/

































































































































































































































Author: Хиггинс И. Джонс Дж. Келли Д.
Tags: общая и теоретическая биология общетехнические дисциплины биология биохимия биотехнологии
ISBN: 5—03—000058—5
Year: 1988




Text
Edited by I. J. Higgins, D. J. Best and J. Jones ,
BIOTECHNOLOGY
PRINCIPLES
AND APPLICATIONS
Blackwell Scientific Publications
Oxford London Edinburgh
Boston Palo Alto Melbourne
БИОТЕХНОЛОГИЯ
Принципы
и применение
Под редакцией
И. Хиггинса, Д. Беста,
Дж. Джонса
Перевод с английского
д-ра биол. наук
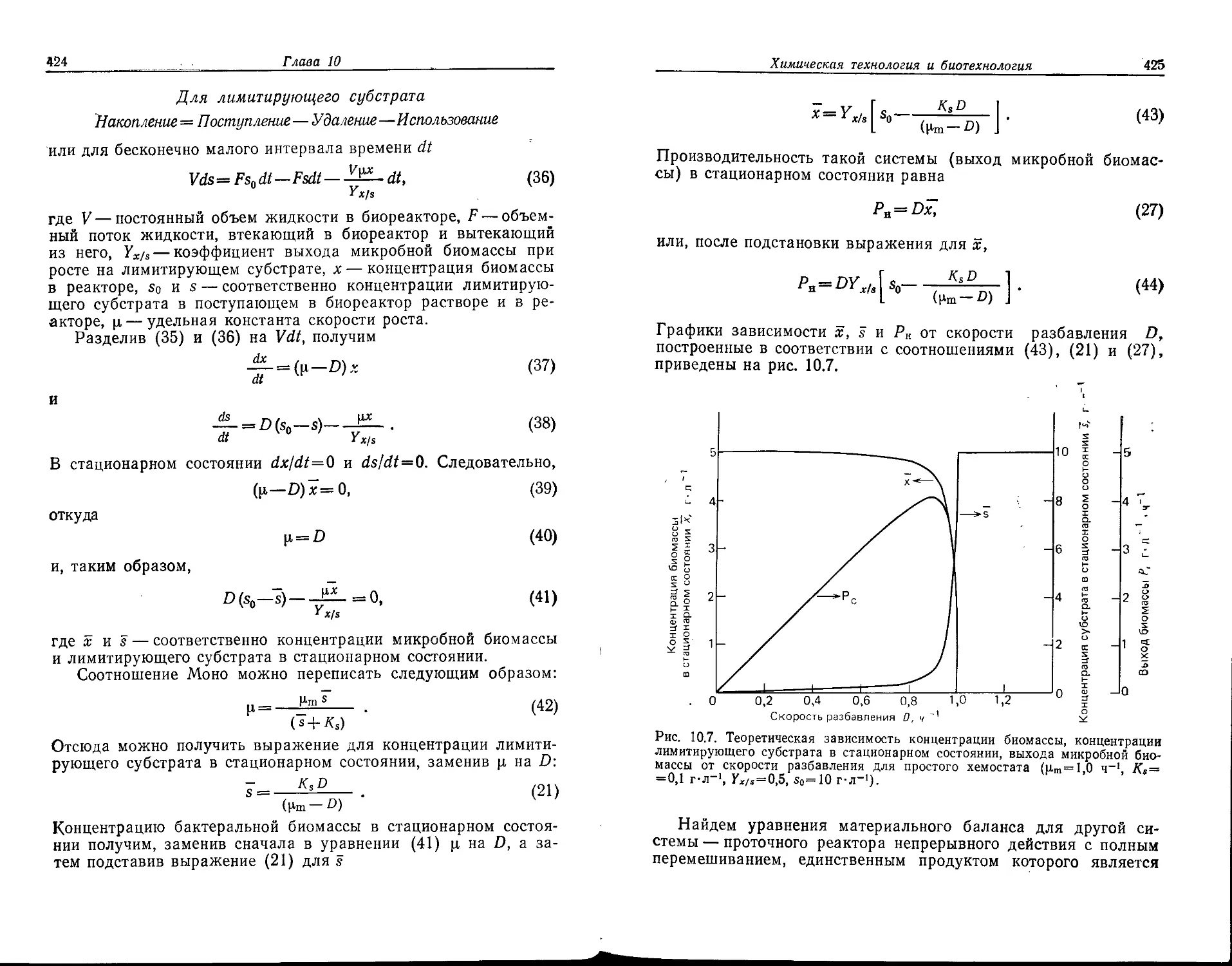
А. С. Антонова
под редакцией
акад. А. А. Баева
Москва «Мир» 1988
ББК 30.16
Б63
УДК 573.6
Бич Г., Бест Д., Брайерли К., Кумбс Дж., Холл Д., Ха-
мер Г., Харди К-, Хиггинс И., Джонс Дж., Келли Д., Мел-
вин М., Оливер С., Пикап Дж., Сил К-, Скиннер Ф., Стаф-
форд Д., Таггарт Дж.
Б63 Биотехнология. Принципы и применение: Пер. с англ./
Под. ред. И. Хиггинса, Д. Беста и Дж. Джонса, — М.: Мир,
1988. — 480 с., ил.
ISBN 5—03—000058—5
Книга, написанная коллективом авторов (Англия, США, Швейцария), пред-
ставляет собой учебник по биотехнологии, освещающий как новые, так и тра-
диционные отрасли промышленности, основанные на применении микроорганиз-
мов. Рассмотрено использование микроорганизмов для получения биотоплива, пи-
щевых продуктов и биоматериалов, а также применение биотехнологии в хими-
ческой промышленности, медицине, сельском хозяйстве и для переработки отхо-
дов. Особое внимание уделено связи биотехнологии и химической технологии.
Для студентов вузов и специалистов, занимающихся исследованиями и раз-
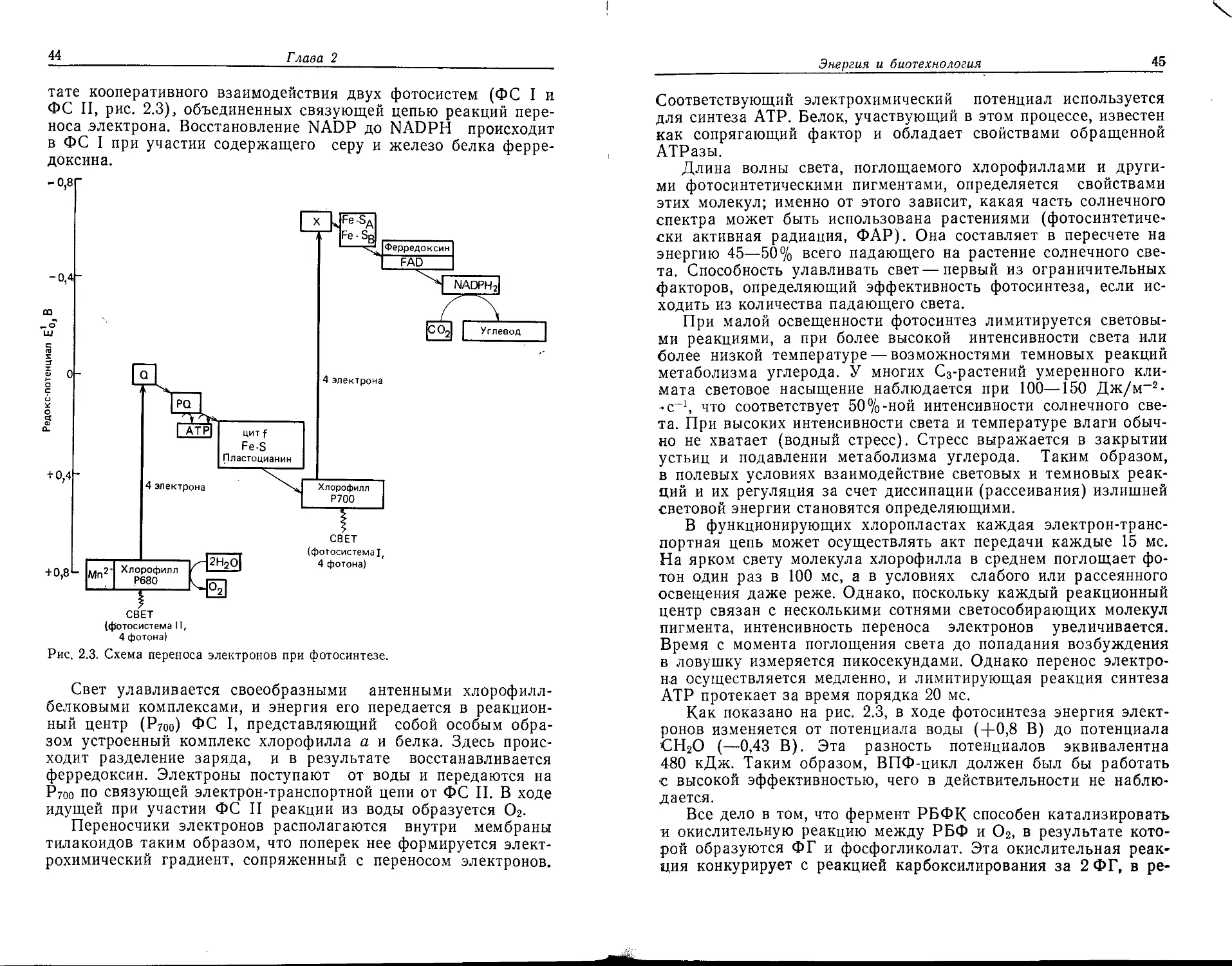
работками в области биотехнологии.
Б
2003000000—285
041 (01)—88
145— 1988,
ч. 1
ББК 30.16
Редакция литературы по биологии
ISBN 5—03—000058—5 (русск.) © 1985 by Blackwell Scientific Publications
ISBN 0—623—01029—0 (англ.) © перевод на русский язык, 1988
Предисловие редактора перевода
Совсем недавно слово «биотехнология» отсутствовало в нашем
языке; вместо него мы употребляли слова «промышленная мик-
робиология», «техническая биохимия» и т. п. Новый термин,
объединивший в себе все прежние названия, появился пример-
но 10 лет назад. Это незначительное на первый взгляд событие
нельзя сводить только к тому, что кому-то посчастливилось при-
думать удачное слово; за ним кроются гораздо более глубокие
причины. Биология, составляющая научную основу любых прак-
тических использований биологических процессов и систем, за
последние несколько десятилетий сделала огромный скачок на
пути познания жизненных явлений, и прежде всего в области
микробиологии, энзимологии, молекулярной биологии и молеку-
лярной генетики. Новые открытия объединили разрозненные
прикладные направления, подвели под них единую фундамен-
тальную основу. В результате биотехнология стала наукой о
практическом использовании биологии в целом, а не отдельных
ее ветвей, как это было прежде. В этом именно и заключается
подлинный смысл явлений, отмеченных введением нового тер-
мина.
Еще с конца прошлого столетия микроорганизмы стали ос-
новой производства целого ряда полезных продуктов (органи-
ческих кислот, этанола для технических целей, ферментов, ви-
таминов, антибиотиков и т. п.). Они нисколько не утратили
своего значения и сейчас, оставаясь главными «машинами» био-
логического производства. Более того, перспективы их дальней-
шего применения вырисовываются все яснее. Достаточно ука-
зать, что из всего огромного разнообразия микроорганизмов,
существующих в природе, используется лишь незначительная
их часть, и притом не самым эффективным образом, поскольку
наши знания о генетике и биохимии микроорганизмов не всегда
достаточны.
Лишь в недавнее время стали использовать для практиче-
ских целей растительные клетки, а животные клетки отчасти
вследствие дороговизны их культивирования нашли лишь огра-
ниченное применение, главным образом для получения моно-
клональных антител. Но методы, основанные на культивирова-
нии и растительных, и животных клеток все же пробили себе
дорогу в биологическую промышленность и со временем, по-ви-
димому, найдут здесь более широкое применение.
Уже используются в биологической промышленности разно-
образные биомолекулы, особенно ферменты, и из них в первую
6 Предисловие редактора перевода *
очередь амилазы и протеазы (они применяются в пивоварении
и производстве моющих средств). Однако нестойкость и относи-
тельная дороговизна ферментов сдерживает внедрение их в про-
мышленность. Крупным успехом стало создание иммобилизо-
ванных ферментов, что позволило решить по крайней мере
часть технологических проблем.
И все же первое место в современной биотехнологии принад-
лежит, конечно, генетической инженерии (технологии рекомби-
нантных молекул ДНК). Она предоставила исследователям но-
вую, исключительно ценную возможность — изменять генетиче-
скую программу бактериальных, растительных и животных кле-
ток, и тем самым как бы завершила формирование биотехноло-
гии. Вероятно, поэтому само название «биотехнология» появи-
лось после того, как возникла генетическая инженерия.
Мы надеемся, что предлагаемый вниманию советских чита-
телей перевод книги «Биотехнология», изданной в Англии в
1985 г., будет весьма полезным, поскольку у нас в стране нет
подобных руководств, а в них испытывается нужда. В книге
намечены исторические корни биотехнологии, что позволяет
проследить весьма своеобразные пути ее развития. Большое
внимание уделено традиционным отраслям биотехнологии, так
как увлечение новыми направлениями не должно приводить к
забвению старых, которые и теперь составляют главную часть
биологического производства.
Книга отражает в значительной степени опыт и подходы
английских биотехнологов (за исключением трех, все авторы
англичане), но это нисколько не умаляет ее достоинств. Главы
несколько неравномерны по объему и содержанию, поскольку
они написаны разными авторами. Можно спорить, следовало
ли включать в 10-ю главу подробные математические выкладки,
касающиеся работы различных биореакторов, но, вероятно, и в
таком виде эта часть книги найдет своего читателя.
В книге часто обсуждаются экономические аспекты приме-
нения биотехнологии, что особенно ценно сейчас, когда в на-
шей стране происходит перестройка экономической системы.
Биотехнология многолика и по своим историческим корням,
и по своей современной структуре, объединяющей элементы
фундаментальных наук и таких прикладных отраслей, как хи-
мическая технология, машиностроение и экономика. Поэтому
по биотехнологии могут быть написаны разные книги, и это ру-
ководство представляет собою одно из возможных.
В заключение хотелось бы поблагодарить аспирантку ка-
федры математического моделирования МХТИ им. Д.' И. Мен-
делеева В. В. Софенину за помощь при редактировании гл. 10.
А. Баев
Предисловие
Цель этой книги — дать читателю полное, глубокое представле-
ние о том, что такое современная биотехнология. Корни ее ухо-
дят в далекое прошлое, а будущее обещает быть блестящим.
Хочется думать, что наш труд окажется полезным как для ис-
следователей в этой области, так и для биотехнологов-практи-
ков, которые хотят расширить свой кругозор. Мы не пытались
написать исчерпывающий обзор, однако считаем, что эта книга
может стать хорошим введением в большинство главных проб-
лем биотехнологии, которые освещены в ней ведущими специа-
листами. Для тех, кто пожелает ознакомиться с этими пробле-
мами более глубоко, мы после каждой главы приводим обшир-
ные списки литературы.
Работа над этой книгой была начата одним из ее редак-
торов за четыре года до выхода издания в свет. Отсюда вовсе
не следует, что авторы пренебрегали своими обязанностями:
просто практические исследования оставляли им слишком ма-
ло свободного времени. За этот период число редакторов книги
выросло от одного до трех, но некоторых рукописей мы так и
не получили! Будем надеяться, что они будут опубликованы во
втором издании.
Редакторы хотели бы выразить свою благодарность некото-
рым авторам этой книги, представившим рукописи за три года
до выхода ее в свет, за проявленное ими долготерпение. Мы
хотели бы также поблагодарить всех авторов и издательство
за их стремление довести наше общее сложное дело до успешно-
го завершения. Мы рады, что нам удалось выступить единым
фронтом, и надеемся, что наши читатели получат от этой книги
и пользу, и удовольствие.
Авторы гл. 3 хотели бы поблагодарить д-ра Дж. Эделмена,
руководителя исследований в RHM PLC, за разрешение опуб-
ликовать результаты, а также администрацию Frish Dites Part-
nership за информацию о микопротеине. Авторы гл. 4 и 6 вы-
ражают свою благодарность г-же Д. Фаулер, выполнившей ри-
сунки. Автор гл. 9 искренне признателен своим коллегам по
Ротамстедской экспериментальной станции за фотографии, ил-
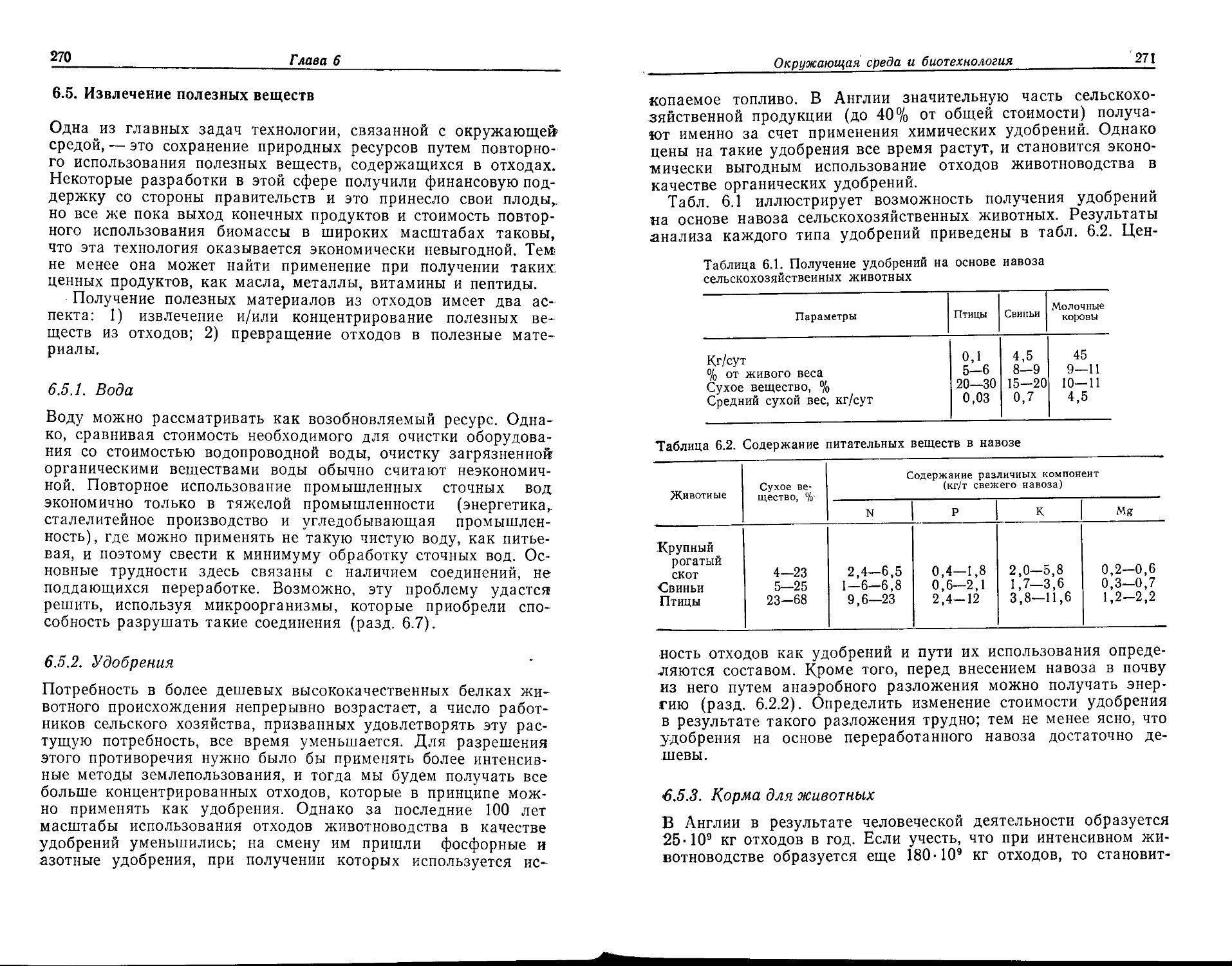
люстрирующие текст: д-ру Барбаре Мосс (рис. 9.1 и 9.2), д-ру
Д. Хеймену (рис. 9.3—9.6), д-ру М. Джонсу (рис. 9.7 и 9.8) и
д-ру Р. Нелсону (рис. 9.9 и 9.10).
Биотехнологический центр
Кренфилдского технологического
института, сентябрь 1984 г.
И. Дж. Хиггинс
Д. Дж. Бест
Дж. Джонс
Глава 1
Что такое биотехнология?
И. Хиггинс
1.1. Введение
Биотехнология — это не просто новомодное, броское название
одной из древнейших сфер деятельности человека; так могут
думать одни только скептики. Само появление этого термина в
нашем словаре глубоко символично. Оно отражает широко рас-
пространенное, хотя и не общепринятое мнение: считается, что
применение биологических материалов и принципов в ближай-
шие десять — пятьдесят лет радикально изменит многие отрасли
промышленности и само человеческое общество. Нетрудно убе-
диться, что интерес к этой науке и темпы ее развития в послед-
ние годы росли очень быстро. Свидетельств тому множество.
Это и появление бесчисленных небольших частных биотехноло-
гических фирм, и образование правительственных комитетов,
призванных оценить возможности нового направления, и чтение
лекций по биотехнологии во многих университетах. Правитель-
ства большинства наиболее развитых стран, как, впрочем, и ря-
да развивающихся, уже вложили значительные средства в раз-
витие биотехнологии. Надо сказать, что как размеры этих вкла-
дов, так и эффективность их использования далеко не одинако-
вы. Специалисты, участвующие в развитии биотехнологии, еди-
нодушно считают, что в масштабах государства успех в этой
области может быть достигнут только при участии правитель-
ственных органов. Их поддержка чрезвычайно важна для раз-
вития этой сложной междисциплинарной технологии. От появ-
ления идеи до ее воплощения в разных отраслях биотехноло-
гии лежит большой путь, и лишь в немногих странах, в частно-
сти в США, действуют сегодня адекватные экономические меха-
низмы, создающие основу для оптимального развития этой тех-
нологии, причем в значительной мере независимого от админи-
страции.
Может быть, правильнее всего называть биотехнологией ис-
пользование в промышленности биологических систем или про-
цессов. В основе ее лежит уникальность биологических систем
в отношении узнавания и катализа. Проявляется это при рас-
познавании других биологических систем или определенных хи-
Что такое биотехнология?
9
мических соединений, а также в удивительной способности фер-
ментов катализировать широкий спектр химических реакций в
мягких условиях. Даже сегодня химикам не удается создать
катализаторы, превосходящие по своей эффективности и специ-
фичности биологические катализаторы, кроме того, наши зна-
ния о механизмах ферментативного катализа остаются весьма
скудными. Несмотря на все достижения химии, которые лежат
в основе многих отраслей промышленности, мы еще очень мало
знаем о природе катализа.
Человек использовал биотехнологию многие тысячи лет:
люди занимались пивоварением, пекли хлеб. Они придумали
способы хранения и переработки продуктов путем ферментации
(производство сыра, уксуса, соевого соуса), научились делать
мыло из жиров, изготавливать простейшие лекарства и перера-
батывать отходы. Однако только разработка методов генетиче-
ской инженерии, основанных на создании рекомбинантных
ДНК (гл. 7), привела к тому «биотехнологическому буму», сви-
детелями которого мы являемся. Эти методы не только откры-
вают возможности улучшения уже освоенных процессов и про-
дуктов, но и дают нам совершенно оригинальные способы полу-
чения новых, ранее недоступных веществ, позволяют осуществ-
лять новые процессы. Сама история этой науки — генетической
инженерии — яркий пример того, как сложно прогнозировать
внедрение в практику достижений фундаментальных наук. Раз-
работка технологии рекомбинантных ДНК—результат значи-
тельных вложений в развитие молекулярной биологии за по-
следние сорок с лишним лет. А ведь не так давно, в конце 60-х
годов, многие биологи сетовали, что слишком уж много вни-
мания уделяется этой престижной области биологии и химии,
которая не дает ничего полезного. Сегодня нам ясно, что от-
крытия молекулярной биологии глубоко скажутся на судьбе че-
ловечества.
Хотя популярность биотехнологии обусловлена главным об-
разом использованием технологии рекомбинантных ДНК, нуж-
но подчеркнуть, что и в других областях науки был сделан ряд
крупных открытий, повлиявших на ее развитие. Наиболее важ-
ные в этом плане достижения отражены в табл. 1.1. Их вклад
в различные отрасли биотехнологии обсуждается в последую-
щих главах этой книги.
Вряд ли найдется иная, помимо биотехнологии, область нау-
ки, к которой так подходят следующие слова: «Нет и еще ты-
сячу раз нет: я не знаю такой науки, которую можно было бы
назвать прикладной. Есть наука и есть области ее применения,
и они связаны друг с другом, как плод с взрастившим его де-
ревом» (Пастер, 1871; цитата взята из Revue Scientifique).
10
Глава 1
Таблица 1.1. Области науки, в которых недавно были получены
новые результаты, важные для развития биотехнологии
Генетическая инженерия (технология рекомбинантных ДНК)
Биокатализ
Ферменты (выделение, иммобилизация, стабилизация)
Целые клетки микроорганизмов и клетки макроорганизмов
(иммобилизация, стабилизация).
Иммунология (особенно моноклональные антитела)
/Производство
Технология ферментации
Переработка отходов
Биоэлектрохимия
Использование научных достижений в биотехнологии тесно
связано с фундаментальными исследованиями и осуществляет-
ся на самом высоком уровне современной науки. Удивительная
научная многоликость биотехнологии, о которой уже шла речь,
отчетливо видна из рис. 1.1. Не все перечисленные на нем от-
Организация/Координация
Биохимия,
Химия
Микробиология
Научные
основы
Электроника
>_ г I1 I у т СМ rl Л
Биотехнология пищевых
продуктов
Биохимическая
технология
Технология
пищевой
технология
Механическая
технология
Рис. 1.1. Междисциплинарная приро-
да биотехнологии.
расли науки вносят свой вклад в осуществление каждого кон-
кретного биотехнологического процесса или в получение того
или иного продукта, но, как правило, таких отраслей несколько.
В наши дни определилась одна важная особенность разви-
тия некоторых перспективных разделов биотехнологии: необхо-
димость тесного международного сотрудничества специалистов,
ученых и технологов. Дело в том, что лишь немногие научные
коллективы в мире обладают достаточным опытом работы в
этой области. Ярким примером тому служит многонациональ-
Что такое биотехнология?
11
ность ряда основанных в последние годы крупнейших междуна-
родных биотехнологических фирм.
В заключительной части этой главы мы вкратце остановим-
ся на проблемах развития и перспективах основных направле-
ний биотехнологии, а также рассмотрим некоторые экономиче-
ские вопросы. Это будет своего рода вступлением к более де-
тальному обсуждению задач биотехнологии в последующих гла-
вах.
1.2. Исторические перспективы
До тех пор, пока всеобъемлющий термин «биотехнология» не
стал общепринятым, для обозначения наиболее тесно связанных
с биологией разнообразных технологий использовали такие на-
звания, как прикладная микробиология, прикладная биохимия,
технология ферментов, биоинженерия, прикладная генетика и
прикладная биология. Если не принимать в расчет производст-
ва мыла, то первая же из числа возникших «технологий» тако-
го рода стала предшественницей прикладной микробиологии.
Наши предки не имели представления о процессах, лежащих в
основе таких технологий. Они действовали скорее интуитивно,
но в течение тысячелетий успешно использовали метод мик-
робиологической ферментации для сохранения пищи (например,
при получении сыра или уксуса), улучшения вкуса (например,
хлеба и соевого соуса) и производства спиртных напитков. Пи-
воварение до сих пор остается наиболее важной (в денежном
исчислении) отраслью биотехнологии. Во всем мире ежегодно
производится около 10й литров пива стоимостью порядка
100 млн. фунтов стерлингов. В основе всех этих производств ле-
жат реакции обмена веществ, происходящие при росте и раз-
множении некоторых микроорганизмов в анаэробных условиях.
В конце XIX в. благодаря трудам Пастера были созданы реаль-
ные предпосылки для дальнейшего развития прикладной (тех-
нической) микробиологии, а также в значительной мере и био-
технологии. Пастер установил, что микробы играют ключевую
роль в процессах брожения, и показал, что в образовании от-
дельных продуктов участвуют разные их виды. Его исследо-
вания послужили основой развития в конце XIX и начале XX вв.
бродильного производства органических растворителей (ацето-
на, этанола, бутанола и изопропанола) и других химических ве-
ществ, где использовались разнообразные виды микроорганиз-
мов. Во всех этих процессах микробы в бескислородной среде
осуществляют превращение углеводов растений в ценные про-
дукты. В качестве источника энергии для роста микробы в этих
условиях используют изменения энтропии при превращениях
веществ. Совсем иначе обстоит дело в аэробных процессах при
контролируемом окислении химических веществ до углекислого
12
Глава 1
газа и воды. При этом организмы извлекают гораздо больше
энергии.
Процессы такого рода, в которых биомасса, т. е. возобно-
вляемый источник сырья, используется для получения химиче-
ских веществ, играли ведущую роль на первом этапе развития
современной биотехнологии. По мере становления нефтехимии
на смену многим из них пришли химические процессы. В тех
случаях, когда некоторые химические соединения, например
цитрат, ацетат и итаконат, широко использовались при произ-
водстве пищевых продуктов, их продолжали получать путем
брожения, самым выгодным с экономической точки зрения пу-
тем. В некоторых странах (например, в Италии) таким спосо-
бом вырабатывали даже технический этиловый спирт. Сегодня
под влиянием энергетического кризиса производство спирта из
растительного сырья получает все более широкое распростране-
ние в США и Бразилии и, видимо, вскоре выйдет на первый
план в странах Дальнего Востока.
Сахара
Фру к тозо-1,6-дифосфат
Дигидрокси-
—NADH
NAD* 1
Глицеральдегид-З-фосфат
Реакции Эмбдена-
Мейергофа
Глицерол-Я-фосфат | г
I । Пируват
’ ; L________со2
Глицерол | f 1
NAD* ) ОбпутьЬ'И Ацетальдегид
Этанол -------------------------1 HSOg
Ацетальдегид бисульфит,
примесное соединение
Рис. 1.2. Переключение спиртового брожения у дрожжей на образование гли-
церола.
Возвращение к производству органических соединений путем
брожения вполне возможно, но в Европе это будут скорее все-
го лишь наиболее дорогостоящие вещества. Очевидным исклю-
чением здесь может быть превращение в технический спирт
винных рек ЕЭС. Это, конечно, странная политика, странная
биотехнология и по меньшей мере необычная экономика, пред-
лагающая химической промышленности нечестные условия со-
ревнования.
На рис. L2 показано, какие биохимические реакции лежат в
основе одного из первых процессов «современной» биотехно-
Что такое биотехнология?
13
логии. Этот рисунок представляет определенный интерес в свя-
зи с тем, что он иллюстрирует «инженерный» принцип модифи-
кации нормального метаболизма дрожжей при образовании ими
спирта. При добавлении бисульфита к сбраживаемой среде аце-
тальдегид не может выступать в роли акцептора восстанови-
тельных эквивалентов, образующихся в реакциях пути Эмбде-
на— Мейергофа. В результате таким акцептором служит фос-
фодиоксиацетон, а вместо этилового спирта образуется глице-
рол. Этот процесс оказался особенно ценным, когда в ходе пер-
вой мировой войны возросли потребности в глицероле (его ис-
пользовали для производства взрывчатых веществ).
Следующим важным этапом в развитии биотехнологии хо-
зяйственно ценных веществ была организация промышленного
производства антибиотиков. Отправной точкой здесь послужи-
ло открытие Флеммингом, Флори и Чейном химиотерапевтиче-
ской активности пенициллина (1940 г.). Сегодня годовой обо-
рот этой отрасли составляет около 2 млрд, фунтов стерлингов.
Как получение химических соединений и пищевых добавок
путем брожения, так и синтез антибиотиков всегда велись в
асептических условиях, но некоторые современные процессы
(например, образование белка одноклеточными организмами)
осуществляют в еще более жестком режиме. Обеспечение таких
особых условий — многоплановая задача. Она решается инже-
нерами-химиками и микробиологами (подробнее об этом будет
рассказано в гл. 10). С другой стороны, использование микро-
организмов при переработке отходов (гл. 6) не требует созда-
ния стерильных условий; напротив, вообще говоря, чем больше
разных микроорганизмов принимает в этом участие, тем лучше.
Впрочем, при планировании и создании заводов по переработке
отходов инженеры-химики и микробиологи столкнулись с про-
блемами иного круга. Процесс минерализации органических
отбросов, основанный на использовании активного ила, был
разработан в 1914 г. С тех пор он был существенно модернизи-
рован, стал более сложным и производительным и используется
сегодня во всем мире для переработки стоков.
В наше время переработка стоков в анаэробных условиях
смешанной микрофлорой, в результате чего попутно образуется
биогаз (который состоит в основном из метана и СОг), приме-
няется все более широко. Такая переработка энергетически вы-
сокоэффективна, так как позволяет сохранять и концентриро-
вать энергию, содержащуюся в различных компонентах стоков
(с газом регенерируется более 80% свободной энергии). В не-
которых случаях, в частности в сельскохозяйственных общи-
нах, с помощью этого процесса можно получать значительную
часть необходимой энергии. Так, в Китае построено более
18 млн. генераторов биогаза. В развитых странах с высоким
14
Глава 1
потреблением энергии превращение отходов в биогаз может
покрыть лишь несколько процентов их энергетических потребно-
стей. Тем не менее на крупных заводах по переработке отходов
биогаз часто сжигают в тепловых машинах, которые приводят
в действие электрогенераторы. В последние годы разработаны
также небольшие установки, предназначенные для переработки
отходов сельского хозяйства.
1.3. Развитие биотехнологической промышленности
после второй мировой войны
Помимо постоянного усовершенствования процессов, о которых
речь шла ранее, в последние сорок лет был разработан ряд
новых; некоторые из них перечислены в табл. 1.2. Мы обратим-
ся к этим процессам в последующих главах книги, но четыре
Таблица 1.2. Некоторые новые направления, развивающиеся
на основе биотехнологии, и продукты, получаемые с ее помощью
Отрасль Примеры
Сельское хозяйство Получение новых штаммов, новые методы селек- ции растений и животных (включая клониро-
Производство химиче- ских веществ вание) Получение органических кислот (например, ли- монной, итаконовой), использование ферментов
Энергетика в составе моющих средств Увеличение потребления биогаза, крупномасштаб-
Контроль за состоянием окружающей среды ное производство этанола как жидкого топлива Улучшение методов тестирования и мониторинга, прогнозирование превращений ксенобиотиков благодаря более глубокому пониманию биохи- мии микроорганизмов, усовершенствованию ме- тодов переработки отходов, особенно промыш-
Пищевая промышлен- ность ленных Создание новых методов переработки и хранения пищевых продуктов, получение пищевых доба- вок (например, полимеров, продуцируемых микроорганизмами аминокислот), использова- ние белка, синтезируемого одноклеточными ор- ганизмами, и ферментов при переработке пище-
Материаловедение вого сырья Выщелачивание руд, дальнейшее изучение и
Медицина контроль биоразложения Применение ферментов для усовершенствования диагностики, создание датчиков на основе фер- ментов, использование микроорганизмов и фер- ментов при производстве сложных лекарств (например, стероидов), синтез новых антибио- тиков, применение ферментов в терапии
Что такое биотехнология?
15
наиболее интересных — производство аминокислот, белка одно-
клеточных организмов (БОО), превращение стероидов и культи-
вирование клеток животных и растений — мы вкратце рассмот-
рим в этом разделе.
За последние тридцать лет производство аминокислот в
аэробных микробиологических процессах получило все более
широкое распространение. В наибольшем количестве выраба-
тывались два продукта — глутамат натрия (ежегодное производ-
ство в мире — около 150 000 т), который служит усилителем
вкуса, и лизин (ежегодное производство в мире—15 000 т),
который используют как пищевую добавку. В мире за год про-
дается аминокислот на сумму 1 млрд, фунтов стерлингов, при-
чем большую часть поставляют японские фирмы. Особую роль
Японии в некоторых областях биотехнологии мы обсудим в этой
главе в разделе «Экономические и коммерческие аспекты био-
технологии».
Микроорганизмы могут превращать растительную биомассу
с низким содержанием белка в пищевые продукты с высоким
его содержанием. В крупных промышленных масштабах этот
процесс использовался в Германии: там в ходе первой мировой
войны выращивали дрожжи Saccharomyces cerevisiae, которые
добавляли главным образом в колбасу и супы. Таким путем
удавалось компенсировать около 60% довоенного импорта пи-
щевых продуктов. Сходные процессы на основе пищевых дрож-
жей Candida arborea и Candida utilis использовались и во вре-
мя второй мировой войны. В 60-х годах ряд нефтяных и химиче-
ских компаний начали исследования и разработки по созданию
новых процессов получения БОО, предназначенного для до-
бавления в пищу людям и животным (гл. 3). В какой-то мере
это было связано с недостатком белковой пищи в мире. В ка-
честве субстратов использовали нефть, метан, метанол и крах-
мал; большинство продуктов вырабатывали для добавления в
корм животным. В целом процессы на основе метанола и крах-
мала оказались наиболее конкурентоспособными. В западных
странах самый крупный завод был построен компанией ICI:
там в одном ферментере при участии метанолпотребляющей бак-
терии Methylophilus methylotrophus получают из метанола около
70 000 т белка прутина (Pruteen) в год. При помощи технологии
рекомбинантных ДНК были модифицированы механизмы асси-
миляции азота этими бактериями, что привело к увеличению
выхода продукта. Это было одним из первых доказательств
практической значимости и потенциальных возможностей гене-
тической инженерии (гл. 7). В СССР ежегодно производится
более 1 млн. т БОО, в основном из углеводородов и отходов
растениеводства. Один из немногих высококачественных продук-
тов из БОО, пригодный для человека, поставляется для проб-
16
Глава 1
ной продажи в Англии фирмой Rank Hovis McDougall. Его вы-
рабатывают из гриба, выращиваемого на содержащем углево-
ды сырье (гл. 3).
Хотя в наше время, и особенно в последние двадцать лет,
интерес к использованию ферментов в промышленности все бо-
лее возрастает, их внедрение в производство происходит мед-
ленно. Набор используемых сегодня ферментов весьма невелик
и применяются они в основном в пищевой промышленности.
Правда, за последние несколько лет значительно возросло чис-
ло ферментов, применяемых в медицине (главным образом для
диагностики). И все же в целом рынок ферментов остается
сравнительно небольшим, порядка 250 млн. фунтов стерлингов
в год. Основные причины такого относительно медленного раз-
вития— нестабильность ферментов, сложность выделения про-
дуктов переработки и проблемы, связанные с добавлением или
заменой кофакторов. Однако в некоторых случаях эти сложно-
сти удается обойти путем использования интактных клеток мик-
роорганизмов. Такой способ стал играть особенно важную роль
в 50-х годах при крупномасштабном производстве лекарствен-
ных препаратов стероидной природы. Было установлено, что
многие микроорганизмы способны строго направленно и стерео-
специфически гидроксилировать сложные молекулы стероидов.
Уже на ранних этапах этого метода было сделано важное на-
блюдение: оказалось, что плесневый гриб Rhizopus arrhizus
способен стереоспецифически, по 11-му положению гидроксили-
ровать женский половой гормон прогестерон. Это существенно
упростило производство кортизона, который применяют для ле-
чения артрита. До внедрения нового метода это соединение по-
лучали с помощью химического синтеза, включавшего 37 ста-
дий, с выходом 0,02%, и поэтому стоимость 1 г его составляла
200 долларов. Благодаря внедрению в этот процесс этапа, ос-
нованного на биотрансформации и существенно упрощающего
синтез, цена упала до 68 центов за 1 г. Впоследствии был выде-
лен еще ряд микроорганизмов, способных специфически гидро-
ксилировать другие углеродные атомы кольца, а недавно мик-
робные системы начали использовать для превращения фитосте-
роидов в С-19-стероидные гормоны с менее громоздкими моле-
кулами. Они находят широкое применение, в частности как пе-
роральные противозачаточные средства.
Химическая промышленность, особенно в США и Японии,
проявляет постоянный интерес к разработке аналогичных про-
цессов получения других дешевых продуктов на основе микро-
биологического окислительного катализа. Считается, что таким
путем удается осуществлять более экономичные процессы, чем
их нынешние аналоги в химической промышленности (гл. 4).
Что такое биотехнология?
17
Ряд производств, например получение вакцин, стал гораздо
эффективнее после освоения методов культивирования расти-
тельных и животных клеток в большом объеме. Был разрабо-
тан также метод слияния клеток различных линий, что позво-
лило, в частности, ученым фирмы Unilever получить новые кло-
ны масличных пальм, как более урожайных, так и дающих
продукцию более высокого качества.
1.4. Перспективы развития биотехнологии
В будущем благодаря расширению сферы своего применения
биотехнология сделает весомый вклад в повышение уровня жиз-
ни (табл. 1.3). Быстрее всего ее применение даст результаты в
Таблица 1.3. Сфера применения методов биотехнологии
Приборы для аналитической химии
Процессы биосинтеза и биодеградации
Углеродсодержащее сырье для химической промышленности
Химическая переработка (очистка продукта)
Химические продукты, использующиеся в быту:
клеи, детергенты, красители, волокна, вкусовые добавки,
желирующие вещества и загустители, камеди и раститель-
ные клеи, душистые вещества, пигменты, пластики, смазки,
воска и пр.
Источники энергии
Контроль за состоянием окружающей среды (воздух, вода и
почва)
Пища и напитки (сельскохозяйственное производство и пере-
работка)
Здравоохранение (диагностика, лечение), борьба с болезня-
ми растений и животных
Добыча минерального сырья на суше и в море
медицине, но в более отдаленном будущем (лет через десять),
по мнению наиболее компетентных специалистов, самый боль-
шой экономичский эффект будет получен от применения биотех-
нологии в сельском хозяйстве и химической промышленности.
Ниже мы вкратце обсудим некоторые последние достижения этой
науки, потенциально важные для медицины, энергетики, про-
изводства пищевых продуктов и напитков, получения химиче-
ских соединений, различных материалов, для защиты окружаю-
щей среды и для сельского хозяйства. Многие из этих аспектов
обсуждаются более подробно в последующих главах.
1.4.1. Медицина
В последнее время все мы имели возможность убедиться, что
благодаря применению технологии рекомбинантных ДНК были
достигнуты крупные успехи в медицине. Многие фирмы, напри-
2—1344
ИМ. Н. И. /i. Ct- ' некого
ШАНСКОГО IDG. ; erCMTETj.
18
Глава 1
мер, весьма преуспели в разработке эффективных методов про-
мышленного производства человеческого интерферона (для это-
го гены человека были клонированы в микроорганизмах). Отме-
тим, впрочем, что многие и сейчас сомневаются в эффективно-
сти интерферона как антивирусного и противоопухолевого
средства. Помимо гена интерферона были клонированы гены
инсулина и гормона роста человека. Эти гены экспрессируются
и в бактериях. В целях крупномасштабного производства были
клонированы гены многих других белков человека, необходи-
мых для диагностики или для лечения. Когда мы писали эту
книгу, уже поступил в продажу полученный при помощи мик-
робов инсулин человека, который используется в медицине.
Большое значение имеет и разработка методов производства
моноклональных антител. Более подробно речь о них пойдет в
гл. 8. Способы их применения сведены в табл. 1.4.
Таблица 1.4. Возможные способы применения
моноклональных антител
Область медицины Способ применения
Анализ Структурные зонды для идентифика- ции специфических особенностей на поверхности клеток
Диагностика Наборы реактивов для диагностики беременности Выявление >строгенных рецепторов для диагностики некоторых форм рака молочной железы
Иммунодиагностика Точное определение количества спе- цифических антигенов
Иммуноочистка Очистка антигенов, например интер-
Терапия Направленный перенос токсинов в раковые клетки, инактивация ядов, пассивная иммунизация, лече- ние аутоиммунных болезней
Не приходится сомневаться, что все шире и шире в диаг-
ностике, терапии и при трансплантации клеток и тканей будут
использоваться ферменты.
К числу активно разрабатываемых, но не вышедших еще из
«детского возраста» сфер применения биотехнологии, которые
окажут, наверное, наибольшее влияние на развитие медицины и
промышленности, относятся биоэлектроника и биоэлектрохимия,
в которых используется взаимодействие биологических, электри-
ческих и электронных систем. В последние годы здесь достиг-
нуты заметные успехи. Так, принципы электронной инженерии
Что такое биотехнология?
19
нашли применение при разработке инфузионных насосов, а так-
же эффективных комплексных устройств, в которых использу-
ются биологические соединения, чаще всего ферменты или анти-
тела, и различные типы электродов. Создан целый ряд чувстви-
тельных датчиков, например контролирующих содержание глю-
козы (для применения в медицине) или же нервных газов
(в военных целях). Действие большинства из разработанных
на сегодня датчиков основано на улавливании продуктов дей-
ствия ферментов. Для этого используются обычные электроды
с иммобилизованной на них биологической системой. Новые
подходы в этой области ставят своей целью создание более чув-
ствительных и эффективных приборов и расширение сферы их
применения. В основе работы таких устройств лежит процесс
прямого переноса электронов между электродами и окисли-
тельно-восстановительными центрами белков. Делаются попыт-
ки подключения белков ко входу полупроводниковых приборов.
Если не вдаваться в детали, то в количественном отношении
наиболее широко в медицине применяются фермент-содержа-
щие датчики нескольких разновидностей для определения со-
держания глюкозы. На их основе будет разработан ряд уст-
ройств, например дешевые, точные и надежные приборы для
проведения анализов in vivo. Считается, что основное приме-
нение они найдут при регуляции содержания сахара в крови у
больных диабетом. Недостаточно точный контроль уровня са-
хара при этой болезни, по-видимому, приводит к развитию от-
даленных, опасных для жизни побочных последствий диабета.
Использование датчиков позволит замкнуть цепь контроля в
аппаратах «искусственная поджелудочная железа».
В ближайшие десять лет, по-видимому, поступят в продажу
и другие датчики, позволяющие, например, определять содержа-
ние отдельных компонентов крови. Появятся и биоэлектронные
иммуносенсоры, причем в некоторых из них будет использо-
ваться полевой эффект транзисторов. На их основе предполага-
ется создать относительно дешевые приборы, способные опре-
делять и поддерживать на заданном уровне концентрацию ши-
рокого круга веществ в жидкостях тела, что может вызвать пе-
реворот в диагностике.
Специфические каталитические свойства биологических си-
стем не нашли пока столь широкого применения, как это про-
гнозировалось в начале 60-х годов; в то же время новые мето-
ды, основанные на способности таких систем к узнаванию, ско-
ро будут внедрены как в медицине, так и в промышленности,
особенно по мере усовершенствования технологии. Объяснение
тому простое: во многих случаях на практике химические ката-
лизаторы при всех их недостатках с экономической точки зре-
ния оказываются выгоднее, чем биокатализаторы. Попытки хи-
2*
20
Глава 1
миков решить проблему специфического узнавания не слишком
обнадеживают, и поэтому, если удастся преодолеть некоторые
трудности, связанные с сопряжением систем и их стабильностью,
биологический подход в целом окажется предпочтительней. Ус-
пешно развивается и направление, связанное с разработкой дат-
чиков, основанных на использовании комбинации иммобилизо-
ванных, стабилизированных микроорганизмов и электродов.
1.4.2. Энергетика
В ходе эволюции в биологических системах сформировался ряд
весьма совершенных механизмов превращения энергии. На рис.
1.3 представлены основные известные их типы, часть которых
Н2 6 — Электричество ---8--- Метан/
Рис. 1.3. Основные пути превращения энергии в живых системах.
используется разными способами, особенно реакции I, 2 и 9.
Более подробно мы обсудим их в гл. 2. Отметим, что биомасса
постоянно используется для получения электрической энергии
за счет небиологических эквивалентов в реакциях 8 и 11.
Что касается возможного вклада биотехнологии в решение
проблем энергообеспечения, то здесь в отличие от других обла-
стей ее применения предсказать что-либо гораздо сложнее.
В последние годы немало говорилось об «энергетическом кризи-
се»: запасы ископаемого топлива ограничены, а население рас-
тет, и потребление энергии per capita все увеличивается. В этом
контексте обсуждаются и перспективы использования ядерной
энергии. Неравномерность распределения запасов ископаемого
топлива, а также наличие ряда сложных политических и эконо-
мических факторов делают любые предсказания особенно
сложными. Наиболее важным является здесь то обстоятельство,
Что такое биотехнология?
21
что около 99,4%, или 1,7-1023 калорий в год, доступной нам не-
ядерной энергии мы получаем от Солнца, и часть ее аккумули-
руется в биомассе, хотя и с малой эффективностью, порядка
1—2%. По этой причине биомасса представляет собой постоян-
но возобновляемый источник’ химической энергии. Ее можно
сжигать или довольно простыми способами превращать при по-
мощи микроорганизмов в жидкое или газообразное топливо
(метан, этиловый спирт или водород). Однако биомасса исполь-
зуется и для других целей: она служит пищей для людей и да-
ет ряд ценных видов сырья.
Со временем биомасса, видимо, будет все более распростра-
ненным исходным продуктом при производстве сырья для хи-
мической промышленности на основе биотехнологических про-
цессов. Примером такого рода может быть превращение лигни-
на в соединения ароматического ряда.
Конкуренция за имеющиеся в наличии запасы биомассы усу-
губляется тем, что площади, пригодные для ее производства,
из-за роста населения постоянно уменьшаются. По этой при-
чине дать точный глобальный прогноз использования энергии,
получаемой из биомассы, довольно трудно. Отметим, однако,
что из-за недостатка ископаемого топлива в некоторых странах
(например, в странах Южной и Северной Америки) производ-
ство этилового спирта путем ферментации становится все более
популярным, особенно для использования в качестве добавки
и/или для замены нефти как горючего на транспорте. Если рас-
сматривать это производство само по себе, то экономическая
его обоснованность оказывается весьма сомнительной, но оно
приемлемо для некоторых стран по политическим соображени-
ям (Бразилия, США).
В последнее время вновь пробудился интерес к разработке
биотопливных элементов, с помощью которых можно с высо-
кой эффективностью и при обычной температуре получать из
ряда видов топлива и биомассы электрическую энергию. Хотя
эти устройства и находят уже применение (например, в качест-
ве специальных датчиков), в большинстве случаев с их помо-
щью сложно получить на электродах ток достаточной плотно-
сти, что позволяло бы использовать их в качестве крупномас-
штабных преобразователей энергии. Возможно, однако, что
уже в ближайшем будущем они найдут применение в специ-
альных областях энергетики. Единственным исключением среди
них является «гибридный» водородный биотопливный элемент:
в нем водород, образующийся при брожении, используется в
обычном водород-кислородном элементе. Впрочем, и другие
биотопливные элементы могут со временем найти применение
для получения дешевой электроэнергии путем переработки сто-
ков, отходов или окиси углерода.
22
Глава 1
Поскольку солнечный свет является мощным источником
энергии, а количество имеющейся биомассы ограничено, неко-
торые биотехнологи, работающие над проблемами энергетики,
занялись разработкой двух проблем, решение которых позволи-
ло бы повысить эффективность использования солнечной энер-
гии. Во-первых, они пытаются найти практические способы
повышения эффективности конверсии солнечного света в био-
массу, например путем выращивания водорослей при высокой
концентрации углекислого газа и ограниченной освещенности в
биореакторах со строго контролируемыми условиями роста. Во-
вторых, они изучают возможность получения водорода путем
расщепления воды при участии фотосистемы фотосинтезирую-
щих организмов, т.е. путем биофотолиза. Технически проще все-
го получать водород, используя интактные сине-зеленые водо-
росли или процессы ферментации (брожения). Надо сказать,
однако, что если биотехнология всерьез намерена внести в бу-
дущем весомый вклад в производство энергии, то ей придется
решить нетривиальную техническую задачу: на основе биофото-
лиза разработать сложный реактор, включающий упорядочен-
ные стабильные биофотосистемы.
В самом ближайшем будущем биотехнология станет играть
все возрастающую роль и при добыче нефти. Поскольку цены
на нефть растут, добыча ее из сложных в эксплуатации зале-
жей становится все более экономически выгодной. Здесь могут
оказаться полезными микроорганизмы. Во-первых, некоторые
образуемые ими полимеры, особенно производные ксантана,
можно использовать в качестве компонентов закачиваемых в
пласт растворов, обладающих нужными реологическими харак-
теристиками, для добычи остаточной нефти (гл. 5). Во-вторых,
в нефтяной промышленности используются поверхностно-актив-
ные вещества микробного происхождения. С экономической точ-
ки зрения производство таких веществ будет особенно выгод-
ным, если их удастся получать путем микробиологической пе-
реработки отходов, содержащих нефть. Как правило, экономи-
ческие характеристики биотехнологических процессов улучша-
ются, если удается совместить переработку отходов с производ-
ством полезного продукта.
Некоторые группы ученых работают сегодня над разработ-
кой более долгосрочных крупномасштабных программ. Предпо-
лагается, например, вводить подходящие микроорганизмы не-
посредственно в нефтяной пласт, чтобы ускорить отток нефти
из пористых пород.
Что такое биотехнология?
23
1.4.3. Пищевые продукты и напитки
Традиционные способы использования микроорганизмов при
производстве различных сортов пива, вина и сброженных про-
дуктов совершенствовались тысячелетиями, и все же до недав-
него времени в них было больше искусства, чем технологии.
Только с развитием микробиологии мы получили возможность
контролировать качество продуктов, добились большей надеж-
ности и воспроизводимости процессов ферментации и научились
получать новые типы продукции (например, БОО и вкусовые
добавки). Сегодня нам еще трудно с уверенностью говорить о
том, каких успехов удастся достичь в этой области с помощью
биотехнологии, но самые общие тенденции вырисовываются до-
вольно ясно. Наиболее успешными представляются два взаимо-
связанных направления. Во-первых, на смену традиционным
способам производства пищи постепенно придут биореакторы,
в которых будут расти клетки животных или растений или же
микроорганизмы. Дело в том, что выход продукции при ис-
пользовании ферментеров или биореакторов может быть суще-
ственно выше, чем в сельском хозяйстве: идущие в них процес-
сы гораздо более интенсивны. Развитию этого направления спо-
собствует и все возрастающая конкуренция за имеющиеся зе-
мельные ресурсы. Во-вторых, эта альтернативная технология
будет становиться все более производительной благодаря ис-
пользованию методов генетической инженерии, которые позво-
ляют получать улучшенные линии клеток и штаммы микроор-
ганизмов.
Такая тенденция в развитии пищевой промышленности пу-
тем интенсификации биотехнологических процессов уже прояви-
лась в полной мере при выработке ценных пищевых добавок.
Наиболее яркий пример — производство лимонной кислоты
(гл. 3). Сегодня ее получают главным образом микробиологи-
ческим методом, а не из цитрусовых. Намечается также рост
использования микробных белков в питании человека.
Новые возможности открываются и в развитии нетрадици-
онных способов выработки пищевых продуктов. Так, в Японии
говядина стоит дорого и производится в недостаточном количе-
стве. Эта страна представляет собой поэтому потенциально
очень емкий рынок для сбыта хороших заменителей говядины,
изготовленных на основе других белковых продуктов.
1.4.4. Химические соединения
Применение биологических систем для производства химических
соединений в принципе дает ряд преимуществ, однако сегодня
лишь малое их число получают с помощью биотехнологиче-
24
Глава 1
ских процессов. К ним относятся сравнительно дешевые, но ис-
пользуемые в больших количествах как топливо этиловый спирт
и метан, а также ряд ценных и довольно дорогих веществ, при-
меняющихся в медицине и для пищевых целей (лимонная кис-
лота, итаконовая кислота, аминокислоты, стероиды и антибио-
тики) .
В принципе производство химических веществ на основе
биокатализа имеет следующие преимущества: специфичность,
легкость контроля, работа при низких температурах, совмести-
мость с окружающей средой и простота. Особенно важны два
первых из них. Так, химическая промышленность органических
соединений базируется сегодня в значительной мере на нефти,
а большинство производимых ею продуктов переработки нефти
получают путем частичного окисления сырья. Достичь специфи-
ческого контролируемого и частичного окисления при помощи
существующих катализаторов довольно сложно, а микроорга-
низмы осуществляют эти типы реакций мастерски, без труда.
По ряду причин биотехнологии еще только предстоит вне-
сти свой вклад в развитие химической промышленности, хотя
уже сегодня мы многое знаем о потенциально полезных в этом
плане биологических системах. Одна из главных проблем заклю-
чается в том, что основанные на биотехнологии отрасли хими-
ческой промышленности будут использовать нетрадиционную
технологию. Реакции, как правило, будут идти при низких тем-
пературах и давлении, в водной среде, хотя ряд эксперимен-
тальных систем работает и в органической фазе. В некоторых
случаях скорость таких процессов невелика, а катализаторы не
очень стабильны. Однако важнее всего, видимо, то обстоятель-
ство, что наибольшее влияние на развитие химической промыш-
ленности по вполне понятным причинам оказывают химики. По
сравнению с другими областями технологии разработка мето-
дов биотехнологического катализа до недавнего времени велась
здесь малыми силами и при скудном финансировании.
Роль биокатализа в химической промышленности сегодня
весьма необычна. С одной стороны, появление генетической ин-
женерии воскресило к нему интерес, поскольку появилась воз-
можность существенно улучшить рабочие характеристики био-
катализаторов, особенно интактных микроорганизмов. С другой
стороны, на разработку таких новых процессов требуется не-
малое время (от десяти до двадцати лет) и это наряду с высо-
кой стоимостью изысканий и внедрения приводит при органи-
зации производства к крупным расходам (10—25% от общей
стоимости осуществления проекта). Новые возможности для
более широкого применения биокатализаторов в химической
промышленности возникли, когда она столкнулась с рядом эко-
номических проблем.
Что такое биотехнология?
25
Все эти соображения, а также тот факт, что недавнее внед-
рение биопроцессов в многотоннажное производство некоторых
химических соединений (например, окиси пропена) привлекли
к себе большое внимание, позволяют думать, что скоро мы ста-
нем свидетелями использования биокатализа, в первую очередь
для синтеза небольших партий веществ с высокой прибавочной
стоимостью. Опыт, накопленный при производстве таких ве-
ществ, поможет развитию многотоннажного производства хи-
мических соединений.
Существуют три главных способа синтеза химических со-
единений на основе биокатализа: 1) путем использования куль-
тур клеток растений или животных, образующих дорогостоя-
щие вещества; 2) путем использования микроорганизмов, при
необходимости измененных методами генетической инженерии,
для биосинтеза или модификации химических веществ; 3) путем
использования измененных методами генетической инженерии
микроорганизмов в качестве «устройств» для экспрессии генов
растений и животных, что позволяет синтезировать в больших
количествах особые, присущие только высшим организмам хи-
мические соединения.
Сделаны первые попытки объединить химическую, биологи-
ческую и электронную технологии, и здесь, по-видимому, будут
достигнуты выдающиеся результаты. Таким путем могут быть
созданы специфические датчики для регуляции процессов в хи-
мической и пищевой промышленности, для медицинской диаг-
ностики, мониторинга и контроля, для наблюдения за состояни-
ем окружающей среды. Особенно заманчивой кажется возмож-
ность создания полупроводниковых биодатчиков, основанных
на микрочипах (micro-chipe). Такие устройства после их усо-
вершенствования будут достаточно дешевы в производстве и
позволят одновременно контролировать многие параметры при
помощи одного крошечного биоэлектронного прибора. Все это
может быть выполнено в течение пяти — двадцати лет. Более
простые датчики, основанные на использовании полупроводни-
ков или технологии тонких слоев, будут разработаны в ближай-
шие пять лет.
В следующем разделе мы обсудим перспективы развития
производства пластмасс, эмульгаторов и загустителей при учас-
тии микроорганизмов.
1.4.5. Материалы
Биотехнология может оказать влияние на получение и исполь-
зование различных материалов по меньшей мере тремя спосо-
бами. Во-первых, она будет способствовать развитию добычи
промышленного сырья, например нефти и других полезных ис-
26
Глава 1
копаемых. Во-вторых, все более широко станут использоваться
продукты микробного происхождения, например для производ-
ства разлагаемых с помощью микроорганизмов пластмасс,
эмульгаторов и загущающих веществ. Наконец, будут усовер-
шенствованы способы защиты различных веществ от разруше-
ния их микроорганизмами.
Подробно перспективы дальнейшего усовершенствования
микробиологических способов выщелачивания руд, а также ис-
пользования полимеров и поверхностно-активных веществ мик-
робного происхождения при добыче нефти обсуждаются в гл. 5.
Наиболее многообещающим сырьем для производства био-
пластмасс является одно из резервных веществ клеток, поли-^-
гидроксибутират (ПГБ). В настоящее время в промышленности
ведутся активные исследования как самого этого вещества и
его производных, так и способов их получения.
Проблема биоповреждений обычно не обсуждается в ряду
проблем биотехнологии, но она, без сомнения, весьма актуаль-
на и сложна с технической точки зрения, а роль ее в экономике
весьма существенна.
Биоповреждения — неизбежное следствие важнейшей роли
микроорганизмов в круговороте элементов в биосфере. Прояв-
ления биоповреждений весьма многообразны: от порчи пище-
вых продуктов до загрязнения смазочных масел и топливных
систем, разрушения бетона и развития электрохимических про-
цессов коррозии под влиянием микроорганизмов. Биотехноло-
гия поможет создать новые методы борьбы с биоповреждениями
благодаря более глубокому пониманию лежащих в их основе
процессов. На этой базе могут быть созданы новые биотехноло-
гические процессы. Примером такого рода служит использова-
ние ферментов в пищевой промышленности.
На основе биотехнологических принципов будут созданы но-
вые типы датчиков для контроля уровня микробного загрязне-
ния сырья.
1.4.6. Окружающая среда
По мере того как увеличивается население Земли и развивает-
ся промышленность, все более серьезной становится проблема
охраны окружающей среды. В решении такого рода задач био-
технология будет играть все возрастающую роль, в частности,
в том, что касается разработки новых или усовершенствования
существующих способов переработки отходов. Такие методы и
системы будут создаваться в значительной мере на основе ис-
следований в области химической инженерии, но свое веское
слово скажут также физиологи микроорганизмов, биохимики и
генетики. Новейшие процессы переработки необычных отходов
Что такое биотехнология?
27
будут основаны на использовании микроорганизмов, обладаю-
щих новыми, неизвестными ранее или искусственно созданны-
ми катаболическими способностями. Более подробно эти две
возможности мы обсудим в гл. 6.
Окружающая среда является, без сомнения, как бы общим
знаменателем для всех видов деятельности, о которых пойдет
речь в этой книге, так как все они оказывают на нее влияние.
Так, расширение использования биотехнологии в химической
промышленности должно привести к созданию новых ее отрас-
лей, лучше совместимых с окружающей средой. Такие же на-
дежды возлагаются и на биоинженерию. Разработка биодатчи-
ков поможет осуществлять мониторинг и контролировать усло-
вия среды. Понятно, что проблемы защиты окружающей среды
неразрывно связаны с сельским хозяйством.
1.4.7. Сельское хозяйство
Точки соприкосновения биотехнологии и сельского хозяйства
весьма многообразны. Продукция сельского хозяйства может
использоваться в промышленности, например для производства
этилового спирта из излишков низкокачественного вина. Такой
подход получил дальнейшее развитие: для выработки спирта
•сельскохозяйственные культуры начали выращивать специаль-
но. Большая часть продукции современного сельского хозяйства
•служит сырьем для развитой пищевой промышленности. В ка-
честве сырья могут использоваться и отходы сельского хозяй-
ства: в частности, большое внимание уделяется возможности
получения топливного газа из навоза с сохранением его ценно-
сти как удобрения. Для усовершенствования процессов в этой
отрасли необходимо иметь более четкое представление о ско-
рости разложения различных субстратов и роли микроорганиз-
мов, в них участвующих.
В ветеринарии биотехнология используется для получения
вакцин и сывороток. Если, как ожидается, вакцины удастся по-
лучать при помощи микроорганизмов, модифицированных мето-
дами генетической инженерии, мы станем свидетелями оконча-
тельного искоренения таких опасных заболеваний,' как ящур и
сонная болезнь. Для увеличения выхода мяса могут использо-
ваться гормоны роста. Современная биотехнология дает нам и
корм для скота, например белково-витаминный концентрат.
Впрочем, нужно еще доказать, что его применение экономичес-
ки целесообразно, по крайней мере в западных странах.
Биотехнология поможет разработать новые способы улуч-
шения сельскохозяйственных культур как по урожайности, так
и по качеству. Можно будет использовать полученные с ее по-
мощью заменители дорогостоящих химических удобрений или
28
Глава 1
пестицидов или же добавки к ним. Так, потребности в азоте
удастся удовлетворить путем внедрения биологической фикса-
ции азота, основанной на симбиозе, а в фосфоре — путем вме-
шательства в процессы, происходящие в микоризах. Задачей от-
даленного будущего является передача способности к фиксации
азота непосредственно отдельным сельскохозяйственным куль-
турам путем введения в них гена нитрогеназы; в результате
такие растения приобретут способность к синтезу фермента, ка-
тализирующего реакцию фиксации азота. Это позволит сэко-
номить энергию, затрачиваемую сегодня при химическом син-
тезе аммиака. Есть надежда, что вместо пестицидов мы в бу-
дущем сможем использовать методы биологического контроля —
как широко известные, так и основанные на применении новых
природных соединений. Так, например, в борьбе с одной из
болезней растений, корончатым галлом, могут помочь бактерии.
С бабочками-вредителями можно бороться с помощью Bacillus
thuringiensis. Разработка таких методов не только принесет
пользу сельскому хозяйству, но и сыграет важную роль в под-
держании на должном уровне условий окружающей среды.
По общему мнению, наибольший вклад биотехнологии в
сельское хозяйство следует ожидать за счет улучшения свойств
самих растений путем использования методов рекомбинантных
ДНК и протопластов растений. Применительно к бобовым и
злакам метод регенерации целых растений из отдельных клеток
не дал пока сколько-нибудь значительных результатов. Однако
работа с люцерной была небезуспешной, и это позволяет на-
деяться, что опыты с бобовыми тоже будут более результатив-
ными по мере разработки все более подходящих условий куль-
тивирования. Применяя подобную технологию, быть может,
удастся получить белки злаков, содержащие незаменимые ами-
нокислоты, которых сейчас в них нет.
1.5. Экономические и коммерческие аспекты биотехнологии
Биотехнология (в широком смысле этого термина) уже сегодня
имеет большое экономическое и социальное значение. Главная
цель этого раздела книги — проанализировать ее возможное
влияние на экономику, описать механизмы новой биотехнологии,
ее быстроразвивающиеся, новые, многообещающие методы. Что-
бы читатель мог составить представление об объеме продукции
биотехнологической промышленности, скажем, что бродильная
промышленность Великобритании дает в год продукции прибли-
зительно на 9 млрд, фунтов стерлингов. В мире производится
антибиотиков на 2 млрд, фунтов, а аминокислот и ферментов —
на 1 млрд, фунтов и 250 млн. фунтов соответственно.
Что такое биотехнология?
29
Если оценить рыночную стоимость отдельных видов биотех-
нологической продукции относительно несложно (например,
в западных странах дешевых, надежных, точных датчиков глю-
козы одноразового пользования за год продается на 800 млн.
фунтов), то предсказать в целом спрос на все виды продукции
и процессы, основанные на применении биотехнологии, гораздо
сложнее. Тем не менее в табл. 1.5 мы приводим относительно
Таблица 1.5. Оценка спроса на мировом рынке
в 2000 г. на различные виды продукции
«новой биотехнологии» (в млрд, фунтов стерлингов)
Химические вещества
Энергия
Пищевые продукты
Медицина
Прочие (например, повышение полноты извлече-
ния нефти, контроль загрязнения окружающей
среды, биопластики, выщелачивание руд)
9
12
12
8
15
56
обоснованные оценки, взятые из разных источников. Предпола-
гается, что во всем мире эта развивающаяся отрасль промыш-
ленности будет давать к 2000 г. продукции на 56 млрд, фунтов
в год, что приблизительно в шесть раз больше, чем дает сейчас
бродильная промышленность Великобритании. Предсказания
относительно дальнейшего развития биотехнологической про-
мышленности сделать еще сложнее, но, вероятно, и в будущем
столетии она будет продолжать развиваться быстрыми темпами.
Интересная особенность данных, приведенных в табл. 1.5,
заключается в следующем: в отличие от общепринятого пред-
ставления, согласно которому биотехнология найдет основное
применение в медицине, они говорят, что к концу века это бу-
дет относительно узкая сфера ее применения. Впрочем, данная
отрасль может оказаться весьма доходной и в ближайшем
будущем развиваться наиболее быстрыми темпами. Так, уже
поступил в продажу человеческий инсулин. Для лечения рака
в 1987 г., видимо, будет широко использоваться интерферон,
а в 1988—1989 гг. он найдет применение как антивирусное и
противовоспалительное средство. Ожидается, что к 1990 г. об-
щедоступными станут вакцина против гепатита В и человече-
ский гормон роста.
1.5.1. Логика развития биотехнологии
Механизмы развития биотехнологии заметно различаются в
разных частях света и зависят от политических и экономических
факторов, характерных для Запада и Востока, Севера и Юга,
30
Глава 1
•стран третьего мира и развитых стран, капитализма и социализ-
ма. Суть технологии также различна: страны третьего мира
обычно развивают альтернативные варианты технологий. Так,
в некоторых странах Дальнего Востока небольшие местные ге-
нераторы биогаза удовлетворяют значительную часть энергети-
ческих потребностей сельских общин. В странах с развитой
экономикой такие установки гораздо менее популярны. В СССР
и других социалистических странах развитие биотехнологии
обычно определяется централизованно. В частности, это позво-
лило осуществить крупную программу по производству белково-
витаминного концентрата. В настоящее время из углеводородов
и растительных отходов там производится за год более миллио-
на тонн этого продукта.
В Японии правительство играет важную роль в координации
развития биотехнологии в университетах и исследовательских
институтах, но разработка и реализация идей осуществляется
главным образом крупными биотехнологическими фирмами.
Правительства большинства западных стран в последние го-
ды выдвинули ряд программ, направленных на содействие раз-
витию биотехнологии. Необходимость их разработки определя-
ется сложной, междисциплинарной природой этой технологии и
продолжительностью подготовительного периода при разработ-
ке способов производства большинства видов продукции. Кроме
того, поскольку многие важные открытия в биотехнологии де-
лаются на переднем крае науки, многие из ведущих ученых,
принимающих участие в таких исследованиях, работают в го-
сударственных учреждениях, чаще всего в системе высшего об-
разования. Правительства должны умело финансировать имен-
но самые нужные исследования, способствовать развитию раз-
личных областей науки и помогать как частному, так и госу-
дарственному сектору в их попытках довести дело до коммерче-
ского продукта, даже если подготовительная работа требует
много времени. Неблагоприятный экономический климат по-
следних лет делает такую «каталитическую функцию» прави-
тельства тем более важной, что расходы на исследования и раз-
работки в учреждениях частного сектора сокращаются.
Что касается США, то правительство этой страны давно
вкладывает значительные средства в развитие различных от-
раслей биотехнологии, а крупные фирмы медицинской, биоло-
гической и энергетической промышленности уже осуществляют
важные программы в области биотехнологии. Кроме того, воз-
никают и частнопредпринимательские биотехнологические
фирмы. Такие скромные по масштабу, новые промышленные
предприятия появляются не только в США; небольшое их чис-
ло основано и в Западной Европе. Некоторые из этих европей-
ских фирм являются «получастными» в том смысле, что они ра-
Что такое биотехнология?
31
ботают в тесном контакте с правительственными или частными
финансовыми учреждениями. Таким образом, участвующие в
развитии биотехнологии фирмы могут быть отнесены к четырем
основным категориям. 1. Специализированные биотехнологиче-
ские фирмы, опирающиеся главным образом на технологию ре-
комбинантных ДНК. 2. Крупные, обычно международные фир-
мы, занятые добычей нефти, получением пищевых продуктов,
химических веществ, переработкой сельскохозяйственной про-
дукции или же работающие в фармакологической промышлен-
ности. Эти фирмы уделяют развитию биотехнологии часть своих
исследовательских и производственных ресурсов. 3. БоЛее мел-
кие фирмы, специализирующиеся в биотехнологическом произ-
водстве; они выпускают, например, препараты для медицины,
продукты ферментации. 4. Фирмы, поставляющие оборудование
для биотехнологической промышленности.
Специализированные биотехнологические фирмы, а их сей-
час в США около двухсот, немало способствовали динамическо-
му развитию технологии. Многие из них, без сомнения, потер-
пят крах. Выживут и добьются успеха те фирмы, которые спо-
собны вести дела на высоком уровне, преданы делу и подходят к
нему творчески; такие качества воспитываются в ходе деятельно-
сти. В более отдаленном будущем некоторые из них, несомненно,
будут поглощены более крупными международными фирмами.
Общеизвестно, что сегодня больше всего новых биотехнологиче-
ских производств организуется в США и это (по меньшей мере
частично) определяется тем, что небольшие фирмы научились
успешно использовать опыт и творческий потенциал ученых и
технологов, которые работают в них в условиях относительной
свободы от множества обязанностей, без перерывов в работе и
конфликтов. Тем не менее ясно, что сегодня существует слиш-
ком много таких фирм, которые пытаются наладить выпуск от-
носительно небольшого набора продуктов, и доля неудачников
будет большой. Недавно Комитет технологических оценок при
конгрессе США провел анализ перспектив производства биотех-
нологических продуктов в мире (см. список рекомендуемой ли-
тературы). Был сделан вывод, что сегодня ведущее положение
занимают США. Они сохранят его и в будущем, при более или
менее выраженной конкуренции со стороны Японии. Из этого
документа следует также, что европейские страны достигнут в
этом деле меньших успехов.
1.5.2. Защита авторских прав в биотехнологии
Подходы к проблеме защиты авторских прав при внедрении
открытий в области биотехнологии в целом сходны с известны-
ми для других видов деятельности. Так, коммерчески важные
32
Глава 1
открытия могут охраняться путем использования «технологии
внутри фирмы» или же обычным патентованием. Патентование
в биологических науках — это весьма специфический вид право-
вой деятельности, но во многих случаях (например, новое уст-
ройство для поточной переработки или новый датчик) сложно-
сти при патентовании сходны с таковыми для любой другой
технологии. Впрочем, появление технологии рекомбинантных
ДНК поставило и ряд новых интересных правовых вопросов.
Упомянем, например, ставшую сегодня широко известной заявку
на патент (патент Коена — Бойера), поданную в 1980 г. Стэн-
фордским и Калифорнийским (Сан-Франциско) университетами.
Этот патент ограничивает право пользования широким кругом
методов генетической инженерии. Хотя многие крупнейшие фир-
мы и купили лицензии на эту технологию, правомочность патен-
та нельзя считать полностью доказанной. Нужно еще посмотреть,
как сложится патентование в столь необычной области создания
новых форм жизни.
1.5.3. Правила техники безопасности
в биотехнологической промышленности и контроль
продукции
Как известно, существуют стандарты безопасности новых видов
продукции. К числу наиболее строгих из них относятся те, ко-
торые касаются медицинских препаратов, а также продуктов,
потребляемых в животноводстве и особенно в пищевой промыш-
ленности. Используемые при этом методы и требования хорошо
обоснованы, и, хотя затраты на контроль велики, это не меша-
ет выработке доброкачественных продуктов биотехнологии, на-
пример грибного белка для питания людей (гл. 3). Тем не ме-
нее сама возможность использования генетической инженерии
вызвала в 70-х годах озабоченность общественности и стала
предметом обсуждения. Правительства стран, в которых воз-
никли такие дебаты, создали подотчетные обществу организа-
ции, призванные контролировать использование методов гене-
тической инженерии. Естественно, что в разных странах для
этой цели были разработаны как разные методы контроля, так
и несколько неодинаковые стандарты. Спустя десять лет в ре-
зультате накопления опыта использования новых методов био-
технологии, мониторинга и контроля стало очевидным, что пер-
воначальные правила безопасности были излишне строгими.
Эти правила для большинства обычных генетических манипуля-
ций постепенно смягчились. В то же время для некоторых ма-
нипуляций правила, касающиеся условий и способов производ-
ства, остались очень строгими и будут по необходимости оста-
ваться таковыми. В этом плане правительство Японии было
Что такое биотехнология?
33
настолько осторожно, что это даже замедлило развитие техно-
логии рекомбинантных ДНК в этой стране. Впрочем, сегодня
работы по биотехнологии в Японии развиваются очень быстро,
так как правила были смягчены, и эта страна уже вышла на
уровень, достигнутый в Западной Европе и Америке.
В Великобритании контроль за работой с рекомбинантными
ДНК осуществляется специальной правительственной Органи-
зацией здравоохранения и безопасности. И эта организация,
и ученые следуют рекомендациям Консультативной группы по
генетическим манипуляциям (GMAG), которая разработала
правила проведения исследований. Эти правила в основном
регламентируют уровни безопасности при проведении различ-
ных исследований. Ученые должны заблаговременно получать
разрешение GMAG для проведения своих опытов, и научные
работники проявляют высокую сознательность при использова-
нии этих правил.
1.6. Заключительные замечания
В этой главе мы попытались дать читателю общее представле-
ние о биотехнологии, ее корнях, нынешнем состоянии и перспек-
тивах. В последующих главах мы более подробно рассмотрим
положение дел в различных отраслях биотехнологии. Эти главы
были написаны специалистами, вклад которых в биотехнологию
и тем самым в улучшение общего уровня жизни не вызывает
сомнений. Разнообразие видов их деятельности до прихода в
биотехнологию убедительно свидетельствует о широте этой
науки и возможностях ее практического применения. Среди ав-
торов есть не только «биотехнологи с академическим уклоном»,
но и лица, тесно связанные с деловым миром и с наукой, со-
трудник международной биотехнологической фирмы, главный
управляющий крупнейшей фирмы продовольственных товаров и
крупный специалист-клиницист. Такова уж природа этой науки!
ЛИТЕРАТУРА
Atkinson В., Mavituna F. (1983). Biochemical Engineering and Biotechnology
Handbook, Macmillan, Byfleet, Surrey.
Brenner S., Hartley B. S., Rodgers P. J. (eds.) (1980). New Horizons in Indust-
rial Microbiology, Royal Society, London.
Bull A. T., Ellwood D. C., Ratledge C. (eds.) (1979). Microbial Technology:
Current Status, Future Prospects, Soc. gen. Microbiol. Symp., 29.
Commercial Biotechnology: An International Analysis (Washington DC: US
Congress Office of Technology Assessment, OTA-BA-218, January 1984).)
Coombs J. (1984). The International Biotechnology Directory, The Nature Press,
New York.
Crespi R. S. (1982). Patenting in the Biological Sciences, John Wiley and Sons,
Chichester.
3—1344
34 Глава 1
Hamdi S. (1983). Biotechnology for Investors, Laing and Cruikshank, London.
Old R. №.. Primrose S. B. (1980). Principles of Gene Manipulation: An Introduc-
tion to Genetic Engineering, Blackwell Scientific Publications, Oxford.
Phelps C. F., Clarke P. H. (eds.) (1983). Biotechnol. Biochem. Soc. Symp., 48.
Rehm H.-J., Reed G. (eds.) (1981). Biotechnology, Vol. 8, Verlag Chemie,.
Weinheim.
Rose A. H. (ed.) (1980). Microbial Enzymes and Biosensors, Economic Microbio-
logy 5, Academic Press, London.
Rothman H., Stanley R., Thompson S., Towalski Z. (1981). Biotechnology: A Re-
view and Annotated Bibliography, Frances Pinter, London.
Science (1983). 219, no. 4585.
Scient. Am. (1982). 247.
Scott C. D. (ed.) (1982). Fourth Symposium in Biotechnology in Energy Produc-
tion and Conservation, Biotechnol. Bioeng. Symp., 12.
Senior P. J., Windass J. (1980). The ICI Single Cell Protein Process, Biotechnol.
Lett., 2, 205—210.
Wiseman A. (ed.) (1983). Principles of Biotechnology, Surrey University Press/'
/Chapman and Hall, New York.
Глава 2
Энергия и биотехнология
Д. Холл, Дж. Кумбс, И. Хиггинс
2.1. Введение
В этой главе мы обсудим роль биотехнологии в производстве
высококачественного топлива («premium fuels») из биологиче-
ского сырья. Начнем с того, что термин «биомасса», который
многими микробиологами понимается в относительно узком
смысле, сегодня при описании самых общих принципов произ-
водства разнообразных видов высококачественного топлива и
веществ специального назначения из растений, выращенных не-
посредственно для этих целей, или из биологических отходов,
образующихся, например, в сельском хозяйстве или пищевой
промышленности, используется в более широком смысле. В ос-
нове как запасания энергии (фотосинтез), так и переработки
сырья (биомассы) в более ценное топливо (путем фермента-
ции) лежат биологические процессы. Особое внимание сегодня
уделяется разработке более изощренных генетических методов:
считается, что они сыграют важную роль как при выведении
улучшенных сортов растений с более высокой урожайностью,
так и новых форм микроорганизмов для осуществления процес-
сов конверсии. Кроме того, вполне возможно создание комбини-
рованных искусственных систем, включающих отдельные ком-
поненты животных и растений. Таким путем можно получить
газообразный водород, связанный С или NH3.
Как видно из рис. 2.1, получение топлива по схеме «био-
масса — биотехнология» основывается на сочетании фотосинте-
за, животноводства, кормопроизводства и ферментации с ис-
пользованием наиболее подходящих организмов. Все это долж-
но быть совмещено с инженерным обеспечением сбора урожая,
его перевозки, обработки и получения конечного продукта.
Единственным поставщиком энергии в такой системе является
солнечный свет (этап фотосинтеза). Соответственно все другие
потребности должны быть удовлетворены за счет комбиниро-
ванных источников энергии (ископаемого топлива, электроэнер-
гии или части самой биомассы). Следовательно, определяющим
фактором является отношение количества солнечной энергии,
запасенной в конечном продукте, к энергии, затраченной на его
3*
36
Глава 2
производство. Очевидно, что в практической жизни вопросы
экономии играют главную роль, однако в большинстве совре-
менных программ по использованию энергии биомассы им от-
водится второстепенное значение. На первом плане стоят зада-
чи самообеспечения, обмена продукцией с другими странами,
увеличения занятости в сельской местности или использования
излишков сельскохозяйственных продуктов. В результате пра-
вительствам приходится субсидировать программы по произ-
водству энергии из биомассы, однако эти вложения не имеют
особого смысла, если весь процесс не дает энергетического вы-
игрыша.
Солнечная энергия
Рис. 2.1. Взаимосвязи между биомассой и биотехнологией.
к сожалению, сегодня многие системы дают относительно
небольшой энергетический «доход», а иногда энергия в них да-
же теряется. Следовательно, с биотехнологических позиций бо-
лее всего нуждаются в усовершенствовании процессы, связан-
ные именно с производством энергии. Прежде всего нужно уве-
личить «урожай энергии» с гектара посевов и уменьшить одно-
временно затраты на сельское хозяйство. Далее, необходимы
энергетически более эффективные способы переработки, особен-
но лигноцеллюлозы. Наконец, мы должны научиться получать
продукцию в более концентрированной форме для уменьшения
энергетических затрат при переработке.
Таким образом, упор в этой области будет делаться на энер-
гетическую сторону процессов производства биомассы и кон-
версии при фотосинтезе, животноводстве и ферментации.
Энергия и биотехнология 37
2.2. Получение биомассы: технология,
основанная на солнечной энергии
Солнце является неиссякаемым источником энергии. Каждый
год на поверхность Земли поступает 3-2024 Дж энергии, в то
время как запасы нефти, природного газа, угля, урана по оцен-
кам эквивалентны 2,5-1022 Дж (8-Ю11 т в «угольном эквивален-
те»), Понятно, что менее чем за неделю Земля получает от
Солнца такое же количество энергии, какое содержится во всех
невозобновляемых ее запасах. Проведем иное сравнение: если
бы только 0,1% поверхности Земли занимали коллекторы, ис-
пользующие солнечную энергию с коэффициентом полезного
действия около 10%, то были бы удовлетворены все текущие
потребности в энергии в мире за год (3-1020 Дж).
Однако у солнечной энергии есть два недостатка: она по-
ступает неравномерно и диффузно. Поэтому необходимо, во-
первых, разработать какие-то системы накопления, так чтобы
энергия была доступна по потребности, а во-вторых, создать
коллекторы большой площади. Оба этих фактора накладывают
жесткие, но все же преодолимые экономические ограничения на
использование систем на основе солнечной энергии. Производ-
ство биомассы путем фотосинтеза решает обе проблемы: во-пер-
вых, коллекторы могут быть выращены из семян и, во-вторых,
получаемый продукт стабилен и может храниться. Впрочем,
и при получении и использовании биомассы для выработки
энергии возникают свои проблемы, которые, однако, уравнове-
шиваются премуществами: ее можно получать во всем мире,
она возобновляется и производится в согласии — в экологиче-
ском смысле — с окружающей средой.
Преимущество использования солнечной энергии, заключен-
ной в биомассе, в том, что она запасается в форме органи-
ческих веществ и поэтому ее можно хранить и перемещать во
времени и пространстве (табл. 2.1). К недостаткам относятся
малая эффективность (обычно менее 1% и редко более 2%)
использования солнечной энергии при фотосинтезе, при образо-
вании продукции растениеводства, диффузный, а часто и сезон-
ный характер продукции и высокое весовое содержание влаги.
По этим причинам для получения высококачественного, богато-
го энергией сырья необходимо осуществить его сбор, перевоз-
ку, удаление воды, концентрирование или же химическую или
биологическую переработку и упаковку. Если же задачей яв-
ляется превращение биомассы в ценные виды топлива, то ду-
мать приходится не только об удалении воды и увеличении
удельного содержания энергии, но и о том, как получить про-
дукт, совместимый с технологией, для которой он предназначен.
Ранее основным путем использования растительного сырья в
38
Глава 2
Таблица 2.1. Некоторые выгоды и проблемы внедрения технологий,
предназначенных для получения энергии из биомассы
Выгоды Проблемы
Экономия энергии
Возобновляемость
Гибкость технологий и разнообразие
продуктов; высокое удельное со-
держание энергии в некоторых из
них
Для внедрения уже разработанных
технологий требуются небольшие
капиталовложения; доступны при
любом уровне доходов
Могут быть внедрены на основе
имеющихся рабочей силы и ресур-
сов
Хорошие перспективы развития био-
логических и инженерных основ
Увеличивается занятость и повы-
шается квалификация
Во многих случаях — умеренная
стоимость
Экологическая безопасность
Не увеличивается содержание СО2
в атмосфере
Конкуренция за земельные и водные
ресурсы
Необходимость отвода земель
На начальных этапах — трудности
с оценкой запасов
Часто — сложности в оценке себе-
стоимости
Нужны удобрения, земельные пло-
щади и вода
Зависимость от существующих мето-
дов ведения сельского и лесного
хозяйства, социальные факторы
Объемистое сырье: сложности с пе-
ревозкой и хранением
Зависимость от изменений климата
Малая эффективность конверсии
Сезонность
качестве топлива во всем мире (а сегодня — во многих разви-
вающихся странах) было прямое сжигание главным образом
древесины и — в меньших масштабах — остатков урожая и на-
воза. В настоящее время на разных стадиях разработки нахо-
дится ряд систем термической модификации такого сырья. Сре-
ди них — установки на основе пиролиза, газификации и гидро-
генизации. Отметим, что для всех этих процессов нужно сырье
с относительно низким содержанием воды, а протекают они при
высокой температуре. Биологические процессы обладают тем
преимуществом, что для них пригодно сырье с высоким содер-
жанием воды, а осуществляются они в интервале температур от
25 до 65 °C. Разнообразие планируемых для использования ви-
дов сырья (биомассы) иллюстрирует табл. 2.2.
Сегодня для этой цели применяются главным образом са-
харный тростник, кукуруза, древесина и стоки, навоз и бытовой
мусор, а также отходы сельского хозяйства и промышленности:
багасса, меласса, побочные продукты производства бумаги и
целлюлозы.
Энергия и биотехнология
39
Таблица 2.2. Источники биомассы для выработки топлива
Отходы Наземные растения Водные растения
Навоз Активный ил Бытовой мусор Пищевые отходы Стоки Отходы древесины, сахарного тростника Солома Шелуха Корки цитрусовых Багасса Меласса Л игноцел люлоза Деревья: эвкалипты тополя ели, сосны Leucaena, Casuarina Культуры, содержащие крахмал Кукуруза Кассава Культуры, содержащие сахар Сахарный тростник Сахарная свекла Водоросли Одноклеточные: Chlorella Scenedesmus Navicula Многоклеточные: бурые водоросли Растения Водяной гиацинт Тростник, камыш
2.2.1. Ресурсы
Основными поставщиками биомассы, идущей на топливо, слу-
жит сельское и лесное хозяйство. Пытаясь оценить их нынешние
возможности, следует, видимо, исходить из наличных земельных
площадей, урожайности современных культур, продуцирующих
сахар и крахмал, и числа работников, занятых в сельском хо-
зяйстве. Ежегодный прирост биомассы во всем мире составляет
около 2-Ю11 т (табл. 2.3), из них приблизительно 1,2-1011 т со-
Таблица 2.3. Годовой прирост биомассы в тоннах
Общая первичная продукция (органические ве- щества) Леса (сухое вещество) Зерновые (урожай) в пересчете на крахмал Корнеплоды (урожай) в пересчете на крахмал Культуры, дающие сахар (урожай) в пересчете на сахар 2-Ю11 9- КР» 1,5-10» 1-Ю9 5,7-108 2,2-108 Ь109 9-107
ставляет древесина (в пересчете на сухое вещество). Примерно
60% вырубаемой древесины используется как топливо. Если
потребление и возобновление леса сбалансированы, то прирост,
не идущий сегодня в дело, составляет 3-1010 т, что эквивалент-
но около 1010 т целлюлозы.
Данные о распределении лесов во всем мире приведены в
табл. 2.4. В ней же указаны земельные площади, используемые
Глава 2
Таблица 2,4. Наличные земельные площади и их использование (в %)
Земельная площадь, млн. га Площади, занятые многолет- ними куль- турами Пашни Паст- бища Леса Орош а- емые земли
Во всем мире 13 078 0,7 10 23 32 1,5
Развитые страны 5 484 0,4 12 23 34 0,9
Развивающиеся страны 7 593 0,9 10 23 30 1,9
США 936 0,2 20 26 31 18,4
Канада 922 — 5 3 36 0,05
Европа 472 3,2 30 19 32 2,8
СССР 2 227 0,2 10 17 41 0,7
Азия 2 757 1,0 16 20 22 4,6
Южная Америка 1 753 1,2 65 25 53 0,4
Африка 2 964 0,5 7 27 22 0,3
Океания 842 0,1 5 56 22 0,2
разными отраслями растениеводства, а также площади оро-
шаемых земель. В табл. 2.5 указаны площади, занимаемые ле-
Таблица 2.5. Площади, занимаемые лесами, и запасы древесины
Площадь лесов, млн. га Запасы древесины
всего, млн. м3 м3/га м3 на душу населения
США 220 20 200 92,1 95,3
Канада 250 17 800 71,2 809,1
Европа 144 14 900 103,5 29,1
СССР 770 81 800 106,2 324,6
Азия и Океания 448 38 700 72,9 18,7
Южная Америка 631 92 000 145,8 362,2
Африка 188 35 200 187,2 90,0
сами, запасы древесины в них и, кроме того, доступное для ис-
пользования ее количество в пересчете на душу населения.
Обращают на себя внимание два обстоятельства: во-первых,
колоссальные запасы древесины в Канаде, СССР и Южной
Америке (в основном в Бразилии и тропиках) и, во-вторых, не-
хватка ее в настояще время в Азии, где быстро растущее насе-
ление расходует биомассу в основном на топливо, используя ее.
с малой эффективностью, без регулярного возобновления.
Нынешний уровень производства сахара и крахмала из ос-
новных сельскохозяйственных культур в мире представлен в
табл. 2.6. Эти данные позволяют еще раз обратить внимание на
хорошо известный факт: в развивающихся странах общая уро-
') Эти цифры отражают чистый урожай, за вычетом потерь, соломы и пр. Данные по зерновым не включают те случаи, когда нх выращи-
вают иа фураж.
42
Глава 2
жайность обычно гораздо ниже, чем в Северной Америке и Ев-
ропе, где она наиболее высока. Особенно низка урожайность в
африканских и многих азиатских странах.
Приведенные здесь данные проливают свет на причину мно-
гих конфликтов при получении энергии за счет биомассы в стра-
нах, где не хватает древесины и продуктов питания и где нуж-
но разрешать дилемму: что важнее — пища или энергия. Отме-
тим, что если удастся повысить продуктивность и сельского, и
лесного хозяйства, то можно решить обе задачи. Применение
агролесных комплексных систем поможет смягчить остроту
многих таких проблем. Для этого нужно оценить пределы про-
дуктивности с учетом эффективности фотосинтеза, пределов
урожайности культур и особенно в зависимости от внешних
источников энергии в сельском хозяйстве и эффективности ее
применения. Еще одна возможность интенсификации заключа-
ется в отборе или выведении новых, дающих высокий выход
биомассы разновидностей растений, которые можно выращивать
на засушливых, болотистых, засоленных или излишне обводнен-
ных землях, продуктивность которых сегодня не идет в сравнение
с достигнутой в сельском или лесном хозяйстве. Рассмотрим все
эти вопросы более подробно.
2.3. Фотосинтез
Фотосинтез является ключевым процессом жизнедеятельности и
осуществляется в основном в растениях. В простейшей форме
он описывается реакцией
Растения
Н2О + СО2 —-------—Органические вещества + О2.
Солнечная энер-
гия
Кроме углерода, водорода и кислорода в ходе светозависи-
мых реакций, протекающих в растениях, в состав органиче-
ских веществ включаются также азот и сера.
Основные процессы фотосинтеза сегодня уже хорошо изве-
стны. Они протекают в хлоропластах (рис. 2.2), которые погло-
щают СОг, поступающий в растение путем диффузии. Первич-
ный процесс карбоксилирования осуществляется в строме
(части хлоропласта, содержащей мало мембран) и катализиру-
ется рибулозобисфосфат-карбоксилазой (РБФК). В результате
образуются две молекулы трехуглеродной кислоты (фосфогли-
церата, ФГ), которые затем восстанавливаются с образованием
молекул трехуглеродного сахара — триозофосфата. Это вещест-
во в хлоропластах служит предшественником крахмала, но мо-
жет поступать и в цитоплазму, где оно используется при синтезе
сахарозы. Часть связанного углерода повторно поступает в вое-
Энергия и биотехнология
43
становительный пентозофосфатный цикл (ВПФ-цикл), поддер-
живая фонд акцептора СО2, рибулозобисфосфата.
ВПФ-цикл «работает» в направлении синтеза, используя
энергию гидролиза АТР и окисления NADPH. АТР и NADPH
Рис. 2.2. Хлоропласты высшего растения.
образуются в ходе так называемых световых реакций фотосин-
теза, которые идут в мембранных (тилакоидных) структурах
хлоропластов. Для восстановления СО2 до СН2О нужно не ме-
нее четырех электронов. Имеющиеся на сегодня данные говорят
в пользу «Z-схемы» фотосинтеза, который протекает в резуль-
44
Глава 2
тате кооперативного взаимодействия двух фотосистем (ФС I и
ФС II, рис. 2.3), объединенных связующей цепью реакций пере-
носа электрона. Восстановление NADP до NADPH происходит
в ФС I при участии содержащего серу и железо белка ферре-
доксина.
-0,8Г
Редокс-потенциал
+ 0,81-
СВЕТ
(фотосистема II,
4 фотона)
Рис. 2.3. Схема переноса электронов при фотосинтезе.
Свет улавливается своеобразными антенными хлорофилл-
белковыми комплексами, и энергия его передается в реакцион-
ный центр (Р700) ФС I, представляющий собой особым обра-
зом устроенный комплекс хлорофилла а и белка. Здесь проис-
ходит разделение заряда, и в результате восстанавливается
ферредоксин. Электроны поступают от воды и передаются на
Р7оо по связующей электрон-транспортной цепи от ФС II. В ходе
идущей при участии ФС II реакции из воды образуется Ог.
Переносчики электронов располагаются внутри мембраны
тилакоидов таким образом, что поперек нее формируется элект-
рохимический градиент, сопряженный с переносом электронов.
Энергия и биотехнология 45
Соответствующий электрохимический потенциал используется
для синтеза АТР. Белок, участвующий в этом процессе, известен
как сопрягающий фактор и обладает свойствами обращенной
АТРазы.
Длина волны света, поглощаемого хлорофиллами и други-
ми фотосинтетическими пигментами, определяется свойствами
этих молекул; именно от этого зависит, какая часть солнечного
спектра может быть использована растениями (фотосинтетиче-
ски активная радиация, ФАР). Она составляет в пересчете на
энергию 45—50% всего падающего на растение солнечного све-
та. Способность улавливать свет — первый из ограничительных
факторов, определяющий эффективность фотосинтеза, если ис-
ходить из количества падающего света.
При малой освещенности фотосинтез лимитируется световы-
ми реакциями, а при более высокой интенсивности света или
более низкой температуре — возможностями темновых реакций
метаболизма углерода. У многих С3-растений умеренного кли-
мата световое насыщение наблюдается при 100—150 Дж/м-2-
-с-1, что соответствует 50%-ной интенсивности солнечного све-
та. При высоких интенсивности света и температуре влаги обыч-
но не хватает (водный стресс). Стресс выражается в закрытии
устьиц и подавлении метаболизма углерода. Таким образом,
в полевых условиях взаимодействие световых и темновых реак-
ций и их регуляция за счет диссипации (рассеивания) излишней
световой энергии становятся определяющими.
В функционирующих хлоропластах каждая электрон-транс-
портная цепь может осуществлять акт передачи каждые 15 мс.
На ярком свету молекула хлорофилла в среднем поглощает фо-
тон один раз в 100 мс, а в условиях слабого или рассеянного
освещения даже реже. Однако, поскольку каждый реакционный
центр связан с несколькими сотнями светособирающих молекул
пигмента, интенсивность переноса электронов увеличивается.
Время с момента поглощения света до попадания возбуждения
в ловушку измеряется пикосекундами. Однако перенос электро-
на осуществляется медленно, и лимитирующая реакция синтеза
АТР протекает за время порядка 20 мс.
Как показано на рис. 2.3, в ходе фотосинтеза энергия элект-
ронов изменяется от потенциала воды (+0,8 В) до потенциала
СН2О (—0,43 В). Эта разность потенциалов эквивалентна
480 кДж. Таким образом, ВПФ-цикл должен был бы работать
«с высокой эффективностью, чего в действительности не наблю-
дается.
Все дело в том, что фермент РБФК способен катализировать
и окислительную реакцию между РБФ и О2, в результате кото-
рой образуются ФГ и фосфогликолат. Эта окислительная реак-
ция конкурирует с реакцией карбоксилирования за 2 ФГ, в ре-
46
Глава 2
зультате чего углерод направляется по С-2-пути фотодыхания,
в ходе которого образуется СО2, утрачиваемый растением.
В результате окислительных процессов и фотодыхания в
Сз-растениях эффективность фотосинтеза уменьшается на 15—
50% в зависимости от вида растений и условий окружающей
среды. Эти изменения обусловлены действием нескольких фак-
торов: 1) потерей субстрата карбоксилирования; 2) потерей уг-
лерода из ВПФ-цикла; 3) потерей СО2 растением; 4) уменьше-
нием диффузионного градиента, способствующего диффузии в
растение.
Скорость фотосинтеза зависит от поступления СО2 в лист, что
в свою очередь зависит от градиента концентрации СО2 и от
ряда противодействующих факторов, связанных с наличием фи-
зических и биологических барьеров для диффузии и ассимиля-
ции:
Р = /<(АСО2)/(г1 + г2 + г3),
где К — коэффициент диффузии; соответствует разным типам
противодействия. Варьирующее противодействие при диффузии
г2 — это сопротивление устьиц. Сила его контролируется све-
том, водой и концентрацией СО2. В частности, при недостатке
воды устьица смыкаются.
Противодействие гз (сопротивление мезофилла) зависит от на-
личия ферментных компонентов, способных к ассимиляции СО2.
Сродство фермента РБФК можно охарактеризовать константой
Михаэлиса (Км). Согласно большинству последних оценок, Км
по СО2 для РБФК составляет 10—20 мкМ, что эквивалентно'
содержанию СО2 в атмосфере, находящемуся в равновесии с
содержанием его в воде. Другими словами, если в клетке нет
механизма концентрирования СО2, то этот фермент будет рабо-
тать со скоростью, равной примерно половине оптимальной.
Механизмы концентрирования СО2 существуют у некоторых
водорослей и ряда высших растений с С4-типом фотосинтеза.
Группа С4-растений включает несколько родов; в основном это
тропические растения. К их числу принадлежат такие коммер-
чески важные культуры, как сахарный тростник, кукуруза и сор-
го, а также тропическая «слоновья трава» (Pennisetum). В опы-
тах с этими растениями была зарегистрирована максимальная
скорость образования сухого вещества: у них имеется дополни-
тельный цикл карбоксилирования (С4-путь), который работает,
как насос, перекачивающий СО2 из атмосферы к месту восста-
новительной реассимиляции за счет ВПФ-цикла в клетках об-
кладки проводящих пучков. Вначале углекислый газ атмосферы
ассимилируется с образованием четырехуглеродных кислот (ма-
лата и аспартата) в наружном (мезофильном) слое фотосин-
тезирующей ткани. Эти кислоты переносятся в слой клеток об-
Энергия и биотехнология
47
кладки проводящих пучков, где СОг высвобождается и заново
ассимилируется. В результате этого процесса фотодыхание в
Сграстениях не так выражено, что увеличивает скорость фото-
синтеза и продуктивность. Впрочем, хотя эти растения и ас-
симилируют углерод более эффективно, они потребляют допол-
нительную энергию для осуществления реакций С4-пути, и по-
этому менее эффективно используют энергию света. С4-растения
также с большим успехом используют воду и меньше страдают
от повышенной температуры. Отметим, однако, что лишь немно-
го видов С4-растений способны хорошо расти в местностях с
умеренным климатом.
2.3.1. Эффективность фотосинтеза
Эффективность фотосинтеза с точки зрения производства био-
массы можно оценить через долю общей солнечной радиации,
попадающей на определенную площадь за определенное время,
которая запасается в органических веществах урожая. Продук-
тивность системы можно оценить по количеству органического
сухого вещества, получаемого с единицы площади за год, и вы-
разить в единицах массы (кг) или энергии (мДж) продукции,
полученной с гектара за год.
Выход биомассы зависит, таким образом, от площади кол-
лектора солнечной энергии (листьев), функционирующих в те-
чение года, и числа дней в году с такими условиями освещенно-
сти, когда возможен фотосинтез с максимальной скоростью, что
определяет эффективность всего процесса. Результаты опреде-
ления доли солнечной радиации (в %), доступной растениям
{фотосинтетически активной радиации, ФАР), и знание основ-
ных фотохимических и биохимических процессов и их термоди-
намической. эффективности позволяют рассчитать вероятные
предельные скорости образования органических веществ в пере-
счете на углеводы.
Растения используют свет с длиной волны от 400 до 700 нм,
т. е. на долю ФАР приходится 50% всего солнечного света.
Это соответствует интенсивности на поверхности Земли 800—
1000 Вт/м2 за обычный солнечный день (в среднем). Усреднен-
ная максимальная эффективность превращения энергии при
фотосинтезе на практике составляет 5—6%. Эти оценки полу-
чены на основе изучения процесса связывания СО2, а также со-
путствующих физиологических и физических потерь. Одному мо-
лю связанного СО2 в форме углевода соответствует энергия
0,47 МДж, а энергия моля квантов красного света с длиной
волны 680 нм (наиболее бедный энергией свет, используемый в
фотосинтезе) составляет 0,176 МДж. Таким образом, минималь-
ное число молей квантов красного света, необходимое для свя-
48
Глава 2
зывания 1 моля СО2, составляет 0,47:0,176 = 2,7. Однако, по-
скольку перенос четырех электронов от воды для фиксации од-
ной молекулы СО2 требует не менее восьми квантов света, тео-
ретическая эффективность связывания равна 2,7:8 = 33%. Эти
расчеты сделаны для красного света; ясно, что для белого све-
та эта величина будет соответственно ниже. В наилучших поле-
вых условиях эффективность фиксации в растениях достигает
3%, однако это возможно лишь в короткие периоды роста и, ес-
ли пересчитать ее на весь год, то она будет где-то между 1
и 3%.
На практике в среднем за год эффективность фотосинтети-
ческого преобразования энергии в зонах с умеренным климатом
составляет обычно 0,5—1,3%, а для субтропических культур —
0,5—2,5%. Выход продукта, который можно ожидать при оп-
ределенном уровне интенсивности солнечного света и разной
эффективности фотосинтеза, легко оценить из графиков, приве-
денных на рис. 2.4.
Эффективность
фотосинтеза
Среднее количество солнечной энергии за год, Вт/м2
(100 Йт/м2- 3150 МДж/м2 за год)
Рис. 2.4. Ожидаемая (хорошая) урожайность однолетнего растения как функ-
ция годичной солнечной энергии при разной эффективности фотосинтеза (Hall,
1982).
2.3.2. Интенсификация фотосинтеза методами биотехнологии
Увеличение выхода биомассы за год в существующих сегодня
системах растениеводства может быть достигнуто двумя путя-
ми: во-первых, за счет увеличения скорости фотосинтеза до пре-
Энергия и биотехнология 49»
делов, возможных в оптимальных условиях, во-вторых, путем,
удлинения периода оптимального фотосинтеза.
Для того чтобы достичь этой цели, необходимо оценить от-
носительную важность различных факторов, ограничивающих
фотосинтез. Действие этих факторов определяется как внутрен-
ними фотобиологическими и физиологическими ограничениями,
так и теми характеристиками окружающей среды, которые ска-
зываются на проявлении этих лимитирующих факторов. К чис-
лу таких важнейших факторов относятся: индекс урожайности,
свет, СО2, вода, температура, питательные вещества, вредители
и болезни, влияние кислорода и фотодыхание, темновое дыха-
ние, ограничение скорости переноса электронов, содержание
ферментов карбоксилирования, светособирающих пигментов,
диссипация энергии в побочных реакциях и скорость переноса
веществ из хлоропластов.
Действие основных лимитирующих факторов можно осла-
бить путем увеличения энергетических затрат при культивиро-
вании, но к их числу относятся и неконтролируемые факторы
внешней среды. Для получения большого количества биомассы
нужны тщательный отбор и выведение сортов растений, спо-
собных эффективно использовать благоприятные условия окру-
жающей среды при минимальных затратах.
Наибольшие успехи в повышении урожайности' сельскохо-
зяйственных культур были достигнуты путем улучшения как
селекционной работы, так и агротехники. Однако в общем
селекция была в основном направлена на увеличение индекса
урожайности культур, а не общего количества собираемой био-
массы.
Приросты урожайности за счет улучшения агротехники обыч-
но связаны с увеличением энергозатрат (в форме горючего для
тракторов, удобрений и средств защиты растений, получаемых
за счет ископаемого топлива). В результате урожай с единицы
площади возрастает, но в пересчете на затраченную энергию
падает. Такие изменения приемлемы при выращивании пище-
вых культур, но нежелательны для «энергетических», когда не-
обходимо получить большой урожай биомассы при малых
энергозатратах. Ясно, что эффективные системы производства
биомассы потребуют дополнительного вклада энергии. Вели-
чину его, а также ожидаемый выигрыш (в смысле увеличения
энергопродуктивности) еще предстоит оценить.
Существенное увеличение выхода продукции было достигну-
то в ходе отбора растений с листьями оптимальном формы, ри-
гидности и расположения, имеющих улучшенный тип кроны и
способных эффективнее улавливать свет. В принципе эффектив-
ность фотосинтеза можно повысить и путем изменения одного
или нескольких составляющих его процессов или факторов:.
4—1344
50
Глава 2
1) улавливания света и переноса электронов; 2) усвоения уг-
лекислого газа и регуляции реакций основного обмена углеро-
да, включающих темновое дыхание и фотодыхание; 3) свойств
мембран, особенно структуры липидов и температурной чувст-
вительности; 4) анатомических признаков для увеличения эко-
номичности использования СОг и воды.
Проведя оценку существующих сортов культур как потен-
циальных продуцентов биомассы, мы можем улучшать их путем
применения новых способов разведения, изучения их фотосин-
тетических возможностей и размножения растений нетрадици-
онными способами. Реализовать эти возможности в будущем
поможет использование технологии рекомбинантных ДНК. Для
продвижения вперед в этой области нам необходимо: 1) разра-
ботать методы выявления положительных изменений в фотосин-
тезе и приспособить сложные лабораторные тесты для работы в
долевых условиях; 2) предложить методы усиления генетиче-
ской изменчивости; 3) понять, как организован геном растений
и хромосома хлоропласта и как регулируется их работа; 4) вы-
явить типы изменений, которые могут быть в них вызваны;
5) разработать новые способы селекции, направленные на уско-
рение размножения и генетическую стабилизацию сортов.
Методы усиления генетической изменчивости многообразны:
для этой цели используют культуру тканей, слияние протоплас-
тов, перенос одиночных генов, гаплоиды., опыление облученной
пыльцой, химический мутагенез, замену митохондриальных и
хлоропластных геномов и т. д. Направленный перенос распоз
наваемых признаков может быть осуществлен методами гене1
тической инженерии. Здесь стоят проблемы выбора вектора,
включение гена в геном и экспрессии нового признака в усло-
виях сложной системы регуляции у растений как на генетиче-
ском, так и на метаболическом уровне.
Таким образом, работы по увеличению продуктивности рас-
тений, выращиваемых для получения энергии (биомассы),
в опытах in vitro будут успешными, если мы: 1) установим ге-
нетические пределы продуктивности; 2) выявим индивидуаль-
ные гены, ответственные за это свойство (признаки); 3) выде-
лим ДНК. или РНК, кодирующие эти признаки; 4) встроим их
при помощи ферментов рестрикции и лигаз в подходящий век-
тор для амплификации в бактериях; 5) введем накопленный та-
ким путем материал в вектор, подходящий для его переноса в
желаемый вид растений; 6) стабилизируем введенную ДНК в
геноме нового хозяина таким образом, чтобы она экспрессиро-
валась как доминантный признак, наследуемый по законам
Менделя; 7) разработаем методы скрининга для выявления та-
ких измененных растений; 8) будем применять альтернативные
подходы, используя для увеличения изменчивости и менее спе-
Энергия и биотехнология 51
цифичные методы, например мутагенез (химический и радиа-
ционный) при работе с культурами тканей и протопластов или
естественную изменчивость, с отбором форм с более эффектив-
ным фотосинтезом.
Хотя сведения о ядерном геноме и пластоме хлоропластов,
а также о механизмах регуляции работы генов при синтезе бел-
ков в хлоропластах и других частях клеток накапливаются
очень быстро, воплощение генно-инженерных возможностей в
практику — дело будущего, отстоящего на десятилетия, Отме-
тим также, что для решения проблемы продуктивности особое
внимание уделяется сегодня развитию фундаментальной фото-
биологии. Предполагается, что в будущем эти знания вопло-
тятся в практику. Пока этого не произошло, производство топ-
лива из биомассы должно опираться на обычное сельское и
лесное хозяйство.
2.4. Сельское и лесное хозяйство
2.4.1. Соотношение видов энергии
При анализе работы любой сельскохозяйственной системы важ-
но учитывать, как соотносится количество энергии, запасенной
в системе, с энергозатратами на ее получение (отношение энер-
гии на входе и выходе). Для систем по производству биомассы
это имеет особое значение. Табл. 2.7 дает представление о ве-
Таблица 2.7. Соотношение энергонакопления и энергозатрат
в сельском хозяйстве Англии
В среднем по фермам Специализированным молочно- товарным 0,38 Т разы При малой эффективности только выпас 9,1
Главным образом молочното- 0,55* выпас и сено 5,6
варным При высокой эффективности
Поставляющим сено 2,4
крупный рогатый скот и овец 0,59 силос 2,4
овец 0,25 Горох свежий 0,9
свиней и птицу 0,32 консервы 0,1
злаковые 1.9 Морковь 1,1
Ячмень (и овес) 2,4 Брюссельская капуста 0,2
Кукуруза (на зерно) 2,3' Тепличный салат 0,002
Пшеница (на зерно) 3,4 Птица (мясо) 0,1
Картофель 1,6 (яйца) 0,1
Сахарная свекла Рыба 0,05
по сахару 3,6 Молоко 0,4
общий выход 4,2
личине этого отношения и иллюстрирует некоторые проблемы
энергетически интенсивного сельского хозяйства, возникающие,
например, при выращивании растений в теплицах. Она иллюст-
4*
52
Глава 2
рирует также малую эффективность превращения энергии в си-
стемах, включающих животных. В доступную для использова-
ния форму превращается лишь одна десятая часть энергии, по-
требляемой животными и растениями, выращиваемыми в теп-
лицах. И тепличные растения, и продукция животноводства —
важные компоненты современного сельского хозяйства. Отме-
ним, однако, что небольшое уменьшение поголовья животных,
выращиваемых для употребления в пищу, если это окажется це-
лесообразным, высвободит большое количество органических
продуктов, которые могут быть использованы как пища, топли-
во и т. д. Сопоставление величин энергетического коэффициента
(выход/вход) при производстве зерна кукурузы в США показа-
ло, что он уменьшился с 3,7 в 1945 г. до 2,8 в 1970 г. Важно
отметить, что удвоение урожая было достигнуто за счет увели-
чения втрое энергозатрат, в основном вследствие более широко-
го применения удобрений. В Великобритании кукурузу выращи-
вают в основном на корма, т. е. растения используются цели-
ком, и поэтому энергетический коэффициент варьирует от 5 до 9.
Если бы удалось уменьшить потребление широко используемых
азотных удобрений (а на их долю приходится часто до 50%
энергозатрат) без уменьшения урожая, например путем биоло-
гической фиксации азота, использования навоза или ила из био-
реакторов, то мы бы сэкономили много энергии. Хотя в прош-
лом практика широкого применения азотных удобрений и кри-
тиковалась, следует отметить, что на каждый джоуль энергии,
затраченный на азотные удобрения для растений, приходится
4—6 Дж запасенной ими энергии. Фотосинтез выступает в роли
катализатора, обеспечивающего «прирост» энергии, но растения
не могут эффективно развиваться в отсутствие оптимального ко-
личества азота (как и других элементов: фосфора, калия и мик-
роэлементов) .
Во многих отраслях сельского хозяйства значительная часть
образовавшейся биомассы остается после уборки в почве или
на ее поверхности. Проблема интегрированной системы топли-
во/пища заключается в том, что, поскольку использоваться мо-
гут все части растений, все их можно и убирать, но такая прак-
тика недопустима. Вынос минеральных питательных веществ
при рубке леса можно компенсировать внесением неорганиче-
ских удобрений, но в тропиках, где почвы обычно более «сла-
бые», эта мера часто оказывается недостаточной, так как глав-
ным фактором устойчивости почвы являются ее органические
компоненты. Они служат источником питательных веществ для
различных микроорганизмов, осуществляющих процессы мине-
рализации и .фиксации атмосферного азота, а также ответствен-
ны за улучшение комковатой структуры почв и их способность
удерживать воду. В целом из экосистем, не получающих под-
Энергия и биотехнология
53
крепления извне, можно изымать не более 10—20% ежегодного
прироста сухого вещества, однако и это относится лишь к слу-
чаям, когда большую часть года существует сомкнутый листо-
вой полог. Если почва оголена, то скорость потери органическо-
го вещества возрастает. Когда целью сельского и лесного хо-
зяйства является производство и топлива, и пищи, следует
совместно выращивать деревья и полевые культуры таким об-
разом, чтобы в системе происходило достаточное накопление
или возврат органического вещества.
2.4.2. Древесина как сырье для производства биотоплива
Как сырье для производства биотоплива древесина обладает
рядом достоинств: выход продукции в пересчете на гектар очень
высок; из древесины мы получаем значительно больше биомас-
сы, чем из любого другого источника; разведение лесов требует
гораздо меньших вложений, чем выращивание других культур.
К числу недостатков нужно отнести длительность роста до зре-
лости, а также тот факт, что главный компонент древесины,
лигноцеллюлоза, очень сложна для переработки. В ближайшем
будущем наиболее удобным и доступным источником сырья бу-
дут отходы деревообрабатывающей промышленности, но впо-
следствии все возрастающее значение будет приобретать «вы-
ращивание топлива». Поскольку основные затраты связаны с
очисткой земли и посадкой, особое внимание уделяется сегодня
выращиванию твердодревесинного быстрорастущего порослево-
го леса. Здесь дело, вероятно, можно организовать так, чтобы
получать продукцию каждые пять лет. Подсчитано, что лесовод-
ческая «энергоферма» может ежегодно давать около 10—30 т
биомассы с гектара.
С начала 50-х годов нашего века в Аргентине и Бразилии на
таких энергофермах на относительно большой площади (8000 га)
выращивали эвкалипт и производили древесный уголь для вы-
плавки стали. В других странах (например, в Южной Америке
и Австралии) производят древесную массу, используемую для
получения ацетатцеллюлозного волокна. В больших масштабах
таким путем производят сырье для выработки бумаги и лесо-
материалов. В зонах умеренного климата пробуют выращивать
Salix и Populus, а в тропиках — Leucaena и Casuarina, но боль-
ше всего продукции (в тех местах, где их можно выращивать)
дают Eucalyptus.
Eucalyptus — это род вечнозеленых растений, в состав кото-
рого входит более 450 видов, разновидностей и гибридных форм.
Большинство их происходит из Австралии, где на их долю при-
ходится до 95% древесины леса. Они растут там и в сухих, и в
заболоченных местах, на почвах самых разных типов. Сегодня
54
Глава 2
они встречаются уже почти по всему земному шару, кроме ре-
гионов, где бывают морозы или же нерегулярно выпадают дож-
ди. При благоприятных условиях они растут быстрее, чем поро-
ды с мягкой древесиной (например, Pinus radiata). Спустя
10 лет после посадки годовой прирост древесины составляет
10 т на гектар. Такие приросты получают лишь в посадках
Pinus radiata двадцатипятилетнего возраста.
Наилучший результат был получен в хозяйстве Aracruz pulp
and fuelwood, расположенном в 500 км к юго-востоку от Рио-де-
Жанейро: при выращивании в отличных погодных условиях от-
борных сортов Eucalyptus на площади 50 000 га удалось полу-
чить за год 45 т древесины с гектара. Посадочный материал по-
лучали путем вегетативного размножения эксплантатов листьев.
Трудности в использовании древесины или лигноцеллюлозы
в качестве сырья для производства жидких энергоносителей за-
ключаются в том, что переработка их сложна: ведь для получе-
ния такого топлива, как этиловый спирт, или же такого химика-
та, как бутанол, необходимы относительно чистые растворы со-
ответствующих сахаров.
2.4.3. Водоросли и водные растения
Потенциальный урожай биомассы у пресноводных и морских
растений весьма велик, но чрезвычайно большое содержание
воды во многих этих растениях при сборе и сложность сушки на
солнце препятствуют использованию их как топлива путем пря-
мого сжигания. По этой причине наиболее подходящей техноло-
гией переработки водных растений и сырых отходов земледелия
в топливо, корма и удобрения является анаэробная фермента-
ция. Эти растения просто процветают в сточных водах. Они ус-
пешно очищают воду и хорошо при этом растут. Таким обра-
зом, они могут играть двойную роль: улучшать состояние окру-
жающей среды и служить важным источником энергии.
В ряде стран из водяных гиацинтов получают биогаз. Их
стали использовать для этой цели, поскольку растения эти ис-
ключительно быстро растут, причем на поверхности воды, и их
легко собирать. Можно использовать и водоросли, растущие в
прудах, в которых перерабатываются сточные воды, содержа-
щие органические вещества (которые загрязняют среду и/или
требуют больших затрат на удаление). Такая технология осо-
бенно пригодна для стран, где много солнца и к тому же неред-
ко возникают проблемы переработки жидких отходов.
Саму идею использовать водоросли и бактерии в биологиче-
ских системах, функционирующих за счет энергии солнечного
света, нельзя считать новой, но в последнее время на нее стали
обращать больше внимания. Одно из достоинств микробиологи-
Энергия и биотехнология
55
ческих систем заключается в том, что в зависимости от условий
они могут быть или технологически сложными, или же просты-
ми. Выбор наиболее подходящей системы определяется местны-
ми «за и против», например соленостью и температурой. Из-
бранные для культивирования виды в этом случае легко приспо-
сабливаются к окружающей среде.
Многие жидкие и полутвердые отходы — идеальная среда
для роста фотосинтезирующих водорослей и бактерий. При хо-
роших условиях они быстро наращивают биомассу и осуществ-
ляют эффективное превращение солнечной энергии (3,5%); вы-
ход продукции составляет 50—80 т с гектара в год. Собранные
водоросли можно прямо скармливать животным, получать из
них метан или сжигать для получения электроэнергии. При этом
одновременно происходит переработка отходов и очистка воды.
По существующим оценкам затраты на такие системы в услови-
ях Калифорнии составляют 50— 75% от затрат на обычные си-
стемы переработки сточных вод. Главная хозяйственная пробле-
ма здесь — затраты на сбор продукции. Ее можно решить, ис-
пользуя иные виды водорослей, которые легче собирать, и новые
технические приемы «сбора урожая». Для полной переработки
жидких стоков сегодня пытаются применять двухступенчатые
водоемы с водорослями. В первом водоеме выращивают водо-
росли, которые собирают путем фильтрации, во втором — азото-
фиксирующие сине-зеленые водоросли (их тоже легко соби-
рать); питательные вещества для их роста поступают из перво-
го водоема. Для увеличения продуктивности можно использо-
вать и промышленные отходы, включая ССЬ. Из собранной био-
массы путем сбраживания можно получать метан (в пересчете
на энергию— 1,1 МДж на килограмм водорослей), причем отхо-
ды от такой переработки будут содержать практически весь
азот и фосфор биомассы водорослей. Это хорошее удобрение
для сельского хозяйства. Один гектар водорослевых прудов мо-
жет давать удобрения для 10—50 гектаров полей.
Путем оптимизации урожаев удается получить в форме мета-
на до 200 КДж с гектара в год с учетом энергозатрат и потерь
при переработке. На широте 30° это соответствует годовой эф-
фективности фотосинтетических процессов, равной 1,5%.
В Калифорнии средний урожай водорослей составляет более
100 кг сухого вещества с гектара в декаду. Летом он бывает
в три раза больше. При урожае 50—60 т сухого вещества с гек-
тара в год из него можно получать 74 000 кВт/ч электроэнергии.
Были сооружены водорослевые пруды объемом 10е л, в которых
фотосинтез идет равномерно и с эффективностью 2—3%. При
крупных предприятиях по откорму скота и птицы водоросли вы-
ращивают в прудах, куда поступают отходы животноводства.
Около 40% азота отходов фиксируется водорослями, которые
56
Глава 2
затем скармливают животным. В культивируемых сегодня зеле-
ных водорослях содержится 50—60% белка, а в сине-зеленых,
с которыми ведутся опыты, содержание экстрагируемого белка
доходит до 60—70%. Водорослевые пруды, в которых перера-
батываются стоки, нашли применение по меньшей мере в десяти
странах, и интерес к таким системам как возможным поставщи-
кам энергии и удобрений все возрастает.
Отметим, что из водорослей могут быть получены некото-
рые углеводороды и редкие химические вещества. Так, у широ-
ко распространенной зеленой водоросли Botryococcus braunii
углеводороды в зависимости от условий роста и разновидности
могут составлять до 75% сухой массы. Длина углеводородной
цепи при этом варьирует от С]7 до С34. Они накапливаются внут-
ри клеток, и водоросли, в которых их много, плавают на по-
верхности. После сбора водорослей эти углеводороды легко от-
делить экстракцией каким-либо растворителем или методом
деструктивной отгонки. Таким путем может быть получено ве-
щество, аналогичное дизельному топливу и керосину. В. braunii
широко распространены в природе; они встречаются в самых
разных местах: от солоноватых озер Австралии (где высохшие
остатки этих водорослей, известные под названием Coorongite,
послужили в прошлом поводом для нефтяной лихорадки) до во-
дохранилищ Лондона. Сходные породы, обнаруженные в других
частях света, называют по месту их обнаружения (N’haugellite
в Мозамбике, балхашит в Казахстане и т. п.).
Оценки продуктивности были получены лишь для лаборатор-
ных культур, выращенных в условиях постоянного освещения,
и в случае непредсказуемых вспышек размножения водорослей
в природных условиях. Для установления продуктивности
В. braunii при массовом культивировании использовать эти оцен-
ки довольно сложно. По мнению группы австралийских ученых,
прежде всего нужно провести следующие исследования: 1) оп-
ределить условия, обеспечивающие максимальную скорость ро-
ста и образования углеводородов в лабораторных и полевых ус-
ловиях; 2) на опытных установках выяснить, можно ли при ин-
тенсивном выращивании добиться скорости роста В. braunii, со-
измеримой с известной для других водорослей; 3) разработать
соответствующие методы выращивания, сбора и переработки;
4) оценить применимость получаемого продукта как альтерна-
тивного источника топлива и смазочных веществ.
В Израиле на опытных установках проводятся эксперименты
с зеленой одноклеточной водорослью Dunaliella bardawil-. она
способна использовать солнечную энергию для синтеза глицеро-
ла, важного органического соединения, широко используемого
промышленностью. Dunaliella может расти и размножаться в
среде с широким диапазоном содержания соли: и в воде океа-
Энергия и биотехнология
57
нов, где она невелика, и в почти насыщенных солевых растворах
Мертвого моря. Она накапливает свободный глицерол, чтобы
противодействовать неблагоприятному влиянию высоких кон-
центраций солей в среде, где она растет. При оптимальных ус-
ловиях и высоком содержании соли на долю глицерола прихо-
дится до 85% сухой массы клеток. Из проведенных опытов сле-
дует, что для роста этим водорослям нужна всего лишь морская
вода, углекислый газ и солнечный свет. Таким образом, они яв-
ляются весьма многообещающими биосинтетическими агентами
для превращения энергии солнечного света в энергию химиче-
ских соединений, запасаемую в молекулах глицерола.
После переработки эти водоросли можно использовать в ка-
честве корма для животных, так как у них нет неперевариваемой
клеточной оболочки, присущей большинству других водорослей.
Кроме того, они содержат значительное количество пигмента
[5-каротина. Таким образом, благодаря возможности получения
трех продуктов — глицерола, пигмента и белка — это опытное
производство представляется вполне перспективным с экономи-
ческой точки зрения, хотя точные оценки продуктивности и за-
трат пока отсутствуют.
Для получения масел или углеводородов не обязательно ис-
пользовать именно водоросли или культуры микроорганизмов.
Подходящим сырьем для прямого получения жидкого углеводо-
родного топлива являются и некоторые высшие растения.
'2.4.4. Масличные растения
Растительные масла могут быть получены из самых разнообраз-
ных растений. Помимо хорошо известных нам подсолнечников,
пальм, кокосовых орехов, оливок и арахиса для этой цели ис-
пользуются и более экзотические виды: джоджоба и гваюла
(оба — растения пустынь), клещевина, рапс (семена), растения,
выделяющие млечный сок, эвкалипты, тыквы, копайба, малме-
лейро, орехи бабассу и др. Масла из этих растений, как и из
многих других, исследуются в разных странах. Однако, прежде
чем принимать решение об использовании этих масел как топ-
лива и о налаживании их производства, необходимо оценить их
стоимость, а также количество энергии, необходимое для выра-
щивания соответствующих растений и переработки сырья. Нет
смысла выращивать растения и получать из них литр раститель-
ного масла, если при этом затрачивается два литра первосорт-
ного горючего. Определяющий фактор здесь — урожайность,
т. е. количество масла, которое фермер может получить с гекта-
ра посевов, а также денежные затраты и трудоемкость. Опыты
с подсолнечником и соей на юге Африки и на Среднем Западе
58
Глава 2
США показали, что можно получить до тонны масла с гектара,
даже если использовать не самый лучший способ отжима. Со-
держание масла в собранном сырье составляет при этом 40—
50% по массе. Выход масла в две тонны с гектара считается
обычным; сообщалось, что он может достигать и пяти тонн. Та-
кое масло стоит около двух долларов за галлон (20 пенсов за
литр). При этом запасенная в продукте энергия относится к
энергозатратам при выращивании и переработке урожая как
3—10 к 1.
Главное препятствие для дальнейшего развития отрасли —
низкая биопродуктивность имеющихся сортов растений. Между
тем опыт осуществления специальных программ (которые долж-
ным образом финансировались) говорит о перспективности воз-
делывания масличных культур. Так, с успехом была выполнена
совместная научная и прикладная программы Великобритании
и Малайзии по выведению высокопродуктивных сортов маслич-
ной пальмы. Были получены рекордные урожаи, до 14 тонн мас-
ла с гектара в год, что в 2—3 раза превосходит обычные; эти
новые сорта быстро размножили путем использования метода
культуры тканей.
Получение топлива из пальмового масла имеет ряд преиму-
ществ перед выработкой этилового спирта из сахара или крах-
малсодержащего сырья. Пальмовое масло получать проще, уро-
жай можно собирать круглый год, при производстве образуется
меньше отходов, загрязняющих окружающую среду, и при этом
одним из «отходов» производства является богатый белком
корм для животных. В идеальном случае можно будет исполь-
зовать выращенное растение целиком и получать набор разных
продуктов. При этом возврат энергии очень велик, и для произ-
водства топлива не требуется воды.
Растительное масло можно использовать в чистом виде или
в смеси с дизельным топливом для применения в двигателях
компрессионного зажигания. Однако эффективность работы и
долговечность таких машин увеличиваются, если масло пере-
работать в метиловые или этиловые эфиры. С другой стороны,
многие виды растений синтезируют углеводороды, которые мож-
но непосредственно использовать как горючее или химическое
сырье. Из последних наиболее известен каучук, получаемый из
культивируемых деревьев Hevea brasilensis. С химической точ-
ки зрения такие углеводороды — это вещества, более восстанов-
ленные, чем углеводы. Их можно использовать непосредственно,
не прибегая к микробиологической или термохимической моди-
фикации, без чего не обойтись в случае углеводов.
В настоящее время предпринимаются серьезные попытки вы-
вести и наладить выращивание сортов растений, синтезирующих
углеводороды более низкой молекулярной массы, чем у каучу-
Энергия и биотехнология
59
ка. Предполагается, что из них можно будет получать жидкое
топливо со свойствами, близкими к таковым у нефти.
