



































































































































































































































































































Author: Андреев М.П. Ахти Т. Гагарина Л.В. Гимельбрант Д.Е.
Tags: lichenes лишайники биологические науки в целом ботаника биология экология флора микология анатомия растений товарищество научных изданий кмк
ISBN: 978-5 -907372-98 -6
Year: 2022




РОССИЙСКАЯ АКАДЕМИЯ НАУК
БОТАНИЧЕСКИЙ ИНСТИТУТ ИМ. В. Л. КОМАРОВА
____ _______ _______
THE RUSSIAN ACADEMY OF SCIENCES
V. L. KOMAROV BOTANICAL INSTITUTE
The Lichen Flora of Russia
Family Parmeliaceae
Edited by
M. P. Andreev, T. Ahti, L. V. Gagarina, D. E. Himelbrant
KMK Scientific Press
Moscow — St. Petersburg 2022
Флора лишайников России
Семейство Parmeliaceae
Ответственные редакторы
М. П. Андреев, Т. Ахти, Л. В. Гагарина, Д. Е. Гимельбрант
Товарищество научных изданий КМК
Москва — Санкт-Петербург 2022
УДК 582.29(470+571)
ББК 28.591(2Рос)
Ф73
Авторы и составители:
М. П. Андреев, Т. Ахти, Л. В . Гагарина, Д. Е . Гимельбрант, И. С. Жданов, Л. А . Конорева,
Е. С. Кузнецова, Т. В. Макрый, Т. Н. Пыстина, Т. Рандлане, А. Сааг, И. Ф. Скирина,
И. С. Степанчикова, И. Н. Урбанавичене, С. В . Чесноков
Contributors:
T. Ahti, M. P. Andreev, S. V. Chesnokov, L. V. Gagarina, D. E . Himelbrant, L. A . Konoreva,
E. S . Kuznetsova, T. V. Makryi, T. N. Pystina, T. Randlane, A. Saag, I. F. Skirina, I. S . Stepanchikova,
I. N. Urbanavichene, I. S. Zhdanov
Флора лишайников России: Семейство Parmeliaceae / Отв. ред. М . П. Андреев, Т. Ахти,
Л. В. Гагарина, Д. Е . Гимельбрант.
—
М.; СПб.: Товарищество научных изданий КМК,
2022. — 187 с.; 1 рис.; 123 цв. фот.; 88 карт.
Настоящий выпуск «Флоры лишайников России» посвящен наиболее широко распространен-
ной в России и в мире группе лишайников, семейству Parmeliaceae. В книге даются краткие
описания анатомического строения и морфологии талломов и репродуктивных структур родов
и видов этой группы лишайников. Приводятся данные о составе содержащихся в них лишайни-
ковых веществ, особенностях распространения таксонов и их экологии. Семейство Parmelia-
ceae — одно из крупнейших во флоре России. Объем книги не позволяет включить в нее все
известные роды. В настоящей, первой части работы приводятся ключи для определения 22
родов и 84 известных для лихенофлоры России видов и 4 подвидов, принадлежащих к этому
семейству. Книга является итогом многолетнего изучения материалов, собранных авторами во
многих областях России, а также многочисленных коллекций, хранящихся в крупнейших гер-
бариях страны и мира. Выпуск иллюстрирован 123 оригинальными цветными фотографиями.
Для всех видов и подвидов приводятся карты распространения на территории России.
Издание рассчитано на студентов, аспирантов и специалистов-лихенологов, ботаников и мико-
логов, а также любителей природы, самостоятельно занимающихся изучением флоры лишай-
ников России.
Рецензенты:
доктор биологических наук А. А. Нотов
доктор биологических наук А. В. Пчёлкин
Издание осуществлено при финансовой поддержке
Российского фонда фундаментальных исследований по проекту 21-14-00013
Издание РФФИ не подлежит продаже
ISBN 978-5 -907372-98 -6
Ф73
© Коллектив авторов, 2022
© Обложка — Е . С . Кузнецова, 2022
© Товарищество научных изданий КМК, издание, 2022
Авторы тома
Андреев Михаил Петрович
Лаборатория лихенологии и бриологии, Ботанический институт им. В. Л . Комарова РАН,
ул. Профессора Попова, д. 2, 197376, Санкт-Петербург.
andreevmp@yandex.ru; andreevmp@binran.ru
Гагарина Людмила Владимировна
Лаборатория лихенологии и бриологии, Ботанический институт им. В. Л . Комарова РАН,
ул. Профессора Попова, д. 2, 197376, Санкт-Петербург.
gagarinalv@binran.ru
Гимельбрант Дмитрий Евгеньевич
Кафедра ботаники, биологический факультет, Санкт-Петербургский государственный уни-
верситет (СПбГУ), Университетская наб., д. 7 –9, 199034, Санкт-Петербург.
Лаборатория лихенологии и бриологии, Ботанический институт им. В. Л . Комарова РАН,
ул. Профессора Попова, д. 2, 197376, Санкт-Петербург.
d_brant@mail.ru; himelbrantde@binran.ru
Жданов Илья Степанович
Лаборатория лихенологии и бриологии, Ботанический институт им. В. Л . Комарова РАН,
ул. Профессора Попова, д. 2, 197376, Санкт-Петербург.
iszhdanov@yandex.ru
Конорева Людмила Александровна
Лаборатория флоры и растительных ресурсов, Полярно-альпийский ботанический сад-
институт им. Н. А. Аврорина КНЦ РАН, 184256, Мурманская обл., Кировск.
Лаборатория лихенологии и бриологии, Ботанический институт им. В. Л . Комарова РАН,
ул. Профессора Попова, д. 2, 197376, Санкт-Петербург.
ajdarzapov@yandex.ru
Кузнецова Екатерина Сергеевна
Кафедра ботаники, биологический факультет, Санкт-Петербургский государственный уни-
верситет (СПбГУ), Университетская наб., д. 7 –9, 199034, Санкт-Петербург.
Лаборатория лихенологии и бриологии, Ботанический институт им. В. Л . Комарова РАН,
ул. Профессора Попова, д. 2, 197376, Санкт-Петербург.
igel_kuzn@mail.ru; kuznetsovaes@binran.ru
Макрый Татьяна Владимировна
Лаборатория низших растений, Центральный Сибирский ботанический сад СО РАН, ул. Зо-
лотодолинская, д. 101, 630090, Новосибирск.
makry@nsu.ru
Пыстина Татьяна Николаевна
Отдел флоры и растительности Севера, Институт биологии Коми научного центра УрО
РАН, ул. Коммунистическая, д. 28, 167982, Сыктывкар.
t.pystina@ib.komisc.ru
Рандлане Тиина (Randlane Tiina)
Тартуский университет, Эстония. Institute of Ecology & Earth Sciences, University of Tartu,
Liivi Str., 2, EE50409, Tartu, Estonia.
tiina.randlane@ut.ee
Сааг Андрес (Saag Andres)
Тартуский университет, Эстония. Institute of Ecology & Earth Sciences, University of Tartu,
Liivi Str., 2, EE50409, Tartu, Estonia.
andres.saag@ut.ee
Скирина Ирина Федоровна
Центр ландшафтно-экологических исследований, Тихоокеанский институт географии ДВО
РАН, ул. Радио, д. 7, 690041, Владивосток.
sskirin@yandex.ru
Степанчикова Ирина Сергеевна
Кафедра ботаники, биологический факультет, Санкт-Петербургский государственный уни-
верситет (СПбГУ), Университетская наб., д. 7 –9, 199034, Санкт-Петербург.
Лаборатория лихенологии и бриологии, Ботанический институт им. В. Л . Комарова РАН,
ул. Профессора Попова, д. 2, 197376, Санкт-Петербург.
stepa_ir@mail.ru; stepanchikovais@binran.ru
Урбанавичене Ирина Николаевна
Лаборатория лихенологии и бриологии, Ботанический институт им. В. Л . Комарова РАН,
ул. Профессора Попова, д. 2, 197376, Санкт-Петербург.
urbanavichen@gmail.com
Чесноков Сергей Владимирович
Лаборатория лихенологии и бриологии, Ботанический институт им. В. Л . Комарова РАН,
ул. Профессора Попова, д. 2, 197376, Санкт-Петербург.
lukinbrat@mail.ru
ПРЕДИСЛОВИЕ
Издание «Флора лишайников России», которое предполагается публиковать в
течение ближайших лет, было задумано как многотомная серия, последователь-
но публикующая результаты монографической обработки тех или иных групп ли-
шайников по мере завершения их изучения на территории России.
Решение об издании серии было принято по инициативе проф. Н. С. Голуб-
ковой в октябре 2006 г. на совещании «Флора лишайников России: состояние и
перспективы исследований», посвященном 120-летию со дня рождения В. П. Са-
вича и проходившем в Ботаническом институте им. В. Л. Комарова в Санкт-
Петербурге. Издание должно было стать логическим продолжением публико-
вавшегося с 1971 по 2008 г. десятитомного «Определителя лишайников СССР»
(«Определителя лишайников России»), у истоков которого стояли крупнейшие
русские и советские ученые-лихенологи — А. Н. Окснер, М. Ф. Макаревич,
В. П. Савич и К. А. Рассадина.
По замыслу участников совещания, каждый из томов серии, подготовленный
специалистами-монографами, должен быть результатом оригинального исследо-
вания, включающего таксономические обработки семейств, порядков, родов или
иных групп лишайников, выявленных на территории России. В основу работы,
кроме оригинальных авторских данных, должна лечь критическая ревизия всех
доступных гербарных материалов, а также современных и исторических литера-
турных источников, как отечественных, так и зарубежных. Таким образом, изда-
ние должно обобщить все имеющиеся к настоящему времени сведения о таксоно-
мическом разнообразии лишайников России на уровне семейств, родов и видов и
явиться базой для дальнейших исследований, как таксономических, так и флори-
стических.
Первый (справочный) том «Флоры лишайников России», опубликованный
в 2014 г., в значительной мере явился руководством для изучения лишайников.
В книгу включены материалы по истории изучения лишайников России и данные
об основных лихенологических гербариях России и мира, где хранятся коллек-
ции лишайников, собранные на территории России. В книге дано описание ана-
томического строения и морфологии талломов и репродуктивных структур ли-
шайников, а также разнообразия их жизненных форм. Самостоятельные разделы
посвящены фотобионтам лишайников, особенностям вторичного метаболизма и
хемосистематике, молекулярно-генетическим методам их изучения, разнообра-
Флора лишайников России Флора лишайников России
8
зию и особенностям распространения лишайников на территории России и их
экологии. Существенное внимание уделено методам сбора и определения и реко-
мендациям по хранению лихенологических коллекций, приведена информация по
номенклатуре и типификации лишайников. В работе дана наиболее актуальная на
тот момент систематическая классификация таксонов и ключи для определения
402 родов лишайников флоры России, включающих более 3.5 тыс. известных на
территории страны видов, а также ключи для определения фотобионтов.
Предыдущий, первый систематический том был посвящен четырем груп-
пам лишайников, достаточно широко распространенным в России и на земном
шаре: роду Protoparmelia, семействам Coenogoniaceae (Fr.) Stizenb., Gyalectaceae
(A. Massal.) Stizenb. и Umbilicariaceae Chevall.
Настоящий выпуск «Флоры лишайников России» посвящен лишайникам ро-
дов Alectoria, Allocetraria, Arctocetraria, Bryocaulon, Bryoria, Cetraria, Cetrariella,
Cetreliopsis, Dactylina, Evernia, Flavocetraria, Gowardia, Letharia, Lethariella,
Masonhalea, Nephromopsis, Oropogon, Platismatia, Tuckermannopsis, Usnocetraria,
Vulpicida семейства Parmeliaceae. Род Protoparmelia семейства Parmeliaceae рас-
сматривался в предыдущем томе.
Семейство Parmeliaceae, одно из крупнейших, насчитывает в своем соста-
ве, по разным оценкам, от 70 до 89 родов и около 2740 видов. Для территории
России известно в настоящее время 54 рода и более 300 видов. Виды семейства
Parmeliaceae широко распространены во всех регионах России и встречаются в
самых разных местообитаниях.
В книге даются описания анатомического строения и морфологии талломов
и репродуктивных структур таксонов, данные о составе содержащихся в них ли-
шайниковых веществ, особенностях их распространения и экологии. Приводятся
ключи для определения 22 родов и 84 известных для лихенофлоры России видов
семейства Parmeliaceae. Книга является итогом многолетнего изучения материа-
лов, собранных авторами во многих областях России, а также многочисленных
коллекций, хранящихся в крупнейших гербариях страны и мира. Выпуск иллю-
стрирован 123 оригинальными цветными фотографиями. Приводятся карты рас-
пространения на территории России всех рассматриваемых видов и подвидов.
Все иллюстрации и карты, включенные в настоящее издание, приведены в автор-
ской редакции.
В работе над книгой приняли участие сотрудники Ботанического института
им. В. Л. Комарова РАН М. П. Андреев, Л. В. Гагарина, И. С. Жданов, И. Н. Ур-
банавичене, С. В. Чесноков, Санкт-Петербургского государственного университе-
та и Ботанического института Д. Е. Гимельбрант, Е. С. Кузнецова, И. С. Степан-
чикова, Полярно-альпийского ботанического сада-института им. Н. А. Аврорина
КНЦ РАН и Ботанического института Л. А. Конорева, Центрального Сибирского
ботанического сада СО РАН Т. В. Макрый, Тихоокеанского института географии
ДВО РАН И. Ф. Скирина, Института биологии Коми научного центра УрО РАН
Т. Н. Пыстина и Тартуского университета (Эстония) Т. Рандлане и А. Сааг.
Предисловие
9
Оригинальные рисунки и обложка выполнены Е. С. Кузнецовой.
Издание рассчитано на студентов, аспирантов и специалистов-лихенологов,
ботаников и микологов, а также любителей природы, самостоятельно занимаю-
щихся изучением лишайников России.
Книга подготовлена и опубликована при финансовой поддержке Российско-
го фонда фундаментальных исследований инициативными и экспедиционными
грантами NoNo 11-04-00901, 11-04-10017, 12-04-10076, 13-04-07028, 13-04-10034,
14-04-00067, 14-04-01411, 14-04-10091, 14-04-31024, 15-44 -05105, 15-04-05971,
15-29-02396 и издательским грантом No 21-14 -00013.
Предисловие
Деление территории Российской Федерации
на регионы
Принятая в настоящем издании схема деления России на физико-географиче-
ские регионы и административные территории (рис. 1) соответствует приведен-
ным в первом и втором томах издания «Флора лишайников России» (2014, 2017).
Авторы стремились максимально приблизить ее к схеме административных еди-
ниц Российской Федерации, хотя в ряде случаев это оказалось невозможным, по-
скольку физико-географические границы рассматривались как более значимые.
Территория Российской Федерации подразделяется на 18 крупных регионов,
присутствие видов в которых указывается в издании отдельно для каждой из ад-
министративных единиц или частей регионов.
Список физико-географических регионов
и административных территорий Российской Федерации
и их частей
Регионы Российской Федерации в работе обозначены сокращениями их рус-
ских названий, приведенными ниже. Названия субъектов Российской Федерации
в текстах видовых очерков приводятся либо полностью (например, области), либо
в сокращенной общепринятой форме (например: Карелия и Башкирия — вместо:
Республика Карелия и Республика Башкортостан или Эвенкия — вместо Эвен-
кийский муниципальный район). Название Республики Алтай приводится цели-
ком, чтобы не путать с горной системой Алтай. Латинские обозначения регионов
предлагаются для возможных иноязычных публикаций.
I. Арктика. — АРКТ. — ARC.
1. Арктика европейская — Аркт. евр. — Ае. Архангельская обл. (архипелаги
Земля Франца-Иосифа и Новая Земля), Ненецкий АО (кроме крайней юго-запад-
ной части, но включая п-ов Канин и острова Колгуев и Вайгач), крайний северо-
восток Республики Коми.
2. Арктика западносибирская — Аркт. зап.-сиб. — Aws. Северная часть Яма-
ло-Ненецкого АО (полуострова Ямал, Тазовский и Гыданский в пределах Ямало-
Ненецкого АО).
3. Арктика таймырская — Аркт. тайм. — At. Север Таймырского Долгано-
Ненецкого района Красноярского края, включая архипелаг Северная Земля.
4. Арктика якутская — Аркт. як. — Aya. Арктические районы Якутии (Ана-
барский, Аллаиховский и Нижнеколымский, тундровые части Булунского и Усть-
Янского административных районов).
5. Арктика чукотская — Аркт. чук. — Ach. Чукотский АО (северная часть).
Д
е
л
е
н
и
е
т
е
р
р
и
т
о
р
и
и
Р
о
с
с
и
й
с
к
о
й
Ф
е
д
е
р
а
ц
и
и
н
а
р
е
г
и
о
н
ы
1
1
Рис. 1. Территориальное деление Российской Федерации (Флора..., 2014, испр.).
1 — Арктика европейская; 2 — Арктика западносибирская; 3 — Арктика таймырская; 4 —
Арктика якутская; 5 — Арктика чукотская; 6 — европейский Север; 7 — Средняя Россия; 8 —
Южная Россия; 9 — Северный Кавказ; 10 — Северный Урал; 11 — Южный Урал; 12 — север
Западной Сибири; 13 — юг Западной Сибири; 14 — Средняя Сибирь; 15 — Южная Сибирь;
16 — Якутия; 17 — север Дальнего Востока; 18 — юг Дальнего Востока.
Д
е
л
е
н
и
е
т
е
р
р
и
т
о
р
и
и
Р
о
с
с
и
й
с
к
о
й
Ф
е
д
е
р
а
ц
и
и
н
а
р
е
г
и
о
н
ы
Флора лишайников России Флора лишайников России
12
II. Европейская Россия. — ЕР. — ER.
6. Европейский Север — Евр. Север — En. Мурманская обл., Республика Ка-
релия, Архангельская обл., юго-западная часть Ненецкого АО, Республика Коми,
Ленинградская и Вологодская области, северо-западная часть Кировской обл. (к
северу примерно от 60° с. ш.).
7. Средняя Россия — Ср. Россия — Em. Калининградская, Псковская, Новго-
родская, Тверская, Ярославская, Костромская, Кировская (кроме северо-западной
части) области, Пермский край (включая Коми-Пермяцкий округ), Смоленская,
Московская, Владимирская, Ивановская, Нижегородская области, Республики
Марий Эл, Чувашия, Татарстан и Удмуртия, Брянская, Калужская, Орловская,
Тульская, Рязанская области, Республика Мордовия, Пензенская, Ульяновская,
Самарская (северная часть) области, Республика Башкортостан, Курская, Липец-
кая, Тамбовская, Белгородская (северная часть), Воронежская (северная часть)
области.
8. Южная Россия — Юж. Россия — Es. Белгородская (юго-восточная часть),
Воронежская (южная часть), Саратовская, Самарская (южная часть), Оренбург-
ская, Ростовская, Волгоградская, Астраханская области, Республика Калмыкия,
Республика Крым.
9. Северный Кавказ — Сев. Кавказ — NC. Краснодарский и Ставропольский
края, Республики Адыгея, Карачаево-Черкесия, Кабардино-Балкария, Северная
Осетия, Ингушетия, Чеченская Республика, Дагестан.
10. Северный Урал — Сев. Урал — Un. Горные районы Республики Коми,
Пермского края (северная часть), Ямало-Ненецкого и Ханты-Мансийского АО и
Свердловской обл. (северная часть). На картах распространения таксонов нахож-
дение видов в частях административных территорий, относящихся к Уралу, от-
дельными точками не показывается, информация об этом приводится в тексте ви-
дового очерка в разделе «Распространение». Например: Коми (горная уральская
часть), Пермский край (внеуральская и горная уральская части).
11. Южный Урал — Юж. Урал — Us. Горные районы Пермского края (юж-
ная часть), Свердловской обл. (южная часть), Республики Башкортостан, Че-
лябинской и Оренбургской областей. На картах распространения таксонов на-
хождение видов в частях административных территорий, относящихся к Уралу,
отдельными точками не показывается, информация об этом приводится в тексте
видового очерка в разделе «Распространение». Например: Пермский край (внеу-
ральская и горная уральская части).
III. Сибирь. — СИБ. — SIB.
12. Север Западной Сибири — Зап. Сиб. сев. — Swn. Южная часть Ямало-
Ненецкого АО, Ханты-Мансийский АО, Свердловская, Тюменская (северная
часть), Томская области.
13. Юг Западной Сибири — Зап. Сиб. юж. — Sws. Челябинская, Курганская,
Тюменская (южная часть), Омская, Новосибирская (кроме горной крайней вос-
точной части) области, Алтайский край (западная, равнинная, степная часть).
Деление территории Российской Федерации на регионы
13
14. Средняя Сибирь — Ср. Сиб. — Sm. Южная часть Таймырского Долгано-
Ненецкого района, север Красноярского края, Эвенкийский муниципальный рай-
он, северная часть Иркутской обл.
15. Южная Сибирь — Юж. Сиб. — Ss. Восточная, горная часть Алтайского
края, крайний восток Новосибирской обл. (Салаирский кряж), Кемеровская обл.,
Республики Алтай, Хакасия, Тыва, Бурятия, Красноярский край (южная часть),
Иркутская обл. (южная часть), Забайкальский край.
16. Якутия — Як. — Sya. Внеарктическая часть Якутии:
Западная — зап. (w) — внеарктическая часть к западу от р. Лены и к северу
примерно от р. Вилюя, или от 64° с. ш. (почти целиком Оленекский эвенкийский
национальный район и западные части Булунского и Жиганского районов).
Центральная — центр. (c) — территории, расположенные к западу и северу
от рек Алдана и Лены и к югу примерно от 64° с. ш. (административные районы
Горный, Сунтарский, Вилюйский, Мирнинский, Вехневилюйский, Нюрбинский и
многочисленные небольшие районы, расположенные вблизи Якутска).
Восточная — вост. (e) — внеарктическая Якутия к востоку от рек Лены и
Алдана в границах Яно-Индигирского и Колымского флористических районов
(Эвено-Бытантайский национальный район, административные районы Верхоян-
ский, Абыйский, Среднеколымский, Верхнеколымский, Момский, Оймяконский,
Томпонский, части Усть-Янского, Кобяйского, Жиганского и Булунского).
Южная — юж. (s) — горные области южной Якутии, расположенные южнее
слияния рек Алдана и Лены, в верховьях рек Амги, Олекмы, Витима и южных
притоков Вилюя (административные районы Усть-Майский, Алданский, Олек-
минский, Ленский и часть Мирнинского).
IV. Дальний Восток. — ДВ. — FE.
17. Север Дальнего Востока — ДВ сев. — Fn. Южная часть Чукотского АО,
Магаданская обл., Камчатский край (Корякский округ и бывшая Камчатская обл.,
включая Командорские о-ва), Хабаровский край (северная часть, включая Шан-
тарские о-ва).
18. Юг Дальнего Востока — ДВ юж. — Fs. Амурская обл., южная часть Ха-
баровского края, Еврейская АО, Сахалинская обл., включая Курильские о-ва,
Приморский край.
Систематическое положение рассматриваемых
таксонов
Систематическое положение родов семейства Parmeliaceae, известных в на-
стоящее время для территории России, приводится в соответствии с первым то-
мом «Флоры лишайников России» (2014) и наиболее актуальными на сегодняш-
ний день системами лихенизированных грибов (Lücking et al., 2016; Divakar et al.,
2017; Wijayawardene et al., 2022), с некоторыми исправлениями и добавлениями
авторов.
Отдел ASCOMYCOTA Caval. Sm. (1998)
Подотдел PEZIZOMYCOTINA O. E. Erikss. et Winka (1997)
Класс LECANOROMYCETES O. E. Erikss. et Winka (1997)
Подкласс LECANOROMYCETIDAE P. M. Kirk, P. F. Cannon,
J. C. David et Stalpers ex Miądl., Lutzoni et Lumbsch (2007)
Порядок LECANORALES Nannf. (1932)
Семейство Parmeliaceae Bercht. et J. Presl (1820)
Alectoria Ach. (1809)
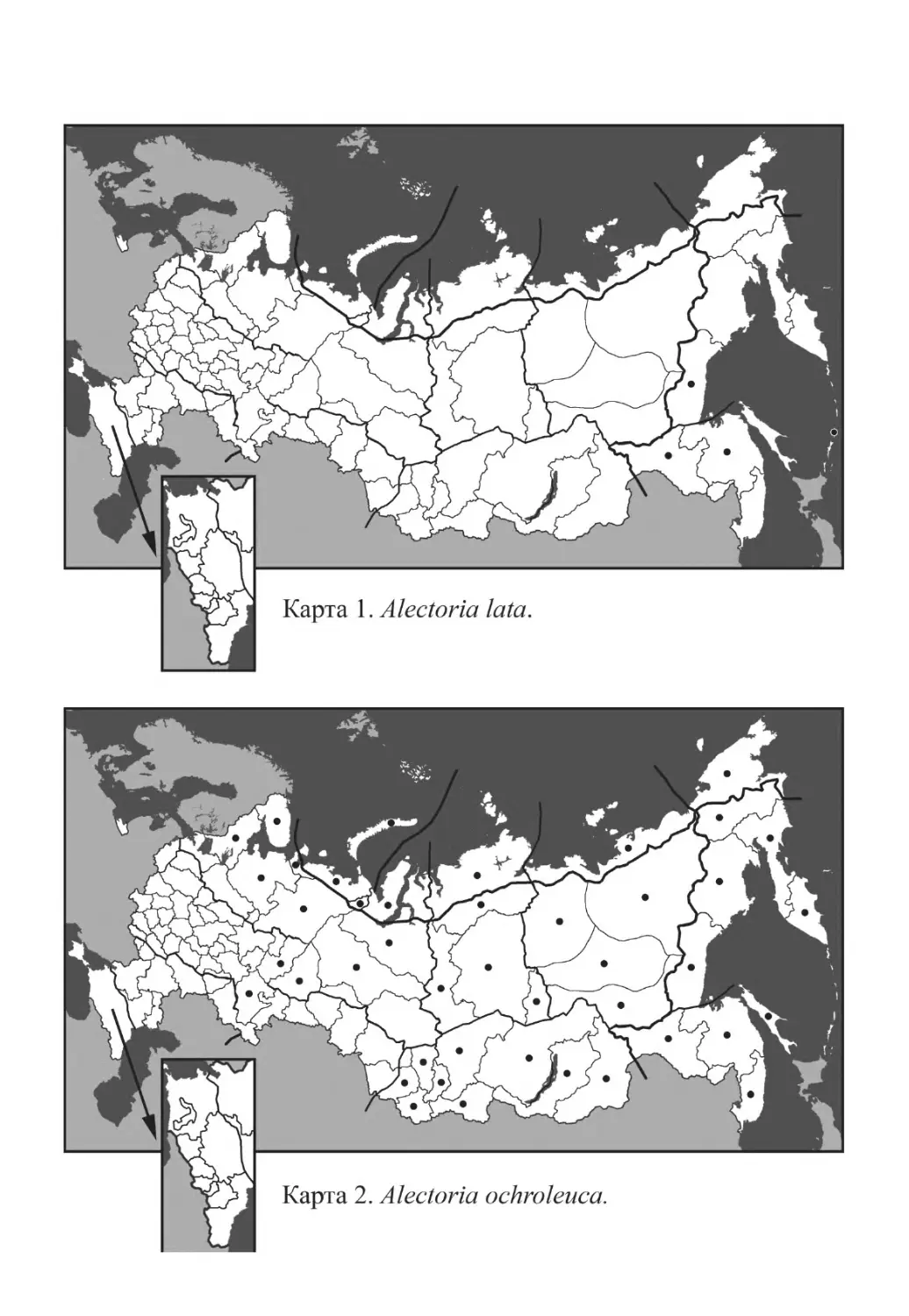
Alectoria lata (Taylor) Linds.
Alectoria ochroleuca (Hoffm.) A. Massal.
Alectoria sarmentosa (Ach.) Ach. subsp. sarmentosa
Alectoria sarmentosa (Ach.) Ach. subsp. vexillifera (Nyl.) D. Hawksw.
Allocetraria Kurok. et M. J. Lai (1991)
Allocetraria madreporiformis (Ach.) Kärnefelt et A. Thell
Allocetraria stracheyi (C. Bab.) Kurok. et M. J. Lai
Arctocetraria Kärnefelt et A. Thell (1993)
Arctocetraria andrejevii (Oxner) Kärnefelt et A. Thell
Arctocetraria nigricascens (Nyl.) Kärnefelt et A. Thell
Asahinea W. L. Culb. et C. F. Culb. (1965)
Asahinea chrysantha (Tuck.) W. L. Culb. et C. F. Culb.
Asahinea scholanderi (Llano) W. L. Culb. et C. F. Culb.
Bryocaulon Kärnefelt (1986)
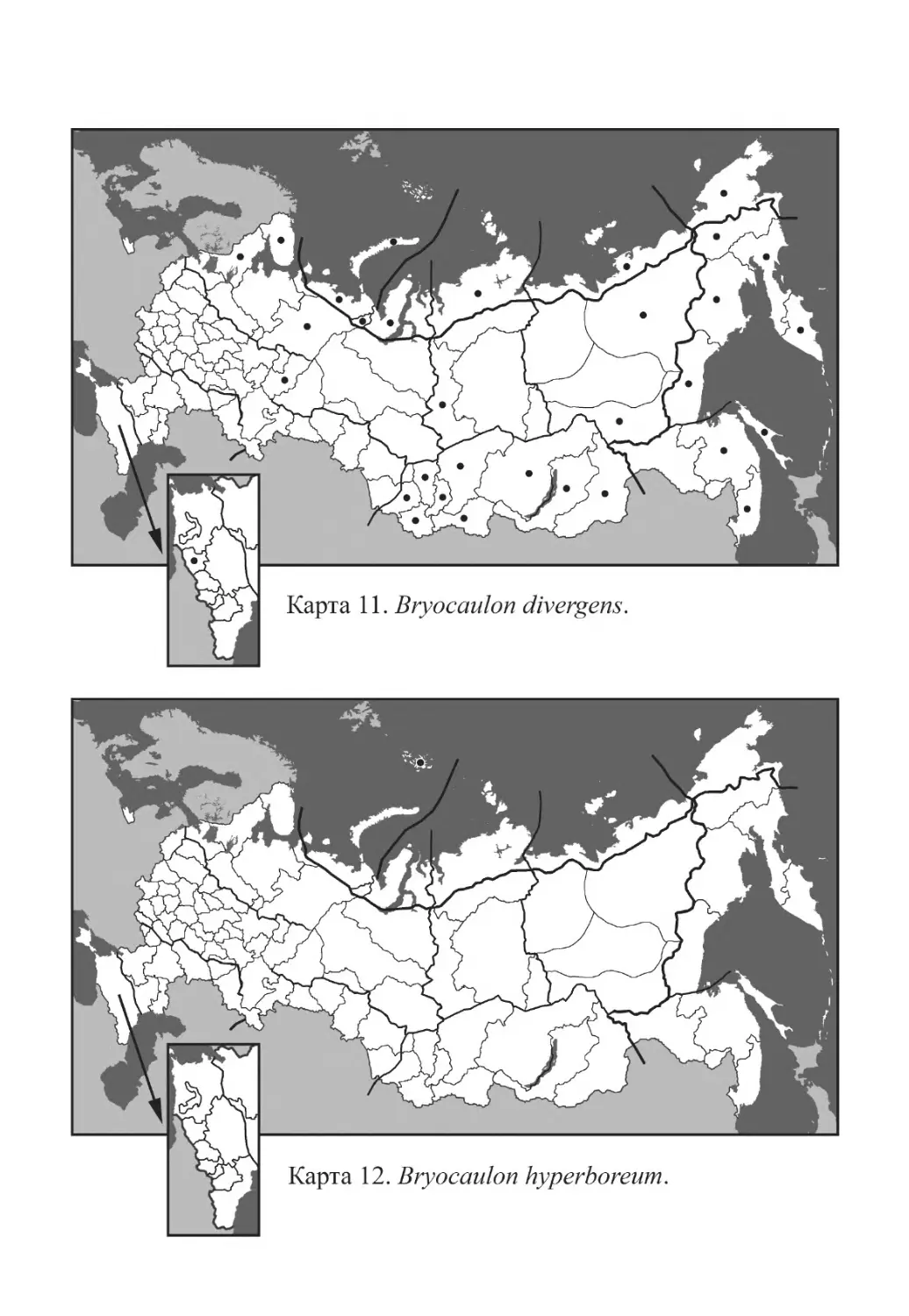
Bryocaulon divergens (Ach.) Kärnefelt
Bryocaulon hyperboreum Øvstedal
Bryocaulon pseudosatoanum (Asahina) Kärnefelt
Систематическое положение рассматриваемых таксонов
15
Bryoria Brodo et D. Hawksw. (1977)
Bryoria americana (Motyka) Holien
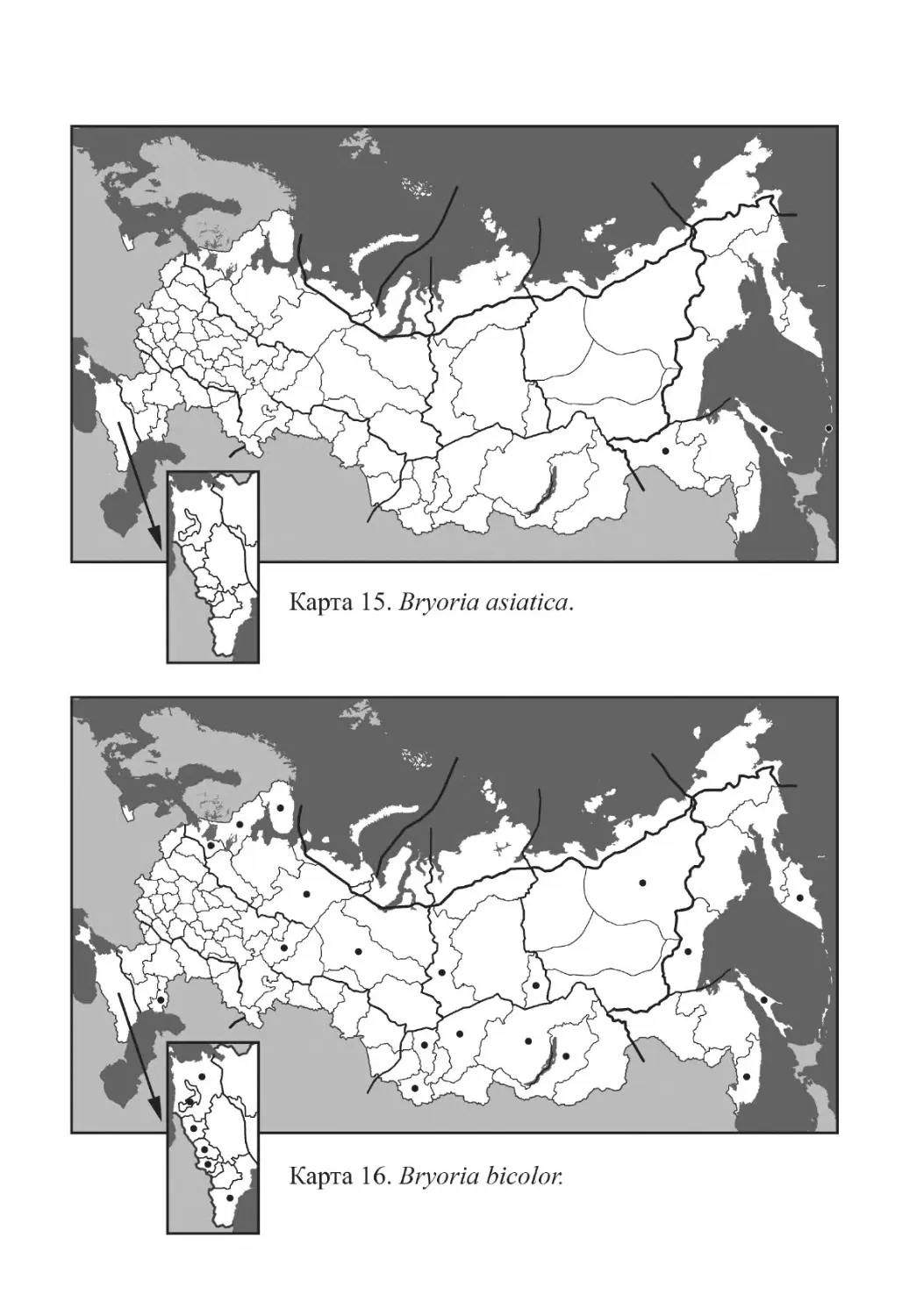
Bryoria asiatica (Du Rietz) Brodo et D. Hawksw.
Bryoria bicolor (Ehrh.) Brodo et D. Hawksw.
Bryoria capillaris (Ach.) Brodo et D. Hawksw.
Bryoria fremontii (Tuck.) Brodo et D. Hawksw.
Bryoria furcellata (Fr.) Brodo et D. Hawksw.
Bryoria fuscescens (Gyeln.) Brodo et D. Hawksw.
Bryoria glabra (Motyka) Brodo et D. Hawksw.
Bryoria implexa (Hoffm.) Brodo et D. Hawksw.
Bryoria kuemmerleana (Gyeln.) Brodo et D. Hawksw.
Bryoria nadvornikiana (Gyeln.) Brodo et D. Hawksw.
Bryoria nitidula (Th. Fr.) Brodo et D. Hawksw.
Bryoria simplicior (Vain.) Brodo et D. Hawksw.
Bryoria smithii (Du Rietz) Brodo et D. Hawksw.
Bryoria tenuis (E. Dahl) Brodo et D. Hawksw.
Bryoria trichodes (Michx.) Brodo et D. Hawksw.
Bryoria vrangiana (Gyeln.) Brodo et D. Hawksw.
Cetraria Ach. (1803)
Cetraria aculeata (Schreb.) Fr. subsp. aculeata
Cetraria aculeata (Schreb.) Fr. subsp. steppae (Savicz) Lutsak, Fernández-
Mendoza et Printzen
Cetraria crispiformis (Räsänen) Makryǐ, comb. et stat. nov.
Cetraria ericetorum Opiz subsp. ericetorum
Cetraria islandica (L.) Ach. subsp. islandica
Cetraria islandica (L.) Ach. subsp. orientalis (Asahina ex M. Satô) Kärnefelt
Cetraria kamczatica Savicz
Cetraria laevigata Rass.
Cetraria muricata (Ach.) Eckfeldt
Cetraria nigricans Nyl.
Cetraria odontella (Ach.) Ach.
Cetraria rassadinae Makryǐ
Cetraria sepincola (Ehrh.) Ach.
«Cetraria» annae Oxner
Cetrariella Kärnefelt et A. Thell (1993)
Cetrariella delisei (Schaer.) Kärnefelt et A. Thell
Cetrariella fastigiata (Nyl.) Kärnefelt et A. Thell
Cetreliopsis M. J. Lai (1980)
Cetreliopsis asahinae (M. Satô) Randlane et A. Thell
Систематическое положение рассматриваемых таксонов
Флора лишайников России Флора лишайников России
16
Dactylina Nyl. (1860)
Dactylina arctica (Richardson) Nyl. subsp. arctica
Dactylina arctica (Richardson) Nyl. subsp. beringica (C. D. Bird et J. W. Thom-
son) Kärnefelt et A. Thell
Dactylina ramulosa (Hook.) Tuck.
Evernia Ach. (1810)
Evernia divaricata (L.) Ach.
Evernia esorediosa (Müll. Arg.) Du Rietz
Evernia mesomorpha Nyl.
Evernia perfragilis Llano
Evernia prunastri (L.) Ach.
Evernia terrestris (Tomin) N. S. Golubk.
Flavocetraria Kärnefelt et A. Thell (1994)
Flavocetraria cucullata (Bellardi) Kärnefelt et A. Thell
Flavocetraria minuscula (Elenkin et Savicz) Ahti, Poryadina et Zhurb.
Flavocetraria nivalis (L.) Kärnefelt et A. Thell
Gowardia Halonen, Myllys, Velmala et Hyvärinen (2009)
Gowardia arctica Halonen, Myllys, Velmala et Hyvärinen
Gowardia nigricans (Ach.) Halonen, Myllys, Velmala et Hyvärinen
Letharia (Th. Fr.) Zahlbr. (1892)
Letharia vulpina (L.) Hue
Lethariella (Motyka) Krog (1976)
Lethariella togashii (Asahina) Krog
Masonhalea Kärnefelt (1977)
Masonhalea inermis (Nyl.) Lumbsch, Nelsen et A. Thell
Masonhalea richardsonii (Hook.) Kärnefelt
Nephromopsis Müll. Arg. (1891)
Nephromopsis endocrocea Asahina
Nephromopsis komarovii (Elenkin) J. C. Wei
Nephromopsis laii (A. Thell et Randlane) Saag et A. Thell
Nephromopsis laureri (Kremp.) Kurok.
Nephromopsis nephromoides (Nyl.) Ahti et Randlane
Nephromopsis ornata (Müll. Arg.) Hue
Nephromopsis pallescens (Schaer.) Y. S. Park var. pallescens
Систематическое положение рассматриваемых таксонов
17
Nephromopsis pseudocomplicata (Asahina) M. J. Lai
Nephromopsis rugosa Asahina
Nephromopsis yunnanensis (Nyl.) Randlane et Saag
Oropogon Th. Fr. (1861)
Oropogon asiaticus Asahina
Platismatia W. L. Culb. et C. F. Culb. (1968)
Platismatia glauca (L.) W. L. Culb. et C. F. Culb.
Platismatia interrupta W. L. Culb. et C. F. Culb.
Platismatia lacunosa (Ach.) W. L. Culb. et C. F. Culb.
Platismatia norvegica (Lynge) W. L. Culb. et C. F. Culb.
Tuckermannopsis Gyeln. (1933)
Tuckermannopsis chlorophylla (Willd.) Hale
Tuckermannopsis ciliaris (Ach.) Gyeln.
Tuckermannopsis gilva (Asahina) M. J. Lai
Tuckermannopsis orbata (Nyl.) M. J. Lai
Tuckermannopsis subalpina (Imshaug) Kärnefelt
Usnocetraria M. J. Lai et J. C. Wei (2007)
Usnocetraria oakesiana (Tuck.) M. J. Lai et J. C. Wei
Vulpicida J.-E. Mattsson et M. J. Lai (1993)
Vulpicida juniperinus (L.) J.-E. Mattsson et M. J. Lai
Vulpicida pinastri (Scop.) J.-E. Mattsson et M. J. Lai
Ниже приводятся остальные роды семейства Parmeliaceae, которые предпола-
гается рассмотреть в следующих томах настоящего издания, и род Protoparmelia,
который рассматривался в предыдущем, 2-м томе издания «Флора лишайников
России» (2017). Все они учитываются в приведенном ниже родовом ключе.
Allantoparmelia (Vain.) Essl.
Anzia Stizenb.
Arctoparmelia Hale
Brodoa Goward
Cetrelia W. L. Culb. et C. F. Culb.
Cornicularia (Schreb.) Hoffm.
Dolichousnea (Y. Ohmura) Articus
Flavoparmelia Hale
Flavopunctelia (Krog) Hale
Hypogymnia (Nyl.) Nyl.
Флора лишайников России Флора лишайников России
18
Hypotrachyna (Vain.) Hale (включая Everniastrum Hale ex Sipman и Parme-
linopsis Elix et Hale)
Imshaugia S. L. F. Mey.
Melanelia Essl.
Melanelixia O. Blanco, A. Crespo, Divakar, Essl., D. Hawksw. et Lumbsch
Melanohalea O. Blanco, A. Crespo, Divakar, Essl., D. Hawksw. et Lumbsch
Menegazzia A. Massal.
Montanelia Divakar, A. Crespo, Wedin et Essl.
Myelochroa (Asahina) Elix et Hale
Neofuscelia Essl.
Nipponoparmelia (Kurok.) K. H. Moon, Y. Ohmura et Kashiw.
Parmelia Ach.
Parmelina Hale
Parmeliopsis (Nyl.) Nyl.
Parmotrema A. Massal.
Pleurosticta Petr.
Protoparmelia M. Choisy
Pseudephebe M. Choisy
Pseudevernia Zopf
Punctelia Krog
Remototrachyna Divakar et A. Crespo
Usnea Dill. ex Adans.
Xanthoparmelia (Vain.) Hale
М. П. Андреев
Семейство Parmeliaceae Bercht. et J. Presl
1820, Prir. Rostlin: 273; Eschw., 1824, Syst. Lich.: 19; Zenker, in Goebel u. Kunze,
1827, Pharmaceutische Waarenkunde (Eisenach) 1 (3): 124. Т и п : Parmelia Ach. —
Пармелиевые
= Alectoriaceae Bercht. et J. Presl, 1820, Prir. Rostlin 1: 273. Т и п : Alectoria Ach.
= Usneaceae Eschw., 1824, Syst. Lich.: 23. Т и п : Usnea Dill. ex Adans.
= Cetrariaceae Schaer., 1850, Enum. Crit. Lich. Eur.: 12. Т и п : Cetraria Ach.
= Corniculariaceae Schaer., 1850, Enum. Crit. Lich. Eur.: 4 («ordo Cornicularii»).
Тип: Cornicularia (Schreb.) Hoffm.
= Everniaceae Tomas., 1949, Arch. Bot. (Forli) 25, ser. 3, 9, 3–4: 238.
Тип: Evernia Ach.
= Anziaceae M. Satô, in Nakai, Honda, 1939, Nov. Fl. Jap. 5: 4. Т и п : Anzia Stizenb.
= Hypogymniaceae Poelt ex Elix, 1980, Brunonia 2: 176. Т и п : Hypogymnia (Nyl.)
Nyl.
Dactylinaceae M. Choisy, 1951, Bull. Mens. Soc. Linn. Lyon 20, 6: 137, nom. inval.
Лихенизированные грибы1
.
ТАЛЛОМ листоватый, с приподнимающимися или
приросшими к субстрату лопастями, либо кустистый, прямостоячий, в виде ку-
стика или свисающий, редко трубчатый (пальцевидный), но может быть умби-
ликатным или пельтатным, до накипного или почти накипного, с развитыми
верхним и нижним коровыми слоями. Лопасти дорсивентральные до изодиаме-
трических. Верхняя поверхность обычно беловато-серая, желтоватая, зеленая или
коричневая до черноватой или разных оттенков этих цветов. Кора параплектен-
химная, иногда прозоплектенхимная, в том числе с антиклинальной (палисадной)
ориентацией гиф. Нижняя поверхность черноватая до светло-коричневой или
одного цвета с верхней поверхностью, обычно с ризинами, редко без них. Серд-
цевина плотная либо часто более рыхлая, белая, у некоторых видов окрашена в
желтоватые или красноватые тона. ПЛОДОВЫЕ ТЕЛА — апотеции зеоринового или
леканорового типа, как правило, с чашевидным талломным краем одного цвета
с талломом, расположены на ножках или непосредственно на поверхности тал-
лома, изредка погруженные, развиваются обычно на поверхности или по краям
лопастей, у некоторых видов — только на концах лопастей. Парафизы простые
или (редко) слегка разветвленные. Сумки обычно булавовидные, реже цилиндри-
ческие, леканорового типа с амилоидным толусом и выраженным аксиальным
телом, реже с сильно амилоидной кольцевой структурой, содержащие обычно 8,
1
Семейство Parmeliaceae включает также ряд родов лихенофильных грибов: Nesolechia
A. Massal., Phacopsis Tul., Raesaenenia D. Hawksw. В публикуемом ниже родовом ключе они не
приводятся.
Флора лишайников России Флора лишайников России
20
реже 2 или 4 аскоспоры. Аскоспоры простые (одноклеточные), эллипсоидные до
круглых, тонкостенные, бесцветные, реже буроватые. КОНИДИАЛЬНЫЕ СПОРОНОШЕ-
НИЯ в форме пикнид. Пикниды погруженные или реже выступающие, темно-ко-
ричневые до черных, образуются на верхней поверхности таллома, могут быть
расположены на концах, по краю и вдоль выростов таллома. Конидии разной
формы, прямые или изогнутые, продолговатые, цилиндрические, палочковидные,
игловидные, нитевидные, обычно с одним или двумя расширенными концами,
либо с расширением в центре (т. е. бутылковидные, гантелевидные или веретено-
видные), иногда с хвостиком. ФОТОБИОНТ Trebouxia-типа, часто Trebouxia jamesii,
у некоторых родов Asterochloris.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Депсиды и депсидоны β-орсинольного типа, жир-
ные кислоты и усниновая кислота.
ЭКОЛОГИЯ. Эпигейные лишайники, эпифиты, эпиксилы, эпибриофиты и эпили-
ты, в тропических широтах также эпифиллы; в разнообразных тундровых, лес-
ных, лесостепных и степных сообществах в горах и на равнине, от полярных пу-
стынь до тропиков.
РАСПРОСТРАНЕНИЕ. Семейство Parmeliaceae, одно из крупнейших и наибо-
лее разнообразное в классе Lecanoromycetes, насчитывает в настоящее вре-
мя по разным оценкам от 70 до 89 (Lücking et al., 2016; Wijayawardene et al.,
2022) родов и около 2740 видов. Наибольшее количество видов известно в ро-
дах Xanthoparmelia (822 вида), Usnea (355 видов), Hypotrachyna (262 вида) и
Parmotrema (250 видов). Представители семейства известны в обоих полушариях,
но центр их разнообразия находится в умеренной и субтропической зонах Юж-
ного полушария. Для территории России известно в настоящее время 54 рода и
более 300 видов. Виды семейства Parmeliaceae широко распространены во всех
регионах России, где встречаются в самых разных местообитаниях.
ПРИМЕЧАНИЕ. Систематика семейства Parmeliaceae в настоящее время не мо-
жет считаться устоявшейся и претерпевает существенные изменения. Несмотря
на то что семейство остается одной из наиболее изучаемых групп лишайников,
некоторые связи внутри него остаются невыясненными. Основываясь на по-
следних филогенетических исследованиях, сем. Parmeliaceae рассматривает-
ся как монофилетическая группа, которая включает ранее описанные семейства
Alectoriaceae, Anziaceae, Hypogymniaceae, Usneaceae и др., которые более не
признаются большинством систематиков. Тем не менее, филогенетический ана-
лиз предполагает существование в семействе от 6 до 9 отдельных клад, напри-
мер Alectorioid (3 рода), Cetrarioid (8 родов), Hypogymnioid (4 рода), Letharioid
(2 рода), Parmelioid (26 родов), Psiloparmelioid (2 рода), причем еще 42 рода пока
не находят в этой системе своего места (Crespo et al., 2010).
ИЗБРАННАЯ ЛИТЕРАТУРА. Голубкова, 1996а; Определитель лишайников России,
1996; Crespo et al., 2007, 2010; Hawksworth et al., 2008; Nordic Lichen Flora, 2011;
Урбанавичюс, 2011; Thell et al., 2012; Lücking et al., 2016; Divakar et al., 2017;
Wijayawardene et al., 2022.
Семейство Parmeliaceae
21
Ключ для определения родов сем. Parmeliaceae России
При составлении ключа для определения родов сем. Parmeliaceae России в ка-
честве вспомогательных использованы ранее опубликованные материалы (Голуб-
кова, 1996а; Nordic..., 2011; Урбанавичюс, 2011; Урбанавичюс и др., 2014; и др.).
1. Таллом накипной, от трещиновато-ареолированного до чешуйчатого или зер-
нистого, от серовато- до темно-коричневого, часто ярко-коричневый, нередко
блестящий ...................................................................................... Protoparmelia.
—
Таллом кустистый или узколопастный (субфрутикозный), прямостоячий или
повисающий, прикрепленный к субстрату основанием с помощью ризоидов,
ризин или псевдогомфа, реже стелющийся, прикрепленный к субстрату во
многих местах гаптерами или ризинами, иногда неприкрепленный ............ 2.
—
—
Таллом листоватый ........................................................................................ 32.
2. Произрастают на почве (в тундрах, болотах, в лесах и степях) .................... 3.
—
Произрастают на древесине и коре (на ветвях и стволах деревьев, на заборах,
крышах и пр.) либо на камнях и скалах ......................................................... 17.
3. Таллом состоит из восходящих, пальцевидно вздутых ветвей, полых внутри:
отдельных или собранных в группы, неразветвленных или (иногда) умеренно
дихотомически разветвленных, коралловидных, гладких, округлых или слегка
уплощенных .......................................................................................... Dactylina.
—
Таллом иной, кустовидно разветвленный; лопасти не полые, плоские до же-
лобчатых или цилиндрические .......................................................................... 4.
4. Таллом неприкрепленный, свободно лежащий на почве поверх мхов и лишай-
ников и способный перемещаться под воздействием ветра ........................... 5.
—
Таллом прикрепленный к субстрату или плотно сидящий в дернинке, прямо-
стоячий из собранных вместе или раздельных лопастей либо в виде сильно
разветвленных кустиков, прямостоячих, реже стелющихся ........................... 9.
5. Таллом желтоватый или зеленоватый ............................................................... 6.
—
Таллом коричневый до черного ......................................................................... 8.
6. Лопасти угловато-округлые до цилиндрических, иногда неравномерной тол-
щины, слегка уплощенные в местах ветвления, одинаковые с верхней и с
нижней стороны ....................................................................................... Evernia.
—
Лопасти плоские, лентовидные, линейные или желобчатые, дорсивентраль-
ные, иногда с заворачивающимися на нижнюю сторону краями .................. 7.
7. Таллом в виде дернинок, 1.5–2.0 (редко до 5) см в диам.; лопасти хрящева-
тые, соломенно-желтые, гладкие, снизу бледнее окрашенные, жилковато- или
сетчато-морщинистые, соралии не образуются; азиатский горностепной и
высокогорный вид ........................................................... Allocetraria stracheyi.
—
Таллом более крупный, 3–6 см дл., лопасти сверху беловато- или серовато-
зеленые, снизу беловатые, иногда одноцветные, соралии округлые; преиму-
щественно в Европейской России ........................................ Evernia prunastri.
Семейство Parmeliaceae
Флора лишайников России Флора лишайников России
22
8. Таллом состоит из одиночных или образующих скопления, неправильно или
дихотомически разветвленных плоских лопастей 0.5–20 мм шир., расходя-
щихся во все стороны подобно рогам оленя, коричневый до темно-бурого;
псевдоцифеллы хорошо выражены, развиваются на нижней поверхности в
виде непрерывной каймы по краям лопастей или покрывают большие участ-
ки нижней поверхности ............................................. Masonhalea richardsonii.
—
Таллом состоит из веточек, объединенных в неприкрепленные шаровидные
пучки 1.5–3(4) см в диам., светло-каштановый до темно-красновато-корич-
невого; веточки 1–2(4) мм в диам., округло- или угловато-цилиндрические,
местами сжатые, иногда ямчатые, относительно мягкие, гибкие, не ломкие,
внутри полые или с рыхлой, паутинистой сердцевиной; боковые веточки ди-
хотомически или неправильно разветвленные, гладкие, обычно матовые или
лишь слегка блестящие; псевдоцифеллы не обильные, но хорошо заметные,
овальные, погруженные, около 0.5 мм шир. и до 2 мм дл. ................................
...........................................................................
Cetraria aculeata subsp. steppae.
9. Таллом бледно-желтый, желтоватый или зеленоватый ................................. 10.
—
Таллом коричневатый или сероватый до черного ......................................... 12.
10. Лопасти плоские, часто желобчатые, не полые, собранные вместе или раз-
дельные, чаще вертикальные, бледно-желтые, в основании красновато-лило-
вые или коричневые ....................................................................... Flavocetraria.
—
Лопасти цилиндрические, неразветвленные или дихотомически разветвлен-
ные, довольно узкие, продолговатые и характерно выпуклые, либо пальце-
видные ................................................................................................................. 11.
11. Веточки цилиндрические, иногда сжатые у основания, желтые или желто-зе -
леные, обычно с чернеющими кончиками, часто с белыми веретеновидными
псевдоцифеллами до 1 мм дл. .......................................... Alectoria ochroleuca.
—
Веточки округлые в сечении, без выраженной дорсивентральности, пальце-
видные, либо желобчатые и трубчато свернутые, до 2–10 мм шир., с волни-
стыми или вырезанными краями, бледно-соломенные, желтые или беловато-
зеленоватые, у основания буроватые, буровато-черные, иногда с красноватым
или лиловым оттенком; псевдоцифеллы отсутствуют ................. Allocetraria.
12. Таллом стелющийся или восходящий; ветви цилиндрические или угловато-
цилиндрические, иногда сжатые у основания и в местах ветвления .......... 13.
—
Таллом, как правило, восходящий; лопасти плоские, желобчатые или трубча-
то свернутые ....................................................................................................... 15.
13. Таллом обычно стелющийся, двухцветный, темно-серый или темно-коричне-
вый до черного, у основания более светлый, матовый или блестящий; ветви
преимущественно цилиндрические; псевдоцифеллы многочисленные, хоро-
шо выраженные, плоские, вытянуто-веретеновидные или овальные, белые;
соралии отсутствуют или образуются очень редко; коровой слой и сердце-
вина от K и P желтеют, от KC краснеют, таллом содержит алекториаловую
и/или барбатоловую кислоты ............................................................. Gowardia.
Семейство Parmeliaceae
23
—
Таллом приподнимающийся или стелющийся, красновато- или каштаново-
коричневый до коричневато-черного, блестящий .......................................... 14.
14. Сердцевина и псевдоцифеллы от C не изменяются; таллом содержит лихе-
стериновую и протолихестериновую кислоты; ветви коричневые до желто-
коричневых, у основания иногда красноватые, многократно разветвленные,
с многочисленными боковыми веточками и шипиками, растущими в разные
стороны; реснички по краям лопастей рассеянные и разветвленные; псевдо-
цифеллы овальные или вытянутые ............................... Cetraria (Coelocaulon).
—
Сердцевина и псевдоцифеллы от C и KC краснеют; таллом содержит оливе-
торовую кислоту; ветви гладкие, местами мелкоямчатые, блестящие, корич-
невые до красно-коричневых, у основания более темные, боковые веточки,
шипики и реснички отсутствуют; псевдоцифеллы веретеновидные, не погру-
женные, приподнятые над поверхностью ветвей или плоские ... Bryocaulon.
15. Верхний и нижний коровые слои двухслойные, 20–130 мкм толщ., наруж-
ный слой более толстый, параплектенхимный, внутренний тонкий, прозо-
плектенхимный; таллом содержит лихестериновую, протолихестериновую,
фумарпротоцетраровую (от P краснеет) и/или норстиктовую (от K краснеет)
кислоты .................................................................................................... Cetraria.
—
Верхний и нижний коровые слои однослойные, 15–40 мкм толщ.; таллом со-
держит другие кислоты ..................................................................................... 16.
16. Сердцевина от C и KC краснеет, содержит гирофоровую и хиасциевую кис-
лоты; конидии бутылковидные; сумки широкобулавовидные, с толусом без
кольцевых структур ............................................................................ Cetrariella.
—
Сердцевина от C и KC не изменяется, содержит рангиформовую и норранги-
формовую кислоты; конидии веретеновидные; сумки узкобулавовидные или
цилиндрические, с кольцевыми структурами в толусе ............. Arctocetraria.
17(2). Произрастают на скалах и камнях ............................................................... 18.
—
Произрастают на деревьях, кустарниках и другом древесном субстрате, но
могут встречаться и на камнях ........................................................................ 21.
18. Сердцевина по периферии паутинистая, в центральной части с плотным
хрящевидным механическим тяжом; таллом чаще однообразно окрашен в
зеленый цвет, либо черная и зеленоватая окраска чередуются в виде полос ..
........................................................................................................................
Usnea.
—
Сердцевина паутинистая, без центрального хрящевидного тяжа ................ 19.
19. Таллом с верхней стороны беловато- или светло-сероватый, с нижней — тем-
но-коричневый; лопасти по краю с хорошо заметными простыми или развет-
вленными ризинами ........................................... Hypotrachyna (Everniastrum).
—
Таллом коричневый до черного; ветви без боковых ризин .......................... 20.
20. Таллом в виде прямостоячих дернинковидных кустиков, прикрепленных
основанием; ветви в некоторых местах и не всегда отчетливо уплощенные;
апотеции развиваются на концах лопастей ..........................................................
..........................................................
Cornicularia normoerica (Gunn.) Du Rietz.
Флора лишайников России Флора лишайников России
24
—
Таллом стелющийся, прикрепленный к субстрату всеми частями с помощью
гаптер; ветви цилиндрические, местами более или менее уплощенные; апоте-
ции боковые ..................................................................................... Pseudephebe.
21. Лопасти более или менее плоские; таллом дорсивентрального строения,
верхняя и нижняя поверхности окрашены в разные цвета .......................... 22.
—
Ветви цилиндрические, угловато-цилиндрические, иногда местами уплощен-
ные; таллом радиального строения, лопасти и ветви одинаково окрашенные
со всех сторон .................................................................................................... 24.
22. Верхняя поверхность зеленоватая, нижняя — светлая до белой ....... Evernia.
—
Верхняя поверхность светлая, сероватая или бежевая, нижняя — темно-ко-
ричневая до черной или пестро окрашенная ................................................. 23.
23. Лопасти по краю с хорошо заметными простыми или разветвленными ризи-
нами; редкий лишайник, встречается на юге Дальнего Востока .......................
................................................................................
Hypotrachyna (Everniastrum).
—
Краевые ризины отсутствуют; широко распространенный лишайник .............
........................................................................
Pseudevernia furfuracea (L.) Zopf.
24. Таллом зеленоватый, желтоватый или серно-желтый ................................... 25.
—
Таллом иначе окрашенный ............................................................................... 29.
25. Ветви таллома нитевидные, сильно удлиненные, тонкие, обычно до 2 мм
толщ., но могут быть и толще, в сечении цилиндрические, реже угловато-
округлые; сердцевина с плотным осевым тяжом, изредка паутинистая, одно-
родная .................................................................................................................. 26.
—
Ветви таллома более толстые, до 5 мм толщ., угловато-округлые, местами
уплощенные; сердцевина паутинистая ............................................................ 28.
26. Сердцевина без осевого тяжа, однородная, ветви легко разрываются; фибрил-
лы отсутствуют; на поверхности ветвей хорошо заметны выступающие вере-
теновидные псевдоцифеллы ............................................ Alectoria sarmentosa.
—
Сердцевина с плотным осевым тяжом, обнажающимся при механическом
разрыве ветвей ................................................................................................... 27.
27. Таллом прямостоячий или свисающий; веточки обычно с боковыми фибрил-
лами, редко без них; осевой тяж от I синеет или не изменяется; псевдоцифел-
лы отсутствуют, если развиваются, то в виде пятен .............................. Usnea.
—
Таллом свисающий; веточки могут быть без коры [напр., D. longissima (Ach.)
Articus], псевдогомф часто отсутствует; осевой тяж от I синеет; псевдоцифел-
лы кольцевые, развиваются между сегментами таллома ........... Dolichousnea.
28. Таллом яркий, серно- или зеленовато-желтый, с мелкими точечными серно-
желтыми соралиями; встречается в горных лесах Крыма и Кавказа ................
...................................................................................................... Letharia vulpina.
—
Таллом зеленоватый; соралии, если образуются, беловатые; широко распро-
страненные лишайники ........................................................................... Evernia.
29. Таллом темно- и пепельно-серый, покрытый короткими сосочкообразными
или удлиненными изидиями; лопасти угловато-цилиндрические, до 5 мм
Семейство Parmeliaceae
25
шир.; сердцевина с центральным цилиндром, внутри полым, пронизанным
отдельными тяжами гиф; редкий причерноморский и средиземноморский
вид ........................................................................................ Lethariella intricata.
—
Таллом коричневатый, коричнево-черный до черного, изредка бледно-серова-
тый, без изидиев; сердцевина паутинистая, однородная .............................. 30.
30. Широко распространенные лесные виды; ветви тонкие, 0.2–0.7 мм в диам.,
волосовидные до нитевидных; коровой слой однослойный с периклинально
(параллельно поверхности) расположенными гифами ....................... Bryoria.
—
Редкие дальневосточные океанические виды; ветви таллома более толстые,
до 1 мм шир. ...................................................................................................... 31.
31. Таллом светло-каштановый до коричневого, обычно повисающий; ветви
гладкие, блестящие, местами мелкоямчатые или с продольными бороздами,
в местах ветвления слегка уплощенные, на концах вильчато разветвленные;
псевдоцифеллы точковидные, слегка приподнятые над поверхностью тал-
лома; аскоспоры одноклеточные, бесцветные; сердцевина белая, от C и KC
краснеет, от K и P не изменяется, содержит оливеторовую кислоту ................
...............................................................................................................
Bryocaulon.
—
Таллом коричневый до темно-коричневого, дернинковидный; ветви гладкие,
более или менее блестящие, на поперечном срезе округлые; псевдоцифеллы в
виде удлиненных беловатых перфораций, открывающихся в центральную по-
лость, реже плоские, закрытого типа; сумки содержат одну темную мураль-
ную аскоспору; сердцевина рыхлая, содержащая желтоватый пигмент, от P
становится золотисто-желтой, от K — слегка пурпурной, от C не изменяется,
содержит псоромовую, иногда также конпсоромовую кислоты .... Oropogon.
32(1). Нижняя кора отсутствует, поверхность лопастей покрыта толстым рыхлым
губчатым слоем черно-коричневого войлока; лопасти более или менее четко-
видные; южные дальневосточные виды ................................................... Anzia.
—
Нижняя поверхность без войлока, покрыта корой, с ризинами или без ризин
............................................................................................................................... 33.
33. Таллом без ризин ............................................................................................... 34.
—
Таллом прикрепляется к субстрату ризинами ................................................ 38.
34. Сердцевина внутри с полостью ....................................................................... 35.
—
Сердцевина компактная, без полости .............................................................. 36.
35. Верхняя поверхность таллома с округлыми или овальными отверстиями ......
..............................................................................................................
Menegazzia.
—
Верхняя поверхность без отверстий .............................................. Hypogymnia.
36. Таллом крупнолистоватый, 10–20 см в диам.; лопасти широкие, округлые,
3–20 мм шир.; верхняя поверхность беловато-сероватая, зеленовато-желтова-
тая, местами оливковая или черно-оливковая .................................... Asahinea.
—
Таллом мелколистоватый, 3–6 см в диам.; лопасти узкие, 0.5–1.5 мм шир.,
вытянутые, линейные, выпуклые; верхняя поверхность обычно темная, ко-
ричневато-черноватая, серовато-черная, черная, реже серебристо-серая ... 37.
Флора лишайников России Флора лишайников России
26
37. Коровой слой от K желтеет, сердцевина от K и P не изменяется, от KC крас-
неет или не изменяется; нижний коровой слой образован палисадной про-
зоплектенхимой; содержит атранорин, хлоратранорин, протоцетраровую,
фумарпротоцетраровую и физодовую кислоты ..................................... Brodoa.
—
Коровой слой от K не изменяется (атранорин отсутствует), сердцевина от K
желтеет, от KC и P становится интенсивно желтой или кирпично-красной;
нижний коровой слой параплектенхимный; содержит оливеторовую, алекто-
риаловую, барбатовую и α-коллатоловую кислоты ............... Allantoparmelia.
38. Таллом очень плотно прилегающий к субстрату; лопасти мелкие, 0.2–15 мм
дл. и 0.1–2.0 мм шир. ........................................................................................ 39.
—
Таллом менее плотно прилегает к субстрату; лопасти более длинные и широ-
кие, по краям нередко приподнимающиеся; иногда таллом неприкрепленный
............................................................................................................................... 40.
39. Таллом с короткоцилиндрическими или зернистыми изидиями; нижняя по-
верхность более или менее светлая до бледно-коричневой со светлыми про-
стыми ризинами; конидии короткие, прямые, бутылковидные, с 1(2) утолще-
нием; сердцевина от K желтеет, содержит тамноловую кислоту (UV–)
...........
................................................................................................................. Imshaugia.
—
Таллом без изидий, с соредиями или без; нижняя поверхность коричневато-
черная, с темными простыми ризинами; конидии серповидные; сердцевина
от K не изменяется, содержит диварикатовую кислоту (UV+ белеет) ..............
............................................................................................................. Parmeliopsis.
40. Таллом неприкрепленный, свободно лежащий на почве поверх мхов и лишай-
ников; произрастают в горных и равнинных степях ..................................... 41.
—
Таллом прикрепленный к субстрату ................................................................ 42.
41. Таллом зеленоватый .................................................................. Xanthoparmelia.
—
Таллом коричневатый до почти черного ........................................ Neofuscelia1
.
42. Таллом неплотно прилегающий к субстрату, с приподнимающимися свобод-
ными краями; ризины немногочисленные, развиваются в центральной части
таллома либо почти достигают края, с голой краевой зоной до 6 мм шир., в
последнем случае края лопастей с ресничками ............................................. 43.
—
Таллом более или менее плотно прилегающий к субстрату всей нижней по-
верхностью; ризины многочисленные, развиваются до самых краев лопастей
............................................................................................................................... 52.
43. Таллом желтый, желтовато-зеленоватый или зеленовато-соломенный ....... 44.
—
Таллом иначе окрашенный, без желтого оттенка .......................................... 47.
44. Сердцевина белая; лопасти широкие, 5–10 мм шир. и 1.5–5.0 см дл., иногда
по краям с узкой черной каймой; апотеции образуются на нижней поверхно-
сти концов лопастей; на краях лопастей иногда развиваются соралии в виде
сплошной или прерывистой каймы ............................................ Nephromopsis.
1
Представители рода Neofuscelia часто рассматриваются в составе рода Xanthoparmelia.
Семейство Parmeliaceae
27
—
Сердцевина желтая, шафраново-желтая или оранжевая ............................... 45.
45. Сердцевина оранжевая, шафраново- или сернисто-желтая, от K интенсивно
желтеет или становится фиолетовой, содержит эндокроцин или секалоновую
кислоту; апотеции развиваются на нижней стороне концов лопастей ..............
..........................................................................................................
Nephromopsis.
—
Сердцевина желтая, от K не изменяется ......................................................... 46.
46. Лопасти часто с восходящими концами, иногда круглые в сечении; сердце-
вина интенсивно желтая, нижняя поверхность светло-желтая, более темная в
центре; апотеции часто с кренулированным краем; сумки широкобулавовид-
ные, 40–50 × 20–27 мкм; пикниды черные, хорошо заметные, конидии 6–8 ×
1–2 мкм; сердцевина и соредии содержат вульпиновую и пинастровую кис-
лоты; виды, широко распространенные в арктических, бореальных и горных
регионах Северного полушария .......................................................... Vulpicida.
—
Лопасти лентовидные, слегка вогнутые и слабо морщинистые; сердцевина
белая до светло-желтой, нижняя поверхность светло-коричневая до бело-
ватой; апотеции с тонким краем, который может быть соредиозным; сумки
узкобулавовидные, 30–40 × 7–12 мкм; пикниды темные или светлые, конидии
9–12 × 1–2 мкм; сердцевина содержит каператовую, лихестериновую, про-
толихестериновую и иногда секалоновую кислоты; довольно редкие лишай-
ники, отмеченные преимущественно в горных районах европейской части
России, на Кавказе и на Дальнем Востоке, но более обычны в горных районах
Юж. Сибири .................................................................................... Usnocetraria.
47. Края лопастей без ресничек .......................................................... Parmotrema1
.
—
Края лопастей с черными ресничками до 1–2 мм дл.; ризины развиваются
почти до краев лопастей, оставляя голой лишь кайму до 6 мм шир. ......... 48.
48. Верхняя поверхность с псевдоцифеллами ...................................................... 49.
—
Верхняя поверхность без псевдоцифелл ........................................................ 51.
49. Таллом некрупный, 3–4 см в диам., с узкими, 1–4 мм шир., слегка желоб-
чатыми лопастями, бледно-коричневый до зеленоватого; псевдоцифеллы
обычно развиты слабо; апотеции многочисленные; содержит алектороновую,
α-коллатоловую и оливеторовую кислоты ........................... Tuckermannopsis.
—
Таллом крупный, 5–25 см в диам., с довольно широкими, 3–25 мм шир., окру-
глыми беловато-сероватыми или зеленовато-сероватыми до коричневатых
лопастями; псевдоцифеллы развиваются в виде округлых, продолговатых или
неопределенной формы пятен .......................................................................... 50.
1
Представители рода Parmotrema, обитающие в горных районах Дальнего Востока и харак-
теризующиеся следующими признаками: лопасти зубчато надрезанные с приподнимающимися
краями; верхняя поверхность серая с сетчатым рисунком псевдоцифелл; нижняя поверхность
черная с густыми, простыми или разветвленными ризинами; споры более мелкие, чем у ти-
пичных Parmotrema, 10–18 × 6–10 мкм, с тонким эписпорием; талломы содержат атранорин,
хлоратранорин, салациновую и каператовую кислоты и норлобаридон, — ранее выделялись в
самостоятельный род Rimelia Hale et A. Fletcher.
Флора лишайников России Флора лишайников России
28
50. Верхняя поверхность таллома часто сильно морщинистая, серая, зеленовато-
коричневая, реже коричневая или беловатая; верхний коровой слой и серд-
цевина от I синеют; аскоспоры мелкие, почти шаровидные, 5–8 × 3–5 мкм;
пикниды образуются редко; содержит атранорин и каператовую кислоту ......
...............................................................................................................
Platismatia.
—
Верхняя поверхность гладкая, беловато-сероватая до почти белой; соредии
иногда образуют белую кайму по краям лопастей, лопастинки отсутствуют;
верхний коровой слой и сердцевина от I не изменяются; аскоспоры эллипсо-
идные, 11–24 × 6–12 мкм; пикниды образуются часто; содержит атранорин,
алектороновую, анциаевую и другие кислоты, каператовая кислота отсутству-
ет ................................................................................................................ Cetrelia.
51. Псевдоцифеллы и апотеции имеются ........................................ Nephromopsis.
—
Псевдоцифеллы отсутствуют; лопасти 3–30 мм шир., часто с соралиями,
прорастающими изидиями; верхняя поверхность таллома зеленовато- или
голубовато-сероватая, реже слегка коричневатая, нижняя светло- или темно-
коричневая, по краю более светлая, блестящая; содержит атранорин и капера-
товую кислоту; широко распространенный вид ................ Platismatia glauca.
52(42). Таллом всегда коричневый, черно-коричневый, серо-коричневый, зелено-
коричневый, верхняя поверхность обычно блестящая (редко матовая и тогда
с налетом), с псевдоцифеллами или без, пикниды погруженные или поверх-
ностные по краю лопастей ............................................................................... 53.
—
Таллом иначе окрашенный ............................................................................... 60.
53. Верхний коровой слой от N окрашивается ..................................................... 54.
—
Верхний коровой слой от N не окрашивается ................................................ 55.
54. Верхний коровой слой от N становится фиолетовым, сердцевина от P стано-
вится желто-оранжевой, от K краснеет; лопасти вогнутые, с приподнимаю-
щимися краями, часто с налетом; на коре деревьев или на скалах ...................
..............................................................................................................
Pleurosticta.
—
Верхний коровой слой от N становится сине-зеленым, сердцевина от P и K не
изменяется; лопасти плоские, с прижатыми к субстрату краями, без налета;
на скалах или почве .......................................................................... Neofuscelia1
.
55. Лопасти вогнутые, с возвышающимися цилиндрическими пикнидами, рас-
положенными по краю; нижняя поверхность светло-коричневая; все реакции
сердцевины негативные ........................... Melanelia commixta (Nyl.) A. Thell.
—
Лопасти обычно плоские, или слегка выпуклые, или вогнутые, пикниды по-
груженные или слегка возвышающиеся; реакции сердцевины позитивные
или негативные .................................................................................................. 56.
56. Сердцевина от C краснеет ................................................................................ 57.
—
Сердцевина от C не изменяется или редко слабо желтеет ........................... 58.
1
Представители рода Neofuscelia часто рассматриваются в составе рода Xanthoparmelia.
Семейство Parmeliaceae
29
57. Преимущественно эпифитные виды; лопасти иногда с тонкими гиалиновыми
волосками, часто с изидиями или соредиями, без псевдоцифелл; содержат
леканоровую кислоту ........................................................................ Melanelixia.
—
Эпилитный вид; лопасти с псевдоцифеллами и соредиями, изидии и гиалино-
вые волоски отсутствуют; содержит гирофоровую кислоту ..............................
.............................................................. Montanelia tominii (Oxner) Divakar et al.
58. Преимущественно лесные эпифитные виды, иногда эпилитные; псевдоци-
феллы слабо выражены, развиваются часто на вершинках бородавочек или
изидий, соредии отсутствуют ......................................................... Melanohalea.
—
Эпилитные, преимущественно высокогорно-арктические виды, псевдоци-
феллы и соралии развиваются на поверхности или по краю лопастей, изидии
отсутствуют ........................................................................................................ 59.
59. Сердцевина от K и P не изменяется, от C не изменяется или краснеет; содер-
жит перлатоловую, стеноспоровую или гирофоровую кислоты .. Montanelia.
—
Сердцевина от K желтеет или не изменяется, от P краснеет или не изменяет-
ся, от C не изменяется; содержит стиктовую, норстиктовую, фумарпротоце-
траровую или протоцетраровую кислоты .......................................... Melanelia.
60(52). Верхняя поверхность таллома с псевдоцифеллами ................................. 61.
—
Верхняя поверхность таллома без псевдоцифелл ......................................... 67.
61. Края лопастей с черными простыми ресничками ....................... Parmotrema.
—
Края лопастей без ресничек ............................................................................. 62.
62. Псевдоцифеллы присутствуют на обеих поверхностях, на верхней поверх-
ности хорошо заметные, окруженные темным ободком или черными воз-
вышениями, на нижней — в виде белых пятен, расположенных в основном
на гребнях ребер таллома, реснички, соредии и изидии отсутствуют; кора
и сердцевина от C не изменяются; редкий эпифитный вид, встречающийся
только на юге Дальнего Востока .................................... Cetreliopsis asahinae.
—
Псевдоцифеллы на верхней поверхности, без темного ободка и без возвыше-
ний; кора и сердцевина от C не изменяются или краснеют ......................... 63.
63. Псевдоцифеллы и макулы пло ские, неправильной формы, штриховидные,
сетчатые и др. ..................................................................................................... 64.
—
Псевдоцифеллы в виде белых точек, от плоских до выпуклых .................. 65.
64. Верхняя поверхность желтовато-зеленоватая до оливково-коричневой; псев-
доцифеллы отсутствуют; в коровом слое, помимо обычно имеющейся ус-
ниновой кислоты, содержатся также стиктовая, протоцетраровая и другие
кислоты; большинство видов обитает в аридных и семиаридных областях ....
....................................................................................................... Xanthoparmelia.
—
Верхняя поверхность зеленовато-сероватая, серая до серовато-коричневой;
псевдоцифеллы неправильной формы (штриховидные, сетчатые и др.), очень
редко точковидные (у трех азиатских видов); содержит в коровом слое атра-
норин и хлоратранорин, усниновая кислота, как правило, отсутствует ...........
...................................................................................................................
Parmelia.
Флора лишайников России Флора лишайников России
30
65. Псевдоцифеллы краевые, очень маленькие (точковидные), расположены на
краях лопастей и лобулей; редкие эпифитные виды хвойно-широколиствен-
ных лесов юга Дальнего Востока .......................................... Nipponoparmelia.
