахар защищает антиген А от расщепле-
ния. Концевой сахар антигена В — галактоза, и это
единственный сахар, защищающий антиген В от
расщепления.
Глава 8
l.N-3nN-7.
2. (5')GCGCAATATTTTGAGAAATATTGCGC(3 );
последовательность содержит палиндром, что
позволит отдельным нитям образовывать шпиль-
ку, а две нити могут образовать крестообразную
структуру.
[304] Приложение Б
3. 9,4 • 10 4 г.
4. а) 40°; б) 0°.
5. Спираль РНК имеет конформацию А, а спираль
ДНК обычно находится в конформации В.
6. В ДНК эукариот около 5% остатков цитозина мети-
лированы. 5-Метилцитозин спонтанным образом
дезаминируется, превращаясь в тимин; образую-
щиеся пары G Т являются одной из самых рас-
пространенных ошибок в ДНК эукариотической
клетки.
7. Кольцо рибозы без основания может раскрываться
и переходить в нециклическую альдегидную фор-
му. Это, а также исчезновение стекинговых взаи-
модействий между основаниями может объяснять
увеличение подвижности ДНК.
8. Стекинговые взаимодействия в нуклеиновых кис-
лотах приводят к уменьшению поглощения в УФ-
области. При денатурации стекинговые взаимодей-
ствия исчезают, и поглощение увеличивается.
9. 0,35 мг/мл
10.
Дезоксирибоза Гуанин Фосфат
Растворимость: фосфат > дезоксирибоза > гуанин
Сильнополяризованпые фосфатные группы и
остатки сахара располагаются на внешней поверх-
ности двойной спирали и находятся в контакте с
водой, тогда как гидрофобные основания распола-
гаются внутри спирали.
11. Без наличия в смеси dCTP на месте первого же
встреченного в матрице остатка G встроится
ddCTP, и полимеризация прекратится. В геле
лишь одна полоса.
13. (5')P-GCGCCAUUGC(3') ОН
(5')P-GCGCCAUUG(3')-OH
(5')P-GCGCCAUU(3')~ ОН
(5')P-GCGCCAU(3> ОН
(5')Р GCGCCA(3') ОН
(5')P-GCGCC(3')-OH
(5')P-GCGC(3') -ОН
(5')Р GCG(3')-ОН
(5')Р GC(3') ОН
и нуклеозид-5'-фосфаты
14. а) Вода обязательный участник большинства хи-
мических реакций, включая те, что вызывают му-
тации. Снижение содержания воды в эндоспорах
ослабляет активность ферментов, вызывающих
мутации, и снижает скорость неферментативных
потерь пурина, имеющих гидролитический меха-
низм. б) УФ-облучение способствует образованию
циклобутановых пиримидиновых димеров. По-
скольку В. subtilis обитает в почве, ее споры могут
подниматься на поверхность почвы или в воздух и
подвергаться длительному УФ-облучению.
15. Концевая группа DMT предотвращает реакцию ос-
нования с самим собой.
16. а) Правая спираль. На одном 5'-конце расположен
аденин, на другом 5'-конце — цитидин, б) Левая
спираль, в) Если вам не удается увидеть стереоизо-
бражение, обратитесь к Интернету.
17. а) Это непростая задача. Образцы из одного и
того же организма достаточно сильно различают-
ся, а выход никогда не достигает 100%. Значения
для С и Т согласуются значительно лучше, чем
для А и G, так что для С и Т проще доказать, что
все образцы из одного организма имеют одина-
ковый состав. Однако даже в случае А и G верно
следующее: (1) диапазоны значений для разных
тканей в значительной степени перекрывают-
ся; (2) различия между разными образцами из
одной и той же ткани примерно такие же, как и
различия между образцами из разных тканей;
(3) в образцах с наиболее высоким выходом зна-
чения согласуются лучше, б) Данный метод недо-
статочно чувствителен, чтобы выявить различия
между здоровой и опухолевой тканями. Опухоль
связана с появлением мутаций, но таких мутаций
всего несколько на миллиард, что слишком мало,
чтобы выявить их данным методом, в) У разных
видов отношения А: G и Т: С очень сильно варьи-
. руют. Например, для бактерии Serratia marcescens
оба отношения составляют 0,4, что говорит о том,
что в последовательности этой ДНК преоблада-
ют G и С. Напротив, для бактерии Haemophilus in-
fluenzae эти соотношения составляют 1,74 и 1,54,
т. е. ее ДНК состоит главным образом из А и Т.
Краткие решения задач и ответы на вопросы |ЗО5]
г) Для справедливости этого утверждения
должны выполняться три условия. (1) А=Т;
данные таблицы показывают, что во всех случа-
ях отношение А : Т близко к единице. Очевидно,
что вариации этого соотношения значительно
слабее, чем вариации отношений А : G и Т : С.
(2) G=C; опять же, отношение G : С близко к
единице, а другие отношения варьируют силь-
нее. (3) (А + G) = (Т + С); это соотношение
пуринов к пиримидинам, которое также близ-
ко к единице, д) Разные фракции «ядра» соот-
ветствуют разным участкам ДНК проростков
пшеницы. Если бы ДНК представляла собой
монотонно повторяющуюся последователь-
ность, нуклеотидный состав всех участков был
бы одинаковым. То, что разные фракции «ядра»
имеют разный состав, говорит о более сложном
характере последовательности.
Глава 9
1,а) (5')--G(3') и (5')ААТТС-------(3')
(3')---СТТАА(5') (3')G---------(5')
б) (5')--GAATT(3') и (5')ААТТС-----(3')
(3')---СТТАА(5') (3')TTAAG-------(5')
в) (5')--GAATTAATTC--------(3')
(3')---CTTAATTAAG--------(5')
г) (5') — — — G(3') и (5')С - - - (3')
(З')---С(У) (3')G---(5')
д) (5')--GAATTC-----(3')
(3')---CTTAAG-----(5')
е) (7)---CAG(3') и (5')CTG-----(3')
(3')---GTC(5') (3')GAC--(5')
ж) (5')----CAGAATTC-------(3')
(3')---GTCTTAAG-------(У)
з) Метод 1: расщепить ДНК рестриктазой EcoRI,
как в пункте (а), обработать, как в пункте (б) или
(г), а затем лигировать синтетический фрагмент
с двумя тупыми концами с последовательностью
узнавания рестриктазы BamHl. Метод 2 (более
эффективный): синтезировать фрагмент ДНК со
следующей структурой:
(5')AATTGGATCC(3')
(3')CCTAGGTTAA(5')
Этот фрагмент эффективно лигируется с лип-
кими концами фрагментов, образующихся в ре-
зультате расщепления рестриктазой EcoRI. Так
можно встроить сайт узнавания для рестрик-
тазы ВяттгН!, но при этом сайт узнавания EcoRI
исчезает.
и) Эти четыре фрагмента (в том же порядке, в кото-
ром они упомянуты в вопросе) следующие (N -
любой нуклеотид):
(5')AATTCNNNNCTGCA(3')
(3')GNNNNG(5')
(5')AATTCNNNNGTGCA(3')
(3')GNNNNC(5')
(5')AATTGNNNNCTGCA(3r)
(3')CNNNNG(5')
(5')AATTGNNNNGTGCA(3')
(3')CNNNNC(5')
2. ДНК фага k может быть упакована в инфекцион-
ные фаговые частицы только в том случае, если ее
размер составляет от 40 000 до 53 000 п. н. Посколь-
ку векторы на основе бактериофагов обычно содер-
жат около 30 000 п. н., они не могут образовывать
фаговые частицы, если в них не встраивается до-
полнительная ДНК необходимой длины (от 10 000
до 23 000 п. н.).
3. а) Восстановленные плазмиды pBR322 без встав-
ки чужеродной ДНК, они сохраняют устойчивость
к ампициллину. Кроме того, две или более молекул
pBR322 могут смыкаться в кольцо как содержащее,
так и не содержащее вставки, б) На дорожках 1 и
2 разделены образцы, содержащие вставку ДНК в
разной ориентации. На дорожку 3 нанесен образец,
содержащий два фрагмента ДНК, встроенных таким
образом, что соединенными оказались проксималь-
ные концы, образующиеся при расщеплении EcoRI.
4. Обратим внимание на аминокислоты, для которых
существует минимальное число кодонов: Met и Тгр.
Лучше всего охватить последовательность от кодона
для первого Тгр до первых двух нуклеотидов кодо-
на для Не. Такой зонд имеет следующую последова-
тельность:
(5')UGGUA(U/C)UG(U/C)AUGGA(U/C)UGGAU
Синтезировать нужно так, чтобы в каждой из ука-
занных позиций стоял либо U, либо С, в результате
чего получится смесь из восьми 20-нуклеотидных
зондов, различающихся только по одной или не-
скольким из указанных позиций.
5. Понадобятся ДНК-праймеры, термостабильная
ДНК-полимераза, дезоксинуклеозидтрифосфаты
и термоциклер (аппарат для проведения ПЦР).
Праймеры нужно синтезировать так, чтобы они
охватывали участок ДНК с повтором CAG. На ри-
сунке изображена кодирующая нить, ориентиро-
ванная слева направо в направлении 5'-*3'. Левый
праймер должен быть идентичен любой последо-
вательности из 25 нуклеотидов, находящихся сле-
ва от повтора CAG. Правый праймер должен быть
[306] Приложение Б
представлен последовательностью комплемен-
тарной и антипар аллельной к последовательно-
сти из 25 нуклеотидов, расположенных справа от
повтора CAG. С помощью этих праймеров можно
амплифицировать участок ДНК, содержащий по-
втор CAG, и определить его размер методом элек-
трофореза путем сопоставления с маркерными
последовательностями. Длина этого фрагмента
ДНК соответствует длине повтора CAG; на этом
основан диагностический тест.
6. Можно использовать ПЦР -праймеры, комплемен-
тарные ДНК из удаленного сегмента, но такие, ко-
торые направляют синтез ДНК не навстречу друг
другу, а в противоположные стороны. Продукт ПЦР
образуется лишь в том случае, если концы удален-
ного сегмента смыкаются с образованием кольцевой
структуры.
7. Чтобы светиться, растению, экспрессирующему лю-
циферазу светляков, нужен субстрат (люциферин),
тогда как растение, экспрессирующее зеленый флу-
оресцентный белок, светится вне зависимости от на-
личия других веществ
8. Нельзя исключить ни одного ребенка, так как каж-
дый содержит фрагменты ДНК, которые он мог
унаследовать от отца
9.
91 5 374 2 68
Н---------1----1------1--1----1---------1--1—1
I-----------А------------1
I--------------В--------------1
I--------------С--------------1
I--------------D--------------1
I------Е-------1
I---------F-----------1
10. Исключительно из соображений удобства; Ti-
плазмида (200 000 п. н.) даже при удалении
Т-ДНК слишком велика, чтобы ее можно было
количественно выделить из клеток и манипули-
ровать ею in vitro. Она также слишком велика для
обратного внедрения в клетки с помощью стан-
дартной процедуры трансформации. Гены vir об-
легчают перенос любой ДНК между повторами
Т-ДНК, даже если они находятся на разных плаз-
мидах. Вторая плазмида в этой двухплазмидной
системе, содержащая только повторы Т-ДНК и
несколько последовательностей для отбора плаз-
миды и ее репликации, имеет сравнительно не-
большой размер; ее легко выделить и ею легко
манипулировать (в частности, вводить и изме-
нять чужеродную ДНК).
11.
АВС
12. Закройте лунку 4, добавьте раствор активирован-
ного Т, осветите, отмойте.
l.A-T 2.G Т З.А Т 4.G-C
Закройте лунки 2 и 4, добавьте раствор активиро-
ванного G, осветите, отмойте.
1. А-Т- G 2. G-T 3. A T-G 4. G-C
Закройте лунку 3, добавьте раствор активирован-
ного С, осветите, отмойте.
l.A T-G С 2.G-T С З.А T-G 4.G-C С
Закройте лунки 1,3 и 4, добавьте раствор активиро-
ванного С, осветите, отмойте.
1.A-T-G С 2.G-T С-СЗ.А T-G 4. G-C-C
Закройте лунки 1 и 2, добавьте раствор активиро-
ванного G, осветите, отмойте.
l.A-T-G-C 2.G-T С-С З.А Т G-C4.GC-C-C
13. Такие векторы вводятся в клетки, инфицирован-
ные вирусом-помощником, который обеспечивает
все необходимое для репликации и упаковки, но
сам не может упаковываться. С помощью этих век-
торов рекомбинантная ДНК встраивается в хро-
мосому хозяйской клетки, причем эта интеграция
очень стабильная, так как отсутствие функции ре-
комбинации и упаковки предотвращает делению и
рекомбинацию интегрированной ДНК.
14. а) Растворы ДНК очень вязкие, так как содержат
очень длинные, переплетенные между собой мо-
лекулы. Чем короче молекулы, тем в меньшей сте-
пени они переплетаются и тем ниже вязкость их
растворов. Поэтому снижение вязкости означает
укорочение нитей полимера, что связано с актив-
ностью нуклеазы б) Это эндонуклеаза. Экзону-
клеаза отщепляет отдельные нуклеотиды от 5'- и
Краткие решения задач и ответы на вопросы [307]
З'-концов цепи, в результате чего образуются рас-
творимые в ТХУ меченные 32Р нуклеотиды. Эндо-
нуклеаза расщепляет внутренние связи в молекуле
ДНК, в результате чего растворимых в ТХУ мечен-
ных 32Р фрагментов образуется мало, в) На 5'-кон-
це. Если бы фосфатная группа осталась на З'-кон-
це, киназа включила бы значительное количество
32Р, поскольку присоединяет фосфатные группы к
5’-концу цепи, и обработка фосфатазой не оказа-
ла бы никакого влияния. В таком случае образцы
А и В содержали бы значительное количество 32Р.
Если же фосфат остается на 5'-конце, киназа не
может включать 32Р, поскольку не может присо-
единять фосфатную группу к уже существующей.
Фосфатаза удаляет фосфат с 5'-конца, и тогда ки-
наза может включать значительное количество
32Р. В таком случае в образце А содержится очень
мало метки, а в образце В - значительно больше,
что и наблюдалось в эксперименте, г) Случайные
разрывы привели бы к образованию фрагментов со
случайным распределением по размеру; образова-
ние специфических фрагментов говорит в пользу
специфического действия фермента, д) Фермент
распознает фрагмент и расщепляет его. В результа-
те на .7-конце этого фрагмента остается специфи-
ческая последовательность. Если бы расщепление
происходило вблизи сайта узнавания, но не в нем
самом, последовательность 5'-концов каждый раз
была бы разной, е) Эти результаты согласуются с
двумя вариантами последовательности (место рас-
щепления показано стрелкой):
I
(7) - - - GTT ААС (3')
(3') - - САА TTG (7)
t
последовательность с фрагментами
(5’)рАрАрС и (З’)ТрТр
I
(7) - - - GTC GAC - - - (3')
(7) - - CAG CTG - - (7)
t
последовательность с фрагментами
(7)pGpApC и (З')СрТр
Глава 10
1. Липидами называют не соединения со специфиче-
ской химической структурой, а те вещества, которые
лучше растворяются в органических растворителях,
чем в воде.
2. а) Число цис-двойных связей. Каждая цис-двойная
связь вызывает изгиб углеводородной цепи, снижая
температуру плавления, б) Можно создать шесть
разных триацилглицеринов, температура плавления
которых повышается в следующем ряду:
ООО < OOP = ОРО < РРО = POP < PPP.
где О = олеиновая кислота, Р = пальмитиновая кис-
лота. в) Жирные кислоты с разветвленной цепью
повышают текучесть мембран, поскольку снижают
плотность упаковки мембранных липидов.
3. Лецитин, являющийся амфифильным соединением,
выступает в качестве эмульгатора, облегчающего со-
любилизацию масла.
5. Мята — это (Я)-карвон, тмин - (5)-карвон.
6.
СООН
Н— С— NH3
СНз
(7?)-2-Аминопропановая
кислота
ОН
Н3С—с—соон
н
(7?)-2-Гидроксипропановая
кислота
СООН
H3N- С-Н
СНз
(8)-2-Аминопропановая
кислота
СООН
н3с—с—он
н
(S) - 2 -Гидроксипропановая
кислота
7. Гидрофобные элементы: (а) две жирные кислоты;
(б), (в) и (г) одна жирная кислота и углеводородная
цепь сфингозина; (д) стероидное ядро и ацильная
боковая группа. Гидрофильные элементы: (а) фос-
фоэтаноламин; (б) фосфохолин; (в) D-галактоза;
(г) остатки сахаров; (д) спиртовые группы (ОН-
группы).
9. Процесс приводит к восстановлению двойных свя-
зей, что повышает температуру плавления липидов,
содержащих соответствующую жирную кислоту.
1308 ] Приложение Б
10. Триацилглицерины в составе животного жира
(сала) гидролизуются под действием NaOH с обра-
зованием мыла (омыление), которые гораздо луч-
ше растворимо в воде, чем исходные соединения.
11. Это может быть только сфинголипид (сфинго-
миелин).
12.
Фосфатидилсерин
13. Длинные цепи насыщенных углеводородов, прак-
тически твердые при температуре окружающей
среды, образуют на поверхности листьев гидро-
фобный слой, через который такие полярные со-
единения, как вода, проникать не могут.
14. а) Свободная ОН-группа у атома С2 и фосфохоли-
новая головка у атома СЗ являются гидрофильны-
ми; жирная кислота у атома С1 лизолецитина явля-
ется гидрофобной, б) Некоторые стероиды, такие
как преднизон, ингибируют действие фосфолипа-
зы А2, блокируя отщепление арахидоновой кисло-
ты от атома С2. Арахидоновая кислота является ис-
ходным соединением для множества эйкозаноидов,
некоторые из них вызывают боль и воспаление,
в) Фосфолипаза А2 высвобождает арахидоновую
кислоту — предшественник других эйкозаноидов,
выполняющих важные защитные функции в орга-
низме. Кроме того, фермент расщепляет поступаю-
щие с пищей глицерофосфолипиды.
15. Группу крови определяет олигосахарид головной
группы мембранного сфинголипида (см. рис. 10-15,
т. 1, с. 503). Этот же олигосахарид соединен с неко-
торыми мембранными гликопротеинами, которые
также служат участками узнавания для антител,
распознающих группу крови.
16. Диацилглицерин — это гидрофобное вещество,
которое остается в мембране. Инозит-1,4,5-три-
фосфат — полярное и очень хорошо растворимое в
воде вещество, которое гораздо легче диффундиру-
ет в цитозоль. Оба вещества выполняют функцию
вторичных посредников.
17. Водорастворимые витамины быстро выводятся с
мочой и не накапливаются в организме. Жирора-
створимые витамины очень плохо растворяются в
воде и запасаются в организме в составе липидов.
18. а) Глицерин и натриевые соли пальмитиновой и сте-
ариновой кислот; б) D-глицерин-З-фосфохолин и
натриевые соли пальмитиновой и олеиновой кислот.
19. Растворимость в воде: моноацилглицерин > ди-
ацилглицерин > триацилглицерин.
20. Вещества сходят с колонки в следующем порядке:
холестерил пальмитат и триацилглицерин; холе-
стерин и н-тетрадеканол; фосфатидилхолин и фос-
фатидилэтаноламин; сфингомиелин; фосфатидил-
серин и пальмитат.
21. а) Следует подвергнуть продукты кислотного ги-
дролиза каждого соединения хроматографическо-
му разделению (газожидкостная хроматография
или тонкослойная хроматография на силикагеле)
и сравнить результаты с известными стандартами.
Гидролизат сфингомиелина: сфингозин, жирные
кислоты, фосфохолин, холин и фосфат; гидролизат
цереброзида: сфингозин, жирные кислоты, саха-
ра, но не фосфаты, б) Сильнощелочной гидролиз
сфингомиелина приводит к появлению сфингози-
на, при гидролизе фосфатидилхолина выделяется
глицерин. Состав продуктов гидролиза можно про-
анализировать методом тонкослойной хроматогра-
фии путем сравнения со стандартными вещества-
ми или с помощью реакции с FDNB (окрашенный
продукт образуется только в реакции со сфинго-
зином). Свободные жирные кислоты выделяются
при обработке фосфолипазой At или А2 только
фосфатидилхолина, но не сфингомиелина.
22. Фосфатидилэтаноламин и фосфатидилсерин.
23. a) GM1 и глобозид. И глюкоза, и галактоза относят-
ся к гексозам, так что «гексоза» соответствует экви-
молярной смеси глюкоза + галактоза. Молярные со-
отношения для четырех ганглиозидов следующие:
GM1 - 1:3:1:1, GM2 - 1:2 :1:1, GM3 - 1:2 :0:1,
глобозид - 1 : 3 : 1 : 0. б) Да, соответствует; при бо-
лезни Тея-Сакса накапливается GM2 (1 : 2 : 1 : 1;
см. т. 1, доп. 1-2, рис. 1). в) Этот метод анализа ана-
логичен тому, что был использован Сенгером для
определения аминокислотной последовательности
инсулина. Анализ каждого фрагмента позволяет
найти только его состав, но не последовательность,
но поскольку каждый фрагмент отличается от пре-
дыдущего лишь на один остаток сахара, можно
определить последовательность звеньев. Структу-
ра нормального асиалоганглиозида: церамид-глю-
коза-галактоза-галактозамин-галактоза (что со-
ответствует последовательности, изображенной в
дополнении 10-2 (т. 1), за исключением удаленного
в результате гидролиза остатка Neu5Ac). г) Асиало-
ганглиозид при болезни Тея-Сакса имеет последо-
вательность церамид-глюкоза-галактоза-галакто-
замин, что соответствует данным, представленным
Краткие решения задач и ответы на вопросы |309|
в доп. 10-2 (т. 1). д) Строение нормального асиа-
логанглиозида GM1 следующее: церамид-глюкоза
(2 ОН-группы участвуют в образовании гликозид-
ных связей, 1 ОН-группа задействована в кольце, 3
ОН-группы (2,3,6) свободны и подвергаются мети-
лированию) — галактоза (2 ОН — в гликозидных
связях, 1 ОН — в кольце, 3 ОН (2,4,6) свободны
для галактозамин (2 ОН — в гли-
козидных связях, 1 ОН — в кольце, 1 NH2 (вместо
ОН) и 2 ОН (4,6) свободны для метилирования)-
галактоза (1 ОН — в гликозидной связи, 1 ОН —
в кольце и 4 ОН (2,3,4,6) свободны для метилиро-
вания). е) Не хватает следующих данных: (1) какой
связью соединены между собой звенья и (2) где
присоединена Neu5Ac?
Глава И
1. Зная количество липида, его молекулярную массу и
площадь поверхности монослоя (определенную, как
показано на рисунке), можно рассчитать площадь
поверхности одной молекулы.
2. Данные указывают на существование двойного
липидного слоя в эритроцитах собаки: единичная
клетка с площадью поверхности 98 мкм2 окружена
монослоем липидов площадью 200 мкм2. В случае
овцы и человека данные указывают на то, что клет-
ки покрыты не двойным, а одинарным липидным
слоем. Эти ранние эксперименты были выполнены
с довольно низкой точностью, более поздние экс-
перименты подтвердили существование двойного
липидного слоя во всех случаях.
3. 63 молекулы SDS на одну мицеллу
4. а) Липиды, образующие двойной слой, являются
амфифильными молекулами: они содержат и ги-
дрофильный, и гидрофобный участки. Для миними-
зации области контакта с водой липиды образуют
двумерные слои, в которых гидрофильные участки
обращены к воде, а гидрофобные сосредоточены
внутри липидного слоя. Кроме того, чтобы границы
слоя не контактировали с водой, двойной липидный
слой замыкается сам на себя, б) Эти слои образуют
замкнутые мембраны, обволакивающие клетки и
внутриклеточные отделы (органеллы).
5. 2 нм; длина двух вытянутых друг за другом молекул
пальмитата составляет примерно 4 нм, что прибли-
зительно соответствует толщине типичного двойно-
го слоя липидов.
6. Снижение скорости диффузии. Перемещение от-
дельных молекул липидов в двойном слое проис-
ходит гораздо быстрее при 37 °C, когда липиды на-
ходятся в «жидкой» фазе, чем при 10 °C, когда они
находятся в «твердой» фазе.
7. 35 кДж/моль, если не учитывать влияние трансмем-
бранного электрического потенциала; 0,60 моль.
8. 13 кДж/моль
9. Большая часть потребляемого тканью кислорода
расходуется на окислительное фосфорилирование,
за счет которого образуется основное количество
клеточного АТР. Следовательно, около 2/3 синте-
зируемого почками АТР расходуется на перекачи-
вание ионов К+ и Na+.
10. Не нарушает. Система симпорта может переносить
более одного эквивалента ионов Na+ при переносе
каждого моля глюкозы.
11. Экстракция солью указывает на периферическую
локализацию белка, а недоступность белка для
протеазы в интактных клетках указывает на его
внутриклеточную локализацию. Судя по всему,
белок X является периферическим белком, находя-
щимся на цитозольной стороне мембраны.
12. Гидрофобные взаимодействия между мембранны-
ми липидами являются нековалентными и обрати-
мыми, что позволяет мембранам самопроизвольно
восстанавливаться при повреждении.
13. Температура конечностей ниже температуры туло-
вища. Поскольку клеточные липиды даже при низ-
кой температуре должны сохранять подвижность,
они должны содержать больше ненасыщенных жир-
ных кислот. Ненасыщенные жирные кислоты сни-
жают температуру плавления липидных смесей.
14. Перемещение полярных и иногда заряженных
групп головок через гидрофобную внутреннюю
среду двойного липидного слоя очень невыгодно
энергетически.
15. При pH 7 триптофан несет и положительный, и от-
рицательный заряды, а индол не заряжен. Переме-
щение менее полярного индола через гидрофобную
среду внутри двойного липидного слоя требует
значительно меньших энергетических затрат.
16 .3 - 1012 с
17. Обработать суспензию клеток немеченым NEM в
присутствии избытка лактозы, затем удалить лак-
тозу и добавить NEM, меченный радиоактивной
меткой. Использовать электрофорез в присутствии
SDS для определения МГ полосы с радиоактивной
меткой (переносчик).
18. Построить график гидропатии; гидрофобные
участки 20 или более остатков могут составлять
трансмембранные сегменты. Определить, взаимо-
действует ли белок из интактных эритроцитов с
проходящими через мембрану реагентами, специ-
фичными к первичным аминам. Если да, то вы име-
ете дело с переносчиком типа I.
[310] Приложение Б
19. Переносчик лейцина специфичен к L-изомеру, од-
нако в его центре связывания может связываться
не только L-лейцин, но и L-валин. Снижение Итах в
отсутствии Na+ указывает на перенос лейцина (или
валина) по механизму симпорта с ионами Na\
20. VmdX снижается, Kt не меняется.
21. Около 1%. Оценка получена путем расчета пло-
щади поверхности клетки и с учетом наличия
10 000 копий молекул переносчика (на основании
размеров молекулы гемоглобина в качестве модели
глобулярного белка, см. с. 233).
22.
10 17
Аминокислоты (V, L, F и С) с наибольшим индек-
сом гидропатии сосредоточены на одной стороне
спирали. Амфифильная спираль как бы погружает-
ся в двойной липидный слой этой своей гидрофоб-
ной поверхностью, тогда как другая поверхность
обращена к воде. В альтернативном варианте не-
сколько спиралей могут сближаться своими гидро-
фобными участками, которые при этом также обра-
щены к двойному липидному слою.
23. Примерно 22. Чтобы оценить долю поверхности
мембраны, покрытой фосфолипидами, нужно
знать (или оценить) среднюю площадь поперечно-
го сечения молекулы фосфолипида в двойном слое
(например, с помощью эксперимента, описанного
в проблеме 1 в данной главе) и среднюю площадь
поперечного сечения белка с молекулярной массой
50 кДа.
24. а) Длина участка а-спирали, приходящаяся на одно
основание (гл. 4), составляет около 1,5 А = 0,15 нм.
Чтобы пересечь двойной липидный слой толщи-
ной 4 нм, спираль должна содержать примерно
27 остатков. Следовательно, чтобы сформировать
семь таких участков, белок должен содержать при-
мерно 190 остатков. Белок с массой Мг 64 000 со-
стоит примерно из 580 остатков, б) Для выявления
трансмембранных участков пользуются индексами
гидропатии, в) Поскольку почти половина этого
участка рецептора адреналина состоит из заряжен-
ных остатков, возможно, он представляет собой
внутриклеточную петлю, соединяющую два сосед-
них трансмембранных участка, г) Поскольку спи-
раль состоит в основном из гидрофобных остатков,
этот участок рецептора, вероятно, образует одну из
трансмембранных спиралей.
25. а) Модель А: согласуется. Две темные полосы это
либо слои белка, либо головки фосфолипидов, а
светлое пространство соответственно либо двой-
ной слой, либо гидрофобное ядро. Модель Б: не
согласуется. Данная модель предполагает более
или менее равномерное окрашивание окружающе-
го клетки слоя. Модель В: согласуется, но с одном
условием. Две темные полосы это головки фос-
фолипидов, светлая зона хвосты. Это означает,
что мембранные белки невидимы, поскольку они
не окрашиваю гея осмием или не содержатся в
данном участке мембраны, б) Модель А: согласу-
ется. «Голый» двойной слой (4,5 нм) плюс два слоя
белка (2 нм) в сумме составляют 6,5 нм, что соот-
ветствует экспериментальной толщине мембраны.
Модель Б: ничего нельзя сказать, поскольку данная
модель не предсказывает толщину мембраны. Мо-
дель В: неясно. Результат трудно интерпретировать
в рамках данной модели, поскольку она утвержда-
ет, что мембрана должна иметь такую же толщину,
как «голый» двойной слой (или быть чуть толще
из-за выступающих концов встроенных белков).
Данные согласуются с моделью только при самых
малых из возможных значений толщины мембра-
ны или только если из двойного слоя выступают
большие участки белков, в) Модель Л: неясно.
Результаты трудно интерпретировать в рамках
модели. Если белки связаны с мембраной ионны-
ми взаимодействиями, в соответствии с моделью
белки должны содержать много заряженных ами-
нокислот, но мы видим обратное. Кроме того, по-
скольку слой белка должен быть очень тонким
(см. (6)), для гидрофобного ядра белка остается
очень мало места, так что гидрофобные остатки
должны выступать наружу. Модель Б: согласует-
ся. В белках есть как гидрофобные остатки (вза-
имодействующие с липидами), так и заряженные
остатки (взаимодействующие с водой). Модель
В: согласуется. Белки содержат как гидрофобные
остатки (связанные с мембраной), так и заряжен-
ные остатки (взаимодействующие с водой), г) Мо-
дель А: неясно. Результаты трудно интерпретиро-
вать в рамках модели, поскольку предсказанное ею
значение соотношения площадей составляет точно
Краткие решения задач и ответы на вопросы [311]
2,0, что довольно трудно себе представить при фи-
зиологическом давлении. Модель Б: ничего нельзя
сказать, нет прогноза о количестве липидов в мем-
бране. Модель В: согласуется. Некоторые участки
поверхности мембраны заняты белками, так что
соотношение площадей составляет меньше 2,0,
что и наблюдается в физиологических условиях,
д) Модель А: неясно. В соответствии с данной мо-
делью белки имеют скорее вытянутую, чем гло-
булярную форму, так что модель согласуется с
реальностью только при предположении, что на-
ходящиеся на поверхности белки содержат спи-
ральные фрагменты. Модель Б: согласуется. В
рамках модели белки имеют глобулярную струк-
туру (содержащую несколько спиральных участ-
ков). Модель В: согласуется. В рамках модели
белки имеют в основном глобулярную структуру,
е) Модель А: неясно. Фосфатные головки защи-
щены слоем белка, но от действия фосфолипа-
зы фосфолипиды защищены лишь в том случае,
если белки полностью покрывают поверхность.
Модель Б: согласуется. Большинство поляр-
ных головок доступно для действия фосфоли-
пазы. Модель В: согласуется. Все полярные го-
ловки доступны для действия фосфолипазы,
ж) Модель А: не согласуется. Белки полностью до-
ступны для расщепления трипсином и практиче-
ски все расщепляются во многих местах, включая
гидрофобные участки. Модель Б: не согласуется.
Практически все белки находятся внутри двой-
ного слоя и недоступны для действия трипсина.
Модель В: согласуется. Участки белков, пронизы-
вающие двойной слой или погруженные в него,
защищены от действия трипсина, а те, что оказыва-
ются на поверхности, подвергаются расщеплению.
Устойчивые к действию трипсина участки отлича-
ются высоким содержанием гидрофобных остатков.
Глава 12
1. Вещество X — это сАМР; его синтез активируется
адреналином, а) При центрифугировании происхо-
дит осаждение аденилатциклазы, катализирующей
образование сАМР. б) Добавление сАМР стимули-
рует гликогенфосфорилазу. в) сАМР устойчив при
нагревании, и его можно получить путем обработки
АТР гидроксидом бария.
2. В отличие от сАМР дибутирил-сАМР легко прони-
кает через плазматическую мембрану.
3. а) Повышает концентрацию сАМР; б) сАМР ре-
гулирует транспорт Na+; в) требуется постоянно
восполнять водный и электролитный баланс в ор-
ганизме.
4. а) В результате мутации R-субъединица теря-
ет способность связывать С-субъединицу, так
что С-субъединица остается постоянно актив-
ной; б) мутация нарушает связывание сАМР с
R-субъединицей, так что С-субъединица ингиби-
руется связанной R-субъединицей.
5. Альбутерол повышает концентрацию сАМР, что
приводит к релаксации и расширению бронхов и
бронхиол. Поскольку p-адренергические рецеп-
торы контролируют и многие другие процессы,
лекарство вызывает ряд нежелательных побоч-
ных эффектов. Для их минимизации нужен спец-
ифический агонист подтипа р-адренергических
рецепторов, содержащихся в гладких мышцах
бронхов.
6. Разрушение гормона, гидролиз GTP, связанного с
G-белком, расщепление, метаболизм или удаление
вторичного посредника, десенсибилизация рецеп-
тора, удаление рецептора от поверхности клетки.
7. Получить слитые белки, состоящие из CFP и
p-аррестина, YFP и цитоплазматического до-
мена p-адренергического рецептора (или на-
оборот). Слитые белки облучить светом с дли-
ной волны 433 нм и измерить пропускание при
476 и 527 нм. Если взаимодействие происходит,
то при добавлении к клеткам, экспрессирую-
щим слитый белок, адреналина должно наблю-
даться увеличение пропускания при 527 нм и
уменьшение пропускания при 476 нм. Если вза-
имодействия нет, проходит только свет с длиной
волны 476 нм. Причин этого может быть несколько:
(1) слитые белки неактивны или не могут взаи-
модействовать; (2) они не попадают к месту своей
обычной локализации; (3) они подвергаются про-
теолитическому расщеплению.
8. Вазопрессин повышает концентрацию Са2+ до
10 6 М, активируя протеинкиназу С. Инъек-
ции ЭГТА блокируют действие вазопрессина,
но не влияют на реакцию на глюкагон, который
использует в качестве вторичного посредника
сАМР, а не Са2+.
9. Амплификация сигнала является результатом
ферментативного каскада, когда один фермент
активирует молекулы другого фермента и т. д.,
например каскада с участием рецептора инсу-
лина, IRS-1, Raf, МЕК, ERK; ERK активирует
транскрипционный фактор, который стимулиру-
ет синтез мРНК.
10. Мутация гена ras, имеющая следствием инак-
тивацию СТРазной активности Ras, приводит
к образованию белка, который после активации
путем связывания GTP продолжает (через Raf)
передавать сигнал инсулина.
13121 Приложение Б
11. Общие свойства Ras и Gs: оба связывают либо GDP,
либо GTP; оба активируются GTP; оба, будучи ак-
тивированными, активируют следующие фермен-
ты каскада; оба обладают собственной GTРазной
активностью, которая отключает их через короткий
период времени. Различия между Ras и Gs: Ras -
маленький мономерный белок, Gs - гетеротример.
Функциональное различие между Gsu (^заключает-
ся в том, что Gs активирует аденилатциклазу, a G!
ее ингибирует.
12. Типы протеинкиназ и факторы их активации
(в скобках): РКА (сАМР), PKG (cGMP), РКС
(Са2+, ДАГ), Са2+/СаМ-киназа (Са2+, СаМ), ци-
клин-зависимая киназа (циклин), тирозинкиназа
(лиганд рецептора, например инсулин), МАРК
(Raf), Raf (Ras), киназа гликогенфосфорилазы
(РКА).
13. При связывании негидролизуемого аналога GTP
Gs остается в активированной форме; таким обра-
зом, аналог продлевает действие адреналина.
14. а) Используйте шарики со связанным «-бунгаро-
токсином для очистки ацетилхолинового рецеп-
тора (см. рис. 3-17, с. 134). Экстрагируйте белки
из электрического органа ската и пропустите
смесь через хроматографическую колонку; рецеп-
тор ацетилхолина связывается с носителем селек-
тивным образом. Смойте белок раствором, осла-
бляющим взаимодействие с а-бунгаротоксином.
б) Используйте связывание [1251]а-бунгаро-
токсина для количественного определения аце-
тилхолинового рецептора при разных методах
очистки. На каждой стадии очистки определяйте
содержание ацетилхолинового рецептора, изме-
ряя связывание [1251]а-бунгаротоксипа с белками
в образце. Оптимизируйте очистку, чтобы добить-
ся максимальной удельной активности рецептора
(имп./мин связанного [1251]а-бунгаротоксина на
мг белка) в конечном образце.
15. а) Нет, не могли. Если бы мембранный потен-
циал Vm определял проницаемость мембраны по
ионам К, то по уравнению Нернста Vin = -90 мВ,
а не -95 мВ, так что мембранный потенциал за-
вит от проводимости других ионов, б) Возможно,
определяющее влияние на Vm оказывает транс-
порт ионов хлора; ЕС] = -94 мВ.
16. а) Vm мембраны ооцита изменяется от -60 до
-10 мВ, т. е. происходит деполяризация мембраны,
б) Влияние КС1 зависит от притока ионов Са2+ из
внеклеточной среды.
17. Гиперполяризация приводит к закрытию потен-
циалзависимых Са2+-каналов в пресинаптическом
отделе палочек сетчатки. Происходящее в резуль-
тате этого снижение концентрации Са2+ приводит
к ослаблению ингибирующего нейромедиатора,
что снижает активность следующего нейрона.
Когда в ответ на световой стимул это ингибиро-
вание снимается, сигнальный путь активируется,
и происходит возбуждение зрительных центров в
головном мозге.
18. а) В этой ситуации нарушается приток ионов Na+
и Са2+ в клетки в ответ на световой сигнал; колбоч-
ки теряют способность передавать в мозг сигнал
о том, что до них дошел свет. Палочки сохраняют
свою функцию, поэтому человек с такой мутацией
сохраняет зрение, но теряет способность различать
цвета, б) В этом случае останавливается отток из
клеток ионов К+, что приводит к деполяризации
мембраны р-клеток и конститутивному высвобож-
дению инсулина в кровь, в) АТР отвечает за закры-
тие этого канала, так что канал остается открытым,
а это мешает деполяризации мембраны р-клеток и
высвобождению инсулина.
19. При болезни Огучи может быть повреждена родоп-
синкиназа или аррестин.
20. Мембранный потенциал палочек больше не из-
меняется в ответ на световой сигнал. Такой экс-
перимент был проделан. Освещение активиро-
вало фосфодиэстеразу, но фермент мало снижал
уровень 8-Br-cGMP, который был намного выше,
чем требуется для открытия ионных каналов. По-
этому свет не вызывал изменения мембранного
потенциала.
21. а) При нагревании каналы TRPV1 открываются,
что вызывает приток Na+ и Са2+ в чувствитель-
ный нейрон. Это приводит к деполяризации мем-
браны и запускает потенциал действия. Когда
потенциал действия достигает окончания аксо-
на, происходит высвобождение нейромедиатора,
сигнализирующего нервной системе о воспри-
ятии тепла, б) Капсаицин имитирует действие
тепла, открывая каналы TRPV1 при низкой тем-
пературе, что вызывает ложное ощущение тепла.
Чрезвычайно низкое значение ЕС50 говорит о
том, что капсаицин оказывает сильнейшее дей-
ствие даже в очень малом количестве, в) В низ-
кой концентрации ментол должен открывать ка-
налы TRPM8, что вызывает ощущение холода; в
более высокой концентрации он открывает кана-
лы и TRPM8, и TRPV3, что вызывает смешанное
ощущение холода и тепла, которое знакомо тем,
кто ел мятные конфеты с очень концентрирован-
ным вкусом.
22. а) Эти мутации могут привести к перманентной
активации рецептора PGE2, что нарушает регуля-
цию клеточного цикла и образованию опухоли,
б) Вирусные гены могут кодировать конститу-
Краткие решения задач и ответы на вопросы [ 3131
тивно активную форму рецептора, заставляя
клетку постоянно делиться, что вызывает обра-
зование опухоли, в) Белок Е1А может связывать-
ся с pRb и препятствовать связыванию E2F, так
что E2F остается в активном состоянии, что спо-
собствует неконтролируемому делению клетки,
г) Обычно клетки легкого не реагируют на PGE2,
поскольку они не экспрессируют соответствую-
щих рецепторов; мутации, вызывающие консти-
тутивную активацию рецептора PGE2, не затра-
гивают легочную ткань.
23. В норме ген опухолевого супрессора кодирует бе-
лок, ограничивающий клеточное деление. Мутант-
ная форма белка не подавляет деление клеток, но
если одна из двух аллелей кодирует нормальный
белок, сохраняется нормальная функция белка.
Онкоген в норме кодирует регуляторный белок,
который запускает клеточное деление, но только в
присутствии правильного сигнала (фактора роста).
Продукт мутантного гена непрерывно заставляет
клетку делиться вне зависимости от присутствия
фактора роста.
24. У ребенка с множественными опухолями в обо-
их глазах с самого рождения все клетки сетчатки
содержат дефектную копию гена Rb. Когда-то в
раннем возрасте в нескольких клетках незави-
симым образом происходят дополнительные му-
тации, повреждающие нормальную копию гена
Rb, что вызывает развитие опухоли. У ребенка
с единственной опухолью при рождении обе ко-
пии гена Rb нормальные; мутации обеих алле-
лей Rb в одной и той же клетке (что происходит
очень редко) вызывают появление единичной
опухоли.
25. Клетки, на поверхности которых экспрессируется
один и тот же рецептор, могут иметь разный набор
белковых мишеней для фосфорилирования.
26. а) Клеточная модель, предсказывающая существо-
вание разных рецепторов на клетках разного типа,
б) Эксперимент подтверждает независимость
разных вкусовых ощущений. Даже если у живот-
ного нет рецепторов сладкого и/или рецепторов
умами, другие вкусовые ощущения сохраняются
в норме; таким образом, приятные и неприятные
ощущения взаимо независимы, в) Да. Потеря
субъединицы T1R1 или T1R3 лишает возмож-
ности воспринимать вкус умами, г) Обе модели.
В соответствии с любой из моделей удаление од-
ного рецептора влечет потерю соответствующего
вкусового ощущения, д) Да. Потеря субъедини-
цы T1R2 или T1R3 приводит почти к полному
исчезновению ощущения сладости; для полного
исчезновения этого ощущения требуется уда-
ление обеих субъединиц, е) При очень высокой
концентрации сахарозы сладкий вкус могут де-
тектировать гомодимерные рецепторы T1R2 и в
меньшей степени T1R3. ж) Результаты согласу-
ются с обеими моделями. Связывание лиганда
может быть отделено от вкусового ощущения.
Если рецептор сладкого связывает любую дру-
гую молекулу, мышь воспринимает ее как слад-
кое вещество.
Глава 13
1. В данном случае система представлена развива-
ющимся цыпленком; питательные вещества, обо-
лочка яйца и внешний мир - окружающая среда.
С одной стороны, превращение единственной
клетки в цыпленка сопровождается чрезвычайно
сильным снижением энтропии системы. С дру-
гой стороны, в начальный момент времени части
яйца вне зародыша (окружающая среда) содержат
сложные молекулы (состояние с низким уровнем
энтропии). В процессе инкубации некоторые из
этих сложных молекул превращаются в простые
молекулы СО2 и Н2О (энтропия повышается). Это
повышение энтропии в окружающей среде боль-
ше, чем снижение энтропии в процессе развития
зародыша (в системе).
2. а) -4,8 кДж/моль; б) 7,56 кДж/моль;
в) -13,7 кДж/моль.
3. а) 262; б) 608; в) 0,30.
4. К\ = 21; \G'° = -7,6 кДж/моль.
5. -31 кДж/моль.
6. а) -1,68 кДж/моль; б) -4,4 кДж/моль; в) При за-
данной температуре &G’° любой реакции имеет
фиксированное значение, определенное в стандарт-
ных условиях (в данном случае при концентрации
фруктозо-6-фосфата и глюкозо-6-фосфата 1 М).
Напротив, АС - величина вариабельная, которая
рассчитывается заново при каждом изменении
концентраций реагирующих веществ и продуктов
реакции.
7. AG'°~0;/Ceq~ 1.
8. Меньше. Суммарный процесс гидролиза АТР мож-
но аппроксимировать следующим уравнением:
АТР4’ + Н2О ADP3 + НРО42 + Н+
Надо понимать, что частицы, несущие заряд, доми-
нируют, но могут присутствовать и другие формы.
При стандартных условиях ([АТР] = [ADP] = [PJ =
1 М) концентрация воды составляет 55 М и не из-
меняется в ходе реакции. Поскольку в этой реакции
образуются ионы Н+, при более высокой концен-
[314] Приложение Б
трации [Н ] (pH 5 0) равновесие сдвигается влево и
выделяется меньше свободной энергии
9. 10.
При низком соотношении [АТР] [ADP] (« 1)
AG гидролиза АТР уменьшается. При данной
концентрации АТР доступное для клетки ко-
личество энергии понижается при уменьшении
отношения | АТР]/[ ADP] и повышается при его
увеличении.
11. а)3 ,85 • 10 3 М; концентрация глюкозо-6-фосфата
составляет 8,9 • 10 8 М; неприемлема, б) ИМ:
поскольку максимальная растворимость глюко-
зы менее 1 М, эта стадия не имеет смысла, в) 837
(АС'0 = - 16,7 кДж/моль); концентрация глюкозы
составляет 1,2 • 10 7 М; да. г) Неразумен. Для это-
го требуется столь высокая концентрация Рр при
которой фосфаты двухвалентных катионов выпа-
дают в осадок, д) Прямой перенос фосфорильной
группы от АТР на глюкозу использует потенциал
переноса фосфорильной группы АТР и не сопро-
вождается образованием высокой концентрации
промежуточных соединений. Ключевым момен-
том этого процесса является ферментативный
катализ.
12. а) -12,5 кДж/моль; б) 14,6 кДж/моль.
13. а) 3-10 4; б) 68,7; в) 7,4 • 104.
14. 13 кДж/моль.
15. 46,7 кДж/моль.
16. В результате изомеризации карбонильная группа
перемещается от атома С1 к атому С2, обеспечи-
вая расщепление связи между атомами СЗ и С4.
Если бы не происходило изомеризации, расще-
плялась бы связь между атомами С2 и СЗ, что
приводило бы к образованию одной молекулы из
двух атомов углерода и одной молекулы из четы-
рех атомов углерода.
17. Механизм аналогичен механизму реакции, ката-
лизируемой алкогольдегидрогеназой (рис. 14-13,
т. 2).
18. Первая стадия — реакция, обратная альдольной
конденсации (рис. 14-5, т. 2), вторая стадия - реак-
ция альдольной конденсации (рис. 13-4, т. 2).
19. а) 46 кДж/моль; б) 46 кг; 68%: в) АТР синтезиру-
ется по мере необходимости, а потом распадается
на ADP и Рь так что в организме его концентрация
поддерживается на стационарном уровне
20. Концентрация АТР находится в динамическом
стационарном состоянии; она остается постоян-
ной, поскольку скорость потребления АТР равна
скорости его синтеза. При гидролизе АТР отще-
пляется концевая (у) фосфорильная группа; при
синтезе АТР из ADP происходит замена этой
группы. Таким образом, концевая фосфорильная
группа находится в быстром обороте. Напротив,
оборот центральной (Р) фосфорильной группы
довольно медленный.
21. а) 1,7 кДж/моль; б) неорганическая пирофосфата-
за катализирует гидролиз пирофосфата и направ-
ляет всю реакцию в целом в сторону образования
ацетил-СоА.
22. 36 кДж/моль.
23. a) NAD NADH; б) нируват/лактат; в) образова-
ние лактата; г) -26,1 кДж/моль; д) 3,63 • 104.
24. а) 1,14 В; б) -220 кДж/моль; в) ~ 4.
25. а) 0,35 В; б) 0,320 В; в) 0,29 В.
26. В порядке усиления тенденции: (а), (г), (б), (в).
27. (в) и (г).
28. а) Состояние с наименьшей энергией и макси-
мальной энтропией достигается при равенстве
концентраций краски в двух клетках. Если меж-
клеточный контакт типа «ловушки для рыбы»
позволяет осуществлять однонаправленный
транспорт, в конечном итоге в олигодендроците
окажется больше краски, чем в астроците. Это
состояние отличается от исходного состояния бо-
лее высокой энергией и более низкой энтропией,
что противоречит второму началу термодинами-
ки, т. е. модель предполагает самопроизвольное
уменьшение энтропии, что невозможно. В энер-
гетических терминах модель предусматривает са-
мопроизвольный переход системы из состояния с
низкой энергией в состояние с высокой энергией
без поступления энергии извне, что опять-таки
невозможно, б) Молекулы, в отличие от рыб, не
проявляют поведенческих реакций, а находятся в
случайном броуновском движении. Диффузия
приводит к суммарному перемещению моле-
Краткие решения задач и ответы на вопросы 13151
кул из области с более высокой концентрацией
в область с более низкой концентрацией по той
простой причине, что при большей численности
молекул (область с высокой концентрацией) ве-
роятность того, что какая-нибудь из них подой-
дет к соединительному каналу, выше. Рассмотрим
процесс с лимитирующей стадией, когда моле-
кулы входят в узкий канал. Узкий конец канала
ограничивает скорость молекул, поскольку при
любом случайном движении молекул вероят-
ность их попадания туда низкая. Широкий конец
канала — это не воронка для молекул (как это мо-
жет быть для рыб), поскольку молекулы не «сдав-
ливаются» стенками сужающегося канала, как
рыбы в ловушке. Узкий конец ограничивает ско-
рость перемещения одинаково в обоих направ-
лениях. При одинаковой концентрации с обеих
сторон выравнивается и скорость перемещения
в обоих направлениях, и концентрация раство-
ренного вещества остается постоянной, в) Пове-
дение рыб не является случайным - оно зависит
от внешних условий. Рыбы, попавшие в широкий
раструб капала, продолжают двигаться вперед по
той причине, что они вообще предпочитают дви-
гаться вперед, и по мере сужения канала они на-
чинают подвергаться «сдавливанию». Им легко
войти в широкий раструб, но выйти из ловушки
гораздо труднее, поскольку трудно пройти узкий
выход, г) Возможных объяснений может быть не-
сколько, и некоторые из них были предложены
критиками данной статьи. Вот два возможных
объяснения. (1) В олигодендроците краситель мо-
жет связываться с другими молекулами. Связы-
вание приводит к тому, что молекулы красителя
перестают быть частью «свободного» раствора,
но продолжают быть видны в флуоресцентный
микроскоп. (2) Краситель может накапливаться
в определенном компартменте (органелле) олиго-
дендроцита либо за счет активного транспорта с
затратой АТР, либо за счет взаимодействия с дру-
гими молекулами в этой органелле.
Глава 14
1. Суммарное уравнение:
Глюкоза + 2АТР -*
2 глицеральдегид-3-фосфат+ 2ADP + 2Н+;
АС'° = 2,1 кДж/моль
2. Суммарное уравнение:
2 Глицеральдегид-З-фосфат + 4ADP + 2Р, -*
2 лактат + 2NAD+;
АС'° = -114 кДж/моль
3. GLUT2 (и GLUT1) содержится в печени и всегда
локализован в плазматической мембране гепато-
цитов. GLUT3 всегда присутствует в плазматиче-
ской мембране некоторых клеток головного мозга.
В норме GLUT4 заключен в везикулы клеток
мышц и жировой ткани и проникает в плазматиче-
скую мембрану только по сигналу инсулина. Таким
образом, печень и головной мозг могут потреблять
глюкозу из крови вне зависимости от выработки
инсулина, тогда как мышцы и жировая ткань по-
требляют глюкозу только при повышении уровня
инсулина, вызванного повышением уровня глюко-
зы крови.
4. СН3СНО + NADH + Н+ СН3СН2ОН + NAD+;
= 1,45 • 104.
5. -8,6 кДж/моль.
6. а) 14СН3СН2ОН; б) [3-14С]глюкоза или [4-14С]глю-
коза.
7. В ходе ферментации выделяется энергия, некото-
рое количество которой запасается в форме АТР,
но значительная часть рассеивается в виде тепла.
Если ферментер не охлаждать, микроорганизмы
могут погибнуть от перегрева.
8. В соевых бобах и в зернах пшеницы содержится
крахмал — полимер глюкозы. Микроорганизмы
расщепляют крахмал до глюкозы, превращают
глюкозу в пируват и, поскольку процесс идет в бес-
кислородной среде (это ферментация), далее пре-
вращают пируват в молочную кислоту и этиловый
спирт. В присутствии О2 пируват окислялся бы до
ацетил-СоА, а затем до СО2 и Н2О. Однако в при-
сутствии кислорода некоторое количество ацетил-
СоА также превращалось бы в уксусную кислоту
(уксус).
9. С1. Эксперимент демонстрирует обратимость аль-
долазной реакции. С1 в глицеральдегид-3-фос-
фате эквивалентен С4 в фруктозо-1,6-бисфосфате
(см. рис. 14-6, т. 2). Исходный глицеральдегид-
3-фосфат содержал меченый атом С1. Атом СЗ в
дигидроксиацетонфосфате становится меченым в
реакции, катализируемой триозофосфатизомера-
зой, в результате образуется меченный по атому СЗ
фруктозо-1,6-бисфосфат.
10. Не принесет. Анаэробный синтез АТР не происхо-
дил бы, а аэробный синтез АТР ослаб бы незначи-
тельно.
11. Нет, не смогли бы. Лактатдегидрогеназа необходи-
ма для рецикла NAD+ из NADH, образовавшегося в
ходе окисления глицеральдегид-3-фосфата.
12. Превращение глюкозы в лактат происходит в мио-
цитах при недостатке кислорода и в этих условиях
[316] Приложение Б
обеспечивает образование АТР. Поскольку лактат
может окисляться до пирувата, глюкоза расходует-
ся не напрасно. При достаточном количестве кис-
лорода пируват окисляется в аэробных реакциях.
Такая гибкость метаболизма позволяет организму
лучше приспосабливаться к изменяющимся усло-
виям внешней среды.
13. Клетка быстро удаляет 1,3-бисфосфоглицерат на
следующей благоприятной стадии процесса, ката-
лизируемой фосфоглицераткиназой.
14. а) Продукт — 3-фосфоглицерат. б) В присутствии
арсената в анаэробных условиях суммарный выход
АТР равен нулю.
15. а) При спиртовом брожении расходуется 2 моль
Pi на 1 моль глюкозы, б) Этанол — это восстанов-
ленный продукт, образующийся в процессе об-
ратного окисления NADH до NAD+, а СО2 — по-
бочный продукт превращения пирувата в этанол.
Пируват должен превращаться в этанол, чтобы
обеспечивать непрерывный источник NAD+ для
окисления глицеральдегид-3-фосфата. Накапли-
вается фруктозо-1,6-бисфосфат, образующийся
в качестве промежуточного продукта гликолиза,
в) Арсенат вытесняет Р, в реакции глицеральдегид-
3-фосфатдегидрогеназы, в результате чего об-
разуется ациларсенат, подвергающийся далее
спонтанному гидролизу. Это препятствует обра-
зованию АТР, но метаболизм 3-фосфоглицерата
продолжается.
16. Ниацин из пищи используется для синтеза NAD+.
Осуществляемые под действием NAD+ окисли-
тельные реакции являются частью циклического
процесса, в которых NAD+ выступает в роли пере-
носчика электронов (восстановительного агента).
Одна молекула NAD+ может окислить несколько
тысяч молекул глюкозы, поэтому потребность в ви-
тамине-предшественнике (ниацине) сравнительно
невысокая.
17. Дигидроксиацетонфосфат + NADH + Н+ -* 3-гли-
церофосфат + NAD+ (катализатор — дегидрогеназа).
18. Недостаточность галактокиназы: галактоза (ме-
нее токсична); недостаточность галактозо-1 -фос-
фат-уридилилтрансферазы: галактозо-1 -фосфат
(более токсичен).
19. Белки расщепляются до аминокислот, которые ис-
пользуются в глюконеогенезе.
20. а) В реакции пируваткарбоксилазы к пирувату
присоединяется 14СО2, но на следующей стадии
ФЕП-карбоксикиназа перемещает тот же СО2.
Таким образом, сначала 14С в глюкозу не включа-
ется.
б)
СН3 с=о 1 сн2 —> с=о — 1 сн2 II С—ОРО3_ 1
14СОО- 14СОО~ 14 000
1-14С-Пируват Оксалоацетат Фосфоенолпирувг
СН2ОРО^
н—с—он сн2орог СН2ОН 1
1 14с-орог н—с—он Н—С—ОРО;
II < 1 < 1
О 14СОО- 14СОО-
1,3-Бисфо- З-Фосфо- 2-Фосфо-
сфоглицерат глицерат глицерат
сн2орог
Н—С—ОН । СН2ОРО|
14 С—н II = с=о 1
II О 14СН2ОН
Глицеральдегид-
3-фосфат
Дигидроксиацетон-
фосфат
2"О3РОН2С СН2ОРОГ
нонс с=о
|| —> —► 3,4-14С-Глюкоза
НО—14 С—14 С—ОН
I I
н н
Фруктозо-1,6-бисфосфат
21. Четыре эквивалента АТР на молекулу глюкозы.
22. Если бы это было так, глюконеогенез был бы очень
высокоэндергоническим процессом и исчезла воз-
можность независимой регуляции гликолиза и
глюконеогенеза.
23. Для превращения пирувата в фосфоенолпируват
клетка затрачивает 1 молекулу АТР и 1 молекулу
GTP.
24. (а), (б) и (г) — глюкогенные, (в) и (д) — нет.
25. Потребление алкоголя усиливает конкуренцию за
NAD+ при метаболизме этанола и глюконеогенезе.
Ситуация усугубляется активной физической на-
грузкой и отсутствием достаточного питания, по-
скольку они также способствуют понижению уров-
ня глюкозы крови.
26. а) Быстрое усиление гликолиза; повышение уров-
ня пирувата и NADH приводит к повышению
Краткие решения задач и ответы на вопросы 13 Ь |
уровня лактата, б) Лактат превращается в глюко-
зу через стадию образования пирувата. Это мед-
ленный процесс, поскольку скорость образования
пирувата ограничена доступностью NAD+, равно-
весие ЛДГ-реакции сдвинуто в сторону образова-
ния лактата, а превращение пирувата в глюкозу
требует энергетических затрат, в) Равновесие ре-
акции, катализируемой ЛДГ, сдвинуто в сторону
образования лактата.
27. В печени лактат превращается в глюкозу в резуль-
тате глюконеогенеза (рис. 14-15 и 14-16, т. 2). Де-
фект фруктозо-1,6-бисфосфатазы в гепатоцитах
препятствует включению лактата в глюконеогенез,
в результате чего лактат накапливается в крови.
28. Сукцинат превращается в оксалоацетат, кото-
рый транспортируется в цитозоль и превраща-
ется в фосфоенолпируват под действием ФЕП-
карбоксикиназы. В соответствии с реакцией,
изображенной на рис. 14-16 (т. 2), для образования
1 моль глюкозы требуется 2 моль ФЕП.
29. Одновременное функционирование анаболиче-
ских и катаболических путей метаболизма глюко-
зы приводит к холостому циклу АТР, сопровожда-
ющемуся дополнительным потреблением О2.
30. Накопление рибозо-5-фосфата, как минимум,
приводило бы к обращению направления реак-
ции в соответствии с законом действующих масс
(уравнение 13-4, т. 2). Кроме того, это могло бы
повлиять и на другие метаболические реакции с
участием рибозо-5-фосфата в качестве субстра-
та или продукта, например на реакции синтеза
нуклеотидов.
31. а) Устойчивость к этанолу, скорее всего, связана
еще со многими другими генами, поэтому та-
кой путь был бы значительно более сложным,
б) L-Арабинозоизомераза (ген агаА) превраща-
ет альдозу в кетозу путем перемещения карбо-
нильной группы в нефосфорилированном сахаре
от атома С1 на атом С2. Подобные ферменты в
данной главе не упоминались; все обсуждавшие-
ся ранее ферменты действуют на фосфорилиро-
ванные сахара. Фермент, осуществляющий ана-
логичные превращения фосфорилированного
фермента, называется фосфогексозоизомеразой.
L-Рибулокиназа (агаВ) фосфорилирует сахар
по атому С5, перенося у-фосфорильную группу
от АТР. В данной главе описано несколько по-
добных реакций, включая реакцию гексокиназы.
Ь-Рибулозо-5-фосфат-эпимераза (araD) пере-
мещает Н- и ОН-группы у хирального атома са-
хара. В данной главе аналогичные реакции не
были описаны, но будут обсуждаться в гл. 20 (см.
рис. 20-1, т. 2). в) Продукты трех генов ага пре-
вращали арабинозу в ксилулозо-5-фосфат в со-
ответствии со следующей последовательностью
реакций: арабиноза (L-арабинозоизомераза) -* L-ри -
булоза (L-рибулокиназа) Ь-рибулозо-5-фосфат
(эпимераза) ксилулозо-5-фосфат. г) Арабиноза
превращается в ксилулозо-5-фосфат, как описа-
но в (в), который включается в метаболический
путь, изображенный на рис. 14-22; образующий-
ся глюкозо-6-фосфат далее ферментируется до
СО2 и этилового спирта, д) 6 молекул арабино-
зы + 6 молекул АТР превращаются в 6 молекул
ксилулозо-5-фосфата, который включается в
путь, изображенный на рис. 14-22 (т. 2), и превра-
щается в 5 молекул глюкозо-6-фосфата, каждая
из которых подвергается ферментации с образо-
ванием 3 молекул АТР — всего 15 молекул АТР.
Таким образом, суммарный выход составляет
15 АТР - 6 АТР = 9 АТР из 6 молекул арабинозы.
Кроме того, в результате процесса образуются 10
молекул этанола и 10 молекул СО2. е) С учетом
более низкого выхода АТР, для такого же роста в
пересчете на эквиваленты АТР бактерия Z. тоЫ-
lis должна ферментировать больше арабинозы
и, следовательно, производить больше спирта,
ж) Один из способов, который заставил бы бак-
терию утилизировать ксилозу, заключается во
введении двух дополнительных генов: аналога
гена araD, продукт которого превращает ксилозу
в рибозу, переставляя Н- и ОН-группы у атома
СЗ, и аналога гена агаВ, продукт которого фос-
форилирует рибозу по атому С5. Образующийся
рибозо-5-фосфат может включаться в существу-
ющий метаболический путь.
Глава 15
1. а) 0,0293; б) 308; в) Нет. Q » К'Ч|, и это означает,
что в клетках реакция PFK-1 далека от равновесия.
Эта реакция протекает медленнее, чем следующие
реакции гликолиза. Поток метаболитов по пути гли-
колиза в значительной степени определяется актив-
ностью PFK-1.
2. а) 1,4-10'9 М; б) Физиологическая концентрация
(0,023 мМ) в 16 000 раз выше равновесной кон-
центрации. Данная реакция в клетках не достигает
равновесия. Многие реакции в клетках далеки от
равновесия.
3. Когда нет кислорода, потребности в АТР удовлет-
воряются путем анаэробного метаболизма глюкозы
(ферментации до лактата). Поскольку при аэроб-
ном окислении глюкозы образуется намного больше
АТР, чем при брожении, для образования того же
количества АТР требуется меньше глюкозы.
[3181 Приложение Б
4. а) Фермент имеет два центра связывания АТР —
активный и регуляторный. Связывание АТР в
регуляторном центре ингибирует PFK-1, снижая
Vinax или повышая Км для связывания АТР в ак-
тивном центре, б) В условиях избытка АТР по-
ток метаболитов по пути гликолиза замедляется,
в) График показывает, что повышение концентра-
ции ADP подавляет ингибирование под действи-
ем АТР. Поскольку пул адениновых нуклеотидов
почти постоянен, потребление АТР вызывает по-
вышение концентрации ADP. Данные показыва-
ют, что активность PFK-1 может регулироваться
соотношением [ATP]/[ADP],
5. При pH 7,0 фосфатная группа глюкозо-6-фосфата
полностью ионизирована, в результате чего моле-
кула в целом приобретает отрицательный заряд.
Поскольку мембраны обычно непроницаемы для
заряженных молекул, глюкозо-6-фосфат не мо-
жет проходить из крови в клетки и, следователь-
но, не может включаться в гликолиз и исполь-
зоваться для синтеза АТР. (По этой же причине
фосфорилированная глюкоза не может выйти из
клетки.)
6. а) В мышцах: распад гликогена в процессе глико-
лиза поставляет клеткам энергию (в форме АТР).
Гликогенфосфорилаза катализирует превращение
запасенного гликогена в глюкозо-1-фосфат, ко-
торый превращается в промежуточный продукт
гликолиза глюкозо-6-фосфат. При активной фи-
зической нагрузке скелетным мышцам требует-
ся больше глюкозо-6-фосфата. В печени: распад
гликогена поддерживает постоянный уровень
глюкозы крови между приемами пищи (глюкозо-
6-фосфат превращается в свободную глюкозу),
б) В активно работающих мышцах потребность в
АТР очень высока, так что глюкозо-1-фосфат дол-
жен образовываться быстро, чем и объясняется
большое значение Vmax.
7. а) [Р]]/[глюкозо-1-фосфат] = 3,3/1; б) и в) Вели-
чина этого отношения в клетке (> 100 : 1) указыва-
ет, что концентрация глюкозо-1-фосфата намного
ниже равновесного значения. Скорость удаления
глюкозо-1-фосфата (путем включения в гликолиз)
выше скорости его образования (в реакции гли-
когенфосфорилазы), так что поток метаболитов
направлен от гликогена к образованию глюкозо-1-
фосфата. Реакция гликогенфосфорилазы, вероят-
но, является регуляторной стадией в процессе рас-
щепления гликогена.
8. а) Повысится; б) понизится; в) повысится
9. В покое\ [АТР] — высокая; [АМР] — низкая;
[ацетил-СоА] и [цитрат] — промежуточная. При
бегстве: [АТР] — промежуточная; [АМР] — вы-
сокая; [ацетил-СоА] и [цитрат] — низкая. Во
время спринта при анаэробном гликолизе поток
глюкозы усиливается по следующим причинам:
(1) ингибирование гликогенфосфорилазы и PFK-1
под действием АТР частично снимается; (2) АМР
стимулирует оба фермента; (3) при снижении кон-
центрации цитрата и ацетил-СоА ослабляется их
ингибирующее действие по отношению к PFK-1 и
пируваткиназе соответственно.
10. Перелетные птицы эффективно используют ана-
эробное окисление жиров (в отличие от бегущего
кролика, использующего анаэробный метаболизм
глюкозы). Птицы берегут мышечный гликоген для
экстренных случаев, когда внезапно может потре-
боваться значительный расход энергии.
И. Случай 1\ е, 3; случай 2: в, 3; случай 3: 3,4;
случай 4: г, 6.
12. а) (1) Жировая ткань: синтез жирных кислот за-
медляется. (2) Мышцы: гликолиз, синтез жирных
кислот и синтез гликогена замедляются. (3) Пе-
чень: гликолиз ускоряется, глюконеогенез, синтез
гликогена и синтез жирных кислот замедляются;
пептозофосфатный цикл происходит без изме-
нений. б) (1) Жировая ткань и (3) печень: синтез
жирных кислот замедляется, поскольку недоста-
ток инсулина приводит к инактивации ацетил-
СоА-карбоксилазы, первого фермента на пути
синтеза жирных кислот. Синтез гликогена инги-
бируется сАМР-зависимым фосфорилированием
гликогенсинтазы. (2) Мышцы: гликолиз замед-
ляется, поскольку GLUT4 неактивен, что делает
невозможным захват глюкозы. (3) Печень: глико-
лиз замедляется, поскольку бифункциональная
PFK-2/FBPa3a-2 переходит в форму с активной
РВРазой-2, в результате чего снижается концентра-
ция фруктозо-2,6-бисфосфата — аллостерического
активатора фосфофруктокиназы и ингибитора
РВРазы-!. Это также вносит вклад в стимуляцию
глюконеогенеза.
13. а) Повышенный; б) повышенный; в) повышенный.
14. а) РКА не может активироваться в ответ на сигнал
глюкагона или адреналина, и гликогенфосфорила-
за остается неактивной, б) РР1 остается активной
и дефосфорилирует гликогенсинтазу (тем самым
активируя ее) и гликогенфосфорилазу (тем самым
ингибируя ее), в) Фосфорилаза остается фосфо-
рилировнной (активной), усиливая расщепление
гликогена, г) Глюконеогенез не стимулируется при
низком уровне глюкозы крови, так что при голода-
нии уровень глюкозы крови падает до опасно низ-
кой величины.
15. Падение концентрации глюкозы в крови вызывает
высвобождение глюкагона поджелудочной желе-
Краткие решения задач и ответы на вопросы [3191
зой. В печени глюкагон активирует гликогенфос-
форилазу через стимуляцию ее сАМР-зависимого
фосфорилирования и стимулирует глюконеоге-
нез путем снижения концентрации фруктозо-2,6-
бисфосфата, таким образом стимулируя РВРазу-1.
16. а) Снижение способности мобилизовать гликоген;
снижение уровня глюкозы крови между приемами
пищи, б) Снижение способности организма пони-
жать уровень глюкозы крови после употребления
в пищу углеводов; повышение уровня глюкозы
крови, в) Снижение концентрации фруктозо-2,6-
бисфосфата в печени, стимуляция гликолиза и
ингибирование глюконеогенеза, г) Снижение кон-
центрации фруктозо-2,6-бисфосфата, стимуля-
ция глюконеогенеза и ингибирование гликолиза,
д) Усиление захвата жирных кислот и глюкозы и
усиленное окисление этих веществ, е) Активиза-
ция превращения пирувата в ацетил-СоА и ускоре-
ние синтеза жирных кислот.
17. а) Учитывая, что каждая частица гликогена содер-
жит около 55 000 остатков глюкозы, эквивалентная
концентрация глюкозы составляет
55 000 • 0,01 мкМ = 550 мМ (0,55 М).
При такой концентрации растворенного вещества в
клетке было бы очень высокое осмотическое давле-
ние (жидкости тела характеризуются намного бо-
лее низкой осмолярностью), б) Чем меньше развет-
влений, тем меньше у полимера свободных концов,
доступных для действия гликогенфосфорилазы, и
тем ниже скорость высвобождения глюкозы. Если
полимер линейный (не имеет разветвлений), фос-
форилаза может действовать всего лишь в одном
положении, в) На внешней поверхности частицы
так много остатков глюкозы, что ферменту трудно
подойти к расщепляемой связи, г) В каждом следу-
ющем ряду число цепей удваивается: ряд 1 содер-
жит одну цепь (2° =1), ряд 2 — две (21 = 2), ряд 3 —
четыре (22 = 4) и т. д. Таким образом, в случае ряда t
число цепей СА в самом верхнем ряду 2Z1. д) Общее
число цепей 2° + 21 + 22 + ...2Z1 = 2' - 1. Каждая цепь
содержит gc звеньев глюкозы, так что общее чис-
ло Ст звеньев глюкозы gc(2t - 1). е) Из цепи дли-
ной gc гликогенфосфорилаза может высвободить
все остатки глюкозы, кроме четырех. Таким обра-
зом, из каждой цепи верхнего ряда фермент может
высвободить (gc - 4) остатков глюкозы. Поскольку
в верхнем ряду имеется 2Z1 цепей, число молекул
глюкозы СРТ, которые может высвободить фер-
мент, (gc - 4)2Z-1. ж) Объем сферы равен Узшг3, где
г - это толщина одного ряда, умноженная на число
рядов, т. е. (0,12gc + 0,35)/" нм. Таким образом, Vs =
4/3w/"3(0,12gc + 0,35)3 нм3, з) Алгебраическим пу-
тем можно показать, что значение gc, при котором
достигается максимальное значение /, не зависит
от t. Пусть t = 3:
gc CA GT GTp V. f
5 4 35 4 11 5,8
6 4 42 8 19 9,7
7 4 49 12 24 12
8 4 56 16 28 14
9 4 63 20 32 15
10 4 70 24 34 16
11 4 77 28 36 16
12 4 84 32 38 17
13 4 91 36 40 17
14 4 98 40 41 17
15 4 100 44 42 16
16 4 110 48 43 16
Оптимальное значение gc (соответствующее мак-
симуму /) равно 13. В природе значение gc варьи-
рует от 12 до 14, что соответствует значениям /,
которые очень близки к оптимуму. Если выбрать
другое значение t, значения будут отличаться, но
оптимальным по-прежнему останется gc = 13.
Глава 16
1. а) Ф Цитратсинтаза:
ацетил-СоА + оксалоацетат + Н2О цитрат + СоА
® Аконитаза-.
цитрат изоцитрат
® Изоцитратдегидрогеназа:
изоцитрат + NAD4 а-кетоглутарат + СО2 + NADH
Ф а-Кетоглутаратдегидрогеназа:
сх-кетоглутарат + NAD+ + СоА
сукцинил-СоА + СО2 + NADH
© Сукцинил-СоА-синтетаза:
сукцинил-СоА + Р; + GDP сукцинат + СоА + GTP
® Сукцинатдегидрогеназа:
сукцинат + FAD фумарат + FADH2
® Фумараза:
фумарат + Н2О малат
® Малатдегидрогеназа:
малат + NAD+ оксалоацетат + NADH + Н+
б), в) Ф СоА, конденсация; © отсутствует, изоме-
ризация; © NAD+, окислительное декарбоксилиро-
вание; © NAD+, СоА и тиаминпирофосфат, окисли-
тельное декарбоксилирование; © СоА, субстратное
фосфорилирование; ® FAD, окисление; ® отсутству-
ет, гидратация; ® NAD+, окисление
г) Ацетил-СоА + 3NAD+ + FAD + GDP + Pj + 2H2O
2CO2 + СоА + 3NADH + FADH2 + GTP + 2H+
[3201 Приложение Б
2. Глюкоза + 4ADP + 4Pj +10NAD+ + 2FAD
4АТР + 10NADH + 2FADH2 + 6СО2
3. а) Окисление; метанол формальдегид + [Н-Н]
б) Окисление; формальдегид формиат + [Н-Н]
в) Восстановление; СО2 + [Н-Н] формиат + Н+
г) Восстановление; глицерат + Н+ + [Н-Н ]
глицеральдегид + Н2О
д) Окисление; глицерин -* дигидроксиацетон + [ Н—Н]
е) Окисление; 2Н2О + толуол бензоат + Н+ + 3[Н-Н]
ж) Окисление; сукцинат фумарат + [Н-Н]
з) Окисление; пируват + Н2О ацетат + СО2 + [ Н—Н ]
4. Из структурных формул видно, что соотношение
связанного с углеродом водорода к углероду (Н/С)
выше для гексановой кислоты (11/6), чем для глю-
козы (7/6). Это означает, что гексановая кисло-
та — более восстановленное соединение и ее полное
окисление до СО2 и Н2О сопровождается выделени-
ем большего количества энергии.
5. а) Окислился; этанол + NAD+
ацетальдегид + NADH + Н+
б) Восстановился;
1,3-бисфосфоглицерат + NADH + Н+ -*
глицеральдегид-3-фосфат + NAD+ + НРО42
в) Не изменил степени окисления; пируват + Н+ -*
-* ацетальдегид + СО2
г) Окислился; пируват + NAD+
ацетат + СО2 + NADH + Н+
д) Восстановился; оксалоацетат + NADH + Н+ -*
малат + NAD+
е) Не изменил степени окисления;
ацетоацетат + Н+ -* ацетон + СО2
6. ТРР: тиазольное кольцо присоединяется к сх-углеро-
ду пирувата и стабилизирует образующийся карб-
анион, выступая в роли ловушки электронов. Липое-
вая кислота, окисляет пируват до состояния окисле-
ния ацетата (ацетил-СоА) и активирует ацетат (тио-
эфир). CoA-SH: активирует ацетат (тиоэфир). FAD:
окисляет липоевую кислоту. NAD+: окисляет FAD.
7. Отсутствие ТРР ингибирует пируватдегидрогеназу;
накапливается пируват.
8. Окислительное декарбоксилирование; NAD+ или
NADP+; сх-кетоглутаратдегидрогеназа
9. Потребление кислорода является мерой активности
двух первых стадий клеточного дыхания — глико-
лиза и цикла лимонной кислоты. Добавление окса-
лоацетата или малата стимулирует цикл лимонной
кислоты и, следовательно, способствует дыханию.
Оксалоацетат или малат выполняет каталитиче-
скую функцию, поскольку регенерируется на после-
дующих стадиях цикла лимонной кислоты.
10. а) 5,6 • 10"6; б) 1,1 • 10 8 М; в) 28 молекул
И. Нужно добавить ADP (или GDP), Рь CoA-SH,
ТРР, NAD+, но не липоевую кислоту, которая кова-
лентно связана с использующими ее ферментами.
12. Не могут синтезироваться флавиновые нуклеоти-
ды FMN и FAD. Поскольку FAD участвует в цикле
лимонной кислоты, недостаточность флавинов
сильно ингибирует цикл.
13. Оксалоацетат может направляться на синтез
аспартата или включаться в глюконеогенез. За-
пас оксалоацетата пополняется в анаплеротиче-
ских реакциях, катализируемых ФЕП-карбокси-
киназой, ФЕП-карбоксилазой, яблочным фермен-
том или пируваткарбоксилазой (см. рис. 16-15,
т. 2).
14. Концевая фосфорильная группа GTP может быть
перенесена на ADP в реакции, катализируемой
нуклеозиддифосфаткиназой с константой равно-
весия 1,0: GTP + ADP -+ GDP + АТР
15. а) ООС-СН2-СН2-СОО (сукцинат), б) Мало-
нат — конкурентный ингибитор сукцинатдегидро-
геназы. в) Остановка цикла лимонной кислоты
приводит к прекращению образования NADH и
переноса электронов и далее к остановке дыхания,
г) Большой избыток сукцината (субстрата) позво-
ляет преодолеть конкурентное ингибирование.
16. а) Добавить равномерно меченную [14С] глюкозу и
следить за выделением 14СО2. б) Равномерно рас-
пределится между атомами С2 и СЗ оксалоацетата;
бесконечное число циклов.
17. Оксалоацетат находится в равновесии с сукцина-
том, в котором атомы С1 и С4 эквивалентны. Ок-
салоацетат, полученный из сукцината, содержит
метку в атомах С1 и С4, а полученный из него ФЕП
содержит метку в атоме С1, откуда метка попадает
в атомы СЗ и С4 в молекуле глюкозы.
18. а) С1; б) СЗ; в) СЗ; г) С2 (метильная группа);
д) С4; е) С4; ж) равномерно распределена между
С2 и СЗ.
19. Тиамин нужен для синтеза ТРР, который высту-
пает в роли простетической группы в пируватде-
гидрогеназном и сх-кетоглутаратдегидрогеназном
комплексах. Недостаточность тиамина снижает ак-
тивность этих ферментных комплексов и вызывает
наблюдающееся накопление метаболитов.
20. Нет. Вхождение в цикл каждых двух атомов в виде
ацетата сопровождается удалением двух атомов в
виде СО2; поэтому суммарного приращения коли-
чества оксалоацетата не происходит. Приток окса-
лоацетата достигается в анаплеротической реакции
карбоксилирования пирувата.
Краткие решения задач и ответы на вопросы [321 ]
21. Да, это приведет к ингибированию цикла лимонной
кислоты. Оксалоацетат содержится в митохондри-
ях в достаточно низкой концентрации, поэтому его
удаление по пути глюконеогенеза сдвинет равнове-
сие реакции цитратсинтазы в сторону образования
оксалоацетата.
22. а) Ингибирование аконитазы; б) фторцитрат; кон-
курирует с цитратом; большим избытком цитрата,
в) Цитрат и фторцитрат ингибируют PFK-1. г) От-
ключаются все катаболические процессы, необхо-
димые для производства АТР.
23. Гликолиз:
Глюкоза + 2Pj + 2ADP + 2NAD+
2 пируват + 2АТР + 2NADH + 2Н+ + 2Н2О
Реакция пируваткарбоксилазы:
2 Пируват + 2СО2 + 2АТР + 2Н2О
-* 2 оксалоацетат + 2ADP + 2Р, +4Н+
Реакция малатдегидрогеназы:
2 Оксалоацетат + 2NADH + 2Н+
-* 2 L-малат + 2NAD+
Так происходит рецикл никотинамидных кофер-
ментов в анаэробных условиях. Суммарную реак-
цию можно записать следующим образом:
Глюкоза + 2СО2 -* 2 L-малат + 4FL
Итак, превращение каждой молекулы глюкозы со-
провождается выделением 4 ионов Н+; в результате
повышается кислотность и терпкость вина.
24. Суммарная реакция:
2 пируват + АТР + 2NAD+ + Н2О
-* а-кетоглутарат + СО2 + ADP + Pj +2NADH + ЗН+
25. Цикл вовлечен и в катаболические, и в анаболиче-
ские процессы. Например, в результате окисления
субстрата в цикле выделяется АТР, но также обра-
зуются предшественники для синтеза аминокислот
(см. рис. 16-15, с. 203).
26. а) Снижает; б) повышает; в) снижает.
27. а) Цитрат образуется из оксалоацетата и ацетил-
СоА под действием цитратсинтазы. Цитратсинтаза
может обеспечивать суммарный прирост цитрата
при следующих условиях: (1) при постоянном при-
токе оксалоацетата и ацетил-СоА и (2) при ограни-
чении синтеза изоцитрата, как в среде с низким со-
держанием Fe3+. Ионы Fe3+ нужны для проявления
активности аконитазы, поэтому в среде с низким
содержанием Fe3+ ее активность низкая.
б) Сахароза + Н2О -* глюкоза + фруктоза
Глюкоза + 2Р; + 2ADP + 2NAD+ ->
2 пируват + 2АТР + 2NADH + 2Н+ + 2Н2О
Фруктоза + 2 Pj + 2ADP + 2NAD+ ->
2 пируват + 2АТР + 2NADH + 2Н+ + 2Н2О
2 Пируват + 2NAD+ + 2СоА -*
2 ацетил-СоА + 2NADH + 2Н+ + 2СО2
2 Пируват + 2СО2 + 2АТР + 2Н2О ->
-* 2 оксалоацетат + 2ADP + 2Р; +4Н+
2 Ацетил-СоА + 2 оксалоацетат + 2Н2О -*
-* 2 цитрат + 2СоА
Суммарная реакция:
Сахароза + Н2О + 2Р; +2ADP + 6NAD+ -*
2 цитрат + 2АТР + 6NADH + 10Н+
в) В суммарной реакции расходуется NAD+. По-
скольку запасы этого окисленного кофермен-
та в клетке ограничены, необходим его рецикл в
электронтранспортной цепи, сопровождающийся
потреблением О2. Итак, превращение сахарозы в
цитрат — аэробный процесс, требующий участия
молекулярного кислорода.
28. Сукцинил-СоА — промежуточное соединение в
цикле лимонной кислоты; его накопление сви-
детельствует о замедлении потока метаболитов
в цикле, требующем сокращения поступления в
цикл ацетил-СоА. Цитратсинтаза, регулирующая
в клетке очень важный окислительный путь, кон-
тролирует наличие NADH и, следовательно, поток
электронов от NADH на О2
29. Катаболизм жирных кислот сопровождается уве-
личением концентрации ацетил-СоА, который сти-
мулирует пируваткарбокси л азу. Происходящий в
результате рост концентрации оксалоацетата сти-
мулирует расход ацетил-СоА в цикле лимонной
кислоты, что приводит к росту концентрации ци-
трата и ингибированию гликолиза на уровне PFK-1.
Кроме того, повышенные концентрации ацетил-СоА
ингибируют пируватдегидрогеназный комплекс, за-
медляя использование пирувата из гликолиза.
30. Кислород нужен для получения NAD+ из NADH, об-
разовавшегося в реакциях цикла лимонной кислоты.
Окисление NADH происходит в реакциях окисли-
тельного фосфорилирования в митохондриях.
31. В соответствии с законом действующих масс при
увеличении отношения [NADH]/[NAD+] цикл ли-
монной кислоты замедляется на трех стадиях, со-
провождающихся восстановлением NAD+. При вы-
сокой концентрации NADH равновесие смещается
в сторону образования NAD+.
32. Реакция направлена в сторону образования цитра-
та; в этих условиях АС реакции цитратсинтазы со-
ставляет примерно -8 кДж/моль.
33. Стадии © и ® необходимы для реокисления липо-
амидного кофактора.
34. Цикл лимонной кислоты играет настолько важную
роль в метаболизме, что серьезные дефекты любого
фермента цикла летальны для эмбриона.
[322] Приложение Б
35. Первый фермент каждого пути находится под
двойным аллостерическим контролем. Ингибиро-
вание одного пути направляет изоцитрат по друго-
му пути
36. а) Единственной реакцией в мышечной ткани, про-
исходящей со значительным потреблением кис-
лорода, является клеточное дыхание, поэтому по
потреблению О2 хорошо оценивать активность ды-
хания. б) В свежеприготовленной мышечной тка-
ни присутствует остаточное количество глюкозы;
потребление О2 связано с окислением этой глюко-
зы. в) Да. Поскольку количество потребленного О2
возрастает при добавлении цитрата или 1-глицеро-
фосфата, в данной системе оба могут служить суб-
стратами в клеточном дыхании, г) Эксперимент I: в
присутствии цитрата затрачивается намного боль-
ше кислорода, чем должно быть использовано на
его полное окисление. Единственным объяснением
может быть то, что каждая молекула цитрата всту-
пает в реакцию более одного раза, т. е. ведет себя
так, как катализатор. Эксперимент II: ответ заклю-
чается в расчете дополнительного О2, использован-
ного каждым образцом по сравнению с контролем
(образец 1).
Избыток
Образец Дополнительный субстрат Потребление потребления
О2, мкл О2, мкл
1 нет 342 0
2 0,3 мл 0,2 М 1 - глицерофосфата 757 415
3 0,15 мл 0,02 М цитрата 431 89
4 0,3 мл 0,2 М 1-глицерофосфата + + 0,15 мл 0,02 М цитрата 1385 1043
Если бы цитрат и 1-глицерофосфат были просто
субстратами, дополнительное количество погло-
щенного кислорода в образце 4 было бы суммой
соответствующих показателей в образцах 2 и 3
(415 + 89 = 504). Однако мы наблюдаем двукрат-
ное превышение этой величины. Таким образом,
цитрат повышает способность ткани метаболизи-
ровать 1-глицерофосфат. Это типичное поведение
катализатора. Для подтверждения этого вывода
необходимы результаты обоих экспериментов (I
и II). Из результатов эксперимента I видно, что
цитрат ускоряет реакцию, но невозможно понять,
способствует ли он метаболизму субстрата или
действует по другому механизму. Из результатов
эксперимента II невозможно установить, какое из
веществ катализатор — цитрат или 1-глицерофос-
фат. Два эксперимента вместе позволяют твердо
сказать, что цитрат выступает в роли катализатора
в процессе окисления 1-глицерофосфата, д) По-
скольку в данном процессе происходит расходова-
ние цитрата (образец 3), действующего в качестве
катализатора, на каком-то этапе он должен реге-
нерировать. Если в последовательности реакций
цитрат сначала расходуется, а потом регенериру-
ется, последовательность реакций не линейная а
циклическая, е) Если процесс останавливается на
а-кетоглутаратдегидрогеназной реакции, цитрат
превращается в а-кетоглутарат, но не претерпевает
дальнейших превращений. Кислород расходуется
на реокисление NADH, образовавшегося в реакции
изоцитратдегидрогеназы.
ж)
Глюкоза-
Оксалоацетат
Цитрат
Малонат
а-Кетоглутарат
Сукцинат
Фумарат
Этот рисунок отличается от рис, 16-7 тем, что здесь
отсутствуют ^wc-аконитат и изоцитрат (между
цитратом и сх-кетоглутаратом), а также сукцинил-
СоА и ацетил-СоА. з) Количественный анализ по-
зволяет исключить разветвленную или другую бо-
лее сложную схему процесса.
Глава 17
1. В составе жирной кислоты; атомы углерода в жирной
кислоте восстановлены сильнее, чем в глицерине.
2. а) 4,0 105 кДж (9,6 104 ккал); б) 18 сут.:
в) 0,48 фунтов/сут.
3. Первая стадия цикла окисления жирных кислот ана-
логична реакции превращения сукцината в фумарат,
вторая стадия — реакции превращения фумарата в
малат, а третья - реакции превращения малата в ок-
салоацетат.
4. Семь циклов; в последнем образуются 2 молекулы
ацетил-СоА.
5. a) R СОО + АТР ацил-АМР + Р^
ацил-АМР + СоА -> ацил-СоА + АМР
б) Необратимый гидролиз PPj до 2РЬ осуществляе-
мый в клетке неорганической пирофосфатазой
Краткие решения задач и ответы на вопросы [3231
6. ф/с-А3-Додеканоил-СоА; он конвертируется в цис-
А2-додеканоил-СоА, а затем в р-гидрокси-
додеканои л - СоА.
