/



















































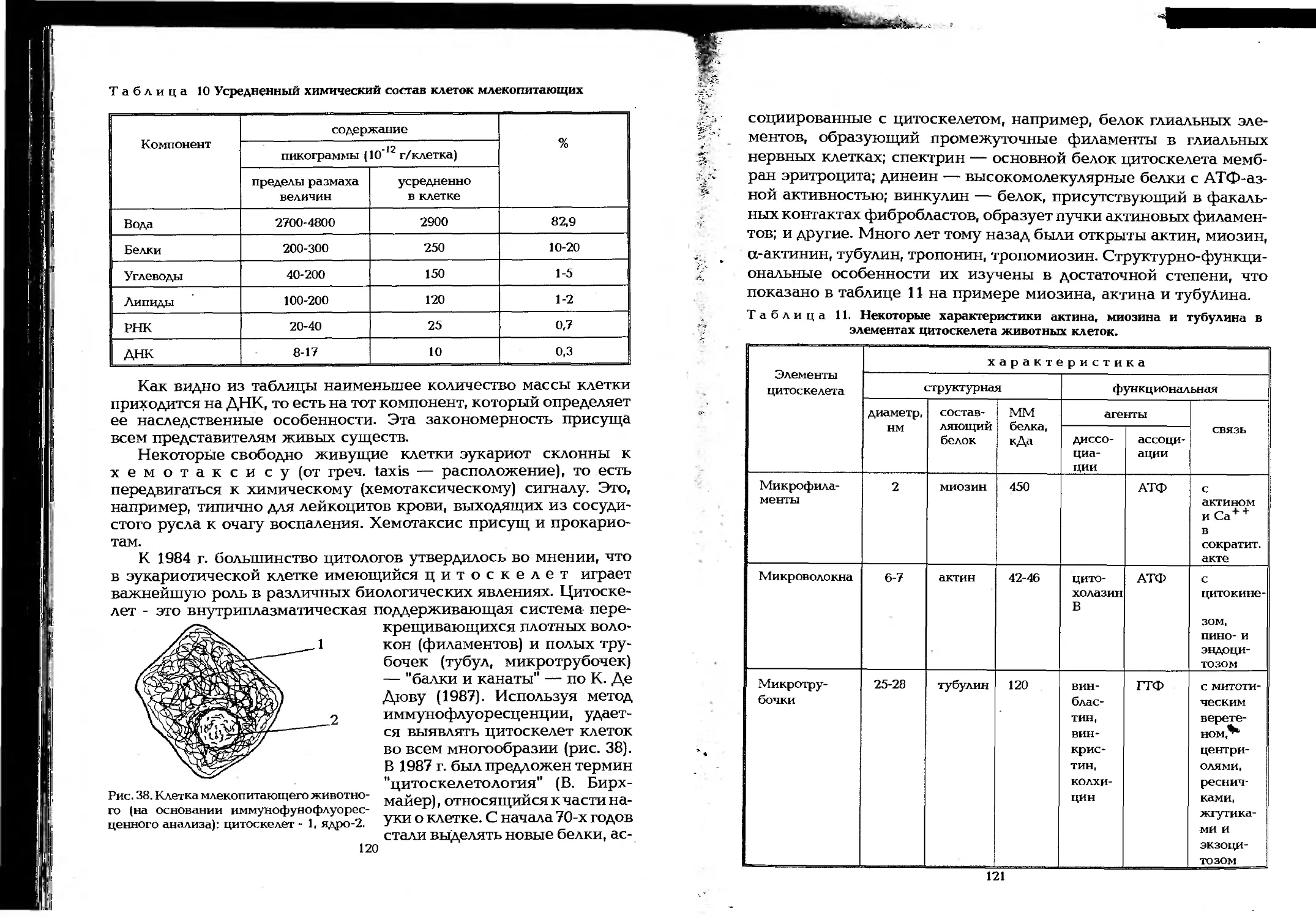


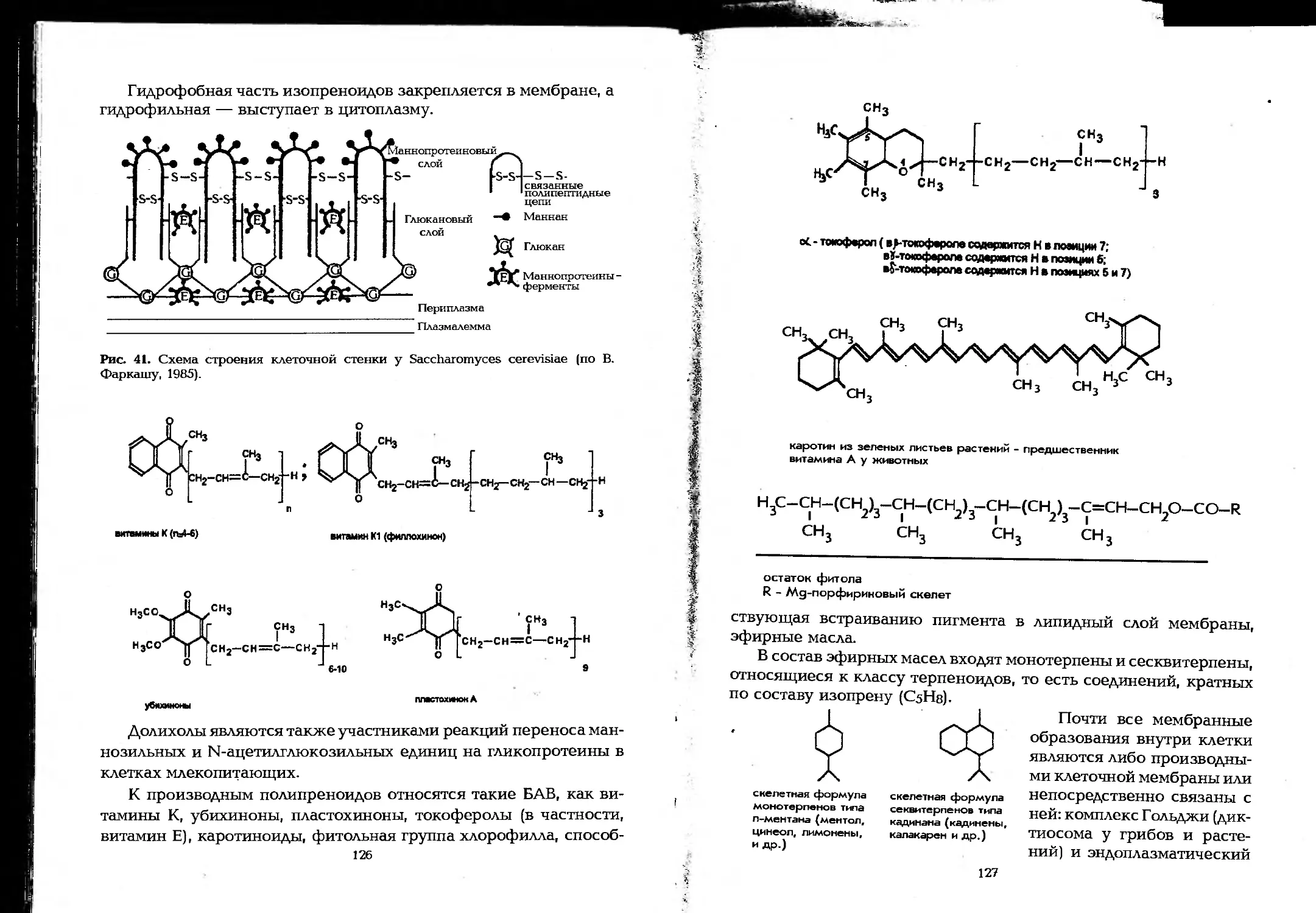




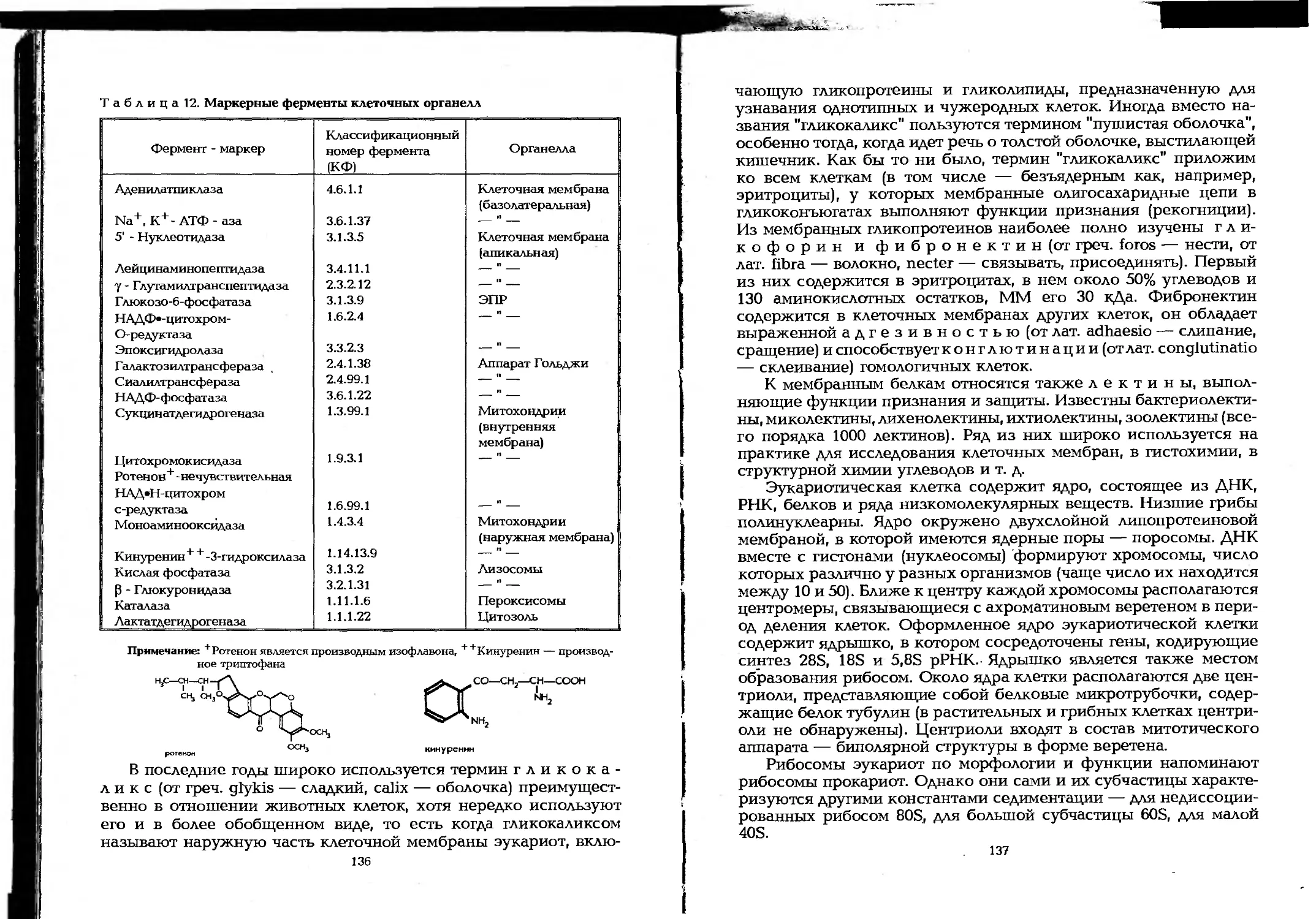

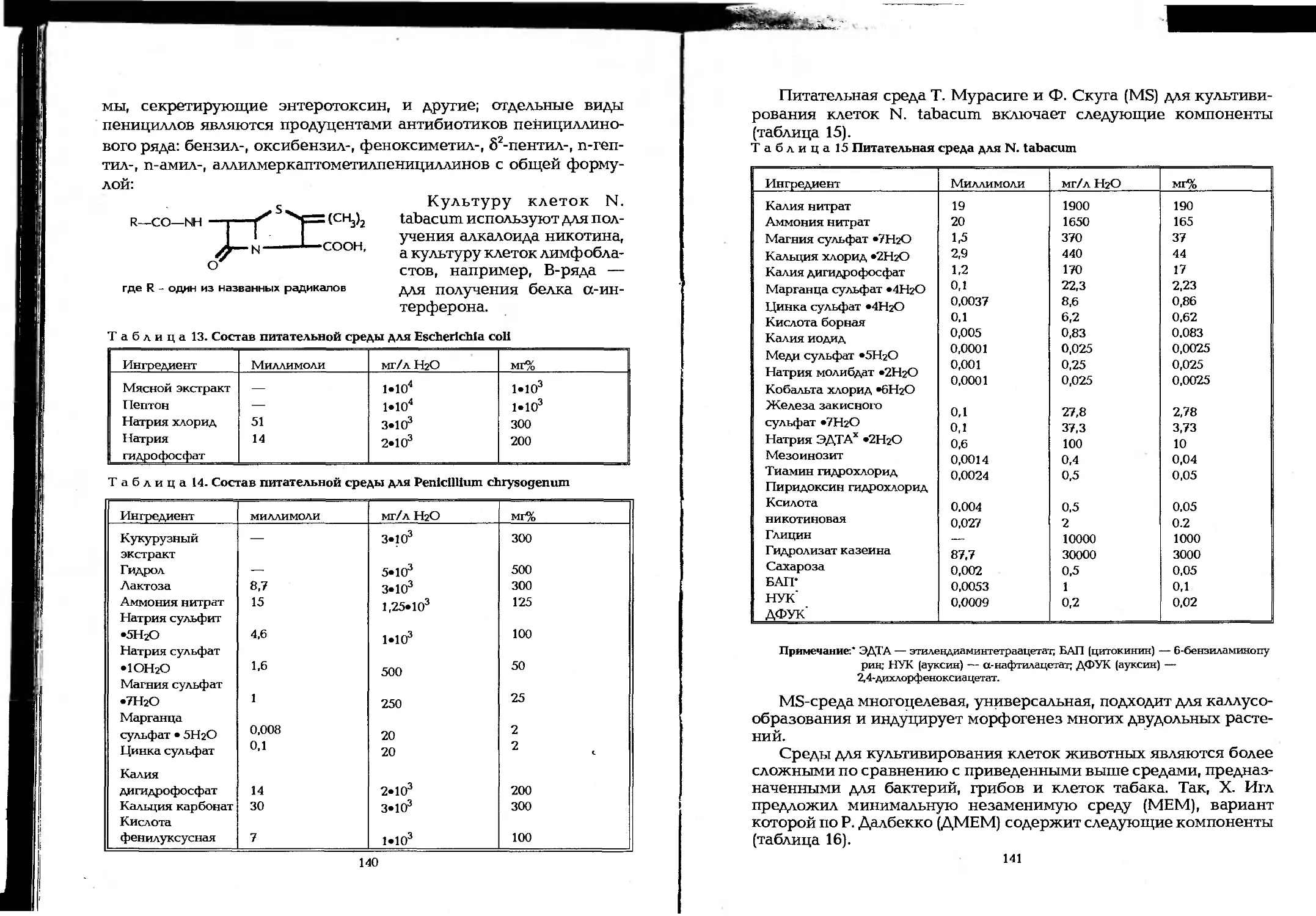
































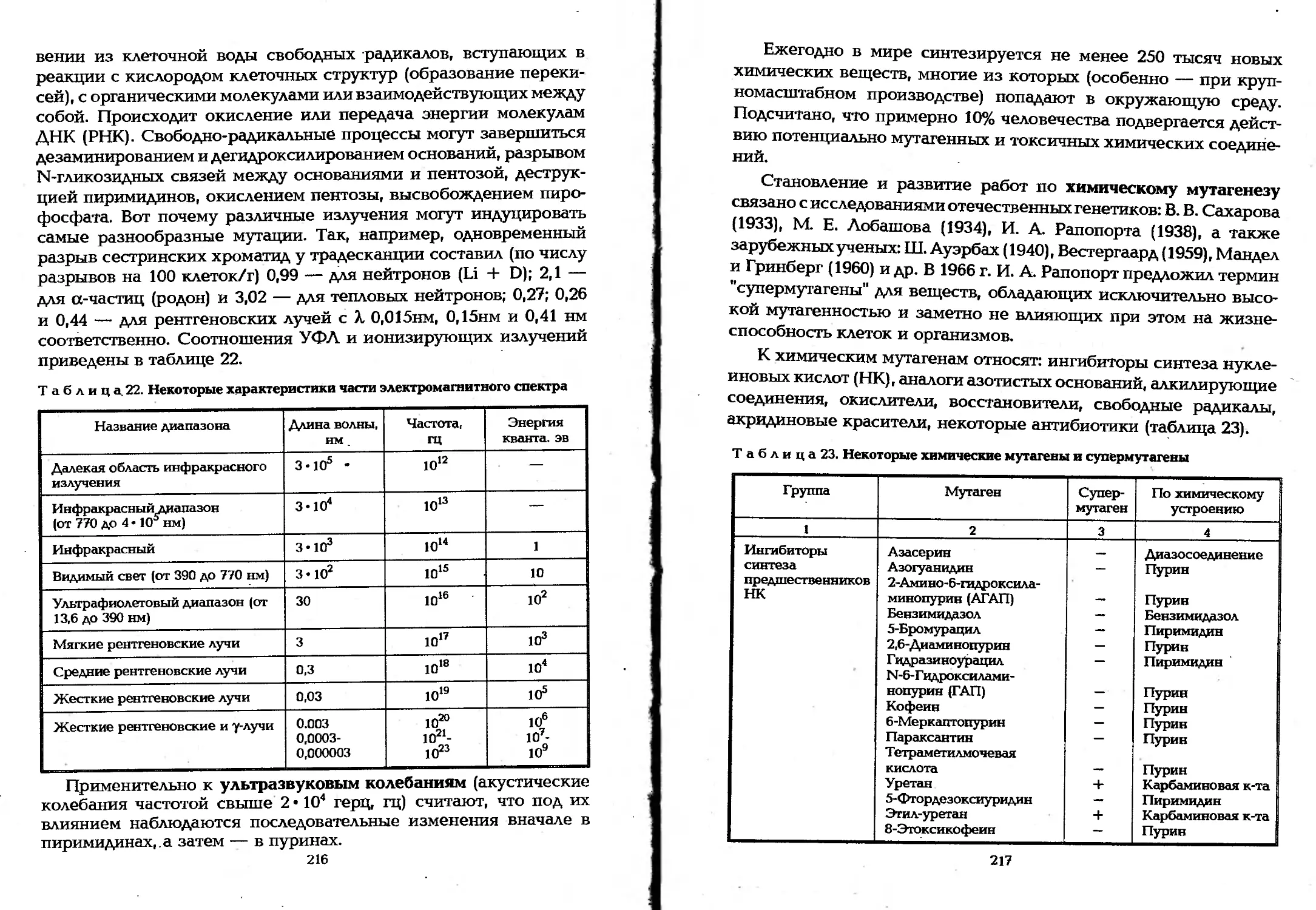



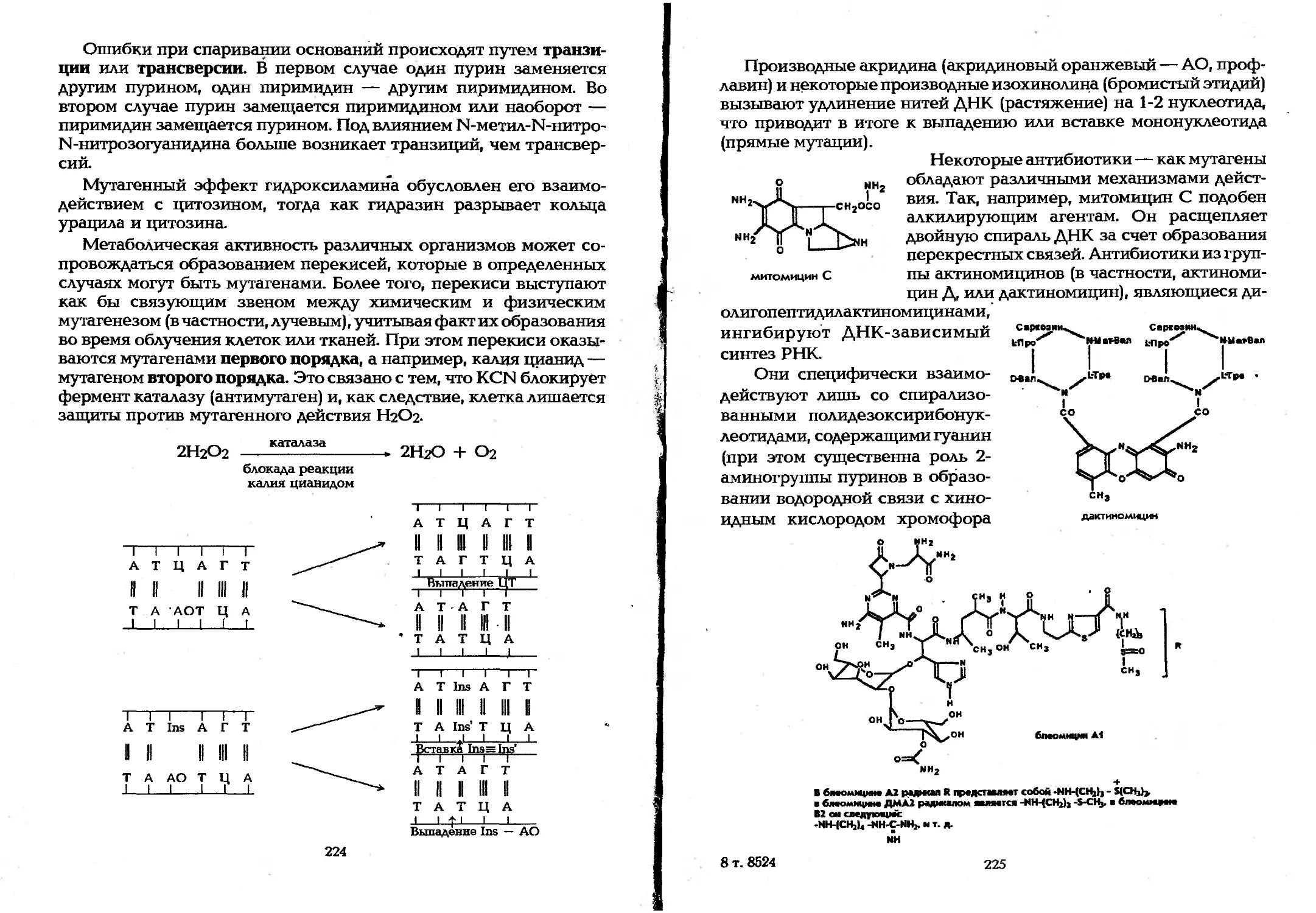



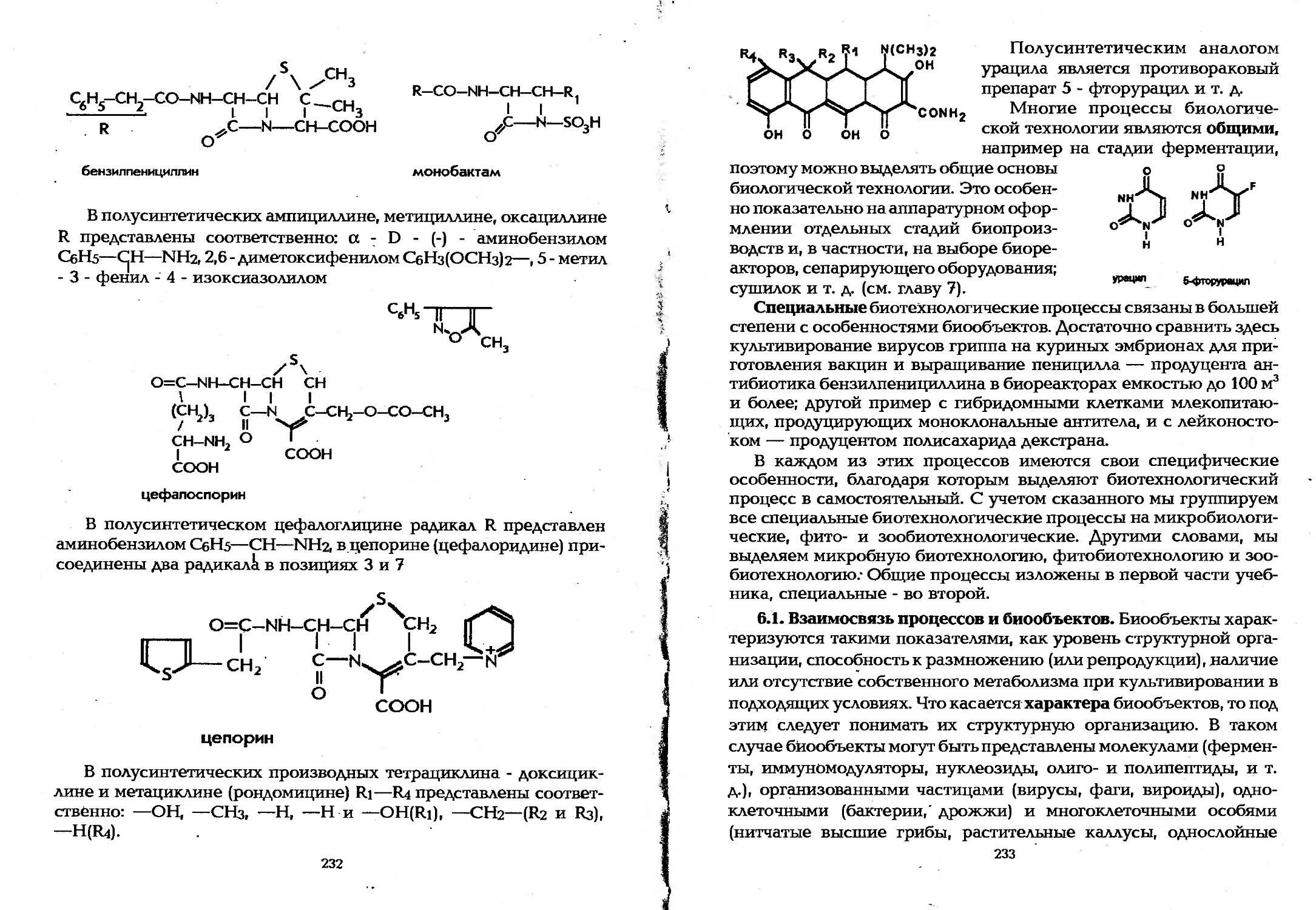





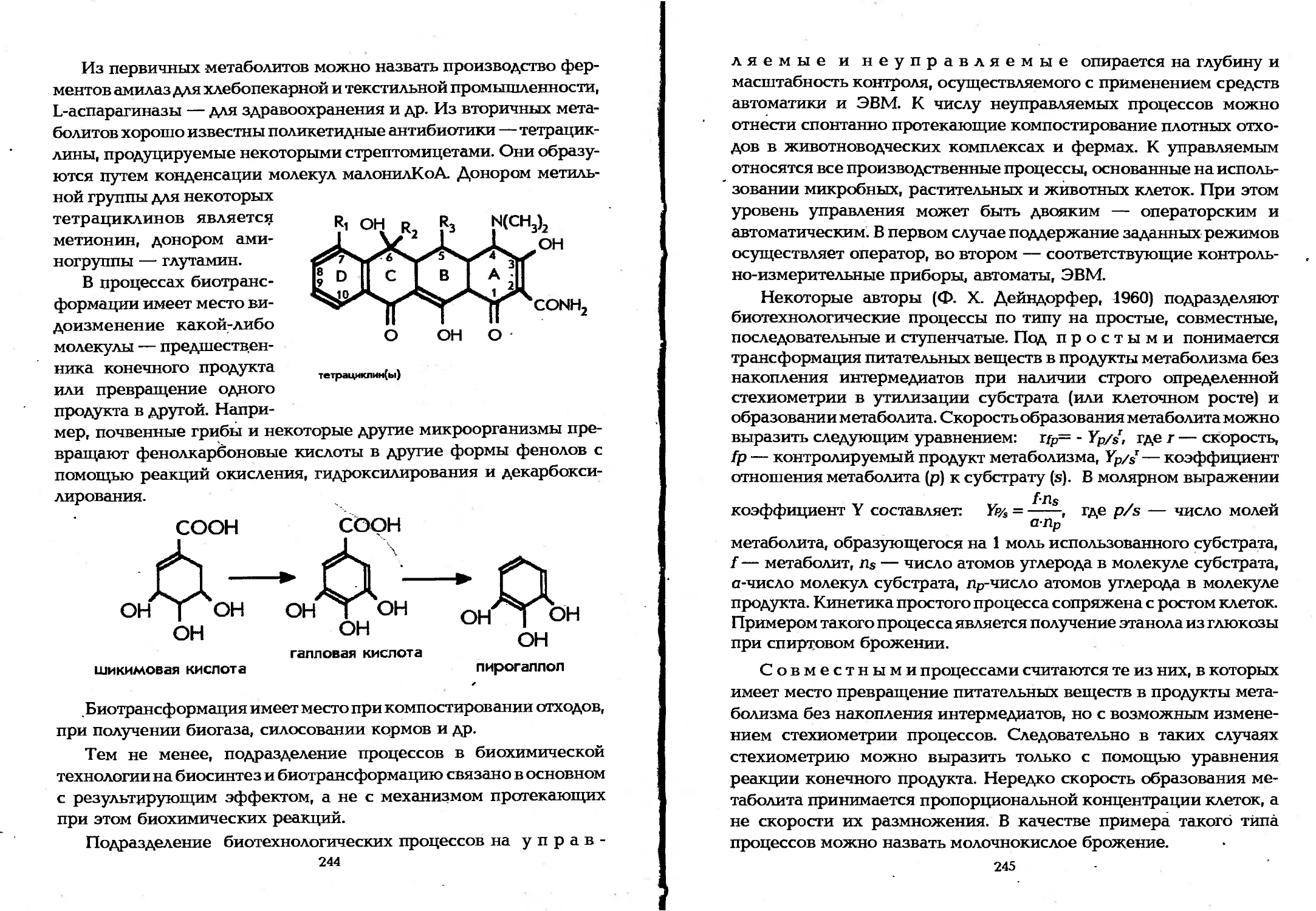

































































































































































Author: Елинов Н.П.