Поиски растений с высоким содержанием углеводородов
периодически проводились и раньше. В 1921 г. была опублико-
вана монография, в которой сообщалось о содержани «каучука»
в растениях Северной Америки. За четыре года до смерти
(1932 г.) американский изобретатель Томас Эдисон изучил в
этом плане около 2000 видов растений. Он нашел, что многие
растения содержат углеводороды, но только у одного-двух из
них молекулярная масса была настолько высока, что они могли
в принципе служить заменителем натурального каучука из He-
vea. Такие поиски велись в основном среди растений, выделяю-
щих млечный сок — латекс. Последний представляет собой по-
хожую на молоко 30%-ную эмульсию углеводородов и является
тем сырьем, из которого на плантациях получают натуральный
каучук. Отметим, что многие виды растений, особенно из семей-
ства Euphorbiaceae, накапливают в латексе углеводороды с су-
щественно меньшей, чем у каучука, молекулярной массой (10—
20 тыс., а не 1—2 млн.). Именно среди веществ с такой молеку-
лярной массой ведутся сегодня поиски заменителей нефти.
Из представителей рода Euphorbia латекс выделяет почти
каждый вид. У дюжины видов Euphorbia он изучен, и в боль-
шинстве случаев в нем обнаружены углеводороды, но молеку-
лярная масса их гораздо ниже, чем у каучука.
Эти растения содержат 10±5% восстановленных органиче-
ских веществ, экстрагируемых ацетоном и бензолом. Если исхо-
дить из содержания «нефти» в 10%, то, по грубой оценке (Cal-
vin, 1976), при ежегодном урожае биомассы в 10 т сухого веще-
ства с акра можно получить 1 т углеводородного сырья. За-
траты на выращивание при этом составляют около 150 долл,
на акр в год, т. е. примерно 20 долл, за баррель. Эти расходы
не включают затрат на переработку, как, впрочем, и стоимость
биомассы отходов. Дополнительные расходы, когда такая
«нефть» используется не как топливо, еще предстоит оценить.
По предварительной оценке стоимость продукта не слишком от-
личается от цены природной нефти, что стимулировало дальней-
шие исследования. Однако проведенные недавно полевые опыты
показали, что получение 1,5 т «нефти» с гектара возможно
только на поливных землях, а при выращивании без полива вы-
ход продукции гораздо ниже.
По-видимому, вопрос о том, получаем ли мы выигрыш в
энергии (отношение запасенной энергии к затраченной более
единицы) при выращивании таких культур, нельзя считать ре-
шенным. Ситуация прояснится, если удастся оценить денежную
стоимость и энергосодержание побочных продуктов производ-
ства.
60
Глава 2
Натуральный каучук можно получить из растущей в пусты-
не гваюлы, что и делали в прошлом в больших масштабах.
Во время второй мировой войны министерство сельского хозяй-
ства США провело большую работу по окультуриванию гваю-
лы, и сегодня это растение — наиболее реальный кандидат для
получения масел разного назначения и углеводородов. Резуль-
таты исследования гваюлы найдут широкое применение и осо-
бенно важны для осуществления перспективных планов получе-
ния углеводородов из новых видов растений.
Чтобы установить возможность интродукции масличных и
углеводород-образующих культур, было изучено большое число
видов растений (Buchanan et al., 1978). Полагают, что вполне
реально при «одомашнивании» этих растений увеличить выход
сухой массы на 50%, а содержание углеводородов и масел —
в два-три раза. Такие прогнозы кажутся вполне оправданны-
ми в свете последних достижений селекционеров растений и аг-
рономов, работающих с такими привычными культурами, как
Hevea. Для того чтобы сравняться с Hevea, необходимо, чтобы
выход масла и углеводородов с гектара в год составлял не ме-
нее двух тонн, а другие показатели были бы сходными с пред-
ставленными в табл. 2.8.
Таблица 2.8. Урожайность и состав продукции модельных масличных
и углеводород-образующих культур (как основа для оценки
вновь осваиваемых культур; Buchanan et al., 1978)
Компонент Каучуконосы Масличные (с побочным образовани- ем каучука) Масличные Гуттаперче- носы
I II 1 1 11 I И
Всего сухого веще- ства 13 500 100 17 900 100 22 500 100 11 500 100
Общий белок 1 485 и 1 610 9 1350 6 1 150 10
Каучук 1 350 10 360 2 — — — —
Гуттаперча — — -— — — 1 380 12
Масло 810 6 2 150 12 2 250 10 920 8
Полифенолы 945 7 1 250 7 4 050 18 805 7
Экстрагированный остаток 10 395 77 14 140 79 16 200 72 8 395 7S
Урожай оценивается после уборки и отражает вес наземной части растений. I — уро-
жай в кг иа 1 га, II — сухое вещество, %.
Лишь немногие из большого числа изученных видов расте-
ний были отобраны для более детального изучения как потенци-
альные продуценты каучука (13 видов), масла и каучука
(11 видов), масла (9 видов) и гуттаперчи (3 вида). В табл. 2.8
Энергия и биотехнология
61
приведены теоретически возможная урожайность и состав
сырья, а также другие интересующие нас характеристики но-
вых культур.
Углеводороды и масла можно использовать после выделения
и химической переработки, например гидрокрекинга и/или эте-
рификации. Отметим, однако, что во всем мире производится се-
годня всего около 40-106 т растительных масел в год, а другие
производства развиты еще недостаточно. По этой причине сбра-
живание сахара в этиловый спирт остается сегодня главным
промышленным способом крупномасштабного производства жид-
кого горючего для нужд транспорта из биомассы растительно-
го сырья. Однако, как уже отмечалось, главная проблема при
использовании сахара или крахмала — малый объем производ-
ства и употребление их в пищу. Единственным потенциально
важным сырьем для бродильной промышленности остается по-
этому лигноцеллюлоза. Создание способов переработки этого
сырья в жидкие полуфабрикаты, пригодные для сбражива-
ния,— главная задача биотехнологии.
2.5. Переработка и различные виды топлива
2.5.1. Производство исходного сырья
Что касается этилового спирта как топлива, то почти все су-
ществующие способы его производства основаны на переработ-
ке мелассы, сока сахарного тростника, кукурузного крахмала
или же в меньшей мере маниока. Сбраживание — это лишь одна
из стадий процесса; помимо него сюда относятся выращивание
растений, их уборка, перевозка на заводы, приготовление сусла,
сбраживание, перегонка, обезвоживание, денатурация, изготов-
ление смесей и реализация продукции. Кроме того, приходится
решать вопрос об удалении или переработке жидких отходов
(кубовых остатков). Понятно, что рассказать обо всем этом
подробно здесь невозможно, и поэтому мы обратим внимание
лишь на те стороны проблемы, которые важны в плане техниче-
ской осуществимости, баланса энергии и экономики, но в то же
время касаются и микробиологических аспектов работы этой си-
стемы, которые могут быть улучшены биотехнологическим пу-
тем. Их можно разделить на две группы. Первая связана с при-
родой сырья, в котором должно содержаться нужное количест-
во сбраживаемого сахара, получаемого дешевым способом и при
малых энергозатратах, а вторая — с отгонкой спирта. В этом
последнем случае можно уменьшить затраты (как энергетиче-
ские, так и экономические), если повысить концентрацию спир-
та в продукте ферментации.
62
Глава 2
При получении этилового спирта из сахар-содержащих куль-
тур отжим содержащего сахар сока ведется стандартными спо-
собами. Простые сахара из сахарного тростника можно полу-
чить механическим отжимом сока, а в случае сахарной свек-
лы— диффузионным методом. Крахмалистое сырье нужно ме-
ханически измельчить до консистенции жидкого теста, а затем
нагреть для разрушения крахмальных зерен. Далее можно при-
менить различные варианты гидролиза, основанного на исполь-
зовании разных сочетаний кислот и/или применении ферментов.
Обычно для разжижения исходного продукта применяют термо-
фильную (90 °C) бактериальную амилазу. На следующем этапе
для осахаривания при 50—60 °C (т. е. гидролиза декстринов до
глюкозы) используют глюкоамилазу. В пересчете на образовав-
шуюся глюкозу выход составляет 51% (по массе). Однако, по-
скольку около 5% сахара расходуется растущими клетками на
энергетические нужды, а также на синтез других органических
соединений (глицерола, уксусной кислоты, ацетальдегида и ряда
других соединений, в основном высших спиртов), предельный
выход составляет около 48% в пересчете на исходный сахар.
Весовой выход продукта зависит также от природы сырья:
1 кг инвертированного .сахара = 0,484 кг этилового спирта = 0,61 л,
1 кг сахарозы = 0.510 кг этилового спирта = 0,65 л,
1 кг крахмала = 0,530 кг этилового спирта = 0,68 л.
Эти различия обусловлены тем, что в реакции гидролиза
олигосахаридов и более высокомолекулярных соединений участ-
вует вода.
При малотоннажном производстве степень очистки исходно-
го сырья, получаемого, например, из кукурузы, может быть
очень небольшой. С другой стороны, на более крупных заводах
использование обычного метода мокрого помола, который при-
меняется при производстве высококачественного сиропа из глю-
козы, дает то преимущество, что при этом удается получить ряд
ценных побочных продуктов, например кукурузное масло и
глютен. Доходы от их продажи позволяют снизить реальные
затраты на производство спирта. Даже при маломасштабном
производстве на основе цельного зерна твердые остатки и дрож-
жи имеют цену как корм для скота. В зависимости от исходного
сырья содержание сахара в сбраживаемом продукте варьирует,
но на практике, чтобы получить достаточно высокую концентра-
цию спирта в сусле, содержание обеспечивающих брожение
твердых веществ должно быть в пределах 16—25%. Это важно,
поскольку потребление пара в обычных системах для перегонки
при производстве 96%-ного спирта из 5%-ного сусла вдвое
больше, чем из 10%-ного.
В настоящее время главные сложности, связанные с произ-
водством спирта как горючего., связаны с тем, что сырье для
Энергия и биотехнология
6»
этого процесса является одновременно и сырьем для! производ-
ства пищевых продуктов и кормов. И'з-за этой конкуренции
стоимость сырья весьма высока. В зависимости от типа сырья
60—85% конечной продажной цены получаемого сегодня спир-
та составляет стоимость сырья. Отметим далее, что, за исклю-
чением спиртовых заводов, перерабатывающих сахарный трост-
ник, на которых волокнистая часть стеблей (багасса) использу-
ется как топливо, в широко распространенных процессах полу-
чения спирта тратится больше энергии, чем ее содержится в по-
лучаемом продукте. Эти проблемы можно будет решить, если
в качестве исходного сырья будет использоваться целлюлоза и
удастся разработать новый энергетически более выгодный ме-
тод выработки спирта.
Сложность использования целлюлозы заключается в том, что
в природном состоянии в клеточных стенках растений она на-
ходится в составе нерастворимого комплекса с гемицеллюлоза-
ми и лигнином. Кроме того, за счет образования водородных
связей отдельные молекулы целлюлозы определенным образом
ориентируются относительно друг друга и образуют микрофиб-
риллы, которые в какой-то мере подобны кристаллам, что препят-
ствует действию гидролитических агентов. Даже после исчерпы-
вающего гидролиза веществ растительных клеток дрожжи, кото-
рые обычно используются при получении спирта, неспособны-
усваивать пятиуглеродные сахара, уроновые кислоты и феноль-
ные соединения, образующиеся из сопутствующих целлюлозе'
веществ клеточных стенок растений.
Чтобы решить эту проблему, работа ведется в нескольких
направлениях. При создании одних методов ставится цель по-
лучить глюкозный сироп, и основное внимание уделяется гидро-
лизу целлюлозы. В других случаях предпринимаются попытки
разрушить ферментами возможно большую часть лигноцеллюло-
зы— либо после разделения клеточных компонентов химически-
ми методами, либо с помощью бактерий, способных непосредст-
венно атаковать как целлюлозу, так и гемицеллюлозу. Размол —
наиболее эффективный физический способ предварительной под-
готовки сырья, но он дорог и энергоемок. Высказывались пред-
положения, что размол в замороженном состоянии обойдется
дешевле.
Для делигнификации можно использовать химическую обра-
ботку такими веществами, как едкий натр, надуксусная кислота
или гипохлорит натрия. Под действием- щелочи волокна набу-
хают и разделяются. Гидролиз ведут либо минеральными кис-
лотами, либо биологическими методами. В последнем случае
используют гидролизующие целлюлозу ферменты грибов, напри-
мер Trichoderma, Aspergillus либо Sporotrichum. После отделе-
ния целлюлозы и гемицеллюлозы для сбраживания глюкозы в*
«4
Глава 2
спирт можно использовать дрожжи. Если для сбраживания ис-
пользуют бактерии, например Klebsiella или Aeromonas, то из
пентоз можно получить бутанол. Другая возможность заключа-
ется в том, что лигноцеллюлозу используют как субстрат для
бактерий Clostridium thermocellum.
Типичный процесс производства глюкозы из целлюлозы
включает механическую первичную обработку, размягчение па-
ром или химическую делигнификацию. Для осуществления всех
этих операций нужна энергия. При использовании химических
методов возникают сложности, связанные с коррозией и нейтра-
лизацией стоков. Процессы, основанные на использовании рас-
творителей, нередко дорогостоящи: приходится регенерировать
растворитель. Что касается свойств продукта, определяющих
саму возможность действия на него ферментов, то к ним отно-
сятся степень кристалличности, лигнификации и размер частиц.
Наиболее хорошо изучена смесь целлюлоз, образуемых
Trichoderma viride и Т. reesei. Детально исследована кинетика
их действия. Получены мутантные штаммы, или разновидности
(например, штамм С-30), обладающие высокой активностью.
Выяснено, что смесь целлюлаз Т. reesei включает три компонен-
та: эндо-фермент (фракция Сх), который атакует субстрат бес-
системно, экзо- 1,4-глюканазу (фракция Ci), в результате дей-
ствия которой образуется целлобиоза, и 1,4-глюкозидазу, кото-
рая превращает целлобиозу в глюкозу. Обычно содержание глю-
козидазы в смеси невелико и его приходится увеличивать, до-
бавляя ферменты других грибов, например Aspergillus. От них
несколько отличается фермент возбудителей бурой гнили: эти
грибы способны образовывать Н2О2, в результате чего процес-
сы первичного разрушения идут быстрее.
При использовании таких ферментных смесей возникают
сложности, связанные с тем, что и активность ферментов, и их
образование ингибируются конечным продуктом катализируе-
мой реакции. Так, 0,01%-ный раствор целлобиозы ингибирует
активность целлюлазы на 75%. Есть надежда, что путем моди-
фикации активности фермента, его регуляции и синтеза с по-
мощью мутагенеза, селекции и методов рекомбинантных ДНК
удастся получать более активные и стабильные ферментные пре-
параты, в меньшей степени ингибируемые конечным продуктом.
Было проведено изучение многих других организмов в плане
возможного их использования для разрушения лигноцеллюлозы.
Некоторые из них перечислены в табл. 2.9. Лигнин деградиру-
ют некоторые штаммы Erwinia, а также дрожжи Trichospora
fermentans; ряд грибов, относящихся к числу хорошо известных
родов Mucor, Penicillium, Trichoderma и Aspergillus, образуют
активную ксиланазу.
Энергия и биотехнология
65
Таблица 2.9. Микроорганизмы, изученные в плане их использования
для расщепления лигноцеллюлозы
Целлюлоза и целлюлазы
Trichoderma viride
Т. reesei
Т. koningii
Coniphora cerebella
Sporotrichum pulverulentum
Palyporus adustus
Myrothesium verrucaria
Penicilliutn funiculosum
Fusarium solani
Aspergillus wentii
Coniphora thermophila
Thielavia terrestris
Phanaerochaete chrysosporium
Clostridium thermohydrosulfuricum
Clostridium thermocellum
Clostridium thermosaccharolyticum
Thermomonospora spp.
Ксиланаза
Chaetomum trilaterale
Глюкозидаза
Aspergillus phoenicus
Лигназа
Erwinia spp.
Trichospora fermentans
2.5.2. Ферментация
Получение метана (биогаза) методом анаэробной переработки
сырья и производство этилового технического спирта путем
сбраживания при участии дрожжей — два главных примера био-
логических технологий, которые могут использоваться для полу-
чения более высококачественного топлива из растений.
Если принять, что исходным сырьем является глюкоза, об-
щую схему осуществляемых превращений можно представить
следующим образом:
Производство биогаза: СбН^Об —► 3(СН4)+3(СО2),
Производство этанола: CsHi2O6 —>- 2(СН3СН2ОН) +2(СО2).
Мы не будем рассматривать здесь все тонкости биохимиче-
ских процессов, лежащих в основе этих превращений. Отметим
только, что в ходе анаэробных превращений часть небольшого
количества энергии, высвобождающейся в реакциях, запасается
в АТР. Это соединение используется как источник энергии для
роста клеток и поддержания жизнедеятельности бактерий или
дрожжей. Поэтому, хотя метаболизм у этих микроорганизмов и
не столь эффективен (в плане использования субстрата для ро-
ста), как у организмов, растущих в присутствии кислорода и
5—1344
66
Глава 2
при дыхании стехиометрически разлагающих глюкозу до угле-
кислого газа и воды, в результате брожения удается получить
достаточно высокий выход продуктов. К тому же удельное со-
держание энергии в них выше, чем в исходном субстрате — био-
массе. В зависимости от фазового состояния (жидкость, твердое
вещество или газ), температуры и давления реагирующих ве-
ществ и продукта теплоты сгорания (калориметрические) со-
ставляют приблизительно 16 кДж/г для глюкозы, 30 кДж/г для
этилового спирта и 56 кДж/г для метана.
Хотя в основе как анаэробных процессов, так и производст-
ва этилового спирта путем брожения лежат микробиологические'
процессы переработки сырья для получения высококачественно-
го топлива из биомассы, только в этом и заключается, навер-
ное, их сходство.
2.5.3. Этиловый спирт
Производство этилового спирта при помощи дрожжей основано
на давно устоявшейся технологии (см. также гл. 3). Для полу-
чения топливного спирта необходимо осуществить ряд процес-
сов (рис. 2.5): подготовить сырье, провести брожение, отгонку
и очистку, обезвоживание (если предполагается использовать
продукт в смеси с нефтью), денатурацию и организовать хране-
ние. Нужно также переработать кубовые остатки (захоронить
или использовать на корм животным и т. п.). На каждой стадии
есть свои узкие места, о которых надо всегда помнить, посколь-
ку это прямо определяет саму возможность использования про-
цесса сбраживания для получения жидкого топлива, его энерго-
баланс и экономичность.
Объем производства крупных спиртовых заводов может быть
очень большим: они ежегодно потребляют тысячи тонн сырья и
выпускают миллионы литров продукции. Наибольший вклад в;
энергобаланс страны производство топливного спирта дает в;
Бразилии. В 1982 г. там было получено 5-Ю9 л спирта, и произ-
водство его по программе Proalcool расширяется. Ожидается,,
что в 1985 г. будет получено более 11*109 л.
Программа эта осуществляется главным образом на заводах,
где проводят прямое сбраживание сока сахарного тростника;
часть спирта производится из мелассы. Вторая крупная нацио-
нальная программа — это производство «бензирта» (gasohol}
в США: оно основано на сбраживании кукурузного крахмала.
В 1984 г. ежегодное производство этого продукта составляло-
5-108 галлонов.
Технический спирт применяют главным образом как горючее
для двигателей внутреннего сгорания. Чаще всего, его испольг
зуют в смесях, но при наличии подходящих машин и в чистом.
Энергия и биотехнология
67
Этанол Рис. 2.5. Схема процесса производ-
5*
68
Глава 2'
виде. Разница между этими двумя способами заключается в том,
что во избежание разделения фаз для смешивания с нефтью
нужно использовать обезвоженный спирт. С другой стороны, спе-
циально сконструированный двигатель может работать и на
95 %-ной смеси спирта с водой, получаемой при первичной от-
гонке. Хотя этиловый спирт можно использовать для приготов-
ления пищи, обогрева, освещения или производства пара и
электричества, особой выгоды получить здесь не удается. Дело
в том, что в процессе превращения биомассы в этанол происхо-
дит значительная потеря энергии.
Энергия потребляется на всех стадиях переработки спирта.
Больше всего тратится ее на концентрирование и обезвоживание
при перегонке. Энергию эту можно получать из отходов сырья
(багассы, соломы и т. д.), сжигая древесину или ископаемое
топливо, газ, нефть или уголь. В целом энергозатраты на пере-
работку сырья близки к количеству энергии, получаемой в фор-
ме спирта. По этой причине энергообеспечение всего процесса
должно идти либо за счет переработки отходов, либо за счет
использования самого дешевого топлива.
Основную массу вырабатываемого на крупных предприятиях
спирта получают сегодня при помощи дрожжей [Saccharomy-
ces, обычно S. cerevisiae, но иногда и S. uvarum (carlsbergensis)
и S. diastaticus]. Первая задача здесь — подобрать дрожжи, под-
ходящие для переработки определенного субстрата. Дрожжи
S. cerevisiae могут расти на глюкозе, фруктозе, мальтозе и маль-
тотриозе, т. е. на сахарах, содержащихся в их обычной «пище»,
которую они получают от сахар- или крахмал-содержащих рас-
тений. S. diastaticus может также использовать, декстрины, а ви-
ды Kluyveromyces fragilis и К. lactus — лактозу.
Образование этилового спирта дрожжами — это анаэробный
процесс, но для их размножения нужен кислород. В следовых
количествах кислород, возможно, нужен и для поддержания
жизнедеятельности клеток, образующих спирт. В ходе метабо-
лизма осуществляется сложная регуляция образования этанола
из глюкозы. Сам процесс метаболизма, жизнеспособность кле-
ток, их рост, деление и образование спирта зависят от концен-
трации субстрата, кислорода и конечного продукта (спирта).
Большую роль в увеличении выхода сыграл отбор штаммов
дрожжей, более устойчивых к повышенным концентрациям как
субстрата, так и спирта.
По возможности содержание твердого сбраживаемого ком-
понента в сырье должно быть 16—25% (вес/объем), тогда ко-,
нечная концентрация этилового спирта составит 6—12%. Для
большинства видов дрожжей оптимальная для их жизнедеятель-
ности температура лежит в пределах от 25 до 33 °C. Осуществ-
ление процесса при pH 4—5 уменьшает опасность микробного
Энергия и биотехнология
69
загрязнения и препятствует развитию так называемых «дрож-
жей-убийц». Требования к дополнительным источникам питания
определяются природой сырья, используемого при подготовке
субстрата. Так, если сырьем служит меласса, то приходится до-
бавлять вещества — источники азота и фосфора, а если в осно-
ве лежит высоко очищенный гидролизат крахмала, то может
понадобиться добавить незаменимые микроэлементы и вита-
мины.
Существуют три основных способа сбраживания сахар-со-
держащего сырья: периодический, периодический с повторным1
использованием клеток и непрерывный. При периодическом про-
цессе субстрат сбраживается после внесения в него свежевыра-
щенной закваски, полученной в аэробных условиях. Брожение
протекает в анаэробных условиях, и весь оставшийся субстрат
превращается при этом в спирт. После завершения брожения
дрожжи удаляют, и для следующего цикла получения спирта
выращивают новую порцию закваски. Размножение дрожжей
является дорогостоящей процедурой, так как расходуется много
субстрата.
При использовании дрожжей по периодической схеме около!
5% сахара расходуется на рост клеток и энергообеспечение син-
теза других соединений: глицерола, уксусной кислоты, ацеталь-
дегида и сивушных масел (в основном высших спиртов). По этой
причине максимальный выход составляет около 48% от суб-
страта по массе. При использовании дрожжей продуктивность
варьирует в пределах 1—2 г этилового спирта в 1 ч на 1 г кле-
ток (сухое вещество). Более значительные выходы (2,5—3,8 г)
получены в опытах с бактериями, например Zymomonas mobilis,
однако такие организмы еще только предстоит внедрить в про-
мышленность. Продуктивность ферментера зависит от режима
работы, штамма дрожжей, плотности клеток и природы суб-
страта. Она варьирует от 1 г-л^'-ч-1 до более 10 г-л-1-ч-1. По-
скольку концентрация спирта при брожении растет, а субстра-
та— уменьшается, за разумные промежутки времени полностью
использовать сахар иногда не удается.
Недостатки процесса (длительное сбраживание и неполное
использование субстрата) можно частично устранить, применяя
периодическую схему с повторным использованием клеток. При
этом в конце цикла дрожжевые клетки отделяют от сброженной
пульпы и сохраняют для использования в следующем цикле.
Следует иметь в виду, однако, что положительным в периоди-
ческом процессе является его устойчивость к нарушениям режи-
ма, поэтому установку может обслуживать персонал не слиш-
ком высокой квалификации. В то же время для него характер-
на малая объемная продуктивность: за 36 часов брожения об-
разуется 5%-ный (вес/объем) раствор спирта, что соответствует
70
Глава 2
образованию в среднем 1,4 г спирта на 1 л за 1 ч. Путем при-
менения схемы с повторным использованием клеток или же
непрерывного брожения выход можно повысить до более чем
10 г-л-1-ч-1. При этом клетки отделяют от барды и использу-
ются заново; тем самым поддерживается высокая концентрация
клеток в среде.
Для работы по этим схемам нужны квалифицированные ра-
ботники, более крупные капиталовложения и сложные контро-
лирующие устройства. При непрерывном процессе степень ис-
пользования субстрата для образования спирта зависит от раз-
ведения. Для достижения высокой продуктивности она должна
быть велика, но при таких условиях уменьшается среднее время
задержки субстрата. Иными словами, задачи оптимизации двух
факторов — объемной продуктивности и полноты использования
сахара — противоречат друг другу. Выбор регламента сбражи-
вания должен строиться исходя из стоимости субстрата, необхо-
димых капиталовложений и производственных расходов. Они
в свою очередь должны определяться в связи с особенностями
всей схемы получения продукции.
По завершении сбраживания концентрация спирта составля-
ет 6—12%. Она зависит от штамма дрожжей и начальной кон-
центрации сахара. Важно достичь наивысшей концентрации
•спирта, так как от этого зависит расход пара на перегонку: он
увеличивается от 2,25 кг на 1 л 96%-ного спирта при отгонке
10%-ного раствора до более чем 4 кг при отгонке 5%-ного. До-
полнительный пар нужен для получения безводного спирта из
азеотропной смеси вода — этиловый спирт, кипящей при посто-
янной температуре. Обычно для этого используют десятикрат-
ный по отношению к количеству удаляемой воды объем бензола.
Сначала при 64—84 °C отгоняется азеотропная смесь бензола,
воды, спирта, а после удаления всей воды, при 68,25 °C, — дру-
гая азеотропная смесь — бензола и спирта. После отгонки всего
бензола остается лишь абсолютный спирт, который собирают,
а бензол используют повторно.
Главными побочными продуктами производства являются
СО2, дрожжи, сивушные масла и кубовые остатки. Каждый из
них обладает определенной ценностью, но переработка жидких
остатков может быть затруднена. В большинстве случаев, по-
скольку за пищевой спирт нужно платить большой налог, алко-
голь денатурируют. Для этого добавляют вещества, придающие
ему горький вкус, или смешивают его с бензином.
2.5.4. Энергобаланс
Общий баланс энергии как при производстве спирта, так и при
анаэробной переработке может быть слабо положительным или
даже отрицательным, поскольку при производстве сырья, его
Энергия и биотехнология . 71
переработке, сортировке, очистке или отжиме потребляется зна-
чительное количество энергии. Эти энергозатраты покрываются
за счет использования остатков растительного сырья (багассы
или соломы), сжигания древесины или ископаемого топлива.
Используются также «тепловые отбросы» электростанций.
Если рассматривать лишь производство этилового спирта, то*
количество энергии, расходуемой на выработку одного литра!
продукта, равно или превышает ее количество, запасенное в дан-
ном объеме спирта. Поэтому, чтобы получить выигрыш, энерго-
обеспечение процесса должно осуществляться за счет возобнов-
ляемых источников либо энергопоставляющих побочных продук-
тов. На сегодняшний день производство на основе сахарного
тростника — это единственный пример процесса с положитель-
ным энергобалансом. Необходимую энергию получают, сжигая
отжатый тростник (багассу). С другой стороны, производства на
основе крахмала или сахарной свеклы требуют дополнительных
энергозатрат. Если для этого используется нефть, то выигрыш в
энергии в пересчете на жидкое топливо бывает либо небольшим,
либо нулевым. С другой стороны, когда топливом служит уголь,
то мы по сути имеем дело с одним из процессов превращения
его в жидкое горючее для транспорта. Это позволяет экономить
энергию, запасенную в нефти. Отметим также, что в производ-
ствах на основе сахара или крахмала стоимость сырья состав-
ляет 60—85% от стоимости получаемой продукции; здесь воз-
можны и снижение цен, и повышение урожайности. В результа-
те с учетом качества энергии, стоимости и экономичности этот
процесс привлекает все большее внимание.
Научные исследования нацелены сегодня на увеличение эф-
фективности использования субстрата, выхода продукта в объ-
емном исчислении, его концентрации и на разработку способов
осуществления процесса при повышенной температуре. Эти усо-
вершенствования помогут экономить дорогой субстрат путем
снижения его затрат на выращивание клеток (повторное их ис-
пользование или иммобилизация) или подавления побочных
реакций, дающих нежелательные продукты (глицерол, ацетат
и т. п.). Кроме того, они помогут экономить энергию: уменьшит-
ся необходимость охлаждения ферментеров, снизятся затраты
на перегонку. Все эти изменения будут плодом совместной рабо-
ты микробиологов и инженеров.
Если рассматривать микробиологическую сторону процесса,
то основной целью здесь является улучшение ответа микроорга-
низмов на изменение концентрации Ог, подбор концентрации
субстрата и специфичности его использования, повышение кон-
центрации спирта. Нужно оптимизировать температуру, соотно-
шение стадий брожения и роста клеток, образование побочных
продуктов и флокуляционные характеристики дрожжей. Послед-
72
Глава 2
нее особенно важно, поскольку самоосаждающиеся клетки легко
отделить простым отстаиванием, не используя дорогие и энерго-
емкие центрифуги. Основной способ улучшения производства
по-прежнему состоит в селекции устойчивых штаммов дрожжей.
Неустойчивость частично обусловлена осмотическими эффекта-
ми, а частично — влиянием накопленного спирта на проницае-
мость мембран. Один из возможных выходов заключается в рас-
ширении набора используемых организмов (путем включения,
например, Z. mobilis, видов Clostridium и т. п.). Другой путь —
использование генетических методов для передачи полезных ка-
честв от различных дрожжей и бактерий дрожжами S. cerevi-
siae. Вторая возможность кажется особенно привлекательной,
так как при работе с этими дрожжами промышленностью на-
коплен большой опыт. Кроме того, хорошо изучен их жизненный
цикл и генетика, получены плазмидные векторы. Следует ска-
зать, что многие используемые промышленностью штаммы
дрожжей являются полиплоидными, не принадлежат к опреде-
ленному типу скрещивания, а жизнеспособность образуемых
ими спор низка. Все это затрудняет их генетический анализ и
улучшение при помощи обычных методов скрещивания, что и
заставляет обратиться к рекомбинантным методам. Сейчас ве-
дутся эксперименты по переносу генов амилазы и целлюлазы
в клетки дрожжей.
Что касается самого брожения, то уже выполнена большая
работа для систем с иммобилизованными клетками, что позво-
лит сделать процесс непрерывным, без повторного использова-
ния клеток. Разрабатываются также системы с повторным ис-
пользованием дрожжей и отгонкой спирта при пониженном дав-
лении (вакуумное брожение). Сложности в разработке обеих
систем связаны с интенсивным выделением СОг, который нуж-
но удалять, и с необходимостью контролировать рост клеток.
Нужна очищенная, не содержащая взвешенных частиц стериль-
ная питательная среда. Далее, немалые затруднения возникают
из-за высокой стоимости среды, поскольку для достижения
большой продуктивности нужны высокие степени разведения,
а при высоком разведении плохо используется субстрат. По
этим причинам, возможно, придется остановиться на серийном
выращивании. В случае микробного заражения потребуется си-
стема дублирования. Учитывая все это, сегодня подобные систе-
мы вряд ли будут экономически конкурентоспособными.
Все эти проблемы придется решать биотехнологам, исполь-
зуя опыт и микробиологов, и инженеров.
Энергия и биотехнология
73
2.5.5. Получение метана в анаэробных условиях
При переработке сырья в анаэробных условиях получается
смесь газов — метана и углекислоты, которые образуются в ре-
зультате разложения сложных субстратов при участии смешан-
ной популяции микроорганизмов разных видов. Поскольку ис-
комый продукт — это газ, сбор его не составляет труда: он про-
сто выделяется в виде пузырьков. Впрочем, иногда при более
сложных способах его использования или распределения по тру-
бам возникает необходимость в очистке от примесей или в ком-
прессии.
В анаэробном реакторе можно перерабатывать самое разно-
образное сырье: отходы сельского хозяйства (испорченные рас-
тительные или пищевые продукты), стоки перерабатывающих
предприятий, содержащие сахар, жидкие отходы, образующие-
ся на сахарных заводах или при отжиме пальмового масла, бы-
товые отходы, сточные воды городов и спиртовых заводов. Мож-
но перерабатывать и специально выращиваемые культуры,
включая и экзотические, растущие в пресной или морской воде
или на бросовых землях: водяной гиацинт, гигантские бурые во-
доросли или слоновью траву.
Весьма важно, что сырье с высоким содержанием целлюлозы
(или лигноцеллюлозы) не так просто использовать для иных
целей: оно дешево или вообще не имеет коммерческой ценности.
Обычно масштабы переработки невелики (в пределах одной
фермы или деревни), хотя были разработаны и проекты более
крупных установок для переработки стоков или же промышлен-
ных отходов. В масштабах государства наибольший вклад в
энергетический бюджет страны вносит такая переработка в Ки-
тае, где построено около семи миллионов небольших (на одну
семью) реакторов.
Неочищенный биогаз обычно используют для приготовления
пищи и освещения. Его можно применять как топливо в стацио-
нарных установках, вырабатывающих электроэнергию. Сжатый
газ в баллонах пригоден как горючее для машин и тракторов.
Его можно подавать в газораспределительную сеть. В послед-
нем случае требуется некоторая очистка биогаза: осушка, удале-
ние углекислоты и сероводорода. Очищенный биогаз ничем не
отличается от метана из других источников, т. е. природного
газа или же SNG (синтетический газ, получаемый из угля или
водородсодержащего сырья). Хотя в большинстве случаев кон-
струкция реакторов рассчитана на получение метана, который
используется как топливо, строить такие установки имеет смысл
не только для этой цели. Нередко, особенно в развитых стра-
нах умеренного пояса, реакторы используют главным образом
для переработки отходов.
74
Глава 2
Установки для производства биогаза по принципу все воз-
растающего объема можно сгруппировать следующим образом:
1) реакторы в сельской местности в развивающихся странах
(обычно имеют объем 1—20 м3); 2) реакторы на фермах раз-
витых стран (объем 50—500 м3) ; 3) реакторы, перерабатываю-
щие отходы промышленности (например, сахарных, спиртовых
заводов и т. п., объем 500—10 000 м3); 4) свалки бытовых и про-
мышленных отходов (объем 1—20-Ю6 м3).
Понятно, что детали технического устройства таких систем
могут сильно различаться. Так, существует несколько конструк-
ций небольших реакторов — от простейшей бродильной ямы в
грунте с фиксированным объемом газа до подземных или полу-
подземных баков с металлическим или резиновым накопителем
газа с изменяющимся объемом. Эти установки могут работать
в режиме полного перемешивания, полного вытеснения, в режи-
ме контактных процессов, как анаэробные фильтры или реакто-
ры с псевдоожиженным слоем. Конструкция таких устройств оп-
ределяется типом перерабатываемого сырья. Задача заключает-
ся в том, чтобы не допустить потери микроорганизмов при ра-
боте системы. Это достигается либо путем повторного их ис-
пользования, либо помещением в реактор поддерживающего
субстрата, на котором и растут клетки. Последний способ осо-
бенно хорош в случае, когда в реактор поступает раствор с низ-
ким содержанием взвешенных частиц.
В развитых странах используется множество разнообразных
установок. Это определяется конкретной задачей — очисткой
стоков или же достижением нужного качества газа. Среди них
есть и небольшие реакторы, мало чем отличающиеся от описан-
ных, и крупные установки с приспособлениями для очистки
газа, электрогенераторами, компрессорами и очистителями воды.
Иногда их строят в составе одного комплекса с другими круп-
ными объектами: канализационными станциями, сахарными и
спиртовыми заводами, животноводческими фермами и молоко-
заводами. В некоторых случаях бывает необходимо провести
предварительную обработку или разведение питательного рас-
твора. Это делается для оптимизации размера частиц во взве-
сях, увеличения глубины переработки или уменьшения токсиче-
ского действия высоких концентраций азота.
В тех случаях, когда главной задачей является переработка
отбросов, приходится прежде всего механическим способом от-
делять крупноразмерную фракцию, например солому и прочее.
При применении такого способа, а также при переработке сточ-
ных вод предприятий пищевой промышленности отношение со-
держания растворенных углеводов к нерастворимым, содержа-
щим лигноцеллюлозу веществам существенно увеличивается,
и обычно доля твердых частиц бывает небольшой. Вследствие
Энергия и биотехнология
75
этого скорость подачи раствора в реактор и время удержания
уменьшаются. Впрочем, с инженерной точки зрения установка
может быть и сложнее, чтобы обеспечить оптимальное взаимо-
действие микробов с субстратом.
Чтобы получить наибольший объемный выход продукции с
небольшой установки, скорость подачи субстрата должна быть
возможно большей, а это в свою очередь связано с поддержа-
нием высокой концентрации жизнеспособных бактерий. При
этом могут возникнуть сложности как в случае субстратов с
высоким содержанием нерастворимых веществ, так и субстра-
тов, содержащих много растворимой органики. В первом случае
в реакторе накапливаются неразрушаемые или медленно раз-
рушаемые вещества, которые в конечном счете будут составлять
более 80% твердых веществ в осадке реактора. Во втором —
переработка растворенных, полностью разрушаемых веществ
приведет к образованию высокоактивного ила, и бактерии бу-
дут составлять до 90% осадка. Удержать такой ил в реакторе
будет сложно. Впрочем, недавно был предложен ряд конструк-
ций, в которых эти проблемы решены. Помимо механического
измельчения сырья с успехом применяется неполный кислотный
или щелочной гидролиз образующих его частиц либо целлюло-
литические ферменты. Еще одно усовершенствование заключа-
ется в механическом перемешивании подаваемого сырья с илом.
Для этой же цели через реактор повторно пропускают выделив-
шийся газ. Перемешивание осадка, содержащего активные бак-
терии, или создание тока жидкости может увеличить количество
выделяющегося газа в зависимости от того, какой вид сырья
используется в процессе.
При решении вопроса об использовании более сложных с ин-
женерной точки зрения, усовершенствованных сооружений или
же каких-то вариантов процесса в первую очередь надо оценить
стоимость их работы. Сегодня из их числа более практичными
могут оказаться системы, перерабатывающие отходы (когда за-
траты на их обезвреживание высоки), а не те крупномасштаб-
ные установки, которые вырабатывают газ как сырье для энер-
гетики. Отметим, однако, что наиболее крупные установки для
получения биогаза всегда очень просто устроены: это могут
быть свалки отходов, в основном бытового мусора. О самой
возможности использования метана, образующегося в таких му-
сорных кучах, задумались, когда стали искать способы предот-
вращения взрывов и пожаров, возникающих в результате выде-
ления в них газа. Кислород, оказавшийся в мусоре при образо-
вании куч, быстро используется аэробными бактериями и гри-
бами, в результате чего условия в них становятся анаэробными.
Влажность поддерживается либо просачивающейся дождевой
водой, либо грунтовыми водами. Если буферная способность ма-
76
Глава 2
териала достаточна для поддержания нейтральных значений
pH, то складываются благоприятные условия для образования
метана. Газ выделяется в смеси с СО2. Собирают его при помо-
щи труб, проложенных в толще мусора под полотнищами
пленки.
Вклад этих технологий в энергообеспечение конкретных ре-
гионов в обозримом будущем (скажем, десять лет) в значитель-
ной мере будет определяться местными условиями. Они будут
зависеть, с одной стороны, от наличия земельных площадей и
сельскохозяйственного сырья, а с другой — от доступности неф-
ти и газа и цен на них. Важную роль сыграют как новые разра-
ботки в области микробиологической технологии, так и инже-
нерные решения, нацеленные на повышение эффективности ра-
боты таких систем при меньших капитальных вложениях и энер-
гопотреблении.
Микробиологические основы процесса
Переработка сырья в метан происходит в ходе сложных взаи-
модействий в смешанных популяциях микроорганизмов. По осо-
бенностям обмена веществ их можно подразделить на три ос-
новные группы: первая осуществляет первичный распад поли-
мерных веществ, вторая образует летучие жирные кислоты,
в частности уксусную, водород и СО2, а третья — метан (метан-
образующие бактерии).
В осуществлении первой стадии процесса принимают участие
разнообразные анаэробные бактерии, превращающие в раство-
римые вещества множество соединений, включая целлюлозу,
жиры и белки. Ключевую роль при этом играют процессы раз-
ложения целлюлозы, так как большинство видов сырья или сточ-
ных вод обогащены лигноцеллюлозой. По оптимальной темпера-
туре жизнедеятельности эти бактерии можно отнести к одной
из трех групп: термофильным организмам, живущим при 50—
60 °C, мезофильным (30—40°) и психрофильным, предпочитаю-
щим комнатную температуру (около 20°C). Большая часть ис-
следований была выполнена для реакторов, работающих на ос-
нове мезофилов. При повышенной температуре скорость распа-
да исходного сырья, особенно целлюлозы, увеличивается, а это —
, важное преимущество. Скорость образования метана лимитиру-
ется интенсивностью процессов разложения сырья. Именно по-
этому время удержания при работе с некоторыми субстратами
бывает так велико.
Время удержания можно уменьшить, если повысить темпе-
ратуру, но это требует энергозатрат. Для получения тепла мож-
но сжигать часть получаемого метана (а в крайних случаях —
и весь газ). Можно использовать и тепловые отбросы сопутст-
Энергия и биотехнология
77
вующих производств (например, воду, использованную в них
для охлаждения). Горячую воду можно получать и с помощью
солнечных батарей. Бактерии, работающие на первом этапе,
лучше всего растут при pH от 6 до 7. В культуре рост многих
разлагающих целлюлозу бактерий подавляется по механизму
обратной связи при накоплении конечных продуктов гидролиза,
однако в смешанной популяции бактерий, существующей в ана-
эробном реакторе, происходит быстрое усвоение этих продуктов
и подавление не так выражено. В результате скорость разруше-
ния полимеров оказывается выше, чем можно было бы ожидать.
Конечные продукты, обладающие свойствами ингибитора, уда-
ляются с помощью бактерий второй группы, которые превраща-
ют различные сахара, аминокислоты и жирные кислоты в лету-
чие жирные кислоты, СО2 и водород.
В ходе этого процесса образуется ряд летучих жирных кис-
лот (молочная, уксусная, пропионовая и др.), но главным суб-
стратом при синтезе метана является уксусная кислота. Метан-
образующие бактерии могут также синтезировать метан из СО2
и Н2. Оптимум pH для них тот же (6—7), что и для бактерий
первой группы, и это важно, поскольку нарушение баланса об-
разования и потребления кислот приведет к падению pH, если
система не обладает достаточными буферными свойствами. Вся-
кое падение pH по этой причине преимущественно сказывается
на активности метанобразующих бактерий, что вызывает даль-
нейшее закисление среды и прекращение образования метана.
С этим можно бороться, добавляя известняк или аммиачную
воду, но при внесении ионов аммония следует соблюдать осто-
рожность. Метанобразующие бактерии могут использовать ам-
монийные ионы как источник азота, но при высоких концентра-
циях они ингибируют их рост. К числу других веществ и соеди-
нений, способных ингибировать процесс, относятся кислород и
окисленные соединения, такие, как нитрат и нитрит, сульфиды,
цианиды, свободные ионы металлов (меди, цинка или никеля),
галогены, формальдегид и сероводород. Система чувствительна
также к резким скачкам температуры.
Выход продукции
При образовании метана, когда субстратом является глюкоза,
весовой выход газа составляет только около 27%, а выход энер-
гии (теоретически) — более 90%. Однако на практике из-за
сложного состава сырья, перерабатываемого в анаэробных ре-
акторах, и низкой эффективности его переработки валовый вы-
ход энергии составляет от 20 до 50%. Определение выхода био-
газа— задача более сложная, чем выхода метана. Прежде все-
го нужно уточнить, что мы хотим измерить: валовый выход.
78
Глава 2
(биогаз-СОг в смеси с метаном) или же один метан. Состав газа-
существенно изменяется в зависимости от условий в реакторе,,
а также от природы подаваемого в него сырья. Теоретически
при переработке углеводов на СОг и метан эти газы должны
образовываться в равных количествах. На самом деле не весь.
СОг выделяется в виде газа, так как он растворяется в воде и
может взаимодействовать с гидроксил-ионами с образованием1
бикарбонатов. Концентрация образующегося бикарбоната будет
зависеть от скорости протока жидкости, pH, температуры и со-
держания в жидкой фазе ионов металлов и других веществ.
Количество образующегося бикарбоната сильно зависит от
содержания белка в сырье: чем оно больше, тем богаче биогаз:
метаном. Обычно биогаз содержит 60—70% метана. Он образу-
ется со скоростью 0,5 м3 на килограмм сухой массы летучих
компонентов; время удержания составляет около 15 суток. Это»
соответствует приблизительно 0,3 м3 метана на килограмм су-
хой массы твердых компонентов. Впрочем, точные оценки дать
сложно, так как выход зависит от условий работы системы,,
сырья и конструкции реактора.
В последних сообщениях об установках, перерабатывающих
биомассу разного качества, приводятся выходы от 0,17 до»
0,4 м3 метана на килограмм сухой массы сырья. Скорость за-
грузки при этом составляла от 1 до более чем 10 кг сырья на»
кубометр реактора в сутки, время удержания 10—40 суток,,
а глубина переработки субстрата от 20 до более чем 70%.
Повышение выхода за счет увеличения содержания энергии
в продукте будет способствовать повышению валового произ-
водства энергии реактором. К сожалению, это не говорит о чис-
том выходе энергии, который будет существенно ниже, так как
при таком ведении процесса энергозатраты возрастут. Количе-
ство энергии, которую необходимо подать в конкретную систему,
будет определяться характером идущего в ней процесса. Он в
свою очередь определяется микробной флорой и природой ис-
пользуемого сырья.
Задачи научных исследований
Сегодня нам еще немного известно о биохимии микробов, участ-
вующих в процессе. Внимания требует и инженерная сторона
дела: нужно усовершенствовать способы закачки сырья, измере-
ния объемов газа, теплообеспечения, использования двигателей
внутреннего сгорания для производства электроэнергии и систе-
мы контроля. Основой существенного улучшения экономических
показателей могло бы стать уменьшение времени удержания,
нужного для эффективной переработки нерастворимых компо-
нентов. Для этого следует в первую очередь улучшить способы
подготовки сырья.
Энергия и биотехнология
79
2.6. Бесклеточные системы
Одна из привлекательных возможностей, предоставляемых тех-
нологией «солнечной энергетики», заключается в использовании
целых организмов как биологических катализаторов при произ-
водстве аммиака и водорода за счет солнечной энергии. Опыты
<с цианобактериями (сине-зелеными водорослями) и зелеными
водорослями показали, что они способны образовывать водород
и кислород путем прямого фотолиза воды. Лежащий в основе
этого явления процесс фотосинтеза сформировался в результате
генно-инженерной деятельности Природы. Фотосинтезирующие
бактерии неспособны разлагать воду, но могут на свету образо-
вывать большие количества водорода (без примесей кислорода)
или аммиака. Для этого им нужны только простые органические
и неорганические субстраты. Такие вещества содержатся в про-
мышленных отходах, и поэтому превращение солнечной энергии
фотосинтезирующими бактериями вполне может быть сопряже-
яо с переработкой отходов.
В ряде лабораторий ведутся на молекулярном уровне иссле-
дования различных процессов образования водорода, а также
механизмов реакции расщепления воды. В образовании водо-
рода принимают участие гидрогеназа и нитрогеназа. Сегодня
активно изучаются свойства этих ферментов из разных организ-
мов, в частности механизмы регуляции их синтеза и активности,
а также стабильность в присутствии кислорода. Предметом
важных исследований является также образование восстанови-
тельных эквивалентов и поток электронов к этим ферментам,
которые при определенных условиях служат факторами, лими-
тирующими активность. Эти опыты позволят понять суть ука-
занных процессов и попытаться оптимизировать выделение во-
дорода имеющимися в нашем распоряжении генетически оха-
рактеризованными организмами. Ряд исследователей-генетиков
занят отбором мутантов с повышенной способностью к образо-
ванию водорода или аммиака. Примерами удачного применения
методов генетической инженерии для создания ферментов с же-
лаемыми свойствами может быть получение устойчивой к кис-
лороду гидрогеназы. Удалось повысить содержание гидрогеназ
в клетках и получить микроорганизмы, способные выделять
фиксированный ими азот в окружающую среду в форме амми-
лка.
В ходе первых исследований была отчасти выяснена физио-
логическая и биохимическая основа процессов образования Нг
и NH3 у фотосинтезирующих организмов, определены условия,
необходимые для максимального проявления этой способности.
В будущем на основе результатов опытов с целыми клетками
мы сможем подобрать условия для максимального и долговре-
80
Глава 2
менного образования Н2 и NH3) оценить техническую возмож-
ность создания лабораторных реакторов с иммобилизованны-
ми клетками фотосинтезирующих организмов.
2.6.1. Комбинированные системы, образующие водород
Около десяти лет назад в области исследований возобновляемых
источников энергии было сделано замечательное открытие. Было
показано, что если взять мембраны, содержащие хлорофилл,
и добавить к окружающему раствору ферменты, действующие
как катализаторы, то на свету будет происходить разложение
воды (фотолиз) на водород и кислород (рис. 2.6). Это открытие
Рис. 2.6. Схема сопряжения процессов поглощения солнечной энергии и обра-
зования водорода на основе стабилизированных хлоропластных мембран и гид-
рогеназных ферментов (Hall, 1982).
было первым шагом на пути к созданию фотореактора, при по-
мощи которого энергия Солнца может запасаться в ценном топ-
ливе— водороде. Фотохимики и фотобиологи проявляют боль-
шой интерес к этим исследованиям, поскольку получение «сол-
нечного водорода» из воды открывает новые перспективы в энер-
гетике. Процессу фотолиза присущ ряд особенностей, которых
нет у каких-либо других систем преобразования энергии. Суб-
стратом в нем является обычная вода, источником энергии с не-
ограниченным запасом — солнечный свет, а продуктом — водо-
род. Его просто хранить, он не загрязняет окружающую среду.
Процесс этот циклический, поскольку при потреблении водорода
регенерируется субстрат-—вода. Система привлекательна и тем,
что она работает при обычной температуре; в ней не образуются
токсичные промежуточные соединения.
Задача создания стабильной биологической системы, способ-
ной работать многие годы, а не часы, чрезвычайно сложна и,
быть может, нерешаема. Если бы мы смогли понять суть био-
логических процессов, нам, возможно, удалось бы их имитиро-
Энергия и биотехнология 81
вать путем создания полной системы синтеза. Такие системы
могли бы использовать свет любой интенсивности в широком
диапазоне температур и этим напоминали бы производящие
электроэнергию солнечные батареи. Надо сказать, что фотобио-
логический способ производства водорода не вышел еще из стен
лабораторий. Целесообразность его дальнейшего развития толь-
ко предстоит доказать в ходе многоплановых исследований.
Впрочем, наши представления о возможностях использования
этой системы в практике могут быстро измениться: достаточно
будет и одного крупного открытия при изучении фотобиологиче-
ских и фотохимических реакций разложения воды.
Несмотря на успешные опыты по стабилизации мембран хло-
ропластов (для этого применялась иммобилизация ферментов и
закрепление их в пленках альгинатного геля и полиуретановых
матрицах), они вряд ли войдут как составная часть в промыш-
ленные системы по улавливанию солнечной энергии. Тем не ме-
нее результаты изучения состава хлоропластов и обмена ве-
ществ в них могут послужить основой для создания таких си-
стем. Чего же мы можем ожидать от исследований в этой об-
ласти? В реакциях фотолиза воды в мембранах хлоропластов„
идущих при участии фотосистемы II, принимает участие комп-
лекс белков с хлорофиллом и магнием. Сегодня нам мало что.
известно и о расположении в нем ионов магния, и о механизмах
фотолиза воды в растениях.
Для практических целей может оказаться необходимым от-
делить активируемую светом стадию фотосинтеза, на которой
образуется кислород, от стадии темновых реакций, где выделя-
ется водород. Одностадийная система будет продуцировать смесь,
кислорода и водорода, и их улавливание и разделение со всей
площади коллектора солнечной энергии могут оказаться невы-
полнимыми. Но можно представить себе и двухстадийный про-
цесс, на первом этапе которого будет функционировать фотока-
талитическая система, в которой образуются неокисляемый пе-
реносчик и кислород. Кислород можно улавливать, а образовав-
шийся переносчик водорода будет использоваться на второй ста-
дии, когда осуществляется темновая реакция образования водо-
рода. Вслед за этим переносчик направляется обратно в первый'
отсек установки, восстанавливается там в ходе световой реак-
ции и может использоваться для повторного цикла реакций.
Переломный момент в проводимых сегодня исследованиях
наступит, когда удастся разработать эффективную стабильную
систему, имитирующую фотосистему II мембран хлоропластов,
т. е. процесс, в ходе которого вода разлагается на кислород
и водород с образованием протонов и восстановительных экви-
валентов. Со вторым этапом, связанным с выделением водорода
в системе, ситуация, видимо, обстоит проще, поскольку в нашем;
6—1344
«2
Глава 2
распоряжении есть большой выбор необходимых переносчиков
электронов и катализаторов образования водорода. Отметим,
однако, что чувствительность многих этих веществ к кислороду
ограничивает возможность их применения. Очень важно отыс-
жать соединение, которое могло бы выполнять функции гидроге-
назы, т. е. служить кислородоустойчивым редокс- и протонным
переносчиком.
Остановимся теперь на водородном фотореакторе. Если бы
нам удалось обойтись без реакции расщепления воды и просто
использовать восстановленные соединения, например аскорбино-
вую кислоту, сульфид натрия, ЭДТА или дитионит в качестве
доноров электронов, то можно было бы без труда получить ста-
бильную систему, интенсивно образующую водород. В роли
-фотокатализатора в ней мог бы выступить какой-нибудь пиг-
мент (например, флавин или даже стабильная фотосистема I
мембран хлоропластов), а в роли переносчиков протонов иэлек-
тронов — красители, гидрогеназа или платина. Можно исполь-
зовать для этого стабильные ферменты и иммобилизованные
системы. Уже созданы небольшие фотореакторы, в которых при
надлежащих условиях образование водорода идет с высокой
скоростью, до нескольких литров Нг в минуту.
Не так-то просто будет получить искусственные мембраны
.для замены нестабильных биологических прототипов, но иссле-
дования в этом направлении уже ведутся. Делаются попытки
применить для этой цели различные решетки с набухающим
слоем, силикаты глин (монтмориллониты и гекториты). Послед-
ние, как известно, способны образовывать стабильные интерка-
ляционные соединения с разнообразными ионными комплекса-
ми и полярными молекулами, например с водой.
.2.6.2. Электроэнергия
Одним из интересных аспектов общей проблемы улавливания
солнечной энергии является использование компонентов биоло-
гических мембран для генерации электропотенциалов. Таким
путем можно попытаться создать солнечную батарею. Так,
в липидные пузырьки или в стопку липидных мембран, содер-
жащих некоторые красители, можно встроить фотосистему пур-
пурных бактерий и один из белков мембран Halobacterium. Всю
эту систему можно поместить на подложку — нитроцеллюлозный
фильтр и установить его между двумя отсеками сосуда. При ос-
.вещении фильтра возникает разность потенциалов. Такую си-
стему можно использовать и для перемещения ионов (например,
Na+) и тем самым для обессоливания. Подобные фотоэлектри-
ческие эффекты уже наблюдались, но сопротивление при этом
было слишком велико для получения достаточно большой силы
тока.
Энергия и биотехнология 83
Недавно было сконструировано устройство, в котором мем-
браны хлоропластов использовались в составе фотогальваниче-
ского элемента для генерирования тока. При освещении объеди-
ненных полуэлементов зарегистрирован потенциал 220 мВ и ток:
800 мкА при плотности тока 16 мкА/см2 (использованы платино-
вые электроды). В одном полуэлементе мембранами хлоропла-
стов осуществляется фотолиз воды, в то время как в другом
идет только частичная реакция (фотосистема I) с образованием
низкого потенциала. Полуэлементы соединены искусственной:
мембраной, проницаемой для ионов, но не для переносчика*
электронов (красителя). Возможно, эту очень интересную фото-
гальваническую батарею удастся модифицировать так, что у
двух ее полюсов будут выделяться Ог и Нг, а тока она давать
не будет.
2.7. Биотопливные элементы
и другие биоэлектрохимические устройства
После того как в конце XIX в. были созданы топливные эле-
менты, появилась возможность эффективно осуществлять пре-
вращение химической энергии в электрическую. Дело в том, что»
на эти элементы не распространяются ограничения, налагаемые?
циклом Карно. Дальнейшее их усовершенствование шло тем не
менее медленно: оказалось, что обеспечить эффективный элект-
рокаталитический перенос электронов от используемого топли-
ва на анод элемента сложно. В результате удалось создать
лишь водородный элемент, дающий достаточную плотность тока.
Он успешно работает при низких температурах и пригоден для
крупномасштабного производства энергии. Схема, объясняющая
принципы работы обычного топливного элемента, приведена на
рис. 2.7. Был предложен ряд элементов, использующих другие
виды топлива (спирты, углеводороды), но они работают лишь
при высоких температурах и дают ток небольшой плотности при
малом коэффициенте полезного действия. Это ограничивает их
применение для производства энергии, но некоторые типы топ-
ливных элементов используются для других целей. Так, один из.
них применяется в качестве датчика в детекторах, выявляющих
наличие спирта в выдыхаемом воздухе.
Важной отличительной чертой биологических систем явля-
ется их способность к контролируемому окислению различных
«топлив» при низкой температуре. Неудивительно, что биоэлек-
трохимики долгое время пытались приспособить участвующие
в этих реакциях катализаторы для работы в топливных элемен-
тах. Ведь в процессах дыхания в живых организмах и реакци-
ях, идущих в биоэлектрохимических элементах, есть много об-
щего.
6*
«4
Глава 2
Анод Мембрана Катод
Рис. 2.7. Схема строения простейшего
кислород-водородного топливного эле-
мента.
нове других организмов и топлив.
Первые опыты с биоло-
гическими топливными эле-
ментами и батареями про-
вел в 1910 г. английский бо-
таник Поттер. Погружая
платиновый электрод в ана-
эробную культуру дрожжей
или Escherichia coli, он об-
наружил, что на нем обра-
зуется потенциал, отрица-
тельный по отношению к
потенциалу такого же элект-
рода, находящегося в аэроб-
ной стерильной среде. На-
пряжение в цепи было при
этом 0,3—0,5 В, а сила сла-
бого тока составляла
0,2 мА. За последующие
50 лет было описано мно-
жество биотопливных эле-
ментов, работающих на ос-
В конце 50-х и начале 60-х
годов интерес к таким устройствам у исследователей, работав-
ших по космическим программам, сильно возрос.
В целом разработанные модификации можно разделить на
три основные группы, описание которых дается далее.
Биотопливный элемент на неэлектроактивных веществах
В таких элементах организмы или ферменты используются
для превращения неэлектроактивных топлив в электроактивные
их производные, которые затем окисляются в обычных элемен-
тах. Вот пример такого процесса:
Сбраживающий
Глюкоза------------
микроорганизм
Н2----> 2Н+ 4- 2е_ (на аноде),
2Н+ + — О2 -J- 2е~--> Н2О (на катоде).
Деполяризирующий биотопливный элемент
В таких элементах микроорганизмы или ферменты применяются
как катализаторы в электрохимической реакции, протекающей
на одном или обоих электродах. Так, можно собрать водород-
ную ячейку с ферментом гидрогеназой, катализирующим реак-
Энергия и биотехнология
85
цию на аноде, и другим ферментом, лакказой, работающим, на
катоде:
Н2____________► 2Н+ + 2е~ (на аноде).
Гидрогенназа
2Н+ 4- —- О2 + 2е"------->• Н2О (на катоде).
2 Лакказа
Регенерируемый биотопливный элемент
В этом устройстве биологический компонент предназначен
для регенерации электрохимически активных веществ, которые
и взаимодействуют с одним или обоими электродами. На рис. 2.8
Рис. 2.8. Биотопливный элемент на основе микробной метанолдегидрогеназы.
МДГ — метанолдегидрогеназа, ФЭС — феназинэтосульфат, ФМС — феназинме-
тосульфат.
приведена схема одного из таких устройств. Это — недавно
созданный биотопливный элемент на основе фермента; в качест-
ве топлива в нем используется метанол.
Хотя некоторые биотопливные элементы стали экономически
выгодными и в них были достигнуты достаточно высокая плот-
ность тока на электроде (до 40 мА/см2) и мощность (около
1 кВт), они еще не доведены до того уровня, чтобы их можно
было широко внедрять в практику.
Отметим, однако, что в самое последнее время технология
биотопливных элементов сильно шагнула вперед. Это касается
и элементов с ферментами, где в качестве топлива используют-
ся спирт и окись углерода, и элементов с клетками микроорга-
низмов. Оба этих типа элементов принадлежат к числу регене-
рируемых. В последних используются редокс-медиаторы, спо-
собные обратимо проникать сквозь стенку и мембрану микроб-
ных клеток. Эти медиаторы реагируют с комцрнентами элект-
рон-транспортной цепи, вызывая нечто вроде короткого замыка-
ния в биохимической электрохимической системе и подсоединяя
86
Глава 2
ее одновременно к аноду топливного элемента (рис. 2.9). В та-
ких устройствах в качестве топлива применяются сахара или.
вещества стоков, но теоретически для этой цели может быть ис-
пользовано любое вещество, пригодное для роста микробов. Та-
кие элементы могут работать довольно длительное время (от не-
скольких недель до месяцев), и наиболее совершенные опытные-
их образцы по эффективности вполне сопоставимы с лучшими
моделями элементов на основе ферментов. Для превращения:
световой энергии в электрическую были созданы и похожие на
них фотобиоэлектрохимические элементы, в которых электроны,,
обладающие высокой энергией, поступают на анод от электрон-
транспортной цепи фотосинтезирующих микроорганизмов.
Рис. 2.9. Принципиальная схема регулируемого топливного элемента, содержа-
щего целые клетки микроорганизма.
Судя по современному состоянию разработок биоэлектрохи-
мических элементов, вполне реально, что их можно будет ис-
пользовать в тех устройствах, где нужна относительно неболь-
шая мощность, например для медицинских или военных целей,,
а также для производства электроэнергии в отдаленных райо-
нах. В ближайшем будущем биотопливные элементы будут ис-
пользоваться как чувствительные датчики, поскольку сила тока
в них пропорциональна количеству переработанного топлива..
В гл. 4 мы обсудим перспективы использования сходных био-
электрохимических элементов в биохимическом синтезе.
ЛИТЕРАТУРА
Общие вопросы
Barnard G. W. (1983). Liquid Fuel Production from Biomass in the developing-
countries: An Agricultural and Economic Perspective. In: Bioconversion Sys-
tems (ed. Wise D. L.), pp. 112—268, CRC Press, Boca Raton, Florida.
Barnard G. W., Hall D. 0. (1982). Energy from renewable resources: ethanol!
fermentation and anaerobic digestion. In: Biotechnology, Vol. Ill (Chapter 7)i
(ed. Dellweg H.), Verlag Chemie, Weinheim,
Энергия и биотехнология 87
Barnett A., Pyle D. L., Subramanian S. К. (eds.) (1978). Biogas Technology in
the Third World: a Multidisciplinary Review, Inti. Devi. Res. Council, 60 Queen
St., Ottawa KIP 5Y7.
BioEnergy Council (1981, 1984). International BioEnergy Directory, BioEnergy
Council, P. O. Box 12807, Arlington, Virginia 22209.
Boland D. L., Turnbull J. W. (1981). Selection of Australian trees other than
Eucaliptus for trials as fuelwood species in developing countries, Aust. For.,
44, 235—246.
Buchanan R. A. et al. (1978). Hydrocarbon and rubber-producing crops, Econ.
Bot., 32, 131—153.
Connell M. G. R. (1982). World Forest Biomass and Primary Production Data,
Academic Press, London.
CBNS (1982). Economic Evaluation and Conceptual Design of Optimal Agricul-
tural Systems for Production of Food and Energy, Center Biol. Natural Sys-
tems report to US-Dept. Energy, Washington DC 20545.
• CIQA (1978) Guayule, Centro Investi. Quimica Appl., Aldama Ote 351, Saltillo,
Mexico.
• Coombs J. (1980). Renewable sources of energy (carbohydrates), Outl. Agric.,
10 235_____245.
• Cote IF. A. (ed.) (1983). Biomass Utilization, Plenum Press, New York.
Bari D. E. (1975). Forest Energy and Economic Development, Clarendon Press,
Oxford.
•Coldman J. C. (1979). Outdoor mass cultures, Water Resour., Wash., 13, 1—19.
•Good N. E. (1981). Fuel from biomass. In: Beyond the Energy Crisis, Vol. II
(ed. Fazzolari R. A. and Smith С. B.), pp. 491—498, Pergamon Press,
Oxford. '
Ball D. O. (1980). Renewalbe resorces (hydrocarbons), Outl. agric., 10, 246—
255; (1981). New Scientist, 89, 524—526.
.Hall D. O. (1981). Solar energy through biology: fuel for the future. In: Advan-
ces in Food Producing Systems for Arid and Semi Acid Lands, (ed. Mannasah
J. T. and Briskey E. J.), Academic Press, New York.
Hall D. O. (1982). Biomass for energy: fuels now and in the future, J. R. Soc.
Arts, 130, 457—471; (1982). Experientia, 38, 3—10.
Hall D. O. (1984). Photosynthesis for energy. In: Advances in Photosynthetic
Research, Vol. II (ed. Sybesma C.), pp. 727—740, Martinus Nijhoff, The
Hague.
Hall D. O., Barnard G. W., Moss P. A. (1982). Biomass for Energy in the
Developing Countries, Pergamon Press, Oxford.
Heden K. (1982). Swedish energy forestry, Biomass, 2, 1—3.
Higgins I. I., Hill H. A. O. (1979). Microbial generation and interconversion of
energy sources. In: Microbial Technology (eds. Bull A. T., Ellwood D. C. and
Ratledge C.), Soc. gen. Microbiol. Symp., 29, 359—377.
Hobson P. N., Bousfield S., Summers R. (1981). Methane Production for Agricul-
tural and Domestic Wastes, Appl. Sci. Publ., London.
Hughes D. E. (ed.) (1982). Anaerobic Digestion (1981). Elsevier, Amsterdam.
ICE (1978). Alcohol Fuels, Inst. Chem. Eng., NSW Branch, Sydney, Australia.
Johnson J., Hinman H. E. (1980). Oil and rubber from arid land plants, Science,
208, 460—464.
Kingsolver В. E. (1982). Euphorbia lathyrus reconsidered: its potential as an
energy crop for arid lands, Biomass, 2, 287—295.
Klass D. L., Emert G. H. (1981). Fuels from Biomass and Wastes, Ann. Arbor
Sci. Publ. Michigan.
tLeach G. (1976). Energy and Food Production. IPC Press, Guildford, UK.
Lewis C. (1983). Biological Fuels, Edward Arnold, London.
Lipinsky E. S. (1981). Chemicals from biomass: petrochemical substitution
options, Science, 212, 1465—1471. •
88
Глава 2
Lipinsky Е. S. (1981). Systems Study of Animal Fats and Vegetable Oils for Use
as Substitute and Emergency Diesel Fuels, Report to US Dept, Energy by
Battelle, Columbus, Ohio 43201.
McLaughlin J., Hoffman J. J. (1982). Survey of biocrude-producing plants from
the southwest, Econ. Bot., 36, 323—339.
Moss P. A., Hall D. 0. (1982). Biomass for energy in Europe, Int. J. Solar
Energy, 1, 239—262.
Nair P. K. (1980). Agroforestry Species, Inti Council Res. Agroforestry, PO
Box 30677, Nairobi.
NAS (1977). Methane Generation from Human, Animal and Agricultural Was-
tes, Natl Acad. Sci. USA, Washington DC 20418.
NAS (1981). Firewood Crops: Shrub and Tree Species for Energy Production,
Natl Acad. Sci. USA, Washington, DC 20418.
NAS (1982). Priorities in Biotechnology R and D for International Develop-
ment, Natl Acad. Sci. USA, Washington DC 20418.
NEPA (1981). Du petrole sur nos terres, Motorisation et Technique Agricole
Suppl. March 1981, NEPA (S. A.) 10 rue Martel, Paris 75493.
OALS (1980). A Technology Assessment of Guayule Rubber Commercialisation,
Office Arid Lands Studies, Univ. Ariz., Tuscon, Arizona.
OTA (1980). Energy from biological Processes, Office of Technology Assessment,
US Congress, Washington DC 20510.
Paiz W., Chartier P., Hall D. O. (eds.) (1981). Energy from Biomass, Appl. Sci.
Publ., London.
Pimentel D., Pimentel M. (1979). Food, Energy and Society, Edward Arnold,
London.
Rabson R., Rogers P. (1981). The role of fundamental biological research in deve-
loping future biomass technologies, Biomass, 1, 17—38.
Richmond A., Preiss K. (1979). The biotechnology of aquaculture, Interdisc. Sci.
Rev., 5, 60—68.
Royal Society London (1982). Industrial and Diagnostic Enzymes, Phil. Trans.
Roy. Soc. Lond.
SAE (1981). Fuels from Crops, Soc. Automative Engin., Natl. Sci. Centre, Mel-
bourne, Australia.
Scurlock J. M. O., Hall D. O. (1984). Energy from Biomass in Europe. In: 5th
Canadian Bioenergy R and D Seminar (ed. Hosnain S.), pp. 56—65, Elsevier,
London.
Shelef G., Soeder C. J. (eds.) (1980). Algae Biomass, Production and Use, Else-
vier, Amsterdam.
Silversides R. (1982). Energy from forest biomass — its effect on forest mana-
gement practices in Canada, Biomass, 2, 29—42.
Slesser M., Lewis C. (1979). Biological Energy Resources, Spon, London.
Sonalysts Inc. (1981). Assessment of Plant Derived Hydrocarbons, Report to
US-Dept. Energy, Washington DC 20545.
Stafford D. A., Wheatley B. L, Hughes D. E. (eds.) (1980). Anaerobic digestion,
Appl. Sci. Publ., London.
Steinbeck K. (1981). Short-rotation forestry as a biomass sourse: an overview.
In: Energy from Biomass (eds. Paiz W., Chartier P. and HallD. O.), pp. 163—
171, Appl. Sci. Publ., London.
Stewart G. A. et al. (1979). The Potential for Liquid Fuels from agriculture and
Forestry in Australia, CSIRO, Div. Chemical Technology, Canberra.
Stewart G. A. et al. (1982). The Potential for Production of Hydrocarbon Fuels
from Crops in Australia, CSIRO Div. Chem. Technol., Canberra, Australia.
Strub A., Chartier P., Schleser G. (eds.) (1983). Energy from Biomass, Vol. 2,
Appl. Sci. Publ., London.
Teri (1982). Biogas Handbook, Tata Energy Res. Inst., Bombay 4000023, India.
_________Энергия и биотехнология ____________________________89
Trindade J. С. (1981). Energy crops—the case of Brazil. In: Energy from Bio-
mass (eds. Paiz W., Chartier P., Hall D. O.), pp. 59—74, Appl. Sci. Publ.,
London.
TRW (1980). Energy Balances in the Production and End-use of Alcohols Deri-
ved from Biomass, TRW Crop. US Dept. Energy, Washington DC 20545.
Zaborsky 0. R. (ed.) (1981). Handbook of Biosolar Resources, Vol. II, Resource
Materials, CRC rPess, Boca Raton, Florida.
Фотосинтез
Beadle C. L„ Long S. P., Hall D. 0., Imbamba S. K„ Olembo R. J. (1985). Photo-
synthesis in Relation to Bioproductivity, UN Environment Programme, PO
Box 30552, Nairobi (publ. Tycooly Inti. Dublin).
Coombs J. (1982). Improving biomass productivity. In: Energy from Biomass,
Vol. II (eds. Strub A., Chartier P., Schleser G.), pp. 105—110, D. Reidel Publi-
shing Co., Dordrecht, Holland.
Coombs J., Hall D. O., Chartier P. (1983). Plants as Solar Collectors, D. Reidel
Publishing Co., Dordrecht, Holland.
Cooper J. P. (ed.) (1975). Photosynthesis and Productivity in Different Environ-
ments, Cambridge University Press.
Ebeling W. et al. (1982). Improving agricultural productivity, Econ. Impact, 39,
6—47.
Edwards G„ Walker D. A. (1983). C-3, C-4: Mechanisms of Cellular and Environ-
mental Regulation of Photosynthesis, Blackwell Scientific Publications, Ox-
ford.
Govindjee (ed.) (1982). Photosynthesis, Vol. 1 and 2, Academic Press, New
York.
Hall D. 0., DaSilva E. J. (1983). Photosynthesis: a bisolar tool for development,
Nature Resour., 19 (2), 2—10.
Hall D. O., Rao К. K. (1981). Photosynthesis, 3rd edn., Edward Arnold, London.
Hall D. 0., Paiz W. (eds.) (1982). Photochemical, photoelectrochemical and pho-
tobiological processes, D. Reidel Publishing Co., Dordrecht, Holland.
Halliwell B. (1981). Chloroplast Metabolism, Clarendon Press, Oxford.
Oswald W. J. (1981). Algae as solar energy converters. In: Energy from Biomass
(eds. Paiz W., Chartier P., Hall D. O), pp. 633—646, Appl. Sci. Publ., London.
Pirson A., Zimmerman M. H. (eds.) (1977). Encyclopaedia of Plant Physiology:
Photosynthesis, Vol. 5, Springer Verlag, Berlin.
UK-ISES (1976). Solar Energy —a UK Assessment, UK Section Inti Solar
Energy Soc., 19, Albemarle St., London Wl.
Бесклеточные системы
Bolton J. R., Hall D. 0. (1979). Photochemical conversion and storage of solar
energy, Ann. Rev. Energy, 4, 353—406.
Braun A. M. (ed.) (1983). Photochemical Conversions, UNESCO, Presses Poly-
technic Romandes, Lausanne.
Calvin M. (1982). Plants can be direct fuel source, Biologist, 29, 145—148;
(1980). Energy, 4, 851—870.
Connolly J. S. (ed.) (1981). Photochemical Conversion and Storage of Solar
Energy, Academic Press, New York.
Этанол
Coombs J. (1981). Ethanol — the process and the technology for production of
liquid transport fuel. In: Energy from Biomass (eds. Paiz W., Chartier P.,
Hall D. O.), pp. 279—291, Appl. Sci. Publ., London.
90
Глава 2
Da Silva J. G. (1978). Energy balance for ethyl alcohol production from crops.
Science, 201, 903—906.
Ferchak J. D., Pye E. D. (1981). Utilization of biomass in the U. S. for the pro-
duction of ethanol fuel as a gasoline replacement, I and II, Solar Energy, 26,
9—25.
Khan A. S., Fox R. W. (1982). Net energy analysis of ethanol production in NE
Brazil, Biomass, 2, 213—232.
Kovarik B. (1982). Fuel Alcohol, IIED/Earthscan, 10, Percy Street, London WL.
Lamed R„ Zeikus J. G. (1980). Ethanol production by thermophilic bacteria, J.
Bacteriol., 144, 569—578.
Lyons T. P. (1981). Gasohol, a step to energy independence, Alltech Publ.,.
Lexington, Kentucky 40503.
SERI (1981). Alcohol Fuels Program Technical Review, Solar Energy Res. Inst.,
Golden, Colorado 80401.
Stone J. E., Marshall H. B. (1980). Analysis of Ethanol Production from Cellu-
losic Feedstocks, Dept Energy, Mines and Resources, Ottawa.
Биотопливные элементы
Aston W. J., Turner A. P. F. (1984). Biosensors and biofuel cell's, BiotechnoL
genet. Eng. Revs, 1, 89—120.
Bennetto H. P. (1984). Microbial biofuel cells, Life Chem. Reps, (in press).
Higgins I. J., Hammond R. C., Plotkin E. V., Hill H. A. O., L/osaki K., Eddo-
wes M. J„ Cass A. E. G.(1980). Electroenzymology and biofuel cells, In: Hyd-
rocarbons in Biotechnology (eds. Harrison D. E. F., Higgins I. J., Watkin-
son R. J.), pp. 181—193, Fleyden, London.
Turner A. P. F., Aston \V. J., Higgins T. J., Davis G., Hitt H. A. O. (1982).
Applied aspects of bioelectrochemistry: Fuel cells, sensors and bioorganic
synthesis, Biotechnol. Bioeng. Symp., 12, 401—412.
Глава 3
Пищевые продукты, напитки
и биотехнология
Г. Бич, М. Мелвин, Дж. Таггарт
3.1. Введение
3.1.1. Микроорганизмы и пищевые продукты
Оставим на время в стороне проблемы, связанные с сельским
хозяйством, о котором речь пойдет в других разделах, и обра-
тимся к производству пищевых продуктов и напитков. Это про-
изводство основано на переработке сырья, в основном постав-
.ляемого сельским хозяйством, или же на использовании опре-
деленных веществ. Все органические вещества, применяемые в
пищевой промышленности, в принципе могут использоваться
микроорганизмами. Уже одно это говорит о ключевой роли
микробиологии при производстве продуктов питания: здесь мик-
робы могут играть и положительную, и отрицательную роль.
Последняя, видимо, более выражена: не случайно меры предо-
сторожности против нежелательной деятельности микробов за-
нимают такое важное место при производстве пищи и ее потреб-
лении. Размножение микробов может вызвать нежелательные
изменения качества пищевых продуктов или их внешнего вида.
При этом нередко образуются вещества, обладающие токсиче-
ским действием. Понятно, что явная порча пищи и связанные с
этим экономические убытки весьма нежелательны, однако наи-
более опасным следствием размножения микробов в пищевых
продуктах является образование токсинов. Некоторые микроор-
ганизмы при подходящих условиях образуют токсины, вызываю-
щие серьезные заболевания или даже смерть.
В биотехнологии микроорганизмы играют положительную
роль, поэтому возможности их применения для сохранения пищи
уже, чем при ее производстве. Ниже мы приведем несколько
примеров того, как используется биотехнология в первом вари-
анте, однако в этой главе мы в основном рассмотрим те про-
цессы, в которых микробы активно участвуют в производстве
пищи. Мы обсудим также применение современных методов и
знаний для интенсификации этих производств.
3.1.2. Две разновидности биотехнологии
Если рассмотреть, чем занимается сегодня биотехнология, то
нетрудно убедиться, что существуют две ее разновидности, раз-
личающиеся по ценности получаемых продуктов и по масштабу
91
92
Глава 3
их производства. Различия эти ясны из табл. 3,1. Биотехнология
маломасштабного производства, дающего дорогую продукцию
специального назначения, сильно отличается от биотехнологии
пищевой промышленности. Определяется это экономикой. Пищу
получают из относительно недорогого сырья и относительно де-
шевыми способами. Стоимость конечного продукта не оправды-
вает дорогостоящих исследований по модернизации производ-
ства, как это имеет место, например, при получении антибиоти-
ков, да это и не требуется. Даже производство микробных фер-
ментов и их использование в пищевой промышленности, связан-
ное с применением сложной технологии, правильно было бы счи-
тать биотехнологией крупномасштабного производства дешевых
продуктов. Каждый, кто знаком с технической биохимией, зна-
Таблица 3.1. Два типа биотехнологий
Маломасштабные Крупномасшта бные
Объем установки Стоимость продукции Тип продукции Основные направления биотехнологических ис- следований и разра- боток Стоимость научных ис- следований и развития 100—1000 л Высокая Для медицины, фарма- цевтической промыш- ленности, высокоспе- циализированная Генетические манипуля- ции Повышенная 10 000 л Невысокая Предметы потребления, малоспециализирован- ная Технология фермента- ции, инженерия про- цессов Пониженная
ет, что получить ферменты высокой чистоты и специфичности —
задача трудоемкая и сложная. Такие препараты дороги, и их не
применяют при выработке пищевых продуктов. Здесь использу-
ют полуочищенные ферменты. Очистку проводят до такой степе-
ни, чтобы их можно было применять в относительно небольшом
объеме. К исключениям относится тот случай, когда фермент
иммобилизуют на твердом носителе, сквозь который пропуска-
ют субстрат. В таких проточных реакторах фермент успешно ис-
пользуется многократно, и это оправдывает применение высоко-
очищенных препаратов. Не следует, впрочем, думать, что пер-
спективы биотехнологии в пищевой промышленности ограниче-
ны. Правильнее говорить о разумных пределах затрат на произ-
водство пищи. Здесь нужен большой выход продукта и простая
Пищевые продукты, напитки и биотехнология
93
технология. По этим причинам главными в биотехнологии пи-
щевой промышленности являются методы крупномасштабного
производства продуктов.
3.1.3. Границы применения биотехнологии в пищевой
промышленности
Спектр продуктов питания, получаемых при помощи микроорга-
низмов, обширен: от вырабатываемых с древних времен за счет
брожения хлеба, сыра, йогурта, вина и пива до новейшего вида
пищевого продукта — грибного белка микопротеина. Микроорга-
низмы при этом играют разную роль: используются продуцируе-
мые ими ферменты или другие метаболиты, с их помощью сбра-
живается пищевое сырье, а некоторые из них выращиваются
для непосредственного потребления. В пищевой промышленно-
сти для осуществления процессов применяют как чистые куль-
туры микроорганизмов, так и дикие формы, содержащиеся в
значительном количестве в сырье, которые начинают размно-
жаться при создании надлежащих условий. Последний способ
особенно характерен для традиционных бродильных произ-
водств, зародившихся во времена, когда о микробах еще ничего
не знали. В мире промышленного производства- такие процессы
обычно ведутся под гораздо более строгим контролем:. Особен-
но это относится к выбору штамма и чистоте культур используе--
мых микроорганизмов.
До недавнего времени биотехнология использовалась в пи-
щевой промышленности с целью усовершенствования освоенных
процессов и более умелого использования микроорганизмов, но-
будущее здесь принадлежит генетическим исследованиям по
созданию более продуктивных штаммов для конкретных нужд*
внедрению новых методов в технологии брожения. Таким путем
мы сможем повысить выход и качество выпускаемой продукции
и освоить производство новых ее разновидностей.
3.2. Молочные продукты
3.2.1. Общие сведения
В пищевой промышленности ферментацию применяют главным
образом для получения молочных продуктов. В сквашивании мо-
лока обычно принимают участие стрептококки н молочнокислые
бактерии; лактоза при этом превращается в молочную кислоту.
Путем использования иных реакций, которые сопутствуют глав-
ному процессу или идут при последующей обработке, получают
и другие продукты переработки молока. Среди них — пахта,
сметана, йогурт и сыр. Свойства конечного продукта зависят при
54
Глава 3
этом от характера и интенсивности реакций ферментации. Те
реакции, которые сопутствуют основному процессу образования
молочной кислоты, и определяют обычно особые свойства про-
дуктов. Так, именно вторичные реакции ферментации, идущие
при созревании сыров, определяют вкус отдельных их сортов.
В некоторых таких реакциях принимают участие пептиды, ами-
нокислоты и жирные кислоты, присутствующие в продуктах.
В молоке при ферментации могут протекать шесть основных
реакций, и в результате образуются молочная, пропионовая или
.лимонная кислота, спирт, масляная кислота или же происходит
колиформное газообразование. Главная из этих реакций — об-
разование молочной кислоты. На ней основаны все способы
-ферментации (сквашивания) молока. Лактоза молока гидроли-
зуется при этом с образованием галактозы и глюкозы. Обычно
-галактоза превращается в глюкозу еще до сквашивания. Имею-
щиеся в молоке бактерии преобразуют глюкозу в молочную кис-
лоту. Образование сгустка казеина происходит в изоэлектриче-
ской точке этого белка (pH 4,6) под действием молочной кисло-
ты. Этот процесс лежит в основе сыроварения (разд. 3.2.2).
При производстве швейцарского сыра ключевую роль играет
маслянокислое брожение с образованием углекислого газа.
Именно оно обусловливает своеобразный вкус (букет) этих сы-
ров и образование глазков. Характерный вкус пахты, сметаны и
сливочного сыра формируется в результате лимоннокислого бро-
жения. Он складывается из составляющих вкусов диацетила,
пропионовой и уксусной кислот и других близких к ним соеди-
нений.
Молочные продукты, полученные на основе спиртового бро-
жения, мало известны в Европе и Америке. Такой тип броже-
ния нашел применение при переработке молока в России, но при
производстве других продуктов он считается нежелательным.
-•Обычно рост вызывающих его дрожжей (Toruld) стараются по-
давить. Нежелательны также маслянокислое брожение и коли-
формное газообразование.
Различные процессы ферментации молока проводят сегодня
.в контролируемых условиях. В течение многих прошедших ты-
сячелетий они осуществлялись при участии бактерий, исходно
присутствующих в молоке. В наше время для этого используют
разнообразные закваски, позволяющие получать молочные про-
дукты нужного качества и типа. Применяющиеся при этом куль-
туры живых бактерий могут представлять либо один какой-то
.штамм определенного вида (культуры моноштаммов), либо не-
сколько штамов и/или видов (многоштаммовые или смешанные
культуры). Коммерческие культуры-закваски состоят из бакте-
рий, образующих молочную кислоту и пахучие вещества.
В табл. 3.2 перечислены некоторые виды бактерий, используе-
Пищевые продукты, напитки и биотехнология
95.
Таблица 3.2. Функциональная роль некоторых бактерий,
используемых при переработке молока
Культура Функция Использование
Propionibacterium Формирование вкуса, об- Производство швейцар-
Р. shermanii Р. petersonil разование глазков Образование кислоты ского сыра
Lactobacillus Созревание, закваска
L. easel для швейцарского сы-
L. helveticus L. bulgaricus L. lactis ра, производство сы- ров типа швейцарского.
Leuconostoc Образование вкусовых Производство сметаны.
L. dextranicum веществ из лимонной сливочного масла, за-
L. citrovorum кислоты (главным об- разом диацетила) квасок
Streptococcus Образование кислоты Производство йогурта и
S. thermophilus швейцарского сыра,.
S. lactis S. cremoris закваски для сыров
мне при производстве молочных продуктов методом фермен-
тации, и указана их роль в этих процессах, а также продукты,,
получаемые с их помощью. Выбор и состав используемых ком-
бинаций из этих штаммов и видов бактерий определяются же-
лаемыми свойствами и условиями получения продуктов, напри-
мер скоростью образования молочной кислоты.
Последующие разделы посвящены разнообразным молочным-
продуктам, вырабатываемым путем ферментации, рассмотрению*
способов их производства и возможных путей их усовершенст-
вования.
3.2.2. Сыр
Сыроварение —один из древнейших процессов, основанных на
ферментации. При производстве сыра сохраняется питательная,
ценность молока. Сыр упоминается в книгах писателей Древней
Греции и Рима. Всего лишь несколько его сортов было получе-
но «по плану», большинство же — результат счастливой слу-
чайности. Известны самые разнообразные сыры — от очень мяг-
ких до твердых. Различие между ними определяется тем, что все-
натуральные мягкие сыры содержат много воды, 50—60%,.
а твердые — всего лишь 13—34%. Хотя свойства сыров чрезвы-
чайно разнообразны, в процессе' выработки всех их есть много*
общего. Первый этап — это подготовка культуры молочнокис-
лых бактерий и засев ею молока. Затем молоко створаживают^
для чего обычно применяют фермент реннин. После отделения!
96
Глава 3
водянистой жидкости (сыворотки) полученную творожистую
массу подвергают термообработке и прессуют в формах. Далее
сгусток солят и ставят на созревание.
Инокуляция
В прошлом сыроделы полагались на бактерии, имеющиеся в
натуральном молоке. Присутствие в нем как нужных, так и не-
желательных микробов приводило к тому, что разные партии
сыра отличались друг от друга.
В наши дни в сыроварении предпочитают использовать мо-
локо с минимальным количеством бактерий и применяют за-
кваски, содержащие различные микроорганизмы в пропорции,
обеспечивающей наилучшее качество. Набор бактерий опреде-
ляется температурой термообработки (табл, 3.3).
Таблица 3.3. Закваски, используемые
при разных температурах обработки сгустка
при производстве сыра
Закваска
Температура
формирования
сгустка, °C
S. lactis или S. cremoris
S. thermophilus +
S. lactis
S. thermophilus +
L. bulgaricus или
L. helveticus или
L. lactis
>38
42—46
49—54
Существуют три причины, по которым образование молочной
кислоты так важно в сыроварении: 1) она способствует образо-
ванию сгустка при добавлении реннина; 2) уменьшает объем
сгустка, что облегчает отделение сыворотки; 3) молочная кис-
лота препятствует развитию нежелательных микроорганизмов в
ходе производства и при созревании сыра; 4) она влияет на
эластичность сгустка и способствует слипанию и затвердению
творожистой массы; 5) сказывается на характере и выражен-
ности ферментативных превращений в ходе созревания, участву-
ет в формировании вкусового букета сыра.
При производстве конкретных сортов сыра скорость образо-
вания кислоты зависит от применения определенных заквасок.
Новинка в этой области — концентрированные, глубоко заморо-
женные или лиофильно высушенные закваски, которые сырова-
ры добавляют прямо в молоко. При получении сухих заквасок
главная сложность — не повредить клетки. Состав заквасок со
Пищевые продукты, напитки и биотехнология
97
временем меняется. В 60-х годах использовали закваски на ос-
нове молока и фосфатного буфера, а затем — на основе сыво-
ротки. Потом стали применять сильно забуференные среды, что
предотвращало падение pH ниже 5,2. Предложено также ис-
пользовать большие объемы заквасок с контролируемыми свой-
ствами, что позволяет поддерживать pH на уровне 6,2.
Створаживание
После внесения закваски для формирования сгустка и сыворот-
ки обычно добавляют сычуг. Роль последнего заключается в
превращении жидкого молока в гель (сгусток). Если этот экст-
ракт сычуга не применяется, то получают творожистую массу
параказеината кальция. Различие между двумя этими способа-
ми створаживания молока в том, что без сычуга оно занимает
до 16 ч, а с ним — всего 15—30 мин. Время, проходящее от мо-
мента добавления закваски до внесения сычуга, называется вре-
менем созревания молока (до 60 мин в зависимости от сорта
сыра). За этот период бактерии успевают выработать достаточ-
но кислоты, чтобы активировать реннин.
Сычуг, применяемый в Сыроварении, —это часть желудка те-
лят. В последние двадцать лет ввиду нехватки этого сырья были
предприняты поиски его заменителей. Функции реннина могут
выполнять большинство протеиназ, однако реннин помимо ство-
раживания молока участвует и в протеолизе, происходящем в
сыре при созревании. Любой заменитель сычуга должен, кроме
створаживания молока, обеспечивать и эту реакцию. Большин-
ство известных заменителей реннина обладают большей, чем у
сычуга, протеолитической активностью при равной способности
к створаживанию. Такая избыточная активность может умень-
шать выход сыра и содержание в нем жиров. Кроме того, неко-
торые продукты протеолиза искажают вкус готового сыра. Так,
горечь сыра определяется некоторыми пептидами, образующи-
мися в сырах при длительной выдержке. Быстро созревающие и
плесневые сыры менее подвержены порче: возможно, в ходе
созревания протеолиз заходит не слишком далеко. Многие про-
теиназы могут створаживать молоко, и поиск возможных заме-
нителей сычуга вели именно среди протеиназ растений, живот-
ных, бактерий и грибов. Как выяснилось, большинство препара-
тов-коагулянтов из растений обладает слишком большой про-
теолитической активностью. Наилучшие заменители были полу-
чены из микробов; они в достаточном количестве поступают на
рынок в Америке и Европе.
Два заменителя сычуга, которые сегодня предлагаются по-
ставщиками, получают довольно сложным путем из плесневого
гриба Mucor miehei. Эти заменители считаются более подходя-
7—1344
98
Глава 3
щими для производства большинства сыров, чем аналогичные
вещества из М. pusillus. В США 60% сыра выделывают с по-
мощью коагулянтов из микробов. Будущее этой отрасли связано-
с применением методов генетической инженерии: реннин будут
синтезировать бактерии. Эта проблема освещена в обзоре Грина
(Green, 1977).
Термообработка и прессование
После коагуляции сгустки измельчают, что способствует пол-
ному отделению сыворотки. Затем эту смесь определенное время
нагревают. После термообработки отделяют сыворотку и созда-
ют условия для накопления молочной кислоты, меняющей хи-
мический состав массы. В ходе этой операции, как и при после-
дующих обработках, создается характерная текстура (строение
массы) сыра. Затем проводят посол и прессование, а также
иную, если требуется, особую обработку.
Такая особая обработка нужна при производстве отдельных
сортов сыра и заключается в следующем: 1) заражении спора-
ми голубых плесневых грибов при производстве рокфора и го-
лубых сыров; 2) нанесении на поверхность спор белых плесне-
вых грибов при производстве камамбера и бри; 3) нанесение
бактерий, нужных для созревания некоторых сыров (Port Salut_
St. Paulin). Примером таких бактерий поверхностного созрева-
ния является Brevibacterium linens.
Созревание
Если необходимо, на следующем этапе сыры отправляют на
созревание или выдержку. К этой группе сыров относятся чед-
дер и швейцарский; сливочные сыры не выдерживают. Созре-
вание происходит в специальных помещениях с контролируемой
температурой и длится до четырех лет. В зависимости от сорта
сыра температура созревания варьирует от 2 до 16 °C. Микро-
организмы и ферменты в ходе этого процесса гидролизуют жи-
ры, белки и некоторые другие .вещества молодого сыра. В ре-
зультате их распада образуются вещества, придающие сырам
характерный вкус. Поскольку процесс этот длительный, сокра-
щение его экономически выгодно. Большинство способов уско-
рения созревания основано на увеличении скорости фермента-
тивного превращения белков или жиров сыра. Сыры с резким
вкусом типа Danish Blue изготавливать проще, чем с более
тонким. Еще сложнее получать ускоренным методом чеддер.
Чтобы добиться естественного сочетания обеспечивающих вкус
веществ, необходимо тщательно подобрать ферменты, добавляе-
мые к исходному продукту. Ранее мы уже говорили об образо-
Пищевые продукты, напитки и биотехнология 99
©ании пептидов бактериями закваски. Эти пептиды могут при-
дать сыру нежелательную горечь, портящую его вкус. Добавле-
ние протеиназ приведет к нужному результату лишь при
определенном уровне активности, иначе пострадает вкус сыра.
Можно надеяться, что эту проблему удастся решить с помощью
новых микробных пептидаз. Обнадеживающие результаты по-
лучены с одной из протеиназ Bacillus subtilis, нейтразой. Для
сыров со средней интенсивностью вкуса время созревания было
сокращено с четырех до двух месяцев, а для сыров с полным
вкусом — с двадцати до шести. Если фермента добавить в ме-
ру, горечь не появляется. Нейтраза удобна и тем, что если сыр
выдерживать при 6 °C, то скорость созревания становится такой
же, как у сыра без добавки, что при неожиданном падении
спроса на сыр дает большие преимущества. Влияние липаз на
созревание менее определенно, хотя указывалось, что они уско-
ряют его при производстве чеддера. При использовании фермен-
тов, добавляемых к молоку, часть их теряется с сывороткой
при отделении ее от сгустка. Это невыгодно и, кроме того, на-
личие ферментов в сыворотке затрудняет ее продажу. Для ре-
шения этих проблем предложено использовать ферменты, за-
ключенные в микрокапсулы. Это один из возможнь(х путей.
Уменьшить время производства и созревания можно, исполь-
зуя молоко с гидролизованной лактозой. Гидролиз лактозы до
глюкозы и галактозы осуществляется ^-галактозидазой (лакта-
зой). Для этого в сырое молоко добавляют S. lactis; при этом
лактоза гидролизуется на 70%. Ускорение производства и созре-
вания сыра происходит, видимо, в результате стимулирующего
влияния глюкозы на рост и образование кислоты бактериями
закваски. Впрочем, ряд ученых иначе трактует влияние молока
с гидролизованной лактозой. Некоторые из них считают, что,
хотя глюкоза и стимулирует рост бактерий закваски, она не ус-
коряет созревания. Механизм действия молока с гидролизован-
ной лактозой не выяснен, но известно, что сыры, полученные на
этой основе, действительно созревают быстрее. Если этот меха-
низм станет нам известен, его можно будет с пониманием ис-
пользовать на практике.
Ускорить созревание можно еще одним способом — увеличив
количество бактерий в закваске. При этом содержание протеи-
наз и пептидаз будет выше, что и приведет к ускорению созре-
вания. Важно, что скорость накопления кислоты останется
прежней, а при производстве определенных сыров это имеет
большое значение. Сегодня для предотвращения излишнего на-
копления кислоты генетически обусловленные свойства бакте-
рий заквасок изменяют методами теплового шока, сенсибилиза-
ции и обработкой растворителями. Некоторые мутантные фор-
мы бактерий образуют больше протеазы или липазы на фоне
у*
100
Глава 3
обычного количества кислоты. Использование заквасок на ос-
нове таких мутантов приводит к более быстрому созреванию;
начальные этапы процесса при этом не меняются. Когда гене-
тика бактерий закваски получит должное развитие, мы сможем
создавать микроорганизмы для определенных целей. Главное
преимущество здесь в том, что в отличие от ферментов моди-
фицированные закваски — это естественный компонент системы.
В табл. 3.4 приведены примеры использования ферментных
Таблица 3.4. Примеры использования ферментных добавок для ускорения
созревания сыров (Law, 1980)
Тип сыра Добавляемые ферменты Стадия добавления
тип ИСТОЧНИК
Cheddar Кислые и нейтраль- ные протеиназы, пептидазы, липазы, декарбоксилазы Различные коммерче- ские препараты и ферменты живот- ных Сгусток
Cheddar Romano Parmesan Липаза, протеиназа Ткани желудка те- ленка Молоко, сгусток
Gouda Протеиназа Aspergillus oryzae То же
Edam Cheddar Протеиназа + пептида- за Pseudomonas fluores- cens Молоко
Rossiiskii Протеиназа Панкреатин
Mozzarella Липазы, эстеразы (микроинкапсулиро- ванные) Секрет из сычуга те- лят
Blue Липаза Aspergillus spp. Сгусток
добавок для ускорения созревания сыров. Более подробные све-
дения содержатся в специальной литературе.
Сыворотка
Сыворотка — это побочный продукт сыроварения; вкратце об
этом уже говорилось ранее. Не так давно ее относили к отходам
промышленности, но в последнее время в этой области намети-
лись изменения: все популярнее становится использование сы-
воротки для производства подсластителей при помощи лактазы.
В Англии получаемые при этом глюкозу и галактозу предпола-
гается использовать в хлебопечении, кондитерской промышлен-
ности и для производства мороженого. Лактазу получают из
Aspergillus niger. Получение сахаров из сыворотки налажено
как в США (Corning/Kroger), так и в Англии (Corning/Dairy
Crest).
Пищевые продукты, напитки и биотехнология 101
3.2.3. Йогурт
Это один из древнейших продуктов, получаемых путем фермен-
тации. После термообработки молоко заквашивают добавлени-
ем 2—3% закваски йогурта. Главную роль здесь играют бакте-
рии Streptococcus thermophillus и Lactobacillus bulgaricus. Для
получения желаемой консистенции продукта, вкуса и запаха
эти организмы должны содержаться в культуре приблизительно
в равных количествах. Кислоту в начале заквашивания обра-
зует в основном S. thermophillus. Смешанные закваски нужно
часто обновлять, поскольку повторные пересевы неблагоприят-
но сказываются на соотношении видов и штаммов бактерий:
в них начинает доминировать L. bulgaricus.
3.2.4. Масло
Из молочных продуктов проще всего получать масло. В зави-
симости от сорта производимого масла используют сливки с
концентрацией от 30—32 до 30—40%. При их сбивании эмуль-
сия масла в воде превращается в эмульсию воды в масле.
При производстве масла для улучшения вкуса и лучшей со-
хранности используют особые культуры бактерий. Улучшение
вкуса было достигнуто путем создания специальных штаммов
бактерий, отобранных по способности синтезировать нужные
вещества, влияющие на вкус. Первыми для этой цели были ис-
пользованы штаммы Streptococcus lactis и близких видов, а за-
тем — смешанные культуры, включающие Streptococcus lactis,
Leuconostoc citrovorum и L. dextranicum.
Помимо улучшения вкуса, таким путем удается устранить
некоторые нежелательные привкусы. Перспективный способ до-
работки масла основан на добавлении липаз. Внедрение его
позволит пускать масло в продажу непосредственно из масло-
бойки.
3.2.5. Сброженная пахта
Сброженный продукт получают из свежей пахты, а чаще из
снятого молока путем добавления закваски, используемой при
производстве масла. Эта закваска представляет собой смесь
молочнокислых стрептококков (S. lactis или S. cremoris) и об-
разующих ароматические вещества бактерий (L. citrovorum и
L. dextranicum). И те и другие бактерии нужны для формиро-
вания полноценного вкуса и запаха пахты; стрептококки при
этом доминируют. Роль молочнокислых стрептококков в заквас-
ке заключается в образовании молочной кислоты (она дает
желаемых кисловатый вкус), свертывании молока и снижении
102 Глава 3
pH до значений, при которых образующие ароматические ве-
щества бактерии синтезируют наибольшее количество летучих
кислот.
3.2.6. Сметана
Ее готовят почти так же, как сброженную пахту. К сливкам
добавляют 0,5—1% закваски, используемой при производстве
масла. Далее продукт выдерживают, пока концентрация кисло-
ты не достигнет 0,6%.
3.2.7. Новые продукты
Известно, что некоторые люди не переносят лактозу; для них
можно выпускать молоко, обработанное ^-галактозидазой —
ферментом, который уменьшает содержание лактозы. Для этой
цели нужно разработать недорогой промышленный способ про-
изводства такого молока. ^-Галактозидазу получают из дрож-
жей, плесеней и бактерий.
3.3. Хлебопродукты =
3.3.1. Хлеб и другие продукты.
В Англии большинство хлебопродуктов производится по техно-
логии Chorleywood Bread Process, но в других странах исполь-
зуется много других технологий хлебопечения. Для производст-
ва хлеба до сих пор применяют в основном дрожжи Saccharo-
myces cerevisiae. Обычно их растят в ферментерах
периодического действия на мелассе (свекловичной или сахар-
ного тростника).
В простейшем случае готовят тесто, смешивая при комнатной
температуре муку, воду, дрожжи и соль. При замесе слои теста
перемещаются, создаются условия для образования пузырьков
газа и подъема теста. Замешанному тесту дают возможность
«подойти», а затем режут на куски нужного веса, формуют (при
этом тесто раскатывают и складывают, чтобы получить нужную
текстуру) и выдерживают во влажной атмосфере. При выдерж-
ке и на первой, следующей за ней стадии выпечки образовав-
шиеся при замесе и формовке «зародыши» газовых пузырьков
наполняются углекислым газом. Он выделяется в ходе анаэроб-
ного сбраживания глюкозы и мальтозы муки. Поднявшееся
тесто выпекают. В ходе этого термического процесса крахмал
желатинизируется, дрожжи погибают и тесто частично обезво-
живается. В результате мы получаем имеющий определенную
форму, плотный по консистенции продукт; с легкой ячеистой
структурой и очень вкусной корочкой.
Пищевые продукты, напитки и биотехнология
103
Помимо углекислого газа при анаэробном брожении обра-
зуются разнообразные органические кислоты, спирты и эфиры.
Все они заметно влияют на формирование вкуса хлеба.
В Германии и США из ржаной муки выпекают хлеб с кис-
линкой. Основа технологии здесь та же, что и при выпечке хле-
ба из пшеничной муки, но при замесе к тесту добавляют пред-
варительно сброженную смесь ржаной муки и воды, заквашен-
ную смешанной культурой лактобацилл. Содержащаяся в этой
закваске (опаре) кислота и придает хлебу особый вкус.
3.3.2. Продукты гидролиза крахмала
Крахмал трех основных поступающих на рынок сортов получа-
ют по обычной технологии. Способы производства кукурузного,
пшеничного и картофельного крахмала были неоднократно и
подробно описаны (см. разд. «Литература»). Гидролиз этих
разновидностейкрахмала в промышленном масштабе осуществ-
ляется разными способами: только кислотой, кислотой и фер-
ментами и только ферментами. В первой четверти нашего века
использовалась простая обработка кислотами, на смену которой
пришли сегодня более совершенные технологические процессы.
История развития. э*гои отрасли может быть хорошим примером
отклика биотехнологии на запросы потребителей и производ-
ства.
В середине 60-х годов на смену кислотному и кислотно-
ферментативному процессам пришел ферментативный способ
переработки крахмала, основанный на последовательном при-
менении а-амилазы В. subtilis и амилоглюкозидазы A. oryzae
или A. niger. Главное его преимущество связано с увеличением
скорости процесса, уменьшением уровня загрязнения продукта-
ми реверсии, получением продукта с высоким декстрозным
эквивалентом (ДЭ). Величина ДЭ гидролизата [в используемой
шкале глюкоза (декстроза) соответствует 100 ед.] отражает
глубину гидролиза крахмала, для которого ДЭ считается рав-
ным нулю.
Следующим важным шагом было внедрение в производство
термостабильных а-амилаз, главным образом из В. lichenifor-
mis и В. amyloliquefaciens, работающих при температуре 100 °C
и позволяющих после обработки аминоглюкозидазой получать
продукцию с ДЭ, близким к 100.
Высокотемпературное ожижение крахмала сегодня стало
обычным промышленным процессом, но следующий этап, оса-
харивание, до сих пор осуществляется при участии «обычной»
глюкоамилазы A. niger путем одноразовой обработки. Предла-
галось использовать для этого иммобилизованные ферменты,
но широкого применения они пока не нашли.
104
Глава 3
Кроме производства глюкозы, наиболее заметным успехом
в этой отрасли промышленности был выпуск смесей глюкозы и
фруктозы. Этот продукт известен под названием изоглюкозы
или кукурузного сиропа с высоким содержанием фруктозы.
Возможности промышленного применения этой технологии осо-
бенно ярко выявились в США, где крахмал-содержащее сырье
производится в избытке, а собственного сахара не хватает. Изо-
глюкоза может заменять сахарозу в большинстве видов пищи.
Что касается Европы, то производство изоглюкозы здесь сдер-
живается по экономическим причинам.
Сырьем для производства изоглюкозы служит деионизован-
ный гидролизат крахмала, получаемый описанным ранее мето-
дом, с ДЭ, возможно более близким к 100. Изомеризация осу-
ществляется ферментами из различных организмов. Выбор их
определяется тем, насколько просто с ними работать, нуждают-
ся ли они в кофакторах и стабильны ли. Источником фермента
чаще всего бывают виды Streptomyces, В. coagulans, Actinopla-
nes missouriensis и виды Arthrobacter. После изомеризации и
дальнейшей обработки образуется продукт, содержащий до
90% фруктозы, хотя исходное сырье, получающееся после уста-
новления равновесия в реакции изомеризации, представляет
собой смесь глюкозы (51%) и фруктозы (42%) с олигосахари-
дами (7%).
Используя разнообразные амилазы, трансглюкозидазы и
ферменты, атакующие разветвленные молекулы, удается полу-
чить множество разновидностей гидролизата крахмала, которые
находят широкое применение в промышленности. На рис. 3.1
сопоставлены данные о сырье, промежуточных и конечных про-
дуктах этого производства.
3.4. Бродильные производства
3.4.1. Производство алкогольных напитков
Получение напитков путем спиртового брожения — одно из
древнейших бродильных производств. Первыми из таких напит-
ков были, видимо, вино и пиво. До появления работ Пастера
в конце XIX в. о сути протекающих при брожении процессов
и их механизмах было известно очень мало. Пастер показал,
что брожение без доступа воздуха осуществляется живыми клет-
ками дрожжей, при этом сахар превращается в спирт и угле-
кислый газ. Тогда же было показано, что брожение осуществ-
ляется под действием каких-то веществ, находящихся внутри
дрожжевых клеток. Одно из главных нововведений в области
микробиологии брожения было предложено Хансеном, работав-
шим в исследовательском центре Карлсберг в Копенгагене с
Пищевые продукты, напитки и биотехнология
1Q5
Процесс
Вещество Действующий агент
Конечный продукт ч
1
6-8
12-14
42
63
94-96
Крахмал
Ожижение
Конверсия
Осахаривание
Деионизация,
изомеризация
| а-Амилаза + нагревание +
Ожиженный _ перемешивание_________________ Декстрин (твердь|й)
крахмал
Ot-Амилаза
Конвертированный а-Амил аза/'кислоты
крахмал “
Пуллуланаза / 3 -амилаза
а -Амилаза/кислота
Аминоглюкозидаза
Хроматографическое
разделение
Декстрозный -
сироп
Изомероэа
Сахар с малым ДЭ
(твердый)
Концентрированный
=► мальтозный сироп
Сахар с ДЭ =42 (сироп)
Сахар с ДЭ=63
(сироп)
Сахар с высоким ДЭ (сироп}
(твердый)
(кристаллы)
>- Изоглюкоза (сироп)
96%-ный сироп фруктозы
Рис. 3.1. Стадии гидролиза крахмала.
дрожжами дикого типа. При производстве пива эти дрожжи
доставляли массу неудобств. Хансен выделил чистые культуры
дрожжей и использовал их в пивоварении; тем самым он стал
пионером применения таких культур при производстве пива.
Алкогольные напитки получают путем сбраживания сахар-
содержащего сырья, в результате которого образуются спирт
и углекислый газ. Сбраживание осуществляется дрожжами ро-
да Saccharomyces. В одних случаях используется природный
сахар (например, содержащийся в винограде, из которого де-
лают вино), в других сахара получают из крахмала (например,
при переработке зерновых культур в пивоварении). Наличие
свободных сахаров обязательно для спиртового брожения при
участии Saccharomyces, так как эти виды дрожжей не могут
гидролизовать полисахариды. Образование этилового спирта из
глюкозы происходит по схеме Эмбдена — Мейергофа—Парна-
са, представленной на рис. 3.2.
В производстве спиртных напитков применяют штаммы
дрожжей Saccharomyces cerevisiae или S. carlsbergensis. Раз-
личие между ними заключается в том, что S. carlsbergensis мо-
гут полностью сбраживать раффинозу, a S. cerevisiae к этому
не способны.
106
Глава 3
СН2ОН
мо\ /он
СН2О®
XZ /г°\к
\он н/
но\1 /он
СН2О©
I СН2ОН
Мн но/
н\___/ОН
н ОН
D -Глюкоза
н он
Г люкозо-6-фосфат
ОН н
Фруктозе- 6-фосфат
!/♦ АТР
сн2о®
снон
ено
Г лицеральдегид-3-фосфат
j * ADP
СН2О®
1/О\СН2О®
—► Гн но>
н\___Ион
он н
Фруктозе-1,6-дифосфат
сн2он
С=О
СН2О®
Дигидроксиацетонфосфат
"/»NAD*
р,
СН2О®
I
снон
СОО"
З-Фосфоглицерат
снон
соо©
АТР ADP
1, З-Дифосфоглицерат
СН2ОН
СНО®:
СОО'
2-Фосфоглицерат
СН2
II
► со®
СОО“
Фосфоенол пируват
ADP АТР
СНЭ
СОО'
Пируват
С = О
СО 2
сн3
СНЭ
н-с-он
с=о
н
н
Этанол
NAD' NADH
Ацетальдегид
Рис. 3.2. Реакции метаболического пути Эмбдена—Мейергофа—Парнаса.
3.4.2. Пиво
Для осуществления спиртового брожения прежде всего необхо-
димо, чтобы в пивоваренном сырье образовался сахар. Тради-
ционным источником нужных для этого полисахаридов всегда
был ячмень, но в качестве дополнительных используются и дру-
гие виды углевод-содержащего сырья. И сегодня ячменный со-
лод составляет основу пива. Ячменный солод и другие компо-
ненты измельчают и смешивают с водой при температуре до
67 °C. В ходе перемешивания природные ферменты ячменного
Пищевые продукты, напитки и биотехнология 107
солода разрушают углеводы зерна. На заключительной стадии
раствор, называемый суслом, отделяют от нерастворимых остат-
ков. Добавив хмель, его кипятят в медных котлах. Для произ-
водства пива с определенным содержанием алкоголя сусло по-
сле кипячения доводят до нужной плотности. Удельная плот-
ность сусла определяется содержанием экстрагированных саха-
ров, подлежащих сбраживанию. По истечении определенного
времени брожение заканчивается, дрожжи отделяют от пива и
выдерживают его некоторое время для созревания. После
фильтрации и других необходимых процедур пиво готово. Схе-
ма производственного процесса дана на рис. 3.3.
Начатое по инициативе Хансена использование индивиду-
альных штаммов дрожжей в пивоварении сегодня стало нормой:
это культуры S. cerevisiae и S. carlsbergensis. Первые представ-
ляют собой дрожжи поверхностного и глубинного брожения:
они применяются в производстве эля. Вторые — дрожжи глу-
бинного брожения, их используют в производстве легкого пива.
Хотя генетика дрожжей развивается уже в течение многих
лет, мы лишь недавно научились осуществлять селекцию дрож-
жей, используемых в производстве пива. По мере углубления
наших знаний о свойствах дрожжей и тех качествах, которые
они придают конечному продукту, все успешнее идет работа по
выведению новых штаммов пивных дрожжей. В конечном счете
мы сможем создать штамм, позволяющий получить идеальный
продукт. Требования к таким идеальным дрожжам будут, ес-
тественно, зависеть от способа сбраживания и желаемых ка-
честв пива. К числу наиболее важных свойств относятся про-
дуктивность, способность формировать осадок, сбраживать
мальтотриозу и т. д. Принимаются во внимание и вкусовые ка-
чества получающегося пива, т. е. образование веществ, ответст-
венных за их формирование. Ранее основным способом получе-
ния штаммов, дающих продукт нужного качества, был их отбор
из существующих пивных дрожжей. Вести отбор было выгоднее,
чем заниматься гибридизацией, отчасти из-за малой способно-
сти пивных дрожжей к спорообразованию и низкой жизнеспо-
собности аскоспор. В каждом аске образуется от одной до че-
тырех спор, но не все они освобождаются при созревании.
Дрожжи из рода Sacharotnyces размножаются в основном веге-
тативно. При этом за счет множественного латерального почко-
вания формируются сферические, эллипсоидные или реже ци-
линдрические дочерние клетки. Поскольку для развития техно-
логии пивоварения могут понадобиться штаммы дрожжей, от-
личающиеся по свойствам от обычно используемых, придется
прибегнуть к гибридизации. Основным вкладом биотехнологии
в пивоваренную промышленность будет создание штаммов
дрожжей, способных давать пиво с желаемыми свойствами.
108
Глава 3
Рис. 3.3. Операции, лежащие в основе пивоварения (с любезного разрешения
д-ра Анны М. Мак-Леод).
Пищевые продукты, напитки и биотехнология 109
3.4.3. Вино
*
Необходимое условие любого спиртового бродильного процес-
са — наличие сахара в сырье. Так, в производстве вина исполь-
зуется сахар виноградного сока. Почти все вино в мире делают
из винограда одного вида, Vitis vinifera. Сок этого винограда —
прекрасное сырье для производства вина. Он богат питательны-
ми веществами, служит источником образования приятных за-
паха и вкуса, содержит много сахара; его природная кислот-
ность подавляет рост нежелательных микроорганизмов.
Виноделие в отличие от пивоварения до самого последнего
времени было основано на использовании диких местных дрож-
жей. Единственная обработка, которой подвергали виноград до
отжима, — окуривание его сернистым газом, чтобы сок не тем-
нел. Кроме того, сернистый газ подавляет деятельность не-вин-
ных дрожжей; это позволяет винным дрожжам, которые менее
чувствительны к нему, осуществлять брожение без помех. При
изготовлении красного вина гребни, косточки и кожица до кон-
ца брожения находятся в виноградном сусле (мусте), а белое
вино делают из чистого сока. Обычно окуривание сернистым
газом проводят до того, как раздавливают ягоды, но иногда его
добавляют и на более поздних стадиях.
В табл. 3.5 перечислены виды дрожжей, наиболее часто
встречающиеся на кожице виноградных ягод. В прошлом имен-
но эти дикие дрожжи и осуществляли спиртовое брожение.
Сегодня в тех районах, где виноделием начали заниматься не-
давно, широко применяются дрожжевые закваски. Связано это
с тем, что желаемая микрофлора может и отсутствовать, а ино-
куляция стандартной культурой дрожжей позволяет получать
вина с нужными свойствами. Кроме того, количество используе-
мого сернистого газа ограничено законом, и это побуждает при-
менять дрожжевые культуры-закваски. Виноделы не очень-то
полагаются на дрожжи дикого типа, если нет уверенности, что
конкуренция со стороны не-винных дрожжей не подавлена.
Использование заквасок дает ряд преимуществ: сокращается
лаг-период размножения дрожжей, образуется продукт с извест-
ными свойствами, уменьшается вероятность появления нежела-
тельного вкуса, поскольку в брожении не участвуют дикие не-
винные дрожжи. В будущем использование специально создан-
ных штаммов будет все более расширяться: это гарантирует
необходимые вкусовые качества вин. Смешанные закваски по-
зволяют получать продукцию с полным букетом, что невозмож-
но при работе с индивидуальными штаммами.
Вкусовые различия между сортами винограда определяются
особыми веществами. Так, в формировании вкуса мускатных
сортов участвуют производные терпенов, линалоол и гераниол.
Таблица 3.5. Дрожжи, обитающие на винограде, а также в виноградном
сусле и вине (Kunkee, Goswell, см. Rose, 1977)
Роды и виды
Источник
Candida pulcherrima (новое назва-
ние — Metschnikowia pulcherrima)
С. scottii (новое название — Leucospo-
ridiurn scottii)
Debaryomyces phaffii
Endomycopsis lindneri
Hansenula saturnus
Kloeckera africana
K. apiculata
Pichia fermentans
Saccharomyces acidifaciens (ныне —
разновидность S. bailii)
S. bayanus (включ. S. oviformis)
S. bisporus
S. carlsbergensis (новое название —
S. uvarum)
S. cerevisiae
S. chodatai (новое название — S. itali-
cus)
S. coreanus
S. elegans (новое название—S. bailii
var. bailii)
S. elegans vaar intermedia (новое на-
звание — S. bailii var. bailii)
S. globosus
S. heterogenicus
S. it aliens
S. oviformis (новое название — S. bay-
anus)
S. pastorianus (новое название —
S. bayanus)
S. rosei
S. rouxii
S, steineri (новое название — S. itali-
cus)
Torulopsis bacillaris (новое назва-
ние — Torulopsis stellata)
Torulopsis burgeffiana (новое назва-
ние — M. pulcherrima)
Обычны для сусла и вина; на вино-
граде
Чехословакия; незрелый виноград,
в Новой Зеландии
Виноград в провинции Коньяк
Виноград и сусло, Бразилия
Вино, полученное из поздно собран-
ного винограда
Весьма обычны для винограда, сусла
и вина
Весьма обычны
Виноград, сусло и вино
Испортившиеся вина, виноград,,
устойчивы к сернистому газу
(фруктофиллы)
Виноград, сусло и вино
Виноград, образуют мало спирта
Виноград, реже — вина (применяют-
ся при производстве легкого пива)
Классические винные дрожжи; по-ви-
димому, наиболее распространен-
ные
Виноград в Грузии (СССР)
Виноград; редкий внд
Виноград и вино (фруктофил)
Виноград в Бразилии
Виноград, виноградный сок и вино;
встречаются редко
Виноград, сусло, виноградный сок;
встречаются редко
Виноград и виноградный сок, выра-
батываемый в местностях с теп-
лым климатом
Виноград, сусло и вино; обычный
вид
Виноград, сусло; обычны для броже-
ния по типу Loire
Виноград и вино; обычный вид
Перезрелый виноград
Вино и виноград
Виноград и вино (фруктофил)
Виноград и сусло
Пищевые продукты, напитки и биотехнология
Ш
Различные вкусовые оттенки появляются при выдержке вина;
хорошо известно, что свой вклад вносит взаимодействие с
древесиной и воздухом при хранении в деревянных бочках.
После завершения спиртового брожения молодое вино хра-
нят в особых условиях, чтобы оно не испортилось. Если вино не
предполагается подвергать яблочно-молочнокислому дображи-
ванию, его обрабатывают сернистым газом, что подавляет окис-
лительные процессы, вызывающие его потемнение. До этого
из вина удаляют дрожжи, чтобы прекратить брожение.
Первосортные вина подвергают выдержке разного рода в за-
висимости от типа вина, а более дешевые разливают, как пра-
вило, в тот же год, когда они получены. Трудности при выра-
ботке дешевых вин обычно связаны с их склонностью к вторич-
ному, яблочно-молочнокислому брожению, которое развивается
ко времени разлива. Если вино склонно к такому брожению,
его искусственно вызывают до разлива, а если нет, то подавля-
ют. При производстве первосортных красных вин такое броже-
ние даже желательно. Оно составляет естественную часть про-
цесса и происходит при хранении. Этот тип брожения осуществ-
ляется молочнокислыми бактериями, в частности Leuconostoc,
Lactobacillus и Pedicoccus. Оно не идет при низких значениях
pH; создав такие условия, его можно подавить. В белых винах
яблочно-молочнокислое брожение происходит реже, так как pH
в них ниже. Среди новшеств в этой области упомянем исполь-
зование для инициации брожения вместо бактерий иммобили-
зованных ферментов.
Некоторые особые сорта вин, например сотерны, получают
при участии гриба Botrytis cinerea. Его развитие на ягодах при-
водит к их обезвоживанию и повышению содержания сахара,
что и определяет сладкий вкус вина. Заражение должно проис-
ходить только перед сбором винограда. Представляет интерес
и еще один процесс, называемый углекислотной мацерацией.
Красные вина, которые должны созреть к 15 ноября в год сбора
винограда, получают особым способом. Виноград не давят,
а помещают целиком в бродильные чаны, где держат в атмо-
сфере углекислого газа. Брожение идет либо прямо в ягодах,
в анаэробных условиях, либо в соке, выделяющемся в резуль-
тате разрушения кожицы углекислым газом. Микробиология
этого процесса пока не исследована.
В крепленых винах часть спирта получается при сбражива-
нии винограда дрожжами, а часть добавляется. К числу таких
вин относятся портвейн, херес и мадера (разд. 3.4.4).
Дальнейшие успехи виноделия будут определяться исполь-
зованием более эффективных штаммов винных дрожжей и ком-
мерческих препаратов дрожжевых заквасок. Это позволит по-
лучать вина особого качества.
112
Глава 3
3.4.4. Спирт
Производство перегнанного спирта моложе, чем неперегнанных
спиртных напитков, но и его корни теряются в веках. Для по-
лучения напитка, содержащего 40% (по объему) спирта, нужна
перегонка. Ее и сегодня осуществляют в перегонных аппаратах,
представляющих собой модификации устройства, предложенно-
го в 1830 г. Коффи и носящего его имя. Различия в сортах спир-
товых продуктов зависят в основном от природы сырья, а также
от того, подвергался ли конечный продукт выдержке. В спирто-
вом производстве используются пригодные для этой цели штам-
мы Saccharomyces. Крупные спиртовые заводы всегда ведут
свою собственную культуру дрожжей в специальных средах.
Выбор штамма дрожжей при производстве спирта определяется
их продуктивностью в особых условиях бродящего сусла. Бро-
жение должно идти активно с образованием спирта в количест-
ве, близком к теоретическому пределу. Хотя в качестве сырья
можно использовать разнообразные продукты, некоторые сорта
спирта обычно производят из вполне определенных его типов.
Так, коньяк, получаемый при перегонке вина, делают из вино-
града, а шотландский виски — из ячменного солода. Другие на-
питки — американский виски, джин и водку, которые обычно’
делают из зерна (например, кукурузы), можно производить и
на основе другого подходящего сырья. Ром обычно получают из
мелассы сахарного тростника или свеклы. Когда сырьем слу-
жит зерно (например, пшеницы или кукурузы), до сбраживания
необходимо гидролизовать крахмал до сахаров. Так, виски —
это продукт перегонки пива без хмеля. Первые стадии процес-
са производства виски такие же, что и при приготовлении сус-
ла в пивоварении. Однако если применяют кукурузу или другие
зерновые, то до приготовления сусла непосредственно в бро-
дильных чанах проводят обработку крахмала в зерне фермен-
тами солода.
Если для производства спирта используют мелассу, такие
предварительные операции не нужны, поскольку углеводы в ней
содержатся в форме, пригодной для сбраживания. Тем не менее
сырье все же приходится подготавливать к процессу: осветлять,
подогревать и разводить водой, чтобы получить концентрацию
сахара, оптимальную для брожения. После подготовки сырья
добавляют культуру подходящих дрожжей и ведут сбражива-
ние. В конце его, когда концентрация спирта достигнет 9—11%
(по объему), отделяют дрожжи путем отстаивания. Остатки их
перед перегонкой можно удалить центрифугированием. Приме-
няются два способа: кубовый и непрерывной перегонки (патент
Коффи). После этого продукт либо отправляют на созревание
(например, виски), либо проводят завершающие операции и
Пищевые продукты, напитки и биотехнология 113
разливают по бутылкам (джин, водка). Для продажи спирто-
продукты обычно разводят до стандартной концентрации в них
спирта (40% по объему).
Остановимся на некоторых важных особенностях производ-
ства отдельных разновидностей спиртопродуктов. Очень важно
использовать правильный штамм дрожжей. При выработке ро-
ма для производства сортов с сильным запахом обычно приме-
няют штаммы дрожжей Schizosaccharomyces, а с менее интен-
сивным — быстро работающие Saccharomyces. Отметим, что
процесс образования спирта ускоряется бактериями Clostridium
saccharobutyricum. Самый лучший ром получают в том случае,
когда соотношение бактерий к дрожжам составляет 1 :5. Куль-
туру бактерий вносят, когда концентрация этанола достигает
3,5—4,5%, а сахара — 6% (вес/объем).
Штаммы дрожжей, используемые в спиртовой промышлен-
ности, должны сохранять жизнеспособность вплоть до концент-
рации этанола 12—15% (по объему). Кроме того, если в каче-
стве сырья используется зерно, дрожжи должны обладать спо-
собностью гидролизовать низкие олигосахариды до глюкозы.
Это необходимо для полного превращения крахмала в этиловый
спирт и углекислый газ.
Перегонка — дорогостоящий этап процесса, так как он энер-
гоемок. Использование дрожжей, выдерживающих повышенные
концентрации спирта, уменьшает энергозатраты на этом этапе,
поскольку для достижения одной и той же концентрации спирта
надо отгонять меньше воды. Ожидается, что работа по созда-
нию новых штаммов дрожжей, устойчивых к еще более высоким
концентрациям спирта, будет успешной.
Пока углеводы не переведены в форму, усваиваемую дрож-
жами, брожение не происходит. Добавление гидролизующих
крахмал ферментов ускоряет этот процесс. Для этого обычно
применяют амилазу из культуральной жидкости штаммов Bacil-
lus subtilis и амилоглюкозидазу, выделяемую из культур грибов
штаммов Aspergillis niger и близких форм. Ферменты исполь-
зуются и при подготовке сусла, и при спиртовом брожении (см.
разд. 3.3.2, где эти вопросы рассматриваются более подробно).
3.4.5. Сидр
Сброженный яблочный сок известен под названием сидр. В тех-
нологии производства сидра и вина есть много сходного.
Когда делают сидр, яблоки прежде всего измельчают в ка-
шицу и отжимают сок. Для этого применяют как прессы, так
и другие устройства, работающие на иных принципах. Способы
подготовки сока перед брожением весьма разнообразны; он мо-
жет использоваться как без обработки, так и после подавления
в нем естественной микрофлоры и замещения ее дрожжами под-
8—1344
a 14
Глава 3
ходящих штаммов. Чаще всего к соку добавляют сернистый газ,
мем подавляют развитие Kloeckera apictilata, неблагоприятно
влияющих на вкус готового сидра. Вслед за этим можно начи-
нать брожение. Оно происходит либо при участии диких дрож-
жей, либо после добавления дрожжевой культуры-закваски.
Требования, предъявляемые к дрожжам при производстве сид-
ра, в общем те же, что и в других бродильных процессах: они
должны обеспечивать полноту сбраживания, высокую его ско-
рость и легко выпадать в осадок. Так как Saccharomyces растут
медленно, при крупномасштабном производстве сидра нередко
к обработанному сернистым газом соку добавляют те или иные
чистые культуры дрожжей. Разные штаммы дрожжей образуют
специфические ароматические вещества. Поэтому при производ-
стве сидра точно так же, как в пивоварении, можно использо-
вать разные штаммы для придания сидру специфического вку-
са. Чтобы получить сидр определенного сорта, добавляемые
дрожжи должны преобладать над дикими, быстрее размножать-
ся и определять конечные свойства продукта. Запуск процесса
доминирующей формой дрожжей — один из наиболее широко
используемых способов, применяемых при крупномасштабном
производстве сидра. Важно, чтобы дрожжи были способны об-
разовывать полигалактуронидазу, необходимую для гидролиза
деэтерифицированных пектинов до галактуроновой кислоты.
В противном случае в конце брожения сидр не просветляется.
Для просветления сидра нередко добавляют гидролизующие
пектин ферменты, включая полигалактуронидазу; их получают
из грибов (разд. 3.7.3).
Для подавления развития всех штаммов дрожжей, кроме
нужных для ферментации, при производстве сидра применяют
сернистый газ по той же схеме, что и в виноделии. Если не за-
ботиться о чистоте аппаратуры и не применять газ, дикие дрож-
жи могут размножиться и вытеснить культурные. При выборе
конкретного штамма дрожжей нужно также учитывать, какие
вкусовые вещества они передадут образующемуся продукту;
можно получить сидр как с небольшим, так и с заметным со-
держанием сивушных масел. Для придания сидру необходимых
качеств нужно использовать определенный штамм дрожжей.
По завершении брожения сидр отделяют от дрожжей и
осветляют. Поскольку в сидре есть яблочная кислота, то, как
и в вине, в нем может начаться яблочно-молочнокислое броже-
ние. Такое брожение не идет, если сидр очень кислый или его
держат на холоде.
Как и в случае пивоварения, развитие производства сидра
будет определяться селекцией новых штаммов дрожжей, позво-
ляющих получать продукты со свойствами, отвечающими вку-
сам разных потребителей.
Пищевые продукты, напитки и биотехнология
115-
3.4.6. Уксус к
Хотя уксус и не принадлежит к алкогольным напиткам, мы ре-
шили остановиться на его производстве в этом разделе, посколь-
ку одна из двух стадий его получения включает спиртовое бро-
жение.
Уксус — это продукт, содержащий не менее 4% (вес/объем)
уксусной кислоты; его получают с помощью двухстадийного-
процесса. Вначале осуществляют спиртовое брожение, в ходе
которого сахар сырья превращается в спирт при участии S. ce-
revisiae. Сырьем может быть любой продукт, который сбражи-
вается с образованием спирта. После завершения этого этапа
дрожжам дают осесть и собирают надосадочную жидкость.