7. 4 ацетил-СоА и 1 пропионил-СоА.
8. Да. Часть трития удаляется из пальмитата в реак-
циях дегидрирования в процессе р-окисления. Уда-
ленный тритий обнаруживается в тритированной
воде.
9. В цитозоле активированные (соединенные с СоА)
жирные кислоты сначала переносятся на карни-
тин, в результате чего высвобождается СоА, а за-
тем транспортируются в митохондрии, где вновь
вступают во взаимодействие с СоА. Таким образом,
цитозольный и митохондриальный пулы СоА со-
держатся раздельно, поэтому радиоактивный СоА
из цитозоля не проникает в митохондрии.
10. а) У голубей преобладает р-окисление, а у фаза-
нов — анаэробный гликолиз гликогена, б) Мыш-
цы голубя потребляют больше О2. в) Жиры содер-
жат больше энергии в пересчете на 1 г вещества,
чем гликоген. Кроме того, анаэробное расщепле-
ние гликогена ограничено толерантностью тка-
ней к лактату. Поэтому голубь, использующий
при полете окислительный катаболизм жиров,
летает на значительно более длинные расстоя-
ния. г) Эти ферменты являются регуляторными
ферментами в соответствующих метаболических
путях и, следовательно, ограничивают скорость
образования АТР.
И. Малонил-СоА больше не ингибирует транспорт
жирных кислот в митохондрии и р-окисление, по-
этому может возникнуть холостой цикл, в котором
одновременно происходит синтез жирных кислот в
цитозоле и их распад в митохондриях.
12. а) Транспорт жирных кислот в митохондрии, опос-
редованный карнитином, определяет скорость
всего процесса окисления жирных кислот. Недо-
статочность карнитина приводит к замедлению
окисления жирных кислот, а добавление карнити-
на повышает скорость окисления, б) Все эти факто-
ры усиливают необходимость окисления жирных
кислот, в) Недостаточность карнитина может быть
следствием недостаточности его предшественника,
лизина, или дефектом одного из ферментов, уча-
ствующих в биосинтезе карнитина.
13. При окислении жиров выделяется метаболическая
вода; 1,4 л воды на 1 кг трипальмитоилглицерина
(небольшой вклад глицерина не учитываем).
14. Бактерии можно использовать для полного окисле-
ния углеводородов до СО2 и Н2О. Однако трудно
обеспечить хороший контакт между углеводоро-
дами и бактериальными ферментами. Кроме того,
может не хватать таких необходимых бактериям
элементов, как азот и фосфор, что может ограничи-
вать их рост.
15. а) Мт 136; фенилуксусная кислота; б) четное
16. Митохондриальный пул СоА не столь большой,
поэтому СоА нужно извлекать из ацетил-СоА в
процессе образования кетоновых тел. Это позво-
ляет осуществлять р-окисление, необходимое для
получения энергии.
17. а) В процессе гликолиза из глюкозы образуется
пируват, а пируват является основным источни-
ком оксалоацетата. При отсутствии глюкозы в
рационе питания концентрация оксалоацетата
падает, и цикл трикарбоновых кислот замедля-
ется. б) С нечетным числом атомов; при превра-
щении пропионата в сукцинил-СоА образуются
промежуточные продукты цикла лимонной кис-
лоты и четырехуглеродные предшественники
для глюконеогенеза.
18. В случае гептановой кислоты, состоящей из не-
четного числа атомов углерода, р-окисление при-
водит к образованию пропионил-СоА, который за
несколько стадий может превращаться в оксало-
ацетат — исходное соединение для глюконеогенеза.
Жирные кислоты с четным числом атомов углеро-
да не могут служить источником исходных соеди-
нений для глюконеогенеза, поскольку полностью
окисляются до ацетил-СоА.
19. При р-окислении со-фторолеата образуется фтор-
ацетил-СоА, который включается в цикл лимонной
кислоты и превращается в фторцитрат, мощный
ингибитор аконитазы. Ингибирование аконитазы
останавливает цикл. Отсутствие восстанавлива-
ющих эквивалентов из цикла лимонной кислоты
делает невозможным окислительное фосфорили-
рование (синтез АТР).
20. Замена Ser на Ala блокирует р-окисление в ми-
тохондриях. Замена Ser на Asp блокирует синтез
жирных кислот и стимулирует р-окисление.
21. Реакция на глюкагон или адреналин стала бы более
длительной, что способствовало бы усилению мо-
билизации жирных кислот в адипоцитах.
22. Комплекс Enz-FAD, имеющий более положитель-
ный стандартный восстановительный потенциал,
является лучшим акцептором электронов, чем
NAD+, и реакция направлена в сторону окисления
ацил-СоА. За это более благоприятное равнове-
сие приходится расплачиваться 1 молекулой АТР:
лишь 1,5 молекулы АТР образуется в расчете на
молекулу FADH2, окисленную в дыхательной
цепи, в отличие от 2,5 молекул АТР на молекулу
NADH.
[324] Приложение Б
23. Девять циклов; арахидоновая кислота (насыщен-
ная кислота, состоящая из 20 атомов углерода) пре-
вращается в 10 молекул ацетил-СоА. из которых
две образуются в девятом цикле.
24. См. рис. 17-11. Образуется [3-14С]сукцинил-СоА,
из которого затем получается меченный по атомам
С2 и СЗ оксалоацетат.
25. Фитановая кислота -> пристановая кислота -> про-
пионил-СоА ->->-> сукцинил-СоА -> сукцинат ->
фумарат -> малат. Все атомы углерода в малате со-
держат метку, но в атомах С1 и С 4 ее в 2 раза мень-
ше, чем в С2 и СЗ.
26. В клеточных реакциях, требующих расхода энер-
гии, гидролиз АТР происходит с участием воды:
АТР + Н2О -> ADP + Рр В суммарной реакции Н2О
нет.
27. Метилмалонил-СоА-мутаза нуждается в кобальт-
содержащем кофакторе, образующемся из вита-
мина В12.
28. Потеря массы за сутки составляет около 0,66 кг,
или 140 кг за 7 месяцев. Кетоза можно избежать
путем деградации неэссенциальных белков в глю-
конеогенезе.
29. а) Жирные кислоты превращаются в СоА-про-
изводные под действием ферментов цитоплазмы,
а затем ацил-СоА переносятся в митохондрии для
окисления. Поскольку в эксперименте использо-
вались изолированные митохондрии, в нем рас-
ходовались производные СоА. б) Стсарил-СоА
быстро превращается в 9 молекул ацетил-СоА в
процессе р-окисления. Все промежуточные со-
единения претерпевают быстрые превращения,
поэтому детектировать их практически пе удаст-
ся. в) Два цикла. В каждом цикле удаляются два
атома углерода, поэтому через два цикла 18-угле-
родная жирная кислота превращается в 14-угле-
родную и 2 молекулы ацетил-СоА. г) Значение
Км больше для транс-изомеров, чем для цис-
изомеров. так что одинаковая скорость расщепле-
ния достигается при более высокой концентрации
трш/с-изомеров по сравнению с ф/с-изомерами.
В общем. трш/с-изомеры связываются хуже цис-
изомеров, вероятно, по геометрическим причи-
нам, пусть даже и непрямо в участке связывания,
д) Накопление каждого конкретного субстрата в
реакциях LCAD VLCAD происходит по-разному,
что и следует ожидать для лимитирующих ста-
дий пути, е) Кинетические данные показывают,
что транс-изомер менее пригоден как субстрат
для LCAD, чем ф/с-изомер, но в случае VLCAD
различие между субстратами небольшое. Будучи
менее пригодным субстратом, трш/с-изомер на-
капливается в большей степени, чем цис-изомер.
ж) Один возможный путь изображен ниже (по-
метки «внутри» и «снаружи» указывают локали
зацию веществ по отношению к митохондриям)
Элаидоил-СоА ___________ элаидоилкарнитин
(снаружи) * (снаружи)
элаидоилкарнитин элаидоил-СоА
(внутри) (внутри)
5-транс-тетрадеценоил-СоА
(внутри)
5-транс-тетрадекановая 5-транс-тетрадекановая
кислота (внутри) кислота (снаружи)
з) Справедливо, что транс-жиры расщепляются
менее эффективно, чем zpc-жиры, и поэтому могут
«вытекать» из митохондрий. Но нельзя сказать, что
транс-жиры совсем не расщепляются клетками:
они расщепляются, но медленнее, чем ^zzc-жиры.
Глава 18
1. о
II
а) ООС—СН2—С—СОО
Оксалоацетат
О
II
б) ООС—СН2—СН2—С—СОО а-Кетоглутарат
О
II
в) СНз—С—СОО
Пируват
Фенилпируват
2. Метод основан на анализе параметров сопряженной
реакции. Продукт медленной реакции трансами-
нирования (пируват) быстро расходуется в следу-
ющей «индикаторной» реакции, катализируемой
лактатдегидрогеназой, которая сопровождается рас-
ходом NADH Таким образом, скорость исчезнове-
ния NADH можно использовать при оценке скорости
аминотрансферазной реакции. За ходом индикатор-
ной реакции следят с помощью спектрофотометра по
уменьшению поглощения NADH при 340 нм.
3. Аланин и глутамин играют особую роль в транспор-
те аминогрупп из мышц и других внепеченочных
тканей (соответственно) в печень.
4. Не будет. В реакции трансаминирования азот из ала-
нина переносится на оксалоацетат, в результате чего
образуется аспартат.
5. 15 моль АТР на моль лактата; 13 моль АТР на моль
аланина, если учитывать удаление азота.
6. а) Голодание приводит к снижению уровня глюко-
зы крови; после приема пищи происходит быстрый
Краткие решения задач и ответы на вопросы [325]
катаболизм глюкогенных аминокислот, б) Окис-
лительное дезаминирование вызывает повышение
уровня NH3, но из-за отсутствия аргинина (проме-
жуточного соединения в цикле мочевины) NH3 не
может включаться в мочевину У кошки аргинин не
синтезируется в достаточном количестве, чтобы пре-
одолеть ситуацию, вызванную экспериментальными
условиями. Таким образом, для этого животного ар-
гинин — незаменимая аминокислота, в) В цикле мо-
чевины орнитин превращается в аргинин.
7. Н2О + глутамат + NAD+ ->
- > а-кетоглутарат + NH4+ + NADH + Н+
NH4+ + 2АТР + Н2О + СО2
- > карбамоилфосфат + 2ADP + Р, + ЗН+
Карбамоилфосфат + орнитин -> цитруллин + Р, + Н+
Цитруллин + аспартат + АТР ->
- > аргининосукцинат + АМР + РР, + Н+
Аргининосукцинат -* аргинин + фумарат
Фумарат + Н2О -> малат
Малат + NAD+ -> оксалоацетат + NADH + Н+
Оксалоацетат + глутамат -> аспартат + а-кетоглутарат
Аргинин + Н2О -> мочевина + орнитин
Итого:
2 Глутамат + СО2 + 4Н2О + 2NAD+ + ЗАТР
-> 2 а-кетоглутарат +2NADH + 7Н+ + мочевина +
+ 2ADP + АМР + PPj + 2Р, (1)
Кроме того, следует учесть дополнительные реакции:
АМР + АТР-> 2ADP (2)
О2 + 8Н+ + 2NADH + 6ADP + 6Р,
2NAD+ + 6АТР + 8Н2О (3)
Н2О + РР1^2Р, + Н+ (4)
Суммируя все приведенные выше реакции, получаем:
2 Глутамат + СО2 + О2 + 2ADP + 2Р, ->
-> 2 а-кетоглутарат + мочевина + ЗН2О + 2АТР
8. Вторая аминогруппа в составе мочевины переносит-
ся туда из аспартата, который образуется в реакции
трансаминирования между глутаматом и оксало-
ацетатом, катализируемой аспартатаминотрансфе-
разой. Примерно половина всех аминогрупп, выде-
ляющихся из организма в составе мочевины, про-
ходит через реакцию аспартатаминотрансферазы,
что и объясняет очень высокую активность этого
фермента по сравнению с другими аминотрансфе-
разами.
9. а) При строгой белковой диете аминокислоты — это
единственный источник метаболической энергии.
Поскольку катаболизм аминокислот сопряжен с вы-
ведением избытка азота в составе мочевины, организм
нуждается в дополнительном количестве воды для
разбавления избытка мочевины в моче. Кроме того,
электролиты в «жидком белке» тоже должны быть
разбавлены и выведены из организма. Если суммар-
ная суточная потеря воды не компенсируется обиль-
ным питьем, происходит потеря воды организмом.
б) При анализе пищевой ценности белка необходимо
учитывать общее количество аминокислот, необхо-
димое для синтеза белка в организме, и аминокислот-
ный состав пищевого белка. Например, содержание
аминокислот в желатине не сбалансировано. При
употреблении в пищу большого количества желатина
и расщеплении избыточного количества аминокис-
лот может быть превышена емкость цикла мочевины,
что приводит к интоксикации аммиаком. Состояние
здоровья человека может усугубляться обезвожива-
нием, вызванным выведением из организма большо-
го количества мочевины. Сочетание этих двух факто-
ров может стать причиной комы и смерти.
10. Лизин и лейцин
11. а) Фенилаланингидроксилаза; диета с понижен-
ным содержанием фенилаланина, б) Блокирован
нормальный путь гидроксилирования фенилала-
нина с образованием тирозина, и поэтому проис-
ходит накопление фенилаланина, в) В результате
трансаминирования фенилаланин превращается в
фенилпируват, который затем восстанавливается
до фениллактата. Для этой реакции трансаминиро-
вания К = 1,0, так что при накоплении фенилала-
нина образуется значительное количество фенил-
пирувата. г) Это связано с низким уровнем синтеза
тирозина — предшественника пигмента меланина.
12. Нарушается катаболизм углеродного скелета ва-
лина, метионина и изолейцина, что связано с от-
сутствием активности мстилмалопил-СоА-мутазы,
которой в качестве кофермента необходим вита-
мин В12. Физиологические последствия недоста-
точности этого фермента описаны в табл. 18-2 и в
дополнении 18-2 (т. 2).
13. В диете веганов недостает витамина В12, что через
несколько лет приводит к тому, что у этих людей
концентрация гомоцистеина и метилмалоната в
крови повышается (что отражает недостаточность
метионинсинтазы и метилмалонилмутазы соответ-
ственно). Лактовегетарианцы получают некоторое
количество витамина В12 в составе молочных про-
дуктов.
14. Генетическая форма пернициозной анемии обычно
возникает как результат нарушения всасывания
пищевого витамина В12 (см. дополнение 17-2, т. 2).
Поскольку соответствующие пищевые добавки
также не всасываются в кишечнике, для лечения
больных применяют внутривенные инъекции пре-
паратов В12
15. Механизм идентичен механизму реакции серинде-
гидратазы (см. рис. 18-20, а, т. 2) с тем только ис-
ключением, что сохраняется дополнительная ме-
тильная группа треонина и поэтому образуется не
пируват, а а-кетобутират.
[326] Приложение Б
16.
a) 16NH2—СО—16NH2
б) ОО14С—СН2—СН2—14С00
15NH О
И 1. II ,,
в) R—NH—С—15NH2 г) R—NH—С— 5NH2
15nh2
I
д) Нет метки, е) ОО14С—С—СН2—14СОО
Н
17. а) Изолейцин II -» IV I -» V III ацетил-
СоА + пропионил-СоА; б) Стадии: (1) трансамини-
рование — аналогичной реакции нет; PLP; (2) окис-
лительное декарбоксилирование — реакция ана-
логична пируватдегидрогеназной реакции; NAD+
ТРР, липоат и FAD; (3) окисление — реакция ана-
логична сукцинатдегидрогеназной реакции; FAD:
(4) гидратация реакция аналогична реакции фу-
маразы; кофактора нет; (5) окисление — реакция
аналогична малатдегидрогеназной реакции; NAD ;
(6) тиолиз (реакция, обратная альдольной конден-
сации) — аналогична реакции тиолазы: СоА.
19. а) Трансаминирование — аналогов нет; PLP.
б) Окислительное декарбоксилирование — анало-
гично окислительному декарбоксилированию пи-
рувата до ацетил-СоА перед вхождением в цикл ли-
монной кислоты или превращению сс-кетоглутарата
до сукцинил-СоА в цикле лимонной кислоты; NAD+,
FAD, липоат и ТРР. в) Дегидратация (окисление)
аналогична дегидратации сукцината с образованием
фумарата в цикле лимонной кислоты или ацил-СоА
с образованием еноил-СоА при [3-окислении; FAD.
г) Карбоксилирование - нет аналогов в цикле ли-
монной кислоты или реакциях [3-окисления; АТР и
биотин, д) Гидратация реакция аналогична гидра-
тации фумарата до малата в цикле лимонной кис-
лоты или еноил-СоА до 3-гидроксиацил-СоА при
(3-окисления; кофакторов нет. е) Реакция, обратная
альдольной реакции, - аналогична обратной ци-
тратсинтазной реакции в цикле лимонной кислоты;
кофакторов нет.
20. а) Лейцин, валин, изолейцин, б) Цистеин (образу-
ющийся из цистина) Если цистеин подвергается
декарбоксилированию, как показано на рис. 18-6,
это приводит к образованию H3N+ CH2-CH2-SH,
который может окисляться до таурина, в) В янва-
18. Вероятный механизм реакции:
оос—С—с—н
I I
NH3 Н
Серин
Н
у
Пиридоксальфосфат
Образующийся на второй стадии формальдегид
(НСНО) в активном центре фермента быстро реа-
гирует с тетрагидрофолатом с образованием N5,N10-
метилентетрагидрофолата (см. рис. 18-17, т. 2).
Краткие решения задач и ответы на вопросы [32 /1
ре 1957 г. в крови отмечается сильно повышенный
уровень изолейцина, лейцина, метионина и валина,
а в моче - повышенный уровень изолейцина, лей-
цина, таурина и валина, г) У всех пациентов высо-
кое содержание изолейцина, лейцина и валина в
крови и моче, что указывает на нарушение процес-
са расщепления этих аминокислот. Учитывая, что в
моче, кроме того, содержится много кетоформ этих
трех аминокислот, метаболический путь блокиро-
ван после стадии дезаминирования, но до дегидри-
рования (как показано на рис. 18-28). д) Модель
не объясняет высокой концентрации метионина в
крови и таурина в моче. Высокий уровень таурина
может быть связан с отмиранием клеток головного
мозга на терминальных стадиях заболевания. Од-
нако причины повышения уровня метионина непо-
нятны. Путь расщепления метионина не связан с
путями расщепления разветвленных аминокислот.
Возможно, повышение уровня метионина является
вторичным эффектом накопления других амино-
кислот. Важно помнить, что образцы января 1957 г.
взяты у умирающего ребенка, так что сравнивать
эти показатели с показателями крови здорово-
го человека, по-видимому, не совсем корректно,
е) Требуется следующая информация (и она впо-
следствии была получена другими исследовате-
лями): (1) у людей с болезнью кленового сиропа
значительно понижена или отсутствует активность
дегидрогеназы; (2) болезнь вызвана дефектом од-
ного единственного гена; (3) этот ген кодирует
часть белка или весь белок с дегидрогеназной ак-
тивностью; (4) генетический дефект приводит к
синтезу неактивного фермента.
Глава 19
1. Реакция 1: (а), (г) NADH; (б), (д) E-FMN;
(в) NAD7NADH и E-FMN/FMNH2
Реакция 2\ (а), (г) E-FMNH2; (б), (д) Fe3+;
(в) E-FMN/FMNH2 и Fe3+/Fe2+
Реакция 3: (а), (г) Fe2+; (б), (д) Q;
(в) Fe3+/Fe2+ и Q/QH2
2. Боковая цепь позволяет убихинону растворяться в
липидах и диффундировать в полужидких мембранах.
3. На основании различия стандартных восстанови-
тельных потенциалов (ДЕ'°) для каждой пары по-
луреакций можно найти значение AG"0. Окислению
сукцината под действием FAD способствует отрица-
тельное стандартное изменение свободной энергии
(AG"° = -3,7 кДж/моль). Окисление NAD+ сопро-
вождалось бы значительным положительным стан-
дартным изменением свободной энергии (AG'° =
= 68 кДж/моль).
4. а) Все переносчики восстановлены; CN блокирует
восстановление О2 под действием цитохромоксида-
зы. б) Все переносчики восстановлены; если нет О2,
восстановленные переносчики не могут окисляться,
в) Все переносчики окислены, г) Переносчики на
ранних стадиях восстановлены, на более поздних
стадиях — окислены.
5. а) Ингибирование NADH-дегидрогеназы ротено-
ном снижает скорость потока электронов в дыха-
тельной цепи, что в свою очередь снижает скорость
образования АТР. Если из-за этого организму пере-
стает хватать АТР, наступает смерть, б) Антими-
цин А сильно ингибирует окисление Q в дыхатель-
ной цепи, снижая скорость переноса электронов и
вызывая все те последствия, что перечислены в (а),
в) Поскольку антимицин А блокирует передачу на
кислород всех электронов, он — более сильный яд,
чем ротенон, который блокирует только передачу
электронов от NADH, но не от FADH2.
6. а) Для удовлетворения потребности в АТР повыша-
ется скорость переноса электронов, следовательно,
соотношение Р/О понижается, б) При высокой кон-
центрации разобщающего агента отношение Р/О
близко к нулю. При снижении этого отношения для
производства того же количества АТР следует окис-
лить больше топливных молекул. Избыток тепла,
выделяющегося в этих окислительных процессах,
повышает температуру тела, в) Повышение актив-
ности дыхательной цепи в присутствии разобщаю-
щего агента требует расщепления дополнительных
топливных молекул. Окисление дополнительных
топливных молекул (включая запасные жиры) для
выработки того же количества АТР приводит к
уменьшению массы тела. При Р/О -> О недостаток
АТР вызывает смерть.
7. Валиномицин выступает в роли разобщающего
агента. Он взаимодействует с ионами К+, образуя
комплекс, проникающий через внутреннюю мем-
брану митохондрий, что нарушает мембранный по-
тенциал. Снижается производство АТР, что вызы-
вает повышение скорости передачи электронов. Это
в свою очередь вызывает повышение протонного
градиента, усиливает потребление кислорода и вы-
деление тепла.
8. а) Ингибируется образование АТР. б) Образова-
ние АТР тесно сопряжено с переносом электронов:
2,4-динитрофенол является разобщающим агентом
для окислительного фосфорилирования, в) С оли-
гомицином.
9. Цитозольная малатдегидрогеназа играет ключевую
роль в транспорте восстановительных эквивалентов
через внутреннюю мембрану митохондрий с помо-
щью малат-аспартатного челночного механизма.
[3281 Приложение Б
10. а) Гликолиз становится анаэробным, б) Потребле-
ние кислорода прекращается, в) Усиливается обра-
зование лактата, г) Выход АТР снижается до 2 АТР
на молекулу глюкозы.
11. Стационарная концентрация Pj в клетке намного
выше концентрации ADP. Рь образующийся при
гидролизе АТР, очень мало влияет на общую кон-
центрацию Pj.
12. Реакция на изменение (а) происходит значительно
быстрее, поскольку реакция на изменение (б) свя-
зана с синтезом белка.
13. а) Вместо молочнокислого брожения происходит
окисление NADH благодаря передаче электронов,
б) Окислительное фосфорилирование более эф-
фективно. в) В условиях высокого отношения дей-
ствующих масс в системе АТР происходит ингиби-
рование фосфофруктокиназы-1.
14. Преимущество в том, что сбраживание древесных
опилок до спирта можно проводить в присутствии
О2 и нет необходимости строго соблюдать анаэроб-
ные условия, что весьма сложно. Эффект Пастера
не наблюдается ввиду неактивности цикла лимон-
ной кислоты и электронтрапспортной цепи.
15. Возможен более эффективный транспорт электро-
нов между комплексами.
16. а) Внешняя среда: 4,0-10 8 М; матрикс: 2,0-10 8 М;
б) энергетический вклад ДрН (градиента концен-
трации Н+) 1,7 кДж/моль; в) 21; г) недостаточно;
д) общий трансмембранный потенциал.
17. а) 0 ,91 мкмоль/(с-г); б) 5,5 с; для поддержания
концентрации АТР на постоянном уровне регуля-
ция его синтеза должна быть слаженной и быстрой.
18. 53 мкмоль/(с-г). При стационарной концентрации
АТР 7,0 мкмоль/г это эквивалентно 10 оборотам
пула АТР в секунду. Этого запаса хватит на 0,13 с.
19. Реактивные формы кислорода взаимодействуют
с макромолекулами, включая ДНК. Если дефект
митохондриального гена приводит к активной про-
дукции РФК, могут повреждаться ядерные гены,
кодирующие протоонкогены, что приводит к об-
разованию онкогенов, неконтролируемому росту
клеток и раку.
20. Различия в степени гетероплазмии по дефектному
гену являются причиной разной степени тяжести
митохондриальных заболеваний.
21. Внутренняя мембрана митохондрий непроницае-
ма для NADH, но восстанавливающие эквивален-
ты NADH переносятся через мембрану косвенным
путем: они передаются на оксалоацетат в цито-
золь, образующийся малат проникает в матрикс
и митохондриальный NAD+ восстанавливается до
NADH.
22. Цикл лимонной кислоты останавливается из-за
отсутствия акцептора электронов, способного
принять их от NADH. Образующийся в процессе
гликолиза пируват не может войти в цикл в виде
ацетил-СоА; накапливающийся пируват в резуль-
тате трансаминирования превращается в аланин и
доставляется в печень.
23. Пируватдегидрогеназа локализована в митохон-
дриях, глицеральдегид-3-фосфатдегидрогеназа — в
цитозоле. Пулы клеточного NAD+ разделены вну-
тренней митохондриальной мембраной.
24. Полное отсутствие глюкокиназы (две дефектные
аллели) делает невозможным проведение гликолиза
с такой скоростью, которая обеспечила бы необхо-
димую для секреции инсулина концентрацию АТР.
25. Дефект комплекса II приводит к усилению синте-
за РФК, повреждению ДНК и мутациям, которые
вызывают нерегулируемое деление клеток (рак).
Пока неизвестно, почему рак возникает именно в
среднем отделе тонкого кишечника.
26. Для достижения максимальной скорости фотосин-
теза ФС1 (которая поглощает свет с длиной волны
700 нм) и ФСП (поглощает свет с длиной волны
680 нм) должны функционировать одновременно.
27. Источником дополнительной массы является вода,
потребленная в суммарной реакции.
28. Пурпурные серные бактерии в качестве донора во-
дорода в процессе фотосинтеза используют H2S.
Кислород в этих реакциях нс участвует, посколь-
ку единственная фотосистема бактерии не имеет
марганецсодержащего расщепляющего воду ком-
плекса.
29. 0,44
30. а) Останавливает; б) замедляет; некоторое коли-
чество электронов продолжает циркулировать по
циклическому пути.
31. При освещении устанавливается градиент протонов.
При добавлении ADP и Pj этот градиент направляет
синтез АТР, но в темноте градиент исчезает.
32. Диурон блокирует перенос электронов между
ФСП и первым участком синтеза АТР.
33. В присутствии вентурицидина блокирован перенос
протонов через комплекс CF0CFb и поток электро-
нов (выделение кислорода) продолжается лишь до
тех пор, пока изменение свободной энергии в про-
цессе перекачки протонов против увеличивающего-
ся протонного градиента не станет равным доступ-
ной свободной энергией фотонов. Динитрофенол,
разрушающий протонный градиент, восстанавли-
вает поток электронов и выделение кислорода.
34. а) 56 кДж/моль; б) 0,29 В.
Краткие решения задач и ответы на вопросы 1329]
35. Из разницы восстановительных потенциалов мож-
но найти, что AG'° реакции составляет 15 кДж/моль.
Рисунок 19-46 показывает, что энергии фотонов из
любой области видимой части спектра более чем
достаточно для проведения этой эндергонической
реакции.
36. 1,35 • 10-77; реакция крайне неблагоприятна! В хло-
ропластах этот барьер преодолевается за счет энер-
гии солнечного света.
37. -920 кДж/моль.
38. Не образуется. Электроны от Н2О поступают на ис-
кусственный акцептор электронов — Fe3+, а не на
NADP+.
39. Примерно 1 раз за 0,1 с; возбуждается 1 молекула
из 108.
40. Свет с длиной волны 700 нм возбуждает ФС1, но не
ФСП. Электроны передаются от Р700 на NADP+,
но от Р680 электроны не поступают. При возбужде-
нии ФСП светом с длиной волны 680 нм электро-
ны начинают перемещаться в направлении к ФС1,
но очень скоро переносчики электронов между
двумя системами полностью восстанавливаются.
41. Не участвует. Возбужденный электрон от Р700
возвращается для заполнения «дырки», возник-
шей при облучении. ФСП не должна участвовать
в передаче электронов, и О2 из Н2О не выделяется.
NADPH не образуется, поскольку возбужденный
электрон возвращается к Р700.
42. а) (1) Наличие Mg2+ согласуется с гипотезой о том,
что хлорофилл непосредственно участвует в ка-
тализе реакции фосфорилирования: ADP + Р, ->
АТР. (2) Многие ферменты (и другие белки), со-
держащие Mg2+, не участвуют в фосфорилирова-
нии, так что наличие Mg2+ в хлорофилле не доказы-
вает его участие в фосфорилировании. (3) Наличие
Mg2+ важно для фотохимических свойств хлоро-
филла — поглощения света и переноса электронов,
б) (1) Ферменты катализируют обратимые реак-
ции, так что изолированный фермент, который в
определенных экспериментальных условиях ка-
тализирует удаление фосфорильной группы, воз-
можно, в других условиях (как в клетке) может
катализировать присоединение фосфорильной
группы. Поэтому возможно, что хлорофилл уча-
ствует в фосфорилировании ADP. (2) Есть два
возможных объяснения: хлорофилл обладает толь-
ко фосфатазной активностью и не катализирует
фосфорилирование ADP в клетке, либо грубый
препарат содержит примесную фосфатазную ак-
тивность, не связанную с реакциями фотосинте-
за. (3) Возможно, препарат содержит примесную
нефотосинтетическую фосфатазную активность,
в) (1) Ингибирования светом следовало бы ожи-
дать, если бы хлорофилловый белок катализировал
реакцию: ADP + Р, + свет АТР. Отсутствие света
благоприятствовало бы протеканию обратной ре-
акции — дефосфорилированию. На свету система
получает энергию и равновесие сдвигается вправо,
снижая фосфатазную активность. (2) Это ингиби-
рование может быть артефактом выделения или
анализа. (3) Вряд ли грубые методы выделения
того времени позволили в целостности сохранить
мембраны хлоропластов, поэтому ингибирование,
скорее всего, является артефактом, г) (1) На све-
ту происходит синтез АТР и потребление других
фосфорилированных веществ. (2) На свету проис-
ходит образование глюкозы и ее метаболизм в про-
цессе клеточного дыхания для производства АТР,
что сопровождается изменением уровня фосфори-
лированных интермедиатов. (3) На свету образует-
ся АТР и расходуются другие фосфорилированные
интермедиаты, д) Световая энергия используется
для образования АТР (как в модели Эмерсона) и
образования восстанавливающей силы (как в мо-
дели Рабиновича), е) Приблизительная стехиоме-
трия процесса фосфорилирования (гл. 19) тако-
ва: 8 фотонов приводят к образованию 2 молекул
NADPH и около 3 молекул АТР. Для восстановле-
ния 1 молекулы СО2 нужны 2 молекулы NADPH и
3 молекулы АТР (гл. 20). Таким образом, для вос-
становления одной молекулы СО2 требуется как
минимум 8 фотонов. Это значение вполне соответ-
ствует модели Рабиновича, ж) Поскольку энергия
света используется для получения как АТР, так и
NADPH, вклад каждого поглощенного фотона за-
ключается больше чем в образовании лишь одной
молекулы АТР. Процесс извлечения энергии света
более эффективен, чем предполагал Рабинович, и
для реакций фотосинтеза годится любой свет, даже
красный.
Глава 20
1. В некоторых органеллах концентрация специфи-
ческих ферментов и метаболитов повышена, и су-
ществуют раздельные пулы кофакторов и интер-
медиатов. Регуляторные механизмы воздействуют
лишь на конкретный пул веществ и конкретную
группу ферментов.
2. Это наблюдение подтверждает, что АТР и NADPH
образуются на свету и необходимы для фиксации
СО2. Конверсия прекращается при исчерпании ре-
зерва АТР и NADPH. Кроме того, в темноте отклю-
чаются некоторые ферменты.
3. X — 3-фосфогл ицерат, Y — рибулозо-1,5-бисфосфат.
4. Рибулозо-5-фосфат-киназа, фруктозе-1,6-бисфос-
фатаза, седогептулозе-1,7-бисфосфатаза и глицер-
1330] Приложение Б
альдегид-3-фосфатдегидрогеназа. Все активируют-
ся в результате восстановления критической дис-
ульфидной связи до пары сульфгидрильных групп.
Иодоацетат необратимо связывается с этими сво-
бодными сульфгидрильными группами.
5. Для осуществления реакций обмена дисульфид-
ных групп, активирующих ферменты цикла Каль-
вина, тиоредоксину нужны обе сульфгидрильные
группы.
6. Восстановительный пентозофосфатный путь приво-
дит к образованию рибулозо-1,5-бисфосфата из три-
озофосфатов, появившихся в процессе фотосинтеза.
Окислительный пентозофосфатный путь обеспечи-
вает NADPH для восстановительного биосинтеза и
пентозофосфаты для синтеза нуклеотидов.
7. Оба типа «дыхания» происходят в растениях и со-
провождаются поглощением О2 и выделением СО2.
(У животных также реализуется механизм ми-
тохондриального дыхания.) Митохондриальное
дыхание осуществляется непрерывно. Электроны
от различных топливных молекул проходят через
цепь переносчиков во внутренней мембране мито-
хондрий и поступают к О2. Фотодыхание проис-
ходит в хлоропластах, пероксисомах и митохон-
дриях. Оно осуществляется в дневное время, когда
происходит процесс фотосинтетической фиксации
углерода. Митохондриальное дыхание происхо-
дит главным образом ночью или в пасмурные дни.
Путь электронов в процессе фотодыхания изобра-
жен на рис. 20-21, а в процессе митохондриального
дыхания — на рис. 19-16.
8. Эта гипотеза подразумевает особую эволюцию, т. е.
эволюцию, направленную на реализацию опреде-
ленной цели. Такие идеи обычно не поддерживают-
ся биологами-эволюционистами. Другие процессы,
такие как сжигание ископаемого топлива и унич-
тожение лесов, сильнее воздействуют на состав
атмосферы. С4-растения, фиксирующие СО2 в ус-
ловиях. когда рубиско предпочитает О2 в качестве
субстрата, также участвуют в установлении отно-
шения СО2/О2 в атмосфере.
9. а) Без поступления NADPH из пентозофосфатного
пути клетки не могли бы синтезировать липиды и
другие восстановленные соединения, б) Без рибу-
лозо-1,5-бисфосфата блокируется цикл Кальвина.
10. Фиксация СО2 кукурузой происходит по С4-пути,
изученному Хетчем и Слеком. В этом процессе
ФЕП быстро карбоксилируется, превращаясь в
оксалоацетат (некоторое количество последнего
подвергается трансаминированию с образованием
аспартата), а затем восстанавливается до малата.
Только вслед за декарбоксилированием СО2 вклю-
чается в цикл Кальвина.
11. Измерить скорость фиксации меченого диоксида
углерода на свету (днем) и в темноте. Если фикса-
ция сильнее в темноте, это CAM-растение. Кроме
того, можно определить титруемую кислотность -
так можно определить количество кислот, запасен-
ных в вакуолях за ночь.
12. Реакция изоцитратдегидрогеназы
13. Запасание 1 моля глюкозо-6-фосфата обходится в
1 моль АТР, что составляет 3,3% всего количества
АТР, образующегося в метаболизме глюкозо-6-
фосфата (т. е. эффективность процесса запасания
глюкозы составляет 96,7%).
14. [PPJ в цитозоле высокая, поскольку там нет неор-
ганической пирофосфатазы.
15. а) Низкая концентрация Р, в цитозоле и высокая
концентрация триозофосфатов в хлоропластах;
б) высокая концентрация триозофосфатов в ци-
тозоле.
16. З-Фосфоглицерат - первый продукт фотосинтеза;
[РД увеличивается при замедлении направляемого
светом синтеза АТР из ADP и Рг
17. а) Сахароза + (глюкоза),, -> (глюкоза)п+1 + фруктоза;
б) Образовавшаяся в синтезе декстрана фрукто-
за быстро захватывается и потребляется бакте-
риями.
18. Вид 1 — С4-растение, вид 2 — С3-растение
19. а) В периферических хлоропластах; (б) и (в) в
центре.
20. а) По аналогии с оксигенным фотосинтезом в рас-
тениях (Н2О + СО2 -> глюкоза + О2) можно запи-
сать реакцию H2S + О2 + СО2 -> глюкоза + Н2О + S.
Это сумма процессов восстановления СО2 под дей-
ствием H2S (H2S + СО2 -> глюкоза + S) и энергети-
ческих затрат (H2S + О2 -> S + Н2О). б) Газы H2S и
СО2 образуются химическим путем в глубоковод-
ных отложениях, но О2 почти повсюду на Земле
образуется в фотосинтезе под действием световой
энергии, в) В экспериментах Робинсона метка 3Н
находится у атома С1 в рибулозо-1,5-бис-фосфате,
так что реакция с СО2 приводит к образованию
1 молекулы [3Н]-3 -фосфоглицерата и 1 молеку-
лы немеченого 3-фосфоглицерата. В реакции с О2
образуется 1 молекула [3Н]-2-фосфогликолата
и 1 молекула немеченого 3-фосфоглицерата. Та-
ким образом, отношение [3Н]-3-фосфоглицерата к
[3Н]-2-фосфогликолату соответствует отношению
скоростей карбоксилирования и оксигенирова-
ния. г) При наличии метки 3Н у атома С5 в обоих
процессах получался бы [3Н]-3-фосфоглицерат, и
нельзя было бы различить, в какой реакции обра-
зуется меченый продукт. Эту реакцию нельзя ис-
пользовать для определения Q.
Краткие решения задач и ответы на вопросы 1331 ]
[COJ , 0,00038
Д) [О2] 0,2 ’
^карбоксилирование _ q [СО2]
^оксигепироваипе [ ^2]
Укарб<>ке.<--»<1>о..а....е =8,6-0,0019 = 0,016
* окснгепироваиие
Таким образом, скорость оксигенирования примерно
в 60 раз выше скорости карбоксилирования! е) Если
для наземного растения Q = 8,6, карбоксилирование
происходит с гораздо меньшей скоростью, чем окси-
генирование. Это очень невыгодно, так что, по всей
вероятности, рубиско наземных растений характери-
зуется значительно более высоким значением Q, чем
8,6. В действительности значения Q для наземных
растений варьируют в диапазоне от 10 до 250. И даже
при таких значениях теоретическая скорость оксиге-
нирования остается достаточно высокой, ж) В реак-
ции рубиско участвует газообразный СО2. При любой
температуре молекулы 13СО2 перемещаются медлен-
нее, чем более легкие молекулы 12СО2, поэтому 13СО2
проникает в активный центр фермента (и включается
в субстрат) медленнее, чем 12СО2. з) В истинно сим-
биотических отношениях трубчатые черви должны
получать значительную долю своего углерода от бак-
терий. Присутствие рубиско в эндосимбионтах про-
сто указывает на их способность осуществлять хемо-
синтез, но ничего не говорит о том, что они снабжают
хозяина значительным количеством собственного
углерода. С другой стороны, тот факт, что соотноше-
ния 13С/12С у хозяина и эндосимбионта значительно
ближе, чем у других морских обитателей, доказывает
бактериальное происхождение значительной части
углерода в клетках трубчатых червей.
Глава 21
1. а) Шестнадцать атомов пальмитата происходят из 8
ацетильных групп 8 молекул ацетил-СоА. Меченый
ацетил-СоА превращается в малонил-СоА, содер-
жащий метку в атомах С1 и С2. б) Метаболический
пул малонил-СоА, источник всех атомов углерода в
пальмитате, за исключением атомов С16 и С15, не
включает метку при введении небольшого количе-
ства меченного 14С ацетил-СоА. Следовательно, об-
разуется только [15,16-14С]пальмитат.
2. В процессе гликолиза в пируват превращается как
глюкоза, так и фруктоза. Пируват конвертируется
в ацетил-СоА под действием пируватдегидроге-
назнорго комплекса. Некоторая часть ацетил-СоА
включается в цикл лимонной кислоты, в котором об-
разуются восстановительные эквиваленты (NADH
и NADPH). В митохондриях перенос электронов на
О2 приводит к образованию АТР.
3. 8 ацетил-СоА + 15АТР + 14NADPH + 9Н2О —
пальмитат + 8СоА + 15ADP +
+ 15Р. + 14NADP+ + 2H+
4. а) Три атома дейтерия на молекулу пальмита-
та, все они локализованы у атома С16; все другие
двухуглеродные звенья образованы из немеченого
малонил-СоА. б) Семь атомов дейтерия в молеку-
ле пальмитата; они расположены у всех четных ато-
мов углерода за исключением атома С16.
5. При использовании трехуглеродного звена
малонил-СоА — активированной формы ацетил-
СоА (вспомните, что для синтеза малонил-СоА
требуется АТР) — метаболизм направлен в сторону
синтеза жирных кислот за счет экзергонического
высвобождения СО2.
6. Скорость синтеза жирных кислот лимитируется
реакцией карбокислирования ацетил-СоА, ко-
торую катализирует ацетил-СоА-карбоксилаза.
Высокая концентрация цитрата и изоцитрата
свидетельствует о том, что условия благоприят-
ствуют синтезу жирных кислот: активный цикл
лимонной кислоты обеспечивает достаточное
количество АТР, восстановленных нуклеотидов
и ацетил-СоА. Цитрат стимулирует (повышает
Vlnax) ацетил-СоА-карбоксилазу. а) Поскольку ци-
трат прочнее связывается с нитчатой формой фер-
мента (активной формой), высокая концентрация
цитрата сдвигает уравнение реакции протомер
нитчатый полимер в сторону образования актив-
ной формы, б) Напротив, пальмитоил-СоА (ко-
нечный продукт синтеза жирных кислот) сдвигает
равновесие реакции в сторону неактивной (прото-
мерной) формы. Следовательно, при накоплении
конечного продукта реакции происходит замедле-
ние биосинтеза.
7. а) Ацетил-СоА (мит) + АТР + СоА (цит)
ацетил-СоА (цит) + ADP + Р, + СоА (мит);
б) 1 АТР на ацетогруппу; в) да.
8. Двойная связь в пальмитолеате возникает в ре-
зультате окислительной реакции, катализируемой
ацил-СоА-десатуразой - оксидазой со смешанной
функцией, использующей в качестве косубстрата
кислород.
9. 3 Пальмитат + глицерин + 7АТР + 4Н2О
трипальмитин + 7ADP + 7Р, + 7Н+
10. В организме взрослых крыс концентрация три-
ацилглицеринов поддерживается на стационарном
уровне путем уравнивания скоростей реакций рас-
щепления и биосинтеза. Поэтому триацилглицери-
ны жировой ткани постоянно находятся в обороте,
что объясняет включение метки 14С из поступив-
шей с пищей глюкозы.
1332] Приложение Б
11. Суммарная реакция:
Дигидроксиацетонфосфат + NADH + пальмитат +
+ олеат + ЗАТР + СТР + холин + 4Н2О
фосфатидилхолин + NAD+ + 2АМР +
+ ADP + Н++ CMP + 5Р;
7 молекул АТР на молекулу фосфатидилхолина
12. Недостаточность метионина снижает уровень ado-
Met, необходимого для de novo синтеза фосфати-
дилхолина. Реутилизационный путь не использует
adoMet, а использует имеющийся холин. Вот поче-
му фосфатидилхолин может синтезироваться даже
при недостатке метионина в пище, пока в организ-
ме имеется холин.
13. Метка 14С появляется в активированном изопрене
в трех местах:
14СН2
^с—14сн2—сн2—
14СНз
14. а) АТР; б) UDP-глюкоза; в) CDP-этаноламин;
г) UDP-галактоза; д) ацил-СоА жирной кислоты;
е) S-аденозилметионин; ж) малонил-СоА; з) А3-
изопренилпирофосфат
15. Линолеат необходим для синтеза простагландинов.
Животные не могут превращать олеат в линолеат,
поэтому линолеат — незаменимая жирная кисло-
та. Растения могут осуществлять это превраще-
ние, поэтому линолеат содержится в растительной
пище (см. рис. 21-12).
16. Лимитирующей стадией в биосинтезе холестери-
на является синтез мевалоната, катализируемый
гидроксиметилглутарил-СоА-редуктазой. Этот
фермент подвержен аллостерической регуляции
мевалонатом и производными холестерина. Кро-
ме того, при высокой внутриклеточной концен-
трации холестерина транскрипция гена HMG-
СоА снижается.
17. При снижении уровня холестерина в результате
приема статинов клетки пытаются компенсировать
синтез путем повышения экспрессии гена HMG-
СоА-редуктазы. Тем не менее статины — хорошие
конкурентные ингибиторы HMG-CoA-редуктазы
и снижают общий уровень синтеза холестерина.
18. Примечание: студент, незнакомый с особенностями
данного фермента, может предложить несколько
возможных альтернатив. Реакция тиолазы начина-
ется с нуклеофильной атаки остатка Cys активного
центра на первую молекулу ацетил-СоА, что при-
водит к вытеснению -S-CoA и образованию кова-
лентной тиоэфирной связи между остатком Cys и
ацетильной группой. Затем основание в молекуле
фермента притягивает протон от метильной груп-
пы второго ацетил-СоА, в результате чего образу-
ется карбанион, атакующий углерод карбонильной
группы тиоэфира, образовавшегося на предыду-
щей стадии. Сульфгидрильная группа остатка Cys
вытесняется, и образуется ацетоацетил-СоА.
Реакция HMG-CoA-синтазы начинается таким же
образом и приводит к образованию тиоэфирной
связи между остатком Cys в молекуле фермента
и ацетильной группой ацетил-СоА с вытеснени-
ем -S-CoA. Далее -S-CoA отщепляется в виде
CoA-SH, а ацетоацетил-СоА связывается с фер-
ментом. От метильной группы связанного с фер-
ментом ацетила отбирается протон, что приво-
дит к образованию карбаниона, который атакует
карбонил в кетогруппе ацетоацетил-СоА. В этой
реакции карбонил превращается в гидроксиль-
ный ион, который протонируется, образуя -ОН.
Затем тиоэфирная связь с ферментом подвергается
гидролитическому расщеплению с образованием
продукта реакции. Реакция HMG-CoA-редуктазы:
один гидрид-ион из NADPH вытесняет -S-CoA, а
затем второй восстанавливает альдегидную группу
до гидроксильной группы.
19. Статины ингибируют HMG-CoA-редуктазу —
фермент на пути синтеза активированных изопре-
нов — предшественников холестерина и многих
изопреноидов, включая кофермент Q (убихинон).
Следовательно, статины могут снижать количество
кофермента Q, доступного для митохондриального
дыхания. Убихинон содержится в пищевых про-
дуктах, а также образуется в результате биосинтеза,
но пока неизвестно, сколько убихинона требуется
организму и насколько полно пищевые источники
могут восполнить недостаток биосинтеза. К по-
бочным эффектам действия статинов относится
уменьшение синтеза некоторых изопреноидов.
Астаксантин
Краткие решения задач и ответы на вопросы 1333]
б) «Голова к голове». Это можно объяснить дву-
мя способами. Во-первых, «хвост» геранилгера-
нилфосфата имеет разветвленную диметильную
структуру, как и оба конца фитоена. Во-вторых, в
результате отщепления PPi не образуется свобод-
ных гидроксильных групп, следовательно, две
-О-(Р)-(Р)-«головки» связываются между со-
бой, образуя фитоен, в) В четырех раундах деги-
дрирования четыре простые связи превращаются
в двойные, г) Нет. Подсчет одинарных и двойных
связей в реакции подсказывает, что одна двойная
связь заменяется двумя одинарными связями,
следовательно, суммарного окисления или вос-
становления в этом процессе не происходит.
д) Стадии Ф-®. Фермент может превращать IPP и
DMAPP в геранилгеранилфосфат, но не катализи-
рует следующие стадии пути, что подтверждается
результатами с другими субстратами, е) Штаммы
1-4 не имеют гена crtE и синтезируют гораздо мень-
ше астаксантина, чем штаммы 5-8, которые харак-
теризуются повышенной экспрессией crtE. Таким
образом, повышенная экспрессия crtE приводит к
значительному усилению синтеза астаксантина. Ди-
кий штамм Е. coli обладает некоторой активностью,
необходимой для реализации стадии ®, но это пре-
вращение фарнезилпирофосфата в геранилгера-
нилпирофосфат является лимитирующим, ж) IPP-
изомераза. Сравнение штаммов 5 и 6 показывает,
что добавление ispA, продукт которого катализиру-
ет стадии Ф и не оказывает большого влияния
на продукцию астаксантина, так что это нелими-
тирующие стадии. Однако из сравнения штаммов
5 и 7 следует, что добавление idi значительно уси-
ливает синтез астаксантина, поэтому реакция IPP-
изомеразы — лимитирующая стадия при повышен-
ной экспрессии crtE. з) Уровень экспрессии был
бы низким (+), сравнимым с уровнем экспрессии в
штаммах 5, 6 и 9. Без повышенной экспрессии idi
синтез астаксантина лимитируется низкой актив-
ностью IPP-изомеразы и, следовательно, низким
уровнем IPP.
Глава 22
1. В условиях симбиоза бактерии поставляют расте-
ниям ион аммония NH4, который получают путем
восстановления атмосферного азота. Этот процесс
требует значительных затрат АТР.
2. Перенос азота от NH3 на углеродный скелет могут
катализировать (1) глутаминсинтетаза и (2) глута-
матдегидрогеназа. Последний образует глутамат,
донор аминогрупп во всех реакциях трансамини-
рования при образовании аминокислот и синтезе
белка.
3. Сначала образуется связь между связанным с фер-
ментом PLP и фосфогомосерином, а затем в резуль-
тате перегруппировки у ct-углерода субстрата образу-
ется кетиминовая группа. Это приводит к активации
p-углерода и передаче протона с последующим вы-
теснением фосфатной группы и образованием двой-
ной связи между р- и у-атомами углерода. Перегруп-
пировка (начинающаяся с удаления протона у атома
углерода, соседнего с азотом аминогруппы субстра-
та) приводит к перемещению двойной связи между
а- и р-атомами углерода, а кетимин превращается в
альдимин. Атака воды на р-атом углерода облегчает-
ся связанным пиридоксалем, после чего происходит
гидролиз иминовой связи между PLP и продуктом с
высвобождением треонина.
133 1] Приложение Б
4. В организме млекопитающих токсичные ионы ам-
мония превращаются в глутамин, что снижает ток-
сическое воздействие на головной мозг.
5. Глюкоза + 2СО2 + 2NH3 -> 2 аспартат + 2Н+ + 2Н2О
6. N-концевой домен глутаминазы примерно такой
же, как у всех глутаминамидотрансфераз. Лекар-
ство, которое связывается с этим доменом, может
ингибировать многие ферменты и, следовательно,
вызывать значительно больше побочных эффек-
тов, чем более специфичный ингибитор, связыва-
ющийся с уникальной С-концевой последователь-
ностью.
7. Если фенилаланингидроксилаза теряет актив-
ность, блокируется путь биосинтеза тирозина, и
организм должен получать тирозин с пищей.
8. В синтезе adoMet происходит выделение трифос-
фата из АТР. Гидролиз трифосфата делает реакцию
термодинамически более выгодной.
9. Если бы ингибирование глутаминсинтазы не было
согласованным, при насыщающей концентрации
гистидина происходило бы отключение фермента-
тивной активности и прекращение синтеза глута-
мина, который нужен бактерии для синтеза других
продуктов.
10. Фолиевая кислота - предшественник тетрагидро-
фолата (рис. 18-16, т. 2), необходимого для синтеза
глицина (рис. 22-12, т. 2), предшественника пор-
фиринов. Это объясняет, почему недостаточность
фолиевой кислоты нарушает синтез гемоглобина.
11. Ауксотрофы по глицину: аденин и гуанин; ауксо-
трофы по глутамину: аденин, гуанин и цитозин;
ауксотрофы по аспартату: аденин, гуанин, цито-
зин и уридин.
12. а) Механизм рацемизации аминокислот отражен
на рис. 18-6 (т. 2), стадия Атом фтора во фтор-
аланине — прекрасная уходящая группа. Фтор-
аланин вызывает необратимое (ковалентное) инги-
бирование аланинрацемазы. На схеме представлен
возможный механизм реакции (Nuc — любая ну-
клеофильная группа в боковой цепи аминокислот-
ного остатка в активном центре фермента):
б) Азасерин (см. рис. 22-48) — аналог глутамина.
Диазоацетильная группа обладает высокой реакци-
онной способностью и образует ковалентные связи
с нуклеофильными группами в активном центре
глутаминамидотрансферазы.
13. а) Как показано на рис. 18-16, пяря-аминобензо-
ат является компонентом М5,М10-метилентетра-
гидрофолата — кофактора, участвующего в перено-
се одноуглеродных фрагментов, б) В присутствии
сульфаниламида, структурного аналога тшря-ами-
нобензоата, бактерии не могут синтезировать тетра-
гидрофолат, который необходим для превращения
AICAR в FAICAR, и в результате происходит на-
копление ATCAR. в) Конкурентное ингибирование
сульфаниламидом фермента, участвующего в био-
синтезе тетрагидрофолата, преодолевается в при-
сутствии избытка субстрата (т?<2р<2-аминобензоата).
14. [14С]Оротат образуется в результате следующей се-
рии реакций (первые три стадии являются частью
цикла лимонной кислоты):
ОО14С - 14СН2-14СН2 - 14соо-
Сукцинат
оо14с н
14С = 14С
Н 14СОО-
Фумарат
ОН Н
I I
оо14с—14с—14с—14соо
I I
н н
Малат
Краткие решения задач и ответы на вопросы [335]
О н
II I
“ оо14с — 14с — 14с — 14СОО"
I
н
Оксалоацетат
| трансаминирование
NH3
оо14с—14с—14сн2—14СОО”
н
Аспартат
О
Нз^° J**2
/С^14СОО-
О N н
Н
О
Н\ 14с
XN/ '''СН.,
I J
,с^14соо~
О N н
Н
гуанина в мочевую кислоту, что вызывает накопле-
ние ксантина, который хуже растворим, чем моче-
вая кислота. Это и является причиной появления
ксантиновых камней. Но поскольку расщепление
GMP происходит в меньшем масштабе, чем рас-
щепление АМР, повреждение почек ксантиновыми
камнями — меньшее зло, чем подагра.
17. 5-Фосфорибозил-1-пирофосфат; это первый ак-
цептор NH3 на пути биосинтеза пуринов.
18. а) Удаляется а-карбоксильная группа, а к у-атому
углерода присоединяется -ОН-группа. б) После-
довательность Btrl гомологична последователь-
ности ацилпереносящих белков. Молекулярная
масса Btrl увеличивается при инкубации в таких
условиях, когда СоА может присоединяться к
белкам. Присоединение СоА к остатку Ser при-
водит к замене -ОН-группы (мол. масса 17) на
4'-фосфопантетеиновую группу (см. рис. 21-5,
т. 2) с формулой CHH21N2O7PS (мол. масса 356).
Таким образом, И 182 - 17 + 356 = 12 151, что
очень близко к экспериментально найденному
значению Мг = 12 153. в) Тиоэфирная связь мо-
жет образовываться с а-карбоксильной группой,
г) В распространенной реакции удаления
а-карбоксила аминокислоты (рис. 18-6, в, т. 2) эта
группа свободна. Более того, трудно себе предста-
вить реакцию декарбоксилирования, начинающу-
юся с карбоксильной группы, находящейся в фор-
ме тиоэфира, д) 12 240 - 12 281 = 41, что близко к
значению Мг для СО2 (44). Поскольку BtrK, скорее
всего, является декарбоксилазой, наиболее вероят-
на следующая структура:
H3N^^y°
S
Btrl
Оротат
15. Клетки не запасают нуклеотиды, чтобы исполь-
зовать их в качестве топливных молекул, и не рас-
щепляют их полностью, но только высвобождают
основания, которые используются в путях реути-
лизации. Из-за низкого значения отношения С : N
нуклеотиды являются бедным источником энергии.
16. Лечение аллопуринолом приводит к следующим
последствиям. (1) Ингибируется превращение
гипоксантина в мочевую кислоту, что вызывает
накопление гипоксантина, который лучше раство-
ряется в воде и легче выводится из организма. Это
смягчает клинические симптомы, связанные с рас-
щеплением АМР. (2) Ингибируется превращение
е) 12 370 - 12 240 = 130. При удалении от глута-
миновой кислоты (C5H9NO4, Мг 147) гидроксиль-
ной группы (-ОН, 17) в процессе глутамилиро-
вания образуется остаток с массой 130. Таким
образом, у-глутамилирование изображенной выше
молекулы приводит к увеличению ее молекуляр-
ной массы па 130. BtrJ способен осуществлять
у-глутамилинование других субстратов, в том чис-
ле и представленной выше молекулы. Скорее всего,
реакция происходит по свободной аминогруппе и
приводит к образованию следующей структуры:
О О
Btrl
1336 | Приложение Б
Антибиотик
Глутамат + Btrl
ОН
Антибиотик
Глава 23
1. Они распознаются двумя типами рецепторов, обыч-
но находящимися в разных типах клеток, и вызыва-
ют различные реакции.
2. Стационарная концентрация АТР поддерживается
за счет переноса фосфорильной группы от фосфо-
креатина на ADP. 1-Фтор-2,4-динитробензол инги-
бирует креатинкиназу.
3. Аммоний чрезвычайно токсичен для нервной ткани,
особенно для клеток головного мозга. Избыток NH3
выводится за счет превращения глутамата в глутамин,
который переносится в печень и впоследствии превра-
щается в мочевину. Дополнительный глутамип возника-
ет в результате превращения глюкозы в а-кетоглутарат,
а-кетоглутарата в глутамат и глутамата в глутамин.
4. Глюкогенные аминокислоты используются в синте-
зе глюкозы для головного мозга; другие окисляются
в митохондриях в цикле лимонной кислоты.
5. Из глюкозы по следующему пути: глюкоза -* диги-
дроксиацетонфосфат (гликолиз); дигидроксиаце-
тонфосфат + NADH + Н+ -> глицерин-3-фосфат +
NAD+ (реакция глицерин-3-фосфатдегидрогеназы).
6. а) Повышенная мышечная активность требует до-
полнительного АТР, который образуется при уси-
ленном потреблении О2. б) После быстрого бега
образовавшийся в анаэробном гликолизе лактат
превращается в глюкозу и гликоген, что также тре-
бует расхода АТР и, следовательно, О2.
7. Глюкоза — основное топливо мозга. ТРР-зависимое
окислительное декарбоксилирование пирувата до
ацетил-СоА — обязательная стадия полного мета-
болизма глюкозы.
8. 190 м
9. а) Инактивация — быстрый способ изменения кон-
центрации гормона, б) Уровень инсулина поддержи-
вается путем выравнивания скоростей его синтеза и
расщепления, в) За счет изменения скорости высво-
бождения из запасных гранул, скорости транспорта
и превращения прогормона в активный гормон.
10. Водорастворимые гормоны связываются с рецеп-
торами на внешней поверхности клетки, запуская
образование вторичных посредников (таких как
сАМР) внутри клетки. Жирорастворимые гормо-
ны проникают через плазматическую мембрану и
непосредственно взаимодействуют с молекулами-
мишенями или с рецепторами.
11. а) В сердце и скелетных мышцах отсутствует
глюкозо-6-фосфатаза. Весь образующийся глюко-
зо-6-фосфат направляется по пути гликолиза, а
при недостатке кислорода превращается в лактат
через пируват, б) При реакции «борьба или бег-
ство» концентрация предшественников гликолиза
должна быть высокой, чтобы обеспечить мышеч-
ную активность. Фосфорилированные соединения
не могут выходить из клетки, поскольку мембрана
непроницаема для заряженных молекул, и поэто-
му глюкозо-6-фосфат не переносится с помощью
переносчика глюкозы. Напротив, печень должна
высвобождать в кровь глюкозу для ее поддержа-
ния на постоянном уровне. Глюкоза образуется из
глюкозо-6-фосфата и переносится в кровоток.
12. а) Усиливается захват глюкозы из крови и ее ме-
таболизм в печени, что приводит к гипогликемии;
отключается катаболизм аминокислот и жирных
Краткие решения задач и ответы на вопросы 133 / ]
кислот, б) Циркулирующих в крови топливных
молекул недостаточно для синтеза АТР. Мозг
повреждается по той причине, что для него глюко-
за — главный источник энергии.
13. Тироксин выступает в роли разобщающего агента в
окислительном фосфорилировании. Разобщающие
агенты снижают отношение P/О, и для удовлетво-
рения потребностей в АТР ткани усиливают про-
цесс дыхания. Термогенез также может объясняться
усиленной утилизацией АТР стимулированной ти-
роксином тканью, поскольку при повышении по-
требности в АТР усиливается окислительное фосфо-
рилирование и повышается интенсивность дыхания.
14. Поскольку прогормоны неактивны, они могут в не-
обходимом количестве храниться в секреторных
гранулах. Быстрая активация прогормонов дости-
гается путем ферментативного расщепления в от-
вет на соответствующий сигнал.
15. В организме животных глюкоза синтезируется из
многих предшественников (см. рис. 1-15). В орга-
низме человека основным предшественником глю-
козы является глицерин из триацилглицеринов и
глюкогенные кислоты из белков.
16. Исходно страдавшая ожирением мышь с геноти-
пом ob/ob начнет худеть, мышь ОВ/ОВ сохранит
свой нормальный вес.
17. ИМТ = 39,3. Человек такого роста имеет нормальный
показатель ИМТ = 25 при массе тела 75 кг. Следова-
тельно, ему нужно похудеть на 43 кг (95 фунтов).
18. Секреция инсулина понизится. Валиномицин
оказывает такое же действие, как открытие К+-
каналов, позволяя ионам К? выходить из клетки,
что вызывает гиперполяризацию.
19. Печень не получает сигнал инсулина и, следова-
тельно, продолжает поддерживать высокий уровень
глюкозо-6-фосфата и глюконеогенеза, повышая
уровень глюкозы крови как при голодании, так и
после приема сахаросодержащей пищи. Повышен-
ный уровень глюкозы крови вызывает высвобожде-
ние инсулина [3-клетками поджелудочной железы,
несмотря на высокий уровень инсулина в крови.
20. Следует учесть несколько аспектов. Какова веро-
ятность возникновения сердечных приступов, свя-
занных с приемом препарата. Как эта вероятность
соотносится с вероятностью сердечных приступов
у людей с хроническими осложнениями диабета 2
типа. Существуют ли столь же эффективные мето-
ды лечения, вызывающие меньше нежелательных
побочных эффектов.
21. При отсутствии глюкозидазной активности кишеч-
ника замедляется всасывание глюкозы из пище-
вого крахмала и гликогена, что снижает обычный
подъем концентрации глюкозы крови после при-
ема пищи. Нерасщепленные олигосахариды фер-
ментируются бактериями в толстой кишке, а вы-
деляющиеся при этом газы вызывают дискомфорт.
22. а) Закрытие ATP-зависимых К+-каналов приводит
к деполяризации мембраны и усилению секреции
инсулина, б) Диабет 2 типа вязан с понижением
чувствительности клеток к инсулину, а не с недо-
статочным синтезом инсулина. Повышение кон-
центрации циркулирующего инсулина ослабляет
симптомы заболевания, в) У больных диабетом
1 типа повреждены [3-клетки поджелудочной желе-
зы, поэтому глибурид таким больным не поможет,
г) Иод, как и хлор, который иод замещает в мече-
ном глибуриде, — галоген, но его размер больше и
он имеет несколько иные свойства. Поэтому воз-
можно, что иодированный глибурид не связыва-
ется с SUR. Если же вместо этого он связывается
с другой молекулой, в результате эксперимента
будет клонирован ген другого белка, д) Хотя белок
очищали, но «очищенный» препарат мог представ-
лять собой смесь нескольких белков, выделяющих-
ся одновременно в одних и тех же эксперименталь-
ных условиях. В таком случае аминокислотная
последовательность могла бы соответствовать
последовательности такого сопутствующего бел-
ка. Вероятность этого события исключили с по-
мощью антител, показавших, что данные пептид-
ные последовательности присутствуют в SUR.
е) Хотя клонированный ген действительно ко-
дирует 25-аминокислотную последовательность,
обнаруженную в SUR, в принципе, возможно, что
этот ген кодирует такую же последовательность
в другом белке. В таком случае этот другой ген с
большой вероятностью должен экспрессироваться
в других клетках, чем ген SUR. Эксперименты по
гибридизации мРНК доказали, что кДНК предпо-
лагаемого гена SUR действительно кодирует SUR.
ж) Избыток немеченого глибурида конкурирует с
меченым глибуридом за связывание с SUR. В ре-
зультате наблюдается связывание гораздо мень-
шего количества меченого глибурида, и поэтому
в белке с массой 140 кДа практически не обнару-
жено радиоактивности, з) В отсутствие избытка
немеченого глибурида меченый белок с массой
140 кДа обнаружен только в присутствии предпо-
лагаемой кДНК SUR. Избыток немеченого глибу-
рида конкурирует с меченым глибуридом, и мечен-
ного 1251 140-кДа белка обнаружено не было. Это
показывает, что кДНК кодирует связывающийся
с глибуридом белок точно такой же массы, как и
SUR — строгое доказательство того, что клони-
рованный ген кодирует белок SUR. и) Возможны
дополнительные эксперименты. (1) Осуществить
1338 ] Приложение Б
экспрессию кДНК предполагаемого SUR в клетках
СНО (клетки яичников китайского хомячка) и по-
казать, что трансформированные клетки обладают
активностью ATP-зависимых К+-каналов. (2) По-
казать, что клетки HIT с мутациями в предполага-
емом гене SUR теряют активность АТР-зависимых
К+-каналов. (3) Показать, что экспериментальные
животные или люди с мутациями в предполагае-
мом гене SUR не способны секретировать инсулин.
Глава 24
1. 6,1 • 104 нм; в 290 раз длиннее головки фага Т2.
2. Число остатков А не равно числу Т, а число G не рав-
но числу С. Это означает, что эта ДНК пе двойная
спираль, образованная при спаривания оснований.
На самом деле фаг М13 имеет однонитевую ДНК.
3. Мг = 3,8-108; длина 200 мкм; LkQ = 55 200; Lk = 51 900.
4. Экзоны содержат 3 п.н./аминокислоту х 192 амино-
кислоты = 576 п. н. Оставшиеся 864 п. н. составляют
последовательности интронов, возможно, лидирую-
щую или сигнальную последовательность, а также
другие некодирующие участки ДНК.
5. 5000 п. н. а) Не изменяется; Lk не может изменяться
без разрыва одних и образования других ковалент-
ных связей в ДНК. б) Становится неопределенной.
По определению кольцевая ДНК с разрывом в од-
ной нити не имеет Lk. в) Снижается. В присутствии
АТР гираза раскручивает ДНК. г) Не изменяется,
если при нагревании в нитях ДНК не произошло
разрывов.
6. Чтобы Lk не изменилось, топоизомераза должна
вводить одинаковое число положительных и отри-
цательных витков.
7.о= -0,067; вероятность >70%.
8. а) Неопределенное; нити никированной ДНК мо-
гут быть разъединены и, следовательно, не имеют
Lk. б) 476; в) 476; ДНК уже релаксирована, так что
топоизомераза не вызывает никаких изменений,
г) 460; каждое действие ДНК-гиразы в присутствии
АТР снижает Lk на две единицы, д) 464; эукариоти-
ческая топоизомераза I повышает Lk слабо закру-
ченной или имеющей отрицательные супервитки
ДНК на одну единицу, е) 460; связывание с нуклео-
сомой не приводит к разрыву нитей ДНК и поэтому
не влияет на значение Lk.
9. Основная структурная единица хроматина повто-
ряется примерно каждые 200 п. н; ДНК доступна
для нуклеазы только через интервалы 200 п. н.
Краткой обработки недостаточно для расщепления
ДНК во всех доступных точках, поэтому возникает
лестница фрагментов ДНК, масса которых кратна
200 п. н. Толщина полос ДНК говорит о некоторой
вариабельности расстояний между участками рас-
щепления. Например, не все фрагменты в самой
нижней полосе имеют размер точно 200 п. н.
10. Правая спираль характеризуется положительным
значением Lk, левая спираль (как в Z-ДНК) - от-
рицательным значением. Снижение Lk замкнутой
кольцевой В-ДНК путем введения отрицательных
витков облегчает образование участков Z-ДНК в
некоторых последовательностях. (О последова-
тельностях, способствующих образованию Z-ДНК,
говорится в гл. 8.)
11. а) Обе нити должны быть замкнуты ковалентны-
ми связями в кольцо, либо ограничены на концах
каким-либо другим образом, б) Образование кре-
стообразных структур, левозакрученной Z-ДНК,
плектонемических или соленоидальных структур,
а также раскрученных молекул ДНК. в) Топоизо-
мераза II или ДНК-гираза Е. coli. г) Фермент свя-
зывается с ДНК в точках пересечения двунитевой
последовательности, расщепляет обе нити одного
из пересекающихся сегментов, протягивает дру-
гой сегмент через разрыв, а затем зашивает разрыв.
В результате значение Lk уменьшается на 2.
12. Центромера, теломеры и автономно реплицирую-
щиеся последовательности или точка начала ре-
пликации.
13. Бактериальный нуклеоид организован в виде доме-
нов длиной примерно по 10 000 п. н. Расщепление
рестриктазой приводит к релаксации ДНК внутри
домена, но не за его пределами. Это повлияет на
экспрессию всех генов домена, зависящую от топо-
логии ДНК, но не на гены в других доменах.
14. а) При соединении концов ДНК с образованием
замкнутого релаксированного кольца, некоторые
типы ДНК полностью релаксированы, тогда как
другие все же имеют некоторое количество левых
или правых витков. Это и объясняет распределе-
ние топоизомеров относительно наиболее рассла-
бленной формы, б) Положительная суперспираль,
в) ДНК, находящаяся в релаксированном состо-
янии даже при добавлении красителя, имеет раз-
рыв в одной или в обеих нитях. В ходе выделения
кольцевой ДНК в некоторых молекулах неизбеж-
но появляется некоторое количество разрывов,
г) Примерно -0,05. Это значение определяется про-
сто путем сравнения нативной ДНК с образцами с
известным значением а. На обоих гелях нативная
ДНК движется ближе всего к образцу с о = -0,049.
15. а) При нерасхождении хромосом одна дочерняя
клетка и все ее потомство имеют две копии син-
тетической хромосомы и окрашены в белый цвет.
Другая дочерняя клетка и ее потомство не имеют
копии синтетической хромосомы и окрашены в
Краткие решения задач и ответы на вопросы [ 339 j
красный цвет. Поэтому половина колонии белая,
а половина красная, б) При потере хромосомы
одна дочерняя клетка и ее потомство имеют одну
копию синтетической хромосомы и окрашены в
розовый цвет. Другая дочерняя клетка и ее потом-
ство не имеют копии синтетической хромосомы и
окрашены в красный цвет. В результате получают-
ся красно-розовые колонии, в) Функциональная
центромера должна иметь размер менее 0,63 т. п. н.,
поскольку все фрагменты больше этого размера
более стабильны при митозе, г) Теломеры нужны
для полной репликации линейных молекул ДНК,
кольцевая ДНК может реплицироваться без них.
д) Чем крупнее хромосома, тем точнее осуществля-
ется процесс сегрегации. Однако представленные
данные не указывают ни на минимальный размер,
ниже которого синтетическая хромосома полно-
стью нестабильна, ни на максимальный размер,
выше которого стабильность уже не меняется.
е) Нормальные хромосомы.
Как показывает график, даже если бы синтетиче-
ские хромосомы имели такую же длину, как нор-
мальные дрожжевые хромосомы, они не были бы
такими же стабильными. Это означает, что на ста-
бильность хромосом влияют еще какие-то другие,
пока неизвестные факторы.
Глава 25
1. При случайной, «дисперсионной» репликации во
втором поколении вся ДНК должна была бы иметь
одинаковую плотность и появляться в виде единой
полосы, а не двух полос, которые наблюдали Ме-
зельсон и Сталь.
2. В этом продолжении эксперимента Мезельсона-
Сталя после трех клеточных делений соотношение
15N-14N ДНК к 14N-14N ДНК составляет 2/6 = 0,33.
3. а) 4,42-105 оборотов; б) 40 минут. В клетках, деля-
щихся каждые 20 минут, цикл репликации начина-
ется каждые 20 минут, так что каждый следующий
цикл начинается до того, как завершится предыду-
щий. в) от 2000 до 5000 фрагментов Оказаки. Эти
фрагменты имеют длину от 1000 до 2000 нуклеоти-
дов и прочно связаны с матрицей путем спаривания
оснований. Каждый синтезированный фрагмент бы-
стро присоединяется к отстающей нити, что обеспе-
чивает правильный порядок фрагментов.
4. А 28,7%, G 21,3%, С 21,3%, Т 27=8,7%. Нить ДНК,
синтезированная на матричной нити: А 32,7%,
G 18,5%, С 24,1%, Т 24,7%; нить ДНК, синтезиро-
ванная на основе комплементарной нити: А 24,7%,
G 24,1%, С 18,5%, Т 32,7%. Предполагается, что обе
матричные нити реплицировались полностью
5. а) Нет. Включение метки 32Р в ДНК происходит в
результате синтеза новой ДНК, что возможно толь-
ко в присутствии всех четырех предшественников,
б) Да. Хотя для синтеза ДНК необходимы все че-
тыре нуклеотидных предшественника, чтобы метка
включилась в новую ДНК, радиоактивным должен
быть лишь один из них. в) Нет. Радиоактивность
включается в ДНК лишь в том случае, если метка
32Р содержится в а-фосфатной группе, поскольку
ДНК-полимераза отщепляет пирофосфат, т. е. р- и
у-фосфатные группы.
6. Механизм 7: З'-ОН присоединяющегося dNTP ата-
кует а-фосфат трифосфата на 5'-конце растущей
нити ДНК, вытесняя пирофосфат. Механизм ис-
пользует нормальные dNTP, и растущая цепь ДНК
всегда имеет трифосфат на 5'-конце.
З'-конец:
134 0] Приложение Б
Механизм 2\ Здесь используются предшественники
нового типа — нуклеотид-З'-трифосфаты. На расту-
щем конце цепи ДНК имеется 5'-ОН-группа, которая
атакует а-фосфат следующего дезоксинуклеотид-3'-
фосфата, вытесняя пирофосфат. Обратите внима-
ние, что для этого механизма требуется эволюция
новых метаболических путей, обеспечивающих син-
тез новых дезоксинуклеотид-З'-трифосфатов.
З'-конец-
3' -конец'
7. Лидирующая нить: предшественники: dATP,
aGTP, dCTP, dTTP (а также матрица ДНК и ДНК-
праймер); ферменты и другие белки: ДНК-гираза,
геликаза, белок, связывающий однонитевую ДНК,
ДНК-полимераза III, топоизомеразы и пирофос-
фатаза. Отстающая нить: предшественники:
АТР, GTP, СТР, UTP, dATP, dGTP, dCTP, dTTP
(а также РНК-праймер); ферменты и другие бел-
ки: ДНК-гираза, геликаза, белок, связывающий
однонитевую ДНК, праймаза, ДНК-полимераза III,
ДНК-полимераза I, лигаза, топоизомеразы и пиро-
фосфатаза. ДНК-лигаза в качестве кофактора ис-
пользует NAD+.
8. Мутанты с дефектной ДНК-лигазой образуют
ДНК-дуплекс, в котором одна из нитей остается
в виде отдельных фрагментов (фрагментов Оказа-
ки). При денатурации такого дуплекса с последую-
щей седиментацией получается одна фракция, со-
держащая интактную нить ДНК (полоса с большой
молекулярной массой), и вторая фракция, содер-
жащая отдельные фрагменты (низкомолекулярная
полоса).
9. Уотсон-криковское спаривание оснований между
матрицей и лидирующей нитью; проверка и уда-
ление неправильно встроенных нуклеотидов под
действием З'-экзонуклеазной активности ДНК-
полимеразы III. Да, это возможно. Поскольку ме-
ханизмы, обеспечивающие точность репликации
функционируют как для лидирующей, так и для
отстающей цепи, отстающая цепь может, в принци-
пе, собираться с такой же точностью. Однако боль-
шое число химических реакций при биосинтезе
этой цепи может быть дополнительным источни-
ком ошибок.
10. ~ 1200 п. н. (600 п. н. в каждом направлении).
11. Небольшая часть (13 из 107 8 9 клеток) ауксотрофных
по гистидину мутантов переживают спонтанную
обратную мутацию и вновь приобретают способ-
ность синтезировать гистидин. 2-Аминоантрацен
повышает вероятность таких мутаций примерно
в 1800 раз и, следовательно, является мутагеном.
Большинство канцерогенных веществ являются
мутагенами, так что 2-аминоантрацен также может
быть канцерогеном.
12. Спонтанное дезаминирование 5-метилцитозина
(см. т. 1, с. 395) приводит к образованию тимина
и аномальной пары G-Т. Это одна из самых рас-
пространенных ошибок в эукариотической ДНК.
Специализированная система репарации находит
дефект и восстанавливает пары G=C.
13. а) Ультрафиолетовое облучение приводит к обра-
зованию пиримидиновых димеров. В нормальных
фибробластах они удаляются путем вырезания
фрагмента поврежденной нити с помощью спе-
циального фермента. Поэтому денатурированная
однонитевая ДНК состоит из множества фраг-
ментов, образовавшихся в результате такого рас-
щепления, и средняя молекулярная масса этой
ДНК меньше. В ДНК больных пигментной ксеро-
дермой такие фрагменты отсутствуют и средняя
молекулярная масса ДНК не изменяется, б) От-
сутствие фрагментов однонитевой ДНК в клетках
больных после УФ-облучения свидетельствует о
дефекте или об отсутствии специфической эксци-
зионной нуклеазы.
14. При гомологичной генетической рекомбинации
между парами гомологичных хромосом практиче-
ски в любых местах могут образовываться струк-
туры Холлидея, и точка ветвления может мигри-
ровать на достаточно большое расстояние. При
сайт-специфической рекомбинации структуры
Холлидея образуются между двумя специфически-
ми участками, и миграция ветви обычно ограниче-
на гетерологичными последовательностями по обе
стороны от участка рекомбинации.
Краткие решения задач и ответы на вопросы [3411
15. При наступлении момента в ходе репликации, ког-
да один сайт рекомбинации уже реплицировался,
а другой еще нет, происходит сайт-специфическая
рекомбинация, сопровождающаяся изменением
направления движения репликативных вилок.
Вилки движутся друг за другом по кольцу ДНК,
создавая множество тандемных копий плазмиды.
Такое мультимерное кольцо может быть разделено
на мономеры в результате дополнительных раун-
дов сайт-специфической рекомбинации.
Точка начала репликации
Точка начала репликации
инициация
репликации
рекомбинация в
сайтах 1 и 2 (инверсия)
Плазмида Частично
с сайтами реплицированная
рекомбинации 1 и 2 плазмида
16. а) Фоновые мутации происходят даже без добав-
ления мутагена - под воздействием радиации, хи-
мических процессов в клетке и других событий,
б) При значительном поражении клеточной ДНК
нарушается функция продуктов многих генов, и
клетка становится нежизнеспособной, в) Клетки
с ослабленной системой репарации ДНК более
чувствительны к действию мутагенов. Поскольку
клетки uvr хуже справляются с репарацией по-
вреждений, вызванных воздействием R7000, в них
выше частота мутаций, в том числе летальных,
г) В штамме uvr+ система эксцизионной репарации
удаляет основания ДНК со связанным [3H]R7000,
снижая содержание 3Н в этих клетках. В штамме
uvr репарация не происходит, и уровень 3Н по-
степенно повышается по мере взаимодействия
[3H]R7000 с ДНК. д) Все перечисленные в табли-
це мутации, за исключением замены А=Т на G=C,
значительно учащаются по сравнению с фоном.
Мутация каждого типа вызвана особым взаимо-
действием между R7000 и ДНК. Поскольку не все
эти взаимодействия равновероятны (из-за раз-
личий в реакционной способности, стерических
ограничений и др.), мутации возникают с разной
частотой, е) Нет, только те, что начинаются с пары
G=C. Таким образом, замены А=Т на C=Cj и А=Т
на Т=А вызваны связыванием R7000 с А или с
Т. ж) R7000-G образует пару с А. Сначала R7000
связывается с G=C с образованием R7000-G=C.
(Сравните с тем, что происходит с CH3-G на
рис. 25-28, б, т. 3). Если не происходит репарации,
одна нить реплицируется как R7000-G=A, что в
результате репарации превращается в Т=А. Дру-
гая нить сохраняется неизменной. Если в результа-
те репликации образуется R7000-G=T, то тот же
самый путь приводит к паре А=Т. з) Нет. Сравни-
те данные двух таблиц, не забывая о том, что раз-
ные мутации происходят с разной частотой.
А=Т на C=G: умеренная в обоих случаях, но в
штамме uvr+ репарация эффективнее
GSC на А=Т: умеренная в обоих случаях, без за-
метной разницы
G=C на C=G: выше в штамме uvr\ безусловно, ре-
парация менее эффективна!
G=C на Т=А: высокая в обоих случаях, без замет-
ной разницы
А=Т на Т=А: высокая в обоих случаях, без замет-
ной разницы
А=Т на G=C: низкая в обоих случаях, без замет-
ной разницы
Некоторые аддукты эффективнее распознаются си-
стемой репарации, чем другие, и поэтому удаляют-
ся быстрее и приводят к меньшему числу мутаций.
Глава 26
1. а) от 60 до 100 с; б) от 500 до 900 нуклеотидов
2. Ошибка в единственном основании при реплика-
ции ДНК приведет к тому, что одна из дочерних
клеток и все ее потомство будут иметь мутирован-
ную хромосому. Ошибка в единственном основа-
нии при транскрипции РНК не влияет на хромосо-
мы, но может привести к синтезу дефектной копии
белка. Но поскольку оборот мРНК осуществляется
достаточно быстро, большинство копий белка это-
го дефекта иметь не будут, и потомство этой клетки
будет нормальным.
3. Блокируется или ингибируется посттранскрипци-
онный процессинг З'-конца (расщепление и поли-
аденилирование).
4. Поскольку матричная РНК не кодирует фермен-
ты, необходимые для инициации вирусной ин-
13\ 21 Приложение Б
фекции, она, скорее всего, окажется инертной или
просто расщепится клеточными рибонуклеазами.
Репликация этой РНК и распространение вируса
возможно лишь в том случае, если в клетки вме-
сте с матричной РНК ввести интактную РНК-
репликазу (РНК-зависимую РНК-полимеразу).
5. (1) В качестве матрицы используют последова-
тельность нуклеиновой кислоты; (2) синтез проис-
ходит в направлении 5'-*3'; (3) в качестве субстра-
тов используют нуклеозидтрифосфаты и образуют
фосфодиэфирные связи с вытеснением РР, Поли-
нуклеотидфосфатаза образует фосфодиэфирные
связи, но отличается от остальных ферментов по
всем остальным пунктам.
6. Обычно две: одна для расщепления фосфодиэфир-
ной связи на одном стыке интрона и экзона, а вто-
рая — для связывания освободившегося конца эк-
зона с экзоном с другой стороны от интрона. Если
в качестве нуклеофила в первой реакции выступает
молекула воды — это стадия гидролиза, и тогда для
осуществления сплайсинга требуется лишь одна
реакция трансэтерификации.
7. В интронах закодированы многие мякРНК, необ-
ходимые для модификаций рРНК. Если не проис-
ходит сплайсинга, мякРНК не образуются.
8. Эти ферменты не имеют корректирующей
3'-*5'-экзонуклеазной активности и поэтому допу-
скают значительное количество ошибок. При этом
вероятность ошибок при репликации, приводящих
к инактивации вируса, в маленьком геноме намно-
го ниже, чем в большом.
9. а) 432 = 1,8-1019; б) 0,006%; в) Для осуществления
«неестественного отбора» используйте хромато-
графическую смолу со связанными молекулами,
которые представляют собой аналоги переходного
состояния в реакции гидролиза сложного эфира
(например, подходящий фосфонат; см. Дополне-
ние 6-3).
10. а-Аманитин останавливает синтез РНК, и когда
через несколько дней все существующие молекулы
мРНК и белков в печени расщепляются, возникает
дисфункция печени и наступает смерть.
11. а) После лизиса клеток и частичной очистки бел-
ковый экстракт следует подвергнуть изоэлектри-
ческому фокусированию. Для идентификации
р-субъединицы можно использовать антитела.
Различие в аминокислотных последовательностях
нормальной р-субъединицы и мутантной формы
(а именно разные заряды аминокислотных остат-
ков) отразится на электрофоретической подвиж-
ности белков в геле, б) Прямое секвенирование
ДНК (по Сенгеру).
12. а) 384 п. н.; б) 1620 п. н.; в) Большинство нукле-
отидов расположено в нетранслируемых участках
на 3'- и 5'-концах мРНК. Кроме того, значительная
часть мРНК кодирует сигнальную последователь-
ность (гл. 27) белков, которая в конечном итоге
отщепляется при формировании зрелого функци-
онального белка.
13. а) кДНК синтезируется в процессе обратной
транскрипции мРНК; таким образом, мРНК, по-
видимому, имела последовательность CGG. По-
скольку геномная ДНК, на основе которой син-
тезирована мРНК, имела последовательность
CAG, первичный транскрипт, вероятно, имел
последовательность CAG, а в процессе посттран-
скрипционной модификации превратился в CGG.
б) Нередактированная последовательность мРНК
совпадает с последовательностью ДНК (за ис-
ключением замены Т на U). Нередактированная
последовательность мРНК имела следующий вид
(* указывает место редактирования):
*
(5')...GUCUCUGGUUUUCCUUGGGUGCCUUUAUGCAGCAAGGAUGCGAUAUUUCGCCAAG...(3')
На стадии 1 праймер 1 отжигается следующим образом:
*
(5')...GUCUCUGGUUUUCCUUGGGUGCCUUUAUGCAGCAAGGAUGCGAUAUUUCGCCAAG...(3')
I I I I I I I I I I I I I I I I I I I I I I I I
Праймер 1: (3')-CGTTCCTACGCTATAAAGCGGTTC-(5')
Последовательность кДНК (подчеркнута) синтезирована справа налево:
*
(5')...GUCUCUGGUUUUCCUUGGGUGCCUUUAUGCAGCAAGGAUGCGAUAUUUCGCCAAG...(3')
I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I
(3/)...CAGAGACCAAAAGGAACCCACGGAAATACGTCGTTCCTACGCTATAAAGCGGTTC~(5/)
На стадии 2 образуется кДНК:
(3')...CAGAGACCAAAAGGAACCCACGGAAATACGTCGTTCCTACGCTATAAAGCGGTTC-(5')
На стадии 3 праймер 2 отжигается на кДНК:
Праймер 2: (5')-CCTTGGGTGCCTTTA-(3')
I I I I I I I I I I I I I I I
(3')..CAGAGACCAAAAGGAACCCACGGAAATACGTCGTTCCTACGCTATAAAGCGGTTC-(5')
*
Краткие решения задач и ответы на вопросы [ 313 ]
ДНК-полимераза добавляет нуклеотиды к З'-концу праймера. Двигаясь слева направо, она встраивает Т, G, С и А.
Однако поскольку А из ddATP не имеет на З'-конце ОН-группы, необходимой для присоединения следующего
нуклеотида, цепь дальше не удлиняется. Это А выделено курсивом, а новая ДНК подчеркнута:
Праймер 2: (5f)~CCTTGGGTGCCTTTATGCA
I I I I I I I I I I I I I I I I I I I
(3')..CAGAGACCAAAAGGAACCCACGGAAATACGTCGTTCCTACGCTATAAAGCGGTTC-(5')
*
В результате образуется нередактированный транскрипт из 19 нуклеотидов. В транскрипте после
редактирования *А заменяется на G; в последовательности кДНК это соответствует С. Начинается стадия 3:
Праймер 2: (5')-CCTTGGGTGCCTTTA-(3')
I I I I I I I I I I I I I I I
(3')...CAGAGACCAAAAGGAACCCACGGAAATACGCCGTTCCTACGCTATAAAGCGGTTC-(5')
*
В данном случае ДНК-полимераза может продвигаться за замененным основанием и останавливается
у следующего Т в последовательности кДНК. Дидезокси-А выделено курсивом, новая ДНК подчеркнута:
Праймер 2: (5')~CCTTGGGTGCCTTTATGCGGCA
I I I I I I I I I I I I I I I I I I I I I I
(3')..CAGAGACCAAAAGGAACCCACGGAAATACGCCGTTCCTACGCTATAAAGCGGTTC-(5')
В результате образуется продукт, состоящий из 22 ну-
клеотидов. в) Факторы, нарушающие функцию белка
(протеазы, тепло), ингибируют редакционный про-
цесс, а те, что не влияют на функцию белков (нукле-
азы), практически никакого действия не оказывают.