—
Псевдоцифеллы ламинальные .......................................................................... 66.
66. Верхняя поверхность желтоватая, желтовато-зеленая, по краям более гладкая,
к центру гофрированная; соралии беловато-серовато-желтоватые, одиночные
или сливающиеся в виде сплошной или прерывистой каймы; нижняя поверх-
ность с немногочисленными ризинами, не доходящими до краев лопастей;
конидии гантелевидные (бифузиформные); содержит в коровом слое уснино-
вую кислоту, в сердцевине — леканоровую ............................. Flavopunctelia.
—
Верхняя поверхность беловато-серая до желтовато-коричневой или к краям
коричневатая; соралии точковидные, ламинальные или краевые; ризины про-
стые, часто скученные; конидии нитевидные или крючкообразные; содержит
в коровом слое атранорин, в сердцевине — гирофоровую или леканоровую
кислоты ................................................................................................... Punctelia.
67(60). Верхняя поверхность таллома сероватая или серовато-зеленоватая ..... 68.
—
Верхняя поверхность таллома зеленовато-желтоватая ................................. 73.
68. Лопасти по краям без ресничек; если же реснички присутствуют, то простые
и ограничены пазухами лопастей; ризины дихотомически разветвленные ......
............................................................................................................................... 69.
—
Лопасти по краям с черными ресничками; ризины простые, неразветвлен-
ные ....................................................................................................................... 70.
69. Лопасти более или менее линейные, с обрезанными или усеченными, иногда
вздутыми концами, с широкоокруглыми пазухами в местах ветвления; содер-
жит в коровом слое атранорин и лихексантон, в сердцевине — гирофоровую,
барбатовую, оливеторовую и другие кислоты .......................... Hypotrachyna.
—
Лопасти узкие, вытянутые, продолговатые и дихотомически разветвленные;
характерная особенность — наружный коровой слой эксципула образован
плектенхимой с очень толстыми клеточными стенками; содержит в коровом
слое атранорин, в сердцевине — гирофоровую, протоцетраровую, салацино-
вую, норстиктовую и стиктовую кислоты; на коре и скалах во влажных от-
крытых местах, преимущественно в горах в тропических районах Юго-Вост.
Азии ............................................................................................ Remototrachyna.
70. Лопасти с простыми редкими или частыми ресничками по краю, иногда
приуроченными только к пазухам; нижняя поверхность черная, с ризина-
ми; сердцевина желто-оранжевая, от C и K желтеет (секалоновая кислота);
кора содержит атранорин, хлоратранорин и секалоновую кислоту; на коре в
лесах в умеренных регионах Вост. Азии и востока Сев. Америки ...................
..............................................................................................................
Myelochroa.
—
Сердцевина белая .............................................................................................. 71.
71. Лопасти широкие, до 4 см шир., с темными ризинами, развивающимися до
самых краев; сердцевина от C краснеет; соредии отсутствуют, изидии могут
Семейство Parmeliaceae
31
развиваться; аскоспоры 6–12 × 4.5–8.0 мкм; на коре; виды океанического и
южного распространения .................................................................... Parmelina.
—
Лопасти до 2 см шир.; ризины часто бледные, не развиваются до краев лопа-
стей, оставляя голую кайму до 6 мм шир.; сердцевина от C краснеет или не
изменяется; соредии и изидии присутствуют; аскоспоры крупнее ............. 72.
72. Лопасти 0.5–5.0 мм шир.; сердцевина от C краснеет; аскоспоры крупные,
12–20 × 8–12 мкм; преимущественно виды Южного полушария .....................
...............................................................................
Hypotrachyna (Parmelinopsis).
—
Лопасти широкие, до 2 см шир.; сердцевина от C не изменяется; апотеции у
северных видов неизвестны; на поросших мхом скалах и основаниях стволов;
преимущественно южные океанические виды ............................ Parmotrema.
73(67). Сердцевина желто-оранжевая (хотя бы местами), от C и K желтеет (сека-
лоновая кислота); кора содержит атранорин, хлоратранорин, зеорин и секало-
новую кислоту; на коре в лесу в умеренных регионах Вост. Азии и востока
Сев. Америки .................................................................................... Myelochroa.
—
Сердцевина белая .............................................................................................. 74.
74. Ризины дихотомически, трихотомически или многократно разветвленные, с
перпендикулярно отходящими ответвлениями .............................................. 75.
—
Ризины простые или слабо разветвленные .................................................... 76.
75. Лопасти линейные, с обрезанными или усеченными, иногда вздутыми конца-
ми, с широкоокруглыми пазухами в местах ветвления ........... Hypotrachyna.
—
Лопасти вытянутые, продолговатые, усеченные и дихотомически разветвлен-
ные; наружный коровой слой эксципула образован плектенхимой с очень тол-
стыми клеточными стенками; на коре и скалах во влажных открытых местах,
преимущественно в горных тропических районах Юго-Вост. Азии ................
......................................................................................................
Remototrachyna.
76. Нижняя поверхность черная или коричнево-черная; ризины черные, не раз-
виваются в зоне около 6 мм шир. у краев лопастей; аскоспоры крупные, более
15 мкм и до 20–24 мкм дл. ............................................................................... 77.
—
Нижняя поверхность светло-коричневая, цвета слоновой кости, редко более
темная в центре; ризины черные или светлые; аскоспоры 6–14 мкм дл. ... 78.
77. Таллом с соредиями, изидии отсутствуют; нижняя поверхность с простыми
ризинами, переходящими в бородавочки; сердцевина от C не изменяется, в
коровом слое, кроме усниновой кислоты, содержатся следы атранорина, в
сердцевине — протоцетраровая и другие кислоты; преимущественно лесные
эпифитные виды умеренных областей ....................................... Flavoparmelia.
—
Таллом с соредиями, изидиями и лобулями или без них; нижняя поверхность
с более светлыми редкими и часто дихотомически разветвленными ризинами;
соредии и изидии присутствуют; сердцевина от C краснеет, содержит в коре
атранорин и хлоратранорин, в сердцевине — гирофоровую и другие кислоты;
преимущественно виды Южного полушария ......................................................
...............................................................................
Hypotrachyna (Parmelinopsis).
Флора лишайников России Флора лишайников России
32
78. Сердцевина от I не изменяется; соралии краевые; растет на деревьях ............
............................................................................................................
Usnocetraria.
—
Сердцевина от I синеет; соралии ламинальные или отсутствуют; виды, ра-
стущие на скалах, почве или свободноживущие в тундрах, северотаежных и
аридных районах ................................................................................................ 79.
79. Таллом в виде концентрических кругов; лопасти до 2 мм шир.; верхняя по-
верхность желто-белая, реже серая; нижняя поверхность цвета слоновой ко-
сти или пурпурно-черно-бархатная; в коровом слое кроме усниновой кислоты
содержится атранорин [за исключением A. aleuritica (Nyl.) Hale]; сердцевина
содержит алектороновую и другие кислоты; арктоальпийские виды ...............
.......................................................................................................... Arctoparmelia.
—
Таллом иной; лопасти до 10 мм шир.; верхняя поверхность желтовато-зеленая
до оливково-коричневой или темно-коричневой; нижняя поверхность бледно-
коричневая до черной; в коровом слое содержатся лишь следы атранорина,
в сердцевине — стиктовая, норстиктовая, иногда фумарпротоцетраровая и
салациновая кислоты; виды, обитающие на скалах и почве по всему земному
шару, но особенно в аридных и семиаридных областях ....... Xanthoparmelia.
Д. Е. Гимельбрант
Alectoria Ach.
in Luyken, 1809, Tent. Hist. Lich.: 95. ≡ Bryopogon Link, 1833, Grundr. Kräuterk. 3:
164, nom. illeg. superfl. Тип: Alectoria sarmentosa (Ach.) Ach. — Алектория
Лихенизированные грибы. ТАЛЛОМ кустистый, дернинковидный или бородо-
видный, обычно средних размеров, реже крупный, состоит из обильно дихото-
мически ветвящихся, восходящих, распростертых по субстрату или повисающих
ветвей, зеленовато-желтый, желтый, реже зеленовато-серый, концы ветвей часто
зачернены. Ветви более или менее цилиндрические, местами уплощенные, глад-
кие или в местах ветвления бороздчато-ребристые, иногда с фовеолами, обычно
с многочисленными хорошо выраженными выпуклыми вытянуто-веретеновид-
ными до яйцевидных белыми псевдоцифеллами, изредка образующими спинулы
или соредии. Соралии отсутствуют или образуются очень редко, бугорковидные,
одного цвета с талломом. Настоящие изидии неизвестны. Коровой слой прозо-
плектенхимный, состоит из периклинально ориентированных гиф, погруженных
в желатинозный матрикс, поверхность таллома матовая или блестящая. Сердце-
вина составлена рыхло расположенными орнаментированными гифами. Перво-
начально таллом прикрепляется к плотному субстрату псевдогомфом, однако
позднее может утратить его и тогда закрепляется на субстрате, цепляясь за него
ветвями. В случае произрастания на почве структуры прикрепления к субстрату
Род Alectoria
33
отсутствуют, талломы погружены отмирающим основанием в растительную дер-
новину. ПЛОДОВЫЕ ТЕЛА — апотеции зеоринового типа, обычны у одних и край-
не редки у других видов, располагаются на ветвях латерально, с возрастом часто
становятся коленчато изогнутыми, край одного цвета с талломом, диск коричне-
вый или красно-коричневый, блестящий, вогнутый, позднее может становить-
ся выпуклым. Эпитеций коричневый, серый или голубовато-зеленый, гимений и
гипотеций бесцветные. Парафизы более или менее разветвленные и анастомози-
рующие, сумки булавовидные, 65–130 × 25–50 мкм, обычно содержат 2–4 аско-
споры. Аскоспоры одноклеточные, эллипсоидные, толстостенные, в зрелости
коричневые с бесцветным эписпорием, 20–48 × 12–28 мкм. КОНИДИАЛЬНЫЕ СПОРО-
НОШЕНИЯ в форме пикнид, шаровидные, погруженные или выступающие, иногда
довольно многочисленные. Конидии палочковидные до веретеновидных, 7–8 ×
0.8 мкм. ФОТОБИОНТ — Trebouxia.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту в значи-
тельной концентрации, сердцевина — алектороновую, диффрактаевую, оливето-
ровую, скваматовую и некоторые другие кислоты.
ЭКОЛОГИЯ. Характерные компоненты эпифитных и напочвенных сообществ,
реже могут поселяться на древесине и каменистых субстратах; в разнообразных
лесных и тундровых сообществах в горах и на равнине.
РАСПРОСТРАНЕНИЕ. Представители рода известны в обоих полушариях, центр
разнообразия находится в Сев. Америке. В России широко распространены, от-
мечены в большинстве северных и горных регионов.
ПРИМЕЧАНИЕ. Включает семь видов, три из которых встречаются на терри-
тории России. Относительно недавно из рода Alectoria выделен род Gowardia
(Halonen et al., 2009) с двумя видами.
Виды рода Alectoria отличаются от видов рода Gowardia наличием желтова-
тых тонов в окраске таллома и составом лишайниковых веществ (не содержат
алекториаловой и барбатоловой кислот). Эпифитные виды рода могут внешне на-
поминать виды рода Usnea, которые легко отличаются не только характером вет-
вления, но и наличием в сердцевине плотного тяжа — стереома. Внешне отда-
ленно похожие представители рода Evernia отличаются более мягким талломом
и отсутствием псевдоцифелл, а наиболее морфологически сходные виды рода
Ramalina Ach. с нитевидными ветвями — крючковидно загнутыми концами ве-
точек, несущими соредиозные зернышки, а также более мелкими точковидными
псевдоцифеллами.
Виды, в норме формирующие многочисленные апотеции, размножаются аско-
спорами. Для остальных представителей рода, особенно напочвенных и бородо-
видно повисающих эпифитных, более характерно вегетативное размножение по-
средством фрагментации талломов, гораздо реже — за счет образования спинул
или соредий.
ИЗБРАННАЯ ЛИТЕРАТУРА: Hawksworth, 1972; Brodo, Hawksworth, 1977; Halonen et
al., 2009; Kärnefelt, Thell, 1992; Thell et al., 2004a; Velmala, Myllys, 2011a.
Род Alectoria
Флора лишайников России Флора лишайников России
34
Ключ для определения видов и подвидов рода Alectoria
1. Таллом с восходящими или распростертыми по субстрату ветвями; преиму-
щественно напочвенные виды ........................................................................... 2.
—
Таллом повисающий или с восходящими ветвями, преимущественно эпифит-
ные виды ............................................................................................................... 3.
2. Таллом с распростертыми по субстрату ветвями, на основных ветвях во мно-
гих местах имеются сильно уплощенные и расширенные участки до 2–4 см
шир.; сердцевина содержит алектороновую кислоту ..........................................
........................................................................
3. A. sarmentosa subsp. vexillifera.
—
Таллом с восходящими ветвями, основные ветви более или менее цилиндри-
ческие, до 2 мм в диам.; сердцевина содержит диффрактаевую кислоту ........
.......................................................................................................
2. A. ochroleuca.
3. Таллом в виде торчащих жестких кустиков с восходящими ветвями или почти
повисающий; апотеции обычно обильные, аскоспоры 35–45 × 18–24 мкм .....
...................................................................................................................
1. A. lata.
—
Таллом бородовидно повисающий, мягкий; апотеции образуются редко, аско-
споры 23–35(48) × 12–20(25) мкм .......... 3. A. sarmentosa subsp. sarmentosa.
1. Alectoria lata (Taylor) Linds.
1859, Trans. Roy. Soc. Edinburgh 22: 135. ≡ Cornicularia lata Taylor, 1847, London J.
Bot. 6: 190. ≡ Bryopogon latus (Taylor) Müll. Arg., 1888, Flora (Regensburg) 71 (9):
130. Т и п : [Мексика] Mexico, ex herb. Hooker (FH ?лектотип, Brodo, Hawksworth,
1977: 61, «holotype»). — Алектория широкая.
ИЗБРАННАЯ ЛИТЕРАТУРА: Brodo, Hawksworth, 1977; Brodo et al., 2001; Ryan, 2002a.
ИЛЛЮСТРАЦИИ: Brodo et al., 2001: 152; Ryan, 2002a: 94.
ТАЛЛОМ кустистый, преимущественно в виде плотных восходящих кустиков
или почти повисающий, редко стелющийся и перепутанный, жесткий, серовато-
зеленый, зеленовато-желтый до желтого, иногда с зачерненными (оливково-чер-
ными) прожилками ближе к основанию ветвей. Основные ветви до 8 (редко до
15) см дл. и 0.5–2.0 мм в диам., обычно ясно не выраженные, более или менее
цилиндрические, неравной толщины, редко угловатые в сечении, иногда в местах
ветвления слегка уплощенные, лишь иногда несут перпендикулярно отходящие
латеральные веточки. Ветвление преимущественно дихотомическое изотомиче-
ское с острыми углами, местами к концам ветвей анизотомическое. Псевдоцифел-
лы многочисленные, хорошо заметные, выпуклые, реже почти плоские, вытяну-
то-веретеновидные или эллипсоидные, белые, 0.2–1.0 мм дл. Соралии и спинулы
отсутствуют. АПОТЕЦИИ встречаются часто, иногда обильны, однако на отдельных
экземплярах могут отсутствовать, расположены на ветвях сбоку или почти терми-
нально, с возрастом могут становиться коленчато изогнутыми, 2–10 мм в диам.,
талломный край цельный или мелко кренулированный, одного цвета с талломом,
Род Alectoria
35
диск коричневый до почти черного. Сумки головчато-овальные, содержат 2 аско-
споры. Аскоспоры эллипсоидные, одноклеточные, в зрелом состоянии могут ста-
новиться коричневыми, 35–45 × 18–24 мкм. ПИКНИДЫ мелкие, до 0.1 мм в диам.,
овальные или округлые, местами выпуклые, темно-коричневые или почти чер-
ные, встречаются редко, однако на некоторых экземплярах обильны. Конидии не
отмечены. — Рис. 2 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K не
изменяется или редко краснеет (неидентифицированное вещество), от C не изме-
няется или редко дает зеленовато-черную окраску, от KC желтеет, от P не изменя-
ется, в UV не флюоресцирует. Сердцевина содержит алектороновую кислоту, от
K, C и P не изменяется, от KC краснеет или не изменяется, в UV флюоресцирует
холодно-голубым цветом. Таллом также может содержать неидентифицированное
вещество, в результате чего от K иногда краснеет.
ЭКОЛОГИЯ. На ветвях хвойных деревьев, на почве и скалах, преимущественно
в высокогорных районах.
РАСПРОСТРАНЕНИЕ. Россия. Горный вид, встречающийся на Дальнем Востоке.
ДВ: ДВ cев.
—
Хабаровский край (сев. часть); ДВ юж.
—
Амурская обл., Саха-
линская обл. (Курильские о-ва), Хабаровский край (юж. часть). — Карта 1.
Общее распространение. Азия (вост.), Сев. и Центр. Америка.
ПРИМЕЧАНИЕ. Эпифитные экземпляры отличаются от Alectoria sarmentosa
subsp. sarmentosa характером роста (в виде плотных торчащих кустиков), часто
обильным развитием апотециев, а также несколько более крупными аскоспорами.
Напочвенные и эпилитные экземпляры отличаются от A. ochroleuca обильным
развитием апотециев и составом лишайниковых веществ сердцевины.
2. Alectoria ochroleuca (Hoffm.) A. Massal.
1856, Sched. Crit. 2: 47. ≡ Usnea ochroleuca Hoffm., 1794, Descr. Pl. Cl. Crypt. 2 (1):
7, Tab. 26. ≡ Lichen ochroleucos Ehrh., 1784, Hannov. Magazin 22 (9): 144, nom.
illeg., non Necker, 1771 et Hoffm., 1784. ≡ Parmelia ochroleuca (Hoffm.) Ach., 1803,
Methodus: 271. ≡ Cornicularia ochroleuca (Hoffm.) DC., in Lam. et DC., 1805, Fl.
Franç., ed. 3, 2: 330. ≡ Evernia ochroleuca (Hoffm.) Fr., 1831, Lichenogr. Eur. Re-
form.: 22. ≡ Bryopogon ochroleucus (Hoffm.) Link, 1833, Grundr. Kräuterk. 3: 164.
Т и п : [Германия] Germany, Fichtelgebirge, Funck, Krypt. Gew. Fichtelgeb. no. 420
(MW-Hoffm. 8562 неотип, Hawksworth, Moore, 1969: 248). — Алектория бледно-
охряная.
= Alectoria ochroleuca (Hoffm.) A. Massal. var. rigida (Fr.) Th. Fr.
ИЗБРАННАЯ ЛИТЕРАТУРА: Hawksworth, 1972; Brodo, Hawksworth, 1977; Крусанова,
1996; Velmala, Myllys, 2011a.
ИЛЛЮСТРАЦИИ: Thomson, 1984: 37; Brodo et al., 2001: 153; Stenroos et al., 2011:
53; Velmala, Myllys, 2011a: 143; Wirth et al., 2013: 148.
Флора лишайников России Флора лишайников России
36
ТАЛЛОМ кустистый, преимущественно с восходящими ветвями или реже сте-
лющийся, жесткий, желтый до зеленовато-желтого, с зачерненными концами вет-
вей. Основные ветви до 10(12) см дл. и 0.5–2.0 мм в диам., ясно выраженные, с
серповидно изогнутыми и поникающими концами, более или менее цилиндриче-
ские, иногда уплощенные от основания и в местах ветвлений, преимущественно
гладкие и матовые, с хорошо заметными и достаточно обычными выпуклыми ве-
ретеновидными белыми псевдоцифеллами около 1 мм дл. Ветвление обильное,
преимущественно анизотомическое дихотомическое. Соралии отсутствуют. АПО-
ТЕЦИИ встречаются редко, боковые, 3–6 мм в диам., одного цвета с талломом, с
желтовато-оранжевым, красновато-коричневым до черного диском, с возрастом
могут становиться коленчато изогнутыми. Сумки головчато-овальные, содержат
2–4 аскоспоры. Аскоспоры эллипсоидные, одноклеточные, в зрелом состоянии
могут становиться темно-коричневыми, 26–42 × 12–28 мкм. ПИКНИДЫ мелкие,
до 0.15 мм в диам., черные, блестящие, располагаются апикально или субапи-
кально, встречаются редко. Конидии палочковидные до веретеновидных, 7–8 ×
0.8 мкм. — Рис. 3 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует. Сердцевина содер-
жит диффрактаевую кислоту, от K, KC, C и P не изменяется, в UV не флюоресци-
рует. Редко сердцевина содержит вместо диффрактаевой алектороновую кислоту
(от KC краснеет), либо крайне редко лишайниковые вещества отсутствуют. Также
таллом в нижней части может содержать неидентифицированное вещество, в ре-
зультате чего от K краснеет.
ЭКОЛОГИЯ. На открытых местах на почве и камнях в тундровых сообществах
на равнинах и в высокогорьях, где может выступать в качестве доминанта напо-
чвенных растительных ассоциаций; очень редко встречается на коре деревьев и
кустарников в равнинных таежных лесах и в субальпийском поясе в горах.
РАСПРОСТРАНЕНИЕ. Россия. Широко распространенный арктоальпийский вид.
АРКТ: все районы. ЕР: Евр. Север — Мурманская обл., Карелия, Архангельская
обл., Ненецкий АО (юго-зап. часть), Коми; Сев. Урал — горные районы Перм-
ского края (сев. часть); Юж. Урал — горные районы Пермского края (сев. часть),
Свердловской обл. и Башкирии. СИБ: Зап. Сиб. сев.
—
Ямало-Ненецкий АО
(юж. часть), Ханты-Мансийский АО; Ср. Сиб. — Таймырский Долгано-Ненецкий
р-н (юж. часть), Красноярский край (сев.), Эвенкия, Иркутская обл. (сев. часть);
Юж. Сиб.
—
горные части всех районов; Як.
—
все районы. ДВ: ДВ сев. — все
районы; ДВ юж. — все районы, кроме Еврейской АО. — Карта 2.
Общее распространение. Европа, Азия, Сев. и Юж. Америка, Гренландия.
ПРИМЕЧАНИЕ. Эпифитные экземпляры отличаются от Alectoria sarmentosa
subsp. sarmentosa характерными зачерненными концами ветвей, а также составом
лишайниковых веществ сердцевины (содержат диффрактаевую кислоту). Об от-
личии от A. lata см. в примечании к этому виду.
Род Alectoria
37
3. Alectoria sarmentosa (Ach.) Ach.
1810, Lichenogr. Universalis: 595. ≡ Lichen sarmentosus Ach., 1795, Kongl. Vetensk.
Aсad. Nya Handl. 16: 212. ≡ Parmelia sarmentosa (Ach.) Ach., 1803, Methodus: 271.
≡ Cornicularia sarmentosa (Ach.) DC. in Lam. et DC., 1815, Fl. Franç., ed. 3, 6: 179.
≡ Evernia sarmentosa (Ach.) Fr., 1826, Nov. Sched. Crit. Lich. 4: 34. ≡ Bryopogon
sarmentosus (Ach.) Link, 1833, Grundr. Kräuterk. 3: 164. ≡ Eualectoria sarmentosa
(Ach.) Gyeln., 1934, Ann. Hist.-Nat. Mus. Natl. Hung. 28: 283. Т и п : [Швеция] Sue-
cia (H-ACH 1818A лектотип, Howe, 1911: 150, pl. 45). — Алектория усатая.
A. Alectoria sarmentosa subsp. sarmentosa. — Алектория усатая подвид усатая.
= Alectoria albida Räsänen; = Alectoria luteola Mont. ex De Not.; = Alectoria
stigmata Bystrek; = Alectoria subsarmentosa Stirt.; = Alectoria sarmentosa (Ach.)
Ach. var. gigantea Räsänen.
ИЗБРАННАЯ ЛИТЕРАТУРА: Hawksworth, 1972; Brodo, Hawksworth, 1977; Крусанова,
1996; Velmala, Myllys, 2011a.
ИЛЛЮСТРАЦИИ: Wirth, 1995: 119; Brodo et al., 2001: 155; Stenroos et al., 2011: 54,
55; Velmala, Myllys, 2011a: 143.
ТАЛЛОМ кустистый, преимущественно повисающий, без явных основных вет-
вей, обычно крупный — до 50 см дл. и более, довольно мягкий, желтый до зе-
леновато-желтого и зеленовато-серого, часто с зачерненными концами ветвей,
иногда с оливково-черными штрихами на отдельных участках. Ветви 0.5–3.0 мм
в диам., более или менее цилиндрические, иногда уплощенные у основания и в
местах ветвлений, гладкие или на отдельных участках перекрученные, местами
с фовеолами, с достаточно обычными и хорошо заметными выпуклыми верете-
новидными или округлыми белыми псевдоцифеллами около 1 мм дл. Ветвление
обильное, преимущественно анизотомическое дихотомическое. Соралии образу-
ются крайне редко, бугорковидные, одного цвета с талломом. АПОТЕЦИИ в россий-
ском материале встречаются достаточно редко, боковые, 2–5 мм в диам., одного
цвета с талломом, с желтовато-оранжевым, красновато-коричневым или почти
черным блестящим или матовым диском, с возрастом могут становиться коленча-
то изогнутыми. Сумки головчато-овальные, содержат 2–4 аскоспоры. Аскоспоры
эллипсоидные, одноклеточные, в зрелом состоянии могут становиться темно-ко-
ричневыми, 23–35(48) × 12–20(25) мкм. ПИКНИДЫ заметные, до 2–3 мм в диам.,
черные, блестящие, располагаются апикально или субапикально, встречаются не-
редко. Конидии неизвестны. — Рис. 4 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K,
C и P не изменяется, от KC желтеет, в UV не флюоресцирует. Сердцевина имеет
довольно вариабельный набор лишайниковых веществ и может содержать в ка-
честве основного компонента алектороновую кислоту, а в качестве дополнитель-
ных — скваматовую, тамноловую, α-коллатоловую, барбатовую, барбатоловую
кислоты, от K, C и P обычно не изменяется (либо редко от K, C и P желтеет), от
Флора лишайников России Флора лишайников России
38
KC краснеет или не изменяется, в UV флюоресцирует холодно-голубым цветом
(иногда флюоресценция не наблюдается). Коровой слой может содержать также
неидентифицированное вещество, в результате чего от K краснеет, а от C приоб-
ретает зеленовато-черную окраску.
ЭКОЛОГИЯ. Эпифитный лишайник, предпочитающий влажные малонарушен-
ные старовозрастные равнинные и горные хвойные леса и являющийся их инди-
катором; реже может поселяться на древесине, почве и каменистых субстратах.
РАСПРОСТРАНЕНИЕ. Россия. Широко распространенный лесной подвид, извест-
ный из многих равнинных и горных территорий России, однако активно сокра-
щающий свой ареал в результате сведения старовозрастных лесов. АРКТ: Аркт.
евр. — Ненецкий АО, Коми (крайний северо-восток). ЕР: Евр. Север — Мурман-
ская обл., Карелия, Архангельская обл., Коми, Ленинградская и Вологодская обл.;
Ср. Россия — Новгородская, Тверская, Ярославская, Костромская обл., Пермский
край, Московская обл.; Сев. Кавказ: Краснодарский край, Адыгея, Карачаево-Чер-
кесия; Сев. Урал — горные районы Пермского края и Свердловской обл.; Юж.
Урал — горные районы Пермского края. СИБ: Зап. Сиб. сев.
—
Ямало-Ненец-
кий АО (юж. часть), Ханты-Мансийский АО; Юж. Сиб. — Алтайский край (вост.
горная часть), Кемеровская обл., Респ. Алтай, Хакасия, Тыва, Красноярский край
(юж. часть); Як.
—
юж.ДВ:ДВюж. —
Хабаровский край (юж. часть), Примор-
ский край. — Карта 3.
Общее распространение. Европа, Азия, Сев. и Юж. Америка.
B. Alectoria sarmentosa subsp. vexillifera (Nyl.) D. Hawksw.
1970, Taxon 19: 241. ≡ Alectoria ochroleuca [subsp.] vexillifera Nyl. in Kihlman,
1891, Meddeland. Soc. Fauna Fl. Fenn. 18: 48. ≡ Alectoria vexillifera (Nyl.) Stizenb.,
1892, Ann. Naturhist. Mus. Wien 7: 122. ≡ Alectoria cincinnata var. vexillifera (Nyl.)
Räsänen, 1931, Ann. Acad. Sci. Fenn. 34 (4): 22. ≡ Alectoria cincinnata f. vexillifera
(Nyl.) Savicz, 1935, Lichenoth. Ross. 4: No. 33. ≡ Bryopogon vexilliferus (Nyl.)
Gyeln., 1935, Feddes Repert. 38: 243. ≡ Alectoria sarmentosa var. cincinnata
f. vexillifera (Nyl.) H. Magn., 1936, Förteckn. Skand. Växt. 4: 68. Т и п : [Россия]
Murmansk Region, Lapponia ponojensis, ad promontorium Orlow, in apertis glareosis,
A. O. Kihlman, 15 May 1889 (H-NYL 35887 лектотип, Brodo, Hawksworth, 1977:
73, «holotype», ZT изолектотип). — Алектория усатая подвид флагоносная.
= Evernia ochroleuca var. cincinnata Fr. ≡ Alectoria cincinnata (Fr.) Lynge ≡
Alectoria ochroleuca var. cincinnata (Fr.) Nyl. ≡ Alectoria sarmentosa var. cincinnata
(Fr.) Nyl. ≡ Cornicularia ochroleuca var. cincinnata (Fr.) Schaer. ≡ Bryopogon
sarmentosus var. cincinnatus (Fr.) Müll. Arg.
ИЗБРАННАЯ ЛИТЕРАТУРА: Hawksworth, 1972; Brodo, Hawksworth, 1977; Thomson,
1984; Крусанова, 1996; Velmala, Myllys, 2011a.
ИЛЛЮСТРАЦИИ: Thomson, 1984: 38; Velmala, Myllys, 2011a: 143.
Род Allocetraria
39
ТАЛЛОМ кустистый, распростертый, обычно достаточно крупный — до 10–
15 см дл., жесткий, желтый до зеленовато-желтого и зеленовато-серого, часто
с зачерненными концами ветвей, с оливково-черными штрихами или целыми
участками таллома, на субстрате лежит свободно, однако иногда способен при-
крепляться к нему гаптерами. Характерной особенностью являются сильно упло-
щенные и расширенные участки основных ветвей (до 2–4 см шир.), бороздчато-
ребристые, с фовеолами, иногда с перфорациями. Такие уплощенные участки
обычно соседствуют с тонкими (0.3–1.0 мм в диам.) цилиндрическими умеренно
или сильно разветвленными частями таллома, иногда напоминающими кисточки.
Соралии не образуются. АПОТЕЦИИ крайне редки. Остальные характеристики, в
том числе состав лишайниковых веществ, соответствуют типовому подвиду.
—
Рис. 5 (вклейка).
ЭКОЛОГИЯ. Напочвенный лишайник, редко переходящий на каменистые по-
верхности, предпочитающий малонарушенные тундровые и тундроидные сооб-
щества, где поселяется на наиболее открытых участках, часто нарушенных крио-
турбацией.
РАСПРОСТРАНЕНИЕ. Россия. Достоверно известен из арктических и субарктиче-
ских регионов России, преимущественно приморских. АРКТ: Аркт. евр.
—
Не-
нецкий АО. ЕР: Евр. Север — Мурманская обл., Карелия, Архангельская обл.
Указания на нахождение в Юж. Сибири нуждаются в проверке. — Карта 4.
Общее распространение. Европа, Африка, Сев. Америка, юж. часть Юж.
Америки (Огненная Земля, Фолклендские о-ва).
ПРИМЕЧАНИЕ. Alectoria sarmentosa subsp. vexillifera — внешне очень необыч-
ный лишайник, таксономический статус которого остается спорным с момента
его описания и до настоящего времени: различные авторы считают его самосто-
ятельным видом, таксоном в ранге подвида либо вовсе не придают отдельного
таксономического статуса. Основные отличия от типового подвида, однако не
только хорошо выражены морфологически, но также проявляются в специфике
экологии и распространения. От другого тундрового напочвенного лишайника —
A. ochroleuca — отличается распростертым талломом с характерно уплощенными
участками ветвей, а также составом лишайниковых веществ сердцевины.
Т. Рандлане, А. Сааг
Allocetraria Kurok. et M. J. Lai
1991, Bull. Natl. Sci. Mus. Tokyo, Ser. B, 17: 60. Т и п : Allocetraria stracheyi
(C. Bab.) Kurok. et M. J. Lai. — Аллоцетрария
Лихенизированные грибы. ТАЛЛОМ листоватый или кустистый, прямостоячий
или почти прямостоячий; верхняя поверхность желтая до желтовато-зеленой или
коричневая, сердцевина белая или светло-желтая до ярко-оранжевой, нижняя по-
Род Allocetraria
Флора лишайников России Флора лишайников России
40
верхность желтая, желтовато-коричневая или коричневая до черной. Лопасти
преимущественно дорсивентральные, обычно распростертые, неразветвленные
или дихотомически разветвленные, довольно узкие, продолговатые и характер-
но выпуклые; у немногих видов лопасти также могут быть цилиндрическими
пальцевидными. Соредии и изидии у всех видов отсутствуют. Псевдоцифеллы
на нижней поверхности, точковидные и нерегулярно разбросанные, либо почти
линейные, расположенные вдоль краев, либо у некоторых видов отсутствуют.
Ризины редкие или отсутствуют. Коровой слой более или менее плектенхим-
ный, состоит из антиклинально ориентированных гиф. ПЛОДОВЫЕ ТЕЛА — апоте-
ции зеоринового типа, до 7–8 мм в диам., с коричневым диском и желтоватым
талломным краем, поверхностные, краевые, субмаргинальные до субламиналь-
ных. Сумки узкобулавовидные до цилиндрических, 30–70 × 8–18 мкм, содержат
8 аскоспор. Аскоспоры простые, округлые, почти округлые, реже широкоэллип-
соидные, бесцветные, 5–10 × 4–8 мкм, обычно расположенные в один ряд. ПИК-
НИДЫ обычно краевые или субмаргинальные, реже поверхностные, погруженные
или расположенные на выступах, пигментированные — темно-коричневые до
черных. Конидии нитевидные, извилистые, с одного конца слегка утолщенные,
10–21 × 0.5–2.0 мкм. ФОТОБИОНТ — Trebouxia.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. В коровом слое содержится усниновая кислота, в
сердцевине — различные жирные кислоты, секалоновая кислота и связанные с
нею пигменты. Один из видов содержит фумарпротоцетраровую кислоту.
ЭКОЛОГИЯ. Эпигейные виды, обитающие в арктических и альпийских обла-
стях, преимущественно на больших высотах (обычно более 3000 м над ур. м.),
некоторые виды в горах могут быть напочвенными и эпифитными на Potentilla
fruticosa, Rhododenron и других кустарниках.
РАСПРОСТРАНЕНИЕ. Представители рода распространены в арктоальпийских и
бореальных регионах Северного полушария, в основном в Юго-Вост. Азии; один
вид имеет более широкое распространение, включая альпийские области Центр.
Европы и Сев. Америки. В России виды отмечены в арктических и/или альпий-
ских сообществах в северных и горных областях.
ПРИМЕЧАНИЕ. Род насчитывает 11 видов, два из которых встречаются на тер-
ритории России. Один вид, который ранее включали в этот род (Allocetraria
oakesiana), сейчас на основе филогенетического анализа (напр., Thell et al., 2012)
выделен в отдельный монотипный род Usnocetraria. Объем рода изменен в одной
из недавних публикаций (Divakar et al., 2017), где на основе современных фило-
генетических методов предлагается разделение всех цетрариоидных лишайников
лишь на три больших рода (Cetraria, Melanelia и Nephromopsis), но эта концепция
авторами не признается.
ИЗБРАННАЯ ЛИТЕРАТУРА: Kurokawa, Lai, 1991; Thell et al., 1995; Kärnefelt, Thell,
1996; Randlane, Thell, 2011a.
Род Allocetraria
41
Ключ для определения видов рода Allocetraria
1. Таллом кустистый, состоит из слегка как бы вздутых цилиндрических вето-
чек; сердцевина в центре веточек паутинистая .......... 1. A. madreporiformis.
—
Таллом листоватый, состоит из дорсивентральных слегка выпуклых лопастей
или иногда почти радиально симметричных цилиндрических веточек; серд-
цевина компактная (не паутинистая) .......................................... 2. A. stracheyi.
1. Allocetraria madreporiformis (Ach.) Kärnefelt et A. Thell
1996, Nova Hedwigia 62: 508. ≡ Dufourea madreporiformis Ach., 1810, Liche-
nogr. Universalis: 525. ≡ Dactylina madreporiformis (Ach.) Tuck., 1862, Proc. Amer.
Acad. Arts Sci. 5: 398. ≡ Cetraria madreporiformis (Ach.) Müll. Arg., 1870, Flora 53:
325. Т и п : [Швейцария] Switzerland, Schleicher, Pl. Crypt. Helv. 2, No. 67 (UPS
лектотип, Randlane, Thell, 2011a: 139). — Аллоцетрария мадрепоровидная.
ИЗБРАННАЯ ЛИТЕРАТУРА: Kärnefelt, Thell, 1996; Randlane et al., 2001; Randlane,
Thell, 2011a.
ИЛЛЮСТРАЦИИ: Brodo et al., 2001: 157; Kärnefelt, Thell, 1996: 493, 496; Randlane,
Thell, 2011a: 143.
ТАЛЛОМ кустистый, пальцевидный, состоящий из слабо разветвленных, слег-
ка вздутых и мелкоямчатых цилиндрических веточек, внутренняя часть которых
заполнена паутинистыми гифами (лопасти никогда не бывают полыми); желтый
до зеленовато-желтого, в освещенных местах коричневатый, без налета. Веточ-
ки растут в виде небольших восходящих пучков, до 3.5 см выс. и 1–3 мм шир.,
обычно цилиндрические и прямые, иногда несколько уплощенные; лопасти за-
полнены плотной белой сердцевиной, в центре паутинистой. Соредии и изидии
отсутствуют. АПОТЕЦИИ редкие, боковые, до 4 мм в диам., со светло-коричневым
диском. Сумки булавовидные, содержат 8 аскоспор, 35–45 × 13–18 мкм; аскоспо-
ры широкоэллипсоидные до почти круглых, 6–10 × 4–8 мкм. ПИКНИДЫ черные,
погруженные или слегка возвышающиеся, разбросаны по всему таллому; кони-
дии нитевидные, извилистые и слегка утолщенные с одного конца, 10–18 × 0.5–
1.5 мкм. — Рис. 6 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует; сердцевина — лихе-
стериновую кислоту, от K, C, KC и P не изменяется, в UV не флюоресцирует.
ЭКОЛОГИЯ. На почве в арктических и альпийских тундровых сообществах.
Предпочитает известковые почвы.
РАСПРОСТРАНЕНИЕ. Россия. Широко распространенный, но довольно редко
встречающийся вид, который в основном известен в северных и горных областях.
АРКТ: Аркт. евр.
—
Ненецкий АО (Архангельская обл.: Вайгач, Земля Фран-
ца-Иосифа, Новая Земля), Коми (крайний северо-восток); Аркт. тайм. (Северная
Земля, о. Визе, Таймыр); Аркт. як. (Новосибирские острова); Аркт. чук. (Чукот-
Флора лишайников России Флора лишайников России
42
ский п-ов, о. Врангеля). ЕР: Евр. Север — Ненецкий АО (юго-зап. часть); Сев.
Кавказ — Адыгея, Карачаево-Черкесия, Кабардино-Балкария, Дагестан. СИБ:
Зап. Сиб. юж. — Алтайский край; Юж. Сиб. — Алтайский край (вост. часть), Ке-
меровская обл., Республика Алтай, Хакасия, Тыва, Бурятия, Красноярский край
(юж. часть), Иркутская обл. (юж. часть); Як. вост.; ДВ: ДВ сев. — Чукотский АО
(юж. часть). — Карта 5.
Общее распространение. Циркумполярный арктоальпийский вид. Европа
(Шпицберген и альпийские регионы Центр. Европы), Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Отличается от видов рода Dactylina отсутствием полости внутри
лопастей.
2. Allocetraria stracheyi (C. Bab.) Kurok. et M. J. Lai
1991, Bull. Natl. Sci. Mus. Tokyo, Ser. B, 17: 62. ≡ Evernia stracheyi C. Bab., 1852,
Hooker’s J. Bot. Kew Gard. Misc. 4: 244 («stracheii»). Т и п : [Индия] India, Uttark-
hand, Kumaon, Gori River; R. Strachey, J. E. Winterbottom (H-NYL 36055 лектотип,
Kurokawa, Lai, 1991: 63). — Аллоцетрария Стрейчи.
= Cetraria everniella (Nyl.) Kremp.; = Cetraria potaninii Oxner.
ИЗБРАННАЯ ЛИТЕРАТУРА: Kurokawa, Lai, 1991; Thell et al., 1995; Randlane et al.,
2001.
ИЛЛЮСТРАЦИИ: Kurokawa, Lai, 1991: 61; Thell et al., 1995: 361.
ТАЛЛОМ листоватый, распростертый до почти прямостоячего; лопасти 2–5 мм
шир. и до 5 см дл., дорсивентральные, характерно выпуклые, с приподнимающи-
мися концами; иногда с радиально симметричными прямыми веточками. Верх-
няя поверхность светло-желтая, гладкая; нижняя поверхность светло-желтая до
коричневой, морщинистая, с редкими краевыми ризинами. Соредии и изидии от-
сутствуют. Сердцевина плотная (не паутинистая), белая до оранжевой. АПОТЕЦИИ
очень редки, концевые или краевые, с коричневым диском, до 5 мм в диам.; зре-
лые сумки и аскоспоры не обнаружены. ПИКНИДЫ краевые, погруженные или на
небольших возвышениях, темноокрашенные; конидии нитевидные, 12–14 × 0.5–
2.0 мкм. — Рис. 7 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует; сердцевина содержит
лихестериновую и протолихестериновую кислоты, иногда также секалоновую
кислоту и сопутствующие пигменты (напр. эндокроцин), придающие ей светло-
желтую до оранжевой окраску, от K, C, KC и P не изменяется, в UV не флюорес-
цирует.
ЭКОЛОГИЯ. На почве в альпийских сообществах выше 2000 м над ур. м.
РАСПРОСТРАНЕНИЕ. Россия. СИБ: Юж. Сиб.
—
Республика Алтай (Кош-
Агачский р-н, плато Укок — Седельникова, 1990). Очень редкий вид, для кото-
рого известна лишь одна хорошо документированная находка в Юж. Сибири. —
Карта 6.
Род Arctocetraria
43
Общее распространение. Азия (Монголия, Китай, Индия, Непал), Сев. Аме-
рика (зап. часть).
ПРИМЕЧАНИЕ. Морфологически очень вариабельный таксон, поскольку тал-
лом может быть дорсивентральным или иногда более или менее радиально сим-
метричным, но в таком случае все же с плотной сердцевиной; дорсивентральные
лопасти Allocetraria stracheyi могут быть распознаны по типичной выпуклой (не
вогнутой) форме.
Т. В. Макрый
Arctocetraria Kärnefelt et A. Thell
1993, Bryologist 96 (3): 402. Т и п : Arctocetraria andrejevii (Oxner) Kärnefelt et
A. Thell. — Арктоцетрария
Лихенизированные грибы. ТАЛЛОМ кустистый или листоватый, вертикальный
или восходящий. Лопасти дорсивентральные, более или менее желобчатые, ме-
стами плоские до почти трубчато свернутых, светло-желтовато-коричневые до
темно-бурых или зеленовато-оливково-серые до черноватых. Псевдоцифеллы
имеются или отсутствуют, на нижней поверхности. Краевые выросты (шипи-
ки), несущие пикниды, редкие, до 0.5 мм дл. Реснички (цилии) имеются или от-
сутствуют. Соралии отсутствуют или очень редкие. ПЛОДОВЫЕ ТЕЛА — апотеции,
зеориновые, субмаргинальные, на концах лопастей, редкие, одного цвета с тал-
ломом. Сумки узкобулавовидные, Lecanora-типа, с 8 аскоспорами. Аскоспоры
одноклеточные, бесцветные, широкоэллипсоидные, 6–8 × 3–5 мкм. КОНИДИАЛЬНЫЕ
СПОРОНОШЕНИЯ — пикниды, обычны, на кончиках шипиков по краям лопастей. Ко-
нидии гантелевидные или веретеновидные. ФОТОБИОНТ Trebouxia-типа.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит жирные кислоты, норранги-
формовую и рангиформовую кислоты.
ЭКОЛОГИЯ. Преимущественно напочвенные лишайники, реже поселяющиеся
на веточках тундровых кустарников, а также на древесине и валежнике; в тундро-
вых сообществах.
РАСПРОСТРАНЕНИЕ. Ареал рода охватывает арктические районы Европы, Азии,
Сев. Америки с центром в Берингии, где представители рода распространены
также в умеренных широтах — в горах.
ПРИМЕЧАНИЕ. Род выделен из рода Cetraria (Kärnefelt et al., 1993) на основании
особого строения сумок и содержания рангиформовой и норрангиформовой кис-
лот. Включает два вида.
ИЗБРАННАЯ ЛИТЕРАТУРА: Рассадина, 1950; Kärnefelt, 1979; Kärnefelt et al., 1993;
Thell, Kärnefelt, 2011a.
Род Arctocetraria
Флора лишайников России Флора лишайников России
44
Ключ для определения видов рода Arctocetraria
1. Лопасти 3–20(30) мм шир., вертикальные, желобчатые до почти трубчато
свернутых, кверху расширяющиеся, с широкоокруглыми отгибающимися
концами, светло-желтовато- или темно-коричневые, в основании светлые, с
многочисленными мелкими точковидными псевдоцифеллами, без ресничек;
сердцевина от I синеет ............................................................... 1. A. andrejevii.
—
Лопасти 1–6 мм шир., восходящие до простирающихся, слабо желобчатые, с
неровно вырезанными краями, зеленовато-оливково-серые до черноватых, с
редкими или отсутствующими точковидными псевдоцифеллами, с ресничка-
ми до 3.5 мм дл.; сердцевина от I не изменяется ............... 2. A. nigricascens.
1. Arctocetraria andrejevii (Oxner) Kärnefelt et A. Thell
1993, Bryologist 96 (3): 402. ≡ Cetraria andrejevii Oxner, 1940, J. Bot. Acad. Sci.
RSS Ukraine 1 (3–4): 44. ≡ Nephromopsis andrejevii (Oxner) Divakar, A. Crespo et
Lumbsch, in Divakar et al., 2017, Fungal Diversity 84: 112. Т и п : [Россия, Респу-
блика Саха (Якутия)] ЯАРСР. Булунський р-н, оленесовхоз, Хараулахские горы,
р. Ухта, верховье, мохово-лишайниковая тундра, 10.06.1938, М. Караваев; р. Кыла
(притока Эбетели), 05.07.1938, М. Караваев; верховье р. Берись, гольцы, дриадо-
во-мохово-лишайниковая тундра, 10.09.1938, М. Караваев; Намський р-н, левый
берег р. Лены у Чукуры, долина ручья, 24.08.1937, Н. Левшина; Вiлюйський р-н,
долина р. Сели Ябыт, притока р. Тюнь (64°30′ с. ш., 121° в. д.), подножье склона к
долине, сфагновое болото, группировка Ledum palustre — Cladoniae, 29.08.1932,
Т. Работнов (KW, LE синтипы). — Арктоцетрария Андреева.
= Cetraria hiascens var. dilatata Vain. ≡ Cetraria hiascens f. dilatata (Vain.) Rass.
≡ Cetraria delisei f. dilatata (Vain.) Rass. ≡ Cetraria kamczatica var. dilatata (Vain.)
Räsänen; = Cetraria simmonsii Krog; = Cetraria simmonsii var. intermedia Krog.
ИЗБРАННАЯ ЛИТЕРАТУРА: Окснер, 1940.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1979: 8, 10, 58–60; Thomson, 1984: 72; Thell, Kärnefelt,
2011a: 144.
ТАЛЛОМ вертикальный, кустистый, до 10 см выс. Лопасти 3–20(30) мм шир.,
более или менее желобчатые до почти трубчато свернутых (или плоских в рас-
ширенных частях), слабо дихотомически разветвленные или почти неразветвлен-
ные, кверху расширяющиеся, с широкоокруглыми отгибающимися концами и
волнистыми краями; края ровные, местами мелкозубчатые благодаря шипикам.
Верхняя поверхность от светло-желтовато-коричневой до темно-коричневой,
обесцвечивающаяся к основанию, неровная — ямчатая, слабо морщинистая и ре-
бристая, глянцевая или матовая; нижняя одинаково с верхней окрашенная, более
морщинистая. Псевдоцифеллы на нижней поверхности, в виде мелких округлых
или узких вдавленных точек или пятнышек, 0.1–0.2(0.5) мм дл. Краевые шипики
редкие, неравномерно развитые, около 0.1–0.2 (редко до 0.5–1.0) мм дл. Соралии
Род Arctocetraria
45
отсутствуют. АПОТЕЦИИ редкие, до 15 мм в диам.; диск коричневый от желтова-
того до темного. Аскоспоры широкоэллипсоидные до почти шаровидных, 6–9 ×
4–5 мкм. ПИКНИДЫ на кончиках краевых шипиков. Конидии от веретеновидных,
утолщенных в центре, до слегка гантелевидных, утолщенных на концах, 4.0–5.5 ×
0.5–1.0 мкм. — Рис. 8 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит рангиформовую и норранги-
формовую кислоты. Коровой слой и сердцевина от K, C, KC и P не изменяются,
сердцевина от I синеет.
ЭКОЛОГИЯ. На почве, обычно во влажных понижениях в арктических и горных
тундрах, а на юге Дальнего Востока на приморских олиготрофных болотах и в
высокогорных ельниках.
РАСПРОСТРАНЕНИЕ. Вид с широким амфиберингийским ареалом, встречающийся
почти циркумполярно по всей Арктике и более широко на Дальнем Востоке, где по
горным системам заходит в умеренные широты. Россия. АРКТ: все районы. ЕР:
Сев. Урал — Коми (Печоро-Илычский заповедник); Ханты-Мансийский АО. СИБ:
Зап. Сиб. сев. — юж. часть Ямало-Ненецкого АО; Ср. Сиб. — юж. часть Таймыр-
ского Долгано-Ненецкого р-на (плато Путорана); Як. — центр. ДВ: ДВ сев. — все
районы; ДВ юж. — Хабаровский край, Сахалинская обл. — Карта 7.
Общее распространение. Европа, Азия, Сев. Америка, Гренландия.
ПРИМЕЧАНИЕ. Слабо полиморфный вид, легко узнаваемый благодаря характер-
ным многочисленным мелким вдавленным псевдоцифеллам, а также ровным кра-
ям лопастей с немногочисленными шипиками, несущими пикниды.
2. Arctocetraria nigricascens (Nyl.) Kärnefelt et A. Thell
1993, Bryologist, 96 (3): 402. ≡ Platysma nigricascens Nyl., in Kihlman, 1891,
Meddeland. Soc. Fauna Fl. Fenn. 18: 50. ≡ Cetraria nigricascens (Nyl.) Elenkin,
1909, Mém. Acad. Imp. Sci. St.-Pétersb., cl. phys.-math. 27: 14. ≡ Nephromopsis
nigricascens (Nyl.) Divakar, A. Crespo et Lumbsch, in Divakar et al., 2017, Fungal
Diversity 84: 113. Т и п : [Россия, Мурманская обл.] Lapponia ponojensis, ad
promontorium Orlov, in lapidosis, 1889, A. O. Kihlman 244 (H9500730 лектотип,
Kärnefelt, 1979: 121). — Арктоцетрария черноватенькая.
= Cetraria rhizophora (Vain.) Rass. ≡ Cetraria hiascens var. rhizophora Vain.;
= Cetraria rhizophora f. arboricola Rass.; = Cetraria arctica H. Magn. ≡ Cetraria
magnussonii Llano; = Cetraria sibirica H. Magn.; = Cetraria elenkinii Krog; =
Cetraria nigricascens var. tominii Rass. ≡ Cetraria elenkinii var. tominii Rass.
ИЗБРАННАЯ ЛИТЕРАТУРА: Еленкин, 1909.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1979: 9, 15, 122–124; Thomson, 1984: 90; Thell,
Kärnefelt, 2011a: 144.
ТАЛЛОМ листоватый, восходящий, вертикальный или простирающийся, 0.5–
5.0 см выс., в виде розеток, пучков, небольших куртинок или бесформенный.
Лопасти дорсивентральные, дихотомически разветвленные, 1–6 мм шир., сла-
Флора лишайников России Флора лишайников России
46
бо желобчатые, местами плоские. Верхняя поверхность ямчато-морщинистая до
сетчато-ребристой или гладкая, бледно-зеленовато-оливково-серая до чернова-
той или буроватой, более светлая в основании, матовая или блестящая; нижняя
одинаково с верхней окрашенная, более морщинистая. Края неровные, нередко с
боковыми лопастинками и тогда с округлыми пазухами между ними. Псевдоци-
феллы на нижней поверхности отсутствуют или редкие, точковидные, до 0.2 мм в
диам. Краевые шипики, несущие пикниды, редкие, до 0.1–0.2(0.5) мм дл. Реснич-
ки обычны, до 3.5 мм дл. Соралии отсутствуют или очень редкие, ламинальные,
до 1 мм в диам. АПОТЕЦИИ редкие, до 8 мм в диам., с кренулированным или почти
ровным краем; диск каштаново-коричневый до темно-бурого. Аскоспоры широ-
коэллипсоидные, 6.0–8.5 × 3.5–4.5 мкм. ПИКНИДЫ на кончиках краевых шипиков.
Конидии гантелевидные, 4.0–5.5 × 1 мкм. — Рис. 9 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит рангиформовую и норранги-
формовую кислоты. Коровой слой и сердцевина от K, C, KC, P и I не изменяются.
ЭКОЛОГИЯ. На веточках тундровых кустарников (Betula nana и др.), иногда
на деревьях (береза, ель), а также на почве, древесине и валежнике; в арктиче-
ских тундрах, лесотундрах, субарктических криволесьях, реже в северотаежных
лесах.
РАСПРОСТРАНЕНИЕ. Арктический вид, распространенный циркумполярно с
центром массовости в Берингии. Россия. АРКТ: все районы. ЕР: Евр. Север —
Мурманская обл., Коми (бассейны рек Колва, Бол. Сыня). СИБ: Ср. Сиб. — юж.
часть Таймырского Долгано-Ненецкого р-на (плато Путорана). ДВ: ДВ сев. —
юж. часть Чукотского АО, Камчатский край (Корякский округ). — Карта 8.
Общее распространение. Европа, Азия, Сев. Америка, Гренландия.
ПРИМЕЧАНИЕ. Весьма полиморфный вид. Узколопастные формы хорошо отли-
чаются от Cetraria nigricans отрицательной реакцией сердцевины с I, а также от-
сутствием красного основания, ямчато-морщинистой поверхностью, более редки-
ми шипиками и ресничками, отсутствием или другим характером псевдоцифелл,
окраской и габитусом. От Tuckermannopsis ciliaris, помимо прочего, отличается
отсутствием цветных реакций. Для юга Дальнего Востока (Чабаненко, 2002) вид
указывался ошибочно.
Е. С. Кузнецова
Asahinea W. L. Culb. et C. F. Culb.
1965, Brittonia 17: 183. Т и п : Asahinea chrysantha (Tuck.) W. L. Culb.
et C. F. Culb. — Асахинея
Лихенизированные грибы. ТАЛЛОМ листоватый, средних размеров или круп-
ный, состоит из распростертых по субстрату слабо прикрепленных коротких и
довольно широких, незначительно перекрывающихся лопастей, приподнимаю-
Род Asahinea
47
щихся по краям. Верхняя поверхность морщинистая или складчатая, матовая или
слегка блестящая, серая или желтая; нижняя — блестящая, черная, лишь по само-
му краю коричневая, без ризин. Псевдоцифеллы на верхней поверхности форми-
руются или отсутствуют. Структуры вегетативного размножения отсутствуют или
представлены хорошо развитыми изидиями. Верхний коровой слой прозоплек-
тенхимный, нижний — параплектенхимный. ПЛОДОВЫЕ ТЕЛА — апотеции зеори-
нового типа, формируются очень редко, располагаются по краям или на поверх-
ности лопастей, талломный край кренулированный, одного цвета с талломом,
диск апотециев коричневый, эксципул, гимений и гипотеций бесцветные, эпите-
ций желто-коричневый. Сумки булавовидные, леканорового типа, парафизы про-
стые с темной апикальной шапочкой. Аскоспоры по 8 в сумке, одноклеточные,
эллипсоидные, бесцветные, 8–14 × 4–8 мкм. КОНИДИАЛЬНЫЕ СПОРОНОШЕНИЯ в фор-
ме погруженных пикнид, располагающихся по краям лопастей или на кончиках
изидий. Конидии гантелевидные, 5–7 × 1 мкм. ФОТОБИОНТ Trebouxia-типа.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит атранорин и усниновую
кислоту, сердцевина — α-алектороновую и α-коллатоловую кислоты.
ЭКОЛОГИЯ. Напочвенные и эпилитные виды, распространены в горных и рав-
нинных тундровых сообществах.
РАСПРОСТРАНЕНИЕ. Арктоальпийский род, представители которого распростра-
нены только в Северном полушарии, в России — в горных и приполярных обла-
стях.
ПРИМЕЧАНИЕ. Род включает два вида, оба известны на территории России.
Ключ для определения видов рода Asahinea
1. Таллом желтый (содержит усниновую кислоту), с псевдоцифеллами на верх-
ней поверхности, без изидий .................................................. 1. A. chrysantha.
—
Таллом белый до желтовато-коричневого (усниновая кислота отсутствует),
без псевдоцифелл, но с многочисленными изидиями на верхней поверхно-
сти ............................................................................................... 2. A. scholanderi.
1. Asahinea chrysantha (Tuck.) W. L. Culb. et C. F. Culb.
1965, Brittonia 17: 184. ≡ Cetraria chrysantha Tuck., V 1858, Amer. J. Sci. Arts, ser.
2, 25 (75): 423. ≡ Platysma chrysanthum (Tuck.) Nyl., fine 1858 («1857»), Mém. Soc.
Imp. Sci. Nat. Cherbourg 5: 334. Т и п : [Россия, Чукотка] Asian coast of the Bering
Strait, C. Wright (FH лектотип, Culberson, Culberson, 1965: 184).
—
Асахинея зо-
лотистая.
= Cetraria saviczii var. candida Oxner et Rass. ≡ Cetraria scholanderi var. candi-
da (Oxner et Rass.) Rass.; = Cetraria chrysantha f. glaucescens Oxner et Rass.; = Ce-
traria chrysantha f. pigmentulata Trass; = Asahinea culbersoniorum Trass; = Platysma
septentrionale Nyl. ≡ Cetraria septentrionalis (Nyl.) Almq.; = Cetraria chrysantha
f. cinerascens Asahina.
Род Asahinea
Флора лишайников России Флора лишайников России
48
ИЗБРАННАЯ ЛИТЕРАТУРА: Savicz, 1926; Рассадина, 1950; Окснер, Рассадина, 1960;
Hakulinen, Ulvinen, 1966; Thomson, 1984; Randlane, Saag, 1989; Gao, 1991; Trass,
1992; Tønsberg et al., 1996.
ИЛЛЮСТРАЦИИ: Thomson, 1984: 44; Brodo et al., 2001: 166; Holien, Tønsberg,
2006: 41; Randlane, Thell, 2011b: 144.
ТАЛЛОМ листоватый, до 27 см в диам., розетковидный или неправильной фор-
мы, состоит из распростертых по субстрату, слабо прикрепленных, широкоокру-
глых лопастей до 3 см шир. Верхняя поверхность желтая, светло-желтоватая,
светлая желтовато-коричневая, изредка серая (хемотип без усниновой кислоты), с
сетчатым рисунком, образованным ребрами и лакунами разного размера. Нижняя
поверхность черная, по краям коричневая, блестящая, без ризин. Псевдоцифеллы
располагаются на верхней поверхности, редкие или многочисленные, желтые с
черным краем, наиболее обычны на ребрах. Изидии отсутствуют, в редких случа-
ях могут формироваться соралии (Savicz, 1926). АПОТЕЦИИ встречаются редко, на
поверхности или по краям лопастей, с красновато-коричневым или коричневато-
черным диском, 3–6 мм в диам. Аскоспоры эллипсоидные, 8–13 × 5–8 мкм. ПИК-
НИДЫ редкие, мелкие, краевые. Конидии гантелевидные, 5–7 × 1 мкм.
—
Рис. 10
(вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит атранорин и усниновую
кислоту (последняя изредка может отсутствовать), от K и KC желтеет, от C и P
не изменяется, в UV не флюоресцирует. Сердцевина содержит α-алектороновую и
α-коллатоловую кислоты (последняя может отсутствовать), от K, C и P не изменя-
ется, от KC розовеет или краснеет, в UV не флюоресцирует.
ЭКОЛОГИЯ. На почве, камнях и растительных остатках в горных и равнинных
тундровых сообществах.
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: Аркт. евр.
—
Архангельская обл. (Новая
Земля), Ненецкий АО; Аркт. зап.-сиб. — Ямало-Ненецкий АО (сев. часть); Аркт.
тайм.
—
Таймырский Долгано-Ненецкий р-н (сев. часть); Аркт. як.
—
арктиче-
ские районы Якутии; Аркт. чук. — Чукотский АО (сев. часть). ЕР: Евр. север —
Мурманская, Архангельская обл. (Новая Земля), Ненецкий АО (юго-зап. часть),
Коми; Сев. Урал — горные районы Коми и Пермского края (сев. часть); Юж.
Урал — горные районы Пермского края (юж. часть) и Свердловской обл. (юж.
часть). СИБ: Зап. Сиб. сев. — Ямало-Ненецкий АО (юж. часть), Ханты-Мансий-
ский АО, Свердловская обл.; Зап. Сиб. юж.
—
Тюменская обл. (юж. часть); Ср.
Сиб. — Таймырский Долгано-Ненецкий р-н (юж. часть), Красноярский край (сев.
часть), Эвенкия, Иркутская обл. (сев. часть); Юж. Сиб. — Кемеровская обл., Ха-
касия, Тыва, Бурятия, Красноярский край (юж. часть), Иркутская обл. (юж. часть),
Забайкальский край; Як.
—
центр., вост. и юж. ДВ: ДВ сев.
—
Чукотский АО
(юж. часть), Магаданская обл., Камчатский край (Корякский округ и бывшая Кам-
чатская обл.), Хабаровский край (сев. часть); ДВ юж. — Амурская обл., Хабаров-
Род Asahinea
49
ский край (юж. часть), Еврейская АО, Сахалинская обл. (Сахалин), Приморский
край. — Карта 9.
Общее распространение. Европа, Азия, Сев. Америка.
ПРИМЕЧАНИЕ. До 1926 г. этот вид не раз приводили для различных регионов
Евразии под ошибочным названием Cetraria lacunosa Ach. (Savicz, 1926). Выявле-
ны три хемотипа вида, различающиеся наличием, кроме атранорина, усниновой
и α-коллатоловой кислот: I — содержит только α-коллатоловую кислоту, II — со -
держит усниновую и α-коллатоловую кислоты, III — содержит только уснино-
вую кислоту. Хемотип I встречается в Приморском и Хабаровском краях, а также
в Японии; II — в Сибири, на юге Дальнего Востока, в Японии и Сев. Америке,
но не представлен на Камчатке и Чукотке; III — встречается в Скандинавии, на
севере Европейской России, в Сибири, на Камчатке, Чукотке и в Сев. Америке
(Randlane, Saag, 1989).
2. Asahinea scholanderi (Llano) W. L. Culb. et C. F. Culb.
1965, Brittonia 17: 187. ≡ Cetraria scholanderi Llano, 1951, J. Washington Acad. Sci.
41: 197. Т и п : [США] Alaska, Lake Schrader, P. F. Scholander, W. Flagg (US голо-
тип, HMAS изотип). — Асахинея Шоландера.
= Cetraria saviczii Oxner et Rass.; = Cetraria saviczii var. nigrita Oxner et Rass.
≡ Cetraria scholanderi var. nigrita (Oxner et Rass.) Rass.; = Cetraria kurodakensis
Asahina ≡ Asahinea kurodakensis (Asahina) W. L. Culb. et C. F. Culb.
ИЗБРАННАЯ ЛИТЕРАТУРА: Окснер, Рассадина, 1960; Hakulinen, Ulvinen, 1966;
Thomson, 1984; Gao, 1991; Tønsberg et al., 1996.
ИЛЛЮСТРАЦИИ: Thomson, 1984: 45; Holien, Tønsberg, 2006: 41.
ТАЛЛОМ листоватый, до 15 см в диам., розетковидный или неправильной
формы, состоит из распростертых по субстрату, слабо прикрепленных, широ-
коокруглых лопастей до 2 см шир. Верхняя поверхность беловатая до желтова-
то-коричневой, с зачерненными пятнистыми краями; нижняя — черная, по кра-
ям коричневая, блестящая. Настоящие псевдоцифеллы отсутствуют, но участки
сердцевины, обнажившиеся на месте отвалившихся изидий, могут напоминать
псевдоцифеллы. Соралии отсутствуют, изидии простые, изредка разветвлен-
ные, одного цвета с талломом, или черные, или с черными кончиками. АПОТЕЦИИ
встречаются очень редко, располагаются на верхней поверхности или по краям
лопастей, с красновато-коричневым или коричневато-черным диском и изидиоз-
ным талломным краем, до 3 мм в диам. Аскоспоры округлые или яйцевидные,
10–13 × 5–8 мкм. ПИКНИДЫ погруженные, располагаются на кончиках изидий. Ко-
нидии палочковидные, простые или слегка изогнутые, 5–10 × 1 мкм.
—
Рис. 11,
12 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит атранорин, от K желтеет, от
C, KC и P не изменяется, в UV не флюоресцирует. Сердцевина содержит алекто-
Флора лишайников России Флора лишайников России
50
роновую и α-коллатоловую кислоты, от K, C и P не изменяется, от KC розовеет
или краснеет, в UV не флюоресцирует.
ЭКОЛОГИЯ. На почве, камнях и растительных остатках в горных и равнинных
тундровых сообществах.
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: Аркт. евр.
—
Ненецкий АО; Аркт. тайм.
—
Таймырский Долгано-Ненецкий р-н (сев. часть); Аркт. як.; Аркт. чук.
—
Чу-
котский АО (сев. часть). СИБ: Юж. Сиб.
—
Бурятия, Красноярский край (сев.
часть), Иркутская обл. (сев. часть), Забайкальский край; Як. — центр., вост. и юж.
ДВ: ДВ сев.
—
Чукотский АО (юж. часть), Магаданская обл., Камчатский край
(Корякский округ и бывшая Камчатская обл.), Хабаровский край (сев. часть); ДВ
юж.
—
Хабаровский край (юж. часть), Сахалинская обл. (Сахалин), Приморский
край. — Карта 10.
Общее распространение. Азия, Сев. Америка.
Л. А. Конорева, С. В. Чесноков
Bryocaulon Kärnefelt
1986, Opera Bot. 86: 17. Т и п : Bryocaulon divergens (Ach.) Kärnefelt. —
Бриокаулон
Лихенизированные грибы. ТАЛЛОМ кустистый, прямостоячий или более или
менее повисающий, неприкрепленный или прикрепленный в основании, обычно
растет пучками или дернинками. Ветви неравномерно дихотомически разветвлен-
ные (до субдихотомических), довольно жесткие, округло- или угловато-цилин-
дрические, гладкие, местами мелкоямчатые или с продольными бороздами, тем-
но- или красновато-коричневые. Соредии и изидии отсутствуют. Псевдоцифеллы
веретеновидные или точковидные, приподнятые или расположенные в плоскости
таллома. Эпикортекс представлен неклеточным слоем 2–5 мкм толщ., поры от-
сутствуют. Кора состоит из двух слоев. Наружная часть представлена 1–3 слоями
плотно упакованных антиклинальных гиф; внутренний коровой слой состоит из
сильно желатинизированных периклинальных гиф. Сердцевина плотная или рых-
лая, включает рассеянные тяжи периклинальных гиф. ПЛОДОВЫЕ ТЕЛА — апотеции
зеоринового типа, латеральные, развиваются на основных ветвях, которые ста-
новятся в этом месте коленчато изогнутыми. Диск апотециев сначала вогнутый,
затем выпуклый, темно-коричневый, с кренулированным краем. Гимений пиг-
ментированный в верхней части. Сумки удлиненно-булавовидные, с сильно ами-
лоидными желатинизированными стенками, Lecanora-типа, толус 4–6 мкм толщ.;
окулярная камера и осевое тело широкие. Аскоспоры простые, бесцветные, эл-
липсоидные, без периспория, расположены по 8 в сумке. Парафизы септирован-
ные, иногда разветвленные, с утолщенными верхушками. Гипотеций бесцветный,
Род Bryocaulon
51
состоит из узких, сильно переплетенных гиф. Эксципул чашевидный, постоян-
ный. КОНИДИАЛЬНЫЕ СПОРОНОШЕНИЯ в форме пикнид, погруженных, грушевидных
или овальных, со стенками, состоящими из сильно желатинизированных гиф.
Пикноконидии гантелевидные. ФОТОБИОНТ Trebouxia-типа, водоросли концентри-
руются под коровым слоем или рассеяны по сердцевине, в виде одиночных кле-
ток или в группах.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Оливеторовая и физодовая кислоты.
ЭКОЛОГИЯ. Виды рода обитают на почве в тундрах (в том числе в горных), на
голых и замшелых скалах, на коре и ветвях деревьев во влажных хвойных и сме-
шанных лесах.
РАСПРОСТРАНЕНИЕ. Род имеет преимущественно арктоальпийское распростра-
нение, но его представители встречаются также в хвойных лесах Вост. Азии и
сев.-зап. части Сев. Америки.
ПРИМЕЧАНИЕ. Морфологически представители рода похожи на виды рода
Bryoria, однако ветви таллома у представителей рода Bryocaulon более толстые,
до 1 мм шир. (у видов рода Bryoria — 0.2–0.7 мм, волосовидные до нитевидных),
кроме того, коровой слой у видов рода Bryoria однослойный, с периклинально
расположенными гифами. Род Nodobryoria Common et Brodo, представленный
обитающим в Гренландии видом N. subdivergens (E. Dahl) Common et Brodo, так-
же морфологически сходен с родом Bryocaulon, но отличается от него формой ро-
ста (повисающий на ветках) и отсутствием вторичных метаболитов. Род включа-
ет в себя 4 вида, из которых в России известно три.
ИЗБРАННАЯ ЛИТЕРАТУРА: Kärnefelt, 1986; Kärnefelt, Thell, 1992; Журбенко, 1996а;
Øvstedal et al., 2009.
Ключ для определения видов рода Bryocaulon
1. Сердцевина от C краснеет (оливеторовая кислота), псевдоцифеллы хорошо
заметные, приподняты над поверхностью таллома ......................................... 2.
—
Сердцевина от C не изменяется (не содержит лишайниковых веществ), псев-
доцифеллы едва заметные, погруженные .......................... 2. B. hyperboreum.
2. Арктоальпийский вид, преимущественно эпигейный ............. 1. B. divergens.
—
Азиатский пацифический вид, преимущественно эпифитный ..........................
.............................................................................................
3. B. pseudosatoanum.
1. Bryocaulon divergens (Ach.) Kärnefelt
1986, Opera Bot. 86: 24. ≡ Cornicularia divergens Ach., 1803, Methodus: 303. ≡
Parmelia divergens (Ach.) Spreng., 1827, Syst. Veg. 4 (1): 276. ≡ Alectoria divergens
(Ach.) Nyl., 1855, Mém. Soc. Imp. Sci. Nat. Cherbourg 3: 171. ≡ Bryopogon divergens
(Ach.) Schwend., 1860, Beitr. Wiss. Bot. 2: 148. ≡ Coelocaulon divergens (Ach.)
R. Howe, 1912, Classif. Usneac. Amer.: 19. Т и п : Lapponia, G. Wahlenberg (H-ACH
1869A лектотип, Brodo, Hawksworth, 1977: 149). — Бриокаулон разветвленный.
Род Bryocaulon
Флора лишайников России Флора лишайников России
52
ИЗБРАННАЯ ЛИТЕРАТУРА: Acharius, 1803; Brodo, Hawksworth, 1977; Kristinsson et
al., 2010; Thell, Kärnefelt, 2011b.
ИЛЛЮСТРАЦИИ: Thell, Kärnefelt, 2011b: 145.
ТАЛЛОМ до 15 см выс., прямостоячий или восходящий, не прикрепленный к
субстрату. Ветви (0.1)0.3–0.5(2.0) мм шир., цилиндрические или слегка уплощен-
ные, гладкие, местами мелкоямчатые, темно-коричневые, часто с красноватым от-
тенком, блестящие. Ветвление дихотомическое анизотомическое. Псевдоцифеллы
обычны, светлоокрашенные, крупные, 0.3 × 0.1 мм, веретеновидные, слегка при-
поднятые над поверхностью таллома. Коровой слой прозоплектенхимный. Серд-
цевина довольно плотная. АПОТЕЦИИ с диском до 10 мм в диам., одного цвета с
талломом, встречаются крайне редко. Гипотеций 15–20 мкм толщ. Эксципул 40–
60 мкм толщ. Гимений 40–55 мкм толщ. Сумки булавовидные. Аскоспоры 6–8 ×
2.5–4.0 мкм. Парафизы 40–55 × 1.0–1.5 мкм, на концах до 2 мкм толщ. ПИКНИДЫ
погруженные, 130 × 110 мкм, с темноокрашенными стенками. Конидии 4.5–5.0 ×
0.8–1.0 мкм. — Рис. 13, 14 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Таллом содержит оливеторовую и 4-O-деметил-
микрофиллиновую кислоты, от C и KC кора и сердцевина краснеют, от K и P не
изменяются, в UV не флюоресцируют.
ЭКОЛОГИЯ. Обитает на почве в тундровых и горно-тундровых сообществах, из-
редка на нижних ветвях деревьев в лесотундре. Повсеместно распространенный
массовый вид, особенно характерный для сухих открытых щебнистых участков.
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: все районы. ЕР: Евр. Север — Мурманская
обл., Карелия, Коми; Ср. Россия — Пермский край (сев. часть); Сев. Кавказ — Ка-
рачаево-Черкесия. СИБ: Ср. Сиб. — Красноярский край (сев. часть); Юж. Сиб. —
горные части Алтайского края и Новосибирской обл., Кемеровская обл., Респу-
блика Алтай, Хакасия, Тыва, Бурятия, Красноярский край (юж. часть), Иркутская
область (юж. часть), Забайкальский край; Як. — вост., юж. ДВ: ДВ сев. — Чукот-
ский АО (юж. часть), Магаданская обл., Камчатский край (Корякский округ и быв-
шая Камчатская обл.), Хабаровский край (сев. часть); ДВ юж. — Хабаровский край
(юж. часть), Сахалинская обл. (Сахалин), Приморский край. — Карта 11.
Общее распространение. Европа, Азия, Сев. Америка, Гренландия.
ПРИМЕЧАНИЕ. Отличается от видов родов Bryoria и Alectoria, растущих в тун-
дре, более толстыми коричневыми блестящими дихотомически разветвленными
ветвями таллома и наличием белых псевдоцифелл. Отличия от других видов рода
указаны в примечаниях к этим видам.
2. Bryocaulon hyperboreum Øvstedal
in Øvstedal, Tønsberg et Elvebakk, 2009, Sommerfeltia 33: 99. Т и п : [Норвегия,
Шпицберген] Svalbard, Kong Karls Land, Hårfagrehaugen, 1936, E. Dahl (O голо-
тип). — Бриокаулон гиперборейский.
ИЗБРАННАЯ ЛИТЕРАТУРА: Øvstedal et al., 2009; Thell, Kärnefelt, 2011b.
Род Bryocaulon
53
ИЛЛЮСТРАЦИИ: Thell, Kärnefelt, 2011b: 145.
ТАЛЛОМ кустистый, до 10 см выс., прямостоячий или распростертый, корич-
невый до коричнево-черного, блестящий, веточки в местах ветвления уплощен-
ные. Ветвление дихотомическое анизотомическое, угол между веточками в верх-
ней части таллома 65 ± 7°. Главные ветви до 0.7 мм шир. Псевдоцифеллы редкие,
веретенообразные, главным образом закрытые (сердцевина не видна), иногда ча-
стично открытые, 0.2–0.3 × 0.1 мм. АПОТЕЦИИ неизвестны. ПИКНИДЫ редкие, по-
груженные. Конидии не обнаружены. — Рис. 15, 16 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Таллом не содержит лишайниковых веществ, кора и
сердцевина от K, KC, C и P не изменяются, в UV не флюоресцируют.
ЭКОЛОГИЯ. Обитает среди мхов на почве в арктических тундрах.
РАСПРОСТРАНЕНИЕ. Россия. Арктический вид. АРКТ: Аркт. евр. — Земля Фран-
ца-Иосифа (о. Гукера, мыс Седова; о. Джексона, мыс Норвегия). — Карта 12.
Общее распространение. Европа (Шпицберген, Земля Франца-Иосифа), Сев.
Америка (Гренландия).
ПРИМЕЧАНИЕ. Отличается от Bryocaulon divergens, также обитающего в Арк-
тике на почве, плохо заметными, закрытыми псевдоцифеллами, отсутствием
в талломе вторичных метаболитов, а также меньшим углом между веточками в
верхней части таллома (78 ± 5° у B. divergens). Bryocaulon hyperboreum морфо-
логически сходен с Nodobryoria subdivergens, однако последний вид отличается
строением корового слоя и отсутствием псевдоцифелл.
3. Bryocaulon pseudosatoanum (Asahina) Kärnefelt
1986, Opera Bot. 86: 28. ≡ Cornicularia pseudosatoana Asahina, 1939, J. Jap. Bot. 15:
356. Т и п : [Япония] Japan, Prov. Suruga, Mt. Fuji, Ohmiyaguchi, 08.1930, Y. Asahi-
na (TNS голотип). — Бриокаулон ложносатоанский.
ИЗБРАННАЯ ЛИТЕРАТУРА: Asahina, 1939; Kärnefelt, 1986; Чабаненко, 2002; Пчёл-
кин, 2008.
ИЛЛЮСТРАЦИИ: Asahina, 1939: 357.
ТАЛЛОМ до 20 см дл., более или менее повисающий, прикрепленный к суб-
страту в основании, светло-каштановый до коричневого. Ветви 0.2–1.0 мм шир.,
гладкие, местами мелкоямчатые или с продольными бороздами, блестящие, вет-
вление дихотомическое. Веточки в местах ветвления слегка уплощенные, на
концах вильчато разветвленные. Коровой слой до 80 мкм толщ. Псевдоцифеллы
точковидные, слегка приподнятые над поверхностью таллома. АПОТЕЦИИ встреча-
ются редко, чашечковидные до бесформенных, до 2 мм в диам., диск без налета.
Гимений 30–35 мкм выс., сумки булавовидные, 35 × 10 мкм. Аскоспоры неокра-
шенные, одноклеточные, эллипсоидные, 7.3–9.0 × 3.5–4.5 мкм. ПИКНИДЫ рассеян-
ные, 80–140 × 70–120 мкм. Конидии 4.9 × 0.7 мкм. — Рис. 17, 18 (вклейка).
Флора лишайников России Флора лишайников России
54
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Таллом содержит оливеторовую кислоту, сердцеви-
на от C и KC краснеет, от K и P не изменяется, в UV не флюоресцирует.
ЭКОЛОГИЯ. Эпифитный лишайник, растет преимущественно на коре и ветвях
хвойных деревьев, реже встречается на лиственных породах в хвойных и сме-
шанных лесах.
