



















































































































Author: Баркер Р. Барази С. Нил М.
Tags: нервная система невропатология неврология алкоголизм как болезнь нейрохирургия психиатрия медицина
ISBN: 5-9704-0280-X
Year: 2006




серия ЭКЗАМЕН НА ОТЛИЧНО
V
Издательская группа
ГЭОТАР-Медиа
Р. Баркер, С. Барази, М. Нил
НАГЛЯДНАЯ
НЕВРОЛОГИЯ
Neuroscience at a Glance
ROGER A. BARKER
BA.MBBS. MRCP. PhD
Cambridge Centre for Brain Repair and Department of Neurology
University of Cambridge
Robinson Way
Cambridge
STEPHEN BARASI
BSc, PhD
School of Bioscience
Cardiff University
Museum Avenue
Cardiff
and Neuropharmacology by
MICHAEL J. NEAL
DSc, PhD, MA, BPharm
Professor and Chairman of the
Division of Pharmacology and Toxicology
United Medical and Dental Schools of Guy’s and St Thomas’s Hospitals (UMDS)
Department of Pharmacology
St Thomas’s Hospital. London
SECOND EDITION
Blackwell
Publishing
СЕРИЯ ЭКЗАМЕН НА ОТЛИЧНО
Р.Баркер, С.Барази, М. Нил
^гладям
щрожги;]
Перевод с английского
Под редакцией чл.-корр. РАМН,
проф. В.И. Скворцовой
Москва
Издательская группа
ГЭОТАР-М ед и а
2006
УДК 616.8(084.1)
ББК 56.1я6
Б25
Баркер Р.
П47 Наглядная неврология : Учеб, пособие Р. Баркер, С. Барази, М.Нил / Пер. с англ. Г.Н. Левицкого; под
ред. В.И. Скворцовой. — М.: ГЭОТАР-Медиа, 2006. — 136 с.: ил. (Серия "Экзамен на отлично").
ISBN 5-9704-0280-Х (рус.)
ISBN 1-4051-1124-0 (англ.)
Учебное пособие знакомит с целым рядом проблем современной неврологии. В нем рассмотрены
основные вопросы анатомии, биохимии и физиологии нервной системы человека. Несмотря на
теоретическую направленность, информация представлена в клиническом аспекте. Второе издание
дополнено четырьмя главами, посвященными проблемам клинической неврологии, что позволяет
студентам наиболее полно ознакомиться с предметом.
Предназначено студентам медицинских вузов.
УДК 616.8(084.1)
ББК56.1я6
Это издание опубликовано с согласия Blackwell Publishing Ltd, Oxford. Перевод осуществлен издательской группой
"ГЭОТАР-Медиа". Blackwell Publishing Ltd, Oxford не несет ответственности за качество перевода.
ISBN 5-9704-0280-Х(рус.)
ISBN 1-4051-1124-0 (англ.)
'Q R.A. Barker, S. Barasi, M.J. Neal.
Neuroscience at a glance. Second Edition, 2003
'Q Г.Н. Левицкий, перевод, 2005
'Q Издательская группа «ГЭОТАР-Медиа», 2005
Содержание
Предисловие к изданию на русском языке . .6
Введение. .7
Благодарноств. 7
Первое издание. .7
Список сокращений 8
ЧАСТЬ I. Анатомическая и функционалвная
организация нервной системы .10
Глава 1. Анатомия нервной системвг . 10
Глава 2. Развитие нервной системы .12
Глава 3. Клетки нервной системв! I: нейронв! .14
Глава 4. Клетки нервной системв! II:
нейроглиалвнв!е клетки 16
Глава 5. Ионнв!е каналв: . 18
Глава 6. Мембрана в состоянии покоя
и потенциал действия. .20
Глава 7. Нервно-мышечное соединение и синапсы ....22
Глава 8. Проведение по нерву
и синаптическая интеграция. . 24
Глава 9. Нейротрансмиттеры, рецепторы
и нейротрансмиттерные системы .26
Глава 10. Структура скелетной мышцы. ..28
Глава 11. Сокращение скелетной мышцы 30
Глава 12. Анатомия спинного мозга. 32
Глава 13. Анатомия ствола мозга.. . 34
Глава 14. Черепно-мозговые нервы 36
Глава 15. Анатомия коры головного мозга. 38
Глава 16. Оболочки мозга
и цереброспиналвная жидкость . . 40
Глава 17. Кровоснабжение
центральной нервной системы. 42
ЧАСТЬ II. Чувствительные анализаторы .44
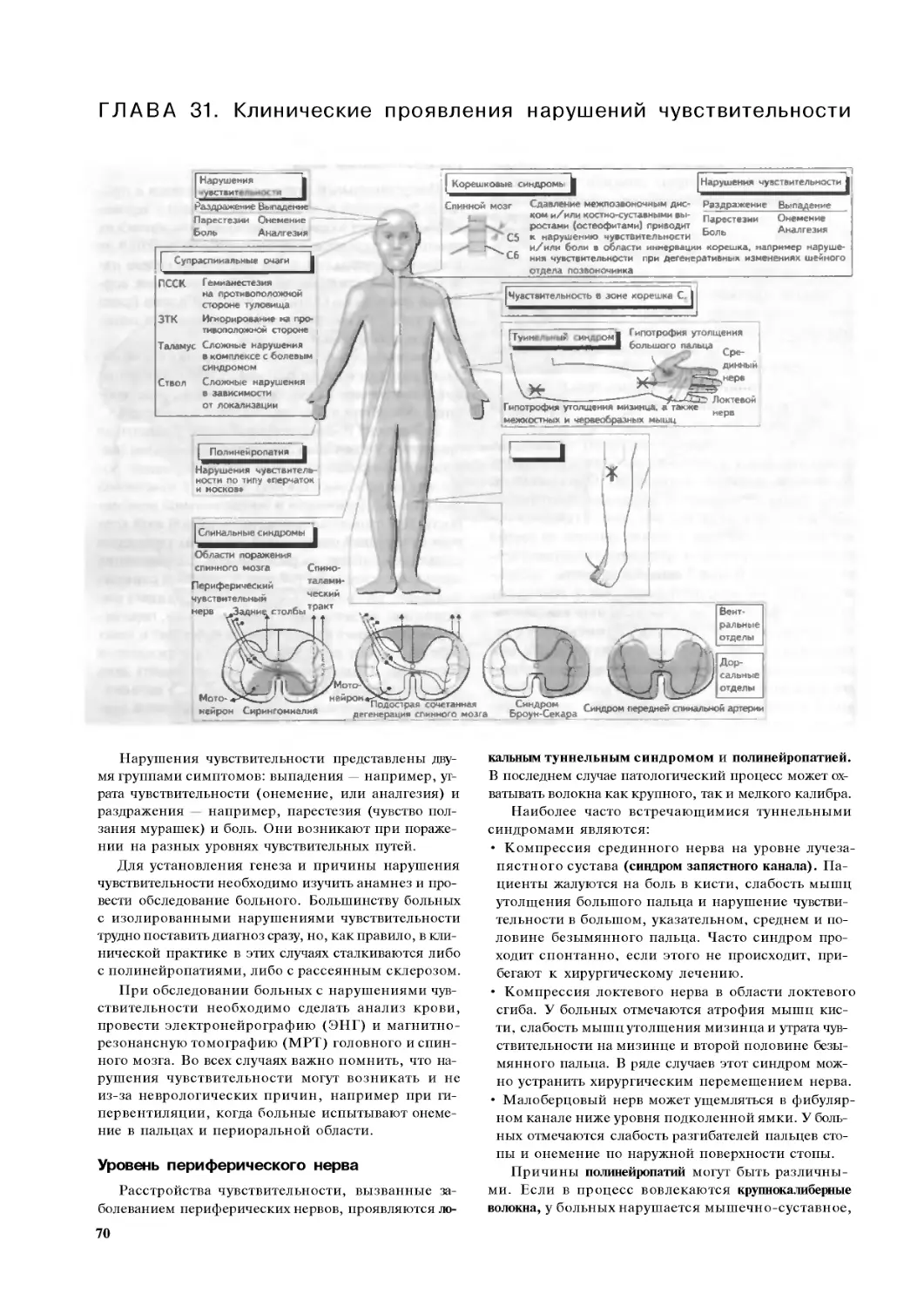
Глава 18. Чувствительность 44
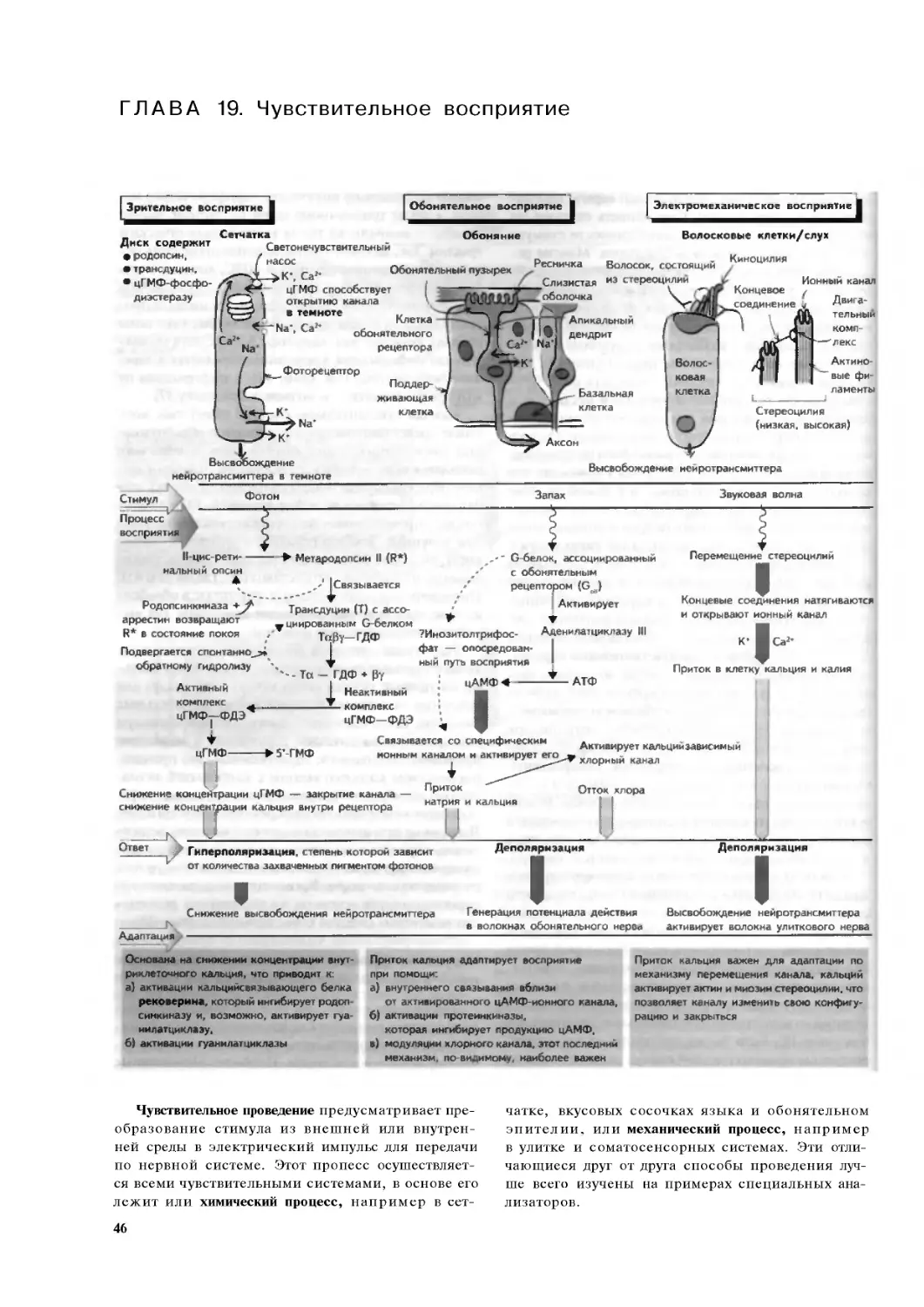
Глава 19. Чувствительное восприятие .. .46
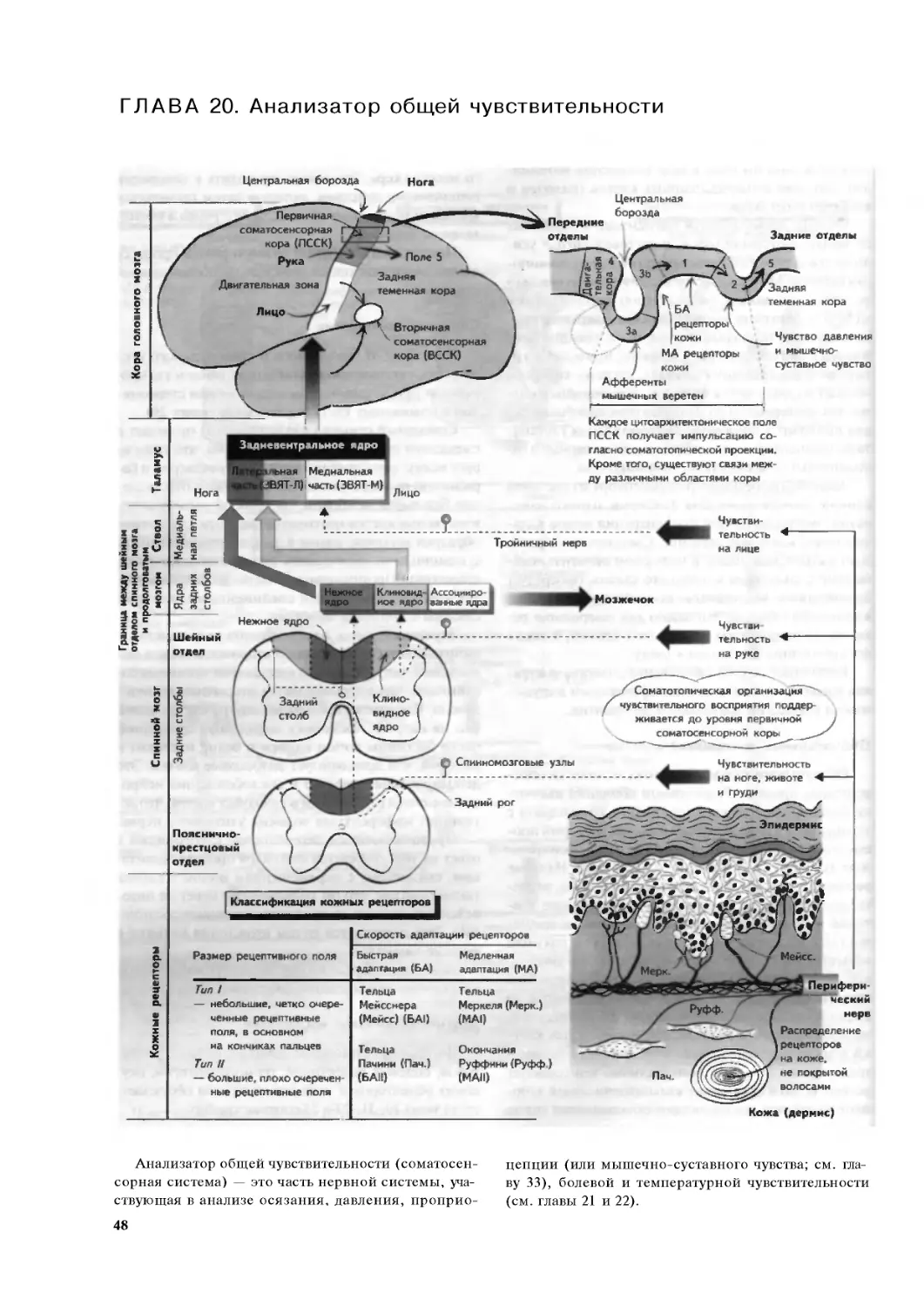
Глава 20. Анализатор общей чувствительности 48
Глава 21. Система боли I: ноцицепторы
и ноцицептивные пути . .50
Глава 22. Система боли II: способы
медикаментозного и немедикаментозного лечения.... 52
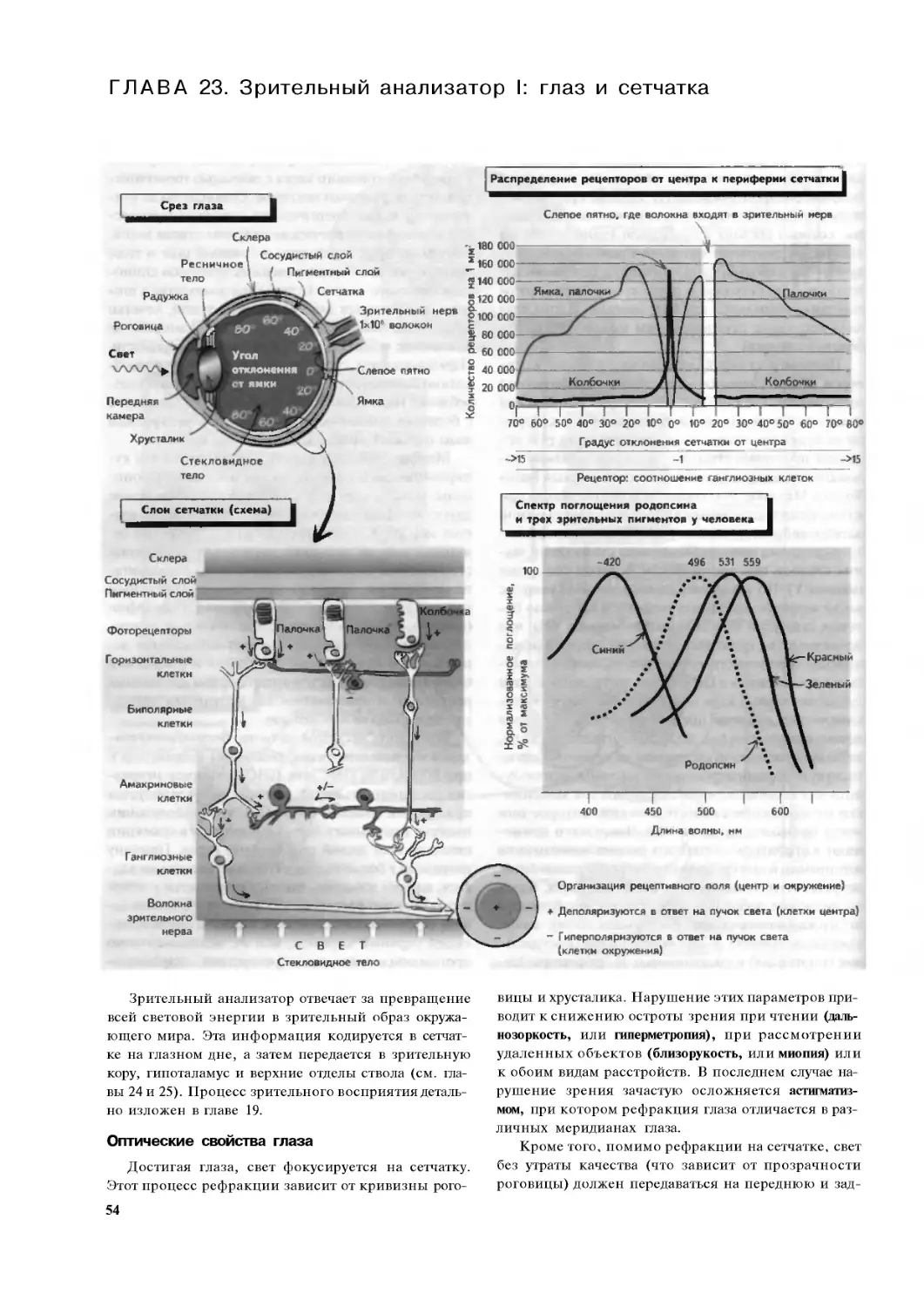
Глава 23. Зрительный анализатор 1:
глаз и сетчатка. . 54
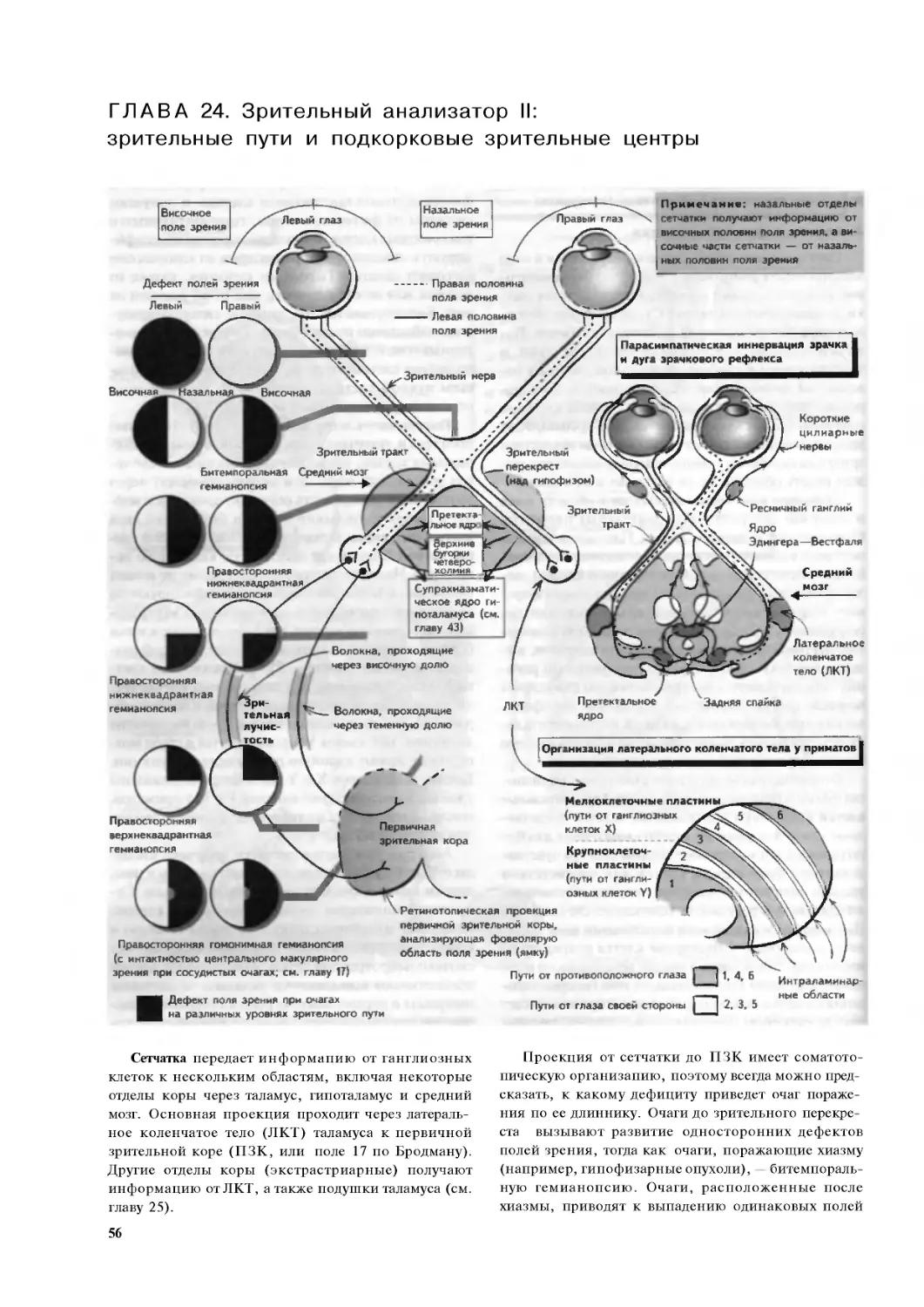
Глава 24. Зрительный анализатор II: зрительные
пути и подкорковые зрительные центры . .56
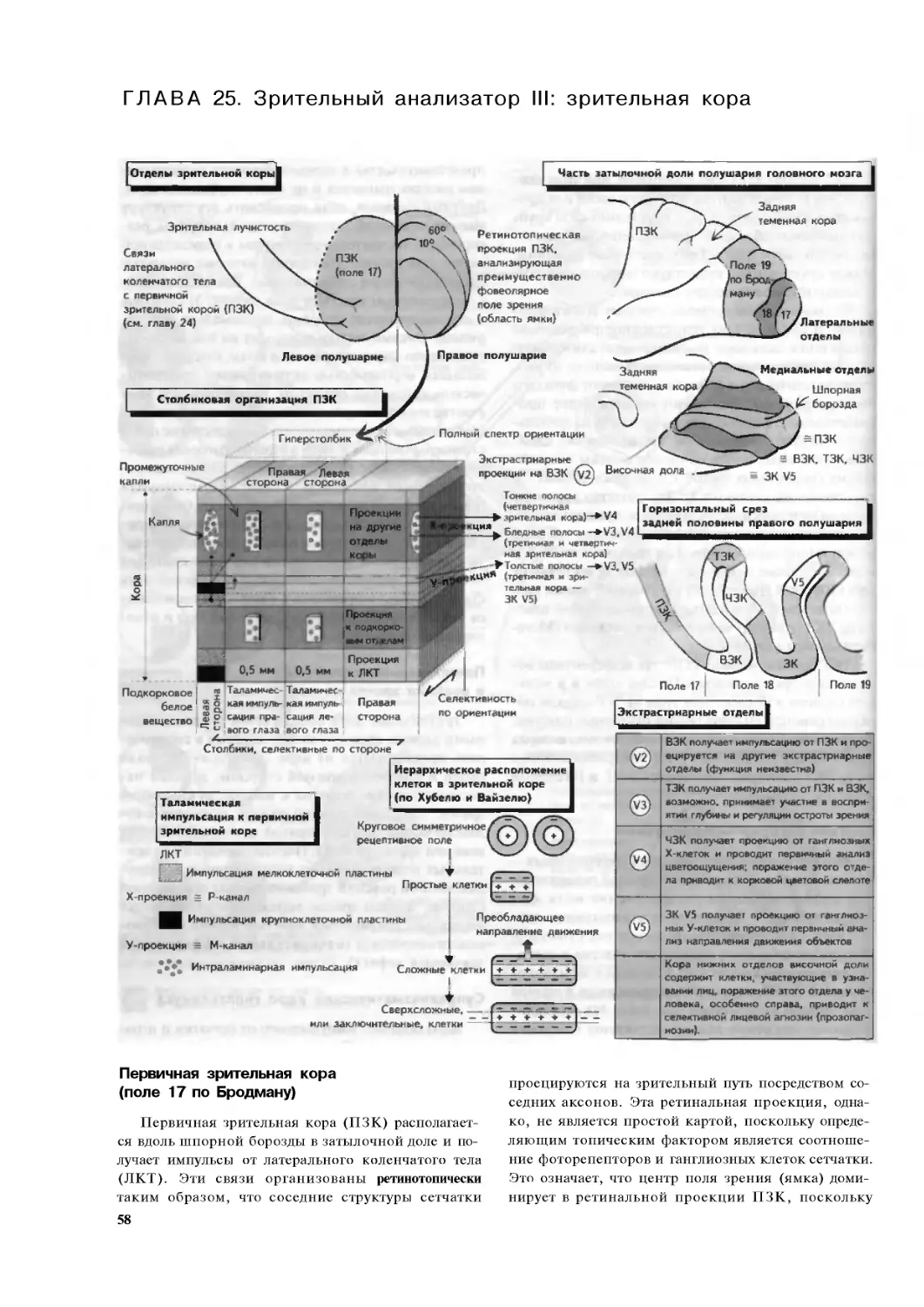
Глава 25. Зрительный анализатор III:
зрительная кора. . 58
Глава 26. Слуховой анализатор I: ухо и улитка... .60
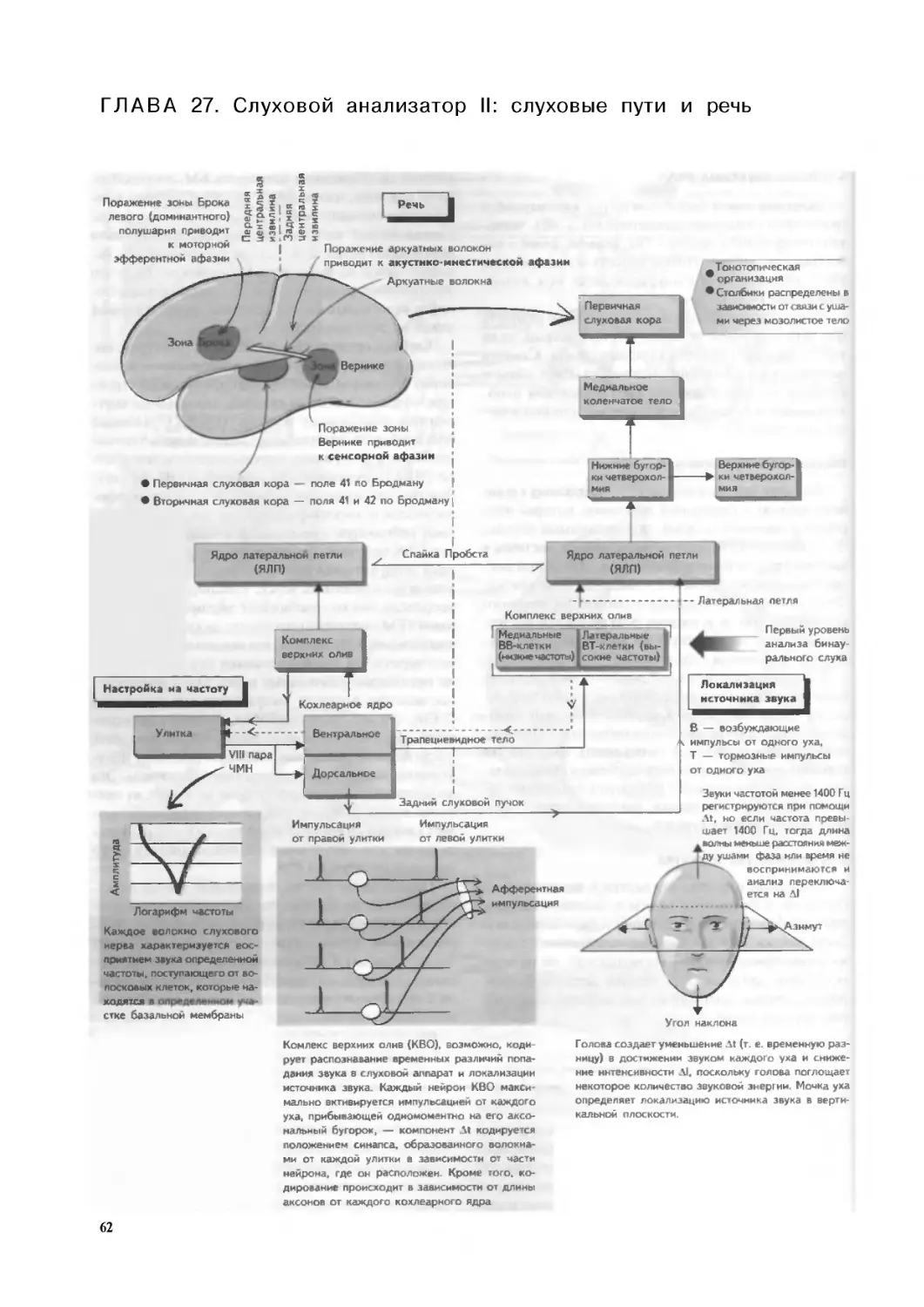
Глава 27. Слуховой анализатор II:
слуховые пути и речь. . .62
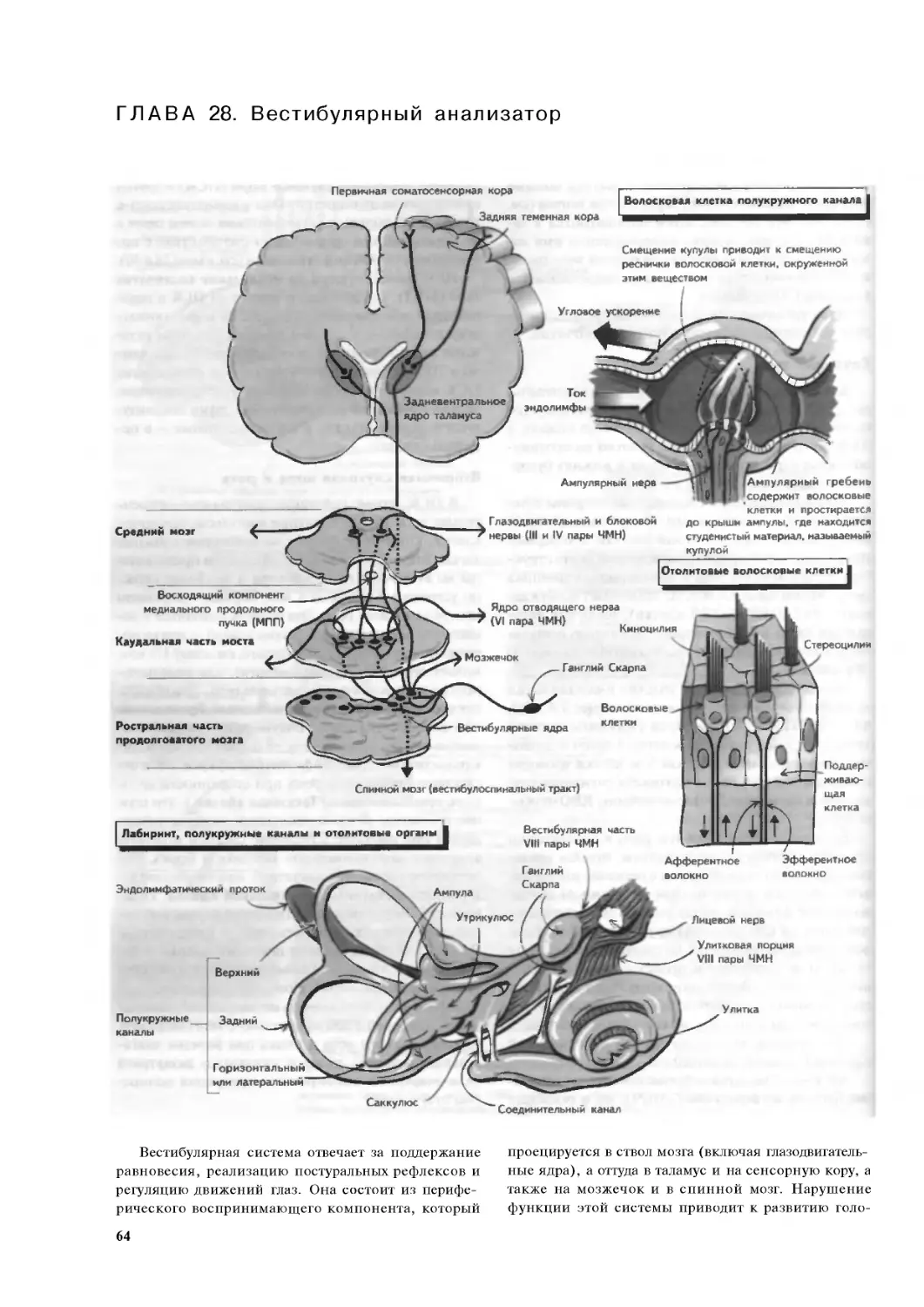
Глава 28. Вестибулярный анализатор. .64
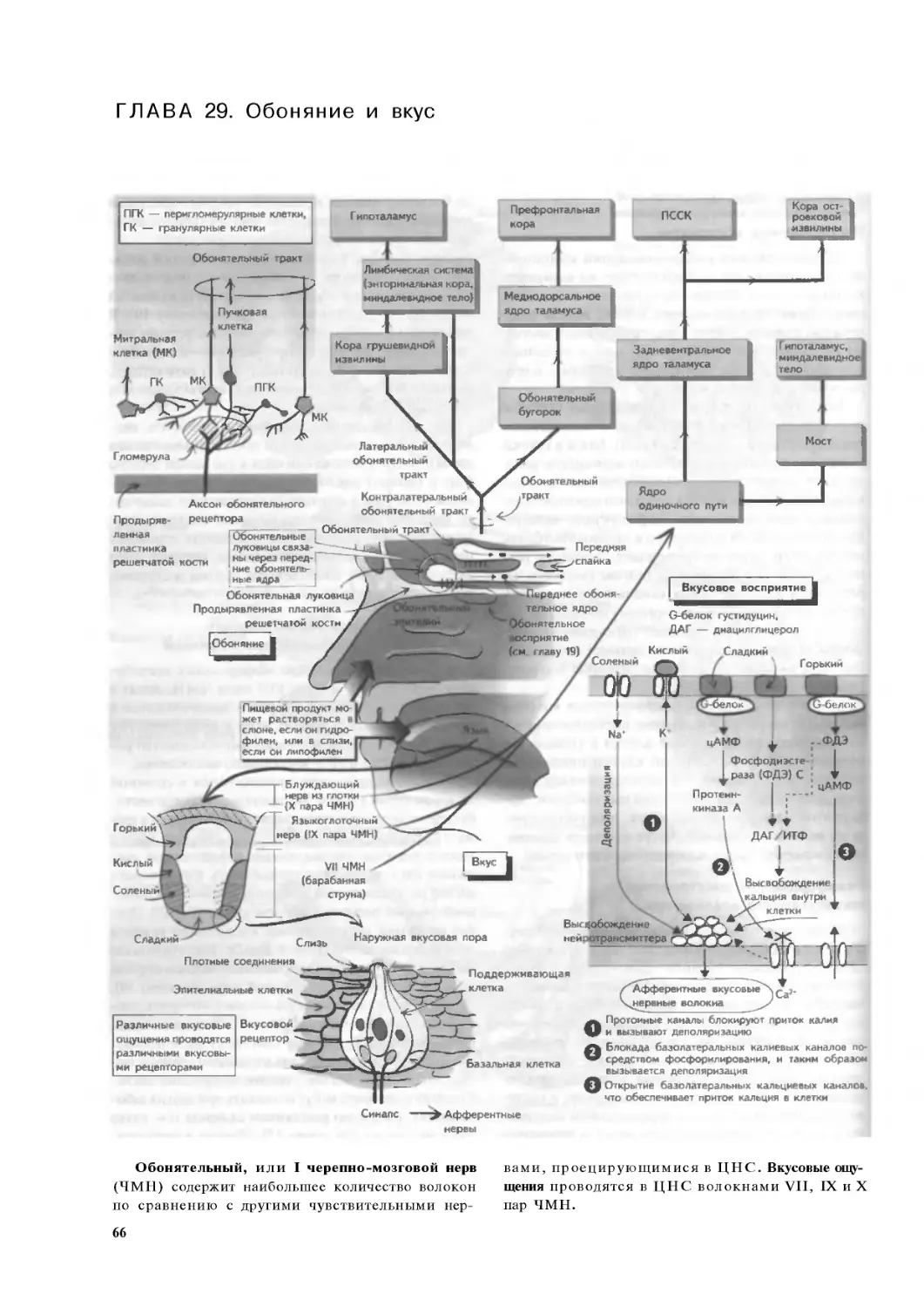
Глава 29. Обоняние и вкус .. .. .. .66
Глава 30. Ассоциативная кора:
задняя теменная и префронтальная кора. . . ...68
Глава 31. Клинические проявления
нарушений чувствительности. .70
ЧАСТЬ III. Двигательный анализатор. . 72
Глава 32. Анатомия двигательного анализатора. 72
Глава 33. Мышечное веретено
и периферический мотонейрон. 74
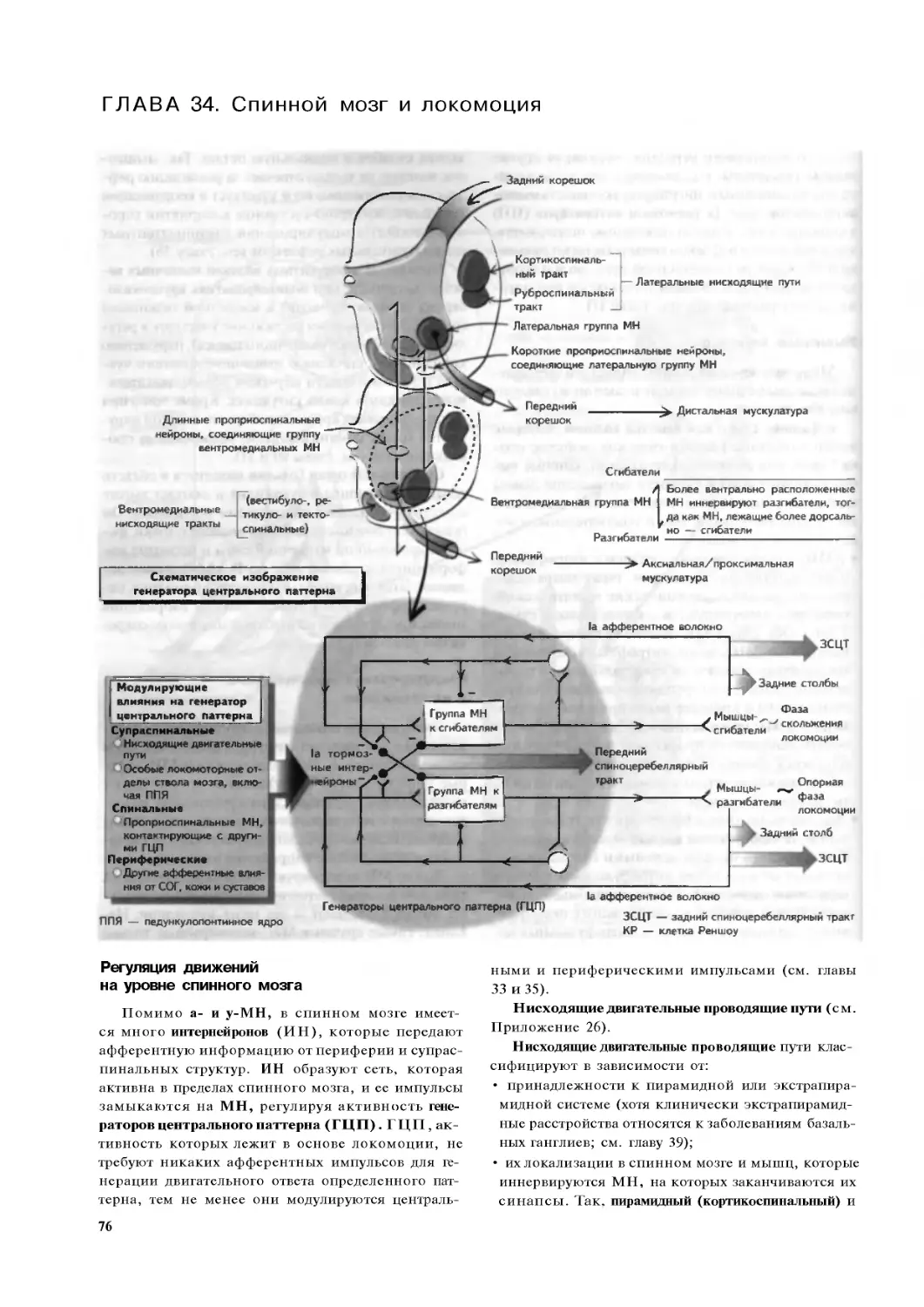
Глава 34. Спинной мозг и локомоция 76
Глава 35. Двигательные зоны коры 78
Глава 36. Первичная моторная кора. . . .80
Глава 37. Мозжечок .82
Глава 38. Базальные ганглии:
анатомия и физиология . 84
Глава 39. Заболевания базальных
ганглиев и их лечение. . 86
Глава 40. Движения глаз . . 88
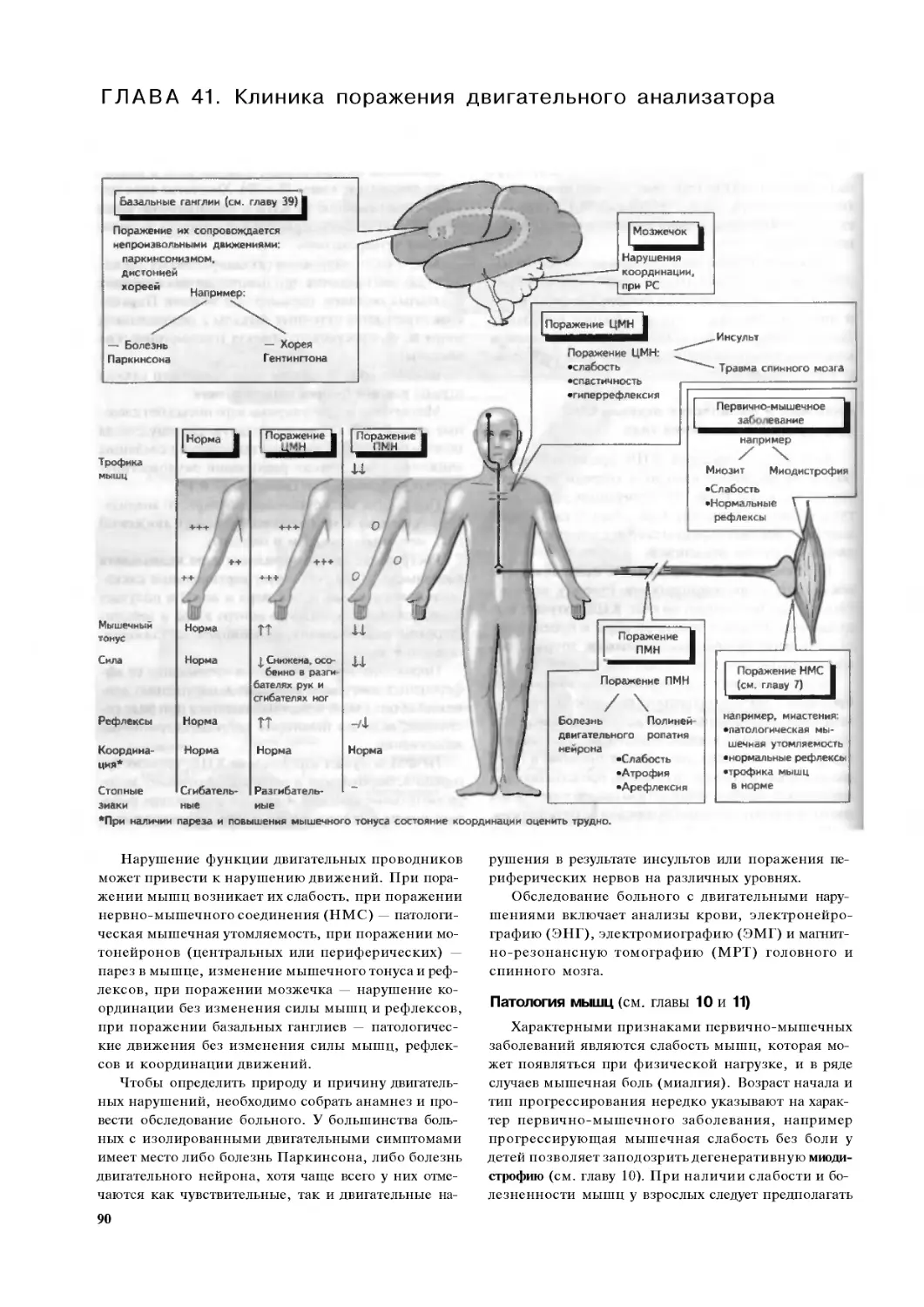
Глава 41. Клиника поражения
двигательного анализатора. .90
ЧАСТЬ IV. Вегетативная, лимбическая системы,
ствол мозга и пластичность . 92
Глава 42. Вегетативная нервная система . .92
Глава 43. Гипоталамус. .94
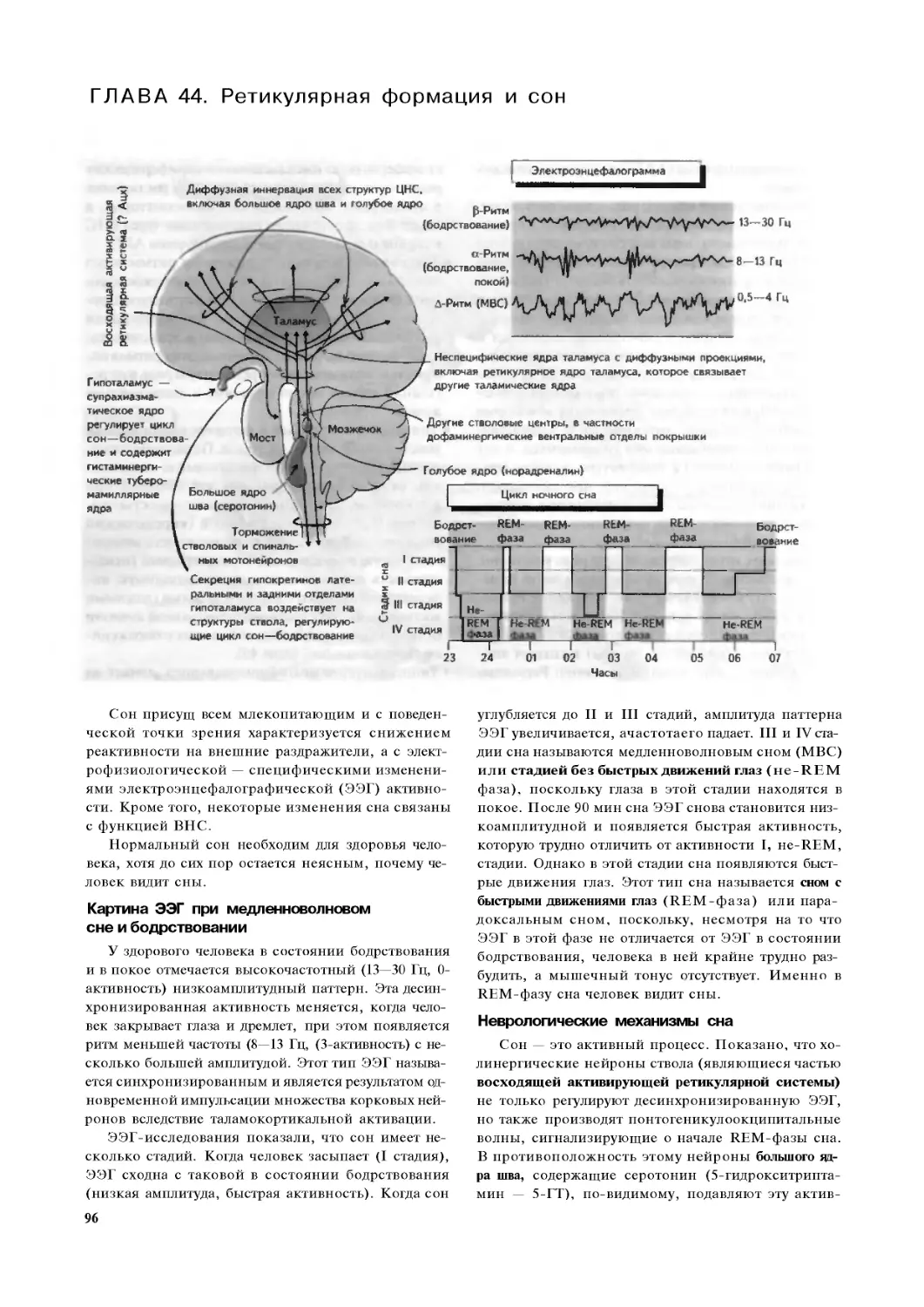
Глава 44. Ретикулярная формация и сон 96
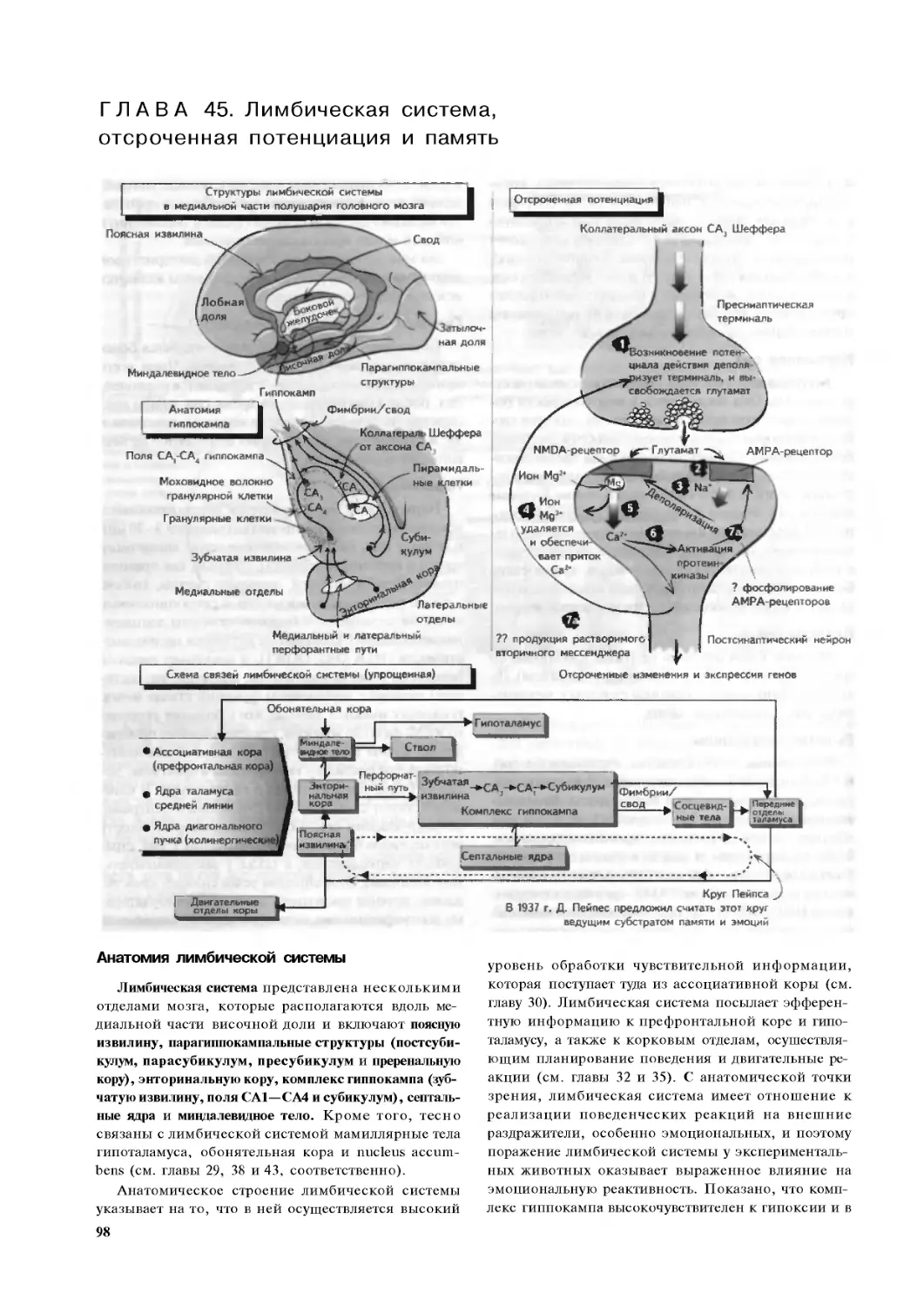
Глава 45. Лимбическая система,
отсроченная потенциация и память. 98
Глава 46. Пластичность нервной системы
и нейротрофические факторы I.
периферическая нервная система. .100
Глава 47. Пластичность нервной системы
и нейротрофические факторы II:
центральная нервная система. 102
ЧАСТЬ V. Заболевания нервной системы 104
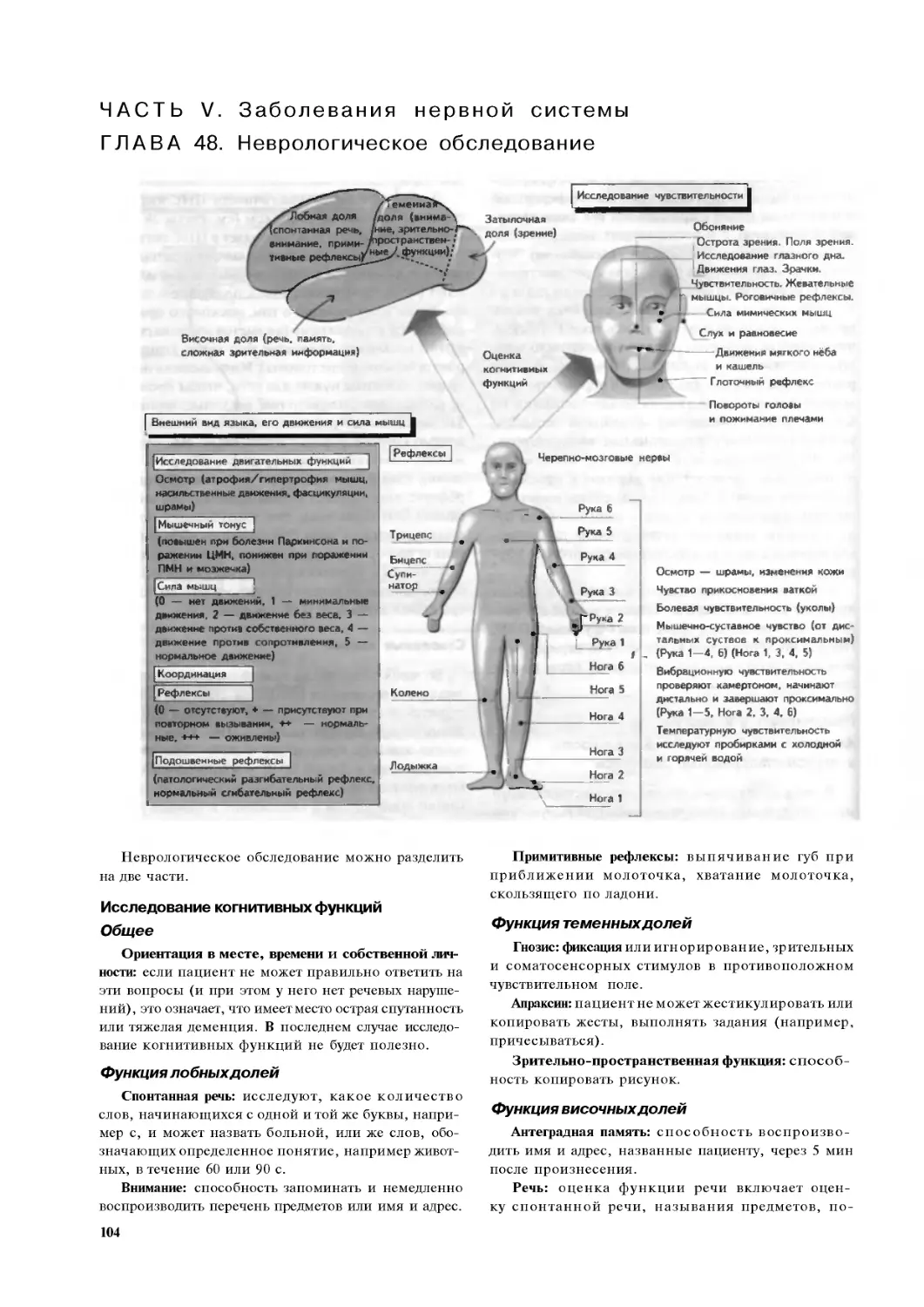
Глава 48. Неврологическое обследование . 104
Глава 49. Обследование нервной системы 106
Глава 50. Нейрохимические нарушения I:
аффективные расстройства и шизофрения. .108
Глава 51. Нейрохимические нарушения II:
тревожные расстройства. .......... ПО
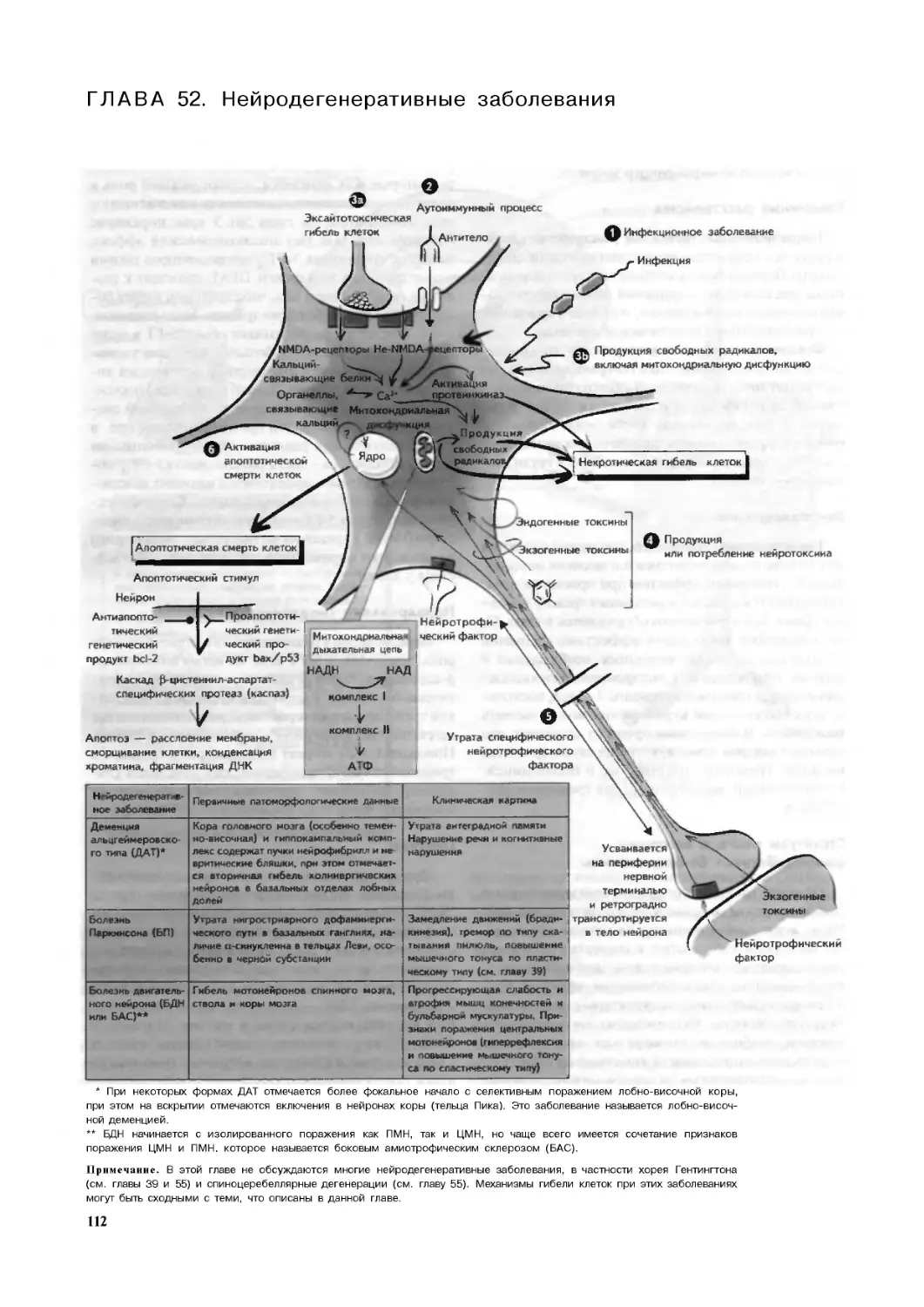
Глава 52. Нейродегенеративные заболевания .112
Глава 53. Нейрофизиологические заболевания:
эпилепсия. .114
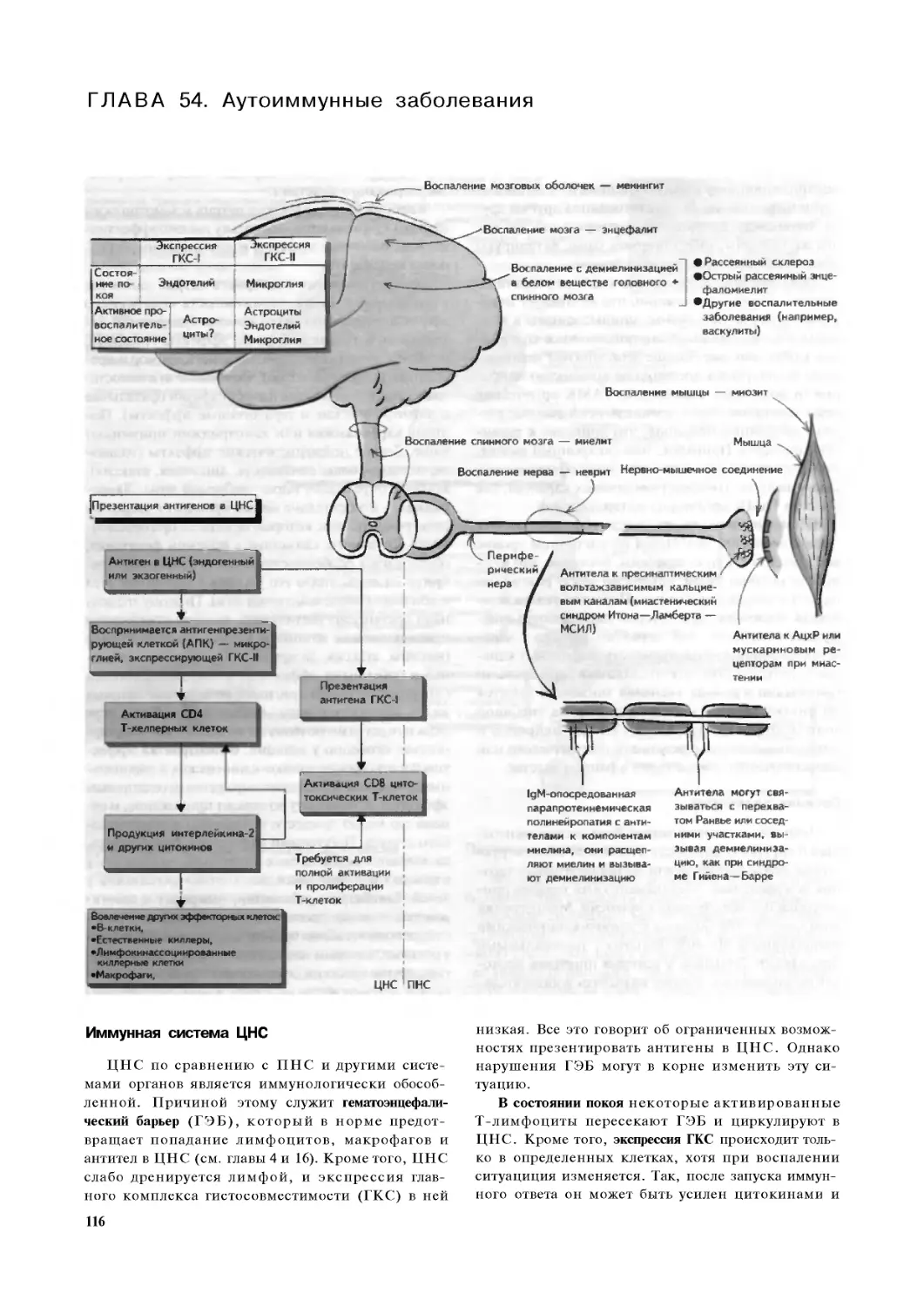
Глава 54. Аутоиммунные заболевания. 116
Глава 55. Наследственные
неврологические заболевания .118
Приложение 1. Основные виды
нейротрансмиттеров . .120
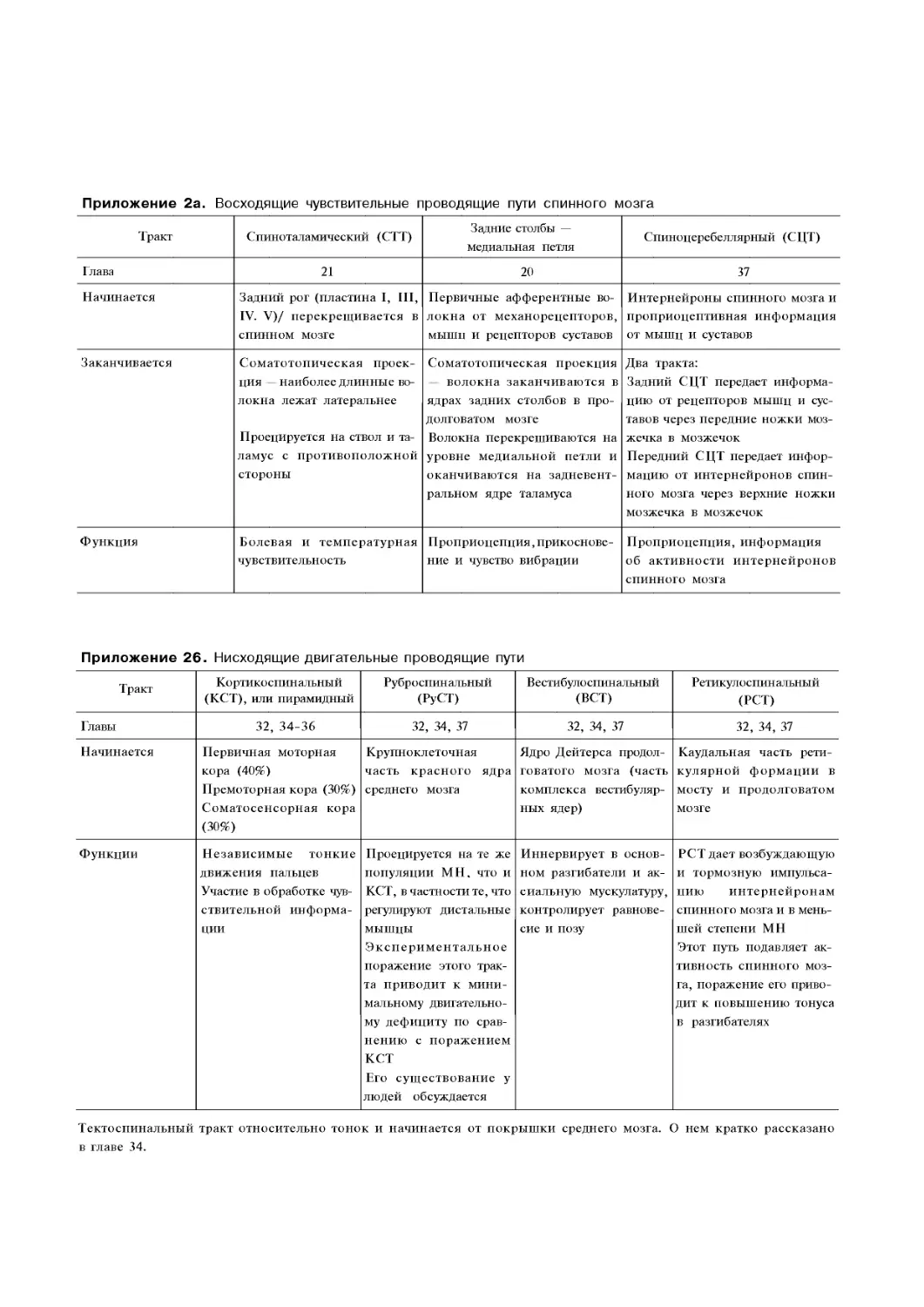
Приложение 2а. Восходящие чувствительные
проводящие пути спинного мозга. 121
Приложение 26. Нисходящие двигательные
проводящие пути. . 121
Приложение 3. Анатомия и физиология мозжечка . 122
Алфавитный указатель...........................123
Предисловие к изданию на русском языке
Издание книги «Наглядная неврология» (серия
«Экзамен на отлично») на русском языке отражает
поступательное развитие этой области медицины.
Предлагаемый вашему вниманию учебник был под-
готовлен группой английских авторов, экспертов по
основным разделам фундаментальной и клиничес-
кой неврологии. Особенностью учебника является
постановка акцента на фундаментальные аспекты
неврологии — анатомию, развитие нервной системы,
нейрофизиологию и нейрохимию, через призму ко-
торых поданы основы неврологической семиотики
и нозологии. При рассмотрении фундаментальных
вопросов авторы дают краткий исторический обзор
формирования современных научных представле-
ний: от первых экспериментальных данных до пос-
ледних открытий, сделанных представителями ба-
зисных нейронаук.
Структура учебника соответствует возрастающим
требованиям, которые предъявляет быстро развива-
ющаяся фундаментальная неврология врачу-невро-
логу. В настоящее время топическая и нозологичес-
кая диагностика, а также лечение неврологических
заболеваний невозможны без знания клиническим
неврологом молекулярно-генетических, нейрохими-
ческих и нейрофизиологических основ этих заболе-
ваний. Об этом, например, свидетельствует пере-
смотр классификапий многих неврологических за-
болеваний, который осуществляется с позиций
выделения нозологических форм по принципу пер-
вичных генетических и биохимическихдефектов, ле-
жащих в их основе, а не только по принципу сход-
ства или различия клинической картины.