Основной недочет этого аргумента заключается в том,
что факторы, нарушающие функцию белка, не вы-
звали полной остановки редактирования. Возможно,
некоторое редактирование или расщепление мРНК
происходит даже без фермента или какая-то часть
фермента сохранила свою активность, г) В полину-
клеотиды включается только а-фосфат NTP. Если бы
исследователи использовали другие типы [32P]NTP,
не было бы меченых продуктов, д) Поскольку мо-
дификация затрагивает только основание А, нас ин-
тересует только судьба всех А в последовательности,
е) Поскольку метка содержалась только в АТР, при
удалении целого нуклеотида из мРНК исчезла бы вся
радиоактивность, и на хроматографической пластин-
ке был бы обнаружен лишь немодифицированный
[32Р]АМР. ж) Если бы основание было удалено или
заменено на другое, исследователи обнаружили бы
только [3Н]АМР. Наличие [3Н]1МР указывает, что
произошла замена А на I без удаления Н в положе-
ниях 2 и 8. Наиболее вероятным механизмом моди-
фикации при замене А на I является гидролитическое
дезаминирование (см. рис. 22-34). з) CAG меняется
на CIG. Этот кодон читается как CGG.
Глава 27
1. a) Gly-GIn-Ser-Leu-Leu-Ile; б) Leu-Asp-Ala-Pro;
в) His-Asp-Ala-Cys-Cys-Tyr; г) Met-Asp-Glu у
эукариот; fMet-Asp-Glu у бактерий.
2. UUAAUGUAU, UUGAUGUAU, CUUAUGUAU,
CUCAUGUAU, CUAAUGUAU, CUGAUGUAU,
UUAAUGUAC, UUGAUGUAC, CUUAUGUAC,
CUCAUGUAC, CUAAUGUAC, CUGAUGUAC
3. Нельзя. Почти все аминокислоты кодируются
несколькими кодонами (например, существует
шесть вариантов кодонов для лейцина), и поэто-
му каждый полипептид может быть закодирован
несколькими способами. И все же для некоторых
аминокислот существует лишь один кодон. Кроме
того, несколько кодонов, кодирующих одну ами-
нокислоту, часто имеют одни и те же нуклеотиды
в двух из трех позиций. Таким образом, некоторые
участки последовательности мРНК, кодирующие
белки с известной аминокислотной последователь-
ностью, можно определить с достаточно высокой
точностью.
4. a) (5')CGACGGCGCGAAGUCAGGGGUGUUAAG(3');
б) Arg-Arg-Arg-Glu-Val-Arg-Gly-Val-Lys; в) Нет.
Комплементарные антипараллельные нити в двой-
ной спирали ДНК не идентичны в направлении
5'-*3'. Из дуплекса РНК считывается лишь с одной
конкретной нити. РНК-полимераза узнает только
«правильную» нить и связывается с ней.
5. Для переноса метионина существует два типа мо-
лекул тРНК: TPHKfMct, инициирующая синтез, и
тРНКМсГ, которая может встраивать остаток Met
внутри полипептидной цепи. Только fMet-TPHKfMet
распознается инициаторным фактором IF-2 и под-
ходит к инициаторному кодону AUG, находящему-
ся в P-сайте рибосомы в инициаторном комплексе.
Кодоны AUG внутри последовательности мРНК
могут связывать только Met-TPHKMet.
[3 'i 11 Приложение Б
6. Взять смесь UDP и UDC, скажем, в соотношении
5 : 1 и подействовать на нее полинуклеотидфосфо-
рилазой. В результате должен получиться полимер,
содержащий много триплетов UUU (Phe), меньше
UUC (Phe), UCU (Ser) и CUU (Leu), значительно
меньше UCC (Ser), CUC (Leu) и CCU (Pro) и со-
всем мало ССС (Pro).
7. Минимум 583 эквивалента АТР (по 4 на каждую
присоединенную аминокислоту, но с учетом того,
что происходит лишь 145 транслокаций). Для ис-
правления каждой ошибки нужно 2 эквивалента
АТР. Для синтеза гликогена нужно 292 эквивален-
та АТР. Дополнительная энергия, затрачиваемая
на синтез 0-глобина, отражает стоимость инфор-
мационного содержания белка. Для синтеза белка
из аминокислот клетке потребуется как минимум
20 активирующих ферментов, 70 рибосомных бел-
ков, 4 рРНК, 32 или более тРНК, одна молекула
мРНК и 10 или более вспомогательных ферментов.
В синтезе цепи (а1->4)-гликогсна из глюкозы уча-
ствуют лишь 4 или 5 ферментов (гл. 15).
8.
Кодоны глицина Антикодоны
(5')GGU (5')ACC, GCC, ICC
(5')GGC (5')GCC, ICC
(5')GGA (5')UCC, ICC
(5')GGG (5')CCC, UCC
а) З'-концевой и средний; б) спаривание с антико-
донами (5’)GCC, ICC и UCC; в) в парах с антико-
донами (5’)АСС и ССС.
9. Только (а), (в), (д) и (ж); (б), (г) и (е) не могут быть
результатом однонуклеотидной замены: для (б) и
(е) требуется замена двух нуклеотидов, а для (г) —
замена всех трех нуклеотидов.
10. Для Glu существует два кодона (GAA и GAG), а для
Vai - четыре кодона (GTT, GTC, GTA и GTG). Одно-
нуклеотидная замена GAA на GTA или GAG на GTG
могла бы привести к замене Glu на Vai в серповидном
гемоглобине. Гораздо менее вероятны замены двух
нуклеотидов: превращение GAA в GTG, GTT и GTC
или превращение GAG в GTA, GTT и GTC.
11. Изолейцин по структуре напоминает некоторые
другие аминокислоты, особенно валин. Чтобы от-
личить изолейцин он валина в процессе аминоаци-
лирования, нужен второй этап проверки. Гистидин
не похож ни на какую другую аминокислоту, и его
структура обеспечивает достаточную специфич-
ность аминоацилирования и связывания тРНК.
12. а) А1а-тРНК-синтетаза распознает пару G3-U70
в аминокислотном плече тРНКА1а. б) Мутантная
тРНКА1а будет вставлять остаток Ala при встрече
с кодонами для Pro. в) Аналогичные последствия
могла бы вызвать мутация тРНКРго, в результа-
те которой она стала бы распознаваться и амино-
ацилироваться Ala-тРНК-синтетазой. г) Такие
мутации вызвали бы инактивацию множества
клеточных белков и привели бы к летальным по-
следствиям для клетки. Этот пример отражает
мощь естественного отбора, препятствующего из-
менениям генетического кода.
13. Лишь последняя присоединенная к полипептид-
ной цепи аминокислота сохраняет ковалентную
связь с тРНК, и, следовательно, она является един-
ственным связующим звеном между полипепти-
дом и кодирующей его мРНК. Исправление ошиб-
ки потребовало бы разрыва этой связи и привело
бы к отрыву пептида от мРНК и преждевременной
остановке синтеза.
14. Белок был бы доставлен в ЭР, а его дальнейший
путь зависел бы от дополнительных сигналов. SRP
связывается с N-концевой сигнальной последова-
тельностью на ранних стадиях синтеза полипеп-
тида и направляет его вместе с рибосомой к ре-
цепторам на ЭР. После синтеза белок переносится
в просвет ЭР, так что NLS недоступна для белков,
ответственных за транспортировку в ядро.
15. Триггерный фактор — это молекулярный шаперон,
стабилизирующий несформированную и готовую к
транслокации форму РгоОшрА.
16. ДНК должна содержать не менее 5784 нуклеоти-
дов; некоторые кодирующие последовательности
вложены друг в друга или перекрываются.
17. а) Спирали удерживаются вместе за счет гидро-
фобных и ван-дер-ваальсовых взаимодействий,
б) R-группы 3, 6, 7 и 10 выступают слева, группы
1, 2, 4, 5, 8 и 9 — справа, в) Одна из возможных по-
следовательностей следующая:
1234 56789 10
A-Phe-Ile-Glu-Vai-Met-Asn-Ser-Ala-Phe-Gln-C
г) Возможная последовательность ДНК-дуплекса
для этой аминокислотной последовательности:
Нематричная цепь
(5')-TTTATTGAAGTAATGAATAGTGCATTCCAG-(3')
I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I
(3')-AAATAACTTCATTACTTATCACGTAAGGTC-(5')
Матричная цепь
д) Phe, Leu, Не, Met и Vai. Все они гидрофобные, но
это не все гидрофобные аминокислоты - здесь не
хватает Trp, Pro, Ala и Gly. е) Туг, His, Gin, Asn, Lys,
Asp и Glu; все они гидрофильные, но Туг меньше
остальных. Однако это не все гидрофильные
аминокислоты — здесь не хватает Ser, Thr и Arg.
Краткие решения задач и ответы на вопросы [345]
ж) Отсутствие в смеси Т позволяет исключить
кодоны, начинающиеся или заканчивающиеся на
Т, т. е. исключить Туг, который обладает слабой
гидрофильностью, и, что более важно, исключить
два стоп-кодона — ТАА и TAG. Никакие другие
аминокислоты при этом из набора NAN не исклю-
чаются. з) Неправильно свернутые белки обычно
уничтожаются. Поэтому, если на основе синтети-
ческого гена синтезируется белок, который при
электрофорезе формирует полосу с определен-
ной молекулярной массой, скорее всего, этот бе-
лок правильно свернут, е) Сворачивание белка
определяется не только водородными связями и
ван-дер-ваальсовыми взаимодействиями. Есть
множество причин, почему синтезированный на
основе случайной нуклеотидной последователь-
ности белок мог бы не сформировать четырехспи-
ральную структуру. Например, структура изме-
нилась бы при наличии водородных связей между
гидрофильными боковыми цепями. Кроме того,
не все последовательности имеют тенденцию об-
разовывать а-спирали.
Глава 28
1. а) Уровень триптофансинтазы остается высоким,
несмотря на присутствие триптофана, б) Уровень
высокий, в) Уровень быстро снижается, предотвра-
щая синтез дополнительного триптофана.
2. а) Конститутивная, слабая экспрессия оперона;
большинство мутаций в операторе с большой веро-
ятностью предотвращает связывание репрессора,
б) Либо конститутивная экспрессия, как в (а), либо
постоянная репрессия, если в результате мутации
нарушена способность связывать лактозу и род-
ственные вещества и, следовательно, отвечать на
присутствие индуктора, г) Повышение или сниже-
ние экспрессии оперона (в условиях индукции) в
зависимости от того, стал ли промотор в результате
мутации больше или меньше похож на консенсус-
ный промотор Е. coli.
3. 7000 копий.
4. 8 -10’9 М, более чем в 105 выше константы диссо-
циации. При наличии в клетке 10 копий активного
репрессора сайт связывания репрессора на опера-
торе всегда занят.
5. (а)-(д): При всех условиях экспрессия генов 1ас-
оперона снижается.
6. а) Ослабление аттенюации транскрипции. Рибосо-
ма, заканчивающая транскрипцию последователь-
ности 1, больше не экранирует последовательность
2 и не блокирует ее транскрипцию. Последователь-
ность 2 всегда доступна для спаривания с последо-
вательностью 3, что предотвращает образование
аттенюатора, б) Усиление аттенюации транскрип-
ции. Последовательность 2 менее эффективно свя-
зывается с последовательностью 3; аттенюаторы
образуются чаще, даже если последовательность
2 не блокирована рибосомой, в) Аттенюация от-
сутствует; единственный механизм регуляции
обеспечивается репрессором Тгр. г) Аттенюация
перестает зависеть от тРНК триптофана и может
стать зависимой от тРНК гистидина, д) Аттеню-
ация происходит редко или не происходит вовсе.
Последовательности 2 и 3 постоянно блокируют
образование аттенюатора, е) Постоянная аттенюа-
ция транскрипции. Аттенюатор образуется всегда,
независимо от наличия триптофана.
7. SOS-ответ не индуцируется, что делает клетку бо-
лее чувствительной к повреждениям ДНК.
8. Каждая клетка бактерии имела бы жгутики, состо-
ящие из флагеллина обоих типов, и была бы чув-
ствительна к антителам, которые вырабатываются
в ответ на оба типа белка.
9. Возможно, в процессе очистки полимеразы были по-
теряны необходимые для активности фермента дис-
социируемые факторы (например, фактор специфич-
ности, аналогичный о-субъединице фермента Е. coli).
10.
Белок Gal4
ДНК-связывающий домен Gal4 Активирующий домен Gal4
Генно-инженерный белок
ДНК-связывающий домен Lac-penpeccopa Активирующий домен Gal4
Генно-инженерный белок не может связываться
с участком связывания Gal4 в гене GAL (UASG),
поскольку он не имеет ДНК-связывающего доме-
на Gal4. Нужно таким образом модифицировать
последовательность ДНК, с которой связывается
Gal4p, чтобы она приобрела нуклеотидную по-
следовательность, с которой обычно связывается
Lac-penpeccop (используя методы, описанные в
гл. 9).
11. Метиламин; реакция включает атаку воды на угле-
род гуанидиновой группы в модифицированном
аргинине.
12. Необходимая для развития яйца ftcrf-мРНК обе-
спечивается матерью. Яйцо нормально развива-
ется даже в том случае, если оно имеет генотип
ЬссГ/ЬссГ, при условии, что мать имеет один нор-
мальный ген bed, а аллель bed является рецессив-
ной. Однако взрослая женская особь с генотипом
1346] Приложение Б
bcd~/bcd~ стерильна, поскольку у нее нет нормальной
копии bed для передачи собственному потомству
13. а) Водородные связи образуются между белком и
скелетом ДНК в позициях ДНК А106, А110, А118,
Т119 и Т122, а также между белком и основаниями
в позициях А106, Т107, А118 и Т119. Четыре по-
следних нуклеотида непосредственно участвуют в
распознавании последовательности ДНК. б) Ске-
лет ДНК: A106-Arg290, АИО-Ser212, A119-Arg199,
T119-Arg204, A122-Ser303. Основания ДНК: A106-
Asn253, T107-Asn253, Al 18-Asn163, T119-Asn163. Asn,
Glu, Gin, Lys и Arg очень часто образуют водород-
ные связи с ДНК. В образовании водородных свя-
зей ТАТА-связывающим белком в основном задей-
ствованы остатки Arg и Asn.
в) ТАТАТАТА (остатки 103-110)
АТАТАТАТ (остатки 122-115)
ТАТА-связывающий белок узнает А106, Т107/Т119
и А118.
г) Гидрофобные взаимодействия очень многочис-
ленны, многие из них происходят с участием ги-
дрофобных поверхностей.
14. а) Для экспрессии 10% (репрессии 90%) 10%
репрессора должно связаться с индуктором, а 90%
находится в свободном виде и может связываться
с оператором. Воспользуемся уравнением 5-8 при
0 = 0,1 иК(1 = IO 4 М:
[IPTG] [IPTG]
[IPTG] + Ха " [IPTGJ+ 10-4М
[IPTG]
’ [IPTG]+10~4M
так что
0,9[IPTG] = 10-5 или [IPTG] = 1,1-10 5М
Для экспрессии 90% с индуктором должно быть
связано 90% репрессора, так что 0 = 0,9. Подстав-
ляя в уравнение 5-8 значения 0 и Kd, получаем
[IPTG] = 910’4 М. Таким образом, при изменении
[IPTG] примерно в 10 раз уровень экспрессии
также изменяется примерно в 10 раз. б) Уровень
белков до индукции низкий, он повышается в про-
цессе индукции и уменьшается при прекращении
синтеза и расщеплении белков, в) Как следует из
пункта (а), экспрессия /яс-оперона не просто вклю-
чается или выключается, следовательно, оперон
не удовлетворяет характеристике А. Как следует
из пункта (б), экспрессия /яс-оперона ослабевает
при удалении индуктора; следовательно, оперон не
удовлетворяет характеристике Б. г) GFP включен'.
уровень экспрессии repts и GFP высокий; repts по-
давляет ОРК, так что Lacl не синтезируется. GFP
выключен: уровень экспрессии Lacl высокий; Lacl
подавляет ОР|ас, так что repts и GFP не синтезиру-
ются. д) Добавление IPTG переключает систему
из состояния «GFP выключен» в состояние «GFP
включен». IPTG действует только в присутствии
Lacl, поэтому он влияет только на состояние «GFP
выключен». Добавление IPTG отменяет репрессию
ОР|ас и приводит к повышению уровня экспрес-
сии repts, что выключает экспрессию Lacl и повы-
шает экспрессию GFP. е) Повышение температуры
переводит систему из состояния «GFP включен» в
состояние «GFP выключен». Тепло оказывает вли-
яние только в присутствии repts, т. е. только в со-
стоянии «GFP включен». Тепло инактивирует repls
и отменяет репрессию ОР, что приводит к высоко-
му уровню экспрессии Lad. Затем Lacl действует
на ОР|ас, подавляя синтез repts и GFP. ж) Харак-
теристика А: система нестабильна в промежуточ-
ном состоянии. В какой-то момент один репрессор
оказывает более сильное влияние, чем другой, про-
сто при случайных флуктуациях экспрессии. Это
отключает экспрессию второго репрессора и фик-
сирует систему в одном из состояний. Характери-
стика Б: экспрессия одного репрессора подавляет
синтез другого, поэтому система остается в одном и
том же состоянии даже после удаления переключа-
ющих стимулов, з) Ни в какой момент времени ни
одна клетка не демонстрировала промежуточного
уровня экспрессии GFP, что подтверждает харак-
теристику А. При промежуточной концентрации
индуктора (X) некоторые клетки перешли в состо-
яние «GFP включен», а другие еще нет и находятся
в состоянии «GFP выключен». Никакого промежу-
точного состояния не существует. Бимодальный
характер экспрессии при [IPTG] = X вызван одно-
временным присутствием в смеси клеток в состо-
янии «GFP включен» и клеток в состоянии «GFP
выключен».
Словарь терминов
АВС-переносчик: представитель большого семей-
ства мембранных переносчиков, содержащих АТР-
связывающие домены (кассеты); перемещают через
плазматическую мембрану различные субстраты,
включая неорганические ионы, липиды и неполярные
молекулы лекарств, используя в качестве источника
энергии молекулы АТР.
ADP (аденозиндифосфат): рибонуклеозид-5'-
дифосфат; акцептор фосфатной группы в энергетиче-
ском метаболизме клетки.
АТР (аденозинтрифосфат): рибонуклеозид-5'-
трифосфат; донор фосфатной группы в энергети-
ческом метаболизме клетки; осуществляет перенос
химической энергии в метаболических путях; общее
промежуточное соединение в сопряженных эндерго-
нических и экзергонических реакциях.
АТРаза: фермент, гидролизующий АТР до ADP и
фосфата; обычно принимает участие в процессах, про-
текающих с затратой энергии.
ATP-синтаза: ферментный комплекс, образующий
АТР из ADP и фосфата в процессе окислительного
фосфорилирования во внутренней мембране мито-
хондрий или в плазматической мембране бактерий,
а также в процессе фотофосфорилирования в хлоро-
пластах.
В-лимфоцит (В-клетка): клетка крови, ответственная
за выработку антител.
сАМР-рецепторный белок: специфический регуля-
торный белок бактерий, контролирующий инициацию
транскрипции генов ферментов, которые в условиях
недостатка глюкозы позволяют клетке переходить на
другие источники углерода; другое название — белок-
активатор катаболизма.
С-концевой остаток: единственный аминокислотный
остаток в полипептидной последовательности, имею-
щий свободную а-карбоксильную группу.
Escherichia coli (Е. coli): распространенная бактерия,
обитающая в тонком кишечнике позвоночных; один
из хорошо изученных организмов.
GLUT: семейство мембранных белков, осуществляю-
щих транспорт глюкозы.
G-белки: семейство гетеротримерных GTP-связы-
вающих белков, участвующих в передаче сигнала
в клетку. Связывание лиганда с рецептором, со-
пряженным с G-белком, приводит к обмену GTP на
GDP, благодаря чему белок получает возможность
активировать следующий фермент сигнального пути.
G-белки обладают собственной СТРазной активно-
стью и, следовательно, инактивируют сами себя.
In situ: на месте, т. е. в естественном положении.
In vitro: «в стекле», т. е. в искусственных условиях, в
пробирке.
In vivo: «в жизни», т. е. в живой клетке или организме.
NAD, NADP (никотинамидадениндинуклеотид
( НАД), никотинамидадениндинуклеотидфосфат
(НАДФ)): никотинамидсодержащие кофакторы, вы-
полняющие функцию переносчиков атомов водорода
и электронов в окисЛительно-восстановительных
реакциях.
Р/О-отношение: количество молей АТР, образую-
щихся в процессе окислительного фосфорилиро-
вания при восстановлении % О2 (т- е- в расчете на
пару электронов, переданных на О2). При передаче
электронов от NADH на О2 Р/О-отношение принято
равным 2,5, а при передаче электронов от FADH на О2
1,5 (в других учебниках 3,0 и 2,0 соответственно).
pH: отрицательный логарифм концентрации ионов
водорода в водном растворе.
рКа: отрицательный логарифм константы диссоциа-
ции кислоты.
R-группа: (1) обозначение алкильной группы;
(2) иногда используется для обозначения заместите-
лей в органических соединениях.
SELEX: метод быстрой идентификации последо-
вательностей нуклеиновых кислот (обычно РНК),
обладающих особыми каталитическими или лиганд-
связывающими свойствами.
8Н2-домен: белковый домен, прочно связывающийся
с остатком тирозинфосфата в некоторых белках, та-
ких как рецепторные тирозинкиназы, что инициирует
образование белкового комплекса, принимающего
участие в передаче сигнала.
Siglec: семейство лектинов клеточной поверхности,
связывающих сиаловую кислоту; содержат множество
иммуноглобулин-подобных доменов.
SOS-ответ: координированная индукция множества
разнообразных генов в клетках бактерий в ответ на
серьезное повреждение клеточной ДНК.
STS-маркер: уникальная для данного локуса ДНК
нуклеотидная последовательность, которую можно
использовать для его идентификации.
Т-лимфоцит (Т-клетка): клетка крови, которая об-
разуется в тимусе и принимает участие в клеточном
иммунном ответе.
Vmax: максимальная скорость ферментативной реак-
ции, которая достигается при насыщении субстратом
центра связывания в молекуле фермента.
[348 ] Словарь терминов
А
Абсолютная конфигурация: конфигурация четырех
разных заместителей у асимметрического атома угле-
рода по сравнению с конфигурацией заместителей в
D- и L-глицеральдегиде.
Автотроф: организм, самостоятельно синтезирующий
сложные молекулы из простейших соединений угле-
рода и азота, таких как диоксид углерода и аммиак.
Агонист: вещество (обычно гормон или нейромедиа-
тор), связывание которого со специфическим рецеп-
тором вызывает физиологический ответ.
S-Аденозилметионин (adoMet): кофактор, участвую-
щий в ферментативном переносе метильных групп.
Адипоцит: клетка животного, специализирующаяся
на запасании жиров (триацилглицеринов).
Активатор: (1) ДНК-связывающий белок, усиливаю-
щий экспрессию (т. е. повышающий скорость транс-
крипции) одного или нескольких генов; (2) положи-
тельный модулятор аллостерического фермента.
Активация аминокислоты: ATP-зависимая фермен-
тативная реакция образования эфирной связи между
карбоксильной группой аминокислоты и З'-гидрок-
сильной группой соответствующей тРНК.
Активный (каталитический) центр: участок на
поверхности фермента, в котором происходит свя-
зывание и каталитическое превращение молекулы
субстрата.
Активный транспорт: перенос растворенных веществ
через клеточную мембрану против градиента концен-
трации, происходящий с затратой энергии.
Актин: белок мышечных волокон, а также важный
компонент цитоскелета многих эукариотических
клеток.
Акцептор протонов: основание, анион, способный
принимать протоны от донора.
Акцептор электронов: вещество, принимающее элек-
троны при окислительно-восстановительной реакции.
Акцепторный контроль: регуляция скорости клеточ-
ного дыхания через изменение концентрации ADP,
который служит акцептором фосфатных групп.
Алкалоз: метаболическое состояние, сопровождаю-
щееся снижением способности организма связывать
гидроксид-ионы; обычно при этом увеличивается pH
крови.
Аллостерический белок: белок (обычно состоящий
из нескольких субъединиц) с несколькими центрами
связывания лиганда, причем связывание лиганда
в одном центре влияет на его связывание в других
центрах.
Аллостерический фермент: регуляторный фермент,
каталитическая активность которого изменяется в ре-
зультате нековалентного взаимодействия со специфи-
ческим метаболитом (эффектором), связывающимся
вне активного центра.
Аллостерический центр: специфический участок на
поверхности молекулы аллостерического фермента, с
которым связывается молекула модулятора (эффек-
тора).
Альдоза: сахар, в молекуле которого карбонильный
углерод находится в составе альдегидной группы на
одном из концов углеродной цепи.
Аминоацил-тРНК: аминоацильное производное
тРНК.
Аминоацил-тРНК-синтетазы: ферменты, катализи-
рующие синтез аминоацил-тРНК с затратой энергии
АТР.
а-Аминокислота: а-аминозамещенная карбоновая
кислота, структурная единица пептидов и белков.
Аминотрансферазы (трансаминазы): ферменты, ка-
тализирующие перенос аминогрупп от а-аминокислот
на а-кетокислоты.
Аммониотелический организм: организм, выделяю-
щий избыток азота в виде аммиака.
Амфиболический путь: метаболическая реакция, во-
влеченная в процессы и катаболизма, и анаболизма.
Амфитропный белок: белок, обратимо связываю-
щийся с клеточной мембраной и поэтому способный
находиться как в цитозоле, так и в мембране.
Амфифильное соединение: соединение, содержащее
полярные и неполярные участки.
Амфолит: вещество, способное выступать в роли и
кислоты, и основания.
Амфотерное вещество: вещество, способное при-
нимать и отдавать протоны и, следовательно, высту-
пать в роли кислоты или основания в зависимости от
условий.
Анаболизм: энергозатратная фаза промежуточного
метаболизма, заключающаяся в биосинтезе клеточных
компонентов из более простых предшественников.
Аналит: анализируемое вещество (например, в масс-
спектрометрии).
Анаммокс: анаэробное окисление аммиака до мо-
лекулярного азота, протекающее с использованием
нитрита в качестве акцептора электронов; осущест-
вляется специализированными хемолитотрофными
бактериями.
Анаплеротическая реакция: ферментативная реак-
ция, поставляющая промежуточные соединения в
цикл лимонной кислоты.
Словарь терминов 13 4 91
Анаэроб: организм, обитающий в бескислородной
среде; облигатные анаэробы в присутствии кислорода
погибают.
Ангидрид: продукт реакции конденсации двух кар-
боксильных или фосфатных групп, протекающей с
выделением молекулы воды;
R—X—О—X—R (X — атом углерода или фосфора).
О О
Ангстрем (А): единица длины. 1 А = 10 8 см. Ис-
пользуется при указании расстояний на атомном и
молекулярном уровне.
Анионообменная смола: полимерная смола с фикси-
рованными катионными группами; используется при
разделении анионов методом ионной хроматографии.
Аномерный углерод: атом углерода в хиральном цен-
тре, возникающем при циклизации молекулы сахара с
образованием полуацеталя; например, карбонильный
углерод в альдегидах и кетонах.
Аномеры: два стереоизомера одного сахара, различа-
ющиеся конфигурацией заместителей у карбонильно-
го (аномерного) атома углерода.
Антагонист: молекула, мешающая физиологическому
действию другой молекулы (агониста), в частности,
связыванию гормона или нейромедиатора с рецепто-
ром.
Антибиотики: большая группа органических со-
единений, которые синтезируются и секретируются
различными микроорганизмами и растениями, где,
по-видимому, выполняют защитную функцию, и ток-
сичные для других организмов.
Антиген: молекула, способная вызывать синтез спец-
ифических антител в организме позвоночных.
Антикодон: специфическая последовательность в
молекуле тРНК, состоящая из трех нуклеотидов и
комплементарная кодону определенной аминокисло-
ты в последовательности мРНК.
Антипараллельная укладка: укладка двух линейных
полимеров с противоположной полярностью, или
ориентацией, цепей.
Антипорт: одновременный перенос (котранспорт)
двух веществ через клеточную мембрану в противопо-
ложных направлениях.
Антитело: защитный белок, синтезируемый иммун-
ной системой позвоночных. См. также Иммуногло-
булин.
Апобелок: белковая часть молекулы белка без орга-
нических или неорганических кофакторов или про-
стетических групп, которые могут быть необходимы
для проявления его активности.
Аполипопротеин: белковая часть липопротеина.
Апоптоз: программируемая гибель клетки в ответ на
внешний сигнал или команду собственных генов, вы-
ражающаяся в планомерном расщеплении собствен-
ного содержимого.
Апофермент: белковая часть фермента без органиче-
ских или неорганических кофакторов или простетиче-
ских групп, которые могут требоваться для проявле-
ния каталитической активности.
Аппарат (комплекс) Гольджи: комплекс мембранных
органелл в эукариотических клетках, участвующий
в п^сттрансляционной модификации белков и их
секреции из клетки или включении в плазматическую
мембрану и мембраны органелл.
Аптамер: олигонуклеотид, связывающийся спец-
ифическим образом с молекулярными мишенями;
получают путем аффинных методов обогащения
(метод SELEX).
Аррестины: семейство белков, связывающихся с
фосфорилированными С-концевыми участками ре-
цепторов, сопряженных с G-белками; предотвращают
взаимодействие рецепторов с G-белками и тем самым
останавливают передачу сигнала посредством этих
рецепторов.
Археи: царства живых организмов; к ним относятся
многие виды организмов, обитающие в экстремаль-
ных условиях повышенной солености и температуры
или низкого значения pH.
Асимметрический атом углерода: атом углерода, свя-
занный четырьмя ковалентными связями с четырьмя
разными химическими группами; может существовать
в двух различных тетраэдрических конфигурациях.
Ассимиляция углерода: последовательность реакций,
в результате которой атмосферный СО2 превращается
в органические соединения.
Аттенюатор: последовательность РНК, участвующая
в регуляции экспрессии некоторых генов; выступает в
роли терминатора транскрипции.
Ауксин: растительный гормон роста.
Ауксотрофный мутант (ауксотроф): мутантный
организм, потерявший способность синтезировать
определенные молекулы и вынужденный получать их
извне.
Аутофосфорилирование: в строгом смысле — фос-
форилирование аминокислотных остатков белка,
катализируемое самим белком; иногда фосфори-
лирование одной субъединицы гомодимера другой
субъединицей.
Ацидоз: метаболическое состояние, сопровождающее-
ся уменьшением способности организма нейтрализо-
вать протоны; обычно при этом уменьшается pH крови.
13501 Словарь терминов
Ацилфосфат: любая молекула с общей формулой
R—С—ОРО? .
II 3
о
Аэроб: организм, который обитает в воздушной
атмосфере и для дыхания использует кислород в каче-
стве конечного акцептора электронов.
Б
Бактериофаг (фаг): вирус, способный реплициро-
ваться в бактериальной клетке.
Бактерии: царство живых организмов; бактерии окру-
жены плазматической мембраной, но не имеют окру-
женных мембранами внутренних органелл и ядра.
Белая жировая ткань: нетермогенная жировая ткань
с высоким содержанием триацилглицеринов, запа-
саемых и высвобождаемых в ответ на гормональные
сигналы. Перенос электронов в дыхательной цепи в
митохондриях белой жировой ткани напрямую связан
с синтезом АТР.
Белковый банк данных (PDB): международная база
данных (www.rcsb.org/pdb), в которой собрана инфор-
мация о трехмерной структуре большинства извест-
ных макромолекул.
Белок: макромолекула, состоящая из одной или не-
скольких полипептидных цепей, каждая из которых
имеет определенную последовательность аминокис-
лот, соединенных пептидными связями.
Бесконкурентный ингибитор: обратимый ингибитор,
который связывается с фермент-субстратным ком-
плексом, но не со свободным ферментом.
Библиотека ДНК: набор клонированных фрагментов
ДНК.
Библиотека кДНК: библиотека ДНК, состоящая из
синтезированных молекул кДНК, комплементарных
всем молекулам мРНК определенного организма или
типа клеток.
Биоанализ: аналитический метод определения коли-
чества биологически активного вещества (например,
гормона) в образце с помощью количественного из-
мерения активности этого вещества в пробирке.
Биоинформатика: компьютерный анализ биологиче-
ских данных с помощью статистических, лингвисти-
ческих, математических, химических и физических
методов; такими биологическими данными часто вы-
ступают аминокислотные или нуклеотидные последо-
вательности, а также структурные данные, результаты
клинических испытаний и т. д.
Биомолекула: органическое соединение, присутству-
ющее в живом организме и выполняющее физиологи-
ческую функцию.
Биоптерин: кофактор на основе птерина, участвую-
щий в ферментативных окислительно-восстанови-
тельных реакциях.
Биосинтез de novo: путь синтеза биомолекулы (на-
пример, нуклеотида) из простых предшественников —
альтернатива повторного использования метаболита.
Биосфера: вся живая материя нашей планеты, в том
числе, в морях и в атмосфере.
Биотин: витамин, кофактор ферментов, участвующих
в реакциях карбоксилирования.
Биоцитин: конъюгированная аминокислота, образу-
ющаяся за счет ковалентной (амидной) связи между
остатком лизина и молекулой биотина.
Брожение (ферментация): неокислительное ана-
эробное расщепление питательного вещества (на-
пример, глюкозы), сопровождающееся выделением
энергии; продуктами брожения являются лактат,
этиловый спирт и другие соединения.
Бурая жировая ткань: термогенная жировая ткань
с большим количеством митохондрий, содержащих
разобщающий белок термогенин, который прерывает
поток электронов между дыхательной цепью и синте-
зом АТР.
Буфер: раствор, который может препятствовать изме-
нению pH системы; готовят из соответствующих осно-
вания и кислоты (кислотно-основная пара, акцептор и
донор протонов), взятых в молярном соотношении ~1.
В
Ван-дер-ваальсовы взаимодействия: слабые межмо-
лекулярные взаимодействия, возникающие благодаря
взаимной поляризации молекул.
Вектор: молекула ДНК, способная независимо репли-
цироваться в клетке хозяина; встраивание чужерод-
ного фрагмента ДНК в вектор позволяет осуществить
репликацию этого фрагмента; например, плазмида
или искусственная хромосома.
Векторный метаболизм: метаболические превра-
щения, в результате которых происходит изменение
локализации (но не химического состава) субстрата
относительно плазматической мембраны или мембра-
ны между двумя клеточными компартментами.
Вирусный вектор: ДНК вируса, измененная таким
образом, что она может использоваться для переноса
рекомбинантной ДНК.
Витамин: органическое соединение, необходимое не-
которым видам организмов в небольшом количестве;
обычно компонент кофермента.
Внеклеточный матрикс: сложная сеть гликозамино-
гликанов, протеогликанов и белков, расположенная
непосредственно на внешней поверхности плазмати-
Словарь терминов |3.> 11
ческой мембраны; обеспечивает адгезию, распознава-
ние и миграцию клеток.
Внепеченочные ткани: все ткани, кроме ткани пече-
ни; термин дополнительно подчеркивает важную роль
печени в метаболизме.
Водородная связь: слабое электростатическое при-
тяжение между электроотрицательным атомом (таким
как кислород или азот) и атомом водорода, ковалентно
связанным со вторым электроотрицательным атомом.
Возбужденное состояние: высокоэнергетическое со-
стояние атома или молекулы; возникает при поглоще-
нии энергии света атомом или молекулой.
Восстанавливающий агент (восстановитель): до-
нор электронов в окислительно-восстановительной
реакции.
Восстанавливающий конец: конец полисахаридной
цепи, на котором находится группа со свободным
аномерным атомом углерода; этот концевой остаток
может выступать в роли восстановителя.
Восстанавливающий сахар: молекула сахара, в кото-
рой есть карбонильный (аномерный) атом углерода,
не участвующий в образовании гликозидной связи и
поэтому подверженный окислению.
Восстановительный эквивалент: общий термин для
обозначения электронов или эквивалентов электро-
нов в форме атома водорода или гидрид-иона.
Восстановление флуоресценции после фотообес-
цвечивания (FRAP): количественный метод анали-
за диффузии компонентов мембраны (липидов или
белков) в двойном слое.
Восстановление (полуреакция): присоединение
электронов к атому или иону.
Время полужизни: время, за которое распадается по-
ловина данного вещества в системе.
Всасывание: процесс переноса веществ при пищева-
рении — из кишечника в кровь.
Вспомогательные пигменты: пигменты растений и
фотосинтезирующих бактерий, под действием света в
видимой области солнечного спектра (каротиноиды,
ксантофиллы и фикобилины), которые дополняют
функцию хлорофиллов по поглощению солнечной
энергии.
Вторичная структура: локальная пространственная
упаковка участка полипептидной или полинуклеотид-
ной цепи.
Вторичный метаболизм: метаболические пути, при-
водящие к образованию специфических продуктов,
содержащихся не во всех живых клетках.
Вторичный посредник (мессенджер): эффекторная
молекула, синтезируемая в клетке в ответ на внешний
сигнал (первичный посредник), такой как гормон.
Второе начало термодинамики: закон, утверждаю-
щий, что любые химические и физические процессы
в замкнутой системе сопровождаются увеличением
энтропии.
Вырожденность кода: возможность обозначения
одного элемента (слова) в языке с помощью несколь-
ких элементов (слов) другого языка; вырожденность
генетического кода заключается в том, что некоторым
аминокислотам соответствует несколько нуклеотид-
ных триплетов.
Высокоэффективная жидкостная хроматография
(ВЭЖХ, HPLC): автоматизированный хромато-
графический метод разделения веществ в условиях
высокого давления; метод позволяет получать вос-
производимые результаты и разделять очень близкие
по свойствам вещества.
Г
Гаметы: репродуктивные клетки с гаплоидным на-
бором генов; сперматозоиды и яйцеклетки.
Ганглиозиды: сфинголипиды, содержащие в головке
сложные олигосахаридные группы; преимущественно
встречаются в нервной ткани.
Гаплоидная клетка: клетка с одним набором генети-
ческой информации, т. е. с одной хромосомой каждого
вида.
Гаптен: небольшая молекула, которая при связывании
с более крупной молекулой может вызывать иммун-
ный ответ.
Гексоза: шестиуглеродный углевод (сахар).
Геликаза: фермент, катализирующий расхождение
нитей ДНК перед репликацией.
Гем: железопорфириновая простетическая группа не-
которых белков.
Гемовый белок: белок, содержащий гем в качестве
простетической группы.
Гемоглобин: гемовый белок эритроцитов; переносчик
кислорода.
Ген: участок хромосомы, кодирующий отдельную функ-
циональную полипептидную цепь или молекулу РНК.
Ген-супрессор опухоли (антионкоген): ген белка, осу-
ществляющего отрицательную регуляцию деления клет-
ки; дефект обеих копий гена приводит к бесконтрольно-
му делению и опухолевой трансформации клетки.
Генетическая информация: наследственная инфор-
мация, закодированная последовательностью нуклео-
тидных оснований в хромосомной ДНК или РНК.
Генетическая карта: схематичное представление рас-
положения специфических последовательностей и
генов на хромосоме.
13521 Словарь терминов
Генетический код: набор триплетов ДНК (мРНК),
кодирующих протеиногенные аминокислоты.
Геном: вся генетическая информация, закодирован-
ная в клетке или вирусе.
Геномика: область науки, изучающая геномы клеток и
организмов.
Геномная библиотека: библиотека ДНК, содержащая
фрагменты ДНК, отражающие всю (или почти всю)
последовательность генома организма.
Генотип: генетическое содержание организма — в от-
личие от его физических характеристик (фенотипа).
Геометрические изомеры (цмс/транс-изомеры):
изомеры, различающиеся расположением заместите-
лей относительно двойной связи.
Гепарансульфат: сульфатированный полимер, состоя-
щий из чередующихся звеньев N-ацетилглюкозамина
и уроновой кислоты (глюкуроновой или идуроновой
кислоты); компонент внеклеточного матрикса.
Гепатоцит: основная клетка печени.
Гетеродуплекс ДНК: дуплекс ДНК, содержащий ком-
плементарные нити, происходящие из двух разных мо-
лекул ДНК с похожими последовательностями; часто
образуется как продукт генетической рекомбинации.
Гетерополисахарид: полисахарид, состоящий из не-
скольких типов моносахаридных звеньев.
Гетеротропный модулятор: аллостерический модуля-
тор, отличный от обычного лиганда.
Гетеротропный фермент: аллостерический фермент,
нуждающийся в модуляторе, отличном от обычного
субстрата.
Гетеротроф: организм, нуждающийся в сложных ор-
ганических молекулах (таких как глюкоза) в качестве
источника питательных веществ и энергии.
Гиалуронат (гиалуроновая кислота): высокомолеку-
лярный кислый полисахарид, обычно построенный из
дисахаридных звеньев GlcUA(pi->3)GlcNAc; основ-
ной компонент внеклеточного матрикса, образующий
крупные комплексы (протеогликаны) с белками и
другими кислыми полисахаридами.
Гидроксония ион (Н3О+): гидратированный протон.
Гидролазы: ферменты (протеазы, липазы, фосфатазы,
нуклеазы), катализирующие реакции гидролиза.
Гидролиз: расщепление связи (например, ангидридной
или пептидной) путем присоединения молекулы воды
с образованием двух или большего числа продуктов.
Гидропатический индекс: шкала, отражающая отно-
сительную гидрофобность и гидрофильность химиче-
ских групп.
Гидрофильность: наличие полярности или заряда,
способствующее взаимодействию молекул или хими-
ческих групп с водой (в том числе их растворению).
Гидрофобность: отсутствие полярности, препятствую-
щее растворению молекул в воде.
Гидрофобные взаимодействия: взаимодействие
неполярных групп молекул в водном растворе, об-
условленное стремлением окружающих молекул воды
перейти в наиболее устойчивое (разупорядоченное)
состояние.
Гиперхромный эффект: значительное усиление
поглощения света при 260 нм, возникающее при рас-
кручивании (плавлении) двойной спирали ДНК.
Гипоксия: метаболическое состояние, вызванное не-
достатком кислорода.
Гистоны: семейство основных белков, прочно связан-
ных с ДНК в хромосомах эукариотических клеток.
Гликан: то же, что полисахарид; полимер, состоящий
из моносахаридных звеньев, соединенных гликозид-
ными связями.
Гликогенез: процесс превращения глюкозы в гликоген.
Гликогенин: белок, инициирующий синтез цепей гли-
когена и катализирующий полимеризацию несколь-
ких первых звеньев в каждой цепи до начала работы
гликогенсинтазы.
Гликогенолиз: ферментативное расщепление запасен-
ного (непищевого) гликогена.
Гликозаминогликан: гетерополисахарид, со-
стоящий из двух чередующихся звеньев, одним
из которых является N-ацетилглюкозамин или
N-ацетилгалактозамин, а вторым — уроновая кислота
(чаще глюкуроновая кислота); прежнее название —
мукополисахарид.
Гликозидная связь: связь между сахаром и другой
молекулой (обычно спиртом, пурином, пиримидином
или сахаром) через атом кислорода (О-гликозидная
связь) или азота (N-гликозидная связь).
Гликоконъюгат: гликопротеин или гликолипид, об-
разованный из углеводного компонента, ковалентно
связанного с белком или липидом.
Гликолиз: катаболический путь, в результате которо-
го молекула глюкозы распадается на две молекулы
пирувата.
Гликолипид: липид с углеводным компонентом.
Гликомика: системный анализ углеводного состава
клетки или ткани, включая компоненты, связанные с
белками или липидами.
Гликопротеин: белок с углеводным компонентом.
Глиоксилатный цикл: у бактерий и некоторых рас-
тений альтернативный вариант цикла лимонной
кислоты, заключающийся в превращении ацетата в
сукцинат и в итоге в новый углевод.
Словарь терминов j 353 j
Глиоксисома: специализированная пероксисома, со-
держащая ферменты глиоксилатного цикла; содер-
жится в клетках прорастающих семян.
Глицерофосфолипиды: амфифильные липиды на
основе глицерина, в которых жирные кислоты при-
соединены сложноэфирной связью к атомам С1 и С2
глицерина, а полярные спирты — фосфодиэфирной
связью к атому СЗ.
Глобулярные белки: растворимые белки глобулярной
(сферической) формы.
Глюкогенные аминокислоты: аминокислоты, угле-
родный скелет которых может превращаться в глюко-
зу или гликоген в процессе глюконеогенеза.
Глюконеогенез: биосинтез углеводов из более про-
стых предшественников (не сахаров), таких как окса-
лоацетат или пируват.
Гомеобокс: консервативная последовательность ДНК
из 180 пар оснований, которая кодирует белковый
домен многих белков, играющих регуляторную роль в
процессах развития.
Гомеодомен: регуляторный белковый домен, опреде-
ляющий сегментацию зародыша; кодируется гомео-
боксом.
Гомеостаз: поддержание динамического стационар-
ного состояния с помощью регуляторных механизмов,
компенсирующих изменения, вызванные внешними
стимулами.
Гомеотические гены: гены, регулирующие развитие
сегментов зародыша дрозофилы; похожие гены обна-
ружены у большинства позвоночных.
Гомологичная генетическая рекомбинация: реком-
бинация между двумя молекулами ДНК с похожей
последовательностью, происходящая во всех клетках;
у эукариот происходит в процессе митоза и мейоза.
Гомологичные белки: белки из разных организмов,
имеющие похожие последовательности и функции
(например, гемоглобины).
Гомополисахарид: полисахарид, состоящий из одно-
типных моносахаридных звеньев.
Гомотропный модулятор: аллостерический модуля-
тор, идентичный нормальному лиганду.
Гомотропный фермент: аллостерический фермент,
для которого субстрат играет одновременно роль
модулятора.
Гормон: химическое соединение, синтезируемое в не-
большом количестве эндокринными железами и пере-
носимое кровью к другим тканям, функцию которых
оно регулирует, выступая в роли посредника.
Грамм-моль: единица количества вещества, численно
равна молекулярной массе этого вещества в граммах.
Грана: стопка тилакоидов (уплощенных мембранных
мешочков или дисков) в хлоропластах.
д
Дальтон: размерность атомной и молекулярной
массы; 1 Да (1,66 • 10 24 г) соответствует массе атома
водорода.
Двойная спираль: естественная конформация двух
комплементарных антипараллельных нитей ДНК.
Двойной (липидный) слой: ориентированное распо-
ложение амфифильных молекул липидов, лежащее в
основе структуры биологических мембран, когда угле-
водородные «хвосты» молекул направлены внутрь
клетки, образуя сплошную неполярную поверхность.
Двойные обратные координатьЦкоординаты Лай-
нуивера-Берка): для более точного определения
значений Vmax и Км вместо координат Vo против [S]
используют координаты 1/VO против 1/[S].
Двухкомпонентная сигнальная система: система
передачи сигнала у бактерий и растений, содержащая
рецепторную гистидинкиназу, которая в результате
связывания с лигандом начинает фосфорилировать
внутренние остатки гистидина, после чего катализи-
рует перенос фосфорильной группы на второй компо-
нент системы — регулятор ответа, который активиру-
ет транскрипцию гена или группы генов.
Двухосновная кислота: кислота, способная при дис-
социации высвобождать два протона.
Дегидрогеназы: ферменты, катализирующие удале-
ние двух атомов водорода из молекулы субстрата.
Дезаминирование: ферментативное удаление амино-
групп из биомолекул, таких как аминокислоты или
нуклеотиды.
Дезоксирибонуклеотиды: нуклеотиды, содержащие в
качестве пентозы 2-дезокси-D-рибозу.
Делеционные мутации: мутации, возникающие в ре-
зультате делении (удаления) одного или нескольких
нуклеотидов из гена или хромосомы.
Денатурация: полное или частичное разворачивание
полипептидной цепи, белка или нуклеиновой кис-
лоты, приводящее к потере биологической функции
молекулы.
Десатуразы: ферменты, катализирующие введение
двойной связи в углеводную часть молекулы жирной
кислоты.
Десенсибилизация: универсальный процесс, за-
ключающийся в ослаблении ответной реакции при
длительном воздействии специфического стимула.
Десольватация: удаление гидратной оболочки, окру-
жающей растворенное вещество в водном растворе.
Диализ: удаление малых молекул (низкомолекуляр-
ныз веществ) из раствора макромолекул путем про-
пускания раствора через полупроницаемую мембрану.
13541 Словарыерминов
Дикий тип: нормальный (немутированный) генотип
или фенотип.
Диплоидная клетка: клетка с двойным набором гене-
тической информации (две хромосомы каждого типа).
Дисахарид: углевод, состоящий из двух моносахарид-
ных звеньев, связанных ковалентной связью.
Дисульфидная связь: ковалентная связь, возникаю-
щая в окислительных условиях между двумя остатка-
ми Cys одной или разных полипептидных цепей; при
этом образуется цистин.
Дифференциальное центрифугирование: разделение
клеточных органелл или других частиц разного раз-
мера, основанное на различии скорости седиментации
при центрифугировании.
Дифференцировка клеток: превращение клеток-
предшественников в специализированные клетки
путем образования новых белков и РНК.
Дифференцировка: приобретение клеткой специфи-
ческих структурных и функциональных особенностей
в процессе роста и развития.
Диффузия: перемещение молекул вещества из обла-
сти с более высокой концентрацией в область с более
низкой концентрацией.
ДНК (дезоксирибонуклеиновая кислота): поли-
нуклеотид со специфической последовательностью
дезоксирибонуклеотидных звеньев, ковалентно свя-
занных между собой 3',5'-фосфодиэфирными связями;
хранилище генетической информации.
ДНК-лигаза: фермент, катализирующий образова-
ние фосфодиэфирной связи между З'-концом одного
фрагмента ДНК и 5'-концом другого.
ДНК-микрочип: набор фрагментов ДНК, располо-
женных определенным образом и иммобилизованных
на твердой подложке, который используется для
гибридизации с анализируемыми последовательно-
стями ДНК.
ДНК-полимераза: фермент, катализирующий
синтез ДНК из молекул дезоксирибонуклеозид-5'-
трифосфатов на основе матрицы.
Домен: четко идентифицируемая структурная едини-
ца полипептида; домены могут выполнять независи-
мые функции и упаковываться независимым образом.
Донор протонов: кислота или другое химическое
соединение, отдающее протоны в кислотно-основной
реакции.
Донор электронов: вещество, отдающее электроны в
ходе окислительно-восстановительной реакции.
Дыхание: любой метаболический процесс, сопрово-
ждающийся потреблением кислорода и выделением
углекислого газа.
Дыхательная цепь: цепь переноса электронов; по-
следовательность белков аэробных клеток, пере-
носящих электроны от субстратов на молекулярный
кислород.
Ж
Жгутик: клеточный отросток, используемый для пе-
редвижения; жгутики бактерий имеют гораздо более
простую структуру, чем жгутики эукариот, которые
напоминают реснички.
Железосерный белок: представитель большого
семейства электронпереносящих белков, в котором
функцию переноса электронов осуществляют ионы
железа, связанные с атомами неорганической серы и
атомами серы в остатках Cys в белке.
Железосерный белок Риске: железосерный белок, в
котором в роли двух лигандов центрального иона же-
леза выступают боковые цепи остатков His; участвует
во многих процессах с переносом электронов, вклю-
чая окислительное фосфорилирование и фотофосфо-
рилирование.
Железосерный центр (кластер): простетическая
группа некоторых белков, задействованная в перено-
се электронов; Fe2+ или Fe3+ связаны с неорганической
серой и атомом серы в остатке Cys в белке.
Желчные соли: амфифильные производные стерои-
дов, обладающие свойствами детергентов и участвую-
щие в расщеплении и всасывании липидов.
Жидкостно-мозаичная модель: модель, рассматрива-
ющая биологическую мембрану в виде жидкого двой-
ного липидного слоя с включенными в него белками;
такой двойной слой обладает как структурной, так и
функциональной асимметрией.
Жирная кислота: алифатическая карбоновая кислота,
присутствующая в природных жирах и маслах; компо-
нент гликолипидов и фосфолипидов мембран.
Жировая ткань: специализированная соединительная
ткань, накапливающая большое количество триацил-
глицеринов; см. также Белая жировая ткань; Бурая
жировая ткань.
3
Закон действия масс: скорость химической реакции
пропорциональна произведению концентраций (или
активностей) реагирующих веществ.
Закрытая система: не обменивается материей и энер-
гией с окружающей средой.
Заменимые аминокислоты: аминокислоты, кото-
рые могут синтезироваться в организме человека и
других позвоночных из простых предшественников
Словарыерминов 1353 j
и поэтому не обязательно должны присутствовать в
составе пищи.
Заместительная мутация: обусловлена заменой одно-
го основания другим.
Зародышевая клетка: клетка животного, образую-
щаяся на ранней стадии эмбриогенеза; может делить-
ся митозом или в результате мейоза давать начало
гаметам (сперматозоидам и яйцеклеткам).
Зеленый флуоресцентный белок (GFP): небольшой
белок из морского организма, ярко флуоресцирую-
щий в зеленой области видимого спектра. Гибридные
белки с GFP часто применяются для определения
внутриклеточной локализации белков методом флуо-
ресцентной микроскопии.
Зимоген: неактивный предшественник фермента; на-
пример, предшественник пепсина — пепсиноген.
Зонд: меченый фрагмент нуклеиновой кислоты, со-
держащий участок, комплементарный искомому гену
или геномной последовательности; используется для
гибридизации.
И
Изменение свободной энергии (AG): количество
свободной энергии, высвобождаемой (АС < 0) или
поглощаемой (АС > 0) в реакции, протекающей при
постоянной температуре и давлении.
Изменение энтальпии (АН): в химической реакции
приблизительно равно разность энергии, затрачива-
емой на разрыв старых связей, и энергии, выделяю-
щейся при образовании новых связей.
Изомеразы: ферменты, катализирующие взаимные
превращения между позиционными изомерами.
Изомеры: молекулы с одинаковой молекулярной
формулой, но разным расположением химических
групп.
Изопрен: 2-метил-1,3-бутадисн, повторяющаяся
структурная единица изопреноидов (терпеноидов).
Изопреноид (терпеноид): представитель многочис-
ленного класса природных веществ, синтезированных
ферментативным путем из двух или большего числа
изопреновых звеньев.
Изотермические условия: постоянная температура.
Изотопы: стабильные или радиоактивные атомы
одного и того же элемента. Изотопы одного эле-
мента имеют одинаковый заряд ядра, но разную
атомную массу, они идентичные по химическим
свойствам. Используются в качестве меток в хими-
ческом анализе.
Изоферменты: множественные формы фермента,
катализирующие одну и ту же реакцию, но разли-
чающиеся по аминокислотной последовательности,
субстратной специфичности, значению Vinax или регу-
ляторным свойствам.
Изоэлектрофокусирование: электрофоретический
метод разделения макромолекул, основанный на раз-
личии значений их изоэлектрических точек.
Изоэлектрическая точка: значение pH, при котором
суммарный электрический заряд молекулы равен
нулю, так что молекула неподвижна в электрическом
поле.
Иммунный ответ: способность организма позвоноч-
ного синтезировать антитела против чужеродных для
организма макромолекул (антигенов).
Иммуноглобулин (антитело): белок, синтезиро-
ванный в ответ на попадание в организм антигена и
способный специфично связываться с ним.
Ингибирование по принципу обратной связи (инги-
бирование конечным продуктом): ингибирование
активности фермента, действующего в начале метабо-
лического пути, конечным продуктом всего пути.
Ингибирующий G-белок (Gj): тримерный GTP-свя-
зывающий белок, который при активации ассоцииро-
ванным рецептором в плазматической мембране на-
чинает ингибировать соседние мембранные фермен-
ты, такие как аденилатциклаза; действие этого белка
противоположно действию Gs.
Индуктор: сигнальная молекула, которая связывается
с регуляторным белком и тем самым вызывает усиле-
ние экспрессии соответствующего гена.
Индукция: усиление экспрессии гена в ответ на из-
менение активности регуляторного белка.
Индуцированное соответствие: изменение конфор-
мации фермента при связывании субстрата, приводя-
щее к переходу фермента в каталитически активное
состояние; модель, описывающая изменение конфор-
мации любой макромолекулы в ответ на связывание
лиганда: центр связывания макромолекулы начинает
лучше соответствовать форме лиганда.
Инициаторный комплекс: комплекс рибосомы, мРНК
и инициаторной Met-TPHKMet или fMet-TPHKfMet,
готовый к стадии элонгации.
Инсерционная мутация: мутация, вызванная инсер-
цией (встраиванием) одного или нескольких основа-
ний в последовательность ДНК.
Инстерционная последовательность (IS-элемент):
мобильные генетические элементы, обычно ограни-
ченные с двух сторон специфическими инвертирован-
ными повторами.
Интегральный белок: белок, прочно связанный с
мембраной благодаря гидрофобным взаимодействиям
(в отличие от периферического белка).
[356] Словарь терминов
Интегрин: представитель большого семейства гетеро-
димерных трансмембранных белков, опосредующих
межклеточные контакты и контакты клеток с внекле-
точным матриксом.
Интеркалирующий агент: вещество (мутаген),
который встраивается в последовательность нукле-
иновых оснований и вызывает сдвиг рамки считы-
вания.
Интеркаляция: встраивание молекулы между арома-
тическими или плоскими кольцевыми молекулами,
например между двумя основаниями нуклеиновой
кислоты.
Интерфероны: класс гликопротеинов, обладающих
противовирусной активностью.
Интрон: последовательность нуклеотидов между
экзонами, которая транскрибируется, но удаляется до
начала трансляции.
Информационные макромолекулы: биомолекулы,
содержащие информацию в форме специфических
последовательностей мономеров, например это могут
быть белки, липиды, полисахариды и нуклеиновые
кислоты.
Ионизирующая радиация: вид излучения (например,
рентгеновское излучение), которое вызывает потерю
электронов некоторыми молекулами, повышая их
реакционную способность.
Ионное произведение воды (Kw): произведение кон-
центраций Н+ и ОН в чистой воде.
Kw = [Н ][ОН ] = 110 14 при 25 °C.
Ионный канал: интегральный белок, обеспечиваю-
щий регулярный транспорт специфических ионов
через клеточную мембрану.
Ионообменная смола: полимерная смола, содержа-
щая иммобилизованные заряженные группы; ис-
пользуется в хроматографии для разделения ионных
соединений.
Ионофор: соединение, которое связывает один или
несколько ионов металла и диффундирует через мем-
брану, перенося связанный ион.
К
Калория: размерность тепловой энергии. 1 кал равна ко-
личеству теплоты, необходимому для повышения темпе-
ратуры 1,0 г воды от 14,5 до 15,5 °C; 1 кал = 4,18 Дж.
Капсид: белковая оболочка вирусной частицы.
Карбанион: атом углерода, несущий отрицательный
заряд.
Карбокатион: атом углерода, несущий положитель-
ный заряд.
Каркасный белок: некаталитический белок, который
способствует образованию полиферментных комплек-
сов, предоставляя ферментам несколько специфиче-
ских участков связывания.
Карнитиновый челнок: механизм переноса жирных
кислот в виде карнитиновых эфиров из цитозоля в
митохондриальный матрикс.
Каротиноиды: жирорастворимые фотосинтетические
пигменты, состоящие из изопреновых звеньев.
Катаболизм: стадия промежуточного метаболизма,
заключающаяся в расщеплении молекул питательных
веществ с выделением энергии.
Катенаны: соединения, в составе которых имеются
два или более цикла, связанных между собой с помо-
щью нековалентных топологических связей, напоми-
нающих механическую связь между звеньями цепи.
Катехоламины: гормоны (например, адреналин),
аминопроизводные катехола.
Катионообменная смола: нерастворимая полимерная
матрица с фиксированными отрицательно заряжен-
ными группами; используется для хроматографиче-
ского разделения катионных соединений.
«Качающиеся» пары: пары, образующиеся при срав-
нительно слабых взаимодействиях между основанием
на З'-конце одного кодона и комплементарным осно-
ванием на 5'-конце антикодана.
Кератины: нерастворимые защитные или струк-
турные белки, состоящие из параллельных поли-
пептидных цепей с а-спиральной или р-кон-
формацией.
Кетогенные аминокислоты: аминокислоты, угле-
родный скелет которых может использоваться для
синтеза кетоновых тел.
Кетоз: аномально высокое содержание кетоновых тел
в крови, тканях и моче.
Кетоза: простой моносахарид, в котором карбонил
представлен кетогруппой.
Кетоновые тела: ацетоацетат, D-p-гидроксибутират и
ацетон; водорастворимые энергетические молекулы,
в норме выделяемые печенью, но присутствующие
в избытке при голодании или некомпенсированном
сахарном диабете.
Киназы: ферменты, катализирующие фосфорилиро-
вание некоторых молекул под действием АТР.
Кинетика: раздел физической химии, изучающий хи-
мическую реакцию во времени, скорость химической
реакции.
Кислородный долг: дополнительное количество кис-
лорода, которое потребляется организмом в период
после интенсивной физической нагрузки, по сравне-
нию с потреблением в состоянии покоя.
Словарыерминов [35/ ]
Клон: потомство одной клетки.
Клонирование: способ получения большого количе-
ства идентичных молекул ДНК, клеток или организ-
мов исходя из одной молекулы ДНК, одной клетки
или одного организма.
Ковалентная связь: химическая связь, возникающая
при обобществлении валентных двух атомов с образо-
ванием общей электронной пары.
Кодон: последовательность из трех нуклеотидов,
кодирующая специфическую аминокислоту.
З'-Конец: в молекуле нуклеиновой кислоты там нахо-
дится свободная ОН-группа рибозы или дезоксирибозы.
5'-Конец: в молекуле нуклеиновой кислоты там на-
ходится свободная фосфатная группа.
Консервативная замена: замена одного аминокислот-
ного остатка в полипептидной цепи другим остатком
с похожими свойствами, не приводящая к изменению
свойств полимера, например, замена Glu на Asp.
Константа неустойчивости (диссоциации) (Kd):
величина, обратная константе комплексообразования;
характеризует способность к диссоциации на отдель-
ные компоненты; например, диссоциация фермент-
субстратного комплекса.
Константа диссоциации кислоты (Ка): характеризует
реакцию диссоциации кислоты на протон и сопряжен-
ное основание.
Константа равновесия (^eq): характеризует любую
химическую реакцию; выражается через равновесные
концентрации всех исходных веществ и всех продук-
тов реакции (реакция в состоянии равновесия) при
заданной температуре и давлении.
Константа скорости: константа пропорциональности,
связывающая скорость химической реакции с концен-
трациями реагирующих веществ.
Конститутивный фермент: фермент, необходимый
клетке непрерывно и присутствующей в постоянной
концентрации, например многие ферменты централь-
ных метаболических путей.
Контиг: набор перекрывающихся клонов или про-
тяженная последовательность, охватывающая целый
фрагмент хромосомы.
Контроль транскрипции: регуляция синтеза белка
путем регуляции образования его мРНК.
Контроль трансляции: регуляция синтеза белка пу-
тем регуляции скорости его трансляции на рибосомах.
Контурная длина: длина спиральной полимерной
молекулы, измеренная вдоль оси спирали.
^-Конформация: растянутая, зигзагообразная
укладка полипептидной цепи; часто реализуется как
вторичная структура белка.
N-Концевой остаток: единственный аминокислот-
ный остаток в составе полипептидной цепи, несущий
свободную а-аминогруппу.
Кооперативность: способность фермента или друго-
го белка изменять сродство к последующим присоеди-
няемым молекулам лиганда после связывания первой
молекулы; при положительной кооперативности
связывание первой молекулы лиганда повышает срод-
ство к следующим молекулам, а при отрицательной
кооперативности — понижает его.
Короткий тандемный повтор (STR): многократный
повтор двух или нескольких пар (обычно от 3 до 6)
примыкающих друг к другу нуклеотидов в последова-
тельности ДНК; используется в судебной медицине и
патологической анатомии.
Кортикостероиды: стероидные гормоны, образу-
ющиеся в коре надпочечников.
Котранспорт: одновременный перенос через мембра-
ну двух растворенных веществ одним и тем же пере-
носчиком. См. также Антипорт; Симпорт.
Коэффициент активности: коэффициент пропор-
циональности между концентрацией растворенного
вещества и его термодинамической активностью в
растворе.
Коэффициент распределения: отношение концен-
траций вещества в двух несмешивающихся фазах,
находящихся в равновесии.
Коэффициент седиментации: показатель скорости
осаждения частицы при центрифугировании.
Кребса цикл: см. Цикл лимонной кислоты
Крестообразная структура ДНК (РНК): вторичная
структура двунитевой РНК или ДНК, образующаяся
в результате денатурации двойной спирали с последу-
ющим спариванием палиндромных повторов, приво-
дящим к образованию крестов или шпилек.
Кривая титрования: зависимость значений pH от
добавленного количества основания при титровании
кислоты.
Кристы: складки внутренней мембраны митохондрий.
Круговорот азота: общий биогеохимический цикл
азота. Разные формы биологически доступного
азота циркулируют в природе, попадая в растения, в
организмы животных, в микроорганизмы, свободный
азот присутствует в атмосфере, а минеральные формы
азота — в геосфере.
Культура ткани: метод длительного сохранения и вы-
ращивания (культивирования) клеток, извлеченных
из многоклеточного организма, на искусственных
питательных средах.
135 <S | Словарыерминов
Л
Лайнуивера-Берка уравнение: алгебраическое пре-
образование уравнения Михаэлиса-Ментен, позволя-
ющее путем экстраполяции более точно определить
значения Vmax и Км.
l/V0 = KM/Vmax[S] + l/Vinax.
Левовращающий изомер: стереоизомер, вращающий
плоскость поляризации плоскополяризованного света
против часовой стрелки.
Лейкотриены: липиды, образующиеся в организме из
арахидоновой кислоты; содержат 20-углеродную цепь.
Относятся к эйкозаноидам.Играют роль в патогенезе
бронхиальной астмы (медиаторы бронхообструкции
при БА).
Лейкоциты: белые кровяные клетки; число лейкоци-
тов характеризует состояние защитных сил организма
(человека и животного).
Лейциновая застежка-молния: структурный бел-
ковый мотив, необходимый для белок-белковых
взаимодействий многих регуляторных белков эука-
риот; состоит из двух взаимодействующих а-цепей,
в которых в каждой седьмой позиции расположен
остаток лейцина.
Лектин: белок, который связывается с углеводом
(обычно олигосахаридом) с очень высоким сродством
и специфичностью и тем самым опосредует межкле-
точные взаимодействия.
Летальная мутация: инактивирует жизненно важную
функцию клетки или организма.
Лиазы: ферменты, катализирующие удаление функ-
циональной (химической) группы, приводящее к
образованию двойной связи, или присоединение по
двойной связи.
Лигазы: ферменты, катализирующие реакцию
конденсации, в которой связь между двумя атомами
образуется за счет использования энергии АТР или
другого макроэргического соединения.
Лиганд: небольшая молекула, специфическим об-
разом связывающаяся с более крупной молекулой;
например, гормон — лиганд специфического белково-
го рецептора.
Лидерная последовательность: короткая N-концевая
последовательность белка или 5’-концевая последова-
тельность РНК со специализированной направляю-
щей или регуляторной функцией.
Лизис: разрушение плазматической мембраны или
клеточной стенки (у бактерий), приводящее к высво-
бождению внутриклеточного содержимого и гибели
клетки.
Лизосома: связанная с мембраной органелла эукари-
отической клетки; содержит разнообразные гидро-
литические ферменты и служит центром деградации
и реутилизации ненужных в данный момент компо-
нентов.
Лимитирующая стадия: (1) в общем случае — фер-
ментативная реакция с максимальной энергией
активации или с максимальной свободной энергией
переходного состояния; (2) в частном случае — самая
медленная стадия метаболического пути.
Лимфоциты: разновидность лейкоцитов; главные
клетки иммунной системы; обеспечивают гумораль-
ный иммунитет.
Липазы: ферменты, катализирующие гидролиз
триацилглицеринов.
Липид: низкомолекулярное соединение, не раствори-
мое в воде, обычно содержит остатки жирных кислот,
стерины или фрагменты изопреноидов.
Липидом: полный липидный состав клетки или ткани
в определенных условиях.
Липкие концы: концы фрагментов ДНК с одинако-
вой или различной последовательностью с корот-
кими комплементарными друг другу однонитевыми
участками, облегчающими лигирование данных
фрагментов.
Липоат (липоевая кислота): промежуточный пере-
носчик атомов водорода и ацильных групп в реакциях
дегидрогеназ а-кетокислот; витамин для некоторых
микроорганизмов.
Липопротеин: липидно-белковый агрегат, с помощью
которого липиды, не растворимые в воде, переносятся
в крови; белковый компонент липопротеина называют
аполипопротеином.
Липосома: ограниченная двойным липидным слоем
небольшая сферическая везикула, спонтанно образу-
ющаяся в суспензии фосфолипидов в водном буфере.
М
Макромолекула: молекулярная масса от нескольких
тысяч до нескольких миллионов.
Макроэргические соединения: группа природ-
ных веществ, молекулы которых содержат богатые
энергией (макроэргические связи); при гидролизе
при стандартных условиях высвобождают энергию
(уменьшается свободная энергия). Присутствуют во
всех живых клетках.
Малая ядерная РНК (мяРНК): небольшая моле-
кула РНК (обычно от 100 до 200 нуклеотидов), содер-
жащаяся в ядре и участвующая в сплайсинге эукарио-
тической мРНК.
Малая ядрышковая РНК (мякРНК): небольшая
молекула РНК (обычно от 60 до 300 нуклеотидов),
участвующая в химической модификации рРНК.
Словарыерминов [3591
Матрикс: водное содержимое клетки или органеллы
(например, митохондрии) с растворенными веще-
ствами.
Матрица: макромолекулярная форма, на основе кото-
рой синтезируется новая информационная макромо-
лекула.
Матричная (информационная) РНК (мРНК): мо-
лекула РНК, комплементарная одной из нитей ДНК;
переносит генетическую информацию от хромосом к
рибосомам.
Матричная нить: нить нуклеиновой кислоты, на
основе которой полимераза синтезирует комплемен-
тарную нить.
Мейоз: способ клеточного деления, в результате
которого диплоидная клетка дает начало гаплоидным
клеткам — гаметам.
Мембранный потенциал (Vm): разность электриче-
ских потенциалов на внутренней и внешней стороне
биологической мембраны, которую можно измерить с
помощью микроэлектрода; типичный мембранный по-
тенциал находится в диапазоне от -25 мВ до -100 мВ
(на мембранах вакуолей некоторых видов растений).
Отрицательный знак потенциала означает, что на
внутренней поверхности мембраны потенциал более
отрицательный, чем снаружи.
Мембранный транспорт: перемещение полярных рас-
творенных веществ через мембрану с помощью специ-
фических мембранных белков (переносчиков).
Метаболизм: ферментативные превращения орга-
нических молекул в живом организме; все процессы
катаболизма и анаболизма.
Метаболит: химическое промежуточное соединение в
реакции метаболизма.
Метаболическая регуляция: механизм, позволяю-
щий клетке противостоять изменению концентрации
отдельных метаболитов при изменении потока по
метаболическому пути в условиях метаболического
контроля.
Метаболический контроль: механизм, позволяющий
изменить поток метаболитов в метаболическом пути в
соответствии с изменением условий.
Метаболом: полный набор всех низкомолекулярных
метаболитов (промежуточных соединений, сигналь-
ных молекул, вторичных метаболитов), присутствую-
щих в данной клетке в конкретных условиях.
Метаболой: надмолекулярный комплекс ферментов,
выполняющих определенные метаболические функ-
ции, и участок клеточной структуры, на котором этот
комплекс локализован.
Металлопротеин: белок с ионом металла в качестве
простетической группы.
Метамерия: сегментарное строение тела, например, у
насекомых.
Миграция точки ветвления: перемещение точки
ветвления разветвленной ДНК, образованной двумя
молекулами ДНК с идентичной последовательно-
стью.
Микро-РНК: малые молекулы РНК (от 20 до 25
нуклеотидов после завершения процессинга), уча-
ствующие в подавлении экспрессии генов путем
ингибирования трансляции и/или расщепления соот-
ветствующей мРНК.
Микросомы: мембранные везикулы, образующиеся
при фрагментации эндоплазматического ретикулума
эукариотической клетки; выделяются методом диф-
ференциального центрифугирования.
Микротельца: связанные с мембраной цитоплазма-
тические везикулы, в которых содержатся ферменты,
участвующие в образовании и разрушении перок-
сидов (перекисей); к ним относятся пероксисомы,
лизосомы и глиоксисомы.
Микротрубочки: тонкие трубочки, собранные из
двух типов глобулярных субъединиц тубулина;
содержатся в ресничках, жгутиках, центросомах и
других двигательных и сократительных структурах
клетки.
Микрофиламенты: тонкие нити актина, содержащие-
ся в цитоплазме эукариотических клеток; выполняют
структурную и двигательную функции.
Миозин: фибриллярный белок, основной компонент
толстых филаментов скелетных мышц и актиномио-
зиновых комплексов.
Миофибрилла: структурный элемент тонких и тол-
стых филаментов мышечных волокон.
Миоцит: клетка мышечной ткани.
Митоз: многостадийный процесс в эукариотической
клетке, приводящий к репликации хромосом и деле-
нию клетки.
Митохондрия: ограниченная двумя мембранами
органелла эукариотической клетки; энергетическая
станция клетки — обеспечивает энергией синтез АТР;
содержит ферментативный аппарат для осуществле-
ния цикла лимонной кислоты, окисления жирных
кислот, переноса электронов и окислительного фос-
форилирования.
Михаэлиса константа (Км): равна концентрации суб-
страта, при которой скорость Vферментативной реак-
ции составляет половину максимальной скорости Vinax.
Михаэлиса-Ментен кинетика: кинетика фермента-
тивной реакции, описываемая гиперболической за-
висимостью начальной скорости реакции от концен-
трации субстрата.
13()() | Словарыерминов
Михаэлиса-Ментен уравнение: уравнение гипер-
болы; описывает зависимость начальной скорости
реакции Vo от концентрации субстрата [S], наблюдаю-
щуюся в случае многих ферментативных реакций:
Vo= Vinax[S]/(XM + [S]).
Мицелла: агрегат амфифильных молекул в воде, в ко-
тором неполярные части молекул направлены внутрь,
а полярные обращены наружу, образуя внешнюю по-
верхность, контактирующую с водной фазой.
Модулятор: метаболит, который связывается с
аллостерическим центром фермента и изменяет его
каталитические свойства.
Молчащая мутация: не приводит к видимым изменени-
ям биологической активности продукта данного гена.
Моль: масса вещества в граммах, численно равная его
молекулярной массе.
Молярный раствор: (водный) раствор, содержащий
1 моль вещества в 1000 мл раствора.
Моноклональные антитела: продуцируются одним
клоном клеток по гибридомной технологии; монокло-
нальные антитела идентичны и направлены против
одного эпитопа целевого антигена.
Моносахарид: углевод, состоящий из единственного
сахарного звена.
Монослой: одинарный слой правильно ориентиро-
ванных липидных молекул.
Моноцистронная мРНК: мРНК, которая может быть
транслирована в один-единственный белок.
Мотив: любой характерный вариант расположения
элементов вторичной структуры белка, встречающий-
ся в одном или нескольких белках; мотив может быть
простым или сложным и может распространяться на
весь белок или только на его часть; то же, что укладка
или супервторичная структура.
Мутазы: ферменты, катализирующие изменение рас-
положения функциональных групп в молекуле.
Мутаротация: изменение оптического вращения,
сопровождающее взаимные превращения а- и
[3-аномерных форм моносахаридов.
Мутация: стойкое, т. е. наследуемое, изменение гено-
типа под влиянием внешней или внутренней среды.
Н
Насыщенная жирная кислота: в углеродном скелете
нет двойных связей.
Нативная конформация: биологически активная
конформация макромолекулы.
Негемовые железосодержащие белки: содержат
железо, но не содержат порфиринового кольца; обыч-
но участвуют в окислительно-восстановительных
реакциях.
Незаменимые аминокислоты: не синтезируются в ор-
ганизме человека (и других позвоночных) и должны
поступать в организм с пищей.
Незаменимые жирные кислоты: полиненасыщенные
жирные кислоты, которые синтезируются растения-
ми, но не синтезируются в организме человека; необ-
ходимый компонент пищевого рациона человека.
Нейромедиатор: низкомолекулярное соединение
(обычно азотсодержащее), секретируемое в аксонных
окончаниях (терминалях) нейрона и связывающееся
со специфическим рецептором следующего нейрона;
осуществляет передачу нервного импульса.
Нейрон: клетка нервной ткани, специализирующаяся
на проведении нервного импульса.
Некодирующая РНК (нкРНК): не кодирует инфор-
мацию для синтеза белкового продукта.
Ненасыщенная жирная кислота: жирная кислота с
одной или несколькими двойными связями.
Неорганическая пирофосфатаза: фермент, гидроли-
зующий молекулу неорганического фосфата с образо-
ванием пирофосфата.
Неполярное вещество: гидрофобное вещество, плохо
растворяется в воде.
Нециклический поток электронов: индуцируемый
светом электронный транспорт от молекулы воды к
NADP+ в кислородзависимых реакциях фотосинтеза;
протекает с участием фотосистем I и II.
Низкоэнергетические фосфатные соединения: фос-
форилированные соединения со сравнительно низкой
стандартной энергией гидролиза.
Нингидриновая реакция: цветная реакция, проис-
ходящая между аминокислотами или пептидами и
нингидрином при нагревании; используется для каче-
ственного и количественного определения аминокис-
лот и пептидов.
Нитрогеназный комплекс: ферментативная система,
способная восстанавливать атмосферный азот до
аммиака в присутствии АТР.
Нонсенс-кодон (бессмысленный кодон или стоп-
кодон): один из триплетов оснований ДНК (UAG,
UAA, UGA), который не кодирует аминокислоты;
терминатор трансляции — поэтому иногда называют
кодном-терминатором.
Нонсенс-мутация: точечная мутация, приводящая
к появлению стоп-кодона и преждевременной оста-
новке синтеза полипептидной цепи.
Нонсенс-супрессор: мутантный ген тРНК, кодирую-
щий молекулу с измененным антикодоном.
Нуклеазы: ферменты, гидролизующие межнуклеотид-
ные (фосфодиэфирные) связи в нуклеиновых кислотах.
Нуклеиновые кислоты: природные полинуклеотиды,
звенья которых образуют специфические последова-
тельности и связаны между собой фосфодиэфирными
связями; ДНК и РНК.
Нуклеозид: соединение, состоящее из пуринового
или пиримидинового основания, ковалентно связан-
ного с пентозой.
Нуклеозиддифосфаткиназа: фермент, катализирую-
щий перенос концевой фосфатной группы нуклеозид-
5'-трифосфата на нуклёозид-5'-дифосфат.
Нуклеозиддифосфатсахара: молекула, напомина-
ющая кофермент и участвующая в ферментативном
синтезе полисахаридов и производных сахаров.
Нуклеозидмонофосфаткиназа: фермент, катализиру-
ющий перенос концевой фосфатной группы АТР на
нуклеозид-5'-монофосфат.
Нуклеоид: означает «подобный ядру»; компартмент
неправильной формы внутри клетки прокариот, в
котором находится генетический материал; геном
бактерий.
Нуклеоплазма (ядерный матрикс): часть содержи-
мого эукариотической клетки, окруженная ядерной
мембраной.
Нуклеосома: структурная единица хроматина у
эукариот; состоит из нити ДНК, обернутой вокруг
гистоновой сердцевины.
Нуклеотид: фосфорный эфир нуклеозида по одной из
гидроксильных групп пентозы.
Нуклеофил: богатая электронами функциональная
группа, способная отдавать электроны электрондефи-
цитным атомам (электрофилам); проявляет актив-
ность в бимолекулярных реакциях замещения.
О
Облегченная диффузия (пассивный транспорт):
диффузия полярных веществ через биологическую
мембрану с помощью белка-переносчика.
Обратимый ингибитор: ингибитор, связывающийся с
ферментом обратимым образом, так что при удалении
ингибитора активность фермента восстанавливается.
Обратная (реверсивная) мутация: возвращающая
организму исходный фенотип.
Обратная транскриптаза: РНК-зависимая ДНК-
полимераза ретровирусов; способна синтезировать
ДНК, комплементарную последовательности РНК.
Общий кислотно-основной катализ: катализ, сопро-
вождающийся переносом протонов от молекул (к
молекулам), отличным от воды.
Словарыерминов [3611
Одноосновная кислота: при диссоциации образуется
один протон.
Окисление (полуреакция): отдача электронов атомом
или ионом.
а-Окисление: реакция окисления р-метилированных
жирных кислот в пероксисомах.
P-Окисление: окислительное расщепление жирных
кислот до ацетил-СоА путем последовательного окис-
ления атома р-углерода.
со-Окисление: альтернативный путь окисления
жирных кислот, при котором сначала происходит
окисление атома углерода, максимально удаленного
от карбоксильной группы.
Окислитель: акцептор электронов в окислительно-
восстановительной реакции.
Окислительно-восстановительная пара (редокс-па-
ра): соединение-донор электронов и соответствующая
ему окисленная форма, например NADH/NAD+.
Окислительно-восстановительная реакция: реакция,
в которой электроны переходят от донора к акцептору.
Оксигеназы: ферменты, катализирующие реакции, в
которых атомы кислорода непосредственно включа-
ются в продукты реакции, образуя гидроксильную
или карбоксильную группу. В реакциях, катализи-
руемых монооксигеназами, происходит встраивание
лишь одного атома кислорода, а другой восста-
навливается до Н2О. В реакциях, катализируемых
диоксигеназами, в продукт встраиваются два атома
кислорода.
Оксидазы со смешанной функцией: ферменты (на-
пример, монооксигеназы), катализирующие окисле-
ние двух восстановителей: обычно один — NADPH,
а другой — субстрат. При этом один атом кислорода
включается в продукт реакции, а другой восстанавли-
вается до Н2О. Для переноса электронов от NADPH
на О2 эти ферменты часто используют цитохром Р450.
Оксидазы: ферменты, катализирующие реак-
ции окисления, в которых в качестве акцептора
электронов выступает молекулярный кислород,
но атомы кислорода не включаются в конечные
продукты.
Олигомер: короткий полимер, состоящий не более
чем из 50 аминокислот, нуклеотидов или сахаров.
Олигомерный белок: состоит из двух или нескольких
идентичных полипептидных цепей.
Олигонуклеотид: короткая нуклеотидная последова-
тельность (обычно не более 50 оснований).
Олигопептид: последовательность из нескольких
аминокислотных остатков, соединенных пептидными
(амидными) связями.
1362 ] Словарыерминов
Олигосахарид: последовательность моносахаридных
звеньев, соединенных гликозидными связями.
Омыление: щелочной гидролиз триацилглицеринов с
образованием жирных кислот.
Онкоген: ген, кодирующий белок, который при нару-
шении регуляции может вызвать быстрое и некон-
тролируемое деление клетки, приводящее к развитию
опухоли. Мутации, вызывающие активацию онкогена,
увеличивают риск превращения клетки в раковую.
Оператор: регулярный участок ДНК, контролирую-
щий экспрессию гена или группы генов путем взаимо-
действия с белком-репрессором.
Оперон: функциональная единица генома прокариот
(участок ДНК), объединенная под общим промотором.
Оптимум pH: диапазон значений pH, в котором фер-
мент демонстрирует максимальную каталитическую
активность.
Оптическая активность: способность вещества вра-
щать плоскость поляризации плоскополяризованного
света.
Органеллы: ограниченные мембранами структуры
эукариотических клеток, содержащие ферменты и
другие компоненты со специфическими функциями.
Ортологи: гены из разных организмов с выраженным
сходством последовательностей и функциональных
связей.
Осмос: диффузия вещества, обычно воды (в общем
случае — растворителя) через полупроницаемую мем-
брану из раствора с меньшей концентрацией раство-
ренного вещества в раствор с большей концентрацией
растворенного вещества.
Осмотическое давление: давление на полупроницае-
мую мембрану, обусловленное диффузией раствори-
теля из раствора с меньшей концентрацией растворен-
ного вещества в раствор с большей концентрацией.
Основная догма молекулярной биологии: основопо-
лагающий принцип молекулярной биологии, заклю-
чающийся в том, что поток генетической информации
направлен от ДНК через РНК к белку.
Основное (невозбужденное) состояние (в отличие
от возбужденного состояния): устойчивое состояние
атома или молекулы.
Основной обмен: один из показателей интенсивности
(скорости) обмена веществ и энергии в организме в
условиях теплового комфорта натощак (в состоянии
полного покоя спустя длительное время после приема
пищи).
Открытая рамка считывания: непрерывный ряд не-
перекрывающихся нуклеотидных кодонов в молекуле
ДНК или РНК, не содержащий стоп-кодона.
Открытая система: система, обменивающаяся с окру-
жающей средой материей и энергией.
Отношение действующих масс (Q): Для реакции
аК + ЬВ * сС + dD
Q=[C]<[Dr/[Ap[Bp.
Отрицательная кооперативность: свойство некото-
рых субъединичных ферментов и белков, заключаю-
щееся в том, что связывание лиганда или субстрата с
одной субъединицей препятствует его связыванию с
другой субъединицей.
Отрицательная обратная связь: способ регуляции
биохимического пути, при котором продукт реакции
ингибирует протекание более ранних стадий пути.
Отстающая нить: нить ДНК, которая в процессе ре-
пликации синтезируется в противоположном направ-
лении к направлению движения репликативной вилки.
Ошибочное спаривание: образование пар между пара-
ми оснований, отличное от уотсон-криковских пар.
п
Палиндром: участок двупитевой ДНК, в котором по-
следовательности оснований в двух нитях симметрич-
ны относительно некоторой оси.
Парадигма: совокупность фундаментальных научных
представлений и терминов, принимаемая научным со-
обществом; в биохимии — экспериментальная модель
или пример.
Патогенный организм: организм, вызывающий за-
болевание.
Пентоза: простой сахар, углеродный скелет которого
состоит из пяти атомов углерода.
Пентозофосфатный (гексозомонофосфатный) путь:
метаболический путь у большинства организмов,
заключающийся во взаимных превращениях гексоз и
пентоз; служит источником восстановительных экви-
валентов (NADPH) и пентоз для реакций биосинтеза;
начинается от глюкозо-6-фосфата и проходит через
образование 6-фосфоглюконата.
Пептид: две или более аминокислоты, соединенные
пептидной связью.
Пептидазы: ферменты, гидролизующие пептидную
связь.
Пептидилтрансфераза: рибозим, синтезирующий
пептидную связь в белках; часть рРНК большой субъ-
единицы рибосомы.
Пептидная связь: амидная связь, образующаяся
между а-аминогруппой одной аминокислоты и
а-карбоксильной группой другой аминокислоты с
удалением молекулы воды.
Словарь терминов [363]
Пептидное картирование: характерная двумерная
картина (на бумаге или в геле) при разделении смеси
пептидов, образовавшихся в результате частичного
гидролиза белка.
Пептидогликан: основной компонент клеточной стен-
ки бактерий; обычно состоит из параллельных цепей
гетерополисахаридов, удерживаемых поперечными
сшивками из коротких пептидных фрагментов.
Первичная структура: последовательность звеньев
в биополимере.
Первичный транскрипт: образующаяся в результате
транскрипции незрелая РНК, подвергающаяся далее
посттранскрипционному процессингу.
Первое начало термодинамики: закон, утверждаю-
щий, что в любом процессе общая энергия системы
остается постоянной.
Передача сигнала (сигнальная трансдукция): усиление
внешнего сигнала (химического, механического или
электрического) и его превращение в клеточный ответ.
Перенос электронов: перемещение электронов от
донора к акцептору, например, от субстратов на кис-
лород через переносчиков в дыхательной (электрон-
транспортной) цепи.
Переносчик электронов: белок (флавопротеин или
цитохром), способный обратимо принимать и от-
давать электроны; участвует в переносе электронов от
органических питательных веществ на кислород или
другие конечные акцепторы.
Переносчики (транспортеры, пермеазы): пронизы-
вающие плазматическую мембрану белки, которые
переносят через нее специфические питательные
вещества, метаболиты, ионы или другие белки.
Перепрограммирование трансляции: программиру-
емое изменение рамки считывания в процессе транс-
ляции мРНК на рибосомах.
Переходное состояние: активированное состояние
молекулы, претерпевшей частичные превращения
в химической реакции; высшая точка координаты
реакции.
Периферические белки: белки, слабо или обратимо
связанные с плазматической мембраной водородными
связями или электростатическими взаимодействия-
ми; обычно при отделении от мембраны растворяются
в водной среде.
Пероксисома: связанная с мембраной органелла эу-
кариотической клетки; содержит ферменты, образую-
щие и разрушающие пероксиды.
Петли ДНК: в результате взаимодействия между бел-
ками, связанными в удаленных друг от друга участках
ДНК, промежуточные участки ДНК способны образо-
вывать петли.
Пираноза: простой сахар, содержащий шестичленное
пирановое кольцо.
Пиридиннуклеотид: нуклеотидный кофермент, со-
держащий производное пиридина никотинамид; NAD
или NADP.
Пиридоксальфосфат: кофермент, содержащий вита-
мин пиридоксин (витамин В6); участвует в реакциях с
переносом аминогрупп.
Пиримидин: азотсодержащее гетероциклическое ос-
нование, содержащееся в нуклеотидах и нуклеиновых
кислотах.
Пиримидиновый димер: димер, образующийся в
результате ковалентного взаимодействия двух со-
седних остатков пиримидина в ДПК; возникает под
действием УФ-излучения; наиболее распространены
тиминовые димеры.
Пищеварение: ферментативное расщепление основ-
ных компонентов пищи до простых веществ в желу-
дочно-кишечном тракте.
Плазмалоген: фосфолипид, в котором в положении
С1 остатка глицерина находится остаток алифати-
ческого спирта, присоединенный простой эфирной
связью.