РАСПРОСТРАНЕНИЕ. Россия. ДВ: ДВ юж.
—
Хабаровский край (юж. часть), Са-
халинская обл. (Сахалин; Курильские о-ва: Кунашир, Итуруп, Шикотан, Уруп). —
Карта 13.
Общее распространение. Пацифический вид, встречается на островах и побе-
режье Тихого океана. Азия (Япония, Тайвань), Сев. Америка (сев.-зап. побережье).
ПРИМЕЧАНИЕ. Близкий вид Bryocaulon satoanum (Gyeln.) Kärnefelt (на террито-
рии России не обнаружен), известный из горных районов Японии, отличается на-
личием физодовой кислоты (от C сердцевина не изменяется), а также более мел-
кими аскоспорами (6.0 × 2.5 мкм). Морфологически сходен с Oropogon asiaticus,
однако последний отличается рыхлой, содержащей желтоватый пигмент сердце-
виной с полостью внутри, а также муральными аскоспорами.
И. Н. Урбанавичене, Т. Н. Пыстина
Bryoria Brodo et D. Hawksw.
1977, Opera Bot. 42: 78. Т и п : Bryoria trichodes (Michx.) Brodo et D. Hawksw. —
Бриория
= Bryopogon Th. Fr., 1860 (prepr.), Nova Acta Regiae Soc. Sci. Upsal., ser. 3, 3: 125,
nom. illeg., non Link, 1833. Т и п : Bryopogon jubatus (L.) Link, nom. rejic.
Лихенизированные грибы. ТАЛЛОМ повисающий, до 10(30) см дл. и более, или
кустисто-дернинковидный или прямостоячий около 5–10 см выс., матовый или
блестящий. Цвет таллома варьирует от беловатого, светло-сероватого, желтоватого
или коричневато-оливкового до темно-, красновато-коричневого или черновато-ко-
ричневого. Ряд видов, особенно обитающих в горных или тундровых местообита-
ниях, имеет отчетливо двухцветный таллом — с темным основанием (примерно на
1⁄2 высоты) главных ветвей и со светлой верхушечной частью. У некоторых видов
с повисающими, однообразно окрашенными талломами ветви имеют небольшие
черноватоокрашенные фрагменты, встречающиеся по всей длине. Таллом в ос-
новном густо, изо- или анизотомически-дихотомически разветвленный, у боль-
шинства видов без четко выраженных главных ветвей, к субстрату прикрепляется
гомфом, у длинных, повисающих морф — часто непосредственно ветвями. Вет-
ви гетеромерные — с прозоплектенхимной корой и очень рыхлой, арахноидной
сердцевиной, часто цилиндрические в сечении до уплощенных; нитевидные, 0.1–
2.0 мм толщ. (до 4 мм у Bryoria fremontii). Основные ветви у ряда видов несут вто-
Род Bryoria
55
ричные, третичные или тонкие, очень короткие боковые веточки — латеральные
спинулы (шипики). Важным диагностическим признаком являются углы в местах
ветвления — тупые (широкие), прямые или острые. На поверхности ветвей ча-
сто развиваются веретенообразные псевдоцифеллы — короткие или длинные (об-
вивающие главные ветви); беловатые, желтоватые или коричневые; плоские или
вдавленные, очень редко — приподнятые. Органами вегетативного размножения
являются соредии, формирующие соралии — беловатые, серовато-зеленые до чер-
новатых или лимонно-желтые. Настоящие изидии отсутствуют, у некоторых видов
(напр., у B. furcellata) развиваются пучки изидиевидных спинул. АПОТЕЦИИ обра-
зуются очень редко, в основном боковые (латеральные); округлые, зеориновые,
край одного цвета с талломом, тонкий, чуть возвышающийся над диском, позднее
у большинства видов становится незаметным. Диск светло-коричневый, коричне-
вый или желтоватый, иногда с небольшим налетом, вогнутый или плоский, выпу-
клый у зрелых апотециев. Сумки Lecanora-типа, узкобулавовидные, битуникатные,
обычно 8-споровые. Аскоспоры округлые до эллипсоидных или широкоэллипсо-
идных, бесцветные, простые, 4–15 мкм дл. КОНИДИАЛЬНЫЕ СПОРОНОШЕНИЯ. Пикниды
образуются редко, встречаются на более тонких веточках; округлые или продолго-
ватые, 0.05–0.10 × 0.05 мм; обычно полностью погружены в веточки, позднее — до
приподнятых. Конидии тонкостенные, веретенообразные, 5–7 × 1 мкм. ФОТОБИОНТ
Trebouxia-типа.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. β-орсинольные депсидоны, атранорин, хлоратра-
норин; фумарпротоцетраровая, протоцетраровая, алекториаловая, барбатоловая,
норстиктовая, коннорстиктовая, гирофоровая, псоромовая и вульпиновая кисло-
ты, иногда присутствуют жирные кислоты.
ЭКОЛОГИЯ. В России наиболее широко встречаются эпифитные и эпибриофит-
ные виды. Довольно редки в производных лесах, предпочитают коренные, где в
достаточно влажных местообитаниях могут густо обрастать стволы и ветви дере-
вьев (рис. 19, вклейка). В горнолесных местообитаниях поселяются на замшелых
скалах и валунах (Bryoria bicolor), в тундровых местообитаниях растут на почве
среди мхов (B. nitidula).
РАСПРОСТРАНЕНИЕ. Произрастают в умеренных и северных, а также горно-боре-
альных регионах Евразии и Сев. Америки, встречаются в горах Юго-Вост. Азии,
Австралазии и Африки.
ПРИМЕЧАНИЕ. Род включает около 50 видов, в России встречается 17 видов.
Изучению экологических, анатомических, морфологических и хемотаксоно-
мических особенностей таксонов рода посвящено большое число публикаций и
монографий (Motyka, 1964; Hawksworth, 1972; Brodo, Hawksworth, 1977; Krog,
1979, 1980; Голубкова, 1996б; и др.). Современные молекулярные исследования
привели к значительным изменениям в понимании межвидовых границ рода
Bryoria (Myllys et al., 2011a; Velmala et al., 2014; Boluda et al., 2019). Например,
C. G. Boluda с соавторами (Boluda et al., 2019) на основе молекулярно-филогене-
тических исследований поставили под сомнение самостоятельность устоявших-
Род Bryoria
Флора лишайников России Флора лишайников России
56
ся, фенотипически выделяемых морфовидов и провели переоценку границ видов
из секции Implexae (Gyeln.) Brodo et D. Hawksw., широко представленных в Го-
ларктике, в том числе произрастающих в России B. capillaris, B. chalybeiformis,
B. implexa, B. kuemmerleana, B. lanestris, B. subcana, B. vrangiana. Все эти виды
предложено считать синонимами B. fuscescens, а североамериканские виды
B. friabilis Brodo et D. Hawksw., B. inactiva Goward, Velmala et Myllys, B. pikei
Brodo et D. Hawksw.
—
синонимами B. pseudofuscescens (Gyeln.) Brodo et
D. Hawksw. Таким образом, перечисленные морфовиды предлагается редуциро-
вать до двух филогенетических видов, а при определении представителей секции
Implexae, в отсутствие молекулярных данных, характеризовать эти таксоны как
Fuscescens- или Pseudofuscescens-комплексы. И только для двух видов секции:
B. kockiana Velmala, Myllys et Goward и B. glabra — этими авторами была под-
тверждена самостоятельность. В качестве доказательства этой точки зрения ав-
торами назван ограниченный набор морфологических признаков и значительная
химическая изменчивость рода Bryoria (Boluda et al., 2019).
Тем не менее, на сегодняшний день вопрос все еще остается дискуссионным
и охарактеризованное выше мнение поддержано далеко не всеми лихенологами.
Опираясь на публикацию T. Spribille с соавторами (Spribille et al., 2020), мы при-
держиваемся более консервативных подходов, в отличие от работы Boluda с соав-
торами (Boluda et al., 2019).
Для характеристики и разграничения видов в пределах секции Implexae
и подготовки ключа мы опирались на исследования S. Velmala с соавторами
(Velmala et al., 2014) и L. Myllys (Myllys et al., 2011a), работу Н. С. Голубковой
(1996), а также на собственные данные. Например, в работах Velmala с соавтора-
ми (Myllys et al., 2011b; Velmala et al., 2014) при изучении проблем таксономии
секции Implexae, идентификации и морфологических особенностей рода в целом
подчеркивается, что хемотипические признаки видов рода Bryoria достаточно
четкие, даже если они не поддержаны генетическими данными. Основываясь на
концепции, подтвержденной молекулярными данными (Velmala et al., 2014), мы
включаем виды B. subcana, B. lanestris и B. chalybeiformis в состав B. fuscescens
s. l. Тем не менее, принимая B. fuscescens в широком объеме, в характеристике
этого вида мы приводим отличительные морфологические и химические призна-
ки B. subcana, B. lanestris и B. chalybeiformis.
Ключ для определения видов рода Bryoria составлен нами с использованием
морфологических признаков и хемотаксономических данных на основе изучения
гербарных коллекций с территории России и других доступных гербариев.
ИЗБРАННАЯ ЛИТЕРАТУРА. Hawksworth, 1972; Brodo, Hawksworth, 1977; Голубкова,
1996б; Velmala et al., 2009, 2014; Myllys et al., 2011a, b.
Род Bryoria
57
Ключ для определения видов рода Bryoria
1. Ветви толстые, 1–2(4) мм в диам., жесткие, неравной толщины, часто пере-
крученные, мелкоямчатые, широко уплощенные в местах ветвления; соралии
бугорчатые, светло- или ярко-желтые (содержат вульпиновую кислоту) .........
.......................................................................................................... 5. B. fremontii.
—
Ветви тоньше, не более 2 мм в диам., варьируют по морфологии; соралии,
если имеются, не бывают ярко-желтыми, лишайниковые вещества иные ... 2.
2. Таллом отчетливо двухцветный, в основании темно-коричневый до черного,
к вершинкам светло-коричневато-оливковый ................................................... 3.
—
Таллом однообразно окрашенный, у ряда видов с единичными черноватыми
фрагментами по всей длине таллома ................................................................ 5.
3. Соралии многочисленные, с изидиоидными шипиками; псевдоцифеллы от-
сутствуют; все химические тесты отрицательные ...................... 14. B. smithii.
—
Соралии отсутствуют; псевдоцифеллы малочисленные, коричневые; внутрен-
няя часть коры от P краснеет ............................................................................. 4.
4. Основные ветви 0.2–0.5 мм в диам., с перпендикулярными спинулами, обиль-
ными вторичными и третичными веточками; углы между ветвями прямые ...
..............................................................................................................
3. B. bicolor.
—
Основные ветви до 0.2 мм в диам., третичные веточки и спинулы слабо
развиты; углы между веточками в верхней части таллома острые, в основа-
нии — тупые .................................................................................... 15. B. tenuis.
5. Таллом короткий (5–7 см выс.), в виде широко разветвленного кустика
(с примерно равными высотой и шириной); псевдоцифеллы отсутствуют, со-
ралии обильные ................................................................................................... 6.
—
Таллом хорошо развитый, в основном длиннее 7 см, повисающий (у B. nitidula
прямостоячий), узко разветвленный, с псевдоцифеллами и соралиями или без
них ......................................................................................................................... 7.
6. Соралии белые, ýже ветвей, с пучками изидиевидных колючек; внутренняя
часть коры от P краснеет (фумарпротоцетраровая кислота) . .............................
....................................................................................................... .. 6. B. furcellata.
—
Соралии зеленовато-черноватые, шире ветвей, без колючек; все химические
тесты отрицательные ................................................................. 13. B. simplicior.
7. Соралии отсутствуют; таллом прямостоячий (до 8 см выс.), псевдоцифел-
лы коричнево-черноватые; главные ветви с тонкими боковыми спинулами;
сердцевина и внутренняя часть коры от P краснеют (фумарпротоцетраровая
кислота); в тундровых или горно-лесных местообитаниях, на почве или зам-
шелых скалах ................................................................................ 12. B. nitidula.
—
Соралии имеются (отсутствуют только у B. asiatica); таллом повисающий
до коротко повисающего, часто более 8 см дл.; псевдоцифеллы имеются или
отсутствуют, их цвет, морфология ветвей и состав лишайниковых веществ
варьируют; эпифиты, редко на замшелых скалах ............................................ 8.
Флора лишайников России Флора лишайников России
58
8. Соралии или таллом от C краснеют (очень редко не изменяются), от KC ста-
новятся розово-красными, от K желтеют или краснеют, реже не изменяются
................................................................................................................................. 9.
—
Соралии и таллом всегда от C, KC и K не изменяются (реже от K кора слабо
желтеет) .............................................................................................................. 12.
9. Таллом серовато-белый, иногда с фиолетовым оттенком, матовый; ветви со
спинулами, темнеющими на кончиках; углы в местах ветвления тупые в ос-
новании таллома и острые в верхушечной части .......... 11. B. nadvornikiana.
—
Цвет таллома варьирует; ветви без укороченных спинул, темнеющих на кон-
чиках; углы в местах ветвления в основном острые ..................................... 10.
10. Таллом светло-серый до беловатого, без фиолетового оттенка; углы в местах
ветвления острые .......................................................................... 4. B. capillaris.
—
Таллом от светло- до темно-коричневого, иногда с оливковым оттенком и
черноватыми фрагментами, углы в местах ветвления варьируют ............... 11.
11. Таллом в основании и местах ветвления часто с небольшим налетом; соралии
и таллом от K краснеют, от P желтеют (норстиктовая, коннорстиктовая кисло-
ты, атранорин) ................................................................... 10. В. kuemmerleana.
—
Таллом без налета, реакции иные (таллом и соралии от P иногда слабо жел-
теют или краснеют — фумарпротоцетраровая кислота или атранорин) ..........
......................................................................................................
17. B. vrangiana.
12. Соралии и коровой слой от P желтеют (псоромовая кислота), кора от K слабо
желтеет ....................................................................................... 9. B. implexa s. l.
—
Соралии и внутренняя часть коры от P краснеют (фумарпротоцетраровая кис-
лота), кора от K не изменяется ......................................................................... 13.
13. Псевдоцифеллы отсутствуют .............................................. 7. B. fuscescens s. l.
—
Псевдоцифеллы имеются .................................................................................. 14.
14. Псевдоцифеллы беловатые ............................................................................... 15.
—
Псевдоцифеллы коричневые или зеленовато-коричневые ............................ 16.
15. Псевдоцифеллы редки, слабо заметные, белые; соралии обильные, белые,
овальные; таллом коричневый или оливково-коричневый, в основном бле-
стящий, изотомически-дихотомически ветвящийся, ветвление регулярное,
все углы в местах ветвления тупые, широкоокруглые; соралии от P краснеют
(фумарпротоцетраровая кислота) .................................................... 8. B. glabra.
—
Псевдоцифеллы беловатые, обильные, приподнятые над поверхностью вет-
вей; соралии редкие; таллом бледно-, красно- или темно-коричневый, часто с
черными фрагментами, с острыми или широкими углами в местах ветвления;
внутренняя часть корового слоя от P краснеет (фумарпротоцетраровая кисло-
та, хлоратранорин) ...................................................................... 16. В. trichodes.
16. Соралии редки, до 1 мм, щелевидные, бугорковидные; псевдоцифеллы ко-
ричневые, веретеновидные и вдавленные; обычны укороченные боковые
спинулы; углы в местах ветвления широкие; таллом коричневый, иногда с
черноватыми фрагментами; сердцевина и соралии от P слабо краснеют (фу-
Род Bryoria
59
марпротоцетраровая кислота, иногда в низкой концентрации); в России не-
редок ............................................................................................. 1. B. americana.
—
Соралии всегда отсутствуют; псевдоцифеллы редкие, слабо заметные, вдав-
ленные, до темно-коричневых; боковые спинулы редкие; углы в местах вет-
вления острые; таллом коричневый, чернеющий в основании; все химические
тесты отрицательные; в России известен только из Амурской и Сахалинской
областей ............................................................................................ 2. B. asiatica.
1. Bryoria americana (Motyka) Holien
1994, Graphis Scripta 6 (1): 40. ≡ Alectoria americana Motyka, 1960, Fragm.
Florist. Geobot. 6 (3): 449. = Bryoria trichodes subsp. americana (Motyka) Brodo
et D. Hawksw., 1977, Opera Bot. 42: 96. Т и п : [Канада] New Brunswick, Grand
Manan, 1879, H. Willey (US голотип). — Бриория американская.
ИЗБРАННАЯ ЛИТЕРАТУРА: Brodo, Hawksworth, 1977; Голубкова, 1996б; Myllys et
al., 2011a, b; Stenroos et al., 2016.
ИЛЛЮСТРАЦИИ: Brodo et al., 2001: 186; Hinds, Hinds, 2007: 139; Myllys et al.,
2011a: 146; Myllys et al., 2011b: 631; Stenroos et al., 2016: 93.
ТАЛЛОМ 7–15(20) см дл., из тонких повисающих веточек, довольно густо и
широко изотомически-дихотомически ветвящийся, коричневый до темно-корич-
невого, часто с черноватыми фрагментами, слегка блестящий, обычно с тупыми
углами в местах ветвления. Главные ветви тонкие, часто с короткими, перпенди-
кулярно отходящими веточками. Соралии отсутствуют или редки, белые, до 1 мм
дл., бугорковидные, приподнятые над поверхностью ветвей или щелевидные,
могут огибать ветви. Псевдоцифеллы коричневатые, редкие, веретенообразные,
вдавленные, 0.5–1.0 мм дл. АПОТЕЦИИ образуются очень редко, боковые, до 2 мм в
диам., диск коричневатый, слегка выпуклый, аскоспоры эллипсоидные, 5.5–7.0 ×
4.0–5.0 мкм. ПИКНИДЫ неизвестны. — Рис. 20–22 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Содержит фумарпротоцетраровую кислоту, часто в
низких концентрациях. Сердцевина и соралии от P краснеют (иногда слабо). Апо-
теции содержат псоромовую кислоту (диски от P желтеют).
ЭКОЛОГИЯ. На стволах и ветвях хвойных, реже лиственных деревьев во влаж-
ных лесах.
РАСПРОСТРАНЕНИЕ. Россия. ЕР: Сев. Кавказ — Адыгея, Краснодарский край.
СИБ: Як. — юж. ДВ: ДВ сев. — Камчатский край (бывшая Камчатская обл.); ДВ
юж. — Сахалинская обл., Хабаровский и Приморский края. — Карта 14.
Общее распространение. Европа, Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Bryoria americana напоминает B. fuscescens и B. implexa, от ко-
торых его отличают очень редко образующиеся соралии, широко расходящиеся,
изотомически-дихотомически ветвящиеся участки таллома, небольшие перпенди-
кулярные боковые веточки, главные ветви с черноватыми фрагментами и корич-
невые вдавленные псевдоцифеллы. От B. trichodes в основном отличается корич-
Флора лишайников России Флора лишайников России
60
невыми вдавленными псевдоцифеллами, более ровными, тонкими, округлыми в
сечении веточками и отсутствием хлоратранорина.
2. Bryoria asiatica (Du Rietz) Brodo et D. Hawksw.
1977, Opera Bot. 42: 155. ≡ Alectoria asiatica Du Rietz, 1926, Ark. Bot. 20A (11):
18. Т и п : [Китай, Сычуань] China, Sichuan prov. (as Sze-chu’an), reg. bor.-
occid., mellan Tsagogamba och Tamba, alt. 4000 m. s. m., på Juniperus, Picea eller
Rhododendron, H. Smith, 5018, 02.10.1922 (UPS голотип). — Бриория азиатская.
ИЗБРАННАЯ ЛИТЕРАТУРА: Wang, Harada, 2001; Wang et al., 2017.
ИЛЛЮСТРАЦИИ: Wang, Harada, 2001: 45–47.
ТАЛЛОМ повисающий, 15–20 см дл., блестящий, темно-коричневый, чернею-
щий в основании, мягкий, анизотомически-дихотомически ветвящийся, к вершин-
кам ветвей ветвление становится изотомически-дихотомическим. Углы в местах
ветвления V-образные, острые — обычно 45–60°. Главные ветви ровные, цилин-
дрические (в сечении округлые, 0.1–0.3 мм в диам.). Боковые спинулы редкие,
0.5–2.0 мм дл., отрастают под острым углом (30–80°), одного цвета с главными
ветвями. Соралии отсутствуют. Псевдоцифеллы иногда слабо заметные, плоские
или слегка вдавленные, коричневые до темно-коричневых. АПОТЕЦИИ образуются
редко, боковые, диск 0.5–1.0 мм в диам., коричневый, блестящий, слегка вогнутый,
с возрастом становится плоским. Аскоспоры почти округлые до эллипсоидных,
6.5–8.0 × 4.0–5.0 мкм. ПИКНИДЫ неизвестны. — Рис. 23, 24 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Методом тонкослойной хроматографии не выявля-
ются. Все химические тесты отрицательные.
ЭКОЛОГИЯ. На ветвях хвойных, реже лиственных деревьев во влажных горных
лесах. Эпифитный вид.
РАСПРОСТРАНЕНИЕ. Россия. ДВ: ДВ юж.
—
Амурская обл., Сахалинская обл.
(Сахалин, о. Кунашир, Курильский заповедник). — Карта 15.
Общее распространение. Азия (Индия, Китай, Тайвань, Япония).
ПРИМЕЧАНИЕ. От морфологически близкого Bryoria americana, также имеюще-
го коричневые псевдоцифеллы, отличается более многочисленными латеральны-
ми спинулами, острыми углами в местах ветвления, отсутствием соралий и фу-
марпротоцетраровой кислоты.
3. Bryoria bicolor (Ehrh.) Brodo et D. Hawksw.
1977, Opera Bot. 42: 99. ≡ Lichen bicolor Ehrh., 1784, Hannover. Mag. 22: 161.
≡ Alectoria bicolor (Ehrh.) Nyl., 1857 («1856»), Actes Soc. Linn. Bordeaux 21 (4):
291. ≡ Bryopogon bicolor (Ehrh.) Stein, 1879, Krypt.-Fl. Schlesien, 2 (2) Flechten: 35.
Т и п : [Германия] Harz («Hercynia»), Ehrhart, Pl. Crypt. Linn. No. 40 (LINN-Sm.
1712.2(3) лектотип, Brodo, Hawksworth, 1977: 99). — Бриория двухцветная.
ИЗБРАННАЯ ЛИТЕРАТУРА: Brodo, Hawksworth, 1977; Голубкова, 1996б; Hinds,
Hinds, 2007; Smith et al., 2009; Myllys et al., 2011a, b.
Род Bryoria
61
ИЛЛЮСТРАЦИИ: Dobson, 2000: 78; Holien, Tønsberg, 2006: 62; Smith et al., 2009:
225; Myllys et al., 2011a: 146; Myllys et al., 2011b: 631.
ТАЛЛОМ отчетливо двухцветный, блестящий, в виде прямостоячих кустиков,
2.0–3.5(4.0) см выc., с явно выраженными главными ветвями, у основания почти
черными и утолщенными до 0.5 мм. В верхней части ветви светлые, оливково-се-
роватые или светло-коричневые, 0.2–0.3 мм в диам., цилиндрические, отходящие
от главных почти под прямым углом. Мелкие колючки (0.03–0.08 мм) многочис-
ленные, перпендикулярные поверхности ветвей. Псевдоцифеллы веретеновид-
ные, 0.2–3.5 мм дл., коричневые, плоские или слегка приподнятые над поверх-
ностью ветвей, не обильные. Соралии отсутствуют. АПОТЕЦИИ образуются очень
редко, до 1 мм в диам., диск темно-коричневый. Аскоспоры округлые до широ-
коэллипсоидных, 6.0–9.0 × 4.0–6.0 мкм. ПИКНИДЫ образуются редко, мелкие, чер-
ные, 0.05–0.10 × 0.05 мм, локализуются ближе к кончикам ветвей, конидии 8.0 ×
0.5 мкм. — Рис. 25 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Содержит фумарпротоцетраровую кислоту. Кора и
сердцевина от P краснеют, от K, C и KC не изменяются.
ЭКОЛОГИЯ. На замшелых скалах и валунах, очень редко как эпифит.
РАСПРОСТРАНЕНИЕ. Россия. ЕР: Евр. Север — Мурманская обл., Карелия, Ле-
нинградская, Архангельская обл.; Сев. Кавказ — Краснодарский край, Адыгея,
Карачаево-Черкесия, Кабардино-Балкария, Северная Осетия; Сев. Урал — Коми
(горная уральская часть); СИБ: Зап. Сиб. сев.
—
Ханты-Мансийский АО; Юж.
Сиб.
—
Кемеровская обл., Красноярский край, Республика Алтай, Бурятия; ДВ:
ДВ сев.
—
Камчатский край (бывшая Камчатская обл.); ДВ юж.
—
Сахалинская
обл., Хабаровский и Приморский края. — Карта 16.
Общее распространение. Европа, Азия, Сев. и Юж. Америка, Африка,
о. Мадейра, о. Сент-Пол.
ПРИМЕЧАНИЕ. Близок по морфологии к Bryoria smithii (двухцветный таллом с
укорачивающимися к вершинке веточками второго порядка и мелкими, перпенди-
кулярно отходящими от них веточками третьего порядка), но у B. smithii образу-
ются соралии и все реакции отрицательные (нет веществ, определяемых методами
тонкослойной хроматографии). Другой близкий по морфологии и набору лишайни-
ковых веществ вид — B. tenuis — характеризуется острыми углами между расходя-
щимися ветвями и отсутствием веточек третьего порядка, типичных для B. bicolor.
4. Bryoria capillaris (Ach.) Brodo et D. Hawksw.
1977, Opera Bot. 42: 115. ≡ Parmelia jubata (L.) Ach. var. capillaris Ach., 1803,
Methodus: 273. ≡ Alectoria jubata (L.) Ach. var. capillaris (Ach.) Ach., 1810,
Lichenogr. Universalis: 593. ≡ Alectoria capillaris (Ach.) Cromb., 1871, J. Bot. 9:
177. ≡ Alectoria implexa var. capillaris (Ach.) Stizenb., 1892, Ann. K. K. Naturhist.
Hofmus. 7: 131. Т и п : [Швеция] Suecia (H-ACH 1799A = H9503990 лектотип,
Hawksworth, 1969: 393). — Бриория волосовидная.
Флора лишайников России Флора лишайников России
62
ИЗБРАННАЯ ЛИТЕРАТУРА: Brodo, Hawksworth, 1977; Thomson, 1984; Holien, 1989;
Purvis et al., 1992; Wirth, 1995; Голубкова, 1996б; Brodo et al., 2001; Myllys et al.,
2011a, b; Velmala et al., 2014; Stenroos et al., 2016.
ИЛЛЮСТРАЦИИ: Hawksworth, 1972: 4; Wirth, 1995: 187; Dobson, 2000: 78; Brodo
et al., 2001: 180; Hinds, Hinds, 2007: 133; Myllys et al., 2011a: 146; Stenroos et al.,
2016: 95.
ТАЛЛОМ до 20(30) см дл., свисающий, беловато-серый до серого, иногда буро-
ватый, матовый до слегка блестящего, нерегулярно ветвящийся, с острыми угла-
ми между ветвями. Ветви преимущественно ровные, иногда скрученные и сжа-
тые, до 0.07–0.40 мм в диам.; колючки и колючкообразные веточки отсутствуют.
Псевдоцифеллы веретеновидные, белые, плоские или слегка вдавленные, мало-
заметные, 0.2–0.7 мм дл., 0.03–0.10 мм шир. Соралии часто обильные, белые, бу-
горчатые, до 0.5 мм в диам., реже слабо развитые. АПОТЕЦИИ образуются редко,
1–2 мм в диам., диск светло-коричневый. Аскоспоры эллипсоидные или почти
округлые, 5–9 × 4–6 мкм. ПИКНИДЫ неизвестны. — Рис. 26 (вклейка).
ЭКОЛОГИЯ. На коре и древесине хвойных и лиственных деревьев в лесных по-
ясах среднегорий и низкогорий, предпочитает влажные темнохвойные или сме-
шанные леса.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Кора и сердцевина от C и KC краснеют (реже от C
не изменяются), от K и P желтеют (атранорин, барбатоловая, алекториаловая кис-
лоты). Соралии от P краснеют (фумарпротоцетраровая кислота).
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: Аркт. евр.
—
Ненецкий АО, Аркт. зап.-
сиб.
—
Ямало-Ненецкий АО; ЕР: Евр. Север — Мурманская обл., Карелия, Ле-
нинградская, Архангельская, Кировская, Вологодская обл., Коми; Ср. Россия —
Калининградская, Псковская, Ярославская обл., Московская, Калужская обл.,
Марий Эл, Мордовия, Татарстан, Чувашия, Башкирия, Ульяновская, Саратовская,
Нижегородская, Пензенская, Смоленская обл.; Юж. Россия — Оренбургская обл.;
Сев. Кавказ — Краснодарский край, Адыгея, Карачаево-Черкесия, Кабардино-
Балкария, Северная Осетия и Ингушетия; Сев. Урал — горные районы Коми,
Пермского края, Свердловской обл.; Юж. Урал — горные районы Свердловской,
Челябинской обл.; СИБ: Зап. Сиб. сев.
—
Ханты-Мансийский АО, Тюменская
обл.; Ср. Сиб.
—
Красноярский край; Юж. Сиб.
—
Алтайский край, Краснояр-
ский край, Республика Алтай, Бурятия, Тыва, Хакасия, Кемеровская обл.; ДВ:
ДВ сев.
—
Камчатский край (бывшая Камчатская обл.); ДВ юж.
—
Сахалинская
обл. — Карта 17.
Общее распространение. Европа, Макаронезия, Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Отличается от близкого вида Bryoria nadvornikiana длинным, ча-
сто повисающим талломом, отсутствием латеральных колючек, более крупными
аскоспорами (4–6 × 5–9 мкм). От других видов со светлым талломом отличается
ярко-желтой реакцией с K (барбатоловая кислота) и розовой реакцией с C (алек-
ториаловая кислота).
Род Bryoria
63
5. Bryoria fremontii (Tuck.) Brodo et D. Hawksw.
1977, Opera Bot. 42: 136. ≡ Alectoria fremontii Tuck., 1858, Amer. J. Sci. Arts, ser. 2,
25 (75): 422. Т и п : [США] California, Sierra Nevada, «camp of Dec. 5–6», 1845,
J. C. Frémont (FH-Tuck. 498 голотип). — Бриория Фримонта.
= Alectoria olivacea Räsänen.
ИЗБРАННАЯ ЛИТЕРАТУРА: Brodo, Hawksworth, 1977; Голубкова, 1996б; Петрова,
2000; Myllys et al., 2006; Velmala et al., 2009; Myllys et al., 2011a, b.
ИЛЛЮСТРАЦИИ: Brodo et al., 2001: 180, 185; Holien, Tønsberg, 2006: 64; Velmala
et al., 2009: 239; Myllys et al., 2011a: 147; Myllys et al., 2011b: 631.
ТАЛЛОМ 10–30(60) см дл., свисающий, бородовидный, желтоватый, краснова-
тый, светло- или темно-коричневый (окраска может варьировать у одного и того
же образца в зависимости от концентрации коричневого пигмента и вульпиновой
кислоты), тусклый или блестящий, анизотомически-дихотомически разветвлен-
ный до почти моноподиального у основания, с острыми углами между расхо-
дящимися ветвями. Ветви (0.2)0.4–1.5(4.0) мм в диам., жесткие, неравной тол-
щины, основные часто перекрученные и мелкоямчатые, в местах ветвления и у
основания уплощенно сжатые, конечные — волосовидные, цилиндрические. От
основных ветвей изредка отходят тонкие, короткие перпендикулярные веточки.
Латеральные колючки отсутствуют. Псевдоцифеллы, если присутствуют, обычно
хорошо заметны, удлиненно-веретеновидные, вогнутые, белые до светло-желтых,
цвет может изменяться в пределах одного и того же экземпляра. Соралии рассе-
янные, часто отсутствуют, бугорчатые, овальные, бледно- или ярко-желтые, рав-
ны ветвям или чуть шире. АПОТЕЦИИ встречаются очень редко, 1.0–2.0(4.0) мм в
диам., латеральные, округлые, молодые вогнутые или плоские, позднее — слегка
выпуклые, диск с желтоватым налетом, эксципул одного цвета с талломом, тон-
кий, позднее становится незаметным. Сумки булавовидные. Аскоспоры 5–8 ×
4–5 мкм, почти округлые, одноклеточные, бесцветные, по 8 в сумке. ПИКНИДЫ не-
известны. — Рис. 27, 28 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Соралии, апотеции, псевдоцифеллы и таллом (ред-
ко) содержат вульпиновую кислоту. В образцах из Сев. Америки отмечены бар-
батовая и норстиктовая кислоты. Все стандартные химические тесты отрицатель-
ные, cоралии и апотеции могут флюоресцировать в UV.
ЭКОЛОГИЯ. На коре крупных деревьев хвойных (обычно сосны, реже ели, ли-
ственницы, кедра), иногда лиственных (береза) пород, часто в кронах деревьев,
в старовозрастных хвойных и смешанных таежных и горных лесах, реже в забо-
лоченных лесах и на болотах. Предпочитает хорошо освещенные местообитания.
РАСПРОСТРАНЕНИЕ. Россия. ЕР: Евр. Север — Мурманская обл., Карелия, Ар-
хангельская обл., Коми, Ленинградская и Вологодская обл.; Ср. Россия — Твер-
ская обл.; Сев. Урал — Коми (горная уральская часть), Пермский край (сев.).
—
Карта 18.
Общее распространение. Европа (сев.), Сев. Америка.
Флора лишайников России Флора лишайников России
64
ПРИМЕЧАНИЕ. От других длинных коричневоокрашенных повисающих видов
рода Bryoria хорошо отличается крупными ярко-желтыми соралиями, иногда
желтоватым цветом таллома и псевдоцифелл. При отсутствии соралий вид можно
узнать по наличию толстых перекрученных, ямчатых, местами уплощенных ос-
новных веточек, желтоватому оттенку псевдоцифелл, отрицательным реакциям
на действие реактивов и наличию вульпиновой кислоты.
6. Bryoria furcellata (Fr.) Brodo et D. Hawksw.
1977, Opera Bot. 42: 103. ≡ Cetraria furcellata Fr., 1825, Syst. Orb. Veg. 1: 283.
Т и п : [США] America borealis (UPS голотип). — Бриория мелковильчатая.
= Alectoria nidulifera Norrl. ≡ Bryopogon niduliferus (Norrl.) Elenkin.
ИЗБРАННАЯ ЛИТЕРАТУРА: Brodo, Hawksworth, 1977; Purvis et al., 1992; Голубкова,
1996б; Петрова, 2000; Hinds, Hinds, 2007; Harada, Wang, 2008; Myllys et al., 2011a;
Stenroos et al., 2016.
ИЛЛЮСТРАЦИИ: Brodo et al., 2001: 181; Hinds, Hinds, 2007: 134; Harada, Wang,
2008: 161, 162; Myllys et al., 2011a: 147; Stenroos et al., 2016: 97.
ТАЛЛОМ в виде прямостоячих, 3–5(8) см выс., сильно разветвленных темно-
коричневых кустиков, изотомически-дихотомически разветвленный, в базальной
части с ветвями, расходящимися под тупым углом. Ветви до 0.5 мм в диам., бле-
стящие, ровные, иногда слега сжатые у основания, с короткими латеральными ко-
лючкообразными веточками, перпендикулярно отходящими от поверхности вет-
вей. Псевдоцифеллы отсутствуют. Соралии многочисленные, белые, щелевидные,
0.3–1.0 мм дл., ýже веточек, на которых развиваются, слегка приподнятые над их
поверхностью, с пучками изидиевидных колючек. АПОТЕЦИИ образуются редко,
до 2 мм в диам., диск светлый до красновато-коричневого, вогнутый, позднее —
выпуклый, с толстым, часто соредиозным краем. Аскоспоры широкоэллипсоид-
ные до почти округлых, 6–7 × 3–4 мкм. ПИКНИДЫ образуются редко, мелкие, чер-
ные, 0.05–0.10 × 0.05 мм, локализуются ближе к кончикам ветвей, конидии 8 ×
0.5 мкм. — Рис. 29, 30 (вклейка).
ЭКОЛОГИЯ. На деревьях хвойных пород, особенно соснах, елях, изредка на по-
верхности валунов, гниющей древесине и почве.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой, сердцевина и cоралии от P краснеют
(фумарпротоцетраровая кислота). От K, C и KC не изменяются.
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: Аркт. евр. — Ненецкий АО, крайний северо-
восток Коми; Аркт. зап.-сиб. — Ямало-Ненецкий АО (п-ов Ямал); Аркт. тайм. —
север Таймырского Долгано-Ненецкого р-на; Аркт. чук.
—
Чукотский АО; ЕР:
Евр. Север — Мурманская обл., Карелия, Ленинградская, Архангельская, Воло-
годская, Кировская обл., Коми; Ср. Россия — Пермский край, Тверская, Ярослав-
ская, Костромская обл.; Сев. Урал — горные районы Коми, Ханты-Мансийского
АО, Свердловской обл.; СИБ: Зап. Сиб. сев.
—
Ямало-Ненецкий и Ханты-Ман-
сийский АО, Свердловская, Томская обл.; Зап. Сиб. юж.
—
Новосибирская обл.;
Род Bryoria
65
Юж. Сиб.
—
Красноярский край, Кемеровская обл., Республика Алтай, Тыва,
Хакасия, Бурятия, Иркутская обл., Забайкальский край; ДВ: ДВ сев.
—
Мага-
данская обл., Камчатский и Хабаровский края; ДВ юж. — Хабаровский край. —
Карта 19.
Общее распространение: Европа, Макаронезия, Азия, Океания, Сев. и
Центр. Америка (Голубкова, 1996б; Myllys et al., 2011a).
ПРИМЕЧАНИЕ. Bryoria furcellata хорошо отличается от других темноокрашен-
ных видов рода наличием соралий с пучками изидиевидных колючек и прямо-
стоячим дернинковидным блестящим талломом. Образцы с плохо развитыми
изидиевидными колючками могут быть спутаны с B. simplicior, который имеет зе-
леновато-черные, более широкие по сравнению с ветвями соралии и негативную
реакцию с P.
7. Bryoria fuscescens (Gyeln.) Brodo et D. Hawksw.
1977, Opera Bot. 42: 83. ≡ Alectoria fuscescens Gyeln., 1932 («1931»), Nyt Mag. Na-
turvidensk. 70: 55, nom. cons. Т и п : [Финляндия] Finland. Tavastia australis, Hol-
lola, 18.10.1882, J. P. Norrlin, in Norrlin, Nylander, Herb. Lich. Fenn. (exs.) No. 446a
(BP 33947 typ. cons.; BM, CANL, H изотипы); [Финляндия] Finland, Etelä-Savo,
Savitaipale, 600 m NW of Mustapää, Tikkioja, 2005, L. Myllys 464 (H 9209715 эпи-
тип, Boluda et al., 2019: 87). — Бриория буроватая.
= Alectoria jubata auct. = Alectoria viridescens Gyeln.; = Bryoria chalybeiformis
(L.) Brodo et D. Hawksw., nom. rejic.; = Bryoria lanestris (Ach.) Brodo et D. Hawksw.;
= Bryoria subcana (Nyl. ex Stizenb.) Brodo et D. Hawksw., nom. rejic.
ИЗБРАННАЯ ЛИТЕРАТУРА: Hawksworth, 1972; Brodo, Hawksworth, 1977; Krog,
1980; Thomson, 1984; Holien, 1989, 1991; Purvis et al., 1992; Wirth, 1995; Голуб-
кова, 1996б; Dobson, 2000; Brodo et al., 2001; Myllys et al., 2011a, b; Hawksworth,
Jørgensen, 2013; Velmala et al., 2014; Boluda et al., 2019.
ИЛЛЮСТРАЦИИ: Hawksworth, 1972: pl. 4E, pl. 5; Thomson, 1984: 59; Wirth, 1995:
183, 185, 188; Goward, 1999: 56, 137; Dobson, 2000: 79; Brodo et al., 2001: 181, 182;
Hinds, Hinds, 2007: 135; McCune, Geiser, 2009: 42; Myllys et al., 2011a: 147–150;
Stenroos et al., 2011: 80; Velmala et al., 2014: 364; Stenroos et al., 2016: 98.
ТАЛЛОМ 5–15(30) см дл., повисающий, серовато-, оливково-коричневый до
коричневато-черного, матовый до слегка блестящего. Ветви 0.07–0.5(0.9) мм в
диам., цилиндрические, в основном ровные, иногда становятся скрученными и
ямчатыми, без выраженных главных ветвей. Ветвление изотомически-дихото-
мическое, иногда, ближе к вершинкам, анизотомически-дихотомическое. Углы в
местах ветвления в основном острые. Псевдоцифеллы не образуются (молодые
узкощелевидные cоралии могут напоминать псевдоцифеллы). Соралии обильные,
реже бедно развиты, бугорчатые или щелевидные, шире ветвей, на которых об-
разуются, белые, иногда с черными вкраплениями и очень редко с короткими спи-
нулами. АПОТЕЦИИ нечастые, до 2 мм в диам., диск коричневый, от плоского до
Флора лишайников России Флора лишайников России
66
выпуклого, аскоспоры эллипсоидные, 6–9 × 4–5 мкм. ПИКНИДЫ образуются редко,
обычно агрегированные, черные и блестящие. — Рис. 31–36 (вклейка).
ЭКОЛОГИЯ. На деревьях хвойных и лиственных пород, сухостое, гниющей дре-
весине. Может поселяться на голых скалах, а также на замшелой почве в примор-
ских и альпийских местообитаниях. Один из наиболее широко распространенных
видов рода в равнинных и горных лесах Северного полушария.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой, сердцевина и cоралии от K, C и KC
не изменяются в окраске, соралии от P краснеют, коровой слой и иногда частично
сердцевина от P краснеют или не изменяются. Содержит фумарпротоцетраровую
кислоту, иногда хлоратранорин. В апотециях содержится псоромовая кислота (от
P диск желтеет).
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: Аркт. тайм. ЕР: Евр. Север — все районы;
Ср. Россия — все районы; Сев. Кавказ — Краснодарский край, Адыгея, Карачае-
во-Черкесия, Кабардино-Балкария, Северная Осетия, Дагестан и Ингушетия; Сев.
Урал — горные районы Коми, Пермского края, Ханты-Мансийского АО, Сверд-
ловской обл.; Юж. Урал — горные районы Пермского края, Свердловской и Че-
лябинской обл.; СИБ: Зап. Сиб. сев.
—
Ханты-Мансийский АО, Тюменская и
Томская обл.; Зап. Сиб. юж.
—
Челябинская, Омская, Новосибирская обл.; Юж.
Сиб.
—
Красноярский край, Новосибирская, Кемеровская и Иркутская обл., Ал-
тайский край, Бурятия, Тыва, Хакасия, Забайкальский край; Як. вост.; ДВ: ДВ
сев. — Магаданская обл., Камчатский и Хабаровский края; ДВ юж. — Амурская
обл., Еврейская АО, Сахалинская обл., Приморский край. — Карта 20.
Общее распространение. Европа, Африка, Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Bryoria fuscescens s. l.
—
очень вариабельный вид, который ха-
рактеризуется в основном коричневоокрашенным талломом, отсутствием псев-
доцифелл, овальными или щелевидными соралиями, которые шире диаметра
ветвей (молодые, узкощелевидные cоралии могут напоминать псевдоцифеллы).
На основании результатов филогенетического анализа, приведенных в ряде ра-
бот (Velmala et al., 2014; и др.), виды B. chalybeiformis, B. lanestris и B. subcana
предложено объединить с B. fuscescens. В результате последующих молекулярных
исследований секции Implexae с использованием пяти генов и 18 микросателлит-
ных маркеров все виды секции предложено свести к четырем (Boluda et al., 2019).
Принимая в целом точку зрения Velmala с соавторами (Velmala et al., 2014), мы,
тем не менее, считаем необходимым привести основные различия между морфо-
логическими видами B. fuscescens s. str., B. chalybeiformis, B. lanestris и B. subca-
na, опираясь на традиционно используемые морфологические, химические и эко-
логические признаки.
Наличие фумарпротоцетраровой кислоты — важный диагностический при-
знак Bryoria fuscescens s. l. (Brodo, Hawksworth, 1977). Bryoria fuscescens и
B. subcana обычно содержат фумарпротоцетраровую кислоту в соралиях, коровом
слое и сердцевине. У B. chalybeiformis и B. lanestris фумарпротоцетраровая кисло-
та содержится только в соралиях.
Род Bryoria
67
Морфология таллома и экологические предпочтения. Таллом Bryoria
chalybeiformis в виде распростертого (рис. 33, вклейка) укороченного кустика;
веточки блестящие, темно-коричневые, 0.1–0.3 мм в диам.; растет в основном на
почве в северных широтах или на ветвях хвойных деревьев в горных районах у
границы леса. Таллом B. lanestris длинный, повисающий, с тонкими, чернова-
тыми, нитевидными и очень хрупкими ветвями (рис. 34, 35, вклейка), распадаю-
щимися на фрагменты; преимущественно эпифит, характерен для горнотаежных
лесов Сев. Урала и Сибири. Таллом B. subcana (рис. 36, вклейка) более светлый,
бледно-буроватый до белого; преимущественно эпифит, приурочен к хвойным,
смешанным и лиственным лесам.
8. Bryoria glabra (Motyka) Brodo et D. Hawksw.
1977, Opera Bot. 42: 86. ≡ Alectoria glabra Motyka, 1960, Fragm. Florist. Geobot. 6:
448. Т и п : [США] USA, Washington, Olympic Peninsula, Clallam Co., Hurricane
Ridge, 5800 ft., 1950, B. I. Brown, W. C. Muenscher 129 (US голотип). — Бриория
оголенная.
ИЗБРАННАЯ ЛИТЕРАТУРА: Brodo, Hawksworth, 1977; Голубкова, 1996б; Myllys et
al., 2011a; Stenroos et al., 2016.
ИЛЛЮСТРАЦИИ: Brodo et al., 2001: 182; Myllys et al., 2011a: 146; Stenroos et al.,
2016: 99.
ТАЛЛОМ повисающий, до 10–15 см дл., коричневый до оливково-коричневого,
иногда с черноватыми участками, блестящий, редко матовый. Ветвление регуляр-
ное, изотомически-дихотомическое, четко U-образное, углы в местах ветвления
тупые или округлые. Тип ветвления (ярусоподобный) хорошо различим в поле-
вых условиях при достаточном увлажнении таллома. Ветви тонкие, 0.2–0.4 мм в
диам., ровные, никогда не становятся извитыми или сжатыми. Латеральные ко-
лючки и веточки отсутствуют. Псевдоцифеллы редкие, незаметные. Соралии об-
разуются часто, белые, мелкие, овальные или щелевидные, обычно ýже ветвей,
на которых развиваются. АПОТЕЦИИ до 1 мм в диам., с бледно-красновато-корич-
невым или темно-бурым диском, развиваются очень редко, в России не найдены.
ПИКНИДЫ неизвестны. — Рис. 37, 38 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой и сердцевина от K, C, KC и P не изме-
няются в окраске, соралии от P краснеют. Содержат фумарпротоцетраровую кис-
лоту и следы протоцетраровой кислоты.
ЭКОЛОГИЯ. На коре хвойных деревьев, а также на березе, в лесных районах с
гумидным климатом.
РАСПРОСТРАНЕНИЕ. Россия. ЕР: Евр. Север — Архангельская обл., Мурманская
обл., Коми; Ср. Россия — Костромская и Нижегородская обл. — Карта 21.
Общее распространение. Европа (в основном на побережье в Фенноскан-
дии), ?Азия, Сев. Америка.
Флора лишайников России Флора лишайников России
68
ПРИМЕЧАНИЕ. Bryoria glabra — не очень вариабельный вид, характеризует-
ся однообразно окрашенным, оливково-черноватым талломом, очень ровными и
гладкими ветвями, строго правильно дихотомически разветвленными, без боко-
вых колючкообразных веточек, мелкими овальными или щелевидными соралия-
ми меньшего, чем ветви, диаметра.
9. Bryoria implexa (Hoffm.) Brodo et D. Hawksw.
1977, Opera Bot. 42: 121. ≡ Usnea implexa Hoffm., 1796, Deutschl. Fl. 2: 134.
≡ Alectoria implexa (Hoffm.) Nyl., 1861, Lich. Scand.: 72. Т и п : sine loco (MW-
Hoffm. 8569 неотип, Hawksworth, 1969: 395). — Бриория переплетенная.
= Bryoria osteola (Gyeln.) Brodo et D. Hawksw.
ИЗБРАННАЯ ЛИТЕРАТУРА: Hawksworth, 1972; Brodo, Hawksworth, 1977; Holien,
1989; Purvis et al., 1992; Голубкова, 1996б; Brodo et al., 2001; Myllys et al., 2011a;
Velmala et al., 2014.
ИЛЛЮСТРАЦИИ: Hawksworth, 1972: pl. 5; Brodo et al., 2001: 135; Myllys et al.,
2011a: 148; Stenroos et al., 2016: 100.
ТАЛЛОМ до 15 см дл., повисающий, светло-коричневый до коричневого, мато-
вый до слегка блестящего, с острыми или тупыми углами между ветвями. Ветви
0.1–0.4 мм в диам., часто сжатые, перекрученные и слегка ямчатые, особенно в
базальной части. Псевдоцифеллы ярко-белые, слегка вдавленные до приподня-
тых над поверхностью ветвей, 1.2 мм дл. и 0.03–0.20 мм шир., удлиненно-вере-
теновидные, на отдельных участках таллома обвивают ветви. Соралии часто от-
сутствуют, бородавчатые до щелевидных. АПОТЕЦИИ образуются редко, до 2 мм
в диам., диск выпуклый, коричневый, со светлым слоевищным краем. Аскоспо-
ры эллипсоидные до почти округлых, 4–6 × 3–5 мкм. ПИКНИДЫ неизвестны.
—
Рис. 39, 40 (вклейка).
ЭКОЛОГИЯ. На стволах и ветвях деревьев хвойных и лиственных пород, иногда
на гниющей древесине, пнях в хвойных и смешанных лесах.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой и соралии от K не изменяются или от
K становятся светло-желтыми, от C и KC не изменяются в окраске, от P коровой
слой, соралии и апотеции желтеют. Содержит псоромовую кислоту, иногда атра-
норин.
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: Аркт. тайм. ЕР: Евр. Север — Архангель-
ская, Вологодская, Мурманская обл.; Ср. Россия — Брянская, Владимирская,
Ивановская, Калужская, Костромская, Московская, Смоленская, Тверская, Ярос-
лавская обл.; Сев. Кавказ — Краснодарский край, Адыгея, Карачаево-Черкесия,
Кабардино-Балкария, Северная Осетия, Дагестан, Ингушетия; Сев. Урал — гор-
ные районы Коми, Пермского края, Ханты-Мансийского АО, Свердловской обл.;
Юж. Урал — Пермский край, Свердловская и Челябинская обл.; СИБ: Зап. Сиб.
сев.
—
Ханты-Мансийский АО, Тюменская обл.; Зап. Сиб. юж.
—
Омская и Но-
Род Bryoria
69
восибирская обл.; Ср. Сиб.
—
Красноярский край, Иркутская обл.; Юж. Сиб.
—
Алтайский и Красноярский края, Республика Алтай, Бурятия, Тыва, Хакасия,
Кемеровская обл., Забайкальский край; Як. вост.; ДВ: ДВ сев.
—
Камчатский и
Хабаровский края; ДВ юж.
—
Амурская обл., Еврейская АО, Сахалинская обл.,
Приморский край. — Карта 22. Распространение и ареал вида недостаточно изу-
чены, часть указаний в литературе может относиться к B. vrangiana.
Общее распространение: Европа, Азия.
ПРИМЕЧАНИЕ. Bryoria implexa отличается от других видов содержанием псоро-
мовой кислоты в вегетативных частях таллома и характерными псевдоцифелла-
ми — белыми и удлиненными (до веретенообразных).
10. Bryoria kuemmerleana (Gyeln.) Brodo et D. Hawksw.
1977, Opera Bot. 42: 155. ≡ Alectoria kuemmerleana Gyeln., 1932 («1931»), Nyt
Mag. Naturvidensk. 70: 49. Т и п : [Словакия] Slovakia. High Tatra Mts., Com.
Szepes, fenyveserdö alomalján, a «Stösschen» lejtőjén a Magas Tátrában, ca. 1380 m,
17.07.1917, G. Timkó 3264 (BP 33952 голотип). — Бриория Кюммерле.
ИЗБРАННАЯ ЛИТЕРАТУРА: Brodo, Hawksworth, 1977; Голубкова, 1996б; Velmala et
al., 2014.
ИЛЛЮСТРАЦИИ: Velmala et al., 2014: 364; Stenroos et al., 2016: 101.
ТАЛЛОМ 15–20 см дл., повисающий, от светло- до темно-коричневого, ино-
гда с черноватыми фрагментами, матовый до слегка блестящего, в основании и
в местах ветвления часто с хорошо заметным налетом. Углы острые до тупых.
Ветвление нерегулярное, в основном изотомически-дихотомическое, ветви 0.07–
0.40(0.50) мм в диам., прямые до скрученных, иногда сплюснутые в местах вет-
вления, без боковых веточек и колючек. Псевдоцифеллы рассеянные, хорошо
заметные, вытянутые, 0.2–0.7(0.9) мм дл., 0.03–0.20 мм шир., беловато-коричне-
ватые. Соралии часто бугорчатые, сероватые, щелевидные, шире ветвей, на ко-
торых образуются. АПОТЕЦИИ с налетом, образуются очень редко.
—
Рис. 41, 42
(вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой и cоралии от K краснеют (рис. 42,
вклейка), от C и KC не изменяются в окраске, от P желтеют. Содержит норстик-
товую кислоту и небольшие количества коннорстиктовой кислоты и атранорина.
Диск апотециев от P желтеет (псоромовая кислота).
ЭКОЛОГИЯ. На коре деревьев хвойных пород, в горных старых лесах.
РАСПРОСТРАНЕНИЕ. Россия. ЕР: Евр. Север — Карелия, Ленинградская обл.;
Ср. Россия — Костромская и Нижегородская обл.; Сев. Кавказ — Краснодарский
край, Адыгея, Дагестан; Сев. Урал — Коми, Пермский край. — Карта 23.
Общее распространение. Европа (в Скандинавии чаще на севере бореальной
зоны), Азия; в Сев. Америке, по-видимому, отсутствует (Velmala et al., 2014). Рас-
пространение изучено недостаточно, поскольку ранее вид рассматривался в каче-
стве хемотипа Bryoria implexa.
Флора лишайников России Флора лишайников России
70
ПРИМЕЧАНИЕ. Bryoria kuemmerleana характеризуется коричневым талломом,
налетом на ветвях, удлиненно-веретеновидными псевдоцифеллами и присутстви-
ем норстиктовой кислоты (Velmala et al., 2014). От других внешне сходных ви-
дов с удлиненными веретеновидными псевдоцифеллами, например от B. implexa
и B. vrangiana, отличается ярко-красной реакцией корового слоя и соралий при
воздействии K (норстиктовая кислота).
11. Bryoria nadvornikiana (Gyeln.) Brodo et D. Hawksw.
1977, Opera Bot. 42: 122. ≡ Alectoria nadvornikiana Gyeln., 1932, Acta Fauna Fl.
Univ., ser. 2, 1: 6. Т и п : [Украина, Закарпатье] Ukraine (Rossia carp.), Turkul, alt.
1850 m, 1930, J. Nádvornik, herb. Servít 17.653 (BP 33953 лектотип, Verseghy, 1964:
123). — Бриория Надворника.
= Alectoria altaica (Gyeln.) Räsänen ≡ Bryopogon altaicus Gyeln.; Alectoria kare-
lica Räsänen, nom. inval.
ИЗБРАННАЯ ЛИТЕРАТУРА: Brodo, Hawksworth, 1977; Thomson, 1984; Purvis et
al., 1992; Голубкова, 1996б; Goward, 1999; Hinds, Hinds, 2007; Smith et al., 2009;
Myllys et al., 2011a, b; Stenroos et al., 2016.
ИЛЛЮСТРАЦИИ: Brodo et al., 2001: 183; Hinds, Hinds, 2007: 138; Myllys et al.,
2011a: 149; Myllys et al., 2011b: 631; Stenroos et al., 2016: 102.
ТАЛЛОМ 4–7(9) см дл., дернинковидный, беловато-сероватый, буроватый, с ро-
зоватым до слегка фиолетового оттенком, редко коричневый; в основании слег-
ка чернеющий, иногда с небольшими черноватыми фрагментами по всей длине;
с тупыми углами в местах ветвления, в верхушечной части с острыми углами.
Ветви (0.1)0.2–0.3 мм в диам., цилиндрические; короткие боковые веточки с тем-
неющими кончиками, в основном многочисленные, перпендикулярно отходят от
поверхности ветвей. Псевдоцифеллы 0.25–0.50 мм дл., плоские, обильные или
слабо развитые, иногда отсутствуют. Соралии до 1 мм дл., бугорчатые, иногда
щелевидные, обычно шире ветвей, на которых образуются, беловатые до светло-
зеленовато-беловатых. АПОТЕЦИИ довольно редки. Сумки узкобулавовидные, 21–
26 мкм дл., аскоспоры бесцветные, 2.5–3.0 × 5.0 мкм, гимений 53–54 мкм выс.,
гипотеций 64 мкм выс. ПИКНИДЫ образуются редко, почти округлые, 0.05–0.10 ×
0.05 мм, локализуются ближе к кончикам ветвей, пикноконидии 8 × 0.5 мкм.
—
Рис. 43 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой, сердцевина и cоралии от K становят-
ся ярко-желтыми, от C — ярко-розовыми (очень редко, при низких концентраци-
ях кислот не изменяются), от KC краснеют. Коровой слой от P желтеет, соралии
краснеют. Диск апотециев от P становится желто-оранжевым. Содержит барбато-
ловую и небольшие количества алекториаловой и фумарпротоцетраровой кислот,
следы атранорина и хлоратранорин.
ЭКОЛОГИЯ. На деревьях хвойных, реже лиственных пород в южных, реже се-
верных бореальных лесах, иногда на вертикальной поверхности скал, особенно
Род Bryoria
71
вдоль берегов озер, рек и в других влажных местообитаниях; на скалах в гипоар-
ктической зоне и субальпийском поясе в горах.
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: Аркт. зап.-сиб.
—
Ямало-Ненецкий АО;
ЕР: Евр. Север — Мурманская обл., Карелия, Ленинградская и Архангельская
обл., сев.-зап. часть Кировской обл., Коми; Ср. Россия — Псковская, Новгород-
ская, Тверская, Ярославская, Московская, Нижегородская, Брянская, Рязанская,
Пензенская, Ульяновская, Самарская обл., Пермский край, Марий Эл, Чувашия,
Татарстан; Сев. Кавказ — Краснодарский край, Адыгея, Карачаево-Черкесия;
Сев. Урал — Коми (горная уральская часть); Юж. Урал — Свердловская и Челя-
бинская обл.; СИБ: Зап. Сиб. сев.
—
Ханты-Мансийский АО, Томская обл.; Ср.
Сиб.
—
Красноярский край; Юж. Сиб.
—
Алтайский и Красноярский края, Ре-
спублика Алтай, Тыва, Бурятия, Иркутская обл., Забайкальский край; Як. ДВ: ДВ
юж. — Сахалинская обл. (Курильские о-ва). — Карта 24.
Общее распространение. Европа, Африка, Азия, Гавайские о-ва, Сев. Аме-
рика.
ПРИМЕЧАНИЕ. Для Bryoria nadvornikiana характерны многочисленные короткие
колючкообразные боковые веточки, темнеющие на кончиках. Таллом в основном
светлоокрашенный, с участками фиолетового оттенка, с тупыми углами между
ветвями в основании таллома и с узкими углами в верхушечной части. Может
быть спутан с B. capillaris, имеющим близкий состав лишайниковых кислот и
характерные реакции, но более светлый до почти белого, повисающий длинный
таллом без колючкообразных боковых веточек.
12. Bryoria nitidula (Th. Fr.) Brodo et D. Hawksw.
1977, Opera Bot. 42: 107. ≡ Bryopogon jubatus var. nitidulus Th. Fr., 1860 (prepr.),
Nova Acta Regiae Soc. Sci. Upsal., ser. 3, 3: 125. = Alectoria nitidula (Th. Fr.) Vain.,
1881, Meddeland. Soc. Fauna Fl. Fenn. 6: 116. Т и п : [Норвегия] Norway, Øst-
Finnmark, Varanger, Klubben, 1857, Th. Fries (UPS лектотип, Brodo, Hawksworth,
1977: 107). — Бриория блестящая.
ИЗБРАННАЯ ЛИТЕРАТУРА: Brodo, Hawksworth, 1977; Purvis et al., 1992; Голубкова,
1996б; Brodo et al., 2001; Myllys et al., 2011a.
ИЛЛЮСТРАЦИИ: Hansen, Andersen, 1995: 17; Brodo et al., 2001: 183; Myllys et al.,
2011a: 149; Stenroos et al., 2016: 103.
ТАЛЛОМ 5–8 см выс., прямостоячий или распростертый, блестящий, красно-
вато- или темно-коричневый, чернеющий у основания, изотомически-дихото-
мически разветвленный в базальной части, в верхушечной — анизотомически-
дихотомически разветвленный, с острыми углами в местах дихотомии. Ветви
0.5–0.6(0.8) мм в диам., ровные, гладкие, прямые. Боковые колючки и колючко-
образные веточки обильные, слегка сдавленные у основания. Псевдоцифеллы
разбросанные, удлиненно-веретеновидные, около 0.5 мм дл., плоские или слегка
Флора лишайников России Флора лишайников России
72
приподнятые, коричневые до темно-коричневых и черных. Соралии отсутствуют.
АПОТЕЦИИ и ПИКНИДЫ неизвестны. — Рис. 44 (вклейка).
ЭКОЛОГИЯ. На скалах и почве в северных и горных тундрах, в лесном поясе гор.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Внутренняя часть корового слоя и сердцевина от K,
C и KC не изменяются в окраске, от P краснеют, хотя бы частично. Содержит фу-
марпротоцетраровую кислоту.
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: все районы. ЕР: Евр. Север — Мурманская
обл., Карелия, Архангельская обл.; Сев. Урал — горные районы Ямало-Ненецко-
го, Ханты-Мансийского АО, Коми; Сев. Кавказ — Карачаево-Черкесия, Адыгея.
СИБ: Юж. Сиб. — Алтайский и Красноярский края, Иркутская обл., Тыва, Буря-
тия; Як. ДВ: ДВ сев.
—
Магаданская обл., Хабаровский и Камчатский края; ДВ
юж. — Сахалинская обл. — Карта 25.
Общее распространение. Сев. Европа, Гренландия, Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Отличается от Bryoria bicolor темноокрашенным, одноцветным
талломом, коричневыми псевдоцифеллами и многочисленными боковыми колюч-
ками и колючкообразными веточками. Обитает главным образом в субарктиче-
ских областях.
13. Bryoria simplicior (Vain.) Brodo et D. Hawksw.
1977, Opera Bot. 42: 109. ≡ Alectoria nidulifera f. simplicior Vain., 1881, Meddeland.
Soc. Fauna Fl. Fenn. 6: 15. ≡ Alectoria simplicior (Vain.) Lynge, 1921, Skr. Vidensk.-
Selsk. Kristiania 1921, Math.-Naturvidensk. Kl. 1, 7: 212 (Stud. Lich. Fl. Norway). Т и п :
[Финляндия] Finland, Lapponia inarensis, Inari, Paatsjoki, 1878, E. A. Vainio (TUR-V
1067 лектотип, Brodo, Hawksworth, 1977: 109). — Бриория простая.
= Alectoria simplicior f. subintricans Vain.; = Bryopogon nitidulus Savicz et Elenkin
f. caespitosus Savicz; = Bryopogon simplicior (Vain.) Gyeln. f. albidosorediosus Gyeln.
ИЗБРАННАЯ ЛИТЕРАТУРА: Brodo, Hawksworth, 1977; Голубкова, 1996б; Goward,
1999; Myllys et al., 2011a; Stenroos et al., 2016.
ИЛЛЮСТРАЦИИ: Thomson, 1984: 65; Myllys et al., 2011a: 150; Stenroos et al., 2016:
103.
ТАЛЛОМ 2–4(5) см дл., дернинковидный, иногда распростертый, красновато-
или темно-коричневый или почти черный, однородно окрашенный, при основа-
нии изотомически-дихотомически разветвленный, с острыми углами в местах
ветвления. Ветви 0.2–0.4 мм в диам., ровные, гладкие, постепенно утончающиеся
к кончикам, с обильными короткими колючками, слегка сжатыми при основании.
Псевдоцифеллы отсутствуют. Соралии обильные, щелевидные, более широкие по
сравнению с ветвями, зеленовато-черные, изредка коричневато-черные, без колю-
чек. АПОТЕЦИИ и ПИКНИДЫ неизвестны. — Рис. 45, 46 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой, сердцевина и cоралии от K, C, KC и P
не изменяются в окраске. Иногда методом тонкослойной хроматографии выявля-
ется содержание жирных кислот.
Род Bryoria
73
ЭКОЛОГИЯ. На хорошо освещенных ветвях и мелких веточках елей, лиственниц
и других деревьев хвойных, а также лиственных пород, изредка на почве, скалах
и гниющей древесине, в северных таежных лесах и в субарктической зоне.
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: Аркт. евр. — Ненецкий АО, крайний северо-
восток Коми; Аркт. зап.-сиб. — Ямало-Ненецкий АО (п-ов Ямал); Аркт. тайм. —
север Таймырского Долгано-Ненецкого р-на; Аркт. чук.
—
Чукотский АО; ЕР:
Евр. Север — Мурманская обл., Карелия, Ленинградская, Архангельская, Киров-
ская обл., Коми; Ср. Россия — Пермский край, Псковская, Тверская, Ярославская,
Кировская, Ивановская, Московская, Рязанская, Ульяновская обл., Чувашия; Сев.
Урал — горные районы Коми, Свердловской обл.; СИБ: Зап. Сиб. сев.
—
Яма-
ло-Ненецкий и Ханты-Мансийский АО, Томская обл.; Зап. Сиб. юж.
—
Ново-
сибирская обл.; Юж. Сиб.
—
Красноярский край, Кемеровская обл., Республика
Алтай, Тыва, Хакасия, Бурятия, Иркутская обл., Забайкальский край; Як. ДВ: ДВ
сев.
—
Магаданская обл., Камчатский и Хабаровский края; ДВ юж.
—
Хабаров-
ский край. — Карта 26.
Общее распространение. Европа, Азия, Сев. Америка, Гренландия.
ПРИМЕЧАНИЕ. Морфологически близок к Bryoria furcellata, от которого отлича-
ется зеленовато-черными соралиями и отсутствием в них пучков изидиевидных
веточек одного цвета с талломом. Соралии у В. furcellata белые, иногда с черны-
ми пятнами, от P краснеют (фумарпротоцетраровая кислота).
14. Bryoria smithii (Du Rietz) Brodo et D. Hawksw.
1977, Opera Bot. 42: 152. ≡ Alectoria smithii Du Rietz, 1926, Ark. Bot. 20A (11): 15.
≡ Alectoria berengeriana var. smithii (Du Rietz) Gyeln., 1935, Ann. Hist.-Nat. Mus.
Natl. Hung. 29: 6. ≡ Alectoria bicolor subsp. smithii (Du Rietz) Räsänen, 1939, Ann.
Bot. Soc. Zool.-Bot. Fenn. Vanamo 12 (1): 32. Т и п : [Китай] China, Prov. Sichuan
(Sze-shu’an), region boreo-occidentalis, inter Tsago-gomba et Tamba, 4000 m,
02.10.1922, H. Smith 5025b (UPS лектотип, Jørgensen, Ryvarden, 1970: 7). — Брио-
рия Смита.
=
Alectoria bicolor var. berengeriana A. Massal. ex Stizenb. ≡ Alectoria
berengeriana (A. Massal. ex Stizenb.) Gyeln.
ИЗБРАННАЯ ЛИТЕРАТУРА: Hawksworth, 1972; Jørgensen, 1972; Brodo, Hawksworth,
1977; Purvis et al., 1992; Wirth, 1995; Голубкова, 1996б; Myllys et al., 2011a;
Stenroos et al., 2016.
ИЛЛЮСТРАЦИИ: Jørgensen, Ryvarden, 1970: 6; Hawksworth, 1972: pl. 8; Myllys et
al., 2011a: 150; Stenroos et al., 2016: 104.
ТАЛЛОМ 4–7(12) см дл., отчетливо двухцветный — у основания темно-корич-
невый до черного, в верхушечной части до светло-оливкового, почти прямостоя-
чий, блестящий. Ветви около 1 мм в диам., цилиндрические или слегка сдавлен-
ные у основания, изотомически-дихотомически разветвленные, с тупыми углами
в местах ветвления. Латеральные колючки 2–7 мм дл. и 0.06–0.10 мм шир., мно-
Флора лишайников России Флора лишайников России
74
гочисленные, перетянутые у основания, часто изогнутые, неразветвленные, рас-
положены перпендикулярно поверхности ветвей. Псевдоцифеллы отсутствуют,
но щелевидные молодые cоралии могут напоминать псевдоцифеллы. Соралии
обильные (в кавказских образцах бывают довольно скудными), щелевидные, во-
гнутые, не шире ветвей, с пучками коротких колючек. АПОТЕЦИИ и ПИКНИДЫ неиз-
вестны. — Рис. 47, 48 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой, сердцевина и cоралии от K, C, KC и
P не изменяются в окраске. Лишайниковые вещества, определяемые с помощью
методов тонкослойной хроматографии, отсутствуют.
ЭКОЛОГИЯ. Произрастает на вертикальной поверхности замшелых скал и валу-
нов, реже на коре деревьев и древесине. Предпочитает влажные местообитания.
Субокеанический вид.
РАСПРОСТРАНЕНИЕ. Россия. ЕР: Евр. Север — Карелия; Сев. Кавказ — Красно-
дарский край, Адыгея, Северная Осетия, Дагестан. — Карта 27.
Общее распространение. Европа, Азия, Гавайские о-ва.
ПРИМЕЧАНИЕ. Морфологически близок к Bryoria bicolor, от которого отли-
чается наличием щелевидных соралий и отсутствием лишайниковых веществ,
определяемых методом тонкослойной хроматографии. Строением соралий (бе-
лых, щелевидных и с густыми пучками изидиевидных колючек) напоминает
B. furcellata, у которого таллом однообразно окрашен, в отличие от отчетливо
двухцветных веточек B. smithii, а также наблюдается красная реакция с P (кора,
сердцевина и соралии содержат фумарпротоцетраровую кислоту).
15. Bryoria tenuis (E. Dahl) Brodo et D. Hawksw.
1977, Opera Bot. 42: 112. ≡ Alectoria tenuis E. Dahl, 1950, Meddel. Grønland 150
(2): 144. Т и п : [Дания, Гренландия] Greenland, Julianehåb (Quaqortoq) District,
Igalikfjord, Eqaluit, 09.08.1937, E. Dahl (O голотип). — Бриория тонкая.
ИЗБРАННАЯ ЛИТЕРАТУРА: Brodo, Hawksworth, 1977; Holien, 1989; Myllys et al.,
2011a; Velmala et al., 2014; Stenroos et al., 2016.
ИЛЛЮСТРАЦИИ: Myllys et al., 2011a: 151; Velmala et al., 2014: 364; Stenroos et al.,
2016: 110.
ТАЛЛОМ прямостоячий или почти повисающий, 4–6(7) см дл., отчетливо двух-
цветный, блестящий. Главные ветви хорошо выражены, цилиндрические, иногда
ближе к основанию слегка сжатые, до 0.4 мм в диам., у основания почти черные,
в верхней части светло-коричневые до коричневых, углы между ветвями обычно
острые, ветви третьего порядка отсутствуют или слабо развиты. Тонкие единич-
ные спинулы перпендикулярно отходят от поверхности ветвей. Псевдоцифеллы
редкие до обильных, обычно темные, малозаметные. АПОТЕЦИИ образуются очень
редко, до 1.5 мм в диам., диск от желтовато- до красновато-коричневого, аскоспо-
ры от почти шаровидных до эллипсоидных, 7–10 × 5–7 мкм. ПИКНИДЫ не обнару-
жены. — Рис. 49, 50 (вклейка).
Род Bryoria
75
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Кора и сердцевина от K, C и KC не изменяются в
окраске, от P краснеют. Содержит фумарпротоцетраровую кислоту.
ЭКОЛОГИЯ. Произрастает на замшелых скалах и ветвях деревьев во влажных
местообитаниях.
РАСПРОСТРАНЕНИЕ. Россия. ДВ: ДВ юж.
—
Сахалинская обл. (о. Кунашир, Ку-
рильский заповедник). — Карта 28.
Возможно нахождение вида у северо-западных границ России, так как он из-
вестен в скандинавских странах.
Общее распространение. Европа, Азия, Сев. Америка, о. Гренландия. Океа-
нический вид. Распространение вида в мире изучено недостаточно.
ПРИМЕЧАНИЕ. От морфологически близкого вида Bryoria bicolor отличается
острыми углами между расходящимися ветвями и отсутствием веточек третьего по-
рядка. У B. smithii образуются часто обильные соралии и все реакции отрицательные
(нет веществ, определяемых с помощью методов тонкослойной хроматографии).
16. Bryoria trichodes (Michx.) Brodo et D. Hawksw.
1977, Opera Bot. 42: 92. ≡ Setaria trichodes Michx., 1803, Fl. Bor.-Amer.
(Michaux) 2: 331, nom. gen. rejic. ≡ Alectoria trichodes (Michx.) D. Hawksw., 1972,
Lichenologist 5 (3–4): 252. Т и п : [Канада] Canada, A. Michaux (PC голотип).
—
Бриория волосистая.
ИЗБРАННАЯ ЛИТЕРАТУРА: Brodo, Hawksworth, 1977; Thomson, 1984; Голубкова,
1996б; Goward, 1999; Thomson, 2003; Harada, Wang, 2008; McCune, Geiser, 2009;
Myllys et al., 2011a, b.
ИЛЛЮСТРАЦИИ: Thomson, 1984: 68; Harada, Wang, 2008: 161, 162; McCune,
Geiser, 2009: 41; Myllys et al., 2011b: 631.
ТАЛЛОМ повисающий, до 15–20 см дл., коричневый, часто с короткими чер-
новатыми фрагментами по всей длине таллома; блестящий, анизо- или изотоми-
чески-дихотомически ветвящийся, с острыми или широкими углами в местах
ветвления. Ветви при основании таллома 0.10–0.30(0.45) мм в диам., цилиндри-
ческие до неровных, изредка с короткими латеральными перпендикулярно отхо-
дящими веточками. Псевдоцифеллы 0.5–1.0 мм дл., обильные, хорошо заметные,
беловатые, щелевидные до овальных, слегка приподнятые (рис. 52, вклейка). Со-
ралии редкие, щелевидные до плоских, с белыми мучнистыми соредиями. АПОТЕ-
ЦИИ образуются редко, диск до 1.5 мм в диам., сначала плоский, затем выпуклый,
светло-коричневый, аскоспоры 7.0–8.5(10) × 4.2–5.0 мкм. ПИКНИДЫ образуются
редко, приподнятые (рис. 52, вклейка), 0.05–0.10 × 0.05 мм, локализуются ближе
к кончикам ветвей, конидии 8 × 0.5 мкм. — Рис. 51–54 (вклейка).
ЭКОЛОГИЯ. Произрастает на хвойных и лиственных деревьях, иногда на скалах.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой и сердцевина от K, C и KC не изменя-
ются, внутренняя часть коры и иногда сердцевина от P частично краснеет. Содер-
жит фумарпротоцетраровую кислоту и хлоратранорин.
Флора лишайников России Флора лишайников России
76
РАСПРОСТРАНЕНИЕ. Россия. ЕР: Ср. Россия — Марий Эл; Сев. Кавказ — Крас-
нодарский край, Карачаево-Черкесия; Сев. Урал — Пермский край (горная ураль-
ская часть); СИБ: Юж. Сиб.
—
Красноярский край, Республика Алтай, Буря-
тия, Хакасия; ДВ: ДВ сев.
—
Камчатский край (бывшая Камчатская обл.), ДВ
юж. — Хабаровский край, Еврейская АО, Сахалинская обл., Приморский край. —
Карта 29.
Общее распространение. Европа, Азия, Сев. Америка.
ПРИМЕЧАНИЕ. От морфологически близкого вида Bryoria americana отличается
белыми псевдоцифеллами (у B. americana они коричневатые, вдавленные и более
длинные), более светлым талломом и присутствием хлоратранорина. Myllys с со-
авторами (Myllys et al., 2011b) отмечают, что у дальневосточных образцов с тер-
ритории России псевдоцифеллы коричневые и довольно длинные — до 0.85 мм,
однако не вдавленные. Принадлежность этих образцов к B. trichodes подтвержде-
на молекулярно-генетическими исследованиями (Myllys et al., 2011b).
17. Bryoria vrangiana (Gyeln.) Brodo et D. Hawksw.
1977, Opera Bot. 42: 97. ≡ Alectoria vrangiana Gyeln., 1932, Magyar Bot. Lapok
31: 46. ≡ Alectoria jubata var. vrangiana (Gyeln.) Räsänen, 1933, Ann. Missouri Bot.
Gard. 20: 10. ≡ Bryopogon vrangianus (Gyeln.) Gyeln., 1935, Feddes Repert. 38: 232.
Т и п : [Швеция] Sweden. Prov. Dalecarlia [Dalarna], par. Leksand [«Loksund»],
Laknäs, 18.07.1931, E. P. Vrang 263 (BP 33.967 голотип; BP 33.968 изотип).
—
Бриория Вранга.
ИЗБРАННАЯ ЛИТЕРАТУРА: Hawksworth, 1972; Brodo, Hawksworth, 1977; Holien,
1989; Velmala et al., 2014; Stenroos et al., 2016.
ИЛЛЮСТРАЦИИ: Hawksworth, 1972: 246; Velmala et al., 2014: 364; Stenroos et al.,
2016: 105.
ТАЛЛОМ 10–30 см, повисающий, ветви ровные, могут становиться извитыми и
приплюснутыми, цвет варьирует, но часто имеет оливково-зеленоватый оттенок,
с бледно- или темно-коричневыми вариациями. Характерный признак — тал-
лом у самого основания от однородно окрашенного до очень светлого (особенно
в тени), часто с чередованием бледных и черноватоокрашенных участков. Кора
матовая до почти блестящей. Ветвление изотомически-дихотомическое, без чет-
ко выраженных главных ветвей, углы в местах ветвления между ветвями тупые.
Ветви 0.07–0.40(0.50) мм в диам., псевдоцифеллы мелкие, узколинейные или
веретенообразные, развиваются на всех зрелых талломах (Velmala et al., 2014),
одного цвета с талломом или немного бледнее у теневых форм. Соралии варьи-
руют в количестве — от обильных до отсутствующих, обычно округлые, выпу-
клые, шире веточек, на которых расположены. АПОТЕЦИИ в некоторых локальных
популяциях довольно обычны, до 1.7 мм в диам., диск коричневый, выпуклый.
Гимений 80 мкм выс. Аскоспоры почти шаровидные до эллипсоидных, 8 × 3.5–
5.0 мкм. ПИКНИДЫ неизвестны. — Рис. 55 (вклейка).