Tags: биологические науки в целом общетехнические дисциплины микробиология биотехнологии
ISBN: 5-02-026027-4
Year: 1995




Text
BBieaertb
F.51
УДК 57«6 +3£^6
Рецензенты:
зав. кафедрой пищевой биотехноло-
гии Санкт-Петербургской государствен-
ной академии холода и пищевых техно-
логий, член-корресповдент академии хо-
лода, профессор Василинец И. М.
зав. кафедрой технологии микробио-
логического синтеза Санкт-Петербург-
ского государственного технологического
института, академик МАНЭБ Яковлев
В. И.
Блинов Н. П.
Е51 Основы биотехнологии. Д ля студентов институтов;
аспирантов и практических работников. Издательская
фирма "Наука” СПБ 1995 г.с., 600 стр. 166 ил.
ISBN 5-02-026027-4
Книга включает общую и специальную биотехнологии. В ней рассмотрены
объекты и методы, фундаментальные и прикладные аспекты микро-, фито- и
зообиотехнологии, приведены основные биотехнологические процессы получения
различных веществ с помощью микробных, растительных и животных клеток, дана
характеристика оборудования, используемого в конкретной биотехнологии, указа-
ны пути утилизации и обезвреживания плотных и жидких отходов в соответству-
ющих производствах.
ББК 30.16
ISBN 5-02-026027-4
© Издательская фирма "Наука" СПб 1995
Учителю и организатору первого
в стране инженерно-микробиологи-
ческого факультета при СПХФИ,
з. д. н. РФ, лауреату Государствен-
ной премии проф. Павлу Николаевичу
Кашкину посвящаю свой труд.
Автор
W Щ A
ISI
ОГЛАВЛЕНИЕ
Предисловие.................................................7
Введение....................................................9
Часть I.
БИОТЕХНОЛОГИЯ — КАК НАУЧНАЯ ДИСЦИПЛИНА . . 11
ГЛАВА 1. Предмет, история развития, цели и задачи биотехнологии ... 11
ГЛАВА 2. Объекты и методы биотехнологии.....................22
2.1. Вирусы......................................25
2.1.1. Вироиды...................................26
2.2. Бактерии....................................27
2.3. Грибы.......................................32
2.4. Растения....................................36
2.4.1. Водоросли.................................36
2.4.2. Клетки высших растений....................37
2.5. Клетки животных •...........................38
Часть II.
РОЛЬ ФУНДАМЕНТАЛЬНЫХ ИССЛЕДОВАНИЙ
В РАЗВИТИИ БИОТЕХНОЛОГИИ.....................................43
ГЛАВА 3. Фундаментальные исследования в области энзимологии ... 45
ГЛАВА 4. Фундаментальные исследования в области структурно-
функциональной организации вирусов, клеток и тканей ... 79
4.1. Акариоты....................................79
4.2. Клетки прокариот............................86
4.3. Клетки эукариот............................. 106
4.4. Некоторые функциональные особенности
клеток и клеточных систем........................ 138
ГЛАВА 5. Фундаментальные исследования в области генетики
и молекулярной биологии вирусов, клеток и клеточных систем 155
5.1. Природа и передача генетической информации.. 157
5.2. Клонирование генов методами генетической
инженерии; рДНК-биотехнология..................... 177
5.3. Изменчивость организмов и ее значение
в биотехнологии...................................212
Часть III.
ПРОЦЕССЫ И АППАРАТЫ В БИОТЕХНОЛОГИИ .... 229
ГЛАВА 6. Процессы в биотехнологии...........................229
6.1. Взаимосвязь процессов и биообъектов.........233
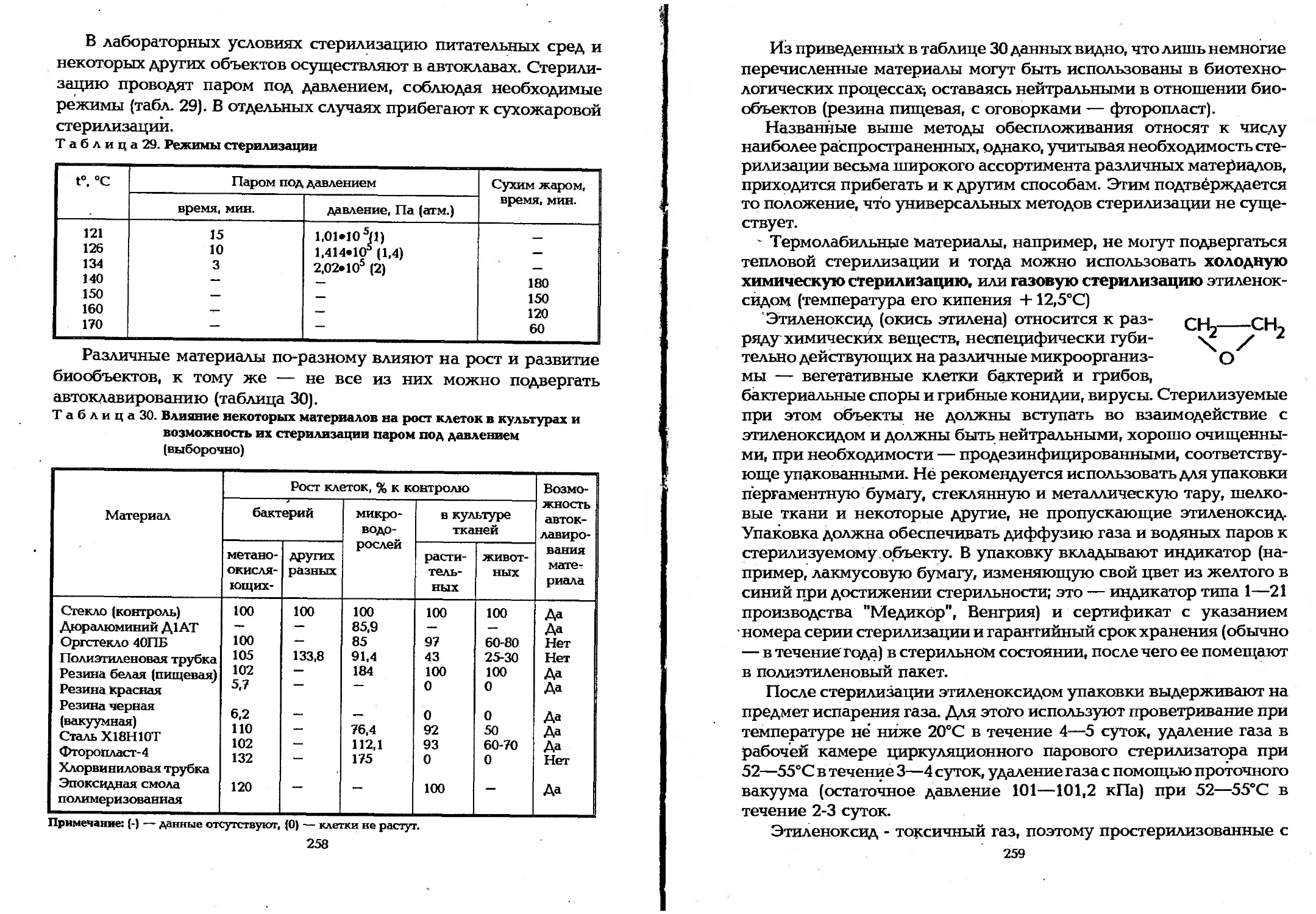
6.2. Значение асептики в биотехнологических процессах . . 246
6.3. Борьба с микробами-контаминантами
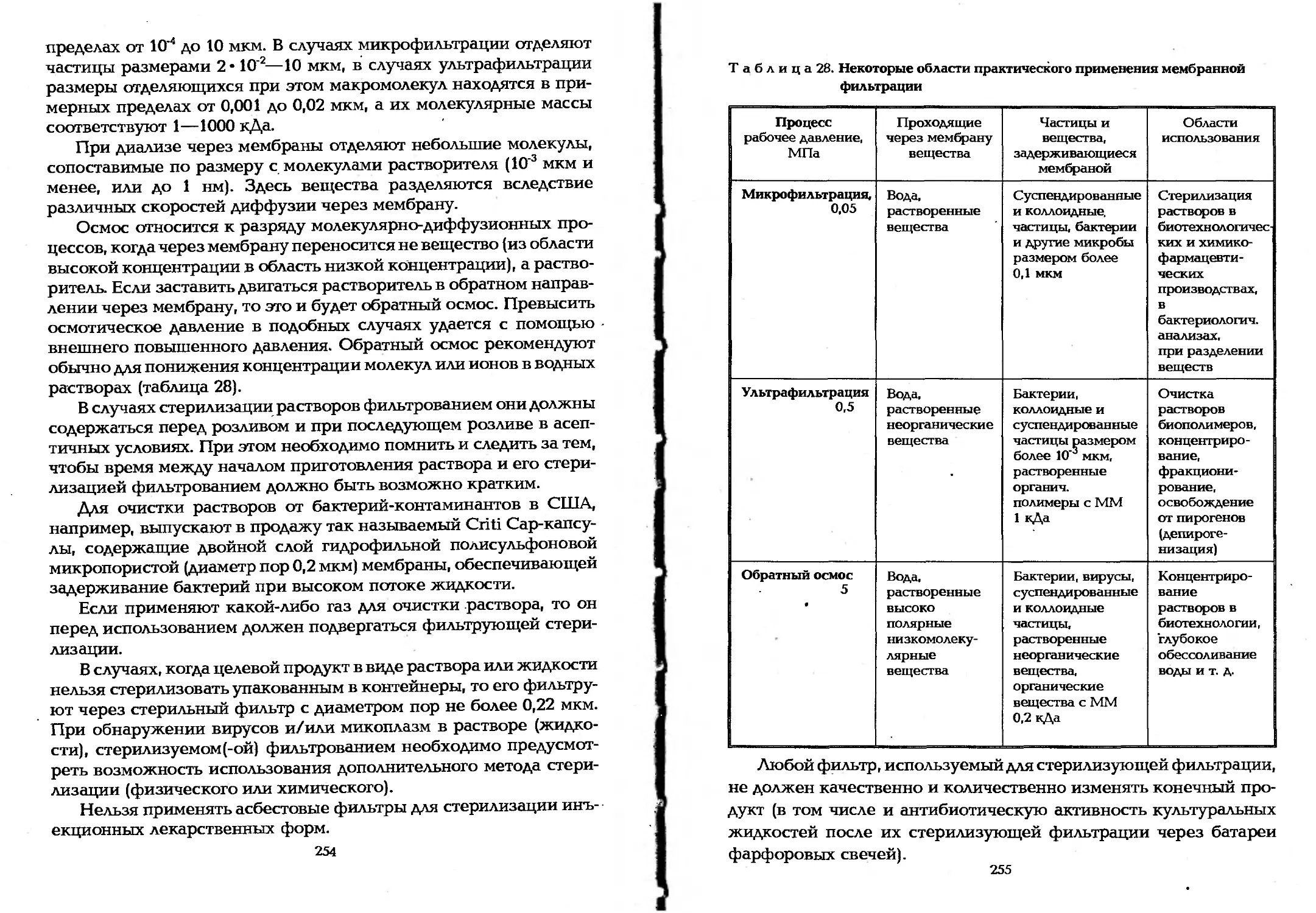
в биотехнологических производствах..........". . . 253
6.4. Биотехнологические процессы в связи с массообменом . 261
6.5. Биотехнологические процессы в связи
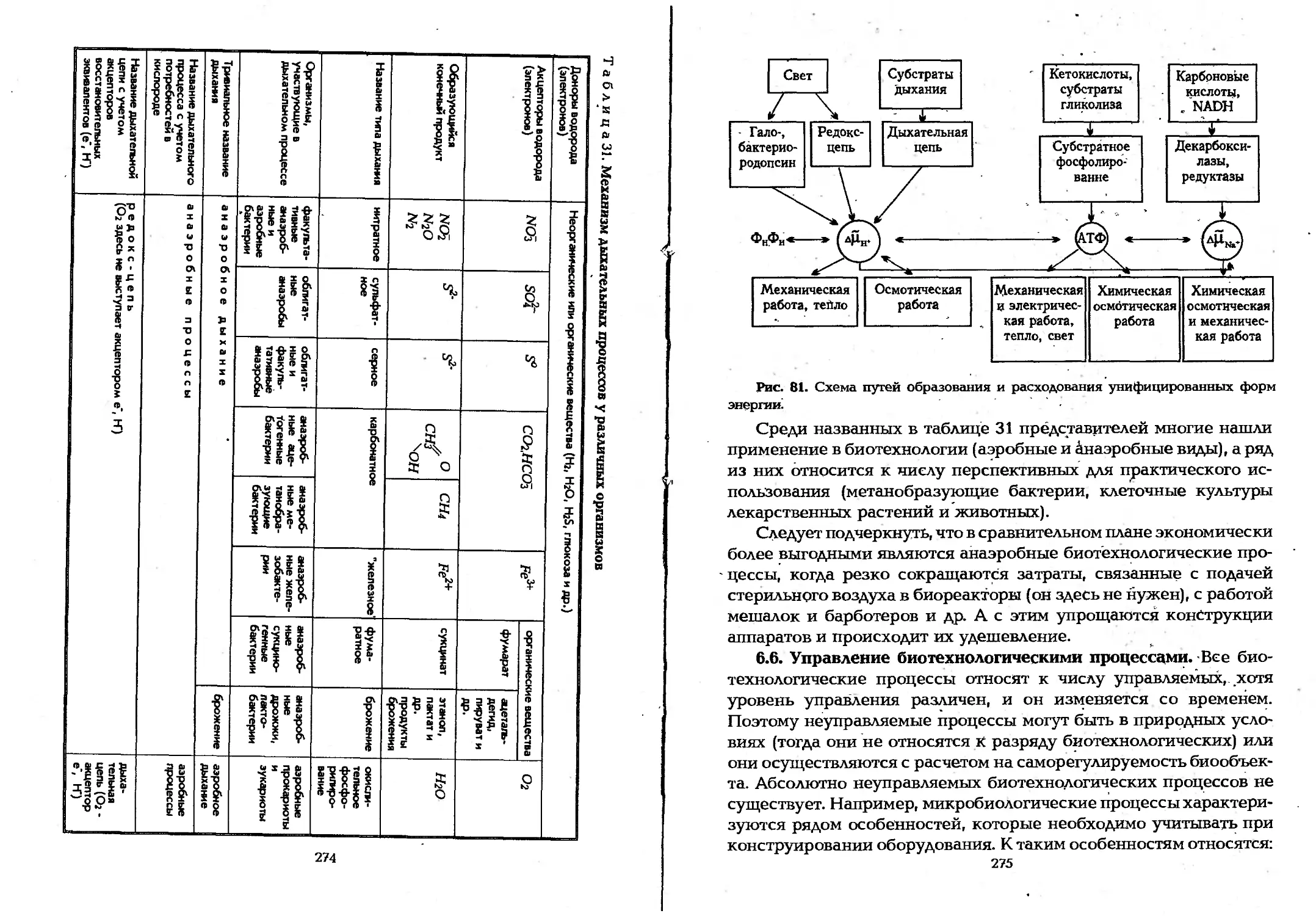
с особенностями метаболизма клеток................269
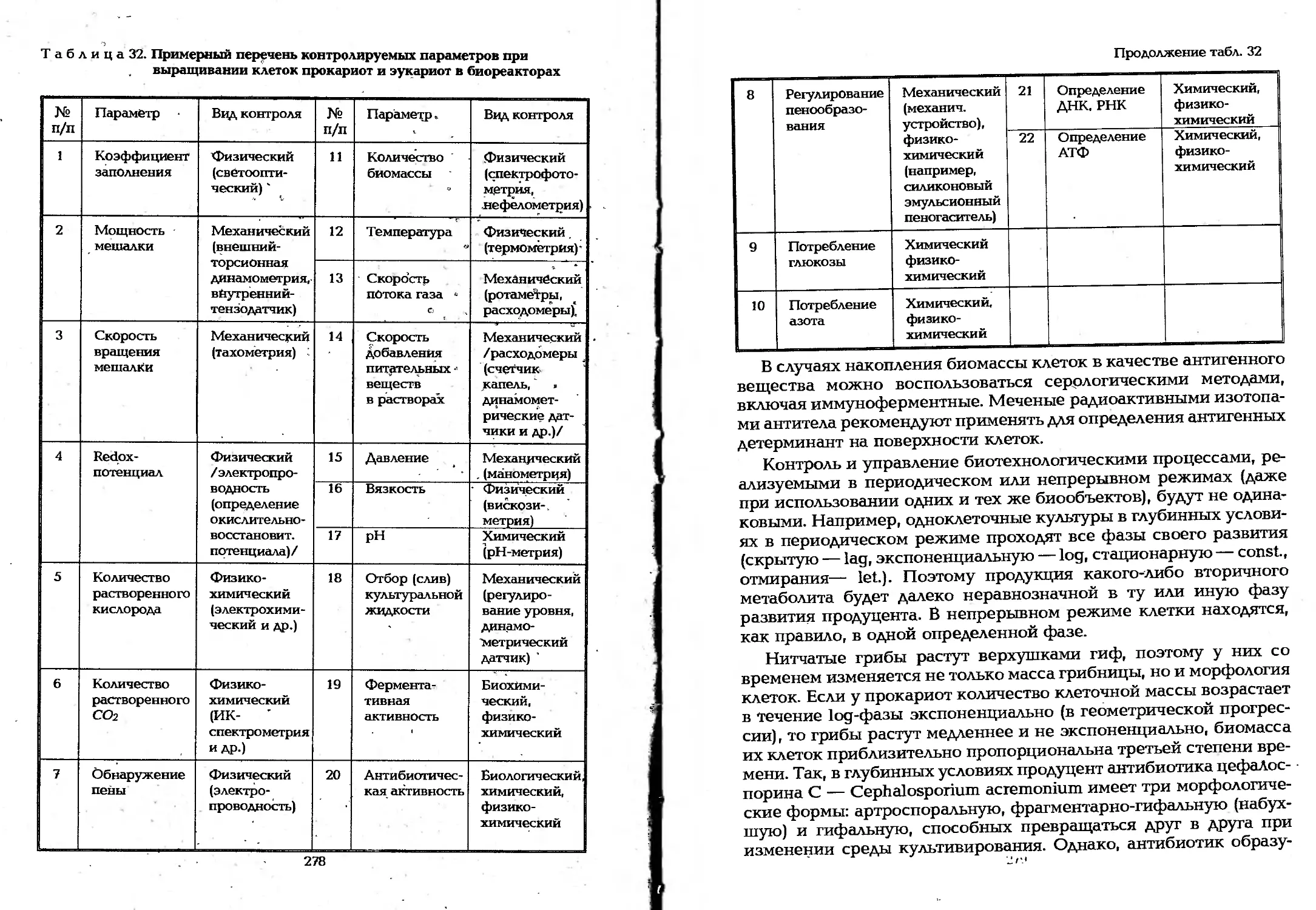
6.6. Управление биотехнологическими процессами...275
6.7. Системы GLP и GMP в связи с качеством
биотехнологических продуктов................ . . 283
ГЛАВА 7. Техническая вооруженность биотехнологических
производств..........................................288
7.1. Аппаратурное оснащение микробиологических
производств......................................297
7.2. Некоторые особенности культивирования биообъектов . 306
5
7.3. Тепловые процессы в ферментаторах . . . • 328
7.4. Аппаратурное оформление процессов выделения
и очистки некоторых продуктов микробного синтеза . . • 331
7.5. Аппаратурное оснащение фитобиотехнологических
производств....................................... 341
7.6. Аппаратурное оснащение зообиотехнологических
производств....................................... 343
ГЛАВА 8. Отходы биотехнологических производств,
их обезвреживание и утилизация............................ .... 349
8.1. Обезвреживание отходов биотехнологических
производств........................................ 353
8.2. Утилизация отходов биотехнологических производств . . 366
Часть IV.