Плазматическая мембрана: внешняя мембрана, окру-
жающая цитоплазму клетки.
Плазмида: внехромосомная, способная реплициро-
ваться автономно небольшая кольцевая молекула
ДНК; используется в генной инженерии.
Пластида: самореплицирующаяся органелла расти-
тельной клетки, может дифференцировать в хлоро-
пласт.
Плектонемическая спираль: структура биополимера,
в которой одна нить обвивается вокруг другой про-
стым и регулярным образом.
Плотность супервитков: количество дополнительных
витков в спиральной молекуле (такой как ДНК) по
сравнению с количеством витков в релаксированной
молекуле.
0-Поворот: участок белковой последовательности,
образованный четырьмя аминокислотными остатка-
ми, заставляющий полипептидную цепь повернуть в
обратном направлении; элемент вторичной структуры
белка.
Поли(А)-«хвост»: последовательность остатков аде-
нозина на З'-конце многих молекул мРНК в эукарио-
тических клетках и некоторых бактериях.
Поликлональные антитела: гетерогенный пул анти-
тел, образованных разными В-лимфоцитами в ответ
на действие антигена; разные антитела в пуле распоз-
нают разные части молекулы антигена (эпитопы).
13641 Словарь терминов
Полилинкер: короткая, часто синтетическим образом
полученная последовательность ДНК, содержащая
сайты узнавания для нескольких рестрикционных
эндонуклеаз.
Полимеразная цепная реакция (ПЦР): лаборатор-
ный метод, заключающийся в многократной ампли-
фикации специфической последовательности ДНК.
Полиморфизм: существование различных, но
полностью функциональных вариантов белка/гена в
популяции организмов.
Полиморфизм длины рестрикционных фрагментов:
вариации длины рестрикционных фрагментов, содер-
жащих специфические последовательности ДНК, среди
индивидуумов в популяции; результат редких измене-
ний в ДНК, создающих или уничтожающих узнаваемые
рестриктазами специфические последовательности.
Полиморфизм последовательности: любые вариации
геномной последовательности (замены нуклеотидов,
инсерции, делении, перестановки), позволяющие вы-
явить одну группы индивидуумов в популяции или
различить организмы разных видов.
Полиморфизм одного нуклеотида (SNP): замена
пары нуклеотидов, позволяющая отличить одну особь
от другой или выделить группу особей в популяции.
Полиненасыщенная жирная кислота: жирная кис-
лота, имеющая несколько двойных связей, обычно
несопряженных друг с другом.
Полинуклеотид: последовательность ковалентно свя-
занных нуклеотидов, в которой З'-гидроксил пентозы
одного нуклеотидного остатка связана фосфодиэ-
фирной связью с 5'-гидроксилом пентозы соседнего
остатка.
Полипептид: длинная последовательность аминокис-
лотных остатков (обычно до 10 000 Да), связанных
пептидными связями.
Полисахарид: линейный или разветвленный поли-
мер, состоящий из моносахаридных звеньев, связан-
ных гликозидными связями.
Полисома (полирибосома): комплекс молекулы
мРНК с двумя или большим числом рибосом.
Полиферментная система (комплекс): группа функ-
ционально взаимосвязанных ферментов в метаболи-
ческом пути.
Полицистронная мРНК: матрица для трансляции
двух или большего числа белков.
Положительная кооперативность: свойство субъе-
диниц белковой молекулы, заключающееся в том, что
связывание первой молекулы лиганда или субстрата
с одной субъединицей облегчает связывание следую-
щих молекул с другими субъединицами.
Полярная молекула: гидрофильное вещество, хорошо
растворяется в воде.
Полярность: (1) в химии — неравномерное распре-
деление электронов в молекуле; полярные молекулы
обычно хорошо растворяются в воде; (2) в молекуляр-
ной биологии — направление от 5'-конца нуклеиновой
кислоты к З'-концу и наоборот.
Порфирин: комплекс азотистых соединений, содержа-
щих четыре замещенных пиррола, объединенных в коль-
цо посредством ковалентных связей; часто в централь-
ной позиции комплекса связывается атом металла.
Порфирия: наследственное заболевание, связанное
с дефицитом одного или нескольких ферментов, уча-
ствующих в синтезе порфиринов.
Порядок зацепления: показатель, определяющий,
сколько раз одна замкнутая кольцевая нить ДНК
пересекается с другой нитью; топологический пара-
метр, отражающий степень скрученности ДНК.
Посттранскрипционный процессинг: ферментатив-
ные модификации первичного транскрипта, приво-
дящие к образованию функциональных мРНК, тРНК,
рРНК и многих других видов РНК.
Посттрансляционная модификация: ферментативная
модификация полипептидной цепи после трансляции
мРНК.
Потенциал переноса групп: характеризует способ-
ность соединения отдавать активированную группу
(например, фосфатную или ацильную); обычно это
стандартная свободная энергия гидролиза.
Потенциал фосфорилирования (ДСр): реальное из-
менение свободной энергии гидролиза АТР в нестан-
дартных условиях, что обычно для клетки.
Правовращающий изомер: стереоизомер, враща-
ющий плоскость плоскополяризованного света по
часовой стрелке.
Праймаза: фермент, катализирующий образование
РНК-олигонуклеотидов, которые используются
ДНК-полимеразами в качестве праймеров.
Праймер: короткий олигомер (например, из сахарных
или нуклеотидных остатков), к которому фермент до-
страивает следующие мономерные звенья.
Праймирование: (1) фосфорилирование амино-
кислотного остатка, который становится центром
связывания и контрольной точкой для фосфорили-
рования других остатков в том же белке; (2) синтез
короткого нуклеотида, к которому ДНК-полимеразы
могут добавлять дополнительные нуклеотиды при
репликации ДНК.
Праймосома: ферментный комплекс, синтезирующий
праймеры для синтеза отстающей нити ДНК при
репликации.
Словарь терминов [365]
Проекционные формулы (формулы Фишера):
способ изображения молекул, позволяющий отразить
конфигурацию заместителей при хиральных центрах.
Проекционные формулы Хеуорса: способ изображе-
ния циклических химических структур, отражающий
конфигурацию всех заместителей; часто применяется
для изображения сахаров.
Прокариоты: исторически принятый термин для обо-
значения всех представителей царств бактерий и архей,
но, поскольку различия между бактериями (прежнее
название — эубактерии) и археями достаточно велико,
данный термин не имеет большого смысла. Достаточно
часто прокариотами называют только бактерии, что
может вызывать путаницу. Кроме того, само слово
«прокариот» отчасти указывает на то, что они были
предками эукариот, что некорректно.
Промежуточное соединение (интермедиат): любое
химическое соединение с ограниченным временем
жизни в реакционном процессе.
Промежуточные структуры Холлидея: образуются
на промежуточной стадии генетической рекомбина-
ции, когда две двунитевые молекулы ДНК удержи-
ваются вместе благодаря взаимодействиям между
отдельными нитями в каждой молекуле.
Промежуточный метаболизм: несколько фермента-
тивных реакций, извлекающих химическую энергию
из питательных веществ и использующих их для
синтеза и сборки клеточных компонентов.
Промотор: последовательность ДНК, с которой
может связываться РНК-полимераза, что инициирует
процесс транскрипции.
Простагландины: жирорастворимые гормонопо-
добные регуляторные молекулы, образующиеся из
арахидоновой кислоты и других полиненасыщенных
жирных кислот.
Простая диффузия: перемещение молекул раство-
ренного вещества через мембрану в область с более
низкой концентрацией без помощи белкового пере-
носчика.
Простетическая группа: ион металла или органиче-
ское соединение (но не аминокислота), ковалентно
связанное с белком и необходимое для проявления
его активности.
Протеасома: сложный ферментный комплекс,
осуществляющий расщепление поврежденных или
ненужных клеточных белков.
Протеинкиназы: ферменты, которые переносят кон-
цевую фосфорильную группу АТР или другого нукле-
озидтрифосфата на боковую цепь остатка Ser, Thr, Туг,
Asp или His в белке-мишени и тем самым регулируют
активность или другие свойства этого белка.
Протеогликан: гибридная макромолекула, состоящая
из гетерополисахарида и полипептида; основной ком-
понент этой молекулы — полисахарид.
Протеом: полный набор белков, экспрессирующихся
в данной клетке, или полный набор белков, который
может экспрессироваться данным геномом.
Протеомика: в широком смысле — изучение белково-
го содержания клетки или организма.
Протомер: общий термин, описывающий любую по-
вторяющуюся единицу одной или нескольких прочно
связанных белковых субъединиц в более крупной
белковой структуре. Субъединицы мультисубъеди-
ничного протомера могут быть как идентичными, так
и разными.
Протондвижущая сила: электрохимический потен-
циал, соответствующий трансмембранному градиенту
концентрации протонов; движущая сила в синтезе
АТР при окислительном фосфорилировании и фото-
фосфорилировании.
Протоонкоген: клеточный ген, обычно кодирующий
регуляторный белок, который в результате мутации
может превратиться в онкоген.
Протоплазма: общий термин, описывающий все со-
держимое живой клетки.
Прохиральная молекула: симметричная молекула,
которая может реагировать несимметричным образом
с ферментом, имеющим хиральный центр, в результа-
те чего образуется хиральный продукт.
Процессивность: способность ферментов, катализи-
рующих синтез биологических полимеров, присоеди-
нять к полимеру множество мономерных звеньев без
диссоциации из комплекса с субстратом.
Пурин: азотсодержащее гетероциклическое основание
в нуклеотидах и нуклеиновых кислотах; состоит из
слитых пиримидинового и имидазольного колец.
Пуромицин: антибиотик, нарушающий синтез по-
липептидов; встраивается в растущую полипептид-
ную цепь и вызывает преждевременную остановку
синтеза.
Р
Равновесие: состояние системы, в котором не про-
исходит никаких суммарных изменений; свободная
энергия такой системы минимальная.
Радикал (свободный радикал): атом или группа
атомов с неспаренным электроном.
Радиоактивный изотоп: изотоп элемента с нестабиль-
ным ядром, который способен испускать ионизирую-
щую радиацию.
13b() | Словарь терминов
Радиоиммуноанализ (РИА): чувствительный
количественный метод определения биологически
активных веществ, основанный на конкурентном
связывании со специфическими антителами.
Разобщающий агент: например, 2,4-динитрофенол,
вещество, разделяющее процессы фосфорилирования
ADP и переноса электронов.
Рамка считывания: непрерывная, неперекрывающая-
ся последовательность тринуклеотидных кодонов
ДНК или РНК.
Рацемическая смесь (рацемат): эквимолярная
смесь D- и L-стереоизомеров оптически активного
вещества.
Реактивные формы кислорода (РФК): высоко
реакционноспособные продукты частичного восста-
новления О2, включая пероксид водорода (Н2О2), су-
пероксидный радикал (-О2) и гидроксильный радикал
(•ОН), образующиеся в качестве побочных продуктов
при окислительном фосфорилировании.
Регулон: группа координированно регулируемых
генов или оперонов, иногда находящихся в удаленных
друг от друга участках хромосомы или генома.
Регуляторная последовательность: последователь-
ность ДНК, участвующая в регуляции экспрессии
гена, например промотор или оператор.
Регуляторный ген: ген, продукт которого регулирует
экспрессию другого гена, например ген белка-репрес-
сора.
Регуляторный фермент: фермент, выполняющий
регуляторную функцию благодаря своей способности
изменять каталитическую активность в результате
аллостерических взаимодействий или ковалентных
модификаций.
Редактирование РНК: посттранскрипционные мо-
дификации мРНК, в результате которых изменяется
смысл некоторых кодонов.
Рекомбинантная ДНК: ДНК, образующаяся путем
соединения генов в новой комбинации.
Рекомбинантная репарация: рекомбинация, на-
правленная на репарацию разрывов или перекрест-
ных сшивок в ДНК, особенно в участках остановки
репликации.
Рекомбинация: любой ферментативный процесс, в
ходе которого происходит расщепление и линейная
перестройка фрагментов хромосом.
Релаксированная ДНК: любая ДНК, существующая
в максимально стабильной и ненапряженной форме,
обычно в В-форме, почти при любых внутриклеточ-
ных условиях.
Ренатурация: свертывание развернутого (денатури-
рованного) глобулярного белка с восстановлением его
нативной структуры и функций.
Рентгеновская кристаллография: определение
трехмерной структуры кристаллов методом рентгено-
структурного анализа.
Репарация: исправление ошибок, допущенных при
синтезе информационных биополимеров, путем уда-
ления неправильных звеньев уже после того, как они
были включены в полимерную цепь.
Репарация ошибочного спаривания: ферментативное
удаление аномально спаренных оснований в ДНК.
Репликазный комплекс: комплекс ферментов и спе-
циализированных белков, осуществляющий реплика-
цию ДНК в клетках.
Репликативная вилка: Y-образная структура, образу-
ющаяся в участке репликации ДНК.
Репликативная форма: полноразмерная структурная
форма вирусной ДНК, образующаяся на промежуточ-
ной стадии репликации некоторых вирусов.
Репликация: синтез дочерних молекул нуклеиновой
кислоты, идентичных родительским молекулам.
Реплисома: полиферментный комплекс, участвую-
щий в синтезе ДНК в репликативной вилке.
Репрессибельный фермент: бактериальный фермент,
синтез которого ингибируется продуктом катализиру-
емой им реакции.
Репрессия: снижение экспрессии гена в ответ на из-
менение активности регуляторного белка.
Репрессор трансляции: репрессор, который связыва-
ется с мРНК и блокирует трансляцию.
Репрессор: белок, который связывается с регулятор-
ной последовательностью или оператором гена, тем
самым блокируя транскрипцию.
Респонсивный элемент: участок ДНК несколько
выше начала гена, который связывает специфический
белок, влияющий на скорость транскрипции гена.
Рестрикционные эндонуклеазы (рестриктазы):
сайт-специфичиые эндодезоксирибонуклеазы, рас-
щепляющие обе нити ДНК вблизи специфического
сайта, узнаваемого ферментом; важный инструмент в
генетической инженерии.
Рестрикционный фрагмент: участок ДНК, вырезае-
мый рестриктазой из более длинной последователь-
ности ДНК.
Ретровирус: РНК-вирус, содержащий обратную
транскриптазу.
Реутилизационный путь: синтез биомолекул, например
нуклеотидов, из промежуточных продуктов расщепле-
ния этих молекул; рецикл в отличие от синтеза de novo.
Словарь терминов 136 / |
Рецептор гормона: белок внутри клетки-мишени
или на ее поверхности, который связывает специфи-
ческий гормон и инициирует клеточный ответ.
Рецепторные тирозинкиназы: большое семейство
белков плазматической мембраны, состоящих из
лигандсвязывающего внеклеточного домена, един-
ственной трансмембранной спирали и цитоплазма-
тического домена с тирозинкиназной активностью,
контролируемой внеклеточным лигандом.
Рецепторы, активируемые пролифераторами
пероксисом (PPAR): семейство ядерных факторов
транскрипции, активируемых липидными лиганда-
ми, которые изменяют экспрессию специфических
генов, включая гены ферментов, участвующих в
синтезе и расщеплении липидов.
Рецепторы, сопряженные с G-белком (GPCR):
большое семейство рецепторных мембранных
белков, состоящих из семи трансмембранных спи-
ральных сегментов, которые образуют ассоцииаты
с G-белками для передачи внеклеточных сигналов
с целью изменения клеточного метаболизма; другое
название — серпентиновые или семиспиральпые
рецепторы.
Рибозимы: молекулы РНК, обладающие каталитиче-
ской активностью.
Рибонуклеаза: нуклеаза, катализирующая гидролиз
некоторых межнуклеотидных связей в РНК.
Рибонуклеотид: нуклеотид, содержащий в качестве
пентозы D-рибозу.
Рибопереключатель: структурный элемент мРНК,
который связывается со специфическим лигандом и
влияет на трансляцию или процессинг мРНК.
Рибосома: надмолекулярный комплекс рРНК
и белков; диаметр от 18 до 22 нм; место синтеза
белка.
Рибосомная РНК (рРНК): тип молекул РНК,
являющихся компонентом рибосом.
Рилизинг-гормоны (либерины): гормоны гипота-
ламуса, способствующие высвобождению других
гормонов гипофизом.
Рилизинг-факторы (факторы терминации): белко-
вые факторы, находящиеся в цитозоле и участвую-
щие в высвобождении готовой полипептидной цепи
из рибосомы.
РНК (рибонуклеиновая кислота): полирибонуклео-
тид со специфической последовательностью, звенья
которого связаны 3',5'-фосфодиэфирными связями.
РНК-полимераза: фермент, катализирующий об-
разование РНК из рибонуклеозид-5'-трифосфатов на
основании матрицы ДНК или РНК.
С
Сайт-направленный мутагенез: генно-инженерный
метод, позволяющий специфическим образом изме-
нять последовательность гена.
Сайт-специфическая рекомбинация: тип генетиче-
ской рекомбинации, затрагивающей только специфи-
ческие последовательности.
Саркомер: функциональная структурная единица со-
кратительного аппарата мышц.
Сателлитная ДНК: нетранслируемый участок эукари-
отической хромосомы, содержащий большое количе-
ство повторов; чаще всего встречается в центромере;
функция неизвестна.
Саузерн-блот: процедура, позволяющая выявить один
или несколько специфических фрагментов ДНК путем
гибридизации с комплементарным меченым зондом.
Сахарный диабет: группа метаболических заболева-
ний, связанных с недостаточным синтезом инсулина
или нарушением его активности; проявляется в на-
рушении транспорта глюкозы из крови в клетки.
Сведберг (S): единица измерения скорости осажде-
ния частицы при центрифугировании.
Суперспираль: спиральная молекула, закрученная
вокруг самой себя.
Суперспиральная ДНК: ДНК, обернутая вокруг
самой себя из-за напряжения, вызванного недоста-
точным или избыточным закручиванием двойной
спирали (по сравнению с В-формой).
Световые реакции: реакции фотосинтеза, которые нс
могут протекать в темноте.
Свободная энергия (G): часть общей энергии систе-
мы, способная превращаться в работу при постоянных
температуре и давлении.
Сдвиг рамки считывания: мутация (инсерция или
деления) нескольких нуклеотидов, изменяющая счи-
тывание кодонов в процессе синтеза белка; начиная
с мутированного кодона, считывается искаженная
аминокислотная последовательность.
Селектины: большое семейство мембранных белков,
прочно и специфично связывающих олигосахариды
на поверхности других клеток, что позволяет переда-
вать сигналы через плазматическую мембрану.
Серповидно-клеточная анемия: заболевание че-
ловека, характеризующееся наличием дефектного
гемоглобина у гомозиготных индивидов с мутантной
аллелью, кодирующей (3-цепь гемоглобина.
Серповидно-клеточный признак: образование эри-
троцитов серповидной формы у гетерозиготных ин-
дивидов с одной мутантной аллелью, ответственной
за серповидно-клеточную анемию; проявляется при
понижении содержания кислорода в воздухе.
13681 Словарь терминов
Сиалоадгезин: лектин на внешней поверхности клет-
ки, который связывается с углеводными лигандами,
имеющими на конце остаток сиаловой кислоты; пред-
ставитель лектинов семейства siglec.
Сигнальная последовательность: аминокислотная
последовательность, расположенная чаще всего на
N-конце, которая указывает назначение или локализа-
цию вновь синтезированного клеточного белка.
Силы Лондона: слабые (недолговечные) силы притя-
жения между молекулами, индуцирующими образо-
вание молекулярных диполей.
Симбионты: два или несколько взаимозависимых орга-
низмов, обычно живущих в непосредственном контакте.
Симпорт: одновременный перенос растворенных ве-
ществ через мембрану в одном и том же направлении.
Синтазы: ферменты, катализирующие реакции кон-
денсации, протекающие без использования нуклео-
зидтрифосфатов в качестве источников энергии.
Синтения: сохранение порядка расположения генов в
хромосомах различных видов организмов.
Синтетазы: ферменты, катализирующие реакции кон-
денсации, протекающие за счет энергии расщепления
АТР или других нуклеозидтрифосфатов.
Системная биология: изучение сложных биохими-
ческих систем, включая все или некоторые функции
клеточных макромолекул (РНК, ДНК, белков).
Складчатый лист: упаковка полипептидных цепей с
(3-конформацией в виде растянутой гармошки, удер-
живаемая водородными связями.
Скрамблазы: мембранные белки, катализирующие
перемещение фосфолипидов в двуслойной мембране,
приводящее к равномерному распределению липидов
между внешним и внутренним слоем.
Следовый элемент (микроэлемент): химический
элемент, необходимый организму лишь в следовых
количествах.
Слитый белок (белок слияния): (1) представитель
семейства белков, облегчающих слияние мембран;
(2) белковый продукт гена, полученного путем соеди-
нения двух разных генов или частей генов.
Соматическая клетка: любая клетка тела, кроме по-
ловой клетки.
Сопряженные реакции: две химические реакции,
имеющие общее промежуточное соединение и способ-
ные обмениваться энергией.
Спаривание оснований: образование водородных свя-
зей между нуклеотидами в цепях нуклеиновых кислот;
например, А образует пару с Т или U, G — пару с С.
Спектроскопия ядерного магнитного резонанса
(ЯМР-спектроскопия): физико-химический метод
анализа, использующий квантово-механические
свойства атомных ядер для изучения структуры и
динамики молекул.
Специфический кислотно-основной катализ: кислот-
ный или основной катализ с участием компонентов мо-
лекулы воды (гидроксид-иона или иона гидроксония).
Специфичность: способность фермента (рецептора)
различать конкурирующие субстраты (лиганды).
а-Спираль: спиральная конформация полипептидной
цепи (обычно закрученная вправо), отличающаяся
максимальным числом водородных связей; одна из са-
мых распространенных вторичных структур в белках.
Спиртовое брожение: анаэробное превращение глю-
козы в этиловый спирт в процессе гликолиза.
Сплайсинг: удаление интронов и сшивание экзонов
первичного РНК-транскрипта.
Сплайсосома: комплекс РНК и белков, осуществляю-
щий сплайсинг мРНК в эукариотических клетках.
Стандартное изменение свободной энергии (AG0):
изменение свободной энергии в реакции, происходя-
щей при стандартных условиях: температура 298 К,
давление 1 атм (101,3 кПа), концентрация всех рас-
творенных веществ 1 моль/л; AG"0 соответствует из-
менению свободной энергии при pH 7,0 в 55,5 М воде.
Стандартный восстановительный потенциал
электродвижущая сила, создаваемая на электроде в
1 М раствора восстанавливающего агента и его окис-
ленной формы при 25 °C и pH 7,0; мера способности
восстановителя отдавать электроны.
Стартовый кодон: AUG (иногда GUG или еще
реже UUG в бактериях и археях); кодирует первую
аминокислоту полипептидной последовательности —
N-формилметиопип в бактериях, метионин в археях и
эукариотах.
Стационарное состояние: состояние системы, при
котором концентрации всех компонентов постоянные.
Стволовые клетки: самообновляющиеся клетки кост-
ного мозга, из которых в результате дифференциров-
ки образуются такие клетки крови, как эритроциты и
лимфоциты.
Стереоизомеры: соединения одного и того же состава
и с одинаковым порядком межатомных связей, но с
разным пространственным расположением атомов.
Стерины (стеролы): липиды, содержащие стероид-
ную структуру.
Стимулирующий G-белок (Gs): тримерный ре-
гуляторный GTP-связывающий белок, который
активируется в результате связывания с рецептором
на плазматической мембране и стимулирует нахо-
дящиеся поблизости мембранные ферменты, такие
как аденилатциклаза; его действие'противоположно
действию Gj.
Словарь терминов 13691
Стоп-кодон (терминирующий кодон): UAA, UAG и
UGA. При синтезе белка служит сигналом для окон-
чания синтеза полипептидной цепи.
Строма: область внутри хлоропласта, окружающая
тилакоиды.
Структурный ген: ген, кодирующий белок или РНК
(в отличие от регуляторного гена).
Субстрат: специфическое соединение, превращение
которого катализирует фермент.
Субстратное туннелирование: перемещение промежу-
точных соединений ферментативного процесса внутри
белкового комплекса напрямую от активного центра
одного фермента в активный центр другого фермента,
катализирующего бедующую стадию процесса.
Субстратное фосфорилирование: фосфорилирова-
ние ADP или другого нуклеозид-5’-дифосфата, сопря-
женное с дегидрированием органического субстрата,
независимо от электрон-транспортной цени.
Суицидный ингибитор: сравнительно малоактивная
молекула, которая связывается с активным центром
фермента, переходит в активную форму и необратимо
инактивирует фермент.
Супрессорная мутация: мутация, которая частично
или полностью восстанавливает функцию, потерян-
ную в результате предыдущей мутации, но проис-
ходит в ином участке, нежели исходная мутация (т. с.
в отличие от обратной мутации, восстанавливает не
генотип, а фенотип).
Сфинголипид: амфифильный липид — сфингозин с
присоединенными к нему остатками жирных кислот и
полярного спирта.
Т
Теломера: специализированная структура на концах
линейных эукариотических хромосом.
Терминальная трансфераза: фермент, катализирую-
щий присоединение нуклеотидных звеньев опреде-
ленного типа к З'-концу цени ДНК.
Терминационная последовательность: последова-
тельность ДНК на конце транскрипционной единицы,
обозначающая окончание транскрипции.
Термогенез: выделение тепла при мышечной актив-
ности, в несопряженном окислительном фосфорили-
ровании или в холостом метаболическом цикле.
Термогенин (разобщающий белок-1): белок вну-
тренней мембраны митохондрий в бурой жировой
ткани, который позволяет протонам проходить через
мембрану, предотвращает использование протонов
для синтеза АТР и рассеивает энергию от окисления
субстратов в виде тепла.
Терпены: органические углеводороды или их произво-
дные, состоящие из повторяющихся изопреновых зве-
ньев; придают вкус и запах растительным продуктам.
Тетрагидробиоптерин: кофермент; восстановленная
форма биоптерина.
Тетрагидрофолат: кофермент; восстановленная, ак-
тивная форма фолата.
Тиаминпирофосфат: кофермент, активная форма
витамина Bf, участвует в реакциях переноса альдегид-
ных групп.
Тилакоиды: замкнутые цистерны или диски, образо-
ванные содержащей пигмент внутренней мембраной
хлоропластов.
Тиоэфир: сложный эфир карбоновой кислоты с тио-
лом или меркаптаном.
Токоферол: форма витамина Е.
Токсины: белки, синтезируемые некоторыми организ-
мами, и токсичные для других организмов.
Топоизомеразы: ферменты, вводящие положитель-
ные или отрицательные витки в замкнутый кольцевой
ДНК-дуплекс.
Топоизомеры: формы кольцевой молекулы ДНК, раз-
личающиеся только порядком зацепления.
Точка начала репликации (он): участок последова-
тельности ДНК, в котором начинается репликация.
Трансаминирование: ферментативный перенос ами-
ногруппы от а-аминокислоты на а-кетокислоту.
Трансгенный организм: организм, в геном которого
с помощью методов рекомбинации включены гены
другого организма.
Трансдукция: (1) превращение одной формы энер-
гии или информации в другую форму; (2) перенос
генетической информации из одной клетки в другую с
помощью вирусного вектора.
Транскриптом: полный набор РНК-транскриптов,
присутствующих в данной клетке или ткани в опреде-
ленных условиях.
Транскрипционный фактор: эукариотический белок,
влияющий на регуляцию и инициацию транскрипции
гена путем связывания с регуляторной последователь-
ностью гена и взаимодействия с РНК-полимеразой
или другими транскрипционными факторами.
Транскрипция: ферментативный процесс, в результа-
те которого генетическая информация, заключенная в
однонитевой ДНК, используется для синтеза компле-
ментарной последовательности мРНК.
Транслоказа: (1) фермент, катализирующий транс-
порт веществ через мембрану; (2) фермент, участву-
ющий в перемещении макромолекул, например в
перемещении рибосом вдоль мРНК.
13/ о | Словарь терминов
Транспирация: испарение воды, прошедшей от кор-
ней растений через сосудистую систему в листья.
Транспозиция: перемещение гена или группы генов
из одного участка генома в другой.
Транспозон: участок ДНК, который может переме-
щаться из одной позиции в геноме в другую.
Транспортная РНК (тРНК): молекулы РНК с массой
от 25 000 до 35 000, каждая из которых связывает одну
специфическую аминокислоту на первом этапе био-
синтеза белка.
Трансформация: введение в клетку чужеродной
ДНК, в результате чего клетка приобретает новый
фенотип.
Триацилглицерины (нейтральные жиры): сложные
эфиры глицерина и трех молекул жирных кислот.
Триоза: простой сахар, остов которого состоит из трех
атомов углерода.
Тромбоксаны: производные арахидоновой кислоты,
участвующие в агрегации тромбоцитов при свертыва-
нии крови.
Тромбоциты: небольшие безъядерные клетки крови,
инициирующие процесс свертывания; образуются из
клеток костного мозга, называемых мегакариоцитами.
Тропный гормон (тропин): пептидный гормон, за-
ставляющий специфическую железу секретировать
ее гормон; например, тиреотропин, синтезируемый
гипофизом, стимулирует секрецию тироксина щито-
видной железой.
Туннелирование: прямой перенос продукта реакции
из активного центра одного фермента в активный
центр другого фермента, катализирующего следую-
щую стадию метаболического пути.
У
Убиквитин: небольшой консервативный белок, свя-
зывающийся с внутриклеточными белками, которые
должны быть направлены на уничтожение в протеа-
сомы. Несколько молекул убиквитина присоединя-
ются к остаткам лизина в белке-мишени ковалентной
связью с помощью специфического фермента.
Удельная активность фермента: количество субстра-
та в микромолях, превращенного ферментом в минуту
в расчете на миллиграмм белка при 25 °C; мера чисто-
ты ферментного препарата.
Удельная теплоемкость: количество энергии (в джо-
улях или калориях), которое нужно затратить, чтобы
повысить температуру 1 г чистого вещества на 1 °C.
Удельное вращение: поворот (в градусах) плоскости
плоскополяризованного света при пропускании через
раствор оптически активного вещества при заданной
длине светового пути при 25 °C; обычно свет с длиной
волны D-линии натрия.
Ультрафиолетовое (УФ) излучение: электромагнит-
ное излучение с длиной волны в диапазоне от 200 до
400 нм.
Унипорт: транспортная система, переносящая лишь
один тип растворенных веществ (в отличие от сим-
порта).
Уреотелический организм: организм, выводящий из-
быток азота в форме мочевины.
Урикотелический организм: организм, выводящий
избыток азота в форме мочевой кислоты (уратов).
Уходящая группа: удаляемая или вытесняемая часть
молекулы в реакции элиминирования или замещения.
Ф
Факторы роста: белки или другие молекулы, дей-
ствующие на клетку извне и стимулирующие ее рост
и деление.
Факторы элонгации: (1) белки, участвующие в транс-
крипции эукариотической ДНК на стадии элонгации;
(2) белки, необходимые для удлинения полипептид-
ной цепи на рибосомах.
Факультативные аэробы/анаэробы: клетки, способ-
ные жить в присутствии/отсутствие кислорода.
Фенотип: видимые признаки организма.
Фермент: белок или РНК, катализирующий специ-
фическую химическую реакцию; фермент не смещает
равновесие реакции, но увеличивает ее скорость,
предлагая новый реакционный путь с более низкой
энергией активации.
Ферментативный каскад: серия реакций, вовлечен-
ных в процесс регуляции, в которых один фермент
активирует другой (часто путем фосфорилирования),
второй активирует третий и т. д. В результате каскада
реакций происходит значительное усиление исходно-
го сигнала.
Ферменты со скрытой функцией: ферменты, вы-
полняющие две различные функции, из которых хотя
бы одна каталитическая; другая функция может быть
регуляторной, каталитической или структурной.
Ферредоксин: железосерный белок (2Fe-2S) хлоро-
пластов, переносящий электроны от железосерных
центров, ассоциированных с фотосистемой I, на
NADP+ в процессе фотофосфорилирования.
Фибриллярные белки: нерастворимые белки, вы-
полняющие защитную или структурную функцию;
состоят из полипептидных цепей, имеющих характер-
ную вторичную структуру.
Словарь терминов [3/11
Фибробласт: клетка соединительной ткани, секрети-
рующая такие белки, как коллаген.
Фиксация азота: превращение атмосферного азота
в восстановленную, биологически доступную форму
под действием азотфиксирующих организмов.
Фиксация углерода: первая реакция при фотосинте-
зе, катализируемая рубиско или другими карбоксила-
зами, в результате которой атмосферный СО2 включа-
ется в состав органического соединения.
Флавинадениндинуклеотид (ФАД; FAD): кофермент
некоторых окислительно-восстановительных фермен-
тов; содержит рибофлавин.
Флавинзависимые дегидрогеназы: дегидрогеназы, ис-
пользующие в качестве кофакторов FMN или FAD.
Флавинмононуклеотид (ФМН, FMN): рибофлавин-
фосфат, кофермент некоторых окислительно-восста-
новительных ферментов.
Флавиновые нуклеотиды: нуклеотидные кофермен-
ты (FMN и FAD), содержащие рибофлавин.
Флавопротеин: фермент, содержащий в качестве про-
стетической группы флавиновый нуклеотид.
Флиппазы: мембранные белки из семейства АВС-
переносчиков, катализирующие перемещение фос-
фолипидов из наружного слоя двойной мембраны в
цитозольный слой.
Флоппазы: мембранные белки из семейства АВС-
переносчиков, катализирующие перенос фосфоли-
пидов из цитозольного слоя двойной мембраны во
внешний слой.
Флуоресценция: испускание света молекулами при их
возвращении из возбужденного в основное состояние.
Фосфатазы: ферменты, расщепляющие фосфоэфир-
ную или фосфоангидридную связь с высвобождением
неорганического фосфата.
Фосфодиэфирная связь: химическая группа, со-
стоящая из двух спиртов, образующих эфиры с одной
молекулой фосфорной кислоты, которая служит
мостиком между ними.
Фосфолипид: липид, содержащий одну или несколь-
ко фосфатных групп.
Фосфорилазы: ферменты, катализирующие фосфо-
ролиз.
Фосфорилирование: образование фосфатного произ-
водного биомолекулы, обычно путем ферментативно-
го переноса фосфорильной группы от АТР.
Фосфоролиз: расщепление связей под действием
фосфатной группы; процесс, аналогичный гидролизу.
Фотовосстановление: индуцированное светом вос-
становление акцептора электронов в фотосинтезиру-
ющих клетках.
Фотодыхание: потребление кислорода, происходя-
щее на свету в клетках растений, главным образом,
как результат окисления фосфогликолата.
Фотон (квант): мельчайшая единица световой энергии.
Фотосинтез: использование энергии света для синте-
за углеводов из углекислого газа (диоксида углерода)
и воды (вода — восстановитель).
Фотосистема: специфические функциональные
группы светопоглощающих пигментов и реакционный
центр в фотосинтезирующих клетках.
Фототроф: организм, способный использовать энер-
гию света для синтеза топливных молекул из таких
простых соединений, как вода, углекислый газ и
кислород (в отличие от хемотрофа).
Фотофосфорилирование: ферментативное образо-
вание АТР из ADP, сопряженное со светозависимым
переносом электронов в фотосинтезирующих клетках.
Фотохимический реакционный центр: часть фото-
синтетического комплекса, в котором энергия по-
глощенного фотона вызывает разделение зарядов,
инициирующее перенос электронов.
Фракционирование: процесс разделения белков или
других сложных смесей на фракции на основании та-
ких свойств, как растворимость, заряд, молекулярная
масса или функция.
Фракция: часть биологического образца после опре-
деленной процедуры, предназначенной для разделе-
ния макромолекул на основании их растворимости,
заряда, молекулярной массы или функций.
Функциональная группа: специфический атом или
группа атомов, придающая биомолекуле специфиче-
ские химические свойства.
Фураноза: простой сахар, содержащий пятичленное
фурановое кольцо.
Футпринтинг: метод идентификации последователь-
ностей нуклеиновых кислот, связанных с ДНК- или
РНК-связывающими белками.
X
Хемиосмотическая теория: теория о запасании энер-
гии в электрон-транспортных реакциях в виде транс-
мембранного потенциала и pH с ее последующим
использованием для образования АТР в реакциях
окислительного фосфорилирования и фотофосфори-
лирования.
Хемиосмотическое сопряжение: сопряжение синтеза
АТР с переносом протонов по трансмембранному по-
тенциалу и по градиенту pH.
Хемотаксис: восприятие клетками специфического
химического сигнала и их движение к источнику
этого сигнала или от него.
[ 3 7 21 Словарь терминов
Хемотроф: организм, получающий энергию за счет
метаболизма органических веществ, синтезированных
другими организмами.
Хендерсона-Хассельбаха уравнение: уравнение,
связывающее pH, рКа и отношение концентраций
акцептора (А ) и донора протонов (НА) в растворе:
pH = pXa + log[A]/[HA],
Хилла коэффициент: характеризует кооперативность
взаимодействия между субъединицами белка.
Хилла реакция: выделение кислорода и восстановле-
ние искусственно введенного акцептора электронов в
образце хлоропластов.
Хиломикрон: липопротеин плазмы крови, состоящий
из триацилглицеринового ядра, покрытого пленкой
белка и фосфолипидов; переносит липиды из кишеч-
ника к тканям.
Химерная ДНК: ДНК, содержащая генетическую
информацию разных видов организмов.
Химотрипсин: хорошо изученная протеаза, расщепля-
ющая полипептидную цепь у ароматических амино-
кислотных остатков со стороны С-конца.
Химотрипсиноген: неактивный предшественник (зи-
моген) химотрипсина.
Хиральное соединение: соединение, содержащее
асимметрический центр (хиральный атом или хираль-
ный центр) и существующее в виде двух несовмести-
мых зеркальных форм (энантиомеров).
Хиральный центр: атом, заместители при котором рас-
положены таким образом, что молекула в целом может
быть совмещена с ее зеркальным отображением.
Хлоропласт: содержащая хлорофилл фотосинтетиче-
ская органелла в некоторых эукариотических клетках.
Холостой цикл: цикл ферментативных реакций, в
котором гидролиз АТР приводит только к выделению
тепловой энергии.
Холофермент: каталитически активный фермент,
содержащий все необходимые для катализа субъеди-
ницы, простетические группы и кофакторы.
Хондроитинсульфат: представитель семейства
сульфатированных гликозаминогликанов, важный
компонент внеклеточного матрикса.
Хроматин: нитчатый комплекс, образованный ДНК,
гистонами и другими белками, составляющий хромо-
сому эукариот.
Хроматография: метод разделения сложных смесей
веществ на основании их различного распределения
между подвижной и неподвижной фазой.
Хромосома: единая крупная молекула ДНК со свя-
занными с ней белками, содержащая в себе множе-
ство генов; форма хранения и передачи генетической
информации.
ц
Цвиттер-ион: биполярный ион с пространственным
разделением положительного и отрицательного за-
рядов.
Центр связывания: щель или карман в молекуле бел-
ка, в котором связывается молекула лиганда.
Центромера: специализированная область хромосо-
мы, где при митозе и мейозе прикрепляется веретено
деления.
Цереброзид: сфинголипид, в головке которого в каче-
стве группы находится один остаток сахара.
Цикл Кальвина: циклический путь метаболизма в
растениях, в результате которого происходит фиксация
диоксида углерода и образование триозофосфатов.
Цикл лимонной кислоты (цикл Кребса или цикл
трикарбоновых кислот): циклический метаболиче-
ский путь окисления ацетильных групп до диоксида
углерода; первая стадия цикла — образование цитрата
(соль лимонной кислоты).
Цикл мочевины: циклический метаболический путь в
печени позвоночных, в результате которого из амино-
групп и диоксида углерода образуется мочевина.
Циклин: представитель семейства белков, регулирую-
щих клеточный цикл путем активации циклин-зави-
симых протеинкиназ.
Циклический АМР (сАМР): вторичный мессенджер;
его образование аденилатциклазой стимулируют не-
которые гормоны и другие сигналы.
Циклический поток электронов: индуцированный
светом поток электронов в хлоронластах, начинаю-
щийся и заканчивающийся в фотосистеме I.
Циклическое фотофосфорилирование: синтез АТР,
направляемый циклическим потоком электронов
через фотосистему I.
Цинковые пальцы: специфический белковый мотив,
участвующий в связывании некоторых белков с ДНК;
в состав входит атом цинка, соединенный координа-
ционными связями с четырьмя остатками лизина или
двумя остатками лизина и двумя остатками гистидина.
Цистрон: участок ДНК или РНК, соответствующий
одному гену.
Цитозоль: водная фаза цитоплазмы, лишенная орга-
нелл, с растворенными в ней веществами.
Цитокин: представитель семейства небольших
секретируемых белков (интерлейкины или интерфе-
роны), активирующих дифференцировку или деление
клетки-мишени путем взаимодействия с рецепторами
на ее плазматической мембране.
Цитокинез: окончательное разделение дочерних кле-
ток после митоза.
Словарь терминов [373]
Цитоплазма: внутриклеточное содержимое между
ядром и плазматической мембраной; содержит раз-
личные органеллы, включая митохондрии.
Цитоскелет: нитчатая сеть, обеспечивающая струк-
турную организацию цитоплазмы; содержит актино-
вые филаменты, микротрубочки и промежуточные
филаменты.
Цитохром Р-450: семейство гем-содержащих фермен-
тов с характерной полосой поглощения при 450 нм,
участвующих в биологических реакциях гидроксили-
рования.
Цитохромы: гемовые белки, выполняющие функцию
переносчиков электронов в процессах дыхания, фото-
синтеза и других окислительно-восстановительных
реакциях.
Ч
Четвертичная структура: трехмерная структура
субъединичного белка, описывающая, в частности,
взаимное расположение его субъединиц.
Число Авогадро (N): число молекул в 1 моль любого
вещества (N= 6,02 • 1023 молекул).
Число оборотов: число молекул субстрата, превра-
щенных молекулой фермента в продукт за единицу
времени, в условиях максимальной активности фер-
мента и при насыщающей концентрации субстрата.
ш
Шайна-Дальгарно последовательность: участок
мРНК, необходимый для связывания с бактериальной
рибосомой.
Шаттл-вектор: векторная ДНК, способная реплици-
роваться в нескольких видах хозяйских клеток.
Шпилька: вторичная структура однонитевой РНК
или ДНК; закрытый с одной стороны антипарал-
лельный дуплекс, образующийся путем спаривания
палиндромных повторов.
э
Эймса тест: способ оценки канцерогенной (мутаген-
ной) активности химических веществ по отношению к
бактериальным клеткам.
Экзергоническая реакция: химическая реакция, про-
текающая с выделением энергии (AG < 0).
Экзон: участок эукариотического гена, кодирующий
часть конечного продукта гена; участок РНК, сохра-
няющийся в зрелой РНК, который транслируется в
белок или входит в структуру РНК.
Экзонуклеаза: фермент, отщепляющий концевые
нуклеотиды в последовательности нуклеиновой
кислоты.
Экзотермическая реакция: химическая реакция, про-
текающая с выделением тепла (АЯ < 0).
Экзоцитоз: слияние внутриклеточных везикул с
плазматической мембраной, в результате которого
содержимое везикул выводится во внеклеточное про-
странство.
Эксклюзионная хроматография (гель-фильтрация):
метод разделения молекул по размерам с помощью
пористых полимеров, удерживающих растворенные
вещества только определенного размера.
Экспрессия гена: образование продукта гена — транс-
крипция и (в случае гена белка) трансляция; сви-
детельством экспрессии гена является наличие его
активного продукта.
Электропорация: введение в клетку макромолекул
при временном повышении проницаемости клеточной
мембраны под действием высокого электрического
напряжения.
Электрофил: электрондефицитная группа с выра-
женной тенденцией принимать электроны от богатой
электронами группы (нуклеофила).
Электрофорез: перемещение ионов в электрическом
поле; часто применяется для разделения смесей ио-
нов, белков или нуклеиновых кислот.
Электрохимический градиент: связан с градиентом
концентрации и электрическим зарядом иона с двух
сторон от мембраны; движущая сила окислительного
фосфорилирования и фотофосфорилирования.
Электрохимический потенциал: энергия, необходи-
мая для поддержания разделения зарядов и концен-
трации иона на разных сторонах мембраны.
Элементы гормонального ответа (HRE): короткие
последовательности ДНК (от 12 до 20 и. н.), которые
связывают рецепторы стероидных, ретиноидных и ти-
реоидных гормонов и витамина D, изменяя экспрес-
сию соответствующих генов. Для каждого гормона
существует своя консенсусная последовательность.
Элюат: жидкая фаза на выходе с хроматографической
колонки.
Энантиомеры: стереоизомеры, различающиеся как
объект и его зеркальное отображение.
Эндергоническая реакция: химическая реакция, со-
провождающаяся расходом энергии (AG > 0).
Эндокринная железа: группа специализированных
клеток, синтезирующих и секретирующих в кровь
гормоны, которые регулируют активность других
клеток.
(3/41 Словарь терминов
Эндонуклеаза: фермент, расщепляющий внутренние
фосфодиэфирные связи в молекулах нуклеиновых
кислот.
Эндоплазматический ретикулум: протяженная
система двуслойных мембран в цитоплазме эукарио-
тической клетки; часть эндоплазматического ретику-
лума, покрытую рибосомами, называют шероховатым
эндоплазматическим ретикулумом.
Эндотермическая реакция: химическая реакция, про-
текающая с поглощением тепла (АН > 0).
Эндоцитоз: захват клеткой внеклеточных веществ
путем заключения их в везикулы (эндосомы),
образующиеся при инвагинации плазматической
мембраны.
Энергетический заряд: соотношение макроэргиче-
ских фосфатных групп в системе ATP/ADP/AMP.
Энергетическое сопряжение: передача энергии
между двумя процессами.
Энергия активации (AG+): количество энергии (в
джоулях), необходимое для перевода 1 моль молекул
реагирующего вещества из основного состояния в
переходное состояние.
Энергия связи: энергия, необходимая для разрыва
химической связи.
Энергия связывания: энергия нековалентного
взаимодействия между ферментом и субстратом или
рецептором и лигандом.
Энтальпия (Я): теплосодержание системы.
Энтропия (У): степень упорядоченности системы.
Энхансер: последовательность ДНК, усиливающая
экспрессию определенного гена; может находиться
на расстоянии нескольких сотен и даже тысяч пар
оснований от этого гена.
Эпигенетика: приобретение живым организмом
некоторого признака не через изменение нуклеотид-
ной последовательности родительских хромосом, а,
например, в результате ковалентной модификации
гистонов.
Эпимераза: фермент, катализирующий обратимые
взаимопревращения двух эпимеров.
Эпимеры: два стереоизомера, различающиеся кон-
фигурацией у одного из асимметрических центров в
молекуле с двумя или большим числом асимметриче-
ских центров.
Эпителиальные клетки: любые покровные клетки
организма или органа.
Эпитоп (антигенная детерминанта): участок на
поверхности макромолекулы (антигена), с которым
связывается конкретное антитело.
Эпитопная метка: белковая последовательность или
домен, связывающийся с хорошо охарактеризован-
ным антителом.
Эритроциты: красные кровяные клетки, содержащие
большое количество гемоглобина и специализирую-
щаяся на переносе кислорода.
Эукариоты: одноклеточные или многоклеточные
организмы, клетки которых имеют заключенное в
мембрану ядро, несколько хромосом и внутренние
органеллы.
Эухроматин: слабо окрашиваемый участок интерфаз-
ной хромосомы — в отличие от конденсированного и
более сильно окрашиваемого гетерохроматина; часто
соответствует участку хромосомы с активной экспрес-
сией генов.
Я
Ядро: содержащая хромосомы и ограниченная мем-
браной органелла эукариотической клетки.
Ядрышко: сильно окрашиваемая структура внутри
ядра эукариотической клетки; участвует в синтезе
рРНК и в образовании рибосом.
Модели молекул
МОЛЕКУЛЯРНАЯ ГРАФИКА. Если не указано иного,
молекулярная графика сделана Jean-Yves Sgro, Ph.D.,
или H. Adam Steinberg из Университета Висконсин-
Мэдисон, биохимический факультет.
АТОМНЫЕ КООРДИНАТЫ. Если не указано иного,
атомные координаты взяты из Protein Data Bank на сайте
Research Collaboratory for Structural Bioinformatics, www.
rcsb.org/pdb/. Cm. Berman, H. M., Westbrook, J., Feng, Z.,
Gilliland, G., Bhat, T. N., Weissig, H., Shindyalov, I. N., &
Bourne, P. E. (2000) The Protein Data Bank. Nucleic Acids
Res. 28, 235-242; Bernstein, F. C., Koetzle, T. E, Williams,
G. J. B., Meyer, E. F.,Jr., Brice, M. D., Rodgers, J. R., Kennard,
O., Shimanouchi, T, & Tasumi, M. (1977) Protein Data Bank:
a computer-based archival file for macromolecular structures.
J. Mo^Biol. 112,535; и Abola, E. E., Bernstein, F. C., Bryant,
S. H., Koetzle, T. E, & Weng,J. (1987) Protein Data Bank. В
Crystallographic Databases — Information Content, Software
Systems, Scientific Applications (Allen, F. H., Bergerhoff, G., &
Sievers, R., eds), Data Commission of the International Union
of Crystallography, Bonn, p. 107.
Некоторые структуры созданы с использованием Sybyl
6.2, Tripos Inc., www.tripos.com или PyMOL, pymol.
sourceforge.net.
Том 1
ГЛАВА 1. C. 15. Francois Jacob (1970) The Logic of Life: A
History of Heredity, Pantheon Books, Inc., New York, 1973;
оригинальная публикация (1970): La logique du vivant:
une histoire de I’heredite, Editions Gallimard, Paris; рис. 1-1,
a. Ron Boardman/Frank Lane Agency/Corbis; рис. 1-1,
6. W. Perry Conway/Corbis; рис. 1-1, в. Karen Tweedy-
Holmes/Corbis; рис. 1-2. The Bridgeman Art Library;
рис. 1-4. По Woese, C. R. (1987) Bacterial evolution.
Microbiol. Rev. 51, 221,
Fig. 4; рис. 1-6 (слева вверху) T. J. Beveridge/Biological
Photo Service; (справа вверху, слева внизу) Biological
Photo Service; (справа внизу) Norma J. Lange/Biological
Photo Service; рис. 1-8. По Alberts, В., Bray, D., Lewis,
J., Raff, M., Roberts, K., & Watson, J. D. (1989) Molecular
Biology of the Cell, 2nd edn, Garland Publishing, Inc.,
New York, pp. 165-166; с изм.; рис. 1-9, а. С любезного
разрешения Invitrogen; рис. 1-9, 6. Dr. Alexey Khodjakov,
Wadsworth Center, New York State Department of Health;
рис. 1-11. По Becker, W. M. & Deamer, D. W. (1991) The
World of the Cell, 2nd edn, Fig. 2-15, Benjamin/Cummings
Publishing Company, Menlo Park, CA; с изм.; рис. 1-16.
Acetyl-CoA из PDB ID 1DM3, Modis, Y. & Wierenga, R.K.
(2000) Crystallographic analysis of the reaction pathway of
Zoogloea ramigera biosynthetic thiolase.
J. Mol. Biol. 297,1171; рис. 1-20. По Carroll, F. (1998)
Perspectives on Structure and Mechanism in Organic
Chemistry, Brooks/Cole Publishing Co., Pacific Grove, CA,
p. 63; с изм.; доп. 1-2 (Пастер). The Granger Collection;
рис. 1-22. PDB ID 1AKX, Brodsky, A. S. & Williamson,
J. R. (1997) Solution structure of the HIV-2 Tar-
argininamide complex./. Mol. Biol. 267, 624; c. 44 (Гиббс)
Historical Pictures Service/Stock Montage; рис. 1-29, a.
Erich Lessing/Art Resource, New York; рис. 1-29, 6. Dr.
Gopal Murti-CNRI/Phototake New York; c. 54. Theodosius
Dobzhansky (1973) Nothing in biology makes sense except
in the light of evolution. The American Biology Teacher
(March) 35,125-129.
ГЛАВА 2. C. 73. Linus Pauling (1939) The Nature of the
Chemical Bond and the Structure of Molecules and Crystals:
An Introduction to Modem Structural Chemistry, Cornell
University Press, Ithaca, NY; рис. 2-9. PDB ID 1A3N,
Tame, J. & Vallone, B. (1998) Deoxy human hemoglobin
(оригинальный источник недоступен); рис. 2-10. По
Nicolls, Р. (2000) Introduction: the biology of the water
molecule. Cell. Mol. Life Sci. 57, 987, Fig. 6a (перерисовано
по данным из PDB и файлам kinemage, опубликованным
в статье Martinez, S.E., Huang, D., Ponomarev, M., Cramer,
W. A., & Smith, J. L. (1996) The heme redox center of
chloroplast cytochrome f is linked to a buried five-water
chain. Protein Sci. 5,1081) с изм.; доп. 2-1. J. В. S. Haldane
(1928) Possible Worlds, Harper and Brothers, New York
and London, pp. 113-126; c. 105. Jon Bertsch/Visuals
Unlimited.
ГЛАВА 3. C. 113. J. J. Berzelius (1838) Letter to G.
J. Mulder. In H. B. Vickery (1950) The origin of the
word protein. Yale Journal of Biology and Medicine 22,
387-393; рис. 3-1, a. Runk/Schoenburger/Grant Heilman
Photography; рис. 3-1, 6. Bill Longcore/Photo Researchers;
рис. 3-1, e. Animals Animals; c. 115 (Дейхофф). C
любезного разрешения Ruth E. Dayhoff; рис. 3-18, 6. Julia
Cox, University of Wisconsin-Madison, Department of
Biochemistry; рис. 3-21, 6. Patrick H. O’Farrell, University
of California Medical Center, San Francisco, Department of
Biochemistry and Biophysics; рис. 3-23. PDB ID 1HGA,
Liddington, R., Derewenda, Z., Dodson, E., Hubbard, R.,
& Dodson, G. (1992) High resolution crystal structures
and comparisons of T-state deoxyhaemoglobin and two
liganded T-state haemoglobins: T(a-oxy) haemoglobin and
T(met)haemoglobin.J. Mol. Biol. 228, 551; c. 143 (Сенгер).
UPI/Corbis-Bettmann; доп. 3-2, рис. 1, 2. По Mann, M.
& Wilm, M. (1995) Electrospray mass spectrometry for
protein characterization. Trends Biochem. Sci. 20, 219; p. 101
(Merrifield) Corbis/Bettmann; с изм.; доп. 3-3, рис. 1.
Информация о последовательности для а - из документа
ID PDGC00017, для б - из документа ID PDGC00018,
www.expasy.org/prosite, Hulo, N., Bairoch, A., Bulliard,
V, Cerutti, L., De Castro, E., Langendijk-Genevaux,
P. S., Pagni, M., & Sigrist, C. J. A. (2006) The PROSITE
database. Nucleic Acids Res. 34, D227; Web Logo из http://
weblogo.berkeley.edu, Crooks, G. E., Hon, G., Chandonia,
J. M., & Brenner, S. E. (2004) WebLogo: a sequence logo
generator, Genome Res. 14, 1188; рис. 3-30,3-31, 3-32.
По Gupta, R. S. (1998) Protein phylogenies and signature
[376] Источники иллюстраций
sequences: a reappraisal of evolutionary relationships among
archaebacteria, eubacteria, and eukaryotes. Microbiol. Mol.
Biol. Rev. 62, 1435, Figs 2, 7, 11, соответственно; с изм.;
рис. 3-33. По Delsuc, E, Brinkmann, H., & Philippe, H.
(2005) Phylogenomics and the reconstruction of the tree of
life. Nat. Rev. Genet. 6, 366; c. 167, задача 21. См. источник
для доп. 3-3, рис. 1., документ ID PDOC00270.
ГЛАВА 4. С. 171. J. С. Kendrew et al. (1958) A three-
dimensional model of the myoglobin molecule obtained by
x-ray analysis. Nature 181, 662-666; рис. 4-1. PDB ID
6GCH, Brady, K., Wei, A., Ringe, D., & Abeles, R. H. (1990)
Structure of chymotrypsin-trifluoromethyl ketone inhibitor
complexes: comparison of slowly and rapidly equilibrating
inhibitors. Biochemistry 29, 7600; координаты для глицина
-- из Sybyl; с. 174. (Полинг) Corbis/Bettmann; (Кори).
AP/Wide World Photos; рис. 4-3. По Creighton, Т. Е.
(1984) Proteins, р. 166. © 1984 by W. Н. Freeman and
Company. Перепечатано с разрешения; рис. 4-4, б, в.
PDB ID 4TNC, Satyshur, К. A., Rao, S. Т, Pyzalska, D.,
Drendel, W, Greaser, M., & Sundaralingam, M. (1988)
Refined structure of chicken skeletal muscle troponin C in
the two-calcium state at 2-angstroms resolution. J. Biol.
Chem. 263, 1628; рис. 4-8, а. См. источник для рис. 4-3;
рис. 4-8, б. С любезного разрешения Hazel Holden,
University of Wisconsin-Madison, Department of
Biochemistry and Enzyme Institute; рис. 4-11. PDB ID
1CGD (с изм.), Bella, J., Brodsky, B., & Berman, H. M.
(1995) Hydration structure of a collagen peptide. Structure
3, 893; рис. 4-12. Science Source/Photo Researchers;
c. 189. Ethel Wedgwood (1906) The Memoirs of the Lord of
Joinville:A New English Version, E. P. Dutton and Company,
New York; (Lind) С любезного разрешения the Royal
College of Physicians of Edinburgh; рис. 4-13, a. PDB ID
1SLK (модель), Fossey, S. A., Nemethy, G., Gibson, K. D., &
Scheraga, H. A. (1991) Conformational energy studies of
p-sheets of model silk fibroin peptides: I. Sheets of poly(Ala-
Gly) chains. Biopolymers 31, 1529; рис. 4-13, 6. Dr. Dennis
Kunkel/Phototake NYC; рис. 4-15. PDB ID 1MBO;
Phillips, S. E. V. (1980) Structure and refinement of
oxy myoglobin at 1.6 angstroms resolution./. Mol. Biol. 142,
531; рис. 4-17, 6. PDB ID 7AHL; Song, L., Hobaugh, M. R.,
Shustak, C., Cheley, S., Bayley, H., & Gouaux, J. E. (1996)
Structure of staphylococcal a hemolysin, a heptameric
transmembrane pore. Science 274, 1859; доп. 4-5, рис. 1, 2,
3. George N. Phillips, Jr., University of Wisconsin-Madison,
Department of Biochemistry; доп. 4-5, рис. 1, г. PDB ID
2MBW; Brucker, E. A., Olson, J. S., Phillips, G. N., Jr., Dou,
Y, & Ikeda-Saito, M. (1996) High resolution crystal
structures of the deoxy-, oxy-, and aquomet- forms of cobalt
myoglobin./. Biol. Chem. 271, 25,419; доп. 4-5, рис. 2, 3, a.
Volkman, В. F, Alam, S. L., Satterlee, J. D., & Markley, J. L.
(1998) Solution structure and backbone dynamics of
component IV-glycera dibranchiata monomeric hemoglobin-
CO. Biochemistry 37, 10,906; доп. 4-5, рис 3, б, в. Созданы
Brian Volkman, National Magnetic Resonance Facility at
Madison, с помощью MOLMOL; PDB ID 1VRF (6) and
1VRE (в), см. источник для доп. 4-5, рис. 2, 3, я; рис. 4-18.
PDB ID 4TNC, см. Источник для рис. 4-4, б, в; рис. 4-19,
в; рис. 4-19, в PDB ID 1DNP, Park, Н. W., Kim, S. Т.,
Sancar, А., & Deisenhofer, J. (1995) Crystal structure of
DNA photolyase from Escherichia coli. Science 268, 1866;
рис. 4-20 PDB ID 1PKN, Larsen, T. M., Laughlin, L. T,
Holden, H. M., Rayment, L, & Reed, G. H. (1994) Structure
of rabbit muscle pyruvate kinase complexed with Mn2+,
K+,and pyruvate. Biochemistry 33, 6301; рис. 4-21 (все a)
PDB ID 1A06, Sugio, S., Kashima, A., Mochizuki, S., Noda,
M., & Kobayashi, K. (1999) Crystal structure of human
serum albumin at 2.5 angstrom resolution. Protein Eng. 12,
439; PDB ID 1BCF, Frolow, E, Kalb (Gilboa), A. J., & Yariv,
J. (1994) The structure of a unique, two-fold symmetric,
haem-binding site. Nat. Struct. Biol. 1, 453; PDB ID 1GAI,
Aleshin, A. E., Stoffer, B., Firsov, L. M., Svensson, B., &
Honzatko, R. B. (1996) Crystallographic complexes of
glucoamylase with maltooligosaccharide analogs:
relationship of stereochemical distortions at the nonreducing
end to the catalytic mechanism. Biochemistry 35, 8319; PDB
ID 1ENH, Clarke, N. D., Kissinger, C. R., Desjarlais, J.,
Gilliland, G. L., & Pabo, С. O. (1994) Structural studies of
the engrailed homeodomain. Protein Sci. 3, 1779; (все 0)
PDB ID 1LXA, Raetz, C. R. H. & Roderick, S. L. (1995) A
left-handed parallel 0-helix in the structure of UDP
N-acetylglucosamine acyltransferase. Science 270, 997; PDB
ID 1PEX, Gomis-Ruth, F. X., Gohlke, U., Betz, M., Knauper,
V, Murphy, G., Lopez-Otin, C., & Bode, W. (1996) The
helping hand of collagenase-3 (Mmp-13): 2.7 A crystal
structure of its C-tenninal haemopexin-like domain./. Mol.
Biol. 264, 556; PDB ID 1CD8, Leahy, D. J., Axel, R., &
Hendrickson, W. A. (1992) Crystal structure of a soluble
form of the human T cell co-receptor Cd8 at 2.6 angstroms
resolution. Cell 68, 1145; (a/0) PDB ID 1DEH, Davis, G.J.,
Stone, C. J., Bosron, W. F., & Hurley, T. D. (1996) X-ray
structure of human 0303 alcohol dehydrogenase: the
contribution of ionic interactions to coenzyme binding./.
Biol. Chem. 271, 17,057; PDB ID 1DUB, Engel, С. K.,
Mathieu, M., Zeelen, J. P, Hiltunen, J. K., & Wierenga, R. K.
(1996) Crystal structure of enoyl-coenzyme A (CoA)
hydratase at 2.5 angstroms resolution: a spiral fold defines
the CoA-binding pocket. EMBOJ. 15, 5135; PDB ID 1PFK,
Shirakihara, Y. & Evans, P. R. (1988) Crystal structure of
the complex of phosphofructokinase from Escherichia coli
with its reaction products./. Mol. Biol. 204, 973; (a + 0)
PDB ID 2PIL, Forest, К. T, Dunham, S. A., Koomey, M., &
Tainer, J. A. (1999) Crystallographic structure of
phosphorylated pilin from Neisseria: phosphoserine sites
modify type IV pilus surface chemistry and morphology. Mol.
Microbiol. 31, 743; PDB ID 1SYN, Stout, T. J. & Stroud,
R. M. (1996) The complex of the anti-cancer therapeutic,
BW1843U89, with thymidylate synthase at 2.0 A resolution:
implications for a new mode of inhibition. Structure 4,
67-77; PDB ID 1EMA, Ormo, M., Cubitt, A. B., Kallio, K.,
Gross, L. A., Tsien, R. Y, & Remington, S. J. (1996) Crystal
structure of the Aequorea victoria green fluorescent protein.
Science 273, 1392; c. 205 (Перутц и Кендрю). Corbis/
Hulton Deutsch Collection; рис. 4-22. PDB ID 2HHB,
Источники иллюстраций [ 3 / / ]
Fermi, G., Perutz, M. E, Shaanan, В., & Fourme, R. (1984)
The crystal structure of human deoxyhaemoglobin at 1.74
angstroms resolution./ Mol. Biol. 175, 159; рис. 4-24, a.
PDB ID 2PLV, Filman, D. J., Syed, R., Chow, M., Macadam,
A. J., Minor, P. D., & Hogle, J. M. (1989) Structural factors
that control conformational transitions and serotype
specificity in type 3 poliovirus. EMBOJ. 8, 1567, как
представлено в базе данных VIPER, Shepherd, С. М.,
Borelli, I. A., Lander, G., Natarajan, P, Siddavanahalli, V,
Bajaj, C., Johnson, J. E., Brooks, C. L. Ill, & Reddy, V. S.
(2006) Nucleic Acids Res. 34, D386; рис. 4-24, 6. Science
Source/Photo Researchers; PDB ID 1VTM, Pattanayek, R.
& Stubbs, G. (1992) Structure of the U2 strain of tobacco
mosaic virus refined at 3.5 angstroms resolution using x-ray
fiber diffraction./ Mol. Biol. 228, 516; рис. 4-25, a. Data
from Sendak, R. A., Rothwarf, D. M., Wedemeyer, W. J.,
Houry, W. A., & Scheraga, H. A. (1996) Kinetic and
thermodynamic studies of the folding/unfolding of a
tryptophan-containing mutant of ribonuclease A.
Biochemistry 35, 12,978; Nishii, I., Kataoka, M., & Goto, Y.
(1995) Thermodynamic stability of the molten globule
states of apomyoglobin./ Mol. Biol. 250, 223; рис. 4-25, 6.
Data from Houry, W. A., Rothwarf, D. M., & Scheraga, H. A.
(1996) Circular dichroism evidence for the presence of
burst-phase intermediates on the conformational folding
pathway of ribonuclease A. Biochemistry 35, 10,125;
рис. 4-27. С любезного разрешения Yong Duan и Peter A.
Kollman, University of California, San Francisco,
Department of Pharmaceutical Science; рис. 4-28. По
Wolynes, P. G., Onuchic, J. N., & Thirumalai, D. (1995)
Navigating the folding routes. Science 267, 1619; рис. 4-30,
6. PDB ID 1AON, Xu, Z., Horwich, A. L, & Sigler, P. B.
(1997) The crystal structure of the asymmetric GroEL-
GroES-(ADP)7 chaperonin complex. Nature 388, 741; c
изм.; рис. 4-31, а. По Selkoe, D. J. (2003) Folding proteins
in fatal ways. Nature 426, 903, Fig. 1; с изм.; рис. 4-31, б.
PDB ID 1IYT, Crescenzi, O., Tomaselli, S., Guerrini, R.,
Salvador!, S., D’Ursi, A. M., Temussi, P. A., & Picone, D.
(2002) Solution structure of the Alzheimer amyloid
P-peptide (1-42) in an apolar microenvironment: similarity
with a virus fusion domain. Eur. J. Biochem. 269, 5642;
рис. 4-31, e. PDB ID 2BEG, Lahrs, T, Ritter, C., Adrian,
M., Riek-Loher, D., Bohrmann, B., Dobeli, H., Schubert, D.,
& Riek, R. (2005) 3D structure of Alzheimer’s amyloid-p
(1-42) fibrils. Proc. Natl. Acad. Sci. USA 102, 17,342;
доп. 4-6, рис. 1. Stephen J. DeArmond; доп. 4-6, рис. 2.
PDB ID 1QM0, Zahn, R., Liu, A., Luhrs, T, Riek, R., von
Schroetter, C., Garcia, F. L., Billeter, M., Calzolai, L., Wider,
G., & Wuthrich, K. (2000) NMR solution structure of the
human prion protein. Proc. Natl. Acad. Sci. USA 97, 145.
ГЛАВА 5. C. 225. Emil Fischer (1906) Untersuchen liber
Aminosauren, Polypeptide und Proteine (Studies of amino
acids, polypeptides and proteins). Berichte derdeutschen
chemischen Gesellschaft zu Berlin 39, 530; рис. 5-1, в. Гем
из PDB ID 1CCR, Ochi, H., Hata, Y, Tanaka, N., Kakudo,
M., Sakurai, T, Aihara, S., & Morita, Y. (1983) Structure
of rice ferricytochrome c at 2.0 angstroms resolution./
Mol. Biol. 166,407; рис. 5-3, 5-5, e, 5-6 (слева). PDB ID
1MBO, Phillips, S. E. V. (1980) Structure and refinement
of oxymyoglobin at 1.6 angstroms resolution./ Mol.
Biol. 142, 531; рис. 5-6 (справа), 5-8, 5-9, a, 5-10
(состояние T), 5-11 (состояние Т). PDB ID 1HGA;
Liddington, R., Derewenda, Z., Dodson, E., Hubbard, R.,
& Dodson, G. (1992) High resolution crystal structures
and comparisons of T state deoxyhaemoglobin and two
liganded T-state haemoglobins: T(a-oxy)haemoglobin
and T(met)haemoglobin./ Mol. Biol. 228, 551; рис. 5-10
(состояние R), 5-11 (состояние R). PDB ID 1BBB,
Silva, M. M., Rogers, P. H., & Arnone, A. (1992) A
third quaternary structure of human hemoglobin a at
1.7-angstroms resolution./ Biol. Chem. 267, 17,248; на
рис. 5-11 состояние R было изменено для того, чтобы
отразить О2 вместо СО; доп. 5-1, рис. 1. По Coburn,
R. Е, Forster, R. Е., & Kane, Р. В. (1965) Considerations
of the physiological variables that determine the blood
carboxyhemoglobin concentration in man./ Clin. Invest.
44,1899; с изм.; доп. 5-1, рис. 2. По Roughton,Е J. W. &
Darling, R. C. (1944) The effect of carbon monoxide on the
oxyhemoglobin dissociation curve. Am. J. Physiol. 141, 17; c
изм.; рис. 5-18. PDB ID 1HGA; см. источник для рис. 5-6
(справа). Координаты для 2,3-бисфосфоглицерата
взяты у Paul Rcisberg, Wellesley College, Department of
Chemistry; рис. 5-19, a. Andrew Syred/Science Photo
Library/Custom Medical Stock Photo; рис. 5-19, 6.
Custom Medical Stock Photo; рис. 5-21, 6. PDB ID 1IGT,
Harris, L. J., Larson, S. B., Hasel, K. W, & McPherson, A.
(1997) Refined structure of an intact IgG2A monoclonal
antibody. Biochemistry 36, 1581; рис. 5-25, a. PDB ID
1GGC, Stanfield, R. L., Takimoto-Kamimura, M., Rini,
J. M., Profy, A. T, & Wilson, I. A. (1993) Major antigen-
induced domain rearrangements in an antibody. Structure
1, 83; рис. 5-25, б, в. PDB ID 1GGI, Rini, J. M., Stanfield,
R. L., Stura, E. A., Salinas, P. A., Profy, A. T, & Wilson,
I. A. (1993) Crystal structure of an human immuno-
deficiency virus type 1 neutralizing antibody, 50.1, in
complex with its V3 loop peptide antigen. Proc. Natl.
Acad. Sci. USA 90, 6325; c. 254 (Кёлер и Милыптейн).
Corbis/UPI/Bettmann; рис. 5-26, 6. State of Wisconsin
Laboratory of Hygiene, Madison, WI; рис. 5-26, e. Son, M.,
Gundersen, R. E., & Nelson, D. L. (1993) A second member
of the novel Ca2+-dependent protein kinase family from
Paramecium tetraurelia\ purification and characterization.
/ Biol. Chem. 268, 5940; рис. 5-27, a. David Shotton,
University of Oxford, Department of Zoology; рис. 5-27,
в. С любезного разрешения Ivan Rayment, University of
Wisconsin-Madison, Enzyme Institute and Department
of Biochemistry (см. также PDB ID 2MYS, Rayment, L,
Rypniewski, W. R., Schmidt-Base, K., Smith, R., Tomchick,
D. R., Benning, M. M., Winkelmann, D. A., Wesenberg, G.,
& Holden, H. M. (1993) Three-dimensional structure of
myosin subfragment-1: a molecular motor. Science 261, 50);
рис. 5-28, a. Eisaku Katayama, Institute of Medical Science,
The University of Tokyo, Department of Fine Morphology;
рис. 5-28, 6. Roger Craig, University of Massachusetts
[ 37s | Источники иллюстраций
Medical School; рис. 5-28, в. См. источник для рис. 5-27,
в; рис. 5-29, б, в. James Е. Dennis/Phototake NYC.
ГЛАВА 6. С. 269. J. В. S. Haldane (1930) Enzymes,
Longmans, Green & Co., London, p. 182; William P. Jencks
(1975) Binding energy, specificity, and enzymic catalysis:
the Circe effect. Advances in Enzymology 43, 219-410;
c. 270 (Бухнер) Science Photo Library/Photo Researchers;
(Самнер и Холдейн) AP/Wide World Photos; рис. 6-1.
PDB ID 7GCH, Brady, K., Wei, A., Ringe, D., & Abeles,
R. H. (1990) Structure of chymotrypsin-trifluoromethyl
ketone inhibitor complexes: comparison of slowly and
rapidly equilibrating inhibitors. Biochemistry 29, 7600;
рис. 6-4. PDB ID 1RA2, Sawaya, M.R. & Kraut, J. (1997)
Loop and subdomain movements in the mechanism of
Escherichia coli dihydrofolate reductase: crystallographic
evidence. Biochemistry 36, 586; c. 287 (Михаэлис)
Rockefeller University Archive Center; (Ментен) C
любезного разрешения Dorothy C. Craig; рис. 6-18, 6, в,
г. PDB ID 7GCH, см. источник для рис. 6-1; рис. 6-22, а.
PDB ID 2YHX, Anderson, С. M., Stenkamp, R. E., & Steitz,
T. A. (1978) Sequencing a protein by x-ray crystallography:
II. Refinement of yeast hexokinase В coordinates and
sequence at 2.1 angstroms./. Mol. Biol. 123, 15; рис. 6-22,
6. PDB ID 1HKG, Steitz, T. A., Shoham, M., & Bennett,
W. S., Jr. (1981) Structural dynamics of yeast hexokinase
during catalysis. Philos. Trans. R. Soc. London Ser. В 293, 43;
координаты для глюкозы (GLC) с изм. взяты из PDB ID
1GLK, St. Charles, R., Harrison, R. W, Bell, G. L, Pilkis,
S. J., & Weber, I. T. (1994) Molecular model of human beta-
cell glucokinase built by analogy to the crystal structure of
yeast hexokinase B. Diabetes 43, 784; рис. 6-23, 6. PDB ID
IONE, Larsen, T. M., Wedekind, J. E., Rayment, L, & Reed,
G. H. (1996) A carboxylate oxygen of the substrate bridges
the magnesium ions at the active site of enolase: structure of
the yeast enzyme complexed with the equilibrium mixture
of 2-phosphoglycerate and phosphoenolpyruvate at 1.8 A
resolution. Biochemistry 35, 4349; рис. 6-24, a. PDB ID
ILZE, Maenaka, K., Matsushima, M., Song, H., Sunada, E,
Watanabe, K., & Kumagai, I. (1995) Dissection of protein-
carbohydrate interactions in mutant hen egg-white lysozyme
complexes and their hydrolytic activity. J. Mol. Biol. 247,
281; рис. 6-25, a, 6. PDB ID 1H6M, Vocadlo, D. J., Davies,
G. J., Laine, R., & Withers, S. G. (2001) Catalysis by hen
egg-white lysozyme proceeds via a covalent intermediate
Nature 412, 835; доп. 6-3, рис. 1. По Fersht, A. (1985)
Enzyme Structure and Mechanism, 2nd edn, p. 318. © 1977,
1985 by W. H. Freeman and Company. Перепечатано c
разрешения, с изм.; рис. 6-32. PDB ID 2AT2, Stevens,
R. C., Reinisch, К. M., & Lipscomb, W. N. (1991) Molecular
structure of Bacillus subtilis aspartate transcarbamoylase at
3.0 angstroms resolution. Proc. Natl. Acad. Sci. USA 88, 6087.
Координаты, зависимые от симметрии, представлены с
любезного разрешения Raymond С. Stevens, University of
California, Berkeley.
ГЛАВА 7. С. 339 E. В. White (1977) Coon Tree. In
Essays of E. B. White, HarperCollins, Perennial Classics
edn, New York, 1999, pp. 47-48; рис. 7-16. С любезного
разрешения Jack Scheper, Floridata.com; рис. 7-17, 6.
Leroy Somon/Visuals Unlimited; рис. 7-19. С любезного
разрешения H.-J. Gabius and Herbert Kaltner, University
of Munich, с иллюстрации, предоставлненной C.-W. von
der Lieth, Heidelberg; рис. 7-20, 6. PDB ID 1C58, Gessler,
K., Uson, L, Takaha, T, Krauss, N., Smith, S. M., Okada,
S., Sheldrick, G. M., & Saenger, W. (1999) V-Amylose at
atomic resolution: x-ray structure of a cycloamylose with 26
glucose residues (cyclomaltohexaicosaose). Proc. Natl. Acad.
Sci. USA 96, 4246; рис. 7-22. PDB ID 1HPN, Mulloy, B.,
Forster, M. J., Jones, C., & Davies, D. B. (1993) N.m.r. and
molecular-modelling studies of the solution conformation
of heparin. Biochem.J. 293, 849; рис. 7-23. PDB ID 1E0O,
Pellegrini, L., Burke, D. E, von Delft, F., Mulloy, B., &
Blundell, T. L. (2000) Crystal structure of fibroblast growth
factor receptor ectodomain bound to ligand and heparin.
Nature 407, 1029; рис. 7-25, а. По Haker, U., Nybakken,
K., & Perrimon, N. (2005) Heparan sulphate proteoglycans:
the sweet side of development. Nat. Rev. Mol. Cell Biol. 6,
532; с изм.; рис. 7-25, 6, 7-26. По Turnbull, J., Powell, A.,
& Guimond, S. (2001) Heparan sulfate: decoding a dynamic
multifunctional cell regulator. Trends Cell Biol. 11, 75; c
изм.; рис. 7-27 врезка. С любезного разрешения Laurel
Ng. Перепечатано с разрешения с Ng, L., Grodzinsky, А.,
Patwari, Р, Sandy, J., Plaas, A. H. K., & Ortiz, C. (2003)
Individual cartilage aggrecan macromolecules and their
constituent glycosaminoglycans visualized via atomic
force microscopy. J. Struct. Biol. 143, 242-257, Fig 7a
left; рис. 7-32. R. M. Genta and D. Y. Graham, Veterans
Affairs Medical Center, Houston, TX; c. 374 (Маршалл,
Уоррен). The Nobel Foundation; рис. 7-33, a, 6. PDB
ID 1M6P, Roberts, D. L., Weix, D. J., Dahms, N. M., &
Kim, J. J. (1998) Molecular basis of lysosomal enzyme
recognition: three-dimensional structure of the cation-
dependent mannose 6-phosphate receptor. Cell 93, 639;
рис. 7-34. По иллюстрации, представленной Dr. C.-W.
von der Lieth, Heidelberg, в Gabius, H.-J. (2000) Biological
information transfer beyond the genetic code: the sugar
code. Naturwissenschaften 87, 108, Fig. 6; с изм.; рис. 7-35.
По Sharon, N. & Lis, H. (1993) Carbohydrates in cell
recognition. Sci. Am. 268 (January), 82; с изм.; рис. 7-37.
С любезного разрешения Anne Dell. Перепечатано с
резрешения из Comelli, Е. М., Head, S. R., Gilmartin, Т,
Whisenant, Т, Haslam, S. М., North, S. J., Wong, N.-K.,
Kudo, T, Narimatsu, H., Esko, J. D., Drickamer, K., Dell,
A., & Paulson, J.C.(2006) A focused microarray approach
to functional glycomics: transcriptional regulation of the
glycome. Glycobiology 16,117-131, Fig. 3.
ГЛАВА 8. C. 391. James D. Watson (1968) The Double
Helix: A Personal Account of the Discovery of the Structure
of DNA, Atheneum, New York; c. 399 (Уотсон и Крик).
Corbis/UPI/Bettmann; рис. 8-12. Science Source/Photo
Researchers; c. 401 (Франклин) Science Photo Library/
Photo Researchers; (Уилкинс) Corbis/UPI/Bettmann;
рис. 8-13, б, e, 8-17. Координаты рассчитаны при
помощи Sybyl; рис. 8-20, б. PDB ID 1ВСЕ, Asensio, J. L.,
Источники иллюстраций 13 / 91
Brown, Т, & Lane, A. N. (1998) Comparison of the solution
structures of intramolecular DNA triple helices containing
adjacent and non-adjacent CG.C+ triplets. Nucleic Acids
Res. 26, 3677; рис. 8-20, г. Теоретическая структура
PDB ID 1QDG, Marathias, V. M., Wang, K. Y, Kumar, S.,
Swaminathan, S., & Bolton, P. H. (1996) Determination
of the number and location of the manganese binding sites
of DNA quadruplexes in solution by EPR and NMR in the
presence and absence of thrombin./ Mol. Biol. 260, 378;
Figure 8-22 Coordinates generated by Sybyl; рис. 8-24.
James, B., Olsen, G. J., Liu, J., & Pace, N. R. (1988) The
secondary structure of ribonuclease P RNA, the catalytic
element of a ribonucleoprotein enzyme. Cell 52, 19-26;
рис. 8-25, a. PDB ID 1TRA, Westhof, E. & Sundaralingam,
M. (1986) Restrained refinement of the monoclinic form of
yeast phenylalanine transfer RNA: temperature factors and
dynamics, coordinated waters, and base-pair propeller twist
angles. Biochemistry 25, 4868; рис. 8-25, 6. PDB ID 1MME,
Scott, W. G., Finch, J. T, & Klug, A. (1995) The crystal
structure of an all-RNA hammerhead ribozyme: a proposed
mechanism for RNA catalytic cleavage. Cell 81, 991;
рис. 8-25, в. PDB ID 1GRZ, Golden, B. L., Gooding, A. R.,
Podell, E. R., & Cech, T. R. (1998) A preorganized active site
in the crystal structure of the Tetrahymena ribozyme. Science
282, 259; рис. 8-27, б. По Marmur, J. & Doty, P. (1962)
Determination of the base composition of deoxyribonucleic
acid from its thermal denaturation temperature./ Mol.
Biol. 5, 109; рис. 8-28. Ross B. Inman, University of
Wisconsin-Madison, Department of Molecular Biology;
рис. 8-33, в. Lloyd Smith, University of Wisconsin-
Madison, Department of Chemistry; рис. 8-34. Данные
предоставлены Lloyd Smith, University of Wisconsin-
Madison, Department of Chemistry.
ГЛАВА 9. C. 433. Emile Zuckerkandl and Linus Pauling
(1965) Molecules as documents of evolutionary history.
Journal of Theoretical Biology 8, 357-366; (Берг) c
любезного разрешения Stanford Visual Art Services;
(Бойер) с любезного разрешения Genentech, Inc.;
(Коэн) с любезного разрешения Stanford Visual Art
Services; рис. 9-4. Elizabeth A. Wood, University of
Wisconsin-Madison, Department of Biochemistry;
рис. 9-15, а. С любезного разрешения Kevin Strange
and Michael Christensen, Vanderbilt University Medical
Center, Department of Pharmacology; доп. 9-1, рис. 1.
Phil Borden/PhotoEdit; доп. 9-1, рис. 2, б. С любезного
разрешения Carol Bingham, Promega Corporation;
с. 458 (Коллинз) Alex Wbng/Newsmakers; (Вентер)
Mike Theiler/Reuters; рис. 9-20. По Wolfsberg, T. G.,
McEntyre, J., & Schuler, G. D. (2001) Guide to the
draft human genome. Nature 409, 824, Fig. 1; с изм.;
рис. 9-23. С любезного разрешения Patrick О. Brown,
Stanford University School of Medicine, Department of
Biochemistry; рис. 9-24. По Pandey, A. & Mann, M. (2000)
Proteomics to study genes and genomes. Nature 405, 837,
Fig. 7; с изм.; рис. 9-29. С любезного разрешения Keith
Wood, University of California, San Diego, Department
of Biology; рис. 9-30 По Fischhoff, D. A. & Bowdish,
K. S. (1987) Insect tolerant transgenic tomato plants.
BioTechnology 5,807. Фотография печатается с любезного
разрешения David Fischhoff, Monsanto Company;
рис. 9-31, а, б. С любезного разрешения Chris Boerboom,
University of Wisconsin-Madison, Department of
Agronomy; рис. 9-33. С любезного разрешения Yorktown
Technologies, LP, www.glofish.com.
ГЛАВА 10. C 487. Michel-Eugene Chevreul (1814)
Chemical inquiries into the nature of several fatty
substances, and particularly on their combinations with the
alkalis. Of a new substance, called margarine, obtained from
the soap made from the fat of pork and potash. Philosophical
Magazine 44, 193; рис. 10-2, a, 6, 10-3. Координаты c
помощью Sybyl; рис. 10-4, a. Biological Photo Service;
рис. 10-4, б. С любезного разрешения Howard Goodman,
Harvard Medical School, Department of Genetics; рис. 10-
6, 6. William J. Weber/Visuals Unlimited; c. 501. Из
Drabkin, D. L. (1958) Thudichum: Chemist of the Brain,
University of Pennsylvania Press, источник: Thudichum,
J. L. W. (1898) Briefe uber offentliche Gesundheitspflege
ihre bisherigen Leistungen und heutigen Aufgaben, F.
Pietzcker, Tiibingen; рис. 10-14. Координаты с помощью
Sybyl; рис. 10-17. PDB координаты предоставлены
Dave Woodcock, Okanagan University College, Kelowna,
British Columbia, Department of Chemistry; доп. 10-2,
рис. 2. Herbert A. Fischler, Isaac Albert Research Institute
of the Kingsbrook Jewish Medical Center; c. 508 (Вейн,
Бергстрём и Самуэльсон). Ira Wyman/Sygma; рис. 10-
20, б. С любезного разрешения Media Center, University
of Wisconsin-Madison, Department of Biochemistry;
c. 512 (Дам и Дойси) AP/Wide World Photos; рис. 10-23
(кардинал) Dr. Dan Suda; (щегол) Richard Day/VIREO;
рис. 10-25. Christie, W. W. (1996) Beginners’ guide to mass
spectrometry of fatty acids: 2. General purpose derivatives.
Lipid Technol. 8, 64.
ГЛАВА 11. C. 525. Robert Frost (1914) Mending Wall. In
North of Boston, David Nutt, London; рис. 11-1.
С любезного разрешения J. D. Robertson; рис. 11-5.
Данные из Zachowski, A. (1993) Phospholipids in
animal eukaryotic membranes: transverse asymmetry and
movement. Biochem. J. 294, 1; рис. 11-7. По Marchesi, V. T,
Furthmayr, H., & Tomita, M. (1976) The red cell membrane.
Annu. Rev. Biochem. 45, 667; с изм.; рис. 11-9. PDB ID
2AT9, Mitsuoka, K., Hirai, T, Murata, K., Miyazawa, A.,
Kidera, A., Kimura, Y, & Fujiyoshi, Y. (1999) The structure
of bacteriorhodopsin at 3.0 A resolution based on electron
crystallography: implication of the charge distribution./
Mol. Biol. 286, 861; рис. 11-10, a. PDB ID 2B6O, Gonen,
T, Cheng, Y, Sliz, P, Hiroaki, Y, Fujiyoshi, Y, Harrison,
S. C., & Walz, T. (2005) Lipid-protein interactions in
double-layered two-dimensional AQP0 crystals. Nature 438,
633; рис. 11-10, 6. PDB ID 2BL2, Murata, T, Yamato, L,
Kakinuma, Y, Leslie, A. G., & Walker, J. E. (2005) Structure
of the rotor of the V-Туре Na+-ATPase from Enterococcus
hirae. Science 308, 654; рис. 11-12. PDB ID 1BL8, Doyle,
D. A., Morais Cabral, J., Pfuetzner, R. A., Kuo, A., Gulbis,
I 3801 Источники иллюстраций
J. M., Cohen, S. L., Chait, В. T, & MacKinnon, R. (1998)
The structure of the potassium channel: molecular basis
of K+ conduction and selectivity. Science 280, 69; PDB ID
1AF6, Wang, Y. E, Dutzler, R., Rizkallah, P. J., Rosenbusch,
J. P, & Schirmer, T. (1997) Channel specificity: structural
basis for sugar discrimination and differential flux rates in
maltoporin.J. Mol. Biol. 272, 56; PDB ID 1QD5, Snijder,
H. J., Ubarretxena-Belandia, L, Blaauw, M., Kalk, К. H.,
Verheij, H. M., Egmond, M. R., Dekker, N., & Dijkstra,
B. W. (1999) Structural evidence for dimerization-regulated
activation of an integral membrane phospholipase. Nature
401, 717; PDB ID 1QJ9, Vogt, J. & Schulz, G. E. (1999)
The structure of the outer membrane protein OmpX from
Escherichia coli reveals mechanisms of virulence. Structure 7,
1301; PDB ID 1PHO, Cowan, S. W., Schirmer, T, Rummel,
G., Steiert, M., Ghosh, R., Pauptit, R. A., Jansonius, J. N.,
& Rosenbusch, J. P. (1992) Crystal structures explain
functional properties of two E. coli porins. Nature 358,
727; рис. 11-13. PDB ID 1FEP, Buchanan, S. K., Smith,
B. S., Venkatramani, L., Xia, D., Esser, L., Palnitkar, M.,
Chakraborty, R., van dor Helm, D., & Dcisenhofer, J. (1999)
Crystal structure of the outer membrane active transporter
FepA from Escherichia coli. Nat. Struct. Biol. 6, 56; PDB ID
1QD5, Snijder, H. J., Ubarretxena-Belandia, I., Blaauw, M.,
Kalk, К. H., Verheij, H. M., Egmond, M. R., Dekker, N., &
Dijkstra, B. W. (1999) Structural evidence for dimerization-
regulated activation of an integral membrane phospholipase.