Род Bryoria
77
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Атранорин, гирофоровая и фумарпротоцетраровая,
протоцетраровая, конфумарпротоцетраровая кислоты. Выявлено три хемотипа
(Velmala et al., 2014). I хемотип: таллом реагирует только с C и KC, дает быстро
проходящее красное окрашивание, содержит гирофоровую кислоту, иногда так-
же атранорин; соралии от P иногда краснеют (фумарпротоцетраровая кислота);
апотеции от P желтеют (псоромовая кислота). II хемотип: таллом и соралии от P
краснеют (фумарпротоцетраровая кислота); апотеции от P желтеют (псоромовая
кислота). III хемотип: таллом иногда содержит атранорин — не реагирует или ре-
агирует с K с окрашиванием в светло-желтый цвет, также может реагировать с P,
давая светло-желтое окрашивание; соралии от P краснеют (фумарпротоцетраро-
вая кислота), иногда также содержат протоцетраровую и конфумарпротоцетраро-
вую кислоты; апотеции от P желтеют (псоромовая кислота).
ЭКОЛОГИЯ. Произрастает на хвойных и лиственных деревьях, иногда на скалах.
РАСПРОСТРАНЕНИЕ. Россия. ЕР: Евр. Север — Мурманская и Архангельская
обл., Карелия, Ленинградская обл., Коми; Ср. Россия — Московская, Ивановская,
Смоленская, Костромская, Нижегородская обл.; Сев. Кавказ — Краснодарский
край, Адыгея, Дагестан, Северная Осетия; Сев. Урал — Коми; СИБ: Юж. Сиб. —
Красноярский край, Бурятия. — Карта 30.
Общее распространение. Европа (Фенноскандия и Европейская Россия),
Азия (Россия), Сев. Америка. Ареал недостаточно изучен; вид, по-видимому, ши-
роко распространен в бореальной зоне.
ПРИМЕЧАНИЕ. Bryoria vrangiana отличается от других повисающих коричнево-
окрашенных имеющих соралии видов рода светло-коричневыми, удлиненно-вере-
тенообразными, часто встречающимися псевдоцифеллами, наличием черноватых
фрагментов на всем протяжении ветвей, а также содержанием более разнообраз-
ных лишайниковых веществ. У довольно близкого вида B. fuscescens всегда от-
сутствуют псевдоцифеллы и состав лишайниковых веществ ограничен только фу-
марпротоцетраровой кислотой; у B. implexa развиваются хорошо заметные белые
псевдоцифеллы и состав лишайниковых веществ ограничен псоромовой кислотой
(таллом от P желтеет); для B. kuemmerleana характерен беловатый налет на ветвях,
а также содержание атранорина, норстиктовой и в небольших количествах коннор-
стиктовой кислот (таллом от K краснеет). Распространение вида не вполне ясно и
требует дополнительных исследований, поскольку ранее он рассматривался в со-
ставе хемотипов B. implexa (Velmala et al., 2014; Stenroos et al., 2016).
Флора лишайников России Флора лишайников России
78
Т. В. Макрый
Cetraria Ach.
1803, Methodus: 292. Т и п : Cetraria islandica (L.) Ach. — Цетрария
= Coelocaulon Link, 1833, Grundr. Kräuterk.: 165. Т и п : Coelocaulon aculeatum
(Schreb.) Link
ТАЛЛОМ кустистый, реже листоватый, вертикальный или восходящий, корич-
невый от светлого, зеленовато-серовато-коричневого до темно-бурого, в основа-
нии обычно красный. Лопасти плоские, дорсивентральные, желобчатые до труб-
чато свернутых, или округло-угловатые; фертильные плоские, значительно шире
стерильных. Края с выростами в виде ресничек (регулярных) или шипиков (более
мелких и разбросанных), несущих пикниды, реже с длинными цилиями (сходны-
ми по структуре с ризинами). Соралии редкие. Псевдоцифеллы развиваются на
нижней поверхности, различной формы, краевые и ламинальные, обычно хорошо
заметны, реже отсутствуют. Коровой слой параплектенхимный, у видов с округ-
ло-угловато-цилиндрическим талломом имеется внутренний прозоплектенхим-
ный слой из периклинально ориентированных гиф. ПЛОДОВЫЕ ТЕЛА — зеориновые
апотеции, расположенные на концах лопастей на верхней поверхности, краевые
или околокраевые; диск коричневый. Сумки булавовидные, Lecanora-типа, с 8
аскоспорами. Аскоспоры одноклеточные, бесцветные, эллипсоидные. КОНИДИАЛЬ-
НЫЕ СПОРОНОШЕНИЯ — пикниды на концах краевых ресничек и шипиков или непо-
средственно на краях лопастей, сидячие, выступающие. Конидии палочковидные
или узковеретеновидные до удлиненно-лимоновидных, у одного вида гантелевид-
ные. ФОТОБИОНТ — Asterochloris или Trebouxia.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит лихестериновую, протолихе-
стериновую, фумарпротоцетраровую и норстиктовую кислоты.
ЭКОЛОГИЯ. В основном напочвенные лишайники, некоторые — эпифиты,
встречаются также на валежнике и замшелых камнях с мелкоземом; обитают в
арктических и горных тундрах, лесотундрах, высокогорных и скально-камени-
стых экотопах, хвойных лесах, степях, на болотах.
РАСПРОСТРАНЕНИЕ. Ареал рода охватывает всю Голарктику, а также горные си-
стемы в палеотропиках (Африка, Малазийская область), неотропиках (Центр. и
Юж. Америка), Австралии, Новой Зеландии и Антарктике.
ПРИМЕЧАНИЕ. Род включает по меньшей мере пять групп видов, достаточ-
но хорошо различающихся по морфологии и анатомии, а также экологии и гео-
графии: Sepincola-группа (C. sepincola), Aculeata-группа (C. aculeata, C. aculeata
subsp. steppae, C. muricata), Islandica-группа (C. islandica s. l., C. crispiformis,
C. laevigata, C. ericetorum), Kamczatica-группа (C. kamczatica), сборная
Rassadinae-nigricans-группа (C. rassadinae, C. odontella, C. nigricans, Cetraria sp.
1
).
1
См. ключ (с. 80) и примечание к Cetraria odontella (с. 94).
Род Cetraria
79
Вид «Cetraria» annae Oxner, имеющий желтоокрашенный листоватый таллом,
лишь формально относится к роду Cetraria и должен быть выделен в самостоя-
тельный род. В ключе для определения он отсутствует. Описание вида размещено
в разделе «Вид, формально относящийся к роду Cetraria» (с. 97).
ИЗБРАННАЯ ЛИТЕРАТУРА: Рассадина, 1950; Kärnefelt, 1979, 1986; Kärnefelt et al.,
1993; Thell, Kärnefelt, 2011c.
Ключ для определения видов и подвидов рода Cetraria
1. Таллом мелкий, листоватый, плотно прикрепленный, с восходящими лопастя-
ми, несущими многочисленные апотеции; эпифит ................ 12. С. sepincola.
—
Таллом кустистый, реже листоватый, слабо прикрепленный; напочвенные
лишайники, реже встречающиеся на других субстратах ................................ 2.
2. Лопасти радиального строения, округло- или угловато-цилиндрические, мес-
тами уплощенные, дихотомически разветвленные, с шипиками и цилиями ...
................................................................................................................................. 3.
—
Лопасти дорсивентральные, от плоских, желобчатых до трубчато свернутых
................................................................................................................................. 5.
3. Таллом образует шаровидные пучки «перекати-поле» 1–3(4) см в диам.; ве-
точки вздутые, широко расходящиеся в разные стороны, почти одинаковые
по толщине на всем протяжении, на концах резко суживающиеся, коротко
разветвленные; псевдоцифеллы крупные, овальные, погруженные, до 2 мм
дл. ........................................................................... 1. С. aculeata subsp. steppae.
—
Таллом образует вертикальные дернинки или подушечки ............................. 4.
4. Таллом образует плотные дернинки 1–3(5) см выс. из вертикальных, густо
многократно коротко разветвленных веточек; псевдоцифеллы мелкие, до
0.3 мм в диам., расположенные на уровне таллома ................. 7. С. muricata.
—
Таллом образует подушковидные куртинки 1–5(8) см выс. из рыхло перепле-
тенных, широко растопыренно разветвленных, торчащих в разные стороны
веточек, в нижней части обычно плоских и бороздчатых; псевдоцифеллы вы-
тянутые, погруженные, до 1 мм дл. ................. 1. C. aculeata subsp. aculeata.
5. Лопасти узкие, 0.1–0.5 мм шир.; таллом мелкий, 0.5–1.5(2.0) см выс. ......... 6.
—
Лопасти более 0.5 мм шир.; таллом более крупный, 1.5–10 (и более) см выс.
................................................................................................................................. 7.
6. Таллом мелкокустистый, из вертикальных лопастей, образующих дернинки;
лопасти в нижней части слегка желобчатые, угловато-округло-сплюснутые,
на концах густо разветвленные на узкие, отчасти вальковатые, направлен-
ные в разные стороны короткие веточки, с шипиками и редкими цилиями до
0.5 мм дл.; основания красные; конидии веретеновидные, 3.6–4.8 × 0.5 мкм;
гипоаркто-монтанный вид ........................................................... 9. С. odontella.
—
Таллом распростертый до восходящего, из отдельных лопастей или их спле-
тений, образующих бесформенные куртинки; лопасти слабо желобчатые
Род Cetraria
Флора лишайников России Флора лишайников России
80
(с ложбинкой в центре, широко V-образные на срезе), неправильно дихото-
мически разветвленные, края с шипиками и немногочисленными цилиями до
0.5 мм дл. ........................................................................................... Cetraria sp.
1
7. Цилии обычны, регулярные, многочисленные, около 2 мм дл.; лопасти
(0.5)1.0–1.5(2.0) мм шир., восходящие, желобчатые (U-образные на срезе)
линейные, правильно дихотомически разветвленные, образующие полушаро-
видные куртинки, подушечки; конидии 6.0–7.5 × 1 мкм; встречается в Аркти-
ке ..................................................................................................... 8. C. nigricans.
—
Цилии отсутствуют или редкие, нерегулярные, до 0.5–1.5 мм дл. ................ 8.
8. Основания лопастей коричневые или желтовато-коричневые, никогда не бы-
вают красными ..................................................................................................... 9.
—
Основания лопастей красные ........................................................................... 10.
9. Лопасти Cucullata-типа, трубчато свернутые, узкие, с волнистыми краями и
отгибающимися концами; краевые реснички (шипики), несущие пикниды,
отсутствуют или редкие, мелкие, до 0.1–0.2 мм дл.; псевдоцифеллы незамет-
ны; cердцевина от K, P и I не изменяется; амфиберингийский вид .................
......................................................................................................
5. C. kamczatica.
—
Лопасти не Cucullata-типа, слегка желобчатые, местами плоские, в верхней
части сильно коротко разветвленные; края конечных веточек неровные, с
многочисленными боковыми лопастинками; краевые реснички, несущие пик-
ниды или с пучком гиф на конце, многочисленные, до 1(2) мм дл.; псевдоци-
феллы ламинальные, в виде мелких мучнистых пятнышек, преимущественно
в краевой зоне; сердцевина от K и P не изменяется, от I синеет; сибирский
кальцефильный вид ................................................................. 11. C. rassadinae.
10. Сердцевина от K и P не изменяется ................................................................ 11.
—
Сердцевина от K становится оранжево-желтой, от P — кирпично-красной ...
............................................................................................................................... 12.
11. Лопасти без предкраевых изгибов (выступов), почти трубчато свернутые, ча-
сто со сходящимися, иногда срастающимися краями; краевые псевдоцифеллы
в виде сплошной узкой линии, ламинальные, как правило, отсутствуют; ам-
фиатлантический подвид, известный также на Аляске и Камчатке ..................
.......................................................................
3. C. ericetorum subsp. ericetorum.
—
Лопасти с предкраевыми изгибами (выступами), желобчатые, с редкими сра-
станиями на уровне предкраевых выступов; ламинальные псевдоцифеллы в
виде небольших пятнышек, краевые иногда образуют узкую неровную кайму
в верхних частях лопастей или незаметны; восточноазиатский подвид ..........
..............................................................................
4. C. islandica subsp. orientalis.
12. Лопасти без предкраевых изгибов (выступов), сильно желобчатые до почти
трубчато свернутых, срастающиеся краями и/или ресничками ................... 13.
1
Таксон требует дополнительного изучения и уточнения статуса, возможно, является формой
вида Cetraria odontella.
Род Cetraria
81
—
Лопасти с предкраевыми изгибами (выступами) и отгибающимися на ниж-
нюю сторону краями ......................................................................................... 14.
13. Нижняя поверхность мелкоямчатая, более или менее сетчато-морщинисто-
ребристая, глянцево блестящая; псевдоцифеллы ламинальные, мелкие, неза-
метные; североамфиатлантический вид ................................ 2. C. crispiformis.
—
Нижняя поверхность не ребристая, матовая, с заметными пятнистыми псев-
доцифеллами ....................................................... 4. C. islandica (var. maculata).
14. Лопасти желобчатые до почти плоских или с ложбинкой в центре, с одним
или несколькими срастаниями на уровне предкраевых выступов; ламиналь-
ные псевдоцифеллы всегда имеются, в виде разнообразных пятен, краевые в
виде неровной каймы или узкой рваной линии, реже отсутствуют ..................
............................................................................... 4. C. islandica subsp. islandica.
—
Лопасти сильно желобчатые до почти трубчато свернутых, с сильно отгиба-
ющимися на нижнюю сторону краями, с многочисленными срастаниями на
уровне предкраевых выступов; ламинальные псевдоцифеллы отсутствуют
или единичные затягивающиеся, краевые в виде сплошной ровной широкой
каймы; азиатско-североамериканский вид ................................. 6. C. laevigata.
1. Cetraria aculeata (Schreb.) Fr.
1852, Syst. Orb. Veg.: 239. ≡ Lichen aculeatus Schreb., 1771, Spic. Fl. Lips.: 125.
≡ Cornicularia aculeata (Schreb.) Ach., 1803, Methodus: 302. ≡ Coelocaulon aculea-
tum (Schreb.) Link, 1833, Grundr. Kräuterk. 3: 165. Т и п : Icon in Dillenius, 1742,
Hist. Musc.: tab. 17, fig. 31 B (лектотип, Laundon, 1984: 216); specimen in Herb. Dil-
lenius (OXF эпитип, Thell, Kärnefelt, 2011c: 139). — Цетрария колючая.
ИЗБРАННАЯ ЛИТЕРАТУРА: Kärnefelt, 1986; Журбенко, 1996б.
A. Cetraria aculeata subsp. aculeata. ― Цетрария колючая подвид колючая.
= Cornicularia tenuissima (L.) Zahlbr. ≡ Cetraria tenuissima (L.) Vain. ≡ Lichen
islandicus var. tenuissimus L.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1986: 48, 55; Thell, Kärnefelt, 2011c: 151.
ТАЛЛОМ кустистый, в виде подушковидных куртинок 2–5(8) см выс. и до
10(15) см в диам. из рыхло переплетенных веточек. Веточки жесткие, ломкие,
внутри с рыхлой паутинистой сердцевиной, местами почти полые, умеренно не-
правильно дихотомически разветвленные, часто односторонние с широко расхо-
дящимися (растопыренными) боковыми веточками, коричневые до темно-бурых,
блестящие, в основании обычно красноватые. Основные веточки 1–2(4) мм шир.,
угловато-цилиндрические, местами вздутые или уплощенные, ямчатые, в ниж-
ней части плоские, бороздчатые; боковые веточки 0.3–1.0 мм в диам., торчащие
в разные стороны. Шипики 0.1–0.3 мм дл. Реснички рассеянные, 0.5–1.0 мм дл.
Соралии редкие (вероятно, как результат повреждений). Псевдоцифеллы обыч-
Флора лишайников России Флора лишайников России
82
ны, хорошо заметные, вытянутые, до 1 мм дл., погруженные. АПОТЕЦИИ редкие, на
концах веточек, до 5 мм в диам., одного цвета с талломом. Аскоспоры эллипсоид-
ные, 5.5–6.5 × 2.5–3.5 мкм. ПИКНИДЫ на кончиках шипиков. Конидии удлиненно-
лимоновидные, 5.0–7.5 × 0.5–1.5 мкм. — Рис. 56 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Содержит лихестериновую и протолихестериновую
кислоты. Коровой слой и сердцевина от K, C, KC и P не изменяются.
ЭКОЛОГИЯ. На кислой дресвянистой, песчаной и торфянистой почве, на кам-
нях и скалах (на кислых и основных породах) — на прослойках мелкозема, очень
редко на коре деревьев и древесине; в сухих, открытых, солнечных местообита-
ниях — в Арктике и высокогорьях, на олиготрофных болотах, в светлых сухих
хвойных лесах, степях, скально-каменистых экотопах.
РАСПРОСТРАНЕНИЕ. Широко распространенный вид. Россия. АРКТ: все районы.
ЕР: Евр. Север — Мурманская обл., Карелия, Ленинградская, Архангельская обл.,
Коми; Ср. Россия — Калининградская, Тверская, Московская обл.; Юж. Россия —
Волгоградская обл., Крым; Сев. Кавказ — Краснодарский край, Адыгея, Карачае-
во-Черкесия, Кабардино-Балкария, Северная Осетия, Дагестан; Сев. Урал — Коми,
юж. часть Ямало-Ненецкого АО, Ханты-Мансийский АО, Свердловская обл. СИБ:
Ср. Сиб. — юж. часть Таймырского Долгано-Ненецкого р-на; Юж. Сиб. — Кеме-
ровская обл., Республика Алтай, Хакасия, Тыва, Бурятия, Красноярский край, Ир-
кутская обл.; Як. — зап. ДВ: ДВ сев. — Камчатский край (Корякский округ и быв-
шая Камчатская обл.); ДВ юж. — Хабаровский край, Сахалинская обл. — Карта 31.
Общее распространение. Европа, Азия, Африка, Сев. и Юж. Америка, Грен-
ландия, Африка, Австралия, Новая Зеландия, Антарктика.
ПРИМЕЧАНИЕ. Довольно полиморфный подвид, как по общему габитусу, так и
по деталям строения. Азиатские образцы сильно отличаются от европейских, что
требует специального изучения. Бывает трудно отличим от Cetraria muricata, осо-
бенно мелкие образцы. Одним из главных отличий, помимо псевдоцифелл, явля-
ется характер ветвления: у C. aculeata subsp. aculeata ветвление большей частью
неправильно дихотомическое, часто одностороннее, веточки относительно длин-
ные, с неодинаковыми расстояниями между ветвлениями, широко расходящиеся
(растопыренные), тогда как у C. muricata ветвление преимущественно правильно
дихотомическое, расстояния между ветвлениями примерно одинаковые, неболь-
шие, веточки всех порядков направлены более или менее вверх.
B. Cetraria aculeata subsp. steppae (Savicz) Lutsak, Fernández-Mendoza et Printzen
in Lutsak et al., 2020, Organisms Diversity Evol. 20 (2): 209. ≡ Cornicularia steppae
Savicz, 1924, Bot. Mater. Inst. Sporov. Rast. Glavn. Bot. Sada R.S.F.S.R. 3 (12): 187.
≡ Coelocaulon steppae (Savicz) Borreno et Vázquez, 1982, Lazaroa 3: 239. ≡ Cetraria
steppae (Savicz) Kärnefelt, 1993, Bryologist 96 (3): 96. Т и п : [Украина, Херсон-
ская обл.] Rossia europaea australis, gub. Jekaterinoslav [Cherson Reg.], «Ascania
Nova» in steppa, 1924, A. N. Oxner (LE L-220 лектотип, Savicz, 1961: 3). — Цетра-
рия колючая подвид степная.
Род Cetraria
83
ИЗБРАННАЯ ЛИТЕРАТУРА: Савич, 1924; Lutsak et al., 2020.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1986: 48, 74.
ТАЛЛОМ неприкрепленный, состоящий из нескольких веточек, образую-
щих шаровидные пучки «перекати-поле» 1.5–3.0(4.0) см в диам., которые мо-
гут переноситься ветром. Веточки относительно мягкие, гибкие, не ломкие (по
сравнению с типовым подвидом), внутри полые или почти полые (с рыхлой па-
утинистой сердцевиной), светло-каштановые до темно-красновато-коричневых,
гладкие, обычно матовые или лишь слегка блестящие. Основные веточки 1–2(4)
мм шир., округло- или угловато-цилиндрические, местами сжатые, иногда ямча-
тые; боковые веточки вздутые, широко расходящиеся в разные стороны преиму-
щественно под прямым или тупым углом, часто с широкоокруглыми пазухами,
почти одинаковой толщины на всем протяжении, (0.2)0.5–1.0(1.5) мм в диам.,
дихотомически или неправильно разветвленные, на концах коротко разветвлен-
ные с растопыренными укороченными конечными веточками, резко заканчиваю-
щимися шипиками 0.1–0.2 мм дл. Реснички рассеянные, разветвленные, до 2 мм
дл. Соралии отсутствуют. Псевдоцифеллы не обильные, но хорошо заметные,
крупные, овальные, погруженные, около 0.5 мм шир., до 2 мм дл. АПОТЕЦИИ неиз-
вестны. ПИКНИДЫ на кончиках шипиков. Конидии удлиненно-лимоновидные, 4.0 ×
0.5 мкм. — Рис. 57 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Содержит лихестериновую, протолихестериновую и
норстиктовую кислоты. Сердцевина и псевдоцифеллы от K и P слегка краснеют;
от C, KC не изменяются.
ЭКОЛОГИЯ. На обнаженных кислых и карбонатных минеральных почвах, голых
песках, выходах мергеля, песчаников, гранитов, сланцев; в песчаных и камени-
стых степях, тимьянниках, на открытых солнечных местах с разреженным траво-
стоем, а также на степных солонцах.
РАСПРОСТРАНЕНИЕ. Вид с узким евразийским ареалом, охватывающим аридную
(степную) зону от Испании до степей Казахстана. Россия. ЕР: Юж. Россия —
Воронежская (юж. часть), Саратовская, Оренбургская, Ростовская, Волгоградская,
Астраханская обл., Калмыкия, Крым; Сев. Кавказ — Краснодарский край, Даге-
стан. СИБ: Юж. Сиб. — Республика Алтай. — Карта 32.
Общее распространение. Европа, Азия.
ПРИМЕЧАНИЕ. Отличается от типового подвида формой таллома «перекати-
поле» с направленными в разные стороны, более мягкими и гибкими, утолщен-
ными веточками с полой сердцевиной, а также гладкой матовой поверхностью,
крупными погруженными псевдоцифеллами. В качестве одного из признаков
указывалось наличие в талломе норстиктовой кислоты (Kärnefelt, 1986). Однако
по некоторым данным (Nadyeina et al., 2013; Lutsak et al., 2017, 2020) содержание
норстиктовой кислоты не строго специфично для Cetraria aculeata subsp. steppae
и не коррелирует с морфологическими признаками.
Флора лишайников России Флора лишайников России
84
2. Cetraria crispiformis (Räsänen) Makryǐ, comb. et stat. nov. MBT 841107.
≡ Cetraria islandica var. crispiformis Räsänen, 1943, Ann. Bot. Soc. Zool.-Bot. Fenn.
Vanamo 18 (1): 19. ≡ Cetraria islandica subsp. crispiformis (Räsänen) Kärnefelt,
1979, Opera Bot. 46: 94. Т и п : [Россия, Мурманская обл.] Lapponia petsamoënsis,
Petsamo, Kervanto, Lupuniemi, 1938, Räsänen (H лектотип, Kärnefelt, 1979: 94). —
Цетрария курчавовидная.
= Cetraria islandica f. castanea Rass.; = Cetraria islandica var. polaris Rass.;
= Cetraria islandica var. polaris f. mediocris Rass.; = Cetraria islandica var. polaris
f. rigidula Rass.; = Cetraria islandica var. polaris f. submaculata Rass.
ИЗБРАННАЯ ЛИТЕРАТУРА: Kärnefelt, 1979.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1979: 11, 14; Thell, Kärnefelt, 2011c: 151.
ТАЛЛОМ кустистый, вертикальный, образующий довольно плотные дернин-
ки (1)3–6(9) см выс. Лопасти узкие, 1–2 (редко более) мм шир. без предкраевых
изгибов (выступов), стройные, сильно желобчатые или трубчато свернутые со
сходящимися и кое-где срастающимися краями и ресничками, дихотомически
разветвленные с немногими короткими боковыми веточками, с дуговидно отги-
бающимися конечными долями; края плоские, с ресничками до 1 мм дл. Нижняя
поверхность мелкоямчатая и более или менее продольно сетчато-морщинисто-
ребристая, от каштаново- до темно-коричневой, иногда зеленовато-коричневая, к
основанию светлеющая, глянцево блестящая, в основании красная или краснова-
то-черная; верхняя одинаково с нижней окрашенная, более гладкая, на конечных
долях поперечно-морщинистая и как бы лакированная. Псевдоцифеллы на нижней
поверхности, ламинальные, многочисленные, плохо различимые без лупы, в виде
мелких пятнышек, точек на дне ямок, краевые отсутствуют, реже в виде узкой рва-
ной линии. АПОТЕЦИИ очень редкие, до 2 см в диам., с коричневым диском. Зрелые
аскоспоры не наблюдались. ПИКНИДЫ на кончиках краевых ресничек. Конидии па-
лочковидные или узковеретеновидные, 6–7 × 1 мкм. — Рис. 58 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит фумарпротоцетраровую, ли-
хестериновую и протолихестериновую кислоты. Коровой слой от K, C, KC и P
не изменяется, сердцевина от K становится оранжево-желтой, от P — оранжево-
красной, от C, KC не изменяется.
ЭКОЛОГИЯ. На кислых и торфянистых почвах, замшелых камнях, в напочвен-
ном покрове, иногда вместе с Cetraria islandica; в арктических тундрах, хвойных
лесах, на болотах. Вид, требовательный к высокой влажности воздуха.
РАСПРОСТРАНЕНИЕ. Вид с североамфиатлантическим ареалом, охватывающим
океанические и субокеанические регионы, включая острова в высокой Арктике.
В России имеет весьма ограниченное распространение. Россия. АРКТ: Аркт.
евр.
—
Ненецкий АО; Аркт. зап.-сиб.
—
сев. часть Ямало-Ненецкого АО; Аркт.
тайм.
—
сев. часть Таймырского Долгано-Ненецкого р-на (о. Уединения). ЕР:
Евр. Север — Мурманская обл., Карелия, Архангельская обл., Коми (Тиманский
кряж); Ср. Россия — Псковская обл. — Карта 33.
Род Cetraria
85
Общее распространение. Европа, Азия, Сев. Америка, Гренландия.
ПРИМЕЧАНИЕ. Не очень полиморфный вид. Наряду с типичными формами со
стройными узкими трубчато свернутыми лопастями встречаются формы со сла-
бо желобчатыми широкими короткими нетипичными лопастями; последние ча-
сто обитают на островах в высокой Арктике (возможно, уродливые экземпляры в
экстремальных условиях существования). Типичные формы, характеризующиеся
сильно морщинисто-ребристой лаково-глянцевой поверхностью, приурочены к
океаническим районам, формы со слабо ребристой поверхностью встречаются в
более континентальных районах. Кроме того, имеются формы с почти гладким, но
с мелкими ямками талломом, которые отмечены на границе ареала, а также формы,
которые, по-видимому, следует рассматривать как гибриды Cetraria crispiformis ×
C. islandica; оба вида обитают в одних и тех же местообитаниях. При этом сраста-
ние талломов и обмен генетическим материалом вполне возможны.
3. Cetraria ericetorum Opiz
1852, Seznam: 173. Т и п : [Швеция] Svecia, O. Swartz (H-ACH 1523B лектотип,
Kärnefelt, 1979: 75). — Цетрария вересковая.
ИЗБРАННАЯ ЛИТЕРАТУРА: Рассадина, 1950; Kärnefelt, 1979.
Cetraria ericetorum subsp. ericetorum. — Цетрария вересковая подвид вере-
сковая.
= Cetraria crispa (Ach.) Nyl. ≡ Cetraria islandica var. crispa Ach.; = Cetraria
tenuifolia (Retz.) R. Howe; = Cetraria crispa var. vagans Mereschk.; = Cetraria
libertina Stuckenb.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1979: 10; Thell, Kärnefelt, 2011c: 151.
ТАЛЛОМ кустистый, вертикальный, образующий плотные дернинки 3–8 см
выс. Лопасти стройные, 1–2(4) мм шир. (фертильные до 10 мм шир.), без пред-
краевых изгибов (выступов), желобчатые или почти трубчато свернутые, часто
со сходящимися, срастающимися ресничками краями, дихотомически развет-
вленные, особенно сильно в верхней части, с узкими линейными конечными до-
лями; края с ресничками до 0.5(1.0) мм дл. Нижняя поверхность коричневая до
темно-коричневой, в средней части лопастей более светлая, в основании красная
до фиолетовой, обычно гладкая и блестящая, реже ямчатая или морщинисто-ре-
бристая; верхняя (внутренняя) поверхность более темная. Соралии отсутствуют
или редкие. Псевдоцифеллы на нижней поверхности, обычны, краевые в виде
узкой белой полоски вдоль краев, иногда также скудные ламинальные. АПОТЕЦИИ
редкие, на сильно расширенных концах лопастей, до 10 мм в диам.; диск корич-
невый. Аскоспоры эллипсоидные, 5–9 × 2–4 мкм. ПИКНИДЫ на кончиках краевых
ресничек. Конидии палочковидные или узковеретеновидные, 6.0–7.5 × 1 мкм. —
Рис. 59 (вклейка).
Флора лишайников России Флора лишайников России
86
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит лихестериновую и протолихе-
стериновую кислоты. Коровой слой и сердцевина от K, C, KC и P не изменяются.
ЭКОЛОГИЯ. В напочвенном покрове, на кислых и торфяных почвах, песках,
вместе с другими лишайниками; в открытых сухих местообитаниях, в сосновых
борах, реже тундрах, на верховых болотах, в верещатниках; от Арктики до боре-
альных лесов, на юге — в горах до высокогорий.
РАСПРОСТРАНЕНИЕ. Вид с амфиатлантическим ареалом, широко распространен-
ный в Европе, заходящий в Зап. Сибирь, а также встречающийся на Камчатке.
Россия. АРКТ: Аркт. евр.
—
Ненецкий АО, северо-восток Коми. ЕР: Евр. Се-
вер — Мурманская обл., Карелия, Архангельская обл., Коми, Ленинградская, Во-
логодская обл.; Ср. Россия — Калининградская, Псковская, Новгородская, Твер-
ская, Московская, Владимирская, Ивановская, Нижегородская обл., Марий Эл,
Татарстан, Брянская, Орловская обл., Мордовия, Пензенская, Липецкая, Тамбов-
ская, Воронежская (сев. часть) обл.; Юж. Россия — Крым; Сев. Кавказ — Крас-
нодарский край, Адыгея, Карачаево-Черкесия, Северная Осетия, Дагестан; Сев.
и Юж. Урал — Коми, Пермский край, Ханты-Мансийский АО, Свердловская и
Челябинская обл. СИБ: Зап. Сиб. сев. — юж. часть Ямало-Ненецкого АО, Ханты-
Мансийский АО; Зап. Сиб. юж. — Новосибирская обл.; Юж. Сиб. — Алтайский
край, Республика Алтай. ДВ: ДВ сев.
—
Камчатский край (бывшая Камчатская
обл.). — Карта 34.
Общее распространение. Европа, Азия, Сев. Америка, Гренландия.
ПРИМЕЧАНИЕ. Слабо полиморфный вид. Помимо кустистой, имеет место фор-
ма с неприкрепленным свободным талломом в виде пучка переплетенных лопа-
стей, приуроченная к открытым песчаным местообитаниям (была описана как
C. crispa var. vagans, C. libertina).
4. Cetraria islandica (L.) Ach.
1803, Methodus: 293. ≡ Lichen islandicus L., 1753, Sp. Pl. 2: 1145. Т и п : sine loco
(LINN 1273.97 лектотип, Howe, 1912: 201).
—
Цетрария исландская, исланд-
ский мох.
ИЗБРАННАЯ ЛИТЕРАТУРА: Рассадина, 1950; Kärnefelt, 1979.
A. Cetraria islandica subsp. islandica. — Цетрария исландская подвид исланд-
ская.
=
Cetraria platyna Ach. ≡ Cetraria islandica var. platyna (Ach.) Ach.;
=
Cetraria islandica var. junipericola Räsänen; = Cetraria islandica var. nuda Vain.;
= Cetraria islandica var. paludosa Räsänen; = Cetraria islandica var. paludosa f.
carelica Räsänen; = Cetraria islandica f. pallida Britzelmayer; = Cetraria islandica
f. ramulicola Savicz; = Cetraria islandica f. vagans Savicz.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1979: 11, 14; Thell, Kärnefelt, 2011c: 151.
Род Cetraria
87
ТАЛЛОМ кустистый или листоватый, вертикальный, образующий рыхлые или
довольно плотные дернинки 3–12 см выс. Лопасти (2)5–10 мм шир. (фертильные
до 45 мм шир.), с заворачивающимися на нижнюю сторону краями и отчетливы-
ми предкраевыми изгибами (выступами), желобчатые до почти плоских, с одним
или несколькими срастаниями на уровне предкраевых выступов, неправильно
дихотомически разветвленные, с короткими, реже длинными боковыми веточка-
ми, с укороченными конечными долями. Края с ресничками до 1 мм дл. Нижняя
поверхность ямчато-неровная («мятая»), обычно светлее верхней; верхняя с вы-
пуклыми участками, соответствующими ямкам нижней поверхности, от серова-
то-зеленой, зеленовато-коричневой до темно-коричневой, в верхних частях лопа-
стей более темная, обычно матовая, реже слегка блестящая, в основании красная.
Соралии отсутствуют или редкие, развиваются из псевдоцифелл. Псевдоцифеллы
на нижней поверхности, ламинальные, обычны, в виде более или менее много-
численных отчетливых пятен неправильной формы, кроме того, нередко имеются
краевые псевдоцифеллы в виде прерывистой неровной линии или сливающихся
пятен. АПОТЕЦИИ более или менее обычны, на концах расширенных лопастей, до
2 см в диам.; диск темно- или светло-коричневый. Споры эллипсоидные, 6–10 ×
3.5–5.0 мкм. ПИКНИДЫ на кончиках краевых ресничек. Конидии палочковидные
или узковеретеновидные, 6–7 × 1 мкм. — Рис. 60 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит фумарпротоцетраровую, ли-
хестериновую и протолихестериновую кислоты. Коровой слой от K, C, KC и P
не изменяется, сердцевина от K становится оранжево-желтой, от P — оранжево-
красной, от C, KC не изменяется.
ЭКОЛОГИЯ. В напочвенном покрове, на дресвянистых, кислых и торфяных по-
чвах, мхах, камнях с прослойками мелкозема, валежнике, реже на стволах и вет-
вях деревьев в комлевой части; в равнинных и горных тундрах, лесотундре, за-
рослях кедрового стланика, хвойных, реже смешанных лесах, верещатниках, на
верховых и прибрежных олиготрофных болотах; от Арктики до лесостепи, на
юге — в горах.
РАСПРОСТРАНЕНИЕ. Вид с широким голарктическим ареалом. В России от запад-
ных границ до Дальнего Востока, обычен в европейской части. Россия. АРКТ:
все районы. ЕР: Евр. Север — Мурманская обл., Карелия, Архангельская обл.,
Коми, Ленинградская, Вологодская обл.; Ср. Россия — все районы, кроме Смо-
ленской обл., Удмуртии и Башкирии; Юж. Россия — Оренбургская обл., Крым;
Северная Кавказ — Краснодарский край, Адыгея, Карачаево-Черкесия, Кабарди-
но-Балкария, Северная Осетия, Чечня, Дагестан; Сев. и Юж. Урал — все райо-
ны. СИБ: Зап. Сиб. сев. — все районы; Зап. Сиб. юж. — Омская, Новосибирская
обл.; Ср. Сиб. — все районы, кроме севера Иркутской обл.; Юж. Сиб. — все рай-
оны; Як. — зап., сев., вост., юж. ДВ: ДВ сев. — все районы; ДВ юж. — все райо-
ны, кроме Еврейской АО. — Карта 35.
Общее распространение. Европа, Азия, Сев. Америка, Гренландия.
Флора лишайников России Флора лишайников России
88
ПРИМЕЧАНИЕ. В понимании I. Kärnefelt (1979) подвид Cetraria islandica subsp.
islandica включает в качестве синонимов все множество описанных форм и
разновидностей, за исключением относящихся к выделенным им подвидам
C. islandica subsp. antarctica Kärnefelt, subsp. crispiformis, subsp. orientalis. Изуче-
ние материалов из России показало, что C. islandica — крайне неоднородный и,
судя по всему, сборный вид, не укладывающийся в четыре подвида, выделенных
Kärnefelt. Кроме них, он включает несколько «форм», различающихся морфоло-
гически и характеризующихся вполне определенной экологией и особенностями
распространения, вследствие чего некоторые из них заслуживают самостоятель-
ного таксономического статуса на уровне видов или подвидов (требуют специ-
ального изучения). Так, C. islandica var. maculata Vain. [≡ C. islandica f. maculata
(Vain.) Savicz], характеризующаяся почти трубчато свернутыми лопастями с не-
отгибающимися или слегка отгибающимися краями, с многочисленными лами-
нальными псевдоцифеллами [рис. 61 (вклейка)], преобладает на большей части
азиатской территории России, а в некоторых регионах северо-восточной Сибири
является единственной «формой». C. islandica subsp. islandica понимается нами, в
отличие от Kärnefelt, более узко, как таксон, включающий формы с желобчатыми
до почти плоских (но не трубчато свернутыми) лопастями с немногочисленными
(1–4) срастаниями на уровне предкраевых выступов. Наряду с кустистыми фор-
мами выделяется f. vagans, отличающаяся неприкрепленным свободным талло-
мом из пучка переплетенных лопастей, обитающая на песчаной почве и примор-
ских дюнах, которую, по-видимому, следует рассматривать как экоморфу.
В. Cetraria islandica subsp. orientalis (Asahina ex M. Satô) Kärnefelt
1979, Opera Bot. 46: 107. ≡ Cetraria islandica var. orientalis Asahina ex M. Satô,
1938, J. Jap. Bot. 14: 786. Т и п : [Япония, Хоккайдо] Hokkaido, prov. Ishikari, Mt.
Ashibetsu, 1935, Asahina (TNS лектотип, Kärnefelt, 1979: 107).
—
Цетрария ис-
ландская подвид восточная.
= Cetraria islandica var. orientalis f. angustifolia Asahina.
ИЗБРАННАЯ ЛИТЕРАТУРА: Satô, 1938, 1959.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1979: 14.
ТАЛЛОМ кустистый, вертикальный, образующий более или менее плотные дер-
нинки 2–6 см выс. Лопасти 2–3(5) мм шир., с выраженными предкраевыми из-
гибами (выступами) и слабо отгибающимися на нижнюю сторону краями, слегка
изогнутые, раскидистые, обычно желобчатые до слабо желобчатых, с немногими
срастаниями на уровне предкраевых выступов, неправильно дихотомически раз-
ветвленные, с удлиненными линейными конечными долями; края с ресничками
0.5–1.0 мм дл. Нижняя поверхность коричневая до темно-коричневой в верхних
частях лопастей, книзу светлеющая, ямчато-неровная («мятая»), матовая; верхняя
более темная, с выпуклыми участками, соответствующими ямкам на нижней по-
верхности, обычно гладкая, слегка блестящая, в основании красная, красновато-
Род Cetraria
89
темно-коричневая, реже желтоватая. Соралии отсутствуют. Псевдоцифеллы на
нижней поверхности, обычны, ламинальные, слегка вдавленные в виде неболь-
ших пятнышек, краевые иногда образуют узкую неровную кайму в верхних ча-
стях лопастей или незаметны. АПОТЕЦИИ редкие, на концах расширенных лопа-
стей, 2–10 мм в диам.; диск светло-коричневый. Аскоспоры эллипсоидные, 6 ×
3.5 мкм. ПИКНИДЫ на кончиках краевых ресничек. Конидии палочковидные или
узковеретеновидные, 6.5–7.0 × 1 мкм. — Рис. 62 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит лихестериновую и протолихе-
стериновую кислоты. Коровой слой и сердцевина от K, C, KC и P не изменяются.
ЭКОЛОГИЯ. В напочвенном покрове; в высокогорьях — в зарослях кедрового
стланика и кустарничков Empetrum и Vaccinium, а также в лесах из березы Эрмана.
РАСПРОСТРАНЕНИЕ. Субокеанический вид с узким восточноазиатско-североаме-
риканским ареалом. Россия. ДВ: ДВ юж. — Курильские о-ва (Satô, 1959). — Кар-
та 36.
Общее распространение. Азия (Япония, Корея, Тайвань, Курилы), Сев. Аме-
рика (восточное побережье).
ПРИМЕЧАНИЕ. Распространение в России плохо изучено.
5. Cetraria kamczatica Savicz
1914, Izv. Imp. Bot. Sada Petra Velikago 14 (1–2): 119. Т и п : [Россия, Камчат-
ский край] Kamtschatka, ad jugum montis Schapoczka non procul pag. Naczika, in
regionibus subalpinis ad terram in tundris crescit, 1909, V. P. Savicz 5873 (LE L-134
лектотип, Kärnefelt, 1979: 109). — Цетрария камчатская.
ИЗБРАННАЯ ЛИТЕРАТУРА: Савич, 1914.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1979: 14, 110.
ТАЛЛОМ кустистый, образующий небольшие дернинки 2–4(5) см выс. или в
виде отдельных лопастей среди мхов и лишайников. Лопасти трубчато свернутые,
узкие, (0.5)1.0–2.0(5.0) мм шир., слабо дихотомически разветвленные, с коротки-
ми боковыми и верхушечными веточками, с волнисто изогнутыми (курчавыми)
краями и отогнутыми наружу верхушками (морфотип Flavocetraria cucullata).
Края цельные, краевые реснички (шипики) отсутствуют или очень редкие, не-
многочисленные, до 0.2 мм дл. Нижняя поверхность тонко ямчато-морщинистая,
каштаново-коричневая до темно-коричневой, глянцево блестящая, одинаковая на
всем протяжении лопастей или в нижней части слегка светлее; верхняя (внутрен-
няя) немного темнее нижней. Соралии отсутствуют. Псевдоцифеллы едва разли-
чимы (в сильную лупу) в виде прерывистой белой линии по самому краю ниж-
ней поверхности лопастей. АПОТЕЦИИ редки (известны у образцов с Аляски), на
концах лопастей, 1–2 мм в диам., со слабо волнистым краем. Аскоспоры широ-
коэллипсоидные, 5–6 × 3.5 мкм. ПИКНИДЫ редкие, очень мелкие, сидячие, в виде
маленьких шипиков, часто отсутствуют. Конидии не наблюдались.
—
Рис. 63
(вклейка).
Флора лишайников России Флора лишайников России
90
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит лихестериновую и протолихе-
стериновую кислоты. Коровой слой и сердцевина от K, C, KC и P не изменяются.
ЭКОЛОГИЯ. На почве среди других лишайников и в моховой подушке; в кустар-
ничково-мохово-лишайниковых тундрах.
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: Аркт. як. (окрестности Тикси); Аркт. чук.
ДВ: ДВ сев. — Камчатский край (Корякский округ и бывшая Камчатская обл.). —
Карта 37.
Общее распространение. Азия, Сев. Америка (США — Аляска).
ПРИМЕЧАНИЕ. Вид с амфиберингийским ареалом и узкой экологической нишей,
морфологически однородный, легко узнаваемый благодаря морфотипу, не свой-
ственному другим видам Cetraria. От похожих мелких форм Cetrariella fastigiata
хорошо отличается отсутствием химических реакций.
6. Cetraria laevigata Rass.
1945, Bot. Mater. Otd. Sporov. Rast. Bot. Inst. Komarova Akad. Nauk S.S.S.R 5 (10–
12): 133. Т и п : [Россия, Бурятия] Russia, Buryatia, ditio Angaro-Sajanensis, Tunka,
1926, V. Smirnov (LE L-140 голотип). — Цетрария сглаженная.
= Cetraria excrispa Savicz; = Cetraria crispa var. japonica Asahina.
ИЗБРАННАЯ ЛИТЕРАТУРА: Рассадина, 1945.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1979: 14, 15.
ТАЛЛОМ кустистый, вертикальный, образующий более или менее плотные дер-
нинки (2)3–6(11) см выс. Лопасти (1)2–3(4) мм шир. (фертильные до 10 мм шир.),
стройные, изящные, с отгибающимися на нижнюю сторону краями и отчетливо
выраженными предкраевыми изгибами (выступами), сильно желобчатые или поч-
ти трубчато свернутые, с многочисленными срастаниями на уровне предкраевых
выступов, в верхней части обычно дихотомически разветвленные, обычно с ред-
кими боковыми веточками; края с ресничками 0.1–1.0 мм дл. Нижняя поверх-
ности гладкая или мелкоямчато-морщинистая, от светло- до темно-коричневой,
книзу постепенно светлеющая, блестящая, реже матовая, в основании красная;
верхняя поверхность немного темнее. Псевдоцифеллы на нижней поверхности,
краевые в виде ровной белой широкой каймы вдоль краев (на отгибах), ламиналь-
ные отсутствуют или единичные, в виде затягивающихся пятнышек. АПОТЕЦИИ
более или менее обычны, до 10 мм в диам., по нескольку на концах расширенных
лопастей; диск коричневый. Аскоспоры эллипсоидные, 6 × 2.5 мкм, часто недо-
развитые. ПИКНИДЫ на кончиках краевых ресничек. Конидии палочковидные или
узковеретеновидные, 6.0–7.5 × 1 мкм. — Рис. 64 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит фумарпротоцетраровую, ли-
хестериновую и протолихестериновую кислоты. Коровой слой от K, C, KC и P не
изменяется, сердцевина от K становится оранжево-желтой, от P оранжево-крас-
ной, от C и KC не изменяется, от I синеет.
Род Cetraria
91
ЭКОЛОГИЯ. В напочвенном покрове, на дресвянистой и торфянистой почве, в
моховой подушке, реже на других субстратах; в горных и арктических тундрах,
редколесьях, зарослях кедрового стланика и ерника, горнотаежных и равнинных
хвойных лесах, на верховых болотах.
РАСПРОСТРАНЕНИЕ. Амфиберингийский вид с обширным ареалом, охватываю-
щим арктические и умеренные области. Широко распространен в азиатской части
России (основная граница ареала проходит по Уралу). Россия. АРКТ: все райо-
ны. ЕР: Евр. Север — Архангельская обл.; Сев. Урал — Коми, Ямало-Ненецкий и
Ханты-Мансийский АО, Свердловская обл.; Юж. Урал — Челябинская обл. СИБ:
Зап. Сиб. сев.
—
все районы; Зап. Сиб. юж.
—
Алтайский край; Ср. Сиб.
—
все
районы; Юж. Сиб.
—
все районы, кроме вост. части Новосибирской обл.; Як. —
все районы. ДВ: ДВ сев. — все районы; ДВ юж. — все районы. — Карта 38.
Общее распространение. Европа, Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Вид легко узнаваем благодаря псевдоцифеллам в виде непрерыв-
ной ровной каймы вдоль краев (на отгибах), почти трубчато свернутым стройным
лопастям с многочисленными срастаниями на уровне предкраевых выступов. Ти-
пичная форма, согласно описанию, имеет гладкую поверхность. Форма с мелкоям-
чато-морщинистой поверхностью одинаково широко распространена на территории
Сибири, иногда гладкие и морщинистые лопасти встречаются в одной дернинке.
7. Cetraria muricata (Ach.) Eckfeldt
1895, Bull. Torrey Bot. Club 22: 240. ≡ Lichen muricatus Ach., 1799 («1798»),
Lichenogr. Suec. Prodr.: 214. ≡ Cornicularia muricata (Ach.) Ach., 1803, Methodus:
302. ≡ Coelocaulon muricatum (Ach.) J. R. Laundon, 1984, Lichenologist 16 (1): 55.
Т и п : [Швеция] Svecia (H-ACH 1865A лектотип, Kärnefelt, 1986: 68).
—
Цетра-
рия остроконечная.
ИЗБРАННАЯ ЛИТЕРАТУРА: Kärnefelt, 1986; Журбенко, 1996б.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1986: 69; Thell, Kärnefelt, 2011c: 152.
ТАЛЛОМ кустистый, в виде плотных дернинок 1–3(5) см выс. из вертикаль-
ных разветвленных веточек. Веточки жесткие, ломкие, с рыхлой или более ком-
пактной сердцевиной, дихотомически густо многократно коротко разветвленные,
коричневые до темно-бурых, блестящие, в основании иногда красноватые. Ос-
новные веточки 0.5–1.0 мм в диам., более или менее цилиндрические, ровные и
гладкие, иногда ямчатые или сжатые, особенно в местах ветвления; боковые и ко-
нечные веточки 0.1–0.3 мм в диам., отходящие большей частью под острым углом
и направленные более или менее вверх. Шипики рассеянные, 0.1–0.2 мм дл.
Реснички рассеянные, 0.5–1.0 мм дл. Соралии редкие. Псевдоцифеллы в основ-
ном на боковых веточках, мелкие, 0.1–0.3 мм в диам., расположенные на уровне
таллома, на основных веточках редкие, до 1 мм дл., слегка погруженные. АПОТЕ-
ЦИИ редкие или обычные, на концах веточек, 2–4(8) мм в диам., с кренулирован-
ным краем, одного цвета с талломом. Аскоспоры эллипсоидные, 5–8 × 2–4 мкм.
Флора лишайников России Флора лишайников России
92
ПИКНИДЫ на кончиках шипиков. Конидии удлиненно-лимоновидные, 5–6 × 1.0–
1.5 мкм. — Рис. 65 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Содержит лихестериновую и протолихестериновую
кислоты. Коровой слой и сердцевина от K, C, KC и P не изменяются.
ЭКОЛОГИЯ. На песчаной почве, камнях и скалах с прослойками мелкозема (на
кислых субстратах и карбонатных); в сухих, открытых, солнечных местообитани-
ях, на альварах, в степях, вересковых пустошах; от умеренных широт до Арктики.
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: все районы. ЕР: Евр. Север — Мурманская
обл., Карелия; Ср. Россия — Калининградская, ?Рязанская обл.; Сев. Кавказ —
Адыгея, Карачаево-Черкесия, Кабардино-Балкария, Дагестан; Сев. Урал — Коми
(Печоро-Илычский заповедник). СИБ: Зап. Сиб. сев.
—
юж. часть Ямало-Не-
нецкого АО; Ср. Сиб.
—
юж. часть Таймырского Долгано-Ненецкого р-на; Юж.
Сиб. — Бурятия, Иркутская обл., Забайкальский край. — Карта 39.
Общее распространение. Европа, Азия, Сев. и Юж. Америка, Гренландия,
Африка, Антарктика.
ПРИМЕЧАНИЕ. О различиях c Cetraria aculeata см. примечания к этому виду.
8. Cetraria nigricans Nyl.
1859, Herb. Mus. Fenn.: 106. ≡ Cetraria odontella var. nigricans (Nyl.) Lynge, 1910,
Bergens Mus. Aarbog 9: 79. Т и п : [ Россия, Мурманская обл.] Rossia, Lapponia
tulomensis, Kola, 1843, F. Nylander (H-NYL 36333 = H9505023 лектотип, Kärnefelt,
1979: 117). — Цетрария черноватая.
= Cetraria capitata Lynge.
ИЗБРАННАЯ ЛИТЕРАТУРА: Рассадина, 1950; Kärnefelt, 1979.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1979: 118; Thell, Kärnefelt, 2011c: 152.
ТАЛЛОМ из сильно переплетенных лопастей, образующих плотные полуша-
ровидные подушковидные куртинки, 1–2(3) см выс., до 10 см в диам. Лопасти
(0.5)1.0–1.5(2.0) мм шир., восходящие до вертикальных, линейные, желобчатые
(U-образные на срезе), многократно правильно дихотомически разветвленные,
с удлиненными веточками между развилками, без боковых ответвлений, с ров-
ными, слегка вздутыми краями. Шипики от рассеянных до многочисленных
0.1(0.2) мм дл. Реснички регулярные, обычно разветвленные, отгибающиеся
книзу, 0.5–2.0 мм дл. Верхняя поверхность коричнево-черная до темно-коричне-
вой, гладкая, блестящая до глянцевой; нижняя более светлая, коричневая, слабо
ребристая, матовая или слегка блестящая; основания лопастей обычно красные.
Соралии отсутствуют (иногда имеются соралиеподобные образования как резуль-
тат инфицирования грибами). Псевдоцифеллы на нижней поверхности, краевые,
очень узкие, слабо различимые, в виде рваной линии. АПОТЕЦИИ редкие, до 10 мм
в диам., с кренулированным (с шипиками) краем и диском одного цвета с талло-
мом. Аскоспоры эллипсоидные, 5 × 2.5 мкм. ПИКНИДЫ на кончиках краевых ши-
пиков. Конидии веретеновидные, 6.0–7.5 × 1 мкм. — Рис. 66 (вклейка).
Род Cetraria
93
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит лихестериновую и протолихе-
стериновую кислоты. Коровой слой и сердцевина от K, C, KC и P не изменяются,
сердцевина от I синеет.
ЭКОЛОГИЯ. На кислой дресвянистой почве и камнях с наносами мелкозема; в
сухих кустарничковых тундрах, на каменных россыпях.
РАСПРОСТРАНЕНИЕ. Арктический вид, заходящий в высокогорья на Сев. Урале и
в Сев.-Вост. Сибири. Россия. АРКТ: все районы. ЕР: Евр. Север — Мурманская
обл., Карелия; Сев. Урал — Коми, Пермский край, Ямало-Ненецкий и Ханты-Ман-
сийский АО, Свердловская обл. СИБ: Зап. Сиб. сев. — юж. часть Ямало-Ненецко-
го АО; Ср. Сиб. — юж. часть Таймырского Долгано-Ненецкого р-на (плато Путора-
на). ДВ: ДВ сев. — все районы, кроме сев. части Хабаровского края. — Карта 40.
Общее распространение. Европа, Азия, Сев. Америка, Гренландия, Новая
Гвинея.
ПРИМЕЧАНИЕ. Слабо полиморфный вид. Восточносибирские образцы отличают-
ся более широкими лопастями. В горах Сибири встречается лишайник, приведен-
ный в ключе как Cetraria sp., который можно спутать с C. nigricans. Основными от-
личительными признаками этого лишайника являются более узкие, почти плоские
или слабо желобчатые лопасти с узкой ложбинкой в центре (широко V-образные на
срезе), неправильно дихотомически разветвленные, а также общий габитус (тал-
лом никогда не образует полушаровидных подушковидных куртинок).
9. Cetraria odontella (Ach.) Ach.
1814, Syn. Meth. Lich.: 230. ≡ Lichen odontellus Ach., 1799 («1798»), Lichenogr.
Suec. Prodr.: 213. ≡ Cornicularia odontella (Ach.) Röhl., 1813, Deutschl. Fl., ed. 2,
3 (2): 141. ≡ Coelocaulon odontellum (Ach.) R. Howe, 1912, Classif. Usneac. Amer.:
20. Т и п : [ Швеция] Suecia (H-ACH 1524 = H9502254 лектотип, Kärnefelt, 1986:
35). — Цетрария зубчиковая.
ИЗБРАННАЯ ЛИТЕРАТУРА: Рассадина, 1950; Kärnefelt, 1986.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1986: 36, 55; Thell, Kärnefelt, 2011c: 152.
ТАЛЛОМ мелкокустистый, вертикальный, образующий плотные подушеч-
ковидные дернинки, 1–3 (редко более) см в диам., 0.5–1.0(1.5) см выс. Лопасти
узкие, 0.1–0.5(1.0) мм шир., плоские, местами слегка желобчатые или округло-
угловатые и перекручивающиеся, правильно или неправильно дихотомически
разветвленные, в основании разделенные на 2(3) основные веточки, на концах
густо разветвленные на узкие, отчасти вальковатые короткие боковые веточки,
направленные в разные стороны. Верхняя и нижняя поверхности более или ме-
нее одинаково окрашенные, в верхней части темно-коричневые, глянцевые, книзу
постепенно светлеющие, матовые, в основании обычно красные. Краевые шипи-
ки около 0.1(0.2) мм дл. Цилии редкие, иногда разветвленные, до 0.5 мм дл. Со-
ралии отсутствуют. Псевдоцифеллы очень мелкие, неотчетливые, в виде точек и
коротких штрихов по краям на нижней поверхности. АПОТЕЦИИ редкие, до 1 мм
Флора лишайников России Флора лишайников России
94
в диам.; диск коричневый, почти одного цвета с талломом. Аскоспоры эллипсо-
идные, 7 × 4.5 мкм. ПИКНИДЫ на кончиках шипиков. Конидии палочковидные или
узковеретеновидные, 3.6–4.8 × 0.5 мкм. — Рис. 67 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит лихестериновую и протолихе-
стериновую кислоты. Коровой слой и сердцевина от K, C, KC и P не изменяются.
ЭКОЛОГИЯ. На замшелых скалах и валунах, камнях с мелкоземом, минеральной
почве (на кислых горных породах), реже на валежнике, корневых лапах деревьев;
в горных хвойных лесах, зарослях кедрового стланика и других высокогорных ку-
старников, на каменных россыпях, в скально-степных экотопах, в открытых су-
хих местообитаниях, в горах и северных широтах до Арктики.
РАСПРОСТРАНЕНИЕ. Голарктический вид, заходящий в горы Юж. Америки. Рос-
сия. АРКТ: все районы, кроме Ненецкого АО. ЕР: Евр. Север — Мурманская
обл., Карелия, Архангельская обл., Коми, Ленинградская обл.; Ср. Россия — Нов-
городская обл.; Сев. и Юж. Урал — все районы. СИБ: Зап. Сиб. сев.
—
Ханты-
Мансийский АО; Ср. Сиб.
—
юж. часть Таймырского Долгано-Ненецкого р-на,
Эвенкия; Юж. Сиб.
—
все районы, кроме востока Новосибирской обл.; Як —
вост., центр., юж. ДВ: ДВ сев.
—
юж. часть Чукотского АО, Магаданская обл.,
Камчатский край (бывшая Камчатская обл.); ДВ юж. — Приморский и Хабаров-
ский края. — Карта 41.
Общее распространение. Европа, Азия, Сев. и Юж. (Венесуэла, Перу) Аме-
рика, Гренландия.
ПРИМЕЧАНИЕ. Легко узнаваемый вид благодаря мелким вертикальным густо
коралловидно разветвленным лопастям с красными основаниями и темно-корич-
невыми глянцевыми конечными веточками. В таких же условиях, что и Cetraria
odontella, часто обитает лишайник (в ключе как Cetraria sp.) с узкими, восходя-
щими или стелющимися, слегка желобчатыми, слабо или густо неправильно ди-
хотомически разветвленными лопастями (см. также примечание к C. nigricans).
Возможно, является формой C. odontella; требует специального изучения.
10. Cetraria rassadinae Makryǐ
1984, Bot. Zhurn. (Moscow et Leningrad) 69 (7): 952. Т и п : [Россия, Иркутская
обл.] Rossia, Sibiria Orientalis, regio Irkutensis, distr. Kazaczinsko-Lenensis, jugum
Bajkalense, prope montem Czerskii, regio subalpina, 1200 m s. m., vallis rivuli Dlinnyi
dicti et fl. Kurkulae (Lenensis), prato-tundra subalpina, in mole erratica (calcarea
dolomitica), 29.07.1979, T. V. Makryi (NSK голотип, LE L-185 изотип).
—
Цетра-
рия Рассадиной.
ИЗБРАННАЯ ЛИТЕРАТУРА: Макрый, 1984; Kärnefelt et al., 1993.
ИЛЛЮСТРАЦИИ: Макрый, 1984: 954; Kärnefelt et al., 1993: 401; Zhurbenko, 2009:
145.
ТАЛЛОМ кустистый, вертикальный, 1.5–2.5(3.0) см выс., образующий более
или менее плотные подушковидные куртинки 3–20 см в диам., иногда сплошной
Род Cetraria
95
покров. Лопасти ломкие, 1–3 мм шир., слабо неравномерно желобчатые, местами
почти плоские, иногда перекрученные, дихотомически разветвленные, особенно
в верхней части, с небольшими боковыми веточками. Края неровные, с много-
численными лопастинками. Верхняя поверхность светло-зеленая, буровато-олив-
ковая до темно-бурой, матовая или слегка блестящая; нижняя светлее верхней,
от беловатой до светло-бурой, более или менее блестящая, с большими пятна-
ми более светлой (до белой) коры, под которыми находятся выпуклые участки
утолщенной сердцевины. Основания лопастей от желтовато-, оранжево-корич-
невых до темно-бурых. Краевые реснички, несущие пикниды, многочисленные,
до 1(2) мм дл., часто с пучком белых гиф на конце, иногда также развиваются на
нижней поверхности. Псевдоцифеллы ламинальные, в верхней части в виде мел-
ких плоских тонкомучнистых пятнышек, преимущественно в краевой зоне (обыч-
но напротив каждой реснички), ближе к основанию вдавленные или выпуклые,
иногда прорывающиеся. Сердцевинный слой неравномерно развит, тонкий до
толстого, иногда прерывистый, вместе с водорослевым слоем 75–210 мкм толщ.
Водоросли часто распределены по всей сердцевине и небольшими скоплениями
под нижней корой. АПОТЕЦИИ редкие, на концах лопастей, до 10 мм в диам., с тем-
но-коричневым выпуклым диском и реснитчатым талломным краем. Аскоспоры
эллипсоидные, 6–7 × 3.0–3.5 мкм. ПИКНИДЫ на кончиках краевых шипиков и рес-
ничек. Конидии узковеретеновидные, 6.5–8.5 × 1 мкм. — Рис. 68 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит лихестериновую и протолихе-
стериновую кислоты. Коровой слой и сердцевина от K, C, KC и P не изменяются;
сердцевина от I синеет (реакция видна через нижнюю кору).
ЭКОЛОГИЯ. Кальцефильный вид. На валунах (доломитовых известняках) с на-
носами мелкозема, реже на почве на близко залегающих известняках. В субаль-
пийских лугах и тундровых луговинах, в зарослях кедрового стланика, кедровых
и лиственничных редколесьях, сосновых лесах, горных степях, на каменных рос-
сыпях. В подгольцовом и верхней части лесного пояса, редко в низкогорьях и на
равнине.
РАСПРОСТРАНЕНИЕ. Вид с ограниченным азиатским ареалом, известный из При-
байкалья, гор Юж. Сибири и сибирской Арктики. Россия. АРКТ: Аркт. зап.-сиб. —
Ямало-Ненецкий АО; Аркт. тайм. — с евер Таймырского Долгано-Ненецкого р-на.
СИБ: Зап. Сиб. сев. — юж. часть Ямало-Ненецкого АО; Ср. Сиб. — юж. часть Тай-
мырского Долгано-Ненецкого р-на (плато Путорана), север Красноярского края,
Эвенкия; Юж. Сиб. — Республика Алтай, Бурятия, юг Красноярского края (Сая-
ны), Иркутская обл., Забайкальский край (Кодар). — Карта 42.
Общее распространение. Азия (Россия).
ПРИМЕЧАНИЕ. Своеобразный вид. При наличии особых характерных при-
знаков, весьма полиморфен по габитусу, поэтому его легко можно спутать
с Cetrariella delisei, отдельными формами Cetraria islandica, а мелкие образцы —
с C. odontella. От первого вида легко отличается отсутствием реакций сердцеви-
ны с C и KC, от второго — желто-коричневым основанием лопастей, отсутствием
Флора лишайников России Флора лишайников России
96
реакций с K и P, от третьего — всегда плоскими, более широкими конечными
веточками с неровными краями.
11. Cetraria sepincola (Ehrh.) Ach.
1803, Methodus: 297. ≡ Lichen sepincola Ehrh., 1783, Hannover. Mag. 2: 206.
≡ Tuckermannopsis sepincola (Ehrh.) Hale, in Egan, 1987, Bryologist 90 (2): 164.
Т и п : [Швеция] Upsaliae, F. Ehrhart, Phytophylacium Ehrhartianum No. 90 (UPS
неотип, Thell, Kärnefelt, 2011c: 139). — Цетрария заборная.
ИЗБРАННАЯ ЛИТЕРАТУРА: Рассадина, 1950.
ИЛЛЮСТРАЦИИ: Thell, Kärnefelt, 2011c: 152.
ТАЛЛОМ мелкий, листоватый, обычно монофилльный, плотно прикрепленный
к субстрату, с восходящими до вертикальных, укороченными лопастями, образу-
ющий выпуклые розетки 0.5–1.5(2.0) см выс., 0.5–2.0(3.0) см в диам. или одно-
сторонне развитый, часто сливающийся с другими талломами. Лопасти плоские
или слегка желобчатые, округлые до удлиненных, 0.3–1.5(2.0) см дл., 1–6(8) мм
шир., слегка неглубоко дихотомически разветвленные, с ровными краями. Верх-
няя поверхность от желтовато-, оливково-, красновато-коричневой до темно-ко-
ричневой, неровная, матовая или блестящая; нижняя светло-коричневая до бело-
ватой у края, морщинистая, с немногочисленными светлыми ризинами. Реснички
редкие. Соралии отсутствуют. Псевдоцифеллы на краях лопастей (слабо замет-
ные) и апотециев. АПОТЕЦИИ всегда присутствуют, многочисленные, обычно на
концах лопастей, латеральные околокраевые, до 8 мм в диам.; диск почти одного
цвета с талломом. Аскоспоры эллипсоидные, 6–10 × 3–6 мкм. ПИКНИДЫ краевые,
погруженные, слегка выдающиеся в виде черных точек. Конидии гантелевидные,
5–7 × 1 мкм. — Рис. 69 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит лихестериновую и протолихе-
стериновую кислоты. Коровой слой и сердцевина от K, C, KC и P не изменяются.
ЭКОЛОГИЯ. На веточках деревьев и кустарников (обычен на березе, ернике,
ольхе), также на валежнике и древесине, редко на камнях; в арктических и гор-
ных тундрах, равнинных и горных хвойных и смешанных лесах, зарослях кедро-
вого стланика и редколесьях, на болотах.
РАСПРОСТРАНЕНИЕ. Широко распространенный вид. Россия. АРКТ: все районы.
ЕР: Евр. Север — Мурманская обл., Карелия, Архангельская обл., Коми, Ленин-
градская, Вологодская обл.; Ср. Россия — все районы, кроме Удмуртии, Ульянов-
ской обл. и Башкирии; Юж. Россия — Самарская, Оренбургская, Волгоградская
обл., Крым; Сев. Кавказ — Краснодарский край, Адыгея, Кабардино-Балкария,
Северная Осетия; Сев. и Юж. Урал — все районы. СИБ: Зап. Сиб. сев.
—
все
районы; Зап. Сиб. юж.
—
Омская, Новосибирская обл., Алтайский край; Ср.
Сиб. —
юж. часть Таймырского Долгано-Ненецкого р-на, Эвенкия; Юж. Сиб.
—
все районы; Як. — все районы. ДВ: ДВ сев. — все районы, кроме юга Чукотского
АО; ДВ юж. — все районы, кроме Еврейской АО. — Карта 43.
Род Cetraria
97
Общее распространение. Европа, Азия, Сев. Америка, Гренландия.
ПРИМЕЧАНИЕ. По анатомо-морфологическому строению (в частности, погру-
женным, слегка выдающимся пикнидам и гантелевидным конидиям), а также со-
гласно результатам молекулярно-филогенетического анализа вид не относится к
роду Cetraria s. str. и должен быть выделен в самостоятельный род. Легко узнава-
емый лишайник, благодаря большому количеству апотециев и ровным краям ло-
пастей без ресничек и шипиков. Полиморфизм не сильно выражен.
Вид, формально относящийся к роду Cetraria
«Cetraria» annae Oxner
1933, Zhurn. Bio-Bot. Tsyklu Vseukraïns’k. Akad. Nauk 2 (7–8): 167. Т и п : [Россия,
Забайкальский край] Oriens Extremus, Provincia Czita, ad saxa in silva prope pag.
Atamanovka, 18.09.1927 [A. M. Oxner]; ad saxa supra thallum Lobariae retigerae ad
flum. Nikischicha in viciniis pag. Atamanovka, 18.09.1927 [A. M. Oxner] (KW синти-
пы). — «Цетрария» Анны.
Избранная литература: Окснер, 1933; Thell et al., 2009.
ИЛЛЮСТРАЦИИ: Thell et al., 2009: 506.
ТАЛЛОМ листоватый, мягкий, более или менее розетковидный, до 3 см шир.,
прикрепленный к субстрату ризоидами. Лопасти 1–3 мм шир., единожды или
дважды разветвленные, около 10(15) мм дл., в основании более узкие, к концам
вееровидно расширяющиеся (за счет ветвления), конечные доли 1–3(5) мм дл.;
края выемчато-волнистые, слегка кренулированные. Верхняя поверхность слабо
сетчато-ямчатая, бледно-желтая или серовато-зеленовато-желтоватая, матовая,
покрытая сизоватым налетом, особенно на концах лопастей; нижняя поверхность
гладкая, бледно-желтовато-серая, с немногочисленными ризоидами. Соралии в
разрывах между верхней и нижней корой, слабо губовидные, соредии зернистые.
Таллом 220–300 мкм толщ.; верхний и нижний коровые слои около 50 мкм толщ.,
параплектенхимные, из нескольких слоев клеток 5–7 мкм в диам.; сердцевина бе-
лая, почти такая же компактная, как коровые слои, из клеток с тонкими оболоч-
ками и широкими просветами. АПОТЕЦИИ неизвестны. ПИКНИДЫ редкие, краевые,
выступающие. Конидии не наблюдались. — Рис. 70 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Содержит усниновую и изонефростериновую кисло-
ты (основные), лихестериновую, скваматовую кислоты и атранорин (в примеси),
а также протолихестериновую и нефростериновую кислоты (следы).
ЭКОЛОГИЯ. На замшелых скалах и камнях, валунах с наносами мелкозема; в
горных лесах и степях, субальпийских экотопах, обычно в долинах рек и по бере-
гам рек и озер.
РАСПРОСТРАНЕНИЕ. Ареал слабо изучен, охватывает Юж. Сибирь и юг Дальне-
го Востока. Россия. СИБ: Юж. Сиб. — Бурятия (Хамар-Дабан, Окинское плато),
Иркутская обл. (Байкальский, Приморский хр.), Забайкальский край (хр. Черско-
Флора лишайников России Флора лишайников России
98
го — долина р. Ингоды). ДВ: ДВ юж.
—
Приморский край (Сихотэ-Алинь).
—
Карта 44.
Общее распространение. Азия (Россия).
Т. В. Макрый
Cetrariella Kärnefelt et A. Thell
in Kärnefelt et al., 1993, Bryologist 96 (3): 402. Т и п : Cetrariella delisei (Schaer.)
Kärnefelt et A. Thell. — Цетрариелла
Лихенизированные грибы. ТАЛЛОМ кустистый, вертикальный. Лопасти более
или менее желобчатые до почти трубчато свернутых, изредка расширяющиеся
кверху, коричневоокрашенные, внизу более светлые, желтоватые. Краевые выро-
сты (шипики), несущие пикниды, редкие, разбросанные, до 0.5(1.0) мм дл. Сора-
лии редкие или отсутствуют. Псевдоцифеллы на нижней поверхности, краевые и
поверхностные. Коровой слой параплектенхимный, сердцевина рыхлая. ПЛОДОВЫЕ
ТЕЛА — апотеции, зеориновые, субмаргинальные, на концах лопастей; диск корич-
невый. Сумки широкобулавовидные, Lecanora-типа, с 8 аскоспорами. Аскоспоры
одноклеточные, бесцветные, эллипсоидные. КОНИДИАЛЬНЫЕ СПОРОНОШЕНИЯ — пик-
ниды, темно-бурые, на кончиках краевых шипиков. Конидии бутылковидные,
утолщенные на одном конце, 7–9 × 0.5–1.5 мкм. ФОТОБИОНТ Trebouxia-типа.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит гирофоровую и хиасциевую
кислоты.
ЭКОЛОГИЯ. Напочвенные лишайники; в арктических и горных тундрах, субаль-
пийских (подгольцовых) сообществах, хвойных бореальных лесах, на болотах.
РАСПРОСТРАНЕНИЕ. Ареал рода охватывает арктические, северотаежные и гор-
ные районы Голарктики, а также высокогорные районы южной оконечности Юж.
Америки и Новой Зеландии.
ПРИМЕЧАНИЕ. Выделен из рода Cetraria (Kärnefelt et al., 1993).
ИЗБРАННАЯ ЛИТЕРАТУРА: Рассадина, 1950; Kärnefelt, 1979; Kärnefelt et al., 1993;
Thell, Kärnefelt, 2011d.
Ключ для определения видов рода Cetrariella
1. Лопасти слабо желобчатые, сильно дихотомически разветвленные, с узкими
рассеченными кончиками ................................................................. 1. С. delisei.
—
Лопасти желобчатые до почти трубчато свернутых, слабо дихотомически раз-
ветвленные, с широкими тупыми закругленными кончиками ...........................
.......................................................................................................... 2. C. fastigiata.
Род Cetrariella
99
1. Cetrariella delisei (Schaer.) Kärnefelt et A. Thell
in Kärnefelt et al., 1993, Bryologist 96 (3): 403. ≡ Cetraria islandica var. delisei
Schaer., 1850, Enum. Crit. Lich. Eur.: 114. ≡ Cetraria delisei (Schaer.) Nyl., in Fries,
1867, Kongl. Svenska Vetensk. Akad. Handl., nov. ser. 7 (2): 11. Т и п : [Канада,
Ньюфаундленд] Terreneuve, 1828, Despréaux (UPS лектотип, Kärnefelt, 1979:
68). — Цетрариелла Делиза.
= Cetraria hiascens (Fr.) Th. Fr. ≡ Cetraria aculeata var. hiascens Fr.
ИЗБРАННАЯ ЛИТЕРАТУРА: Kärnefelt, 1979; Kärnefelt et al., 1993; Thell, Kärnefelt,
2011d.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1979: 10, 68; Kärnefelt et al., 1993: 401; Hansen,
Andersen, 1995: 19; Brodo et al., 2001: 219; Hinds, Hinds, 2007: 148; Thell, Kärnefelt,
2011d: 152.
ТАЛЛОМ кустистый, вертикальный, образующий плотные дернинки. Лопасти
5–8 см выс., сильно дихотомически разветвленные, с сильно рассеченными на
узкие доли кончиками, в основании слабо желобчатые, 2–8 мм шир., на концах
плоские, 0.5–1.0 мм шир., темно-коричневые до бурых, к основанию постепен-
но светлеющие до желтоватых. Верхняя поверхность гладкая, слегка блестящая;
нижняя — с продольными ребрами, матовая; сердцевина рыхлая. Края ровные,
с редкими шиповидными выростами, 0.1–0.5 мм дл. Соралии редкие. Псевдоци-
феллы отчетливые, на нижней поверхности. АПОТЕЦИИ редкие, на расширенных
лопастях, до 17 мм в диам.; диск одного цвета с талломом. Аскоспоры эллипсо-
идные, 6.0–8.5 × 2.5 мкм. ПИКНИДЫ темно-бурые, на кончиках краевых шипиков.
Конидии бутылковидные, 7–9 × 0.5–1.5 мкм. — Рис. 71 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит гирофоровую и хиасциевую
кислоты. Коровой слой от K, C, KC и P не изменяется, сердцевина и псевдоцифел-
лы от K и P не изменяются, от C слабо краснеют, от KC становятся ярко-красными.
ЭКОЛОГИЯ. В напочвенном покрове на торфянистой и песчаной почве, обыч-
но во влажных понижениях рельефа; в равнинных и горных кустарничковых тун-
драх, ерниках, верещатниках, на верховых болотах, в местах наледей и снежни-
ков в долинах рек и ручьев, а также на каменных россыпях с залеживающимся
снегом.
РАСПРОСТРАНЕНИЕ. Арктоальпийский вид, широко распространенный в Ар-
ктике и на севере бореальной зоны, а также в горах. Россия. АРКТ: все райо-
ны. ЕР: Евр. Север — Мурманская обл., Карелия, Архангельская, Ленинградская
обл., Коми; Ср. Россия — Псковская, Новгородская, Тверская обл.; Сев. Урал —
Коми, Пермский край, Ямало-Ненецкий и Ханты-Мансийский АО, Свердловская
обл.; Юж. Урал — Свердловская обл. (Висимский заповедник). СИБ: Зап. Сиб.
(сев.) — Ямало-Ненецкий АО, Ханты-Мансийский АО, Томская обл.; Ср. Сиб. —
все районы, кроме сев. части Иркутской обл.; Юж. Сиб.
—
все районы, кроме
востока Новосибирской обл.; Як. — центр., вост., юж. ДВ: ДВ сев. — все районы;
ДВ юж. — Амурская обл., Хабаровский край, Сахалинская обл. — Карта 45.
Род Cetrariella
Флора лишайников России Флора лишайников России
100
Общее распространение. Европа, Азия, Сев. и Юж. Америка, Гренландия,
Новая Зеландия.
ПРИМЕЧАНИЕ. Довольно полиморфный вид. Различаются мелкие и крупные
формы, с узкими или довольно широкими лопастями, слабо или сильно развет-
вленными, с широкими и короткими или узкими удлиненными верхними долями.
2. Cetrariella fastigiata (Nyl.) Kärnefelt et A. Thell
in Kärnefelt et al., 1993, Bryologist 96 (3): 403. ≡ Cetraria delisei subsp. fastigiata
Nyl., in Norrl., 1873, Not. Sällsk. Fauna Fl. Fenn. Förh. 13: 325. ≡ Cetraria hiascens
var. fastigiata (Nyl.) Vain., 1881, Meddeland. Soc. Fauna Fl. Fenn. 6: 119. ≡ Cetraria
fastigiata (Nyl.) Kärnefelt, 1977, Bot. Not. 130 (2): 128. Т и п : [Финляндия]
Finland, Enontekiö Lapland, Enontekiö («Enontekis»), Hetta, 1867, J. P. Norrlin
(H9500705 лектотип, Kärnefelt, 1977b: 128). — Цетрариелла равновершинная.
ИЗБРАННАЯ ЛИТЕРАТУРА: Kärnefelt, 1979; Kärnefelt et al., 1993; Thell, Kärnefelt,
2011d.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1979: 11, 86; Hansen, Andersen, 1995: 19; Thell et al.,
2004b: 439; Thell, Kärnefelt, 2011d: 153.
ТАЛЛОМ кустистый, вертикальный, образующий плотные дернинки. Лопасти
около 3–6 см выс., 2–5 мм шир., желобчатые до почти трубчато свернутых, сла-
бо дихотомически разветвленные, с короткими боковыми веточками, с закруглен-
ными отогнутыми верхушками (морфотип Flavocetraria cucullata), коричневые
до темно-бурых, к основанию постепенно светлеющие до желтоватых. Верхняя
поверхность гладкая или ямчатая, матовая или блестящая; нижняя обычно бле-
стящая. Края неровно вырезанные, волнистые. Краевые шипики редкие или от-
сутствуют, 0.1–0.5(1.0) мм дл. Соралии отсутствуют. Псевдоцифеллы преиму-
щественно краевые, очень узкие, неразличимые, поверхностные редкие, слабо
развитые. АПОТЕЦИИ редкие, на расширенных лопастях, до 10 мм в диам.; диск
одного цвета с талломом. Аскоспоры эллипсоидные, 6.0–8.5 × 2.5–3.5 мкм. ПИК-
НИДЫ темно-бурые, на кончиках краевых шипиков. Конидии бутылковидные, 7–9
× 0.5–1.5 мкм. — Рис. 72 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит гирофоровую и хиасциевую
кислоты. Коровой слой от K, C, KC и P не изменяется, сердцевина и псевдоци-
феллы от K и P не изменяются, от C слабо краснеют, от KC становятся ярко-крас-
ными.