Содержание спирта доводят до 10—13%, и, если в ходе броже-
ния для подавления роста бактерий добавлялся сернистый газ,,
его перед дальнейшими операциями удаляют. На следующем
этапе этиловый спирт превращается в уксусную кислоту (про-
межуточным продуктом является ацетальдегид). Все процессы
получения уксуса идут при участии смешанных культур Aceto-
bacter; в некоторых случаях применяют закваски. На этой стадии
основную роль играют бактерии A. schuetzenbachii, A. curvim,
A. orleanense и близкие к ним микроорганизмы. Брожение про-
исходит в аэробных условиях с потреблением больших коли-
честв кислорода и выделением тепла. Ранее этот второй процесс
шел медленно, но его удалось усовершенствовать и увеличить
конверсию этилового спирта в уксусную кислоту. Метод глубин-
ной ферментации в этом производстве применить долгое время
не удавалось. Проблему удалось решить лишь после второй
мировой войны: этому помогли исследования по аэрации куль-
тур при производстве антибиотиков. Сегодня эта технология
нашла широкое применение.
3.5. Белковые продукты
3.5.1. Традиционные белковые продукты,
получаемые путем ферментации
Микроорганизмы начали использовать в производстве белковых
продуктов задолго до возникновения микробиологии. Достаточ-
но упомянуть всевозможные разновидности сыра, а также про-
дукты, получаемые путем ферментации соевых бобов. И в пер-
вом, и во втором случае питательной основой является белок.
Способы производства этих продуктов рассмотрены в разд. 3.2
и 3.7. При выработке их при участии микробов происходит глу-
бокое изменение свойств белоксодержащего сырья. В результа-
те получают пищевые продукты, которые можно дольше хранить
S*
116
Глава 3
(сыр) или удобнее потреблять (соевый творог). Микробы игра-
ют роль и в производстве некоторых мясных продуктов, пред-
назначенных для хранения. Так, при изготовлении некоторых
сортов колбасы (bologna, salami) используется кислотное бро-
жение, обычно при участии комплекса молочнокислых бактерий.
Образовавшаяся кислота способствует сохранности продукта
и вносит вклад в формирование его особого вкуса. Кислотооб-
разующие бактерии используются и при засоле мяса — еще од-
ном способе консервации. Ряд блюд восточной кухни получают
путем ферментации рыбы: для этого применяют плесневые гри-
бы и дрожжи.
Этим, пожалуй, и ограничивается использование микроорга-
низмов в переработке белков. Возможности современной биотех-
нологии в этих производствах невелики, за исключением сыро-
делия (разд. 3.2.2). Другое дело — выращивание и сбор
микробной массы, перерабатываемой в пищевые продукты:
здесь биотехнология может проявить себя во всей полноте.
3.5.2. Белок одноклеточных организмов
По многим важным показателям биомасса микроорганизмов
может обладать весьма высокой питательной ценностью. В не-
малой степени эта ценность определяется белками: у большин-
ства видов они составляют значительную долю сухой массы
клеток. На протяжении десятилетий активно обсуждаются и ис-
следуются перспективы увеличения доли белка микроорганиз-
мов в общем балансе производимого во всем мире белка. Такое
увеличение возможно как в косвенной форме, путем введения
белковых добавок в корм животным (что уменьшает потреб-
ность в таких продуктах, как соевая и рыбная мука), так и в
прямой, путем получения продуктов питания. О белках, идущих
на корм скоту, речь идет в гл. 9. Здесь мы рассмотрим те слу-
чаи, когда биомасса микроорганизмов непосредственно исполь-
зуется в пищу.
Чтобы отличать такой тип продуктов от белков высших мно-
гоклеточных животных и растений, для микробного белка при-
думано специальное название — белок одноклеточных организ-
мов (БОО)1. Производство его связано с крупномасштабным
выращиванием определенных микроорганизмов, которые соби-
рают и перерабатывают в пищевые продукты. В основе лежит
технология ферментации — ветвь бродильной промышленности
и производства антибиотиков. Чтобы осуществить возможно
более полное превращение субстрата в биомассу микробов, тре-
1 В отечественной литературе обычно употребляется термин «белково-ви-
таминный концентрат», но он уже по смыслу. — Прим, перев.
Пищевые продукты, напитки и биотехнология 117
буется многосторонний подход. Выращивание микробов в пи-
щевых целях представляет интерес по двум причинам. Во-пер-
вых, они растут гораздо быстрее, чем растения или животные:
время удвоения их численности измеряется часами. Это сокра-
щает сроки, нужные для производства определенного количест-
ва пищи. Во-вторых, в зависимости от выращиваемых микроор-
ганизмов в качестве субстратов могут использоваться разнооб-
разные виды сырья. Что касается субстратов, то здесь можно
идти по двум главным направлениям: перерабатывать низкока-
чественные бросовые продукты или ориентироваться на легко-
доступные углеводы и получать за их счет микробную биомассу,
содержащую высококачественный белок. И в том и в другом
случае технология ферментации играет ключевую роль. Здесь
важно подобрать оптимальный состав среды, создать опреде-
ленные условия для роста, разработать конструкции ферменте-
ров, правила их эксплуатации и системы контроля, выработать
методы отделения биомассы от культуральной среды. Промыш-
ленное производство белка одноклеточных организмов всегда
осуществляется методом глубинного культивирования в жидких
средах; применяются как одноэтапное, так и непрерывное куль-
тивирование. Непрерывное культивирование сложнее, чем одно-
этапное, но более экономично: производительность ферментеров
выше. Именно этот метод был избран для промышленного про-
изводства БОО.
Непрерывное культивирование
Метод непрерывного культивирования основан на поддержании
в системе динамического равновесия. Для перемешиваемой глу-
бинной культуры постоянного объема это означает постоянство
скорости роста микроорганизмов, которое обеспечивается путем
равномерного ее разбавления свежей питательной средой (при
сохранении объема). Среды, используемые при непрерывном
культивировании, всегда составляют таким образом, чтобы один
из субстратов (обычно это источник углерода) лимитировал
рост, поэтому его содержание в культуральной жидкости мини-
мально. Такой способ широко применяется в экспериментах по
физиологии микроорганизмов. Даже в микробиологических
лабораториях, где работа с чистыми культурами в асептических
условиях — обычное дело, опыты по непрерывному культивиро-
ванию требуют особого внимания. Специальное устройство ап-
паратуры, строгое соблюдение правил работы — все направлено
на то, чтобы избежать загрязнения посторонней микрофлорой.
Важность асептики при непрерывном культивировании стано-
вится особенно ясной, если учесть, что метод этот представляет
интерес как для лабораторий, так и для промышленности толь-
118
Глава 3
ко в том случае, если стационарное состояние культуры удается
поддерживать в течение нескольких дней, недель или даже ме-
сяцев. Все это время культуру надо перемешивать, подавать
непрерывно стерильную питательную среду и воздух и часть ее
постоянно удалять. Температура должна поддерживаться по-
стоянной; обычно регулируется и pH среды. Нужно периодиче-
ски отбирать пробы для контроля постоянства условий и чисто-
ты культуры. При промышленном производстве условия асеп-
тики могут быть соблюдены только в специальных сложных
установках. В их конструкции должна быть учтена необходи-
мость стерилизации перед началом каждого производственного
цикла. Это касается также работы любого датчика или пробо-
отборника, встроенного в ферментер для непрерывного наблю-
дения и контроля всех параметров культуры.
О мерах безопасности при производстве белка
одноклеточных организмов
Микроорганизмы, традиционно используемые в пищевой про-
мышленности, часто входят в состав конечного продукта (хотя
доля их там обычно невелика). Как показывает опыт, безопас-
ность этих продуктов не вызывает сомнений. Особенность белка
одноклеточных организмов заключается в том, что этот продукт,
во-первых, практически целиком состоит из микробной биомас-
сы и, во-вторых, в его производстве нередко принимают участие
микробы, опыт использования которых мал и которые ранее
в пище отсутствовали. Понятно, что государственные учрежде-
ния, контролирующие качество пищевых продуктов, требуют,
чтобы выходу на рынок БОО предшествовали испытания на
безопасность нового продукта. Такие испытания всегда дорого-
стоящи, и это сдерживает развитие производства, в частности
производства продуктов на основе БОО, особенно предназна-
ченных в пищу. По этой причине крен в развитии производства
БОО был сделан в сторону выработки кормов для животных,
а не белков, непосредственно идущих в пищу. Правила оценки
безвредности и способы тестирования продуктов, идущих на
приготовление кормов, менее жесткие, а роль эстетического
фактора не столь значительна. По этой причине для производ-
ства таких белков можно использовать более широкий круг суб-
стратов, в том числе и органические вещества отходов. К числу
БОО-продуктов, производимых промышленностью на корм жи-
вотным, относятся Pruteen фирмы ICI (биомасса бактерий, вы-
ращенных на метаноле), Toprina фирмы ВР (дрожжи, выра-
щенные на н-алканах) и грибная масса, получаемая по техно-
логии фирмы Finnish Pekilo. При ее производстве в качестве
субстрата используется сульфитный щелок, отход бумажной
Пищевые продукты, напитки и биотехнология 119
промышленности. Все эти БОО выпускаются в виде слабо окра-
шенных порошков.
БОО-продукты, предназначенные в пищу, немногочисленны.
Из их числа давно применяется дрожжевой экстракт (гидроли-
зат пекарских дрожжей). Вообще говоря, это не белковая пища,
так как используется он в небольшом количестве как вкусовая
И витаминная приправа. Кроме того, это скорее побочный про-
дукт бродильной промышленности, а не главный продукт про-
цесса производства БОО. Во время второй мировой войны в пи-
щевых целях в Германии выращивали дрожжи Candida, но это
производство не получило дальнейшего развития. Относительно
недавно фирма Hoechst стала выпускать на основе бактерий,
растущих на метаноле, продукт, содержащий 90% белка. Он
представляет собой одно из веществ, получаемых при фракцио-
нировании клеток выращенных бактерий. Сообщалось, что этот
белок обладает нужными функциональными свойствами и его
можно использовать в пищу. Процесс, разработанный фирмой
Hoechst, позволяет получат^ ряд продуктов лишь на основе зна-
чительных вложений и поэтому относится скорее к группе мало-
масштабных биотехнологий по градации, используемой в табл.
3.1. Единственный новый официально разрешенный вид белко-
вой пищи микробного происхождения — это микопротеин, про-
изводство которого налажено в Англии фирмой Ranks Hovis Me
Dougall.
3.5.3. Грибной белок (микопротеин)
Микопротеин — это пищевой продукт, состоящий в основном
из мицелия гриба. При его производстве используется штамм
Fusarium gratninearum, выделенный из почвы. И процесс, и про-
дукт — это результат осуществления программы по их интен-
сивному развитию, изучению и испытанию. Микопротеин про-
изводят сегодня на опытной установке методом непрерывного
выращивания. В качестве субстрата используется глюкоза и
другие питательные вещества, а источниками азота служат
аммиак и аммонийные соли. Общая схема функционирования
установки приведена на рис. 3.4. После завершения стадии фер-
ментации культуру подвергают термообработке для уменьшения
содержания рибонуклеиновой кислоты, а затем отделяют мице-
лий методом вакуумного фильтрования.
Если сопоставить производство микопротеина с процессом
-синтеза белков животных, то выявится ряд его преимуществ.
Помимо того что здесь выше скорость роста (это характерно
для производства всех БОО-продуктов), превращение субстра-
та в белок происходит несравненно эффективнее, чем при ус-
воении пищи домашними животными. Это отражено в табл. 3.6.
120
Глава 3
Рис. 3.4. Схематическое изображение производства микопротеина.
Работа ведется при соблюдении
правил гигиены
Работа ведется в асептических
условиях
Таблица 3.6. Эффективность конверсии при образовании белка для различных
животных и Fusarium graminearum
Исходный продукт Продукция
белок, г общая, г
Корова 1 кг корма 14 68 говядины
Свинья То же 41 200 свинины
Курица » 49 240 мяса
Fusarium grami- 1 кг углеводов + неор- 136 1080 клеточной
nearum ганический азот массы, сырая масса
Нелишне напомнить, что корма для животных должны содер-
жать некоторое количество белка, до 15—20% в зависимости
от вида животных и способа их содержания. Положительным
фактором является и волокнистое строение выращенной куль-
туры (рис. 3.5); текстура массы мицелия близка к таковой у
естественных продуктов, поэтому у продукта может быть ими-
тирована текстура мяса, а за счет добавок — его вкус и цвет.
Плотность продукта зависит от длины гиф выращенного гриба,
которая определяется скоростью роста. На рис. 3.6 показано,
как выглядит готовый сформованный продукт. Для хранения
его обычно замораживают, но иногда и высушивают до порош-
ка путем распыления.
После проведения всесторонних исследований питательной
ценности и безвредности микопротеина министерство сельского
Пищевые продукты, напитки и биотехнология
121
Рис. 3.5. Строение гиф Fusarium graminearum. Микрофотография получена с
помощью сканирующего электронного микроскопа.
Рис. 3.6. Так выглядит искусственная пища из грибного белка микопротеина.
122
Глава 3
хозяйства, рыболовства и пищевых продуктов дало разрешение
на его продажу в Англии. Содержание основных питательных
веществ в нем указано в табл. 3.7.
Таблица 3.7. Средний состав микопротеина
и сравнение его с составом говядины
Состав, % (на сухой вес)
Компоненты микопротенн постный сы- рой бифштекс
Белки 47 68
Жиры , 14 30
Пищевые волокна 25 Следы
Углеводы 1 10 0
Зола \ 3 2
РНК (рибонуклеино- вая кислота) 1 Следы
3.6. Пищевые добавки и ингредиенты
3.6.1. Подкислители v
Подкислители применяются в основном как вкусовые добавки
для придания продуктам «острого» вкуса. В практику они во-
шли скорее всего в результате широкого использования орга-
нических кислот для сохранения пищи.
По-видимому, самым популярным подкислителем в пищевой
промышленности сегодня является лимонная кислота. Сначала
этот важный продукт получали, отжимая сок из лимонов
(в Италии): таким путем до начала 20-х годов удовлетворялось
три четверти мировой потребности в лимонной кислоте. Сегодня
ее получают при участии A. niger, сбраживая мелассу и содер-
жащие глюкозу гидролизаты. Отметим, что процесс сбражива-
ния нужно строго контролировать, так как лимонная кислота
в отличие от других вторичных метаболитов играет важную
роль в регуляции обмена веществ. При консервировании поми-
доров широко используют яблочную кислоту; ее образует A. fla-
vus. К числу других кислот, находящих широкое применение в
пищевой промышленности, относятся итаконовая (продуцент —
A. terreus), глюконовая, используемая в форме глюконолактона
(продуцент — A. niger), и фумаровая (виды Rhizopus).
3.6.2. Аминокислоты (см. также гл. 4)
Сегодня во всем мире производится более 200 000 т аминокис-
лот в год; их используют главным образом как добавки к кор-
мам и пищевым продуктам. Основную часть аминокислот полу-
Пищевые продукты, напитки и биотехнология 123
чают методами ферментации, но все большее значение приобре-
тают химический и ферментативный синтез.
Главными продуктами, получаемыми по технологии фермен-
тации, являются глутаминовая кислота (продуцент — Согупе-
bacterium glutamicum) и лизин (В. flavum). В большом коли-
честве производят еще две аминокислоты, глицин и метионин,
но до сих пор только методом химического синтеза.
3.6.3. Витамины и пигменты
Основные потребности промышленности в этих соединениях
удовлетворяются за счет природных источников и химического
синтеза, но два из них, ^-каротин и рибофлавин, традиционно
получают методами биотехнологии. Последний образуется при
глубинном культивировании Eremothecium ashbyii или Ashbya
gossypii. Возможности микробиологического производства не-
токсичных пищевых красителей исчерпаны далеко не полно-
стью; р-каротина сегодня вырабатывают очень мало, а важную
группу красителей красного цвета до сих пор получают из вино-
градных выжимок (экстракта).
3.6.4. Усилители вкуса .
Расщепляя с помощью нуклеазы Penicillum citrinum нуклеино-
вые кислоты, получают в промышленном масштабе б'-нуклео-
тиды (содержащие главным образом инозин и гуанин), которые|
находят применение как усилители вкуса. Вещества, усиливаю-
щие оттенки вкуса, содержатся и в природных пищевых продук-
тах. Главным усилителем вкуса считается натриевая соль
глутаминовой кислоты: ее можно получить при помощи Micro-
coccus glutamicus. Пионером использования усилителей вкуса
является Япония, но сам принцип применялся при создании ре-
цептов многих блюд во всем мире.
3.6.5. Жиры и масла
Анализ перспектив использования жиров и масел, продуцируе-
мых бактериями (в особенности содержащих необычные поли-
ненасыщенные жирные кислоты, а потому дорогостоящих), по-
казал, что в нынешних экономических условиях использовать
эти вещества вместо жиров растительного и животного проис-
хождения нецелесообразно.
3.6.6. Растительные клеи и загустители
Натуральные растительные клеи, получаемые из камедей рас-
тений или морских водорослей, принадлежат к числу давно из-
вестных пищевых добавок; в Китае, например, они широко
124
Глава 3
употреблялись еще в первом столетии до нашей эры, а об их
использовании упоминается даже в Ветхом завете. Раститель-
ные клеи относятся к группе пищевых добавок — гидроколлои-
дов. Их применяют для изменения консистенции пищевых про-
дуктов: эти вещества способствуют их загущению и образова-
нию студней. Структура пищевых продуктов при этом стабили-
зируется, улучшаются их внешний вид и вкус.
Большое значение для этой отрасли имеет разработка про-
мышленного производства полисахаридов из Pseudomonas spp.
Их предполагается использовать вместо глюкоманнозы, одного
из наиболее широко применяемых загустителей. Свойства этого
нового продукта детально изучены. В кондитерской промышлен-
ности и при производстве мороженого в качестве стабилизатора
сегодня широко используется декстран из Leuconostoc mesen-
teroides (гл. 5).
3.7. Фрукты и овощи
3.7.1. Консервированные овощи
Как и в случае многих других разновидностей пищевого сырья,
необходимость сохранения овощей для употребления их в тече-
ние всего года привела к созданию ряда новых пищевых про-
дуктов. До того как в практику вошли консервирование в бан-
ках и замораживание, для сохранения овощей использовалась
главным образом соль. Низкие концентрации соли (2—2,5%)
при переработке содержащих сахара овощей с малой долей
белков не препятствуют брожению с образованием кислот, иду-
щему при участии бактерий. Этот способ дает хорошие резуль-
таты, но если белка в овощах много (горох, фасоль),то продукт
портится. Если такие овощи засаливают, то соли добавляют
столько, чтобы полностью подавить брожение. Когда соли до-
бавляют мало, основную роль в консервации играют молочно-
кислые бактерии. Образование молочной кислоты из сахаров
препятствует развитию бактерий кишечной группы, протеолити-
ческих бактерий, анаэробных и спорообразующих видов.
Кислую капусту готовят из свежей измельченной капусты.
После добавления соли на первых стадиях брожения доминиру-
ют бактерии Leuconostoc mesenteroides, которые в анаэробных
условиях превращают сахара в молочную и уксусную кислоты,
этиловый спирт, маннитол, эфиры и СО2. В дальнейшем обра-
зование молочной кислоты из сахаров и маннитола осуществ-
ляется при участии Lactobacillus plantarum. Разложение ман-
нитола — важный этап, так как он придает продукту горький
вкус. Хотя при получении кислой капусты условия сбраживания
Пищевые продукты, напитки и биотехнология 125
в какой-то степени контролируются, применять закваски нет не-
обходимости, так как никаких преимуществ они не дают.
Пикули делают из засоленных огурцов. Конечный продукт
получают путем полного или частичного сбраживания либо без
такой обработки. При брожении важную роль опять-таки игра-
ют молочнокислые бактерии, осуществляющие ферментацию
сахаров. Соли обычно добавлят немного, и рассол с самого на-
чала подкисляют уксусной кислотой. Если к нему добавляют
укроп и другие пряности, то получают укропные пикули.
Оливки перерабатывают путем засолки и обработки щело-
ком. Когда консервируют зеленые плоды, молочнокислое бро-
жение осуществляется при участии Leuconostoc mesenteroid.es,
а затем Lactobacillus planarum и продолжается от шести до
десяти месяцев. Спелые оливки либо не сбраживают вовсе,
либо сбраживают недолго. И в том и в другом случае большое
значение имеет обработка щелоком, так как при этом удаляется
олеуропеин (глюкозид, имеющий горький вкус).
Микроорганизмы играют важную роль и на определенных
стадиях выработки некоторых других продуктов, особенно кофе
и какао. При замочке плодов на них развиваются молочнокис-
лые бактерии и дрожжи, что способствует отделению кожуры
от зерен; влияние микробов на качество конечного продукта
незначительно.
Приведенные примеры использования микроорганизмов в пе-
реработке овощей — это процессы с простой технологией. Хотя
в случае засолки деятельность микробов и определяет основные
свойства конечного продукта, суть ее проста — это в основном
образование кислоты. Сходную продукцию можно получить за
более короткое время — для этого достаточно добавить к све-
жим овощам соответствующие кислоты и тем самым обойтись
без брожения. Отметим, что, хотя для сохранения овощей ис-
пользуются главным образом консервирование в банках и за-
мораживание, спрос на заквашенные продукты сохраняется, так
как они обладают специфическим вкусом.
3.7.2. Продукты из сои
Соя издавна принадлежит к числу главных пищевых культур
в странах Азии, особенно в Китае и Японии. В восточной кухне
она служила главным поставщиком белка и масла задолго до
того, как ее стали использовать как источник этих веществ в
странах Запада. На основе соевых бобов на Востоке вырабаты-
вают множество традиционных пищевых продуктов; их особый
вкус определяется деятельностью микроорганизмов. Это глав-
ным образом грибы, в частности представители рода Aspergil-
lus.
•126
Глава 3
Соевый соус готовят на основе кашицы из набухших и отва-
ренных бобов сои. В нее вносят закваску, содержащую разнооб-
разные микроорганизмы, главным образом Aspergillus oryzae.
В ходе выдержки в течение 3—5 сут при 25—30° гриб активно
разрастается на поверхности. Затем в смесь добавляют соль
(до 20%) и оставляют ее созревать на 0,5—2 года. В настоя-
щее время применяют чистые культуры A. oryzae, поэтому срок
.выдержки сокращается до одного-трех месяцев. В завершение
жидкость сливают с массы или отделяют под прессом и полу-
чают соевый соус. Помимо ускорения процесса путем исполь-
зования чистых культур разработаны и сугубо химические спо-
собы получения соевых гидролизатов. Так готовят несброжен-
ный соевый соус.
В Японии из размельченных соевых бобов (иногда с добав-
кой риса) делают похожую приправу — тамари. Продолжитель-
ность сбраживания при этом меньше, чем в случае китайского
соевого соуса, а ведущую роль в процессе играет Aspergillus
tamari. Еще одну приправу, мизо, готовят в Японии из отварен-
ных соевых бобов, которые заражают A. oryzae,. добавляют к
ним соль и воду и ставят на созревание на несколько дней, по-
сле чего тщательно измельчают. Когда делают натто, сваренные
соевые бобы укутывают в рисовую солому и ставят на созрева-
ние на 1—2 сут. Главную роль здесь играет Bacillus subtilis,
-образующая слизистую пленку.
В восточной кухне часто используется соевый сыр, или курд.
Соевые бобы сначала замачивают и измельчают в пасту, а за-
тем отжимают ее через полотно. Для створаживания применяют
соли кальция и магния и полученный продукт формуют в виде
кубиков. В ходе выдержки в течение месяца при 14 °C активно
развиваются белые плесневые грибы. Окончательное созревание
происходит в рассоле или особым образом сделанном вине.
Современная биотехнология, видимо, лишь в малой степени
может способствовать развитию этих традиционных производств.
Все произошедшие здесь изменения заключались обычно в
уменьшении времени ферментации; в отдельных случаях про-
цессы стали безмикробными. Впрочем, возможно дальнейшее
усовершенствование процессов путем улучшения штаммов и
контроля за ферментацией. Кроме того, соевые бобы могут
стать тем сырьем, из которого на основе традиций восточной
кухни для рынка как Запада, так и Востока можно будет полу-
чать совсем новые продукты ферментации. В этих случаях пере-
рабатываются целые бобы, однако известны уже и полученные
с помощью биотехнологии новые продукты из белков сои. Их
вырабатывают путем контролируемого гидролиза белков сои
•ферментами микроорганизмов. Свойства таких продуктов (рас-
творимость, вкус и т. д.) можно в определенной мере регулиро-
Пищевые продукты, напитки и биотехнология 127
вать. Наиболее привлекательным из них представляется изо-
электрический растворимый гидролизат белков сои. Как заме-
нитель мяса он лучше, чем блюда из соевых бобов. В странах,,
где население получает с пищей недостаточно белка, им обога-
щают безалкогольные напитки.
3.7.3. Применение ферментов при выработке фруктовых соков
Применение4ферментов из микроорганизмов — один из главных,
путей, которые биотехнология использует и будет использовать-
для обновления пищевой промышленности. Наибольшие успехи
были достигнуты при производстве фруктовых соков: здесь ис-
пользуют такие ферменты, как пектиназы, целлюлазы, гемицел-
люлазы, амилазы и протеиназы. Эти ферменты применяются
не только в давно освоенных производствах; с их помощью
удалось расширить ассортимент и добиться большего выхода
продукции из сырья. Ферменты используются на следующих
основных стадиях переработки фруктов. 1. Обработка мезги:
разрушение мякоти при выработке фруктовой кашицы или нек-
таров; увеличение выхода сока; лучшее отделение веществ, от-
ветственных за цвет и вкус. 2. Обработка сока: уменьшение
вязкости; облегчение изготовления концентратов; упрощение
процедур осветления, фильтрования и стабилизации сока.
Выбор ферментов и способов их применения для получения
наилучших результатов при выработке соков производится с
учетом трех обстоятельств. 1. Активности фермента. Для полу-
чения желаемого действия при производстве фруктовых соков-
необходимо подобрать правильную концентрацию, температуру
и продолжительность обработки. 2. Необходимости деградации
пектина. Качество продукции сильно зависит от типа пектина
и свойств конкретного фермента. 3. Механизмов осветления.
Этот процесс состоит из трех последовательных стадий. Пони-
мание их особенностей исключительно важно при производстве
соков: это стадии дестабилизации, коагуляции и осаждения.
При производстве соков из фруктов большую роль играют
пектины. Если их слишком много, то соки получаются излишне
вязкими и мутными. Контролируя содержание пектинов, удает-
ся существенно повысить выход сока и его качество. Пектино-
вые вещества, или пектины, представляют собой гетерополиса-
хариды, производные галактуроновой кислоты. Они являются
субстратами для ряда ферментов: пектинлиаз, пектатлиаз, по-
лигалактуронидаз или пектинэстераз. Действие их заключается
в деполимеризации путем расщепления глюкозидных связей;,
пектиноэстеразы атакуют этерифицированные карбоксильные
группы.
128
Глава 3
3.8. Заключение
3.8.1. Ближайшие перспективы
По оценкам примерно 15% реализуемой продукции пищевой
промышленности вырабатывается на основе биотехнологии, но
влияние ее на эту промышленность сегодня не больше, чем 25
лет назад (Tonge, Jarman, 1981). Определяется это тремя при-
чинами. 1. Производство пищевых продуктов и сегодня являет-
ся трудоемкой отраслью промышленности с большим объемом
ручного труда и низким уровнем технологии. Многие производ-
ственные процессы в ней есть не что иное, как увеличенные
копии кулинарных приемов. Наука, изучающая их основы, сла-
бо развита, а суть самих процессов не до конца понята.
2. В «табеле о рангах» биотехнологии, о котором речь шла в
начале этой главы, изготовление пищевых продуктов относится
к крупнотоннажным производствам с малой прибавочной стои-
мостью. В сущности именно малая прибавочная стоимость пи-
щевых продуктов мешает вкладывать средства в изучение и
развитие производства и методов определения безопасности
продуктов, а это необходимо для того, чтобы сделать биотехно-
логические процессы и продукты конкурентоспособными.
3. Главные усилия при модернизации производства пищевых
продуктов должны быть направлены на уменьшение или полное
подавление нежелательных изменений, вызываемых биологиче-
ски активными побочными продуктами. Сегодня для описания
таких процессов применяется термин «отрицательная биотехно-
логия»; по сути здесь решаются задачи сохранения пищи. Вне
зависимости от термина такие исследования все же не относятся
к числу важнейших в биотехнологии. В ближайшем будущем
новой технологии основные успехи будут достигнуты, по-види-
мому, при производстве продуктов с высокой прибавочной стои-
мостью: пищевых и вкусовых добавок, пищевых красителей и
функциональных агентов. Выпуск таких продуктов в чем-то
сходен с производством новых лекарственных препаратов в фар-
мавцевтической промышленности: обязательны испытания на
безвредность; необходимы надежные источники сырья с высо-
кой, но приемлемой стоимостью.
3.8.2. Отдаленные перспективы
Затраты на организацию многотоннажных биотехнологических
производств столь велики, что лишь фирмы, способные осуще-
ствлять долгосрочные стратегические программы, могут решить-
ся на внедрение такой технологии. Главное преимущество новой
технологии заключается в том, что выход продукции с фермен-
Пищевые продукты, напитки и биотехнология
129
тера или биореактора несравненно выше, чем от растения или
животного, так что производство предметов потребления таким
способом всегда оказывается более выгодным.
Как это ни парадоксально, именно продуктивность оказалась
тем важным фактором, который повлиял на внедрение этой тех-
нологии. Планируемая замена привычных предметов потребле-
ния сказалась на финансовой сфере, и это повредило экономике.
Подобные неблагоприятные явления относительно часто проис-
ходят при производстве продуктов с малой прибавочной стои-
мостью. В последние годы они сопутствовали производству
изоглюкозы. Развитие этой технологии в(странах ЕЭС сдержи-
валось системой квот и налогов. В то же время в США про-
грамма по производству спирта-горючего получила поддержку
правительства (Economic Development Administration, 1981)
в форме дотаций и налогов в масштабах всей страны и отдель-
ных регионов. По-видимому, искусственно привносимые влияния
на экономическую жизнеспособность той или иной программы
будут иметь место и впредь: нужно учитывать, что сельское хо-
зяйство играет в экономике стран особую роль.
Однако в будущем, когда потребности в той или иной про-
дукции превысят возможности традиционных технологий, пре-
имущества, связанные с большей продуктивностью, все же про-
явятся. Примером того, как биотехнология помогает решать за-
дачи, стоящие перед пищевой промышленностью, может быть
такой факт: лимонную кислоту сегодня получают с помощью
грибов, и в результате земельные площади в Европе не при-
шлось занимать посадками лимонных деревьев.
ЛИТЕРАТУРА
Arima К,. (1977). Recent developments and future directions of fermentations in
Japan, Devs ind. Microbiol., 18, 78—117.
Aunstrup K,. (1979). Production of extracellular enzymes. In: Applied Bioche-
mistry and Bioengineering, Vol. 2 (eds. Wingard L. B., Katchalski-Katzir E,
and Goldstein L.), pp. 27—69, Academic Press, London.
Birch G. G., Blakebrough N., Parker K. J. (eds.) (1981). Enzymes and Food
Processing, Appl. Sci. Publ., London.
Beech F. W. (1972). Cider making and cider research: a review, J. Inst. Brew.,
78, 477—490.
Beauchat L. R. (ed.) (1978). Food and Beverage Mycology, AVI Publishing,
Westport, Connecticut.
Collins T. H. (1968). A Summary of Breadmaking Processes, Flour Milling and
Baking Research Association Report Number 13.
Economic Development Administration (1981). Alternative Utilization of Wheat
Starch, US Department of Commerce, Washington DC.
Glicksman M. (1969). Gum Technology in the Food Industry, Academic Press,
London.
Green M. L. (1977). Review of the progress of dairy science: milk coagulants.
J. Dairy Res., 44, 159—188.
9—1344
130
Глава 3
Greenshields R. N. (1978). Acetic Acid: vinegar. In: Primary Products of Meta-
bolism (ed. Rose A. H.), pp. 121—186, Academic Press, London.
Herbert D., Elsworth R., Telling R. C. (1956) . The continuous culture of bacteria;
a theoretical experimental study, J. gen. Microbiol., 14, 601—622.
Hersorn A. C., Hulland E. D. (1980). Canned Foods. Thermal Processing and:
Microbiology, p. 43, Churchill Livingstone, Edinburgh.
Hirose У., Sano K., Shibai H. (1978). Amino acids. In: Annual Reports on Fer-
mentation Processes, Vol. 2 (ed. Perlman D.), pp. 155—189, Academic Press,
London.
Howling D. (1979). The general science and technology of glucose syrups. In:
Sugar: Science and Technology (eds. Birch G. G. and Parker K. J.), pp. 259—
285, Appl. Sci. Publ., London.
Jarman T. R. (1979). Bacterial alginate synthesis. In: Microbial Polysaccharides-
and Polysaccharases (eds. Berkeley R. C. W., Gooday G. W. Ellwood D. C.),
pp. 35—50, Academic Press, London.
Jarvis B., Holmes A. W. (1982). Biotechnology in relation to the food industry,
J. Chem. Tech. BiotechnoL, 32, 224—232.
Jarvis B., Paulus K. (1982). Food preservation: an example of the application of
negative biotechnology, J. Chem. Tech. BiotechnoL, 32, 233—250.
Kooi E. R., Armbruster F. C. (1967). Production and use of dextrose. In: Starch:
Chemistry and Technology, Vol. II (eds. Whistler R. L. and Paschall E. F.),
pp. 553—568, Academic Press, London.
Kunkee R. E. (1967). Malo-lactic fermentation. In: Advances in Applied Micro-
biology, Vol. 9 (ed. Umbreit W. W.), pp. 235—279, Academic Press, London.
Lancrenon X. (1978). Recent trends in the manufacturing of natural red colours,
Process Biochem., 13, no. 10, 16.
Law B. A. (1980). Accelerated ripening of cheese, Dairy Inds, Inti, May 1980, 17,
Law B. A. (1981). Short cuts to faster flavour, Food, March 1981, 17—19.
Lorenz K. (1981). Sourdough processes — methodology and biochemistry, Ba-
kers’ Dig., 55, 32—36.
MacLeod G„ Seyydain-Ardebili M. (1981). Natural and simulated meat flavors,
Crit. Rev. Food Sci. Nut., 14, 309—437.
Maga J. A. (1974). Bread Flavor, Crit. Rev. Food Technol., 5, 55—142.
Radley J. A. (ed.) (1976). Starch Production technology, Appl. Sci. Publ.,
London.
Rankine В. C. (1972). Influence of yeast strain and malo-lactic fermentation ore
composition and quality of table wine, Am. Enol. Vitic., 23, 152—158.
Ratledge C. (1979). Resources conservation by novel biological processes. 1.
Grow fats from wastes, Chem. Soc. Rev., 8, 283—295.
Reed G. (ed.) (1975). Enzymes in Food Processing, Academic Press, London.
Reed G. (ed.) (1982). Prescott and Dunn’s Industrial Microbiology, 4th end.
Macmillan, London.
Robinson R. K. (1981). Dairy Microbiology, Vol. 2, Appl. Sci. Publ., London.
Rose A. H. (ed.) (1977). Alcoholic Beverages, Academic Press, London.
Rose A. H. (ed.) (1982). Fermented Foods, Academic Press, London.
Schierholt J. (1977). Fermentation processes for the production of citric acid.
Process Biochem., 12, no. 9, 20—21.
Schlingmann M., Faust U., Scharf U. (1981). Bioproteins for human nutrition,
FIE Conference, 1981, London.
Scharma S. C. (1981). Gums and hydrocolloids in oil-water emulsions, Food
Technol., 35, no. 1, 59—67.
Sittig W. (1982). The present state of fernentation reactors, J. Chem. Tech. Bio-
technol., 32, 47—58.
Sodeck G., Modi J., Kominek J., Salzbrunn W. (1981). Production of citric acid
according to the submerged fermentation process, Process Biochem., 16 no- 6,
9—11.
Пищевые продукты, напитки и биотехнология 131
Spinks A. (Chairman) (1980), Biotechnology: Report of a Joint Working Party,
p. 35, HMSO, London.
Stevens T. J. (1966). A method of selecting pure yeast strains for ale fermenta-
tions, J. Inst. Brew., 72, 369—373.
Tonge G. M., Jarman T. R. (1981). Opportunities for biotechnology in the food
industry, FIE Conference, 1981, London.
Whitworth D. A., Ratledge C. (1974). Microorganisms as a potential source of
oils and fats, Process Biochem., 9, no. 9, 14—22.
Wierzbicki L. E., Kosikowski F. V. (1972). Food syrups from acid whey treated
with beta-galactosidase of Aspergillus niger, J. Dairy Sci., 56, 1182—1184.
Williams A. (1975). The history of the Chorleywood bread process. In: Bread-
making: The Modern Revolution (ed. Williams A.), pp. 25—39, Hutchinson
Benham, London.
Yamada K., Kinoshita J., Tsunoda T., Aida K. (1972). The Microbial Production
of Amino Acds by Fermentation, John Wiley and Sons, New York.
9‘
Глава 4
Химия и биотехнология
Д. Бест
4.1. Введение
Во всех главах этой книги, так или иначе, затрагивается вопрос
об использовании удивительной способности биологических си-
стем к узнаванию и выполнению каталитических функций. Быть
может, наиболее эффективное применение такие системы могут
найти в современной химической промышленности. Мы знаем,
что биомасса тоже представляет собой сложную химическую
систему, а большинство процессов и продуктов биотехнологии
имеют биохимическую природу, будь то производство веществ,
используемых как горючее (гл. 2), получение путем брожения
продуктов питания и напитков (гл. 3), синтез биополимеров
(гл. 5), использование организмов, участвующих в круговороте
химических веществ на Земле (гл. 6), применение сложных
химических соединений в медицине (гл. 8) или сельском хозяй-
стве (гл. 9). В этой главе речь пойдет главным образом о прин-
ципах, перспективах и технологии получения химической про-
дукции на базе биотехнологии. Основное внимание будет уде-
лено химическим процессам и соединениям, которые в других
главах не обсуждаются.
Корни современной прикладной микробиологии и соответст-
венно биотехнологии уходят в химическую промышленность на-
чала нынешнего века: именно тогда были разработаны основы
промышленного производства ряда химических веществ (напри-
мер, ацетона, этилового спирта, бутандиола, бутанола и изопро-
панола) из углеводов растений (об истории развития таких про-
изводств рассказано в гл. 1). На смену этой важной отрасли
промышленности пришла быстро развившаяся нефтехимическая
промышленность. Однако сейчас запасы ископаемого сырья
стали предметом конкуренции, так как оно требуется для про-
изводства химических веществ, энергии и даже пищевых про-
дуктов; все это усугубляется повышением цен на нефть и уголь.
В таких условиях применение процессов нового типа при про-
изводстве химических веществ из возобновляемой биомассы
становится все более перспективным. Идет переоценка возмож-
Химия и технология
133
ностей использования модернизированных процессов, по сути
своей подобных разработанным в начале века.
В этом направлении уже сделаны первые шаги. Так, глице-
рол получают с помощью водорослей; активно ведется работа
по изучению возможности использования лигнина древесины.
С технической точки зрения это сложная проблема: решение ее
откроет новый путь получения многих ценных ароматических
соединений, играющих столь важную роль в современной хи-
мической промышленности.
| Помимо новых способов получения химических веществ из
биомассы биотехнология дает нам также более эффективные и
производительные катализаторы для осуществления химических
। взаимопревращений. Они могут использоваться двояким путем.
Во-первых, специфичность действия фермента (или ферментов)
! может применяться при катализе in vivo или in vitro простых,
1 но технически трудно осуществимых превращений. Примером
такого рода служит гидроксилирование стероидов при синтезе
лекарственных веществ. Во-вторых, с их помощью можно осу-
ществлять полный биосинтез сложных и дорогостоящих тонких
химических продуктов из простых составных частей, например
образование антибиотиков в ходе вторичного метаболизма у гри-
бов. В табл. 4.1 приведены различные варианты использования
биокаталитической активности. Многообещающей областью
дальнейшего развития представляется производство ценных ве-
ществ из растений [среди которых упомянем терпены (арома-
тические вещества) и алкалоиды (используемые при производ-
стве лекарств)] путем массового культивирования клеток рас-
тений.
4.2. Развитие современной химической биотехнологии
4.2.1. Бродильное производство растворителей
О бродильном производстве глицерола шла речь в гл. 1, а о по-
лучении этанола — растворителя, пищевого продукта, промежу-
точного вещества в реакциях химического синтеза и горючего —
в гл. 2. К числу других важных бродильных производств отно-
сится получение ацетона и бутанола. Впервые в промышленном
масштабе они были осуществлены в Манчестере Вейсманном в
ходе первой мировой войны. Ацетон был необходим для произ-
водства кордита и как метательное взрывчатое вещество в тя-
желой артиллерии. До начала военных действий его импорти-
ровали из Германии. Ацетон низкого качества получали путем
сухой перегонки древесины, но для упомянутых целей нужен
был высококачественный растворитель.
134
Глава 4
Таблица 4.1. Биотехнология и химическая промышленность
Биокатализ на стадии роста
Нерегулируемый
Изменение среды с целью модифика-
ции метаболизма
Получение мутантов, способных
к сверхпродукции промежуточных
продуктов метаболизма
Индукция определенных фермента-
тивных процессов
Ингибированная ферментация
Направленный синтез из предшест-
венников в обход метаболического
контроля
Биокатализ по завершении роста
Одностадийные превращения, позво-
ляющие обойтись без очистки фер-
меита или принятия мер по сохра-
нению его стабильности
Многостадийные процессы фермента-
тивной конверсии
Биокатализ in vitro
Использование очищенного фермен-
та для одностадийной конверсии
какого-либо природного субстрата
Одностадийное образование химиче-
ских промежуточных продуктов из
неприродных субстратов с исполь-
зованием очищенных ферментов
с широким спектром действия
Многостадийные полусинтетические
метаболитические процессы
Химический катализ на основе био-
логических принципов
Создание химических аналогов фер-
ментативных процессов
Получение химических катализаторов
с биологической специфичностью
(образование «биоорганических
комплексов»)
Белок одноклеточных организмов
Органические кислоты, аминокислоты
Аминокислоты
Антибиотики
Аминокислоты, антибиотики
Изомеризация глюкозы, получение
глюконата, трансформация стерои-
дов
Трансформация стероидов
Глюкозоизомер аза, аминокислоты
Получение гидрохинона с помощью
глюкозооксидазы
Бродильный процесс (ферментация) был основан на перера-
ботке крахмала, концентрация которого составляла до 3,8%
(вес/объем), анаэробными спорообразующими бактериями
Clostridium acetobutylicum. Превращению подвергалось до 30%
субстрата, в результате чего получалась смесь растворителей
(60 /о бутанола, 30% ацетона, 5—10% этанола, изопропанола и
мезитилоксида). Остальная часть субстрата в ходе процесса,
представленного на рис. 4.1, превращалась в водород и угле-
кислый газ.
Поскольку образовывались большие объемы газов, при круп-
номасштабном производстве перемешивания не требовалось,
Химия и технология
135
Крахмал
Глюкоза
Г люкозо-6-фосфат
Триозофосфат
Рис. 4.1. Схема реакций ацетон-бутанольного брожения.
а главная сложность заключалась в гашении пены. В зависи-
мости от штаммов отношение ацетон : спирт несколько варьиро-
вало. Многие микробы, разрушающие крахмал и способные
образовывать растворители, могут также сбраживать мелассу
при содержании сахара в среде до 6% (вес/объем). Фактором,
определяющим количество использованного субстрата, оказа-
лась чувствительность организмов, участвующих в процессе,
к н-бутанолу (верхний предел — около 1,2%, по объему) и аце-
тону (0,4%). Заражения бродильных емкостей аэробными бак-
териями обычно не происходило, и главной проблемой была
инфекция бактериофагами. Впоследствии выяснилось, что уча-
ствующие в процессе микроорганизмы можно «иммунизировать»
путем нескольких пересевов в присутствии бактериофага. Было
установлено, что фаговая инфекция является штамм-специфич-
ной.
Растворители отделяли от среды отгонкой. В конце первой
мировой войны главную роль стало играть производство бута-
нола: он нашел применение при получении широкого круга
136
Глава 4
веществ, включая мочевиноформ альдегидные пластмассы, пла-
стификаторы и тормозные жидкости. Побочный продукт, водо-
род, стал использоваться в производстве синтетического метано-
ла и для гидрогенизации пищевых масел; углекислый газ либо
сжижали, либо превращали в сухой лед. Твердые вещества
отходов содержали большое количество рибофлавина (витами-
на Вг), и их можно было использовать как богатую белком
добавку к кормам.
После второй мировой войны бродильное производство этих
растворителей, однако, сильно сократилось, так как относитель-
ная стоимость нефтехимических продуктов по сравнению с по-
лимерами сахаров уменьшилась. Производство н-бутанола
путем ферментации продолжалось лишь в ЮАР. Однако в на-
стоящее время получение бутанола с помощью ферментации
становится все более выгодным, и очень может быть, что мето-
дами генетической инженерии удастся создать такие микроор-
ганизмы-продуценты, применение которых перетянет чашу весов
в сторону этого способа. Главный недостаток существующих
штаммов — низкая устойчивость к конечным продуктам и отно-
сительно низкий выход растворителей.
В ходе второй мировой войны активно исследовалась воз-
можность получения бутилен-2,3-гликоля; в последнее время
к этому процессу микробиологической конверсии также прояв-
ляется интерес. Одной из проблем молочной промышленности
является использование сыворотки. Между тем она может быть
источником углерода при образовании бутиленгликоля бакте-
риями Klebsiella pneumoniae или Enterobacter aerogenes, кото-
рый превращают затем в сырье для производства синтетическо-
го каучука.
4.2.2. Производство органических кислот
Среди органических кислот самая важная — уксусная. На ры-
нок США ее ежегодно поступает около 1,4 млн. т общей стои-
мостью до 500 млн. долл, (без учета уксуса). В прошлом ос-
новную часть уксусной кислоты получали путем микробиологи-
ческого окисления этанола, но сегодня, за исключением
производства уксуса, этот процесс по экономическим соображе-
ниям не применяется. Впрочем, в результате ведущихся иссле-
дований термофильных бактерий, способных превращать цел-
люлозу в уксусную кислоту, а также штаммов Acetobacter и
Clostridium, способных синтезировать ее из водорода и угле-
кислого газа, этот метод, может быть, восстановит свои позиции.
Техническая уксусная кислота используется при выработке
многих химических веществ, включая каучук, пластмассы, во-
локна и инсектициды. При обычном способе производства мик-
Химия и технология
137
робиологическая конверсия этанола в уксусную кислоту при
участии штаммов Acetobacter и Gluconobacter идет в аэроб-
ных условиях и поэтому, строго говоря, не является процессом
брожения. Уксус по праву считается важнейшим продуктом
микробиологической промышленности (см. гл. 3).
В конце XIX в. началось промышленное производство мо-
лочной кислоты при участии молочнокислых бактерий Lactoba-
cillus delbrueckii, L. leichmannii и L. bulgaricus. Это был один
из первых процессов, где применялась частичная стерилизация
среды нагреванием. Этот микроаэрофильный процесс осуществ-
ляется при высокой температуре (45—50°). В нем используют
содержащее крахмал сырье, которое предварительно обраба-
тывают ферментами или подвергают кислотному гидролизу
(разд. 4.2.6). L. bulgaricus активно сбраживает лактозу и может
поэтому использовать молочную сыворотку в качестве питатель-
ного субстрата. В других случаях конверсии подвергается саха-
роза (концентрация 12—18%, вес/объем). Процесс идет 3—
4 суток; при этом в больших количествах выделяется углекис-
лый газ, что облегчает создание в среде оптимальных полуаэроб-
ных условий. Описаны также способы конверсии 1,2-пропандио-
ла в молочную кислоту при помощи Arthrobacter oxydans, Alca-
ligenes faecalis или Fusarium solani. Эти микроорганизмы в
основном образуют L( + ) -изомер молочной кислоты, но некото-
рые штаммы L. leichmanii синтезируют D(—)-изомер. Было
изучено образование молочной кислоты при непрерывном куль-
тивировании. В одностадийном процессе выход в случае L. del-
brueckii составлял 89 г/л в сутки. При использовании препара-
тов молочнокислых бактерий, иммобилизованных в альгинатных
гелях, степень конверсии достигала 97%. Доля L( + )-изомера
составляла 90%, а время полужизни—100 суток. В этих про-
цессах молочную кислоту получают в форме кальциевой соли;
чтобы выделить конечный продукт, ее обрабатывают серной
кислотой. Молочную кислоту используют в качестве добавки к
безалкогольным напиткам, эссенциям, фруктовым сокам, дже-
мам и сиропам, для декальцификации кож в дубильной про-
мышленности, а также при производстве пластмасс, когда
L( + )-форму полимеризуют в полилактат, применяемый для
производства пластиковых оберток. Соли молочной кислоты ис-
пользуются в медицине.
Производство лимонной кислоты методом ферментации при
участии грибов (рис. 4.2) также принадлежит к числу давних
биотехнологических процессов; оно было налажено в 1893 г.
Его развитие шло в тесной связи с разработкой многих фунда-
ментальных аспектов микробиологии. Вначале основные проб-
лемы были связаны с микробным загрязнением. В поисках их
решения было найдено, что процесс можно вести при очень
138
Глава 4
Г люкоза
- Стимуляция потока углерода
при гликолизе
Стимуляция
(2?) Подавление
Рис. 4.2. Производство лимонной кислоты.
низких pH, и это почти не сказывается на образовании кислоты
грибами. В таких условиях создавать и поддерживать стериль-
ность гораздо проще. За 1—2 недели ферментации при высоких
концентрациях сахара в сырье выход достигал 60%. Наиболь-
ший выход получали, когда тем или иным способом ограничи-
вали рост мицелия. Первоначальный вариант процесса основы-
вался на поверхностной ферментации, но в 1950 г. было внесено
важное изменение — освоено глубинное культивирование. Было
Химия и технология
139
показано, что стабильный процесс глубинной ферментации воз-
можен только в том случае, £сли он осуществляется в две
стадии: на первой идет рост мицелия, а на второй (в несодер-
жащей фосфор среде)—образование лимонной кислоты. За ко-
роткий срок были разработаны схемы, основанные на исполь-
зовании дешевого углеводного сырья: мелассы, крахмала и
глюкозного сиропа. Наличие ионов металлов в исходном сырье
приводит к резкому падению выхода; их нужно удалять либо
путем осаждения гексацианоферратом, либо пропусканием через
ионообменные смолы, либо применением солей четвертичного
аммония. Для устранения вредного влияния этих примесей ши-
роко используется также метанол и другие низшие спирты.
Механизм их действия неизвестен. Возможно, они как-то влия-
ют на цитоплазматическую мембрану. В 60-х годах для произ-
водства лимонной кислоты был предложен новый процесс на
основе н-парафинов (С9_3о) и штаммов Corynebacterium, Arthro-
bacter и Brevibacterium, но рыночной продукции с его помощью
получено не было. Изучалось также образование лимонной кис-
лоты дрожжами Candida. Они синтезируют смесь лимонной и
изолимонной кислот в соотношении, зависящем как от генети-
ческих факторов, так и от условий ферментации. Было найдено,
что ключевую роль здесь играет аконитат-гидролаза: мутанты
с малой активностью этого фермента продуцировали больше
лимонной кислоты. Растущие на углеводородах дрожжи также
способны синтезировать лимонную кислоту из глюкозы. Гриб
Trichoderma viride образует большое количество цитрата из
глюкозы; это позволяет вырабатывать лимонную кислоту из
целлюлозы. С помощью некоторых видов Penicillium можно
вести ферментацию с образованием Ь5-алло-изолимонной кис-
лоты, диастереомера изолимонной кислоты.
В промышленном производстве лимонной кислоты в основ-
ном используется Aspergillus niger, но применяется также и
A. went И. Процесс ферментации очень сложен, так как лимон-
ная кислота является продуктом первичного метаболизма этих
грибов, и любое сколько-нибудь существенное выделение этого
промежуточного соединения обмена веществ в окружающую
среду свидетельствует о сильном нарушении метаболизма, воз-
никающем вследствие его дисбаланса или генетических нару-
шений. Рост грибов обычно регулируют путем изменения соста-
ва среды (Р, Мп, Fe, Zn). Субстрат должен легко усваиваться;
негидролизованные полимеры обычно не используют, так как
в этом случае внеклеточный гидролиз будет лимитировать ско-
рость всего процесса.
Сверхпродукция лимонной кислоты является ответной реак-
цией на недостаток фосфата, но при выраженной нехватке ме-
таллов лимитирующим фактором не обязательно является фос-
140
Глава 4
фат. Роль металлов при этом до конца еще не понята. Оптимум
pH составляет 1,7—2,0; в более щелочной среде происходит об-
разование заметных количеств щавелевой и глюконовой кислот.
Таким образом, тщательный контроль за культуральной средой
позволяет обойти регуляторные системы обмена и создает оп-
тимальный фон для образования лимонной кислоты. Видимо,
в этих условиях стимулируется гликолиз и обеспечивается не-
ограниченное поступление углерода в реакции промежуточного
метаболизма. Уровень накопления цитрата зависит при этом
от поступления оксалоацетата.
При недостатке марганца активность ферментов цикла три-
карбоновых кислот уменьшается, что в свою очередь подавляет
анаболизм. Такое нарушение обмена приводит к повышению
концентрации аммонийных ионов внутри клеток, и они могут
смягчать ингибирующее влияние цитрата на фосфофруктокина-
зу. Кроме того, марганец, видимо, как-то влияет на биохимиче-
ские свойства поверхности клеток и морфологию гиф. Посколь-
ку в процессе потребляется много кислорода, возможно повтор-
ное окисление цитоплазматического NADH без образования
АТР. В нем участвует альтернативная, а не основная цепь ды-
хательных реакций. В результате без сколько-нибудь выражен-
ного изменения обмена возникает метаболическая «утечка»
(flux) через гликолиз. Эта утечка, происходящая при участии
конститутивной пируваткарбоксилазы и некоторых ферментов
цикла трикарбоновых кислот, а также необычная кинетика
действия ферментов, участвующих в метаболизме оксалоацета-
та, приводят к увеличению внутриклеточной концентрации цит-
рата. Последний способствует дальнейшему накоплению цитра-
та путем ингибирования изоцитратдегидрогеназы.
В промышленном производстве лимонной кислоты применя-
ется несколько вариантов процесса. Традиционным твердофаз-
ным вариантом является процесс Коджи; он имеет много обще-
го с процессом поверхностной ферментации. Глубинная фермен-
тация с технической точки зрения сложнее, чем поверхностная,
но возможна в разных вариантах: периодическом с подпиткой
и непрерывном. Периодическая ферментация используется при
работе с глюкозосодержащими субстратами, а ее вариант с
подпиткой чаще применяется при переработке мелассы. Непре-
рывное культивирование, дающее наибольший выход продукта,
также возможно, но применение этого способа в промышленно-
сти в обозримом будущем маловероятно. Для процесса харак-
терно два максимума скорости: роста и образования продукта.
На первом этапе образуется значительное количество продукта,
зависящее от скорости роста. На втором этапе рост отсутствует,
а предельное количество образующегося продукта определяется
концентрацией биомассы. В конце ферментации массу мицелия
Химия и технология
141
отделяют фильтрованием и промывают. Затем при рН<3,0
осаждают щавелевую кислоту в форме оксалата кальция. Бо-
гатый белком мицелий можно использовать на корм скоту.
Лимонную кислоту осаждают из жидкой фазы в форме каль-
циевой трехзамещенной соли в комплексе с четырьмя молеку-
лами воды. Осадок отфильтровывают, промывают и свободную
кислоту получают путем обработки сульфатом кальция. Далее
ее очищают при помощи активированного угля и ионообменных
смол. Можно также экстрагировать кислоту растворителем.
У лимонной кислоты приятный кислый вкус, она хорошо
растворима в воде. Ее широко используют в пищевой, фарма-
цевтической и косметической промышленности. Эфиры лимон-
ной кислоты применяются в производстве пластмасс. Поскольку
лимонная кислота связывает (хелатирует) металлы, ее исполь-
зуют для их очистки. В составе детергентов она легко разруша-
ется живыми организмами, и ею заменяют фосфаты.
4.2.3. Другие органические кислоты
Процессы, основанные на микробиологической ферментации,
разработаны и для получения ряда других органических кислот.
Среди них — глюконовая кислота и ее производные, яблочная,
виннокаменная, салициловая, янтарная, пировиноградная и кое-
вая кислоты. Хотя некоторые из них и поступают на рынок,
в нынешних условиях в большинстве случаев такое производст-
во экономически невыгодно.
D-глюконовая кислота и ее 6-лактон представляют собой
простые продукты окисления (дегидрогенизации) глюкозы
(рис. 4.3). Еще в начале 20-х годов было налажено промышлен-
ное производство этой кислоты из глюкозы при участии Asper-
gillus niger. Нейтрализация кислоты позволяла получать боль-
шой выход продукта. В погруженных культурах за 48 ч
конверсия субстрата составляла 90%. Исследования на полу-
промышленных установках показали, что если ферментацию
вести при повышенном давлении, то выход кислоты за 24 ч
составляет 95% от теоретического при использовании раствора
глюкозы с концентрацией 150—200 г/л. Процесс можно вести
в полунепрерывном режиме, заново используя мицелий (до де-
вяти раз подряд). Более того, концентрацию глюкозы можно
довести до 350 г/л, если для удаления кислоты использовать
комплексообразование с соединениями бора и получать боро-
глюконат кальция. Однако для осуществления этого процесса
нужны особые, устойчивые штаммы. От него отказались после
того, как было выяснено, что эта соль неблагоприятно влияет
на кровеносные сосуды животных. Контроль за pH осуществля-
ли путем добавления углекислого кальция либо едкого натра.
142
Глава 4
он
D Гпюкоза
! Ферментация
| Спонтанный гидролиз
СН2ОН
I
неон
неон
I
носн
I
неон
1 .
соон
D-Глюконовая кислота Ферментативный
। процесс
;
•неон
I
неон
I
носн
I
с=о
I
соон
2-Оксоглюконовая
коспот а
ен2он
с=о
!
неон
I
носн
неон
I
соон
5-Оксоглюконовая
кислота
Ферментативный
------1 | Химический
L-Идоновая Ферментация
кисло та 1---——г----------
9=0 Синтез
неон —► аскорбиновой
I кислоты
носн
I
С^=0
!
соон
2,5-Диоксоглюконовая
кислота
Ферментация
СООН СН2ОН
носн неон
I I
СООН НОСН
D (+) -Винная кислота с
СООН
2-Оксоглюконовая кислота
Синтез аскорбиновой кисоты
Рис. 4.3. Органические кислоты, получаемые из глюкозы.
Было найдено, что образование глюконата прямо связано с ко
личеством глюкозооксидазы в среде. Этот фермент в промыш
ленном масштабе получают методом ферментации.
Натриевая соль глюконовой кислоты используется для из
влечения металлов. Когда pH процесса контролируют с по
Химия и технология
143
мощью едкого натра, получают именно эту соль. Был разрабо-
тан непрерывный процесс ее производства, при котором выход
<натриевой соли из 35%-ного (вес/объем) раствора глюкозы
•составляет 95%. Предпринимались попытки применить при бро-
жении иммобилизованные системы (как целые клетки, так и
глюкозооксидазу). Натриевая соль глюконовой кислоты в при-
сутствии едкого натра играет роль ловушки кальция и поэтому
используется в составе щелочных средств для мытья бутылок.
Она также способна связывать ионы железа в широком диапа-
зоне pH и как препятствующий отложению железа агент при-
меняется в составе щелочных препаратов для борьбы со ржав-
чиной. Кальциевые и железные соли глюконовой кислоты
применяются как пероральные и внутривенные препараты в ме-
дицине, а чистая кислота — как моющее средство в молочной
промышленности. Глюконолактон находит применение как мед-
ленно действующий подкислитель в составе пекарских порош-
ков, при переработке мяса и в других отраслях пищевой про-
мышленности.
Процесс образования глюконовой кислоты при участии бак-
терий нашел ограниченное применение лишь в некоторых стра-
нах Востока. Например, так называемый «чайный гриб» (ассо-
циация дрожжей, уксуснокислых и глюконовокислых бактерий)
^превращает подслащенный заваренный чай в напиток, содер-
жащий смесь этих кислот. Малая эффективность реакций окис-
ления глюкозы, идущих при участии бактерий, определяется
тем, что одновременно с главным процессом идет образование
2-оксоглюконовой, 5-оксоглюконовой и диоксоглюконовой кис-
лот. Для выработки этих кислот, которые могут служить субст-
ратами для дальнейшей биологической или химической перера-
ботки, были разработаны специальные процессы. Так, 5-оксо-
глюконовую кислоту можно гидрогенизировать в ходе химиче-
ской реакции и получить L-идоновую кислоту, которая в свою
очередь служит субстратом в ферментативной реакции синтеза
2-оксогулоновой кислоты (рис. 4.3). Такие превращения можно
•осуществить и чисто биологическим путем: при помощи видов
Acetobacter глюкозу переводят в 2,5-диоксоглюконовую кисло-
ту, которая превращается в 2-оксогулоновую кислоту при уча-
стии Corynebacterium или Brevibacterium в ходе двух- или
одностадийного процесса на основе смешанных культур этих
бактерий. Выход продукта, правда, невелик. 2-Оксогулоновая
кислота — весьма ценный продукт, так как ее метиловый эфир
в щелочных условиях легко превращается в аскорбиновую кис-
лоту.
Виннокаменная кислота является обычным побочным про-
дуктом виноделия. Однако ее можно получать и путем микроб-
ной трансформации 5-оксоглюконовой кислоты. Штаммы, спо-
144
Глава 4
собные превращать глюкозу в 5-оксоглюконат через глюконат,
могут путем дальнейшей ферментации образовывать тартрат.
Для этой цели обычно используют мутанты Acetobacter и Glu-
conobacter. Виннокаменную кислоту можно вырабатывать так-
же из транс- или qwc-эпоксиянтарной кислоты. Соли ее (тартра-
ты) находят широкое применение в пищевой промышленности,
но методы биотехнологии в ее производстве обычно не исполь-
зуются.
Яблочную кислоту, которая применяется в качестве подкис-
лителя в пищевой промышленности, можно получать из фума-
ровой либо путем ферментации при участии видов Paracolo-
bactrum, либо с помощью иммобилизованной фумаразы. Описа-
ны также способы ее получения из н-парафинов при помощи
дрожжей и из этанола при участии Schizophyllum commune.
Итаконовую кислоту, идущую на производство пластмасс
и красителей, получают с высоким выходом путем ферментации
глюкозы с участием грибов из рода Aspergillus. Совсем недавно
на основе биотехнологии из углеводных субстратов, а также
С12-14-парафинов при участии Candida hydrocarbofurmarica по-
лучали 2-оксоглутаровую кислоту, но на смену этому способу
пришло каталитическое окисление бензола.
Большинство органических кислот, вырабатываемых с по-
мощью микробов, является продуктом переработки пищевого
сырья; исключение составляют кислоты, производимые из н-па-
рафинов. О возможности использования других видов углеводо-
родного сырья как потенциального источника более ценных
органических соединений говорится уже давно, но лишь немно-
гие процессы используются сегодня для получения промышлен-
ной продукции. Так, из нафталина при помощи микробов вы-
рабатывают салициловую кислоту и другие окисленные его
производные. Об этом в последние двадцать лет писали не раз
(Cain, 1980; Tangnu, Ghose, 1980, 1981).
К числу бактерий, способных вырабатывать салициловую
кислоту (рис. 4.4) при росте в средах с нафталином, принадле-
жат многие виды Pseudomonas, Achromobacter и Corynebacte-
rium. Запатентован способ выработки о-гидроксибензальпиро-
виноградной кислоты и 1,2-дигидро-1,2-дигидроксинафталина
при участии видов Nocardia. Большинство диких штаммов бак-
терий, расщепляющих нафталин, при хорошей аэрации в про-
стых солевых средах редко образует салицилат в концентрации,
превышающей 1%, но путем изменения сред и отбора подходя-
щих штаммов могут быть получены и более высокие выходы.
Одним из основных факторов, влияющих на выход, является
доступность субстрата, и накопление салицилата происходит
лишь при постоянном присутствии нафталина: это угнетает
дальнейшие окислительные превращения. Механизм деградации
Химия и технология
145-
зависит от относительной концентрации нафталина и салицило-
вой кислоты. Сложность заключается в том, что полиаромати-
ческий нафталин плохо растворим в воде, в среде ферментации
он обычно присутствует в виде тонкой взвеси. Добавление
эмульгаторов типа Span 80, Span 20, лецитина, кефалина и дру-
гих поливиниловых спиртов существенно увеличивает накоп-
Нафталин
О2
I ,
Нафталин-оксигеназа
Ключ
Накопление мутантами
Химическое производство
Накопление в результате
изменения среды или
применения методов
генетической инженерии
н
G-Г идроксибензаль'
пировиноградная
_____кислота______
Дальнейшая
деградация
Декарбоксилаза
салициловой
кислоты
Рис. 4.4. Микробиологическая конверсия нафталина в полезные органические
вещества.
Химическая
дегидратация
1,2-Дигидро-
ксинафталин-
оксигеназа
Катехол
ления салицилата, так как при этом повышается доступность
субстрата. Обычно используют чистый нафталин, но салицилат
можно получать и из неочищенных нафтафракций. Примеси
(алкилнафталины, тиофен, бензотиофен и крезолы) этому не
мешают.
В ходе ферментации pH быстро падает, так что нужно ис-
пользовать сильно забуференные среды с высокой концентраци-
ей фосфата либо добавлять мочевину или углекислый кальций.
Для максимального накопления салицилата необходимы ионы
10—1344
446
Глава 4
различных металлов. Сообщалось, что выход можно еще более
повысить, если внести в среду особые добавки: органические и
неорганические производные алюминия или бора, пантотеновую
кислоту и ряд других веществ. Ферментация регулируется на-
капливающимся продуктом, а не субстратом; удаление салици-
лата из среды снимает его ингибирующее влияние на рост и
приводит к дальнейшему образованию салициловой кислоты.
Продукт отделяют двумя способами. Для этого используют ани-
онообменную смолу (типа амберлит IRA-400), которую либо
вносят прямо в среду, либо помещают в диализный мешок; при
этом салициловая кислота адсорбируется смолой. Культураль-
ную жидкость можно пропускать через колонку с ионообменной
смолой, смонтированную около ферментера. При этом концент-
рация продукта в ферментере все время поддерживается на
низком уровне, что многократно увеличивает выход (до 6 раз);
возрастает полнота извлечения продукта. Альтернативный спо-
соб удаления продукта — диализная ферментация. Применение
этого процесса на небольшой опытной установке позволило уве-
личить выход салицилата от 10 до 206 г/л. Преимущество ме-
тода состоит в том, что удается избежать неблагоприятного
воздействия ионообменных смол; с другой стороны, приходится
использовать большие объемы жидкой среды, что снижает кон-
центрацию в ней продукта. Применяются и другие, более тра-
диционные способы отделения продукта, например экстракция
растворителем. Производство салицилата путем ферментации
также страдает от фаговой инфекции, и приходится вести рабо-
ту по селекции устойчивых к фагам мутантов.
Показано, что деградация нафталина и салицилата микро-
организмами нескольких родов детерминируется плазмидами.
Так, за превращение нафталина в салицилат ответственна плаз-
мида NAH: она несет гены ферментов, осуществляющих этот
процесс (нафталиноксигеназы, 1,2-диоксинафталиноксигеназы,
дегидрогеназы салицилового альдегида). Таким образом, у мно-
гих микроорганизмов, использующих нафталин, генетическая
информация для осуществления этого процесса закодирована
в плазмиде, но это бывает не всегда. В ходе использования та-
ких плазмид создаются предпосылки для встраивания соответ-
ствующего генетического материала в хромосомы клеток хо-
зяина, что превращает штаммы в продуценты салицилата.
Способность использовать или окислять нафталин обычно за-
крепляется при росте на нафталине, салицилате или его анало-
гах, таких как бензоат или аминобензоат. Описаны интересные
процессы сопутствующего окисления: мутантные штаммы Pseu-
domonas putida, выращенные на среде с глюкозой, которая слу-
жит единственным источником углерода и энергии, способны
окислять нафталин до дигидрокси-1,2-дигидронафталина и 1,2-
Химия и технология
147’
гидроксинафталина на основе индукции ферментов нафталином
или другими соединениями-индукторами, происходящей после-
завершения роста. Первое из этих соединений после кислотной-
дегидратации превращается в а-нафтол — важное моноокислен-
ное производное нафталина (рис. 4.4).
4.2.4. Аминокислоты (см. также гл. 3)
Производство аминокислот при помощи бактерий
и их мутантов
Все аминокислоты, из которых состоят белки, являются-
L-a-амино- (или имино-) кислотами. Они находят применение-
как пищевые добавки, приправы, усилители вкуса, как сырье в
парфюмерной и фармацевтической промышленности и при про-
изводстве других веществ. Их можно получать как из природ-
ных продуктов (главным образом при гидролизе белков рас-
тений), так и путем химического, микробиологического или
ферментативного синтеза. Если химический синтез дает продукт-
рацемат, который требует дальнейшей обработки (разд. 4.2.6),
то последние два метода позволяют получить оптические чистые
аминокислоты.
Секретом большинства производственных процессов с уча-
стием микроорганизмов, о которых говорится в этой главе, яв-
ляется изменение условий среды: именно за счет этого достига-
ется синтез избыточных количеств желаемого продукта. Необ-
ходимого дисбаланса метаболизма можно добиться путем
эмпирического изменения таких факторов, как концентрация
субстрата, pH, концентрация продукта, или же путем установ-
ления критических уровней содержания других веществ (ионов
металлов, органических добавок) в среде. При переводе биоло-
гических процессов образования аминокислот на коммерческую
основу были выработаны новые способы желаемых изменений
метаболизма у организмов-продуцентов, направленных на уве-
личение выхода промежуточных продуктов, образование кото-
рых в иных условиях находится под строгим метаболическим
контролем.
Для производства аминокислот бактерии стали использо-
ваться с начала 50-х годов. Штаммы их постоянно улучшали
генетическими методами, выделяя ауксотрофные мутанты и му-
танты с измененными регуляторными свойствами. Чтобы обес-
печить образование аминокислот в больших количествах, в лю-
бом случае необходимо изменить систему регуляции обмена.
Для этого можно либо стимулировать потребление субстрата
в некоторых путях биосинтеза и выделение аминокислот в среду,
J0*
148
Глава 4
либо подавить побочные реакции и процессы деградации амино-
кислот.
Производство таких аминокислот, как L-глутамат, L-валин,
DL-аланин, L-глутамин и L-пролин, при участии диких штам-
мов бактерий основано либо на использовании присущих этим
бактериям особенностей метаболизма, либо на стимуляции об-
разования аминокислот в ответ на изменение условий внешней
среды. Образовывать аминокислоты способны бактерии многих
родов (например, Corynebacterium, Brevibacterium, Bacillus,
Aerobacter, Microbacterium, Escherichia), причем они настолько
продуктивны, что производство становится рентабельным. Так,
вцды Corynebacterium или Brevibacterium, выращиваемые на
углеводном сырье (гидролизат крахмала, мелассы из сахарного
тростника и свеклы), на этаноле или ацетате при наличии до-
статочного количества биотина в среде способны синтезировать
до 30 г/л глутамата. Для накопления этой аминокислоты важ-
ным условием является полное или частичное подавление ак-
тивности а-кетоглутаратдегидрогеназы. Образование продукта
увеличивается при добавлении ^-лактамных антибиотиков (пе-
нициллина, цефалоспорина С), поверхностно-активных веществ
и жирных кислот. Влияние двух последних агентов обусловлено
увеличением проницаемости клеточных мембран для глутамата,
которая зависит от внутриклеточной концентрации жирных кис-
лот и липидов. Путем изменения условий среды процесс фер-
ментации, в ходе которого образуется L-глутамат, может быть
переключен на синтез L-глутамина или L-пролина. При высокой
концентрации биотина и ионов аммония создаются благоприят-
ные условия для образования L-пролина, а при больших кон-
центрациях аммония и ионов цинка в слабо кислой среде уси-
ливается синтез L-глутамина. DL-аланин, видимо, образуется
в реакции трансаминирования при участии пирувата; затем он
подвергается рацемизации ферментом аланинрацемазой.
Ауксотрофные мутанты не могут образовывать ингибиторы
соответствующего метаболического пути, работающие по прин-
ципу отрицательной обратной связи, так как у них отсутствует
определенная ключевая ферментативная реакция. Поэтому при
выращивании мутантного штамма в среде с минимальной кон-
центрацией необходимого питательного ингредиента они способ-
ны образовывать избыточные количества вещества-предшест-
венника или близких к нему метаболитов блокированной реак-
ции. Так, первые реакции в цепи биосинтеза L-аргинина
(рис. 4.5) у Corynebacterium glutamicum ингибируются по ме-
ханизму обратной связи самим конечным продуктом, и образо-
вание соответствующих ферментов подавляется L-аргинином.
У цитруллинового ауксотрофа этой бактерии отсутствует фер-
мент, называемый орнитин-карбамоилтрансферазой, который
Химия и технология
149
Перенос
ацетильной
группы
Глюкоза
I
I
Глутамат
N -Ацетиглутамат
N 'Ацетилглутамилфосфаг
Орнитин
Глутамат
М -Ацетилорнитин ।---Ф---------
а-Кетоглутарат
Карбамилфосфа т (орни тин-карбамоил трансфераза)
Цитруллин
Аспартат, АТР
Фумарат
Аргинин
^Ацетилглутамат-
семиальдегид
Ингибирование аргинином по механизму
обратной связи
Репрессия аргинином по механизму
обратной связи
Рис. 4.5. Биосинтез L-аргинина.
катализирует превращение орнитина в цитруллин на одном из
промежуточных этапов биосинтеза L-аргинина. Синтез аргинина
не идет, что приводит к снятию ингибирования по принципу
обратной связи со всех ферментов этого пути и накоплению
избытка орнитина. Ауксотрофиые мутанты находят применение
и в тех случаях, когда необходимо синтезировать соединения,
являющиеся конечными продуктами разветвленных цепей мета-
болических реакций. Так, L-аспартат является общим предшест-
венником для L-лизина, L-треонина, L-метионина и L-изолей-
цина. Первая реакция в процессе образования всех этих амино-
кислот катализируется аспартокиназой, активность которой
может быть ингибирована по механизму обратной связи при
150
Глава 4
совместном действии L-лизина и L-треонина. У мутантов, аук-
сотрофных по гомосерину или треонину/метионину, существен-
но снижается внутриклеточная концентрация L-треонина, что
снимает блокаду с аспартокиназы и позволяет клеткам накап-
ливать L-лизин,
Получить ауксотрофные мутанты, способные накапливать
конечные продукты неразветвленных цепей биосинтеза, напри-
мер L-аргинин, невозможно. В таких случаях приходится отби-
рать мутанты с частично нарушенной регуляцией биосинтеза,
что позволяет получить повышенный выход конечного продукта.
Такие мутанты известны под названием регуляторных; их отби-
рают по устойчивости к аналогам аминокислот или же среди
ревертантов ауксотрофов. В основе использования аналогов
аминокислот лежит их сходство с природными аминокислотами.
Они ингибируют рост бактерий, но этот эффект может быть
уменьшен путем добавления соответствующей природной ами-
нокислоты. Таким образом, аналоги выступают в роли искусст-
венных, работающих по принципу обратной связи, ингибиторов
ферментов, обеспечивающих биосинтез природных аминокислот,
и одновременно подавляют процесс их включения в белки.
Появление мутантов, устойчивых к этим ложным ингибиторам,
означает, что регуляторные ферменты соответствующего пути
обмена становятся у них нечувствительными к аналогу. Так,
серусодержащий аналог лизина 5-(2-аминоэтил)^-цистеин яв-
ляется у Brevibacterium flavum ложным, действующим по прин-
ципу обратной связи ингибитором аспартокиназы. Устойчивые
к его действию мутанты вырабатывают фермент, который в 150
раз менее чувствителен к ингибированию по механизму обрат-
ной связи, чем фермент исходного штамма, и в результате про-
дуцируют до 33 г/л лизина. Для увеличения выхода можно
воспользоваться как ауксотрофией, так и дефектами регуляции
одновременно. Так, у Corynebacterium glutamicum и Brevibac-
terium flavum сверхпродукции L-треонина не наблюдается, по-
скольку происходит сочетанное ингибирование по механизму
обратной связи аспартокиназы и L-треонином, и L-лизином,
а L-треонин ингибирует и гомосериндегидрогеназу. Мутант, ус-
тойчивый к аналогу треонина, сс-амино-^-оксивалериановой кис-
лоте, синтезирует в избыточном количестве треонин, так как
ферменты его, ингибированные этой аминокислотой, десенсиби-
лизированы. Принимающие участие в синтезе треонина гомо-
сериндегидрогеназа и киназа также «выключаются» L-метиони-
ном, и поэтому ауксотрофы по метионину образуют L-треонин
с еще большим выходом.
Регуляторные мутанты можно получить путем трансдукции,,
проводя при этом отбор отдельных мутаций, вызывающих пол-
ное рассогласование регуляторных механизмов, а затем объеди-
Химия и технология
151
.яяя эти признаки путем ко-трансдукции. Таким способом у од-
ного штамма последовательно может быть закреплена устойчи-
вость к нескольким аналогам (гл. 7).
Производство аминокислот из биосинтетических
предшественников
Использование предшественников при производстве аминокис-
лот позволяет успешно «обходить» метаболический контроль,
осуществляющийся по механизму обратной связи и репрессии.
Рассмотрим процесс синтеза L-лейцина из L-треонина через
а-кетобутират. Первый фермент в этом пути биосинтеза, гидра-
таза, у Serratia marcescens ингибируется L-изолейцином по ме-
ханизму обратной связи. При добавлении в среду D-треонина
происходит индукция D-треонингидратазы, которая L-изолейци-
ном не ингибируется, и поэтому синтез L-изолейцина из D-трео-
нина может миновать метаболический контроль. С другой сто-
роны, для того чтобы обойти контроль, можно использовать
предшественники, превращаемые в ходе обмена в кетобутират
(например, а-амино-, а-бром- и а-гидроксибутират); точно так
же при участии гидроксиметилтрансферазы и в присутствии до-
статочного количества метилентетрагидрофолата из предшест-
венников глицина можно получать L-серин. В качестве постав-
щиков С-1 могут выступать глицин, формальдегид, формиат,
саркозин, холин или метионин. При наличии в среде глицина,
глюкозы и метанола метиониновый ауксотроф Artrobacter glo-
biformis образует до 5,2 г/л L-серина. Для такой конверсии,
а также для образования L-глутамата, L-метионина и аромати-
ческих аминокислот могут использоваться и другие штаммы
растущих на метаноле бактерий.
Синтез аминокислот с помощью ферментов
Об использовании чистых ферментов для нужд химии подробно
будет говориться в разд. 4.2.6, а здесь мы остановимся на том,
какова роль и ожидаемые преимущества применения ферментов
при синтезе аминокислот. Эти процессы бывают одно- и много-
стадийными, а используемые в них методы весьма разнообраз-
ными от применения in situ интактных, но не растущих орга-
низмов до иммобилизованных препаратов. В этой связи целе-
сообразно рассмотреть пять классов ферментов.
1. Гидролитические ферменты (или гидролазы), например
L-a-амино-е-капролактам-лиаза (синтез L-лизина) или 2-амино-
тиазолин-4-карбоксилатгидролаза (синтез L-цистеина) (рис. 4.6).
Чтобы можно было использовать неочищенные ферменты, целые
клетки обрабатывают поверхностно-активными веществами, вы-
152
Глава 4
нгс^с^сн2
н2
Рацемическая смесь
D- И к-<Х-Дми«о- е-капролактама
Г идролаза
Рацемаза
H2N(CH2)4CH(NH2)COOH
L -Лизин
+
D* Об -Амино- S-капролактам
сн,—сн—соон СН2—ОН—СООН сн2—СН—СООН-
II II .11
q к]---------------------► S NH? * SH NH2
Гидролаза! |, г Гидролаза II
2 ' . ^Н2 L-Цистеин
L-2 -Амино-^-тиазолин-4-карбоксилат s .Kap6aM0UJ,.L.Qucrei,H
Место гидролитической атаки
Рис. 4.6. Применение гидролитических ферментов при производстве аминокис-
лот.
зывающими изменение проницаемости. Кроме того, могут быть
получены мутанты, у которых искомый продукт не вовлекается
более в обмен веществ.
2. Лиазы. Эти ферменты часто используются в реакциях
дезаминирования. Так, для образования L-аспарта из фумарата
аммония может использоваться (в обратной реакции) аспарта-
за, или L-аспартат — аммиак-лиаза. В качестве доноров аммо-
ния могут, кроме того, выступать гидразин или гидроксиламин.
Сходным образом L-фенилаланин — аммиак-лиаза может ката-
лизировать распад L-фенилаланина с образованием транс-ко-
ричной кислоты и аммиака. Хотя обычно равновесие в этих
реакциях сдвинуто в сторону реакций распада, при высоких
концентрациях аммонийных ионов начинают преобладать про-
цессы синтеза.
3. Ферменты, содержащие пиридоксальфосфат. Это обычные
коферменты, участвующие в метаболизме аминокислот. Они ка-
тализируют множество реакций: рацемизацию, трансаминирова-
ние, декарбоксилирование, реакции замещения и элиминации
и являются своего рода универсальными. По-видимому, роль
этих коферментов состоит в активации аминокислот, что облег-
чает их взаимодействие с апоферментом. Мы рассмотрим здесь
лишь несколько ферментов из этой группы. Так, L-тирозин —
фенол-лиаза (p-тирозиназа) катализирует реакцию р-элимина-
ции, в которой тирозин распадается с образованием пирувата,
фенола и аммиака. При оптимальных условиях Erwinia herbi-
cola может синтезировать очень много этого фермента (до 10%
Химия и технология
153
,0) 0-Тирозиназа
4а) Процесс, обратный элиминации
+ СНзСОСООН
Пируват
+ NH3
А ммиак
Н20
CH2CH(NH2)COOH
Тирозин
СН2ОНСН(1ЧН2)СООН
Серин
4.2) Триптофаназа; процесс, обратный -элиминации
+ СНзСОСООН + NH3
Пировиноградная Аммиак
CH2CH(NH2)COOH
Триптофан
кислота
(3) L-Метионин - у • лиаза
CH3SeH + СН3СН2СОСООН + NH3
Метилселенол а -Кетобутират Аммиак
CH3 — Se — (CH2)2CH(NH2)COOH + Н20
Селен метионин
Рис, 4.7. Реакции синтеза, катализируемые пиридоксалевыми ферментами.
растворимого белка). Его используют для синтеза тирозина:
в иммобилизованной форме он применялся для непрерывного
его производства. Субстратная специфичность этого фермента
такова, что он может также осуществлять реакцию ^-замещения
между DL-серином и пирокатехолом, в результате которой об-
разуется L-ДОФА (рис. 4.7). Примером широкого распростра-
ненного в природе фермента, осуществляющего дезаминирова-
ние, может служить L-триптофан — индол-лиаза (триптофаназа).
154
Глава 4
Она катализирует реакции аф-элиминации и (3-замещения и.
также характеризуется широкой субстратной специфичностью
(L-триптофан, L-цистеин, З-метил-Ь-цистеин, (З-хлор-Ь-аланин,,
L-серин). Действие ее может быть обращено, и тогда она будет
способствовать синтезу L-триптофана из индола, пирувата и
аммиака. Примером индуцибельного фермента является L-ме-
тионин — г/-лиаза: она катализирует реакцию элиминации, где
субстратами могут быть разнообразные аминокислоты, включая
производные L-метионина и L-цистеина. L-метиопин расщепля-
ется с образованием метантиола, а-кетобутирата и аммиака.
Когда фермент «работает на синтез», его можно применять для
производства новых серусодержащих аминокислот на основе-
алкантиолов и арилтиоспиртов. Фермент может также расщеп-
лять селенметионин. Использование селенолов в обратных ре-
акциях замещения, идущих с образованием гомоцистеинов,
содержащих селен вместо серы, — это первый случай, когда,
селен удалось включить в состав аминокислот (рис. 4.7).
Эту лиазу можно применять для синтеза меченых аминокис-
лот.
4. Дегидрогеназы аминокислот, например лейцин- и аланин-
дегидрогеназы. Эти ферменты катализируют обратимые реак-
ции дезаминирования. Их применяют в непрерывных процессах,
синтеза аминокислот из соответствующих кето-аналогов.
В мембранном реакторе дегидрогеназы аминокислот удержива-
ются ультрафильтрующей мембраной и используют в своей ра-
боте один и тот же пул NADH, который сохраняется в реакторе,
так как ковалентно связан с полиэтиленгликолем. Регенерация
его в ходе процесса осуществляется с помощью формиатдегид-
рогеназы (разд. 4.4.2).
5. Глутаминсинтаза. Этот фермент катализирует АТР-зави-
симую реакцию аминирования глутамата, которая была сопря-
жена со сбраживанием сахара дрожжами. Высвобождающаяся
при брожении энергия используется для синтеза глутамина.
При распаде фруктозо-1,6-дифосфата, образовавшегося при
сбраживании глюкозы, продуцируется АТР, которая необходи-
ма для энергоснабжения эндергонической реакции, катализи-
руемой синтазой. При использовании бесклеточного экстракта
пекарских дрожжей и глутаминсинтазы Gluconob act er suboxy-
dans из глюкозы, глутамата и ионов аммония в качестве суб-
стратов с высоким выходом (92 мол. %) был получен глутамин.
Применение аминокислот
Аминокислоты находят применение во многих сферах.
1. Их используют в качестве пищевых добавок. Так, лизи-
ном, триптофаном и треонином обогащают растительные белки,
а метионин включают в блюда из сои.
Химия и технология
155
2. При выработке пищевых продуктов аминокислоты находят
применение в роли усилителей вкуса и добавок. Благодаря вы-
раженному мясному вкусу широко используется L-энантиомер
мононатриевой соли глутаминовой кислоты. Глицин добавляют
как подсластитель, бактериостатическое вещество и антиокси-
дант.
3. Аминокислоты применяются в медицине (вливания),
.а некоторые их аналоги используются для лечения психических
заболеваний.
4. В химической и фармацевтической промышленности ами-
нокислоты широко используются как предшественники в произ-
водстве детергентов, полиаминокислот (из них делают синтети-
ческие волокна и пленки), полиуретана и химикатов для сель-
ского хозяйства.
4.2.5. Антибиотики и стероиды
Антибиотики (группа веществ микробного происхождения)
играют большую роль в нашей жизни. В медицине и ветерина-
рии они с успехом применяются как противомикробные и про-
тивоопухолевые препараты; с их помощью контролируется рост
растений и ведется борьба с болезнями. Все антибиотики были
выделены в ходе систематического скрининга микроорганизмов;
число их было существенно увеличено путем химической моди-
фикации, цель которой состоит в 1) расширении спектра дейст-
вия и повышении эффективности; 2) снижении токсичности и
устранении нежелательных побочных эффектов; 3) создании
аналогов, устойчивых к разрушению микробами и обладающих
поэтому большим временем полужизни; 4) усовершенствовании
способов их введения.
Со времени открытия и описания первых антибиотиков полу-
чили путевку в жизнь целое поколение полусинтетических пени-
циллинов, цефалоспоринов, аминогликозидов, тетрациклинов,
рифамицинов, макролидов и линкозаминидов. Эти соединения
очень сложны, и метод полного химического синтеза не может
конкурировать с их производством методом ферментации. По-
скольку микробы могут разрушать антибиотики, возникла
мысль модифицировать природные антибиотики ферментами
микроорганизмов (вспомним о превращениях стероидов),
но лишь немногие из таких веществ оказались экономически
приемлемыми (6-аминопенициллановая кислота, 6-диметилтет-
рациклины). Существует три основных способа получения но-
вых антибиотиков.
1. Прямая ферментация. В этом случае используются обра-
зующие антибиотик микроорганизмы, которые синтезируют но-
вые биологически активные соединения в присутствии подходя-
156
Глава 4
Таблица 4.2
Тетрациклин
NH
NH ||
ОН о
он
он
Стрептомицин
щих предшественников или ингибиторов метаболизма. Так,
Penicillium chrysogenum не только синтезирует пенициллин, на
и включает фенилуксусную кислоту в бензилпенициллин, а дру-
гие предшественники — в аналоги пенициллина. Этот принцип
находит широкое применение, например, при получении новых
блеомицинов путем добавления аминов к культуре S. verticillub
Химия и технология
157
и новых актиномицинов — путем добавления 4-метилпролина к
среде для выращивания S. parvulus. При подавлении синтеза
антибиотиков также иногда образуются полезные вещества.
Так, при подавлении процесса присоединения хлора S. aureofa-
ciens образует тетрациклин, а не хлортетрациклин. Если в сре-
ду ферментации добавить L-метионин, то ингибируются реакции
метилирования и синтезируется 6-деметил-7-хлортетрациклин
(табл. 4.2).
2. Мутанты организмов-продуцентов иногда образуют био-
логически активные промежуточные продукты какого-то опре-
деленного пути биосинтеза антибиотиков либо соединения, ко-
торые могут оказаться полезными как предшественники при
создании новых аналогов. «Блокированные» мутанты этого ти-
па не способны образовать нужный антибиотик, если в среде
отсутствует метаболитический предшественник, который в нор-
ме образуется при участии фермента, действующего вслед за
-ОН СН3СО------СН3
в -сн2соон -Н
Рис. 4.8. Структура рифамицина.
блокированным звеном метаболизма. Поскольку ферменты,,
участвующие во вторичном метаболизме, нередко обладают от-
носительно низкой субстратной специфичностью, аналоги пред-
шественников антибиотиков могут быть относительно легко
превращены мутантом в аналоги самого антибиотика в ходе
процесса, известного как мутационный биосинтез, или мутасин-
тез. Nocardia medlterranei синтезирует около двадцати разных
рифамицинов (рис. 4.8). Путем добавления барбитала ход фер-
ментации и как следствие спектр образуемых антибиотиков
существенно меняется. Мутанты этого организма, у которого-
158
Глава 4
подавлена способность к ацилированию, образуют предшествен-
ник рифамицина В — рифамицин SV, который служит исход-
ным веществом для получения многих синтетических рифами-
цинов (например, рифампицина, препарата для лечения
туберкулеза, который действует и на возбудителя проказы).
.Другой мутант, с блокированным метилированием, синтезирует
27-деметилрифамицин SV, ценный субстрат для синтезов, кото-
рый исключительно сложно получить химическими методами.
3. Особенно успешно модификация антибиотиков микроба-
ми идет в следующих двух процессах.
а. При ферментативном гидролизе пенициллина с образо-
ванием 6-аминОпенициллановой кислоты (6-АП К, рис. 4.9), ко-
торая является ценным исходным продуктом при производстве
некоторых полусинтетических, важных для медицины аналогов
Пенициллин
О Н
Пенцциллоиновая кислота
Пенициллин • ацилаза
Ъ-Аминопенициллановая кислота
RCOOH
Рис. 4.9. Гидролиз пенициллина с образованием 6-АПК (ядра молекулы пени-
циллина) или пенициллиновой кислоты.
пенициллина. В промышленности 6-АПК, ядро молекулы пени-
циллина, получают путем гидролиза пенициллина или бензил-
пенициллина при участии штаммов, с высоким выходом обра-
зующих в ходе ферментации пенициллинацилазу; для этой же
цели используют также иммобилизованную пенициллинацилазу.
Исходя из типа пенициллина, который ацилазы предпочти-
тельно гидролизуют, их подразделяют на группы. Некоторые из
них способны катализировать и обратные реакции. На основе
•6-АПК было получено более 40 000 полусинтетических пеницил-
линов. В некоторых случаях не было необходимости выделять
•6-АПК: примером может быть превращение бензилпенициллина
в ампициллин. Гидролиз бензилпенициллина осуществляют при
участии мутанта Kluyvera citrophila, после чего в ферментер
Химия и технология
15»
вносят мутант Pseudomonas melanogenum и метиловый эфир-
DL-фенилглицина. Условия процесса изменяют таким образом,
чтобы они способствовали образованию ампициллина (рис. 4.10).
В роли катализатора выступает ацилаза, образуемая вторым
мутантным организмом, которая не способна в этих условиях
гидролизовать или синтезировать бензилпенициллин. В ходе
этого двухстадийного процесса образуется только ампициллин
[D (—) -а-аминобензилпенициллин].
6-АПК
Рис. 4.10. Синтез ампициллина.
б. В клинике широко применяются аминогликозидные анти-
биотики (стрептомицин, неомицин, канамицин, гентамицин).
Бактерии, способные их инактивировать, были выделены не-
только от больных, но и как самостоятельные, образующие ан-
тибиотики штаммы. Их ферментативная активность может быть-
частью механизма детоксикации, при помощи которого организ-
мы-продуценты защищают себя от неблагоприятного воздейст-
вия образуемых ими же веществ. К числу модификаций, проис-
ходящих при инактивации антибиотиков, относятся N-
ацетилирование, О-фосфорилирование, О-аденилирование и
О-нуклеотидилирование. Установление механизма модификации
позволило планировать и осуществлять химический синтез но-
вых аналогов, устойчивых к такой инактивации.
Инактивация антибиотиков по другому механизму, включая1
гидролиз, гидроксилирование, эпоксидирование, сульфоокисле-
ние, фосфорилирование или восстановление, обычно приводит
!60
Глава 4
к образованию или полностью, или частично неактивных про-
изводных. Их изучение позволит синтезировать новые аналоги,
выявить те участки молекул, которые ответственны за антибио-
тическую активность, а также создать рациональные основы
«конструирования» антибиотиков и усовершенствования произ-
водства.
Пенициллиназы (р-лактамазы) гидролизуют р-лактамное
кольцо молекулы субстрата и являются основой устойчивости
болезнетворных бактерий к пенициллинам и цефалоспоринам.
В результате гидролиза амидной связи кольца образуется пе-
нициллиновая кислота (рис. 4.9), которая полностью лишена
антимикробной активности. Эти ферменты синтезируются мно-
гими бактериями; они могут различаться по строению и специ-
Химический гидролиз
7-Аминоцефалоспорановая кислота
Рис. 4.11. Гидролиз цефалоспорина С.
фичности. Глубокое изучение механизмов их действия позволи-
ло наладить производство устойчивых аналогов антибиотиков,
таких как ампициллин и карбенициллин, а также найти при-
родные ингибиторы лактамазы — клавулановую и оливановую
кислоты. р-Лактамазы используются для оценки количества
пенициллинов в пищевых продуктах и биологическом сырье,
а также для инактивации пенициллина в молоке, что предот-
вращает аллергические реакции у его потребителей.
В некоторых случаях получить полезные предшественники
с помощью микробов не удается. Так, при выработке цефало-
спорина в основном образуется цефалоспорин С, который при-
ходится затем химическим методом гидролизовать до 7-амино-
цефалоспорановой кислоты (рис. 4.11) и уже ее использовать
Химия и технология
161
как субстрат для получения новых полусинтетических цефало-
споринов.
Способность микроорганизмов выступать в роли химических
катализаторов впервые удалось использовать в полной мере для
синтеза промышленно важных стероидов. В последние тридцать
лет субстратная и стереоспецифичность ферментов нашла ши-
рокое применение в производстве стероидов при осуществлении
разнообразных реакций: гидроксилирования, дегидроксилирова-
ния, эпоксидирования, окисления, восстановления, гидрогениза-
ции, дегидрогенизации, этерификации, гидролиза эфиров и изо-
меризации. Целью всеобъемлющих исследований в этой обла-
сти было осуществление специфических структурных пере-
строек стероидов при мягких условиях. Специфич-
ность таких реакций определяется либо выбором оп-
ределенного вида микроорганизмов, либо химической модифи-
кацией субстрата, стереохимически исключающей другие реак-
ции. Понимание зависимости между строением молекул
субстрата и характером его перестройки, осуществляемой мик-
роорганизмами, позволило сформулировать требования для
каждой конкретной реакции, например для гидроксилирования.
В определении скорости и направления реакции главную роль,
как выяснилось, играют положение и ориентация замещающих
групп в молекулах стероидов. История развития методов мик-
робиологического преобразования стероидов представляет собой
прекрасный пример сочетания химического подхода со специ-
фичностью и разнообразием биологических систем. Кроме того,
на этой основе может быть осуществлен синтез новых стероидов,
обладающих лучшими фармакологическими свойствами.
Первый запатентованный процесс микробной трансформации
стероидов был разработан в 1937 г., но внедрить его в промыш-
ленность удалось лишь в 1952 г. [процесс 11-«-гидроксилирова-
ния прогестерона некоторыми видами грибов (рис. 4.12)].
С технологической точки зрения этот процесс не потерял зна-
чения и теперь. Сегодня в нем используются виды грибов с
весьма высокой субстратной специфичностью относительно мес-
та гидроксилирования. Дальнейшее усовершенствование процес-
са может быть основано на использовании спор грибов или же
на изменении состава культуральных сред. Упомянутая выше
трансформация может быть выполнена с высоким выходом при
концентрации субстрата 20—50 г/л. Сходным образом по поло-
жению 7 и 14 может быть гидроксилирован дезоксикортикосте-
рон. Если провести ба-метилирование ядра молекулы стероида,
то нежелательного гидроксилирования по 7а-положению не
произойдет. Направленное гидроксилирование путем химиче-
ской модификации широко используется для повышения эффек-
тивности процесса.
11-1344
162
Г пава »
СН3
11 а Гидроксипрогестерон
Рис. 4.12. Схема применяемого в промышленности метода микробной транс-
формации прогестерона.
Большинство поступающих в продажу стероидов, обладаю-
щих противовоспалительным действием, — это производные
преднизолона, и именно этим определяется важная роль про-
цессов микробного гидроксилирования кортикостерона (веще-
ства S Рейхштейна) и его производных. В промышленном мас-
штабе производство гидрокортизона путем гидроксилирования
кортикостерона осуществляется при участии некоторых видов-
грибов (например, Cunninghamella blakesleeana) с начала 50-х
годов. За это время процесс был неоднократно усовершенство-
ван. Проблемы, связанные с деградацией субстрата, которая
происходит при обычных условиях производства, можно решить-
путем регулярного его добавления или использования других
микроорганизмов, например Thieghemella orchidis. Кроме того,
ход синтеза можно контролировать, применяя метод химической
модификации. Так, метилирование по 16а-положению подавля-
ет нежелательное восстановление кетогруппы при С-20, а обра-
зование уксуснокислого эфира по С-17 стереохимически препят-
ствует другим побочным реакциям. Конечное превращение гид-
Химия и технология
163
рокортизона в коммерческие
продукты со структурой пред-
низолона также осуществляет-
ся с помощью микробов. Хи-
мические методы здесь явно
проигрывают по сравнению с
микробиологическими. Одной
из главных реакций в этом
процессе является образова-
ние 1,2-двойной связи; для де-
гидрогенизации по положению
1 используют главным обра-
зом Mycobacterium globiforme
и Arthrobacter simplex. Выход
этой реакции зависит от того,
® какой форме подается суб-
страт. Если он поступает в
среду в микрокристаллах, то
концентрацию субстрата мож-
но довести до 400 г/л и полу-
чить выход 80—90%. При мо-
дификации прегнана получают
многие фармакологически цен-
ные кортикоиды, гестагены и
.анестезирующие средства сте-
роидной природы. Их произ-
водство основано на проведе-
нии широкого спектра превра-
щений, осуществляемых мик-
робами.
Другой важной группой со-
единений, модифицируемых с
помощью микробов, являются
.андростаны и эстраны. Их
используют в промышленном
синтезе половых гормонов и
Вещество S Рейхштейна
Curvutaria lunata
Arthrobacter simplex
Рис. 4.13. Превращение вещества S
Рейхштейна в преднизолон.
минералокортикоидных соеди-
нений. Примером такого рода
служит превращение дрожжа-
ми 4-андростен-3,17-диона в
тестостерон. Все возрастающее значение приобретает процесс
окислительного расщепления боковой цепи Сщ-стероидов: он
позволяет использовать дешевые стероиды для производства
предшественников, идущих на синтез стероидов, крайне нуж-
ных фармакологам. И в этом случае приходится проводить
химическую модификацию, чтобы предотвратить разрушение
ап
164
Глава 4
«скелета» молекул стероидов. Она заключается в гидроксили-
ровании по положению С-9 и направляет процесс микробной
перестройки структур на разрушение боковых цепей. Так, холе-
стерол или его соли превращаются в 4-холестен-З-он и 1,4-анд-
ростадиен-3,17-дион. Для более глубокого знакомства с проб-
лемой микробной трансформации стероидов мы отсылаем чи-
тателя к литературе, приведенной в конце главы.
Дальнейшие успехи в области применения методов микро-
биологии в химии стероидов будут основаны на применении
иммобилизованных клеток и органических растворителей, что
позволит расширить масштабы производства.
4.2.6. Коммерческие аспекты применения ферментов
Применение ферментов в химической технологии обычно бывает
обусловлено их высокой избирательностью и стереоспецифич-
ностью, однако, как отмечалось ранее, эти их свойства не всегда
оказываются желательны. Примером такого рода могут служить
случаи использования широкой субстратной специфичности фер-
мента для производства аналогов основного продукта. Второе
важное преимущество технологии на основе ферментов перед
химическим катализом заключается в том, что при относительно
мягких условиях удается достичь более высоких скоростей пре-
вращений. Об использовании отдельных ферментативных реак-
ций для получения аминокислот и антибиотиков мы уже гово-
рили; в этом разделе будет описано сегодняшнее положение дел
в сфере использования ферментных препаратов в промышлен-
ности (табл. 4.3).
Протеиназы
Протеиназы давно применяются в пищевой промышленности.
Ранее ферменты для этих целей выделяли из животных и рас-
тений; сегодня их частично замещают протеазы микробов. Пер-
вым ферментом, нашедшим применение в промышленности, бы-
ла а-амилаза (такадиастаза) из Aspergillus oryzae, производ-
ство которой началось в 1890 г. Эти препараты содержали
значительную примесь протеазы, и их рекомендовали исполь-
зовать как средство, способствующее пищеварению. Отметим,
что производство и поступление на рынок такого рода продук-
тов было весьма ограниченным вплоть до начала 60-х годов,
когда их стали использовать в составе детергентов. Впрочем,
о такой возможности было известно за пятьдесят лет до этого;
средство для замачивания белья, содержащее соду и панкреа-
тические ферменты, продавалось еще в 1913 г. Крупный успех
в производстве и продаже таких средств был достигнут лишь
Химия и технология
165
Таблица 4.3
после того, как стали использоваться протеазы Bacillus subtilis,
в особенности субтилизин Carlsberg. В конце 60-х годов при-
близительно 50% всех детергентов, выпускавшихся в Европе
и США, уже содержали протеазы. Постоянно ведется работа
по увеличению активности ферментов (особенно их способности
удалять пятна) и стабильности их в моющих растворах. Еще
одна отрасль, где наблюдалось бурное развитие технологий на
основе протеаз, — это производство сыра. Здесь все усилия бы-
ли направлены на поиск микробных заменителей сычуга телят.
Для выработки протеаз в промышленном масштабе нужны
штаммы микроорганизмов, синтезирующие внеклеточные про-
теазы с высоким выходом. Эти ферменты подразделяют сегодня
на три группы: сериновые, кислые и металлопротеазы.
Среди сериновых протеаз на первом месте стоит субтилизин
Carlsberg. При участии Bacillus licheniformis ежегодно произ-
водится около 500 т очищенного фермента. Накопление про-
теазы начинается в конце логарифмической фазы роста микро-
бов, и выход фермента увеличивается при использовании аспо-
рогенных штаммов (они удобны еще и тем, что при этом удается
исключить технически сложную операцию по отделению спор
от фермента). Сериновые протеазы не гидролизуют белки до
аминокислот. В стиральные порошки обычно добавляют 0,5%
166
Глава 4
(по весу) препарата, содержащего 3% активного фермента.
Хотя содержание фермента в них и мало, при стирке он кон-
центрируется на пятнах белковой природы из-за сродства к
субстрату. В меньшей степени применяются протеазы алкало-
фильных видов Bacillus-, они активны при pH 9—12 и темпера-
туре выше 50 °C.
В состав металлопротеаз входит атом металла, обычно цин-
ка, без которого фермент не активен. В промышленности ме-
таллопротеазы получают с помощью Bacillus amyloliquefaciens
и В. thermoproteolyticus. Специфичность действия этих фермен-
тов выше, чем у сериновых протеаз, но их нельзя использовать
как «микробный сычуг», так как уровень неспецифического про-
теолиза у них все же весьма высок. Они применяются в пиво-
варении, при гидролизе белков ячменя, так как сериновые про-
теазы ингибируются веществами солода. Удаление с их помощью
белков позволяет избежать помутнения пива при охлаждении,
которое происходит в результате взаимодействия белков с тан-
нинами при хранении пива на холоде. С той же целью часто
используются протеазы растений папаин и бромелаин.
Кислые протеазы синтезируются грибами. По свойствам они
похожи на пищеварительные ферменты животных пепсин и рен-
нин. Применяют их для гидролиза соевого белка при производ-
стве соевого соуса, в хлебопекарной промышленности (здесь
с их помощью видоизменяют свойства клейковины муки так,
чтобы получить мягкое, пластичное тесто, из которого делают
бисквиты), как средства, способствующие пищеварению или же
предотвращающие помутнение пива при охлаждении. Большин-
ство протеаз вызывает свертывание молока, но творог получа-
ется невкусным, из-за глубокого гидролиза казеина. Субстрат-
ная специфичность кислых протеаз термофильных грибов Мисог
pusillus и Мисог miehei уже. Они похожи на ферменты сычуга
и широко применяются для створаживания молока. Менее ши-
роко используются протеазы Enthothia parasitica, так как они
обладают большей, чем «сычужные» ферменты Мисог, протео-
литической активностью; они применяются, например, при вы-
делке эмментальского сыра. Разрабатывается и другой подход:
на основе технологии рекомбинантных ДНК проведено клониро-
вание и получена экспрессия гена реннина телят в микроорга-
низмах (например, Е. coli). Синтезированный таким способом
фермент успешно прошел испытания при опытном производстве
сыра (гл. 3).
Протеазы находят применение и в кожевенной промышлен-
ности, при удалении шерсти и умягчении кож. Такая обработка
делает кожи мягкими и эластичными.
Химия и технология
167
Глюкозоизомераза
«Королевой» иммобилизованных ферментов в промышленности
можно считать глюкозоизомеразу, которая катализирует пре-
вращение глюкозы во фруктозу. Коммерческие препараты ее
известны под фирменным названием «Sweetzyme» или «Маха-
zyme». Их появление послужило толчком для развития круп-
ного производства фруктозного сиропа. При высоких концент-
рациях субстрата и нейтральных pH несладкая глюкоза с вы-
ходом 42—47% изомеризуется ферментом в более сладкую
фруктозу. Такие фруктозные сиропы (Isomerose, Isosyrup, Corn-
sweet, Isosweet) сегодня широко потребляются пищевой про-
мышленностью. Запатентовано множество способов иммобили-
зации и использования как самой изомеразы, так и содержащих
ее клеток. Процесс идет при 60—65 °C при pH 7,0—8,5 в при-
сутствии ионов магния. При производстве насыщенного фрук-
тозного сиропа из кукурузы в качестве субстрата используется
либо глюкоза, либо продукт комплексной ферментативной пе-
реработки, заключающейся в ожижении и осахаривании крах-
мала.
Амилазы и амило глюкозид азы
Использование ферментов в производстве крахмала позволяет
контролировать глубину его гидролиза и получать продукцию
с желаемыми свойствами: вязкостью, сладостью, осмотическим
давлением и устойчивостью к кристаллизации. Гидролиз ката-
лизируется ферментами трех разновидностей: эндоамилазами,
экзоамилазами и а-1,6-глюкозидазами.
Эндоамилазы — это а-амилазы, они расщепляют а-1,4-глю-
козидные связи в амилозе и амилопектине с образованием оли-
госахаридов с разной длиной цепи и «-конфигурацией при
Ci-атоме глюкозы, способной служить восстановителем. Для
ожижения крахмала при высокой температуре используют тер-
мостабильные а-амилазы. Так, температурный оптимум у фер-
мента из Bacillus amyloliquefaciens лежит при 70 °C, а у амила-
зы В. licheniformis — при 92 °C. Непродолжительное время по-
следняя может работать при температуре до НО °C. В этих
условиях активность фермента стабилизируется ионами каль-
ция и высокой концентрацией субстрата в среде. Для ожиже-
ния крахмал диспергируют в воде при нагревании; для умень-
шения вязкости и во избежание оседания крахмала при охлаж-
дении проводят частичный его гидролиз. При одностадийном
ожижении фермент добавляют в самом начале, до приготовле-
ния суспензии, которое проводят при 150 °C в течение 5 мин,
после чего гидролиз ферментом ведут 2 ч при 95°C .При кис-
168
Глава 4
лотно-ферментном ожижении фермент вносят после желирова-
ния крахмала, вызванного нагреванием.
При осахаривании используются термостабильные а-амила-
зы, особенно мальтогенные ферменты из грибов. Лучше всего
они работают при 55 °C и концентрации субстрата 30—40%.
Процесс обычно продолжается более 48 ч. Получаемые из крах-
мала сиропы содержат много мальтозы (40—50%); они находят
применение при производстве карамели и замороженных десерт-
ных блюд.; Для получения сиропов с очень высоким содержани-
ем мальтозы (80%) мальтогенные экзоамилазы используются
вместе с а-1,6-глюкозидазами.
Экзоамилазы расщепляют а-1,4-глюкозидные связи, а глю-
когенные экзоамилазы гидролизуют а-1,6-глюкозидные связи в
разветвленных молекулах олигосахаридов, в то время как маль-
тогенные экзоамилазы (например, р-амилаза злаков) не спо-
собны проходить эти точки ветвления. Последовательное уда-
ление низкомолекулярных компонент (глюкозы или мальтозы)
с нередуцирующего конца полимерной молекулы крахмала при-
водит к постепенному уменьшению вязкости; образующийся
продукт имеет p-конфигурацию при Ci-атоме восстанавливаю-
щей молекулы глюкозы. Для промышленных целей глюкоами-
лазы получают из Aspergillus niger или Rhizopus spp. Это низ-
коспецифичные ферменты, которые гидролизуют связи ct-1,3 и
а-1,6 медленнее, чем а-1,4. Они стабильны в широком диапазо-
не pH и более активны при 75 °C, хотя обычно им приходится
функционировать при 65 °C. Эти ферменты могут также ката-
лизировать полимеризацию глюкозы с образованием мальтозы
и изомальтозы, но осахаривание никогда не достигает равно-
весия, и эта реакция особого значения не имеет. В препаратах
этих ферментов присутствует загрязняющая примесь трансглю-
козилаза, катализирующая образование несбраживаемых олиго-
меров глюкозы за счет реакции переноса молекул глюкозы при
осахаривании. Это может существенно снижать выход глюкозы.
Глюкоамилазы применяются в основном в производстве кон-
центрированного сиропа (90—97% D-глюкозы), из которого вы-
рабатывают кристаллическую глюкозу или же концентрирован-
ные фруктозные сиропы (см. предыдущий раздел). Для полу-
чения максимального выхода продукта при осахаривании клю-
чевое значение имеет процесс ожижения; при ферментативном
ожижении выход продукта больше, чем при кислотном, так как
в этом случае образуется меньше побочных продуктов. Глюко-
амилазы применяются также при выработке сиропов с высоким
декстрозным эквивалентом или с высокой степенью конверсии
(35—43% глюкозы, 30—37% мальтозы и 8—13% мальтотриозы)
для пищевой промышленности. Сиропы, полученные кислотным
гидролизом, обрабатывают мальтогенными и глюкогенными ами-
Химия и технология
169
лазами. Состав продукта при этом зависит от соотношения ис-
пользованных в производстве ферментов.
Большинство разновидностей крахмала, важных в промыш-
ленном отношении, содержит 80% амилопектина, который лишь
частично расщепляется в ходе ферментативного гидролиза. Под
действием а-амилаз из него образуются производные декстри-
нов, которые далее уже гидролизоваться не могут. Так как по
точкам ветвления гидролиз под действием глюкоамилаз идет
медленно, используют а-1,6-глюкозидазы, из них в промышлен-
ном масштабе — пуллуланазу (пуллулан-6-глюканогидролазу).
Ферментативная переработка крахмала проводится сегодня
в широком диапазоне pH. Задачей будущего является выделе-
ние термостабильных ацидофильных ферментов, что позволит
вести переработку при одном и том же pH (4—5).
Другие ферменты, имеющие коммерческое значение
(см. также гл. 3)
Сегодня ферменты применяются наиболее широко для превра-
щения углеводов, играющих особую роль в пищевой и молоч-
ной промышленности. Так, р-галактозидазу (лактазу) применя-
ют для гидролиза лактозы в снятом молоке. Получаемый без-
лактозный продукт используется теми людьми, организм кото-
рых не способен синтезировать лактазу. Этот фермент приме-
няется, кроме того, для переработки лактозы сыворотки в глю-
козогалактозные сиропы, а также подкисленной сыворотки,
образующейся при получении творога.
Под действием инвертазы из сахарозы получают глюкозу.
Для определения количества глюкозы в жидкостях тела в кли-
никах в последнее время стала широко использоваться глюкозо-
оксидаза — как в свободной форме, так и иммобилизованная.
На коммерческий уровень поставлено ферментативное раз-
деление рацемических смесей аминокислот и эфиров терпенов.
Такие смеси аминокислот образуются при химическом синтезе,
и разделение их по оптическим свойствам составляющих имеет
важное практическое значение. Известно, что для этого можно
использовать соответствующие физико-химические методы (ме-
ханическое разделение, избирательная кристаллизация, хрома-
тографическое разделение) и химические подходы (фракцион-
ная кристаллизация солей диастереомеров), но гораздо более
эффективными и удобными оказываются процессы, основанные
на стереоспецифичности ферментов. Укажем некоторые из ис-
пользуемых здесь приемов.
1. Получение асимметричных производных, например обра-
зование амидов П-ацил-Б-аминокислот, катализируемое папаи-
ном, бромелином или фицином. Так; взяв анилин и папаин, мы
170
Глава 4
получаем нерастворимые L-анилиды аминокислот. Их можно
гидролизовать химическим способом и получить L-изомер.
2. Асимметричный гидролиз. В этом случае осуществляется
стереоспецифический гидролиз производных аминокислот, в ре-
зультате которого образуется лишь один энантиомер. Эфиры
L-аминокислот гидролизуют протеолитическими ферментами
(например, химотрипсином); нужный продукт и не вступающие
в реакцию D-эфиры разделяют по растворимости.
3. Использование амидаз. Эти ферменты из почек и подже-
лудочной железы млекопитающих или из микроорганизмов сте-
реоспецифически гидролизуют амиды L-аминокислот, которые
затем отделяют методом дифференциальной растворимости.
Амиды менее склонны к спонтанному гидролизу, чем эфиры,
и с помощью этого метода получают оптически более чистые
препараты аминокислот.
4. Стереоспецифический гидролиз М-ацил-Ь-аминокислот.
В этом случае используются аминоацилазы или карбоксипеп-
тидазы, и в результате образуются L-аминокислоты. Ацилазы
получают из самых разных микроорганизмов. Иммобилизован-
ная аминоацилаза одного из грибов успешно использовалась
в установке для непрерывного ферментативного разделения.
В конце 60-х годов в промышленности стали применять ме-
тод разделения рацемических смесей эфиров терпенов при по-
мощи гидролаз: было обнаружено, что ферментативный гидро-
лиз этих эфиров идет только в том случае, если эфирная связь
экваториальна или же легко принимает эту конформацию. Раз-
деление по оптическим свойствам зависело также от положения
диастереомерных центров; оно шло успешно в случае эпимеров
вторичных спиртов. В качестве катализатора применяли дрож-
жи, иммобилизованные в полиуретане. Опыт проводили с
DL-ментолсукцинатом, для которого в водонасыщенном н-геп-
тане степень конверсии составила 72,6%. В результате был по-
лучен оптически чистый L-ментол. Этот метод разделения с про-
изводительностью 800 кг был запатентован. Рацематы терпенов
можно разделять и путем обращенного гидролиза: сначала при
участии липазы получают эфиры, а затем гидролизуют их хи-
мическим методом.
Будущее технологии иммобилизованных ферментов
Сегодня в промышленности реализовано всего четыре крупно-
масштабные технологии на основе иммобилизованных фермен-
тов (глюкозоизомеразы, аминоацилазы, пенициллинацилазы и
лактазы). Последнюю иммобилизовали на частицах кремнезема
и применяли для конверсии лактозы сыворотки в глюкозу и га-
Химия и технология
171
лактозу. В обозримом будущем иммобилизованные ферменты
могут быть использованы для следующих целей.
1. Холинэстераза может применяться для определения пести-
цидов. Степень ингибирования этого фермента в присутствии
пестицидов оценивают электрохимическими или колориметриче-
скими методами.
2. Аналогичным образом другие ферменты могут использо-
ваться для определения токсических веществ. Так, карбоангид-
раза очень чувствительна даже к малым концентрациям хлор-
производных углеводородов, гексокиназа — к хлордану, линдану
и токсафену.
3. Иммобилизованная диизопропилфторфосфатаза нервных
клеток кальмара может найти применение для обезвреживания
фосфоорганических нервных газов (зомана, зарина).
4. Иммобилизованная гепариназа может применяться для
предотвращения тромбообразования в аппаратах искусственно-
го кровообращения.
5. Иммобилизованная билирубиноксидаза может использо-
ваться для удаления билирубина из крови новорожденных, стра-
дающих желтухой.
6. Предложен новый способ применения иммобилизованного
гемоглобина. Суть его состоит в том, что включенный в поли-
уретановую матрицу белок образует «гемогубку», способную
поглощать кислород прямо из воды с эффективностью 80%.
Далее кислород высвобождается из полимера под действием
слабого электрического разряда или в вакууме. Предполагается,
что такая система может снабжать кислородом водолазов либо
работающие под водой двигатели.
7. Возможно, вскоре удастся создать системы из нескольких
иммобилизованных ферментов. Так, если заключить в микро-
капсулы три фермента — уреазу, глутаматдегидрогеназу и глю-
козодегидрогеназу, то их можно будет использовать для удале-
ния мочевины из крови больных с почечной недостаточностью.
8. Иммобилизованные ферменты найдут дальнейшее приме-
нение в молочной промышленности. При производстве сыра мо-
гут использоваться иммобилизованные свертывающие молоко
белки — реннин и пепсин. Для гидролиза жира в молоке можно
использовать иммобилизованные липазы и эстеразы.
9. Разнообразные иммобилизованные ферменты со временем
найдут применение и в датчиках для быстрого анализа. Сегодня
в таком качестве используются лишь несколько ферментов, но
когда будет решена проблема стабилизации, их число увеличит-
ся. Особенно полезными из-за их высокой стабильности могут
оказаться ферменты термофилов.
172
Глава 4
4.3. Получение химических веществ из биомассы
4.3.1. Биотехнология на основе растительных клеток
Растения издавна являются поставщиками химических соедине-
ний для самых разных отраслей химической промышленности.
Это не только такое сырье, как сахара, но и целый набор слож-
ных вторичных метаболитов, например каучук, кокаин, вещест-
ва, использующиеся в качестве красителей, вкусовых добавок
и пряностей. Получить такие вещества методом химического син-
теза часто бывает невозможно из-за сложности их строения.
Сегодня, воодушевленные успехами биотехнологии, ученые вновь
обращаются к царству растений. Они не только пытаются отыс-
кать пути к улучшению способов выработки уже освоенной про-
дукции (например, аймалина и кодеина), но и разработать
новые принципы биотрансформации и получить новые продук-
ты. Нам предстоит в ближайшие годы заставить гены растений
работать в бактериальных клетках; сложность этой задачи со-
стоит в том, что мы плохо знаем, как они работают даже в соб-
ственных клетках. Кроме того, вторичные метаболиты образу-
ются в результате многоступенчатых процессов, о регуляции
которых нам тоже почти ничего не известно. Можно думать, что
путем использования культур растительных тканей мы сможем
разработать новые подходы к получению ценных химических
продуктов, особенно лекарственных веществ, а также улучшить
сорта растений. Работая с культурами тканей растений, мы
сможем контролировать образование таких веществ и при этом
не зависеть от капризов погоды и не думать о вредителях рас-
тений, которые так сильно влияют на образование нужных нам
веществ.
Культуры растительных тканей можно получить из любого
вида растений. При этом используются разнообразные среды.
Познание особенностей физиологии и биохимии таких культур
позволили значительно повысить их урожайность и выход био-
массы. Однако многие такие культуры не могут считаться истин-
ными автотрофами, так как для роста им необходимы внешние
источники углерода (в форме глюкозы или сахарозы), азот, ми-
неральные вещества и факторы роста. Сегодня мы умеем полу-
чать большое количество биомассы (20—30 г/л), но повысить
выход интересующих нас веществ обычно удается лишь за счет
снижения выхода биомассы и подавления роста. Выход различ-
ных веществ в культуре может быть в 10 раз выше, чем в слу-
чае растения, но для этого необходимо разработать новые стра-
тегии скрининга и селекции, особенно если искомый продукт
образуется в малых количествах.
Химия и технология
173
С проблемами биотехнологии растительных клеток можно
познакомиться на примере организации промышленного произ-
водства первого вещества, полученного из культуры тканей рас-
тения. Известно, что корни растения Lithospermum erytrorhizon
содержат шиконин и его производные. Это растение издавна
используется в Японии как лекарственное, поскольку оно обла-
дает антибактериальной и противовоспалительной активностью.
•Шиконин, ярко-красное вещество, производное нафтохонина,
применяли и как краситель. Для того чтобы концентрация ши-
конина в корнях достигла 1—2%, возраст растения должен со-
-ставлять 5—7 лет. Так как выращивать Lithospermum erythror-
hizon в промышленном масштабе в Японии невозможно, его
приходилось ввозить из Кореи и Китая, а поэтому стоимость
чистого природного вещества составляла 4500 долл, за 1 кг.
Представлялось заманчивым наладить его промышленное про-
изводство на основе культуры тканей растений. Клетки растений
не выделяют вторичные продукты в среду, а запасают их в ва-
куолях и органеллах, что затрудняет их получение. Однако на-
копление шиконина облегчает отбор наиболее продуктивных
линий, поскольку содержащие этот краситель клетки имеют яр-
ко-красный цвет. Удалось выделить линию, накапливающую до
15% шиконина на сухую массу клеток. Последующая оптими-
зация среды позволила достичь тринадцатикратного увеличения
продуктивности. Был разработан двухступенчатый процесс куль-
тивирования, в котором на первой стадии создавались оптималь-
ные условия для наращивания биомассы, а на втором —для
образования вторичных продуктов. Клетки растят в суспензион-
ной культуре; для этого лучше всего подходят эрлифтные фер-
ментеры, так как в них осуществляются одновременно как ин-
тенсивное перемешивание, так и разделение клеток, а вероят-
ность повреждения относительно хрупких клеток минимальна.
Процесс производства шиконина начинается с наращивания кле-
ток в ферментере объемом 200 л. Затем его содержимое пере-
носят в аппарат объемом 750 л и проводят экстракцию обыч-
ными химическими методами. Выход продукта за один цикл
составляет 5 кг, поэтому стоимость его гораздо ниже, чем при
получении упомянутым выше способом.
Шиконин можно выделять и иным способом — путем изме-
нения pH среды или же добавления некоторых влияющих на
проницаемость клеток веществ, например диметилсульфоксида.
Такой подход может оказаться полезным при работе с иммоби-
лизованными клетками: в этом случае благодаря селективной
проницаемости можно будет постоянно удалять вторичные мета-
болиты из внутриклеточных вакуолей без глубокого нарушения
первичного обмена веществ. Необходимо отметить, что иммо-
билизация, осуществленная наиболее мягким, щадящим мето-
174
Глава 4
дом, может способствовать внутриклеточному накоплению со-
единения в ответ на создание микроокружения, имитирующего
условия в продуктивных частях растения. Рост можно замед-
лить, направив питательные вещества на образование вторич-
ных продуктов метаболизма. Так, клетки Catharanthus, иммо-
билизованные в альгинат-акриламидной матрице, в больших
количествах выделяют в среду аймалин и серпентин. Для полу-
чения ряда алкалоидов, производных индола [аймалина, вин-
бластина и винкристина (два последних соединения применя-
ются при лечении рака)], можно, по-видимому, использовать-
барвинок Catharanthus roseus. Необходимо отметить, что кон-
центрация этих алкалоидов обычно крайне мала, и их прихо-
дится выделять из смеси других образуемых растением алка-
лоидов. Кроме того, в культурах тканей винбластин и винкри-
стин не образуются, но разработан альтернативный способ их
производства. Эти алкалоиды представляют собой асимметрич-
ные димеры катарантина и виндилина, причем катарантин мож-
но получать методом культивирования в большом количестве
с относительно небольшой примесью других алкалоидов. Диме-
ры затем можно синтезировать химическим путем.
Культивируемые ткани растений применяются также для
проведения биотрансформации. Примером такого рода может
служить образование стимулирующего работу сердца дигоксина
путем 12-р-гидроксилирования дигитоксина в суспензионных
культурах или в иммобилизованных клетках наперстянки Digi-
talis lanata. На основе этих систем, а также иммобилизованных
клеток моркови удалось осуществить и синтез ряда новых кар-
денол идов.
Методы культивирования тканей растений применялись и для
улучшения сортов сельскохозяйственных культур: повышения их
устойчивости к болезням и неблагоприятным условиям среды,,
увеличения содержания сахарозы и крахмала, повышения уро-
жайности. Эти методы используют для выведения более продук-
тивных древесных пород—источников лигноцеллюлозы, а так-
же растений — продуцентов эластомеров (гваюла, молочай) или
масла (масличная пальма, соя).
Таким образом, перспективы развития биотехнологии на ос-
нове растительных клеток представляются весьма многообещаю-
щими. Будет налажено производство новых лекарственных пре-
паратов, подсластителей, средств защиты растений, веществ для
косметической и парфюмерной промышленности. Возможности
этой быстро развивающейся технологии будут еще более рас-
ширены в результате дальнейшей разработки способов получе-
ния гибридов путем слияния протопластов.
Химия и технология
175
4.3.2. Химические вещества, получаемые из биомассы
Известно, что после окончания второй мировой войны химиче-
ская промышленность получала из природного газа и нефти
разнообразные виды сырья высокой чистоты в большом коли-
честве и по относительно стабильным ценам. Сегодня многое
в этой сфере изменилось, кроме того, за последние пятнадцать
лет больших успехов достигла биотехнология. Все это побудило
переоценить возможности использования биомассы для произ-
водства химических веществ. До развития нефтехимии из био-
массы получали многие виды сырья для химической индустрии,
например жиры и масла (для производства мыла и детергентов),
метанол (путем сухой перегонки древесины) и растворители
(за счет сбраживания крахмала и сахаров). Цены и доступность
этого сырья (биомассы) существенно менялись. Как уже гово-
рилось в этой главе и в гл. 5, большую роль в производстве
химических веществ и полимеров, которые находят применение
во многих отраслях промышленности, играет биомасса в виде
сбраживаемых сахаров. Но в этом разделе мы уделим особое
внимание использованию главных запасов биомассы — лигно-
целлюлозы. Утилизация биомассы основана на процессе ее кон-
версии в сбраживаемые субстраты, которые в свою очередь
служат сырьем для многих отраслей микробиологической про-
мышленности, производящих химические вещества, горючее и
продукты питания. При этом необходимо эффективно исполь-
зовать все компоненты биомассы: целлюлозу, гемицеллюлозу и
лигнин. Лигнин защищает полиглюкан от действия ферментов,
так что для успешного использования полимерной целлюлозы
необходимо прежде всего удалить гемицеллюлозы, далее раз-
рушить комплекс лигнина с целлюлозой, а затем —и кристал-
лическую структуру самой целлюлозы. Эти задачи могут быть
решены с применением разнообразных химических, физических
и микробиологических методов.
Гемицеллюлозы легко удаляются путем растворения в сла-
бой кислоте в ходе первичной обработки биомассы. При произ-
водстве глюкозы из древесины довольно успешно применяется
гидролиз слабым раствором кислоты при высоких давлении и
температуре. Этот способ нашел применение в ряде стран. Го-
раздо эффективнее гидролиз концентрированной кислотой, но
для осуществления этого процесса требуются большие капита-
ловложения. Особенно успешно применяется плавиковая кисло-
та, причем безводную кислоту можно регенерировать. Лигноцел-
люлозу можно также размалывать и затем подвергать радиа-
ционному облучению в больших дозах. Получаемый такими
способами лигнин различается по своим свойствам. Очищенные
лигнины используются при выработке клеев, смол, адгезивов,
176
Глава 4
типографской краски, для диспергирования красителей; они
применяются как адсорбенты, изоляторы, используются при до-
быче нефти, в асфальтовых смесях и при нанесении полимеров.
После удаления лигнина экстракцией растворителями получают
кристаллическую целлюлозу, которая идет на нужды бумажной
промышленности. Рентабельность этого процесса определяется
главным образом стоимостью регенерации растворителя. При-
мером одностадийного процесса, протекающего в мягких усло-
виях, является экстракция фенолом при 100 °C, когда лигнин-
и гемицеллюлозы растворяют в феноле, а высвободившуюся
после фильтрования целлюлозу используют для выделки бумаж-
ной массы. По мере остывания фенольного экстракта гемицел-
люлоза концентрируется главным образом в водной фазе, от-
деляясь от масел и лигнина, которые можно использовать для
производства новых порций фенола методом гидрокрекинга.
Таким образом, в отношении фенола и энергетических потреб-
ностей— это самообеспечивающийся процесс. При «паровом
взрыве» лигноцеллюлоза претерпевает превращения, в резуль-
тате которых она становится более чувствительной к фермен-
тативному гидролизу (как в рубце животных, так и вне его).
При этом увеличивается и выход реакционноспособных лигни-
нов, высвобождаемых под действием слабой щелочи. При раз-
рушении замораживанием используется жидкий аммиак. В слу-
чае умеренно твердой древесины метод автогидролиза на основе
парового взрыва дает отходы, пригодные для дальнейшей пере-
работки микробами; древесина хвойных и тропических пород,
более богатая лигнином, плохо поддается такой обработке.
Для ферментативного превращения целлюлозы в глюкозу
необходимы дорогостоящие целлюлазы и, возможно, для их по-
лучения придется провести отбор мутантов с повышенной цел-
люлолитической активностью. Достигнутая на сегодня скорость,
гидролиза целлюлозы все еще низка,, но такую переработку
можно объединить с процессом брожения при участии дрожжей,
так что глюкоза будет и образовываться, и потребляться в од-
ном и том же ферментере. Достоинства и недостатки такого-
процесса были изучены на опытных установках. Проведены ис-
следования по клонированию генов целлюлаз и включению их
в геном подходящих организмов-хозяев, которые тем самым
приобретают способность к деградации содержащих целлюлозу
субстратов. Такой прием позволяет осуществлять более широ-
комасштабное и эффективное производство химических веществ
из биомассы с помощью ферментов. Альтернативой прямой мик-
робной конверсии является применение высокотемпературных
анаэробных' процессов при участии Clostridium thermocellum^
когда одновременно идут и гидролиз, й ферментация.
Химия и технология
\П
При помощи ферментов ксиланаз гемицеллюлозы могут быть
разрушены с образованием пентозных сахаров. Найдены виды
дрожжей, способные осуществлять ферментацию пентоз, но они
чувствительны к спирту. Порог этой чувствительности может
быть изменен генетическими методами; кроме того, путем кло-
нирования гена ксилоизомеразы дрожжам Saccharomyces может
быть передана способность потреблять ксилозу. Это позволит
им осуществлять конверсию ксилозы (субстрата, на котором
они не растут) в ксилулозу (субстрат, который они сбражи-
вают).