Nature 401, 717; PDB ID 1MAL, Schirmer, T, Keller, T. A.,
Wang, Y. E, & Rosenbusch, J. P. (1995) Structural basis for
sugar translocation through maltoporin channels at 3.1 A
resolution. Science 267, 512; рис. 11-15. Heller, H., Schaefer,
M., & Schulten, K. (1993) Molecular dynamics simulation
of a bilayer of 200 lipids, in the gel and in the liquid-crystal
phases./. Phys. Chem. 97,8343; рис. 11-18. С любезного
разрешения Takahiro Fujiwara, Ken Ritchie, Hideji
Murakoshi, Ken Jacobson, and Akihiro Kusumi; доп. 11-
1, рис. 2, а. С любезного разрешения Daniel J. Mueller,
Ueli Aebi и Andreas Engel, M. E. Muller Institute (MSB),
Biozentrum, University of Basel; доп. 11-1, рис. 2, б. С
любезного разрешения S. Scheuring, D. Fotiadis, С. Moller,
S. A. Muller, A. Engel и D. J. Mueller; доп. 11-1, рис. 2, в.
С любезного разрешения Daniel J. Mueller, Max-Planck
Institute of Molecular Cell Biology and Genetics; рис. 11-
20, б. По микрофотографии, любезно предоставленной
J. М. Edwardson, University of Cambridge, Department
of Pharmacology; с изм.; рис. 11-21, а. С любезного
разрешения R. G. Parton. Перепечатано с разрешения
из Parton, R. G. & Simons, K. (2007) The multiple faces
of caveolae. Nat. Rev. Mol. Cell Biol. 8, 185-194, Fig. la;
рис. 11-23. По Zimmerberg, J. & Kozlov, M. M. (2006)
Nat. Rev. Mol. Cell Biol. 7, 15; с изм.; рис. 11-24. По Chen,
Y. A. & Scheller, R. H. (2001) SNARE-mediated membrane
fusion. Nature 2, 98; с изм.; рис. 11-29, а, в. По Mueckler,
M. (1994) Facilitative glucose transporters. Eur. J. Biochem.
219, 713; с изм.; доп. 11-2, рис. 1. По Lienhard, F. E., Slot,
J. W, James, D. E., & Mueckler, M. M. (1992) How cells
absorb glucose. Sci. Am. 266 (January), 86; с изм.; рис. 11-
35. PDB ID 1SU4, Toyoshima, C., Nakasako, M., Nomura,
H., & Ogawa, H. (2000) Crystal structure of the calcium
pump of sarcoplasmic reticulum at 2.6 angstrom resolution.
Nature 405, 647; рис. 11-36. По Kiihlbrandt, W. (2004)
Biology, structure and mechanism of P-type ATPases.
Nat. Rev. Mol. Cell Biol. 5, 291; с изм.; c. 563 (Скоу). C
любезного разрешения Information Office, University
of Aarhus, Denmark; рис. 11-41. PDB ID 1L7V, Locher,
К. P, Lee, A. T, & Rees, D. C. (2002) The E. coli BtuCD
structure: a framework for ABC transporter architecture
and mechanism. Science 296, 1091; доп. 11-3, рис. 2. Tom
Moninger, University of Iowa, Ames; рис. 11-43, a. PDB
ID 1PVL, Abramson, J., Smirnova, L, Kasho, V, Verner, G.,
Kaback, H. R., & Iwata, S. (2003) Structure and mechanism
of the lactose permease of Escherichia coli. Science 301, 610;
рис. 11-43, 6. PDB ID 2CFQ, Mirza, O., Guan, L., Verner,
G., Iwata, S., & Kaback, H. R. (2006) Structural evidence foi
induced fit and a mechanism for sugar/H+ symport in LacY.
EMBOJ. 25, 1177; рис. 11-45. Координаты рассчитал
для The Virtual Museum of Minerals and Molecules, www.
soils.wisc.edu/virtual_museum Phillip Barak, University
of Wisconsin-Madison, Department of Soil Science,
используя данные из Neupert-Laves, К. & Dobler, M.
(1975) The crystal structure of a K+ complex of valinomycin
Helv. Chim. Acta 58, 432; c. 571 (Эгр). С любезного
разрешения Royal Swedish Academy of Sciences; рис. 11-
46, a. PDB ID 2B5F, Tornroth-Horsefield, S., Wang, Y,
Hedfalk, K., Johanson, U., Karlsson, M., Tajkhorshid, E.,
Neutze, R., & Kjellbom, P. (2006) Structural mechanism
of plant aquaporin gating. Nature 439, 688; рис. 11-46, 6.
По PDB ID 1J4N, Sui, H., Han, B.-G., Lee, J. K., Walian,
P, & Jap, В. K. (2001) Structural basis of water-specific
transport through the AQP1 water channel. Nature 414,
872; с изм.; c. 575 (Heep, Закман) The Nobel Foundation;
(Маккиннон) С любезного разрешения Royal Swedish
Academy of Sciences; рис. 11-48 a, 6. PDB ID 1BL8,
Doyle, D. A., Cabral, J. M., Pfuetzner, R. A., Kuo, A., Gulbis,
J. M., Cohen, S. L., Chait, В. T, & Mackinnon, R. (1998)
The structure of the potassium channel: molecular basis
of K+ conduction and selectivity. Science 280, 69; рис. 11-
48, в. По Yellen, G. (2002) The voltage-gated potassium
channels and their relatives. Nature 419, 37; с изм.; рис. 11-
49. PDB ID 1J95, Zhou, M., Morais-Cabral, J. H., Mann, S.,
& MacKinnon, R. (2001) Potassium channel receptor site
for the inactivation gate and quaternary amine inhibitors.
Nature 411, 657; рис. 11-50, а, б, г. PDB ID 2A79, Long,
S. B., Campbell, E. B., & MacKinnon, R. (2005) Crystal
structure of a mammalian voltage-dependent Shaker family
K+ channel. Science 309,897; рис. 11-50, в. По Gandhi,
C. S. & Isacoff, E. Y. (2005) Shedding light on membrane
proteins. Trends Neurosci. 28,476.
ГЛАВА 12. C. 591. Earl W. Sutherland (1971) Nobel
Address, www.nobel.se/medicine/laureates/1971 /
sutherland-lecture.html; доп. 12-2, рис. 1 (Гилман)
Office of News and Publications, The University of Texas
Southwestern Medical Center at Dallas; (Родбелл) C
любезного разрешения Andrew Rodbell; доп. 12-2,
Источники иллюстраций [ 3s 1 ]
рис. 2. PDB ID 5Р21, Pai, Е. Е, Krengel, U., Petsko,
G. A., Goody, R. S„ Kabsch, W, & Wittinghofer, A. (1990)
Refined crystal structure of the triphosphate conformation
of H-ras p21 at 1.35 A resolution: implications for the
mechanism of GTP hydrolysis. EMBOJ. 9, 2351; доп. 12-
2, рис. 3. По Vetter, I. R. & Wittinghofer, A. (2001) The
guanine nucleotide-binding switch in three dimensions.
Science 294, 1300, Fig. 3; с изм.; рис. 12-6, 6. PDB ID
1U7E, Kim, C., Xuong, N.-H., & Taylor, S. S. (2005)
Crystal structure of a complex between the catalytic
and regulatory (Ria) subunits of PKA. Science 307, 690;
доп. 12-3, рис. 1. Chris Parks/ImageQuest Marine;
доп. 12-3, рис. 2. PDB ID 1GFL, Yang, E, Moss, L. G., &
Phillips, G. N., Jr. (1996) The molecular structure of green
fluorescent protein. Nat. Biotechnol. 14, 1246; рис. 12-11,
a. PDB ID 1CLL, Chattopadhyaya, R., Meador, W. E.,
Means, A. R., & Quiocho, F. (1992) A calmodulin structure
refined at 1.7 angstroms resolution./ Mol. Biol. 228, 1177;
рис. 12-11, б, в. PDB ID 1CDL, Meador, W. E., Means,
A. R., & Quiocho, F. A. (1992) Target enzyme recognition
by calmodulin: 2.4 angstroms structure of a calmodulin-
peptide complex. Science 257, 1251; рис. 12-12, a. C
любезного разрешения Michael D. Cahalan, University
of California, Irvine, Department of Physiology and
Biophysics; рис. 12-12, 6. Rooney, T. A., Sass, E. J., &
Thomas, A. P. (1989) Characterization of cytosolic calcium
oscillations induced by phenylephrine and vasopressin in
single fura-2-loaded hepatocytes./ Biol. Chem. 264, 17,131;
рис. 12-13. По Foskett, J. K., White, C., Cheung, K.-H.,
& Mak, D.-O. D. (2007) Inositol trisphosphate receptor
Ca2+ release channels. Physiol. Rev. 87, 593; рис. 12-14, 6.
(Рецептор инсулина) с изм. из PDB ID 2DTG, McKern,
N. М., Lawrence, М. С., Streltsov, V. A., Lou, M.-Z., Adams,
T. E., Lovrecz, G. O., Elleman, T. C., Richards, К. M.,
Bentley, J. D., Pilling, P. A., Hoyne, P. A., Cartledge, K. A.,
Pham, T. M., Lewis, J. L., Sankovich, S. E., Stoichevska, V,
Da Silva, E., Robinson, С. P, Frenkel, M. J., Sparrow, L. G.,
Fernley, R. T, Ера, V. C., & Ward, C. W. (2006) Structure
of the insulin receptor ectodomain reveals a folded-over
conformation. Nature 443, 218; (инсулин) PDB ID 2CEU,
Whittingham, J. L., Zhang, Y., Zakova, L., Dodson, E. J.,
Turkenburg, J. P, & Dodson, G. G. (2006) 1222 crystal
form of despentapeptide (B26-B30) insulin provides new
insights into the properties of monomeric insulin. Acta
Crystallogr. D Biol. Crystallogr. 62, 505-511; рис. 12-14,
e. PDB ID 1IRK, Hubbard, S. R., Wei, L., Ellis, L., &
Hendrickson, W. A. (1994) Crystal structure of the tyrosine
kinase domain of the human insulin receptor. Nature 372,
746; рис. 12-14, г. PDB ID 1IR3, Hubbard, S. R. (1997)
Crystal structure of the activated insulin receptor tyrosine
kinase in complex with peptide substrate and ATP analog.
EMBOJ. 16, 5572; рис. 12-21. PDB ID 1SHC, Zhou,
M. M., Ravichandran, K. S., Olejniczak, E. E, Petros, A. M.,
Meadows, R. P, Sattler, M., Harlan, J. E., Wade, W. S.,
Burakoff, S. J., & Fesik, S. W. (1995) Structure and ligand
recognition of the phosphotyrosine binding domain of She.
Nature 378, 584; рис. 12-23. По Pawson, T, Gish, G. D.,
& Nash, P. (2001) SH2 domains, interaction modules and
cellular wiring. Trends Cell Biol. 11, 504, Fig. 5; с изм.;
рис. 12-26. По Taylor, R. (1994) Evolutions: the voltage-
gated sodium channel./ NIH Res. 6, 112; с изм.; рис. 12-27,
я, б. По Changeux, J. P. (1993) Chemical signaling in the
brain. Sci. Am. 269 (November), 58; с изм.; рис. 12-33. По
Quaked, E, Rozhon, W, Lecourieux, D., & Hirt, H. (2003)
А МАРК pathway mediates ethylene signaling in plants.
EMBOJ. 22, 1282; с изм.; рис. 12-34. По Tichtinsky, G.,
Vanoosthuyse, V, Cock, J. M., & Gaude, T. (2003) Making
inroads into plant receptor kinase signalling pathways.
Trends Plant Sci. 8, 231, Fig. 1; с изм.; рис. 12-37. PDB ID
1BAC, Chou, К.-C., Carlacci, L., Maggiora, G. M., Parodi,
L. A., & Schulz, M. W. (1992) An energy-based approach
to packing the 7-helix bundle of bacteriorhodopsin.
Protein Sci. 1, 810; рис. 12-39. По Nathans, J. (1989) The
genes for color vision. Sci. Am. 260 (February), 42; с изм.;
доп. 12-4, рис. 1. С любезного разрешения Professor
J. D. Mollon, Cambridge University, Department of
Experimental Psychology; рис. 12-44, a. PDB ID 1HCK,
Schulze-Gahmen, U., De Bondt, H. L., & Kim, S.-H. (1996)
High-resolution crystal structures of human cyclin-
dependent kinase 2 with and without ATP: bound waters
and natural ligand as guides for inhibitor design./ Med.
Chem. 39, 4540; рис. 12-44, 6. PDB ID 1FIN, Jeffrey, P. D.,
Russo, A. A., Polyak, K., Gibbs, E., Hurwitz, J., Massague,
J., & Pavletich, N. P. (1995) Mechanism of Cdk activation
revealed by the structure of a cyclin a-Cdk2 complex.
Nature 376, 313; рис. 12-44, в. PDB ID 1JST, Russo, A. A.,
Jeffrey, P. D., & Pavletich, N. P. (1996) Structural basis of
cyclin-dependent kinase activation by phosphorylation.
Nat. Struct. Biol. 3, 696; рис. 12-45. Данные из Pines, J.
(1999) Four-dimensional control of the cell cycle. Nat. Cell
Biol. 1, E73; доп. 12-5, рис. 1. CNRI/Photo Researchers;
доп. 12-5, рис. 2. PDB ID 1S9I, Ohren, J. E, Chen, H.,
Pavlovsky, A., Whitehead, C., Zhang, E., Kuffa, P, Yan,
C., McConnell, P, Spessard, C., Banotai, C., Mueller,
W. T, Delaney, A., Omer, C., Sebolt-Leopold, J., Dudley,
D. T, Leung, I. K., Flamme, C., Warmus, J., Kaufman,
M., Barrett, S., Tecle, H., & Hasemann, C. A. (2004)
Structures of human MAP kinase kinase 1 (MEK1) and
MEK2 describe novel noncompetitive kinase inhibition.
Nat. Struct. Mol. Biol. 11, 1192; доп. 12-5, рис. 3, a. PDB
ID IIEP, Nagar, B., Bornmann, W, Pellicena, P, Schindler,
T, Veach, D. R., Miller, W. T, Clarkson, B., & Kuriyan, J.
(2002) Crystal structures of the kinase domain of c-Abl
in complex with the small molecule inhibitors PD173955
and imatinib (STI-571). Cancer Res. 62, 4236; доп. 12-5,
рис. 3, 6. PDB ID 1M17, Stamos, J., Sliwkowski, M. X.,
& Eigenbrot, C. (2002) Structure of the epidermal growth
factor receptor kinase domain alone and in complex with a
4-anilinoquinazoline inhibitor./ Biol. Chem. 277, 46,265;
доп. 12-5, рис. 3, в. PDB ID 1S9I, см. источник для
доп. 12-5, рис. 2; доп. 12-5, рис. 3, г. PDB ID 2A4L, De
Azevedo, W. E, Leclerc, S., Meijer, L., Havlicek, L., Strnad,
M., & Kim, S. H. (1997) Inhibition of cyclin-dependent
kinases by purine analogues: crystal structure of human
13821 Источники иллюстраций
cdk2 complexed with roscovitine. Eur. J. Biochem. 243,
518; рис. 12-50. По Kinzler, К. W. & Vogelstein, B. (1996)
Lessons from hereditary colorectal cancer. Cell 87, 159;
с изм.
Tom 2
ГЛАВА 13. C. 11. Rudolf Clausius (1865) Uber
Verschiedene far die Anwendung bequeme Formen der
Hauptgleichungcn der mechanischen Warmetheorie
(On several convenient forms of the second fundamental
equations of the mechanical theory of heat). In The
Mechanical Theory of Heat with Its Applications to the Steam-
Engine and to the Physical Properties of Bodies (Tyndall, J.,
trans.), John van Voorst, London, p. 365; Francois Jacob
(1970) The Logic of Life: A History of Heredity, Pantheon
Books, Inc., New York, 1973; оригинальная публикация
(1970) La logique du vivant: une histoire de I’hertdite,
Editions Gallimard, Paris; (Лавуазье) The Granger
Collection; c. 12. © Sidney Harris; рис. 13-7. По Layer,
G., Heinz, D. W, Hahn, D., & Schubert, W.-D. (2004)
Structure and function of radical SAM enzymes. Curr. Opin.
Chem. Biol. 8, 472, Fig. 4; с изм.; доп. 13-1, рис. 1. E. R.
Degginger/Photo Researchers; рис. 13-25. PDB ID 1LDH,
White, J. L., Hackert, M. L., Buehner, M., Adams, M. J.,
Ford, G. C., Lentz, P. J.,Jr., Smiley, I. E., Steindel, S. J., &
Rossmann, M. G. (1976) A comparison of the structures
of apo dogfish M4 lactate dehydrogenase and its ternary
complexes./. Mol. Biol. 102, 759; c. 53 (Стронг, Эльвейем)
С любезного разрешения University of Wisconsin-
Madison, Department of Biochemistry; (Вули) Rockefeller
Archive Center.
ГЛАВА 14. C. 65. Arthur Harden (1923) Alcoholic
Fermentation, 3d edn, Longmans, Green and Co., London;
c. 66 (Эйлер-Челпин) Austrian Archives/Corbis;
(Эмбден) С любезного разрешения Institut far Biochemic
I: Molekulare Bioenergetik, Universitaetsklinikum
Frankfurt, ZBC; (Мейергоф) Hulton-Seutsch Collection/
Corbis; c. 71 (Гарден) Hulton Getty/Liaison; (Янг) C
любезного разрешения Medical History Museum, The
University of Melbourne; c. 80 (Варбург) Hulton Archive
by Getty Images; доп. 14-1, рис. 3. ISM/Phototake;
доп. 14-2. Animals Animals; доп. 14-3, рис. 1. Charles
O’Rear/CORBIS.
ГЛАВА 15. C. 121. C. F. Cori and G. T. Cori (1929)
Glycogen formation in the liver from d- and 1-lactic acid.
Journal of Biological Chemistry 81, 389-403; Alfred A.
Knopf, New York; рис. 15-1. www.genome.ad.jp/kegg/
pathway/map/mapOl 100.html; c. 132 (Бухнер) The Nobel
Foundation; рис. 15-7. Данные из Torres, N. V, Mateo,
E, Melendez-Hevia, E., & Kacser, H. (1986) Kinetics of
metabolic pathways: a system in vitro to study the control
of flux. Biochem.J. 234, 169; доп. 15-1, рис. 2. Fell, D.
(1997) Understanding the Control of Metabolism, Portland,
London, p. 103; рис. 15-14, a. PDB ID 1PFK, Shirakihara,
Y. & Evans, P. R. (1988) Crystal structure of the complex
of phosphofructokinase from Escherichia coli with its
reaction products./. Mol. Biol. 204, 973; рис. 15-18, a.
PDB ID 2NPP, Xu, Y., Xing, Y, Chen, Y, Chao, Y, Lin, Z.,
Fan, E., Yu, J. W, Strack, S., Jeffrey, P. D., & Shi, Y. (2006)
Structure of the protein phosphatase 2A holoenzyme. Cell
127, 1239; рис. 15-23. По Chakravarty, К., Cassuto, H.,
Reshef, L., & Hanson, R. W. (2005) Crit. Rev. Biochem. Mol.
Biol. 40, 133, Fig. 2; с изм.; рис. 15-24. BCC Microimaging,
перепечатано с разрешения; с. 158 (Лелуар) The Nobel
Foundation; доп. 15-4 (Кори) АР; рис. 15-32. PDB ID
1LL2, Gibbons, В. J., Roach, Р. J., & Hurley, T. D. (2002)
Crystal structure of the autocatalytic initiator of glycogen
biosynthesis, glycogenin./. Mol. Biol. 319,463; c. 165
(Сазерленд) The Nobel Foundation.
ГЛАВА 16. C. 182. H. A. Krebs and W. A. Johnson
(1937) The role of citric acid in intermediate metabolism
in animal tissues. Enzymologia 4, 148; c. 183 (Кребс) C
любезного разрешения Public Relations Office, University
of Sheffield; рис. 16-5, а, б. С любезного разрешения
Dr. Z. Hong Zhou, University of Texas-Houston Medical
School, Department of Pathology and Laboratory Medicine;
рис. 16-8, a. PDB ID 5CSC, Liao, D.-L, Karpusas, N^.,
& Remington, S. J. (1991) Crystal structure of an open
conformation of citrate synthase from chicken heart at 2.8-A
resolution. Biochemistry 30, 6031; рис. 16- 8, 6. PDB ID
5CTS, Karpusas, M., Branchaud, B., & Remington, S. J.
(1990) Proposed mechanism for the condensation reaction
of citrate synthase: 1.9-angstroms structure of the ternary
complex with oxaloacetate and carboxymethyl coenzyme
A. Biochemistry 29, 2213; рис. 16-9. По Remington, J. S.
(1992) Mechanisms of citrate synthase and related enzymes
(triose phosphate isomerase and mandelate racemase).
Curr. Opin. Struct. Biol. 2, 730; с изм.; доп. 16-1, рис. 1. По
Eisenstein, R. S. (2000) Iron regulatory proteins and the
molecular control of mammalian iron metabolism. Annu.
Rev. Nutr. 20, 637, Fig. 1; с изм.; доп. 16-1, рис. 2, a. PDB
ID 2B3Y, Dupuy, J., Volbeda, A., Carpentier, P, Darnault,
C., Moulis, J. M., & Fontecilla-Camps, J. C. (2006) Crystal
structure of human iron regulatory protein 1 as cytosolic
aconitase. Structure 14, 129; Box доп. 16-1, рис. 2, б.
PDB ID 2IPY, Walden, W. E., Selezneva, A. L, Dupuy, J.,
Volbeda, A., Fontecilla-Camps, J. C., Theil, E. C., & Volz, K.
(2006) Structure of dual function iron regulatory protein
1 complexed with ferritin IRE-RNA. Science 314, 1903;
рис. 16-12, 6. PDB ID 1SCU, Wolodko, W. T, Fraser,
M. E., James, M. N. G., & Bridger, W. A. (1994) The crystal
structure of succinyl-Co A synthetase from Escherichia
coli at 2.5 angstroms resolution./. Biol. Chem. 289, 10,883;
рис. 16-21. Richard N. Trelease, Arizona State University,
Department of Botany.
ГЛАВА 17. C. 225. John Clarke (1639) Paroemiologia
Anglo-Latina (Proverbs English and Latin), imprinted by F.
Kyngston for R. Mylbourne, London; доп. 17-1.
Stouffer Productions/Animals Animals; доп. 17-2
(Ходжкин) The Nobel Foundation.
ГЛАВА 18. C. 261. Hans Krebs (1970) The history of
Источники иллюстраций [3831
the tricarboxylic acid cycle. Perspectives in Biology and
Medicine 14, 154-170, 161-162; рис. 18-5, в, г, d. PDB
ID 1AJS, Rhee, S., Silva, M. M., Hyde, С. C., Rogers,
P. H., Metzler, С. M., Metzler, D. E., & Arnone, A. (1997)
Refinement and comparisons of the crystal structures of pig
cytosolic aspartate aminotransferase and its complex with
2-methylaspartate./ Biol. Chem. 272, 17,293.
ГЛАВА 19. C. 305. Claude Bernard (1865) An Introduction
to the Study of Experimental Medicine (H. C. Greene,
trans.), Harry Schumann, Paris. Reprinted (1957) by
Dover Publications, New York; Peter Mitchell (1979)
Keilin’s respiratory chain concept and its chemiosmotic
consequences. Science 206, 1148-1159; c. 306 (Ленинджер)
Alan Mason Chesney Medical Archives of The Johns
Hopkins Medical Institutions; рис. 19-5, г. PDB ID
1FRD, Jacobson, B. L., Chae, Y. K., Markley, J. L., Rayment,
I., & Holden, H. M. (1993) Molecular structure of the
oxidized, recombinant, heterocyst (2Fe-2S) ferredoxin from
Anabaena 7120 determined to 1.7 angstroms resolution.
Biochemistry 32, 6788; рис. 19-9. PDB ID 2FUG, Sazanov,
L. A. & Hinchliffe, P. (2006) Structure of the hydrophilic
domain of respiratory complex I from Thermus thermophilus.
Science^, 1430; рис. 19-10. PDB ID 1ZOY, Sun, E, Huo,
X., Zhai, Y., Wang, A., Xu, J., Su, D., Bartlam, M., & Rao,
Z. (2005) Crystal structure of mitochondrial respiratory
membrane protein Complex II. Cell 121, 1043; рис. 19-11,
a. PDB ID 1BGY, Iwata, S., Lee, J. W., Okada, K., Lee,
J. K., Iwata, M., Rasmussen, B., Link, T. A., Ramaswamy, S.,
& Jap, В. K. (1998) Complete structure of the 11-subunit
bovine mitochondrial cytochrome bcl complex. Science
281, 64; рис. 19-13. PDB ID 1OCC, Tsukihara, T,
Aoyama, H., Yamashita, E., Tomizaki, T, Yamaguchi, H.,
Shinzawa-Itoh, K., Nakashima, R., Yaono, R., & Yoshikawa,
S. (1996) The whole structure of the 13-subunit oxidized
cytochrome c oxidase at 2.8 A. Science 272, 1136; рис. 19-
14. По Williams, R. J. P. (1995) Nature 378, 235; с изм.,
исправлено по сравнению с Williams, R. J. P. (1995)
Purpose of proton pathways. Nature 376, 643; рис. 19-
15, а, б. С любезного разрешения Egbert Boekema.
Перепечатано с разрешения из Heinemeyer, J., Braun,
H.-Р., Boekema, E. J., & Kour, R. (2007) A structural model
of the cytochrome c reductase/oxidase supercomplex from
yeast mitochondria./ Biol. Chem. 282, 12,240-12,248, Figs
4A and 5A; c. 327 (Митчелл) AP/Wide World Photos;
доп. 19-1, рис. 1. D. Cavagnaro/Visuals Unlimited; c. 331
(Рэкер) С любезного разрешения E. Racker; рис. 19-16.
PDB ID 2FUG, 1ZOY, 1BGY и 1OCC; см. источники
для рис. 19-9, 19-10, 19-11 л и 19-13; PDB ID 1HRC,
Bushnell, G. W., Louie, G. V., & Brayer, G. D. (1990)
High-resolution three-dimensional structure of horse heart
cytochrome c.J. Mol. Biol. 214, 585; рис. 19-23, 6. PDB ID
1BMF, Abrahams, J. P, Leslie, A. G., Lutter, R., & Walker,
J. E. (1994) Structure at 2.8 A resolution of Fl-ATPase from
bovine heart mitochondria. Nature 370, 621; рис. 19-25, 6,
в. PDB ID 1BMF, см. источник для рис.19-23, б; с. 333
(Уокер) The Nobel Foundation; рис. 19-25 г, д. PDB ID
1QO1, Stock, D., Leslie, A. G. W, & Walker, J. E. (1999)
Molecular architecture of the rotary motor in ATP synthase.
Science 286, 1700; белковая основа рассчитана из данных
Са PDB с помощью MIDAS-PLUS и Sybyl; рис. 19-25,
е. По Fillingame, R. Н. (1999) Protein structure: molecular
rotary motors. Science 286,1687; c. 336 (Бойер) The
Nobel Foundation; рис. 19-27 (слева). По Sambongi, Y,
Iko, Y, Tanabe, M., Omote, H., Iwamoto-Kihara, A., Ueda,
L, Yanagida, T, Wada, Y., & Futai, M. (1999) Mechanical
rotation of the c subunit oligomer in ATP synthase (F0Ft):
direct observation. Science 286, 1722-1724; с изм.; рис. 19-
27 (справа). С любезного разрешения Ryohei Yasuda
и Kazuhiko Kinosita из Yasuda, R., Noji, H., Kinosita, K.,
Jr., & Yoshida, M. (1998) FrATPase is a highly efficient
molecular motor that rotates with discrete 120° steps.
Cell93, 1117; рис. 19-31. PDB ID 1OHH, Cabezon, E„
Montgomery, M. G., Leslie, A. G. W., & Walker, J. E. (2003)
The structure of bovine FrATPase in complex with its
regulatory protein I Fl. Nat. Struct. Biol. 10, 744; рис. 19-
32. По Harris, D. A. (1995) Bioenergetics at a Glance,
Blackwell Science, London, p. 36; с изм.; рис. 19-35. Don
W. Fawcett/Photo Researchers; рис. 19-37. По Riedl,
S. J. & Salvesen, G. S. (2007) The apoptosome: signaling
platform of cell death. Nat. Rev. Mol. Cell Biol. 8, 409, Fig. 3;
с изм; рис. 19-38, a. Morris, M.A. (1990) Mitomutations
in neuro-ophthalmological diseases: a review./ Clin.
Neuroophthalmol. 10, 159; рис. 19-38, б. Из Wallace, D.,
Zheng, X., Lott, M. T, Shoffner, J. M., Hodge, J. A., Kelley,
R. L, Epstein, С. M., & Hopkins, L. C. (1988) Familial
mitochondrial encephalomyopathy (MERRF): genetic,
pathophysiological, and biochemical characterization of
a mitochondrial DNA disease. Cell 55, 601; рис. 19-40.
Michael W. Davidson, Florida State University; рис. 19-41,
б. С любезного разрешения Rob Taylor. Перепечатано
с разрешения из Taylor, R. W. & Turnbull, D.M. (2005)
Mitochondrial DNA mutations in human disease. Nat.
Rev. Genet. 6, 389-402, Fig. 2a; рис. 19-45, 6. Biological
Photo Service; рис. 19-49. PDB ID 2BHW, Standfuss,
J., Terwisscha van Scheltinga, A. C., Lamborghini, M., &
Kuhlbrandt, W. (2005) Mechanisms of photoprotection
and nonphotochemical quenching in pea light-harvesting
complex at 2.5 A resolution. EMBOJ. 24, 919; рис. 19-50,
19-54, a, 6,19-60, 6,19-61,19-65. По Heldt, H.-W. (1997)
Plant Biochemistry and Molecular Biology, Oxford University
Press, Oxford, pp. 57, 62, 63, 100, 101, 133; рис. 19-55. PDB
ID 1PRC, Deisenhofer, J., Epp, O., Sinning, L, & Michel,
H. (1995) Crystallographic refinement at 2.3 angstroms
resolution and refined model of the photosynthetic reaction
center from Rhodopseudomonas viridis./ Mol. Biol. 246,
429; рис. 19-57. По Rutherford, A.W. & Faller, P. (2001)
The heart of photosynthesis in glorious 3D. Trends Biochem.
Sci. 26,341, Fig. 1; с изм.; рис. 19-58, а. По Kuhlbrandt,
W. (2001) Structural biology: chlorophylls galore. Nature
411, 896, Fig. 1; рис. 19-58, б, в. PDB ID 1JBO, Nield,
J., Rizkallah, P. J., Barber, J., & Chayen, N. E. (2003) The
1.45 A three-dimensional structure of c-phycocyanin from
the thermophilic cyanobacterium Synechococcus elongatus.
J. Struct. Biol. 141, 149; рис. 19-59, a, 6 PDB ID 1UM3,
[ 38\ | Источники иллюстраций
Kurisu, G., Zhang, H., Smith, J. L., & Cramer, W. A. (2003)
Structure of the cytochrome b6 f complex of oxygenic
photosynthesis: tuning the cavity. Science 302 1009;
c. 382 (Арнон) University of California, Berkeley; c. 383
(Ягендорф) Cornell University; рис. 19-60, a. PDB ID
2AXT, Loll, B., Kern, J., Saenger, W., Zouni, A., & Biesiadka,
J. (2005) Towards complete cofactor arrangement in the 3.0
A resolution structure of photosystem II. Nature 438, 1040;
PDB ID 2E74, Yamashita, E., Zhang, H., & Cramer, W. A.
(2007) Structure of the cytochrome b6 f complex: quinone
analogue inhibitors as ligands of heme cn.J. Mol. Biol. 370,
39; PDB ID 1A70, Binda, C., Coda, A., Aliverti, A., Zanetti,
G., & Mattevi, A. (1998) Structure of the mutant E92K
of [2Fe-2S] ferredoxin I from Spinacia oleracea at 1.7 A
resolution. Acta Crystallogr. D Biol. Crystallogr. 54, 1353;
PDB ID 1AG6, Xue, Y, Okvist, M., Hansson, O., & Young,
S. (1998) Crystal structure of spinach plastocyanin at 1.7
A resolution. Protein Sci. 7, 2099; PDB ID 2001, Amunts,
A., Drory, O., & Nelson, N. (2007) The structure of a plant
photosystem I supercomplex at 3.4 A resolution. Nature
447, 58; PDB ID 1QG0, Deng, Z., Aliverti, A., Zanetti,
G., Arakaki, A. K., Ottado, J., Orellano, E. G., Calcaterra,
N. B., Ceccarelli, E. A., Carrillo, N., & Karplus, P A.
(1999) A productive NADP4 binding mode of ferredoxin-
NADP+ reductase revealed by protein engineering and
crystallographic studies. Nat. Struct. Biol. 6, 847; PDB ID
1QO1, см. источник для рис. 19-25, z, д; рис. 19-66, а.
PDB ID 1C8R, Luecke, Н., Schobert, В., Richter, Н.-Т,
Cartailler, J.-P, & Lanyi, J. К. (1999) Structural changes in
bacteriorhodopsin during ion transport at 2 A resolution.
Science 286, 255; рис. 19-66, б. По Gennis, R. B. & Ebrey,
T. G. (1999) Proton pump caught in the act. Science 286,
252; с изм.
ГЛАВА 20. C. 403. Melvin Calvin (1961) Nobel Address,
www.nobel.se/chemistry/laureates/1961/calvin-lecture.
html; рис. 20-1. PhotoDisc; c. 405 (Кальвин) The Nobel
Foundation; рис. 20-2. Ken Wagner/Visuals Unlimited;
рис. 20-5, a. PDB ID 8RUC, Andersson, I. (1996)
Large structures at high resolution: the 1.6 angstroms
crystal structure of spinach ribulose-1,5-bisphosphate
carboxylase/oxygenase complexed with 2-carboxyarabinitol
bisphosphate.J. Mol. Biol. 259,160; рис. 20-5, 6. PDB
ID 9RUB, Lundqvist, T. & Schneider, G. (1991) Crystal
structure of activated ribulose-1,5-bisphosphate carboxylase
complexed with its substrate, ribulose-1,5-bisphosphate.
J. Biol. Chem. 266,12,604; рис. 20-6. PDB ID 1RCX,
Taylor, T. C. & Andersson, I. (1997) The structure of
the complex between rubisco and its natural substrate
ribulose 1,5-bisphosphate. J. Mol. Biol. 265,432; рис. 20-18.
Halliwell, B. (1984) Chloroplast Metabolism: The Structure
and Function of Chloroplasts in Green Leaf Cells, Clarendon
Press, Oxford, p. 97; рис. 20-23, a. Ray Evert, University
of Wisconsin-Madison, Department of Botany; рис. 20-29
(целлюлоза) Ken Wagner/Visuals Unlimited; рисунок
no Becker, W. M. & Deamer, D. W. (1991) The World of
the Cell, 2nd edn, The Benjamin/Cummings Publishing
Company, Inc., Redwood City, CA, p. 60, Fig. 3-20; рис. 20-
30, врезка. С любезного разрешения Mark J. Grimson,
Texas Tech University, and Candace H. Haigler, North
Carolina State University; c. 443 (задача 19). С любезного
разрешения Elena V. Voznesenskaya, Vincent R. Franceschi,
Olavi Kiirats, Elena G. Artyusheva, Helmut Freitag, и
Gerald E. Edwards.
ГЛАВА 21. C. 445. Konrad Bloch (1987) Summing up.
Annual Review of Biochemistry 56,10; рис. 21-3, a.
PDB ID 2CF2, Maier, T, Jenni, S., Ban, N. (2006)
Architecture of mammalian fatty acid synthase at 4.5 A
resolution. Science 311,1258; рис. 21-3, 6. PDB IDs
2UV9, 2UVA, 2UVB, and 2UVC, Jenni, S., Leibundgut,
M., Boehringer, D., Frick, C., Mikolasek, B., Ban, N. (2007)
Structure of fungal fatty acid synthase and implications
for iterative substrate shuttling. Science 316, 254; рис. 21-
11, 6. Daniel Lane, The Johns Hopkins University, School
of Medicine; c. 472 (Кеннеди) Harvard Medical School;
c. 484 (Блох, Линеен, Корнфорт) AP/Wide World
Photos; (Попьяк) Corbis/UPI/Bettmann; рис. 21-39,
б. С любезного разрешения Robert L. Hamilton and
the Arteriosclerosis Specialized Center of Research,
University of California, San Francisco; рис. 21-40, 6. Alan
Attic, University of Wisconsin-Madison, Department of
Biochemistry; c. 491 (Браун и Голдштейн) Corbis/UPI/
Bettmann; доп. 21-3 (Эндо) С любезного разрешения
Akira Endo, Ph.D.; (Альберте) С любезного разрешения
Alfred W. Alberts; (Вагелос) С любезного разрешения Р.
Roy Vagelos.
ГЛАВА 22. С. 505. Gertrude Elion, цит. no S. В. McGrayne
(2002) Damn the torpedoes. Full speed ahead. Science 296,
851-852; доп. 22-1, рис. 1, 2. По van Niftrik, L. A., Fucrst,
J. A., Damste,J. S. S., Kuenen,J. G.Jetten, M. S. M., &
Strous, M. (2004) The anammoxosome: an intracytoplasmic
compartment in anammox bacteria. FEMS Microbiol. Lett.
233, 10, Figs 4 and 3; с изм.; доп. 22-1, рис. 3. С любезного
разрешения John Fuerst. Перепечатано с разрешения
из Lindsay, М. R., Webb, R. L, Strous., М., Jetten, М. S.,
Butler, М. К., Forde, R. J., & Fuerst, J. А. (2001) Cell
compartmentalisation in planctomycetes: novel types of
structural organisation for the bacterial cell. Arch. Microbiol.
175, 421, Fig. 6A; рис. 22-3. PDB ID 1N2C, Schindelin, H.,
Kisker, C., Schlessman, J. L., Howard, J. B., & Rees, D. C.
(1997) Structure of ADP X Alf4 -stabilized nitrogenase
complex and its implications for signal transduction. Nature
387, 370; рис. 22-4, a. С. P. Vance/Visuals Unlimited;
(врезка) Ric Ergenbright/Corbis; рис. 22-4, 6. Jeremy
Burgess/Photo Researchers; рис. 22-5. PDB ID 2GLS,
Yamashita, M. M., Almassy, R. J., Janson, C. A., Cascio, D.,
& Eisenberg, D. (1989) Refined atomic model of glutamine
synthetase at 3.5 angstroms resolution. J. Biol. Chem. 264,
17,681; доп. 22-3, рис. 1. John Mansfield, University of
Wisconsin-Madison, Department of Veterinary Science;
c. 545 (Бьюкенен) Massachusetts Institute of Technology
Museum Collection; рис. 22-37. PDB ID 1M6V, Thoden,
J. B„ Huang, X., Raushel, F. M., & Holden, H. M. (2002)
Carbamoyl-phosphate synthetase: creation of an escape
Источники иллюстраций 13 85]
route for ammonia./. Biol. Chem. ТП, 39,722; Jim Thoden
and Hazel Holden, University of Wisconsin-Madison,
Department of Biochemistry and Enzyme Institute,
provided preliminary data for the channel path; рис. 22-
40, a. Thelander, L. & Reichard, P. (1979) Reduction of
ribonucleotides. Annu. Rev. Biochem. 48, 133; рис. 22-
40, 6. PDB ID 1PFR, Logan, D. T, Su, X. D., Aberg, A.,
Regnstrom, K., Hajdu, J., Eklund, H., & Nordlund, P. (1996)
Crystal structure of reduced protein r2 of ribonucleotide
reductase: the structural basis for oxygen activation at
a dinuclear iron site. Structure 4, 1053; c. 557 (Элайон
и Хитчингс) с любезного разрешения Kathy Bendo
Hitchings.
ГЛАВА 23. С. 567. Charles Edouard Brown-Sequard and
J. d’Arsonval (1891) Additions a une note sur 1’injection
des extraits liquides de divers organes, comme methode
therapeutique (Additions to a note on the injection of
aqueous extracts of various organs, as a therapeutic method).
Comptes Rendus de la Societe de Biologie (9)3, 267; c. 569
(Гиймен) Corbis/UPI/Bettmann; (Шалли) AP/Wide
World Photos; (Ялоу) с любезного разрешения Rosalyn
S. Yalow, Veterans Affairs Medical Center, New York;
доп. 23-1. (Бантинг, Маклеод, Бест, Коллип) Corbis/
UPI/Bettmann; рис. 23-3. По Orth, D. N. (1975) General
considerations for radioimmunoassay of peptide hormones.
Methods Enzymol. 37, 22; с изм.; рис. 23-16, a, Fred
Hossler/Visuals Unlimited; рис. 23-19. Перепечатано с
разрешения из Blei, М. L., Conley, К. Е., & Kushmerick,
М. J. (1993) Separate measures of АТР utilization and
recovery in human skeletal muscle./. Physiol. 465, 210, Fig.
4; рис. 23-21. D. W. Fawcett/Photo Researchers; рис. 23-
23. С любезного разрешения M. L. Thomas, H. C. Sing,
G. Belenky, Walter Reed Army Institute of Research, U.S.
Army Medical Research Materiel Command, Division of
Neuropsychiatry; рис. 23-29, б. Координаты с любезного
разрешения Frances М. Ashcroft, Oxford University,
использованы с разрешения S. Haider and М. S. Р. Sansom,
чтобы заново построить модель, опубликованную
в Antcliff, J. Е, Haider, S., Proks. Р, Sansom, М. S. Р,
Ashcroft, Е М. (2005) Functional analysis of a structural
model of the АТР-binding site of the KATP channel Kir6.2
subunit. EMBOJ. 24, 229-239; рис. 23-34. John Sholtis,
The Rockefeller University, New York; рис. 23-35. По
Ezzell, C. (1995) Fat times for obesity research: tons of new
information, but how does it all fit together?/. NIH Res. 7,
39; с изм.; рис. 23-36. По Schwartz, M. W. & Morton, G. J.
(2002) Obesity: keeping hunger at bay. Nature 418, 595,
Fig. 1; с изм.; рис. 23-37. По Auwerx, J. & Staels, B. (1998)
Leptin. Lancet 351, 737; с изм.; рис. 23-40. По http://web.
indstate.edu/thcme/mwking/ampk.html и Steinberg, G. R.
& Kemp, В. E. (2007) Adiponectin: starving for attention.
Cell Metab. 6, 4, Fig. 1; с изм.; рис. 23-42. По Evans, R. M.,
Barish, G. D., & Wang, Y.-X. (2004) PPARs and the complex
journey to obesity. Nat. Med. 10, 355-361, Fig. 3; с изм.;
рис. 23-43. По Cummings, D. E., Purnell, J. Q., Frayo,
R. S., Schmidova, K., Wisse, В. E., & Weigle, D. S. (2001)
A preprandial rise in plasma ghrelin levels suggests a role in
meal initiation in humans. Diabetes 50, 1714, Fig. 1; с изм.;
рис. 23-44. По Nadler, S. T. & Attie, A. D. (2001) Please
pass the chips: genomic insights into obesity and diabetes./.
Nutr. 131,2080.
Tom 3
ГЛАВА 24. C. 7. N. R. Cozzarelli (1993) The structure
and function of DNA supercoiling and catenanes. Harvey
Lectures 87,35-55; рис. 24-1. Из Kleinschmidt, A. K., Land,
D., Jackerts, D., & Zahn, R. K. (1962) Darstellung und
Langenmessungen des gesamten Desoxyribonucleinsaure
Inhalter von T2-Bakteriophagen. Biochim. Biophys. Acta
61, 857-864; c. 8 (Бидл) Archive Photos; (Татум) Corbis/
UPI/Bettmann; рис. 24-4. Huntington Potter and
David Dressier, Harvard Medical School, Department of
Neurobiology; рис. 24-5, a, 6. G. F. Bahr/Biological Photo
Service; рис. 24-6. D. W. Fawcett/Photo Researchers;
рис. 24-11. По Cozzarelli, N. R, Boles, T. C., & White,
J. H. (1990) Primer on the topology and geometry of DNA
supercoiling. In DNA Topology and Its Biological Effects
(Cozzarelli, N. R. & Wang, J. C., eds), Cold Spring Harbor
Laboratory Press, Cold Spring Harbor, NY, pp. 139-184;
с изм.; рис. 24-12. По Saenger, W. (1984) Principles of
Nucleic Acid Structure, Springer-Verlag, New York, p. 452;
с изм.; рис. 24-13. Laurien Polder, из Kornberg, A. (1980)
DNA Replication, W. H. Freeman & Company, New York,
p. 29; рис. 24-14, 24-15. См источник для рис. 24-11;
рис. 24-20. Keller, W. (1975) Characterization of purified
DNA-relaxing enzyme from human tissue culture cells.
Proc. Natl. Acad. Sci. USA 72, 2553; рис. 24-21, 24-22. По
Champoux, J. J. (2001) DNA topoisomerases: structure,
function, and mechanism. Annu. Rev. Biochem. 70, 369, Figs
3, 11; с изм.; рис. 24-23, a. James H. White, T. Christian
Boles, and N. R. Cozzarelli, University of California,
Berkeley, Department of Molecular and Cell Biology;
рис. 24-26, 6. Ada L. Olins and Donald E. Olins, Oak
Ridge National Laboratory; рис. 24-27. PDB ID 1AOI,
Luger, K., Maeder, A. W., Richmond, R. K., Sargent, D. E, &
Richmond, T. J. (1997) Crystal structure of the nucleosome
core particle at 2.8 A resolution. Nature 389, 251; доп. 24-
2, рис. 1. По Sarma, К. & Reinberg, D. (2005) Histone
variants meet their match. Nat. Rev. Mol. Cell Biol. 6, 140;
доп. 24-2, рис. 2, б. Данные с любезного разрешения
Steve Henikoff. Перепечатано с разрешения из Mito, Y,
Henikoff, J. G., & Henikoff, S. (2005) Genomescale profiling
of histone НЗ.З replacement patterns. Nat. Genet. 37, 1092;
рис. 24-30, 6. Barbara Hamkalo, University of California,
Irvine, Department of Molecular Biology and Biochemistry;
рис. 24-31. D. W. Fawcett/Visuals Unlimited; рис. 24-34,
а, б. По Hirano, T. (2002) The ABCs of SMC proteins: two-
armed ATPases for chromosome condensation, cohesion, and
repair. Genes Dev. 16, 399; с изм.; рис. 24-34, e. Thomas E.
Melby, Charles N. Ciampaglio, Gina Briscoe, and Harold
P. Erickson; рис. 24-34, г, д. По Nasmyth, К. & Haering,
С. Н. (2005) The structure and function of SMC and kleisin
complexes. Annu. Rev. Biochem. 74, 611, 614, Figs 4, 5; c
1386 ] Источники иллюстраций
изм.; рис. 24-35. По Bazett-Jones, D. Р., Kimura, К., &
Hirano, Т. (2002) Efficient supercoiling of DNA by a single
condensin complex as revealed by electron spectroscopic
imaging. Mol. Cell 9,1183, Fig. 5; с изм.; c. 40 (задача 9)
Roger Kornberg, MRC Laboratory of Molecular Biology;
(задача 14) Bowater, R. P. (2005) Supercoiled DNA:
structure. In Encyclopedia of Life Sciences, doi: 10.1038/npg.
els.0006002, John Wiley & Sons, Inc./Wiley InterScience,
www.els.net.
ГЛАВА 25. C. 43. Arthur Kornberg (1975) For the love
of enzymes. In In Honour of Severo Ochoa (A. Kornberg
et al., eds), Pergamon Press, New York, 1976, pp. 243-
251; рис. 25-3, б. С любезного разрешения Dr. R. G.
Wake, University of Sydney, Australia. Перепечатано c
разрешения из Gyurasits, E. B. & Wake, R. G. (1973)
Bidirectional chromosome replication in Bacillus subtilis. J.
Mol. Biol. 73, Plate 1(f); c. 49 (Корнберг) AP/Wide World
Photos; рис. 25-8. PDB ID 3BDP, Kiefer, J. R., Mao, C.,
Braman, J. C., & Beese, L. S. (1998) Visualizing DNA
replication in a catalytically active Bacillus DNA polymerase
crystal. Nature 391, 304; рис. 25-10, 6. PDB ID 2POL,
Kong, X.-P, Onrust, R., O’Donnell, M., & Kuriyan, J. (1992)
Three-dimensional structure of the p subunit of Escherichia
coli DNA polymerase III holoenzyme: a sliding DNA clamp.
Cell 69, 425; рис. 25-12. По рисункам в Erzberger, J. P,
Mott, M. L., & Berger, J. M. (2006) Structural basis for
АТР-dependent DnaA assembly and replication-origin
remodeling. Nat. Struct. Mol. Biol. 13, 676-683; с изм.;
рис. 25-14. По анимации, любезно предоставленной Mike
O’Donnell, The Rockefeller University; рис. 25-18. Wake,
R. G. & King, G. E (1997) A tale of two terminators: crystal
structures sharpen the debate on DNA replication fork arrest
mechanisms. Structure 5, 1; рис. 25-18, б. По Lemon, К. P.
& Grossman, A. D. (2001) The extrusion-capture model for
chromosome partitioning in bacteria. Genes Dev. 15, 2031; c
изм.; рис. 25-19. Bruce N. Ames, University of California,
Berkeley, Department of Biochemistry and Molecular
Biology; рис. 25-20. По иллюстрации в Sivaprasad, U.,
Dutta, A., & Bell, S. P. (2006) Assembly of pre-replication
complexes. In DNA Replication and Human Disease
(DePamphilis, M. L., ed.), Cold Spring Harbor Laboratory
Press, Cold Spring Harbor, NY, pp. 141-152; с изм.; рис. 25-
21. По иллюстрации, предоставленной Paul Modrich;
с изм.; рис. 25-22. Grilley, М., Griffith, J., & Modrich, Р.
(1993) Bidirectional excision in methyl-directed mismatch
repair./. Biol. Chem. 268,11,830; рис. 25-23. Watson,
J. D., Hopkins, N. H., Roberts, J. W., Steitz, J. A., & Weiner,
A. M. (1987) Molecular Biology of the Gene, 4th edn, The
Benjamin/Cummings Publishing Company, Menlo Park,
CA, p. 350; рис. 25-24. По иллюстрации, предоставленной
Aziz Sancar; с. 81 (МакКлинток) AP/Wide World
Photos; рис. 25-32, 6. Bernard John, The Australian
National University; рис. 25-33, 6. Huntington Potter
and David Dressier, Harvard Medical School, Department
of Neurobiology; рис. 25-36, a. John Heuser, Washington
University Medical School, Department of Biochemistry;
рис. 25-36, 6. PDB ID 2REB, Story, R. M., Weber, I. T, &
Steitz, T. A. (1992) The structure of the E. coli RecA protein
monomer and polymer. Nature 355, 318; рис. 25-38. Roca,
A. I. & Cox, M. M. (1990) The RecA protein: structure and
function. Crit. Rev. Biochem. Mol. Biol. 25,415; рис. 25-40,
а. По Guo, E, Gopaul, D. N., & Van Duyne, G. D. (1997)
Structure of Cre recombinase complexed with DNA in a
site-specific recombination synapse. Nature 389, 40; с изм.;
рис. 25-40, 6. PDB ID 3CRX, Gopaul, D. N„ Guo, E, &
Van Duyne, G. D. (1998) Structure of the Holliday junction
intermediate in Cre-Loxp site-specific recombination.
EMBOJ. 17,4175.
ГЛАВА 26. C. 105. Francis H. C. Crick (1958) The
biological replication of macromolecules. Symposium of
the Society of Experimental Biology 12, 146; рис. 26-4.
PDB ID 1IW7, Vassylyev, D. G., Sekine, S., Laptenko, O.,
Lee, J., Vassylyeva, M. N., Borukhov, S., & Yokoyama, S.
(2002) Crystal structure of a bacterial RNA polymerase
holoenzyme at 2.6 A resolution. Nature 417, 712; рис. 26-
6, б. По координатам, предоставленным Seth Darst,
The Rockefeller University; рис. 26-8. Landick, R. (1997)
RNA polymerase slides home: pause and termination site
recognition. Cell 88, 741; доп. 26-1, рис. 2. Carol Gross,
University of California, San Francisco, Department of
Stomatology; рис. 26-10, 6. PDB ID 1TGH, Juo, Z. S.,
Chiu, T. K., Leiberman, P. M., Baikalov, L, Berk, A. J.,
& Dickerson, R. E. (1996) How proteins recognize the
TATA-box.J. Mol. Biol. 261, 239; рис. 26-10, в. По Klug,
A. (2001) A marvelous machine for making messages.
Science 292, 1844-18Й6; рис. 26-11, 6. PDB ID 1DSC,
Lian, C., Robinson, H., & Wang, A. H.-J. (1996) Structure
of actinomycin D bound with (GAAGCTTC)2 and
(GATGCTTC)2 and its binding to the (CAG)N:(CTG)
N triplet sequence by NMR analysis. J. Am. Chem. Soc.
118, 8791; рис. 26-13, a. Pierre Chambon, Laboratorie
de Genetique Moleculaire des Eucaryotes, Faculte de
Medecine (CNRS); рис. 26-13, б, в. Chambon, P. (1981)
Split genes. Sci. Am. 244 (May), 60; c. 125 (Чек) Corbis/
UPI/Bettmann; Figure 26-16 Cech, T. R. (1986) RNA as
an enzyme. Sci. Am. 255 (November), 64; рис. 26-17, a.
Kramer, A. (1996) The structure and function of proteins
involved in mammalian pre-mRNA splicing. Annu. Rev.
Biochem. 65, 367; рис. 26-22. По Blencowe, В. J. (2006)
Alternative splicing: new insights from global analyses. Cell
126, 38, Fig. 2; с изм.; рис. 26-26. По Kiss, T. (2002) Small
nucleolar RNAs: an abundant group of noncoding RNAs
with diverse cellular functions. Cell 109,146; рис. 26-28.
По Wienholds, E. & Plasterk, R. H. A. (2005) MicroRNA
function in animal development. EEBS Lett. 579, 5914; c
изм.; рис. 26-29, 6. PDB ID 1MME, см. Источник для
рис. 8-25, 6; рис. 26-30. Cech, Т. R., Damberger, S. H.,
& Gutell, R. R. (1994) Representation of the secondary
and tertiary structure of group I introns. Struct. Biol. 1,
273; рис. 26-31. Cech, T. R. (1986) RNA as an enzyme.
Sci. Am. 255 (November), 64; c. 143 (Грюнберг-Манаго)
С любезного разрешения Marianne Grunberg-Manago;
(Очоа) AP/Wide World Photos; c 145 (Темин) Corbis/
UPI/Bettmann; (Балтимор) AP/Wide World Photos;
Источники иллюстраций [387 ]
рис. 26-36. Haseltine, W. А. & Wong-Staal, Е (1988) The
molecular biology of the AIDS virus. Sci. Am. 259 (October),
52; рис. 26-37. Kingsman, A. J. & Kingsman, S. M. (1988)
Ту: a retroelement moving forward. Cell 53, 333; рис. 26-39,
a. Boeke, J. D. (1990) Reverse transcriptase, the end of the
chromosome, and the end of life. Cell 61, 193; рис. 26-39, в.
Jack Griffith, University of North Carolina at Chapel Hill,
Comprehensive Cancer Center; c. 152 (Вёзе, Крик, Оргел)
AP/Wide World Photos; доп. 26-3, рис.З PDB ID 1RAW,
Dieckmann, T, Suzuki, E., Nakamura, G. K., & Feigon, J.
(1996) Solution structure of an АТР-binding RNA aptamer
reveals a novel fold. RNA 2, 628.
ГЛАВА 27. c. 165. Gerald Joyce, цит. по M. M. Waldrop
(1992) Finding RNA makes proteins gives “RNA World”
a big boost. Science 256,1396-1397; c. 166 (Замечник)
News Office, Massachusetts General Hospital; рис. 27-1.
D. W. Fawcett/Visuals Unlimited; c. 167 (Ниренберг)
AP/Wide World Photos; c. 170 (Корана) С любезного
разрешения Archives, University of Wisconsin-Madison;
c. 180 (Номура) с любезного разрешения Masayasu
Nomura; рис. 27-13, а, б, в. PDB ID 2OW8, 1VSA, 1GIX,
Korostelev, A., Trakhanov, S., Laurberg, M., & Noller, H. F.
(2006) Crystal structure of a 70S ribosome-tRNA complex
reveals functional interactions and rearrangements. Cell 126,
1065; рис. 27-13, г. PDB ID 1Q7Y, Hansen, J. L., Schmeing,
T. M., Moore, P. B., & Steitz, T. A. (2002) Structural insights
into peptide bond formation. Proc. Natl. Acad. Sci. USA 99,
11,670; доп. 27-2, рис. 1. PDB ID 1NKW, Harms, J. M.,
Schluenzen, F, Zarivach, R., Bashan, A., Gat, S., Agmon,
I., Bartels, H., Franceschi, F, & Yonath, A. (2001) High
resolution structure of the large ribosomal subunit from
a mesophilic eubacterium. Cell 107, 679; c. 185 (Холли)
Corbis/UPI/Bettmann; рис. 27-18, 6. PDB ID 4TRA,
Westhof, E., Dumas, P, & Moras, D. (1988) Restrained
refinement of two crystalline forms of yeast aspartic acid
and phenylalanine transfer RNA crystals. Acta Crystallogr.
A 44, 112; рис. 27-22, a. PDB ID 1QRT, Arnez, J. G. &
Steitz, T. A. (1996) Crystal structures of three misacylating
mutants of Escherichia coli glutaminyl-tRNA synthetase
complexed with tRNA(Gln) and ATP. Biochemistry 35,
14,725; рис. 27-22, 6. PDB ID 1ASZ, Cavarelli, J., Eriani,
G., Rees, B., Ruff, M., Boeglin, M., Mitschler, A., Martin, E,
Gangloff, J., Thierry, J. C., & Moras, D. (1994) The active
site of yeast aspartyl-tRNA synthetase: structural and
functional aspects of the aminoacylation reaction. EMBO
J. 13, 327; доп. 27-3, рис. 2. По Xie, J. M. & Schultz, P. G.
(2006) Innovation: a chemical toolkit for proteins — an
expanded genetic code. Nat. Rev. Mol. Cell Biol. 7, 778;
рис. 27-30, б (слева). PDB ID 1B23, Nissen, P, Thirup, S.,
Kjeldgaard, M., & Nyborg, J. (1999) The crystal structure
of Cys-tRNACys-EF-Tu-GDPNP reveals general and
specific features in the ternary complex and in tRNA. Struct.
Fold. Des. 7, 143; рис. 27-30, б (справа). PDB ID 1DAR,
al-Karadaghi, S., Aevarsson, A., Garber, M., Zheltonosova,
J., & Liljas, A. (1996) The structure of elongation factor
G in complex with GDP: conformational flexibility and
nucleotide exchange. Structure 4, 555; рис. 27-32, 6. Steven
L. McKnight and Oscar L. Miller, University of Virginia,
Department of Biology; c. 210 (Блобел) с любезного
разрешения Gunter Blobel, The Rockefeller University;
c. 211 (Паладе) AP/Wide World Photos; рис. 27-42,
6. D. W. Fawcett/Photo Researchers; рис. 27-45. По
Mayor, S. & Pagano, R. E. (2007) Pathways of clathrin-
independent endocytosis. Nat. Rev. Mol. Cell Biol. 8, 604; c
изм.; рис. 27-46, «.John Heuser, Washington University
Medical School, Department of Biochemistry; рис. 27-48,
a. PDB ID 1IRU, Unno, M., Mizushima, T, Morimoto, Y,
Tomisugi, Y, Tanaka, K., Yasuoka, N., & Tsukihara, T. (2002)
The structure of the mammalian 20S proteasome at 2.75 A
resolution. Structure 10, 609.
ГЛАВА 28. C. Francois Jacob and Jacques Monod
(1961). Genetic regulatory mechanisms in the synthesis of
proteins. Journal of Molecular Biology 3, 318-356; p. 1119
(Жакоб, Моно) Corbis/Bettmann; рис. 28-7, б. С
любезного разрешения М. Thomas Record and Oleg
Tsodikov, University of Wisconsin-Madison, Department
of Biochemistry; рис. 28-7, в, г. PDB ID 1LBG, 1LBH,
Lewis, M., Chang, G., Horton, N. C., Kercher, M. A.,
Pace, H. C., Schumacher, M. A., Brennan, R. G., & Lu, P.
(1996) Crystal structure of the lactose operon repressor
and its complexes with DNA and inducer. Science 271,
1247. Боковые цепи добавлены только для вводных
Cot-only PDB с помощью программ MIDAS-PLUS и
Sybyl; рис. 28-11, а, в, г. PDB ID 1LCC, Chuprina, V .Р,
Rullmann, J. А. С., Lamerichs, R. М. J. N., Van Boom,
J. H., Boelens, R., & Kaptein, R. (1993) Structure of the
complex of Lac repressor headpiece and an 11 base-pair
half-operator determined by nuclear magnetic resonance
spectroscopy and restrained molecular dynamics./ Mol. Biol.
234,446; рис. 28-11, 6. PDB ID 1LBG, см. источник для
рис. 28-7, в, г\ рис. 28-12. PDB ID 1A1L, Elrod-Erickson,
М., Benson, Т. Е., & Pabo, С. О. (1998) High-resolution
structures of variant Zif268-DNA complexes: implications
for understanding zinc finger-DNA recognition. Structure 6,
451; рис. 28-13. PDB ID 1B8I, Passner, J. M., Ryoo, H.-D.,
Shen, L., Mann, R. S., & Aggarwal, A. K. (1999) Structure
of a DNA-bound ultrabithorax-extradenticle homeodomain
complex. Nature 397, 714; рис. 28-14, a. McKnight, S. L.
(1991) Molecular zippers in gene regulation. Sci. Am. 264
(April), 54-64; рис. 28-14, 6. PDB ID 1YSA, Ellenberger,
T. E., Brandl, C. J., Struhl, K., & Harrison, S. C. (1992) The
GCN4 basic region leucine zipper binds DNA as a dimer of
uninterrupted a helices: crystal structure of the protein-
DNA complex. Ce//71, 1223; рис. 28-15. PDB ID 1HLO,
Brownlie, P, Ceska, T. A., Larners, M., Romier, C., Theo,
H., & Suck, D. (1997) The crystal structure of an intact
human max-DNA complex: new insights into mechanisms of
transcriptional control. Structure 5, 509; рис. 28-16. PDB
ID 1RUN, Parkinson, G., Gunasekera, A., Vojtechovsky,
J., Zhang, X., Kunkel, T. A., Berman, H., & Ebright, R. H.
(1996) Aromatic hydrogen bond in sequence-specific
protein-DNA recognition. Nat. Struct. Biol. 3,837; рис. 28-
20. PDB ID 1TRO, Otwinowski, Z., Schevitz, R. W, Zhang,
R.-G., Lawson, C. L., Joachimiak, A. J., Marmorstein, R.,
[3881 Источники иллюстраций
Luisi, В. Е, & Sigler, Р. В. (1988) Crystal structure of Trp
repressor operator complex at atomic resolution. Nature 335,
321; рис. 28-21, a. Watson, J. D., Hopkins, N. H., Roberts,
J. W, Steitz, J. A., & Weiner, A. M. (1987) Molecular Biology
of the Gene, 4th edn, The Benjamin/Cuminings Publishing
Company, Menlo Park, CA, p. 487; рис. 28-23. Nomura,
M., Course, R., & Baughman, G. (1984) Regulation of the
synthesis of ribosomes and ribosomal components. Annu.
Rev. Biochem. 53, 75; рис. 28-25. По Szymanski, M. &
Barciszewski, J. (2002) Beyond the proteome: non-coding
regulatory RNAs. Genome Biol. 3, 6; с изм.; рис. 28-26.
По Winkler, W. C. & Breaker, R. R. (2005) Regulation
of bacterial gene expression by riboswitches. Annu. Rev.
Microbiol. 59, 493; рис. 28-27. John D. Cunningham/
Visuals Unlimited; рис. 28-31. Schwabe, J. W. R. & Rhodes,
D. (1991) Beyond zinc fingers: steroid hormone receptors
have a novel structural motif for DNA recognition. Trends
Biochem. Sci. 16, 291; рис. 28-37. E R. Turner, University
of Indiana, Bloomington, Department of Biology; p.
1149 (Nusslein-Volhard) AP/Wide World Photos/Mark
Lennihan; рис. 28-39. Wolfgang Driever and Christiane
Nusslein-Volhard, Max-Planck-Institut; рис. 28-40. По
рисунку, предоставленному Marv Wickens, University of
Wisconsin-Madison, Department of Biochemistry; с изм.;
рис. 28-41, б. С любезного разрешения Phillip Ingham,
Imperial Cancer Research Fund, Oxford University;
рис. 28-43, a, 6. F. R. Turner, University of Indiana,
Bloomington, Department of Biology; рис. 28-43, в, г.
E. B. Lewis, California Institute of Technology, Division of
Biology; доп. 28-1, рис. 1. По Abzhanov, A., Kuo, W. P,
Hartmann, C., Grant, B. R., Grant, P. R., & Tabin, C. J.
(2006) The calmodulin pathway and evolution of elongated
beak morphology in Darwin’s finches. Nature 442, 565, Fig.
4; с изм.
f
Предметно-именной указатель
А
АВС-нуклеазы 3: 74, 78
ABC-транспортеры 1: 565; 2: 489
АСАТ (ацил-СоА-холестеринацил-
трансфераза) 2:485,492
ADAR3:176-178
AdoMet, см. Аденозинметионин
ADP (аденозиндифосфат)
- в синтезе жирных кислот 2: 451,
452
- в фотосинтезе 2: 357,358
- перенос фосфатной группы
2: 73-76
- синтез 1: 45,46
ADP-глюкоза 2: 426-428
ADP-глюкозопирофосфорилаза 2:430
ADP-рибозилирование ферментов
1:324,602
Aequorea victoria, флуоресцентные
белки 1: 451, 612
Agrobacterium tumefaciens, в клониро-
вании 1: 469-471
AlkB белок, в репарации ДНК 3: 68,
75, 77
АМР (аденозинмонофосфат) 1: 44,
394; 2:33, 546, 547, 550
- варианты формы 1: 427
- концентрация 1: 608, 614, 615
- модификация аллостерических
ферментов 1: 329, 600, 604
АМРК (AMP-активируемая про-
теинкиназа) 2: 131, 243
- влияние адипонектина 2: 612
- стимуляция лептином 2: 609, 610
AMP-активируемая протеинкиназа,
см. АМРК
АР-эндонуклеазы 3: 68, 72, 73, 79
Apaf-1 фактор апоптоза 2: 350
АРС, мутации 1: 671
АроВ-100 2: 487-490
- редактирование РНК 3: 177
АроВ-48, редактирование РНК 3: 177
АРОВЕСЗ: 176, 177
ароЕ
- в болезни Альцгеймера 2: 487
- связывание с рецепторами 2:485,
487
Ар-сайт в репарации ДНК 3: 71
Arabidopsis thaliana
— аквапорины 1: 572
— сигнальные пути 1: 648, 649
— синтез целлюлозы 2: 432
Aspergillus niger, синтез лимонной
кислоты 2: 204
АТР (аденозинтрифосфат) 1: 45, 46,
424,425
- в активном транспорте 2: 40
- в биолюминесценции у светляков
2: 39
- в гликолизе 2: 75-80
- в глюконеогенезе 2: 103, 104
- в инициации репликации 3: 56, 57
- в протеолизе 3: 220, 221
- в регуляции гликолиза и глюко-
неогенеза 2: 143,144
- в регуляции метаболизма 2: 129,
130
- в сердечной мышце 2: 589
- в сокращении мышц 2: 40, 589
- в фиксации азота 2: 507, 510
- в фотосинтезе 2:404, 405,415,416
- в цикле Кальвина 2: 415, 416
- выход при окислении глюкозы
2: 201,202
- гидролиз 1: 425; 2: 18,31, 35,37
— и функция моторных белков
1: 256-262
— биохимическое/химическое урав-
нение 2: 27
— константа равновесия 2: 18
— изменение свободной энергии
2: 16, 29,31
- ингибирование пируваткиназы
2:147
- комплексы Mg2+ 1: 308; 2: 27, 30
- концентрация 2: 30
- перенос фосфорильных групп
2: 28-43
- при гипоксии 2: 344
- регуляция в окислительном фос-
форилировании 2: 343-346
- синтез 2: 327-342
— в галофильных бактериях 2: 387,
388
— при окислительном фосфорили-
ровании 2:343-346
— при фотофосфорилировании
2:381-384
— регуляция 2:347
- синтез сАМР 1: 427
АТ Разы
- ААА+ 3: 56
- F-типа 1: 563; 2: 331; см. также
АТР-синтаза
- Na+ /К+ в поляризации мембраны
1: 563, 633, 634, 635
-Р-типа 1:540, 541, 560, 561
- V-типа 1: 564
- в мембранном транспорте 1: 564,
565
- в нейронах 2: 590
- в сетчатке 1: 651
АТР-синтаза 2:327,334, 351, 385
- в хлоропластах 2: 384
- конформация субъединиц 2: 333
- модель действия 2: 333-338, 384
- функциональные домены 2: 331
АТР-синтасома 2: 340
ATP-управляемые К+-каналы 2: 596
Azotobacter vinelandii, фиксация азота
2:512
AZT (азидотимидин) 3: 147
А-ДНК 1: 405
А-сайт 3: 197
В
ВАС (искусственная бактериальная
хромосома) 1: 439, 442
BRCA1/2, и рак молочной железы
3: 80
В-ДНК 1:405
В-клетки (В-лимфоциты) 1:170
- рекомбинация 3: 96, 97
bicoid 3: 273-275
Втр4 3: 280
С
С2-путь 2: 422
С3-растения 2: 423
С4-путь 2: 422-425
С4-растения 2: 423
Са2+
- концентрация в цитозоле/ внекле-
точных жидкостях 1: 634, 635
- осцилляции 1: 615, 618
Са2+/кальмодулин-зависимые про-
теинкиназы 1: 614-616
Са2+-каналы 1: 573, 580
- в метаболизме глюкозы 2: 596
- в передаче сигнала 1: 611, 616
- дефекты 1: 579,581
Са2+-насос 1: 560, 611, 616
-SERCA 1:560, 561
сАМР (циклический аденозинмоно-
фосфат) 1: 427
- гормональная регуляция 1: 599
- измерение методом FRET 1: 614,
615
- как вторичный мессенджер 1: 427,
607
- расщепление 1: 605, 609
- синтез под действием аденилатци-
клазы 1: 598, 599
- структура 1: 427
сАМР-зависимая протеинкиназа А
(РКА) 1:599, 604
сАМР-чувствительный элемент
(CRE)3: 268
[3901 Предметно-именной указатель
сАМР-рецепторный белок (CRP)
3: 116
САМ-растения 2: 426
С ASP, предсказание структуры белка
1:212
caudal 3: 275, 276
CDK9 3: 120
CDP (цитидиндифосфат) 2: 471
CDP-диацилглицерин 2: 471-473
Celera Corporation 1: 458
CFP (голубой флуоресцентный
белок) 1:612-515
cGMP (циклический гуанозинмоно-
фосфат)
- роль в зрительной функции
1:651-654
- структура 1: 627
cGMP-зависимая протеинкиназа
(протеинкиназа G) 1: 627
cGMP-фосфодиэстераза (PDE)
1: 628, 653
ChREBP-транскрипционный фактор
2: 150,151
Clostridium
- acetobutyricum 2: 96
- botulinum 1 549
- perfringens 1: 222
CMP (цитидинмонофосфат) 1: 394,
424; 2: 548
CO2, см. Диоксид углерода
СоА (кофермент А) 1: 426; 2: 183
CODIS база данных 1: 455,457
СОХ (циклооксигеназа) 2: 461, 462
- ингибиторы 2: 461, 462
СОХ4 2: 345,346
Crassulaceae, фотосинтез 2: 426
CREB 1:608; 3: 268
CTF1 3: 265, 266
С-белок 1: 260
С-концевой остаток 1: 128
— посттрансляционные модифика-
ции 3: 206
D
Dam-метилаза 3: 58
- в репарации 3: 68
dAMP (дезоксиаденозинмоно-
фосфат) 1: 394
dCMP (дезоксицитидинмоно-
фосфат) 1: 394
DDI3: 147
dGMP (дезоксигуанозинмоно-
фосфат) 1: 393, 394
Dicer 3: 138, 270
DnaA-белок, в инициации реплика-
ции 3: 56-60
DnaB-хеликаза 3: 57-60
DnaC-белок, в инициации реплика-
ции 3: 57-60
DnaG-белок 3: 58
DnaK/DnaJ, в фолдинге белка 1: 212,
213
Drosha 3: 138, 270
Drosophila melanogaster
— жизненный цикл 3: 272
— регуляция генов 3: 272-278
dTMP (дезокситимидинмоно-
фосфат) 1: 393, 394
D-плечо, в тРНК 3: 185
Е
Е. coli, см. Escherichia coli
El, Е2, ЕЗ ферменты в расщеплении
белков 3: 220, 221
е1Е4-связывающие белки 3: 198, 199
ELISA (ИФА) 1: 255, 256; 2: 570
епсЪ. 145,146,148,176
erbB2 онкоген 1: 670
ERK киназа 1: 327, 619, 621, 622, 625
Escherichia coli 1: 20-24
— F^o-комплекс 2: 331, 335-338
- - lac оперон 3: 235-245
— ДНК 1:51
— карта хромосомы 3: 44
— клонирование 1: 434-480
— липополисахариды 1: 370
— метилирование ДНК 1: 419
— молекулярный состав 1: 33
— переносчик лактозы 1: 567, 568
— плазмидные векторы 1: 439, 440
— репликация 3: 50-65
— рибосомы 3: 180,184
— РНКаза Р 1: 410; 3: 136,141, 142
— секвенирование генома 1: 61
— сигнальные пути 1: 644, 645
— синтез фосфолипидов 2: 472
— транскрипция 3: 106-115
— цикл лимонной кислоты и глиок-
силатный цикл 2: 212, 214
— шапероны 1: 213, 214
— экспрессирующие векторы 1: 444,
445
EST (метка экспрессируемой после-
довательности) 1: 451
Е-сайт, на рибосоме 3: 197
F
F^o-комплекс Е. coli 2: 331, 335-338
Fi-компонент АТР-синтазы 2: 331,
332,334-338
Fab-фрагмент иммуноглобулина
1:251,254
FAD (флавинадениндинуклеотид)
1:426; 2: 54,308
FADD-белок 1:672, 673
Fas-рецептор, в апоптозе 1: 672, 673
Fc-фрагмент иммуноглобулина
1:251
Fe-S-центр 2: 311, 368; см. также
Железосерные кластеры
fMet-тРНК, в синтезе белка
3: 195-198
FMN (флавинмононуклеотид) 2: 54,
308
FOXO1-фактор 2: 151,152
Е0-компонент АТР-синтазы 2: 331,
335-338
FRAP (метод обесцвечивания флуо-
ресценции) 1: 541, 542
FRET (флуоресцентный резонанс-
ный перенос энергии) 1:609,
613-615
F-актин 1: 258, 260, 261
G
gagb. 144,145
- сдвиг рамки считывания
3:175-177
gag-pol белок, и сдвиг рамки считы-
вания 3: 175-177
GAL-гены, регуляция 3: 264, 265
Са14р-белок 1: 467
— кислый активирующий домен
3: 265
GAP-белки (активаторы СТРаз)
1:601,602,658
gap-гены 3: 273, 276
GATC-последовательность
— в репарации 3: 68-70
— в репликации 3: 58
GDP (гуанозиндифосфат)
- в |3-адренергическом пути
1:599-605
- при зрительном процессе 1: 652,653
- при обонянии 1: 656
GEF (факторы обмена гуаниновых
нуклеотидов)1:601
Gemmata obscuriglobus 2: 509
GLUTl-переносчик 1: 553-555
СЕиТ2-переносчик 1: 556, 569
СЕиТ4-переносчик 1: 556
GMP (гуанозинмонофосфат) 1: 394,
424
GPI (гликозилфосфатидилинозит)
1:530, 536, 537, 545
GPR14 1:478
Grb2 1: 619, 625, 626, 629
GRK-киназы 1: 606, 607
GroEL/GroES 1: 19,159
- в фолдинге 1: 213, 214
GSH, см. Глутатион
Предметно-именной указатель [391 ]
GTP (гуанозинтрифосфат) 1: 68, 424,
598, 599
- в |3-адренергическом пути
1: 597-605
- и восприятие запаха и вкуса 1: 656,
657
- и зрительный процесс 1: 652-654
- синтез cGMP 1: 627
СТРазу активирующие белки (GAP)
1:601,602
СТРазы 1: 599, 601, 602, 605, 658
G-актин 1: 258
G-белки 1: 600-605, 658
- G, 1: 598, 608
-GoH 1:656
- Gq 1: 611
- Gs 1: 598, 599, 608
- Ras-типа 1: 600, 619
- гастдуцин 1: 656
G-тетраплекс 1: 407
н
Halobacterium salinarum 2: 387, 388
— бактериородопсин 1: 532, 533, 544
HCR репрессор 3: 269
Helicobacter pylori, лектины 1: 374,
375
HIF-l (индуцируемый гипоксией
транскрипционный фактор-1)
2:81,211,345
HIRA, в ремоделировании хромати-
на 3: 259
/zzs-оперон 3: 247
HMG-CoA2: 252
- в синтезе холестерина 2: 480, 481
Н MG-Co А-редуктаза
- в синтезе холестерина 2: 480, 481,
491,492
- ингибиторы 2: 494,495
HMG-CoA-синтаза, в синтезе
холестерина 2: 480, 481
HOY-гены 3: 276, 277, 279
HRE (элементы гормонального
ответа) 1: 643; 3: 267, 268
Hsp70, в фолдинге 1: 212, 213
HU, в репликации 3: 56-58
hunchback 3: 276
I
IgA 1: 252
IgD 1:252
IgE 1: 252, 253
IgG 1:251, 252, 254
- рекомбинация генов 3: 96, 97
IgM 1: 252
Inr (инициаторная последователь-
ность) 3: 117, 119, 261,264
IP3 (инозит-1,4,5-трифосфат) 1: 507,
610,611
ISWI семейство, в ремоделировании
хроматина 3: 259
J
JAK (Янус-киназа) 1: 624; 2: 608
JAK-STAT-путь 1: 624, 625; 2: 608
J-сегмент легкой цепи 3: 96, 97
L
L-19 IVS-рибозим 3: 140,141
/ас-оперон 3: 235-245
/ас-промотор 3: 235-245
Lac-penpeccop 3: 235-245
Lactobacillus bulgaricus, в фермента-
ции 2: 96
/ew-оперон 3: 247
LexA-penpeccop, в SOS-реакциях
3: 249, 250
LINE (длинные рассеянные повто-
ры) 3: 14
Lon-протеаза 3: 220
LRP-рецептор 2: 491
М
MALDI-масс-спектрометрия 1: 149
МАРК-киназы 1: 622, 632
— каскад 1: 622
МСМ-белки 3: 64, 65
MDRl-переносчик 1: 564
МЕК-протеинкиназа 1: 619, 622
MFP-белок 2: 248
MFS-факторы, облегчающие транс-
порт 1: 568
MjTyrRS3:192-194
МутРНКт>'гЗ: 192-194
MODY (сахарный диабет взрослого
типа у молодых) 2: 152, 153, 355
MutH в репарации ДНК 3: 68-71
MutL в репарации ДНК 3: 68-71
MutS в репарации ДНК 3: 68-71
М-белок 1: 260
N
Na+/K+-ATPa3a 1: 560, 562, 563; 2: 40
- в нейронах 2: 590
- в поляризации мембраны 1: 633, 634
- в сетчатке 1: 651
Иа+-глюкозный симпорт 1: 569, 570
NAD+ (никотинамидаденинди-
нуклеотид) 1: 426; 2: 50, 454
- в цикле Кальвина 2: 415
NADH
- в гликолизе 2: 80
- дегидрогеназные реакции 2: 51, 52
- окисление в митохондриях рас-
тений 2: 326
- поглощение света 2: 50
- шаттл-системы 2:340-342
NADH-дегидрогеназа 2:308, 313, 314
NADH-убихинон-оксидоредуктаза
(комплекс I; NADH-
дегидрогеназа) 2: 314
NADP+ 2: 50
- дегидрогеназные реакции 2: 51, 52
- недостаточность витаминной фор-
мы 2: 53, 54
NADPH 2: 307,308,454
- в защите клеток от активных форм
кислорода 2: 107,108,324
- в пентозофосфатном пути 2:107-111
- в распределении глюкозо-6-
фосфата 2: 112
- в реакциях анаболизма 2:447
- в синтезе глицеральдегид-3-
фосфата 2: 410, 411
- в синтезе жирных кислот
2:447-455
- в фотосинтезе 2: 358,359,415,416
- в цикле Кальвина 2: 415,416, 418
- дегидрогеназные реакции 2: 51, 52
- синтез 2: 454
nanos 3: 276
Neu5Ac (сиаловая кислота) 1: 350,
372, 373, 376, 502
NLS (сигнал ядерной локализации)
3:216,217
NO-синтаза 1: 628; 2: 576
NS-домен 1: 364-366
NuA4, в ремоделировании хроматина
3: 259
NURF, в моделировании хроматина
3: 259, 260
о
OmpLA, структура 1: 536
ОтрХ, структура 1: 536
ORC-комплекс 3: 64
oriC (точка начала репликации)
3: 44, 56-58
Р
р53 1: 665
- мутации 1: 670
Р/2е_ коэффициент 2: 338
P/О отношение 2: 338
Paramecium 1: 526
PCNA 3: 65
PDK1 1:622
pH 1:92
- буферов 1: 97
- в биологических системах Г. 96
[3921 Предметно-именной указатель
- в связывании кислорода гемогло-
бином 1: 244
- оптимум для ферментов 1: 102, 303
p/ze-оперон 3: 247
pl (изоэлектрическая точка) 1: 125,
126
- аминокислот 1: 116
- определение 1: 136
рХа 1: 94-98
- R-групп аминокислот 1: 116, 125
- определение по кривой титрования
1: 124, 126
Plasmodium falciparum 2: 107
Pol I, см. РНК-полимераза I
Pol II, см. РНК-полимераза II
Pol III, см. РНК-полимераза III
ро/-ген 3: 145, 146,148
PPAR (активируемые пролиферато-
рами пероксисомные рецепто-
ры) 2: 612, 613
pRb (белок ретинобластомы) 1: 596,
665, 670
Propionibacterium freudenreichii, в
ферментации 2: 96
pumilio 3: 275, 276
Р-сайт 3: 197
R
R2C2 комплекс протеинкиназы
1:599, 604,615
Raf-1 1:619, 622
RAG-белки 3: 97, 98
Ran СТРаза 3:216,217
Ra-белок 1:600, 601,619
— мутации 1: 670
— переключатели 1: 600
ras-онкоген 1: 670, 671
/М-ген 1:670, 671
— белок 1: 665
RecA 3: 86-89
- в SOS-ответе 3: 78
RecBCD фермент 3: 85,86, 89
RF-1 3:179, 202, 204
RF-2 3: 179, 202, 204
RFA 3: 65
RFC 3: 65
RGS-белки 1: 601, 658
Rhodobacter sphaeroides 2: 368,369
Rhodopseudomonas viridis 2:368,369
Rhodospirillum rubrum 2:367, 408
RS-номенклатура стереоизомеров
1:36, 37,39
RU486 (мифепристон) 1: 644
RuvA-белок 3: 85, 89
RuvB-белок 3:85, 89
RuvC-белок 3: 85, 89,93
R-состояние гемоглобина 1: 235
s
AS (изменение энтропии) 1: 44, 212,
282
Saccharomyces cerevisiae, см. Дрожжи
Salmonella typhimurium 3: 254
— липополисахариды 1: 370
SCOP, база данных 1: 202-205
SDS, см. Додецилсульфат натрия
SecA/B-шапероны 3: 218
ЗЕЬЕХЯйетод 3: 154, 155
SERCA-насос 1: 560, 561
5Н2-домен 1: 619, 622, 624, 625
SINE3: 177, 178
SN1 -механизм 1: 312, 313
SN2-MexaHH3M 1: 312, 313
SNAP25 1:548
SNARE-белки 1: 548
SNP (полиморфизм одного нуклео-
тида) 1: 460
SOS-ответ 3: 249
Spl, домены 3: 265, 266
Src 1: 619, 630
SREBP, транскрипционный фактор
2: 151
SRP (сигнал-распознающая частица)
3:211,212
STAT, транскрипционные факторы
1:624-627
Streptococcus pneumoniae 1: 400
Streptomyces lividans 1: 536, 575
SWI/SNF, в ремоделировании хро-
матина 3: 259, 260, 265
SWR1, в ремоделировании хромати-
на 3: 259, 260
Т
Т3 (трииодтиронин) 2: 575
Т4 (тироксин)2: 575
ТАТА-бокс 3: 112,118, 119
ТАТА-связывающий белок (ТВР)
3: 118-120
ТуС-плечо тРНК 3: 185
Тег-последовательность 3: 62, 63
Tetrahymena thermophila 3: 127, 139,
140,151
TFII3: 118-121
Ti-плазмида, в клонировании рас-
тений 1:469-471
TRF1 3: 150,151
TRF2 3: 150,151
7ф-оперон, аттенюация транскрип-
ции 3: 246-249
Тгр-репрессор 3: 247
Trypanosoma brucei rhodesiense 2: 541
t-SNARE 1: 548
TUFPHK3: 157
Tus-Ter комплекс 3: 63
Т-ДНК, в клонировании растений
1:471
Т-клетки 1: 249, 250
- рецепторы 1: 250
Т-петля 3:150,151
Т-состояние гемоглобина 1: 235
Т-хелперы (Тц-клетки) 1: 250
и
UAS (активаторная последователь-
ность) 3: 261
иЬхЪ. 277
UDP (уридиндифосфат) 2: 89,435
UDP-глюкоза 2: 16, 89
- в синтезе гликогена 2: 161-163
— сахарозы 2: 428,429
— целлюлозы 2: 432-434
UDP-глюкозопирофосфорилаза
2: 161,429
ultrabithorax 3: 277
UMP (уридин-5'-монофосфат)
1:394; 2: 547
Umu белки, в репарации ДНК 3:77,78
UTP (уридинтрифосфат) 1: 48
UvrA, в репарации ДНК 3: 74, 78
UvrB, в репарации ДНК 3: 74, 78
UvrC, в репарации ДНК 3: 74, 78
uvrD, в репарации ДНК 3: 78
V
v-SNARE 1: 548
X
XerCD система 3: 93
Х-сцепленная адренолейкодистро-
фия (XALD) 2: 246
Z
Z-диск 1: 259-261
Z-ДНК!: 405
Z-cxeMa2: 372, 373
Предметно-именной указатель [ 3931
А
Абзимы, см. Каталитические анти-
тела
Абиотический синтез 1: 56
Абсолютная конфигурация 1: 117
Абсолютная температура, единицы
1:276
Авандия 2: 470, 612, 617
Авидин 2: 207, 338
Авогадро число 2: 13
Автономно реплицирующиеся по-
следовательности (АРП) 3: 64
Автотрофы 2: 5-7
Агар 1: 359
Агароза 1: 359, 362
Аггрекан 1: 366, 367
Агонисты рецепторов 1: 597
Адаптерная гипотеза 3: 166, 167
Адаптерные белки, в сигнальных
путях 1: 617, 629
Адгезионные молекулы 1: 372, 549
Аденилаткиназа 2: 41, 550, 610
Аденилатциклаза 1: 326,427, 599,
602-604,608
Аденилилтрансфераза 2: 515, 516
Аденилирование 2: 38, 515, 516
- ферментов 1: 324
Аденин, 1:392,399; 2: 544
- биосинтез 2: 557, 574
- дезаминирование 1: 416
- клеточная концентрация 2: 30
- регуляция метаболизма 2: 129-131
- транслоказа 2: 339
- эволюционное значение 3: 152, 153
S-Аденозилгомоцистеин 2: 284
S-Аденозилметионин (AdoMet)
2: 284
- в качестве мутагена 1: 323
- синтез 2: 294
Аденозин 1: 394
- анти-форма 1: 404
- как кофактор ферментов 1: 425,
426
- метилирование 1: 419
- син-форма 1: 404
- эволюционное значение 1: 425
Аденозин-3’,5’-циклический моно-
фосфат, см. сАМР
Аденозин-2’,З’-циклический моно-
фосфат 1:394
Аденозин-2’-монофосфат 1: 395
Аденозин-З’-монофосфат 1: 394
Аденозин-5’-монофосфат 1: 394; см.