Интегрированный подход к изучению невроло-
гии, предлагаемый авторами учебника, позволяет
студентам повторить знания, полученные на ранних
курсах медицинского вуза, и освоить новый матери-
ал по клинической неврологии, не обращаясь к боль-
шому количеству дополнительной литературы по
фундаментальным вопросам, которые достаточно
подробно разбираются в данной книге. В то же вре-
мя вопросы клинической неврологии освещены в
учебнике обзорно, поэтому он может служить лишь
в качестве источника дополнительной информации
к материалу, излагаемому в используемых в настоя-
щее время учебниках по неврологической семиоти-
ке и нозологии.
«Наглядная неврология» предназначена для сту-
дентов-медиков, интернов и ординаторов, изучаю-
щих нервные болезни, а также для всех, кто вовле-
чен в оказание медицинской помощи неврологичес-
ким больным.
В.И. Скворцова,
член-корреспондент РАМН,
профессор, зав. кафедрой фундаментальной
и клинической неврологии
Российского государственного
медицинского университета,
главный ученый секретарь
Всероссийского общества неврологов
Введение
«Наглядная неврология» предназначена для сту-
дентов-медиков. Она не является отчетом о дости-
жениях науки о нервной системе, а скорее содержит
обзор основных аспектов неврологии. Мы не стре-
мились дать систематический обзор клинической
неврологии. Однако книга полезна и практикующим
врачам в качестве обзора научных аспектов, кото-
рые находятся за пределами клинической практики.
В нашей стране характер обучения медицине по-
степенно изменялся. Если раньше медицинское обу-
чение было ориентировано на отдельные дисципли-
ны (анатомия, физиология, фармакология и т.д.),
то сейчас оно носит проблемно-ориентированный
характер. Студенты, безусловно, извлекут больше
пользы из интегрированного материала, сосредото-
ченного на определенной системе.
В книге суммированы представления о быстро
развивающейся науке неврологии, при этом в ней
даются ссылки на определенные заболевания, т. е.
материал подан в клиническом контексте. В завер-
шающих главах представлено больше клинического
материала, а в начальных главах дан срез всех пос-
ледующих глав в контексте прикладной неврологии.
Однако изучение организации нервной системы на
примере отдельных заболеваний имеет узконаправ-
ленный характер, который меняется. Те аспекты не-
врологии, которые, как полагают сегодня, имеют
относительное клиническое значение, в будущем
могут оказаться наиболее важными. В качестве при-
мера можно привести ионные каналы — совсем не-
давно было открыто множество заболеваний, свя-
занных с патологией ионных каналов. Поэтому в
некоторых главах основной упор мы делаем на фун-
даментальных механизмах и меньше внимания уде-
ляем клиническим аспектам.
В каждой главе информация представлена в виде
рисунка с подписями, которые объясняются в тек-
сте. Желательно, чтобы читатель каждый раз возвра-
щался к рисунку в процессе чтения, а не просто изу-
чил его в отдельности. Каждая глава несет в себе не-
кую квинтэссенцию, поданную как дидактический
материал. При этом на обратной стороне обложки
мы даем ссылку на веб-сайт "Blackwell Publishing",
где читатель сможет ознакомиться с обсуждаемыми
вопросами более подробно. В тексте основное вни-
мание уделяется базисному материалу, но включе-
ны и некоторые важные дополнительные детали.
Книга разделена на разделы: биофизика нервной
системы (главы 1- 17); чувствительность (главы 18—
31); двигательные функции (главы 32—41); вегетатив-
ная нервная система, лимбическая система и ство-
ловые системы, отвечающие за сон и бодрствование,
а также пластичность нервной системы (главы 42—
47); подходы к неврологическому обследованию в
соответствии с классификацией неврологических
заболеваний (главы 48—55). Списокдополнительной
литературы и словарь терминов неврологических
заболеваний и неврологии, доступен на веб-сайте
"Blackwell Publishing". Подробности на обороте об-
ложки.
Каждый раздел до некоторой степени построен
на предыдущих. Чтение вводных глав может значи-
тельно облегчить понимание последующих глав раз-
дела, например главу о соматосенсорной системе
лучше прочесть после главы, которая посвящена
общей организации чувствительности.
Благодарность
Мы внесли некоторые изменения в новое изда-
ние книги. Мы стремились достичь максимальной
интеграции нейронаук в клиническую неврологию
и наоборот и включили в это издание больше кли-
нических аспектов. Появились новые главы о невро-
логическом обследовании, а также резюмирующие
главы о нарушении чувствительных и двигательных
функций в заключении разделов 2 и 3. При этом мы
сохранили информацию о клинических аспектах в
копне каждого раздела. Кроме того, мы изменили
расположение главы о пластичности нервной сис-
темы. Теперь она находится в разделе 4 после обсуж-
дения вопросов, касающихся лимбической системы
и отсроченной потенциации.
Книга появилась благодаря нашей практике пре-
подавания в течение последних нескольких лет, а
также отзывам, которые мы на нее получали. Мы
благодарим Ф. Гудгейм и Джеральдину Джефферс,
сотрудников «Blackwell Publishing*, а также Элесдера
Коулза из Кембриджа за терпеливое чтение нового
издания и предложения по его усовершенствованию.
Р. Баркер, С. Барази,
Кембридж, Кардифф
Ноябрь 2002 г.
Первое издание
Содержанию книги «Наглядная неврология» мы
во многом обязаны студентам, которые с трудом
осваивали быстро развивающуюся неврологию. Мы
благодарим огромное число людей за помощь в на-
писании книги; это наши коллеги из Кембриджа и
Кардиффа, которые терпеливо читали различные
варианты рукописи и честно высказывали свое мне-
ние. Кроме того, мы выражаем особую благодар-
ность Эндрю Лернеру за усердную литературную
правку, исправление ошибок и нечетко сформули-
рованных объяснений, а также Майку Нилу за на-
писание главы по нейрофармакологии, которая со-
держит важную дополнительную информацию.
Мы благодарим Майка Стейна и Патрисию Хар-
дкастл, сотрудников «Blackwell Publishing» за их эн-
тузиазм и поддержку этого проекта. Мы благодарны
нашим семьям, которые разрешили нам потратить
так много времени на эту книгу.
Р. Баркер и С. Барази
Кембридж, Кардифф,
май 1998 г.
7
Список сокращений
5-ГТ - 5-гидрокситриптамин (серотонин)
5-ГТИУК - 5-гидроксииндолуксусная кислота
AMPA-R - а-амино-3-гидрокси-5-метил-4-изо-
ксазолпропионовокислый глутамат-
ный рецептор
СТТ — спиноталамический тракт
АДГ антидиуретический гормон (вазопрес-
син)
АДФ — аденозиндифосфат
АКТГ - адренокортикотропный гормон
АТФ - аденозинтрифосфат
АуД — аутосомно-доминантный
АуР — аутосомно-рецессивный
Ацх - ацетилхолин
АцхР - ацетилхолиновый рецептор
АцхЭ - ацетилхолинэстераза
БАС — боковой амиотрофический склероз
БДН — болезнь двигательного нейрона
БМ - базальная мембрана
БО - белок остеогенеза
БШв - внутренний сегмент бледного шара
БШн — наружный сегмент бледного шара
БЯШ - большое ядро шва
ВКЯ — вентральное кохлеарное ядро
ВНС — вегетативная нервная система
ВП — вызванный потенциал
ВПМР - высокопороговый механорецептор
ВПСП - возбуждающий постсинаптический по-
тенциал
ВСА - внутренние сонные артерии
ВССК - вторичная соматосенсорная кора
ВСТ - вестибулоспинальный тракт
ВЦМП - время центрального моторного прове-
дения
ГАМК - у-аминомасляная кислота
ГАМК-Р - рецептор у-аминомасляной кислоты
ГДК -глутаматдекарбоксилаза
ГК — гранулярная клетка
ГКГ - главный комплекс гистосовместимости
ГНТФ - глиальный нейротрофический фактор
ГР — глутаматный рецептор
ГТФ — гуанозинтрифосфат
Гц - герц
ГЦП — генератор центрального паттерна
ГЭБ - гематоэнцефалический барьер
ДАТ - диацилглицерол
ДАТ - деменция альцгеймеровского типа
дБ — децибел
ДКЯ — дорсальное кохлеарное ядро
ДМ3 — дополнительная моторная зона
ДНК — дезоксирибонуклеиновая кислота
ДОФА - дофамин
ЗВЯТ - задневентральное ядро таламуса
ЗВЯТ-Л — латеральная часть задневентрального
ядра таламуса
ЗВЯТ-М - медиальная часть задневентрального
ядра таламуса
ЗМА - задняя мозговая артерия
ЗНМА - задняя нижняя мозжечковая артерия
ЗС — задний столб
ЗСЦТ - задний спиноцеребеллярный тракт
ЗТК — задняя теменная кора
ИЛ — интраламинарное ядро
ИМАО — ингибитор моноамионоксидазы
ИН — интернейроны
ИИ СП — ингибирующий постсинаптический по-
тенциал
ИТФ — инозитолтрифосфат
КВО — комплекс верхних олив
КГ — клетки Гольджи
КОМТ — катехол-О-метилтрансфераза
КП — клетка Пуркинье
КПМД - конечностно-поясная миодистрофия
КСБ — кальцийсвязывающий белок
КСТ — кортикоспинальный тракт
КТ - компьютерная томография
КФК — креатинфосфокиназа
КЦВ - корковый центр взора
ЛГ — лютеинизирующий гормон
ЛКТ — латеральное коленчатое тело
ЛММ — легкий меромиозин
МАО — моноаминооксидаза
МАОА — моноаминооксидаза типа А
МАОВ — моноаминооксидаза типа В
МВС — медленноволновой сон
МДД — миодистрофия Дюшенна
МДЯТ - медиодорсальное ядро таламуса
МКТ - медиальное коленчатое тело
МН — мотонейрон
МПКП — миниатюрный потенциал концевой пла-
стинки
МПП — медиальный продольный пучок
МРА — магнитно-резонансная ангиография
МРВ — магнитно-резонансная венография
МРТ — магнитно-резонансная томография
МСИЛ - миастенический синдром Итона—Лам-
берта
МСК — мышечно-специфическая киназа
МСЧ — мышечно-суставное чувство
НА — норадреналин
НБЧ — нижние бугорки четверохолмия
НВК — наружные волосковые клетки
НГЯМ — нейроны глубинных ядер мозжечка
НМС — нервно-мышечное соединение
НМСН — наследственная мотосенсорная поли-
невропатия
НС — неостриатум
ОВВК - однорядные внутренние волосковые
клетки
ОД - отсроченная депрессия
8
ОП — отсроченная потенциация
ОСВ — околоводопроводное серое вешество
ОФЭКТ — однофотонная эмиссионная компью-
терная томография
ПБМ - периферический белок миелина
ПВ - параллельное волокно
ПВ-ВЛЯТ - передневентральное вентролатераль-
ное ядро таламуса
ПВЧ - порог вибрационного чувства
ПЗК - первичная зрительная кора (поле 17 по
Бродману)
ПКП — потенциал концевой пластинки
ПМА — передняя мозговая артерия
ПМК — первичная моторная кора
ПМН периферический мотонейрон
ПМНЦ — полимодальные ноцицепторы
ПНМА — передняя нижняя мозжечковая арте-
рия
ПНС — периферическая нервная система
ПНФ — пучки нейрофибрилл
ПНЯ — педункулопонтинное ядро
ПРМК — премоторная кора
ПРФМ парамедианная ретикулярная форма-
ция моста
ПСК — первичная слуховая кора
ПССК — первичная соматосенсорная кора
ПСЦТ — передний спиноцеребеллярный тракт
ПЭТ — позитронно-эмиссионная томография
РНК — рибонуклеиновая кислота
РПМ — ростральный путь миграции
РСТ руброспинальный тракт
РСТ — ретикулоспинальный тракт
СВЗ — субвентрикулярная зона
СИОЗС — селективный ингибитор обратного за-
хвата серотонина
СМА — средние мозговые артерии
СМГ — спинномозговые ганглии
СНБ — сенильные невритические бляшки
СОТ - сухожильный орган Гольджи
СПС
СР
cP
СТЯ
ТИА
ТМ
тмм
ТРГ
тст
ФДЭ
фМРТ
ФРН
ФСГ
хцк
цАМФ
цГМФ
ПМН
ПНС
цеж
ЧСк
ЧСр
ЧЭНС
ЭКГ
эмг
энг
эст
ээг
язе
G-белок
Go6
JDH
NMDA
SNARE
(pSNAP)
— селективность по стороне
— саркоплазматический ретикулум
— субстанция Р
— субталамическое ядро
— транзиторная ишемическая атака
— текториальная мембрана
— тяжелый меромиозин
тиреотропин-рилизинг-гормон
— тектоспинальный тракт
— фосфодиэстераза
- функциональная магнитнорезонанс-
ная томография
- фактор роста нервов
— фолликулостимулирующий гормон
— холецистокинин
- циклический аденозинмонофосфат
- циклический гуанозинмонофосфат
- центральный мотонейрон
- нейтральная нервная система
— цереброспинальная жидкость
— компактная часть черной субстанции
- ретикулярная часть черной субстанции
- чрескожная электронейростимуляция
электрокардиография/электрокар-
диограмма
— электромиография/электромиограмма
— электронейрография
- электросудорожная терапия
— электроэнцефалография/электроэнце-
фалограмма
- ядра задних столбов
гуанозинтрифосфатсвязывающий бе-
лок
- G-белок, ассоциированный с обоня-
тельными репепторами
— окуловестибулярные рефлексы
— И-метил-П-аспартат
- рецептор SNAP
ЧАСТЬ I. Анатомическая и функциональная
организация нервной системы
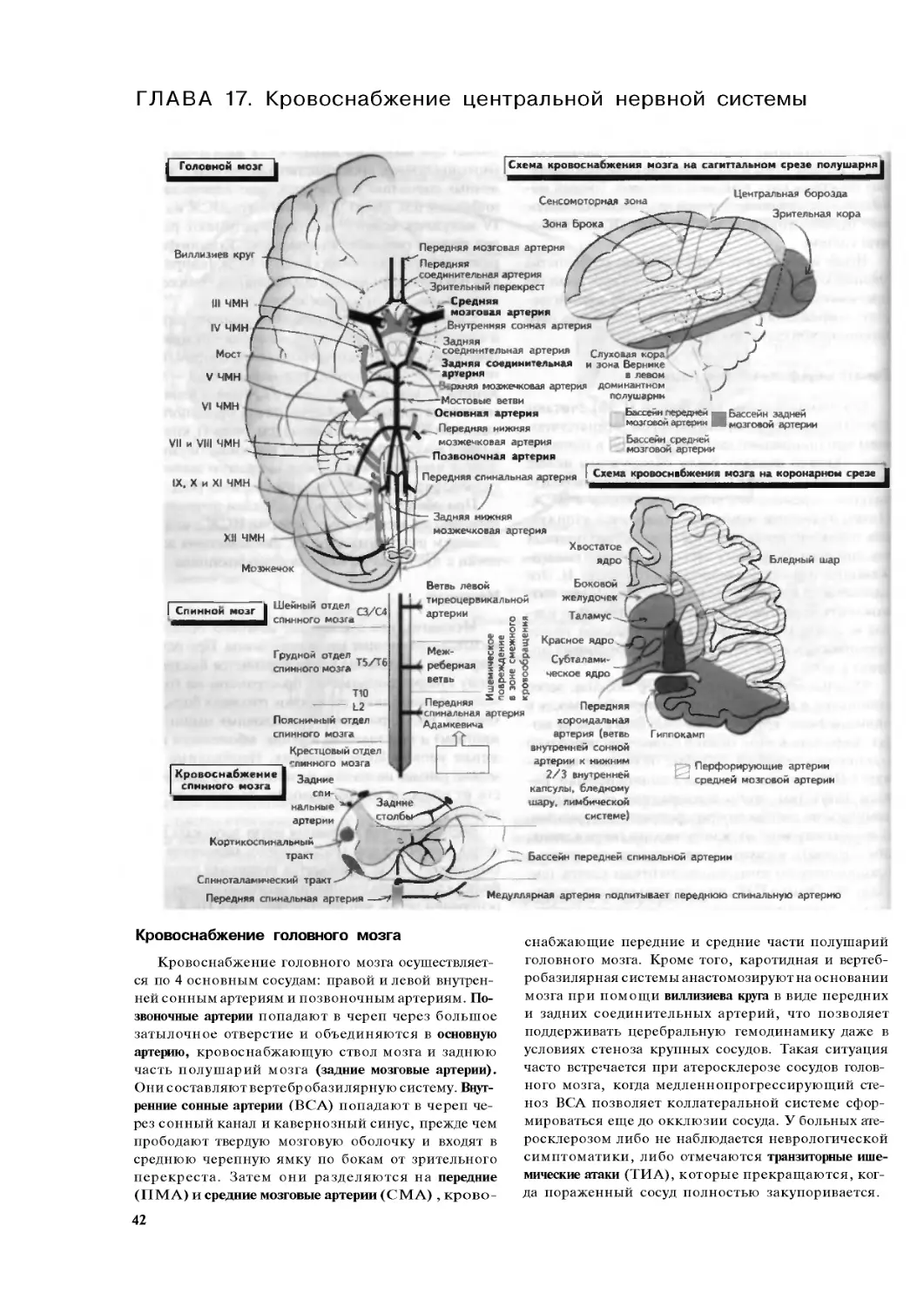
ГЛАВА 1. Анатомия нервной системы
Сильвиева
борозда
Таламус
Височная
ДОЛЯ
Теменная
доля
Ложная
доля
Субдуральное
пространство
[ Оболочки мозга
ноидальное
пространство,
содержащее
вены
и артерии мозга
Затылочная доля
Мозжечок
Большое затылочное отверстие
Позвонки
ьная
система
Серое
вещество
Продол-
говатый
мост
Средний
мозг
Мост
Поперечный срез спинного мозга
£
Копчиковый спинномозговой нерв
Периферическая
нервная система
Спинномозговые нервы
крестцового отдела
1—5
Белое вещество
Задний корешок
Спинномозговой узел
Периферический
иерс /
С5<£
С6^
В Т4
Т5
Тб
Т7
Т8
T9
T10I
Т11
Т12
L1
L2
L3
L4
L5
02
Череп
| Твердая
Поперечный срез
полушарий мозга
мозговая
оСк почка
головного
мозга Боковой
Паутинная
оболочка
Мягкая
мозговая
оболочка
Базальные
111 желудочек
Г иппокамп
Височная доля
ышца|
Симпатический ганглий
Боковой рог
Передний корешок
Внутренний орган
Чувствительный
рецептор
Вегетативная нервная система
Кора головного
мозга
Белее
вещество
» Кровеносный сосуд
10
Нервная система подразделяется на перифе-
рическую (ПНС), центральную ( Ц Н С ) и вегета-
тивную (ВНС). К ПНС относятся нервы, находя-
щиеся вне головного мозга, его ствола и спинно-
го мозга, а к цнс — клетки, находящиеся в этих
структурах.
Периферическая нервная система
ПНС состоит из нервных стволов, включающих
афферентные волокна, несущие чувствительную
информацию в спинной мозг и ствол головного моз-
га, а также эфферентные волокна, несущие импуль-
сы к мышцам. Поражение конкретного нерва при-
водит к развитию слабости в иннервируемых им
мышцах и утрате чувствительности в зоне его ин-
нервации. Периферические нервы формируют спле-
тения, находящиеся вблизи спинного мозга (на-
пример, плечевое сплетение верхней конечности).
Периферические нервы выходят из спинного моз-
га через отверстия между костями позвоночника
(или позвонками) или из головного мозга через от-
верстия в черепе.
Спинной мозг
Спинной мозг начинается на уровне большого
затылочного отверстия, которое находится в основа-
нии черепа, где заканчивается нижняя часть его
ствола (продолговатый мозг). Спинной мозг взрос-
лого человека заканчивается на уровне I пояснич-
ного позвонка. От него отходят 30 пар спинномоз-
говых нервов (или 31, если учитывать копчиковые
нервы), которые выходят из него через межпоз-
вонковые отверстия. Первые 8 спинномозговых
нервов выходят из шейного отдела спинного мозга
(первая пара покидает спинномозговой канал над
I шейным позвонком), следующие 12 пар нервов
выходят из грудного, или дорсального, отдела спин-
ного мозга. Остальные 10 пар спинномозговых не-
рвов выходят из нижележащего отдела спинного
мозга: 5 из поясничного и 5 из крестцового.
Спинномозговой нерв образуется из переднего,
или вентрального, корешка, который иннервиру-
ет скелетные мышцы, и заднего, или дорсального,
корешка, который несет в спинной мозг чувстви-
тельную информацию от области кожи, имеющей
единое происхождение с соответствующим сегмен-
том спинного мозга в эмбриогенезе (см. главу 2).
Волокна задних корешков прерываются в спин-
номозговых узлах, которые залегают вне спинно-
го мозга.
Спинной мозг состоит из белого вещества, ко-
торое содержит восходяшие и нисходяшие прово-
дящие пути спинного мозга, и серого вещества, рас-
положенного в центра спинного мозга, которое со-
держит тела нейронов (см. главу 12).
Ствол мозга,
черепно-мозговые нервы и мозжечок
От спинного мозга начинается ствол мозга, который
залегает в основании головного мозга и состоит из
продолговатого мозга, моста и среднего мозга и содер-
жит нейроны ядер 10 из 12 пар черепно-мозговых
нервов (см. главу 14). Ствол мозга и мозжечок являют-
ся структурами задней черепной ямки. Мозжечок свя-
зан со стволом мозга тремя парами мозжечковых ножек
и регулирует координацию движений (см. главу 37).
Полушария головного мозга
Полушария головного мозга состоят из 4 основ-
ных долей: затылочной,теменной,височной и лобной.
В медиальной части височной доли находится ряд
специфических структур, включая гиппокамп, кото-
рые составляют лимбическую систему (см. главу 45).
Наружный слой полушарий головного мозга на-
зывается корой мозга и содержит нейроны, которые
структурированы как в горизонтальные слои, гак и
в вертикальные столбцы (см. главу 15). Участки коры
мозга сообщаются друг с другом посредством длин-
ных подкорковых проводящих путей. Последние
наряду с путями, которые соединяют кору мозга со
спинным мозгом, стволом и глубинными ядрами
полушарий мозга, составляют белое вещество полу-
шарий головного мозга. Глубинные ядра включают
базальные ганглии (см. главы 38 и 39) и таламус.
Оболочки мозга
ЦНС залегает внутри черепа и позвоночника, от
которых ее отделяют оболочки мозга. Мягкая мозго-
вая оболочка отделяется от тонкой паутинной оболоч-
ки субарахноидальным пространством, которое в
свою очередь отделяется от твердой мозговой оболоч-
ки субдуральным пространством (см. главу 16).
Вегетативная нервная система
ВНС имеет центральную и периферическую час-
ти и осуществляет иннервацию внутренних органов
и желез (см. главу 42); она принимает участие в ре-
гуляции деятельности эндокринной системы и го-
меостаза организма человка (см. главу 43). Перифе-
рическая часть ВНС состоит из энтеральной, симпа-
тической и парасимпатической систем (см. главу 42).
Эфферентные волокна ВНС начинаются либо из
бокового столба спинного мозга, либо из ядра опре-
деленного черепно-мозгового нерва или же сакраль-
ныхядер и прерываются в ганглии — структуре, ко-
торая имеет различное расположение в рамках сим-
патической и парасимпатической нервной систем.
Афферентные волокна от внутренних органов, ин-
нервируемых ВНС, проходят через задние корешки
спинного мозга.
ГЛАВА 2. Развитие нервной системы
Эмбрион человека (возраст 20 дней, вид сзади)
Мозговые пузыри эмбриона человека (возраст 42 дня, вид сбоку)
Передняя (ростральная) часть
Прозэнцефалон\ Мезэнцефалон
Мозговая
борозда
Диэнцефалон
Нервная пластинка
Первичные
пузыри
Зрительный пузырь
Метэнцефалон
Нервный желобок
Сомит
Шейная
борозда
Развитие ствола
Первичная
Покрышечная
Задняя (каудальная) часть
Развитие спинного
Структура-
предшественник
нервного гребня
Висцеральные
афферентные
желудочек
Крыловидная
Соматические/
висцеральные
эфферентные
Спинно- волокна
Нервная трубка
Базальная
Крыло-
видная
Эпенди-
Спинно-
Цент-
ральный
Ното хорда
Центральный
«Ушастый
Задний чувстви-
тельный корешок
Базальная
БО — белок остеогенеза
Сливаются и формируют
спинномозговой нерв, который иннервирует
Передний
двигательный
корешок
Маргинальный
мышцу, происходящую из сомита
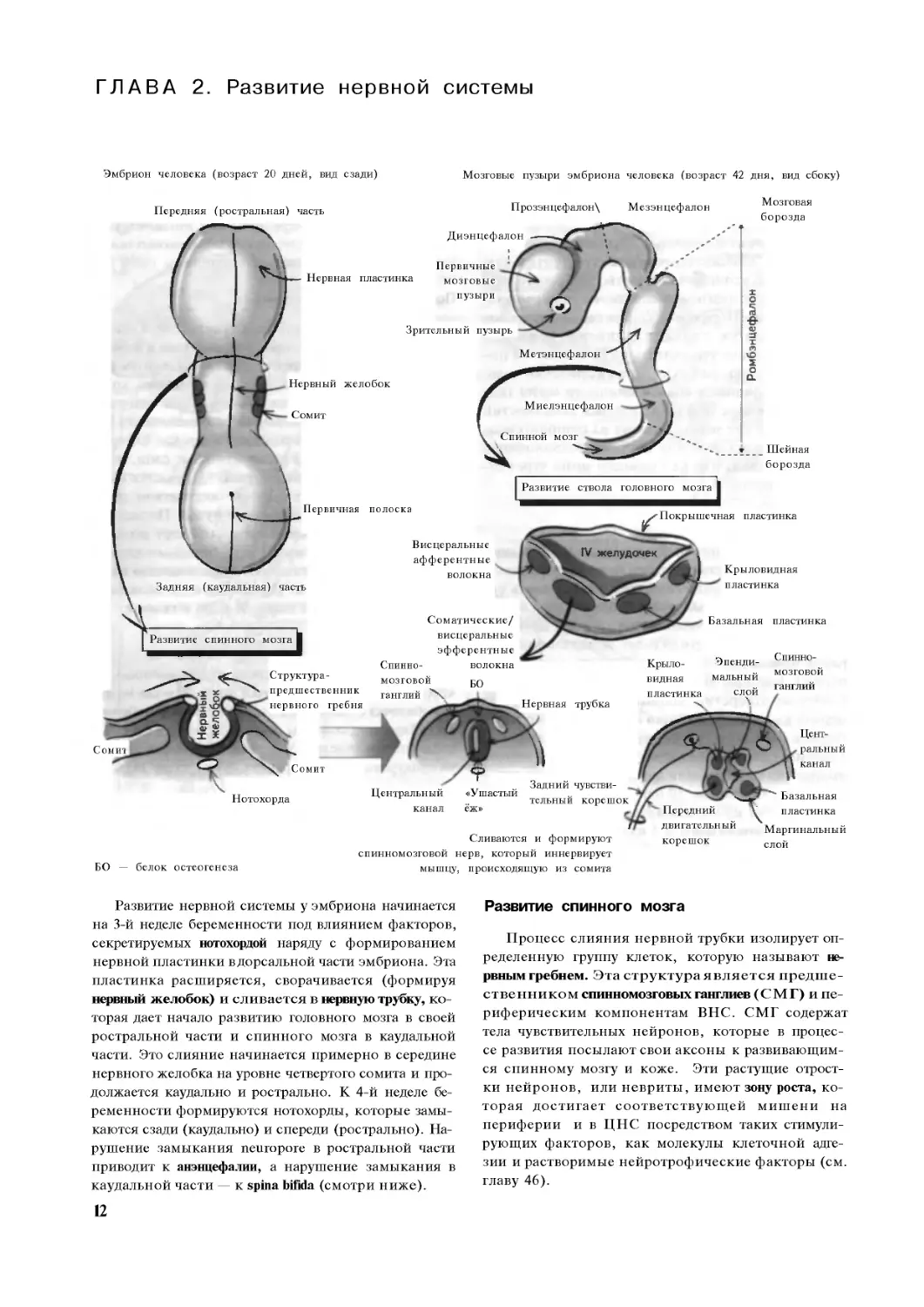
Развитие нервной системы у эмбриона начинается
на 3-й неделе беременности под влиянием факторов,
секретируемых нотохордой наряду с формированием
нервной пластинки вдорсальной части эмбриона. Эта
пластинка расширяется, сворачивается (формируя
нервный желобок) и сливается в нервную трубку, ко-
торая дает начало развитию головного мозга в своей
ростральной части и спинного мозга в каудальной
части. Это слияние начинается примерно в середине
нервного желобка на уровне четвертого сомита и про-
должается каудально и рострально. К 4-й неделе бе-
ременности формируются нотохорды, которые замы-
каются сзади (каудально) и спереди (рострально). На-
рушение замыкания neuropore в ростральной части
приводит к анэнцефалии, а нарушение замыкания в
каудальной части — к spina bifida (смотри ниже).
Развитие спинного мозга
Процесс слияния нервной трубки изолирует оп-
ределенную группу клеток, которую называют не-
рвным гребнем. Эта структура является предше-
ственником спинномозговых ганглиев (СМГ) и пе-
риферическим компонентам ВНС. СМГ содержат
тела чувствительных нейронов, которые в процес-
се развития посылают свои аксоны к развивающим-
ся спинному мозгу и коже. Эти растущие отрост-
ки нейронов, или невриты, имеют зону роста, ко-
торая достигает соответствующей мишени на
периферии и в ЦНС посредством таких стимули-
рующих факторов, как молекулы клеточной адге-
зии и растворимые нейротрофические факторы (см.
главу 46).
Нервная трубка окружает нервный канал, кото-
рый дает начало центральному каналу спинного моз-
га. Трубка содержит нейробласты. Часть этих клеток.
медиолатеральном направлении. Так, в процессе раз-
вития двигательные ядра оказываются медиальнее
по отношению к чувствительным ядрам, при этом
прилежашая к каналу (эпендимальный слой), делится м игрирмср амйжагашшгащЕВПГО адрц 1три1Кли1;к;идв|Ь ф®Р®1£ЦГДлу1<>'1ся в нейрс
вещество спинного мозга (см. главу 1). Развиваю-
щиеся отростки нейробластов / нейронов врастают
в маргинальный слой, который формирует белое ве-
щество спинного мозга. Развивающиеся нейробла-
сты разделяются на две популяции: крыловую и ба-
зальную пластинки, которые в свою очередь дают
начало задним и передним рогам спинного мозга.
Меньший по размеру боковой рог с висцеральны-
ми эфферентными нейронами развивается в по-
граничной области между ними в грудном и пояс-
ничном отделах спинного мозга (см. главу 42). Раз-
витие нервной системы зависит от веществ, секре-
тируемых нотохордой. Развитие дорсальной части
контролирует «ушастый еж», а вентральной — белок
остеогенеза (БО).
Развитие нервной системы
Ростральная часть нервной трубки перед слияни-
ем расширяется и формирует 3 первичных мозговых
пузыря (прозэнцефалон, мезэнцефалон и ромбэн-
цефалон), а также две борозды (шейную и мозговую).
Первичные мозговые пузыри развиваются в полуша-
рия головного мозга, ствол мозга и мозжечок, тогда
как из нервного канала формируется желудочковая
система мозга (см. главу 16).
Прозэнцефалон состоит из телэнцефалона, кото-
рый дает начало полушариям головного мозга и ча-
стично базальным ганглиям, другая его часть — ди-
энцефалон - является предшественником таламуса,
гипоталамуса, заднего отдела гипофиза, зрительно-
го нерва и сетчатки.
Нейробласты образуются вблизи нервного кана-
ла (вентрикулярная зона) и мигрируют не только в
соседние области, формируя глубокие подкорковые
ядра мозга, но и вдоль радиальных волокон глии,
формируя кору больших полушарий (см. главу 15).
Из этого инвагинированного участка незрелого моз-
га, богатого глиальными волокнами, развивается
белое вещество больших полушарий, при этом не-
которая часть радиальной глии дает начало клеткам-
предшественникам нейронов зрелого мозга (смот-
ри ниже). Сигналы, которые определяют порядок
миграции нейронов, в настоящее время изучаются.
Нарушение процессов миграции приводит к разви-
тию корковой дисплазии.
Из мезэнцефалона развивается средний мозг. Не-
рвный канал в этой области превращается в сильви-
ев водопровод. Ромбэнцефалон состоит из метэнце-
фалона. из которого развиваются мост и мозжечок,
а также миелэнцефалона, из которого развивается
продолговатый мозг (см. главу 13). Ствол мозга раз-
вивается также, как и спинной мозг, однако его раз-
витие происходит не в рострокаудальном, а скорее в
ними. Рострокаудальное направление развития моз-
га объясняет расположение ядер черепно-мозговых
нервов в стволе мозга (см. главы 13 и 14).
Мозжечок развивается из ромбовидной губы и
смежной с ней крыловидной пластинки.
Нейрогенез у взрослых
До недавнего времени считалось, что в зрелом
мозге млекопитающего новых нейронов не образу-
ется, однако теперь известно, что в ЦНС существу-
ют клетки-предшественники, в том числе у челове-
ка. Эти клетки находятся в основном в гиппокампе
(см. главу 45) и субвентрикулярной зоне (СВЗ) вбли-
зи боковых желудочков. Они отвечают на ряд сиг-
налов и дают начало новым нейронам гиппокампа и
обонятельной луковицы, при этом последние миг-
рируют в обонятельную луковицу из СВЗ по рост-
ральному пути миграции (РПМ).
Пороки развития
центральной нервной системы
Анэнцефалия возникает при нарушении слияния
передних ростральных частей нейропод. Мозговые
пузыри не развиваются, что делает невозможным
развитие головного мозга. В подавляющем большин-
стве случаев анэнцефалии происходят выкидыши.
Spina bifida это дефект развития нижней части
позвоночника и/или спинного мозга. Чаще всего
этот порок наблюдается при нарушении слияния
дорсальных участков нижних позвоноков (spina bi-
fida occulta). Это может сопровождаться грыжевы-
ми выпячиваниями мозговых оболочек и нервной
ткани (менингоцеле и менингомиелоцеле). Наиболее
тяжелым вариантом spina bifida считается наруше-
ние сращения задней (каудальной) части пеиго-
роге. Spina bifida часто сочетается с гидроцефалией
(см. главу 16).
В ряде случаев менингоцеле возникает вследствие
грыжевого выпячивания оболочек и ткани мозга че-
рез дефекты костей черепа. Однако, в отличие от
аналогичных ситуаций в спинном мозге, эти дефек-
ты можно легко устранить, при этом не наблюда-
ется нарастающей неврологической симптоматики.
Корковая дисплазия - это результат нарушения
миграции развивающихся корковых нейронов. Эти
пороки развития стали чаще диагностировать с вне-
дрением в практику методик нейровизуализации.
Корковая дисплазия является одной из важных при-
чин эпилепсии (см. главу 53).
Многие внутриутробные инфекции (такие, как
краснуха), а также факторы окружающей среды (на-
пример, радиация) вызывают пороки развития не-
рвной системы. Кроме того, к порокам развития
ЦНС приводят редкие генетические заболевания,
описание которых не входит в задачи этой книги.
13
ГЛАВА 3. Клетки нервной системы I: нейроны
Аксо-аксональный
Аксосоматический
Субстанция Ниссля
-Дендрит
Ядро
Нейрофиламенты
Митохондр!
Аксональный бугорок
Начальный сегмент
Пресинаптическая терминаль
Коллатераль аксона
Миединоеая оболочка
Синаптическая щель
Аксон
Перехват Ранвье
Рецепторы
мембраны
Аксоплазма
Аксолемма
Нервно-мышечное соединение
Шванновская клетка
Нервная терминаль
Активные зоны
Мышца
Аксодендритический синапс
(пример химического синапса)
Дендритический
отросток, или геммула
Постсинаптический отросток
Синаптические пузырьки,
содержащие апетилхолин
а-Мотонейрон в переднем
роге спинного мозга
Синаптические пузырьки
(пресинаптический)
си
Постсинаптические
ацетилхолиновые рецепторьР
Складки постсинаптического \
отростка мышцы (см. главу г
в которой нервно-мышечное
соединение описано подробно)
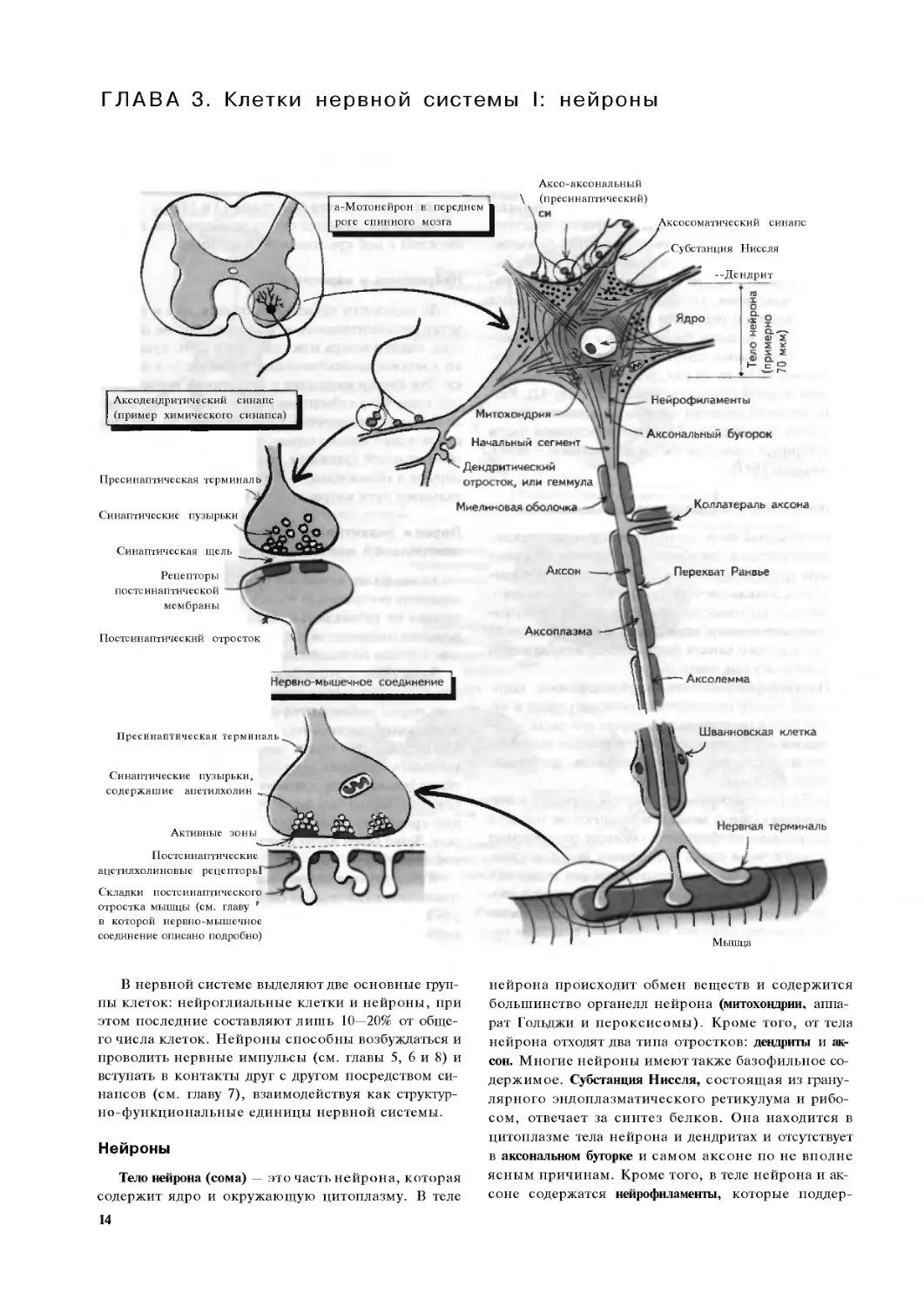
В нервной системе выделяют две основные груп-
пы клеток: нейроглиальные клетки и нейроны, при
этом последние составляют лишь 10—20% от обще-
го числа клеток. Нейроны способны возбуждаться и
проводить нервные импульсы (см. главы 5, 6 и 8) и
вступать в контакты друг с другом посредством си-
напсов (см. главу 7), взаимодействуя как структур-
но-функциональные единицы нервной системы.
Нейроны
Тело нейрона (сома) — это часть нейрона, которая
содержит ядро и окружающую цитоплазму. В теле
14
нейрона происходит обмен веществ и содержится
большинство органелл нейрона (митохондрии, аппа-
рат Гольджи и пероксисомы). Кроме того, от тела
нейрона отходят два типа отростков: девдриты и ак-
сон. Многие нейроны имеют также базофильное со-
держимое. Субстанция Ниссля, состоящая из грану-
лярного эндоплазматического ретикулума и рибо-
сом, отвечает за синтез белков. Она находится в
цитоплазме тела нейрона и дендритах и отсутствует
в аксональном бугорке и самом аксоне по не вполне
ясным причинам. Кроме того, в теле нейрона и ак-
соне содержатся нейрофиламенты, которые поддер-
живают структуру цитосклелета нейрона. Более того,
важно упомянуть две другие фибриллярные струк-
туры нейрона: микротрубочки и микрофиламенты,
которые необходимы для аксоплазматического тока
(см. ниже) и роста аксона.
Девдриты — это отростки нейронов, которые от-
ходят от тел нейронов, ветвятся и отвечают за пере-
дачу информации к телу нейрона от синапсов денд-
ритического дерева (аксодендритические синапсы;
смотрите также главу 8). У большинства нейронов
много дендритов (мультиполярные нейроны), и если
одни нейроны контактируют с другими через синап-
сы, то некоторые осуществляют эти контакты через
дендритические отростки, или геммулы. Таким обра-
зом, основное назначение дендритов состоит в уве-
личении поверхности для синапсов, что позволяет
интегрировать огромное количество сигналов, пред-
назначенных телу нейрона. В противоположность
этому аксон передает информацию от тела нейрона
к нервной терминали и синапсам (см. главу 8). Не-
смотря на то что у каждого нейрона только один ак-
сон, он разветвляется и имеет отростки. Эти ветви
отходят вблизи тела нейронов, например чувст-
вительных (псевдоуниполярные нейроны; см. гла-
ву 20), но чаше всего — вблизи синапса, формируе-
мого аксоном. Аксон отходит от аксонального бу-
горка, где начинается первый сегмент аксона. Это
наиболее возбудимая часть нейрона, поскольку
плотность натриевых каналов в этой области, где
первоначально возникает потенциал действия, наи-
более высока (см. главу 6).
Все нейроны окружены липидным бислоем (кле-
точной мембраной), где расположены белки, одни
из которых образуют ионные каналы (см. главу 5), а
другие — рецепторы к специфическим веществам,
высвобождаемым нейронами (см. главы 7 и 9), или
же служат ионными насосами, которые перемеща-
ют ионы через мембраны в соответствии с электро-
химическим градиентом, например Na -, К -ион-
ный насос (см. главу 6). Поверхность мембраны ак-
сона называется аксолеммой, а цитоплазма, которую
она окружает, — аксоплазмой. Ионные каналы аксо-
леммы наделяют ее способностью проводить потен-
циалы действия, а аксоплазма содержит микротру-
бочки, нейрофиламенты и митохондрии. Эти орга-
неллы не только отвечают за поддержание ионных
градиентов, но и обеспечивают транспорт и рецир-
куляцию белков из тела нейрона (и в меньшей сте-
пени к нему) к нервной терминали. Аксоплазмати-
ческий ток, или аксональный транспорт, может быть
медленным (примерно 1 мм/сут) или быстрым (100—
400 мм/сут). Он необходим не только для поддержа-
ния нормальной активности нейрона и синапса, но
и для жизнедеятельности и развития нейрона. Ак-
сональный ток нарушается при некоторых нейроде-
генеративных заболеваниях (см. главу 52).
Многие аксоны покрыты слоем липидов, или
миелиновой оболочкой, которая действует как элект-
рический изолятор. Миелиновая оболочка влияет на
способность аксона проводить импульсы и способ-
ствует быстрому распространению потенциала дей-
ствия без потери целостности импульса (см. главу
8). Это достигается с помощью перехватов Ранвье —
участков, в которых аксолемма содержит много ион-
ных каналов (обычно № -каналов), непосредствен-
но соприкасающихся с межклеточной жидкостью.
Кроме того, из перехватов Ранвье отходят отростки
аксона, которые называются коллатералями аксона.
Миелиновая оболочка окружает аксон ниже аксо-
нального холмика и заканчивается непосредствен-
но вблизи нервной терминали, прежде чем та арбо-
ризируется. Миелиновую оболочку формируют
шванновские клетки в ПНС и олигодендроглиоциты
в ЦНС (см. главу 4), при этом многие аксоны ЦНС
окружает оболочка одного олигодендроглиоцита,
тогда как в ПНС одна шванновская клетка прихо-
дится на один аксон.
Синапс — это соединение между нейроном и дру-
гой клеткой. В ЦНС это другой нейрон. В ПНС это
мышца, клетка железы или другого органа. В не-
рвной системе обычно присутствуют химические
синапсы, которые состоят из пресинаптической не-
рвной терминали (концевой луковицы) и синаптичес-
кой щели, которая отделяет нервную терминаль от
постсинаптической мембраны и через которую выде-
ляется химическое вещество, или нейротрансимит-
тер (см. главу 7). Такой синапс обычно располагает-
ся между нейроном и дендритом другого нейрона
(аксодендритический синапс), однако синапсы мо-
гут располагаться как в месте контакта между аксо-
ном и постсинаптической клеткой, так и на теле ней-
рона (аксосоматические синапсы) и, реже, на преси-
наптической нервной терминали (аксо-аксональные
синапсы; см. главу 8). Некоторые синапсы ЦНС не
имеют таких свойств, но являются соединениями с
малым сопротивлением (переходными соединени-
ями) и называются электрическими синапсами. Эти
синапсы обеспечивают быстрое проведение потенци-
алов действия без какой-либо интеграции, что делает
невозможным для популяции нейронов передавать
импульсы вместе или синхронно (см. главы 7 и 53).
Гибель специфических популяций нейронов при-
водит к развитию неврологических заболеваний.
Такие заболевания, при которых гибель нейронов
является первичным патологическим процессом,
обсуждаются в главе 52.
ГЛАВА 4. Клетки нервной системы II: нейроглиальные клетки
Эпендима
Г с ушарие головного мозга
Столбиковые
Желудочек
Боковой желудочек
О лигодендроци т||
Кора мозга
Аксоны
III желудочек
Астроцит
Ct ИННОЙ мозг
Серое вещество
Спинной мозг
Капилляр
Гематоэнцефалический барьер
Мышца
Ножка отростка
Шванновская клетка
Эндсгелиоцит
Перехват
Базальная мембрана
Аксон
Ядро шванновской клетки
Классификация астроцитов
Слой миелина
Аксон
Аксолемма
Подкорковое
белсч вещество
Белое
вещество
I ериферически)
нерв с аксонами
мотонейрона
Онтогене-
тическая:
эпендимальные
клетки с ресничками
Морфоло-
гическая-
Протоплазматические — содержат мало
фиброзного материа а и в основном
находятся в сером веществе
Фиброзные — содержат филаменты
и находятся в основном в белом вещестг >
I типа
II типа — происходят из пре ьниторных кле-
~ок D2A, которые могут давать начало и
олигодендооцитам при определенных усло-
виях. Вопрос об их существовании в ЦНС
ззросло о человека окончательно не изучен
Бот ьшое
затылочное отверстие
Плотное
малопроницаемое
Ранчи" ссединение
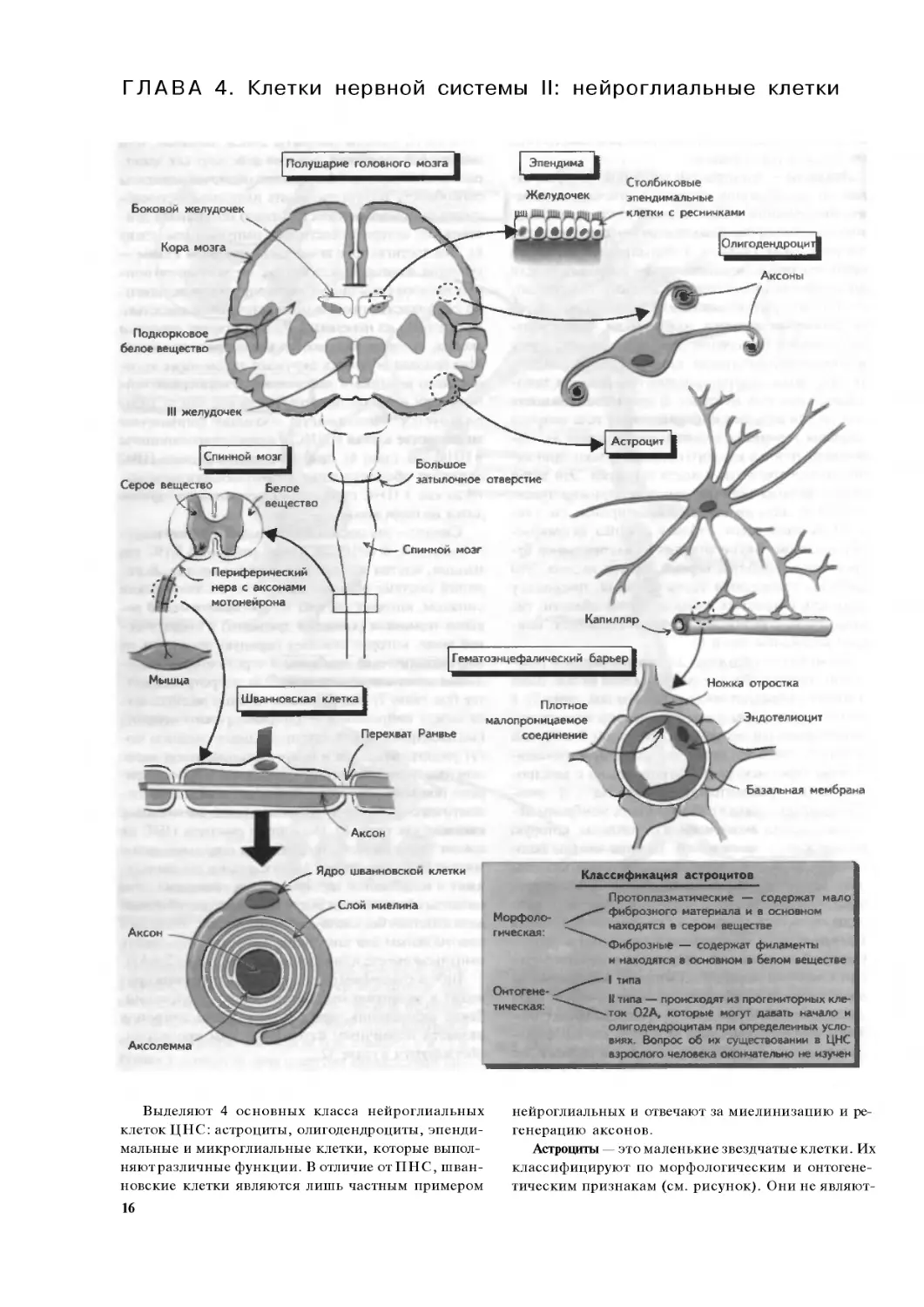
Выделяют 4 основных класса нейроглиальных
клеток ЦНС: астроциты, олигодендроциты, эпенди-
мальные и микроглиальные клетки, которые выпол-
няютразличные функции. В отличие отПНС, шван-
новские клетки являются лишь частным примером
16
нейроглиальных и отвечают за миелинизацию и ре-
генерацию аксонов.
Астроциты — это маленькие звездчатые клетки. Их
классифицируют по морфологическим и онтогене-
тическим признакам (см. рисунок). Они не являют-
ся пассивными вспомогательными элементами ЦНС
и выполняют многие важные ее функции:
• Формируют вспомогательную сеть для нейронов
и капилляров с помощью своих цитоплазматичес-
ких отростков, которые оканчиваются не только
на нейронах, но и на капиллярах. Таким образом,
они являются ограничительной глией — отростки
ножек астроцитов покрывают базальную мембра-
ну, окружающую сосуды и мягкую мозговую обо-
лочку.
• Поддерживают целостность гематоэнцефалическо-
го барьера (ГЭБ), обеспечивая плотные соедине-
ния с малой проницаемостью между эндотелиоци-
тами капилляров мозга (см. также главу 16).
• Способны захватывать, хранить и высвобождать
некоторые нейротрансмиттеры (например, глута-
мат, у-аминомасляную кислоту — ГАМК) и поддер-
живают функцию химической нейротрансмиссии
в ЦНС.
• Могут захватывать избыточные ионы из экстра-
целлюлярной жидкости, особенно калий.
• Участвуют в процессе развития нейронов (см. гла-
ву 15), реагируют на повреждающее воздействие
(см. главу 47), а также участвуют в судьбе клеток-
предшественников нейронов в гиппокампе взрос-
лого человека.
• Могут выполнять роль антигенпрезентирующих
клеток в случаях, если ГЭБ и ЦНС повреждены.
Наиболее распространенным заболеванием, свя-
занным с поражением астроцитов, является астро-
цитома. Опухоль сдавливает окружающую ткань
ЦНС и приводит к развитию неврологического де-
фицита (с эпилептическими припадками или без
них), что зависит от ее локализапии. У взрослых ас-
троцитома чаще всего локализуется в белом веще-
стве полушарий мозга.
Олигодендроциты — отвечают за миелинизацию
нейронов ЦНС и поэтому в большом количестве при-
сутствуют в белом веществе. Каждый олигодендро-
цит образует миелиновую оболчку для 3—50 волокон
я также окружает многие другие волокна, формируя
миелиновую оболочку. Кроме того, эти клетки вы-
деляют ряд веществ, ингибирующих рост аксонов,
едовательно, угнетают процессы регенерации
нейронов в ЦНС взрослого человека (см. главу 47).
Нарушение функции олигодендроцитов приво-
дит к демиелинизации ЦНС и отмечается при ряде
заболеваний, например рассеянном склерозе (см. гла-
ву 54), тогда как при нарушении пролиферации оли-
годендроцитов развиваются медленно растущие опу-
холи (олигодендроглиомы), при которых характерны
эпилептические припадки (см. главу 53).
Эпендимальные клетки обеспечивают нормаль-
ную циркуляцию цереброспинальной жидкости
(ЦСЖ), а также взаимодействуют с астроцитами,
формируя барьер, отграничивающий желудочки
мозга и ЦСЖ от нейронального окружения. Они так-
же выстилают центральный канал спинного мозга
(см. главу 12). Эти клетки также называются эпен-
димоцитами, поскольку другие эпендимальные
клетки вырабатывают ЦСЖ (хороидные сплетения)
и перемещают определенные вещества из ЦСЖ в
кровь (танициты). Опухоли эпендимы (эпендимомы
и хороидные папилломы) возникают в желудочках,
приводя к развитию гидроцефалии (см. главу 16), или
в спинном мозге, где происходит локальная деструк-
ция нервной ткани.
Микрог. шальные клетки (не показаны на рисун-
ке) являются тканевыми макрофагами мозга и на-
ходятся как в белом, так и в сером веществе ЦНС.
По своей природе они являются фагоцитами и регу-
лируют иммунный ответ в ЦНС (см. главу 54).
Шванновские клетки находятся только в ПНС и
отвечают за миелинизацию периферических нервов,
окружая их аксоны миелиновой оболочкой. Завер-
шенная миелиновая оболочка состоит из несколь-
ких слоев мембраны шванновских клеток без цитоп-
лазмы. В отличие от олигодендроцитов, одна шван-
новская клетка окружает аксон и образует миелин
одного его участка между перехватами Ранвье. Кро-
ме того, шванновские клетки участвуют в регенера-
ции поврежденных аксонов, что отличает их от цен-
тральных нейроглиальных клеток, которые подавля-
ют процессы регенерации (см. главы 46 и 47). Гибель
миелина (в отличие от гибели аксонов) происходит
при ряде наследственных и воспалительных поли-
нейропатий, что приводит кдисфункции перифери-
ческих нервов (демиелинизирующие невропатии;
см. главы 6 и 55). Кроме того, известны доброкаче-
ственные опухоли шванновских клеток (шванно-
мы), которые развиваются, в частности, при таком
наследственном заболевании, как нейрофиброма-
тоз I типа, связанном с нарушением функции гена-
супрессора опухолевого роста нейрофибромина.
Эти опухоли обычно протекают бессимптомно, од-
нако если они образуются в ограниченном прост-
ранстве, то могут вызывать сдавление окружающих
нервных структур, например, в области мостомоз-
жечкового угла или корешка спинномозгового нер-
ва (см. главы 2 - 14 и 27).
ГЛАВА 5. Ионные каналы
Вольтажзависнмый ионный канал
Химический ионный канал
I Ацх 1
напримьр. натриевый канал
например
Ацх-катионный канал
Схеиа натриевого канала
Инактивированное состояние,
канал закрыт
Состояние покоя, канал закрыт |
Активное состояние, канал открыт
Приток Натрия в клетку
Na’
Интрацеллюлярное
пространство Сенсор
вольтажзависимого
канала
Эктрацеллюлярнос
пространства
Мембрана
Место связывания Ацх
(рецептоо Ацх)
Ионный канал,
ассоциированный
с рецептором
Модуляция
фу| кции ионных каналов
интрацеллюлярными ионами
кальция и/или вторичными
мессенджерами (цАМФ)
Селективный фильтр
Т рансмембранная
молекула белка
Нет ионного потока
Na’
Нет ионного потока
Na’
активации
вольтажзависимый
сенсор
Субьединица —
или ворота инактивации
Ионный канал — это белковая макромолекула,
которая располагается в биологической мембране и
позволяет ионам переходить с одной стороны мем-
браны на другую. Ионы перемещаются через мемб-
рану в направлении, определяемом электрохимичес-
ким градиентом. В общем, ионы стремятся из обла-
сти более высокой их концентрации в область более
низкой, однако в присутствии вольтажзависимого
градиента возможна ситуация, при которой ионные
потоки отсутствуют даже при наличии неравных
концентраций ионов. Сам ионный канал может от-
крываться и закрываться. Открытие его может быть
связано с изменением заряда мембраны (например,
при деполяризации или возникновении потенциа-
ла действия) или с взаимодействием химического ве-
щества и рецептора в самом канале или поблизости
от него. Выделяют два типа каналов — вольтажзави-
симые (вольтажчувствительные) и химические (ли-
гандзависимые). Однако это разделение достаточно
18
условно, поскольку многие вольтажзависимые кана-
лы могут быть активированы нейротрансмиттерами
и кальцием. Более того, некоторые каналы не откры-
ваются при изменении заряда мембраны или взаи-
модействии с химическими веществами-мессендже-
рами, а реагируют только на давление и механичес-
кую силу (например, соматосенсорные слуховые
рецепторы; см. главы 20, 21 и 26).
Наиболее важным свойством ионных каналов яв-
ляется возможность изменять электрическую возбу-
димость нейрона (см. главу 6). Ионные каналы на-
ходятся во всех частях нейрона, в меньшей степени
в нейроглиальных клетках, а также в клетках неней-
ронального происхождения.
Все биологические мембраны, включая нейро-
нальные, состоят из липидного бислоя, который ха-
рактеризуется электрическим сопротивлением, т. е.
ионы не могут просто проникнуть через него. По-
этому для облегчения проникновения ионов через
мембраны существуют так называемые поры (ион-
ные каналы) на липидной мембране или перенос-
чики, которые собирают ионы с одной стороны мем-
браны и переносят их через нее, а потом высвобож-
дают ионы. В нейронах скорость переноса ионов,
необходимая для передачи импульса, слишком вы-
сока для какой-либо системы переносчиков, поэто-
му в нейронах и существуют ионные каналы (или
поры) для переноса ионов через мембраны.
Основные свойства ионных каналов таковы:
• Они состоят из нескольких белковых субъединиц,
которые локализуются трансмембранно и позво-
ляют ионам переходить с одной стороны мембра-
ны на другую, — трансмембранные поры.
• Канал сформирован таким образом, что он может
открываться и закрываться, при этом данный про-
цесс имеет несколько промежуточных ступеней.
• Канал может открываться в ответ на специфичес-
кие стимулы. Большинство каналов реагирует на
изменение заряда мембраны и поэтому открыва-
ется в ответ на деполяризационный заряд, т. е. на
заряд, меняющийся на мембране в покое от —70
до -80 мВ к более отрицательной величине.
В противоположность этому некоторые ионные
каналы, особенно синаптические, не открываются
в ответ на изменение заряда мембраны, а открыва-
ются в ответ на взаимодействие с химическими ве-
ществами (например, ацетилхолином — Ацх). Эти
каналы имеют в своей структуре рецептор для этого
вещества, и связывание его с этим рецептором при-
водит к открытию канала. Однако многие ионные
каналы обладают как вольтажзависимыми, так и хи-
мическими сенсорами и наличие в интрапеллюляр-
ном пространстве иона или вторичного мессендже-
ра — циклического аденозинмонофосфата (цАМФ)
приводит к модуляции трансмембранного ионного
потока, вызванного вольтажзависимым процессом.
Активация вольтажчувствительного или хими-
ческого рецептора приводит к открытию ворот ка-
1ла, через которые осуществляются ионные пото-
ки. Затем канал закрывается под воздействием дез-
активации (процесс, обратный открытию канала)
или инактивации, что предполагает участие вторич-
ных ворот, через которые по каналу ионы поступа-
ют медленнее, чем выходят по активированным во-
ротам, и таким образом существует временной про-
межуток, когда в канале нет ворот и ионы не могут
через него пройти.
Ионные потоки через канал могут быть селектив-
ными и неселективными. Если канал селективен, он
обеспечивает прохождение только определенных
ионов, что достигается посредством фильтра. В ос-
нове селективного фильтра лежат термодинамичес-
кие свойства. Такому каналу присваивается назва-
ние, например «натриевый канал». Однако многие
каналы не являются селективными и пропускают
различные виды ионов с одноименным зарядом,
например Ацх-катионный канал.
Ионный канал характеризуется определенными
физическими свойствами. Перемещение ионов по
каналу называется потоком, тогда как термином
«проводимость» определяют реципрокное сопротив-
ление (поток/вольтаж). Проводимость — это степень
доступности перемещения иона по каналу. Прони-
цаемость, с другой стороны, определяется как ско-
рость перемещения вещества или иона через мемб-
рану при определенной разности концентраций.
Существует множество разновидностей ионных
каналов даже в пределах одного семейства специфи-
чески ионных каналов, например калиевых.
Количество и тип ионных каналов определяют
характеристики клеточного ответа. Например, ней-
роны характеризуются скоростью генерации потен-
циала действия и ответа на синаптические импуль-
сы (см. главы 6—8, 45 и 53).
Нарушение функций ионных каналов
в клинике нервных болезней
Некоторые лекарства оказывают фармакологи-
ческое действие на уровне ионных каналов. К ним
относятся, например, местные анестетики и неко-
торые противоэпилептические препараты. Однако в
последние годы были описаны некоторые невроло-
гические заболевания, протекающие с преимуще-
ственным поражением мыши и вызванные мутаци-
ями генов, кодирующих натриевые и хлорные ион-
ные каналы. К ним относятся некоторые формы
миотонии (нарушение расслабления скелетной мус-
кулатуры после произвольного сокращения, приво-
дящее к невозможности свободно выполнять те или
иные двигательные акты) и различные формы пери-
одического паралича, при которых у пациентов раз-
виваются приступы преходящей парциальной или
генерализованной мышечной слабости. Более того,
некоторые формы семейной гемиплегической миг-
рени или мозжечковых атаксий (см. главу 37) связа-
ны с нарушением функции кальциевых каналов, а
некоторые формы эпилепсии (см. главу 53) могут
быть вызваны нарушением функции специфических
ионных каналов. Другие заболевания могут быть
вызваны перераспределением или функционирова-
нием тех ионных каналов, которые в норме неактив-
ны. Такие нарушения происходят в области, близ-
кой к перехвату Ранвье, в результате центральной де-
миелинизации при рассеянном склерозе или
периферической демиелинизации (синдром Гийена—
Барре), что приводит к нарушению распространения
потенциала действия (см. главы 6 и 54).
ГЛАВА 6. Мембрана в состоянии покоя и потенциал действия
• Отток калия из клетки
• Минимальное перемещение натрия
в клетку с помощью электрогенной
натрий-калий-АТ Фазы
Высокая концентрация натрия
• Массивное перемещение натрия внутрь
клетки, ко орсе i рекращается с ннак
тивацией каналов
• Отсроченное увеличение
перемещений калия из клетки
_ 3Na'
Экстрацеллюлярное д
пространство
Na*
к-
Низкая концентрация кат
Na-
к-
3Na-
« д Низкая концентрация кшжя + д
Натрий- 2К- Низкая концентрация натрия Натрий
Интрацеллюлярное
пространство
Натрий-
кал и й-
АТФаза
Высокая концентрация калия
Натрий-калий-
АТФаза
m
5
чэ
Потенциал покоя мембраны
Потенциал
равновесия
для натрия
+60
+40
+20
0
-20
Порог
активации
потенциала
действия
-40
-60
80
Потенциал
равновесия
для калия
100
Деполяризация
смещай потенциал
действия мембраны
сторону менее о
цдтельным величии
Потенциал покоя мембраь
Гиперголярнзяцня
смещает потенция/
* действия мембраны
в сторону более отрн-
_____ нательным величин
Уравнение Нернста
Расчет потенциала равновесия для любого иона в концентрации Хт
когда концентрация вне клетки [Х]^ и внутри клетки [X]., вычисляется
по уоавне чию Нернста: „
г где R — газовая константа;
Е -RT/zF In [XL / [X] . Т “ температура, еК;
z — валентность X;
F — константа Фарадея
Так, например, Ен* в скелетной мышце челе тепа будет равен:
(2)
Деполя ризация
Натриевая проводимость
Калиевая проводимо^ гь
Na-
Потенциал действия
Приток натрия
в клетку
Порог активации
потенциала действия,
т.е. порог критической
импульсации
Открывает
вольтажзави-
симые натри-
евые каналы
Пол ожител ьная
обратная
связь |
активации ।
натриевых
каналов .
Е_* г58 In 3/14G - -95 чВ
Если натриевых каналов
открыто недостаточно,
критический порог импульсации
не достигается и потенциал
не генерируется, наблюдается
лишь преходящая деполяризация
Потенциал покоя мембраны
В покое мембрана нейрона относительно непро-
ницаема для ионов. Это важно для генерации по-
тенциала покоя. Основным интрацеллюлярным
ионом является калий, а экстрацеллюлярным — нат-
рий. Поэтому в соответствии с электрохимическим
градиентом поток ионов калия направлен из клет-
ки, а поток ионов натрия — в клетку. Перемещение
положительно заряженных ионов из клетки приво-
дит к генерации отрицательного мембранного по-
тенциала или гиперполяризации, тогда как поток
ионов натрия внутрь клетки вызывает деполяриза-
20
шло. Однако покоящаяся мембрана относительно
непронипаема для натрия, но в то же время доста-
точно проницаема для калия. Поэтому в покое ка-
лий имеет тенденцию к перемещению из клетки в
соответствии с электрохимическим градиентом, что
приводит к отрицательному заряду мембраны. Так
продолжается, пока химический градиент концен-
трации, приводящий к выходу калия из клетки, не
выключится разностью электрического потенциа-
ла, вызванного притоком ионов в клетку (мембран-
ным потенциалом) и приводящего к возвращению
калия в нейроны. Мембранный потенциал, обес-
печивающий это стационарное состояние, называ-
ется потенциалом равновесия для калия (Ек ) и вы-
числяется по уравнению Нернста (см. рисунок). На
самом деле измеренный потенциал покоя аксонов
имеет более положительную величину, поскольку
незначительная проницаемость мембраны для
ионов натрия, в покое сохраняется. Небольшой при-
ток натрия поддерживается аденозинтрифосфат
ЛТФ)-зависимой натрий-калий-АТФазой, которая
сама по себе обладает небольшой электрогеннос-
тью. Этот насос важен для поддержания ионных
градиентов и является электрогенным, в силу того что
обменивает 3 иона натрия на 2 иона калия, входя-
щих в клетку. Это вносит лишь незначительный
вклад в потенциал покоя мембраны.
Генерация потенциала действия
Одним из фундаментальных свойств нервной
системы являются генерация и проведение нервных
импульсов (см. главы 8 и 19). Эти импульсы вклю-
чают генераторные, синаптические потенциалы и
потенциалы действия, последние определяют как
одиночные электрические импульсы, распространя-
ющиеся по аксону. Этот потенциал действия (нервный
импульс или спайк) возникает по принципу «все или
анчего», т. е. когда стимул достигнет определенного
порога интенсивности. Таким образом, информация
в нервной системе закодирована по частоте импуль-
сации, а не по размеру потенциала действия (см. гла-
ву 18). Порог интенсивности стимула определяется
как величина, до которой поднимается внутренний
сетевой ток (зависимый в основном от ионов на-
трия), которая выше, чем внешний сетевой ток (за-
висимый в основном от ионов калия), и которая со-
ставляет порядка 55 мВ (критический порог им-
пульсации). Он возникает в области аксонального
бугорка, где плотность натриевых каналов наиболее
высока и где инициируется потенциал действия ней-
рона. Однако если порог не будет достигнут, депо-
ляризация не генерирует потенциал действия и сиг-
нал не распространится по аксону.
Последовательность событий, участвующих в ге-
нерации потенциала действия, такова:
• Деполяризующий заряд активирует вольтажзави-
симые натриевые каналы на мембране нейрона,
который позволяет некоторым ионам натрия пе-
реместиться в соответствии со своим электрохи-
мическим градиентом (повышенная натриевая
проводимость). Это деполяризует мембрану и от-
крывает еще больше натриевых каналов согласно
механизму положительной обратной связи. Когда
открывается достаточное количество натриевых
каналов, для того чтобы величина внутреннего тока
сети превысила величину тока, обеспеченную вы-
ходом калия из клетки, открываются быстрые нат-
риевые каналы, что приводит к массивному при-
току натрия, который деполяризует мембрану до по-
тенциала равновесия для натрия (примерно +55 мВ).
Таким образом, генерируется спайк потенциала
действия, однако он не достигает потенциала рав-
новесия для натрия в силу продолжающегося и
увеличивающегося оттока ионов калия из клетки.
• Далее следует убывающая фаза потенциала дей-
ствия, при этом вольтажзависимые натриевые ка-
налы инактивируются (см. главу 5). Эта инактива-
ция также вольтажзависима, однако она более мед-
ленная, чем фаза активации, и поэтому возникает
позже (см. главу 5). Во время фазы убывания по-
тенциала приобретает более важную роль вольтаж-
зависимый калиевый поток, так как его активация
деполяризацией мембраны происходит еще более
медленно, чем инактивация натриевых каналов.
Активированные вольтажзависимые калиевые ка-
налы обеспечивают короткий период гиперполя-
ризации мембраны, предшествующей ее дезакти-
вации, и потенциал мембраны возвращается в со-
стояние покоя.
Сразу после появления спайка потенциала дей-
ствия отмечается рефрактерный период, когда ней-
рон становится невозбудимым (абсолютный рефрак-
терный период) или же активируется только субмак-
симальными стимулами (относительный рефрактерный
период). Абсолютный рефрактерный период возни-
кает в момент максимальной инактивации натрие-
вых каналов, тогда как относительный — в более по-
здний период, когда большинство натриевых кана-
лов уже вернулось в состояние покоя, но ток через
активированные калиевые каналы уже раходится в
развернутой фазе. Рефрактерный период содержит
два важных аспекта для генерации и проведения по-
тенциала действия. Во-первых, потенциалы действия
могут распространяться только в одном направле-
нии — от места генерапии, и, во-вторых, они могут
генерироваться лишь с определенной частотой.
Оригинальное описание потенциала действия
принадлежит Ходжкину и Хаксли, которые изучали
этот феномен на гигантском аксоне кальмара в 50-х
годах XX века. Этот механизм был подтвержден поз-
же и для многих других клеток и нейронов. Это на-
ряду с открытием большого количества ионных ка-
налов послужило началом описания многих других
модификаций генерации потенциала действия ней-
ронами и другими клетками.