СПЕЦИАЛЬНЫЕ БИОТЕХНОЛОГИИ......................................373
ГЛАВА 9. Микробиотехнология............................ . . 374
9.1. Принципы культивирования микроорганизмов . ... 378
9.2. Выделение конечных продуктов ферментации ... 386
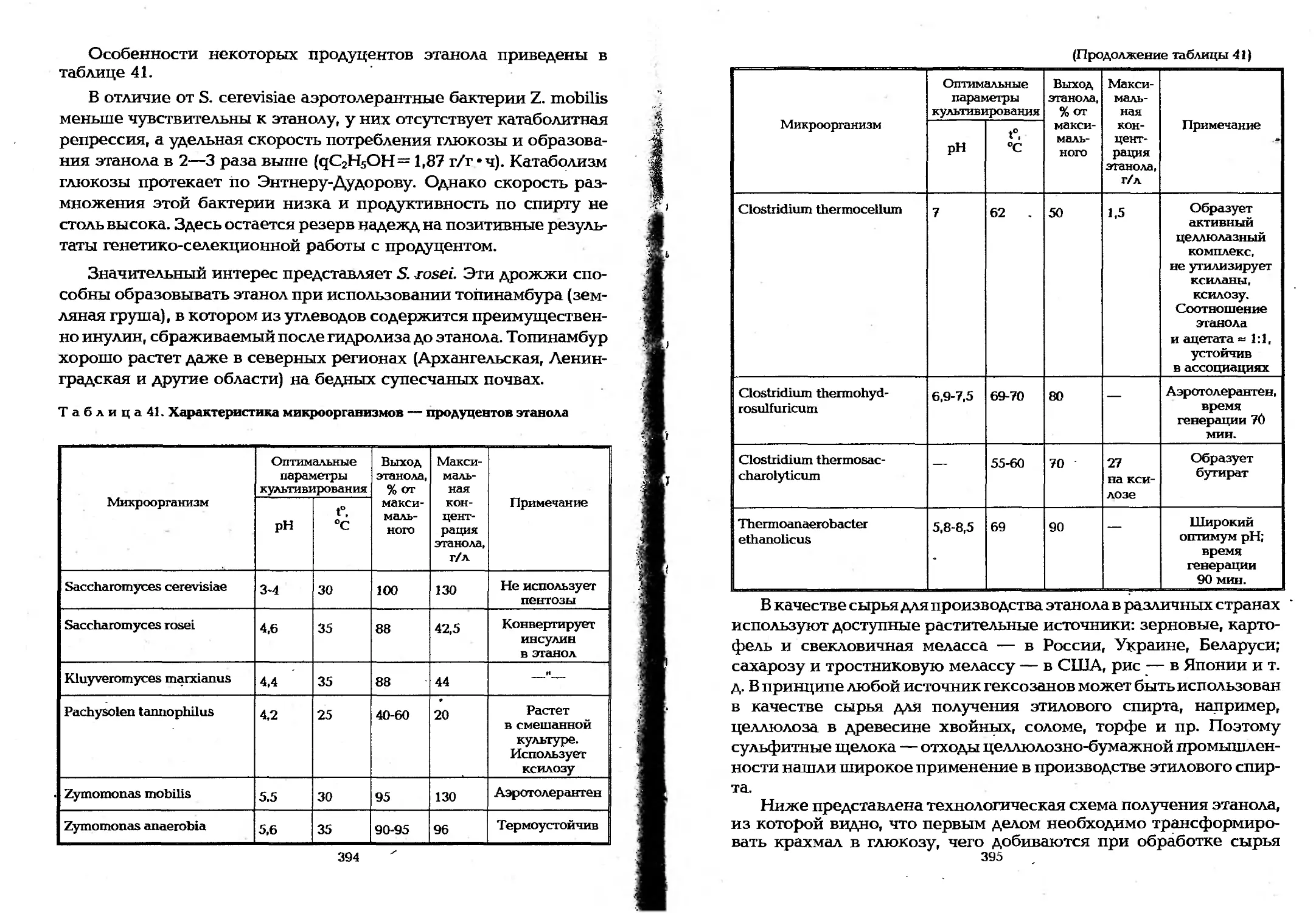
9.3. Микробиотехнологические процессы .... ... 392
9.3.1. Получение продуктов брожения ................392
9.3.2. Получение органических кислот................411
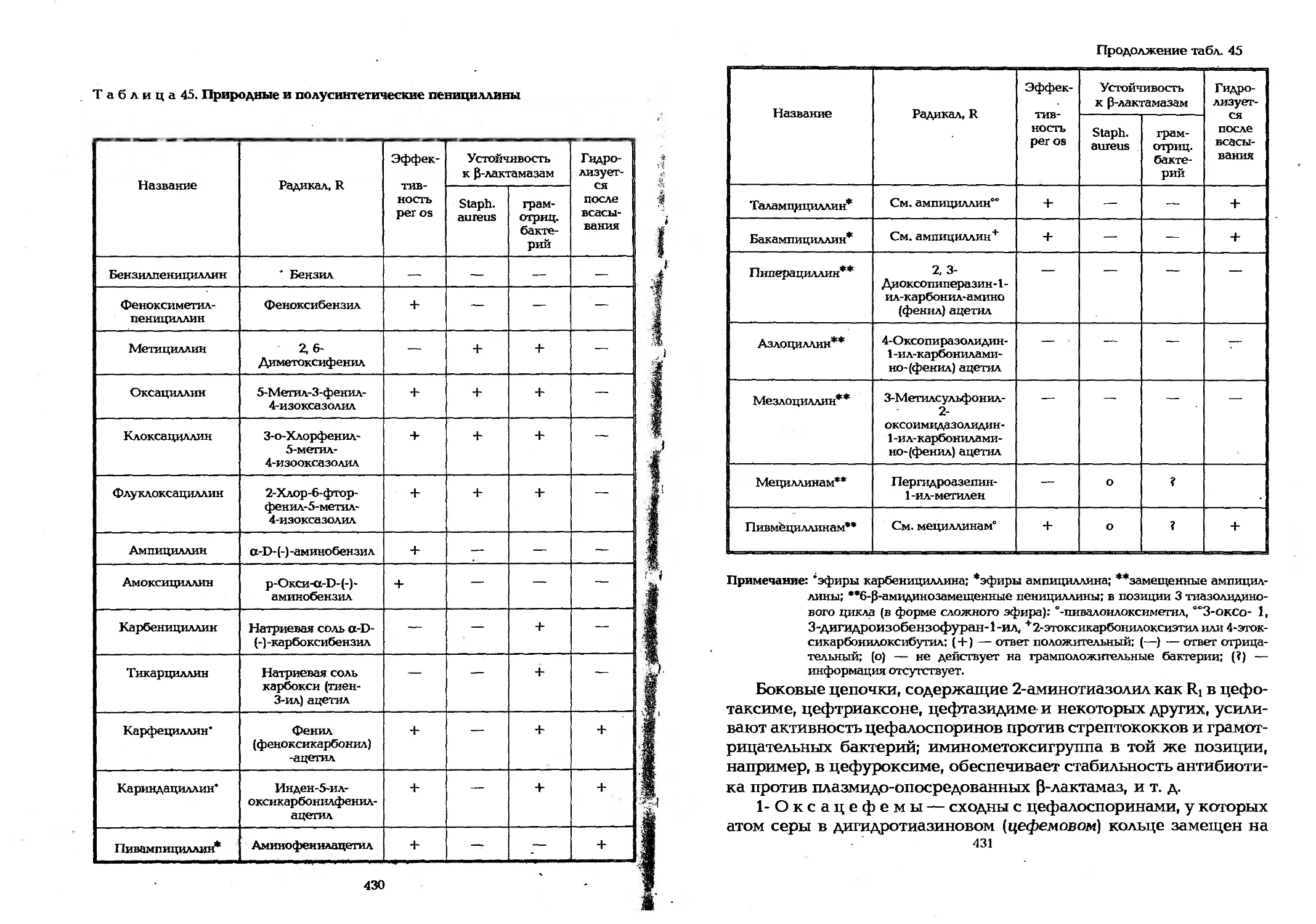
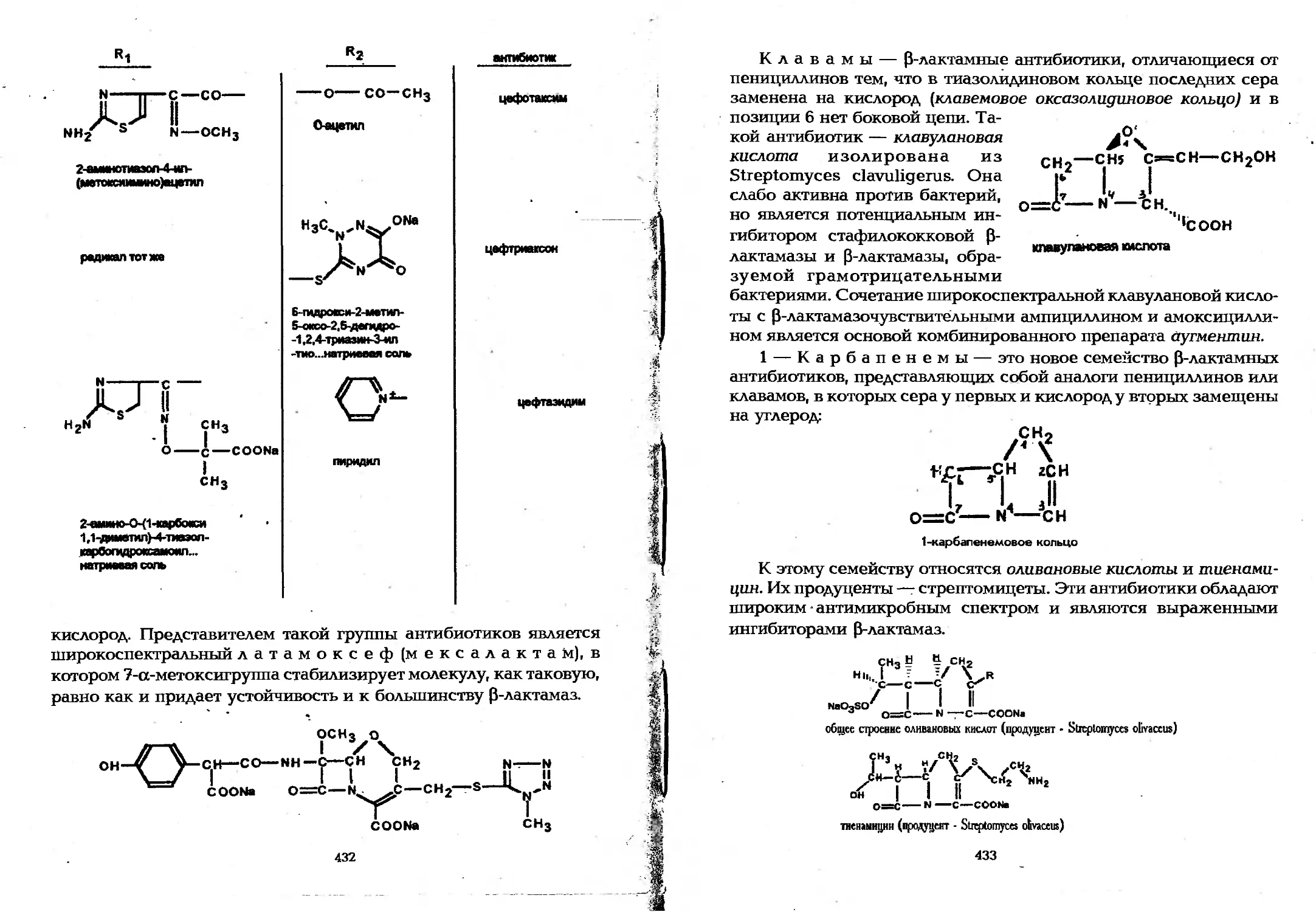
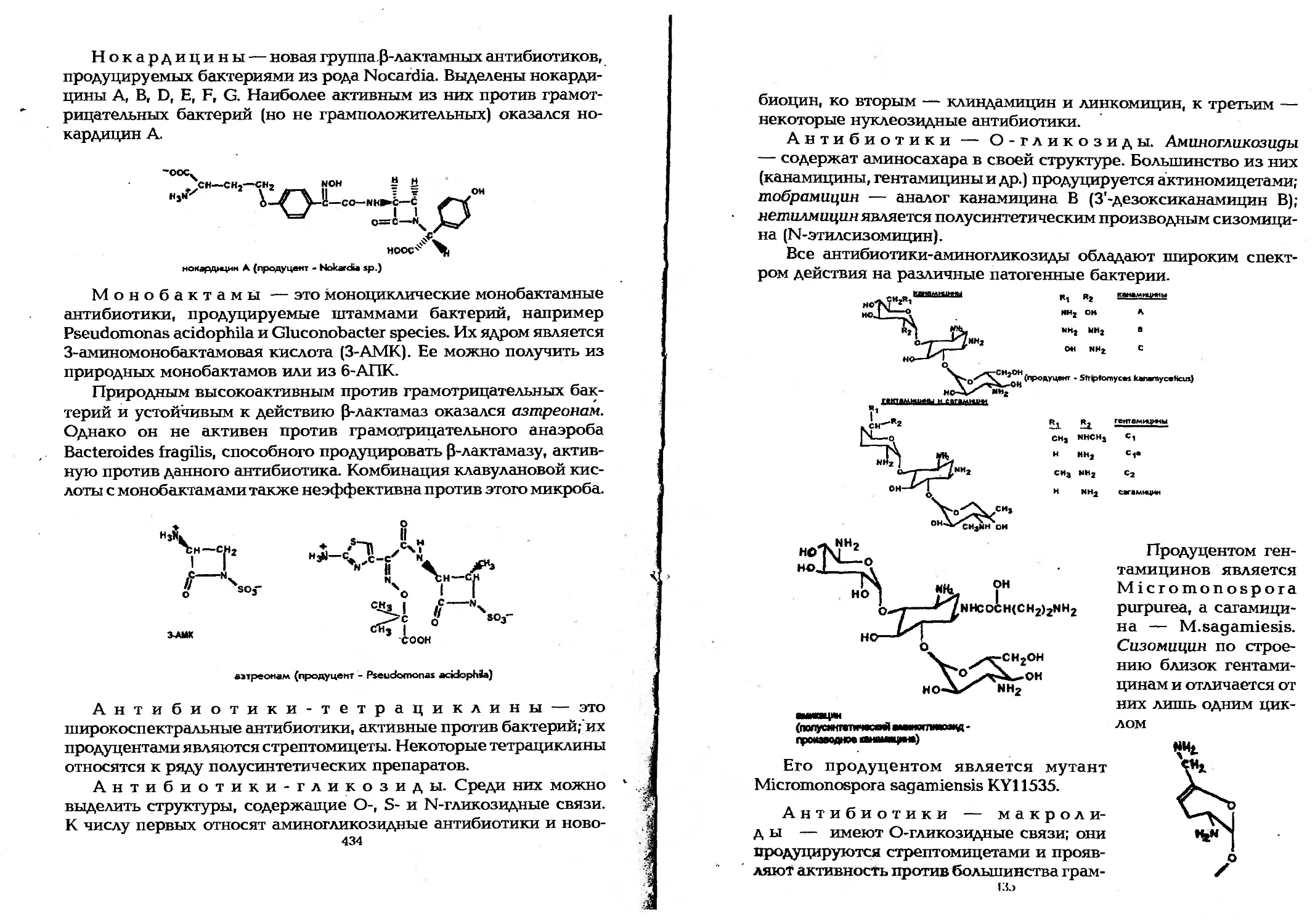
9.3.3. Получение антимикробных веществ..............428
9.3.4. Получение аминокислот........................444
9.3.5. Получение витаминов .........................479
9.3.6. Получение микробных препаратов —
удобрителей почв, стимуляторов и регуляторов
роста растений.....................................455
9.3.7. Получение микробных полимеров ... 459
ГЛАВА 10. Фитобиотехнология ...................................489
10.1 Термины, используемые в фитобиотехнологии .....492
10.2. Вегетативное размножение растений методом
культур тканей.....................................500
10.3. Культивирование клеток растений
в глубинных условиях ..............................507
10.4. Использование методов генетической инженерии,
или рДНК в фитобиотехнологии........................510
10.5. Получение биогаза и удобрений на основе
использования растений............................. 520
10.6. Коллекционные центры сохранения генофонда
растений............................................526
ГЛАВА 11. Зообиотехнология.....................................532
11.1. Способы выращивания клеток животных...........534
11.2. Эмбриональные и другие ткани для репродукции
вирусов и получения вирусных препаратов............544
11.3. Получение инсектопатогенных вирусов
в клеточных культурах...............................555
11.4. Интерфероны...................................556
11.5. Получение и использование гомо-,
гетеро- и синкариотических гибридов ................562
11.6. Трансгенные животные......................... 581
11.7. Иммуномодуляторы..............................587
11.8. Коллекционные центры клеточных культур,
их роль в сохранении генофонда животных организмов 597
Литература ..................................'.................599
6
ПРЕДИСЛОВИЕ
Первоначальный вариант этой книги был утвержден Госкомоб-
разованией СССР как учебник для студентов, изучающих биотех-
нологические производства на соответствующих факультетах вы-
сших учебных заведений. Его структура сохранена в прежнем виде.
Весь материал книги подразделен на четыре части. В первой из
них рассмотрены предмет, история развития, цели и задачи био-
технологии; объекты и методы биотехнологии; во второй — роль
фундаментальных исследований в развитии биотехнологии; в
третьей - процессы и аппараты в биотехнологии; техническая
вооруженность биотехнологических производств; отходы биотех-
нологических производств, их обезвреживание и утилизация. В
четвертой части представлено основное содержание микробиотех-
нологии, фитобиотехнологии и зообиотехнологии.
В основу книги положен курс лекций, читаемый автором
студентам биотехнологического факультета Санкт-Петербургского
химико-фармацевтического института в течение последних 8 лет
по утвержденной программе. За это время издан ряд монографи-
ческих работ и пособий биотехнологической направленности, од-
нако до сих пор не опубликован учебник, в котором были бы
объединены микро-, фито- и зообиотехнологии. Первая попытка
такого рода всегда сопряжена с возможными недочетами и несо-
вершенством, поэтому все критические и конструктивные замеча-
ния будут приняты нами с признательностью.
В подготовке 7-й главы учебника активное участие приняли
доценты Р. М. Бакаева и Н. А. Силуянова. При окончательном
оформлении книги учтены пожелания и рекомендации профессо-
ров, доцентов и ассистентов СПХФИ в составе: проф. Г. А.
Битовской, проф. Н. А. Заикиной, доц. В. К. Грековой, доц. М.А.
Кашкиной, доц. Ю. Б. Марюхта, доц. И. П. Соколовой, к. б. н. асе.
С. В. Гуриной; в подготовке рукописи и иллюстративного материала
большую помощь оказали С. В. Волобуева, С. Л. Воротынская, А.
О. Григорянц, Н. Н. Блинова, А. В. Караваева, О. А. Кудряшова, Н.
В. Разукрантова, М. А Савинова, О. М. Тихомирова, Л. Г. Федорова,
В. В. Южанина. Всем названным лицам выражаю свою искреннюю
благодарность.
Автор также признателен рецензентам проф. И. М. Василинецу
и проф. В. И. Яковлеву.
Издание учебника было бы просто невозможным без велико-
душной спонсорской поддержки директора АП "Новая Заря" Юрия
Федоровича Назарова, чье понимание социальных задач и вера в
7
успех подготовки молодых кадров инженеров-биотехнологов в
России по новейшим направлениям развития науки и техники
явились основным побудительным мотивом содействовать нашим
усилиям.
Публикация книги в настоящем виде осуществлена в сжатые
сроки благодаря умелой организации и благосклонности генераль-
ного директора акционерного объединения "Роспечать" Белгород-
ской области Виктора Васильевича Гончарова, директора Белго-
родской областной типографии Галины Николаевны Левиной,
технического редактора Валентины Степановны Просековой, кор-
ректора Светланы Сергеевны Кухаревой. 7
Заслуженный деятель науки России,
академик СПбИА, проф. Н. П. Блинов—
8
ВВЕДЕНИЕ
Современный уровень развития биотехнологии обусловлен
общим прогрессом науки и техники, особенно — в течение послед-
них 50 лет. Достаточно отметить лишь такие события, как установ-
ление структуры и функций нуклеиновых кислот, обнаружение
ферментов рестрикции ДНК и выявление их значения в жизни
клеток с последующим использованием в генно-инженерных ра-
ботах, создание гибридом и получение моноклональных антител,
внедрение ЭВМ и компьютерной техники в биотехнологические
процессы и т. д.
Изучение биотехнологии невозможно без знания основ хими-
ческих дисциплин и, прежде всего, органической, биологической
и коллоидной химии, ряда биологических дисциплин (общей био-
логии, микробиологии, ботаники, генетики, иммунологии, эколо-
гии), а также процессов и аппаратов химической и биологической
технологии и ряда других общеинженерных дисциплин.
Биотехнологию относят к числу приоритетных наук, где можно
прогнозировать более быстрые и важнейшие достижения для
социально-экономического прогресса общества.
Ведущее положение в области внедрения биотехнологических
разработок ныне занимают США, где к концу 80-х. годов биотех-
нологическими проблемами было занято 349 компаний, включая
296 — чисто биотехнологических, расходовавших на эти цели,
например, в 1987 г. 1 млрд. 200 млн. долларов. Среди них можно
назвать Genentech Inc., Alza corp., Amgen Inc., Cetus corp., Biogen,
Ceptocor Inc., Chiron corp., Xoma corp., Immunex corp., DNA Plant
Technology, Celgene corp., Damon Biotech Inc., и др. При этом
области исследования включали: создание лекарств и диагности-
ческих средств (467 млн. долларов), специальных химических
соединений (144 млн. долларов); животноводство и растениеводст-
во (168 млн. долларов); оборудование (48 млн. долларов) и т. д.
Food and Drug Administration (FDA) в 1982 г. одобрила и
разрешила применение первого биотехнологического продукта —
генно-инженерного инсулина; в последующие годы было разреше-
но применять еще 8 биотехнологических препаратов: два варианта
гормона роста человека, два варианта 2а-интерферона для лечения
волосатоклеточной лейкемии, активатор тканевого плазминогена
(ТРА) для лечения тромбоза коронарных сосудов, вакцину против
гепатита В, фактор VIII для лечения гемофилии, мышиные моно-
клональные антитела для предупреждения отторжения почечных
трансплантатов.
9
К 1988 г. в США свыше 80 лекарственных веществ и вакцин
были объектами биотехнологических разработок, 14 из которых
были представлены на утверждение в FDA: тромболитические
агенты; ферменты дисмутазы, предупреждающие клеточные по-
вреждения, наступающие при реперфузии гипоксических тканей;
гормон эритропоэтин, стимулирующий эритропоэз; эпидермаль-
ный ростовой фактор для ускоренного заживления ран; монокло-
нальные антитела для трансплантации костного мозга, лечения
септического шока, рака и патологических состояний от передо-
зировок лекарств и др. (к 1988 г. было одобрено 300 наборов
моноклональных антител). К 1995 г. стоимость поступающих на
рынок только моноклональных антител прогнозировалась в 6,4
биллиона долларов.
Из научных учреждений России ведущее место по биотехно-
логии занимает институт биохимии и физиологии микроорганиз-
мов РАН (ИБФМ), сотрудники которого, например, совместно с
учеными научно-исследовательского вычислительного центра
(НИВЦ) в 1985 г. создали автоматизированный комплекс "Фермен-
тер-ЭВМ", что обеспечивает возрастание эффективности управле-
ния процессом биосинтеза; существенный вклад в решение био-
технологических проблем в России внесли коллективы ВНИИ
"Синтез-белок”, ВНИИ биотехнологии, ВНИИА, Сибирское отде-
ление РАН, институт молекулярной биологии РАН, институт гене-
тики и селекции промышленных микроорганизмов и др.
В России давно и хорошо известны предприятия, выпускающие
биотехнологическую продукцию (антибиотики, ферменты, различ-
ные дрожжи и т. д.). Это же можно сказать и о предприятиях в
странах СНГ.
Основной вклад в подготовку кадров инженеров-биотехнологов
с 1945 г. вносил и продолжает вносить Санкт-Петербургский
химико-фармацевтический институт; с начала 80-х годов подготов-
ка таких специалистов была расширена за счет открытия новых
биотехнологических отделений и факультетов в других вузах
страны. Сегодня изучение биотехнологии является "велением вре-
мени” для высших и средних специальных учебных заведений, в
которых подготавливаются молодые специалисты для соответству-
ющих отраслей производства. Объем и программы курсов могут
варьировать в зависимости от профилизации учебного заведения.
В настоящем издании, по возможности, затронуты все основные
аспекты биотехнологии, и поэтому эта книга может быть исполь-
зована также и сотрудниками учреждений, призванных выпускать
биотехнологическую продукцию.
ю
Часть I
Биотехнология —
как научная дисциплина
Глава 1
ПРЕДМЕТ, ИСТОРИЯ РАЗВИТИЯ,
ЦЕЛИ И ЗАДАЧИ БИОТЕХНОЛОГИИ
Биотехнология — это наука об использовании биологических
процессов в технике и промышленном производстве. Название ее
происходит от греческих слов bios — жизнь, teken — искусство,
logos — слово, учение, наука. К числу биологических процессов
п
относят те из них, в которых применяют биологические объекты
различной природы (микробной, растительной или животной),
например, производство ряда продуктов медицинского, пищевого
и другого назначения — антибиотики, вакцины, ферменты, кор-
мовой и пищевой белки, полисахариды, гормоны, гликозиды, ами-
нокислоты, алкалоиды, биогаз, удобрения и пр.
В соответствии с определением Европейской Федерации Био-
технологов (ЕФБ, 1984) биотехнология базируется на интегральном
использовании биохимии, микробиологии и инженерных наук в
целях промышленной реализации способностей микроорганизмов,
культур клеток тканей и их частей. Уже в самом определении
предмета отражено его местоположение как пограничного, благо-
даря чему результаты фундаментальных исследований в области
биологических, химических и технических дисциплин приобрета-
ют выраженно прикладное значение. Биотехнология непосредст-
венно связана с общей биологией, микробиологией, ботаникой,
зоологией, анатомией и физиологией, биологической, органиче-
ской, физической и коллоидной химией, иммунологией, биоинже-
нерией, электроникой, технологией лекарств, генетикой и другими
научными дисциплинами.
Человек, рождающийся для познания мира (в том числе — и
самого себя), давным-давно освоил на практике различные процес-
сы биотехнологии, не зная по существу, что они относились к
такому разряду. В самом деле, с библейских времен известно
виноделие, тысячелетия насчитывает хлебопечение и т. д.
Познавательная деятельность людей непосредственно сказыва-
лась на уровне социального развития общества. Недаром вторую
половину XX столетия мы называем периодом научно-технической
революции. Наука сегодня имеет огромное значение в жизни
людей, и научный подход к решению любой задачи — веление и
требование времени.