Биодеградация лигнина — это окислительный процесс, осуще-
ствляемый в первую очередь грибами. Высвобождающейся при
этом энергии, по-видимому, недостаточно для обеспечения роста
микроорганизмов. Сегодня изучению микробиологических и био-
химических аспектов деградации лигнина уделяется все больше-
внимания. Показано, что в эксперименте внеклеточные фермен-
ты гриба Phanerochaete chrysosporium могут осуществлять де-
градацию лигниновых модельных субстратов, при этом в про-
цессе окислительного расщепления пропильных боковых цепей
лигнина участвует перекись водорода.
Обнаружено, что Acetobacter xylinum может синтезировать
целлюлозу из сахаров и крахмала. Это создает предпосылки
для получения чистого полимера, необходимого для производ-
ства вискозы, целлофана и других целлюлозных полимеров, вы-
рабатываемых сегодня на основе лигноцеллюлозы, из которой
удален лигнин.
Дискутируется вопрос о том, смогут ли конкурировать с та-
ким сырьем, как целлюлоза, хитин (высокомолекулярный по-
лимер N-ацетилглюкозамина) или же хитозан (сходный полимер-
с небольшим числом ацетилированных N-групп), особенно если
иметь в виду, что запасы их ограничены. Хитин можно получать
из антарктического криля (рачков), отходов при переработке-
водных животных, имеющих панцирь, или же из грибного ми-
целия-отхода бродильной промышленности. Оба полимера мо-
гут иметь разнообразное применение: как адгезивы, коагулянты,
переносчики лекарственных веществ, а также как добавки при
выдёлке бумаги и тканей.
4.4. Перспективы развития
В будущем влияние биотехнологии на развитие химической про-
мышленности будет определяться возможностью объединения:
принципов микробиологии, биохимии и химической технологии.
Основной предпосылкой использования биологического катализа
в химии является способность ферментов катализировать энан-
тиомерно однозначные, стереохимически определенные реакции
12—1344
178
Глава 4
синтеза. Сегодня многие биологические процессы имеют такие
характеристики, что с экономической точки зрения они не могут
конкурировать с альтернативными химическими процессами.
Принято считать, что для успешного осуществления крупно-
масштабного биологического процесса, направленного на полу-
чение некоего химического соединения, необходимо, чтобы при-
бавочная стоимость продукта составляла 500—1000 долл, за
тонну сверх стоимости сырья. Впрочем, все же существуют об-
ласти, в которых применение биотехнологии обещает быть пер-
спективным.
1. Тип реакции: прямые реакции окисления — восстановле-
ния; использование обычных ферментов для проведения нетри-
виальных реакций; белковая инженерия, нацеленная на изме-
нение свойств катализатора; использование биокатализа в не-
водных средах.
2. Конфигурация реактора: оптимизация биокатализаторов
методами генной инженерии; выбор термостабильных систем
или использование иммобилизованных конфигураций; разработ-
ка дешевых, эффективных и общеудобных методов повторного
использования кофактора; химическая инженерия в связи с про-
блемами крупномасштабных биокаталитических систем (см.
гл. 10).
4.4.1. Тип реакции
Реакции прямого окисления и оксигенации
Для преобразования сложных молекул в ходе органического
синтеза используются оксидоредуктазы со строгой структурной,
сайт- и стереоспецифичностью. В случае более широкой суб-
стратной специфичности эти ферменты могут использоваться
как катализаторы типовых реакций. В реакциях превращения
спиртов в карбонилы находят применение нуклеотид-зависимые
дегидрогеназы. Так, хорошо изучена алкогольдегидрогеназа из
печени лошади: известны ее субстратная специфичность и сте-
реохимия катализируемых реакций. Она атакует моно-, ди- и
тетрациклические структуры. Построена модель ее активного
центра, что позволяет прогнозировать активность этого фермен-
та в отношении новых субстратов (см. разд. 4.1.1). Отметим,
что ациклические вторичные спирты — плохой субстрат для это-
го фермента, и если ставится задача осуществления синтеза на
их основе, то целесообразно попытаться использовать другие
дегидрогеназы (возможно, термофильные). Особенность окси-
геназ состоит в их способности с высокой эффективностью и
специфичностью включать кислород прямо в органические суб-
страты; проведение такого прямого окисления неактивирован-
Химия и технология
17»
ных органических субстратов — вечный камень преткновения в
работе химика-синтетика. Реакции этого типа весьма разнооб-
разны: они лежат в основе превращения алканов в спирты,
олефинов — в эпоксиды, сульфидов — в сульфоксиды; это реак-
ции гидроксилирования ароматического кольца и его раскры-
тия, а также окислительного деметилирования. Вполне возмож-
но, что результаты глубокого изучения механизма активации и
присоединение кислорода ферментами найдут применение для
создания новых химических каталитических систем. В последнее
время большое внимание уделялось использованию различных
микробных оксигеназных систем в стереоспецифическом эпокси-
окислении олефинов, в результате которого образуются «строи-
тельные блоки» для синтеза полимеров. В микроорганизмах,
окисляющих алканы и алкены, функционируют соответствую-
щие монооксигеназы, но если исходить из нынешнего уровня
технологии, то вряд ли хоть один процесс такого типа целесо-
образен с экономической точки зрения. Поскольку оксигеназы
нестабильны, принято считать, что эти процессы основаны на
биокаталитической активности целостных организмов. Предло-
жен интересный альтернативный путь таких превращений: трех-
ступенчатый ферментативный процесс с участием оксидаз-
(рис. 4.14). На первом этапе пиранозо-2-оксидаза из гриба-ба-
зидиомицета катализирует образование перекиси водорода, ис-
пользуя в качестве субстрата и источника энергии глюкозу.
Далее галопероксидаза осуществляет реакцию перекиси водо-
рода с алкеном и ионом какого-либо галогена с образованием
алкенгалогидрина, от которого затем отщепляются атомы во-
дорода и галогена и образуется соответствующий алкеноксид.
Побочными продуктами процесса являются фруктоза и глюко-
новая кислота. Стереоспецифичность окислительного катализа
может использоваться в процессе синтеза диэпоксимономеров
для производства стереорегул яр ных полимеров. Мономерные
единицы могут быть получены и путем превращений бензола
под действием оксигеназ (гл. 5) или в результате двойного мо-
ноокисления дифенила с образованием 4,4-дигидроксидифенила.
Использование обычных ферментов в нетривиальных
химических реакциях
В органическом синтезе ферменты применяются пока не очень
широко в основном потому, что большинство интересующих ис-
следователей веществ не являются их природными субстратами.
Приходится специально изучать субстратную специфичность
ферментов и условия протекания реакций, катализируемых наи-
более освоенными промышленностью ферментами, с тем чтобы
выяснить возможность вовлечения этих ферментов в каталити-
12»
<80
Глава 4
А
сн2=сн—сн3 + 02
Пропан
NADH2 NAD
/°\
сн2— сн —СН3 + Н2О
1,2-Эпоксипропан
Например, метанмонооксигеназы,
алканмонооксигеназы,
алкенмонооксигеназы
Г люкозв
СН2 —СН — СН3
Г люкозо-2-
оксидаза
> Н2О;
--- Вг” *-------
Г алопероксидаза
2-Г ексулоза
Вг ОН
СН2 —СН —СН3
Химический
катализ
Фруктоза
Основная или
галогидрин-эпоксидаза
--> Вг“
/°\
сн2—сн—СНз
Рис. 4.14. Осуществляемое при участии микроорганизмов образование мономе-
ров эпоксидов. А — прямое окисление с помощью монооксигеназ с широким
спектром действия; Б — многоступенчатый процесс.
ческие процессы, интересующие химика-синтетика. Так, промыш-
ленностью производится фермент глюкозооксидаза, проявляю-
щий высокую специфичность к донору электронов D-глюкозе.
С другой стороны, специфичность его к акцепторам электронов
гораздо шире, и природный акцептор, кислород, может быть
заменен некоторыми синтетическими красителями. Это послу-
жило основой для использования фермента в реакции восста-
новления бензохинона в гидрохинон (рис. 4.15), причем реакция
шла в три раза быстрее, чем в присутствии кислорода. Фермент
может восстанавливать и другие хиноны (некоторые из них
применяются в биологии и медицине), а также ряд ароматиче-
ских нитросоединений. Было показано, что галактозооксидаза
катализирует реакцию стереоспецифического превращения али-
фатических спиртов (не сахаров) в альдегиды; так, глицерол
превращается в S-(—)-глицеральдегид. Для разделения [}-арил-
Химия и технология
181
А
D -Глюкоза + О2
D Тлюконолактон + Н2О
E.FAD + Глюкоза
E.FADH2 + О2
t Е FADH2 + Тлюконолактон
♦ E.FAD + Н2О2
Рис. 4.15. Использование глюкозооксидазы и неприродных акцепторов электро-
нов при химическом синтезе органических веществ. А — обычная реакция; Б —
£
D-Глюкоза + Бензохинон .--------k D -Тлюконолактон + Гидрохинон
реакция с неприродным акцептором электронов.
акролеинов может быть использована ксантиноксидаза, так
как она окисляет транс-изомер в сто раз быстрее, чем цис.
Изменяя условия, в которых действует фермент, мы также
можем существенно повлиять на химические особенности ката-
лизируемой реакции. Так, пероксидаза из хрена гидроксилирует
ароматические соединения с небольшой специфичностью и ма-
лым выходом. Понижение температуры с 25 до О °C подавляет
побочные реакции, в результате чего в основном гидроксилиру-
ются пара-замещенные ароматические соединения по мета-поло-
жению и vice versa. Карбоксиэстераза из печени свиньи способ-
на осуществлять как реакцию трансэтерификации, так и гидро-
лиз эфиров, причем последняя реакция доминирует, поскольку
плохо растворимые спирты не могут эффективно конкурировать
с молекулами воды. Фермент можно заставить работать по пер-
вому механизму, применив двухфазные системы, в которых фер-
мент растворен в минимальном объеме воды, а основная орга-
ническая среда образована спиртом и эфиром.
182
Глава 4
Инженерия белка
Белковая инженерия может быть основана на химической мо-
дификации готового белка или на методах генетической инже-
нерии, позволяющих получать модифицированные варианты
природных белков.
Конструирование определенного биологического катализато-
ра ведется с учетом как специфичности белка, так и каталити-
ческой активности металлоорганического комплекса. Вот при-
меры такой модификации, проведенной для получения «полу-
синтетических биоорганических комплексов». Миоглобин каша-
лота способен связывать кислород, но не обладает биокатали-
тической активностью. В результате объединения этой биомо-
лекулы с тремя электрон-переносящими комплексами, содержа-
щими рутений, которые связываются с остатками гистидина на
поверхности молекул белка, образуется комплекс, способный
восстанавливать кислород при одновременном окислении ряда
органических субстратов, например аскорбата, со скоростью-
почти такой же, как для природной аскорбатоксидазы. В прин-
ципе белки можно модифицировать и другими способами. Рас-
смотрим, например, папаин. Он относится к числу хорошо изу-
ченных протеолитических ферментов, для которого определена
трехмерная структура. Поблизости от остатка цистеина-25 на
поверхности белковой молекулы располагается протяженный
желобок, в котором протекает реакция протеолиза. Этот уча-
сток может быть алкилирован производным флавина без изме-
нения доступности участка связывания потенциальных субстра-
тов. Такие модифицированные флавопапаины использовались-
для окисления Ы-алкил-1,4-дигидроникотинамидов, и каталити-
ческая активность некоторых из этих модифицированных белков
была существенно выше, чем у природных флавопротеин-NADH-
дегидрогеназ. Таким образом удалось создать очень эффектив-
ный полусинтетический фермент. Использование флавинов с вы-
сокоактивными, находящимися в определенном положении элек-
трон-оттягивающими заместителями, возможно, позволит разра-
ботать эффективные катализаторы для восстановления никотин-
амида.
Крупные успехи, достигнутые за последнее время в химиче-
ском синтезе ДНК, открыли перед белковой инженерией прин-
ципиально новые возможности: конструирование уникальных,,
не встречающихся в природе белков. Для этого необходимо и
дальнейшее развитие технологии, так чтобы изменение генов
методами генетической инженерии приводило к предсказуемым
изменениям белков, к улучшению вполне определенных функ-
циональных их характеристик: числа оборотов, Дм для конкрет-
ного субстрата, термостабильности, температурного оптимума.
Химия и технология
183
стабильности и активности в неводных растворителях, субстрат-
ной и реакционной специфичности, потребности в кофакторах,
оптимуме pH, устойчивости к протеазам, аллостерической ре-
гуляции, молекулярной массы и субъединичного строения. Обыч-
но такого улучшения достигали с помощью мутагенеза и отбора,
л в последнее время — путем химической модификации и иммо-
билизации. Для успешного конструирования конкретного типа
молекул белка необходимо выявить ряд основополагающих за-
кономерностей, связывающих структурные особенности белков
и их желаемые свойства. Так, зная точную кристаллическую
структуру молекулы изучаемого белка, мы можем идентифици-
ровать те ее участки, которые следует направленно модифици-
ровать для увеличения его каталитической активности. Такая
модификация может состоять в изменении аминокислотной по-
следовательности белка.
Первая контролируемая модификация белка была проведена
в середине 60-х годов Кошландом и Бендером. Для замены гид-
роксильной группы на сульфгидрильную в активном центре
протеазы — субтилизина они применили метод химической мо-
дификации. Однако, как выяснилось, такой тиолсубтилизин не
сохраняет протеазную активность. Вообще говоря, методы хи-
мической модификации не только жестки и неспецифичны; они
плохи еще и тем, что с их помощью невозможно вызвать мно-
жественные желаемые изменения, особенно если модифицируе-
мые аминокислотные остатки погружены в глубь третичной
структуры белка. Для этого нужна белковая инженерия, осно-
ванная на генетической инженерии. Сегодня она осуществля-
ется при помощи двух хорошо освоенных методов (гл. 7). Так,
сайт-специфический мутагенез осуществляется следующим об-
разом. Клонируют ген того белка, который интересует исследо-
вателя, и встраивают его в подходящий генетический носитель.
Затем синтезируют олигонуклеотидную затравку с желаемой
мутацией, последовательность которой из десяти — пятнадцати
нуклеотидов в достаточной степени гомологична определенному
участку природного гена и поэтому способна образовывать с
ним гибридную структуру. Эта синтетическая затравка исполь-
зуется полимеразами для начала синтеза комплементарной ко-
пии вектора, которую затем отделяют от оригинала и исполь-
зуют для контролируемого синтеза мутантного белка. Альтер-
нативный подход основан на расщеплении цепи, удалении под-
лежащего изменению сайта и замещении его синтетическим
аналогом с желаемой последовательностью нуклеотидов.
Тирозил-тРНК—синтетаза катализирует реакцию аминоаци-
лирования тирозиновой тРНК, которая включает активирование
тирозина с помощью АТР с образованием тирозиладенилата.
Ген этого фермента, выделенный из Bacillus stearothermophilus,
184
Глава 4
был встроен в бактериофаг М13. Затем каталитические свойст-
ва фермента, особенно его способность связывать субстрат, бы-
ли изменены путем сайт-специфической модификации. Так, трео-
нин-51 был заменен на аланин. Это привело к двукратному
увеличению связывания субстрата, видимо, из-за невозможности
образования водородной связи между этим остатком и тирозил-
аденилатом. При замене аланина пролином нарушается конфи-
гурация молекулы фермента, но способность к связыванию суб-
страта увеличивается в сто раз, так как облегчается его взаимо-
действие с гистидином-48. Сходные сайт-специфичные изменения,
были получены в р-лактамазе, и обычно они сопровождались
инактивацией фермента. Замена серина-70 на цистеин приводиг
к образованию р-тиоллактамазы, константа связывания у кото-
рой не отличается от таковой для природного фермента, но ак-
тивность по отношению к пенициллину составляет всего 1—2%..
Тем не менее активность этого мутантного фермента в отноше-
нии некоторых активированных цефалоспоринов не меньше ис-
ходной активности или даже превышает ее; эти белки также-
более устойчивы к действию протеаз.
Мутации, вызываемые путем сайт-специфичного воздействия^
используют сегодня для проверки адекватности результатов
структурных исследований. В некоторых случаях с их помощью-
удалось показать, что структурная стабильность белка и его
каталитическая активность могут быть разобщены. Накопив
достаточное количество информации о взаимосвязи между ста-
бильностью структуры белка и его функцией, мы, возможно,
сумеем осуществлять тонкую регуляцию активности биологиче-
ских катализаторов и создавать полностью синтетические их
аналоги. Недавно появилась работа, в которой сообщалось о
клонировании первого синтетического гена фермента, кодирую-
щего активный фрагмент молекулы рибонуклеазы.
Биологический катализ в неводных средах
Известно, что биологический катализ осуществляется в природе
в водной среде, но сфера применения ферментов в биотехнологии
не может ограничиваться только этими условиями. Нередко
нужно подвергнуть изменению структуры липофильных или во-
донерастворимых веществ, например стероидов или углеводоро-
дов. Применение органических растворителей может не только
увеличить каталитическую активность определенного фермента
путем повышения доступности субстрата, но и сместить равно-
весие соответствующей химической реакции.
Главным фактором здесь является стабильность биокатали-
затора в органическом растворителе. Активность ферментов во
многих не смешивающихся с водой органических растворителях.
Химия и технология
185
нередко сильно падает. Так, в качестве катализаторов реакции
дегидрирования стероидов в среде бензола и гептана использо-
вали разные виды Nocardia. Для увеличения их стабильности
применяли иммобилизацию; при этом нужно было учитывать
гидрофобность носителя. Бактерии, заключенные в гидрофобные
гели, обладали большей каталитической активностью, чем на-
ходящиеся в гидрофильном окружении, что определялось ха-
рактером распределения субстрата между гелем и окружающим
его растворителем. Сегодня мы умеем конструировать носители
е нужной степенью гидрофобности: для этого используются фо-
тосшиваемые преполимеры смол и уретановые преполимеры.
Иммобилизация фермента на поверхности или внутри твердого
'носителя повышает его устойчивость к действию органических
растворителей, препятствуя разрушению третичной структуры
белка в таких средах. Многие ферменты, катализирующие ре-
акции превращения липофильных субстратов in vivo, локализу-
ются в мембранах, и иммобилизация как бы имитирует их ста-
билизированное связанное состояние в мембранах. Так, для син-
теза этилового эфира М-ацетил-Ь-триптофана из этанола и
К[-ацетил-Б-триптофана использовали химотрипсин в хлороформе.
•Липаза, иммобилизованная на гидрофобном полимере, ка-
тализирует в н-гексане реакцию переэтерификации. Таким путем
из оливкового масла и насыщенных высших жирных кислот
.получают заменитель кокосового масла.
Комбинируя разные описанные в этой главе способы, мы
сможем существенно расширить границы применения биологи-
ческих катализаторов в производстве и анализе химических ве-
ществ, применяемых в промышленности.
4.4.2. Конфигурация реактора
Оптимизация биокатализатора
Особенности конфигурации биореактора, используемого в био-
технологическом процессе, определяются биохимическими и био-
физическими свойствами избранного биокатализатора. От его
природы зависит также и способ дальнейшей переработки по-
лученного продукта. В этой связи при разработке процесса
особое значение приобретает улучшение свойств катализатора
методами генетической инженерии, например изменения тех фи-
зических параметров, которые определяют его способность ра-
ботать в определенной среде, специфичность и производитель-
ность, а также локализацию синтезируемого продукта (вспом-
ним о внеклеточном образовании некоторых веществ клетками
растений). Более подробно роль генетических методов в решении
задач биотехнологии разбирается в гл. 7.
186
Глава 4
Регулировать особенности протекания процесса можно так-
же путем разумного выбора катализатора. Сегодня большие
усилия затрачиваются на выделение, изучение свойств и спосо-
бов использования термофильных микроорганизмов и их фер-
ментов во многих уже применяющихся в промышленности био-
катализируемых реакциях. Получаемые при этом преимущества
определяются увеличением стабильности белков и их более
высокой каталитической активностью. Модернизировать процес-
сы можно и путем применения организмов, обитающих в экс-
тремальных условиях (например, при низких pH или высоких
концентрациях солей). Так, для производства органических кис-
лот имеет смысл попытаться использовать ацидофильные гало-
филы; галофильные виды уже сейчас применяются при выра-
ботке полиолов (образование глицерола при участии Duniella).
Применение иммобилизованных биокатализаторов как в со-
ставе целых организмов, так и в виде изолированных фермен-
тов будет играть все большую роль в разработке и проведении
биотехнологических процессов, поскольку это облегчает работу
с ними, повторное их использование и отделение продукта ре-
акции. Сама иммобилизация тоже может благоприятно сказы-
ваться на стабильности и рабочих характеристиках катализато-
ра, что в конечном счете может создать основу для кардиналь-
ного пересмотра экономической конкурентоспособности опреде-
ленного типа биопроизводства.
Повторное использование кофактора
Для проявления каталитической активности 30% известных фер-
ментов нужен один из пяти кофакторов (NAD, NADP, АТР,
FAD либо СоА). Само применение этих ферментов в биоката-
лизе будет определяться тем, удастся ли повторно использовать
эти дорогие вещества либо вообще обойтись без них. Существу-
ет три подхода к решению этой проблемы: ферментативный,
химический и электрохимический.
Ферментативную регенерацию кофакторов, например, можно
осуществить при участии ряда имеющихся в продаже фермен-
тов, которые в качестве восстановителя используют дешевые
субстраты. В этом направлении была проведена большая работа
по использованию формиатдегидрогеназы — фермента, который
катализирует восстановление NADH при превращении формиа-
та в углекислый газ. Так как продукт реакции является газом,
он легко удаляется из реакционной смеси и не мешает выделе-
нию из нее продукта реакции. Основная трудность здесь — по-
теря самого кофактора из реакционной смеси. Предпринимались
попытки повторного использования регенерированного кофакто-
ра в такой системе путем присоединения его к более крупным
Химия и технология
187
молекулам с сохранением его каталитической активности. Такой
комплекс можно удерживать в реакторе при помощи ультра-
4)ильтрующей или гидрофобной жидкой мембраны. NAD вклю-
чали в полиэтиленгликоль (с мол. массой 10000—40 000) и ис-
пользовали в мембранном реакторе. К сожалению, каталитиче-
ская активность таких иммобилизованных кофакторов обычно
сильно падает и составляет всего несколько процентов от их
активности в свободной форме. Коферменты и соответствующие
регенерирующие ферменты стабилизировали также путем им-
мобилизации в структуре гелей.
Химическое окисление NADH можно осуществить в реакции
присоединения к нему кислорода с использованием акцепторов
электронов вроде PMS/DCPIP или FMN.
Несомненный интерес представляет и электрохимическое
•окисление NAD(P)H и восстановление NAD(P)+. Для окисле-
ния восстановленных коферментов был предложен ряд электро-
дов. Осуществить стереоспецифическое электрохимическое вос-
становление оказалось гораздо сложнее. Недавно были разра-
ботаны соответствующие процессы с применением электродов
с ферментами, которые участвуют в реакции стереоспецифиче-
ского восстановления за счет электрохимического восстановле-
ния редокс-центров ферментов. Это удалось осуществить бла-
годаря разработке методов переноса электронов между элект-
родами и этими редокс-центрами. В результате в некоторых
случаях каталитический процесс протекает в отсутствие кофак-
тора. Такое замещение кофактора модифицированным электро-
дом, видимо, легче осуществить для реакций восстановления,
чем окисления.
4.4.3. Будущий вклад биотехнологии в химическую
промышленность
Источником сырья для различных отраслей химической промыш-
ленности в обозримом будущем будут нефть и ее производные.
Получаемые из них с малыми затратами продукты вряд ли по-
требуется производить при помощи какой-то другой технологии.
Факторами, которые могут оказать сильное влияние на внедре-
ние биотехнологии в эту область, являются истощение источни-
ков сырья, повышение стоимости энергии и постоянная необхо-
димость эффективной переработки отходов. Уменьшение доступ-
ных источников горючего приведет к тому, что все более широко
будут использоваться ресурсы биомассы. Бродильные производ-
ства и технологии на основе ферментов будут и далее дополнять
спектр обычных химических технологий. Что же касается при-
менения биотехнологии в крупномасштабных производствах хи-
мических веществ или полимеров, то перспективы здесь весьма
188
Глава 4
ограничены. С экономической точки зрения наиболее целесооб-
разным представляется использование специфических преиму-
ществ биокаталитических процессов в малообъемных производ-
ствах редких химических веществ с высокой прибавочной стои-
мостью.
ЛИТЕРАТУРА
Современная химическая биотехнология
Atkinson В., Maviiuna F. (1983). Biochemical and Bioengineering Handbook,
Nature Press, UK.
Cain R. B. (1980). Transformation of aromatic hydrocarbons. In: Hydrocarbons
in Biotechnology (eds. Harrison D. E. F., Higgins I. J. and Watkinson R.),
pp. 99—133, Heyden, London.
Dellweg H. (ed.) (1983). Biotechnology, Vol. 3: Biomass, Microorganisms for
Special Applications, Microbial Products 1, Energy from Renewable Sources,
Verlag Chemie, Weinheim.
Evelegh D. E. (1981). The microbiological production of industrial chemicals,
Scient. Am., 245, 154—178.
Godfrey T„ Reichelt J. (1983). Industrial Enzymology: The Application of Enzy-
mes in Industry, The Nature Press, Macmillan, UK.
Kieslich K., Sebek О. K. (1979). Microbial transformation of steroids. In: Annual
Reports on Fermentation Processes, Vol. 3 (ed. Perlman D.), pp. 267—297,
Academic Press, New York.
Peppier H. J., Perlman D. (eds.) (1979). Microbial Technology, Vol. 1 and 2:
Microbial Processes, Academic Press, London.
Rose A. H. (ed.) (1978—1980). Economic Microbiology Series, Vol. 2: Primary
Products of Metabolism, Vol. 3: Secondary Products of Metabolism, Vol. 5:
Microbial Enzymes and Bioconversions, Academic Press, London.
Schindler Schmid R. D. (1982). Fragrance or aroma chemicals: microbial
synthesis and enzymatic transformation — a review, Process Biochem., 17(5),
2—8.
Scott C. D. (ed.) (1983). Fifth Symposium on Biotechnology for Fuels and Che-
micals, Biotechnology and Bioengineering Symposium Number 13, John Wiley
and Sons, New York.
Sebek 0. K, Kieslich K. (1977). Microbial transformation of organic compounds:
alkanes, alicyclics, terpenes and alkaloids. In: Annual Reports on Fermentation
Processes, Vol. I (ed. Perlman D.), pp. 267—297, Academic Press, New York.
Smith J. E., Berry D. R,, Kristiansen B. (eds.) (1983). The Filamentous Fumgi:
Volume IV, Fungal Technology, Edward Arnold, London.
Получение химических веществ из биомассы
Curtin М. Е. (1983). Harvesting profitable products from plant tissue culture.
Biotechnology, 1(8), 649—657.
Eveleigh D. E. (1983). Biological routes for cellulose utilisation. In: Biotech’83,
Proceedings of the International Conference on the Commercial Applications
and Implications of Biotechnology, pp. 539—548, Online Conferences Ltd.,
Midlesex, UK.
Flinn J. E„ Lipinsky E. S. (1983). Production of chemicals via advanced biotech-
nological processes. In: Biotech’83, Proceedings of the International Confe-
rence on the Commercial Applications and Implications of Biotechnology,
pp. 523—538, Online Conferences Ltd., Middlesex, UK.
Fowler M. IF. (1981). Plant cell biotechnology to produce desirable substances
Chem. Ind., 7, 229—233.
Химия и технология
189
Jones L. Н. (1983). Plant cell cloning and culture products. In: Biotechnology
(eds. Phelps C. F. and Clarke P. H.), pp. 221—232, Biochemical Society Sym-
posium Number 48, Biochemical Society, London.
Staba E. 1. (ed.) (1980). Plant Tissue Culture as a Source of Biochemicals, CRC
Press, Boca Raton, Florida.
Перспективы развития
Brown G. B., Manecke G., Wingard L. B. (eds.) (1978). Enzyme Engineering,.
Vol. 4, Plenum Press, New York.
Klibanov A. M. (1983). Unconventional catalytic properties of conventional"
enzymes: applications in organic chemistry, Basic Life Sci., 25, 497—518.
Maugh T. H. (1984). Semisynthetic enzymes are new catalysts, Science, 223,.
154—156.
Maugh T. H. (1984). Need a catalist? Design an enzyme, Science, 223, 269—
271.
Neidleman S. L., Geigert J. (1983). Biological galogenation and epoxidation. In:
Biotechnology (eds. Phelps C. F. and Clarke P. H.), pp. 3&—52, Biochemical'
Society Symposium Number 48, Biochemical Society, London.
Omata T., lida T., Tanaka A., Fukui S. (1979). Transformation of steroids by gel-
entrapped Nocardia rhodochrous cells in organic solvents, Eur. J. appl. Micro-
biol. Biotechnol., 8, 143—155.
Pye E. K., Weetail H. H. (eds.) (1978). Enzyme Engineering, Vol. 3, Plenum
Press, New York.
Ulmer К. M. (1983). Protein engineering, Science, 219, 666—671.
Wingard L. B., Berezin I. V., Klyosov A. A. (eds.) (1980). Enzyme Engineering,
Future Directions, Plenum Press, New York.
Wiseman A. (ed.) (1977—1983). Topics in Enzyme and Fermentation Biotechno-
logy, Vols. 1—7, Fillis Horwood, Chichester.
Глава 5
Материалы и биотехнология
К. Брайерли, Д. Келли, К. Сил, Д. Бест
-5.1. Введение
.Вероятно, из всех аспектов микробиологической технологии
меньше всего рекламируется и больше всего недооценивается
применение микроорганизмов для экстракции металлов из ми-
нералов, для концентрирования и извлечения драгоценных ме-
таллов из растворов, а также для получения новых промыш-
ленных биоматериалов.
Еще за 1000 лет до нашей эры римляне, финикийцы и люди
других ранних цивилизаций извлекали медь из рудничных вод
или вод, просочившихся сквозь рудные тела. В XVII в. валлий-
цы в Англии (графство Уэльс) и в XVIII в. испанцы на место-
рождении Рио-Тинто применяли такой процесс «выщелачива-
ния» для получения меди из содержащих ее минералов. Эти
древние горняки и не подозревали, что в подобных процессах
экстракции металлов активную роль играли бактерии. В на-
стоящее время этот процесс, известный как бактериальное вы-
щелачивание, применяется в широких масштабах во всем мире
.для извлечения меди из бедных руд, содержащих этот и другие
ценные металлы в незначительных количествах. Биологическое
выщелачивание применяется также (правда, менее широко) для
высвобождения урана. Проведены многочисленные исследования
природы организмов, участвующих в процессах выщелачивания
металлов, их биохимических свойств и возможностей примене-
ния в данной области. Результаты этих исследований показы-
вают, в частности, что бактериальное выщелачивание может
гшироко использоваться в горнодобывающей промышленности и,
по всей видимости, сможет полностью удовлетворить потребно-
сти в энергосберегающих, не оказывающих вредного влияния на
•окружающую среду технологиях.
Несколько менее известно, но столь же важно использование
микроорганизмов в горнодобывающей промышленности для из-
влечения металлов из растворов. Некоторые прогрессивные тех-
нологии уже включают биологические процессы для получения
•металлов в растворенном состоянии или в виде твердых частиц
«из моечных вод, остающихся от переработки руд. О способности
Материалы и биотехнология
19В
микроорганизмов накапливать металлы известно уже давно,
и энтузиасты издавна мечтали об использовании микробов для
получения ценных металлов из морской воды. Проведенные
исследования рассеяли некоторые надежды и в значительной
степени определили области применения микроорганизмов. Из-
влечение металлов при их участии остается многообещающим
способом дешевой обработки загрязненных металлами промыш-
ленных стоков, а также экономичного получения ценных ме-
таллов.
Давно известно и о способности микроорганизмов синтези-
ровать полимерные соединения; в самом деле, большинства
компонентов клетки — это полимеры. Однако на сегодняшний
день менее 1% всего количества полимерных материалов про-
изводит микробиологическая промышленность; остальные 99%
получают из нефти. Пока биотехнология не оказала решающего
влияния на технологию полимеров. Возможно, в будущем с по-
мощью микроорганизмов удастся создавать новые материалы
специального назначения.
Биоповреждение обычно не рассматривают как один из ас-
пектов биотехнологии, но тем не менее оно имеет огромное
экономическое значение. Микробы способны разрушать мате-
риалы, находящиеся в земле и под водой; они размножаются
в промышленных водах, нефтепродуктах и эмульсиях, вызывая
коррозию промышленных сооружений или образование слизи,,
а также повреждают топливные баки. Последнее создает осо-
бенно серьезные проблемы в авиации и заставляет проводить-
дорогостоящие контрольные измерения.
Цель этой главы состоит в том, чтобы рассмотреть роль
микроорганизмов в коррозии, в извлечении металлов и получе-
нии биоматериалов. Описаны отдельные микроорганизмы и их
использование в современной технологии, а также их значение-
для дальнейшего развития технологии на нашей планете.
5.2. Микробное выщелачивание
Методы извлечения меди из пород, содержащих минералы, пу-
тем обработки их кислыми растворами используются уже много-
веков. Однако лишь в 50-е и 60-е гг. нашего столетия выясни-
лось, что в получении металлов из минералов решающую роль
играют бактерии. В 1947 г. Колмер и Хинкл выделили из шахт-
ных дренажных вод бактерию Тiobacillus ferrooxydans. Этот
организм окислял двухвалентное железо и восстанавливал се-
русодержащие соединения, а также, возможно, и некоторые;
металлы. Вскоре оказалось, что он участвует и в переводе медиа
из рудных минералов в раствор.
1.92
Глава 5
Сейчас известны и другие микроорганизмы, активно участ-
вующие в извлечении металлов из минералов; большинство ми-
нералов сульфидной природы разрушается именно этим путем.
.Хотя технология бактериального выщелачивания используется
в основном для извлечения меди и урана, она находит доста-
точно широкое применение и в переработке минерального сырья.
В данном разделе рассматриваются организмы, участвующие
в выщелачивании металлов, и механизмы их действия. Описаны
сферы применения микробного выщелачивания в настоящем и
возможное будущее биоэкстрактивной металлургии.
5.2.1. Выщелачивающие микроорганизмы
В бактериальном выщелачивании участвуют следующие микро-
организмы.
Thiobacillus ferrooxidans
Этот наиболее изученный из всех выщелачивающих организ-
мов почти всегда можно выделить из среды, в которой проис-
ходит окисление железа или минералов. Т. ferrooxidans, вероят-
но, представлен в различных природных средах штаммами с
температурными оптимумами от 10 до 30 °C. Максимальная
.переносимая температура равна 37°C (или ниже).
Leptospirillum ferrooxidans
Этот организм, который, видимо, лучше называть Ferrovibrio,
впервые был выделен в Армении, однако теперь известно, что
юн встречается во многих местах, где осуществляется выщела-
чивание. Он может расти при 40 °C и pH 1,2 на пирите (FeS2)
и, по-видимому, окисляет только железо, не затрагивая серу.
Этим он отличается от Т. ferrooxidans, который окисляет серу
так же хорошо, как железо.
Thiobacillus thiooxidans, Т. acidophilus и Т. organoparus
Эти ацидофильные организмы окисляют только серу и ее
соединения, а также действуют на серу пирита совместно с Lep-
tospirillium ferrooxidans. Они могут участвовать в окислении
серы, образующейся в результате химической реакции между
ионами трехвалентного железа и сульфидами меди. Т. thiooxi-
,dans изменяет pH среды, в которой он растет, до значительно
меньших величин ( — 0,65), чем те, которые переносит хорошо
охарактеризованный штамм Т. ferrooxidans, и способен таким
Материалы, и биотехнология
193
образом повышать эффективность прямого кислотозависимого
выщелачивания сульфидов (например, PbS, CdS, NiS) н присут-
ствии элементарной серы.
Умеренные термофилы
Обнаружены различные термофильные, окисляющие пирит, же-
лезо и серу бактерии, которые лучше всего растут при темпе-
ратурах около 50 °C. Эта группа умеренных термофилов вклю-
чает факультативных гетеротрофов, хемолитотрофных гетеро-
трофов и автотрофов, причем обнаруживаются все новые и но-
Рис. 5.1. Микрофотография Sulfolobus brlerleyi, растущего на элементарной
сере при 60 °C. Получена с помощью сканирующего электронного микроскопа
(увеличение 10 000).
вые организмы этого класса. Данные организмы могут играть
существенную роль в выщелачивании саморазогревающихся ми-
нералов и угольных отвалов.
Крайние ацидотермофилы
Лучше всего изучен род Sulfolobus (рис. 5.1). Все его виды
окисляют серу, а некоторые (например, S. brierleyi) способны
к окислению железа и таких минералов, как халькопирит. Эти
13-1344
194
Глава 5
организмы переносят температуру до 85 °C и выделены в основ-
ном из горячих источников. Впрочем, недавно из дренажных
вод угольных отвалов был получен организм, сходный с Sulfo-
lobus. Род Sulfolobus относится к Archebacteriaceae — отдельной
группе бактерий, которую предлагают считать третьим царством
живых организмов. Представители этого рода, вероятно, играют
важную роль в выщелачивании минералов при повышенных
температурах.
Все упомянутые выщелачивающие бактерии переводят ме-
таллы в раствор различными путями. Соответствующие методы
были названы «прямыми» и «непрямыми». Окислительным про-
цессом, «прямо» катализируемым бактериями, является окис-
ление железа,
4FeSO4 + О2 + 2Н2 SO4-> 2Fe2 (SO4)3 + 2Н3 О, (1 >
и окисление серы,
S8 + 12О2 + 8Н2 О-> 8Н2 SO4. (2>
Ряд минералов непосредственно окисляется некоторыми выще-
лачивающими организмами. Примерами такого рода могут быть
окисление пирита,
4FeS2 + 15О2 + 2НаО-> 2Fe2 (SO4)3 + 2Н2 SO4, (3>
и сфалерита,
ZnS + 2О2---> ZnSO4 (4>
Ион трехвалентного железа служит сильным окисляющим аген-
том, переводящим в раствор многие минералы, например халь-
коцит,
Cu2 S + 2Fe2 (SO4)3 > 2CuSO4 4- 4FeSO4 + S°, (5>
и уранинит,
UO2 + Fe2 (SO4)3--> UO2 SO4 + 2FeSO4. (6>
Выщелачивание, происходящее при участии иона трехвалент-
ного железа, который образуется в результате жизнедеятельно-
сти бактерий, называют «непрямой» экстракцией. Нередко в хо-
де этой химической реакции образуется элементная сера [реак-
ция (5)], которая может непосредственно окисляться бактерия-
ми до серной кислоты [реакция (2)].
5.2.2. Промышленное применение биоэкстрактивной
металлургии
В настоящее время бактериальное выщелачивание, известное-
также как биогидрометаллургия или биоэкстрактивная метал-
лургия, применяется в промышленных масштабах для перевода
в растворимую форму меди и урана.
Материалы и биотехнология 195
Выщелачивание медных отвалов
Методы, использовавшиеся в XVIII в. на месторождении Рио-
Тинто (Испания) для извлечения меди из груд выветрившейся
породы, в основном сохранились до наших дней. В нынешнем
столетии выщелачивание отвалов, как называют этот процесс,
развивалось в США; оно используется для получения меди из
бедных руд [содержащих менее 0,4% меди (по весу)], а также
из отвального материала с очень низким содержанием меди.
Такие отвальные материалы накапливаются при крупномас-
штабной открытой разработке руды. Бедную руду или отваль-
ную породу перевозят из карьера куда-либо поблизости (обыч-
но в долину), где естественный уклон дает возможность соби-
рать используемые растворы. Во избежание загрязнения под-
почвенных и поверхностных вод выбирают непроницаемые для
воды участки. Отвалы, образующиеся в результате работы зем-
леройной техники, имеют огромные размеры, достигая в высоту
300 и более метров. Самым большим в мире отвалом является
Бингхэм-Каньон («Кеннекотт Коппер Корпорэйшн»), Он вмеща-
ет около 3,6-1012 кг породы.
Для начала процесса выщелачивания отвал смачивают во-
дой, подкисленной серной кислотой до pH 1,5—3,0, путем ее
распыления, полива или инъекции через трубы, помещенные
вертикально внутри породы. Этот кислый раствор, или «выще-
лачиватель», просачивается сквозь бедную руду или отвальные
материалы. Он содержит кислород и углекислый газ и создает
благоприятную среду для размножения ацидофильных гиоба-
цилл, широко распространенных в сульфидных рудах. В неко-
торых случаях содержание Thiobacillus ferrooxidans превышает
106 клеток на 1 кг породы и на 1 мл выщелачивающего раство-
ра. Этот организм активно окисляет растворимые ионы двух-
валентного железа и воздействует на серу- и железосодержащие
минералы. Активность Т. ferrooxidans необходима для оптималь-
ного выщелачивания. При pH ниже 3,5 окисление железа пере-
стает зависеть от pH:
d (Fe+2)
- \.... -=К' (Fe+2) (О2),
dt
где К/=1,0-10-7 атм-1-мин-1 при 25 °C. Следовательно, при кис-
лых значениях pH, необходимых для выщелачивания отвалов,
и в отсутствие Т. ferrooxidans железо оставалось бы в двухва-
лентном состоянии и экстракция меди из сульфидных минералов
была бы минимальной. Т. ferrooxidans ускоряет окисление двух-
валентного железа в 106 раз. При окислении медно-сульфидных
минералов нередко образуется элементарная сера. Эта сера
маскирует частицы минералов, ограничивая воздействие на
аз*
Т96
Глава 5
них со стороны трехвалентного железа. Т. ferrooxidans, присут-
ствующая в количестве 103—105 клеток на 1 г породы и на 1 мл
выщелачивающего раствора, окисляет некоторые растворимые-
соединения серы и элементарную серу. Разрушение серы этим
организмом приводит к удалению маскирующего слоя серы,,
окружающего некоторые частицы минералов, и усиливает про-
цесс выщелачивания. Таким путем Thiobacillus thiooxidans и
Thiobacillus ferrooxidans совместно разлагают минералы суль-
фидной природы и являются мощным окислителем для раство-
рения медно-сульфидных минералов и образования серной кис-
лоты. Эта последняя создает благоприятную среду для деятель-
ности микроорганизмов и удерживает ионы двухвалентной меди
в растворе.
Поскольку при выщелачивании отвалов в среде развиваются
природные тиобациллы, никакого засева не проводят. Проявле-
нию необходимой активности микроорганизмов способствуют
обеспечение кислотности отвала и обилие кислорода. Последнее
достигается путем аэрирования выщелачивающего раствора;
циркуляции воздуха внутри породы способствует и особая фор-
ма отвалов (с гребнями или ребрами). Иногда вертикально’
внутри отвала помещают трубы с отверстиями и через них про-
дувают сжатый воздух, способствующий протеканию биологи-
ческих и химических реакций.
В выщелачиваемых отвалах происходит также много важ-
ных небиологических реакций. Самой ценной с экономической
точки зрения является окисление медно-сульфидных минералов
образующимися биологическим путем ионами трехвалентного
железа [например, реакция (5)]. К другим небиологическим
реакциям, протекающим в выщелачиваемых отвалах, относятся
гидролиз солей трехвалентного железа с последующим осажде-
нием основного сульфата трехвалентного железа,
Fe2 (SO4)s + 2Н2О-> 2Fe (ОН) (SO4) + Н2 SO4, (7)
растворение карбонатных минералов,
СаСО3 + Н2 SO4 + Н2О > CaSO4 + 2Н2О + СО2, (8)
и твердофазные превращения, приводящие к образованию вто-
ричных минералов. Все эти реакции стабилизируют pH отваль-
ных пород и выщелачивающего раствора на нужном уровне.
Выщелачиваемые отвалы имеют значительные размеры. Это
создает много инженерных проблем и может препятствовать
деятельности бактерий и протеканию важных химических ре-
акций. К таким проблемам относятся, во-первых, уплотнение
отвалов и образование осадков, которые затрудняют взаимо-
действие раствора с минералами, во-вторых, попадание внутрь
отвалов крупных минерализованных глыб, мало подверженных
Материалы и биотехнология
197
разрушению, и в-третьих, повышение температуры отвалов за
счет протекания в них экзотермических реакций. Так, в Бинг-
хэм-Каньоне было отмечено повышение температуры выше 80 °C.
При таких температурах тиобациллы инактивируются, но может
повышаться активность термофильных выщелачивающих бакте-
рий. Из отвалов были выделены ацидотермофильные штам-
мы TH, близкие к Thiobacillus, однако крайне термофильные
штаммы Sulfolobus не обнаружены. Правда, это не исключает
их существования в подобных средах.
Из выщелачиваемых отвалов вытекают растворы, содержа-
щие 0,75—2,2 г меди в 1 л. Эти растворы направляют в отстой-
ники; медь из них получают путем осаждения с использованием
железа или экстракцией растворителями. В первом случае соз-
дают условия, при которых растворы контактируют с железом
и протекает следующая реакция:
CuSO4 + Fe° Cu° + FeSO4. (9)
«Отработанные» выщелачивающие растворы вновь поступают в
отвал. В последние годы для получения меди из раствора нача-
ли применять экстракцию растворителями. Ионы меди из вод-
ной фазы экстрагируют органическими жидкостями, только ча-
стично растворимыми в воде. Затем медь извлекают из орга-
нического растворителя.
Выщелачивание урана
Для экстракции урана бактерии применяются реже. Для того
чтобы при выщелачивании урана можно было использовать
микробиологическую технологию, руда и/или связанные с ней
породы должны быть богаты сульфидными минералами и не
слишком интенсивно поглощать кислоту. Бактериальное выще-
лачивание урана применяли в восточных районах Канады для
извлечения остаточного урана на уже выработанных площадях,
а также из отвалов. В первом случае стенки и крыши забоев
(при подземной выработке) промывали обычной или подкис-
ленной водой. Для роста бактерий достаточно 3—4 месяцев, за
это время Т. ferrooxidans окисляет железо до трехвалентного
состояния. Затем трехвалентное железо окисляет восстановлен-
ный уран до растворимого окисленного состояния в соответствии
с реакцией (6). По прошествии этого периода забои снова про-
мывают. Промывные воды, содержащие уран, собирают и из-
влекают из них уран с помощью ионного обмена либо экстра-
гируют растворителями.
Бактериальное выщелачивание применялось в Канаде и в ка-
честве первичного средства для получения урана. Рудное тело
разрушали взрывом и осуществляли выщелачивание in situ.
198
Глава 5
Как и при выщелачивании меди в отвалах, при выщелачивании
in situ часто приходится осуществлять инженерные мероприя-
тия, что отрицательно сказывается на активности бактерий.
5.2.3. Возможности применения бактериального
выщелачивания
Инженерные проблемы
Из-за огромных масштабов операций по выщелачиванию отва-
лов активность бактерий, развивающуюся в ходе процесса, мож-
но контролировать только в ограниченной степени. Для наибо-
лее эффективного использования бактериального выщелачива-
ния необходимо создавать такие инженерные схемы, которые
позволяли бы осуществлять определенный контроль за актив-
ностью микробов. Помимо выщелачивания отвалов в горноруд-
ной промышленности существуют и другие средне- и высокотех-
нологичные процедуры, при которых для экстракции металлов
используются гидрометаллургические процессы (реакции, про-
исходящие в воде). Эти технологии (выщелачивание in situ,
чановое выщелачивание, кучное выщелачивание) применимы и
к процессам бактериальной экстракции металлов.
Выщелачивание in situ с успехом используют для извлечения
урана из песчаниковых формаций с низким содержанием руд-
ного минерала. Выщелачивающие растворы вводят в неразру-
шенное урансодержащее рудное тело через инъекционную сква-
жину. Эти растворы, содержащие химический окислитель (на-
пример, перекись водорода), взаимодействуют с минералом,
окисляя уран и переводя его в растворимую форму. Далее
урансодержащие растворы выкачивают из минерализованной
зоны через выходные скважины (рис. 5.2). На западе США и
в южном Техасе, где широко практикуется выщелачивание
in situ, применяют карбонатные растворы с нейтральным pH.
По всей видимости, бактерии в этом процессе экстракции не
участвуют. Технология in situ находит применение не только
при выщелачивании урана. Тот же подход возможен и при
экстракции других металлов, присутствующих в низких кон-
центрациях в глубоко залегающих месторождениях. Бактерии
могут использоваться для выщелачивания сульфидных минера-
лов или для разрушения жильных минералов при извлечении
металлов с помощью других гидрометаллургических техноло-
гий. Большинство исследований по использованию бактерий для
выщелачивания in situ было проведено в лаборатории при вы-
соких давлении и температуре. Эти исследования показали, что
гидростатическое давление в 30,4 МПа (эквивалентное давлению
на глубине в 3000 м) не сказывается на жизнедеятельности
Материалы и биотехнология
199
Рис. 5.2. Схематическое изображение процесса выщелачивания in situ. А. Кон-
трольные скважины для отбора проб с целью наблюдения за передвижением
выщелачивающего раствора из рудного тела. Б. При обычно используемой для
выщелачивания in situ схеме «пяти точек» объем выщелачивающего раствора,
выкачиваемого из четырех рабочих скважин, больше, чем объем вводимого
раствора. Это сводит к минимуму возможность загрязнения подпочвенных
вод.
железоокисляющих бактерий. Однако при закачивании раство-
ров на большую глубину ограничивающим фактором для ис-
пользования бактерий может оказаться гипербарический кис-
лород. Влияние высокой температуры, существующей на боль-
шей глубине, недостаточно изучено. Не исключено также, что
из-за более низкой проницаемости для бактерий монолитных,
сильно уплотненных включений в месторождениях эффектив-
ность выщелачивания резко уменьшается. Эта возможность так-
же не изучалась детально. Несмотря на все эти пробелы в на-
ших знаниях, можно утверждать, что достоинством технологии
in situ является то, что она представляет собой систему, в ко-
торой можно контролировать много факторов, в том числе со-
держание кислорода и питательных веществ, pH т. д.
200
Глава 5
Чановое выщелачивание используется в горнорудной про-
мышленности для извлечения урана, золота, серебра и меди из
окисных руд. Медные и урановые руды сильно измельчают и
смешивают с растворами серной кислоты в больших емкостях
(обычно размером 30X50X6 м) для перевода металла в раство-
римую форму. Время выщелачивания, как правило, составляет
несколько часов. Медь получают из кислого раствора электро-
лизом, уран — ионообменным путем или экстракцией раствори-
телем. Ферментация в чанах, а также в отстойниках с постоян-
ным или предварительным перемешиванием может с успехом
применяться для бактериального выщелачивания потому, что
при этом легко контролировать факторы, влияющие на актив-
ность микроорганизмов. К этим факторам относятся: размер
частиц руды, ее качество, плотность пульпы (масса руды на
единицу объема раствора), pH, содержание углекислого газа,
кислорода, время удержания (время нахождения частиц в ре-
акторе), температура и содержание питательных веществ. Хотя
руда и не стерилизуется, возможен строгий контроль за видо-
вым составом и количеством микроорганизмов. Чановое выще-
лачивание создает предпосылки для использования специфиче-
ских штаммов микроорганизмов (например, ацидотермофиль-
ных бактерий) или микробов-выщелачивателей, полученных ме-
тодами генетической инженерии. Вначале чановое выщелачива-
ние применяли для руд с очень высоким содержанием металлов,
однако эта технология может использоваться и в случае мате-
риалов более низкого качества. При этом следует учитывать
экономические и технологические факторы.
Кучное выщелачивание применяют для химической экстрак-
ции урана, меди, золота и серебра. При выщелачивании урана
и меди руду измельчают и помещают на специальные водоне-
проницаемые поверхности. При извлечении меди и урана кучи
могут содержать 10—50-108 кг руды и в высоту достигать
4,5—5,5 м. Вершины куч выравнивают и наносят на них раствор
серной кислоты. Новые кучи часто помещают поверх уже суще-
ствующих. Такой способ выщелачивания урана и меди сходен
с выщелачиванием отвалов; однако здесь используются более
концентрированные растворы серной кислоты, частицы породы
меньше по размеру, а качество породы (содержание металла
в ней) выше. Кучное выщелачивание длится несколько месяцев,
а для выщелачивания отвалов требуются годы. Этот метод при-
меним также для экстракции золота и серебра из руд и даже
из отходов, подобных шламу (пустая порода, остающаяся после
извлечения руды и размельчения). Чтобы обеспечить эффектив-
ное протекание выщелачивающего раствора, тонко измельчен-
ный шлам должен быть подвергнут агломерации (спекание в
шарики). В щелочных растворах цианидов серебро и золото
Материалы и биотехнология
201
образуют комплексы, которые затем отделяют от раствора с по-
мощью активированного угля. Кучное выщелачивание требует
относительно небольших капиталовложений и технического обес-
печения и представляет собой легко реализуемую систему. Бла-
годаря небольшим масштабам операций при таком выщелачи-
вании величину частиц породы, размеры куч, контакт между
раствором и частицами, аэрацию, pH, состав растворов можно
строго контролировать; с таким выщелачиванием хорошо со-
прягаются процессы биоэкстракции. Их использование расши-
ряет возможности методов кучного выщелачивания, позволяя
осуществлять извлечение других металлов или проводить пред-
варительную биологическую обработку материала до начала
обычных процессов экстракции.
Экономическая значимость
Хотя процессы биологического выщелачивания и представляют
собой альтернативу обычным процессам экстракции, маловеро-
ятно, что микробиологическая технология в ближайшем буду-
щем заменит такой издавна существующий процесс, как вы-
плавка металлов. Тем не менее, подобно другим гидрометал-
лургическим процессам типа кислотного кучного выщелачивания
урановых и медных окисных руд и выщелачивания золотонос-
ных и серебряных руд с помощью цианидов, эффективные ме-
тоды бактериального выщелачивания, несомненно, могут ока-
зать заметное влияние на технологию переработки минераль-
ного сырья.
Предполагается, что микробиологическая технология позво-
лит перерабатывать руды и отходы, использование которых
обычными методами неэкономично. Примерами такого рода
являются переработка огромных количеств шламов и отходов
с небольшим, но все же заметным содержанием драгоценных
или стратегических металлов, а также экстракция металлов,
заключенных в минеральных матриксах. Бактерии легко разла-
гают пирит, арсенопирит и другие минералы с освобождением
металлов.
Еще одна область применения биологического выщелачива-
ния, которая обещает быть экономически выгодной, — избира-
тельное выщелачивание некоторых металлов; при этом один из
металлов какого-либо минерала переходит в раствор, а осталь-
ные остаются нерастворенными. Некоторые минералы для уве-
личения их способности к флотации можно подвергнуть пред-
варительному биологическому выщелачиванию. Флотация пред-
ставляет собой метод разделения минералов, состоящий в том,
что образуемая реагентами пена поднимает на поверхность во-
202
Глава 5
ды некоторые тонко измельченные минералы, тогда как другие
тонут.
Последствия для окружающей среды, к которым ведет ис-
пользование многих обычных методов добычи руд, их обогаще-
ние, а также выплавка металлов, весьма существенны. Обще-
принятые способы добычи и обогащения руд часто ведут к на-
рушению поверхностных слоев Земли из-за создания гигантских
открытых карьеров и гор пустой породы и шламов. Это может
создавать экологические проблемы. При выплавке металлов из
сульфидных руд и сжигании углей с высоким содержанием серы
образуются зола и двуокись серы, представляющие потенциаль-
ную опасность для окружающей среды. Биогидрометаллургия
в сочетании с технологией выщелачивания in situ способна
уменьшить разрушение земной поверхности, вызываемое разра-
боткой и обогащением руд общепринятыми методами, а также
устранить необходимость в выплавке металлов из сульфидных
минералов. С помощью этого подхода можно будет извлекать
сульфидные минералы при небольшом их содержании и большой
глубине залегания. Разработка подобных месторождений обще-
принятыми подземными методами или открытым способом не-
рентабельна.
Одна из самых многообещающих возможностей бактериаль-
ного выщелачивания — использование его для удаления серы
из угля перед сжиганием последнего. Выщелачивающие бакте-
рии легко катализируют растворение неорганической (пирит-
ной) серы, содержащейся в каменном угле; однако на органи-
ческую серу эти бактерии не действуют. Были исследованы и
другие бактерии, способные эффективно удалять серусодержа-
щие органические вещества из каменного угля.
Прежде чем проводить точный экономический анализ техно-
логии бактериального выщелачивания, необходимо тщательно
изучить все ее достоинства и недостатки, сравнив их с таковы-
ми для существующих пирометаллургических и гидрометаллур-
гических процессов. По сравнению с обычными методами добы-
чи и обогащения руд и выплавки металлов бактериальное вы-
щелачивание может оказаться вполне конкурентоспособным
благодаря меньшим энергозатратам, снижению расхода реаген-
тов при экстракции металлов, а также меньшему влиянию на
окружающую среду.
Недостатки метода
В предыдущих разделах в общих чертах говорилось о практи-
ческом использовании бактериального выщелачивания в настоя-
щее время и в перспективе. Однако немедленное практическое
применение бактериального выщелачивания сдерживается по
Материалы и биотехнология 203
ряду причин. Главное препятствие заключается в том, что про-
цесс еще плохо исследован как на опытных установках, так и
в полевых условиях. Большинство экспериментов было прове-
дено в полупроизводственных условиях. Поэтому трудно судить
об экономической значимости процессов бактериального выще-
лачивания и оценить технологические трудности, которые могут
возникнуть при широкомасштабном промышленном использова-
нии микробиологических процессов. Обычно применяемые про-
цессы бактериального выщелачивания страдают также от не-
достатка хорошей техники. Вполне вероятно, что создание спе-
циальной системы оптимизации биологической активности силь-
но расширило бы .использование бактерий при выщелачивании.
К параметрам, которые должны при этом учитываться, отно-
сятся температура, питательные вещества, содержание кисло-
рода и углекислого газа, размер частиц, качество минерала,
плотность пульпы (масса частиц на единицу объема выщела-
чивающего раствора), скорость протекания выщелачивающего
раствора и pH.
Процессы бактериального выщелачивания нередко протека-
ют медленнее, чем химические процессы, в которых мелкие
частицы обрабатывают сильными реагентами при повышенных
температуре и давлении. При экстракции металлов из ряда
минеральных концентратов быстрые химические процессы эко-
номически более выгодны. Однако в тех случаях, когда при
обработке минералов стоимость химических реагентов и энер-
гозатраты очень высоки, а скорость переработки не имеет ре-
шающего значения, правильно выбранные, технологичные спо-
собы бактериального выщелачивания могут оказаться предпо-
чтительными.
При бактериальном выщелачивании сульфидных минералов
в системе должно образовываться достаточное количество кис-
лоты и не содержаться слишком много минералов, поглощаю-
щих кислоты. Для бактерий, окисляющих железо и серу, тре-
буется кислая среда. Поэтому для переработки непригодны
руды и отходы, поглощающие кислоты в большом количестве.
Для выщелачивания металлов in situ бактерии применялись
мало. При подземном выщелачивании с помощью растворов
следует принимать во внимание такие факторы, как влияние на
активность бактерий повышенного гидростатического давления
и гипербарической оксигенации. Гидростатическое давление воз-
никает в результате введения выщелачивающих растворов под
давлением, а также за счет веса столба жидкости, а гиперба-
рическая оксигенация обусловлена введением кислорода под
давлением в рудное тело in situ для регенерации окисляющего
агента. Однако при использовании бактерий кислород в такой
концентрации не нужен, поскольку эти организмы сами регене-
204
Глава 5
рируют окислитель. Кислород требуется лишь в концентрациях,
необходимых для обеспечения жизнедеятельности микроорганиз-
мов. Первоначально технология выщелачивания in situ приме-
нялась для пород типа песчаников; при этом размельчать поро-
ду не было необходимости, поскольку она обладала высокой
проницаемостью для выщелачивающего раствора. Для выще-
лачивания такого типа микроорганизмы не использовались. При
бактериальном же выщелачивании встает вопрос: не будет ли
рост бактерий на ненарушенном массиве пород ограничивать
проницаемость и не уменьшится ли при этом циркуляция рас-
твора? Такая проблема вряд ли возникнет при выщелачивании
рудных тел in situ, предварительно разрушенных взрывом или
иным способом.
Если какой-то микробиологический процесс отработан для
выщелачивания металла (металлов) из определенного рудного
тела, то вряд ли его удастся применить без изменений для по-
лучения оптимальных результатов при выщелачивании подоб-
ного металла из другого рудного тела. Даже если металлы
сходны, тип рудного минерала и содержащей его породы впол-
не могут различаться. Выщелачивающие бактерии действуют
на разные минералы неодинаковым образом. Известно, напри-
мер, что некоторые халькопириты огнеупорнее других, и такие
огнеупорные руды устойчивее к прямому действию микроорга-
низмов или продуктов их жизнедеятельности.
В связи с огромными объемами перерабатываемого материа-
ла выщелачивание проводят под открытым небом, а не в поме-
щениях со строго контролируемыми условиями. Поэтому микро-
организмам приходится «работать» при разных погодных усло-
виях, существенно различающемся насыщении минеральными
солями и неодинаковых pH. Ни система в целом, ни рудное
тело не бывают стерильными: в них всегда присутствуют при-
родные бактерии. Специально подобранные или мутантные
штаммы выщелачивающих бактерий должны хорошо сочетаться
с природной микрофлорой. Несомненно, что с помощью генети-
ческих манипуляций могут быть получены штаммы с повышен-
ной способностью окислять железо или минералы, а также пе-
реносить высокие концентрации металлов или кислот. Работы
в этом направлении ограничиваются неполнотой наших знаний
обо всех интересующих нас микроорганизмах и о деталях ме-
ханизма разложения сульфидных минералов; кроме того, мы
почти ничего не знаем о генетических особенностях выщелачи-
вающих микроорганизмов (например, об их хромосомных кар-
тах, наличии и функциях плазмид, способности к трансформа-
ции или переносу плазмид). Здесь имеется широкое поле для
исследований, очень важных с точки зрения биотехнологии.
Материалы и биотехнология
225
5.3. Превращение, накопление
и иммобилизация металлов микроорганизмами
Побуждаемая строгими законами об охране окружающей сре-
ды, необходимостью извлечения ценных металлов и очистки
промышленных вод для их повторного использования, горно-
рудная промышленность все шире применяет новые физико-хи-
мические технологии для очистки сточных вод. Слишком часто
эти технологии оказываются крайне дорогостоящими и неэф-
фективными. Все больше фирм приходят к убеждению, что для
•очистки сточных вод можно использовать биологические про-
цессы, причем эти процессы могут быть более экономичными и
эффективными, чем обычно применяемые методы. Некоторые
промышленные предприятия широко используют эти процессы
для удаления из рудничных сточных вод примесей неорганиче-
ских ионов. Применяемые системы обычно представляют собой
‘большие отстойники или проточные пруды с медленным тече-
нием, в которых растут водоросли и микроорганизмы. Эти ор-
ганизмы накапливают растворенные металлы и их частицы или
•образуют продукты, переводящие примеси в нерастворимую
форму. Обычно используемые процессы биологической очистки
сточных вод мало подвержены прогрессу, эту технологию мож-
но рассматривать как несложную. Исследования последних лет
.показывают, что многие микроорганизмы способны накапливать
металлы в больших концентрациях и содержат структурные
компоненты, которые могут избирательно связывать специфи-
ческие ионы. Селекция микроорганизмов, способных накапли-
вать металлы, и создание технически более совершенных систем
в целях использования этих организмов для удаления всех или
отдельных загрязняющих ионов, присутствующих в малых ко-
личествах в больших объемах сточных вод, могли бы получить
широкое применение в горнодобывающей промышленности и
в других отраслях индустрии, где образуются сточные воды.
Извлекать металлы из окружающей среды способны все
микроорганизмы, поскольку такие металлы, как железо, маг-
ний, цинк, медь, молибден и многие другие входят в состав
ферментов или пигментов, подобных цитохромам или хлоро-
филлам. В ряде случаев металлы накапливаются микроорга-
низмами в значительных количествах; в бактериальной клетке
могут содержаться ионы калия в концентрации 0,2 М, даже
если в среде калий присутствует в концентрациях 0,0001 М и
ниже. В ходе эволюции у микроорганизмов сформировались
системы поглощения, специфичные к определенным металлам
и способные к значительному их концентрированию. В резуль-
тате метаболических реакций, протекающих у микроорганизмов,
могут происходит различные превращения металлов: выделяе-
206
Глава 5
мне в окружающую среду продукты метаболизма способны
образовывать комплексы с металлами или осаждать их из рас-
творов; некоторые металлы могут переводиться с их помощью-
в летучие формы и удаляться из раствора; металлы могут окис-
ляться или восстанавливаться. Так, в разделе, посвященном
выщелачиванию металлов, мы уже рассматривали окисление
железа [Fe(II)—>Fe(III) +е~]. Fe(III) может также восста-
навливаться до Fe(II) при дыхании ряда бактерий. За счет
использования восстановительных эквивалентов дыхания может
происходить восстановление селената, селенита, теллурата и
теллурита до свободных металлоидов; таким путем осуществля-
ется иммобилизация этих металлоидов.
Рис. 5.3. Возможные взаимодействия между металлами и микробной клеткой:
поверхностная адсорбция, внутриклеточное накопление, перевод в летучую
форму, комплексообразование с выделяемыми органическими соединениями в
внеклеточное осаждение.
Ниже перечислены основные механизмы иммобилизации,
комплексообразования или других способов удаления метал-
лов из растворов микроорганизмами: 1) перевод в летучую
форму; 2) внеклеточное осаждение; 3) внеклеточное комплексо-
образование и последующее накопление; 4) связывание клеточ-
ной поверхностью; 5) внутриклеточное накопление.
Все эти реакции изображены на рис. 5.3. Понятно, что ука-
занные процессы могут перекрываться, поскольку один из них
способен давать начало другому или быть его составной частью.
Материалы и биотехнология 207
5.3.1. Перевод в летучую форму
В настоящее время твердо установлено, что многие микроорга-
низмы способны метилировать ртуть. Это приводит к превра-
щению ионов Hg(II) из осадка или раствора в метилртутные
•соединения (например, диметилртуть), которые уходят в атмо-
сферу. Такое превращение может быть важным этапом в при-
родном круговороте ртути. Возможно также микробиологиче-
ское метилирование других металлов, например мышьяка, тел-
лура и селена, которые таким способом удаляются из почвы и
воды. Подобные процессы могут играть важную роль в при-
родных циклах этих металлов и иметь значение, например, при
образовании обедненных селеном почв или при удалении ток-
сичных металлов при обработке сточных вод. Как бы то ни
'было биотехнологические исследования, направленные на умень-
шение или увеличение подобной микробной активности, пред-
ставляются весьма перспективными.
5.3.2. Внеклеточное осаждение
Металлы могут иммобилизовываться и накапливаться в почвах
и в осадочных породах за счет связывания с продуктами мета-
болизма микробов или с накапливающимися органическими
•остатками. Эти процессы издавна использовались человеком
при очистке сточных и промышленных вод. При обычной очист-
ке сточных вод образующийся ил содержит целый набор ме-
таллов, перешедших из воды. Живые клетки и органические
остатки, присутствующие в отстойниках или проточных прудах,
будут накапливать эти металлы, которые впоследствии оказы-
ваются в осадках. Для удаления металлов из промышленных
•стоков или из рудничных вод в горнорудной промышленности
используют пруды, в которых «цветут» водоросли (их усилен-
ный рост стимулируется органическими или минеральными пи-
тательными веществами, которые содержатся в воде).
Один из наиболее удачных примеров внеклеточного осажде-
ния — перевод металлов в осадок путем осаждения сероводо-
родом, образуемым сульфатредуцирующими бактериями. Эти
бактерии обитают в анаэробных средах во всех частях земного
шара (в озерных, океанических и некоторых речных осадках,
бескислородных почвах, болотах и т. п.), и с их помощью про-
исходит сопряжение окисления органических веществ с восста-
новлением сульфатов до сульфидов:
H2SO4+ 8Н = Н2 S + 4Н2О. (10)
Эта реакция может приводить к осаждению металлов в почвах
и осадках, например в виде пирита (FeS2). В масштабе геоло-
.гического времени таким способом могут накапливаться боль-
208
Глава 5
шие массы сульфидов металлов, например CuS. К образованию
сульфидов ведет разложение биомассы водорослей в осадочных
породах. Схематически такой процесс представлен на рис. 5.4.
Оба этих процесса осуществляются в природе и используются
человеком в относительно небольших масштабах, если говорить
об общем количестве загрязненной металлами воды, которая
нуждается в очистке. Потенциальные возможности применения
биотехнологии для бактериального восстановления сульфатов в
Стадия 1: рост и отмирание водорослей и осаждение биомассы
Стадия 2: отложение и накопление сульфидов металлов в осадках
Поступление воды,
содержащей металл~\^
______________________Металл + Сульфид
H2S
Отток очищенной водьи
Сульфиды металла
* Сульфат редуцирующие х
бактерии в осадках используют простые! ♦
органические соединения из разлагаемых1
микроорганизмами водорослей
Рис. 5.4. Существующая в природе система осаждения металлов. Отмершие
водоросли оседают на дно водоема (стадия 1) и при их последующем разло-
жении под действием микроорганизмов образуется сероводород, осаждающий
металлы (стадия 2).
целях очистки вод от металлов, а также для извлечения метал-
лов из разбавленных растворов совершенно очевидны. На
рис. 5.5 показаны две высокотехнологичные системы такого ро-
да. В одноступенчатой системе в хемостат с культурой сульфат-
редуцирующих бактерий непрерывно поступает раствор, загряз-
ненный каким-либо металлом и содержащий сульфат и пита-
тельные вещества типа лактата или в зависимости от природы
используемой бактерии углеводород либо органическую кисло-
ту. Для одновременного разложения сложных органических
остатков можно применять смешанные культуры бактерий.
Материалы и биотехнология
209
функционирующих в анаэробных условиях. При более строго
контролируемом процессе для получения сероводорода следует
использовать раздельные культуры сульфатредуцирующих бак-
терий; при этом сероводород необходимо прокачивать сквозь
загрязненную металлом воду, протекающую через осадочный чаи .
Сульфид металла будет осаждаться, а поток свободной от ме-
талла (или по крайней мере сильно очищенной) воды — выте-
кать из резервуара. Потери избыточного сульфида можно конт-
ролировать, меняя соотношение между поступающими сульфа-
том и металлом. Однако некоторое количество избыточного
растворимого сульфида все-таки неизбежно образуется. Его
можно удалять путем спонтанного химического окисления, если
поддерживать на соответствующем уровне количество раство-
ренного сульфата. Альтернативным решением является окисле-
Одностадийная система
Рис. 5.5. Две высокотехнологичные системы извлечения металлов из растворов
в виде сульфидов. В одностадийной системе применяют сульфатредуцирующие-
бактерии; в двустадийной системе образуется сероводород, который в дальней-
шем используется для осаждения металлов в отдельном резервуаре.
ние сульфида серуобразующими фотосинтезирующими бактерия-
ми в трехступенчатом процессе. Далее возможно экономически:
выгодное получение элементарной серы. Излишки газообразного.-
сероводорода легко вновь использовать в цикле.
5.3.3. Внеклеточное комплексообразование
Некоторые микроорганизмы синтезируют специфические хими-
ческие соединения, обладающие высоким сродством к опреде-
ленным металлам. Наиболее известны соединения, образующие
комплексы с железом. Молибден, ванадий и другие микроэле-
менты, участвующие в метаболизме бактерий, также способны
поступать в клетку в форме внеклеточных комплексов. Хотя*
14—1344
210
Глава 5
эти комплексообразующие соединения не осаждают металлы,
на их основе может быть создана новая технология извлечения
•отдельных металлов из растворов. Для иммобилизации раство-
ренных металлов пригодны комплексообразующие агенты, вы-
деленные из культур микроорганизмов, а также химически син-
тезированные соединения, имитирующие эти агенты. Эффектив-
ность связывания металлов с подобными соединениями можно
увеличить путем повышения их емкости по отношению к данным
металлам, увеличения скорости соответствующих реакций и спе-
цифичности комплексообразующих агентов. Для этого можно
попытаться синтезировать химическим путем такие агенты, для
которых в качестве моделей служили бы природные продукты.
Перспективным представляется и создание методами генетиче-
ской инженерии микроорганизмов, синтезирующих нужные ве-
щества в большом количестве.
5.3.4. Внутри- и внеклеточное накопление металлов
микроорганизмами
О прямом накоплении металлов микроорганизмами уже шла
речь в предыдущих разделах. Теперь мы рассмотрим лежащие
в основе этого явления биохимические процессы и возможности
их использования в прикладной микробиологии. Идея примене-
ния микроорганизмов для извлечения металлов из растворов
издавна представлялась привлекательной не только для очистки
.воды, но и для получения ценных или экономически важных
металлов. Возможность эксплуатации такого процесса концент-
рирования несомненна, поскольку хорошо известна способность
живых организмов извлекать металлы из разбавленных раство-
ров и накапливать их. Многие растения и животные концентри-
руют элементы из окружающей их среды в миллионы раз.
Возможные пути биотехнологического использования накоп-
ления металлов микробами иллюстрирует рис. 5.6. Существуют
два альтернативных механизма: накопление металлов организ-
мами в растущей культуре и суспензиями нерастущих организ-
мов. Конечно, можно использовать и проточные колонки с им-
мобилизованными микроорганизмами или другие биофильтрую-
щие устройства. Если предполагается применение растущих
культур, то потребуется довольно сложная технология. Необхо-
димо учесть и привести в соответствие друг с другом следую-
щие факторы: 1) скорость роста организма (проточная культу-
ра); 2) способность к накоплению металла и, следовательно,
•скорость его включения; 3) возможная токсичность металла
для данных организмов; 4) конкуренция за извлекаемый ме-
талл между данным организмом и возможными комплексооб-
разующими агентами среды (органическими и неорганически-
Материалы и биотехнология
211
ми); 5) конкуренция между данным металлом и другими не-
обходимыми металлами за включение в организм. Будучи
высокотехнологичным, процесс извлечения металлов с по-
мощью растущих культур требует учета слишком многих пара-
метров и создает много проблем. Более привлекательным пред-
ставляется накопление металлов нерастущими или иммобили-
зованными микроорганизмами. При этом легче определять и
контролировать физико-химические условия среды, в которой
находятся клетки, и их способность к накоплению металла.
Кроме того, металл может быть переведен в раствор, относи-
Раствор металла
Культура
микроорганизмов
Суспензия
микроорганизмов
Микроорганизм + Связанные металлы
Деструктивное
извлечение металла
Металл Повторное
использование
Недеструктивное
извлечение металла
микроорганизма
Рис. 5.6. Возможные способы извлечения металлов из раствора с помощью»
микроорганизмов в качестве биосорбентов.
тельно свободный от комплексообразующих агентов, в кон-
центрациях, нелетальных для живых клеток.
После концентрирования металла в микроорганизме возни-
кает проблема извлечения из него металла. Для этого можно
использовать либо недеструктивный способ высвобождения ме-
талла из микроорганизма, либо экстракцию путем разрушения.
В последнем случае микроорганизмы подвергают пирометал-
лургической обработке либо разрушают их концентрированной
щелочью или кислотой. Выбор метода определяется тем, на-
сколько легко высвобождается металл из микроорганизма и
насколько ценен сам микроорганизм для того, чтобы имело
смысл повторно его использовать. Ясно, что, если стоимость
получения микроорганизма высока, его повторное использова-
ние весьма желательно. С другой стороны, если данный орга-
14*
212
Глава 5
низм представляет собой дешевый побочный продукт иных
производств (например, дрожжи) и/или извлекаемые металлы
являются крайне ценными (например, металлы платиновой
группы), то экономически предпочтителен деструктивный про-
цесс. В любом варианте выбор организма и процесса экстрак-
ции должен определяться основными свойствами аккумулирую-
щих металл микробных систем; самое главное — четкое пони-
мание биохимических процессов, характерных для данного ми-
кроорганизма.
Микроорганизмы способны концентрировать металлы одним
из следующих способов: 1) внеклеточное накопление участву-
ющих или не участвующих в метаболизме металлов путем свя-
зывания или осаждения их на клеточной стенке или мембранах;
2) внутриклеточное накопление нужных для метаболизма ме-
таллов (например, К, Fe, Mg, Мо, следы Си, Ni); 3) внутри-
клеточное накопление относительно больших количеств несу-
щественных для метаболизма металлов (например, Со, Ni, Си,
Cd, Ag) в основном с помощью механизмов, служащих для
накопления существенных для метаболизма металлов.
Поглощение некоторых металлов дрожжами и бактериями
осуществляется почти исключительно за счет поверхностного
связывания; примером служит накопление урана дрожжами
или свинца у Micrococcus. Внутриклеточное накопление может
сопровождаться незначительным поверхностным связыванием;
пример тому — накопление калия. Процесс накопления метал-
лов нередко характеризуется двухфазной кинетикой (рис. 5.7).
Рис. 5.7. Обычно наблюдаемая кинетика накопления металлов бактериями: ста-
дия 1 —энергонезависимая, стадия 2—энергозависимая.
Материалы и биотехнология 213
Сразу после введения металла в среду последний быстро свя-
зывается с клеточной поверхностью за счет независимого от
энергии процесса, а затем происходит медленный перенос ме-
талла в цитоплазму клетки. Последний процесс часто является
энергозависимым и протекает лишь при активном дыхании. Он
может блокироваться дыхательными ядами и анаэробиозом,
ингибирующими аэробное дыхание или запасание энергии.
У бактерий связывание металлов с клеточной поверхностью
происходит интенсивнее, чем у большинства дрожжей; так,
скорость связывания металлов у Saccharomyces cerevisiae на-
много ниже, чем у Escherichia coli или Bacillus (рис. 5.8). Свя-
занные с клеточной поверхностью металлы легко отделяются
от нее хелатирующими агентами или разбавленными кислота-
ми; например, кобальт, связанный с поверхностью Bacillus,
легко удаляется при помощи ЭДТА (рис. 5.8). На внутрикле-
точное накопление металла ЭДТА не влияет. Бактериальные
системы представляются предпочтительными для удаления ме-
таллов с последующим быстрым связыванием; однако энерго-
зависимое поглощение металлов в случае дрожжей часто ока-
зывается более эффективным, чем для бактерий. У различных
штаммов родственных бактерий уровень поверхностного связы-
вания существенно различается. Например, Bacillus megate-
rium КМ (при концентрации 1 г сухой массы на литр) при 20 °C
связывает 43 мг кадмия на 1 г сухой массы из раствора, со-
держащего Cd в концентрации 112 мг/л (в то время, как
В. polymyxa — всего 10 мг Cd на 1 г сухой массы). Сильно свя-
зывающий штамм В. megaterium (при концентрации 1 г сухой
массы клеток на литр) извлекает 38 и 68% кадмия из раство-
ров, содержащих соответственно 112 и 11 мг Cd в 1 л.
Последующее накопление металлов внутри клетки, как пра-
вило, требует специфических транспортных систем. Четко уста-
новлено, что поглощение никеля или кобальта происходит при
участии системы транспорта магния, а поглощение рубидия,
вероятно, при участии системы транспорта калия. Различные
металлы могут конкурировать за карбоксилы, гидроксилы
и другие участки связывания на поверхности клетки или за
транспортные системы. При поступлении в клетку иона какого-
либо металла из цитоплазмы выходят одноименно заряженные
ионы. В зависимости от организма это могут быть протоны,
ионы магния или калия (рис. 5.9).
Над применением микроорганизмов в биотехнологии в ка-
честве биосорбентов металлов еще предстоит поработать. Не-
давно было показано, что уран может сорбироваться из мор-
ской воды или из растворов водорослями, дрожжами или
Pseudomonas. Pseudomonas накапливает уран в цитоплазме
(рис. 5.10), а для Saccharomyces характерно поверхностное
214
Глава 5
Рис. 5.8. Поглощение кобальта дрожжами и бациллами из 0,002 М PIPES.
(Norris, Kelly, 1977; Norris, Kelly, 1979.) 1 —Saccharomyces cerevisiae, 2 —
Bacillus megaterium, 3 — кобальт, удерживаемый В. megaterium после промыв-
ки 1 мМ ЭДТА.
связывание (рис. 5.11). Этот связанный уран легко удаляется,,
и дрожжи можно использовать повторно. Потенциальные воз-
можности развития технологии экстракции определяются мно-
гими факторами. 1. Специфичность связывания должна быть
достаточной для удаления определенного металла или смеси
металлов из разбавленного раствора. 2. Специфичность био-
аккумулятора по отношению к металлу должна быть эквива-
лентной таковой для физико-химических методов (ионный об-
мен, экстракция растворителями и т. п.). 3. Биоаккумулятор
должен обладать способностью к извлечению из среды боль-
ших количеств металла. 4. Чтобы процесс был экономически
Материалы и биотехнология
215
Выход эквивалентного иона (заряда)
Рис. 5.9. Схематическое изображение ионного равновесия, поддерживаемого
микроорганизмами, которые накапливают металлы внутри клетки.
выгоден, стоимость извлекаемых металлов должна быть экви-
валентна затратам на получение и, возможно, повторное ис-
пользование микроорганизмов (или превышать их). 5. Другие
вещества окружающей среды не должны оказывать вредного
воздействия на биоаккумуляторы.
Широкое разнообразие микробных систем и возможность
существенного изменения микроорганизмов генетическими ме-
тодами позволяют надеяться на создание биотехнологии, отве-
чающей всем этим требованиям. Однако к спекуляциям по по-
воду разработки такой технологии следует относиться с осто-
рожностью. Дело в том, что извлечение металлов с помощью
микробиологических процессов подчиняется тем же химиче-
ским законам, что и получение их обычными физическими и
химическими методами. По мере изучения способности микро-
216
Глава 5
Рис. 5.10. Электронная микрофотография Pseudomonas aeruginosa, демонстри-
рующая внутриклеточное накопление урана (увеличение 27 000). Фотография
любезно предоставлена Дж. У. Страндбергом (G. W. Strandberg, Oak Ridge
National Laboratory, Oak Ridge, TN).
организмов к накоплению металлов может обнаружиться, что
микробные системы способны служить лишь моделями для
разработки технологий искусственного извлечения металлов.
5.4. Биополимеры
5.4.1. Введение
Термин «биополимеры» относится ко многим высокомолекуляр-
ным соединениям (например, к нуклеиновым кислотам, полиса-
харидам и липидам), синтезируемым самыми разными организ-
мами. В этом разделе мы особенно подробно рассмотрим обра-
зование микроорганизмами полисахаридов и поли-В-гидрокси-
бутирата. Эти биополимеры часто синтезируются в ответ на
Материалы и биотехнология
217
Рис. 5.11. Электронная микрофотография Saccharomyces cerevisiae NRRLY2574,
демонстрирующая накопление урана на поверхности клеток (увеличение
35 000). Фотография любезно предоставлена Дж. У. Страндбергом (G. W. Strand-
berg, Oak Ridge National Laboratory, Oak Ridge, TN).
специфические условия среды в тех случаях, когда соединения
углерода не являются фактором, лимитирующим рост, и, следо-
вательно, могут служить резервным источником углерода и/или
энергии (разд. 5.4.7).
5.4.2. Полисахариды
Полисахариды служат источником энергии и структурными
компонентами клеточных стенок и внеклеточных капсул. Мно-
гие из этих полимеров, имеющие коммерческую ценность как
промышленные клеи, были получены из растительных тканей
(экстракты семян и морских водорослей, древесные экссудаты
и т. п.). Способность таких полисахаридов изменять реологиче-
ские свойства воды, вызывая образование геля и влияя на
218
Глава 5
свойства водных растворов в потоке, привели к их широкому
промышленному использованию в самых различных ситуациях.
Полисахаридные гидроколлоиды часто применяются в пищевой,,
фармацевтической, парфюмерно-косметической, нефтяной, бу-
мажной и текстильной промышленности. Например, из красных,
водорослей производят в промышленных масштабах карраге-
нан и агар, а из бурых — альгинаты. Однако получение полиса-
харидов из растений и водорослей обладает своими недостат-
ками.
1. Химический состав полисахаридов зависит от метаболи-
ческих потребностей синтезирующих их организмов, связанных,
в свою, очередь с изменениями внешних условий (например, се-
зонные изменения, разные циклы развития растений, время их
сбора и т. д.). Поэтому при производстве сырья невозможно,
обеспечить контроль за его качеством.
2. При переработке происходят изменение и разрушение-
продукта, поскольку такая переработка нередко включает гру-
бые воздействия (щелочная экстракция, приводящая к р-эли-
минации и разрушению; выщелачивание горячей водой; отбели-
вание). При этом конечный продукт может приобрести неже-
лательный запах или цвет.
3. Количество получаемого растительного продукта зависит
от урожайности, погодных условий, заболеваний растений или'
загрязнения окружающей среды.
При получении полисахаридов из микроорганизмов обеспе-
чивается контролируемый синтез полимеров и постоянство про-
дукции. Кроме того, микробные полисахариды часто обладают
уникальными физическими и химическими свойствами, улуч-
шенными функциональными характеристиками; биологическая)
потребность в кислороде при их образовании невелика. Микро-
организмы синтезируют множество полисахаридов в форме-
внеклеточных капсул или слизей, не связанных с клеточной;
стенкой. Как правило, в их состав входит небольшой набор мо-
носахаридов (нейтральные гексозы, метилпентозы, кетосахара,,
аминосахара, уроновые кислоты), однако разное их сочетание-
дает полимеры с разнообразными физическими свойствами.
Отметим, что получение микробных полисахаридов — относи-
тельно дорогой процесс: для его осуществления требуются-
большие капиталовложения и энергетические затраты и необ-
ходим квалифицированный персонал. Видимо, микробные поли-
меры не вытеснят окончательно крахмал и его производные из-
всех сфер их использования. Оценивая целесообразность про-
мышленного производства того или иного полисахарида, сле-
дует учитывать следующие факторы: 1) потенциальный объем
годового производства продукта и спрос на него как в настоя-
щее время, так и в будущем; 2) уникальность свойств данного
Материалы и биотехнология
219
продукта по сравнению с другими микробными и растительны-
ми полисахаридами; 3) экономичность производства и предпо-
лагаемую длительность применения продукта.
Микроорганизмы, образующие полисахариды, найдены в
самых разных средах, но вряд ли существует определенная
стратегия их выделения. Было показано, что синтез полимеров
идет как в аэробных, так и в анаэробных культурах. Установ-
лено, что психрофильные микроорганизмы синтезируют полиса-
хариды, относительно же активности такого рода у термофилов
мало что известно.
5.4.3. Образование полисахаридов при брожении
Для образования большого количества полимера требуется
легкодоступный и дешевый источник углерода. Ферментация
позволяет культивировать организм-продуцент в строго опреде-
ленных условиях среды, контролируя, таким образом, процесс
биосинтеза и влияя на тип продукта и его свойства. Специфи-
чески изменяя условия роста, можно менять молекулярную
массу и структуру образующегося полимера. В ряде случаев
максимальная скорость синтеза полисахарида достигается в
логарифмической стадии роста, в других — в поздней логариф-
мической или в начале стационарной. Обычно углеводными
субстратами служат глюкоза и сахароза, хотя полисахариды
могут образовываться и при росте микроорганизмов на н-алка-
нах( С12-61), керосине, метаноле, метане, этаноле, глицероле и
этиленгликоле. Недостатком проведения процесса в ферменте-
рах является то, что среда часто становится очень вязкой, по-
этому культура быстро начинает испытывать недостаток кис-
лорода; мы все еще не умеем рассчитывать соотношение меж-
ду скоростью перемешивания неньютоновских жидкостей и по-
дачей кислорода. Необходимо также контролировать быстрые
изменения pH среды. И все же упомянутый метод позволяет
быстро синтезировать полимер для того, чтобы определить его
физические свойства, а также дает возможность оптимизиро-
вать состав среды, главным образом в отношении эффективно-
сти различных углеводных субстратов. Часто в качестве лими-
тирующего фактора применяют азот (соотношение углерод:
азот—Ю:1), хотя можно использовать и другие (серу, маг-
ний, калий и фосфор). Природа лимитирующего фактора спо-
собна определять свойства полисахарида, например его вяз-
костные характеристики и степень ацилирования. Так, многие
полисахариды, синтезируемые грибами, фосфорилированы. При
недостатке фосфора степень фосфорилирования может умень-
шаться или становиться равной нулю; в этих условиях может
даже измениться соотношение моносахаридов в конечном по-
220
Глава 5
лимере. Недостаток калия приводит к снижению синтеза поли-
сахарида из-за уменьшения поступления питательных веществ.
После удаления микробных клеток центрифугированием
продукт извлекают из культуральной жидкости относительна
мягким и простым способом, который включает осаждение ор-
ганическими растворителями (спиртами, ацетоном). Таким обра-
зом уменьшается вероятность разрушения или модификации:
полимера. Крупные клетки грибов лучше всего удалять центри-
фугированием. При крупномасштабном производстве эффектив-
ное удаление продуцента часто бывает затруднено, и культу-
ральную жидкость пастеризуют нагреванием или гомогениза-
цией. Чтобы при отделении не возникало лишних сложностей,
нужно разумно сбалансировать степень конверсии углеводного-
сырья и вязкость среды при завершении процесса. Нередко,
осаждению полимера в присутствии смешивающихся с водой
органических растворителей способствует добавление какого-
либо электролита. Другой вариант этого метода предусматри-
вает добавление растворителя в меньшем количестве, чем;
требуется для осаждения полимера. После удаления твердых
компонентов фильтрованием при 100 °C добавляют еще одну
порцию растворителя для осаждения полисахарида. Можна
также высушивать отфильтрованную культуральную жидкость,
распылением, осаждать полимер путем добавления поливалент-
ных катионов или с помощью четвертичных аммониевых соеди-
нений (цетилтриметиламмонийхлорида) в сочетании с метано-
лом. Последний метод используется нечасто.
Недостатки, присущие культивированию в ферментерах, для
которого характерны постоянные изменения концентрации пи-
тательных веществ, микроорганизмов и синтезируемых про-
дуктов, можно устранить, применяя непрерывные культуры.
Исходно такие культуры использовались при изучении физио-
логии синтеза полисахаридов, однако они оказались весьма
перспективными для получения стабильно высокой продуктив-
ности. Рост микроорганизмов можно регулировать с помощью-
только одного лимитирующего компонента и непрерывно кон-
тролировать остальные параметры. Таким образом можна
изучать влияние разных лимитирующих питательных веществ,
и разных скоростей роста при постоянных прочих условиях
среды (например, регулировать количество растворенного кис-
лорода путем изменения скорости вращения мешалки и интен-
сивности аэрации). У некоторых микроорганизмов, например у
Xanthomonas campestris и Azotobacter vineland.il, скорость об-
разования полисахарида зависит от разведения, причем при-
более низком разведении наблюдается увеличение выхода по-
лимера. У других организмов синтез не зависит от разведения.
В случае непрерывных культур возникает проблема стабиль.
Материалы и биотехнология 22)
ности штаммов, т. е. появления вариантов, не образующих по-
лисахарид. Все здесь определяется природой используемого-
организма и условиями его культивирования. Так, Xanthomo-
nas juglandis, специфичный по отношению к хозяину вариант
X. campestris, в культурах с ограниченным содержанием азота
не меняется на протяжении более чем 900-часового непрерыв-
ного синтеза полисахарида; при этом наблюдается высокий
выход полимера. Параметром, способствующим возникновению
мутантов с измененной способностью к синтезу полисахаридов;
иногда служит ограничение по углероду в непрерывных куль-
турах. Ферментацию с образованием полисахаридов удается
контролировать многими способами, но постоянного выхода
продукта можно достичь путем регуляции вязкости культуры..
Однако при культивировании в ферментерах с перегородками
иногда образуются области с повышенной вязкостью из-за при-
стеночного роста или накопления полисахарида в других участ-
ках. При таких неоднородных условиях роста контроль за вяз-
костью среды осуществлять не удается.
К нерешенным биоинженерным проблемам относятся: боль-
шой расход энергии на перемешивание вязких сред, недостаток
кислорода из-за неидеального перемешивания и необходимость,
постоянной обработки культуральной среды, включающей уда-
ление клеток и использование больших количеств растворите-
лей для осаждения. В связи с низкой концентрацией продукта
(часто ниже 5%, вес/объем) требуются большие ферментеры
(50—200 м3). Для оценки реологических характеристик куль-
туральных сред проводят исследования на опытных установках..
Полученные данные используют для изменения конструкции
ферментеров и мешалок с целью увеличения их эффективности
при работе с псевдопластичными культуральными средами.
Проблемы последующей обработки конечного продукта при
синтезе полисахаридов связаны прежде всего с удалением ми-
кроорганизмов, что крайне важно, если этот продукт приме-
няется в пищевой промышленности. Для разрушения бактерий
используют литические и протеолитические ферменты, что
в свою очередь приводит к дальнейшему загрязнению среды„
поскольку при этом добавляется потенциальный субстрат для
роста микробов. При удалении бактериальных остатков фильт-
рованием применяют обработку полимера протеолитическими
ферментами при щелочном pH с последующим добавлением
кремниевого адсорбента. Во избежание деградации полимера
при щелочном pH можно при необходимости подкислять среду.
Для осветления растворов полисахаридов их пропускают чере»
иммобилизованные эндоглюканазы, что приводит к незначи-
тельному уменьшению вязкости вследствие гидролиза глико-
зидных связей.
222
Глава 5
В настоящее время осуществляется промышленное произ-
водство ряда микробных полисахаридов [декстран, ксантан,
геллановая смола, занфло (Zanflo) и политран (Polytran)].
Получение многих других находится на стадии разработки.
В следующем разделе описаны основные свойства полисахари-
дов микробного происхождения и перспективы их применения.
5.4.4. Микробные полисахариды:
свойства, применение и коммерческая ценность
Ксантан [келтрол (Keltrol), келзан (Kelzan), Родогель
(Rhodogel) ]
Ксантан синтезируется Xanthomonas campestris при росте на
-глюкозе, сахарозе, крахмале, кукурузной декстрозе и барде.
В качестве источников углерода могут использоваться промыш-
ленные отходы, например сыворотка, образующаяся при выра-
ботке творога. Этот полимер, имеющий аллюлозный остов, по-
строен из повторяющихся пятичленных блоков, содержащих
D-глюкозу, D-маннозу, D-глюкуроновую кислоту; к некоторым
из них присоединены остатки уксусной и пировиноградной кис-
лот. Мол. масса его варьирует от 2-Ю6 до 15-Ю6. Точная тре-
тичная структура ксантана еще не определена, однако резуль-
таты конформационного анализа позволяют предположить, что
полимер имеет форму спирали. Остается выяснить, одиночная
она или двойная. Ксантан был первым микробным полисаха-
ридом, который начали производить в промышленном масшта-
бе (1967 г.). Он нашел широкое применение в промышленности.
Это вещество обладает высокой вязкостью при малых концен-
трациях и низкой скоростью сдвига. Вязкость его остается
постоянной в широком диапазоне pH, не зависит от темпера-
туры и присутствия солей, например хлористого калия. В соче-
тании с растительным полисахаридом из семян лжеакации вод-
ные растворы ксантана образуют стабильные гели. Ксантан
получают обычным способом в ферментере с использованием
•коммерческой D-глюкозы в качестве источника углерода. Пе-
ред осаждением полимера метанолом или изопропанолом в при-
сутствии хлористого калия клетки обычно не удаляют. Изуча-
лась возможность непрерывного получения ксантана, но про-
мышленное внедрение этого метода осуществлено не было.
Уникальные свойства ксантана предопределили его широкое
применение в самых разных отраслях промышленности в каче-
стве стабилизатора и средства для контроля за состоянием
•суспензий, гелеобразованием и вязкостью. Псевдопластические
текучие свойства этого полимера в сочетании с устойчивостью
ж нагреванию, кислотам, щелочам и присутствию катионов
Материалы и биотехнология 22$
обеспечивают ему преимущества над другими смазками в со-
ставе бентонитовых шламов, и ксантан широко используется
при добыче нефти. Он применяется также для повышения вы-
хода нефти, где в сочетании с поверхностно-активными веще-
ствами и углеводородами служит в качестве агента, контроли-
рующего вязкость жидкости, закачиваемой в нефтеносные
пласты. Молекулы полимера, использующегося для повышения
выхода нефти, должны быть достаточно малого размера, что-
бы они могли проникать в поры породы, образуя растворы,
с высокой вязкостью, которая не изменяется при высоких кон-
центрациях солей, высоких температурах и давлении, а также-
под действием сдвиговых напряжений, создаваемых при работе-
насосов. Ксантан, очищенный от бактериальных остатков, об-
ладает более низкой степенью удержания, чем широко ис-
пользуемый полиакриламид. При добыче нефти применяются
водные растворы сульфатированного полисахарида, так как
они подвержены меньшему разжижению под действием сдвиго-
вых напряжений. В комплексах с бурой ксантан применяют
в качестве гелеобразующего агента при производстве взрывча-
тых веществ.
В 1969 г. было разрешено использовать ксантан в пищевой
промышленности для улучшения вкусовых свойств консерви-
рованных и замороженных продуктов, приправ, соусов, быст-
ро приготовляемых продуктов, заправок, кремов и фруктовых
напитков. Благодаря синергическому взаимодействию между
ксантаном и растительными галактоманнанами вместе с ка-
медью из семян лжеакации он нашел применение в производст-
ве кормов, например консервированного корма для домашних
животных, где он конкурирует с агаром; в качестве суспенди-
рующего агента он используется также в производстве кормов
с низким содержанием твердых компонентов. Простые и слож-
ные эфиры ксантана применяют в косметике и в текстильной
промышленности.
Декстран
ДексТран — это a-D-глюкан, синтезируемый самыми разными
грамположительными и грамотрицательными бактериями, та-
кими как Aerobacter spp., Streptococcus bovis и S. viridans,.
а также Leuconostoc mesenteroides. В промышленности этот
полимер получают выращиванием последнего из перечисленных
микроорганизмов на сахарозе. Большинство полисахаридов яв-
ляются продуктами внутриклеточного синтеза, однако при об-
разовании декстрана субстрат не проникает в клетку. Декстра-
ны классифицируют в зависимости от относительного содержа-
ния каждого из трех имеющихся типов связей (а-1->3„
224
Глава 5
•а-1->4 и а-1—-6), а также по растворимости в воде. Высокомо-
лекулярный полимер осаждают органическими растворителями,
а затем разрушают ферментативным путем (используя экзо-
и эндодекстраназы) с помощью гидролиза слабой кислотой
либо нагреванием до получения продукта с нужной молекуляр-
ной массой. Степень такой деградации контролируется по изме-
нению вязкости. В качестве альтернативы можно использовать
низкомолекулярные полисахариды как затравку для получения
декстранов с желаемой степенью полимеризации. Бактериаль-
ные инокуляты часто неоднократно промывают солевым раст-
вором для удаления примесей высокомолекулярных полиме-
ров, которые в противном случае смогли бы послужить затрав-
ками при полимеризации. Помимо природы акцептора глюко-
зы, молекулярная масса декстрана определяется также кон-
центрацией сахарозы и температурой реакции. При высокой
концентрации сахарозы (70% по весу) образуется в основном
однородный набор низкомолекулярных декстранов. При ис-
пользовании низкомолекулярной декстрановой затравки
(10 000—25000) и небольших концентраций сахарозы (10% по
весу, pH 5, 15 °C) значительная часть получаемого продукта
(50%) имеет мол. массу в пределах 50 000—100 000; при этом
отпадает необходимость в его дальнейшем фракционировании
для применения в медицине.
Внеклеточный фермент, катализирующий синтез декстрана,
декстрансахараза (а-1,6-глюкан : О-фруктозо-2-глюкозил-
трансфераза, К.Ф.2.4.1.5), является индуцибельным у Leuconos-
toc mesenteroides\ у Streptococcus mutans или S. sanquis он син-
тезируется в течение логарифмической фазы роста. Этот фер-
мент освобождает фруктозу и переносит глюкозные остатки на
связанную с ферментом молекулу акцептора. Растущая цепь
декстрана остается прочно связанной с ферментом. Исследова-
ния иммобилизованной декстрансахаразы показали, что она
образует продукты с узким спектром молекулярных масс, тогда
как в растворимой системе синтезируются очень высокомолеку-
лярные декстраны. Синтезирующие декстран организмы проду-
цируют большие количества этого фермента в растворимой
форме или в связанном с клеткой состоянии. Производство дек-
странов микробной природы остается, однако, ограниченным,
несмотря на успехи в разработке бесклеточных систем.
Прежде всего декстраны используют в качестве замените-
лей плазмы (для увеличения объема крови); кроме того, они
применяются в медицине для создания гидрофильного слоя на
обожженных поверхностях в целях поглощения жидких экссу-
датов. Для разделения и очистки биологических молекул на-
ходят широкое применение производные декстранов с попереч-
ными сшивками, в которых функциональные группы (например,
Материалы и биотехнология
225
карбоксиметильные или диаминогруппы) соединены с глюкоз-
ными остатками эфирными связями. Сульфатированные декст-
раны используют в качестве полиэлектролитов.
Микробный альгинат
Источником альгинатов издавна служили морские водоросли
(например, Laminaria spp.), однако по природе своей этот ис-
точник непостоянен. Среди бактерий близкие к альгинату гете-
рополисахариды образуют из D-маннуроновой и L-глюкуроно-
вой кислот Pseudomonas aeruginosa и Azotobacter vinelandii.
Этот процесс осуществляют в промышленном масштабе, выра-
щивая Azotobacter в условиях избытка углерода. Микробный
альгинат отличается от соответствующего продукта из водорос-
лей наличием О-ацетильных групп, связанных с остатками D-
маннуроновой кислоты. Тип получаемого альгината можно из-
менять, варьируя различные параметры. Например, при низком
содержании фосфата образуется в основном высокомолекуляр-
ный полимер с хорошим выходом, а при разной концентрации
ионов кальция можно получать разное соотношение между
маннуроновой и галактуроновой кислотами; это соотношение
в свою очередь влияет на эпимеризацию одной кислоты в дру-
гую. Было изучено также образование микробного альгината
в непрерывной культуре, где его выход при выращивании на
сахарозе увеличивается до 50% по сравнению с 25% в фермен-
тере. При высокой интенсивности дыхания могут происходить
большие потери углеродного субстрата вследствие его превра-
щения в углекислый газ. Максимальная скорость синтеза поли-
сахарида в непрерывной культуре была достигнута при недо-
статке в среде молибдена. При ограниченном содержании фос-
фора и низкой плотности клеток в культурах Azotobacter vine-
landii происходит дополнительная потеря углерода из-за
синтеза поли-р-гидроксибутирата.
В настоящее время альгинаты из растительных источников
используются в основном в пищевой промышленности в каче-
стве загустителей или гелеобразующих агентов. Их применяют
для стабилизации йогурта, для предотвращения образования
кристаллов льда при получении мороженого; подобно производ-
ным пропиленгликоля, их добавляют в содержащие кислоту
продукты, например в приправы для салатов, поскольку эти
соединения образуют гели только при pH ниже 3. Альгинатные
полисахариды превращаются в гель в присутствии поливалент-
ных катионов (Са+2, Sr+2) и поэтому применяются для мягкой
иммобилизации микроорганизмов. В других условиях подобное
гелеобразование может быть нежелательным и его пытаются
предотвратить, получая соответствующие производные поли-
мера.
15—1344
226
Глава 5
Геллановая камедь
Геллан — полисахарид, состоящий из остатков глюкозы, рамно-
зы, глюкуроновой кислоты и содержащий О-ацетильные группы
(3—4,5%), — получают методом аэробной ферментации при
участии Pseudomonas elodea АТСС 31461 на каком-либо угле-
водном источнике углерода. Этот продукт существует в трех
формах: нативной, низкоацетильной и низкоацетильной/освет-
ленной. Низкоацетильная форма, легко получаемая из натив-
ной нагреванием при pH 10, образует при нагревании и охлаж-
дении твердые хрупкие гели. Прочность геля зависит от кон-
центрации камеди и солей, а также от природы присутствующих
катионов. Наиболее прочный гель образуется при более низких
концентрациях двухвалентных катионов (Са2+, Mg2+) по срав-
нению с одновалентными. Хотя этот полимер еще не был ре-
комендован для применения в пищевой промышленности, воз-
можно, в будущем он заменит каррагенан и агар. Его уже ис-
пользуют в микробиологии в качестве компонента различных
сред, где он известен под коммерческим названием гелрит
(Gelrite). Здесь его преимуществами перед агаром являются
более высокая прозрачность, та же прочность геля при вдвое
меньшей концентрации полимера, меньшая токсичность и вы-
сокая устойчивость к действию ферментов.
Занфло (Zanflo)
Полисахарид занфло, получаемый из Erwinia tahitica, облада-
ет сходными с ксантаном свойствами; единственное отличие со-
стоит в том, что его вязкость претерпевает обратимые термиче-
ские изменения (при температуре выше 60°C). Этот полисаха-
рид, растворы которого обладают высокой вязкостью, состоит
из остатков фукозы, галактозы, глюкозы и уроновой кислоты
и содержит некоторое количество этерифицированных О-аце-
тильных групп. В качестве источника углеводов при его выра-
ботке применяют лактозу, гидролизованный крахмал или их
смеси. Этот полимер был создан специально для разметки до- ।
рожных покрытий, поскольку он устойчив к замораживанию— I
оттаиванию и действию ферментов, а также обладает хорошей
текучестью и наносится ровным слоем на поверхность. Благо-
даря этим качествам он может найти применение в лакокра-
сочной промышленности. Erwinia tahitica синтезирует также
целлюлазу; это налагает ограничения на использование занф-
ло при производстве красок, содержащих целлюлозные загус-
тители.
Материалы и биотехнология
227
Политран (склероглюкан)
Политран представляет собой линейный р-1,3-глюкан, выделяе-
мый грибом Sclerotium glucanicum и близкими к нему видами
при выращивании в глубинной культуре на среде с кукурузным
экстрактом. К каждому третьему остатку в цепи связью р-1->6
присоединена одна D-глюкопиранозильная группа. Политран
обладает псевдопластическими свойствами в широком диапазо-
не pH и температуры и нечувствителен к различным солям.
Его применяют для стабилизации бентонитовых шламов при
«бурении и для повышения нефтедобычи; он используется также
в керамических глазурях, латексных и типографских красках
и при дражировании семян. Этот нейтральный полисахарид
разрушается экзоглюканазами до глюкозы и гентобиозы.
В настоящее время намечается возможность промышленного
получения и многих других микробных полисахаридов. За по-
следние пять лет в выделении и производстве различных поли-
меров наблюдается быстрый прогресс.
Микробные полисахариды, синтезируемые Alcaligenes spp.
Компанией Kelco в США за последнее время доведено до про-
мышленных масштабов получение нескольких полисахаридов
при участии различных видов Alcaligenes.
В глубинных культурах образуется с большим выходом по-
лимер SI30 неизвестной структуры. Он обладает высокой вяз-
костью при низкой концентрации, прекрасной растворимостью,
большой вязкостью в морской воде и в солевых растворах и не
утрачивает этих свойств при высоких температурах (~ 149°C).
Большинство растворов полисахаридов теряют свою вязкость
при температурах свыше 93 °C, а вязкость растворов полимера
S130 при 149 °C остается такой же, как при комнатной темпе-
ратуре. Кроме того, при таких высоких температурах он не
разрушается в течение длительного времени, особенно в при-
сутствии малых количеств кислорода. Будучи очень вязкими
при низких скоростях сдвига, его растворы при высоких скоро-
стях сдвига становятся жидкими почти как вода. Благодаря
таким свойствам данный полимер может найти широкое при-
менение в нефтяной промышленности в качестве хорошего су-
спендирующего агента и вяжущего компонента буровых раство-
ров с низким содержанием твердых частиц.
Сходными с S130 свойствами обладает полимер S194, кото-
рый, кроме того, нечувствителен к сдвиговым напряжениям.
В сочетании с высокой устойчивостью к солям эти качества
обусловливают его применение в роли суспендирующего агента
в суспензиях пестицидов (концентрированные суспензии нера-
15»
228
Глава 5
створимого в воде материала), а также в суспензиях удобре-
ний, используемых в сельском хозяйстве.
Полисахарид S198 содержит О-ацетильные и О-сукциниль-
ные остатки; его получают при аэробной ферментации с ис-
пользованием глюкозы в качестве источника углерода. Его ра-
створы тоже обладают высокой вязкостью при низкой концен-
трации и достаточно стабильны в широком интервале pH и
температур. Кроме того, благодаря устойчивости к сдвиговым
напряжениям он является прекрасным суспендирующим аген-
том. S198 остается стабильным в среде, загрязненной тяжелы-
ми металлами. Все это позволяет думать, что водорастворимые
смеси на основе этого полисахарида могут быть использованы
вместо смазочных материалов для гидравлических систем, из-
готавливаемых на основе нефти.
Курдлан
Курдлан —это а-1,3-глюкан, синтезируемый Alcaligenes faeca-
lis, var. myxogenes, штамм 10C3. При нагревании до темпера-
туры выше 54 °C происходит необратимое гелеобразование это-
го полимера; прочность геля зависит от температуры: она по-
стоянна в интервале 60—80 °C и возрастает в интервале 80—
100 °C. При температуре выше 120 °C молекулярная структура
этого полисахарида изменяется: одиночная спираль переходит
в тройную. Курдлан нерастворим в холодной воде, и его гели
можно получать также путем диализа щелочных растворов
против воды. Этот полисахарид может найти применение в ка-
честве гелеобразователя в кулинарии, он может использоваться
как молекулярное сито, как подложка при иммобилизации фер-
ментов и как связующий агент.
Пуллулан
Пуллулан представляет собой a-D-глюкановый полисахарид,
состоящий из a-1-’-б-мальтотриозных и небольшого числа маль-
тотетраозных единиц. Он синтезируется Aureobacidium pullulans
и образует прочные, упругие пленки и волокна, которые можно
формовать. По сравнению с целлофаном и полипропиленом эти
пленки мало проницаемы для кислорода. Пуллулан, возможно,
найдет применение в качестве упаковочного материала или
флюккулирующего агента в суспензиях глин в горной промыш-
ленности. Он устойчив к амилазам, но разрушается ферментом
пуллуланазой.
Было описано также образование полисахаридов видами
Arthrobacter, Beijerinckia и бактериями, использующими метан
и метанол.
Материалы и биотехнология
229
5.4.5. Биосинтез полисахаридов
Хотя у некоторых бактерий синтез полисахаридов (например,
декстранов) осуществляется вне клетки, в большинстве случа-
ев полисахариды синтезируются внутри нее, а для этого необ-
ходимо, чтобы соответствующий субстрат проник через клеточ-
ную мембрану. Почти во всех работах, касающихся биосинтеза
полисахаридов, изучались организмы, имеющие небольшое про-
мышленное значение или не имеющие его вовсе. Эти результа-
ты можно, видимо, экстраполировать и на промышленные
организмы.
Поглощение субстрата осуществляется путем облегченной
диффузии, активного транспорта (при этом субстрат проникает
в клетку в неизмененном состоянии) или групповой транслока-
ции (при этом субстрат подвергается фосфорилированию).
Скорость синтеза полисахарида, по-видимому, зависит от ско-
рости поступления субстрата, которая, таким образом, может
быть первым из факторов, лимитирующих синтез полисахари-
да. В то же время скорость конверсии углерода обычно очень
высока, поэтому она, видимо, не может быть фактором, лими-
тирующим синтез. Специфические потребности в углероде для
образования полисахарида иногда связаны со специфическими
механизмами поглощения субстрата.
Субстрат, поступающий в клетку, подвергается превраще-
ниям в ходе анаболических или катаболических процессов; пер-
вые включают его превращение в полисахариды, липополиса-
хариды или гликоген. Гликоген редко синтезируется в проли-
ферирующих бактериях, но он является возможным источником
утечки углерода в условиях, неблагоприятных для роста мик-
роорганизма. Примером такого рода может служить двуста-
дийный процесс, при котором вслед за стадией роста микробов
наблюдается синтез полисахарида. Контроль за синтезом гли-
когена осуществляется путем аллостерической регуляции обра-
зования ADP-глюкозы без участия изопреноидных липидов.
Синтез экзополисахаридов зависит от наличия нуклеотидди-
фосфатмоносахаридов, например UDP-глюкозы. Ключевую роль
в этом процессе играет UDP-глюкозо — пирофосфорилаза, ка-
тализирующая образование предшественников для синтеза по-
лимеров клеточной стенки, например тейхоевых кислот и
липополисахаридов, а также экзополисахаридов. Направление
предшественников на синтез того или иного полимера осуще-
ствляется путем строгой регуляции активности таких активи-
рующих ферментов. Одни нуклеотиддифосфатмоносахара
(UDP-галактуроновая и GDP-маннуроновая кислоты) являются
предшественниками в синтезе только экзополисахаридов, дру-
гие служат также активированными предшественниками при
230
Глава 5
образовании моносахаридов (например, UDP-глюкоза являет-
ся предшественником D-галактозы и D-глюкуроновой кисло-
ты). Чтобы какой-либо моносахарид включился в полисахарид,
организм должен синтезировать активированный нуклеотидный
предшественник. Единственным исключением является L-гулу-
роновая кислота, присутствующая в бактериальном альгинате.
Она образуется из GDP-маннозы и GDP-маннуроновой кисло-
ты, но сама по себе не активирована. Альгинат синтезируется
с использованием активированных маннуронозильных остатков,
которые затем подвергаются действию эпимеразы in situ. Сте-
пень специфичности этой эпимеразы точно не известна, но она
действует только на неацилированные остатки. L-гулуроновая
кислота была найдена также в некоторых полисахаридах, не
содержащих остатков маннуроновой кислоты. Отметим, что
в полисахаридах были обнаружены моносахариды (например,
амино- и метилуроновые кислоты, а также метилгексозы), для
которых вообще не идентифицировано соответствующее активи-
рованное углеводное производное.
Дальнейший контроль за пулом активированных моносаха-
ридов может осуществляться с помощью гидролаз UDP-саха-
ров, хотя эти ферменты и являются периплазматическими. Кро-
ме того, некоторые ферменты, участвующие в синтезе нуклео-
тиддифосфатуглеводов, связаны с мембраной и неизвестно,
могут ли продукты этих реакций находиться в свободном со-
стоянии в цитоплазме.
Затем углеводные остатки последовательно переходят от
нуклеотида на липидный переносчик, которым обычно служит
изопреноидный алкогольфосфат (С50—С6о), активированный
при переносе сахарофосфата. Так образуются олигосахаридные
блоки (от тетра- до октасахаридов), из которых путем после-
довательного их присоединения к редуцирующему концу цепи
и образуется полисахарид. Точный механизм удлинения цепи
и высвобождения' полимера неизвестен. Однако участие изо-
преноидных липидов не является обязательным: например,
они не обнаружены у Azotobacter, синтезирующего альгинат.
В тех организмах, где эти липиды присутствуют, контроль за
синтезом экзополисахаридов может осуществляться за счет из-
менения доступности таких липидных переносчиков, которая
в свою очередь может регулироваться изменением соотношения
между свободным спиртом и фосфатом. Регуляция путем де-
фосфорилирования требует наличия ATP-зависимой киназы для
реактивации свободного спирта. Сзб-изопреноид-алкоголь-кина-
за ингибируется антибиотиком моеномицином. По-видимому,
большинство клеток обладает достаточным количеством изопре-
ноидных липидов для одновременного осуществления всех не-
обходимых реакций синтеза, но если их не хватает, то полиса-
Материалы и биотехнология 231
хариды будут синтезироваться только в позднюю логарифмиче-
скую фазу или при низкой^температуре. Антибиотик бацитра-
цин эффективно связывается с изопреноидными липидами, так
что они не могут участвовать в биосинтезе. У мутантов с повы-
шенной резистентностью к этому антибиотику часто наблюда-
ется более интенсивный синтез полисахаридов. Отмечено, что
у мутантов, неспособных синтезировать пептидогликаны, обра-
зуется больше липидов, нужных для синтеза экзополисахари-
дов. По-видимому, потребность в изопреноидных липидах при
биосинтезе полисахаридов можно представить в виде следую-
щего ряда: пептидогликан>липополисахарид>экзополисаха-
рид.
В синтезе экзополисахаридов могут участвовать различные
типы липидных переносчиков, а для синтеза гетерополисахари-
дов иногда требуется не один вид переносчиков. Модификации
полимера, например О-ацетилирование или присоединение пи-
руваткеталей, возможны до тех пор, пока он еще связан с ли-
пидным переносчиком. Ацетилирование не является обязатель-
ным этапом; его отсутствие может быть связано с тем, что в
системе нет соответствующих ацетилаз либо достаточного ко-
личества предшественника, предположительно ацетил-СоА.
При синтезе бактериального альгината ацетильные остатки,
связанные в его молекулах с остатками D-маннуроновой кис-
лоты, должны вводиться до эпимеризации; было высказано
предположение, что эти ацетильные группы способны предот-
вращать эпимеризацию. Соотношение пируват/ацетат в ксанта-
новых камедях сильно различается, и процесс модификации по-
лимера должен включать механизм, обеспечивающий эти раз-
личия. Неизвестно, является ли присоединение пирувата
одно- или многоступенчатым процессом; неизвестно также, за-
висит ли этот процесс от внутриклеточной концентрации фос-
фоенолпирувата. Полимер ксантан может к тому же состоять
из содержащих и не содержащих остатки пирувата цепей в раз-
личных соотношениях.
Механизм высвобождения полимера из комплекса с липид-
ным переносчиком до сих пор подробно не изучен. Видимо, ка-
кую-то роль здесь играет лигазная реакция, в ходе которой
полимер освобождается и связывается с клеточной поверх-
ностью. Обычно после экскреции полисахариды остаются свя-
занными с клеточной стенкой; местом присоединения может
служить какой-либо наружный мембранный белок. Очевидно,
существует определенное число мест связывания, после насы-
щения которых избыток полисахарида выделяется уже в виде
слизи. Возможно также, что места связывания «приспособле-
ны» к полимеру определенного размера. По-видимому, слизи-
стые мутанты образующих капсулы бактерий либо не имеют
232
Глава 5
фермента киназы, либо лишены самих мест связывания. Точ-
ная локализация мест связывания неизвестна, однако процесс
экскреции должен включать перемещение полимера от места
его синтеза — цитоплазматической мембраны — к месту его
конечной внеклеточной локализации; для этого гидрофильная
молекула должна пройти сквозь гидрофобную мембрану. Вы-
сказывалось предположение, что у грамотрицательных организ-
мов местом выхода полисахаридов могут быть участки адгезии
(сайты Байера), где внутренняя и наружная мембраны соеди-
няются друг с другом. Однако ни механизм этого явления, ни
способы его регуляции до сих пор не известны.
5.4.6. Подходы к усовершенствованию производства
микробных полисахаридов
Использование микроорганизмов для получения промышленно
ценных полисахаридов можно сделать более эффективным с
помощью следующих усовершенствований: 1) увеличения ско-
рости образования полисахаридов и повышения их выхода;
2) модификации получаемых полисахаридов; 3) изменения по-
верхностных свойств микроорганизмов-продуцентов для об-
легчения отделения клеток на последующих этапах переработ-
ки; 4) устранения ферментативных активностей, способных вы-
звать нежелательные модификации полисахаридов; 5) переноса
генетических детерминант синтеза полисахаридов в технологи-
чески более удобные организмы-продуценты.
Скорость или степень превращения углеводного субстрата
в полимерный продукт можно увеличить путем повышения
удельной активности участвующих в синтезе ферментов, изме-
нения механизмов регуляции синтетического процесса или уве-
личения доступности предшественников полисахарида. Число
ферментативных стадий биосинтетического процесса зависит от
сложности данного полимера, и любая попытка увеличить вы-
ход полимера должна быть основана на ясном представлении
о данном биосинтетическом пути и механизмах его метаболиче-
ского контроля. В настоящее время выход увеличивают путем
отбора случайных мутантов. Скорость потребления субстрата
можно повысить путем дупликации генов, продукты которых
участвуют в формировании механизмов поглощения, но такой
способ не обязателен, если у данного организма имеется не-
сколько путей транспорта для каждого субстрата.
Для модификации полисахарида в целях улучшения того
или иного его свойства также необходимо располагать некото-
рыми сведениями об особенностях данного пути синтеза. Ксан-
тан в водных растворах образует микрогели за счет взаимо-
действия пируваткеталей полимера с катионами. Эти группы
Материалы и биотехнология
233
можно удалить путем обработки полимера щавелевой или
трифторуксусной кислотой. Существуют также мутанты, обра-
зующие ксантаны, лишенные остатков пирувата или ацетиль-
ных групп, а в остальном имеющие неизмененную углеводную
структуру и более низкую вязкость. Однако отбор таких му-
тантов затруднен тем, что модифицированные продукты часто
неотличимы от исходных полисахаридов, хотя колонии и могут
различаться по внешнему виду и иметь тенденцию к прикреп-
лению к поверхности культуральной среды. Другие способы
решения данной проблемы предусматривают выращивание
бактерий дикого тйпа в присутствии ингибиторов ацилирования
или при недостатке калия либо магния. Можно также выде-
лить мутанты, образующие полисахариды с большей молеку-
лярной массой и, следовательно, с большей вязкостью, но в ос-
тальном не отличающиеся от обычных по химическому составу.
Длина полимерных цепей иногда зависит от условий роста.
Например, в непрерывных культурах Xanthomonas juglandis
при малом разведении образуются более длинные неразветв-
ленные молекулы.
Процесс выделения полисахаридов можно облегчить путем
изменения поверхностных свойств микроорганизма-продуцента
(например, за счет удаления поверхностного полимерного ма-
териала типа липополисахаридов). В подобных мутантных
культурах происходит аутоагглютинация и спонтанная флоку-
ляция, что уменьшает число необходимых операций центрифу-
гирования. Однако нужно внимательно следить за тем, чтобы
у таких мутантов клеточный материал, например белки, не
«утекал» из периплазматического пространства или не проис-
ходил лизис с загрязнением конечного продукта. К другим из-
менениям относятся мутации капсулообразующих организмов,
приводящие к появлению стабильных, образующих слизи бак-
терий, а также получение устойчивых к фагам мутантов, что
уменьшает риск заражения фагом в процессе производства.
Некоторые микроорганизмы образуют экзополисахариды,
впоследствии разрушаемые гидролитическими ферментами. На-
пример, Azotobacter vinelandii синтезирует альгинат и альгина-
зу (альгинат — лиазу). Продуценты ксантана часто выделяют
активную целлюлазу, которая способна вызвать деградацию
при последующем добавлении ксантана к продуктам, содержа-
щим целлюлозу. Устранить такую нежелательную фермента-
тивную активность можно либо путем умелого подбора условий
культивирования, либо за счет использования мутантов, неспо-
собных к образованию подобных гидролитических ферментов.
Штаммы, продуцирующие полисахариды, синтезируют так-
же другие полимерные продукты, например другие полисахари-
ды, поли-р-гидроксибутират или гликоген. При этом из нужного
234
Глава 5
биосинтетического процесса может изыматься значительное ко-
личество углерода. Следует заняться поисками мутантов, ли-
шенных подобного рода альтернативных синтетических путей.
Образование гликогена у прокариот отличается от других пу-
тей синтеза полисахаридов, и получение мутантов, дефектных
по ключевым ферментам этого пути (ADP-глюкозопирофосфо-
рилазе и гликоген-синтетазе), является эффективным решением
данной проблемы. Однако для реализации этого подхода необ-
ходимо иметь представление о регуляторных механизмах обоих
путей. Некоторые виды продуцентов курдлана синтезируют
также значительные количества сукциноглюкана. Были полу-
чены мутанты Alcaligenes spp., Agrobacterium radiobacter и
Rhizobium trifolii, не обладающие этой активностью. Выделены
четыре типа мутантов: продуценты сукциноглюкана, продуцен-
ты курдлана, продуценты обоих полимеров и мутанты, не син-
тезирующие курдлан и образующие небольшие количества сук-
циноглюкана. Сукциноглюкан является гетерополисахаридом
глюкозы и галактозы. Оба полимера содержат ацетильные
группы и остатки пировиноградной кислоты. Механизм этих
изменений в синтезе полимеров неизвестен.
В некоторых случаях предпочтительным является перенос
генетических детерминант синтеза полисахаридов. Например,
перенос детерминант синтеза альгината из штамма Pseudomo-
nas aeruginosa, выделенного исходно от больного цистофибро-
зом, в непатогенные виды Pseudomonas создает предпосылки
для промышленного использования этого процесса. Положение
генов, ответственных за синтез альгината, уже установлено.
Представляет интерес и перенос генов, детерминирующих син-
тез ксантана, в хозяина, непатогенного для растений. Детерми-
нанты синтеза полисахаридов можно ввести и в более быстро-
растущие бактериальные штаммы. Выявлена также возмож-
ность превращения штаммов, не способных к синтезу
экзополисахаридов, в штаммы — продуценты этих полимеров
при помощи отбора на резистентность к карбенициллину. Эта
методика с успехом применялась в опытах с некоторыми вида-
ми Pseudomonas с целью получения тнс-мутантов, синтезиру-
ющих полисахариды. По всей вероятности, в подобных случаях
у организмов дикого типа синтез альгината был супрессирован,
а мутация по соответствующим генам создала условия для
экспрессии.
5.4.7. Поли-^-гидроксибутират
Поли-р-гидроксибутират (ПГБ) — это термопластичный по-
лиэфир, состоящий из повторяющихся блоков
—СН (СН3)—СН2—СО—О— и, как было установлено более
Материалы и биотехнология
235
50 лет назад, являющийся резервным энергозапасающим со-
единением. Он накапливается самыми разнообразными микро-
организмами (например, видами Alcaligenes, Azotobacter,. Ba-
cillus, Nocardia, Pseudomonas и Rhizobium). В некоторых ус-
ловиях отдельные виды, в частности Alcaligenes eutrophus и Azo-
tobacter beijerinckii, способны аккумулировать этот полимерный
материал в таком количестве, что он составляет до 70% их су-
хой массы. Накоплению ПГБ внутри клеток при росте на уг-
леводах или на других источниках углерода, например на ме-
таноле, способствует недостаток фосфора или азота. Поэтому
образование данного полимера может быть одностадийным
процессом, при котором лимитирующим фактором является
с самого начала роста определенное питательное вещество.
Процесс может быть и двухстадийным, когда микроорганизмы
сначала растят в среде, содержащей достаточное количество
питательных веществ, для накопления биомассы, а затем пере-
водят в условия, где лимитирующим рост фактором служит
какой-либо компонент среды, кроме углерода, что вызывает
значительное повышение уровня внутриклеточного ПГБ. К на-
коплению ПГБ способны многие виды, но выход продукта и
молекулярная масса синтезируемого полимера у них обычно
недостаточны для использования этих организмов в промыш-
ленном производстве данного полимера. Необходимо, чтобы
содержание ПГБ в биомассе составляло не менее 30—40% (по
весу), а мол. масса полимера была порядка 200 000—300 000.
Поскольку ПГБ образуется в виде внутриклеточных гранул,
его экстрагируют из микроорганизмов после разрушения кле-
ток. Предпочтительным методом его очистки служит экстрак-
ция растворителями типа галогенированных углеводородов, на-
пример 1,2-дихлорэтаном. Альтернативный подход к получе-
нию полимера в пригодном к использованию виде состоит
в прессовании в формах или экструзии высушенных клеток са-
мого микроорганизма с содержанием ПГБ по весу не менее
50%. Для получения композиции, пригодной к созданию пред-
метов определенной формы, клетки разрушают и при необхо-
димости смешивают с нужным количеством экстрагированного
полимера. Один из технологических подходов к изменению по-
лимерных свойств ПГБ предусматривает изменение состава го-
мополимера in situ. Всегда считалось, что этот резервный энер-
гетический материал является простым полиэфиром мономера
p-гидроксибутирата. Однако сейчас установлено, что во многих
случаях он представляет собой более сложный гетерополимер
разных жирных p-гидроксикислот. Следовательно, состав этого
полимера можно менять в процессе роста микроорганизма, на-
пример добавлением пропионовой кислоты к культуральной
среде: таким способом в образующийся сополимер вводятся
236
Глава 5
остатки p-гидроксивалерата. Это в свою очередь изменяет рео-
логические свойства полимера.
Появление на рынке синтезируемых микробами гетерополи-
меров ПГБ вряд ли окажет существенное влияние на процве-
тающую индустрию пластмасс; однако полезные свойства этого
полимера, особенно его подверженность биодеградации, могут
способствовать его применению в ряде областей.
5.4.8. Другие полимеры, образуемые микроорганизмами
Все описанные до сих пор биополимеры полностью синтезиру-
ются определенными микроорганизмами в процессе роста на
том или ином источнике углерода. Существуют и иные способы
получения новых полимерных материалов, предусматривающие
использование микроорганизма лишь на отдельных этапах син-
теза. При этом микроорганизм служит только микробиологиче-
ским катализатором для проведения химического превращения,
которое в иных условиях затруднено. В гл. 4 уже упоминалось
о реакциях гидроксилирования с участием микробов для полу-
чения фенолов и дигидродиолов в целях последующей химиче-
ской полимеризации. Речь идет о биотехнологическом процес-
се, предложенном для получения нового полимера, полифени-
лена, который представляет интерес для специалистов по
материаловедению благодаря своей термостабильности и высо-
кой электропроводности. Этот полимер можно синтезировать
из бензола и кислорода, используя генетический вариант Pseu-
domonas putida для осуществления начального превращения
бензола в дигидродиол. Этот дважды окисленный продукт вы-
деляется клетками в окружающую среду; его экстрагируют
растворителями, а затем получают производные, необходимые
для химического синтеза полифенилена. Существуют и иные
возможности использования подобной технологии для получе-
ния полимеров нафталина и бифенила.
5.5. Биоповреждение материалов
5.5.1. Введение
Термин «биоповреждение» вошел в наш язык лишь в послед-
нее время, но обозначаемые им процессы известны человеку
издавна, с тех пор, как он начал перерабатывать природное
сырье и заботиться о сохранности пищевых продуктов. Орга-
низмы, ответственные за процессы биоповреждения, сопровож-
дали нас и в еще более отдаленном прошлом, будучи сущест-
венным звеном в круговороте элементов биосферы. Человек ра-
но осознал необходимость защиты сырья и пищевых продуктов
Материалы и биотехнология 237
от возвращения их в этот круговорот и стал использовать для
замедления жизнедеятельности микроорганизмов консервирова-
ние или другие защитные способы. Консервирование как эмпи-
рический процесс существует несколько тысячелетий, и имеется
множество описаний его использования.
Однако для более полного понимания принципов этого
процесса и его научного осмысления потребовалось появление
микроскопа и возможностей селективного культивирования и
идентификации организмов, вызывающих биоповреждения.
Начало исследованиям процессов биоповреждения положи-
ла вторая мировая война; это было связано с поступлением
чувствительных к биологическим воздействием материалов на
театры военных действий, где высокая температура и влаж-
ность ускоряли их порчу. С тех пор мы все более глубоко
осознаем те проблемы, которые связаны с биоповреждениями
продуктов, изготовленных из природного, а позднее — из син-
тетического сырья. Для полного понимания природы этих про-
цессов требуются совместные усилия экологов, . физиологов,
биохимиков и ученых других, небиологических направлений,
таких как технология и материаловедение.
5.5.2. Определение биоповреждений
Под биоповреждением понимают «любое нежелательное изме-
нение свойств какого-либо материала, вызванное жизнедея-
тельностью различных организмов». В широком смысле это
процесс, приводящий к уменьшению ценности любого матери-
ала. При этом имеются в виду те свойства данного материала,
которые обусловливают его использование в определенных це-
лях. По своей природе эти изменения могут быть механически-
ми, физическими или касаться эстетических свойств материала
и не обязательно приводят к его химическому разрушению.
Последний момент важен для определения различий между
биоповреждением и биоразложением (биодеградацией). «Био-
повреждение» — термин более широкий, «биоразложение» —
ограниченный, относящийся только к разрушению какого-либо
продукта (часто сырья), попавшего в окружающую среду (на-
пример, нефтепродуктов, пестицидов или детергентов). В об-
щем смысле биоповреждение — процесс нежелательный, а био-
разложение обычно рассматривается как желательный.