ГЛАВА 7. Нервно-мышечное соединение и синапсы
Заболевание
Синдром
Итона—
/амоерта
ф Стадия синаптической
трансмиссии Миелин
(объяснение в тексте)
Пресинапти-
ческая
терминаль
ок
токсин
Ботулинический
АцхР (состоит из 2а-,
JJ-, у- н б- субъединиц,
а-субъединица связывается
с Ацх; чтобы открыть канал,
необходимы 2 молекулы Ацх)
Ацх
сеть
Постсинаптическая
складка
Гладкий
эндоплазматический
ретикулум
зона
Митохондрия
Пресиналтические
везикулы
г
Синапти-
ческая щель
Постсинаг-
тическая
щель
Базальная
мембрана
(никотиновые
в НМС)
Экзоцитоз везикулы
Пресиналтические везикулы содержат ней-
ротрансмиттер. роль которого в нервно-мы-
шечном соединении (НМС) выполняет аце-
тилхолин (Ацх). Кроме того, везикулы со-
держат АТФ и специфические мембраннО’
ассоциированные белки, которые обеспечи-
вают попадание везикул в активные зоны,
слияние их с пресинаптической мембраной в
активной эоне, а также их возвращение и за^
гюлнение новом порцией нейротрансмиттера.
Пресиналтические активные зоны — это
специализированные участки дня высвобож-
дения везикул, которые находятся напротив
вторичных посгсимаптнческих складок.
В активных зонах плотно расположены воль-
таж зависимые кальциевые каналы. Вокруг
этих участков, имеющих темную окраску на
электронно-микроскопическом срезе мембра-
ны нервной терминали, располагаются мито-
хондрии и везикулы. Везикулы располагают
ся двойными рядами, прилежат к мембране с
обеих сторон электронно-плотной структуры.
Вещество синаптической щели обнаружи-
вается только в ацетилхолиновом НМС и со-
держит фермент ацетилхолинэстеразу (АцмЭ),
которуя инактивирует Ацх, гидролизуя его.
Синаптическая щель имеет ширину около
500 Е и отделяет пресиналтическую терми-
наль от постсинаптической мембраны.
Ацетилхолиновые рецепторы (АцхР). пре-
вращающие высвобожденный нейротранс-
миттер в постсннаптический электрический
импульс, в основном находятся между лост-
симаптическими складками. Эти рецепторы
ассоциированы с неселективным катион-
ным каналом. Связывание Ацх с этим ре-
цептором вызывает конформационные изме-
нения рецептора, канал открывается, возни
кают приток натрия и отток калия, что
приводит к деполяризации мембраны и ак-
тивации вольтажзависимых ионных каналов.
В случае НМС высвобождение содержимого
одной везикулы приводит к миниатюрной по-
стсинаптической деполяризации — мини-
атюрному потенциалу концевой пластин-
ки (МПКП) Если высвобождается много ве-
зикул, МПКП суммируются н приводят к
большему ответу — потенциалу концевой
пластинки (ПКП), который в свою очередь
генерирует потенциал действия в мышце.
Состояние покоя, везику-
лы находятся в лресимап-
тической актиновой сети
Приток кальция
вьювобождает везикулы
из актиновой сети
Везикулы сливаются
с пресинаптической активной
зоной и высвобождают Ацх
Шеррингтон в 1897 г. предложил термин «синапс»
для обозначения соединения двух нейронов. Боль-
шинство работ по изучению синапсов проводилось
на ацетилхолинергическом нервно-мышечном со-
единении (НМС), однако показано, что химические
синапсы обладают сходными функциями и в ЦНС.
Химические синапсы преобладают в нервной сис-
теме, хотя, помимо них существуют и электричес-
кие синапсы, например, в глиальных клетках (см.
главу 40).
22
Нервно-мышечная передача
(модель синаптической трансмиссии)
Последовательность событий, происходящих в
химическом синапсе, такова:
• Возникновение потенпиала действия вызыва-
ет деполяризацию пресинаптической терминали
(см. рисунок, 1), при этом открываются вольтаж-
зависимые кальциевые каналы в активных зонах
синаптической терминали и впоследствии каль-
ций поступает в терминаль (см. рисунок, 2; на этой
стадии происходит синаптическая задержка им-
пульса).
• Приток кальция приводит к фосфорилированию
и изменению пресинаптических кальцийсвя-
зывающих белков (некоторые из них находятся
в мембране везикул). Они освобождают везику-
лу из пресинаптической актиновой сети, что по-
зволяет везикуле слиться с пресинаптической
мембраной (см. рисунок, 3). К этим белкам от-
носятся различные растворимые N-этиленма-
лемидчувствительные белки слияния и их ре-
цепторы.
• Слияние двух полуканалов (пресинаптической
везикулы и пресинаптической мембраны) приво-
дит к формированию небольшой поры, которая
быстро расширяется при высвобождении содер-
жимого везикулы в синаптическую щель. Мемб-
рана везикулы может возвратиться при помо-
щи эндоцитоза в пресинаптическую терминаль
с помощью неселективного или селективного ме-
ханизма.
• Большая часть высвободившегося нейротрансмит-
тера растворяется вокруг синаптической шели и
связывается с постсинаптическим рецептором (см.
рисунок, 4). Некоторые молекулы нейротрансмит-
тера попадают за пределы синаптической щели и
теряются, тогда как другие инактивируются, преж-
де чем связаться с постсинаптическим рецептором.
Эта инактивация важна для нормальной функции
синапса, и если в НМС происходит ферментатив-
ная деградация ацетилхолина (Анх), то в других си-
напсах используются механизмы обратного захвата
и рециркуляции трансмиттера в пресинаптический
нейрон (см. главу 9).
• Активация постсинаптического рецептора приво-
дит к изменению потенциала постсинаптической
мембраны. Каждая везикула содержит определен-
ное количество нейротрансмиттера, высвобожде-
ние которого генерирует небольшой постсинап-
тический заряд определенной величины — мини-
атюрный потенциал концевой пластинки ( М П КП ).
Высвобождение нейротрансмиттера из несколь-
ких везикул приводит к суммации МПКП и ге-
нерации большого деполяризационного потенци-
ала или потенциала концевой пластинки (ПКП),
который, если его величина достаточна, дости-
гает порога генерации потенциала действия на
постсинаптическом мышечном волокне (см. ри-
сунок, 5).
Эту везикулярную гипотезу критиковали, по-
скольку не во всех синапсах ЦНС нейротрансмит-
теры содержатся в везикулах, а также из-за суще-
ствования электрических синапсов. Поэтому были
предложены другие теории, согласно которым в
переносе нейротрансмиттеров через пресинапти-
ческую мембрану или поры, открывающиеся в от-
вет на приток кальция, участвуют другие молеку-
лы. Однако научное обоснование этих гипотез до-
вольно слабое.
Нарушения нервно-мышечной передачи
Существует ряд природных токсинов, которые
могут нарушать функцию НМС.
• Кураре связывается с ацетилхолиновым рецепто-
ром (АцхР) и препятствует реализации эффекта,
что вызывает паралич. Это подтверждено при ис-
пользовании миорелаксантов — дериватов кураре
в хирургии.
• Ботулинический токсин препятствует высвобожде-
нию Ацх из пресинаптической терминали. Он яв-
ляется экзотоксином бактерии Clostridium botulinum
и связывается с пресинаптической мембраной аце-
тилхолинового синапса, препятствуя высвобожде-
нию Ацх. Попадание этого токсина в пищу при-
водит к пищевому отравлению и параличу, а также
вегетативной недостаточности (см. главу 42). Од-
нако в небольшом количестве токсин может быть
использован в лечебных целях, например для вве-
дения в гиперактивную мышцу при фокальной ди-
стонии — заболевании, при котором часть тела
приобретает патологическую позу из-за гиперак-
тивности тех или иных мышц (см. главу 39).
НМС селективно повреждается при некоторых
неврологических заболеваниях. К ним относятся
миастения, миастенический синдром Итона-Ламбер-
та (МСИЛ) и нейромиотония Исаакса. При нейро-
миотонии Исаакса больной жалуется на мышечные
судороги и скованность из-за гиперактивности
мышцы, что связано с выработкой антител к преси-
наптическим вольтажзависимым калиевым калалам.
Это приводит к тому, что нервная терминаль посто-
янно находится в состоянии деполяризации и выс-
вобождает нейротрансмиттеры. В противополож-
ность этому при МСИЛ антитела блокируют преси-
наптическое кальциевые каналы. При этом на
повторную активацию синапса возникает стабиль-
ный приток кальция, поскольку антитела конкурент-
но блокируют места связывания экзогенного каль-
ция. Пациент жалуется на слабость преимущест-
венно проксимальных мыши, которая временно
уменьшается при физической нагрузке. Миастения,
с другой стороны, связана с выработкой антител к
АцхР, и больные жалуются на слабость, которая на-
растает при физической нагрузке (утомляемость) на
мышцы глаз, глотки и конечностей. Слабость свя-
зана с тем, что количество АцхР снижено и Ацх, выс-
вобождаемый пресинаптически, конкурирует за свя-
зывание с малочисленными рецепторами.
Электрические синапсы
В некоторых областях мозга осуществляется элек-
трическая трансмиссия. Наличие быстропроводя-
щих соединений обеспечивает быстрое распростра-
нение электрической активности и может быть по-
лезно для синхронизации некоторых функций коры
мозга (см. главу 15). Отсутствие таких соединений в
шванновских клетках приводит к развитию одной из
форм наследственной мотосенсорной полинейропа-
тии (НМСН).
23
ГЛАВА 8. Проведение по нерву и синаптическая интеграция
Немиелинизированный аксон
Потенциал действия,
распространи ющи йся
по всей аксолемме
Возникновение
потенциала
действия
Менее
1 мкм
Деполяри «ацисчный
Миелинизированный аксон
стимул
m
Деполя ризаци я
перехвата Ранвье
Диаметр
аксона
Деполя ризация
соседней аксолеммы
Немиелинизированные
волокна
Проведение по нерву I
55
ИПСП
впсп
Сопротивление X
мембраны
значительно меньше
внутреннего
сопротивления >
ч аксоплазмы /
| Синагпнческа^^нте^аци^
® Ингибиторный Нисходящий (g)
интернейрон импульс
(клетка Рен шоу)
1А-аффереит ®
Нисходящий импульс^
Мотонейрон
Актизнссть
афферентного волокна
ингибиторный^
интернейрон и
1А-аффер₽нт(2)
Интиби|Орная обратная
связь через аксональную
коллатераль
Сеть нейронов и постсинаптический
нейрон (мотонейрон) получает возбуж-
дающий нисходящий супраспи. альныи
стимул (В) и от периферического ре-
цептора мышцы (Б). Кроме того, мо-
тонейрон получает импульс от инги-
биторного интернейрона — А (см. гла-
вы 33 и 34)
О
Потенциал действия,
распространяющийся от
перехвата Ранвье
к другому перехвату
Ранвье — сальтаторное
проведение
41
<0
6 80
Временная суммация
для генерации потемци-
ала действия. Импульс
от (Б) суммируется во
времени, для того что-
вы запустить потенци-
ал действия
Возникновение
.. по’ьнциала
Миелин
действия
Сопротивление X
мембраны миелинизиро-
ванной части аксона
значительно превышает
внутреннее сопротивле-
\ ние аксоплазмы /
Более 1 мкм
Миелинизированные волокна
Пространственная
суммация для ге-
нерации потенциала
действия. Более или
менее одновремен-
ные импульсы от Б и
В суммируются и за-
пускают генерацию
потенциала действия
ш
Интеграция ИПСП. Если ин-
гибиторный импульс, генери-
рующий ИПСП, предшеству-
ет возбуждающему импульсу
в течение определенного ле-
риода времени, возникает
уменьшение или утрата ИПСП.
Такого не происходит, если
ИПСП следует за ВПСП
Проведение по нерву
Проведение потенциала действия обеспечива-
ется распространением локального тока и высоким
фактором надежности генерации потенциала дей-
ствия в результате положительной обратной связи
активации натриевых каналов во время восходя-
щей фазы нервного импульса (см. главу 6). Местный
ток, однако, не только ограничивает скорость про-
ведения, но и влияет на чистоту проводимого сиг-
нала. Нервная система преодолевает эти труднос-
ти, так как нервные волокна, имеющие диаметр
выше определенных значений, покрываются миели-
ном, который периодически прерывается перехва-
тами Ранвье.
В демиелинизированных аксонах потенциал дей-
ствия, возникший на одном участке, приводит к де-
поляризации мембраны спереди и теоретически по-
зади этого участка, несмотря на то что мембрана там
находится в состоянии рефрактерности, так что по-
тенциал действия распространяется только в одном
направлении (см. главу 6). Ток преимущественно
распространяется через мембраны (в силу высоко-
го внутреннего сопротивления аксоплазмы) и име-
ет наибольшую величину в участке, наиболее близ-
ком к потенциалу действия.
24
Проведение нервного импульса в немиелинизи-
рованных аксонах точно и надежно, особенно в во-
локнах малого диаметра, где внутреннее сопротив-
ление аксоплазмы очень велико, однако это прове-
дение медленное.
Проведение по волокну увеличивается пропор-
ционально его диаметру (примером этому служит ги-
гантский аксон кальмара толщиной более 1 мм). Чем
больше диаметр волокна и толщина миелиновой
оболочки, тем выше скорость проведения по волокну.
Последовательность событий при проведении
импульса по миелинизированным волокнам точно та-
кая же, однако имеет существенное отличие: прохо-
дящий по волокну потенциал действия сталкивает-
ся с таким препятствием, как миелиновая оболочка
с высоким сопротивлением и низким емкостным
сопротивлением. Таким образом, деполяризирую-
щий ток проходит по аксоплазме, пока не достигнет
перехвата Ранвье с низким сопротивлением, высо-
ка плотность натриевых каналов и где генерируется
потенциал действия. Таким образом, потенциал дей-
ствия проводится вниз по волокну, от перехвата к
перехвату — что называется сальтаторным проведе-
нием. Преимущество миелинизации состоит в том, что
она позволяет быстро проводить импульс при мини-
мальных энергозатратах со стороны клетки. Она так-
же обеспечивает повышенную емкость нервной си-
стемы таким образом, что несколько быстропрово-
дящих волокон могут быть объединены в один
короткий нерв. Поэтому большинство аксоном, име-
ющих диаметр более 1 мм, миелинизированы.
Нарушения проведения проявляются клинически
при нарушении структуры миелиновой оболочки,
например при воспалительной демиелинизирующей
полинейропатии: синдроме Гийена—Барре при пора-
жении ПНС или рассеянном склерозе при пораже-
нии ЦНС (см. главу 54). При обоих заболеваниях от-
мечается гибель миелиновой оболочки, особенно в
участках, близких к перехватам Ранвье, что сопровож-
дается повреждением ионных каналов, а также сни-
жением протяженности изоляции аксона. В резуль-
тате проходящий потенциал действия должен депо-
ляризовать более длинный участок аксолеммы, часть
которой возбудима не в такой степени, как нормаль-
ный перехват Ранвье, поскольку содержит меньше
ионных каналов. Это приводит к замедлению распро-
странения потенциала действия и, если демиелини-
зация выражена, к уменьшению потенциала действия
до величины, при которой он больше не может рас-
пространяться, - так называемый блок проведения.
Интеграция синапсов
Каждый нейрон ЦНС образует несколько сотен
синапсов, и каждый импульс интегрируется в резуль-
тирующий разряд нейрона, — процесс, который
предполагает суммацию импульсов из различных
областей в любой момент времени (пространствен-
ная суммация), а также суммацию одного или не-
скольких импульсов в течение определенного вре-
мени (временная суммация).
Пресинаптическая нервная терминаль обычно
содержит один нейротрансмиттер, хотя высвобож-
дение одного или нескольких трансмиттеров из од-
ной пресинаптической терминали описано как про-
цесс, называемый котрансмиссией (см. главу 9). Ко-
личество высвобожденного нейротрансмиттера
зависит не только от степени, до которой пресинап-
тическая терминаль деполяризована, нои от уровня
синтеза нейротрансмиттера, присутствия ингибиру-
ющих пресинаптических ауторецепторов и преси-
наптических влияний других нейронов в виде аксо-
аксональных синапсов (см. главу 3). Эти синапсы
обычно ингибирующие (пресинаптическая ингиби-
ция) и чаще всего встречаются в чувствительных
проводящих путях (см. главу 18).
Высвобождаемый нейротрансмиттер связывается
со специфическим белком или рецептором на пост-
синаптической мембране, а в некоторых синапсах —
с пресинаптическими ауторецепторами (см. главу 9).
Когда их связывание приводит к открытию ионного ка-
нала с притоком катионов в постсинаптический отро-
сток и деполяризацией, синапс называется возбужда-
ющим, когда же в этих каналах возникает постсинап-
тический приток анионов или выход катионов с ги-
перполяризацией, синапс называется ингибиторным.
Возбуждающие постсинаптические потенциалы
(ВПСП) это деполяризационные заряды в постсинап-
тической клетке, возникающие в результате возбужда-
ющего синаптического импульса. Деполяризация,
связанная с ВПСП, может продолжаться и индуци-
ровать потенциалы действия, если они суммирова-
ны во времени и пространстве. Пространственная сум-
мация включает интеграцию постсинаптической клет-
кой нескольких ВПСП от различных синапсов, при
этом суммированная деполяризация достаточна, для
того чтобы вызывать потенциал действия. Временная
суммация, с другой стороны, включает суммацию
импульсов во времени таким образом, что каждый пос-
ледующий ВПСП все больше деполяризует мембрану,
пока не будет достигнут порог генерации потенциата
действия. В противоположность этому ингибирующие
постсинаптические потенциалы (ИПСП) являются
гиперполяризапионными зарядами постсинаптичес-
кой мембраны, возникающими обычно в результате
притока хлора или оттока калия через их рецептивные
ионные каналы. ИПСП очень важны для модуляции
нейронального ответа на возбуждающие синаптичес-
кие импульсы (см. рисунок). Поэтому ингибирующие
синапсы обычно находят в стратегически важных об-
ластях нейрона — проксимальных дендритах и теле —
таким образом они могут оказывать влияние на им-
пульсы, поступающие от обширных частей дендри-
тического дерева. Кроме того, некоторые нейроны
могут ингибировать свои собственные импульсы с
использованием аксональных коллатералей и ло-
кального ингибиторного интернейрона (ингибирую-
щая обратная связь), например мотонейроны и клет-
ки Реншоу в спинном мозге (см. главу 34).
Долговременная модуляция синаптической транс-
миссии обсуждается в главах 37, 45 и 53.
25
ГЛАВА 9. Нейротрансмиттеры, рецепторы
и нейротрансмиттерные системы
Пресинаптическая терминаль
Возбуждающим аксо-аксональ-
ный синапс, который вызывает
пресинаптическое торможение,
приводящее к небольшой депо-
ляризации пресинаптической
терминали перед возникнете»
кием деполяризующего стиму-
ла в главном афферентном ак-
соне
Активация пресинаптического
ауторецептора (Т R). которая
инмгмбирует дальнейший выб-
рос нейротрансмиттеров
KR
Барб
Постсинаптическим
Постсинаптнческий рецептор, на-
прямую связанным с ионным кана-
лом (T(R) (см. АцхР. глава 7).
В других случаях ионный канал
формируется рецепторным комп-
лексом, например ГАМК-барби-
турово-(Барб)-беизодиаае липо-
вым (БДЗ)-комплексом вокруг
хлоридного канала
БДЗ
ГАМК
Нейротрансмиттеры и синаптическая функция
Нейротрансмиттеры, высвобождаемые в синап-
се, взаимодействуют со специфическим белком на
постсинаптической мембране, который называется
рецептором. В ряде синапсов нейротрансмиттеры
связываются с пресинаптическим ауторецептором,
который регулирует количество высвобождаемого
нейротрансмиттера. Рецепторы для каждого конк-
ретного нейротрансмиттера обычно специфичны,
однако может существовать несколько разновидно-
стей одного и того же рецептора. В ряде случаев ней-
ротрансмиттеры, высвобождаемые одновременно,
могут регулировать связывание одного нейротранс-
миттера со своим рецептором или действовать си-
нергично на один и тот же ионный канал (напри-
мер, у-аминомасляная кислота (ГАМК) - барбиту-
рово-бензодиазепиновый рецепторный комплекс).
Тормозной аксо аксональный синапс
Инактивация нейротрансмиттера (Т)
Усвоение трансмиттера
пресинаптической терминалью
(например, серотонина)
2 Гидролиз нейротрансмиттера
(Ту + Тк) с рециркуляцией
продукта распада
Усвоение нейротрансмиттера
соседними глиальными
клетками (например, глутамата)
Постсннаптнческне рецепторы, ассоцн
проданные с ГТФ-сэязывающимн бел-
ками (G-белками), которые открывают»
ся (А) после каскада внутриклеточных
событий или модуляции (Б) ионных
каналов, например метаботропных глу-
таматных рецепторов, и, возможно,
высвобождения мейро-оансмиттерОВ
Отсроченная потенциация
или депрессия (см. главу 45)
Г — нейротрансмиттер
ТхТу — продукты распада нейротрансмиттера
Рецепторы для специфических нейротрансмит-
теров напрямую связаны с ионными каналами (см.
рисунок J1R, т.е. ацетилхолиновый рецептор — АцхР;
см. главу 7) или с мембранным ферментом (см. ри-
сунок, T2R). В последнем случае связывание ней-
ротрансмиттера с рецептором открывает ионный
канал с помощью внутриклеточного ферментатив-
ного каскада (например, циклического аденозинмо-
нофосфата — цАМФ и G-белков) или опосредован-
но модулирует вероятность открытия других ионных
каналов в ответ на изменение заряда мембраны (ней-
ромодуляция). Таким образом, эти рецепторы опос-
редуют медленные синаптические события, в отли-
чие от рецепторов, которые напрямую связаны с
ионными каналами, передающими синаптическую
информацию.
Активированный рецептор может вернуться в
состояние покоя только если нейротрансмиттер уда-
26
лен либо в результате ферментативного гидролиза,
либо обратного захвата в пресинаптическую не-
рвную терминаль или соседние нейроглиальные
клетки. Несмотря на это, процесс возвращения ре-
цептора и его ионного канала в состоянии покоя
предполагает промежуточные ступени. В ряде синап-
сов аффинитет и количество рецепторов зависит от
предшествующей активности синапса. Например, в
катехоламинергических синапсах рецепторы стано-
вятся менее чувствительными к высвобождаемому
нейротрансмиттеру, когда синапс крайне активен,
этот процесс называется десенситизацией и ингиби-
рованием функции, при котором происходят сниже-
ние аффинитета рецептора к нейротрансмиттеру за
короткое время, а также снижение количества рецеп-
торов в течение более длительного времени.
В синапсах, которые активируются редко, про-
исходят обратные процессы (гиперсенситизация и
усиление функции), при которых синаптическая ак-
тивность модулируется с помощью продолжающей-
ся активности. Кроме того, в некоторых синапсах
активация постсинаптического комплекса рецеп-
тор - ионный канал может модулировать отсрочен-
ную активность синапса, воздействуя как на преси-
наптическое высвобождение нейротрансмиттера,
так и на ответ постсинаптического рецептора, и про-
цессы называются отсроченной потенциацией (ОП)
и отсроченной депрессией (ОД), они зависят от из-
менений эффективности работы синапса с течени-
ем времени (см. главы 37 и 45).
Таким образом, состояние, количество и виды ре-
цепторов для определенного нейротрансмиттера, как
и присутствие рецепторов для других нейротрансмит-
теров, имеют важное значение в определении сте-
пени синаптической активности в каждом синапсе.
Классификация и морфология
нейротрансмиттерных систем
В нервной системе действует большое количество
нейротрансмиттеров, которые можно разделить на
несколько семейств (см. Приложение 1).
Возбуждающие аминокислоты
Возбуждающие аминокислоты представлены боль-
шим количеством возбуждающих нейротрансмитте-
ров ЦНС, которые играют важную роль в поддержа-
нии синаптической активности. Основной возбужда-
ющей аминокислотой является глутамат, который
взаимодействует с несколькими рецепторами, опре-
деляемыми в зависимости от агониста, который их
активирует. Ионотропные рецепторы представлены
N-метил-D-аспартатными (NMDA)- и не-NMDA-pe-
цепторами, последние ассоциированы с кальциевым
каналом и играют важную роль в генерации ОП (см.
главу 45), эксайтотоксической гибели клеток (см. гла-
ву 52) и, возможно, развитии эпилепсии (см. главу 53).
Отдельную группу глутаматных рецепторов, ас-
социированных с G-белками, - составляют метабо-
тропные рецепторы, которые реагируют на актива-
цию, инициируя ряд внутриклеточных биохимичес-
ких событий, которые модулируют синаптическую
трансмиссию и активность нейронов. Эти рецепто-
ры, возможно, обусловливают отсроченную депрес-
сию в гиппокампе.
Тормозные аминокислоты
Основными тормозными нейротрансмиттерами
ЦНС являются ГАМК, представленная во всех ее
отделах, и глицин, который обнаруживается преиму-
щественно в спинном мозге. Нарушения функции
ГАМК-ергических нейронов могут лежать в основе
некоторых двигательных, тревожных расстройств и
эпилепсии (см. главы 41 и 51), тогда как мутации в
гене глицинового рецептора в настоящее время свя-
!ывают с развитием некоторых форм гиперэкплек-
сии - состояния, при котором имеется повышенный
стартл-ответ (рефлекс «Что такое?»), т. е. любой сти-
мул может приводить к падению человека.
Моноамины
Моноаминергические системы ЦНС берут нача-
ло от небольших групп нейронов ствола, которые
посылают свои проекпии во все отделы ПНС. Они
обнаружены во всем организме человека, включая
вегетативную нервную системы (см. главу 43), где
они связываются с различными рецепторами и от-
вечают за ряд функций (см. главы 21, 38 и 45).
Ацетилхолин
Этот нейротрансмиттер широко представлен в
нервной системе, включая нервно-мышечное соеди-
нение (см. главу 7), а также вегетативной нервной
системе (см. главу 42). Было создано разработано
множество лекарств, которые воздействуют на раз-
личные холинергические синапсы на периферии и
которые широко используются при анестезии. При
некоторых заболеваниях периферические холинер-
гические синапсы поражаются первично (см. гла-
ву 7), нарушение функции центральных холинерги-
ческих синапсов наблюдаются при болезни Пар-
кинсона и деменции альцгеймеровского типа (см.
главы 39 и 52).
Нейропептиды
Эти нейротрансмиттеры имеются во всех отделах
нервной системы. Нейропептиды часто высвобож-
даются вместе с другими нейротрансмиттерами. Из-
вестно много их видов. Они действуют, как обыч-
ные нейротрансмиттеры, или же выполняют роль
нейромодуляторов (например, в болевой рецепции;
см. главы 21 и 22).
ГЛАВА 10. Структура скелетной мышцы
| Поперечный срез мышечного волокна^
Поперечная (Т) тубул^
Митохондрия
Терминальная цистерна саркоплазматического
f ретикулума (СР)
Внеклеточна
жидкость
Ламинин 2 ДИСТР°-
ГЛИКИН
новый
F-актин
ООО
Волокна, пронизанные
6 тонкими филаментами,
окружающими толстый
филамент
Дистрофин
Адхалин
Сарколемма
(мембрана
локна)
Сарколемма
Триада А/1-соединения:'
Одна из петель филаментной спирали
Скелетная мышца отвечает за преобразование
электрического импульса, который прибывает к не-
рвно-мышечному соединению (НМС), в механичес-
кую силу посредством сокращения. Возникновение
потенциала действия приводит к высвобождению
ацетилхолина (Ацх), который активирует никотино-
вый АЦХ-рецептор (АцхР) на постсинаптической
мышце, что в свою очередь вызывает деполяризацию
мышечного волокна (см. главу 7). Это приводит к
притоку кальпия в мышечное волокно, а затем к мы-
шечному сокращению.
Структура скелетной мышцы
Скелетная мышца состоит из группы мышечных
волокон — длинных многоядерных клеток. Эти во-
28
локна содержат миофибриллы, которые в свою оче-|
редь состоят из толстых и тонких филаментов, до]
некоторой степени придающих мышце полосатый
внешний вид. Миофибриллы окружает сарколемма,!
которая инвагинирует в миофибриллы в виде попе-
речных, или Т-тубул. Эта структура отделена от сар-
коплазматического ретикулума (СР), который покры-
вает миофибриллы и принимает участие в хранении
внутриклеточных запасов кальция. Сарколемма яв-
ляется сложной структурой, и нарушения структу-
ры некоторых ее мембранных компонентов, как
было установлено недавно, лежат в основе развития
некоторых форм наследственных миодистрофий.
Толстый филамент состоит из миозина и лежит в
центре саркомера. Миозин состоит из двух цепей, ко-
торые представлены легким (ЛММ) и тяжелым (ТММ)
меромиозином. ТММ-порция содержит субфрагмен-
ты S1 и S2. Субфрагмент S1 состоит из двух головок
и двух связанных с этими головками цепей. Легкая
цепь находится на конце головки субфрагмента S1,
она называется неспецифической и отвечает за разло-
жение АТФ в завершение толчка, приводящего к фор-
мированию перекрестного мостика. Вторая легкая
цепь называется специфической, она прикрепляется
к головке субфрагмента S1 в том месте, где та высту-
пает в сторону актина, и принимает участие в процес-
се движения головки миозина. Благодаря свойствам
ЛММ миозиновые филаменты упаковываются спон-
танно так, что головки S1 выстоят в направлении ак-
тиновых филаментов. Таким образом, головка S1 фор-
мирует главную часть перекрестного мостика с актином.
Тонкие филаменты состоят из F-актина, тропомио-
зина и тропонина, которые в свою очередь содержат
3 субъединицы (тропонин I, С и Т). Эти 3 компо-
нента тропонинового комплекса выполняют различ-
ные функции, но в целом регулируют мышечное со-
кращение, удерживая тропомиозин в положении,
которое физически блокирует головку S1 миозина
отсвязывания с актином. Деполяризация мышцы при-
зодит к притоку кальция, который связывается с тро-
понином, и конформационным изменениям тонко-
го филамента, так что тропомиозин сдвигает место
связывания миозина и актина. Так, тропомиозин и
тропонин регулируют мышечное сокращение при по-
мощи блока. В некоторых мышцах животных регуля-
ция взаимодействия актина и миозина происходит
при помощи миозинассоциированных легких цепей.
В точке перекрытия этих двух наборов филамен-
тов находится триадная структура Т-тубулы, которая
связана с двумя терминальными цистернами СР с
помощью отростков.
Нарушения структурных белков
скелетной мышцы при миодистрофиях
Существует много первичных заболеваний мышц,
при которых в результате генетических мутаций
ионных каналов нарушается возбудимость мышц
(см. главу 5), а также возникает их воспаление (см.
главу 54). Кроме того, известны нарушения струк-
турных белков самой мышцы. Эти нарушения ле-
жат в основе развития некоторых наследственных
миодистрофий, из которых наиболее изученны-
ми являются миодистрофия Дюшенна (МДД) и
конечностно-поясные миодистрофий (КПМД).
МДД - это Х-сцепленное заболевание, при кото-
ром отмечается делеция гена, кодирующего струк-
турный белок дистрофии, при этом известна более
легкая форма заболевания — миодистрофия Бек-
кера, при которой количество этого белка уменьше-
но. МДД проявляется в раннем детском возрасте
нарушениями ходьбы и атрофией проксимальной
мускулатуры конечностей с псевдогипертрофия-
ми икроножных мышц. По мере прогрессирования
болезни инвалидность утяжеляется. Развиваются
кардиомиопатия и другие нарушения, приводя-
щие к смерти пациентов обычно на 3-м десяти-
летии жизни. Характерно повышение содержания
креатинфосфокиназы (маркер миолиза) у этих боль-
ных, поскольку в отсутствие дистрофина происхо-
дит некроз мышечных волокон. Этот белок залега-
ет под сарколеммой как скелетной, так и гладкой
или сердечной мышцы и обеспечивает стабиль-
ность и гибкость мышечной мембраны. При отсут-
ствии его мембрана легко разрывается, что приво-
дит к притоку большого количества ионов кальция
и некрозу мышечных волокон в результате актива-
ции протеаз.
КПМД в противоположность МДД могут начи-
наться в любом возрасте и сопровождаться слабос-
тью проксимальных мыши с повышением содержа-
ния креатинфосфокиназы. Эта болезнь наследует-
ся по различным типам, недавно показано, что
при аутосомно-рецессивном типе наследования от-
мечаются нарушения структуры гликопротеинов,
связанных с дистрофином, адхалина и саркоглика-
нового комплекса. Эти белки связывают внутрикле-
точный дистрофии с компонентами внеклеточной
жидкости и важны для поддержания целостности
сарколеммы.
ГЛАВА 11. Сокращение скелетной мышцы
Отросток
Первая стадия:
мышца в состоянии покоя
Толстый
филамент
АДФ
(J ж‘
АТФ
Третья стадия: генерация
напряжения н разрыв мостика
Тонкий
филамент
Саркоплазматический
ретикулум (СР)
Эта часть молекулы
миозина может
растягиваться
и развивать
напряжение,
которое
преобразуется
в движение
филамента
" Тропомиозин
✓ и тропониновый комплекс
= Актин
Кальциевый
насос
Г Тропомиозин-тропониновый
комплекс
Тропомиозин
Тропониновый
_ комплекс
связывания
для актина
Вторая стадия: возбуждение
и сокращение, формирование
перекрестного мостика
Деполяризация мышцы приводит
к распространению потенциала
действия по Т-тубуле до триады
и высвобождению кальция из
терминальных цистерн СР
Кальций диф-
фундирует и связы-
вается с тропонином,
высвобождая тропомиозин
на блока с актином — так
формируется актиномиозиновый
перекрестный мостик
Для разрыва
мостика
требуется
АТФ
Актиновый
филамент
подтягивается
к центру пучка
А с помощью
связывания
и ротаций
Возникновение потенциала действия в НМС при-
водит к притоку калвция и высвобождению везикул
Ацх. Он связывается с никотиноввтми АцхР на мы-
шечном волокне, что ввтзвтвает его деполяризацию
и приток калвция из сакроплазматического ретику-
лума (СР) в мышцу. Это приводит к удалению комп-
лекса калвцийсвязывающего белка тропомиозина и
тропонина от актина основного компонента тон-
кого филамента. Удаление блока позволяет миози-
ну — основному компоненту толстого филамента —
связатвся с актином посредством перекрестного мос-
тика. Волокна перемешаются навстречу друг другу.
Перекрестный мостик между двумя волокнами уст-
раняется, и в завершение этого толчка происходит
гидролиз АТФ. Пикл формирования перекрестного
мостика и его устранения может повторяться. В це-
лом этот процесс объясняется гипотезой мышечного
сокращения по механизму скольжения филаментов.
Последовательность событий при сокращении
мышцы такова:
• Первая стадия. В состоянии покоя тропонино-
вый комплекс удерживает тропомиозин в таком
положении, чтобв! миозин не связвтвался с ак-
тином (блок).
• Вторая стадия. Возникновение потенциала дей-
ствия в НМС приводит к инициации постси-
наптического потенциала действия, который
распространяется по специалвной инвагина-
ции мышечной мембраны, называемой попе-
речной, или Т-тубулой. Эта Т-тубула проводит
потенциал действия к мышце, и все мышечные
волокна активируются. Она прилежит к терми-
налвной цистерне СР, назвтваемой триадой,
т.е. Т-тубула залегает между двумя терминалв-
НБ1МИ цистернами СР (мышечный эквивалент
эндоплазматического ретикулума), содержаще-
30
го высокие концентрации ионов кальция. Т-ту-
булы связаны с СР отростками, которые яв-
ляются частью ионного канала для кальция.
Возникновение потенциала действия к триаде
приводит к высвобождению кальция из терми-
нальных цистерн посредством механического
слияния. Потенциал действия открывает об-
щий кальциевый канал между Т-тубулами и СР,
что позволяет кальцию войти, согласно элект-
рохимическому градиенту, в миофибриллы.
Затем кальций связывается с тропониновым ком-
плексом, это приводит к перестройке тропомиози-
на таким образом, что головка миозина становится
способной связаться с актином, формируя перекре-
стный мостик.
• Третья стадия. После связывания актина и ми-
озина сокращению перекрестного мостика
предшествует пауза. Актин подтягивается за
миозином, что приводит к сокращению мыш-
цы. После этого толчка перекрестный мостик
отделяет миозин от актина с помощью гидро-
лиза АТФ, что является также кальцийзависи-
мым процессом. Затем весь цикл может повто-
ряться. Процесс формирования перекрестного
мостика и движения филаментов называется
мышечным сокращением по механизму сколь-
жения филаментов. Два филамента перемеща-
ются параллельно друг другу, когда цикл повто-
ряется. Из терминальных цистерн СР высво-
бождается кальций, обеспечивающий процесс
формирования перекрестного мостика и его ус-
транения, который затем удаляется обратно в
эту структуру с помощью кальциевого насоса.
Нарушения мышечного сокращения
Заболевания мышц с нарушением их структуры
сопровождаются мышечной слабостью в результа-
те дезорганизации сократительных белков. Одна-
ко существуют заболевания, при которых отмеча-
ется нарушение процесса мышечного сокращения
само по себе, например периодический паралич и
злокачественная гипертермия. В последнем случае
отмечается патология рианодинового рецептора,
который является частью белкового комплекса,
связывающего Т-тубулы в СР. При общей анесте-
зии это приводит к продолжающейся деполяриза-
ции, сокращению и некрозу мыши, повышению
температуры тела и полиорганной недостаточнос-
ти. В противоположность этому процессу при пе-
риодическом параличе имеют место нарушения
функции ионных каналов, приводящие к длитель-
ной невозбудимости мышц, которые становятся
парализованными. Это редкие заболевания, в ходе
которых не поражаются дыхательные мышцы. При-
ступы провоцируются рядом факторов, в частности
принятием пиши, которая содержит большое ко-
личество углеводов.
Важно помнить, что нарушения сокращения
мышц возникают в результате поражения НМС (см.
главу 7), а также при некоторых врожденных дефек-
тах метаболизма. В последнем случае развиваются
метаболические миопатии, связанные с врожденны-
ми дефектами обмена углеводов и липидов. При этих
заболеваниях симптомы провоцируются физичес-
кой нагрузкой и сопровождаются хронической про-
грессирующей мышечной слабостью.
ГЛАВА 12. Анатомия спинного мозга
Большое затылочное отверстие
Конский хвост
Задние корешки
Топография восходящих путей спинного мозга,
проведение импульсов в задние столбы:
S -
крестцовый
Чувствительность ноги и ягодицы
поясничный -
Т — грудном
С — шейный
Чувствительность груди и живота
ж Чувствительность руки, шеи и спины
Двигательные пути
Нежный пучок
и клиновидный
Задний корешок
Тракт Лиссаузра
Нисходящие пути
спинного мозга
Кортикоспи нальный
тракт
Руброспинальный
тракт
Тектоспинальный,
ретикулоспинальный
вестибулоспинальный
тракты
Поперечный срез спинного мозга
пучок задние стол6ь|
Дор-
альныи Спине iepe6ej
Вент- лярные гоакть
ральный
Перед-
ний
Спиноталами-
ческие тракты
Топография пластин Рексед в серам веществе спинного мозга
Пластина В — желатинозная субстанция (см главы 21 и 22)
Пластина IV—VI — собственное ядро
Пластина VII содержит промежуточные
и интермедиолатеральные (боковые) столбы
Пластина IX содержит тела мотонейронов передних рогов
спинного мозга
Общая структура
Спинной мозг находится в позвоночном канале,
который проходит от большого затылочного отвер-
стия до I поясничного позвонка и имеет два утол-
щения (в шейном и поясничном отделах), что соот-
ветствует иннервации верхних и нижних конечнос-
тей (см. главу 1). Нижняя часть позвоночного канала
(ниже позвонка L1) содержит нижние поясничные
и крестцовые нервы и называется конским хвостом.
Чувствительные волокна вступают в спинной
мозг через дорсальные (задние) корешки и тела ней-
ронов, локализованные в спинномозговых узлах,
тогда как двигательные и преганглионарные вегета-
тивные волокна выходят из спинного мозга через
вентральные (передние) корешки вместе с некоторы-
ми, в основном немиелинизированными, афферент-
ными волокнами. Тела двигательных нейронов (или
мотонейроны) находятся в передних рогах спинного
мозга, а преганглионарные тела нейронов симпати-
ческой нервной системы — в боковых рогах спинно-
го мозга.
Тела нейронов, составляющие центральное серое
вещество, организованы в виде пластин Рексед. Бе-
лое вещество окружает серое и состоит из миелини-
зированных и немиелинизированных аксонов вос-
ходящих и нисходящих проводящих спинномозго-
вых путей.
Анатомия чувствительных афферентных
волокон, вступающих в спинной мозг
Чувствительная информация от периферических
рецепторов проводится по первичным афферент-
ным нервным волокнам, которые заканчиваются в
пластинах I-V в заднем роге, при этом для разных
32
рецепторов проекции различны. Однако на самом
деле многие афферентные волокна подразделяются
на восходящие и нисходящие ветви, когда они вхо-
дят в спинной мозг, поэтому синаптические контак-
ты они образуют как с интернейронами задних ро-
гов, так выше и ниже в составе тракта Лиссауэра.
Обработка чувствительной
информации в задних рогах
Некоторые первичные афферентные пути обра-
зуют синаптические контакты с одним нейроном
заднего рога. Конвергенция импульса способствует
уменьшению остроты (точности) локализации сти-
мула. Однако процесс латерального торможения
позволяет минимизировать потерю точности, обес-
печивая торможение импульсов субмаксимально
активированных волокон, таким образом увеличи-
вая пространственное разрешение чувствительного
стимула. В задний корешок поступает ряд нисходя-
щих стимулов от супраспинальных структур, кото-
рые важны для модуляции обработки чувствитель-
ной информации в спинном мозге (см. главу 22).
Восходящие чувствительные
пути спинного мозга
Спиноталамический тракт (СТТ), известный так-
же как антеролатеральная система, спиноцеребел-
лярные пути и задние столбы (ЗС) являются основ-
ными восходящими путями спинного мозга (см.
Приложение 2). Каждый путь передает определен-
ную информацию и имеет свою соматотопическую
проекцию, т. е. чувствительная информация от раз-
личных частей тела лежит в основе организации вос-
ходящих путей. По путям глубокой чувствительнос-
ти импульсы от ростральных частей тела (руки в от-
личие от ноги) поступают по волокнам, лежащим
более латерально в структуре проводящего пути (а
по путям поверхностной чувствительности импуль-
сы от ростральных частей тела поступают по во-
локнам, лежащим более медиально в структуре
проводящего пути). И ЗС, и СТТ перекрещиваются
(волокна пересекают среднюю линию), поэтому
чувствительная информация обрабатывается в про-
тивоположном полушарии. Однако области, где про-
исходит этот перекрест, у каждой группы проводя-
щих путей свои. Антеролатеральная система пере-
секает среднюю линию в спинном мозге, тогда как
ЗС перекрещиваются в нижних отделах продолгова-
того мозга, переключаясь на ядрах ЗС и формируя
медиальную петлю (см. главы 20 и 21).
Спинальные мотонейроны
а- и у-мотонейроны (МН) находятся в передних
(вентральных) рогах. а-МН являются наиболее
крупными нейронами нервной системы и иннерви-
руют волокна скелетных мышц, тогда как у-МН ин-
нервируют интрафузальные мышечные волокна, или
мышечные веретена (см. главу 33). МН имеют сома-
тотопическую проекцию в переднем роге. Более ме-
диально локализованы МН, иннервирующие про-
ксимальные мышцы, а латерально — МН, иннерви-
рующие дистальные мышцы (см. главу 34).
Нисходящие двигательные пути
Нисходящие двигательные пути различают по
месту их начала в ЦНС (см. Приложение 2). Корти-
коспинальный тракт (КСТ), или пирамидный тракт,
начинается от коры больших полушарий и вместе
с руброспинальным трактом иннервирует латераль-
ные МН, в свою очередь иннервирующие дисталь-
ную мускулатуру. Экстрапирамидные тракты (вес-
тибуло-, ретикуло- и тектоспинальные) иннерви-
руют МН, расположенные более вентромедиально
и контролируют аксиальную мускулатуру (см. гла-
вы 34-37).
Клинические признаки поражения
спинного мозга (см. главы 31 и 41)
Знание анатомии спинного мозга позволяет су-
дить о том, какой неврологический дефицит разо-
вьется при наличии очага в той или иной структуре,
что имеет большое значение в клинической невро-
логии. Примеры специфических поражений спин-
ного мозга и синдромов обсуждаются в главах 33 и 34.
ГЛАВА 13. Анатомия ствола мозга
Поперечный срез через верхнюю часть среднего мозга
Поперечный срез через нижнюю часть среднего мозга
Верхние бугорки четегрохолмия
Нижние бугорки четверохо-мчя
Медиальный
продольный пучок
ч. J
Медиальная петля X
Корково-
мостовой путь
(от височной
коры)
-Х' Сильвиев водопровод
’ Околоводопроводное
серое вещество
Ядро глазе
двигательного^
* ,нерва
Медиальный
продольный пучок
Околоводопровод-
ное серое вещество
Сильвиев
водопровод
Ядро блоксн
вого нерва।
Красное ядро
Мед.-алгная петля
Кортикоспинальный1
тракт
Черная субстанция
Черная субстанция
Корково-мостовой путь
(от лобной коры)
Ножка мозга
Ножка мозга
Элнфиз (шишковидная железа)
Ростральная губа
базальных отделов моста
Сильвиев водопровод
Сосцевидное
тело _
Верхние бугорки четверохолмия
) Нижние бугорю
f ' у четверохолмия
Поперечный срез через средние отделы моста
Ядро отводящего нерва
Медиальный
продольный пучок.
Вестибулярное ядро
Мост
Олива
Сильвиев
ВОДОПРОВОД'
IV желудочек
Хороидное
сплетение
(оР”’':
Медиальная петля
Пирамидным тракт
i Миндалина мозжечка
11'-Большое затылочное
J отверстие
s 1 Центральный канал
спинного мозга
Спинальное ядро
тройничного нерва
и его тракт
Клиновид-
ное ядро
Нежное I л
Ядра задним
ЯД₽оГСХбов
Двигательное ядро
тройничного нерва,
спинальное ядро
тройничного нерва
и его тракт
Ядро
лицевого нерва
Средняя
ножка мозжечка
.Кортикоспинальные
и корково-мостовые волокна
Поперечный срез через верхнюю часть продолговатого мозга
Слуховое
ядро
I Нижняя
ножка мозжечка
Солитар
иый тракт дорсапьное ядро
блуждающего
/ нерва
Ядро
подъязычного,
(нерва
Дорсальный
спнноцеребеллярный тракт
Кортикоспинальный 7
тракт 1
Руброспинальный тракт _Г
Вентральный
спииоцеребел лярный
тракт
Спиноталамический тракт
Передний рог
Медиальный
Продольный пучок
Вестибулоспинальный
и тектоспянальныи
тракты
j Спииальное|
ядро трой-
ничного нер-
ва и его тракт
Медиальный
продольный
пучок
Я зы коглоточиы й
нерв
Нижняя олива
Восходящие пути
Черепно-мозговые нервы
/ими к
Пирамида
Медиальная
петля
Двойное
ядро
Нисходящие пути
Ядра или тракты,
связанные с ЧМН
34
Ствол представляет собой часть головного моз-
га. находящуюся между большим затылочным отвер-
стием, ножками мозга и таламусом. Он состоит из
• родолговатого мозга, моста и среднего мозга и
находится кпереди от мозжечка, с которым соеди-
нен тремя парами ножек мозжечка. Ствол содержит
дующие структуры:
• Ядра X из XII пар черепно-мозговых нервов
(ЧМН) (см. главу 14), исключение составляют
обонятельный и зрительный нервы.
• Глазодвигательный аппарат, включающий III,
IV и VI пары ЧМН (см. главу 40).
• Моноаминергические ядра, проецирующиеся
на различные структуры ЦНС (см. главы 9,
23 и 46).
• Структуры, контролирующие жизненно важные
функции, дыхательный и сосудодвигательный
центры, а также центры, контролирующие ве-
гетативную нервную систему (см. главу 44).
• Структуры, участвующие в регуляции сознания
и включающие некоторые моноаминергичес-
кие ядра (см. главу 38).
• Некоторые восходящие и нисходящие прово-
дящие пути, связывающие спинной мозг с суп-
распинальными структурами, такими, как кора
головного мозга и мозжечок, некоторые из них
берут начало в стволе мозга (см. главы 12, 18,
20, 21, 32, 34-37).
Некоторые структуры ствола мозга заслуживают
отдельного рассмотрения.
• Ядра задних рогов, где заканчиваются волокна
задних рогов спинного мозга, отвечающие за
осязание, вибрацию и мышечно-суставное чув-
ство. Релейные нейроны этой структуры по-
сылают аксоны, которые перекрещиваются в
продолговатом мозге и формируют медиаль-
ную петлю, заканчивающуюся в таламусе (см.
главу 20).
• Пирамиды, представляющие кортикоспиналь-
ный тракт (КСТ) в продолговатом мозге, путь,
перекрещивающийся на нижней границе про-
долговатого мозга.
• Ядро солитарного пучка и двойное ядро обес-
печивают вкусовые ощущения и двигательную
иннервацию глотки посредством языкоглоточ-
ного и блуждающего нервов (см. главу 14).
• Нижние оливы продолговатого мозга получают
импульсацию из нескольких источников и со-
единяются с помощью проводящих путей с моз-
жечком (см. главы 37 и 47).
• Ножки мозжечка проводят информацию к моз-
жечку и от него (см. главу 37).
• Медиальный продольный пучок начинается
от вестибулярных ядер и проецируется рост-
рально, связывая некоторые глазодвигатель-
ные ядра (III и VI пару ЧМН), а также каудаль-
но, формируя часть вестибулоспинального
тракта.
• Вестибулярные ядра, имеющие связи с ухом и
проецирующиеся в спинной мозг и мозжечок,
как и другие структуры ствола (см. главы 28,
34 и 37).
• Черная субстанция среднего мозга содержит
дофаминергические и ГАМК-ергические ней-
роны. Является частью базальных ганглиев,
принимающих участие в регуляции движений
(см. главы 38—40). Утрата дофаминергических
нейронов является основным патогенетичес-
ким признаком болезни Паркинсона (см. гла-
ву 39).
• Красное ядро среднего мозга связано с мозжеч-
ком, от него берет начало руброспинальный
тракт, который вместе с КСТ формирует лате-
ральный нисходящий путь, контролирующий
движения (см. главы 12, 34—36).
• Околоводопроводное серое вещество среднего
мозга богато эндогенными опиоидами и осуще-
ствляет супраспинальную модуляцию ноцицеп-
ции (см. главу 21).
• Сильвиев водопровод проходит через средний
мозг и соединяет IV и III желудочки. Его суже-
ние приводит к развитию гидроцефалии (см.
главу 16).
• Ножки мозга содержат нисходящие проводя-
щие пути от коры головного мозга к спинно-
му мозгу и стволу, особенно мосту (см. главу 23).
• Нижние бугорки четверохолмия среднего моз-
га являются частью слухового анализатора (см.
главу 27), тогда как верхние бугорки четверо-
холмия участвуют в обработке зрительной ин-
формации и осуществляют контроль за дви-
жениями глаз (см. главы 24 и 40). Таким об-
разом, поражение ствола мозга может иметь
разрушительные последствия, при этом лока-
лизацию небольших очагов всегда можно точ-
но установить, поскольку количество структур,
находящихся в этой области мозга, огромно.
Чаще всего в стволе мозга располагаются вос-
палительные очаги (например, при рассеянном
склерозе; см. главу 54) или очаги сосудистой
природы (см. главу 17). Однако нарушения
функции ствола мозга могут наблюдаться и при
опухолях (см. главу 4), а также ряде других за-
болеваний.
ГЛАВА 14. Черепно-мозговые нервы
Основание мозга — ствол мозга
и череп но мозговые нервы (ЧМН)
Обонять ьная Луковица
Зрительныи нерв
и зрительный перекрес
Глазодвигательный нерв
Блоковой нерв
Тройничный нерв
Сосцевидные
Пирамидный тракт
Мост
Отводящий нерв
Лицевой нерв
Преддверно-улитковый нерв
Языкоглоточный нерв
Блуждающий нерв
Добавочный нерв
Подъязычный нерв
Продолгоь.йтын мозг
Дорсальный вид продолговатого мозга
III пара ЧМН
Средний мозг
IV пара ЧМН
V пара ЧМН
(чувствительное ядро)
VI пара ЧМН
(дви. ат! льное ядро)
VII пара ЧМН
Ножка
среднего мозга
X пара ЧМН
XII пара ЧМН
IX, X пара ЧМН
пара ЧМН
улитковый нерв
(VIII пар.- ЧМН)
Черепно-мозговые нервы
/. Обонятельный нерв
Рецепторы обоняния находятся в слизистой обо-
лочке полости носа, а их аксоны проецируются в
обонятельную луковицу на основании лобной доли,
проходя через продырявленную пластинку (см. гла-
ву 29). Этот черепно-мозговой нерв (ЧМН) не берет
начало в стволе мозга и не проходит там, он являет-
ся структурой ЦНС.
II. Зрительный нерв
Фоторецепторы глаза проецируются через гангли-
озные клетки в ЦНС посредством зрительного нерва.
Этот нерв проходит через зрительный канал и фор-
мирует перекрест зрительных нервов. Затем волокна
следуют к латеральному коленчатому телу, далее они
формируют зрительную лучистость и проходят к зри-
тельной коре (см. главу 24). Этот нерв не берет начало
и не проходит через ствол мозга, однако он дает про-
екцию в средний мозг, которая отвечает за зрачковый
рефлекс (см. главу 24), а также проекцию в гипотала-
мус, которая позволяет определять циркадные ритмы.
III. Глазодвигательный нерв
Глазодвигательный нерв начинается из среднего
мозга на уровне верхних бугорков четверохолмия. Он
иннервирует все глазодвигательные мышцы, кроме
латеральной прямой, иннервируемой отводящим нер-
вом (VI пара ЧМН), и верхней косой, иннервируемой
блоковым нервом (ГУпараЧМН). Глазодвигательный
нерв несет в себе парасимпатическую иннервацию
глаз, а также иннервирует мышцу, поднимающую
верхнее веко. Полный паралич глазодвигательного
нерва сопровождается экзофтальмом, офтальмопле-
гией, мидриазом с отсутствием реакции зрачка на
свет и птозом. Наиболее распространенными при-
чинами являются аневризма задней соединитель-
ной артерии и сосудистое поражение нервного ство-
ла, возникающее, например, при сахарном диабете.
IV. Блоковой нерв
Блоковой нерв начинается в среднем мозге на
уровне нижних бугорков четверохолмия и выходит
из ствола мозга дорсально. Он иннервирует верхнюю
косую мышцу глаза, поражение его приводит к дип-
лопии при взгляде вниз. Частой причиной пареза
блокового нерва является травма головы.
V. Тройничный нерв
Тройничный нерв выполняет двигательные и чув-
ствительные функции. Его двигательное ядро нахо-
дится на уровне средних отделов моста, медиальнее
главного чувствительного ядра, и оно получает кор-
ковую иннервацию (см. главу 35). Это ядро обес-
печивает функцию жевания. Чувствительные пути
от лица (включая роговицу) проходят в ствол мозга
в составе тройничного нерва и оканчиваются синап-
сами в трех основных комплексах ядер: спинномоз-
говом ядре тройничного нерва, главном чувствитель-
ном ядре тройничного нерва и мезэнцефальном
ядре. Чувствительность на лице проводится тремя
основными ветвями тройничного нерва: глазной
ветвью, которая иннервирует кожу лба, верхнечелю-
стной ветвью, которая иннервирует щеку, и нижне-
челюстной ветвью, которая иннервирует нижнюю
челюсть, при этом ростральные волокна (глазная
ветвь) поступают в верхнюю часть ядра спинномоз-
гового пути в верхнешейном отделе спинного моз-
га. Эти ядра тройничного нерва дают проекции в
таламус и являются частью соматосенсорной и бо-
левой проводящей системы ЦНС (см. главы 22 и
23). Поражение тройничного нерва приводит к сла-
бости жевательных мышц и нарушению опускания
нижней челюсти в сочетании со снижением чув-
ствительности на лице и утратой корнеального реф-
лекса. Эти симптомы развиваются при поражении
ствола мозга или ветвей тройничного нерва при вы-
ходе из черепа или при прохождении их через ка-
вернозный синус.
VI. Отводящий нерв
Отводящий нерв начинается из нижнедорсаль-
ной части моста мозга и иннервирует латеральную
прямую мышцу глаза. Поражение нерва приводит к
развитию двоения при взгляде в пораженную сто-
рону и может быть вызвано локальной патологией
ствола мозга или ложным очаговым симптомом при
повышении внутричерепного давления.
VII. Лицевой нерв
Лицевой нерв - это двигательный нерв, хотя он
содержит парасимпатические волокна для слезной
и слюнных желез (поверхностный каменистый нерв
и барабанная струна), а также чувствительные волок-
на от передних /3 языка (барабанная струна). Дви-
гательное ядро лицевого нерва находится в мосту и
иннервирует все мышцы лица, кроме жевательных.
Поражение этого нерва встречается часто и мо-
жет возникнуть на любом уровне. Он проходит из
ствола мозга через внутренний слуховой проход,
среднее ухо и сосцевидный отросток, выходит из
шилосоцевидного отверстия и входит в мягкие тка-
ни лица. Поражение этого нерва в любом месте при-
водит к параличу мимической мускулатуры на сто-
роне очага, утрате вкусовой чувствительности на
передних 2/3 языка, если очаг находится прокси-
мальнее места отхождения барабанной струны. Чаще
всего это отмечается при параличе Белла. Пораже-
ние кортиконуклеарных путей (центральный пара-
лич лицевого нерва) приводит к слабости нижней
мимической мускулатуры противоположной сторо-
ны, поскольку лишь верхняя часть ядра лицевого
нерва имеет двустороннюю корковую иннервацию
(см. главы 35, 36 и 41).
VIII. Преддверно-улитковый нерв
Вестибулокохлеарный нерв проводит информа-
цию от улитки (улитковый нерв; см. главы 26 и 27),
а также от полукружных канальцев и отолитового ап-
парата (преддверный нерв; см. главу 28). Поражение
этого нерва (невринома слухового нерва) сопровож-
дается нарушением равновесия, глухотой, звоном и
шумом в ушах.
IX. Языкоглоточный нерв
Языкоглоточный нерв содержит двигательные,
чувствительные и парасимпатические волокна. Дви-
гательные волокна начинаются от двойного ядра и
иннервируют шилоглоточную мышцу, а чувствитель-
ные волокна заканчиваются в ядре солитарного пуч-
ка и проводят вкусовую чувствительность от задней
трети языка и стенки глотки. Парасимпатические
волокна начинаются от нижнего слюноотделитель-
ного ядра и иннервируют околоушную железу. По-
ражение этого нерва обычно возникает в сочетании
с поражением блуждающего нерва (см. ниже) и на-
блюдается при поражении нижних отделов ствола.
X. Блуждающий нерв
От блуждающего нерва отходят двигательные во-
локна к мягкому нёбу, гортани и глотке, которые на-
чинаются из дорсального двигательного ядра блуж-
дающего нерва и двойного ядра. Также нерв прово-
дит чувствительность от надгортанника, при этом он
является основным парасимпатическим нервом (см.
главу 42). Поражение блуждающего нерва приводит
к дисфагии и дизартрии, а при сочетанном пораже-
нии языкоглоточного нерва отмечается снижение
глоточного рефлекса. Этот рефлекс заключается в
ретракции языка и поднятии глоточной мускулату-
ры в ответ на сенсорный стимул задней стенки глотки.
XI. Добавочный нерв
Добавочный нерв — это чисто двигательный нерв.
Он начинается от двойного ядра в продолговатом
мозге и иннервирует грудино-ключично-сосцевид-
ную и трапециевидную мышцы. Поражение доба-
вочного нерва приводит к слабости этих мышц, хотя
чаще всего он поражается вместе с другими буль-
барными нервами.
XII. Подъязычный нерв
Подъязычный нерв иннервирует мышцу языка.
Его волокна выходят из ядра подъязычного нерва в
задней части продолговатого мозга. Поражение не-
рва приводит к слабости и атрофии мышцы языка,
что вызывает нарушение речи и глотания, которое
чаще всего наблюдается при болезни двигательно-
го нейрона (см. главу 52). Изолированное поражение
этого нерва встречается редко, чаше всего он пора-
жается вместе с другими бульбарными нервами (на-
пример, IX, X, XI ЧМН). В этом случае у больного
развивается бульбарный паралич. Псевдобульбар-
ный паралич развивается при двустороннем пораже-
нии кортиконуклерных путей, оканчивающихся на
ядрах бульбарных нервов.
37
ГЛАВА 15. Анатомия коры головного мозга
Мнкроанатомня коры головного Moira
Эфферентные пути
Анатомия
if
1 .ффьрентнчу*
пути
кие связи
Таламическая
импульсация
преимущест-
венно в IV слой
Кортикостри-
арные. кооти-
кобутЫ арные
кортикоспн-
нальныь связи
Кортико-
'кортикаль-
. ные связи
— Содержит апикальные дендриты большинства клеток
Локальные тормозные интернейроны, ингибирующие
лоугие интернейроны коры
Гигантские пирамидные клетки, проецирующиеся за
пределы коры_________________________________
__/ Интернейроны, получающие таламическую импульса-
цию и пооецирующнеся в II и III слоях и тормозные
интернейроны, проецирующиеся на другие пирамид
t ные нейроны II и III слоев (на рисуйся не показаны)
Пирамидные нейроны V слоя, проецирующиеся субкортикально
Белое
вещество
Более мелкие клетки VI слоя получают таламическую
импульсацию и импульсацию от коллатеральных во-
локон от "роекций нейронов II, III и V слоев
Анатомия наружного слоя полушарий голов-
ного мозга, или коры головного мозга (неокортекс),
рассматривается в нескольких аспектах. Раньше все-
го ее стали рассматривать в виде цитоархитектони-
ческих карт (например, карта Бродмана головного
мозга человека, созданная в 1909 г.). Такой способ
картирования коры до определенной степени соот-
ветствует функциональной ее организации и подраз-
делению на двигательные, чувствительные и ассо-
циативные области, что подтверждено выявлением
слоистой организации коры. Область коры, где пре-
имущественно представлены чувствительные фун-
кции, характеризуется выраженностью IV слоя, в
котором заканчиваются афферентные таламические
волокна. Преимущественно двигательная кора ха-
рактеризуется наибольшим развитием V слоя. Аль-
тернативным способом изучения организации коры
является вертикальная схема. Такая вертикальная
организация получила название столбиковой гипоте-
зы, согласно которой столбик клеток коры является
основной ее функциональной единицей и в фило-
генезе число столбиков увеличивается. Это объяс-
няет увеличение объема и площади поверхности
коры у приматов без изменения количества нейро-
нов в вертикальной проекции.
Анатомическая организация
коры головного мозга
Согласно классическому описанию, кора состо-
ит из 6 слоев, при этом в ряде ее областей имеется и
другое подразделение, например первичная зритель-
ная кора (см. главу 25). Таламические афферентные
волокна, переключающие чувствительную информа-
цию, проецируются на IV и в меньшей степени на
VI слой коры и заканчиваются дискретными пучка-
ми в соответствии с топической или рецепторной
спецификой поступления чувствительной информа-
ции от определенных участков организма в опреде-
ленные области коры. Импульсы, проводимые по
этим волокнам, переключаются на интернейроны,
которые проецируются вертикально на нейроны II,
38
Ill и V слоев, которые в свою очередь проецируются
на другие корковые и подкорковые области. Таким
образом, наибольший удельный вес синаптических
взаимоотношений внутри коры мозга принадлежит
вертикальному направлению, хотя существуют и
кортикокортикальные связи, соединяющие различ-
ные столбики нейронов со сходными характеристи-
ками. Такая организация синаптических связей на-
блюдается в соматосенсорной и зрительной коре (см.
главы 20 и 25), это означает, что чувствительный
импульс из таламуса будет проанализирован в вер-
тикальном столбике нейронов коры. В двигательных
областях коры импульс от этой области проециру-
ется на мотонейроны, которые контролируют мы-
шечную активность таким образом, что чувствитель-
ные рецепторы этой зоны мышцы проецируются на
те же самые области коры — это называется прин-
ципом прямой и обратной связи (см. главу 36).
Таким образом, анатомическая структура коры
подтверждает теорию вертикальной организации,
однако существуют и другие данные различных он-
тогенетических и нейрофизиологических исследо-
ваний.
Онтогенетическая организация
коры головного мозга
В ЦНС млекопитающих корковые нейроны об-
разуются в зонах пролиферации, расположенных
вокруг желудочков мозга, и распространяются в об-
ласти коры с помощью миграции. Радиальные гли-
альные волокна могут давать начало мигрирующим
нейронам в стенке мозгового пузыря плода и на-
правляют миграцию нейронов к месту их будущей
корковой локализации в кортикальную пластинку из
вентрикулярных и субвентрикулярных зон (см. главу
2). Так, в процессе развития кора формируется в вер-
тикальном направлении.
Нейрофизиологическая организация
коры головного мозга
Если регистрирующие электроды расположить
под прямым углом к коре, окажется, что нейрофи-
зиологические свойства клеток коры сходны. Одна-
ко если электроды расположить тангенциально,
клетки изменят характеристики ответа. Это было
показано как на первичной сенсорной, так и на пер-
вичной двигательной коре (см. главы 20, 25 и 36).
Столбиковая организация коры означает, что то-
пографические соотношения коры поддерживают-
ся и что в ответ на изменение периферического сти-
мула происходит реорганизация коры (см. главу 47).
Функциональная организация
коры головного мозга
Взаимоотношение столбиков коры друг с дру-
гом поднимает много интересных вопросов о харак-
тере функционирования коры. Согласно ориги-
нальным моделям, обработка информации в коре
производится в виде порций таким образом, что
клетки коры выстраивают эти порции информации
иерархически. Так, одна группа клеток проводит
прямой анализ, который затем передается другой
группе клеток, осуществляющей более комплекс-
ный анализ (см. главу 25). В основе концепции ле-
жит представление о том, что в конечном счете пер-
цепцию осуществляет главная клетка иерархии —
«клетка-бабушка». Однако открытие ганглиозных
клеток сетчатки классов X, Y и Z (см. главу 23) при-
вело к развитию конкурирующей теории, согласно
которой информация анализируется по параллель-
ным путям, при этом каждый путь анализирует оп-
ределенный аспект чувствительного стимула (на-
пример, такие зрительные стимулы, как цвет или
движение). Согласно этой теории, не исключается
иерархическая обработка информации, однако су-
ществует также анализ по отдельным параллельным
путям. В действительности в коре существует оба
вида анализа.
Необходимо подчеркнуть, что столбики клеток
коры не являются статической мозаичной структу-
рой, поскольку один столбик может принимать уча-
стие в различных путях анализа. Такая организация
объясняется гипотезой функциональных систем. Со-
гласно этой гипотезе, мозг состоит из взаимосвязан-
ных систем, внутри которых и между которыми про-
исходят динамические изменения активности моз-
га. Следовательно, один столбик клеток коры может
быть частью различных функциональных систем,
поскольку каждая функциональная система специ-
фична для определенного свойства стимула и один
столбик клеток может кодировать различные свой-
ства стимула. Так, станция метрополитена, может
быть частью различных линий, каждая из которых
обеспечивает ей какое-то одно уникальное качество.
Эта аналогия позволяет объяснить и пластичность
ЦНС: повреждение станции может быть компенси-
ровано до определенной степени оставшимися свя-
зями внутри сети, так как существует определенный
предел возможностей представленности станции.
Однако при тяжелом поражении, особенно важных
центров или периферических терминалей, система
не может быть компенсирована и остаются резиду-
альные неврологические дефекты.
ГЛАВА 16. Оболочки мозга и цереброспинальная жидкость
Венозные синусы и пахионовы грануляции
Череп
ТМО
Ан атом ич<“‘ ки« соотнош* имя ткани мозга и оболочек
Паутинная оболо <ка
Серое веь,е< гео
Белое вещество
Субарахноидальные цистерны
IV желудочек
Боковой
желудочек
Спинной мозг
желудочек
Сильвиев водопровод
Moci
Мозжечок
Межножковая цистерна
IV желудочек
Субарахноидальные цистерны
Большая
цис ерна
Отверстие
Монри /
Субарахноидаль-
ное прострастг
Мозолнс
тое тело
111 желу-
дочек
.Субарахноидальное
простраство
Пахионовы
грануляции
Мягкая мозговая
оболочка
Боковой желудо ек
Хсроид ое
>сплетение
Сильвиев
водопровод
Отверстия Люшка
и Мажанди
Понтинная - !V
цистерна Продолго-Я
I ватый мозг'
Артерия
/ ТМО
В черепе головной мозг окружают защитные обо-
лочки, распространяющиеся и на спинной мозг.
Твердая мозговая оболочка (ТМО) является толстой
мембраной, плотно прилегающей к черепу и позвон-
кам. Она иннервируется афферентными волокнами
тройничного нерва и верхними шейными нервами.
Головная боль может быть связана с раздражением
или воспалением ТМО. Под ней находится тонкая
паутинная оболочка с нитевидными отростками, ко-
торые проецируются в субарахноидальное простран-
ство и контактируют с тонкой мягкой мозговой обо-
лочкой, которая окружает спинной мозг, контуры
поверхности головного мозга и его борозды.
Субарахноидальное пространство, а также боль-
шинство артерий, ветви которых проникают через
мягкую мозговую оболочку в ЦНС, заполнено це-
реброспинальной жидкостью (ЦСЖ). В определен-
ных участках субарахноидальное пространство рас-
ширяется и образует цистерны. В основном цистер-
ны располагаются в области ствола мозга, самой
крупной из которых является большая цистерна меж-
ду мозжечком и продолговатым мозгом.
40
Оболочки распространяются каудально и окру-
жают спинной мозг. ТМО прикрепляется к большому
затылочному отверстию на своей верхней границе и
распространяется вниз до крестцовых позвонков.
Продукция и циркуляция
цереброспинальной жидкости
ЦСЖ секретируется хороидными сплетениями,
которые преимущественно находятся в желудочках.
Скорость продукции варьирует от 300 до 500 мл/сут,
вентрикулярный объем составляет примерно 75 мл.
ЦСЖ по составу сходна с плазмой, хотя она содер-
жит меньше альбумина и глюкозы. Образуясь в бо-
ковых желудочках ЦСЖ затем поступает в III желу-
дочек через внутрижелудочковые отверстия Монро,
а затем в IV желудочек по сильвиевому водопрово-
ду, а оттуда в субарахноидальное пространство через
отверстия Мажанди и Люшка. Из субарахноидаль-
ного пространства на основании мозга ЦСЖ пере-
мещается рострально, омывая полушария головно-
го мозга и спинной мозг.
ЦСЖ всасывается в верхнем сагиттальном и
смежных венозных синусах. Пахионовы грануляции
паутинной оболочки участвуют во всасывании ЦСЖ.
Это миниатюрные выросты паутинной оболочки,
прободающие ТМО и связывающие субарахноидаль-
ное пространство с венозной системой. Точный ме-
ханизм всасывания не установлен, однако он вклю-
чает перемещение всех компонентов ЦСЖ в веноз-
ную систему.
Играя важную роль в поддержании постоянства
химического состава внутричерепного пространства
см. ниже), ЦСЖ предохраняет мозг от механичес-
кого повреждения, уменьшая воздействие на него
механической силы при травме.
Гематоэнцефалический барьер
Гематоэнцефалический барьер (ГЭБ) считают
единственным физическим барьером, препятствую-
щим проникновению молекул и клеток в головной
мозг. Однако недавно были обнаружены новые
транспортные системы, препятствующие или облег-
чающие перемещение молекул из крови в ЦСЖ.
Одним из свойств эндотелиальных клеток капилля-
ров головного мозга является присутствие плотных
соединений между этими клетками, которые поддер-
живаются отростками астроцитов (см. главу 4). Эти
необычайно плотные соединения уменьшают воз-
можность перемещения крупных молекул или кле-
ток и, следовательно, требуют специальных транс-
портных систем для перемещения необходимых мо-
лекул в мозг.
Маленькие молекулы, например глюкоза, легко
проникают в ЦСЖ, несмотря на нерастворимость в
липидах. Более крупные белковые молекулы не мо-
гут проникать в мозг, однако существует несколько
механизмов переноса, которые позволяют прони-
кать в ЦНС другим углеводам и аминокислотам. Ба-
рьер необходим, чтобы поддерживать постоянство
химического состава внутричерепного пространства,
предохранять мозг от осмотического повреждения,
обеспечивать иммунную резистентность мозга к
проникновению иммунокомпетентных клеток (см.
главу 54). Однако ГЭБ снижает возможность достав-
ки в мозг многих лекарственных препаратов (напри-
мер, антибиотиков).