ЭКОЛОГИЯ. В напочвенном покрове на торфянистой и песчаной почве, обычно
во влажных понижениях рельефа, а также на наносах мелкозема на камнях; в ар-
ктических и горных тундрах, ерниках, на болотах, а также на каменных россыпях
с залеживающимися снежниками.
РАСПРОСТРАНЕНИЕ. Циркумполярный, в основном арктический вид с североам-
фиатлантическим и амфиберингийским центрами массовости, проникающий в
умеренные широты до 60(55)° с. ш. и по высокогорьям заходящий в горы Юж.
Род Cetreliopsis
101
Сибири (Становое нагорье). Россия. АРКТ: все районы. ЕР: Евр. Север — Мур-
манская обл., Карелия, Архангельская обл., Коми, Ленинградская обл.; Ср. Рос-
сия — Псковская обл.; СИБ: Юж. Сиб.
—
Забайкальский край (хребты Кодар и
Каларский). ДВ: ДВ сев. — Магаданская обл., Камчатский край (Корякский округ
и бывшая Камчатская обл.). — Карта 46.
Общее распространение. Европа, Азия, Сев. Америка, Гренландия.
ПРИМЕЧАНИЕ. На Становое нагорье вид, вероятно, проник по Верхоянскому и
Становому хребтам, что позволяет предполагать, что в Восточной Сибири, а точ-
нее в горах Якутии, он распространен значительно шире.
Т. Рандлане, А. Сааг
Cetreliopsis M. J. Lai
1980, Quart. J. Taiwan Mus. 33: 218. Т и п : Cetreliopsis rhytidocarpa (Mont.
et Bosch) M. J. Lai. — Цетрелиопсис
Лихенизированные грибы. ТАЛЛОМ листоватый или почти кустистый; верхняя
поверхность зеленовато-желтая до соломенно-желтой, сердцевина белая, нижняя
поверхность или полностью черная, или черная лишь в центральной части и ко-
ричневатая у края. Лопасти дорсивентральные, часто с приподнимающимися кра-
ями. Псевдоцифеллы присутствуют на обеих поверхностях — хорошо заметные,
окруженные черным ободком, на возвышениях, развивающихся на верхней по-
верхности, либо в виде небольших белых точек на нижней поверхности. У неко-
торых видов могут развиваться реснички и соредии; изидии отсутствуют. Ризины
редкие или многочисленные, черные. Верхняя и нижняя кора параплектенхимная.
ПЛОДОВЫЕ ТЕЛА — апотеции зеоринового типа, с коричневым диском и таллом-
ным краем, краевые или субмаргинальные. Сумки широкобулавовидные, 35–60 ×
11–20 мкм, содержат 8 аскоспор. Аскоспоры простые, эллипсоидные, бесцветные,
6–12 × 4–7 мкм. ПИКНИДЫ часто многочисленные, краевые и поверхностные, чер-
ные, в основном на выступающих возвышениях, хорошо заметные. Конидии ган-
телевидные, 5 × 1–2 мкм. ФОТОБИОНТ Trebouxia-типа.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Кора содержит усниновую кислоту, сердцевина —
фумарпротоцетраровую кислоту и другие сходные депсидоны β-орсинольного
типа вместе с жирными кислотами.
ЭКОЛОГИЯ. Эпифитные лишайники, растущие на хвойных и лиственных дере-
вьях в лесных сообществах; один вид напочвенный, в горных лугах.
РАСПРОСТРАНЕНИЕ. Довольно редкие виды, распространенные в Вост. и Юго-
Вост. Азии и на некоторых островах Азии (Тайвань, Филиппины, Борнео, Ява,
Новая Гвинея), В России очень редок, встречается только на Дальнем Востоке.
Род Cetreliopsis
Флора лишайников России Флора лишайников России
102
ПРИМЕЧАНИЕ. Род включает семь видов, из которых лишь один известен в
России. В одной из недавних публикаций (Divakar et al., 2017), в которой на ос-
нове современных филогенетических методов предлагается разделение всех
цетрариоидных лишайников лишь на три больших рода (Cetraria, Melanelia и
Nephromopsis), виды рода Cetreliopsis вновь отнесены к роду Nephromopsis, но та-
кая концепция авторами не признается.
ИЗБРАННАЯ ЛИТЕРАТУРА: Lai, 1980; Randlane et al., 1995.
Cetreliopsis asahinae (M. Satô) Randlane et A. Thell
in Randlane et al., 1995, Cryptog. Bryol. Lichénol. 16: 49. ≡ Cetraria asahinae
M. Satô, 1936, Res. Bull. Saito Ho-on-kai Mus. 11: 22. ≡ Nephromopsis asahinae
(M. Satô) Räsänen, 1952, Kuopion Luonnon Ystäväin Yhdistyksen Julkaisuja, B, 2, 6:
50. Т и п : [Россия, Курильские острова, Кунашир] (Russia) Kuril Islands, Kunashi-
ri, Tomari-mura, 16.08.1923, A. Yasuda (TNS голотип). — Цетрелиопсис Асахины.
ЛИТЕРАТУРА: Красная книга Приморского края..., 2008; Чабаненко, 2002; Rand-
lane et al., 1995.
ИЛЛЮСТРАЦИИ: Randlane et al., 1995: 50.
ТАЛЛОМ листоватый, дорсивентральный, с довольно широкими (8–15 мм) и
приподнимающимися лопастями; верхняя поверхность желтовато-зеленая, ниж-
няя в центральной части черная, с коричневыми или светло-коричневыми краями.
Псевдоцифеллы присутствуют на обеих поверхностях, на верхней поверхности
хорошо заметные, окруженные темным ободком или черными возвышениями,
на нижней — в виде белых пятен, расположенных в основном на гребнях ребер
таллома. Реснички, соредии и изидии отсутствуют. Ризины редкие или многочис-
ленные, черные. АПОТЕЦИИ обычны, краевые или субмаргинальные, с коричневым
диском и желтовато-зеленым талломным краем, до 15 мм в диам. Сумки 35–40 ×
12–17 мкм, содержат 8 аскоспор. Аскоспоры 8–12 × 4–7 мкм. ПИКНИДЫ часто мно-
гочисленные и хорошо заметные, краевые и поверхностные, черные, на образую-
щихся возвышениях. Конидии гантелевидные, 4 × 1 мкм. — Рис. 73, 74 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту (редко
кислота отсутствует), от K, C и P не изменяется, от KC желтеет или не изменяет-
ся, в UV не флюоресцирует; сердцевина содержит протоцетраровую (основная),
фумарпротоцетраровую и физодаловую (±) кислоты, от K, C и KC не изменяется,
от P краснеет, в UV не флюоресцирует.
ЭКОЛОГИЯ. Эпифитный вид, растущий на хвойных (Abies, Picea) и лиственных
(Betula, Quercus, Tilia) деревьях в смешанных и лиственных лесах на равнине, а
также в горах на малых и средних высотах.
РАСПРОСТРАНЕНИЕ. Россия. Редкий вид, встречающийся только на Дальнем
Востоке. ДВ: ДВ юж. — Амурская обл., Хабаровский край (юж. часть), Сахалин-
ская обл. (Курильские о-ва: Кунашир, Шикотан), Приморский край. — Карта 47.
Общее распространение. Вост. и Юго-Вост. Азия. Имеет более широкое рас-
пространение, чем другие таксоны этого рода.
Род Dactylina
103
ПРИМЕЧАНИЕ. Таксон, морфологически легко распознаваемый по наличию на
верхней поверхности характерных псевдоцифелл, окруженных темным ободком
и/или выступающим возвышением.
Д. Е. Гимельбрант, И. С. Степанчикова
Dactylina Nyl.
1860, Syn. Meth. Lich. 1 (2): 286. Т и п : Dactylina arctica (Richardson) Nyl. —
Дактилина
Лихенизированные грибы. ТАЛЛОМ кустистый, средних размеров, состоит из
отдельных или собранных в группы восходящих неразветвленных пальцевид-
но вздутых или умеренно дихотомически разветвленных коралловидных ветвей
зеленовато-желтого до желтовато-коричневого, иногда почти золотистого цвета,
более темных к основанию. Ветви округлые или слегка уплощенные, гладкие,
без макул, псевдоцифелл, соралий и изидий, полые внутри или местами с рыхло
рассеянными пучками толстостенных гиф сердцевины. Коровой слой перекрыт
эпикортексом 5 мкм толщ., прозоплектенхимный, состоит из антиклинально ори-
ентированных гиф, 25–40 мкм толщ., поверхность таллома матовая или блестя-
щая, голая или покрыта тонким светлым налетом. Альгальный слой не выражен,
клетки фотобионта расположены группами в периферической части сердцевины.
Структуры прикрепления к субстрату отсутствуют, талломы погружены нижней
отмирающей частью в растительную дернину и таким образом закреплены в ней.
ПЛОДОВЫЕ ТЕЛА — апотеции зеоринового типа, образуются редко, располагают-
ся на концах боковых ветвей таллома, до 5 мм в диам. Талломный край одного
цвета с талломом, диск коричневый, блестящий, эксципул, гимений и гипотеций
бесцветные, эпитеций коричневый. Парафизы септированные, неразветвленные
или местами слабо разветвленные, головчатые, со светло-коричневыми верхуш-
ками апикальных клеток, сумки широкобулавовидные, 25–65 × 10–18 мкм, со-
держат 8 аскоспор. Аскоспоры одноклеточные, округлые или слегка эллипсоид-
ные, бесцветные, толстостенные, 4–6 мкм в диам. КОНИДИАЛЬНЫЕ СПОРОНОШЕНИЯ в
форме погруженных или выступающих черных округлых пикнид. Конидиеносцы
3–4 -клеточные, до 20 мкм дл. Конидии веретеновидные до почти палочковидных,
прямые или слегка изогнутые, 5–7 × 1 мкм. ФОТОБИОНТ Trebouxia-типа.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую и гирофоровую
кислоты, сердцевина — гирофоровую, физодовую и физодаловую кислоты, а так-
же леканоровую, хиасциевую и орселлиновую кислоты в качестве дополнитель-
ных веществ.
ЭКОЛОГИЯ. Напочвенные виды; в различных типах тундровых сообществ в го-
рах и на равнине.
Род Dactylina
Флора лишайников России Флора лишайников России
104
РАСПРОСТРАНЕНИЕ. Широко распространенные в Северном полушарии аркто-
альпийские виды с циркумполярным характером распространения, южнее зоны
тундр встречаются в высокогорьях. В России также широко распространены в по-
лярных, приполярных и горных регионах.
ПРИМЕЧАНИЕ. Включает два вида, оба встречаются на территории Рос-
сии. Виды рода Dactylina морфологически наиболее сходны с Allocetraria
madreporiformis, однако последний отличается наличием паутинистой сердцеви-
ны, занимающей всю внутреннюю часть таллома, а также иным составом лишай-
никовых веществ (в коровом слое содержится только усниновая, а в сердцеви-
не — лихестериновая кислота).
Поскольку апотеции формируются весьма редко, основным типом размно-
жения является вегетативное посредством фрагментации талломов (особенно за
счет мелких боковых выростов-веточек).
ИЗБРАННАЯ ЛИТЕРАТУРА: Thomson, Bird, 1978; Thomson, 1984; Журбенко, 1996в;
Kärnefelt, Thell, 1996; Thell, Kärnefelt, 2011e.
Ключ для определения видов и подвидов рода Dactylina и Allocetraria
madreporiformis
1. Таллом большей частью неразветвленный, во всех частях полый, обычно бо-
лее 2–3 см выс. и более 3 мм шир., без налета ............................................... 2.
—
Таллом явственно дихотомически разветвленный, в большей или меньшей
степени заполнен рыхлой паутинистой сердцевиной, обычно до 2–3 см выс. и
до 3 мм шир., без налета или с тонким светлым налетом на концах ветвей ...
................................................................................................................................. 3.
2. Сердцевина содержит гирофоровую кислоту, от C краснеет, от P не изменяет-
ся ................................................................................ 1. D. arctica subsp. arctica.
—
Сердцевина содержит физодовую и физодаловую кислоты, от C не изменяет-
ся, от P становится оранжево-красной ............. 1. D. arctica subsp. beringica.
3. Таллом большей частью полый (особенно ближе к центру), желтовато-корич-
невый до коричневого, концы ветвей, как правило, покрыты тонким светлым
налетом; сердцевина от P обычно краснеет ............................. 2. D. ramulosa.
—
Таллом полностью заполнен рыхлой паутинистой сердцевиной, желтый до
зеленовато-желтого, ветви без налета; сердцевина от P не изменяется ...........
...............................................................................
Allocetraria madreporiformis.
1. Dactylina arctica (Richardson) Nyl.
1860, Syn. Meth. Lich. 1 (2): 286. ≡ Dufourea arctica Richardson, 1823, Bot. App. in
Franklin, Narr. Journey Polar Sea: 762. ≡ Evernia arctica (Richardson) Tuck., 1848,
Proc. Amer. Acad. Arts Sci. 1: 203. ≡ Cladonia arctica (Richardson) Th. Fr., 1861
(1860 prepr.), Nova Acta Regiae Soc. Sci. Upsal., ser. 3, 3: 260. ≡ Cetraria arctica
(Richardson) Tuck., 1882, Syn. N. Amer. Lich. 1: 30. Т и п : [Канада] Canada,
Род Dactylina
105
Northwest Territories, [Great] Bear Lake, J. Franklin (FH-Tuck лектотип, Kärnefelt,
Thell, 1996: 502). — Дактилина арктическая.
A. Dactylina arctica subsp. arctica. — Дактилина арктическая подвид арктиче-
ская.
= Dufourea polaris Rupr.; = Cetraria arctica var. minor Elenkin; = Dufourea
arctica var. papillata Savicz ≡ Cetraria arctica var. papillata (Savicz) Zahlbr.
ИЗБРАННАЯ ЛИТЕРАТУРА: Thomson, Bird, 1978; Thomson, 1984; Журбенко, 1996в;
Kärnefelt, Thell, 1996; Thell, Kärnefelt, 2011e.
ИЛЛЮСТРАЦИИ: Thomson, 1984: 203; Kärnefelt, Thell, 1996: 489, 495; Brodo et al.,
2001: 294; Thell, Kärnefelt, 2011e: 153.
ТАЛЛОМ кустистый, состоит из отдельных или собранных в группы восходя-
щих неразветвленных или единично разветвленных пальцевидно вздутых ветвей
до 8 см выс. и до 15 мм толщ., светло-желтого до коричневато-желтого, иногда
почти золотистого цвета, темнеющих к основанию (коричневатый оттенок силь-
нее проявляется в более освещенных местообитаниях). Ветви округлые, иногда
слегка уплощенные, гладкие, местами с фовеолами, притупленные на концах,
полые внутри, редко с единичными округлыми перфорациями. Поверхность тал-
лома блестящая, к основанию становится матовой, без налета. АПОТЕЦИИ образу-
ются очень редко (известны по материалам из Сибири), располагаются на концах
небольших боковых веточек, до 5 мм в диам., диск коричневый, блестящий. Сум-
ки широкобулавовидные, 25–65 × 10–18 мкм. Аскоспоры одноклеточные, окру-
глые, 4–6 мкм в диам. ПИКНИДЫ погруженные, круглые, черные. Конидии верете-
новидные, 5–6 × 1 мкм. — Рис. 75 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую и гирофоровую
кислоты, от K и P не изменяется, от C становится розовым (изредка не изменяет-
ся), от KC желтеет, в UV не флюоресцирует. Сердцевина содержит гирофоровую
кислоту, в качестве дополнительных веществ может также содержать леканоро-
вую, хиасциевую и орселлиновую кислоты, от K и P не изменяется, от C и KC
краснеет, в UV не флюоресцирует.
ЭКОЛОГИЯ. Обитает на почве в открытых местообитаниях, предпочитает ув-
лажненные места; в различных типах тундровых сообществ в горах и на равнине.
РАСПРОСТРАНЕНИЕ. Россия. Широко распространенный арктоальпийский ли-
шайник. АРКТ: все районы. ЕР: Сев. Урал — горные районы Коми и Свердлов-
ской обл.; СИБ: Зап. Сиб. сев. — Ямало-Ненецкий АО (юж. часть), Ханты-Ман-
сийский АО; Ср. Сиб.
—
Таймырский Долгано-Ненецкий р-н (юж. часть); Юж.
Сиб.
—
Кемеровская обл., Республика Алтай, Хакасия, Тыва, Бурятия, Красно-
ярский край (юж. часть), Иркутская обл. (юж. часть), Забайкальский край; Як. —
все районы. ДВ: ДВ сев.
—
все районы; ДВ юж.
—
Сахалинская обл. (Саха-
лин). — Карта 48.
Флора лишайников России Флора лишайников России
106
Общее распространение. Европа, Азия, Сев. Америка. Арктоальпийский
лишайник с циркумполярным распространением в Северном полушарии, южнее
зоны тундр встречается преимущественно в высокогорьях.
ПРИМЕЧАНИЕ. Dactylina arctica — весьма полиморфный вид, включающий
три подвида, отличающиеся исключительно составом лишайниковых веществ
сердцевины. Два из них известны с территории России. Отличия D. arctica
subsp. beringica от типового подвида приведены ниже. D. ramulosa отличается от
D. arctica небольшим размером таллома, как правило, развитием налета на кон-
цах ветвей, присутствием хотя бы местами рыхлой сердцевины и составом ме-
дуллярных лишайниковых веществ.
B. Dactylina arctica subsp. beringica (C. D. Bird et J. W. Thomson) Kärnefelt et
A. Thell
1996, Nova Hedwigia 62 (3–4): 503. ≡ Dactylina beringica C. D. Bird et J. W. Thomson,
1978, Canad. J. Bot. 56 (14): 1612. Т и п : [Канада] Canada, British Columbia, Summit
Lake, Mile 292.5 Alaska Highway, 58°40′ N, 124°43′ W, on summit tundra, 5300 ft,
13.08.1967, J. W. Thomson et T. Ahti 16553 (WIS голотип; CANL, COL, DUKE, H,
UAC, US изотипы). — Дактилина арктическая подвид берингийская.
ИЗБРАННАЯ ЛИТЕРАТУРА: Thomson, Bird, 1978; Thomson, 1984; Журбенко, 1996в;
Kärnefelt, Thell, 1996; Thell, Kärnefelt, 2011e.
От типового подвида отличается исключительно составом лишайниковых ве-
ществ сердцевины. — Рис. 76 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Содержит физодовую и физодаловую кислоты, в ка-
честве дополнительных веществ может содержать также леканоровую, хиасцие-
вую, орселлиновую кислоты и неидентифицированное вещество; гирофоровая
кислота в сердцевине не обнаружена. От K и C сердцевина не изменяется, от KC
становится розовой, от P — оранжево-красной, в UV не флюоресцирует.
ЭКОЛОГИЯ. Обитает на почве в открытых местообитаниях, часто совместно
с типовым подвидом; в различных типах тундровых сообществ в горах и на рав-
нине.
РАСПРОСТРАНЕНИЕ. Россия. Арктоальпийский лишайник, вероятно, широко рас-
пространенный в арктических и субарктических районах Ср. и Вост. Сибири, а
также на севере Дальнего Востока, в высокогорьях Юж. и Вост. Сибири. АРКТ:
Аркт. як.; Аркт. чук.
—
Чукотский АО (сев.). Ср. Сиб.
—
Таймырский Долга-
но-Ненецкий р-н (юж. часть), Красноярский край (сев. часть); Юж. Сиб.
—
Бу-
рятия, Иркутская обл. (юж. часть), Забайкальский край; Як.
—
все районы. ДВ:
ДВ сев. — Чукотский АО (юж.), Магаданская обл., Камчатский край (Корякский
округ). — Карта 49.
Общее распространение. Азия, Сев. Америка. Арктоальпийский лишайник,
возможно, с циркумполярным распространением в Северном полушарии, однако
ареал его изучен явно недостаточно.
Род Dactylina
107
ПРИМЕЧАНИЕ. Какие-либо морфологические или анатомические отличия
Dactylina arctica subsp. beringica от типового подвида не обнаружены, оба под-
вида отмечены в одних и тех же местообитаниях. Несколько отличающийся от
типового подвида характер распространения может объясняться недостаточным
вниманием исследователей к составу лишайниковых веществ.
Еще один подвид Dactylina arctica — subsp. chinensis (Follmann) Kärnefelt et
A. Thell, — известный только из типового местонахождения в высокогорье про-
винции Шаньси (Shaanxi) в Китае, по набору морфологических и анатомических
характеристик также достоверно не отличается от типового подвида, однако со-
держит в коровом слое только усниновую кислоту, в сердцевине обнаружена фи-
зодаловая кислота, а в качестве дополнения — орселлиновая кислота и неиденти-
фицированное вещество (коровой слой от K желтеет, сердцевина от P становится
оранжево-красной, остальные реакции негативные). Не исключена возможность
обнаружения этого подвида в высокогорьях Юж. Сибири и юга Дальнего Востока.
2. Dactylina ramulosa (Hook.) Tuck.
1862, Proc. Amer. Acad. Arts Sci. 5: 397. ≡ Dufourea ramulosa Hook., 1825, App.
Parry J. Sec. Voy.: 414. ≡ Evernia ramulosa (Hook.) Tuck., 1848, Proc. Amer. Acad.
Arts Sci. 1: 203. ≡ Cetraria ramulosa (Hook.) Tuck., 1882, Syn. N. Amer. Lich. 1: 30.
Т и п : [Канада] Canada, Northwest Territories, Melville Peninsula, Igloolik, Parry
Expedition (BM лектотип, Kärnefelt, Thell, 1996: 505). — Дактилина мелковетви-
стая.
= Dufourea muricata Laurer.
ИЗБРАННАЯ ЛИТЕРАТУРА: Thomson, Bird, 1978; Thomson, 1984; Журбенко, 1996в;
Kärnefelt, Thell, 1996; Thell, Kärnefelt, 2011e.
ИЛЛЮСТРАЦИИ: Thomson, 1984: 207; Kärnefelt, Thell, 1996: 491, 495; Brodo et al.,
2001: 294; Stenroos et al., 2011: 200; Thell, Kärnefelt, 2011e: 153; Wirth et al., 2013:
446.
ТАЛЛОМ кустистый, хрупкий, обычно напоминает более или менее рыхлую
подушечку, состоящую из восходящих или почти распростертых, явственно вет-
вящихся пальцевидных элементов, реже состоит из одиночных ветвей. Основ-
ные ветви почти цилиндрические, местами уплощенные, до 2 см выс. и до 3 мм
толщ., гладкие или изредка с отдельными фовеолами, матовые или блестящие,
желтовато-коричневые или соломенного цвета (в хорошо освещенных местооби-
таниях коричневые), темнеющие к основанию, в верхней части покрыты отчет-
ливым тонким беловатым до фиолетового налетом, реже голые, могут нести ко-
роткие боковые веточки, обычно более темные. Элементы таллома внутри полые,
однако местами заполнены рыхло расположенными гифами сердцевины, менее
плотными в центральной части. АПОТЕЦИИ образуются редко (известны по мате-
риалам из Сибири), располагаются на концах небольших боковых веточек, обыч-
но до 3 мм в диам., диск коричневый, блестящий. Сумки широкобулавовидные,
Флора лишайников России Флора лишайников России
108
30–40 × 13–18 мкм. Аскоспоры одноклеточные, округлые, 4.5–6.5 мкм в диам.
ПИКНИДЫ несколько выступающие, округлые, черные. Конидии веретеновидные
до почти палочковидных, прямые, 5–7 × 1 мкм. — Рис. 77, 78 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, а затем становится розовым, в UV не флюо-
ресцирует. Сердцевина содержит физодаловую и физодовую кислоты, а также не-
идентифицированное вещество, от C и K не изменяется, от KC и P краснеет, в
UV не флюоресцирует (иногда физодаловая кислота отсутствует или содержится
в следовых количествах, в таком случае сердцевина от P не изменяется).
ЭКОЛОГИЯ. Обитает на почве в открытых местообитаниях; в различных типах
тундровых сообществ в горах и на равнине.
РАСПРОСТРАНЕНИЕ. Россия. Широко распространенный арктоальпийский ли-
шайник, встречающийся в Арктике, на Урале, в Сибири и на Дальнем Востоке.
АРКТ: все районы, кроме крайнего северо-востока Коми. ЕР: Сев. Кавказ —
Адыгея; СИБ: Ср. Сиб. — Таймырский Долгано-Ненецкий р-н (юж. часть); Юж.
Сиб.
—
Кемеровская обл., Республика Алтай, Хакасия, Тыва, Бурятия, Красно-
ярский край (юж. часть), Иркутская обл. (юж. часть), Забайкальский край; Як. —
вост., юж. ДВ: ДВ сев. — Магаданская обл., Камчатский край (Корякский округ и
бывшая Камчатская обл.). — Карта 50.
Общее распространение. Европа, Азия, Сев. Америка. Арктоальпийский
лишайник с циркумполярным распространением в Северном полушарии, южнее
зоны тундр встречается в высокогорьях.
ПРИМЕЧАНИЕ. Об отличиях от филогенетически наиболее близкого вида
Dactylina arctica см. в примечаниях к нему. Анатомо-морфологически D. ramulosa
более сходен с Allocetraria madreporiformis, однако отличается от него значитель-
но более слабым развитием паутинистой сердцевины, занимающей внутреннюю
часть таллома лишь местами, желтовато-коричневой или коричневой окраской, а
также другим составом лишайниковых веществ (в сердцевине содержатся физо-
даловая и физодовая кислоты).
И. С. Степанчикова, Д. Е. Гимельбрант
Evernia Ach.
1810, Lichenogr. Universalis: 84, 441. Т и п : Evernia prunastri (L.) Ach. — Эверния
Лихенизированные грибы. ТАЛЛОМ кустистый, разветвленный, иногда листо-
ватый с узкими разветвленными лопастями (Evernia prunastri), средних размеров
или крупный, в отдельных случаях до 40 см дл., желтоватый, серовато-желтова-
тый, реже серовато-зеленоватый, иногда с коричневатыми или чернеющими кон-
цами ветвей (в гербарии часто темнеет). Ветви таллома восходящие, полупови-
Род Evernia
109
сающие, повисающие или распростертые, в большинстве случаев мягкие, вялые,
гибкие, иногда более жесткие, приподнятые, редко твердые, торчащие и ломкие
(E. perfragilis). Деление на верхнюю и нижнюю поверхности у большинства ви-
дов выявить невозможно. Поверхность таллома матовая, без выраженного нале-
та, часто ямчатая, с широкими морщинами или складками, иногда с кольцевыми
трещинами, реже гладкая. Настоящие псевдоцифеллы отсутствуют, на поверхно-
сти таллома могут располагаться соредии и/или изидии. Таллом прикрепляется к
субстрату псевдогомфом, иногда в развитом состоянии может не иметь органов
прикрепления. Коровой слой тонкий, состоит из антиклинально ориентирован-
ных разветвленных гиф, погруженных в матрикс. Гифы корового слоя ориенти-
рованы антиклинально, внешняя часть коры представлена параплектенхимой,
внутренняя — прозоплектенхимой. Альгальный слой тонкий, расположен под ко-
ровым слоем. Сердцевина толстая, белая, волокнистая, рыхлая ватообразная или
относительно компактная, состоит из периклинально ориентированных широких
ровных гиф. ПЛОДОВЫЕ ТЕЛА — апотеции леканорового типа, с хорошо развитым
талломным краем, у большинства видов крайне редки, располагаются на ветвях
латерально, маргинально или терминально, сидячие или формируются на корот-
ких талломных «ножках», диск коричневый или красновато-коричневый, иногда
светлый, гладкий, часто блестящий. Эпитеций коричневый, гимений и гипотеций
бесцветные. Сумки Lecanora-типа, содержат 8 аскоспор. Аскоспоры бесцветные,
одноклеточные, эллипсоидные до широкоэллипсоидных, с тупыми концами, тон-
костенные, 6–11 × 4–6 мкм. КОНИДИАЛЬНЫЕ СПОРОНОШЕНИЯ в форме пикнид, окру-
глые, погруженные, черные вблизи остиол, у многих видов редки. Конидии па-
лочковидные, часто заостренные с обоих концов. ФОТОБИОНТ — Trebouxia.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит атранорин, хлоратранорин,
усниновую кислоту, сердцевина — диварикатовую или эверновую кислоты.
ЭКОЛОГИЯ. Эпифитные и напочвенные виды, также могут произрастать на дре-
весине и каменистых субстратах; в различных типах сообществ в горах и на рав-
нинах.
РАСПРОСТРАНЕНИЕ. Род широко представлен в различных природных зонах Се-
верного полушария. Виды Evernia встречаются в Европе, Азии, Сев. Америке,
Африке.
ПРИМЕЧАНИЕ. Включает 8 видов, шесть из которых известны на территории
России. Не обнаруженный в России вид Evernia illyrica (Zahlbr.) Zahlbr., возмож-
но, является редким хемотипом вида E. divaricata, в талломе которого не содер-
жится усниновой кислоты. Эпифитные виды рода Evernia морфологически сход-
ны с некоторыми представителями рода Ramalina, которые отличаются более
плотными талломами (что обусловлено наличием под коровым слоем элементов
стереома), часто более светлым матовым диском апотециев, а также двуклеточны-
ми аскоспорами и строением сумок. Lethariella togashii отличается присутстви-
ем плотного тяжа в сердцевине и составом вторичных метаболитов. Hypotrachyna
cirrhata (Fr.) Divakar, в отличие от представителей рода Evernia, имеет темную
Род Evernia
Флора лишайников России Флора лишайников России
110
нижнюю поверхность и многочисленные ризины по краям и на нижней поверх-
ности лопастей. Виды рода Evernia могут также отдаленно напоминать виды рода
Alectoria Ach., которые отличаются наличием хорошо развитых псевдоцифелл,
характером ветвления и составом вторичных метаболитов.
ИЗБРАННАЯ ЛИТЕРАТУРА: Bird, 1974; Голубкова, 1981, 1996в; Thomson, 1984;
Ryan, 2002b; Moberg, Thell, 2011a.
Ключ для определения видов рода Evernia
1. Таллом листоватый, нижняя поверхность светлее верхней; соредии присут-
ствуют, изидии не развиваются; содержит эверновую кислоту .........................
.........................................................................................................
5. E. prunastri.
—
Таллом кустистый, поверхность окрашена равномерно; соредии и изидии
присутствуют или не развиваются; содержит диварикатовую кислоту ........ 2.
2. Изидии присутствуют, часто совместно с соредиями ....... 3. E. mesomorpha.
—
Изидии отсутствуют, соредии крайне редки .................................................... 3.
3. Таллом очень мягкий и легкий, с кольцевыми растрескиваниями корового
слоя и рыхлой сердцевиной, часто длиннее 10 см, может быть легко разорван
на части ................................................................................................................. 4.
—
Таллом более плотный, часто короче 10 см ..................................................... 5.
4. Лесной эпифитный лишайник, таллом повисающий; коровой слой тонкий
(менее 10 мкм толщ.)
.................................................................
1. E. divaricata.
—
Напочвенный вид открытых местообитаний, таллом распростертый; коровой
слой более толстый (более 10 мкм толщ.)
.................................
6. E. terrestris.
5. Таллом прикреплен к субстрату псевдогомфом, часто несет многочисленные
апотеции; обитает, как правило, на коре деревьев ................. 2. E. esorediosa.
—
Таллом обычно не имеет органов прикрепления, апотеции неизвестны; напо-
чвенные виды открытых местообитаний .......................................................... 6.
6. Таллом жесткий, хрупкий, восходящий, ветви относительно тонкие (1–2 мм
шир.), гладкие или продольно-ребристые; сердцевина компактная ..................
........................................................................................................
4. E. perfragilis.
—
Таллом мягкий, распростертый по субстрату; ветви неравномерной толщины
(у основания до 1 см шир., на концах до 1–2 мм шир.), сетчато-лакунозные;
сердцевина рыхлая ....................................................................... 6. E. terrestris.
1. Evernia divaricata (L.) Ach.
1810, Lichenogr. Universalis: 85, 441. ≡ Lichen divaricatus L., 1767, Syst. Nat., ed.
12, 2: 713. ≡ Letharia divaricata (L.) Hue, 1899, Nouv. Arch. Mus. Hist. Nat., sér. 4,
1: 59. Т и п : [Германия, Майсен] Germany, Saxony, Missniae (Meissen), J. Schreber
(LINN 1273.277 лектотип I, Howe, 1912: 201; лектотип II, Jørgensen et al., 1994:
375). — Эверния растопыренная.
= Evernia altaica Florovsk.
Род Evernia
111
ИЗБРАННАЯ ЛИТЕРАТУРА: Ryan, 2002b; Moberg, Thell, 2011a.
ИЛЛЮСТРАЦИИ: Thomson, 1984: 228; Brodo et al., 2001: 312; Moberg, Thell,
2011a: 154; Stenroos et al., 2011: 215; Wirth et al., 2013: 472.
ТАЛЛОМ кустистый, повисающий, мягкий, светлый серовато-зеленый до жел-
товато-зеленого, исходно прикреплен к субстрату узким псевдогомфом, позднее
свободно свисает с ветвей или цепляется за неровности субстрата. Основные вет-
ви до 30 (редко до 40) см дл. и 1–4 мм в диам., неравной толщины, матовые, сет-
чато-лакунозные, угловатые в сечении, иногда в местах ветвления слегка упло-
щенные, несут редко расположенные перпендикулярно отходящие латеральные
веточки. Ветвление преимущественно неравно-дихотомическое, с острыми угла-
ми. Коровой слой очень тонкий (менее 10 мкм) и мягкий, с многочисленными по-
перечными, часто кольцевыми трещинами, обнажающими белую рыхлую сердце-
вину. Соралии очень редки; изидии и псевдоцифеллы отсутствуют. Размножается
преимущественно вегетативно посредством фрагментации. Дочерние талломы,
образовавшиеся в результате фрагментации, не имеют специализированных орга-
нов прикрепления и фиксируются, зацепляясь за неровности коры и иногда нама-
тываясь на ветви; могут частично прирастать к субстрату отдельными участками.
АПОТЕЦИИ редки, 1.5–8.0 мм в диам., талломный край цельный или мелко крену-
лированный, одного цвета с талломом, диск красно-коричневый. Аскоспоры 7–8
× 4–5 мкм. ПИКНИДЫ крайне редки, формируются на кончиках боковых веточек,
мелкие, до 0.1 мм в диам. — Рис. 79, 80 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту (часто в
небольших количествах), от K и C не изменяется, от KC не изменяется или дает жел-
товатую окраску, от P не изменяется, в UV не флюоресцирует. Сердцевина содержит
диварикатовую кислоту, от K, C, KC и P не изменяется, в UV флюоресцирует.
ЭКОЛОГИЯ. На ветвях преимущественно хвойных деревьев, предпочитает ме-
стообитания с высоким уровнем влажности, наиболее обилен в малонарушенных
лесных массивах.
РАСПРОСТРАНЕНИЕ. Россия. Широко распространенный лесной вид, известный
из многих равнинных и горных территорий России, однако за последнее столе-
тие значительно сокративший свой ареал в результате сведения и фрагмента-
ции старовозрастных лесов, поэтому из некоторых регионов известен только по
историческим данным. ЕР: Евр. Север — Мурманская обл., Карелия, Архангель-
ская обл., Коми, Ленинградская, Вологодская, Кировская (сев.-зап. часть) обл.;
Ср. Россия — Псковская, Новгородская, Тверская, Костромская, Кировская обл.,
Пермский край (равнинная часть), Смоленская, Московская, Нижегородская обл.,
Марий Эл, Татарстан, Удмуртия, Мордовия; Юж. Россия — Саратовская, Вол-
гоградская обл., Крым; Сев. Кавказ — Краснодарский край, Адыгея, Карачаево-
Черкесия, Кабардино-Балкария, Ингушетия, Дагестан; Сев. Урал — горные райо-
ны Пермского края и Свердловской обл.; Юж. Урал — горные районы Пермского
края, Башкирии, Челябинской и Оренбургской обл. СИБ: Зап. Сиб. сев.
—
Хан-
Флора лишайников России Флора лишайников России
112
ты-Мансийский АО, Томская обл.; Юж. Сиб.
—
Алтайский край (вост. горная
часть), Новосибирская (крайний восток), Кемеровская обл., Республика Алтай,
Хакасия, Тыва, Бурятия, Красноярский край (юж. часть); Як. — центр., вост., юж.
ДВ: ДВ юж. — Хабаровский край (юж. часть), Приморский край. — Карта 51.
Общее распространение. Европа, Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Evernia divaricata отличается от других представителей рода
наиболее мягким и длинным повисающим талломом, обычно лишенным ве-
гетативных пропагул. Видом, наиболее похожим внешне, является Lethariella
togashii, обитающий в лесных сообществах юга Дальнего Востока и отличаю-
щийся присутствием плотного тяжа в сердцевине и составом вторичных метабо-
литов.
В некоторых регионах Северной Америки и Европы на песчаной почве в от-
крытых местообитаниях встречается лишайник, обычно идентифицируемый как
Evernia divaricata. Таксономический статус напочвенных популяций вида, на наш
взгляд, неоднозначен и требует дополнительных филогенетических исследова-
ний, в том числе сравнения с E. terrestris.
2. Evernia esorediosa (Müll. Arg.) Du Rietz
1924, Svensk Bot. Tidskr. 18: 390. ≡ Evernia mesomorpha f. esorediosa Müll. Arg.,
1891, Flora 74 (1): 110. ≡ Evernia thamnodes f. esorediosa (Müll. Arg.) Zahlbr., 1930,
Cat. Lich. Univ. 6: 362. ≡ Letharia thamnodes f. esorediosa (Müll. Arg.) Hue, 1899,
Nouv. Arch. Mus. Hist. Nat., sér. 4, 1: 58. Т и п : [Япония] Japan, Honshu, ad Nikko in
Japonia, M. Miyoshi [M0205337 синтип («isotype»)]. — Эверния несоредиозная.
ИЗБРАННАЯ ЛИТЕРАТУРА: Голубкова, 1981, 1996в.
ИЛЛЮСТРАЦИИ: Голубкова, 1996в: 51.
ТАЛЛОМ кустистый, относительно мягкий, зеленовато-желтоватый, до
12(15) см дл., прикреплен к субстрату псевдогомфом. Ветви торчащие или по-
висающие и приподнятые от основания, дихотомически равно или неравно раз-
ветвленные, 1.5–2.0 мм в диам., угловато-округлые в сечении, уплощенные и рас-
ширенные в местах ветвления, к концам постепенно утончающиеся, с матовой,
продольно-лакунозной поверхностью. Соредии и изидии отсутствуют. АПОТЕЦИИ
обычно многочисленные, до 17 мм в диам., округлые, на талломных «ножках».
Диск коричневый, гладкий до блестящего, сначала вогнутый, позднее становится
плоским. Талломный край апотециев складчатый. ПИКНИДЫ иногда очень много-
численные, черные, располагаются на поверхности ветвей. — Рис. 81 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K
и C не изменяется, от KC желтеет, от P не изменяется, в UV не флюоресцирует.
Сердцевина содержит диварикатовую кислоту, от K, C, KC и P не изменяется, в
UV флюоресцирует.
ЭКОЛОГИЯ. На коре преимущественно хвойных деревьев, реже на веточках ку-
старничков в горнотаежных лесах.
Род Evernia
113
РАСПРОСТРАНЕНИЕ. Россия. Вид, широко распространенный в Сибири и на Даль-
нем Востоке. СИБ: Зап. Сиб. сев. — Томская обл.; Зап. Сиб. юж. — Омская, Ново-
сибирская обл.; Ср. Сиб. — Таймырский Долгано-Ненецкий р-н (юж. часть), Крас-
ноярский край (сев. часть), Эвенкия; Юж. Сиб. — все районы, кроме вост. горной
части Алтайского края; Як. — центр., вост., юж. ДВ: ДВ сев. — Магаданская обл.,
Хабаровский край (сев. часть); ДВ юж. — Хабаровский край (юж. часть), Амурская
обл., Еврейская АО, Сахалинская обл. (Сахалин), Приморский край. — Карта 52.
Общее распространение. Азия (Россия, Монголия, Китай, Япония).
ПРИМЕЧАНИЕ. Вид морфологически наиболее близок к Evernia mesomorpha, от
которого отличается отсутствием изидий и соредий и наличием апотециев.
3. Evernia mesomorpha Nyl.
1861, Lich. Scand.: 74. ≡ Letharia mesomorpha (Nyl.) Du Rietz, 1913, Svensk Bot.
Tidskr. 9: 427. Т и п : [Финляндия] Savolaxia (Etelä-Savo), Taipalsaari, 1852,
E. Nylander (H-NYL 35772 = H9505060 лектотип, Moberg, Thell, 2011a: 139;
H9504281, 9504282 изолектотипы). — Эверния мезоморфная.
= Evernia prunastri var. thamnodes Flot. ≡ Evernia thamnodes (Flot.) Arnold
≡ Letharia thamnodes (Flot.) Hue; = Evernia thamnodes f. arenicola Savicz; = Evernia
thamnodes f. tecticola Savicz.
ИЗБРАННАЯ ЛИТЕРАТУРА: Ryan, 2002b; Moberg, Thell, 2011a.
ИЛЛЮСТРАЦИИ: Thomson, 1984: 229; Brodo et al., 2001: 312; Moberg, Thell,
2011a: 154; Stenroos et al., 2011: 216.
ТАЛЛОМ кустистый, мягкий, зеленовато-желтоватый, до 8(10) см дл., прикре-
плен к субстрату псевдогомфом. Ветви у основания часто приподнятые, ближе
к концам повисающие, иногда торчащие, дихотомически равно или неравно раз-
ветвленные, 1–3 мм в диам., угловато-округлые в сечении, уплощенные и рас-
ширенные в местах ветвления, к концам постепенно утончающиеся, с матовой,
продольно-лакунозной поверхностью. Изидии, как правило, многочисленные,
диффузно распределены по поверхности таллома, часто совместно с мелкими
округлыми соралиями; соредии тонкие, мучнистые. АПОТЕЦИИ редки, до 6(9) мм
в диам., округлые. Диск красно-коричневый, гладкий, чаще всего блестящий. Тал-
ломный край апотециев тонкий, неровный, может быть покрыт соредиями и изи-
диями. ПИКНИДЫ располагаются на поверхности ветвей, черные, встречаются не-
часто. — Рис. 82 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K
и C не изменяется, от KC желтеет, от P не изменяется, в UV не флюоресцирует.
Сердцевина содержит диварикатовую кислоту, от K, C, KC и P не изменяется, в
UV флюоресцирует.
ЭКОЛОГИЯ. На коре и древесине преимущественно хвойных деревьев, иногда
на ветвях кустарников, очень редко на почве, в лесных сообществах и на болотах,
в основном в таежной зоне.
Флора лишайников России Флора лишайников России
114
РАСПРОСТРАНЕНИЕ. Россия. Наиболее широко распространенный на территории
России вид рода. АРКТ: Аркт. евр. — Ненецкий АО, Коми (крайний северо-вос-
ток); Аркт. зап.-сиб. — Ямало-Ненецкий АО (сев. часть); Аркт. як. — арктические
районы Якутии. ЕР: Евр. Север — все районы; Ср. Россия — все районы, кроме
Калининградской обл.; Юж. Россия — Саратовская, Самарская (юж. часть), Орен-
бургская, Волгоградская обл., Калмыкия, Крым; Сев. Кавказ — Краснодарский
край, Северная Осетия; Сев. Урал — все районы; Юж. Урал — все районы. СИБ:
Зап. Сиб. сев. — все районы; Зап. Сиб. юж. — все районы, кроме зап. части Ал-
тайского края; Ср. Сиб. — все районы; Юж. Сиб. — все районы; Як. — все райо-
ны. ДВ: ДВ сев. — все районы; ДВ юж. — все районы. — Карта 53.
Общее распространение. Евразия, Сев. Америка.
ПРИМЕЧАНИЕ. Единственный представитель рода, образующий изидии.
4. Evernia perfragilis Llano
1951, J. Washington Acad. Sci. 41: 199. ≡ Alectoria arctica Elenkin et Savicz, 1912,
Acta Horti Petropol. 32: 73. ≡ Evernia arctica (Elenkin et Savicz) Lynge, 1928, Lich.
Nov. Zemlya: 209, nom. illeg., non (Richardson) Tuck., 1823. Т и п : [Россия] Но-
вая Земля, Крестовая губа, 1901, И. В. Палибин (LE L-40 лектотип, hic designatus,
MBT 10004194; LE L-15305 изолектотип MBT 10004195).
—
Эверния очень
ломкая.
= Evernia esorediosa var. tundrae Oxner.
ИЗБРАННАЯ ЛИТЕРАТУРА: Голубкова, 1981, 1996в.
ИЛЛЮСТРАЦИИ: Thomson, 1984: 230; Голубкова, 1996в: 53.
ТАЛЛОМ кустистый, жесткий и ломкий, с торчащими восходящими ветвями,
бледно-желтоватый или зеленоватый, иногда чернеющий ближе к концам вет-
вей, не прикрепленный к субстрату, лежащий на поверхности почвы или закре-
пленный в мохово-лишайниковой дернине. Ветви дихотомически разветвленные,
2–5 см дл., 1–2 мм шир. (в местах ветвления до 3–4 мм), в сечении угловато-окру-
глые до округлых, слегка уплощенные в местах ветвления. Поверхность про-
дольно-лакунозная или почти гладкая, матовая, местами с матовым блеском, ино-
гда ближе к концам ветвей покрыта тонким светлым налетом. Коровой слой до
55 мкм толщ. Сердцевина компактная. Соредии и изидии не формируются. АПО-
ТЕЦИИ неизвестны. ПИКНИДЫ в развитом состоянии не отмечены, в некоторых слу-
чаях встречаются на концах ветвей, недоразвитые, коричнево-черные; конидии не
обнаружены. — Рис. 83 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит в небольших количествах
усниновую кислоту (иногда в очень малых концентрациях), от K и C не изменяет-
ся, от KC не изменяется или слабо желтеет, от P не изменяется, в UV не флюорес-
цирует. Сердцевина содержит диварикатовую кислоту, от K, C, KC и P не изменя-
ется, в UV флюоресцирует.
ЭКОЛОГИЯ. На почве в арктических регионах и высокогорьях.
Род Evernia
115
РАСПРОСТРАНЕНИЕ. Россия. Арктоальпийский вид, встречающийся в Аркти-
ке, в горных регионах Юж. Сибири и на севере Дальнего Востока. АРКТ: Аркт.
евр. — Архангельская обл. (Новая Земля); Аркт. зап.-сиб. — Ямало-Ненецкий АО
(сев. часть); Аркт. тайм. — Таймырский Долгано-Ненецкий р-н (сев. часть); Аркт.
як.
—
арктические районы Якутии; Аркт. чук.
—
Чукотский АО (сев. часть).
СИБ: Ср. Сиб. — Таймырский Долгано-Ненецкий р-н (юж. часть); Юж. Сиб. —
Кемеровская обл., Республика Алтай, Хакасия, Тыва, Бурятия, Красноярский край
(юж. часть). ДВ: ДВ сев. — Чукотский АО (юж. часть). — Карта 54.
Общее распространение. Европа (Россия), Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Отличается от других видов рода Evernia плотным, жестким,
ломким талломом с более толстым коровым слоем и компактной сердцевиной.
Морфологически близок к некоторым видам родов Alectoria и Ramalina, от кото-
рых отличается более хрупким талломом, отсутствием настоящих псевдоцифелл,
а также составом вторичных метаболитов. По характеру роста и окраске может
также отдаленно напоминать Cladonia amaurocraea (Flörke) Schaer.
5. Evernia prunastri (L.) Ach.
1810, Lichenogr. Universalis: 85, 442. ≡ Lichen prunastri L., 1753, Sp. Pl. 2: 1147.
Т и п : [Швеция] (LINN 1273.125 лектотип, Jørgensen et al., 1994: 380).
—
Эвер-
ния сливовая.
= Evernia arenaria (Fr.) Fr.; = Evernia arenaria Elenkin, nom. illeg. ≡ Evernia
elenkiniana Zahlbr. («elenkiana»).
ИЗБРАННАЯ ЛИТЕРАТУРА: Ryan, 2002b; Moberg, Thell, 2011a.
ИЛЛЮСТРАЦИИ: Brodo et al., 2001: 313; Moberg, Thell, 2011a: 154; Stenroos et al.,
2011: 217; Wirth et al., 2013: 471.
ТАЛЛОМ узколопастный, листоватый, кустовидный, мягкий, зеленоватый, до
10 см дл., прикреплен к субстрату псевдогомфом. Лопасти торчащие или к кон-
цам повисающие, дихотомически правильно или неправильно разветвленные,
1–6 мм шир., дорсивентральные, плоские, иногда слегка выпуклые. Верхняя по-
верхность таллома часто неровная, ямчатая, серо-зеленоватая или зеленоватая,
нижняя ямчатая или сетчатая, иногда ровная, белая. Разница в окраске верхней
и нижней поверхностей обусловлена расположением альгального слоя непо-
средственно под верхним коровым слоем (иногда альгальный слой распределен
неравномерно, тогда верхняя поверхность таллома слегка пятнистая — макулоз-
ная). При хранении в гербарии различия в окраске верхней и нижней поверхно-
стей могут становиться незаметными. Соралии беловатые, округлые, часто сли-
вающиеся, расположены преимущественно по краям лопастей, реже на верхней
поверхности таллома; соредии тонкие, мучнистые. Иногда помимо соредий на-
блюдается формирование напоминающих изидии лопастинок, однако настоя-
щие изидии не формируются. АПОТЕЦИИ редки, до 1.5 см в диам., округлые. Диск
светлый. Аскоспоры 6–11 × 4–6 мкм. ПИКНИДЫ иногда встречаются на краях и на
Флора лишайников России Флора лишайников России
116
поверхности лопастей, погруженные; конидии палочковидные, 6–7 × 0.5 мкм. —
Рис. 84 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту (редко
отсутствует), хлоратранорин и атранорин, от K желтеет, от C не изменяется, от
KC желтеет, от P не изменяется, в UV не флюоресцирует. Сердцевина содержит
эверновую кислоту, от K, C, KC и P не изменяется, в UV не флюоресцирует.
ЭКОЛОГИЯ. На коре и древесине деревьев различных пород, иногда на почве и
каменистых субстратах; преимущественно в лесной зоне.
РАСПРОСТРАНЕНИЕ. Россия. Один из наиболее широко распространенных видов
лишайников на территории Европейской России, крайне редко встречающийся
за Уралом. Указания на нахождение вида в Юж. Сибири нуждаются в проверке.
АРКТ: Аркт. евр. — Ненецкий АО, Коми (крайний северо-восток). ЕР: все райо-
ны. СИБ: Зап. Сиб. сев. — Ханты-Мансийский АО, Свердловская обл.; Зап. Сиб.
юж. — Челябинская и Курганская обл. ДВ: ДВ юж. — Сахалинская обл. (Куриль-
ские о-ва: Итуруп, пос. Лесозаводский). — Карта 55.
Общее распространение. Европа, Азия, Африка, Макаронезия, Сев. Америка.
ПРИМЕЧАНИЕ. Единственный листоватый представитель рода Evernia. От эпи-
фитных видов рода Ramalina отличается выраженной дорсивентральностью тал-
лома, более мягкими веточками (что вызвано отсутствием элементов стереома) и
более тонким коровым слоем. Морфологически крайне вариабелен, в связи с чем
было описано множество форм, в настоящее время не считающихся таксономиче-
ски значимыми.
6. Evernia terrestris (Tomin) N. S. Golubk.
1981, Konspekt Flory Lishaynikov Mong. Nar. Resp.: 107. ≡ Evernia esorediosa
f. terrestris Tomin, 1933, Animadvers. Syst. Herb. Kryloviano Univ. Tomsk. 5–6:
9. ≡ Evernia thamnodes f. terrestris (Tomin) Zahlbr., 1940, Cat. Lich. Univ. 10:
551. Т и п : [Монголия] in vicinitate lacus Kholbi-Nor, Mongolia, W. I. Baranov,
04.08.1926 (LE L-15306 лектотип, hic designatus, MBT 10004196; LE L-15307 изо-
лектотип MBT 10004197); [Казахстан] alt. ca. 2700 m, prope flum. Kokdshar, Ka-
zakstania, W. Sapozhnikov, B. Schischkin, 26.07.1912 (синтип, местонахождение об-
разца неизвестно). — Эверния наземная.
= Evernia crispata Florovsk.; = Evernia taurica Florovsk.
ИЗБРАННАЯ ЛИТЕРАТУРА: Голубкова, 1981, 1996в.
ИЛЛЮСТРАЦИИ: Голубкова, 1996в: 56.
ТАЛЛОМ кустистый, мягкий, зеленовато-желтоватый, 5–7 см дл., не прикреплен
к субстрату, распростерт по поверхности почвы в виде беспорядочных скоплений.
Ветви различной толщины и формы, от широких, почти плоских, лентовидных
ветвей первого порядка (могут достигать 0.5–1.2 см шир., в местах ветвления до
1.5 см шир.) до узких (до 1.0–1.5 мм) концевых ветвей, многократно дихотоми-
Род Evernia
117
чески правильно или неправильно разветвленных, угловато-округлых в сечении.
Поверхность мятая, ясно сетчато-лакунозная, матовая, с нерегулярными, часто
кольцеобразными трещинами. Сердцевина рыхлая. Соредии и изидии не форми-
руются. АПОТЕЦИИ неизвестны. ПИКНИДЫ неизвестны. — Рис. 85, 86 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит в небольших количествах
усниновую кислоту (иногда в очень малых концентрациях), от K и C не изменяет-
ся, от KC не изменяется или слабо желтеет, от P не изменяется, в UV не флюорес-
цирует. Сердцевина содержит диварикатовую кислоту, от K, C, KC и P не изменя-
ется, в UV флюоресцирует.
ЭКОЛОГИЯ. На почве среди мхов и других растений, в горных тундрах и высо-
когорных степях, преимущественно на высотах 2600–3000 м.
РАСПРОСТРАНЕНИЕ. Россия. Редкий горный лишайник. ЕР: Юж. Россия —
Крым. СИБ: Юж. Сиб. — Республика Алтай, Хакасия, Тыва, Бурятия, Краснояр-
ский край (юж. часть). — Карта 56.
Общее распространение. Азия (Россия, Казахстан, Киргизия, Монголия).
ПРИМЕЧАНИЕ. Морфологически вид наиболее близок к Evernia divaricata, эко-
логически — к E. perfragilis. По окраске и характеру произрастания сходен с
Alectoria sarmentosa subsp. vexillifera, от которого отличается отсутствием насто-
ящих псевдоцифелл и более мягким талломом с рыхлой сердцевиной; по имею-
щимся данным, ареалы этих видов не пересекаются.
Т. В. Макрый
Flavocetraria Kärnefelt et A. Thell
in Kärnefelt et al., 1994, Acta Bot. Fenn. 150: 81. Т и п : Flavocetraria cucullata
(Bellardi) Kärnefelt et A. Thell. — Флавоцетрария
Лихенизированные грибы. ТАЛЛОМ кустистый, вертикальный, из собранных
вместе или раздельных лопастей, бледно-желтый, в основании красновато-лило-
вый или коричневый. Лопасти слабо дихотомически разветвленные, более или
менее желобчатые или трубчато свернутые, с более или менее ровными или вол-
нисто вырезанными краями. Соралии отсутствуют или редкие. Псевдоцифеллы
на нижней поверхности, в виде мелких пятен или реже — узкой прерывистой
линии, проходящей по самому краю. Коровой слой тонкий; сердцевина рыхлая.
ПЛОДОВЫЕ ТЕЛА — зеориновые апотеции, расположенные на концах расширен-
ных лопастей на верхней поверхности вблизи края; диск красновато-коричневый.
Сумки узкобулавовидные, Lecanora-типа, с 8 аскоспорами. Аскоспоры однокле-
точные, бесцветные, эллипсоидные, 5–10 × 3.0–5.5 мкм. КОНИДИАЛЬНЫЕ СПОРОНО-
ШЕНИЯ — пикниды, черные, расположенные преимущественно по краям лопа-
стей, слегка выступающие в виде коротких шипиков. Конидии гантелевидные и
палочковидные, 6–7 × 0.8–1.0 мкм. ФОТОБИОНТ Trebouxia-типа.
Флора лишайников России Флора лишайников России
118
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Кора содержит усниновую кислоту; сердцевина —
лихестериновую, протолихестериновую кислоты и антрахиноны.
ЭКОЛОГИЯ. Напочвенные лишайники на минеральных и торфянистых почвах,
мхах, реже на валежнике и стволах деревьев; в горных и арктических тундрах,
лесотундрах, хвойных лесах, на сфагновых болотах и марях, в зарослях ерника,
кедрового стланика, на высокогорных мелкотравных лугах, в редколесьях, на ка-
менных россыпях и в других горных экотопах.
РАСПРОСТРАНЕНИЕ. Ареал рода охватывает северную часть Голарктики, а также
отдельные горные регионы Южного полушария (Юж. Америка, Индонезия, Па-
пуа Новая Гвинея).
ПРИМЕЧАНИЕ. Полифилетический род (Thell et al., 2009; Nelsen et al., 2011;
Divakar et al., 2017). Был выделен из рода Cetraria s. l. (Kärnefelt et al., 1994) на
основании таких признаков, как кустистый таллом, бледно-желтая окраска и ган-
телевидные конидии. Согласно последним молекулярно-филогенетическим ис-
следованиям, два вида отнесены к роду Nephromopsis, один — к роду «Cetraria».
ИЗБРАННАЯ ЛИТЕРАТУРА: Рассадина, 1950; Kärnefelt et al., 1994; Randlane, Thell,
2011c.
Ключ для определения видов рода Flavocetraria
1. Лопасти желобчатые до почти плоских, лакунозно-ямчатые, морщинистые ..
...............................................................................................................
3. F. nivalis.
—
Лопасти трубчато свернутые или сильно желобчатые .................................... 2.
2. Лопасти крупные, до 10 см выс. и 3–5 мм шир., сильно желобчатые до почти
трубчатых, свернутые верхней поверхностью внутрь, с несливающимися кра-
ями, с отогнутыми верхушками ................................................... 1. F. сucullata.
—
Лопасти мелкие, 1–4 см выс., 1–2(3) мм шир., свернутые в узкую трубочку
нижней поверхностью внутрь, с заворачивающимися внутрь, налегающими
один на другой, иногда сливающимися краями и шлемовидно загнутыми вер-
хушками ........................................................................................ 2. F. minuscula.
1. Flavocetraria cucullata (Bellardi) Kärnefelt et A. Thell
in Kärnefelt et al., 1994, Acta Bot. Fenn. 150: 81. ≡ Lichen cucullatus Bellardi,
1788, Osserv. Bot.: 54. ≡ Cetraria cucullata (Bellardi) Ach., 1803, Methodus: 293.
≡ Allocetraria cucullata (Bellardi) Randlane et Saag, 1992, Mycotaxon 44 (2): 492.
≡ Nephromopsis cucullata (Bellardi) Divakar, A. Crespo et Lumbsch, in Divakar et al.,
2017, Fungal Diversity 84: 112. Т и п : [Италия, Южный Тироль] Al monte Ritten
presso Bolzano, nel Tirolo meridionale l’esempl. inferiore, il superiore dalla Valdobbia,
1862, F. von Hausmann et A. Carestia, Erbar. Crittogam. Ital. no. 926 (FI неотип,
Randlane, Thell, 2011c: 139). — Флавоцетрария клобучковая.
ИЗБРАННАЯ ЛИТЕРАТУРА: Рассадина, 1950; Randlane, Thell, 2011c.
ИЛЛЮСТРАЦИИ: Randlane, Thell, 2011c: 154.
Род Flavocetraria
119
ТАЛЛОМ кустистый, вертикальный, из плотно собранных или раздельных ло-
пастей, бледно-желтый или зеленовато-беловатый, в основании красновато-лило-
вый до красновато-бурого. Лопасти крупные, 4–10 см выс., (1)2–5 мм шир., сла-
бо неправильно дихотомически разветвленные, трубчато свернутые или сильно
желобчатые, с несливающимися, извилисто вырезанными, волнистыми краями
и отогнутыми верхушками. Верхняя (внутренняя) поверхность ровная или слег-
ка ямчатая, гладкая, глянцевая; нижняя (наружная) более или менее продольно-
морщинистая и ребристая. Псевдоцифеллы на нижней (наружной) поверхности,
в виде мелких пятен. Соралии отсутствуют. АПОТЕЦИИ расположены на концах
плоских, расширенных до 10 мм (иногда более) лопастей; диск красновато-корич-
невый. Аскоспоры эллипсоидные, 5–10 × 3.0–5.5 мкм. ПИКНИДЫ многочисленные,
черные, расположенные по краям лопастей, выступающие, иногда в виде шипи-
ков. Конидии гантелевидные, 6 × 1 мкм. — Рис. 87 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Кора содержит усниновую кислоту, от KC желтеет,
от K, C и P не изменяется. Сердцевина содержит лихестериновую и протолихе-
стериновую кислоты, от K, C, KC и P не изменяется. В базальной части таллома
содержатся антрахиноны.
ЭКОЛОГИЯ. На дресвянистой и пеcчаной почве, торфе, мхах, реже на валежни-
ке, иногда на стволах лиственницы (на комле); в горных и арктических тундрах, в
хвойных лесах, зарослях кедрового стланика, на высокогорных лугах, в верещат-
никах, скально-степных сообществах, на каменных россыпях, торфяных болотах,
приморских дюнах.
РАСПРОСТРАНЕНИЕ. Широко распространенный арктоальпийский лишайник.
Россия. АРКТ: все районы. ЕР: Евр. Север — Мурманская обл., Карелия, Архан-
гельская обл., Коми; Ср. Россия — Кировская обл.; Сев. Кавказ — Краснодарский
край, Адыгея, Карачаево-Черкесия, Кабардино-Балкария, Северная Осетия, Даге-
стан; Сев. и Юж. Урал: Коми, Ямало-Ненецкий, Ханты-Мансийский АО, Перм-
ский край, Свердловская и Челябинская обл., Башкирия. СИБ: Зап. Сиб. сев.
—
Ямало-Ненецкий и Ханты-Мансийский АО; Зап. Сиб. юж.
—
Алтайский край;
Ср. Сиб.
—
все районы; Юж. Сиб.
—
все районы; Як.
—
все районы. ДВ: ДВ
сев. — все районы; ДВ юж. — все районы, кроме Еврейской АО. — Карта 57.
Общее распространение. Европа, Азия, Сев. и Юж. Америка (Перу), Грен-
ландия, Океания (Папуа Новая Гвинея).
ПРИМЕЧАНИЕ. Слабо полиморфный вид. Выделена форма с сильно отгибающи-
мися, курчавыми верхушечными лопастями (f. crispata Rass.), а также широколо-
пастные лакунозные формы (var. wainioi Räsänen, f. major Rass.).
2. Flavocetraria minuscula (Elenkin et Savicz) Ahti, Poryadina et Zhurb.
in Zhurbenko et al., 2005, Mycotaxon 92: 202. ≡ Cetraria cucullata f. minuscula
Elenkin et Savicz, 1910, Trudy Bot. Muz. Imp. Akad. Nauk 8: 43. ≡ Cetraria
minuscula (Elenkin et Savicz) McCune, in McCune et al., 2018, Mycosphere 9 (4):
867. Т и п : [Россия, Хабаровский край] Rossia, Regio Jakutensis [Chabarovsk], [fl.]
Род Flavocetraria
Флора лишайников России Флора лишайников России
120
Czelasin, ad turfam siccam in palude turfosa, 21.05.1903, I. M. Sczegolev (LE L-117
лектотип, Савич, 1964: 224, «typus»; L-118 изолектотип). — Флавоцетрария мел-
коватая.
= Cetraria cucullata var. rabotnovii Oxner.
ИЗБРАННАЯ ЛИТЕРАТУРА: Еленкин, Савич, 1911; Савич, 1964; Zhurbenko et al.,
2005; Thell et al., 2009.
ИЛЛЮСТРАЦИИ: Савич, 1964: 224; Zhurbenko et al., 2005: 203; Thell et al., 2009: 507.
ТАЛЛОМ кустистый, вертикальный, из собранных в рыхлые куртинки лопа-
стей, бледно-желтый или зеленовато-белый, в основании темно-коричневый. Ло-
пасти стройные, изящные, 2–4 см выс., 0.5–2.0(3.0) мм шир., слабо неправиль-
но дихотомически разветвленные, узко трубчато свернутые на всем протяжении
(включая конечные веточки), с заворачивающимися внутрь, налегающими один
на другой, иногда сливающимися, извилисто вырезанными курчавыми краями,
с шлемовидно загнутыми верхушками. Верхняя (наружная) поверхность более
или менее ровная, матовая, иногда со слабо заметными участками белого нале-
та; нижняя (внутренняя) немного светлее верхней. Псевдоцифеллы в виде узкой
прерывистой линии по самому краю нижней (внутренней) поверхности. Соралии
отсутствуют. АПОТЕЦИИ неизвестны. ПИКНИДЫ редкие, черные, расположенные по
краям лопастей, выступающие в виде бугорков. Конидии палочковидные, 6–7 ×
0.8 мкм. — Рис. 88 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Кора содержит усниновую кислоту, от KC желтеет,
от K, C и P не изменяется. Сердцевина содержит протолихестериновую кислоту,
от K, C, KC и P не изменятся.
ЭКОЛОГИЯ. На дресвянистой почве, в мохово-лишайниковой подушке, неболь-
шими дернинками среди других видов Flavocetraria и Cladonia P. Browne; в гор-
ных экотопах, тундрах, зарослях кедрового стланика и ерника, в сосновых, ли-
ственничных и еловых горных лесах, на сфагновых болотах. Встречается редко.
РАСПРОСТРАНЕНИЕ. Бореально-монтанный вид с широким амфиберингийским
ареалом, в Азии простирающимся от Тихого океана (Чукотка — Эгвекинот) по
горным системам Сев.-Вост. Сибири до гор Прибайкалья (Байкальский хр.). Рос-
сия. АРКТ: Аркт. чук. СИБ: Юж. Сиб.
—
Бурятия, Иркутская обл., Забайкаль-
ский край; Як. — центр. (Амгинский р-н), вост. (Оймяконский р-н), юж. (Алдан-
ский р-н). ДВ: ДВ сев. — Магаданская обл., Камчатский край (Корякский округ и
бывшая Камчатская обл.), Хабаровский край (сев. часть). — Карта 58.
Общее распространение. Азия, Сев. Америка (Аляска, Канада — Северо-За-
падные территории и Лабрадор).
ПРИМЕЧАНИЕ. Слабо полиморфный вид; имеются формы с более или менее
ровными и, наоборот, с сильно изрезанными курчавыми краями лопастей. Хо-
рошо отличается от мелких форм Flavocetraria cucullata более узкими, трубчато
свернутыми верхней поверхностью наружу лопастями, шлемовидно загнутыми
верхушками, а также более длинными и узкими конидиями. В природе издалека
Род Flavocetraria
121
лишайник выглядит как слабо разветвленная Cladonia. Указания для европейско-
го и таймырского секторов Арктики и Зап. Сибири ошибочны; в высокой Аркти-
ке вид отсутствует. Результаты молекулярно-филогенетического анализа (Thell et
al., 2009; Divakar et al., 2017) показывают отдаленное родство таксона с другими
представителями рода, с чем можно не соглашаться.
3. Flavocetraria nivalis (L.) Kärnefelt et A. Thell
in Kärnefelt et al., 1994, Acta Bot. Fenn. 150: 81. ≡ Lichen nivalis L., 1753, Sp. Pl.:
1145. ≡ Cetraria nivalis (L.) Ach., 1803, Methodus: 294. ≡ Allocetraria nivalis (L.)
Randlane et Saag, 1992, Mycotaxon 44 (2): 492. ≡ Nephromopsis nivalis (L.) Divakar,
A. Crespo et Lumbsch, in Divakar et al., 2017, Fungal Diversity 84: 113. Т и п :
[Швеция] Sweden (LINN-HL 1273.101 лектотип, Jørgensen et al., 1994: 378).
—
Флавоцетрария снежная.
ИЗБРАННАЯ ЛИТЕРАТУРА: Рассадина, 1950; Randlane, Thell, 2011c.
ИЛЛЮСТРАЦИИ: Randlane, Thell, 2011c: 154.
ТАЛЛОМ вертикальный, из более или менее плотно собранных или раздельных
лопастей, бледно-желтый или зеленовато-беловатый, в основании желтовато-
коричневый или буроватый. Лопасти 3–8 см выс., 3–10(15) мм шир., слегка же-
лобчатые или плоские, слабо неправильно разветвленные, с короткими боковы-
ми и конечными долями и ровными (в промежутках между веточками) краями,
в верхней части обычно веерообразно разветвленные. Верхняя (внутренняя) по-
верхность ямчато-лакунозная, морщинистая, гладкая, глянцевая; нижняя — мор-
щинистая и ребристая. Псевдоцифеллы на нижней поверхности, в виде мелких
пятен. Соралии отсутствуют. АПОТЕЦИИ на концах расширенных лопастей; диск
красновато-коричневый. Аскоспоры эллипсоидные, 5–10 × 3.0–5.5 мкм. ПИК-
НИДЫ многочисленные, черные, расположенные по краям лопастей и на ребрах,
выступающие в виде бугорков и коротких шипиков. Конидии гантелевидные,
6 × 1 мкм. — Рис. 89 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Кора содержит усниновую кислоту, от KC желтеет,
от K, C и P не изменяется. Сердцевина не содержит никаких веществ, от K, C, KC
и P не изменяется. В базальной части таллома содержатся антрахиноны.
ЭКОЛОГИЯ. На дресвянистой и пеcчаной почве, торфе, камнях с наносами мел-
козема; в горных и арктических тундрах, лесотундре, в горных сухих сосново-ли-
ственничных лесах, в зарослях кедрового стланика, высокогорных кобрезиевых
скально-степных сообществах, на каменных россыпях, верховых болотах, при-
морских дюнах; в малоснежных местообитаниях.
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: все районы. ЕР: Евр. Север — Мурманская
обл., Карелия, Архангельская обл., Ненецкий АО, Коми, Ленинградская обл.; Ср.
Россия — Калининградская и Кировская обл.; Сев. Кавказ — Краснодарский
край, Адыгея, Карачаево-Черкесия, Кабардино-Балкария, Северная Осетия, Чеч-
ня, Дагестан; Сев. и Юж. Урал — Коми, Ямало-Ненецкий и Ханты-Мансийский
Флора лишайников России Флора лишайников России
122
АО, Свердловская обл., Пермский край, Башкирия. СИБ: Зап. Сиб. сев.
—
Яма-
ло-Ненецкий и Ханты-Мансийский АО; Ср. Сиб. — все районы, кроме сев. части
Иркутской обл.; Юж. Сиб.
—
все районы, кроме крайнего востока Новосибир-
ской обл.; Як. — все районы. ДВ: ДВ сев. — все районы; ДВ юж. — все районы,
кроме Еврейской АО. — Карта 59.
Общее распространение. Европа, Азия, Сев. и Юж. Америка (Аргентина),
Гренландия.
ПРИМЕЧАНИЕ. Довольно полиморфный вид. Наибольшее разнообразие форм
наблюдается в горах и Арктике, где в одном и том же экотопе можно наблюдать
одновременно до четырех мозаично произрастающих форм, различающихся по
окраске, размеру талломов, форме и разветвленности лопастей. Заслуживают
специального изучения разновидность var. saviczii Rass. с широкими (10–15 мм
шир.), слабо разветвленными, сильно морщинисто-лакунозными лопастями и
форма f. suffruticosa Rass. с узкими, почти изодиаметрическими конечными доля-
ми сильно кустисто разветвленных верхушек лопастей.
Д. Е. Гимельбрант
Gowardia Halonen et al.
2009, Bryologist 112 (1): 141. Т и п : Gowardia nigricans (Ach.) Halonen et al. —
Говардия
Лихенизированные грибы. ТАЛЛОМ кустистый, с восходящими или стелю-
щимися по субстрату ветвями, сероватый до черного, обычно более или менее
двухцветный, в нижней части более светлый, поверхность таллома матовая или
блестящая. Ветви преимущественно цилиндрические, местами неравной тол-
щины или уплощенные, иногда с фовеолами, с редкими или многочисленными
хорошо выраженными плоскими вытянуто-веретеновидными или овальными бе-
лыми псевдоцифеллами. Соралии отсутствуют или образуются очень редко. Изи-
дии неизвестны. Коровой слой прозоплектенхимный, состоит из периклинально
ориентированных гиф. Сердцевина составлена рыхло расположенными орнамен-
тированными гифами. Таллом, в случае произрастания на почве, не имеет струк-
тур прикрепления к субстрату, погружен отмирающим основанием в раститель-
ную дерновину, на плотном субстрате закрепляется, цепляясь ветвями. ПЛОДОВЫЕ
ТЕЛА — апотеции зеоринового типа, талломный край одного цвета с талломом,
диск плоский до выпуклого, желтовато-коричневый до красновато-коричневого.
Эпитеций коричневатый или серый, гимений и гипотеций бесцветные. Парафизы
многократно разветвлены, сумки булавовидные, обычно содержат 2–4 аскоспоры.
Аскоспоры одноклеточные, эллипсоидные, толстостенные с выраженным гиали-
новым эписпорием, в зрелости красновато-коричневые, 20–41 × 12–28 мкм. КОНИ-
ДИАЛЬНЫЕ СПОРОНОШЕНИЯ неизвестны. ФОТОБИОНТ — Trebouxia.
Род Gowardia
123
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Содержит алекториаловую, часто барбатоловую
кислоты. При длительном хранении в гербарии бумага, контактирующая с талло-
мом лишайника, может приобретать розовато-охристую или розовато-коричневую
окраску и обесцвечиваться. При изучении состава лишайниковых веществ сле-
дует иметь в виду, что алекториаловая кислота со временем способна дегради-
ровать с образованием других соединений, в том числе гематоммовой кислоты,
5,7-дигидрокси-6-метилфталида и алекториалина.
ЭКОЛОГИЯ. Напочвенные лишайники, реже могут поселяться на каменистых
субстратах или коре кустарников; в сообществах равнинных и горных тундр, ино-
гда в северной тайге.
РАСПРОСТРАНЕНИЕ. Представители рода известны в обоих полушариях, для од-
ного характерно циркумполярное распространение, для другого — биполярное.
В России один вид встречается преимущественно в высокоширотной Арктике,
другой широко распространен в арктических и субарктических регионах, а также
в высокогорьях.
ПРИМЕЧАНИЕ. Относительно недавно описанный род, выделенный из рода
Alectoria (Halonen et al., 2009). Включает три вида, между которыми, возможно,
существуют переходные формы; два вида встречаются на территории России, а
недавно описанный вид Gowardia zebrina Goward et Myllys — в зап. Канаде (Go-
ward, Myllys, 2020). От близкого рода Alectoria отличается окраской таллома, со-
ставом лишайниковых веществ, а также формой и цветом псевдоцифелл.
ИЗБРАННАЯ ЛИТЕРАТУРА: Hawksworth, 1972; Brodo, Hawksworth, 1977; Kärnefelt,
Thell, 1992; Thell et al., 2004a; Halonen et al., 2009; Velmala, Myllys, 2011b; Zakeri
et al., 2017; Goward, Myllys, 2020.
Ключ для определения видов рода Gowardia
1. Таллом черный или коричнево-черный, иногда более светлый у основания,
преимущественно блестящий, обычно без выраженных основных ветвей; не
содержит барбатоловой кислоты ................................................... 1. G. arctica.
—
Таллом ясно двухцветный, преимущественно матовый, часто с выраженными
основными ветвями; содержит барбатоловую кислоту ........... 2. G. nigricans.
1. Gowardia arctica Halonen et al.
2009, Bryologist 112 (1): 141. ≡ Alectoria gowardii Lumbsch, 2010, Fieldiana, Bot.
1: 48. Т и п : [Канада] Canada, Northwest Territories, Inuvaluit. Banks Island, Swan
Lake, 100 m, mainly mesic mountain heath, 28.07.1999, J.-E. Mattsson 5255 (UPS го-
лотип; H, OULU изотипы). — Говардия арктическая.
ИЗБРАННАЯ ЛИТЕРАТУРА: Halonen et al., 2009.
ИЛЛЮСТРАЦИИ: Halonen et al., 2009: 144.
Род Gowardia
Флора лишайников России Флора лишайников России
124
ТАЛЛОМ кустистый, с восходящими до распростертых по субстрату ветвями,
до 5 см выс., черный до коричнево-черного, иногда светло-коричневый или свет-
ло-серый в нижней части, непосредственно контактирующей с субстратом. Ветви
таллома неравной толщины, часто в местах ветвления слегка уплощенные, ино-
гда с фовеолами, 0.3–0.5(1.0) мм в диам., большей частью блестящие, местами с
матовыми участками. Ветвление анизотомическое дихотомическое, ближе к кон-
цам ветвей изотомическое, основные ветви обычно явно не выражены. Псевдо-
цифеллы единичные или многочисленные, хорошо заметные, большей частью
плоские, вытянуто-веретеновидные или нитевидные, до 0.5 мм дл., белые, кон-
трастно выделяющиеся на фоне темных ветвей. Соралии, изидии и спинулы от-
сутствуют. АПОТЕЦИИ неизвестны. ПИКНИДЫ неизвестны. — Рис. 90 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Содержит алекториаловую кислоту и два неиденти-
фицированных вещества, от K и KC краснеет, от C приобретает зеленовато-чер-
ную окраску, от P становится желтоватым, в UV не флюоресцирует.
ЭКОЛОГИЯ. На почве в различных типах тундровых сообществ высокоширот-
ной Арктики.
РАСПРОСТРАНЕНИЕ. Россия. Вероятно, широко распространен на островах и ма-
териковом побережье Арктики. АРКТ: Аркт. евр.
—
Архангельская обл. (Земля
Франца-Иосифа); Аркт. зап.-сиб.
—
Ямало-Ненецкий АО (Ямал); Аркт. тайм.
—
Таймырский Долгано-Ненецкий р-н (Северная Земля: о. Большевик); Аркт. як. —
арктические районы Якутии; Аркт. чук. — Чукотский АО (сев.). — Карта 60.
Общее распространение. Европа (Россия), Азия (Россия), Сев. Америка (Ка-
нада).
ПРИМЕЧАНИЕ. Отличается от близкородственного вида Gowardia nigricans, с
которым связан комплексом переходных форм, слабой выраженностью основных
ветвей, более темной и более однотонной окраской таллома, преимущественно
блестящей поверхностью ветвей, характерной зеленовато-черной окраской при
действии C и отсутствием барбатоловой кислоты.
2. Gowardia nigricans (Ach.) Halonen et al.
2009, Bryologist 112 (1): 145. ≡ Cornicularia ochroleuca var. nigricans Ach., 1810,
Lichenogr. Universalis: 615. ≡ Alectoria ochroleuca var. nigricans (Ach.) Körb.,
1859, Parerga Lichenol. 1: 5. ≡ Alectoria nigricans (Ach.) Nyl., 1861, Flora 44: 372.
≡ Bryopogon nigricans (Ach.) Gyeln., 1935, Feddes Repert. 38: 239. Т и п : [Шве-
ция] Lapponia (H-ACH 1859B = H9502960 лектотип, Howe, 1911: 150, pl. 45).
—
Говардия черноватая.
= Alectoria ochroleuca subsp. thulensis Th. Fr. ≡ Alectoria thulensis (Th. Fr.)
Th. Fr. ex Kremp. ≡ Alectoria nigricans var. thulensis (Th. Fr.) Vain. ≡ Bryopogon
nigricans f. thulensis (Th. Fr.) Gyeln.; = Alectoria nigricans var. tschuktschorum
Vain.; = Alectoria nigricans var. nipponica Asahina; = Alectoria nigricans f. sorediata
E. Dahl; = Alectoria furcellata R. Sant.; Alectoria boryana Delise, nom. inval.
Род Gowardia
125
ИЗБРАННАЯ ЛИТЕРАТУРА: Hawksworth, 1972; Brodo, Hawksworth, 1977; Крусанова,
1996; Velmala, Myllys, 2011b.