Употребление слова «организм» предусматривает участие
в этом процессе представителей животного и растительного
мира; так оно и есть на самом деле. Микроорганизмы как фак-
торы биоповреждения широко изучались и хорошо представле-
ны в литературе. Однако нельзя недооценивать роль насеко-
мых, грызунов, зеленых растений (в том числе водорослей) и
даже птиц.
238
Глава 5
5.5.3. Классификация процессов биоповреждения
Использование термина «материалы» в определении биопов-
реждений означает, что речь идет о «неживом», в отличие от
патологии, изучающей различные нежелательные процессы
в живой материи. Различия эти зачастую очень тонки, иногда
наблюдается перекрывание в том смысле, что организмы, об-
наруживаемые в живой материи или органических остатках,
сохраняют свою жизнедеятельность и в неживой материи,
уменьшая ценность данного продукта в процессе хранения.
Однако во многих случаях со смертью организма-хозяина из-
меняются условия питания и клеточные компоненты, что при-
водит к изменению и типа организма, «колонизирующего» дан-
ный материал. Так, говоря о действии грибов на хлебные зла-
ки, мы разграничиваем «полевые грибы» и «грибы хранилищ».
Условно можно выделить три типа биоповреждений
(табл. 5.1), которые часто в значительной мере перекрываются.
Таблица 5.1. Классификация типов биоповреждений
Процесс Примеры
1. Механический
2. Химический
а) ассимиляционный
б) диссимиляционный
3. Засорение и загрязне-
ние
Повреждение «несъедобных» материалов (напри-
мер, свинцовых труб, пластмассовых покрытий)
грызунами и насекомыми (погрызы, сквозные
отверстия). Повреждения дорожных покрытий,
и стен, вызванные ростом растений
Использование в качестве источника питатель-
ных веществ субстратов, содержащихся в тех.
или иных материалах (например, целлюлозы,
древесины или кератина шерсти)
Продуцирование организмом какого-либо продук-
та (например, кислоты или токсичного веще-
ства), вызывающего коррозию материала ил»
другие повреждения, в результате которых ма-
териал становится непригодным для использо-
вания
Засорение трубопроводов; обрастание ракушкам»
и водорослями корпуса судов; коррозия и по-
тускнение декоративных покрытий и пластико-
вых занавесей в результате роста грибов ие-
на самом материале, а на поверхностных за-
грязнениях
Об их наличии очень полезно знать при выборе направления
исследований или при разработке рекомендаций, касающихся
условий хранения продуктов и материалов.
Материалы и биотехнология 239
3.5.4. Материалы, подверженные биоповреждениям
При описании биоповреждений легче всего проводить их клас-
сификацию по типу продукта. Однако это оказывается затруд-
нительным, если мы имеем дело со сложными продуктами, на-
пример с красками, где встречаются комбинации исходного
сырья, такого как целлюлоза и синтетический полимер; здесь
классификация по типу продукта невозможна. Было показано,
что среда, в которой хранится и используется данный продукт,
часто оказывает заметное влияние на организмы, которые в
нем обитают, и на активности этих организмов. В следующих
разделах мы вкратце рассмотрим продукты, подвергающиеся
биоповреждениям. Для более полного знакомства с этими
процессами читателю следует обратиться к списку рекомендуе-
мой литературы в конце главы.
Пищевые продукты
В тех странах, где наиболее остро стоит продовольственная
проблема, особенно велики и потери сырья после уборки уро-
жая. В развитых странах продукты различными способами
защищают от грибов, насекомых и грызунов, так что потери
сводятся к минимуму. При хранении зерна необходимо исполь-
зовать различные химические и физические способы защиты,
например пестициды и высушивание. Много неприятностей при-
чиняет присутствие микотоксинов в продуктах, которые были
заражены грибами, часто на ранних стадиях хранения. Это мо-
жет приводить к браковке крупных партий зерна, тем более
если оно используется в качестве корма. Особенно тщательной
должна быть защита от заражения готовых продуктов. Упаков-
ка может приводить как и подавлению роста микроорганизмов,
так и к его стимулированию. Использование немногочисленных
химических консервантов регулируется в соответствии с их хи-
мической природой законодательным путем.
Целлюлоза
Целлюлоза в своей исходной форме, в виде различных волокон
и древесины, столетиями служила сырьем для получения мно-
гих материалов и продуктов. Специалисты по защите материа-
лов постоянно занимались вопросами сохранности изделий из
материалов на основе целлюлозы, и сегодня результаты этих
исследований широко используются в деревообрабатываю-
щей и текстильной промышленности. Организмы, расщепляю-
щие целлюлозу, составляют лишь небольшой процент от обще-
го числа известных видов грибов и бактерий; несмотря на это,
240
Г шва 5
разрушение материалов на основе целлюлозы прецставляет
собой весьма распространенное явление и в соответствующих
условиях может происходить очень быстро. В земле при 25 °C
хлопчатобумажная ткань полностью теряет сг.ою прочность за
10 сут. Насколько нам известно, за разрушение целлюлозы
ответственны скорее всего грибы. Условия внутри целлюлоз-
ных материалов (относительная влажность меньше 90%, очень
низкое содержание азота, кислый pH) часто оказываются бла-
гоприятными для их развития. Разветвленные гифы грибов
с легкостью проникают сквозь клеточные стенки, ближе к цел-
люлозе, обычно тесно связанной с лигниновой и гемицеллюлоз-
ной матрицей. Определенную роль, несомненно, играют и бак-
терии: они наверняка участвуют в разрушении пектиновых
слоев и углублений в древесине мягких пород, что приводит
к проникновению внутрь древесины воды и бактерий.
Подверженность древесины биоповреждениям обусловлена
характером распределения в ней питательных веществ. Если
древесина соответствующим образом не просушена, а заболонь
не срезана для предотвращения миграции питательных веществ,
то во влажных условиях может развиться ее голубое окраши-
вание и начаться поверхностное повреждение.
Для повышения качества или улучшения тех или иных
свойств древесины ее можно химически модифицировать. По-
вышенной устойчивостью к микроорганизмам (видимо, из-за
уменьшения гидролизуемости) обладают ацетат целлюлозы,
вискоза и различные замещенные производные целлюлозы (на-
пример, карбокси- и этоксицеллюлоза). При введении в эмуль-
сии замещенные производные целлюлозы действуют как напол-
нители и агенты, влияющие на вязкость.
Продукты животного происхождения
Большинство продуктов животного происхождения, чувстви-
тельных к биоповреждениям, имеет белковую природу. К ним
относятся шкуры, шерсть и клеи. Бактерии и грибы часто ока-
зывают неблагоприятное воздействие на шкуры и шерсть уже
на ранних этапах их обработки. Более того, из-за большой за-
грязненности свежих шкур или шерсти их порча может начать-
ся в течение 48 ч, еще на бойне или в помещении для стрижки.
Для предотвращения этого процесса шкуры дубят, а шерсть
обезжиривают. Однако при некоторых способах дубления шку-
ры вымачивают вначале в воде. Если такое вымачивание про-
водится при повышенных температурах и продолжается дли-
тельное время, происходит размножение бактерий. В коже,
используемой для изготовления книжных переплетов, нередко
бывает повышено содержание углеводов, что способствует раз-
Материалы и биотехнология 241
витию плесени при хранении в условиях повышенной влаж-
ности.
В целом шерсть наиболее чувствительна к бактериям во
время обработки, а клеи могут разрушаться как при хранении
в различных емкостях, так и после высыхания и образования
пленки.
Поверхностные покрытия
Поверхностные покрытия (краски, различные типы лаков) иг-
рают двоякую роль: они выполняют декоративную функцию и
защищают покрываемую поверхность от вредных воздействий
среды, в том числе и от микроорганизмов. Из-за постепенного
отказа от введения свинца в состав красок и широкого рас-
пространения эмульсионных покрытий возникла проблема био-
повреждения самих красок. Такое повреждение происходит как
при хранении красок в емкостях, так и после нанесения их на
поверхность и высыхания с образованием пленки. Большинст-
во исследований в этой области направлено на создание эф-
фективных защитных систем, которые действовали бы все то
время, пока существует данное покрытие. Краски содержат
пигменты, связывающие вещества, эмульгаторы, масла, смолы
и смачивающие агенты; они могут быть растворены в воде или
в специальных растворителях. Некоторые из этих ингредиен-
тов, например казеин, крахмал, целлюлоза и пластификаторы,
могут разрушаться микробами, а применение альтернативных,
устойчивых к микробному разрушению компонентов зачастую
невозможно. Развитие микроорганизмов в пленках очень силь-
но зависит от факторов окружающей среды: температуры,
влажности, наличия на поверхности питательных веществ (на-
пример, удобрений, приносимых ветром). Повреждения в ем-
костях часто связаны с жизнедеятельностью бактерий, но могут
быть обусловлены и развитием грибов. Кроме того, в жидких
эмульсионных красках могут оставаться внеклеточные фермен-
ты, например входящие в состав целлюлазной системы; эти
ферменты способны снижать вязкость эмульсии.
Резины и пластмассы
Резины и пластмассы представляют собой материалы, содер-
жащие каучук или какой-либо синтетический полимер. До 50%
их состава может приходиться на долю добавок, используемых
в качестве пластификаторов, антиоксидантов и веществ, защи-
щающих данный материал от гидролиза и УФ-света. Кроме то-
го, добавки служат наполнителями и пигментами. Многие из
них чувствительнее к повреждениям, чем сам полимерный «ске-
16—1344
242
Глава 5
лет». Так, поливинилхлорид (ПВХ) в непластифицированной
форме очень устойчив к биоповреждениям, однако применение
его как такового ограничивается тем, что он не пластичен.
Для придания такому материалу пластичности в него вводят
пластификатор, которым часто служит сложный эфир органи-
ческой кислоты, однако этот же пластификатор повышает чув-
ствительность материала к биоповреждениям.
Природные каучуки состоят из регулярно повторяющихся
изопреновых остатков, подверженных окислению, в результате
которого возрастает вероятность разрушения их под действием
микроорганизмов. Введение поперечных сшивок и добавление
антиоксидантов замедляют этот процесс. Были созданы синте-
тические каучуки с улучшенными свойствами, в том числе ус-
тойчивые к биоповреждениям. Очень устойчивы к действию
микробов силиконовые, нитрильные и неопреновые резины.
В целом синтетические полимеры, используемые в качестве
пластмасс (такие, как полиэтилен, полистирол и ПВХ), сами
по себе устойчивы к микроорганизмам. Однако некоторые из
полимеров, применяемых все более широко, оказались чувстви-
тельными к ним. Таковы полиуретаны —нечетко ограниченное
семейство полимеров, выделяемое по признаку наличия урета-
новой группировки; молекула их содержит также замещенные
мочевинные, биуретовые, амидные и аллофанатные группиров-
ки. Кроме того, для придания такому полимеру эластичности
в него химическим путем вводят простые и сложные эфирные
группировки. Большая часть всех этих группировок в соответ-
ствующих условиях чувствительна к химическому, а также, ве-
роятно, к ферментативному гидролизу. Видимо, подвержен-
ность гидролизу определяется конфигурацией молекулы; из-
вестно также, что сложноэфирные связи особенно легко
гидролизуются микроорганизмами. По крайней мере в одном
случае было показано, что вероятность биоповреждения умень-
шается в результате использования соединений, препятствую-
щих гидролизу. Тем не менее наши знания относительно путей
расщепления полиуретанов остаются далеко не полными. Высо-
кая износоустойчивость полиуретанов и их стойкость к истира-
нию гарантировали им прочное место на рынке пластмасс, и ма-
ловероятно, что в ближайшем будущем им будет найдена за-
мена.
Топлива и смазочные материалы
К этой группе веществ относятся прежде всего фракции нефти
с четко выраженными гидрофобными свойствами. При их кон-
такте с водой может происходить целый ряд процессов с уча-
стием микроорганизмов. Подобно пластмассам, эти продукты
Материалы и биотехнология
243
содержат многочисленные добавки, улучшающие их свойства,
которые повышают чувствительность такого рода продуктов
к биоповреждениям.
В качестве топлива для реактивных самолетов широко ис-
пользуют керосин. В емкостях для его хранения и в топливных
баках самолетов неизбежно присутствует вода, поэтому на по-
верхностях раздела топливо — вода и вода — металл начина-
ется рост грибов. При отсутствии контроля развивающиеся
грибы отделяются от поверхности и попадают в топливную си-
стему. Это приводит к засорению фильтров и топливопроводов
и тем самым к искажениям показаний приборов. Из сред по-
добного рода неоднократно выделяли особый гриб Cladosporium
resinae. Перемещение топлива внутри топливных систем само-
лета увеличивает аэрацию, а в ультразвуковом самолете топли-
во служит тепловым амортизатором. Внутри некоторых топлив-
ных баков температура доходит до 55°C. По-видимому, все это-
способствует росту термофильных грибов.
Более тяжелая фракция нефти (масла) используется про-
мышленностью в гидравлических системах для их смазки, а
также применяется для облегчения механической обработки
металлов на токарных станках и при шлифовании. Попадание
воды в такие системы приводит к их повреждению либо в ре-
зультате заклинивания из-за снижения эффективности смазоч-
ного материала, либо в связи с началом коррозии, вызываемой
кислыми промежуточными продуктами жизнедеятельности ми-
кробов. В металлообрабатывающей промышленности важней-
шие проблемы связаны с микробным заражением рабочих жид-
костей; оно вызывает расслоение эмульсий, коррозию обраба-
тываемых деталей и повышенный износ токарных станков.
Относительная роль отдельных микроорганизмов в биоповреж-
дениях масел подобного типа до конца не изучена. Однако
в подходящих условиях, видимо, можно легко выделить орга-
низмы, способные к прямому использованию данного масла,
разрушению эмульгатора и образованию сероводорода.
Металлы и камни
Строгих доказательств связи между активностью определенных
микроорганизмов и процессами коррозии не существует. Воз-
можны три механизма коррозии: образование корродирующих
веществ (кислоты, сероводород, аммиак); образование ячеек
с различной аэрацией; катодная деполяризация. Все это при-
водит к появлению ржавчины, а затем к разъеданию стенок
емкостей для хранения горючего, имеющих водный отстой, и
баков для самолетного топлива, а также к образованию в во-
допроводных трубах пробок, затрудняющих ток воды. Для за-
16*
244
Глава 5
щиты от коррозии применяются обычные методы, в том числе
механические ингибиторы, поверхностные покрытия и анодная
защита.
Камни подвержены воздействиям многочисленных химиче-
ских и иных факторов окружающей среды, способствующих их
эрозии. Проблема биоповреждений камня возникает в связи
с применением его в строительстве или для изготовления па-
мятников, когда необходимо сохранить данную конструкцию.
Как и в случае металлов, продемонстрировать непосредствен-
ное участие микроорганизмов в подобных процессах разруше-
ния крайне трудно. Тем не менее было предложено несколько
механизмов. Первый из них — механическое воздействие: раз-
витие микроорганизмов способствует накоплению воды, замер-
зание и оттаивание которой приводит к разрушению поверхно-
стей. Второй механизм состоит в расширении и сжатии
микробных клеток, а третий — в образовании хелатных комп-
лексов между минералами и органическими кислотами, выде-
ляемыми микробами. Было показано, что бактерии могут пере-
водить в раствор нерастворимые фосфаты и силикаты за счет
образования 2-кетоглутаровой кислоты. Видимо, в разрушении
камней существенную роль играют не упоминавшиеся до сих
пор лишайники, что может быть обусловлено их способностью
к сжатию и расширению (изменение влажности от 15 до 300%
за 2—3 ч) при высушивании или увлажнении и к проникнове-
нию внутрь пород. Недавно были изучены спилы пород, насе-
ленных лишайниками; показано, что гриб-симбионт способен
глубоко в них проникать и избирательно растворять минераль-
ные компоненты.
ЛИТЕРАТУРА
Микробное выщелачивание
Brierley С. L. (1978). Bacterial leaching, Crit. Rev. Microbiol., 6, 207—262.
Brierley C. L. (1982). Microbiological mining, Scient. Am., 247, 42—51.
Fenchel T., Blackburn T. H. (1979). Bacteria and Mineral Cycling, Academic
Press, London.
Murr L. E., Torma A. E., Brierley J. A. (eds.) (1978). Metallurgical Applications
of Bacterial Leaching and Related Microbiological Phenomena, Academic Press,
New York.
Potter G. M. (1981). Design factors for heap leaching operations, Mining Eng.,
33, 277—281.
Иммобилизация металлов
Kelly D. P„ Norris P. R., Brierley C. L. (1979). Microbiological methods for the
extraction and recovery of metals. In: Microbial Technology: Current State,
Future Prospects (eds. Bull. A. T., Ellwood D. C. and Ratledge C.), Cambridge
University Press, Cambridge.
Материалы и биотехнология
245
Norris Р. R., Kelly D. Р. (1979). Accumulation of metals by bacteria and yeast,
Devs ind. Microbiol., 20, 299—308.
Norris P. R., Kelly D. P. (1977). Accumulation of cadmium and cobalt by Sac-
charomyces cerevisiae, J. gen. Microbiol., 99, 317—324.
Norris P. R., Kelly D. P. (1982). The use of mixed microbial culture in metal
recovery. In: Microbial Interactions and Communities, Vol. I (eds. Bull А. Т.»
Slater J. H.), Academic Press, London.
Strandberg G. Shumate S. E„ Parrot J. R. (1981). Microbial cells as biosor-
bents for heavy metals: accumulation of uranium by Saccharomyces cerevisiae
and Pseudomonas aeruginosa, Appl. Environ. Microbiol., 41, 237—245.
Tuovinen О. H., Kelly D. P. (1974). Use of microorganisms for the recovery of
metals, Int. Metall. Revs, 19, 21—31.
Биополимеры
Baird J. K-, Sandford P. A., Cottrell I. W. (1983). Industrial applications of some
new microbial polysaccharides, Biotechnology, 1, 778—783.
Berkely R. C. W., Gooday G. W., Ellwood D. C. (eds.) (1979). Microbial Poly-
saccharides and Polysaccharases, Society for General Microbiology and Aca-
demic Press, London.
Dawes E. A., Senior P. J. (1973). Energy reserve polymers in microorganisms,
Adv. Microbial. Physiol., 10, 136—266.
Sutherland I. W. (1982). Biosynthesis of microbial exopolysaccharides, Adv. Mic-
robial Physiol., 24, 79.
Sutherland I. W. (1983). Extracellular polysaccharides, In: Biotechnology, Vol. 3,
Biomass, Microorganisms, Products 1, Energy (ed. Dellweg H.), Verlag Che-
mie, Weinheim.
Биоповреждение материалов
Eggins H. 0. W., Oxley T. A. (1980). Biodeterioration and Biodegradation Bull.,
16, 53—56.
Gilbert L. J., Lovelock D. W. (eds.) (1975). Microbial Aspects of the Deteriora-
tion of Materials, Academic Press, London.
Onions A. H. S., Allsopp D., Eggins H. O. W. (1981). Smith’s Introduction to
Industrial Mycology, 7th edn, Edward Arnold, London.
Rose A. H. (ed.) (1981). Microbial Biodeterioration, Economic Microbiology,
Vol. 6, Academic Press, London.
Seal K. J., Eggins N. 0. W. (1981). The bioterioration of materials. In: Essays in
Applied Microbiology (eds. Norris J. R. and Richmond M. H.), John Wiley
and Sons, Chichester.
Глава 6
Окружающая среда и биотехнология
Д. Бест, Дж. Джонс, Д. Стаффорд
6.1. Введение
С момента возникновения цивилизованного общества перед
ним все время стояла проблема охраны окружающей среды.
Из-за промышленной, сельскохозяйственной и бытовой деятель-
ности человека постоянно происходили изменения физических,
химических и биологических свойств окружающей среды, при-
чем многие из этих изменений были весьма неблагоприятны.
Мы ожидаем, что биотехнология будет оказывать многообраз-
ное и все возрастающее влияние на способы контроля за ок-
ружающей средой и на ее состояние. Хорошим примером та-
кого рода служит внедрение новых, более совершенных методов
переработки отходов, однако этим применение биотехнологии
в данной сфере отнюдь не ограничивается. Она будет играть
все большую роль в химической промышленности и сельском
хозяйстве и поможет хотя бы отчасти решить многие из су-
ществующих здесь проблем.
В этой главе мы рассмотрим проблему переработки отхо-
дов как в исторической перспективе, так и в плане той роли,
которую биотехнология уже играет и будет играть в этой сфе-
ре. Мы остановимся также на применении биотехнологии
в борьбе против недавно возникшей для окружающей среды
угрозы — распространения ксенобиотиков и нефтяных загряз-
нений.
Сегодня быстро развиваются разнообразные отрасли про-
мышленности, в которых процессы жизнедеятельности микроор-
ганизмов используются для создания замкнутых систем, для
контроля за загрязнением сточных вод, для использования
альтернативных энергоресурсов и химического сырья в про-
мышленности; эти процессы широко используются и в сельском
хозяйстве. В развивающихся странах подобная деятельность,
известная под названием «целенаправленная технология»,
могла бы привести к значительному повышению жизненного
уровня и — что еще более важно — повышению качества жиз-
ни многих миллионов людей. Масштабы некоторых из этих но-
вых биотехнологических процессов и их применение для реше-
Окружающая среда и биотехнология 247
ния задач охраны окружающей среды могут быть совершенно
ошеломляющими. Так, для переработки отходов уже построены
огромные биореакторы емкостью 4000—5000 м3. Поскольку
концентрация бактерий в таком реакторе может быть порядка
108—Ю9 клеток в 1 мл, биотехнологи получают в свое распоря-
жение достаточно мощный источник «биологической энергии».
Биологическая переработка отходов опирается на целый
ряд дисциплин — биохимию, генетику, химию, микробиологию,
химическую технологию и вычислительную технику. Усилия
всех этих дисциплин концентрируются на трех основных на-
правлениях: 1) деградация органических и неорганических ток-
сичных отходов; 2) возобновление ресурсов для возврата
в круговорот веществ углерода, азота, фосфора и серы; 3) по-
лучение ценных видов органического топлива.
6.2. Переработка отходов
Тысячелетиями отходы деятельности человека перерабатыва-
лись естественным путем, при участии соответствующих микро-
организмов. В наиболее широко распространенных установках
для очистки сточных вод выполняются четыре основные опе-
рации (рис. 6.1).
1. При первичной обработке удаляются твердые частицы,
которые либо отбрасываются, либо направляются в реактор.
2. На втором этапе происходит разрушение растворенных
органических веществ при участии природных аэробных микро-
организмов. Образующийся ил, состоящий главным образом из
микробных клеток, либо удаляется, либо перекачивается в ре-
актор. По технологии, использующей активный ил, часть его
возвращается в аэрационный тэнк.
3. На третьем этапе (необязательном) производится хими-
ческое осаждение и разделение фосфора и азота.
4. Для переработки ила, образующегося на первом и вто-
ром. этапах, обычно используется процесс анаэробного разло-
жения. При этом уменьшается объем осадка и количество па-
тогенов, устраняется запах, а кроме того, образуется ценное
органическое топливо — метан.
Сходные процессы применяют при переработке промышлен-
ных сточных вод, особенно в химической, пищевой, целлюлоз-
но-бумажной промышленности. Поэтому совершенно очевидно,
что любые биотехнологические усовершенствования этих про-
цессов найдут немедленное применение в промышленности. Та-
кие усовершенствования могут быть направлены на увеличение
мощности перерабатывающих установок, повышение выхода
полезных побочных продуктов, на замену обычно применяе-
Первичная переработка Вторичная переработка
Рис. 6.1. Стадии переработки отходов путем анаэробного разложения.
Окружающая среда и биотехнология
249
мых синтетических химических добавок, устранение запаха и
удаление металлов, а также не поддающихся переработке со-
единений.
£.2./. Аэробная переработка отходов (см. также гл. 9)
Аэробная переработка стоков — это самая обширная область
контролируемого использования микроорганизмов в биотехно-
логии. Она включает следующие стадии: 1) адсорбция субстра-
та на клеточной поверхности; 2) расщепление адсорбированно-
го субстрата внеклеточными ферментами; 3) поглощение рас-
творенных веществ клетками; 4) рост и эндогенное дыхание;
5) высвобождение экскретируемых продуктов; 6) «выедание»
первичной популяции организмов вторичными потребителями.
В идеале это должно приводить к полной минерализации от-
ходов до простых солей, газов и воды. Эффективность пере-
работки пропорциональна количеству биомассы и времени
контактирования ее с отходами.
Системы аэробной переработки можно разделить на систе-
мы с перколяционными фильтрами и системы с использовани-
ем активного ила.
Перколяционные фильтры
Перколяционный фильтр был самой первой системой, приме-
ненной для биологической переработки отходов, причем его
конструкция фактически не изменилась со временени создания
в 1890 г. Эта система используется в 70% очистных сооруже-
ний Европы и Америки и обладает рядом преимуществ, кото-
рые состоят в простоте, надежности, малых эксплуатационных
расходах, образовании небольших излишков биомассы и воз-
можности длительного использования установки (в течение
30—50 лет).
Основной недостаток перколяционного фильтра — избыточ-
ный рост на нем микроорганизмов; это ухудшает вентиляцию,
ограничивает протекание жидкости и приводит в конечном сче-
те к засорению фильтра и выходу его из строя. Одна из недав-
них модификаций установки состоит в использовании чередую-
щегося двойного фильтрования (ЧДФ), когда фильтры, на
которые сначала поступает поток жидкости, периодически ме-
няют местами с другими фильтрами. ЧДФ особенно ценно при
очистке промышленных стоков. Для разрушения слоев грязи
в толще фильтров используют также обратную циркуляцию и
пульсирующую подачу. Это улучшает величину биохимической
потребности в кислороде (ВПК), но снижает нитрифицирую-
щую активность. Другие модификации в конструкции и работе
250
Глава 6
установок с перколяционными фильтрами состоят в уменьше-
нии скорости поступления жидкости для более равномерного
распределения биомассы, а также в использовании прямой
двойной фильтрации с большим объемом среды, когда на
первый фильтр поступает больший объем среды и тем самым
увеличивается его загрузка.
В 1970 г. на смену клинкеру, камню или гравию в системах
с перколяционными фильтрами пришли пластмассы. Это позво-
лило Применять такие системы для переработки некоторых про-
мышленных стоков высокой концентрации. Другим важным
преимуществом явилось то, что пластмассы — легкий материал,
и это позволяет строить высокие, не занимающие много места
очистные сооружения. Для создания оптимальной поверхност-
ной площади, вентиляции и пористости пластмассы размалы-
вают.
Основное изменение в конструкцию очистных сооружений
в Англии было внесено в 1973 г., когда был создан вращаю-
щийся биологический реактор. Он представляет собой враща-
ющиеся «соты» из пластиковых полос, попеременно погружае-
мые в сточные воды и поднимаемые на поверхность. При таком
способе увеличивается площадь поверхности, с которой может
контактировать биомасса, и улучшается аэрация (разд. 6.6).
Среда, в которой находится перколяционный фильтр, не
является водной в прямом смысле, поскольку это всего лишь
тонкая водная пленка над слоем биомассы. Определить, какие
именно микроорганизмы присутствуют в среде, довольно труд-
но из-за сложности и гетерогенности биомассы. По-видимому,
основной активной группой бактерий, участвующих в перера-
ботке сточных вод, служат ZoOgloea, хотя определенную роль
играет и ряд других бактерий. В очистных сооружениях отме-
чается также активный рост некоторых видов нитчатых бакте-
рий и грибов. Из водорослей чаще всего присутствуют сине-зе-
леные (Cyanophyceae) и Chlorophyceae. Встречаются и много-
численные Metazoa, в том числе земляные черви, насекомые и
ракообразные. Мухи и черви очень важны для регуляции раз-
вития пленок.
Активный ил
Переработка отходов с помощью активного ила, осуществляе-
мая сложной смесью микроорганизмов, была предложена в
1914 г. Этот процесс более эффективен, чем фильтрация, и по-
зволяет перерабатывать сточные воды в количестве, в десять
раз превышающем объем реактора. Однако он обладает рядом
недостатков: более высокими эксплуатационными расходами
из-за необходимости перемешивания и аэрации; большими
Окружающая среда и биотехнология 251
трудностями в осуществлении и поддержании процесса; образо-
ванием большого избытка биомассы. Несмотря на все это,
процесс, использующий активный ил, остается наиболее рас-
пространенным методом переработки сточных вод в густонасе-
ленных районах, поскольку требует меньших площадей, чем
эквивалентная фильтрационная система.
Как и в фильтрационные системы, в систему с активным
илом были внесены некоторые изменения. Следующие из них
связаны с аэрацией.
1. Градиентная аэрация, приводящая интенсивность аэрации
в соответствие с потребностью в кислороде, которая на входе
больше, чем на выходе.
2. Ступенчатая аэрация, при которой по всей длине тэнка
сточные воды поступают с интервалами.
3. Констактная стабилизация, при которой повторно исполь-
зуемый ил аэрируется, что способствует более полной утилиза-
ции микроорганизмами любых доступных питательных компо-
нентов. Это приводит к более полной ассимиляции отходов при
возврате в основные рабочие тэнки. В результате объем ила
на стадии аэробного разложения уменьшается, и поэтому
в принципе это аналогично увеличению аэрации.
4. Использование чистого кислорода в закрытых тэнках, ко-
торые поэтому могут работать при более высоких концентраци-
ях биомассы; таким образом уменьшается время пребывания
сточных вод в тэнке и, кроме того, решается проблема «разбу-
хания» (избыточного роста нитчатых бактерий и грибов, пре-
пятствующего оседанию ила).
5. Разработка колонного эрлифтного ферментера компани-
ей ICI в 1974 г. (рис. 6.2). Он более экономичен, чем обыч-
ный, благодаря уменьшению времени пребывания сточных вод
в тэнке и снижению эксплуатационных расходов.
Активный ил — это истинно водная среда. Как и в перко-
ляционных фильтрах, основная группа бактерий, участвующих
в процессе переработки, — это Zoogloea. Считается, что актив-
но растет только небольшая часть флокуляционного ила. По
сравнению с перколяционными фильтрами в активном иле на-
блюдается меньшее экологическое разнообразие. Рост водорос-
лей ограничивается недостатком света, а виды и разнообразие
присутствующих в иле простейших определяются степенью пе-
реработки отходов (рис. 6.3).
Для успешной переработки бытовых и промышленных отхо-
дов необходимо точно знать состав и концентрацию стоков.
Это служит «руководством к действию»: зная качественные и
количественные характеристики среды, можно сразу устано-
вить, какой микробный посевной материал необходим для ини-
циации работы системы. Часто бывает трудно показать, что
Рис. 6.2. Колонный эрлифтный ферментер.
Окружающая среда и биотехнология
253
Рис. 6.3. Микроорганизмы, присутствующие в активном иле. Микрофотографии
получены с помощью сканирующего электронного микроскопа и любезно пре-
доставлены Дж. Моррисом. A. Epistylis (Х1000). Б. Стебельчатые бактерии
(Х16 000). В. Жгутиковые бактерии (Х5000). Г. Vaginocolidae (Х750). Д. Ко-
лония Epistylis sp. (Х350). Е. Vorticella sp. (Х1000).
именно те микроорганизмы, которых выделяют из систем био-
логической переработки отходов, осуществляют окисление при-
сутствующих соединений.
Микробиологическое изучение любой системы, использую-
щей активный ил, включает: 1) идентификацию микроорганиз-
мов и определение их численности; 2) оценку микробиологиче-
ской активности как популяции в целом, так и отдельных ви-
дов; 3) оценку соотношения между (1) и (2), с одной сторо-
ны, и количеством вводимых питательных веществ и продуктов
переработки — с другой.
Микробиологическую активность активных илов можно
оценивать по приросту биомассы или по интенсивности общего
метаболизма; последний включает изменения, происходящие
254
Глава 6
в среде. Измерения могут проводиться и для какой-то отдель-
ной популяции микроорганизмов. Можно показать, что актив-
ность ила связана с определенными бактериями, точно подсчи-
тать их число и определить метаболическую активность. Далее
можно выяснить, в какой мере та или иная специфическая ак-
тивность ила определяется конкретными видами бактерий с из-
вестными свойствами, и установить, какое влияние оказывают
на них неблагоприятные условия, в которых они оказываются
из-за поступления в среду тех или иных питательных веществ
или продуктов метаболизма других микроорганизмов. Для
-сточных вод, поступающих в емкость с активным илом, харак-
терны высокие концентрации органических соединений и, сле-
довательно, наличие больших количеств хемоорганотрофных
видов, например Achromobacter, Flavobacterium, Pseudomonas
и Moraxella, а также многих других бактерий. При высоких
концентрациях неорганических соединений в стоках обнаружива-
ются бактерии Thiobacillus, Nitrosomonas, Nitrobacter и Ferro-
bacillus spp. окисляющие соответственно серу, аммиак и желе-
зо. Эти организмы были выделены из систем для переработки
-отходов и идентифицированы с помощью методов селективных
культур. В ходе этих работ важно установить, играют ли ка-
кие-либо виды главенствующую роль в тех процессах, которые
протекают в активном иле. Этот аспект часто недооценивает-
ся, особенно небиологами. Нередко бывает трудно однозначно
установить роль того или иного микроорганизма. Например, ес-
ли из системы по переработке отходов выделены Tbiobacillus,
окисляющие соединения серы, то это еще не означает, что вся
активность такого рода определяется именно этими микроорга-
низмами: частичное окисление ряда соединений серы осущест-
вляют и виды Pseudomonas.
Взаимосвязи между организмами, участвующими в катабо-
лизме органических и неорганических субстратов, имеют важ-
ное значение для регуляции процессов, происходящих в актив-
ном иле. Промежуточные продукты метаболизма у одного вида
бактерий способы оказывать влияние на процессы деградации
у другого. Например, известно, что фенол подавляет активность
•организмов, окисляющих аммиак: он может ингибировать этот
-окислительный процесс даже при столь малых концентрациях,
как 3—4 м. д.
Промежуточные продукты расщепления бензойной кислоты
до катехола, сукцината и ацетата ингибируют образование
-ферментов, участвующих в начальных этапах расщепления. Ка-
техол и сукцинат подавляют синтез ферментов, разрушающих
-бензоилформиат и бензальдегид, по механизму обратной связи,
а ацетат действует как катаболитный репрессор: наличие про-
стого органического соединения подавляет расщепление более
Окружающая среда и биотехнология 255
сложных молекул до тех пор, пока это более простое соедине-
ние не будет использовано. Когда ингибирование снимается,,
синтезируются новые ферменты, ответственные за расщепление
более сложных ароматических структур. На практике при на-
личии в отходах гомологичных рядов каких-либо соединений
необходимо образование ферментов, способных справиться с
расщеплением самой сложной молекулы данного ряда. Полное
расщепление таких соединений должно происходить в течение
определенного минимального времени удержания (нахожде-
ния отходов в реакторе) в процессе переработки. Следователь-
но, можно предсказать, какая обработка потребуется для
окисления фенольных соединений; например, чем сложнее бо-
ковая цепь молекулы, тем больше времени необходимо для:
ферментативного разрушения этого вещества.
Эффективность данного процесса можно повысить, изучив;
механизмы регуляции метаболизма в микрофлоре систем с ак-
тивным илом. Регуляция биодеградации — это сложная зада-
ча. Однако, зная биохимию соответствующих процессов, мы„
по-видимому, сможем вмешиваться и в их регуляцию. Напри-
мер, добавление к илу промежуточных продуктов цикла три-
карбоновых кислот в низких концентрациях (2—5 мг/л), глю-
козы, аминокислот и витаминов (в частности, аланина и нико-
тиновой кислоты) приводит к ускорению окисления ряда
соединений. Введение этих промежуточных продуктов в состав-
биомассы увеличивает энергетические потребности системы,,
стимулирует синтез АТР за счет усиленного окисления неорга-
нических веществ типа серы или аммиака. Понимание биохи-
мии подобных процессов, видимо, даст возможность вмеши-
ваться в процессы регуляции метаболизма.
Задача микробиолога-биотехнолога при разработке методов;
очистки сточных вод состоит в более полном изучении и учете-
взаимосвязи между активностью микроорганизмов, образовани-
ем хлопьев ила и производительностью установки по перера-
ботке отходов. В этом смысле превращения в системе активно-
го ила следует рассматривать в основном как окислительные-
процессы во влажной среде, сопровождающиеся увеличением
объема ила, которое можно расценивать как вредное или по-
лезное (последнее — когда ил используется повторно). Совер-
шенно очевидно, что биологический способ переработки приго-
ден для множества различных органических и неорганических
соединений и устраняет их вредное воздействие на окружаю-
щую среду.
Акт о защите среды от загрязнений от 1974 г. по мере прет-
ворения его в жизнь будет оказывать все возрастающее влия-
ние на технологию очистки сточных вод. Термин «биодеграда-
ция» используется сейчас очень широко, но имеет множество:
256
Глава 6
толкований. Иногда под биодеградацией понимают полную ми-
нерализацию какого-либо соединения микроорганизмами с об-
разованием углекислого газа, сульфата, нитрата и воды; это
одна крайность. Другая крайность состоит в том, что данный
термин используют применительно к незначительным измене-
ниям соединений, приводящим к утрате некоторых характер-
ных их свойств. Стандартные методы оценки деградации по-
зволяют определить термин «биодеградация» следующим обра-
зом: 1) первичная деградация, при которой характерные свой-
ства исходного соединения утрачиваются и перестают выяв-
ляться специфическими химическими тестами; 2) допустимая
для окружающей среды биодеградация, при которой происхо-
дит минимальное изменение исходного соединения, необходимое
для утраты его свойств (оба этих определения основаны на
произвольных критериях и поэтому неточны); 3) окончательная
биодеградация, включающая полное превращение исходного
соединения в неорганические конечные продукты и связанная
с нормальными процессами метаболизма микробов.
В ходе изучения деградации широкого круга органических
веществ были выделены микроорганизмы, способные к разру-
шению весьма необычных соединений (рис. 6.4 и 6.5).
Принцип «псевдоожиженного слоя»
Данная технология, введенная в практику в 1980 г., во многих
отношениях представляет собой сочетание систем перколяци-
онных фильтров и активного ила. Она весьма экономична бла-
годаря использованию высоких концентраций микроорганиз-
мов и отсутствию необходимости в осаждении конечных про-
дуктов. Существуют два основных типа установок.
1. Уловитель Саймона Хартли. Он был разработан на ос-
новании исследований, проведенных в научно-техническом ин-
ституте Манчестерского университета. Биомассу наращивают
в пустотах внутри прокладок из пористого полиэфира, которые
удерживаются внутри реактора с помощью сеток. Прокладки
периодически удаляют из реактора, густую биомассу (до 15 кг
на каждый кубометр обводненного носителя) отжимают и пу-
етые прокладки возвращают в реактор.
2. Оксигенатор Дорра — Оливера. Здесь в качестве подлож-
ки используется песок; его периодически выпускают из реакто-
ра, очищают и используют снова.
Возможное нежелательное последствие интенсификации
аэробной обработки — это излишнее образование ила. Стои-
мость его удаления может составить до 50% расходов на пере-
работку сточных вод. Альтернативные решения состоят или
Окружающая среда и биотехнология
257
Лекарственные вещества,
А продуцируемые живыми системами
Хинин
Никотин
£ Окислитель и ускоритель вулканизации
В Противокоррозионные
присадки
2Меркаптобензо тиазол
Морфолин
Рис. 6.4. Основные группы соединений, которые обнаруживаются в отходах
производства лекарственных препаратов, синтетических гербицидов и других
нефтехимических продуктов и подвержены биодеградации.
Тиофен-2-карбоноеая кислота
Тиофен-2-карбоксил-СоА
НООС HSC — СОСоА
Сложный эфир СоА-2'меркапто-
пентендиоевой кислоты
5 -Гидрокси тиофен-2-карбоксил-СоА
H2S
НООС О=С —СОСоА
а 'Оксоглутарил-СоА
Рис. 6.5. Промежуточный метаболизм аналога тиофена в одном из видов Fla-
vobacterium.
в последующем использовании этого ила (разд. 6.5), или
в разобщении анаболической и катаболической активности для
неполного превращения субстрата в биомассу (этого можно
достичь, создав условия постоянной нехватки микроэлементов
или делая перерывы в подпитке).
17—1344
258
Глава 6
6.2.2. Анаэробное разложение (см. также гл. 2 и 9)
Все возрастающая стоимость переработки отходов с помо-
щью аэробного разложения и энергетический кризис, с одной
стороны, и новые достижения микробиологии и технологии —
с другой, возродили интерес к анаэробной переработке. Самая
распространенная технология анаэробной переработки — разло-
жение ила сточных вод. Эта хорошо разработанная технология
с успехом используется с 1901 г. Однако здесь существует ряд.
проблем, обусловленных малой скоростью роста облигатных
анаэробных метанобразующих бактерий, которые использу-
ются в данной системе. К ним относятся также чувствитель-
ность к различным воздействиям и неприспособленность к из-
менениям нагрузки. Конверсия субстрата также происходит
довольно медленно и поэтому обходится дорого. Некоторые-
проблемы связаны с неудачными инженерными решениями. Тем
не менее этот подход представляется перспективным с точки
зрения биотехнологии; например, можно добавить к отходам
ферменты для повышения эффективности процесса или попы-
таться усилить контроль за переработкой путем изменения тех
или иных биологических параметров (разд. 6.3).
Анаэробная ферментация отходов или растительных куль-
тур, специально выращиваемых для получения энергии, очень-
перспективна для экономичного получения газообразного топ-
лива при умеренных температурах (30—35°C). Эта новая от-
расль биотехнологии была развита микробиологами в сотруд-
ничестве с инженерами-химиками и механиками, работниками
сельского хозяйства и экономистами.
При выращивании сообщества различных бактерий на смеси
органических соединений происходят сложные биохимические-
реакции (рис. 6.6) Метанобразующие бактерии способны к син-
тезу энергоносителя непосредственно из водорода и углекисло-
го газа. Микроорганизмы, расщепляющие целлюлозу, синтези-
руют жирные кислоты, которые могут подвергаться восстанови-
тельному расщеплению до метана и углекислого газа; некото-
рые бактерии способны даже образовывать молекулярный
водород. Описано сложное, взаимозависимое микробное сообще-
ство, в котором можно выделить три группы бактерий: бакте-
рии, осуществляющие гидролиз и брожение, бактерии, образую-
щие водород и уксусную кислоту, а также водородотрофные,:
метанобразующие бактерии. Метанобразующие бактерии рас-
тут медленно и очень чувствительны к резким изменениям за-
грузки реактора и накоплению водорода. Можно надеяться,,
что усовершенствование конструкции реактора и контроль за
процессом помогут уменьшить колебания загрузки реактора и
позволят контролировать ее, определяя содержание водорода а
Окружающая среда и биотехнология
259
Липиды Лигнины Белки
МЕТАН + УГЛЕКИСЛЫЙ ГАЗ
Рис. 6.6. Биохимическое расщепление отдельных соединений до метана и угле-
кислого газа при анаэробном разложении отходов.
промежуточных продуктов типа пропионовой и масляной кис-
лот. Проблемы перегрузки, особенно существенные в случае
промышленных стоков, можно обойти, увеличивая скорости
оборота и применяя в качестве буферных систем сточные воды
химических предприятий и бытовые сточные воды. Для увели-
чения метаногенной активности бактерий можно использовать
обычные методы отбора или методы генетической инженерии.
Оценить возможность использования данного процесса при пе-
реработке смешанных отходов, а также охарактеризовать по-
требности в питательных веществах и усовершенствовать на-
чальный этап процесса за счет уменьшения количества необ-
ходимого микробного посевного материала поможет
дальнейшее изучение физиологии и экологии участвующих в
процессе микроорганизмов.
Для получения энергии и полезных побочных продуктов
можно использовать самые разнообразные отходы и сырье.
87'
260
Глава 6
К культурам, выращиваемым специально в целях конверсии,
энергии в газообразное топливо, относится кассава; конечными
продуктами служат метанол или этанол. Некоторые страны,,
например Бразилия, Австралия и Новая Зеландия, намерены
к 2000-му году использовать подобные вещества, получаемые
биологическим путем, в качестве основного источника топлива.
Сходные проекты обсуждаются и в некоторых европейских
странах, например в Финляндии, Швеции и Ирландии.
В Англии работа по биоконверсии энергии проводится в;
рамках Программы по использованию солнечной энергии (ми-
нистерства энергетики); за счет этой программы финансиру-
ются и проекты ЕЭС по получению энергии биологическими
способами. В США используется множество подходов; так, од-
но очистное сооружение за счет биологической конверсии бы-
тового мусора позволяет получить газ в количестве, достаточ-
ном для обеспечения им 12 тыс. домов. Основные микробиоло-
гические и технологические проблемы этой технологии и пер-
спективы ее применения в развивающихся странах были рас-
смотрены на Первой международной конференции по анаэроб-
ному разложению, состоявшейся в Кардиффе в 1979 г. Анаэроб-
ные ферментеры могут применяться также в целях получения
промежуточных продуктов для химической промышленности
(например, уксусной, молочной и акриловой кислот в качестве
химического сырья, идущего на переработку; гл. 4).
Однако широкое использование анаэробных реакторов в
целях получения газообразного топлива сдерживается рядом
причин. Традиционно в конструкцию реакторов входили тэнки
с мешалками, рассчитанные на длительное пребывание перера-
батываемого материала. В целях сокращения этого времени
были созданы реакторы, в которых переработанные отходы от-
деляются от биомассы, используемой повторно. Чтобы процесс
был экономически выгодным, должны быть разработаны не-
дорогие конструкции, которые не засоряются и включают про-
стые в эксплуатации устройства для отвода тепла. Основные
усилия в области анаэробной ферментации должны быть на-
правлены на изучение этапов, лимитирующих скорость процес-
са. На первом из них происходит гидролиз целлюлозы и крах-
мала с образованием растворимых органических кислот и спир-
та. Вторым лимитирующим этапом может быть образование
метана из этих жирных кислот с короткой цепью. Моделирова-
ние процесса разложения осложняется тем, что трудно ска-
зать, какие микроорганизмы доминируют на том или ином
этапе, и установить, какие именно этапы лимитируют скорость
процесса. Возможно, в условиях реактора лимитирующими
окажутся другие стадии. Крайне важно определить количество
образуемых микроорганизмами газов, особенно водорода, уг-
Окружающая среда и биотехнология
261
Вход жидкости
Выход жидкости
Выход теплоносителя
Биогаз
В
Углекислый газ
Радиальная опорная балка
Емкость из полимерной пленки
Изолирующая панель
Стальная сетка
Арматура
Бетонный кольцевой
Фундамент
Изолирующая пленка
Теплообменник
Рис. 6.7. Три типа установок, использующихся при очистке сточных вод пище-
вой промышленности. А. Анаэробный фильтр. 5. Упрощенная схема установки,
в которой используется перемешивание с помощью винтового насоса и вытяж-
ной трубы. Образование пены контролируется диспергированием содержимого
реактора над поверхностью. В. Высокоскоростной реактор Коулзэрда.
лекислого газа и сероводорода, который ингибирует активность
метанобразующих бактерий. Недавно было проведено исследо-
вание анаэробного превращения ряда субстратов культурами
известных микроорганизмов. Это очень сложный процесс; из
262
Глава 6
природных систем было выделено много новых участвующих
в нем типов бактерий.
Промышленное применение систем анаэробного разложения
неуклонно возрастает; они используются при переработке от-
ходов животноводческих ферм и промышленных, в том числе
пищевых, отходов, а также для переработки культур, специаль-
но выращиваемых для получения энергии. На рис. 6.7, 6.8 и 6.9
Анаэробный реактор с перегородками (дефлекторами)
Реактор со слоем ила и потоком, неправленым вверх
Рис, 6.8. Типы реакторов для переработки отходов животноводческих ферм
(Л) и различных стоков (5).
схематически представлены некоторые из имеющихся в продаже
установок. Конструкция реакторов была существенно усовер-
шенствована, что увеличило их эффективность на 300 %. Многие
новые модели еще не вышли из стен лабораторий или находят-
ся на стадии производственных испытаний, однако некоторые
Окружающая среда и биотехнология
263
полномасштабные системы уже работают и имеются в про-
даже.
Получение энергии из отходов представляет несомненный
интерес для развивающихся стран, поскольку эту энергию мож-
но извлекать и из природных продуктов. Сотрудничество меж-
ду развитыми и развивающимися странами постоянно возрас-
тает. В развивающихся странах созданы учерждения для
А
Б
Предохранительный
Выход газа перегрузочный Крышка
патрубок люка Рабочий уровень
------13,5м ----------
Реактор поршневого типа
Рис. 6.9. Анаэробное разложение ила, образовавшегося в сточных водах (А),
и отходов животноводческих ферм (Б).
практического использования технологий, разработанных глав-
ным образом в Европе и Америке. Некоторые развивающиеся
страны ведут самостоятельные исследования в этой сфере и
сегодня лидируют в области фундаментальных разработок по
возобновляемым источникам энергии.
264
Глава 6
6.3. Биологический контроль
за системами микробиологической переработки отходов
Основным условием применения биологической переработки
сточных вод является постоянный контроль за возможным ток-
сическим действием на установку со стороны поступающих сто-
ков, с тем чтобы предотвратить серьезные повреждения систе-
мы или даже выход ее из строя. При эксплуатации установок
по переработке отходов и промышленных сточных вод важно
следить за тем, чтобы не возникали перегрузки. Обычные ме-
тоды проведения анализов (например, измерение потребности
в кислороде или определение pH) часто и недостаточно быст-
ры, и малочувствительны. Однако в метаболической активности
микроорганизмов существуют ключевые моменты, анализ кото-
рых дает возможность улавливать малейшие изменения в их
состоянии. Например, благодаря внутриклеточному контролю
метаболизма содержание АТР в популяции микроорганизмов
сохраняется на относительно постоянном уровне, около 2 мкг
на 1 мг сухой массы клеток. Изменения доступности субстрата
или введение токсических веществ быстро сказываются на кон-
центрации АТР внутри клеток. Мгновенная гибель клеток ведет
к полной потере АТР за счет автолиза. Время оборота для
АТР обычно не превышает 1 с.
Следовательно, о состоянии популяции (наступление стрес-
са или периода покоя) можно судить по изменению уровня
внутриклеточного АТР. Измерение концентрации АТР в актив-
ном иле нормально функционирующей системы позволяет опре-
делить «активную биомассу» данной системы. В процессе про-
изводства клеткой энергии участвует кислород, и при его недо-
статке отношение между его поступлением и потребностью
в нем отразится на уровне АТР (рис. 6.10).
По технологии, основанной на применении активного ила,
большая его часть используется повторно, поступая из послед-
него седиментационного тэнка в аэрационный тэнк, а некоторая
его часть поступает в реактор аэробного или анаэробного раз-
ложения. Доля повторно используемого ила и скорость его кру-
гооборота определяют время контакта между поступающими
сточными водами и микроорганизмами ила, а следовательно,
и скорость очистки сточных вод. Эти факторы влияют на воз-
раст ила, который в свою очередь определяет его способность
к оседанию (рис. 6.11). Производительность активного ила
в значительной степени зависит от скорости нарастания новой
биомассы в аэрационном тэнке, которая в свою очередь зави-
сит от изменений скорости поступления сточных вод и их со-
става. Поэтому контроль за процессом с целью его нормализа-
ции и получения на выходе конечного продукта, отвечающего
Окружающая среда и биотехнология
265
определенным критериям, сопряжен с трудностями. Такой кон-
троль можно проводить, определяя уровень АТР, регулируемый
по принципу обратной связи. Действительно, измеряя концен-
трацию АТР, удалось заметно улучшить контроль за переработ-
кой сточных вод с применением активного ила. Содержание
АТР можно определять вручную; однако для постоянного кон-
троля за процессом требуются автоматические измерительные
устройства. Пробы жидкости, обработка которой закончена,
отбирают через определенные промежутки времени на протя-
жении суток. Анализ уровня АТР в этих пробах позволяет най-
ти отношение количества суспендированных в жидкой смеси
твердых частиц к содержанию АТР, что в свою очередь пока-
зывает, какое количество активного ила, содержащего АТР,
нужно использовать повторно и какова подходящая скорость.
Рис. 6.10. Влияние введения в отходы токсичного вещества на активную био-
массу (по результатам определения АТР). Показаны также изменения актив-
ности возвращаемого активного ила (/) и содержания суспендированных твер-
дых частиц в смешанной жидкости (II).
сброса. Содержание АТР определяется в течение 10 мин после
отбора пробы, следовательно, контрольные измерения можно
проводить каждые 15 мин. Использование микропроцессоров
позволяет автоматически поддерживать скорость сброса с по-
мощью клапанов или насосов, направляющих избыток активно-
го ила в тэнк для разложения, а также менять скорость сброса
ила в течение суток в ответ на изменение внешних условий.
При таком методе контроля получают гораздо более освет-
ленный конечный продукт с меньшим содержанием суспенди-
266
Глава 6
Рис. 6.11. Изменение количества активной биомассы (///), объема ила (//) и пе-
ремешиваемого удельного объема (/) для отходов, перерабатываемых в аэра-
ционном тэнке, в зависимости от возраста ила.
рованных твердых частиц и меньшей величиной ВПК. Преиму-
щества данного метода огромны. В сочетании с системой кон-
троля за поступлением кислорода он позволяет существенно
повысить экономичность процесса переработки.
Окружающая среда и биотехнология
267
6,4. Контроль за патогенностью.
Одно из основных достоинств процесса микробного анаэробно-
го разложения состоит в элиминации с его помощью патоген-
ных микроорганизмов, в особенности агентов, вызывающих
порчу пищи (главные образом Salmonella). Для анализа вы-
живаемости этих организмов, а также микробов группы Е. coll
в системах лабораторного масштаба использовали чувстви-
тельный метод наиболее вероятных чисел (НВЧ). К числу ко-,
нечных продуктов неметаногенного разложения относятся на-
сыщенные жирные кислоты, и есть данные, что к молекуле
жирной кислоты — перед деградацией за счет р-окисления —
присоединяется водород. Образующаяся октановая кислота
особенно эффективно убивает патогенные микроорганизмы. На
рис. 6.12 представлены данные о выживаемости Salmonella в
Время, ч
Рис. 6.12. Кривые выживаемости Salmonella в отстое, образующемся в реакторе
при анаэробном разложении.
супернатанте, образующемся при анаэробном разложении.
Видно, что жирные кислоты подавляют рост этих бактерий и
приводят к их гибели.
Процессы анаэробного разложения оказывают ингибирую-
щее действие и на микроорганизмы, патогенные для растений.
268
Глава 6
Недавние исследования показали, что численность вида Fusa-
rium, Corynebacterium и Globodera в ходе разложения умень-
шается и падает почти до нуля через 10 сут после начала очи-
стки сточных вод (рис. 6.13 — 6.15). Это позволяет надеяться
на возможность использования процессов анаэробного разло-
жения для переработки растительного сырья, пораженного воз-
будителями различных заболеваний {Fusarium oxysporum —
дикариотическим грибом — возбудителем вилта и корневой гни-
ли томатов, гвоздики и риса; Corynebacterium michiganese —
Рис. 6.13. Среднее число нематод Globodera pallida, вышедших из цист, на
100 цист в зависимости от времени воздействия на них содержимого анаэроб-
ного реактора при 35 °C. Контрольная величина (7 сут в воде) —8739±444.
возбудителем сосудистого вилта, рака и листовой пятнистости
томатов, картофеля и табака; Globodera pallida — широко из-
вестной корневой нематодой картофеля, разрушающей корни
растения-хозяина).
В сточных водах, образующихся в других условиях, напри-
мер в регионах с тропическим климатом, могут присутствовать
и другие патогенные микроорганизмы, например Entamoeba
hystolytica, вызывающая амебную дизентерию. Это необходимо
учитывать при разработке способов очистки отходов. Все био-
технологические подходы, столь полезные с точки зрения здра-
воохранения, следует развивать в самых широких масштабах.
Окружающая среда и биотехнология
269
Рис. 6.14. Численность популяции Fusarium oxysporum до (ф) и после (О)
введения ее в анаэробный реактор.
Время, сут
Рис. 6.15. Изменение численности популяции Corynebacterium michiganese
при 35 °C в течение 5 сут нахождения в анаэробном реакторе. Приведены ре-
зультаты трех экспериментов.
270
Глава 6
6.5. Извлечение полезных веществ
Одна из главных задач технологии, связанной с окружающей
средой, — это сохранение природных ресурсов путем повторно-
го использования полезных веществ, содержащихся в отходах.
Некоторые разработки в этой сфере получили финансовую под-
держку со стороны правительств и это принесло свои плоды,,
но все же пока выход конечных продуктов и стоимость повтор-
ного использования биомассы в широких масштабах таковы,
что эта технология оказывается экономически невыгодной. Тем-
не менее она может найти применение при получении таких,
ценных продуктов, как масла, металлы, витамины и пептиды.
Получение полезных материалов из отходов имеет два ас-
пекта: 1) извлечение и/или концентрирование полезных ве-
ществ из отходов; 2) превращение отходов в полезные мате-
риалы.
6.5.1. Вода
Воду можно рассматривать как возобновляемый ресурс. Одна-
ко, сравнивая стоимость необходимого для очистки оборудова-
ния со стоимостью водопроводной воды, очистку загрязненной
органическими веществами воды обычно считают неэкономич-
ной. Повторное использование промышленных сточных вод
экономично только в тяжелой промышленности (энергетика,
сталелитейное производство и угледобывающая промышлен-
ность), где можно применять не такую чистую воду, как питье-
вая, и поэтому свести к минимуму обработку сточных вод. Ос-
новные трудности здесь связаны с наличием соединений, не
поддающихся переработке. Возможно, эту проблему удастся
решить, используя микроорганизмы, которые приобрели спо-
собность разрушать такие соединения (разд. 6.7).
6.5.2. Удобрения
Потребность в более дешевых высококачественных белках жи-
вотного происхождения непрерывно возрастает, а число работ-
ников сельского хозяйства, призванных удовлетворять эту рас-
тущую потребность, все время уменьшается. Для разрешения
этого противоречия нужно было бы применять более интенсив-
ные методы землепользования, и тогда мы будем получать все
больше концентрированных отходов, которые в принципе мож-
но применять как удобрения. Однако за последние 100 лет
масштабы использования отходов животноводства в качестве
удобрений уменьшились; на смену им пришли фосфорные и
азотные удобрения, при получении которых используется ис-
Окружающая среда и биотехнология
271
копаемое топливо. В Англии значительную часть сельскохо-
зяйственной продукции (до 40% от общей стоимости) получа-
ют именно за счет применения химических удобрений. Однако
цены на такие удобрения все время растут, и становится эконо-
мически выгодным использование отходов животноводства в
качестве органических удобрений.
Табл. 6.1 иллюстрирует возможность получения удобрений
на основе навоза сельскохозяйственных животных. Результаты
анализа каждого типа удобрений приведены в табл. 6.2. Цен-
Таблица 6.1. Получение удобрений на основе навоза
сельскохозяйственных животных
Параметры Птицы Свиньи Молочные коровы
Кг/сут 0,1 4,5 45
% от живого веса 5—6 8—9 9—11
Сухое вещество, % 20—30 15-20 10-11
Средний сухой вес, кг/сут 0,03 0,7 4,5
"Таблица 6.2. Содержание питательных веществ в навозе
Животные Сухое ве- щество, %• Содержание различных компонент (кг/т свежего навоза)
N р К Mg
Крупный рогатый скот •Свиньи Птицы 4—23 5—25 23—68 2,4—6,5 1-6—6,8 9,6—23 0,4—1,8 0,6—2,1 2,4-12 2,0—5,8 1,7—3,6 3,8—11,6 0,2-0,6 0,3—0,7 1.2-2,2
ность отходов как удобрений и пути их использования опреде-
ляются составом. Кроме того, перед внесением навоза в почву
из него путем анаэробного разложения можно получать энер-
гию (разд. 6.2.2). Определить изменение стоимости удобрения
в результате такого разложения трудно; тем не менее ясно, что
удобрения на основе переработанного навоза достаточно де-
шевы.
6.5.3. Корма для животных
В Англии в результате человеческой деятельности образуется
25-10® кг отходов в год. Если учесть, что при интенсивном жи-
вотноводстве образуется еще 180-10® кг отходов, то становит-
272
Глава 6
Рис. 6.16. Получение белка из активного ила, образующегося в сточных водах.
ся ясно, что при переработке всех этих отходов мы можем по-
лучить многие тонны активного ила. В процессе переработки
отходов при участии микроорганизмов образуется много мик-
робного белка, который можно повторно использовать как
корм для скота, поскольку 30—40% сухой массы выросших
клеток — это неочищенный белок. На рис. 6.16 описан метод
экстракции белка из активного ила, а в табл. 6.3 приведен со-
став белка одноклеточных организмов (БОО) из того же ис-
точника. Тяжелые металлы, обнаруженные в отстое сточных
вод (например, медь из отходов свиноводства, где ее присут-
ствие обусловлено применением концентратов меди для корм-
Окружающая среда и биотехнология
273
Таблица 6.3. Состав белка одноклеточных
организмов, обитающих в активном иле
Компонент
Вода
Общий белок (NX6,25)
Истинный белок
Жиры
Зола
Кислотонерастворимая зола
Углеводы (в пересчете на глюко-
зу)
Волокнистые вещества
Валовое содержание энергии
(ккал/г)
Вес. %
7,85
37,80
31,25
1,73
25,16
12,33
9,80
10,30
16,20
ления животных), необходимо удалять. Идеальным результа-
том такого повышения качества ила сточных вод путем экст-
ракции белка должен быть безвредный, чистый, экономичный
корм для скота. Кроме того, в иле образуются и другие цен-
ные биологические соединения, например аминокислоты и ви-
тамины. В табл. 6.4 представлена концентрация аминокислот
Таблица 6.4. Концентрация аминокислот
в белковом продукте из активного ила
Аминокислота г/100 г пробы
Аспарагиновая кислота Серин Глутаминовая кислота Пролин Глицин Аланин Цистеин Тирозин Треонин Валин Метионин Изолейцин Фенилаланин Гистидин Лизин Аргинин Триптофан’’ 2,86 1,4 3,62 1,28 2,26 2,95 0,25 0,72 1,86 2,19 0,25 1,42 2,07 0,69 1,68 1,59
') Разрушается при кислотном гидролизе.
18—1344
274
Глава 6
в белковом продукте из активного ила, применяемом в качест-
ве корма для рыб. Однако бытовые и сельскохозяйственные от-
ходы, по-видимому, непригодны для промышленного получения
БОО. Это связано с их низкой пищевой ценностью и с необ-
ходимостью получения продукта, свободного от токсичных ве-
ществ и патогенных микроорганизмов. Существуют и экономи-
ческие проблемы, определяемые расходами на ферментацию и
оборудование, необходимое для высушивания. Незаряженные
отходы производства пищевых продуктов и напитков, а также
бродильной промышленности можно использовать в качестве
добавок к корму. Однако экономическая целесообразность их
применения еще не доказана.
Министерство сельского хозяйства, рыболовства и пищевой
промышленности опубликовало инструкцию «О порядке пере-
работки белка» от 1981 г., касающуюся использования белко-
вых продуктов, получаемых из ила, в качестве корма. В ней
правительство ограничилось предложениями по ужесточению
•стандартов, указанных в Актах о болезнях животных от 1950
я 1975 гг. В них особое внимание уделено контролю за таки-
ми возбудителями заболеваний, как Salmonella.
6.6. Биологическая переработка промышленных отходов
Промышленные отходы можно в первом приближении разде-
лить на две категории: 1) отходы производств, основанных на
использовании биологических процессов (производство пище-
вых продуктов, напитков, ферментация); 2) отходы химической
промышленности. В первом случае отходы имеют различный
состав и обычно перерабатываются путем биологического
окисления, как это делалось традиционно в случае бытового
мусора. Однако такой способ экономически невыгоден, и в на-
стоящее время широко обсуждается вопрос о возможности
уменьшения объема разбавленных сточных вод либо их непо-
средственного использования — трансформации (для получения
биомассы или других ценных продуктов) или же путем извле-
чения из них ценных соединений.
В многочисленных и разнообразных отраслях химической
промышленности образуется большое количество отходов, при-
чем многие из них с трудом поддаются разрушению и длитель-
ное время присутствуют в среде. Поэтому часто перед обыч-
ной биологической переработкой отходов бывает необходимо
провести их предварительную химическую или физическую об-
работку. Использование специфических микроорганизмов для
расщепления ксенобиотиков при переработке отходов еще не
нашло широкого применения в промышленности, и тем не ме-
нее подобный подход представляется весьма перспективным.
Окружающая среда и биотехнология 27!>
Это может быть: 1) деградация отдельных видов отходов in
situ с помощью специализированных культур микроорганизмов-
или их сообществ; 2) введение специально подобранных куль-
тур в обычные системы переработки отходов; 3) ликвидация и
Обезвреживание разливов нефти; 4) извлечение металлов^
(гл. 5); 5) биологическая очистка газов от пахучих и вредных
соединений (меркаптанов, сероводорода, цианида, хлорзаме-
щенных углеводородов и т. д.); 6) получение биомассы из от-
ходов; 7) превращение отходов в метан (гл. 2).
В результате широкого применения человеком продукции
химической промышленности в окружающую среду попадают
различные типы ксенобиотиков: пластмассы (пластификаторы),
взрывоопасные вещества, добавки, полимеры, красители, по-
верхностно-активные вещества, пестициды и органические со-
единения — производные нефти. Что касается бытового мусо-
ра; то для его переработки созданы широко применяемые си-
стемы, использующие активный ил и оросительные фильтры..
Сточные же воды химической промышленности, как правило,,
не соответствуют возможностям подобных систем. Интенсив-
ность переноса кислорода в ходе процессов, обычно протекаю-
щих в таких системах, бывает недостаточна для поддержания:
максимальной скорости окисления при участии микрофлоры.
Эти процессы чувствительны также к колебаниям в загрузке-
реактора, особенно если токсичные вещества и ингибиторы по-
ступают в систему в высоких и непостоянных концентрациях.
Проблему недостатка кислорода, возникающую при перера-
ботке отходов химической промышленности в обычно исполь-
зуемых системах на основе активного ила, пытались решить
несколькими способами. В двух случаях (распределитель с
пробулькиванием и система «Аноке») для увеличения скоро-
сти переноса газа использовали чистый кислород. В одной и»
новых систем переработки отходов — колонном эрлифтном фер-
ментере, разработанном фирмой ICI, — пошли по пути уве-
личения количества растворенного кислорода (рис. 6.2).
В центральной части колонны имеется не доходящая до дна-
вертикальная секция, в которую сверху поступают отходы и;
повторно используемый активный ил; туда же вводится воз-
дух. Когда смесь выходит из ферментера вверх по наружной
секции колонны, давление в системе падает, что вызывает про-
булькивание пузырьков воздуха. Благодаря высокому содер-
жанию растворенного кислорода и турбулентности биомасса
поддерживается в высокоактивном состоянии и становится бо-
лее устойчивой по отношению к перегрузкам, а также к умень-
шению аэрации и времени нахождения отходов в ферментере,
особенно в случаях высоко концентрированных отходов.
- Такие процессы с повышенной аэрацией устойчивы к рез-
18
276
Глава 6
ким перегрузкам отходами, не оказывающими токсического или
ингибирующего действия. В случае же токсичных отходов
более пригодными оказываются системы, в которых использу-
ются микроорганизмы, растущие в пленках. Такие популяции
микробов не вымываются из системы, даже если на их рост
и метаболизм оказывают неблагоприятное воздействие посту-
пающие сточные воды. Кроме того, внутри пленки из-за огра-
ничения диффузии создаются градиенты концентрации. Это
приводит к понижению концентраций токсичных продуктов
внутри пленки, а следовательно, к повышению скорости их
усвоения и окисления. Пленка создает также экологическую ни-
шу для организмов, рост которых в присутствии высоких
концентраций отходов при перегрузках существенно за-
медляется. Самая простая форма пленочной системы — это
перколяционный фильтр (разд. 6.2.1), однако подобного рода
пленки разрушаются, если они становятся очень тонкими, при
уменьшении концентрации субстрата на поверхности подлож-
ки. В таком случае клетки погибают и пленка отпадает, за-
соряя фильтры внутри системы переработки отходов. При
слишком высоких концентрациях субстрата происходит быст-
рый рост микроорганизмов, что приводит к образованию тол-
стой пленки и к ее периодическому отслоению. Интенсивность
подобных процессов можно снизить, разбавив поступающий
раствор с питательными веществами осветленными сточными
водами. Разработка новых методов сохранения толщины плен-
ки представляет безусловный интерес. Так, при помощи мед-
ленного вращения диска из полистирола внутри протекающих
сточных вод толщина пленки поддерживается постоянной за
счет гидродинамических сил и аэрации при выходе пленки из
воды. Такая эффективная и простая система была предложена
для очистки стоков с низкой величиной ВПК. Еще один эффек-
тивный метод переработки токсичных отходов in situ может
быть основан на использовании реакторов с ожиженной под-
ложкой, где микроорганизмы растут на поверхности неболь-
ших инертных частиц (песок, стекло, антрацит), через слой ко-
торых пропускают с контролируемой скоростью сточные воды
и воздух.
Отходы, не содержащие азота или фосфора, не способны
поддерживать рост микроорганизмов. В подобных случаях для
окисления токсичных соединений до двуокиси углерода можно
использовать покоящиеся клетки при условии, что активность
их гидролитических и окислительных ферментов не подавляет-
ся. Поскольку среда при переработке отходов в колонных реак-
торах периодически меняется, микроорганизмы оказываются в
условиях голодания и в это время их рост прекращается. При
поступлении источника углерода на короткое время включает-
Окружающая среда и биотехнология
217
ся несопряженный метаболизм, когда организмы дышат, но не
растут. Это дает то преимущество, что уменьшается общий вы-
ход биомассы (ила).
Рассмотрим методы биологической переработки промышлен-
ных отходов на примерах молочной, бумажной промышленности
и производства красителей.
€.6.1. Отходы молочной промышленности;
сыворотка (см. также гл. 3)
Сыворотка является побочным продуктом сыроварения. Ее со-
став зависит от типа используемого молока и вырабатываемо-
го сыра. В высушенном или концентрированном виде сыворот-
ка применялась в качестве корма для животных; однако ее не-
достатком является то, что она несбалансирована с точки зре-
ния содержания питательных веществ: в ней слишком высока
концентрация минеральных веществ и лактозы. Разработаны
способы извлечения из сыворотки белков путем ультрафильтра-
ции, осаждения или выделения с помощью ионного обмена. Из
таких белков можно получать белковые гидролизаты, исполь-
зуя для этого ферментеры. После извлечения белков получают
большие объемы фильтратов с высокими концентрациями лак-
тозы (35—50 г/л), минеральных веществ, витаминов ,и молочной
кислоты, и встает проблема дальнейшего их использования.
Если превратить лактозу в молочную кислоту при участии мо-
лочнокислых бактерий, то мы получим источник углерода, ко-
торый может сбраживаться дрожжами (например, смешанными
культурами Lactobacillus bulgaricus и Candida krusei). Возмож-
но и прямое сбраживание лактозы дрожжами Kluyveromyces
jragilis или Candida intermedia. После подобного сбраживания
не обязательно отделять микроорганизмы от среды, объем кото-
рой можно уменьшить и получить обогащенную белком сыво-
ротку.
Из сыворотки получают не толко белковые продукты, но
и— путем ферментации — сырье для химической промышлен-
ности (например, этанол). Путем химического гидролиза лак-
тозы с последующим удалением глюкозы из раствора с по-
мощью ферментации можно получать галактозу. Альтернатив-
ный биологический путь — использование мутантных дрожжей,
лишенных р-галактозидазы. Такие мутанты сохраняют способ-
ность к гидролизу лактозы и используют образующуюся глю-
козу в качестве источника углерода. В результате гидролиза
лактозы фильтрат становится более сладким; на опытных ус-
тановках такой гидролиз осуществляют с помощью иммобили-
зованной р-галактозидазы (гл. 4). Гидролизованный фильтрат
не только находит применение в пищевой промышленности, но
278
Глава 6
может оказаться полезным и при решении проблем, связанных
с недостатком ферментов у некоторых животных-отъемышей и
с непереносимостью лактозы у человека. Из сыворотки полу-
чают и другие химические соединения: лактозу, лактулозу,
лактитол и лактобионовую кислоту.
6.6.2. Отходы целлюлозно-бумажной промышленности
Волокнистый материал, применяющийся при производстве бу-
маги и других продуктов, получают как из древесных, так и:
из травянистых растений после химического расщепления лиг-
нина. Однако этот процесс сопровождается потерей большого-
количества древесины и образованием огромного количества
отходов. Все это должно стимулировать разработку альтерна-
тивной химической технологии.
В настоящее время применяют два процесса получения дре-
весной пульпы. Основной из них — это щелочная варка (суль-
фатный процесс), в результате которой образуется темная
сульфатная варочная жидкость. Эти отходы содержат трудна
перерабатываемые ароматические продукты расщепления лиг-
нина и низкомолекулярные органические кислоты (глюкоизоса-
хариновую, молочную, уксусную и муравьиную). При получе-
нии пульпы из смолистой древесины сосны образуются талло-
вое масло и терпены, широко использующиеся в промышлен-
ности. Сульфатную варочную жидкость не удается перерабаты-
вать биологическими способами, которые могли бы применять-
ся в промышленном масштабе; гораздо экономичнее упаривать-
эту жидкость и сжигать ее, получая таким образом энергию из
отходов.
Сульфатная варка целлюлозы применяется реже; она дает
отходы следующего состава: лигносульфонаты с ароматически-
ми элементами (60%), сахара (манноза, галактоза, глюкоза,,
ксилоза, арабиноза; 36%), уксусная кислота, метанол и фур-
фураль. Эти жидкие отходы — хорошее сырье для ферментации
благодаря высокому содержанию в них углеводов. Их фермен-
тация в широких масштабах начата в 1909 г. В настоящее
время традиционным методом удаления пентоз, гексоз и уксус-
ной кислоты из таких отходов служит их ферментация при
участии дрожжей. Помимо этих традиционных методов вскоре
будут использоваться и новые процессы превращения отходов
в грибной белок с помощью Paecilomyces variotii, Sporotrichum
pulverulentum и Chaetomicum cellulolyticum. Неподдающиеся
переработке соединения можно концентрировать и сжигать.
Лигносульфонаты применяют в качестве связывающих ве-
ществ и вспомогательных средств при бурении; щелочным окис-
лением при повышенном давлении их можно превращать в ва-
Окружающая среда и биотехнология 279
нилин. Вообще говоря, главное в переработке отходов целлю-
лозно-бумажной промышленности — это понижение энергоза-
трат, а какой химический принцип при этом используется —
менее существенно.
Основная экологическая проблема, порождаемая целлюлоз-
но-бумажной промышленностью, — это очистка сточных вод, а
также обработка конденсатов, образующихся в испарителях и
реакторах. Сточные воды осветляют путем нейтрализации и от-
стаивания, окисления в одно- и двухстадийных установках с ак-
тивным илом, в аэрируемых отстойниках или путем сочетания
биологических и химических способов окисления. Эти методы
пригодны для эффективного удаления соединений, подвержен-
ных биодеградации, а также токсичных производных фенола,
однако они оказываются дорогими и неэффективными в слу-
чае производных лигнина, с трудом поддающихся переработ-
ке. Отбеливатели, содержащие хлорпроизводные бифенилов,
можно обесцвечивать с помощью грибов — возбудителей белой
гнили.
Среди побочных продуктов сульфитного процесса получения
целлюлозы преобладают химически модифицированные лигни-
ны, образующиеся во многих реакциях между активным суль-
фитом и каким-либо сложным природным полимером. Структу-
ра лигносульфонатов в деталях неизвестна. Они представляют
собой гетерогенную смесь соединений с широким спектром мо-
лекулярных масс (300—100000); состав смесей определяется
природой перерабатываемой древесины. Образование сульфо-
натов приводит к частичной солюбилизации лигниновых фраг-
ментов. Сложность структуры лигносульфонатов затрудняет
изучение их биодеградации. Для упрощения задачи обычно ис-
пользуют модельные соединения, например дегидрополимеры
кониферилового спирта или другие низкомолекулярные продук-
ты. Низкомолекулярные лигносульфонаты чувствительнее к
биодеградации, чем высокомолекулярные; с другой стороны,
производные лигнина, видимо, устойчивее к разрушению, чем
сам лигнин. Следовательно, образование сульфопроизводных
затрудняет переработку. В таких сопряженных окислительно-
деградативных процессах почвенные грибы и бактерии более
эффективны, чем гнилостные грибы; для осуществления этих
процессов требуется также дополнительный источник углерода.
Распад лигносульфонатов нередко сопровождается полимери-
зацией, в результате чего наблюдается сдвиг в распределении
полимеров по молекулярным массам. Эти изменения могут кор-
релировать с присутствием внеклеточных фенолоксидаз (на-
пример, лакказы), физиологическая роль которых остается не-
известной. Фенолы превращаются в соответствующие хиноны и
фенокси-радикалы, которые спонтанно полимеризуются. Таким
280
Глава 6
образом, полимеризация и деградация происходят одновремен-
но. Однако в случае некоторых грибов реакции полимеризации
не протекают в присутствии целлюлозы. Целлюлоза распада-
ется до целлобиозы, являющейся субстратом для целлобио-
за : хинон оксидоредуктазы, которая одновременно окисляет
целлобиозу и восстанавливает хиноны и фенокси-радикалы.
Может существовать и другая оксидоредуктазная система, в ко-
торой легко доступные источники углерода используются для
восстановления хинонов. Возможная роль подобной биологиче-
ской полимеризации состоит в облегчении осаждения лигно-
сульфонатов. Лигносульфонаты применяются как связывающие
вещества при производстве отдельных видов картона, где в ка-
честве катализатора полимеризации используют содержащие
лакказу культуральные фильтраты. Фенолоксидазы могут играть-
важную роль в определении судьбы многих ксенобиотиков в
окружающей среде, участвуя в полимеризации фенолов и в об-
разовании органических полимеров почвы.
Чувствительность лигносульфонатов к биодеградации увели-
чивается после их химической или физической модификации.
Под действием УФ-облучения и озонирования происходит фраг-
ментация этих молекул, а удаление остатков сульфоновой кис-
лоты, хотя и снижает растворимость лигносульфонатов, одно-
временно уменьшает их устойчивость к биодеградации. Пред-
принимались попытки использовать для микробного десульфи-
рования анаэробные сульфатредуцирующие бактерии и неко-
торые виды Pseudomonas. Большими потенциальными возмож-
ностями в этом смысле обладают смешанные культуры. Исполь-
зование таких культур для разрушения лигносульфонатов мо-
жет оказаться более эффективным, чем применение отдельных,
штаммов, поскольку при этом могут быть созданы сообщества
с широким спектром активностей, например способные к де-
сульфированию, расщеплению прочных связей, метилированию-
и деполимеризации. В результате может быть получена высо-
коэффективная биоокислительная система. В одной из опыт-
ных установок для получения БОО из углеводов, содержа-
щихся в отходах целлюлозно-бумажной промышленности, ис-
пользуют Candida utilis, а для разрушения остаточных лигно-
сульфонатов — смешанную культуру. Биомасса, образующаяся
на второй стадии этого процесса, не находит сбыта, поэтому ее
повторно используют после термообработки для ускорения ро-
ста Candida.
6.6.3. Отходы от производства красителей
Текстильная промышленность и производство красителей от-
правляют в отходы устрашающее количество красителей и-
лигментов, единственным общим структурным свойством ко-
Окружающая среда и биотехнология
281
торых является наличие хромофорной группировки. Они посту-
пают в окружающую, среду со сточными водами; с количест-
венной точки зрения эти соединения не относятся к числу ос-
новных ее загрязнителей. Кроме того, эти отходы обычно не
рассматриваются как токсичные или канцерогенные для рыб
или млекопитающих (за исключением бензидина и катионных
красителей).
Для очистки окрашенных сточных вод применяют химиче-
ские методы; удаление красителей и пигментов с помощью мик-
робов весьма ограничено. Устранение этих продуктов из отхо-
дов с помощью активного ила заключается в основном в ад-
сорбции, а не в деградации. Степень их разложения микроор-
ганизмами определяется растворимостью, ионными свойства-
ми, а также природой заместителей и их числом. Оказалось,
что микроорганизмы способны разлагать красители, но только
после адаптации к значительно более высоким концентрациям,
чем те, которые обычно обнаруживаются в сточных водах. По-
скольку микроорганизмы могут использоваться для контроля
за загрязнением окружающей среды этими соединениями, был
проведен скрининг, направленный на выявление тех микроор-
ганизмов, которые способны к расщеплению модельных ве-
ществ типа n-аминобензолов, а также скрининг и селекция ор-
ганизмов из дренажных канав предприятий по производству
красителей. Были сделаны попытки найти взаимосвязь между
подверженностью соединения биодеградации и его химической
структурой.
Компоненты азокрасителей могут разлагаться в аэробных
или анаэробных условиях. Анаэробная биодеградация осу-
ществляется относительно легко, поскольку многие организмы
синтезируют неспецифические ферменты, катализирующие вос-
становительное расщепление азогруппы (рис. 6.17). Однако
для дальнейшей деградации необходим двухстадийный процесс,
продукты которого — амины — могут быть потенциально вред-
ными и их ' подвергают окислительному расщеплению. Азоре-
дуктазы, выделенные из аэробных систем, видимо, обладают
большей субстратной специфичностью и не находят широкого
применения при переработке отходов. Процесс разложения
азокрасителей изучался на примере простого модельного соеди-
нения 4,4'-дикарбоксиазобензола (ДКАБ). Выделены орга-
низмы, использующие это соединение в качестве единственного
источника углерода, азота и энергии. Ранее считалось, что
азосвязь в природе не образуется. Однако среди продуктов,
синтезируемых одним из грибов — патогеном насекомых — бы-
ло идентифицировано азосоединение, так что бактериям при-
ходилось встречаться с этим типом связей in vivo. Был выде-
лен один из видов Flavobacterium, разрушающий только
282
Глава 6
Рис. 6.17. Анаэробное восстановитель-
ное расщепление модельных азокра-
сителей.
трсшс-ДКАБ; если использо-
вать ингибиторы ферментов,,
раскрывающие кольцо, то это
приводит к накоплению ами-
нобензоата. Тем не менее дан-
ный организм не расщепляет
промышленные красители, а
попытки ввести его в непре-
рывную культуру для дли-
тельной адаптации оказались
безуспешными. Flavobacteriunt
не выдерживала конкуренции
со стороны другого организ-
ма — одной из псевдомонад^
способной расти при высоких
концентрациях (750 мг/л) ис-
пользуемого ею синтетического
красителя и, следовательно,
эффективно разрушать его-
При повышенной температуре
эта способность утрачивалась.
Возможно это связано с тем, что упомянутая функция кодиру-
ется плазмидой. Следует отметить, что полученные штаммы об-
ладают высокой субстратной специфичностью и едва ли выжи-
вут в установках для очистки сточных вод.
6.6.4. Биологическая очистка газов
Очистка отходов от вредных, токсичных и пахучих газов — это
серьезная экологическая проблема. Во многих промышленных
производствах (в фотопромышленности, при перегонке нефти,,
очистке природного газа и в целлюлозно-бумажной промыш-
ленности) образуются восстановленные соединения серы (тио-
сульфат, сероводород, метилмеркаптаны, диметилсульфид) .
Эти соединения являются также побочными продуктами ана-
эробного разложения отходов животноводства с высоким со-
держанием органических веществ. Большинство неорганических:
восстановленных соединений серы служат источником энергии
для целого ряда микроорганизмов, растущих в аэробных или
анаэробных условиях (рис. 6.18). Могут быть созданы очист-
ные системы, основанные, например, на использовании тйоба-
цилл: в таких системах анаэробное десульфурирование сопря-
жено с денитрификацией. Один из методов очистки от серово-
дорода состоит в пропускании газа через солевой раствор, на-
пример раствор сульфата меди. В результате происходит
осаждение нерастворимого сульфида металла, который затем
Окружающая среда и биотехнология
283
может быть окислен при участии микроорганизмов (см. также
тл. 5, где рассматривается переработка отходов горнодобываю-
щей промышленности). Количество отходов с неприятным за-
пахом увеличивается и в результате интенсификации животно-
водства. Для устранения этого запаха из отходов удаляют,
в частности, восстановленные соединения серы; такое удале-
ние происходит с потерей азота или без потери (путем образо-
вания аммиака) в зависимости от того, какие микроорганизмы
при этом используются. Органические сульфиды часто бывают
токсичными для микроорганизмов. Например, обогащение от-
Аэробный процесс
H2S + 2О2 --► H2SO4
(CH3)2S + 5О2 -► 2СО2 + H2SO4 + 2Н2О
Анаэробный процесс
5H2S + 8NaNO3 -► 4Na2SO4 + H2SO4 + 4H2O + 4N2
(CH3)2S + 4NaNO3 -► 2CO2 + Na2SO4 + 2NaOH + 2H2O + 2N2
Рис. 6.18. Разрушение восстановленных соединений серы.
ходов микроорганизмами, способными использовать диметил-
сульфид, затруднено, хотя в принципе можно выделить сооб-
щество микробов, растущее на очень близком по структуре суб-
страте диметилсульфоксиде. Преобладающий в таком сообще-
стве организм Hyphotnicrobiutn spp. быстро окисляет диметил-
сульфид, так что есть основания надеяться на создание отно-
сительно простого микробиологического способа переработки
таких ядовитых отходов.
Для детоксикации цианида в промышленных отходах были
предложены различные биологические методы: от использова-
ния активного ила до применения специфических ферментов,
разрушающих цианид. Так, роданаза, обнаружена у Bacillus
stearothermophilus, катализирует превращение цианида в тио-
цианат. Альтернативная иммобилизованная система основана
на гидролизе цианида до формамида, катализируемом инду-
цибельным ферментом цианидгидратазой. Этот фермент был
обнаружен в грибах, паразитирующих на растениях-цианоге-
нах. Таким образом, мы имеем ряд микробиологических мето-
дов очистки промышленных стоков, однако нам еще предстоит
определить их эффективность по степени очистки на выходе.
284
Глава 6
6.7. Биодеградация ксенобиотиков в окружающей среде
Биодеградация органических соединений, загрязняющих окру-
жающую среду, оправдана только в том случае, если в резуль-
тате происходит их полная минерализация, разрушение и де-
токсикация; если же биохимическая модификация этих соеди-
нений приводит к повышению их токсичности или увеличивает
время нахождения в среде, она становится не только нецеле-
сообразной, но даже вредной. Детоксикация загрязняющих сре-
ду веществ может быть достигнута путем всего одной модифи-
кации структуры. Судьба ксенобиотика зависит от ряда слож-
ным образом взаимосвязанных факторов как внутреннего ха-
рактера (устойчивость ксенобиотика к различным воздействи-
ям, растворимость его в воде, размер и заряд молекулы, лету-
честь), так и внешнего (pH, фотоокисление, выветривание).
Все эти факторы будут определять скорость и глубину его пре-
вращения. Скорость биодеградации ксенобиотика данным со-
обществом микроорганизмов зависит от его способности прони-
кать в клетки, а также от структурного сходства этого синтети-
ческого продукта и природного соединения, который подверга-
ется естественной биодеградации. В удалении ксенобиотиков
из окружающей среды важную роль играют различные меха-
низмы метаболизма.
В большинстве случаев при исследовании биодеградации
использовался традиционный подход, основанный на выделе-
нии и анализе свойств чистых изолятов из окружающей среды.
С другой стороны, из-за гетерогенности среды в ней формиру-
ются местообитания для множества разных микроорганизмов
с самыми разнообразными метаболическими свойствами. Эти
местообитания не могут не быть взаимосвязанными друг с
другом. Ксенобиотики подвергаются действию смешанных по-
пуляций микроорганизмов, т. е. сообществ, для которых ха-
рактерны отношения кооперации, комменсализма и взаимопо-
мощи.
6.7.1. Участие микробных сообществ в биодеградации
ксенобиотиков
Можно выделить стабильные сообщества, в которых взаимо-
действия между отдельными его членами дает им ряд преиму-
ществ, в результате чего такая ассоциация становится более-
эффективной, чем отдельно взятые виды. Классификация мик-
робных сообществ основана на характере взаимосвязей между
отдельными видами.
1. Сообщества, в которых один или несколько членов не
способны к синтезу тех или иных компонентов, необходимых
Окружающая среда и биотехнология 285>
для роста; этот дефицит покрывается за счет метаболической
активности других членов сообщества. Сложность различных
взаимоотношений в подобных сообществах может варьировать,
но в любом случае взаимосвязи такого типа, по всей видимо-
сти, играют роль в деградации многих соединений. Например,
сообщество из двух микроорганизмов, растущее на циклогекса-
не, включает один из видов Nocardia, способный окислять цик-
логексан, но не способный к росту на нем одном, а также один
из видов Pseudomonas, который поставляет другим видам не-
обходимые для роста соединения, возможно биотин. Такие-
взаимосвязи могут быть облигатными для роста определенных
видов или поставлять дополнительные питательные вещества,
способствующие росту первично окисляющего организма и
увеличению скорости деградации. Это со всей очевидностью
показывает, что истинную скорость деградации ксенобиотиков;
в окружающей среде можно оценить, только учитывая воз-
можности микробных сообществ, а не отдельных видов микро-
организмов.
2. Сообщества, в которых метаболиты, в какой-то мере по-
давляющие рост организма-продуцента или других организмов;
в том же местообитании, удаляются остальными членами сооб-
щества.
3. Сообщества, в которых параметры роста одного или не-
скольких его членов изменяются так, что получается более
конкурентоспособное сообщество, устойчивое к изменениям
среды. Такое сообщество может стать менее чувствительным к
ингибированию субстратом и приобрести способность справ-
ляться с перегрузками, возникающими в очистных системах.
4. Крайне важная роль микробных сообществ в деградации
ксенобиотиков состоит в том, что они способны осуществлять-
совместную «метаболическую атаку» на субстрат. Отдельные
члены такого сообщества в отличие от сообщества в целом не
обладают метаболической активностью, необходимой для пол-
ной деградации данного соединения. Так, в лаборатории иног-
да не удается добиться минерализации какого-либо вещества,
но происходит это потому, что деградацию тщетно пытаются
осуществить с помощью одного вида микроорганизмов. Отме-
чалось также, что такого рода комбинированная метаболиче-
ская система может возникать в результате синтеза отдельны-
ми видами разных компонентов ферментного комплекса, с про-
явлением ферментативной активности только у целого сооб-
щества (например, синтез активной лецитиназы двумя видами;
Pseudomonas).
5. Ранее подчеркивалась важность сопряженного метаболиз-
ма для расщепления ксенобиотиков. Микроорганизмы, расту-
щие на одном субстрате, превращают другой в ходе одной или
-286
Глава 6
нескольких ферментативных реакций. Эти реакции не связаны
с ростом микроорганизма в том смысле, что в них не образуются
промежуточные продукты, которые данный организм использо-
вал бы для роста. Однако эти промежуточные продукты могут
•служить источником углерода для других членов сообщества.
Выделение или создание подобных сообществ для решения
проблем загрязнений среды ксенобиотиками вполне реально,
хотя о стабильности таких ассоциатов в условиях переработки
отходов известно немного.
6. Были выявлены сообщества, в которых осуществляется
передача восстановительных эквивалентов от одной популяции
другой. В анаэробных условиях классическим ферментативным
способом получения избытка восстановительных эквивалентов
было восстановление конечных продуктов обмена. Однако в
упомянутых сообществах используется второй, акцепторный ор-
ганизм. На сегодняшний день уже выделено немало таких со-
обществ. Анаэробное разрушение подобными сообществами аро-
матических соединений может иметь экономическое значение.
7. При изучении непрерывных культур были обнаружены со-
общества, в которых к полному использованию лимитирующего
рост субстрата способна не одна, а несколько первичных попу-
ляций, хотя можно было бы ожидать, что наиболее конкуренто-
способный организм окажется доминирующим. Следовательно,
существующие в сообществе взаимодействия должны стабили-
зировать свободную конкуренцию между его членами.
В стабильных сообществах микроорганизмов создаются ус-
ловия для свободного обмена генетической информацией меж-
ду популяциями. Важным эволюционным механизмом появле-
ния новых штаммов in vivo, способных взаимодействовать с но-
выми компонентами окружающей среды, может служить пере-
нос плазмид между сообществами микроорганизмов (гл. 7).
Частота таких событий для природных сообществ неизвестна,
•однако в условиях лаборатории они действительно происходят.
Так, в одном из опытов в течение многих поколений выращива-
ли смешанные непрерывные культуры разных видов Pseudomo-
nas. Один из видов мог расти на хлоркатехолах, а другой об-
ладал плазмидой TOL, несущей ген фермента бензолдиоксиге-
назы. На 4-хлорбензоате не мог расти ни один из штаммов,
•однако, использовав принцип обогащения, удалось выделить
мутант, способный к росту на этой хлорсодержащей аромати-
ческой кислоте. По всей видимости, здесь произошел перенос
плазмиды, в результате которого мутантный штамм приобрел
•способность к окислительному декарбоксилированию 4-хлор-
бензойной кислоты с помощью приобретенной диоксигеназы с
широкой специфичностью, а следовательно, и способность к ро-
•сту на образующемся хлоркатехоле.
Окружающая среда и биотехнология 287
В свете взаимодействий подобного рода, существенных для
окружающей среды, мы рассмотрим еще несколько специфи-
ческих примеров деградации ксенобиотиков. Потенциальная'
возможность использования микроорганизмов при биотехно-
логической переработке промышленных отходов обсуждается1
в разд. 6.6. Мы знаем, что в промышленных сточных водах со-
держатся стабильные побочные продукты реакций, часто из-
вестного состава. Вполне понятно, что технология контроля за
ними хорошо разработана. Однако в окружающую среду могут
попадать и сложные смеси промышленных продуктов, например
при разнообразных неспецифических выбросах на любом из
этапов технологического процесса (утечки и т. п.). Деградация
ксенобиотиков будет рассмотрена именно в этом контексте.
6.7.2. Хлорпроизводные углеводородов
С-1- и С-2-хлорпроизводные углеводородов широко использу-
ются в качестве растворителей и представляют собой важный
фактор загрязнения окружающей среды. Тем не менее о мик-
робной деградации этих соединений известно немного. Были
выделены организмы, способные к использованию дихлорме-
тана, однако механизм его деградации до конца не выяснен.
По-видимому, в результате первичного дегалогенирования, ка-
тализируемого какой-то галоидгидролазой, образуется хлорме-
танол, спонтанно разлагающийся до формальдегида.
Дегалогенирование галогензамещенных ароматических со-
единений происходит в основном после расщепления аромати-
ческих групп системы под действием диоксигеназ. Связь угле-
род—галоген становится лабильной после присоединения атома,
кислорода к углероду, связанному с галогеном, с образовани-
ем галогензамещенных катехолов. Относительно широкая спе-
цифичность ферментов начальных этапов катаболизма может
обусловливать довольно быстрое превращение разных галоген-
замещенных ароматических соединений, хотя работа ферментов
и осложняется стерическими и негативными индуктивными эф-
фектами боковых групп-заместителей. Это пример участия
обычных катаболических ферментов в первичной атаке на ксе-
нобиотики. Галогензамещенные катехолы представляют собой
ключевые метаболиты в деградации арилгалогенов. Они разру-
шаются путем орто-расщепления, поскольку ферменты мета-
расщепления необратимо инактивируются веществами, образу-
ющимися в ходе реакции. Обычные пирокатехазы расщепляют
галоген-замещенные катехолы с относительно низкой эффектив-
ностью, поэтому основное значение в деградации этих соедине-
ний приобретает раскрытие кольца. На последующих этапах,
метаболизма, включая циклоизомеризацию, происходит от-
288
Глава 6
щепление оставшихся замещающих групп кольца. На работе
обычно применяемых очистных сооружений плохо сказывают-
ся перегрузки их отходами, содержащими хлорзамещенные
ароматические соединения: путь мета-расщепления нарушается,
д неэффективность орто-пути приводит к накоплению токсич-
ных фенолов и темноокрашенных продуктов их самоокисления.
6.7.3. Другие замещенные простые ароматические
соединения
При деградации арилгалогенов замещающие группы часто от-
щепляются на последних этапах катаболизма после разруше-
ния ароматических колец системы. В случае сульфонированных
ароматических соединений связь углерод — заместитель высо-
кополярна и должна стать лабильной на первых же этапах,
в противном случае всем последовательно работающим фермен-
там придется «иметь дело» с этой замещающей группой. Суль-
фонированные нафталины широко используются в качестве
эмульгаторов и смачивающих агентов, а также при производ-
стве азокрасителей; в обычных очистных сооружениях эти со-
единения не разлагаются микроорганизмами. Однако в непре-
рывной культуре одно из использующих нафталин микробных
сообществ, выделенное из сточных вод, приобрело способность
к расщеплению нафталинсульфоновых кислот. Первые этапы
катаболизма включают диоксигенацию, удаление замещающей
группировки и реароматизацию — ключевой этап, для которо-
го необходима диоксигеназа с широкой специфичностью, спо-
собная к специфическому гидроксилированию по положению
1,2 кольца (рис. 6.19).
Спонтанная
элиминация
Рис. 6.19. Элиминация остатка сульфоновой кислоты из молекулы нафталин-
сульфоновой кислоты.
Окружающая среда и биотехнология
289
Иитротолуолы представляют собой токсичные компоненты
сточных вод военных заводов, которые в природе быстро раз-
рушаются. Однако они минерализуются не полностью, посколь-
ку продукты их деградации (ароматические амины) конденси-
руются с карбоксильными группами биологических макромо-
лекул, образуя полиамиды, которые, по-видимому, устойчивы
к последующему действию микробов.
6.7.4. Полиароматические углеводороды
Полихлорбифенилы (ПХБ) — это очень устойчивые соедине-
ния, которые долго остаются в окружающей среде и прочно ад-
сорбируются биологическими и осадочными материалами.
В почвах они практически не мигрируют, а микроорганизмы не
могут их глубоко деградировать. ПХБ в пробах из окружаю-
щей среды отличаются по составу от ПХБ, получаемых в про-
мышленности, поскольку они модифицируются природными
системами. Микробная деградация бифенила осуществляется
при участии систем катаболизма, сходных с известными для
других ароматических углеводородов. С увеличением степени
хлорирования скорость метаболизма падает. Сообщалось, что
смешанные культуры микроорганизмов осуществляют деграда-
цию промышленных ПХБ до неохарактеризованных углеводоро-
дов, при этом расщепляются преимущественно молекулы с бо-
лее низкой степенью хлорирования. Если замещающих группи-
ровок больше, чем в тетрахлор-ПХБ, то молекула становится
полностью резистентной. В большинстве работ исследовали пре-
вращения чистых изомеров ПХБ.
Бензпирен представляет собой устойчивое полиароматиче-
ское соединение, при деградации которого образуются канце-
рогенные гидрокси- и эпоксипроизводные. Он не минерализу-
ется в системах активного ила, хотя было описано несколько
культур микроорганизмов, способных частично метаболизиро-
вать этот продукт с помощью индуцибельных и неиндуцибель-
ных гидроксилазных систем. При этом образуется сложная
смесь конъюгированных производных.
Полистирол очень устойчив к биодеградации. Был описан
процесс, в котором тонко измельченные автомобильные шины,
изготовленные из стирол-бутадиеновой резины, частично разла-
гались микроорганизмами после добавления какого-либо по-
верхностно-активного вещества. Содержащиеся в шинах анти-
озонаты, антиоксиданты и ускорители вулканизации замедля-
ют этот процесс; конечный продукт можно использовать для
улучшения почвы. Имеются сообщения о росте сообщества мик-
роорганизмов на стироле, в результате чего удаляется ингиби-
тор полимеризации 4-трет-бутилкатехол. В результате свобод-
19—1344
290
Глава 6
норадикальной полимеризации стирола происходит осаждение
полистирола. Впоследствии этот полимер исчезает, что свиде-
тельствует о способности данного микробного сообщества к раз-
рушению полистирола.
6.7.5. Биодеградация нефтяных загрязнений
Рассмотрим теперь процессы биодеградации сложных смесей
углеводородов и их производных в средах, загрязненных
нефтью. Речь пойдет как о сточных водах нефтяной промыш-
ленности, так и о загрязнении нефтью окружающей среды. Ис-
точники таких загрязнений могут быть самые разнообразные:
промывка корабельных бункеров для горючего, аварии на Дан-
керах в открытом море (основная причина нефтяных загрязне-
ний окружающей среды), утечки в нефтехранилищах и сброс
отработанных нефтепродуктов. 1
' ' Сточные воды нефтяной промышленности обычно очищают
биологическим способом после удаления большей части нефти
• физическими способами или с помощью коагулянтов. Токсиче-
ское воздействие» компонентов таких сточных вод на системы
активного ила можно свести к минимуму путем постепенной
«акклиматизации» очистной системы к повышенной скорости
поступления стоков и последующего поддержания скорости по-
тока и его состава на одном уровне. Однако загрузка этих'си-
стем 1 может значительно варьировать и, видимо, лучше ис-
пользовать более совершенные технологии, например системы
с илом, аэрированным чистым кислородом, или же колонные
биореакторы.
Самые большие утечки нефти в окружающую среду проис-
ходят в море, где она затем подвергается различным физиче-
ским превращениям, известным как выветривание. В ходе этих
абиотических процессов, включающих растворение, испарение
и фотоокисление, разлагается — в зависимости от качества
нефти и от метеорологических условий — 25—40% нефти. На
этой стадии разрушаются многие низкомолекулярные алканы.
Степень микробиологической деградации выветрившихся нефтя-
ных рйзлйвов определяется рядом факторов. Весьма важен со-
став нефти: относительное содержание насыщенных, аромати-
ческих, содержащих азот, серу и кислород соединений, а так-
же асфальтенов в различных типах нефти различно. Опреде-
ленную устойчивость нефти придают разветвленные алканы,
серусодержащие ароматические соединения и асфальтены.
-Кроме того, скорость роста бактерий, а следовательно, и Ско-
рость биодеградации определяются доступностью питательных
веществ, в частности азота и фосфора. Оказалось, что добав-
ление таких веществ увеличивает скорость биодеградации.
Окружающая среда и биотехнология • 291
Количество разных организмов, способных расти на компонен-
тах нефти, зависит от степени загрязненности углеводородами.
Например, больше всего их находят поблизости от крупных
лортов или нефтяных платформ, где среда постоянно загряз-»
лена нефтью. Полная деградация нефти зачастую не происхо-
дит даже при участии богатых по видовому составу микроб-
ных сообществ. Наиболее биологически инертные компоненты,
например асфальтены, содержатся в осадочных породах и неф-
тяных залежах. Основные физические факторы, влияющие на
скорость разложения нефти,—это температура, концентрация
(кислорода, гидростатическое давление и степень дисперсности
нефти. Наиболее эффективная биодеградация осуществляется >
тогда, когда нефть эмульгирована в воде.
Особую проблему представляют выбросы или . ^случайные
разливы нефти на поверхности почвы, поскольку они могут
привести к загрязнению почвенных вод и источников питьевой
:воды. В почве» содержится очень много микроорганизмов, спо-
собных разрушать углеводороды. Однако даже их активность не
•всегда достаточна, если образуются растворимые производные .>
или поверхностно-активные соединения, увеличивающие рас-
пространение остаточной нефти.
*6.7.6. Пестициды
’Слив отходов производства пестицидов сегодня'строго контро- -
лируётся; технология очистки сточных вод или их детоксика-
ции хорошо разработана, хотя остается сложной и многообраз-
ной. Она включает сначала экстракцию пестицидов растворите-1
лями, а затем обычную биологическую обработку. Для ликви-
дации непредусмотренных выбросов, происходящих при утечках
пли при промывке и замене контейнеров с пестицидами, подхо-
дящая технология пока отсутствует. Пестициды попадают в
окружающую среду и в результате использования их для об-
работки сельскохозяйственных культур. Большинство пестици-
дов расщепляется бактериями и грибами. Превращение исход-
ного пестицида в менее сложные соединения нередко осуществ-
ляется при участии сообществ микроорганизмов. Были описа-
ны различные стадии и промежуточные продукты- процессов
деградации-ДДТ, идущей, например, ’в ходе сопряженного ме-
тоболизма и приводящей к полной минерализации этого стой-
'кого пестицида. Часто из среды, содержащей ксенобиотик,
можно выделить сообщества такого рода, в которых он служит
не основным источником углерода, а источником фосфора, се-
ры или азота. Чрезвычайно высокая токсичность пестицидов1
зачастую утрачивается на первой же стадии их модификации.
Это позволяет разработать относительно несложные микробио-
d9*
292
Глава 6
логические способы их детоксикации. Например, в результате
гидролиза может значительно уменьшиться токсичность пести-
цида или увеличиться вероятность биодеградации. Для этого
хорошо было бы использовать внеклеточные ферменты, способ-
ные функционировать в отсутствие коферментов или специфи-
ческих факторов и осуществлять детоксикацию разнообразных
пестицидов. Это могут быть такие гидролазы, как эстеразы,.
ациламидазы и фосфоэстеразы. Чтобы выбранный фермент
можно было применять in situ, он должен обладать подходя-
щей кинетикой в широком диапазоне температур и pH, быть
нечувствительным к небольшим количествам растворителей
или тяжелых металлов, не ингибироваться субстратом при кон-
центрациях, характерных для содержимого очистных систем,
а также хорошо храниться. В ряде случаев в качестве биоло-
гического агента детоксикации была испробована паратионгид-
ролаза, выделенная из Pseudomonas sp. С ее помощью удалось
удалить 94—98% остаточного паратиона (около 75 г) из кон-
тейнера с пестицидом за 16 ч при концентрации субстрата 1 %,
(по весу). Забуференные растворы паратионгидролазы исполь-
зовали также для детоксикации паратиона в разливах на поч-
ве, где его концентрация, по-видимому, была весьма высока.
Скорость разложения паратиона в этом случае зависела от ти-
па почвы, влажности, буферной емкости раствора и концент-
рации фермента. При этом значительные количества пестицида
были обезврежены всего за 8 ч. Как показали лабораторные
эксперименты, еще одна возможная сфера применения иммоби-
лизованных ферментов — это очистка сточных вод. Были опи-
саны гидролазы для детоксикации других пестицидов. Многие
из них обладают широкой субстратной специфичностью, что
открывает большие возможности для создания других простых
систем детоксикации пестицидов. В будущем подобные систе-
мы смогут применяться при промывке промышленных химиче-
ских установок и реакторов, ферменты в виде аэрозолей — для
удаления пестицидов с поверхностей, а ферменты в сочетании
с пестицидами — для быстрого разрушения пестицидов после
их использования.
6.7.7. Биодеградация поверхностно-активных веществ
По чувствительности к биодеградации синтетические поверх-
ностно-активные соединения, применяемые в быту и в промыш-
ленности как моющие средства, можно разделить на «жест-
кие» и «мягкие». Анионные соединения этой группы, такие как
алкилбензолсульфонаты, в конце 50-х гг. привлекли к себе
внимание тем, что они загрязняют окружающую среду: это
проявлялось в образовании пены в водоемах. Сначала в прода-
Окружающая среда и биотехнология
293
жу поступали «жесткие» детергенты, устойчивость которым
придавали их разветвленные алкильные боковые цепи. Чтобы
предотвратить их накопление в природе, промышленность доб-
ровольно перешла к производству подверженных биодеграда-
ции, линейных неразветвленных алкилбензолсульфонатов. Раз-
рушение этих поверхностно-активных соединений начинается
с окисления концевых метильных групп, после чего за счет
р-окисления идет расщепление линейных боковых цепей. Коль-
цевые структуры молекул обычно разрушаются только после
полной деградации боковой цепи. Данный процесс осуществля-
ется только в аэробных условиях, поскольку для начального
окислительного этапа требуется кислород. Разветвленные мо-
лекулы не всегда оказываются устойчивыми, хотя процесс их
р-окисления и затруднен. Механизм разрушения разветвлен-
ной цепи до конца не установлен. Связь углерод — сера явля-
ется очень прочной, и это увеличивает биологическую инерт-
ность молекулы детергента. Реакции десульфирования деталь-
но не изучены, но скорее всего в них участвуют гидроксилазы
или монооксигеназы. По-видимому, далее сульфонатный оста-
ток превращается в сульфат, возможно, с образованием суль-
фита в качестве промежуточного продукта. Метаболизм арил-
сульфонатов уже был рассмотрен нами в разд. 6.7.3. Есть ос-
нования считать, что десульфирование и лета-расщепление
ароматического кольца детерминируются плазмидами.
. Алкилсульфаты все еще используются как моющие средст-
ва на фабриках-прачечных и в косметической промышленно-
сти; основную их массу составляют первичные алкилсульфа-
ты. Линейные сульфаты легко разрушаются, но этот процесс
замедляется из-за наличия разветвленных участков. В первич-
ной атаке этих молекул участвуют сульфатазы, образующие
соответствующие спирты, которые подвергаются затем даль-
нейшему метаболизму. Для этого процесса кислород не ну-
жен, и он может идти в анаэробных условиях. Необычность
алкилсульфатаз по сравнению с сульфатазами вообще состоит
в том, что они атакуют связь С—О в группах С—О—S. Пер-
вичные и вторичные алкилсульфаты индуцируют образование
сложного комплекса сульфатаз.
Неионные детергенты применяются в быту; поскольку они
облегчают смачивание и способствуют образованию эмульсий,
их с успехом используют при производстве аэрозолей для
сельского хозяйства и в косметической промышленности. Ли-
нейные первичные алкогольэтоксилаты минерализуются быстро
и до конца, но более высокомолекулярные гомологи более ус-
тойчивы. Деградация осуществляется путем окисления конце-
вых метильных групп и последующего р-окисления с образова-
нием низкомолекулярных алканоатэтоксилатов, лишенных по*
?94 Глава 6
верхностно-активных свойств. У вторичных алкогольэтоксила-
тов, гидрофобная алкильная цепь разрушается с обоих концов
за,, счет со- и р-окисления. Имеющиеся в этих соединениях •
эфирные связи увеличивают их устойчивость к биодеградации. •
Были предложены возможные способы расщепления этих свя-
зей, частности монооксигеназное расщепление, гидролиз, уча*
стие углерод: кислород—лиазы и окисление а-атома угле-?
рода эфирной связи с последующим гидролизом сложного
эфира. -
Имеющиеся в продаже детергенты редко содержат по весу
более 30% поверхностно-активного соединения. К числу осталь-
ных компонентов относятся оптические отбеливатели, отбели-
ватели-окислители, вспомогательные пенообразователи, анти-
коррозийные добавки и (в ряде случаев) ферменты. Основную
массу составляет носитель (наполнитель). Наполнители нужны
для: 1) уменьшения концентрации свободного кальция и маг- -
ния с целью предотвращения образования неорганических осад-
ков; в противном случае выпадут в осадок соли щелочнозе-
мельных металлов и аниона детергента; 2) диспергирования
агрегатов почвенных частиц и стабилизации почвенных суспен-
СООН.СН2.О.СН(СООН).СН2СООН кмс
СООН.СН2.О.СН2.СООН ОДА
СООН.СН2.О.СН2.СН2.О.СН2.СООН ЭГТА
N(CH2COOH)3 н га
Рис. 6.20. Структура некоторых органических; соединений, ,прцгодных_для по,- ;
вышения эффективности детергентов. , ,ч.
зий. Наполнители должны иметь хорошую буферную емкость
и не вступать в нежелательные реакции с другими компонен-
тами смеси. В течение многих лет для этих целей использова-
ли тринатрийфосфат, но он ускоряет зарастание внутренних
водоемов. Процесс эвтрофикации водоемов может ускоряться
в результате биодеградации и минерализации любого азоти-
стого Или фосфорного соединения. В поисках кандидатов на •
роль свободных от азота и фосфора наполнителей были выбра-
ны синтетические карбоксиэфиры (КМС, карбоксиметилсукци-
нат; ОДА, окисидиацетат; ЭГДА, этиленгликольдиацетат; рис.
6.20). Изучение метаболизма КМС и его аналогов позволило
понять, каким путем микроорганизмы приобретают способность
к деградации новых ксенобиотиков. Сам КМС быстро разруша-
ется в природе; в.начальной реакции участвует индуцибельная,
•лиаза, расщепляющая молекулу КМС с образованием гликола.-.
Окружающая средй и биотехнология
295
та и фумарата. Аналоги КМС оказались более стойкими, види-
мо, вследствие субстратной специфичности индуцируемых
лиаз. Устойчивость этих аналогов не связана с процессом их
поглощения клетками, поскольку было п’оказано, что они по-
ступают в них при участии конститутивной системы транспор-
та цитрата. КМС-лиаза оказалась удивительно сходной с по-
лигалактуронатлиазой; организмы, развивающиеся на КМС,
так же хорошо растут на полигалактуронате. Это весьма пока-
зательный пример появления способнобти к деградации путем
приобретения новой функции уже существующими ферментами.
Возможно, что эта лиаза кодируется плазмидой. В качестве на-
полнителя широко использовался нитрилтриацетат (НТА), по-
скольку он подвержен быстрой биодеградации в системах ак-
тивного ила и в речной воде.
ЛИТЕРАТУРА
Callely A. G., Forster С. F., Stafford D. A. (eds.), 1977. Treatment of Industrial
Surfactants, pp. 283—327, Hodder ans Stoughton, London.
Chater K. №. A., Somerwille M. J. (eds.), 1978. The Oil Industry and Microbial
Ecosystems, Heyden and Son Ltd. (on behalf of the Institute of Petroleum),
London.
Gibson D. T. (ed.), 1984, Microbial Degradation of Organic Compounds, Micro-
biology Series, Vol. 13, Marcel Dekker, New York.
Higgins I. J., Burns R. G., 1975. The Chemistry and Microbiology of Pollution,
Academic Press, London. y
Lei'singer T., Cook A. M., Hutter R., .N'uesch 7., 1981. Microbial Degradation of
Xenobiotics and Recalcitrant Compounds, FEMS Symp., 12, Academic Press
(on behalf of the Federation of European Microbiological Societies), London.
Monlin G., Galzy P., 1984. Whey, a Potential Substrate for Biotechnology. In:
Biotechnology and Engineering Reviews, Vol. I, (ed. Russel G. E.), pp. 347—
374, Intercept Ltd., Newcastle-upon-Tyne-. ‘ ’
Stafford D. A., 1982. Biological Treatment.of Organic Compounds and Solvents.
In: Safe Use of Solvents (eds. Collings A. J., Luxton S. G.), pp. 293—304,
Academic Press, London.
Stafford D. A., Etheridge S. P. 1982. Farm Wastes, Energy Production and the
Economics of Farm Anaerobic Digesters. In: Anaerobic Digestion, pp. 255—
268, Elsevier Biomedical Press, Amsterdam.
Stafford D. A., Hawkes D. L., Horton R., 1980. Methane Production from Waste
Organic Matter, CRC Press, Boca Raton, Florida.
Turner ]„ Stafford D. A., Hughes D. E., Clarkson J. (1983). The Reduction of
Three Plant Pathogens (Fusarium, Corynebacterium and Globodera) in
. Anaerobic Digestion, Agric. Wastes, 6, 1—11. ! -
Wheatley A. D., 1984. Biotechnology and Effluent Treatment. In: Biotechnology
and Genetic Engineering Reviews, Vol. I (ed. Russel G. E.), pp. 261—310,
Intercept Ltd., Newcastle-upon-Tyne.
Глава 7
Генетика и биотехнология
К. Харди, С. Оливер
7.1. Введение
При оптимизации любого промышленного процесса, протекаю-
щего с участием живых организмов, основные усилия бывают
направлены на улучшение их генетически обусловленных
свойств. Традиционно для повышения продуктивности штаммов
использовали мутагенез с последующим скринингом и отбором
подходящих вариантов. В «допастеровский период» отбор при
проведении наиболее «древних» процессов ферментации (на-
пример, в пивоварении или сыроделии) осуществлялся, очевид-
но, бессознательно. В последнее время для объединения же-
лаемых свойств разных штаммов в одном организме стали ис-
пользовать гибридизацию.
Необходимой предпосылкой создания более продуктивных
штаммов путем отбора мутантов и рекомбинации является глу-
бокое знание биохимии и физиологии процесса ферментации.
Нам нужна также информация о том, какие стадии метаболиз-
ма являются лимитирующими, каковы термодинамические пре-
делы повышения выхода продукта. Обычно эффективность про-
цесса удается поднять, используя приемы как физиологии, так
и генетики: при этом возможности двух наук дополняют друг
друга, а не противопоставляются.
Сегодня в этой области произошли явные перемены: если
раньше единственным используемым генетическим методом
был отбор улучшенных штаммов, то сегодня предлагаются сов-
сем новые подходы, основанные на технологии рекомбинантных
ДНК (генетическая инженерия). С их помощью путем фермен-
тации можно получать новые виды продукции: белки и пепти-
ды человека, антигены вирусов. Надо сказать, что большой ин-
терес к биотехнологии в значительной мере обусловлен именно
появлением генетической инженерии. В этой главе мы рассмот-
рим, какое применение могут найти методы модификации гене-
тического материала как in vivo, так и in vitro для разработки
новых и модернизации существующих биотехнологических про-
цессов.
Генетика и биотехнология
297
7.2. Способы увеличения продуктивности штаммов
7.2.1. Мутагенез и отбор
В прошлом для увеличения продуктивности штаммов обычно
использовали мутагенез и отбор: именно таким путем удалось
повысить выход антибиотиков, синтезируемых грибами и акти-
номицетами. Рис. 7.1 иллюстрирует применение этих методов
для повышения выхода пенициллина. Было последовательно
отобрано свыше двадцати штаммов, продуцирующих все боль-
ше пенициллина, и в конечном счете продуктивность увеличи-
лась в 55 раз. Как в этом случае, так и во многих других пря-
мого отбора не происходило, поскольку не удавалось создать
условия, при которых росли бы только искомые штаммы. Вме-
сто этого пришлось применять метод скрининга: клетки, выжив-
шие после воздействия больших доз мутагенов, размножали в
колбах на качалках, после чего в фильтратах культуральной
среды определяли количество антибиотика.
Скрининг требует много времени и напряженного труда. Ча-
сто штаммы, для которых был получен большой выход при
выращивании в колбах, оказывались непродуктивными в усло-
виях роста в ферментере большого объема. Нередко успеху
способствовало знание «секретов мастерства»: так, оказывает-
ся, что большая продуктивность обычно свойственна особым
морфологическим вариантам.
Скрининг с использованием чашек с агаром вместо колб на
качалках позволяет обследовать гораздо больше мутантов. Об-
разование антибиотиков или метаболитов нередко оценивается
методом биопроб, когда в агар вводят индикаторные организ-
мы. Разработаны устройства для автоматического скрининга
чашек. В них используются механические приспособления для
инокуляции и производится периодическое фотографирование
и последующий компьютерный анализ изображений.
Можно создать особые условия, при которых мутанты с
нужными свойствами не растут. В этом случае применяется
так называемый метод обогащения, когда активно растущие
клетки дикого типа погибают, а нерастущие мутантные выжи-
вают. Так, например, пенициллин или нистатин вызывает ги-
бель растущих клеток бактерий или грибов соответственно.
При работе с грибами недостаток в среде инозитола приводит
к автолизу растущих, но не покоящихся клеток.
Наиболее подходящим способом является, конечно, прямой
отбор, когда создаются условия для роста только мутантных
клеток. Этот подход применялся для выделения штаммов —
сверхпродуцентов некоторых метаболитов, например аминокис-
лот или цитрата. Обработанную мутагеном культуру выращи-
|uv
Wis. B13-D10 (не пигмент.и-
। рованный)
IS
Wis. 47-638
Is
Wis. 47-1564
- |s
Wis. 48-701
| NM
Wis. 49-133
|s ‘
Wis. 51-20
E-1
|nm.
E-3
|NM
E-4.
| NM
-E-6
| NM
E-S
| NM
E-9
| NM
£-10
|nm
£-12 •
|nm
£-13
| NM
Е-14
| NM
£-15
E.15.1 |
University
of Wisconsin
Обозначения
При получении каждого последу-
ющего штамма были использованы
следующие методы:
S - естественный отбор;
UV ' ультрафиолетовое облучение;
X - рентгеновское облучение;
1 NM' обработка азотистым ипритом
[метил-бис-(р-хлорэтил) -амином]
SA -обработка сарколизином
[ л-ди- (2-хлорэтил)-аминофенил-
аланином]
Lilly -
Industries
Ltd.
Последний штамм
A
Генетика и биотехнология
299
Б
100000 г
I
Рис. 7.1. А. Последовательный отбор штаммов, образующих все большее коли-
чество пенициллина, с помощью генетических методов (Impacts of Applied
Genetics, Office of Technology Assessment, US Government Printing Office, Wa-
shington DC, USA). Б. Иллюстрация тех изменений, которые произойди в про-
изводстве пенициллина. Показано, как росла конечная концентрация пеницил-
лина в культуральной среде при его промышленном производстве в период
с 1940 по 1975 г.
вают в присутствии ингибитора (например, аналога аминокис-
лоты) , в результате него отбираютсй мутанты, преодолевающие
нарушение обмена за счет образования избытка желаемого со-
единения.
Отбор на чашках с агаром основан на том, что часть орга-
низмов отвечает на- воздействие по принципу «всё или ничего».
Колонии мутантов растут, а диких клеток — нет. Однако при
получении промышленных штаммов, особенно предназначенных
для использования в длительных процессах ферментации, не-
редко стремятся получить для конкретных условий относитель-
но небольшие различия в скорости роста. В этом случае отбор
происходит при непрерывном культивировании. В хемостатах
при длительном выращивании культура все время находится
в экспоненциальной фазе роста, и это позволяет выделять да-
же те мутанты, у которых сродство к субстрату, удельная ско-
рость роста или устойчивость к токсическому действию высо-
ких концентраций субстрата или продукта лишь немного пре-
вышает исходный уровень.
300
Глава 7
7.2.2. Гибридизация путем скрещивания
Наиболее простой путь создания организмов с желаемым комп-
лексом генетически обусловленных признаков — это скрещива-
ние штаммов, принадлежащих к противоположным половым
типам. Как про-, так и эукариотические микроорганизмы скре-
щиваются при контакте клеток, и этот процесс используется
для получения рекомбинантов.
Конъюгация у бактерий
У бактерий половой процесс называют конъюгацией. Контакт
между двумя клетками осуществляется за счет образования
длинного конъюгационного мостика, который служит для пере-
носа ДНК из одной клетки в другую. Способность к формиро-
ванию мостика закодирована во многих плазмидах (см. далее);
по нему они переносят свои гены, а в некоторых случаях и гены
клетки-хозяина в клетку-реципиент. Плазмиды, осуществляю-
щие перенос генов клетки-хозяина, обладают, таким образом,
способностью к мобилизации хромосом. У Escherichia coli та-
кие F-плазмиды выступают в роли фактора пола. Они способ-
ны мобилизовать хромосому не только Е. coli, но и родствен-
ных энтеробактерий (Shigella, Klebsiella, Salmonella, Erwinia)
и поэтому могут использоваться для переноса генов между
бактериями разных родов. Некоторые плазмиды, выделенные
из Pseudomonas aeruginosa, еще более «неразборчивы». Так,
плазмида R68.45 может переносить хромосомные гены между
видами Pseudomonas, в том числе и Pseudomonas putida, но
кроме этого, обладает способностью к мобилизации хромосом
и таких родов, как Rhizobium, Rhodopseudomonas, Azospiril-
lum, Agrobacterium и Escherichia.
К числу наиболее широко используемых в промышленности
микроорганизмов относятся разнообразные виды Streptomy-
ces-. с их помощью получают более 60% разновидностей при-
меняющихся сегодня антибиотиков. У этих организмов хорошо
развиты системы скрещивания, которые обнаружены и у их
близких родственников, видов Nocardia (они синтезируют ан-
тибиотики рифамицины). Способность к конъюгации была от-
крыта при изучении Streptomyces coelicolor (этот вид примене-
ния в промышленности не нашел и используется в лабораториях).
В клетках этого организма было найдено два типа плазмид,
SCP1 и SCP2, несущих фактор пола. SCP2 продуцирует ре-
комбинантные формы с очень высокой частотой. SCP1 по сво-
им свойствам близка к F-плазмиде Е. coli. Она может встраи-
ваться в хромосому Streptomyces, а также включать фрагмен-
ты хромосомной ДНК. Это довольно крупная плазмида (Afr =
Генетика и биотехнология
301
= 18-106), несущая все шестнадцать генов, нужных для обра-
зования антибиотика метиленомицина.
Многие используемые в промышленности виды Streptomy-
ces, видимо, не обладают собственной системой скрещивания,
«о у Streptomyces rimosus, продуцента окситетрациклина, най-
дена плазмида SRP1, которая несет активный фактор пола, и
при скрещивании SRP1+XSRP1- рекомбинантные формы обра-
зуются с частотой 1СГ4—10~3.
У других грамотрицательных бактерий (помимо Streptomy-
ces) конъюгационные плазмиды встречаются относительно ред-
ко. Отметим, что недавно система скрещивания была обнару-
жена у бактерий такого важного в промышленном отношении
рода, как Bacillus. Существенно, что эти бактерии способны
.легко акцептировать «голую» ДНК, и поэтому рекомбинант-
ные формы несложно получить путем трансформации. Такая
возможность была использована при создании штаммов Bacil-
lus, у которых около 50% синтезируемого ими белка составля-
ет гидролизующий крахмал фермент а-амилаза. При этом бы-
ли отобраны несколько мутантов по разным генам с повышен-
ной способностью к образованию фермента. Эти разные мута-
ции были затем сведены воедино у одного штамма путем по-
следовательной трансформации хромосомной ДНК и отбора на
повышенную способность гидролизовать крахмал.
Системы скрещивания у грибов
У грибов существуют разнообразные типы скрещивания, кото-
рые используются в генетических исследованиях. Многие гри-
бы-аскомицеты и базидиомицеты обладают сложноорганизован-
ными системами скрещивания, препятствующими самооплодо-
творению и другими формами инбридинга. Половой процесс
.контролируется системой несовместимости. У некоторых грибов
'Система несовместимости биполярна; при этом процесс скрещи-
вания контролируется всего одним локусом, который существу-
ет в двух альтернативных аллельных формах. Это дает два ти-
па спаривания, например а и а у Saccharomyces cerevisiae,
причем разрешена лишь комбинация а/а. У других грибов (на-
пример, Schizophyllum commune) система несовместимости
тетраполярна. У них тип спаривания определяется двумя гена-
ми, каждый из которых имеет множество аллельных форм. Для
успешного скрещивания два партнера должны обладать разны-
ми аллелями каждого из двух локусов типа спаривания (на-
пример, АхВуХАуВх— это полностью совместимое спарива-
ние, а комбинации АхВуХАхВх или же АхВуХАуВу и АхВуХ
ХАхВу невозможны, так как приводят к гибели клеток на раз-
ных стадиях процесса скрещивания).
302
Глава 7
Парасексуальный цикл у грибов
Многие мицелиальные формы грибов, применяющиеся в про-
мышленности, не имеют истинного полового цикла, во времж
которого можно было бы провести скрещивание с целью кон-
струирования более продуктивных штаммов. Однако в этом
случае для осуществления рекомбинации можно использовать-
парасексуальный цикл. Именно этот подход применялся при-
работе с1 грибами таких промышленно важных родов, как Л$-
pergillus, Penicillium, Cephalosporium и Fusarium. Суть- пара-
сексуального цикла отражена на рис. 7.2.
I
Родители-гаплоиды
и рекомбинантные
гаплоиды
Гаплоид
Гаплоид
Плазмогамия
Гетерокарион.
Вторичное
Митотическая
рекомбинация
Кариогамия-----
Первичное
нерасхождение
хромосом
Митотическая
рекомбинация
Соматический
диллоид.
Рекомбинантные
диплоиды
Рис. 7.2. Обычный парасексуальный цикл. [С. Е. Caten (1981) Parasexual pro-
cesses in. fungi. In: The Fungal Nucleus (eds. K. Gull, S. G. Oliver), pp. 191—
214. Cambridge University Press.) Тонкие линии-—гаплоидные или анеуплоид-
ные ядра; жирные линии — диплоидные ядра.
Гетерокарионы, у которых в одной клетке сосуществуют ге-
нетически разные ядра, могут быть получены либо путем слия-
ния ' (анастомоза) гиф разных мицелиев, либо в результате-
образования мутантного ядра в мицелии. Слияние ядер в гете-
рокарионе происходит с низкой частотой (10-5—10~4), но этот
процесс1 ускоряется под действием (+) -камфоры или УФ-све-
та. Митотическую рекомбинацию в диплоидном ядре можно ин-
дуцировать с помощью химических и физических воздействий,
которые либо вызывают мутации, либо ингибируют синтез-
ДНК. Возврат к гаплоидному состоянию после рекомбинации,,
видимо, происходит в результате постепенной утраты отдель-
ных хромосом. Этому способствуют такие вещества, как п-фтор-
фенилаланин и карбаматы бензимидазола, препятствующие об-
разованию микротрубочек.
Генетика и биотехнология
303
По сравнению с истинными системами пола парасексуаль-
лый цикл — это малоэффективный механизм образования ре-
комбинантов. Впрочем, его эффективность можно повысить,
применяя интенсивный отбор и разнообразные вещества. Эта
система была использована для повышения выхода антибиоти-
ков, синтезируемых Penicillum и Cephalosporium.
7.3. Генетические перестройки in vivo
7.3.1. Плазмиды
.Многие свойства бактерий, интересные с точки зрения биотех-
нологии, кодируются плазмидами. Плазмиды — это кольцевые
молекулы ДНК, которые стабильно передаются потомству бак-
териальных клеток независимо от хромосомной ДНК- В гене-
тической инженерии плазмиды используются для клонирования
нужных генов. Мы опишем в этом разделе некоторые важней-
шие свойства.плазмид и обсудим те из них, которые особенно
ценны для биотехнологии.
Структура и репликация
Мрл. масса плазмид составляет от 1-Ю6 до 200-106. Таким об-
разом, самые мелкие плазмиды кодируют один-два белка сред-
него размера, тогда как более крупные — 300 или более бел-
ков. Крупные плазмиды могут кодировать множество фермен-
тов, необходимых для работы целой последовательности биохи-
мических реакций, например для превращения толуола в кате-
хол. Плазмиды с мол. массой более 100-106 обнаружены толь-
ко у грамотрицательных бактерий, в частности у видов Pseudo-
monas и Agrobacterium. Мол. масса плазмид грамположитель-
ных бактерий лишь в отдельных случаях превышает 40-106.
В. бактериальных клетках плазмиды существуют в виде
кольцевых двухцепочечных ДНК, которые, кроме того, находят-
ся в сверхспиральном состоянии. Некоторые из плазмид могут
существовать лишь в клетках одного или двух близких видов.
К числу плазмид с широким кругом хозяев относятся RP4,
R68.45, RK2 и сходные с ними плазмиды, несущие маркер ус-
тойчивости к лекарственным препаратам (R-плазмиды), кото-
рые принадлежат P-группе несовместимости (члены одной и
той же группы несовместимости не могут сосуществовать в бак-
териальной клетке). Эти плазмиды были перенесены во многие
виды грамотрицательных бактерий; создается впечатление, что
их хозяевами могут быть все штаммы грамотрицательных бак-
терий.
Таким образом, R-плазмиды, относящиеся к группе Р, осо-
бенно подходят для генетических экспериментов с грамотрица-
304
Глава 7
тельными бактериями, используемыми в промышленности. При
помощи этих плазмид мы можем осуществлять перенос хромо-
сомных генов между неродственными видами. Небольшие по
размеру производные плазмид RP4 и RK2 используются как
векторы для клонирования молекул ДНК, которая затем мо-
жет быть перенесена в клетки самых разных видов.