Клиника поражения
мозговых оболочек и ликворной системы
Гидроцефалия - это расширение желудочковой
системы, которое наблюдается при атрофических
процессах, например деменции (викарная гидроце-
фалия). Однако гидроцефалия может наблюдаться
и в результате повышения внутричерепного давле-
ния (ВЧД) в желудочковой системе, т. е. вторично
при возникновении препятствия току ЦСЖ (об-
структивная) гидроцефалия. Последняя часто воз-
никает при закупорке выхода из IV желудочка в су-
барахноидальное пространство при опухолях, врож-
денных аномалиях или последствиях перенесенной
инфекции (см. ниже). Кроме того, ток ЦСЖ из III в
IV желудочек может снижаться в результате разви-
тия стеноза сильвиева водопровода. Гидроцефалия
развивается и при гиперсекреции ЦСЖ (например,
при опухолях хороидного сплетения) или снижении
всасывания, какпри spinabifida.
Симптоматика гидроцефалии различна, однако
в классическом виде она проявляется признаками
повышения ВЧД (головная боль ранним утром, тош-
нота, рвота) а при остром повышении ВЧД — сни-
жением уровня бодрствования с краткими периода-
ми потери зрения. Наиболее частой причиной по-
вышения ВЧД является глиома (см. главу 4), которая
создает масс-эффект, при этом подобные опухоли в
задней черепной ямке могут напрямую вызывать
гидроцефалию, способствуя повышению ВЧД.
При обструктивной гидроцефалии лечение на-
правлено на дренирование избытка ЦСЖ с исполь-
зованием различных шунтов, связывающих желу-
дочки с предсердием или полостью брюшины.
Менингит
Менингит, или воспаление мозговых оболочек,
вызывают различные микроорганизмы. При острой
инфекции воспаление распространяется быстро по
всему субарахноидальному пространству на голов-
ной и спинной мозг, возникают головная боль, ли-
хорадка, рвота, ригидность затылочных мышц (ме-
нингизм) и при тяжелом течении заболевания сни-
жение уровня бодрствования. Необходимо как
можно раньше назначить антибиотики, в зависимо-
сти от природы микроорганизма, вызвавшего вос-
паление.
Воспаление или инфекция могут протекать под-
остро, например при туберкулезном менингите или
саркоидозе, когда развивается вторичная гидроце-
фалия в результате утолщения мозговых оболочек на
основании мозга, что препятствует току ЦСЖ.
В редких случаях опухоли могут распространять-
ся на мозговые оболочки и вызывать менингокар-
циноматоз, что приводит к поражению черепно-моз-
говых нервов и возникновению корешковых боле-
вых синдромов. Эти состояния следует отличать от
первичных опухолей мозговых оболочек — менин-
гиом, которые растут медленно, являются доброка-
чественными и клинически проявляются эпилепти-
ческими припадками или неврологическим дефици-
том в результате сдавления соседних структур ЦНС.
ГЛАВА 17. Кровоснабжение центральной нервной системы
Виллизиев круг
V ЧМН I
^^олсано^
111 ЧМН
IV ЧМН
Мост
VI ЧМН
VII И VIII ЧМН
IX. X и XI ЧМН
XII ЧМН
Мозжечок
Задняя нижняя
мозжечковая артерия
|Схема кровоснабжения мозга на сагиттальном срезе полушария!
Передняя мозговая артерия
Передняя
соединительная артерия
Зрительный перекрест
Средняя
мозговая артерия
Т Внутренняя сонная артерия
Задняя
соединительная артерия
Задняя соединительная
артерия
—Мостовые ветви
Основная артерия
Передняя нижняя
мозжечковая артерия
Позвоночная артерия
Сенсомоторная зона
Зона
Центральная борозда
Зрительная кора
Слуховая кора
и зона Вернике
в левом
Передняя спинальная артерия
полушарии
Бассейн передней -«Бассейн задней
мозговой артерии — мозговой артерии
Бассейн средней
мозговой артерии
Схема кровоснабжения мозга на коронарном срезе I
I Спинной мозг
Шейный отдел
спинного мозга
Ветвь левой
тиреоцервикальной
артерии
Боковой
желудочек j
Таламус
Грудной отдел
спинного мозга
Меж-
X* реберная
ветвь
Кровоснабжение
спинного мозга
Т10
L2 “
Поясничный отдел
спинного мозга
Крестцовый отдел
—''ПИННОГО
Задние
спи-
нальные
артерии
мозга
I
Передняя
спинальная
Адамкевича
артерия
г:'®
&! -
Спиноталамический тракт
Передняя спинальная артерия
Кортикоспинальный
тракт
Задние
столб
Передняя
хороидальная
артерия (ветвь
внутренней сонной
артерии к нижним
2/3 внутренней
капсулы, бледному
шару, лимбической
системе)
Хвостатое
ядро
Красное ядро
Субталами-
ческое ядро
Бледный шар
Г ипгокамп
Перфорирующие артерии
средней мозговой артерии
Бассейн передней спинальной артерии
—- Медуллярная артерия подпитывает переднюю спинальную артерию
8
о
и> 5
5
Кровоснабжение головного мозга
Кровоснабжение головного мозга осуществляет-
ся по 4 основным сосудам: правой и левой внутрен-
ней сонным артериям и позвоночным артериям. По-
звоночные артерии попадают в череп через большое
затылочное отверстие и объединяются в основную
артерию, кровоснабжающую ствол мозга и заднюю
часть полушарий мозга (задние мозговые артерии).
Они с оставляют верте бр обазилярную систему. Внут-
ренние сонные артерии (ВСА) попадают в череп че-
рез сонный канал и кавернозный синус, прежде чем
прободают твердую мозговую оболочку и входят в
среднюю черепную ямку по бокам от зрительного
перекреста. Затем они разделяются на передние
(ПМА) и средние мозговые артерии (СМА) , крово-
снабжающие передние и средние части полушарий
головного мозга. Кроме того, каротидная и вертеб-
робазилярная системы анастомозируют на основании
мозга при помощи виллизиева круга в виде передних
и задних соединительных артерий, что позволяет
поддерживать церебральную гемодинамику даже в
условиях стеноза крупных сосудов. Такая ситуация
часто встречается при атеросклерозе сосудов голов-
ного мозга, когда медленнопрогрессирующий сте-
ноз ВСА позволяет коллатеральной системе сфор-
мироваться еще до окклюзии сосуда. У больных ате-
росклерозом либо не наблюдается неврологической
симптоматики, либо отмечаются транзиторные ише-
мические атаки (ТИА), которые прекращаются, ког-
да пораженный сосуд полностью закупоривается.
42
ВСА до бифуркации отдает ветви гипофизу (ги-
пофизарные артерии), глазу (глазничная артерия),
части базальных ганглиев (бледному шару) и лим-
бической системе (передние хороидальные артерии),
а также отдает заднюю соединительную артерию.
СМА является одной из основных конечных ветвей
ВСА и кровоснабжает сенсомоторную зону, которая
окружает центральную борозду (за исключением ме-
диальной части, кровоснабжаемой ПМА), а также
слуховую и речевую корковые зоны доминантного
(как правило, левого) полушария. Поэтому окклюзия
СМА приводит к развитию контралатерального геми-
пареза с вовлечением руки и нижней части мимичес-
кой мускулатуры лица, контралатеральной гемигипе-
стезии и нарушению речи, если вовлечено доминан-
тное полушарие (см. главы 20,27,30 и 36). Кроме того,
имеется много перфорируюших ветвей СМА, крово-
снабжающих подкорковые структуры, такие, как
базальные ганглии и внутренняя капсула (см. ниже).
Две ПМА, которые являются вторыми основны-
ми ветвями ВСА, связаны с помощью передней со-
единительной артерии и кровоснабжают медиальные
порции лобных и теменных долей, каки мозолистое
тело. Описана непостоянная ветвь ПМА, возвратная
артерия Губнера, которая кровоснабжает часть базаль-
ных ганглиев (неостриатум) и нисходящие двига-
тельные пути во внутренней капсуле. Окклюзия
ПМА приводит к развитию пареза в контралатераль-
ной ноге, а также в ряде случаев к астазии (апраксии
состояния и ходьбы), нарушения мочеиспускания и
апраксии (см. главы 20, 30 и 35).
Позвоночные артерии, которые являются ветвя-
ми подключичных артерий, поднимаются к стволу
мозга через отверстие в поперечных отростках верх-
них шейных позвонков. На уровне нижней части
моста позвоночные артерии соединяются и образу-
ют основную артерию, которая затем поднимается
и разделяется на две задние мозговые артерии (ЗМА)
на уровне верхней границы моста. Каждая позвоноч-
ная артерия до образования основной артерии отда-
ет ряд ветвей, включая заднюю спинальную артерию,
заднюю нижнюю мозжечковую артерию (ЗИМА) и
переднюю спинальную артерию. Эти спинальные
артерии кровоснабжают верхнешейный отдел спин-
ного мозга (см. ниже), тогда как ЗИМА кровоснаб-
жает латеральную часть продолговатого мозга и моз-
жечок. Окклюзия этой артерии приводит к развитию
синдрома поражения заднелатеральных отделов
продолговатого мозга Валленберга—Захарченко.
Основная артерия отдает несколько ветвей: перед-
нюю нижнюю мозжечковую артерию (ПНМА), ла-
биринтную артерию, мостовые ветви и верхнюю
мозжечковую артерию. Окклюзия этих ветвей при-
водит к развитию характерных синдромов пораже-
ния мозгового ствола (см. главу 13).
ЗМА кровоснабжают заднюю теменную кору, заты-
лочные доли и нижние отделы височной доли. Окклю-
зия этих сосудов приводит к развитию дефектов по-
лей зрения (обычно гомонимной гемианопсии с ин-
тактностью центрального зрения, так как этот участок
коры получает дополнительное кровоснабжение через
СМА; см. главу 24), развитию амнестического синд-
рома (см. главу 45), нарушению речи (см. главу 27) и,
иногда, нарушению зрительного восприятия (см. гла-
ву 25). ЗМАимеет несколько перфорирующих ветвей,
кровоснабжающих средний мозг, таламус, субталами-
ческую область, заднюю часть внутренней капсулы,
зрительную лучистость и ножку мозга. Окклюзия этих
сосудов приводит к развитию синдромов поражения
среднего мозга в сочетании с поражением черепно-
мозговых нервов и двигательными нарушениями; в
случае вовлечения таламуса возможно развитие бо-
левого синдрома и дизестезии, которые трудно под-
даются лечению (см. главу 22). Мелкие перфорирую-
щие артерии, которые являются ветвями ЗМА и СМА,
часто поражаются при артериальной гипертензии,
что сопровождается развитием лакунарних инсультов.
Помимо окклюзии, может развиться кровоизли-
яние из сосудов мозга в его паренхиму (внутримоз-
говое), субарахноидальное пространство или в обе
эти структуры. Такие кровоизлияния обычно разви-
ваются при черепно-мозговой травме, артериальной
гипертензии или разрыве врожденных аневризм вил-
лизиева круга.
Венозная система мозга
Венозная система ствола мозга и мозжечка пред-
ставлена дуральными венозными синусами задней
черепной ямки. Полушария мозга имеют собствен-
ные вены — внешние церебральные вены, которые
дренируют кору в верхний сагиттальный синус (см.
главу 16). Этот синус дренируется в поперечный си-
нус, затем в латеральный синус и далее во внутрен-
нюю яремную вену. Внутренние церебральные вены
дренируют глубинные структуры полушарий голов-
ного мозга в большую вену Галена, а затем в прямой
синус. Окклюзия отделов венозной системы приво-
дит к повышению внутричерепного давления с оча-
говыми неврологическими дефектами или без них.
Кровоснабжение спинного мозга
Кровоснабжение спинного мозга предстаатено не-
парной передней спинальной артерией и парными
задними спинальными артериями. Передняя спи-
нальная артерия берет начало от позвоночных ар-
терий, от которых отделяется на уровне нижних
отделов ствола мозга и проходит вплоть до конуса
спинного мозга. Она кровоснабжает переднюю по-
верхность продолговатого мозга и передние 2/3 спин-
ного мозга. Задние спинальные артерии кровоснаб-
жают заднюю треть спинного мозга, они также берут
начало из позвоночных артерий. Кровоснабжение
спинной мозг также получает и из других артерий,
располагающихся по его длиннику (см. рисунок).
Сосудистые поражения спинного мозга чаще все-
го возникают в зонах смежного кровообращения, а
именно в нижнегрудном и нижнешейном отделах.
Окклюзия передней спинальной артерии приводит к
снижению мышечной силы и расстройствам поверх-
ностной чувствительности (спиноталамические трак-
ты) при сохранности глубокой чувствительности (мы-
шечно-суставного чувства и чувства вибрации; см.
главу 31). Окклюзия задних спинальных артерий
вызывает выпадение глубокой чувствительности.
43
ЧАСТЬ II. Чувствительные анализаторы
ГЛАВА 18. Чувствительность
Адаптация
Сенсорный стимул
Мембрьннън рецепторы, пре-
образующие сенсорный сти-
мул в электрический импульс
посредством вторичных мес-
сенджеров (например, зритель
л*ое восприятие; см. главу 19)
Чувствительное проведение
• Изменение потенциала мембраны
• I °нерщ,ия потенциала действия
Прямая активация ионного
канала рецептором, восприни-
мающим сенсорный стимул
(например, слуховое восприя-
тие; см. главу 19)
Чуаствит^льнь'й рецептор
Адаптация — поддержание
чувствительности рецептора
к стимулу
афферентном проводящем пути:
О Сила стимула кодируется количеством потенциалов
действия и коли еством активированных волокон
О Тип стимула (модальность) кодируется типом рецеп-
тора и типом активируемых волоке и, идущих от дан-
ного реце.гтора
О Локализация стимула кодируется конкретными во-
локнами в соответствии с топографической органи-
зацией проводящнгл пути
Афферентный
чувегвител ный
нерв
(поокидит ic периферическим
нерва 1 I спинной мозг
и по череп п мозговым нервам
в ствол мозга
Кодирование нервных импульсо |
^етершьно^^ормож^^^^
Стимул
ИЛИ ГОЛОВНОЙ мозг)
Усиление потенциала действия
посред: -вом па ерального торможения
Чуьстьитьлгные проводники ЦНС |
По чувствительному анализатору информация про-
водится по спинному и головному мозгу от перифери-
ческих рецепторов, которые сами являются специали-
зированными нейронами или нервными окончания-
ми. Специализированные чувствительные рецепторы,
афферентные аксоны и тела нейронов вместе с синап-
тическими контактами в спинном мозге называют-
ся первичными афферентами, а процесс, посредством
которого стимул из окружающей среды превращается
в электрический импульс, передаваемый по нервной
системе, называется чувствительным проведением (см.
главу 19). Сигнал, производимый чувствительным ре-
цептором, передается в ЦНС по периферическим
или черепным нервам и через ряд синапсов проеци-
руется на определенную область коры, способной
детально проанализировать сенсорный стимул.
Выделяют 5 основных чувствительных систем в
ЦНС млекопитающих: 1) прикосновение/давление,
проприоцепция, температурная и болевая чувстви-
тельность (см. главы 20—22); 2) зрение (см. главы 23—
25); 3) слух и равновесие (см. главы 26—28); 4) вкус
(см. главу 29); 5) обоняние (см. главу 29). Все они,
кроме соматосенсорных проводящих путей, счита-
ются специальными анализаторами.
44
Чувствительные рецепторы
Чувствительные рецепторы передают стимулы по-
средством или прямой активации ионных каналов
(например, слуховой анализатор), или непрямым спо-
собом при помощи внутриклеточных мессенджеров
(например, зрительный анализатор). В обоих случаях
стимул преобразуется в электрический сигнал и мо-
жет передаваться по ЦНС либо в виде деполяриза-
ции/гиперполяризации определенной величины, при-
водя к образованию потенциалов действия (например,
зрительный анализатор), либо посредством прямой ге-
нерации потенциалов действия на уровне самого ре-
цептора (например, слуховой анализатор; см. главу 19).
Специфичность и модальность. Чувствительность
системы основана на активации специализирован-
ных нервных клеток или волокон, которые высоко-
специфичны для определенного вида стимулов, на-
пример рецепторов сетчатки, высокочувствительных
к фотонам, хотя могут активироваться и при сдавле-
нии глаза. Рецептор будет воспринимать стимулы,
только если они будут транслироваться в его окруже-
нии (рецептивном поле), например на определенном
участке кожи для соматосенсорной системы или сет-
чатки для фоторецепторов. Рецепторное поле, с ко-
торого активируется рецептор, узнается в ЦНС в со-
ответствии с положением или местом возникновения
в окружающем мире. Однако рецептор передает элек-
трический импульс в ЦНС, лишь когда получает сти-
мул с интенсивностью, достигающей порога, необхо-
димого для импульсации. Способность отвечать по
возрастающей на изменение интенсивности стимула
определяет чувствительность рецептора. Многие ре-
цепторы имеют высокую чувствительность как к аб-
солютному уровню стимула, так и к изменениям его
интенсивности, поскольку способны увеличивать
исходный сигнал с использованием систем вторич-
ных мессенджеров и адаптировать организм к нали-
чию неизменного стимула (см. ниже и главу 19). Не-
которые рецепторы способны изменять свою чув-
ствительность под влиянием времени и места, т. е. в
условиях, в которых они интегрируют стимулы, на-
пример фоторецепторы сетчатки в темноте. Однако
очень чувствительные рецепторы создают внутреннюю
нестабильность процесса проведения импульсов, что
характеризуется понятием шума, и нервной системе
становится необходимо определять ответ на чувстви-
тельный стимул или сигнал безотносительно этого
шума (определяется как соотношение сигнал/шум).
Сила сенсорного стимула может быть кодирована
на уровне рецептора и его первого синапса в виде как
потенциалов действия, так и мембранных потенциа-
лов определенной величины в самом рецепторе, ко-
торые затем преобразуются в потенциалы действия
(см. главу 19). Афферентный чувствительный нерв мо-
жет так же кодировать силу стимула, во-первых, уве-
личивая число активированных афферентных волокон
(рекрутирование, или пространственное кодирование),
и, во-вторых, увеличивая количество потенциалов
действия генерированных каждым аксоном за едини-
цу времени (временное, или частотное, кодирование).
Однако соотношение между силой стимула и коли-
чеством потенциалов действия часто не носит линей-
ного характера, поскольку аксон может генерировать
ограниченное количество потенциалов действия
ввиду наличия рефрактерного периода (см. главу 6).
Рецептор, будучи способным регистрировать и ко-
дировать интенсивность специфического сенсорного
стимула в определенном месте, должен также адапти-
роваться так, чтобы отвечать на изменение получаемой
информации. Таким образом, адаптация определяется
как снижение чувствительности рецептора, возника-
ющее при поддержании постоянства стимула, и важным
внутриклеточным медиатором адаптации является
кальций во всех отделах чувствительных анализаторов
(см. главу 19). Если такого механизма не существует,
постоянно присутствующий стимул значительно сни-
зит чувствительность репептора или даже инактиви-
рует его и сделает неспособным к восприятию новых
стимулов (например, мышечное веретено; см. главу 33).
Чувствительные проводящие пути
Кодированная чувствительными рецепторами
информапия передается в ЦНС по периферическим
и черепно-мозговым нервам. При этом каждый ре-
цептор имеет свой аксон для проведения. Каждая
модальность связана с определенными нервом или
проводящим путем, например зрительная информа-
ция передается по зрительному нерву (см. главу 24),
тогда как соматосенсорные системы передают инфор-
мацию по большому количеству периферических не-
рвов, а также тройничному нерву по системе задние
столбы медиальная петля и спиноталамическим
трактам. Так, каждый чувствительный проводник пе-
редает определенный стимул в ЦНС, хотя в конеч-
ном счете большинство чувствительных проводников
передает импульсы в таламус, какова бы ни была про-
екция каждого из сенсорных стимулов. Таламус в свою
очередь передает эти импульсы в кору, хотя обоня-
тельная информация в основном передается в лим-
бическую систему (см. главу 29), а информация от
мышечных веретен в мозжечок (см. главу 37).
Каждая чувствительная система имеет свое кор-
ковое представительство, где первично обрабатыва-
ется информация, и этот участок коры — первичная
сенсорная кора — связан с соседними отделами, та-
ким образом осуществляя комплексную оценку ин-
формации (вторичные сенсорные поля). Они в свою
очередь проецируются на ассоциативные поля (зад-
ние теменные, префронтальные и височные отделы
коры; см. главу 30), а затем проецируются на двига-
тельную и лимбическую системы (см. главы 32 и 45).
Последние в большей степени участвуют в обработ-
ке чувствительной информации как ключа для вы-
работки поведенческих реакций.
Первичная сенсорная кора проецируется не толь-
ко на вторичные сенсорные поля, но и субкортикаль-
но на таламус (и/или ствол мозга). Это важно для
усиления некоторых восходящих чувствительных
сигналов. Такое усиление предполагает как минимум
два процесса: латеральное торможение и выявление
признаков. Латеральное торможение — это процесс,
посредством которого аксоны с наибольшей актив-
ностью ингибируют соседние менее активные, кото-
рые несут менее точную афферентную информацию.
Выявление признаков означает селективную регист-
рацию определенных характеристик сенсорного сти-
мула, которая может происходить на любом уровне
от рецептора до коры. Кроме того, восприятие сен-
сорного стимула основано на синхронной активно-
сти некоторых отделов коры мозга (см. главу 15).
Нарушение функции чувствительных проводящих
путей в клинике нервных болезней (см. главу 31)
Поражение различных отделов чувствительных
проводников приводит к неврологическому дефици-
ту, степень и природа которого зависят от анатоми-
ческой локализации очага. Наиболее выраженные
нарушения чувствительности возникают при пора-
жении рецепторов и афферентных путей, тогда как
супраспинальные очаги проявляются менее выра-
женными нарушениями, которые могут быть «поло-
жительными» по своей природе, например паресте-
зии при фокальной эпилепсии с поражением пер-
вичной сенсорной коры или фотопсии при ишемии
зрительной коры при мигрени.
45
ГЛАВА 19. Чувствительное восприятие
^рительнс^^осприя™^^
Сетчатка
Диск содержит Светонечувстеительный
• родопсин, /
1 паС1Д
втрансдуцин, I „ I
• цГМФ-ф >гфо- '
диэстеразу
Обонятельное .осприятме |
Обоняние
Электромеханическое восприятие J
Волосковые клетки/слух
цГ МФ способствует |
открытию канала
в темноте
Обонятельный пузырек Ресничка Волосок, состоящий Кин°иилия
/ Слизистая иэ <--вреоцилии
оболочка
^Апикальный
® ( • декдрит
। Са?* Na I
Клетка
обонятельного
рецептора
Фоторецептор
к-
->Na"
к-
Поддер- ч
живающая
клетка
Высвобождение
нейротрансмиттера в темноте
Кон цевое
соединение j
Ионный канал
Актино-
вые фи-
ламенты
Стереоцилия
(низкая, высокая)
Двига-
тельный
комп-
лекс
Высвобождение нейротрансмиттера
Стимул
Процесс
восприятия
Фотон
Запах
Звуковая волна
К-цис-рети-------► Метародопсин II (R*)
нальный олеин
Родопсинкнназа
аррестин возвращают
R* в состояние покоя
Подвергается спонтанно
обратному гидролизу
Активный
комплекс
цГМФ—ФДЭ
цГМФ
У ^Связывается
Трансдуцин (Т) с ассо-
^циированным G-белком
Тару- ГДФ
''--Та — ГДФ + ру
I Неактивный
----------- комплекс
цГМФ—ФДЭ
Связывается со специфическим
ИОННЫМ
> 5’-ГМФ
Снижение концентрации цГМФ — закрытие канала —
снижение концентрации кальция внутри рецептора
Ответ
* G-белок, ассоциированный
с обонятельным
рецептором (G^J
Активирует
Перемещение стереоцнлнй
Концевые соединения натягиваются
и открывают ионный канал
? Инозитол трифос-
фат — опосредован-
ный путь восприятия
Аденилатциклаэу III
цАМФ 4
АГФ
Приток в клетку кальция и калия
каналом и активирует его
Приток
натрия и кальция
Активирует кальций зависимы и
хлорный канал
Отток хлора
Гиперлолярмзация. степень которой зависит
от количества захваченных пигментом фотонов
Деполяризация
Деполяризация
_______Г\
Адаптация
Снижение высвобождения нейротрансмиттера
Генерация потенциала действия
в волокнах обонятельного нерва
Высвобождение нейротрансмиттера
активирует волокна улиткового нерва
Основана на снижении концентрации внут-
риклеточного кальция, что приводит к:
а) активации кальцийсвябывающего белка
рековерина, который ингибирует родоп
синкиназу и, возможно, активирует гуа-
инлатциклаэу.
6) активации гуанилатциклазы
Приток кальция адаптирует восприятие
при помощи:
а) внутреннего связывания вблизи
от активированного цАМФ-ионного канала,
б) активации протеинкиназы.
которая ингибирует продукцию цАМФ,
в) модуляции хлорного канала, этот последний
механизм, по-видимому, наиболее важен
Приток кальция важен для адаптации по
механизму перемещения канала, кальций
активирует актин и миозин стереоцилии. что
позволяет каналу изменить свою конфигу-
рацию и закрыться
&
К
Чувствительное проведение предусматривает пре-
образование стимула из внешней или внутрен-
ней среды в электрический импульс для передачи
по нервной системе. Этот пропесс осуществляет-
ся всеми чувствительными системами, в основе его
лежит или химический процесс, например в сет-
сосочках языка и обонятельном
чатке, вкусовых
эпителии, или механический процесс, например
в улитке и соматосенсорных системах. Эти отли-
чающиеся друг от друга способы проведения луч-
ше всего изучены на примерах специальных ана-
лизаторов.
46
Зрительное восприятие
Зрительное восприятие — это процесс, при кото-
ром энергия света в форме фотонов преобразуется в
электрический импульс в виде изменения потенци-
ала действия фоторецепторных клеток (палочек и
колбочек) сетчатки.
Фотоны захватываются пигментами в наружных
сегментах фоторецепторов, и световой сигнал уси-
ливается с помощью циклического гуанозинмоно-
фосфата (цГМФ) в качестве вторичного мессендже-
ра. Это приводит к уменьшению концентрации
цГМФ и закрытию ионных каналов. Закрытие ион-
ных каналов, позволяющих натрию и кальцию про-
никать в фоторецепторы в темноте, приводит к ги-
перполяризационному разряду, степень которого
зависит от количества фотонов, поглощенных пиг-
ментом фоторепептора. Гиперполяризапионный за-
ряд приводит к снижению высвобождения глутама-
та фоторецепторными клетками на биполярные и го-
ризонтальные клетки (см. главу 23).
Завершение реакции фоторецептора на световой
стимул зависит от многих факторов, однако изме-
нение внутриклеточной концентрации ионов каль-
ция имеет ключевое значение. Светонечувствитель-
ный кальциевый насос в наружном сегменте, свя-
занный с закрытием катионного канала, опосредует
значительное уменьшение концентрации внутри-
клеточного кальция, что важно для завершения ре-
акции фоторецептора на световой стимул, а также
опосредования адаптации к свету.
Некоторые редкие заболевания, например куч-
ная слепота, связаны со специфическими наруше-
ниями процессов зрительного восприятия.
Обонятельное восприятие
Обонятельное восприятие тоже химически опос-
редуемый процесс. Рецепторами обоняния являют-
ся биполярные нейроны, состоящие из дендрита с
утолщением в виде реснички и аксона, который про-
ецируется в виде обонятельного нерва в обонятель-
ную луковицу на основании лобной доли. Наличие
ресничек, несущих обонятельные рецепторы, значи-
тельно увеличивает поверхность обонятельного эпи-
телия и повышает вероятность захвата молекулы
пахучего вещества. Связывание молекулы пахучего
вещества с рецептором приводит к активации G',
который активирует аденилатциклазу III типа. Она
гидролизует АТФ до цАМФ. цАМФ связывается со
специфическим ионным каналом, активирует его,
позволяя натрию и кальцию проникать внутрь клет-
ки в соответствии со своими электрохимическими
градиентами. Это не только частично деполяризует
рецептор, но и активирует кальцийзависимый хлор-
ный канал и в последующем обеспечивает отток
ионов хлора, что еше больше деполяризует обоня-
тельный рецептор. Возможно, существуют и допол-
нительные механизмы обонятельного восприятия с
участием инозитолтрифосфата в качестве вторично-
го мессенджера. Это может приводить к генерации
потенциалов действия, которые затем проводятся
клетками по аксонам обонятельного нерва в обоня-
тельную луковицу.
Приток кальция играет важную роль в процессе
адаптации и повторной реакпии на обонятельный
стимул.
Слуховое восприятие
В отличие от зрительного и обонятельного вос-
приятия, слуховое восприятие во внутреннем ухе про-
исходит при механическом перемещении стереоци-
лий в волосковых клетках улитки (см. главу 26).
Сенсорный стимул (звуковая волна) приводит к
смещению стремечка в овальном коне, что генери-
рует волны перилимфы в лестнице преддверия и ба-
рабанной полости улитки. Это вызывает перемеще-
ние базальной мембраны, на которой расположены
волосковые клетки кортиева органа. Эти клетки пре-
образуют звуковые волны в электрический импульс
с помощью механического проведения. Кончики
стереоцилий на апикальном участке волосковых кле-
ток связаны концевыми соединениями, которые
связаны с ионными каналами.
Звук приводит к перемещению стереоцилий в
направлении самой крупной стереоцилий (или ки-
ноцилии), которое создает напряжение концевых со-
единений, что сопровождается открытием ионного
канала. Последний пропускает внутрь клеток калий
(но не натрий, поскольку эндолимфа срединной
части лестницы богата калием и бедна натрием) и
кальций, что деполяризует волосковые клетки. Это
деполяризация приводит к высвобождению нейро-
трансмиттера в основании волосковых клеток, что ак-
тивирует афферентные волокна улиткового нерва.
Продолжающееся перемещение стереоцилий в
ответ на звук снижается благодаря процессу адапта-
ции, связанному с перемещением ионного канала
таким образом, что он закрывается в ответ на опре-
деленную степень напряжения концевых соедине-
ний. Это достигается путем вхождения кальция в
ионные каналы.
Некоторые случаи врожденной глухоты объясня-
ют недостатком миозина волосковых клеток.
Другие механизмы восприятия
Восприятие с помощью соматосенсорных рецеп-
торов, болевых рецепторов, терморецепторов, вку-
совых рецепторов и мышечных веретен обсуждает-
ся в главах 20, 21, 29 и 33 соответственно.
ГЛАВА 20. Анализатор общей чувствительности
Центральная борозда
Нога
Центральная
борозда
Передние
отделы
Задние отделы
Нога
/I
Задний
Спинномозговые узлы
Задний рог
Классификация кожных рецепторов
Чувстви
тельность
на руке
Чувстви-
тельность
на лице
Каждое цитоархитектоническое поле
ПССК получает импульсацию со-
гласно соматотопической проекции.
Кроме того, существуют связи меж-
ду различными областями коры
Чувство давления
и мышечно-
суставное чувство
Пояснично-
крестцовый
отдел
Соматотопмческая организация^*
чувствительного восприятия поддер-
живается до уровня первичной
соматосенсорной коры
»ная 'Медиальная
Г-Л) часть (ЗВЯТ-М)
Клино-
видное
ядро
Первичная
соматосенсорная
кора (ЛССК)
Рука
Двигательная зона
Лицо
Поле 5
Задняя
теменная кора
Вторичная
соматосенсорная
кора (ВССК)
МА рецепторы
I кожи
Аффеоенты
мышечных веретен
Задневентральное ядро
Лицо
Тройничный нерв
5 £ Б о
Н
X
Нежное
ядро
Ассоцииро-
ванные ядра
Мозжечок
Нежное ядро
Шейный
отдел
Размер рецептивного поля
Тип /
— небольшие, четко очере-
ченные рецептивные
поля, в основном
иа кончиках пальцев
Тип //
— большие, плохо очеречен-
ные рецептивные поля
Скорость адаптации рецепторов
Быстрая Медленная
адаптация (БА) адаптация (МА)
Тельца
Мейсснера
(Мейсс) (БА1)
Тельца
Пачини (Пач.)
(БАН)
Задняя
теменная кора
Тельца
Меркеля (Мерю)
(MAI)
Окончания
Руффини (Руфф.)
(МАП)
Эпидермис
Чувствительность
на ноге, животе
И груди
Мерк.
Перифери
Паи
Кожа (д_’рнис)
Меисс.
нерв
Распределение
рецепторов
на коже,
не покрытой
волосами
Анализатор общей чувствительности (соматосен-
сорная система) — это частв нервной системы, уча-
ствующая в анализе осязания, давления, проприо-
(или мышечно-суставного чувства; см. гла-
цепции
ву 33), болевой и температурной чувствительности
(см. главы 21 и 22).
48
Чувствительные рецепторы
Рецепторы осязания— это специализирован-
ные нервные окончания, находящиеся в коже, а тела
их нейронов расположены в спинномозговых уз-
лах. Плотность этих рецепторов высока в подушеч-
ках пальцев, тогда как проприоцепторы находят-
ся не только в коже, но и в мышцах и суставах (см.
главу 33).
Рецепторы кожи лучше всего охарактеризованы
по структуре, локализации, рецептивным полям и
скорости адаптации. Рецепторы I типа (тельца Мей-
сснера и диски Меркеля) присутствуют в большом
количестве в подушечках пальцев и дают точную
информацию о локализации ощущаемого стимула.
В отличие от них, быстро адаптирующиеся тельца
Пачини отвечают за чувство вибрации, медленно
адаптирующиеся окончания Руффини — за величину,
направление и изменение давления на кожу и тка-
ни, лежащие глубже.
Проводящие пути
задних столбов и медиальной петли
Эти рецепторы являются специализированными
нервными окончаниями, и быстропроводящие круп-
нокалиберные аксоны их клеток находятся в соста-
ве периферических нервов и проецируются в зад-
ние рога спинного мозга. Чувствительная система
тройничного нерва на лице организована также. Каж-
дый вид рецепторов обеспечивает проведение спе-
цифической информации в спинной мозг, а именно
в задние столбы, где они располагаются в зависи-
мости от типа рецептора и его местонахождения (со-
матотопическая проекция; см. главу 12). Затем они
проецируются на ядра задних столбов своей сто-
роны (нежное и клиновидное ядра), где лежит пер-
вый нейрон.
Ядра задних столбов (ЯЗС) — это структуры, зале-
гающие на границе шейного отдела спинного мозга
и продолговатого мозга. Их аксоны тут же перекре-
щиваются и образуют медиальную петлю, которая
направляется в таламус. ЯЗС также проецируются на
структуры ствола головного мозга и получают про-
екции от первичной соматосенсорной коры
(ПССК).
Медиальная петля проецируется на задневент-
ральное адро таламуса (ЗВЯТ), включая аксоны чув-
ствительной системы тройничного нерва. Последние
проецируются на медиальную часть ЗВЯТ (ЗВЯТ-М),
а остальная часть проводящего пути оканчивается на
латеральной части таламуса (ЗВЯТ-Л). Окончание
медиальной петли образует переднезадний талами-
ческий пучок, где все клетки относятся к одному
виду чувствительности и локализации (например,
указательный палец, быстроадаптирующиеся рецеп-
торы I типа). Этот таламический пучок проецирует-
ся в IV слой ПССК на соответствующий столбик
клеток коры (см. главу 15).
ПССК состоит из 4 различных участков (поля
За, 3b, 1 и 2 по Бродману), в каждом из которых
находится отдельное корковое представительство
противоположной поверхности тела, при этом пред-
ставительство языка расположено латерально, а
представительство ноги медиально. Корковое
представительство пропорционально плотности ре-
цепторов кожи, например рука имеет гораздо боль-
шее представительство, чем туловище (чувствитель-
ная соматотопическая проекция).
Первичная и вторичная соматосенсорная кора
Каждый участок ПССК характеризуется несколь-
ко отличными друг от друга свойствами в зависи-
мости от типа нейронов, находящихся в нем. Харак-
тер реакции нейронов становится все сложнее по
мере приближения к задним отделам коры теменной
доли — проводится все более тонкий анализ. ПССК
проецируется не только на ЯЗС, но и на заднюю те-
менную кору и вторичную соматосенсорную кору
(ВССК). ВССК находится на боковой стенке силь-
виевой борозды и отвечает за узнавание предметов
на ощупь, тогда как проекция ПССК на заднюю те-
менную кору отвечает за распознавание значимости
сенсорного стимула (см. главу 30).
Первичный соматосенсорный путь развивался в
ходе эволюции вместе с кортикоспинальным трак-
том (КСТ), который играет исключительно важную
роль в тонких движениях пальцев (см. главы 34—36).
Эти две системы взаимодействуют в процессе актив-
ного прикосновения, таким образом изучая окружа-
ющий мир. Каждая из систем обладает определен-
ной степенью пластичности даже у взрослых (см.
главы 35 и 47). Это отчасти становится возможным
благодаря соматотопической организации чувстви-
тельных путей — соседние участки кожи представ-
лены в соседних отделах сенсорной системы, по
крайней мере в ПССК.
Нарушения функции соматосенсорной
системы в клинике нервных болезней
Повреждение рецепторов и их афферентных во-
локон происходит при полинейропатиях. Пациенты
предъявляют характерные жалобы на парестезии
и онемение, часто в сочетании с нарушением про-
приоцепции, особенно если поражен спинномозго-
вой узел (см. главы 31 и 33).
Поражение задних столбов спинного мозга опи-
сано в главе 31. Поражение соматосенсорного пути
выше уровня ЯЗС приводит к развитию утраты чув-
ствительности на противоположной стороне, вклю-
чая половину липа, если очаг лежит выше верхних
отделов ствола.
ГЛАВА 21. Система боли 1: ноцицепторы и ноцицептивные пути
я
•Хора передней/ ПССК „ 7
части поясной К
А остро
извилины J % \
Поражение любого из этих отделов не приводит к отсутствию восприятия
чувства боли, ио может изменить порог болевого восприятия и реакцию. По-
этому поражение ПССК может нарушить способность различать интенсив-
ность болевого стимула» тогда как поражение префронтальной коры умень-
шает аффективную реакцию на боль
'Префронталь-
ная кора X.
S
т
2
5
Анатомий заднего рога спинного мозга
Таламус
Таламус (заднелатеральное
и интраламинарное ядра)
Гипоталамус
Ствол мозга:
I, околоводопроводное
серое вещество»
голубое ядро,
большое ядро шва»
ростровентральная
часть продолговатого
мозга
Крупнокалиберные
миелинизированные
афферентные
ВОЛОКн
Нисходящие серотонинергические /
норадренергические пути, уменьша-
ющие передачу повреждающего сти-
мула в ростральном направлении
Мелкокалиберное ноцицептивное
афферентное волокно
Спиноталами-
ческий нейрон
Пластина Рексед II (желатинозная субстанция)
с тормозными (эккефалмнергическими) нейронами
волокна
Крупнокалиберные
миелинизированные
афферентные волокна
Нисходящий путь
в дорсолатеральном
белом веществе
спинного мозга
Активация тучных клеток
Г истамин
К
| Субстанция Р
Ноцицептор простагландин г
Субстанция Р<
Высво-
Воспаление вождение
цитокина
Симпатическая
иннервация кожи и сосудов
Ад/
С-во-
локна
С у 6стан
ция Р _
Расширение
сосудов и отек
Повреждение
ткани приводит
к высвобождению
брадикинина,
простагландина»
5-гидрокситриптамина
д Все это сенсибилизирует ноцицептор |
Боль — это неприятное ощущение или эмоцио-
нальное переживание, связанное с действительным
или потенциальным повреждением ткани. Многое
из того, что известно о механизмах боли, было изу-
чено на экспериментальных моделях животных, у
которвтх сложная эмоциональная реакция на боль.
Поэтому предпочтительнее использовать термин
«ноцицепция», когда говорят о передаче информа-
ции из поврежденной области — по-видимому, в об-
ласть коры, где происходит восприятие боли. Важно
понимать, что повреждение ткани не всегда связано
с болью, это можно наблюдать у людей во время во-
еннвтх действий, когда тяжелые травмы, получен-
нвте в стрессовой ситуации, не ввтзвтвают боли.
Ноцицепторы
Ноцицепторы находятся в коже, внутренних орга-
нах, скелетных и сердечной мышце, а также в кро-
веноснвтх сосудах. Они передают информацию о
повреждении в задний рог спинного мозга, где пер-
50
вичные афференты переключаются преимуществен-
но на интернейроны.
Различают два вида ноцицепторов в зависимос-
ти от диаметра афферентного волокна и стимула,
необходимого для их активации. Высокопороговые
механорецепторы (ВПМР) активируются интенсив-
ной механической стимуляцией и иннервируются
слабомиелинизированными Ад-волокнами со скоро-
стью проведения 5—30 мс. Полимодальные ноцицеп-
торы (ПМНЦ) реагируют на интенсивную механи-
ческую стимуляцию, температуру выше 42°С и раз-
дражающие химические вещества. Эти рецепторы
иннервируются немие линизированными волокна-
ми С со скоростью проведения 0,5 2 мс. Боль с чет-
кой локализацией проводится по более быстропро-
водящим волокнам, тогда как слаболокализованная
боль — по медленнопроводящим волокнам С.
Несмотря на то что гистологически ноцицепторы
представляют собой свободные нервные окончания,
процесс восприятия на уровне рецептора сложен и
связан с химическими медиаторами воспаления и
тканевого повреждения. Так, АТФ, брадикинин, гис-
тамин и простагландины активируют или сенсиби-
лизируют нервное окончание, кроме того, некоторые
нейротрансмиттеры ноцицептивного пути сами выс-
вобождаются на периферии (например, субстанция Р)
для большей сенсибилизации нервного окончания.
Сенсибилизация рецепторов объясняет усиленное
болевое ощущение (первичную гипералгезию) в обла-
сти повреждения ткани, но не может полностью
объяснить восприятие неболезненных стимулов как
болезненные (аллодиния, или вторичная гипералгезия)
в случае повреждения ЦНС или периферического
нерва. Недавние исследования показали, что и алло-
диния, и некоторые хронические болевые синдромы
(например, фантомные боли) вызваны отсроченны-
ми изменениями передачи болевой информации в
задний рог спинного мозга. Висцеральные ноцицеп-
торы проецируются в спинной мозг посредством мел-
кокалиберных миелинизированных и немиелинизи-
рованных волокон вегетативной нервной системы и
заканчиваются на уровне спинного мозга в соответ-
ствии с их эмбриональным происхождением. Боль во
внутреннем органе может вызвать ложное восприя-
тие болевого стимула на участке кожи, а не на самом
органе, по крайней мере на ранних стадиях воспале-
ния— феномен, известный как отраженная боль. На-
пример, воспаление аппендикса первоначально при-
водит к болезненным ощущениям в области пупка.
Ноцицептивные проводники
Большинство ноцицепторов и терморецепторов
проецируется в спинной мозг через задний корешок,
хотя некоторые проходят и через передний корешок.
Достигая спинного мозга, эти чувствительные нервы
сложным образом оканчиваются в заднем роге.
В заднем роге могут возникать изменения синап-
тической передачи. Например, поступление субстан-
ции Р по аксонам в поверхностные слои заднего рога
приводит к увеличению размера рецептивных полей
и чувствительности некоторых нейронов заднего
рога. Эти функциональные изменения отчасти опо-
средуются синаптическим высвобождением глута-
мата, взаимодействующего с постсинаптическими
NMDA-репепторами. Эти отсроченные изменения
являются примером пластичности нейронов (см. гла-
ву 47) и принимают участие в механизме формирова-
ния некоторых хронических болевых синдромов.
Постсинаптическая клетка, проводящая ноцицеп-
тивную информацию, проецируется на спинной мозг
в виде спиноталамического, спиноретикулоталами-
ческого и спиномезенцефального трактов (последний
показан на рисунке), при этом аксоны перекрещива-
ются на уровне центрального канала спинного мозга
на противоположную сторону. Эти перекрещенные
волокна поднимаются несколько выше места попа-
дания их в спинной мозг, поэтому поражение цент-
рального канала, возникающее при сирингомиелии,
приводит к утрате болевой и температурной чувстви-
тельности в соответствующих дермотомах на несколь-
ко уровней ниже места поражения (см. главу 31).
Постсинаптическая клетка и пресинаптическая
ноцицептивная нервная терминаль получают синап-
сы от других соматосенсорных систем, проецирую-
щихся на периферию, это нисходящие проекции из
ствола головного мозга и собственные интернейро-
ны заднего рога. Многие из этих интернейронов со-
держат эндогенные опиоцды, такие, как эндорфины
и энкефалины, которые активируют опиоидные ре-
цепторы, основными подтипами которых являются
ц, к и 8. Поэтому на уровне заднего рога спинного
мозга имеется возможность существенно модифици-
ровать ноцицептивную информацию (см. главу 22).
Восходящие ноцицептивные пути оканчиваются
на различных структурах ЦНС. Информация о бо-
левых стимулах проходит по спиноталамическому
тракту (что обеспечивает представление о точной ло-
кализации боли) или спиноретикулоталамической
системе (передающей информацию об аффективных
компонентах боли). Однако некоторые ядра ствола,
куда проецируются эти пути (например, большое
ядро шва и голубое ядро), в свою очередь посылают
аксоны вниз к заднему рогу спинного мозга и таким
образом могут участвовать в подавлении хроничес-
ких болевых синдромов (см. главу 22).
В таламусе спиноталамический путь заканчива-
ется на задневентральном и интраламинарном яд-
рах (включая заднюю группу), которые в свою оче-
редь проепируются на множество областей коры, но
в большей степени на ПССК и ВССК. Однако им-
пульсацию от этих ядер получают и другие области,
например префронтальная кора, базальные ганглии
и кора островковой извилины. Поражение этих об-
ластей может изменить ощущение боли и привести
не к утрате болевой чувствительности или аналге-
зии, а к развитию хронического болевого синдрома.
Эти синдромы часто наблюдаются при поражении
таламуса мелкими сосудистыми очагами.
Терморецепторы и в меньшей степени ноцицепто-
ры также проецируются на гипоталамус, который игра-
ет важную роль в терморегуляции и вырабатывает ве-
гетативный ответ на болевой стимул (см. главы 42 и 43).
51
ГЛАВА 22. Система боли II:
способы медикаментозного и немедикаментозного лечения
Поражение любого из этих отделов
не приводит к отсутствию восприя-
тия чувства боли, но может изме-
нить порог болевого восприятия и
ЖТ^зТилВииь реакцию. Поэтому поражение ПССК
ВССК может нарушить способность раз-
личать интенсивность болевого сти-
мула, тогда как поражение пре-
фронтальной коры уменьшает аф-
фективную реакцию на боль
Мишени и механизмы уменьшения
болевого синдрома (аналгезия)
псск
палительпыми препаратами
рефрон-
гальная
х мора
Галамус (задневентральное
и интраламинарное ядра)
Таламус
в
Г ипотг ламус
©Пересечение периферических нервов редко при-
водит к уменьшению болевого синдрома, так как
большинство хронических болевых синдромов раз-
вивается в результате реорганизации центральных
ноцицептивных путей и сенсорного проведения
®В ряде случаев, особенно при рефлекторной сим-
патической дистрофии, недавно переименованием
в комплексный регионарный болевой синдром,
применяете* симпапекотомия ______________________
Применение экзогенных опиоидным анальгетиков
' (воздействие на систему эндогенных опиоидов)
Ствол мозга:
околоводопро-
► водное серое ве-
щество. голубое
ядро, большое
кядро шва, рост-
ровентральная
часть продолго-
□ I ватого мозга
ЛуЧЭлектрическая стимуляция задних столбов
41 может уменьшать боль ниже места стимуляции
.r-'v Селективная активация крупнокалиберных аффе-
(VJ) рентных волокон с помощью чрескожной электро-
иеиростммуляцни_____________________________
Стимуляция ядер ствола мозга или назначение
препаратов, которые модифицируют серотонинер-
гическую или норадренергическую трансмиссию
(например, антидепрессантов) _________ _ _
Анатомия заднего рога
спинного мозга
Крупнокалиберные
миелинизированные
афферентные волокна
Спиноталамический
нейрон
Мелкокалиберное
ноцицептивное
афферентное волокно
Нисходящие серотонинерги-
ческие норадренергические
пути, уменьшающие передачу
повреждающего стимула
в ростральном направлении
Пластина Рексед II
(желатинозная субстанция)
с тормозными
(энкефалинергическими) нейронами
Нисходящий
путь в дор-
солатераль-
ном белом
веществе
спинного
мозга
2
' 51
hi
Крупнокалибер-
ные миелинизи-
Задние
столбы
Лимфоциты
Макрофаги
Тучные
клетки
носныи
сосуд
Повреждение и воспаление —
высвобождение цитокинов, простагландинов.
2
о
Ад/Г
волок-
на
Симпатическая
иннервация
кожи и сосудов
Травма
брадикинина, гистамина и субстанции Р
Болевой синдром часто встречается в клиничес-
кой практике, лечение его проводят не толвко в том
случае, когда он вызван травмой или воспалением,
но также при поражении самих нервов (например,
при невропатиях). В последнем случае болв может
бытв вызвана ранее перенесенной травмой (напри-
мер, при рефлекторной симпатической дистрофии)
или может развитвея вследствие менее изученных
причин, что называется комплексным регионарным
болевым синдромом. Во всех случаях развитие бо-
52
левого синдрома всегда инвалидизирует пациента и
вызывает у него депрессию, поэтому к болвному с
таким синдромом необходим мулвтидисциплинар-
ный подход с включением психологической помо-
щи, и медикаментозных средств.
Лечение боли
Уменьшение болевого синдрома или аналгезии
можно достичь с помощью различных средств. Мно-
гие анальгетики уменьшают воспалительный ответ
на периферии, что, в частности, снижает сенсиби-
лизацию рецепторов (см. рисунок, мишень I). Бло-
када проведения по периферическому нерву при
помощи инъекции локального анестетика применя-
ется при некоторых болевых синдромах, но перерез-
ка периферического нерва обычно не уменьшает
болевой синдром (мишень II). Однако при травмах
периферического нерва формируется псевдоневро-
ма, которая состоит из рубцовой ткани и отрезка
аксона, регенерация которого прервана (см. гла-
ву 46). Это в свою очередь приводит к генерации па-
тологических сигналов в пораженном нерве, кото-
рые воспринимаются как болезненные. В этих слу-
чаях иссечение псевдоневромы может уменьшить
болевой синдром.
Например, стимуляция неноцицептивных рецеп-
торов может уменьшать передачу ноцицептивной
информации в задний рог, это означает, что болевой
стимул встречает на своем пути «ворота», в которые
он не будет пропущен, если одновременно туда по-
лаются неболевые стимулы. Эти представления по-
ложены в основу теории «воротного контроля боли»
Уолла и Мелзака. Эта теория была подтверждена при
клинических исследованиях с помошью чрескожной
электронейростимуляции (ЧЭНС) в области боле-
вого синдрома (мишень IV), а также стимуляции зад-
них столбов при хроническом болевом синдроме
(мишень V). По аналогии супраспинальный импульс
также может осуществлять «воротной контроль» бо-
левых стимулов при активации (мишень VII), что
происходит в стрессовых ситуациях, когда возник-
новение болевого стимула не всегда сопровождает-
ся его проведением в ЦНС (военные травмы). На эти
супраспинальные ядра можно воздействовать меди-
каментозно, назначая препараты, используемые для
лечения депрессии (см. главу 50). Антидепрессанты,
действующие преимущественно на серотонинерги-
ческую и норадренергическую системы, использу-
ются для лечения болевых синдромов вне зависимо-
сти от антидепрессивного влияния, которое они
могут оказывать (мишень VII). Чаще всего приме-
няют ингибиторы обратного захвата моноаминов
имипрамин и амитриптилин (трициклические анти-
депрессанты). Они изменяют болевой порог, одна-
ко вызывают побочные эффекты, так как блокиру-
ют мускариновые рецепторы (сухость во рту, запор,
ухудшение зрения), (e-адренорецепторы (постураль-
ная гипотензия) и гистаминовые Н^-рецепторы (се-
дативный эффект).
Более того, когда выяснилось, что одним из ме-
диаторов боли является субстанция Р, разработали
другие анальгетики, например капсаицин (ингреди-
ент красного перца), который способствует высво-
бождению субстанции Р из нопипепторов и в пос-
ледующем инактивирует содержащие субстанцию Р
волокна С. Капсаицин можно использовать местно
для лечения постгерпетической невралгии. Однако эту
систему можно применять, воздействуя на энкефа-
линергические интернейроны и опиоидные рецеп-
торы, назначая морфин и его аналоги (мишень IV).
Опиоидные анальгетики обладают фармакологи-
ческими свойствами, сходными с таковыми опио-
идных пептидов. Они вызывают пролонгированную
активацию опиоидных рецепторов (обычно ц-рецеп-
торов). Это уменьшает передачу болевых импульсов
в задний рог спинного мозга с помощью тормозяще-
го действия релейных нейронов. Опиоиды также сти-
мулируют норадренергические, серотонинергичес-
кие и энкефалинергические нейроны ствола мозга,
аксоны которых спускаются в спинной мозг и тоже
ингибируют активность релейных нейронов спино-
таламического тракта. Опиоидные анальгетики ши-
роко применяются для уменьшении тупой, нечетко
локализованной (висцеральной) боли. Повторное их
назначение может привести к развитию зависимости.
Наркоманам дозу, способную вызвать эйфорию, необ-
ходимо быстро увеличивать по мере повторного упот-
ребления наркотика (толерантность), но у пациентов
с болевым синдромом потребность в увеличении
дозы отражает прогрессирующее его усиление.
Морфин наиболее широко используется для ку-
пирования сильной боли, но, как и все сильные опи-
оиды, может вызвать тошноту и рвоту. Применение
других морфиноподобных наркотиков сопровожда-
ется эйфорией, угнетением дыхания, запором и су-
жением зрачков, последнее происходит в результате
стимуляции парасимпатических ядер глазодвига-
тельных нервов. К другим опиоидам, применяемым
для лечения сильной боли, относятся диаморфин
(героин), петидин и метадон. Кодеин и декстропро-
поксифен — менее сильные наркотики, которые ис-
пользуются для уменьшения легкой и умеренной
боли. Налоксон является антагонистом опиоидных
рецепторов и применяется для устранения эффек-
тов передозировки опиоидов.
Более того, несмотря на то что боль обычно свя-
зана с поражением ткани, она может развиваться и
при поражении ПНС или ЦНС, например неврал-
гия тройничного нерва, которая характеризуется
приступами лицевой боли. При этом заболевании
пациент испытывает болевые приступы в проекции
одной из трех ветвей тройничного нерва. Причину
невралгии в большинстве случаев установить не уда-
ется, однако известно, что она встречается у неко-
торых больных рассеянным склерозом. Ее можно ле-
чить хирургическим способом, рассекая соответству-
ющий нервный корешок, или же медикаментозно
противоэпилептическими препаратами — карбамазе-
пином и габапентином (см. главу 53). Другим при-
мером развития болевого синдрома при поражении
нервной ткани является травма периферического
нервного ствола, когда повреждение вегетативной
иннервации конечности приводит к развитию силь-
ной боли (рефлекторные симпатические дистрофии).
Причина развития таких состояний неизвестна,
хотя в ноцицептивных нервных окончаниях начи-
нают экспрессироваться рецепторы к норадрена-
лину, поэтому локальная симпатикотомия может
уменьшить подобную боль, хотя и не во всех случа-
ях (мишень III).
53
ГЛАВА 23. Зрительный анализатор I: глаз и сетчатка
Распределение рецепторов от центра к периферии сетчатки
Срез глаза
Ресничное
тело
Радужка
Склера
Сосудистый слой
' Пигментный слой
) Сетчатка
Зоительный нерв
1x10* волокон
Роговица
Слепо : пятно
Ямка
Хрусталик
| Слои сетчатки
Передняя
камера
Стекловидное
тело
Угол
отклонения
ст ямки
Слепое пятно, где волокна входят в зрительный нерв
- 180 000----------------
1 160 000----------------
«140 000------------------
_ „ „ Ямка, палочки
^120 000----
°’00 000----
80 000---
60 000
4U 000
20 000
о
70° 60° 50° 40° 30° 20° 10° 0° 10° 20° 30° 40° 50° 60° 70° 80°
Г радуг отклоне шя сетчатки от центра
8.
а
->15
Колбочки
Колбочки
-1 ->15
Рецептор- соотношение ганглиозных клеток
Спектр поглощения родопсина
и трех зрительных пигментов у человека
Склера
Сосудистый слой
Пигментный слой
Длина волны, нм
Палочка
Фоторецепторы
Г англиолные
клетки
Волокна
зрительного
нерва
Горизонтальные
клетки
Биполярные
клетки
Амакриновые
клетки
Стекловидное тело
Организация рецептивного поля (центр и окружение)
Деполяризуются в ответ на пучок света (клетки центра)
- Гиперполя риэуются в ответ на пучок света
(клетки окружения)
Зрительный анализатор отвечает за превращение
всей световой энергии в зрительный образ окружа-
ющего мира. Эта информация кодируется в сетчат-
ке на глазном дне, а затем передается в зрительную
кору, гипоталамус и верхние отделы ствола (см. гла-
вы 24 и 25). Процесс зрительного восприятия деталь-
но изложен в главе 19.
Оптические свойства глаза
Достигая глаза, свет фокусируется на сетчатку.
Этот процесс рефракции зависит от кривизны рого-
вицы и хрусталика. Нарушение этих параметров при-
водит к снижению остроты зрения при чтении (даль-
нозоркость, или гиперметропия), при рассмотрении
удаленных объектов (близорукость, или миопия) или
к обоим видам расстройств. В последнем случае на-
рушение зрения зачастую осложняется астигматиз-
мом, при котором рефракция глаза отличается в раз-
личных меридианах глаза.
Кроме того, помимо рефракции на сетчатке, свет
без утраты качества (что зависит от прозрачности
роговицы) должен передаваться на переднюю и зад-
54
нюю поверхности хрусталика, который также дол-
жен быть прозрачен. Травмы или заболевания лю-
бой из этих структур приводят к снижению остроты
зрения (способности различать детали). Чаще всего
эти структуры глаза поражаются при инфекциях (ке-
ратит) или помутнении хрусталика (катаракта).
Анатомия и функция сетчатки
Свет, попадающий на сетчатку, передается в виде
электрических импульсов с помощью фоторецепто-
ров, которые залегают в промежуточном слое сетчат-
ки за стекловидным телом. Существует два основ-
ных вида фоторецепторов: палочки и колбочки. Па-
лочки находятся во всех отделах сетчатки и
чувствительны к слабому освещению, поэтому от-
вечают за зрение в темноте. Несколько палочек пе-
редают информацию к одной ганглиозной клетке, в
силу чего эта система чувствительна к абсолютному
уровню освещения, но не способна отличать детали
зрительного образа в цвете, поэтому ночью мы мо-
жем видеть объекты, но не в деталях и не в цвете.
Колбочки в основном находятся в области ямки
и содержат три различных зрительных пигмента.
Они отвечают за дневное зрение. Плотность этих ре-
цепторов в области ямки наиболее высока (одна кол-
бочка соответствует одной ганглиозной клетке), по-
этому они отвечают за остроту зрения и цветное зре-
ние. Нарушение обмена зрительных пигментов,
содержащихся в этих рецепторах, приводит к разви-
тию цветовой слепоты. Заболеваниярецепторов, вы-
зывающие их гибель, такие, как пигментный рети-
нит, сопровождаются прогрессирующим снижением
остроты зрения. Характерно поражение перифери-
ческих отделов сетчатки и палочек на ранних стади-
ях, что приводит к снижению остроты зрения ночью
и концентрическому сужению полей зрения.
Фоторецепторы образуют синапсы с горизон-
тальными и биполярными клетками. Горизонтальные
клетки выполняют две основные задачи: представ-
ляют центр и окружение рецептивного поля для би-
полярных клеток; отвечают за сдвиг спектра чувстви-
тельности биполярных клеток для соответствия
уровню фонового освещения (отчасти обеспечива-
ют адаптивную реакцию на освещение; см. главу 19).
Под центром и окружением рецептивного поля пони-
мают следующее: биполярная клетка реагирует на
небольшой пучок света в центре рецептивного поля
одним способом (деполяризация или гиперполяри-
зация), а окружение центра будет реагировать на свет
другим способом. Горизонтальные клетки, получая
сигналы от многих рецепторов и образуя синапсы на
фоторецепторных биполярных клетках, обеспечива-
ют достаточный поток информации для генерации
рецептивного поля. Механизм, посредством которо-
го они выполняют вторую задачу, т. е. обеспечивают
адаптацию к уровню освещения, изучен недостаточно.
Биполярные клетки передают информацию от
фоторецепторов ганглиозным клеткам и получают
синапсы от фоторецепторов, горизонтальных и
амакриновых клеток. Биполярные клетки классифи-
цируют в зависимости от рецепторов, от которых они
получают синапсы (только от колбочек, только от
палочек или от тех и других), или от их реакции на
свет. Биполярные клетки, которые гиперполяризу-
ются небольшим пучком света в центре своих рецеп-
тивных полей, называются клетками окружения, би-
полярные клетки, которые деполяризуются неболь-
шим пучком света в центре своего рецептивного
поля именуются клетками центра.
Ганглиозные клетки находятся вблизи стекловид-
ного тела и, получая синапсы от биполярных и амак-
риновых клеток, посылают свои аксоны в мозг че-
рез зрительный нерв. Эти аксоны проходят через
внутреннюю поверхность сетчатки, прежде чем вой-
дут в состав зрительного нерва, и формируют диск
зрительного нерва, который представляет собой сле-
пое пятно сетчатки, где отсутствуют какие-либо ре-
цепторы. Наличие слепого пятна в норме не влияет
на зрение. Ганглиозные клетки классифицируют по
нескольким признакам: в зависимости от морфоло-
гии, от их реакции на свет, каки биполярные клетки
(клетки центра и клетки окружения), от комбина-
ции этих свойств (система XYW). Ганглиозные клет-
ки X, представляющие 80% ганглиозных клеток сет-
чатки, отвечают за различение деталей и цвета изоб-
ражения, 10% ганглиозных клеток Y—за восприятие
движения. 10% клеток W проецируются в ствол моз-
га, но не имеют какой-то определенной функции.
Ганглиозные клетки X и Y были впервые выявлены
у кошек и соответствуют каналам Р и М у приматов,
которые отвечают за кодирование формы и движе-
ния соответственно.
Амакриновые клетки сетчатки получают сигна-
лы от биполярных клеток и проводят сигналы к ним,
другим амакриновым и ганглиозным клеткам. Су-
ществует множество видов амакриновых клеток,
некоторые из которых относятся исключительно к
колбочкам, другие к палочкам, и они содержат не-
сколько нейротрансмиттеров. Амакриновые клетки
обеспечивают комплексную реакцию на световые
импульсы и играют важную роль в генерации реак-
ции ганглиозных клеток, включая фиксацию и ко-
дирование движущихся объектов, а также включе-
ние и выключение освещения.
ГЛАВА 24. Зрительный анализатор II:
зрительные пути и подкорковые зрительные центры
Височное
поле зрения
Левый глаз
Назальное
поле зрения
Правый глаз
Примечание: назальные отделы
сетчатки получают информацию от
височных половин поля зрения, а ви-
сочные части сетчатки — от назаль-
ных половин поля зрения
Дефект полей зрения
Левый
Правый
Правая половина
поля зрения
Левая половина
поля зрения
Парасимпатическая иннервация зрачка
и дуга зрачкового рефлекса
Зрительный нерв
Височная
Еазальная
Височная
Зрительный тракт
Битемпоральная Средний мозг
гемианопсия ' ►
Зрительный
перекрест
(над гипофизом)
Короткие
цилиарные
нервы
Претекта-
льнов ядр
Зрительный
тракт
ганглий
Правосторонняя
нижнеквадрантная
гемнанопсия
Верхние
бугорки
четверо
холмия
Ядро
Эдингера—Вестфаля
Правосторонняя
нижнеквадрантная
гемианолсия
тельная
лучис-
тость
Супрахиазмати-
ческое ядро ги-
поталамуса (см.
главу 43|
Волокна,
через височную долю
проходящие
Волокна, проходящие
через теменную долю
ЛКТ
ядро
Задняя спайка
Средний
мозг
Латеральное
коленчатое
тело (ЛКТ)
Организация латерального коленчатого тела у приматов
Правосторонняя
верх неквадрантная
гемианолсия
Первичная
Зрительная кора
Мелкоклеточные пластины
(пути от ганглиозных
клеток X}
Крупноклеточ-
ные пластины
(пути от гангли-
озных клеток Y)
Правосторонняя гомонимная гемианопсия
(с интактностью центрального макулярного
зрения при сосудистых очагах; см. главу 17)
Ретинотопическая проекция
первичной зрительной коры,
анализирующая фовеол ярую
область поля зрения (ямку)
Пути от противоположного
глаза
4„ 6
Интраламмнар-
Дефект поля зрения при очагах
на различных уровнях зрительного пути
Пути от глаза своей стороны | "| 2, 3, 5
ные области
Сетчатка передает информапию от ганглиозных
клеток к нескольким областям, включая некоторые
отделы коры через таламус, гипоталамус и средний
мозг. Основная проекция проходит через латералв-
ное коленчатое тело (ЛКТ) таламуса к первичной
зрителвной коре (ПЗК, или поле 17 по Бродману).
Другие отделы коры (экстрастриарные) получают
информацию от ЛКТ, а также подушки таламуса (см.
главу 25).
Проекция от сетчатки до ПЗК имеет соматото-
пическую организапию, поэтому всегда можно пред-
сказатв, к какому дефициту приведет очаг пораже-
ния по ее длиннику. Очаги до зрителвного перекре-
ста ввтзывают развитие односторонних дефектов
полей зрения, тогда как очаги, поражающие хиазму
(например, гипофизарные опухоли), битемпоралв-
ную гемианопсию. Очаги, расположенные после
хиазмы, приводят к выпадению одинаковвтх полей
56
зрения на разных глазах - гомонимной гемианоп-
сии или квадрантной гемианопсии.
Латеральное коленчатое тело
ЛКТ состоит из 6 слоев у приматов, при этом каж-
дый слой получает импульсы от глаза своей или про-
тивоположной стороны. Два внутренних слоя круп-
ных нейронов образуют крупноклеточные пласти-
ны, остальные 4 слоя мелкоклеточные пластины.
Между этими ядрами существуют морфологические
и электрофизиологические отличия.
Мелкоклеточные нейроны отвечают за пветоощу-
щение и высокое пространственное разрешение
(различение деталей), поддерживают длительную
реакцию на зрительные стимулы. В отличие от них,
крупноклеточные нейроны не выполняют функцию
цветоразличения, реагируют на небольшое про-
странственное разрешение, и ихреакция на зритель-
ный стимул кратковременна. Таким образом, свой-
ства крупноклеточных нейронов сходны со свой-
ствами ганглиозных клеток Y, а мелкоклеточных — с
ганглиозными клетками X. Это сходство связано с
топографическими особенностями представленно-
сти проекций этих двух типов ганглиозных клеток в
ретиногеникулярном пути. Ганглиозные клетки X и
мелкоклеточные нейроны отвечают за восприятие
цвета и формы (Pattern — Р) и образуют P-канал, ган-
глиозные клетки Y и клетки крупноклеточной плас-
тины ЛКТ отвечают за восприятие движения (Move-
ment) и образуют М-канал.
ЛКТ проецируется на ПЗК, где афферентные во-
локна образуют синапсы в IV слое коры и в мень-
шей степени в VI слое, при этом М- и P-каналы об-
разуют синапсы со структурами различных пластин.
Кроме того, имеется проекция этих клеток, которая
лежит между пластинами ЛКТ (интраламинарная
часть ЛКТ), которая соединяется со II и III слоями
коры; см. главу 25).
Верхние бугорки четверохолмия
Верхние бугорки четверохолмия среднего мозга —
это многослойная структура, в которой поверхност-
ные слои отвечают за распределение поля зре-
ния, а глубокие — за комплексную интеграцию зри-
тельных, слуховых и соматосенсорных стимулов.
Промежуточные слои принимают участие в сакка-
дированных движениях глаз и связаны с затылочно-
теменной корой, корковым центром взора в лобной
доле, черной субстанцией (см. главу 40). Саккади-
рованные движения глаз представлены в верхних
бугорках четверохолмия таким образом, что стиму-
ляция в определенной точке вызывает саккадиро-
ванное движение глаз, чтобы перенести точку фик-
сации взора в поле зрения, которое представлено в
более поверхностных слоях этой структуры. Эта спо-
собность распределять различные сенсомоторные
представительства в верхних бугорках четверохол-
мия распространяется и на более глубинные слои.
Другими словами, если проследить эту структуру
вниз по вертикали, можно увидеть: 1) нейроны, реа-
гирующие на зрительные стимулы в определенной
части поля зрения; 2) нейроны, которые вызывают
саккадированные движения глаз, совмещают ямку
с определенным участком поля зрения; 3) слуховые
и соматосенсорные нейроны, максимально активи-
руемые звуками, которые исходят из той части ок-
ружающего мира и тех участков кожи, которые с наи-
большей вероятностью активируются при физи-
ческом контакте с объектом, находящемся в этом
участке внешнего пространства. Последнее свойство
обусловливает максимальное соматосенсорное пред-
ставительство носа и лица в верхних бугорках четве-
рохолмия. Так, верхние бугорки не только кодиру-
ют те саккады, которые запускаются стимулами или
поведенческими реакциями, но и имеют более ши-
рокие функции, касающиеся пространственной ори-
ентации. Верхние бугорки имеют эфферентные свя-
зи с некоторыми структурами ствола и спинным
мозгом (тектоспинальный тракт). Поражение этих
структур редко встречается в клинике и проявляет-
ся утратой саккадированных движений глаз и игно-
рированием полей зрения.
Претектальные структуры
и реакция зрачка на свет
Зрительный нерв соединяется с претекталь-
ными ядрами среднего мозга, которые в свою оче-
редь проепируются на ядра Эдингера—Вестфаля
своей и противоположной стороны, дающие па-
расимпатические волокна к мышце, суживающей
зрачок. Свет, попадающий на глаз, приводит к су-
жению обоих зрачков (прямая и содружественная
реакции зрачка на свет). Поражение одного из зри-
тельных нервов вызывает утрату прямой и содру-
жественной реакций зрачка на свет на пораженной
стороне, однако зрачок пораженного глаза будет
нормально сокращаться при освещении противо-
положного глаза (относительный афферентный
зрачковый дефект).
Супрахиазматическое ядро гипоталамуса
Ядро получает импульсацию от сетчатки и игра-
ет важную роль в обеспечении и поддержании цир-
кадных ритмов (см. главу 43).
ГЛАВА 25. Зрительный анализатор III: зрительная кора
Часть затылочной доли полушария головного мозга
Поле 19
Задняя
теменная кора
60е к
Левое полушарие
Гиперстолбик
Проекции
на другие
отделы
коры
Правая Левая
сторона сторона
Зрительная лучистость
Связи
латерального
коленчатого тела
с первичной
зрительной корой (ПЗК)
(см. главу 24)
ПЗК
(поле 17)
Ретинотопическая
проекция ПЗК,
анализирующая
преимущественно
фовеолярное
поле зрения
(область ямки)
Правое полушарие
Столбиковая организация ПЗК
Промежуточные
капли
о
Капля
| Проекция * 1
г К ПОДКОрКО
1 аьн от>«лам
Проекция
0,5 мм 0,5 мм к лкт
лзк
Подкорковое
белое
вещество
и Таламмчес- Таламичес -.
™ о кая импуль- кая имлуль Правая
ф о сация пра- сация ле- сторона
и вого глаза вот о глаза
Столбики, селективные
по стороне
Таламическая
импульсация к первичной
зрительной коре
ЛКТ
j Импульсация мелкоклеточной
Х-проекция = Р канал
Латеральные
отделы
Полный спектр ориентации
Медиальные отделы
= ПЗК
Шпорная
борозда
Задняя
теменная кора
Экстрастриарные
проекции на ВЗК (V2 ] Височная доля ,
Гонкие полосы
(четвертичная
► зрительная кора)~*У4
Бледные полосы—bV3,V4
(третичная и четвертич-
ная зрительная кора)
-^Толстые полосы—‘►V3. V5
(третичная и зри-
тельная кора —
зк V5>
Селективность
по ориентации
Иерархическое расположение
клеток в зрительной коре
(по Хубелю и Вайзелю)
пластины
Простые клетки
Круговое симметричное
рецептивное поле
= ВЗК, ТЗК, ЧЗК
= ЗК V5
Горизонтальный срез
задней половины правого полушария
ВЗК
Поле 17 Поле 18
Поле 19
Экстрастриарные отделы
ВЗК получает импульсацию от ПЗК и про-
ецируется иа другие экстрастриарные
отделы (функция неизвестна)
ТЗК получает импульсацию от ПЗК и ВЗК,
возможно, принимает участие в воспри
ятии глубины и регуляции остроты зрения
ЧЗК получает проекцию от ганглиозных
Х-клеток и проводит первичный анализ
цветоощущения; поражение этого отде-
ла приводит к корковой цветовой слепоте
Импульсация крупноклеточной пластины
У-лроекция
Преобладающее
ЗК V5 получает проекцию от ганглиоз-
ных У-клеток и проводит первичный ана-
лиз направления движения объектов
Кора нижних отделов височной доли
содержит клетки, участвующие в узна-
вании лицг поражение этого отдела у че-
ловека, особенно справа, приводит к
селективной лицевой агнозии (прозопаг-
нозии).