также АМР
Аденозиндезаминаза 2: 556
- генная терапия при недостаточно-
сти 1: 475
Аденозиндифосфат, см. ADP
Аденозинтрифосфат, см. АТР
Аденозинфосфорибозилтрансфераза
2: 556
Адипозная ткань, см. Белая жировая
ткань, Бурая жировая ткань
Адипокины 2: 605
Адипонектин 2: 610-612, 617
Адипоциты 1:491,492; 2:585-587,605
- синтез NADH 2:454
Адреналин 1: 597,598; 2: 173,539, 602
- в метаболизме глюкозы 2: 539
— липидов 2: 539
- как нейромедиатор и гормон 2: 566
- как усилитель сигнала 2: 570,578
- каскадный механизм 1: 606; 2: 166
- регуляция 1: 597, 598
- синтез 2: 539
- синтетические аналоги 1: 597
Адренергические рецепторы 1: 597
— в рафтах 1: 632
— десенсибилизация 1: 606, 607
— киназа (ARK) 1:327
Адренолейкодистрофия 2: 246
Адсорбционная хроматография
1:515,516
Азасерин 2: 558
Азидотимидин (AZT) 3: 147
Азот
- включение в аминокислоты 2: 513
- восстановление до аммиака
2:510-513
- выведение 2: 273
- круговорот в биосфере 2: 6
- метаболизм 2: 510-517
- оксид, см. Монооксид азота
- растворимость в воде 1: 79
- фиксация нитрогеназным ком-
плексом 2: 510-513
- цикл 2: 506
Азотистая кислота как мутаген
1:418
Азотистые основания 1: 392; см. так-
же Пурины; Пиримидины
— спаривание 1: 399
Азотистый иприт 1: 418
Аквапорины 1: 556, 571-573
Аконитаза (аконитатгидратаза)
2:192-195
- «скрытая функция» 2: 193
цис- Аконитат 2: 192
Акридин 3: 121, 122
Активаторная последовательность
(UAS) 3:261
Активаторы транскрипции 3: 232
Активационный барьер 1: 47, 48
Активный транспорт 1: 558, 567
— вторичный 1: 558
— первичный 1: 558
— роль АТР 2: 339, 593
Активный центр 1: 273
— комплементарность переходному
состоянию 1: 278
Активные формы кислорода (АФК)
2:324,345
Актин 1: 258-261
- филаменты 1: 24, 258-261
а-Актинин 1: 259
Актиномицин D 3:121
Актос (пиоглитазон) 2: 612, 617
Акцепторный контроль 2: 343
— коэффициент 2: 343
Аланил-глутамил-глицил-лизин
1:129
Аланин 1: 34; 2: 70
- в транспорте аминогрупп в печень
2: 271
- расщепление до пирувата 2: 70,
100, 584
- свойства 1: 179; 2: 581
- стереоизомеры 1: 37, 115, 117
- структура 1:34
Аланинаминотрансфераза (АЛТ)
2: 269, 271
Алкалоз 1: 93
Алкаптонурия 2: 292
Алкены 2: 46
Алкилирующие агенты как мутагены
1:418
Алкогольдегидрогеназа 1: 297; 2: 53,
250
Аллантоин 2: 556
Аллели 1: 246,455, 670
- АРОЕ, при болезни Альцгеймера
2: 487
Аллигаторы, анаэробный метабо-
лизм 2: 92
Аллоза 1: 342
Аллопуринол 2: 557
Аллостерическая регуляция
— аспартаткарбамоилазы 2: 548, 549
— биосинтеза аминокислот
2: 529-531
— глутаминсинтетазы 2: 529, 530
— метаболизма жиров 2: 173
— метаболизма углеводов
2:171-173
— образования ацетил-СоА 2: 208,
209
Аллостерические белки 1: 237
Аллостерические модуляторы
1:319-322
13941 Предметно-именной указатель
Аллостерические ферменты
1:319-323
Аллостерические эффекторы 1: 319
Алпер Тиква 1: 218
АЛТ, см. Аланинаминотрансфераза
Альберте Альфред 2: 494, 495
Альбинизм 2: 287
Альбумин
- в сыворотке 2: 230,536, 584, 592
-ген 3: 131
Альдегиддегидрогеназа 2: 250
Альдегиды 1: 31, 340
- и полуацетали 1: 343, 344
- и полукетали 1: 343,344
Альдогексозы 1: 341,342
Альдозы 1: 341-344
Альдолазы 2: 73, 74, 128
- в цикле Кальвина 2: 412, 413
- классы 2: 73, 74
Альдольная конденсация 1: 23 <
Альдостерон 1: 509
Альтроза 1: 342
Альцгеймера болезнь
— аполипопротеины 2: 487
-- отложение амилоида 1: 215, 216
а-Амантин 3: 122
Амарил 2: 597
Амиды, стандартное изменение сво-
бодной энергии 2: 16
Амилаза 1: 355 2: 86, 87
Амилоза 1: 353-355,358; см. также
Крахмал
Амилоид 1: 215-217
Амилоидоз 1: 215-217
Амилопектин 1: 353-355, 358; 2: 87,
428; см. также Крахмал
Амилопласты 2: 405, 406
Аминоацил-связывающий (А) сайт
рибосомы 3: 197
Аминоацил-тРНК 3: 179, 186
Аминоацил-тРНК-синтетазы 3: 179,
186
а-Аминогруппы, перенос на
а-кетоглутарат 2: 263, 266
Аминокислотная последователь-
ность 1:33, 53, 142-148
— в эволюционном анализе
1: 156-160
— гомологии 1: 158, 462, 483
— консенсусная 1: 155, 156
— определение 1: 142-152
--методами масс-спектрометрии
1: 148-151
--по Сенгеру 1: 145
--по Эдману 1: 145
— синтез коротких пептидов 1: 152,
153
— соответствие последовательности
ДНК1:151,152
Аминокислотные остатки 1: 33, 38
— С-концевые 1: 128
— N-концевые 1: 128, 144, 145
— изопренелирование 3: 207
--и продолжительность жизни
белка 3: 221, 222
— последовательность 1: 142-148
— фарнезелирование 3: 207
— фосфорилирование 1: 324-327
Аминокислоты 1: 113-127; 2: 278; см.
также названия аминокислот
- 20 основных 1: 119
-R-группы 1:114,118-120
- активация, в синтезе белка 3: 179
- биосинтез 2: 518-532
- в а-спиралях 1: 177-181
- в |3-структурах 1: 181-183
- глюкогенные 2: 104, 105, 280, 281
- заменимые 2: 278
- как амфолиты 1: 123
- катаболизм 2: 261-298
— генетические нарушения 2: 287,
290-292
- кетогенные 2: 280, 281
- кислотно-основные свойства
1: 126, 127
- кодоны 3: 167, 191-194
- кривые титрования 1: 123-126
- незаменимые 2: 278
- нестандартные 1: 120-123
- образование пептидной связи
1: 127
- окисление 2:261-294
- полярность 1: 118
- с разветвленной цепью, отсутствие
расщепления в печени 2: 296,
297
- сокращенные названия 1:115
- стереоизомеры 1: 38, 115-118
- цвиттер-ионы 1: 123
6-Аминолевулинат, биосинтез 2: 533
у-Аминомасляная кислота (ГАМК)
2: 272, 539
— рецепторы как ионные каналы
1: 579, 640
Аминопептидазы 2: 264
Аминоптерин 2: 558
Аминосахара 1: 346, 360
Аминотрансферазы 2: 266, 267; см.
также Трансаминазы
- пиридоксальфосфат как простети-
ческая группа 2: 266, 267
Амины 1: 31
- как продукты декарбоксилирова-
ния аминокислот 2: 268, 538, 539
Аммиак
- в метаболизме азота 2: 6, 506-509
— аминокислот 2: 262-264
- в образовании мочевины 2: 273,
274
- высвобождение глутаматом 2: 269
- перенос аланином 2: 270, 271
- токсичность 2: 270, 272, 278
Аммониотелические виды 2: 273
Аммония цианид 3: 152
Амоксициллин 1: 317
Амплификация ДНК методом ПЦР
1:452
Амфиболические пути 2: 203
Амфитропные белки 1: 530, 531
Амфифильные соединения 1: 78, 81,
496
Амфолиты 1: 123,127
Амфотерные аминокислоты 1: 123
Анаболизм 1: 48, 49; 2: 7
- цикл лимонной кислоты 2: 182, 203
Анаболические пути 2: 8,9, 582, 610
— в метаболизме углеводов
2: 403, 582
— перенос энергии 2: 7, 404, 454
Аналит 1: 149
Анаммокс 2: 507-509
Анаммоксосомы 2: 508, 509
Анандамид 1: 622
Анаплеротические реакции, в цикле
лимонной кислоты 2:188,
203-205
Анаэробные бактерии, неполный
цикл лимонной кислоты 2: 202
Анаэробный метаболизм у целакан-
тов 2:92
Ангидриды кислот, стандартное
изменение свободной энергии
2: 16, 29
Ангстрем (А) 1: 177
Андерсена болезнь 2: 159
Андрогены 2: 495
- синтез 2: 575
Анемия
- межобластная 2: 284
- пернициозная, роль витамина В12
2: 284
Анионобменивающий (АЕ) белок
1:557
Анионообменник 1: 132; см. также
Ионообменная хроматография
Аничков Н. Н. 2: 493
Анкирин 1: 543
Аннотирование генов 1: 62
Аномальные трихроматы 1: 656
Аномерный углерод Г. 344,350,351
Аномеры 1: 344
Предметно-именной указатель [ 395 ]
Анорексигенные нейроны 2: 607-609
Анри Виктор 1: 286
Антагонисты рецепторов 1: 597, 644
Антеннапедии 3: 278, 279
Антенны хлорофиллов 2: 304, 362,
365,366,369-377
Антибиотики
- ингибирование гликозилирования
белков 3: 213, 214
- ингибиторы топоизомераз 3: 24
- ингибиторы транскрипции 3: 121,
122
- ингибиторы трансляции 3: 208, 209
- механизм действия 1: 314-317, 570;
2: 560; 3: 208, 209
- перенос генов устойчивости на
плазмидах 1: 21, 439, 471; 3: 10
- устойчивость бактерий 1: 157
Антигенная вариабельность 3: 256
Антигенная детерминанта (эпитоп)
1:250
Антигены 1: 250
- взаимодействие с антителами
1:249-251
Аптидиуретический гормон (вазо-
прессин) 2: 576-578
Антикодон 3: 173
Антипорт 1: 370, 380; 2: 411, 416
- Na+/K+-ATPa3a 2: 590
— в мембранном транспорте в ней-
ронах 2: 590
— в сетчатке 1: 651-653
— роль в поляризации мембраны
2: 339, 340
- для триозофосфатов 2: 417,418
Антитела 1: 249, 251; см. также
Иммуноглобулины
- в анализе 1: 254-256
- как результат рекомбинации 3: 96,
97
- каталитические 1: 307,308
- моноклональные 1: 254
- поликлональные 1: 254
- разнообразие 1: 250
- специфичность связывания 1: 253
Анфинсен Кристиан 1: 209, 210
Апоаконитаза 2: 193
Аполипопротеины 2: 228,485-489
- в болезни Альцгеймера 2: 487, 488
- редактирование РНКЗ: 176-178
Апопротеин 1: 272
Апоптоз 1: 672, 673
- липоапоптоз 2: 616
- роль митохондрий 2: 347, 350
- роль цитохрома Р-450 2: 347
- фактор активации Apaf-1 2: 350
Апоптосома 2:350
Апофермент 1: 272
Аппетит, гормональный контроль
2: 605-610
Аптамеры 3: 154,155, 254
Арабиноза 1: 343
Арахидовая кислота 1: 489
Арахидонат 1: 508; 2: 461-463
Арахидоновая кислота 1: 508; 2: 572
Арбер Вернер 1: 435
Аргиназа 2: 273, 520
Аргинин 1: 116
- биосинтез 2: 519-521
- в синтезе оксида азота 2: 542, 576
- превращение в а-кетоглутарат
2: 292
- свойства 1: 116
Аргининемия 2: 287
Аргининосукцинат 2: 273, 275
Аргининосукцинатная ацидемия
2: 287
Аргининосукцинатсинтетаза 2: 273,
275
Арнон Даниэль 2: 382
Ароматические аминокислоты,
см. также Аминокислоты
— в биосинтезе веществ в растениях
2: 523, 526
Аррестины
- 1 1: 654
- 2 (0-аррестин) 1: 606, 607, 654
Археи 1: 18-20
- мембранные липиды 1: 496, 500,
501
Асимметрия молекул 1: 184
Аскорбат 1: 190,191
Аскорбиновая кислота, см. Витамин
С
Аспарагин 1: 114, 116; 2: 278, 304,
518, 523
- расщепление до оксалоацетата
2:104,297
- свойства 1: 116, 120
Аспарагиназа 2: 523
Аспарагиновая кислота 1: 116; 2: 304,
518, 523
Аспартам 1: 39, 69, 129
Аспартат 1: 78, 116, 120; 2: 274, 523,
531
- в С4-пути 2: 424, 425
- расщепление до оксалоацетата 2: 297
- свойства 1: 116, 120
- синтез пиримидинов 2: 544, 547,
548
Аспартатаминотрансфераза (ACT)
2: 267, 269, 274
Аспартат-аргининосукцинатный
шунт 2: 276
Аспартаттранскарбамоилаза 1: 321;
2: 548, 549
Аспирин 2: 461-463; см. также Аце-
тилсалициловая кислота
Астбери Уильям 1: 177
Атеросклероз 2: 493, 494
- роль тирш/с-жирных кислот 1: 494
Атом
- электроотрицательность 1: 70, 75,
76, 304
- водорода, в переносе электронов
2: 46
Атомная масса, единицы 1: 32
Атомно-силовая микроскопия 1: 544
Атриальный натрийуретический
фактор (ANF) 1: 627
Аттрактанты 1: 645
Ауксины 1: 470; 2: 538
Аутокринные гормоны 2: 571, 605
Аутосплайсирующиеся интроны
3: 125,140,153
Аутофосфорилирование, см. также
Фосфорилирование
- в бактериях 1: 645
- в проведении сигнала инсулина
1:621
- в растениях 1: 648, 649
- в сигнальных путях 1: 625
Африканская сонная болезнь (три-
паносомоз) 1: 540-542
Аффинная хроматография 1: 135,
379, 447
Ахиральные молекулы 1: 35
Ацеталь 2: 343, 348
Ацетальдегид 2: 49
Ацетат
- активированный 2: 199-203
- в синтезе жирных кислот 2: 213,
214
- в синтезе холестерина 2: 480, 481
- как источник фосфоенолпирувата
2:212
- транспорт 2: 213, 214
Ацетил-СоА 1: 426
- в метаболизме глюкозы 2: 582
- в синтезе жирных кислот
2: 446-456
- в синтезе холестерина 2: 480
- гидролиз 2: 34
- метаболизм в печени 2: 252, 582,
583, 584
- образование при 0-окислении
2: 235
- окисление в цикле лимонной кис-
лоты 2: 237
- окисление пирувата 2: 183
Ацетил-СоА-АСР-трансфераза 2: 450
13961 Предметно-именной указатель
Ацетил-СоА-карбоксилаза
- в бактериях 2: 457
- в растениях 2: 457
- в синтезе жирных кислот 2: 151,
243, 446, 457
N-Ацетилгалактозамин 1: 390
N-Ацетилглутамат в синтезе мочеви-
ны 2: 277
N-Ацетилглутаматсинтаза 2: 277, 279
N-Ацетилглюкозамин 1: 311, 313, 490
- в синтезе пептидогликанов 2: 435
Ацетилен 2: 45
Ацетил-кофермент А, см. Ацетил-
СоА
N-Ацетилмурамовая кислота 1: 311,
313,347
— в синтезе пептидогликанов 2: 435
N-Ацетилнейраминовая кислота
1:311,313,376 *
— в ганглиозидах 2: 502
Ацетилсалициловая кислота (аспи-
рин) 2: 461,462
Ацетилхолиновый рецептор 1: 578,
579, 638
— в передаче сигнала 1: 635, 636
— дефекты 1: 581
— открытая/закрытая конформация
1:579
— синаптическая агрегация 1: 542
— структура 1: 638
Ацетоацетат 2: 63, 85, 251-252
Ацетоацетатдекарбоксилаза 2: 252
Ацетоацетил-АСР 2: 450, 451
Ацетоацетил-СоА 1: 425; 2: 252, 253
Ацетогруппа
- в синтез жирных кислот 2: 213,
214
- транспорт 2: 213, 214
Ацетон 2: 466, 603
- при диабетическом кетоацидозе
2: 603
Ацивин 2: 558
Ацивицин 2: 558
Ацидемия
- аргининосукцинатная 2:287
- метилмалоновая 2: 287, 295
Ацидоз 1:93, 102; 2: 254, 604
- диабетический 2: 603, 604
Ацикловир 3: 66
Ацил-СоА
- в превращениях жирных кислот
2: 231
- производные жирных кислот
2: 230, 231
— в синтезе триацилглицеринов
2: 464-466
Ацил-СоА-ацетилтрансфераза 2: 235
Ацил-СоА-ацилтрансфераза (тиола-
за) 2: 235
Ацил-СоА-дегидрогеназы 2: 234, 237,
316
- генетические дефекты 2: 245
- средней цепи (MCAD) 2: 245
Ацил-СоА-десатураза 2: 458
Ацил-СоА-синтетазы 2: 230, 231
- в синтезе триацилглицеринов
2: 465
Ацил-СоА-холестеринацил-
трансфераза (АСАТ) 2: 485, 492
Ацилкарнитиновый переносчик
2: 231,232
Ацилпереносящий белок (АСР)
2: 449, 450
N-Ацилсфинганин 2: 476, 478
N-Ацилсфингозин 2: 476, 478
Ацилферменты 1: 302, 305
Ацил фосфат 2: 33, 76
Аэробные организмы, эволюция
1:60
Аэробный метаболизм, у мелких по-
звоночных 2: 92
Аэролизин k 1: 150
Б
Базы данных
— белковых структур 1: 431, 463
— геномные 1: 444, 449, 457, 459,
462, 466
Бактериальный геном 3: 10, 11
— секвенирование 1: 458, 459
Бактерии 1: 19-24; см. также
Escherichia coli
- аминокислотный состав 2: 538
- анаэробные, фиксация азота
2: 507-509
- галофильные, синтез АТР 2:
387-389
- гетерополисахариды клеточной
стенки 1: 358
- грамотрицательные 1: 21
- грамположительные 1: 21
- зеленые серные 2: 368, 386
- калиевые каналы Г. 575, 576
- лектины 1: 374
- липополисахариды 1: 370
- пурпурные 1: 60, 532, 544; 2: 367,
369,386
- регуляция экспрессии 3: 243-256
- репликация ДНК 3: 45-66
- синтез гликогена 2: 427
- синтез пептидогликанов 2: 433-435
- синтез целлюлозы 2: 433
- устойчивость к антибиотикам
1:316,317
- эндосимбиотические 1: 59, 60
— эволюция хлоропластов 2: 386
— эволюция митохондрий 2: 352
Бактериородопсин 1: 222; 2: 388,389
- как световой протонный насос
1: 84,2: 389
- микроскопический анализ 1: 544
- структура 1:532
Бактериофаг k 1: 435,440-442; 3:91,
92
Баллард Джон 2: 468
Балтимор Дэвид 3: 144, 145
Бантинг Фредерик 2: 568
Бевацизумаб (авастин) 1: 669
Бекстра 2: 462,463
Белая жировая ткань 2: 585-588
Белки 1: 132-262
- аллостерические 1: 237
- аполипопротеины 2: 485-487
- базы данных 1: 193, 203-205
- биосинтез 3: 178-209
- время полужизни 2: 125, 582, 512
- вторичная структура 1: 141,
177-183
- глобулярные 1: 185, 192-205
- гомологичные 1: 61
- денатурация 1: 208, 209
- домены 1: 200
- железо-серные 2: 309, 310
— Риске 2:310,317, 376
- заякоривающие киназу А (АКАР)
1:604, 608, 609,616,617
- зимогены 1: 328, 329
- иммунной системы, см. Иммуно-
глобулины
- каталитические, см. Ферменты
- кислород-связывающие 1: 226-246
- классификация по структурным
признакам 1: 203, 204
- конформация 1: 172
- конъюгированные 1: 363-369
- липопротеины 2: 485-487
- мембранные, см. Мембранные
белки
- методы выделения и анализа
1:132-140,149-152,184,196-199
- мотивы укладки 1: 200-205;
3: 239-242
- моторные 1: 256-262
- мультимерные 1: 205
- мультисубъединичные 1: 130
- мультифункциональный 2: 248
- нативные 1: 172
- олигомернные 1: 130
- ортологичные 1: 61
- паралогичные 1: 61
- первичная структура 1: 141-148
Предметно-именной указатель [3971
- плазмы крови 2: 485, 591, 592
- полиморфные 1: 142
- посттрансляционный процессинг
3:180, 208
- пробелки 1: 328
- протеолитическая активация
1:319, 328, 329
- протомерные 1: 130
- разобщающий, см. Термогенин
- регуляторные 3: 237-242
- ренатурация 1: 209
- рибосомные 3: 180, 250, 251
- с высокой электрофоретической
подвижностью (HMG) 3: 262
- свертывание (фолдинг) 1: 210-215
— нарушения 1: 215-218
- семейства и суперсемейства 1: 203
- слияния 1: 547
- сложные 1: 131
- стабильность 1: 172
- третичная и четвертичная структу-
ра 1: 141, 184-208
- трифункциональный 2: 235
- фибриллярные 1: 185-192
- флавопротеины 2: 54, 55
- фосфорилирование/дефосфорили-
рование 2: 127
Белые клетки крови (лейкоциты)
2: 592
Белые мышцы 2: 587
Бензоаты 2: 278
Бензоил-СоА 2: 278
Берг Пол 1:433,438
Бергстрем Суне 1: 508
Берсон Соломон 2: 569
Бест Чарлз 2: 568
Бидл Джордж 2: 193; 3: 8
Бикарбонат 1: 79
- буферные свойства 1: 99-101
- образование 1: 242,556
Биливердин 2: 535
Билирубин 2: 535, 536
Биоинформатика 1: 115, 156, 212
Биолюминесценция, цикл у светля-
ков 2: 39
Биомолекулы
- амфифильные 1: 78, 81
- асимметричные 1: 35
- конформация 1: 38
- макромолекулы 1: 31
- молекулярная масса 1: 32
- происхождение 1: 55, 56
- стереоспецифичность 1: 37, 38
- функциональные группы 1: 29, 30
- хиральные 1: 35
Биотин 2: 205, 207, 281
- в пируваткарбоксилазной реакции
2: 100, 101,205-207
- в реакции ацетил-СоА-
карбоксилазы 2: 446, 447
- в синтезе фосфоенолпирувата из
пирувата 2: 100
- недостаточность 2: 207
Биохимические реакции
— внутримолекулярные перегруп-
пировки 2: 23
— изомеризация 2: 23, 24
— и химические реакции 2: 27, 28
— нуклеофильные 2: 21
— окислительно-восстановительные
2: 26; см. также Окислительно-
восстановительные реакции
— распространенные типы 2: 20-26
— расщепление связи 2: 20, 21
— свободнорадикальные 2: 24, 25
— с переносом групп 2: 25
— электрофильные 2: 21
— элиминирование 2: 23, 24
Биоцитин 2: 63
Биоэнергетика 1: 44; 2: 13
Бисульфиты, мутагенные свойства
1:418
Бисфосфаты 2: 73
1,3-Бисфосфоглицерат 2: 202
- гидролиз 2: 32
- синтез 2: 76, 79
2,3-Бисфосфоглицерат, в связывании
гемоглобина с кислородом 1: 245
Бишоп Майкл 3: 146
Блобел Гюнтер 3: 210
Блох Конрад 2: 445, 484
Богатый глутамином домен 3: 265,
266
Бойер Герберт 1: 433
Бойер Поль 2: 336, 338
Больцмана константа 2: 13
Большая бороздка 1: 401
Бор Кристиан 1: 243
Бор Нильс 1: 243
Бора эффект 1: 243
Борьба или бегство, реакция 1: 173,
602, 603
а/|3-Бочонок 1: 451, 200, 202
Брассинолид 1: 509, 647
Брассиностероиды 1: 647
Браун Майкл 2: 490, 493
Брожение, при пивоварении 2: 94-96;
см. также Ферментация
Бунгаротоксин 1: 581
Бурая жировая ткань 2: 585-588
--митохондрии 2: 348
Буферные системы 1: 96-98
Бухнер Эдуард 1: 270; 2: 66, 132
Быстро сокращающиеся мышцы
2: 587, 588
Бычья губчатая энцефалопатия
1:217,218
Бьюкенен Джон 2: 544
В
Вагелос П. Рой 2: 494
Вазопрессин 2: 576, 577
Вакцины, на основе вирусов 3: 146
Валин 1: 118
- биосинтез 2: 523, 524
- в гемоглобине S 1: 247
- катаболизм 2: 288, 294, 296, 297
Валиномицин 1: 571
Ванадат 1:560
Ван-дер-Ваальса радиус 1: 34, 35
Ван-дер-ваальсовые взаимодействия
1: 82, 83
Вант-Гоффа уравнение 1: 85
Варбург Отто 2: 60, 66, 80-83
Варбурга эффект 2: 80
Варм ус Харольд 3: 146
Варфарин 1: 512
Везикулы 1: 529
Вейн Джон 1: 508
Векторные реакции, в окислитель-
ном фосфорилировании 2: 315,
323
Векторы, см. Клонирующие векторы
Вентер Дж. Крэйг 1: 458
Вернике-Корсакова синдром 2: 112
Вёзе Карл 1: 19, 156; 3: 152
Виагра (силденафил) 1: 628
Видимый свет
-- поглощение фотопигментами
2: 361-365
— электромагнитное излучение
2: 359, 360
Виментин 1: 259
Винтовая (спиральная) симметрия
1:207
Виокс 2: 462, 463
Вирусная инфекция
— вакцины 3: 146
— и онкогены 1: 666
— и рак 3: 146
— проникновение в клетку 1: 374
Вирусные векторы 1: 473, 474
Вирусоид 3: 139, 142
Вирусы, см. также ВИЧ; Ретрови-
русы
- в генной терапии 1: 475, 476
- геном 3: 145
- ДНК 3:9
- ДНК-полимераза 3: 66
[398] Предметно-именнойу ка зател ь
- и лектины 1: 374
- и селектины 1: 374
- капсид 1: 207
- простого герпеса, ДНК-полимераза
3: 66
- РНКЗ: 151
- саркома Рауса 3: 146
Витамины 1: 510-512
- А (ретинол) 1: 511, 652
- В12 (кобаламин) 2: 240-242
- С 1:189-191
- D 2: 575
- D2 (эргокальциферол) 1: 511
- D3 (холекальциферол) 1: 510, 511
- Е (токоферол) 1: 512
- К 1: 512
- К] (филлохинон) 1: 512
- К2 (менахинон) 1: 5i£
- изопреноиды 1: 512, 513
— биосинтез 2: 497
- ниацин 2: 51, 53
ВИЧ (вирус иммунодефицита
человека)
- как пример ретровируса 1: 317;
3: 146, 147
- лечение 1: 307, 318; 3: 147
Вкус, сигнальные механизмы 1: 656,
657
Внеклеточный матрикс
— гликозаминогликаны 1: 359, 360
— протеогликаны 1: 363, 364
Внутренняя адаптерная последова-
тельность 3: 139
Внутренние петли, РНК 1: 409, 410
Внутренний фактор 2: 242
Вода 1: 73-79
- мембранный транспорт 1: 556,
571-573
- расщепление 2: 379-381
Водные растворы, см. также Вода
-pH 1:88-91
- амфифильные соединения 1: 78,
81,496
- буферные системы 1: 97, 98
- гипертонические 1: 86
- гипотонические 1: 86
- изотонические 1: 85-87
- коллигативные свойства 1: 84
- осмолярность 1: 85
- слабые кислоты и основания
1: 94-96
Водород, перенос гемоглобином
1: 242, 244
Водородные связи 1: 75
— в белках!: 173, 174, 178
— в воде 1: 73-75
— в нуклеиновых кислотах 1: 52,
399,402, 406
— в полисахаридах 1:353, 355, 358
— типы в биологических системах
1:76
Водоросли, гетерополисахариды
клеточной стенки 1: 359
Возбужденное состояние молекулы
2: 360
Волосы
- а-кератин 1: 177, 185, 186
- химическая завивка 1: 187
Воск 1: 493, 495
- пчелиный 1: 495
Восстанавливающие концы 1: 347,
354
Восстанавливающие сахара 1: 347
Восстановительный пентозофосфат -
ный путь 2: 112,406
Восстановительный потенциал 2: 311
- - стандартный 2: 47, 48, 312, 322
Восстановительный эквивалент
2: 46, 309
Вращательная (поворотная) симме-
трия 1: 207
Вращательный катализ 2:306, 336,
337
Врожденная ночная слепота 1: 581
Врожденная парамиотония 1: 579,
581
Врожденные дефекты метаболизма
1:505
Врожденный миастенический син-
дром 1: 581
Вспомогательные пигменты 2: 361
Вторичная структура 1: 141,
177-184,211
— коллагена 1: 188
— мотивы 1: 200
— супервторичная 1: 200
— ДНК/РНК 1:406, 410
Вторичный мессенджер (посредник)
1:427, 595-515, 607, 609, 658
— в регуляции экспрессии 3: 252,
268
— нуклеотиды 1: 391, 427, 627
Вторичный метаболит 1: 32
Вторичный системный амилоидоз
1:215
Вули Д. Уэйн 2: 53
Вырожденность генетического кода
3: 171
Высокоэффективная жидкостная
хроматография (ВЭЖХ) 1: 135
Вюрц Шарль-Адольф 1: 273, 286
Г
Г6ФДГ, см. Глюкозо-6-фосфат-
дегидрогеназа
Газовая постоянная (R) 1: 47, 85, 634;
2: 47
Газожидкостная хроматография
(ГЖХ) 1:516, 517
Газы, растворимость 1: 79
D-Галактитол 2: 90
Галактоза 1: 343, 345, 359, 377
- окисление 1: 347
- превращение в глюкозо-1-фосфат
2: 89
- регуляция метаболизма 3: 263, 264
Галактозамин 1: 345
Галактоземия 2: 90, 117
0-Галактозидаза 1: 441; 3: 234, 235
Галактозидпермеаза 3: 234, 235
0-Галактозиды, и /яс-оперон 3: 235
Галактокиназа 2: 89, 90
Галактолипиды 1: 496, 500, 526
Галапагосские вьюрки, эволюция
3: 278-280
Галофильные бактерии, синтез АТР
2: 387-389
ГАМК (у-аминомасляная кислота)
2: 272, 539
- рецепторы 1: 579, 640
Ганглиозидоз 1: 505
Ганглиозиды 1: 502, 503; 2: 477
-GD3 1:376
-GM1 1:375
- GM2 1: 505
- в болезни Тея-Сакса 1: 505
- функции 1: 369
Гаптены 1: 250
Гарден Артур 2: 71
Гастдуцин 1: 656
Гастрин 2: 264
Гашиш 1: 622
ГДГТ (глицериндиалкилглицерин-
тетраэфиры) 1: 501
Гексадекановая кислота 1: 488
Гексозомонофосфатный путь 2: 107,
108; см. также Пентозофосфат-
ный путь
Гексозы 1: 340-342
- производные 1: 345, 346
- фосфорилированные, в гликолизе
2: 70
Гексокиназы 1: 293, 308, 309; 2: 71
-12: 141
-II2: 140, 141
-IV 2:142, 581,595
- изоформы 2: 71, 72, 140
Предметно-именной указатель [399]
- ингибиторы, в противоопухолевой
терапии 2: 82,83
- регуляция 2: 141-143
Гель-электрофорез, см. Электро-
форез
Гем 1: 227
- А 2:310
- Ь,в комплексе III 2: 317
- С 2: 310
- в синтезе d-аминолевулината
2: 532, 533
- как источник желчных пигментов
2: 534
- структура 1: 227, 228
Гемагглютенин (белок НА) 1: 374
Гемовая группа 1: 205, 206
Гемовые кофакторы цитохромов
2: 309,310,317
Гемоглобин
- А 1: 247
- R/T-формы 1: 235, 236
- S 1: 247
- генетические вариации 1: 264, 265
- гликированный 1: 348, 349
- олигомерная структура 1: 130
- при серповидно-клеточной анемии
1: 246-248
- связывание кислорода 1: 233, 237
— эффект Бора 1: 243
— влияние БФГ 1: 245
- связывание монооксида углерода
1:240, 241
- структура 1: 141, 205, 206
- субъединицы 1: 83, 233-235
- транспорт водорода 1: 242, 244
- транспорт диоксида углерода
1:242
- транспорт кислорода 1: 233
Гемоцитобласты 1: 233
Генерализованная эпилепсия с фе-
брильными судорогами 1: 581
Генерализованный ганглиозидоз
1:505
Генетика 1: 50-52
Генетическая карта Е. coli 3: 44
Генетическая рекомбинация
3: 81-89; см. также Рекомбина-
ция ДНК
Генетические заболевания
— генная терапия 1: 475, 476
— нарушение фолдинга 1: 215-218
Генетические мутации, см. Мутации
Генетические нарушения метаболиз-
ма аминокислот 2: 287
-----ацил-СоА-дегидрогеназы
2: 245
-----в цикле мочевины 2: 277, 278
Генетический код 3: 166-178
Генная инженерия 1: 434 479;
см. также Клонирование
Генная терапия 1: 475,476
Геном 1: 60
- аннотированный 1: 61, 62
- бактерий 3: 10
- вирусов 3: 9
- дрожжей 3: 11
- картирование 1: 450-452,458, 459
- проект «Геном человека» 1: 458,
459
- ПЦР в анализе 1: 452
- секвенирование 1: 421, 458,459
- синтения 1: 462
- человека 1: 460
- эволюция 1: 62
- эукариот 3: 11
Геномика 1: 434, 449
- сравнительная 1: 62, 452, 467
- функциональная 1: 62
Геномные библиотеки 1: 449-451
Гены 3: 8
- гомологичные 1: 61
- домашнего хозяйства 3: 230
- иммуноглобулинов, рекомбинация
3: 96,97
- материнские 3: 273
- ортологичные 1: 61
- паралогичные 1: 61
- парного правила 3: 274
- полярности сегментов 3: 274
- пробела 3: 273, 276
- сегментации 3: 273
- смотрители 1: 670
Гепарансульфат 1: 361, 364-367
Гепарин 1: 360, 361
Гепатоциты 2: 580
- гранулы гликогена 2: 154, 155
- каскад адреналина 1: 606
- метаболизм
— аминокислот 2: 582-584
— глюкозы 2: 581, 582
— жирных кислот 2: 584
— питательных веществ 2: 580-582
— углеводов 2: 581,582
- реутилизация триацилглицеринов
2: 467
-синтез NADPH 2: 454, 455
- синтез жирных кислот 2: 455-457
Гептозы 1:341,370
Геранилгеранильная группа 1: 573
Ге ран ил пирофосфат, в синтезе
холестерина 2: 23, 483, 496, 502
Гербициды, устойчивость растений
1:471-473
Герса болезнь 2: 159
Гетерозиготы, с геном серповидных
эритроцитов 1:247,248
Гетеролитическое расщепление кова-
лентной связи 2: 20, 21
Гетероплазмия 2: 354
Гетерополисахариды 1: 352, 359;
см. также Полисахариды
Гетеротропная регуляция 1: 245,322
Гетеротропные ферменты 1: 321,323
Гетеротрофы 1: 20, 58
Гетерохроматин 3: 257
Гиалуронат 1: 361, 367
Гиалуронидаза 1: 361
Гиалуроновая кислота 1:360
Гиббс Дж. Уиллард 1: 44
Гиббса энергия 2: 13; см. также Сво-
бодная энергия
Гибридизация, см. ДНК, гибридиза-
ция; Клонирование
Гибридные дуплексы 1: 414, 415
Гигантские поры 2: 350
Гидрид-иоп 2: 21,183, 193,308, 309
0-Гидрокси-0-метилглутарил-СоА,
см. HMG-CoA
0-Гидроксиацил-АСР-дегидратаза
2:448, 452
0-Гидроксиацил-СоА
(3-гидроксиацил-СоА) 2: 234
0-Гидроксиацил-СоА-дегидрогеназа
2: 234
0-Гидроксибутират 2: 251, 253
- в головном мозге 2: 590
0- Гидроксибутиратдегидрогеназа
2: 252
Гидроксилазы 2: 460
5-Гидроксилизин 1: 120
- в коллагене 1: 188
5-Гидроксиметилцитидин 1: 395
4-Гидроксипролин 1: 120
- в коллагене 1: 187, 189, 190
2-Гидроксипропановая кислота,
см. Молочная кислота
Гидролазы 1: 104
Гидролиз 1: 104
- АТР 2: 29, 35
- ацетил-СоА 2: 34
- 1,3-бисфосфоглицерата 2: 32
- гликозидной связи 2: 86, 87
- глюкозо-6-фосфата 2: 103
- дисахаридов 1: 351
- и фосфоролиз 2: 87, 88,155
- изменение свободной энергии
2: 16,33,34
- креатинфосфата 2: 33
- переходное состояние 1: 307
- сложных эфиров 2: 34
- тиоэфиров 2: 33, 34
| \ 001 Предметно-именной указатель
- фосфоенолпирувата 2: 32
- фосфорилированных соединений
2: 32, 33
Гидропатический индекс, см. Индекс
гидрофобности
Гидрофильные соединения 1: 77
Гидрофобные взаимодействия 1: 81,
211; см. также Слабые взаимо-
действия
— в глобулярных белках 1: 194,195,
199
— и стабильность белков 1: 173, 209
Гидрофобные соединения 1: 77, 81
Гиймен Роже 2: 567
Гилман Альфред 1: 598, 600
Гильберт Вальтер 1: 419, 420
Гипераммониемия 2: 278
Гипергликемия, секреция инсулина
2: 85,153
Гиперглицинемия некротическая
2: 287
Гиперинсулинемия 2: 598
Гиперинсулинизма/гипераммо-
ниемии синдром 2: 270
Гиперкалиемический паралич
(врожденная парамиотония)
1:579, 581
Гипертонический раствор 1: 86
Гиперфенилаланинемия 2: 291
Гиперхолестеринемия наследствен-
ная 2: 493-495
Гиперхромный эффект 1: 413
Гипогликемия 2: 592, 597, 598
Гипоксантин 1: 394, 395; 2: 555
Гипоксантин-гуанинфосфорибозил-
трансфераза 2: 556, 557
Гипоксия 1: 245; 2: 69, 81
- адаптационные реакции 2: 345
- активные формы кислорода 2: 345
- гидролиз АТР 2: 344
Гипоталамо-гипофизарная система
2: 577
Гипоталамус 2: 576-578
- в регуляции массы тела 2: 606, 607
Гипотонический раствор 1: 86
Гипофиз, гормоны 2: 576-568
Гипохромный эффект 1: 413
Гиппурат 2: 278
Гирке болезнь 2: 159
Гистамин 2: 530,539, 542
Гистидин 1: 98, 119, 120
- биосинтез 2: 529, 530
- буферные свойства 1: 98, 127
- кривая титрования 1: 126
- превращение в а-кетоглутарат
2: 292, 293
Гистидинкиназа 1: 645
Гистонацетилтрансферазы 3: 258, 259
Гистондеацетилазы 3: 258, 259
Гистоны 3: 28, 258, 259
Глибурид 2: 597
Гливек (иматиниб) 1: 669
Гликаны 1: 352-354; см. также По-
лисахариды
Гликированный гемоглобин 1: 349
Гликобиология 1: 339-390
Гликоген 1: 353; 2: 121
- биосинтез в бактериях 2: 428
- болезни накопления 2: 159, 160
- в мышцах 2: 92,155
- в цикле Кори 2: 92, 97
- гидролиз 1: 325, 326
- гранулы, в гепатоцитах 2: 154, 155,
164
- как форма запасания глюкозы
1: 352-354
- метаболизм 2: 154-165
— регуляция 2: 165-173
- отщепление глюкозы 2: 156
- расщепление 2: 87, 88, 155-158
- роль гликогенина 2: 163, 164
- сахаронуклеотиды 2: 158
- синтез 2: 158, 162
— регуляция 2: 137, 138
- трехмерная структура 1:358
Гликогенез 2: 155
Гликогенин2: 163, 164
- и гранулы гликогена 2: 164
Гликогенозы 2: 159
Гликогенолиз 2: 155
Гликогенсвязывающий белок GM
2: 171
Гликогенсинтаза 1: 327; 2: 162, 163,
168
- а 2: 168
- (3 2:168
- киназа (GSK3) 1: 622, 630; 2: 168
- праймер 2: 163, 164
регуляция 2: 168
- фосфорилирование 1: 168
Гликогенфосфорилаза 2: 87, 155, 156,
160, 167
- регуляция 1: 325, 326; 2: 165, 167
Гликозаминогликаны 1: 359-361,
363-365
Гликозидазы 1:379, 515
Гликозидная связь 1: 348, 350, 355,
357, 368
— в нуклеотидах 1: 392, 404
— гидролиз 1: 311, 313, 416
— изменение свободной энергии
2:16
Гликозилирование белков 3: 207,
213,214
Гликозилфосфатидилинозит (GPI)
1: 530, 536, 537, 545
Гликоконъюгаты 1: 339, 363
- гликолипиды 1: 364, 369
- гликопротеины 1: 363, 367-369,
3: 207,213
- протеогликаны 1: 363-367
Гликолатный путь 2: 421, 422
Гликолиз 2: 66-90, 144, 149
- в опухолевых тканях 2: 81-83
- в хлоропластах 2: 430
- взаимопревращения триозофосфа-
тов 2: 74, 75
- и глюконеогенез 2: 99, 105
— согласованная регуляция
2:139-153
— роль фруктозо-2,6-бисфосфата
2: 144-146
- изменение свободной энергии в
эритроцитах 2: 98
- образование АТР 2: 80
- подготовительная стадия 2: 67, 68,
71-75
- при пониженном содержании кис-
лорода 2: 92
- пути, питающие 2: 86-90
- регуляция 2: 80, 105
- роль глюкозо-6-фосфатдегидро-
геназы 2: 106, 107
- стадия выплаты процентов 2: 67,
68, 75-79
- участие глюкозо-6-фосфата 2: 112,
140
Гликолипиды 1: 364, 369, 496; см.
также Липиды
- нейтральные 1: 502, 515
- синтез 2: 476, 477
- транспорт 2: 478
Гликомика 1: 369
Гликопротеины 1: 363,367-369;
3: 207,213
- анализ олигосахаридной составля-
ющей 1: 379, 380
- мембранные 1: 526, 527
- связывание с лигандами 1: 373
- топология 1: 530-532
Гликосфинголипиды 1: 503, 543
Гликофорин 1: 369, 527, 531, 532
Глиоксилат 2: 212, 288
Глиоксилатный цикл 2: 212-215
— в растениях 2: 247, 436
Глиоксисомы 2: 213, 214, 436
- (3-окисление 2: 246, 247, 249
Глипизид 2: 597
Глипиканы 1: 364, 365
Глифосат, устойчивость растений
1: 472, 473
Предметно-именной указатель [401 ]
Глицеральдегид 1: 37, 117, 342
Глицеральдегид-З-фосфат 1: 281;
2: 58, 75
- в гликолизе 2: 67-69, 74, 75, 98
- в цикле Кальвина 2: 406, 407,
410-416
- окисление до 1,3-бисфосфоглице-
рата 2: 76
- синтез 2: 406, 410-412
Глицеральдегид-З-фосфатдегидро-
геназа 2: 53, 68, 69, 76
- активация светом 2: 418, 419
- механизм реакции 2: 77
Глицерин 1: 491
- в мембранных липидах архей
1:500, 501
- в синтезе триацилглицеринов
2: 465
- в триацилглицеринах 1: 491
- в фосфолипидах 1: 496, 497
- структура 1: 491
- хиральность 1: 497
Глицерин-З-фосфат 1: 497
- в синтезе липидов 2: 465, 477
- в синтезе углеводов 2: 436, 437
- синтез 2: 468
Гл и церин-3-фосфатдегидрогеназа
2: 342, 465
Глицерин-З-фосфатный челнок
2: 341,342
Глицериндиалкилглицеринтетра-
эфиры (ГДГТ) 1: 501
Глицеринкиназа 2: 465
Глицеропеогенез 2: 468, 469; см. так-
же Метаболизм глюкозы
Глицерофосфатацилтрансфераза
2:611
Глицерофосфолипиды 1: 497, 498;
см. также Триацилглицерин
- действие фосфолипаз 1: 504
- номенклатура 1: 518, 519
- полярные головки 1: 496
- присоединение головки 2: 470, 471,
473
- синтез 2: 464, 465
- транспорт 2: 478
Глицин 1: 93,114, 115,125, 126, 180;
2: 532, 537
- биосинтез 2: 519, 521
- буферные свойства 1: 109, 125
- в а-спиралях 1: 176-181
- в |3-поворотах 1: 182,183
- в (3-слоях 1: 181, 182
- в коллагене 1: 188
- в синтезе пуринов 2: 544
- в фотосинтезе 2: 326, 423
- как предшественник порфиринов
2: 532, 533
- кривая титрования 1: 123, 124
- расщепление до пирувата 2: 285
- рецепторы 1: 579, 639
Глициндекарбоксилазный комплекс
2: 421,423
Глицинрасщепляющий фермент
(глицинсинтаза) 2: 287, 521
Глицинсинтаза 2: 287, 521
Глобины 1: 203, 228; см. также Гемо-
глобин; Миоглобин
- структура 1: 198, 228
Глобозиды 1: 502, 505
Глобулярные белки 1: 185 192-207
— в модели липопротеиновых ком-
плексов 1: 589
— денатурация и фолдинг 1: 208
— методы анализа структуры
1: 196-199
— (3-повороты 1: 182
Глутамат (глутаминовая кислота)
1: 114; 2: 17, 274, 279,514
- биосинтез 2: 513, 524
- в биосинтезе пролипа и аргинина
2:519, 520
- в метаболизме азота 2: 263, 264
- включение аммония 2: 513
- высвобождение аммония 2: 269
- как усилитель вкуса 1:681
- катаболизм 2: 292, 293
- кривая титрования 1: 126
Глутамат-у-полуальдегид 2: 519-521
Глутамат-аспартатная транспортная
система 2:341
L-Глутаматдегидрогеназа 2: 269-271
Глутамат-оксалоацетаттрансаминаза
(ГОТ) 2: 269
Глутамат-пируват-трансаминаза
(ГПТ) 2: 269
Глутаматсинтаза 2: 513, 514
Глутамин 1: 120; 2: 105, 278, 517, 519
- биосинтез 2: 513
- в метаболизме азота 2: 263, 513
- как переносчик аминогрупп 2: 263,
264, 270
- катаболизм 2: 274, 292, 293
Глутаминаза 2: 270, 271,513
Глутаминамидотрансфераза 2: 517
- ингибиторы 2: 589
- механизм реакции 2: 549
Глутаминсинтетаза 2: 35, 270, 272,
513-516
- аллостерическая регуляция 1: 329,
330; 2:514,515
Глутаредоксин 2: 537, 550, 551
Глутатион 2:48, 326, 536, 551
- биосинтез 2: 536, 536
- защита клетки от окислителей
2: 106-108
Глутатионпероксидаза 2: 324, 326,
537
Глутатионредуктаза 2: 324, 326
Глюкагон
- в мобилизации жирных кислот
2: 228, 229, 467
- в регуляции метаболизма глюкозы
2:146,167,170, 595, 598, 599
- в регуляции метаболизма холесте-
рина 2: 491, 492
- каскадный механизм 2: 166
Глюканотрансфераза 2: 156
Глюкогенные аминокислоты 2: 98,
104, 280
Глюкоза 1: 26,343
- UDP-глюкоза 2: 16, 89
- а-форма 1: 344
- (3-форма 1: 344
- в анализе крови 1: 348, 349
- в дисахаридах 1: 350, 351
- в целлюлозе 1: 355
- запасные формы 1: 353
--гликоген 1: 353, 354
--крахмал 1: 353, 354
- и /яс-оперон 3: 235-245
- использование в клетке 2: 65
- как восстанавливающий сахар
1:347
- катаболизм 2: 581, 582
— в опухолевой ткани 2: 81-83
- метаболизм, см. также Брожение;
Гликолиз; Глюконеогенез
— в головном мозге 2: 590, 591
— в жировой ткани 2: 585
— в мышцах 2: 587-589
— в печени 2: 581, 582
— в поджелудочной железе 2: 595,
596
— при голодании 2: 598, 599, 601
— при сахарном диабете 2: 601, 603
— функция адреналина 2: 602
— функция глюкагона 2:598, 599
— функция инсулина 2: 594, 595
— функция кортизола 2: 602
- окисление 1: 46, 347
— выход АТР 2: 201, 202
— в нейронах 2: 590-591
— пентозофосфатный путь
2:109-111
— в клетках до углекислого газа 2: 49
- реакция гексокиназы 1:308
- транспорт 1:553-556; 2: 581
— симпорт с Na+ 1: 569, 570
14 021 Предметно-именной указатель
- уровень в крови 2: 592, 593
-----при диабете 1: 348; 2: 603
-----регуляция 2: 593-604
- фосфорилирование 2: 18, 19
- эпимеры 1: 343
Глюкозамин 1: 346
Глюкозилцереброзид 1: 501
Глюкозо-1-фосфат 2: 89, 157, 160
- в гликолизе 2: 165
- в синтезе крахмала 2: 426-428
- образование из галактозы 2: 89, 90
Глюкозо-6-фосфат 1:45,46; 2: 98, 99,
112, 122, 158
- в гликолизе 2: 112
- в пентозофосфатном пути 2: 107,
108, 110
- в печени 2: 99, 161, 598
- в синтезе гликогена 2: 161-163
- гидролиз 2: 157
- превращение в глюкозу 2: 99, 104,
598, 599
- превращение в фруктозо-6-фосфат
2: 72
- реакция гексокиназы 1: 308, 309
- регуляция функции инсулина
2: 595
Глюкозо-6-фосфатаза 2: 103, 128, 143
- в печени 2: 157, 158
- гидролиз глюкозо-6-фосфата 2: 158
Глюкозо-6-фосфатдегидрогеназа
(Г6ФДГ) 2: 109
- активация светом 2: 419
- недостаточность 2: 106
Глюкозофосфатизомераза (фосфо-
глюкоизомераза) 2: 72, 111
Глюкозо-аланиновый цикл 2: 271
Глюкозурия 2: 603
Глюкокиназа 2: 60, 141, 154, 161
- дефекты 2: 355
- кинетические свойства 2: 142, 581
- регуляция 2: 142,143
Глюкокортикоиды 2: 468, 469, 495,
574, 575
- синтез 2: 495, 496
Глюконат 1: 346
Глюконеогенез 2: 97-105
- в мышцах 2: 588, 589
- в печени 2: 583, 584, 589
- в прорастающих семенах 2:436, 437
- в хлоропластах 2: 430
- глюкогенные аминокислоты 2: 104,
280
- и гликолиз 2: 99, 105
-----согласованная регуляция
2: 139-153
-----роль фруктозо-2,6-бисфосфата
2: 144-146
- и глиоксилатный цикл 2: 212-215
- и глицеронеогенез 2: 468, 469
- и цикл лимонной кислоты 2: 104,
105
- обходные пути 2: 99-103
- при голодании 2: 253, 598, 601
- энергетические затраты 2: 103
Глюконолактон 1: 346
Глюкопираноза 1: 344
Глюкотрол 2: 597
Глюкуронат 1: 347, 361
Голдбергер Джозеф 2: 53
Голдштейн Джозеф 2: 490,491, 493
Головной мозг
— метаболизм 2: 590, 599-601
— потребление глюкозы 2: 97, 590,
591
Голодание
- метаболизм глюкозы 2: 599-601
- синтез кетоновых тел 2: 252, 253
Голубой флуоресцентный белок
(CFP) 1:612-515
Гольджи комплекс
— лектины 1: 375, 376
— сортировка белков 3: 214
— транспортные везикулы, см.
Транспортные везикулы
Гомеобокс 3: 241
- гены 3: 276
Гомеодомен 3: 241
Гомеостаз 2: 123
Гомеотическис гены 3: 276
Гомогентизатдиоксигеназа 2: 287, 292
Гомозиготы, с геном серповидных
эритроцитов 1: 247, 248
Гомолитическое расщепление кова-
лентной связи 2: 20, 21
Гомологи 1: 61, 157
Гомологическая рекомбинация
3:81-88
Гомоплазмия 2: 354
Гомополисахариды 1: 352; см. также
Гликоген; Крахмал; Полисаха-
риды
Гомоцистинурия 2: 287
Гормон роста, у растений 1: 470; см.
также Ауксины
Гормональная регуляция
- гликогенфосфорилазы 1: 326;
2: 172
- массы тела 2: 604-614
- метаболизма жиров 2: 173, 457,
466, 469
- метаболизма углеводов 2: 144, 173,
228, 593-600
- транскрипции 1: 643, 644
- уровня глюкозы 2: 593-600
- уровня холестерина 2: 491
- экспрессии генов 3: 263, 267, 268
Гормональный каскад 2: 570, 577
Гормон-чувствительная липаза
2: 599, 602,611
Гормон-чувствительные элементы
(HRE) 1:643; 3: 267, 268
Гормоны 2: 566-618
- адипокины 2: 605
- активация триацилглицеринов
2: 228, 229
- анализ 1: 593; 2: 567-569
- аутокринные 2: 571
- в качестве сигнальных молекул
1: 608, 609
- в пищеварении 2: 264, 265
- витамин D 2: 575
- время реакции 2: 571
- ингибирующее действие 1: 608
- каскады 2: 570, 577, 578
- катехоламины 2: 574
- классификация 2: 571, 572
- коры надпочечников 1: 509; 2: 496,
576
- механизм действия 2: 570, 571
- монооксид азота 2: 576
- открытие 2: 567-569
- паракринные 2: 571
- пептидные 2: 572
- половые 1: 509; 2: 495
- растительные 1: 470
- регуляция высвобождения
2: 576-579
- рекомбинантные 1: 479
- ретиноидные 2: 575
- рецепторы 2: 570
- связывание лектинов 1: 372
- синтез в митохондриях 2: 347, 348
- стероидные 1: 509; 2: 574
— рецепторы 3: 263, 267, 268
— синтез 2: 495, 496
- ткани-мишени 2: 577
- щитовидной железы 2: 575
- эйкозаноиды 1: 507; 2: 574
- эндокринные 2: 571
ГОТ (глутамат-оксалоацетат-транс-
аминаза) 2: 269
Гоше болезнь 1: 505
ГПТ (глутамат-пируваттранс-
аминаза) 2: 269
Градиент концентрации 1: 550, 551,
553, 559, 567
— в поляризации мембраны 1: 633
Граны 2: 358
Грелин 2: 607,613,614
Гринберг Дж. Роберт 2: 544
Предметно-именной указатель [ 4 03]
Грипп
- методы лечения 1: 374
- функция лектинов 1: 374
Грубый экстракт 1: 132
Грюнберг-Манаго Марианна 3: 142,
143
Гуанилат 1: 393; 2: 544
Гуанилилциклазы 1: 595, 627, 628
- и зрение 1: 655
Гуанилин 1: 627
Гуанин 1: 399
- дезаминирование 1: 416; 2: 555
- метилирование 1:418,419
Гуаниновые нуклеотиды, регуляция
биосинтеза 2: 547
Гуанозин 1: 393, 400,407
- в сплайсинге 3: 126, 153, 154
Гуанозиндифосфат, см. GDP
Гуанозинмонофосфат, см. GMP
Гуанозинтетрафосфат (ppGpp) 1: 427
Гуанозинтрифосфат, см. GTP
Губчатая энцефалопатия 1: 217, 218
Гу л оз а 1: 342
Гуморальный иммунитет 1: 249
д
Дабсилхлорид 1: 144
Дальгарно Линн 3: 196
Дальтон (Да) 1: 32
Дальтон Джон 1: 655
Дальтонизм 1: 655
Дам Хенрик 1: 512
Дансилхлорид 1: 144
Двойная спираль
— в транскрипции 3: 106, 107
— варианты 1:403-406
— ДНК 1:52, 391, 403,413
---раскручивание 3: 15-18
— РНК 1:406,410,413
— суперскрученная 3: 15-18
Двойной липидный слой 1: 496
---диффузия флип-флоп 1: 540
---кавеолы 1: 546
---латеральная диффузия 1:540,
541
---расположение аминокислотных
остатков белков 1: 535
---рафты 1: 545
Двугибридные системы в дрожжах
1:467, 468
Двумерная ЯМР-спектроскопия
1: 198,199
Двумерный электрофорез 1: 139, 149,
463
Двухкомпонентная сигнальная
система 1: 645-648
Двухплазмидная система 1: 470, 471
ДГДГ (дигалактозилдиацилглице-
рин) 1: 499
Де Дюв Кристиан 2: 160
Девсона-Даниелли- Робертсона
модель 1: 589
Дегидрирование 2: 46
Дегидроаскорбат 1: 191
Дегидрогеназы 2: 27, 46, 51
- стереоспецифичность в использо-
вании NAD+ и NADP+ 2: 51, 52
Дегидрогидроксилизинонорлейцин
1: 188
7-Дегидрохолестерин 1: 510; 2 575
Деградация по Эдману 1: 145
Дезаминирование нуклеотидных
оснований 1: 416
---окислительное 2: 269
---трансдезаминирование 2: 270
Дезокси-О-рибоза 1: 340,393,424
Дезокси-О-рибофураноза 1: 393
Дезоксиаденилат 1: 393, 394
Дезоксиаденозилкобаламин 2: 240,
242, 553
Дезоксиаденозильная группа 2:240,
242
Дезоксиаденозин 1: 393, 394
Дезоксиаденозинмонофосфат
(dAMP) 1:394
Дезоксигемоглобин 1: 206, 235, 246
Дезоксигуанилат 1: 393, 394
Дезоксигуанозин 1: 292, 294
Дезоксигуанозинмонофосфат
(dGMP) 1:393,394
Дезоксинуклеозидтрифосфаты, регу-
ляция рибонуклеотидредуктазы
2: 549-551
Дезоксирибонуклеотиды 1: 393, 394
- рибонуклеотиды как предшествен-
ники 2:550
Дезоксисахара 1: 346
Дезокситимидилат 1:393,394
Дезокситимидин 1: 393, 394
Дезокситимидинмонофосфат
(dTMP) 1:393,394
Дезоксицитидилат 1:393,394
Дезоксицитидин 1: 393,394
Дезоксицитидинмонофосфат
(dCMP) 1:394
Дейхофф Маргарет 1:115
Декарбоксилирование аминокислот
2: 538,539
— а-кетокислот 2: 22
— окислительное 2: 182
— с инициацией свободными ради-
калами 2: 24, 25
Дексаметазон 2:468
Декстран 1:354,357
Декстроза 1: 339
Деления 3: 67, 68, 168
Денатурирующее картирование 3: 47
Денатурация
- белков 1: 184, 208-210, 271, 332
-гибридов ДНК-РНК 1:414
-ДНК 1:413-415
Дендротоксин 1: 581, 640
Денитрификация 2: 507-509
Дерматансульфат 1: 361
Десатуразы 2:458, 461
Десенсибилизация рецепторов
1: 605, 606
Десмин 1: 259
Десмозин 1: 122,123
Джонсон В. А. 2: 181, 222
Диабет несахарный 1:556, 572
Диабет сахарный
— анализ на сахар 1: 102, 347-349
— ацидоз 1:93, 102
— взрослого типа у молодых
(MODY) 2:152,153,355
— диагностика 1: 348; 2: 604
— кетоз/кетоацидоз 2: 84, 85, 254,
603,604
— лечение 2:469, 568, 597, 617
---инсулином 2: 568, 569
---производными сульфонилмоче-
вины 2: 597
— метаболизм жиров 2: 84
— метаболизм углеводов 2: 84
— митохондриальные мутации
2: 355,356
— нарушение синтеза жирных кис-
лот 2: 466
— нарушение транспорта глюкозы и
воды 1: 556
— патофизиология 2: 598, 601,603
— редкие формы 2: 153
- - связь с ожирением 2: 604
— теория «липидной нагрузки»
2:616
— тип 1 2: 603
— тип 2 2: 603,615,616
Диализ, для очистки белков 1: 132,
165,168
Диастереомеры 1: 36
Диацилглицерин 1: 498, 507, 610, 611
- в синтезе триацилглицеринов
2: 471
Диацилглицерин-З-фосфат, в синте-
зе триацилглицеринов 2: 465
Дигалактозилдиацилглицериды
(ДГДГ) 1:499
Дигидробиоптеринредуктаза 2: 292
Дигидроксиацетон 1: 340, 341
| \ () 4 ] Предметно-именной указатель
Дигидроксиацетонфосфат
- в гликолизе 2: 67, 68
- в глицеронеогенезе 2: 468
- в цикле Кальвина 2: 415-417
- обмен на неорганический фосфат
2:417,418
1, 25-Дигидроксихолекальциферол
1:510, 511; 2: 572,575
Дигидролипоилдегидрогеназа 2: 184;
см. также Пируватдегидроге-
назный комплекс
Дигидролипоилтрансацетилаза
2: 184; см. также Пируватдеги-
дрогеназный комплекс
Дигидрофолатредуктаза 2: 554,
558-560
- связывание субстрата 1: 278
Дигидрофосфат, буферные свойства
1:96
Дидезоксиинозин (DDI) 3:147
Дидезокси-метод секвенирования
ДНК (метод Сенгера) 1: 420
Дикого типа клетки 1: 54
Димерные хромосомы, репарация
3: 93, 94
Диметилаллилпирофосфат, в синте-
зе холестерина 2: 482
Диметилнитрозамин, как мутаген 1:418
Динамическое стационарное состоя-
ние 1: 40
Динцис Говард 3: 195
Динциса эксперимент 3: 195
Динеины 1: 257
Динитрогеназа 2: 511, 512
- редуктаза 2:511,512
Диоксигеназы 2: 459
Диоксид углерода
— ассимиляция 2: 404-417
— в С2-растениях 2: 423
- - в С4-растениях 2: 423-425
— в САМ-растениях 2: 426
— в окислении
--глюкозы 2: 49
--а-кетоглутарата 2: 194
--пирувата 2:183
--цитрата 2: 192
— в фотосинтезе, см. Цикл Каль-
вина
— влияние на связывание гемогло-
бина с кислородом 1: 243, 244
— круговорот 2: 6
— парциальное давление 1: 100, 101
— перенос гемоглобином 1: 242, 243
— растворимость в воде 1: 79
Дисахариды 1: 339,340, 348,351
- в составе гликозаминогликанов
1: 359, 360
- восстанавливающие 1: 350
- гидролиз до моносахаридов 1: 86,
88
Дистальный гистидин 1: 232
Дисульфидные связи, при секвени-
ровании белка 1:143-145
Дифтерии токсин 3: 209
Дифференцировка клеток 2:124
Диффузионный предел 1: 292
— растворенных веществ и размеры
клетки 1: 18
Диффузия
- мембранных липидов 1: 540-543
- облегченная 1: 552-554, 556; см.
также Переносчики
- простая 1: 551
- скачкообразная 1: 543
- флип-флоп 1: 540
Дихроматы 1: 655
Диэлектрическая проницаемость
1:77
ДНК
- А-форма 1: 404
- В-форма 1: 404; 3: 17
- G-тетраплекс 1: 407
- Z-форма 1:404
- амплификация, методом ПЦР
1:452, 453
- бактериальная 3: 10,11
- библиотеки 1:449-452
- большая и малая бороздки 1: 401
- в хлоропластах 3: 12
- в эукариотах 3: 11-15
- вирусная 3: 9
- гибридизация 1: 443, 463, 465
— в клонировании 1: 451
— в создании библиотек 1: 451
- гибриды 1: 414, 415
- гликозидазы, в репарации 3: 68,
71,72
- двойная спираль 1: 400
- двухцепочечные разрывы 3: 77,84,
90, 97
- денатурационное картирование
3: 47, 92,168
- денатурация 1: 413, 414
- дрожжевая 3: 11
- зонды 1: 443, 444, 454, 455
- и гистоны 3: 257, 258
- клонирование 1:434-443
- кольцевая 3: 17,44-48
- комплементарная, см. Комплемен-
тарная ДНК
- комплементарность нитей 1: 402
- линкерная, в нуклеосомах 3: 258
- матричная цепь 3: 108
- метилирование 1: 419; 3: 58, 68, 69
- микроинъекции 1: 473
- митохондриальная 2: 351-354;
3:171-173
- мусорная 3: 156
- мутации 1: 415; 3: 67
- нематричная (кодирующая) цепь
3: 108
- нуклеосомы 3: 258
- нуклеотидный состав 1: 394, 395
- однонуклеотидный полиморфизм
(SNP) 1:460
- отжиг 1: 413
- палиндром 1: 404
- плазмиды 1: 439
- повреждение 3:66, 67
- повторы 3: 56, 58, 94,118, 145, 148,
178
— инвертированные 1: 404
— короткие тандемные (STS) 1: 455,
456
- порядок зацепления 3: 18
- регуляторные последовательности
3:8
- рекомбинантная 1: 434
— применение в медицине 1: 479
- релаксированная 3: 16
- рестрикция 1: 434-438
— липкие концы 1: 436
— тупые концы 1: 438
- сателлитная 3: 13
- сверхспирализация 3: 15-17
— и топоизомеразы 3: 20
— плектонемическая 3: 24
— плотность 3: 19, 20
— причины 3: 15-17
— раскручивание 3: 15-17
— соленоидная 3: 25
- секвенирование 1: 419-422, 458,
459
- синтез, см. Репликация ДНК
- спаривание оснований 1: 399
- температура плавления 1: 403
- типирование1:454
- топоизомеры 3: 20
- топология 3: 18
- транспозиция 3: 81, 94
- транспозоны 3: 94
- трехцепочечная 1: 405
- упаковка 3:16
- фингерпринтинг 1: 454
- фосфодиэфирные связи 1: 396
- химический синтез 1: 422-424
- Чаргаффа правило 1: 400, 401
- человеческая 3: 11-15
- шпильки 1: 405
- элементы DUE 3: 56, 57
ДНКазы 3: 48
Предметно-именной указатель [4 05 ]
ДНК-хеликаза II, в репарации 3: 68,
70, 74
ДНК-гираза 3: 57, 61
ДНК-зависимая РНК-полимераза
3: 108,109, 121
ДНК-зависимая транскрипция
3: 106-122; см. также Транс-
крипция ДНК
ДНК-лигазы 1: 434, 435; 3: 48, 55, 62
- в ник-трансляции 3: 53
- в репарации 3: 68, 72-74
ДНК-маркирующий сайт (STS)
1:450
ДНК-микрочипы (ДНК-чипы)
1:463-465
ДНК-полимеразы 1: 272, 420,435;
3: 52
- I 3: 49, 52
— в ник-трансляции 3: 53
— открытие 3: 49
— фрагмент Клёнова 3: 53
- II 3:52
- ШЗ: 52-55
— в репарации 3: 68
— субъединицы 3: 54, 55
- IV 3:79
- V3:78
- а, 6 3: 64, 65
- е 3: 65
- т], (3, i и ХЗ: 79
- в ник-трансляции 3: 53
- в репликации
— бактерий 3: 56-62
— эукариот 3: 64-66
- вирусная 3: 66
- коррекция ошибок 3: 52
- матрица 3: 45
- процессивность 3: 50
- РНК-зависимая 3: 144
- 3'-*5'-экзонуклеазная активность
3:51,52
- 5'-*3'-экзонуклеазная активность
3: 52, 53
ДНК-праймазы 3: 55, 58-60
ДНК-репликативная система
(реплисома) 3: 54
ДНК-связывающие белки 3: 54,58,
111
ДНК-связывающий домен/мотив
3: 237-241
ДНК-фотолиазы 3: 68, 74, 80
Добжанский Феодосий 1: 54
Додекановая кислота 1: 488
Додецилсульфат натрия (SDS)
1: 136, 529
Дойси Эдвард 1: 512
Долихол 1: 513, 514, 518
Домен (группа организмов) 1: 18, 19
Домен (надмолекулярная структура)
1: 154,200, 201
- смерти 1: 672, 673
Доминантная тугоухость 1: 581
Доноры водорода, в фотосинтезе
2:386
Дофамин 2: 538, 539
Дочерние хромосомы 1: 403, 660
Дрожжи
- двугибридная система 1: 467, 468
- искусственные хромосомы 1: 442,
443
- переключение типов спаривания
3: 256
- регуляция метаболизма галактозы
3: 263, 264
- репликация 3: 64
- РНК-полимераза II 3: 118
- секвенирование генома 1: 459
- тРНК аланина 3:184, 185
- тРНК фенилаланина 1: 411; 3:186
- факторы спаривания 1: 658
- ферментация 2: 66,67, 69
- хромосомы 3: И, 14
- центромеры 3: 14
Дыхательная цепь (цепь переноса
электронов) 2: 182,306-326
Е
Еноил-АСР-редуктаза 2: 448,452
Еноил-СоА-гидратаза 2: 234, 235, 248
Еноил-СоА-изомераза 2: 238,239, 248
Енолаза 2: 79, 128
- каталитическая активность 1: 309,
310
Ж
Жакоб Франсуа 1: 15, 408; 3: 234
Жасмонат 1: 647
Жгутики 1: 656, 657
- движение 1: 257, 645, 648, 649
Железо
- в протопорфирине IX 1: 227, 228;
2:310
- гемовое 1: 194,195, 227, 228;
см. также Гем
- гомеостаз 2: 193
- двухвалентное, окисление 2: 44,
46, 48
- железо-чувствительные элементы
(IRE) 2: 194
- потребность организма 1: 29
- регуляторные белки (IRP) 2: 193,
194
Железосерные белки 2: 309, 310
— Риске 2: 310
Железосерные кластеры 2: 192, 311
— в аконитазе 2:192,193
— реакции 2: 370,371
Желтый флуоресцентный белок
(YFP) 1:612-615
Желудочно-кишечный тракт 2: 264
Желчные кислоты 1: 504, 519; 2: 484,
494
Желчные пигменты, как продукт
распада гема 2: 535
Жеттен Майк 2: 508
Жидко-мозаичная модель мембраны
1: 528, 589
Жизнь, происхождение 1: 56, 57, 156;
3: 172; см. также Эволюция
Жирные кислоты 1: 487-519; см.
также Липиды
— активация и транспорт 1: 580;
2: 38, 226-23
— амфифильная природа 1: 80
— анализ 1: 515-518
— в бактериальных липополисаха-
ридах 1: 370
— в восках 1: 495
— в глицерофосфолипидах
1:496-498; 2: 464
— в клетках Е. coli 1: 539
— в пищевых жирах 1: 494
— в триацилглицеринах 1: 491,492;
2: 464
---активация 2: 229
— двойные связи 1: 489
— десатурация 2: 458-461
— как липидные якоря 1: 536,545
— метаболизм
---в головном мозге 2: 590
---в жировой ткани 2: 468, 585,586
---в мышцах 2: 587, 588
---в печени 2: 584
---координированная регуляция
синтеза и распада 2: 244
---при диабете 1: 102; 2: 84, 85
---роль AMP-зависимой протеин-
киназы 2: 131
---роль инсулина 2: 150
---у позвоночных в сравнении с
другими организмами 2: 105, 211
— мобилизация 2: 467, 599, 602
— незаменимые 2: 459
— номенклатура 1: 488
— окисление 2: 225-250
— а 2: 250
— р 2: 225, 233-236
---в пероксисомах 2: 245, 246
---ферменты из разных органелл
2: 247, 248
---полное 2: 232, 233
[ 4 061 Предметно-именной указатель
---со-, в эндоплазматическом рети-
кулуме 2: 249
---ненасыщенных жирных кислот
2: 237-239
---с нечетным числом атомов угле-
рода 2: 239
---регуляция 2: 241-244
— омега-3 1:489
— омега-6 1: 489
— переэтерификация 1: 517; 2: 230
— полиненасыщенные 1: 489
— превращение в ацил-СоА-
производные 2: 230, 231
— свободные 1: 490; 2: 229
— синтез 2: 445-464
---в хлоропластах растений 2:453,
454
---десатурация 2: 458,461
---длинноцепочечных насыщен-
ных жирных кислот 2: 457
---регуляция 2: 244,455-457
---синтаза жирных кислот 2:
447-453
---система элонгации 2: 457
---эйкозаноидов 2: 461-463
— температура плавления 1: 490
— транс 1: 489,492,494
— упаковка 1:490
Жировые клетки, см. Адипоциты
Жиры; см. также Жировая ткань;
Триацилглицериды
- прогоркшие 1:493
- производство тепла 2: 236, 585
3
Закман Берт 1: 575
Закон действующих масс 2: 17,77,
128
---для определения энергетическо-
го состояния клетки 2: 344
Закрытая система 1:41
Заменимые аминокислоты 2: 278;
см. также Аминокислоты
Заместительная мутация 3: 67
Замечник Пол 1:10; 3: 166
Занамивир (реленза) 1: 374
Запасные липиды 1:487-495
Запасные полисахариды 1: 353
Запахи, пути передачи 1: 650-656
Заряд аминокислоты 1:120,125
Звездчатые эндотелиоциты 2: 580
Зеленый флуоресцентный белок
(GFP) 1:451,478, 609,612,
613-615
Зеркальный повтор, в ДНК 1: 406
Зимоген 1: 328
Змеиный яд 1: 522,581
Зокор (симвастатин) 2: 494
Зонды
- флуоресцентные 1: 421, 541,542,
609
- ДНК/РНК 1:443,444,454,455
Зрение 1: 650-656
И
Ибупрофен 2:462
Ивата Coy 1: 568
Иглобрюх, яд 1: 579
Иди-Хофсти уравнение 1: 335
Излучение электромагнитное 2: 359,
360
— повреждение ДНК 1: 417, 418
— поглощение ДНК 1: 413
Изменение биохимической свобод-
ной энергии 1: 274
Изолейцин 1:49,118
- катаболизм 2: 289
- превращение в сукцинил-СоА
2: 288, 289, 294
-синтез 2: 523,524,531
Изолированная система 1: 41
Изомеразы 2: 87, 109
Изомеризация 2: 23, 24
Изомеры
- геометрические 1: 34,35
- конфигурационные 1: 35
- стереоизомеры 1: 35,36, 39
Изопентенилпирофосфат, в синтезе
холестерина 2: 23,482
Изопрен, в синтезе холестерина
2:480-483
Изопреноиды
- как липидные якоря 1: 536, 537,
545
- синтез 2: 496,497
Изопропилтиогалактозид (ИПТГ)
3:236, 285
Изопротеренол 1: 67
Изоферменты 2: 71, 72
Изоцитрат 2: 192, 200,212
Изо цитрат дегидрогеназа 2: 192,193,
210,214
Изоцитратлиаза 2: 212
Изоэлектрическая точка (pl) 1: 125,
126,137
Изоэлектрофокусирование 1:
137-139
Икосаэдрическая симметрия 1: 207
Иматиниб (гливек) 1: 669 .
Иммунизация вирусными вакцина-
ми 3: 14
Иммунитет
- гуморальный 1: 249
- клеточный 1: 249
Иммунная система 1: 249
— интегрины 1: 642
— клетки 1: 249, 250
— клональная селекция 1: 250, 256
— коррекция дефектов методами
генной терапии 1: 475-477
— селектины 1: 373,374
— у растений/животных 1: 648,649
— эволюция 3: 97, 98
Иммуноблоттинг 1: 256
Иммуноглобулиновая складчатость
1:252
Иммуноглобулиноподобные белки
1:549
Иммуноглобулины (Ig) 1: 249, 251;
см. также Антитела
- А 1: 252
- D 1:252
- Е 1:252,253
- G 1: 252-254
-Ml: 252
- рекомбинация 3: 96,97
Иммунодефициты
- лечение 3: 147
— генная терапия 1: 475-477
Иммунопреципитация, для выделе-
ния белков 1: 467
Иммуноферментный анализ, см.
ИФА
Импортины 3: 216, 217
Инактивация генов, метод анализа
1:461
Инвертированные повторы ДНК
1:405,406
Ингибирование
- кинетические методы анализа
1:296
- кооперативное 2: 529,531
- последовательное с отрицательной
обратной связью 2: 532
Ингибиторный G-белок (Gi) 1: 608
Ингибиторы
- бесконкурентные 1: 295-298
- в медицине 1: 314-318
- действующие по принципу обрат-
ной связи 1: 49,321
- кинетические методы определения
типа 1: 296
- конкурентные 1: 295-298
- неконкурентные 1: 297
- необратимые 1: 298
- обратимые 1: 295
- смешанные 1: 295-298
- суицидные 1: 298
Индекс гидрофобности 1: 116,534
Индирубин 1: 669
Индолилацетат (ауксин) 1:470, 647
Предметно-именной указатель [407]
Индольное кольцо, в триптофане
1:118, 119, 377
Индукторы 3: 235, 236
Индукция 3: 230, 235, 236
Индуцибельные гены 3: 230
Индуцированное соответствие 1: 282
— модель 1: 243
— при связывании антигена с анти-
телом 1: 252
— при связывании фермента с суб-
стратом 1: 278
Индуцируемый гипоксией фактор
HIF-1 2: 81, 211,345, 346
Инициаторная последовательность
(Inr) 3: 117
Инициаторный (стартовый) кодон
3:170,199
Инициаторный комплекс
- бактерий 3: 196, 198
- эукариот 3: 198,199
Инициация трансляции 3:195-198
Инман Росс 3:47
Инозин 1: 395; 3: 176
Инозинат (IMP) 2: 545-547
- в антикодонах 3: 174,185
Инозит, в синтезе липидов 2: 474
Инозит-1,4,5-трифосфат (IP3) 1: 507,
610,611
Инсерционные мутации 3: 67
Инсерционные последовательности
(IS) 3: 94
Инсулин 1: 143; 2: 572
- аминокислотная последователь-
ность 1: 143, 168, 169
- в метаболизме
--жиров 2: 244
--углеводов 2: 84,137,146, 170
--холестерина 2: 84,492
- в регуляции массы тела 2: 607-614
- в сигнальных путях 1: 619-622,
624,625
- в синтезе
--гликогена 2: 137, 168,170, 171
--триацилглицеринов 2: 466
- как пептидный гормон 2: 572
- нечувствительность 2: 469, 612,
615
- открытие 2: 568, 569
- отсутствие при нарушении окис-
лительного фосфорилирования
2: 356
- при диабете 2: 348, 349, 556; 2: 84,
85,153,603
- регуляция уровня глюкозы 1: 556,
557; 2: 85,593-596
- регуляция экспрессии 1: 621; 2: 149
- рецептор 1: 170, 619, 624, 626
— субстрат-1 1: 619
— тирозинкиназная активность
1:619, 620, 624
- сигнальная последовательность
2: 572
- синтез 2: 572, 573
Инсулиннезависимый сахарный диа-
бет 2: 612, 615
Инсулинозависимый сахарный диа-
бет, см. Сахарный диабет I типа
Интеграза 1: 317, 474; 3: 91-93, 145
Интегральные мембранные белки
1:531-533
Интегрины 1: 367,549, 641, 642
Интермедиат (промежуточное со-
единение) 1: 275
Интерференция РНК 3: 270
Интроны 3:12, 122,123
- сплайсинг 3: 125-129, 131, 133,139,
140, 148
Инфаркт миокарда 1: 140,141
Информационная РНК, см. мРНК
Информационные макромолекулы
1:33, 391
— сахара 1:363,371
Ион гидроксония 1: 89
Ионизация
- аминокислот 1: 119, 124
- воды 1: 89
- зависимость от pH 1: 96
- кислот 1: 98
- константа равновесия 1: 89,94
- пептидов 1: 128, 299
Ионизирующая радиация, поврежде-
ние ДНК 1: 417,418
Ионная помпа, см. АТРазы; Пере-
носчики
Ионное произведение воды (Кхх)
1:91
Ионные взаимодействия 1: 77, 78; см.
также Слабые взаимодействия
— и стабильность белков 1: 96, 173,
174,235, 236
Ионные каналы 1: 553, 573, 595, 633
— а-спирального типа 1: 638
— Са2+ 1:561, 611, 618, 635
---дефекты 1: 581
— Cl" 1: 633-635, 639
---при кистозном фиброзе 1: 565,
566
— К+ 1: 536, 575-578; см. также
Калиевые каналы
---дефекты 1: 581
---в сигнальных путях 1: 635
— Na+1:635
---дефекты 1: 579, 581
---в сигнальных путях 1: 635, 637
— анализ функции 1: 574
— ацетилхолиновый рецептор 1: 638
— в нейронах 1:639
— действие токсинов 1: 579, 602
— лигандзависимые 1: 574, 579, 636
— отличие от переносчиков 1:574
— потенциалзависимые 1: 574, 578,
579,636
Ионообменная хроматография
1: 132, 133
Ионофоры 1:550, 570
Ионы водорода, концентрация 1: 89,
96; см. также pH
— градиент 1: 567
— как внутриклеточные мессендже-
ры 1: 633
— концентрация в цитозоле/внекле-
точной жидкости 1: 634, 635
— содержание в крови 2: 591, 592
Иринотекан 3: 25
Искусственные хромосомы 1: 439,
442,443; 3: 15
— бактериальные 1: 439, 442
— дрожжевые 1: 442, 443; 3:15
— человеческие 3: 15
ИФА (ELISA) 1: 255, 256; 2: 570
Ишемия, гидролиз АТР 2: 344, 345
К
Кавеолин 1: 545, 546, 632
Кавеолы 1: 545, 546, 609, 632
Казеинкиназа II 2: 168
Кайзер Дейл 1: 438
Калиевые каналы 1: 575-578, 635
— АТР-зависимые 2: 597
— в метаболизме глюкозы 2: 595,
596
— дефекты 1: 581
— структура 1: 575-578
Калий
- концентрация в клетке 1: 634
- транспорт с помощью Na+/K+-
АТРазы 1: 562
- уровень в крови 2:591, 592
Калнексин 1: 376
Кальвин Мелвин 2: 403-406
Кальвина цикл 2: 404, 407
— реакции фиксации углерода
2:406 407
— рубиско 2: 407-413
— стехиометрия 2: 415
— участие АТР 2: 415
— участие NADPH 2: 415
Кальмодулин 1: 614-616
Кальретикулин 1: 376
[ 4OS | Предметно-именной указатель
Кальций
- в зрительной функции 1: 651, 659
- в мышечном сокращении 1: 261
- регуляция 2: 576
- содержание в крови 2: 592
Кальцитонин
- альтернативный процессинг гена
3: 131,132
Кальцитриол 2: 576
Каннабиноиды 1: 622
Капсид вируса, симметрия 1: 207
Карбаминогемоглобин 1: 244
Карбамоилглутамат 2: 279
Карбамоилфосфат, образование
2: 547, 548
Карбамоилфосфатсинтетаза
- I 2: 273, 275, 277, 287, 548
— недостаточность 2: 279
- II 2: 548,549
Карбанион 2:21-23,95, 111, 198
Карбоангидраза 1: 105, 242,336
Карбокатион 2: 21-23
Карбоксибиотин 2: 100, 101
Карбоксигемоглобин 1: 240
у-Карбоксиглутамат 1: 123
Карбоксипептидаза
- А 2: 264
- В 2: 264
Карбонаты, переходное состояние
при гидролизе 1: 307
Карбонильная группа 2: 22-24
Карбоновые кислоты 1: 31, 32; 2: 181
Карвон, стереоизомеры 1: 39
Кардиолипин 1: 498, 527; 2: 321
- синтез 2: 472,474
Карнитин 2: 230
Карнитинацилтрансфераза I/II
2: 230-232,610
Карнитиновый переносчик 2:230-232
Карнубский воск 1: 495
Р-Каротин 1: 511, 512, 652; 2: 362
Каротиноиды 2: 361
Картирование
- Е. coli 3: 44
- геномов 1: 451, 458
- денатурацией 3: 47
Карутерс Марвин 1: 423
Каспазы, в апоптозе 1: 672, 673; 2: 350
Катаболизм 1: 48, 49; 2: 7-9, 69, 581
- аминокислот 2: 261-272, 280-298
- белков, жиров и углеводов при
клеточном дыхании 2: 181, 182
- глюкозы, в опухолевых тканях
2:81-83
- жирных кислот 2: 226-250
- пиримидинов 2: 556,557
- пуринов 2: 555,556
Каталаза 2: 146, 246, 247
Катализ
- в движении липидов через двой-
ной слой 1: 540, 541
- вращательный 2: 306
- ионами металла 1: 284
- кислотно-основной 1: 283, 284
- ковалентный 1: 284
- регуляторные ферменты 1:319, 320
- специфичность 1: 281
Каталитическая константа (число
оборотов) (&cat) 1: 291
Каталитическая триада 1: 303
Каталитические антитела 1: 250, 307,
308
Катенаны 3: 63
Катехоламины 2: 358, 359, 574
Катионообменная хроматография
1: 132,133
Качающегося банана модель 1: 568
Качающиеся пары нуклеотидов
3: 174,175
Кашалот, запасные жиры 1: 493
Квант 2: 359, 360
кДНК (комплементарная ДНК)
1:451
- библиотеки 1: 450
- клонирование 3: 156, 192
Келер Георг 1: 254
Кендрю Джон 1: 171, 194, 197, 205
Кеннеди Юджин 2: 188, 230, 306, 472
Кератансульфат 1: 360, 367
а-Кератин 1: 177, 178, 185, 186
Кернс Джон 3: 46
Кетали 1: 343
Кетоацидоз, при диабете 2: 84, 85,
604
Кетоацидурия (болезнь кленового
сиропа) 2: 287
(3-Кетоацил-АСР-редуктаза 2:448,
452
р-Кетоацил-АСР-синтаза 2: 448, 450
р-Кетоацил-СоА 2: 234, 235
p-Кетоацил-СоА-трансфераза 1: 425;
2: 253
Кетогексозы 1: 343, 344
Кетогенные аминокислоты 2: 105,
280, 281
а-Кетоглутарат (оксоглутарат)
1: 190,191; 2: 104, 188, 193, 199,
200
- в малат-кетоглутаратной транс-
портной системе 2: 341, 324
- в метаболизме глюкогенных ами-
нокислот 2: 104, 280, 292, 293
- образование из изоцитрата 2: 192,
201
- окисление до сукцинил-СоА 2: 194,
201
- перенос а-аминогруппы 2: 266
а-Кетоглутаратдегидрогеназа 2: 194,
195,210
Кетоз 2: 204, 466, 603
Кетозы 1: 341
- D- и L-изомеры 1: 341
Кетокислоты
- а, дегидрогеназный комплекс
2: 296
- р, декарбоксилирование 2: 22
Кетоновые тела 2: 251-253
— в печени 2: 580, 584
— образование из аминокислот
2: 280
— при голодании 2: 598, 601
— при диабете 2: 253, 254, 466, 603
— при мышечном сокращении
2: 587-589
Кетоны 1: 31, 340, 343; 2: 251
Килби Б. А. 1:302
Киназы 2: 26,197; см. также Про-
теинкиназы
- казеинкиназа 2: 168
- креатинкиназа 2: 41,141, 588
- пируваткиназа 2: 79, 128, 148
- триозокиназа 2: 89
- фосфоглицераткиназа 2: 77
- фосфорилазы р 1:325,604,605;
2:166
Кинезины 1: 257
Кинетика 1:47
- Михаэлиса-Ментен 1: 287, 289,
290; 2: 126, 135, 136
- стационарная 1: 287, 288
- предстационарная 1: 294, 302
- ферментативных реакций 1: 285
Кинетопласт 3: 12
Кислород
- круговорот 2: 6
- парциальное давление 1: 232
- растворимость в воде 1: 79
- связывание с гемоглобином
1: 235-238
- транспорт в крови 2: 591, 592
Кислородобразующий комплекс
2:380
Кислородсвязывающие белки 1: 228
Кислотно-основное титрование
1: 123,124
Кислотно-основной катализ
— общий 1: 283, 284
— специфический 1: 283, 284
Кислотно-основные пары 1: 93,94
— как буферные системы 1: 96, 97
Предметно-именной указатель [ 4 091
Кислоты
- аминокислоты 1: 116,125
- в качестве буферов 1: 97-99
- константа диссоциации 1: 94-97
- кривая титрования 1:95
- сильные 1: 94-97
- слабые 1: 94-97
- Хендерсона-Хассельбалха уравне-
ние 1: 98
Кислые сахара 1: 346
Кистозный фиброз
— дефекты ионных каналов 1: 565
— нарушение фолдинга белка
1:215,217
Клавулановая кислота 1: 316,317
Кларк А. Р. 1:337, 338
Кларк Джон 2: 225
Кленового сиропа болезнь 2: 287, 297
Клетки 1: 17
- бактериальные 1: 20, 21, 33
- киллеры (цитотоксические
Т-клетки) 1:249, 250
- Лангерганса 2: 595
- основные химические компоненты
1:26,32
- поджелудочной железы
— а 2: 595
— р 2:568,573, 595, 596, 603, 617
- размер 1: 18
- структура 1: 17, 18
- эукариотические 1: 22-24
Клеточная оболочка 1: 20, 21
Клеточная смерть программируемая,
см. Апоптоз
Клеточная стенка, синтез полисаха-
ридов 2: 431-433
Клеточное дыхание 2: 181, 182
— альтернативные пути в растениях
2: 420
— перенос электронов в митохон-
дриях 2:327-329
Клеточный иммунитет 1: 249;
см. также Иммунная система
Клеточный цикл
— мейоз 3: 82
— регуляция 1: 664, 665
— роль циклинзависимых протеин-
киназ 1:664
— стадии 1: 660
— хромосомные изменения 3: 27
Клёнова фрагмент 3: 53
Клод Альбер 1: 24
Клон 1: 254, 434
Клональная селекция 1: 250, 256
Клонирование 1:434-448
- белковые тэги для хроматографии
1:447
- в бактериях 1: 434-448
- в животных 1: 473,475, 479
- в растениях 1: 469-473
- векторы, см. Клонирующие век-
торы
- гибридизация 1: 443
- ДНК-лигазы 1:434; 3: 55, 62
- кДНК 1:450; 3: 156-158
- линкеры/полилинкеры 1: 438
- микроинъекция 1: 473
- мутагенез 1:446
- получение большого количества
белка 1: 444
-ПЦР 1:452, 453
- рестриктазы 1: 434,436,437
- слитые белки 1:447
- трансформация 1:449, 470, 473,
476, 477
- ферменты 1: 435
- флуоресцентных маркеров 1: 612
- электропорация 1:439
Клонирующие векторы 1: 434,435,
437-440
— бактериофаги 1: 441
— вирусные 1: 473,474
— для растений 1: 469-472
— искусственные бактериальные
хромосомы 1:439, 441, 442
— искусственные дрожжевые хро-
мосомы 1: 442,443
— плазмиды 1: 434,439, 440
— Til: 469,470
— ретровирусные 1: 473, 474
— фаг XI: 441; 3: 92,93, 285
— шаттлы 1: 443
— экспрессирующие 1: 444, 445
Клубеньки, фиксация азота 2:510,
512,513
Кляйзена конденсация 2: 22, 23
Коагуляция
- протеолиз 1:329
- роль селектинов 1: 373, 374
Коактиваторы 3: 260-262
Кобаламин, см. Витамин В12
Кобротоксин 1: 581, 640
Ковалентная связь 1: 27, 29, 75, 82,
114,172
— в соединениях фосфора 2: 35
— в ферментативных реакциях
1: 277, 284
— гетеролитическое расщепление
2: 20,21
— гомолитическое расщепление
2: 20,21
Ковалентные модификации
регуляторных ферментов 1:319,
323-328; 2:127,515,516
Ковалентный катализ 1: 283,284
Когезины 3:36
Кодирующая цепь ДНК 3: 108, 109
Кодоны 1: 445,483; 3: 167-180,
191-204
Коинтеграт 3: 95
Коклюшный токсин 1: 602
Колбочки 1: 650-656
Коллаген 1: 187, 188
- аминокислотный состав 1: 187
- и аскорбиновая кислота 1: 189-191
- и протеогликаны 1: 367
- нарушения структуры 1:191, 192
Коллигативные свойства 1: 84
Коллинз Френсис 1: 457, 458
Коллип Дж. Б. 2: 568
Колоночная хроматография 1:132,
133
Кольцевая ДНК 3: 44-48
Кольцевые липиды 1: 534
Компактин 2: 494
Комплекс
- I (NADH/убихинон) 2:313,314,
322
- II (сукцинат/убихинон) 2: 313,
316,322
- III (убихинон/цитохром с) 2: 313,
317,318, 321,322
- IV (цитохром с/О2) 2: 313,318,
320-322
- лиганда с рецептором 1: 592-594
- транслокации 3: 212, 218
- фермент-субстратный, см. Фер-
мент-субстратный комплекс
Комплементарная ДНК (кДНК)
1:451; 3:146
— библиотеки 1: 451; 3: 156-158
— для гибридизации 1:463,465;
3:158
Конденсация 1: 104, 272, 510
- при образовании пептидной связи
1: 127
- сложноэфирная (Кляйзена) 2: 22,
23
- хромосом 3: 36
Конденсины 3: 36
Конкурентный ингибитор 1: 295-298
Консенсусные последовательности
1: 155, 156
— oriC 3: 56
— промоторов 3: 231, 232, 244
— узнаваемые протеинкиназами
1:325, 327, 603,615
Консенсусное древо жизни 1: 19
Константа
- ассоциации (Ка) 1: 229
— в анализе по Скэтчарду 1: 593
[410] Предметно-именной указатель
- Больцмана (k) 2:13
- диссоциации комплекса (К(|)
---антигена с антителом 1: 253
---лиганда с рецептором 1: 231,
592, 593
---фермента с субстратом 1: 290
- кислотной диссоциации (Ка)
1: 93, 94
- Михаэлиса (Км) 1: 288, 290, 291
— кажущаяся 1: 295, 296
- равновесия (Kcq) 1: 276
- скорости (k) 1: 229, 276, 277
Конститутивная экспрессия 3: 230
Контиги 1:449, 458
Конфигурация 1: 345
Конформация
- al: 181,187,192, 200-202
— небольших глобулярных белков
1: 199
- ванна 1: 354
- и конфигурация 1: 354
- кресло 1: 345
- нативная 1: 53
Кооперативное ингибирование 2: 530
Кооперативное связывание
1: 237-239, 242-244
Корана X. Гобинд 1: 423; 3: 170
Кори (Форбса) болезнь 2: 159
Кори Герти 2: 92, 121,159,160, 165
Кори Карл 2: 92,121,159,160,165
Кори Роберт 1: 35,174,177,181, 186
Корнберг Артур 2: 160; 3: 49
Корнберг Ханс 2: 212
Корнфорт Джон 2: 484
Коровьего бешенства болезнь, см.