Рис. 7.3. Плазмиду pBR322 получают путем объединения плазмид ТпЗ,
pSClOl и pMBl. [J. G. Sutcliffe (1979) Cold Spring Harbor Simposia on Quanti-
tative Biology, 43, 77—90.] Плазмида pMBl сходна с плазмидой ColEl. Устой-
чивость к ампициллину определяется геном р-лактамазы. Указано направление
транскрипции этого гена. Стрелками отмечены уникальные сайты расщепления
pBR322 некоторыми ферментами.
Репликация некоторых типов плазмид тщательно изучена,
и нам известны сегодня многие детали этого процесса, особен-
но для небольшой плазмиды ColEl. Эта информация помогает
при использовании плазмид для клонирования генов. Чаще все-
го применяют производные плазмиды pBR322 длиной 4362 пар
нуклеотидов, несущей гены устойчивости к пенициллину и тет-
рациклину. Эти гены были перенесены из R-плазмид (рис. 7.3)
Генетика и биотехнология
305
в фрагмент небольшой плазмиды pMBl, которая близка кочень
хорошо изученной небольшой плазмиде ColEl.
Плазмиды pBR322 и ColEl могут реплицироваться в Е. со-
И и близких видах Enterobacteriaceae. В бактериальных клет-
ках они представлены несколькими копиями; так,для ColEl чис-
ло плазмидных копий на хромосому равно примерно 20, а для
pBR322 — 40. Это обстоятельство очень важно для клонирова-
ния. Чем больше число копий клонированного гена, тем выше
выход кодируемого белка. Кроме того, многокопийные плазми-
ды не так легко утрачиваются при культивировании. С другой
стороны, малое число копий плазмиды иногда имеет свои пре-
имущества, например при клонировании таких генов, продукты
которых гибельны для бактериальной клетки в большом коли-
честве.
Изучению механизмов контроля репликации и числа копий
бактериальных плазмид посвящено множество работ. Было
обнаружено, что число копий плазмид примерно одинаково в-
клетках, размножающихся с разной скоростью. Из этого сле-
дует, что репликация плазмид как-то связана с ростом бакте-
рий. Такая координация достигается при участии механизмов,,
контролирующих начало репликации. Если уж репликация на-
чалась, то она идет с относительно постоянной скоростью при
любом темпе размножения бактерий. При высокой скорости
размножения репликация индуцируется чаще в случае много-
копийных плазмид, чем малокопийных.
Начало репликации ColEl и pBR322 контролируется неболь-
шими молекулами РНК, которые взаимодействуют с особым
участком плазмиды, расположенным поблизости от точки ини-
циации репликации. Начавшись, репликация идет в одном нап-
равлении и заканчивается вблизи точки инициации. Генетики,
стремящиеся максимально увеличить выход белков, кодируе-
мых клонированными в pBR322 генами, получали мутанты с
модифицированными РНК. У некоторых таких мутантов число
копий плазмиды составляет более 100, что приводит к суще-
ственному увеличению выхода продуктов, кодируемых клониро-
ванными генами. Интересная особенность репликации ColEl
и pBR322 заключается в том, что она полностью обеспечива-
ется ферментами хозяина. Сами плазмиды не несут информа-
ции, нужной для кодирования ферментов репликации.
Процессы репликации pBR322 и ColEl, с одной стороны, и.
хромосомы Е. coli — с другой, различаются тем, что первый не
подавляется хлорамфениколом, который блокирует синтез бел-
ка. В присутствии этого соединения репликация хромосом пре-
кращается по завершении уже начавшегося раунда реплика-
ции. Такого подавления репликации pBR322 не происходит, и:
плазмиды накапливаются в обработанных хлорамфениколом;
20-1344
306-
Глава 7
клетках, так что в конечном счете они составляют до 50% всей
ДНК. При выделении плазмид для увеличения выхода плаз-
мидной ДНК к культуре за несколько часов до сбора клеток
иногда добавляют хлорамфеникол.
Инициация репликации плазмиды R1 Е. coli регулируется
белком-репрессором, кодируемым одним из генов R1. Были по-
лучены мутантные плазмиды, которые кодируют аномальный
репрессор или же имеют несовершенный механизм регуляции
его образования. Некоторые из этих мутантов т—температуро-
чувствительные (репрессор неактивен при 43°C). При 30°C
репликация идет нормально, и на клетку образуется 1—2 плаз-
мидные копии, а при 43 °C репликация инициируется гораздо
чаще, так что в клетке накапливается по нескольку сот плаз-
мид. Небольшой сегмент R1, содержащий участок начала реп-
ликации и связанные с ними регулирующие элементы, был ис-
пользован для создания плазмид-векторов, применяемых при
клонировании разнообразных генов. -Преимущество таких век-
торов заключается в том, что при повышенной температуре с
их помощью можно получать множества копий клонируемого
гена, а следовательно, и много белка, кодируемого этим геном.
Важнейшие гены плазмид
Для биотехнологии особенно интересны те гены плазмид, в ко-
торых закодирована способность к фиксации азота и деграда-
ции органических соединений, а также факторы вирулентности
патогенных бактерий. ...
По-видимому, одним из крупных достижений генной инже-
нерии может оказаться использование бактерий или их генов
для передачи растениям способности фиксировать азот. На
корнях бобовых растений при участии бактерии Rhizobium об-
разуются клубеньки, в которых и происходит фиксация азота —
перевод его в форму, доступную растениям. Недавно было ус-
тановлено, что по крайней мере некоторые гены бактерий, де-
терминирующие образование клубеньков и фиксацию азота, ло-
кализованы в плазмидах. Впервые указания на это были полу-
чены в опытах по передаче плазмид при конъюгации от одного
штамма Rhizobium другому. Каждый конкретный вид из рода
Rhizobium обычно вступает в симбиотические отношения только
с одним определенным бобовым растением. Так, R. phaseoli
участвует в образовании клубеньков у фасоли, a R. legumino-
sarurn — у гороха. Было найдено, что варианты R. leguminosa-
гит, утратившие способность индуцировать образование клу-
беньков у гороха, вновь ее обретали, если путем конъюгации им
передавалась плазмида от штамма, обладающего такой способ-
ностью. Более того, если подобная конъюгативная плазмида
. Генетика и биотехнология- 307
передавалась от R. leguminosarum к R. phaseoli, то эти бакте-
рии с большей эффективностью индуцировали образование клу-
бщщца у гороха, чем у фасоли. Крупные плазмиды с мол. мас-
сой более 90-106, которые имеются у Rhizobium trifolii и R. me-
. liloti, также несут по меньшей мере часть генов, нужных для.
образования клубеньков и фиксации азота.
Плазмиды могут обусловливать увеличение вирулентности
патогенных бактерий. В некоторых случаях идентификация свя-
. занных с плазмидами факторов вирулентности помогает в борь-
бе с патогенными микроорганизмами. К числу наиболее изу-
ченных в этом отношении плазмид принадлежат плазмиды ви-
рулентности патогенных .штаммов Е.. coli.. Энтеротоксигенные'
штаммы Е. coli часто вызывают острую диарею у молодняка
животных, а у людей — «диарею путешественников». Плазми-
ды этих штаммов нередко несут ген температурочувствительно'-
го токсина, близкого к токсину холеры, и ген температуроус-
стойчивого токсина. - Энтеротоксигенные штаммы обычно со-
держат и плазмиды с генами фимбрий- (pili), которые называ-
ют также «факторами колонизации». Эти факторы находятся
на поверхности бактериальных, клеток, которые с их помощью»
прикрепляются к стенке тонкого кишечника. Штаммы, инфи-
цирующие определенный вид животных, обладают обычно оди-
наковым фактором колонизации. Так, у штаммов Е. coli, вызы-
вающих, диарею у свиней, в состав этих липких выростов обыч-
но входит антиген К88 или 987Р, в то время как у штаммов,,
выделенных от телят, образуется антиген К89. Потеря энт^ро-
токсигенными штаммами плазмид, кодирующих антигены —
факторы колонизации, сильно снижает их вирулентность, хотя»
- штаммы и продолжают образовывать токсины.
Одним из способов борьбы с заболеванием свиней, вызывае-
- мым энтеротокоигенными штаммами, была вакцинация живот-
- ных препаратом, содержащим антиген К88. Предпринимаются
попытки повысить выход применяемого в вакцинах антигена
К88, используя технодогию рекомбинантных ДНК- Эти методы-
нашли применение также и в производстве некоего токсоида,,
который можно не опасаясь вводить животным для образова-
ния у них антител к токсинам, продуцируемым энтеротоксиген-
ными бактериями. Другой подход в борьбе с диареей в живот-
новодстве состоит в выведении пород, устойчивых к кодируе-
мым плазмидами антигенам — факторам колонизации.
Транспозоны
. Транспозоны и вставочные последовательности — это сходные-
элементы в хромосомных ДНК бактерий, ДНК бактериофагов,
и плазмид. В опытах- с. бактериями транспозоны используются»
20
308
Глава 7
для получения мутантов и для клонирования нужных бакте-
риальных генов. Открыты они были Хеджесом и Жакобом, ко-
торые показали, что небольшой фрагмент плазмиды RP4, опре-
деляющий устойчивость к пенициллину, может встраиваться
в другие плазмиды, последовательность которых не гомологич-
на RP4. Обычный механизм рекомбинации в клетках бактерий,
•основанный на взаимодействии гомологичных молекул ДНК и
зависимый от продукта бактериального гена rec А+, в переносе
этого гена устойчивости к антибиотику не используется. В по-
следующие годы было описано множество других транспозонов.
Большинство из них было обнаружено в R-плазмидах грамот-
рицательных бактерий; обычно они придают им устойчивость
к антибиотикам. Лучше всего изучен транспозон ТпЗ длиною
4957 пар нуклеотидов, обеспечивающий устойчивость к пени-
циллину. Последовательности из 38 пар нуклеотидов на концах
ТпЗ одинаковы, но противоположно направлены. В транспозо-
не закодирована информация о ферменте транспозазе, играющем
важную роль в его перемещениях (транспозициях). Этот транс-
позон может встраиваться во множество сегментов молекулы
ДНК, но предпочитает АТ-богатые участки.
Транспозоны могут служить маркером гена, предназначен-
ного для клонирования. Как известно, при клонировании хро-
мосомных генов бактерий иногда возникают сложности, свя-
занные с тем, что нет простого метода, позволяющего выявить,
какая из плазмид, несущих встроенный фрагмент хромосомной
ДНК, содержит интересующий исследователя ген. Иногда эту
проблему удается решить, выделив сначала мутант по этому
гену с включенным в него или расположенным поблизости
транспозоном.
7.3.2. Слияние протопластов
С целью преодоления преград для генетического обмена, су-
ществующих в обычных системах скрещивания, был разработан
метод слияния протопластов (клеток с удаленными клеточны-
ми оболочками). Этот метод пригоден для получения межвидо-
вых и даже межродовых гибридов. Его можно использовать
при гибридизации клеток одного вида, которые принадлежат к
несовместимым группам спаривания или же когда природная
система скрещивания малоэффективна в плане образования ге-
нетических рекомбинантов.
Слияние протопластов бактерий
Метод слияния протопластов был впервые использован в опы-
тах с бактериями для получения межвидовых гибридов Bacillus
imegateriurnKBacillus subtilis. В природе эти бактерии не
Генетика и биотехнология
309
конъюгируют, но легко трансформируются изолированной
ДНК. Путем слияния протопластов, индуцируемого либо поли-
этиленгликолем, либо СаСЬ, получить их гибриды несложно.
При этом частота рекомбинации может достигать 10~5—10-4;
процесс нечувствителен к ДНКазе, следовательно, он осу-
ществляется без трансформации.
Этот метод нашел широкое применение при гибридизации
видов Streptomyces. Так, Streptomyces coelicolor обладает плаз-
мидозависимой системой фертильности или скрещивания
(с. 300), однако эта система малоэффективна, а многие исполь-
зуемые в промышленности виды Streptomyces вообще не спо-
собны к обычному скрещиванию. Если же применить метод
слияния протопластов, индуцируемого полиэтиленгликолем, то
'частоту рекомбинации удается повысить примерно на четыре
порядка по сравнению с наблюдаемой при обычном скрещива-
нии. Максимальная частота рекомбинации, достижимая при ра-
боте этим методом, составляет 10—20%• Штаммы не обяза-
тельно должны нести селективные генетические маркеры, по-
скольку при использовании этого метода возможен прямой от-
бор гибридов с нужным свойством. Это дает большие преиму-
щества при конструировании промышленно важных штаммов,
поскольку работа по введению подходящих маркеров нередко
превращается в утомительное трудоемкое занятие.
Слияние протопластов грибов
Образование гибридов грибов с помощью слияния протопластов
изучалось очень активно; этот метод нашел применение в про-
мышленности при создании штаммов Cephalosporium acremo-
nium, для которых характерны высокая скорость роста и боль-
шой выход цефалоспорина. Первые опыты по слиянию были
выполнены с ауксотрофными мутантами Geotrichum candidum,
у которых не обнаружено ни половой, ни парасексуальной си-
стем. Слияние протопластов осуществляли путем центрифугиро-
вания, причем прототрофные гетерокарионы образовывались с
низкой частотой. Впоследствии слияние обычно осуществляли,
добавляя ионы Са2++ или ПЭГ.
Межвидовые слияния удалось провести как в опытах с раз-
личными мицелиальными грибами, так и с дрожжами, среди
которых были и такие промышленно важные организмы, как
Penicillium chrysogenum, Aspergillus niger, Candida tropicalis,
Yarrowia lipolytica, Kduyveromyces lactis и Saccharomyces ce-
revisiae. Слияние обычно происходит с частотой 0,2—2%. Ве-
роятность слияния ядер, приводящего после елияния клеток к
образованию диплоидов, зависит от вида организма. Здесь
вскрыта определенная закономерность: мицелиальные грибы
310
Глава 7
дают гетерокарионы, диморфные дрожжи — нестабильные
диплоиды, а истинные дрожжи — стабильные диплоиды (рис.
7.4).
Межвидовые слияния также происходят с достаточно высо-
кой частотой, особенно у относительно близкородственных ви-
дов Aspergillus или Penicilliutn. Если сливаются ядра, то за
этим обычно следует потеря хромосом. Нередко после выра-
Г етерокарионы
Geotrichum candidum
Phycomyces blakesleeanus
Mucor racemosus
Гетерокарионы нечасто образующиеся
нестабильные диплоиды
Cephalosporium acremonium
Гетерокарионы и редкие стабильные диплоиды
Aspergillus
Penicillum
Гетерокарионы и часто образующиеся
стабильные диплоиды
Candida tropicalis
Нестабильные диплоиды
Yarrowia lipolytica
Стабильные диплоиды
Saccharomyces cerevisiae
Kluyveromyces lactis
Рис. 7.4. Результаты внутривидовых слияний протопластов.
щивания в неселективных условиях образуются споры с гапло-
идными ядрами лишь одного из двух родительских видов.
Предпринимались попытки осуществить слияние протопла-
стов дрожжей разных родов (например, CandidaxSaccharomy-
copsis, Уarrow lax Pichia, YarrowiaXKluyveromyces, Saccharo-
mycesXSchizosaccharomyces). Однако такие гибриды оказа-
лись нестабильными, а частота их образования очень низкой
(10-5—10-4). В последнем варианте обычно формировались
многоядерные клетки. Сообщалось об успешном слиянии про-
топластов мицелиальных грибов (целлюлолитический организм
Генетика и биотехнология
311,
Trichoderma reesii) и дрожжей (5. cerevisiae), но эти наблюде-
ния нуждаются в подтверждении.
При многих слияниях образуются нестабильные гетерока-
рионы, в которых обычно доминирует ядро одной из родитель-
ских форм. Впрочем, далеко не всегда это препятствует гене-
тическому обмену. Так, убедительно показана возможность
межъядерных генетических обменов в гетерокарионах Schizop-
hilum commune n Saccharomyces cerevisiae. Таким образом,
слияние протопластов может быть полезным методом установ-
ления гетерокариотического состояния, при котором возможен
в ограниченном объеме перенос генов.
Образование гибридов растений путем слияния протопластов
Протопласты растений можно получить путем механического
или ферментативного разрушения клеточных стенок. Такие про-
топласты— ценный инструмент в: руках генетика растений. Они
довольно быстро и эффективно регенерируют' Клеточную стен-
ку, у них легко индуцировать деление с образованием каллус-
ной ткани. Из каллуса можно вырастить растения, способные
цвести и давать всхожие семена. Доступность жизнеспособных
протопластов позволила генетикам растений проводить опыты
по мутагенезу и селекции на уровне единичной клетки, т. е.
примерно так, как это делают микробиологи. В результате, на-
пример, были получены растения табака, устойчивые к болезни
«wild-fire»: мутации у протопластов вызывали этилметансуль-
фонатом, а затем отбирали мутанты, устойчивые к действию
метионинсульфоксима. Дело в том, что токсин, образуемый
Pseudomonas tabaci (организмом, вызывающим упомянутое за-
болевание), — это тоже аналог метионина.
Слияние протопластов растений осуществляют с помощью
ряда агентов, включая азотнокислый натрий, гидроксид каль-
ция и ПЭГ. Наибольшая частота слияния наблюдается при
действии ПЭГ, но гибриды, полученные в присутствии кальция,
проще регенерировать в целое растение. Таким путем удалось
получить межсортовые внутривидовые гибриды, гибриды между
видами одного рода и даже разных родов. Опыты эти проводи-
лись на таких важных культурах, как картофель, табак и
спаржа.
Результатом слияния может быть простое сращение клеток
одной и той же родительской линии или образование гибридов
двух типов клеток. Как и для большинства других разновидно-
стей скрещивания, основная трудность здесь состоит в выборе
метода отбора гибридов; кроме того, работа с растениями силь-
но осложняется недостатком подходящих генетических марке-
ров. В большинстве тех случаев, когда гибриды удавалось по-
312
Глава 7
лучить путем слияния протопластов, гибридизация уже была
осуществлена обычными генетическими методами. Таким об-
разом, фенотип гибрида был уже известен, и его можно было
принять за стандарт при отборе. В этой новой области селек-
ции растений предстоит еще большая работа, и успех ее опре-
деляется тем, сможем ли мы использовать обширный набор-
маркеров (например, устойчивости к химическим веществам и
токсинам) или ауксотрофных мутантов растений.
7.3.3. Слияние клеток животных
Осуществить слияние клеток и получить внутри- или межви-
довые гибриды клеток млекопитающих методически проще, чем
в случае микроорганизмов или растений, так как клетки млеко-
питающих не имеют клеточной стенки, которую необходимо
удалять перед слиянием. Первый метод контролируемого слия-
ния клеток млекопитающих был основан на использовании ин-
активированного вируса Сендай. Этот вирус относится к группе
парамиксовирусов; его вирион покрыт липидсодержащей обо-
лочкой, сливающейся с мембраной клетки-хозяина и обеспечи-
вающей проникновение вируса в клетку. Вирус способствует
слиянию, связываясь одновременно с мембранами двух клеток.
В опытах по слиянию клеток млекопитающих используются
также некоторые химические вещества (ионы кальция, лизоле-
цитин, ПЭГ).
Отбор гибридов из смеси продуктов слияния лучше всего-
вести в селективных условиях, хотя для этого можно использо-
вать и трудоемкий метод скрининга. Так, отбор может происхо-
дить в результате комплементации ауксотрофных и температу-
рочувствительных мутаций, но чаще всего для этого использу-
ются маркеры устойчивости к химическим веществам. Послед-
ний метод был предложен Литтлфилдом, который провел опы-
ты по слиянию устойчивой к бромдезоксиуридину линии
(BUdRr) L-клеток мыши и еще одной линии, устойчивой к аза-
гуанину (AGr). В клетках BUdRr отсутствует фермент тимидин-
киназа (ТК), а в клетках AGr — инозинпирофосфорилаза
(IPP). Ни клетки ТК_1РР+, ни клетки TK+IPP- не растут на
среде, содержащей гипоксантин, аминоптерин и тимидин. Ами-
ноптерин ингибирует биосинтез нуклеотидов; трифосфаты пури-
новых и пиримидиновых нуклеозидов могут синтезироваться
только клетками, способными использовать и тимидин, и гипо-
ксантин. Для жизнедеятельности этим клеткам необходимы ак-
тивные ферменты ТК и IPP. Таким образом, среди продуктов
слияния TK+IPP-, ТК~1РР+ и TK+IPP+ способностью к росту
будут обладать лишь клетки, у которых активны оба фермента,.
Генетика и биотехнология
313
Использование таких приемов отбора позволило получить
гибриды не только путем слияния линий клеток одного вида,
но и межродовые гибриды клеток человека с клетками мыши
в крысы. Эти линии клеток грызунов/человека нестабильны,
'причем легче они теряют хромосомы человека, так что после
тридцати делений в процессе выращивания у них остается все-
то семь из 24 хромосом человека, изначально присутствовавших
в гибридной клетке. Этот процесс элиминации хромосом был
применен при картировании генома человека, поскольку та-
ким путем удается быстро локализовать определенные гены на
.хромосомах.
Моноклональные антитела
Юдин из результатов использования метода слияния клеток
млекопитающих чрезвычайно быстро нашел применение в био-
технологии: это линии клеток, полученных при гибридизации с
участием клеток миеломы (так называемые «гибридомы»), с
помощью которых могут вырабатываться моноклональные ан-
титела. Этот метод, разработанный в Кембридже Мильштейном
и его сотр., основан на создании «бессмертных» клеток, произ-
водящих антитела, за счет слияния их с клетками миеломы.
Большинство антигенов вызывает в организме образование
целого набора антител, но каждый отдельно взятый лимфоцит
цродуцирует лишь одно из них. Миеломы — это злокачествен-
ные новообразования иммунной системы; они развиваются в ре-
зультате неконтролируемой пролиферации одной линии лимфо-
цитов. При этом в больших количествах синтезируется один тип
белков-антител. Иными словами, миеломы являются природны-
ми продуцентами моноклональных антител. По методу Миль-
штейна проводят слияние нормальных (неопухолевых) лимфо-
цитов и клеток миеломы. Вначале полученные гибриды синте-
зируют смесь антител, включающую два типа антител роди-
тельских клеток, а также гибридные их формы, образующиеся
путем ассоциации тяжелых и легких цепей антител двух роди-
тельских форм. Впоследствии в результате элиминации хромо-
сом образуются клетки, способные к синтезу антител лишь од-
ного типа. Это могут быть либо антитела, закодированные в ге-
номе лимфоцита, либо антитела, характерные для клеток мие-
ломы. (Скрининг и обнаружение клона, синтезирующего иско-
мое антитело, ведут при помощи биохимических и иммунохими-
ческих методов. Схема этого метода дана на рис. 7.5.
Область возможного применения моноклональных антител
весьма обширна. Их используют в диагностике, например для
быстрого и точного типирования тканей и органов, предназна-
ченных для пересадки. Они находят применение при очистке
белков и других биологических соединений методом иммуноад-
314
Глава 7
Введение антигена
в организм мыши
Культура клеток
миеломы
Клетки
селезенки
Слияние
В полиэтиленгликоле
Клетки
миеломы.
Скрининг для отбора клонов,
образующих специфические антитела
против введенного антигена
Размножение нужных клонов
Крупномасштабное производство моноклональных-
антител с помощью:
1) выращивания большого количества клеток или
2) индукции опухолей,мышей путем инъекции
Рис. 7.5. Метод Мильштейна’.
Генетика'и биотехнология
315
сорбции. Секер и Берке использовали моноклональные антите-
ла для очистки больших количеств интерферона из лимфоцитов
человека (гл. 8). Очевидно, что такой подход пригоден и для
очистки интерферона, синтезируемого бактериями-рекомбинан-
тами. Возможности применения моноклональных антител для
пассивной Иммунизации ограничены, во всяком случае, у боль-
ных людей, поскольку пока не удалось получить стабильные
клоны на основе клеток человека.
7.4. Опыты по генетической инженерии in vitro
7,4.1. Технология
Для получения разнообразных белков эукариот и вирусов
животных широко применяются бактерии и дрожжи Saccharo-
myces cerevisiae. При этом используются самые разные методы,
но наиболее широко, те из дих, которые описаны ниже.
. Прежде всего необходимо изолировать нужный ген. Если
ген животного должен экспрессироваться в клетках бактерий
или дрожжей, то обычно вначале выделяют, соответствующую
мРНК. Как правило, осуществить прямую экспрессию генов
животных в бактериальных клетках не удается, поскольку эти
гены содержат интроны. Интроны — это декодирующие участки
гена; вся информация об аминокислотной последовательности
белка содержится, в других участкахэкзонах. Зрелые моле-
кулы мРНК клеток млекопитающих не содержат последова-
тельностей, комплементарных интронам: они удаляются фер-
ментами в процессе, называемом сплайсингом. Следовательно,
такие молекулы можно транскрибировать с помощью фермен-
тов ревертазы (обратной транскриптазы) и ДНК-полимеразы
с образованием ДНК.,Некоторые гены животных, например ко-
дирующие а-лейкоцитарные интерфероны, не содержат интро-
нов и могут прямо экспрессироваться в бактериальных или
дрожжевых клетках. Если в качестве исходного объекта прихо-
дится использовать мРНК, то прежде всего следует найти ее
источник — ткань, в которой этот продукт образуется в наи-
большем количестве. Иногда таким источником служат опухо-
левые ткани или же клетки в культуре, образующие необходи-
мую мРНК.
После транскрипции данной мРНК.,и получения комплемен-
тарной ДНК (кДНК) ‘ необходимо определить, какие из мно-
жества молекул этой кДНК содержат ген, подлежащий выде-
лению и экспрессии. На этом этапе работы молекулы кДНК
обычно вводят в состав плазмид или генома бактериофагов, ко-
торые служат векторами. Затем рекомбинантные плазмиды со
встроенными молекулами кДНК. переносят в клетки подходя-
щих бактерий-хозяев.
316
Глава 7
Идентификация искомых клонов занимает иногда много»
времени. Если же клонирование гена проведено, выявить сход-
ные гены в банках кДНК относительно несложно, если приме-
нить метод Грюнштейна и Хогнесса. В этом случае колонии
бактерий, содержащих рекомбинантные плазмиды, выращива-
ют на нитроцеллюлозных фильтрах, помещенных на поверх-
ность питательной среды. Затем бактериальные клетки разру-
шают, а высвободившуюся ДНК денатурируют. При этом она)
остается связанной с фильтром. Далее на фильтр наносят ра-
диоактивный зонд — денатурированную ДНК или РНК, кото-
рая связывается с гомологичной или частично гомологичной
бактериальной ДНК. Избыток радиоактивных молекул зонда,,
которые не связались специфически с гомологичной ДНК, уда-
ляют промыванием и выявляют лизированные колонии, связав-
шие зонд, методом радиоавтографии. Бактерии, для которых
наблюдалась положительная реакция в этом тесте, можно вы-
делить из дубликата — реплики набора колоний.
Если специфический зонд отсутствует, то его можно синте-
зировать химическими методами, если известна полностью или
частично аминокислотная последовательность белка — продук-
та интересующего нас гена. Конечно, определить точную нук-
леотидную последовательность, исходя из аминокислотной по-
следовательности, нельзя, поскольку каждая аминокислота мо-
жет кодироваться разными триплетами (от 1 до 4). Однако,
зная аминокислотную последовательность, можно синтезиро-
вать набор зондов и идентифицировать с их помощью плазмиды
с частично гомологичными последовательностями.
Другие методы идентификации рекомбинантных плазмид ос-
нованы на экспрессии включенного в них гена. При этом иден-
тифицируется белковый продукт, для чего используются имму-
нологические методы или методы определения его специфиче-
ской активности. Для обнаружения антигенов в бактериях пу-
тем скрининга, на основе специфического их взаимодействия с
антителами, часто используется метод, разработанный Брумом:
и Джилбертом. Для этого пластиковый диск покрывают слоем
специфических радиоактивных антител и помещают его на
предварительно лизированные колонии. Антигены из лизиро-
ванных бактерий связываются с антителами на диске. Положе-
ние антигенов, адсорбированных на диске, определяют с по-
мощью радиоавтографии.
Включение ДНК в плазмидные и фаговые векторы
Обычно выбор вектора определяется штаммом хозяина, кото-
рый используется для экспрессии клонированной ДНК. Если в
роли хозяина выступает Е. coli, плазмидный вектор скорее все-
Генетика и биотехнология
317
го представляет собой произ-
водное pBR322, а если хозяи-
ном является Bacillus spp., то
векторные плазмиды получают
из различных видов Bacillus
или Staphylococcus. Вектор для
Saccharomyces cerevisiae кон-
струируют либо на основе
плазмиды длиной 2 мкм, либо
из фрагментов хромосомы
дрожжей, способных реплици-
роваться подобно плазмидам.
Нередко используются чел-
ночные векторы, которые мо-
гут реплицироваться в одном
или нескольких организмах-
хозяевах. Применение таких
векторов оказывается весьма
....AGCT____
. ...TCGA....
t
... .GGATCC....
.. . .CCTAGG....
t
Рис. 7.6. Нуклеотидные последователь-
ности, узнаваемые и расщепляемые эн1
донуклеазами рестрикции А1и\ (ввер-
ху) и BamHl (внизу).
перспективным в связи с тем,
что нередко для наработки большого количества плазмид и для
трансформации ДНК удобнее в роли хозяина использовать
Е. coli. Строение плазмиды pBR322 показано на рис. 7.3. Она
несет гены устойчивости к тетрациклину и ампициллину. На
карте указаны места расщепления этой молекулы эндонуклеа-
зами рестрикции (рестриктазами).
Используемые при клонировании рестриктазы выделяют из-
различных прокариот. Всего было получено более ста пятидеся-
ти их разновидностей. Они отличаются друг от друга тем, что
узнают и расщепляют разные нуклеотидные последовательно-
сти. Сайты узнавания для большинства ферментов, используе-
мых в генетической инженерии, представляют собой палиндро-
мы из 4, 5 или 6 нуклеотидов. Расщепление происходит с обра-
зованием либо «тупых» (как в случае Alul), либо липких (спо-
собных к комплементарному связыванию) концов (как в случае-
5amHI; см. рис. 7.6). Неспаренные нуклеотиды липкого конца
оканчиваются З'-гидроксильной или 5'-монофосфатной группи-
ровкой в зависимости от фермента.
Обычный метод клонирования ДНК основан на расщепле-
нии плазмидной и встраиваемой ДНК одной и той же рестрик-
тазой. При этом получаются молекулы с липкими концами, ко-
торые затем отжигают, получают кольцевую рекомбинантную-
молекулу и сшивают ДНК-лигазой фага Т4. В случае плазми-
ды pBR322 встраивание обычно осуществляют по генам устой-
чивости к антибиотикам, поскольку такие рекомбинантные мо-
лекулы легко распознать по их неспособности обеспечить устойг
чивость.
318
ГлЬва 7 '
ДНК-лигазу фага Т4 можно применять и для сшивания «ту-
пых» концов молекул ДНК, хотя этот процесс осуществляется
труднее, чем в случае липких концов. Липкие концы можно
.превратить в тупые путем отщепления одноцепочечных высту-
пающих участков специфичными только к- таким участкам нук-
.леазами. Альтернативный способ основан на использовании
ДНК-полимеразЫ: она присоединяет к короткой Цепи недо-
стающие нуклеотиды, комплементарные б'-оДйоцепойечной вы-
ступающей части молекулы. Лигаза применяется также для
встраивания в плазмиды синтезированных фрагментов ДНК с
целью получения на стыке ДНК-вставки и вектора нужной по-
следовательности. Комплементарные одноЦепочечные концы у
вектора и встраиваемой ДНК могут быть образованы при по-
мощи концевой ДНК—‘НуклеотидйЛтрансферазы: она присоеди-
няет нуклеотиды к З'-кбнцам молекуй ДНК-?С ’её помощью к
З'-концам вектора ’могут быть добавлены «хвосты» из несколь-
ких остатков дезоксигуаниловой кислоты [oligo(dG)], а к кон-
;цам встраиваемой молекулы — комплементарная последова-
тельность oligo(dC). Смысл этой процедуры состоит в том, что
повторное лигирование вектора становится невозможным.
Трансформация
Полученные in vitro рекомбинантные шлазмиды необходимо пе-
ренести в подходящую клетку-хозяина, Природа которой и оп-
ределяет особенности способа трансформации. Так, клетки
Е. coli становятся компетентными ,(т. е. способными захваты-
ва.ть очищенную ДНК) после обработки их на холоду СаСЬ,
а цлетки Bacillus subtilis. приобретают компетентность на опре-
деленной фазе клеточного цикла при их разовом культивирова-
нии в условиях дефицита питательных веществ. Многие дру-
гие бактерии, включая виды Streptomyces, можно трансформи-
ровать только в форме протопластов, когда удалены клеточные
стенки. Для облегчения проникновения ДНК в дрожжевые
клетки после обработки их СаС12 или ПЭГ из этих клеток так-
же часто получают протопласту. .Интактные, дрожжевые клет-
ки становятся компетентными после обработки ионами щелоч-
ных металлов, например Ц+. .....
7.4.2. Экспрессия клонированных генов
Дель многих опытов по клонированию состоит в наработке в
большом количестве какого-либо белка эукариот. Именно для
этого и встраивают гецы эукариот, в плазмиды. бактерий. Что-
бы достичь высокого уровня экспрессии гена, эукариотическую
ДНК нужно встроить вблизи от активного промотора транск-
Генетика я биотехнология
31»
рипции; образующаяся мРНК при этом должна эффективно
транслироваться. Промоторы, использующиеся для обеспечения
активной экспрессии, обычно не принадлежат к числу изна-
чально присутствующих в плазмидах. Как правило, это бывают
промоторы фаговых или хромосомных генов, включаемые в
плазмидные векторы. Типичным примером такого рода служит
промотор PL бактериофага %. ДНК с таким промотором эффек-
тивно транскрибируется, и образуется большое количество
мРНК. Контроль за работой Pl осуществляется белком-репрес-
сором, продуктом гена Q этого бактериофага. Ген Ci857 кодиру-
ет температурочувствительный белок-репрессор, который акти-
вен при 30°С, но не работает при 42°С. Если внести этот ген
в составе плазмиды или фага в ту же клетку, что и промотор
Pl, то экспрессию клонированных генов, контролируемую PL,
можно регулировать, изменяя температуру.
У дрожжей Saccharomyces cerevisiae векторы с высоким-
уровнем экспрессии конструируют на основе промоторов генов,
которые кодируют важнейшие клеточные белки (например, ал-
когольдегидрогеназу и фосфоглицераткиназу). Так, с помощью-
промотора алкогбльдегидрогеназы С1 удалось добиться прямой
транскрипции в дрожжах гена инсулина человека, а в опытах
с использованием промотора фосфоглицераткиназы получен
высокий уровень экспрессии генов интерферонов.
С помощью бактерий были получены с высоким выходом не-
которые белки — продукты генов животных и их вирусов. Так,
были созданы штаммы Е. coli, у которых 20% всего клеточно-
го белка составляли коровый антиген вируса гепатита В, гор-
мон роста человека или главный капсидный антиген вируса
ящура. У одного из сконструированных штаммов В. subtilis
последний составлял около 1% синтезируемого этой бактерией
белка. Однако добиться экспрессии в бактериальных клетках
генов некоторых белков животных или их вирусов совсем не-
просто, даже если эти гены сопряжены с сигналами инициации
транскрипции и трансляции, которые обеспечивают в норме-
высокий уровень экспрессии генов прокариот. Причины такой
неэффективной экспрессии не всегда ясны, но в некоторых слу-
чаях удалось установить, что протеазы бактерий быстро разру-
шают белки животных и вирусов. В подобных ситуациях мож-
но повысить выход, применяя несодержащие протеаз мутанты.
При выработке проинсулина, предшественника инсулина, неко-
торая защита от протеаз обеспечивается тем, что полипептид
секрётируетсй в периплазматичёскбе пространство у клеточной
стенки Е. coli. На N-конце молекулы препроинсулина нахо-
дится последовательность гидрофобных аминокислот, с по-
мощью которой (с одновременным ее отщеплением) осуществ-
ляется транспорт этой' молекулы через мембрану в- перипла!з-
320
Глава 7
матическое пространство. Время полужизни проинсулина в
клетке 2 мин, а в периплазматическом пространстве 20 мин.
Другой способ получения чувствительных к протеазам бел-
ков основан на выделении их бактериями Bacillus в окружаю-
щую среду. Эти бациллы в отличие от Е. coll способны экскре-
тировать некоторые белки. Так, получены штаммы Bacillus,
осуществляющие экскрецию проинсулина. Выделенный в среду
€елок обычно проще очищать, чем внутриклеточный. Помимо
этого, использование штаммов Bacillus имеет то преимущество,
что они не образуют эндотоксинов. Выяснилось, однако, что
клетки Bacillus эффективно экскретируют, видимо, лишь неко-
торые типы полипептидов. В опытах с дрожжами проблема сек-
реции была решена путем создания векторов, содержащих сиг-
нальные последовательности генов a-фактора гормона спарива-
ния или токсина-«убийцы».
7.4.3. Прикладные аспекты генетической инженерии
Не вызывает сомнения, что методы генетической инженерии бу-
дут играть ведущую роль в развитии биотехнологии и найдут
в ней самое широкое применение. Уже сегодня с помощью бак-
терий и дрожжей мы получаем в больших количествах белки
эукариот и вирусов, которые применяются в медицине и вете-
ринарии. В табл. 7.1 перечислены некоторые белки, для синте-
Таблица 7.1. Белки эукариот и вирусов,
получаемые методом клонирования генов
в микроорганизмах
Инсулин человека
a-, [3- и 7-интерфероны
Коровый антиген вируса гепатита В
Поверхностный антиген вируса гепатита В
Гормон роста человека
Капсидный белок вируса ящура
Реннин теленка
за которых используется этот подход. Образование больших
количеств ненужного бактериальной клетке белка ставит ее в
неблагоприятное положение при отборе, так что приходится
решать сложные проблемы стабилизации таких штаммов. Их
удается решить — по крайней мере в случае выработки очень
ценных продуктов — путем создания жестких условий отбора,
направленных на сохранение плазмидного вектора, а иногда и
клонируемого в нем гена. Использование индуцибельных про-
моторов типа XPl препятствует постоянному синтезу продукта,
и он образуется лишь в результате индукции в определенное
.время клеточного цикла.
Генетика и биотехнология
321
Проблема стабилизации не возникает, если вместе с клони-
руемым геном клетка-хозяин получает определенные преиму-
щества при отборе. Так, методы генетической инженерии по-
зволяют расширить спектр используемых микроорганизмом
субстратов. Пивоварение, например, заинтересовано в создании
штаммов дрожжей, способных к секреции ферментов, разруша-
ющих крахмал и белки. Появление у организма ряда новых
ферментативных активностей может увеличить эффективность
его обмена веществ. Так, фирме ICI удалось повысить эффек-
тивность усвоения азота клетками Methylophilus methylotrop-
hus, продуцента БОО, введя в них ген фермента глутаматде-
гидрогеназы Е. coll (рис. 7.7). Это позволило увеличить выход
БОО примерно на 7% при росте бактерий на метаноле и ам-
миаке. Сегодня мы можем уже конструировать искусственные
линейные хромосомы дрожжей Saccharomyces cerevisiae, что в
принципе позволяет осуществлять стабильную модификацию
клеток путем формирования в них новых путей метаболизма.
Введение чужеродного гена в клетки организма-проду-
цента— это отнюдь не единственный генно-инженерный способ
конструирования новых штаммов. Иногда оказывается полез-
ным клонирование собственных генов организма. Так, если
известно, что определенная ферментативная реакция лимитиру-
ет скорость какого-то метаболитического процесса, то введе-
ние многих копий соответствующего гена в рекомбинантные
плазмиды может ликвидировать это «узкое место» благодаря
образованию большего числа молекул фермента. Главной об-
ластью применения «самоклонирования», видимо, может стать
направленный мутагенез. При обычном методе получения но-
вых штаммов с помощью мутагенеза и отбора действию мута-
гена подвергается весь геном организма-продуцента. При этом,
естественно, не гарантируется, что полезные мутации произой-
дут именно в интересующих нас генах. Мутируют также и дру-
гие гены, и некоторые из таких мутаций неблагоприятно по-
влияют на жизнеспособность организма-продуцента. Если же
провести клонирование нужных генов, то их обработку мута-
геном можно провести in vitro, а затем вернуть эти гены в ор-
ганизм. Это гарантирует получение только желаемых мутаций.
Основой развития фармацевтической промышленности до
недавнего времени был скрининг микроорганизмов и синтети-
ческих химических веществ. Появление новейших методов хи-
мического синтеза генов, возможно, позволит пойти другим
путем: конструирования случайных нуклеотидных последова-
тельностей, их клонирования и оценки биологической активно-
сти соответствующих белков и пептидов. Хотя занятие это мож-
но уподобить написанию сонета Шекспира группой печатающих
21—1344
322
Глава 7
А
(a) GDH путь-Е. соП Глутамат
2-Оксоглутарат + NHJ + NAD(P)H дегидр2ген.!^ Глутамат + NAD(P)+
Б
Лизогенизация донорного штамма gdh+ Е. соН транспозоном RP4.Mu
Скрещивание с реципиентным штаммом gdh Е. со// и отбор gdh*~
рекомбинантов'
Выделение плазмид RP4gdh
I
Субклонирование гена gdh в рТВ70-плазмидном векторе,
способном экспрессироваться и реплицироваться в Methy/ophUus
Перенос рТВ70 gdh в Methy/ophUus с помощью конъюгации при участии RP4
Выключение GOGAT-пути у Methy/ophilus с помощью мутагенеза:
при этом весь аммиак ассимилируется при участии фермента
глутаматдегидрогеназы, кодируемого геном gdhE. co/i
Рис. 7.7. Увеличение эффективности ассимиляции азота бактериями Methylo-
philus methylotrophus, используемыми для получения БОО. А. Пути ассимиля-
ции аммиака у бактерий. Путь (а) эффективнее, чем (б), так как в нем не
потребляется АТР. Б. Клонирование гена gdh Е. coll (гена глутаматдегидроге-
назы) и его введение в Methylophilus.
на машинке обезьян, случайная крупная находка поможет убе-
диться, что и такой стратегией поиска не следует пренебре-
гать.
Генетика и биотехнология
323
7.5. Итоги и перспективы
Новые разработки в области генетической теории и технологии
послужили основой для создания целого арсенала методов по-
лучения новых штаммов и путей модернизации технологиче-
ских процессов. Хорошо освоенные мутагенез и отбор будут
использоваться и далее, но специфичность этого метода станет
гораздо выше благодаря введению мутаций в клонированные
гены in vitro. Усовершенствования оборудования и способов
контроля за работой ферментеров существенно облегчат посто-
янный отбор, нацеленный на улучшение количественных харак-
теристик организма-продуцента.
Определяющую роль в развитии генетики клеток растений
и млекопитающих должно сыграть внедрение метода слияния
клеток. Уже сегодня важные средства диагностики — монокло-
нальные антитела — получают с помощью линий клеток-гибри-
дом; возможно, они внесут свой вклад и в развитие терапии.
Обычным способом скрещивания у видов Streptomyces стало
слияние протопластов, но при работе с грибами этот метод
имеет пока второстепенное значение. Открытие межъядерного
переноса генов у грибов позволит более сознательно использо-
вать метод слияния протопластов. Есть основания считать, что
генетическая инженерия привнесет важные изменения в меди-
цину и сельское хозяйство, и в немалой степени потому, что
технология рекомбинантных ДНК позволит нам глубже по-
нять главные молекулярно-биологические особенности клеток
растений и животных. Этот подход уже позволяет получать ин-
формацию о процессах, лежащих в основе таких сложных для
лечения болезней, как малярия и болезнь Чагаса.
С помощью методов генетической инженерии можно будет
получать продукты, которые не могли ранее применяться из-
за того, что выделять их из тканей животных было сложно и
дорого. К числу их относятся интерфероны. В клиниках сейчас
очень широко изучается эффективность синтезируемых бакте-
риями интерферонов при лечении вирусных заболеваний и раз-
ных форм рака. С внедрением биотехнологических подходов су-
щественно снизится стоимость уже используемых медицинских
средств, а инсулин человека, образуемый бактериями или
дрожжами, со временем, вероятно, заменит инсулин свиньи
(гл. 8).
Генетические свойства многих промышленных микроорга-
низмов изучены еще очень слабо. Нередко тому есть объектив-
ные причины, например отсутствие у них пола. Новые методы
генетики позволят быстрее и на более рациональной основе
улучшать такие штаммы.
21*
324
Глава 7
ЛИТЕРАТУРА
Beggs J. D., 1981. Gene cloning in yeast. In: Genetic Engineering, Vol. 2 (ed.
Williamson R.), pp. 175—203, Academic Press, London.
Bolivar F., Rodriguez R. L„ Greene P. J., Betlach M. C., Heynecker H. L„
Boyer H. W. (1977). Construction of new cloning vehicles II. A multipurpose
cloning system, Gene, 2, 95—113.
Broda P., 1979. Plasmids, W. H. Freeman and Co., Oxford.
Broome S., Gilbert W. (1978). Immunological screening method to detect specific
translation products, Proc. natn. Acad. Sci. USA, 75, 2746—2749.
Brown S. W., Oliver S. G. (1982). Isolation of ethanol-tolerant mutants of yeast
by continuous selection, Eur. J. appl. Microbiol. Biotechnol., 16, 119—122.
Caten С. E., 1981. Parasexual processes in fungi. In: The Fungal Nucleus (eds.
Gull K., Oliver S. G.), pp. 191—214, Cambridge University Press, Cambridge.
Ferenczy L., 1981. Microbial protoplast fusion. In: Genetics as a Tool in Micro-
biology (21st Symposium of the Society for General Microbiology; (eds. Glo-
ver S. W. and Hopwood D. A.), pp. 187—218, Cambridge University Press,
Cambridge.
Fincham J. R. S„ Day P. R., Radford A., 1979. Fungal Genetics, 4th edn., Black-
well Scientific Publications, Oxford.
Grunstein M., Hogness D. S. (1975). Colony hybridisation: A method for the
isolation of cloned DNAs that contain a specific gene, Proc. natn. Acad. Sci,
USA, 72, 3961—3965.
Hardy K. G., 1981. Bacterial Plasmids, Van Nostrand Reinhold, London.
Holloway B. W. (1979). Plasmids that mobilise the bacterial chromosome, Plas-
mid, 2, 1—19.
Hopwood D. A., 1981. Genetic studies of antibiotics and other secondary meta-
bolites. In: Genetics as a Tool In Microbiology (21st Symposium of the Society
for General Microbiology; eds. Glover S. W. and Hopwood D. A.), pp. 187—
218, Cambridge University Press, Cambridge.
Klechner N. (1981). Transposable elements in prokaryotes, Ann. Rev. Genet.,
15, 341—404.
Kohler F., Milstein C. (1975). Continuous culture of fused cells secreting anti-
body of predefined specificity, Nature, 256, 495—497.
Kubitschek H. E., 1970. Introduction to Research with Continuous Culture, Pren-
tice-Hall, Englewood Cliffs, New Jersey.
Maxam A., Gilbert W. (1980). Sequencing end-labelled DNA with base-specific
chemical cleavages, Meth. Enzym., 65, 499—560.
Milstein C. (1980). Monoclonal antibodies, Scient. Am., 243, no. 4, 56—64.
Old R. W., Primrose S. W., 1985. Principles of Gene Manipulation: An Introduc-
tion to Genetic Engineering, Blackwell Scientific Publications, Oxford.
Sanger F., Coulson A. R., Barrell B. G., Smith A. J. H., Roe B. A. (1980). Clo-
ning in single-stranded bacteriophage as an aid to rapid DNA sequencing, J.
Mol. Biol., 143, 161—178.
Shortle D., DiMaio D., Nathans D. (1981). Directed mutagenesis, Ann. Rev.
Genet., 15, 265—294.
Struhl K. (1983). The new yeast genetics, Nature, 305, 391—397.
Vasil I. K„ Ahuja M. R., Vasil V. (1978). Plant tissue cultures in genetics and
plant breeding, Adv. Genet., 20, 127—215.
Windass J. D., Worsey M. J., Pioli E. M., Pioli D., Barth P. T., Atherton К. T.,
Dart E. C., Byrom D., Powell K„ Senior P. J. (1980). Improved conversion of
methanol to single-cell protein by Methylophilus methylotrophus, Nature, 287,
396—401.
Williams B. G., Blattner F. R., 1980. Bacteriophage lambda vectors for DNA
cloning. In: Genetic Engineering, Principles and Methods, Vol. 2 (eds. Set-
low J. K. and Hollaender A.), Plenum Press, New York.
Глава 8
Медицина и биотехнология
Дж. Пикап
8.1. Введение
Особенно ярко новейшие успехи биотехнологии проявляются в
практической медицине главным образом потому, что их рас-
пространение из лабораторий в промышленность, а затем и в
клинику происходит в последние годы удивительно быстро. Так,
первые публикации об экспрессии гена инсулина человека в
Е. coll появились в 1979 г. Полученный на основе рекомбинант-
ных ДНК инсулин человека был испытан на добровольцах, не
страдающих диабетом, в 1980 г., а уже в 1981 г. велись раз-
вернутые клинические испытания.
Крупные открытия в науке обычно делаются при разработ-
ке фундаментальных проблем. Мы разделяем мнение большин-
ства врачей о том, что последние достижения биотехнологии,
нашедшие применение в самых важных отраслях медицины,
оказывают и будут оказывать революционизирующее воздейст-
вие на диагностику, лечение и понимание основ патологии мно-
гих тяжелых заболеваний. Ориентируясь на читателей, не име-
ющих медицинской подготовки, мы расскажем о том, какую
важную роль играют в клинической практике некоторые новые
подходы, а также широко используемые методы диагностики.
Мы по необходимости ограничимся лишь немногими примера-
ми, но читатель может без труда дополнить их множеством
других: использованием в терапии белков, которые можно син-
тезировать при помощи видоизмененных методами генетической
инженерии микроорганизмов, применением моноклональных
антител, ферментов и т. д. Мы не обсуждаем использующиеся
при этом технологические процессы сколько-нибудь подробно
(о них речь идет в других главах); исключение составляет лишь
раздел о синтезе инсулина человека: дело в том, что ин-
сулин был первым белком, полученным с помощью технологии
рекомбинантных ДНК и испытанным на людях, а также пер-
вым (или одним из первых) препаратом такого рода, нашед-
шим применение в клинике.
326
Глава 8
8.2. Немодифицированные и мутантные клетки
и синтезируемые ими соединения (см. также гл. 7)
8.2.1. Антибиотики
Можно считать, что клиническая биотехнология зародилась
с началом промышленного производства пенициллина в 40-х гг.
и его использования в терапии. По-видимому, применение этого
первого природного пенициллина повлияло на снижение забо-
леваемости и смертности больше, чем какого-либо другого пре-
парата, но, с другой стороны, поставило ряд новых проблем,
которые удалось решить опять-таки с помощью биотехнологии.
Во-первых, успешное применение пенициллина вызвало боль-
шую потребность в этом лекарственном препарате, и для ее
удовлетворения нужно было резко повысить выход пеницилли-
на при его производстве. Во-вторых, первый пенициллин —G
(бензилпенициллин) — действовал главным образом на грампо-
ложительные бактерии (например, Streptococci и Staphylococ-
ci), а нужно было получить антибиотики с более широким
спектром действия и/или активностью, поражающие и грамот-
рицательные бактерии типа Е. coli и Pseudomonas. В-третьих,
поскольку антибиотики вызывали аллергические реакции (чаще
всего незначительные, вроде сыпи на коже, но иногда и тяже-
лые, угрожающие жизни проявления анафилаксии), необходи-
мо было иметь целый набор антибактериальных средств, с тем
чтобы можно было выбрать из равноэффективных препаратов
такой, который не вызывал бы у больного аллергию. В-четвер-
тых, пенициллин нестабилен в кислой среде желудка и его
нельзя назначать для приема внутрь. Наконец, многие бакте-
рии приобретают устойчивость к антибиотикам. Классический
пример тому — образование стафилококками фермента пени-
циллиназы (правильнее, fi-лактамазы), который гидролизует
амидную связь в р-лактамном кольце пенициллина с образова-
нием фармакологически неактивной пенициллоиновой кислоты
(рис. 8.1).
Увеличить выход пенициллина при его производстве удалось
в основном благодаря последовательному использованию серии
мутантов исходного штамма Penicillium chrysogenum (получен-
ных в результате воздействия УФ- и рентгеновского облучения,
азотистого иприта и в результате спонтанного мутагенеза),
а также путем изменения условий выращивания. Были выделе-
ны новые антибиотики, эффективные в случае грамотрицатель-
ных бактерий: стрептомицин, синтезируемый нитчатыми бакте-
риями (Actinomycetes) рода Streptomycetes, и цефалоспорин,
продуцируемый плесневым грибом Cephalosporium. Хотя цефа-
лоспорины и относятся к р-лактамным антибиотикам, содержа-
щим р-лактамное ядро, аналогичное таковому у пенициллинов,
Медицина и биотехнология
327
Б ензилпенициллин
Ампициллин
H2N Н
С
/ \
НООС СН2.СН2.СН2.СО— NH — сн— сн сн2
I I I
со —N С —СН2.О.СО.СН3
хс^
соон
Цефалоспорин С
Рис. 8.1. Структурные формулы природного бензилпенициллина и сопоставле-
ние с полусинтетическими пенициллинами (ампициллином и клоксациллином)
и другим природным р-лактамным антибиотиком — цефалоспорином. А. Бен-
зилпенициллин; (а) — место отщепления боковой цепи от 6-аминопеницилла-
новой кислоты, (б) —место расщепления fl-лактамного кольца пенициллиназа-
ми. Б. Боковая цепь ампициллина. В. Боковая цепь клоксациллина. Г. Цефало-
спорин С.
по своему строению они существенно отличаются от последних,
что позволяет назначать их больным, у которых пенициллин
вызывает аллергию (рис. 8.1).
Путем замещения боковой цепи природной р-лактамной мо-
лекулы получено множество полусинтетических антибиотиков
с новыми свойствами: с другим спектром действия, чувстви-
'328
Глава 8
тельностью к пенициллиназе и содержимому желудочно-кишеч-
ного тракта и т. д. Вначале бензильную группу пенициллина G
удаляли химическими методами с образованием G-аминопени-
циллановой кислоты, но если в культуральную среду ввести
бактерии, образующие амидазы, то это превращение можно
осуществить и биологическим путем. Так, ампициллин (рис. 8.1)
является лолусинтетическим производным бензилпенициллина,
отличающимся от него всего лишь наличием добавочной ами-
ногруппы в боковой цепи. Тем не менее он активен при перо-
ральном введении и действует на широкий круг бактерий, в том
числе на некоторые грамотрицательные, вызывающие заболева-
ния органов дыхания (Haemophilus influenzae), пищеварения
(Schigella и Salmonella) и выделения (Е. coli, Proteus). Устой-
чив к кислоте и клоксациллин (рис. 8.1), и к тому же он не
разрушается (J-лактамазами. Его часто назначают вместе с ам-
пициллином тем больным, у которых обнаружены стафилокок-
ки, синтезирующие пенициллиназу, или же при «госпитальных
инфекциях», когда наличие таких ферментов весьма вероятно.
8.2.2. Культуры других клеток
Другими примерами использования интактных клеток с неиз-
мененным геномом для производства лекарственных препаратов
могут быть синтез интерферона культивируемыми клетками
лимфобластомы (процесс Wellcome Foundation, см. разд. 8.3.2)
и вирусных антигенов для выработки вакцин при выращивании
клеточных культур на инертных микроносителях.
8.2.3. Различные варианты биоконверсии (см. гл. 4)
Микроорганизмы используются и на отдельных стадиях синте-
за лекарственных веществ, который ранее осуществлялся пу-
тем многоступенчатых и дорогостоящих химических реакций.
Так, один из штаммов хлебной плесени, Rhizopus arrhizus, на
начальном этапе синтеза производного стероида, кортизона,
может гидроксилировать прогестерон по 11-му положению.
Применение подобной стратегии биоконверсии наряду с тради-
ционными химическими превращениями позволило получать
многие стероиды более простыми и дешевыми способами на ос-
нове стеролсодержащего растительного сырья. Именно благода-
ря этому такие стероиды, как преднизон, дексаметазон, тесто-
стерон и эстрадиол, могут сегодня широко применяться в кли-
нике. Некоторое представление об исключительной важности
этих веществ в терапии можно получить, ознакомившись с
табл. 8.1, где перечислены основные области применения сте-
роидов.
Медицина и биотехнология
329
Таблица 8.1. Примеры использования в клинике стероидных
препаратов
Стероид Заболевание
Глюкокортикоиды кортизон гидрокортизон преднизон преднизолон дексаметазон Заболевания крови: гемолитическая анемия, ост- рый лейкоз, идиопатическая тромбоцитопениче- ская пурпура, угнетение костного мозга Замещение: аддисонова болезнь Аллергии: астма, экзема (очаговая), анафилаксия Иммуносупрессия: последствия пересадки органов Аутоиммунные заболевания: системная красная волчанка, узелковый полиартериит, ревматоид- ный артрит, височный артериит Прочие: язвенный колит, саркоидоз, нефротиче-
Половые стероидные гор- моны тестостерон эстрадиол прогестерон ский синдром Замещение: гипогонадизм Гинекологические заболевания: симптомы мено- паузы, дисменорея, эндометриоз, избыточное кровотечение при менструации (менорагия), оральные контрацептивы Прочие: рак молочной железы, остеопороз
8.3. Модифицированные клетки и образуемые ими вещества
8.3.1. Моноклональные антитела
Еще до разработки технологии гибридом, позволившей полу-
чать гомогенные антитела, большое влияние на развитие кли-
нической медицины оказали «обычные» антитела. Отметим, что
наработка подходящих антител всегда осложнялась непредска-
зуемостью иммунного ответа, и их титр, как и перекрестные ре-
акции, варьировали от животного к животному и от одной пар-
тии сыворотки к другой. Это определялось тем, что данный ан-
тиген вызывает образование целого набора антител. В основу
метода получения практически неограниченного количества го-
могенных антител, которые находят самое широкое применение,
легла фундаментальная работа Кёлера и Мильштейна, выпол-
ненная в начале 70-х г. Рассмотрим несколько примеров.
Иммунологический анализ
Разработка Ялоу и Берсоном метода радиоиммунологического
анализа (РИА) оказала глубочайшее влияние на многие обла-
сти клинической медицины и науку вообще. Он позволяет оп-
ределять очень небольшие количества вещества путем вытес-
330
Глава 8
нения меченного радиоактивным изотопом антигена из комп-
лекса со специфическим антителом при добавлении все возра-
стающего количества немеченого испытуемого или стандартно-
го антигена. Особенно ярко достоинства метода проявились в
эндокринологии, так как «рабочая» концентрация гормонов
обычно очень невелика, а определение их при помощи биологи-
ческих методов анализа — долгая, утомительная, иногда попро-
сту неосуществимая процедура. Анализируемые вещества бы-
вают нестабильны даже вне условий анализа, а при анализе
их нередко приходится концентрировать; кроме того, они со-
держат примеси, которые могут обладать биологической ак-
тивностью, сходной с таковой у изучаемого гормона. Так, ин-
сулиноподобные факторы роста, соматомедины, можно опреде-
лять биологическими методами, но они не инактивируются ан-
тисыворотками против инсулина. Приведем лишь один пример
гормона, определение которого с помощью радиоиммунологиче-
ского анализа открыло принципиально новые возможности: это
пролактин. Применяя быстрый и относительно простой способ
его определения при помощи РИА в плазме большого числа
пациентов и здоровых добровольцев, удалось установить, что
повышенное содержание пролактина в плазме ассоциировано
с наличием наиболее распространенного типа опухолей перед-
него отдела гипофиза (пролактиномы), а нередко и с индуци-
руемыми пролактином галактореей и аменореей (отсутствием
менструаций), а также иными нарушениями менструального
цикла. Сегодня определение содержания пролактина стало
важной частью исследований бесплодия и аменореи. Приблизи-
тельно у 20% женщин с аменореей обнаруживается повышен-
ное содержание пролактина в плазме, причиной которого мо-
жеть быть опухоль гипофиза, прием лекарств, вызывающих
повышение уровня пролактина, и т. д.
Моноклональные антитела уже используются в коммерчес-
ских наборах для проведения радиоиммунологического анали-
за, и их доступность позволила предложить альтернативные
методы анализа, когда используется избыток меченых антител
(иммунорадиометрический анализ). Эти методы иногда обла-
дают большей чувствительностью.
Иммуногистохимия
Меченые антитела могут использоваться для изучения распре-
деления антигенов в срезах тканей с помощью как светового,
так и электронного микроскопа. При работе по общепринятому
«сэндвич-методу» на срез наносят немеченые специфические
антитела, отмывают их избыток, а затем добавляют антитело
против иммуноглобулина (обычно несущее флуоресцентную
Медицина и биотехнология 331
метку или связанное с ферментом). Таким образом осуществ-
ляется непрямое мечение изучаемого антигена. В биологиче-
ских исследованиях и в медицине применение этого метода ока-
залось исключительно плодотворным. Одним из примеров ис-
пользования обычных поликлональных антисывороток может
быть топографическое картирование различных типов клеток
в тканях, например островков Лангерганса. Иммуногистохими-
ческие опыты с применением различных антисывороток пока-
зали, что содержащие инсулин В-клетки располагаются в цент-
ральной части островков, а секретирующие глюкагон А-клет-
ки — на их периферии. На стыке двух слоев располагаются
D-клетки, образующие соматостатин. Возможно, такое взаимо-
расположение клеток имеет функциональное значение.
Примером современного метода использования монокло-
нальных антител для гистопатологической диагностики заболе-
ваний человека может быть анализ биоптатов (образцов тка-
ни) лимфатических узлов как способ определения типа опухо-
лей лимфатической системы (например, болезни Ходжкина,
различных лимфом и др.).
Методы иммуногистохимии находят также применение в
диагностике для определения природы клеток, давших начало
опухоли, по типу содержащегося в них антигена.
Типирование подлежащих пересадке тканей
Гуморальные и клеточные реакции, ответственные за оттор-
жение тканей и органов при межвидовой их пересадке или же
при пересадке пациенту, не состоящему в родстве с донором,
направлены в основном против так называемых антигенов ги-
стосовместимости, расположенных на поверхности клеток.
Впервые эти белки были открыты при изучении антигенов лей-
коцитов человека (HLA). Кодирующие их гены расположены
на 6-й хромосоме. Эти поверхностные маркеры играют ключе-
вую роль в узнавании организмом «своих» и «чужих» клеток.
Предварительный подбор HLA-гаплотипов донора и реципиента
существенно улучшил результаты операций по пересадке орга-
нов. Четыре из пяти главных локусов (А, В, С и Dr) комплек-
са генов HLA (рис. 8.2) кодируют серологически определенные
)г?
Рис. 8.2. Основные локусы HLA (А, В, С, D и Dr) на 6-й хромосоме. Гипоте-
тический ген иммунного ответа Dr, видимо, близок к локусу D. Связь между
HLA и подверженностью определенным заболеваниям может опосредоваться
именно этим локусом.
332
Глава 8
антигены, т. е. типируемые при помощи антител. В отличие от
них локус D отвечает за пролиферативный ответ культуры при
смешивании лимфоцитов, хотя он может быть таким же, как
в случае локуса Dr. Гены каждого локуса исключительно поли-
морфны: одному антигену соответствует до тридцати аллелей.
Типирование HLA-комплекса у пациентов можно существенно
улучшить путем применения моноклональных антител против
различных антигенов комплекса HLA, так как это повышает
надежность и воспроизводимость результатов.
Антигены HLA связаны также с некоторыми заболевания-
ми. Изучение частоты встречаемости их у населения позволило
выявить, что некоторые болезни возникают чаще в группах с
определенными специфическими гаплотипами HLA. Так, сте-
пень риска возникновения целиакии составляет около 73 для
гаплотипа D3, а ревматоидного артрита — 4 для D4, т. е. эти
заболевания встречаются соответственно в 73 или 4 раза чаще
у лиц, обладающих этими антигенами, чем у тех, кто этих ан-
тигенов не имеет. Таким образом, HLA-типирование позволяет
оценить вероятность возникновения определенного заболевания
в конкретной популяции и способствует осуществлению мер по
идентификации и предотвращению болезней (там, где такие ме-
ры предпринимаются).
Диагностика злокачественных новообразований
и наблюдение за ними
Известны несколько специфических опухолевых маркеров, ко-
торые с успехом используются в диагностике, прогнозировании
и выявлении распространения опухолей (т. е. метастазов). Не-
которые из них обнаруживаются в крови (обычно с помощью
метода радиоиммунологического анализа), а другие находят в
препаратах опухолей. Так, а-фетопротеин является главным
белком сыворотки плода; его содержание уменьшается в те-
чение первого года жизни. Определяя содержание а-фетопро-
теина в плазме при помощи метода РИА, удалось установить,
что оно повышается у многих больных с гепатомой (рак пече-
ни) и при раке семенников (тератоме). У многих больных,
страдающих раком прямой кишки, в плазме отмечается повы-
шенное содержание карциноэмбрионального антигена. Дальней-
шее его увеличение может служить указанием на неэффектив-
ность химио- или лучевой терапии.
Помимо того что моноклональные антитела применяются
как специфические реагенты в стандартных тестах, они могут
использоваться и для идентификации новых, более специфиче-
ских маркеров опухолей. Так, Ритсу и его сотр. (Ritz et al.,
1980) удалось получить моноклональные антитела к антигену
клеток при остром лимфолейкозе человека. Были также выде-
Медицина и биотехнология
333
-лены гомогенные антитела к клеткам злокачественной мелано-
мы человека (одна из разновидностей рака кожи), которые не
давали перекрестной реакции с нормальными клетками кожи.
Введение радиоактивных и флуоресцентных меток в опухоле-
специфичные антитела, вероятно, облегчит выявление метаста-
зов и оценку первичных реакций опухолей в ходе лечения как
in vivo, так и при анализе препаратов, полученных при биоп-
сии. Развитию новых способов лечения (см. далее) может спо-
собствовать направленное введение лекарственных препаратов,
присоединенных к антителам против данных опухолей.
Направленное введение лекарственных препаратов
Моноклональные антитела могут найти применение для вве-
дения лекарственных веществ и токсинов в определенную часть
тела (например, опухоль) либо путем их непосредственного
присоединения к таким веществам, либо путем связывания с
поверхностью липосом, содержащих внутри такие вещества.
Были получены комплексы антител к поверхностным анти-
генам раковых клеток со многими неспецифичными противора-
ковыми средствами, но далеко не всегда они оказывались эф-
фективными. Вероятно, наиболее многообещающим является
использование сильнодействующих растительных и бактериаль-
ных токсинов, одна молекула которых, как принято считать,
может вызвать гибель клетки. Молекулы токсина дифтерии и
рицина образованы двумя полипептидными цепями, связанны-
ми дисульфидными мостиками. Цепь В связывается с клеточ-
ной поверхностью, а цепь А, обладающая ферментативной ак-
тивностью, проникает внутрь клетки и нарушает работу ме-
ханизма биосинтеза белка. Были предприняты попытки заме-
нить В-цепь токсинов специфическими антителами, преимуще-
ственно гомогенными. Недавно был получен препарат моно-
клональных антител к антигену раковых клеток прямой кишки,
ассоциированных с A-цепью токсина, который избирательно
действует на эти клетки в культуре.
Получен ассоциат липосом, содержащих метатрексат (инги-
битор дигидрофолатредуктазы, противораковое средство), и мо-
ноклональных антител к антигенам гистосовместимости мышей.
Было показано, что он оказывает специфическое влияние на
клетки селезенки. Надо сказать, что исследования по направ-
ленному введению лекарственных веществ, содержащихся в
липосомах, только начинаются.
Передозировка лекарств
Антитела против лекарственных препаратов, например дигокси-
ага, могут оказаться полезными для снятия последствий его
334
Глава 8
передозировки, хотя сегодня и не ясно, нужно ли применять
их экстракорпорально, связанными с твердым носителем, через;
который циркулирует кровь, или же вводить в кровоток.
Получение важных для медицины веществ
Примером использования моноклональных антител для очист-
ки и крупномасштабного производства важных для медицины
веществ может быть работа Секера и Берке (Secher, Burke,.
1980). Им удалось ковалентно пришить мышиное моноклональ-
ное антитело против лейкоцитарного интерферона человека к.
твердому носителю — сефарозе. С помощью полученного таким
образом специфического иммуноадсорбента удалось за один
цикл очистить в 5000 раз лимфобластоидный интерферон. Важ-
ное преимущество этого способа заключается в том, что для
производства антител требуется очень немного частично очи-
щенного антигена, поскольку разработаны методы отбора ш
клонирования искомого гибрида.
8.3.2. Технология рекомбинантных ДНК. (см. также гл. 7)
Общеизвестно, что разработка методов изменения генетическо-
го аппарата клеток, позволяющих вводить в них чужеродные
гены, клонировать их, экспрессировать и получать нужные про-
дукты, совершила настоящую революцию в биологии. Эти до-
стижения находят самое широкое применение и в медицине.
Белки и пептиды, доступные совсем недавно лишь в очень не-
большом количестве, сегодня предполагается производить в.
массовом масштабе и использовать для лечения многих боль-
ных. Вот несколько тому примеров.
Инсулин
Сегодня 1—2% населения Англии, Европы и США страдают
диабетом, и приблизительно 20% этих больных не могут су-
ществовать без инъекций инсулина. Со времени проведения
первых опытов по использованию инсулина для лечения диабе-
та в 1922 г. этот гормон выделяли из поджелудочной железы
животных (коров или свиней). Существует несколько причин,
по которым целесообразна разработка методов крупномасштаб-
ного производства и выпуск на рынок инсулина человека: среди
них и коммерческие соображения, и эмоциональные факторы,
и потребности развития науки, и возможные преимущества при
лечении.
Инсулин как коров, так и свиней немного отличается па-
аминокислотной последовательности от инсулина человека. Осо-
Медицина и биотехнология
335
бенно близки инсулины человека и свиньи: у последнего лишь
С-концевой треонин В-цепи заменен на аланин. Инсулины ко-
ровы и человека различаются по трем аминокислотным остат-
кам, и именно этими различиями определяется повышенная
•иммуногенная активность инсулина коровы по сравнению с ин-
сулином свиньи. Почти у всех больных, которых лечат обычным
методом, т. е. вводят им инсулин коровы, в крови появляются
антитела к инсулину. Антигенные свойства инсулина частично
определяются и примесями в его препаратах. Скорее всего
именно образованием антител к инсулину объясняются некото-
рые незначительные побочные эффекты при инъекциях инсу-
лина коровы, например атрофия подкожной жировой прослой-
ки в месте повторного введения. В случае высокоочищенного
инсулина свиньи эти эффекты отсутствуют. Далее, антитела к
инсулину инактивируют его в кровотоке, так что больному
приходится вводить большие дозы, чем это необходимо. Анти-
тела, вероятно, влияют и на продолжительность биологическо-
го действия введенного инсулина. Надежных данных о том, что
антитела способны вызвать более долгосрочные и серьезные
нарушения, нет. Можно думать, что инсулин человека будет
еще менее иммуногенным, чем инсулин свиньи (это естествен-
но, поскольку он не является чужеродным белком).
Так как число больных диабетом и соответственно потреб-
ность в инсулине растут, то вполне возможно, что Доступность
его в будущем уменьшится из-за нехватки исходного сырья —
поджелудочной железы животных. Столь же очевидно, что из-
за чисто эмоциональных факторов лучше использовать инсулин
человека, чем каких-либо животных.
Ряд производителей инсулина во главе с фирмой Eli Lilly
and Со. использовали как основу для производства инсулина
человека технологию рекомбинатных ДНК. Согласно схеме
процесса, разработанного Eli Lilly в сотрудничестве с фирмой
Genenetech Inc. (рис. 8.3), на первом этапе воссоздают последо-
вательность ДНК по аминокислотной последовательности инсу-
лина, раздельно синтезируя искусственные гены его А- и В-це-
пей. На 5'-конце каждого из них располагается кодон метиони-
на (который в синтезированном белке оказывается N-конце-
вым), а на З'-конце— терминирующие последовательности.
Каждый из генов встраивают затем в ген р-галактозидазы
плазмид, а их в свою очередь вводят в клетки Е. coll. Посколь-
ку бактерии растят на среде с галактозой, а не с глюкозой, то
в них индуцируется синтез р-галактозидазы, а вместе с ней —
А- и В-цепей инсулина, присоединенных через остаток метиони-
на. После лизиса бактерий и обработки бромцианом, который
специфически расщепляет белки по остатку метионина, цепи
инсулина отделяют от р-галактозидазы (инсулин метионина не
336
Глава 8
Синтез гена А-цепи
Встраивание в плазмиду
Синтез гена В-цепи
Встраивание в плазмиду
Введение в Е. coli Введение в Е. СоП
Очистка
Лизис клеток
Расщепление бромцианом
A-цепь инсулина
В-цепь инсулина’
I I
S S
Инсулин
Рис. 8.3. Получение инсулина человека на основе технологии рекомбинантных
ДНК (Miller, Baxter, 1980).
содержит). Затем проводят очистку цепей и их объединение
(с низким выходом), в результате чего образуется нативный
двухцепочечный инсулин. Было показано, что он не содержит
белков Е. coli, эндотоксинов и пирогенных веществ, по физиче-
ским свойствам неотличим от инсулина из поджелудочной же-
лезы человека и в тест-системе (гипогликемия у животных)
проявляет полную биологическую активность.
Медицина и биотехнология
337
Содержание глюкозы В плазме, ммоль/л
Рис. 8.4. Изменение содержания глюкозы в плазме здоровых добровольцев под
действием малых и больших доз инсулина человека, полученного на основе
технологии рекомбинантных ДНК, и инсулина из поджелудочной железы
свиньи (Keen et al., 1980). Вверху — биологический ответ при подкожном вве-
дении, внизу — при внутривенном введении в течение 1 ч.
Впоследствии был испытан альтернативный метод: синтези-
ровали ген молекулы-предшественника, проинсулина, который
и вводили в Е. coli. После очистки проинсулина его расщепля-
ли трипсином и p-карбоксипептидазой и получали нативный
инсулин.
Инсулин человека, полученный с помощью Е. coli, оказался
первым «генно-инженерным» белком, испытанным на людях.
В опытах со здоровыми добровольцами было установлено, что
он безопасен, во всяком случае при краткосрочном применении
(не вызывает аллергических и иных нежелательных реакций)
и обладает практически одинаковой с инсулином свиньи спо-
собностью снижать уровень глюкозы в крови при введении его
под кожу или внутривенно (рис. 8.4). В настоящее время такой
22—1344
'338
Глава 8
инсулин человека получают множество диабетиков во всем ми-
ре. Этому предшествовали клинические испытания, в ходе ко-
торых изучались изменения метаболизма и иммунологические
эффекты. Сегодня это уже обычный метод лечения.
Интерферон
Интерфероны — это группа белков, открытых в ходе изучения
веществ, вырабатываемых клетками, зараженными вирусами.
•Они индуцируют как локальные, так и системные противови-
русные реакции в других клетках. Кроме того, интерфероны об-
ладают еще двумя важными свойствами: подавляют пролифе-
рацию клеток (и, таким образом, потенциально являются про-
тивоопухолевым средством) и модулируют иммунную систему.
Недавно была упорядочена классификация интерферонов. Их
делят на группы, включающие по нескольку белков: а (ранее —
лейкоцитарные интерфероны), р (интерфероны фибробластов)
и у (иммунные интерфероны).
До недавнего времени интерфероны были доступны лишь
•в небольшом количестве. Частично очищенные препараты полу-
чали главным образом из лейкоцитов человека. Из-за высокой
видоспецифичности для клинического применения необходимо
было синтезировать именно интерферон человека, но его нель-
зя было всесторонне исследовать в опытах на животных. Про-
ведение должным образом спланированных и широкомасштаб-
ных испытаний интерферона на больных вирусными заболева-
ниями и раковых больных было затруднено, но мы уже имеем
какое-то представление о потенциальных возможностях этого
препарата. Так, лейкоцитарный интерферон существенно
уменьшает распространение пузырьков и продолжительность
их формирования, а также снимает боль у больных с одной из
форм герпеса (herpes zoster, опоясывающий лишай). Гепа-
тит В также поддается лечению интерфероном (см. ниже). Сре-
ди онкологических больных были проведены испытания на па-
циентах с метастазирующим раком молочной железы, и у две-
надцати из сорока трех диаметр опухоли уменьшился по мень-
шей мере на 50%. Изучалось его действие на лимфомы (иные,
чем лимфома Ходжкина), остеосаркомы и злокачественные ме-
ланомы. Однако здесь нужно проделать большую работу по
подбору оптимальных доз, частоты и способа введения интер-
ферона и выяснению эффективности комплексного лечения с
применением других противоопухолевых средств. Неоднократно
сообщалось о побочных эффектах (лихорадка, общее недомога-
ние, потеря веса) при применении интерферона.
Понятно, что в изучении, клинических испытаниях и внед-
рении интерферона в практику будет достигнут большой про-
Медицина и биотехнология 339*
гресс, если удастся получать в большом количестве чистый
препарат на основе технологии рекомбинатных ДНК. Уже
синтезирован ген лейкоцитарного интерферона человека длиной:
514 пар нуклеотидов; его включали в плазмиду и клонирова-
ли затем в Е. coli. Удалось достичь экспрессии гена интерферо-
на человека в клетках дрожжей, а ген фибробластного интер-
ферона был включен в Е. coli. За его экспрессией можно было
проследить по образованию продукта с противовирусной ак-
тивностью. Интересен сам факт проявления биологической ак-
тивности, так как этот природный интерферон гликозилирован,,
а получаемый по технологии рекомбинантных ДНК — нет..
Производство интерферона на основе рекомбинантных ДНК ос-
воили уже по меньшей мере две фармацевтические компании.
Начались и клинические испытания этих препаратов.
Гормон роста
Гормон роста человека — это белок, состоящий из 191 амино-
кислотного остатка и имеющий мол. массу 22 000. Он образу-
ется и секретируется передней долей гипофиза и необходим для
роста костей. Выяснено, что у 7—10 людей на 1 млн. этот гор-
мон образуется в недостаточном количестве, что приводит к за-
держке роста (карликовости). Хотя это заболевание обычно
врожденное, задержка роста становится заметной лишь в более
позднем, детском возрасте, так как гормон не нужен для внут-
риутробного развития. Строение гормона роста видоспецифич-
но, и в клинике можно применять лишь гормон роста человека.
До недавнего времени его получали из гипофиза трупов, но
по вполне понятным причинам этот способ имеет свой ограни-
чения. Фармацевтическая компания Kabi Vitrum в сотрудниче-
стве с Genentech Inc. предприняла попытки наладить произ-
водство гормона роста на основе технологии рекомбинантных
ДНК с использованием Е. coli. Можно надеяться, что с по-
мощью такого подхода мы вскоре не будем испытывать в нем
недостатка. Очищенный препарат гормона из бактерий по био-
логической активности подобен гормону из гипофиза. Сейчас
ведутся клинические испытания: гормон роста вводят детям,,
страдающим от его недостатка.
Вакцины
Гепатит В — заболевание, при котором трудно получить под-
ходящий антиген для иммунизации. Исход этой вирусной бо-
лезни печени трудно прогнозировать, особенно у пожилых и ос-
лабленных больных. Заражение происходит при переливании
крови и плазмы, при использовании плохо стерилизованных
22*
340
Глава 8
шприцев, игл и других медицинских инструментов, а также при
половых сношениях. К числу групп с высокой степенью риска,
тде может помочь вакцинация, относятся больные с синдромом
приобретенного иммунодефицита, раковые больные, которым
необходимо переливать кровь в большом количестве, а также
лица, проводящие гемодиализ, осуществляющие транспланта-
ции, персонал онкологических учреждений, наркоманы, умст-
венно отсталые и обслуживающие их, проститутки и гомосек-
суалисты.
Вирус гепатита В не размножается в культуре тканей. Вак-
цины против него получали путем тепловой обработки сыво-
ротки бессимптомных носителей. Впоследствии была предло-
жена вакцина на основе инактивированных сферических части-
чек его поверхностного антигена (HBsAg), присутствовавших в
сыворотке носителей. Эти вакцины эффективны и безопасны,
однако для их получения необходимо большое количество
смешанной сыворотки, осуществление дорогостоящих защитных
мер и крупные затраты на оборудование.
Альтернативным способом получения антигена может быть
наработка белков вируса гепатита В клетками прокариот, ко-
торый и был разработан Эдманом и др. (Edman et al., 1981).
Другой подход заключается в инокуляции пептидов синтетиче-
ского поверхностного антигена. Исходя из нуклеотидной после-
довательности вирусного генома была реконструирована ами-
нокислотная последовательность HBsAg, и многие исследова-
тели попытались с помощью ЭВМ выявить в ней антигенные
последовательности. Так, Дрисману и др. (Dreesman et al.,
1982) удалось синтезировать два циклических пептида, которые
при одноразовом введении вызывали образование антител у
мышей.
Ферменты
Ферменты составляют основу многих тестов, используемых в
клинической медицине. Они все чаще применяются при авто-
матизированном анализе и биохимическом скрининге жидко-
стей тела, которые ведутся в биохимических лабораториях сов-
ременных клиник. Примером таких ферментов могут быть глю-
козооксидаза, гексокиназа, холестеролоксидаза и эстераза, уре-
аза, уриказа, лактатдегидрогеназа и алкогольдегидрогеназа.
Иногда ферменты применяют и в терапии (например, стрепто-
киназу или урокиназу; они оказывают мощное фибринолитиче-
ское действие при тяжелом тромбозе сосудов).
В будущем ферменты (скорее всего иммобилизованные)
найдут, вероятно, значительно более широкое применение в
составе фермент-содержащих электродов, используемых для
Медицина и биотехнология
341
мониторинга in vivo и in vitro. Много подобных устройств уже
сконструировано, но в клинике не применяется. Так, многими
учеными был разработан электрод с глюкозооксидазой. В уст-
ройстве Апдайка и Хикса глюкозооксидаза наслоена непосред-
ственно на поверхность обычного платинового электрода
(рис. 8.5). Чем больше кислорода потребляется в реакции
Платиновый катод
Рис. 8.5. Устройство ферментного электрода с глюкозооксидазой (Updike,
Hicks, 1967).
Глюкоза-)-Кислород = Глюконовая кислота+Перекись водоро-
да, тем меньше количество его регистрируется внутренней
частью электрода. К сожалению, это устройство все еще рабо-
тает недостаточно надежно, что не позволяет использовать его
как имплантируемый аппарат для постоянной регистрации со-
держания глюкозы. Возникающие здесь проблемы связаны с
наличием конкурентных отношений между глюкозой и кислоро-
дом в жидкостях тела, инактивацией фермента in vivo, слож-
ностью калибровки и дрейфом характеристик электрода. Веду-
щиеся интенсивные исследования позволяют надеяться, что,
усовершенствовав такие электроды с ферментами, удастся со
временем создать датчик глюкозы для автономно работающего,
полностью автоматического и небольшого по размеру протеза
поджелудочной железы, нужного для лечения больных диабе-
том. В этой связи особенно интересны последние достижения в
области разработки фермент-содержащих электродов. Исследо-
ватели, работающие в Крэнфилдском технологическом инсти-
туте Оксфордского университета и в госпитале Гая в Лондоне,
разрабатывают глюкозный электрод, в котором для переноса
342
Глава 8
электронов от простетической группы (ФАД) глюкозооксидазы
на графитовый электрод используется органический медиатор-
(например, ферроцен), т. е. процесс идет без участия кислоро-
да, который обычно выступает в роли конечного акцептора
электронов. Работа этого электрода, таким образом, не связана
с кислородом. Он может оказаться особенно полезным при соз-
дании имплантируемого, способного работать in vivo устройст-
ва для больных диабетом.
По-видимому, основные усилия в ближайшие несколько
лет будут направлены на развитие технологии биодатчиков.
Ферменты (и другие лиганды, например антитела; см. разд.
8.5) могут оказаться весьма полезными для контроля за кон-
центрацией (мониторинга) разнообразных веществ, интересую-
щих клиницистов: промежуточных метаболитов, лекарственных:
препаратов и гормонов. Свою роль сыграет здесь биотехноло-
гия: она предоставит и обычные, и редкие ферменты микробов,,
полученные как путем крупномасштабного их выращивания^,
так и, возможно, с помощью технологии рекомбинантных.
ДНК.
В качестве примера возможного применения ферментов в
терапии укажем на предложение вводить инкапсулированные
в липосомах ферменты для лечения некоторых болезней, свя-
занных с накоплением лизосом. Такой способ был испытан на
нескольких больных болезнью Гоше, с врожденной недостаточ-
ностью по глюкоцереброзид : р-глюкозидазе. Полученные резуль-
таты не позволяют сделать однозначных выводов.
8.4. Применение методов молекулярной генетики
и технологии рекомбинантных ДНК в медицинской
диагностике и патологии
8.4.1. Пренатальная диагностика наследственных болезней
Многие методы молекулярной генетики начинают широко при-
меняться в пренатальной диагностике наследственных болезней,,
например гемоглобинопатий. Так, в 1978 г. Кен и Доузи раз-
работали метод диагностики серповидноклеточной анемии путем-
анализа ДНК из клеток околоплодной жидкости. Это несрав-
ненно более безопасный метод, чем взятие для анализа крови
плода, когда вероятность аборта доходит до 7%. Серповидно-
клеточная анемия — это одно из наиболее часто встречающихся
нарушений синтеза гемоглобина. Она развивается в результате
замены глутаминовой кислоты в 6-м положении p-цепи гемогло-
бина на валин. При дезоксигенации эритроциты, содержащие
аномальный HbS (две обычные а- и две аномальные р-цепи),
приобретают форму полумесяца (серповидную). Такие негибкие
Медицина и биотехнология
343
серповидные клетки затрудняют кровообращение в капиллярах
л быстро разрушаются. Клиническими проявлениями болезни
являются анемия, прекращение роста, приступы острой боли
(кризы), а также острые или хронические заболевания органов
вследствие закупорки сосудов (например, инсульт).
В работе Кена и Доузи (рис. 8.6) ДНК выделяли из лейко-
Фрагменты ДН К
Л-----1 Н НИН (один из них с геном глобина)
Разделение с помощью
электрофореза
Ретикулоциты
Выделение РНК
Глобиновая мРНК
Обратная 32
транскриптаза +
Е —{ Глобиновая 32Р-кДНК
*
Г ибридизация
Глобинспецифичный участок ДНК,
локализованный методом
радиоавтографии
Рис. 8.6. Схема, иллюстрирующая использование генных зондов и анализ по-
лиморфизма длины рестрикционных фрагментов в пренатальной диагностике
гемоглобинопатий, например серповидноклеточной анемии (Emery, 1981).
щитов крови двух родителей, их ребенка, страдающего серпо-
видноклеточной анемией, и здорового добровольца (контроль).
.ДНК была выделена также из клеток околоплодной жидкости,
отобранной в ходе третьей беременности матери. Вошедший не-
давно в практику метод картирования с помощью эндонуклеаз
рестрикции (гл. 7) позволяет сопоставлять разные ДНК; мече-
ную Р32-кДНК, комплементарную глобиновому гену, можно по-
дучить путем выделения глобиновой мРНК из ретикулоцитов
щ использования обратной транскриптазы, с помощью которой
'из Р32-нуклеотидов синтезируется кДНК. При расщеплении со-
шоставляемых ДНК эндонуклеазой (в данном случае —Нра!)
344
Глава 8
образуются фрагменты разной длины, которые разделяют ме-
тодом гель-электрофореза. Затем фрагменты переносят на нит-
роцеллюлозный фильтр и проводят гибридизацию с радиоактив-
ной кДНК-зондом (метод саузерн-блотинг). Нуклеотидные
последовательности, связавшие зонд, локализуют методом ра-
диоавтографии. Именно они содержат гены глобина. У здоро-
вого человека ген (3-глобина локализован во фрагменте длиной
7,6 kbp. У больного ребенка он был обнаружен во фрагменте
длиной 13,0 kbp, а анализ распределения фрагментов расщеп-
ленной ДНК у обоих родителей и в клетках плода выявил
участки длиной как 7,6, так и 13,0 kbp (т. е. они являются ге-
терозиготами и несут локус серповидноклеточности). Полимор-
физм определяется вариациями в локализации сайта узнавания
эндонуклеазой, расположенном приблизительно на расстоянии
5000 нуклеотидов от З'-конца гена.
Сходный метод был применен для пренатальной диагностики
(3-талассемии. При этой гемоглобинопатии (3-цепь гемоглобина
синтезируется либо в недостаточном количестве, либо не син-
тезируется вообще. При использовании метода определения по-
лиморфизма рестрикционных фрагментов по длине (RFLP) глу-
боких знаний о молекулярных основах патологических измене-
ний при данном заболевании не требуется. Это по сути «метод,
отпечатков пальцев», при помощи которого выявляются анома-
лии, сопутствующие клиническим симптомам, и поэтому он мо-
жет применяться при лечении многих наследственных болезней..
8.4.2. Роль генетических факторов в патологии
Примером генетически обусловленного заболевания может быть,
и диабет, но механизм наследования и молекулярная основа,
его остаются неясными. У пациентов группы 1, страдающих
юношеским диабетом, наблюдается полная или почти полная
гибель (3-клеток островков Лангерганса, и инсулин у них не об-
разуется. Такая разновидность диабета чаще всего встречается
у гаплотипов Dr3 и Dr4 HLA. В группе 2 (диабет взрослых)
уровень инсулина в крови больных близок к норме или повы-
шен; аномалии у них иные, и среди них — нечувствительность
рецепторов к инсулину. Они и приводят к недостатку инсулина.
У больных диабетом группы 2 взаимосвязи с определенными
типами HLA не выявлено. Ротвейн и др. (Rotwein et al., 1981)
использовали метод RFLP для анализа ДНК 35 здоровых лю-
дей, 17 больных диабетом из группы 1 и 35 — из группы 2..
У 26% здоровых людей в последовательности ДНК, прилегаю-
щей к 5'-концу гена инсулина, были обнаружены вставки дли-
ной 1,5—3,4 kbp. Такие же вставки присутствуют и в ДНК
35% больных группы 1 и 66%—группы 2 (рис. 8.7). Была
Медицина и биотехнология
345
определена полная нуклеотидная последовательность этого по-
лиморфного участка, прилегающего к гену инсулина, и показа-
но, что различия определяются числом и расположением повто-
ряющихся последовательностей. Из ряда опытов следует, что
последовательность прилежащего к 5'-концу гена участка может
сказываться на экспрессии гена. В этой связи существенно
<5олее высокий полиморфизм этого участка при диабете типа 2
представляет особый интерес (следует, впрочем, отметить, что
некоторым исследователям не удалось подтвердить наличия та-
кой взаимосвязи). Сегодня мы еще не можем утверждать, что
эти изменения достаточны или обязательны для развития бо-
Рис. 8.7. Полиморфизм длины рестрикционных фрагментов б'-фланкирующего
участка гена инсулина человека. Указано место вставок длиной 1,5 и 3,4 kbp.
Pre — участок, кодирующий аминокислотную последовательность, примыкаю-
щую к N-концу проинсулина, которая, видимо, способствует его секреции в эн-
доплазматический ретикулум; В, С и А — последовательности, кодирующие
В-цепь, кониекторный пептид и А-цепь инсулина. Заштрихованный участок —
интрон (гл. 7).
лезни, и трактовку результатов нельзя считать однозначной.
Еще два наследственных заболевания, точная генетическая
основа которых нам не известна, но для которых недавно была
установлена взаимосвязь между особым типом полиморфизма
ДНК и наличием «больного» гена — это мышечная дистрофия
Дюшена и хорея Хантингтона. Последнее заболевание неизле-
чимо, наследуется по аутосомному доминантному механизму и
выражается в прогрессирующем слабоумии и параличе, насту-
пающем на тридцатом — сороковом году жизни. К сожалению,
до недавнего времени мы не располагали методом выявления
носителей такого гена. Поскольку здесь налицо определенная
связь с маркерами ДНК, ее следует иметь в виду при генети-
ческом консультировании, и возможно, что со временем на этой
основе будут идентифицированы и сам ген хореи Хантингтона,
и соответствующий продукт.
346
Глава 8
8.5. Перспективы развития
Развитие новых направлений биосенсометрии, видимо, будет за-
висеть от успехов микроэлектроники, основанной на применении
продуктов биотехнологии, например ферментов и антител. Не-
давно были созданы ион-селективные транзисторы на основе-
полевого эффекта (ЕЕТ’ы): в таких устройствах на изолирую-
щий слой транзистора помещается мембрана с избирательной
проницаемостью. На поверхность транзистора можно было бы
нанести ферменты и антитела так, чтобы он «чувствовал» свя-
зывание белка и/или возможные его конформационные измене-
ния и/или реакцию с субстратом. Первым шагом здесь явля-
ется разработка чувствительного к пенициллину FET’a, в кото-
ром применен фермент пенициллиназа (Caras, Janata, 1980).
Зададимся теперь вопросом, можно ли лечить болезни чело-
века путем трансплантации нормальных и измененных генети-
ческими методами культивируемых клеток либо тканей орга-
низма-хозяина, видоизмененных методами клеточной инженерии?
Напомним, что интерес к лечению наследственных аномалий
обмена веществ методом трансплантации недавно вновь возрос
в результате изучения эпителиальных клеток амниона человека.
Эти клетки легко получить из плаценты и культивировать. Они
синтезируют и выделяют в среду ферменты, отсутствующие при
некоторых заболеваниях, связанных с накоплением лизосом,,
а кроме того, по-видимому, не образуют поверхностных антиге-
нов и поэтому не будут отторгаться реципиентом.
Надо сказать, что лечение заболеваний, развивающихся из-за
недостатка соединений, образуемых в норме определенным ти-
пом клеток (например, гормонов, синтезируемых эндокринными
железами, или молекул глобинов, формируемых клетками кост-
ного мозга), — задача архисложная. Несингенные клетки чело-
века (взятые от трупа, плода или от неидентичного донора),,
образующие нужное вещество, будут неизменно отторгнуты, ес-
ли только поверхностные антигены HLA или иммунологический
ответ на них не модифицировать или подавить каким-то обра-
зом. Можно также попытаться изменить геном одной из клеточ-
ных линий самого пациента (например, фибробластов) путем
встраивания генов (с использованием ДНК-трансформации или
рекомбинантных вирусных векторов), дерепрессии или же ме-
тодом слияния клеток. Здесь сразу же возникают опасения, свя-
занные с возможностью реимплантации людям генетически не-
полноценных, зараженных вирусами клеток, риском неопроли-
ферации. Следует учитывать, что вместе с синтетическим геном
клетке, видимо, должна быть передана способность и к осуще-
ствлению других необходимых функций. Например, для лечения
диабета клетку нужно «научить» не только вырабатывать ин-
Медицина и биотехнология
347
сулин, но и упаковывать его в секреторные гранулы, регулиро-
вать его синтез и экзоцитоз в соответствии с содержанием
глюкозы в крови (т. е. нужен еще и некий датчик глюкозы).
Механизм генетического контроля этого процесса, видимо, очень
сложен, и сегодня мы о нем ничего не знаем.
Высказывалось мнение, что методы цитогенетической моди-
фикации в первую очередь следовало бы применить для лечения
таких потенциально летальных болезней, как серповидноклеточ-
ная анемия или талассемия. В этих случаях болезнь возникает
яз-за аномалий в одном гене или комплексе генов, а экспрессия
происходит в одном типе тканей. Плюрипотентные кроветворные
стволовые клетки костного мозга продолжают реплицироваться
всю жизнь, и, если модифицировать их генетическими методами
а реимплантировать в костный мозг, они смогут стать важным
компонентом синтизирующей гемоглобин ткани. Впрочем, если
сегодня такие исследователи, как Меркола и Клайн, уже ставят
вопрос о том, не пришло ли время экспериментов по трансплан-
тации генов человека, то многие в противовес им указывают,
что, прежде чем двигаться дальше, нам нужно провести мно-
жество опытов на животных, оценить степень риска и т. д. Нет
сомнения, что в будущем, кроме того, придется решать пробле-
му совмещения научных и медицинских возможностей биотех-
нологии и связанных с их использованием вопросов этики.
ЛИТЕРАТУРА
Антибиотики и биоконверсия
Aharonowitz У., Cohen G. (1981). The microbiological production of pharmaceuti-
cals, Scient. Am., 245, 106—118.
Finter N. B., Fantes К. H., 1980. The purity and safety of interferons prepared
for clinical use: the case for lymphoblastoid interferon. In: Interferon 2 (ed.
Gresser I.), pp. 65—80, Academic Press, London, New York.
Van Wesel A. L„ van Steenis G., Hannick C. A., Cohen H. (1978). New approach
to the production of concentrated and purified inactivated polio and rabies tis-
sue culture vaccines, Devi Biol. Standard, 41, 159—168.
Моноклональные антитела
Belchetz P. E., Braidman I. P., Cralwy J. C. W., Gregoriadis G. (1977). Treat-
ment of Gaucher’s disease with liposome-entrapped glucocerebroside: P-glucosi-
dase, Lancet, ii, 116—117.
Buckman R. (1982). Tumour markers in clinical practice, Br. J. Hosp. Med., 27,
9—20.
Coons A. H., Kaplan M. H. (1950). Localisation of antigen in tissue cells. Impro-
vements in a method for the detection of antigen by means of fluorescent anti-
body, J. exp. Med., 91, 1—13.
Daussef J., Svejgaard A. (1977). HLA and Disease, pp. 316, Munksgaard, Co-
penhagen.
Diamond B. A., Yelton D. E., Scharff M. D. (1981). Monoclonal antibodies. A new
technology for producing serological reagents, New Engl. J. Med., 304, 1344—
1349.
348
Глава 8
Gilliland D. G., Streplewski Z., Collier R. J., Mitchell K. F., Chang T. H., Koprow-
ski H. (1980). Antibody directed cytotoxic agents: use of monoclonal antibody
to direct the action of toxin A chains to colorectal carcinoma cells, Proc, natn,
Acad. Sci. USA, 77, 4539.
Gregoriadis G. (1981). Targeting of drugs: implications in medicine, Lancet, if,
241—246.
Kohler G., Milstein C. (1975). Continuous cultures of fused cells secreting anti-
body of predefined specificity, 256, 495—497.
Koprowski H., Steplewski Z., Herlyn D., Herlin M. (1978). Study of antibodies
against human melanoma produced by somatic cell hybrids, Proc. natn. Acad.
Sci. USA, 75, 3405—3409.
Leserman L. D., Machy P., Barbet J. (1981). Cell-specific drug transfer from
liposomes bearing monoclonal antibodies, Nature, 293, 226—228.
Nairn R. C. (1976). Fluorescent Protein Tracing, 4th edn., pp. 648, Livingstone,
Edinburgh.
Olsnes S. (1981). Directing toxins to cancer cells, Nature, 290, 84.
Ritz J., Pesando J. M., N otis-McConarty J., Lasarus H., Schlossman S. F. (1980).
A monoclonal antibody to human acute lymphoblastic leukaemia antigen, Natu-
re, 283, 583—585.
Secher D. S., Burke D. C. (1980). A monoclonal antibody for large-scale purifi-
cation of human leukocyte interferon, Nature, 285, 446—450.
Yalow R. S., Berson S. (1960). Immunoassay of endogenous insulin in man, J.
clin. Invest., 39, 1157—1175.
Yeh M. Y., Hellstrom I., Brown J. P., Warner G. A., Hansen J. A., Hellstom К. E.
(1979). Cell surface antigens of human melanoma identified by monoclonal
antibody, Proc. natn. Acad. Sci. USA, 76, 2927—2931.
Лекарственные препараты, получаемые с помощью технологии рекомбинатных
ДНК
Bloom В. R. (1980). Interferons and the immune system, Nature, 284, 593—
595.
Crea R., Kraszewski A., Hirose T., Itakura K. (1978). Chemical synthesis of
genes for human insulin, Proc. natn. Acad. Sci. USA, 75, 5764—5769.
Derynck R., Remant E., Saman E., Stanssens P., De Clerq E., Content J., Fiers W.
(1980). Expression of human fibroblast interferon gene in Escherichia coli.
Nature, 287, 193—197.
Dreesman G. R„ Sancheq Y„ lonescu-Matia I., Sparrow J. T„ Six H. R„ Peter-
son D. L., Hollinger F. B., Melnick J. L. (1982). Antibody to hepatitis В sur-
face antigen after a single inoculation of uncoupled synthetic HBsAg peptides,
Nature, 295, 158—160.
Edge M. D., Greene A. R., Heathcliffe G. R., Meacock P. A., Schuch W., Scan-
lon D. B., Atkinson T. C., Newton C. R., Markham A. F. (1981). Total synthesis
of a human leukocyte interferon gene, Nature, 292, 756—762.
Edman J. C„ Hallewell R. A., Valenzuela P„ Goodman H. M„ Rutter W. J.
(1981). Synthesis of hepatitis В surface and core antigens in E. coll. Nature,
291, 503—506.
Goeddel D. V., Kleid D. G., Bolivar F., Heyneker H. L„ Yansura D. G., Crea R.,
Hirose T„ Kraszewski A., Itakua K„ Riggs A. D. (1979). Expression in Esche-
richia coli of chemically synthesized genes for human insulin, Proc natn Acad
Sci. USA, 76, 106—110.
Goeddel D. V. et al. (1979). Direct expression in Escherichia coli of a DNA
sequence coding for human growth hormone, Nature, 281, 544—548.
Hitzeman R. A., Hagie F. E., Levine H. L„ Goeddel D. V., Ammerer G„ Hall B. D.
(1981). Expression of a human gene for interferon in yeast, Nature 293
717—722.
Медицина и биотехнология
349
Keen Н., Glynne A., Pickup J. С., Viberti G. С., Bilous R. W., Jarrett R. J.,
Marsden R. (1980). Human insulin produced by recombinant DNA technology:
safety and hypoglycaemic potency in healthy men, Lancet, ii, 398—401.
Olson K-, Fenno J., Lin N., Harkins R. N., Snider C„ Kohr W. H., Ross M. J.,
Fodge D., Prender G„ Stebbing N. (1981). Purified human growth hormone
from E. coli is biologically active, Nature, 293, 408—411.
Miller W. L„ Baxter J. D. (1980). Recombinant DNA — A new source of insulin,
Diabetologia, 18, 431—436.
Southern E. M. (1975). Detection of specific sequences among DNA fragments-
separated by gel electrophoresis, J. Mol. Biol., 98, 503—517.
Stiehm R., Kronenberg L. H., Rosenblatt H. M., Bryson Y., Merlgan T. C. Inter-
feron: immunobiology and clinical significance, Ann. Intern. Med., 96, 80—93.
Turner A. P. F., Pickup J. C. (1985). Diabetes mellitus: biosensors for research
and management, Biosensors, 1, 85—115.
Updike S. J., Hicks G. P. (1967). The enzyme electrode, Nature, 214, 986—988.
Zuckerman A. J. (1982). Developing synthetic vaccines, Nature, 295, 98—99.
Молекулярная генетика болезней человека
Anonymous (1984). Molecular genetics for the clinician, Lancet, i, 257—259.
Bell G. J., Selby M. J., Rutter IF. J. (1982). The highly polymorphic region near
the human insulin gene is composed of simple randomly repeating sequences,
Nature, 295, 31—35.
Davies К. E., Pearson P. L., Harper P. S., Murray J. M., O’Brien I., Sarfarazi M.r
Williamson R. (1983). Linkage analysis of two cloned DNA franking the
Duchenne muscular dystrophy locus on the short arm of the human X chro-
mosome, Nucleic Acids Res., 11, 2303—2312.
Emery A. E. H. (1981). Recombinant DNA technology, Lancet, ii, 1406—1409.
Gusella J. F., Wexler N. S., Conneally P. M. et al. (1983). A polymorphic DNA
marker genetically linked to Huntington’s disease, Nature, 306, 234—238.
Kay Y. W., Dozy A. M. (1978). Antenatal diagnosis of sickle-cell anaemia by DNA
analysis of amniotic cells, Lancet, ii, 910—912.
Little P. F. R., Annison G., Darling S., Williamson R., Camba L., Model B.
(1980). Model for antenatal diagnosis of B-thalassaemia and other monogenic
disorders by molecular analysis of linked DNA polymorphisms, Nature, 285,
144—147.
Rotwein P., Chyn R., Chirgwin J., Cordell B., Goodman H. M. (1981). Polymor-
phism to the 5' flanking region of the human insulin gene and its possible
relation to type 2 diabetes, Science, 213, 1117—1120.
Weatherall D. J. (1979). Mapping haemoglobin genes, Br. med. J., 2, 352—354.
Перспективы развития: биосенсоры, трансплантация и генная терапия
Adinolphi М., Ackle С. A., McColl I., Fenson А. Н., Tansley L., Connolly Р.,
Hsi В. L., Faulk W. Р., Traves Р., Bodmer W. F. (1982). Expression of HLA
antigens, p2-microglobulin and enzymes by human amniotic epithelial cells,
Nature, 295, 325—327.
Caras S., Janata J. (1980). Field effect transistor sensitive to penicillin, Analyt.
Chem., 52, 1935—1937.
Mercola К. E., Cline M. J. (1980). The potentials of inserting new genetic infor-
mation, New Engl. J. Med., 303, 1297—1300.
Turner A. P. F., Pickup J. C. (1985). Diabetes mellitus: biosensors for research
and treatment, Biosensors, 1, 85—115.
Глава 9
Сельское хозяйство и биотехнология
Ф. Скиннер
“ЭЛ. Введение
Зрелище возделанных полей стало для нас настолько привычно,
что мы не замечаем искусственности такого пейзажа. Между
тем очевидно, что деятельность живущих на Земле людей очень
сильно сказывается на ее облике, ибо с незапамятных времен
•они выращивают на ней растения и содержат домашних живот-
ных. Фермеров можно считать предшественниками современных
биотехнологов, так как они постоянно занимались улучшением
пород животных. Впрочем, поскольку делалось это в основном
эмпирически, называть фермеров биотехнологами было бы все
же неправильно. Ведь биотехнология — это целенаправленное и
научно обоснованное использование биологических процессов
при производстве, переработке и использовании сырья. На прак-
тике многие из таких процессов идут при участии микроорга-
низмов.
Сегодня ферма — это центр по производству пищевого сырья
за счет земельных ресурсов, но ранее ее функции были шире,
так как на ней осуществлялась и его переработка. Навыки хле-
бопечения, пивоварения, сыроделия и производства сброженных
продуктов были естественным дополнением к умению выращи-
вать зерно, получать мясо и молоко. Всем этим нужно было за-
ниматься на месте, чтобы не допустить порчи продуктов под
действием микробов. Сегодня многие из этих вторичных про-
изводств, в которых ведущую роль играет жизнедеятельность
соответствующих микроорганизмов, перенесены с ферм на заво-
ды (так, во всяком случае, обстоит дело в развитых странах).
Здесь они были усовершенствованы с целью повышения каче-
ства продукции и эффективности процесса.
Нередко на заводах осуществляется и более глубокая пере-
работка пищевых продуктов, обычно путем ферментации, когда
из них производится сырье для химической промышленности.
Хорошо известным примером может быть выработка техниче-
ского спирта из избытков низкокачественного вина; менее из-
вестно, что спирт в больших количествах получают из специаль-
Сельское хозяйство и биотехнология
351
но выращиваемых для этих целей культур (гл. 2). Промышлен-
ная биотехнология оказывает фермеру и прямую помощь,,
снабжая его вакцинами и антибиотиками, а также добавками
к кормам. Таким образом, взаимосвязь между сельским хозяй-
ством и биотехнологией весьма многообразна, и поэтому описать
в одной статье все ее аспекты просто невозможно. Быть может,
имеет смысл остановиться лишь на том, какое содействие может
она оказать фермеру-земледельцу и садоводу в деле повышения
урожайности и улучшения сортовых особенностей растений.
Необходимость увеличения объема сельскохозяйственной*
продукции и ее качества (питательной ценности, вкусовых до-
стоинств и рыночных качеств) общеизвестна. При этом оно-
должно осуществляться экономически приемлемыми способами,,
а также с учетом влияния на окружающую среду.
Развитые страны могут позволить себе в больших масшта-
бах применять химические удобрения, но многим другим стра-
нам это недоступно, и они вынуждены искать иные пути. Основ-
ным необходимым для роста элементом является азот. Он от-
нюдь не принадлежит к числу редких, но чтобы перевести азот
в форму, доступную растениям, его надо фиксировать. К счастью,,
в ходе эволюции выработался и закрепился биологический ме-
ханизм фиксации азота при симбиозе — весьма эффективный*
процесс, на котором мы остановимся подробнее. Сходен с ним
и процесс несимбиотической фиксации азота, который сейчас ак-
тивно изучается, но на практике применяется пока в ограничен-
ном масштабе.
Большое внимание привлекают к себе биологические способы-
снабжения растений фосфором, а также контроля за вредителя-
ми и болезнями растений. В этой области намечается переход
от лабораторных экспериментов к практике. Разрабатываются
способы выращивания ценных культур в контролируемых усло-
виях— это новый и многообещающий подход. Быть может, са-
мый большой вклад, который может внести биотехнология в
сельское хозяйство, — это улучшение сортов растений; сущест-
венный прогресс здесь будет достигнут благодаря использова-
нию методов генетической инженерии и технологии слияния про-
топластов.
9.2. Метод питательной пленки
9.2.1. Основы метода
Для полноценного роста растения нуждаются в воде и кисло-
роде, но эти жизненно важные вещества редко имеются в опти-
мальном количестве при выращивании растений в почве или:
других твердых средах. Излишняя вода заполняет промежутки’.
352
Глава 9
между частицами почвы и лишает корни кислорода. С другой
стороны, хорошая аэрация обычно достигается лишь при недо-
статке воды. При использовании метода питательной пленки
эти недостатки удается преодолеть.
У растений, выращиваемых таким способом, корни развива-
ются в тонком слое постоянно возобновляемого питательного
раствора, содержащего все необходимые вещества. Уровень
раствора поддерживается на строго определенной высоте, так
что нижняя часть развивающейся подушки корней постоянно
находится в растворе, а верхняя выступает над ним, но оказы-
вается покрытой тонкой пленкой жидкости. Очень важно, чтобы
эта пленка постоянно существовала, так как, если бы все корни
сказались погруженными в раствор, им недоставало бы кисло-
рода.
Рис. 9.1. Подушки корней кукурузы, выращенной по методу питательной плен-
ки (Elmes, Mosse, 1984). В лоток (емкость) или в мешочек из муслина, рас-
положенный ниже выходного патрубка, была помещена фосфатсодержащая
порода (вид сверху). Слева — вид на подушку из корней сбоку. Корневые си-
стемы так сильно переплетаются, что растения не нуждаются в опоре.
Основной отличительной особенностью метода питательной
пленки является то, что корневая система каждого растения
развивается настолько интенсивно и так тесно переплетается
с корнями соседей, что растения не нуждаются в опоре (рис.9.1).
Таким образом, метод питательной пленки существенно от-
личается от метода водных культур (гидропоники), когда вода
и питательные вещества имеются в избытке, но аэрация корне-
Сельское хозяйство и биотехнология
353
вой системы обычно недостаточна. Трудности закрепления рас-
тений при выращивании по этому методу остаются и в том слу-
чае, когда корни развиваются не в водной среде, а в толще
инертного материала (например, песка, гравия или пемзы),
смачиваемого питательным раствором за счет капиллярных сил.
При таком культивировании условия выращивания очень близ-
ки к таковым для почв и обладают всеми свойственными им
недостатками.
Установки, использующиеся в методе питательной пленки,
их эксплуатация и применение метода подробно описаны в кни-
ге Купера (Cooper, 1979), откуда мы и взяли большинство при-
водимых в данной главе сведений.
9.2.2. Основные требования к оборудованию
Прямоугольная выровненная поверхность, на которой предпо-
лагается выращивать растения, должна иметь двухсторонний
уклон. По всей длине самой низкой ее части проходит канавка-
ловушка. Ее выстилают непромокаемой полимерной пленкой и
закрывают крышкой, чтобы туда не попадал свет и не проис-
ходило испарение. Если питательный раствор подогревается, то
и желобок, и крышка должны быть изолированы. В наиболее
низкой части установки помещаются отстойник и насос для пе-
рекачки раствора. Жесткие емкости (ячейки), в которых будут
выращиваться растения, можно помещать прямо на выровнен-
ную поверхность, начиная с верхней части установки и вплоть
до канавки-ловушки, в которую из них стекают излишки рас-
твора. Другой способ — покрыть всю поверхность бетоном или
же сделать цементную полоску вокруг каждой емкости (ячей-
ки), а затем поместить на поверхность с небольшим ровным
наклоном гибкие емкости с плоским дном.
Питательный раствор подается насосом через полиэтилено-
вую трубку в самую верхнюю часть установки, откуда через
узкие полиэтиленовые же трубочки распределяется по емкостям.
Под действием силы тяжести раствор стекает по ним очень
тонким слоем к канавке-ловушке и возвращается к насосу. Около
насоса к трубке подачи подсоединена возвратная трубка с ре-
гулируемым клапаном, так что часть раствора перекачивается
прямо в канавку и не протекает через емкости.
При этом можно, регулируя клапан, легко контролировать
скорость протекания раствора через емкости; для перемешива-
ния и аэрации подаваемый в систему раствор можно разбрыз-
гивать в воздухе; при обслуживании установки можно опорож-
нять кюветы-желобки, не прерывая циркуляции раствора в ем-
костях: для этого следует просто переключить возвратный канал
на слив. При этом в отстойнике будет достаточно раствора для
23—1344
354
Глава 9
обеспечения потребностей растений. На случай аварии нужно
иметь запасной насос, а при отключении электричества может
понадобиться аварийный генератор.
Рис. 9.2, Небольшая экспериментальная установка, применяющаяся для выра-
щивания фасоли (Phaseolus vulgaris) по методу питательной пленки. Пита-
тельный раствор подается с помощью насоса по пластмассовой трубке к даль-
нему краю лотка и затем стекает из передней части лотка через широкую труб-
ку в пластмассовую емкость (на переднем плане).
По размерам установки, использующиеся в методе питатель-
ной пленки, сильно различаются: от очень больших, промыш-
ленных, для массового выращивания, например, помидоров или
салата до небольших установок для научных целей. На рис. 9.2
показана опытная установка.
9.2.3. Фитотоксичность
Ясно, что контактирующие с питательным раствором части уста-
новки не должны выделять токсичные вещества, которые могли
бы замедлить рост растений или даже вызвать их гибель. Без
опаски можно использовать полиэтилен и, по-видимому, такие
материалы, как полипропилен и акрилонитрилбутадиенстирол.
Твердый поливинилхлорид фитотоксического действия не ока-
зывает, но гибкий поливинилхлорид лучше не применять, так
как иногда он оказывается фитотоксичным. В установках не
следует использовать металлы, которые относятся к числу мик-
Сельское хозяйство и биотехнология
355
роэлементов, поскольку в питательном растворе они могут на-
капливаться до токсических концентраций. Так, не нужно при-
менять трубки с гальваническим покрытием: цинк с их поверх-
ности постепенно выщелачивается. Если о токсичности конструк-
ционных материалов ничего не известно, то их следует предва-
рительно выдерживать в аэрируемом питательном растворе, на
котором выращиваются проростки.
$.2.4. Устройство емкостей, используемых в методе
питательной пленки
Ширина емкостей варьирует от 10 до 23 см в зависимости от
вида выращиваемых растений. Устройство их может существен-
но различаться, но основной принцип, которого необходимо при-
держиваться, состоит в том, что конструкция должна быть пло-
ской и жесткой. Если поверхность, на которую помещают
емкости, неровная, они должны быть жесткими, если же осно-
вание твердое и плоское, можно применять более дешевые гиб-
кие емкости. Емкость из темного полиэтилена, которую при
необходимости помещают в твердый каркас, должна быть до-
статочно глубокой, чтобы у нее можно было загнуть внутрь
края, что уменьшает испарение и укрывает корни от света, но
не препятствует росту стебля. При другой конструкции в осно-
вании при его изготовлении заранее делают углубления, но та-
кой способ годится только для выращивания растений с раски-
дистыми листьями, например салата. Они закрывают емкость
и предотвращают испарение. Известны многие варианты таких
конструкций.
$.2.5. Опора для молодых растений
Для молодых растений, пересаженных в емкости устройства
метода питательной пленки, нужна опора, так чтобы листья их
находились на свету, а корни — в питательном растворе. Обыч-
но проростки еще до образования корней высаживают в кубик
из адсорбента, например из торфа, или же помещают в сосуды,
содержащие среду для роста корней. За счет капиллярных сил
корни в достатке получают и воду и питательные вещества из
тонкого слоя протекающего раствора.
$.2.6. Болезни
Конструкция установки, использующейся в методе питательной
пленки, как нельзя лучше способствует быстрому распростране-
нию в ней патогенных микроорганизмов, однако связанные с
этим проблемы оказались не столь серьезными, как представ-
23*
356
Глава 9
лялось вначале. Возможно, устойчивость выращиваемых по ме-
тоду питательной пленки растений к возбудителям различных
заболеваний, распространяемым вместе с питательной средой,
обусловлена тем, что корни их не повреждаются механическим
или иным способом, как это бывает при выращивании растений
в почве, и тем самым отсутствуют «ворота инфекции».
9.2.7. Перспективы
Метод питательной пленки уже применяется для выращивания
растений как в открытом грунте, так и в теплицах в Англии и
ряде других стран. Кроме того, он может получить широкое
распространение как метод исследования, поскольку позволяет
легко контролировать условия выращивания. Можно поддержи-
вать температуру питательной среды с помощью термостата,
осуществлять постоянный контроль за pH как вручную, так и
автоматически, оптимизировать концентрацию питательных ве-
ществ. Для борьбы с заболеваниями и подавления нежелатель-
ной микрофлоры в систему можно добавлять фунгициды, пести-
циды и антибиотики. В опытных теплицах можно развертывать
небольшие установки. Сделаны первые попытки создать устрой-
ство для работы в стерильных условиях. В работе Купера
(Cooper, 1979) обсуждаются интересные проекты использования
метода питательной пленки в растениеводстве закрытого грун-
та, а также для получения дерна, кормов для скота и каучука.
В целом направление это представляется весьма перспективным.
9.3. Бобовые культуры и фиксация азота при симбиозе
9.3.1. Из истории вопроса
Семейство Leguminoseae включает около 625 родов и 18 000 ви-
дов. Это одно из самых многочисленных и наиболее важных
с экономической точки зрения семейств цветковых растений.
Семена многих видов бобовых богаты белком. Они являются
ценным сырьем для производства кормов и пищевых продуктов.
Из растений умеренного климата можно назвать горох и фасоль.
В тропических и субтропических зонах возделываются самые
разнообразные культуры, включая сою, вигну китайскую, чече-
вицу и арахис. Многие бобовые — ценные пастбищные культуры
(например, различные виды клевера, Stylosanth.es и др.). Они
могут служить зеленым удобрением (например, люцерна), дают
нам лесоматериалы, фрукты, овощи, клеящие вещества, волок-
на, лекарства и пряности. Роль бобовых определяется еще и
тем, что они способны фиксировать азот атмосферы в корневых
клубеньках, которые формируются при участии почвенных бак-
Сельское хозяйство и биотехнология
357
терий из рода Rhizobium. Образующиеся в них азотистые ве-
щества необходимы для роста растений.
Если об участии клубеньковых бактерий в симбиогенной
фиксации атмосферного азота стало известно сравнительно не-
давно (менее 100 лет назад), то о многих преимуществах вы-
ращивания бобовых люди узнали гораздо раньше (по меньшей
мере две тысячи лет тому назад). Культура, из которой можно
получать богатые белком пищевые продукты и которая не тре-
бует (или требует очень мало) азотных удобрений, несомненно,
выгодна, и неудивительно, что образующие клубеньки бобовые
стали важной сельскохозяйственной культурой в Европе, Север-
ной Америке и на других континентах.
Установлено, что среди бобовых, играющих важную роль
в сельском хозяйстве (в основном из Papilionoideae), приблизи-
тельно 98% видов могут образовывать клубеньки. У диких ви-
дов они чаще всего формируются при участии особых разновид-
ностей бактерий-ризобиумов, существующих в их природном
местообитании. Если какой-то вид бобовых (дикий или куль-
тивируемый) долго растет на одном месте, это приводит к по-
степенному накоплению в почве сожительствующей с ним раз-
новидности ризобиумов. Урожай, особенно если в почве недо-
стает азотистых соединений, нередко зависит от того, образова-
лась ли в данном месте эффективная (т. е. способная к фикса-
ции азота) ассоциация из растения-хозяина и соответствующей
разновидности (штамма) Rhizobium. Нужный штамм не всегда
присутствует в том месте, где предполагается выращивать дан-
ную культуру, и в таком случае его приходится вносить в почву.
Как только стала ясна роль симбиоза бактерий рода Rhizo-
bium и бобовых, были разработаны способы внесения этих бак-
терий в почву для улучшения условий культивирования. Их ста-
ли добавлять также и к семенам. Затраты на внедрение этого
способа инокуляции невелики, транспортные расходы незначи-
тельны, а сами методы достаточно несложны, чтобы их можно
было внедрить в сельское хозяйство развивающихся стран, где
высокая стоимость удобрений ложится тяжелым грузом на пле-
чи фермеров. Выращивание бобовых с применением метода ино-
куляции семян нередко благоприятно сказывается на состоянии
окружающей среды: оно помогает бороться с наступающей пу-
стыней, облегчает борьбу с эрозией почв, уменьшает перенос
почвы ветрами и позволяет с большим успехом восстанавливать
истощенные земли. Большинство образующих клубеньки бобо-
вых способны полностью удовлетворить свои потребности в азо-
те. Это, конечно, бывает только тогда, когда и прочие условия
благоприятствуют росту растений, т. е. растения получают до-
статочно воды и других питательных веществ.
358
Глава 9
9.3.2. Инокуляция бактериями Rhizobium
Наиболее простой способ инокуляции основан на использовании
почвы, взятой с полей, где выбранная для выращивания куль-
тура бобовых растет хорошо. Этот способ вполне пригоден и
широко применялся в конце XIX в.
Недостаток его заключается в том, что при этом приходится
перемещать много почвы, так как Rhizobium составляют лишь
малую часть микрофлоры и ничтожную часть самой почвы.
В Америке обычно вносили 100—1000 кг почвы на 1 га. Брали
эту почву с расположенного где-нибудь недалеко поля, где был
получен хороший урожай нужной бобовой культуры. Еще один
недостаток этого метода — возможность распространения с поч-
вой болезней растений.
Гораздо меньше почвы необходимо вносить при помощи сея-
лок, когда бактерии попадают прямо к семенам, где они нужнее
всего. На основе этого простого способа был разработан метод
прямой инокуляции семян. Сначала применяли измельченную
почву (всего около 0,5 кг почвы на 1 кг семян), а затем стали
использовать методы, основанные на введении чистых культур
бактерий.
Первая разновидность коммерческой культуры для инокуля-
ции была запатентована Ноббе и Хилтнером в 1896 г. Она по-
ступала на рынок под названием «Nitragin». Для разных бобо-
вых выпускалось 17 ее вариантов. Уже в 20-х гг. на рынок
поступало много других разновидностей инокулятов. Некото-
рые из них представляли собой чистые культуры бактерий, сме-
шанные с почвой, песком, торфом, навозом или измельченной
породой, другие — культуры, выращенные на агаре или в жид-
кой среде. Далеко не в каждом случае качество таких иноку-
лятов было высоким, не каждый раз их правильно применяли,
так что и высокие урожаи получали не всегда.
9.3.3. Инокуляция сем.ян
Суть этого метода заключается в том, что на семена наносят
большое число клеток Rizobium, соответствующих определенно-
му виду растения-хозяина, что увеличивает вероятность быстро-
го образования клубеньков у проростков при участии данных
микроорганизмов. Для этого нужно достаточно много бактерий,
которые должны сохранять жизнеспособность до тех пор, пока
в почве они не проникнут в корневые волоски.
Культуры на агаре
Для прямой инокуляции семян пригодны культуры Rhizobium,
выращенные в пробирках или колбах на среде с агаром. Обыч-
но для наращивания бактерий применяют агаровую среду с ман-
Сельское хозяйство и биотехнология
359
нитолом и дрожжевым экстрактом следующего состава: 0,5 г
К2НРО4, 0,2 г MgSO4-7H2O, 0,1 г NaCl, 10 г маннитола, 0,4 г
дрожжевого экстракта (например, Difco, Oxoid), 15 г агара,
до 1 л дистиллированной воды, pH 6,8—7,0; среду автоклави-
руют при 121 °C 15 мин. В 10 мл такой среды может содержать-
ся ~1010 клеток ризобиумов. На холоде их можно хранить
несколько недель.
В промышленном масштабе пробирочные культуры Rhizobi-
um meliloti для инокуляции люцерны (альфальфы, Medicago
sativa) производились в Англии с 1929 по 1963 г. За их качест-
вом следил отдел микробиологии почвы Ротамстедской экспе-
риментальной станции. Для инокуляции семян бактерии со-
скабливали с поверхности косяка и суспендировали в снятом
молоке, к которому добавляли немного фосфата кальция. Эту
смесь выливали на кучу зерна и хорошо все перемешивали,
а затем давали просохнуть в тени. При этом бактерии прили-
пали к поверхности семян, а пленка высохшего молока надежно
их защищала. В идеальном случае семена вскоре высевали.
Этот метод пригоден для выработки небольших партий иноку-
лята и соответствовал потребностям потребителей, выращиваю-
щих люцерну в Англии, но сегодня такие культуры здесь боль-
ше не производят. В каждой пробирке содержалось достаточно
клеток Rhizobium, чтобы обработать семена для —0,5 га посе-
вов люцерны. К середине 40-х г. ежегодно продавалось около
40 000 таких препаратов, но к концу 50-х г. число их упало до
9000—10 000, а к 1964 г. до 1000—2000. Это было связано, по
крайней мере частично, с уменьшением площадей, засеваемых
люцерной. Сегодня в Англии коммерческие препараты-инокуля-
ты не производятся, но из Австралии и США импортируются
высококачественные препараты на торфяной основе.
9.3.4. Современные инокуляты на твердых носителях
Rhizobium, выращенные на агаре или в жидкой среде, после
высушивания на поверхности семян быстро погибают, да и са-
ми культуры их нежизнестойки. Этих недостатков лишены ино-
куляты на торфяной основе, которые были созданы в США,
а сегодня применяются повсеместно.
При получении культуру нужного штамма Rhizobium выра-
щивают обычным способом в ферментере объемом в несколько
литров в жидкой среде (например, содержащей маннитол и
дрожжевой экстракт). Установка может быть очень несложной,
лишь бы ее устройство обеспечивало защиту от микробного
загрязнения. Температуру среды поддерживают в интервале
25—28 °C, и осуществляют интенсивную аэрацию, продувая че-
рез среду стерильный воздух. Контроль за pH осуществлять не
360
Глава 9
нужно, особенно если в среду добавлено немного СаСОз. К мо-
менту смешивания с торфом (носителем) —стерильным или не-
стерильным— плотность культуры должна быть достаточно вы-
сокой, 5-Ю8—4-Ю9 клетка/мл. В случае медленно растущих
штаммов (например, R. lupini} со средним временем удвоения
числа клеток около 10 ч обычно бывает сложно получить куль-
туру с высокой плотностью, и нередко даже размножение более
быстро растущих штаммов (со средним временем удвоения
3 ч) подавляется видами-загрязнителями. Для обеспечения быст-
рого роста культуры на жидкой среде полезно вносить сразу
много бактерий.
Культуры на торфяном носителе
Торф высушивают либо при обычной температуре, либо при
осторожном нагревании до влажности около 10% (на сухую
массу), а затем измельчают при помощи молотковой мельницы
до порошка, который просеивают через сито (200 меш) и дово-
дят pH до 6,5—7,0, добавляя тонко измельченный СаСОз. Такой
торф можно использовать как стерильный или нестерильный
носитель. О пригодности партии торфа для получения инокуля-
тов Rhizobium нельзя судить по общим его свойствам, и на каж-
дой из них проводят пробные выращивания именно тех штам-
мов, которые предполагается использовать. Рост и выживае-
мость бактерий в торфе зависят от многих факторов.
Нестерильные инокуляты на торфе
К подготовленному измельченному торфу добавляют до 40%
(по массе) жидкой культуры, тщательно перемешивают смесь
и оставляют ее для созревания в неглубоких лотках. Через не-
сколько суток смесь опять перемешивают, а затем фасуют,
обычно в заплавленные мешочки из полиэтиленовой пленки
(толщиной 0,0375 мм). За первые несколько недель после при-
готовления смеси число жизнеспособных клеток Rhizobium
обычно увеличивается в 2—5 раз. Если держать фасовки при
низкой температуре, бактерии обычно долгое время сохраняют
жизнеспособность. Возможное время хранения оценивают ис-
ходя из продолжительности периода, за который погибает 90%
клеток (время уменьшения численности до '/ю от первоначаль-
ной). Это время варьирует от 90 недель (при 5 °C) до 8 (при
25°C). При благоприятных условиях культуры можно хранить
до года (время уменьшения численности до ’/ю >16 недель),
но срок этот заметно меньше, когда они содержатся при ком-
натной температуре. Точно предсказать возможную продолжи-
тельность хранения нельзя, и ее приходится определять для
каждой партии отдельно.
Сельское хозяйство и биотехнология
361
Стерильные инокуляты на торфе
Заплавленные полиэтиленовые или полиамидные мешочки с под-
готовленным измельченным влажным торфом стерилизуют в ав-
токлавах (например, при 115 °C в течение 4 ч) или же путем
у-облучения (примерно 4,5-106 рад). Затем при помощи шпри-
ца для инъекций в них вводят нужный объем жидкой культуры
(порядка 10 мл). Прокол запаивают и мешочки ставят на ин-
кубацию, в ходе которой происходит существенное размножение
бактерий.
Носители для Rhizobium
Твердый инокулят состоит из бактерий и носителя, роль кото-
рого заключается в поддержании жизнеспособности клеток, по-
скольку он частично защищает их от пересыхания. Кроме того,
носитель способствует более равномерному распределению бак-
терий в массе семян и помогает им прикрепиться к их поверх-
ности. Хотя в случае бактериальных суспензий нередко получа-
лись хорошие результаты, считается, что применение торфа как
носителя более эффективно: при использовании жидких культур
или же суспензий' клетки Rhizobium после инокуляции и при-
креплении их к поверхности семян оказываются практически
беззащитными. Поэтому при производстве коммерческих ино-
кулятов вначале чаще всего использовали именно торф. Однако
торф есть далеко не везде, а если он и имеется, то сказать
заранее, пригоден ли он как носитель, невозможно. В связи
с этим были предприняты поиски альтернативных носителей с
такими же защитными свойствами, как у торфа.
С неодинаковым успехом для этой цели были испробованы
всевозможные композиции: разнообразные смеси почвы и торфа,,
в которые были добавлены молотая люцерна, измельченная со-
лома, дрожжи или сахар; нильский ил с добавками питательных
веществ; почва, смешанная с кокосовыми охлопьями или дре-
весным углем. Опробованы также вермикулит, перегнившие
опилки, бентонит, кизельгур и каменный уголь. В Африке удач-
ный носитель был получен путем компостирования кукурузных
кочерыжек в течение 30 недель вместе с молотым известняком,
суперфосфатом и азотнокислым аммонием. Полученный ком-
пост высушивали на воздухе, измельчали, просеивали, разве-
шивали по 100 г в плотные полиэтиленовые мешочки и авто-
клавировали.
Сегодня для поддержания жизнеспособности Rhizobium по-
лучают носители из самых разнообразных веществ, но лучшим
носителем все же является торф. В некоторых регионах, напри-
мер в развивающихся странах, определенную ценность могут
представлять дешевые местные заменители торфа.