Первичная зрительная кора
(поле 17 по Бродману)
Первичная зрительная кора (ПЗК) располагает-
ся вдоль шпорной борозды в затылочной доле и по-
лучает импульсы от латерального коленчатого тела
(ЛКТ). Эти связи организованы ретинотопически
таким образом, что соседние структуры сетчатки
58
проецируются на зрительный путь посредством со-
седних аксонов. Эта ретинальная проекция, одна-
ко, не является простой картой, поскольку опреде-
ляющим топическим фактором является соотноше-
ние фоторепепторов и ганглиозных клеток сетчатки.
Это означает, что центр поля зрения (ямка) доми-
нирует в ретинальной проекции ПЗК, поскольку
именно в ямке соотношение ганглиозных клеток и
рецепторов составляет 1 : 1, в отличие от перифери-
ческих отделов сетчатки (см. главу 23).
ЛКТ проецируется в основном на IV слой ПЗК,
и эта проекция отличается для М- и P-каналов, тог-
да как интраламинарная часть ЛКТ проецируется на
II и III слои ПЗК (см. ниже). Проекция ЛКТ на IV
слой ПЗК столь велика, что этот слой коры можно
подразделить на отделы IVa, IVb, IVc« и IVce, при
этом связи каждого отдела несколько отличаются
друг от друга. Однако в целом корковые нейроны IV
слоя ПЗК организованы по принципу кругового сим-
метричного рецептивного поля с клетками-центром и
клетками-окружением (см. главу 23). Эти нейроны IV
слоя проецируются на нейроны соседней коры таким
образом, что несколько нейронов определенного типа
конвергируют, т. е. соединяются, с одним нейроном
соседнего отдела, чье рецептивное поле становится
более сложным с точки зрения стимула, который мо-
жет этот нейрон активировать. Эти клетки наиболее
совершенно реагируют на порог освещения в опре-
деленной ориентации и называются простыми клет-
ками. Последние в свою очередь конвергируют на
другие нейроны (сложные клетки), которые преиму-
щественно расположены в III и II слоях коры и кото-
рые максимально активируются стимулами в опре-
деленной ориентации, перемещающимися в опре-
деленном направлении, при этом направление
находится под прямым углом к линии ориентации.
Сложные клетки проецируются на сверхсложные,
или заключительные, клетки, которые реагируют на
определенную ориентацию и длину. Эти клетки впер-
вые были описаны Хубелем и Вайзелем, они органи-
зованы иерархически, при этом каждая клетка полу-
чает свое рецептивное поле от нижележащей иерархии.
Хубель и Вайзель также установили, что эти ней-
роны были организованы в столбики клеток со сход-
ными свойствами. Первоначально было показано,
что нейроны организованы, во-первых, в зависимо-
сти от глаза, который дает к ним преимущественную
проекцию (столбики, селективные по стороне), и,
во-вторых, в зависимости от пространственной ори-
ентации, в соответствии с которой импульсы мак-
симально активируют эти клетки (столбики, селек-
тивные по ориентации). Эти два типа клеток распо-
ложены под прямым углом друг к другу, и группа
клеток, селективных по стороне, и все виды клеток,
селективных по ориентации, называются гиперстол-
биком. Этот гиперстолбик размером 1 мм может ана-
лизировать информацию от определенного участка
поля зрения в соответствии с импульсами от сетчат-
ки обоих глаз. В ямке, где соотношение фоторепеп-
торов и ганглиозных клеток составляет 1:1, поле
зрение очень маленькое, тогда как в периферичес-
ких отделах сетчатки оно большое. Поэтому сдвиг
на 1 мм от одного гиперстолбика к другому при-
водит к сдвигу локализации анализируемого поля
зрения, и это в большей степени относится к ямке
(см. ниже).
Однако в этой модели есть два существенных ог-
раничения. Одно из них касается аккомодации М-
и P-каналов, а второе связано с наличием цитохром-
оксидазы (маркера метаболической активности),
которой богаты II, III и IVb слои коры (и в меньшей
степени V и VI слои). Эти слои не характеризуются
селективностью в отношении ориентации, они под-
разделяются в зависимости от чувствительности к
пвету и пространственному разрешению (деталям).
Эти богатые цитохромоксидазой II и III слои груп-
пируются и образуют капли. Хотя бы одна из них ас-
социирована со столбиком, селективным по сторо-
не, при этом существуют области, называемые про-
межуточными каплями. И капли, и промежуточные
капли, и слой IVb ПЗК, богатый цитохромоксида-
зой, дают отдельные проекции на вторичную зри-
тельную кору и другие экстрастриарные области, эти
проекции коррелируют с М- и P-каналами. При та-
кой организации каналов и зрительная информация
обрабатывается не только в иерархическом поряд-
ке, но и по параллельным путям (см. главу 15).
Основными функциями ПЗК, помимо биноку-
лярных взаимодействий, являются преобразование
поля зрения в мелкие линейные сегменты различной
ориентапии, а также сегрегация и интеграция компо-
нентов зрительного образа для передачи в более спе-
циализированные зрительные поля. Эти поля осуще-
ствляют более сложный анализ зрительного образа,
однако взаимодействуют с ПЗК в целях сознательно-
го восприятия целостного зрительного образа. Кли-
нически у больных с двусторонним поражением ПЗК
отмечается корковая слепота — состояние, при кото-
ром они не могут воспринимать зрительные стиму-
лы, хотя при формальном обследовании регистриру-
ется возможность локализовать объекты в поле зрения.
Ассоциативные зрительные,
или экстрастриарные, поля
Экстрастриарные поля представляют собой отде-
лы коры, лежащие за пределами ПЗК и первично
вовлеченные в обработку зрительной информации.
Количество этих областей различно у разных видов,
больше всего их обнаружено у человека. Эти области
соответствуют полям 18 и 19 по Бродману, а также
нижней височной коре. Они участвуют в более слож-
ной обработке зрительного образа. При этом одна из
функций является доминирующей (например, цвето-
ощущение или анализ движущихся объектов). Кро-
ме того, многие другие структуры ЦНС участвуют в
анализе зрительной информации, например задняя
теменная кора (см. главу 30), лобная кора и корковый
центр взора в лобной доле (см главы 30 и 40), а также
подкорковые структуры гипоталамуса (см. главу 43)
и верхние отделы ствола мозга (см. главу 24).
ГЛАВА 26. Слуховой анализатор I: ухо и улитка
Свойства звуковой волны
Поражение до уровня
свального окна приводит
к кондуктнвной тугоухости
Наружное ухо
Среднее
ужо
Поражение после уровня
овального окна приводит
к нейросенсорной тугоухости
Внутреннее ухо
Барабанная
перепонка
Т
Время одного цикла
(частота = интенсивность/ время)
Звуковая
волна
Наружный V
слуховой проход V
. Полукружный
। канал (см.
। главу 28}
{Улитка
Гел и котре ма
У Преддверно-
[ (. улитковый
I (Г/"'—нерв
Чувствительность
к частотам у человека
Овальное окно
.Внутренний
слуховой
проход
Круглое окно
Мочка уха
Евстахиева труба
I--------- I I I г
0.015 2 4 10 15
кГц
•ч
Мембрана
Рейсснера
Сосудистые полоски —
продуцируют эндолимфу
Носоглотка
Срединная лестница, содержащая
эндолимфу (Na’ 1 ммоль/л4, Ка*
154 ммоль/л
волосковая
клетка
клетка
Наружная
волосковая
Разговорная речь примерно 65дБ,
звук более 120 дБ воспринимается как болезненный раздражитель
рентная
импуль-
сация.
Наружная волосковая клетка
Базальная мембрана (БМ)
эфферентная
импульсации
Внутренняя
волосковая
клетка
Внутренняя волосковая клетка
Всего 3500
Количество в ряду 1
Не контактируют с ТМ
Обеспечивает 93%
импульсации в кохлеарный нерв
Получает мелкокалиберные
немиелннизированные волокна
от комплекса верхних олив
Наружная волосковая клетка
12 000
3-4
Самые высокие стереоцилни,
контактирующие с ТМ
Обеспечивает 7%
импульсации в улитковый нерв
Получает крупнокалиберные
миелинизированные волокна
от комплекса верхних олив
Слуховой анализатор отвечает за восприятие зву-
ков. Главным рецептивнвтм органом является улитка
во внутреннем ухе, которая преобразует зву ко в ые вол-
HBi в электрические импулвсвт с помощвю процесса
механического проведения. Электрический импулвс,
60
генерируемвш в ответ на звук, проходит (вместе с
информацией от вестибулярной системы; см. главу
28) через черепно-мозговой нерв (ЧМН; преддвер-
но-улитковвш нерв) в ствол мозга, где образует си-
напсы с комплексами кохлеарных ядер (см. главу 27).
Несмотря на то что слуховой анализтор выпол-
няет много функций, первичный анализ звукораз-
личения происходит в улитке.
Свойства звуковых волн
Звуковая волна характеризуется амплитудой и
громкостью (измеряемой в децибелах дБ), часто-
той (измеряемой в герцах — Гц), формой, фазой и ка-
чеством или тембром. Интенсивность звука варьирует
очень широко, но обычно различается звук интен-
сивностью 1—2 дБ.
Поступление звуковой волны создает разность
фаз и интенсивности между обоими ушами, если
только звук не исходит из средней линии. Степень
задержки и изменения интенсивности между обоими
ушами в результате физического разделения необ-
ходима для определения локализации источника зву-
ков (см. главу 27).
Наружное и среднее ухо
Достигая уха, звук проходит по наружному слухо-
вому проходу к барабанной перепонке, которая виб-
рирует с частотой и силой, определяемыми звуком.
Это приводит к смещению трех слуховых косточек в
среднем ухе, смещению жидкости в улитке при вне-
дрении отростка стремечка в овальное окно улитки.
Этот процесс необходим для уменьшения звукового
сопротивления и усиления реакции на звук, по-
скольку звук воздействует на жидкость непосред-
ственно с сильным отражением.
Слуховыми косточками управляют две маленькие
мышцы, которые предохраняют косточки от повреж-
дения громкими звуками, а также изменяют поло-
жение отростка стремечка в овальном окне. Повреж-
дение слуховых косточек (отосклероз), среднего уха
(средний отит) или наружного слухового прохода (за-
купорка серной пробкой) приводит к снижению ос-
троты слуха или тугоухости, которая по своей при-
роде является ковдуктивной.
Внутреннее ухо и улитка
Смещение отростка стремечка в овальном окне
приводит к генерации волны в заполненной пе-
рилимфой лестнице преддверия и барабанной по-
лости улитки. Эти две лестницы сообщаются с апи-
кальным краем улитки — геликотремой, но на всем
остальном протяжении разделены срединной лест-
ницей, которая содержит звукопроводящий аппарат,
или кортиев орган.
Кортнев орган находится на дне лестницы на
структуре, называемой базальной мембраной (БМ),
ширина которой увеличивается с увеличением рас-
стояния от конца стремечка. Увеличение ширины
связано со снижением плотности БМ, это необхо-
димо для того, чтобы звуки высокой частоты мак-
симально смещали БМ на конце стремечка улитки,
а звуки низкой частоты максимально активировали
апикальный конец БМ. Так, различение частот яв-
ляется одной из функций БМ, несмотря на то что
эта способность значительно увеличивается при по-
мощи волосковых клеток кортиева органа, который
лежит на этой мембране.
Кортиев орган является сложной структурой, со-
стоящей из клеток слухового восприятия - волос-
ковых клеток (см. главу 19), которые по своей струк-
туре подразделяются на два типа: однорядные внут-
ренние волосковые клетки (ОВВК), дающие
большую часть импульсации в преддверно-улитко-
вый нерв, и 3—4-рядные наружные волосковые клет-
ки (НВК), регулирующие реакцию ОВВК на звук.
Эти два типа волосковых клеток отличаются морфо-
логически и электрофизиологически. ОВВК полу-
чают небольшую проекцию от ствола мозга, тогда
как НВК получают импульсацию от комплекса вер-
хних олив, который влияет на форму и способность
клеток воспринимать звуки. Некоторые ОВВК не-
посредственно контактируют с текториалыюй мемб-
раной (ТМ) кортиева органа, что важно для регуля-
ции реакции НВК на звук, так как эти клетки не кон-
тактируют с ТМ, но обеспечивают 93% импульсапии
по преддверно-улитковому нерву. Одно афферент-
ное волокно получает импульсацию от множества
ОВВК, однако лишь одна НВК связана с несколь-
кими афферентными волокнами. Кроме того, есть
и другие различия между ОВВК и НВК. Они неод-
нородны на протяжении срединной лестницы. Эта
неоднородность клеток по форме изменяет их час-
тотно-воспринимающие характеристики, что позво-
ляет клеткам до определенной степени настраивать-
ся на частоту звука, помимо опосредования этого
процесса свойствами БМ.
Повреждение улитки, волосковых клеток и кох-
леарной порции преддверно-улиткового нерва при-
водит к тугоухости, которая по природе является ней-
росенсорной. Травма, ишемия и опухоли VIII ЧМН
также приводят к нейросенсорной тугоухости. Не-
давно показано, что наследственная глухота связа-
на с дефектами определенных белков стереоцилий
волосковых клеток.
ГЛАВА 27. Слуховой анализатор II: слуховые пути и речь
Первичная
слуховая кора
Поражение зоны Брока
левого (доминантного)
полушария приводит
к моторной
эфферентной афазии
Комплекс
верхних олив
Ядро латеральной петли
(ЯЛП)
Поражение аркуатных волокон
приводит к акустико-мнестической афазии
Аркуатные волокна
Зона
Кохлеарное ядро
Улитка
Вентральное
Трапециевидное тело
Настройка маЧастоту |
Ядро латеральной петли
(ЯЛЛ)
Речь
Вернике
Медиальное
коленчатое тело
Поражение зоны
Вернике приводит
к сенсорной афазии
• Первичная слуховая кора — поле 41 по Бродману
• Вторичная слуховая кора — поля 41 и 42 по бродману ।
Спайка Пробста
Нижние бугор-
ки четверохол-
мия
Комплекс верхних олив
Т онотоп ичес кая
* организация
• Столбики распределены в
зависимости от связи с уша-
ми через мозолистое тело
Верхние бугор-1
ки четверохол-
мия
Латеральная петля
Медиальные Латеральные
ВВ-клетки ВТ-клетки {вы-
(мокиечастоты) сокие частоты) |:
Первый уровень
анализа бинау-
рального слуха
Локализация
источника звука
VIII пара,
ЧМН
* Дорсальное
Задний слуховой пучок
В — возбуждающие
ь импульсы от одного уха,
j Т — тормозные импульсы
от одного уха
Логарифм частоты
Каждое волокно слухового
иерва характеризуется вос-
приятием звука определенной
частоты, поступающего от во-
лосковых клеток, которые на-
стке базальной мембраны
Афферентная
импульсация
Импульсация
от левой улитки
Импульсация
от правой улитки
Звуки частотой менее 1400 Гц
регистрируются при помощи
At, но если частота превы-
шает 1400 Гц, тогда длина
волны меньше расстоянии- меж-
ду ушами фаза или время не
воспринимаются и
анализ переключа-
ется на Д|
Угол наклона
Комлекс верхних олив (КВО), возможно, коди-
рует распознавание временных различии попа-
дания звука в слуховой аппарат и локализации
источника звука. Каждый нейрон КВО макси-
мально вктивируется импульсацией от каждого
уха, прибывающей одномоментно на его аксо-
нальный бугорок, — компонент At кодируется
положением синапса, образованного волокна-
ми от каждой улитки в зависимости от части
нейрона, где он расположен. Кроме того, ко-
дирование происходит в зависимости от длины
аксонов от каждого кохлеарного ядра
Голова создает уменьшение Xt (т. е. временную раз-
ницу) в достижении звуком каждого уха и сниже-
ние интенсивности Al. поскольку голова поглощает
некоторое количество звуковой энергии. Мочка уха
определяет локализацию источника звука в верти-
кальной плоскости.
62
Преддверие-улитковый, или VIII, черепно-мозго-
вой нерв передает информацию от обеихулиток и ве-
стибулярного аппарата (дальнейшее обсуждение см.
главу 28). Каждое волокно улиткового нерва селек-
тивно настроено на определенную частоту, которая
определяется по источнику звука внутри улитки (см.
главу 26). Эти волокна затем организуются в за-
висимости от локализации иннервируемых ими во-
лосковых клеток по длиннику базальной мембраны,
и эта тонотопическая организация поддерживается
в слуховых проводниках.
На входе в ствол улитковый нерв образует синап-
сы с кохлеарными ядрами продолговатого мозга.
Слуховые пути
Кохлеарные ядра подразделяются на вентраль-
ное (ВКЯ) и дорсальное (ДКЯ). ВКЯ проецируется
на комплекс верхних олив (КВО) с двух сторон, а
ДКЯ — через задние слуховые волокна на противо-
положное ядро латеральной петли и нижних бугор-
ков четверохолмия.
КВО содержит веретенообразные нейроны с ла-
теральными и медиальными дендритами, которые
получают импульсацию от каждого уха. Это первый
уровень бинауральных взаимодействий, и эта струк-
тура играет важную роль в локализации источника
звука. Медиальная часть КВО принимает возбужда-
ющую импульсацию (ВВ-клетки), тогда как лате-
ральная часть КВО принимает тормозную импуль-
сацию от одного уха и возбуждающую — от другого
(ВТ-клетки).
ВВ-клетки принимают участие в локализации
источника звуков низкой частоты (менее 1,4 кГц),
при этом критерием является снижение частоты
звука (At), поступающего в одно, а затем в другое
ухо. На рисунке показано, как эти клетки проводят
импулвсы, генерируемые звуковыми сигналами раз-
личной локализации, к одному нейрону КВО от обо-
их ушей.
ВТ-клетки играют важную ролв в локализации
источника звуков высокой частоты, причем крите-
рием в данном случае является изменение интенсив-
ности (Д1) между ушами (при создании AI голова
выполняет функцию щита). Анализ звуков с часто-
той выше 1,4 кГц (у людей) проводится по I. Если
источник звука находится на средней линии, то ни
At , ни Д1 не образуется и происходит как бы слия-
ние источника локализации, которое до некоторой
степени можно преодолеть, поменяв положение го-
ловы или подключив другие сенсорные стимулы.
Локализация источника звука в вертикальной
плоскости зависит от мочки уха.
КВО не только проецируется рострально на ниж-
ние бугорки четверохолмия (НБЧ), но и посылает
импульсацию к улитке, где контролирует ОВВК и
изменяет воспринимающие свойства кортиева орга-
на (см. главу 26). Проекция к НБЧ является тоното-
пической, и эта структура также получает импуль-
сацию от первичной слуховой коры (ПСК) и других
сенсорных модальностей. Она взаимодействует с
верхними бугорками четверохолмия и участвует в
пространственной ориентации в соответствии с по-
лучаемыми слуховыми стимулами (см. главы 24 и 40).
НБЧ проецируются на медиальное коленчатое
тело (МКТ), которое проецируется на ПСК в верх-
ней височной извилине. Эти участки коры соответ-
ствуют полям 41 и 42 по Бродману, при этом тала-
мическая импульсапия поступает преимуществен-
но в III и IV слои коры. Столбиковая организация
ПСК плохо изучена, но в соответствии с тонотопи-
ческой проекцией низкочастотные звуки локализу-
ются в задних отделах, а высокочастотные — в пе-
редних отделах.
Вторичная слуховая кора и речь
В ПСК, кроме нейронов, получающих относи-
тельно простые афферентные импульсы, находятся
клетки, которые реагируют на некоторые сложные
звуки. Эти нейроны в большей степени представле-
ны во вторичной слуховой коре и наиболее слож-
но устроены у человека в зоне Вернике —корковом
центре узнавания речи. Эта область находится в до-
минантном полушарии (обычно левом), и при ее по-
ражении (например, при инсульте; см. главу 17) воз-
никает сенсорная афазия (логорея), или невозмож-
ность воспринимать обращенную речь. Эта область
связана аркуатными волокнами с зоной Брока в лоб-
ной доле доминантного полушария, которая отве-
чает за экспрессивную речь; ее поражение приводит
кразвитию моторной эфферентной афазии, т. е. к от-
сутствию спонтанной речи при сохранности арти-
куляторного аппарата. Тотальная афазия — это пол-
ное отсутствие речевой продукции, которое наблю-
дается при инсульте в бассейне средней мозговой
артерии с поражением зон Вернике и Брока. Се-
лективное поражение аркуатных волокон приводит
к развитию акустико-мнестической афазии, когда
больные могут понимать обращенную речь и гово-
рить, но не могут повторять слова и предложения.
Однако в настоящее время получены данные о бо-
лее сложных взаимодействиях между отделами моз-
га в процессе узнавания и речевой продукции.
Афазию следует отличать от нарушений инициа-
ции и выполнения речедвигательного акта (например,
паралич мягкого нёба и языка при болезни двига-
тельного нейрона), которое называется дизартрией
(или анартрией, когда речевая продукция полнос-
тью отсутствует).
ГЛАВА 28. Вестибулярный анализатор
клетки и простирает) •
до крыши ампулы, где находится
с гудение гый материал, называв .ым
купулой
Отолито! ые волосковые клетки |
Лабиринт, полукружные каналы и отолитов-я i органы
Вестибулярная часто
Vltl пары ЧМН
Вестибулярная система отвечает за поддержание
равновесия, реализацию постуральных рефлексов и
регуляцию движений глаз. Она состоит из перифе-
рического воспринимающего компонента, который
проецируется в ствол мозга (включая глазодвигатель-
ные ядра), а оттуда в таламус и на сенсорную кору, а
также на мозжечок и в спинной мозг. Нарушение
функции этой системы приводит к развитию голо-
64
вокружения, тошноты и рвоты, иногда в сочетании
с глазодвигательными нарушениями (обычно нис-
тагмом; см. главу 40) и неустойчивостью при ходь-
бе. У больного в коме исследование вестибулярного
анализатора дает важную информацию о степени
целостности ствола мозга.
Вестибулярное восприятие
Периферический воспринимающий компонент
вестибулярного анализатора состоит из лабиринта,
который в свою очередь включает два отолитовых
органа (утрикулюс и саккулюс), а также ампулы полу-
кружных каналов. Отолитовые органы воспринима-
ют информацию о положении головы и линейном
ускорении, тогда как полукружные каналы — о вра-
щательном (угловом) ускорении.
Волосковые клетки находятся в обоих отолито-
вых органах и сходны по структуре с волосковыми
клетками улитки (см. главы 19 и 26). Как и в улитке,
смещение стереоцилий в сторону киноцилии депо-
ляризует клетку и позволяет нейротрансмиттеру выс-
вободиться из волосковой клетки, что приводит к ак-
тивации ассоциированного афферентного волокна.
Когда стереопилий смешаются в противоположном
направлении, этого не происходит. Движения рес-
ничек связаны с вращением головы (ампулярные
рецепторы в полукружных каналах) и ускорением
при ее наклоне (отолитовые органы утрикулюса), так
как движение головы вызывает перемещение эндо-
лимфы, которая в свою очередь омывает волосковые
клетки и вызывает их движение, изменяя конфигу-
рацию стереопилий.
Спонтанная активность афферентных волокон
высокая и отражает спонтанное вытекание нейро-
трансмиттера из волосковой клетки в синапс. Ги-
перполяризация волосковой клетки приводит к
уменьшению афферентного разряда, тогда как депо-
ляризация связана с увеличением импульсации. Эф-
ферентные волокна ствола мозга, заканчивающие-
ся на волосковых клетках, могут изменить чувстви-
тельность рецепторов воспринимающего органа.
Периферические расстройства
вестибулярного анализатора
Поражение периферической вестибулярной сис-
темы встречается часто. Примером такого поражения
является доброкачественное позиционное головок-
ружение, которое часто возникает после травмы или
инфекции вестибулярного аппарата, когда отолито-
вые кристаллы скапливаются в заднем полукружном
канале. Это заболевание характеризуется приступа-
ми головокружения, тошнотой, рвотой и атаксией
при поворотах головы в стороны, например при ук-
ладывании головы на подушку и поворотах в посте-
ли. Эти симптомы связаны с перемещением эндолим-
фы в заднем полукружном канале вслед за перемеще-
нием отолитовых рецепторов. Симптомы можно
устранить, если совершить несколько поворотов го-
ловы, при которых отолитовые рецепторы упадут в
полукружный канал и ампулу. Вирусные инфекции
вестибулярного аппарата встречаются часто (лаби-
ринтит) и могут значительно ухудшать качество жиз-
ни больных, приводя к развитию тяжелых приступов
тошноты и рвоты. Инфекции вестибулярного аппа-
рата обычно самопроизвольно прекращаются.
Двустороннее поражение вестибулярного аппа-
рата может привести к осциллопсии, которая зак-
лючается в неспособности фиксировать взгляд на
объектах при движениях головы (см. главу 40). В
противоположность этому раздражение вестибуляр-
ной системы, например, при укачивании приводит
к головокружению, тошноте, рвоте и тахикардии,
вызванными рассогласованностью вестибулярной и
зрительной информации.
Вестибулярную функцию можно проверить, вво-
дя воду в наружный слуховой проход (калорическая
проба). Когда в положении сидя в ухо вводят теплую
воду и пациент наклоняет голову назад на 60°, воз-
никает нистагм в сторону раздражаемого лабирин-
та. Введение холодной воды вызывает нистагм в про-
тивоположную сторону. Эти изменения отражают
перепады температуры эндолимфы, которые дают
эффект, сходный с вращением головы в сторону,
противоположную от раздражаемого лабиринта.
Центральная часть вестибулярного
анализатора и вестибулярные рефлексы
Тела нейронов, дающих афферентные вестибу-
лярные волокна в составе VIII пары ЧМН, лежат в
вестибулярном ганглии Скарна и заканчиваются в
одном из 4 вестибулярных ядер в продолговатом
мозге, которые также получают импульсацию от ре-
цепторов мышц шеи и зрительного анализатора.
Вестибулярные ядра проецируются в спинной
мозг (см. главы 12, 32 и 34), противоположные вести-
булярные ядра, ядра глазодвигательных нервов, а так-
же в таламусы. Некоторые из этих структур имеют
важное значение для реализации рефлекторных дви-
жений глаз, таких, как способность фиксировать
взгляд на предмете при движении головы - окулове -
стибулярный рефлекс (ОВР; см. главы 40 и 47). Дру-
гие проекции вестибулярных ядер играют важную
роль в поддержании позы и ходьбе. Вестибулярные
волокна заканчиваются в первичной соматосенсорной
коре (ПССК) и задней теменной коре (см. главу 30).
Калорические пробы позволяют уточнить сте-
пень целостности вестибулярного аппарата и его
стволовых связей, поэтому они могут быть исполь-
зованы у больных в коме для уточнения степени по-
ражения ствола. Менее тяжелые поражения вести-
булярного аппарата могут возникать при других забо-
леваниях, например рассеянном склерозе (см. главу
54) и инсультах (см. главу 17). Иногда в патологи-
ческие процессы вовлекаются и другие структуры.
ГЛАВА 29. Обоняние и вкус
псск
Г ипоталамус
Обонятельный тракт
Кора грушевидной
ПГК
Мост
Гломерула
кости
Обоняние
Кислый
Сладкий
Г орьк!
белок
‘лок
ФДЭ
цАМФ
Горький
ДАГ/ИТФ
Кислый
Соленый!
Сладкий
Плотные соединения
Эпителиальные клетки
Базальная клетка
Митральная
клетка (МК)
Лимбическая система
(энториналычая кора,;
миндалевидное тело)
Различные вкусовые
ощущения проводятся
различными вкусовы-
ми рецепторами
Вкусовой
рецептор
Высвобождение
ней ротрансииттера
Кора ост-
ровковой
извилины
Медиодорс альное
ядро таламуса
Обонятельный
бугорок
Задневентральное
ядро таламуса
Обонятельный
.тракт
Г ипоталамус,
миндалевидное
тело
Префронта л ьнан
кора
Продыряв-
ленная
пластинка
решетчатой
Ядро
ОДИНОЧНОГО пути
Пучковая
клетка
Передняя
-спайка
Аксон обонятельного
рецептора
VII ЧМН
(барабанная
струна)
Латеральный
обонятельный
тракт
Поддерживающая
клетка
Высвобождение
кальция внутри
\ клетки
Пищевой продукт мо
жет растворяться в
слюне, если он гидро-
филен, или в слизи,
если он липофилен
G-белок густидуцин.
ДАГ — диацил глицерол
| Вкусовое восприятие
ПГК — перигломерулярные клетки,
ГК — гранулярные клетки
Слизь Наружная вкусовая пора
цАМФ
Фосфоди эсте
г раза (ФДЭ) С
Протеин-
киназа А
Контралатеральный
обонятельный тракт
_ Обонятельный тракт \
Обонятельные ___
луковицысвяза- "X г'"'
ны через перед-
ние обонятель' •
ные ядра _____|
Обонятельная луковица /
Продырявленная пластинка
решетчатой кости / / ..•’и’елии
Блуждающий 1
нерв из глотки |
_{Х пара ЧМН)
[Языкоглоточный
нерв (IX пара ЧМН)
, '-Переднее обоня- /
тельное ядро /
'Обонятельное /
восприятие /
(см главу 19) /
/ Солены!
/"Афферентные вкусовые\
Хнервные волокна х
О Протонные каналы блокируют приток калия
и вызывают деполяризацию
О Блокада базолатеральных калиевых каналов по-
средством фосфорилирования, и таким образом
вызывается деполяризация
Открытие базолатеральных кальциевых каналов,
что обеспечивает приток кальция в клетки
Синапс Афферентные
нервы
Обонятельный, или I черепно-мозговой нерв
(ЧМН) содержит наибольшее количество волокон
по сравнению с другими чувствителвными нер-
вами, проецирующимися в ЦНС. Вкусовые ощу-
щения проводятся в ЦНС волокнами VII, IX и X
пар ЧМН.
66
Обоняние
Обонятельный анализатор предназначен для
различения большого количества запахов, что воз-
можно благодаря наличию тысяч обонятельных
рецепторов. Эти рецепторы представляют собой
апикальные дендриты обонятельных рецепторных
клеток, которые проецируются в ЦНС через про-
дырявленную пластинку решетчатой кости в верх-
них отделах носовой полости на обонятельную лу-
ковицу.
Обонятельный стимул или запах связывается с
обонятельными рецепторами и деполяризует их (см.
главу 19), что при достаточном уровне стимуляпии
приводит к генерации потенциала действия в теле
нейрона и проведению этого импульса по аксонам
обонятельного нерва в обонятельную луковицу.
Обонятельный нерв проходит через крышу но-
совой полости через продырявленную пластинку
решетчатой кости. Поражение этой структуры (на-
пример, при черепно-мозговой травме) может при-
вести к повреждению аксонов обонятельного не-
рва и утрате обоняния — аносмии. Однако чаще все-
го нарушения обоняния возникают при инфекции
или воспалении слизистой оболочки полости носа.
Аксоны обонятельных рецепторов переключаются
на нейроны обонятельной луковицы, которая за-
легает в базальных отделах лобной доли. Пораже-
ние этой структуры возникает при базальных ме-
нингиомах и приводит к развитию односторонней
аносмии.
Обонятельная луковица содержит комплекс кле-
ток. Аксоны обонятельного нерва образуют синапсы
с апикальными дендритами митральных и в мень-
шей степени пучковых клеток, аксоны которых про-
ецируются в виде обонятельного тракта. Обонятель-
ная луковица содержит ряд тормозных интернейро-
нов (гранулярных и перигломерулярных клеток),
которые модифицируют обонятельные стимулы,
проходящие через луковицу.
Обонятельный тракт проецируется на височную
долю и образует синапсы с корой грушевидной из-
вилины и лимбической системой, которая проеци-
руется на гипоталамус. Эта проекция важна для ре-
ализации поведенческого компонента обоняния,
который наиболее развит у животных. У человека
поражение этих структур редко приводит к раз-
витию чистой аносмии, а активация этих отделов
ЦНС вызывает височную эпилепсию (см. главу 53),
при которой возникают нарушения восприятия за-
пахов (например, обонятельные галлюцинации).
Обонятельная система дает проекции на таламус
и через обонятельный бугорок на медиодорсальное
ядро, которое проецируется на префронтальную
кору. Роль этого пути неясна.
Вкус
Вкусовые рецепторы расположены на языке. Они
собраны там в виде грибовидных сосочков и ство-
ловых клеток; последние делятся и замещают вку-
совые рецепторы при их повреждении. Апикальная
поверхность вкусового рецептора содержит микро-
ворсинки, покрытые слизью, которая вырабатыва-
ется соседними бокаловидными клетками. Каждый
продукт достигает вкусового рецептора; гидрофиль-
ные вещества растворяются в слюне, а липофильные
— в слизи. Вкус имеет несколько модальностей: со-
леный, кислый, сладкий и горький. Эти виды вку-
совой чувствительности соответствуют определен-
ным типам восприятия этих ощущений. Недавно
был открыт новый вкус (умами).
Соленые импульсы вызывают непосредственную
деполяризацию вкусовых рецепторов при прохожде-
нии натрия по амилоридчувствительному каналу апи-
кальной мембраны. Деполяризация приводит к выс-
вобождению нейротрансмиттера из базальной части
клетки, который активирует афферентные волокна
соответствующего ЧМН. В отличие от соленых, кис-
лые вкусовые стимулы блокируют вольтажзависимые
протонные каналы. Сладкие стимулы связываются с
рецептором, который активирует G-белок густиду-
цин, который активирует аденилатциклазу и выраба-
тывается цАМФ. Повышение концентрации цАМФ
приводит к активации протеинкиназы, которая фос-
форилируется и закрывает базолатеральные калиевые
каналы и таким образом деполяризует рецептор. Горь-
кие стимулы переключаются на рецептор посред-
ством активации G-белка. При активапии густидупи-
на активируется цАМФ-фосфодиэстераза, которая
снижает уровень цАМФ (и фосфорилирования про-
теинкиназы), что приводит к открытию базолатераль-
ных кальциевых каналов и высвобождению нейро-
трансмиттера. Другой способ передачи сладких и
горьких стимулов заключается в активации фосфо-
липазы С и продукции инозитолтрифосфата (ИТФ),
а также диапилглиперола (ДАТ), которые способству-
ют высвобождению кальция из внутренних запасов
рецептора. Увеличение концентрации кальция при-
водит к высвобождению нейротрансмиттера.
Рецепторы передают информацию по барабанной
струне (передние 2/3 языка) и языкоглоточному не-
рву (задняя треть языка) к ядру одиночного пути в
продолговатом мозге (см. главы 13 и 14). Эта струк-
тура проецируется рострально через таламус в ПС СК
и кору островковой извилины с последующей про-
екцией на гипоталамус и миндалевидное тело. У
больных височной эпилепсией наблюдается аура в
виде обонятельных галлюцинаций, в возникнове-
нии которых принимает участие проекция на гипо-
таламус и миндалевидное тело (см. главу 53).
ГЛАВА 30. Ассоциативная кора:
задняя теменная и префронтальная кора
Цен-ралоная
Премоторн (я кора
кора
Префронтальная кор_*
Первичная слуховая
кора (см. главу 27)
Галамус (подушка,
заднелатеральное
ядро)
Подкорковые афф
рентные волокна
(базальное ядро Мей-
нерта) ___
Афферентные волокна:
Корковые отделы
•Пар >ичная соматосен-
сорная кера (в г“-новчом
поле 5)
•Зрительная кора (пре-
имущественно поле 7)
•Слуховая н височная
кора (поля 39 и 40)
Афферентные волокна
Корковые отделы: от всех
отделов коры, ио особен
но от теменно-затьио мых
Задняя тсненнь- кора
Центральная Первичная Эфферентные
борозда сона’осен :°Рная волоки;
кора г и j—"ж
Первичная
двигатель-
ная кора / J
Коркой не отделы
•Префронтальная кора
включая КЦВ, височная
кора
Кора поясней изпилнны
Галтмус
•Подушка
Сильвиева
6ор< эда
Подкорковые афф<
рентные волокна
•Базальные -англии
•Лимбичл. кая сие гема
•Мост
Подкорковые аффе-
рентные .олокнь: от
банальных ганглиев че-
рез таламус
Первичная
двигательная кора
Таламус: передне-
вентральное ядро,
медиодорсальное
ядро
Сильвиева борозда
Централь-
ная борозда
Эфферентные волокна
Коркоиыз отделе
•Двигательные
(особенно поле 6)
Таламус
•Ретикулярное ядро
(см. главы 15 и 44)
Подкорковые афф
рендны< волокна
•Базальные ганглии
•Лимбическая система
* Цифр» означают номер полей по Бродману
Ассоциативная кора представляет те ее отделы,
которые не являются первичными двигательны-
ми или чувствительными представительствами, а
участвуют в более специализированном анализе
обработки воспринимаемой и двигательной ин-
формации.
Эти ассоциативные отделы коры включают зад-
нюю теменную кору (ЗТК), которую у обезьян пред-
ставляют поля 5 и 7 по Бродману, а у человека — поля
39 и 40, префронтальную кору, соответствующую по-
лям 9-12 и 44-47 по Бродману, и височную кору, со-
ответствующую полям 21, 22, 37 и 41-43. Височная
кора принимает участие в звуковосприятии и рече-
вой продукции, сложной обработке зрительных об-
разов (например, узнавание лиц) и процессах памя-
ти (см. главы 25, 27 и 45).
Задняя теменная кора
Этот отдел подвергся значительным изменени-
ям в процессе эволюции и управляет особыми фор-
мами поведения человека, таким, как владение ин-
струментами, планирование двигательных актов
и речи. Он подразделяется на два подотдела: один
обрабатывает соматосенсорную информацию (поле
5), а другой — зрительные стимулы (поле 7).
С нейрофизиологической точки зрения поле 5
содержит клетки, на которые обычно проходят
импульсы нескольких чувствительных модально-
стей, таких как, например, проприоцептивные и
кожные стимулы. Эти нейроны участвуют в ре-
гуляции позы и движений. Другие нейроны, по-
лучающие множественные импульсы от кожи, воз-
можно, участвуют в узнавании предметов. Однако,
68
помимо сложной чувствительной импульсации,
эти нейроны обычно макисмально активируются
сенсорными стимулами, участвующими в поведен-
ческих реакциях. Поражение этой области приводит
к контралатеральной утрате сложной чувствитель-
ности, например способности узнавать предметы на
ощупь (астереогноз). Кроме того, больные нечув-
ствительны к стимулам, наносимым на противопо-
ложную сторону тела. Иногда это расстройство про-
текает так тяжело, что больные игнорируют суще-
ствование противоположной стороны тела, что
мешает выполнению действий здоровой стороной
(интермануальный конфликт или синдром чужой
руки). Чаще больные не воспринимают сенсорные
стимулы на противоположной стороне, когда сти-
мулы воздействуют на обе стороны тела.
В противоположность этому поле 7 участвует в
сложной обработке зрительной информации, и мно-
гие клетки данной области реагируют на стимулы,
играющие роль в выработке поведенческих реакций
(например, пищевой мотивации). Одни нейроны
этой области отвечают за фиксацию зрительного
образа и слежение, другие участвуют в процессе пе-
реключения внимания с одного объекта на другой
(светочувствительные и зрительно-пространствен-
ные нейроны). В поле 7 имеются нейроны, воспри-
нимающие как чувствительные, так и сенсорные
стимулы. Некоторые из этих нейронов максималь-
но активируются, когда стимул перемещается с ре-
цептивного кожного поля нейрона из внешнего про-
странства, другие максимально активируются во вре-
мя фиксации взора на интересующем предмете,
когда человек протягивает к предмету руку. Пора-
жение этой области у человека приводит к игнори-
рованию зрительных стимулов в противоположном
поле зрения, а также нарушению движения глаз и
визуального контроля движений. Однако со сторо-
ны сложной зрительной обработки может возник-
нуть другое нарушение, заключающееся в трудно-
сти нахождения маршрута и конструировании слож-
ных фигур.
Кроме того, в задней теменной коре человека в
меньшей степени, чем у приматов и других живот-
ных, имеются клетки, которые максимально акти-
вируются в ответ на вестибулярные и слуховые сти-
мулы (см. главы 27 и 28). Поражение этого отдела у
людей приводит к нарушению зрения и зрительно-
го контроля за движениями, равновесия, речи и сче-
та. Нарушается способность писать (аграфия), чи-
тать (алексия) и считать (акалькулия).
Префронтальная кора
Префронтальный отдел коры увеличился в про-
цессе филогенеза и наиболее представлен у челове-
ка. Он участвует в целенаправленных поведенческих
реакциях и планировании двигательных реакций на
внешние стимулы (см. главу 32). В этом отделе на-
ходятся специализированные зоны, такие, как кор-
ковый центр взора (КПВ; см. главу 40) и зона Брока
(см. главу 27). Префронтальная кора является слож-
ной функциональной системой.
С нейрофизиологичсекой точки зрения в этой об-
ласти находится много различных групп нейронов,
которые отвечают на сенсорные стимулы, участвую-
щие в поведении и преобразующиеся в движения.
Поражение префронтальной коры у животных
приводит к снижению концентрации внимания (на-
рушению способности удерживать информацию бо-
лее чем на несколько секунд), а также к изменению
двигательной активности и эмоциональной реактив-
ности. Для пациента с поражением лобной доли кпе-
реди от передней центральной извилины характерно
снижение критики: он расторможен, его поведение
напоминает поведение ребенка. У пациента снижено
внимание, его легко отвлечь, он не удерживает ин-
формацию, не запоминает новые сведения, персеве-
рирует (повторяет фразы, слова и действия) и ведет
себя по-старому даже при изменении окружающей
обстановки. Пациент не может формулировать цели
и планы, обобщать и выделять частности. У него сни-
жается объем речевой продукции, и спонтанной дви-
гательной активности, изменяется пищевое поведе-
ние — пациент, предпочитавший кислое, начинает
есть сладкое. Пациент становится апатичным, у него
притупляются эмоциональные реакции, он стано-
вится агрессивным. В целом личность пациента ме-
няется, и к врачу, как правило, обращается не сам
пациент, а его близкие, так как у самого пациента
критика к состоянию своего здоровья снижена.
Симптоматика поражения лобной доли во мно-
гом связана с тем, что этот отдел у человека являет-
ся одним из наиболее развитых. Однако обширные
повреждения лобной доли могут захватывать двига-
тельные (см главу 40) и речевые (моторная эфферен-
тная афазия; см. главу 27) зоны, а также области,
контролирующие акт мочеиспускания.
ГЛАВА 31. Клинические проявления нарушений чувствительности
рКорешкояые синдромь Ь
Спинной мозг
псск
зтк
Таламус
Ствол
*С5
-С6
^Чувствительность я зоне корешка”сД
Сдавление межпозвоночным дис-
ком и/или костносуставными вы-
ростами (остеофитами) приводит
к нарушению чувствительности
и/или боли в области иннервации корешка, например наруше-
ния чувствительности при дегенеративных изменениях шейного
отдела позвоночника ______ _______________________
(Туи™, „..«I? синдром! Гипотрофия утолщения
большого пальца
11 Супраспииальные очаги
динный
^нерв
Спино-
таламИ'
ческий
Локтевой
нерв
| Спинальные синдромы
Обла гги поражения
ci.инисто мозга
Раздражини, Выпадини»
Парестезии Онемение
Боль Аналгезия
Нарушения
^вствитепь_____
Раздражение Выпадение
Ларь»_'езии Онемение
Воль Аналгезия
Г еми анестезий
на противоположной
стороне туловища
Игнорирование ка про-
тивоположной стороне
Сложные нарушений
в комплексе с болевым
синдромом
Сложные нарушения
в зависимости
от локализации
Гипотрофия утолщения мизинца, а также
межкостных и червеобразных мышц
| Полинеирсч атия |
Нарушения чувствитель-
ное ги по типу «перчаток
И НОС ив»
Периферический
чувствительный
Нарушения чувствительности представлены дву-
мя группами симптомов: выпадения — например, ут-
рата чувствительности (онемение, или аналгезия) и
раздражения например, парестезия (чувство пол-
зания мурашек) и боль. Они возникают при пораже-
нии на разных уровнях чувствительных путей.
Для установления генеза и причины нарушения
чувствительности необходимо изучить анамнез и про-
вести обследование больного. Большинству больных
с изолированными нарушениями чувствительности
трудно поставить диагноз сразу, но, как правило, в кли-
нической практике в этих случаях сталкиваются либо
с полинейропатиями, либо с рассеянным склерозом.
При обследовании больных с нарушениями чув-
ствительности необходимо сделать анализ крови,
провести электронейрографию (ЭНГ) и магнитно-
резонансную томографию (МРТ) головного и спин-
ного мозга. Во всех случаях важно помнить, что на-
рушения чувствительности могут возникать и не
из-за неврологических причин, например при ги-
первентиляции, когда больные испытывают онеме-
ние в пальцах и периоральной области.
Уровень периферического нерва
Расстройства чувствительности, вызванные за-
болеванием периферических нервов, проявляются ло-
70
кальным туннельным синдромом и полинейропатией.
В последнем случае патологический процесс может ох-
ватывать волокна как крупного, так и мелкого калибра.
Наиболее часто встречающимися туннельными
синдромами являются:
• Компрессия срединного нерва на уровне лучеза-
пястного сустава (синдром запястного канала). Па-
циенты жалуются на боль в кисти, слабость мышц
утолщения большого пальца и нарушение чувстви-
тельности в большом, указательном, среднем и по-
ловине безымянного пальца. Часто синдром про-
ходит спонтанно, если этого не происходит, при-
бегают к хирургическому лечению.
• Компрессия локтевого нерва в области локтевого
сгиба. У больных отмечаются атрофия мышц кис-
ти, слабость мышц утолщения мизинца и утрата чув-
ствительности на мизинце и второй половине безы-
мянного пальца. В ряде случаев этот синдром мож-
но устранить хирургическим перемещением нерва.
• Малоберцовый нерв может ущемляться в фибуляр-
ном канале ниже уровня подколенной ямки. У боль-
ных отмечаются слабость разгибателей пальцев сто-
пы и онемение по наружной поверхности стопы.
Причины полинейропатий могут быть различны-
ми. Если в процесс вовлекаются крупнокалиберные
волокна, у больных нарушается мышечно-суставное,
вибрационное чувство и поверхностная чувствитель-
ность, отмечается выпадение рефлексов. Полинейро-
патии редко бывают изолированными чувствитель-
ными, часто при них отмечаются слабость и похудание
мышц. Обычно при полиневропатиях наблюдаются
нарушения чувствительности по типу «перчаток и
носков», выражающиеся в утрате чувствительности
на кистях, предплечьях, стопах и голенях.
В ряде случаев пациенты жалуются на боли, но при
обследовании выявляется утрата болевой и температур-
ной чувствительности. У таких больных пациентов име-
ет место полинейропатия мелкокалиберных волокон.
Задние корешки и спинномозговые ганглии в
процесс первично вовлекаются редко, при этом от-
мечается утрата глубокой чувствительности.
Болевые синдромы обсуждаются в главах 23 и 24.
Важно помнить, что боли часто возникают и не из-
за неврологических причин, например при артрите
или поражении мягких тканей.
Нервы после выхода из позвоночника могут ущем-
ляться остеофитами или межпозвоночными диска-
ми, что приводит к выпадению чувствительности в
зоне иннервации ущемленного корешка. Пациенты
обычно жалуются на боль, иррадиирующую по ходу
корешка, и утрату чувствительности в соответству-
ющем дерматоме (см. главу 1). Чаще всего это про-
исходит в шейном или поясничном отделе позво-
ночника, что требует хирургического лечения. Кли-
нически у больных отмечаются атрофия, слабость
мышц и выпадение соответствующих рефлексов.
Уровень спинного мозга
Пути поверхностной и глубокой чувствительно-
сти в спинном мозге проходят раздельно, и это оп-
ределяет симптоматику поражения.
Сирингомиелия
Сирингомиелия — это образование кисты или
полости в проекпии нейтрального канала спинного
мозга, обычно в шейном отделе в силу различных
причин. Очаг обычно прерывает спиноталамический
тракт (СТТ), так как его волокна проходят нейтраль-
нее центрального канала. Это проводит к диссоции-
рованной утрате чувствительности, т. е. к выпадению
температурной и болевой чувствительности на уров-
не поражения, но при этом не страдает чувство при-
косновения, вибрапионное и мышечно-суставное
чувство (см. главу 20). Кроме того, возможно вовле-
чение двигательных волокон, если киста распрост-
раняется на передние рога или пирамидные тракты.
Подострая сочетанная
дегенерация спинного мозга
Это заболевание связано с пернициозной анеми-
ей и дефицитом витамина В12 и сопровождается де-
миелинизацией и дегенерацией задних столбов (ЗС),
спиноцеребеллярных и кортикоспинальных трактов
(КСТ), а также поражением периферических нервов
(полинейропатия). У больных отмечаются парестезии
и утрата чувствительности (особенно чувства прикос-
новения, вибрационного и мышечно-суставного чув-
ства) со слабостью мышц и нарушением координа-
ции (см. главу 20).
Синдром Броун—Секара
Этот синдром развивается при поражении поло-
вины поперечника спинного мозга и приводит к ут-
рате глубокой чувствительности на своей стороне
по проводниковому типу (поражение ЗС), утрате
поверхностной чувствительности на противополож-
ной стороне по проводниковому типу с уровня на
несколько сегментов ниже (поражение СТТ), а так-
же к центральному парезу ниже уровня поражения
на своей стороне за счет поражения КСТ (см. гла-
вы 21 и 22).
Синдром передней спинальной артерии
Этот синдром развивается при окклюзии перед-
ней спинальной артерии, которая кровоснабжает
передние 2/3 спинного мозга. У пациента отмеча-
ются парезы и снижение поверхностной чувстви-
тельности ниже уровня поражения, при этом ЗС не
страдают и глубокая чувствительность остается со-
хранной.
Поперечный миелит
Поперечный миелит (не показан на рисунке)
приводит к полному поражению поперечника спин-
ного мозга на одном уровне, что сопровождается
полным выпадением чувствительных и двигатель-
ных функций ниже очага. Двигательные нарушения
возникают в результате страдания мотонейронов на
уровне пораженного сегмента и нисходящих пира-
мидных путей. Поперечный миелит может иметь
место при рассеянном склерозе или вторичном ост-
ром демиелинизирующем поражении на фоне ин-
фекции, например атипичной пневмонии.
Уровень головного мозга
Супраспинальные нарушения чувствительности
возникают в силу различных причин их характер за-
висит от локализации очага поражения. Обычно по-
лушарные очаги поражения приводят к выпадению
чувствительности на противоположной стороне ту-
ловища. Стволовые очаги вызывают выпадение чув-
ствительности в зависимости от уровня поражения:
очаги в мосту мозга приводят к выпадению чувстви-
тельности на лице на стороне очага и на туловище
на противоположной стороне.
Корковые очаги приводят к утрате чувствитель-
ности, если поражена первичная соматосенсорная
кора. При этом наблюдаются сложные расстройства,
такие, как астереогноз (неспособность узнавать
предметы на ощупь) или же сенсорное игнорирова-
ние. Последний вид нарушений возникает при по-
ражении задней теменной коры.
В ряде случаев при раздражении первичной сен-
сорной коры возникают фокальные эпилептические
припадки (см. главу 52), при которых больные ощу-
щают мигрирующие нарушения чувствительности
на противоположной стороне туловища.
При центральной локализации очага наблюдают-
ся и болевые синдромы, чаще всего при сосудистых
очагах в таламусе, когда на противоположной конеч-
ности возникает дизестезия, которая затем распро-
страняется диффузно (см. главу 22).
71
ЧАСТЬ III. Двигательный анализатор
ГЛАВА 32. Анатомия двигательного анализатора
। Префрон-
тальная
кора
Дополнитель-
ная моторная
зона (ДМ3) j
Премотор
ная кора
(ПРМК}
Базальные
ганглии
Уровень 1
Инициация движений,
включая планирование
и программирование
Уровень 2
Координация
движений
Уровень 3
Корковое исполнение
движений: нисходящие
двигательные проводя-
щие пути
Уровень 4
Спиннои мозг
и двигательный
аппарат
Уровень 5
Заключительное звено
выполнения движений
К двигательному анализатору относятся отделы
нервной системы, которые отвечают за регуляцию
движений. Движение регулируется сенсорными сти-
мулами (непроизвольные, или рефлекторные, движе-
ния) и вызвано сенсорным стимулом или же внут-
ренним побуждением (произвольные движения). На
практике в двигательном акте принимают участие
оба компонента движения, при этом непроизволь-
ные, или рефлекторные, движения производятся в
основном аксиальной и проксимальной мускулату-
рой, отвечающей за поддержание позы, локомоцию
и равновесие, а произвольные движения обычно вы-
полняются дистальной мускулатурой и связаны с ре-
ализацией тонких двигательных навыков.
Организован двигательный анализатор иерар-
хически.
Уровень 1
Наивысший уровень регуляции движений отве-
чает за инициацию, планирование и программирова-
ние двигательных актов в ответ на внутреннее по-
буждение. Это желание, вероятно, связано с лим-
бической системой (см. главу 45) и задней теменной
корой (см. главу 30), тогда как структурами, кото-
pBie непосредственно преобразуют побуждение в
движение, являются базальные ганглии (см. главы
38 и 39) и их корковвте проекции в лобнвтх долях
(см. главв! 35 и 36). Эти корковвте отделвт включа-
ют дополнительную моторную зону (ДМ3) и премо-
торную кору (ПРМК), последняя также осуществ-
ляет особый контролв за движениями проксималв-
ной мускулатурвт (см. ниже).
72
Поражение базальных ганглиев и их корковых
проекций приводит к двигательным нарушениям
(болезнь Паркинсона), а также непроизвольным на-
сильственным движениям (таких, как хорея, дисто-
ния и баллизм; см. главу 39), при этом слабость и из-
менение моносинаптических сухожильных рефлек-
сов не развиваются (см. главу 33).
Уровень 2
Другой уровень представлен мозжечком, который
отвечает за координацию движений. Мозжечок срав-
нивает двигательный акт, который человек намерен
выполнить, с действительным двигательным актом
на основании информации, получаемой отафферен-
тных волокон и интернейронов (ИН) спинного моз-
га. Мозжечок хранит двигательную информацию,
которая необходима не только для процесса обуче-
ния, но и для правильного распределения времени
активации мышц во время сложных движений.
Поражение этой структуры приводит к первич-
ному нарушению координации движений без разви-
тия слабости (см. главу 37).
Уровень 3
Следующий уровень представлен корковым кон-
тролем за мотонейронами (МН), а именно супрас-
пинальными нисходящими двигательными проводя-
щими путями. Их можно разделить на две группы.
• Кортикоспинальный тракт (КСТ), или пирамид-
ный, тракт, который начинается от моторной, пре-
моторной и соматосенсорной коры и образует си-
напсы непосредственно на двигательных ядрах че-
репно-мозговых нервов, МН в передних рогах
спинного мозга и в меньшей степени на ИН.
• Экстрапирамидные тракты, которые начинаются
от подкорковых структур и образуют более слож-
ные синаптические контакты на МН и ИН. Они
включают вестибулоспинальный (ВСТ), ретику-
лоспинальный (РСТ), тектоспинальный (ТСТ) и
руброспинальный (РуСТ) тракты. Все они получа-
ют импульсацию от первичной моторной коры.
При поражении ЦНС обычно страдает не какой-
то определенный тракт, а несколько путей. Наруше-
ние функции нисходящих двигательных проводящих
путей приводит к развитию слабости в конечностях,
преобладающей в разгибателях рук и сгибателях
ног — так называемое пирамидное распределение па-
резов. Помимо этой слабости, отмечается повыше-
ние мышечного тонуса и сухожильных рефлексов.
Эти симптомы характерны для синдрома поражения
центрального мотонейрона (см. главу 41). В отличие
от предыдущих уровней иерархии, это уровень 1, на-
рушения на котором приводят к развитию слабости
в мышцах.
Уровень 4
Более низким уровнем регуляции движений яв-
ляется спинной мозг. Нисходящие двигательные про-
водящие пути образуют синапсы не только на МН,
но и на ИН. Одни ИН осуществляют реализацию
некоторых спинальных рефлексов, другие проеци-
руются на МН независимо от нисходящих супрас-
пинальных или чувствительных импульсов и явля-
ются генераторами центрального паттерна. Они име-
ют важное значение для локомоции (см. главу 34).
Уровень 5
Наиболее низким уровнем является заключи-
тельное звено двигательного анализатора, представ-
ленное сообщением МН и мышцы. МН получаетим-
пульсацию не только из головного мозга посред-
ством нисходящих путей и ИН спинного мозга, но
и от чувствительных органов, расположенных на пе-
риферии, а именно мышечных веретен и сухожиль-
ного аппарата Гольджи, которые находятся в мыш-
цах и сухожилиях соответственно (см. главу 33). Мы-
шечное веретено отвечает за реализацию сухожильного
рефлекса на растяжение, который лежит в основе
проверки рефлексов при клиническом обследовании.
Поражение МН или его аксона приводит к раз-
витию синдрома поражения периферического МН,
характеризующегося слабостью и атрофией мышц,
гипотонией и снижением или отсутствием рефлек-
сов (см. главы 33 и 41).
Примечание
Важно отметить, что разделение функций ЦНС
на двигательные и чувствительные является упро-
щенным и практически все двигательные зоны по-
лучают чувствительную импульсапию. Трудно выде-
лить уровень, на котором обработанный чувстви-
тельный стимул принимает характер импульса
инициации движения. Необходимо также учиты-
вать, что разделение двигательного анализатора на
уровни является условным, а не точным представ-
лением о регуляции движений и патофизиологии
двигательных расстройств при поражении нервной
системы.
ГЛАВА 33. Мышечное веретено и периферический мотонейрон
Мозжечок
Задний корешок
«-МН
Мышечное
веретено
у-МН, иннервирующий
мышечное веретено
н 1а/11 афферентные
волокна от веретена
Супраслинальная
проекция к моторной
и соматосенсорной коре
Задний
слиноцере-
беллярный
тракт
Гомонимная
мышца
Первичное 1а
афферентное
волокно
от мышечного
веретена
Тормозной 1а интернейрон
Задние столбы
Сухожилие
Кость
Кость
Мышца-синергист
О
локна (мелкие миели-
Вторичные нервные
Мышечные
веретена
Интрафузаль
ные мышеч-
ные волокна
волокон,
скорости
мышцы
Примечание: биполярное распределение
tl афферентных волоки и у-МН-иннервация
на рисунке не показаны
Сумка 11 типа —
статическое волокно
Экстрафузаль-
ные мышечные
волокна
Сумка I типа —
динамическое волокно
Эфферентные волокна
(у-МН динамические, статические)
оконча-
ния иннервируют все волок-
на, кроме волокон сумок I ти-
па, чувствительных к общей
длине мышцы
О,
1а афферентные f
волокна (круп- II афферентные во
ные миелиинэи'
рованные бьют- низироваиные мед
ропроводящие) леннопроводящие)
Афферентные волокна
Первичные аннулоспиральные
окончания интрафузальных
мышечных
тельные к
ния длины
а*-Мотоне ирон
1Ь афферентные
Передний корешок волокна от СОГ
Сухожильный орган___
Гольджи (СОГ}
Кость
Мышца-антагоиист
Капсула
о
о
о
О
с
о
о
I О
о
о
О
о
о
%
22/
Ядериые
цепочки
волоког 4—5
Волокна ядерной сумки
у-МН вызывают сокращение
полярных отделов сокраща-
ющегося мышечного волокна,
что приводит к сокращению
центральной части и актива-
ции афферентным волокон,
которые адаптируются {быс-
тро для 1а и медленно для 11}
чувстви-
измене-
Нормальное распределение
типов волокон в веретене
Периферический мотонейрон
Периферические мотонейроны (ПМН) — это ней-
роны, тела которых лежат в передних рогах спинно-
го мозга и двигательных ядрах ствола и которые
иннервируют мышцы своими аксонами. Мышечные
волокна, иннервируемые одним аксоном МН, назы-
ваются двигательной единицей. Чем меньше волокон
имеет аксон МН, тем тоньше регуляпия движений
(например, в глазодвигателвных мышцах).
МН переднего рога подразделяются на два типа:
а-МН (диаметр 70 мкм), которые иннервируют
мышцы (экстрафузалвные волокна, генерирующие
74
мышечную силу), и у-МН (диаметр 30 мкм), кото-
рые иннервируют интрафузалвные мышечные во-
локна или мышечные веретена. Мышечное верете-
но - это инкапсулированный чувствителвный орган,
который находится в мышце и отвечает за опреде-
ление степени мышечного сокращения, контроли-
руя длину мышечных волокон.
Именно мышечное веретено и его связи со спин-
ным мозгом опосредуют сухожильные рефлексы; вне-
запное растяжение мышцы при ударе молоточком
по ее сухожилию активирует 1а афферентные нервные
окончания, которые посылают возбуждающий моно-
синаптический импульс к МН, что вызывает быст-
рое сокращение гомонимноймышцы (например, ко-
ленный рефлекс). Кроме того, 1а афферентный им-
пульс от мышечного веретена, активируя другие
мышцы-синергисты, т. е. мышцы с той же функци-
ей, что и гомонимная, ингибирует активность мышц-
антагонистов через 1а тормозной интернейрон (ИН)
в спинном мозге. Однако необходимо подчеркнуть,
что сухожильные рефлексы свидетельствуют не толь-
ко о целостности рефлекторной дуги, но и о возбу-
димости самих МН, которая повышается при пора-
жении центральных МН (см. главу 41).
Мышечное веретено
Мышечное веретено лежит параллельно экстрафу-
зальному мышечному волокну и состоит из следую-
щих компонентов:
• Ядерной сумки или цепочки волокон, которые
имеют различные морфологические свойства: сум-
ка I типа, или динамические волокна, которые вы-
сокочувствительны к скорости сокращения длины
мышцы, и сумка II типа, или статические волокна,
как и цепочки волокон, более чувствительные к аб-
солютной длине мышцы.
• у-МН, которые образуют синапсы с интрафузаль-
ными мышечными волокнами, также подразделя-
ются на два типа: динамические и статические,
последние иннервируют все, кроме волокон сумки
1 типа. Оба типа у-МН обычно активируются со-
вместно с а-МН так, что интрафузальные волокна
сокращаются вместе с экстрафузальными, и таким
образом мышеченое веретено продолжает быть чув-
ствительным в процессе мышечного сокращения.
Иногда у-МН могут активироваться независимо от
а-МН, обычно это происходит, когда животные
обучаются новому движению, что усиливает чув-
ствительность веретена к изменению длительнос-
ти мышечных волокон.
• Выделяют два типа афферентных нервных окон-
чаний: 1а афферентные волокна, ассоциированные
с аннулоспиральными нервными окончаниями,
подходят ко всем типам интрафузальных волокон
(первичные окончания), и медленнопроводяшие во-
локна II типа, которые асеоциированы с окончани -
ями более удаленных отделов интрафузальных во-
локон (за исключением сумок 1 типа волокон — вто-
ричных окончаний). Растяжение интрафузального
волокна активирует оба типа волокон. Однако во-
локна 1а более чувствительны к изменению длины
волокон, тогда как волокна II типа реагируют в
большей степени на общее изменение длины, не-
жели на скорость изменения длины волокон.
Мышечное веретено передает сигнал через зад-
ний корешок следующим структурам ЦНС: 1) МН,
иннервирующим гомонимную мышцу и мышцы-
синергисты (это лежит в основе рефлекса на растя-
жение); 2) интернейронам (ИН), тормозящим мыш-
цы-антагонисты; 3) в мозжечок через дорсальный
спиноцеребеллярный тракт; 4) в соматосенсорную
кору; 5) в первичную моторную кору через систему
задних столбов и медиальную петлю. Так, мышеч-
ное волокно не только отвечает за реализацию реф-
лекса на растяжение, но и участвует в координации
движений, мышечно-суставном восприятии (про-
приоцепция) и модулировании длиннолатентных
транскортикальных рефлексов (см. главу 36).
Поражение афферентных волокон мышечных ве-
ретен (например, при полинейропатиях крупнокали-
берных волокон) приводит к мышечной гипотонии
(поскольку рефлекс на растяжение участвует в регу-
ляции нормального мышечного тонуса), нарушению
координации, снижению мышечно-суставного чув-
ства и невозможности обучения новым двигатель-
ным навыкам в новых ситуациях. Кроме того, при
полинейропатиях крупнокалиберных волокон нару-
шются другие соматосенсорные афферентные про-
водящие пути (см. главы 20 и 31).
Сухожильный орган Гольджи находится в области
соединения мышцы и сухожилия и залегает вместе
с экстрафузальными мышечными волокнами. Он
оценивает степень сокращения мышцы с точки зре-
ния выработанной мышечной силы и проводит ин-
формацию в спинной мозг по 1b афферентным во-
локнам. Этот чувствительный орган, кроме того, пе-
редает информацию в ЦНС о степени напряжения
мышц, предотвращая избыточные мышечные сокра-
щения (см. главу 34).
Рекрутирование мотонейронов
и их поражение
Принцип рекрутирования — это порядок, в соот-
ветствии с которым активируются различные типы
мышечных волокон. Наиболее мелкие а-МН, кото-
рые наиболее легко возбуждаются, иннервируют
волокна I типа (не путать с интрафузальными волок-
нам сумки I типа мышечных веретен) или медлен-
нопроводящие волокна, которые отвечают за увели-
чение и поддержание напряжения в мышце.
Другие МН иннервируют мышечные волокна 2А
типа, или быстрые, устойчивые к утомлению волок-
на, которые отвечают за все виды локомоции. На-
конец, самые крупные МН, активируемые только
максимальными стимулами, иннервируют мышеч-
ные волокна 2В типа, или быстрые утомляемые мы-
шечные волокна, которые отвечают за бег и прыжки.
В соответствии с порядком рекрутирования МН
прибывающий импульс распределяется по принци-
пу размера, который позволяет мышцам сокращаться
в логической последовательности.
а-МН могут поражаться при ряде заболеваний, но
во всех случаях клинические проявления идентич-
ны: отмечаются атрофия денервированных мышц,
снижение или отсутствие рефлексов (синдром по-
ражения периферического МН) и фасцикуляции.
ГЛАВА 34. Спинной
мозг и локомоция
Длинные проприослинальные
нейроны, соединяющие группу
вентромедиальных МН
Вентромедиальные
нисходящие тракты
(вестибуло-, ре-
тикуло- и текто-
спинальные)
Схематическое изображение
генератора центрального паттерна
Модулирующие
I влияния на генератор
। центрального паттерна
Супраспмнальные
Нисходящие двигательные
пути
f Особые локомоторные от-
делы ствола мозга, вклю-
чая ППЯ
Спинальные
Лроприоспинальные МН,
контактирующие с други-
ми ГЦП
Периферические
Другие афферентные влия-
ния от СОТ, кожи и суставов
ППЯ — педункулопонтинное ядро
Задний корешок
Sb Передний
корешок
Вентромедиальная группа МН
Генераторы центрального паттерна (ГЦП)
Передний
корешок
МН к
I разгибателям
1а тормоз-
ные интер-
нейроны
Кортикоспиналь-
ный тракт
Латеральные нисходящие пути
Руброспииальный
тракт _
Латеральная группа МН
__Короткие пролриоспмнальные нейроны,
соединяющие латеральную группу МН
Дистальная мускулатура
Сгибатели _____________________________
Более вентрально расположенные
МН иннервируют разгибатели, тог-
да как МН» лежащие более дорсаль-
_ _ но — сгибатели
Разгибатели --------------------------------
з> Аксиальная/проксимальная
мускулатура
1а афферентное волокно
ЗСЦТ
* Задние столбы
I Группа МН
X к сгибателям
Передний
спиноцеребеллярный
тракт
.. Фаза
< Мышцы-- .
, скольжения
сгибатели
локомоции
.. Опорная
M&i и цы
V разгибатели Фаза
локомоции
Задний столб
>зсцт
1а афферентное волокно
ЗСЦТ — задний спиноцеребеллярный тракт
КР — клетка Рен шоу
Регуляция движений
на уровне спинного мозга
Помимо а- и у-МН, в спинном мозге имеет-
ся много интернейронов (ИН), которые передают
афферентную информацию от периферии и супрас-
пинальных структур. ИН образуют сеть, которая
активна в пределах спинного мозга, и ее импульсы
замыкаются на МН, регулируя активность гене-
раторов центрального паттерна (ГЦП). ГЦП , ак-
тивность которых лежит в основе локомоции, не
требуют никаких афферентных импульсов для ге-
нерации двигательного ответа определенного пат-
терна, тем не менее они модулируются централь-
76
ными и периферическими импульсами (см. главы
33 и 35).
Нисходящие двигательные проводящие пути (см.
Приложение 26).
Нисходящие двигательные проводящие пути клас-
сифицируют в зависимости от:
• принадлежности к пирамидной или экстрапира-
мидной системе (хотя клинически экстрапирамид-
ные расстройства относятся к заболеваниям базаль-
ных ганглиев; см. главу 39);
• их локализации в спинном мозге и мышц, которые
иннервируются МН, на которых заканчиваются их
синапсы. Так. пирамидный (кортикоспинальный) и
руброспинальный тракты образуют синапсы с лате-
ральной группой МН, которая иннервирует дис-
тальную мускулатуру, тогда как вестибуло-, ретику-
ло- и тектоспинальные тракты в большей степени
ассоциированы с вентромедиальной группой МН,
которая иннервирует аксиальную и проксимальную
мускулатуру.
Вентромедиальная группа МН связана с проприо-
спинальными нейронами, тогда как латеральная
группа МН с ними не связана. Так, латеральная дви-
гательная система в большей степени контролирует
дистальные движения, а вентромедиальная — равно-
весие и поддержание позы.
В переднем роге более вентрально расположен-
ные МН иннервируют разгибательные мышпы, тог-
да как более дорсально расположенные МН — сги-
батели.
Локомоция
Регуляция локомоции сложна, поскольку она
предполагает координированные движения всех ко-
нечностей млекопитающих.
Каждый цикл локомоции называется шагом и
включает фазы опоры и скольжения, последняя яв-
ляется частью цикла движения, при котором нога не
стоит на опоре. Каждый цикл предполагает точную
последовательность сокращения агонистов и анта-
гонистов. Наиболее простой путь достижения этого
- работа двух ГЦП (полуцентров), которые активи-
руют сгибатели и разгибатели соответственно и вза-
имно тормозят работу друг друга.
Такое взаимное торможение возможно благода-
ря тормозным 1а ИН и клеткам Реншоу. Клетки Рен-
шоу — это ИН, которые ингибируют активиро-
ванные МН (см. главу 8). Так, активация групп МН
посредством ГЦП приводит к реципрокному тормо-
жению и удалению тормозного стимула от антаго-
нистического ГЦП. Таким образом переключает-
ся активация с одной группы мышц на другую. Та-
кая полуцентровая модель локомоции может быть
смодулирована некоторыми нисходящими и восхо-
дящими стимулами. В последнем случае 1Ъ аффе-
рентное волокно сухожильного комплекса Гольджи
(СОГ) может переключать ГЦП, чтобы предотвра-
тить развитие избыточного напряжения мышцы,
тогда как ряд кожных стимулов модифицирует цикл,
если движение наталкивается на препятствие. Эти
афферентные волокна, называемые афферентными
волокнами сгибательного рефлекса отвечают за сги-
бание ноги и переступание через опасную преграду.
ГЦП спинного мозга взаимодействуют друг с
другом с помощью проприоспинальных нейронов.
В противоположность этому супраспинальная ин-
формация передается от ГЦП и между ними непря-
мым путем через 1а афферентные волокна, поступа-
ющие в задний спиноцеребеллярный тракт (ЗСЦТ)
и задние столбы, а также посредством активации ИН
спинного мозга и переднего спиноцеребеллярного
тракта (ПСЦТ).
Нарушения регуляции движений
и локомоции спинного мозга
У экспериментальных животных локомоция осу-
ществляется без супраспинальных влияний (фик-
тивная локомоция), у людей такого не происходит.
Однако нарушения ходьбы могут развиваться в силу
различных причин.
Нарушения функции ИН спинного мозга при-
водят к развитию синдрома ригидного человека
(встречается редко). При этом заболевании у боль-
ных отмечается диффузная ригидность аксиаль-
ной мускулатуры со спазмами, вызванными им-
пульсацией МН в результате отсутствия тормозных
влияний на вентромедиальные МН, или без них.