Бычья губчатая энцефалопатия
Коронарный тромбоз 1: 512, 628;
2: 493, 495, 590
Корончатый галл 1: 470
Короткие тандемные повторы (STR)
1:455
Корректирующая активность
— в репликации 3: 50-52
— в транскрипции 3: 110, 146
— в трансляции 3: 188,189
Корриновая кольцевая система
2: 240
Кортизол 1: 509; 2: 468,576,578
- в метаболизме глюкозы 2: 602, 603
Кортикостероиды 1: 509; 2: 468, 574,
602, см. также Стероидные
гормоны
- синтез 2: 495
Котранспортные системы 1: 556, 558,
562, 567
Кофакторы
- ферментов Г. 271
- нитрогеназного комплекса
2:510-512
Коферменты 1: 271; 2: 201, 281
- А (СоА) 1:271,426; 2: 183, 184
- В122: 240, 242, 284
- NAD+ и NADP+1: 426
— стереоспецифичность дегидроге-
наз 2: 50
- Q (убихинон) 1:512, 513 2: 309,
314,322
- биотин 2: 100, 101
- в пируватдегидрогеназном ком-
плексе 2: 183,184
- как универсальные переносчики
электронов 2: 49,307,460
- липоевая кислота 2: 183
- пиридоксальфосфат 2: 266, 267
- простетические группы 1: 271
- тиаминпирофосфат 2: 94, 95
- флавиннуклеотиды 2: 426
Кошланд Даниэль 1: 242, 282
Коэн Стенли 1: 433
Коэффициент
- специфичности (kCM/KM) 1: 292
- эластичности (е) 2: 134-136
Красные кровяные клетки,
см. Эритроциты
Красные мышечные волокна 2: 587
--синдром МЭРКМВ 2: 355
Крахмал 1: 353-358
- биосинтез 2: 426-428
- в амилопластах 2: 405, 406
- расщепление 2: 86-88
- фосфорилаза 2: 87
Крахмаленнтаза 2: 427,428
Креатин 2: 536, 537
- в мышечном сокращении 2: 588,
589
Креатинкиназа 2: 588, 589
Кребс Ханс 2: 83, 181, 182, 222, 223,
261,273
Кребс Эдвин 2: 160
Кребса бицикл 2: 275, 276
Кребса цикл (цикл трикарбоновых
кислот, цикл лимонной кисло-
ты) 2: 181,182,188-207
Крейцфельда-Якоба болезнь 1:217,
218
Крестоносцы 1: 189
Крестообразные структуры, ДНК
1:406
Крик Френсис 1: 9, 398-403; 3: 45, 50,
152,167, 174
Криптохромы 2: 55
Кровь
-pH 1:92, 93,102
- буферные свойства 1: 98-101
- группы, роль сфинголипидов 1: 503
- липопротеины 2: 485-488
- метаболические функции 2:591,592
- перенос аммиака 2: 270
- перенос кислорода 1: 228-233
- плазма 1: 86; 2: 591, 592
- содержание глюкозы 1: 553-556;
2: 592
- состав 2: 591
- транспортные функции 2: 591, 592
- электролиты 2: 591
Крокодилы 2: 92
Кроссинговер 3: 83
Круговой дихроизм 1: 590
Кручение (ДНК) 3: 20
Ксантин 1: 395; 2: 555
Ксантиноксидаза 2: 555, 557
Ксенобиотики 2: 349
Ксилоза 1: 108, 364
Ксилулоза1:342
Ксилулозо-5-фосфат 2:81,82,109,111
- в регуляции метаболизма углево-
дов и жиров 2: 146,150, 151
- в цикле Кальвина 2: 412,413
Куенен Гижс 2: 508
Культура ткани 1: 473
Кунитц Мозес 1: 270
Кэбак Рон 1: 568
5'-Кэп 3: 123
5'-Кэп-связывающий комплекс
3: 123, 124
Кюне Фредерик 1: 270
Л
Лавуазье Антуан 2: И, 12
Ладдсраны 2: 508, 509
Лазерная десорбционноиониза-
ционная масс-спектрометрия
(MALDI) 1: 149
Лайнуивера-Берка уравнение 1: 289
Лактальбумин 1: 369
(З-Лактамаза 1: 316, 317
(З-Лактамные антибиотики, устойчи-
вость бактерий 1: 314-316; 3:10
Лактат 2: 27, 300, 386
- в синтезе глюкозы 2: 102
- в сокращении мышц 2: 118, 588,
589
Лактатдегидрогеназа (ЛДГ) 1: 333,
337; 2: 27, 52,91
- изоформы 2: 140, 141
Лактовегетарианцы 2: 301
Лактоза 1: 348, 351; 2: 86, 88
- /ac-оперон 3: 235-245
- непереносимость 2: 88
- переносчик (лактопермеаза) 1: 567,
568, 588
Предметно-именной указатель | \ 111
Лактозилцерамид 1: 501
Лактоназа 2: 109
Лактоны 1: 346, 347
Лактопермеаза 1: 567, 568, 588
Ламберта-Бера закон 1: 121
Ламеллы 2: 358, 378
Ланолин 1: 495
Ланостерин 2: 484
Латеральная диффузия 1: 540-543
Латеральный (горизонтальный)
перенос генов 1:157
Лауриновая кислота 1: 488
ЛДГ, см. Лактатдегидрогеназа
Леви тельца 1: 217
Левинталь Сайрус 1:210
Леггемоглобин 2: 512, 513
Легкие цепи
— иммуноглобулина 1: 251; 3: 96
— миозина 1: 257
Ледер Филипп 3: 169
Лейкотриены 1: 507, 508; 2: 463, 574;
см. также Эйкозаноиды
Лейкоциты 1: 249, 250, 475; 2: 592
Лейцин 1:116,119; 2:105, 278,518
- биосинтез 2: 523, 524
- катаболизм 2: 280, 281, 288, 296
— нарушения 2: 287, 297
Лейциновая молния (застежка)
3: 241,242
Лекарства
- вклад геномики и протеомики в
создание 1: 478-480
- метаболизм, роль цитохрома Р-450
2: 349,460
Лекарственная устойчивость
— роль АВС-транспортеров 1:
564-566
— роль плазмид 3: 10
Лектины 1:372-378
ЛелуарЛуис 2: 158, 160
Ленинджер Альберт 2: 188, 230, 306
Лептин 2: 605, 606, 608-610
- рецептор 2: 606
Лернер Ричард 1: 308
Летсингер Роберт 1: 423
Лецитин, см. Фосфатидилхолин
Лецитин-холестеринацилтранс-
фераза (LCAT) 2: 489
Лёша-Нихена синдром 2: 557
Лиазы 2: 197
Лигазы 1: 197
Лигандзависимые каналы
1: 574-579
— в передаче сигнала 1: 634-640
Лиганды 1: 135
- в иммунных реакциях 1: 249-254
- взаимодействие с рецепторами
1:592, 593
— количественный анализ 1:
592-594
- связывание с белками 1: 225-248
— кооперативное 1: 237-242
Лигнин 2: 538
Лигноцеровая кислота 1: 488
Лидирующая цепь 3: 48
Лизин 1: 115-117
- биосинтез 2: 518, 523, 524, 531
- карбамоилирование 2: 407
- катаболизм 2: 280, 288
Лизолецитин 2: 489
Лизосомы
- в заместительной терапии 1: 375
- мечение белков для доставки
1:375, 378; 3:210, 214, 220
- ферменты 1: 503, 505
Лизофосфолипазы 1: 504
Лизоцим!: 129, 139, 200, 272
- каталитическая активность
1:310-313
- структура 1: 311
Ликсоза 1: 342
Лимфоциты 1: 249, 250
- В 1:249, 250, 252; 2 592
- рекомбинация 3:97
- Т 1:249, 250
- и селектины 1: 373
- при иммунодефицитах 2: 556
- хоминг 1: 373, 374
Линд Джеймс 1: 189
Линен Феодор 2: 484
Линия М (М-диск) 1: 259
Линкер 1: 437, 438
Линолеат 2: 458, 459, 461
- синтез 2: 459
Линолевая кислота 1: 499, 521; 2: 238
Линоленат 2: 458,459, 461
Линоленовая кислота 1: 489,521
Липазы 1: 379, 491
- гормон-чувствительная 1: 605;
2: 229, 599
Липидная гипотеза 2: 493, 494
Липидные якоря 1: 536, 537
— и рафты 1: 543, 545
Липидный слой, см. Двойной липид-
ный слой
Липидом 1: 519
Липидомика 1: 517
Липиды 1: 33, 487,496-517, см.
также Воски; Гликолипиды;
Глицерофосфолипиды; Жирные
кислоты; Липополисахариды;
Сфинголипиды; Фосфолипиды
- архей 1: 500
- биосинтез
— глицеронеогенез 2: 468
— мембранных фосфолипидов
2: 470-476
— плазмалогенов 2: 476, 477
— стероидов в митохондриях 2: 348,
349
— сфинголипидов 2: 476-478
— триацилглицеринов 2: 464-467
— функция инсулина 2: 466
— холестерина, стероидов и изопре-
ноидов 2: 479-484, 497
--регуляция 2: 491-493
— эфиров холестерина 2: 485
— эйкозаноидов 2: 461-463
- выделение 1: 515, 516
- галактолипиды тилакоидных мем-
бран 1: 499, 500
- ганглиозиды 1: 369
- гидролиз 1:517
- запасные 1: 487, 496
- запасы организма 2: 600
- как окислительно-восстановитель-
ные кофакторы 1:512
- как пигменты 1:514
- как сигнальные вещества
1:507-512
- кольцевые 1: 533, 534
- мембранные 1: 26, 369, 370,
496-506
— болезни накопления 1: 505
— микродомены 1: 543, 545
— рафты 1: 543, 545
— у разных организмов 1: 526, 527
- метаболизм
— в жировой ткани 2: 228, 229, 468,
585-587
— в печени 2: 584, 585, 594
— внутриклеточная локализация
2:454
— и ожирение 2: 605-614
— функция адреналина 2: 602
— функция кортизола 2: 602
— функция ксилулозо-5-фосфата
2: 146, 147
- методы анализа 1: 515-517
- основные категории 1: 519
- перемещение через двойной ли-
пидный слой 1: 540, 541, 543
— измерение скорости 1: 542
- пищевые, всасывание 2: 226, 227
- плазмалогены 1: 498, 499
- при зимней спячке у животных
2:236
- различные функции 1: 513
- разрушение в лизосомах 1: 503
| \ 12 ] Предметно-именной указатель
- расщепление, мобилизация и
транспорт 2: 226
- связывание с мембранными белка-
ми 1: 536, 537
- стерины 1: 504
- структурные, см. Липиды мембран-
ные
- транспорт 2: 485-489
Липкие концы 1: 437, 438
Липоапоптоз 2: 616
Липоат (липоевая кислота)
2: 183-186, 207
Липоиллизиновая группа 2: 184,186,
187
Липополисахариды 1: 370, 649;
см. также Полисахариды
Липопротеинлипаза2: 228
Липопротеиновых субъединиц мо-
дель (модель Бенсона) 1: 589
Липопротеины 2: 485-490
- высокой плотности (ЛПВП)
2: 488-491
- классификация 2: 486
- низкой плотности (ЛПНП)
2: 486-491
- очень низкой плотности
(ЛПОНП)2: 486-491
- простетические группы 1: 131
- транспорт 2: 485
- хиломикроны 2: 227, 228
Липосомы 1: 529
- в клонировании животных 1: 473
Литотрофы 1: 20
Ли-Фраументи раковый синдром
1:671
Лобан Петер 1: 438
Ловастатин (мевакор) 2: 494
Лондона (Ван-дер-Ваальса) силы
1:82
ЛПВП, см. Липопротеины высокой
плотности
ЛПНП, см. Липопротеины низкой
плотности
ЛПОНП, см. Липопротеины очень
низкой плотности
Лютеин 2: 361-363
Лютеинизирующий гормон 1: 369
Люцифер желтый 2: 63
Люцифераза, в генетических экс-
периментах 1: 471,472
Люциферин 2: 39
м
Магния ионы
— в комплексе с АТР 2: 27, 28,30
— в цикле Кальвина 2: 405, 408, 418
Мак-Ардля болезнь 2: 159
Маккарти Маклин 1: 400
Маккинкон Родерик 1: 575
Макклинток Барбара 3: 81
Маклеод Дж. Дж. Р. 2: 568
Маклеод Колин 1: 400
Макромолекулы 1: 32
- информационные 1: 33
- слабые взаимодействия 1: 82, 83
- энергия для сборки 2: 40
Макрофаги 1: 249, 250
Макроциты 2: 284
Максам Алам 1: 419, 420
Максама-Гильберта метод секвени-
рования 1: 420
Максимальная скорость (Vmax) 1: 286,
288,289
Макэлрой Уильям 2: 39
Малат 2: 101,198,199
- окисление до оксалоацетата 2: 199
- транспорт 2:340,455, 456
Малат-а-кетоглутаратная система
2: 341,455,456
Малат-аспартатный челнок 2: 340,
341
Малатдегидрогеназа 2: 199, 425,454,
455
-синтез NADPH 2: 454,455
Малатсинтаза 2: 212
Малая бороздка 1: 401
Малеиновая кислота (малеат) 1: 34;
2: 199
Малик-фермент, см. Малатдегидро-
геназа
Маллис Кэри 1: 10, 452
Малонил/ацетил-СоА-АСР-
трансфераза 2: 450, 451
Малонил-СоА 2: 241, 242, 244, 611
- в синтезе жирных кислот 2:447-463
- синтез 2: 446
Малые РНК
— временные (мвРНК) 3: 270
— интерферирующие (миРНК)
3: 270
— ядерные (мяРНК) 3:127, 128, 138,
157
— ядрышковые (мякРНК) 3: 135,
136,157
Мальтоза 1: 350, 351
Мальтозосвязывающий белок 1: 447
Мальтопорин 1: 536,357
Малярия, и серповидно-клеточная
анемия 1: 248
Манноза 1: 343
- окисление 1:347
Маннозамин 1: 345
Маннозо-6-фосфат 1: 375
- рецептор 1:378
Маннозосвязывающий белок 1: 372
Марихуана 1: 622
Маршалл Барри 1: 374
Масса тела, регуляция 2: 604-614
— PPAR 2: 612, 613
---адипонектином 2: 610-617
---грелином 2: 607, 613, 614
---инсулином 2: 609-612
---лептином 2: 605-610
---термогенином 2: 606, 609
---теория «липидной нагрузки»
2:615-617
---функция гипоталамуса 2: 131,
605-614
Масс-спектрометрия 1: 32,149-151
-MALDI 1:149,381
- в анализе
---аминокислотной последователь-
ности 1: 148, 149
---липидов 1: 517, 518
---сахаров 1: 380,381
- тандемная 1: 150, 151
- электроспрей 1: 149
Материнская мРНК 3: 273
Материнские гены 3: 273
Матричная цепь
— в репликации ДНК 3: 49
— в репликации РНК 3: 144,154
— в транскрипции 3: 108
Матричная РНК, см. мРНК
Маттеи Генрих 3: 167
МГДГ (моногалактозилдиацилгли-
церин) 1: 499
Мевакор (ловастатин) 2: 494
Мевалонат 2: 494
- в синтезе холестерина 2: 480, 481
Мегалобластная анемия 2: 284
Мегалобласты 2: 284
Медиатор 3: 261-266
Медленно сокращающиеся мышцы
2: 587
Медузы, флуоресцентные белки
1:451
Медь, в комплексе IV 2: 320-322
Межклеточные взаимодействия/
адгезия
— интегрины 1: 549
— кадгерины 1: 549
— лектины 1: 372, 73
— протеогликаны/олигосахариды
1: 364-370
— селектины 1: 373,374, 549
Мезельсон Мэтью 3: 46
Мезельсона-Сталя эксперимент
3: 46
Мезофилл листьев, в С4-растениях
2:424,425
Предметно-именной указатель [ \ 13 ]
Мейергоф Отто 2: 66, 83
Мейоз 3: 82-84
Меланокортин 2: 608
Меланоцитстимулирующий гормон,
см. Меланокортин
Мембранные белки 1:363,364; см.
также Гликопротеины; Протео-
гликаны
— амфитропные 1: 530, 531
— анализ методом атомно-силовой
микроскопии 1: 544
— в рафтах 1: 545
— гидропатический индекс 1: 534
— дефекты, как причина заболева-
ний 1:217
— интегральные 1: 530-532
---связь с липидными кольцами
1:532
— остатки Тгр и Туг 1: 535, 536
— периферические 1: 530
— порины 1: 535
— связь с липидными компонента-
ми 1: 536, 537
— а-спирали 1: 533-535
— [3-цилиндр (бочонок) 1: 535, 536
Мембранные гликопротеины 1: 369
Мембранные липиды 1: 26, 369, 370,
496-506; 2: 470-476
— архей 1: 500
-- биосинтез, см. Липиды, биосинтез
— болезни накопления 1: 505
— ганглиозиды 1: 369
— диффузия 1: 540, 541, 543
---измерение скорости 1: 542
— кольцевые 1: 533, 534
— микродомены 1: 543, 545
— плазмалогены 1: 497, 499
— разрушение в лизосомах 1: 503
— растений 1: 500
— рафты 1: 543, 545
— связывание с мембранными бел-
ками 1: 536, 537
— стерины 1: 504
— сфинголипиды 1: 501-504
— тилакоидных мембран 1: 499, 500
— у разных организмов 1: 526, 527
— фосфолипиды 1: 497
— эфирные 1: 499
Мембранные липополисахариды
1:369,270
Мембранные переносчики, в мито-
хондриях 2: 307-321
Мембранные рафты 1: 543-546, 632
— в передаче сигнала 1: 632
Мембранный потенциал (Vm) 1: 551,
559, 562,574, 577,651
Мембранный транспорт 1: 550-581
— ABC-транспортеры 1: 564, 566
— АТР-зависимый 1: 563
— АТР-синтаза 1: 63,564
— Са2+-насос 1: 560
— SERCA 1:560,561
— Ыа+К?-АТРаза 1: 562
— аквапорины 1: 570-573
— активный 1: 558
---первичный 1:558
---вторичный 1: 558,567
— антипорт 1:558
— в нейронах 2: 578; 2: 590
— глюкозы 1: 553-556
— изменение свободной энергии
1:552
— ионный градиент 1: 567
— ионофоры 1: 570
— котранспортные системы 1: 558
— липидов 2: 478
— мембранный потенциал 1: 551
— облегченная диффузия (пассив-
ный транспорт)1: 552
— пассивный 1: 552
— порины 1: 535
— симпорт 1: 558
— энергетические затраты 1: 569
— транспортеры, см. Ионные кана-
лы; Переносчики; Транспортеры
— унипорт 1: 558
— факторы, облегчающие транспорт
(MFS) 1:569
— хлоридно-бикарбонатный обмен-
ник 1: 556,557
— электрогенный 1: 559
— электронейтральный 1: 558
— электрохимический градиент
1:551
Мембраны 1: 525-549
- асимметрия 1: 528, 530
- динамика 1: 538-540
- диффузия молекул 1: 540
- двойной липидный слой 1:528, 529
— основные состояния 1: 539
- кавеолы 1: 546
- модель строения 1: 527, 528
- плазматическая
— белки 1: 530, 537, 549
— липидный состав 1: 527, 530
— нервных клеток 2: 590
— проницаемость 1: 85, 527
— рафты 1: 543, 545
— связывание белков 3: 207,214,
218
— состав у разных организмов 1: 526
— трансмембранный потенциал
1: 633
- поляризация 1: 633, 634
- слияние 1: 546-548
— в синапсах 1: 548
Менахинон (витамин К2) 1: 512
Ментен Мод 1: 287
Меринг (фон) Йозеф 2: 568
Меромиозин 1: 257
Меррифилд Р. Брюс 1: 152, 423
Мерцающие кластеры 1: 75
Метаболизм 1: 48; 2: 7, 579
- анаэробный, у целакантов 2: 92
- аэробный, у позвоночных 2: 92
- в головном мозге 2: 590, 591
- в жировой ткани 2: 585-587
- в мышцах 2: 587-590
- в печени 2: 579-585
- внутренние перегруппировки
2: 23, 24
- и биоэнергетика 2: 12-19
- окислительно-восстановительные
реакции 2: 26, 27
- промежуточный 2: 7
- реакции с переносом групп 2: 25,26
- регуляция по принципу обратной
связи 1: 49
- роль АТР 1: 49
- свободно-радикальные реакции
2: 25
- функция крови 2: 591, 592
Метаболиты 1: 18, 32; 2: 7, 122
- вторичные 1: 32
- изменение концентрации
2: 128-131
- пул, в растениях 2: 437, 438
Метаболическая вода 1: 104, 105
Метаболические пути 1: 48; 2: 7-9
- анаболические 2: 7
- близость к равновесию 2: 128, 129
- катаболические 2: 7
- конвергентные 2:8
- дивергентные 2: 8
- циклические 2: 8
Метаболический алкалоз 1: 93
Метаболический ацидоз 1: 93, 102
Метаболический контроль 2: 127,
132
— анализ 2: 134-138
Метаболический синдром 2: 615-618
Метаболом 1:32; 2: 126
Метаболой 2: 210, 211
Металлопротеины 1: 131
Металлы
- как катализаторы 1: 284, 285
- как кофакторы 1: 271,309,310
- как переносчики кислорода 1: 227
Метамерия 3: 272
Метан 2: 46
141 \ | Предметно-именной указатель
Метанол
- в экстракции липидов 1: 515-517
- отравления 1: 297
Метиладенин, деметилирование
3: 75, 77
Метиладенозин 1: 395
Метиламин, рКа 1: 124,125
7-Метил гуанозин 1: 395;3: 75, 77
Метилирование
- в анализе сахаров 1: 379
- в репарации ДНК 3: 74-77
- нуклеиновых оснований 1: 418,
419, 435, 436
- ферментов 1: 323
6-М-Метиллизин 1: 120
Метилмалонил-СоА 2: 239
Метилмалонил-СоА-мутаза 2: 240,
243
- дефекты 2: 287, 295
- при пернициозной анемии 2: 284
Метилмалонил-СоА-эпимераза
2: 240
Метилмалонилполуальдегид 2: 556
Метилмалоновая ацидемия 2: 287,
295, 296
5-Метилцитидин 1: 395
Метилцитозин, деметилирование
3: 68, 72, 75
Метионин 1: 118; 2: 523, 538, 542
- биосинтез 2: 524, 531
- превращение в сукцинил-СоА
2: 294
Метионинаденозилтраисфераза
2: 283
Метионинсинтаза 2: 283, 284
Метка экспрессируемой последова-
тельности (EST) 1: 451
Метотрексат 2: 558, 559
Метэнкефалин 1: 68
Миелиновая оболочка 1: 502, 526
Микродомены мембраны 1: 543, 545
Микроинъекции ДНК 1: 473
Микроназ 2: 597
МикроРНКЗ: 138, 270
Микроскопия атомно-силовая 1: 544
Микротрубочки 1: 24, 25, 257
Миллера-Юри эксперимент 1: 55, 56
Мильштейн Сезар 1: 254
Минералокортикоиды 2: 496,575
- синтез 2: 348
Минихромосомы, белки МСМ 3: 64,
65
Минковски Оскар 2: 568
Минус-зеленые дихроматы 1: 655
Минус-красные дихроматы 1: 655
Миоглобин 1:129,139, 194, 200, 234
- гемовая группа 1: 195
- изучение методом ЯМР 1:197-199
- рентгеноструктурный анализ
1: 194,196,197
- связывание кислорода 1: 226,
228-233
Миозин 1: 257
- в мышечном сокращении
1: 260-262
- взаимодействие с актином 1: 257,
260-262
- суперспираль 1: 257
- толстые нити 1: 258
- фосфорилирование 1: 665
Миокарда инфаркт 2: 140,141
Миоклоническая эпилепсия с разо-
рванными красными мышечны-
ми волокнами (МЭРКМВ)
2: 351,356
Миофибриллы 1: 258, 259
Миоцит 2: 140,141, 166, 229, 587
- контроль синтеза гликогена 2: 137
Миристиновая кислота 1: 488
Миристоильная группа 1: 323, 537,
545
миРНК 3: 270
мир РНК, гипотеза 1: 57; 3: 153-159
Митоз 1: 660, 662, 663
Митохондриальная ДНК, см. мтДНК
Митохондриальная энцефаломиопа-
тия 2: 354
Митохондриальное дыхание 2: 420
Митохондриальные гены, мутации
2: 353-356
Митохондрии
- ATP-синтазный комплекс
2: 331-336
--механизм вращательного ката-
лиза 2: 336-338
--при гипоксии 2: 344, 345
- апоптоз 2:350
- в растениях, альтернативный путь
окисления NADH 2: 325, 326
- вариации генетического кода
3: 172,177
- гены 2: 351
— и диабет 2: 355, 356
— мутации 2: 353-355
- гетероплазмия 2: 354
- гомоплазмия 2: 354
- ДНК, см. мтДНК
- доставка NADH из цитозоля
2: 340-342
- доставка белков 3: 210, 215
- и ксенобиотики 2: 349
- и старение 2: 346, 353
- и хемиосмотическая теория 2: 306,
327, 328
- матрикс 2: 307
- мембранные переносчики 2: 309
- мембраны 2: 307
- образование активных форм кис-
лорода 2: 324, 345
- р-окисление 2: 232-237
— ферменты, отличие от ферментов
пероксисом и глиоксисом 2: 248
- отношение концентраций NADH/
NAD+ 2: 102
- происхождение 2: 352, 353
- реакции переноса электронов
2: 306-320
-----разобщение 2: 329, 330
- респирасомы 2: 320, 321
- синтез АТР 2: 237, 327-338
--выход 2: 343
--регуляция 2: 346, 347
- синтез стероидов 2: 348, 349
- строение 2: 307
- термогенез 2: 348
- типы цитохромов 2: 309, 310
- транспорт жирных кислот
2: 230-232
Митчелл Питер 2: 305, 306, 327
Мифепристон 1: 644
Михаэлис Леонор 1: 287
Михаэлиса кинетика 1: 287-290
Михаэлиса константа (Км) 1: 288,
290, 291
— кажущаяся 1: 295, 296
Михаэлиса-Ментен уравнение 1: 288
— интерпретация параметров 1: 290
— преобразования 1: 289
Мицеллы 1: 80, 81, 529
Мишер Фридрих 1: 400
Млекопитающие
- аквапорины 1: 571
- жировые запасы 1: 491-495
- клонирование 1:473-479
- концентрация глюкозы в крови
1:354
- повреждение ДНК 1: 416, 417
- протеогликаны 1: 364
- сигнальные пути 1: 646, 647
Мобильные элементы 3: 94, 148
Модуляторы, в связывании белков с
лигандами 1: 320
Мозаичная модель мембран 1: 589
Молекулярная масса 1: 32
Молекулярные паразиты 1: 460;
3:94,155
Молотоголовый рибозим 3: 138, 139
Молочная кислота 1: 522
Молочнокислое брожение 2: 69, 91
— при сокращении мышц 2: 92
— роль пирувата 2: 91
Предметно-именной указатель 1415 ]
Молчащие мутации 3: 67
Моно Жак 1: 9, 28, 242, 408; 3: 234
Моногалактозилдиацилглицерин
(МГДГ) 1:499
Моноклональные антитела 1: 254,
308, 479
— в противораковой терапии 1: 669
— каталитические 1: 308
Монооксигеназы 2: 460
Монооксид азота (NO) 1: 628; 2: 576
— в роли гормона 2: 572, 576
— при стенокардии 1: 628
— синтез из аргинина 2: 542, 543
Монооксид углерода
— отравление 1: 240, 241
— связывание с гемоглобином
1:228, 240, 241
— физиологическое действие 1: 240,
241
Моносахариды 1: 339, 340, 347
- D- и L-изомеры 1: 341
- альдоза 1: 341, 342
- в гликолизе 2: 87
- восстанавливающие свойства
1:347
- гексоза Г. 341,345
— производные 1: 346, 347
- гептоза 1: 341
- кетозы 1: 341, 342
- конформация 1:345
- мутаротация 1: 344
- номенклатура 1: 341-343
- образование при гидролизе дисаха-
ридов и полисахаридов 2: 86
- окисление 1: 347
- пентоза 1:341
- пиранозы 1: 344
- полуацеталь 1: 343
- полукеталь 1: 343
- тетроза 1: 341
- триоза 1: 341
- фосфорилирование 1: 347
- фураноза 1: 344
- хиральные центры 1: 341
- циклические формы 1: 344
---формулы Хеуорса 1: 345
- энантиомеры 1: 341
- эпимеры 1: 343
Моно-Уаймена-Шанже модель
1:242,243
Моноцистронная мРНК 1: 408
Морфогены 3: 273, 275
Мотивы белковые 1: 200-203
- ДНК-связывающие 3: 237-242
Моторные белки 1: 256-262
Мочевая кислота 2: 555
Мочевина 2: 273, 601
- в метаболизме аминокислот 2: 263,
583, 584
- в метаболизме пиримидинов
2: 555, 556
мРНК (матричная РНК) 1: 391, 408;
3: 105
- в эволюции 1: 56
- время полужизни 3: 142
- длина 1: 408
- искусственная 3: 167
- 5’-кэп 3: 123
- материнская 3: 273
- моноцистронная 1: 408
- поли(А)-«хвост» 3: 122, 123, 130
- полицистронная 1: 408
- процессинг 3: 122-133
- расщепление 3: 142
- редактирование 3:176
- скорость синтеза 2:124
- спаривание оснований с тРНК
3:173,174
- сплайсинг 3: 122,125
- стабильность 2:124
- функции 1: 408
- шпильки 1: 406; 3: 142
МСГ (мслапоцитстимулирующий
гормон), см. Меланокортин
мтДНК (митохондриальная ДНК)
3: 172
- вариации генетического кода
3: 172, 173
Мультимер 1: 205
Мультисубъединичные белки 1: 130,
240
Мультифункциональный белок
(MFP) 2: 248
Муравьиная кислота 2: 45
Мурамовая кислота 1: 346
Мусорная ДНК 3:156
Мутагенез
- олигонуклеотид-направленный
1:446
- сайт-направленный 1: 446, 447
Мутазы 2:87
Мутаротация 1: 344
Мутации 1: 54, 415; 3: 67
- ацил-СоА-дегидрогеназ 2: 245
- в цикле лимонной кислоты, при
раке 2: 211
- в эволюции 1: 54, 55; 3: 278-280
- глутаматдегидрогеназы 2: 270
- дегидрогеназы СоА-производных
жирных кислот 2: 245
- делеции 3: 67
- замены 3: 67
- и апоптоз 2: 666
- и окислительный стресс 1:419; 2:316
- инсерции 3: 67
- канцерогенные 1: 601, 666-671;
3: 67, 79
- митохондриальных генов,
см. Митохондрии
- молчащие 3: 67
- нонсенс 3: 203
- под действием излучения 1:417
- при амилоидозе 1: 215
- при кистозном фиброзе 1: 565, 579
- при ретинобластоме 1: 670
- при сахарном диабете 2: 152, 153,
355, 597
- при синдроме Вернике-Корсакова
2: 112
- при тромбастении 1: 642
- роль алкилирующих агентов 1:418,
419
- супрессоров 3: 203
Муцины 1: 368
Мышечные белки 1: 258-262
— актин 1: 257
— миоглобин 1: 194
— миозин 1: 257
— тропонин 1: 201, 261
Мышечные волокна 1: 258, 259
Мышечные сокращения 1: 260-262
— быстрые 2: 587
— источники энергии 2: 588, 589
— медленные 2: 587
— функция АТР 1: 260, 261
Мыши, трансгенные 1: 477, 478
Мышцы
- белки, см. Мышечные белки
- белые 2:587
- выведение аммиака 2: 272
- источники энергии 2: 588, 589
- красные 2: 587
— МЭРКМВ2: 351,356
- метаболизм 2: 588-590
— углеводов 2: 173
- сердечные 2: 589
- структура 1: 258, 259
Мэдсен Нейл 2: 212
МЭРКМВ (миоклоническая эпилеп-
сия с разорванными красными
мышечными волокнами) 2: 351,
356
н
Надмолекулярные комплексы 1:18,
27,33
Направляющая РНКЗ: 176
Натриевые каналы 1: 578, 579,
635-637
— в сигнальных путях 1: 627, 628
| \ 16 ] Предметно-именной указатель
Натрий
- содержание в крови 2: 592
- транспортная АТРаза 1: 569, 633
Натрий-глюкозный симпорт 1: 569
Надпочечники 2: 495
Налидиксовая кислота 3: 24
Напроксен 2: 462
Наследственная (семейная) гиперхо-
лестеринемия 2: 493-495
Наследственная нейропатия зри-
тельного нерва Лебера 2: 351,
354
Наследственный неполипозный рак
толстой кишки 3: 80
Натансон Даниэль 1: 436
Нативная конформация (белка)
1:53, 172
Начальная скорость (г0) 1: 286
Небулин 1: 259
Невосстанавливающие концы 1: 347,
354
Неер Эрвин 1: 575
Незаменимые аминокислоты 2: 278,
523, 524; см. также Аминокис-
лоты
Незаменимые жирные кислоты
2: 459
Нейромедиаторы
- биосинтез из аминокислот 2: 538,
539
- в роли гормонов 2: 566
- высвобождение 1: 635, 636
— при слиянии мембран 1: 548
- рецепторы 1: 638, 639
Нейроны
- №+-каналы 1: 578, 635-637
- анорексигенные 2: 607-609
- вкусовые 1: 656
- зрительные 1: 650-656
- мембранный транспорт 2: 590
- обонятельные 1: 656
- орексигенные 2: 607, 608
- светочувствительные 1: 650
Нейропептид Y (NPY) 2: 607
Нейроэндокринная система 2: 566
Нейтральное значение pH 1: 91, 92
Нейтральные гликолипиды 1: 502
Нейтральные жиры, см. Триацилгли-
церины
Нековалентное связывание, см. Сла-
бые взаимодействия
Некодирующая РНК (нкРНК) 3: 271
Неконкурентный ингибитор 1: 297
Нексавар (сорафениб) 1: 669
Необратимые ингибиторы 1: 295, 298
Неокислительная стадия пентозо-
фосфатного пути 2: 108, 109
Неравновесные стадии метаболиче-
ских путей 2: 128, 129
Нерв Лебера, наследственная нейро-
патия 2: 351, 354
Нервная сигнализация 2: 566
Нервный импульс, передача 1: 636
Несовершенный остеогенез 1: 191
Нестероидные противовоспалитель-
ные средства (НПВС) 1: 508;
2: 462, 463
Ниацин (никотиновая кислота)
2: 53, 54
Никотин 2: 53
Никотинамид 2: 53
Никотинамидадениндинуклеотид,
см. NAD+
Никотинамидадениндинуклеотид-
фосфат, см. NADP+
Никотинамиднуклеотидсодержащие
дегидрогеназы 2: 307, 308
Никотиновая кислота, см. Ниацин
Никотиновый ацетилхолиновый
рецептор 1: 578, 579 638
---агрегаты в синапсах 1: 542
---дефекты 1: 579
---открытая/закрытая конформа-
ция 1: 638, 639
Ник-трансляция 3: 53
Нимана-Пика болезнь 1: 505
Ниренберг Маршалл 3: 167
Нитрификация 2: 507
Нитрогеназный комплекс 2: 510-513
Нитроглицерин 1: 628
нкРНК (некодирующая РНК) 3: 271
Номура Масайасу 3: 180
Нонсенс-кодоны 3: 170, 203
Нонсенс-мутации 3: 203
Нонсенс-супрессоры 3: 203
Норадреналин 2: 538, 567, 574, 602
Нортроп Джон 1: 270
НПВС (нестероидные противовос-
палительные средства) 1: 508;
2: 462, 463
Нуклеазы 3: 48
Нуклеин 1: 400
Нуклеиновые кислоты 1: 33, 391-399
— гибриды 1: 414, 415
— гидрофильный остов 1: 396
— ДНК 1:393, 394,400-403;
см. также ДНК
---З'-конец 1: 396
---5'-конец 1: 396
---пространственное строение
1: 403-408
---зонды 1: 443
---неферментативные превраще-
ния 1: 415-419
---определение последовательно-
сти 1: 419-422
---полярность 1: 396
— РНК 1: 393, 394, 408; см. также
РНК
---пространственное строение
1:409-412
---состав 1: 392-396
— синтез, см. Репликация;
Транскрипция
— фосфодиэфирные связи 1: 396
— функции 1: 391
— химический синтез 1: 422-424
Нуклеозиддифосфаткиназа 2: 41,
198, 550
- механизм пинг-понг 2: 41
Нуклеозиддифосфаты 1: 424
Нуклеозидмонофосфаткиназы 2: 550
Нуклеозидмонофосфаты 1: 424, 550
- превращение в нуклеозидтрифос-
фаты 2: 550
Нуклеозидтрифосфаты 1: 424
- образование из нуклеозидмоно-
фосфатов 2: 550
Нуклеозиды 1: 393, 394; 3: 49, 51
Нуклеоид 1: 18, 21; 3: 36, 37
Нуклеосомы 3: 27, 28
- ацетилирование 3: 258
- сборка хроматина 3: 28-31
- фибриллы 3: 31
5'-Нуклеотидаза 2: 555
Нуклеотидная последовательность
— в эволюционных исследованиях
1: 158, 159
— и аминокислотная последова-
тельность 1: 148, 152; 3: 171
— определение 1: 419-424
Нуклеотидсвязывающий мотив 1: 425
Нуклеотиды 1: 391-394
- биосинтез 2: 543-554
— реутилизация 2: 556
- гидролиз N-p-гликозидной связи
1:416,417
- депуринизация 1: 416, 417
- как кофакторы и коферменты
1: 425, 426; 2: 52, 54
- как сигнальные молекулы 1: 427
- как универсальные переносчики
электронов 2: 49, 50
- метаболизм 2: 543-557
— образование мочевины и мочевой
кислоты 2: 555-557
- неферментативные реакции
1:415-419
- сахаронуклеотиды 2: 158, 428
- сокращенные обозначения 1: 392,
394
Предметно-именной указатель [417]
- состав 1: 392
- спектр поглощения 1:398
- структура 1: 392
- трансфосфорилирование 2: 40,41
- триплеты, см. Кодоны
- ферменты биосинтеза как мишени
противоопухолевой терапии
2: 558-560
- флавиновые 1: 426; 2: 54, 55
- фосфаты, как переносчики химиче-
ской энергии 1:424,425
- фосфодиэфирные связи 1: 396
--и фосфоэфирные связи 1: 425
- эволюция 1: 54, 56; 3: 151-155
Нуклеофил 2: 21
Нуклеофильное замещение в реак-
циях АТР 2: 37-39
Нюсляйн-Фольхард Кристиана
3: 273
о
Обесцвечивание флуоресценции
(FRAP) 1:541, 542
Облегченная диффузия 1: 552, 553;
см. также Переносчики
Обмен головками, в синтезе фосфо-
липидов 2: 472
Обратимые ингибиторы 1: 295
Обратная транскриптаза 3: 144
Обходные реакции глюконеогенеза
2:98-103
Общий кислотно-основной катализ
1: 283, 284
Один ген - один белок, гипотеза 3: 8
Ожирение 2: 600, 604-614
Оказаки фрагменты 3:48
Окаймленные пузырьки 3: 219
Окисление
- аминокислот, см. Аминокислоты,
окисление
- ацетата 2: 202
- в пентозофосфатном пути 2: 107
- в цикле лимонной кислоты
2: 188-199
— изоцитрата до а-кетоглутарата и
СО2 2: 192
— а-кетоглутарата до сукцинил-
СоА и СО2 2:194,195
— малата до оксалоацетата 2: 199
— пирувата до ацетил-СоА и СО2
2: 69, 182-187
— сукцината до фумарата 2: 198
- глицеральдегид-3-фосфата до
1,3-бисфосфоглицерата 2: 76
- глюкозы 2:49
— выход АТР 2: 80
- жирных кислот
— а-окисление 2: 250
— (3-окисление 2: 232-236, 245-248
---образование ацетил-СоА и АТР
2: 232-235
— а)-окисление 2: 249
- углерода в биологических процес-
сах 2:21,27,45
Окислительно-восстановительные
реакции 1:43; 2: 26, 27
— восстановительный потенциал
2: 47,48
— и дегидрирование 2: 45
— полуреакции 2: 44
— электродвижущая сила 2: 44
Окислительное дезаминирование
2: 270
Окислительное декарбоксилирова-
ние 2: 183
Окислительное фосфорилирование
2: 305-346
— в сердечной мышце 2: 589
— выход АТР 2: 343
— и активные формы кислорода
2: 324, 345
— и субстратное фосфорилирование
2: 78
— ингибирование гидролиза АТР
2: 344
— мембранные переносчики
2: 309-313
— полиферментные комплексы
2:313-320
— разобщение 2: 329, 330, 348
---термогенин 2: 348
— регуляция 2: 343-347
— респирасомы 2: 320
— синтез АТР 2: 327-342
------вращательный катализ
2: 336-339
------регуляция 2: 346, 347
— термогенез в бурой жировой
ткани 2: 347,348
— функция митохондрий 2: 306, 307,
347; см. также Митохондрии
— хемиосмотическая теория 2: 306,
327
— электрохимический потенциал
2: 322,323
Окислительные стадии пентозофос-
фатного пути 2: 108,109
Окислительный пентозофосфатный
путь 2: 111,419,435,438
Окислительный стресс, роль мито-
хондрий 2: 355
Оксалоацетат 2:100
- в С4-пути 2:424,425
- в глиоксилатном цикле 2: 212-214
- в синтезе аминокислот 2: 518,523,
524
- в цикле лимонной кислоты 2:190,
191,199, 205
- как продукт окисления малата
2:102,199
- как продукт превращения глюко-
генных аминокислот 2: 104, 280
- как продукт расщепления аспара-
гина и аспартата 2: 274, 297, 298
Оксигеназы 2: 659, 560
- со смешанной функцией 2:460
Оксидазы 2: 27, 250
- со смешанной функцией 2: 291,
458-460,477,478, 483, 496
Оксидоредуктазы 2: 51
Окситоцин 2: 577
Оксоглутарат, см. а-Кетоглутарат
Октадекадиеновая кислота 1: 488
Октадекановая кислота 1: 488
Октадекатриеновая кислота 1: 488
Октадеценовая кислота 1: 488
Олеат 2: 238
- синтез 2:458,459
Олеиновая кислота 1: 488
Олигомеры 1: 32, 130
Олигонуклеотид-направлспный
мутагенез 1: 446
Олигонуклеотиды 1: 397, 423
Олигопептиды 1: 128
Олигосахариды 1: 33,339; см. также
Дисахариды; Полисахариды;
Углеводы
- анализ 1: 379-381
- в ганглиозидах 1: 502
- в гликопротеинах 1: 363, 368-371;
3: 207,213
- в гликофорине 1: 527, 531
- взаимодействие с лектинами
1: 372-378
- конформация 1: 357
- номенклатура 1: 351
- N-связанные 1:368; 3: 207, 213, 214
- О-связанные 1: 368; 3: 207, 214
- синтез 2: 403-438
- структура 1: 371
- химический синтез 1: 381
Олтман Сидней 3: 141
Онкогены 1: 666-670
- мутации 1: 671; 3: 146,207
Оператор 3: 232
Оперон 3:234
- his3: 247
-1асЗ: 234-236
- leu 3:247
- phe 3: 247
| \ 1 <s | Предметно-именной указатель
- trp 3: 246-249
- регуляция 3: 234-236
Опины Г. 469, 470
Опсины 1: 652, 655; см. также Ро-
допсин
Оптическая активность 1: 37, 117
Опухоль 1: 666, 671
- анализ на микрочипах 1: 465, 466
- в растениях 1: 470
- гены супрессоров 1: 666, 667, 670
- методы лечения 1: 476,477,
667-669
- промоторы 1: 611
- роль р53 1: 671
- транскрипционный профиль 1: 466
- фактор некроза (TNF) 1: 477, 672
Органеллы 1: 22-25
- разделение 1: 23
- растений 2: 358,405
Органические растворители, экс-
тракция липидов 1: 510, 515, 516
Органотрофы 1: 20
Оргел Лесли 3:152
Ориджин 3:48
Орексигенные нейроны 2: 607, 608
Ориджин-независимое начало ре-
пликации 3: 89
Орнитин 1: 122, 123; 2: 519
- в цикле мочевины 2: 273-275
Орнитин-6-аминотрансфераза 2: 519,
521
Орнитиндекарбоксилаза 2: 540-542
Орнитинтранскарбамоилаза 2: 273
Оротат 2: 543
Ортологи 1: 61, 157,462
Осельтамивир (тамифлю) 1: 374
Осмолярность 1: 85
Осмос 1: 85, 86
Осмотический лизис 1: 86
Основания
- азотистые (в нуклеотидах) 1: 26,
392; см. также Пиримидины;
Пурины
— алкилированные, репарация
3: 74-77
— анти-форма 1: 403,404
— в кодонах 3: 171
— водородные связи 1: 399
— дезаминирование 1: 416-418
— и азотистая кислота 1: 418
— качающиеся пары 3: 172-175
— метилирование 1: 419
— минорные 1: 395
— син-форма 1: 403, 404
— спаривание 1: 399
— таутомерные формы 1: 398
— функциональные группы 1: 398
— химические свойства 1: 397, 398
— Чаргаффа правила 1: 400
- аминокислот 1: 123
- в буферных системах 1: 96-98
- слабые 1: 89,93
Основное состояние 1: 274; 2: 361
Основные транскрипционные факто-
ры 3: 118, 120
Островковые клетки (клетки Лан-
герганса) 2: 595
Острый лимфобластный лейкоз
(ОЛЛ) 2: 523
Острый миелоидный лейкоз 1: 668
Отжиг ДНК 1: 410, 413,414, 420
Открытая рамка считывания 3: 171
Открытая система 1: 41
Относительная молекулярная масса
(Mr) 1: 32
Отравление моллюсками 1: 581
Отстающая цепь 3: 48
Очоа Северо 2: 83, 160, 359; 3: 142,
143
п
Паладе Джордж 3:211
Палиндромная ДНК 1: 405
Пальмитат
- в синтезе жирных кислот
2: 452-454,458
- десатурация 2: 458
- синтез 2:449,452-454
Пальмитиновая кислота 1: 488
Пальмитоильная группа
— окисление 2: 235, 236
— пальмитоил-СоА 2: 458
— связывание с мембраной 1: 543,
545
Пальмитолеиновая кислота 1: 488
Пальмитоолеат, синтез 2: 458
Панкреатит острый 2: 266
Панкреатические клетки
— а 2: 595
— 02:595, 596,615
Панкреатический ингибитор трипси-
на 2: 264
Паразиты молекулярные, эволюция
3: 94, 155
Паракринные гормоны 2: 571, 574
— эйкозаноиды 1: 507
Паралоги 1: 61, 157,462
Парамиозин 1: 260
Паращитовидная железа 2: 576
Паркинсона болезнь, аномалия фол-
динга 1: 215, 217
Парного правила гены 3: 274
Пассивный транспорт 1: 552, 553
Пастер Луи 1: 36, 37; 2: 80
Пейс Норман 3: 141
Пеллагра 2: 63
Пенициллин
- ген устойчивости на плазмидах
3:10
- механизм действия 1: 314
Пентозофосфатный (фосфоглюко-
натный, гексозомонофосфат -
ный) путь 2: 107,406,438
— восстановительный 2: 112, 406
— и гликолиз 2: 112
— неокислительный этап 2: 108,109,
406-415
— общая схема 2: 108
— окислительный этап 2: 108,109
— окислительный 2: 108, 111
— синдром Вернике-Корсакова
2: 112
— синтез NADPH 2: 107
— функция глюкозо-6-фосфата
2: 109, 112,582
--дефект Г6ФДГ 2: 106
Пентозофосфаты 2: 82, 109
- превращения 2: 412
- синтез в цикле Кальвина 2: 406
Пентозы 1: 341
- в нуклеиновых кислотах 1: 396, 404
- в нуклеотидах 1: 392
Пепсиноген 2: 264
Пептидилпролил-г^с-тирш/с-
изомераза (PPI) 1: 214
Пептидилтрансфераза 3: 200
Пептидильный (Р) сайт рибосомы
3: 197-202
Пептидная группа 1: 174
Пептидная связь 1: 127, 175
— в а-спирали 1: 177
— образование в синтезе белка
3: 200
— транс-кон фигурация 1: 182
— ^wc-конфигурация 1: 182
Пептидные гормоны 2: 567, 569-574
Пептидогликаны 1: 362; 2: 433
- ингибирование синтеза пеницил-
лином 1: 314, 315
- синтез в бактериях 2: 433, 434
Пептиды 1: 127; см. также Белки;
Полипептиды
- аминокислотный состав 1: 127, 128
- ионизация 1: 128
- номенклатура 1: 128
- определение последовательности
1: 143-145, 147
- химический синтез 1: 152-154
Первичная структура белка 1: 141
--анализ 1: 143-151
Предметно-именной указатель [419]
Первичный активный транспорт
1:558
Первичный бульон 1: 56-58
Первичный системный амилоидоз
1:215
Первичный транскрипт 3:122
— процессинг 3: 122-143
— сплайсинг 3:122,125
Передняя доля гипофиза 2: 566, 567
Переключатели, в G-белках
1: 599-602
Переключение типа спаривания у
дрозофил 3: 256
Перенос генов, латеральный 1: 157
Переносчики 1: 271, 525, 540, 552,
558, 560 см. также Ионные
каналы; Транспортеры
-АВС 1:565; 2:489
- Са2+-насос 1: 560, 561
- F-типа АТРаза 1: 563
- GLUT 1: 553-556
- Ма+/К+-АТРазы 1: 562
- №+/глюкозный симпорт 1: 569,
570
- P-типа АТРазы 1: 560
- SERCA насос 1: 560, 561
- V-типа АТРазы 1: 564
- активные 1: 553
- аланин как переносчик аммиака
2: 271,272
- ацилкарнитиновый/карнитиновый
2: 230-232
- глутамин как переносчик аммиака
2: 270
- глюкозы 2: 553-570
— Ма+/глюкозный симпорт 1: 569,
570
— в кишечнике (GLUT2) 1: 556, 569
— в мышцах (GLUT4) 1: 556
— в эритроцитах (GLUT1) 1:
553-555
— дефекты при диабете 1: 556
- жирных кислот 2: 229-232
- и ионные каналы 1: 552, 553
- ионофоры 1: 570
- котранспортные системы 1: 567
- коферменты как переносчики
электронов, атомов и функцио-
нальных групп 1: 271
- лактозы 1: 567, 568, 588
- липидов 2:485-487
- мальтозы 1: 536
- мультилекарственные 1: 564
- пассивные 1: 553
- скрамблазы 1: 541
- флиппазы 1: 541
- флоппазы 1: 541
- фосфолипидов 1: 540
- цитрата 2: 455
Переходное состояние
— аналоги 1: 307
— в ферментативной реакции 1: 47,
48, 268, 274, 306
— комплементарность ферменту
1: 279, 280, 306-308
Перилипин 2: 228, 229
Пермеазы 1: 552, 567
Пернициозная анемия 2: 284
Пероксисома 2: 213, 245, 246
-PPAR2: 244,612
- метаболизм липидов 2: 421,422
- растений 2: 247
Перутц Макс 1: 205
Петли
- в белковых структурах 1: 200, 201
— р-а-р 1: 202
-ДНК 1:414,417; 3: 37,47
-РНК 1:409,410
Печень
- высвобождение аммиака глутама-
том 2: 263, 264, 269
- гликоген 1: 353
- глицеронеогенез 2: 469
- детоксикация 2: 585
- доставка аммония аланином 2: 271,
272
- каскад адреналина 1: 606
- метаболизм 2: 579, 580
— аминокислот 2: 582-584
--с разветвленной цепью 2: 296,
297
— гликогена 2: 154-164
— глутамина 2: 269, 270
— глюкозы 2: 581, 582
— жирных кислот 2: 584
— углеводов 2: 581,582
- образование кетоновых тел
2: 251-253
- рецикл триацилглицеринов 2: 467,
468
- синтез холестерина 2:483, 484
- ферменты 2: 579, 589
Пигментная ксеродерма 3: 79, 80
Пигменты 2: 535, 536, см. также
Фотопигменты желчи
- зрительные 1: 511, 512, 514
- светопоглощающие Halobacterium
salinarum 2: 388
Пили 1: 21
Пинг-понг (механизм реакции)
1: 293, 294
Пиоглитазон (актос) 2: 612, 617
Пиранозы 1: 344, 345
Пиридиннуклеотиды 2: 51
Пиридоксальфосфат 2: 266, 267
- в реакции гликогенфосфорилазы
2: 155
Пиримидиновые димеры
— образование 1:417
— репарация фотолиазой 3: 74
Пиримидиновые нуклеотиды,
см. Пиримидины
Пиримидины 1: 392, 397, 398, 416
- биосинтез 2: 543
- катаболизм 2: 555
- реутилизация 2: 556
- синтез de novo 2: 548
Пирофосфатаза неорганическая
2: 38, 39,161
Пирофосфорильная группа 2: 37, 73
Пирролизин 3:191
Пируват
- альтернативные пути превращений
2: 69, 149
- в анаплеротических реакциях
2: 204, 205
- в биосинтезе аминокислот 2: 518,
523
- в гликолизе 2: 66-70
---энергетический выход 2: 202,
343
- в глицеронеогенезе 2: 468
- в глюконеогенезе 2: 97, 99
- в молочнокислом брожении 2: 69
- в спиртовом брожении 2: 69, 93
декарбоксилирование и дегидриро-
вание под действием ПДГ 2: 186
- как продукт расщепления амино-
кислот 2: 285, 288
- метаболизм в печени 2: 582-584
- окисление до ацетил-СоА и СО2
2:182, 183
- перенос из цитозоля в митохон-
дрии 2: 456
- превращение в фосфоенолпируват
2: 100-103
---регуляция 2:148, 149
---в растениях 2: 425
- при фенилкетонурии 2: 291
- таутомеризация 2: 79
Пируватдегидрогеназа 2:184
Пируватдегидрогеназный комплекс
2:182-187
— синтез ацетил-СоА 2: 186
Пируватдекарбоксилаза 2: 93, 95
Пируваткарбоксилаза 2: 100,101,
149, 205
Пируваткиназа 2: 32, 79, 128
- регуляция 2: 147, 148
Пируват-фосфатдикиназа, в С4-пути
2: 425, 426
[420] Предметно-именной указатель
Питание, гормональный контроль
2: 607-615
Питательные вещества
— перенос кровью 2: 591, 592
— расщепление в печени 2: 579
Плазма крови 2: 591, 592
— белки 2: 592
— глюкоза 2: 592
— транспорт липопротеинов 2:
485-488
Плазмалогены 1: 497, 499; 2: 476, 477
Плазматическая мембрана
— глипиканы 1: 364, 365
— ионные каналы 1: 633-639
— липидные рафты 1: 632
— липополисахариды 1: 369, 370
— нейронов, транспорт веществ
1:639
— проницаемость 1: 85, 573, 574
— связывание белков 3: 214
- связывание ДНК при репликации
3: 58
— синдекан 1:364, 365
Плазмидные вектора 1: 434, 436, 437,
439-443
— pBR322 1:439
— Til: 469-471
Плазмиды 1: 21, 439; 3: 10
Плазмодесмы 2:424, 425
Планктомицеты 2: 508, 509
Пластиды 2: 405,406
- эволюция 1: 59
Пластохинон 1: 512, 513
- PQA 2: 374,376
Пластоцианин 2: 373, 376, 373
Плектонемическая спираль 3: 24-26
Поворотная симметрия 1: 206
|3-Повороты 1: 182, 183
Повторы ДНК 3: 56, 58,94, 148, 238
Поглощение света (А) 1: 121
— NAD+nNADH2: 50
— опсинами 1: 655
— спектр цитохрома с 2: 310
Подагра 2: 557, 563
Поджелудочная железа 2: 568
— в регуляции метаболизма глюко-
зы 2: 595, 596
Полани Майкл 1: 280
Поли(А)-«хвост» 3:130, 156
Полиаденилатполимераза 3: 130
Полиакриламид, для электрофореза
1: 136
Поликлональные антитела 1: 254
Полилинкер 1: 437, 438
Полимеразная цепная реакция
(ПЦР) 1:452, 453
---в методе отпечатков 1: 455-457
Полимеразы, см. ДНК-полимеразы;
РНК-полимеразы
Полиморфизм 1: 142
- длины рестрикционных фрагмен-
тов 1: 455
- одного нуклеотида 1: 460
Полиморфные белки 1: 142
Полиненасыщенные жирные кисло-
ты 1: 489
Полинуклеотидкиназа 1: 435
Полинуклеотидфосфатаза 3: 142
Полинуклеотиды 1: 397
- синтетические 3: 142
Полиовирус, симметрия 1: 207
Полипептидная цепь, элонгация
3: 179, 180
Полипептиды 1: 127, 128, см. также
Пептиды
Полисахариды 1: 340, 352-362,
см. также Углеводы
- во внеклеточном матриксе 1: 359
- конформация 1: 357-358
- функции 1: 362
Полисомы 3: 204
Полифермснтный комплекс в окис-
лительном фосфорилировании
------комплекс I 2: 313-315
------комплекс II 2: 316-318
------комплекс III 2: 318
------комплекс IV 2: 318-320
— туннелирование субстрата 2: 187
Полифосфат неорганический 2: 42
Полифосфаткиназы 1 и 2 2: 42
Полицистронная мРНК 1: 408
Полностью тиранс-ретиналь 1: 511
Половые гормоны 1: 509; 2: 574
— синтез 2: 348, 495, 574
Полоса
- А 1: 259
- 11: 259
Полуацеталь 1: 434
Полукеталь 1: 434
Полуконсервативная репликация
3: 45
Полуреакция 2: 44, 45
- стандартный восстановительный
потенциал 2: 47
Поляризация мембран, в передаче
сигнала 1: 633-640
Полярность
- аминокислот 1:118
— определение 1: 534
- в эмбриональном развитии 3: 271,
273
- сегментов, гены 3: 274
Полярные липиды, разделение 1:515
Помпе болезнь 2: 159
Попьяк Джордж 2: 484
Порины 1: 535, 536; 2: 306
Портер Родни 1: 251
Порфирин 2: 532
Порфирия 2: 534
Порфириновое кольцо 1: 227
Порфобилиноген 2: 532
Порядок зацепления (Lk) 3: 18
Последовательность ядерной лока-
лизации (NLS) 3:216, 217
Посттрансляционная модификация
белков 3: 179, 180, 206
Потенциал мембранный (Vm) 1: 551
Потенциалзависимые ионные кана-
лы 1:574-581
--в сигнальных путях 1: 635-639
Поток метаболитов (/) 2: 123, 128,
133-137
— коэффициент управления (С) 2:135
Поток электронов
— в антеннах хлорофиллов 2: 374,
375
— в бактериях 2: 367, 368
— в хлоропластах 2: 372
— индуцированный светом 2: 367
— протонный градиент 2: 382, 383
— расщепление воды 2: 379, 380
— роль комплекса цитохрома Ь6/
2: 376, 377
Почки
- аквапорины 1: 556, 571-573
- ионный баланс 2: 592
- метаболизм глутамина 2: 270, 271
- расщепление разветвленных ами-
нокислот 2: 296
- эндокринная функция 2: 576
Правастатин (правахол) 2: 494
Правахол (правастатин) 2: 494
Прадера-Вилли синдром 2: 614
Праймаза 3: 55, 58, 59
Праймер
- в репликации ДНК 3: 55
- в репликации РНК 3: 146
Праймирование 2: 168
Праймосома 3: 89
Пребиотическая химия 1: 55, 56
Преднизолон 1: 508, 509
Преднизон 1: 508, 509
Предстационарная кинетика 1: 287,
294
Пренилирование 2: 496
Пренилтрансфераза 2: 23
Препроинсулин 2: 572
Пререпликативный комплекс 3: 64
Прерибосомная рРНК (пре-рРНК)
3: 117, 134,135
Предметно-именной указатель 14 211
Прионные болезни 1: 217, 218
Прионы 1: 217, 218
Пробела гены 3: 273, 276
Пробелки 1: 328
Прогестерон, синтез 2: 495
Прогормоны 2: 572
Программируемая клеточная смерть,
см. Апоптоз
Проинсулин 2: 572, 573
Прокарбоксипептидаза
- А 2: 264
- В 2: 264
Прокариоты 1: 18, см. также Бак-
терии
Пролил-4-гидроксилаза 1: 190, 191
Пролин 1: 118; 2: 292
- биосинтез 2: 519, 520
- в активационных доменах 3:
- в коллагене 1: 187-191
- в p-поворотах 1: 182
- в р-слоях 1: 183
- в а-спиралях 1: 180
- превращение в а-кетоглутарат
2: 293
Промежуточное соединение 1: 305
Промежуточные филаменты 1: 24
— белки 1: 185, 186
Промотор 3: 110, 231
- в экспрессирующих векторах
1:444,445
- опухоли 1: 611
- специфичность 3: 118, 232
Проницаемость мембран 1: 525, 527
Проопиомеланокортин (ПОМК)
1:573
Пропионат 2: 239
Пропионил-СоА 2: 239, 240
Пропионил-СоА-карбоксилаза 2: 239
Пропластиды 2: 405
Пропранолол 1: 597
Прорастающие семена, глюконеоге-
нез 2: 105,436
Простагландин-Н2-синтаза (цикло-
оксигеназа) 2: 461
Простагландины 1: 508
- Е1 1:508, 647
- G2 2: 461
- Н2 2:463
- синтез 2: 461,462, 574
Простая транспозиция 3: 95
Простетическая группа 1: 131, 271
— гем 1: 227
— посттрансляционное присоедине-
ние 1: 227; 3: 207
Протеазы
- аспартильные 1: 318
- в секвенировании белков Г. 146
- ингибиторы 1: 307, 314
- металлопротеиназы 1: 318
- сериновые 1: 303,318
- цистеиновые 1: 318
Протеасома 1: 18, 663; 3: 220, 222
Протеинкиназы 1: 324; 2: 127
- А (РКА) 1:599
— активация 1: 599, 604
— анализ методом FRET 1: 612-614
— в p-адренергических путях 1: 605,
606
— и АКАР 1: 608, 609
— инактивация 1: 605-608
— регуляция ферментов 1: 326, 565,
566, 605
- АМР-зависимая 2: 130,131
- В (РКВ) 1: 622, 623
- С (РКС) 1:507, 610, 611
— и форболовые эфиры 1:611
- G (PKG) 1: 627, 628
- Са2+/кальмодулин-зависимые
1:614,616
- сАМР-зависимые, см. Протеинки-
наза А
- cGMP-зависимые, см. Протеинки-
наза G
- аутоингибирование 1: 599, 630
- в регуляции клеточного цикла
1: 660-665
- в сигнальных путях 1: 595, 646
- в транскрипции 3: 120
- гистидиновые 1: 646-648
- как мишени в противоопухолевой
терапии 1: 667-670; 2: 82
- консенсусные последовательности
1: 325, 327, 603
- рецепторные тирозинкиназы
1:619-624
— система JAK-STAT 1: 624, 625
- рецептороподобные, в растениях
1:648, 649
- сопряженные с G-белком 1: 606,
607
- субстратная специфичность
1: 325-327
- фосфорилирующая активность
1: 325-328
- циклинзависимые 1: 660-665
Протеинфосфатазы 1: 606; 2: 127
Протеогликаны 1: 363-366
- агрегаты 1: 366
Протеолитический процессинг
3: 207, 221
- в активации белков 1: 328
Протеолитические ферменты, регу-
ляция 1: 328, 663
Протеом 1: 461; 2:126
Протеомика 1: 434
Противовирусные препараты 1: 374
Протомеры 1: 130, 205
Протондвижущая сила 2: 323, 327,
339
— во вращении жгутиков 2: 352
Протонная помпа, см. АТРаза
Протоны
- градиент 2: 322-326, 382, 383
- перенос при кислотно-основном
катализе 1: 283, 284
- прыжки 1: 84, 90, 572
Протоонкогены 1: 666
Протопорфирин 1: 227; 2: 310, 534
Протопорфириноген 2: 24
Протромбин 1: 512
Проферменты 1: 328
Прохиральные молекулы 2: 200
Процессивность ДНК-полимеразы
3: 50
Процессинг РНК 3: 122-143
Прузинер Стенли 1:218
Прыгающие гены 3: 81
Прямая транспозиция 3: 95
Псевдоуридин 1: 395; 3: 128,134, 135
Пуриновое кольцо, происхождение
атомов 2: 544
Пуриновые нуклеотиды, см. Пурины
Пуриновые основания 1: 392, 393
— анти-форма 1: 403, 404
— водородные связи 1: 399
— дезаминирование 1: 416-418
— минорные 1: 395
— потеря 1: 416-418
— реутилизация 2: 556
— син-форма 1: 403,404
— таутомерные формы 1: 398, 419
— Чаргаффа правила 1: 400
Пурины 1:392, 397, 398, 416
- биосинтез 2: 543
- расщепление 2: 555
- регуляция синтеза 2: 547
- реутилизация 2: 556
- синтез de novo 2: 544, 545
Пуромицин 3: 208
Пурпурные бактерии 1: 60
- бактериородопсин 1: 533, 544
ПЦР (полимеразная цепная реак-
ция) 1: 10, 452
Пэтч-кламп метод 1: 574, 575
Р
Равновесие 1: 46, 47, 274; 2: 14
- константа (Keq) 1: 47, 90, 94, 276;
2:14-16
— в метаболизме углеводов 2:128,
129
[ 4 221 Предметно-именной указатель
— ионизации воды 1: 90,91
— и изменение свободной энергии
1:47; 2:14-17
— расчет 2:18,19
— двух связанных реакций 2: 19
— гидролиза АТР 2: 18,19, 27
Радикал 2: 21
- гидроксильный 2: 324
- свободный 2: 24, 25, 324
- супероксидный 2: 316, 324
- тирозиновый 2: 551
Радиоиммуноанализ 2: 569
Разветвляющий фермент хлоропла-
стов 2: 428
--в метаболизме гликогена 2: 156
Развитие, регуляция генов 3:
271-280
Разобщающий белок (термогенин)
2: 348,586, 609
РайзингЗ: 20
Рак
- ABC-транспортеры 1: 564,565
- генная терапия 1: 475-477
- и гены супрессоров 1: 667, 670, 671
- и система репарации ДНК 3: 79, 80
- интегрины 1: 641, 642
- как осложнение генной терапии
1: 475-477
- как результат повреждений ДНК
1:419
- кожи, при ксеродерме 3: 79
- лечение
— генная терапия 1: 475-477
— ингбиторы топоизомераз 3: 24, 25
— ингибиторы протеинкиназ
1: 667-670; 2:82
— ингибиторы ферментов биосин-
теза нуклеотидов 2: 558,559
— стероидные препараты 1: 644
— химиотерапия 1: 82; 2: 523,558,
559; 3: 24, 25, 204
- метаболизм глюкозы 2: 81-83
- микрочипы в анализе 1: 466
- молочной железы 1: 644; 3: 80
- мутации 1: 601, 666-671; 2: 211;
3: 145, 207
- онкогены 1: 666-670
- ПЭТ-сканирование 2: 83
- ретровирусы 3: 146
- селектины 1: 374
- толстой кишки 1: 667; 3: 80
— мутации 1: 670, 671
Рамачандрана карта 1: 176
Рамка считывания 3: 167
- открытая 3: 171
- сдвиг 3: 175
Рамноза 1: 345
Раскручивание ДНК 3: 21-24
Расплавленная глобула 1:211
Растения
-С32: 422-425
- С4 2: 422-425
- САМ 2: 426
- аквапорины 1: 572, 573
- амилопласты 2: 405
- брассинолид 1: 509
- гликолатный путь 2: 421, 422
- глиоксисомы 2: 213, 246-248
- гормоны роста 1: 470
- десатуразы 2: 459,461
-ДНК 3:173, 270
- иммунные реакции 1: 649
- клонирование 1:469-473
- летучие сигнальные вещества
1:509,510
- мембранные липиды 1: 500
- метаболизм липидов 2: 454
- метаболизм углеводов 2: 426-433;
см. также Цикл Кальвина
— интеграция 2: 435-438
- митохондриальное дыхание 2: 420
- митохондрии, альтернативный
путь окисления NADH 2: 326
- органеллы 2: 22, 23, 213, 246, 358,
405
- осмотическое давление 1: 87
- пентозофосфатный путь 2: 406
- пероксисомы 2: 246-248
- пластиды 2: 405,406
- пропластиды 2: 405
- пулы метаболитов 2: 437
- рецептор этилена 1: 647, 648
- сигнальные пути 1: 646-649
- синтез NADPH 2: 453-455
- синтез клеточной стенки
2: 431-433
- сосудистые 1: 646
- структура клетки 1: 22, 27
- фиксация азота 2: 512, 513
- фотодыхание 2: 420, 422
- фотосинтез, см. Фотосинтез
- хлоропласты, см. Хлоропласты
Раус Ф. Пейтон 3: 146
Рауса саркома, вирус 3: 146
Рахит 1:510,511
Рацемическая смесь 1: 36, 37
Регулон 3: 245
Регуляторные белки 3: 233, 237, 241
Регуляторные последовательности
3: 8, 234
Регуляторные ферменты 1: 319-322
Регуляция экспрессии генов
3: 229-280
--SOS-ответ 3: 76, 249
----ТАТА-связывающий белок
3: 118
----активаторы 3: 232
----транскрипции 3: 266
----активирующая последователь-
ность UAS 3: 261
----антигенные вариации 3: 256
----аттенюация транскрипции
3: 245-249
----богатый глутамином домен
3:265
----богатый пролином активацион-
ный домен 3: 266
----в бактериях 3: 243-256
----в развитии 3: 271-280
----в эукариотах 3: 257-280
----вторичные мессенджеры 3: 252,
268
----гормональная 1: 619, 621;
3: 266-268
----ДНК-связывающие домены
3: 237
----индукторы 3: 235
----индукция 3: 230, 236
----катаболитная репрессия 3: 244
----кислый активирующий домен
3: 265
----коактиваторы 3: 262
----медиатор 3: 262
----нарушение в опухолевых клет-
ках 2: 81-84
----операторы 3: 232
----опероны, см. Опероны
----отрицательная 3: 232
----переключение типа спаривания
у дрожжей 3: 256
----положительная 3: 232
----принципы 3: 229-243
----регулоны 3: 245
----рекомбинационная 3: 256
----ремоделирование хроматина
3:257-260
----репрессия 3: 231
----репрессоры 3: 232
----рибопереключатели 3: 253
----РНК-интерференция 3: 270
----сайленсинг 3: 270
----сайт-специфическая рекомбина-
ция 3: 89, 256
----строгий ответ 3: 251
----фазовая вариация 3: 256
----факторы специфичности 3: 232
----функция инсулина 1: 619, 621;
2:149,150
----энхансеры 3: 233
----эффекторы 3: 232
Редактирование РНКЗ: 175,176
Предметно-именной указатель [ 423]
Рекомбиназа 3: 91-95
Рекомбинантная ДНК 1: 434
— в клонировании животных
клеток 1: 473-477
— в медицине 1: 479, 480
— в секвенировании геномов
1:449-461
— в сельском хозяйстве 1: 469-473
— для создания библиотек ДНК
1:449-451
— методы 1: 434-461
Рекомбинационная репарация ДНК
3: 78,81,89, 90
Рекомбинация ДНК 3: 43, 81
— в репарации 3: 77, 78
— генов иммуноглобулинов
3: 95-98
— гомологичная 3: 81
— и мейоз 3: 82, 84
— кроссинговер 3: 83
— миграция точки ветвления 3: 85
— реакция обмена цепей ДНК 3: 88
— сайт-специфическая 3: 81, 89-94
— сигнальные последовательности
3: 97
— транспозиция 3:81, 94, 95
— ферменты 3: 85-89
Релаксированная ДНКЗ: 16-21
Реленза (занамивир) 1: 374
Ренатурация белков 1: 209
Рентгеновская дифракция, метод
анализа
-----белков 1: 532, 533,573
-----ДНК 1:400-402
-----полисахаридов 1: 357, 358
Репарация ДНК 1: 446, 670, 671;
3: 66-80
-SOS-ответ 3: 76,78, 249
— TLS-полимеразы 3: 76, 79
— в димерных хромосомах 3: 93
— ДНК-полимеразы 3: 52, 79, 89
— и рак 3: 79, 80
— корректирующая активность
3:51-53
— ник-трансляция 3: 53
— ошибочно спаренных оснований
3: 68, 99
— прямая 3: 68, 74
— рекомбинационная 3: 78, 81, 89,
90
— с эксцизией нуклеотидов 3: 68,
73, 74
— с эксцизией оснований 3: 68,
71-73
— функция циклинзависимых про-
теинкиназ 1: 661-664
— через повреждение 3: 76
Репликативная вилка 3: 47, 48
— восстановление 3: 89
— остановка 3: 63, 77, 80, 89
Репликативная транспозиция 3: 95
Репликативные формы 3: 9
Репликатор 3: 64
Репликация ДНК: 1: 52; 3: 43-66
— Тег-последовательность 3: 63
— Tus-Ter комплекс 3: 63
— в бактериях 3: 52-63
— в эукариотах 3: 64, 65
— ДНК-полимеразы 3: 52, 61, 64, 65
— ДНК-связывающие белки 3:58
— инициация 3: 56-58
— корректирующая активность 3: 50
— лидирующая цепь 3: 48, 58-60
— матрица 3: 50
— направленность 3: 48
— ник-трансляция 3: 53
— нуклеофильная атака 3: 49
— отстающая цепь 3: 48, 58-62
— ошибки 3: 50-52
— полуконсервативная 3: 45
— праймер 3: 49, 50, 55
— репликатор 3: 64
— реплисомы 3: 54
— скорость
---в бактериях 3: 50, 52
---в эукариотах 3: 64
— терминация 3: 62, 63
— топоизомеразы 3: 63
— точка начала (ориджин)
---в бактериях 3: 56
---в эукариотах 3: 64
— точность 3: 50
— удлинение цепи 3: 58
— ферменты 3: 61, 64
— фрагменты Оказаки 3:48, 59-61,
65
— хеликазы 3: 54, 57-60, 64
— через повреждение 3: 76
— эволюционное значение 1: 54, 55
Репликация РНКЗ: 144
— интроны 3: 122, 124,125
— обратная транскриптаза
3: 144-146
— ретротранспозоны 3: 148
— РНК-репликазаЗ: 151
— саморепликация 1: 56; 3:152
— теломеразы 3: 149, 152
— хоминг 3: 148
— эволюционное значение 1:56-58;
3: 152
Реплисома 3: 54
Репрессия 3: 231
- катаболитная 3: 244
Репрессоры 3: 232, 233
-1асЗ: 235, 236, 244, 245
-trp3: 246-249
- в эукариотах 3: 263, 269, 274
- и SOS-ответ 3: 249, 250
- трансляции 3: 250, 251
Респирасомы 2:320
Респонсивные элементы 2: 124
Рестрикции-модификации система
1:435
Ретикулоциты, регуляция трансля-
ции 3: 269
Ретиналь полностью транс 1: 34,
511,652
- 11-цис 1:34, 511, 652
Ретинобластома 1: 670
- pRb 1: 665, 670
Ретиноевая кислота 1:511; 3: 267
Ретиноидные гормоны 3: 266, 267
Ретинол (витамин А) 1: 511
Ретровирусы 1: 317; 3: 144
- векторы 1: 473-476
- лекарства 1: 317
Ретротранспозоны 3: 148
Ретрохоминг 3: 148
Реутилизации путь 2: 543, 556
Рефсума болезнь 2: 250
Рецепторные каналы Г. 579, 639
Рецепторные киназы 1: 606, 624, 667,
668
— JAK-STAT 1:624
Рецепторные ферменты
— гистидинкиназа 1: 645
— гуанилилциклаза 1: 627, 528
— тирозинкиназа 1: 619, 624, 632
Рецепторный потенциал 1: 656
Рецепторы
- ANF 1: 627, 628
- Fas 1: 673
- LRP 2:
- NPY 2: 607
- PYY 2: 607
- Т-клеточный 1: 250
- агонисты 1: 597
- p-адренергические 1: 597, 598
- активируемые пролифераторами
пероксисом (PPAR) 2: 612, 613
- антагонисты Г. 597
- в эндоцитозе 2: 490; 3: 219
- глицина 1: 579, 639, 640
- гормонов 2: 570, 571; 3: 266, 267
- грелина2: 614
- гуанидина 1: 627
- десенсибилизация 1: 606
- инсулина 1: 170, 619, 624, 626
- как усилители сигнала 2: 570
- каннабиноидов 1:622
[424] Предметно-именной указатель
- лептина 2: 605, 606, 608, 610
-ЛПНП 2:490
- меланокортина 2: 607
- никотиновые ацетилхолиновые
1: 578, 579
- обонятельные 1: 656-659
- родопсин 1: 654, 656
- семиспиральные 1: 597
- серпентиновые 1: 597
- сладкого вкуса 1: 681
- сопряженные с G-белками
1:595-610, 650, 658
- сродство к лигандам Г. 592
- стероидов 1: 509, 643, 644; 2: 570
- фактора роста тромбоцитов 1: 624
- эволюционное значение 1: 658
- эпидермального фактора роста
1: 595, 624
-----мутации 1: 667
- эритропоэтина 1: 624, 625
- этилена 1: 647, 648
- ядерные 1: 643, 644
Решеф Леа 2:468
Рибоза 1:340, 341,392
- конформация 1: 393
Рибозимы Г. 411; 3: 122, 139-142,
154, 182, 200
Рибозо-5-фосфат 2: 108, 109,438
Рибозо-5-фосфатизомераза 2:412
Рибозофосфат-пирофосфокиназа
2:519
Рибонуклеаза 1: 129, 200
- ренатурация 1: 209
Рибонуклеиновая кислота, см. РНК
Рибонуклеозидмонофосфаты цикли-
ческие 1: 395
Рибонуклеотидредуктаза 2: 550-553
Рибонуклеотиды 1: 392-394, 424
- восстановление 2: 550
Рибопереключатели 3: 253, 254
Рибосомная РНК, см. рРНК
Рибосомные белки 3: 180, 250-252
Рибосомы 3: 180
- бактериальные 3: 180-183
- белки 3: 250-252
- как место синтеза белка 3: 180
- коррекция ошибок 3: 202
- образование 3: 180, 250
- открытие 3: 169
- полисомы 3: 204, 205
- рецикл 3: 203, 204
- сайты связывания аминоацил-
тРНК (А, Р и Е) 3: 197
- структура 3:180
- субъединицы 3: 180, 181
- эукариотические 3:183, 184
Риботимидин Г. 393
Рибофураноза 1: 393
Рибулоза 1:343
Рибулозо-1,5-бисфосфат 2: 406-416
Рибулозо-1,5-бисфосфаткарбокси-
лаза/оксигеназа, см. Рубиско
Рибулозо-5-фосфат 2: 109,415,438
Рибулозо-5-фосфаткиназа 2:415,418
Ринальдо Пьеро 2: 295, 296
Риске Джон 2:310
Риске железо-серный белок 2: 310,
317
Рифампицин 3: 122
Рицин 3: 209
РНК 1: 33, 53; см. также Нуклеино-
вые кислоты
-TUF3: 157
- аптамеры 3: 154, 155, 254
- вирусная, см. Ретровирусы; РНК-
вирусы
- вторичная структура 1: 406,410
- гидролиз 1: 397
- интерференция 3: 270
- каталитические 3: 122, 139-142,
154,182,200
-5'-кэпЗ: 122-124
- малая 3: 270
— временная 3: 270
— интерферирующая 3: 270
— ядерная 3: 127, 128, 138, 157
— ядрышковая 3: 135, 136, 157
- матричная, см. мРНК
- микро 3: 138, 270
- направляющая 3: 176
- некодирующая 3: 271
- нуклеотидный состав 1: 392-395
- оборот 3: 123
- паразитарная 3: 155, 156
- прерибосомная, процессинг 3: 134,
135
- процессинг 3: 122-143
— в бактериях 3: 134
— в эукариотах 3: 122-134
— миРНК 3: 270
— мРНК, см. мРНК
— мякРНК 3: 135, 136, 157
— мяРНК 3: 127, 128, 138, 157
— поли(А)-«хвост» 3: 130, 142, 156
— полиаденилатполимераза 3: 130
— полинуклеотидфосфорилаза
3: 142
— редактирование 3: 139,175-177
— рРНКЗ: 134, 135,183
— сплайсинг 3: 122,125
- расщепление 3: 123, 142
- редактирование 3: 175, 176
- репликация, см. Репликация РНК
- рибозимы, см. Рибозимы
- рибосомная, см. рРНК
- самореплицирующаяся 1: 56; 3: 152
- синтез 3: 106; см. также Транс-
крипция
- создание, метод SELEX 3: 154, 155
- спаривание оснований 1: 399
- сплайсинг 3:122-133
- транскрипты
— первичные 3: 122, 123
— с неизвестной функцией
3: 156-158
— сложные 3: 132
- транспортная, см. тРНК
- трехмерная структура 1: 408-412
- фосфодиэфирные связи 1: 396
- г^с/тиряис-механизм регуляции
3: 252-254
- шпильки 3: 116, 142, 190, 247, 252,
270
- эволюционная роль 1: 56, 57
РНКаза Р 1:410; 3: 134, 136, 138,
141-143
РНК-вирусы 3: 144, 270
РНК-ДНК гибриды, денатурация
1:414
РНК-зависимая ДНК-ролимераза
3: 144, 145, 151
РНК-зонд 1:443
РНК-полимеразы 3: 117
-13:117
-113:117, 261
— связывание с промотором 3: 117,
119
- III 3: 117
- в транскрипции 3: 106-108
— бактерий 3: 108, 109
— эукариот 3: 117-122
- ДНК-зависимая 3: 108
- ингибиторы 3: 121, 122
- РНК-зависимая 3: 151
- связывание с промотором 3: 231
- факторы специфичности 3: 232
РНК-репликаза 3: 151
Робертс Ричард 3: 125
Родбел Мартин 1: 598, 600
Родопсин 1: 651, 652, 655
Родопсинкиназа 1: 654
Розетки
- гликогена 2: 155
- целлюлозы 2: 432,433
Розиглитазон (авандия) 2: 470
Россмана укладка 2: 52, 163
рРНК (рибосомная РНК) 3: 134, 135,
183
- интроны 3: 139, 140
- процессинг 3: 133-136
- синтез 3:135
Предметно-именной указатель [ 4 25]
Рубиско 2:407-410,417-420
Рубиско-активаза 2:410
Рэкер Эфраим 2: 331
С
Сазерленд Эрл 2: 160, 165, 166
Сайт-направленный мутагенез 1: 446
Сайт-специфическая рекомбинация
3:81,89
Сакситоксин 1: 579
Салициловая кислота 2: 461
Самнер Джеймс 1: 270
Самуэльсон Бенгт 1: 508
Сандхофа болезнь 1: 505
Саркомер 1: 259, 260
Саркоплазматический ретикулум
1:259, 261,560, 561
Сателлитная ДНК 3: 13
Саузерн Джереми 1: 454
Саузерн-блоттинг 1: 454, 455
Сахара 1: 69, 339, см. также Моно-
сахариды, Полисахариды,
Углеводы
- аминосахара 1: 346
- восстанавливающие 1: 347, 354
- и долихолы 1: 513
- кислые 1: 346
- невосстанавливающие 1: 347, 354
- нуклеотидные 2: 158-161
- эпимеры 1: 343
Сахарный диабет, см. Диабет сахар-
ный
Сахарный код 1: 371, 372
Сахароза 1: 23, 68, 69, 87, 275, 371;
2: 94
- в прорастающих семенах 2: 436,
437
- синтез 2: 426, 428, 429
Сахарозо-6-фосфат 2: 428, 429
Сахарозо-6-фосфатсинтаза 2: 428,
429, 431
Сахарозо-6-фосфатфосфатаза 2: 428,
429
Сахарозосинтаза 2: 433
Сахаронуклеотиды 2: 158, 428
Свертывание крови
— протеолитическая активация
факторов 1: 329
— рекомбинантные продукты 1: 479
— роль интегринов 1: 641, 642
Сверхспирализация ДНК, плотность
3: 16, 19
Свет; см. также Зрение; Флуорес-
ценция; Фотосинтез
- как стимул для сигнальной систе-
мы растений 1: 646-650
- плоскополяризованный, вращение
1:36, 37, 117
- поглощение
— аминокислотами 1: 120
-ДНК 1:397,413
— закон Ламберта-Бера 1: 121
— липидными пигментами 1: 514
- флуоресцентными белками
1:612-614
Светляки, цикл биолюминесценции
2: 39
Световые реакции 2: 358
Светопоглощающие комплексы
2 : 361
Светособирающие (антенные) пиг-
менты 2: 362, 365
Светочувствительные нейроны
1:650
Светящиеся растения 1: 471, 472
Свободная энергия (С) (энергия
Гиббса) 1:44; 2: 13
— в стабилизации структуры белка
1: 173, 174
— в ферментативном катализе
1: 274, 276, 280
— в фолдинге белка 1: 211, 212
— гидролиза 2: 16
— изменение (АС) 1: 44, 47, 48;
2: 14-16
Свободноживущие азотфиксирую-
щие бактерии 2: 510
Свободные жирные кислоты 1: 488,
489; 2: 229; см. также Жирные
кислоты
Свободные радикалы 2: 20
— реакции 2: 20-25
Связь; см. также Слабые взаимодей-
ствия
- водородная, см. Водородная связь
- гликозидная 1: 343
— N-гликозидная 1: 348
— О-гликозидная 1: 348
— расщепление лизоцимом 1: 311,
312
- дисульфидная 1: 120
— расщепление 1: 145
- ковалентная 1: 27
— С-Н2:21
— О-Н1:75
— О-Р2: 25, 26
— расщепление 2: 20, 21
- нековалентная 1: 27; см. также
Слабые взаимодействия
- пептидная, см. Пептидная связь
- расщепление при секвенировании
пептидов 1:145
- углерода 1: 29, 31, 75; 2: 21-23
- фосфодиэфирная 1: 396
- энергия диссоциации 1: 75
Связывание
- антител 1: 254
- кооперативность 1: 237, 242
- субстрата в активном центре фер-
мента 1: 273
- центры 1: 239 320, 367
- энергия 1: 48, 280, 281
Сдвиг рамки считывания 3: 175
Сегментации гены 3: 273
Седогептулозо-1,7-бисфосфат 2:415
Седогептулозо-7-фосфат 2: 414, 415
Секвенатор 1: 145
Секвенирование
- белков и пептидов 1: 143-148
-----методом масс-спектрометрии
1: 151
-ДНК 1:420-422, 430
— автоматическое 1: 421, 422
— и определение белковой последо-
вательности 1: 148, 152
— по Сенгеру 1: 144,420-422
Секретин 2: 264
Селектины 1: 373, 374, 549
Селеноцистеин 1: 123; 3: 191
Семенники 2: 575
Семиспиральные рецепторы 1: 597
Сенгер Фредерик 1: 143, 145, 168,
169,419
Сердечная мышца 2: 589, 590
— изоферменты ЛДГ2: 140,141
Сердечно-сосудистые заболевания
— амилоидозы 1: 215
— антикоагулянты 1: 479
— атеросклероз 2: 493-495
— гликогенозы 2: 159
— как осложнение диабета 1: 348
— нитросоединения 1: 628
— приступы 2: 141, 344, 463
— роль тиряис-жирных кислот
1: 493, 494
— статины 2: 494
Серин 1: 120
- биосинтез 2: 326, 519, 521
- в активности транспептидазы
1:314
- в активности химотрипсина 1:303
- в гликолатном пути 2: 421,422
- в протеогликанах 1: 364
- в синтезе липидов 2: 472-478
- расщепление до пирувата 2: 285
- фосфорилирование 1: 325
- циклосерин 2: 538
Серингидроксиметилтрансфераза
2: 285, 288,421, 423, 519, 554
Сериндегидратаза 2: 285
[ 4261 Предметно-именной указатель
Сериновые протеазы 1: 303
Сероводород 1: 79
Серотонин 1: 500, 647; 2: 290, 539
- рецепторы 1: 579, 608
Серповидно-клеточная анемия
1: 246-248
Сестринские хроматиды 3: 82, 83
Сетчатка 1: 650-652, 655
- колбочки 1: 651, 652, 655
- палочки 1: 651, 652
Сиалоадгезин 1: 376
Сиаловая кислота 1: 347
— в ганглиозидах 1: 502, 503, 531
Сигнал-распознающая частица
(SRP) 3:211,212
Сигнальная последовательность
3: 131,206,210-212
— бактериальных белков 3: 217, 218
— инсулина 2: 572, 573
— ядерных белков 3: 216
Сигнальные белки 1: 629-632
Сигнальные системы 1: 591-672
— в микроорганизмах 1: 644, 645
— в нейроэндокринной системе
2: 566
— в протеолизе 3: 221
-- в растениях 1: 509; 646-650;
2: 463
— в регуляции генов эукариот 3: 266
— в регуляции клеточного цикла
1: 660-665
— гормональные 2: 566, 570, 571
— гормональные каскады 2: 570
— зрение, обоняние и вкус
1: 650-659
— ионные каналы 1: 633
— кавеолы 1: 632
— лиганд-рецепторные взаимодей-
ствия 1: 592-594
— механизмы 1: 595, 596
— рафты 1: 632
— роль липидов 1: 506-514
— роль углеводов 1: 33, 363,375
— роль фосфатидилинозита 1: 622;
2: 507
— роль эйкозаноидов 2: 508
— стероидные гормоны 1: 643, 644
— ферментативные каскады 1: 592
— функция инсулина 2: 149, 151