Наука формировалась и эволюционировала по мере формиро-
вания и развития человеческого общества. Это, в частности, не-
посредственно относится и к биотехнологии. Ее возникновение,
становление и развитие условно можно подразделить на 4 периода:
эмпирический, этиологический, биотехнический и генотехниче-
ский. Эмпирический (от греч. empeirikos — опытный) или доисто-
рический период — самый длительный, охватывающий примерно
8000 лет, из которых более 6000 лет — до' нашей эры и около 2000
лет — нашей эры. Древние народы того времени интуитивно
использовали приемы и способы изготовления хлеба, пива и
12
некоторых других продуктов, которые теперь мы относим к раз-
ряду биотехнологических. Кризис охотничьего промысла (хозяй-
ства) стал побудительным мотивом революции в изготовлении
продуктов питания. Эта революция началась около 8000 лет назад
и привела к изобретению техники земледелия — началу произво-
дительного ведения хозяйства (неолит и бронзовый века.) Стали
формироваться так называемые приречные цивилизации Месопо-
тамии, Египта, Индии и Китая. Шумеры — первые жители Месо-
потамии (на территории современного Ирака) создали цветущую
в те времена цивилизацию. Они выпекали хлеб из кислого теста,
владели искусством готовить пиво. В этом следовали им ассирийцы
и вавилоняне, жившие также в Месопотамии, египтяне и древние
индусы. В течение нескольких тысячелетий известен уксус, издрев-
ле приготавливавшийся в домашних условиях, хоуя о микробах —
индукторах этого процесса мир узнал в 1868 г. благодаря работам
Пастера, и это несмотря на существование с XIV в. так называемого
"Орлеанского способа" приготовления уксуса; первая дистилляция
вина осуществлена в XII в.; водку из хлебных злаков получили в
XVI в.; шампанское известно с XVIII в., но получение почти
абсолютного этанола впервые удалось в XIV в. испанцу Раймунду
Луллию (ок. 1235 — 1315) благодаря перегонке вина с негашеной
известью.
В те древние времена продукты питания растительного и
животного происхождения использовались не только в пищу, но
и для лечебных целей. Например, в ассирийской столице Ниневии
(8 — 7 века до н. э.) была царская библиотека, насчитывавшая
более 30 000 клинописных табличек, из которых в 33 имелись
сведения о лекарственных средствах и их рецептуре, и в самом
городе размещался сад лекарственных растений.
тому же эмпирическому периоду относятся: получение кис-
ломолочных продуктов, квашеной капусты, медовых алкогольных
напитков, силосование кормов, мочка лубоволокнистых растений.
Длительное накопление фактов происходило и в области ми-
кологии (от греч. го у Kes — гриб). Сведения о грибах можно найти
в писаных источниках древности, а Луций Лициний Лукулл (106
— 56 гг. до н. э.), славившийся богатством, роскошью и пирами ("лу-
куллов пир”), предпочитал всем съедобным грибам кесарев гриб
(Amanita cesarea, L.). Древние евреи хорошо знали ржавчину
хлебных злаков и головню. В IV — I веках до н. э. были собраны
интересные материалы о грибах, нашедшие отражение в работах
Аристотеля, Диоскорида, Плиния Младшего, Теофраста. В после-
13
дующие века нашей эры микология стала самостоятельной наукой
— велика роль в этом Д. Персоона и Э. М. Фриза, по праву
считающихся отцами систематической микологии.
Таким образом, народы исстари пользовались на практике
микробиологическими процессами, ничего не зная о микробах.
Эмпиризм также был характерен и в практике использования
полезных растений и животных.
Второй, этиологический (от греч. aitia — причина) период в
развитии биотехнологии охватывает вторую половину XIX века и
первую треть XX века (1856 — 1933 гг.). Он связан с выдающимися
исследованиями великого французского ученого Луи Пастера (1822
— 1895) — основоположника научной микробиологии и ряда
микробиологических дисциплин (промышленной, медицинской,
химической, санитарной). С аналитической микробиологией не-
посредственно связано открытие Пастером молекулярной ассимет-
рии (стереоизомерии). Это, по существу, бриллиантовый век мик-
робиологии. Пастер вскрыл микробную природу брожений, дока-
зал возможность жизни в бескислородных условиях, эксперимен-
тально опроверг ходячее тогда представление о самопроизвольном
зарождении живых существ, создал научные основы вакцинопро-
филактики и вакцинотерапии; предложил метод стерилизации,
называемый по его имени пастеризацией и т. д.
Немеркнущая слава Пастера не затмила имен его выдающихся
учеников и сотрудников: Э. Дюкло, Э. Ру, Ш. Э. Шамберлана, Ж.
А. Вильемена, И. И. Мечникова. В этот же период творили Р. Кох,
Д. Листер, Ш. Китазато, Г. Т. Риккетс, Д. И. Ивановский, А. Лаверан
и другие.
Параллельно с Пастером трудился в Германии, а позднее — во
Франции, выдающийся миколог А де Бари (1831 — 1888) —
основоположник физиологической микологии. Изучив стадии раз-
множения и историю индивидуального развития грибов (онтоге-
нетический метод), с учетом их взаимоотношений с другими
видами, а также цитологических и биологических особенностей,
де Бари создал классификацию, которая и сегодня лежит в основе
современных классификационных схем микро- и макромицетов.
Де Бари — основоположник микофитопатологии — науки о
грибных болезнях растений (от греч. fiton — растение, pathos —
болезнь), под его руководством сформироваласьплеяда выдающих-
ся ученых (в том числе — из России): Ф. М Бальфур, И. В.
Баранецкий, М. Бейеринк, О. Брефельд, М. С. Воронин, А. Кох, А
С. Фаминицин и др.
14
В биотехнологии важными являются питательные среды для
культивирования ряда биообъектов. Уже Л. Пастер приготовил
первую жидкую питательную среду в 1859 году, метод выращива-
ния грибов на желатине предложил О. Брефельд в 1864 г., Ж. Ролен
сообщил о жидких средах для выращивания нитчатых грибов в
1870 г., Р. Коху в 1876 г. удалось вырастить бациллы сибирской
язвы в капле водянистой влаги, извлеченной из глаза погибшей
коровы. В 80-е годы XIX столетия Р. Кох предложил метод культи-
вирования бактерий на стерильных ломтиках картофеля и затем
— на агаризованных питательных средах.
В настоящее время, предлагая самые сложные и необычные в
каком-либо отношении среды для выращивания биообъектов, мы
опираемся на основополагающие результаты этих выдающихся
ученых. Аналогичным образом можно сказать и о вариантах
способов стерилизации питательных сред, имея в виду тиндализа-
цию, кипячение, дробную стерилизацию и др. Все они основыва-
лись на необходимости уничтожения посторонней микрофлоры,
которая попадала в среды в процессе их изготовления.
В ряду открытий всемирного значения стоит обнаружение в
1892 г. вируса мозаичной болезни табака Д. И. Ивановским (1864
— 1920). Последовавшие за этим обнаружения других вирусов
обеспечили становление новой научной дисциплины — вирусоло-
гии: Ф. Леффлер и П. Фрош в 1898 г. открыли вирус ящура, Д.
Кэррол в 1901 г. — вирус желтой лихорадки, ф. Туорт в 1915 г. и
Ф. д'Эрелльв 1917 г. — вирусы бактерий (бактериофаги). Большой
вклад в вирусологию был внесен отечественными и зарубежными
учеными — Л. А. Зильбером, А. А. Смородинцевым, М. П. Чума-
ковым, А. Борелем, К. Левадити, К. Ландштейнером, В. Стэнли, П.
Лейдлоу, П. Руа, П. Ф. Эндерсом и многими другими.
Этиологический период знаменателен тем, что удалось доказать
индивидуальность микробов и получить их в чистых культурах.
Более того, каждый вид мог быть размножен на питательных средах
и использован в целях воспроизведения соответствующих процес-
сов (бродильных, окислительных и др.). Например, маслянокислые
бактерии и вызываемое ими маслянокислое брожение, лактобак-
терии и молочнокислое брожение, дрожжи — сахаромицеты и
спиртовое брожение, уксуснокислые бактерии и окисление этано-
ла до уксусной кислоты и т. д. В этот период было начато
изготовление прессованных пищевых дрожжей, а также некото-
рых продуктов обмена (метаболизма) — ацетона, бутанола, лимон-
ной и молочной кислот; во Франции приступили к созданию
15
биоустановок для микробиологической очистки сточных вод.
Знание причин биологических процессов еще не исключало
нестерильные операции, хотя и стремились к использованию
чистых культур микроорганизмов.
Для всестороннего изучения морфолого-физиологических
свойств и продуктов обмена, прежде всего, микробов все ранее
предложенные способы их выращивания оказались малопригод-
ными. Более того, накопление однородной по возрасту большой
массы клеток оставалось исключительно трудоемким процессом.
Вот почему требовался принципиально иной подход для решения
многих задач в области биотехнологии. В 1933 году А. Клюйвер и
Л. X. Ц. Перкин опубликовали работу "Методы изучения обмена
веществ у плесневых грибов", в которой изложили основные
технические приемы, а также подходы к оценке и интерпретации
получаемых результатов при глубинном культивировании грибов.
С этого времени начинается третий период в развитии биологи-
ческой технологии — биотехнический. Началось внедрение в
биотехнологию крупномасштабного герметизированного оборудо-
вания, обеспечившего проведение процессов в стерильных усло-
виях. Особенно мощный толчок в развитии промышленного био-
технологического оборудования был отмечен в период становления
и развития производства антибиотиков (время второй мировой
войны 1939 — 1945 гг., когда возникла острая необходимость в
противомикробных препаратах для лечения больных с инфициро-
ванными ранами). Все прогрессивное в области биологических и
технических дисциплин, достигнутое к тому времени, нашло свое
отражение в биотехнологии. Следует отметить, что уже в 1869 г..
Ф. Мишер получил "нуклеин" (ДНК) из гнойных телец (лейкоци-
тов); В. Оствальд в 1893 г. установил каталитическую функцию
ферментов; Т. Леб в 1897 г. установил способность к выживанию
вне организма (в пробирках с плазмой или сывороткой крови)
клеток крови и соединительной ткани; Г. Хаберланд в 1902 г.
показал возможность культивирования клеток различных тканей
растений в простых питательных растворах; Ц. Нейберг В 1912 г.
раскрыл механизм процессов брожения; Л Михаэлис и М. Л.
Ментен в 1913 г. разработали кинетику ферментативных реакций,
а А. Каррел усовершенствовал способ выращивания клеток тканей
животных и человека и впервые применил экстракт эмбрионов
для ускорения их роста; Г. А. Надсон и Г. С. Филлипов в 1925 г.
доказали мутагенное действие рентгеновских лучей на дрожжи, а
в 1937 г. Г. Кребс открыл цикл трикарбоновых кислот (ЦТК); в 1960
16
г. Ж. Барски и др. впервые обнаружили соматические гибриды
опухолевых клеток мыши. Следовательно, накопленные научные
факты стали побудительным мотивом для разработки способов
крупномасштабного культивирования клеток различного проис-
хождения. Это необходимо было д ля получения различных клеточ-
ных продуктов и самих клеток для нужд человека, и, прежде всего,
в качестве или в составе лечебных и профилактических средств:
пенициллина, стрептомицина, тетрациклинов, декстрана, ряда ами-
1 нокислот и многих других веществ. К 1950 г. Ж. Моно (Франция)
разработал теоретические основы непрерывного управляемого
культивирования микробов; в 50-е годы вопросам практической
реализации непрерывного культивирования микроорганизмов по-
святили свои исследования М. Стефенсон, И. Малек, Н. Д. Иеру-
салимский и др.
Примерно за 40 лет третьего периода были решены основные
задачи по конструированию, созданию и внедрению в практику
необходимого оборудования, в том числе главного из них —
биореакторов. Это оборудование используют и в настоящее время.
Четвертый период в биотехнологии — генотехнический (от
греч. genesis — происхождение, возникновение, рождение) начал-
ся с 1972 г. В этом году П. Берг со своими сотрудниками в США
создали первую рекомбинантную молекулу ДНК. Однако следует
отметить, что в 1969 г. Дж. Бекуит с коллегами выделил в химически
чистом виде лактозный 1ен из кишечной палочки, показав тем
самым возможность направленных манипуляций с генетическим
материалом бактерий.
Естественно, что без фундаментальной работы Ф. Крика и Дж.
Уотсона (1953) по установлению структуры ДНК было невозмож-
ным достигнуть современных результатов в области биотехноло-
гии. Выяснение механизмов функционирования и регуляции ДНК,
выделение и изучение специфичных ферментов привело к фор-
мированию строго научного подхода к разработке биотехнологи-
ческих процессов на основе генно-инженерных работ. В этом суть
генотехнического периода.
Уже в 1982 г. поступил в продажу человеческий инсулин,
выработанный кишечными палочками, несущими в себе искусст-
венно встроенную генетическую информацию об этом гормоне.
На таком же уровне или с близким к тому заделом находятся
следующие генно-инженерные препараты: интерфероны, фактор
некротизации опухоли (TNF), интерлейкин-2, соматотропный гор-
мон человека и аналог его соматомеднп Ц и другие^ _ -----~~~
17 I Г' Г'’" '’ ’' Л
i v - <. н эй
. u i м С К О И < А Д Е У :';И
Зная строение аппарата наследственности у разных организ-
мов, удается манипулировать не только нуклеиновыми кислотами,
но и целыми хромосомами (хромосомная инженерия) и клетками
(клеточная инженерия).
Для генотехнического периода характерны: разработка интен-
сивных процессов (вместо экстенсивных) на основе направленных
фундаментальных исследований (с продуцентами антибиотиков,
ферментов, аминокислот, витаминов), получение суперпродуцен-
тов; создание продуцентов, несущих в себе бессмысленную гене-
тическую информацию (например, гены интерферона человека в
клетках Pseudomonas aeruginosa); создание необычных организ-
мов, ранее не существовавших в природе (неклубеньковых расте-
ний, несущих гены азотобактерий, ответственные за способность
фиксировать молекулярный азот из воздуха); разработка и внед-
рение экологически чистых и, по возможности, безотходных тех-
нологий; разработка и внедрение в практику специальной аппара-
туры блочного (сменного) типа для различных биотехнологических
схем; автоматизация и компьютеризация биотехнологических про-
цессов; создание экономически оптимальных производственных
процессов при максимальном использовании сырья и минималь-
ном потреблении энергии. Вот почему инженер — биотехнолог в
современном пониманикгдолжен быть широко и глубоко подготов-
ленным специалистом, в распоряжении которого оказываются
сложнейшие биологические системы (или аналоги их), синхронно
работающие в заданном направлении. В любом биотехнологиче-
ском процессе наиважнейшим звеном является биообъект, "кап-
ризы" которого по любому поводу могут пагубно сказаться на
результатах этого процесса.
В течение последних 10 — 15 лет текущего столетия происхо-
дило бурное развитие биотехнологии, определились сферы при-
оритетного внедрения конкретных результатов биотехнологиче-
ских разработок, и, как следствие, появились такие названия, как
медицинская биотехнология, иммунобиотехнология (от лат.
immunus — невосприимчивый), биогеотехнология (от греч. geo —
земля), инженерная энзимология (от греч. еп — в, zyme — заква-
ска) . Одни из них прочно входят в лексикон специалистов, напри-
мер, иммунобиотехнология, инженерная энзимология, другие на-
звания приживаются плохо или с трудом (медицинская биотехно-
логия, биогеотехнология). К медицинской биотехнологии
относили те производственные процессы, которые завершались
созданием с помощью биообъектов средств или веществ медицин-
ского назначения (прежде всего профилактического или лечебного
18
действия на организм человека). Это — антибиотики, некоторые
витамины, коферменты и ферменты, отдельные микробные пол-
исахариды — как самостоятельные препараты или вспомогатель-
ные вещества при создании различных лекарственных форм,
аминокислоты, нуклеозиды и др.
Иммунобиотехнология объединяет производства
вакцин, иммуноглобулинов крови, иммуномодуляторов, иммуно-
медиаторов, моноклональных антител и некоторых других. Можно
заметить, что на основе иммунобиотехнологических процессов
создаются также профилактические и лечебные средства, объеди-
няемые под эгидой медицинской биотехнологии. Следовательно
иммунобиотехнология представляется здесь частным случаем ме-
дицинской биотехнологии. Вместе с тем, иммунобиотехнологиче-
ские процессы по целевым продуктам вышли за пределы медицин-
ского назначения (например, в иммуноферментном анализе, им-
муноблотинге). В равной мере большинство ферментов (как и
аминокислот или некоторых других продуктов) производится не
для целей здравоохранения. Поэтому термин "Медицинская био-
технология" представляется во многом искусственным и, очевидно,
отсюда плохо приживающимся. Напротив, вычленение иммуноби-
отехнологии в качестве самостоятельной научной субдисциплины
является обоснованным, и производственные процессы здесьчетко
ограничены использованием иммунной системы того или иного
макроорганизма или отдельных компонентов ее (макрофаги, лим-
фоциты, различные иммуноглобулины).
Биогеотехнология — это субдисциплина, ранее
называвшаяся геологической микробиологией. Сущность ее сво-
дится к использованию микроорганизмов для добычи полезных
ископаемых, например, цветных металлов, нефти; для окисления
метана в угольных шахтах и пр. Спектр биогеотехнологических
производственных процессов невелик, но очень важен для жизне-
обеспечения народного хозяйства. Необходимо подчеркнуть, что
не все процессы, относимые ныне к биогеотехнологическим, яв-
ляются сугубо технологическими (например, окисление метана с
помощью специальных микроорганизмов). Обычно принято счи-
тать, что в результате биотехнологического процесса образуется
какой-то целевой продукт, используемый на практике. В приведен-
ном примере окисления метана преследуется иная цель — сниже-
ние концентрации метана до безопасного (например, для шахте-
ров) уровня. Хотя и в этой реакции образуются конечные продук-
ты, которые могут быть использованы на практике:
СН4 + О2—• СО2 + 2Н2
19
Инженерная энзимология — это отрасль
биотехнологии, базирующаяся на использовании каталитических
функций ферментов (или ферментных систем) в изолированном
состоянии или в составе живых клеток для получения соответст-
вующих целевых продуктов. Биообъект здесь — фермент (или
комплекс ферментов). На практике обычно используют иммоби-
лизованные ферменты (реже — иммобилизованные клетки), бла-
годаря чему стабилизируется и пролонгируется их ферментативная
-активность. Иногда инженерную энзимологию отождествляют с
биотехнологией. В этом содержится большая доля истины, так как
все реакции в клетках катализируются ферментами. Однако слово
"инженерная" привносит свою специфику, заключающуюся в
акценте на создание конструкции (от франц, engin — машина), в
данном случае — на конструирование биокатализаторов с задан-
ными свойствами с последующим использованием в биотехноло-
гическом процессе.
В научной литературе можно встретить и другие названия
биотехнологических процессов, например, "Биотехнология живо-
тной клетки", "Экономическая микробиология", "Ферментация и
биоинженерия”, "Промышленная микробиология", "Сельскохозяй-
ственная биотехнология", "Биохимическая инженерия" и др. Более
того, в принципе можно говорить и писать о биотехнологии
каждого индивидуального продукта, образуемого каким-либо мик-
робом, какими-либо клетками растений и животных, или появля-
ющегося под каталитическим воздействием фермента; в равной
мере речь может идти и о биотехнологии продуцентов каких-либо
веществ, например, о Panax ginzeng (жень-шень), о Rauwolfia
serpentina (Раувольфия змеевидная), об Actinomyces spp. (актино-
мицеты) и т. д. Поэтому наиболее рациональным является подраз-
деление биотехнологии на микробную биотехнологию, раститель-
ную, или фитобиотехнологию, и животную, или зообиотехноло-
гию, включающую также и биотехнологические процессы, осно-
ванные на использовании клеток человека.
Из приведенной схемы видно, что наибольшее число реализо-
ванных процессов имеет место в микробной биотехнологии. Мно-
гие микроорганизмы обладают заметным преимуществом перед
растительными и животными объектами по таким показателям,
как скорость размножения, лабильность и быстрота адаптации к
изменяющимся условиям среды обитания. Этим и определяется
диапазон применения микробов в различных отраслях производ-
ственной деятельности человека, то есть все то, что составляет
20
микробиотехнология
Биотехнология —
фитобиотехнология
зообиотехнология
использование
в легкой
промышленности, в
пищевой
промышленности,
в медицинской
промышленности,
в химической
промышленности,
в металлургии,
в нефтедобывающей
промышленности,
в водном хозяйстве,
в защите окружающей
среды,
в энергетике,
в косметической
промышленности
в агропромышленном
комплексе,
в медицинской и
косметической
промышленности
в животноводстве,
в медицинской
промышленности,
в пищевой
промышленности
предмет "Промышленная микробиология". Это можно представить
в следующем виде (таблица 1).
Какие же основные цели и задачи стоят перед биотехнологами?
Во-первых, поддержание и активация путей обмена клеток, веду-
щих к накоплению целевых продуктов при заметном подавлении
других реакций обмена у культивируемого организма; во-вторых,
получение клеток или их составных частей (преимущественно —
ферментов) для направленного изменения сложных молекул (на-
пример, рестриктазы, изомеразы и т. д.); в-третьих, углубление и
совершенствование рДНК-биотехнологии и клеточной инженерии
на предмет получения особо ценных результатов в фундаменталь-
ных и прикладных разработках; в-четвертых, создание безотход-
ных и экологически безопасных биотехнологических процессов;
в-пятых, совершенствование и оптимизация аппаратурного офор-
мления биотехнологических процессов с целью достижения мак-
симальных выходов конечных продуктов при культивировании
естественных видов с измененной наследственностью методами
клеточной и генной инженерии; в-шестых, повышение технико-
экономических показателей биотехнологических процессов по
сравнению с существующими.
21
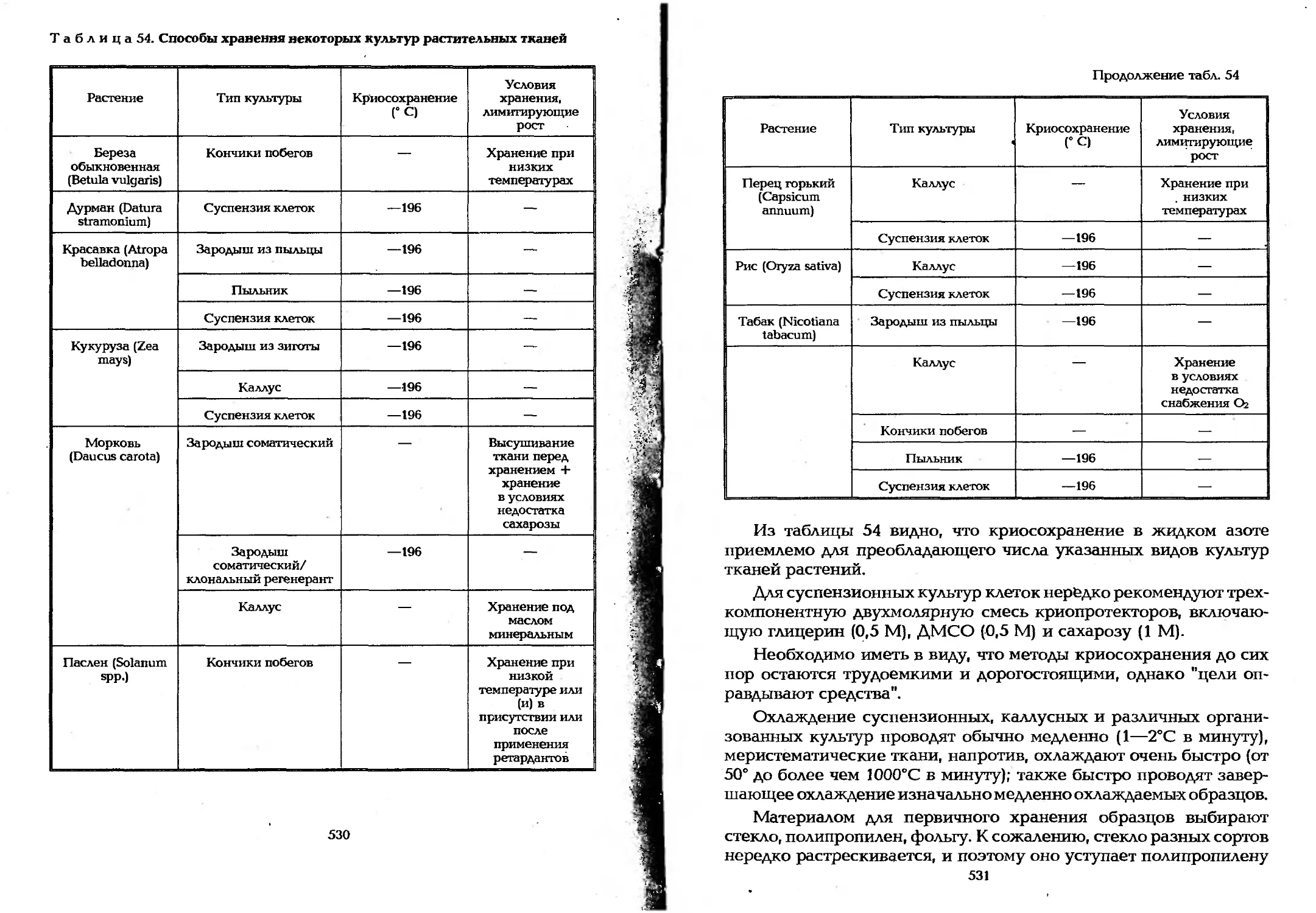
Таблица 1 Процессы в промышленной микробиологии
Характеристика процесса Целевые продукты Названия целевых [ продуктов или процессов
Биосинтез Метаболиты: преметаболиты Аминокислоты ?
Нуклеозиды j
Нуклеотиды ||
первичные Нуклеиновые кислоты ||
Ферменты
вторичные Алкалоиды
Антибиотики
Гиббереллины j
Гликаны и гликоконъюгаты в
Органические кислоты, кетоны, спирты,
Липиды
Аминокислоты, пептидные гормоны
Клеточная масса Пекарские и пивные дрожжи 1
Кормовой и пищевой белок
Вакцины и антигенные вещества
Трансформация Неорганические вещества Обнаружение металлов
Обогащение металлов !