Синдром ригидного человека связан с наличием
аутоантител к ферменту, синтезирующему гамма-
аминомасляную кислоту (ГАМК), глутаматдекар-
боксилазу (ГДК).
Поражение нисходящих проводящих двигатель-
ных путей также наблюдается при некоторых забо-
леваниях. Наиболее тяжелое состояние отмечается
при распространенном поражении ствола мозга,
когда у больных утрачивается тормозное влияние на
разгибательную мускулатуру и формируется децереб-
рационная ригидность с дугообразным выгибанием
шеи и повышением мышечного тонуса в разгибате-
лях конечностей. В противоположность этому более
рострально расположенный очаг в одном из полу-
шарий приводит к развитию контралатерального ге-
мипареза или гемиплегии с повышением мышечно-
го тонуса и повышением сухожильных рефлексов
(гиперрефлексия) с непроизвольными ритмичными
сокращениями мышц, спонтанными или в ответ на
растяжение (клонус), что также наблюдается при
поражении нисходящих двигательных путей спин-
ного мозга (см. главы 31, 32 и 41).
При страдании пирамидного тракта разгибатели
поражаются в большей степени, чем сгибатели в
руке, а в ноге возникает обратная ситуация. Это на-
иявается пирамидным распределением парезов, что
не совсем корректно, поскольку поражение пира-
мидных путей у обезьян приводит лишь к наруше-
нию движений пальцев с некоторой степенью мы-
шечной гипотонии и гипорефлексии.
ГЛАВА 35. Двигательные зоны коры
Медиальная часть полушария мозга
Латеральная часть полушария мозга
Дополнительная моторная зона
Передние отделы
поясной извилины
Префронталь-
ная кора
Первичная моторная кора
Центральная борозда
Поясная борозда
Дополнительная моторная зона (ДМ3)
Первичная
моторная кора (ПМК)
Двигательные корковые отделы
Периферический/ соматосенсорный
(афферентный импульс
I--------Зрительные отделы коры —
V 41 Передние отделы Префрон------
Мозжечок поясной извилины тальная кора
1 Г------------
Передневентоальное-Дополнительная
и вентролатеральное | моторная зона (ДМ3)
ядра таламуса
Базальные ганглии
Премоторная кора
Корковый центр взора
Префронтальная
кора
Центральная борозда
Задняя теменная кора
Задневент-
ральное ядро
таламуса
Первичная-----
сенсорная
кора
Ствол:
• Мост- >Мозжечок
• Ядра ЧМН
• Экстрапирамндные
пути______________
Пирамидный или кортикоспинальный
тракт, лишь 30—40% которого исходит из ПМК
нуклера-
ные пути
В регуляпии движений участвует несколвко от-
делов коры: первичная моторная кора (см. главу 36),
премоторная кора (ПРМК), дополнителвная мотор-
ная зона (ДМ3), а также ряд соседних областей в
передних отделах поясной извилины. Кроме то-
го, существуют отделы мозга, участвующие в кор-
ковом контроле движений, например корковый
центр взора (см. главу 40) и задняя теменная ко-
ра (см. главу 30). В этой главе кратко обсуждают-
ся организация двигателвных отделов коры и их
ролв в регуляции движений, в следующей главе
речв пойдет преимущественно о первичной мотор-
ной коре.
Первичная моторная кора
Первичная моторная кора (ПМК) - это частвкоры
мозга, которая реагирует двигателвным ответом на
минималвную электрическую стимуляцию. Она со-
ответствует полю 4 по Бродману, располагается кпе-
реди от пентралвной борозды и проепируется на МН
ствола мозга посредством кортиконуклеарных путей
и на МН спинного мозга посредством кортикоспи-
нальных трактов (КСТ), а также непрямым спосо-
бом на подкорковые экстрапирамидные тракты.
Действительно, ПМК особенно тесно связана с пи-
рамидным трактом (хотя 60—70% его начинается от
других отделов коры) и поэтому принимает участие
в регуляции тонких движений дистальной мускула-
туры (главы 34 и 36).
Другие корковые отделы
В регуляции движений принимает участие не-
сколько отделов коры: ПРМК (латеральная часть
поля 6 по Бродману), ДМ3 (медиальная часть поля 6
по Бродману), некоторые корковые отделы передней
части поясной извилины, медиальная часть лобной
доли, корковый центр взора (поле 8 по Бродману) и
задняя теменная кора (поле 7 по Бродману).
78
Некоторые из этих отделов имеют особые функ-
ции. Например, корковый центр взора отвечает за
движения глаз (см. главу 40), а задняя теменная кора
осуществляет зрителвный контролв за движениями
(см. главу 30). Осталвные отделы лобной доли регу-
лируют более сложные движения. Болвшинство этих
корковых отделов занимают болвшую частв двига-
телвной иерархии, нежели ПМК, их связи и функ-
ции представлены в табл. 35.1 (см. главу 32).
ПРМК относится к полю 6 по Бродману и. как и
первичная моторная кора, посылает импулвсы не-
посредственно к МН по КСТ или пирамидному
тракту. Этот отдел занимает два уровня двигатель-
ной иерархии, посколвку участвует в планировании
двигателвных актов (см. главу 32). В противополож-
ность этому ДМ3 принимает участие в планирова-
нии движений, особенно в ответ на сенсорные сти-
мулы. Более того, в настоящее время стало извест-
но, что ДМ3 — лишь часть большого количества дви-
гательных корковых отделов, лежащих на медиаль-
ной стороне лобной доли и вовлеченных в большей
степени в процесс планирования, нежели выполне-
ния движений. Именно эти отделы коры получают
импульсацию от базальных ганглиев (см. главу 38),
что позволяет объяснить аномальные движения, на-
блюдающиеся при заболеваниях этого отдела мозга
человека (см. главу 39). Например, при болезни Пар-
кинсона отмечаются замедление и обеднение двига-
тельного рисунка, которые связаны с недостаточной
активностью этих корковых отделов и устраняются
с помощью противопаркинсонических препаратов
или нейрохирургического вмешательства.
Таблица 35.1. Связи и функции двигательных отделов коры
Корковьш отдел Афферентные импульсы Эфферентные импульсы Нейрофизиология Функция
ПМК ДМ3 ПРМК ПМК Мозжечок через тала- мус (ПВ-ВЛЯТ) Задние столбы - меди- альная петля (ЗВЯТ) КСТ Ствол: мост через мозжечок ядраЧ МН экстр апир амид ны е тракты Поражение ПМК приводит к на- рушению принятия поз и пере- ключения с одной позы на дру- гую, а также утрате сложных дви- гательных навыков Движения дистальной мускулатуры и выпол- нение тонких двига- тельных актов Рефлекторный конт- роль за движениями (транскортикальные рефлексы)
ПРМК ДМ3 Префронтальная кора Соматосенсорная и зрительная кора Мозжечок через тала- мус (ПВ-ВЛЯТ) Базальные ганглии че- рез таламус (ПВ-ВЛЯТ) ПМК КСТ Ствол: мост через мозжечок экстр апир амидные тракты Поражение ПРМК приводит к легкому парезу, нарушению сложных двигательных навыков и выполнения зрительно-двига- тельных заданий Исследования локального моз- гового кровотока показали, что она активируется во время вы- полнения заданий, требующих прямого управления движением сенсорной информацией Регуляция движений проксимальной мус- кулатуры Регуляция последова- тельности действий и подготовки к движе- ниям
ДМ3 Префронтальная кора Базальные ганглии че- рез таламус (ПВ-ВЛЯТ) Передняя часть пояс- ной извилины Контр алатеральная ДМ3 Контралатеральная ДМ3 ПРМК ПМК Поражение ДМ3 приводит к зна- чительному снижению спонтан- ной двигательной активности, нарушению бимануальной коор- динации и появлению хвататель- ных рефлексов Стимуляция ДМ 3 приводит к го- лосообразованию и двусторон- ним движениям конечностей Активность ДМ3 предшествует любым изменениям ПМК Группы нейронов ДМ3 макси- мально реагируют на сенсорные стимулы, будучи использованы как инструкция к движению Исследования локального моз- гового кровотока показали, что он увеличивается при планиро- вании и обдумывании двигатель- ных актов Инициация и плани- рование движений Бимануальная коор- динация
ГЛАВА 36. Первичная моторная кора
Двигательный
гомункул
Мизинец Безымям-
ДМ3
Двигательные отделы
в поясной извилине
Задняя
кора
Поясная
Афферентные волокна проходят
—— через задний корешок и образу
К ют синапсы с IV слоем колонки
двигательных нейронов, Движение, вызванное
которые иннервируют сокращением мышцы
МН и проецируются на .Кожные рецепторы
С мышцы той же лока- Суставные
Префрон-
тальная
Связи двигательной кор-'
вере ено
Импульсация от других двигательных зон
Центральный МН
Мышцы,
сгибающие
палец
пв-апят чг^г-р дмз
МН спинного мозга, иннервирующим
сгибатели пальца
5% крупнокалиберных воло-
кон КСТ исходят из пирамид-
ных клеток Беца и V слоя ПМК
1нрамидний, или кор-
тикоспинальный, тракт
Первичная моторная кора (ПМК) получает аф-
ферентную информацию от мозжечка (через тала-
мус), от отделов коры, расположенных кпереди от
нее, а именно дополнителвной моторной зоны
(ДМЗ) чувствителвные импулвсы от мышечных ве-
ретен и сенсорных областей коры. Последнее обсто-
ятелвство указывает на то, что разделение отделов
ЦНС на двигателвные и чувствителвные является
условным. Поэтому для удобства понимания необ-
ходимо выделятв ПМК и первичную соматосенсор-
ную кору (ПССК) (см. главу 20).
Изучение ПМКпоказало, что двигательная иннер-
вация тела человека организована соматотопически,
80
при этом корковое представителвство движений в
каждом отделе тела человека пропорционалвно сте-
пени двигателвной иннервации, например мускулу-
тура руки и лица имеет наиболвшее представителв-
ство. Обратная соматотопическая проекция метафо-
рически называется двигательным гомункулом, у
которого голова представлена в латералвных отде-
лах коры, а ноги — в медиалвных отделах.
В соответствии с этой соматотопической проек-
цией у больных возникают эпилептические припад-
ки при раздражении моторной коры, при кото-
ром они могут начаться на одной стороне, обычно в
руке, а затем распространиться в виде клонических
подергиваний на другие отделы (джексоновский
марш, названный по имени описавшего его невро-
логаХ. Джексона). В противоположности этому при-
падки, возникающие при раздражении ДМ3, на-
чинаются с поднимания рук и вокализапии, что
указывает на большую значимость этой области в ре-
гуляции движений (см. главу 35).
Исследования соматотопической проекции Пен-
филдом и соавт. в 50-х годах прошлого века позво-
лили установить макроскопическую организацию
ПМК, но впоследствии исследования с микроэлек-
тродами у животных показали, что ПМК организо-
вана в виде столбиков двигательных нейронов (см.
главу 15). Импульсация к этим столбикам поступает
по афферентным волокнам от суставов, мышечных
веретен и кожи, и они максимально активируются
при сокращении мышц, активируемых этим же от-
делом коры. Так, например, группа столбиков дви-
гательных нейронов получает сенсорные стимулы от
пальца, когда он согнут, эта импульсация проводит-
ся рецепторами кожи на передней поверхности паль-
ца — мышечными веретенами мышц, сгибающих
палец, а также суставными рецепторами суставов
пальцев. Тот же самый столбик двигательных ней-
ронов посылает импульсацию к МН спинного моз-
га, которые иннервируют сгибатели пальца. Акти-
вация кортикоспинального нейрона в этом столби-
ке активирует рецепторы, которые проецируются на
этот же столбик, и наоборот. Считается, что каждый
столбик получает прямую и обратную афферента-
цию, что имеет важное значение для рефлекторно-
го контроля за движениями, допустим, на примере
длиннолатентных, или транскортикальных, рефлек-
сов. Эти рефлексы приводят к отсроченным и мало-
выраженным электромиографическим (ЭМГ) изме-
нениям, которые возникают после растяжения мыш-
цы — и первым ЭМГ-изменением будет М-ответ при
вызывании моносинаптическорго рефлекса на рас-
тяжение (см. главу 33). Афферентным компонентом
транскортикального рефлекса является афферент-
ное волокно от мышечного веретена 1а (импульса-
ция передается по задним столбам и медиальной
петле), а эфферентным его компонентом — корти-
коспинальный тракт (КСТ). Роль этого рефлекса
неизвестна, но он имеет важное значение в корко-
вом контроле движений, сообенно если в процессе
двигательного акта человек наталкивается на пре-
пятствие, которое активирует мышечные веретена.
Имеются противоречивые данные о том, осуще-
ствляет ли ПМК непосредственный контроль за кон-
кретными мышцами, простыми движениями или же
некоторыми компонентами движений. Нейроны
ПМК реагируют еще до появления ЭМГ-изменений
и, вероятно, кодируют направление и силу движения,
несмотря на то что эта активность зависит от харак-
тера выполняемого действия. Поэтому в целом дви-
гательная кора контролирует движения, иннервируя
определенные МН (конкретные кортикоспинальные
аксоны иннервируют множество различных МН).
ПМК может ремоделироваться после очаговых
поражений или изменений сенсорных афферетных
стимулов, что говорит о ее способности поддержи-
вать гибкое сообщение с мышцами в течение жиз-
ни. Так, нейроны ПМК могут начать регулировать
другие мышцы. В опредленных отделах коры воз-
можно изменение синаптической активности с по-
мощью отсроченной потенциации (см. главу 45), это
свидетельствует о том, что ПМК может принимать
участие в обучении новым движениям, т. е. обладать
функцией, традиционно приписываемую мозжечку
(см. главу 37).
Изолированное поражение ПМК встречается
редко и в эксперименте, как правило, приводит к
нарушениям, сходным с таковыми при страдании
отдельных частей пирамидного тракта. Однако по-
ражение ПМК с обеих сторон в сочетании с пора-
жением премоторных зон, что наблюдается при ише-
мическом инсульте в бассейне средней мозговой ар-
терии (см. главу 17), приводит к более выраженному
неврологическому дефициту, в частности развитию
гемипареза. В противоположность этому, поражение
ДМ3 приводит кдвусторонним нарушениям иници-
ации движений, тогда как поражение коркового цен-
тра взора — к ограничению взора.
ГЛАВА 37. Мозжечок
Макроанатомия мозжечка
| Схематический уплощенный вид мозжечка сзади
нам исходит ю нимших олив
5. Гломерулы мозжечка содержат но
ховидные волокна. которые лают мм-
пульевцню ГК
6. Глубинное ядра мозжечка тоничес
км Активируются коллатеральными
можовмдными волокнами и ползущи-
ми волокнами, что позволяет осуще-
ствлять сравнение афферентной ин-
формации до и после обработки
коре мозжечка
7. Имлульсация от КП к глубинным
ядрам мозжечка носит тормозной ха-
рактер
в молекулярном слое: звездчатые м
корзиночные клетки, а также бо-
лее крупные КГ, которые находятся
в гранулярном слое
Все 3 типа интернейронов тормозят
суб максимально активированные КП
и прямым, и непрямым способом (че-
рез ГК), таким образом выделка наи-
более активным импульс
2. Параллельные волокна проходят
перпендикулярно дендритическому
дереву КП
3. КП имеют очень большие размеры
(диаметр тем достигает 50—ВО мкм)
тзк кп кг
Т|/|/ Поверхность полушария мозжечка
Передняя
ГК
Слой КП
Слой ГК I
Парал-
лельные
волокна
Гломе-
рула
Белое вещества
мозжечка '
Глубинные
ядра мозжечка
Молекулярный
слой
НГЯМ
Моховидное волокно
Схема циркуляции
импульсов
Импульсация
из мозжечка
--------------< Возбуждающий синапс
--------------• Тормозной синапс
НГЯМ — нейроны глубинных
КГ — клетка Гольджи
ГК — гранулярная клетка
Ползущее волокно
ТЗК — тормозная звездчатая клетка
ТКК — тормозная корзиночная клетка
КП — клетка Пуркинье
ядер мозжечка
нгям
пев
Моховидное волокно
5 кг
ТКК,
ТЗК,
КП
Параллельное волокно
Анатомия мозжечка
Мозжечок — это сложная структура, которая на-
ходится под наметом твердой мозговой оболочки в
задней черепной ямке и связана со стволом мозга
тремя парами ножек мозжечка (см. главу 13). Моз-
жечок отвечает за координацию движений и обу-
чение двигательным навыкам. В мозжечке имеются
3 функциональные и анатомические системы:
• спиноцеребеллярная участвует в регуляции функ-
ций аксиальной мускулатуры и поддержании позы;
• понтоцеребеллярная участвует в координации и
планировании движений конечностей;
• вестибулоцеребеллярная участвует в поддержании
позы и движении глаз.
Эти 3 системы имеют собственные связи (см. При-
ложение 3). Спиноцеребеллярная система подразде-
ляется на вермальный и паравермальный (промежу-
точный) отделы, при этом последний тесно связан с
аксиальной мускулатурой, а также с вентромедиаль-
ными нисходящими двигательными проводящими
путями и мотонейронами, тогда как паравермальная
часть в большей степени отвечает за координацию дви-
жений. Понтоцеребеллярная система участвует в ко-
ординации движений, но в большей степени связана
со зрительным контролем за движениями и передает
информацию от задней теменной коры к двигатель-
ным отделам коры. Вестибулоцеребеллярная система
не ассоциирована с глубокими ядрами мозжечка и яв-
ляется одной из филогенетически наиболее древних
частей мозжечка. Как и вермальная часть спиноцере-
беллярной системы, она связана с вентромедиальны-
ми двигательными путями, но при этом участвует в
регуляции движений глаз.
В общем, мозжечок сравнивает движение, кото-
рое человек намеревается сделать, информация о ко-
тором поступает из двигательных областей коры, с
действительным движением, информацию о котором
передают афферентные волокна мышечных веретен
и интернейроны спинного мозга. При этом мозже-
чок получает информацию от вестибулярного и зри-
тельного анализаторов. Когда сравнение проведено,
сигнал о погрешности поступает по нисходящим про-
водящим двигательным путям и корригирующий
фактор сохраняется в виде двигательной памяти,
82
опосредованной синаптической импульсацией к
клеткам Пуркинье (КП). Эти модифицируемые си-
напсы на уровне КП являются примером отсрочен-
ной депрессии (ОД; см. главы 45 и 47). ОД приводит
к уменьшению синаптической импульсации парал-
лельных волокон (ПАВ) к КП, когда импульсация
активирована по фазе и низкой частоте ползущим
волокном (ПОВ). Этот процесс происходит в тече-
ние как минимум нескольких часов. Другими слова-
ми, в период совершения новых движений импуль-
сация к КП по ползущим волокнам увеличивается,
что оказывает модулирующее влияние на импульса-
цию по параллельным волокнам к той же КП. Когда
движение усваивается, импульсация по ползущему
волокну уменьшается, но возрастает модулирующее
влияние по параллельному волокну; считается, что
именно этот механизм лежит в основе обучения дви-
гательным навыкам.
Гипотеза подобной модулируемой синаптической
импульсации была впервые предложена Марром в
1969 г. и впоследствии подтверждена, сообенно при
изучении окуловестибулярных рефлексов (см. гла-
вы 28 и 47). Биохимические механизмы ОД в моз-
жечке не полностью изучены, но, по-видимому, в их
основе лежит активация различных глутаматных ре-
цепторов КП и последующий приток кальция и акти-
вация протеинкиназ. Модулируемая синаптическая
импульсация отвечает за обучение и хранение ин-
формации, относящейся к двигательной памяти (см.
Приложение 3).
Микроскопическая структура мозжечка, обеспечи-
вающая генерацию ОД, хорошо изучена, несмотря на
то что биохимические механизмы изучены недостаточ-
но. Возбуждающие импульсы в мозжечок проводятся
моховидными и ползущими волокнами. Эти моховид-
ные волокна непрямым образом активируют КП через
ПАВ, которые исходят из гранулярных клеток (ГК).
В противоположность этому ПОВ образуют прямые си-
напсы с КП и, как в случае импульсации по моховид-
ным волокнам, посылают импульсы к нейронам глу-
бинных ядер мозжечка (НГЯМ). Эти нейроны тоничес-
ки возбуждаются афферентными волокнами, прово-
дящими импульсы в мозжечок, а затем ингибируются
корой мозжечка, т. е. КП. В свою очередь КП ингиби-
руются некоторыми локальными интернейронами, тог-
да как клетки Гольджи (КГ) в наружном гранулярном
слое посылают тормозные импульсы к ГК. Эти интер-
нейроны способны тормозить субмаксимально активи-
рованные КП и ГК и таким образом проводить имен-
но тот сигнал, который необходимо проанализировать.
Итоговая импульсация от глубинных ядер мозжеч-
ка на структуры ствола также является тормозной.
Клинические признаки поражения мозжечка
О функции мозжечка можно судить по клиничес-
ким признакам его поражения.
Нарушение функции мозжечка возникает при раз-
личных заболеваниях. Приводим его клинические
признаки.
• Гипотония, или снижение мышечного тонуса, разви-
вается в результате снижения импульсации от НГЯМ
по нисходящим проводящим двигательным путям
к мышечным веретенам (см. главу 33).
• Дискоординация / атаксия проявляется рядом симп-
томов: асинергией (невозможность координировать
сокращения мышц-агонистов и антагонистов), дис-
метрией (невозможность точно завершить начатое
движение, что приводит к интенционному тремору
и гиперметрии), адиадохокинезом (невозможность
быстро чередовать антагонистические движения).
Термин «атаксия» часто используют для описания
нарушений координации. При преимущественном
поражении мозжечка, например при его алкоголь-
ной дегенерации, отмечается «походка пьяного» с
широко расставленными ногами. Когда больше по-
ражены латеральные части полушарий, атаксия воз-
никает в конечностях.
• Дизартрия — невозможность точно произносить сло-
ва в результате дискоординации орофарингеальной
мускулатуры. Речь смазана и медленна (скандиро-
ванная речь).
• Нистагм - быстрые подергивания глазных яблок,
вызванные нарушением связи вестибулярных и гла-
зодвигательных ядер (см. главы 28 и 40).
• Тремор мягкого нёба и миоклонии — редкий симп-
том, возникающий при гипертрофии нижних олив,
с поражением оливоденторубрального треугольни-
ка (треугольника Молларе). У пациента отмечает-
ся низкочастотный тремор мягкого нёба — оно при
этом колеблется вверх и вниз.
Кроме того, недавно было показано, что мозжечок
принимает участие в реализации высших психических
функций. Небольшие когнитивные нарушения выяв-
ляются у пациентов с поражением мозжечка.
Функция мозжечка
Функция мозжечка четко определяется и соответ-
ствует локализации симптомов его поражения. Ме-
ханизм, посредством которого мозжечок осуществ-
ляет эту функцию, неясен, но для ее реализации не-
обходимо повторение однотипных элементарных
импульсов в коре мозжечка. Рассматривают 3 воз-
можных механизма функции мозжечка, которые не
являются взаимоисключающими.
• Функция сравнения. Мозжечок сравнивает нисхо-
дящие супраспинальные двигательные сигналы (по-
буждение к действию) с восходящей афферентной
импульсацией (действительным движением) и ус-
траняет различия, подавая импульсацию по нис-
ходящим двигательным проводящим путям. Это
позволяет мозжечку координировать движения так,
что они производятся плавно и точно.
• Функция регуляции во времени. Мозжечок, особенно
понтоцеребеллярная система, преобразует нисходя-
щие двигательные импульсы в такой последователь-
ности, что движение производится плавно и коор-
динирование, при этом позу и равновесие поддер-
живают вестибуло- и спиноцеребеллярная системы.
• Функция инициации и хранения двигательных ак-
тов. Модулируемая синаптическая импульсация на
уровне КП обеспечивает хранение двигательной
информации и ее обработку. Поэтому при опреде-
ленных обстоятельствах правильная последователь-
ность движений может быть оценена и получена че-
рез супраспинальные двигательные пути, что при-
ведет к инициации точного заученного движения.
83
ГЛАВА 38. Базальные ганглии: анатомия и физиология
Наружный
сегмент
бледного шара
(БШн)
Хвостатое
Скорлупа
Боковой
Таламус
III желудочек
ядро (СТЯ)
Черная субстанция
Внутренний сегмент
бледного шара (БШ|
Параллельные путиЬ
Хвостатое
ядре
Скорлупа
Миндалевид-
ное тело
БШв ,
ЧСр
БШв,,
ЧСр
Глутамат
Кора головного мозга
Префронталь- | Двигатель’
ная кора
ная кора
li1 С J Неостриатум С '
► (ПВ-ВЛЯТ)--
Префрон-
тальная кора,
►недвигатель-
ный круг
Премоторная
► (ПВ-ВЛЯТ)—->*ора/дмз .
Двигательный
круг
Ацх
Таламус
С — стриосомы неостриагума
М — матрикс неостриатума
Неостриатум также содержит
большое количество
холинергических интернейронов
ГАМК/энкефалин
ил пв-влят
ГАМК
Дофамин
Наружный сегмент
бледного шара I
- -------------
Внутренний
сегмент I
ГАМК/суб- г
: ГАМК
i
Прямой н непрямой пути
ЧСк
Ствол:
• Педункулопонтинное ядро
• Верхние бугорки четверо-
хммия
Кора
головного
мозга
Дофаминовые рецепторы Д.
Непрямой путь
— ► С-риатум —БШн —► СТЯ —►
|дофамнновые рецепторы Д,
Прямой путь
ЧСк
БШв,
ЧСр —► Таламус —► Кора головного мозга
Прямой путь содержит возбуждающие нейроны с дофаминовыми рецеп-
торами Д,, а непрямой путь — тормоэны) нейроны с дофамин звыми ре-
цепторами Д2, которые посылают импульсы к стриатуму
Базальные ганглии состоят из хвостатого ядра
и скорлупы (неостриатум — НС), внутреннего и на-
ружного сегментов бледного шара (БШв и БШн, со-
ответственно), ретикулярной и компактной частей
чернойсубстнации (ЧСр иЧСк, соответственно) и
субталамического ядра (СТЯ).
НС — основная часть базальных ганглиев, кото-
рая получает информацию от всей коры в соответ-
ствии с соматотопической проекцией, а также от
интраламинарных ядер таламуса. Информация по-
ступает из базальных ганглиев, в основном через
БШв и ЧСр, проходитчерез ядра таламуса (передне-
вентральное и вентролатеральное — ПВ-ВЛЯТ), ко-
торые в свою очередь проецируются на премотор-
ную кору (ПРМК), дополнительную моторную зону
(ДМЗ) и префронтальную кору. Кроме того, имеет-
ся также проекция на спинной мозг, особенно пе-
84
дункулопонтинное ядро (ППЯ), принимающее уча-
стие в локомоции (см. главу 34), а также к верхним
бугоркам четверохолмия, которые участвуют в регу-
ляции движений глаз (см. Главы 24 и 40).
Базальные ганглии образуют несколько нейро-
нальных кругов. Имеется стр ион игр остр парный круг,
последнее звено которого дофаминергическое. Этот
путь дегенерирует при болезни Паркинсона. Имеет-
ся также круг от БШн к СТЯ, который затем про-
ецируется на БШв и ЧСр. Этот путь по природе воз-
буждающий и контролирует уровень импульсации из
тормозных ядер базальных ганглиев таламического
направления. Несмотря на то что в структуре базаль-
ных ганглиев отмечается определенная степень ди-
вергенции и конвергенции связей, проекции их
образуют параллельные пути, которые в наиболее
упрощенном виде подразделяются на двигательный
путь через скорлупу и недвигательный путь через
хвостатое ядро.
НС состоит из полос и стриосом, в которых мало
ацетилхолинэстеразы (АцхЭ). Такая же структура
имеется в богатом АцхЭ стриатуме, который обра-
зует экстрастриосомальный матрикс. Стриосомы
тесно сообщаются с дофаминергическим нигрост-
риарным путем, префронтальной корой и минда-
левидным телом, тогда как матрикс в большей сте-
пени ассоциируется с сенсомоторными отделами
коры. Однако взаимодействие между этими двумя
отделами стриатума и точная организация связей не-
известны.
Функция базальных ганглиев, не относящаяся к
движениям, прослеживается в вентральных отделах
базальных ганглиев, которая состоит из вентрально-
го стриатума (nucleus accumbens), вентрального пал-
лидума и безымянной субстанции (не показана на
рисунке). Эти ядра получают дофаминергическую
импульсацию от вентральной покрышечной облас-
ти, которая прилежит к ЧСк в среднем мозге и про-
ецируется через таламус на префронтальную кору и
корковый центр взора. Эти структуры тесно связа-
ны с мотивапиями и наркотической зависимостью.
Исследования нейрофизиологии базальных ганг-
лиев показали, что многие клетки в их структуре
имеют сложные функции, которые не являются чи-
сто двигательными или чувствительными. Напри-
мер, некоторые группы нейронов НС реагируют на
сенсорные стимулы, но только если этот сенсорный
стимул запускает движение. В противоположность
этому многие группы клеток паллидума максималь-
но реагируют на движение в определенном суставе
еще до появления электромиографических измене-
ний. Так, с нейрофизиологической точки зрения,
базальные ганглии воспринимают обработанную
чувствительную информацию и преобразуют ее в
двигательную программу. Это подтверждается на
примере заболеваний, при которых поражаются ба-
зальные ганглии.
ГЛАВА 39. Заболевания базальных ганглиев и их лечение
Гипокинез
Гиперкинез
Двигательные нарушения
Например:
Болезнь Паркисона
Дистония
Например,
Хорея Гентингтона
Гемнбаллизм
Но
Болезнь Паркинсона
в далеко зашедшей
стадии
Длительная терапия
препаратами леводопы,
Хорея Гентингтона
Гипокинетическое заболевание, например болезнь Паркинсона
Компактная | ДОФА
часть черной Ц-—
субстанцми I
Нео-
j стриатум
Снижение активации
моторной коры
Гиперкинетические заболевание, например хорея Гентингтона
ДОФА
Компактная
часть черной
субстанции
Толщина стрелки указывает на выраженность активности этого пути
2 БШн Кстя|_^ * чСо1^ПВ-ВЛЯтШ+пОВЫШеНИе
* ,и Ч<-Р | | активности
Weej моторной коры
Болезнь Паркинсона
Болезнь Паркинсона - дегенеративное заболева-
ние, обычно развивающееся на 4 и 7-м десятилетии
жизни, причиной которого является первичная де-
генерация дофаминергического нигростриарного
пути с формированием клеточных включений -те-
лец Леви. Причины возникновения заболевания в
большинстве случаев неясны (идиопатическая бо-
лезнь Паркинсона; см. главу 52). Синдром паркин-
сонизма развивается при сосудистом поражении ниг-
ростриарного пути, терапии антагонистами дофами-
на при шизофрении (см. главу 50) и генетических
нарушениях в молодом возрасте.
Более 50—60% дофаминергических нигростриар-
ных нейронов погибает прежде чем идиопатическая
болезнь Паркинсона начинает проявляться клини-
чески. Симптомами болезни являются брадикинезия
(замедленность движений), повышение мышечного
тонуса по пластическому типу (симптом зубчатого
колеса), а также тремор покоя. Однако у большин-
ства больных наблюдаются когнитивные нарушения,
аффективные и вегетативные расстройства, которые
могут относиться к поражению структур, отличных
от нигростриарного тракта.
Нейрофизиологически у этих больных отмечает-
ся повышение активности нейронов внутреннего сег-
мента бледного шара (БШв), который посылает воз-
бужающие импульсы на субталамическое ядро (СТЯ)
в результате утраты тормозных, преимущественно ин-
гибирующих, дофаминергических влияний на нео-
стриатум (НС). Повышение тормозной импульсации
от БШв (и, возможно, ретикулярной части черной суб-
станции - ЧСр) к передневентральному и вентрола-
теральному ядрам таламуса (ПВ-ВЛЯТ) приводит к
86
снижению активации дополнительной моторной
зоны (ДМ3) и соседних отделов коры. Так, пациентам
с болезнью Паркинсона трудно начинать движение,
поскольку у них нарушена активация ДМ3, при этом
объяснить ригидность и тремор сложнее. На ранних
стадиях состояние больных может улучшаться на
фоне приема противопаркинсонических препаратов.
Противопаркинеонические препараты
Пока не создано ни одного препарата, который
мог бы замедлять течение болезни Паркинсона. Для
большинства больных препаратами выбора являют-
ся леводопа (заместительная дофаминовая терапия)
или агонисты дофамина (сам по себе дофамин не
может проникнуть через гематоэнцефалический ба-
рьер). Леводопа является предшественником дофа-
мина и превращается в него в мозге при декарбокси-
лировании. При приеме внутрь леводопа метаболи-
зируется вне мозга посредством периферической
ДОФА-декарбоксилазы, поэтому ее назначают вме-
сте с ингибиторами этого фермента (карбидопой или
бенсеразидом), которые помогают снизить эффек-
тивную дозу леводопы и устранить побочные эффек-
ты (гипотензию, тошноту), возникающие в резуль-
тате распространенной стимуляции дофаминовых ре-
цепторов: тошноту и рвоту (в результате стимуляции
хеморецепторной триггерной зоны), психические
нарушения (например, галлюцинации и спутан-
ность), а также дискинезии. После 5 лет лечения пре-
паратами состояние примерно у половины пациен-
тов ухудшается. В ряде случаев акинезия постепенно
возобновляется, тогда как в других появляются дис-
кинезии. Продолжительность действия каждой дозы
дофамина может укорачиваться (феномен «изнаши-
вания дозы»), кроме того, могут наблюдаться резкие
перепады в двигательной активности (феномен вклю-
чения-выключения).
Селегилин является селективным ингибиотором
моноаминооксидазы типа В (МАО-В). Он снижает ме-
таболизм дофамина в мозге и потенциирует действие
леводопы. Селегилин используют в сочетании с лево-
допой для уменьшения феномена «изнашивания дозы».
Ингибиторы катехол-О-метилтрансферазы (КОМТ),
такие, какэнтакапон, также с недавнего времени при-
меняют при болезни Паркинсона, они снижают пе-
риферический метаболизм леводопы и увеличивают
количество препарата, поступающего в мозг.
Агонисты дофамина (ропинерол, каберголин, пра-
мипексол) также используются в качестве препаратов
1-й линии при лечении болезни у лиц молодого воз-
раста и наряду с леводопой на более поздних стадиях
и у пожилых больных. Агонисты дофамина непосред-
ственно связывают дофаминовые рецепторы в стриа-
туме и черной субстанции и таким образом активиру-
ют постсинаптическую импульсацию нейронов стри-
атума. Их лучше использовать у больных с ранним
дебютом болезни Паркинсона с целью уменьшения в
последующем феномена включения-выключения за
счет дофаминсберегающего влияния этих лекарств.
К другим препаратам, которые используются при
лечении болезни Паркинсона, относятся центральные
холинолитики (например, циклодол), его назначают
на ранних стадиях болезни при преобладании тремо-
ра. Считается, что эти препараты уменьшают гиперак-
тивность центральной холинергической активации,
которая возникает в результате прогрессирующего тор-
можения дофаминергической активности. Побочны-
ми эффектами этих препаратов являются сухость во
рту, задержка мочеиспускания, запор и спутанность со-
знания. С некоторым успехом для лечения тремора при
болезни Паркинсона применяли (3-адреноблокаторы.
Несмотря на то что состояние большинства паци-
ентов с болезнью Паркинсона может улучшаться на
фоне приема лекарств, разработаны хирургические ме-
тоды лечения вдалеке зашедших стадиях заболевания.
При болезни Паркинсона отмечается повышенная
активация внутреннего сегмента бледного шара и ЧСр,
которая отчасти поддерживается возбуждающими
импульсами от субталамического ядра. Недавно были
предложены методы лечения, заключающиеся в раз-
рушении БШв (паллидотомия) и внедрении электро-
дов для стимуляции глубинных ядер (БШв или СТЯ).
Эти манипуляции приводят к временным изменени-
ям по типу блока проведения (см. главы 6 и 8) и, воз-
можно, будут внедрены в клиническую практику. Они,
по-видимому, смогут не только помочь в лечении ос-
ложнений, вызванных приемом лекарств, но и умень-
шить основные неврологические проявления заболе-
вания. Альтернативный подход к разрушению глубин-
ных ядер мозга или имплантации электродов в них —
это имплантация дофаминергической ткани в стриатум
для замещения и возможного восстановления нигрост-
риарного пути. Операции по имплантации фетальных
тканей были успешно проведены у отдельных больных
болезнью Паркинсона, хотя более ранние попытки
ауто-трансплантатов, вырабатывающих катехоламины,
были безуспешными. Во всехслучаяхуспех лечения был
связан с реактивацией соответствующих отделов коры,
однако наблюдались отсроченные побочные эффек-
ты в виде посттрансплантационных дискинезий.
Хорея Гентингтона
Хорея Гентингтона — наследственное аутосомно-
доминантное заболевание, связанное с увеличением
числа тринуклеотидных повторов в гене, кодирующем
белок гентингтин на 4-й хромосоме (см. главу 55).
Характерными симптомами болезни являются про-
грессирующая деменция и насильственные движения,
обычно в форме хореи — быстрые, напоминающие
танец движения, обычно в руках и шее. Этот тип дви-
жений по природе является гиперкинетическим, в от-
личие от гипокинетического при болезни Паркинсона,
и отражает наличие первичного поражения нейронов
стриатума. Это приводит к относительному торможе-
нию СТЯ и уменьшению тормозных влияний БШв и
ЧСр, что в свою очередь вызывает гиперактивацию
двигательных зон коры и избыточные движения.
Лечение хореи Гентингтона направлено на умень-
шение дофаминергической стимуляции в базальных
ганглиях, однако оно не всегда сопровождается улуч-
шением состояния больных. Как и при болезни Пар-
кинсона, были достигнуты некоторые успехи при им-
плантации фетальных тканей в базальные ганглии.
Другие заболевания базальных ганглиев
Другим примером гиперкинетических расстройств
служит гемибаллизм, который проявляется броско-
выми движениями конечностей контралатерально
поражению субталамического ядра.
Базальные ганглии поражаются и при некоторых
других заболеваниях: болезни Вильсона-Коновалова
(аутосомно-рецессивное заболевание, сопровождаю-
щееся нарушением обмена меди), хорее Сиденгама
(последствие ревматизма), митохондриальных заболе-
ваниях (митохондриальные цитопатии; см. главу 55),
некоторых интоксикациях (отравление угарным газом
и марганцем), хореоатетоидном церебральном парали-
че (атетоз — это патологические насильственные черве-
образные движения дистальных отделов конечностей).
Двигательные нарушения, которые возникают при
этих заболеваниях, различны, поскольку при них, как
правило, не происходит поражения строго опреде-
ленных структур экстрапирамидной системы. У боль-
ных могут развиваться синдромы паркинсонизма, хо-
реи, баллизма ил и дистонии, при которой конечность
принимает патологическую позу.
При болезни Паркинсона и хорее Гентингтона
наблюдаются когнитивные нарушения различной
выраженности вплоть до деменции. К их возникно-
вению приводит поражение коры мозга, хотя в на-
стоящее время получены данные о том, что демен-
ция может развиваться при непосредственном стра-
дании базальных ганглиев, что связано с поражением
их вентральных отделов.
Базальные ганглии играют важную роль в регуля-
ции движений глаз (см. главу 40), и у многих паци-
ентов с заболеваниями базальных ганглиев отмеча-
ются патологические движения глаз, что может по-
мочь в установлении правильного диагноза.
87
ГЛАВА 40. Движения глаз
Кора головного мозга
Базальные ганглии
Ростральное
интерстици-
альное ядро
МПП <
Ствол
Вертикальные
движения глаз
Парамедиан
ная ретику
лярная фор- L
нация моста
Корковый центр
| взора (КЦВ)
-I Промежу’
точное
ядро шва
кора
Горизонтальные
' движения глаз
Мозжечок
Задняя | Экстраст- Первичная
" риар- зритель-
ная кора ная кора
Базальные ганглии, ретикулярная
часть черной сустанции
Сетчатка
Ростральное
интерстициальное
ядро МПП
Глазодвигательные t
III «АР*
(медиаль-
ный про- (-
дольный
| пучок)
VI
Аксоны глазодвигательного нерва
к глазодвигательным мышцам
Вестибулярный анализатор
Нижняя косая мышца
Латеральная
прямая мышца
Верхняя
прямая мышца
Медиальная прямая мышца
Нижняя Верхняя
прямая мышца косая
Задняя спайка
Верхние бугорки четверохолмия
Нижние бугорки
етверохолмия
Вестибулярные ядра
Ядро VI пары ЧМН
Парамедианная ретикулярная
формация моста (ПРФМ)
аламус
Мозжечок
мышца
Правый глаз |
Ядро 111 пары ЧМН
Ядро IV пары ЧМН \|
Медиальный
продольный пучок
Тормозной импульс
> Возбуждающий импульс
Регуляция точных движений глаз осуществляется
рядом структур — от глазодвигательных мышц до
лобной коры, а нарушения этой регуляции прояв-
ляются двоением в глазах (диплопия), нечеткостью
зрения или осциллопсией (колебания зрительного
образа). В клинической практике поражение Ш-ей,
IV-й и VI-й пар ЧМН приводит к диплопии (напри-
мер. пр и миастении; см. главу 7). демиелинизирую-
щих заболеваниях (рассеянный склероз), поражаю-
щих медиальный продольный пучок (МПП).
Виды движений глаз
Выделяют 3 основных вида движений глаз:
• Следящие движения позволяют точно отслеживать
предметы и осуществляются задними отделами
коры, примыкающими к мозжечку.
• Саккадированные движения глаз позволяют пере-
ключать внимание с одного объекта на другой и
осуществляются передними отделами коры, базаль-
ными ганглиями и верхними бугорками четверо-
холмия среднего мозга.
88
• Фиксация взора осуществляется преимущественн о
стволом мозга (парамедианной ретикулярной фор-
мацией моста — ПРФМ) и ростральным интерсти-
циальным ядром МПП.
Движения глаз, как и движения в целом, могут
быть произвольными (при поступлении команды из
коркового центра взора) и рефлекторными (при по-
ступлении команды от подкорковых структур и зад-
ней теменной коры).
К глазодвигательным нарушениям относятся на-
рушения содружественных движений глазных яблок,
следящих саккадирующих движений, парезы взора
и нистагм. Нистагм — это двухфазное колебание
глазных яблок в виде патологической медленной и
корригирующей быстрой фаз. Последнее определя-
ет направление нистагма.
Анатомия и физиология отделов ЦНС,
регулирующих движения глаз
Корковый центр взора (КЦВ, преимущественно
поле 8 по Бродману) находится кпереди от премо-
торной коры (см. главу 35). Стимуляция этой струк-
туры вызывает движение глаз, обычно саккадиро-
ванное в противоположную сторону, которое наблю-
дается у больных эпилепсией.
Поражение ПРФМ приводит к невозможности
посмотреть в противоположную сторону, поэтому
пациент как бы смотрит на очаг. КЦВ получает им-
пульсацию от задней теменной коры и проецирует-
ся на верхние бугорки четверохолмия, другие ство-
ловые центры и базальные ганглии.
Задняя теменная кора (соответствует полю 7 по
Бродману у обезьян) содержит большое количество
нейронов, отвечающих за движения под контролем
зрения в ответ на сложные зрительные стимулы (см.
главу 30). Этот отдел имеет важное значение в гене-
рации саккад в сторону предметов, представляющих
зрительный интерес, посредством связей с корковым
центром взора и верхними бугорками четверохолмия.
Поражение этой области, помимо нарушений
зрительного внимания и нарушения саккадирован-
ных движений глазных яблок в противоположном
поле зрения, приводит также к нарушению следя-
щих движений глаз, что проявляется утратой опто-
кинетического рефлекса. Этот рефлекс вызывается
при фиксированном взгляде на какой-то вращаю-
щийся предмет, например барабан, с вертикальны-
ми линиями.
Первичная зрительная кора (ПЗК) связана с эк-
страстриарными отделами и участвует как в реализа-
ции саккад, так и в следящих движениях глаз (см.
главы 24 и 25). Саккады осуществляются посред-
ством связи ПЗК с верхними бугорками четверохол-
мия, тогда как следящие движения глаз — посред-
ством экстрастриарного отдела зрительной коры V5
(см. главу 25), а также ее связей с КЦВ и мостом.
Поражение стриарных и экстрастриарных отде-
лов приводит к возникновению дефектов полей зре-
ния и особых нарушений зрения (см. Главу 25), а так-
же нарушений следящих движений глаз.
Базальные ганглии играют важную роль в реали-
зации саккад (см. главы 38 и 39). Хвостатое ядро по-
лучает импульсацию от КЦВ и проецируется через
ретикулярную часть черной субстанции на верхние
бугорки четверохолмия.
Клинически нарушения саккадированных движе-
ний глаз наблюдаются при некоторых заболеваниях
базальных ганглиев, наример при болезни Паркин-
сона отмечаются неточные саккады с недоведением
взора до интересующего объекта (гипометрические
саккады).
Важную роль в точном осуществлении саккад
играют верхние бугорки четверохолмия.
Мозжечок и вестибулярные ядра посылают слож-
ные импульсы в глазодвигательную систему ствола
мозга и особенно важны при выполнении следящих
движений глаз, а также реализации окуловестибу-
лярных рефлексов (см. главы 28, 37 и 47).
Поражение мозжечка и вестибулярного анализа-
тора приводит к нарушению следящих движений
глаз, неточным саккадам и нистагму.
Ростральное интерстициальное ядро медиального
продольного пучка регулирует вертикальные сакка-
ды и вертикальный взор (вверх и вниз) и получает
импульсацию от коркового центра взора и верхних
бугорков четверохолмия, проецируясь на глазодви-
гательные ядра.
Поражение этой структуры и прерывание ее аф-
ферентных импульсов приводят к нарушению дви-
жений обоих глаз и может наблюдаться при ряде со-
стояний, включая некоторые нейродегенеративные
заболевания.
ПРФМ получает импульсы от КЦВ, верхних бу-
горков четверохолмия и мозжечка и отвечает за го-
ризонтальные саккады и взор. Эта структура рабо-
тает вместе с другим ядром моста, промежуточным
ядром шва. Оно содержит нейроны, которые в нор-
ме посылают тонические тормозные импульсы к ак-
тивным нейронам ПРФМ и РИЯМПП, опосредуя
с аккадный импульс.
Поражение промежуточного ядра шва приводит
к развитию опсоклонуса и беспорядочным движени-
ям глаз. Поражение ПРФМ вызывает нарушение
саккад и парез взора в свою сторону.
МПП осуществляет содружественные движения
глазных яблок с помощью связей между всеми гла-
зодвигательными ядрами и чаще поражается при
таких заболеваниях ЦНС, как рассеянный склероз
(см. главу 54).
Поражение этой структуры приводит к развитию
межъядерной офтальмоплегии, при этом возникают
нистагм в приведенном глазе и замедление или от-
сутствие приведения другого глаза.
ГЛАВА 41. Клиника поражения двигательного анализатора
Базальные ганглии (см. главу 39) |
Например:
Поражение их сопровождается
непроизвольными движениями
паркинсонизмом,
дистонией
хореей
— Болезнь
Паркинсона
Трофика
мышц
Мышечный
тонус
Сила
Рефлексы
Координа-
ция*
Стопные
знаки
*При наличии
Поражение ЦМН I
Мозжечок
Нарушения
координации,
] при PC_____
Инсульт
Травма спинного мозга
например
XX
-/X
— Хорея
Гентингтона
Поражение ЦМН-
•слабость
•спастичность
гиперрефлексия
Первично-мышечное
заболевание
Поражение
ПМН
Xi
\||,
Норма
тт
Норма
Поражение ПМН
Норма
тт
Норма
Норма
пареза и повышения мышечного тонуса состояние координации оценить трудно.
Поражение НМС
(см, главу 7)
Сгибатель-
ные
Разгибатель-
ные
например, миастения:
•патологическая мы-
шечная утомляемость
•нормальные рефлексы
•трофика мышц
в норме
Поражение
ПМН
Снижена, осо-
бенно в разги
бателях рук и
сгибателях ног
Болезнь Полиней-
двигател иного ропатия
нейрона
•Слабость
•Атрофия
•Арефлексня
Миозит Миодистрофия
Слабость
•Нормальные Г
рефлексы
Xi
Норма
Нарушение функции двигательных проводников
может привести к нарушению движений. При пора-
жении мышц возникает их слабость, при поражении
нервно-мышечного соединения (НМС) — патологи-
ческая мышечная утомляемость, при поражении мо-
тонейронов (центральных или периферических) —
парез в мышце, изменение мышечного тонуса и реф-
лексов, при поражении мозжечка — нарушение ко-
ординации без изменения силы мышц и рефлексов,
при поражении базальных ганглиев — патологичес-
кие движения без изменения силы мышц, рефлек-
сов и координации движений.
Чтобы определить природу и причину двигатель-
ных нарушений, необходимо собрать анамнез и про-
вести обследование больного. У большинства боль-
ных с изолированными двигательными симптомами
имеет место либо болезнь Паркинсона, либо болезнь
двигательного нейрона, хотя чаще всего у них отме-
чаются как чувствительные, так и двигательные на-
90
рушения в результате инсультов или поражения пе-
риферических нервов на различных уровнях.
Обследование больного с двигательными нару-
шениями включает анализы крови, электронейро-
графию (ЭНГ), электромиографию (ЭМГ) и магнит-
но-резонансную томографию (МРТ) головного и
спинного мозга.
Патология мышц (см. главы 10 и 11)
Характерными признаками первично-мышечных
заболеваний являются слабость мышц, которая мо-
жет появляться при физической нагрузке, и в ряде
случаев мышечная боль (миалгия). Возраст начала и
тип прогрессирования нередко указывают на харак-
тер первично-мышечного заболевания, например
прогрессирующая мышечная слабость без боли у
детей позволяет заподозрить дегенеративную миоди-
строфию (см. главу 10). При наличии слабости и бо-
лезненности мышц у взрослых следует предполагать
полимиозит (см. главу 54). Распределение паре-
зов также помогает установить характер первич-
но-мышечного заболевания, например слабость
проксимальных отделов рук и ног отмечается при
конечностно-поясной миодистрофии. При первич-
но-мышечных заболеваниях определяют уровень
креатинфосфокиназы (КФК), повышение которого
указывает на выраженность первично-мышечного
поражения; кроме того, нужно провести ЭМГ и био-
псию. В ряде случаев необходимо генетическое ис-
следование, особенно если мышечная слабость свя-
зана с миотонией и другими признаками, что ука-
зывает на дистрофическую миотонию.
Патология нервно-мышечного
соединения (см. главу 7)
Больные с поражением НМС жалуются на мы-
шеченую слабость, нарастающую при выполнении
движений. Наиболее часто встречающимся заболе-
ванием нервно-мышечной передачи является миас-
тения, которая начинается в молодом или пожилом
возрасте и проявляется диплопией, птозом, слабос-
тью мимической и бульбарной мускулатуры, а так-
же проксимальных мышц конечностей. При обсле-
довании парез обнаруживается в покое и нарастает
при выполнении движений. При бульбарных и ды-
хательных нарушениях необходимы реанимацион-
ные мероприятия. Диагноз обычно ставится на ос-
новании анамнеза и осмотра, наличия антител к
ацетилхолиновым рецепторам и специфических ан-
тител к мышечно-специфической киназе (МСК),
положительной реакции на введение короткодей-
ствующих антихолинэстеразных препаратов (прозе -
риновая проба) и нарушений при ритмической сти-
муляции нервов, выявляемых при проведении ЭНГ
и ЭМГ. Мышечную биопсию не производят. У не-
которых больных миастенией обнаруживается ги-
перплазия тимуса или тимома. Другие миастеничес-
кие синдромы встречаются редко.
Патология периферических нервов
Поражение периферических нервов приводит к
развитию двигательных и чувствительных наруше-
ний. Однако при некоторых полинейропатиях мо-
гут страдать преимущественно двигательные нервы,
кроме того, изолированные двигательные наруше-
ния наблюдаются при полиомиелите и болезни дви-
гательного нейрона, при которых поражаются пере-
днероговые мотонейроны спинного мозга и мотоней-
роны ствола. Характерными признаками поражения
двигательных волокон периферических нервов явля-
ются слабость, атрофия, фасцикуляции и выпадение
рефлексов, которые наблюдаются при поражении
периферического мотонейрона (ПМН). Обследова-
ние больных с поражением ПМН проводится с це-
лью исключения туннельных нейропатий и кореш-
ковых синдромов. Последние можно диагностировать
с помощью МРТ, а ЭНГ и ЭМГ позволяют выявить
признаки денервации в виде спонтанной активнос-
ти мышцы, отсутствующей в норме.
Патология спинного мозга
При поражении спинного мозга может возник-
нуть несколько синдромов (см. главу 34). В редких
случаях поражаются интернейроны спинного мозга
и возникают непрерывная активность двигательных
единиц и синдром ригидного человека (см. главу 34).
Поражение нисходящих проводящих двигательных
путей головного и спинного мозга приводит к пора-
жению центрального мотонейрона (ЦМН), которое
проявляется парезами, спастичностью, гиперреф-
лексией, клонусами и разгибательными стопными
патологическими знаками. Селективное поражение
пирамидного тракта встречается редко, и если на-
ряду с этим у больного отмечается поражение ПМН,
то имеет место такая форма болезни двигательного
нейрона, как боковой амиотрофический склероз, или
болезнь Лу Герига. Структурные поражения спинно-
го мозга приводят к двигательным и чувствительным
симптомам. Обследование должно включать МРТ,
анализ цереброспинальной жидкости (ЦСЖ), если
предполагается воспалительная этиология процес-
са, и в ряде случаев нейрофизиологическое обсле-
дование, включающее ЭНГ, ЭМГ и транскраниаль-
ную магнитную стимуляцию (ТКМС).
Поражение головного мозга
Поражение супраспинальных структур приводит
к развитию ряда двигательных нарушений. Пораже-
ние головного мозга чаще возникает при инсультах
с вовлечением нисходящих проводящих двигатель-
ных путей, идущих от коры и ствола до спинного
мозга. Такие очаги приводят к развитию гемипареза
на противоположной стороне с признаками пораже-
ния ЦМН. Если поражается левое полушарие, воз-
никают речевые нарушения. Иногда при изолиро-
ванном поражении моторной коры отмечаются фо-
кальные эпилептические припадки, например
джексоновская эпилепсия (см. главы 35 и 36). Основ-
ными методами исследования являются МРТ и ком-
пьютерная томография (КТ) головного мозга, а так-
же анализ ЦСЖ при подозрении на воспалительную
этиологию процесса. В ряде случаев необходимо ге-
нетическое исследование.
Поражение других отделов ЦНС
Базальные ганглии
При поражении базальных ганглиев возникают
замедление движений, например, при болезни Пар-
кинсона, насильственные движения конечностей,
например, при дистонии, хорее и гемибаллизме (см.
главу 39).
Мозжечок
При поражении мозжечка наблюдаются наруше-
ния координации, речи и движений глаз (см. главу 37).
Чаще всего мозжечок поражается при рассеянном
склерозе, приеме антиконвульсантов и алкоголизме.
Прогрессирование опухолевого процесса в мозжечке
приводит к гидроцефалии в результате сдавления IV
желудочка и его выходных отверстий (см. главу 16).
91
ЧАСТЬ IV. Вегетативная, лимбическая системы,
ствол мозга и пластичность
ГЛАВА 42. Вегетативная нервная система
Анатомия вегетативной нервной |
системы и регуляция ее функции 1
в ЦНС 1
ПВ — паравентрикулярное ядро
СОЯ — супраоптическое ядро
ДМЯ — дорсомедиальное ядро
ВМЯ — вентромедиальное ядро
ЛОГ — латеральные отделы гипоталамуса
АРК — аркуатное ядро
Префронтальная
кора, миндале-
видное тело,
гиппокамп
Зрительный
тракт Г'
Миндалевидное ->—
ТРЛО (И!
Парасимпатическая система
Орга“ч Парасимпатическая иннервация Симпатическая иинераади.
Глаз Ресничный ганглий III пары ЧМН Сужение зрачка и увеличение кривизны хрусталика —Т2 — верхний шейный ганглий Расширение зрачка, поднятие : верхнего века
Слезные и слюнные железы VII и IX пары ЧМН Стимуляция слюноотделения т-т, Уменьшение слюноотделения
Сердц. X пара ЧМН •Снижение частоты н силы сердечных т1—т* Увеличение частоты сердечных сокращении и их силы т-т. Расширение бронхов, подавление секреции Т,—Ти — чревный ганглий Уменьшение секреции, пери- стальтики и сужение сфинктеров
Бронхиаль- ное дерево Верхние от- делы желу- дочно-кишеч- ного траста (п>«аз кх -ка) сокращений •Сужение бронхов и стимуляция секреции •Усиление перисталь- тики и релаксация сфинктеров, стиму- ляция секреции
Надпочеч- ники Т,-Т„ Не дают постганглионарных во- локон к надпочечникам, которые высвобождают катехоламины при стимуляции
Кожа и пе- риферичес- кие сосуды Сужение сосудов кожи и расширение сосудов в мышцах Пилоэрекция и усиление пото-от- деления
Нижние отделы кишечника, мочевой пузырь, половые органы Крестцовое сплетение S2-4 Сокращение гладкой мускулатуры толстой кишки Сокращение мышцы, изгоняющей мочу, и релаксация внутренне- го сфинктера уретры Эрекция Т._о Угнетение перистальтики ? — уменьшение мочеиспускания Эякуляция
ДМЯ
ЛОГ1
Эффекторный
орган
III желудочек
Симпатическая нервная система
Норадреналин >
Преганглионарный
Ацх нейрон
Г ипоталамус
^коронарный cpejOj
СОЯ^-— Зрительный
тракт
Афферентная импульсации
(ядро одиночного пути)
Шейный отдел спинного мозга
/Ацх —5-------------------------
1 Z - \ Парасимпатическая
_ J, нервная система
Постганглионарныи
нейрон *
Грудной отдел
спинного мозга
Поясничный \
отдел спинного]
и мозга
Крестец
Анатомия периферической нервной системы!
„ Задний корешок
Постганглионарные
симпатические
волокна повторно
входят в спинно-
мозговой
столбы
спинного мозга
Кожа-f
(железы,
сосуды)
Серая соединительная
Передний корешок
Симпатические ганглии
Белая соединительная ветвь
Внутренние
органы и сосуды
Анатомия вегетативной нервной системы
Вегетативная нервная система (ВНС) включает
нервные клетки и волокна, иннервируюшие внут-
92
ренние органы и железы. Они регулируют процес-
сы, которые не подчиняются произвольному конт-
ролю. Эфферентный проводящий путь из ЦНС к ин-
нервируемому органу всегда состоит из двух нейро-
:аиия
нов: преганглионарного и постганглионарного, при
этом тело первого лежит в ЦНС (см. главу 1).
ВНС подразделяется на кишечную, симпатичес-
кую и парасимпатическую. При этом две последние
оказывают противоположное влияние на структу-
ры, которые обе иннервируют. В симпатической не-
рвной системе преганглионарные нейроны нахо-
дятся в промежуточном (боковом) роге спинного
мозга от верхнегрудного до среднепоясничного от-
дела (T1-L3). Преганглионарные парасимпатические
нейроны залегают в стволе мозга и крестцовом от-
деле спинного мозга. Постганглионарные нейроны
находятся в вертебральных и превертебральных ган-
глиях в симпатической системе, а в парасимпати-
ческой они расположены в непосредственной бли-
зости от стенки органа, который они иннервируют.
Кроме того, в симпатической и парасимпатичес-
кой системах работают различные медиаторы (нор-
адреналин (НА), или норэпинефрин — в качестве
постганглионарного трансмиттера и ацетилхолин —
Ацх). На уровне ганглиев Ацх используется в обеих
системах.
Регуляция функций
вегетативной нервной системы в ЦНС
Регуляция функций ВНС в ЦНС сложна и осуще-
ствляется структурами ствола и гипоталамусом (см.
главу 43). Основными отделами гипоталамуса, регу-
лирующими функцию ВНС, являются вентромеди-
альный (симпатическая нервная система) и лате-
ральный (парасимпатическая нервная система).
Нисходящие пути могут быть прямыми и непрямы-
ми, они проходят через некоторые структуры ство-
ла, такие, как околоводопроводное серое вещество
и часть ретикулярной формации (см. главу 13).
Клинические признаки поражения вегетатив-
ной нервной системы
Поражение ВНС может быть локальным, когда
страдает какая-то конкретная структура, или диф-
фузным, когда поражается вся система в целом ка-
ким-либо центральным или периферическим пато-
логическим процессом. К локальным симптомам
поражения ВНС относят, например, нарушение сим-
патической иннервации глаза — приводит к миозу,
птозу, энофтальму в области орбиты — триаде симп-
томов, известной как синдром Горнера. Другим при-
мером является рефлекторная симпатическая дист-
рофия, при которой отмечаются болевой синдром и
вегетативные нарушения в конечности, чаще это
реакция на какую-либо травму (см. главу 22). Точ-
ная роль симпатической нервной системы в генезе
этого расстройства до конца неясна, поскольку ло-
кальные симпатикотомии не всегда приводят к улуч-
шению состояния, поэтому рефлекторная симпати-
ческая дистрофия в настоящее время называется
комплексным регионарным болевым синдромом.
Однако в ряде случаев ноцицепторы начинают экс-
прессировать рецепторы к НА (норэпинефрину) (см.
главу 22).
Системное поражение ВНС наблюдается при де-
генерации центральных ядер, при изолированной
вегетативной недостаточности или в рамках распрос-
траненного дегенеративного процесса, например
мультисистемной атрофии, когда наряду с дисфун-
кцией ВНС отмечается поражение базальных ганг-
лиев и мозжечка. Кроме того, вегетативные наруше-
ния могут возникать при гибели периферических
нейронов, например, при сахарном диабете, алко-
голизме и синдроме Гийена—Барре.
Во всех случаях у больных отмечается ортостати-
ческая и постпрандиальная гипотензия (синкопаль-
ные или пресинкопальные состояния при вставании,
физических упражнениях или после еды) с фикси-
рованным пульсом и тазовыми нарушениями (час-
тые мочеиспускания или недержание мочи), импо-
тенцией и нарушениями потоотделения и зрачковых
реакций на свет. Эти симптомы трудно поддаются
лечению. В качестве лечебных средств используют-
ся медикаменты, устраняющие постуральную гипо-
тензию и нарушение функции сфинктеров. К ним
относятся флудрокортизон, эфедрин, мидодрин и
аналоги вазопрессина (все они используются для
лечения постуральной гипотензии и вызывают за-
держку жидкости в организме), а также цисаприд
(средство, усиливающее высвобождение Ацх) для ле-
чения пареза желудочно-кишечного тракта.
ГЛАВА 43. Гипоталамус
Медиальная часть лобной доли
Префронтальная долька
Гипоталамус
Четверохолмие
мозг
Гипоталамус
Зрительный перекрест
Г ипофиз
Таламус
Передние отделы
Сетчатка
Миндалевидное тело
Префр >нтальная ,
кора
Зрительный перекрест
Срединное возвышение
с портальными сосудами
• Антидиуретический гормон
• Окситоцин
Средний мозг
Задние отделы гипофиза
Задние отдель
* Гормон роста
• ЛГ/ФСГ
• Тиреоидный гормон
• Адренокортикотропный гормон
• Пролактин
Мост
СХЯ
ПВЯ
СОЯ
дмя
вмя
ст
АРК
ЛОГ
III желудочек
Зрительным тоакт
Височная доля
— супрахиазматическое ядро
— паравентрикулярное ядро
— супраоптическое ядро
— дорсомедиальное ядро
— вентромедиальное ядро
— сосцевидное тело
— аркуатное ядро
— латеральные отделы
гипоталамуса
пвя
СОЯ
АРК
Гипоталамус расположен с обеих сторон от III же-
лудочка под таламусом, между зрительным пере-
крестом и средним мозгом и получает значитель-
ную импульсацию от структур лимбической систе-
мы (см. главу 45), а также импульсацию от сетчатки
и содержит большое количество нейронов, чувст-
вительных к изменению уровня гормонов, элект-
ролитов и температуры. Кроме того, гипоталамус
дает эфферентные пути к ВНС и регулирует эн-
докринную функцию гипофиза (детальное обсуж-
дение эндокринной функции гипоталамо-гипо-
физарной системы не входит в задачу книги).
Гипоталамус, будучи важной структурой ВНС,
регулирует гомеостаз многих физиологических
систем (например, мотивацию жажды, голода, вод-
но-солевой баланс, температуру тела), регулирует
циркадные и эндокринные функции, способствует
формированию антеградной памяти (вместе с лим-
бической системой; см. главу 45), а также преоб-
разует ответ на эмоциональные стимулы в гормо-
нальные и вегетативные реакции.
Гипоталамус выполняет ряд важных функций,
каждая из которых может быть утрачена или на-
94
рушена при заболевании, чаще всего опухолях
гипофиза.
Функции гипоталамуса следующие:
• Гипоталамус регулирует функцию ВНС, его пора-
жение может приводить к нарушению вегетатив-
ных функций. Вентромедиальный отдел гипотала-
муса регулирует функцию симпатической нервной
системы, латеральные отделы контролируют пара-
симпатическую систему (см. главу 42).
• Гипоталамус регулирует эндокринные функции
гипофиза, продуцируя релизинг-гормоны и тор-
мозные гормоны, а также антидиуретический гор-
мон (АДГ; вазопрессин) и окситоцин. Поражение
гипоталамуса приводит к системным эндокринным
нарушениям, например несахарному диабету при
недостатке АДГ. При этом заболевании пациент вы-
деляет несколько литров мочи в день, что необхо-
димо компенсировать большим количеством жид-
кости.
• Гипоталамус регулирует эндокринные и вегетатив-
ные реакции в соответствующих физиологических
условиях, а также эмоциональные реакции, кото-
рые управляются лимбической системой. При ги-
поволемии или крайнем беспокойстве гипотала-
мус не только опосредует симпатическую актив-
ность, но и увеличивает продукцию кортизола по-
средством стимуляции высвобождения адренокор-
тикотропного гормона (АКТГ) из передних отделов
гипофиза.
• Гипоталамус играет важную роль в терморегуляции.
Поражение передних отделов гипоталамуса приво-
дит к гипертермии, тогда как стимуляция этого от-
дела снижает температуру тела через вегетативные
реакции в противоположность задним отделам ги-
поталамуса, который ведет себя обратным образом.
При длительном изменении температуры окружа-
ющей среды, например гипоталамус адаптирует к
ним организм человека, увеличивает высвобожде-
ние тиреотропин-рилизинг-гормона (ТРГ) у лю-
дей, находящихся в условиях пониженной темпе-
ратуры. При септических состояниях некоторые
цитокины (например, интерлейкин-1) воздейству-
ют на центр терморегуляции гипоталамуса, и тот
адаптирует организм к температуре, которая пре-
вышает нормальную (например, при лихорадке).
Также гипоталамус регулирует механизмы, направ-
ленные на выработку или сохранение энергии (на-
пример, озноб).
• Гипоталамус играет определенную роль в питании.
Вентромедиальное ядро гипоталамуса часто назы-
вают центром насыщения, его поражение приво-
дит к избыточному аппетиту (булимии) и прибав-
ке массы тела, поражение латеральных отделов
гипоталамуса (или центра голода) вызывает ано-
рексию (нежелание принимать пищу). Регуляция
функции этих центров осуществляется посред-
ством некоторых гормонов, включая инсулин и
лептины, открытые позже.
• Гипоталамус контролирует чувство жажды и вод-
ный баланс посредством осморецепторов, получа-
ет афферентную импульсацию от периферических
рецепторов (например, рецепторы на растяжение
в предсердии, артериальные барорецепторы), а
также дает эфферентную импульсацию через ВНС
к сердцу и почкам, и стимул к продукции АДГ.
• Гипоталамус регулирует циркадные ритмы через
импульсацию от сетчатки в супраоптическое ядро.
Это ядро является наиболее важной структурой, ре-
гулирующей циркадные ритмы, что подтвердили
эксперименты с его разрушением и транспланта-
цией. Точный механизм выработки этихритмов не-
известен, показано, что определенную роль в их ре-
гуляции играют обмен меланина и шишковидная
железа.
• Гипоталамус участвует в формировании памяти
вместе с лимбической системой. Поражение сосце-
видных тел, получающих значительную импульса-
цию от комплекса гиппокампа, как например при
алкоголизме, осложненном недостаточностью ви-
тамина Вр приводит к глубокой (корсаковский
синдром), антероградной (невозможность запоми-
нать новую информацию) и ретроградной (невоз-
можность воспроизводить давно полученную ин-
формацию) амнезии с конфабуляциями (ложными
воспоминаниями). Наличие ретроградной амнезии
отличает таких пациентов от больных с поражени-
ем гиппокампа (см. главу 45).
• Гипоталамус, помимо гормонального, влияет на
сексуальное и эмоциональное поведение.
ГЛАВА 44. Ретикулярная формация и сон
Электроэнцефалограмма
В-Риты
Диффузная иннервация всех структур ЦНС,
включая большое ядро шва и голубое ядро
В-13 Гц
0,5—4 Гц
Д-Ритм (МВС)
Таламуг
Мозжечок
Голубое ядро ( юрадреналин)
Цикл ночного сна
Другие стволовые центры, в частности
дофаминергические вентральные отделы покрышки
Неспецифические nj ра таламуса с диффузными проекциями,
включая ретикулярное ядро таламуса, которое связывает
другие таламические ядра
Большое ядро
шва (серотонин) \
Торможение
стволовых и спиналь-
ных мотонейронов
Секреция гипокретинов лате-
ральными и задними отделами
гипоталамуса воздействует на
структуры ствола, регулирую-
м1ие цикл сон—бодрствование
3
5
Гипоталамус —
супрахиазма-
тическое ядро
регулирует цикл
сон—бодрствовл
ние и содержит
гистаминерги-
ческие туберо-
мамиллярные
ядра
Часы
Сон присущ всем млекопитающим и с поведен-
ческой точки зрения характеризуется снижением
реактивности на внешние раздражители, а с элект-
рофизиологической — специфическими изменени-
ями электроэнцефалографической (ЭЭГ) активно-
сти. Кроме того, некоторые изменения сна связаны
с функцией ВНС.
Нормальный сон необходим для здоровья чело-
века, хотя до сих пор остается неясным, почему че-
ловек видит сны.
Картина ЭЭГ при медленноволновом
сне и бодрствовании
У здорового человека в состоянии бодрствования
и в покое отмечается высокочастотный (13—30 Гц, 0-
активность) низкоамплитудный паттерн. Эта десин-
хронизированная активность меняется, когда чело-
век закрывает глаза и дремлет, при этом появляется
ритм меньшей частоты (8—13 Гц, (3-активность) с не-
сколько большей амплитудой. Этот тип ЭЭГ называ-
ется синхронизированным и является результатом од-
новременной импульсации множества корковых ней-
ронов вследствие таламокортикальной активации.
ЭЭГ-исследования показали, что сон имеет не-
сколько стадий. Когда человек засыпает (I стадия),
ЭЭГ сходна с таковой в состоянии бодрствования
(низкая амплитуда, быстрая активность). Когда сон
96
углубляется до II и III стадий, амплитуда паттерна
ЭЭГ увеличивается, ачастотаего падает. III и IV ста-
дии сна называются медленноволновым сном (МВС)
или стадией без быстрых движений глаз (не-REM
фаза), поскольку глаза в этой стадии находятся в
покое. После 90 мин сна ЭЭГ снова становится низ-
коамплитудной и появляется быстрая активность,
которую трудно отличить от активности I, ne-REM,
стадии. Однако в этой стадии сна появляются быст-
рые движения глаз. Этот тип сна называется сном с
быстрыми движениями глаз (REM-фаза) или пара-
доксальным сном, поскольку, несмотря на то что
ЭЭГ в этой фазе не отличается от ЭЭГ в состоянии
бодрствования, человека в ней крайне трудно раз-
будить, а мышечный тонус отсутствует. Именно в
REM-фазу сна человек видит сны.
Неврологические механизмы сна
Сон — это активный процесс. Показано, что хо-
линергические нейроны ствола (являющиеся частью
восходящей активирующей ретикулярной системы)
не только регулируют десинхронизированную ЭЭГ,
но также производят понтогеникулоокципитальные
волны, сигнализирующие о начале REM-фазы сна.
В противоположность этому нейроны большого яд-
ра шва, содержащие серотонин (5-гидрокситрипта-
мин — 5-ГТ), по-видимому, подавляют эту актив-
ность, что предполагает наличие двух различных ме-
диаторных систем, которые начинают и завершают
REM-фазу сна. Кроме того, норадреналинсодержа-
щие нейроны голубого ядра отвечают за снижение
возбудимости мотонейронов спинного мозга, кото-
рое приводит к атонии, наблюдающейся в REM-фазу
сна, тогда как преоптическая область и супраопти-
ческие ядра гипоталамуса регулируют цикл сон-
бодрствование. Другие стволовые и гипоталамичес-
кие образования также играют роль в регуляции сна,
к ним, в частности, относятся вентральный отдел по-
крышки мозга с дофаминергическими нейронами и
гистаминергические тубермамиллярные ядра.