ИЛЛЮСТРАЦИИ: Thomson, 1984: 35; Brodo et al., 2001: 153; Stenroos et al., 2011:
226; Velmala, Myllys, 2011b: 155; Wirth et al., 2013: 148.
ТАЛЛОМ кустистый, с восходящими до распростертых по субстрату ветвями,
подушковидный, до 10 см выс., обычно явно двухцветный — серый до розовато-
серого в нижней части и темно-серый до почти черного в верхней. Ветви таллома
обычно спутанные, неравной толщины, часто в местах ветвления и у основания
слегка уплощенные, иногда местами с фовеолами, 0.5–1.5 мм в диам., большей
частью матовые, местами в верхней части с блестящими участками. Ветвление
анизотомическое дихотомическое, основные ветви хорошо выражены. Псевдоци-
феллы более или менее многочисленные, погруженные, плоские или слегка вы-
пуклые, вытянуто-веретеновидные до щелевидных, до 1 мм дл., белые, заметные
на фоне темных ветвей. Соралии образуются крайне редко («Alectoria nigricans
f. sorediata»), изидии и спинулы отсутствуют. АПОТЕЦИИ образуются редко, рас-
положены на ветвях сбоку, до 2–9 мм в диам., талломный край цельный, одного
цвета с талломом, со временем исчезает, диск вогнутый или позднее выпуклый,
желтовато-коричневый до коричневого. Сумки содержат 2, редко 4 аскоспо-
ры. Аскоспоры эллипсоидные, одноклеточные, в зрелом состоянии становятся
красновато-коричневыми, 20–41 × 12–28 мкм. ПИКНИДЫ неизвестны.
—
Рис. 91
(вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Содержит алекториаловую и часто барбатоловую
кислоту, от K слабо желтеет, от KC и C краснеет, от P желтеет, в UV не флюорес-
цирует.
ЭКОЛОГИЯ. На почве, иногда на скалах, реже на ветвях кустарничков в различ-
ных типах тундровых сообществ в высокогорьях, арктических и субарктических
регионах Северного полушария, а также в соответствующих регионах Южного.
РАСПРОСТРАНЕНИЕ. Россия. Широко распространенный арктоальпийский вид,
встречающийся в Арктике, на Европейском Севере, на Урале, в Сибири и на
Дальнем Востоке. АРКТ: все районы. ЕР: Евр. Север — Мурманская обл., Каре-
лия, Архангельская обл., Ненецкий АО (юго-зап. часть), Коми; Сев. Урал — гор-
ные районы Пермского края (сев. часть). СИБ: Зап. Сиб. сев. — Ямало-Ненецкий
АО (юж. часть), Ханты-Мансийский АО; Ср. Сиб.
—
Таймырский Долгано-Не-
нецкий р-н (юж. часть), Иркутская обл. (сев. часть); Юж. Сиб.
—
Кемеровская
обл., Республика Алтай, Хакасия, Тыва, Красноярский край (юж. часть), Иркут-
ская обл. (юж. часть), Забайкальский край; Як. — все районы. ДВ: ДВ сев. — все
районы; ДВ юж. — Сахалинская обл. (Сахалин). — Карта 61.
Общее распространение. Европа, Азия, Сев. и Юж. Америка, Австралия,
Тасмания, Новая Зеландия, Антарктида и субантарктические острова.
ПРИМЕЧАНИЕ. Об отличиях от близкородственного вида Gowardia arctica см.
в примечаниях к нему.
Флора лишайников России Флора лишайников России
126
Л. В. Гагарина
Letharia (Th. Fr.) Zahlbr.
1892, Hedwigia 31: 36. ≡ Evernia [unranked] Letharia Th. Fr., 1871, Lichenogr.
Scand. 1: 322. Т и п : Letharia vulpina (L.) Hue. — Летария
Лихенизированные грибы. ТАЛЛОМ кустистый, радиальный, ярко-желто-зеле-
ный, прикрепляется к субстрату псевдогомфом. Ветви угловато-округлые, места-
ми сдавленные, дихотомически ветвящиеся. Соралии и изидии могут образовы-
ваться. Коровой слой параплектенхимный. Сердцевина рыхлая, гифы собраны в
пучки. ПЛОДОВЫЕ ТЕЛА — апотеции зеоринового типа, до 2 см в диам., с темно-
коричневым диском, терминальные, образуются редко. Сумки овальные, содер-
жат по 8 аскоспор. Аскоспоры одноклеточные, эллипсоидные, бесцветные, 5–8 ×
4–5 мкм. КОНИДИАЛЬНЫЕ СПОРОНОШЕНИЯ в форме пикнид, слегка выпуклые, черные.
Конидии игольчатые, 8–9 × 1 мкм. ФОТОБИОНТ — Trebouxia.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит вульпиновую кислоту и
атранорин, в апотециях обнаружена норстиктовая кислота.
ЭКОЛОГИЯ. Эпифитные лишайники, реже могут поселяться на древесине и ка-
менистых субстратах; в горных лесах.
РАСПРОСТРАНЕНИЕ. Представители рода известны только в Северном полуша-
рии, центр разнообразия находится в Сев. Америке; в России обитают в горных
лесах Кавказа.
ПРИМЕЧАНИЕ. Включает 3 вида, один из них встречается на территории России.
ИЗБРАННАЯ ЛИТЕРАТУРА: Котлов, 1996а; Brodo et al., 2001; Moberg, Thell, 2011b;
Altermann et al., 2016.
Letharia vulpina (L.) Hue
1899, Nouv. Arch. Mus. Hist. Nat., sér. 4, 1: 57. ≡ Lichen vulpinus L., 1753, Sp. Pl.
2: 1155. ≡ Usnea vulpina (L.) Hoffm., 1796, Deutschl. Fl. 2: 138. ≡ Parmelia vulpina
(L.) Ach., 1803, Methodus 2: 267. ≡ Evernia vulpina (L.) Ach., 1810, Lichenogr. Uni-
versalis: 443. ≡ Chlorea vulpina (L.) Nyl., 1855, Mém. Soc. Imp. Sci. Nat. Cherbourg
3: 170. ≡ Alectoria vulpina (L.) Mudd, 1861, Man. Brit. Lich.: 71. ≡ Rhytidocaulon
vulpinum (L.) Elenkin, 1916, Izv. Imp. Bot. Sada Petra Velikago 16 (2): 264. Т и п :
[Европа] Europe (LINN 1273.298 лектотип I, Howe, 1912: 383; лектотип II,
Jørgensen et al., 1994: 383). — Летария лисья.
ИЗБРАННАЯ ЛИТЕРАТУРА: Hue, 1899; Котлов, 1996а; Brodo et al., 2001; Ryan, 2002c.
ИЛЛЮСТРАЦИИ: Arup et al., 1997: 184; Brodo et al., 2001: 413; Holien, Tønsberg,
2006: 45.
ТАЛЛОМ кустистый, в виде торчащего, реже повисающего кустика, ярко-жел-
то-зеленый, до 15(20) см дл., прикрепляется к субстрату псевдогомфом. Лопасти
до 2 мм шир., угловато-округлые, одинаково окрашенные со всех сторон. Поверх-
Род Letharia
127
ность лопастей складчато-лакунозная, покрытая мелкоточечными серо-желтыми
соралиями, на которых развиваются цилиндрические изидии. АПОТЕЦИИ редки,
до 10 мм в диам., с вогнутым темно-коричневым диском и более светлым краем.
Аскоспоры 5–8 × 4–5 мкм, одноклеточные, эллипсоидные, бесцветные. ПИКНИДЫ
слегка выпуклые, черные. Конидии игольчатые, 8–9 × 1 мкм. — Рис. 92 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит вульпиновую кислоту и
атранорин, от KC краснеет; в апотециях обнаружена норстиктовая кислота.
ЭКОЛОГИЯ. Обитает на стволах и ветвях деревьев хвойных пород, реже на гни-
ющей древесине в горных сосновых и других хвойных и смешанных лесах в суб-
альпийском поясе.
РАСПРОСТРАНЕНИЕ. Россия. ЕР: Сев. Кавказ — Краснодарский край, Адыгея,
Карачаево-Черкесия, Кабардино-Балкария, Северная Осетия. — Карта 62.
Общее распространение. Европа, Азия, Сев. Африка, Сев. Америка.
ПРИМЕЧАНИЕ. Вид характеризуется ярким желто-зеленым талломом, серо-жел-
тыми соралиями и более или менее многочисленными цилиндрическими изидия-
ми. Вероятно, представляет собой комплекс видов. Чувствителен к загрязнению
атмосферного воздуха, включен в Красную книгу России и региональные Крас-
ные книги.
И. Ф. Скирина
Lethariella (Motyka) Krog
1976, Norweg. J. Bot. 23 (2): 88. ≡ Usnea subgen. Lethariella Motyka, 1936, Lich.
Gen. Usnea Stud. Monogr. Pars Syst. 1: 39. Т и п : Lethariella intricata (Moris)
Krog. — Летариелла
Лихенизированные грибы. ТАЛЛОМ кустистый, жесткий или повисающий, ди-
хотомически или нерегулярно ветвящийся, прикрепляется к субстрату хорошо
выраженным псевдогомфом. Коровой слой тонкий, рыхлый, пористый, у боль-
шинства видов 20–40 мкм толщ., часто сетчато-складчатый и морщинистый или
с продольными бороздками. Центральный цилиндр хрупкий или эластичный, по-
лый или сплошной. Сосочки, псевдоцифеллы и фибриллы никогда не образуются.
АПОТЕЦИИ развиваются очень редко, леканориновые, с матовым, коричневым до
черного диском, талломный край без фибрилл. Аскоспоры одноклеточные, эллип-
соидные, бесцветные, по 8 в сумке. ПИКНИДЫ неизвестны.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой у всех видов содержит атранорин,
у видов Lethariella canariensis (Ach.) Krog, L. cashmeriana Krog, L. cladonioides
(Nyl.) Krog и L. zahlbruckneri (Du Rietz) Krog — также канарионовую кислоту,
сердцевина — орсинолтридепсидгирофоровую, псоромовую, конпсоромовую, ме-
рохлорофаевую, 4-О-метилкриптохлорофаевую и вульпиновую кислоты.
Род Letharia
Флора лишайников России Флора лишайников России
128
ЭКОЛОГИЯ. Виды рода являются эпифитами, но могут произрастать на камнях.
Приурочены к областям с холодным, умеренным или тропическим климатом.
РАСПРОСТРАНЕНИЕ. Род имеет европейско-азиатское распространение: Европа
(Средиземноморье, Макаронезия), Азия (горы Центр. Азии, Дальний Восток Рос-
сии, Япония).
ПРИМЕЧАНИЕ. Род включает шесть видов: Lethariella canariensis, L. cashmeri-
ana, L. cladonioides, L. intricata (Moris) Krog, L. togashii, L. zahlbruckneri. На тер-
ритории России произрастает один вид — L. togashii.
ИЗБРАННАЯ ЛИТЕРАТУРА: Motyka, 1936–1938; Krog, 1976; Котлов, 1996б.
Lethariella togashii (Asahina) Krog
1976, Norweg. J. Bot. 23 (2): 95. ≡ Letharia togashii Asahina, 1952, J. Jap. Bot. 27:
295. Т и п : [Япония] Japan, Honshu, Prov. Kai, Yamanaka, Mt. Fuji, 11.08.1952, M.
Togashi (TNS голотип). — Летариелла Тогаши.
ИЗБРАННАЯ ЛИТЕРАТУРА: Asahina, 1952; Krog, 1976; Ohmura, Kashiwadani, 1997;
Чабаненко, 2002; Скирина, 2006; Kurokawa, Kashiwadani, 2006; Скирина и др.,
2016; Каганов, Ёжкин, 2019.
ИЛЛЮСТРАЦИИ: Скирина и др., 2016; Скирина, 2019.
ТАЛЛОМ свисающий, до 20 см дл., дряблый, с дихотомическим или нерегуляр-
ным ветвлением, зеленовато-беловато-серый до бледно-коричневого, в молодых
частях с желтым оттенком ветвей, прикрепляется к субстрату псевдогомфом. Ве-
точки почти цилиндрические, но чаще сильно сплюснутые, продольно-бороздча-
тые, 1.2–1.4 мм в диам., в местах ветвления 1.5–2.5 мм шир., постепенно утон-
чающиеся к кончикам, часто с раздвоенными вершинками. Коровой слой тонкий
(20–30 мкм толщ.), мягкий. Образцы с Дальнего Востока в основном имеют неж-
ный зеленовато-беловато-серый оттенок и местами прерывистые бороздки на по-
верхности таллома. Сердцевина паутинистая, с желтоватым оттенком, особенно
в более молодых частях ветвей. Центральный цилиндр 0.4–2.0 мм в диам., твер-
дый, тонкий (занимает менее половины диаметра ветви), эластичный. Соредии,
изидии и фибриллы отсутствуют. АПОТЕЦИИ неизвестны. — Рис. 93 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит атранорин, сердцевина —
вульпиновую кислоту и тритерпеноиды. Отмечены также два неидентифициро-
ванных лишайниковых вещества (тритерпеноид и алифатическое соединение).
Коровой слой и частично сердцевина от K слабо желтеют (со временем окраска
становится более яркой, особенно в молодых частях), от KC, C и P не изменяют-
ся, в UV не флюоресцируют.
ЭКОЛОГИЯ. Растет в пихтово-еловых лесах, чаще на ветвях и стволах хвойных,
реже лиственных деревьев. Требователен к повышенной влажности воздуха, за-
тененности и невысоким температурам.
РАСПРОСТРАНЕНИЕ. Россия. ДВ: ДВ юж.
—
Сахалинская обл. (Сахалин: Мака-
ровский р-н, хр. Жданко; окрестности пос. Горное; Холмский р-н, гора Мицул),
Род Lethariella
129
Хабаровский край (Ульчский и Николаевский районы: мысы Каменный и Давы-
дова, бухта Табо, бассейн р. Татарка). — Карта 63.
Общее распространение. Азия (Россия; Япония: острова Хонсю и Хоккай-
до).
ПРИМЕЧАНИЕ. Lethariella togashii отличается от других видов рода твердым
эластичным центральным цилиндром, отсутствием в коровом слое оранжевого
пигмента — канарионовой кислоты, наличием вульпиновой кислоты в сердцеви-
не.
Редкий субокеанический вид с охотско-сахалино-японским ареалом, в России
произрастает на побережье Татарского пролива в окрестности бухты Табо (Хаба-
ровский край), где находится основная часть ареала, в пределах которой он встре-
чается спорадически, но со значительной численностью. В остальных частях аре-
ала встречается единичными экземплярами.
Т. В. Макрый
Masonhalea Kärnefelt
1977, Bot. Not. 130 (1): 101. Т и п : Masonhalea richardsonii (Hook.) Kärnefelt. —
Мэйсонхэйлея
Лихенизированные грибы. ТАЛЛОМ листоватый, жесткий, из одиночных или
образующих небольшие дернинки, неправильно или дихотомически разветвлен-
ных лопастей, коричневый до темно-бурого. Псевдоцифеллы на нижней поверх-
ности, хорошо выражены, в виде непрерывной каймы по краям лопастей или
покрывающие большие участки нижней поверхности, оставляя лишь островки,
покрытые коровым слоем. ПЛОДОВЫЕ ТЕЛА — апотеции, зеориновые, латеральные,
почти на ножке, одного цвета с талломом. Аскоспоры одноклеточные, бесцвет-
ные, широкоэллипсоидные или почти шаровидные. КОНИДИАЛЬНЫЕ СПОРОНОШЕ-
НИЯ — пикниды, краевые, редкие или многочисленные, слегка выступающие в
виде шипиков, бурые или черные; под стенкой пикниды находится слой коровой
плектенхимы. Конидии палочковидные. ФОТОБИОНТ Trebouxia-типа.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит алектороновую или лихесте-
риновую и протолихестериновую кислоты.
ЭКОЛОГИЯ. Напочвенные и эпифитные (на веточках тундровых кустарников)
лишайники; обитают в арктических и горных тундрах, подгольцовых сообще-
ствах.
РАСПРОСТРАНЕНИЕ. Ареал рода охватывает арктические районы Европы, Азии,
Сев. Америки, с центром в Берингии, где представители рода встречаются также
в умеренных широтах — в высокогорьях.
ПРИМЕЧАНИЕ. Выделен из рода Cetraria (Kärnefelt, 1977a) как монотипный род.
В настоящее время на основании молекулярно-филогенетического и анатомо-
Род Lethariella
Флора лишайников России Флора лишайников России
130
морфологического анализов в него включено два вида (Thell, 2011a; Nelsen et al.,
2013), с чем можно не соглашаться.
ИЗБРАННАЯ ЛИТЕРАТУРА: Kärnefelt, 1977a, 1979; Nelsen et al., 2013.
Ключ для определения видов рода Masonhalea
1. Лопасти крупные, разветвленные в виде оленьих рогов, распростертые с при-
поднимающимися концами; на почве .................................. 2. M. richardsonii.
—
Лопасти мелкие, в виде язычков, слабо разветвленные или неразветвленные,
отогнутые кверху или вертикальные; на веточках тундровых кустарников,
реже на почве .................................................................................. 1. M. inermis.
1. Masonhalea inermis (Nyl.) Lumbsch, Nelsen et A. Thell
in Thell, 2011, Nordic Lichen Flora, 4: 67. ≡ Cetraria crispa f. inermis Nyl., 1887,
Bull. Soc. Linn. Normandie, sér. 4, 1: 214. ≡ Cetraria inermis (Nyl.) Krog, 1973,
Bryologist 76 (2): 299. ≡ Tuckermannopsis inermis (Nyl.) Kärnefelt, 1993, Bryologist
96 (3): 403. ≡ Nephromopsis inermis (Nyl.) Divakar, A. Crespo et Lumbsch, in Divakar
et al., 2017, Fungal Diversity 84: 112. Т и п : [ Россия, Чукотка] Sibiria septentriona-
lis, sinus Konyam ad fretum Bering, 64°50′ lat. bor., 173° long. occid., 1879, E. Alm-
quist (S лектотип, Krog, 1973: 299; H-NYL 36339 = H9505022 изолектотип).
—
Мэйсонхэйлея безоружная.
ИЗБРАННАЯ ЛИТЕРАТУРА: Krog, 1973; Kärnefelt, 1979; Thell et al., 2009; Thell,
2011a; Nelsen et al., 2013.
ИЛЛЮСТРАЦИИ: Krog, 1973: 300; Thell et al., 2009: 508; Thell, 2011a: 158; Nelsen
et al., 2013: 730.
ТАЛЛОМ листоватый, мелкий, редко более 2 см выс., из одиночных или собран-
ных в небольшие плотные дернинки (на почве), вертикальных или отгибающихся
кверху лопастей, в основании прикрепленный. Лопасти плоские или слегка же-
лобчатые, 0.5–3.0 мм шир., неразветвленные или в основании слабо разветвлен-
ные, в верхней части в виде язычков, иногда с маленькими краевыми лопастин-
ками, с более или менее ровными до слегка зубчатых (благодаря выступающим
пикнидам) краями. Верхняя поверхность гладкая или слегка поперечно-ямча-
то-волнистая, коричневая, матовая до лоснящейся; нижняя — светлая до белой,
в центральной части выпуклая (соответствует желобку верхней поверхности), с
плоскими или вогнутыми бортиками по краям, со слегка загибающимися на ниж-
нюю сторону краями, несущими кое-где редкие реснички (ризины). Псевдоци-
феллы на нижней поверхности в виде непрерывной каймы вдоль краев. АПОТЕЦИИ
обычны, редко отсутствуют, краевые, 0.5–4.0 мм в диам., с коричневым блестя-
щим диском и слегка кренулированным краем. Аскоспоры шаровидные, 3.5–5.0 ×
3.5–6.0 мкм в диам. ПИКНИДЫ краевые, иногда переходящие на верхнюю поверх-
ность, выступающие, в виде шипиков (зубчиков), темно-бурые или черные. Ко-
Род Masonhalea
131
нидии палочковидные до слегка утолщенных на одном или обоих концах, 8–10 ×
1.0–1.3 мкм. — Рис. 94 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит лихестериновую и протолихе-
стериновую кислоты. Коровой слой и сердцевина от I, K, C, KC и P не изменяются.
ЭКОЛОГИЯ. На веточках тундровых кустарников (Salix spp., Betula nana и др.),
реже на почве в мохово-лишайниковой дернине и на камнях с наносами мелкозе-
ма; в тундровых сообществах и на каменных россыпях.
РАСПРОСТРАНЕНИЕ. Амфиберингийский вид, широко распространенный в Ар-
ктике и в горных районах Дальнего Востока. Россия. АРКТ — все районы. ЕР:
Сев. Урал — Коми (Печоро-Илычский заповедник). СИБ: Ср. Сиб. — юж. часть
Таймырского Долгано-Ненецкого р-на (Путорана). ДВ: ДВ сев.
—
Магаданская
обл., Камчатский край (бывшая Камчатская обл.). — Карта 64.
Общее распространение. Европа, Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Вид отнесен к роду Masonhalea на основании молекулярного
анализа и анатомо-морфологических признаков (Thell, 2011a; Nelsen et al., 2013).
Однако близкое родство его с M. richardsonii недостаточно убедительно, по мно-
гим признакам (в частности, по строению сумок) виды сильно различаются. По
результатам последних исследований (Divakar et al., 2017) таксон отнесен к роду
Nephromopsis, что также не представляется корректным.
Вероятно, в Сев.-Вост. Азии вид распространен значительно шире, по край-
ней мере, в некоторых районах Магаданской области это массовый лишайник.
2. Masonhalea richardsonii (Hook.) Kärnefelt
1977, Bot. Not., 130 (1): 104. ≡ Cetraria richardsonii Hook., in Richardson, 1823, Bot.
App. in Franklin, Narr. Journey Polar Sea: 761. ≡ Evernia richardsonii (Hook.) Nyl.,
1858 («1857»), Mém. Soc. Imp. Sci. Nat. Cherbourg 5: 99. ≡ Parmelia richardsonii
(Hook.) Nyl., 1860, Flora 43: 42. ≡ Platysma richardsonii (Hook.) Nyl., 1860, Syn.
Meth. Lich. 1 (2): 306. ≡ Nephromopsis richardsonii (Hook.) Divakar, A. Crespo et
Lumbsch, in Divakar et al., 2017, Fungal Diversity 84: 113. Т и п : [Канада] Barren
grounds from Point Lake to the Arctic Sea, 1820, J. Franklin, N. American Land
Expedition 1820 No. 43 (BM001096797 лектотип, Kärnefelt, 1977a: 104).
—
Мэй-
сонхэйлея Ричардсона.
ИЗБРАННАЯ ЛИТЕРАТУРА: Савич, 1923; Рассадина, 1950; Kärnefelt, 1977a.
ИЛЛЮСТРАЦИИ: Kärnefelt, 1977a: 104; Макрый, 2002: 230; Nelsen et al., 2013: 730.
ТАЛЛОМ листоватый, из не прикрепленных к субстрату, крупных, около 10–
15 см дл., хрящеватых, распростертых, отгибающихся кверху лопастей, в сухом
состоянии шарообразно скручивающихся и переносимых ветром. Лопасти снизу
слабо желобчатые со слегка заворачивающимися на нижнюю сторону краями,
сверху слегка выпуклые, 2–8(15) мм шир., неправильно дихотомически развет-
вленные, с широко расходящимися веточками первого порядка. Верхняя поверх-
ность от светло- до темно-коричневой (влажная оливково-зеленая до зелено-
Род Masonhalea
Флора лишайников России Флора лишайников России
132
вато-коричневой), гладкая, матовая; нижняя — серовато-белая с коричневыми
пятнами. Ризины и реснички отсутствуют. Соралии отсутствуют (отмечены со-
ралиеподобные образования). Псевдоцифеллы на нижней поверхности, образуют
широкие неровные каймы вдоль краев лопастей, местами сливающиеся, оставляя
лишь посередине лопастей участки, покрытые корой. АПОТЕЦИИ редки (извест-
ны из Сев. Америки), 4–8 мм в диам., с блестящим диском почти одного цвета с
талломом и кренулированным краем. Аскоспоры широкоэллипсоидные, 6.0–6.5 ×
3–6 мкм. ПИКНИДЫ краевые, немногочисленные до редких, слегка выступающие в
виде шипиков, черные. Конидии палочковидные, 5 × 0.75 мкм. — Рис. 95 (вклей-
ка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит алектороновую кислоту. Ко-
ровой слой от K, C, KC и P не изменяется. Сердцевина от I, K, C и P не изменяет-
ся, от KC краснеет.
ЭКОЛОГИЯ. На почве поверх мхов и других лишайников; в арктических и гор-
ных тундрах, на нивальных луговинах и болотах, в гольцовых лугово-степных со-
обществах, подгольцовых зарослях кедрового стланика и лиственничных редко-
лесьях.
РАСПРОСТРАНЕНИЕ. Вид с широким амфиберингийским ареалом, в Азии дале-
ко проникающий в высокогорья умеренной области до Байкала и гор Юж. Сиби-
ри (Рассадина, 1950; Макрый, 2008). Россия. АРКТ: Аркт. як., Аркт. чук. СИБ:
Юж. Сиб.
—
Кемеровская обл. (Кузнецкий Алатау), Хакасия (Вост. Саян), Буря-
тия, Красноярский край (Зап. Саян), Иркутская обл., Забайкальский край; Як. —
центр., вост., юж. ДВ: ДВ сев.
—
юж. часть Чукотского АО, Магаданская обл.,
Камчатский край (бывшая Камчатская обл.), сев. часть Хабаровского края.
—
Карта 65.
Общее распространение. Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Хорошо отграниченный от всех коричневых цетрариоидных ли-
шайников, слабо полиморфный вид.
Род Nephromopsis
133
Т. Рандлане, А. Сааг
Nephromopsis Müll. Arg.
1891, Flora 74: 374. Т и п : Nephromopsis stracheyi (C. Bab.) Müll. Arg. —
Нефромопсис
= Cetrariopsis Kurok., 1980, Mem. Natl. Sci. Mus. Tokyo 13: 140. Т и п : Cetrariop-
sis wallichiana (Taylor) Kurok.
= Tuckneraria Randlane et A. Thell, in Randlane et al., 1994, Acta Bot. Fenn. 150: 144.
Тип: Tuckneraria pseudocomplicata (Asahina) Randlane et Saag [≡ Nephromopsis
pseudocomplicata (Asahina) M. J. Lai]
Лихенизированные грибы. ТАЛЛОМ листоватый, может быть очень крупным,
до 20 см в диам., часто сильно морщинистый или сетчато-складчатый; верхняя
поверхность зеленовато- или серо-желтая до желтой, сердцевина белая или слег-
ка желтоватая до ярко-оранжевой, нижняя поверхность желтовато-коричневая
или коричневая до черной. Лопасти дорсивентральные, до 2.5 см шир., округ-
лые или продолговатые до удлиненных, с прижатыми или приподнимающимися
краями, у некоторых видов с редкими краевыми ресничками. Соредии имеются
у одного, изидии — у двух видов, известны также филлидии (лобули). Псевдо-
цифеллы всегда образуются на нижней поверхности, в виде либо маленьких бе-
лых точек, либо более крупных правильной формы пятен; часто расположены на
гребнях ребер на нижней поверхности или на особых пробкообразных выростах.
Ризины редкие или многочисленные, того же цвета, что и нижняя поверхность.
Коровой слой параплектенхимный. ПЛОДОВЫЕ ТЕЛА — апотеции зеоринового типа,
от мелких (1.0–2.5 мм) до очень крупных (около 30 мм в диам.), краевые на ниж-
ней стороне таллома, при этом часто повернуты вверх, так что можно неверно
оценить их положение, или реже на верхней поверхности; с коричневым диском,
круглые или почковидные, иногда на короткой ножке, с желтоватым талломным
краем. Сумки узкобулавовидные до булавовидных, 30–70 × 8–14 мкм, содер-
жат 8 аскоспор. Аскоспоры простые, эллипсоидные, почти круглые до круглых,
бесцветные, 6–12 × 2.5–6.0 мкм. КОНИДИАЛЬНЫЕ СПОРОНОШЕНИЯ — пикниды, часто
встречающиеся, краевые и поверхностные, часто на черных выступающих возвы-
шениях, но могут быть и погруженными в таллом; темноокрашенные до черных.
Конидии гантелевидные, 4–5 × 1.0–1.5 мкм. ФОТОБИОНТ Trebouxia-типа.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту; серд-
цевина — различные жирные кислоты, орсинол или β-орсинольные депсиды и
депсидоны; у некоторых видов в сердцевине содержится секалоновая кислота и
сопутствующие пигменты, окрашивающие ее в желтый или оранжевый цвет.
ЭКОЛОГИЯ. Представители рода растут преимущественно на коре лиственных или
хвойных деревьев в горных лесах; один вид растет на скалах и валунах, два напо-
чвенных вида встречаются в альпийских областях на высотах 3500–4800 м над ур. м.
Род Nephromopsis
Флора лишайников России Флора лишайников России
134
РАСПРОСТРАНЕНИЕ. Большинство видов распространены в Вост. и Юго-Вост.
Азии; один вид (Nephromopsis laureri) имеет более широкое распространение,
включающее, кроме Азии, горные леса Центр. Европы и Юж. Америки. Боль-
шинство видов, отмеченных в России, распространены на Дальнем Востоке и/или
в Юж. Сибири.
ПРИМЕЧАНИЕ. Род включает 19 видов, 10 из которых встречаются в России.
В недавней публикации (Divakar et al., 2017) на основе современных фило-
генетических методов предлагается разделение всех цетрариоидных лишайников
лишь на три больших рода (Cetraria, Melanelia и Nephromopsis). В соответствии
с этой работой род Nephromopsis должен включать более 50 видов, но такая кон-
цепция авторами не признается.
ИЗБРАННАЯ ЛИТЕРАТУРА: Randlane et al., 1995; Randlane, Saag, 1998; Thell et al.,
2005.
Ключ для определения видов рода Nephromopsis
1. Таллом соредиозный ........................................................................ 4. N. laureri.
—
Таллом несоредиозный ....................................................................................... 2.
2. Эпилитный вид ........................................................................... 2. N. komarovii.
—
Эпифитные виды, растущие на коре (реже на древесине) ............................. 3.
3. Сердцевина желтая или оранжевая, от K становится ярко-желтой или лило-
вой ......................................................................................................................... 4.
—
Сердцевина белая, от K не изменяется ............................................................. 5.
4. Сердцевина светло-желтая, от K становится ярко-желтой (секалоновая кисло-
та) ....................................................................................................... 6. N. ornata.
—
Сердцевина оранжевая, от K становится лиловой (эндокроцин) .......................
......................................................................................................
1. N. endocrocea.
5. Апотеции мелкие и многочисленные, в основном ламинальные ......................
........................................................................................................
7. N. pallescens.
—
Апотеции разного размера и в разном числе (редко отсутствуют), преимуще-
ственно краевые ................................................................................................... 6.
6. Сердцевина от C или KC краснеет (оливеторовая, физодовая или алектороно-
вая кислоты) ......................................................................................................... 7.
—
Сердцевина от C и KC не изменяется (жирные кислоты) .............................. 9.
7. Таллом правильно и отчетливо сетчатый ...................................... 9. N. rugosa.
—
Таллом гладкий или только слегка морщинистый ........................................... 8.
8. Таллом равномерно зеленовато-желтый, часто с краевыми филлидиями,
краевые реснички всегда отсутствуют; аскоспоры продолговатые, 7–10 ×
3–5 мкм .................................................................................................... 3. N. laii.
—
Таллом зеленовато-желтый, но часто с коричневой полосой вдоль краевой
области, краевые филлидии отсутствуют, иногда с краевыми ресничками;
аскоспоры почти круглые, 5–6 × 4–5 мкм .................. 8. N. pseudocomplicata.
Род Nephromopsis
135
9. Нижняя поверхность сильно ребристая; псевдоцифеллы расположены на ре-
брах и пробковидных выростах .......................................... 10. N. yunnanensis.
—
Нижняя поверхность ровная или умеренно ребристая; псевдоцифеллы или на
ребрах, или на поверхности, но не на особых выростах .............................. 10.
10. Таллом гладкий или слегка морщинистый, без краевых филлидий; псевдоци-
феллы средние до крупных, плоские или вогнутые, на поверхности нижней
стороны ................................................................................. 5. N. nephromoides.
—
Таллом умеренно ребристый, часто с краевыми филлидиями; псевдоцифеллы
мелкие и плоские, в основном на ребрах ............................................ 3. N. laii.
1. Nephromopsis endocrocea Asahina
1935, J. Jap. Bot. 11: 24. ≡ Cetraria endocrocea (Asahina) M. Satô, in Nakai, Hon-
da, 1939, Nov. Fl. Jap. 5: 37. Т и п : [Япония] Japan, insula Nippon (Honshu), Na-
suzan, 1897, U. Faurie 339 (KY лектотип, Satô, 1939: 37).
—
Нефромопсис
внутришафранный.
ЛИТЕРАТУРА: Randlane, Saag, 1998; Чабаненко, 2002; Красная книга Приморско-
го края..., 2008.
ИЛЛЮСТРАЦИИ: Yoshimura, 1979: pl. 19.
ТАЛЛОМ листоватый, до 15 см в диам., с удлиненными приподнимающими-
ся лопастями до 8 мм шир.; верхняя поверхность желтовато-зеленая, кожистая,
гладкая или слегка сетчатая; сердцевина темно-желтая или оранжевая; нижняя
поверхность светло- или темно-коричневая до почти черной, регулярно сетчатая.
Соредии и изидии отсутствуют. Псевдоцифеллы в виде небольших белых точек
на нижней поверхности: краевые, поверхностные или на гребнях ребер. Ризины
редкие или многочисленные. АПОТЕЦИИ краевые на нижней поверхности талло-
ма, круглые или почковидные, до 12 мм в диам., диск коричневый, повернутый
вверх. Сумки 45 × 12 мкм; аскоспоры продолговатые, 9–10 × 4–5 мкм. ПИКНИДЫ
черные, краевые и поверхностные, на выступающих возвышениях, иногда очень
многочисленные; конидии гантелевидные, 5 × 1.5 мкм. — Рис. 96 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует; сердцевина — лихе-
стериновую и протолихестериновую кислоты вместе с эндокроцином, от C и P не
изменяется, от K приобретает лиловый цвет, в UV не флюоресцирует.
ЭКОЛОГИЯ. Растет на коре хвойных (Abies, Larix, Tsuga) или лиственных
(Betula) деревьев в лесных сообществах в нижнем и среднем поясах гор.
РАСПРОСТРАНЕНИЕ. Россия. Очень редкий вид, известный только на Дальнем
Востоке. ДВ: ДВ юж.
—
Хабаровский край (юж. часть), Сахалинская обл. (Ку-
рильские о-ва: Итуруп, Кунашир, Шикотан), Приморский край. — Карта 66.
Общее распространение. Азия (Китай, Япония).
ПРИМЕЧАНИЕ. Морфологически сходен с Nephromopsis ornata, но у последнего
светло-желтая сердцевина и несколько более грубый общий внешний вид.
Флора лишайников России Флора лишайников России
136
2. Nephromopsis komarovii (Elenkin) J. C. Wei
1991, Enum. Lich. China: 158. ≡ Cetraria komarovii Elenkin, 1903, Izv. Imp. S.-Pe-
terburgsk. Bot. Sada 3: 51. Т и п : [Россия] Russia, Irkutsk, Nilova Pustyn, Mt. Chon-
goldoi, 1902, Elenkin 155 (LE голотип, FH изотип). — Нефромопсис Комарова.
= Cetraria perstraminea Zahlbr.
ЛИТЕРАТУРА: Randlane, Saag, 1998; Randlane et al., 2001; Чабаненко, 2002;
Красная книга Приморского края..., 2008.
ТАЛЛОМ листоватый, до 15 см в диам., с удлиненными лопастями до 2 см
шир.; верхняя поверхность от лимонно-желтой до желтовато-зеленой, сильно
морщинисто-складчатая, с образованием своего рода концентрических узоров;
сердцевина белая; нижняя поверхность светло-коричневая, гладкая или слегка
морщинистая. Соредии и изидии отсутствуют. Псевдоцифеллы ламинальные на
нижней поверхности, в виде правильных белых пятен разного размера. Ризины
многочисленные до редких, светло-коричневые до беловатых. АПОТЕЦИИ много-
численные, краевые на нижней поверхности таллома, иногда поверхностные,
круглые или почковидные, до 15 мм в диам., диск коричневый, обращенный
вверх, часто имеются только молодые апотеции. Сумки 30 × 9 мкм; аскоспоры
продолговатые, 6–8 × 3–5 мкм. ПИКНИДЫ неизвестны. — Рис. 97, 98 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует; сердцевина — лихе-
стериновую и протолихестериновую кислоты, от K, C, KC и P не изменяется, в
UV не флюоресцирует; дополнительно в сердцевине некоторых образцов обнару-
жены фумарпротоцетраровая и стиктовая кислоты вместе с констиктовой, в та-
ких случаях сердцевина от K приобретает коричневую окраску или желтеет, от P
краснеет или становится оранжевой.
ЭКОЛОГИЯ. Эпилитный вид, растет на скалах и валунах в нижнем и среднем
поясах гор (200–1500 м над ур. м.); часто во влажных экотопах, например в доли-
нах горных речек, но также в тенистых местах в горных лесах.
РАСПРОСТРАНЕНИЕ. Россия. Редкий вид, известный только в Юж. Сибири и на
Дальнем Востоке, местонахождения в России находятся на северной границе аре-
ала. СИБ.: Юж. Сиб.
—
Республика Алтай, Тыва, Бурятия, Иркутская обл. (юж.
часть), Забайкальский край. ДВ: ДВ юж. — Хабаровский край (юж. часть), При-
морский край. — Карта 67.
Общее распространение. Азия (Монголия, Китай).
ПРИМЕЧАНИЕ. Легко распознается по интенсивному лимонно-желтому или зе-
леному цвету и характерному складчато-морщинистому листоватому таллому с
концентрическим рисунком. Единственный эпилитный вид в роде Nephromopsis.
3. Nephromopsis laii (A. Thell et Randlane) Saag et A. Thell
1997, Bryologist 100 (1): 111. ≡ Cetrariopsis laii A. Thell et Randlane, in Rand-
lane et al., 1995, Cryptog. Bryol. Lichénol. 16: 46. Т и п : [Россия] Russia, Pri-
Род Nephromopsis
137
morye, Kedrovaya Padj Nature Reserve, 1961, U. Pärn (TU голотип; LD изотип). —
Нефромопсис Лая.
ЛИТЕРАТУРА: Randlane et al., 1995, 2001; Randlane, Saag, 1998; Чабаненко, 2002;
Красная книга Приморского края..., 2008.
ИЛЛЮСТРАЦИИ: Randlane et al., 1995: 45.
ТАЛЛОМ листоватый, до 8 см в диам., с округлыми приподнимающимися ло-
пастями до 10 мм шир.; верхняя поверхность зеленовато-желтая, умеренно мор-
щинистая; сердцевина белая; нижняя поверхность светло- или темно-коричневая
с беловатыми краями, густосетчатая. У многих образцов по краю лопастей раз-
виты краевые филлидии в виде бахромы. Соредии, изидии и краевые реснички
отсутствуют. Псевдоцифеллы на нижней поверхности в виде небольших белых
круглых или овальных пятен, иногда окруженных коричневым ободком, располо-
жены большей частью на гребнях ребер. Ризины редкие, коричневые. АПОТЕЦИИ
многочисленные, большей частью краевые, иногда субмаргинальные, часто рас-
положены на филлидиях, до 5 мм в диам., продолговатые или почковидные, диск
коричневый, обращенный вверх, часто имеются только молодые апотеции в виде
краевой коричневой линии. Сумки 40–45 × 8–10 мкм; аскоспоры продолговатые,
8 × 3 мкм. ПИКНИДЫ неизвестны. — Рис. 99 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует; сердцевина — лихе-
стериновую и протолихестериновую кислоты, от K, C, KC и P не изменяется, в
UV не флюоресцирует; дополнительно в сердцевине может присутствовать алек-
тороновая кислота, в таких случаях сердцевина от KC краснеет.
ЭКОЛОГИЯ. Растет на коре хвойных (Abies, Picea, Pinus) или лиственных
(Betula, Quercus, Sorbus) деревьев в лесных сообществах в нижних и средних по-
ясах гор (600–3000 м над ур. м.).
РАСПРОСТРАНЕНИЕ. Россия. Редкий вид, известный только на Дальнем Востоке,
местонахождения в России находятся на северной границе ареала. ДВ: ДВ юж. —
Хабаровский край (юж. часть), Еврейская АО, Сахалинская обл. (Курильские
о-ва: Кунашир), Приморский край. — Карта 68.
Общее распространение. Вост. и Юго-Вост. Азия.
ПРИМЕЧАНИЕ. В некоторой степени сходен с Nephromopsis nephromoides, но эти
два вида имеют разные псевдоцифеллы на нижней поверхности: у N. laii псев-
доцифеллы мелкие и плоские, преимущественно расположенные на коричнево-
окрашенных складках в целом более светлоокрашенной нижней поверхности, в
то время как у N. nephromoides — среднего размера или крупные, плоские или
вогнутые и в основном развиваются прямо на нижней поверхности.
4. Nephromopsis laureri (Kremp.) Kurok.
1991, J. Jap. Bot. 66: 156. ≡ Cetraria laureri Kremp., 1851, Flora 34: 673. ≡ Tuckneraria
laureri (Kremp.) Randlane et A. Thell, in Randlane et al., 1994, Acta Bot. Fenn. 150:
Флора лишайников России Флора лишайников России
138
149. Т и п : [Германия, Бавария] Germany, Bavaria (Bayern), Inzell, Appichl, A. von
Krempelhuber (WU лектотип, Obermayer, 2004: 513). — Нефромопсис Лаурера.
= Cetraria straminea Kremp. ex Schwend.
ЛИТЕРАТУРА: Randlane et al., 1994; Чабаненко, 2002; Thell et al., 2005; Красная
книга Приморского края..., 2008.
ИЛЛЮСТРАЦИИ: Randlane et al., 1994: 145–146.
ТАЛЛОМ листоватый, до 7 см в диам., с округлыми лопастями до 5–10 мм шир.
с приподнимающимися краями; верхняя поверхность светло-желтая или желто-
вато-зеленая, кожистая, гладкая; сердцевина белая; нижняя поверхность белая до
светло-коричневой. Соредии беловатые, многочисленные, в краевых соралиях;
изидии отсутствуют. Псевдоцифеллы на нижней поверхности в виде плоских,
круглых или неправильной формы белых пятен, часто окруженных светло-ко-
ричневым ободком. Ризины разбросанные, редкие, довольно длинные, светло-ко-
ричневые до беловатых. АПОТЕЦИИ очень редки, краевые, до 8 мм в диам., с ко-
ричневым диском и соредиозным талломным краем. Сумки 35–45 × 10–12 мкм;
аскоспоры почти круглые, 5–6 × 4–5 мкм. ПИКНИДЫ краевые на выступающих воз-
вышениях; конидии гантелевидные, 5 × 1.0–1.5 мкм. — Рис. 100 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует; сердцевина — лихе-
стериновую и протолихестериновую кислоты, от K, C, KC и P не изменяется, в
UV не флюоресцирует.
ЭКОЛОГИЯ. Растет преимущественно на коре хвойных (Abies, Juniperus, Picea,
Pinus), реже лиственных (Betula, Quercus, Salix, Sorbus) деревьев в горных лесах.
РАСПРОСТРАНЕНИЕ. Россия. Широко распространенный, но довольно редко
встречающийся вид, известный во многих регионах, преимущественно в гор-
ных областях. ЕР: Евр. Север — Карелия, Коми, Ленинградская обл.; Ср. Рос-
сия — Новгородская, Костромская, Кировская обл., Пермский край, Марий Эл,
Татарстан, Башкирия; Сев. Кавказ — Краснодарский край, Северная Осетия; Сев.
Урал; Юж. Урал. СИБ: Зап. Сиб. сев.
—
Ханты-Мансийский АО, Свердловская
обл.; Зап. Сиб. юж.
—
Челябинская, Новосибирская обл., Алтайский край (зап.
часть); Юж. Сиб.
—
Алтайский край (вост. часть), Кемеровская обл., Республи-
ка Алтай, Хакасия, Тыва, Бурятия, Красноярский край (юж. часть), Иркутская
обл. (юж. часть), Забайкальский край; Як. — вост., юж.; ДВ: ДВ юж. — Амурская
обл., Хабаровский край (юж. часть), Еврейская АО, Сахалинская обл., Примор-
ский край. — Карта 69.
Общее распространение. Европа (горы Центр. Европы), Азия (Монголия,
Китай, Япония, Непал), Юж. Америка (Колумбия, Венесуэла).
ПРИМЕЧАНИЕ. Единственный соредиозный вид в роде Nephromopsis; имеет
сходство с другим лишайником, имеющим краевые соредии и желтый листоватый
таллом, — Usnocetraria oakesiana, — но лопасти последнего более узкие и ленто-
видные, и псевдоцифеллы на нижней поверхности отсутствуют.
Род Nephromopsis
139
5. Nephromopsis nephromoides (Nyl.) Ahti et Randlane
in Randlane, Saag, 1998, Cryptog. Bryol. Lichénol. 19: 183. ≡ Platysma nephromoi-
des Nyl., 1869, Flora 52: 442. ≡ Cetraria nephromoides (Nyl.) D. D. Awasthi, 1982,
Bull. Bot. Surv. India 24: 11. Т и п : [Индия] India, West Bengal, Darjeeling District,
J. D. Hooker fil. et T. Thomson 2080 (H-NYL 36068 лектотип, Awasthi, 1982: 11). —
Нефромопсис нефромовидный.
= Nephromopsis ectocarpisma (Hue) Gyeln.
ЛИТЕРАТУРА: Randlane, Saag, 1998; Randlane et al., 2001; Чабаненко, 2002.
ИЛЛЮСТРАЦИИ: Yoshimura, 1979: pl. 19.
ТАЛЛОМ листоватый, до 20 см в диам., с округлыми широкими лопастями, ко-
торые могут закручиваться; верхняя поверхность серовато-желтая или зеленая, ко-
жистая и толстая, гладкая или слегка сетчатая; сердцевина белая; нижняя поверх-
ность светло-коричневая, гладкая или немного сетчатая у краев. Соредии и изидии
отсутствуют. Псевдоцифеллы на нижней поверхности белые, крупные и хорошо
заметные, овальные или круглые, плоские до вогнутых, расположены преимуще-
ственно поверхностно, но и в краевой зоне могут встречаться на гребнях ребер.
Ризины редкие. АПОТЕЦИИ обычно многочисленные, краевые, довольно мелкие (до
8 мм в диам.), с коричневым диском, круглые или овальные, обращенные вверх.
Сумки 35–40 × 10 мкм; аскоспоры эллипсоидные, 7–8 × 3 мкм. ПИКНИДЫ редкие,
краевые, погруженные; конидии неизвестны. — Рис. 101, 102 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует; сердцевина — лихе-
стериновую и протолихестериновую кислоты, а также некоторые другие допол-
нительные жирные кислоты (например, каператовую кислоту), от K, C, KC и P не
изменяется, в UV не флюоресцирует.
ЭКОЛОГИЯ. Эпифит, растет на коре живых и упавших деревьев в лесных сооб-
ществах на средних высотах в горах (до 3600 м над ур. м.).
РАСПРОСТРАНЕНИЕ. Россия. Очень редкий вид, известный только на Дальнем
Востоке. ДВ: ДВ юж. — Приморский край. — Карта 70.
Общее распространение. Вост. и Юго-Вост. Азия.
ПРИМЕЧАНИЕ. Отличия от Nephromopsis laii см. в примечании к этому виду.
6. Nephromopsis ornata (Müll. Arg.) Hue
1900, Nouv. Arch. Mus. Hist. Nat., sér. 4, 2: 90. ≡ Cetraria ornata Müll. Arg., 1891,
Nuovo Giorn. Bot. Ital. 23: 122. Т и п : [Япония] Honshu, Mt. Ontake, 1890,
M. Miyoshi 109 (G00291076 голотип). — Нефромопсис украшенный.
ЛИТЕРАТУРА: Randlane, Saag, 1998; Randlane et al., 2001; Чабаненко, 2002; Thell
et al., 2005; Красная книга Приморского края..., 2008; Скирина и др., 2010.
ИЛЛЮСТРАЦИИ: Yoshimura, 1979: pl. 19; Скирина и др., 2010: тaбл. IX, 4.
Флора лишайников России Флора лишайников России
140
ТАЛЛОМ листоватый, до 15 см в диам., с приподнимающимися лопастями до
15 мм шир.; верхняя поверхность желтовато-зеленая, кожистая, гладкая или слег-
ка сетчатая; сердцевина светло-желтая; нижняя поверхность светло- или темно-
коричневая до почти черной, полностью сетчатая. Соредии и изидии отсутствуют.
Псевдоцифеллы в виде небольших белых точек на нижней поверхности: краевые,
поверхностные и на гребнях ребер. Ризины редкие. АПОТЕЦИИ краевые, располо-
жены на нижней поверхности таллома, круглые, до 3 см в диам., с коричневым
диском, обращенные вверх. Сумки 40–45 × 10 мкм; аскоспоры продолговатые,
7–9 × 4–5 мкм. ПИКНИДЫ черные, преимущественно краевые, но также и поверх-
ностные, на выступающих возвышениях, иногда очень многочисленные; конидии
гантелевидные, 5 × 1 мкм. — Рис. 103 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует; сердцевина — лихе-
стериновую и протолихестериновую кислоты вместе с секалоновой кислотой, от
C и P не изменяется, от K становится ярко-желтой; дополнительно в сердцевине
могут быть следы эндокроцина и фумарпротоцетраровой кислоты.
ЭКОЛОГИЯ. Эпифит, растет на коре хвойных (Abies, Larix, Picea, Pinus, Ta x u s ) и
лиственных (Alnus, Betula, Populus, Salix) деревьев, иногда также на валунах, по-
крытых мхами, в лесных сообществах на низких и средних высотах в горах (900–
3200 м над ур. м.).
РАСПРОСТРАНЕНИЕ. Россия. Редкий вид, известный только на Дальнем Востоке;
местонахождения в России находятся на северной границе ареала. ДВ: ДВ юж. —
Амурская обл., Хабаровский край (юж. часть), Еврейская АО, Сахалинская обл.
(Курильские о-ва: Итуруп, Кунашир), Приморский край. — Карта 71.
Общее распространение. Азия (Китай, Южная Корея, Япония).
ПРИМЕЧАНИЕ. Мофологически довольно близок к Nephromopsis endocrocea,
но у последнего ярко-желтая до оранжевой сердцевина и более изящный общий
внешний вид. N. ornata — филогенетически наиболее обособленный вид внутри
рода, формирующий сестринскую монофилетическую группу по отношению к
другим видам рода Nephromopsis.
7. Nephromopsis pallescens (Schaer.) Y. S. Park var. pallescens
1990, Bryologist 93 (2): 122. ≡ Cetraria pallescens Schaer., in Moritzi, 1846 (1845–
1846), Syst. Verz.: 129. ≡ Cetrariopsis pallescens (Schaer.) Randlane et A. Thell, in
Randlane et al., 1995, Cryptog. Bryol. Lichénol. 16: 42. Т и п : [Индонезия] Indone-
sia, Java, Mt. Pangerango, W. Zollinger 449 (G00291082 голотип). — Нефромопсис
бледнеющий.
= Cetraria wallichiana (Taylor) Müll. Arg. ≡ Cetrariopsis wallichiana (Taylor)
Kurok. ≡ Ahtia wallichiana (Taylor) M. J. Lai.
ЛИТЕРАТУРА: Randlane et al., 1995, 2001; Чабаненко, 2002; Thell et al., 2005;
Красная книга Приморского края..., 2008.
ИЛЛЮСТРАЦИИ: Yoshimura, 1979: pl. 19; Randlane et al., 1995: 45.
Род Nephromopsis
141
ТАЛЛОМ листоватый, крупный, до 16 см в диам., с круглыми лопастями; верх-
няя поверхность зеленовато-желтая, обычно морщинистая или сетчатая; серд-
цевина белая; нижняя поверхность белая до желтой, слегка или чаще сильно
морщинистая. Соредии и изидии отсутствуют. Псевдоцифеллы на нижней по-
верхности в виде небольших белых пятен, расположены на особых пробкообраз-
ных выростах, иногда окруженные светло-коричневым ободком. Ризины редкие,
разбросанные, белые или светло-коричневые. АПОТЕЦИИ до 3 см в диам., с корич-
невым плоским или слегка выпуклым диском, в основном поверхностные, но мо-
гут быть также субмаргинальные и краевые, временами очень многочисленные и
покрывающие весь таллом, но иногда расположены в основном в маргинально-
субмаргинальной части нижней поверхности и только очень маленькие апотеции
возникают по-настоящему ламинально; молодые апотеции с тонким талломным
краем. Сумки 35–40 × 10 мкм; аскоспоры продолговатые, 7–10 × 3–4 мкм. ПИКНИ-
ДЫ неизвестны. — Рис. 104, 105 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует; сердцевина — алекто-
роновую и/или лихестериновую и протолихестериновую кислоты, от K, C и P не
изменяется, от KC краснеет или не изменяется, в UV не флюоресцирует.
ЭКОЛОГИЯ. Эпифит, растет преимущественно на коре хвойных (Larix, Pinus)
и лиственных (Carpinus, Populus, Salix, Sorbus, Quercus) деревьев и кустарников
(Rhododendron) в горных лесах.
РАСПРОСТРАНЕНИЕ. Россия. Редкий вид, известный только на Дальнем Востоке.
ДВ: ДВ юж. — Еврейская АО, Приморский край. — Карта 72.
Общее распространение. Вост. и Юго-Вост. Азия.
ПРИМЕЧАНИЕ. Наиболее характерным признаком этого вида является наличие
многочисленных поверхностных апотециев, хотя образцы с преимущественно
маргинально-субмаргинальными апотециями тоже встречаются, и тогда для иден-
тификации могут быть использованы признаки нижней поверхности таллома —
псевдоцифеллы, расположенные на особых пробковидных выростах морщини-
стой нижней поверхности.
8. Nephromopsis pseudocomplicata (Asahina) M. J. Lai
1981 («1980»), Quart. J. Taiwan Mus. 33: 224. ≡ Cetraria pseudocomplicata Asahi-
na, 1936, J. Jap. Bot. 12: 804. ≡ Tuckneraria pseudocomplicata (Asahina) Randlane
et Saag, in Randlane et al., 1994, Acta Bot. Fenn. 150: 150. Т и п : [Япония] Japan,
Shikoku, Prov. Iyo, Mt. Ishizuchi, 26.8.1933, F. Fujikawa (DUKE лектотип, Lai, 1980:
224; TNS изолектотип). — Нефромопсис ложноскладчатый.
= Nephromopsis nipponensis (Asahina) M. J. Lai.
ЛИТЕРАТУРА: Randlane et al., 1994; Чабаненко, 2002; Красная книга Приморско-
го края..., 2008.
ИЛЛЮСТРАЦИИ: Yoshimura, 1979: pl. 19; Randlane et al., 1994: 145.
Флора лишайников России Флора лишайников России
142
ТАЛЛОМ листоватый, крупный; лопасти округлые, до 7 мм шир., с приподни-
мающимися краями, иногда несущие рассеянные краевые реснички; верхняя по-
верхность зеленовато-желтая, часто с коричневой полосой вдоль краевой зоны,
гладкая или слегка морщинистая; сердцевина белая; нижняя поверхность бело-
ватая, желтоватая, светло-коричневая или коричневая, также гладкая или слегка
морщинистая. Соредии и изидии отсутствуют. Псевдоцифеллы на нижней по-
верхности, немногочисленные, в виде небольших круглых белых пятен, иногда
окруженные светло-коричневой каймой. Ризины разбросанные, белые или светло-
коричневые, простые, длинные (3–4 мм). АПОТЕЦИИ краевые, округлые, до 6 мм
в диам., с коричневым диском; иногда отсутствуют. Сумки 30–35 × 12–14 мкм;
аскоспоры почти круглые, 5–6 × 4–5 мкм. ПИКНИДЫ многочисленные, на крае-
вых черных выступающих возвышениях; конидии гантелевидные, 5 × 1 мкм.
—
Рис. 106 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует; в отношении химиче-
ского состава сердцевины выделяются два хемотипа: (1) с алектороновой и кол-
латоловой кислотами и (2) с физодовой и конфизодовой кислотами; оба хемотипа
дополнительно содержат лихестериновую и протолихестериновую жирные кис-
лоты и дают одинаковые цветные реакции: от K, C и P сердцевина не изменяется,
от KC краснеет.
ЭКОЛОГИЯ. Эпифитный лишайник, растущий на коре хвойных деревьев (Abies,
Larix, Pinus, Tsuga) в горных лесах.
РАСПРОСТРАНЕНИЕ. Россия. Редкий вид, известный только на Дальнем Востоке.
ДВ: ДВ юж. — Амурская обл., Хабаровский край (юж. часть), Сахалинская обл.,
Приморский край. — Карта 73.
Общее распространение. Азия (Япония, Тайвань).
ПРИМЕЧАНИЕ. Характерные признаки Nephromopsis pseudocomplicata включают
довольно гладкий зеленовато-желтый таллом, часто с коричневой полосой вдоль
краевой зоны, красную реакцию сердцевины с KC и почти круглые аскоспоры.
9. Nephromopsis rugosa Asahina
1935, J. Jap. Bot. 11: 12. ≡ Cetraria rugosa (Asahina) M. Satô, in Nakai, Honda,
1939, Nov. Fl. Jap. 5: 46. Т и п : [Япония] Japan, Honshu, Prov. Musashi, Mt. Ko-
bushi, 02.07.1933, Y. Asahina (TNS голотип, DUKE изотип).
—
Нефромопсис
морщинистый.
ЛИТЕРАТУРА: Randlane, Saag, 1998; Чабаненко, 2002; Красная книга Приморского
края..., 2008.
ИЛЛЮСТРАЦИИ: Yoshimura, 1979: pl. 19.
ТАЛЛОМ листоватый, крупный, до 20 см в диам., с округлыми приподнимаю-
щимися лопастями до 2.5 см шир.; верхняя поверхность желтоватая или мато-
во-оливковая, часто с заметным зеленым оттенком, отчетливо правильно сетча-
Род Nephromopsis
143
тая; сердцевина белая; нижняя поверхность светло-коричневая, желтоватая или
беловатая, сильно сетчатая. Соредии и изидии отсутствуют. Псевдоцифеллы на
нижней поверхности в виде небольших белых пятен, расположены в основном
на гребнях ребер. Ризины разбросанные. АПОТЕЦИИ краевые, на нижней поверх-
ности таллома, округлые или почковидные, до 20 мм в диам., с коричневым об-
ращенным вверх диском. Сумки 35–40 × 10 мкм; аскоспоры продолговатые, 7–9 ×
3–5 мкм. ПИКНИДЫ многочисленные, краевые и поверхностные, на черных высту-
пающих возвышениях, которые часто расположены вдоль краев лопастей и ребер
на верхней поверхности; конидии гантелевидные, 5 × 1.0–1.5 мкм.
—
Рис. 107
(вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует; в отношении хими-
ческого состава сердцевины выделяются два хемотипа: (1) с оливеторовой кис-
лотой: сердцевина от C краснеет, от K и P не изменяется — и (2) с физодовой и
конфизодовой кислотами: сердцевина от K, C и P не изменяется, от KC краснеет;
оба хемотипа дополнительно содержат лихестериновую и протолихестериновую
жирные кислоты.
ЭКОЛОГИЯ. Эпифитный лишайник, в основном растущий на коре хвойных де-
ревьев (Abies, Larix, Picea) в горных лесах на небольшой высоте (до 1700 м над
ур. м.).
РАСПРОСТРАНЕНИЕ. Россия. Очень редкий вид, известный только на Дальнем
Востоке. ДВ: ДВ юж. — Приморский край. — Карта 74.
Общее распространение. Азия (Монголия, Япония).
ПРИМЕЧАНИЕ. Наиболее характерные признаки этого вида — отчетливо сетча-
тая верхняя и нижняя поверхность лопастей с характерным зеленоватым оттен-
ком в комбинации с красной реакцией сердцевины с C и KC.
10. Nephromopsis yunnanensis (Nyl.) Randlane et Saag
1992, Mycotaxon 44: 488. ≡ Platysma yunnanense Nyl., 1888, Lich. Nov. Zeland.:
150. ≡ Cetraria yunnanensis (Nyl.) Zahlbr., 1911 («1909»), Trudy Troitskos.-Kyak-
htinsk. Otd. Priamursk. Otd. Imp. Russk. Geogr. Obshch. 12: 89. Т и п : [Китай]
China, Yunnan, J. M. Delavay 1602 (H-NYL 36134 лектотип, Randlane et al., 1995:
39). — Нефромопсис юннаньский.
ЛИТЕРАТУРА: Randlane et al., 1995, 2001; Randlane, Saag, 1998.
ТАЛЛОМ листоватый, крупный, до 7 см в диам., с приподнимающимися лопа-
стями; верхняя поверхность светло-зеленая или желтая, сильно морщинистая и
сетчатая; сердцевина белая; нижняя поверхность белая до светло-коричневой,
сильно сетчатая с характерными выростами. Соредии и изидии отсутствуют.
Псевдоцифеллы многочисленные на нижней поверхности в виде небольших бе-
лых пятен, расположены на гребнях ребер и на особых выростах. Ризины разбро-
санные, светлые, простые. АПОТЕЦИИ краевые, округлые, до 13 мм в диам., с ко-
Флора лишайников России Флора лишайников России
144
ричневым диском. Сумки 40–45 × 12–14 мкм; аскоспоры продолговатые, 8–9 ×
4–5 мкм. ПИКНИДЫ чрезвычайно многочисленные, краевые и поверхностные (ино-
гда ламинально и на нижней поверхности), на выступающих возвышениях, ко-
торые бывают коричневатыми или чаще — цвета таллома и чернеют только на
верхушке, легко отламываются; конидии гантелевидные, 4 × 1 мкм.
—
Рис. 108
(вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует; сердцевина — лихе-
стериновую и протолихестериновую жирные кислоты, от K, C, KC и P не изменя-
ется, в UV не флюоресцирует.
ЭКОЛОГИЯ. Эпифитный лишайник, растущий на коре хвойных (Picea) и ли-
ственных (Quercus, Sorbus) деревьев в горных лесах на средних высотах (до
3200 м над ур. м.).
РАСПРОСТРАНЕНИЕ. Россия. Очень редкий вид, известный только на Дальнем
Востоке. ДВ: ДВ юж.
—
Еврейская АО (заповедник «Бастак» — Скирина, 2015:
57). — Карта 75.
Общее распространение. Азия (Китай).
ПРИМЕЧАНИЕ. Наиболее характерные признаки этого вида — обилие ламиналь-
ных пикнид, многочисленных и расположенных на светлых возвышениях, и при-
сутствие псевдоцифелл на складках и выростах очень морщинистой нижней по-
верхности таллома.
И. C. Жданов
Oropogon Th. Fr.
24 I 1861, Gen. Heterolich. Eur.: 49. ≡ Atestia Trevis., 28 I 1861, Flora 44: 49, nom.
illeg. superfl. Т и п : Oropogon loxensis (Fée) Th. Fr. [≡ Atestia loxensis (Fée) Trevis.,
nom. illeg. superfl.]. — Оропогон
Лихенизированные грибы. ТАЛЛОМ кустистый, бородовидный, дернинковид-
ный, распростертый по субстрату или повисающий, коричневый, сероватый, серо-
вато-черный до черного. Ветвление изотомическое дихотомическое, реже анизото-
мическое, редко с боковыми сегментами. Ветви большей частью цилиндрические,
реже слегка сплющенные, особенно в местах ветвления, более или менее гладкие,
изредка с соредиями или спинулами, без изидий, с псевдоцифеллами. Последние
представляют собой отверстие, открывающееся в центральную полость, реже они
плоские, закрытого типа. Коровой слой прозоплектенхимный. Сердцевина обыч-
но представляет собой слой, выстилающий изнутри центральную полость, реже
сердцевина плотная, без полости внутри. ПЛОДОВЫЕ ТЕЛА — апотеции, обычно при-
сутствуют, образуются латерально, иногда на окончаниях ветвей, до 5 мм в диам.
Род Oropogon
145
Диск первоначально вогнутый, впоследствии становится плоским или выпуклым.
Край ровный или кренулированный, иногда со спинулами. Гимений до 240 мкм
выс. Сумки 90–200 × 30–70 мкм, амилоидные. Аскоспоры муральные, темные, 74–
157 × 23–52 мкм, по одной в сумке. ПИКНИДЫ иногда присутствуют, погруженные
или слегка выступающие, конидии 4–7 × 0.5–1.0 мкм.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Основные лишайниковые вещества, характерные
для рода: алекториаловая, барбатовая, барбатоловая, диффрактаевая, каператовая,
норстиктовая, оливеторовая, плакодиоловая, протоцетраровая, псоромовая, сала-
циновая, физодаловая, фумарпротоцетраровая и эверновая кислоты, атранорин,
жирные кислоты.
ЭКОЛОГИЯ. Представители рода произрастают на коре деревьев, почве, камени-
стом субстрате, обычно на значительной высоте над уровнем моря.
РАСПРОСТРАНЕНИЕ. Род имеет отчетливое амфипацифическое распространение,
приурочен к горным районам Юж. и Вост. Азии, Центр. и Юж. Америки и Вест-
Индии.
ПРИМЕЧАНИЕ. Род насчитывает около 40 видов. Бóльшая часть видов известна в
Новом Свете; в азиатской части ареала насчитывается лишь восемь видов. Только
два вида являются общими для обеих частей ареала. Морфологически виды рода
Oropogon напоминают представителей рода Bryoria; их можно легко спутать. Ос-
новное отличие рода Oropogon — сумки с одной темной муральной аскоспорой
(у Bryoria аскоспоры одноклеточные, бесцветные, по 8 в сумке). В стерильном со-
стоянии большинство видов Oropogon отличается наличием отверстий, открываю-
щихся в центральную полость. На территории России известен один вид.
ИЗБРАННАЯ ЛИТЕРАТУРА: Satô, 1967; Esslinger, 1980, 1989, 2016; Chen, 1996; Wei
et al., 2017.
Oropogon asiaticus Asahina
in M. Satô, 1937, J. Jap. Bot. 13: 596. Т и п : [Япония] Japan, Honshu, Prov. Musashi,
Nippara prope Tokyo, 16.10.1922, Y. Asahina (TNS голотип). — Оропогон азиатский.
ИЗБРАННАЯ ЛИТЕРАТУРА: Satô, 1937; Asahina, 1952; Esslinger, 1989; Красная книга
Российской Федерации..., 2008.
ИЛЛЮСТРАЦИИ: Красная книга Российской Федерации..., 2008: 734.
ТАЛЛОМ кустистый, дернинковидный, до 10(13) см дл., из дихотомически раз-
ветвленных, расходящихся, спутанных между собой веточек, большей частью ко-
ричневый до темно-коричневого, веточки до 0.8 мм толщ., гладкие, более или ме-
нее блестящие. Соредии отсутствуют. Перфорации имеются, слегка удлиненные,
беловатые. Веточки на поперечном срезе округлые. Коровой слой до 130 мкм
толщ., наружная часть коричневая, тонкая, до 10 мкм толщ., внутренняя белая,
из плотно расположенных гиф, параллельных оси веточек. Сердцевина рыхлая,
содержит желтый пигмент, внутри с полостью. Апотеции блюдцевидные, около
2 мм в диам., приросшие к боковой поверхности веточек, со слегка городчатым
Род Oropogon
Флора лишайников России Флора лишайников России
146
краем и диском, окрашенным в каштановый цвет. Гимений бесцветный, около
200 мкм выс. Гипотеций бесцветный, около 80 мкм выс. Сумки субцилиндриче-
ские. Аскоспоры первоначально бесцветные, впоследствии буроватые, эллип-
соидные, прямые, 100–150 × 28–45 мкм. Парафизы нитевидные, разветвленные,
септированные, на вершине с незначительным утолщением или вовсе не утол-
щенные. — Рис. 109 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Содержит псоромовую, иногда также конпсоромо-
вую кислоты. Сердцевина от P становится золотисто-желтой, от K — слегка пур-
пурной, от C не изменяется, в UV не флюоресцирует.
ЭКОЛОГИЯ. Произрастает на коре деревьев.
РАСПРОСТРАНЕНИЕ. Россия. ДВ: ДВ юж.
—
Сахалинская обл. (южные острова
Курильской гряды — Итуруп, Кунашир и Шикотан). — Карта 76.
Общее распространение. Вост. Азия (Япония, Китай, Тайвань).
ПРИМЕЧАНИЕ. Наиболее существенный отличительный признак вида — содер-
жание псоромовой кислоты. Помимо Oropogon asiaticus, это вещество характерно
только для еще одного американского представителя рода — O. colibor Essl. При-
мечательно, что местонахождения O. asiaticus на Курильских о-вах являются са-
мыми северными для всего рода Oropogon.
Е. С. Кузнецова, И. С. Степанчикова
Platismatia W. L. Culb. et C. F. Culb.
1968, Contr. U.S. Natl. Herb. 34: 524.
Тип: Platismatia glauca (L.) W. L. Culb. et C. F. Culb. — Платизмация
Лихенизированные грибы. ТАЛЛОМ листоватый, средних размеров или круп-
ный, состоит из распростертых по субстрату слабо прикрепленных лопастей,
частично перекрывающихся и приподнимающихся над субстратом. Верхняя по-
верхность матовая или блестящая, пепельно-серая, серо-зеленая, желтовато-бе-
лая, желтовато-коричневая или зеленовато-коричневая; нижняя — блестящая,
кремово-белая, коричневая или черная, более светлая по краям, c немногочислен-
ными светлыми или черными ризинами. Лопасти или короткие, довольно широ-
кие, по краю рассеченные, или вытянутые, узкие, с гладкой или складчатой по-
верхностью, часто с рельефным сетчатым рисунком, образованным выпуклыми
ребрами. Псевдоцифеллы располагаются вдоль ребер или отсутствуют; у неко-
торых видов псевдоцифеллы формируются также на нижней поверхности. Сора-
лии и/или изидии хорошо развиты или отсутствуют, краевые или поверхностные,
часто развиваются по ребрам, иногда на месте псевдоцифелл. Верхний коровой
слой прозоплектенхимный. ПЛОДОВЫЕ ТЕЛА — апотеции зеоринового типа, сидя-
чие или слегка приподнятые, располагаются по краям лопастей или на верхней
Род Platismatia
147
поверхности у краев лопастей, до 4 см в диам., у большинства видов редки. Тал-
ломный край кренулированный, одного цвета с талломом, часто очень тонкий,
диск плоский до выпуклого, коричневый, иногда перфорированный. Сумки ле-
канорового типа, широкобулавовидные, содержат 8 аскоспор. Аскоспоры одно-
клеточные, эллипсоидные, бесцветные, (3.5)5.0–8.0(10) × 3–5 мкм. КОНИДИАЛЬНЫЕ
СПОРОНОШЕНИЯ в форме мелких погруженных или полупогруженных пикнид, рас-
полагающихся по краям лопастей. Конидии палочковидные, бутылковидные или
кеглевидные, 5–8 × 1.0–1.5 мкм. ФОТОБИОНТ Trebouxia-типа.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Атранорин, хлоратранорин, фумарпротоцетраровая
и каператовая кислоты. В качестве дополнительных веществ могут встречаться
хлоратранорин, джекиевая, паннаровая и псевдоплакодиоловая кислоты, редко —
неидентифицированный желтый пигмент.
ЭКОЛОГИЯ. Обитают на коре деревьев (преимущественно хвойных), изредка на
древесине, почве и скалах; в разнообразных лесных и тундровых сообществах в
горах и на равнине.
РАСПРОСТРАНЕНИЕ. Виды рода распространены преимущественно вокруг Ти-
хого океана, однако известно несколько видов, ареалы которых ограничены Сев.
Америкой или Вост. Азией.
ПРИМЕЧАНИЕ. Род включает 11 видов, из них на территории России достоверно
известны четыре. Platismatia herrei (Imshaug) W. L. Culb. et C. F. Culb. и P. erosa
W. L. Culb. et C. F. Culb. ранее ошибочно приводились для юга Дальнего Восто-
ка России (Скирина, 1995; Чабаненко, 2002; Скирин, Скирина, 2012; Яковченко
и др., 2013), все изученные образцы относятся к широко распространенному и
весьма морфологически изменчивому виду P. interrupta.
ИЗБРАННАЯ ЛИТЕРАТУРА: Culberson, Culberson, 1968; Obermayer, Randlane, 2012.
Ключ для определения видов рода Platismatia
1. Таллом состоит из широких лопастей и сильно разветвленных тонких выро-
стов, псевдоцифеллы отсутствуют .................................................. 1. P. glauca.
—
Таллом состоит только из широкоокруглых или лентовидных лопастей ..... 2.
2. Соредии и/или изидии присутствуют ................................................................ 3.
—
Соредии и изидии отсутствуют ......................................................................... 7.
3. Соредии присутствуют (часто в дополнение к изидиям) ............................... 4.
—
Соредии отсутствуют, образуются только изидии ........................................... 5.
4. Псевдоцифеллы на верхней поверхности лопастей округлые или неправиль-
ной формы, вытянутые или сливающиеся ............................... 2. P. interrupta.
—
Псевдоцифеллы на верхней поверхности отсутствуют ................ 1. P. glauca.
5. Верхняя поверхность с явно выраженным сетчатым рисунком из ребер .........
.........................................................................................................
4. P. norvegica.
—
Верхняя поверхность гладкая или со слабо выраженным сетчатым рисунком
................................................................................................................................. 6.
Род Platismatia
Флора лишайников России Флора лишайников России
148
6. Верхняя поверхность лопастей с псевдоцифеллами .............. 2. P. interrupta.
—
На верхней поверхности псевдоцифеллы отсутствуют ................ 1. P. glauca.
7. Верхняя поверхность покрыта рельефной сетью ребер и глубоких лакун,
сердцевина от P становится оранжево-красной ......................... 3. P. lacunosa.
—
Верхняя поверхность иная, сердцевина от P не изменяется ..............................
........................................................................................................
2. P. interrupta.
1. Platismatia glauca (L.) W. L. Culb. et C. F. Culb.
1968, Contr. U.S. Natl. Herb. 34 (7): 530. ≡ Lichen glaucus L., 1753, Sp. Pl. 2: 1148.
≡ Lobaria glauca (L.) Hoffm., 1796, Deutschl. Fl. 2: 149. ≡ Cetraria glauca (L.) Ach.,
1803, Methodus: 296. ≡ Physcia glauca (L.) DC., in Lam. et DC., 1805, Fl. Franç.,
ed. 3, 2: 401. ≡ Platysma glaucum (L.) Frege, 1812, Deutsch. Bot. Taschenbuch 2:
167. ≡ Parmelia glauca (L.) Hepp, 1824, Lich.-Fl. Würzburg: 23. Т и п : [Европа]
(LINN-HL 1273.139 лектотип I, Howe, 1912: 201; лектотип II, Jørgensen et al.,
1994: 377). — Платизмация сизая.
ИЗБРАННАЯ ЛИТЕРАТУРА: Kärnefelt, 1987; Thell, 2011b.
ИЛЛЮСТРАЦИИ: Brodo et al., 2001: 575; Holien, Tønsberg, 2006: 40; Randlane et
al., 2011: 234; Thell, 2011b: 164; Sharnoff, 2014: 113; Stenroos et al., 2016: 531.
ТАЛЛОМ листоватый, до 15 см в диам., неправильной формы, состоит из рас-
простертых по субстрату или приподнимающихся, восходящих, слабо прикре-
пленных, широкоокруглых или слегка вытянутых, по краю рассеченных лопа-
стей до 2 cм шир. Некоторые лопасти или части лопастей могут разрастаться в
обильно ветвящиеся коралловидные выросты. Верхняя поверхность лопастей
матовая или блестящая, голубовато-зеленая, серовато-зеленая или коричневатая,
гладкая до сетчато-морщинистой. Простые или коралловидные изидии, а также
зернистые соредии обычно развиваются каймой по краям лопастей, однако ино-
гда могут формироваться и на их верхней поверхности. Соралии светло-серые до
почти белых. Псевдоцифеллы отсутствуют. Нижняя поверхность блестящая, кре-
мово-белая или коричневая до черной, часто более светлая по краям, гладкая или
морщинистая, несет рассеянные, светлые до черных ризины. АПОТЕЦИИ образуют-
ся редко, краевые, с коричневым, иногда перфорированным диском, до 10 мм в
диам. Сумки и субгимений от I дают, соответственно, синее до сине-зеленого и
фиолетовое до ярко-пурпурного окрашивание. Аскоспоры эллипсоидные до яйце-
видных, 3.5–8.5 × 3–5 мкм. ПИКНИДЫ краевые, погруженные, очень редки. Кони-
дии бутылковидные, 5.6–7.8 мкм дл. — Рис. 110 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит атранорин, от K желтеет, от
KC, C и P не изменяется. Сердцевина содержит каператовую кислоту, от K, KC, C
и P не изменяется, от I становится фиолетовой, в UV не флюоресцирует.
ЭКОЛОГИЯ. Эпифитный вид, растет на стволах и ветвях хвойных деревьев и бе-
резы, реже на других лиственных деревьях; изредка произрастает на древесине, а
Род Platismatia
149
также на почве и камнях. Встречается в лесных сообществах различных типов, а
также в зональных и горных тундрах.
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: Аркт. евр.
—
Ненецкий АО; Аркт. зап.-
сиб. — Ямало-Ненецкий АО (сев. часть); Аркт. як. ЕР: Евр. Север — Мурманская
обл., Карелия, Архангельская обл., Ненецкий АО (юго-зап. часть), Коми, Ленин-
градская, Вологодская, Кировская обл. (сев.-зап. часть); Ср. Россия — Калинин-
градская, Псковская, Новгородская, Тверская, Ярославская, Костромская, Киров-
ская обл., Пермский край, Смоленская, Московская, Владимирская, Ивановская,
Нижегородская обл., Марий Эл, Чувашия, Татарстан, Калужская, Орловская,
Тульская, Рязанская, Пензенская, Ульяновская, Самарская обл. (сев. часть), Баш-
кирия, Курская, Белгородская обл.; Юж. Россия — Саратовская обл., Крым; Сев.
Кавказ — Краснодарский край, Адыгея, Карачаево-Черкесия; Сев. Урал — Коми,
Пермский край (сев. часть), Свердловская обл. (сев. часть); Юж. Урал — Перм-
ский край (юж. часть), Свердловская обл. (юж. часть), Башкирия, Челябинская
обл. СИБ: Зап. Сиб. сев. — Ханты-Мансийский АО, Свердловская, Томская обл.;
Зап. Сиб. юж. — Челябинская, Новосибирская обл., Алтайский край (зап. часть);
Юж. Сиб. — Кемеровская обл., Хакасия, Тыва, Бурятия, Красноярский край (юж.
часть), Алтайский край (вост. часть), Республика Алтай; Як. — юж. — Карта 77.
Общее распространение. Европа, Азия, Африка, Сев. и Юж. Америка, Ан-
тарктида и субантарктические острова.
ПРИМЕЧАНИЕ. Указания этого вида для юга российского Дальнего Востока
(Чабаненко, 2002; Чабаненко и др., 2002; Скирина, 2007, 2016) не подтвержде-
ны гербарными образцами. Все доступные образцы были переопределены как
P. interrupta (Kuznetsova et al., 2021).
В 2011 г. из Platismatia glauca был выделен вид P. wheeleri Goward, Altermann
et Björk (Lumbsch et al., 2011), отличающийся сильно волнистыми краевыми со-
ралиями, прорывающимися из сердцевины, и отсутствием изидий. В настоящее
время вид известен из Сев. Америки и Словакии, может быть обнаружен в России.