Преимущественно органические вещества Компостирование отходов, получение биогаза [
Детоксикация, дезодорация « и обезвреживание, например, ! ПАВ (поверхностно-активных в-в) |
Определение (анализ) веществ по | продуктам трансформации |
Кисломолочные продукты и сыры
Хлебно-булочные изделия
Квашение и соление овощей ||
Силосование кормов
Мочка льна и джута J
Ферментация чая, табака, кофе, [ какао, маслин [
Пивоварение, виноделие, | винокурение j
Глава 2
ОБЪЕКТЫ И МЕТОДЫ БИОТЕХНОЛОГИИ
Объектами биотехнологии являются вирусы, бактерии, грибы
— микромицеты и макромицеты, протозойные организмы, клетки
22
(ткани) растений, животных и человека, некоторые биогенные и
функционально сходные с ними вещества (например, ферменты,
простагландины, лектины, нуклеиновые кислоты и др.). Следова-
тельно, объекты биотехнологии могут быть представлены органи-
зованными частицами (вирусы), клетками (тканями) или их мета-
болитами (первичными, вторичными). Даже при использовании
биомолекулы как объекта биотехнологии исходный биосинтез ее
осуществляется в большинстве случаев соответствующими клет-
ками. В этой связи можно сказать, что объекты биотехнологии
относятся либо к микробам, либо к растительным и животным
организмам. В свою очередь организм можно образно характери-
зовать как систему экономного, сложнейшего, компактного, само-
регулируемого и, следовательно, целенаправленного биохимиче-
ского производства, устойчиво и активно протекающего при оп-
тимальном поддержании всех необходимых параметров. Из такого
определения следует, что вирусы не являются организмами, но по
содержанию молекул наследственности, приспособляемости, из-
менчивости и некоторым другим свойствам они относятся к пред-
ставителям живой природы.
Как видно из приводимой схемы, объекты биотехнологии
исключительно разнообразны, диапазон их распространяется от
организованных частиц (вирусов) (рис. 1А) до человека.
Вирусы занимают положение между живой и неживой приро-
дой, у них нет ядра, хотя имеется наследственный ядерный мате-
риал — рибонуклеиновая кислота (РНК) или дезоксирибонуклеи-
новая кислота (ДНК).
В отличие от микробов клеточной организации РНК и ДНК в
вирусных частицах вместе никогда не обнаруживаются.
Из сказанного следует, что названия "биологическая техноло-
гия, или биотехнология" и "биохимическая технология" тождест-
венны, так как биологические процессы, используемые в технике
и промышленном производстве, базируются на биохимической
основе.
В настоящее время большинство объектов биотехнологии со-
ставляют микробы, относящиеся к трем надцарствам (безъядер-
ные, предъядерные, ядерные) и пяти царствам (вирусы, бактерии,
грибы, растения и животные). Причем первые два надцарства
состоят исключительно из микробов, тогда как третье — преиму-
щественно из растений и животных.
23
Микробами среди растений являются микроскопические водо-
росли (AJgae), а среди животных — микроскопические простейшие
(Protozoa). Из эукариот к микробам относятся грибы и, при опре-
деленных оговорках, лишайники, которые являются природными
симбиотическими ассоциациями микроскопических грибов и мик-
роводорослей или грибов и цианобактерий.
В первой половине XIX в. было сделано одно из самых основных
обобщений биологии — клеточная теория (М. Шлейден, Т. Шванн,
Р. Вирхов), которая стала общепризнанной. Она же оказалась
фундаментом науки — цитология (от греч. kitos — полость). Из
всех объектов биотехнологии лишь вирусы, вироиды и биомоле-
кулы не имеют клеточной организации. Однако вирусы, находясь
в клетках, ведут себя как живые существа — они реплицируются
("размножаются") и их генетический материал функционирует, в
24
основном^ по общим законам, присущим клеткам любого проис-
хождения. По мере совершенствования методов и техники цито-
логических исследований ученые глубже проникают в сущность
организованных частиц и клеток, а в результате такого проникно-
вения удается обосновать принадлежность всех живых существ к
трем надцарствам: Acaryotae — безъядерные, Procaryotae — предъ-
ядерные и Eucaryotae — ядерные (от греч. а — нет, pro — до, ей
— хорошо, полностью, кагуоп — ядро). К первому относятся
организованные частицы — вирусы и вироиды, ко второму —
бактерии, к третьему — все другие организмы (грибы, водоросли,
растения, животные).
Несмотря на то, что представители всех надцарств содержат
генетический материал, различные акариоты лишены какого-либо
одного типа нуклеиновой кислоты РНК или ДНК. Они не способны
функционировать (в том числе — реплицироваться) вне живой
клетки, и, следовательно, правомочно именовать их безъядерными.
Бактерии имеют клеточную организацию и у них имеются
нуклеиновые кислоты обоих типов — РНК и ДНК, из которых ДНК
представлена в виде одиночной (кольцевидной) хромосомы. Боль-
шинство из них размножается на питательных средах (вне орга-
низма), а если среди бактерий и есть безусловные (облигатные)
паразиты, приближающиеся по данному признаку к вирусам (хла-
мидии, спироплазмы, риккетсии), то паразитизм их отличается по
своему механизму — его можно назвать клеточным. Паразитизм
вирусов развивается на генетическом уровне. Таким образом,
бактерии — это организмы, состоящие из функционально связан-
ных структур, в том числе, генетических. Несмотря на то, что
генетические структуры бактериальной клетки функционируют
полноценно, они не сгруппированы в форме отграниченного ядра,
и поэтому бактерии отнесены к предъядерным (прокариотическим)
организмам.
Клетки грибов, водорослей, растений и животных имеют на-
стоящее, отграниченное от цитоплазмы, ядро и поэтому их относят
к эукариотам.
2.1. Вирусы. Среди микробов вирусы характеризуются
наименьшей величиной — они измеряются в нанометрах (нм.), и
облигатным паразитизмом. Последний признак положен в основу
классификации их на вирусы бактерий, или бактериофаги, вирусы
растений и вирусы животных; имеются также и вирусы грибов.
Как уже было сказано, структурно вирусы представляют собой
организованные частицы, содержащие один какой-либо тип нук-
леиновой кислоты (РНК или ДНК), не обладающие собственным
25
обменом веществ, но способные к репликации в клетках организ-
ма-хозяина или интеграции с его геномом, ведя при этом "скрытое
существование”. Под организованностью вирусной частицы пони-
мают специфическое построение, или архитектонику (от греч. archi
— начальный, главный, первый, tecton — искусник, мастер) струк-
турных блоков, характерную для того или иного вируса, сущест-
вующего вне организма — в и р и о н (см. рис. 22а). Каждый вирион
в очищенном виде представляет собой истинный кристалл, кото-
рый построен из нуклеиновой кислоты и белка, не связанных друг
с другом ковалентными связями. Понятие "вирион" относится к
интактной вирусной частице (от лат. intactus — нетронутый,
неповрежденный), способный к инфицированию или заражению
(от лат. infectiosus — заразный).
Нуклеиновые кислоты — вещества наследственности вирусов.
По типу нуклеиновой кислоты их подразделяют на РНК-содержа-
щие вирусы и ДНК-содержащие вирусы. К первым относят все
вирусы растений, ко вторым — большинство бактериофагов, ряд
вирусов человека и животных (аденовирусы, вирусы герпеса,
осповакцины и др.).
Белок структурируется вокруг вирусной нуклеиновой кислоты
(генома) в виде оболочки и называется капсидом. Форма
вириона определяется его капсидом. Вместе с нуклеиновой кисло-
той капсид образует н у к л е о к а п с и д.
Примерный перечень вирусов включает 17 семейств вирусов
позвоночных и 7 семейств вирусов беспозвоночных животных, 10
семейств вирусов бактерий. Описаны 20 родов вирусов растений
и 5 родов вирусов грибов. Классификационные схемы вирусов до
конца еще не устоявшиеся, к тому же открывают новые для науки
вирусы (пример с вирусами эбола, иммунодефицита человека —
ВИЧ). Представителями ДНК-содержащих вирусов являются ви-
русы контагиозного моллюска, оспы, герпеса, большинство фагов
бактерий; РНК-содержащими являются вирусы растений, вирусы
гриппа человека, бешенства, полиомиелита и др.
2.1.1. Вироиды. В 1971 г. Т. О. Динер (США) впервые описал
субвирусный возбудитель (патоген) веретеновидности клубней
картофеля (ВВКК), названный вироидом. К 1984 г. было известно
10 болезней культурных растений (в том числе — зерновых),
вызываемых вироидами. По молекулярной структуре вироиды
представляют собой одноцепочечные, ковалентно замкнутые, коль-
цевые молекулы РНК, лишенные капсидов. Число нуклеотидов в
таких РНК находится в пределах 240 — 400. По форме вироиды
26
могут быть линейные и кольцевидные, они способны принимать
шпилечную, квазидвухцепочечную конформацию (от лат. quasi —
якобы, как-будто, почти, близко; conformatio — форма, расположе-
& >: а
iW
Рис. 1. Бактериофаг Т2(А): 1 — капсомеры, по-
крывающие головку, 2— воротничок, 3 — шей-
ка, 4 — полый стержень, 5 — чехол, 6 — нити,
7 — икосаэдрическая головка; мендсикуты Б (а-
д): галобактерии (а), метанобактерии (б), вегета-
тивные формы (в) и покоящиеся формы (г);
термоацидофилы (д); тенерикуты (е): микоплаз-
мы (ж), спироплазмы (з).
ние). Каждый тип вироида, например ВВКК или вироид экзокор-
тиса цитрусовых (ВЭЦ), содержит уникальный, только ему прису-
щий, особый вид низкомолекулярной РНК. Размеры вироидов
находятся в пределах 15 нм. В чувствительных клетках растений-
хозяев они сосредоточиваются в ядре, ассоциируясь с ядрышком
в виде белково-нуклеинового комплекса, и реплицируются авто-
номно-целиком при помощи предсуществутощих или активирован-
ных ферментов хозяина. Вироиды не транслируются! Это подтвер-
ждается структурным сходством их между собой и отсутствием у
ряда вироидов кодонов-инициаторов. В то же время репликация
происходит благодаря транскрипции последовательностей вироид-
ных РНК с РНК-матриц при участии РНК-полимераз.
2.2. Бактерии — существа клеточной организации, у
которых ядерный материал не отделен от цитоплазмы элементар-
ными мембранами и не связан (!?) с какими-либо основными
белками. Цитоплазма в них с нерегулярно разбросанными рибо-
сомами (705-типа) неподвижна, клетки не обладают способностями
к эндо- и экзоцитозу. В большинстве своем бактерии одноклеточны,
наименьший диаметр их 0,2 — 10,0 мкм.
Все бактерии составляют единое царство Bacteria, хотя одни из
них — археобактерии (Archaeobacteria) заметно отличаются от
других, названных эубактериями (Eubacteria) (От греч, ей — хоро-
шо). Очевидно, археобактерии являются более древними предста-
вителями прокариот, чем эубактерии. Они обитают в средах с
экстремальными условиями (от лат. extremus — крайний) — высо-
кие концентрации неорганических солей, повышенные темпера-
туры, оксид и диоксид углерода — как единственные источники
углерода. К археобактериям относятся галобактерии, термоацидо-
фильные бактерии и метанобразующие, или метаногенные бакте-
рии (рис. 1Б). Дендрограмма (от греч. dendron — дерево, gramma
— описание) прокариот может быть изображена следующим об-
разом:
Галофильные Термоацидофиль— Метаногенные фототрофные Хемотрофные
бактерии ные бактерии бактерии бактерии бактерии
Эубактерии
Предковые формы
Фототрофными бактериями являются оксигенные цианобакте-
рии, аноксигенные пурпурные и зеленые бактерии; хемотрофными
— грамположительные и грамотрицательные бактерии и бациллы,
миксобактерии, стебельковые и почкующиеся бактерии, вибрио-
ны, спириллы, спирохеты, актиномицеты, коринебактерии, мико-
бактерии, риккетсии, хламидии, микоплазмы и спироплазмы.
Галобактерии включают роды Haloarcula, Halobacterium,
Halococcus, Natrobacterium, Natrococcus. Они обнаруживаются в
морских солеварнях (оптимум натрия хлорида для них 3,5 — 5 М).
Термоацидофильные бактерии обитают в кислых горячих ис-
точниках при pH 2 — 3 и температуре 70° — 90°С (Sulfolobus
acidocoldarius), в самонагревающихся терриконах угольных шахт
при pH 1 — 2 и температуре 59°С (Thermoplasma acidophilum), в
горячих источниках на дне морей и на склонах вулканов при
температуре 85 — 105°С (Thermoproteus tenax, Т. neutrophilus и
ДР-)-
Метаногенные бактерии, к которым относятся кокки
(Methanococcus vannielli), сарцины (Methanosarcinabarkeri), палоч-
ки (Methanobacteriumformicicum, Methanobrevibacterruminantium),
спириллы (Methanospirillum hungatei) и другие формы, являются
анаэробными микроорганизмами. Они обитают в отстойниках
сточных вод городов и населенных пунктов, в навозе, в осадках на
дне прудов и озер, в рисовых полях, в лиманах и эстуариях (от лат.
aestuarium — берег, заливаемый приливом), в рубце жвачных
28
животных. Приуроченность их к широкому диапазону температур
естественна, учитывая среды обитания. Тем не менее, например в
рубце жвачных животных температура достаточно постоянная. .
Согласно определителю Д. X. Берги (1984, 1986) археобактерии
относятся к отделу мендосикутов, все другие бактерии, или эубак-
терии — к отделам: грациликутов, фирмикутов и тенерикутов (от
лат. mendosus—ложный, фальшивый, gracilis—стройный, тонкий,
firmus — прочный, tener—чувствительный, нежный, cutis—кожа).
Грациликуты объединяют грамотрицательные бактерии с двух-
слойной клеточной стенкой (они, как правило, содержат фосфо-
липидную мембрану во внешнем слое клеточной стенки). Форма
клеток у них различная — от сферической до палочковидной, от
прямых до искривленных (изогнутых); подвижные или неподвиж-
ные; не образуют эндоспор, размножаются делением (некоторые
— почкованием); многие обладают пилями (волосками, или фимб-
риями). По типам питания к грациликутам относятся фототрофы
(включая цианобактерии) и хемотрофы, по типам дыхания —
аэробы, анаэробы и факультативные анаэробы, по патогенности
— сапрофиты и паразиты.
Микроорганизмы с многослойным муреиновым каркасом от-
носятся к фирмикутам. Все они грамположительные, спорообра-
зующие или не образующие спор; сюда же включены актиноми-
цеты и родственные им бактерии. Форма клеток различная —
круглые, палочковидные, ветвящиеся, нитевидные, неветвящиеся.
По типу питания — преимущественно хемогетеротрофы, по типу
дыхания — аэробы, анаэробы и микроаэрофилы, патогенные и
сапрофиты.
К тенерикутам относятся микоплазмы и спироплазмы (от греч.
тукев — гриб, plasma — вязкая, эластичная; speira — завиток,
спираль, кольцо) (рис. 1Б). Это мельчайшие свободноживущие
полиморфные бактерии, без клеточной стенки, сгруппированные
в классе Mollicutes (от лат. mollis — мягкий). Воспроизводятся они
почкованием, делением, сегментацией ветвистых структур; цитоп-
лазматическая мембрана трехслойна; все известные виды микоп-
лазм патогенны. Диаметр их 0,15 — 0,25 мкм, хотя полиморфизм
по длине клеток весьма широкий. Микоплазмы полностью устой-
чивы к пенициллину; растут на специальных агаризованных сре-
дах.
К мендосикутам относится единственный уже рассмотренный
класс археобактерий, заметно отличающийся от других микроор-
ганизмов.
Известны еще так называемые мини- и макси-клетки бактерий,
29
в частности, Е. coli. Установление, что клетки кишечной палочки,
несущие две мутации (min А и min В) делятся ассиметрично и в
каждое второе деление образуется круглая безъядерная мини-клет-
ка (примерно в 3 раза меньше родительской клетки по размеру).
Мутации в генах гес А и uvt А сопровождаются инактивацией
основных систем репарации и существенным возрастанием чувст-
вительности клеток к ультрафиолетовым лучам. Клетки Е. coli при
этом увеличиваются в размерах (макси-клетки). Мини- и макси-
клетки используются для включения многокопийных плазмид при
постановке генно-инженерного эксперимента.
Бактерии подразделяют на группы по источникам энергии,
углерода и донорам электронов (таблица 2), благодаря чему можно
выделить типичных представителей в каждой из них.
Так, цианобактерии относят к первой подгруппе, зеленые
несерные бактерии — к второй, нитрифицирующие бактерии —
к третьей, водородные бактерии — к четвертой и, наконец, бациллы
и другие микроорганизмы — к пятой. Для сравнения можно
указать, что дрожжи и нитчатые грибы из эукариот также отно-
сятся к хемогетероорганотрофам. '
В учебной и научной литературе можно встретить и такие
термины, как прототрофы, метатрофы, паратрофы, миксотрофы.
Следует подчеркнуть, что все эти термины были предложены в
начале XX в. в связи с разделением бактерий на группы по типам
Таблица 2. Деление микробов на группы по источникам энергии, углерода
и доноров электронов.
Группа Источник Номер и название подгруппы 1
энергии углерода доноров электронов (водорода)
Фотот- рофные бактерии Свет Неоргани- ' Неорганические 1. Фотоавтолито-
чески й Органи- ческий вещества Органические вещества трофы 1 2. Фотогетероор- ган отрофы+
Хемот- рофные бактерии Химичес- кие реакции окисления - восста- новления Неоргани- Неорганические 3. Хемоавтолито-
ческий Органи- ческий вещества Неорганические трофы 4. Хемогетероли-
вещества Органические вещества тотрофы 5. Хемогетероор- ганотрофы
Примечание: От греч. photos — свет, autos — сам, lythos — камень, trophia —
питание, etheros — иной, другой, organicos — органический.
30
питания. Так А. Фишер в 1903 г. предложил различать прототрофы
(автотрофы —' по В. Пфеферу), не нуждающиеся в готовом орга-
ническом веществе для питания, метатрофы (гетеротрофы — по
В. Пфеферу), нуждающиеся в органическом веществе — это
большинство сапрофитных микроорганизмов, и паратрофы, пита-
ющиеся живым белком и обитающие лишь в организме других
существ — это все облигатные болезнетворные микробы; те виды,
которые могут расти на питательной среде в пробирке (in vitro) и
в организме чувствительного животного (in vivo), были названы В.
Пфефером миксотрофами (от лат. mixtus — смешанный). Приве-
денные названия (прототрофы, метатрофы и миксотрофы) теперь
имеют больше исторический смысл. В 1946 г. была предложена
классификация микробов по способам питания, или трофики,
которой пользуются и в настоящёе время (таблица 3).
Таблица 3. Деление микробов по трофике
Признак Тип питания
Источник энергии 1) живой организм 2) химическая реакция 3) фотохимическая реакция Источник углерода 1) диоксид углерода 2) органические вещества Донор электрона 1) неорганический 2) органический Паратрофия Хемотрофия Фототрофия Автотрофия Гетеротрофия Литотрофия Органотрофия
Применительно к характеристике способностей микроорганиз-
мов питаться живым белком in vitro или, чаще in vivo, используют
термин патогенность (от греч. pathos — болезнь, genesis — воз-
никновение, происхождение), то есть способность вызывать забо-
левание. Поэтому все бактерии по признаку патогенности подраз-
деляют на две большие группы — сапрофитные (неболезнетвор-
ные, от лат. saprotes — гниение, гниль) и патогенные (болезнетвор-
ные). Между ними находятся промежуточные виды — облигатные
и факультативные паразиты (от лат. obligatus — обязательный,
безусловный, facultativus — факультативный, возможный в изве-
стных условиях). Облигатными являются, например, спирохеты
сифилиса, гонококки, факультативными — синегнойная палочка,
вульгарный протей. Следовательно, облигатные, или безусловные
паразиты не развиваются вне организма или развиваются с трудом
на питательных средах с нативными белками; факультативные
31
паразиты растут и размножаются во внешней среде, но при
некоторых обстоятельствах (например, при снижении защитных
сил макроорганизма) они могут быть причиной инфекционного
(заразного) заболевания.
У фототрофных и хемотрофных бактерий, различающихся по
источникам потребляемой энергии, существенно различны и ре-
акции, лежащие в основе энергетического и конструктивного
обменов. В любом случае разнообразие бактериальных видов
настолько велико, что даже ничтожно малая доля их, используемых
в биотехнологических целях, требует к себе исключительного
внимания. Достаточно назвать из числа эубактерий такие далеко
отстоящие микроорганизмы, как Micromonospora sp. из группы
актиноплан, Hydrogenomonas (Alcaligenes) eutropha из группы во-
дородных бактерий, Lucibacterium harveyi из группы светящихся
бактерий, палочку туберкулеза — Mycobacterium tuberculosis, не-
которых псевдомонасов (например, Pseudomonas aeruginosa), эше-
рихии (Escherichia coli) и многие другие, то становится понятной
исключительная важность бережного (можно сказать — любовно-
го) обращения с каждым биообъектом для сохранения стабильно-
сти его морфо-физиологических свойств и продуктивности в со-
ответствующих условиях.
2.3. Г р и б ы. К царству низших эукариот — Mycota относятся
микромицеты, то есть микроскопические грибы (например, дрож-
жи, пенициллы, аспергиллы и др.) и макромицеты, формирующие
в процессе своего роста и развития визуально наблюдаемые пло-
довые тела — трутовики, агариковые грибы и др. Микро- и
макромицеты могут быть объектами биотехнологии.
Примечательно, что грибы имеют сходство и с растениями
(верхушечный, или апикальный рост, прочная клеточная стенка,
наличие вакуолей и поперечных перегородок у многих из них) и
с животными (гетеротрофный тип питания, большая или мёныпая
потребность в витаминах, наличие хитина или хитозана, синтез
гликогена). Следовательно, грибы эволюционно произошли рань-
ше — до дивергенции растений и животных в самостоятельные
царства. В то же время лишь грибам присуще мицелиальное
строение и, как следствие, абсорбционный способ питания (осмот-
рофия); для них известны явления дикариозиса (раздельное на-
хождение двух ядер в одной клетке, способных к одновременному
делению и имитирующих диплоидное ядро) и гетерокариозиса
(нахождение разнокачественных ядер в одной клетке).
Основные таксономические группы грибов (от греч. taxis —
32
приведение в порядок, устройство, nomos — закон) являются
достаточно устоявшимися, однако предлагаемые разными автора-
ми классификационные схемы весьма многочисленны и, порой, во
многом различны. В этой связи целесообразно и научно оправданно
придерживаться следующей схемы. Царство грибов включает два
отдела — Myxomycota и Eumycota, то есть грибы-слизевики (от
греч. пгуха — слизь) и настоящие грибы (от греч. ей — хорошо, в
смысле — типичный, хорошо развитый). Первые из них немного-
численны и представлены "голой" плазменной массой — плазмо-
дием. Они претерпевают своеобразный цикл развития и образуют
половые аттрактанты (от лат. attractio — притяжение, тяготение).
Отдел эумицетов включает 7 классов: Chytridiomycetes,
Hyphochytridiomycetes, Oomycetes, Zygomycetes, Ascomycetes,
Basidiomycetes и Deuteromycetes. К хитридиевым (от греч.
chytridion — капелька, что отражает содержание капли жира в
зооспорах) относится более 500 видов грибов, в основном пред-
ставленных плазмодиальными образованиями, то есть у них пол-
ностью отсутствует мицелий, а если он есть, то находится лишь в
зачаточном состоянии. Зооспоры и планогаметы (клетки размно-
жения) располагают только одним задним бичевидным (плетевид-
ным) жгутиком, что имеет определенное таксономическое значе-
ние.
Гифохитридиевые имеют зооспоры с одним передним биче-
видным жгутиком, нити их не имеют перегородок, класс представ-
лен немногими видами.
Оомицеты также включают свыше 500 видов. Это водные
грибы, объединяемые на основании оогамии — полового процесса.
Бесполые зооспоры их обладают двумя разными жгутиками, один
из которых передний сверкающий (блестковидный, перистый),
другой — задний бичевидный.
Зигомицеты, включающие свыше 500 видов, полностью утра-
тили подвижные стадии в циклах развития. Половой процесс у них
— зигогамия. Мицелий, как правило, хорошо развит и, в основном,
без перегородок.
Нередко, в отечественной и зарубежной учебной и научной
литературе все, преимущественно водные, грибы (включая и "вы-
шедших на сушу" зигомицетов) объединяют в один класс
Phycomycetes (от греч. phycos — водоросль). Все водные грибы
либо лишены мицелия, либо он в зачаточном или развитом состо-
янии, но не имеет перегородок (септ) или они редкие — такие
грибы относят к низшим. Нитчатые грибы с перегородками в
2 т. 8524 33
мицелии относят к высшим. Не путать эти понятия с понятиями
совершенные и несовершенные грибы, из коих первые обладают
половым процессом размножения, вторые — не обладают им.
Например, Mucor rouxii является низшим совершенным грибом, а,
например, Stilbella aurantica является высшим несовершенным
грибом.
Следовательно, к высшим грибам относят аскомицеты, или
сумчатые грибы; базидиомицеты, или базидиальные грибы и дей-
теромицеты, или несовершенные грибы (Fungi imperfecti).
Сумчатые грибы — наиболее обширный класс, включающий
свыше 15 000 видов самых различных строения, формы и место-
обитания. Отличительным признаком их являются сумки, образу-
ющиеся на аскогенных гифах в результате полового процесса. В
сумках формируются половые споры, с помощью которых они
размножаются. Чаще образуется 8 спор, хотя нередки и исключе-
ния из этого правила. Мицелий у них септированный (от лат. septum
— перегородка), в септах имеются центральные поры, обеспечи-
вающие сообщения между клетками и обмен клеточным содержи-
мым.
Сформировавшиеся сумки образуют плодовые тела, которые
могут быть закрытыми (клейстотеции, или клейстокарпии, от лат.
cleistos — могущий закрываться, смыкающийся; teke — капсула,
покрышка, мешок; carpos — плод), грушевидными с круглой по-
рой-остиолой наверху —перитеции (от греч. peri — вокруг) и
открытыми блюдцевидными или чашевидными апотециями (от
греч. приставки аро — со значением удаления или отделения,
обратности или возвращения и др.). Для сумчатых грибов харак-
терно и бесполое размножение с помощью конидий, образующих-
ся на гаплоидном мицелии. Следовательно, в цикле развития
аскомицетов имеются половые и бесполые стадии (рис. 2).