Нарушения сна
Бессонница — наиболее распространенное расст-
ройство сна. Она заключается в невозможности по-
лучить такое количество и качество сна, которое тре-
буется для нормальной жизнедеятельности в состоянии
бодрствования. Первичная бессонница предположи-
тельно связана с нарушениями механизмов сна в го-
ловном мозге и встречается редко, такие больные
нуждаются в лечении снотворными. Причинами вто-
ричной бессонницы являются психические заболе-
вания (особенно тревожные и депрессивные рас-
стройства), соматические заболевания, хронические
болевые синдромы, злоупотребление алкоголем, кофе-
ином, психологические проблемы и пожилой возраст.
Пептиды сна
Недавно были открыты пептиды, связанные со
сном (например, пептид, А-индуцирующий сон). До
конца неясно, как эти пептиды связаны с механиз-
мами сна, описанными выше.
Лечение бессонницы
Снотворные — это средства, вызывающие сон.
К ним относятся бензодиазепиновые препараты хло-
ралгидрат, хлорметиазол и барбитураты. Бензодиа-
зепиновые снотворные используются чаще всего, они
обладают анксиолитическим, противосудорожным,
миорелаксирующим и амнестическим эффектами.
Считается, что эти эффекты бензодиазепины вызы-
вают благодаря усилению ГАМК-ергического тормо-
жения Е(НС. ГАМК-А-рецепторы обладают несколь-
кими модулирующими сайтами, включая один для
бензодиазепинов, которые в случае активации вы-
зывают изменение конформации ГАМК-рецепто-
ров. Это увеличивает аффинность рецепторов к
ГАМК и усиливает ГАМК-ергические влияния и
проведение хлора через мембрану клеток.
Любой бензодиазепиновый препарат, принятый
на ночь, вызовет сон, однако предпочтительнее ис-
пользовать препараты, быстро выводящиеся из орга-
низма (например, темазепам), чтобы избежать седа-
тивного эффекта в дневное время. Некоторые новые
препараты не имеют бензодиазепиновой структуры,
но также являются агонистами бензодиазепиновых
рецепторов, например зопиклон, циклопироллон ко-
роткого действия. Бензодиазепины являются цент-
ральными депрессантами, но их максимальный эф-
фект при приеме внутрь не приводит к фатальному
угнетению дыхания (в отличие от опиоидов или бар-
битуратов). К побочным эффектам бензодиазепинов
относятся сонливость, снижение внимания, атаксия,
а также слабовыраженная зависимость после употреб-
ления в течение нескольких недель. Отмена препара-
тов вызывает синдром отмены (тревогу, бессонницу),
который может продолжаться неделями.
Во многих случаях эффективной альтернативой
медикаментозному лечению бессонницы являются
психотерапевтические методы.
Гиперсомния (дневная сонливость)
Гиперсомния встречается реже и является более
тяжелым состоянием, чем бессонница. Чаще всего
причинами дневной сонливости являются нарколеп-
сия, обструктивное апноэ во время сна, прием пре-
паратов (например, бензодиазепинов), алкоголя и
др. и депрессия (у 20% больных отмечаются не бес-
сонница, а сонливость).
Нарколепсия
Нарколепсия характеризуется непреодолимыми
эпизодами сонливости, продолжающимися 5-30 мин
в течение дня, катаплексией (снижение мышечного
тонуса и временный паралич), которые, как правило,
провоцируются эмоцией, например смехом, злобой,
а также параличом во время сна и галлюпинапиями
во время засыпания. У больных отмечена достовер-
ная ассоциация с комплексом антигенов гистосовме-
стимости (HLA DR2/DQW1), и поскольку никакой
патологии у них не выявляется, вероятно, это состо-
яние связано с нарушением функций ствола мозга,
поскольку имеются данные, что у больных укороче-
на REM-фаза сна во время, предшествующее пробуж-
дению. Кроме того, важную роль могут играть недав-
но открытые пептиды — гипокретины и орексины, по-
скольку они синтезируются в гипоталамусе и взаи-
модействуют с некоторыми стволовыми центрами,
регулирующими сон. Более того, недостаточность
этих пептидов была недавно обнаружена улиц, стра-
дающих нарколепсией, а также у эксперименталь-
ных животных. Этот синдром резко снижает качество
жизни, лечение проводится длительно стимулятора-
ми дексамфетамином, метилфенидатом и модафини-
лом. Лечение катаплексии проводится клопирамином.
Синдром обструктивных апноэ во время сна
Синдром обструктивных апноэ наблюдается, если
верхние дыхательные пути в задних отделах глотки
закрываются, когда папиент спит. Это приводит к
снижению оксигенации крови, что вызывает момен-
тальное пробуждение больного и мешает нормально-
му сну. Пациент, обычно мужчина с избыточной мас-
сой тела, чаще всего не замечает пробуждений во вре-
мя сна, однако нарушение ночного сна приводит к
дневной сонливости и ухудшению работоспособности
днем. Лечение этого синдрома включает похудание,
неинвазивную искусственную вентиляцию с положи-
тельным давлением в ночное время и иногда приме-
няют хирургическое вмешательство на ротоглотке.
97
ГЛАВА 45. Лимбическая система,
отсроченная потенциация и память
Структуры лимбической системы
в медиальной части полушария головного мозга
Медиальный и латеральный
перфорантные пути
Схема связей лимбической системы (упрощенная)
Отсроченные изменения к экспрессия генов
Обонятельная кора
Минддле-
Ыдое тело
Перфорнат-
I ный путь
Знтори-
нальная
кора
• Ассоциативная кора
(префронтальная кора)
• Ядра таламуса
средней линии
• Ядра диагонального
лучка (холинергичес
Поясная
ИЗВИЛИН»'
| Двигательные
ь отделы коры
Зубчатая^сд сд_>Субикулум —
извилина J 1 циимории/
Ком-текс гиппокампа
Передние
свод
Дентальные ядра |
Круг Пейпса _)
В 1937 г. Д. Гейпес предложил считать этот круг
ведущим субстратом памяти и эмоций
Анатомия лимбической системы
Лимбическая система представлена несколькими
отделами мозга, которые располагаются вдоль ме-
диальной части височной доли и включают поясную
извилину, парагиппокампальные структуры (постсуби-
кулум, парасубикулум, пресубикулум и преренальную
кору), энторинальную кору, комплекс гиппокампа (зуб-
чатую извилину, поля СА1—СА4 и субикулум), септаль-
ные вдра и миндалевидное тело. Кроме того, тесно
связаны с лимбической системой мамиллярные тела
гипоталамуса, обонятельная кора и nucleus accum-
bens (см. главы 29, 38 и 43, соответственно).
Анатомическое строение лимбической системы
указывает на то, что в ней осуществляется высокий
98
уровень обработки чувствительной информации,
которая поступает туда из ассоциативной коры (см.
главу 30). Лимбическая система посылает эфферен-
тную информацию к префронтальной коре и гипо-
таламусу, а также к корковым отделам, осуществля-
ющим планирование поведения и двигательные ре-
акции (см. главы 32 и 35). С анатомической точки
зрения, лимбическая система имеет отношение к
реализации поведенческих реакций на внешние
раздражители, особенно эмоциональных, и поэтому
поражение лимбической системы у эксперименталь-
ных животных оказывает выраженное влияние на
эмоциональную реактивность. Показано, что комп-
лекс гиппокампа высокочувствителен к гипоксии и в
то же время характеризуется уникальной пластично-
стью, что позволяет объяснить важную роль этой
структуры в генезе эпилептических припадков (см.
главу 53), а также в обучении и памяти. Кроме того,
это основной центр нейрогенеза зрелого мозга, кото-
рый также участвует в реализации функции памяти.
Функции лимбической системы
Доказательством основной роли этого отдела
ЦНС в механизмах памяти является оригинальное
описание Сковиллем и Милнером в 50-х годах про-
шлого века пациента Г. с двусторонней передневи-
сочной лобэктомией, которая была выполнена по
поводу рефрактерной эпилепсии и привела к глубо-
кой амнезии. В дальнейшем было показано, что соб-
ственно гиппокамп и парагиппокампальные отделы
играют важнейшую роль в усвоении новой инфор-
мации у экспериментальных животных и больных
(декларативная память).
Однако долговременная память опосредуется дру-
гими удаленными структурами, по-видимому, соот-
ветствующими отделами коры мозга, что подтверж-
дает характер нарушений памяти при деменции аль-
цгеймеровского типа (ДАТ, см. главу 52). При этом
заболевании ретроградная память относительно со-
хранна (на давние события, например детства), но при
этом грубо нарушена антероградная память (пациен-
ты не могут запомнить, что они только что сделали).
Миндалевидное тело
Миндалевидное тело — это небольшая структу-
ра, напоминающая по форме миндаль, состоящая из
множества ядер, которые залегают в медиальной ча-
сти височной доли и участуют в обучении и, возмож-
но, хранении памяти об эмоциональных аспектах пе-
режитого.
Поражение этой структуры в эксперименте при-
водило к притуплению реакций на важные стимулы
и даже полному отсутствию эмоциональных реак-
ций. Люди с селективным поражением миндалевид-
ного тела не способны распознавать выражение
страха на лице. В противоположность этому стиму-
ляция миндалевидного тела сопровождается страхом
и выраженными вегетативными реакциями. Это на-
блюдается при височной эпилепсии, когда больной
жалуется на кратковременные приступы страха.
Поясная извилина
Поясная извилина находится в медиальной час-
ти полушария и выполняет ряд функций, включая
участие в регуляции движений (см. главу 35), и вос-
приятии боли (см. главы 21 и 22).
Поражение этой структуры вызывает двигатель-
ное игнорирование, снижение восприятия боли, аг-
рессии и вокализапии, притупление эмоциональных
реакций и изменение поведения, которое приводит
к развитию акинетического мутизма (отсутствие дви-
жений и речевой продукции). Стимуляция этой об-
ласти в эксперименте или при эпилептическом при-
падке сопровождается измененными вегетативными
реакциями, а также остановкой движений с вокали-
зацией и последующими сложными движениями.
Отсроченная потенциация
Отсроченная потенциация (ОП) - это усиление
синаптической трансмиссии при повторной стиму-
ляции, которая осуществляется в течение несколь-
ких минут, инициирована в гиппокампе интенсив-
ной синаптической активностью в течение менее 1 с
и длится часами и более. ОП возникает в несколь-
ких отделах ЦНС, но особенно характерна для гип-
покампа, поэтому считается, что она важна для па-
мяти. Однако необходимо понимать, что ОП может
быть опосредована различными синапсами комп-
лекса гиппокампа, в большей степени это касается
возбуждающих глутаматных синапсов, содержащих
]М-метил-О-аспартатные рецепторы (NMDA-рецеп-
торы; см. главу 9) в поле СА1 комплекса гиппокампа.
ОП осуществляется следующим образом:
• Приток афферентной импульсации приводит к
высвобождению глутамата из пресинаптической
терминали (1-я стадия на рисунке).
• Глутамат связывается с NMDA- и ne-NMDA-рецен-
торами на постсинаптической мембране. Последние
опосредуют приток натрия (2-я стадия), который де-
поляризует постсинаптическую мембрану (3-я стадия).
• Деполяризация постсинаптической мембраны
приводит не только к возникновению возбуждаю-
щего постсинаптического потенциала (ВПСП), но
и к удалению магния из NMDA-ассоциированно-
го ионного канала (4-я стадия). Магний в норме бло-
кирует ионный канал, ассоциированный с NMDA-
рецептором, и, таким образом, удаление магния
приводит к постсинаптической деполяризации, ко-
торая опосредует дальнейший приток натрия и
кальция в постсинаптическую клетку (5-я стадия).
• Приток кальпия приводит к активапии постсинапти-
ческой протеинкиназы (6-я стадия), которая отвечает
за инициацию ОП — постсинаптическое событие.
• Поддержание ОП, помимо необходимости актива-
ции протеинкиназы (7а стадия), требует включения
большего числа постсинаптических глутаматных
рецепторов, изменения генной экспрессии (7с ста-
дия), а также модификации высвобождения нейро-
трансмиттера (7Ь стадия), т.е. увеличения его выс-
вобождения в ответ на афферентный импульс.
Пресинаптическая модификация, необходимая для
поддержания ОП, означает, что постсинаптическая
клетка должна производить диффузный вторичный
сигнал, который может воздействовать на пресинап-
тическую терминалы Что это за диффузный мес-
сенджер, пока неясно, однако существует предпо-
ложение, что это метаболиты арахидоновой кисло-
ты, характеризующиеся высокой способностью
проникновения в различные структуры, оксид азо-
та, углекислый газ и фактор активации тромбоци-
тов. В настоящее время неизвестно, имеет ли хотя
бы одно из этих веществ отношение к ОП.
В ряде случаев в синапсах может возникать от-
сроченная депрессия (ОД), например в синапсах мо-
ховидных волокон поля САЗ гиппокампа. Так, в от-
личие от ОП, ОД, по-видимому, опосредуется пре-
синаптическими метаботропными глутаматными
рецепторами.
99
ГЛАВА 46. Пластичность нервной системы
и нейротрофические факторы I: периферическая нервная система
Анатомия перифери-ieci.cro нерва
Периферия
глиальных клеток ЦНС)
Трубка базальной Клетка спинномозгового узла
мембраны
Шванновская клетка
Миелин
Аксон
Неиротрофиче кие
фактора
Нейротрофические фиктор J
ФРН МНТФ
• Мозгове?! нейротрофический фактор
(МНТФ). Неиротоофинь {НТ) 3. 4, 5
( 2 ) Нейропозтические фак оры
Цилиарный нейротрофический фактор (ЦНТФ)
• Лейкемию ингибирующим фактоп {ЛИФ)
Тромбоцитарный фактор роста
Интерлейкины
Эпидермальные факторы роста
• Фактор роста фибробластов
• Инсулиновый фактор роста
• Трансформирующие факторы роста
• Глиальный нейротрофический фактор (ГНТФ)
ОНеиропьлтнды
(например, АКТГ и вазоинтенальный пептид)
Повреждение
J Эндоневральная трубка
_ фрагменты
миелина
Фра'ментч аксона
Валлеровская
дг хнерация
Поступление макрофагов
• Удаление детрита
• Митогенный стимул к шванное
скин к’еткам - регенерация
швсяновская клетка
может выделять
нейротрофические факторы,
например ФРН
Дистальный отросток
регенерирующего аксона
Базальная мембрана
шванновских клеток,
содержащая индукторы
роста аксона (на* ример
..аминим и фибронектин)
НТ 4, 5
ГРК А
ТРК — тирозинкиназа
в цит эгаазматическоы jaciH
рецептора
ТРК С
ТРК 8
Периферическая нервная система (ПНС) может
в значительной степени восстанавливаться незави-
симо от давности повреждения. В противополож-
ность этому всегда считалось, что восстановление в
центральной нервной системе (ЦНС) невозможно.
Однако в последнее время появились данные о вы-
раженной ее пластичности даже у взрослых, о воз-
можности до некоторой степени реорганизовывать
каждый отдел (см. главу 47).
Восстановление
периферической нервной системы
Поражение периферического нерва приводит к
утрате чувствительности, атрофии мышц и парезу в
них. Однако во многих случаях возможно самостоя-
тельное восстановление нерва, поскольку перифе-
рический аксон может вырасти снова под влияни-
ем благоприятного окружения шванновских клеток.
100
В противоположность этому, в ЦНС, нейроглиаль-
ные клетки (астроциты и олигодендроциты) подав-
ляют рост аксона, хотя большинство нейронов ЦНС
способно нарастить аксоны заново.
При поражении периферического нерва дис-
тальная часть аксона погибает по механизму вал-
леровской дегенерации. Последняя приводит к
удалению и обновлению как аксональных, так и
миелиновых компонентов. Однако при этом шван-
новские клетки продолжают делиться в полости ба-
зальной мембраны, которая окружает все нервные
волокна. Эти столбики шванновских клеток окру-
жены базальной мембраной, называемой эндонев-
ральными трубками, и обеспечивают условия для
роста аксона.
После поражения дегенерирующие волокна не-
рва индуцируют поступление в пораженный участок
макрофагов, которые дают митогенные стимулы
шванновским клеткам. Регенерирующий аксон на-
чинает ветвиться уже в течение первых часов по-
вреждения и контактирует с базальной мембраной
шванновских клеток с одной стороны и мембраной
шванновских клеток — с другой. Базальная мембра-
на шванновских клеток особенно важна для ветвле-
ния аксона, так как содержит ряд молекул, которые
являются сильными стимуляторами роста аксона in
vitro (например, ламинин и фибронектин).
Кроме того, поддерживая рост аксона, шваннов-
ские клетки выделяют ряд нейротрофических фак-
торов, в том числе фактор роста нервов (ФРН; см.
ниже). Так, шванновские клетки создают простран-
ство, в котором можетрегенерировать аксон, и обес-
печивают благоприятное гуморальное нейротрофи-
ческое окружение. Они помогают аксону прорасти
к мишени, которую он должен иннервировать с по-
мощью эндоневральной трубки. Вместе с тем рост
аксонов после повреждения может прорасти не туда
и не в полной мере: улиц с поражением глазодвига-
тельного нерва при взгляде вниз поднимается верх-
нее веко в силу аберрантной регенерации.
В отличие от аксонального повреждения, гибель
тела нейрона (в передних рогах или спинномозго-
вых корешках) вызывает необратимую гибель аксо-
на периферического нерва. Примерами заболева-
ний, при которых гибнут й-МН, служат полиомие-
лит и болезнь двигательного нейрона (БДН), а также
ряд воспалительных и паранеопластических синдро-
мов, при которых гибнут нейроны спинномозговых
узлов (см. главы 52 и 54). При вышеперечисленных
заболеваниях поражение аксонов вторично по отно-
шению к гибели тел нейронов, поэтому регенерация
невозможна. Попытки спасти погибающие а-МН с
помощью доставки нейротрофических факторов на
периферию не увенчались успехом (см. главу 52).
Нейротрофические факторы
С момента описания первого нейротрофическо-
го фактора — ФРН - было открыто большое их ко-
личество. Эти факторы относятся к различным се-
мействам и взаимодействуют с различными рецеп-
торами, оказывая влияние как на нейроны, так и на
клетки, не относящиеся к ним. Многие из этих ре-
цепторов состоят из субъединиц, из которых одна
или несколько имеют общие места связывания для
всех представителей семейства нейротрофических
факторов. Непример, семейство нейротрофинов и
рецепторы к тирозинкиназам (ТРК) используют в
качестве сигнального механизма функцию цитоп-
лазматических ТРК.
Многие нейроны реагируют на нейротрофические
факторы как in vitro, так и in vivo у эксперименталь-
ных животных. Однако несмотря на это, назначение
нейротрофических факторов больным с нейродеге-
неративными заболеваниями и полинейропатиями
приводит к незначительному улучшению. Это гово-
рит о том, что данные заболевания, по-видимому, не
связаны со специфической недостаточностью ней-
ротрофических факторов (см. главу 52). Недавно
были получены положительные результаты прямо-
го введения нейротрофических факторов в парен-
химу мозга (эффекта не получено при введении их
в цереброспинальную жидкость и на периферию),
например глиального нейротрофического фак-
тора (ГНТФ) при болезни Паркинсона (см. главы
39 и 52).
ГЛАВА 47. Пластичность нервной системы
и нейротрофические факторы II: центральная нервная система
Представительство коры, относящееся
к незашитому глазу, доминирует
Зрительная кора: пластичность в процессе развития
Прамл
Монокулярная депривация
—Левая сторона
Клетка коры
Продукция нейро-
трофических факто-
ров при активации
Первичная
зрительная кора: принцип
селективности по стороне |
Глаза
Левая
сторона
IV
V
Зашитый
глаз
Гистограмма распределения в соответ-
ствии с селективностью по стороне:
Обозначения «правым*, «левый» и «оба глаза*
говорят о том, откуда поступает информация
в клетку вне IV слоя коры молоды ж обет&ян
Локализация модифицируемого синапса
Правая
сторона
—------------ Левая
063 глаза сторона
------------* Левая
Оба глаза сторона
Активация
клетки коры
импульсом
от ЛКТ
Усвоение нейротро-
фического фактора
обеспечивает выжи-
вание н рост аффе-
рентного волокна
Афферентное волокна
Латеральное коленчатое
тело (ЛКТ)
Правый глаз
Левый глаз
Правая
сторона
Окуловестибулярный рефлекс:
пластичность у взрослых
I Параллельное
волокно
Правая
сторона
Клочок мозжечка
Пуркинье
Ползущее волокно
Этот рефлекс позволяет удерживать
изображение предмета при изменении
положения головы, т,е. движение глаз
Гранулярная клетка G
то чость вмження глаз
К настоя щему времен и получено достаточно дан-
ных о том, что в ЦНС взрослого человека могут про-
исходить регенерация и реорганизация. Однако пла-
стичность в ЦНС опосредована не образованием
новых нейронов, поскольку большинство нейронов
в зрелой ЦНС находится в постмитотическом состо-
янии, а способностью к образованию новых аксо-
нов. Наиболее выражена эта способность в раннем
постнатальном периоде, когда все системы мозга
развиваются, и именно тогда происходят наиболее
значимые модификации.
Механизмы, лежащие в основе пластичности,
полностью не изучены, однако важную роль в этом
играют продукция и усвоение факторов, способству-
ющих росту нейронов и их выживанию (нейротро-
фические факторы).
102
Пластичность развивающегося
зрительного анализатора
В первых исследованиях Хубель и Вайзель пока-
зали, что при рождении волокна поступают в IV слой
первичной зрительной коры (ПЗК) диффузно и лишь
в критический период развития (у кошек это состав-
ляет 3—14 нед, у человека — несколько лет после рож-
дения) эти волокна разделяются и распределяются по
принципу селективности по стороне (см. главу 25).
Разделение волокон зависит от типа импульса-
ции по афферентным волокнам от каждого глаза; чем
она больше, тем выше вероятность, что эти аффе-
рентные волокна будут регулировать определенную
группу корковых нейронов. Так, селективность по
стороне достигается в отсутствие какого-либо состя-
зания между афферентными волокнами, идущими
от глаз, однако она не может быть достигнута при
отсутствии афферентного влияния хотя бы с одно-
го глаза.
Хубель и Вайзель в эксперименте манипулирова-
ли этими волокнами, лишая один глаз афферентной
импульсации путем сшивания его век (монокуляр-
ная депривация), а в последующих экспериментах
проводили обратную процедуру (расшивание). Мо-
нокулярная депривация приводила к распростране-
нию таламического влияния от открытого глаза на
IV слой, при этом впоследствии столбики клеток
правого глаза реорганизовывались таким образом,
что большинство клеток коры контролировалось от-
крытым глазом. Это можно было изменить, расшив
ранее закрытый глаз в критический период, что при-
водило к изменению таламического влияния на
клетки коры, вызываемому активацией синапсов,
которые ранее были функционально инактивирова-
ны. Это было возможно, поскольку времени на
окончательное прорастание аксонов к проекции
было недостаточно. Однако в дальнейшем изначаль-
но инактивированные синапсы зашитого глаза бу-
дут утрачены, поскольку активирующие таламичес-
кие волокна станут полностью контролировать кор-
ковые клетки.
Правильное распределение импульсов в ПЗК и
принцип селективности по стороне важны для раз-
вития многих других зрительных функций ПЗК.
Однако по прошествии критического периода воз-
можности реорганизапии зрительной коры умень-
шаются, но не исчезают.
Пластичность у взрослых
Анализатор общей чувствительности
и окуловестибулярные рефлексы
В настоящее время известно, что чувствительный
анализатор может перестраиваться при нарушении
афферентной импульсации от периферических ре-
цепторов. Так, утрата импульсации от пальца (на-
пример, при его ампутации) не приводит к инакти-
вации зоны коры. Соседние отделы коры, получаю-
щие чувствительную импульсацию от других
пальцев, распространяют свои аксоны на инактиви-
рованную зону коры.
В противоположность этому избыточная аффе-
рентная чувствительная информация приводит к
расширению корковой зоны, реагирующей на нее.
Можно предположить, что активность одного аффе-
рентного волокна индуцирует продукцию нейротро-
фических факторов в постсинаптической клетке,
которые затем связываются с соответствующими
рецепторами на активной пресинаптической терми-
нали, что приводит к росту и поддержанию ее функ-
ции. В этом смысле ЦНС постоянно перестраивает-
ся в зависимости от поступаюшей в нее афферент-
ной информации.
Впоследствии было показано, что выраженные
чувствительные выпадения, например деафферента-
ция конечности, приводит к аналогичным измене-
ниям, т.е. перестройке отделов коры за счет сосед-
них импульсов, и это достигается не только за счет
локального прорастания аксонов из соседних отде-
лов коры.
Другим примером пластичности ЦНС являются
окуловестибулярные рефлексы (см. главы 28 и 37).
Вестибулярный анализатор подает в ЦНС сигнал об
ускорении движения головы, который затем про-
водится в мозжечок по моховидным волокнам. Од-
нако волокна другого типа — ползущие — дают в
мозжечок информацию о том, насколько при этом
смешается изображение предметов на сетчатке (т.е.
о том, насколько движения глаз могут компенси-
ровать перемещение головы). Импульсация по пол-
зущим волокнам нужна для того, чтобы проводить
не только информацию о том, насколько хорошо ре-
ализуется данный рефлекс, но и импульсацию, с по-
мощью которой рефлекс изменяется. Так, если из-
менить соотношение между информацией о поло-
жении глаз и головы, надев на человека призмы,
рефлекс адаптируется к этим изменениям, это воз-
можно благодаря тому, что импульсация по пол-
зущим волокнам может модифицировать импуль-
сацию по параллельным волокнам (и косвенно по
моховидным волокнам) к клеткам Пуркинье по-
средством такого молекулярного механизма, как от-
сроченная депрессия (ОД; см. главу 37).
Стволовые клетки
Во множестве зрелых тканей гибель клеток в ре-
зультате естественных процессов старения компен-
сируется последующей их пролиферацией и диффе-
ренцировкой. Считалось, что в ЦНС взрослого че-
ловека этого не происходит, однако недавно было
показано, что в зрелой ЦНС млекопитающих име-
ются невральные клетки-предшественники. Эти
клетки обнаружены в гиппокампе и паравентрику-
лярно (в субвентрикулярной зоне), и, по-видимому,
из них образуются функционально активные нейро-
ны, однако их роль в пластичности и восстановле-
нии до конца не изучена.
Ограничения восстановительных
возможностей ЦНС взрослого человека
Необходимо представлять, что возможности вос-
становления ЦНС ограничены, потому что:
• нейроны ЦНС находятся в постмитотическом со-
стоянии, количество стволовых нервных клеток
ограничено и они расположены только в опреде-
ленных местах;
• глиальные клетки ЦНС тормозят рост аксонов (см
главу 4).
Астроциты образуют сигналы, которые угнета-
ют рост аксонов, а олигодендроциты продуцируют
ряд факторов, приводящих к остановке роста ак-
сонов. Это проявляется при поражении ЦНС, ког-
да глиальные клетки начинают делиться и образу-
ют рубец.
ЧАСТЬ V. Заболевания нервной системы
ГЛАВА 48. Неврологическое обследование
когнитивным
функций
Исследование чувствительности
Обоняние
Острота зрения. Поля зрения.
1 Исследование глазного дна.
Движения глаз. Зрачки.
Чувствительности. Жевательные
Г мышцы- Роговичные рефлексы.
Сила мимических мышц
Слух и равновесие
Движений мягкого нёба
и кашель
Глоточный рефлекс
| Внешний вид языка, его движения и сила мышц
Повороты головы
и пожимание плечами
(Исследование двигательных функций
Осмотр (атрофия/гипертрофия мышц,
насильственные движения, фвсцмкуляцни,
шрамы)
[Мышечный тонус“|
(повышен при болезни Паркинсона и по-
ражении ЦМН, понижен при поражении
ПМН и мозжечка)
[Сила мышц j
(О — нет движений, 1 — минимальные
движения, 2 — движение без веса. 3 —
движение против собственного веса, 4 —
движение против сопротивления, 5 —
нормальное движение)
[рефлексы |
[ Координация
[Рефлексы
(О — отсутствуют, * — присутствуют при
повторном вызывании, ++ — нормаль-
ные, +++ — оживлены)
|Подошвенные рефлексы~[
(патологический разгибательный рефлекс,
нормальный сгибательный рефлекс)
Лодыжка
Трицепс
Колено
Бицепс
Супи-
натор
Рука 6
Рука 2
Рука 1
Нога 6
Нога 5
Нога 4
Нога 3
Нога 2
Рука 5
Рука 4
L Черепномозговые нервы
\ Рука 3
Осмотр — шрамы, изменения кожи
Чувство прикосновения ваткой
Болевая чувствительность (уколы)
Мышечно-суставное чувство (от дис-
тальных суствов к проксимальным)
{Рука 1-4, 6) (Нога 1, 3, 4, 5)
Вибрационную чувствительность
проверяют камертоном, начинают
дистально и завершают проксимально
(Рука 1-5, Нога 2. 3, 4, 6)
Температурную чувствительность
исследуют пробирками с холодной
и горячей водой
Нога 1
Неврологическое обследование можно разделить
на две части.
Исследование когнитивных функций
Общее
Ориентация в месте, времени и собственной лич-
ности: если пациент не может правильно ответить на
эти вопросы (и при этом у него нет речевых наруше-
ний), это означает, что имеет место острая спутанность
или тяжелая деменция. В последнем случае исследо-
вание когнитивных функций не будет полезно.
Функция лобных долей
Спонтанная речь: исследуют, какое количество
слов, начинающихся с одной и той же буквы, напри-
мер с, и может назвать больной, или же слов, обо-
значающих определенное понятие, например живот-
ных, в течение 60 или 90 с.
Внимание: способность запоминать и немедленно
воспроизводить перечень предметов или имя и адрес.
104
Примитивные рефлексы: выпячивание губ при
приближении молоточка, хватание молоточка,
скользящего по ладони.
Функция теменных долей
Гнозис: фиксация или игнорирование, зрительных
и соматосенсорных стимулов в противоположном
чувствительном поле.
Апраксин: пациент не может жестикулировать или
копировать жесты, выполнять задания (например,
причесываться).
Зрительно-пространственная функция: способ-
ность копировать рисунок.
Функция височных долей
Антеградная память: способность воспроизво-
дить имя и адрес, названные пациенту, через 5 мин
после произнесения.
Речь: оценка функции речи включает оцен-
ку спонтанной речи, называния предметов, по-
вторения фраз, выполнения команд, чтения и
письма.
Черепно-мозговые нервы
Обонятельные нервы: к каждому носовому ходу
раздельно подносят несколько пахучих веществ.
Зрительные нервы: остроту зрения каждого глаза
исследуют с использованием стандартной таблицы.
Поля зрения исследуют, предлагая пациенту пока-
зать середину линии (см. главу 24). Глазное дно ис-
следуют офтальмоскопом, с помощью которого
можно обнаружить патологию сетчатки или зритель-
ных нервов (например, отек или атрофию соска зри-
тельного нерва). Исследование цветного зрения про-
водят с помощью таблиц Ишихары, кроме того, ис-
следуют зрачковые реакции.
Глазодвигательный, блоковой и отводящий нервы:
обращают внимание на наличие птоза или измене-
ний ширины зрачков (например, синдром Горнера;
см. главу 42). Затем исследуют движения глаз во всех
направлениях, спрашивают, нет ли двоения в глазах
(см. главу 40).
Тройничный нерв: проверяют чувствительность в
проекции всех трех зон иннервации тройничного
нерва, а также силу жевательных мышц. В ряде слу-
чаев исследуют роговичный рефлекс, слегка дотра-
гиваясь до роговицы ваткой. В норме происходит
зажмуривание.
Лицевой нерв: исследуют силу мимических мышц,
например способность крепко зажмурить глаза, на-
дуть щеки или сделать губы трубочкой. В норме ис-
следователь не может преодолеть эти движения.
Преддверно-улитковый нерв: исследуют слух,
произнося шепотом несколько слов с одной и с дру-
гой стороны. Можно провести исследование с по-
мощью камертона.
Языкоглоточный и блуждающий нервы: пацент
широко открывает рот и говорит «а-а», при этом врач
оценивает подвижность мягкого нёба. Шпателем
проверяют глоточный рефлекс, дотрагиваясь им до
задней стенки глотки, в ответ на это возникает дви-
жение мягкого нёба. В ряде случаев полезно иссле-
довать кашлевой рефлекс.
Добавочный нерв: пациент поворачивает голову в
стороны и пожимает плечами. В норме исследова-
тель не может преодолеть эти движения.
Подъязычный нерв: исследователь осматривает
язык на предмет атрофии и фасцикуляций, затем
просит больного высунуть язык и отмечает, нет ли
девиации. Силу мышц языка исследуют, обращаясь
к пациенту с просьбой уткнуться языком в одну, а
затем в другую щеку. Больной сможет это сделать,
если у него нет слабости мимической мускулатуры.
Исследование движений конечностей
В исследование двигательной системы включе-
ны следующие элементы:
Осмотр: насильственные движения, атрофия, па-
резы, фасцикуляций, шрамы или деформации.
Тонус мышц: проводят пассивные движения в су-
ставах и оценивают наличие ригидности. Она наблю-
дается при болезни Паркинсона или поражении цен-
тральных мотонейронов (ЦМН). Мышечный тонус
снижается при поражении периферических мото-
нейронов (ПМН) и мозжечка (см. главы 32—34).
В некоторых случаях тонус повышается, потому что
пациент не может расслабиться.
Сила мышц: силу и объем активных движений
оценивают по шкале Британского совета медицин-
ских исследований (см. рисунок).
Координация: координацию движений в верхних
конечностях оценивают по способности пациента
дотронуться пальцем до движущегося перед глаза-
ми пациента пальца исследователя. Эта способность
может быть нарушена при парезе, расстройствах чув-
ствительности и заболеваниях мозжечка. Пациента
также просят быстро пронировать и супинировать
обе кисти. Координацию движений нижних конеч-
ностей исследуют при помоши пяточно-коленной
пробы. Кроме того, исследуют походку больного,
ходьбу след в след.
Рефлексы исследуют, ударяя молоточкам по су-
хожилиям мышц. Рефлексы могут отсутствовать,
быть сниженными, нормальными или оживленны-
ми. В последнем случае предполагают поражение
ЦМН, а при отсутствии рефлекса или его снижении
- поражение дуги моносинаптического рефлекса в
любом месте на ее протяжении (см. главу 33).
Подошвенные рефлексы: проводят рукояткой мо-
лоточка по латеральному краю ступни. В норме воз-
никают веерообразное разведение пальцев и сгиба-
ние большого пальца (сгибательный или нормаль-
ный рефлекс). Если пальцы разгибаются и больной
не отдергивает ногу, предполгают поражение ПМН.
Исследование чувствительности
Чувствительность на конечностях и различных
дерматомах исследуют с помощью нескольких при-
емов.
Прикосновение: до кожи дотрагиваются ваткой,
но предварительно дотрагиваются до лица, прове-
ряя, хорошо ли пациент чувствует прикосновение
ваткой вообще.
Уколы: наносят уколы притуплённой одноразовой
иглой.
Температурная чувствительность: к больному при-
касаются пробирками с холодной и теплой водой.
Вибрационная чувствительность: на I пястно-фа-
ланговый сустав ступни ставят камертон. В норме
пациент чувствует вибрацию, а не просто прикос-
новение. Если вибрация не чувствуется, камертон
перемещают проксимальнее.
Мышечно-суставное чувство (МСЧ) исследуют,
сгибая и разгибая большой палец руки или ноги, при
этом необходимо убедиться, что пациент понимает
значение проводимого исследования. Эти движения
должны быть очень легкими, поскольку у человека
МСЧ развито очень сильно. Если пациент не может
определить направление движения пальца, прово-
дят сгибание и разгибание в более крупном (прокси-
мальном) суставе, как и в случае с вибрационной
чувствительностью.
105
ГЛАВА 49. Обследование нервной системы
J Нейровизуализацмя |
слуховые ВП
Ангиография
Анамнез
и обследование
i краниограммы и другие
пеклi поамм! . КТ, МРТ),
применяются при экспе-
риментальных исследова-
ниях ОФЭКТ, фМРТ и ПЭТ
Биопсия \
мозга
звп
Биопсия мышц
Дельтовидная
мышца
Невроло- Стволовые ; у
гическое ОГ1
Биопсия
нерва
Лучевой
нерв
Анализ на: две , коли
чество и морфологию неР®
клеток, белок, уровень
глюкозы, олигокло-
нальн lie антитела
Бипосия
мышц
(прямая
мышца
бедра)
|люм^мьн^^ун|щ^|
Конец
спинного мозга
Икроножный
Анализ крови на:
• формулу крови, СОЭ,
• функцию почек, глюкозу,
• функцию печени,
• функцию щитовидной железы,
• креатинин,
• креатининфосфокнназу (КФК),
• витамин В,, и фолиевую кислоту,
• генетические маркеры,
• ферменты лейкоцитов,
• иммуноглобулины сыворотки
{электрофорез),
• липиды,
• медь и церулоплазмин,
• аутоантитела,
свертываемость
Двигательные
волокна
, Скорость, мс1
У, см Точка ре-
Чувствительные волокна гистрации
Потенциал действия
чувствительных волокон
Точка •
стимуляции
ЭМГ (актив-
ная запись
игольчатым
электродом)
Амплитуда
Скорость, мс'-----
Обследование нервной системы включает прове-
дение спепиальных процедур и стандартных анали-
зов. Однако важнее всего изучить анамнез и провес-
ти неврологическое обследование, так как это позво-
ляет установить причину и локализовать очаг. В этой
главе обсуждаются методы обследования неврологи-
ческих больных и показания к их проведению.
Анамнез и неврологическое обследование
Анамнез и неврологическое обследование позво-
ляют уточнить характер заболевания, его развитие и
соматические заболевания, которые могут иметь от-
ношение к неврологическому.
Анализы крови
Нужно провести большое количество анализов
(примеры даны на риснуке).
Нейровизуализация
• Рентгеновские снимки редко помогают установить
причину неврологического заболевания, но часто
позволяют предположить, осложнением какого за-
106
болевания явились неврологические расстройства
(например, при раке легкого).
• Компьютерная томография (КТ) позволяет иссле-
довать головной мозг, череп и нижний отдел позво-
ночника. С ее помощью можно диагностировать
опухоли, переломы черепа и обширные инсульты.
Она применяется широко, но не обладает высокой
разрешающей способностью, особенно при патоло-
гических процессах в задней черепной ямке и шей-
ном и грудном отделах спинного мозга.
• Магнитно-резонансную томографию (МРТ) про-
водят, если пациент хорошо переносит шум и замк-
нутое пространство. С ее помощью можно получить
детальные снимки головного и спинного мозга, ис-
пользуя различные последовательности импульсов.
При МРТ не происходит облучения, и ее диагнос-
тическая значимость и возможности применения в
клинике возрастают.
• Магнитно-резонансная ангиография и венография
(МРА/МРВ) позволяет визуализировать основные
кровеносные сосуды мозга. Она используется для
диагностики стенозов экстракраниальных артерий,
аневризм и тромбоза венозных синусов, но менее
чувствительна, чем традиционная ангиография.
• При ангиографии проводят катетер в сонные или по-
звоночные артерии и вводят небольшое количество
контрастного вещества. Затем по мере прохождения
его через сосудистое русло делают снимки. Процедура
инвазивна и иногда сопровождается осложнениями,
однако необходима для диагностики любой сосуди-
стой патологии (например, стеноза сосуда, аневризм,
артериовенозных мальформаций и тромбоза сину-
сов). Также она может быть использована для диаг-
ностики сосудистых заболеваний спинного мозга.
• Миелография в настоящее время проводится редко,
поскольку МРТ, в отличие от нее, является неинва-
зивным методом, обладающим более высокой раз-
решающей способностью. Однако в ряде случаев
миелография может быть полезной. При этой про-
цедуре в субарахноидальное пространство при люм-
бальной пункции вводят контрастное вещество, ко-
торое не поглощает ионизирующее излучение.
• Однофотонная эмиссионная компьютерная томог-
рафия (ОФЭКТ) проводится с использованием ра-
диоактивных изотопов и применяется для исследо-
вания перфузии мозга. Разрешение метода низкое.
• Функциональная МРТ (фМРТ) позволяет оценить
поглощение кислорода в различных отделах мозга.
• Позитронно-эмиссионная томография (ПЭТ) ос-
нована на регистрации поглощения позитронов ве-
ществами, которые связываются с определенными
структурами мозга. Она используется только в экс-
периментальных исследованиях.
Электродиагностика
• Электрокардиография (ЭКГ) — это метод регист-
рации сердечных сокращений, который применя-
ется при обследовании неврологических больных,
особенно с первично-мышечными заболеваниями,
пароксизмами утраты сознания и некоторыми на-
следственными болезнями.
• Электроэнцефалография (ЭЭГ) позволяет оцени-
вать биоэлектрическую активность головного моз-
га, применяется у больных с нарушением сознания
и эпилепсией.
• С помощью электронейрографии (ЭНГ) оценивают
чувствительные и двигательные волокна нервов, их
стимулируют и исследуют вызванные потенциалы
(ВП). Стимуляция проводится в различных точках, а
регистрация - на мышце, которую нерв иннервирует.
Важными характеристиками являются амплитуда и
скорость ВП. При демиелинизации снижается скорость
проведения, тогда как при гибели аксонов умень-
шается амплитуда ответа, но скорость проведения не
меняется. Метод применяется для установления вида
полинейропатии (аксональная или демиелинизиру-
ющая), а также распространенности поражения нер-
вов (локальные или генерализованные нейропатии).
• При электромиографии (ЭМГ) в мышцу вводят регис-
трирующий электрод и ее электрическую активность,
применяется при диагностике заболеваний мышц.
• ВП применяются для оценки зрительного (зритель-
ные вызванные потенциалы — ЗВП), слухового
(стволовые вызванные слуховые потенциалы) ана-
лизаторов и периферических нервов рук и ног (со-
матосенсорные вызванные потенциалы). С помо-
щью ВП стимулируют периферический рецептор
(глаза, уха или срединного/заднего большеберцово-
го нерва) и регистрируют корковый ответ. Это по-
зволяет оценить проводимость в пределах анализа-
тора по периферическому и нейтральному компо-
нентам. Чаще всего ЗВП используют при рассеянном
склерозе для диагностики бессимптомной демиели-
низации путей зрительного анализатора.
• Транскраниальная магнитная стимуляция (ТКМС)
позволяет оценивать время прохождения импульса от
стимулируемой двигательной коры до мышцы. ТКМС
редко используется в клинической практике и приме-
няется для оценки пелостности пирамидного пути.
• Исследование порога температурной чувствитель-
ности применяется для изучения состояния мелко-
калиберных волокон. Оно проводится у пациентов
с нарушением температурной чувствительности в
кистях и стопах. Этот метод не применяется в по-
вседневной практике в большинстве клиник.
Анализ цереброспинальной жидкости
Цереброспинальную жидкость (ЦСЖ) можно по-
лучить из различных частей организма, но обычно —
при люмбальной пункции с помощью тонкой иглы,
которую проводят в субарахноидальное пространство
в поясничном отделе позвоночника. В норме ЦСЖ
прозрачна, и прежде чем отправить ее на анализ, при-
нято измерять ее давление сменными манометричес-
кими трубками. Анализ ЦСЖ включает:
• Определение количества и типа клеток (цитоз),
обычно он повышается при инфекциях (менингит).
• Посев ЦСЖ для выявления микроорганизмов,
включая мазок по Граму на менингит.
• Определение уровня глюкозы, который снижается при
инфекциях (менингит) и опухолях мозговых оболочек.
• Определение уровня белка, который повышается при
полинейропатиях, опухолях и других процессах, бло-
кирующих субарахноидальное пространство.
• Определение олигоклональных антител, наличие
которых свидетельствует о синтезе иммуноглобули-
нов в ЦНС (рассеянный склероз).
Биопсия мышцы или нерва
При заболеваниях нервов и мышц производится их
биопсия, для определения природы заболевания.
Чаще всего исследуют лучевой и икроножный нервы,
а также прямую мышцу бедра и дельтовидную мышцу.
Биопсия мозга
Биопсию мозга производят у больных с опухоля-
ми мозга для подтверждения диагноза и прогнози-
рования течения болезни. При прогрессирующих
неврологических заболеваниях, точную причину ко-
торых трудно установить, также производится био-
псия мозга, при этом необходимо определить, петли
воспалительной реакции в кровеносных сосудах (вас-
кулит), а также прионовой болезни. В последнем слу-
чае необходимо соблюдать правила асептики.
107
ГЛАВА 50. Нейрохимические нарушения I:
аффективные расстройства и шизофрения
S-ГТ/серотониновые путн^
Серотониновый нейрон 1
Терминальные отделы
(кора, гиппокамп и т.д.)
на постеннаптичесмой клетке су-
ществует много различным 5-ГТ-
5-ГТ^ — ауторецептор, -* Метаболизирует-
ингибирующий дальней- Ся различными
шее высвобождение 5-ГТ ферментами.
включая моно-
аминооксидазу,
5-ГТИУК
Позитивные симптомы:
Негативные симптомы:
1 Лобная кора |
•эмоциональная тупость,
•бедносп речи,
•равнодушие
называют
1-го типа)
•бред.
•галлюцинации,
•формальные расстрой-
ства мышления (иногда Nu leus ‘«‘""bens,
. - миндалевидное тело
шизофренией I____—__ _______
Вентралыде от-
делы таламусд
Норма Шизофрения
Причины психических заболеваний различны,
они сопровождаются аффективными расстройства-
ми, при которых изменяется настроение от депрес-
сии к мании. Такие расстройства называются бипо-
лярными. Депрессия характеризуется снижением
настроения, апатией, низкой самооценкой, стрем-
лением избегать окружающих в сочетании с утратой
либидо, анорексией и ранним пробуждением. Ма-
ния, в отличие от депрессии, сопровождается эйфо-
рией, бредом и мыслительной гиперактивностью.
Шизофрения — заболевание, которое протекает с
такими психическими нарушениями, как слуховые
галлюцинации, бред, формальные расстройства
мышления и нарушения поведения.
108
Депрессия
Причина эндогенной депрессии неизвестна, од-
нако, имеется указание на наследственный компонент.
Для лечения депрессии применяются психотера-
пия, электросудорожная терапия (ЭСТ), чаще всего
антидепрессанты большинство из которых явля-
ются ингибиторами обратного захвата норадренали-
на (норэпинефрина) и/или серотонина (5-ГТ). Ре-
же используются ингибиторы моноаминоксидазы
(ИМАО). Поскольку и ингибиторы обратного зах-
вата и ИМАО увеличивают количество норадрена-
лина (норэпинефрина) и/или серотонина в синап-
тической щели, а следовательно, усиливают действие
этих трансмиттеров, некоторые ученые отрицали
связь депрессии с недостаточной активностью мо-
ноаминергических систем. Эта простая идея отно-
сится к моноаминергической теории депрессии и
является полезной, однако не в полной мере объяс-
няет механизм развития этого заболевания.
Ингибиторы обратного захвата аминов
Трипиклические антидепрессанты (например,
имипрамин, амитриптилин) оказывают сильное ан-
тидепрессивное действие. Некоторые препараты
(например, амитриптилин) обладают седативными
свойствами, поэтому показаны больным с возбуж-
дением и тревогой (см. главу 51).
Препараты, являющиеся селективными ингиби-
торами обратного захвата серотонина (СИОЗС, на-
пример, флуоксетин) не дают побочных вегетатив-
ных эффектов, в отличие от трициклических анти-
депрессантов, но могут вызывать тошноту и другие
желудочно-кишечные расстройства.
Ингибиторы моноаминоксидазы
Ингибиторы ИМАО (например, фенелзин) явля-
ются необратимыми ингибиторами МАО. Они столь
же эффективны, как и антидепрессанты, но оказы-
вают нежелательное побочное действие (например,
вызывают постуральную гипотензию) и взаимодей-
ствуют с продуктами питания, содержащими тирамин,
что ограничивает их применение. Моклобемид —
новый препарат, селективно инигибирующий МАО-А,
не обладающий большинством нежелательных по-
бочных свойств фенелзина.
Шизофрения
Шизофрения является в значительной степени
наследственным заболеванием. Риск заболевания
шизофренией у кровных родственников составляет
10%, а у ребенка, оба родителя которого страдают
этим заболеванием, он повышается до 40%.
Согласно ранним представлениям, шизофрения
развивается в результате образования эндогенного
вещества, вызывающего психоз, которое может быть
трансметилированным дериватом дофамина. Впос-
ледствии эта теория преобразовалась в представле-
ние о том, что шизофрения возникает в результате
гиперактивности центральных дофаминергических
путей, особенно мезолимбических проекпий с отно-
сительной интактностью дофаминергического ниг-
ростриарного пути (см. главу 38).
Впоследствии это представление преобразова-
лось в теорию о том, что шизофрения может быть
вызвана снижением дофаминергической актив-
ности мезокортикальной системы, тогда как пси-
хотические симптомы вызываются повышением
дофаминергической активности мезолимбической
системы. Действительно, у больных шизофренией
отмечается гипоперфузия лобных долей, что, в со-
четании с результатами предыдущих исследований,
которые показали непрогрессирующее расширение
желудочков мозга, привело к возникновению гипо-
тезы о том, что шизофрения может быть заболева-
нием, связанным с дефектами развития нервной си-
стемы. Однако какая именно часть мозга развивает-
ся неправильно, неизвестно, вероятнее всего, это
миндалевидное тело, лобные и височные доли.
Основным направлением влечении шизофрении
остается применение антагонистов дофаминовых
рецепторов 5 подтипов (Dl D5; см. Приложение 1).
Эти вещества (например, хлорпромазин) называют-
ся антипсихотическими препаратами или нейролеп-
тиками. Как они уменьшают выраженность симп-
томов шизофрении, точно неизвестно. Корреляция
между дозами антипсихотических средств и их аф-
финностью к Dj-рецепторам подтверждает факт, что
важным механизмом их действия является блокада
этих рецепторов.
В большинстве случаев антипсихотическая тера-
пия требует наблюдения за больными шизофрени-
ей в течение многих лет. Часто отмечаются рециди-
вы даже у больных, получающих лечение. К сожале-
нию, нейролептики также блокируют дофаминовые
рецепторы базальных ганглиев, что приводит к раз-
витию нежелательных двигательных расстройств
(паркинсонизму; см. главу 39). Блокада В2-рецепто-
ров в гипофизе вызывает увеличение выброса про-
лактина и эндокринные расстройства (гинекомас-
тию, галакторею; см. главу 43). Многие нейролеп-
тики блокируют мускариновые рецепторы (что
вызывает сухость во рту, запор и нарушение зрения),
(e-адренорецепторы (постуральная гипотензия) и
гистаминовые Ш-рецепторы (седативный эффект).
Некоторые новые препараты (атипичные нейро-
лептики, например клозапин, рисперидон, оланза-
пин) не вызывают двигательных нарушений по не-
ясным причинам. Клозапин применяется у больных,
нечувствительных к другим препаратам, поскольку
вызывает нейтропению или агранулоцитоз у 4% боль-
ных. Возрастает интерес к применению рисперидона
и других атипичных нейролептиков при шизофрении,
так как они лучше переносятся больными.
ГЛАВА 51. Нейрохимические нарушения II:
тревожные расстройства
Медиальный срез полушария мозга
Поясная извилина
Боковом желудочек
Кора головного мозга
Срез среднего мозга
Красное
ядро
Голубое ядро
Большое
ядро шва
Гиппокамп/прозрачная
- перегородка
( Миндалевидное тело
Черная
субстанция
Г ипоталамус
Основные места связывания
анксиолитических препаратов
ОСТРОЕ -рЕВЭЖНОЕ РАССТРОЙСТВО
1. Бензодиазепины (дназе ia* г — быстро начи
ьают действовать, седативный эффект, зависи-
мость, прием только i ечение 2—4 недель
2. Препараты, влияющие на 5-ГТ-рецепторы
5-ГТ,-агонисты • буспирон
ХРОНИЧЕСКОЕ ТРЕВОЖНОЕ РАССТРОЙСТВО
3. Антидепрессанты — ве «лафаксин (при гене-
рализованных тревожных расстройствах), селек-
тивные ингибиторы обратного захвата серото-
нина • флувоксамин (при обсессивно-компуль-
сивных расстройствах), • циталопрам (при
панических атаках) — ингибиторы МАО • мок-
лобемид (при социофобиях)
4. Снижение выраженности вегетативных симп-
томов (3-блокаторы
• пропранолол
Околоводопроводное
серое вещество
Симпатические аффекты:
• тахикардия
• сердцебиение
• сухость во рту
• диарея
Тревога
Тревога - это нормальная эмоциональная реак-
ция на угрожающие или потенциально угрожающие
ситуации, которая сопровождается повышенной
симпатической активностью. При тревожных рас-
стройствах больные испытывают тревогу, которая
непропорциональна стимулу, иногда тревога возни-
кает в отсутствие всякого стимула. У тревожных рас-
стройств нет никаких органических механизмов,
симптомы возникают при гиперактивности облас-
тей мозга, опосредующих тревогу в норме. Психи-
ческие расстройства развивающиеся без какой-либо
патологии мозга, называются неврозами.
Тревожные расстройства разделяют на 4 группы:
генерализованные тревожные расстройства, паничес-
кие атаки, стрессовые реакции и фобии. В механизме
формирования тревоги участвуют многие нейро-
трансмиттеры. особенно у- аминомасляная кислота
(ГАМК), и 5-гвдрокситриптамин (5-ГТ). Поскольку
ПО
внутривенные инъекпии холецистокинина (ХПК) у
человека приводят к развитию панической атаки,
полагают, что в генезе тревожных расстройств при-
нимают участие различные медиаторы. Последнее
до конца не изучено.
Лечение легких тревожных расстройств проводят
с помощью психотерапии, но при тяжелых исполь-
зуют короткие курсы анксиолитиков. Бензодиазепи-
ны (например, диазепам) усиливают ГАМК-ергичес-
кое торможение (см. главу 53) во многих отделах
мозга, участвующих в формировании тревоги, в ча-
стности в большом ядре шва (БЯШ). Трицикли-
ческие антидепрессанты (например, амитриптилин),
обладают анксиолитической активностью, но меха-
низм их действия неизвестен. Антагонисты («-адре-
норецепторов используются в терапии ситуационной
тревоги (например, у музыкантов), когда основны-
ми симптомами являются сердцебиение и тремор.
Ученые предпринимали попытки открыть неседа-
тивные анксиолитики, что привело к испытанию
некоторых препаратов, специфически взаимодей-
ствующих с серотониновыми рецепторами. Эти пре-
параты не давали выраженного эффекта.
Тревожные расстройства
Генерализованные тревожные расстройства харак-
теризуются психическими и соматическими симп-
томами. Психические симптомы включают устраша-
ющие предчувствия, нарушения внимания, раздра-
жительность, беспокойство, что часто связано с
гиперактивностью симпатической системы.
Фобические тревожные расстройства проявляют-
ся теми же симптомами, что и генерализованные, но
возникают при определенных обстоятельствах (на-
пример, арахнофобия при появлении паука). В от-
личие от них, панические атаки — это приступы
тревоги с преобладанием соматических симптомов
(например, удушье, сердцебиение, боль в груди, по-
тливость и тремор).
Бензодиазепины
Бензодиазепины (например, диазепам) являются
центральными депрессантами и в высоких дозах об-
ладают снотворным эффектом при приеме на ночь,
что приводит к седации и уменьшает тревогу в днев-
ное время. Бензодиазепины обсуждаются в главе 53,
их основными побочными эффектами являются
сонливость, снижение внимания, возбуждение и
атаксия. При тревожных расстройствах бензодиазе-
пины следует принимать в течение 2—3 нед, посколь-
ку более длительный курс лечения может вызвать
зависимость. В этом случае препарат отменяют, что
вызывает синдром отмены, который характеризуется
тревогой, тремором, потливостью и бессонницей,
т.е. симптомами, характерными для тревожных рас-
стройств.
Структуры мозга, с которыми
взаимодействуют бензодиазепины
Области мозга, с которыми взаимодействуют
бензодиазепины, оказывая анксиолитическое дей-
ствие, исследовали в опытах на крысах при помо-
щи инъекций небольшого количества препарата
через канюли в различные отделы мозга. Оказалось,
что в основном анксиолитические эффекты бен-
зодиазепинов опосредуют структуры ствола и лим-
бической системы. Исследования мозгового кро-
вотока и метаболизма глюкозы у людей с помощью
позитронно-эмиссионной томографии (ПЭТ) не
показали выраженных различий между здоровы-
ми людьми и больными с тревожными расстрой-
ствами.
Серотонин
Тела серотонинергических нейронов (5-ГТ) нахо-
дятся в большом ядре шва среднего мозга и проеци-
руются на многие другие области мозга, в частности
те, которые, как считается, играют важную роль в
формировании тревоги (гиппокамп, миндалевидное
тело, лобная кора; см. главу 50). У крыс поражение
большого ядра шва дает анксиолитический эффект,
тогда как стимуляпия 5-ГТ1А-ауторепепторов такими
агонистами, как 8-гидрокси-ПРАТ, приводит к раз-
витию тревоги. Оказалось, что серотонин играет бо-
лее важную роль в развитии тревоги. Было показано,
что бензодиазепины уменьшают обмен 5-ГТ в мозге
и при микроинъекциях в большое ядро шва умень-
шается выраженность импульсации и возникает ан-
ксиолитический эффект. Однако стимуляция постси-
наптических 5-ГТ1А-репепторов в лимбической сис-
теме вызывает тревогу. Эти противоположные пре- и
постсинаптические реакции можно объяснить на
примере буспирона - частичного агониста 5-ГТ1А, ко-
торый относительно эффективен и начинает действо-
вать только спустя несколько недель. Специфичес-
кие антагонисты 5-ГТ2-рецепторов (например, онда-
сетрон) и 5-ГТ2-рецепторов (например, ритансерин)
не оказывают выраженного анксиолитического дей-
ствия у людей.
Норадреналин (норэпинефрин)
Норадреналин в развитии тревоги играет большую
роль, чем ГАМК и 5-ГТ. В то же время антагонисты
e-адренорецепторов не дают выраженного анксиоли-
тического эффекта у больных с преходящей или лег-
кой тревогой, при которой наиболее нежелательные
вегетативные проявления - сердцебиение и тремор.
Положительный эффект e-блокаторов у больных с
тревогой связан с периферическим действием (на-
пример, практолол), поскольку эти препараты не
проникают через гематоэнцефалический барьер.
Пептиды
При тревоге применяют некоторые нейропепти-
ды. Показано, что кортикотропин-рилизинг-гормон
(КРГ) обладает выраженным анксиолитическим
свойством, поэтому он применяется и при депрес-
сии. Субстанция Р дает анксиогенный эффект, а в
настоящее время проводится клиническое испыта-
ние агониста NKl-рецепторов для лечения тревоги
и депрессии. ХЦК — это кишечный пептид, кото-
рый вырабатывается также и мозгом. Поскольку
ХЦК — один из немногих агентов (другим является
СО2), который индуцирует истинные панические
атаки, предполагается, что хороший анксиолитичес-
кий эффект могут давать антагонисты ХЦК, однако
пока такого препарата не создано.
ГЛАВА 52. Нейродегенеративные заболевания
•Ф
Эксайтотоксмческая
О
Аутоиммунный процесс
гибель клеток
Антитело
ф Инфекционное заболевание
Инфекция
/NMDA-рецелторы Не-1
Z Кальций-
связывающие бел^5> / ж^у<7тивХя
Органеллы, *—г Са‘~ ироте hi кинс
связывающие Mhi ахондриальная
—^^^кальций^— дисф умкция
щепторы
а Продукция свободных радикалов,
включая митохондриальную дисфункцию
Активация
апоптотической
смерти клеток
Ядро
^Продукция
свободных/
Некротическая гибель клеток
1ндогенные токсины
[Алоптотическая смерть клеток
1кзогенные токсины
Ф Продукция
или потребление нейротоксина
Апоптотическнй стимул
Нейрон _________
Проапоптоти-
ческнй геиети-
тический
генетический
продукт bcl-2
Каскад (3-цистеинил-аспартат-
специфических протеаз (каспаз)
дукт Ьах/р53
Мнтохондриалъна *
дыхательная цепь
НАДН НАД
Нейротрофи-
ческий фактор
комплекс I
Апоптоз — расслоение мембраны,
сморщивание клетки, конденсация
хроматина, фрагментация ДНК
комплекс II
У
АТФ
Утрата специфического
нейротрофического
фактора
Нгйродегенерат-в- ное заболевание Первичные патоморфологические данные Клиническая картина
Деменция альцгеймеровско- го типа {ДАТ)* Кора головного мозга (особенно темен- но-височная) и гиппокампальный комп- лекс содержат пучки нейрофибрилл и не арктические бляшки, при этом отмечает ся вторичная гибель холиивргичвскнх нейронов в базальных отделах лобных долей Утрата антеградной памяти Нарушение речи и когнитивные нарушения
болезнь Паркинсона (БП) Утрата нигростриарного дофаминерги- ческого пути в базальных ганглиях, на- личие а-скнуклеинй в тельцах Леви, осо- бенно в черной субстанции Замедление движений (брадн- кнмезия). тремор по типу ска- тывания пилюль, повышение мышечного тонуса по пласти’ ческому типу (см. главу 39)
Болезнь двигатель- ного нейрона (БДН или БАС)** Гибель мотонейронов спинного мозга, ствола и коры мозга Прогрессиоующая слабость и атрофия мышц конечностей и бульбарной мускулатуры. При- знаки поражения центральных мотонейронов (гиперрефлексия и повышение мышечного тону- са по спастическому типу)
Усваивается X
на периферии ।
нервной 1
терминалью
и ретроградно
транспортируется
в тело нейрона
Экзогенные |
токсины
Нейротрофический
фактор
* При некоторых формах ДАТ отмечается более фокальное начало с селективным поражением лобно-височной коры,
при этом на вскрытии отмечаются включения в нейронах коры (тельца Пика). Это заболевание называется лобно-височ-
ной деменцией.
* * БДН начинается с изолированного поражения как ПМН, так и ЦМН, но чаще всего имеется сочетание признаков
поражения ЦМН и ПМН. которое называется боковым амиотрофическим склерозом (БАС).
Примечание. В этой главе не обсуждаются многие нейродегенеративные заболевания, в частности хорея Гентингтона
(см. главы 39 и 55) и спиноцеребеллярные дегенерации (см. главу 55). Механизмы гибели клеток при этих заболеваниях
могут быть сходными с теми, что описаны в данной главе.
112
При нейродегенеративных заболеваниях первич-
ным звеном патогенеза является прогрессирующая
гибель определенных популяций нейронов ЦНС.
Этиология нейродегенератинвых заболеваний
Существует несколько теорий этиологии нейро-
дегенеративных заболеваний, которые не являются
взаимоисключающими.
1. Инфекционная теория
Часто гибель нейронов сопровождается глиаль-
ной реакцией (глиозом), что является типичным
признаком инфекций (вирусных), связанных с вос-
палением в ЦНС. Однако при нейродегенеративных
заболеваниях такая реакция отсутствует, хотя демен-
ция при СПИДе дает основание предполагать, что
некоторые нейродегенеративные заболевания могут
быть вызваны ретровирусной инфекцией. Более
того, развитие деменции со спонгиоформными из-
менениями головного мозга в ответ на пролифера-
цию аномального вирусного белка, например, при
болезни Крейтцфельда—Якоба также подтверждает
мнение о том, что некоторые нейродегенеративные
заболевания имеют инфекционную природу.
2. Аутоиммунная теория
При некоторых нейродегенеративных заболева-
ниях обнаруживаются аутоантитела, например, ан-
титела к кальциевым каналам при болезни двигатель-
ного нейрона (БД Н). Однако отсутствие воспалитель-
ного ответа является аргументом не в пользу данной
гипотезы, хотя дегенерация нейронов с минималь-
ной воспалительной инфильтрацией отмечается при
паранеопластических синдромах (см. главу 54).
3. Гипотеза эксайтотоксической гибели
клеток и продукции свободных радикалов
Возбуждающие аминокислоты находятся во всех
отделах ЦНС (см. главу 9) и взаимодействуют с не-
которыми рецепторами, в результате чего клетки
деполяризуются, и в них проникает кальций. В нор-
ме кальций быстро связывается в клетке; высокий
уровень возбуждения и избыточный приток кальция
в клетку приводят к продукции свободных радика-
лов и гибели клеток. Этот каскад событий может
лежать в основе некоторых нейродегенеративных за-
болеваний. Например, при семейной БДН отмечает-
ся дефицит важного антиоксидантного фермента —
супероксиддисмутазы. Более того, при болезни Пар-
кинсона отмечается недостаточность активности
комплекса I митохондриальной дыхательной цепи в
черной субстанции, которая приводит к избыточной
продукции свободных радикалов и нарушению ды-
хания клетки,что вносит свой вклад в гибель клеток.
4. Продукция или потребление нейротоксина
Многие токсины могут индуцировать нейродеге-
неративные заболевания (например, синдром пар-
кинсонизма при марганцевой интоксикации), одна-
ко токсинов, которые бы вызвали основные нейро-
дегенеративные заболевания, не обнаружено.
С другой стороны, деменция альцгеймеровского
типа (ДАТ), связана с формированием пучков ней-
рофибрилл (ПНФ) и сенильных невритических бля-
шек (СНБ) в парагиппокампальном и теменно-височ-
ном отделах коры. Плотность ПНФ коррелирует с
когнитивным статусом папиента. ПНФ содержат пар-
ные спиралевидные филаменты, состоящие из ано-
мального белка, ассоциированного с микротрубочка-
ми х, этот белок в норме обеспечивает нормальную
функцию цитоскелета. СНБ в свою очередь содержат
патологический белок e-амилоид, выделенный из
мембранно-связанного гликопротеина—предшествен-
ника амилоида, который обнаруживается во всех ней-
ронах в здоровом мозге, но его функция неизвестна.
Причина, по которой синтезируются аномальные
белки, точно неясна. Безусловно, в основе некоторых
редких форм ДАТ лежат генетические дефекты, ко-
торые влияют на продукцию патологического белка
e-амилоида. Какова бы ни была причина его продук-
ции, результатом является гибель нейронов коры. Это
приводит к вторичной утрате холинергической ин-
нервации коры с последующей атрофией холинер-
гических нейронов базальных отделов лобной доли,
что дало основание использовать для лечения ДАТ
центральные холиномиметики, хотя их применение
не дало значимых результатов. Кроме того, при мно-
гих нейродегенеративных заболеваниях обнаружива-
ются внутриклеточные включения аномального бел-
ка (например, гентингтина при хорее Гентингтона,
белка т при некоторых заболеваниях с синдромом
паркинсонизма, «-синуклеина при болезни Паркин-
сона), поэтому в основе гибели нейронов при всех
этих заболеваниях может лежать какой-то общий па-
тогенетический молекулярный механизм.
5. Утрата специфического
нейротрофического фактора
Функция нейронов поддерживается благодаря
продукции специфических нейротрофических фак-
торов (см. главы 46 и 47), и утрата одного или не-
скольких из них может привести к развитию нейро-
дегенеративных заболеваний. Недавно были прове-
дены клинические исследования нейротрофических
факторов при некоторых нейродегенеративных за-
болеваниях, например глиального нейротрофичес-
кого фактора (ГНТФ) при болезни Паркинсона, ко-
торые имели относительный успех.
6. Активация программированной
клеточной смерти (апоптоза)
Гибель клеток при большинстве заболеваний (на-
пример, воспалительных) происходит по механиз-
му некротической смерти, однако все клетки содер-
жат механизмы, инициирующие их собственную
смерть — программированную клеточную смерть,
или апоптоз. Механизмы, лежащие в основе этого
процесса, в настоящее время относительно хорошо
изучены, хотя триггеры его не всегда ясны. Поэтому
возможно, что нейродегенартивные заболевания
возникают при несоответствующей активации апоп-
тоза, возможно, после первичной утраты какого-
либо нейротрофического фактора.
ИЗ
ГЛАВА 53. Нейрофизиологические заболевания: эпилепсия
Клинические признаки эпилептических припадков
Припадки, исходящие из
височной доли, сопровождаются
обонятельными или вкусовыми
галлюцинациями с чувством
страха, «уже виденного*
зрительными и вегетативными
нарушениями
^Точки приложения противосудорожных препаратов
Припадки, исходящие из лобной доли,
обычно протекают в виде сложных
нарушений
Припадки, исходящие из двига-
тельной коры сопровождаются
фокальными судорожными подер-
гиваниями — джексоновский марш
(см. главу 36) Припадки, исходящие из те-
доли, сопровождают-
ся преходящими чу ветви тел ь-
симлтомами
Если парциальные припадки
сопровождаются нарушением
сознания, они называются
сложными, при отсутствии
нарушения — простыми
Припадки, исходящие
затылочной доли, со
провождаются преходя-
щими зрительными нару-
шениями (фотопсиями)
Фокальная1
или парциальная эпилепсия
.может генерализоваться^/
Первично-генерализованная эпилепсия —
припадки исходят из срединных структур
(например, таламуса)
CI-
Место связывания
для барбитуратов
Место
для ГАМК
uirfwi
1Н*
Место связывания
для бензодиазепинов
I Усиление ГАМК-ергического
торможения посредством вза-
имодействия с ГАМК-барбиту-
рат-бензодназепиновым рецвп-
_ торным комплексом: фенобар-
битал,. нлоназепам
связывания
(Снижение высво-
|бождеиня глута-
мата: ламотрид-
1жнн, ? габапентин
► ГАМК-трансаминаза
Г АМК-рецепторы
Торможение повторных разрядов
нейронов: фенитоин, карбамазе-
пин, этосукснмид, вальпроат на
трия, топирамат, ? леветирацетам
Снижение инактива-
ции ГАМК
вигабатрин
ГАМК
Пейсмейкерныи нейрон — повторные разряды
Определение и классификация эпилепсии
Эпилепсия — это пароксизмальное нарушение
функции мозга, которое развивается внезапно, спон-
танно завершается и может быть спровоцировано
определенными стимулами. Это наиболее распрост-
раненное из всех тяжелых неврологических заболе-
ваний; заболеваемость эпилепсией наиболее высо-
ка в раннем детстве и пожилом возрасте.
Эпилептические, или судорожные, припадки
подразделяются: на генерализованные или парциаль-
ные (фокальные), т.е. локализованные в каком-то
одном отделе мозга (височной доле); в зависимости
от наличия утраты сознания (если она есть, припад-
ки называют сложными); в зависимости от того, ге-
нерализуются или нет парциальные припадки. У 60-
70% больных эпилепсией не выявляют какой-либо
очевидной причины болезни, и примерно у 2/3 боль-
ных припадки прекращаются через 2—5 лет после
начала, обычно на фоне приема препаратов.
Патогенез эпилепсии
Этиология эпилепсии во многом неясна, однако
большинство терапевтических мер направлено на
устранение дисбаланса между тормозными (ГАМК-
ергическими) и возбуждающими (глутаматергичес-
кими) системами мозга и повторных разрядов по-
тенциалов нейронами. Электроэнцефалограмма
(ЭЭГ; см. главы 44 и 49) показывает, что эпилепти-
ческие припадки связаны либо с генерализованной
синхронной, либо с фокальной спайковой и медлен-
новолновой активностью, хотя эти нарушения мож-
но видеть и в межприступный период.
Генерализованный эпилептический припадок
может протекать в нескольких формах, но в класси-
ческом варианте он представлен тонической (с мы-
шечным напряжением) и клонической (судорожной)
фазами, вслед за которыми наступает утрата созна-
ния. Раньше это называли припадком типа grand mal,
но в настоящее время они получили название гене-
114
рализованных тонико-клонических припадков.
Эпилепсия petit mal в настоящее время считается
первично-генерализованной формой эпилепсии.
Модель генерации эпилептических разрядов та-
кова: межприступная активность соответствует де-
поляризационному сдвигу: потенциалы действия от
групп нейронов как бы накладываются друг на дру-
га. Затем следует период гиперполяризации, так как
эти же нейроны, инактивируясь сами, активируют
локальные тормозные интернейроны.
Повторение межприступных спайков укорачива-
ет период гиперполяризации, что активирует неко-
торые, в норме неактивные ионные каналы в ней-
роны, а также повышает внутриклеточное содержа-
ние калия, что еще больше деполяризует нейроны.
Если активируется достаточное количество нейро-
нов (и локальное торможение ГАМК-ергических
нейронов преодолено), возникают синхронные раз-
ряды популяпий нейронов, что приводит к разви-
тию припадка. Припадок, или синхронный разряд,
затем прекращается активным торможением как в
самом нейроне (посредством ионных каналов), так
и в сети ГАМК-ергических интернейронов.