2. Platismatia interrupta W. L. Culb. et C. F. Culb.
1968, Contr. U.S. Natl. Herb. 34 (7): 539. Т и п : [Япония] Japan, Prov. Shinano
(Pref. Nagano), Mt. Norikura, 06.08.1952, Y. Asahina (TNS голотип). — Платизма-
ция прерывистая.
ИЗБРАННАЯ ЛИТЕРАТУРА: Culberson, Culberson, 1968.
ТАЛЛОМ листоватый, до 12 см в диам., розетковидный или неправильной фор-
мы, состоит из распростертых по субстрату, слабо прикрепленных, восходящих,
широкоокруглых лопастей до 2 см шир. Верхняя поверхность лопастей серая,
светло-коричневая по краям, гладкая до слабо морщинистой, с прерывистым
сетчатым узором. Псевдоцифеллы крупные, округлые или неправильной фор-
мы, вытянутые или сливающиеся, у некоторых образцов грубо соредиозные или
изидиозные. Изидии простые, палочковидные или округлые, распадающиеся на
Флора лишайников России Флора лишайников России
150
соредии, развиваются на поверхности лопастей. Нижняя поверхность угольно-
черная в центре и более светлая — орехово-коричневая до белой — по краям, с
редкими черными ризинами. АПОТЕЦИИ не обнаружены. ПИКНИДЫ неизвестны.
—
Рис. 111, 112 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит атранорин, от K желтеет, от
KC, C и P не изменяется, от I становится светло-фиолетовым или не изменяется,
в UV не флюоресцирует. Сердцевина содержит каператовую кислоту и неиденти-
фицированный желтый пигмент, от K, KC, C и P не изменяется, в UV не флюорес-
цирует.
ЭКОЛОГИЯ. На коре и древесине деревьев разных пород, редко на почве. Встре-
чается в лесных сообществах различных типов.
РАСПРОСТРАНЕНИЕ. Россия. СИБ: Юж. Сиб.
—
Бурятия, Иркутская обл. (юж.
часть). ДВ: ДВ сев. — Хабаровский край (сев. часть); ДВ юж. — Амурская обл.,
Хабаровский край (юж. часть), Еврейская АО, Сахалинская обл. (Сахалин, Ку-
рильские о-ва), Приморский край. — Карта 78.
Общее распространение. Азия (Россия, Япония).
3. Platismatia lacunosa (Ach.) W. L. Culb. et C. F. Culb.
1968, Contr. U.S. Natl. Herb. 34 (7): 541. ≡ Cetraria lacunosa Ach., 1803, Methodus:
295. ≡ Platysma lacunosum (Ach.) Nyl., 1855, Bot. Not.: 137. Т и п : [Сев. Америка]
North America (S L2515 неотип, Culberson, Culberson, 1968: 541).
—
Платизма-
ция ямчатая.
= Cetraria lacunosa f. cavernosa G. Merr.
ИЗБРАННАЯ ЛИТЕРАТУРА: Culberson, Culberson, 1968; Kuznetsova et al., 2021.
ИЛЛЮСТРАЦИИ: Brodo et al., 2001: 576; Kuznetsova et al., 2021: 186.
ТАЛЛОМ листоватый, до 16 см в диам., розетковидный или неправильной фор-
мы, состоит из распростертых по субстрату, слабо прикрепленных, широкоокру-
глых лопастей до 1.5 см шир. Верхняя поверхность лопастей серовато-белая, зеле-
новато-серая, серая, светло-коричневая по краям, с хорошо выраженным сетчатым
узором из приподнимающихся ребер и глубоких лакун. Псевдоцифеллы иногда
присутствуют на гребнях ребер, однако плохо заметны на общем светлом фоне
таллома. Соредии и изидии отсутствуют. Нижняя поверхность угольно-черная в
центре и более светлая — орехово-коричневая до белой — по краям, местами с
сетчатым узором, с редкими или многочисленными черными ризинами. АПОТЕЦИИ
редки, неправильной формы, располагаются на верхней поверхности ближе к кра-
ям лопастей, диск коричневый, без перфораций, до 2 см в диам. Аскоспоры эллип-
соидные, 7–10 × 3.0–4.5 мм. ПИКНИДЫ краевые или поверхностные, располагаются
на ребрах. Конидии палочковидные, 1 × 5–7 мкм. — Рис. 113 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит атранорин, от K желтеет, от
KC, C и P не изменяется, в UV не флюоресцирует. Сердцевина содержит капе-
Род Platismatia
151
ратовую и фумарпротоцетраровую кислоты, от K, KC, C и I не изменяется, от P
окрашивается в оранжево-красный цвет, в UV не флюоресцирует.
ЭКОЛОГИЯ. На камнях в лишайниковых и кустарничковых тундрах. На западе
Сев. Америки вид встречается на коре, древесине, скалах и мхах в прибрежных
лесах.
РАСПРОСТРАНЕНИЕ. Россия. ДВ: ДВ сев.
—
Камчатский край (Командорские
о-ва). — Карта 79.
Общее распространение. Азия (Россия, Командорские о-ва), Сев. Америка
(тихоокеанское побережье, Алеутские о-ва).
ПРИМЕЧАНИЕ. От всех прочих видов рода отличается положительной реакцией
сердцевины с P (фумарпротоцетраровая кислота). Отличается от внешне наибо-
лее сходного вида Platismatia norvegica отсутствием изидий. До 1926 г. (Savicz,
1926) этот вид ошибочно указывали для различных регионов Сев. Евразии, путая
его с Asahinea chrysantha (≡ Cetraria chrysantha). Указания Platismatia lacunosa
для Новой Земли, Земли Франца-Иосифа и п-ова Таймыр (Kristinsson et al., 2010)
являются ошибочными и относятся к Asahinea chrysantha (образцы в LE).
4. Platismatia norvegica (Lynge) W. L. Culb. et C. F. Culb.
1968, Contr. U.S. Natl. Herb. 34 (7): 543. ≡ Cetraria lacunosa f. norvegica Lynge,
1921, Skr. Vidensk.-Selsk. Kristiania 1921, Math.-Naturvidensk. Kl. 1, 7: 196 (Stud.
Lich. Fl. Norway). ≡ Cetraria norvegica (Lynge) Du Rietz, 1924, Bot. Not.: 59. Т и п :
[Норвегия] Norway, Hordaland, Store Kalsø, 1920, B. Lynge (O лектотип, Culberson,
Culberson, 1968: 543). — Платизмация норвежская.
= Cetraria norvegica f. sorediata Degel. ≡ Cetraria tuckermanii Oakes var. sore-
diata (Degel.) Räsänen.
ИЗБРАННАЯ ЛИТЕРАТУРА: Thell, 2011b.
ИЛЛЮСТРАЦИИ: Brodo et al., 2001: 577; Holien, Tønsberg, 2006: 40; Thell, 2011b:
165; Stenroos et al., 2016: 531.
ТАЛЛОМ листоватый, до 23 см в диам., розетковидный или неправильной фор-
мы, состоит из распростертых по субстрату, слабо прикрепленных, восходящих,
широкоокруглых, рассеченных по краям лопастей до 2.5 см шир. Верхняя по-
верхность лопастей серая, желтовато-серая, зеленовато-серая, изредка слегка ко-
ричневатая по краю, с ясно выраженным рельефным сетчато-ямчатым рисунком.
Псевдоцифеллы мелкие, неправильной формы, приурочены преимущественно к
гребням ребер. Изидии располагаются как на верхней поверхности лопастей (на
ребрах), так и по их краям, тонкие, палочковидные или коралловидные, обычно
ассоциированы с псевдоцифеллами. Нижняя поверхность коричневая до черной,
по краям более светлая — коричневая, цвета верхней поверхности или почти бе-
лая. Ризины немногочисленные, черные. АПОТЕЦИИ очень редки, неправильной
формы, располагаются у краев лопастей, с коричневым диском, перфорирован-
Флора лишайников России Флора лишайников России
152
ные, до 1.5 см шир. Аскоспоры эллипсоидные, 6.0–8.5 × 3–4 мкм. ПИКНИДЫ редки,
краевые, погруженные. — Рис. 114 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит атранорин, от K желтеет, от
KC, C и P не изменяется, в UV не флюоресцирует. Сердцевина содержит капера-
товую кислоту, от K, KC, C и P не изменяется, от I становится фиолетовой или не
изменяет окраску, в UV не флюоресцирует.
ЭКОЛОГИЯ. Океанический вид, обитает на скалах и на еловой коре в хвойных и
смешанных лесах, во влажных местообитаниях.
РАСПРОСТРАНЕНИЕ. Россия. ЕР: Евр. Север — Архангельская обл. — Карта 80.
Общее распространение. Европа, Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Указания этого вида для юга российского Дальнего Востока
(Толпышева, 1990; Чабаненко, 2002; Чабаненко и др., 2002; Скирина, 2007, 2016)
не подтверждены гербарными образцами. Все доступные образцы были пере-
определены как Platismatia interrupta (Kuznetsova et al., 2021).
Platismatia norvegica может быть спутан с P. glauca, от которого отличается
ясно выраженным рельефным сетчато-ямчатым рисунком на верхней поверхно-
сти лопастей, более широкими лопастями, наличием псевдоцифелл и поверхност-
ных изидий (соредии не формируются). У P. glauca вегетативные пропагулы при-
урочены преимущественно к краям лопастей.
Е. С. Кузнецова
Tuckermannopsis Gyeln.
1933, Acta Fauna Fl. Universali, ser. 2, Bot. 1 (5–6): 6. Т и п : Tuckermannopsis cilia-
ris (Ach.) Gyeln. — Тукерманнопсис
Лихенизированные грибы. ТАЛЛОМ листоватый, средних размеров или круп-
ный, состоит из приподнимающихся над субстратом лопастей, слабо прикре-
пленных в центральной части. Лопасти вытянутые, узкие, слабо каналовидные,
изредка разветвленные, с ровными или волнистыми краями, у некоторых видов
несущими реснички. Верхняя поверхность лопастей зеленовато-коричневая, сла-
бо морщинистая, без псевдоцифелл или с редкими белыми псевдоцифеллами.
Нижняя поверхность от светло-коричневой до черной, с мелкими белыми псев-
доцифеллами и немногочисленными ризинами. Соредии формируются или от-
сутствуют, изидии не формируются. Верхний и нижний коровые слои параплек-
тенхимные. ПЛОДОВЫЕ ТЕЛА — апотеции леканоринового типа, диск апотециев
коричневый, блестящий. Сумки Lecanora-типа, узкобулавовидные, с небольшим
толусом и аксиальным телом 3–5 мкм выс., содержат 8 аскоспор. Аскоспоры од-
ноклеточные, округлые, 3.5–6.0 мкм в диам. КОНИДИАЛЬНЫЕ СПОРОНОШЕНИЯ в форме
коричневых или черных пикнид, располагающихся на поверхности или по краям
лопастей. Конидии гантелевидные или палочковидные. ФОТОБИОНТ —Trebouxia.
Род Tuckermannopsis
153
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой может содержать атранорин, сердце-
вина — алектороновую, оливеторовую, физодовую и α-коллатоловую кислоты,
орсинол-депсиды и депсидоны.
ЭКОЛОГИЯ. Виды рода поселяются преимущественно на стволах и ветвях хвой-
ных пород и берез, изредка на скалах. В разнообразных лесных и тундровых со-
обществах в горах и на равнине.
РАСПРОСТРАНЕНИЕ. Виды рода распространены в Европе, Азии, Африке, Сев. и
Юж. Америке, Австралии.
ПРИМЕЧАНИЕ. Включает семь видов, шесть из которых встречаются на тер-
ритории России. До недавнего времени в этот род включали Cetraria sepincola
и Masonhalea inermis. В 2017 г. коллектив авторов (Divakar et al., 2017) предло-
жил отнести род Tuckermannopsis к роду Nephromopsis, однако в международную
практику это новшество к настоящему времени не вошло.
ИЗБРАННАЯ ЛИТЕРАТУРА: Kärnefelt, Thell, 2001; Saag et al., 2002; Divakar et al.,
2017.
Ключ для определения видов рода Tuckermannopsis
1. Соралии краевые .................................................................................................. 2.
—
Соралии отсутствуют .......................................................................................... 3.
2. Верхняя поверхность от светло-коричневой до коричневой, матовая, от KC не
изменяется ............................................................................... 1. T. chlorophylla.
—
Верхняя поверхность темно-коричневая, местами блестящая, от KC розовеет
..................................................................................................................
3. T. gilva.
3. Лопасти вытянутые, лентовидные, с вытянутыми псевдоцифеллами вдоль
краев .............................................................................................. 5. T. subalpina.
—
Лопасти более или менее округлые, псевдоцифеллы не развиваются или ко-
роткие, не вытянутые .......................................................................................... 4.
4. От KC сердцевина не изменяется; содержит протолихестериновую кислоту ..
............................................................................................................... 4. T. orbata.
—
От KC сердцевина розовеет или краснеет; не содержит протолихестериновой
кислоты .............................................................................................. 2. T. ciliaris.
1. Tuckermannopsis chlorophylla (Willd.) Hale
in Egan, 1987, Bryologist 90 (2): 164. ≡ Lichen chlorophyllus Willd., in Humb., 1793,
Fl. Friberg.: 20. ≡ Platysma chlorophyllum (Willd.) Vain., 1881, Meddeland. Soc.
Fauna Fl. Fenn. 6: 121 ≡ Cetraria chlorophylla (Willd.) Vain., 1896, Acta Soc. Fauna
Fl. Fenn. 13: 7. Т и п : [Дания] Denmark, Jylland, Vendsyssel, W of Råbjerg Mile,
1994, S. C. Christensen 11056 in Hansen, Lich. Danici Exs. no 34 (C неотип, Ahti,
Thell, 2011: 140). — Тукерманнопсис хлорофилловый.
= Cetraria scutata auct.; = Cetraria ulophylla (Ach.) Rebent.
ИЗБРАННАЯ ЛИТЕРАТУРА: Kärnefelt, 1987; Ahti, Thell, 2011.
Род Tuckermannopsis
Флора лишайников России Флора лишайников России
154
ИЛЛЮСТРАЦИИ: Wirth, 1995: 269; Brodo et al., 2001: 693; Holien, Tønsberg, 2006:
36; Ahti, Thell, 2011: 166; Randlane et al., 2011: 260; Sharnoff, 2014: 123; Stenroos et
al., 2016: 690.
ТАЛЛОМ листоватый, до 6 см в диам., неправильной формы, состоит из слабо при-
крепленных в центральной части, приподнимающихся или повисающих лопастей
до 1 см шир. Верхняя поверхность лопастей оливковая до коричневой, гладкая или
слегка морщинистая, обычно матовая. Нижняя поверхность оливковая до коричне-
вой, несколько светлее верхней, темнеющая к краям, с немногочисленными свет-
лыми ризинами. Псевдоцифеллы отсутствуют. Серовато-белые мучнистые соредии
формируют кайму по краям лопастей, среди них изредка присутствуют немногочис-
ленные краевые изидии. АПОТЕЦИИ очень редки, располагаются на краях лопастей,
с коричневым диском и соредиозным краем, до 4 мм в диам. Сумки узкобулавовид-
ные, 30–40 × 7–10 мкм. Аскоспоры округлые, 4–7 мкм в диам. ПИКНИДЫ краевые,
погруженные. Конидии гантелевидные, 5–6 × 1.5 мкм. — Рис. 115 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой от K, KC, C и P не изменяется, в UV
не флюоресцирует. Сердцевина содержит протолихестериновую и рангиформо-
вую кислоты, от K, KC, C и P не изменяется, в UV не флюоресцирует.
ЭКОЛОГИЯ. На коре ветвей или стволов хвойных и лиственных деревьев, изред-
ка на древесине и скалах.
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: Аркт. евр. —
Ненецкий АО, Коми (крайняя
сев.-вост. часть); Аркт. зап.-сиб.
—
Ямало-Ненецкий АО (сев. часть). ЕР: Евр.
Север — Мурманская обл., Карелия, Архангельская обл., Коми, Ленинградская,
Вологодская, Кировская обл. (сев.-зап. часть); Ср. Россия — Калининградская,
Псковская, Новгородская, Тверская, Ярославская, Костромская, Кировская обл.,
Пермский край, Смоленская, Московская, Владимирская, Ивановская, Нижего-
родская обл., Марий Эл, Чувашия, Татарстан, Калужская, Орловская, Тульская,
Рязанская обл., Мордовия, Пензенская, Ульяновская, Самарская обл. (сев. часть),
Башкирия, Курская, Белгородская, Воронежская обл.; Юж. Россия — Саратов-
ская, Самарская, Оренбургская, Волгоградская обл., Крым; Сев. Урал.
—
Коми,
Пермский край (сев. часть), Свердловская обл. (сев. часть); Юж. Урал — Перм-
ский край (юж. часть), Свердловская обл. (юж. часть), Башкирия, Челябинская
обл. Сев. Кавказ — Краснодарский край, Адыгея. СИБ: Зап. Сиб. сев.
—
Яма-
ло-Ненецкий АО (юж. часть), Ханты-Мансийский АО, Свердловская, Томская
обл.; Зап. Сиб. юж. — Челябинская, Курганская, Новосибирская обл.; Ср. Сиб. —
Красноярский край (сев. часть), Эвенкия; Юж. Сиб. — Кемеровская обл., Респу-
блика Алтай, Хакасия, Тыва, Бурятия, Красноярский край (юж. часть), Иркутская
обл. (юж. часть); Як.
—
центр. и юж. ДВ: ДВ сев.
—
Камчатский край (бывшая
Камчатская обл.); ДВ юж.
—
Хабаровский край (юж. часть), Амурская обл., Ев-
рейская АО, Приморский край. — Карта 81.
Общее распространение. Европа, Азия, Юж. Африка (ЮАР), Сев. и Юж.
Америка, Австралия, Новая Зеландия.
Род Tuckermannopsis
155
2. Tuckermannopsis ciliaris (Ach.) Gyeln.
1933, Acta Fauna Fl. Universali, ser. 2, Bot. 1 (5–6): 6. ≡ Cetraria ciliaris Ach., 1810,
Lichenogr. Universalis: 508. ≡ Nephromopsis ciliaris (Ach.) Hue, 1899, Nouv. Arch.
Mus. Hist. Nat., sér. 4, 1: 216. Т и п : [США] USA, America borealis, G. H. E. Müh-
lenberg (H-ACH 1511 = H9502237 лектотип, Culberson, Culberson, 1967: 159,
«holotype»). — Тукерманнопсис реснитчатый.
= Cetraria halei W. L. Culb. et C. F. Culb. ≡ Cetraria ciliaris var. halei
(W. L. Culb. et C. F. Culb.) Ahti ≡ Tuckermannopsis halei (W. L. Culb. et C. F. Culb.)
M. J. Lai; = Tuckermannopsis americana (Spreng.) Hale.
ИЗБРАННАЯ ЛИТЕРАТУРА: Randlane, Saag, 1992; Ahti, Thell, 2011.
ИЛЛЮСТРАЦИИ: Brodo et al., 2001: 692; Hinds, Hinds, 2007: 475; Ahti, Thell, 2011:
166; Stenroos et al., 2016: 691.
ТАЛЛОМ листоватый, до 5–8 см в диам., розетковидный или неправильной
формы, состоит из распростертых по субстрату или приподнимающихся, слабо
прикрепленных в центральной части, широкоокруглых, рассеченных лопастей до
4–6 мм шир., с длинными ресничками, рассеянными по краям. Верхняя поверх-
ность лопастей от светло- до темно-коричневой, более или менее морщинистая.
Нижняя поверхность коричневая, несколько светлее верхней, темнеющая к краям,
с немногочисленными ризинами. Псевдоцифеллы, соредии и изидии отсутству-
ют. Верхний коровой слой 12–50 мкм толщ. АПОТЕЦИИ краевые или концевые, с
коричневым диском и тонким бородавчатым талломным краем, до 12 мм в диам.
Сумки узкобулавовидные. Аскоспоры округлые, 5–7 × 4–6 мкм в диам. ПИКНИДЫ
краевые, иногда поверхностные, бочонковидные, черные. Конидии гантелевид-
ные, 5 × 1 мкм. — Рис. 116, 117 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит атранорин, от K желтеет, от
C, KC и P не изменяется, в UV не флюоресцирует. Сердцевина содержит оливето-
ровую и физодовую кислоты, от C и KC розовеет или краснеет, от K и P не изме-
няется, в UV не флюоресцирует.
ЭКОЛОГИЯ. Преимущественно на коре деревьев хвойных пород и берез, редко
на древесине.
РАСПРОСТРАНЕНИЕ. Россия. ЕР: Евр. Север — Мурманская обл., Карелия, Ар-
хангельская обл., Коми, Ленинградская обл.; Ср. Россия — Новгородская, Ко-
стромская обл., Пермский край (Коми-Пермяцкий округ). СИБ: Зап. Сиб. сев. —
Ямало-Ненецкий АО (юж. часть), Ханты-Мансийский АО, Свердловская обл.; Ср.
Сиб. — Красноярский край (сев. часть), Эвенкия; Юж. Сиб. — Кемеровская обл.,
Республика Алтай, Хакасия, Тыва, Бурятия, Красноярский край (юж. часть), Ир-
кутская обл. (юж. часть), Забайкальский край. Як. — центр., вост. и юж. ДВ: ДВ
сев.
—
Чукотский АО (юж. часть), Магаданская обл., Камчатский край (Коряк-
ский округ и бывшая Камчатская обл.); ДВ юж. — Амурская обл., Еврейская АО,
Сахалинская обл. (Курильские о-ва), Приморский край. — Карта 82.
Флора лишайников России Флора лишайников России
156
Общее распространение. Европа, Азия, Сев. Америка (США), Юж. Америка
(Аргентина).
ПРИМЕЧАНИЕ. Некоторые авторы рассматривают хемотип, содержащий алекто-
роновую и α-коллатоловую кислоты вместо оливеторовой и физодовой кислот, в
качестве самостоятельного вида Tuckermannopsis americana.
3. Tuckermannopsis gilva (Asahina) M. J. Lai
1980, Quart. J. Taiwan Mus. 33: 225. ≡ Cetraria gilva Asahina, 1953, J. Jap. Bot.
28: 139. Т и п : [Япония] Japan, Honshu, Prov. Shinano, Mt. Norikura, 06.08.1952,
Y. Asahina (TNS голотип). — Тукерманнопсис зубчатый.
ИЗБРАННАЯ ЛИТЕРАТУРА: Lai, 1980.
ТАЛЛОМ листоватый, состоит из слабо прикрепленных к субстрату, приподни-
мающихся, неправильно разветвленных лопастей до 5 мм шир. Верхняя поверх-
ность темно-коричневая или оливковая, местами блестящая. Псевдоцифеллы и
краевые реснички отсутствуют. Серовато-белые мучнистые соредии формируют
кайму по краям лопастей. Нижняя поверхность светло-коричневая, с рассеянны-
ми редкими светлыми ризинами. АПОТЕЦИИ не обнаружены. ПИКНИДЫ неизвест-
ны. — Рис. 118 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Содержит алектороновую кислоту. Коровой слой от
KC окрашивается в розовый цвет, кора и сердцевина от K, C и P не изменяются, в
UV не флюоресцируют.
ЭКОЛОГИЯ. На тонких ветвях березы.
РАСПРОСТРАНЕНИЕ. Россия. СИБ: Юж. Сиб. — Иркутская обл. (юж. часть). ДВ:
ДВ юж. — Амурская обл., Приморский край. — Карта 83.
Общее распространение. Азия (Япония, Северная Корея).
ПРИМЕЧАНИЕ. Вид близок к Тuckermannopsis chlorophylla и T. ulophylloides
(Asahina) M. J. Lai (неизвестен в России), от первого отличается розовой реакци-
ей коры с KC, а от второго — отсутствием ресничек.
4. Tuckermannopsis orbata (Nyl.) M. J. Lai
1981, Quart. J. Taiwan Mus. 33: 226. ≡ Platysma orbatum Nyl., 1869, Flora 52: 442.
Т и п : [США, Калифорния] U.S.A., California, Oakland Hills, H. N. Bolander
(H-NYL 36066 = H9505046 голотип). — Тукерманнопсис лишенный.
ИЗБРАННАЯ ЛИТЕРАТУРА: Степанчикова, Гимельбрант, 2011.
ИЛЛЮСТРАЦИИ: Brodo et al., 2001: 694–695; Sharnoff, 2014: 123.
ТАЛЛОМ листоватый, довольно мелкий, неправильной формы, состоит из рас-
простертых по субстрату или приподнимающихся, слабо прикрепленных, ши-
рокоокруглых или неправильных, рассеченных, слегка вогнутых лопастей, кре-
нулированных по краю, до 1 см дл. и 6 мм шир. Верхняя поверхность лопастей
светло-серо-коричневая, оливково-коричневая, коричневая до почти черной, бле-
Род Tuckermannopsis
157
стящая по краям, морщинистая. Псевдоцифеллы мелкие, расположены по краю
лопастей и по краю апотециев, матовые, сероватые, очень плохо заметные. Со-
редии и изидии отсутствуют, но иногда разветвленные филлидии на краях лопа-
стей могут напоминать изидии. Нижняя поверхность светло-коричневая, сильно
морщинистая. Ризины немногочисленные, светлые. Верхний и нижний коровые
слои параплектенхимные. АПОТЕЦИИ очень обычны и многочисленны, с краем и
диском цвета таллома, располагаются на поверхности и по краям лопастей, до
4 мм в диам. Сумки узкобулавовидные, 30–40 × 8–10 мкм, парафизы слабо раз-
ветвленные, с расширенными концами. Аскоспоры округлые, 3.5–6.0 мкм в диам.
ПИКНИДЫ многочисленные, сидячие или слегка погруженные основанием, крае-
вые или поверхностные, бочонковидные, черные. Конидии гантелевидные, 5–6 ×
1–2 мкм. — Рис. 119 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Сердцевина содержит протолихестериновую кисло-
ту. Кора и сердцевина от K, KC, C и P не изменяются, в UV не флюоресцируют.
ЭКОЛОГИЯ. На коре хвойных пород и берез, изредка на сухостое.
РАСПРОСТРАНЕНИЕ. Россия. ДВ: ДВ сев.
—
Камчатский край (бывшая Камчат-
ская обл.). — Карта 84.
Общее распространение. Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Вид со стабильным составом вторичных метаболитов, но в
остальном весьма полиморфный. Например, образцы, лишенные филлидий, мо-
гут быть ошибочно определены как Tuckermannopsis ciliaris / T. americana. Не-
которые образцы могут напоминать Cetraria sepincola, который отличается мень-
шим количеством пикнид, обычно погруженных или слегка выступающих над
поверхностью и расположенных только по краю лопастей, почти плоскими восхо-
дящими лопастями, более контрастными псевдоцифеллами, а также более обиль-
ным развитием апотециев. По составу лишайниковых веществ эти два вида не
различаются.
5. Tuckermannopsis subalpina (Imshaug) Kärnefelt
1993, Bryologist 96 (3): 403. ≡ Cetraria subalpina Imshaug, 1950, Mycologia 42: 746.
Т и п : [США, Вашингтон] U.S.A., Washington, Pierce Co., Mt. Rainier National
Park, Mt. Wow, 6000 ft, 21.08.1948, H. A. Imshaug 1876 (MICH00062631 голотип,
H9500223, MSC, US изотипы). — Тукерманнопсис субальпийский.
= Cetraria arborialis (G. Merr.) G. E. Howard.
ИЗБРАННАЯ ЛИТЕРАТУРА: Kärnefelt, 1979.
ТАЛЛОМ листоватый, неправильной формы, состоит из восходящих, слабо
прикрепленных к субстрату, лентовидных, дихотомически разветвленных, пере-
путанных желобчатых лопастей 150–220 мкм толщ. и до 6.5 мм шир., с редкими
ресничками по краям (реснички могут отсутствовать). Верхняя поверхность ло-
пастей блестящая, желтовато-, красновато- или зеленовато-коричневая, гладкая,
нижняя — более светлая, коричневая, коричнево-желтая или оливковая, без ри-
Флора лишайников России Флора лишайников России
158
зин. Псевдоцифеллы плохо заметные, расположены на верхней поверхности или
вытянуты в тонкую линию вдоль краев лопастей. Соредии и изидии отсутствуют.
АПОТЕЦИИ обычны, формируются на верхней поверхности по краям лопастей, с
зубчатым краем, исчезающим с возрастом, оливковые или светло-коричневые, до
8 мм в диам. Гимений 50–80 мкм выс., бесцветный, желтоватый в верхней части,
субгимений 68–82 мкм выс., бесцветный, сумки 30–54 × 8.0–12.7 мкм, цилин-
дрические или узкобулавовидные, парафизы простые, неветвящиеся. Аскоспоры
округлые, 3.5–6.4 мкм в диам. ПИКНИДЫ формируются по краям лопастей. Кони-
дии палочковидные, 7–10 × 1.0–1.3 мкм. — Рис. 120 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой от K, KC, C, I и P не изменяется, в UV
не флюоресцирует. Сердцевина содержит протолихестериновую и лихестерино-
вую кислоты, от K, KC, C, I и P не изменяется, в UV не флюоресцирует.
ЭКОЛОГИЯ. На коре тонких веточек хвойных пород, а также на почве среди
мхов.
РАСПРОСТРАНЕНИЕ. Россия. АРКТ: Аркт. чук. — Чукотский АО (сев. часть). —
Карта 85.
Общее распространение. Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Не полиморфный, морфологически четко выраженный вид.
Т. Рандлане, А. Сааг
Usnocetraria M. J. Lai et J. C. Wei
in Lai et al., 2007, J. Natl. Taiwan Mus. 60 (1): 57–58.
Тип: Usnocetraria oakesiana (Tuck.) M. J. Lai et J. C. Wei. — Усноцетрария
Лихенизированные грибы. ОПИСАНИЕ — см. Usnocetraria oakesiana.
ПРИМЕЧАНИЕ. Недавно описанный род Usnocetraria первоначально включал 11
видов (Lai et al., 2007); девять из них были опубликованы недействительно, по-
скольку базионимы не цитировались (Thell et al., 2009). Систематическое поло-
жение еще одного вида, Nephromopsis kurokawae (Shibuichi et K. Yoshida) Kurok.,
внутри рода Usnocetraria не подтверждается данными филогенетического анали-
за. Таким образом, род можно считать монотипным, содержащим только один вид
U. oakesiana (Thell et al., 2009; Thell, 2011c). Последний не является родственным
ни одному из первоначально (недействительно) включенных в род Usnocetraria
видов. В другой новейшей публикации (Divakar et al., 2017), в которой на осно-
ве современных филогенетических методов пересмотрены границы родов сем.
Parmeliaceae, предлагается разделение цетрариоидных лишайников лишь на три
крупных рода (Cetraria, Melanelia и Nephromopsis). Такая концепция не признает-
ся авторами настоящей публикации.
ИЗБРАННАЯ ЛИТЕРАТУРА: Lai et al., 2007; Thell et al., 2009; Thell, 2011c.; Divakar et
al., 2017.
Роды Usnocetraria,
159
Usnocetraria oakesiana (Tuck.) M. J. Lai et J. C. Wei
in Lai et al., 2007, J. Natl. Taiwan Mus. 60 (1): 58. ≡ Cetraria oakesiana Tuck., 1841,
Boston J. Nat. Hist. 3: 445. ≡ Tuckermannopsis oakesiana Hale, in Egan, 1987, Bryo-
logist 90 (2): 164. ≡ Allocetraria oakesiana (Tuck.) Randlane et A. Thell, in Thell et
al., 1995, Flechten Follmann: 363. Т и п : [США] USA, New Hampshire, White Mts,
1839, W. Oakes (FH лектотип, Thell et al., 1995: 363). — Усноцетрария Оукса.
ЛИТЕРАТУРА: Thell et al., 1995; Thell, 2011c.
ИЛЛЮСТРАЦИИ: Brodo et al., 2001: 157; Thell, 2011c: 172.
ТАЛЛОМ листоватый, дорсивентральный, прижатый; верхняя поверхность
желтовато-зеленая; сердцевина белая до светло-желтой в нижних частях; нижняя
поверхность светло-коричневая до беловатой, с разбросанными светло-коричне-
выми ризинами; лопасти лентовидные, редко разветвленные, слегка вогнутые и
слабо морщинистые, 1–4 мм шир. Псевдоцифеллы и изидии отсутствуют; сора-
лии краевые, многочисленные, светло-желтые, с зернистыми соредиями. Верх-
ний и нижний коровые слои параплектенхимные, иногда ориентированы анти-
клинально. ПЛОДОВЫЕ ТЕЛА — апотеции зеоринового типа, немногочисленные,
субмаргинальные, до 7 мм в диам., со светло-коричневым диском и тонким тал-
ломным краем, который может быть соредиозным. Сумки узкобулавовидные, со-
держат 8 аскоспор, 30–40 × 7–12 мкм; аскоспоры круглые, около 5 мкм в диам.
ПИКНИДЫ краевые, темные или светлые, погруженные до слегка выступающих;
конидии бутылковидные, 9–12 × 1–2 мкм. ФОТОБИОНТ Trebouxia-типа. — Рис. 121
(вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет; сердцевина — каператовую, лихестериновую и
протолихестериновую кислоты, от K, C, KC и P не изменяется; дополнительно в
сердцевине может присутствовать секалоновая кислота, в таких случаях сердце-
вина светло-желтая, от K желтеет, от KC становится оранжевой.
ЭКОЛОГИЯ. Эпифитный вид, встречается на коре деревьев хвойных и листвен-
ных пород в горных лесах, чаще всего на Betula и Picea.
РАСПРОСТРАНЕНИЕ. Россия. Довольно редкий вид, отмеченный в европейской
части России, на Кавказе и на Дальнем Востоке, но наиболее обычен в горных
районах Юж. Сибири. ЕР: Ср. Россия — Марий Эл; Сев. Кавказ — Адыгея.
СИБ: Юж. Сиб.
—
Республика Алтай, Хакасия, Бурятия, Красноярский край
(юж. часть), Иркутская обл. (юж. часть); Як. — юж. ДВ: ДВ юж. — Приморский
край. — Карта 86.
Общее распространение. Европа (преимущественно горные районы Центр.
Европы, единственное местонахождение в Скандинавии), Вост. Азия, Сев. Аме-
рика (вост. районы).
ПРИМЕЧАНИЕ. Морфологически сходен с Nephromopsis laureri, но отличается от
последнего отсутствием псевдоцифелл на нижней поверхности.
Роды Usnocetraria,
Флора лишайников России Флора лишайников России
160
Т. Рандлане, А. Сааг
Vulpicida J.-E. Mattsson et M. J. Lai
1993, Mycotaxon 49: 427. Т и п : Vulpicida juniperinus (L.) J.-E. Mattsson et
M. J. Lai. — Вульпицида
Лихенизированные грибы. ТАЛЛОМ листоватый, образующий неправильной
формы розетки или почти кустистый; верхняя поверхность зеленовато-желтая
до ярко- или темно-желтой; сердцевина интенсивно желтая; нижняя поверхность
полностью светло-желтая с темной центральной частью. Лопасти в основном
дорсивентральные, часто с восходящими концами, иногда круглые в сечении.
Соредии ярко-желтые, имеются у одного вида. Изидии отсутствуют. Ризины ред-
кие, светлые или черные. Верхний коровой слой параплектенхимный. ПЛОДОВЫЕ
ТЕЛА — апотеции зеоринового типа с коричневым диском и часто с кренулирован-
ным краем, краевые или поверхностные. Сумки широкобулавовидные, 40–50 ×
20–27 мкм, содержат 8 аскоспор. Аскоспоры простые, широкоэллипсоидные до
почти сферических, бесцветные, 5–6 × 5 мкм. КОНИДИАЛЬНЫЕ СПОРОНОШЕНИЯ —
пикниды, часто многочисленные, преимущественно краевые и субмаргинальные,
черные, погруженные или на возвышениях, хорошо заметные. Конидии бутыл-
ковидные, 6–8 × 1–2 мкм (у североамериканских видов конидии лимоновидные).
ФОТОБИОНТ Trebouxia-типа.
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, сердце-
вина и соредии — вульпиновую и пинастровую кислоты.
ЭКОЛОГИЯ. Встречаются преимущественно как эпифиты на хвойных деревьях,
эпиксильно на древесине и как эпигейные лишайники на известковой почве; в
лесных и тундровых сообществах, иногда в травяных сообществах на известня-
ках.
РАСПРОСТРАНЕНИЕ. Два вида широко распространены в арктических, бореаль-
ных и горных регионах Северного полушария, в то время как два других вида —
эндемики Сев. Америки. В России широко распространены в большинстве реги-
онов.
ПРИМЕЧАНИЕ. Род легко отличается от других родов желтоватым талломом и
интенсивно желтой сердцевиной. Недавние филогенетические исследования
(Saag et al., 2014) показали, что род включает четыре вида (вместо ранее при-
знававшихся шести), два из которых встречаются в России. В другой новейшей
публикации (Divakar et al., 2017), где на основе современных филогенетических
методов пересмотрены границы родов сем. Parmeliaceae и предлагается разделе-
ние цетрариоидных лишайников лишь на три крупных рода (Cetraria, Melanelia и
Nephromopsis), виды рода Vulpicida отнесены к роду Cetraria. Такая концепция не
признается авторами настоящей публикации.
ИЗБРАННАЯ ЛИТЕРАТУРА: Mattsson, 1993; Mattsson, Lai, 1993; Thell et al., 2011;
Saag et al., 2014; Stenroos et al., 2016.
Vulpicida
161
Ключ для определения видов рода Vulpicida
1. Таллом всегда листоватый; лопасти с краевыми соралиями ...... 2. V. pinastri.
—
Таллом часто листоватый, иногда почти кустистый; соралии отсутствуют .....
......................................................................................................
1. V. juniperinus.
1. Vulpicida juniperinus (L.) J.-E. Mattsson et M. J. Lai
1993, Mycotaxon 49: 427. ≡ Lichen juniperinus L., 1753, Sp. Pl.: 1147, nom. cons.
≡ Cetraria juniperina (L.) Ach., 1803, Methodus: 298. ≡ Tuckermannopsis juniperina
(L.) Hale, in Egan, 1987, Bryologist 90 (2): 164. Т и п : [Швеция] Sweden, Härje-
dalen, Storsjö, Flatruet W of Falkvålen, 02.08.1991, J.-E. Mattsson 2340 (LD typ.
cons., Mattsson, 1993: 35). — Вульпицида можжевельниковая.
=
Cetraria alvarensis (Wahlenb.) Vain. ex Lynge; = Cetraria tilesii Ach.
≡ Vulpicida tilesii (Ach.) J.-E. Mattsson et M. J. Lai; = Cetraria tubulosa (Schaer.)
Zopf ≡ Vulpicida tubulosus (Schaer.) J.-E. Mattsson et M. J. Lai.
ЛИТЕРАТУРА: Mattsson, 1993; Randlane, Saag, 2005; Saag et al., 2014.
ИЛЛЮСТРАЦИИ: Randlane et al., 2011: 287, 291; Stenroos et al., 2016: 733; Рандла-
не и др., 2019: 291.
ТАЛЛОМ листоватый до почти кустистого, образует розетки неправильной
формы, свободно прикрепленные до плотно прижатых, до 5 см в диам. Лопасти
1–5 мм шир., дорсиветральные и немного желобчатые или круглые в сечении,
восходящие до почти вертикальных. Верхняя поверхность интенсивно желтая в
открытых местообитаниях, до зеленовато-серо-желтой — в затененных; сердце-
вина ярко-желтая; нижняя поверхность светло-желтая до коричневато-белой, с
жилками. Соредии и изидии отсутствуют. Ризины коричневато-белые, оттопырен-
ные. АПОТЕЦИИ редкие до частых, субмаргинальные до почти поверхностных, до 6
мм в диам., с кренулированным талломным краем и коричневым диском. Сумки
широкобулавовидные, содержат 8 аскоспор; аскоспоры широкоэллипсоидные до
почти круглых, 5–6 × 5 мкм. ПИКНИДЫ редкие и разбросанные или многочислен-
ные, краевые до поверхностных, погруженные или на коротких черных возвыше-
ниях; конидии бутылковидные, 6–8 × 1–2 мкм. — Рис. 122, 123 (вклейка).
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует; сердцевина — вуль-
пиновую и пинастровую кислоты, от K, C, KC и P не изменяется, в UV не флюо-
ресцирует.
ЭКОЛОГИЯ. Встречается в арктических, альпийских, горных и бореальных со-
обществах в Северном полушарии. Эпигейный лишайник на известковых почвах
и эпифит на коре, преимущественно на Juniperus communis (в Европе) и на Pinus
pumila (в Азии).
РАСПРОСТРАНЕНИЕ. Россия. Широко распространенный вид, встречающий-
ся в Арктике, на Европейском Севере, в Ср. России, в горах Кавказа, в Сибири
Vulpicida
Флора лишайников России Флора лишайников России
162
и на Дальнем Востоке. АРКТ: Аркт. евр.
—
Ненецкий АО (Архангельская обл.:
Новая Земля, Канин); Аркт. зап.-сиб. (Ямал); Аркт. тайм. (Северная Земля, Тай-
мыр); Аркт. як. (Новосибирские о-ва, Анабарский р-н); Аркт. чук. (Чукотский
п-ов, о. Врангеля). ЕР: Евр. Север — Мурманская обл., Карелия, Ненецкий АО
(юго-зап. часть), Коми, Ленинградская и Вологодская обл.; Ср. Россия — Твер-
ская обл., Пермский край, Башкирия; Сев. Кавказ — Кабардино-Балкария, Север-
ная Осетия; Сев. Урал; Юж. Урал. СИБ: Зап. Сиб. сев.
—
Ямало-Ненецкий АО
(юж. часть); Зап. Сиб. юж. — Алтайский край (зап. часть); Ср. Сиб. — Краснояр-
ский край (сев. часть); Юж. Сиб.
—
Алтайский край (вост. часть), Кемеровская
обл., Республика Алтай, Хакасия, Тыва, Бурятия, Красноярский край (юж. часть);
Як. — вост. ДВ: ДВ сев. — Магаданская обл., Камчатский край (Корякский округ
и бывшая Камчатская обл.); ДВ юж. — Амурская, Сахалинская обл. (Курильские
о-ва: Кунашир), Приморский край. — Карта 87.
Общее распространение. Европа (Скандинавия и прилегающие территории
и альпийские регионы Центр. Европы), Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Два ранее разделявшихся вида: Vulpicida tilesii и V. tubulosus (по-
следний обычно с кустистым талломом и растущий преимущественно на поч-
ве) — были объединены с V. juniperinus из-за сходства молекулярных признаков и
постепенных переходов в морфологии (Saag et al., 2014).
2. Vulpicida pinastri (Scop.) J.-E. Mattsson et M. J. Lai
1993, Mycotaxon 49: 428. ≡ Lichen pinastri Scop., 1772, Fl. Carn., ed. 2, 2: 382. ≡ Cetra-
ria pinastri (Scop.) Gray, 1821, Nat. Arr. Brit. Pl. 1: 432. ≡ Tuckermannopsis pinastri
(Scop.) Hale, in Egan, 1987, Bryologist 90 (2): 164. Т и п : [Италия] Italy, Friuli Ve-
nezia Giulia, Prov. Udine, Carnic Alps, road between Sauris and Casera Razzo, 1750 m,
1993, P. L. Nimis (LD неотип, Mattsson, 1993: 39). — Вульпицида сосновая.
= Cetraria caperata auct. non (L.) Vain.: Vainio, p. p., excl. typo.
ЛИТЕРАТУРА: Mattsson, 1993; Randlane, Saag, 2005; Saag et al., 2014.
ИЛЛЮСТРАЦИИ: Randlane et al., 2011: 289; Stenroos et al., 2016: 734.
ТАЛЛОМ листоватый, образует свободно прикрепленные розетки до 3 см в
диам., лопасти 1–5 мм шир., дорсивентральные, с приподнятыми краями и окру-
глыми концами. Верхняя поверхность интенсивно желтая в открытых место-
обитаниях, до зеленовато-серо-желтой — в затененных; сердцевина ярко-желтая;
нижняя поверхность светло-желтая до коричневато-белой, с жилками. Соралии
ярко-желтые, краевые, иногда поверхностные. Ризины коричневато-белые, от-
топыренные. АПОТЕЦИИ очень редки, субмаргинальные, с гладким талломным
краем и коричневым диском. Сумки широкобулавовидные, содержат 8 аскоспор;
аскоспоры широкоэллипсоидные до почти круглых, 5–6 × 5 мкм. ПИКНИДЫ ред-
кие, краевые, на коротких черных возвышениях; конидии бутылковидные, 6–8 ×
1–2 мкм. — Рис. 124 (вклейка).
Общий ключ для коричневых цетрариоидных лишайников
163
ЛИШАЙНИКОВЫЕ ВЕЩЕСТВА. Коровой слой содержит усниновую кислоту, от K, C
и P не изменяется, от KC желтеет, в UV не флюоресцирует; сердцевина — вуль-
пиновую и пинастровую кислоты, от K, C, KC и P не изменяется, в UV не флюо-
ресцирует.
ЭКОЛОГИЯ. Встречается в бореальных и альпийских регионах Северного полу-
шария. Преимущественно эпифит, растущий на коре Betula, Pinus и других хвой-
ных деревьев, но также и на древесине.
РАСПРОСТРАНЕНИЕ. Россия. Широко распространенный вид. АРКТ: Аркт.
евр.
—
Ненецкий АО (Архангельская обл.: Новая Земля, Канин); Аркт. зап.-сиб.
(Ямал); Аркт. тайм. (Таймыр); Аркт. як. (Анабарский р-н); Аркт. чук. (Чукотский
п-ов). ЕР: Евр. Север — Мурманская обл., Карелия, Архангельская обл., Коми,
Ленинградская обл.; Ср. Россия — Псковская, Новгородская, Тверская, Ярослав-
ская, Костромская, Кировская обл., Пермский край, Московская, Нижегородская
обл., Чувашия, Татарстан, Рязанская обл., Мордовия, Пензенская, Ульяновская
обл., Самарская обл. (сев. часть), Башкирия, Белгородская обл. (сев. часть); Юж.
Россия — Саратовская обл., Самарская обл. (юж. часть), Оренбургская, Волго-
градская обл.; Сев. Кавказ — Адыгея, Карачаево-Черкесия, Дагестан; Сев. Урал;
Юж. Урал. СИБ: Зап. Сиб. сев.
—
Ямало-Ненецкий АО (юж. часть), Свердлов-
ская обл., Тюменская обл. (сев. часть), Томская обл.; Зап. Сиб. юж.
—
Омская
обл., Новосибирская обл., Алтайский край (зап. часть); Юж. Сиб.
—
Алтайский
край (вост. часть), Кемеровская обл., Республика Алтай, Хакасия, Тыва, Бурятия,
Красноярский край (юж. часть), Забайкальский край; Як. — вост.; ДВ: ДВ сев. —
Камчатский край (Корякский округ и бывшая Камчатская обл.); ДВ юж. — Амур-
ская обл., Хабаровский край (юж. часть), Еврейская АО, Сахалинская обл. (Ку-
рильские о-ва: Итуруп, Кунашир), Приморский край. — Карта 88.
Общее распространение. Европа, Азия, Сев. Америка.
ПРИМЕЧАНИЕ. Легко распознается по комбинации светло-желтого листоватого
таллома и интенсивно желтых краевых соралий.
Т. В. Макрый
Общий ключ для коричневых цетрариоидных лишайников
Группа коричневых цетрариоидных лишайников объединяет представителей
рода Cetraria s. str. и родов Arctocetraria, Cetrariella и Masonhalea, выделенных
из рода Cetraria s. l. По своему габитусу коричневые цетрариоидные лишайники
даже внутри одного отдельно взятого рода довольно разнообразны. В то же время
для них характерны некоторые общие признаки, что позволяет (а в ряде случаев и
более удобно) определять их по общему ключу.
В этой группе выделяется несколько морфотипов.
Общий ключ для коричневых цетрариоидных лишайников
Флора лишайников России Флора лишайников России
164
Cucullata-тип (Cetraria kamczatica, Cetrariella fastigiata) характеризует-
ся почти трубчато свернутыми лопастями с округлыми отогнутыми верхушка-
ми (схожий с Flavocetraria cucullata). Masonhalea-тип (Masonhalea richardsonii)
имеет крупные дорсивентральные неприкрепленные лопасти и характерные псев-
доцифеллы, покрывающие бóльшую часть нижней поверхности. Aculeata-тип
(Cetraria aculeata, C. aculeata subsp. steppae, C. muricata) отличается почти ради-
альным строением лопастей. Все эти типы характеризуются наличием редких,
разрозненных коротких краевых выростов (шипиков), несущих пикниды.
Большая группа видов, относящихся к морфотипам Islandica-типа (с желобча-
тыми до почти плоских лопастями, с отгибающимися на нижнюю сторону несхо-
дящимися краями), Ericetorum-типа (с почти трубчато свернутыми лопастями, со
сходящимися краями) и Laevigata-типа (с почти трубчато свернутыми лопастями,
отгибающимися краями, с многочисленными срастаниями на уровне предкрае-
вых выступов), характеризуется наличием регулярных реснитчатых краевых вы-
ростов (ресничек), несущих пикниды.
Самостоятельные морфотипы характерны для видов рода Arctocetraria,
Cetrariella delisei, Masonhalea inermis и других. При этом Cetraria nigricans и
Arctocetraria nigricascens имеют сходные морфотипы (желобчатые лопасти и на-
личие длинных краевых цилий).
Ключ для определения коричневых цетрариоидных лишайников
1. Сердцевина от KC краснеет ............................................................................... 2.
—
Сердцевина от KC не изменяется ...................................................................... 3.
2. Таллом кустистый, из вертикальных лопастей ............................... Cetrariella.
—
Таллом крупный, распростертый, неприкрепленный, из дорсивентральных,
неправильно дихотомически разветвленных лопастей .......................................
........................................................................................
Masonhalea richardsonii.
3. Лопасти радиального строения, округло- или угловато-цилиндрические ........
........................................................................................ Cetraria aculeata-группа.
—
Лопасти не радиального строения ..................................................................... 4.
4. Лопасти узкие, 0.1–0.5 мм шир., таллом мелкий, 0.5–1.5(2.0) см выс. ......... 5.
—
Лопасти более широкие, 0.5 мм шир.; таллом более крупный, 1–10 (и более)
см выс. .................................................................................................................. 6.
5. Таллом мелкокустистый, из вертикальных лопастей, образующих дернинки;
лопасти в основании угловато-округло-сплюснутые, на концах более или
менее густо коралловидно разветвленные на тонкие, отчасти вальковатые,
направленные в разные стороны короткие веточки с шипиками и редкими ци-
лиями до 0.5 мм дл.; основания красные; конидии веретеновидные, 3.6–4.8 ×
0.5 мкм; гипоаркто-монтанный вид ..................................... Cetraria odontella.
—
Таллом распростертый до восходящего, из отдельных лопастей или их спле-
тений, образующих бесформенные куртинки; лопасти слабо желобчатые
Общий ключ для коричневых цетрариоидных лишайников
165
(с ложбинкой в центре, широко V-образные на срезе), неправильно дихото-
мически разветвленные, края с шипиками и немногочисленными цилиями до
0.5 мм дл. ........................................................................................... Cetraria sp.
1
6. Основания лопастей красные; края с ресничками до 0.5–1.0(1.5) мм дл.; ци-
лии обычно отсутствуют ........................................... Cetraria islandica-группа.
—
Основания лопастей не красные, а если красные, то имеются цилии; шипики
или реснички имеются или отсутствуют .......................................................... 7.
7. Цилии имеются, длинные, 2.0–3.5 мм дл. ........................................................ 8.
—
Цилии отсутствуют или они редкие и короткие, до 0.5–1.5 мм дл. .............. 9.
8. Таллом коричнево-черный в виде округлых подушковидных куртинок; ло-
пасти 0.5–2.0 мм шир., линейные, правильно дихотомически разветвленные,
желобчатые, с краевыми узкими псевдоцифеллами в виде рваной линии;
края ровные, слегка вздутые, с шипиками и цилиями до 2 мм дл.; конидии
веретеновидные, 6.0–7.5 × 1 мкм; сердцевина от I синеет; напочвенный вид в
Арктике ................................................................................... Cetraria nigricans.
—
Таллом зеленовато-оливково-серый до черноватого или буроватого в виде
куртинок или розеток; лопасти 1–6 мм шир., неправильно дихотомически
разветвленные, слабо желобчатые, ямчато-морщинистые, обычно без псев-
доцифелл; края неровные, с шипиками и цилиями до 3.5 мм дл.; конидии
гантелевидные, 4.0–5.5 × 1 мкм; сердцевина от I не изменяется; эпифитный и
напочвенный вид в Арктике ................................... Arctocetraria nigricascens.
9. Эпифиты, редко встречающиеся на других субстратах ................................ 10.
—
Напочвенные или обитающие на каменистом субстрате виды .................... 11.
10. Таллом листоватый, плотно прикрепленный, с восходящими короткими ло-
пастями 0.5–2.0 см дл., несущими многочисленные апотеции; пикниды не-
многочисленные, выступающие, незаметные; гипоаркто-бореальный вид ......
...................................................................................................
Cetraria sepincola.
—
Таллом из одиночных, реже собранных в плотные дернинки (на почве), вер-
тикальных или отгибающихся кверху лопастей, до 2 см выс.; лопасти в виде
язычков; псевдоцифеллы образуют сплошную кайму вдоль краев; края мел-
козубчатые благодаря выступающим темным пикнидам; апотеции обычны;
арктический широко-амфиберингийский вид ................. Masonhalea inermis.
11. Краевые шипики или реснички (несущие пикниды) многочисленные, часто с
пучком гиф на конце; основания лопастей желтовато-коричневые до бурых;
лопасти слабо желобчатые до плоских; конечные веточки с многочисленны-
ми боковыми долями и лопастинками; сердцевина от I синеет; сибирский
кальцефильный вид ............................................................. Cetraria rassadinae.
—
Краевые шипики отсутствуют или редкие, немногочисленные, разбросанные
............................................................................................................................... 12.
1
Таксон требует дополнительного изучения и уточнения статуса, возможно, является формой
вида Cetraria odontella.
Флора лишайников России Флора лишайников России
166
12. Лопасти крупные, 3–20(30) мм шир., желобчатые до почти трубчато сверну-
тых, кверху расширяющиеся, с широкоокруглыми отгибающимися концами;
края ровные; псевдоцифеллы в виде мелких погруженных точек на нижней
поверхности; арктический широко-амфиберингийский вид ..............................
.........................................................................................
Arctocetraria andrejevii.
—
Лопасти узкие, 1–2 мм шир., трубчато свернутые, Cucullata-типа, с волнисты-
ми краями и отгибающимися концами; псевдоцифеллы едва заметные, распо-
ложены по самому краю нижней поверхности; узкий амфиберингийский вид
...............................................................................................
Cetraria kamczatica.
Номенклатурные новинки
Cetraria crispiformis (Räsänen) Makryǐ — comb. et stat. nov.
................................
84
Evernia perfragilis Llano — лектотип, изолектотип ............................................. 114
Evernia terrestris (Tomin) N. S. Golubk. — лектотип, изолектотип .................... 116
Литература
Голубкова Н. С. 1981. Конспект флоры лишайников Монгольской Народной Республики.
Л.: Наука. 200 с.
Голубкова Н. С. 1996а. Таблица для определения родов семейств Alectoriaceae и
Parmeliaceae // Определитель лишайников России. Вып. 6. СПб.: Наука. С. 7–15.
Голубкова Н. С. 1996б. Род Bryoria // Определитель лишайников России. Вып. 6. СПб.: На-
ука. С. 18–32.
Голубкова Н. С. 1996в. Род Evernia // Определитель лишайников России. Вып. 6. СПб.: На-
ука. С. 49–56.
Еленкин А. А. 1909. Лишайники полярного побережья Сибири // Записки Имп. Акад.
наук. Физ.-мат. отд-ние. Т. 27, No 1. С. [1–4]+1–53; ил.
Еленкин А. А., Савич В. П. 1911. Список лишайников, собранных Ир. М. Щеголевым в
Якутской и Приморской областях по хребту Джугджуру (Становому) и его отрогам
между Нельканом и Аяном в 1903 г. // Труды Ботан. музея Имп. Акад. наук. Вып. 8.
С. 26–49.
Журбенко М. П. 1996а. Род Bryocaulon // Определитель лишайников России. Вып. 6. СПб.:
Наука. С. 39–41.
Журбенко М. П. 1996б. Род Cetraria // Определитель лишайников России. Т. 6. СПб.: На-
ука. С. 41–45.
Журбенко М. П. 1996в. Dactylina // Определитель лишайников России. Вып. 6. СПб.: На-
ука. С. 45–49.
Каганов В. В., Ёжкин А. К. 2019. Новые местонахождения редких и охраняемых лишайни-
ков с острова Сахалин // Бюл. Ботан. сада-ин -та ДВО РАН. Вып. 21. С. 1–8.
Котлов Ю. В. 1996а. Род Letharia // Определитель лишайников России. Вып. 6. СПб.: На-
ука. С. 58.
Котлов Ю. В. 1996б. Род Lethariella // Определитель лишайников России. Вып. 6. СПб.:
Наука. С. 58–59.
Красная книга Приморского края: Растения. Редкие и находящиеся под угрозой исчезно-
вения виды растений и грибов. 2008 / Биолого-почвенный институт ДВО РАН. Влади-
восток: АВК «Апельсин». 688 с.
Красная книга Российской Федерации (растения и грибы). 2008 / гл. редколл.: Ю. П. Трут-
нев и др.; сост. Р. В. Камелин и др. М.: Т-во науч. изд. КМК. 855 c.
Крусанова З. Г. 1996. Alectoria // Определитель лишайников России. Вып. 6. СПб.: Наука.
С. 16–18.
Макрый Т. В. 1984. Cetraria rassadinae — новый вид лишайника из Прибайкалья // Ботан.
журн. Т. 69, No 7. С. 952–957.
Макрый Т. В. 2002. Мэйсонхэйлеа Ричардсона // Красная книга Читинской области и
Агинского Бурятского автономного округа. Растения. Чита. С. 230.
Макрый Т. В. 2008. Мэйсонхэйлеа Ричардсона: Masonhalea richardsonii (Hook in Richards.)
Kärnefelt // Красная книга Российской Федерации (растения и грибы). М.: Т-во науч.
изд. КМК. С. 729–730.
Окснер А. М. 1933. Нови види обрiсникiв з Азii // Журн. бiо-ботан. циклу Всеукр. Акад.
Наук. No 6–7. С. 167–172.
Окснер А. М. 1940. Лишайники басейну рік Індігірки, Яни, Лени і південного Прибайкал-
ля. IV // Ботан. журн. АН УРСР. 1940. Т. 1, No 3–4. С. 31–57.
Окснер А. Н., Рассадина К. А. 1960. Новые представители рода Cetraria в СССР // Ново-
сти систематики низших растений. Т. 13. С. 5–14.
Определитель лишайников России. Вып. 6. Алекториевые, Пармелиевые, Стереокаулоно-
вые. 1996 / отв. ред. Н. С. Голубкова. СПб.: Наука. 213 с.
Литература
169
Петрова О. В. 2000. Род Bryoria в Мурманской области. Препринт. Апатиты: Изд-во КНЦ
РАН. 28 с.
Пчёлкин А. П. 2008. Бриокаулон псевдосатоанский: Bryocaulon pseudosatoanum (Asahina)
Kärnefelt // Красная книга Российской Федерации (растения и грибы). М.: Т-во науч.
изд. КМК. С. 721–722.
Рандлане Т., Сааг А., Мартин Л., Дегтяренко П., Оя Э. 2019. Эпигейные макролишайники
Эстонии. Тарту: Изд-во Тартуского ун-та. 319 c.
Рассадина К. А. 1945. О новом виде и новых формах Cetraria // Ботан. материалы Отд.
споровых растений Ботан. ин-та им. В. Л. Комарова Акад. наук СССР. Т. 5, вып. 10–
12. С. 133–137.
Рассадина К. А. 1950. Цетрария (Cetraria) СССР // Труды Ботан. ин-та им. В. Л. Комарова
Акад. наук СССР. Сер. 2. Споровые растения. Вып. 5. М.; Л. С. 171–304.
Савич В. А. 1914. Новые виды и формы лишайников Камчатки // Известия Ботан. сада Пе-
тра Великого. Т. 14, вып. 1–2. С. 111–128.
Савич В. П. 1923. О лишайнике Cetraria Richardsonii Hook. // Ботан. материалы Ин-та спо-
ровых растений Гл. ботан. сада РСФСР. Т. 2, вып. 12. С. 189–191.
Савич В. П. 1924. О новом почвенном лишайнике Cornicularia steppae mihi и лишайнике
Cornicularia tenuissima // Ботан. материалы Ин-та споровых растений Гл. ботан. сада
РСФСР. Т. 3, вып. 12. С. 185–188.
Савич В. П. 1964. О форме Cetraria cucullata (Bell.) Ach. f. minuscula Elenk. et Savicz // Но-
вости систематики низших растений. С. 123–125.
Седельникова Н. В. 1990. Лишайники Алтая и Кузнецкого нагорья: Конспект флоры. Но-
восибирск: Наука. 175 с.
Скирин Ф. В., Скирина И. Ф. 2012. Эколого-субстратная приуроченность эпифитных ли-
шайников пихтово-еловых и кедрово-широколиственных лесов южного Сихотэ-Алиня
// Turczaninowia. Т. 15, No 1. С. 70–79.
Скирина И. Ф. 1995. Лишайники Сихотэ-Алинского биосферного района. Владивосток:
Дальнаука. 132 с.
Скирина И. Ф. 2006. Новый для лихенофлоры России вид Lethariella togashii
(Parmeliaceae) с юга Дальнего Востока // Ботан. журн. Т. 91, No 7. С. 1143–1145.
Скирина И. Ф. 2007. Лишайники // Флора, микобиота и растительность заповедника «Ба-
стак». Владивосток: Дальнаука. С. 209–260.
Скирина И. Ф. 2015. Список лишайников заповедника «Бастак» // Биoта и среда
заповедникoв Дальнегo Вoстока. No 4. С. 28–87.
Скирина И. Ф. 2016. Лишайники // Растения, грибы и лишайники Сихотэ-Алинского запо-
ведника. Владивосток: Дальнаука. С. 458–525.
Скирина И. Ф. 2019. Лишайники // Красная книга Хабаровского края: Редкие и находящи-
еся под угрозой исчезновения виды растений, грибов и животных: официальное из-
дание / Министерство природных ресурсов Хабаровского края, Ин-т водных и экол.
проблем ДВО РАН. Воронеж: ООО «МИР». С. 310–332.
Скирина И. Ф., Коженкова С. И., Родникова И. М. 2010. Эпифитные лишайники Примор-
ского края и использование их в экологическом мониторинге. Владивосток: Дальнау-
ка. 134 c.
Скирина И. Ф., Салохин А. В., Царенко Н. А., Скирин Ф. В. 2016. Новые местонахождения
редких и охраняемых лишайников острова Сахалин // Turczaninowia. Т. 19, No 2. C. 54–63.
Степанчикова И. С., Гимельбрант Д. Е. 2011. Первые находки Tuckermannopsis orbata
(Parmeliaceae) и Chaenothecopsis parasitaster (Mycocaliciaceae) на Камчатке // Ново-
сти систематики низших растений. Т. 54. С. 219–222.
Литература
Флора лишайников России Флора лишайников России
170
Толпышева Т. Ю. 1990. Platismatia norvegica (Lynge) Culb. et C. Culb. — новый вид лишай-
ника для СССР // Новости систематики низших растений. Т. 27. С. 117–118.
Урбанавичюс Г. П. 2011. Роды листоватых и кустистых лишайников семейства Parmelia-
ceae России и ключ для их определения // Новости систематики низших растений.
Т. 45. С. 242–256.
Урбанавичюс Г. П., Макрый Т. В., Кузнецова Е. С., Гагарина Л. В., Конорева Л. А., Андре-
ев М. П., Гимельбрант Д. Е. 2014. Ключи для определения родов лишайников флоры
России // Флора лишайников России: Биология, экология, разнообразие, распростра-
нение и методы изучения лишайников / под ред. М. П. Андреева, Д. Е. Гимельбранта.
М.: Т-во науч. изд. КМК, 2014. С. 292–364.
Флора лишайников России: Биология, экология, разнообразие, распространение и методы
изучения лишайников / под ред. М. П. Андреева, Д. Е. Гимельбранта. М.; СПб.: Т-во
науч. изд. КМК, 2014. 392 с.
Флора лишайников России: Род Protoparmelia, семейства Coenogoniaceae, Gyalectaceae и
Umbilicariaceae / отв. ред. М. П. Андреев, Д. Е. Гимельбрант. М.; СПб.: Т-во науч. изд.
КМК, 2017. 195 с.
Чабаненко С. И. 2002. Конспект флоры лишайников юга российского Дальнего Востока.
Владивосток: Дальнаука. 232 с.
Чабаненко С. И., Скирина И. Ф., Княжева Л. А. 2002. Список лишайников Приморского
края и обитающих на них грибов. Южно-Сахалинск: Сахалинский ботан. сад; СахНЦ;
ДВО РАН. 89 с.
Яковченко Л. С., Галанина И. А., Малашкина Е. В., Бакалин В. А. 2013. Мохообразные
и лишайники малонарушенных лесных сообществ в нижнем Приамурье (Российский
Дальний Восток) // Комаровские чтения. Т. 60. С. 9–68.
Acharius E. 1803. Methodus, qua omnes detectos lichenes secundum organa carpomorpha ad
genera, species et varietates redigere atque observationibus illustrate tentavit. Stockholmiae:
Typis C. F. Marquard. 393 p.
Ahti T., Thell A. 2011. Tuckermannopsis // A. Thell, R. Moberg (eds). Nordic lichen flora.
Vol. 4: Parmeliaceae. Göteborg: Nordic Lichen Society. P. 105–106.
Altermann S., Leavitt S. D., Goward T. 2016. Tidying up the genus Letharia: introducing L. lu-
pina sp. nov. and a new circumscription for L. columbiana // Lichenologist. Vol. 48, pt. 5.
P. 423–439.
Arup U., Ekman S., Kärnefelt I., Mattsson J.-E. (eds). 1997. Skyddsvärda lavar i sydvästra Sve-
rige. Lund: SBF-förlaget. 276 p.
Asahina Y. 1939. Cornicularia-Arten aus Japan // J. Jap. Bot. Vol. 15. P. 353–358.
Asahina Y. 1952. Lichenologische Notizen (85–87) // J. Jap. Bot. Vol. 27. P. 293–296.
Awasthi D. D. 1982. Lichen genus Cetraria in India and Nepal // Bull. Bot. Surv. India. Vol. 24,
No. 1–4. P. 1–27.
Bird C. D. 1974. Studies on the lichen genus Evernia in North America // Canad. J. Bot. Vol. 52,
No. 11. P. 2427–2434.
Boluda C. G., Rico V. J., Divakar P. K., Nadyeina O., Myllys L., McMullin R. T., Zamora J. C.,
Scheidegger C., Hawksworth D. L. 2019. Evaluating methodologies for species delimita-
tion: the mismatch between phenotypes and genotypes in lichenized fungi (Bryoria sect. Im-
plexae, Parmeliaceae) // Persoonia. Vol. 42. P. 75–100.
Brodo I. M., Hawksworth D. L. 1977. Alectoria and allied genera in North America // Opera
Bot. No. 42. P. 1–164.
Brodo I. M., Sharnoff S. D., Sharnoff S. 2001. Lichens of North America. New Haven; London:
Yale Univ. Press. 795 p.
Литература
171
Chen J. 1996. The lichen genus Oropogon from China // Acta Mycol. Sinica. Vol. 15, No. 3.
P. 173–177.
Crespo A., Kauff F., Divakar P. K., del Prado R., Pérez-Ortega S., de Paz G. A., Ferencova Z.,
Blanco O., Roca-Valiente B., Núñez-Zapata J., Cubas P., Argüello A., Elix J. A., Esslin-
ger T. L., Hawksworth D. L., Millanes A., Molina M. C., Wedin M., Ahti T., Aptroot A.,
Barreno E., Bungartz F., Calvelo S., Candan M., Cole M., Ertz D., Goffinet B., Lindb-
lom L., Lücking R., Lutzoni F., Mattsson J.-E., Messuti M. I., Miadlikowska J., Piercey-
Normore M., Rico V. J., Sipman H. J. M., Schmitt I., Spribille T., Thell A., Thor G., Up-
reti D. K., Lumbsch H. T. 2010. Phylogenetic generic classification of parmelioid lichens
(Parmeliaceae, Ascomycota) based on molecular, morphological and chemical evidence //
Taxon. Vol. 59, No. 6. P. 1735–1753.
Crespo A., Lumbsch H. T., Mattsson J. E., Blanco O., Divakar P. K., Articus K., Wiklund E.,
Bawingan P. A., Wedin M. 2007. Testing morphology-based hypotheses of phylogenetic re-
lationships in Parmeliaceae (Ascomycota) using three ribosomal markers and the nuclear
RPB1 gene // Molec. Phylogen. Evol. Vol. 44, No. 2. P. 812–824.
Culberson W. L., Culberson C. F. 1965. Asahinea, a new lichen genus in the Parmeliaceae //
Brittonia. Vol. 17, No. 2. P. 182–190.
Culberson W. L., Culberson C. F. 1967. A new taxonomy for the Cetraria ciliaris group // Bry-
ologist. Vol. 70. P. 158–166.
Culberson W. L., Culberson C. F. 1968. The lichen genera Cetrelia and Platismatia (Parmeli-
aceae) // Contr. U.S. Natl. Herb. Vol. 34, No. 7. P. 449–558.
Divakar P. K., Crespo A., Kraichak E., Leavitt S. D., Singh G., Schmitt I., Lumbsch H. T. 2017.
Using a temporal phylogenetic method to harmonize family and genus-level classification in
the largest clade of lichen-forming fungi // Fungal Diversity. Vol. 84. P. 101–117.
Dobson F. S. 2000. Lichens: An illustrated guide to the British and Irish species. Slough, Eng-
land: Richmond Publ. Co. Ltd. 431 p.
Esslinger T. L. 1980. Typification of Oropogon loxensis and description of two related species //
Bryologist. Vol. 83, No. 4. P. 529–532.
Esslinger T. L. 1989. Systematics of Oropogon (Alectoriaceae) in the New World // Syst. Bot.
Monogr. Vol. 28. P. 1–111.
Esslinger T. L. 2016. Oropogon in Mexico // Biblioth. Lichenol. Vol. 110. P. 289–307.
Gao X. 1991. Studies in species of the lichen genus Asahinea // Nordic J. Bot. Vol. 11, No. 4.
P. 483–485.
Goward T. 1999. The lichens of British Columbia, illustrated keys. Pt. 2: Fruticose species / Brit-
ish Columbia Ministry of Forests. Victoria (BC): Crown Publ. 319 p. (Spec. Rep. Ser. No. 9).
Goward T., Myllys L. 2020. Gowardia zebrina sp. nov., a new species in a little-known genus of
arctic-alpine lichens (Parmeliaceae) // Pl. Fungal Syst. Vol. 65, No. 1. P. 219–226.
Hakulinen R., Ulvinen T. 1966. Asahinea chrysantha (Tuck.) Culb. et Culb. in Fennoskandien //
Ann. Univ. Turku. Ser. A.II. Vol. 36. P. 101–105.
Halonen P., Myllys L., Velmala S., Hyvärinen H. 2009. Gowardia (Parmeliaceae) — a new
alectorioid lichen genus with two species // Bryologist. Vol. 112, No. 1. P. 138–146.
Hansen E. S., Andersen J. 1995. Greenland lichens. Copenhagen: Rhodos; Danish Polar Centre.
124 p.
Harada H., Wang L.-S. Taxonomic study on Bryoria (lichenized Ascomycota, Parmeliaceae) of
east Asia (4). External morphology and anatomy of pycnidia // Lichenology. 2008. Vol. 7,
No. 2. P. 159–168.
Hawksworth D. L. 1969. Chemical and nomenclatural notes on Alectoria (Lichenes) I // Taxon.
Vol. 18. P. 393–399.
Флора лишайников России Флора лишайников России
172
Hawksworth D. L. 1972. Regional studies in Alectoria (Lichenes) II. The British species // Li-
chenologist. Vol. 5. P. 181–261.
Hawksworth D. L., Blanco O., Divakar P. K., Ahti T., Crespo A. 2008. A first checklist of par-
melioid and similar lichens in Europe and some adjacent territories, adopting revised ge-
neric circumscriptions and with indications of species distributions // Lichenologist. Vol. 40,
pt. 1. P. 1–21.
Hawksworth D. L., Jørgensen P. M. 2013. (2196) Proposal to conserve the name Alectoria fusce-
scens (Bryoria fuscescens) against Lichen chalybeiformis and Alectoria subcana (Ascomy-
cota: Lecanorales: Parmeliaceae) // Taxon. Vol. 62, No. 5. P. 1057.
Hawksworth D. L., Moore D. M. 1969. Some lichens from Tierra del Fuego with notes on their
chemical constituents // Bryologist. Vol. 72, No. 2. P. 247–251.
Hinds J. W., Hinds P. L. 2007. The macrolichens of New England. Bronx; New York: New York
Bot. Gard. Press. 584 p. (Mem. New York Bot. Gard. No. 96).
Holien H. 1989. The genus Bryoria sect. Implexae in Norway // Lichenologist. Vol. 21, pt. 3.
P. 243–258.
Holien H. 1991. Bryoria subcana in Norway // Graphis Scripta. Vol. 3. P. 95–96.
Holien H., Tønsberg T. 2006. Norsk lavflora. Trondheim: Tapir Akad. Forlag. 206 p.
Howe R. H. Jr. 1911. American species of Alectoria occurring north of the fifteenth parallel //
Mycologia. Vol. 3. P. 106–150.
Howe R. H. Jr. 1912. The lichens of the Linnean herbarium, with remarks on Acharian material
// Bull. Torrey Bot. Club. Vol. 39. P. 190–203.
Hue A. M. 1899. Lichenes extra-europaei a pluribus collectoribus ad Museum Parisiense missi
(Suite) // Nouv. Arch. Mus. Hist. Nat. Sér. 4. T. 1. P. 27–220.
Jørgensen P. M. 1972. Further studies in Alectoria sect. Divaricatae DR // Svensk Bot. Tidskr.
Vol. 66. P. 191–201.
Jørgensen P. M., James P. W., Jarvis C. E. 1994. Linnaean lichen names and their typifications //
Bot. J. Linn. Soc. Vol. 115. P. 261–405.
Jørgensen P. M., Ryvarden L. 1970. Contributions to the lichen flora of Norway // Årbok Univ.
Bergen, Mat.-Naturvitensk. Ser. 1969. No. 10. P. 1–24.
Kärnefelt I. 1977a. Masonhalea, a new lichen genus in the Parmeliaceae // Bot. Not. Vol. 130.
P. 101–107.
Kärnefelt I. 1977b. Three new species of brown fruticose Cetraria // Bot. Not. Vol. 130. P. 125–
129.
Kärnefelt I. 1979. The brown fruticose species of Cetraria // Opera Bot. Vol. 46. P. 1–150.
Kärnefelt I. 1986. The genera Bryocaulon, Coelocaulon and Cornicularia and formerly associ-
ated taxa // Opera Bot. Vol. 86. P. 1–90.
Kärnefelt I. 1987. Cetraria (Parmeliaceae) and some related genera on the African continent //
Bothalia. Vol. 17. P. 45–49.
Kärnefelt I., Mattsson J.-E., Thell A. 1993. The lichen genera Arctocetraria, Cetraria and Ce-
trariella (Parmeliaceae) and their presumed evolutionary affinities // Bryologist. Vol. 96,
No. 3. P. 394–404.
Kärnefelt I., Thell A. 1992. The evaluation of characters in lichenized families, exemplified with
the alectorioid and some parmelioid genera // Pl. Syst. Evol. Vol. 180, No. 3–4. P. 181–
204.
Kärnefelt I., Thell A. 1996. A new classification for the Dactylina / Dufourea complex // Nova
Hedwigia. Vol. 65. P. 487–511.
Kärnefelt I., Thell A. 2001. Delimitation of the lichen genus Tuckermannopsis Gyeln. (Asco-
mycotina Parmeliaceae) based on morphology and DNA sequences // P. M. McCarthy,
Литература
173
G. Kantvilas, S. H. J. J. Louwhoff (eds). Lichenological contributions in honour of Jack
Elix. P. 193–209. (Biblioth. Lichenol. Bd. 78).
Kärnefelt I., Thell A., Randlane T., Saag A. 1994. The genus Flavocetraria Kärnefelt & Thell
(Parmeliaceae, Ascomycotina) and its affinities // Acta Bot. Fenn. Vol. 150. P. 79–86.
Kristinsson H., Zhurbenko M., Hansen E. S. 2010. Panarctic checklist of lichens and lichenicolous
fungi // CAFF Technical Report No. 20. Akureyri (Iceland): CAFF Int. Secretariat. P. 1–120.
Krog H. 1973. Cetraria inermis (Nyl.) Krog, a new lichen species in the Amphi-Beringian flora
element // Bryologist. Vol. 76. P. 299–300.
Krog H. 1976. Lethariella and Protousnea, two new genera in Parmeliaceae // Norweg. J. Bot.
Vol. 23. P. 83–106.
Krog H. 1979. On the identity of Bryoria vrangiana // Norweg. J. Bot. Vol. 26. P. 179–181.
Krog H. 1980. On Bryoria chalybeiformis and some related species // Lichenologist. Vol. 12.
P. 243–245.
Kurokawa S., Kashiwadani H. 2006. Checklist of Japanese lichens and allied fungi // Natl. Sci.
Mus. Monogr. Vol. 33. P. 1–157.
Kurokawa S., Lai M.-J. 1991. Allocetraria, a new genus in the Parmeliaceae // Bull. Natl. Sci.
Mus. Tokyo. Ser. B. Botany. Vol. 17. P. 59–65.
Kuznetsova E. S., Stepanchikova I. S., Skirina I. F., Chesnokov S. V., Himelbrant D. E. 2021.
A revision of the lichen genus Platismatia (Parmeliaceae) in Russia, with a key to the spe-
cies // Novosti sistematiki nizshikh rastenii = Новости систематики низших растений.
Vol. 55, pt. 1. P. 179–194.
Lai M. J. 1980. Studies on the cetrarioid lichens in Parmeliaceae of East Asia // Quart. J. Taiwan
Mus. Vol. 33. P. 215–229.
Lai M. J., Qian Z. G., Xu L. 2007. Synopsis of the cetrarioid lichen genera and species (Parme-
liaceae, lichenized Ascomycotina) in China // J. Natl. Taiwan Mus. Vol. 60. P. 45–62.
Laundon J. 1984. The typification of Withering’s neglected lichens // Lichenologist. Vol. 16,
No. 3. P. 211–239.
Lücking R., Hodkinson B. P., Leavitt S. D. 2016. The 2016 classification of lichenized fungi
in the Ascomycota and Basidiomycota — Approaching one thousand genera // Bryologist.
Vol. 119, No. 4. P. 361–416.