Базидиальные грибы. Насчитывается свыше 30 000 видов ба-
зидиомицетов (от греч. basis — основание), различающихся боль-
шим разнообразием по строению, форме и размерам.-Для базиди-
омицетов так же характерны половая и бесполая стадии развития.
Первая завершается формированием базидии — репродуктивного
органа, на котором образуются, как правило, по 4 споры (базиди-
оспоры), сидящих на стеригмах. Бесполая стадия кратковременна
— она представлена прорастающими трубками спор и позже —
дикариотическим мицелием. Из этого последнего возникает пло-
довое тело — базидиокарп, на котором затем образуются базидии.
Для многих базидиомицетов характерны пряжки (рис. 3), об-
разующиеся на мицелии и участвующие в синхронном процессе
деления ядер, а в мицелиальных септах — долипоры (см. рис. 32).
34
Рис. 2. Цикл развития аскомицетов на примере Pyronema species: 1-мицелий, 2-ко-
нидии, образующиеся на гаплоидном мицелии, 3-( + ) и (-) типы спаривания (соот-
ветственно мужские и женские клетки), 4-аскогенные гифы (а-д), 5-сумки со спо-
рами, 6-аскоспоры.
Дейтеромицеты (от греч. _______
deiteros — второй) — сбор- < Т*"
ный класс грибов, поскольку
в него включены все те пред-
ставители микромицетов, у
которых не обнаружен поло- ________
вой процесс развития. Если
же половая стадия выявляет-
ся, ТО такой гриб сразу же Рис. 3. Пряжки у базидиальных грибов,
переносят в соответствую-
щий ему класс.
Насчитывают многие тысячи видов несовершенных грибов. Им
присущ хорошо развитый септированный мицелий и конидиальное
(бесполое) спороношение. У несовершенных грибов могут иметь
место гетерокариоз и парасексуальный цикл.
Лишайники (от лат. Lichenes) — это естественные симбионты
грибов (микобионтов) и водорослей (фикобионтов) или грибов и
цианобактерий (бактериобионтов). Лишайники выделяют в само-
стоятельную группу организмов, изучаемую в специальной науч-
35
ной дисциплине—лихенология. В настоящее время известно около
30 000 видов лишайников. Микобионтами у них выступают пре-
имущественно аскомицеты (исключительно редко — базидиоми-
цеты), фикобионтами и бактериобионтами — зеленые и желто-зе-
леные водоросли, цианобактерии. Называют лишайники по мико-
бионту. Лишайники подразделяют по форме на листоватые (в том
числе — кочующие), кустистые и корковые (накипные). Размно-
жаются они бесполым (кусочками, конидиями) и половым путем
за счет микобионта (рис. 4).
Рис. 4. Лишайники: корковый или накипной - 1, листоватый - 2, кустистый - 3,
кочующий - 4.
Из сказанного можно составить представление о том, что
потенциальных биообъектов для использования в производстве
самых различных веществ обильное множество и большинство из
них лежит пока нетронутым кладом для будущих поколений кла-
доискателей.
2.4 Растения. Царство растений Plantae включает подцарства
багрянок (Rhodophyta), водорослей (Phycophyta) и высших расте-
ний (Embryophyta). У первых двух нет дифференциации тела на
органы и ткани — они представляются слоевищем (thallus) и
обитают главным образом в воде. Тело высших растений расчле-
нено на органы и ткани. К настоящему времени насчи-
тывают сотни тысяч видов растений, многие из которых исполь-
зуют в различных отраслях народного хозяйства.
Для растений характерны: способность к фотосинтезу, наличие
целлюлозы, биосинтез крахмала.
2.4.1. Водоросли. К водорослям относятся: багрянки,
пиррофитовые, золотистые, желто-зеленые, эвгленовыеихаровые.
36
Как правило, водоросли являются водными организмами, их на-
считывают около 100 000 видов. Все они пигментированы за счет
хлорофилла, каротиноидов, ксантофиллов, фикобилинов. Водорос-
ли — важный источник различных полисахаридов и других био-
логически активных веществ. Размножаются они вегетативно,
бесполым и половым путями. Как биообъекты используются недо-
статочно, хотя, например, ламинария под названием морской
капусты производится промышленностью различных стран. Хоро-
шо известны агар-агар и альгинаты, получаемые из водорослей.
2.4.2. Клетки высших растений. Высшие растения
(порядка 300 000 видов) — это дифференцированные многоклеточ-
ные, преимущественно наземные организмы. Способы их беспо-
лого и полового размножения хорошо описаны в учебниках бота-
ники. В процессе дифференциации и специализации клетки рас-
тений группировались в ткани (простые — из однотипных клеток,
и сложные — из разных типов клеток). Ткани, в зависимости от
функции, подразделяют на образовательные, или меристемные (от
греч. meristos—делимый), покровные, проводящие, механические,
основные, секреторные (выделительные). Из всех тканей лишь
меристематические способны к делению и за их счет образуются
все другие ткани. Это важно для получения клеток, которые затем
должны быть включены в биотехнологический процесс (см. спе-
циальную часть),
Клетки меристемы, задерживающиеся на эм-
бриональной стадии развития в течение всей
жизни растения, называются инициальными,
другие постепенно дифференцируются и превра-
щаются в клетки различных постоянных тканей
— конечные клетки.
В зависимости от топологии в растении мери-
стемы подразделяют на верхушечные, или апи-
кальные (от лат. apex — верхушка), боковые, или
латеральные (от лат. lateralis — боковой) и про-
межуточные, или интеркалярные (от лат.
intercalaris — промежуточный, вставной) (рис. 5).
В 1902 г. Г. Хаберландт впервые сделал попыт-
ки культивировать клетки растений. В середине
текущего столетия промышленное производство
декоративных и плодоовощных культур в ряде
стран мира базировалось преимущественно на
Рис. 5. Меристемы
растительные: вер-
хушечные — 1. бо-
ковые — 2, проме-
жуточные — 3.
37
методе культур тканей и органов растений — были получены линии
с заданными характеристиками. Здесь следует отметить, что Г.
Хаберландт впервые выдвинул гипотезу о тотипотент-
ности любой живой клетки растения. Тотипотентность — это
свойство соматических клеток растений полностью реализовать
свой потенциал развития вплоть до образования целого растения.
Любой вид растения может дать в соответствующих условиях
неорганизованную массу делящихся клеток — каллус (от лат. callus
— мозоль), особенно при индуцирующем влиянии растительных
гормонов. Массовое производство каллусов с дальнейшей регене-
рацией побегов пригодно для крупномасштабного производства
растений. Вообще каллус представляет собой основной тип куль-
тивируемой на питательной среде растительной клетки. Каллусная
ткань из любого растения может длительно рекультивироваться.
При этом первоначальные растения (в том числе и меристемати-
ческие), дедифференцируются и деспециализируются, но индуци-
руются к делению, формируя первичный каллус.
Кроме выращивания каллусов удается культивировать клетки
некоторых растений в суспензионных культурах.
Важными биообъектами представляются также и протопласты
растительных клеток. Методы их получения принципиально сход-
ны с методами получения бактериальных и грибных протопластов.
Последующие клеточно-инженерные эксперименты с ними заман-
чивы по возможным ценным результатам.
2.5. Клетки животных. Из царства Animalia биообъектами
могут быть простейшие организмы — Protozoa и высшие животные.
И если сегодня о биотехнологии Protozoa мало что-либо известно,
то применительно к биотехнологии животных имеются внедрен-
ные развитые технологические процессы, написаны соответству-
ющие монографии в нашей стране и за рубежом. Тем не менее,
высокие дифференциация и специализация эукариотических кле-
ток животных объясняют те трудности, с которыми приходится
сталкиваться исследователям и практическим работникам, имея
дело с подобным материалом.
Простейшие (Protozoa) — это одноклеточные микроскопиче-
ские животные. В данном отделе различают классы жгутиковых
(Flagellata, или Mastigofora, от лат. flagellum — бич, жгут, masticatus
— жевательный, от греч. foros — нести), саркодовые (Sarcodina, от
греч. sarcos — мясо), споровики (Sporozoa) и реснитчатые (Ciliofora,
38
или Ciliata). Простейшие широко распространены в природе,
некоторые из них обитают и в теле человека. По своему строению
клетки их напоминают клетки животных и содержат все основные
структурные элементы (органоиды и включения). Многие протозоа
активно передвигаются с помощью ложноножек, жгутиков и ре-
сничек. По типу питания они являются гетеротрофами, обладаю-
щими специальными структурами для захвата пищи или поглощают
ее посредством фагоцитоза.
Культивирование протозоа in vitro не простое, однако доступно
при необходимом усердии, а препарат "круцин" из Trypanosoma
kruzi является свидетельством удачного использования этого орга-
низма в биотехнологии.
В начале XX в. Р. Гаррисон и А. Каррель установили факт
возможного культивирования клеток животных in vitro, то естьони
доказали способность животных клеток к независимой жизни в
питательной среде вне живого организма.
С учетом всех особенностей животных клеток (в отличие,
например, от растительных и бактериальных) порой невозможно
отказаться от них во избежание каких-то чрезвычайных затрудне-
ний, так как лишь только с их помощью можно получить какой-то
особый продукт (вещество). Для примера назовем моноклональные
антитела, получение трансгенных животных и т. д.
Для реализации биотехнологических процессов важными па-
раметрами биообъектов являются: чистота, скорость размноже-
ния клеток и репродукции вирусных частиц, активность и ста-
бильность биомолекул или биосистем.
Следует иметь в виду, что при создании благоприятных условий
для избранного биообъекта биотехнологии эти же условия, могут
оказаться благоприятными, например, и для микробов — контами-
нантов, или загрязнителей (от лат. contaminatio — заражение,
загрязнение). Представителями контаминирующей микрофлоры
оказываются вирусы, бактерии и грибы, находящиеся в культурах
растительных или животных клеток. В этих случаях микробы-кон-
таминанты выступают вредителями производств в биотехнологии.
При использовании ферментов в качестве биокатализаторов воз-
никает необходимость предохранения их в изолированном или
иммобилизованном состоянии от деструкции банальной сапрофит-
ной (не болезнетворной) микрофлорой, которая может проникнуть
в сферу биотехнологического процесса извне вследствие несте-
39
рильности системы, например, из-за негерметичности в каком-ли-
бо узле.
Скорости размножения клеток и репродукции вирусных час-
тиц прямо пропорционально сказываются на возрастании клеточ-
ной массы и образовании метаболитов или, применительно к
фагам, на возрастании массы лизирующихся клеток. В этом смысле
подавляющее большинство микроорганизмов выгодно отличается
от клеток растений и животных. Однако не следует упускать из
виду важность конечного продукта. Например, уже упомянутые
моноклональные антитела можно получить лишь с помощью жи-
вотных клеток, когда длительность культивирования их не приоб-
ретает самодовлеющего значения.
Активность и стабильность в активном состоянии биообъектов
— одни из важнейших показателей их пригодности для длительного
использования в биотехнологии.
Таким образом, независимо от систематического положения
биообъекта, на практике используют либо природные организо-
ванные частицы (фаги, вирусы) и клетки с естественной генети-
ческой информацией, либо клетки с искусственно заданной гене-
тической информацией, то есть в любом случае используют клетки,
будь то микроорганизм, растение, животное или человек. Для
примера можно назвать процесс получения вируса полиомиелита
на культуре клеток почек обезьян в целях создания вакцины против
Рис. б. Биореактор фир-
мы Inceltech для выращи-
вания бактерий и грибов
(общий вид).
Рис. 7. Биореактор для
растений (общий вид).
40
этого опасного заболевания. Хотя мы заинтересованы здесь в
I накоплении вируса, репродукция его протекает в клетках живо-
!тного организма. Другой пример с ферментами, которые будут
использованы в иммобилизованном состоянии. Источником фер-
ментов также являются изолированные клетки или специализиро-
г ванные ассоциации их в виде тканей, из которых изолируют
- нужные биокатализаторы.
Биотехнологии присущи свои специфические методы — это
крупномасштабное глубинное культивирование биообъектов в пе-
риодическом, полунепрерывном или непрерывном режиме; выра-
щивание клеток растительных и животных тканей в особых усло-
виях. Биотехнологические методы культивирования биообъектов
выполняются в специальном оборудовании, например, в фермен-
таторах выращивают бактерии и грибы при получении антибио-
тиков, ферментов, органических кислот, некоторых витаминов
(рис. 6);
в подобных же ферментаторах выращивают некоторые клетки
человека (бласты) для получения белка— интерферона. Раститель-
ные клетки чаще выращивают в стационарных условиях на среде
. с уплотненной (например, агаризованной) подложкой в стеклян-
ных или полиэтиленовых емкостях, хотя некоторые виды расти-
тельных клеток можно культивировать в специальных фермента-
торах (рис. 7).
В стеклянных роллерах культивируют и большинство живо-
тных клеток или, например, в куриных эмбрионах.
Другие методы, используемые в биотехнологии, являются об-
щими, например, с методами в микробиологии, биохимии, биоин-
женерии, органической химии и других науках.
Тем не менее, следует особо выделить методы клеточной и
генной инженерии, когда в экспериментальных условиях удается
создавать клетки с заведомо известными свойствами. Так осуще-
ствлены соматическая гибридизация клеток картофеля и томата
(гибрид назван "помато”), перенос генетической информации о
синтезе человеческого или животного гормона инсулина в бакте-
риальные клетки (кишечной палочки), способных затем продуци-
ровать полипептидные цепи инсулина.
Эти методы и, прежде всего, генно-инженерные легли в основу
современной биотехнологии. Более того, некоторые ученые стре-
: мятся отождествить генную инженерию и биотехнологию, а это
41
неправильно, поскольку генная инженерия представляет методи-
ческую сущность биотехнологии. Наука не может быть наукой,
если в ее определении отсутствуют специфичные, только ей при-
сущие, объекты и методы.
Однако даже в случае реализации генно-инженерных разрабо-
ток измененная наследственная информация на уровне молекул
инкорпорируется затем в клетках, с которыми и приходится иметь
дело в биотехнологическом процессе. Из этого можно вывести
представление об уровнях биотехнологии: клеточном и молеку-
лярном. Тот и другой определяются биообъектами. В первом случае
дело имеют с клетками, например, актиномицетов при получении
антибиотиков, микромицетов при получении лимонной кислоты,
животных при изготовлении вирусных вакцин, человека при из-
готовлении интерферона. Во втором случае дело имеют с молеку-
лами, например, с нуклеиновыми кислотами в так называемой"ре-
комбинантной ДНК-биотехнологии” (рДНК-биотехнология), бази-
рующейся на генной инженерии и составляющей сущность пред-
мета "Молекулярная биотехнология"; или на использовании отдель-
ных ферментов (ферментных систем), например, протеаз в мою-
щих средствах, липаз для модификации вкуса молочных продуктов
и т. д. Однако необходимо помнить, что в начальной или конечной
стадии молекулярный уровень трансформируется в клеточный.
Так, ферменты продуцируются клетками, а при генно-инженерных
разработках реципиентом новой генетической информации стано-
вится также клетка.
Особенность методов, используемых в биотехнологии, заклю-
чается в том, что они должны выполняться, как правило, в асеп-
тических условиях (от греч. а — не, septicos — гнилостный), то
есть при исключении возможности попадания в среду культиви-
рования биообъекта болезнетворных (патогенных) и неболезнет-
ворных (сапрофитных) микроорганизмов. Патогенные виды пред-
ставляют непосредственную опасность для занятых в производстве
людей и для потребителей конечных продуктов; сапрофитные виды
могут выступать конкурентами за питательные субстраты, антаго-
нистами, продуцентами токсических веществ, включая пирогены.
42
Часть IL
Роль фундаментальных исследований
в развитии биотехнологии
Подразделение научных исследований (равно как и наук) на
фундаментальные и прикладные в какой-то мере условно, хотя
фактически эти термины в достаточно широком ходу. Более того,
фундаментальные и прикладные исследования включают в перс-
пективные планы научных разработок в соответствующих акаде-
мических и отраслевых учреждениях.
Принято считать, что фундаментальные (от лат. fundamentum-
основа, опора, основание) исследования изначально нацелены на
решение основополагающих проблем, а конкретные результаты,
вытекающие из них, составляют суть прикладных исследований.
Это можно показать на следующих примерах (таблица 4).
43
Таблица 4. Соотношение фундаментальных и прикладных исследований
Научные исследования
фундаментальные прикладные
Открытие Г. Менделем единиц наследственности у садового гороха; исследования Т. Моргана и сотрудников с мушкой дрозофилой, приведшие к всеобщему признанию законов Менделя. Вскрытие Л. Пастером причины болезни шелковичных червей во Франции. Обезвреживание болезнетворных микробов при сохранении их антигенности. Выведение М. Фарадеем законов электролиза на основании изучения действия электрического тока на водные растворы различных веществ. Открытие радиоактивных элементов Марией и Пьером Кюри. Формулирование проблемы антагонизма между микробами и ее экспериментальное подтверждение. На основании принципов Менделя - Моргана были созданы рациональные методы селекции в растениеводстве и животноводстве, в микробиологии. С помощью этих методов были выведены новые сорта сельскохозяйственных растений, новые породы домашних животных, высокопродуктивные штаммы микроорганизмов; выявлены наследственные болезни у человека, определены пути и созданы методы рациональной их профилактики и терапии. Разработаны и осуществлены меры по спасению производства шелка. Создание профилактических и лечебных иммунопрепаратов (вакцин, сывороток, иммуноглобулинов). Создание методов и приборов для выделения веществ при электролизе, для определения электропроводности, для профилактической и терапевтической электростимуляции при некоторых патологических состояниях у людей и т. д. Создание и применение рентгеновских методов исследования, радиотерапия раковых заболеваний, эксплуатация атомных электростанций и др. Создание промышленности антибиотиков и применение антибиотических препаратов в здравоохранении.
Подобного рода примеры весьма многочисленны и множество
их убедительно доказывает скорость и глубину социального про-
44
гресса человечества на долгом пути его эволюции. И наука здесь
выступает решающим фактором, так как наука - это отрасль
человеческой деятельности, направленная на познание мира (ми-
роздания).
Фундаментальные исследования завершаются, как правило,
установлением или доказательством новых фактов, закономерно-
стей в соответствующей отрасли знаний. Рано или поздно эти
факты (закономерности) находят практическое воплощение. Тем
не менее, нельзя противопоставлять фундаментальные и приклад-
ные исследования, так как на определенном этапе нередко можно
видеть трансформацию их в противоположном направлении. На-
пример, на основании обобщения полученных сведений о нукле-
иновых кислотах Дж. Уотсон и Ф. Крик смогли предложить
природную модель двойной спирали ДНК и, тем самым, подвели
материальный фундамент под ранее известный якобы "мифиче-
ский" ген. На этой основе возникли многочисленные практические
разработки по молекулярной биологии гена и, в том числе, напри-
мер, по мутагенезу. С другой стороны, практические разработки
по выделению и манипулированию с ДНК различного происхож-
дения стали основой для постулирования общегенетических зако-
нов, присущих всему живому. Вот почему и сегодня справедливы
слова великого Л. Пастера, сказанные по этому поводу: "Нет,
тысячу раз нет, не существует ни одной категории наук, которой
можно было бы дать название прикладых наук. Существуют науки
и приложение наук, связанные между собой как плод и породившее
его дерево".
Биологическая технология - как наука является воплощением
симбиоза теоретических исследований и практической реализации
предварительно полученных фактов в работах с организованными
частицами (вирусами), клетками и тканями, с первичными и вто-
ричными метаболитами, с генетическим материалом.
Глава 3
Фундаментальные исследования
в области энзимологии
Любая клетка - целостная система, составные части которой
структурно и функционально взаимозависимы. Эта зависимость
выражается прежде всего в генетически обусловленном синтезе
белковых молекул - преимущественно ферментов. В хронологиче-
ском порядке только белки - продукты матричного синтеза должны
45
относиться к разряду первичных; все другие молекулы, возника-
ющие под каталитическим .действием ферментов, оказываются
вторичными. Выдающийся шведский химик И. Я. Берцелиус,
предложивший в 1836 г. термин "катализ" (за 35 лет до выявления
М. Манасеиной каталитических функций дрожжевого сока в 1871
г.), пророчески писал: "У нас есть все основания предполагать, что
в тканях и жидкостях растений и животных происходят тысячи
каталитических процессов”. И это в то время, когда ферменты еще
не были известны науке. Теперь, например, утверждается, что в
мельчайшей клетке (0,1 мкм в диаметре), относящейся к Mollicutes,
содержится более, чем 100 ферментов, то есть столько, чтобы
клетка могла самостоятельно функционировать как организм. Ес-
тественно, что в клетке других прокариот и эукариот насчитывают
более 1000 биокатализаторов. Большинство микробов лишено спе-
циализированных структур, хотя бы отдаленно напоминающих
органы дыхания, пищеварения и другие, присущих высшим эука-
риотам. Поэтому основная метаболическая нагрузка (метаболизм
- это биологический обмен веществ и энергии) в процессах их
роста, развития и размножения падает на ферменты. В любой
живой клетке находятся малые и большие (полимерные) молекулы
различных веществ. Они должны синтезироваться в клетке, а
некоторые секретироваться из нее (нередко, распадаясь перед этим
на более мелкие составные части или их производные). Во всех
таких процессах участвуют ферменты. Уже сегодня известно около
2000 индивидуальных ферментов и это не предел. В клетках
прокариот и эукариот ферменты распределены и локализованы
целесообразно, например, в цитоплазме находятся почти все фер-
менты гликолиза, в матриксе митохондрий - ферменты цикла
трикарбоновых кислот и р-окисления жирных кислот, ферменты
окислительного фосфорилирования - во внутренней мембране
митохондрий.
В клетке E.coli нить ДНК имеет размеры 1,4«106хЗ,0 нм, а массу
1»1014 г. В разомкнутом состоянии длина ее составит примерно 1,4
мм, то есть подобная ДНК приблизительно в 500 раз длиннее
бактериальной клетки, вмещающей эту ДНК. Такая хромосома
кишечной палочки заключает в себя- информацию, достаточную
для кодирования 4500 белков, значительная часть которых будет
представлена ферментами.
Ферменты составляют основную массу клеточных белков. На
долю одного фермента приходится от сотых долей процента (у
некоторых вирусов) до 10 -12% (у ряда микроорганизмов клеточной
организации, например у E.coli). В то же время хромосомная ДНК
не есть простая последовательность множества генов. Поэтому
46
нельзя считать, что количество хромосомной ДНК, например у
представителей эукариотических организмов пропорционально
уровню их эволюционного развития. В таком случае некоторые
ля1ушки и рыбы были бы развиты более, чем животные и человек,
поскольку уних размер генома достигает 10'°-10” пар нуклеотидов
(пн), а у млекопитающих и человека он на 1 - 2 порядка ниже (109
- 1О10 пн). Суть здесь в том, что у высокоразвитых существ
значительная часть ДНК оказываетсяя молчащей, так как она
образована не генными последовательностями (у человека к этому
типу относится 80 - 90% всей ДНК) или повторяющимися огромное
число раз идентичными последовательностями. Все это не может
не сказываться на ферментном наборе в клетках, органах и тканях,
равно как и на их функциональной активности.
Ферменты давно являются объектами биотехнологии - их ин-
дустрия зародилась в начале XX в., и объемы ферментного произ-
водства продолжают нарастать, а что касается публикаций о био-
катализаторах, то в мире появляется более 10 000 статей ежегодно.
Ферменты присущи любой живой клетке и, в небольшом ассорти-
менте - организованным частицам (вирусами). Наука, изучающая
ферменты, называется энзимологией, а инженерная энзимология
- это ветвь или субдисциплина биологической технологии, изуча-
ющая биотехнологические процессы, в которых используется ка-
талитическое действие ферментов. Главная задача инженерной
энзимологии заключается в практической реализации фермента-
тивных процессов в целях экономичного получения различных
веществ и энергии для нужд народного хозяйства.
Чтобы выйти на уровень инженерной энзимологии, необходи-
мо решать многие основополагающие проблемы и задачи, к кото-
рым можно отнести:
1) способы выделения и очистки ферментов в зависимости от
их топологии в биообъекте или при выделении в среду обитания;
2) определение состава и строения фермента;
3) особенности кинетики ферментативного действия в зависи-
мости от внешних условий;
4) оценку активности ферментов в зависимости от их чистоты
или, напротив, от наличия естественных примесей или искусствен-
ных добавок;
5) механизмы и пути регуляции синтеза и активности фермен-
тов;
6) моделирование действия ферментов в иммобилизованном
состоянии, учитывая при этом иммобилизацию индивидуальных
ферментов и полиферментных систем, каковыми являются живые
клетки;
47
7) аппаратурное оформление ферментативных процессов;
8) экономичность ферментативных процессов, рекомендуемых
к внедрению в производство.
Некоторые из этих проблем и задач будут рассмотрены в
данном разделе, другие (6-8) - во второй части учебника.
Распределение ферментов в клеточных структурах неравноз-
начное, и независимо от того, что биосинтез их осуществляется в
элементах ядерного аппарата, часть ферментов секретируется
внеклеточно - преимущественно гидролазы. Это обусловлено не-
обходимостью расщепления полимерных веществ до такого уров-
ня, чтобы продукты гидролиза могли поступить в клетку для
конструктивного и/или энергетического обменов.
К ферментам внеклеточного типа можно отнести микробные
амилазы, липазы и пептид-гидролазы, катализирующие реакции
гидролиза соответственно крахмала; жиров и белков. Животная
протеаза (пепсин) условно также может быть причислена в разряд
внеклеточных, так как она поступает из соответствующих клеток
(главных клеток слизистой оболочки желудка) в полость желудка;
то же можно сказать и о ферментах поджелудочной железы,
поступающих в просвет двенадцатиперстной кишки.
Следует иметь в виду, что многие ферменты синтезируемые в
клетках, являются зимогенами, или неактивными
ферментами, и для трансформации их в активные формы необхо-
дим ограниченный (специфический) посттрансляционный проте-
олиз.