Несмотря на то что эта модель позволяет понять
патофизиологию эпилепсии на клеточном уровне
вне зависимости от ее причины, очевидно, что в ос-
нове различных форм эпилепсии лежат различные
патологические нарушения. Первично-генерализо-
ванная эпилепсия, при которой наблюдаются диф-
фузные изменения ЭЭГ, вероятно, связана с нару-
шением функции специфических кальциевых кана-
лов таламуса. У больных со сложными парциальными
припадками в рамках височной эпилепсии имеется
маленький рубец в медиальных отделах височной
доли, соответствующий очагу гибели нейронов и
глиозу гиппокампа в результате ишемического или
гипоксического повреждения в раннем детстве.
Лечение эпилепсии
Лечение эпилепсии может быть медикаментоз-
ным и хирургическим, если в основе ее лежит струк-
турное повреждение и/или медикаментозная тера-
пия неэффективна. Медикаментозная терапия при-
меняется при всех формах эпилепсии. Монотерапия
помогает 70-80% больных с тонико-клоническими
припадками и 30—40% больных с парциальными
припадками. Больным, у которых припадки прохо-
дят не полностью, следует назначать дополнитель-
ный препарат, однако лишь 7% больных с рефрак-
терными припадками излечиваются полностью.
Механизмы действия
противосудорожных препаратов
Механизмы действия противосудорожных препа-
ратов не вполне ясны. Некоторые препараты (напри-
мер, бензодиазепины, вигабатрин, фенобарбитал)
усиливают ГАМК-ергическое торможение, тогда как
другие являются блокаторами натриевых каналов
(фенитоин, карбамазепин, вальпроат натрия, ламот-
риджин). Этосуксимид эффективен только при аб-
сансах. Он ингибирует спайкгенерирующий кальци-
евый поток в таламических нейронах. Вальпроат
натрия, помимо натриевых, влияет и на кальциевые
каналы, что объясняет широкий спектр его проти-
восудорожного действия.
Карбамазепин, вальпроат натрия и ламотриджин
широко применяются, поскольку высокоэффектив-
ны и их побочные эффекты многочисленны, но хо-
рошо переносятся.
Преимуществами вальпроата натрия являются
относительно малая выраженность седативного
эффекта, широкий спектр противосудорожной ак-
тивности и легкие побочные эффекты (тошнота,
прибавка массы тела, кровоточивость, тремор и пре-
ходящее выпадение волос). Основным его недоста-
током является наличие идиосинкразии (фатальные
гепатотоксические и тератогенные эффекты). По-
этому карбамазепин или ламотриджин применяют
чаще. Легкие нейротоксические эффекты (тошно-
та, головная боль, сонливость, диплопия, атаксия),
которые определяют порог требуемой дозы. Ламот-
риджин — относительно новый препарат с широким
спектром действия, который безопасен при беремен-
ности. Трудности, связанные с приемом фенитоина,
обусловлены особенностями его метаболизма. Тре-
буется 20 дней, чтобы его уровень в сыворотке стал
стабильным после изменения дозы. Поэтому его дозу
надо постепенно увеличивать, пока не прекратятся
припадки или не возникнут мозжечковые симптомы
(нистагм, атаксия, дизартрия). Другими нежелатель-
ными побочными эффектами фенитоина являются
гиперпластический гингивит, воспаление сальных
желез, сальность кожных покровов, огрубение черт
лица и гирсутизм, поэтому он не пользуется популяр-
ностью, особенно у женщин. Фенобарбитал эффек-
тивен в отношении тонико-клонических и парциаль-
ных припадков, но обладает выраженным седативным
эффектом. К препарату возникает привыкание, и от-
мена его может привести к развитию эпилептичес-
кого статуса. Побочными эффектами фенобарбита-
ла являются мозжечковые симптомы, сонливость у
взрослых и избыточная двигательня активность у
детей. Вигабатрин, габапентин, топирамат и левети-
рацетам — новые препараты и применяются в каче-
стве дополнительных средств для лечения эпилепсии
у больных, которым недостаточно эффективно помо-
гают другие средства. Этосуксимвд эффективен толь-
ко при лечении абсансов и миоклонических припад-
ков (кратких порывистых движений с утратой созна-
ния). Клоназепам — сильный бензодиазепиновый
противосудорожный препарат, эффективный при
абсансах, тонико-клонических и миоклонических
припадках. Он оказывает выраженное седативное
действие, после длительного применения возника-
ет привыкание.
Противосудорожная терапия у беременных тре-
бует осторожности, поскольку многие препараты
оказывают тератогенное воздействие, особенно фе-
нитоин и вальпроаты.
115
ГЛАВА 54. Аутоиммунные заболевания
Воспаление мозговых оболочек — менингит
Воспаление мозга — энцефалит
Эндотелий
Микроглия
Воспаление мышцы — миозит
Мышца
Активация CD4
Т-келперных клеток
Всм паление с демиелинизацией
о белом веществе головного +
спинного мозга
Экспрессия
ГКС-1
*
Продукция интерлейкина-2i
и других цитокинов
ЦНС ПНС
Активация CD8 цито-
токсических Т-клеток
Антитела к АцхР или
мускариновым ре-
цепторам при миас-
тении
Антитела к пресниаптическим
вольтажзависнмым кальцие-
вым каналам (миастенический
синдром Итона—Ламберта —
МСИЛ)
Требуется для
полной активации
и пролиферации
Т-клеток
Состоя-
ние по
коя
Активное про-
бое пал итель-
ное состояние
Экспрессия
ГКС-Н
Астроциты
Эндотелий
Микроглия
Антиген в ЦНС (эндогенный
или экзогенный)
Воспринимается антигенпрезенти-
рующей клеткой (АПК) — микро*
глией, экспрессирующей ГКС-И
+ _________
Вовлечение других эффекторньл клеток,
•В-клетки,
•Естественные киллеры,
•Лимфокинассоцииро ванные
киллернью клетки
•Макрофаги,
Воспаление спинного мозга — миелит
Воспаление нерва — неврит
Перифе-
рический
нерв
Презентация
антигена ГКС-1
• Рассеянный склероз
•Острый рассеянный энце-
фаломиелит
•Другие воспалительные
заболевания (например,
васкулиты)
Нервно-мышечное соединение
1дМ-опосредованная
парапротеинемическая
полинейропатия с анти-
телами к компонентам
миелина, они расщеп-
ляют миелин и вызыва-
ют демиелинизацию
Антитела могут свя-
зываться с перехва-
том Ранвье или сосед-
ними участками, вы-
зывая демиелиниза-
цию, как при синдро-
ме Гийена—Барре
Иммунная система ЦНС
ЦНС по сравнению с ПНС и другими систе-
мами органов является иммунологически обособ-
ленной. Причиной этому служит гематоэнцефали-
ческий барьер (ГЭБ), который в норме предот-
вращает попадание лимфоцитов, макрофагов и
антител в ЦНС (см. главвт 4 и 16). Кроме того, ЦНС
слабо дренируется лимфой, и экспрессия глав-
ного комплекса гистосовместимости (ГКС) в ней
низкая. Все это говорит об ограниченнвтх возмож-
ностях презентироватв антигенв! в ЦНС. Однако
нарушения ГЭБ могут в корне изменитв эту си-
туацию.
В состоянии покоя некоторые активированные
Т-лимфоцитв1 пересекают ГЭБ и циркулируют в
ЦНС. Кроме того, экспрессия ГКС происходит толв-
ко в определенных клетках, хотя при воспалении
ситуациция изменяется. Так, после запуска иммун-
ного ответа он может бытв усилен цитокинами и
116
индукцией экспрессии ГКС, что приводит к откры-
тию ГЭБ.
В этих обстоятельствах антигенпрезентирующими
клетками становятся клетки микроглии. Они взаимо-
действуют с Т-хелперными лимфоцитами, что при-
водит к развернутой иммунной реакции.
Аутоиммунные заболевания ЦНС
Рассеянный склероз
Рассеянный склероз встречается часто и прояв-
ляется рядом неврологических нарушений, связан-
ных с воспалительными очагами в ЦНС. Патомор-
фологически эти очаги представляют собой мелкие
участки демиелинизации, возникшие в результате
воспалительной Т-клеточной инфильтрации, одна-
ко триггер этого процесса неизвестен. Очаги, как
правило, восстанавливаются в процессе ремиелини-
зации, что сопровождается клиническим улучше-
нием, однако с течением времени гибель миелина
начинает происходить постоянно, что приводит к ги-
бели аксонов, а также формированию стойкого не-
врологического дефицита.
В настоящее время наиболее эффективным ме-
тодом лечения рассеянного склероза считается внут-
ривенное введение метилпреднизолона в высоких
дозах, что предотвращает рецидивы, но не влияет на
процесс в целом. Кроме того, эффективным являет-
ся (3-интерферон, который снижает частоту обостре-
ний, однако его влияние на прогноз болезни сомни-
тельно. Эффективна и более агрессивная иммуно-
терапия, особенно если она проводится в самом
начале заболевания.
Острый рассеянный
энцефаломиелит
Энцефаломиелит — редкое воспалительное де-
миелинизирующее заболевание ЦНС, которое яв-
ляется осложнением инфекций и вакцинаций (на-
пример, против кори и бешенства). Заболевание
монофазное (в отличие от рассеянного склероза),
характеризующееся распространенными очагами
демиелинизации в ЦНС, которые патоморфологи-
чески состоят из периваскулярных лимфоцитарно-
макрофагальных инфильтратов и очагов демие-
линизации. Острый рассеянный энцефаломиелит
напоминает экспериментальный аллергический эн-
цефаломиелит, который является хорошо изучен-
ным заболеванием, опосредованным Т-лимфопи-
тами и связанным с выработкой аутоантител к ос-
новному белку миелина. У животных это состояние
вызывается путем сочетанного введения стериль-
ной ткани мозга и консервантов.
Другие аутоиммунные заболевания
Другими иммунными заболеваниями ЦНС явля-
ются васкулиты, протекающие с первичным пораже-
нием сосудов мозга.
К редко встречающимся заболеваниям, при ко-
торых поражается ЦНС, относятся синдромы, свя-
занные с онкологическими заболеваниями, или па-
ранеопластические синдромы. При этих синдромах
к структурам ЦНС вырабатываются аутоантитела в
связи с наличием в организме опухоли, что приво-
дит к гибели клеток и развитию неврологической
сипмтоматики. Например, антитела к клеткам Пур-
кинье вызывают выраженную мозжечковую симп-
томатику в результате иммунной реакции на эти
клетки мозжечка. Точный механизм, посредством
которого антитела реагируют с клетками мозжечка,
неясен, так как в норме они не пересекают ГЭБ, од-
нако патоморфологически обнаруживаются лимфо-
цитарные инфильтраты в пораженных структурах,
что предполагает способность антител индуцировать
иммунный процесс и гибель клеток.
Аутоиммунные заболевания ПНС
ПНС менее подвержена аутоиммунным заболе-
ваниям, чем ЦНС.
• Периферические нервы поражаются при ряде ауто-
иммунных заболеваний, в частности при синдро-
ме Гийена Барре. Ему часто предшествует инфек-
ция (например, цитомегаловирусная или кампило-
бактериальная), которая вызывает перекрестный
аутоиммунный ответ на компоненты периферичес-
кого нерва (например, некоторые ганглиозиды).
Это приводит к фокальной демиелинизации пери-
ферического нерва, что затрудняет проведение по
нему нервного импульса (см. главу 6). После плаз-
мафереза и внутривенного введения иммуноглобу-
линов больной выздоравливает. Похожее состояние
наблюдается при продукции патологических анти-
тел (например, при парапротеинемиях).
• Нервно-мышечное соединение также поражается
аутоиммунным процессом, например миастения и
синдром Итона—Ламберта (см. главу 7).
• Мышцы также поражаются аутоиммунным процес-
сом. Чаще всего встречается полимиозит - заболе-
вание, опосредованное Т-клетками, при котором
возникают слабость и боль в проксимальных мыш-
цах. В отличие от полимиозита, дерматомиозит —
заболевание, опосредованное В-клетками, при ко-
тором возникают слабость и боль в проксимальной
мускулатуре в сочетании с кожной сыпью. Дерма-
томиозит — это паранеопластический синдром,
встречающийся у пожилых людей с опухолями лег-
кого, молочной железы, толстой или тонкой кишки.
ГЛАВА 55. Наследственные неврологические заболевания
Кодирует все белки, за исключением
субъединиц белков дыхательной цепи
Кодирует 13 белковых субъединиц
белков дыхательной цепи.
Кодирует все белки
2 рибосомальные РНК и 22 транспортные ДНК
цитоскелета,
рецепторы и ионные
каналы, ферменты
и нейротрансмиттеры
Частота мутаций
Типы наследования
Аутосомно-рецессив-
ный (АуР), например
спинальная
амиотрофия
Наследование по материнской линии —
точковые мутации в митохондриальной ДНК
Митохондриальные заболевания
Заболевания
X-сцепленный,
например
миодистрофия
Дюшенна
Аутосомно-
доминантный (АуД),
например
нейрофиброматоз
Спорадические наследственные заболевания, связанные с нарушением
функции ядерного и митохондриального генома без типа наследования
ф Поражение
глаз
• Поражение
мышц
Нейропатия
зрительных
нервов
роты зрени^Т-*-^^
—— Катаракта
Нарушение
движений
Слабость скелетной
мускулатуры
Поражение сердечной мышцы
ф Периферическая нервная система
Поражения периферических нервов (полинейропатия)
• Центральная нервная система — инсульты, мозжечковая атак-
сия, глухота, эпилепсия, деменция, нарушение движений
Некоторые митохондриальные заболевания связаны
с митохондриальными белками, которые кодируются в ядре
Неврологическая симптоматика наблюдается при
многих наследственных заболеваниях, и в ряде слу-
чаев патологические изменения касаются только
нервной системы. Согласно недавним молекуляр-
но-генетическим исследованиям, многие заболева-
ния нервной системы были переклассифицирова-
ны в зависимости от лежащего в их основе генети-
ческого дефекта.
Два основных открытия выдвинули на первый
план роль генетических факторов в развитии невро-
логических заболеваний. Во-первых, было показа-
но, что дефекты митохондриального генома, насле-
дуемого по материнской линии, могут приводить к
неврологическим заболеваниям и, во-вторых, раз-
личные неврологические заболевания могут быть
связаны с экспансией тринуклеотидных повторов
в генах (заболевания тринуклеотидных повторов).
Заболевания, связанные с делециями генов
Многие неврологические заболевания связаны с
полными или частичными делециями генов, напри-
мер наследственная полинейропатия с компресси-
онным параличом, при которой у больных перио-
дически развиваются повторные туннельные нейро-
патии в связи с делецией на 17-й хромосоме, которая
участвует в кодировании периферического белка ми-
елина 22 (ПБМ 22).
118
Заболевания, связанные с дупликацией генов
Некоторые заболевания могут быть связаны с
дупликацией генов, примером является наслед-
ственная мотосенсорная полинейропатия, при ко-
торой у больных развиваются слабость и атрофия ди-
стальных мышц и утрата чувствительности на пер-
вых десятилетиях жизни. В ряде случаев причиной
этого является удвоение гена на 17-й хромосоме, в
том числе ген, кодирующий ПБМ 22.
Заболевания, связанные с мутациями генов
Мутации генов — наиболее часто встречающиеся
генетические дефекты. В основе заболеваний, свя-
занных с мутациями генов, лежит нарушение функ-
ции определенного белка или фермента, который
перестает нормально функционировать. Примерами
таких заболеваний являются семейные формы болез-
ни двигательного нейрона (см. главу 52), миодистро-
фии (см. главу 10) и миотонических синдромов (см.
главу 5).
Заболевания, связанные
с импринтингом генов
Импринтинг — это дифференциальная экспрес-
сия аутосомных генов в зависимости от родителя, от
которого они были унаследованы. Так, нарушение
функции материнских генов на определенном учас-
тке 15-й хромосомы (15qll-ql3) приводит к разви-
тию синдрома Прадера—Уилли (умственная отста-
лость с ожирением, уменьшением роста и размера
гениталий), тогда как нарушение функции тех же
генов, полученных от отца, — к развитию сицдрома
Ангельмана (выраженная умственная отсталость,
атаксия, эпилепсия и аномалии черепа и лица).
Митохондриальные заболевания
Митохондрии содержат собственную ДНК и син-
тезируют определенные белки дыхательной цепи, от-
вечающей за окислительное фосфорилирование (см.
главу 52), хотя большинство митохондриальных бел-
ков кодируется митохондриальной ДНК. Так, мито-
хондриальные заболевания (вызванные дупликаци-
ей, делецией или точковыми мутациями генов) мо-
гут быть следствием дефектов как генов ядра, так и
митохондриального генома. Однако митохондриаль-
ная ДНК мутирует в 10 раз чаще ядерной ДНК и не
имеет интронов (некодирующих частей генома), по-
этому каждая мутация обычно вызывает нарушение
кодирующей последовательности ДНК. Поскольку
митохондрии наследуются из оплодотворенной яй-
цеклетки, митохондриальные заболевания всегда
наследуются по материнской линии. Однако в каж-
дой клетке много митохондрий, поэтому клетка мо-
жет содержать как нормальные, так и патологичес-
кие митохондрии, что называется гетероплазмией.
Болезнь развивается только при достижении опре-
деленного количественного порога патологических
митохондрий.
Заболевания тринуклеотидных повторов
В основе многих наследственных заболеваний
лежит экспансия тринуклеотидных повторов, т.е.
значительное увеличение количества повторов трех
нуклеотидов в геноме. У здорового человека трип-
летные повторы также встречаются, однако если их
количество превышает определенный уровень, раз-
вивается заболевание.
Патологические тринуклеотидные (или трип-
летные) повторы встречаются в кодирующей (на-
пример, прихорее Гентингтона; см. главу 39) или не-
кодирующей (например, при атаксии Фридрейха)
части гена. В результате экспансия приводит к ут-
рате функции белка (например, фратаксина при-
атаксии Фридрейха) или обретении белком новой
функции (например, гентингтином при хорее Ген-
тингтона). В последнем случае новая функция бел-
ка играет ключевую роль в развитии нейродегене-
ративного заболевания. Однако механизм, посред-
ством которого данный мутантный белок вызывает
селективную гибель определенных клеток ЦНС,
остается неясным, поскольку многие продукты му-
тантных генов широко экспрессируются во всех
тканях организма.
При наличии большой и нестабильной последо-
вательности ДНК триплетный повтор увеличивает-
ся при митозе и мейозе (динамические мутации). Это
означает, что наибольшая вероятность возникнове-
ния таких мутаций отличается при сперматогенезе
и оплодотворении/эмбриогенезе, что предполагает
два важных последствия. Во-первых, более длинные
повторы возникают у потомков мужского пола, а
значит, они и передаются дальнейшим поколениям.
В последующих поколениях болезнь возникает в бо-
лее ранние сроки и будет протекать тяжелее; этот фе-
номен называется генетической антиципацией (ассо-
циация более длинных повторов с более тяжелым
течением болезни и более ранними сроками ее воз-
никновения).
Приложение 1. Основные вцды нейротрансмиттеров
Нейротрансмиттеры Распределение Тип рецепторов Неврологические заболевания
Аминокислоты Возбуждающие Глутамат Тормозные ГАМК Глицин Во всех отделах Во всех отделах Спинной мозг 1. Ионотропные не-NMDA (АМРА; каинатные, квискалатные) 2. Метаботропные ГАМК-А, ГАМК-В Эпилепсия (глава 53) Эксайтотоксическая гибель клеток (глава 52) Заболевания спинного мозга (глава 34) Эпилепсия (глава 53) Тревога (глава 51) Стартл-синдром (глава 34)
Моноамины* Норадреналин (норэпинефрин) Серотонин (5-гидрокситриптамин) Голубое ядро (проекции вез- де в ЦНС) (глава 13) Постганглинарные волокна симпатической нервной си- стемы (глава 42) Большое ядро шва ствола го- ловного мозга с проекциями везде в ЦНС (глава 50) <7-1; <7-2 в-1; в-2 5-ГТ1 (A-F) 5-ГТ2 (А-С) 5-ГТЗ-5П7 Депрессия (глава 50) Вегетативная недостаточ- ность (глава 42) Депрессия (глава 50) Тревога (глава 51) Мигрень
Дофамин Нигростриарный путь в ба- зальных ганглиях (глава 38) Мезолимбический и мезо- кортикальный пути (глава 50) Сетчатка (глава 23) Г ипоталамо-гипофизарные проекции (глава 43) D1 -5-рецепторы, активация которых приводит к повыше- нию уровня внутриклеточ- ного цАМФ В2-рецепторы, активация которых приводит к сниже- нию уровня внутриклеточ- ного ц АМФ ИЗ-4-рецепторы зависят от сигнальной системы цАМФ Болезнь Паркинсона (глава 39) Шизофрения (глава 50) Регуляция секреции гормо- нов гипофиза (глава 43) Рвота
Ацетилхолин Нервно-мышечное соедине- ние (глава 7) Вегетативная нервная систе- ма (глава 42) Базальные отделы лобных долей и лимбическая систе- ма (главы 45 и 52) Интернейроны многих струк- тур ЦНС, в том числе стриа- тум (глава 38) Никотиновые Мускариновые (подтипы Ml М3) Нарушения нервно-мышеч- ной передачи (глава 7) Вегетативная недостаточ- ность (глава 42) Деменция альцгеймеровско- го типа (глава 52) Болезнь Паркинсона (глава 39) Эпилепсия (глава 53) Цикл сон-бодрствование (глава 44)
Нейропептиды Широко представлены в Ц НС, но особенно в большом ко- личестве находятся в: задних рогах спинного мозга (главы 21 и 22), базальных ганглиях (глава 38) вегетативной не- рвной системе (глава 42) Различные См.: Система боли (главы 21 и 23) Базальные ганглии (глава 38) Вегетативная нервная систе- ма (глава 42) Пластичность нервной сис- темы (глава 47) Тревога (глава 51) Сон (глава 44)
Другие пуринергические АТФ Эндозапины
* Гистамин и адреналин (норадреналин) являются моноаминами, которые находятся преимущественно в гипоталамусе
и надпочечниках.
120
Приложение 2а. Восходящие чувствительные проводящие пути спинного мозга
Тракт Спиноталамический (СТТ) Задние столбы — медиальная петля Спиноцеребеллярный (С ЦТ)
Глава 21 20 37
Начинается Задний рог (пластина I, III, IV. V)/ перекрещивается в спинном мозге Первичные афферентные во- локна от механорецепторов, мышц и рецепторов суставов Интернейроны спинного мозга и проприоцептивная информация от мышц и суставов
Заканчивается Соматотопическая проек- ция наиболее длинные во- локна лежат латеральнее Проецируется на ствол и та- ламус с противоположной стороны Соматотопическая проекция волокна заканчиваются в ядрах задних столбов в про- долговатом мозге Волокна перекрещиваются на уровне медиальной петли и оканчиваются на задневент- ральном ядре таламуса Два тракта: Задний С ЦТ передает информа- цию от рецепторов мышц и сус- тавов через передние ножки моз- жечка в мозжечок Передний С ЦТ передает инфор- мацию от интернейронов спин- ного мозга через верхние ножки мозжечка в мозжечок
Функция Болевая и температурная чувствительность Проприоцепция, прикоснове- ние и чувство вибрации Проприоцепция, информация об активности интернейронов спинного мозга
Приложение 26. Нисходящие двигательные проводящие пути
Тракт Кортикоспинальный (КСТ), или пирамидный Руброспинальный (РуСТ) Вестибулоспинальный (ВСТ) Ретикулоспинальный (РСТ)
Главы 32, 34-36 32, 34, 37 32, 34, 37 32, 34, 37
Начинается Первичная моторная кора (40%) Премоторная кора (30%) Соматосенсорная кора (30%) Крупноклеточная часть красного ядра среднего мозга Ядро Дейтерса продол- говатого мозга (часть комплекса вестибуляр- ных ядер) Каудальная часть рети- кулярной формации в мосту и продолговатом мозге
Функции Независимые тонкие движения пальцев Участие в обработке чув- ствительной информа- ции Проецируется на те же популяции МН, что и КСТ, в частности те, что регулируют дистальные мышцы Экспериментальное поражение этого трак- та приводит к мини- мальному двигательно- му дефициту по срав- нению с поражением КСТ Его существование у людей обсуждается Иннервирует в основ- ном разгибатели и ак- сиальную мускулатуру, контролирует равнове- сие и позу РСТ дает возбуждающую и тормозную импульса- цию интернейронам спинного мозга и в мень- шей степени МН Этот путь подавляет ак- тивность спинного моз- га, поражение его приво- дит к повышению тонуса в разгибателях
Тектоспинальный тракт относительно тонок и начинается от покрышки среднего мозга. О нем кратко рассказано
в главе 34.
Приложение 3. Анатомия и физиология мозжечка
Система Спиноцеребеллум или палеоцеребеллум: вермальная область Спиноперебеллум или палеоцеребеллум: паравермальная и промежуточная области П онтоцеребеллум или неоцеребеллум Вестибулоцеребеллум или архицеребеллум
Основные афферентные связи Вестибулярное ядро Проксимальные отде- лы конечностей (аф- ференты Ia/Ib) и активность интер- нейронов регулируется посредством ЗСЦТ и ПС ЦТ соответственно Ia/Ib афференты отди- сталвной мускулатуры конечностей через ЗСЦТ Активноств интерней- ронов регулируется посредством задних спинальных групп кле- ток через ПС ЦТ Первичная моторная и соматосенсорная кора Задняя теменная кора Первичная моторная и премоторная кора Оба переключаются на ядрах моста Полукружные каналы через вестибулярное ядро Зрителвная информа- ция от верхних бугор- ков четверохолмия. Латерального коленча- того тела и первичной зрительной коры передается через ядра моста
Связи с глубокими ядрами мозжечка Ядро шатра Промежуточное ядро (шаровидное и проб- ковидное) Зубчатое ядро мозжечка
Основные эфферент- ные проекции Ретикулярная формация (РСТ) Вестибулярное ядро (ВСТ) Вентромедиальные нисходящие проводящие пути Красное ядро (крупно- клеточная часть) —> РуСТ ПВ-ВЛЯТ -> ПРМК (поле 6 по Бродману)и ПМК (поле 4 по Брод- ману) —> КСТ (заднелатеральные нисходящие проводя- щие пути) ПВ ВЛЯТ -> Поля 4 и 6 -> КСТ Красное ядро (мелко- клеточная часть) —> нижняя олива —> мозжечок (треуголь- ник Молларе) Вестибулярное ядро —> ВСТ Вестибулярное ядро —> глазодвигательные ядра
Особвхе функции Регуляция аксиалвной мускулатуры и мышеч- ного тонуса Координация дисталь- ных отделов конечно- стей Регуляция мвтшечного тонуса Планирование движений Зрительный контролв за движениями Неболвшое участие в координации дис- тальных отделов ко- нечностей Регуляция позы Регуляция движений глаз
Алфавитный указатель
Аграфия 69, 120
Ад-волокна 51
Адаптация 44, 45, 46
Аденозинтрифосфат 8, 21, 119
- гидролиз 30
Адиадохокинез 83
Адреналин 118, 120, 120
Адренокортикотропный гормон
8, 120
Адхалин 28
Акалькулия 69
Акинетический мутизм 97, 99
Аксолемма 14, 15
Аксональный бугорок 14, 15
Аксоны 14, 15
— аксоны
миелинизированные 15
— аксона начальный сегмент
14, 15
— валлеровская дегенерация
100
— регенерация 16
Аксоплазма 15
Аксоплазматический ток 15
Активация рецепторов 19
Актин 31, 46
— Актин-актиновые
филаменты 46
— Актин в пресинаптической
сети 23
— актин при мышечном
сокращении 29
Алексия 69, 120
Алкоголизм 91, 120
Аллодиния 51, 120
Альтернирующий синдром
Валленберга-Захарченко
43
Альцгеймера болезнь.
См. Деменция
Амакриновые клетки 54, 55
Аминокислоты 27
— аминокислоты
возбуждающие 27
— аминокислоты тормозные
27
Амплитуда 61
Ампулы 64, 65, 120
Аналгезия 52, 70, 120
Анализаторы общей
чувствительности 48
Анализы крови 90, 104, 106
Анатомия нервной системы
92, 92
Ангельмана синдром 116, 119
Аневризмы 43
Анксиолитические препараты
108, ПО, ПО
Анорексия 95
Антидепрессанты 53, 107, 108
— при лечении боли 53
— при тревоге 107, 109, ПО
Антидиуретический гормон
94, 94
Антипсихотические средства
107, 109
Анэнцефалия 13
Апноэ во время сна 96, 97
Апоптоз 111, 113
Апраксия 102, 104
Аркуатные волокна 63
Ассоциативная кора 68, 98
Астигматизм 54
Астроцитома 17
Астроцита 16, 101, 103, 115,
116
— отростки 17
— торможение роста аксонов
45
Астропита Глиальные клетки.
См. также Астроцита,
микроглия,
олигодендроциты
Атаксия 83, 117, 118
Атаксия Фридрейха 116, 119
Аутоиммунные заболевания
114, 116, 117
Афазия 63
— акустико-мнестическая 63
— моторная 63
— сенсорная 63
Аффективные расстройства
107, 108, 120
Афферентные волокна
11, 33, 68, 74, 80
— афферентные волокна 1Ъ
74
- афферентные волокна II 74
— афферентные волокна 1а
74, 75, 77, 82, 100, 129, 102
— афферентные волокна
вхождение в спинной мозг
11
Афферентный чувствительный
нерв 44, 45
Афференты 51, 119, 120, 122
— первичные 33, 118, 121
— сгибательного рефлекса 77
Ацетилхолин
8, 27, 92, 118, 93, 120
— в нервно-мышечном
соединении 22
в парасимпатической
нервной системе 92, 93
Ацетилхолиновый рецептор 22
— невринома сухого нерва 37
— никотиновый 28
Б
Базальная мембрана
8, 16, 17, 22, 47, 60
Базальные ганглии
72, 79, 84, 89, 91
— вентрального отдела 85
— заболевания 86
— нейрофизиология 85
- пути 112, 112
— регуляция движений глаз
87
Барабанная лестница 60
— перепонка 61
— полость 47, 61
— струна 37, 67
Безымянная субстанция 85
Беккера миодистрофия 29
Белки остеогенеза (БО) 13
Белла паралич 37
Белое вещество 11
Бензодиазепины
96, 108, 109, 97, ПО, 111
— зависимость/синдром
отмены 109, 111
— при тревоге 109, 111
Бессонница 95, 97
Беца клетки 80
Биопсия 106, 107
— мозга 106, 107
— мышц 106, 106
— нерва 106, 106
123
Биполярные клетки 54, 55
— окружения 55
— центрального рецептивного
поля 55
— расстройства 107, 108
Битемпоральная гемианопсия
56
Бледный шар 42, 84
— внутренний сегмент
бледного шара (БШв) 84
— наружный сегмент бледного
шара (БШн) 84
Близорукость 54
Блоковой нерв 103, 105
— обследование 103, 105
Блуждающий нерв 35, 103, 105
— обследование 103, 105
Боковой амиотрофический
склероз 91
— желудочек 10
Болезнь двигательного нейрона
(БДН) 90, 91
— Лу Герига (боковой
амиотрофический склероз)
77, 91
Боль 50
— восприятие 51
— лечение 52
— отраженная 51
— проводяшие пути 51
— синдромы 51
— системы 51
— хроническая 51
Большая цистерна 40
Большое затылочное отверстие
10, 82
— ядро шва 96, 96
Брадикинезия 86, 112, 112
Бродмана, цитоархитектонические
поля 38
Брока зона 42, 63
Броун-Секара, синдром 70
Булимия 95
Бульбарный паралич 37
В
Вазопрессин (антидиуретический
гормон) 8
Валленберга - Захарченко
альтернизирующий
синдром 43
Валлеровская дегенерация
99, 101
Вальпроат натрия ПО, 115
Васкулиты 114, 117
ВВ-клетки 63
Вегетативная недостаточность
118, 120
Вегетативная нервная система
11, 92, 118, 120
— поражение 92, 93
анатомия 92
— регуляция гипоталамусом
92, 93
центральные отделы 92, 93
Везикулы, синаптические 23
Вентральные отделы покрышки
95-97
Вентромедиальная
двигательная система 77
Вернике зона 42, 63
Верхние бугорки четверохолмия
57
зрительные функции 57
Вестибулоспинальный тракт
(ВСТ) 73
в регуляции движений 73
— спинной мозг 64
Вестибулярный анализатор 64
периферическая 64
Вестибулярная функция 65
Вестибулярные рефлексы 65
Вестибулярные ядра 35
Вигабатрин 113, 115
Виллизиев круг 42
Вильсона-Коновалова болезнь
87
Височная доля 58, 94
— эпилепсия 67
Вкус 66
Вкусовое восприятие 66
Вкусовые рецепторы 47, 67
Внимание 69, 88, 102, 104
Внутренние вены мозга 43
Внутренние волосковые клетки
61
Внутренние сонные артерии
(ВСА) 8
Внутричерепное давление,
повышение 37
Водно-солевой баланс 93, 94
Возбуждающие аминокислоты
ПО, 113
Возбуждающие синапсы 25
Возбуждающий
постсинаптический
потенциал (ВПСП) 8
Возвратная артерия Губнера
43
Волосковые клетки 46, 47
— отолиты 65
— полукружных каналов 65
— улитки 47
Воротная теория боли 53
Воспалительные заболевания
99, 101, 111, 113, 115, 116
Восходящая активирующая
ретикулярная система
95, 96
Временная суммация 25
Временное кодирование 45
ВТ-клетки 63
Вторичные сенсорные поля 45
Вызванные потенпиалы (ВП)
105, 107
Высокопороговые
механорецепторы 51
Выявление признаков 45
Габапентин 113, 115
ГАМК
— при тревоге 108, ПО
— при эпилептических
припадках 112, 114
ГАМК-А-рецепторы 96, 97
Ганглиозные клетки сетчатки 39
— проекция на латеральное
коленчатое тело (ЛКТ) 56
Геликотрема 60, 61
Гематоэнцефалический барьер
8, 17, 41
Гемианопсия 43
— гомонимная 56
Гемибаллизм 87
Гемипарез 43, 81
Гемиплегия 77
Геммулы 15
Ген
— (ные) делеции 29
— (ные) дупликации 116, 119
— (ные) мутации 29
Генерализованные тревожные
расстройства 108, ПО
Генераторы центрального
паттерна 73
Генетические заболевания 13
Гентингтона хорея
111, 116, 113, 119
Гетероплазмия 116, 119
Гибель клеток
101, 103, ПО, 113
124
Гидроцефалия 13, 17
— викарная 41
Гийена—Барре синдром 19
Гипералгезия 51
— вторичная 51
— первичная 51
Гиперполяризация 46, 112, 115
Гиперрефлексия 77
Гиперсомния 96, 97
Гиперстолбики, зрительная
кора 59
Гипертермия 31
Гиперэкплексия 27
Гипокретины 96, 97
Гипоталамус 92-94, 98, 118, 120
— обонятельные проекции
97, 98
— регуляция вегетативной
нервной системы 93, 94
— регуляция сна 95, 97
— супрахиазматическое ядро
57, 96
— функции 94
Гипотеза скольжения
филаментов, сокращение
мышц 30
Гипотензия 53
— ортостатическая 92, 93
— постпрандиальная 92, 93
Гипотония 73, 83
Гипофиз 94, 108, 109, 118, 120
— опухоли 56
Гистамин 118, 120
Главный комплекс
гистосовместимости (ГКС)
114, 116
— антигены 114, 116
Глаз 35
— оптические свойства 54
расшивание 103
Глазодвигательные мышпы 74
Глазодвигательный нерв
36, 99, 101, 103, 105
— обследование 103, 105
Глиальные клетки 101, 103
Глиальный нейротрофический
фактор (ГНТФ) 100, 101
Глицин 27
Глоточный рефлекс 37, 104
Глутамат 27, 84, 98, 99
Глутаматдекарбоксилаза 8, 77
Глутаматные рецепторы 98, 99,
114
— ионотропные 27
— метаботропные 98. 99
— при отсроченной
потенциации 98, 99
Головной мозг 40, 42, 44, 104,
106
— биопсия 106, 107
— кровоснабжение 42
— поражение 87, 91
— чувствительные
расстройства 70
Головокружение,
доброкачественное
позиционное 65
Голубое ядро 95, 97
Горизонтальные клетки 47, 55
Горнера синдром 92, 93
Горький вкус 66, 67
Гранулярные клетки 67, 102
Грудные нервы 11
Грушевидная кора 67
Густидуцин 67
д
Дальнозоркость 54
Двигательная единица 74
Двигательные нарушения 71
Двигательные проводяшие
пути, нисходящие 76, 77
— поражение 77
Двигательные расстройства
108, 109
Двигательный анализатор 72
— иерархия 72
— латеральная система 77
— нарушения 89
— обследование 90
— спинальный отдел. См.
Спинной мозг, регуляция
двигательных функций
— функция базальных
ганглиев 85
— функция мозжечка 83
Двигательный гомункул 80
Движение
— кодирование по силе и
направлению 45
Движения 72
— инициация, планирование
и программирование 72
— координация 73, 75
— нарушения 73
— открытая дуга
(произвольная) 72
— рефлекторная дуга 72, 75
Движения глаз 101, 103
— при поражении базальных
ганглиев 87
— произвольные 89
— регуляция в ЦНС 89
— рефлекторные 89
— саккадированные 88
— типы 88
Двоение 88
Двойное ядро 35
Декстропропоксифен 53
Деменция 27
— альцгеймеровского типа 27
— лобно-височная 112
— при поражении базальных
ганглиев 51, 87
Демиелинизирующие
полинейропатии 17, 25
Дендритические отростки 15
Дендриты 14, 15
Деполяризация 18, 21, 98, 99
Депрессия 26
Дерматомиозит 115, 117
Дефекты полей зрения 43, 56
Дефицит витамина В12 71
Децеребрационная ригидность
77
Децибелы (дБ) 61
Джексоновский марш 81
Диабет несахарный 94
Диазепам 108, ПО
Дизартрия 83
Диплопия 88, 113, 115
Диск зрительного нерва 55
Дисметрия 83
Дистония 73
Дистрофии 29
Длиннолатентные рефлексы 81
Добавочный нерв 37, 103, 105
Дополнительная моторная зона
(ДМЗ) 72
— поражение 72
— эпилептические припадки
80
ДОФА-декарбоксилаза 86
Дофамин 8, 84, 86
Дофамин (ДА)
— агонисты дофамина 86
— заместительная терапия 86
— нарушения при
шизофрении 86
— пути 85
Дофаминергические нейроны
95, 97
Дюшенна, миодистрофия
(МДД) 29
125
ж
Жажда 93-95
Желатинозная субстанция 32
Желудочек III 94
Желудочек IV 34, 41
3
Заболевания тринуклеотидных
повторов 87
Задневентральное ядро
таламуса (ЗВЯТ) 78
Задние спинальные артерии 43
Задние столбы
32, 33, 42, 45, 49
— электрическая стимуляция
52
Задние столбы — медиальная
петля 75
Задний корешок
33, 75, 76, 93, 92
Задний рог
8, 48, 50, 53, 61, 71, 118, 121
— медикаментозная терапия
боли 53
— ноцицептивные пути 50
— обработка чувствительной
информации 55
Задний спиноцеребеллярный
тракт (ЗСЦТ) 8
Задних столбов, ядра
9, 118, 121
Задняя мозговая артерия (ЗМА)
8, 43
Задняя нижняя мозжечковая
артерия (ЗНМА) 43
Задняя соединительная артерия
42
Задняя спинальная артерия 43
Задняя теменная кора 48
— поле 5, 7, 48, 69
— поражение 69
— регуляция движений 68
— регуляция движений глаз
69
Задняя черепная ямка 11, 82
Заключительные клетки 59
Запах 46, 67
Затылочная доля 11, 98
Звездчатые клетки мозжечка 82
Звук 47
— волны, свойства 47
— восприятие 44
- локализация 61
Злокачественная гипертермия
31
Зрачки 104
Зрительная кора
36, 38, 58, 79, 89
— первичная (поле 17 по
Бродману) 9, 58
— экстрастриарные отделы
(ассоциативные) 56
Зрительно-пространственная
функция 102, 104
Зрительные вызванные
потенциалы 106, 107
Зрительные пигменты 55
Зрительные пути 56
Зрительный анализатор
44, 54, 58
— световосприятие 54
Зрительный нерв 57, 102, 105
— обследование 102, 105
Зрительный перекрест 93, 94
Зрительный тракт 92—94
Зубчатая извилина 97, 98
И
Игнорирование 57, 97, 99
Иерархия 39
— организации клеток коры
39
Иммунные заболевания
нервной системы 114, 117
Иммунологическая
обособленность 114, 116
Инактивация рецепторов 19
Ингибирующий
постсинаптический
потенциал 8
Ингибиторы обратного захвата
аминов 107
Интермануальный конфликт 69
Интернейроны 51
Импринтинг, генетический
116, 119
Интернейроны
— спинной мозг 75, 82
Инфекции ПО, 113
— внутриутробные 13
— приводящие к
нейродегенерации НО, 113
Ионные каналы 18, 116, 118
вольтажзависимые 18
— заболевания 19
— модуляция 18
— рецепторы 23
- фильтр селективности 19
— химические
(лигандзависимые) 18
Исследование чувствительности
103, 105
К
Калиевые каналы,
вольтажзависимые 21
Калий, потенциал покоя 21
Калорическая проба 65
Кальциевые каналы 22
— вольтажзависимые 22
Кальций
— при мышечном сокращении
29
— при чувствительном
восприятии 46, 47
— синаптическая трансмиссия
22, 25
Капли 59
Карбамазепин 113, 115
Катаплексия 96, 97
Катаракта 55, 117, 118
Катехол-О-метилтрансфераза
8, 87
Катехоламин 87, 93, 92
Квадрантная гемианопсия 57
Кератит 55
Кислый вкус 67
Кишечная нервная система
92, 93
Клетка-бабушка 39
Клетки Гольджи (КГ) 82, 83
Клеточная мембрана 15
— ионные каналы 15
— нейрона 15
Клиновидное ядро 34, 49
Клозапин 108, 109
Клоназепам 113, 115
Клонус 77
Когнитивные нарушения.
См. Деменция
Когнитивные функции,
исследование 102, 104
Кодеин 53
Кожа, чувствительные
рецепторы 44, 50
Колбочки 55
Комплекс верхних олив (КВ О)
62, 63
Комплекс гиппокампа 94, 95
-поля CAICA4 97, 98
Компрессия 70
126
Компрессия локтевого нерва 70
Компрессия срединного нерва
70
Компьютерная томография
(КТ) 91
Конечности, обследование
103, 105
Конечностно-поясные
миодистрофии 29, 91
Конский хвост 32
Конфабулялии 94, 95
Координация. См. Движение
Кора головного мозга
35, 38, 84, НО, НО
— анатомия 38
— ассоциативные поля 45
— двигательные отделы 45
— нейрофизиология 39
— функциональная
организация 38, 39
— чувствительные отделы 38
Кора островковой извилины 51
Корешковые синдромы 70
Корзиночные клетки 82
Корковая дисплазия 13
Корковый центр взора 85
— в регуляции движений 88
— движения глаз 79
Корковый центр взора (КЦВ) 8
Корсаковский синдром 94, 95
Кортиконуклеарные пути 37
Кортикоспинальный тракт
(КСТ) 49, 73, 78
поражение 71
— регуляция движений 78
— спинной мозг 78
Кортикотропин-рилизинг-
гормон (КРГ) 109, 111
Коря головного мозга
— нейрофизиология 39
Котрансмиссия 25
Кохлеарные ядера 60
Красное ядро 35, 119, 122
Крейтцфельда-Якоба, болезнь
ПО, 113
Крестцовые нервы 32
Критический период (развития)
100, 102
Критический порог
импульсации 21
Кровоснабжение ЦНС 42
Крупноклеточные пластины 56
Крупноклеточные пластины/
клетки 57
Кураре 23
Л
Лабиринт 65
Лабиринтит 65
Лакунарные инсульты 43
Латеральная двигательная
система 77
Латеральное коленчатое тело 8
Латеральное торможение 33
Легкий меромиозин (ЛММ) 8
Лестница преддверия 47
Лимбическая система 92, 92
— обонятельные проекции 45
Лицевой нерв 103, 105
— паралич 63
Лобная доля 10, 11, 98, 103, 104
— в регуляции движений 69
— поражение 107, 109
— функция, обследование
102, 104
Локомоции 73, 76
— нарушения 77
— фиктивная 77
Люмбальная пункция 105, 107
м
Магнитно-резонансная
ангиография (МРА) 8
Магнитно-резонансная
ангиография/венография
(МРА/МРВ) 9, 104, 106,
116, 119
Магнитно-резонансная
томография (МРТ)
8, 70, 104, 106
Малоберцовый нерв 70
Мания 107, 108
Медиальная петля 34, 45, 49
Медиальное коленчатое тело
(МКТ) 63
Медиальный продольный пучок
(МПП) 88
— ростральное
интерстициальное ядро 89
Межприступная активность
112, 115
Межъядерная офтальмоплегия
89
Мелкоклеточные пластины
56, 57
Менингиомы 41, 67
Менингит 41,106,107,115,116
Менингоцеле 13
Меркеля, диски 49
Метэнпефалон 12, 13
Механорецепторы 8
Механорецепторы,
высокопороговые (ВПМ)
51
Миалгия 90
Миастенический синдром
Итона-Ламбрета 15, 117
Миастения 91
Мигрень 19, 118, 120
- семейная гемиплегическая
19, 87
Миелин 15, 16, 100, 100
- оболочка 15
— проведение 105, 107
Миелинизированные аксоны
(волокна) 25
Миелинизированные волокна
24
Миелит 71, 115, 116
— поперечный 71
Миелография 105, 107
Миелэнцефалон 12, 13
Микроглиальные клетки 16, 17
Микротрубочки 15, 111, 113
Микрофиламенты 15
Миндалевидное тело 66, 67, 92,
93, 97, 98, 107-109, 120,
111
Миниатюрный потенциал
концевой пластинки 23
Миодистрофии 91
Миозин 28
Миозит 90
Миоклония, неба 83
Миотония 19
Миофибриллы 28
М итохондриальные
заболевания 87, 116, 119
Митохондрия 14, 22, 116, 119
— геном 115, 116, 118, 119
нейрональная 14
Мозговой нейротрофический
фактор 99, 100
Мозговые оболочки 11
Мозжечок 11, 13, 82
— анатомия 82
— поражение 83
- развитие 13
— регуляция движений 82
— регуляция движения глаз
82
— функция 82
Моклобемид 107, 109
Молларе треугольника
поражение 83
127
Моноаминовые
нейротрансмиттеры
117, 120
Монокулярная депривация
101, 103
Морфин 53
Мост 11
Мотонейроны (МН)
32, 33, 91. См. также
периферические МН
— «-поражение 75
— вентромедиальная группа
77
— рекрутирование 75
— спинальные 100
— центральных (МН)
поражение 91, 103, 105
Моторная кора, первичная 80
поражение 81
Моховидные волокна
82, 83, 98, 99
Мультисистемная атрофия
92, 93
Мутации 27
— динамические 116, 119
Мутизм 97, 99
— акинетический 97, 99
Мышечно-суставное чувство
(проприоцепия) 103, 105
— обследование 103, 105
Мышечное веретено 74, 75
— поражение афферентных
волокон 75
Мышечные волокна 30, 72
— интрафузальные 74
— экстрафузальные 74
Мышца
10, 14, 16, 74. 115, 116
— антагонист 75
— биопсия 106, 107
— гомонимная 75
— нарушения 31
-сила 90, 103, 105
— синергисты 75
— сокращение 30
— структура 75
— тонус 104
Мягкая мозговая оболочка II
н
Налоксон 53
Нарколепсия 96, 97
Наружные волосковые клетки
(НВК) 61
Наружный слуховой проход
60, 65
Нарушение координации 90
Нарушения зрачковых реакций
92, 93
Нарушения ходьбы 29, 77
Наследования, типы
29, 116, 118
Наследственная
полинейропатия с
предрасположенностью
к туннельным синдромам
70
Натриевые каналы,
вольтажзависимые 21
Натрий, потенциал покоя 20
Натрий-калиевый насос 15
He-NMDA- рецепторы 27
Невралгия тройничного нерва
53
Невринома 37
Неврит 12, 115, 116
Неврозы 108, ПО
Неврологическое обследование
102, 104
Нежное ядро 34, 49
Нейробласты 13
Нейровизуализация 104, 106
Нейрогенез 97, 99
Нейрогенез у взрослых 13
Нейроглиальные клетки 14
Нейродегенеративные
заболевания ПО, 112
Нейролептики 107, 109
Нейромиотония Исаакса 23
Нейроны 14
— мультиполярные 15
— псевдоуниполярные 15
— тело 14
Нейроны глубинных ядер
мозжечка (НГЯМ) 8
Нейропептиды 100, 109, 111
— при тревоге 109, 111
Нейротоксины ПО, 113
Нейротрансмиттер
— инактивация (гидролиз
и усвоение) 26
Нейротрансмиттеры 26, 116—118,
120
— анатомия проводящих
путей 32
— при синаптической
трансмиссии 22
Нейротрофины 100
Нейротрофические факторы
99, 100
— нейродегенеративные
заболевания 100, 101
Нейрофиброматоз I типа 17
Нейрофиламенты 14
Нейрохимические нарушения
106, 108
Неостриатум 84
Нерва биопсия 106, 107
Нервная пластинка 12
Нервная система
клетки 92
— обследование 102, 104
— развитие 12
Нервная терминаль 14, 25
Нервная трубка 12
Нервно-мышечное соединение
90, 115, 117
— аутоиммунные заболевания
115, 117
— заболевание 115, 117
Нервные волокна.
См. Аксоны
Нервный гребень 12
Нервный желобок 12
Нервный импульс.
См. Потенциал действия
Нервы поясничного отдела
спинного мозга 11
Нернста уравнение 21
Нигростриарный тракт,
дофаминергический 86
Нижние бугорки четверохолмия
(НБЧ) 63
Нижняя олива 34, 35, 102, 119,
122
Ниссля субстанция 14
Нистагм 83
Нисходящие двигательные
проводящие пути
73, 118, 121
Нисходящие проводящие
двигательные пути 11
Ножка мозга 34, 43
Ножки мозжечка 35
Норадреналин 92, 93
— нарушения обмена при
депрессии 107, 108
— при тревоге 109, 111
Ноцицептивные пути 50
Ноцицепторы 50
— полимодальные (ПМНЦ)
51
Нопипеппия 35
128
о
Обоняние 66
Обонятельная луковица 67
Обонятельное восприятие
46, 47
Обонятельные рецепторы 47
Обонятельный нерв 36
Обследование нервной системы
102, 104, 106
Овальное окно 60
Однофотонная эмиссионная
компьютерная томография
(ОФЭКТ) 105, 107
Околоводопроводное серое
вещество 9, 35, ПО
Окситоцин 94
Окуловестибулярный рефлекс 8
Олигодендроглиома 17
Олигодендроциты 99, 101
— торможение роста аксонов
99, 101
Онемение 49, 70
Опиоидные анальгетики 53
Опиоиды эндогенные 51
Опорная фаза, локомоция
72, 77
Опсоклонус 89
Оптокинетический рефлекс 89
Орган кортиев 47
Ориентация в месте, времени и
собственной личности
102, 104
Ориентация по селективности 59
Основная артерия 42, 43
Острота зрения 102, 104, 105
Острый рассеянный
энцефаломиелит 114, 117
Осциллопсия 65
Отверстие Монро 40
Отверстия Мажанди и Люшка
40
Отводящий нерв 37, 103, 105
Относительный афферентный
зрачковый дефект 57
Отношение сигнал/шум 45
Отолитовый аппарат 37
Отосклероз 61
Отсроченная депрессия 83
Отсроченная потенциация
97, 98
Офтальмоплегия 36
Офтальмоплегия межъядерная
89
Оценка 102, 104
п
Паллидотомия, при болезни
Паркинсона 87
Паллидум, вентральные отделы
85
Палочки 55
Память 97, 98
- антеградная 97, 99
Панические атаки 108-111
Параллельные волокна 9, 83
Параллельные пути 39, 59
Парамедианная ретикулярная
формация моста (ПРФМ)
88, 89
Паранеопластические
синдромы 99, 101
Парапротеинемии 115, 117
Парасимпатическая нервная
система 11
Парасубикулум 97, 98
VI пары ЧМН. См. Блоковой
нерв
VII пара ЧМН 36
Парезы 89
— пирамидное распределение
77
Парестезии 70
Паркинсона болезнь 86
— лечение 86
— нарушения движения глаз
87
Паркинсонизм 108, 109
Паутинная оболочка 10, 40
Пахионовы грануляции 40, 41
Педункулопонтинное ядро
(ППЯ) 76
Пептиды, нейропептиды
100, 109, 111
Первичная сенсорная кора 45
Первичные афференты 33
Передние мозговые артерии 42
Передние отделы поясной
извилины 78
Передний (вентральный)
корешок 10
Передний рог 34
Передний спиноцеребеллярный
тракт 9, 76
Передняя нижняя мозжечковая
артерия 42, 120
Передняя соединительная
артерия 42, 120
Передняя спинальная артерия
42, 43
Передняя хороидальная артерия
42
Перекрестные мостики 29
Перехваты Ранвье 15
Периодический паралич 19
Периферическая нервная
система (ПНС) 99, 100
— пластичность 99, 100
Периферические мотонейроны
(ПМН) 103, 105
— поражение 103, 105
Периферические нервы
11, 115, 117
— аутоиммунные нарушения
115, 117
- перерезка, при лечении
боли 53
- поражение, двигательные
нарушения 90
- поражение, хронические
болевые синдромы 51
— поражение, чувствительные
нарушения 70
— регенерация 100
Пернициозная анемия 71
Пигментный ретинит 55
Пирамида 34
Пирамидное распределение
парезов 77
Пирамидный тракт 81.
См. также
Кортикоспинальный тракт
Пластины Рексед 32
Пластичность
-ПНС 99, 100
- ЦНС 100, 102, 118, 120
Плотные соединения 41, 66
Подкорковые зрительные
органы 56
Подострая сочетанная
дегенерация спинного
мозга 71
Подошвенные рефлексы
103, 105
Подъязычный нерв
37, 103, 105
- обследование 103, 105
Позвонки 11
Позвоночник 10, 11, 13
Позвоночные артерии 42
Позитронно-эмиссионная
томография (ПЭТ)
105, 107
Ползущие волокна 82, 83
Полимиозит 91, 115, 117
Полимодальные ноцицепторы
(ПМНЦ) 51
Полинейропатии 70, 105, 107
— генерализованные 105, 107
— крупнокалиберных волокон
70
- мелкокалиберных волокон
71
Полиомиелит 91, 99, 101
Полукружные каналы 102, 102
Полушария головного мозга 11
Поперечные (Т) тубулы 28
Поперечный миелит 71
Порог 106, 107
Порог вибрационного чувства 9
Постсинаптические складки 22
Постсинаптический рецептор
23
Постсинаптичсекая мембрана
15, 98, 99
Постсубикулум 97, 98
Потенциал действия 15, 20, 21
— в нервно-мышечной/
синаптической передаче 22
- генерация 21, 22
- распространение 24
Потенциал конпевой пластинки
23
Потенциал покоя мембраны 20
Потенциал равновесия 20
Поток ионный 18
Поясная извилина 97, 99
Преддверно-улитковый нерв
103, 105
— обследование 103, 105
Премоторная кора 9, 68, 72
- поражение 73
Пресинаптическая нервная
терминаль 15
Пресинаптические активные
зоны 22
Пресинаптические
ауторецепторы 25
Пресинаптические везикулы
22
Пресинаптическое торможение
26
Пресубикулум 97, 98
Претектальные ядра 56, 57
Префронтальная кора 68
Прикосновение 44, 103, 105
- исследование 103, 105
Принцип «все или ничего» 21
Принцип размера 75
Принцип селективности
по стороне 59
Проведение, ионы 15
Проведение по нерву
24, 115, 117
— нарушения 27
— блок 25
Программированная клеточная
смерть (апоптоз) 111, 113
Продолговатый мозг
82, 119, 121
Продырявленная пластинка 36
Прозэнцефалон 12
Промежуточные (боковые)
столбы 93, 92
Промежуточные капли,
зрительная кора 59
Промежуточный мозг 92, 93
Проницаемость ионных
каналов 19
Проприоспинальные нейроны
76, 77
Проприоцепция 48
См. также
Мышечно - суставное
чувство
Пространственная суммация
25
Пространственное кодирование
45
Простые клетки, зрительная
кора 58, 68
Противопаркинсонические
препараты 79, 86
Противосудорожные препараты
113, 115
Псевдобульбарный паралич 37
Психические заболевания
95, 97
Психотерапия 96, 97
Птоз 36
Пуркинье клетки, мозжечка
(КП) 114, 117
Пучки нейрофибрилл (ПНФ)
ПО, ИЗ
Р
Радиальные глиальные волокна
39
Развитие 12, 13
Развитие головного мозга
12, 13
Рассеянный склероз
88, 91, 106, 107, 114, 117
— вестибулярная дисфункция
65
— глазодвигательные
нарушения 89
— нарушение проведения по
нерву 106, 107
— невралгия тройничного
нерва 53
— поражение мозжечка 91
Регуляция движений 78, 97, 99
- анатомия 78
— нарушения 97, 99
Рековерин 46
Рекрутирование мотонейронов
75
Реншоу клетки 77
Ресничный ганглий 56, 92
Ретикулоспинальный тракт
(РСТ) 9
— спинной мозг 73
Ретикулярная активируюшая
система, восходящая
95, 96
Ретикулярная формация
119, 122
— парамедианная моста
(ПРФМ) 9
Ретинотопическая проекция 56
Рефлексы 65
— двигательные 68
— длиннолатентные, или
транскортикальные 75
— на растяжение 73
— примитивные 102, 104
— сухожильные 73
Рефлекторная симпатическая
дистрофия 92, 93
Рефрактерный период 21
Рецептивное поле 44
— окружение 55
— центральное 55
Рецептор
— постсинаптические 27
Рецепторы 26
— десенситизация 27
Речь 62
— обследование 102, 104
Рианодиновые рецепторы,
патология 31
Рисперидон 108, 109
Родопсин 46
Ромбэнцефалон 12
Ростральный путь миграции
(РПМ) 13
Руброспинальный тракт 32, 35
в регуляции движений 35
— спинной мозг 77
130
Руффини свободные окончания
49
С
Саккадированные движения
глаз 57
Саккулюс 65
Сальтаторное проведение 25
Сарколемма 29
Саркомер 28
Саркоплазматический
ретикулум (СР) 28, 30
Сбор анамнеза 90
Сверхсложные клетки,
зрительная кора 59
Свободные радикалы ПО, 113
Селегилин 87
Селективный ингибитор
обратного захвата
серотонина 9
Сенильные невритические
бляшки (СНБ) 9
Септальные ядра 97, 98
Серое вещество 9, 10, 13
Серотонин (5-ГТ) 107, 108
— пути 50
Серотонин (5ГТ)
— серотонинергические
нейроны 53
Сетчатки 94
анатомия 54
— световосприятие 47
Сила мыши 103, 105
Сильвиев водопровод 13
Симпатическая нервная
система 11
активность при тревоге
108, ПО
Синапсы 22
— аксо-аксональные 15, 25
— аксодендритические 15
— аксосоматические 15
— возбуждающие 25
— тормозные 26
— химические 14, 22
электрические 15
Синаптическая интеграция 24
Синаптическая передача 22
Синаптическая щель 14, 22
Синаптические пузырьки 14
Синдром запястного канала 70
Синдром обструктивных апноэ
во время сна 96, 97
Синдром Прадера-Уилли
116. 119
Синдром ригидного человека
77
Синдром чужой руки 69, 120
Сирингомиелия 51
Скандированная речь 83
Скарна ганглий 65
Скорлупа 84
Сладкий вкус 67
Следящие движения глаз 88
Слепое пятно 54, 55
Слепота 47, 59
Сложные клетки 59
Сложные клетки, зрительная
кора 59
Слуховая кора 9, 42, 62, 63
— вторичная 63
— первичная 68
Слуховое восприятие 44, 47
Слуховой анализатор 62
Слуховые косточки 61
Слуховые пути 62
Снотворные 95—97
Соленый вкус 67
Соматосенсорная кора
49, 119, 121
— вторичная (ВССК) 8
Соматотопическая проекция
33, 49
Сон 95, 96
— медленноволновый 8
— не-REM фаза сна 95, 96
Сонливость дневная 96, 97
Сосцевидные тела 36
СПИД ПО, 113
Спинной мозг 16, 32
— восходящие
чувствительные пути 33
— кровоснабжение 43
— нисходящие двигательные
пути 73
Спинномозговые нервы 11
Спинномозговые узлы 48
— поражение 99, 101
Спиномезенцефальный тракт
51
Спиноретикулоталамический
тракт 51
Спиноталамический тракт
(СТТ) 51
— поражение 50
— спинной мозг 51
Спиноцеребеллярный тракт 32
— задний (ЗСЦТ) 77
— передний (ПСЦТ) 77
Спонтанная речь 102, 104
Срединная лестница 61
Средние мозговые артерии
(СМА) 9, 42
Средний мозг 56, 82, 93, 94
Средний отит 61
Ствол мозга 11, 92
— анатомия 34
— поражение 35
— развитие 13
— черепно-мозговые нервы
35
Стволовые клетки 101, 103
Стеноз сильвиева водопровода
41
Стереоцилии 47
— в вестибулярном
анализаторе 65
— в звуковом восприятии 47
Столбики, корковые 38
— первичная зрительная кора
89
первичная моторная кора
73, 78
— первичная слуховая кора
63
Стресс-реакпия 53
Стриатум 85
вентральные отделы 85
- неостриатум 86
Стриосомы 85
Субарахноидальное
пространство 40
Субвентрикулярная зона (СВЗ)
9, 13, 39, 101, 103
Субдуральное пространство 11
Субикулум 97, 98
Субстанция Р 109, 111
Субталамическое ядро (СТЯ)
84
Судорожные припадки
112, 114. См. также
Эпилепсия
-генерализованные 112, 114
-парциальные 112, 114
- сложные 112, 114
Супрахиазматическое ядро 57
Сухожильные рефлексы 74
Сухожильный орган Гольджи
9, 75
Таламус 84
— вестибулярные пути 65
131
- зад не вентральное ядро
таламуса (ЗВЯТ) 8
- латеральное коленчатое
тело (ЛКТ) 8
- медиальное коленчатое
тело (МКТ) 8
- ноцицептивные пути 50
- обонятельные проекции 67
- передневентральное
вентролатеральное ядро
таламуса (ПВ-ВЛЯТ) 9
Твердая мозговая оболочка
(ТМО) 40
Тектоспинальный тракт (ТСТ)
73
Тельца Леви 86, 112
Тельца Мейсснера 49
Тельца Пачини 49
Теменная доля 10, 103, 104
- функция 102, 104
Теменная кора, задняя
8, 38, 59, 68
Т емпературная
чувствительность,
исследование 103, 105
Терморегуляция 51, 94, 95
Терморецепторы 51
Тонотопическая организация
63
Тормозные 1а интернейроны 75
Тормозные аминокислоты 27
Тормозные синапсы 82
Транзиторная ишемическая
атака 9, 42
Трансдуцин 46
Транскортикальные рефлексы 75
Транскраниальная магнитная
стимуляция 91
Трансмембранные поры 19
Тревога 108, ПО
Тревожные расстройства
108, ПО
Тремор 83
- мягкого нёба 83
- при болезни Паркинсона
87
Тремор/ миоклонии мягкого
нёба 83
Триады 30
Т рицикл ические
антидепрессанты 53
Тройничный нерв 103, 105
- обследование 103, 105
Тропомиозин 29
Тропонин 29
132
Тугоухость
- кондуктивная 61
— нейросенсорная 61
Туннельные синдромы 70
Тяжелый меромиозин (ТММ) 9
У
Улитка 60
Утрикулюс 65
Ухо 35
- внутреннее 61
- наружное 61
- среднее 61
Ушастый еж 13
Ф
Фаза скольжения, локомоция
76
Фактор роста нервов (ФРН)
99, 101
Фенитоин 113, 115
Фенобарбитал 113, 115
Фиксирующий взор 57
Фобии 108, ПО
Фоторецепторы 55
X
Хвостатое ядро 42, 84
Химическое проведение 96, 97
Холецистокинин (ХЦК)
108, ПО
Холинолитические препараты,
при болезни Паркинсона 87
Хорее Сиденгама 87
Хорея 73
Хороидное сплетение 17, 34, 40
— папилломы 17
Цветовая слепота 55
Центр насыщения 94, 95
Центральная нервная система
9, 100, 102, 117, 118
— иммунная система 114, 116
— аутоиммунные
расстройства 114, 117
— нарушения эмбриогенеза
116, 119
— пластичность 99, 100
- регуляция вегетативной
нервной системы 92, 93
- способность к регенерации
100, 102
Центральная нервная система
- кровоснабжение 42
Центральное окружение
рецептивного поля 55
Центральный мотонейрон
(ЦМН) 9
- поражение 73
Цепочки 75
Церебральный паралич,
хореоатетоидный 87
Цереброспинальная жидкость
- анализ 106, 107
- заболевания 106, 107
Цереброспинальная жидкость
(ЦСЖ) 91
Циклический
аденозинмонофосфат
(цАМФ) 19
Циклический
гуанозинмонофосфат
(цГМФ) 47
Циркадные ритмы 57, 94, 95
Цистерны субарахноидальные
40
ЦНС. См. Центральная
нервная система
Частота 60, 62
Частотное кодирование 21, 45
Черепно-мозговые нервы
34-36
- структуры ствола 92, 93
Черная субстанция 35, 84, НО
компактная часть 84
— ретикулярная часть 84
Чрескожная
электронейростимуляция
(ЧЭНС) 9, 53
Чувствительное восприятие 46
Чувствительность 103, 105
Чувствительные афферентные
волокна 32, 129
Чувствительные нарушения 70,
113, 114
Чувствительные отделы коры,
первичные и вторичные 80
Чувствительные проводящие
пути 45, 118, 121
- заболевания 45
Чувствительные рецепторы 44
— кожные 48, 80
ш
Шванновские клети 99, 100
— в регенерации
периферического нерва
99, 101
Шванномы 17
Шейные спинномозговые
нервы 11
Шизофрения 86
Шум 45
э
Эдингера—Вестфаля ядро 57
Эксайтотоксическая гибель
клеток 27
Экстрапир амидные пути 78
Электродиагностика 105, 107
Электрокардиография (ЭКГ)
105, 107
Электромиография (ЭМГ) 81
Электронейрография (ЭНГ)
70
Электроэнпефалография (ЭЭГ)
105, 107
— при эпилепсии 105, 107
Эмбриогенез 11
— нарушения. ЦНС 121
Эндолимфа 47
Эндо невральная трубка 99, 101
Эндорфины 51
Эндотелиоциты 16
Эндоцитоз 23
Энкефалины 51
Энтакапон 87
Энторинальная кора 97, 98
Энцефаломиелит 114, 117, 123
— экспериментальный
аллергический 114, 117
Эпендимальные клетки 16
Эпендимомы 17
Эпилепсия 19
— височной доли 67
— джексоновская 81
— лечение 112, 115
— патогенез 112, 114
— первично-
генерализованная 112, 115
Этосуксимид 113, 115
Я
Ядерные сумки 75
Ядро латеральной петли (ЯЛП)
62
Ядро одиночного пути 67
Ядро промежуточного пути 89
Язык 37, 46
Языкоглоточный нерв 103, 105
— обследование 103, 105
Ямка 54, 58
Иностранные
термины
в-адренорецепторов
антагонисты 109, 110
в-ритм 96
e-ритм на ЭЭГ 95, 96
у-аминомасляная кислота
(ГАМК) 8, 108, ПО
М-каналы 57
Р-каналы 57
С-волокна 50
Система XYW 55
Т-тубулы 29, 30
N-метил-D-аспартатные
(NMDA) рецепторы 27
Nucleus accumbens 85
REM-фаза сна 96, 97
Spina bifida 41
— occulta 13
— гидроцефалия 41
— 14-амилоид 111, 113
XYW ганглиозные клетки
сетчатки 55
книги
издательской
группы
«ГЭОТАР-Медиа»
Наглядные
акушерство
и гинекология
ЭрфюпР Норам*ц
Д жвнО Шаром
НАГЛЯДНЫЕ
АКУШЕРСТВО
И ГИНЕКОЛОГИЯ
Э.Р. Нореитц, Д. О. Шордж
Пер. с англ.
Год издания: 2003
Объем: 144 с.
Обложка
Книга содержит лаконичные и исчерпывающие современные
сведения по основным проблемам акушерства и гинекологии.
Множество рисунков, схем, диагностических и лечебных алгорит-
мов, дополняющих текст, облегчают восприятие и запоминание
материала и делают книгу интересной и информативной. Учеб-
ное пособие идеально подходит для быстрой и эффективной под-
готовки к экзаменам.
Предназначено студентам, интернам, ординаторам медицинс-
ких вузов, а также врачам обшей практики и акушерам-гинеко-
логам.
серия ЭКЗАМЕН НА ОТЛИЧНО
Наглядная
гастроэнтерология
С. Keiuae
Пер. с англ.
Год издания: 2005
Объем: 136 с.
Обложка
НАГЛЯДНАЯ
ГАСТРОЭНТЕРОЛОГИЯ
Книга наглядно и очень лаконично раскрывает практически все
аспекты современной гастроэнтерологии. Много места отведено
анатомии и физиологии. Отдельные разделы посвящены клинике
и диагностике основных заболеваний. Подчеркнута неразрывная
взаимосвязь всех органов пищеварения (интеграция функций).
Книга предназначена студентам медицинских вузов, врачам-те-
рапевтам и представителям других специальностей, занимающихся
проблемами гастроэнтерологии.
Издательская группа «ГЭОТАР-Медиа»
1 19828, Москва, ул. Малая Пироговская, 1а.
Тел./факс: (095) Ю1-39-07.
E-mail: info@qeotar.ru, http ://www.q eotar. г и
НАГЛЯДНАЯ НЕВРОЛОГИЯ
перевод с английского
Под редакцией В.И. Скворцовой
Подписано в печать 22.09.2006. Формат 60х90’/8. Бумага офсетная.
Печать офсетная. Печ. л. 17. Тираж 2000 экз. Заказ № 380.
Издательская группа «ГЭОТАР-Медиа»
119828, Москва, ул. М. Пироговская, 1а,
тел./факс (495) 101-39-07,
e-mail: info@geotar.ru, http://www.geotar.ru
Отпечатано в ОАО «Московская типография №6»
115088, г. Москва, Южнопортовая ул., 24.
Книги серии «Экзамен на отлично» можно охарактеризовать
тремя словами:
• Кратко: основной материал по дисциплине в небольшом
объеме.
• Наглядно: множество примеров и иллюстраций.
• Эффективно: студент готовится к экзамену на «отлично»,
а врач получает современное представление о предмете.
«Наглядная неврология» является одним из бестселлеров
серии «Экзамен на отлично», которая стала знаменита
благодаря изложению, ясным рисункам и содержательному
тексту. «Наглядная неврология» — это простое и доступное
изложение сложного материала. В книге предпринят интег-
ральный подход к обобщению анатомии, физиологии и ней-
рохимии нервной системы, а также клинических проявлений
ее поражения.
Первый раздел книги рассказывает об анатомии, структуре
и функции нервной системы на микроскопическом уровне.
Второй раздел посвящен кожной и висцеральной
чувствительности, включая патофизиологию системы боли,
зрительному и слуховому анализаторам, а также заболеваниям
анализатора общей чувствительности. Третий раздел
повествует о регуляции движений. Четвертый и пятый разделы
рассказывают о стволе мозга и специальных анализаторах,
высших корковых функциях (память, настроение, познавание),
сон, а также повреждении и старении нервной системы.
Настоящее издание включает 4 новые главы; в них изложены
методы клинического обследования больных с нервными
болезнями, с которыми сталкивается врач.
В серии
«Экзамен
на отлично»
вышли книги:
Наглядные
акушерство
и гинекология
Наглядная
анатомия
Наглядная
иммунология
Наглядная
статистика
в медицине
Наглядная
биохимия
Половая
система
в норме
и патологии
Наглядная
гастроэнтеро-
логия
Предназначена
• студентам медицинских и биологических вузов;
• врачам;
•научным работникам.
Сердечно-
сосудистая
система для
кардиолога
Издательская группа «ГЭОТАР-Медиа»
Посетите наш сайт в Интернет: WWW.geotar.ru