362
Глава 9
9.3.5. Методы инокуляции
Самый простой, но, наверное, наименее эффективный метод ино-
куляции — смешивание сухого инокулята и семян перед посе-
вом. При этом к семенам прикрепляется мало бактериальных
клеток, большая их часть теряется, и необходимое условие на-
несения достаточного числа клеток Rhizobium на семена не вы-
полняется. Поэтому лучше вносить инокулят в виде водной
кашицы. Хорошо прилипает к семенам инокулят на торфе, осо-
бенно если добавить к нему водорастворимый клей (карбокси-
метилцеллюлозу или гуммиарабик). К тому же при этом по-
вышается выживаемость бактерий после высушивания семян.
Дражирование семян
Цель этой обработки состоит в интенсификации образования
клубеньков; при этом вместе с бактериями к семенам бобовых
добавляют относительно малорастворимые неорганические по-
рошкообразные вещества (фосфат кальция, измельченный из-
вестняк и т. п.). Хороший эффект при внедрении культуры кле-
вера (Trifolium subterraneum) на кислых почвах Австралии
дало дражирование известью. Сначала семена дражировали в
тонко измельченном известняке, а затем непосредственно перед
высевом смешивали с влажным инокулятом на торфе. Такой
способ позволяет нанести больше инокулята и увеличивает вы-
живаемость бактерий на семенах. К инокуляту на торфе при
этом добавляют раствор гуммиарабика ( — 40%), и все это тща-
тельно перемешивают с семенами (например, в небольшой, чи-
стой бетономешалке), добиваясь равномерного их покрытия.
Затем за один прием добавляют необходимое количество кар-
боната кальция (но не гидроокиси кальция, которая является
слишком сильной щелочью) и продолжают перемешивание до
тех пор, пока инокулированные семена не покроются ровным
слоем известковой муки. Количество добавок и время обработки
нужно строго контролировать, так чтобы образовались плотные
гранулы, содержащие большое число жизнеспособных бактери-
альных клеток. Семена после дражирования следует высевать
как можно быстрее, пока число жизнеспособных клеток на них
максимально.
Используемые в настоящее время инокуляты
на основе почвы
Хотя сегодня в нашем распоряжении есть хорошие инокуляты
семян, их не всегда удается использовать с полной эффектив-
ностью. Дело в том, что часто семена бывает необходимо об-
Сельское хозяйство и биотехнология
363
рабатывать фунгицидами и пестицидами, а некоторые из них
либо детальны для Rhizobium, либо оказывают на них небла-
гоприятное воздействие. Кроме того, в оболочке семян ряда
бобовых содержатся природные антибиотики. Проблемы возни-
кают и при высеве семян в нагретую сухую почву: при этом
бактерии могут погибнуть до того, как пройдет дождь и семена
прорастут.Поэтому разумно было бы вносить Rhizobium в почву
так, чтобы они находились поблизости от семян, в пределах
досягаемости для развивающихся корней, но все же на некото-
ром расстоянии, чтобы свести к минимуму влияние токсинов,
содержащихся в семянах. Сегодня вновь привлекает к себе вни-
мание инокуляция почвы, а не семян, с использованием улуч-
шенных инокулятов.
Так, очень удобен инокулят на основе пористых гранул; да-
же после нескольких месяцев хранения в нем остается доста-
точно много жизнеспособных бактерий. Такие гранулы можно
изготовить из увлажненного гипса с добавлением 0,2% натрие-
вой карбоксиметилцеллюлозы, замедляющей схватывание. Бак-
терии выращивают в жидкой питательной среде, к которой до-
бавляют сформированные гранулы, затем их высушивают на
воздухе, фасуют и в таком виде хранят.
Можно приготовить и гранулированный инокулят на основе
торфа, в котором создаются условия для размножения Rhizo-
bium. Его, как и гипсовые гранулы, можно высевать вместе с
семенами или же помещать в почву под ними, если нужно за-
щитить бактерии от вредного воздействия жары и сухости.
9.3.6. Улучшение симбиоза между бобовыми и Rhizobium
Симбиотические отношения, приводящие к фиксации азота,—
это наиболее эффективный способ биологического образования
аммиака, потребляемого сельскохозяйственными культурами.
Влияя на них, мы сможем достичь значительного прогресса
в использовании биологической фиксации азота для производ-
ства пищевых продуктов. Для расширения масштабов и эффек-
тивности систем фиксации азота необходимо глубже понять ге-
нетику бактерий Rhizobium, чтобы не зависеть столь сильно от
природных систем симбиоза, а формировать их с участием
любого желаемого вида растений, употребляемых в пищу.
Генетические эксперименты
с азотфиксирующими организмами
В последние десять лет для изучения биологической азотфикса-
ции стали применять методы генетики микробов и молекулярной
биологии. После того как в 1971 г. Штейхер и др. (Steicher et
364
Глава 9
al., 1971) показали, что колифаг Pi можно размножать в Kleb-
siella pneumoniae М.5 (свободноживущая азотфиксирующая бак-
терия) и трансдуцировать с ее помощью ntf-гены (гены азот-
фиксации), появилась надежда, что генетическими методами
удастся повысить способность к азотфиксации и получать более
высокие урожаи.
До недавнего времени мы не знали достаточно определенно,
какой из партнеров в симбиотической системе Rhizobium — бо-
бовое несет nif-гены, ответственные за фиксацию азота, но
в 1975 г. несколько групп ученых обнаружили, что чистым куль-
турам некоторых медленно растущих видов Rhizobium присуща
нитрогеназная активность. Из этих опытов следовало, что
nif-гены содержатся в геноме бактерий, а не растения-хозяина.
В подтверждение этого было показано, что с помощью плазми-
ды возможен перенос nif-генов от R. trifolii к неспособному
фиксировать азот штамму Klebsiella aerogenes. Было установ-
лено, что некоторые из плазмид, несущих гены, ответственные
за формирование клубеньков и фиксацию азота, относительно
легко передаются от штамма к штамму Rhizobium при конъ-
югации.
9.4. Везикулярно-арбускулярная микориза
9.4.1. Роль везикулярно-арбускулярной микоризы
Корни растений в природных условиях никогда не бывают сте-
рильными. На их поверхности всегда присутствует типичная
для данного местообитания микрофлора, причем нередко корни
бывают заражены почвенными патогенными микробами. Под
действием некоторых непатогенных грибов формируются осо-
бые структуры; одни грибы, размножаясь, образуют вокруг
мелких корешков чехол — эктомикоризу, другие, проникая в
кортикальную ткань корней, образуют эндомикоризу. В резуль-
тате устанавливаются симбиотические отношения, выгодные
растениям-хозяевам. Эктомикоризы, образованные грибами-ба-
зидиомицетами и аскомицетами, обнаружены у некоторых де-
девьев и кустарников, в частности тех, которые произрастают
в умеренном поясе северного полушария, а две из трех извест-
ных разновидностей эндомикоризы встречаются только в пре-
делах семейств Ericaceae и Orchidaceae. Третья из них известна
под названием «везикулярно-арбускулярная микориза» (ВА):
при ее образовании в покровных тканях формируются харак-
терные структуры. Эта разновидность микориз особенно важна
для сельского хозяйства (рис. 9.3—9.5). Микориза ВА, обра-
зуемая грибом-фикомицетом из семейства Endogonaceae, встре-
чается особенно часто в большинстве почв всех климатических
Сельское хозяйство и биотехнология
365
Рис. 9.3. Интактные корни
лука, инфицированные вези-
кулярно-арбускулярным ми-
лоризным грибом (Phillips,
Hayman, 1970). После освет-
ления препарат был окра-
шен КОН-трипановым си-
ним. Корни ииокулированы
штаммом Endogone с жел-
тыми вакуолярными спора-
ми. Видны выходящие нару-
жу гифы и несколько зон
проникновения, но инфекция
в глубине ткани слабая.
Рис. 9.4. Корень лука (см.
рис. 9.3) с везикулами и ги-
фами, располагающимися в
коре продольно (Phillips,
Hayman, 1970).
зон: она присуща большей части покрытосеменных, многим го-
лосеменным, а также некоторым папоротникам и печеночникам.
Микориза ВА найдена у большинства важнейших видов сель-
скохозяйственных растений, она обычно встречается у важней-
ших культур из семейств Gramineae и Leguminoseae, но не най-
дена, у видов семейства Cruciferae и сахарной свеклы.
Значение BA-микоризы в сельском хозяйстве определяется
тем, что она помогает растениям поглощать из почвы фосфаты.
Фосфат-ионы в почве не очень подвижны, и поэтому вокруг
корней их часто недостает. Гифы микоризы, вырастающие из
366
Глава 9
Рис. 9.5. Корень лука (см. рис. 9.3) (Phillips, Hayman, 1970). Видны арбуску-
лы, ответвляющиеся от расположенных продольно гиф.
корневого мицелия и распространяющиеся за пределы области,
где недостает фосфата, переносят его прямо в клетки хозяина.
Растения со слабо развитой корневой системой, имеющие ко-
роткие торчащие корни с небольшим числом волосков (напри-
мер, лук и цитрусовые), обычно не очень успешно поглощают
фосфаты из почвы и «полагаются» в этом на ВА-микоризу.
Сельское хозяйство и биотехнология
367
Благодаря микоризам нередко существенно улучшается рост
растений на почвах, бедных фосфором. При этом также увели-
чивается поступление микроэлементов, например цинка и меди.
Выявлено, что концентрация гормонов роста в растениях с ми-
коризой бывает выше, чем в ее отсутствие. Есть и другие пре-
имущества; когда образование микоризы происходит в присут-
ствии подходящих видов Rhizobium, у бобовых усиливаются
образование клубеньков и азотфиксация.
Поскольку фосфатные удобрения приходится расходовать
экономно, большое внимание сегодня уделяется тому, как эф-
фективнее использовать преимущества BA-микоризы в растение-
водстве. Хотя микоризные грибы встречаются повсеместно,
в почве, на которой предполагается выращивать новую куль-
туру, может и не быть нужных их разновидностей. Они навер-
няка отсутствуют в стерилизованной почве теплиц и средах для
выращивания растений в горшках. Частичная стерилизация,
необходимая для уничтожения патогенов и вредителей расте-
ний, убивает и полезные грибы-эндофиты. Чтобы размножить
в почве эндофиты или ввести их туда заново, нужна инокуляция,
но здесь возникает немало сложностей, поскольку гриб не раз-
вивается без растения-хозяина. Осыпающиеся с корней крупные
покоящиеся споры гриба-эндофита (около 0,15 мм в диаметре)
можно отсеять от почвы и поместить на среду с агаром. Они
прорастут, но в отсутствие растения-хозяина, в которое прони-
кает мицелий, вскоре перестанут расти. Единственный способ
получения больших количеств эндофита — это выращивание его
на подходящей линии растений. На самом деле инокулятом
должна служить смесь корней мицелия и спор. На рис. 9.6 по-
казано, какое благотворное действие на растения оказывает
внесение в почву грибов, вызывающих образование ВА-мико-
ризы.
9.4.2. Инокуляция эндофитом
Выделение спор эндофита из почвы
Почву, содержащую тонкие корешки, замачивают в воде и раз-
мешивают до разделения почвенных частиц, а затем процежи-
вают через сито с отверстиями диаметром 500—600 мкм. Про-
шедшую сквозь сито киселеобразную массу перемешивают,
встряхивают и дают постоять, чтобы осели грубые минеральные
частицы, после чего пропускают через другое сито, с диаметром
отверстий 70—100 мкм. Осадок на сите изучают под микроско-
пом. Такой способ отсеивания дает вполне хорошие результаты,
поскольку покоящиеся споры эндофита диаметром 100—200 мкм
превышают по размеру крупнейшие среди грибов. Этот метод
368
Глава 9
Рис. 9.6. Проростки белого клевера, растущие на нестерильном торфе, взятом
с горного луга в среднем Уэльсе. Обе группы растений были инфицированы
одним и тем же местным микоризным грибом, но росли плохо (слева) до тех
пор, пока не была проведена инокуляция микоризным грибом, взятым с бли-
жайшего горного луга той же местности (справа). Во все горшки был добав-
лен суперфосфат (40 кг/га).
особенно хорош для легких песчаных почв, так как при избытке
органики в почве обнаружить споры бывает сложно. Морфоло-
гические признаки таких спор весьма своеобразны, и на них
основана систематика этих грибов.
Другой метод заключается в выделении спор из обогати-
тельной культуры, которую получают путем инокуляции опре-
деленных сортов растений, выращиваемых в горшках, кусочка-
ми корней с микоризой. В результате заражения в пересчете
на горшок образуется гораздо больше спор (несколько тысяч),
чем в полевых условиях (несколько сот на 100 г почвы), по-
этому выделить их намного проще.
Получение одноразовых культур
Для получения одноразовой культуры на растениях определен-
ного сорта для инокуляции корней проростков в стерильной
почве берут 10—20 идентичных спор. В случае успешного зара-
жения через 3—4 мес в горшке развивается гомогенная попу-
ляция спор. Их можно выделить одним из описанных выше спо-
собов и использовать для заражения новых партий опытных
растений.
Крупномасштабное производство инокулятов
Технология, предложенная Менге и др. (Menge et al., 1977)\
представляет собой по сути дела модификацию только что опи-
санного маломасштабного способа культивирования в горшках.
Сельское хозяйство и биотехнология
369
Корни растения с BA-микоризой или же инфицированную почву
используют для инокуляции растения-хозяина. При этом у рас-
тения-донора и растения-хозяина, для которого предназначен
вырабатываемый инокулят, не должно быть общих заболеваний
корневой системы. Так, инокулят для цитрусовых можно полу-
чать, размножая эндофит на суданской траве; использовать для
этого цитрусовые не следует.
Споры, полученные при таком способе культивирования,
можно подвегнуть поверхностной стерилизации и использовать
для инокуляции других линий растений той же разновидности.
Убедившись, что горшечные растения свободны от болезней и
вредителей, которые могли бы поразить выбранные для иноку-
ляции растения-хозяева, можно провести инокуляцию послед-
них содержимым горшков. При получении инокулята рекомен-
дуется использовать подходящие пестициды и фунгициды, не
действующие на BA-микоризу. Методы, подобные описанному,
позволяют получать инокуляты в количестве, достаточном для
промышленного выращивания растений с хорошо развитой
BA-микоризой в горшечной культуре. И все же метод горшечной
культуры малоэффективен, и таким путем трудно получить ино-
куляты в количестве, необходимом для полевых опытов. Не
удивительно поэтому, что наряду с ним изучаются и другие
методы. Кроме того, есть надежда, что исследование физиоло-
гии эндофита позволит разработать метод его выращивания в
чистой культуре, без растения-хозяина, в лабораториях. Это*
существенно упростило бы производство инокулята, позволило-
бы получать более гомогенный, свободный от патогенов препа-
рат. Сегодня же основой для производства инокулята остаются
несущие микоризу растения.
Получение чистых инфицированных корней
В настоящее время изучается возможность получения чистых
корней, инфицированных эндофитом ВА (с помощью метода
питательной пленки; разд. 9.2). Опубликованы результаты пер-
вых опытов, в которых при производстве инокулята использо-
вали фасоль (Phaseolus vulgaris) и кукурузу (Zea mays). Спо-
ры или корешки с микоризой, завернутые в небольшие кусочки
гигроскопичного материала, помещали между корнями, где цир-
кулировал питательный раствор, и получали гораздо более чи-
стые инфицированные корни, чем при выращивании в почве,
но эндофит распространялся в корневой массе медленно. Когда
такие корни с микоризой применяли для инокуляции в опытах
с проростками салата, лука, фасоли и кукурузы, наблюдалось
нормальное образование BA-микориз. При инфицировании про-
ростков фасоли и салата культурами трех разных вадов эндо-
24—1344
370
Глава 9
фитов рода Glomus, полученными с помощью метода питатель-
ной пленки, во всех случаях образовывались видоспецифичные
микоризы. Каждый препарат данного инокулята был не менее
эффективен, чем препарат такой же массы, полученный методом
горшечной культуры. При высушивании на воздухе инфекцион-
вость уменьшалась, но инактивации препарата не происходило
(Elmes, Mosse, 1980).
Метод питательной пленки хорош тем, что можно строго
контролировать условия роста и видоизменять их так, чтобы
микориза развивалась успешно, без помех со стороны коллои-
дов почвы. Так, опыты, о которых речь шла выше, показали,
что для хорошего развития проростков кукурузы и микоризы
необходимо присутствие 5,6 мг/л железа в форме NaFe-ЭДТА;
при концентрации азота в циркулирующем растворе 11 мг/л
(95% в форме МОз~ и 5% в виде NH4+) наблюдалось подав-
ление заражения эндофитом, но если концентрация азота в рас-
творе была 1 мг/л, то заражение протекало нормально. Таким
образом, этот метод весьма перспективен как для изучения ми-
коризных заражений, так и при крупномасштабном производ-
стве инокулятов для полевых опытов и на продажу.
9.4.3. Методы инокуляции
Инокулировать BA-эндофитами молодые многолетние растения,
например сеянцы деревьев или черенки, относительно просто,
особенно если выращивать их в контейнерах. Несколько грам-
мов неочищенного инокулята, полученного из горшечной куль-
туры растения-хозяина, либо добавляют в среду, либо помеща-
ют его поблизости от молодых корней так, чтобы до пересадки
при участии эндофита успела образоваться мощная микориза.
Те, кто занимается разведением лесов и цитрусовых, все более
убеждаются, что это очень эффективный метод.
В практику сельского хозяйства инокуляция полевых куль-
тур BA-эндофитами не внедрена, но в опытах используется ши-
роко. Этот метод хорош для выращивания многолетних культур
в контейнерах, но не нашел применения для инокуляции в по-
левых условиях, так как препарата нужно очень много. На
гектар приходится вносить около 2—3 т неочищенного инокуля-
та, а получить такие количества методом горшечной культуры
растения-хозяина практически невозможно. Даже при работе
на небольших опытных делянках для получения хорошего ре-
зультата следует вносить ценный инокулят очень аккуратно.
Рассмотрим один пример такого рода.
В опытах с красным клевером (Trifolium pratense) на Ротам-
стедской станции в 1979 г. было проведено сопоставление че-
тырех способов инокуляции. Для заражения использовали не-
Сельское хозяйство и биотехнология
371
очищенные препараты (растение-хозяин с почвой), содержащие
эндофиты двух типов. Инокулят вносили: 1) под семена в бо-
розды; 2) разбрасыванием, с заделкой граблями вместе с се-
менами; 3) в форме многосеменных гранул объемом около 1 см3.
По четвертому методу, называемому «мокрым высевом», неочи-
щенный инокулят обогащали, отмывая его на сите, суспендиро-
вали в 4 %-ном растворе метилцеллюлозы вместе с пророщенны-
ми семенами и вносили эту кашицу в борозды. Через девять
месяцев выяснилось, что второй метод дает плохие результаты,
так как BA-микориза образовывалась всего у 5—10% расте-
ний— примерно у такого же их числа, как и в контроле без.
инокуляции, когда она формировалась при участии природных
эндофитов почвы. При внесении гранул микориза образовыва-
лась чаще, приблизительно в 25% случаев, но наилучшие ре-
зультаты были получены при внесении инокулята в борозды
пли при «мокром высеве». И в том и в другом случае микориза
формировалась у большинства растений (65—70%).
Мокрый высев, по-видимому, наиболее удобен для проведе-
ния инокуляции в полевых условиях, так как при этом нужно1
меньше препарата, чем при его внесении другими способами.
Кроме того, в метилцеллюлозный гель можно добавлять и дру-
гие микроорганизмы, например Rhizobium. Использование гра-
нул также имеет свои преимущества, поскольку с семенами и:
инокулятом можно вносить одновременно большие количества'
малорастворимых веществ, например извести (Hayman et al.,.
1981).
9.5. Биологический контроль
9.5.1. Основы биологического контроля
Уже в самом начале развития микробиологии стало известно,
что одни микроорганизмы могут подавлять рост других. Наи-
более важным результатом интенсивных исследований в этой
области было, наверное, открытие антибиотиков и разработка
способов их применения в клинике. Большое внимание привлек-
ла к себе и сама возможность использования одних микроорга-
низмов для регуляции численности популяции других благодаря
действию антагонистических или конкурентных механизмов.
К сожалению, в ходе этих исследований почти ничего важного
для сельского хозяйства открыто не было. Тем не менее сама
идея такого биологического широкомасштабного контроля про-
должает привлекать внимание ученых.
Биологический контроль осуществляется в природе и помо-
гает предотвращать болезни растений, но мы далеко не всегда
понимаем, каков его механизм и как им можно управлять с
24*
372
Глава 9
пользой для сельского хозяйства. Успехи в этой прикладной
области исследований весьма незначительны, вне всякого со-
мнения потому, что слишком мало усилий предпринималось для
изучения поведения смешанных популяций микроорганизмов в
почве и на поверхности растений. Есть, однако, и примеры си-
стем биологического контроля, которые разработаны до такого
уровня, что их не без основания можно считать биотехнологи-
ческими.
3.5.2. Примеры биологического контроля
Антагонистическое действие Trichoderma
Об антагонистической активности гриба Trichoderma известно
давно. Если внести во влажную почву значительное количество
Trichoderma lignorum, то он подавит выпревание проростков
(болезнь «черная ножка»), главным образом благодаря дейст-
вию токсина, который можно выделить из фильтратов культур
гриба. Было показано, что к этому эффекту причастен и прямой
паразитизм гриба. Известно, что другие виды Trichoderma всту-
пают в антагонизм или прямо паразитируют на многих грибах
и способны существенно снижать заболеваемость, вызываемую
рядом почвенных патогенов растений. Некоторые виды Tricho-
derma вполне могут использоваться в системах биологического
контроля. В работе Уэлса и др. (Wells et al., 1972) сообщалось,
что в полевых условиях Trichoderma harzianum подавляет забо-
левания стеблей и корней, вызываемые Sclerotium rolfsii, если
вносить его в почву в больших количествах (1 : 10, по объему).
Бэкмен и Родригес-Кабана (Backman, Rodriguez-Kabana, 1975)
предложили использовать для инокуляции гранулированный но-
ситель из диатомовой земли, пропитанный 10%-ным раствором
мелассы: он способствовал росту Trichoderma harzianum и его
было легко рассеивать. Применение этого инокулята позволяет
бороться с болезнями, вызываемыми Sclerotium rolfsii в поле-
вых условиях. На 1 га вносили 140 кг гранул, и это было вы-
годней, чем обработка пестицидами.
В условиях теплиц Trichoderma harzianum сдерживает вы-
превание проростков фасоли и редьки, но целесообразность его
применения в этом качестве в полевых условиях не установ-
лена. Trichoderma harzianum оказывает ингибирующее действие
на Rhizoctonia solani и защищает проростки инокулированных
.им семян гороха и редьки от выпревания, вызываемого Rhizoc-
.tonia solani или Руthium spp.
Сельское хозяйство и биотехнология
373
Грибная гниль
В Европе базидиомицет Fames annosus является основным воз-
будителем сердцевинной гнили хвойных, особенно ели Picea
rabies. Он поражает также лиственные породы и лесоматериалы.
Заражение этим грибом сосновых пней тоже нежелательно, так
как инфекция распространяется на их корни, а затем и на кор-
ни соседних, здоровых деревьев. Заселение пней этим грибом
можно предотвратить, засеяв их спорами другого гриба-бази-
диомицета, Peniophora gigantea. Гриб разрастается на пнях,
и, конкурируя с Fames annosus, подавляет развитие сердцевин-
ной гнили. Этот способ применяется в Англии и США, где нуж-
ный инокулят имеется в продаже.
Скорее всего любой организм, избранный в будущем для
осуществления биологического контроля, будет действовать на
патогены двояким способом: либо образуя вещества-ингибито-
ры, либо конкурируя за питательные вещества, либо проявляя
и тот и другой эффект. Многие сообщения на эту тему пред-
ставляют сегодня лишь научный интерес; оценить значение их
для практики нет возможности.
9.5.3. Корончатые галлы и борьба с ними
Природа заболевания
Некоторые разновидности почвенной бактерии Agrobacterium
tumefaciens способны внедряться в ткани двудольных растений
через раневые поверхности и вызывать образование опухолей —
корончатых галлов. Хотя об этом явлении известно с 1907 г.,
механизм патогенности удалось установить совсем недавно. Бы-
ло показано, что клетки патогенных штаммов этой бактерии со-
держат крупную плазмиду (Ti) с мол. массой от 90-10® до
150-10®, зависящей от того, какие иные свойства, кроме пато-
генности, в ней закодированы. Имеются данные, что превраще-
ние нормальных клеток растения в опухолевые обусловлено
переносом в них плазмиды Ti и включением ее в хромосомную
днк хозяина.
Клетки опухоли растений могут расти на питательной среде
в форме недифференцированной массы, но иногда из них обра-
зуются тератомы, которые порождают внешне нормальные рас-
тения. Эти тератомы все еще содержат плазмидную ДНК и син-
тезируют опины, но обычно опухоли у них отсутствуют и могут
развиваться либо спонтанно, либо после повреждений.
Было показано, что плазмиду Ti методами генетической ин-
женерии можно модифицировать, в результате чего она приоб-
374
Глава 9
ретает способность переносить чужеродную ДНК в растение-
хозяина. Существование жизнеспособных тератом позволяет
надеяться, что на основе такого плазмидного переноса ДНК
можно получить стабильно трансформированные растения.
Любопытно, что хотя тератомы могут цвести и образовывать
семена, выращенные из таких семян растения не несут плазми-
ды Ti. Видимо, она каким-то образом утрачивается при мейозе.
Сегодня содержащие ДНК Ti-плазмид растения размножают
только вегетативно.
Agrobacterium tumefaciens не инфицирует однодольные рас-
тения, и поэтому векторы на основе плазмиды Ti нельзя исполь-
зовать в генетических экспериментах с культивируемыми зла-
ками.
Биологический контроль
В Австралии корончатые галлы очень часто встречаются у мин-
даля, персиков и роз, и в 1973 г. садоводы-профессионалы раз-
работали метод защиты этих культур, состоящий в том, что
саженцы (проростки, трансплантаты и черенки) смачивают в
суспензии бактерий, которые подавляют рост патогена. Такая
обработка почти полностью предотвращает заболевание и яв-
ляется первым примером практического использования бакте-
рий для борьбы с корончатыми галлами.
Не все агробактерии являются патогенами. Многие их штам-
мы, выделенные из почвы питомников, где выращивали косточ-
ковые культуры, были непатогенными, но отличить их от пато-
генных по какому-либо иному, чем степень патогенности, приз-
наку было невозможно. Для почвы вокруг больных растений
отношение патогенных штаммов к непатогенным было высоким,
а для почвы вокруг здоровых растений — низким.
Изучение взаимоотношений между двумя формами в лабо-
ратории, когда в качестве типичной непатогенной формы был
взят штамм 84, показало, что если в опытах с проростками
помидоров или персиков отношение числа патогенных штаммов
к непатогенным было меньше или равно 1, то галлы не разви-
вались. Конечная цель этих экспериментов заключалась в соз-
дании условий, при которых указанное отношение для поверх-
ности корней не превышало бы единицы, для чего предприни-
мались попытки повысить содержание непатогенных форм на
семенах или корнях молодых деревьев при пересадке. В опытах
с персиками путем инокуляции корней удалось защитить около
95% растений, росших год на почве без патогенов, и около»
99%, когда инокулировались и семена, и корни.
Сельское хозяйство и биотехнология
375
Способ обработки
Сама обработка крайне проста: семена, черенки или корни мо-
лодых растений погружают в суспензию бактерий, а затем не-
медленно высаживают. Хорошие результаты получают при плот-
ности суспензии порядка 107—108 бактерий/мл; при этом забо-
леваний практически не наблюдается. В Австралии, Новой Зе-
ландии и США такие инокуляты имеются в продаже; некоторые
из них делают на основе торфа, как и в случае Rhizobium,
а другие представляют собой культуры, выращенные на агаре.
Механизм контроля
Штамм 84 синтезирует высокоспецифичный антибиотик, кото-
рый подавляет рост большинства патогенных агробактерий. Это
нуклеотидный бактериоцин, известный под названием агро-
цин 84. В клетках этого штамма содержится небольшая плаз-
мида с мол. массой 30-106, детерминирующая синтез агроцина,
и крупная плазмида с мол. массой 124-10®, в которой закоди-
рована способность использовать нопалин — необычную амино-
кислоту типа опина, присутствующую только в тканях корон-
чатых галлов. Крупная плазмида Ti, которая несет гены пато-
генности и чувствительности к агроцину, в этих клетках отсут-
ствует. Когда патогенный штамм выращивают в присутствии
агроцина 84, образуется небольшое количество устойчивых к не-
му колоний, как правило непатогенных. При этом, видимо, либо
теряется плазмида Ti, либо происходит мутация (делеция), ко-
торая проявляется не только в форме устойчивости к агроци-
ну 84, но и в потере патогенности.
Проблемы борьбы с корончатыми галлами
К сожалению, метод борьбы с корончатыми галлами с помощью
штамма 84 не удается распространить на все подверженные
этой болезни культуры, так как некоторые из них, например
-винный виноград, инфицируются штаммами, устойчивыми к аг-
роцину 84. Кроме того, в некоторых полевых опытах из галлов
были выделены патогенные, образующие агроцин 84, штаммы.
Все известные синтезирующие агроцин 84 штаммы устойчивы
к этому бактериоцину, так что бороться с ними с помощью
штамма 84 невозможно.
Опыты показали, что такие устойчивые патогенные штаммы
могут появляться в результате конъюгации между штаммом 84
и патогенными штаммами. В ходе конъюгации две плазмиды
штамма 84 передаются независимо друг от друга, в то время
как плазмида Ti патогена не обязательно проникает в клетку-
376
Глава 9
реципиент. Из шести возможных трансконъюгатов три содержат
Ti-плазмиду, а два из них к тому же и небольшую плазмиду,
в которой закодирована способность к синтезу агроцина 84.
В лаборатории конъюгация происходит только в присутствии
нопалина, а в природе, по-видимому, непосредственно в самой
ткани корончатого галла либо вблизи него. Можно предполо-
жить, что иногда при этом появляются патогенные штаммы,
образующие агроцин. Ясность в этот вопрос будет внесена лишь
тогда, когда будет обнаружен или создан методами генетической
инженерии мутант штамма 84, неспособный к конъюгации с па-
тогеном или же к переносу плазмиды с маркером агроцин 84.
9.6. Аэробная переработка отходов в сельском хозяйстве
(см. также гл. 6)
9.6.1. Проблема хранения и переработки отходов
Если сельское хозяйство ведется традиционными способами, то>
отходов животноводства образуется немного, и их несложно'
использовать для удобрения ^расположенных поблизости пахот-
ных земель. Сегодня, когда й животноводстве применяется ин-
тенсивная технология, жидкие и твердые отходы образуются
в большом количестве, и площадь близлежащих земель может
оказаться недостаточной для их использования. Хранить и пе-
рерабатывать такие отходы весьма непросто. Кроме того, в по-
следнее время проблемы использования отходов животновод-
ства привлекают пристальное внимание специалистов по охране
окружающей среды и органов здравоохранения, которых тре-
вожит возможность проникновения загрязнений в водоемы и
распространения таким путем возбудителей заболеваний. Не-
обходимо научиться наиболее полно и экономично использовать
ту часть отходов, которую можно применять как удобрения,
и в то же время решить проблемы, связанные с возможным
загрязнением среды из-за большого объема таких отходов. Важ-
ность задачи потребовала разработки и внедрения различных
систем переработки отходов.
Отличительной особенностью аэробной биологической систе-
мы, которая столь успешно применяется для переработки бы-
товых сточных вод, является свободный доступ воздуха к аэроб-
ным микроорганизмам, участвующим в превращении различных
веществ, содержащихся в отходах, в относительно стабильные
конечные продукты. Было показано, что контролируемая пере-
работка отходов сельского хозяйства в аэробных условиях
вполне возможна. Ряд научных центров в Европе и Северной
Америке приняли участие в исследованиях и разработке про-
грамм использования различных систем переработки. Главными
Сельское хозяйство и биотехнология
377
особенностями системы, используемой на фермах, должны быть
долговечность, простота в эксплуатации и желательно автома-
тизация всех процессов. Сегодня уже предложено несколько
таких маломасштабных систем.
9.6.2. Системы переработки отходов в аэробных условиях
Для переработки твердых отходов необходимо много времени
и средств, поэтому на фермах с интенсивной технологией для
их удаления стали широко использовать воду. Образующуюся
взвесь закачивают в хранилища или в системы переработки.
Водоем для окисления
Это наиболее простая установка, представляющая собой неглу-
бокую (не более 150 см) емкость, наполненную отходами и
имеющую достаточную площадь поверхности для обеспечения
аэрации. На поверхности этого водоема растут фотосинтезирую-
щие водоросли, которые повышают эффективность системы бла-
годаря выделению кислорода. Скорость поступления отходов
не должна быть слишком высокой. Недостатки таких установок
состоят в том, что 1) переработка отходов требует много вре-
мени; если объем отходов велик, необходимы водоемы большой
площади; 2) накапливаются твердые отходы, которые разлага-
ются в анаэробных условиях; 3) создаются условия для раз-
множения насекомых. Но есть и два преимущества: не нужна
механизация и обслуживающий персонал.
Аэрируемый водоем сходен с водоемом для окисления и от-
личается лишь тем, что в нем установлено механическое устрой-
ство для усиления аэрации, перемешивания и сохранения твер-
дых частиц в суспендированном состоянии. При одинаковой
загрузке такой водоем может быть меньше по площади и глуб-
же, чем водоем для окисления. Для него проще оценить время
переработки и качество стоков.
Такие системы особенно удобны для сбора и переработки
отходов, когда жидкость нельзя сливать в природные водоемы,
а также для сохранения воды для целей ирригации.
Каскадные бассейны
Каскадные бассейны — это тоже простая немеханизированная
система, применяемая в Англии. В отличие от водоемов для
окисления отходы поступают в нее постоянно, и время удержа-
ния их в системе невелико. Они включают первичный отстойник
(желательно с дефлекторами, позволяющими контролировать
скорость протекания), в котором осаждаются крупные частицы,
378
Глава 9
а также каскад мелких бассейнов, разделенных перегородками
или плотинами, через которые перетекает вода. Переливаясь
из бассейна в бассейн, вода аэрируется. Если время удержания
подобрано правильно, то глубина переработки оказывается не
меньше, чем в водоеме для окисления. Как и в неаэрируемых
водоемах, перемешивание осадка идет плохо, а активность мик-
роорганизмов может подавляться из-за недостатка кислорода
или же слишком медленн-ой диффузии конечных продуктов об-
мена.
Канава Пасвира (Pasveer ditch)
Эта система для окисления отходов является модификацией
установки с активным илом и представляет собой одну непре-
рывную, вытянутую в длину емкость, которую часто располагают
под полом хлева, откуда поступают отходы; в ней они и окис-
ляются. Жидкость (толщина слоя 0,3—0,6 м) аэрируют и пере-
мешивают с помощью ротора, помещенного в емкость. Имеются
приспособления для добавки новой порции отходов и удаления
отработанной жидкости. Эта и ряд других систем, основанных
на принудительной аэрации, во многом сходны с реакторами
непрерывного действия, в которых развивается особая микро-
флора (аналогичная таковой в хлопьях активного ила), мета-
болизирующая субстрат. Сходство заключается в том, что от-
ходы (субстрат) поступают в объеме, небольшом по сравнению
с общим их объемом, перерабатываемым в реакторе. Этот тип
перерабатывающего устройства активно изучался для выясне-
ния его пригодности для переработки отходов животноводства
в контролируемых условиях.
Основной недостаток всех этих систем — невозможность увя-
зать поступление кислорода с его потреблением. Их эффектив-
ность можно будет повысить, если удастся точнее оценить ско-
рость расщепления разных субстратов и лучше изучить микро-
организмы, участвующие в процессе.
9.7. Переработка отходов сельского хозяйства
в анаэробных условиях (см. также гл. 6)
9.7.1. Главные особенности процесса
При переработке органических отходов в анаэробных условиях
образуется горючий газ, на 60% состоящий из метана, и твер-
дый остаток, содержащий весь или почти весь азот и все другие
питательные вещества, содержащиеся в исходном растительном
материале. В природе такой процесс развивается при недостат-
ке кислорода в местах скопления веществ растительного или
Сельское хозяйство и биотехнология
379
животного происхождения: в болотах, осадках на дне озер,
а также в желудке травоядных. Он может протекать и в закры-
той емкости, наполненной подходящим органическим веществом,
куда не поступает воздух. Метанобразующие бактерии и неко-
торые другие микроорганизмы, продуцирующие нужные этим
бактериям субстраты, формируют в таких условиях систему
прочных симбиотических отношений, которая может функцио-
нировать неопределенно долгое время, если в нее в подходящем
количестве поступают все новые порции отходов.
Температурный оптимум лежит в интервале 30—35 °C, и для
его поддержания нужен подогрев. Этот процесс начали изучать
еще во времена Пастера; ему посвящено множество работ, а сам
процесс был значительно модифицирован, в частности примени-
тельно к очистке сточных вод. Это весьма удачный пример дей-
ствующей биотехнологии, поскольку она может применяться в
больших масштабах, оправдывающих капиталовложения и те-
кущие расходы по внедрению. Переработка отходов при участии
метанобразующего ила применяется на ряде крупных установок
в Англии и других странах, что существенно уменьшает ее стои-
мость. Она осуществляется, например, на Магденской станции
по переработке сточных вод в западной части Лондона (Thames
Water Authority). Там установлено двадцать тэнков для пере-
работки жидких отходов, диаметр каждого из них составляет
21,3 м, а глубина—10,4—16,0 м, с общим объемом 79000 м3.
Эта станция продолжает действовать, хотя переработка отходов
непрерывно производилась на ней в течение почти пятидесяти
лет, и вырабатывает метан в количестве, достаточном для ра-
боты двухтопливных машин, постоянно подающих сжатый воз-
дух для перемешивания содержимого тэнков и вырабатываю-
щих электроэнергию, потребляемую всей станцией.
9.7.2. Переработка отходов сельского хозяйства
Еще в начале века было выявлено, что из навоза можно полу-
чать горючий газ, а отходы использовать как удобрение. Пред-
принимались попытки найти практическое применение этому про-
цессу, но в целом интерес к нему был невелик до тех пор, пока
нехватка горючего во время войны не заставила обратить на
него внимание. С тех пор было сконструировано множество
установок для производства метана, но все они имели две основ-
ные части: 1) герметичный тэнк или реактор, в котором осуще-
ствлялась ферментация; 2) емкость для газа, обычно накопи-
тельный плавающий колокол с емкостью, близкой к таковой
у реактора.
В 40-х г. простейшие установки этого типа с реакторами
емкостью около 10 м3 использовались французами в Северной
380
Глава 9
Африке, и многие из них продолжали работать как там, так и
во Франции до начала 50-х г. В это время в Англии тоже стали
выпускать такие небольшие установки для продажи фермерам,
но, по-видимому, лишь немногие из них нашли применение.
Простейшие установки периодического действия имеют ряд
серьезных недостатков. Основной из них — трудность загрузки
реактора полутвердыми отходами и его опорожнение, которые-
обычно проводят вручную. Неудобство заключается и в том,
что надо иметь несколько реакторов, процесс в которых нахо-
дится на разных стадиях, чтобы сгладить неравномерность в
интенсивности образования газа, а также иметь возможность
подогревать реакторы до 30—35 °C. Все это, без сомнения, пре-
пятствовало широкому использованию генераторов метана на
фермах, и от них практически отказались, когда в 60-х годах
было налажено снабжение баллонами с пропаном и бутаном.
Совсем иначе обстояло дело в послевоенной Германии, где
были предприняты разработки крупных высокомеханизирован-
ных установок. Классическим примером может быть установка.
«Bihugas», построенная Ф. Шмидтом в Аллерхопе на ферме
площадью 220 акров, где в 1950 г. содержалось 55 голов круп-
ного рогатого скота, 180 свиней и 650 кур. По расчетам навоза
от этих животных, если смешать его с соломой, было достаточ-
но для производства 4,5-104 м3 газа в год. Установка включала
несколько цилиндрических подогреваемых наземных реакторов,
в которые насосами подавалась взвесь из навоза и резаной со-
ломы. Ферментацию проводили непродолжительное время, в те-
чение 5—21 сут, когда газ образовывался наиболее интенсивно.
Увеличить выход газа путем продолжительной неэкономичной
переработки не пытались. Безусловно, работа такой установки
была весьма эффективной, но ее размеры и стоимость были
таковы, что она могла работать только на самых крупных жи-
вотноводческих фермах. Такие крупные установки, несмотря на
их эффективность, нередко не выдерживали конкуренции с бо-
лее обычными и дешевыми источниками энергии, и ими пере-
ставали пользоваться.
Резкий подъем цен на нефть в 70-х гг. вынудил фермеров
к экономии, и неудивительно, что при этом вновь пробудился
интерес к генераторам метана и эффективным методам перера-
ботки навоза. Нефтяной кризис послужил толчком к публика-
ции в общедоступной прессе множества статей и заметок, ко-
торые призывали наладить производство метана в масштабах
всей страны, на каждой ферме, в каждом доме и даже машине.
Такие предложения нередко были сущей чепухой и свидетель-
ствовали о неправильном понимании сути дела. Интерес к про-
блеме сохраняется до сих пор, и в области метаногенной фер-
ментации были достигнуты серьезные успехи.
Сельское хозяйство и биотехнология
381
9.7.3. Микробиологические основы процесса
Метанобразующие бактерии являются строгими анаэробами,,
и при их исследовании ученые столкнулись с рядом проблем.
Это касалось прежде всего их выделения, но сегодня эта зада-
ча облегчается тем, что сконструированы удобные установки,
где созданы анаэробные условия. В настоящее время изучено-
множество таких бактерий, и мы многое узнали об особенностях
их метаболизма.
На первой стадии процесса ферментации из растительной
и фекальной массы, образуются летучие жирные кислоты (ук-
сусная, масляная). Важную роль при этом играют клостридии.
Кислоты (за исключением уксусной) служат далее субстратом
для группы уксуснокислых бактерий. В конечном счете в ре-
зультате совместного действия этих групп бактерий образуются
уксусная кислота, водород и углекислый газ, которые являются
подходящим субстратом для метанобразующих бактерий. Эти
бактерии относятся либо к ацетокластикам (например, Metha-
nosarcina barkeri, которая превращает уксусную кислоту в ме-
тан и СОг), либо к группе бактерий, использующих водород
(например, Methanospirillum hungatei, которая синтезирует ме-
тан из водорода и СО2). Взаимоотношения бактерий этих двух
групп, а также способы регуляции самого процесса переработки
подробно рассматриваются Мозеем (Mosey, 1982).
9.7.4. Современные разработки
Основная проблема, которая возникает на фермах, где содер-
жится много животных, заключается в хранении навоза и ис-
пользовании его наиболее выгодным образом. Если при этом
в качестве побочного продукта будет образовываться метан и
затраты на хранение навоза не увеличатся, то фермеры, безу-
словно, отнесутся к такой возможности положительно. Однако
осуществить эту возможность на практике вряд ли удастся, по-
скольку эффективные механизированные установки, предназна-
ченные для использования в развитом сельском хозяйстве, весь-
ма дороги. В Англии сегодня выпускают реакторы улучшенной
конструкции для переработки отходов ферм, но затраты на них
едва ли можно компенсировать доходами от производства ме-
тана. Для переработки разбавленных промышленных отходов
также сконструировано несколько интересных новых типов ана-
эробных реакторов, в которых используется принцип псевдо-
ожиженного слоя, но здесь основная цель состоит в очистке сто-
ков, а не в получении горючего газа.
Многие тысячи простейших реакторов были построены в Ин-
дии, Африке, на Дальнем Востоке и в Китаем. Здесь есть кому
382
Глава 9
их обслуживать из-за избытка рабочих рук. Очень трудно, од-
нако, оценить, насколько они на самом деле полезны и как дол-
го прослужат.
$.8. Использование протопластов в селекции растений
9.8.1. Вегетативное размножение
Изменчивость высших растений определяется в основном пере-
распределением генов при половом размножении. Хотя такая
изменчивость и важна с эволюционной точки зрения, она слу-
жит помехой при разведении сортов растений с желаемым на-
бором признаков. К счастью, не все цветковые растения раз-
множаются только половым путем. У многих из них образуются
особые органы вегетативного размножения, например видоиз-
мененные стебли—известные всем усы земляники и клубни кар-
тофеля. Многие растения обладают ценной особенностью: они
способны к полной регенерации из небольшого отрезка стебля,
черенка. Эта особенность широко используется для разведения
растений с ценными сортовыми признаками. Только таким спо-
собом можно разводить растения, неспособные давать потомство
половым путем. Некоторые растения полностью регенерируют
из целых листьев (многие суккуленты) или их частей (напри-
мер, бегонии).
Полноценное растение может развиться и из каллусной тка-
ни, выращенной из меристемы in vitro. Этот метод позволил
получить безвирусные сорта картофеля. Вот уже несколько лет
метод культуры тканей используется для выведения оздоров-
ленных и устойчивых к болезням растений, для создания сортов
с определенным генотипом, а также для сохранения и разведе-
ния сортов ценных растений.
Сегодня мы знаем, что многие изолированные клетки, если
их культивировать в соответствующих условиях, могут регене-
рировать в целое растение. Клетка, обладающая такой способ-
ностью к росту и формированию ткани, из которой затем раз-
вивается полноценное растение, называется тотипотентной. В се-
редине 60-х гг. тотипотентность клеток была обнаружена у мно-
гих видов растений.
Из тотипотентных клеток легко получить лишенные клеточ-
ной стенки протопласты. Разработаны методы их культивиро-
вания для получения каллусной ткани и затем — небольших
растеньиц, которых можно размножать обычными способами.
Протопласты получены из тканей большинства органов рас-
тений, в том числе из корней, листьев, лепестков, плодов, коле-
оптил, запасающих органов, тетрад микроспор (из пыльников),
корневых клубеньков бобовых и каллусных тканей. Чаще всего
Сельское хозяйство и биотехнология
385
для этого берут мезофильную ткань листа: из нее образуется
много единообразных протопластов. Нередко из протопластов
получают каллусные культуры, из которых можно регенериро-
вать целое растение. К сожалению, до последнего времени не
удавалось вырастить целые растения из изолированных прото-
пластов бобовых и злаков — двух важнейших групп культиви-
руемых растений. Нередко в относительно простых условиях
выращивания и с высоким выходом была осуществлена реге-
нерация в целое растение протопластов Medicago sativa (лю-
церна, альфальфа). Это позволяет надеяться, что подобрав со-
ответствующие условия, удастся регенерировать из протопла-
стов и бобовые. Сегодня мы еще не умеем регенерировать
растения из протопластов хлебных злаков и пищевых бобовых,
но в 1980 г. Вернике и Бреттел (Werniche, Brettell, 1980) доби-
лись образования каллуса и регенерации растений из листьев
злаков, так что можно ожидать, что и в случае протопластов
злаков будет достигнут прогресс.
Особая ценность протопластов для селекции растений опре-
деляется целым рядом их свойств. Во-первых, протопласты
можно получать в большом количестве и отбирать из них раз-
новидности с полезными свойствами. Хотя сами протопласты
генетически единообразны, формирующиеся из них каллусы да-
ют растения, существенно различающиеся по внешним призна-
кам. Во-вторых, отсутствие клеточной стенки облегчает слияние
протопластов и образование гибридов. Поскольку при этом сли-
ваются соматические, как минимум диплоидные, клетки, селек-
ционер растений получает в свои руки мощный инструмент для
отбора. В-третьих, в отсутствие клеточной стенки облегчается
захват чужеродной ДНК — фрагментов молекул или же бакте-
риальных плазмид, в результате чего формируются растения
с совершенно новым набором признаков.
Изменчивость
Хотя растения, полученные в результате размножения вегета-
тивным путем (клоны), обычно похожи на родительское расте-
ние, из этого вовсе не следует, что все эти клоны генетически
одинаковы. Иногда возникают клоны, которые существенно от-
личаются от исходной формы. Их называют соматическими ва-
риантами или «спортами», и появляются они в результате ге-
нетических изменений в клетках меристемы, которые дают на-
чало всему новому растению или его части. В ряде случаев
такие спорты были размножены и привели к созданию новых
сортов. Примером могут быть апельсины «Навель» и персики-
нектарины. В основе соматической изменчивости лежит ряд ге-
нетических механизмов: изменение числа хромосом в ядре, му-
•384
Глава 9
тация отдельных генов, модификация внеядерных генов клеточ-
ных органелл (хлоропластов и митохондрий). Хотя изменчивость
растений картофеля, полученных из протопластов, обычно не-
велика, для протоклонов тетраплоидного сорта Russet Burbank
она оказалась существенной. Изучение изменчивости у регене-
рированных растений картофеля сорта Maris Bard показало,
что она возникает при росте каллусов из генетически едино-
образных протопластов и не является следствием экспериментов
с самими протопластами.
Изменчивость протоклонов, наблюдающаяся в отсутствие
мутагенов, весьма важна для растениеводства: благодаря ей
селекционер растений получает богатый исходный материал.
Так, среди регенерантов картофеля были выявлены растения
с потенциально полезными признаками: низкорослые, с изме-
ненными клубнями и с разными сроками созревания.
9.8.2. Регенерация растений из протопластов
Шеферд и Тоттен (Shephard, Totten, 1977 г.) в опытах с кар-
тофелем, у которого регенерация растений из культуры тканей
затруднена, разработали метод, позволяющий достаточно успеш-
но регенерировать растения из протопластов. Их способ был
несколько необычным, поскольку предусматривал особые усло-
вия выращивания растений, из которых потом получали прото-
пласты. Кроме того, на каждой стадии развития растений-реге-
нерантов они применяли особую среду.
Выращивание растений для опытов
Растения картофеля (американский сорт Russet Burbank) вы-
ращивали из клубней, свободных от Х-вируса, при высокой
освещенности (продолжительность светового дня 12 ч), при 24 °C
и относительной влажности 70—75%. Перед началом опыта рас-
тения выдерживали 4—10 сут при той же температуре и влаж-
ности, но более слабом (7000 люкс) и менее продолжительном
освещении (6 ч в сутки). Содержание растений в условиях сла-
бого и непродолжительного освещения приводило к существен-
ному увеличению выхода жизнеспособных протопластов из тка-
ней листьев (до 2—3-106 на 1 г ткани).
Среды для выделения и размножения
При выделении и размножении протопластов применяли среды
пяти типов (А —Е), которые представляли собой варианты
среды, предложенной Лэмом (Lam, 1975). В их состав входили
основные питательные вещества растений, микроэлементы, ами-
Сельское хозяйство и биотехнология
385
нокислоты, факторы роста, фитогормоны и регуляторы осмоти-
ческого давления. Содержание веществ подбиралось так, чтобы
обеспечить оптимальный рост на каждом этапе регенерации.
Все среды содержали 1,5—2,0% агара, кроме полужидкой сре-
ды А, где его концентрация составляла только 0,5%. Среды
D и Е, использованные на заключительном этапе, содержали
1650 мг/л NH4NO3.
Выделение протопластов
4 г ткани, вырезанной из стерилизованных с поверхности моло-
дых листьев, инкубировали в 200 мл среды А (без сахарозы и
агара) в темноте при 4 °C в течение 16—24 ч. Затем среду за-
меняли на 100 мл забуференного (pH 5,6) раствора с фермен-
тами, чтобы отделить мезофильные клетки и удалить их клеточ-
ные оболочки. При встряхивании смеси процесс заканчивался
за 4 ч при 28 °C. Жизнеспособные протопласты после процежи-
вания смеси собирали центрифугированием, промывали сре-
дой А, снова осаждали и хранили в жидкой среде А при кон-
центрации 6-Ю5 клетка/мл.
Рост каллуса
При инкубации протопластов на твердой среде В или на моди-
фицированной твердой среде происходит регенерация клеточных
стенок и ограниченное деление клеток, в результате чего обра-
зуются микрокаллусы. После переноса в среду С они растут
далее до стадии, когда становится возможным перенос в сре-
ду D, где индуцируется образование (морфогенез) побегов. На
заключительной стадии каллусы с побегами длиной около 1 мм
переносят на среду Е для завершения образования побегов и
формирования корней. Укорененные растения пересаживают в
небольшие горшочки с вермикулитом, где они и растут далее.
Здесь изложена суть метода, который следует использовать
для регенерации целых растений из одиночных протопластов,
и проиллюстрировано, как экспериментатору приходится варь-
ировать условия на разных стадиях развития, чтобы добиться
успеха. На рис. 9.7—9.10 показано, как выглядят свежевыде-
ленные протопласты, молодые каллусы и крошечные растения,
регенерированные из ткани каллуса. Особенно важно в этой
работе строго выдерживать надлежащие условия, однако почти
наверняка их придется специально подбирать для каждого изу-
чаемого вида растений.
Томасу (Thomas, 1981) удалось получить культуры побегов
тетраплоидного картофеля сорта Maris Bard, которые использо-
вались как исходный материал для выделения протопластов^
25—1344
386
Глава 9
Рис. 9.7, Свежевыделенные протопласты Solanum brevidens.
Пазушные почки стерилизовали с поверхности раствором гипо-
хлорида натрия и культивировали затем на среде Мурасиге
и Скуга (Murashige, Skoog, 1962), содержащей, кроме того,
сахарозу (30 г/л), агар (6 г/л) и 6-бензиламинопурин (6-БАП,
0,5 мг/л), pH 5,8. В ходе инкубации при 26 °C при освещенности
~400 люкс (16 ч днем, 8 ч ночью) за 3—5 нед образовывались
ростки длиной около 3 см. Их можно было использовать для
дальнейшего размножения на той же среде, но без 6-БАП, для
чего брали короткие (3—8 мм) отрезки стебля с листом и па-
зушной почкой. Рост продолжался три недели, после чего по-
беги можно было опять использовать для размножения или же
выделения протопластов. Цельные проростки (но не взятые от-
дельно листья) давали жизнеспособные протопласты, которые
могли длительное время делиться в культуре.
9.8.3. Методы слияния протопластов
Хотя о возможности слияния протопластов растений было из-
вестно уже давно, метод контролируемого слияния с воспроиз-
водимыми результатами был разработан лишь в 1970 г. (Power
et al., 1970). Тем самым был сделан первый шаг к соматической
гибридизации растений. При суспендировании протопластов
в 0,25 М растворе азотнокислого натрия слияние происходило
очень быстро. Впоследствии для индукции слияния протопластов
Parthenocissus tricuspidata и Petunia hybrida использовали
10,2%-ный раствор сахарозы, содержащий 5,5% азотнокислого
натрия. Для той же цели применяли хлористый кальций.
Сельское хозяйство и биотехнология
387
Рис. 9.8, Каллусы, образовавшиеся из протопластов Solatium brevidens на ага-
ре. Снимок сделан примерно через месяц после получения протопластов. Для
выращивания использована чашка Петри диаметром 9 см.
Было показано (Као, Michayluk, 1974), что полиэтиленгли-
коль с большой молекулярной массой также вызывает неспе-
цифическое слипание, а затем и слияние протопластов растений.
При инкубации в самом полиэтиленгликоле слияний почти не
происходило, но при его разведении гибриды образовывали до
10% протопластов. ПЭГ способствовал также захвату хлоро-
пластов водоросли Vaucheria dichotoma протопластами морко-
ви. Методы слияния протопластов подробно описаны в руковод-
стве Пауэра и Дэйви (Power, Davey, 1979).
Получение соматических гибридов
Для осуществления гибридизации соматических клеток методом
слияния протопластов необходимо выполнить следующие опе-
рации: 1) выделить протопласты; 2) осуществить слияние;
3) регенерировать клеточные стенки; 4) осуществить слияние
ядер так, чтобы получить полноценное гибридное ядро; 5) раз-
множить гибридные клетки; 6) регенерировать целое растение.
Осуществить, слияние протопластов растений, вообще гово-
25'
388
Глава 9
Рис. 9.9. Регенерация мини-растений Solarium tuberosum из каллусной ткани,
полученной из протопластов (снимок сделан примерно через 100 сут после пе-
ренесения на агаровую среду, способствующую регенерации).
ря, несложно, но из этого не следует, что столь же просто раз-
множить гибридные клетки или же регенерировать гибридное
растение. Прежде всего нужно отобрать гибридные протопла-
сты, которые составляют лишь малую часть всех клеток, а за-
тем создать им условия для развития. Примером работы такого
рода может быть эксперимент по соматической гибридизации
Petunia hybrida и Petunia parodii. Здесь отбор был основан на
различиях в способности к росту протопластов из листьев этих
двух видов растений. Дело в том, что протопласты Petunia pa-
rodii в использованной в этом опыте среде могли формировать
лишь микрокаллусы, состоящие не более чем из пятидесяти
клеток, в то время как из протопластов Petunia hybrida можно
получать растущие каллусы. Было использовано также и то
обстоятельство, что протопласты Petunia hybrida более чувстви-
тельны к актиномицину D, чем протопласты Petunia parodii.
Гетерокарион, образующийся в результате слияния двух раз-
ных протопластов, может после слияния ядер превратиться в
гибридную клетку. Имеется в виду, что происходит объединение
всех несущих гены, самореплицирующихся органелл обоих про-
Сельское хозяйство и биотехнология
389
Рис. 9.10. Регенерация мини-растений Solatium brevidens из каллусной ткани,
полученной из протопластов (снимок сделан приблизительно через 100 сут
после перенесения на агаровую среду, способствующую регенерации).
топластов, в то время как при обычном половом скрещивании
от каждой родительской клетки поступает по ядру, несущему
хромосомные гены (кариомы), но гены, передаваемые с пласти-
дами (пластидомы) и митохондриями (хондриомы), передаются
обычно лишь от женской клетки. Таким образом, методы на
основе слияния протопластов позволяют получать комбинации
двух полных родительских геномов. Слияние протопластов двух
скрещивающихся половым путем видов нередко приводит к об-
разованию стабильных амфиплоидных гибридных клеток, из ко-
торых могут быть регенерированы растения — соматические гиб-
риды. Надо сказать, что метод слияния протопластов пригоден
для работы не только со столь близкими видами или гибридны-
ми клетками.
Томас и др. (Thomas et al., 1979) перечисляет ряд работ,
в которых была осуществлена регенерация в экспериментах по
слиянию протопластов растений, принадлежащих к одному и
тому же (7 случаев) и разным (14 случаев) видам. В четырех
экспериментах по межвидовому слиянию были получены линии
клеток, а в четырех других — осуществлен захват клеточных
390
Глава 9
органелл протопластами. В двух опытах в протопласты растений
была введена чужеродная ДНК, которая экспрессировалась,
а в трех из протоклонов были получены мутантные растения.
Способы применения метода слияния протопластов весьма мно-
гообразны.
Возможность образования соматических гибридов создает
предпосылки для изменений внеядерного генного компонента
гибридных клеток. Для этого, например, можно использовать
протопласты-альбиносы, в которых нет хлоропластных генов,
или же протопласты, у которых во время слияния проводится
инактивация ядерного генома одного из родителей. Донорами
цитоплазматических генов, вводимых путем слияния в полно-
ценные протопласты, могут быть безъядерные субпротопласты
или микропласты.
Захват протопластами микроорганизмов и ДНК
Дэйви и Пауэр (Davey, Power, 1975) обнаружили, что ПЭГ
способствует захвату клеток и протопластов дрожжей и клеток
сине-зеленых водорослей протопластами Parthenocissus tricuspi-
data. При этом микроорганизмы локализуются в ограниченных
мембранами пузырьках (везикулах) цитоплазмы протопластов.
Значительный интерес представляет возможность введения в
протопласты интактных микроорганизмов с полезными свойст-
вами, например таких, которые способны фиксировать азот.
Другой вариант — это введение в высшие растения генетическо-
го материала микробов с последующей его экспрессией. В прин-
ципе методы слияния протопластов вполне могут использовать-
ся для этой цели, но пока успехи и в этой области весьма скром-
ны (Thomas et al., 1979).
При введении в протопласты нового генетического материа-
ла в виде чистой ДНК последняя не всегда сохраняется и экс-
прессируется в хозяйских клетках. Весьма перспективным пред-
ставляется использование в качестве векторов Ti-плазмид Agro-
bacterium tumefaciens (раздел 9.5.3), так как они непосредст-
венно включаются в ДНК хозяина и легко экспрессируются при
образовании опухолей. Ti-плазмиды, содержащие или не содер-
жащие чужеродной ДНК, могут вызвать трансформацию про-
топластов растений. По-видимому, такие исследования будут
проводиться все более интенсивно.
ЛИТЕРАТУРА
Allen М. F., Moore Т. S. jun., Christensen М. (1980). Phytohormone changes in
Bouteloua gracilis infected by vesicular-arbuscular mycorrizae. I. Cytokinin
increases in the host plant, Canad. J. Bot., 58, 371—374.
Сельское хозяйство и биотехнология
391
Aviv D., Fluhr R., Edelman M., Galun E. (1980). Progeny analysis of the inter-
specific hybrids: Nicotiana tabacum (CJAS>)+Nicotiana sylvestris with respect
to nuclear and chloroplast markers, Theor. appl. Genet., 56, 145—150.
Backman P. A., Rodriguez-Kabana R. (1975). A system for the growth and deli-
very of biological control agents to the soil, Phytopathology, 65, 819—821.
Beringer J. E., 1982. Microbial genetics and biological nitrogen fixation. In:
Advances in Agricultural Microbiology (ed. Subba Rao N. S.), pp. 2—23, Ox-
ford and IBM Publishing Co., New Delhi.
Bilkey P. C., Cocking E. C. (1980). Isolation and properties of plant microplasts:
newly identified subcellular units capable of wall synthesis and division into
separate microcells, Eur. J. Cell Biol., 22, 502.
Bonnett H. T., Eriksson T. (1974). Transfer of algal chloroplasts of higher plants,
Planta, 120, 71—79.
Brown M. E., 1982. Nitrogen fixation by free-living bacteria associated with
plants — fact or fiction? In: Bacteria and Plants (eds. Rhodes-Roberts M. E.
and Skinner F. A.), pp. 25—41, Academic Press, London.
Burton J. C. (1980). New developments in inoculating legumens. In: Recent
Advances in Biological Nitrogen Fixation (ed. Subba Rao N. S.), pp. 380—
405, Edward Arnold, London.
Burton J. C., Curley R. L. (1965). Comparative efficiency of liquid and peat-
base inoculants of field-grown soybeans (Glycine max), Agron. J., 57, 379—
381.
Carr J. G. (1982). The production of foods and beverages from plant materials
by micro-organisms. In: Bacteria and Plants (ed. Rhodes-Roberts M. E. and
Skinner F. A.), pp. 155—167, Academic Press, London.
Cocking E. C. (1981). Opportunities from the use of protoplasts, Phil. Trans. Roy.
Soc. Lond., B292, 557—568.
Coombs J. (1981). Biogas and power alcohol, Cherny Ind. 4 April, 223—229.
Cooper A., 1979. The ABC of NET, Grower Books, London.
Corby H. D. L., 1975. A method of making a pure-culture, peat-type, legume
inoculant, using a substitute for peat. In: Symbiotic Nitrogen Fixation in
Plants (ed. Nutman P. S.), pp. 169—173, Cambridge University Press, Cam-
bridge.
Corby H. D. L., 1980. The systematic value of leguminous root nodules. In:
Advances in Legume Systematics, Part 2 (eds. Polhill R. M. and Raven P. H.),
pp. 657—669, Royal Botanic Gardens, Kew.
Davey R. M., Power J. B. (1975). Polyethylene glycol-induced uptake of micro-
organisms into higher plant protoplasts. An ultrastructural study, Plant Sci.
Lett., 5, 269—274.
Deacon J. IF., 1983. Microbial Control of Plant Pests and Diseases, Aspects of
Microbiology 7, Van Nostrand Reinhold (UK) Co., Ltd., Nottingham, England.
Dunican L. K., O’Gara F., Tierney A. B., 1976. Plasmid control of effectiveness in
Rhizobium. Transfer of nitrogen fixing genes on a plasmid from Rhizobium
trifolii to Klebsiella aerogenes. In: Symbiotic Nitrogen Fixation in Plants (ed.
Nutman P. S.), pp. 77—90, Cambridge University Press, Cambridge.
Dye M.. 1979. The Rothamsted Rhizobium culture collection and inoculant use in
the UK, Rothamsted Report for 1978, Part 2, 119—130.
Elmes R. P„ Mosse B-, 1980. Nutrient film technique, Rothamsted Report for
1979, Part I, 188.
Elmes R. P., Mosse B. (1984). Vesicular-arbuscular endomycorrhizal inoculum
production. II. Experiments with maize (Zea mays) and other hosts in nutrient
flow culture, Canad. J. Bot, 62, 1531—1536.
Fraser M. E. (1966). Pre-inoculation of lucerne seed, J. appl. Bacterio!., 29,
587—595.
Fraser M. E. (1975). A method of culturing Rhizobium meliloti on porous granu-
les to form a pre-inoculant for lucerne seed, J. appl. Bacteriol., 39, 345—351.
392
Глава 9
Fred Е. В., Baldwin I. L., McCoy E., 1932. Root Nodule Bacteria and Legumi-
nous Plants, University of Wisconsin Press, Madison, Wisconsin.
Hadar Y., Chet I., Henis Y. (1979). Biological control of Rhizoctonia solani
damping-off with wheat bran culture of Trichoderma harzianum. Phytopatho-
logy, 69, 64—68.
Hayman D. S. (1977). Mycorrhizal effects on white clover in relation to hill land
improvement, ARC Res. Rev., 3, 82—85.
Hayman D. S. (1980). Mycorrhiza and crop production, Nature, 287, 487—488.
Hayman D. S., Mosse B. (1979). Improved growth of white clover in hill gras-
slands by mycorrhizal inoculation, Ann. appl. Biol., 93, 141—148.
Hayman D. S., Morris E. J., Page R. J. (1981). Methods for inoculating field
crops with mycorrhizal fungi, Ann. appl. Biol., 99, 241—253.
Hepburn A. G., 1982. The biology of the crown gall — a plant tumour induced
by Agrobacterium tumefaciens. In: Bacteria and Plants (eds. Rhodes-Ro-
berts M. E. and Skinner F. A.), pp. 101—113, Academic Press, London.
Hobson P. N., Feilden N. E. H. (1982). Production and use of biogas in agricul-
ture, Prog. Energy Combust. Sci., 8, 135—158.
Kao K- N., Michayluk M. R. (1974). A method for high-frequency intergeneric
fusion of plant protoplasts, Planta, 115, 355—367.
Kerr A. (1972). Biological control of crown gall: seed inoculation, J. appl. Bac-
terid., 35, 493—497.
Kerr A. (1980). Biological control of crown gall through production of agro-
cin 84, Pl. Disease, 64, 25—30.
Khin Htay., Kerr A. (1974). Biological control of crown gall: seed and root
inoculation, J. appl. Bacteriol., 37, 525—530.
Kumar A., Wilson D., Cocking E. S. (1981). Polypeptide composition of Frac-
tion 1 protein of the somatic hybrid between Petunia parodii and Petunia par-
viflora, Biochem. Genet., 19, 255—261.
Lam S. (1975). Shoot formation in potato tuber discs in tissue culture, Am. Po-
tato J„ 52, 103—106.
Lawrie J. (1961). Natural Gas and Methane Sources, pp, 82—88, Chapman and
Hall, London.
Liu S., Baker R. (1980). Mechanism of biological control in soil suppressive to
Rhizoctonia solani, Phytopathology, 70, 404—412.
Menge J. A. (1983). Utilization of vesicular-arbuscular mycorrhizal fungi in
agriculture, Canad. J. Bot., 61, 1015—1024.
Menge J. A., Lembright H., Johnson E. L. V. (1977). Utilization of mycorrhizal
fungi in citrus nurseries, Proc. Int. Soc. Citriculture, 1, 129—132.
Mosey F. E. (1982). New developments in the anaerobic treatment of industrial
wastes, Wat. Pollut. Contr., 81, 540—552.
Munns D. N., Mosse B., 1980. Mineral nutrition of legume crops. In: Advances in
Legume Science (eds. Summerfield R. J. and Bunting A. H.), pp. 115—125,
Royal Botanic Gardens, Kew.
Murashige T., Skoog F. (1962). A revised medium for rapid growth and bioassay
with tobacco tissue cultures, Physiologia Pl., 15, 473—497.
Nemec S. (1980). Effects of 11 fungicides on endomycorrhizal development in
sour orange, Canad. J. Bot., 58, 522—526.
New P. B., Kerr A. (1972). Biological control of crown gall: field measurements
and glasshouse experiments, J. appl. Bacteriol., 35, 279—287.
Noack W., 1955. Biogas in der Landwirtschaft, Otto Elsner Verlagsgesellschaft,
Darmstadt.
Pasveer A. (1959). A contribution to the development of the activated sludge
process, J. Proc. Inst. Sewage Purif., 4, 436.
Phillips J. M., Hayman D. S. (1970). Improved procedures for clearing roots and
staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid asses-
sment of infection, Trans. Br. mycol. Soc., 55, 158—161.
Сельское хозяйство и биотехнология
393
Polhill R. М., Raven Р. Н. (eds.), 1980. Advances in Legume Systematics, Royal
Botanic Gardens, Kew.
Power J. B., Cummins S. E., Cocking E. C. (1970). Fusion of isolated plant pro-
toplasts, Nature, 225, 1016—1018.
Power J. B., Davey M. R., 1979. Laboratory manual: Plant Protoplasts (isolation,
fusion, culture, genetic transformation), Department of Botany, University of
Nottingham, Nottingham.
Power J. B., Frearson E. M., Hayward C., Cocking E. C. (1975). Some conse-
quences of the fusion and selective culture of Petunia and Parihenocissus
protoplasts, Plant Sci. Lett., 5, 197—207.
Power J. B., Frearson E. M„ Hayward C., George D., Evans P. K-, Berry S. F.,
Cocking E. C. (1976). Somatic hybridisation of Petunia hybrida and P. parodii,
Nature, 263, 500—502.
Rishbeth J. (1963). Stump protection against Fames annosus. III. Inoculation
with Peniophora gigantea, Ann. appl. Biol., 52, 63—77.
Roberts IF. P., Tate M. E., Kerr A. (1977). Agrocin 84 is a 6-N-phosphoramidate
of an adenine nucleotide analogue, Nature, 265, 379—381.
Robinson K-, 1971. Aerobic treatment of agricultural wastes. In: Microbial
Aspects of Pollution (eds. Sykes G. and Skinner F. A.), pp. 91—102, Academic
Press, London.
Robinson K- (1974). The use of aerobic processes for the stabilization of animal
wastes, CRC Crit. Rev. Environ. Contr., 4, 193—220.
Robinson K- (1979). Developments in aerobic stabilization of animal wastes in
Northeast Scotland: a review, Environ. Protec. Eng., 5, 127—143.
Roughley R. J. (1968). Some factors influencing the growth and survival of
root nodule bacteria in peat culture, J. appl. Bacteriol., 31, 259—265.
Roughley R. J. (1970). The preparation and use of legume seed inoculants, Pl.
Soil., 32, 675—701.
Roughley R. J., Date R. A., Walker M. H. (1966). Inoculating and lime pelleting
legume seed, Agric. Gaz. N. S. W., March, 142—146.
Roughley R. J., Vincent J. M. (1967). Growth and survival of Rhizobium spp. in
peat culture, J. appl. Bacteriol., 30, 362—376.
Santos A. V. P. Dos, Outka D. E., Cocking E. C„ Davey M. R. (1980). Organo-
genesis and somatic ebbryogenesis in tissues derived from leaf protoplasts and
leaf explants of Medicago sativa, Z. Pfl Physiol., 99, 261—270.
Schroth M. N., Hancock J. G. (1981). Selected topics in biological control, Ann.
Rev. Microbiol., 33, 453—476.
Shanmugam К- T„ Hennecke H., 1980. Microbial genetics and nitrogen fixation.
In: Recent Advances in Biological Nitrogen Fixation (ed. Subba Rao N. S.),
pp. 227—256, Edward Arnold, London.
Shephard J. F., Bidney D., Shahin E. (1980). Potato protoplasts in crop improve-
ment, Science, 208, 17—24.
Shephard J. F., Totten R. E. (1977). Mesophyll cell protoplasts of potato. Isola-
tion, proliferation and plant regeneration, Pl. Physiol., 60, 313—316.
Steinhorn J., Roughley R. J. (1974). Sodium chloride as a cause of low numbers
of Rhizobium in legume inoculants, J. Appl. Bacteriol., 37, 93—99.
Stewart W. D. P. (ed.), 1975. Nitrogen Fixation by Free-living Microorganisms,
IBP 6, Cambridge University Press, Cambridge.
Streicher S., Gurney E., Valentine R. C. (1971). Transduction of the nitrogen-
fixation genes in Klebsiella pneumoniae. Proc. natn. Acad. Sci USA 68,
1174—1177.
Strijdom B. W., Deschodt С. C., 1975. Carriers of rhizobia and the effects on the
survival of rhizobia. In: Symbiotic Nitrogen Fixation in Plants (ed. Nut-
man P. S.), pp. 151—168, IBP 7, Cambridge University Press, Cambridge.
Thomas E. (1981). Plant regeneration from shoot culture-derived protoplasts of
tetrapioid potato (Solanum tuberosum cv. Maris Bard), Plant Sci. Lett., 23,
81—88.
394 Глава 9
Thomas Е., Bright S. №. J., Franklin J., Lancaster V. A., Miflin B. J., Gib-
son R. (1982). Variation amongst protoplast-derived potato plants (Solanum
tuberosum cv. «Maris Bard»). Theor. appl. Genet., 62, 65—68.
Thomas E., King P. J., Potrykus I. (1979). Improvement of crop plants via
single cells in vitro — an assessment, Z. Pfl Ziicht, 82, 1—30.
Weindling R. (1946). Microbial antagonism and disease control, Soil. Sci., 61,
23____30.
Wells H. D., Bell D. K-, Jaworksi C. A. (1972). Efficacy of Trichoderma har-
zianum as a biocontrol for Sclerotium rolfsii, Phytopathology, 62, 442—447.
Wenzel G., Schieder O., Przewozny T., Sopory S. K., Melchers G. (1979). Compa-
rison of single cell culture-derived Solanum tuberosum L. plants and a model
for their application in breeding programs, Theor. appl. Genet., 55, 49—55.
Wernicke W., Brettell R. (1980). Somatic embryogenesis from Sorghum bicolor
leaves, Nature, 287, 138—139.
Vincent J. M. (1970). A Manual for the Practical Study of the Rootnodule Bac-
teria, IBP Handbook, No. 15, Blackwell Scientific Publications, Oxford.
.Zaenen I., van Larebeke N., Teuchy H., van Montagu M., Schell J. (1974). Super-
coiled circular DNA in crown gall inducing Agrobacterium strains, J. Mol.
Biol., 86, 109—127.
Глава 10
Химическая технология и биотехнология
Г. Хамер
10.1. Введение
Основные точки соприкосновения между химической технологи-
ей и биотехнологией лежат в области изучения, разработки,
проектирования, создания и осуществления процессов микро-
биологического производства продуктов, качественно превосхо-
дящих сырье, из которого они получены. Как и всякая другая
технология, биотехнология впитала в себя достижения целого
ряда наук, и круг решаемых с ее помощью задач определяется
относительным вкладом в нее отдельных дисциплин. Сегодня
основная проблема во взаимосвязях между биотехнологией и
химической технологией — это четкое определение сфер прило-
жения. В случае микробиологических процессов мы видим две
возможности: подход, основанный на биохимической технологии,
и подход, основанный на физиологии микроорганизмов. Привер-
женцы химической технологии часто отдавали предпочтение
первому из них, но надо сказать, что за последние 25 лет таким
путем не удалось разработать способы коренного улучшения
микробиологических процессов, которые принесли бы такой же
успех, какой был достигнут химическим производством в целом
или в нефтеперерабатывающей промышленности на основе прин-
ципов химической технологии. Разумеется, по крайней мере
частично это можно объяснить консерватизмом ряда произ-
водств, основанных на микробиологических процессах, а также
несогласованностью и непоследовательностью в их развитии.
Основной же причиной является относительно небольшой мас-
штаб упомянутых процессов. Чтобы получить право на незави-
симое существование, биохимическая инженерия должна раз-
работать гораздо более эффективные методы решения проблем,
связанных с осуществлением микробиологических процессов и
переработкой, что в свою очередь должно выразиться в значи-
тельном экономическом эффекте. Из самой сути биотехнологии,
опирающейся главным образом на физиологию микроорганиз-
мов, следует, что оптимизацией процессов микробиологического
396
Глава 10
производства должны заниматься узкие специалисты, т. е. мик-
робиологи, изучающие именно данный процесс, и инженеры-хи-
мики. При этом последние не обязательно должны детально
разбираться в микробиологической стороне проблемы. Такой
подход означает, что микробиолог или биотехнолог, помимо
глубоких знаний физиологических проблем, связанных с исполь-
зованием различных микроорганизмов, и навыков их решения,
должен обладать опытом сотрудничества с химиками-техноло-
гами. Можно надеяться, что благодаря плодотворному союзу
между биологами-исследователями и химиками-технологами в
ближайшее десятилетие будут созданы предпосылки как для
технологических нововведений, так и для экономического про-
гресса в области производств, основанных на микробиологиче-
ских процессах. Главная цель этой главы — способствовать раз-
витию более эффективных взаимосвязей между биологами и хи-
миками-технологами, и следовательно, повышению экономич-
ности как уже существующих, так и новых биотехнологических
производств.
Многие из биотехнологических производств по природе своей
строго традиционны и служат человечеству уже много столетий.
К их числу относятся производство спиртных напитков, фермен-
тированных молочных, соевых и рыбных продуктов. В этих от-
раслях промышленности качество и ценность продукта чаще
оцениваются по тонким различиям во вкусе и запахе, чем по
какому-либо иному легче поддающемуся определению критерию.
Для многих традиционных продуктов подобные критерии каче-
ства останутся важными и в будущем, однако маловероятно,
чтобы так же оценивались и новые продукты. Спрос на какой-
либо продукт будет зависеть главным образом от его функцио-
нальных качеств, общей стоимости его производства и ее воз-
можных неконтролируемых колебаний (например, из-за изме-
нений стоимости сырья и энергии).
Обычно расширение сферы применения биотехнологии про-
исходит в ответ на неотложные запросы практики, а не в ре-
зультате открытия нового способа синтеза некоего нового со-
единения при участии какого-то вновь выделенного микроорга-
низма. Вот ряд проблем, необходимость решения которых
привела к развитию исследований основных микробиологиче-
ских процессов и к созданию программ технических разработок.
1. Начиная с конца 60-х гг. Европа и Америка столкнулись
бы с катастрофическим уменьшением содержания белковых ком-
понентов в кормах для животных, если бы не были найдены
альтернативные способы их получения.
2. Начиная с середины 70-х гг. истощение запасов или сокра-
щение добычи жидкого ископаемого топлива привели к повы-
шению цен на него. Эти цены так возросли, что в большинстве
Химическая технология и биотехнология
397
развитых и развивающихся стран пришлось получать жидкое
топливо из альтернативных местных источников.
3. В будущем возможность максимального извлечения нефти
из разведанных месторождений сырой нефти будет зависеть от
синтеза новых химических соединений, позволяющих увеличить
нефтедобычу.
Попытки решения тех проблем, которые указаны в первом
примере, привели к интенсивному развитию исследований и раз-
работке методов получения белка одноклеточных организмов.
Для решения второй задачи потребовалось создание эффектив-
ных способов превращения возобновляемых местных источников
биомассы в этанол и/или метан. Для решения третьей из упо-
мянутых проблем сначала попытались использовать синтетиче-
ские полимеры, но не получили желаемого результата; тогда
основные усилия были направлены на поиски и производство
биополимеров, по своим свойствам пригодных для повышения
нефтедобычи.
Экономический эффект любой из этих программ определялся
и продолжает определяться тем, как долго будет существовать
спрос на тот или иной продукт, а также тем, не изменится ли
политическая ситуация и, следовательно, те предпосылки, кото-
рые использовались для оценки спроса.
Приведем три примера микробиологических производств, по-
явившихся в результате открытия нового пути синтеза новых
продуктов и никак не связанных с проблемами спроса. Речь
идет о предложениях по использованию для производства пласт-
масс поли-р-гидроксимасляной кислоты, синтезируемой Alcali-
genes eutrophus; о получении эмульгаторов при участии многих
бактерий, окисляющих углеводороды; об инсектицидах, проду-
цируемых Bacillus thuringiensis.
Взаимоотношения между биотехнологами и химиками-техно-
логами лучше всего прослеживаются на примере производства
основных продуктов промышленных микробиологических про-
цессов. В химической промышленности различают продукты
тонкого химического синтеза, продукты маломасштабного и ва-
лового синтеза. При создании производственных установок для
получения продуктов, относящихся к каждой из этих трех ка-
тегорий, используются совершенно разные подходы. Продукты
тонкого химического синтеза получают обычно в реакторах пе-
риодического действия, используемых также и для производ-
ства многих сходных веществ. Чаще всего производительность
одной установки лежит в интервале 100 кг/г.— 100 т/г. Значи-
тельную часть стоимости продукта составляют затраты на
очистку и анализы, направленные на то, чтобы по своему ка-
честву продукт соответствовал предъявляемым к нему требо-
ваниям. Согласно принятой классификации, к продуктам тон-
398
Глава 10
кого химического синтеза микробного происхождения относятся
вакцины, витамины, б'-нуклеотиды, некоторые аминокислоты»
антибиотики и ряд ферментов, предназначенных для использо-
вания в медицине. Химические продукты маломасштабного син-
теза обычно производятся в количестве 100 т/г. — 20 000 т/г. Ча-
ще всего к их качеству предъявляются менее строгие требова-
ния, чем к качеству продуктов тонкого химического синтеза.
Обычно их получают на установках, предназначенных только
для данного продукта, однако непрерывный процесс при этом
используется не всегда. К продуктам данной категории, произ-
водимым микробиологическим способом, относятся глутамино-
вая кислота, широко использующаяся как усилитель вкуса, ан-
тибиотики, применяющиеся для защиты сельскохозяйственных
культур, ферменты, предназначенные для использования в про-
мышленности, органические кислоты, в частности лимонная, мо-
лочная и глюконовая; раньше сюда же относили такие раство-
рители, как ацетон и бутанол. В эту же категорию можно вклю-
чить многие пищевые продукты и напитки, полученные путем
ферментации. И наконец, продукты третьей категории произво-
дят обычно на установках непрерывного действия, производи-
тельность которых превышает 20000 т/г. В этом случае рыноч-
ная стоимость продукта определяется простотой его получения
и общей производительностью установки, а не жесткими тре-
бованиями к качеству. К этой категории продуктов микробного
происхождения принадлежат БОО, различные виды топлива,
например биогаз (метан), спирт, использующийся как горючее
(этанол), и биополимеры, применяющиеся для более полного
извлечения нефти. Кроме того, валовым продуктом можно, ко-
нечно, считать и воду, сбрасываемую из установок для биоло-
гической очистки сточных вод и отходов, где идут самые круп-
номасштабные непрерывные микробиологические процессы.
В этих случаях конечный продукт не имеет обычно прямой
экономической ценности, но крайне важен для благополучия
и процветания общества. С увеличением масштабов производ-
ства расширяются и взаимосвязи между биотехнологией и хи-
мической технологией. Однако даже при получении продуктов
тонкого химического синтеза эти взаимосвязи должны стать
еще более глубокими, если мы хотим разработать особо эффек-
тивные технологии.
Основная задача практической химической технологии со-
стоит в создании производственных систем, с помощью которых,
используя достижения науки и техники, из сырья и материалов
можно экономичным путем получать продукты, пользующиеся
спросом. В биотехнологических процессах применяется' специ-
альное оборудование для превращения сырья в готовую продук-
Химическая технология и биотехнология
399
цию с помощью биохимических реакций, фазовых переходов,
агломерации, разделения, экстракции, высушивания и т. д.
Предприниматель, вкладывая капитал в какую-либо отрасль,
ставит своей целью получить максимальную прибыль. Если для
производства какого-то продукта предполагается использовать
микробиологический процесс, то прежде всего необходимо срав-
нить его с альтернативными процессами, а в некоторых случа-
ях— ис другими источниками получения этого продукта (на-
пример, если речь идет о белковых ингредиентах кормов, то это
может быть сельское хозяйство).
Экономическая оценка промышленного производства и про-
изводственных взаимосвязей, имеющая важнейшее значение, со-
пряжена со многими трудностями. Методика при этом, как пра-
вило, такова, что оценки занижаются по сравнению с требова-
ниями, выдвигаемыми лицами, которые желают оценить то или
иное производство или каналы перемещения капиталов. Однако
излишняя жесткость в оценке перспективности нового производ-
ства может привести к тому, что эта сфера деятельности пере-
станет представлять для конкурентов коммерческий интерес.
В зависимости от степени финансового риска общую сумму
капиталовложений в любое производство можно разделить на
три части: 1) основной капитал в сфере непосредственной обра-
ботки сырья, т. е. капиталовложения внутри отрасли; 2) капи-
таловложения во вспомогательные подразделения, т. е. инвести-
ции за пределами отрасли; 3) оборотный капитал.
Взятые вместе, эти капиталовложения составляют сумму,
которой рискует предприниматель, осваивающий новую сферу
производства. Наибольшая степень риска соответствует основ-
ному капиталу, наименьшая — оборотному.
Накладными расходами называются затраты, связанные с
бесперебойным функционированием производственного процес-
са. Их тоже можно разделить на три части: 1) расходы, про-
порциональные основным капиталовложениям, т. е. затраты на
эксплуатацию, страхование и управление; они не зависят от
производительности труда и могут быть выражены в процентах
от основного капитала; 2) расходы на сырье, коммунальные
службы, химические реактивы и прочие материалы; расходы,
связанные с контролем за качеством, финансовыми операциями,
покупкой патентов, лицензий и технологий; эти затраты зависят
от производительности; 3) прямые затраты на содержание
управленческого персонала, включающие помимо выплаты жа-
лования административные накладные расходы и расходы на
надзор.
Валовая прибыль, получаемая в результате функционирова-
ния конкретного предприятия, исчисляется как разность между
чистым годовым доходом (за вычетом издержек обращения,
400
Глава 10
премий и издержек сбыта продукции) и годовой суммой себе-
стоимости. Чистая прибыль — это ожидаемая выручка за год
за вычетом амортизации и налогов. Амортизационный регла-
мент порой очень сложен, его условия оговариваются государ-
ственными инстанциями и зависят от местоположения пред-
приятия; зачастую налоговые стимулы требуют оптимизации
разрешенного амортизационного регламента.
Оценивая возможность увеличения рентабельности производ-
ства путем интеграции и оптимизации производственного про-
цесса, необходимо отчетливо понимать, какие технологические
факторы наиболее важны с точки зрения экономики. В случае
микробиологического производства это могут быть либо соб-
ственно микробиологические, либо производственно-технологиче-
ские факторы. Прежде чем мы перейдем к их рассмотрению,
попытаемся вначале пояснить, что понимается под интеграцией
производства.
В рамках традиционной практики проектирования производ-
ства при разработке путей оптимизации отдельных процессов
почти не уделяется внимание их интеграции в единое производ-
ство. За последнее десятилетие во многих странах несколько
раз резко повышались цены на сырую нефть — главный источ-
ник энергии для промышленности — и нефтепродукты. До этих
акций — в значительной степени политических — в основе стра-
тегии нефтеперерабатывающей промышленности лежала пред-
посылка, согласно которой стоимость продукта должна снижать-
ся с увеличением масштабов производства. Рост цен на топливо
и сырье, а также связанное с этим удорожание строительства
предприятий привели к тому, что этот принцип едва не рухнул;
становилась все более очевидной необходимость интеграции
производства для снижения капиталовложений и экономии энер-
гии и сырья. Интеграция производства может охватывать как
выпуск изделий в рамках внутриотраслевой технологии, так и
взаимосвязь между производствами внутри отрасли и вне ее.
Мы остановимся подробнее на первом варианте. Однако вна-
чале сделаем несколько замечаний по поводу взаимосвязей вто-
рого типа, особенно характерных для промышленного микро-
биологического производства: в чем трудность развития таких
взаимосвязей на предприятиях, первоначально созданных для
валового химического производства, на основе обычной техно-
логии?
Промышленные микробиологические процессы протекают в
основном при низких температурах, обычно ниже 40 °C. Для
этого необходимо много воды (как в качестве хладагента, так
и в качестве высокочистого рабочего тела), а также значитель-
ные количества высокочистого рабочего пара и воздуха. Сырье
и готовые продукты должны храниться в условиях, при которых
Химическая технология и биотехнология
401
их загрязнение было бы минимальным. На предприятиях мик-
робиологической промышленности предъявляются повышенные
требования к гигиене производства. Необходимо особым спосо-
бом перерабатывать отходы и осуществлять серьезный контроль
за качеством продуктов. Все эти условия очень трудно выпол-
нить на типовых предприятиях для валового химического син-
теза, в частности на нефтехимических заводах. Для этого при-
ходится— в основном в пределах отрасли — проводить сущест-
венную перестройку производства, что требует новых капитало-
вложений и приводит к увеличению финансового риска. В целом
экономика микробиологического производства явно улучшается,
когда несколько таких производств обслуживаются централи-
зованными мощностями, должным образом оптимизированными
и ориентированными не только на узкие внутриотраслевые
нужды.
Наиболее важными для промышленного микробиологическо-
го производства являются следующие микробиологические фак-
торы: 1) коэффициенты выхода продукта или продуктов; 2) ско-
рости роста и/или скорости получения конечного продукта;
3) сродство культуры-продуцента к углеродным энергетическим
субстратам; 4) стабильность и неприхотливость культуры про-
дуцента.
К наиболее важным технологическим факторам относятся:
1) конверсия сырья и степень конверсии; 2) производительность;
3) концентрации продукта.
В высокотехнологичных и рентабельных процессах эти мик-
робиологические технологические факторы тесно взаимосвя-
заны.
Принципы и методология изучения рентабельности химиче-
ских предприятий обсуждались Раддом и Уотсоном (Rudd, Wat-
son, 1968).
10.2. Микробиологические факторы,
влияющие на производительность и экономичность процесса
В любом микробиологическом процессе ключевую роль играет
культура используемых микроорганизмов. В промышленной мик-
робиологии широко применяются плесневые грибы, дрожжи,
актиномицеты, бактерии и водоросли в виде чистых или сме-
шанных культур. В традиционных процессах ферментации пред-
почтение обычно отдается смешанным культурам, а в большин-
стве современных ферментационных процессов — монокультурам,
выращиваемым в асептических условиях. Говорить об идеаль-
ных культурах можно лишь в самых общих чертах. Следует
отдавать себе отчет в том, что идеального микроорганизма не
существует, а разнообразие продуктов, которые можно получать
26-1344
402
Глава 10
с помощью микроорганизмов, определяется разнообразием са-
мих микроорганизмов и их субстратов. Впрочем, возможно, ког-
да-нибудь мы и сумеем создавать такие идеальные микроорга-
низмы с помощью генетической инженерии и тем самым опро-
вергнем высказанное выше утверждение. Большинство исполь-
зуемых сегодня культур получено из природных источников;
однако эти культуры были затем улучшены или путем выращи-
вания в условиях, характерных для данного процесса (для по-
вышения выхода биомассы и первичных метаболитов), или с по-
мощью мутагенеза (для производства вторичных метаболитов).
Приведем основные уравнения, описывающие рост микроор-
ганизмов, в частности одноклеточных, в ферментерах и в других
микробиологических реакторах. Для выращивания любой куль-
туры необходимы: 1) жизнеспособный посевной материал;
2) источники энергии и углерода; 3) питательные вещества для
синтеза биомассы; 4) отсутствие ингибиторов роста; 5) соответ-
ствующие физико-химические условия.
Если все эти требования выполнены, то скорость роста (уве-
личения биомассы) одноклеточных микроорганизмов с бинар-
ным делением, размножающихся в условиях хорошо перемеши-
ваемой периодической культуры, будет пропорциональна кон-
центрации микробной биомассы, т. е.
dx
~Г=^’ (1)
где dxldt— скорость роста, ц— коэффициент пропорционально-
сти, обычно называемый удельной скоростью роста, х — концент-
рация биомассы (на сухой вес). Если ц— постоянная величина,
уравнение (1) можно преобразовать и проинтегрировать:
1пх = 1пх0+р/, (2)
где Хо — концентрация микробной биомассы в момент времени
/=0, t — время. Уравнение (2) можно переписать в виде
In (х/х0) = рЛ (3)
откуда
х = х0 е^. (4)
Если построить график зависимости 1пх от времени, то мы по-
лучим прямую с наклоном ц. Такой рост называют экспонен-
циальным или логарифмическим; он имеет место тогда, когда
состав микробной биомассы и условия окружающей среды оста-
ются постоянными. Приведенные выше уравнения применимы
и к смешанным культурам, в которых одноклеточные организмы
равномерно распределены в культуральной среде.
Химическая технология и биотехнология
403
При описании роста микроорганизмов с бинарным делением
в периодических культурах широко используется величина, ха-
рактеризующая время удвоения биомассы. Связь между удель-
ной скоростью роста р, и временем удвоения биомассы ty можно
найти из уравнения (3), заменив х на 2х0, a t — на ty:
(5>
10.2.1. Эффективность роста: коэффициент выхода биомассы
При любой количественной оценке роста микроорганизмов и/или
синтеза конечного продукта необходимо связать образование
микробной биомассы и продуктов с расходованием субстратов
и питательных веществ. Для описания соотношения между ро-
стом микроорганизмов (в расчете на сухой вес) и потреблением
углеродного энергетического субстрата были введены понятия
урожая бактериальной культуры и выхода биомассы (Monody
1942). В математической форме коэффициент выхода микроб-
ной биомассы Yx/s можно представить следующим образом:
Ухц--~, (6)
ds
где ds — бесконечно малое уменьшение концентрации субстрата,
соответствующее бесконечно малому увеличению концентрации
микробной биомассы dx. Знак минус указывает, что знаки из-
менения х и s противоположны. При постоянных условиях роста
коэффициент выхода остается постоянным, т. е. если хо и So —
концентрации субстрата и биомассы в момент времени to, а х
и s — в момент I, то
(х—x0) = Yx/s(s0—s). (7)
В общем случае коэффициент выхода биомассы является пере-
менной величиной, зависящей от условий культивирования. Ва-
риабельность этого коэффициента очень важна с точки зрения
оптимизации процесса. Понятие коэффициента выхода можно
распространить на все разнообразие питательных веществ, ис-
пользуемых растущими микроорганизмами, и на все получаю-
щиеся продукты. Кроме того, при оценке относительной эффек-
тивности биохимических путей, лежащих в основе роста и ме-
таболизма микроорганизмов, широко применяют понятия мо-
лярного урожая биомассы и выхода, когда учитывается число
электронов, поступающих от субстратов, и суммарное количе-
ство потребляемой энергии.
Когда микроорганизмы растут на хорошо растворимых суб-
стратах, коэффициент выхода и коэффициент конверсии совпа-
26*
404
Глава 10
дают; однако при росте на плохо растворимых и несмешиваю-
щихся субстратах они могут заметно различаться. Коэффициент
конверсии можно определить как количество микробной био-
массы, образующейся на единицу массы субстрата.
Для описания процесса образования какого-либо продукта
микроорганизмом, растущим на данном субстрате, можно вос-
пользоваться уравнениями, аналогичными (6) и (7):
^/.= —(8)
(Р—Po) = Yp/Asa—s)> (9)
где Yp/S — коэффициент выхода продукта, dp — бесконечно ма-
лое увеличение концентрации продукта, ро и р — соответственно
исходная концентрация продукта и концентрация его в момент
времени t.
/
Поток углерода
_____Поток энеогии
Рис. 10.1. Потоки углерода и энергии при гетеротрофном росте микробов и об-
разовании продукта.
Для роста любого микроорганизма необходимы источники
углерода и энергии. В случае гетеротрофных микроорганизмов
это одно соединение или смесь углеродсодержащих соединений,
которые удовлетворяют обе эти потребности. У автотрофных
микроорганизмов углеродные и энергетические субстраты раз-
личаются. Для гетеротрофных микроорганизмов и коэффициент
выхода биомассы, и коэффициент выхода продукта зависят от
распределения энергии и углерода между процессами анаболиз-
ма и катаболизма. На рис. 10.1 представлена диаграмма рас-
пределения потоков энергии и углерода при росте гетеротроф-
ных микроорганизмов и образовании ими соответствующего
Химическая технология и биотехнология
405
продукта. Коэффициенты выходов зависят от природы источ-
ника углерода и энергии, а также от биохимических особенно-
стей данного микроорганизма; когда источник энергии и угле-
рода подавляет рост, коэффициенты выхода зависят также и от
его концентрации. В случае источника углерода и энергии с от-
носительно высоким содержанием углерода (по сравнению с
другими элементами) и, следовательно, с высокой энергетиче-
ской константой при аэробном окислении наблюдается наруше-
ние равновесия между высвобождением энергии и потреблением
углерода; это ограничивает максимальный коэффициент выхода
биомассы.
К этой категории углеродных энергетических субстратов от-
носятся углеводороды. Избыточная энергия, высвобождаемая
при их использовании, рассеивается в виде тепла и приводит
к необходимости охлаждения микробиологических реакторов.
Скорость потребления определенного субстрата растущими
культурами часто представляют в виде
(10>
dt
где q — метаболический коэффициент, который очень широко
используется в микробиологии, особенно для кислорода и угле-
родных энергетических субстратов. Подставив в это уравнение
выражение для х из уравнения (1), получим
dx р,
ds q
откуда с помощью уравнения (6) имеем
(Н)
(12)
Yxis
10.2.2. Кинетика роста и образования продукта
Экспоненциальная модель, описываемая уравнением (1), пред-
ставляет собой лишь одну из нескольких относительно широко
распространенных моделей роста микроорганизмов; каждая из
таких моделей применима к специфическим физическим усло-
виям или той или иной морфологии биомассы. Другими альтер-
нативными моделями роста являются линейная модель и модель
кубического корня. Первая из них относится к случаю, когда
рост лимитируется диффузией субстрата или какого-либо пита-
тельного вещества. Вторая модель часто применяется при опи-
сании образования осадков (в случае плесневых грибов) или
бактериальных хлопьев, которые разрастаются только по пери-
ферии. На самом деле она представляет собой комбинацию
406
Глава 10
модели экспоненциального роста и модели, в которой лимити-
рующим фактором является перенос вещества. Модель кубиче-
ского корня адекватно описывает образование мицелиальных
осадков в периодической культуре, однако малопригодна для
случая непрерывной культуры, поскольку не учитывает разру-
шение хлопьев в биореакторе под действием гидродинамических
сил.
Если для периодической культуры наблюдается экспоненци-
альный рост, то это означает, что для широкого круга субстра-
тов и в широком диапазоне их концентраций скорость роста
фактически не зависит от концентрации субстрата. В этом слу-
чае процесс роста следует кинетике нулевого порядка и можно
ожидать, что потребление субстрата будет подчиняться следую-
щему уравнению ферментативной кинетики:
7m 8
s + Ks
(13)
где <?m — максимальный метаболический коэффициент при
Ks — константа насыщения, эквивалентная константе Михаэли-
са— Ментен в ферментативной кинетике. Подставив вместо q
и qm выражения, полученные из (12), мы придем к хорошо из-
вестному соотношению Моно:
(14)
„ __ Pms
И (s+Ks)
График этой функции представлен на рис. 10.2. Соотношение
Моно широко используется не только потому, что дает универ-
Рис. 10.2. Графическое представление соотношения Моно, описывающего за-
висимость удельной константы скорости роста от концентрации лимитирую-
щего субстрата.
Химическая технология и биотехнология
407
сальное описание кинетики роста микроорганизмов, но и бла-
годаря своей простоте. Для оценки цт и Ks (последняя из этих
величин обратно пропорциональна сродству микроорганизмов к
определенному субстрату) уравнение (14) можно преобразовать
следующим образом:
—=-^-4—- . (15)
[A S[lm p,m
Таким образом, график зависимосоти 1/р, от 1/s (т. е. график
Лайнуивера — Бэрка) является прямой и пересекает ось абс-
цисс в точке —1/Ks, а ось ординат — в точке 1/цт. Данных о
значениях константы насыщения Ks очень немного. Для угле-
родных энергетических субстратов и для минеральных пита-
тельных веществ значение К? обычно составляют —10~5 М,
а для кислорода ~ 10-6—10-5 М.
Рост микроорганизмов (прирост биомассы) в промышленной
установке можно описать с помощью уравнений (1), (14) и (7),
к которым нужно еще добавить уравнения, описывающие по-
токи в реакторе:
dx
----= их,
dt
„__ Рта s
И (s + ^s) ’
(х—x0) = Yx/s(s0—s).
(1)
(Н)
(7)
Если при этом происходит образование какого-то продукта, то
нужно использовать еще и уравнение (9):
(Р—Po) = ^p/S(so—s)- (9)
Однако образование микробных продуктов отнюдь не обязатель-
но связано с ростом. Поэтому, прежде чем применять те или
иные кинетические уравнения для анализа процессов в реакторе,
необходимо установить, какая именно взаимосвязь существует
между образованием продукта и ростом микроорганизмов.
Пожалуй, наиболее полезная классификация процессов об-
разования продукта в периодических культурах была предло-
жена более двадцати пяти лет назад Гэйденом (Gaden, 1959).
Согласно этой классификации, все процессы можно разделить
на три основные группы, которые обозначают как процессы ти-
па I, II, III.
В процессах типа I основной продукт образуется в ходе ре-
акций первичного энергетического метаболизма, когда между
потреблением субстрата и образованием продукта существует
постоянное стехиометрическое соотношение, метаболические пу-
ти представляют собой цепь последовательных реакций, а ско-
408
Глава 10
рость любого процесса имеет один максимум, обычно совпадаю-
щий с максимумом для других процессов. Типичными примерами
процессов типа 1 служат образование этанола, глюконовой и
молочной кислот из глюкозы, а также, разумеется, образование
микробной биомассы.
В процессах типа II основной продукт образуется в ходе
реакций энергетического метаболизма непрямым путем: либо
в ходе побочных реакций, либо в результате последующего вза-
имодействия между прямыми продуктами метаболизма. Кине-
тические кривые имеют сложный вид, причем максимумы ско-
ростей различных процессов не совпадают. Типичными приме-
рами процессов типа II являются образование лимонной и ита-
коновой кислот, а также некоторых аминокислот.
К процессам типа III относятся те, в которых основной про-
дукт образуется не в результате энергетического метаболизма,
а независимым путем. В таких процессах рост и метаболическая
активность достигают максимума на ранних этапах периодиче-
ского процесса, а максимум образования интересующего нас
продукта приходится на более поздние стадии, когда окисли-
тельная активность снижается. Продукты, образующиеся в ходе
таких процессов, называются вторичными метаболитами; это,
например, антибиотики пенициллин и стрептомицин.
Три этих типа процессов схематически представлены на
рис. 10.3. Совершенно очевидно, что подобная классификация
была принята только из соображений удобства и не является
ни совершенной, ни всеобъемлющей. Существуют процессы
с совсем иными характеристиками, протекающие в средах раз-
ного состава и при разных условиях. Отметим, что замена штам-
ма мало сказывается на общем характере кинетических кривых.
Впрочем, известны и исключения, особенно в процессах типа III.
Это наводит на мысль, что по мере усложнения биосинтетиче-
ских реакций, в ходе которых образуется данный продукт, дан-
ный тип процессов можно подразделить на более мелкие классы.
Обратимся теперь к непрерывным микробиологическим про-
цессам. Чтобы обеспечить протекание процесса в оптимальных
условиях стационарного состояния, необходимо располагать со-
ответствующими кинетическими данными. Если в условиях пе-
риодической культуры процесс -относился к типу I, то необходи-
мые условия для непрерывного культивирования можно теорети-
чески определить прямо из данных для периодического процесса.
В целом процессы такого типа лучше всего протекают в стацио-
нарных условиях в одностадийном биореакторе с полным пере-
мешиванием. Те процессы, для которых в условиях периодиче-
ской культуры характерны кинетики типа II или III, при не-
прерывном культивировании, по всей видимости, лучше всего
осуществлять в многостадийных последовательных биореаКто-
Тип I
Рис. 10.3. Кинетические кривые для периодического процесса ферментации,
предложенные Гэйденом (Gaden, 1959). Штриховые кривые —рост, пунктир-
ные— использование субстрата, сплошные — образование продукта.
410
Глава 10
pax. В этом случае можно так подобрать условия, чтобы ско-
рость роста и связанный с ним энергетический метаболизм до-
стигали максимума на одной стадии, а образование продукта —
на другой. Прежде чем переводить такие процессы из периоди-
ческих в непрерывные, очень важно выяснить соотношения меж-
ду скоростями отдельных процессов и основными контролируе-
мыми переменными.
В настоящее время некоторые процессы, характеризующиеся
кинетикой типа I в условиях периодической культуры, осущест-
вляются как непрерывные; для процессов же с кинетикой ти-
па II и III известны лишь единичные примеры такого рода.
Впрочем, эти процессы находят в последнее время все более
широкое применение именно при периодическом поступлении
питательных веществ.
10.2.3. Сродство к субстрату и рост микроорганизмов
При анализе роста культур в биотехнологических процессах
необходимо учитывать остаточную концентрацию питательных
веществ (субстратов), конкурентоспособность микроорганизмов
и особенности их роста на смешанных углеродных энергетиче-
ских субстратах. Первый из этих факторов имеет два важных
аспекта. Один из них экономический, связанный с полнотой ис-
пользования субстрата; второй касается качества продукта, его
чистоты и нетоксичности, поскольку остатки субстрата попада-
ют в конечный продукт. Полнота использования субстрата, ко-
торый является лимитирующим в периодической или непрерыв-
ной культуре, определяется сродством к нему данных микро-
организмов.
По-видимому, наиболее известной кинетической моделью
ферментативных процессов является модель Михаэлиса — Мен-
тен. Реакция фермента с субстратом описывается следующим
уравнением:
E-j-S^E —S—Д-E-f-P, (16)
где Е — фермент, S — субстрат, Е—S — фермент-субстратный
комплекс, Р — продукт, fe+i — константа скорости первой реак-
ции в прямом направлении, k-i — константа скорости первой
реакции в обратном направлении, k2 — константа скорости вто-
рой реакции, которая считается необратимой. Изменение кон-
центрации фермент-субстратного комплекса описывается сле-
дующим уравнением:
~ = k+i(e—c)s—k_lc—k2c, (17)
Химическая технология и биотехнология
411
где е — концентрация фермента, с — концентрация фермент-суб-
стратного комплекса, s — концентрация субстрата. Если s^e,
то для стационарного состояния
Для ферментативной реакции, описываемой уравнением (16),
скорость образования продукта v равна
v = k, с =-----------------------v-^-------= —max- - , (19)
[(fe_i + ^г)/^+11 + s + (k2/k+1) + s Л’м—s
где Umax—максимальная скорость образования продукта, т. е.
скорость, при которой весь фермент входит в состав фермент-
субстратного комплекса, Ks — константа равновесия для реак-
ции диссоциации фермент-субстратного комплекса, Лм=Л« +
+/г2/^-ц — константа Михаэлиса. Когда скорость образования
продукта определяется константой k2, т. е. &2-С&+1, то
Лм = ^- (20)
При выполнении условия (20) величина Км обратно пропорцио-
нальна сродству фермента к субстрату: чем меньше Км, тем вы-
ше сродство. Если МЫ заменим величины V, Umax и Км в (19)
на у,, pirn и Ks соответственно, то получим соотношение Моно
(14), которое уже было введено нами ранее и графически пред-
ставлено на рис. 10.2. Микроорганизмы, для которых кривая
зависимости между ц и s имеет большой начальный наклон,
обладают высоким сродством к лимитирующему субстрату.
И наоборот, микроорганизмы, для которых эта кривая пологая,
обладают низким сродством, и в таких культурах будут наблю-
даться более высокие остаточные концентрации субстрата.
При непрерывном культивировании в хемостате конкурен-
ция между двумя видами микроорганизмов за один лимитирую-
щий субстрат определяется обычно именно их сродством к этому
субстрату, а не максимальной удельной константой скорости
роста цт. При стационарном режиме в простом хемостате без
повторного использования биомассы концентрация лимитирую-
щего субстрата равна
K*D- , (21)
Pm В
где D — скорость разбавления. Из этого соотношения видно, что
концентрация лимитирующего субстрата в стационарном состоя-
нии зависит от скорости образования клеток за счет роста, ко-
торая в точности равна скорости разбавления, и от параметров,
412
Глава 10
характеризующих рост данного микроорганизма. Ясно, что па-
раметры, характеризующие рост разных микроорганизмов на
одном и том же субстрате, вряд ли будут одинаковы. Предпо-
ложим, что мы имеем культуру микроорганизмов А и В. По-
строив график зависимости удельной константы скорости роста ц
от концентрации лимитирующего субстрата s, мы получим два
разных случая (рис. 10.4,А и Б). В ситуации, изображенной
Б
Рис. 10.4. Соотношение Моно для культуры, представляющей собой смесь мик-
роорганизмов А и В, которые растут на одном лимитирующем субстрате (Har-
der et al., 1977). А. ц.тл>Ртв, Ksa<Asb. Г. РтА<1Хтв, Asa<1'Csb. В хемостате
при обеих степенях разбавления А и Рь А будет задерживаться, а В — вымы-
ваться.
на рис. 10.4, А, для микроорганизма А при любой данной ско-
рости разбавления (скорости роста) будет наблюдаться более
низкая концентрация лимитирующего субстрата, чем для мик-
роорганизма В при той же скорости разбавления (скорости
роста). Следовательно, в хемостате, где рост двух видов мик-
роорганизмов А и В лимитируется одним и тем же субстратом,
А всегда будет вытеснять В, поскольку последний не способен
сохранять достаточно высокую скорость роста при той концент-
рации субстрата, которая наблюдается для вида А, даже если
максимальная удельная скорость роста для В не была превы-
шена. Напротив, в ситуации, изображенной на рис. 10.4, Б, отно-
сительная конкурентоспособность двух видов микроорганизмов,
растущих в хемостате на одном лимитирующем субстрате, будет
зависеть от степени разбавления. Подобный анализ конку-
рентного роста в хемостатах можно распространить и на много-
компонентные смешанные микробные популяции, однако такая
экстраполяция не всегда оказывается возможной, поскольку в
Химическая технология и биотехнология
413
сложных смешанных культурах могут существовать несколько
другие взаимоотношения. Кроме того, в смешанных культурах,
применяющихся в биотехнологических процессах, использовать
основной субстрат обычно может только один вид микроорга-
низмов из всех присутствующих, а другие лишь модифицируют
среду с тем, чтобы оптимизировать рост основного вида, осо-
бенно если этот последний слишком прихотлив.
Вообще говоря, любой гетеротрофный микроорганизм спосо-
бен расти на самых разных субстратах — источниках углерода
и энергии. Большинство углеводов служат субстратом для очень
широкого спектра микроорганизмов; для углеводородов этот
спектр уже; на некоторых синтетических органических соедине-
ниях способны расти лишь очень немногие микроорганизмы,
а большинство таких соединений вообще не могут служить
субстратами. Один из основных вопросов, касающихся исполь-
зования смешанных субстратов данным гетеротрофным штам-
мом, состоит в том, как расходуются эти субстраты — одновре-
менно (конкурентным образом) или последовательно.
Для периодической чистой культуры микроорганизмов, рас-
тущей на смеси двух субстратов, обычно наблюдается так назы-
ваемый диауксичный рост. При этом на кинетической кривой
выявляются две четко различимые экспоненциальные фазы,
разделенные интервалом, на котором скорость роста проходит
через минимум. Такой характер роста обусловлен последова-
тельным использованием субстратов. Диауксичный рост на са-
харах впервые наблюдал Моно в 1942 г., однако подобная кар-
тина имеет место и для других субстратов. Диауксичный рост
характеризуется тем, что микроорганизмы не способны синте-
зировать ферменты, необходимые для метаболизма второго
углеродного субстрата, несмотря на его наличие, т. е. индукции
не происходит даже в присутствии индуктора. Это явление
получило название катаболитной репрессии.
При росте периодических культур на двухкомпонентном
субстрате наблюдаются и другие явления. Субстраты могут
использоваться независимо, так что потребление одного не
сказывается на потреблении другого. В таком случае они могут
метаболизироваться одновременно, и при этом иногда наблю-
дается даже стимуляция роста. Возможны ситуации, когда
ферменты, необходимые для метаболизма второго субстрата,
синтезируются, но их активность ингибируется, и тогда суб-
страты используются последовательно, но рост, строго говоря,
не является диауксичным. В случаях двухкомпонентных смесей
последовательно используемых субстратов тот субстрат, кото-
рый метаболизируется быстрее, обычно расходуется первым,
хотя известны и исключения. Однако в каждой конкретной двух-
компонентной смеси субстратов быстрее может использоваться
414
Глава 10
то один, то другой субстрат в зависимости от того, какой микро-
организм растет на этой смеси.
В связи со всем сказанным выше важно отметить, что мно-
гие культуральные среды, применяющиеся в промышленных
биотехнологических процессах, представляют собой сложные
смеси субстратов и их состав влияет на производительность
процесса. Характер потребления смешанных субстратов в не-
прерывных проточных культурах не всегда можно предсказать
исходя из результатов, полученных на экспериментальных
установках с периодическими культурами, растущими на таких
же смесях субстратов. В непрерывных проточных системах
компоненты смеси могут потребляться одновременно, даже если
в периодической культуре того же микроорганизма они исполь-
зовались последовательно. Особый интерес представляет сле-
дующее наблюдение, сделанное на хемостатной культуре. Ока-
залось, что в двухкомпонентной смеси субстратов критическая
скорость разбавления для полного использования того компо-
нента, который обеспечивает более медленный рост периодиче-
ской культуры, может значительно превышать нормальную
скорость вымывания. Кроме того, применяя двухкомпонентные
смеси субстратов в непрерывной культуре, можно контролиро-
вать поступление углерода в те или иные метаболические пути,
изменяя состав субстратной смеси или условия протекания про-
цесса, и регулировать тем самым количество образующейся
энергии или какого-либо продукта.
10.2.4. Стабильность культуры и требования,
предъявляемые ею
Пригодность выбранной культуры для использования ее в дан-
ном промышленном микробиологическом процессе часто не
оценивается и становится критическим фактором, когда дело
доходит до практического ее применения. Совершенно очевидно,
что, вообще говоря, самое главное — это генетическая стабиль-
ность культуры, но обсуждение вопросов этого круга выходит
за рамки данной главы. Здесь в общих чертах мы рассмотрим
физические, химические и пищевые факторы, а также требова-
ния, предъявляемые микроорганизмами. Основные условия,
необходимые для роста микроорганизмов, были перечислены
выше, но всегда остается вопрос: обеспечивают ли выбранные
условия оптимальный рост и/или образование продукта? К ка-
ким последствиям могут привести изменения этих условий,
в частности, как это скажется на продуктивности процесса?
Теплопередача и перенос веществ в биореакторах увеличи-
ваются за счет диссипации энергии в системе, обычно в резуль-
тате интенсивного механического перемешивания содержимого
Химическая технология и биотехнология
415
реактора вращающимися мешалками. Такие мешалки оказыва-
ют двоякое действие: способствуют диспергированию мелких
пузырьков воздуха в толще культуральной среды и обеспечива-
ют перемешивание, необходимое для минимизации концентраци-
онных градиентов, эффективного охлаждения реактора и умень-
шения выделения пузырьков воздуха. Однако возникающие при
таком перемешивании гидродинамические силы могут повреж-
дать клетки, особенно в случае волокнистых плесневых грибов,
актиномицетов, бактерий, имеющих жгутики, фимбрии, а так-
же хрупких клеток тканей, выращиваемых в суспензионной куль-
туре. Образование специфического продукта может зависеть от
морфологии плесневого гриба; по этой причине явно непригод-
ны для использования в промышленных процессах те штаммы,
морфология которых значительно изменяется при изменении
условий культивирования. Кроме того, следует избегать таких
условий проведения процесса, когда мицелий плесневых грибов;
повреждается.
Ранее считалось, что вероятность повреждений бактериаль-
ных клеток определяется скоростью вращения лопастей мешал-
ки. Однако, согласно наиболее правдоподобной современной
гипотезе о механизме повреждений, они обусловлены кавитаци-
онными явлениями в вихрях, возникающих сразу за лопастями
мешалок. Вопрос о повреждении бактериальных клеток в интен-
сивно перемешиваемых культурах все еще не решен. Если
анализировать свойства суспензий бактериальных клеток в по-
токе, используя для определения гидродинамических характе-
ристик любой отдельной бактериальной клетки предположение,
что она представляет собой сферическую частицу соответствую-
щего диаметра, то оценить силы, вызывающие механическое
повреждение клетки, не удается. Подобный подход не учиты-
вает наличие жгутиков или фимбрий: они — особенно это каса-
ется жгутиков — могут в несколько раз превосходить по длине
самые крупные несущие их бактериальные клетки. Вполне воз-
можно, что именно разрыв этих структур и последующее выте-
кание клеточного содержимого ответственны за разрушение
клеток при интенсивном перемешивании бактериальных куль-
тур. Заметим, что значительная часть исследований по генети-
ческой инженерии направлена на осуществление передачи
специфических свойств от различных бактерий к Escherichia
coli, которая не особенно пригодна для использования ее в
процессах с интенсивным перемешиванием именно из-за нали-
чия фимрий и жгутиков и, следовательно, подверженности ме-
ханическим повреждениям.
Если среди плесневых грибов и бактерий, вероятно, можно
отобрать выносливые микроорганизмы, более или менее при-
годные для использования их в условиях, характерных для
416
Глава 10
интенсивных процессов, то культуры тканей, в частности куль-
туры клеток млекопитающих, нуждаются в особых биореакто-
рах, конструкция которых учитывает присущую этим клеткам
хрупкость.
Если говорить о других условиях среды, то совершенно
очевидно, что наиболее пригодны для использования в промыш-
ленности микроорганизмы с широкими диапазонами оптимумов
pH, концентрации растворенного кислорода и температуры.
Впрочем, частично эти трудности позволяют обойти техниче-
ское усовершенствование установок и улучшение контроля за
условиями в них. Отметим, однако, что в некоторых новых
конструкциях промышленных биореакторов, в частности в ко-
лонных аппаратах и в циклических системах под давлением,
создаются значительные градиенты как физических параметров,
так и концентраций питательных веществ. Если экономические
преимущества подобных систем над более привычными доста-
точно велики, то для достижения высокой экономичности про-
цесса в целом крайне важно, чтобы в нем принимали участие
микроорганизмы, сохраняющие высокую продуктивность в быст-
ро меняющихся условиях.
В заключение следует отметить, что в последнее время не
уделялось достаточно внимания использованию в промышлен-
ных биотехнологических процессах смешанных, неприхотливых
культур определенного состава. Впрочем, даже в процессах,
осуществляемых при участии таких культур, значительные из-
менения условий могут приводить к нарушению равновесия в
соотношении между разными микроорганизмами, составляющи-
ми эти смешанные популяции.
10.3. Технологические факторы, влияющие
на производительность и экономичность процесса
Любой процесс, в котором участвуют микроорганизмы, незави-
симо от способа его осуществления можно отнести к одной из
двух категорий, определяемой характером поставленной задачи:
1) получение нужных микроорганизмов; 2) осуществление опре-
деленных химических превращений.
Последний тип процессов можно подразделить еще на два
класса: а) получение нужного конечного продукта; б) перера-
ботка сырья.
Все процессы, осуществляемые с целью химического превра-
щения, обладают одной общей особенностью, резко отличающей
их от процессов, цель которых — получение микробной биомас-
сы: микроорганизмы, осуществляющие данное превращение, не
обязательно должны при этом находиться в состоянии роста.
Этот аспект нередко недооценивается, хотя общеизвестно, что
Химическая технология и биотехнология
417
химические превращения могут осуществляться даже микроор-
ганизмами, уже потерявшими жизнеспособность. За такие
превращения ответственны клеточные ферменты. Совершенно
очевидно, что для контролируемого осуществления многих пре-
вращений с самым высоким выходом лучше всего использовать
иммобилизованные покоящиеся клетки или иммобилизованные
ферменты, выделенные из этих клеток. Роль таких систем при
проведении некоторых превращений в промышленных масшта-
бах становится все больше, однако при этом возникают про-
блемы, связанные со стабильностью подобных биокаталитиче-
ских систем, регенерацией кофакторов и с диффузией. Наиболее
широко применяемой в промышленных масштабах техноло-
гией остается химическое превращение, осуществляемое расту-
щими культурами микроорганизмов. С биохимической точки
зрения микробиологические процессы можно подразделить на
две основные категории: катаболические, в ходе которых слож-
ные соединения расщепляются до более простых, и биосинтети-
ческие, в ходе которых сложные соединения образуются из бо-
лее простых.
В основе любого процесса, протекающего при участии мик-
роорганизмов, лежит та или иная реакция или последователь-
ность реакций. Тем не менее при реализации этого процесса
в промышленном масштабе лимитирующим фактором, как пра-
вило, оказывается не кинетика этой реакции или последователь-
ности реакций, а другой, на первый взгляд менее существенный
фактор, который и определяет возможности использования дан-
ного процесса в промышленности.
Ключевыми факторами, влияющими на экономичность про-
цесса в биореакторе, являются производительность и степень
превращения, а они в свою очередь определяются теми физиче-
скими факторами, от которых зависит теплопередача и перенос
массы. К этим факторам относятся: 1) гидродинамические
свойства суспендированных микроорганизмов; 2) реологические
свойства культуральных сред; 3) электрокинетические свойства
микроорганизмов; 4) давление; 5) поверхностные и пристеноч-
ные эффекты, а также эффекты, возникающие на границе раз-
дела фаз; 6) эффекты, связаные с наличием нескольких фаз
в потоке; 7) флотационные, седиментационные и сегрегацион-
ные эффекты.
Интересно было бы обсудить, какую же реальную информа-
цию о любом из этих факторов и/или их влиянии на рост мик-
роорганизмов или на образование продукта могут дать микро-
биологические исследования процессов, протекающих с участием
микроорганизмов.
27—1344
418
Глава 10
10.3.1. Классификация биореакторов и их производительность
Химические реакции можно классифицировать по-разному.
Если иметь в виду выбор конструкции реактора, то, вероятно,,
наиболее полезной будет классификация, основанная на том,,
в однофазной или в многофазной среде протекает данная реак-
ция. Реакции, идущие в однофазной среде, называются гомоген-
ными, а в многофазных — гетерогенными. Для большинства
ферментативных реакций и многих реакций, протекающих при
участии микроорганизмов, подобная классификация не являет-
ся однозначной, поскольку они занимают весьма обширную
область между гомогенными и гетерогенными реакциями. Чтобы
упростить классификацию биохимических реакций, условились
считать реакционные системы, в которых существуют только
микроградиенты концентраций, гомогенными, а системы с мак-
роградиентами— гетерогенными. Следовательно, для биохими-
ческих реакций понятия гомогенности и гетерогенности не совпа-
дают с таковыми для химических реакций.
Любую систему, у которой существует ограничивающая ее
поверхность и в которой протекают биохимические реакции,
можно назвать биореактором. Промышленный биореактор —
это емкость, в которой осуществляются рост микроорганизмов
и/или различные химические превращения.
Промышленные биореакторы могут работать в периодиче-
ском режиме, периодическом режиме с доливом субстрата,
полунепрерывном (полупериодическом) и непрерывном проточ-
ном режимах. Исторически в промышленности утвердился
периодический способ работы при осуществлении химических
превращений и полунепрерывный — при получении микробной
биомассы. В последнее время для химических превращений ста-
ли применять реакторы с периодическим режимом и с доливом
субстрата, а для получения микробной биомассы — реакторы,
работающие в непрерывном проточном режиме. Традиционно
биореакторы, работающие в непрерывном проточном режиме,
использовались в промышленном масштабе только для аэробной
переработки сточных вод и отходов (т. е. в процессах с самой
большой пропускной способностью среди всех технологических
операций), а также при производстве уксуса. За исключением/
этих двух случаев, биологическая промышленность проявляла
исключительный консерватизм в том, что касалось перехода на
непрерывную проточную технологию, причем без достаточных
на то оснований.
Детальный анализ некоторых потенциальных режимов рабо-
ты биореакторов указывает на превосходство проточного непре-1
рывного режима. При работе по периодическому режиму в ре-!
актор загружают все необходимые компоненты (за исключени-
Химическая технология и биотехнология
419
ем кислорода для аэробных процессов), ведут процесс до
конца и собирают конечный продукт. Периодический режим с
добавлением субстрата предусматривает периодическое или не-
прерывное введение субстрата без удаления конечного продукта,
который собирают только по завершении процесса. При полу-
непрерывном ведении процесса, который называют также
полупериодическим, по завершении начальной стадии в перио-
дическом режиме реактор наполовину опорожняют, чтобы час-
тично собрать продукт, а затем снова заполняют таким же
объемом свежей среды и доводят процесс до конца; затем
снова осуществляют ту же последовательность операций. По-
добный подход направлен на лучшее использование производ-
ственной установки.
Сравним производительность реактора, работающего в
периодическом и непрерывном проточном, с идеальным переме-
шиванием, режимах. Периодический процесс включает несколь-
ко этапов: подготовительный, лаг-период, фазу экспоненциаль-
ного роста и период удаления продукта; с точки зрения
производительности реактора важен только период экспонен-
циального роста. Время, необходимое для завершения цикла
/ц, можно представить как сумму продуктивного времени t и
общего непродуктивного времени ts, т. е.
k = i + iK- (22)
Из уравнения (3), имея в виду, что для экспоненциального рос-
та ц=цт, получаем
t = -L In ( —V (23)
Р-П1 \ *
так что
= + (24)
Pm \ хо /
Для периодического процесса общая продуктивность равна
суммарной продукции, деленной на полное время цикла. Сум-
марная продукция задается соотношением (7), но для таких
периодических процессов, которые мы рассматриваем, остаточ-
ная концентрация субстрата s фактически равна нулю. Следо-
вательно, суммарная продукция равна
(x—x0) = Yx/ss0 (25)
и производительность периодического процесса Рл составляет
Рп =-------------------------yx/ssoPm--- . (26)
In I ~ I + С pm
\ хо /
27*
420
Глава 10
По определению производительность реактора (выход биомас-
сы), работающего в непрерывном проточном режиме с идеаль-
ным перемешиванием, равна
Pn = Dx, (27)
где х — концентрация биомассы в реакторе в стационарном
состоянии. Если принять, что коэффициент выхода биомассы
Уж/s и концентрация субстрата в свежей среде so для периоди-
ческого и непрерывного процессов одинаковы, a s — остаточная
концентрация лимитирующего субстрата, то уравнение (27)
имеет вид
Pa=DYx/,(s0-l), (28)
откуда при $<с$о получаем
Pn = DYx/ss0. (29)
Следовательно, отношение производительностей равно
^=-5-[1п(х/х0) + /яИт]. (30)
Рп Нт
В реакторе, работающем в проточном непрерывном режиме с
полным перемешиванием, максимальная скорость разбавления
равна Z) = p,m. Однако во избежание нестабильности на самом
деле поддерживают скорость разбавления на уровне 0,8 р,т,
и тогда
-^- = О,8[1п(х/хо) + /яИт]. (31)
Графическое сопоставление продуктивности реактора в периоди-
ческом и непрерывном режимах приводится на рис. 10.5.
Рабочие характеристики проточных биореакторов непрерыв-
ного действия лучше всего оценивать исходя из расчета мате-
риального баланса по биомассе, лимитирующему субстрату и
продукту. Используя самую приближенную классификацию
проточных биореакторов непрерывного действия с суспензион-
ными культурами, можно выделить два типа реакторов: реак-
торы с идеальным перемешиванием и проточные биореакторы в
режиме полного вытеснения (реакторы поршневого типа). Биоре-
акторы с идеальным перемешиванием могут работать как хемо-
статы или как турбидостаты. В хемостате поддерживается
постоянная плотность микробной культуры за счет потребления
лимитирующего субстрата или какого-либо иного питательного
вещества, а в систему турбидостата входит светочувствительное
устройство, которое измеряет оптическую плотность культуры
и обеспечивает ее постоянство. В промышленности, как правило.
Химическая технология и биотехнология
421
Рис. 10.5. Соотношение между образованием микробной биомассы в реакторе,
работающем в непрерывном и периодическом режимах, для различных коли-
честв посевого материала, максимальных удельных констант скорости роста и
продолжительностей непродуктивного этапа периодического цикла; D=Q,8 pm
для непрерывной системы (Aiba et al., 1973).
применяются реакторы, работающие в режиме хемостата, поэто-
му о турбидостатах мы далее говорить не будем.
Любой анализ производственных характеристик биореактора
включает изучение свойств существующих в нем потоков, а так-
же кинетики роста микроорганизмов и осуществляемых с их
участием превращений. Для описания потока в проточном ре-
акторе непрерывного действия удобнее всего использовать
422
Глава 10
распределение частиц, проходящих через реактор, по времени
1их пребывания в реакторе. В случае проточного реактора пол-
ного вытеснения все элементы жидкости проходят через реактор
строго упорядоченно, так что любой данный элемент никак не
перемешивается с элементами, поступающими в реактор до или
после него, т. е. отсутствует осевое перемещение. Следователь-
но, для стационарного состояния проточного реактора с полным
вытеснением все порции жидкости, поступающие в реактор,
находятся в нем одинаковое время. Среднее время пребывания
задается соотношением
где V — объем реактора, F — объемный поток жидкости, посту-
пающий в реактор и вытекающий из него. Напротив, содержи-
мое проточного реактора непрерывного действия с идеальным
перемешиванием является полностью однородным, а потому
его состав идентичен составу вытекающей из реактора жидко-
сти. Питательные вещества, поступающие в реактор, немедлен-
но перемешиваются с его содержимым и находятся в реакторе
разное время. Тем не менее среднее время пребывания для тако-
го реактора тоже определяется соотношением (32), хотя кривые
распределения времени пребывания для этих двух типов реак-
торов совершенно различны.
Кривые вымывания для данных реакторов также сильно
различаются. Представим себе, что реактор каждого типа на-
полнен жидкостью, в которой до начала поступления новых ее
порций дискретно и равномерно распределены инертные, несе-
грегирующие частицы, причем объемные потоки, выходящие из
каждого реактора, одинаковы. Тогда число частиц в каждом
реакторе будет равно
Qo = Vxo, (33)
где V—объем жидкости, х0— начальная концентрация. При
поступлении в реактор новых порций чистой жидкости из него
будет вытекать поток (обеспечивающий постоянство объема
содержимого реактора), концентрация твердых частиц в кото-
ром будет неодинакова для разных типов реакторов. При этом
число частиц, остающихся в реакторах, будет непрерывно умень-
шаться.
Для идеального проточного реактора с полным вытеснением
концентрация частиц в вытекающей жидкости будет равна х0
до тех пор, пока время вытекания t не станет равно времени
пребывания т; в этот момент х станет равным нулю. Число
частиц, остающихся в реакторе, будет линейно уменьшаться во
времени и при t=x станет равным нулю. Для реальных проточ-
Химическая технология и биотехнология
423
ных реакторов с полным *
вытеснением наблюдает- so-------*------ г---------*------s
ся несколько иная карти- F F
на из-за осевого переме-
шивания и диффузии.