В любом случае получение экзо- и эндоферментов на опреде-
ленных этапах как бы унифицируется, когда все стадии выделения
и очистки будут определяться лишь их физико-химическими ха-
рактеристиками. Так, при выделении экзофермента клетки проду-
цента становятся отходом, а культуральная жидкость или, в другом
случае, желудочный сок - целевым продуктом-сырцом. Если речь
идет о необходимости получения эндоферментов, то содержащие
их клетки и ткани измельчают (дезинтегрируют) и экстрагируют
подходящим растворителем. Полученный раствор также представ-
ляет собой полупродукт — сырец. И если речь здесь идет об одном
и том же ферменте,- но разного происхождения и топологии (экзо-
и эндо-), то, начиная с сырца, технологические схемы их выделения
будут во многом тождественными. В этом случае можно использо-
вать такие подходы, как высаливание, сепарирование в градиентах
плотности каких-либо веществ, мембранную фильтрацию,- гель-
хроматографию, афинную хроматографию, ионный обмен и дру-
48
гие. При необходимости выделют ферменты, локализованные в
каких-либо структурах клетки. Тогда, с учетом физико-химических
характеристик таких структур (размер, плотность, форма), приме-
няют дифференциальное центрифугирование после дезинтегра-
ции клеток (тканей). При этом сферические частицы различной
плотности и/или размера будут перемещаться в центрифужной
пробирке на одно и то же расстояние за разное время. Время
прохождения частицы из одного положения (и) в другое (гг) можно
определить, исходя из следующего уравнения:
t 9 Я г Г2
2 со R (Pp-pf) Л
где t — время, ц — вязкость центрифугируемой жидкости, со
— угловая скорость, R— радиус частицы, рр — плотность частицы,
р/ — плотность центрифугируемой жидкости, — коэффициент
силы тяжести.
Форма выделяемых частиц не всегда сферическая, и тогда в
используемые методы вносят необходимые поправки, или выбира-
ют какие-либо другие способы изоляции и расчета. Ферменты —
глобулярные белки, поэтому при их разделении играют роль, в
основном, молекулярные массы (ММ) молекул, а не число субъ-
единиц (С) в них. В качестве примера можно назвать химотрипсин
(ММ = 24500 Да, С = 3), щелочную фосфатазу (ММ = 80000 Да,
С = 2), лактатдегидрогеназу (ММ = 140000 Да, С = 4), триптофаназу
(ММ = 220000 Да,С = 8), и др. Более того, известны ферменты,
катализирующие одну и ту же реакцию в организме одного вида,
но представляющие собой различные молекулярные формы. Их
называют изоферментами, выделение которых по высказанным
причинам заметно осложняется.
Большие по размеру компоненты (интактные клетки, неразру-
шенные частицы ткани, ядра) осаждаются при дифференциальном
центрифугировании быстрее — их собирают в осадок при срав-
нительно невысоких скоростях; супернатант (от. лат. supematans
— всплывающий), или надосадок подвергают последующему цен-
трифугированию при более высоких скоростях. При этом в осадке
могут быть митохондрии. В случае дальнейшего центрифугирова-
ния при еще более высоких скоростях вращения ротора центри-
фуги можно собрать рибосомы.
Во время вращения ротора с большой угловой скоростью со
развивается центробежное ускорение (G), во много раз превосхо-
дящее силу тяжести и способное достигать величин 600—600000 g
49
(g=ускорение силы тяжести). G = со2г (г — расстояние, пройден-
ное частицей). Так, для выделения ядер из дезинтеграта эукарио-
тических клеток требуется 10 минут центрифугирования при 600
д, для изоляции лизосом и митохондрий — 5 мин. при 15000 д, для
выделения рибосом — 1 час при 100000 д. Скорость продвижения
частицы в направлении г (Vr) описывается уравнением
Vr = (t — время)
Центрифугирование дезинтеграта клеток или тканей при боль-
ших скоростях в водной суспензии представляет собой турбулен-
тный поток и сила сопротивления движению различных частиц
("твердых тел") определяется не вязкостью жидкости (ц), а ее
плотностью (р), тогда сила сопротивления называется гидравличе-
ской и описывается формулой F = CfSpv2, где F — гидравлическая
сила сопротивления, Cf — коэффициент, зависимый от формы
твердого тела, S — площадь поперечного сечения тела. Для сфери-
ческого тела значения Cf находятся в интервале 0,05 — 0,2 в
зависимости от характера поверхности (гладкая, шероховатая); для
тонкого диска, перпендикулярного потоку, Cf равна примерно 0,55.
Относительная роль динамического сопротивления и вязкого
трения сказывается на поведении движущейся жидкости и харак-
теризуется безразмерным числом Рейнольдса:
Re=^pf =
T]7v т]
где 1 — характерный линейный размер. При обтекании тел 1 —
это длина или поперечный размер, при течении в длинных трубках
1 — это диаметр трубки. В условиях центрифугирования скорости
частиц и, следовательно, их числа Рейнольдса очень малы. Поэтому
для определения силы сопротивления среды, действующей на
частицу, пользуются законом Стокса. Согласно этому закону сила
сопротивления, испытываемая твердым шаром при его медленном
поступательном движении в неограниченно вязкой жидкости,
описывается выражением Р = 6тгцРу, где R — радиус шара, т}
—коэффициент вязкости жидкости, v — скорость движения шара.
Параметры движения частицы можно вычислить по следующе-
му уравнению, исключив из него коэффициент силы тяжести,
поскольку направление г перпендикулярно направлению силы
тяжести:
G(pp-pp),
где Vr скорость частицы в направлении г, G — центробежное
ускорение, рр — плотность частицы, рр— плотность жидкости, т]
— вязкость жидкости, R — радиус частицы.
50
При подходе к выбору биообъекта — продуцента того или иного
энзима важно иметь хорошую (по скорости роста, продуктивности
и активности фермента) культуру — штамм (не клон), так как
поддерживать его в лабораторных и производственных условиях
легче, чем клон. К тому же клоновые культуры используют, как
правило, для генетических целей.
Необходимо иметь в виду и такие факты, как полифермент-
ность культуральных жидкостей или тканевых экстрактов, из
которых предстоит выделить те или иные ферменты в индивиду-
альном виде. Это особенно относится к гликансинтетазам и глика-
назам, что является своеобразным отражением разветвленности
углеводных полимеров (участие в синтезе более, чем одной синте-
тазы) и их полидисперсности (опосредованность действия генети-
ческого кода на процесс синтеза полисахарида во время роста и
развития культуры клеток или ткани, то есть здесь имеет место
нематричный синтез углеводного полимера). В таких случаях це-
лесообразно применить афинную хроматографию (от англ, affinity
— сродство).
Афинная хроматография основана на специфичности биомо-
лекул. Принципиально этим методом возможно получение абсо-
лютно чистых веществ. В нативном состоянии ферменты за счет
своего активного центра и регуляторного участка (одного или
более) взаимодействуют с небольшим числом лигандов, которые
представляют собой субстраты, либо эффекторы. Благодаря высо-
кому сродству активного участка фермента к лиганду, их обрати-
мому связыванию и отсутствию какой-либо другой реакции между
лигандом и матрицей возможно эффективное проведение афинной
хроматографии.
В качестве лиганда обычно используют конкурентный обрати-
мый ингибитор (см.), его ковалентно сшивают с соответствующей
нерастворимой матрицей так, чтобы он не терял своей способности
связываться с ферментом. Матрицей с лигандом в соответствую-
щем буферном растворе заполняют колонку, а затем через нее
пропускают раствор фермента, подлежащий очистке. В колонке
задерживается лишь специфический фермент, а все другие при-
меси уходят. После этого проводят элюцию специфического фер-
мента, используя раствор субстрата в среде с другим pH и (или)
ионной силой. При разделении ферментов методом афинной хро-
матографии необходимо знать все кинетические параметры целе-
вого фермента, располагать хорошей матрицей и удачно подобран-
ным лигандом. В качестве примера приводим схему путей сшива-
ния лиганда с одной из возможных гидроксильных групп полиса-
51
харида агарозы (или сефарозы) через удлиняющие мостики, на-
пример диаминов типа H2N(CH?)xNH2 (х = 2 — 6) и £-аминокапро-
новой кислоты:
н,с—сн—сн,—сн,—сн,—с ООН
3 I Z Z Z
NH2
Е- -аминокапроновая кислота
агарозная матрица
I I " I— —Г
он он
ОН ОН
NH
R - ЛИГАНД I
R
,CNBr/R(CH,),NH
бромциан
C=NH
(СН2)Х
R
производные
изомочееины
Хорошей матрицей для афинной хроматографии является три-
сакрил марки GF 2000 (Франция), получаемый сополимеризацией
1Ч-акрилоил-2-амино-2-гидроксиметил-1,3-пропандиола и гидро-
ксилированного акрилового бифункционального мономера. Три-
52
сакрил — высоко гидрофильный сополимер за счет вторичных
амидных и первичных гидроксиметильных групп. Последние на-
ходятся на поверхности молекулы, тогда как полиэтиленовый кор
(от англ, core — сердцевина) полностью скрыт внутри.
HN—С—СН2ОН HN——СН20Н
со сн,он с0 сн2он
I I
--сн—СН2—сн—СН2---сн— СН2—сн—сн2—
со-----------------со
I сн9он I сн^он
I I 2 I I 2
HN-C—CH2OH HN—С—СН2ОН
сн2он сн2он
трисакрип GF2000
Рис. 8. Получение некоторых сорбентов для а финной хроматографии на основе
трисакрила GF 2000 (КДА — карбонилдиимидазол, НФХФ — нитрофенилхлоро-
формиат, БЭ — бисэпоксироны, ГА — глутаралгдегид, М-Э-2ЭХ — N-этоксикарбо-
нил -2-ЭТОКСИ-1, 2-дигидрохинолин, АЯК — ангидрид янтарной кислоты).
Трисакрил стабилен при разных температурах — до 121°С и
при рН1— 11, равно как устойчив к диссоциирующим агентам и
поверхностно-активным веществам. В целях использования его для
53
афинной хроматографии проводят активацию такими веществами,
как глутаральдегид, эпихлоргидрин, дивинилсульфон, р-нитрофе-
нил-хлороформиат, карбонилдиимидазол, этилендиамин и др. (рис.
8).
Активированный трис акрил используют затем для сшивания с
лигандом. В качестве примера можно привести иммобилизацию
лиганда на трисакриле, активированном глутаральдегидом (рис. 9).
о сно сно
> -C-NH”CH-CH-(CH2)2-CH=C-(CII2)2-C.HO + H^N-j лиганд!-*
(«Уз
СНО
о сно сно
II I I
-С-КН-СН-СННСН^-СН-СННСН^-СНО
(СН2)3 NH
СНО | ЛИГАНД |
Рис. 9. Схема иммобилизации лиганда с трисакрилом, активированным глутараль-
дегидом.
В любом случае выделение и очистка ферментов — это наука,
граничащая с искусством. В самом деле, если химическая реакция
протекает с выходом 0,1% и с сотней побочных продуктов, то вряд
ли химик-органик проявит интерес к процессу получения (и, тем
более, очистки) целевого продукта. Но именно такие задачи встают
перед биотехнологами, желающими получить индивидуальный бе-
лок. В работе с биологическим материалом необходимы и важны
знания предварительных характеристик по содержанию белка в
различных биообъектах (даже в органах и тканях одного и того же
вида животного). От этого зависит выбор материала, его предва-
рительная подготовка для включения в технологический процесс.
Молекулы ферментов чувствительны к различным воздействи-
ям, поэтому при их выделении и очистке требуется соблюдать
необходимые меры предосторожности: контролировать темпера-
туру (нередко поддерживать ее близкой к точке замерзания ис-
пользуемого растворителя), pH (как правило — с помощью буфер-
54
ных растворов), концентрацию белка, ионную силу раствора,
концентрацию ионов тяжелых металлов (удаляют их с помощью
комплексообразователей, например ЭДТА), значение окислитель-
но-восстановительного потенциала (Ео), который должен поддер-
живаться на низком уровне (например, с помощью меркаптоэта-
нола). Следует иметь в виду и тот факт, что многие белки в
разбавленных растворах проявляют склонность к денатурации —
устойчивость возрастает в присутствии их субстратов.
Из вспомогательных приемов очистки ферментов часто поль-
зуются диализом, лиофилизацией, пропусканием растворов через
соответствующие молекулярные сита (гелевые, мембранные). В
качестве примера очистки гипотетического фермента приведена
таблица 5. Из данных в таблице обращают на себя внимание
сокращение объемов раствора, содержащего целевой продукт (в
200 раз), уменьшение содержания белка в 5 раз при заметном
возрастании активности фермента в 100 раз (в расчете на 1 мг
белка).
Т а б л и ц а 5. Сравнительная характеристика методов очистки фермента "X”
Содер- жание белка, мг/мл Активность Сум- марный выход, % Сте- пень (крат- ность) очист- ки
Стадии очистки Объем раствора, л
Ед/мл удель- ная, Ед/мг белка
Экстракт белков - сырец 200 2,5 100 100 100 1
Высаливание аммония сульфатом 80 1,25 200 160 80 4
Хроматография на ДЭАЭ-целлюлозе 20 1,0 500 500 37,5 12,5
Препаративный электрофорез 5 1,0 1500 1500 37,5 37,5
Изоэлектрическое осаждение 3 0,3 2000 6700 30 167
Кристаллизация 1 0,5 5000 10000 25 250
55
Суммарный выход достаточно высокий, а степень очистки
превосходная - в 250 раз. Но такие показатели далеко не типичны
для разных ферментов.
В качестве критериев чистоты ферментов используют данные
электрофореза, ультрацентрифугирования (седиментограмма), оп-
ределения молекулярной массы различными методами, по раство-
римости (кривые растворения), оценки полидисперсности с опре-
делением специфической активности каждой фракции, хроматог-
рафирования на различных носителях и в нескольких системах,
аминокислотного состава (особенно — при обнаружении белковых
примесей), включая секвенирование (от англ, sequence — после-
довательность) на автоматизированных приборах - секвенаторах,
и т. д.
В последние годы продолжает расти интерес к внеклеточным
ферментам микроорганизмов, поскольку микробы — продуценты
относительно легко культивируются в контролируемых условиях
и подвергаются направленному изменению в сторону получения
высокопродуктивных вариантов.
У грамположитёльных бактерий экзоферменты топологически
могуг быть строго внеклеточными (многие протеазы, гликозидазы
и др.) и локализованными с наружной стороны клеточной мемб-
раны, например, ос-глюкозидаза у Вас. licheniformis (это можно
доказать при протопластировании клеток, когда ферменты пере-
ходят в окружающую среду после удаления клеточной стенки).
Грибы в отношении секреции экзоферментов уподобляют грам-
положительным бактериям. У грамотрицательных бактерий экзо-
ферменты дополнительно могут находиться в периплазм атическом
пространстве. Казалось бы, что такие ферменты должны рассмат-
риваться внутриклеточными, однако они достаточно легко осво-
бождаются при осмотическом шоке или в результате протоп-
ластирования клеток.
Строго внеклеточных ферментов у грамположительных бакте-
рий больше, чем у грамотрицательных, однако среди последних
имеются выраженные продуценты их (аэромонасы, псевдомонасы,
некоторые энтеробактерии и др.).
Каким же образом секретируемые белки транспортируются
через структуры клеточной оболочки?
В конце 70-х годов XX в. была предложена так называемая
сигнальная гипотеза (Г. Блобелидр., 1979). Согласно этой гипотезе
56
секретируемый белок через ЬтН?-конец удлиняется на 15—30
аминокислот, вкупе составляющих лидерный, или сигна-
льный пептид, направляющий рибосому, из которой он
появляется, к мембране. В мембране лидерный пептид вместе с
другими мембранными белками формирует канал, стабилизирую-
щийся при присоединении рибосомы. Рибосома синтезирует бе-
лок, и он тут же экспортируется через пору. Этот механизм
называется котрансляционной секрецией. После частичного или
полного экспорта белка сигнальный пептид удаляется с помощью
специфической сигнальной эндопептидазы. Решающую роль в
секреции играет гидрофобная центральная часть сигнальной по-
следовательности размером около 5,5 нм (от 15 до 19 аминокислот)
и находящаяся в высокоупорядоченной конформации. NH?-Конец
в составе гидрофильного домена (от англ, domain — регион,
область) присоединяется к отрицательно заряженной внутренней
поверхности цитоплазматической мембраны. В ходе трансляции
белка гидрофобный участок сигнальной последовательности по-
степенно внедряется в мембрану до появления участка расщепле-
ния в форме петли на ее внешней стороне. После этого сигнальная
эндопептидаза отщепляет сигнальную последовательность от рас-
тущей цепи полипептида, способствуя тем самым котрансляцион-
ной секреции белка (рис. 10).
К настоящему времени доказано, что котрансляционная секре-
ция присуща прокариотам и эукариотам. Таким путем секретиру-
ются, например, а-амилаза у Bac.subtilis, 0-лактамаза у
Bac.licheniformis, экзотоксин у Corynebacterium diphtheriae.
После завершения котрансляционного транспорта рецептор-
ные белки мембраны освобождаются и могут функционировать в
плоскости мембраны.
У прокариот и эукариот имеется также другой тип секреции
—посттрансляционный, когда через мембрану транспортируется
завершенный белок. Механизм транспорта объясняется либо с
привлечением триггерной гипотезы У. Уикнера (1979), либо ранее
рассмотренной сигнальной гипотезы (рис. И). Согласно обеим
гипотезам роль белка - предшественника, обладающего сигнальной
последовательностью, весьма велика. В первом случае эта после-
довательность обеспечивает скручивание полипептида так, что
гидрофобные области его связываются с мембраной. Затем белок
может освобождаться из мембраны благодаря отделению сигналь-
57
Рис. 10. А - экспорт белка через мембрану путем котрансляционной секреции
(1 —рибосомы, 2 — мембрана. 3 — пора в мембране, 4 — сигнальная последо-
вательность, 5 — сигнальный пептид, 6 — рецептор сигнального пептида, 7 —
сайт действия сигнальной эндопептидазы, 8 — рецептор рибосомы); Б —
строение сигнального пептида белка 1am В Escherichia coli (А — гидрофильный
сегмент, Б — гидрофобный сегмент, p-сайт расщепления; последовательность
а инокислот; 1—метионин, 2 — изолейцин, 3 — треонин, 4 — лейцин, 5 —
аргинин, 6 — лизин, 7 — аланин, 8 — валин, 9 — глицин, 10 — серин, И —
глутамин, 12 — пролин); С — секреция белка через мембрану (М) по типу петли
(1 — NH? - конец, 2 — гидрофобный участок, СП — сигнальная пептидаза).
58
ной последовательности за счет
эндопротеолиза. Во втором слу-
чае сигнальная последователь-
ность полной полипептидной це-
пи вместе с рецепторными бел-
ками мембраны формирует по-
ры. Бедок развертывается и про-
исходит перемещение (трансло-
кация), сопровождаемая удале-
нием сигнальной последователь-
ности.
Для эукариот доказано, что
их эндоплазматический ретику-
лум содержит для работающей
рибосомы своеобразные "при-
чальные белки", к которым при-
соединяется комплекс рибосомы
с частицей, узнающей сигналь-
Рис. 11. Схема посттрансляционного
транспорта белков через мембрану со-
гласно "триггерной гипотезе" по Уикне-
ру (1979) — I и "сигнальной гипотезе" по
Блобелу и др. (1979) — П.
ную последовательность. Частица включает шесть полипептидных
цепей и небольшую молекулу РНК с константой седиментации 7S.
Причаливание рибосомы с частицей индуцирует трансляцию при
последующей секреции растущего белка (рис. 12) до момента
появления сигнального пептида из рибосомы, когда происходит
блокада трансляции.
На основе рассмотренной
"сигнальной гипотезы" мож-
но сформулировать следую-
щие основные положения:
1) процесс экспорта бел-
ков эволюционно консерва-
тивен (прокариотические
клетки узнают сигнальную
последовательность эукариот
и наоборот);
2) в мембране существует
некий механизм экспорта,
или аппарат секреции;
I п
Рис. 12. Начальные этапы векторного пере-
носа белков в эукариотических клетках: I —
блокирование трансляции, II — снятие блока
трансляции после “причаливания” рибосомы
— 1, сигнальная последовательность — 2;
частица, узнающая сигнал — 3, "причаль-
ный" белок — 4, мембрана — 5, сигнальный
пептид — 6.
3) свободные рибосомы в
цитоплазме клетки и рибосомы, связанные с мембраной, не раз-
личаются между собой;
59
4) у эукариот на эндоплазматическом ретикулуме имеются так
называемые причальные белки для функционирующих рибосом,
соединенных с частицами, узнающими сигнальную последователь-
ность;
5) секретируемые белки синтезируются обычно в форме более
длинного предшественника;
6) белки, как правило, секретируются котрансляционно (реже
— посттрансляционно), то есть растущий полипептид переносится
через мембрану по мере трансляции;
7) в структуре подлежащего секреции белка может существо-
вать "стоп"-последовательность, останавливающая секрецию и да-
ющая начало интегральному мембранному белку.
Следует вспомнить, что молекулярные массы (ММ) ферментов
достаточно большие и находятся в пределах примерно от 12 до
нескольких тысяч килодальтон (кДа). Ферменты с ММ более 100
кДа обычно представлены субъединицами (двумя и более).
Нелегко себе вообразить, например, что кишечная палочка,
размножающаяся в течение 20—30 минут, реализует множество
направленных и строго избирательных биокаталитических процес-
сов. Прямыми и косвенными доказательствами в опытах с фер-
ментом трансаминазой аспарагиновой кислоты установлено, что
отдельные стадии каталитической реакции протекают в простран-
стве около нескольких сотен кубических миллимикрометров (нм)
за время от десятитысячных до миллионных долей секунды.
Для того, чтобы управлять действием ферментов, необходимы
фундаментальные исследования по расшифровке молекулярных
основ ферментативного катализа. Многое уже сделано в данном
направлении, но это лишь начало, а желательно знать как можно
больше о всех группах ферментов.
Согласно классификации (КФ) ферменты подразделяются на
шесть главных классов, каждый из которых включает подклассы
и подподклассы:
I. Оксидоредуктазы (КФ1.) — катализируют окислительно-вос-
становительные реакции;
II. Трансферазы (КФ2.) — катализируют реакции переноса
функциональных групп (фосфатных, ацильных, гликозильных, аль-
дегидных и др.);
III. Гидролазы (КФЗ.) — катализируют реакции гидролиза
60
(перенос функциональных групп на молекулу воды);
IV. Лиазы (КФ4.) — катализируют реакции присоединения по
двойным связям и обратные реакции;
V. Изомеразы (КФ5.) — катализируют реакции изомеризации,
то есть перенос групп внутри молекулы с образованием изомерных
форм;
VI. Лигазы (КФ6.) — катализируют реакции образования связей
С — С, С — S, С — ОиС — N, сопряженных с расходованием
энергии АТФ или других нуклеозидтрифосфатов.
Под класс и подподкласс ферментов соответственно обозначают
вторым и третьим числами; порядковый номер фермента - четвер-
тым числом в его подподклассе. Например, бетаин-гомоцистеин-
метилтрансфераза имеет кодовый номер (шифр) — КФ 2.1.1.5, то
есть согласно классификации (КФ) этот фермент относится к
трансферазам (П-й главный класс), переносящим одноуглеродные
остатки (первый подкласс) в форме метила — метилтрансферазы
(первый подподкласс), номер конкретного фермента 5 (бетаин:
L-гомоцистеин S-метилтрансфераза), катализируемая реакция
здесь следующая:
coo" сн,—SH
I I
CHj-N+=(CHj)3 + CHj
, НС-NH,
бетаин | *
соон
COOH CHj-S—CHj
CH2-N=(CH3)2 + CH2
CH-NHj
диметилглицин |
соон
L-гомоцистеин
L- метионин
Классификация и номенклатура ферментов имеют универсаль-
ное значение, то есть они распространены на все ферменты,
независимо от источника их получения.
Со времени выделения Дж. Б. Самнером в 1926 г. уреазы в
кристаллическом виде несомненно признавалось, что все фермен-
ты — это простые или сложные белки. В 1981—1982 гг. Т. Р. Чех
с сотрудниками открыл каталитическую активность у рибонукле-
иновой кислоты. Этим опровергается универсальность принципа
"фермент — это белок", а кроме того привносятся новые концепции
в ранние этапы эволюции живой материи. Некоторые исследова-
тели (Дж. Дарнелл-младший) считают, что первым веществом
наследственности была РНК, а не ДНК, поскольку ей присуща
61
большая функциональная универсальность — она способна хра-
нить информацию, воспроизводиться, направлять синтез белка и
вести себя как фермент. Вместе с тем следует оговориться, что
диапазон реакций, катализируемых РНК, еще недостаточно велик,
чтобы указанное выше опровержение принималось сегодня в
"конкурентный расчет".
В отличие от небиологических катализаторов ферменты —
высоко специфичны, они имеют активные центры, многие из них
проявляют свою активность в присутствии коферментов (коэнзи-
мов) небелковой природы, то есть в таких случаях ферменты
являются сложными белками. К сожалению, до сих пор нет
достаточной четкости в определениях коферментов и кофакторов.
Одни авторы отождествляют эти понятия, другие используют лишь
один термин — коферменты, третьи выделяют ионы некоторых
металлов в разряд активаторов, прямо не связывая их с кофермен-
тами, и т. д. Накопленный фактический материал служит веским
основанием подразделять белки-ферменты на две группы — про-
стые ферментные белки и сложные ферментные белки, из которых
сложные содержат в своем составе неорганические и/или органи-
ческие небелковые структуры — коферменты. Коферменты могут
обратимо или необратимо (простетические группы) связываться с
белковой частью — апоферментом. Полный ферментный комплекс
называют холоферментом. Если же действие некоторых фермен-
тов лишь активируется неорганическими ионами или атомами
металлов и не снимается полностью в их отсутствии, то такие
активаторы следует называть кофакторами.
Функции контакта с субстратом и катализа у простых фермен-
тных белков выполняют боковые радикалы аминокислот в пол-
ипептидной молекуле; в сложных ферментных белках эти функции
выполняют преимущественно коферменты.