Lumbsch H. T., Ahti T., Altermann S., Amo De Paz G., Aptroot A., Arup U., Bárcenas Peña A.,
Bawingan P. A., Benatti M. N., Betancourt L., Björk C. R., Boonpragob K., Brand M., Bun-
gartz F., Cáceres M. E. S., Candan M., Chaves J. L., Clerc P., Common R., Coppins B. J.,
Crespo A., Dal-Forno M., Divakar P. K., Duya M. V., Elix J. A., Elvebakk A., Fankhaus-
er J. D., Farkas E., Ferraro L. I., Fischer E., Galloway D. J., Gaya E., Giralt M., Goward T.,
Grube M., Hafellner J., Hernández M. J. E., Herrera Campos M. A., Kalb K., Kärnefelt I.,
Kantvilas G., Killmann D., Kirika P., Knudsen K., Komposch H., Kondratyuk S., Law-
rey J. D., Mangold A., Marcelli M. P., McCune B., Messuti M. I., Michlig A., Miranda Gon-
zalez R., Moncada B., Naikatini A., Nelsen M. P., Øvstedal D. O., Palice Z., Papong K.,
Parnmen S., Pérez-Ortega S., Printzen C., Rico V. J., Rivas Plata E., Robayo J., Rosabal D.,
Ruprecht U., Salazar Allen N., Sancho L., Santos de Jesus L., Vieira T. S., Schultz M.,
Seaward M. R. D., Sérusiaux E., Schmitt I., Sipman H. J. M., Sohrabi M., Søchting U.,
Søgaard M. Z., Sparrius L. B., Spielmann A., Spribille T., Sutjaritturakan J., Thammatha-
worn A., Thell A., Thor G., Thüs H., Timdal E., Truong C., Türk R., Umaña Tenorio L.,
Upreti D. K., Van den Boom P., Vivas Rebuelta M., Wedin M., Will-Wolf S., Wirth V.,
Wirtz N., Yahr R., Yeshitela K., Ziemmeck F., Wheeler T., Lücking R. 2011. One hundred
new species of lichenized fungi: a signature of undiscovered global diversity // Phytotaxa.
Vol. 18. P. 1–127.
Флора лишайников России Флора лишайников России
174
Lutsak T., Fernández-Mendoza F., Kirika P., Wondafrash M., Printzen C. 2020. Coalescence-
based species delimitation using genome-wide data reveals hidden diversity in a cosmopoli-
tan group of lichens // Organisms Diversity Evol. Vol. 20, iss. 2. P. 189–218.
Lutsak T., Fernández-Mendoza F., Nadyeina O., Şenkardeşler A., Printzen C. 2017. Testing the
correlation between norstictic acid content and species evolution in the Cetraria aculeata
group in Europe // Lichenologist. Vol. 49, pt. 1. P. 39–56.
Mattsson J.-E. 1993. A monograph of the genus Vulpicida (Parmeliaceae, Ascomycetes) // Opera
Bot. Vol. 119. P. 1–61.
Mattsson J.-E., Lai M. J. 1993. Vulpicida, a new genus in Parmeliaceae (lichenized Ascomyce-
tes) // Mycotaxon. Vol. 46. P. 425–428.
McCune B., Geiser L. 2009. Macrolichens of the Pacific Northwest. Ed. 2. Corvallis: Oregon
State Univ. Press. 464 p.
Moberg R., Thell A. 2011a. Evernia // A. Thell, R. Moberg (eds). Nordic lichen flora. Vol. 4:
Parmeliaceae. Göteborg: Nordic Lichen Society. P. 49–51.
Moberg R., Thell A. 2011b. Letharia // A. Thell, R. Moberg (eds). Nordic lichen flora. Vol. 4:
Parmeliaceae. Göteborg: Nordic Lichen Society. P. 65–66.
Motyka J. 1936–1938. Lichenum generis Usnea studium monographicum. Pars systematica.
Vol. 1, 2. Leopoli. 651 p.
Motyka J. 1964. The North American species of Alectoria // Bryologist. Vol. 67, No. 1. P. 1–44.
Myllys L., Halonen P., Velmala S. 2006. Notes on some rare species of Bryoria from Finland //
Graphis Scripta. Vol. 18. P. 23–26.
Myllys L., Velmala S., Holien H. 2011a. Bryoria // A. Thell, R. Moberg (eds). Nordic lichen
flora. Vol. 4: Parmeliaceae. Göteborg: Nordic Lichen Society. P. 26–37.
Myllys L., Velmala S., Holien H., Halonen P., Wang L.-S., Goward T. 2011b. Phylogeny of the
genus Bryoria // Lichenologist. Vol. 43, pt. 6. P. 617–638.
Nadyeina O., Lutsak T., Blum O., Grakhov V., Scheidegger C. 2013. Cetraria steppae Savicz is
conspecific with Cetraria aculeata (Schreb.) Fr. according to morphology, secondary chem-
istry and ecology // Lichenologist. Vol. 45, pt. 6. P. 841–856.
Nelsen M. P., Chavez N., Sackett-Hermann E., Thell A., Randlane T., Divakar P. K., Rico V. J.,
Lumbsch H. T. 2011. The cetrarioid core group revisited (Lecanorales: Parmeliaceae) // Li-
chenologist. Vol. 43, pt. 6. P. 537–551.
Nelsen M. P., Thell A., Leavitt S. D., Hampton-Miller C. J., Lumbsch H. T. 2013. A reappraisal
of Masonhalea (Parmeliaceae, Lecanorales) based on molecular and morphological data //
Lichenologist. Vol. 45, pt. 6. P. 729–738.
Nordic Lichen Flora. Vol. 4: Parmeliaceae. 2011 / A. Thell, R. Moberg (eds). Göteborg: Nordic
Lichen Society. 184 p.
Obermayer W. 2004. Additions to the lichen flora of the Tibetan region // P. Döbbeler, G. Ram-
bold (eds). Contributions to Lichenology. Festschrift in honour of Hannes Hertel. P. 479–
526. (Biblioth. Lichenol. Vol. 88).
Obermayer W., Randlane T. 2012. Morphological and chemical studies on Platismatia erosa
(Parmeliaceae) from Tibet, Nepal and Bhutan // Bryologist. Vol. 115, No. 1. P. 51–60.
Ohmura Y., Kashiwadani H. 1997. Lichens of Mt. O-akan and its adjacent areas, Hokkaido, Ja-
pan // Bull. Natl. Sci. Mus., Tokyo. Ser. B. Vol. 23, No. 1. P. 1–24.
Øvstedal D., Tønsberg T., Elvebakk A. 2009. The lichen flora of Svalbard // Sommerfeltia.
Vol. 33. P. 3–393.
Purvis O. W., Coppins B. J., Hawksworth D. L., James P. W., Moore D. M. (eds). 1992. The Li-
chen flora of Great Britain and Ireland. London: Nat. Hist. Mus. Publ. 710 p.
Randlane T., Saag A. 1989. Chemical variation and geographical distribution of Asahinea chry-
santha (Tuck.) Culb. et C. Culb. // Lichenologist. Vol. 21, pt. 4. P. 303–311.
Литература
175
Randlane T., Saag A. 1992. Tuckermannopsis americana contra Cetraria ciliaris in Russia // Fo-
lia Cryptog. Estonica. Fasc. 29. P. 33–36.
Randlane T., Saag A. 1998. Synopsis of the genus Nephromopsis (fam. Parmeliaceae, lichenized
Ascomycota) // Cryptog. Bryol. Lichénol. Vol. 19, No. 2–3. P. 175–191.
Randlane T., Saag A. 2005. Distribution patterns of primary and secondary species in the genus
Vulpicida // Folia Cryptog. Estonica. Fasc. 41. P. 89–95.
Randlane T., Saag A., Martin L., Timdal E., Nimis P. L. 2011. Eesti puudel kasvavad suursamb-
likud. Tartu: Tartu Ülikooli Kirjastus. 326 lk.
Randlane T., Saag A., Obermayer W. 2001. Cetrarioid lichens containing usnic acid from the Ti-
betan area // Mycotaxon. Vol. 80. P. 389–425.
Randlane T., Saag A., Thell A., Kärnefelt I. 1994. The lichen genus Tuckneraria Randlane &
Thell — a new segregate in the Parmeliaceae // Acta Bot. Fenn. Vol. 150. P. 143–151.
Randlane T., Thell A. 2011a. Allocetraria // A. Thell, R. Moberg (eds). Nordic lichen flora.
Vol. 4: Parmeliaceae. Göteborg: Nordic Lichen Society. P. 128–130.
Randlane T., Thell A. 2011b. Asahinea // A. Thell, R. Moberg (eds). Nordic lichen flora. Vol. 4:
Parmeliaceae. Göteborg: Nordic Lichen Society. P. 21–22.
Randlane T., Thell A. 2011c. Flavocetraria // A. Thell, R. Moberg (eds). Nordic lichen flora.
Vol. 4: Parmeliaceae. Göteborg: Nordic Lichen Society. P. 52–53.
Randlane T., Thell A., Saag A. 1995. New data about the genera Cetrariopsis, Cetreliopsis and
Nephromopsis (Fam. Parmeliaceaee, lichenized Ascomycotina) // Cryptog. Bryol. Lichénol.
Vol. 16, No. 1. P. 35–60.
Ryan B. D. 2002a. Alectoria // T. H. Nash III, B. D. Ryan, C. Gries, F. Bungartz (eds). Lichen flora
of the Greater Sonoran Desert Region. Vol. 1. Tempe: Arizona State University. P. 92–95.
Ryan B. D. 2002b. Evernia // T. H. Nash III, B. D. Ryan, C. Gries, F. Bungartz (eds). Lichen flora
of the Greater Sonoran Desert Region. Vol. 1. Tempe: Arizona State University. P. 188–190.
Ryan B. D. 2002c. Letharia // T. H. Nash III, B. D. Ryan, P. Diederich, C. Gries, F. Bungartz
(eds). Lichen flora of the Greater Sonoran Desert Region. Vol. 1. Tempe: Arizona State
Univ. P. 267–270.
Saag A., Randlane T., Thell A., Obermayer W. 2002. Phylogenetic analysis of cetrarioid lichens
with globose ascospores // Proc. Estonian Acad. Sci., Biol., Ecol. Vol. 51, No. 2. P. 103–123.
Saag L., Mark K., Saag A., Randlane T. 2014. Species delimitation in the lichenized fungal ge-
nus Vulpicida (Parmeliaceae, Ascomycota) using gene concatenation and coalescent-based
species tree approaches // Amer. J. Bot. Vol. 101, No. 12. P. 2169–2182.
Satô M. 1937. Enumeratio lichenum insulae Formosae. III // J. Jap. Bot. Vol. 13. P. 595–599.
Satô M. 1938. Enumeratio lichenum ins. Formosae (V) // J. Jap. Bot. Vol. 14. P. 783–791.
Satô M. 1939. Parmeliales (I) // T. Nakai, M. Honda (eds). Nova Flora Japonica. Vol. 5. Tokyo;
Osaka: Sanseidō. P. 1–87.
Satô M. 1959. Range of the Japanese lichens (VIII) // Bull. Fac. Arts Sci. Ibaraki Univ., Nat. Sci.
Vol. 13. P. 39–51.
Satô M. 1967. Distribution and substratum of the Japanese lichens (2). The genus Oropogon
Th. Fr. // Misc. Bryol. Lichenol. Vol. 4. P. 129–131.
Savicz V. P. 1926. Notes on Cetraria chrysantha Tuck. and C. lacunosa Ach. in Russia // Bry-
ologist. Vol. 29, No. 2. P. 26–28.
Savicz V. P. 1961. Lichenotheca Rossica (regionibus confinibus completa). Edidit Institutum Bo-
tanicum nomine V. L. Komarovii Academiae Scientiarum URSS. Decas XI (1961) // Bot.
Mater. Otd. Sporov. Rast. Bot. Inst. Komarova Akad. Nauk S.S.S.R. = Ботан. материалы
Отд. споровых растений Ботан. ин-та им. В. Л. Комарова АН СССР. T. 14. P. 1–6.
Sharnoff S. 2014. A field guide to California lichens. New Haven: Yale University Press. 405 p.
Флора лишайников России Флора лишайников России
176
Smith C. W., Aptroot A., Coppins B. J., Fletcher A., Gilbert O. L., James P. W., Wolseley P. A.
(eds). 2009. The lichens of Great Britain and Ireland. London: British Lichen Society. 1046 p.
Spribille T., Fryday A. M., Pérez-Ortega S., Svensson M., Tønsberg T., Ekman S., Holien H.,
Resl P., Schneider K., Stabentheiner E., Thüs H., Vondrák J., Sharman L. 2020. Lichens
and associated fungi from Glacier Bay National Park, Alaska // Lichenologist. Vol. 52, pt. 2.
P. 61–181.
Stenroos S., Ahti T., Lohtander K., Myllys L. (eds). 2011. Suomen jäkäläopas Luonnontieteelli-
nen keskusmuseo. Helsinki. 534 p. (Norrlinia. Vol. 21).
Stenroos S., Velmala S., Pykälä J., Ahti T. 2016. Lichens of Finland. Helsinki: Finnish Museum
of Natural History LUOMUS. 896 p. (Norrlinia. Vol. 30).
Thell A. 2011a. Masonhalea // A. Thell, R. Moberg (eds). Nordic lichen flora. Vol. 4: Parmelia-
ceae. Göteborg: Nordic Lichen Society. P. 66–67.
Thell A. 2011b. Platismatia // A. Thell, R. Moberg (eds). Nordic lichen flora. Vol. 4: Parmelia-
ceae. Göteborg: Nordic Lichen Society. P. 97–98.
Thell A. 2011c. Usnocetraria // A. Thell, R. Moberg (eds). Nordic lichen flora. Vol. 4: Parmelia-
ceae. Göteborg: Nordic Lichen Society. P. 127–128.
Thell A., Ahti T., Randlane T. 2011. Vulpicida // A. Thell, R. Moberg (eds). Nordic lichen flora.
Vol. 4: Parmeliaceae. Göteborg: Nordic Lichen Society. P. 128–130.
Thell A., Crespo A., Divakar P. K., Kärnefelt I., Leavitt S. D., Lumbsch H. T., Seaward M. R. D.
2012. A review of the lichen family Parmeliaceae — history, phylogeny and current taxo-
nomy // Nordic J. Bot. Vol. 30. P. 641–664.
Thell A., Feuerer T., Kärnefelt I., Myllys L., Stenroos S. 2004a. Monophyletic groups within the
Parmeliaceae identified by ITS rDNA, beta-tubulin and GAPDH sequences // Mycol. Progr.
Vol. 3, No. 4. P. 297–314.
Thell A., Högnabba F., Elix J. A., Feuerer T., Kärnefelt I., Myllys L., Randlane T., Saag A., Sten-
roos S., Ahti T., Seaward M. R. D. 2009. Phylogeny of the cetrarioid core (Parmeliaceae)
based on five genetic markers // Lichenologist. Vol. 41, pt. 5. P. 489–511.
Thell A., Kärnefelt I. 2011a. Arctocetraria // A. Thell, R. Moberg (eds). Nordic lichen flora.
Vol. 4: Parmeliaceae. Göteborg: Nordic Lichen Society. P. 18–19.
Thell A., Kärnefelt I. 2011b. Bryocaulon // A. Thell, R. Moberg (eds). Nordic lichen flora:
Vol. 4: Parmeliaceae. Göteborg: Nordic Lichen Society. P. 24–26.
Thell A., Kärnefelt I. 2011c. Cetraria // A. Thell, R. Moberg (eds). Nordic lichen flora. Vol. 4:
Parmeliaceae. Göteborg: Nordic Lichen Society. P. 37–43.
Thell A., Kärnefelt I. 2011d. Cetrariella // A. Thell, R. Moberg (eds). Nordic lichen flora. Vol. 4:
Parmeliaceae. Göteborg: Nordic Lichen Society. P. 43–45.
Thell A., Kärnefelt I. 2011e. Dactylina // A. Thell, R. Moberg (eds). Nordic lichen flora. Vol. 4:
Parmeliaceae. Göteborg: Nordic Lichen Society. P. 48–49.
Thell A., Randlane T., Kärnefelt I., Gao X.-Q., Saag A. 1995. The lichen genus Allocetraria (Asco-
mycotina, Parmeliaceae) // F. J. A. Daniels, M. Schulz, J. Peine (eds). Flechten Follmann: Con-
tributions to lichenology in honour of Gerhard Follmann. Cologne: Univ. Cologne. P. 353–370.
Thell A., Randlane T., Saag A., Kärnefelt I. 2005. A new circumscription of the lichen genus
Nephromopsis (Parmeliaceae, lichenized Ascomycetes) // Mycol. Progr. Vol. 4, No. 4.
P. 303–316.
Thell A., Westberg M., Kärnefelt I. 2004b. Biogeography of the lichen family Parmeliaceae in the
Nordic countries with taxonomic remarks // Symb. Bot. Upsal. Vol. 34, No. 1. P. 429–452.
Thomson J. W. 1984. American Arctic lichens. 1: The macrolichens. New York: Columbia Univ.
Press. 504 p.
Литература
177
Thomson J. W. 2003. Lichens of Wisconsin. Madison: Wisconsin State Herbarium, Univ. of
Wisconsin. 386 p.
Thomson J. W., Bird C. D. 1978. The lichen genus Dactylina in North America // Canad. J. Bot.
Vol. 56, No. 14. P. 1602–1624.
Tønsberg T., Gauslaa Y., Haugan R., Holien H., Timdal E. 1996. The threatened macrolichens of
Norway — 1995 // Sommerfeltia. Vol. 23. P. 1–258.
Trass H. 1992. A new species of Asahinea (Ascomycotina, Parmeliaceae) // Folia Cryptog. Es-
tonica. Fasc. 29. P. 31–32.
Velmala S., Myllys L. 2011a. Alectoria // A. Thell, R. Moberg (eds). Nordic lichen flora. Vol. 4:
Parmeliaceae. Göteborg: Nordic Lichen Society. P. 14–16.
Velmala S., Myllys L. 2011b. Gowardia // A. Thell, R. Moberg (eds). Nordic lichen flora. Vol. 4:
Parmeliaceae. Göteborg: Nordic Lichen Society. P. 55.
Velmala S., Myllys L., Goward T., Holien H., Halonen P. 2014. Taxonomy of Bryoria section
Implexae (Parmeliaceae, Lecanoromycetes) in North America and Europe, based on chemi-
cal, morphological and molecular data // Ann. Bot. Fenn. Vol. 51, No. 6. P. 345–371.
Velmala S., Myllys L., Halonen P., Goward T., Ahti T. 2009. Molecular data show that Bryoria fre-
montii and B. tortuosa (Parmeliaceae) are conspecific // Lichenologist. Vol. 41. P. 231–242.
Verseghy K. 1964. Typen-Verzeichnis der Flechtensammlung in der Botanischen Abteilung des
Ungarischen Naturwissenschaftlichen Museums. Budapest: Term. Muz. 149 p.
Wang L. S., Harada H. 2001. Taxonomic study of Bryoria asiatica-group (lichenized Ascomy-
cota, Parmeliaceae) in Yunnan, southern China // Nat. Hist. Res. Vol. 6, No. 2. P. 43–52.
Wang L. S., Wang X. Y., Liu D., Myllys L., Shi H. X., Zhang Y. Y., Yang M. X., Li L. J. 2017.
Four new species of Bryoria (Lichenized Ascomycota: Parmeliaceae) from the Hengduan
Mountains, China // Phytotaxa. Vol. 297, No. 1. P. 29–41.
Wei X.-L., Leavitt S. D., Huang J.-P., Esslinger T. L., Wang L.-S., Moncada B., Lücking R.,
Divakar P. K., Lumbsch H. T. 2017. Parallel Miocene-dominated diversification of the li-
chen-forming fungal genus Oropogon (Ascomycota: Parmeliaceae) in different continents //
Taxon. Vol. 66, No. 6. P. 1269–1281
Wijayawardene N. N., Hyde K. D., Dai D. Q., Sánchez-García M., Goto B. T., Saxena R. K.,
Erdoğdu M., Selçuk F., Rajeshkumar K. C., Aptroot A., Błaszkowski J., Boonyuen N.,
da Silva G. A., de Souza F. A., Dong W., Ertz D., Haelewaters D., Jones E. B. G., Karunara-
thna S. C., Kirk P. M., Kukwa M., Kumla J., Leontyev D. V., Lumbsch H. T., Maharachchi-
kumbura S. S. N., Marguno F., Martínez-Rodríguez P., Mešić A., Monteiro J. S., Oehl F.,
Pawłowska J., Pem D., Pfliegler W. P., Phillips A. J. L., Pošta A., He M. Q., Li J. X.,
Raza M., Sruthi O. P., Suetrong S., Suwannarach N., Tedersoo L., Thiyagaraja V., Tib-
promma S., Tkalčec Z., Tokarev Y. S., Wanasinghe D. N., Wijesundara D. S. A., Wimalase-
ana S. D. M. K., Madrid H., Zhang G. Q., Gao Y., Sánchez-Castro I., Tang L. Z., Stadler M.,
Yurkov A., Thines M. 2022. Outline of Fungi and fungus-like taxa — 2021 // Mycosphere.
Vol. 13, No. 1. P. 53–453.
Wirth V. 1995. Die Flechten Baden-Württembergs. Teil 1–2. Stuttgart: Ulmer. 1006 p.
Wirth V., Hauck M., Schultz M. 2013. Die Flechten Deutschlands. Bd. 1. Stuttgart: Ulmer. 672 p.
Yoshimura I. 1979. Lichen flora of Japan in colour. Osaka: Hoikusha Publ. 349 p.
Zakeri Z., Elix J. A., Otte V. 2017. Degradation of alectorialic acid in the lichen genus Usnea //
Lichenologist. Vol. 49, pt. 5. P. 539–543.
Zhurbenko M. P. 2009. Lichenicolous fungi and lichens from the Holarctic. Part II // Opusc.
Philolichenum. Vol. 7. P. 121–186.
Zhurbenko M. P., Laursen G. A., Walker D. A. 2005. New and rare lichenicolous fungi and li-
chens from the North American Arctic // Mycotaxon. Vol. 92. P. 201–212.
Указатель латинских названий таксонов
лишайников и грибов1
1
Курсивом набраны синонимы, прописными буквами — названия таксонов в ранге выше се-
мейства; полужирные цифры — страницы, на которых приведено основное описание.
Ahtia wallichiana (Taylor) M. J. Lai 140
Alectoria Ach. 8, 14, 19, 32–34, 52, 110, 115,
123
—
albida Räsänen 37
—
altaica (Gyeln.) Räsänen 70
—
americana Motyka 59
—
arctica Elenkin et Savicz 114
—
asiatica Du Rietz 60
—
berengeriana (A. Massal. ex Stizenb.)
Gyeln. 73
—
—
var. smithii (Du Rietz) Gyeln. 73
—
bicolor (Ehrh.) Nyl. 60
—
—
subsp. smithii (Du Rietz) Räsänen 73
—
—
var. berengeriana A. Massal. ex Sti-
zenb. 73
—
boryana Delise 124
—
capillaris (Ach.) Cromb. 61
—
cincinnata (Fr.) Lynge 38
—
—
var. vexillifera (Nyl.) Räsänen 38
—
—
f. vexillifera (Nyl.) Savicz 38
—
divergens (Ach.) Nyl. 51
—
fremontii Tuck. 63
—
furcellata R. Sant. 124
—
fuscescens Gyeln. 65
—
glabra Motyka 67
—
gowardii Lumbsch 123
—
implexa (Hoffm.) Nyl. 68
—
—
var. capillaris (Ach.) Stizenb. 61
—
jubata (L.) Ach. var. capillaris (Ach.) Ach.
61
—
—
var. vrangiana (Gyeln.) Räsänen 76
—
jubata auct. 65
—
karelica Räsänen 70
—
kuemmerleana Gyeln. 69
—
lata (Taylor) Linds. 14, 34, 36
—
luteola Mont. ex De Not. 37
—
nadvornikiana Gyeln. 70
—
nidulifera Norrl. 64
—
—
f. simplicior Vain. 72
—
nigricans (Ach.) Nyl. 124
—
—
var. nipponica Asahina 124
—
—
var. thulensis (Th. Fr.) Vain. 124
—
—
var. tschuktschorum Vain. 124
—
—
f. sorediata E. Dahl 124, 125
—
nitidula (Th. Fr.) Vain. 71
—
ochroleuca (Hoffm.) A. Massal. 14, 22, 34,
35, 39
—
—
subsp. thulensis Th. Fr. 124
—
—
[subsp.] vexillifera Nyl. 38
—
—
var. cincinnata (Fr.) Nyl. 38
—
—
var. nigricans (Ach.) Körb. 124
—
—
var. rigida (Fr.) Th. Fr. 35
—
olivacea Räsänen 63
—
sarmentosa (Ach.) Ach. 24, 32, 37
—
—
subsp. sarmentosa 14, 34–37
—
—
subsp. vexillifera (Nyl.) D. Hawksw.
14, 34, 38, 39, 117
—
—
var. gigantea Räsänen 37
—
—
var. cincinnata (Fr.) Nyl. 38
—
—
—
f. vexillifera (Nyl.) H. Magn. 38
—
simplicior (Vain.) Lynge 72
—
—
f. subintricans Vain. 72
—
smithii Du Rietz 73
—
stigmata Bystrek 37
—
subsarmentosa Stirt. 37
—
tenuis E. Dahl 74
—
thulensis (Th. Fr.) Th. Fr. ex Kremp. 124
—
trichodes (Michx.) D. Hawksw. 75
—
vexillifera (Nyl.) Stizenb. 38
—
viridescens Gyeln. 65
—
vrangiana Gyeln. 76
—
vulpina (L.) Mudd 126
Alectoriaceae Bercht. et J. Presl 19, 20
Allantoparmelia (Vain.) Essl. 17, 26
Allocetraria Kurok. et M. J. Lai 8, 14, 22, 39,
41
—
cucullata (Bellardi) Randlane et Saag 118
—
madreporiformis (Ach.) Kärnefelt et A.
Thell 14, 41, 104, 108
—
nivalis (L.) Randlane et Saag 121
—
oakesiana (Tuck.) Randlane et A. Thell 40,
159
Указатель латинских названий таксонов лишайников и грибов
179
—
stracheyi (C. Bab.) Kurok. et M. J. Lai 14,
21, 39, 41, 42, 43
Anzia Stizenb. 17, 19, 25
Anziaceae M. Satô 19, 20
Arctocetraria Kärnefelt et A. Thell 8, 14, 23,
43, 44, 163, 164
—
andrejevii (Oxner) Kärnefelt et A. Thell
14, 43, 44, 166
—
nigricascens (Nyl.) Kärnefelt et A. Thell
14, 44, 45, 164, 165
Arctoparmelia Hale 17, 32
—
aleuritica (Nyl.) Hale 32
Asahinea W. L. Culb. et C. F. Culb. 14, 25,
46, 47
—
chrysantha (Tuck.) W. L. Culb. et
C. F. Culb. 14, 46, 47, 151
—
culbersoniorum Trass 47
—
kurodakensis (Asahina) W. L. Culb. et
C. F. Culb. 49
—
scholanderi (Llano) W. L. Culb. et
C. F. Culb. 14, 47, 49
ASCOMYCOTA Caval. Sm. 14
Atestia Trevis. 144
—
loxensis (Fée) Trevis. 144
Brodoa Goward 17, 26
Bryocaulon Kärnefelt 8, 14, 23, 25, 50, 51
—
divergens (Ach.) Kärnefelt 14, 50, 51, 53
—
hyperboreum Øvstedal 14, 51, 52, 53
—
pseudosatoanum (Asahina) Kärnefelt 14,
51, 53
—
satoanum (Gyeln.) Kärnefelt 54
Bryopogon Link 32
Bryopogon Th. Fr. 54
—
altaicus Gyeln. 70
—
bicolor (Ehrh.) Stein 60
—
divergens (Ach.) Schwend. 51
—
jubatus (L.) Link 54
—
—
var. nitidulus Th. Fr. 71
—
latus (Taylor) Müll. Arg. 34
—
niduliferus (Norrl.) Elenkin 64
—
nigricans (Ach.) Gyeln. 124
—
—
f. thulensis (Th. Fr.) Gyeln. 124
—
nitidulus Savicz et Elenkin f. caespitosus
Savicz 72
—
ochroleucus (Hoffm.) Link 35
—
sarmentosus (Ach.) Link 37
—
—
var. cincinnatus (Fr.) Müll. Arg. 38
—
simplicior (Vain.) Gyeln. f. albidosoredio-
sus Gyeln. 72
—
vexilliferus (Nyl.) Gyeln. 38
—
vrangianus (Gyeln.) Gyeln. 76
Bryoria Brodo et D. Hawksw. 8, 15, 25, 51,
52, 54–57, 64, 145
—
sect. Implexae (Gyeln.) Brodo et
D. Hawksw. 56, 66
—
americana (Motyka) Holien 15, 59, 60, 76
—
asiatica (Du Rietz) Brodo et D. Hawksw.
15, 57, 59, 60
—
bicolor (Ehrh.) Brodo et D. Hawksw. 15,
55, 57, 60, 61, 72, 74, 75
—
capillaris (Ach.) Brodo et D. Hawksw. 15,
56, 58, 61, 71
—
chalybeiformis (L.) Brodo et D. Hawksw.
56, 65–67
—
fremontii (Tuck.) Brodo et D. Hawksw.
15, 54, 57, 63
—
friabilis Brodo et D. Hawksw. 56
—
furcellata (Fr.) Brodo et D. Hawksw. 15,
55, 57, 64, 65, 73, 74
—
fuscescens (Gyeln.) Brodo et D. Hawksw.
15, 56, 58, 59, 65, 66, 77
—
glabra (Motyka) Brodo et D. Hawksw. 15,
56, 58, 67, 68
—
implexa (Hoffm.) Brodo et D. Hawksw.
15, 56, 58, 59, 68, 69, 70, 77
—
inactiva Goward, Velmala et Myllys 56
—
kockiana Velmala, Myllys et Goward 56
—
kuemmerleana (Gyeln.) Brodo et D.
Hawksw. 15, 56, 58, 69, 70, 77
—
lanestris (Ach.) Brodo et D. Hawksw. 56,
65–67
—
nadvornikiana (Gyeln.) Brodo et D.
Hawksw. 15, 58, 62, 70, 71
—
nitidula (Th. Fr.) Brodo et D. Hawksw. 15,
55, 57, 71
—
osteola (Gyeln.) Brodo et D. Hawksw. 68
—
pikei Brodo et D. Hawksw. 56
—
pseudofuscescens (Gyeln.) Brodo et
D. Hawksw. 56
—
simplicior (Vain.) Brodo et D. Hawksw.
15, 57, 65, 72
—
smithii (Du Rietz) Brodo et D. Hawksw.
15, 57, 61, 73–75
—
subcana (Nyl. ex Stizenb.) Brodo et
D. Hawksw. 56, 65–67
Указатель латинских названий таксонов лишайников и грибов
Флора лишайников России Флора лишайников России
180
—
tenuis (E. Dahl) Brodo et D. Hawksw. 15,
57, 61, 74
—
trichodes (Michx.) Brodo et D. Hawksw.
15, 54, 58, 59, 75, 76
—
—
subsp. americana (Motyka) Brodo et
D. Hawksw. 59
—
vrangiana (Gyeln.) Brodo et D. Hawksw.
15, 56, 58, 69, 70, 76, 77
Cetraria Ach. 8, 15, 19, 23, 40, 43, 78, 79, 90,
97, 98, 102, 118, 129, 134, 158, 160, 163
—
aculeata (Schreb.) Fr. 78, 81, 92, 164
—
—
subsp. aculeata 15, 79, 81, 82
—
—
subsp. steppae (Savicz) Lutsak,
Fernández-Mendoza et Printzen 15, 22,
78, 79, 82, 83, 164
—
—
var. hiascens Fr. 99
—
alvarensis (Wahlenb.) Vain. ex Lynge 161
—
andrejevii Oxner 44
—
annae Oxner 15, 79, 97
—
arborialis (G. Merr.) G. E. Howard 157
—
arctica H. Magn. 45
—
arctica (Richardson) Tuck. 104
—
—
var. minor Elenkin 105
—
—
var. papillata (Savicz) Zahlbr. 105
—
asahinae M. Satô 102
—
caperata auct. 162
—
capitata Lynge 92
—
chlorophylla (Willd.) Vain. 153
—
chrysantha Tuck. 47, 151
—
—
f. cinerascens Asahina 47
—
—
f. glaucescens Oxner et Rass. 47
—
—
f. pigmentulata Trass 47
—
ciliaris Ach. 155
—
—
var. halei (W. L. Culb. et C. F. Culb.)
Ahti 155
—
crispa (Ach.) Nyl. 85
—
—
var. japonica Asahina 90
—
—
var. vagans Mereschk. 85, 86
—
—
f. inermis Nyl. 130
—
crispiformis (Räsänen) Makryǐ 15, 78, 81,
84, 85, 167
—
cucullata (Bellardi) Ach. 118
—
—
var. rabotnovii Oxner 120
—
—
f. minuscula Elenkin et Savicz 119
—
delisei (Schaer.) Nyl. 99
—
—
subsp. fastigiata Nyl. 100
—
—
f. dilatata (Vain.) Rass. 44
—
elenkinii Krog 45
—
—
var. tominii Rass. 45
—
endocrocea (Asahina) M. Satô 135
—
ericetorum Opiz 78, 85
—
—
subsp. ericetorum 15, 80, 85
—
everniella (Nyl.) Kremp. 42
—
excrispa Savicz 90
—
fastigiata (Nyl.) Kärnefelt 100
—
furcellata Fr. 64
—
gilva Asahina 156
—
glauca (L.) Ach. 148
—
halei W. L. Culb. et C. F. Culb. 155
—
hiascens (Fr.) Th. Fr. 99
—
—
var. dilatata Vain. 44
—
—
var. fastigiata (Nyl.) Vain. 100
—
—
var. rhizophora Vain. 45
—
—
f. dilatata (Vain.) Rass. 44
—
inermis (Nyl.) Krog 130
—
islandica (L.) Ach. 78, 81, 84–86, 88, 95, 165
—
—
subsp. antarctica Kärnefelt 88
—
—
subsp. crispiformis (Räsänen) Kärnefelt
84, 88
—
—
subsp. islandica 15, 81, 86, 88
—
—
subsp. orientalis (Asahina ex M. Satô)
Kärnefelt 15, 80, 88
—
—
var. crispa Ach. 85
—
—
var. crispiformis Räsänen 84
—
—
var. delisei Schaer. 99
—
—
var. junipericola Räsänen 86
—
—
var. maculata Vain. 81, 88
—
—
var. nuda Vain. 86
—
—
var. orientalis Asahina ex M. Satô 88
—
—
—
f. angustifolia Asahina 88
—
—
var. paludosa Räsänen 86
—
—
—
f. carelica Räsänen 86
—
—
var. platyna (Ach.) Ach. 86
—
—
var. polaris Rass. 84
—
—
—
f. mediocris Rass. 84
—
—
—
f. rigidula Rass. 84
—
—
—
f. submaculata Rass. 84
—
—
f. castanea Rass. 84
—
—
f. maculata (Vain.) Savicz 88
—
—
f. pallida Britzelmayer 86
—
—
f. ramulicola Savicz 86
—
—
f. vagans Savicz 86, 88
—
juniperina (L.) Ach. 161
—
kamczatica Savicz 15, 78, 80, 89, 164, 166
—
—
var. dilatata (Vain.) Räsänen 44
Указатель латинских названий таксонов лишайников и грибов
181
—
komarovii Elenkin 136
—
kurodakensis Asahina 49
—
lacunosa Ach. 49, 150
—
—
f. cavernosa G. Merr. 150
—
—
f. norvegica Lynge 151
—
laevigata Rass. 15, 78, 81, 90
—
laureri Kremp. 137
—
libertina Stuckenb. 85, 86
—
madreporiformis (Ach.) Müll. Arg. 41
—
magnussonii Llano 45
—
minuscula (Elenkin et Savicz) McCune
119
—
muricata (Ach.) Eckfeldt 15, 78, 79, 82,
91, 164
—
nephromoides (Nyl.) D. D. Awasthi 139
—
nigricans Nyl. 15, 46, 78, 80, 92–94, 164,
165
—
nigricascens (Nyl.) Elenkin 45
—
—
var. tominii Rass. 45
—
nivalis (L.) Ach. 121
—
norvegica (Lynge) Du Rietz 151
—
—
f. sorediata Degel. 151
—
oakesiana Tuck. 159
—
odontella (Ach.) Ach. 15, 78–80, 93–95,
164, 165
—
—
var. nigricans (Nyl.) Lynge 92
—
ornata Müll. Arg. 139
—
pallescens Schaer. 140
—
perstraminea Zahlbr. 136
—
pinastri (Scop.) Gray 162
—
platyna Ach. 86
—
potaninii Oxner 42
—
pseudocomplicata Asahina 141
—
ramulosa (Hook.) Tuck. 107
—
rassadinae Makryǐ 15, 78, 80, 94, 165
—
rhizophora (Vain.) Rass. 45
—
—
f. arboricola Rass. 45
—
richardsonii Hook. 131
—
rugosa (Asahina) M. Satô 142
—
saviczii Oxner et Rass. 49
—
—
var. candida Oxner et Rass. 47
—
—
var. nigrita Oxner et Rass. 49
—
scholanderi Llano 49
—
—
var. candida (Oxner et Rass.) Rass. 47
—
—
var. nigrita (Oxner et Rass.) Rass. 49
—
scutata auct. 153
—
sepincola (Ehrh.) Ach. 15, 78, 79, 96, 153,
157, 165
—
septentrionalis (Nyl.) Almq. 47
—
sibirica H. Magn. 45
—
simmonsii Krog 44
—
—
var. intermedia Krog 44
—
steppae (Savicz) Kärnefelt 82
—
straminea Kremp. ex Schwend. 138
—
subalpina Imshaug 157
—
tenuifolia (Retz.) R. Howe 85
—
tenuissima (L.) Vain. 81
—
tilesii Ach. 161
—
tubulosa (Schaer.) Zopf 161
—
tuckermanii Oakes var. sorediata (Degel.)
Räsänen 151
—
ulophylla (Ach.) Rebent. 153
—
wallichiana (Taylor) Müll. Arg. 140
—
yunnanensis (Nyl.) Zahlbr. 143
Cetraria sp. 78, 80, 93, 94, 165
Cetrariaceae Schaer. 19
Cetrariella Kärnefelt et A. Thell 8, 15, 23, 98,
163, 164
—
delisei (Schaer.) Kärnefelt et A. Thell 15,
95, 98, 99, 164
—
fastigiata (Nyl.) Kärnefelt et A. Thell 15,
90, 98, 100, 164
Cetrariopsis Kurok. 133
—
laii A. Thell et Randlane 136
—
pallescens (Schaer.) Randlane et A. Thell
140
—
wallichiana (Taylor) Kurok. 133, 140
Cetrelia W. L. Culb. et C. F. Culb. 17, 28
Cetreliopsis M. J. Lai 8, 15, 101, 102
—
asahinae (M. Satô) Randlane et A. Thell
15, 29, 102
—
rhytidocarpa (Mont. et Bosch) M. J. Lai
101
Chlorea vulpina (L.) Nyl. 126
Cladonia P. Browne 120, 121
—
amaurocraea (Flörke) Schaer. 115
—
arctica (Richardson) Th. Fr. 104
Coelocaulon Link 23, 78
—
aculeatum (Schreb.) Link 78, 81
—
divergens (Ach.) R. Howe 51
—
muricatum (Ach.) J. R. Laundon 91
—
odontellum (Ach.) R. Howe 93
—
steppae (Savicz) Borreno et Vázquez 82
Coenogoniaceae (Fr.) Stizenb. 8
Cornicularia (Schreb.) Hoffm. 17, 19
—
aculeata (Schreb.) Ach. 81
Флора лишайников России Флора лишайников России
182
—
divergens Ach. 51
—
lata Taylor 34
—
muricata (Ach.) Ach. 91
—
normoerica (Gunn.) Du Rietz 23
—
ochroleuca (Hoffm.) DC. 35
—
—
var. cincinnata (Fr.) Schaer. 38
—
—
var. nigricans Ach. 124
—
odontella (Ach.) Röhl. 93
—
pseudosatoana Asahina 53
—
sarmentosa (Ach.) DC. 37
—
steppae Savicz 82
—
tenuissima (L.) Zahlbr. 81
Corniculariaceae Schaer. 19
Dactylina Nyl. 8, 16, 21, 42, 103, 104
—
arctica (Richardson) Nyl. 103, 104, 106,
108
—
—
subsp. arctica 16, 104, 105
—
—
subsp. beringica (C. D. Bird et
J. W. Thomson) Kärnefelt et A. Thell
16, 104, 106, 107
—
—
subsp. chinensis (Follmann) Kärnefelt et
A. Thell 107
—
beringica C. D. Bird et J. W. Thomson 106
—
madreporiformis (Ach.) Tuck. 41
—
ramulosa (Hook.) Tuck. 16, 104, 106, 107,
108
Dactylinaceae M. Choisy 19
Dolichousnea (Y. Ohmura) Articus 17, 24
—
longissima (Ach.) Articus 24
Dufourea arctica Richardson 104
—
—
var. papillata Savicz 105
—
madreporiformis Ach. 41
—
muricata Laurer 107
—
polaris Rupr. 105
—
ramulosa Hook. 107
Eualectoria sarmentosa (Ach.) Gyeln. 37
Evernia Ach. 8, 16, 19, 21, 24, 33, 108–110,
115, 116
—
[unranked] Letharia Th. Fr. 126
—
altaica Florovsk. 110
—
arctica (Elenkin et Savicz) Lynge 114
—
arctica (Richardson) Tuck. 104
—
arenaria Elenkin 115
—
arenaria (Fr.) Fr. 115
—
crispata Florovsk. 116
—
divaricata (L.) Ach. 16, 109, 110, 112, 117
—
elenkiniana Zahlbr. 115
—
esorediosa (Müll. Arg.) Du Rietz 16, 110,
112
—
—
var. tundrae Oxner 114
—
—
f. terrestris Tomin 116
—
illyrica (Zahlbr.) Zahlbr. 109
—
mesomorpha Nyl. 16, 110, 113
—
—
f. esorediosa Müll. Arg. 112
—
ochroleuca (Hoffm.) Fr. 35
—
—
var. cincinnata Fr. 38
—
perfragilis Llano 16, 109, 110, 114 , 117,
167
—
prunastri (L.) Ach. 16, 21, 108, 110, 115
—
—
var. thamnodes Flot. 113
—
ramulosa (Hook.) Tuck. 107
—
richardsonii (Hook.) Nyl. 131
—
sarmentosa (Ach.) Fr. 37
—
stracheyi C. Bab. 42
—
taurica Florovsk. 116
—
terrestris (Tomin) N. S. Golubk. 16, 110,
112, 116, 167
—
thamnodes (Flot.) Arnold 113
—
—
f. arenicola Savicz 113
—
—
f. esorediosa (Müll. Arg.) Zahlbr. 112
—
—
f. tecticola Savicz 113
—
—
f. terrestris (Tomin) Zahlbr. 116
—
vulpina (L.) Ach. 126
Everniaceae Tomas. 19
Everniastrum Hale ex Sipman 18, 23, 24
Flavocetraria Kärnefelt et A. Thell 8, 16, 22,
117, 118, 120
—
cucullata (Bellardi) Kärnefelt et A. Thell
16, 89, 100, 117, 118 , 120, 164
—
—
var. wainioi Räsänen 119
—
—
f. crispata Rass. 119
—
—
f. major Rass. 119
—
minuscula (Elenkin et Savicz) Ahti,
Poryadina et Zhurb. 16, 118, 119
—
nivalis (L.) Kärnefelt et A. Thell 16, 118, 121
—
—
var. saviczii Rass. 122
—
—
f. suffruticosa Rass. 122
Flavoparmelia Hale 17, 31
Flavopunctelia (Krog) Hale 17, 30
Gowardia Halonen et al. 8, 16, 22, 33, 122,
123
—
arctica Halonen et al. 16, 123, 125
Указатель латинских названий таксонов лишайников и грибов
183
—
nigricans (Ach.) Halonen et al. 16,
122–124
—
zebrina Goward et Myllys 123
Gyalectaceae (A. Massal.) Stizenb. 8
Hypogymnia (Nyl.) Nyl. 17, 19, 25
Hypogymniaceae Poelt ex Elix 19, 20
Hypotrachyna (Vain.) Hale 18, 20, 23, 24, 30,
31
—
cirrhata (Fr.) Divakar 109
Imshaugia S. L. F. Mey. 18, 26
LECANORALES Nannf. 14
LECANOROMYCETES O. E. Erikss. et
Winka 14, 20
LECANOROMYCETIDAE P. M. Kirk et al.
14
Letharia (Th. Fr.) Zahlbr. 8, 16, 126
—
divaricata (L.) Hue 110
—
mesomorpha (Nyl.) Du Rietz 113
—
thamnodes (Flot.) Hue 113
—
—
f. esorediosa (Müll. Arg.) Hue 112
—
togashii Asahina 128
—
vulpina (L.) Hue 16, 24, 126
Lethariella (Motyka) Krog 8, 16, 127
—
canariensis (Ach.) Krog 127, 128
—
cashmeriana Krog 127, 128
—
cladonioides (Nyl.) Krog 127, 128
—
intricata (Moris) Krog 25, 127, 128
—
togashii (Asahina) Krog 16, 109, 112, 128,
129
—
zahlbruckneri (Du Rietz) Krog 127, 128
Lichen aculeatus Schreb. 81
—
bicolor Ehrh. 60
—
chlorophyllus Willd. 153
—
cucullatus Bellardi 118
—
divaricatus L. 110
—
glaucus L. 148
—
islandicus L. 86
—
—
var. tenuissimus L. 81
—
juniperinus L. 161
—
muricatus Ach. 91
—
nivalis L. 121
—
ochroleucos Ehrh. 35
—
odontellus Ach. 93
—
pinastri Scop. 162
—
prunastri L. 115
—
sarmentosus Ach. 37
—
sepincola Ehrh. 96
—
vulpinus L. 126
Lobaria glauca (L.) Hoffm. 148
Masonhalea Kärnefelt 8, 16, 129–131, 163
—
inermis (Nyl.) Lumbsch, Nelsen et A. Thell
16, 130, 153, 164, 165
—
richardsonii (Hook.) Kärnefelt 16, 22,
129–131, 164
Melanelia Essl. 18, 29, 40, 102, 134, 158, 160
—
commixta (Nyl.) A. Thell 28
Melanelixia O. Blanco et al. 18, 29
Melanohalea O. Blanco et al. 18, 29
Menegazzia A. Massal. 18, 25
Montanelia Divakar et al. 18, 29
—
tominii (Oxner) Divakar et al. 29
Myelochroa (Asahina) Elix et Hale 18, 30, 31
Neofuscelia Essl. 18, 26, 28
Nephromopsis Müll. Arg. 8, 16, 26–28, 40,
102, 118, 131, 133, 134, 136, 138, 140,
153, 158, 160
—
andrejevii (Oxner) Divakar, A. Crespo et
Lumbsch 44
—
asahinae (M. Satô) Räsänen 102
—
ciliaris (Ach.) Hue 155
—
cucullata (Bellardi) Divakar, A. Crespo et
Lumbsch 118
—
ectocarpisma (Hue) Gyeln. 139
—
endocrocea Asahina 16, 134, 135, 140
—
inermis (Nyl.) Divakar, A. Crespo et
Lumbsch 130
—
komarovii (Elenkin) J. C. Wei 16, 134, 136
—
kurokawae (Shibuichi et K. Yoshida)
Kurok. 158
—
laii (A. Thell et Randlane) Saag et A. Thell
16, 134–136, 137, 139
—
laureri (Kremp.) Kurok. 16, 134, 137, 159
—
nephromoides (Nyl.) Ahti et Randlane 16,
135, 137, 139
—
nigricascens (Nyl.) Divakar, A. Crespo et
Lumbsch 45
—
nipponensis (Asahina) M. J. Lai 141
—
nivalis (L.) Divakar, A. Crespo et Lumbsch
121
—
ornata (Müll. Arg.) Hue 16, 134, 135, 139,
140
Флора лишайников России Флора лишайников России
184
—
pallescens (Schaer.) Y. S. Park var. pal-
lescens 16, 134, 140
—
pseudocomplicata (Asahina) M. J. Lai 17,
133, 134, 141, 142
—
richardsonii (Hook.) Divakar, A. Crespo et
Lumbsch 131
—
rugosa Asahina 17, 134, 142
—
stracheyi (C. Bab.) Müll. Arg. 133
—
yunnanensis (Nyl.) Randlane et Saag 17,
135, 143
Nesolechia A. Massal. 19
Nipponoparmelia (Kurok.) K. H. Moon,
Y. Ohmura et Kashiw. 18, 30
Nodobryoria Common et Brodo 51
—
subdivergens (E. Dahl) Common et Brodo
51, 53
Oropogon Th. Fr. 8, 17, 25, 144–146
—
asiaticus Asahina 17, 54, 145, 146
—
colibor Essl. 146
—
loxensis (Fée) Th. Fr. 144
Parmelia Ach. 18, 19, 29
—
divergens (Ach.) Spreng. 51
—
glauca (L.) Hepp 148
—
jubata (L.) Ach. var. capillaris Ach. 61
—
ochroleuca (Hoffm.) Ach. 35
—
richardsonii (Hook.) Nyl. 131
—
sarmentosa (Ach.) Ach. 37
—
vulpina (L.) Ach. 126
Parmeliaceae Bercht. et J. Presl 8, 14, 17,
19–21, 158, 160
Parmelina Hale 18, 31
Parmelinopsis Elix et Hale 18, 31
Parmeliopsis (Nyl.) Nyl. 18, 26
Parmotrema A. Massal. 18, 20, 27, 29, 31
PEZIZOMYCOTINA O. E. Erikss. et Winka 14
Phacopsis Tul. 19
Physcia glauca (L.) DC. 148
Platismatia W. L. Culb. et C. F. Culb. 8, 17,
28, 146, 147
—
erosa W. L. Culb. et C. F. Culb. 147
—
glauca (L.) W. L. Culb. et C. F. Culb. 17,
28, 146–148, 149, 152
—
herrei (Imshaug) W. L. Culb. et C. F. Culb.
147
—
interrupta W. L. Culb. et C. F. Culb. 17,
147–149, 152
—
lacunosa (Ach.) W. L. Culb. et C. F. Culb.
17, 148, 150, 151
—
norvegica (Lynge) W. L. Culb. et
C. F. Culb. 17, 147, 151, 152
—
wheeleri Goward, Altermann et Björk 149
Platysma chlorophyllum (Willd.) Vain. 153
—
chrysanthum (Tuck.) Nyl. 47
—
glaucum (L.) Frege 148
—
lacunosum (Ach.) Nyl. 150
—
nephromoides Nyl. 139
—
nigricascens Nyl. 45
—
orbatum Nyl. 156
—
richardsonii (Hook.) Nyl. 131
—
septentrionale Nyl. 47
—
yunnanense Nyl. 143
Pleurosticta Petr. 18, 28
Protoparmelia M. Choisy 8, 17, 18, 21
Pseudephebe M. Choisy 18, 24
Pseudevernia Zopf 18
—
furfuracea (L.) Zopf 24
Punctelia Krog 18, 30
Raesaenenia D. Hawksw. 19
Ramalina Ach. 33, 109, 115, 116
Remototrachyna Divakar et A. Crespo 18, 30,
31
Rhytidocaulon vulpinum (L.) Elenkin 126
Rimelia Hale et A. Fletcher 27
Setaria trichodes Michx. 75
Tuckermannopsis Gyeln. 8, 17, 27, 152, 153
—
americana (Spreng.) Hale 155–157
—
chlorophylla (Willd.) Hale 17, 153, 156
—
ciliaris (Ach.) Gyeln. 17, 46, 152, 153,
155, 157
—
gilva (Asahina) M. J. Lai 17, 153, 156
—
halei (W. L. Culb. et C. F. Culb.) M. J. Lai
155
—
inermis (Nyl.) Kärnefelt 130
—
juniperina (L.) Hale 161
—
oakesiana (Tuck.) Hale 159
—
orbata (Nyl.) M. J. Lai 17, 153, 156
—
pinastri (Scop.) Hale 162
—
sepincola (Ehrh.) Hale 96
—
subalpina (Imshaug) Kärnefelt 17, 153,
157
—
ulophylloides (Asahina) M. J. Lai 156
Указатель латинских названий таксонов лишайников и грибов
185
Tuckneraria Randlane et A. Thell 133
—
laureri (Kremp.) Randlane et A. Thell 137
—
pseudocomplicata (Asahina) Randlane et
Saag 133, 141
Umbilicariaceae Chevall. 8
Usnea Dill. ex Adans. 18–20, 23, 24, 33
—
subgen. Lethariella Motyka 127
—
implexa Hoffm. 68
—
ochroleuca Hoffm. 35
—
vulpina (L.) Hoffm. 126
Usneaceae Eschw. 19, 20
Usnocetraria M. J. Lai et J. C. Wei 8, 17, 27,
32, 40, 158
—
oakesiana (Tuck.) M. J. Lai et J. C. Wei
17, 138, 158, 159
Vulpicida J.-E. Mattsson et M. J. Lai 8, 17,
27, 160, 161
—
juniperinus (L.) J.-E. Mattsson et M. J. Lai
17, 160, 161, 162
—
pinastri (Scop.) J.-E. Mattsson et M. J. Lai
17, 161, 162
—
tilesii (Ach.) J.-E. Mattsson et M. J. Lai
161, 162
—
tubulosus (Schaer.) J.-E. Mattsson et M. J.
Lai 161, 162
Xanthoparmelia (Vain.) Hale 18, 20, 26, 28,
29, 32
Содержание
Авторы тома....................................................................................................................5
Предисловие ................................................................................................................... 7
Деление территории Российской Федерации на регионы........................................ 10
Систематическое положение рассматриваемых таксонов ....................................... 14
Семейство Parmeliaceae (М. П. Андреев) .................................................................. 19
Род Alectoria (Д. Е. Гимельбрант) .............................................................................. 32
Род Allocetraria (Т. Рандлане, А. Сааг) ....................................................................... 39
Род Arctocetraria (Т. В. Макрый) ................................................................................. 43
Род Asahinea (Е. С. Кузнецова).................................................................................... 46
Род Bryocaulon (Л. А. Конорева, С. В. Чесноков) ...................................................... 50
Род Bryoria (И. Н. Урбанавичене, Т. Н. Пыстина) .................................................... 54
Род Cetraria (Т. В. Макрый)......................................................................................... 78
Род Cetrariella (Т. В. Макрый)..................................................................................... 98
Род Cetreliopsis (Т. Рандлане, А. Сааг) ..................................................................... 101
Род Dactylina (Д. Е. Гимельбрант, И. С. Степанчикова) ....................................... 103
Род Evernia (И. С. Степанчикова, Д. Е. Гимельбрант) .......................................... 108
Род Flavocetraria (Т. В. Макрый) .............................................................................. 117
Род Gowardia (Д. Е. Гимельбрант) ........................................................................... 122
Род Letharia (Л. В. Гагарина) .................................................................................... 126
Род Lethariella (И. Ф. Скирина) ................................................................................ 127
Род Masonhalea (Т. В. Макрый)................................................................................. 129
Род Nephromopsis (Т. Рандлане, А. Сааг) ................................................................. 133
Род Oropogon (И. С. Жданов).................................................................................... 144
Род Platismatia (Е. С. Кузнецова, И. С. Степанчикова).......................................... 146
Род Tuckermannopsis (Е. С. Кузнецова) .................................................................... 152
Род Usnocetraria (Т. Рандлане, А. Сааг) ................................................................... 158
Род Vulpicida (Т. Рандлане, А. Сааг) ......................................................................... 160
Общий ключ для коричневых цетрариоидных лишайников (Т. В. Макрый)........ 163
Номенклатурные новинки ......................................................................................... 167
Литература .................................................................................................................. 168
Указатель латинских названий таксонов лишайников и грибов............................ 178
Содержание
187
Contents
Contributors ..................................................................................................................... 5
Preface.............................................................................................................................. 7
Regional subdivision of the Russian Federation............................................................ 10
Systematic position of taxa ............................................................................................ 14
Family Parmeliaceae (M. P. Andreev) ........................................................................... 19
Genus Alectoria (D. E. Himelbrant) .............................................................................. 32
Genus Allocetraria (T. Randlane, A. Saag) ................................................................... 39
Genus Arctocetraria (T. V. Makryi)................................................................................ 43
Genus Asahinea (E. S. Kuznetsova) ............................................................................... 46
Genus Bryocaulon (L. A. Konoreva, S. V. Chesnokov) .................................................. 50
Genus Bryoria (I. N. Urbanavichene, T. N. Pystina) ..................................................... 54
Genus Cetraria (T. V. Makryi) ....................................................................................... 78
Genus Cetrariella (T. V. Makryi) ................................................................................... 98
Genus Cetreliopsis (T. Randlane, A. Saag).................................................................. 101
Genus Dactylina (D. E. Himelbrant, I. S. Stepanchikova) .......................................... 103
Genus Evernia (I. S. Stepanchikova, D. E. Himelbrant).............................................. 108
Genus Flavocetraria (T. V. Makryi) ............................................................................. 117
Genus Gowardia (D. E. Himelbrant)........................................................................... 122
Genus Letharia (L. V. Gagarina) ................................................................................. 126
Genus Lethariella (I. F. Skirina) .................................................................................. 127
Genus Masonhalea (T. V. Makryi) ............................................................................... 129
Genus Nephromopsis (T. Randlane, A. Saag) .............................................................. 133
Genus Oropogon (I. S. Zhdanov) ................................................................................. 144
Genus Platismatia (E. S. Kuznetsova, I. S. Stepanchikova)......................................... 146
Genus Tuckermannopsis (E. S. Kuznetsova) ................................................................ 152
Genus Usnocetraria (T. Randlane, A. Saag)................................................................ 158
Genus Vulpicida (T. Randlane, A. Saag)...................................................................... 160
Generic key for brown cetrarioid lichens (T. V. Makryi) ............................................. 163
Nomenclatural novelties .............................................................................................. 167
References .................................................................................................................... 168
Index of Latin names ................................................................................................... 178
Содержание
Рис. 2. Alectoria lata. 1 — таллом с апотециями; 2 — участок таллома с псевдоцифелла-
ми; 3 — апотеции. Масштабная линейка — 5 мм. Фото: И. С. Степанчикова.
Рис. 3. Alectoria ochroleuca. Общий вид лишайника: 1 — в естественной среде обитания
(тундра); 2 — гербарный образец. Масштабная линейка — 5 мм. Фото: И. С. Степанчикова.
Рис. 4. Alectoria sarmentosa subsp. sarmentosa. 1, 2 — общий вид в естественной среде
обитания; 2 — участок таллома с апотециями. Масштабная линейка — 5 мм. Фото:
И. С. Степанчикова.
Рис. 5. Alectoria sarmentosa subsp. vexillifera. 1, 2 — общий вид лишайника. Масштаб-
ная линейка — 5 мм. Фото: И. С. Степанчикова.
Рис. 6. Allocetraria madreporiformis. Общий вид лишайника (TU-76713). Масштабная
линейка — 1 см. Фото: А. Сааг.
Рис. 7. Allocetraria stracheyi. Общий вид лишайника (TU-88234). Масштабная линей-
ка — 1 см. Фото: А. Сааг.
Рис. 8. Arctocetraria andrejevii. Общий вид лишайника. Масштабная линейка — 1 см.
Фото: Т. В. Макрый.
Рис. 9. Arctocetraria nigricascens. Общий вид лишайника. Масштабная линейка —
5 мм. Фото: Т. В. Макрый.
Рис. 10. Asahinea chrysantha. 1, 2 — общий вид лишайника. Масштабная линейка —
1 см. Фото: Д. Е. Гимельбрант.
Рис. 11. Asahinea scholanderi. Общий вид лишайника. Масштабная линейка — 1 см.
Фото: Д. Е. Гимельбрант.
Рис. 12. Asahinea scholanderi. 1
—
апотеции; 2 — изидии. Масштабная линейка —
5 мм. Фото: И. С. Степанчикова.
Рис. 13. Bryocaulon divergens. Общий вид лишайника в естественной среде обитания
(каменистая тундра). Фото: С. В. Чесноков.
Рис. 14. Bryocaulon divergens. Гербарный образец с псевдоцифеллами. Масштабная
линейка — 2 мм. Фото: С. В. Чесноков.
Рис. 15. Bryocaulon hyperboreum. Общий вид лишайника, гербарный образец (общий
вид). Фото: С. В. Чесноков.
Рис. 16. Bryocaulon hyperboreum. Гербарный образец с псевдоцифеллами. Масштаб-
ная линейка — 2 мм. Фото: С. В. Чесноков.
Рис. 17. Bryocaulon pseudosatoanum. Общий вид лишайника в естественной среде
обитания. Фото: С. В. Чесноков.
Рис. 18. Bryocaulon pseudosatoanum. Гербарный образец с псевдоцифеллами. Мас-
штабная линейка — 2 мм. Фото: С. В. Чесноков.
Рис. 19. Стволы и ветви деревьев в старовозрастном лесу, густо обросшие лишайни-
ками рода Bryoria. Фото: И. Н. Урбанавичене.
Рис. 20. Bryoria americana. Фрагмент таллома. Масштабная линейка — 5 мм. Фото:
И. Н. Урбанавичене.
Рис. 21. Bryoria americana. Углы в местах ветвления и шипики. Масштабная линей-
ка — 3 мм. Фото: И. Н. Урбанавичене.
Рис. 22. Bryoria americana. Коричнево-черные псевдоцифеллы. Масштабная линей-
ка — 2 мм. Фото: И. Н. Урбанавичене.
Рис. 23. Bryoria asiatica. Боковые спинулы (шипики) и углы ветвления. Масштабная
линейка — 2 мм. Фото: И. Н. Урбанавичене.
Рис. 24. Bryoria asiatica. Коричневые псевдоцифеллы. Масштабная линейка — 2 мм.
Фото: И. Н. Урбанавичене.
Рис. 25. Bryoria bicolor. Двухцветный таллом с обильными третичными веточками.
Масштабная линейка — 5 мм. Фото: И. Н. Урбанавичене.
Рис. 26. Bryoria capillaris. Апотеции и углы ветвления. Масштабная линейка — 2 мм.
Фото: И. Н. Урбанавичене.
Рис. 27. Bryoria fremontii. Ветви и углы в местах ветвления. Масштабная линейка —
5 мм. Фото: И. Н. Урбанавичене.
Рис. 28. Bryoria fremontii. Желтые соралии. Масштабная линейка — 2 мм. Фото:
И. Н. Урбанавичене.
Рис. 29. Bryoria furcellata. Соралии с изидиоидными спинулами (шипиками). Мас-
штабная линейка — 2 мм. Фото: И. Н. Урбанавичене.
Рис. 30. Bryoria furcellata. Апотеции и соралии. Масштабная линейка — 2 мм. Фото:
И. Н. Урбанавичене.
Рис. 31. Bryoria fuscescens s. str. Повисающий таллом. Масштабная линейка — 10 см.
Фото: И. Н. Урбанавичене.
Рис. 32. Bryoria fuscescens s. str. Таллом с соралиями. Масштабная линейка — 5 мм.
Фото: И. Н. Урбанавичене.
Рис. 33. Bryoria fuscescens s. l. (B. chalybeiformis). Фрагмент таллома и соралии. Мас-
штабная линейка — 2 мм. Фото: И. Н. Урбанавичене.
Рис. 34. Bryoria fuscescens s. l. (B. lanestris). Фрагмент таллома с соралиями. Мас-
штабная линейка — 2 мм. Фото: И. Н. Урбанавичене.
Рис. 35. Bryoria fuscescens s. l. (B. lanestris). Ветви и углы ветвления. Масштабная ли-
нейка — 3 мм. Фото: И. Н. Урбанавичене.
Рис. 36. Bryoria fuscescens s. l. (B. subcana). Фрагмент таллома с соралиями. Масштаб-
ная линейка — 3 мм. Фото: И. Н. Урбанавичене.
Рис. 37. Bryoria glabra. Соралии, псевдоцифеллы и широкие, U-образные углы вет-
вления. Масштабная линейка — 2 мм. Фото: И. Н. Урбанавичене.
Рис. 38. Bryoria glabra. Повисающий таллом и U-образный тип ветвления. Масштаб-
ная линейка — 10 мм. Фото: И. Н. Урбанавичене.
Рис. 39. Bryoria implexa. Фрагмент таллома и соралии. Масштабная линейка — 5 мм.
Фото: И. Н. Урбанавичене.
Рис. 40. Bryoria implexa s. l. (B. osteola). Ветви и углы ветвления. Масштабная линей-
ка — 3 мм. Фото: И. Н. Урбанавичене.
Рис. 41. Bryoria kuemmerlleana. Соралии и углы ветвления. Масштабная линейка —
5 мм. Фото: И. Н. Урбанавичене.
Рис. 42. Bryoria kuemmerlleana. Реакция таллома с K. Масштабная линейка — 2 мм.
Фото: И. Н. Урбанавичене.
Рис. 43. Bryoria nadvornikiana. Веточки, спинулы и соралии, острые углы ветвления.
Масштабная линейка — 2 мм. Фото: И. Н. Урбанавичене.
Рис. 44. Bryoria nitidula. Острые углы в местах ветвления, веточки с латеральными
спинулами и коричневыми псевдоцифеллами. Масштабная линейка — 2 мм. Фото:
И. Н. Урбанавичене.
Рис. 45. Bryoria simplicior. Веточки и черноватые соралии. Масштабная линейка —
2 мм. Фото: И. Н. Урбанавичене.
Рис. 46. Bryoria simplicior. Фрагмент таллома с соралиями. Масштабная линейка —
10 мм. Фото: И. Н. Урбанавичене.
Рис. 47. Bryoria smithii. Веточки с соралиями и изидиоидными шипиками. Масштаб-
ная линейка — 2 мм. Фото: И. Н. Урбанавичене.
Рис. 48. Bryoria smithii. Соралии с изидиоидными шипиками. Масштабная линейка —
2 мм. Фото: И. Н. Урбанавичене.
Рис. 49. Bryoria tenuis. Двухцветные веточки и редкие спинулы. Масштабная линей-
ка — 10 мм. Фото: И. Н. Урбанавичене.
Рис. 50. Bryoria tenuis. Фрагмент таллома, углы ветвления. Масштабная линейка —
15 мм. Фото: И. Н. Урбанавичене.
Рис. 51. Bryoria trichodes. Участок таллома с латеральными шипиками (спинулами) и
псевдоцифеллами, с острыми углами в местах ветвления. Масштабная линейка — 2 мм.
Фото: И. Н. Урбанавичене.
Рис. 52. Bryoria trichodes. Псевдоцифеллы и пикниды. Масштабная линейка — 5 мм.
Фото: И. Н. Урбанавичене.
Рис. 53. Bryoria trichodes. Апотеции и псевдоцифеллы. Масштабная линейка — 3 мм.
Фото: И. Н. Урбанавичене.
Рис. 54. Bryoria trichodes. Аскоспоры. Масштабная линейка — 20 мкм. Фото:
И. Н. Урбанавичене.
Рис. 55. Bryoria vrangiana. Ветви, углы ветвления, соралии и псевдоцифеллы. Мас-
штабная линейка — 3 мм. Фото: И. Н. Урбанавичене.
Рис. 56. Cetraria aculeata. Конечные веточки лопастей. Масштабная линейка — 2 мм.
Фото: Т. В. Макрый.
Рис. 57. Cetraria aculeata subsp. steppae. Общий вид таллома и отдельная веточка.
Масштабная линейка — 5 мм. Фото: Т. В. Макрый.
Рис. 58. Cetraria crispiformis. Общий вид лишайника и отдельные лопасти: формы с
ямчатой и выраженной сетчато-ребристой поверхностью с многочисленными краевы-
ми срастаниями. Масштабная линейка — 5 мм. Фото: Т. В. Макрый.
Рис. 59. Cetraria ericetorum subsp. ericetorum. Общий вид лишайника и верхняя часть ло-
пастей с многочисленными краевыми срастаниями. Масштабная линейка — 1 см. Фото:
Т. В. Макрый.
Рис. 60. Cetraria islandica subsp. islandica. Общий вид стерильных и фертильных ло-
пастей. Масштабная линейка — 1 см. Фото: Т. В. Макрый.
Рис. 61. Cetraria islandica var. maculata. Общий вид стерильных лопастей. Масштаб-
ная линейка — 1 см. Фото: Т. В. Макрый.
Рис. 62. Cetraria islandica subsp. orientalis. Общий вид стерильных лопастей. Мас-
штабная линейка — 1 см. Фото: Т. В. Макрый.
Рис. 63. Cetraria kamczatica. Общий вид стерильных лопастей. Масштабная линей-
ка — 5 мм. Фото: Т. В. Макрый.
Рис. 64. Cetraria laevigata. Общий вид лопастей с многочисленными срастаниями на
уровне предкраевых выступов и псевдоцифеллами в виде ровной каймы. Две формы:
типичная (гладкая) и с мелкоямчато-морщинистой поверхностью. Масштабная линей-
ка — 1 см. Фото: Т. В. Макрый.
Рис. 65. Cetraria muricata. Конечные веточки лопастей (край дернинки). Масштабная
линейка — 2 мм. Фото: Т. В. Макрый.
Рис. 66. Cetraria nigricans. Общий вид лишайника. Масштабная линейка — 5 мм.
Фото: Т. В. Макрый.
Рис. 67. Cetraria odontella. Общий вид лишайника (край дернинки). Масштабная ли-
нейка — 5 мм. Фото: Т. В. Макрый.
Рис. 68. Cetraria rassadinae. Общий вид отдельных лопастей: верхняя и нижняя по-
верхности (голотип). Масштабная линейка — 5 мм. Фото: Т. В. Макрый.
Рис. 69. Cetraria sepincola. Общий вид лишайника. Масштабная линейка — 5 мм.
Фото: Т. В. Макрый.
Рис. 70. «Cetraria» annae. Общий вид (участок таллома). Масштабная линейка —
5 мм. Фото: Т. В. Макрый.
Рис. 71. Cetrariella delisei. Общий вид лишайника. Масштабная линейка — 5 мм.
Фото: Т. В. Макрый.
Рис. 72. Cetrariella fastigiata. Общий вид стерильных лопастей. Масштабная линей-
ка — 5 мм. Фото: Т. В. Макрый.
Рис. 73. Cetreliopsis asahinae. Общий вид лишайника (TU-88057). Масштабная линей-
ка — 1 см. Фото: А. Сааг.
Рис. 74. Cetreliopsis asahinae. Ламинальные псевдоцифеллы на верхней поверхности,
окруженные пикнидиальными выступающими повышениями (TU-88057). Масштаб-
ная линейка — 1 см. Фото: А. Сааг.
Рис. 75. Dactylina arctica subsp. arctica. Общий вид лишайника. Масштабная линей-
ка — 5 мм. Фото: И. С. Степанчикова.
Рис. 76. Dactylina arctica subsp. beringica. Общий вид лишайника. Масштабная линей-
ка — 5 мм. Фото: И. С. Степанчикова.
Рис. 77. Dactylina ramulosa. Общий вид лишайника. Масштабная линейка — 5 мм.
Фото: И. С. Степанчикова.
Рис. 78. Dactylina ramulosa. Общий вид лишайника. Масштабная линейка — 5 мм.
Фото: И. С. Степанчикова.
Рис. 79. Evernia divaricata. 1, 2 — общий вид лишайника. Масштабная линейка —
1 см. Фото: И. С. Степанчикова, Д. Е. Гимельбрант.
Рис. 80. Evernia divaricata. Общий вид лишайника. Масштабная линейка — 1 см.
Фото: И. С. Степанчикова, Д. Е. Гимельбрант.
Рис. 81. Evernia esorediosa. 1, 2 — общий вид лишайника с апотециями. Масштабная
линейка — 1 см. Фото: И. С. Степанчикова, Д. Е. Гимельбрант.
Рис. 82. Evernia mesomorpha. 1 — общий вид лишайника; 2 — апотеций. Масштабная
линейка — 1 см. Фото: И. С. Степанчикова, Д. Е. Гимельбрант.
Рис. 83. Evernia perfragilis. Общий вид лишайника. Масштабная линейка — 1 см.
Фото: И. С. Степанчикова.
Рис. 84. Evernia prunastri. 1, 2 — общий вид лишайника. Масштабная линейка —
1 см. Фото: И. С. Степанчикова, Д. Е. Гимельбрант.
Рис. 85. Evernia terrestris. Общий вид лишайника. Масштабная линейка — 1 см. Фото:
И. С. Степанчикова.
Рис. 86. Evernia terrestris. 1, 2 — общий вид лишайника. Масштабная линейка —
1 см. Фото: И. С. Степанчикова.
Рис. 87. Flavocetraria cucullata. Общий вид стерильных и фертильных лопастей. Мас-
штабная линейка — 1 см. Фото: Т. В. Макрый.
Рис. 88. Flavocetraria minuscula. Общий вид лишайника: формы с более или менее
извилисто вырезанными краями. Масштабная линейка — 5 мм. Фото: Т. В. Макрый.
Рис. 89. Flavocetraria nivalis. Общий вид стерильных и фертильных лопастей. Мас-
штабная линейка — 1 см. Фото: Т. В. Макрый.
Рис. 90. Gowardia arctica. Общий вид лишайника. Масштабная линейка — 5 мм.
Фото: И. С. Степанчикова.
Рис. 91. Gowardia nigricans. 1, 2 — общий вид лишайника. Масштабная линейка —
5 мм. Фото: И. С. Степанчикова.
Рис. 92. Letharia vulpina. 1 — общий вид лишайника. Масштабная линейка — 1 см.
Фото: З. М. Ханов. 2 — цилиндрические изидии. Масштабная линейка — 1 мм. Фото:
И. С. Степанчикова.
Рис. 93. Lethariella togashii. Общий вид лишайника. Масштабная линейка — 5 см.
Фото: А. А. Смирнов.
Рис. 94. Masonhalea inermis. Общий вид лишайника: верхняя и нижняя поверхности
лопастей, многочисленные апотеции. Масштабная линейка — 5 мм. Фото: Т. В. Ма-
крый.
Рис. 95. Masonhalea richardsonii. Общий вид лишайника: верхняя и нижняя поверхно-
сти лопастей. Масштабная линейка — 1 см (по: Макрый, 2002).
Рис. 96. Nephromopsis endocrocea. Общий вид лишайника (TU-85986) и оранжевая
сердцевина (TU-88065). Масштабная линейка — 1 см. Фото: А. Сааг.
Рис. 97. Nephromopsis komarovii. Общий вид лишайника (TU-88189). Масштабная ли-
нейка — 1 см. Фото: А. Сааг.
Рис. 98. Nephromopsis komarovii. Многочисленные краевые апотеции (TU-74076).
Масштабная линейка — 1 см. Фото: А. Сааг.
Рис. 103. Nephromopsis ornata. Общий вид лишайника (Япония, Хонсю, Националь-
ный парк Никко) и светло-желтая сердцевина (TU-53847). Масштабная линейка —
1 см. Фото: Т. Рандлане и А. Сааг.
Рис. 104. Nephromopsis pallescens. Общий вид лишайника (TU-88230). Масштабная
линейка — 1 см. Фото: А. Сааг.
Рис. 105. Nephromopsis pallescens. Нижняя беловатая сторона таллома (TU-88230).
Масштабная линейка — 1 см. Фото: А. Сааг.
Рис. 106. Nephromopsis pseudocomplicata. Общий вид лишайника (TU-88330). Мас-
штабная линейка — 1 см. Фото: А. Сааг.
Рис. 107. Nephromopsis rugosa. Общий вид лишайника с явно выраженным сетчатым
рисунком из ребер на верхней поверхности (TU-88327). Масштабная линейка — 1 см.
Фото: А. Сааг.
Рис. 108. Nephromopsis yunnanensis. Общий вид лишайника с пикнидиальными высту-
пами на сильно морщинисто-ребристой нижней поверхности (TU, s. n.). Масштабная
линейка — 1 см. Фото: А. Сааг.
Рис. 109. Oropogon asiaticus. Общий вид лишайника с апотециями. Масштабная ли-
нейка — 2 мм. Фото: И. С. Жданов.
Рис. 110. Platismatia glauca. 1, 2 — общий вид лишайника. Масштабная линейка —
1 см. Фото: И. С. Степанчикова, Д. Е. Гимельбрант.
Рис. 111. Platismatia interrupta. 1 — общий вид лишайника; 2 — верхняя поверх-
ность с псевдоцифеллами. Масштабная линейка — 1 см. Фото: И. С. Степанчикова,
Д. Е. Гимельбрант.
Рис. 112. Platismatia interrupta. Таллом: 1 — верхняя поверхность с псевдоцифелла-
ми; 2 — нижняя поверхность без псевдоцифелл. Масштабная линейка — 1 см. Фото:
И. С. Степанчикова, Д. Е. Гимельбрант.
Рис. 113. Platismatia lacunosa в естественной среде обитания (Командорские о-ва,
о. Медный, скальный останец). Таллом с апотециями. Масштабная линейка — 1 см.
Фото: И. С. Степанчикова.
Рис. 114. Platismatia norvegica. Таллом: 1 — верхняя поверхность с изидиями;
2 — нижняя поверхность. Масштабная линейка — 1 см. Фото: И. С. Степанчикова,
Д. Е. Гимельбрант.
Рис. 115. Tuckermannopsis chlorophylla. 1 — общий вид лишайника. Масштабная ли-
нейка — 1 см. 2 — соралии. Масштабная линейка — 5 мм. Фото: И. С. Степанчикова.
Рис. 116. Tuckermannopsis ciliaris. Общий вид лишайника с апотециями и развитыми
ресничками. Масштабная линейка — 1 см. Фото: И. С. Степанчикова.
Рис. 117. Tuckermannopsis ciliaris. Участок таллома с апотециями и пикнидами. Мас-
штабная линейка — 1 см. Фото: И. С. Степанчикова, Д. Е. Гимельбрант.
Рис. 118. Tuckermannopsis gilva. Общий вид лишайника. Масштабная линейка — 1 см.
Фото: И. С. Степанчикова.
Рис. 119. Tuckermannopsis orbata. Общий вид лишайника с апотециями и пикнидами.
Масштабная линейка — 1 см. Фото: И. С. Степанчикова.
Рис. 120. Tuckermannopsis subalpina. Общий вид лишайника с апотециями. Масштаб-
ная линейка — 1 см. Фото: И. С. Степанчикова.
Рис. 121. Usnocetraria oakesiana. Общий вид лишайника (TU-58915). Масштабная ли-
нейка — 1 см. Фото: А. Сааг.
Рис. 122. Vulpicida juniperinus. Общий вид таллома листоватого морфотипа (Эстония,
округ Сааре). Масштабная линейка — 1 см. Фото: А. Сааг.
Рис. 123. Vulpicida juniperinus. Общий вид таллома полукустистого морфотипа (Эсто-
ния, округ Сааре). Масштабная линейка — 1 см. Фото: А. Сааг.
Рис. 124. Vulpicida pinastri. Общий вид таллома с краевыми соралиями (Эстония,
округ Тарту). Масштабная линейка — 1 см. Фото: А. Сааг.
Íàó÷íîå èçäàíèå
Ôëîðà ëèøàéíèêîâ Ðîññèè: Ñåìåéñòâî Parmeliaceae.
Ì.; ÑÏá.: Òîâàðèùåñòâî íàó÷íûõ èçäàíèé ÊÌÊ.
2022. 187 ñ., 44 ÷/á âêë., 62 öâ. âêë.
Ïðè ó÷àñòèè ÈÏ Ìèõàéëîâà Ê.Ã.
Ðåäàêòîð: È. . Ñîêîëîâà
Îðèãèíàë-ìàêåò: Ò.À . Ãîðëèíà
Ïîäïèñàíî â ïå÷àòü 12.12.2021. Çàêàç 1
Ôîðìàò 70́100/16. Îáú̧ì 12 + 7 ï.ë. Óñë. ïå÷.ë. 24,7.
Áóìàãà îôñåòí. è ìåëîâ. Òèðàæ 300 ýêç. (1-é çàâîä)