Для реакторов с пол-
ным перемешиванием
концентрация частиц в
вытекающей жидкости
всегда будет равна кон-
центрации частиц в со-
держимом реактора. Кри-
вая вымывания для та-
ких реакторов, как и кри- Рис. ю.б. Проточный биореактор не-
вая распределения вре- прерывного действия с полным пере-
мени пребывания, будет мешиванием (см. текст),
представлять собой отри-
цательную экспоненту. Изменение концентрации частиц и их
числа в реакторе описываются уравнением
---*1 =----dQ = Dx (34)
dt dt
Используя соотношения для скорости вымывания, уравнения
роста микроорганизмов и образования продукта, а также вы-
ражение для коэффициентов выхода биомассы, можно получить
уравнения материального баланса, которые описывают работу
идеального проточного реактора непрерывного действия. Для
этого нужно найти соотношение, в котором накопление какого-
либо продукта выражено через увеличение его количества бла-
годаря входящему потоку, уменьшение из-за вытекающего
потока, увеличение за счет роста или образования продукта
в реакторе, уменьшение за счет использования в реакторе и
увеличение за счет рециркуляции потока.
Уравнение материального баланса для биореактора, рабо-
тающего в проточном режиме с идеальным перемешиванием
без рецикла, т. е. обычного хемостата (рис. 10.6), где продуктом
служит только микробная биомасса, выглядят следующим об-
разом.
Для микробной биомассы
Накопление = Рост — Удаление
или для бесконечно малого интервала времени
Vdx = V pxdt—Fxdt. (35)
424
Глава 10
Для лимитирующего субстрата
Накопление= Поступление—Удаление—Использование
или для бесконечно малого интервала времени dt
Vds=Fsodt-Fsdi—-^-dt, (36)
Kt/S
где V — постоянный объем жидкости в биореакторе, F — объем-
ный поток жидкости, втекающий в биореактор и вытекающий
из него, Ух/s — коэффициент выхода микробной биомассы при
росте на лимитирующем субстрате, х — концентрация биомассы
в реакторе, So и s — соответственно концентрации лимитирую-
щего субстрата в поступающем в биореактор растворе и в ре-
акторе, у, — удельная константа скорости роста.
Разделив (35) и (36) на Vdt, получим
(37)
at
И
£ = D(s0-s)---(38)
at Yx/s
В стационарном состоянии dx/dt=Q и ds!dt=Q. Следовательно,
(у.—£>)х=0, (39)
откуда
р = £> (40)
и, таким образом,
D(s0-7)—(41)
‘ X/S
где х и s — соответственно концентрации микробной биомассы
и лимитирующего субстрата в стационарном состоянии.
Соотношение Моно можно переписать следующим образом:
и = _,_Нд-!- . (42)
(s + *s)
Отсюда можно получить выражение для концентрации лимити-
рующего субстрата в стационарном состоянии, заменив у на D:
Концентрацию бактеральной биомассы в стационарном состоя-
нии получим, заменив сначала в уравнении (41) у, на D, а за-
тем подставив выражение (21) для s
Химическая технология и биотехнология
425
X=Yxts So
KSD
(Rm—О)
(43)
Производительность такой системы (выход микробной биомас-
сы) в стационарном состоянии равна
/’я=1?х,
(27)
или, после подстановки выражения для х,
— ^Yx/3 s0-
KSD
(Pon
(44)
Графики зависимости х, s и Рп от скорости разбавления D,
построенные в соответствии с соотношениями (43), (21) и (27),
приведены на рис. 10.7.
Рис. 10.7. Теоретическая зависимость концентрации биомассы, концентрации
лимитирующего субстрата в стационарном состоянии, выхода микробной био-
массы от скорости разбавления для простого хемостата (р.т=1,0 ч-1, Ks=
=0,1 г-л-1, Ух/5=0,б, so= 10 г-л-1).
Найдем уравнения материального баланса для другой си-
стемы— проточного реактора непрерывного действия с полным
перемешиванием, единственным продуктом которого является
426
Глава 10
микробная биомасса; в таком реакторе для интенсификации
процесса часть концентрированной микробной биомассы ис-
пользуется повторно, поступая из сепаратора, расположенного
на выходе из реактора (это может быть седиментационный
тэнк, применяемый при очистке сточных вод; центрифуга или
система ультрафильтрации). В подобной системе, изображенной
на рис. 10.8, скорость разбавления D равна F/V, где F — по-
F
so
s
X
Рис. 10.8. Проточный реактор непрерывного действия с полным перемешива-
нием и повторным использованием концентрированной микробной биомассы
(см. текст).
ток жидкости через систему в целом, V — постоянный объем
жидкости в биореакторе. Поток, выходящий из биореактора (но
не из системы в целом), можно представить в виде
Fs^P+aF,, (45)
или
где а — доля вытекающего из реактора содержимого, которая
используется повторно. Если принять, что микробная биомасса
становится более концентрированной в g раз, то количество
биомассы, повторно используемой в биореакторе, будет равно
aFsgx.
Уравнение материального баланса по биомассе будет в
этом случае иметь вид
Химическая технология и биотехнология
427
Накопление = Рост—Удаление 4- Повторное использование,
или для бесконечно малого интервала времени dt
Vdx = V yxdt—Fs xdt+aFs gxdt, (47)
а соответствующее уравнение по лимитирующему субстрату —
Накопление — ПоступлениеПовторное использование — Удаление—
— Расходование,
или для малого интервала времени
Vds = Fsodt~VaFssdt—Fssdt—'^—dt. (48)
Yx/s
Подставив в (47) выражение для Fs и разделив на Vdt, мы по-
лучим для стационарного состояния (при котором dx!dt = ty:
[и---^-as)D 1~=0 (49)
L (\-а) ]
или
(l-ag)Z) . (50)
Г (1-а)
В предположении, что выполняется соотношение Моно, полу-
чаем
_ (1 —ag)Z>
;T—KS (1 —“)
или
Г.. DKt(\ — ag)
Ит(1 — а)~ 0(1— ag) *
Подставив в (48) выражение для Fs и разделив на Vdt, мы по-
лучим для стационарного состояния (при котором ds/dt=Q)
уравнение, которое представляет собой уравнение для обычного
хемостата (41). Подставив в него выражение для $, будем
иметь
77 Yx/s. (5о s) (1 а)
Для определения концентрации микробной биомассы hx в ос-
ветленном потоке, вытекающем из сепаратора, нужно найти
уравнение материального баланса по биомассе в сепараторе.
В предположении, что сепаратор работает в стационарном ре-
жиме, т. е. биомасса в нем не накапливается, уравнение ма-
428
Глава 10
териального баланса для биомассы можно записать следую-
щим образом:
Поступление = Повторное использование-{-Концентрированные от-
ходы + Осветленные отходы
или
Fg х = aFa gx+cFgx + (\—c)Fhx, (54)
где с — доля потока F, вытекающего из системы в виде кон-
центрированных отходов, h — концентрационный коэффициент
по биомассе в осветленном потоке, вытекающем из сепаратора.
Подставив сюда выражение для Fs [соотношение (46)], по-
лучим
д = (1 —ag) — eg (1 — а)
(1-а) (1-е)
(55)
Если реактор работает в стационарном режиме, то его произво-
дительность (но не производительность системы в целом) по
биомассе будет равна
’ — Fsx aFsSx
н V V
(56)
или, после замены Fs,
р, __ Dx(l-ag)
Н (1-а)
(57)
Зависимость между производительностью Р'я, концентрацией
микробной биомассы в стационарном состоянии х, концентра-
цией лимитирующего субстрата в стационарном состоянии s и
скоростью разбавления представлена на рис. 10.9. Из этих со-
отношений с очевидностью следует потенциальная возможность
увеличения рабочей скорости разбавления или благодаря пов-
торному использованию биомассы, как в рассматриваемом на-
ми случае, или путем удержания активной биомассы.
Вернемся теперь к анализу работы проточного биореактора
с полным вытеснением. В этом случае в каждом малом эле-
менте жидкости dv, проходящем через биореактор, имеет ме-
сто в основном экспоненциальный рост (ц. = цт), если только
концентрация первого лимитирующего субстрата значительно
превышает константу насыщения Ks- Рост биомассы в таком
биореакторе без рецикла описывается следующим уравнением:
In х = In х0 + pm t, (58)
где Хо — концентрация биомассы в среде, поступающей в био-
реактор, х — концентрация в момент времени t. Если о — объ-
Химическая технология и биотехнология
429
Рис. 10.9. Теоретическая зависимость концентрации биомассы и лимитирующе-
го субстрата в стационарном состоянии, а также выхода микробной биомассы
в биореакторе от скорости разбавления для хемостата с концентрированием
микробной биомассы и ее повторным использованием ([Хт=1,0 ч-1, Ks =
= 0,1 г-л-1, Ух/з=0,5, 5о=Ю г-л-1, а=0,2, g=4).
ем культуральной среды, вышедшей из биореактора за время t,
то t~vlF = vlVd. Подставив это выражение для t в уравнение
(58), получаем
1пх = 1пх0+^. (59)
При рециркуляции микробной биомассы в проточном биореак-
торе с полным вытеснением никакой внешней инокуляции не
требуется. В подобной системе общий поток, вытекающий из
биореактора (но не из всей системы), описывается уравнением
(46), как и в случае рассмотренного выше реактора с рецирку-
ляцией. Если биомасса концентрируется в сепараторе в g раз,
то доля ее, повторно используемая в биореакторе, будет равна
ag. Если xw— концентрация биомассы в потоке, вытекающем
из биореактора, то в стационарном состоянии xa = agxv. Время
t, за которое из реактора вытечет объем V, равно
/= . (60)
Приняв v = Vn — объему, при котором происходит исчерпание
лимитирующего субстрата, и заменив х и t в уравнении (59),
для стационарного состояния получим
430
Глава 10
Va
=-------------- (In —
(1 — a) |xm \ ag
(61>
При Va = V (объем жидкости в биореакторе) это соотноше-
ние принимает вид
— ------------(In —\ . (62>
Р (1 — а) |лт \ ag )
Максимальная величина xw, хт, равна
хт=У$0 + х0.
(63>
В случае суспензионных культур проточные реакторы с полным
вытеснением используются редко. Как правило, реакторы рабо-
тают в режиме, близком к режиму полного перемешивания,
разумеется, применительно к жидкой фазе. Тем не менее не
следует считать, что в системе действительно осуществляется>
идеальное перемешивание, пока не проанализированы все воз-
можные причины появления неоднородности (например, из-за
пристеночного роста и т. п.). Вероятно, самым лучшим прибли-
жением к реактору с полным вытеснением был бы каскад по-
следовательных реакторов с идеальным перемешиванием без
дополнительных поступлений питательных веществ, однако для
этого число биореакторов, составляющих такой каскад, долж-
но быть бесконечным. Неидеальный поток с полным вытеснени-
ем можно получить, лишь когда работает более шести после-
довательных биореакторов.
10.3.2. Анализ размерностей и масштабирование
Анализ размерностей и моделирование с помощью безразмер-
ных величин широко применяются в химической технологии и
не менее важны для конструирования биореакторов и их мас-
штабирования. С анализом размерностей и с безразмерными
величинами хорошо знакомы лишь немногие микробиологи.
Это прежде всего метод, с помощью которого можно получить
какую-то информацию о взаимосвязях между переменными,
характеризующими определенные физические системы. Цен-
ность такого подхода состоит в том, что его можно применять
в тех случаях, когда мы располагаем неполной информацией о
системе, и тогда получаемая частичная информация нередко1
имеет большую ценность, позволяя уменьшить число экспери-
ментов, необходимых для получения полной информации.
Рассмотрим, например, физическую абсорбцию кислорода'
в аэрируемом биореакторе. Очевидно, что для полного описа-
ния системы с физической точки зрения необходимо вывести
сложные гидродинамические уравнения, описывающие переме-
Химическая технология и биотехнология
431
щение газообразной и жидкой фаз и диффузию в жидкой фазе.
Количество поглощенного кислорода будет сложным образом
зависеть от скорости поступления газа и от всех других пере-
менных, входящих в уравнения, которые описывают перемеще-
ние фаз. Определить точный вид зависимости между скоростью
подачи газа и другими переменными практически невозможно;
однако анализ размерностей позволяет установить ряд важных
соотношений, которые должны выполняться для любой такой
зависимости. Это связано с тем, что корректные способы изме-
рений удовлетворяют требованию, согласно которому число,
равное отношению между результатами измерений двух объ-
ектов, не зависит от того, какая единица используется для из-
мерений, а корректные системы уравнений имеют такой вид,
который не зависит от системы единиц.
Решение той или иной технологической проблемы обычно
начинают с очень приближенного анализа, учитывая как мож-
но меньше деталей. В результате исходные характеристики про-
цессов, протекающих в реакторе, могут быть представлены в
виде простого соотношения между двумя или тремя безразмер-
ными величинами и соответствующими эмпирическими констан-
тами. Существует около трехсот безразмерных величин, пред-
ставляющих интерес с точки зрения технологии, но на самом
деле большинство биотехнологов имеют дело только с двумя
десятками.
Наиболее широко используемая безразмерная величина —
это число Рейнольдса NKe, которое применяется при описании
потока жидкости:
NRe = -^ , (64)
п
где L — характеристический размер, и — скорость потока, р —
плотность жидкости, ц — ее вязкость. При конструировании
биореакторов числа Рейнольдса используются при описании по-
тока жидкости в трубопроводах, движения капель, частиц и
пузырьков газа, а также вращающихся мешалок. Для потока
жидкости в трубопроводе
NRe=-^-, . (65)
Л
где d — диаметр трубопровода, и — скорость потока. Для ча-
стиц, пузырьков или капель, погружающихся в жидкости или
всплывающих в ней,
N = _^Р£_, (66)
Т]с
где d3 — эквивалентный диаметр частиц, пузырька или капли,
432
Глава 10
т. е. диаметр сферы, равной по объему частице, пузырьку или
капле, U — скорость частицы, всплывающей или погружающей-
ся в жидкость, ,рс — плотность среды, т]С — ее вязкость. Для
вращающихся мешалок
NRe=-^^-, (67)
Чс
где D — диаметр перемешивающего устройства, N — скорость
вращения.
Перечислим некоторые другие безразмерные величины,
обычно используемые химиками-технологами: число Нуссель-
та Nnu, число Прандтля Npr, число Гразгофа Nor, число Фруда
NFr, число Пекле NPe, число Стэнтона Nst, число Шмидта NSc,
число Шервуда Nsh, число Вебера NWe. Как и число Рейнольд-
са, некоторые из этих величин имеют по нескольку определе'-
ний, а, например, числа Пекле, Прандтля и Нуссельта записы-
ваются в одинаковом виде в случае теплопередачи и переноса
массы. Кроме того, одни из этих безразмерных величин, на-
пример число Стэнтона, представляют собой комбинации дру-
гих величин, а некоторые имеют более одного названия.
В качестве примера использования безразмерных величин
можно рассмотреть процесс теплопередачи. Когда имеет место
только естественная конвекция, теплопередача зависит исклю-
чительно от структуры потока, определяемой числом Гразгофа,
а число Рейнольдса может быть исключено из безразмерных
уравнений, описывающих этот процесс. В условиях же принуди-
тельной конвекции эффектами структуры потока обычно мож-
но пренебречь и исключить из уравнений число Гразгофа. Сле-
довательно, уравнение, использующееся в случае естественной
конвекции, имеет вид
NNu = /(NGr,NPr), (68)
а в случае принудительной конвекции —
NNu = f(NRe,NPr). (69)
При увеличении масштаба возникают проблемы, связанные с
экстраполяцией данных, полученных в лаборатории или на по-
лупроизводственной установке, на случай промышленной уста-
новки. Оборудование для биотехнологической промышленности
можно успешно создавать и эксплуатировать только при усло-
вии правильного масштабирования. Ясно, что с проблемой уве-
личения масштаба мы сталкиваемся при конструировании всех
установок, используемых в биотехнологических процессах,
включая теплообменники, центрифуги, испарители, сушилки и
т. д., но здесь мы рассмотрим в основном биореакторы.
Химическая технология и биотехнология 43&
Любой материальный объект или физическая система ха-
рактеризуются тремя параметрами: размером, формой и со-
ставом. Все эти параметры меняются независимо друг от дру-
га, так что два объекта могут различаться по размеру, но<
иметь одинаковую форму и состав или быть одинаковыми
только по форме, но существенно различаться по размерам и
составу. Принцип подобия устанавливает взаимосвязь между
физическими системами р-азных размеров; на нем основано
увеличение или уменьшение масштабов процессов и оборудо-
вания. Согласно этому принципу, пространственная и времен-
ная конфигурации системы определяются отношением между
соответствующими величинами внутри систем и не зависят от
того, в каких единицах эти величины измеряются. Принцип
подобия нередко путают с методом анализа размерностей, хотя
логически они совершенно различны. Первый из них — это об-
щий принцип природы, тогда как второй — всего лишь один из
методов его реализации.
Биотехнолог имеет дело со сложными системами, включаю-
щими твердые тела и жидкости, в которых происходит пеоенос
энергии и вещества и протекают различные реакции; для таких
систем принцип подобия связан в основном с понятием формы..
Когда понятие формы применяется к сложным биотехнологи-
ческим системам, оно включает помимо геометрических про-
порций и такие факторы, как структура потока жидкости, тем-
пературные градиенты, зависимость концентрации от времени
и т. д. Системы, обладающие одинаковой конфигурацией по
одному или более из этих параметров, можно рассматривать,
как подобные.
Подобие можно определить двумя способами: устанавливая?
отношение результатов различных измерений для одного и то-
го же объекта, т. е. внутренние пропорции, или отношения меж-
ду результатами соответствующих измерений разных объектов,,
т. е. масштаб. Геометрическое подобие лучше всего определя-
ется через масштабные соотношения, а подобие по таким пе-
ременным, как скорость, сила или температура, — через внут-
ренние соотношения для каждой системы. Эти внутренние со-
отношения представляют собой безразмерные величины, харак-
теризующие подобие в различных условиях.
Для систем, в которых протекают биотехнологические про-
цессы, существуют четыре типа подобия: 1) геометрическое,.
2) механическое, 3) тепловое и 4) химическое, причем каждый
из этих типов подобия связан со всеми остальными. Для полно-
го химического подобия необходимо наличие геометрического,
механического и теплового подобия, но часто приходится огра-
ничиваться приближенным химическим подобием, например,,
когда имеет место лишь приближенное механическое подобие-.
28—1341
434
Глава 10
На самом деле такой элемент приближения имеется всегда,
поскольку в реальных системах обязательно существуют возму-
щения и идеальное подобие становится недостижимым. Так,
рассматривая потоки жидкости в геометрически подобных си-
стемах, мы не можем считать их идентичными, поскольку нет
гарантии, что все неровности поверхностей геометрически по-
добны.
Из рассмотрения четырех типов подобия, перечисленных вы-
ше, следует, что геометрическое подобие между двумя объектами
имеет место тогда, когда каждой точке одного из них соответ-
ствует определенная точка другого. Однако каждой точке пер-
вого объекта может соответствовать более одной точки второ-
го; кроме того, масштаб может быть одинаковым не по всем
•осям. О подобии объектов с такими различиями в масштабе
говорят как об искаженном.
Механическое подобие включает подобие статическое, ки-
нематическое и динамическое; каждое из них можно рассмат-
ривать как распространение понятия геометрического подобия
на стационарные или движущиеся системы, на которые действу-
ют силы. Статическое подобие относится прежде всего к де-
формации структур и представляет для биотехнологов лишь не-
большой интерес. Напротив, кинематическое и динамическое
подобие очень важны и касаются систем, для которых харак-
терно движение. В случае геометрического подобия использу-
ется декартова система координат, при кинематическом подо-
бии вводится дополнительная переменная — время. О геомет-
рически подобных движущихся системах говорят как о кине-
матически подобных в тех случаях, когда соответственные ча-
стицы описывают за соответственные интервалы времени по-
добные траектории. Когда две геометрически подобные жидкие
системы подобны кинематически, свойства потоков геометри-
чески подобны, а процессы переноса массы и тепла в этих двух
системах связаны друг с другом простыми соотношениями. Ки-
нематическое подобие в жидкостях влечет за собой геометриче-
ское подобие как турбулентных систем, так и пограничных
(пристеночных) слоев жидкости. Динамически подобны силы
(гравитационные, центробежные и т. п.), под действием кото-
рых осуществляется ускоренное или замедленное движение тел
в динамических системах. В жидких и дисперсионных системах
кинематическое подобие предусматривает и динамическое подо-
бие, поскольку характер движения в таких системах определя-
ется приложенными силами. Динамически подобные системы —
это геометрически подобные движущиеся системы, в которых
соотношения между всеми соответственными силами одинако-
вы. Динамическое подобие в потоке жидкости очень важно
.для предсказания изменения давления или потребления энер-
Химическая технология и биотехнология
43б>
гии; в случае переноса массы и теплопередачи оно имеет только
косвенное значение как способ установления кинематического
подобия.
Понятие теплового подобия применяется в случае техноло-
гических систем, в которых происходит теплопередача; в этом:
случае к единицам длины, силы и времени добавляется темпе-
ратура. Тепло может передаваться от одной точки системы к
другой с помощью разных механизмов: излучения, теплопровод-
ности, конвекции или переноса вещества под действием гради-
ента давления. Первые три процесса осуществляются за счет
разницы в температурах, а четвертый определяется характе-
ром потока в системе. Тепловое подобие имеет место в геомет-
рически подобных системах, когда отношения соответственных
разностей температур постоянны, а когда такие системы нахо-
дятся в движении, они также и кинематически подобны. Теп-
ловое подобие должно учитываться при разработке режима сте-
рилизации биореакторов, особенно в плане теплового поврежде-
ния материала, а также при охлаждении биореакторов.
Понятие химического подобия относится к системам, в кото-
рых протекают различные реакции и состав которых меняется
от точки к точке либо — в случае периодических или цикличе-
ских процессов — во времени. При этом никаких новых фунда-
ментальных единиц не вводится, однако должны быть заданы
один или более концентрационных параметров в зависимости от
числа независимо изменяющихся химических компонентов, по»
которым должно быть установлено подобие. Химическое подо-
бие имеет место в геометрически подобных системах и систе-
мах с тепловым подобием, когда отношение между соответст-
венными разностями концентрации остается постоянным и ког-
да эти системы, если они находятся в движении, кинематиче-
ски подобны. Внутренним критерием, который характеризует-
химическое подобие, в дополнение к тем критериям, которые-
необходимы для кинематического и теплового подобия, яв-
ляется отношение скорости образования продукта к скорости
всего потока или к скорости молекулярной диффузии.
Механическое, тепловое или химическое подобие между гео-
метрически подобными системами определяется такими крите-
риями, как соотношения размеров, сил или скоростей внутри1
этих систем. Поскольку эти критерии представляют собой от-
ношение между одинаковыми величинами, они безразмерны и
могут быть получены с помощью анализа размерностей.
Безразмерные критерии подобия представляют собой отно-
шения физических величин, являющихся функциями или раз-
личных движущих сил, или сил сопротивления, которые лими-
тируют процесс. В реальных системах часто существуют мно-
жественные лимитирующие факторы. Например, сопротивле-
28*
436
Глава IV
ние движению жидкости может быть обусловлено вязкостью,
гравитационными силами или силами поверхностного натяже-
ния, для которых соответствующими безразмерными величина-
ми являются числа Рейнольдса, Фруда и Вебера. Для самого
простого случая гомологичных систем с разными абсолютными
размерами эти три критерия несовместимы, поскольку для них
имеет место разная функциональная зависимость скорости
движения жидкости от линейных размеров; так, для одинако-
вых чисел Рейнольдса скорость«длине-1, для одинаковых чисел
Фруда скорость «длине0,5, для одинаковых чисел Вебера ско-
рость «дли не-0’5. При увеличении масштаба сложного биотех-
нологического процесса лучше всего было бы найти такие усло-
вия, при которых скорость процесса в целом определялась в
основном каким-то одним безразмерным критерием, но, как
правило, приходится использовать как минимум два критерия.
Для биореакторов с вращающимися мешалками, благодаря
которым воздух диспергируется в толще культуральной среды,
очень часто основным экономическим фактором при масштаби-
ровании является потребление энергии при перемешивании.
В общем виде безразмерное уравнение для двухфазного пото-
ка жидкости в емкости с вращающейся мешалкой выглядит
следующим образом:
Np=f(NRe,NFr,NWe), (70)
или
Р DN2 О8№ри \ ,71)
рн№П6 \ л ’ G и )’ { '
где Р — потребляемая энергия, ,р„— плотность жидкой (не-
прерывной) фазы, N — скорость вращения мешалки, D — диа-
метр мешалки, т]н — вязкость жидкой (непрерывной) фазы,
G — гравитационная постоянная, о — поверхностное натяже-
ние.
Для систем газ — жидкость зависимость от чисел Ройнольд-
са и Вебера в уравнении потока жидкости можно с достаточной
точностью представить как степенные функции этих чисел, хо-
тя показатели степени и не являются постоянными для всех
условий. Далее, зависимость от числа Фруда возникает только
при наличии вихрей на поверхности жидкости, а в биореакто-
рах с вращающимися мешалками образование вихрей предот-
вращается с помощью перегородок. Следовательно, для аэри-
руемых биореакторов с вращающимися мешалками
Np=/(NRe)“(NWe)v, (72)
тде показатели степени а и у зависят от условий процесса, в
частности от структуры потока.
Химическая технология и биотехнология
437
Анализ размерностей массопередачи в аэрируемых геомет-
рически подобных биореакторах с мешалками дает
Nsh = /(NRe)“(NSc)v, (73)
или
ky,D = х / Д2Урн \ а / Г)н у <74.
<© \ Пн ) \ PaD / ’
где — коэффициент массопередачи, 3) — коэффициент моле-
кулярной диффузии кислорода в жидкой (непрерывной) фазе.
Хотя у нас нет оснований сомневаться в применимости уравне-
ний (72) и (73) к случаю биореакторов с вращающимися ме-
шалками, следует помнить о тех трудностях, которые возника-
ют при оценке влияния перемешивания на биологическую ак-
тивность; эти трудности вполне могут привести к несостоятель-
ности обсуждаемого подхода. Скорость вращения лопастей ме-
шалки nND задает максимальную скорость сдвига в аэрируе-
мых биореакторах, от которой зависит вероятность поврежде-
ния жизнеспособных клеток, особенно нитевидных бактерий
или образующих хлопья микробов. При увеличении размеров
биотехнологических установок необходимо уделять особое вни-
мание связям между механическими и физическими факторами,
с одной стороны, и биологическими — с другой. В противном
случае задача увеличения масштаба вряд ли будет решена кор-
ректно.
10.3.3. Перенос кислорода
Одна из основных задач, которые приходится решать химикам-
технологам при конструировании биореакторов, — это обеспе-
чение эффективного переноса кислорода в промышленных био-
реакторах, где осуществляются аэробные микробиологические
процессы. Независимо от того, в каком режиме осуществляется
аэробный микробиологический процесс — периодическом, полу-
непрерывном или непрерывном, — в установку должен непре-
рывно подаваться кислород для достижения достаточно высо-
кой производительности. Потребность аэробной культуры в
кислороде зависит от концентрации микроорганизмов в реакто-
ре, скорости их роста и соответствующего коэффициента вы-
хода. В некоторых случаях могут играть роль и другие факто-
ры, например необходимость удаления из культуры двуокиси
углерода, особенно для процессов, в которых в качестве источ-
ника кислорода используется воздух, который как раз и слу-
жит источником кислорода в подавляющем большинстве аэроб-
ных микробиологических процессов. При тех температурах, при
которых обычно протекают микробиологические процессы (10—
438
Глава 10
40°C), и при атмосферном давлении равновесная концентрация
растворенного кислорода крайне мала по сравнению с концент-
рациями в растворе почти всех основных питательных веществ,
или субстратов, хотя именно концентрация последних лимити-
рует рост микроорганизмов в непрерывном проточном хемоста-
те. Такой недостаток кислорода приводит к снижению произво-
дительности, к ухудшению качества продукта или к образова-
нию побочных продуктов. При реализации большинства аэроб-
ных микробиологических процессов расходы на снабжение-
кислородом составляют значительную часть общей стоимости,
производства.
В равновесии растворимость газа в жидкости представляет
собой весьма небольшую, вполне определенную величину, зави-
сящую от природы этих газа и жидкости, от температуры и
давления в системе. В случае газовых смесей, например возду-
ха, растворимость компонентов зависит от парциального дав-
ления каждого из них. При прочих равных условиях, чем выше
температура, тем меньше растворимость, и чем выше давле-
ние (или парциальное давление), тем растворимость выше. За-
висимость между растворимостью газа и давлением известна-
как закон Генри, согласно которому при постоянной температу-
ре масса газа, растворенного в данном объеме жидкости, про-
порциональна давлению газа, с которым эта жидкость находит-
ся в равновесии. Большинство культуральных сред представля-
ют собой разбавленные водные растворы, и для простоты их
можно считать сходными с водой. Концентрация кислорода,
растворенного в воде, находящейся в равновесии с воздухом,,
т. е. в состоянии насыщения, при постоянной температуре и по-
стоянном суммарном давлении равна
х — Р°2
01 Но,
где /?о2 —парциальное давление кислорода в воздухе, Но2—
постоянная Генри для данной системы при соответствующей
температуре. Значения этих постоянных приводятся в физических
и химических справочниках, и нужно только правильно вы-
брать единицы измерения.
Для описания переноса (абсорбции) газов в жидкостях
обычно используются три гидродинамические модели: 1) мо-
дель двух пленок Льюиса — Уитмена; 2) модель проницаемо-
сти с систематически обновляемой поверхностью (модель Хиг-
би); 3) модель проницаемости со случайно возобновляемой по-
верхностью (модель Данквертса).
Для описания физической абсорбции газов системами аэроб-
ных микробных культур обычно применяют модель Льюиса —
(75>
Химическая технология и биотехнология
439
Уитмена, поэтому только ее мы здесь и рассмотрим. Перенос
кислорода из жидкой фазы в бактериальную клетку также ча-
сто рассматривают с позиций, аналогичных модели двух пле-
нок.
Согласно модели Льюиса — Уитмена, на поверхности разде-
ла фаз газ — жидкость образуются две тонкие пленки, тесно
прилегающие к этой поверхности. Пленки являются в основном
неподвижными, а в толще газовой и жидкой фаз происходит
турбулентное движение. Абсорбция газа осуществляется в ходе
стационарных процессов молекулярной диффузии в этих двух
неподвижных пленках. Считается, что на поверхности раздела
фаз газ — жидкость мгновенно устанавливается равновесие, а
Направление транспорта кислорода
Рис. 10.10. Модель двух (ламинарных) пленок, предложенная Уитменом для
процесса физической абсорбции (транспорта) кислорода из воздуха в водную
среду (обозначения см. в тексте).
скорость абсорбции определяется относительными скоростями
диффузии в пленках. Предположим, что абсорбция осуществля-
ется из пузырьков воздуха, диспергированных в жидкости; ра-
диус кривизны пузырьков на несколько порядков превышает
толщину гипотетической пленки, и транспорт кислорода из пу-
зырьков воздуха в жидкость можно представить так, как это
сделано на рис. 10.10. В газовой пленке Парциальное давление
кислорода меняется от ро2 — парциального давления во всем
объеме газовой фазы до Ро2 i — парциального давления в газо-
440
Глава 10
вой пленке непосредственно у границы раздела фаз; соответст-
венно в жидкой пленке концентрация растворенного кислорода-
меняется от Co2i — равновесной концентрации при насыщении,,
соответствующей ро2 ь до Со2 — концентрации во всем объеме-
жидкости.
Поскольку кислород на поверхности раздела не накаплива-
ется, скорость переноса кислорода на единицу площади в газо-
вой и жидкой пленках будет одинакова и равна
^о2 = kc (Ро2 —Рол) = (Со21~Со2). (76>
где &g и — коэффициенты массопереноса соответственно для
газовой и жидкой пленок. Для воздуха и других газовых сме-
сей, содержащих кислород, ро2; и Co2i определить нельзя;
в этих случаях используются коэффициенты переноса Kg для
газа и kL для жидкости:
^о2 — (Ро2—Р*о2) — К-ъ (с*о2 со2)> (77)
где р*о2 — равновесное парциальное давление, соответствующее
Со2, а с*о2 — равновесная концентрация, соответствующая пар-
циальному давлению ро2. Тогда, согласно закону Генри, полу-
чаем
Ро2 = Но2с*02, (78).
Р*о2 = НО2СО2, (79)
Poai= Н02 co2t • (80)
Сделав подстановку в уравнения (75) и (76), имеем
= (81)
Kg
Величины, обратные коэффициентам массопереноса, называют-
ся коэффициентами сопротивления транспорту, т. е. 1/Pg и
1/йь — соответственно коэффициенты сопротивления в газовой
и жидкой пленках, a \/Kg и 1//Cl — соответственно коэффици-
енты сопротивления для всего объема газа и всего объема
жидкости. Их относительные значения зависят от растворимо-
сти газа. При абсорбции очень хорошо растворимого газа, на-
пример аммиака в воде, постоянная Генри Hnh3 очень мала,
и последним членом в уравнении, аналогичном (81), можно
пренебречь, так что коэффициент массопереноса для газовой
Химическая технология и биотехнология
441
фазы Ка практически равен коэффициенту для газовой пленки
абсорбция лимитируется процессами, происходящими в га-
зовой пленке. При абсорбции (транспорте) кислорода, который
растворяется в воде слабо, постоянная Но2 очень велика, так
что в уравнении (82) член 1/Ho2&g очень мал и им можно пре-
небречь. Коэффициент массопереноса для жидкой фазы Лц
практически равен коэффициенту массопереноса для жидкой
пленки &L; абсорбция кислорода лимитируется процессами, про-
текающими в пленке жидкости. Когда газовая фаза содержит
кислород, po2i = po2 и kG — бесконечно большая величина, так
что абсорбция в этом случае также лимитируется процессами
в жидкой пленке.
В большинстве аэробных биореакторов кислород поступает
в культуральную среду из пузырьков воздуха, диспергирован-
ных в ней; в этом случае необходимо оценить скорость массо-
передачи кислорода в единице объема, и уравнение (77) при-
нимает следующий вид:
#O2 — а (С*О2 со2)> (83)
где Ко2 — скорость переноса кислорода на единицу объема
жидкости, а — удельная площадь поверхности раздела фаз
газ — жидкость, т. е. площадь поверхности раздела фаз на еди-
ницу объема жидкости. При насыщении с*о2=Со2 и перенос
кислорода не происходит. Однако находящиеся в среде мик-
роорганизмы будут потреблять кислород в процессе своей жиз-
недеятельности, так что с*о2 станет больше Со2 и начнется пе-
ренос. В случае модели двух пленок Кь определяется следую-
щим образом:
^=4"’ (84)
о
где — коэффициент диффузии, б — толщина пленки жидко-
сти. Для другой модели поглощения газа KL определяется иным
способом. В модели Хигби
(85)
а в модели Данквертса
KL=/%, (86)
где t* — это время нахождения элементарного объема жидко-
сти на поверхности раздела, a tD — эквивалентное время диф-
фузии.
Транспорт кислорода из пузырька в жидкую среду — это
лишь часть общего процесса переноса кислорода к бактериаль-
442
Глава 10
ным клеткам. Как было показано для переноса кислорода из;
пузырьков в жидкость, транспорту препятствуют разного рода
факторы, один из которых является лимитирующим. Понятие
сопротивления переносу можно распространить и на последую-
щие стадии процесса. Кислород, поступив в культуральную сре-
ду, должен затем транспортироваться к месту его использова-
ния в микроорганизме. Сопротивление переносу складывается
Длина пути переноса (масштаб не соблюден)
Рис. 10.11. Различные типы сопротивления транспорту кислорода из пузырька
газа к месту его использования микроорганизмом (обобщение модели Уитме-
на; обозначения см. в тексте).
из сопротивления со стороны жидкой фазы (1/&в), которым1
для невязких, ньютоновских жидкостей в условиях полной тур-
булентности можно пренебречь, из сопротивления со стороны
жидкой пленки, прилегающей к поверхности бактериальной
клетки, хлопьям, осадку или к поверхности мицелия (1/&м) и
либо внутриклеточного сопротивления в случае дисперсионных
культур, либо межклеточного (включая и внутриклеточное) в
случае хлопьев, осадков и мицелия (1/&с). Все типы сопротив-
ления переносу кислорода представлены на рис. 10.11. В слу-
Химическая технология и биотехнология
443
чае дисперсионных культур перенос кислорода лимитируется
•сопротивлением в жидкой пленке пузырька (1/£l). Когда же
микроорганизмы растут в виде хлопьев, осадков или мицели-
альной массы, в некоторых случаях роль лимитирующего фак-
тора играет не сопротивление в жидкой пленке газового пузырь-
•ка, а внутри-/межклеточное сопротивление или сопротивление
конгломерата.
В связи с очевидностью роли переноса кислорода из пузырь-
ков газа в жидкость попытаемся с помощью уравнения (83)
оценить возможности увеличения скорости транспорта кисло-
рода. В принципе можно варьировать все три члена этого урав-
нения: KL, а и (с*оа—Соа) (последний член характеризует дви-
жущую силу процесса). Однако KL является характеристикой
•системы в целом и не может изменяться независимо, поэтому
обычно ее считают константой, хотя под действием поверхност-
но-активных веществ, присутствующих в микробиологических
установках, она все-таки изменяется. Удельную площадь по-
верхности раздела фаз газ — жидкость и член, характеризую-
щий движущую силу, легко увеличить, изменяя контролируе-
мым образом условия проведения процесса. Обычно первая
из этих величин возрастает при интенсификации перемешива-
ния, а вторая — при увеличении суммарного или парциального
давления.
Удельная площадь поверхности раздела фаз газ — жид-
кость определяется следующим образом:
где а — задержка газа на единицу диспергируемого объема,
D — средний диаметр пузырьков (в случае пузырьков неодина-
кового размера). Обычно для определения среднего диаметра
пузырька используют формулу Саутера:
п.
У\пО3
D= —------, (88)
п
^nD2
1
где п — число пузырьков, D — диаметр эквивалентной сферы.
Ясно, что для интенсификации транспорта кислорода путем
увеличения а необходимо либо повысить содержание газа в
диспергируемой среде (т. е. е), либо уменьшить средний диа-
метр пузырьков. /
Диспергирование пузырьков газов в жидкостях — сложный
процесс. Независимо от способа получения пузырьков для них
444
Глава 10
всегда наблюдается некое распределение по размерам. Размер
любого диспергированного пузырька определяется равновеси-
ем между двумя основными типами напряжений: напряжением
сдвига и поверхностным натяжением. На размер пузырьков за-
метно влияет присутствие в среде растворенных неорганических;
и органических веществ, при этом изменяется либо коалесцен-
ция, либо поверхностное натяжение.
В биореакторах коалесценция и поверхностное натяжение1
не могут изменяться независимо друг от друга и произвольным-
образом, однако несомненно они изменяются в ходе процесса
или в результате добавления соединений, контролирующих це-
нообразование. Теоретически уменьшение среднего размера пу-
зырьков при диспергировании газов в жидкостях может проис-
ходить при интенсификации перемешивания. Однако на самом1
деле при обычных удельных мощностях перемешивания сред-
ний размер пузырьков остается примерно постоянным, зато за-
метно увеличивается содержание газа, т. е. другой компонент
величины а; это связано с рециркуляцией жидкости, благодаря
которой в свою очередь снижается потеря пузырьков газа.
Механизм разрушения пузырьков в изотопическом турбулент-
ном потоке мы обсуждать не будем. Отметим лишь, что в ре-
зультате рассмотрения этих механизмов для случая отсутствия
коалесценции было получено уравнение, удовлетворяющее мно-
жеству имеющихся экспериментальных данных для систем с
очень высокой энергоемкостью:
T) = J
L р°.2 (p/v)m ]
где J — константа, о — поверхностное натяжение, р— плот-
ность жидкости, P/V— энергоемкость на единицу объема в ус-
ловиях насыщения газом. При наличии коалесценции уравнение
(89) принимает вид
D = J
a°>e
(89>
аМ
р0,2 (Р/У)О,4
(90)
е“.
Здесь показатель степени а, в который возводится величина е,.
характеризует степень коалесценции.
Итак, очевидно, что задержка газа и средний размер пу-
зырьков прямо связаны между собой; при этом задержка газа
зависит от интенсивности перемешивания и от объемной скоро-
сти потока газа. Соответствующее эмпирическое уравнение для
жидкости с постоянными реологическими характеристиками
имеет вид
e = J'(P/y)₽(Vn)v, (91)
где Г, р и у — константы, зависящие от системы и оборудова-
Химическая технология и биотехнология
445
ния, а Уп — поверхностная скорость газа, т. е. отношение объ-
емной скорости газового потока к площади поперечного сечения
биореактора. В биореакторах с вращающимися мешалками и
высокой энергоемкостью происходит рециркуляция значитель-
ной части газовой фазы и соответственно уменьшается влияние
поверхностной скорости газа на величину е. В то же время в;
барботажных биореакторах относительная роль Vn сильно воз-
растает.
Оценивая транспорт кислорода из газовой фазы в жид-
кость в микробиологических реакторах, Эндрью (Andrew, 1982}
высказал мнение, что при конструировании любых промышлен-
ных аэробных биореакторов, в том числе больших танков с ме-
шалками, их можно рассматривать как аппараты колонного
типа со свободным прохождением пузырьков газа по высоте
аппарата. Крупные биореакторы-тэнки с мешалками относят к
категории вышеупомянутых биореакторов, основываясь на яв-
лении массопереноса, а также в связи с тем, что при обычной
промышленной энергоемкости мешалка не способна создавать
рециркуляцию жидкости, достаточную для возврата пузырьков,
стремящихся выйти на поверхность жидкости.
Рассмотрим теперь вопрос о возможности изменения скоро-
сти транспорта кислорода путем изменения того члена уравне-
ния (83), который характеризует движущую силу процесса,
т. е. (с*о2—Соа). Здесь имеется несколько возможностей.
В первую очередь уменьшение до минимума рабочей концент-
рации растворенного кислорода в среде, т. е. Со2. Однако, ког-
да эта величина приближается к нулю, дальнейшее увеличение
движущей силы становится невозможным. Максимально дости-
жимая, постоянная скорость транспорта кислорода /?о2т равна
Яоат = *ьас*о2. (92>
Уменьшать содержание растворенного кислорода в культураль-
ной среде следует с осторожностью, поскольку микроорганиз-
мы при этом вынуждены будут расти в условиях недостатка
кислорода, что может отрицательно сказаться на их росте и/или
образовании продукта. Если микроорганизмы растут в виде
хлопьев, образуют осадок или мицелий и величина 1/&с стано-
вится фактором, лимитирующим весь процесс транспорта кис-
лорода, концентрация Со2 должна поддерживаться на уровне
значительно выше нуля, если мы хотим, чтобы во всем объеме
микробной биомассы, находящейся в реакторе, поддерживался
аэробный метаболизм.
Прежде чем говорить об изменении с*о2, следует сначала
определить, какое именно парциальное давление кислорода со-
ответствует с*о2 в тех или иных типах биореакторов. Для био-
446
Глава 10
реакторов с высокой интенсивностью перемешивания, т. е. ре-
акторов, которые обычно используются в лаборатории, было
показано, что газовая и жидкая фазы полностью перемешива-
ются. Следовательно, парциальное давление кислорода в дис-
пергированных пузырьках равно парциальному давлению кис-
лорода в газе, выходящем из жидкости, т. е. в свободном про-
странстве биореактора. Для биореакторов колонного типа со
свободным прохождением пузырьков по высоте биореактора, в
том числе и для промышленных биореакторов с перемешивани-
ем, работающих в условиях обычной энергоемкости, в первом
приближении можно считать, что жидкая фаза перемешивает-
ся полностью, а газовая находится в режиме идеального вытес-
нения, т. е. существует градиент парциального давления кисло-
рода в направлении от дна биореактора к его верху. Для этих
.двух крайних случаев уравнение (83) можно переписать сле-
дующим образом:
(93)
~-2 Но2
(для случая полного перемешивания) и
Яо = KLaPCp Г-----(У1-У2)--1 (94)
\ Уз Уз /
(для случая полного вытеснения). Здесь Р — давление в сво-
бодном пространстве, Рср — среднее рабочее давление в реакто-
ре (давление в рабочем пространстве+гидростатическое давле-
ние) ух, У2 и уз — мольные доли кислорода во входящем и вы-
ходящем газе и в газовой смеси, соответствующей равновесной
концентрации растворенного кислорода.
Из уравнений (93) и (94) с очевидностью следует, что с
увеличением давления в свободном пространстве биореактора
повышается скорость транспорта кислорода в обоих рассмот-
ренных случаях, а с увеличением парциального давления кис-
.лорода путем обогащения им газовой смеси, продуваемой че-
рез биореактор, или полной замены ее кислородом повышается
скорость транспорта кислорода в случае полного вытеснения.
В реакторах с идеальным перемешиванием парциальное давле-
ние кислорода в пузырьках, а следовательно, и скорость транс-
порта кислорода должны увеличиваться с уменьшением кон-
версии кислорода; однако это вряд ли можно считать экономи-
чески целесообразным.
Поскольку микроорганизмы имеют тенденцию скапливаться
у поверхности раздела между воздухом и жидкостью, видимо,
Химическая технология и биотехнология
447
правильнее было бы записать уравнение (82) следующим обра-
зом:
/?о2=-Е'^ьа(с*о2—Со.)- (?5>
Ясно, что в отсутствие такого накопления Е'=\, однако обычно,,
особенно в случае бактерий значительно меньшего размера,,
чем толщина пленки (~20 мкм), концентрация их у поверхно-
сти раздела фаз будет гораздо больше, чем в толще жидкости.
Влияние этого явления на транспорт кислорода, вероятно, в
какой-то степени сходно с эффектами, наблюдаемыми при по-
глощении газа в условиях одновременного протекания химиче-
ской реакции; тем не менее прямая аналогия здесь вряд ли воз-
можна, поскольку концентрация, бактерий внутри пленки меня-
ется нелинейно, а текучесть пленки уменьшается из-за присут-
ствия в ней бактериальных клеток.
10.3.4. Теплопередача и охлаждение биореактора
К числу наименее разработанных проблем биотехнологии отш>-
сятся разнообразные проблемы осуществления процессов тепло-
передачи, которые обеспечивают поддержание температуры-
аэрируемой культуральной среды на уровне, соответствующем
оптимуму анаболической и катаболической активности данных
микроорганизмов. Для большинства из них диапазон темпера-
тур, оптимальных для роста или образования продукта, очень-
узок, нередко всего несколько градусов. При температурах ни-
же оптимальной скорость роста любого микроорганизма повы-
шается в ответ на повышение температуры относительно мед-
ленно, а при температурах, лишь на несколько градусов превы-
шающих этот оптимум, происходит резкое снижение скорости
роста до нуля. Подавляющее большинство микроорганизмов,,
используемых в биотехнологических процессах, — мезофилы:,
температурный оптимум для них лежит между 15 и 40 °C. Тем-
пературный оптимум термофильных микроорганизмов значи-
тельно превышает 40°C, а у облигатных психрофильных микро-
организмов он лежит ниже 10 °C.
При производстве любого валового микробиологического-
продукта аэробным способом охлаждение биореактора предс-
тавляет собой серьезную технологическую проблему. Если тем-
пературный оптимум, при котором достигается максимальный
выход продукта и производительность, лежит ниже 40 °C, на
охлаждение приходится основная часть производственных расхо-
дов. Обычно в практике биотехнологических процессов промыш-
ленного масштаба используют в качестве охлаждающей среды
воду, а не какой-либо хладагент. При определении размеров-.
448
Глава 10
охлаждающей поверхности, необходимой для поддержания
температуры любого процесса, особое значение имеет макси-
мальная температура охлаждающей воды, а производственные
расходы на системы охлаждения определяются средней темпе-
ратурой охлаждающей воды, используемой на разных этапах
процесса. Следует подчеркнуть, что при очистке валового про-
дукта требуется значительно меньшее охлаждение, чем при
осуществлении обычных биологических процессов. Однако не-
которые из недавно разработанных физических методов разде-
ления протекают при температурах, близких к обычным, и
представляются весьма перспективными в плане использования
их в биотехнологии.
Температура охлаждающей воды зависит от многих факто-
ров. К ним относятся происхождение этой воды, климатические
условия и различия в местоположении установки, а также тип
используемой системы охлаждения: это может быть проточная
система или система с рециркуляцией. В будущем для охлаж-
дения, вероятно, все больше будут применяться замкнутые си-
стемы (с рециркуляцией), с тем чтобы исключить тепловое и
возможное химическое загрязнение окружающей среды в ре-
зультате сброса воды из проточных систем охлаждения. В ус-
ловиях умеренного климата и при использовании мезофильных
организмов в крупных, интегрированных производствах опти-
мальная температура процесса будет лишь немного отличаться
ют температуры охлаждающей воды.
Хотя калориметрические методы измерения количества теп-
ла, выделяющегося при росте и дыхании микроорганизмов,
разработаны уже давно, использованию этого процесса посвя-
щено сравнительно немного работ. Измерения количества выде-
ляющегося тепла производили обычно для оценки интенсивно-
сти роста микроорганизмов. В аэробных микробиологических
процессах количество тепла, выделяющегося в единицу време-
ни, прямо пропорционально скорости потребления кислорода,
которая в свою очередь связана с ростом с помощью соответст-
вующего коэффициента выхода. Считается, что это эмпириче-
ское соотношение между скоростью потребления кислорода и
количеством выделяющегося тепла можно применять для оцен-
ки последней величины, и прямые ее измерения проводят толь-
ко в редких случаях. Это соотношение имеет вид
Q = ^o2- (96)
Здесь Q — количество тепла, выделяющегося при росте аэроб-
ных микробов в единицу времени на единицу объема, Ro2 —
скорость потребления кислорода на единицу объема (равная
скорости транспорта кислорода), k—константа, равная 124,
если Q выражено в ккал-м_3-ч-1, a Ro 2—в моль-м~3-ч-1. Оцен-
Химическая технология и биотехнология
449
ки, получаемые с помощью этого уравнения, близки к таковым
при расчете количества тепла исходя из теплоты сжигания суб-
стратов и микробной биомассы соответствующего состава.
Большинство промышленных биореакторов снабжено ох-
лаждающим кожухом и/или внутренними охлаждающими
змеевиками. В последнее время для интенсификации процессов
в биореакторах как обычной конструкции, так и новых конфи-
гураций стали использовать наружные охлаждающие контуры.
В некоторых реакторах новых конфигураций внутренняя ох-
лаждающая система настолько нарушает основные гидродина-
мические рабочие характеристики, что исключает такое охлаж-
дение. При увеличении размера реактора создание необходимой
внутренней охлаждающей поверхности становится все более
трудной задачей. Для геометрически подобных емкостей увели-
чение объема пропорционально кубу линейных размеров, при
этом площадь увеличивается только пропорционально квадра-
ту линейных размеров. Эффективность охлаждения аэробных
биореакторов зависит от следующих факторов: 1) переноса
тепла от отдельных клеток в культуральную среду; 2) перено-
са тепла от культуральной среды, содержащей диспергирован-
ные пузырьки газа и микроорганизмы, к охлаждающим по-
верхностям; 3) поступления тепла за счет рассеяния мсханиче-—
ской энергии, затрачиваемой на перемешивание среды с целью
ее аэрации; 4) охлаждения при испарении, выравнивания тем-
ператур и работы при расширении, связанной с прохождением
воздуха через среду; 5) изменения коэффициента теплопереда-
чи вследствие загрязнения охлаждающих поверхностей накап-
ливающимися на них микроорганизмами. Потенциальную зна-
чимость этих факторов необходимо рассматривать отдельно
для каждой технологической системы.
Когда микроорганизмы диспергированы в толще водной сре-
ды, как бывает обычно в процессах, где культивируют бакте-
рии и дрожжи, скорость теплопередачи лимитируется перено-
сом тепла от культуральной среды к охлаждающим поверхно-
стям.
Содержимое аэробного биореактора, работающего в услови-
ях интенсивного перемешивания, можно представить как массу
пузырьков, но нужно еще учесть, что жидкая фаза содержит
суспендированные микроорганизмы, оказывающие влияние на
ее вязкость. Эта последняя зависит от размера микроорганиз-
ма, его морфологии и концентрации, а также от способности
синтезировать внеклеточные полимерные продукты.
Одна из основных задач аэрации содержимого биореакто-
ра— это получение мелких пузырьков газа и тем самым мак-
симальное увеличение площади поверхности, через которую
происходит перенос газа в среду. Во многом благодаря свойст-
29—1344
450
Глава 10
вам культуральной среды образующиеся в ней пузырьки имеют
сферическую форму и относительно слабо деформируются. Фак-
тически они представляют собой упругие сферы, гидродинами-
ческие свойства которых сходны со свойствами твердых сфери-
ческих частиц. Хотя плотность и теплопроводность твердых ча-
стиц, с одной стороны, и пузырьков — с другой, явно различны,,
поведение пузырьков в некоторых аспектах сходно с поведени-
ем частиц в псевдоожиженном слое, хотя в последнем случае-
содержание диспергированных частиц обычно выше.
Плотность клеток микроорганизмов очень близка к плотно-
сти водной среды, в которой они растут, и при анализе тепло-
передачи в дисперсионных культурах бактерий и дрожжей,
обычно имеющих диаметр эквивалентной сферы меньше-
10 мкм, культуральную среду можно рассматривать как одно-
родную водную фазу. В аэрируемых биореакторах перенос теп-
ла от культуральной среды к охлаждающим поверхностям осу-
ществляется путем принудительной конвекции. Чтобы судить
о потенциальной эффективности охлаждения, необходимо оце-
нить соответствие характеристик водно-газовых смесей, образу-
ющихся при оптимальном транспорте кислорода, факторам,
способствующим увеличению теплопередачи от этой смеси к
поверхности.
Благодаря хорошим характеристикам теплопередачи реак-
торы с псевдоожиженным слоем применяются для проведения
реакций, которые требуют тщательного контроля температуры.
Из-за ожижения частиц увеличивается коэффициент теплопере-
дачи между образуемым ими слоем и теплопередающими по-
верхностями. Это увеличение определяется в основном тремя
факторами: 1) высокой теплоемкостью на единицу объема ча-
стиц, благодаря чему они выполняют функцию теплоперенося-
щих агентов; 2) разрушением частицами ламинарного слоя на
теплопередающей поверхности и вследствие этого уменьшением
эффективной толщины этого слоя; 3) нестационарностью про-
цесса теплопередачи, обусловленной тем, что частицы движут-
ся к теплопередающей поверхности и от нее в виде «пакетов».
Проанализируем значимость этих факторов в применении
к процессу диспергирования газа в жидкости. Если главную
роль играет первый фактор, то коэффициент теплопередачи
между пузырьками и теплопёредающей поверхностью будет
существенно меньше из-за низкой теплоемкости пузырьков. Ес-
ли основным является второй фактор, то коэффициент тепло-
передачи будет больше, а если третий, то может наблюдаться
повышенная коалесценция и образование сгустков. В псевдо-
ожиженных системах задержка твердых частиц нередко дости-
гает 0,4, тогда как в реакторах с обычным потреблением энер-
Химическая технология и биотехнология
451
сии задержка газа вряд ли превышает 0,12. При задержке вы-
ше 0,2 начинается образование газовых «пробок».
В барботажных колоннах пробулькивание газа через жид-
кость приводит к двум типам перемещений: 1) циркуляции
всего содержимого, вызываемой всей массой пузырьков; 2) ло-
кальным микроперемещениям, вызываемым отдельными пу-
зырьками. Экспериментальные данные для суммарного коэф-
фициента теплопередачи в условиях охлаждения жидкости, в
которой диспергированы пузырьки газа, за счет теплопередачи
к стенке колонны описываются уравнением
КРг«’33Мж)-0’14, (97)
Пж /
а за счет теплопередачи к охлаждающему змеевику — уравне-
нием
(О-РП2 \ 0 33
^) NPr0’33 (Tlw/Пж)-0’14 (4/dT)°’33> (98)
где h — суммарный коэффициент теплопередачи, В и В' — кон-
станты для системы, kiK — теплопроводность жидкости, g — гра-
витационная постоянная, е — задержка газа, рж — плотность
жидкости, т]ж — вязкость жидкости, Npr — число Прандтля,
<r]w — вязкость жидкости при температуре теплопередающей
поверхности, dK — диаметр колонны, — диаметр трубки ох-
лаждающего змеевика. Ни одно из этих уравнений нельзя пря-
мо использовать в случае биореактора с интенсивным переме-
шиванием с помощью мешалки. Из-за значительной рецирку-
ляции в реакторах подобного типа показатели степеней этих
уравнений должны быть другими.
Для интенсификации переноса кислорода в биореакторах
системы аэрации должны обеспечивать образование как можно
более мелких пузырьков. Однако при уменьшении размера пу-
зырьков уменьшается теплопередача. С другой стороны, тепло-
передача увеличивается при увеличении задержки газа. Из
этого следует, что в биореакторах с высокой скоростью перено-
са кислорода будет, при прочих равных условиях, наблюдаться
и более интенсивная теплопередача.
10.3.5. Конверсия и рециркуляция
Как мы уже говорили, основным технологическим фактором,
ют которого зависит экономичность любого микробиологическо-
го процесса, является производительность биореактора. Одна-
ко в случае нерастворимых или несмешивающихся с водой суб-
стратов и нелимитирующих рост питательных веществ сущест-
венную роль начинает играть и величина конверсии. Удельную
29*
452
Глава 10
конверсию субстрата или какого-либо питательного вещества
можно определить как отношение массы использованного суб-
страта к массе субстрата, исходно введенного в систему. Для
микробиологических процессов удельная конверсия полностью
растворимого, лимитирующего рост субстрата практически рав-
на единице, т. е. остаточная концентрация субстрата, завися-
щая от сродства участвующих в процессе микроорганизмов к
лимитирующему субстрату, близка к нулю. Для нерастворимых
или несмешивающихся с водой субстратов и для нелимитирую-
щих рост питательных веществ удельная конверсия часто быва-
ет значительно меньше единицы. Для таких систем коэффици-
ент конверсии, определяемый как вес сухой микробной био-
массы или продукта, получаемых на единицу веса введенного
субстрата или питательного вещества, будет меньше соответст-
вующего коэффициента выхода. В одних случаях величина
конверсии кислорода и газообразных углеродных энергетиче-
ских субстратов, несмешивающихся с водой и нерастворимых
в ней углеродных энергетических субстратов, а также раство-
римых нелимитирующих рост питательных веществ определя-
ется режимом работы биореактора, в других более важными
факторами являются массоперенос, геометрия биореактора и
технологические параметры.
Хотя стоимость сырья при аэробной переработке и не повы-
шается из-за использования воздуха в качестве источника кис-
лорода, капитальные и эксплуатационные расходы значительно
возрастают в связи с необходимостью получения сжатого воз-
духа и его стерилизации. В процессах промышленного масшта-
ба важно снизить такие расходы до минимума, максимально
увеличивая степень конверсии кислорода, при условии, что рас-
ходы на это не приведут к снижению экономичности процесса
в целом.
Если в качестве источника кислорода используется воздух,
то удельная конверсия кислорода в высокоэффективных лабо-
раторных биореакторах редко превышает 0,2, а в обычных про-
мышленных биореакторах она обычно составляет —0,1. Даже
при таких низких величинах конверсии кислород нередко явля-
ется лимитирующим рост фактором, поскольку скорость его
транспорта не соответствует скорости потребления. Из рассмот-
рения уравнения (83), описывающего физическую абсорбцию
кислорода культуральной средой, следует, что в системах с пол-
ностью перемешиваемой газовой фазой высокие значения
удельной конверсии кислорода несовместимы с увеличением
скорости транспорта кислорода до максимума. Очевидно, при
решении этой проблемы следует учитывать технические и эко-
номические факторы. Что касается биореакторов, в которых га-
зовая фаза перемещается в режиме полного вытеснения, то по
Химическая технология и биотехнология
453
сравнению с полностью перемешиваемыми системами для них
следует ожидать значительно более высоких величин конвер-
сии кислорода при данной скорости транспорта.
Для увеличения удельной конверсии в технологических си-
стемах чаще всего используют рециркуляцию. В случае мик-
робиологических процессов такой подход применим в равной
мере и к газовой, и к жидкой фазам, впрочем, с некоторыми
модификациями. В случае повторного использования газовой
фазы их можно рассмотреть на примере двух схем потоков,
приведенных на рис. 10.12, — работы в однократном проточном
Без повторного
использования
Рис. 10.12. Повторное использование газовой фазы для увеличения удельной
конверсии кислорода при низких степенях конверсии (обозначения см, в тек-
сте).
режиме и при рециркуляции газовой фазы в предположении,
что переносимый компонент газовой фазы (например, кислород
воздуха) является минорным и поэтому газовые потоки, нап-
равленные внутрь биореактора и из него, практически одинако-
вы. Без рециркуляции удельная конверсия кислорода в возду-
хе Ло2 может быть представлена в виде
О)21 — б’р22 _ Qo2
°2 CO21 gco2i
(99)
454
Глава 10
В условиях рециркуляции соответствующая величина Х'о2 рав-
на
Q'o2
%, _ cO2i~ С'ог2________________
°2 Co2i “ (G-G')Co21 ’
(ЮО)
где Co2i —концентрация кислорода в потоке газа, поступающее
го в обе системы, Со22 и С'о22 — концентрации кислорода в
потоке газа, выходящем из систем без рециркуляции и с ре-
циркуляцией соответственно, Qo2 и Q'o2 — скорости утилиза-
ции кислорода в системе без рециркуляции и с рециркуляцией
соответственно, G и G' — скорости газового потока, проходяще-
го через биореактор и используемого повторно. Когда кислород
не является фактором, лимитирующим рост, т. е. Qo2 = Q'o2.
отношение величин удельной конверсии при рециркуляции и
без нее задается уравнением
X О2 G
Ло2 (G— G')
(101)
Если же кислород служит лимитирующим фактором, то из-за
изменения в движущей силе транспорта кислорода Qo2>Q'o2>
и для полностью перемешиваемой газовой фазы уравнение
(101) принимает вид
. (102)
*о2 Q(G-G')
Как бы мы ни увеличивали удельную конверсию кислорода,
при условии постоянства режима работы при рециркуляции га-
зовой фазы производительность биореактора будет неизбежно
уменьшаться в результате снижения движущей силы транспор-
та кислорода. Ясно, что конверсию нельзя оптимизировать не-
зависимо от производительности.
Когда в биореакторы добавляют газообразные или летучие
углеродные энергетические субстраты, конверсия рассматрива-
ется с совершенно иных позиций. В отличие от кислорода воз-
духа подобные субстраты представляют собой дорогостоящее
сырье, и процессы, в которых они используются, должны быть
направлены на достижение очень высокой конверсии либо на
полное их использование в системах безотходной технологии
с привлечением других процессов. По-видимому, из газообраз-
ных углеродных энергетических субстратов наиболее известен
метан, хотя в качестве промышленных субстратов можно ис-
пользовать и другие газообразные алканы. При условии, что
метан как сырье для микробиологических процессов можно
Химическая технология и биотехнология
455
использовать непосредственно в виде природного газа и вер-
нуть стоимость неутилизованного метана, используя его в та-
ких энергоемких процессах, как сушка, в биореакторе могут
быть достигнуты относительно высокие значения удельной кон-
версии — 0,7—0,8. При этом необходимы реакторы с полным
вытеснением газовой фазы и высоким давлением, а также, воз.
можно, с повторным использованием газовой фазы. В качестве
альтернативы можно использовать несколько последователь-
ных реакторов высокого давления с полным перемешиванием.
Обратимся теперь к жидкой фазе и рассмотрим потенци-
альные возможности ее повторного использования. Следует
подчеркнуть, что все микробиологические процессы протекают
при относительно низких концентрациях микроорганизмов,
субстратов, питательных веществ и продуктов в культуральной
среде. В них расходуется большое количество воды, в которой
эти микроорганизмы, субстраты, питательные вещества и про-
дукты диспергированы или растворены. Концентрация микробов
обычно лимитируется такими факторами, как ингибирование
субстратом, питательными веществами или продуктами, а в
аэробных процессах — скоростью транспорта кислорода. Одна-
ко в технологических системах с рециркуляцией биомассы мо-
гут иметь место и очень высокие концентрации микроорганиз-
мов.
При микробиологическом получении продуктов с высокой или
средней стоимостью имеет смысл повторное использование или
регенерация среды только для процессов, в которых задейство-
вано уже частично использованное очень дорогостоящее сырье.
В случае крупномасштабного производства повторное исполь-
зование среды очень выгодно как в плане экономии большого
количества высококачественной воды и питательных веществ,
так и уменьшения объема сточных вод.
В любой технологической схеме рециркуляции воды/среды
ключевым моментом является установление механизма взаимо-
действия между различными элементарными операциями, обра-
зующими данный цикл. При повторном использовании среды
возникают следующие проблемы: 1) накопление в среде инги-
биторов из-за метаболической активности, сопровождающейся
образованием побочных продуктов, или за счет лизиса в про-
цессе рециркуляции; 2) накопление ингибиторов за счет мета-
болической активности, лизиса или разрушения клеток в био-
реакторе; 3) накопление в сырье нежелательных, неиспользуе-
мых примесей; 4) рециркуляция неиспользованных субстратов
или питательных веществ, вызывающих нарушение физическо-
го и химического равновесия и динамическую неустойчивость
в биореакторе.
456
Глава 10
В случае смешанных культур рециркуляция может приво-
дить к избирательному возвращению в биореактор микроорга-
низмов, составляющих эту смешанную культуру, с неизбежным
нарушением популяционного равновесия в биореакторе.
Повторное использование микробной биомассы — это ско-
рее способ интенсификации микробиологических процессов, чем
средство увеличения конверсии. В отличие от немногих приме-
ров повторного использования газовой или жидкой фазы в
промышленных микробиологических процессах повторное ис-
пользование микробной биомассы представляет собой хорошо
разработанную процедуру, применяемую уже около 70 лет при
очистке сточных вод с помощью активного ила.
В математическом представлении непрерывный микробио-
логический процесс с повторным использованием микроорганиз-
мов эквивалентен непрерывному микробиологическому процес-
су, в котором происходит преднамеренная или случайная за-
держка микробов в реакторе. Возможно, такая операция най-
дет широкое применение в промышленности. Повторное исполь-
зование и задержка микроорганизмов дают существенное пре-
имущество в тех случаях, когда участвующие в процессе мик-
роорганизмы медленно растут или синтезируют не связанный с
ростом продукт. Отметим, что при использовании такого спосо-
ба интенсификации процесса производительность реактора
нельзя будет повысить до тех пор, пока его характеристики, от-
носящиеся к теплопередаче и массопереносу, не будут соответ-
ствовать рассчитываемому повышению производительности.
Если теплопередача или массоперенос в биореакторе в рабочем
режиме без рециркуляции или задержки микробов являются
лимитирующими факторами, то производительность реактора
можно повысить, только улучшив тем или иным способом эти
параметры.
10.3.6. Характеристики микробных и мицелиальных суспензий
Состав микробов обычно выражают в виде процентного содер-
жания углерода, водорода, кислорода, азота и золы в расчете
на сухую массу. На самом деле жизнеспособные микроорганиз-
мы состоят в основном из воды: на ее долю приходится при-
мерно 70% массы клеток.
Форма и размеры разных микроорганизмов описаны в так-
сономической литературе, однако следует помнить, что они мо-
гут изменяться в зависимости от условий культивирования. При
анализе массопереноса и теплопередачи в биореакторах, а так-
же процессов отделения микроорганизмов от конечного продук-
та часто пользуются понятием «эквивалентный размер» («диа-
метр эквивалентной сферы»). Этот термин применяется для
Химическая технология и биотехнология
культур, в которых клетки растут в виде дискретно дисперги-
рованных суспензий. Понятие эквивалентного размера является
полезным упрощением, позволяющим в более простой форме
описывать динамическое поведение клеток в суспензии. Кроме
того, следует принимать во внимание и плотность суспензии.
Обычно она составляет 1,07—1,09 г-см-3, хотя иногда бывает
и ниже—1,03 г-см-3. Таким образом, плотность водных куль-
туральных жидкостей обычно несколько выше, чем плотность
воды.
Единственный случай, для которого было получено выраже-
ние для силы сопротивления движению сферических частиц в
жидкости, — это погружение частицы с малой скоростью в жид-
кость бесконечного объема. Для такой частицы Стокс получил
уравнение
F =3 (TVC\ud), (103)
где F—-сила сопротивления, ц— вязкость жидкости, d —
диаметр сферической частицы, и — скорость жидкости относи-
тельно частицы. На самом деле условия, в которых выполняет-
ся закон Стокса, реализуются редко. В большинстве систем ча-
стицы непрерывно испытывают ускорение или замедление; кро-
ме того, даже в условиях ламинарного потока частицы редко
бывают изолированы друг от друга, обычно они взаимодейству-
ют между собой.
Конечную скорость погружения частицы и0 можно полу-
чить, приравняв силу сопротивления к эффективной силе тяже-
сти, т. е.
3(лф0Д = -^-(рч—pm)g, (104)
о
где рч и рж — соответственно плотности частицы и жидкости.
Отсюда
(рч Рж) g /1
“°---------------• (105)
Закон Стокса для скорости оседания цс суспензий мелких ча-
стиц одинакового размера имеет вид
, (Ю6)
Чс
где рс — средняя плотность суспензии, т]с — кажущаяся вяз-
кость. Это уравнение учитывает влияние концентрации частиц
как на вязкость, так и на плотность жидкости. В случае неод-
нородных частиц в уравнение (106) можно ввести член, учиты-
вающий изменение их формы и размера.
458
Глава 10
Что касается биологических процессов, то для них наибо-
лее важным параметром является вязкость соответствующих
жидкостей. Она определяется как мера внутреннего трения в
жидкости, и в условиях ламинарного потока, т. е. потока с низ-
кими числами Рейнольдса, равна отношению напряжения сдви-
га к соответствующей скорости сдвига, т. е.
Т = т] — , (107)
dy
где Т — напряжение сдвига, ц— вязкость жидкости, du/dy —
скорость сдвига, выраженная через градиент скорости в жид-
кости.
Вязкость можно представить либо как произведение силы
на время и на величину, обратную квадрату длины, либо как
произведение массы на величины, обратные времени и длине.
Во всех системах единиц численные значения абсолютной вяз-
кости одинаковы. В технической литературе нередко использу-
ется термин «кинематическая вязкость», которая представляет
собой произведение абсолютной вязкости на величину, обрат-
ную плотности жидкости.
Любую жидкость, вязкость которой при постоянных давле-
нии и температуре постоянна и не зависит от скорости сдвига,
называют ньютоновской. Свойствами ньютоновской жидкости
обладают все газы и многие гомогенные жидкости. Однако
большинство жидкостей не являются гомогенными; так, у мно-
гих культуральных жидкостей вязкость меняется с изменени-
ем скорости сдвига. Подобные жидкости называют неньюто-
новскими. Для них отношение напряжения сдвига к скорости
сдвига равно кажущейся вязкости, которая, разумеется, зави-
сит от скорости сдвига. Неньютоновские жидкости можно клас-
сифицировать по форме соотношения между напряжением сдви-
га и скоростью сдвига.
Очень многие культуральные среды для выращивания бак-
терий и дрожжей по существу являются ньютоновскими жидко-
стями даже при очень высокой плотности клеток, если только
в среду не экскретируются в значительном количестве синтези-
руемые этими микроорганизмами биополимеры. Напротив,
культуральные жидкости, в которых растут плесневые грибы
или актиномицеты, часто являются неньютоновскими, причем
степень отклонения их от поведения ньютоновских жидкостей
зависит как от концентрации мицелия, так и от его морфоло-
гии. При удалении мицелия из большинства культуральных
жидкостей плесневых грибов и актиномицетов они становятся
ньютоновскими.
Многие культуральные среды для выращивания плесневых
грибов и актиномицетов ведут себя как псевдопластические
Химическая технология и биотехнология
459
жидкости, т. е. с увеличением
скорости сдвига их кажущаяся
вязкость сначала уменьшает-
ся. График зависимости напря-
жения сдвига от скорости
сдвига для псевдопластиче-
ской жидкости сначала имеет
форму выпуклой кривой, а за-
тем становится линейным. При
экстраполяции линейного
участка до пересечения с осью
напряжений сдвига мы полу-
чим точку, отвечающую пре-
дельному напряжению сдвига.
Для пластических жидкостей
предельное напряжение сдвига
соответствует предельной точ-
ке. На рис. 10.13 приведены
Рис. 10.13. Связь между напряжени-
ем сдвига и скоростью сдвига для
жидкостей с разными реологическими
свойствами.
типы зависимостей между напряжением сдвига и скоростью
сдвига. Иногда мицелиальные культуральные жидкости оказы-
ваются тиксотропными, т. е. кажущаяся вязкость уменьшается
не только с увеличением скорости сдвига, но и по мере переме-
шивания при данной скорости сдвига.
В общем виде зависимость между напряжением сдвига и
скоростью сдвига для неньютоновских жидкостей имеет вид
/ би \
\ dy /
(Ю8)
где К и п — константы. Псевдопластическую жидкость можно в
первом приближении охарактеризовать величиной предельного
напряжения сдвига и пластической вязкости, т. е. наклоном
прямолинейного участка зависимости между напряжением
сдвига и скоростью сдвига в линейных координатах. Однако, ес-
ли построить зависимость напряжения сдвига от скорости сдви-
га в логарифмических координатах, то линейный участок с на-
клоном п и точка пересечения К будут определяться начальным
участком графика. В истинных бингхэмовских пластических
жидкостях движение начинается только после того, как напря-
жение сдвига превышает предельное значение. В псевдопласти-
ческих жидкостях некоторое движение возникает даже при
низких напряжениях сдвига. При одинаковой энергоемкости
и одинаковой геометрии реакторы с псевдопластической жид-
костью заметно отличаются от реакторов с ньютоновской сре-
дой по задержке газа, распределению пузырьков по размеру
и времени перемешивания.
460
Глава 10
10.4. Вспомогательное оборудование,
использующееся в биотехнологических процессах
Основное внимание в предыдущей части этой главы было уде-
лено вопросам, связанным с центральным звеном любого био-
технологического процесса — биореактором. Однако, каким бы
ни был процесс, это основное звено не может существовать
изолированно. При получении высокоценных продуктов тонко-
го химического синтеза в биореакторах периодического дейст-
вия иногда удается осуществлять некоторые дополнительные
процессы, например приготовление и стерилизация сред, кон-
центрирование и экстрагирование продуктов, хотя и не обяза-
тельно в оптимальных условиях. Несмотря на то, что концеп-
ция использования одного и того же элемента технологической
установки для осуществления нескольких процессов представ-
ляет значительную ценность при получении некоторых продук-
тов, в целом она изжила себя. Исключение составляют приго-
товление и стерилизация сред. Все чаще, несмотря на возмож-
ное увеличение капитальных затрат, для каждой стадии техно-
логического процесса применяется специально созданное обо-
рудование. Это облегчает осуществление процесса в целом и
повышает его эффективность.
При получении продуктов валового синтеза биотехнологиче-
скими методами из экономических соображений независимо от
сырья или технологической схемы в промышленном масштабе
обычно реализуют непрерывный процесс производства. Такие
производства включают ряд последовательных или параллель-
ных элементарных процессов, обеспечивающих надежное полу-
чение продукта на протяжении 300—330 дней в году. Основная
задача состоит в оптимальном использовании ресурсов. К сожа-
лению, подобный путь параллельного проведения элементарных
процессов обычно затрудняет управление производством в це-
лом и поддержание больших скоростей потребления сырья и
образования продукта. Перечислим стадии, обычно составляю-
щие полный непрерывный биотехнологический процесс: 1) хра-
нение сырья, его предварительная обработка и смешивание;
2) проверка степени загрязнения; 3) переработка сырья с
целью получения продукта; 4) концентрирование продукта;
5) отделение продукта; 6) конечная обработка продукта;
7) хранение продукта. Если говорить о капитальных затратах
и эксплуатационных расходах, то на долю ключевого этапа
производства приходится не более 30—40% от общего их ко-
личества. Отсюда сразу становится ясно, насколько значимы
вспомогательные технологические этапы.
Разумеется, не все перечисленные выше стадии присущи ис-
ключительно биотехнологическим процессам. Некоторые из них
Химическая технология и биотехнология
461
часто встречаются и в других технологиях. Однако есть опера-
ции, характерные только для биотехнологии: это, например,
стерилизация сред и воздуха. При осуществлении некоторых
других операций (например, концентрирование, сушка и хране-
ние) должны учитываться особенности биологических материа-
лов.
10.4.1. Стерилизация среды и воздуха
Обязательным условием осуществления многих биотехнологи-
ческих процессов является работа в асептических условиях,
поэтому решающее значение имеет стерилизация среды, а в
случае аэробных процессов — и воздуха. Тем не менее для
разработки и оптимизации эффективных методов стерилизации
сред и воздуха было сделано удивительно мало. В случае био-
технологических процессов жидкие среды стерилизуют исклю-
чительно нагреванием до высоких температур, а воздух, как
правило, лишь фильтрованием; мы обсудим только эти два ме-
тода стерилизации.
Рациональное конструирование стерилизаторов культураль-
ных сред основывается на данных о кинетике гибели микроор-
ганизмов. Нагревание вызывает потерю их жизнеспособности,
но не физическое разрушение. Скорость инактивации микроор-
ганизмов при данной температуре описывается уравнением
— = — kN, (109)
dt
где k — зависящая от температуры константа скорости реак-
ции, N — число потенциально жизнеспособных микробных кле-
ток, t — время. Важно отметить, что в это уравнение входит
число клеток или спор, а не их масса, фигурировавшая раньше
в уравнениях роста. При стерилизации ключевую роль играет
именно число клеток и спор как потенциальных центров ин-
фекции.
Для вегетативных клеток кинетика гибели первого порядка
[уравнение (109)] является хорошим приближением, хотя на
нее может влиять и множество дополнительных факторов. Од-
нако в случае спор наблюдаются отклонения от этого уравне-
ния. И для вегетативных клеток, и для спор влияние темпера-
туры на константу скорости реакции К, входящую в уравнение
(109), описывается формулой Аррениуса:
k = ae~E^T, (ПО)
где а — эмпирическая константа, Е — энергия активации, R —
газовая постоянная, Т — абсолютная температура. Преобразо-
462
Глава 10
вав уравнение (109) и подставив в него выражение для k, по-
лучим
-^- = ae~E^Tdt ,
N
(111).
или в интегральной форме, приняв, что N=N0 при / = 0,
In ~ = — a j е-^rf/. (112>
° о
Цель всех процессов стерилизации состоит в уменьшении числа
выживших потенциально жизнеспособных клеток и спор до-
величины, меньшей единицы. Поэтому такие процессы лучше
всего рассматривать с вероятностной точки зрения. Стерилиза-
цию среды можно проводить как в периодическом, так и в не-
прерывном режимах. В периодическом режиме тепловая обра-
ботка среды обычно бывает излишне продолжительной, что
приводит к ухудшению качества среды. При непрерывном ре-
жиме эти недостатки можно устранить.
Цикл периодической стерилизации состоит из трех стадий:
нагревание, выдерживание (обычно при 121 °C) и охлаждение.
Инактивация клеток и спор происходит на всех трех стадиях;,
поэтому, если время нагревания равно Л, время выдержива-
ния — t2, время охлаждения — t%, а общая продолжительность
цикла — t, то
In А- = In +In + In =
N Л\ N2 N
Г h ti ts
= a J е-£/«г dt + j“ e-£/«r dt + f e~£/«r dt
[o 6 6
(113>
где Ni и N2 — общее число выживших клеток и спор после на-
гревания и выдерживания соответственно.
При стерилизации в периодическом режиме стадии нагре-
вания и охлаждения нередко бывают весьма продолжительны-
ми, а время выдерживания обычно составляет 0,3—0,5 ч при»
температуре 121 °C.
Существует два разных типа систем для стерилизации сре-
ды в непрерывном режиме: системы с прямой инжекцией пара,,
требующие сверхчистого пара, и системы с косвенным нагрева-
нием с помощью теплообменников. Ясно, что системы обоих
типов работают при повышенном давлении. Системы с прямой
инжекцией включают инжектор пара, секцию для выдержива-
ния, расширительный клапан и секцию для быстрого охлажде-
ния. В установках с косвенным' нагревом среда обычно подвер-
Химическая технология и биотехнология
463
гается предварительному нагреванию уже стерилизованной
средой для возврата отработанного тепла. В систему входят
также: безынжекционный теплообменник, в котором среда ме-
нее чем за 1 мин нагревается до 135°C; секция, в которой
среду выдерживают 3—5 мин при 1351ОС; секция для быстрого
охлаждения, где стерилизованную среду сначала охлаждают
вновь поступающей средой, а затем — охлаждающей водой.
Можно применять плоские теплообменники либо змеевики, од-
нако предпочтение часто отдается первым: у них легче очищать
поверхности теплообмена, которые быстро загрязняются при
стерилизации многих сред.
При проведении непрерывной стерилизации следует пом-
нить, что в среде, протекающей через теплообменники, всегда
существуют градиенты скорости, и режим полного вытеснения
не достигается. Поэтому при оценке вероятности выживания
клеток и спор необходимо учитывать зависимость распределе-
ния времени пребывания среды в теплообменниках от режима
потока в них.
Стерилизация среды имеет ключевое значение во многих
биотехнологических процессах; не менее важна и стерилизация
воздуха, используемого для аэрации. При этом из огромных
объемов воздуха, часто необходимых для осуществления аэроб-
ных процессов, следует удалить микроорганизмы размером
до 0,5 мкм. Хотя выделяющееся при сжатии воздуха тепло спо-
собствует эффективному уничтожению содержащихся в нем
микробов, воздух обычно необходимо также фильтровать через
волокнистые спеченные или ацетилированные плоские фильтры
из поливинилового спирта (ПВС). Удаление микроорганизмов
с помощью воздушных фильтров (кроме фильтров из ПВС)
происходит не благодаря малому размеру пор, а вследствие
того, что воздух проходит через фильтр очень сложным путем,
из-за чего увеличивается вероятность соударения увлекаемых
им микроорганизмов с фильтром. В подобных системах часто
происходит значительное падение давления. Материал, из ко-
торого изготавливают фильтры и подводящие устройства, не
должны разрушаться при стерилизации. Для обеспечения вы-
сокой производительности фильтры следует держать сухими.
10.4:2. Выделение продукта и завершающие операции
В биотехнологической промышленности применяют множество
способов выделения конечного продукта и его завершающей об-
работки. Те же способы, за исключением предназначенных для
разрушения клеток, широко используются и в других отраслях
промышленности, поэтому здесь мы не будем рассматривать
отдельные элементарные технологические приемы. Обсуждая
464
Глава 10
применение этих методов в биотехнологических производствен-
ных процессах, всегда следует помнить два момента: во-первых,
необходимо предотвратить денатурацию продукта, которая мо-
жет происходить из-за недостаточного охлаждения в ходе про-
цесса, химического или биологического загрязнения, а также
из-за разного рода биологических воздействий, если продукт
получен в нестабильном состоянии. Во-вторых, всегда нужно
учитывать потенциальную взрывоопасность и опасность для
здоровья пыли белковой природы, которая скапливается после
высушивания биологических продуктов. Обе эти проблемы дол-
жны учитываться при разработке эффективной технологии.
10.5. Перспективы развития промышленных
биотехнологических процессов
В предыдущих разделах данной главы относительно подробно
обсуждались ключевые аспекты применения химической техно-
логии к биотехнологическим, преимущественно микробиологиче-
ским, процессам, а также теория и экономика промышленных
технологических систем. К сожалению, данных, необходимых
для дальнейшего развития биотехнологии, удручающе мало.
Так, нам практически ничего не известно о физических свойст-
вах микроорганизмов. Вместе с тем за последние сто лет ог-
ромных успехов достигли гидродинамика, химия поверхностей
и процессов на границе раздела фаз, а также молекулярная
биология, имеющие решающее значение для разработки био-
технологических процессов. Такое несоответствие совершенно
недопустимо, особенно если учесть, что существует много мето-
дов, позволяющих получить необходимые физические данные
о микроорганизмах, но ими просто не пользуются.
Напротив, кинетика биотехнологических процессов в боль-
шинстве случаев хорошо изучена и в последние годы широко
использовалась для развития технологии. Но даже в этом слу-
чае еще не решены проблемы, возникающие при использовании
полученных в лаборатории данных для анализа промышленных
процессов и создания технологических схем. Остаются спорны-
ми вопросы, связанные с вариабельностью данных и с теми
предельными значениями величин, которые можно использо-
вать для систем, отличных от тех, для которых эти величины
были получены. Например, вариация в коэффициентах выхода
или конверсии в 15% может привести как к значительным’
коммерческим выгодам, так и к полному краху. Однако лишь
немногие микробиологи, занимающиеся биотехнологией, могут
дать более точные оценки этих коэффициентов, особенно если
анализ процесса или разработка технологии должны быть осу-
ществлены за короткие сроки. Другие типичные примеры того»
Химическая технология и биотехнология
465
же рода — это влияние давления и количества растворенного
углекислого газа, а также условий роста на размер микробов.
Что касается таксономических аспектов, то слишком многие
микробиологические данные получены при условиях, весьма
далеких от тех, при которых протекают интересующие нас про-
цессы. Экстраполяция подобных данных нередко приводит к
таким результатам, которые можно назвать мистикой или
фольклором в микробиологии. Для разработки эффективных
биотехнологических процессов крайне важны два момента: по-
строение как умозрительных, так и основанных на реальных
данных гипотез, их тщательная проверка в соответствующих
условиях с целью их подтверждения или опровержения и одно-
временная разработка строгих теорий и установление законов,
которым подчиняется поведение микробов в той среде, в кото-
рой их предполагается использовать. Только при таких услови-
ях можно надеяться на успешное применение биотехнологии
во многих областях промышленности в ближайшие десятиле-
тия.
ЛИТЕРАТУРА
Aiba S„ Humphrey А. Е„ Millis N. F., 1973. Biochemical Engineering (2nd edn.),
p. 434, Academic Press, New York.
Andrew S. P. S. (1982). Gas-liquid mass-transfer in microbiological reactors,
Trans. Inst. Chem. Engrs, 60, 3—13.
Atkinson B., Sainter P. (1982). Development of downstream processing, J. Chem.
Tech. Biotechnol., 32, 100—108.
Bailey J. E., Ollis D. F., 1977. Biochemical Engineering Fundamentals, p. 753,
McGraw-Hill, Kogakusha, Tokyo.
Calderbank P. H. (1967). Gas absorption from bubbles, Chem. Engr. bond.,
No. 212, Oct., 209—233.
Cooney C. L., Wang D. I. C., Mateles R. I. (1968). Measurement of heat evolu-
tion and correlation with oxygen consumption during microbial growth, Bio-
technol. Bioengng, 11, 269—281.
Gaden E. L. (1959). Fermentation process kinetics, J. biochem. microbial. Technol
Engng, 1, 413—429.
Harder W., Kuenen J. G., Matin A. (1977). Microbial selection in continuous
culture, J. appl. Bacteriol., 43, 1—24.
Herbert D., 1961. A theoretical analysis of continuous culture systems. In: Conti-
nuous Culture of Micro-organisms, S. С. I., Monograph No. 12, pp. 21—53,
Soc. Chem. Industry, London.
Johnstone R. E., Thring M. IF., 1957. Pilot Plants, Models, and Scale-up Methods
in Chemical Engineering, p. 307, mcGraw-Hill, New York.
Monod J., 1958. Recherches sur la croissance des cultures bacteriennes 2nd edn.,
p. 210, Herman, Paris.
Payne W. J. (1970). Energy yields and growth of heterotrophs, Ann. Rev. Micro-
biol., 24, 17—52.
Pirt S. J., 1975. Principles of Microbe and Cell Cultivation, p. 274, Blackwell
Scientific Publication, Oxford.
Rudd D. F., Watson С. C., 1968. Strategy of Process Engineering, p. 466, John
Wiley and Sons, New York.
Richards J. W. (1961). Studies in aeration and agitation, Prog. ind. Microbiol,
3, 141—172.
30—1344
Предметный указатель
Лгроци!. 375
Азокрасители, биодеградация -281, 2S2
Азотные удобрения 52
-Актиномицины 157
DL-аланин 148
Алкилсульфаты 293
.Алкогольные напитки, производство 104—
114
Альгинат 225
Амидазы 170
а-Амнлаза (такадиастаза) нз Aspergillus
oryzae 164
Амилазы 167—169
'«-Амилазы термостабильные из Bacil-
lus 167
Амилоглюкозидазы 167—169
— из Aspergillus 103
-Амилопектин 169
-Аминокислоты 14—15, 122—123, 147—155
— в качестве пищевых добавок 154—155
— образование у бактерий 147—150
— применение 154—155
— производство из биосинтетических пред-
шественников 151
— рацемические смеси 169—170
— синтез с помощью ферментов 151—154
Ампициллин 327
-Анаэробное разложение, образование ме-
тана 65—66, 73—78
Анаэробные реакторы 260, 261, 262, 263
Антагонистическое действие гриба Trie bo-
iler та на патогенные организмы 372
Антибиотики 13, 155—161, 326—328
— аминогликозидные 159
— и конъюгация у Strepiomyces 300—301
— получение с помощью Strepiomyces
156—157
Антигены лейкоцитов человека (HLA)
331—332, 344, 346
Антитела моноклональные 16, 313—315,
329—333
Аргинин, биосинтез 148—149
Арилгалогены 288
Аскомицеты 301, 362
•L-аспартат 149
Ацетон 133—135
Ацидотермофилы крайние 193—194
АТР, концентрация в активном иле 264
Базидиомицеты 301, 364
Бактерии, механические повреждения в
биореакторах 415—416
— слияние протопластов 308—309
— характеристики суспензий 457—459
— экспрессия генов животных 315—316
(Безразмерные величины 430, 431—432,
435-436
Белковые продукты 115—122
.Белок одноклеточных организмов (БОО)
15, 116—119, 397
-------из активного ила, концентрации
аминокислот 273
-------состав 272, 273
-------отходов целлюлозно-бумажной
промышленности 280
-------—--------получение при участии
Candida 280
Бензилпенициллин 156, 158, 326, 327
Бензохинон 180, 181
Бензпирен 289
Бесклеточные системы 79—83
Биллирубнноксидаза иммобилизованная 171
Биогаз см. Метан
Биодатчики полупроводниковые 25
Биодеградация нефтяных загрязнений
290—291
— поверхностно-активных веществ 292—295
Биокатализаторы, оптимизация 185—186
Биологическая очистка газов 282—283
Биологический катализ в неводных средах
184—185
Биомасса 12, 20, 21, 22, 35—42, 47—52, 54
— коэффициент выхода 403—405
— получение химических веществ 172—177
Биоповреждения, авиационное топливо 243
— зерно 239
— камни 244
— клен 240
— масла нефтяные 243
— материалы 26, 236—244
— металлы 243—244
— определение и классификация 437—438
— пищевые продукты 239
— пластмассы 241—242
— .поверхностные покрытия (краски, лаки)
241
— шолиуретаны 242
— продукты животного происхождения
240—241
— резина и пластмассы 241
— целлюлоза 239—240
— шерсть 240
Биополимеры 216—236
Биореактор вращающийся 250
Биореакторы, анализ размерностей и мас-
штабирование 430—437
— классификация и производительность
4.18—430
— конверсия газовой и жидкой фаз 451—
454
— перенос кислорода 437—447
— .периодического действия 418, 419, 421
-------с доливом субстрата 418
— полунепрерывного (полупериодического)
действия 418, 419
— проточные непрерывного действия с пол-
Предметный указатель
467
ным вытеснением (реакторы поршневого
типа) 263, 420, 422, 428—430, 452, 455
--------------(идеальным) перемешивание
ем 260. 419, 420, 422, 423, 425—426, 453
— рециркуляция газовой и жидкой фа»
453—455
— с вращающимися мешалками 436
— теплопередача и охлаждение 447—45-1
Бнотопливные элементы 21, 83—86
----деполяризующие 84—85
----на основе неэлектроактивных ве-
ществ 84
•--------ферментов 85
---------Escherichia coli 84
----регенерируемые 85—86
Бнофотолиз 22
Блеомицины 156
Бобовые культуры 28, 356—364
Болезни растений, биологический контроль
371—376
----и Trichoderma 372
Болезнь Гоше 342
Брожение, образование полисахаридов
219—222
Бумажная масса, промышленное произ-
водство 176
Бутанол 133—136
Бутиленгликоль 136
Вакуумное брожение 72
Вакцины 339—340
Вещество S Рейхштейна 162, 163
Винибластин 174
Винкристии 174
Вино 109—111
Виноделие, использование Botrytis cinerea
111
----Hansenula saturnus 110
Вирус гепатита В, антиген 319, 320, 340
— ящура, капсидный белок 319, 320
Витамины 123
Водные растения как источник биомассы
39, 54—57
Водород, получение путем биофотолиза
22, 80—82
Водоросли 22, 54—57, 250, 251
Водяной гиацинт, получение биогаза 54
Восстановительный пентозофосф атный цикл
.(ВП-цикл) 42—43, 45, 46
Восстановленные соединения серы в отхо-
дах, разложение 282—283
Вредители сельскохозяйственных культур,
борьба с помощью Bacillus 28, 397
Выщелачивание кучное 200
— чановое 200
— In situ 198—199, 203—204
Галактозооксидаза 180
Галофилы ацидофильные 186
Геллан 226
Гемоглобинопатии, диагностика 342—344
Генетическая инженерия, опыты in vitro
315—322
— — прикладные аспекты 320—321
Генри закон 438
Геи тирозил-тРНК—синтетазы нз Bacillus
stearothermophilus 183—184
Гетерокарионы 302, 310, 388—389
Гибридизация путем скрещивания 300—303
— соматических клеток 387—390
Гибриды растений, образование путем
слияния протопластов 311—312
Гндроколлоиды как пищевые добавки 124
Гидролазы 151
30*
Гидрохинон 180, 181
Гликоген, синтез 229, 233, 234
Глицин 123, 155
Глутамат, ATP-зависимое аминирование
154
L-глутамнн 148
Глутаминовая кислота 123
Глюкоамилазы из Aspergillus niger 168
Глюкозооксидаза, ферментный электрод 341
Глюконовая кислота 122, 141—143
--- натриевая соль 142
D-глюконовая кислота, производство пр»
участии Aspergillus niger 141
Говядина и микопротенн, сравнение сред-
него состава 122
Гормоны половые стероидные 163, 328—329*
Грибная гниль 373
Грибы, парасексуальный цикл 302—303
— плесневые, механические повреждения
в- биореакторах 415—416
— системы скрещивания 301
------типы спаривания у Saccharomyces
cerevisiae 301
— слияние протопластов 309—311
— характеристика суспензий 457—459
Датчики 19, 25, 340—342
— контролирующие содержание глюкозы
19, 29
ДДТ, биодеградация 291
Декстран 124, 223—225
Декстрозный эквивалент (ДЭ) ЮЗ, 169
Детергенты, разложение 293—294
Диабет 19, 334—338, 344—345
Диарея 307
Дигоксин 174, 333
Диплоиды 309—310
ДНК, включение в векторные плазмиды
Bacillus 317
-----------Saccharomyces 317
— химический синтез 182
ДНК-лигаза фага Т4 317, 318
Древесина 39—40, 53—54
Древесная пульпа 278
------ щелочная варка (сульфатный про-
цесс) 278
Дрожжи Torula 94
L-DOPA 153
Железо, окисление 191, 192, 194
— трехвалентное 192, 194
Жирные кислоты полиненасыщенные 123
Жиры и масла 123
Загустители 26, 124, 225
Занфло 226
Зерновые как продуценты крахмала 40, 41
Изменчивость 383—384
Изоглюкоза, получение с помощью Sirep-
totnyces 104
L-изолейцин 149
Иммуногистохимия 330—331
Инвертаза 169
Ингибирование по принципу обратной
связи в биосинтезе аминокислот 148—150
Инженерия белка 182—184
Инокуляция растений везикулярно-арбу-
скулярной микоризой 367—371
— семян бобовых бактериями Rhizobium
358—363
Инсулин 320, 334—338
468
Предметный указатель
— человека, получение с помощью Esche-
richia 335—338
Интерферон 18, 29, 315, 334, 338—339
Ион-селективный транзистор на основе по-
левого эффекта 346
Итаконовая кислота, получение при учас-
тии Aspergillus 122, 144
Йогурт 101
Каллус, образование из протопластов 385—
386
Капуста, квашение 124—125
Карбоангидраза 171
Карбоксиметнлсукцинат (КМС) 294—295
Карденолиды 174
^-Каротин 123
Каучук 58—59, 60
Кинетика образования продукта в перио-
дических культурах микроорганизмов
407—410
— роста микроорганизмов 405—407
Кислород, перенос в биореакторах 437—447
----из воздуха в жидкость, модель Дан-
квертса 438, 441
----------------Льюиса—Уитмена 438—
447
----------------Хигби 438, 441
Клеи 123—124
Клокациллин 327
Кобальт, поглощение микроорганизмами
213, 214
Кожевенная промышленность 166, 240
Конверсия в биореакторах 45.1—454
Консервирование мясных продуктов, роль
микроорганизмов 116
Конъюгация у бактерий 300—301
Корм для животных 15, 271—274
Корончатые галлы 373—376
Кортизон 16
Кортикостерон (вещество S Рейхштейна)
162
Кофакторы, повторное использование 186—
187
Коэффициент выхода биомассы 403—405
----продукта 404—405
Крахмал, продукты гидролиза 103—104, 105
— уровень производства 40—42
Ксантан 22, 222—223, 232—233
Ксантиноксидаза 180
Ксенобиотики, биодеградация 274—275,
284—295
----роль Pseudomonas 285, 286, 292
Кукурузный сироп с высоким содержанием
фруктозы 104, 167
Культуры тканей 172—Г74, 415, 416
Курдлан 228
<Лайнуивера—Бэрка график 407
Лактоза, сбраживание дрожжами 277
Латекс, образование в растениях 59
Лекарственные препараты, направленное
введение 333
----передозировка 333—334
Лиазы, использование в реакциях дезами-
нирования 152
Лигнин 63, 64, 175—177
Лигносульфонаты 278, 279—280
— десульфирование при участии Pseudo-
monas 280
Лигноцеллюлоза 175—177
— разложение 63—65
Лизин 15, 23, 149, 150, 151, 154
Лимонная кислота 122, 129, 137—141
-----образование дрожжами Candida 139
---получение из целлюлозы 139
---процесс Коджи 140
Лимоннокислое брожение 94
Лишайники, разрушение камней 244
Масла растительные 57—58, 60, 61
Масличные растения 57—61
Масло, использование специальных штам-
мов бактерий 101
Медицина 14, 17—19, 325—347
Медь, выщелачивание 195—197, 200
Меласса 61, 112
Мембрана с избирательной проницаемо-
стью 346
Мембраны хлоропластов 80—81
Металлопротеазы 166
Металлы, бактериальное выщелачивание
191—204
— внеклеточное комплексообразование 206,
209—210
---осаждение 206, 207—209
— внутриклеточное накопление микроорга-
низмами 206, 210—216
— перевод в летучую форму 206, 207
— связывание с клеточной поверхностью
206, 212—213
— удаление из растворов микроорганиз-
мами 205—216
Метан, анаэробная переработка сырья 65—
66, 73—78
— получение 13—14, 65—66, 73—78, 258—
259, 378—381
Метионин 123, 149—151, 153, 154
Метод водных культур (гидропоника)
352—353
— Мильштейна, получение моноклональ-
ных антител 313, 314
— питательной пленки 351—356, 369—370
— слияния протопластов 386—390
Микропротеин (грибной белок), получение
с помощью Fusarium graminearum 119—
122
Микориза везикулярно-арбускулярная 364—
371
Микробиологическая переработка отходов,
биологический контроль 264—266
------контроль за патогенностью 267—268
Микробиологическое производство, выделе-
ние конечного продукта 463—464
---культуры микроорганизмов 401—403
---процессы н их экономичность 12—20,
28—31, 346—401, 416, 417
--- требования к условиям культивиро-
вания микроорганизмов 400—401, 414—416
Михаэлиса—Ментен модель ферментатив-
ных процессов 410—412
Молоко с гидролизованной лактозой 99
— сквашивание 93—95
— с низким содержанием лактозы 102, 169
Молочная кислота, образование 137, 277
------при участии Lactobacillus 137
— промышленность, применение Leucono-
stoc dextranicum 95, 101
Молочные продукты 93—102, 169, 171
Моно соотношение 406, 412, 424, 427
Морфолин 257
Мутагенез 183—184, 297—299
— сайт-специфический 183—184
Навоз, содержание питательных веществ
271
Нафтализ, превращение в салициловую
кислоту 144—147
Нафталинсульфоновая кислота 288
Предметный указатель
469
Нейтраза, ее роль в созревании сыров 99
Нервные газы фосфоорганическне, обез-
вреживание 171
Нефть, биодеградация загрязнений 290—
291
Никотин 257
Нитрилтриацетат (НТА) 294—295
Ннтротолуолы 289
Окисление и оксигенация 178—179
Окислитель и ускоритель вулканизации 257
Окружающая среда, влияние биотехноло-
гии 14. 17, 26—27, 202, 246—295
Оксигенатор Дорра—Оливера 256
Оксидиацетат (ОДА) 294
2-оксоглутаровая кислота, получение при
участии Candida 144
Оливки 125
Опухолевые маркеры 332
Органические кислоты, производство 136—
147
— растворители, бродильное производство
(ферментация) 133—136
Осахаривание 103, 105
Отходы, извлечение полезных веществ
270—274
— переработка анаэробная 13, 14, 27, 258—
263, 378—382
------- получение метана из навоза 27,
379—380, 381
----аэробная 249—257, 376—378
-------с помощью активного ила 250—256
— превращение в микопротеин (грибной
белок) 278
— промышленные, переработка биологиче-
скими методами 13—14, 26, 274—283
----------- использование пленок микро-
организмов 276
Папаин 182
Парасексуальный цикл у грибов 302—303
Паратионгидролаза из Pseudomonas sp.,
детоксикация паратиона 292
Патогенные микроорганизмы при анаэроб-
ном разложении отходов 267—268
Пектины, расщепление ферментами 114,
127
Пенициллин 155—158, 326—328
— мутагенез и отбор 297, 298, 299
Пенициллиназы (ft-лактамазы), их инги-
биторы 160
Пенициллиновая кислота 158, 160
Пепсин 166, 171
Периодическая культура микроорганизмов,
днауксичный рост 413
-------образование продукта 407—408, 409
Перколяционные фильтры 249—250, 276
Пестициды, детоксикация 291—292
Пивоварение 11, 106—108
Пигменты 123, 280—281
Пикули 125
Пиридоксальфосф ат 152—154
Пирит, окисление 192, 194
Пищевые добавки 122—124, 223
— продукты и напитки, промышленное
производство 14, 15, 23, 91—127
----использование дрожжей Candida 15,
119
Плазма крови, заменители 224
Плазмида R1 Escherichia coli 306
— Ti 373—374, 390
Плазмидные гены Rhizobium, участие в
фиксации азота 305—307
Плазмиды вирулентности патогенных
штаммов Escherichia coli 307
— включение ДНК 316—318
— генетические перестройки in vivo 300—
301, 303—308
— гены устойчивости к лекарственным
препаратам 304—305
— перенос генов из клеткн-хозяина 300,
303
— репликация в клетках Escherichia coll
304—306
Пластмассы 26, 236
Подкислители 122
Подсолнечник, выход масла 57—58
Поли-Р-гидроксибутират (ПГБ) 26, 234—236
Полисахариды 124, 217—234
— биосинтез 229—232
— образование при брожении 219—222
----у Pseudomonas 226, 234
----Rhizobium 234
Полистирол 289
Политран (склероглюкан) 227
Полифенилен 236
Полихлорбифеннлы (ПХБ) 289
Полиэтиленглнколь (ПЭГ) 309, 311, 312,
387, 390
Половые гормоны 163
Преднизолон 162, 163
Предшественники аминокислот 151
— антибиотиков 157—158
Пренатальная диагностика наследственных
болезней 342—344
Прогестерон 16, 161, 162
Проинсулин, синтез в клетках Escherichia
coli 319—320
L-пролин 148
Протеазы 165—166
— Bacillus 165—166
Протопласты, использование в селекции
растений 382—390
— методы слияния 386—390
— слияние у бактерий и грибов 308—311
Прутин 15, 118
«Псевдоожиженный слой», переработка
отходов 256—257
Пуллулан 228
Пуллуланаза 169
Радионммунологический анализ 329—330
Размножение вегетативное 382—386
Рак 332, 338
Реакторы см. Биореакторы
Регенерация растений из протопластов
384—386
Редокс-меднаторы 85
Рестрнктазы 317
Рестрикционные фрагменты, полиморфизм
по длине 344, 345
Рециркуляция газовой и жидкой фаз в
биореакторах 453—455
Рибофлавин 123
Рибулозобифосф ат-карбоксилаза (РБФК)
42, 45. 46
Рифамицнны 157—158
Рост микроорганизмов, модели 405—407
----сродство к субстрату 410—414
Салициловая кислота 144—147
----образование у Pseudomonas 144
Самоклонирование 321
Сахар, получение этилового спирта 58, 61,
62—63, 66—72
— уровень производства 40—42
Сахара, образование прн сульфатной вар-
ке целлюлозы 278
470
Предметный указатель
Селен в составе аминокислот 153, 154
Сельскохозяйственные культуры, пронзвод-
ство сахара и крахмала 40—42
Семена бобовых, дражирование 362
Сера, окисление 192, 193, 194
Серповидноклеточная анемия, диагностика
н лечение 342—344, 347
Сидр 113—114
Симбиоз бактерий Rhizobium и бобовых
356, 357, 363, 364
Слияние клеток животных 312—313, 314
—------маркеры устойчивости к химиче-
ским веществам 312
— протопластов, гибридизация соматиче-
ских клеток 387—390
---грибов 309—311
---межвидовое 309, 310
— — образование гибридов растений 311—
312, 386-390
— — у Bacillus 308
-------Saccharotnyces 309, 310
-------Streptomyces 309
Сметана, использование культуры бакте-
рий 102
Соевые бобы, получение белковых про-
дуктов 115, 125—127, 166
Соевый соус, использование Aspergillus
oryzae 126
Солнечная энергия, использование в бес-
клеточных системах 79—82
— — технология получения биомассы 35—
42
Солод 106—107
Соя, выход масла 57—58
Стокса закон 457
Сточные воды 54, 56, 247, 249, 252, 264
— — анаэробное разложение в реакторах
различных типов 261, 262, 263
---------ила в тэнке Имхоффа 263
— — очистка 205, 247, 255, 261, 279, 290—
291
--- повторное использование
Стерилизация воздуха 463
— сред 460, 461—463
Стероиды 16, 161—164, 328—329
— андростаны 163
— эстраны 163
Стрептомицин 156
Субтилизин Carlsberg 165
Сукциноглюкан 234
Суспензии клеток, их характеристики 457—
459
Сыворотка 100, 169, 277—278
Сыроварение, использование протеаз Ел-
thothia parasitica 166
— — протеиназы (нейтразы) Bacillus 99
---Aspergillus oryzae 100
— — Brevibacterium linens 99
— — Propionibacterium 95
— получение заменителей сычуга нз Mucor
miehel 97—98
Сыры, производство 95—100, 116, 165, 166
— созревание при участии Pseudomonas
100
Сычуг и его заменители 97—98, 165, 166
Э-Талассемия, пренатальная диагностика
344
Термофилы умеренные 193
Терпены 170
Тетрациклин 155, 156
Техника безопасности и контроль продук-
ции 32—33
Технология рекомбинантных ДНК, приме-
нение в медицине 334—347
Тиофея-2-карбоновая кислота 257
Тнпирование тканей человека 331—332
L-тирозин—феиол-лиаза (В-тирозиназа>
152—153
Топливный элемент кислород-водородный
83, 84
Топливо, использование биологических ис-
точников 21—22, 61—78, 258—263, 378—382
Тотипотентность растительных клеток 382
Трансглюкозилаза 168
Транспозоны 307—308
Трансформация бактерий 318
Треонин 149, 150, 151
L-триптофан—индол-лиаза (триптофаназа>
152—153
Углеводороды, образование в растениях
58, 59—61
— полиаром этические 289—290
— получение из водорослей 56
— хлорпроизводные 287—288
Углекислый газ, образование в анаэробных
условиях 258, 259
Удобрения 270—271
Уксус 115, 136—137
— получение прн участии культур Aceto-
bacter 115
Уксусная кислота 136—137
--- образование при получении метана 77
Уран, выщелачивание 197—198, 200
— накопление в микроорганизмах 213—214,
216, 217
Усилители вкуса 123
Фаговые векторы, включение ДНК 316
Фенилглицин 159
Ферментация, получение белковых продук-
тов 115—116
---этанола 65—70
Ферментер колонный эрлифтный 251, 252,
275
Ферменты 9, 16
— гидролитические (гидролазы) 151—152
— использование в коммерческих целях
164—171
------медицине 340—342
— производство фруктовых соков 127
— синтез аминокислот 151—154
Ферменты, созревание сыров 100
а-Фетот.ротеин 332
Фиксация азота 28
---и плазмидиые геиы 306—307
---плазмиды Rhizobium 306—307
---при участии клубеньковых бактерий
и бобовых 356—364
---симбиоз Rhizobium и бобовых 357,
363, 364
Фотосинтез 42—51
— интенсификация 48—51
— эффективность 47—48
Фотосинтезирующие бактерии 79
Фотосинтетически активная радиация
(ФАР) 45, 47
Фруктовые соки 127
Фруктозные сиропы 167
Фрукты и овощи 124—127
Халькопирит 193
Химические соединения 23—25, 132—164,
173, 175-177
--- определение токсичности 171
--- получение с помощью биокатализа-
торов 133, 134
Предметный указатель
471
Хммотрипсин 185
Хинин 257
Хитин 177
Хлебопечение 102—103
— использование Saccharomyces cerevisiae
102
Холинэстераза 171
Хорея Хантингтона 345
Целлюлоза 63—64, 175—176
— образование у Acetobacter xylinum 177
— — — Trichoderma 64
— сульфатная варка 278
Цефалоспорин 309, 326
- С 160, 327
Цианид, детоксикация 283
---с помощью родоназы Bacillus 283
Цистеин 151, 152, 154
С<-растения 46, 47
Шиконин 173
Экзоамилазы 168
Экспрессия генов животных у Saccharo-
myces cerevisiae 315
— клонированных генов 318—320
------у Bacillus 319, 320
Электроэнергия 21, 82—83
Эмбдена—Мейергофа—Парнаса метаболиче-
ский путь 106
Эмульгаторы 25, 26, 397
Эндоамилазы 167—168
Этнленгликольдиацетат (ЭГДА) 294
Этиловый спирт, образование из глюкозы
105
— — перегонка 112—113
— — периодический способ получения 69
Этиловый спирт, производство 58, 61, 62—
63, 66—72, 112—113
------использование Saccharomyces 68,
72, 105, 107, 110, 112, ИЗ
Челночные векторы 317
Яблочная кислота 114, 122, 144
Указатель латинских названий
Acetobacter curvim 115
— orleanense 115
— schuetzenbachii 115
— xylinum 177
Achromobacter 254
Actinoplanes missouriensis 104
Aerobacter 148
Aeromonas 64
Agrobacterium 300, 303
— radiobacter 324
— tumefaciens 373, 374, 390
Alcaligenes 227—228, 235
— eutrophus 397
— faecalis 137
Arthrobacter 104, 139, 228
— globijormis 151
— oxydans 137
— simplex 163
Ashbya gossypii 123
Aspergillus 144, 302, 310
— flavus 122
— niger 103, 122, 139, 141, 168, 309
— oryzae 103, 126, 164
— tamarii 126
Aureobasidium pullulans 228
Azospirillum 300
Azotobacter beijertnck.il 235
— vinelandii 220, 233
Bacillus 148, 235, 301, 317, 320
— amyloliquefaciens 166, 167
— licheniformis 165, 167
— megaterium 213, 318
— polymyxa 213
— stearothermophilus 183, 283
— subtilis 99, 113, 126, 165, 308, 318
— thermoproteolyticus 166
— thuringiensis 28, 397
Beijerinckia 228
Botryococcus braunii 56
Botrytis cinerea 111
Brevibacterium 139, 148
— flavum 123, 150
— linens 98
Chorophyceae 250
Cladosporium resinae 243
Clostridium 136
— aceiobutylicum 134
— saccharobutyricum 113
— thermocellum 64, 176
Corynebacterium 139, 144
— glutamicum 123, 148, 150
— michiganese 268
Crucifarae 365
Cunninghamblla blakesleeana 162
Curvularia lunata 163
Cyanophyseae 250
Debaryomyces phaffii 110
Dunaliella bardawil 56
Endogonaceae 364—365
Endomycopsis lindneri 110
Entamoeba hystolitica 268
Enterobacter aerogenes 136
Enterobacteriaceae 305
Enthothia parisilica 166
Epistylis 253
Eremothecium ashbyii 123
Ericaceae 364
Erwinia 300
— herbicola 152—153
— tahitica 226
Escherichia 148
— colt 300, 318, 319, 322, 335, 337, 415
Eucalyptus 53—54
Euphorbia 59
Ferrobccillus 254
Ferrovibrio 192
Flavobacterium 254, 257, 281, 282
Fames annosus 373
Fusarium 268, 269, 302
— graminearum 119—121
— oxysporum 268, 269
— solani 137
Candida 119, 139, 280
— arborea 15
— hydrocarbofumarica 144
— intermedia 277
— krusei 277
— pulcherrima 110
— scottii 110
— tropicalis 309, 310
— utiUs 15, 280
Catharantus roseus 174
Cephalosporium 303, 326
— acremonium 309, 310
Chaetomicum cellulolyiicum 278
Geotrichum candidum 309, 310
Globodera 268
Gluconobacter 144
— suboxydans 154
Gramineae 365
Halobacterium 82
Hansenula saturnus 110
Hevea 58, 60
Hyphomicrobium 283
Указатель латинских изданий
473
Klebsiella 64, 300
— aerogenes 364
— pneummoniae 136, 364
Kloeckera ПО, 114
Kluyveromyces 68
— fragilis 277
— lactis 309, 310
Lactobacillus bulgaricus 95, 96, 101, 277
— casei 95
— helveticus 95, 96
— lactis 95, 96
Leguminoseae 365
Leptospirillum ferrooxidans 192
Leuconostoc citrovorum 95, 101
— dextranicutn 95, 101
— mesenteroides 124, 223, 224
Lithospertnutn erythrorhizon 173
Medicago sativa 359, 383
Metazoa 250
Methanosarcina barkeri 381
Methanospirillum hungatei 381
Methylophilus methylotrophus 15, 321, 322
Microbacterium 148
Micrococcus 212
— glutamicus 133
Mucor tniehei 97—98
— racemosus 310
Mycobacterium globiforme 163
— fluorescens 100
— putida 236, 300
Rhizobium 300, 357—362, 364
— leguminosarutn 306, 307
— meliloti 307. 359
— phaseoli 306, 307
— trifolii 234, 307, 364
Rhizopus 122, 168
— arrhizus 16
Saccharomyces 112, 113, 177, 213
— carlsbergensis 105, 107, 110
— cerevisiae 15, 68, 72, 102, 105, 107, 110,
115, 213, 214, 217, 301, 309, 310, 315, 317
— diastaticus 68
— uvarum 68, 110
Salmonella 267, 274, 300
Schizophyllum commune 144, 301, 311
Sclerotium glucanicum 227
— rolfsii 372
Serratia marcescens 151
Sporotr'chum 65
— pulcerulentum 278
Streptococcus 95, 96. 101, 224
Strepiomyces 104, 300, 301, 318
— coellcolor 300, 309
— parvulus 157
— rimosus 301
— verticillus 156
Nitrobacter 254
Nitrosvmonas 254
Mocardia 144, 185, 300
— tnediterranei 157
Crchidaceae 364
Paecilcmyces variotii 278
Paracolobactrum 144
Parthenocissus tricuspidata 386, 390
Penicillium 139, 302, 310
— chrysogenum 156, 298, 309, 326
— citrinum 123
Peniophora gigantea 373
Petunia hybrida 386, 388
— parodii 388
Phanerochaete chrysosporiutn 177
Phaseolus vulgaris 369
Phycomyces blakesleeanus 310
Pichia 310
— fermentans 110
Pinus radiata 54
Propionibacterlum 95
Pseudomonas 144, 213, 234, 254, 280, 286, 292,
300, 303
— aeruginosa 216, 234, 300
— elodea 226
Thieghemella orchidis 162
Thiobacillus 254
— acidophilus 192
— ferrooxidans 191, 192, 195, 197
— organoparus 192
— thiooxldans 192, 197
Torula 94
Torulopsis 110
Trichoderma 372
— reesei 64, 311
— viride 64, 139
Trichospora fermentans 64
Trifolium subterraneum 362
Vaginocolidae 253
Vaucheria dichotomy 387
Vitis vinifera 109
Vorticetta 253
Xanthamonas 220—222, 233
Yarrowia 309, 310
Zea mays 369
Zoogloea 250, 251
Zymomonas mobilis 69
Оглавление
Предисловие редактора перевода ..................................... 5
Предисловие......................................................... 7
Глава 1. Что такое биотехнология? И, Хиггинс........................ 8
1.1. Введение.................................................... 8
1.2. Исторические перспективы...............................11
1.3. Развитие биотехнологической промышленности после второй ми-
ровой войны......................................................14
1.4. Перспективы развития биотехнологии..........................17
1.4.1. Медицина...............................................17
1.4.2. Энергетика.............................................20
1.4.3. Пищевые продукты и напитки.............................23
1.4.4. Химические соединения..................................23
1.4.5. Материалы..............................................25
1.4.6. Окружающая среда.......................................26
1.4.7. Сельское хозяйство.....................................27
1.5. Экономические и коммерческие аспекты биотехнологии ... 28
1.5.1. Логика развития биотехнологии..........................29
1.5.2. Защита авторских прав в биотехнологии..................31
1.5.3. Правила техники безопасности в биотехнологической про-
мышленности и контроль продукции ........................... 32
1.6. Заключительные замечания ...................................33
Литература....................................................33
Глава 2. Энергия и биотехнология. Д. Холл, Дж. Кумбс, И. Хиггинс 35
2.1. Введение....................................................35
2.2. Получение биомассы: технология, основанная на солнечной энер-
гии .............................................................37
2.2.1. Ресурсы................................................39
2.3. Фотосинтез..................................................42
2.3.1. Эффективность фотосинтеза..............................47
2.3.2. Интенсификация фотосинтеза методами биотехнологии 48
2.4. Сельское и лесное хозяйство.................................51
2.4.1. Соотношение видов энергии..............................51
2.4.2. Древесина как сырье для производства биотоплива . . 53
2.4.3. Водоросли и водные растения............................54
2.4.4. Масличные растения.....................................57
2.5. Переработка и различные виды топлива........................61
2.5.1, Производство исходного сырья...........................61
Оглавление
475
2.5.2. Ферментация......................................65
2.5.3. Этиловый спирт...................................66
2.5.4. Энергобаланс.....................................70
2.5.5. Получение метана в анаэробных условиях .... 73
2.6. Бесклеточные системы........................................79
2.6.1. Комбинированные системы, образующие водород ... 80
2.6.2. Электроэнергия...................................82
2.7. Биотопливные элементы и другие биоэлектрохимические устрой-
ства ............................................................83
Литература...................................................86
Глава 3. Пищевые продукты, напитки и биотехнология. Г. Бич, М. Мел-
вин, Дж. Таггарт..........................................................91
3.1. Введение.........................................-
3.1.1. Микроорганизмы и пищевые продукты.....................
3.1.2. Две разновидности биотехнологии.......................
3.1.3. Границы применения биотехнологии в пищевой промыш-
ленности ....................................................
3.2. Молочные продукты . ...................................
3.2.1. Общие сведения........................................
3.2.2. Сыр...................................................
3.2.3. Йогурт................................................
3.2.4. Масло . . . ...................................
3.2.5. Сброженная пахта......................................
3.2.6. Сметана...............................................
3.2.7. Новые продукты........................................
3.3. Хлебопродукты..............................................
3.3.1. Хлеб и другие продукты................................
3.3.2. Продукты гидролиза крахмала...........................
3.4. Бродильные производства....................................
3.4.1. Производство алкогольных напитков ......
3.4.2. Пиво..................................................
3.4.3. Вино..................................................
3.4.4. Спирт.................................................
3.4.5. Сидр..................................................
3.4.6. Уксус.................................................
3.5. Белковые продукты..........................................
3.5.1. Традиционные белковые продукты, получаемые путем фер-
ментации ...................................................
3.5.2. Белок одноклеточных организмов........................
3.5.3. Грибной белок (микопротеин)...........................
3.6. Пищевые добавки и ингредиенты..............................
3.6.1. Подкислители..........................................
3.6.2. Аминокислоты..........................................
3.6.3. Витамины и пигменты...................................
3.6.4. Усилители вкуса.......................................
3.6.5. Жиры и масла . ...................................
3.6.6. Растительные клеи и загустители.......................
3.7. Фрукты и овощи.............................................
3.7.1. Консервированные овощи................................
3.7.2. Продукты из сои.......................................
3.7.3. Применение ферментов при выработке фруктовых соков
3.8. Заключение.................................................
3.8.1. Ближайшие перспективы ................................
3.8.2. Отдаленные перспективы................................
Литература...................................................
91
91
91
93
93
93
95
101
101
101
102
102
102
102
103
104
104
106
109
112
115
115
116
119
122
122
122
123
123
123
123
124
124
125
127
128
128
128
129
476
Оглавление
Глава 4. Химия и технология. Д. Бест................... 132
4.1. Введение....................................................132
4.2. Развитие современной химической биотехнологии .... 133-
4.2.1. Бродильное производство растворителей.................133
4.2.2. Производство органических кислот......................136
4.2.3. Другие органические кислоты........................141
4.2.4. Аминокислоты..........................................147
4.2.5. Антибиотики и стероиды................................155
4.2.6. Коммерческие аспекты применения ферментов . . 164?
4.3. Получение химических веществ из биомассы...........172
4.3.1. Биотехнология на основе растительных клеток . . . 172
4.3.2. Химические вещества, получаемые из биомассы . . 175
4.4. Перспективы развития........................................177
4.4.1. Тип реакции...........................................178
4.4.2. Конфигурация реактора.................................185
4.4.3. Будущий вклад биотехнологии в химическую промышлен-
ность .......................................................187
Литература.........................................188
Глава 5. Материалы и биотехнология. К. Брайерли, Д. Келли, К. Сил,
Д. Бест....................................................190
5.1. Введение ...................................................190
5.2. Микробное выщелачивание...........................191
5.2.1. Выщелачивающие микроорганизмы...............192
5.2.2. Промышленное применение биоэкстрактивной металлургии 194
5.2.3. Возможности применения бактериального выщелачивания 198
5.3. Превращение, накопление и иммобилизация металлов микроор-
ганизмами ....................................................205
5.3.1. Перевод в летучую форму............................207
5.3.2. Внеклеточное осаждение.............................206
5.3.3. Внеклеточное комплексообразование..................209
5.3.4. Внутри- и внеклеточное накопление металлов микроорга-
низмами .....................................................210
5.4. Биополимеры.................................................216
5.4.1. Введение..............................................216
5.4.2. Полисахариды..........................................217
5.4.3. Образование полисахаридов при брожении .... 219
5.4.4. Микробные полисахариды: свойства, применение и коммер-
ческая ценность .............................................222
5.4.5. Биосинтез полисахаридов...............................229
5.4.6. Подходы к усовершенствованию производства микробных
полисахаридов .............................................. 232
5.4.7. Поли-р-гидроксибутират................................234
5.4.8. Другие полимеры, образуемые микроорганизмами . 236
5.5. Биоповреждение материалов...................................236
5.5.1. Введение..............................................236
5.5.2. Определение биоповреждений............................237
5.5.3. Классификация процессов биоповреждения .... 238
5.5.4. Материалы, подверженные биоповреждениям .... 239
Литература...................................................244
Глава 6. Окружающая среда и биотехнология. Д. Бест, Дж. Джонс,
Д. Стаффорд........................................................246
6.1. Введение....................................................246
6.2. Переработка отходов.........................................247
6.2.1. Аэробная переработка отходов..........................249
Оглавление
477
6.2.2. Анаэробное разложение.................................258
6.3. Биологический контроль за системами микробиологической пере-
работки отходов.................................................264
6.4. Контроль за патогенностью...................................267
6.5. Извлечение полезных веществ.................................270
6.5.1. Вода..................................................270
6.5.2. Удобрения.............................................270
6.5.3. Корма для животных..................................271
6.6. Биологическая переработка промышленных отходов . . . 274
6.6.1. Отходы молочной промышленности; сыворотка . . . 277
6.6.2. Отходы целлюлозно-бумажной промышленности . . . 278
6.6.3. Отходы от производства красителей.....................280
6.6.4. Биологическая очистка газов...........................282
6.7. Биодеградация ксенобиотиков в окружающей среде .... 284
6.7.1. Участие микробных сообществ в биодеградации ксенобио-
тиков ......................................................284
6.7.2. Хлорпроизводные углеводородов....................287
6.7.3. Другие замещенные простые ароматические соединения 288
6.7.4. Полиароматические углеводороды...................289
6.7.5. Биодеградация нефтяных загрязнений ...............290
6.7.6. Пестициды........................................291
6.7.7. Биодеградация поверхностно-активных веществ . . . 292
Литература...................................................295
Глава 7. Генетика и биотехнология. К. Харди, С. Оливер .... 296
7.1. Введение....................................................296
7.2. Традиционные способы увеличения продуктивности штаммов 297
7.2.1. Мутагенез и отбор.....................................297
7.2.2. Гибридизация путем скрещивания........................300
7.3. Генетические перестройки in vivo............................303
7.3.1. Плазмиды..............................................303
7.3.2. Слияние протопластов................................. 308
7.3.3. Слияние клеток животных...............................312
7.4. Опыты по генетической инженерии in vitro...................315
7.4.1. Технология ...........................................315
7.4.2. Экспрессия клонированных генов............318
7.4.3. Прикладные аспекты генетической инженерии .... 320
7.5. Итоги и перспективы .......................................323
Литература...................................................324
Глава 8. Медицина и биотехнология.Дж. Пикап........................325
8.1. Введение................................................. 325
8.2. Немодифицированные и мутантные клетки и синтезируемые ими
соединения.....................................................326
8.2.1. Антибиотики......................................... 326
8.2.2. Культуры других клеток............................. 328;
8.2.3. Различные варианты биоконверсии......................328
8.3. Модифицированные клетки и образуемые ими вещества . . 329*
8.3.1. Моноклональные антитела..............................329
8.3.2. Технология рекомбинантных ДНК.............334
8.4, Применение методов молекулярной генетики и технологии реком-
бинантных ДНК в медицинской диагностике и патологии . . 342
8.4.1. Пренатальная диагностика наследственных болезней . . 3421
8.4.2. Роль генетических факторов в патологии.............. 344
8.5. Перспективы развития.......................................346
Литература................................................ 347
478
'Оглавление
Глава 9. Сельское хозяйство и биотехнология. Ф. Скиннер , . . . 350
9.1. Введение....................................................350
9.2. Метод питательной пленки....................................351
9.2.1. Основы метода.........................................351
9.2.2. Основные требования к оборудованию....................353
9.2.3. Фитотоксичность.......................................354
9.2.4. Устройство емкостей, используемых в методе питательной
пленки.......................................................355
9.2.5. Опора для молодых растений............................355
9.2.6. Болезни ..............................................355
9.2.7. Перспективы...........................................356
9.3. Бобовые культуры и фиксация .азота при симбиозе .... 356
9.3.1. Из истории вопроса....................................356
9.3.2. Инокуляция бактериями Rhizobium.......................358
9.3.3. Инокуляция семян......................................358
9.3.4. Современные инокуляты на твердых носителях . . . 359
9.3.5. Методы инокуляции.....................................362
9.3.6. Улучшение симбиоза между бобовыми и Rhizobium . . 363
9.4. Везикулярно-арбускулярная микориза.........................364
9.4.1. Роль везикулярно-арбускулярной микоризы .... 364
9.4.2. Инокуляция эндофитом..................................367
9.4.3. Методы инокуляции.....................................370
9.5. Биологический контроль.....................................371
9.5.1. Основы биологического контроля........................371
9.5.2. Примеры биологического контроля.......................372
9.5.3. Корончатые галлы и борьба с ними......................373
9.6. Аэробная переработка отходов в сельском хозяйстве . . . 376
9.6.1. Проблема хранения и переработки отходов .... 376
9.6.2. Системы переработки отходов в аэробных условиях . . 377
9.7. Переработка отходов сельского хозяйства в анаэробных услови-
ях (см. также гл. 6).............................................378
9.7.1. Главные особенности процесса..........................378
9.7.2. Переработка отходов сельского хозяйства...............379
9.7.3. Микробиологические основы процесса....................381
9.7.4. Современные разработки................................381
9.8. Использование протопластов в селекции растений .... 382
9.8.1. Вегетативное размножение..............................382
9.8.2. Регенерация растений из протопластов..................384
9.8.3. Методы слияния протопластов...........................386
Литература...................................................390
Глава 10. Химическая технология и биотехнология. Г. Хамер . . 395
10.1. Введение....................................................395
10.2. Микробиологические факторы, влияющие на производительность
и экономичность процесса........................................401
10.2.1. Эффективность роста: коэффициент выхода биомассы 403
10.2.2. Кинетика роста и образования продукта...............405
10.2.3. Сродство к субстрату и рост микроорганизмов . . . 410
10.2.4. Стабильность культуры...............................414
10.3. Технологические факторы, влияющие на производительность и
экономичность процесса..........................................416
10.3.1. Классификация биореакторов и их производительность 418
10.3.2. Анализ размерностей и масштабирование .... 430
10.3.3. Перенос кислорода...................................437
.10.3.4. Теплопередача и охлаждение биореактора .... 447
Оглавление 479'
10.3.5. Конверсия и рециркуляция.......................... 451
10.3.6. Характеристики микробных и мицелиальных суспензий 456'
10.4. Вспомогательное оборудование, использующееся в- биотехноло-
гических процессах............................................ 460'
10.4.1. Стерилизация среды и воздуха.....................461
10.4.2. Выделение продукта и завершающие, операции . . 463;
10.5. Перспективы развития промышленных биотехнологических про-
цессов ......................................................464
Литература............................................... 465'
Предметный указатель.............................................466
Указатель латинских названий.......................................472
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее оформле-
нии, качестве перевода и другие просим присы-
лать по адресу;
129820, Москва,
1-й Рижский пер., д. 2,
издательство «Мир»
Учебное издание
Г. Бич, Д. Бест., К. Брайерли и др.
БИОТЕХНОЛОГИЯ. ПРИНЦИПЫ И ПРИМЕНЕНИЕ
Под редакцией И. Хиггинса, Д. Беста и Дж. Джонса
Заведующий редакцией чл-корр. АН СССР Т. М. Турлаев
Зам. зав. редакцией М. Д. Гроздова
Ст. научный редактор Н. Н. Шафрановская
Мл. научный редактор 3. В. Соллертинская
Художник В. Е. Карпов
Художественный редактор А. Я. Мусин
Технический редактор Е. В. Алехина
Корректор Л. Д. Панова
ИБ № 6351
Сдано в набор 14.10.87. Подписано к печати 8.04.88.
Формат 60Х901/1в. Бумага книжно-журнальная. Печать вы-
сокая. Гарнитура литературная. Объем 15,00 бум. л. Усл.
печ. л. 30,00. Усл. кр.-отт. 30,00. Уч.-изд. л. 30,76. Изд.
№ 4/5325. Тираж 10,000 экз. Зак. 1344. Цена 2 р. 80 к.
ИЗДАТЕЛЬСТВО «МИР».
129820, ГСП, Москва, И-110, 1-й Рижский пер., 2.
Московская типография № 11 Союзполиграфпрома при
Государственном комитете СССР по делам издательств,
полиграфии и книжной торговли. 113105, Москва, Нага-
тинская ул., д. 1.