— функция лептина 2: 606, 609, 610
Силденафил (виагра) 1: 628
Симбиоз
- в фиксации азота 2: 513
- в эволюции митохондрий 2: 352
- в эволюции хлоропластов 2: 386
Симбионты 2: 6, 510
Симвастатин (зокор) 2: 494
Симметричная модель, см. Модель
Моно-Уаймена-Шанже
Симметрия в белках
--вращательная (поворотная)
1: 206, 207
--икосаэдрическая 1: 207
--спиральная (винтовая) 1: 207
Симпорт 1: 558
- №+/глюкоза 1: 569, 570
Синапс
- передача сигнала 1: 635, 636
- слияние мембран 1: 548
Синдекан 1: 364, 365
Синдром X (метаболический син-
дром) 2: 615
Синий флуоресцентный белок
(BFP) 1:613-615
Синтаза 2: 197
- NO 2: 576
- жирных кислот 2: 150, 447-453
Синтения 1: 462
Синтетаза 2: 197
а-Синуклеин 1: 217
Системная биология 1: 462
Ситостерол 2: 433
Сквален 2: 480-484
Сквален-2,3-эпоксид 2: 483, 484
Скваленмонооксигеназа 2: 483, 484
Скользящий зажим 3: 60, 61
Скорость реакции
- константа (k) 1: 229, 277
- лимитирующая стадия 1: 287, 290,
291
- уравнение 1: 277, 292
Скоу Йенс 1: 562
Скрамблаза 1: 540, 541
Скрепи (почесуха) 1: 217
Скэтчарда анализ 1: 592-594
Слабые взаимодействия 1: 73-84
— в стабилизации белков 1: 172,
173,210
— в структуре полисахаридов 1: 356
— между ферментом и субстратом
1: 277, 279, 280, 282
Слабые кислоты 1: 93-96
— в составе буферов 1: 98
Слабые основания 1: 93-96
Слай Уильям 2: 295
Следовые элементы 1: 29
Сложноэфирная конденсация Кляй-
зена 2: 22, 23
р-Слои 1: 181
- в крупных глобулярных белках
1:201
- скрученные 1: 201
Слэк Роджер 2: 424
Смешанное ингибирование 1:295-298
Смит Хамилтон 1: 436
Соленоидная спираль 3: 25, 26
Солнечная энергия 2: 357-363
Сольватная оболочка 1: 173
Соматостатин 2: 572, 595
Сопряженные с G-белками рецептор-
ные киназы (GRK) 1: 606, 607
Сорафениб (нексавар) 1: 669
Сорбоза 1: 343
Сосудистые растения 1: 32, 646
Спаривание оснований
— аномалии 3: 51, 68
— ДНК 1:9,402
-ДНК-РНК 1:409
— кодонов и антикодонов 3: 174
— при ДНК-белковых взаимодей-
ствиях 3: 237, 238
— при репликации 3: 51
— РНК-РНК 3: 139
— тРНКЗ: 185
Спектрин 1: 542, 543
Спектрофотометрия 1: 121
Спермацетовое масло 1: 493
Спермидин 2: 540, 542, 543
Спермин 2: 540, 542, 543
Специфический кислотно-основной
катализ 1: 284
Специфичности константа (kCM/KM)
1: 292
СПИД, см. Вирус иммунодефицита
человека
Спираль
-al: 177, 178, 200
- актина 1: 258
- в мембранных белках 1: 531—533,
561,564-568, 572
- в полисахаридах 1: 354, 358
- в трансмембранных каналах 1: 552,
553, 576
- двойная ДНК 1: 52, 399-402
- a-кератина 1: 185, 186
- миоглобина 1: 194, 195
- миозина 1: 257
-РНК 1:409, 410
- родопсина 1:652
- суперскрученная ДНК 1: 186
---в транскрипции 3: 107
---варианты 1: 404,405
---раскручивание 1: 413
- тройная коллагена 1: 187, 188, 190
Спираль-петля-спираль мотив
1: 616; 3: 242
Спираль-поворот-спираль мотив
3: 239, 240
Спирты
- в экстракции липидов 1: 515
- и полуацетали 1: 343,344
Предметно-именной указатель [427]
- и полукетали 1:343, 344
- ферментация 2: 69,93,94
Сплайсинг 3: 122-133
Сплайсосома 3: 127
Сплайсосомные интроны 3: 127
Спудич Джеймс 1: 267
Сравнительная геномика 1: 462, 467
Сталь Франклин 3:
Стандартное изменение свободной
энергии (АС0) 1: 274; 2: 15
-----аддитивность 2: 18, 19
-----гидролиза АТР 2: 29
-----гликолиза 2: 70, 98
-----зависимость от концентрации
2: 17
-----переноса электронов 2: 322
-----расчет 2: 16
-----связь с константой равновесия
1:276; 2: 14, 15
Стандартные приведенные параме-
тры 2: 15
Стандартные условия 2: 15
Стандартный восстановительный
потенциал (Е°) 2: 47, 48
Старение
- и митохондриальная ДНК 2: 346
- и теломеры 3: 151
Стартовый кодон 3: 195, 196
Статины 2: 493, 494
Стационарная кинетика 1: 287-294
Стационарное состояние 1: 287
— клеточного роста 2: 123
Стеарат 2: 457,458
Стеариновая кислота 1: 490
Стекинговые взаимодействия 1: 398,
409,413
— при репликации 3: 50
Стенокардия, прием вазодилатато-
ров 1: 628
Стереоизомеры 1: 33-40
- аминокислот 1: 115-118
- липидов 1: 497
- сахаров 1: 340-344
Стерины 1: 504
- в мембранах 1: 496
- как липидные якоря 1: 527, 532, 545
Стероидные гормоны 1: 509
— регуляция транскрипции 1: 643
— роль митохондрий 2: 348
— синтез 2:495, 574
Стероиды 1: 509
- синтез 2: 480-485
Стоп-кодоны 3: 170
Стрептомицин 3: 209
Стресс
- окислительный 2: 324
— и малярия 2: 107
- функция адреналина 2: 602
- функция кортизола 2: 602
Строма 2: 359
Стронг Фрэнк 2: 53
Субстрат 1: 226
- инсулинового рецептора 1: 624
— IRS-1 1:619, 622; 2: 170
— IRS-2 2: 610
- туннелирование 2: 187
Субстратный цикл 2: 140
Судебная медицина, анализ ДНК
1:454,455
Суицидный ингибитор 1: 298
Сукцинат 2: 188, 189
- в глиоксилатном цикле 2: 212
- окисление до фумарата 2: 198
Сукцинат дегидрогеназа 2: 198, 211
Сукцинаттиокиназа 2: 196, 197
Сукцинил-СоА
- в распаде аминокислот 2: 279
- превращение в сукцинат 2: 196
Сукцинил-СоА-синтетаза, см. Сук-
цинаттиокиназа
Сульфатирование, в протеогликанах
1:360,361,364,365
Сульфодезокси-а-В-глюкопирано-
зилдиацилглицерин 1: 499, 500
Сульфолипиды 1: 496,500
Сульфонилмочевина 2: 597, 617
Сунитиниб (сутент) 1: 669
Супероксиддисмутаза 2: 326
Супероксидный радикал 2: 106, 316
Суперспираль
- ДНК, см. ДНК, сверхспирализация
- а-кератина 1: 185, 186
- коллагена 1: 187,188
Супрессорная тРНК 3: 203
Сутент (сунитиниб) 1: 669
Сфинганин 2: 476,478
Сфингозин 1: 501, 502
Сфинголипиды 1: 496, 501, 503
- расщепление 1: 503, 505
- синтез 2: 478
Сфингомиелин 1: 502, 505, 506;
2:476
- распределение в мембране 1:
528-546
Сфингомиелиназа 1: 505
Сывороточный альбумин 2: 229, 230,
592
Сывороточный амилоид А 1: 215
т
Табачной мозаики вирус 1: 207
Тагатоза 1: 342
Талоза 1: 342
Тамифлю (осельтамивир) 1: 374
Тамоксифен 1: 644
Тандемная масс-спектрометрия
1: 150, 151
Таруи болезнь 2: 159
Тарцева (эрлотиниб) 1: 669
Таурохолевая кислота 1: 506
Тейтем Эдвард Л. 2: 193; 3: 8
Теломераза 3: 149-151
Теломеры 3: 14, 15, 149-151
Темин Говард 3: 144, 145
Температура абсолютная, размер-
ность 2: 13
Температура кипения
— воды 1: 74
— некоторых растворителей 1: 74,
76
Тепло, образование см. Термогенез
Теплового шока белки
--в фолдинге 1: 167, 212
--промоторы генов 3: 115, 232
Теплота испарения 1: 74
— воды 1: 74, 76
Терминальная трансфераза 1: 435
Терминальный комплекс, в целлюло-
зе 2: 432
Терминаторы транскрипции
3: 116, 247-249
Терминация транскрипции
- р-зависимая 3: 117
- р-независимая 3: 116, 142
Термиты 1: 355
Термогенез 2: 325, 586
- в митохондриях 2: 348
- сократительный 2: 589
Термогенин 2: 348, 586
Термодинамика
- второй закон 1: 551; 2:12
- и биоэнергетика 2: 12-19
- первый закон 1: 41; 2: 12
- предотвращение рассеивания
энергии 2: 371
Тест на толерантность к глюкозе
2:604
Тестостерон 1: 509; 2:495, 575
Тетрагидробиоптерин 2: 284, 292
- в реакции фенилаланин-
гидроксилазы 2: 291
Тетрагидроканнабинол 1: 622
Тетрагидрофолат 2: 281, 282, 517, 554
- связывание с ферментом 1: 278
Тетрадекановая кислота 1: 488
Тетракозановая кислота 1: 488
Тетрануклеотидная гипотеза, опро-
вержение 1: 432
Тетраплексная ДНК 1:406, 407
Тетрациклины 3: 208, 209
Тетродотоксин 1: 640
| \ 28 | Предметно-именной указатель
Тетрозы 1: 341
Тея-Сакса болезнь 1: 505
Тиазолидиндионы 2: 469, 470
Тиаминпирофосфат 2: 93, 112, 183
- в цикле Кальвина 2: 413
Тилакоиды 2: 358, 359
- поток протонов и электронов
2: 367-385
Тимидилат, биосинтез 2: 554, 558,
559
Тимидилатсинтаза 2: 554, 559
Тимин 1: 392
- эволюционное значение 1: 416, 417
Тиолаза 2: 235, 252, 253, 480
Тиоредоксин 2:419, 550, 551
Тиоредоксинредуктаза 2:419,550,551
4-Тиоуридин 1: 395
Тиоэфиры 2: 34, 183
- свободная энергия гидролиза 2: 35
Типирование ДНК 1: 454-456
Тиреоглобулин 2: 575
Тиреоидные гормоны 2: 571, 575
Тиреотропин-высвобождающий
гормон 2: 567
Тиреотропный гормон (ТТГ) 1: 130
Тирозин 1: 118; 2: 280
- биосинтез 2: 523-529
- в мембранных белках 1: 535,536
- в синтезе гормонов 2: 574, 575
- поглощение света 1: 120
- расщепление 2: 280
- фосфорилированный 1: 608
— в регуляции клеточного цикла
1:662
Тирозиназа 2: 287
Тирозинкиназы
- как мишени для противоопухоле-
вой терапии 1: 667-669; 2: 81, 82
- растворимые 1: 624, 625
- рецепторные 1: 595, 619-624
Тирозиновый радикал 2: 551-553
Тироксин (Т4) 2: 575
Титин 1: 130, 260
Титрование
- аминокислот 1: 123-126
- кривая 1: 94
- пептидов 1: 128, 129
- слабых кислот 1:95, 97
- уравнение Хендерсона-Хассель-
баха 1: 98
Токоферолы 1: 512
Токсины 1: 579, 581, 640
- ингибирование трансляции
3: 207-209
Толбутамид 2: 597
Толстые филаменты 1: 258
Тонкие филаменты 1:258
Тонкий кишечник, всасывание жи-
ров 2: 226,227
Тонкослойная хроматография 1: 169,
515-517
Топоизомеразы 3: 23, 54, 61, 63
- ингибиторы 3: 24, 35
Топоизомеры 3: 20, 21
Топология 3: 16, 18
Топотекан 3: 25
Торсионный угол 1: 176, 177, 357
Точка начала репликации (oriC)
3: 44, 46,48, 56, 58, 64, 65
Трансальдолаза 2: 111
- в цикле Кальвина 2: 412, 413
Трансаминаза 2: 266
Трансаминирование 2: 266
Трансгенная мышь 1: 477
Трансдезаминирование 2: 270
Трансдуцин 1: 652, 653
Транс-жирные кислоты 1: 493, 494
Транскетолаза 2: 111, 112
- в цикле Кальвина 2: 412-415
- дефект при синдроме Вернике-
Корсакова 2: 112
Транс-конфигурация пептидной
связи 1: 176, 182
Транскрипт
- первичный 3: 122, 123
- сложный 3: 132
- сплайсинг 3:122-133
Транскриптом 2: 126; 3: 106, 156-158
Транскрипционные факторы 2: 124;
150-152; 3: 120
— основные 3: 120
Транскрипционный профиль 1: 466
Транскрипционный пузырек
3: 106-108
Транскрипция 1: 408, 409; 3: 105
- активаторы 3: 232
- аттенюация 3: 245-249
- ингибирование лекарствами 3: 121,
122
- инициация 3: 112-115, 232-234
- интронов 3:131
- корректирующая активность 3: 110
- обратная 1: 317; 3: 144
- промоторы 3: 110, 112, 260
- регуляция
— в бактериях 3: 234-236
— в эукариотах 3: 257-263, 271
— отрицательная 3: 232, 233, 235
— положительная 3: 232, 233
- репрессоры 3: 232
— стероидными гормонами 1: 643
- сопряжение с трансляцией 3: 204,
205
- терминация 3: 116
Транслокация
- в перемещении белка 3: 212, 218
- в синтезе белка 3: 200, 201
Трансляция 3: 179-181
- регуляция 3: 250, 268
- репрессия 3: 250, 268
- сдвиг рамки 3: 175
- сопряжение с транскрипцией
3: 204, 205
- стадии 3: 179, 181
Транспептидаза 1: 314, 315
Транспозиция 3:94
- прямая 3: 95
- репликативная 3: 95
Транспозоны 1: 460; 3: 94
- инсерционные последовательности
3: 95
- сложные 3: 94
Транспортеры 1: 552, 553; см. также
Ионные каналы; Переносчики
Транспортная РНК, см. тРНК
Транспортные везикулы 1: 375
— доставка белков 3: 214, 215
— доставка липидов 2: 265
Трансферрин 2: 193
- рецепторы 2: 193, 194
Трансформация, при клонировании
1: 439,449
Трансфосфорилирование, в нуклео-
тидах 2: 40
Трастузумаб (герцептин) 1: 669
Трегалоза 1: 351
Треоза 1: 342
Треонин!: 114, 120; 2: 523
- биосинтез 2: 523
- превращение в пропионил-СоА
2: 295
- превращение в сукцинил-СоА
2: 294
- расщепление до пирувата 2: 285,
288
- фосфорилированный 1: 632
Треониндегидрогеназа 1: 291
- ингибирование 1: 321; 2: 531
Треонинсинтаза 2: 524, 525
Триацилглицеринлипаза 2: 586
Триацилглицерины 1: 491,492; 2: 584;
см. также Жирные кислоты
- в прорастающих семенах 2: 247
- всасывание 2: 226-228
- гормональная мобилизация 2: 228,
229
- запасание в адипоцитах 2: 585, 586
- накопление при ожирении
2:615-617
- реутилизация 2: 466, 467
- синтез 2: 464-469
Предметно-именной указатель [429]
- содержание в организме 2: 600
- транспорт 2:485-487
- цикл 2: 467
Триглицериды, см. Триацилглице-
рины
Трииодтиронин (ТЗ) 2: 575
Трикарбоновых кислот цикл (цикл
лимонной кислоты, цикл Креб-
са) 2: 181
Триметоприм 2: 558, 560
Триозокиназа 2:89
Триозофосфатизомераза 1: 281; 2: 58,
75, 89,112, 195,411
Триозофосфаты
- антипорт 2: 416-418
- в цикле Кальвина 2: 406-416
- взаимные превращения 2: 412
- превращение в сахарозу/крахмал
2: 426-430
- синтез 2: 410-415
Триозы 1: 340, 341
Трипаносомоз (африканская сонная
болезнь) 1: 541, 542
Триплетный код 3: 167-171
Трипсин 1: 102, 146, 257, 272; 2: 264
- ингибитор 1: 328; 2: 264
- синтез 1: 328, 329
- фрагменты 1: 147
Трипсиноген 1: 328, 329; 2: 264
Триптофан 1:69, 116, 118
- биосинтез 2: 523, 527, 528
- в гидрофобных взаимодействиях
1:377
- в мембранных белках 1: 535, 536
- поглощение света 1: 120
- расщепление до ацетил-СоА
2: 288-290
- расщепление до пирувата 2: 285
Триптофансинтаза 2: 527, 528
Трисахаридный мостик 1: 364-366
Трихроматы 1: 656
тРНК (транспортная РНК) 1: 391;
3:136
- D-плечо 3: 184
- TipC-плечо 3: 185
- аминоацилирование 3: 179,
185-188
- аминокислотное плечо 3: 184
- антикодоновое плечо 3: 184
- в трансляции 3: 185
- минорные основания 1: 395
- процессинг 3: 137
- спаривание с мРНК 3: 136
- структура 1: 411; 3: 184-186
- супрессоры 3: 203
Тройная связь углерода 1: 29, 31
Тройная спираль
— ДНК 1:407
— коллагена 1: 187-192
Тройной комплекс, в двухсубстрат-
ной реакции 1: 293, 294
Тромбин 1: 329, 361
Тромбоз
- протеолиз 1: 329
- селектины 1: 373, 374
Тромбоксансинтаза 2: 463
Тромбоксаны 1: 507-509
- синтез 2: 461-463
Тромбоциты 2: 592
- интегрины 1: 642
- фактор активации 1:499; 2: 476
- фактор роста (PDGF) 1: 624
Тропные гормоны 2: 576
Тропоколлаген 1:188
Тропомиозин 1: 258, 261
Тропонин 1: 258, 261, 614
Тубокурарин 1: 581
Тудикум Йохан 1: 501, 503, 523
Туникамицин 3: 213, 214
Тупые концы 1: 437, 438
Тяжелая цепь
— иммуноглобулина 1: 251, 252
---рекомбинация 3: 97
— миозина 1: 257
Тяжелый комбинированный имму-
нодефицит 1:475
У
Уаймен Джеффри 1: 242
Убиквитин 1: 323, 662, 663;
3: 220-222
- в протеолизе 3: 221, 222
Убихинон (кофермент Q) 1: 512, 513;
2:309,313-318
Убихинон: цитохром-с-оксидо-
редуктаза2: 313, 318
Углеводы 1: 339-381; см. также
Дисахариды; Моносахариды;
Олигосахариды; Полисахариды
- анализ 1: 379-381
- активация долихолами 1: 513
- гликоконъюгаты 1: 363-370
- гликолиз 2: 66-84
- код сахаров 1: 371
- метаболизм 66-112
— анаболизм 2: 581
— в печени 2: 172, 581, 582
— в растениях и бактериях
2: 403-445
— катаболизм 2: 581
— метаболический контроль 2: 137,
138
— при сахарном диабете 2:84,85
— регуляция 2: 105, 112, 139-152
---аллостерическая и гормональ-
ная 2: 170-173
---на уровне экспрессии генов
2: 149-152
— ферменты 2: 124-127
- номенклатура 1: 339-343
- окисление 1: 347
- связывание с лектинами 1: 372-378
- синтез, см. также Глюконеогенез
— С4-путь2: 420-426
— гликолатный путь 2: 421, 422
— интеграция процессов 2: 435-438
— крахмала 2: 426-428
— пентозофосфатный путь
2: 107-112,438
— пептидогликанов 2: 433-435
— сахарозы 2: 428-430
— фотодыхание 2: 420, 421
— фотосинтез 2: 404-419
— целлюлозы 2: 431-433
- формулы Фишера 1: 341
- химический синтез 1: 381
Углерод
- аномерный 1: 344
- асимметрический 1: 35, 341
- окисление 2: 21, 26, 27, 45
Углеродная связь 1: 29-31
— гидролиз 2: 32, 33
— реакции 2: 20-24
Уилкинс Морис 1: 400, 401
У итерс Стефен 1: 313
Уксусная кислота 1: 93-95; 2: 116
— pXd 1: 95
— в буферной системе 1: 97
— кривая титрования 1: 95
Унипорт 1: 558
Уокер Э. Джон 2: 333, 334
Уоррен Дж. Робин 1: 374
Уотсон Джеймс Д. 1:9, 142, 400-405,
457; 3:45, 50
Уотсона-Крика правило спаривания
оснований 3: 108
Уратоксидаза 2: 556
Урацил 1: 392, 393
- дезаминирование цитозина 1: 416
- при заболеваниях 2: 554
- таутомерные формы 1: 397, 398
Урацил-ДНК-гликозилаза 3: 72
Уреотелические организмы 2: 273
Уридилат 1: 393
Уридилилирование 2:516
Уридилилтрансфераза 2: 515, 516
Уридин 1: 393
Уридин-5'-монофосфат (UMP)
1: 394; 2: 547
Уридиндифосфат (UDP) 2: 89,435
[ 4301 Предметно-именной указатель
Урикотелические организмы 2: 273
Уроновая кислота 1: 347
— в гликозаминогликанах 1: 360,
361
Уротензин II1: 478
Усай Бернардо 2: 160
Усиление сигнала 1: 575, 592, 604,
606,618, 626, 659
Ф
Фабри болезнь 1: 505
Фаг к, как вектор 1: 438, 440,441;
3: 92
Фагоцитоз 1: 253
Фактор роста сосудистого эпителия,
рецептор 1: 624, 667
Факторы высвобождения (термина-
ции) 3: 202-204
Факторы обмена гуаниновых нукле-
отидов (GEF) 1: 601
Факторы роста 1: 664, 666-668
Факторы специфичности 3: 114, 115,
232
Фанкони-Бикель болезнь 2: 159
Фарадея постоянная 1: 559; 2: 13, 47
Фелинга реакция 1: 347, 348
Фенилаланин 1: 78, 116, 118; 2: 291
- биосинтез 2: 518, 523, 528, 529
- катаболизм 2: 280, 288
-тРНК 1:411
Фенилаланингидроксилаза 2: 287,
291,528
Фенилацетат 2: 279, 291, 292
Фенилацетил-СоА 2: 279
Фенилацетилглутамин 2: 279
Фенилкетонурия 2: 287, 291
- скрининг у новорожденных 2: 292
Фенилпируват 2: 291, 292
Фенотип 3: 8
Феофитин 2: 367-371
Феофитин-хиноновая система
2: 367-371
Ферментативная кинетика 1: 285
— двухсубстратных реакций
1: 293,294
— для регуляторных ферментов
1:319-322
— зависимость от pH 1: 299,303
— константа Михаэлиса 1: 288, 290
— лимитирующая стадия 1: 275
— максимальная скорость 1: 285,
290
— начальная скорость 1: 285, 289
— переходное состояние 1: 274, 306,
307
— предстационарная 1: 287, 294, 302
— стационарная 1: 287, 294
— уравнение скорости реакции
1: 276, 287
— уравнение Михаэлиса-Ментен
1:288
---преобразования 1: 289, 290
Ферментативные реакции 1: 300-330
— второго порядка 1: 229
— двухсубстратные 1: 293
— ингибирование 1: 295-299; см.
также Ингибиторы ферментов
---по принципу обратной связи
1:321
---суицидными ингибиторами
1:298
— интермедиаты 1: 275
— константа диссоциации 1: 290
— константа скорости 1: 276
— координата реакции 1: 274
— лимитирующая стадия 1: 275
— механизм
---гексокиназы 1: 308, 309
---енолазы 1: 309, 310
---катализа ионами металлов
1:284,309,310
---кислотно-основного катализа
1: 283, 284
---ковалентного катализа 1: 284
---лизоцима 1: 310-313
---пинг-понг 1: 293, 294
---Филлипса, для лизоцима 1: 312,
313
---химотрипсина 1: 301-305
— основное состояние 1: 274
— первого порядка 1: 229
— переходное состояние 1: 274
— прсдстационарное состояние
1: 287, 294
— принципы катализа 1: 277-281
— равновесие 1: 276
---константа 1: 276
— разрушение сольватной оболочки
1:282
— скорость 1:274
---максимальная 1: 286
---начальная 1: 286
— уменьшение энтропии 1: 282
— стационарное состояние 1: 287
— уравнение скорости 1: 276
— энергия активации 1: 274, 275,
280
— энергия связывания 1: 277, 280
Ферментативный каскад 1: 592
— МАРК 1:619-621
— в растениях 1: 648, 649
Ферментация (брожение) 2: 66, 96
- в пивоварении 2: 93
- молочнокислое брожение 2: 69, 91,
581
- полезные продукты 2: 96
- производство спирта 2: 65, 69, 93
Фермент-субстратный комплекс
1: 278, 280, 285-287
— индуцированное соответствие
1:282
— ковалентные/нековалентные
взаимодействия 1: 277
Ферменты 1: 270-330
- активность 1: 140; 2: 125
— удельная 1: 140
- активный (каталитический) центр
1: 226, 273
— комплементарность переходному
состоянию 1: 278, 279, 306, 307
- аллостерические 1: 319-323
- апоферменты 1: 272
- зимогены 1: 328
- классификация 1: 272
- кофакторы 1: 271
- коферменты 1: 271
- коэффициент управления 2: 135,
136
- нитрогеназного комплекса 2: 510,
511
- очистка 1: 132-140
- печени 2: 579
- пируватдегидрогеназного комплек-
са 2: 183, 184
- простетические группы 1: 271
- протеолитические, активация
1: 328, 329
- проферменты 1: 328
- регуляторные 1: 319
— аллостерические 1: 319-323
— гомотропные 1: 320
- регуляция 1: 319-330
— и состояние равновесия 2: 128
— ковалентная модификация
1: 323-328
— на уровне экспрессии генов
2: 124-126
— протеолитическое расщепление
1: 328, 329
— фосфорилирование 1: 322-328
- рестрикционные, см. Эндонуклеа-
зы рестрикции
- рецепторные, см. Рецепторные
ферменты
- специфичность 1: 281
— коэффициент 1: 292
- субстраты 1: 226, 273
Предметно-именной указатель [ 431 ]
- холоферменты 1: 271
- число оборотов 1: 291
- эластичность 2: 134
Ферредоксин Fd 2:48, 311, 374, 375,
419
Ферредоксин-НАОР+-оксидо-
редуктаза 2: 374, 378
Ферредоксин-тиоредоксинредуктаза
2:419
Ферритин 2: 193, 194
Фибриллярные белки 1: 185-192
— а-кератин 1: 185-187
— коллаген 1: 187-192
— фиброин 1: 192
Фибрин 1: 192
Фибриноген 1: 329, 512
Фибробласты, фактор роста (ФРФ)
1:361,365, 624
Фиброин шелка 1: 192
Фибронектин, и протеогликаны
1: 359, 367, 549
Фикобилин 2: 361, 362
Фикобилипротеин 2: 361
Фикобилисома 2: 364
Фикоцианобилин 2: 361-364
Фикоэритробилин 2: 361-364
Фиксация азота 2: 6, 510
— бактерией Azotobacter vinelandii
2:512
Фиксация углерода 2: 358, 406, 407;
см. также Диоксид углерода,
ассимиляция
Филаменты цитоскелета 1: 24
Филлипс Дэвид 1: 310
Филлипса механизм 1: 312, 313
Филлохинон (витамин КД 1: 512;
2: 374, 375
Фитановая кислота, а-окисление
2: 250
Фитол 2:361
Фишер Эмиль 1: 117, 225, 278
Фишера проекционные формулы
1:341,344
Флавинадениндинуклеотид, см. FAD
Флавинмононуклеотид, см. FMN
Флавиновые нуклеотиды 2:48,54,
55, 308
Флаводоксин 2:511,512
Флавопиридол 1: 669
Флавопротеины 2: 55, 308, 314
- электронпереносящий (ЭПФ)
2:314
Флиппазы 1: 540,541
Флип-флоп диффузия 1: 540
Флоппазы 1:540, 541
Флуоресцентный резонансный
перенос энергии (FRET) 1: 609,
613-615
Флуоресценция 2: 361
- FRAP 1:541, 542
- FRET 1:613-615
Фолат
- Н4, см. Тетрагидрофолат
- метаболизм, в химиотерапии 2: 558
- недостаточность 2: 284
Фолдинг белка 1: 200-215; 3: 179,
180
— in vitro 1: 440, 442
— белковые домены 1: 200
— мотивы 1: 200-202
— нарушения 1: 215-218
— парадокс Левинталя 1: 210
— правила 1: 201
— расплавленная глобула 1: 211
— роль дисульфидизомеразы 1: 213
— супервторичная структура 1: 200
— термодинамика 1:211,212
— шаперонины 1: 212-214
— шапероны 1: 212-214
Форболовые эфиры 1: 611
Форбса (Кори) болезнь 2: 159
Формальдегид 2: 45
N-Формилметионин 3: 195, 196, 200
- посттрансляционные модифика-
ции 3: 206
N-Формилметионин-тРНК 3: 195,
196
Фосфагены 2: 42
Фосфат
- в качестве буфера 1: 93,95, 97
- в нуклеотидах 1: 392, 394
- неорганический
— в окислении глюкозы 1: 46
— в синтезе крахмала 2: 427-430
— в фотосинтезе 2: 415-417
Фосфатаза 2:197
- фосфорилазы а (фосфопротеин-
фосфатаза-1) 2:127, 167, 168,
170
Фосфатидилглицерин 1: 527, 533;
2:472
Фосфатидилинозит 1: 507, 527, 530;
2: 472, 474
Фосфатидилинозит-3,4,5-трифосфат
1:507, 622; 2: 170
Фосфатидилинозит-4,5-бисфосфат
1:507, 609, 622; 2: 170
Фосфатидилинозиткиназа 2: 170,472
Фосфатидилинозитная система
1:610
Фосфатидилсерин 1: 497, 530, 541;
2:475
Фосфатидилсеринсинтаза 2: 475
Фосфатидилхолин (лецитин) 1: 78,
497, 502, 527; 2: 461,472,474,
475, 478
Фосфатидилэтаноламин 1: 497, 530;
2: 472, 474-476
Фосфатидная кислота 1: 497; 2: 465,
471
— фосфатаза 2: 465
Фосфатная связь, энергетическое
содержание 2: 35, 36
Фосфаттранслоказа 2: 159, 340
Фосфоангидридная связь 1: 44, 45,
104,425
Фосфогексозоизомераза 2: 24, 430
2-Фосфогликолат 2: 420-422
2-Фосфоглицерат 1: 309, 310; 2: 78,
79,98, 104
З-Фосфоглицерат 2: 33, 76, 78, 98,
104
- в фиксации углерода 2: 406-418
З-Фосфоглицераткиназа 2: 76, 411
Фосфоглицератмутаза 2: 78,128
Фосфоглицериды, см. Глицерофос-
фолипиды
З-Фосфоглицериповая кислота
2: 32, 33
Фосфоглюкоизомераза (глюкозо-
фосфатизомераза) 2: 72, 111
Фосфоглюкомутаза 2: 87, 157
Фосфоглюконат дегидрогеназа 2: 109
Фосфоглюконатный путь, см. Пенто-
зофосфатный путь
Фосфодиэстеразы циклических
нуклеотидов 1: 606
--и зрение 1: 602, 653, 654
Фосфодиэфирная связь, в нуклеино-
вых кислотах 1: 396
Фосфоенол пиру ват (ФЕП) 1: 309,
310; 2: 79, 100
- в глиоксилатном цикле растений
2:211,212
- в глицеронеогенезе 2: 468
- в глюконеогенезе 2: 98, 104
- в переносе фосфорильных групп
2:36, 37, 79
- гидролиз 2: 32
- образование
— из оксалоацетата 2: 212
— из пирувата 2: 100
— из 2-фосфоглицерата 2: 79
Фосфоенолпируваткарбоксикиназа
(ФЕП-карбоксикиназа) 2: 101
Фосфоенолпируваткарбоксилаза
(ФЕП-карбоксилаза) 2: 424
Фосфоинозитид-З-киназа 1: 616, 622
14 321 Предметно-именной указатель
Фосфокреатин 2: 536, 537
- в мышцах 2: 588, 589
Фосфолипаза 1: 504, 517
- А: 1 503, 504, 536; 2: 461
- С 1: 504, 507, 530, 609, 659
Фосфолипиды 1: 496
- в составе мембран 1: 527, 528, 530,
533, 534
- группы головки 2: 470, 471, 473
- двойной слой 1: 527, 528
- инозита 1: 507
- методы анализа 1: 517
- переносчики 1: 540, 541
- расщепление в лизосомах 1: 503
- синтез 2: 470,472, 476
- транспорт 2: 485,486
Фосфоманноизомераза 2: 90
Фосфопантетеин 2: 450-452
Фосфопентозоизомераза 2: 109
Фосфопорин Е 1: 536
Фосфопротеинфосфатаза
- 1 (фосфатаза фосфорилазы а)
2: 127, 167, 168,170
-2А2: 127, 146, 150
- в регуляции цикла лимонной кис-
лоты 2: 208, 214
Фосфопротеины 1: 131
Фосфор, ковалентные связи 2: 25, 26
Фосфорамидитный метод синтеза
ДНК 1:422,423
5-Фосфорибозил-1 -пирофосфат
2:518
5-Фосфорибозиламин 2: 545, 547
Фосфорибозилпирофосфат (PRPP)
2:544
Фосфорилаза, см. также Гликоген-
фосфорилаза 1: 325; 2: 165, 166
- крахмала 2: 87
Фосфорилирование; см. также Ауто-
фосфорилирование
- АТРаз 1: 560-562
- аминокислотных остатков 1: 122,
123
- ацетил-СоА-карбоксилазы 2: 243
- белка ретинобластомы 1: 664, 665
- в метаболизме углеводов 1: 347
- в передаче сигнала 1: 619-627
-----сАМР 1: 599, 602-604
-----cGMP 1: 627
-----в |3-адренергическом пути
1: 606, 607
-----в бактериях 1: 644-646
-----в поливалентных адаптерных
белках 1: 629-631
-----в пути 1Р3/диацилглицерина
1:611
-----в пути JAK-STAT 1: 624, 625
----в пути PI-3K-PKB 1: 622, 623
----в растениях 1: 644-646
----киназой р-адренергического
рецептора 1: 606
----протеинкиназой А 1: 599,
602-604
----рецепторными протеинкиназа-
ми 1: 619, 624
- в процессинге РНКЗ: 143
- в регуляции
--активности ферментов 1: 324,
327, 605
--экспрессии 3: 268, 269
- в репликации ДНК 3: 64
- в сердечной мышце 2: 589, 590
- в синтезе сахарозы 2: 428,429
- в транскрипции 3: 120
- в ферментативных каскадах 1: 606,
619-626
- гликогенфосфорилазы 1: 326
- глюкозы 2:71
- консенсусные последовательности
1: 327, 603, 605
- множественное 1: 325, 328
- обратимость 1: 328
- окислительное 2: 78, 305-346
- посттрансляциоппос 3: 206
- потенциал (DGp) 2: 30
- протонный градиент 2: 382
- родопсина 1: 654
- субстратное 2: 78
- фруктозо-6-фосфата 2: 72, 73
- циклинзависимых протеинкиназ
1:661-663; 3: 64,120
Фосфорилированные соединения,
энергия гидролиза 2: 29-33
Фосфорильная группа
— и АТР 2: 25, 26, 35-38,41
— перенос 2: 25, 26
--от неорганических полифосфа-
тов 2: 42
— от ФЕП 2: 37
— функция в гликолизе 2: 71-80
Фосфоролиз 2: 87, 155,197
Фосфотирозинсвязывающий домен
1:630
Фосфотирозинфосфатазы 1: 632
Фосфофруктокиназа
- 1 (ФФК-1) 2: 72, 73, 99,103,430
— регуляция 2: 143, 346
- 2 (ФФК-2) 2: 146, 429,430, 611
Фотодыхание 2: 420,422
Фотолиазы 2: 55
- в репарации ДНК 3: 74, 75
Фотолитография 1: 463, 464
Фотон 2: 359
- энергия 2: 359-370
Фотопигменты 1: 655
- главные и вспомогательные 2: 362,
363
6,4-Фотопродукт 1: 417, 418
Фотосинтез 2: 356-366
- С2-путь 2: 422
- Р,-триозофосфатный антипорт
2:416,417
- ассимиляция СО2 2: 404
- в С4-растениях 2: 420, 423, 424
- в САМ-растениях 2: 420
- восстановительный пентозофос-
фатный путь 2: 406
- гликолатный путь 2: 420-422
- и фотофосфорилирование 2: 357
- поглощение энергии света хлоро-
филлом 2: 359-366
- световые реакции 2: 358
- синтез крахмала 2: 426-428
- синтез сахарозы 2: 428-430
- спектр действия 2: 362, 364
- темновые реакции 2: 358
- фиксация углерода 2: 358;
см. также Цикл Кальвина
- фотодыхание 2: 420
- функция АТР 2: 415
- функция NADPH 2: 415
- эволюция 2: 385-387
Фотосистема 2: 365
- I/II 2: 367-375
- связь с цитохромом Ь6/2: 376-379
Фототрофы 1: 20
Фотофосфорилирование 2: 357
- в хлоропластах 2: 358, 359
- и окислительное фосфорилирова-
ние 2: 382
- поглощение света 2: 359-365
- расщепление воды 2: 379, 380
- реакция Хилла 2: 359
- синтез АТР 2: 381, 382
- стехиометрия 2: 383, 384
- хемиосмотическая теория 2: 328
Фотохимические реакционные цен-
тры 2: 365, 366
— Fe-S2: 370-373
----бактерий 2: 367, 368
----растений 2: 372
Фракционирование
- белков 1: 132
- клеток 1: 24, 87
Франклин Розалинд 1: 400, 401
Фруктоза 1: 340, 341, 343, 344
Фруктозо-1,6-бисфосфат 2: 72, 73
- в цикле Кальвина 2: 411-413
- расщепление 2: 73
- регуляция 2: 143, 144
Предметно-именной указатель [433]
Фруктозо-1,6-бисфосфатаза 2: 103,
139,412,418,429
- активация светом 2:418, 419
Фруктозо-1-фосфат 2: 88, 89
Фруктозо-2,6-бисфосфат 2: 144, 146
- в синтезе сахарозы 2:429, 430
Фруктозо-2,6-бисфосфатаза 2: 146
Фруктозо-6-фосфат 2: 146
- в синтезе сахарозы 2:411, 430
- в цикле Кальвина 2:411, 413
Фруктозодифосфатальдолаза (аль-
долаза) 2: 73
Фруктозофосфатальдолаза 2: 89
Фруктокиназа 2: 88
L-Фтораланин 2: 538
Фтордезоксиглюкоза 2: 82, 83, 591
Фторурацил 2: 558, 559
X
Хансон Ричард 2:468
Хантингтин 1: 217
Хантингтона болезнь 1:484
— аномальный фолдинг 1: 215, 217
Хартли Б. С. 1: 302
Хеликазы 3: 54
- в репарации 3: 70, 73, 74
- в репликации 3: 54, 55, 57-65
Хемиосмотическая теория (теория
Митчелла) 2: 305, 327-329, 339,
382
Хемогетеротрофы 1: 58
Хемотаксис, двухкомпонсптпая
передача сигнала 1: 644-646
Хемотрофы 1: 20
Хендерсона-Хассельбаха уравнение
1: 98, 99
Хенселейт Курт 2: 273
Херши Альфред 1: 400
Херши-Чейза эксперимент 1: 400
Хеуорса проекционные формулы
1:344, 345,351
Хилл Арчибальд 1: 239
Хилл Роберт 2: 359
Хилла график 1: 242
Хилла коэффициент 1: 239; 2: 127,
136
Хилла реагент 2:359
Хилла уравнение 1: 239
Хиломикроны 2: 228,485-487
Химиотерапевтические препараты
— ингибиторы протеиназ 1: 667, 668
— ингибиторы топоизомеразы
3 : 24, 25
— мишени в биосинтезе нуклеоти-
дов 2: 558-560
— транскрипционный профиль
1:465, 466
Химическая реакция
— механизм 1: 304, 305
— отличие от биохимический реак-
ции 2: 27
Химические модели молекул
— СРК 1: 33-35
— шаро-стержневая 1: 33, 34
— перспективная 1: 33, 34
Химические элементы
— незаменимые 1: 29
— следовые (микроэлементы) 1: 29
Химический синтез ДНК 1: 643
--фосфорамидитный метод 1: 422
Химотрипсин 1: 129, 130, 200, 273,
298; 2: 25, 264,568
- каталитическая активность 1: 291,
302
- комплементарность переходному
состоянию 1: 306
- механизм реакции 1: 283, 301-305
- синтез 1: 328,329
- структура 1:301
Химотрипсиноген 1: 131, 139; 2: 264
Хиральные молекулы 1: 35-38
— оптическая активность 1: 117
Хиральный центр 1: 35
Хитин 1: 356
Хитчингс Джордж 2: 557, 560
хлДНК (ДНК хлоропластов) 3: 12
Хлорамфеникол 3: 209
Хлоридно-бикарбонатный обменник
1: 556-558
Хлориды
- в плазме крови 2: 591
- в цитозоле и внеклеточной жидко-
сти 1: 634
Хлорные каналы
— в сигнальных путях 1: 633-635
— при кистозном фиброзе 1: 565,
566, 579
Хлоропласты 1: 26,159; 2:405,406
- АТР-синтаза 2: 384
- ДНКЗ: 12
- интеграция фотосистем I и II
2: 367,372,373, 377, 379
- мембранные липиды 1: 500, 526,
527
- метаболизм липидов 2: 453, 454
- синтез
— АТР 2:377
— NADPH 2:375, 454, 455
— жирных кислот 2:453, 454
— крахмала 2: 426-430
- транспорт белков 3: 210, 215
- фотосинтез 2: 366
- эволюция 2:386,387
Хлорофилл 2: 359,361-363
- антенны 2: 365
- передача возбуждения 2:364-366
- типы 2: 361, 363
Хлороформ, для экстракции липидов
1:515,516
Хогланд Малой 1: 10; 3: 166
Ходжкин Дороти Кроуфут 2: 240
Холдейн Дж. Б. С. 1: 269, 270, 280, 287
Холдейн Хейзел 1: 258
Холекальциферол (витамин D3)
1:510,511
Холерный токсин 1: 324, 677
Холестерин 1:504
- биосинтез 2: 479-484
- в мембранах 1: 526, 527, 545, 632
- в синтезе изопреноидов 2: 479-484,
497
- в синтезе стероидных гормонов
2:495,496
- и транс-жирные кислоты 1: 494
- избыточный синтез 2: 492
- липопротеиновые комплексы
2: 227, 228
- медикаментозное снижение содер-
жания 2: 493,494
- опосредованный рецепторами
эндоцитоз 2:490,491
- регуляция синтеза 2: 491, 492
- структура 1:504
- транспорт 2:485-489
— обратный 2: 488, 489
- этерификация 2: 484, 485
- эфиры 2: 484, 485
Холецистокинин 2: 264
Холидея структуры 3: 84, 85,89,93
Холли Роберт 3: 166
Холофермент 1: 271
Хондроитинсульфат 1: 364, 365
Хоризмат, в биосинтезе аминокислот
2:524, 526-528
Хоминг 3:148,149
Хроматиды сестринские 3:82, 83
Хроматин 3: 27
- активный/неактивный 3: 257, 258
- ацетилирование/деацетилирова-
ние 3: 257, 258
- в 30-нм волокнах 3: 31, 34
- гетерохроматин 3: 257
- и гистоны 3: 27-29, 258
- изменения при транскрипции
3: 257, 258
- конденсированный 3: 257
- нуклеосомы 3: 27-35
- ремоделирование 3: 258
- сборка 3: 27-35
- эухроматин 3:32, 257
| \ 3 \ ] Предметно-именной указатель
Хроматография
- адсорбционная 1: 515, 516
- аффинная 1: 134,135
- высокоэффективная жидкостная
1: 135
- газожидкостная 1: 516, 517
- ионообменная 1: 132-134
- колоночная 1: 132,135, 254
- тонкослойная 1: 169, 515, 516
- эксклюзионная 1: 134
Хроматофор 2: 382
Хромосомы 3: 27
- бактериальные 1:439,442; 3:10,
36,37
- димерные, репарация 3: 93,94
- дочерние 1: 403
- дрожжевые искусственные 1: 442,
443; 3: 15
- конденсация 3: 27
- структура 3: 27-35
- эукариотические 3: 11, 27-35
Хугстин Карст 1: 406
Хугстиновские пары 1: 406
Хэтч Маршалл 2: 424
ц
Цветовое зрение 1: 654-656
Цвиттер-ион 1: 123
Целакант, анаэробный метаболизм
2: 92
Целебрекс 2: 462,463
Целлюлаза 1: 355
Целлюлоза 1: 355,357; 2: 431
- синтез 2: 432-434
Целлюлозосинтаза 2: 432-434
Цельвегера синдром 2: 246
Центральная догма молекулярной
биологии 3: 6, 144
Центрифугирование
- дифференциальное 1: 23, 24
- изопикническое 1: 23, 24
Центромера 3: 14
а-Цепь коллагена 1: 187,188
Церамид 1:502,505
- регуляторная функция 1: 507
Цереброзиды 2:478-478
Церулоплазмин 1: 372, 373
Цетуксимаб (эрбитукс) 1: 669
Цианобактерии 1: 21, 61; 2: 203
- фиксация азота 2: 6, 510
- фотофосфорилирование 2: 372, 373
- эволюция 1: 58-60; 2: 386,387
Цианокобаламин 2: 240
Цикл лимонной кислоты (цикл
трикарбоновых кислот, цикл
Кребса) 2:181-207
---активация ацетата 2: 182
----анаплеротические реакции
2: 204
---в метаболизме глюкозы 2: 582
----в метаболизме липидов 2: 584
----в печени 2: 582-584
----и глиоксилатный цикл
2:211-213
----компоненты 2: 203
----неполный, в анаэробных бакте-
риях 2: 202
---образование изоцитрата 2: 192
----образование цитрата 2: 190
---- окисление
----ацетата 2: 202
----изоцитрата до а-кетоглутарата
и СО2 2:192
----малата до оксалоацетата 2: 199
----сукцината до фумарата 2: 198
----онкогенные мутации 2: 211
----открытие 2: 222
----пируватдегидрогеназный ком-
плекс 2: 182-187
----пируваткарбоксилазная реакция
2: 205, 206
----превращение сукцинил-СоА в
сукцинат 2: 196
---продукты 2: 201
----реакции 2: 188,189
----регуляция 2: 208-211
----связь с циклом мочевины 2: 275,
276
----стадии 2: 190
----энергия окисления 2: 201
Цикл мочевины 2: 273-279
— генетические дефекты 2: 277-279
— связь с циклом лимонной кисло-
ты 2: 275, 276
Циклинзависимые протеинкиназы
(CDK) 1:661-664
— ингибирование 1:664
— продукты онкогенов 1: 668, 669
Циклины 1: 661-664; 3: 64
Циклический АМР, см. сАМР
Циклический GMP, см. cGMP
Циклогексимид 3: 209
Циклооксигеназа (СОХ) 2: 461,462
- ингибиторы 2: 461, 462
Циклосерин 2: 538
Циметидин 2: 542
Цинга 1: 189-191
Цинковые пальцы 3: 240, 241
Ципрофлоксацин 3: 24
Цис-конфигурация пептидной связи
1: 182,183
Цистатионин-|3-синтаза 2: 522
- дефект 2: 287
Цистатионин-у-лиаза 2: 522
Цистеин!: 120, 121,146
- биосинтез 2: 519, 521, 522, 532
- в железосерных кластерах 2: 311
- расщепление до пирувата 2: 285
Цистин 1:120, 121,146
Цг/с-тиране-изомеры 1: 34
Цитидилат 1: 393
Цитидилатсинтаза 2: 549
Цитидин 1: 393,400, 419
Цитидиндифосфат (CDP) 2: 471, 472
Цитидинмонофосфат (CMP) 1: 547;
2: 471
Цитидинтрифосфат (СТР) 1: 48
Цитозин 1: 392,393, 399,424
- дезаминирование 1: 416,417
- метилирование 1:419
Цитозоль 1: 18
- метаболизм липидов 2: 454
- синтез сахарозы 2: 436
- синтез целлюлозы 2: 433
Цитокинины 1: 470
Цитокины 1: 624,475
- в регуляции клеточного цикла
1:664
Цитоплазма 1: 18, 20, 24
- филаменты 1: 24
Цитоскелет 1: 24, 25
Цитотоксические Т-клетки
(Тс-клетки) 1: 250
Цитохромоксидаза 1: 271; 2: 313,
318, 320, 460; см. также Ком-
плекс IV
Цитохромы 2: 48,309, 310,312
- Ь52: 459
— цитохром-65-редуктаза 2: 459
- Ь6/2: 365,372, 373,376-379,387
- Ьс 1 2: 317, 318, 368, 370; см. также
Комплекс III
- с 1: 129,130,139, 200; 2: 309,310,
313,318,350,387
- /1:84; 2: 376
- Р-450 2: 348,349,459,460,496
- связь фотосистем I и II 2: 372, 373,
376
Цитрат 2: 192, 200
- асимметрические реакции 2: 199,
200
- в синтезе жирных кислот 2: 455,
456
- образование в цикле лимонной
кислоты 2: 188,190
- переносчик 2: 455,456
Цитратлиаза 2: 456
Цитратсинтаза 2: 190,191,204,455,
456
Цитруллин 1: 122,123,162
Цукеркандл Эмиль 1: 156, 433
Предметно-именной указатель [435]
Ч
Чаргафф Эрвин 1: 400
Чаргаффа правила 1: 400, 401
Чейз Марта 1: 400
Четвертичная структура белков
1: 141, 185-214
Чек Томас 3: 126
Число оборотов (&са1) 1: 291
Чувствительные нейроны 1: 633-659
ш
Шайн Джон 3: 196
Шайна-Дальгарно последователь-
ность 3: 196, 197
Шалли Эндрю 3: 567
Шанже Жан-Пьер 1: 242
Шаперонины 1: 212-214
Шапероны 1: 53,158, 212, 213,487;
3: 253, 258
Шаро-стержневая модель 1: 178, 188
Шарп Филипп 3: 125
Шаттл-вектор Г. 443
Шафрир Элеазар 2: 468
Шпильки
- в аттенюаторе 3: 247
- в ДНК 1:406
- в РНК 1:409,410; 3: 116, 142, 190,
247,252, 270
Шульц Петер 1: 308
Шумейкер Джеймс 2: 295
щ
Щелочная фосфатаза 1: 435,485
Щитовидная железа 2: 571
— гормоны 2: 571, 575; см. также
Тиреоидные гормоны
э
Эволюционное древо 1: 19, 159, 160
Эволюция 1: 19, 20, 54
- in vitro (SELEX) 3: 154-158
адаптация к водной среде 1: 73
- бактерий 1: 21; 2: 508, 509
- галапагосских вьюрков 3: 278-280
- горизонтальный перенос генов
1: 157
- и гипотеза «мира РНК» 1: 55, 56;
3: 153-159
- и гомологии в белках и генах
1:60-62,155-161
- и семейства белков 1: 157-159, 203
- иммунной системы 3: 97,98
- интронов 3: 148
- Миллера-Юри эксперимент 1: 55
- митохондрий 2: 352
- молекулярных паразитов 3: 97, 155
- ретротранспозонов 3: 148
- роль аденина 3:152,153
- роль мутаций 1: 54; 3: 278-280
- роль эндосимбиоза 1: 59, 60; 2:352
- фотосинтеза 2: 385-387
- хлоропластов 2:386
- цианобактерий 2:181,386,387
- эукариотических клеток 1: 59, 60
Эгр Питер 1: 571
Эдельман Джеральд 1: 251
Эдман Пер 1: 145
Эйвери Т Освальд 1: 400
Эйвери-Маклеода-МакКарти экс-
перимент 1: 400
Эйкозановая кислота 1: 488
Эйкоаноиды 1: 507-510; 2: 458, 574;
см. также Лейкотриены; Про-
стагландины; Тромбоксаны
- сигнальная функция 2: 572, 574
- синтез 2:458, 461, 574
Эйкозатриеноат и эйкозатетраеноат,
синтез 2:461
Эйлер-Челпин Ганс 2: 66
Эймс Брюс 3: 67
Эймса тест 3: 67
Экзергонические реакции 1: 44-46,
558
— сопряжение с эндергоническими
реакциями 1: 44-46, 558
Экзонуклеазы 3: 48
-III 1:435
- бактериофага X 1: 435
- в репарации ДНК 3: 50, 51, 70, 71
- корректирующая активность
3:51-54,63
Экзоны 3: 12, 13,122
- транскрипция 3: 123,131-133
Экзоцитоз 1: 25
Экситон 2: 361
- перенос 2: 361
Экспрессия генов 3: 230
- индукция 3: 230
- конститутивная 3: 230
- регуляция 3: 229-280
- репрессия 3: 230, 231
Эксцизионная нуклеаза 3: 74
Эксцизионная репарация 3: 68
- нуклеотидов 3: 73, 74
- оснований 3: 71-73
Элайон Гертруда 2: 505, 557
Эластин, и протеогликаны 1: 359,367
Эластичности коэффициент
2: 134-136
Электрический заряд
— аминокислот 1: 120,125
— влияние pH 1: 120,125
Электрогенный транспорт 1: 559
Электродвижущая сила 2: 44
Электролиты, содержание в плазме
2: 591,592
Электромагнитное излучение 2: 359
Электронейтральный транспорт
1:558
Электронпереносящий флаво-
протеин (ETF) 2: 234
Электроны
- акцепторы универсальные 2: 307
- в молочнокислом брожении 2: 90,
91
- переносчики 2: 44,45
— NADH и NADPH 2: 49, 50
— специализированные 2: 49
— универсальные 2: 49, 50
- стандартный восстановительный
потенциал 2: 47
Электроотрицательность атомов
1: 75,76
Электропорация 1: 439
Электроспрей (ESI-масс-
спектрометрия) 1: 149
Электростатические взаимодействия
с водой 1: 75,77
Электрофилы, в ферментативных
реакциях 1: 304; 2: 21-24
Электрофорез
- белков 1: 136-139
- в клонировании 1: 449
- в секвенировании ДНК 1: 420-422
- двумерный 1: 139,463; 2: 126
-ДНК 1:454-456
- пульсирующий 1: 443
Электрохимический градиент 1: 551;
2:323
— в поляризации мембраны 1: 634
— изменение свободной энергии
1: 560; 2:323
Элементы гормонального ответа
(HRE) 1:643; 3:267, 268
Элерса-Данлоса синдром 1: 191
Элиминирования реакции 2: 20, 23,
25
Эллиптицин 3: 25
Элонгация 3: 179, 180, 199-202
- в транскрипции 3:106, 112, 118
Эльвейм Конрад 2: 53
Эмбден Густав 2: 66
Эмбриональное развитие, регуляция
генов 3: 271-279
Энантиомеры 1: 35, 36
- моносахариды 1: 341
Эндергонические реакции 1:44-46,
558
— сопряжение с экзергоническими
реакциями 1:44-46,558
Эндо Акиро 2: 493,494
[ 4 361 Предметно-именной указатель
Эндокринные железы 2: 576,577
— секреция гормонов 2: 566,574,578
Эндонуклеазы 3:48
- АР-эндонуклеаза 3: 73
- в репарации ДНК 3: 69, 73, 74
- рестрикции 1:434-438
— в создании библиотек ДНК 1: 449
Эндоплазматический ретикулум
— а-окисление 2: 249
— посттрансляционная модифика-
ция белков 3: 211
— синтез жирных кислот 2: 457, 458
— ферменты семейства Р-450 2: 349
Эндосимбиоз 1: 59,60; 2: 352,386
Эндосимбиотические бактерии, эво-
люция хлоропластов 1: 60; 2: 386
Эндоцитоз 1: 25
- опосредованный рецепторами
2: 490; 3:219
Энергетический метаболизм 2: 6-10
Энергетическое сопряжение реакций
1:45
Энергия активации 1: 47,48, 274
— белковых конформаций 1: 172,
173
— Гиббса (0 1:44,47; 2:13-15
— диссоциации 1: 75
— запасенная в АТР 1: 45,425 2: 29,
587,588
— и константа скорости 1: 277
— источники в организме 2: 600
— конформации сахаров 1: 357
— мембранного транспорта
1: 550-559
— протондвижущая сила 2: 323
— растворения 1: 79
— света, поглощение хлорофиллом
2: 359, 360
— связывания 1: 277
— солнечная 1: 105; 2: 6,356-358
— термодинамические законы
преобразования в организме
2: 12-20
— ферментативной реакции
1:274-282
— фолдинга 1: 173,174,179, 211, 212
Энергия диссоциации 1: 75
Энтальпия (Я) 1 44; 2: 13
- изменение (АЯ) 1: 76; 2: 14
Энтеропептидаза 2: 264
Энтропия (5) 1: 44; 2: 13
- и стабильность белков 1:211,212
- изменение (А5) 1: 44, 212, 282;
2: 14
- снижение в ферментативных реак-
циях 1: 282
Энхансеры 3: 233, 260
Энцефалопатия, митохондриальная
2: 354
Эпигенетика 3: 32
Эпимеры 1: 343,361
Эпитоп 1: 250, 254
- в очистке белковых комплексов
1:467
- каталитические антитела 1: 308
Эргокальциферол (витамин D2)
1:511
Эритроза 1: 342
Эритрозо-4-фосфат, в цикле Кальви-
на 2:413,414
Эритропоэтин 1: 624
- рекомбинантный 1: 479
- рецептор 1: 625
Эритроциты 1: 233, 373,479; 2: 591,
592
-JAK-STAT-путь 1:624
- аквапорины 1: 570,571
- гликофорин 1: 369,527, 531
- изменение свободной энергии в
реакциях гликолиза 2: 98
- мембранные белки 1: 530, 531
- образование 1: 233
- транспорт глюкозы 1: 553, 554
- форма 1: 246, 247
- хлоридно-бикарбонатный обмен-
ник 1: 556-558
Эритрулоза 1: 342
Эрлотиниб (тарцева) 1: 669
Эстрадиол 1: 509,647; 2: 495
Эстрогены 1: 643,644; 2: 495
- синтез 2: 575
Этан 2:46
Этанол, см. Спирты
Этаноламинкиназа, мутации 2: 474
Этилен, рецепторы в растениях
1:647
Этопозид 3: 25
Эубактерии 1: 19, 21
Эукариоты
- ДНКЗ: 11-15
- клеточный цикл 1: 660
- структура клеток 1:18, 22, 24
- эволюция 1: 19,59, 60
Эухроматин 3: 257
Эфиры, гидролиз
— изменение стандартной свобод-
ной энергии 2: 16
— переходное состояние 1: 307
— сложных эфиров и тиоэфиров
2: 33,34
Эффекторы 3: 232
Я
Яблочный фермент, см. Малатдеги-
дрогеназа
Ягендорф Андрэ 2: 382
Ядерные белки 3: 216, 217
Ядерные рецепторы 1: 595
Ядерные факторы транскрипции
3: 268
Ядерный магнитный резонанс
(ЯМР)
--в анализе углеводов 1: 380
--в определении структуры белка
1: 197-199
Ядро 1: 18
- транспорт белков 3: 216, 217
Яды
- и ионные каналы 1: 579,581
- ингибирование трансляции
3: 207-209
Язва желудка 1: 61, 374
Яичники 2: 575, 576
Ялоу Розалин 2: 568,569
ЯМР, см. Ядерный магнитный резо-
нанс
Янг Уильям 2: 71
Янус-киназа (JAK) 1: 624; 2: 608
Оглавление
III ПУТИ ПЕРЕДАЧИ ИНФОРМАЦИИ
24 Гены и хромосомы 7
24.1. Элементы хромосом 7
Гены — это участки молекул ДНК,
кодирующие полипептиды
и молекулы РНК 8
Молекулы ДНК гораздо крупнее, чем
клеточные или вирусные структуры,
в которые они упакованы 9
Гены и хромосомы эукариот
очень сложно организованы 12
Краткое содержание 15
24.2. Сверхспирализация ДНК 15
Большинство клеточных ДНК
раскручены 17
Степень скручивания ДНК
определяется топологическим
параметром — порядком зацепления 18
Топоизомеразы катализируют
изменение порядка
зацепления в ДНК 21
Для компактной упаковки ДНК нужна
особая форма сверхспирализации 23
Дополнение 24-1. Медицина. Лечение заболеваний
путем ингибирования топоизомераз 24
Краткое содержание 26
24.3. Структура хромосом 27
Хроматин состоит из ДНК и белков 27
Гистоны — небольшие основные белки 28
Нуклеосомы — основные структурные
единицы хроматина 28
Нуклеосомы образуют структуры
с более сложной организацией 31
Дополнение 24-2. Медицина. Эпигенетика,
структура нуклеосом
и варианты гистонов 32
Структура конденсированных хромосом
поддерживается SMC-белками 35
Бактериальная ДНК тоже сложно
организована 36
Краткое содержание 37
Ключевые термины 38
Дополнительная литература для дальнейшего
изучения 38
Вопросы и задачи 39
Анализ экспериментальных данных 41
25 Метаболизм ДНК 43
25.1. Репликация ДНК 45
Основные принципы репликации ДНК 45
ДНК разрушается нуклеазами 48
ДНК синтезируется
ДНК-полимеразами 49
Репликация — очень точный процесс 50
У Е. coli не менее пяти ДНК-полимераз 52
В репликации ДНК участвует
множество ферментов
и белковых факторов 54
Репликация хромосомы Е. coli
происходит постадийно 56
Репликация в эукариотических клетках
происходит по похожей схеме,
но сложнее 64
Вирусные ДНК-полимеразы являются
мишенями для противовирусной
терапии 66
Краткое содержание 66
25.2. Репарация ДНК 66
Онкологические заболевания
связаны с мутациями 67
Все клетки имеют несколько систем
репарации ДНК 68
Взаимодействие репликативных вилок
с повреждением в ДНК может
запустить подверженный ошибкам
синтез ДНК через повреждение 76
Дополнение 25-1. Медицина. Репарация ДНК
и рак 79
Краткое содержание 80
25.3. Рекомбинация ДНК 81
Гомологичная генетическая
рекомбинация выполняет
несколько функций 81
Рекомбинация в ходе мейоза
начинается с двухцепочечных
разрывов 84
В рекомбинации участвует множество
ферментов и других белков 85
Для репарации заблокированных
репликативных вилок
используются все возможности
метаболизма ДНК 89
Сайт-специфическая рекомбинация
приводит к точным
перестройкам ДНК 89
|zi38| Оглавление
Для полной репликации хромосомы
может потребоваться сайт-
специфическая рекомбинация 93
Подвижные генетические элементы
перемещаются из одного
участка ДНК в другой 94
Сборка генов иммуноглобулинов
происходит путем рекомбинации 95
Краткое содержание 98
Ключевые термины 98
Дополнительная литература
для дальнейшего изучения 98
Вопросы и задачи 100
Анализ экспериментальных данных 102
26 Метаболизм РНК 105
Молекулы рРНК и тРНК также
подвергаются процессингу 133
РНК со специализированными
функциями подвергаются
различным вариантам процессинга 137
Каталитические РНК осуществляют
некоторые реакции
метаболизма РНК 138
Ферментативные свойства интронов
группы I 139
мРНК в клетке разрушаются
с разной скоростью 142
Полинуклеотидфосфорилаза
создает случайные РНК-подобные
полимеры 142
Краткое содержание 143
26.1. ДНК-зависимый синтез РНК 106
РНК синтезирует РНК-полимераза 108
Синтез РНК начинается с промоторов 110
Дополнение 26-1. Практическая биохимия.
РНК-полимераза оставляет
свой след на промоторе 111
Транскрипция регулируется
на нескольких уровнях 115
Специфические последовательности
подают сигнал прекращения
синтеза РНК 116
В клетках эукариот содержатся
РНК-полимеразы трех типов 117
Для проявления активности
РНК-полимеразы II требуются
другие белковые факторы 118
Возможно селективное ингибирование
ДНК-зависимой РНК-полимеразы 121
Краткое содержание 122
26.2. Процессинг РНК 122
К 5'-концу эукариотической мРНК
присоединяется кэп 123
Из ДНК в РНК транскрибируются
и интроны, и экзоны 125
РНК катализирует сплайсинг
интронов 125
На З'-конце молекулы мРНК
имеются характерные структуры 130
Альтернативный процессинг РНК
приводит к образованию
нескольких продуктов одного гена 131
26.3. РНК-зависимый синтез РНК и ДНК 144
Обратная транскриптаза синтезирует
ДНК с матрицы вирусной РНК 144
Некоторые ретровирусы вызывают
рак и СПИД 146
Дополнение 26-2. Медицина. Борьба со СПИДом
с помощью ингибиторов обратной
транскриптазы 147
Многие транспозоны, ретровирусы
и интроны могут иметь общее
эволюционное происхождение 148
Теломераза — специализированная
обратная транскриптаза 148
Некоторые вирусные РНК
реплицируются РНК-зависимой
РНК-полимеразой 151
Синтез РНК открывает важный
подход к изучению биохимической
эволюции 151
Дополнение 26-3. Практическая биохимия.
Метод SELEX для получения РНК
с заданными свойствами 154
Дополнение 26-4. Расширяющийся мир РНК,
или транскрипты с неизвестной
функцией 156
Краткое содержание 159
Ключевые термины 159
Дополнительная литература
для дальнейшего изучения 159
Вопросы и задачи 161
Биохимия в Интернете 162
Анализ экспериментальных данных 162
Оглавление [439]
27 Метаболизм белка 165
27.1. Генетический код 166
Генетический код был расшифрован
с помощью искусственных мРНК 167
Дополнение 27-1. Исключение, подтверждающее
правило: природные вариации
генетического кода 172
«Качание» позволяет некоторым
молекулам тРНК распознавать
более одного кодона 172
Считывание последовательности
зависит от сдвига рамки
и редактирования РНК 175
Краткое содержание 178
27.2. Синтез белков 178
Синтез белка происходит
в пять стадий 179
Рибосома — сложная
надмолекулярная машина 180
Дополнение 27-2. Из мира РНК в мир белка 182
Транспортные РНК имеют
специфическую структуру 184
Стадия 1: аминоацил-тРНК-синтетазы
присоединяют определенные
аминокислоты к соответствующим
молекулам тРНК 186
Дополнение 27-3. Естественное и искусственное
расширение генетического кода 191
Стадия 2: синтез белка инициирует
определенная аминокислота 195
Стадия 3: пептидные связи
образуются на стадии элонгации 199
Стадия 4: для прекращения синтеза
полипептида нужен специальный
сигнал 202
Дополнение 27-4. Индуцированные вариации
генетического кода: нонсенс-
супрессия 203
Стадия 5: вновь синтезированные
полипептиды сворачиваются
и процессируются 205
Многие антибиотики и токсины
ингибируют синтез белка 207
Краткое содержание 209
27.3. Транспорт и расщепление белков 210
Посттрансляционная модификация
многих эукариотических белков
начинается в эндоплазматическом
ретикулуме 211
Гликозилирование играет ключевую
роль в транспорте белка 212
Сигнальные последовательности
ядерных белков не отщепляются 216
Бактерии тоже используют
сигнальные последовательности
для транспорта белков 217
Белки проникают в клетки путем
опосредованного рецепторами
эндоцитоза 219
Расщепление белков во всех
клетках осуществляется
специализированными системами 220
Краткое содержание 223
Ключевые термины 223
Дополнительная литература
для дальнейшего изучения 224
Вопросы и задачи 225
Анализ экспериментальных данных 227
28 Регуляция экспрессии генов 229
28.1. Принципы регуляции генов РНК-полимераза связывается с ДНК 230
в области промоторов Инициация транскрипции регулируется белками, которые связываются с промоторами или 231
недалеко от них Многие бактериальные гены собраны в кластеры и регулируются в виде 232
оперонов Отрицательная регуляция лактозного 234
оперона Регуляторные белки содержат специальные ДНК-связывающие 235
домены Регуляторные белки содержат также домены, ответственные 237
за взаимодействия белка с белком 241
Краткое содержание 243
28.2. Регуляция экспрессии генов
у бактерий Положительная регуляция 243
лактозного оперона Многие гены ферментов биосинтеза аминокислот регулируются путем 243
аттенюации транскрипции 245
[4 4()| Оглавление
При индукции SOS-ответа
происходит разрушение
репрессорных белков 249
Синтез рибосомных белков
происходит координированно
с синтезом рРНК 250
Функция некоторых мРНК
регулируется малыми РНК
по цис- или ттфбшс-механизму 252
Некоторые гены регулируются
путем генетической
рекомбинации 254
Краткое содержание 256
28.3. Регуляция экспрессии генов
у эукариот 257
Транскрипционно активный хроматин
по структуре отличается
от неактивного хроматина 257
Хроматин ремоделируется путем
ацетилирования и перемещения
нуклеосом 258
Многие эукариотические промоторы
подвергаются положительной
регуляции 260
ДНК-связывающие активаторы
и ко-активаторы способствуют
сборке основных факторов
транскрипции 260
Гены метаболизма галактозы
в дрожжах подвергаются и
положительной, и отрицательной
регуляции 263
Активаторы транскрипции
имеют модульное строение 265
Экспрессия эукариотических
генов может регулироваться
внеклеточными
и внутриклеточными сигналами 266
Регуляция может осуществляться
путем фосфорилирования
ядерных факторов транскрипции 268
Трансляция многих эукариотических
мРНК подавляется 268
Посттранскрипционный сайленсинг
гена опосредован РНК 270
У эукариот реализуется несколько
вариантов РНК-опосредованной
регуляции экспрессии генов 271
Развитие контролируется
каскадами регуляторных белков 271
Дополнение 28-1.0 плавниках, крыльях
и клювах 278
Краткое содержание 281
Ключевые термины 281
Дополнительная литература
для дальнейшего изучения 282
Вопросы и задачи 283
Биохимия в интернете 284
Анализ экспериментальных данных 285
Приложение А. Принятые в биохимии
сокращения и аббревиатуры 287
Приложение Б. Краткие решения задач
и ответы на вопросы 291
Словарь терминов 347
Источники иллюстраций 375
Предметно-именной указатель 389
Оглавление тома 1
Предисловие к русскому изданию Краткое содержание трех томов Об авторах книги Несколько слов о науке Предисловие Благодарности 1 Основы биохимии 1.1. Принципы организации клетки 1.2. Химические основы биохимии 1.3. Физические основы биохимии 1.4. Генетические основы биохимии 1.5. Эволюционные основы биохимии I СТРОЕНИЕ И КАТАЛИЗ 2 Вода 2.1. Слабые взаимодействия в водных средах 2.2. Ионизация воды, слабые кислоты и слабые основания 2.3. Роль буферных систем в поддержании pH в биологических системах 2.4. Участие воды в реакциях 2.5. Живые организмы приспособлены к водной среде 3 Аминокислоты, пептиды и белки 3.1. Аминокислоты 3.2. Пептиды и белки 3.3. Как работать с белками 3.4. Структура белка: первичная структура 4 Трехмерная структура белков 4.1. Обзор белковых структур 4.2. Вторичная структура белка 4.3. Третичная и четвертичная структуры белка 4.4. Денатурация и фолдинг белка 5 Функции белков 5.1. Обратимое связывание белков с лигандами: белки, связывающие кислород 5.2. Комплементарное взаимодействие между белками и лигандами: иммунная система и иммуноглобулины 5.3. Энергозависимые взаимодействия белков:актин, миозин и молекулярные моторы 6 Ферменты 6.1. Введение 6.2. Как работают ферменты 6.3. Ферментативная кинетика как подход к пониманию механизма действия ферментов 6.4. Примеры ферментативных реакций 6.5. Регуляторные ферменты 7 Углеводы и гликобиология 7.1. Моносахариды и дисахариды 7.2. Полисахариды 7.3. Гликоконъюгаты: протеогликаны, гликопротеины и гликолипиды 7.4. Углеводы как информационные молекулы: код сахаров 7.5. Методы анализа углеводов 8 Нуклеотиды и нуклеиновые кислоты 8.1. Основные понятия 8.2. Структура нуклеиновых кислот 8.3. Химия нуклеиновых кислот 8.4. Другие функции нуклеотидов 9 Технологии на основе информации из ДНК 9.1. Клонирование ДНК: основные понятия 9.2. От генов к геномам
[442] Оглавление тома 1
9.3. От геномов к протеомам
9.4. Изменения генома и новые продукты
биотехнологии
10 Липиды
10.1. Запасные липиды
10.2. Структурные липиды в мембранах
10.3. Липиды как сигнальные вещества, кофакторы и пигменты
10.4. Методы анализа липидов
И Биологические мембраны и транспорт
11.1. Состав и строение мембран
11.2. Динамика мембран
11.3. Транспорт веществ через мембраны
12 Биосигнализация
12.1. Общие свойства систем передачи
сигналов
12.2. Рецепторы, сопряженные с G-белком,
и вторичные мессенджеры
12.3. Рецепторные ферменты
12.4. Рецепторные гуанилатциклазы,
cGMP и протеинкиназа G
12.5. Мультивалентные адаптерные
белки и мембранные рафты
12.6. Регулируемые ионные каналы
12.7. Интегрины: двунаправленные
рецепторы, ответственные
за клеточную адгезию
12.8. Регуляция транскрипции
стероидными гормонами
12.9. Сигнализация у микроорганизмов
и растений
12.10. Сенсорная передача сигнала в процессах
зрения,обоняния и вкуса
12.11. Регуляция клеточного цикла
протеинкиназами
12.12. Онкогены, гены опухолевых супрессоров
и программируемая гибель клетки
Оглавление тома 2
II БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ
13 Основы биоэнергетики. Типы химических реакций
13.1. Биоэнергетика и термодинамика
13.2. Химические основы биохимических реакций
13.3. Перенос фосфатных групп и АТР
13.4. Окислительно-восстановительные реакции в биологических системах
14 Гликолиз, глюконеогенез и пентозофосфатный путь
14.1. Гликолиз
14.2. Метаболические пути, питающие гликолиз
14.3. Превращение пирувата в анаэробных условиях: брожение
14.4. Глюконеогенез
14.5. Пентозофосфатный путь окисления глюкозы
15 Принципы регуляции метаболизма
15.1. Регуляция метаболических путей
15.2. Теория контроля метаболизма
15.3. Согласованная регуляция гликолиза и глюконеогенеза
15.4. Метаболизм гликогена в клетках животных
15.5. Согласованная регуляция синтеза и распада гликогена
16 Цикл лимонной кислоты
16.1. Образование ацетил-СоА — активированного ацетата
16.2. Реакции цикла лимонной кислоты
16.3. Регуляция цикла лимонной кислоты
16.4. Глиоксилатный цикл
17 Катаболизм жирных кислот____________
17.1. Расщепление, активация
и транспорт жиров
17.2. Окисление жирных кислот
17.3. Кетоновые тела
18 Окислительное
расщепление аминокислот
и образование мочевины
18.1. Метаболические пути аминогрупп
18.2. Выделение азота и цикл мочевины
18.3. Пути деградации углеродного
скелета аминокислот
19 Окислительное
фосфорилирование
и фотофосфорилирование
ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ
19.1. Реакции переноса электронов
в митохондриях
19.2. Синтез АТР
19.3. Регуляция окислительного
фосфорилирования
19.4. Роль митохондрий в термогенезе,
синтезе стероидов и апоптозе
19.5. Митохондриальные гены:
происхождение и мутации
ФОТОСИНТЕЗ: ПОГЛОЩЕНИЕ
СОЛНЕЧНОЙ ЭНЕРГИИ
19.6. Основные особенности
фотофосфорилирования
19.7. Поглощение света
19.8. Основной фотохимический процесс —
это индуцированный светом
перенос электронов
19.9. Синтез АТР в процессе
фотофосфорилирования
19.10. Эволюция оксигенного
фотосинтеза
[444] Оглавление тома 2
20 Биосинтез углеводов у растений и бактерий 20.1. Фотосинтез углеводов 20.2. Фотодыхание, С4- и САМ-пути 20.3. Биосинтез крахмала и сахарозы 20.4. Синтез полисахаридов клеточной стенки: целлюлоза растений и пептидогликан бактерий 20.5. Интеграция углеводного метаболизма в растительной клетке 21 Биосинтез липидов 21.1. Биосинтез жирных кислот и эйкозаноидов 21.2. Биосинтезтриацилглицеринов 21.3. Биосинтез мембранных фосфолипидов 21.4. Биосинтез холестерина, стероидов и изопреноидов 22 Биосинтез аминокислот, нуклеотидов и родственных соединений 22.1. Общий обзор метаболизма азота 22.2. Биосинтез аминокислот 22.3. Производные аминокислот 22.4. Биосинтез и деградация нуклеотидов 23 Гормональная регуляция и интеграция метаболизма млекопитающих 23.1. Гормоны: различные структуры для различных функций 23.2. Тканеспецифичный метаболизм: разделение функций 23.3. Гормональная регуляция энергетического метаболизма 23.4. Ожирение и регуляция массы тела 23.5. Ожирение, метаболический синдром и диабет П типа
Замеченные опечатки
Том 1
Страница,строка, колонка Напечатано Следует читать
5, справа в сноске jehninger5e lehninger5e
10, 2, справа Мюллисом Маллисом
65, 27, справа (4/3)0)^ (4/3)лг3
66,4 слева О)Г2Й nr2h
66,12, справа 4сог2 4лг2
68, слева формулы фосфатидилхолина и метэнкефалина перепутаны местами
109,35, слева концентраций концентрации
217,17, справа извест-но известно
224, справа, таблица, 3-й столбец L-протеаза L-субстрат
239, справа; 2 раза в формуле (5-13) (5-16)
248, 20, слева последст-вием последствием
265,4 слева Proviodence Providence
265, 22, слева а2р2-димеров аР-димеров
266,1, слева в Интернете программ в Интернете и программ
267, справа, подпись Частота связывания миозина Плотность связывания миозина
под рисунком или фрагментов миозина с шариками или фрагментов миозина на шариках
288, последнее уравнение справа 0o = MEt] [S]/(KM + [S]) »0 - г'тах [S] / (Км + [S])
289, последнее 1 Км , [S] 1 = + 1
— 1 Т 7 ГО1 Т 7 ГО1
уравнение слева ?’О ^niax [S] ^max [S] ^0 Knax [S] Кпах
333, И, слева происходящую находясь
337, 3, слева приведенная приведенную
338, 6, справа более крупной аминокислоты на более крупную аминокислоту
338, 9, справа Объясните этот результат. Объясните, что в этом удивительного.
388,12, слева 160 г/м ОЛЬ 162 г/моль
430, 20, слева гидролизиует гидролизует
431, 33, слева на расстояние на расстоянии
442, 28, слева электропореза электропорации
450,3, слева последовательно-меченные сайты ДНК-маркирующие сайты
450,34, слева последовательно-меченным сайтом ДНК-маркирующим сайтом
470,3, справа стимулирующих стимулирующие
482, 24, справа селектируемого селективного
482, 26, справа Потерю своей функции нарушенным геном Потерю функции гена
484, 2 слева ...рекомбинации, ДНК на обоих концах сегмента, которые скрепляются вместе, а удаленная... ...рекомбинации, при которой участки на концах сегмента слипаются, и удаленная...
485,19, слева привнесенной чужеродной
488, табл. 10-1, 4-й столбец, 7-я строка Аальмитолеиновая Пальмитолеиновая
[ \ \ б ] Замеченные опечатки
Страница,строка, колонка Напечатано Следует читать
494, 3, справа ЛВВП ЛПВП
500, подпись под рисунком, 4-я строка дифтаниловые дифитанильные
506, рисунок Таурохолиновая Таурохолевая
508, рис.10-18 в изображении простагландина Е1 пятичленный цикл должен быть замкнут
521,45, слева могут быть образовываться могут быть образованы
545,18, справа предпочений предпочтений
564, 41,44 слева, 3 справа MDRN1 MDR1
567, рис.11-42, б вертикальная ось - [Лактоза]всреде, штриховая линия - [Лактоза]внутри вертикальная ось - [Лактоза] внутри, штриховая линия - [Лактоза]всреде
587, 33, справа человечка человека
589,16, слева адреналин адреналина
599,1, слева гетеродимер гетеротример
602, доп. 12-2, 19, слева находящуюся находящейся
617, рис.12-12, а [Са2+] (нм) [Са2+] (мкМ)
617, рис.12-12, б [Са2+]в цитозоле (мМ) [Са2+]в цитозоле (нм)
630, 7, слева в то числе в том числе
640, 4, справа сильные усилители эффективные усилители
655,13, слева их из
658, 6, слева G-белки G-белков
677,10, слева пионерные пионерские
678 8 2 раза ЭДТА ЭГТА
679, 8, слева - 95 мВ -95 мВ
681, 44 слева белокм белок
681, под графиком аменорид амелорид
Замеченные опечатки [ \ \ 7 ]
Том 2
Страница, строка, колонка Напечатано Следует читать
17,7, справа закон действующих масс отношение действующих масс
37,43, справа (с. 861) (с. 518)
60,7 слева Приемлима Приемлема
60,17, справа Сопряжение разрыва АТР Сопряжение гидролиза АТР
61,7, слева «Химическая» Химическая
62,19, слева масса человека тело человека
62, 29, справа Энергетический спектр Энергетика
63,10, слева Электронное сродство соединений Сродство соединений к электронам
63, рисунок (В) (Б)
81,20, слева вначале в начале
101,1, слева молочной кислоты яблочной кислоты
117, 21, слева гексозобисфосфата, та, даже гексозобисфосфата, даже
117,29, слева 3-фосфоглицерат 3-глицерофосфат
159, табл. 1, 1-й столбец, 5-я строка болезнь Помпа Болезнь Помпе
205, табл. 16-2, 4-я строка малат-фермент малик-фермент
208,15, слева малат-фермент малик-фермент
219,13, слева был в 7 раз выше необходимо было в 7 раз выше необходимого
221, 21, справа значения в 0,5 мг/л значения 0,5 мг/л
221, 35, справа описывается S-образной описывается S-образной кривой
222, 25, справа В организмах Существуют организмы
234,40, справа Р-нидроксиацил Р-гидроксиацил
235,30, слева трехфункциональным трифункциональным
256,39, слева кофермент А-эфиры эфиры кофермента А
257, 20, слева одинаково хороши были бы так же хороши
257,1, справа окисления карнитина равна окисления олеата равна
258, 24, справа использует ферментативно связанный использует связанный
264,31, справа специфический ингибитор — белок ингибитора трипсина специфический ингибитор трипсина
291, И, слева фенилаланингидролазы фенилаланингидроксилазы
300,25, слева Объясните, принцип Объясните принцип
300,30, слева но не в различных но в различных
300,33, слева аминных групп в аминогруппах
300,34, слева збыток избыток
301,15, слева у ребенка бы частая рвота у ребенка частая рвота
301,17, слева отставало нормы отставали от нормы
302, 6, справа (образовании глицина) (образования глицина)
302,10, справа аналогичных некоторым в цикле аналогичных некоторым реакциям в цикле
350, рис. 19-37, внизу к гибели и утилизации к гибели и расщеплению
350,32, слева апоптозоме апоптосоме
[ zi \ (S ] Замеченные опечатки
Страница,строка, колонка Напечатано Следует читать
397,18, справа мутантных агентов мутагенов
398,13, слева которая потенциально соответствует которая соответствует
400, 5, справа фотосинтезирующего процесса процесса фотосинтеза
401, 3 слева Образуется ли NADPH. Образуется ли NADPH?
438, подпись под рис. 20-35 в каждом пуле соединения легко в каждом пуле легко
440,43, справа в которых совместно используются в которых используются
450,1, слева 4'-фосфопантатеин 4'-фосфопантетеин
450,4, слева 4'-фосфопантатеиновая 4'-фосфопантетеиновая
450, на рисунке и в подписи фосфопантатеин, фосфопантотеин фосфопантетеин
450,31, справа фосфопантатеина фосфопантетеина
452,15,21,28, справа фосфопантатеина фосфопантетеина
453, подпись к рисунку фосфопантатеиновая фосфопантетеиновая
462, 2, слева НПВП НПВС
463,16, справа НПВП НПВС
464,14, слева фосфопантатеин фосфопантетеин
468, И, слева Леа Решеф Леей Решеф
495,45, селва минералкортикоиды минералокортикоиды
500,17, справа жирноацил-СоА СоА-производных жирных кислот
501, 31, слева волосы шерсть
502,1, слева Изобразите Найдите
515, 7, справа (уридилирования) (уридинилирования)
515,12, справа уридилирование уридинилирование
515,13, справа деуридилирование деуридинилирование
519, уравнение слева Рибоза-5-фосфат Рибозо-5-фосфат
540, 3, справа орнитинкарбоксилазой орнитиндекарбоксилазой
544, 8, слева Фосфорибозил пирофосфат (PRPP) Фосфорибозилпирофосфат (PRPP)
важную роль играет важную роль
567, подпись под рис. 23-2 ТТГ ТРГ
568,10, справа молодой ученый, работающий молодой ученый, работавший