Коферменты классифицируют по каталитическим и структур-
но-функциональным признакам. Например, коферментами окси-
до-редуктаз являются: НАД, НАДФ, ФАД, ФМН, липоевая кислота
и др.; коферментами трансфераз-пантотенат, УДФ-глюкоза, ЦДФ-
холин, тетрагидрофолиевая кислота и др. Ряд коферментов явля-
ются производными витаминов: тиамина (декарбоксилазы а-кето-
кислот, транскетолазы), рибофлавина (ФМН, ФАД), пантотеновой
кислоты (кофермент А), никотинамида (НАД, НАДФ) и т. д. Отсюда
62
понятна необходимость отдельных витаминов в питательных сре-
дах для биообъектов.
Достаточно много ферментов (порядка 25%) относится к метал-
лоферментам, представляющим собой одну из групп металлобио-
молекул.
Такие ферментные белки без металлов не проявляют фермен-
тативной активности. В тех же случаях, когда ионы металлов не
выполняют роли коферментов (металлы-кофакторы), а лишь акти-
вируют ферментативные реакции, то такие ферменты по сути
Металлоби о молекулы
Ферментные белки
Оксщщредуктазы
Оксидазы
(Fe.Cu.Mo)
Дегидрогеназы
(Fe,Cu.Mo)
Гидроксилазы
(Fe,Cu,Mo)
Оксигеназы
(Fe)
Супероксид—
дисмутаза
(Cu,Zn,Mn)
Нитрогеназы
(Fe,Mo)
Гидрогеназы
(Fe)
Транспортные и
аккумулирующие белки
4
Переносчики
электронов
Цитохромы (Fe)
Fe — S(Fe)— белки
Си—белки
Аккумулирующие,
уравсреррцыг It
Структурные
Ферритин (Fe)
Трансферрин (Fe)
Церулоплазмин (Fe)
Небелковые молекулы
Фотовосстанав —
_лйвающй_е."
Хлорофилл (Мд)
и Фотосистема II
(Мп, Мд)
Транспортирующие
металлы и структур —
.мгё.
Сидерофоры (Fe)
Скелетные (Ca,Si)
Гидролазы
Карбоксипепти—
даза (Zn)
Аминопептидазы
(Мд, Мп)
Фосфатазы
(Mg,Zn,Cu)
Связывающие О2
Миоглобин (Fe)
Гемоглобин (Fe)
Гемеритрин (Fe)
Гемоцианин (Fe)
Изомердзы _и_синтетазы
Витамин В12—белок
Коэнзимы (Со)—белки
63
своей не являются металлоферментами. Их следует называть ме-
таллоактивйрованными ферментами.
Можно утверждать, что в металлоферментах ионы металлов
выполняют и каталитическую и структурную роли, тогда как в
металлоактивированных ферментах — преимущественно катали-
тическую. Тем не менее металлы в качестве коферментов и
кофакторов в какой-то части сходны в своих функциональных
проявлениях. Для металлоактивированных ферментов возможны
4 схемы трехчленных комплексов, участниками которых являются
фермент (Ф), субстрат (С) и металл (М) в стехиометрическом
соотношении 1:1:1.
М
1) Ф—С—М, 2) Ф—М—С, 3) М—Ф—С, 4) Ф [
С
В первой схеме мостиковое положение занимает субстрат, во
второй — металл, в третьей — фермент и в четвертой — металл в
циклическом комплексе. Металлоферменты, например, не форми-
руют комплекс Ф—С—М, так как при очистке они выявляются в
форме Ф—М.
Металлоферментами являются так называемые геминовые
ферменты (каталаза, пероксидаза, цитохромоксидаза), в составе
которых имеется простетическая железопорфириновая группи-
ровка, или гем; трансферазы, у которых коферментная форма
витамина В12 представлена 5'-дезоксиаденозилкобаламином.
Все ферменты с коферментом В12 катализируют реакции об-
мена водорода, связанного с углеродом, на какую-либо иную
64
Витамин Bi2
группу (алкильную, аминную, гидроксильную, карбоксильную).
Другая коферментная форма витамина В12 — метил-кобаламин
(группа CN в коррине замещена метилом) участвует в реакциях
переноса метильных групп. Кофермент В12 в определенной степени
сходен с гемом, однако вместо железа в нем содержится кобальт.
В настоящее время стали известны и новые коферменты.
Например, корфин (фактор 430) представляет собой никельтетра-
пиррол и напоминает рассмотренные выше макроциклические
структуры гема и коррина. Корфин выступает коферментом окси-
доредуктазу метанобразующих бактерий, связанных с окислитель-
но-восстановительным превращением одноуглеродного субстрата.
3 т. 8524 65
В метанообразующих бактериях найдены также фактор 420,
оказавшийся гидридным донором, и фактор, восстанавливающий
диоксид углерода.
У метилотрофных бактерий, окисляющих метан, обнаружен
кофермент метоксатин, содержащий в своей молекуле ортохино-
новую простетическую группу. Оксидоредуктаза — глюкозодегид-
рогеназа Acinetobacter calcoaceticus содержит сходную ортохино-
новую простетическую группу. Вот почему такие ферменты пол-
учили название хинобелки.
66
фактор, восстанавливающий COj
Механизм действия этих коферментов точно пока неизвестен.
Кофакторами в металлоактивированных ферментах часто вы-
ступают ионы магния, цинка, кальция, то есть металлы с постоян-
ной валентностью. Например цинк активирует алкогольдегидроге-
назу, щелочную фосфатазу; кальций - а-амилазу; магний — АТФ-
азу, гексокиназу и многие другие ферменты; марганец (или магний)
— енолазу; калий — пируваткиназу, и т. д.
Из сказанного о металлоферментах и металлоактивированных
ферментах следует, что наличие определенных ионов в среде
культивирования биообъектов строго обязательно.
Кинетические характеристики биокатализаторов, определяю-
щие активность и длительность работы ферментов в различных
условиях, являются важными показателями и при промышленном
получении ферментов. Все эти данные подробно рассматриваются
в курсе биохимии. Здесь же мы останавливаемся лишь на важней-
ших параметрах, без которых биотехнология ферментов будет
недостаточно охарактеризованной.
В процессе биокатализа активность ферментов и, следователь-
но, их каталитические функции снижаются. Кажущаяся структур-
ная громоздкость их в нативном виде (от лат. nature — природа) с
лихвой компенсируется эффективностью катализа, если сравни-
вать их с неорганическим ("простыми") катализаторами. Однако в
изолированном виде ферменты достаточно легко инактивируются,
что приобретает важное значение для практики. Отсюда возникает
проблема стабилизации активности ферментов в случаях их про-
мышленной эксплуатации. Следует заметить, что in vivo инактива-
ция и активация ферментов, равно как и синтез их de novo или
denuo (по-лат. — заново), регулируется самим организмом. К тому
же в организме (клетке, ткани) многие ферменты находятся в
связанном состоянии. Очевидно, что при этом их конформации
больше или меньше изменяются, что не безразлично для клетки с
67
функциональной точки зрения. Из практической работы известно
немало случаев, когда неполностью очищенные ферменты были
какое-то время активнее и стабильнее чистых образцов биоката-
лизаторов. На этом основании можно предполагать, что так назы-
ваемые примеси создавали благоприятное окружение ферменту и,
очевидно, выгодное потребителю. Стабилизация ферментов могла
быть обусловлена разными факторами, например, иммобилиза-
цией на мембранных структурах, соединением с высокополимер-
ными углеводами или гликоконъюгатами и пр. Подобная стабили-
зация сродни консервации белковой молекулы, так как здесь речь
идет о сохранении фермента какое-то (обычно более или менее
длительное) время со всеми присущими ему параметрами.
В любом случае при получении ферментов в производстве
необходимо опираться на воспроизводимые результаты, заложен-
ные в регламенте.
Активность и стабильность — как параметры важны и для вновь
внедряемых в производство ферментов, поскольку предполагаемая
их "выходная активность” приобретает существенное значение с
точки зрения выбора (или проектирования) конструкции реактора,
оснащении его необходимой контрольно-измерительной аппарату-
рой и т. д. (в частности, для исключения быстрой инактивации
фермента).
Активность биокатализаторов измеряют в международных еди-
ницах (ME) — 1 ME соответствует количеству фермента, превра-
щающему 1 микромоль (мк/моль; 10’6 моль) субстрата в 1 мин. в
стандартных условиях. Единицы активности могут быть выражены
в мкЕ, нЕ и пЕ, то есть соответственно в микро-, нано- и пикоеди-
ницах с учетом превращения 10'6, 10'9 или 10'12 моля субстрата в 1
мин. Удельную активность выражают в единицах на 1 мг белка. С
1973 г. введена единица ферментативной активности катал (кат.),
соответствующая количеству катализатора, способного превра-
щать 1 моль субстрата в продукт за 1 секунду. Тогда соотношения
катал и ME будут следующими: 1 кат= 1 моль субстрата • С'1 = 60
моль» мин1 =60»106 мкмоль» мин-1 = 6»107 ME, 1 МЕ= 1 мкмоль*
мин ‘= 1/60 мкмоль* с’* = 1/60 мккат= 16,67 нкат= 16670 пкат. Все
это косвенно подтверждает факт трудности выделения ферментов
в чистом виде и на практике приходится чаще иметь дело с группой
белков в продукте — сырце или в исходном материале, или,
наконец, в других партиях материала, подлежащих разделению.
В лабораторных и производственных условиях нередко прихо-
68
дится иметь дело с денатурацией ферментов в зависимости от ряда
факторов, например, биологических — гидролиз целевого продукта
какой-либо пептид-гидролазой, находящейся в исходном сырье или
привнесенной случайно, например, при микробном загрязнении;
физических — неадекватное температурное, механическое, ради-
ационное воздействия; химических — pH, комплексообразовате-
лей, растворителей, ионогенных поверхностно-активных веществ,
тяжелых металлов и т. д.
Естественно, что от качества выделения и очистки фермента
зависит надежность анализа его структуры. Тем не менее, следует
помнить, что одинаковые ферменты, выделяемые из различных
источников, могут иметь разные аминокислотные последователь-
ности и, следовательно, они не тождественны — их каталитическая
активность и свойства будут различными. В качестве примера
можно назвать вид Вас. coagulans, образующий глюкозоизомеразу;
этот фермент активируется ионами Со2+ , а такой же фермент,
продуцируемый мутантом данного вида при pH более 8, не нуж-
дается в ионах кобальта. Вот почему точность имеющейся инфор-
мации об источнике фермента, его технологии, паспортных данных
приобретает исключительную важность при сравнении фермента,
выпущенного разными фирмами-изготовителями.
М. Левитт и К. Чотиа в 1976 г. подразделили структурно
известные белки по наличию и расположению сс-спиралей и
(3-слоев на пять классов:
I — белки, состоящие только из ос-спиралей, уложенных вместе
в глобулярную структуру;
И — белки, состоящие только из (3-слоев и формирующие
глобулу в форме бочонка;
III — белки, включающие ос-спирали и (3-слои (а + (3-белки), но
разобщенные в третичной структуре;
IV — белки сс/(3, в которых а- и ^-структурные участки чере-
дуются в первичной структуре, формируя третичную структуру, в
центре которой находятся обычно параллельно уложенные (3-слои,
окаймленные с обеих сторон ос-спиралями;
V — обычно малые молекулы с многими -S—S-мостиками или
ассоциированные с крупным кофактором "неупорядоченные бел-
ки", обладающие слабой вторичной структурой.
На рис. 13 представлены схемы Дж. Ричардсон (1981) для
различных белков, отнесенных к какому-либо одному из пяти
вышеназванных классов.
69
Рис. 13. Пять классов белков по Дж. Ричардсон
(1981):
А — миохемеритин (класс I),
Б -— Си, Zn—супероксиддисмутаза (класс II),
В -— лизоцим (а+Р-структура, III класс),
Г и Д — две ортогональные проекции НАД—свя-
зывающего домена лактатдегидрогеназы (IV
класс),
Е — тризофосфатизомераза (IV класс),
Ж — нейраминидаза вируса гриппа (V класс).
ж
Более или менее длинные цепи белков, как правило, формируют
домены, представляющие собой свернутую в пространстве струк-
туру, имитирующую маленькую белковую молекулу. Доменам
присущи функции связывания, и в ферментных белкюс активный
центр располагается преимущественно на границе между двумя
или большим числом доменов. К настоящему времени установлено,
что домены способны перемещаться друг относительно друга в
процессе функционирования содержащей их молекулы. В трипси-
ногене домен из неупорядоченного состояния переходит в упоря-
доченное в ходе активации зимогена.
70
Заметный прогресс в изучении структуры белков начался с
конца 70-х годов текущего столетия. Если до этого структуры их
выводили на основании метода диффракции нейтронов, карт
распределения электронных плотностей с введением в молекулы
белков тяжелых атомов, то в последующие годы были развиты
методы синхронной радиации, методы с использованием компью-
терной техники, что сослужило огромную службу кристаллогра-
фии данных биополимеров. В этой связи статичный метод рентге-
ноструктурного анализа одиночного кристалла способен дать ди-
намичную информацию. На основании указанных методов пока-
зано, например, что фермент лизоцим имеет оболочку из 33—35
прочно связанных молекул воды и менее упорядоченную область
из 95—105 молекул воды, соединенных с белком лишь одной
водородной связью. Около 60—80% остальной воды распределено
в промежутках между кристаллами и не влияет на электронную
плотность белка.
Молекулы белков не являются жесткими структурами — они
подвижны л, следовательно, достаточная лабильность молекул
ферментов (молекулярная вибрация) играет важную роль в узна-
вании субстрата и стабилизации переходного состояния.
Чтобы увидеть ферменты в действии стали использовать метод
рентгеноструктурного анализа при низких температурах (криоэн-
зимология) — можно сделать "стоп-кадры" в кинетически значи-
мых точках катализируемой реакции.
Теперь хорошо известно, что ферменты, внедряемые в про-
мышленность выгодно отличаются от неорганических (или синте-
тических) катализаторов рядом преимуществ, существенным из
которых следует назвать низкую энергоемкость ферментативных
процессов, то есть сравнительно небольшие затраты внешней
энергии. На каталитические процессы в промышленности прихо-
дится 80—90% и доля их продолжает возрастать. В неорганическом
синтезе, например, получают контактным способом десятки мил-
лионов тонн серной кислоты в год. При контактном процессе
окисление SO2 в SO3 протекает в присутствии катализатора V2O5
с добавлением К2О и других оксидов.
В органическом синтезе на никелевых катализаторах осущест-
вляют многие реакции гидрирования соединений, включающих
С = С, С= С, С = О, NO2 — группы. Для примера назовем алюмо-
молибденовые, алюмохромовые и алюмоплатиновые катализато-
ры, применяемые для дегидрирования линейных и разветвленных
алканов в олефины. Так, катализаторы на основе оксида хрома,
применяемые в реакции превращения бутана в бутен, работают
при температуре около 600°С, а регенерация их протекает при
640—650°С. Оксидные катализаторы окисления С03О4, СщОз, NiO
71
эффективны также при температурах 250—600°С. Лишь отдельные
катализаторы полимеризации (ВЁз) проявляют свое действие при
—80°—100°С, например, при получении полиизобутилена. Однако
чтобы добиться понижения температуры до —80° или —100° С,
также необходима затрата энергии.
Напротив, все биохимические реакции в живых системах
потекают при сравнительно низких температурах. Например, фер-
менты микроорганизмов — сапрофитов, живущих в почве средних
широт нашей страны, относятся к мезофилам, то есть их ферменты
эффективно действуют в пределах от + 10 до +50°С (с преимуще-
ственным интервалом от + 20 до + 30°С). Нечто аналогичное можно
сказать и о ферментах растений. Ферменты животных более
эффективны при нормальной температуре тела.
Тем не менее, следует помнить, что для каждого фермента есть
свои так называемые кардинальные точки температуры, pH и др.
(minimum, optimum, maximum).
Необходимо также отметить и тот факт, что скорости фермен-
тативных реакций при оптимальных условиях многократно превы-
шают скорости реакций, в которых используются неорганические
(синтетические) катализаторы.
Ферменты, с точки зрения энергетики, заметно снижают энер-
гию активации химических реакций. Под энергией активации
понимают количество энергии в Джоулях, необходимое для при-
ведения всех молекул 1 моля вещества при определенной темпе-
ратуре в состояние критического энергетического уровня (пере-
ходный уровень), при котором начинает происходить химическая
реакция. Если не все молекулы вещества достигают переходного
уровня, то скорость химической реакции снижается.
Основные пути повышения скоростей реакции — это либо
использование температурного фактора, либо катализаторов. В
случае применения неорганических или синтетических катализа-
торов необходимо использовать в большинстве случаев высокие
температуры. Как уже было сказано, биокатализаторы эффективно
работают при сравнительно низких температурах. Скорости реак-
ций зависят от концентраций субстратов. Когда фермент насыщен
субстратом, он не может функционировать быстрее — это и есть
максимальная скорость реакции (Vmax), то есть, когда доля свобод-
ного фермента оказывается предельно низкой.
Общая скорость ферментативной реакции должна быть про-
порциональной концентрации фермент-субстратного комплекса
ES (Е — энзим, S— субстрат). Зависимость скорости фермента-
тивной реакции от концентрации субстрата графически представ-
ляется гиперболической кривой (рис. 14). Км на рисунке представ-
ляет собой константу Михаэлиса—Ментен, то есть концентрацию
Рис. 14. Зависимость скорости
(V) ферментативной реакции
от концентрации субстрата
специфического субстрата, при которой конкретный фермент
обеспечивает скорость реакции, равную половине Vmax- До насто-
ящего времени анализ кинетики всех ферментативных реакций
проводят на основе следующего математического уровнения Ми-
хаэлиса—Ментен:
v _ Vmax [-S]
° Км + [S]
где Vo — начальная скорость при концентрации S, Vmax —
максимальная скорость и Км — константа Михаэлиса—Ментен для
данного фермента, соответствующая определенному субстрату.
Нередко прибегают к преобразованиям уравнения Михаэли-
са—Ментен для получения каких-либо преимуществ при изучению
кинетики ферментативных реакций. В частности, к таким преоб-
разованиям относится уравнение Лайнуивера-Бэрка:
1 _ Км 1 1
Vo Vmax [S] Vmax
Согласно этому уравнению, по графику, построенному в двой-
ных обратных координатах, можно более точно определить Vmax
(рис. 15).
На основании взаимодействия фер-
ментов и специфичных им субстратов, а
также учитывая множество фермента-
тивных реакций в клетке или в организ-
ме, и, наконец, принимая во внимание
другие взаимодействия в биосистемах
(антигены-антитела, токсины и их рецеп-
торы и т. д.), можно говорить о компле-
ментарности как основе биологической
специфичности. В большинстве случаев
имеет место молекулярная комплемен-
тарность. Такие взаимодействия проте-
кают согласно законам термодинамики.
Однако известны взаимодействия фер-
ментов с другими веществами, кото-
рые сопровождаются снижением J ш
полным подавлением активности био-
катализаторов. Вещества, подавляю-
щие ферментативные реакции, назы-
вают ингибиторами (от англ, inhibit —
препятствовать, сдерживать). Они из-
бирательно взаимодействуют с фер-
ментами, например, цианиды, оксид
углерода, азид натрия ингибируют оп-
73
Рис. 15. График двойных обрат-
ных координат.
ределенные окислительно-восстановительные ферменты; иодаце-
тамид, ртутные и мышьяковистые соли блокируют тиоловые фер-
менты; амины, гидразиды и др. взаимодействуют с карбонильной
группой ферментов и, как следствие, ингибируют их активность.
Очевидно, что в живой клетке на принципе "активация-ингибиро-
вание" проявляется и требуемая клетке функциональная актив-
ность в соответствующих условиях существования.
Все ингибиторы подразделяют на обратимые и необратимые:
среди обратимых выделяют конкурентные и некон-
курентные. Обратимые ингибиторы образуют непрочные
комплексы с ферментами [EI], которые способны распадаться на
исходные компоненты.
Е+1 <-> EI
Мерой ингибирования является та концентрация ингибитора,
которая вызывает угнетение фермента наполовину (I50). Отсюда,
чем меньше величина I, тем сильнее ингибитор, а чем больше I,
тем слабее. В случае взаимодействия необратимых ингибиторов
комплексы [EI] не распадаются и реакция протекает лишь вправо
Е + I —► EI
Конкурентный ингибитор связывается с активным центром
фермента и далее, в отличие от фермент-субстратного комплекса,
не подвергается ферментативной трансформации. Следовательно,
конкурентный ингибитор выступает конкурентом субстрату за
связывание с активным центром. Изменяя концентрации субстра-
та, можно вытеснить из комплекса ингибитор. Примером конку-
рентного ингибитора сукцинатдегидрогеназы является малонат (в
норме — субстратом выступает сукцинат).
Неконкурентный ингибитор связывается где-то в другом месте
фермента, а не с активным центром. При этом изменяется кон-
формация всей молекулы фермента, сопровождающаяся обрати-
мой инактивацией его каталитического центра. Неконкурентные
ингибиторы взаимодействуют и со свободным ферментом и его
фермент-субстратным комплексом:
Е + 1**Е1
ES+I о ESI
Комплексы EI и ESI становятся неактивными.
Из сказанного можно сделать вывод о том, что в работу
ферментов может вмешиваться множество различных веществ, с
помощью которых регулируется их биосинтез и активность. Био-
синтез ферментов будет рассмотрен позднее в этой главе. Что же
74
ПВ
отходы
Рис. 16. Идеализированная
клетка в устойчивом состоянии
(ПВ — питательные вещества).
касается регуляции активности биокатализаторов, то, наряду с
большим накопленным материалом в этом-направлении, открыва-
ются новые пути обмена, в регуляции которых участвуют фермен-
ты, и познание которых становится насущно необходимым; давно
возникла потребность полнее изучить
взаимодействие между известными ме-
таболическими циклами с определением
роли и механизмов энзиматического ре-
гулирования такого взаимодействия, а
также исследовать глубину участия раз-
личных ферментов в поддержании гоме-
остаза организмов (от греч. oemoios —
подобный, одинаковый, stasis — состоя-
ние, неподвижность), то есть устойчи-
вость их внутренней среды.
Любая живая клетка представляется
устойчивой системой, поддерживаю-
щейся однонаправленным потоком ме-
таболитов (рис. 16).
В зрелых клетках концентрации ме-
таболитов относительно постоянны.
Гибкость устойчивой системы, каковой
является зрелая клетка, подтверждается
слабыми изменениями и уравновешива-
нием сдвигов, благодаря чему организм
поддерживает постоянство внутренней среды вопреки широкому
разнообразию питательных веществ, качества воды, внешней тем-
пературы и т. д.
При переходе от одноклеточных организмов к многоклеточным
регуляторные механизмы ферментативных процессов существен-
но усложняются, хотя основные концепции приложимы к сущест-
вам любой организации.
В лабораторных или производственных условиях трудно или
невозможно увязать изменение ферментативной активности в
клетке (ткани) с каким-либо одним (из двух) процессом — возра-
станием количества фермента или возросшей эффективностью
биокатализатора. Более определенно можно говорить о повышении
или понижении активности в тех случаях, когда имеют дело с
ферментом в системе (-ах) in vitro, тем более, что большинство
результатов по оценке регуляции их активности получены в таких
системах.
В механизмах регуляции ферментативной активности у эука-
риотических клеток (в отличие от прокариот) существенна роль
физического разграничения (компартментализация) внутреннего
75
содержимого (протопласта). Организация набора ферментов, ка-
тализирующих соответствующую последовательность метаболиче-
ских реакций, в виде макромолекулярного комплекса координи-
рует ферменты и "канализует" интермедиаты (промежуточные
продукты) по данному метаболическому пути. Более того, конфор-
мационные изменения в одном компоненте комплекса может
передаться благодаря бёлок-белковому взаимодействию другим
ферментам комплекса. Вследствие этого возможно приумножение
регуляторных эффектов.
Велика роль ионов металлов в металлоферментах и металлоак-
тивированных ферментах, в которых аденозин-трифосфат (АТФ)
выступает субстратом. Когда комплекс "АТФ-ион металла" оказы-
вается субстратом реакции, то обычно максимальная активность
наблюдается при молярном соотношении АТФ и металла близким
к единице. При избытке того или иного из этих ингредиентов
отмечается ингибиторный эффект. Поскольку нуклеозидди- и три-
фосфаты образуют стабильные комплексы с дивалентными кати-
онами, постольку внутриклеточные концентрации нуклеотидов
могут влиять на внутриклеточное содержание свободных ионов
металлов и, как следствие, на активность определенных ферментов.
Например, конфигурация глутаматсинтетазы кишечной палочки в
отсутствие ионов металлов релаксирует ("расслабляется") и стано-
вится каталитически неактивной; добавление Мо2+ или Мп2+ вос-
станавливает активность фермента.
Каталитическая активность и регуляция активности ферментов
могут протекать в форме мгновенного обратимого ингибирования
по типу обратной связи (Feedback inhibition) с использованием
аллостерических эффекторов небольшой молекулярной массы, не
имеющих структурного сходства с субстратами или коферментами
и связывающимися с аллостерическими сайтами (не активными
центрами) ферментов.
д Ф1 _ Ф2 _ ФЗ г Ф4 ..
А------>Б------>В----->Г------>Д
Это — механизм грубого контроля регуляции активности фер-
ментов. В указанной схеме продукт Д способен связываться с
первым ферментом (Ф1) или ингибировать его. Следовательно, Д
— это отрицательный аллостерический эффектор или Feedback-
ингибитор Ф1 и таким образом может регулироваться синтез Д. В
качестве примера такой регуляции можно назвать ингибирование
триптофаном антранилат-синтетазы у микроорганизмов на пути
76
синтеза триптофана, где промежуточным продуктом является ан-
траниловая кислота.
Различают кумулятивную, мультивалентную (согласованную)
и кооперативную Feedback-ингибиции. В первом случае два или
более конечных продукта ингибируют строго аддитивно один
регуляторный фермент; во втором — два и более конечных про-
дукта находятся в избытке и только при этом условии происходит
полная ингибиция реакции; при кооперативной ингибиции един-
ственный конечный продукт, присутствующий в избытке, тормо-
зит действие регуляторного фермента, но когда, при наличии двух
и более конечных продуктов, торможение заметно превышает
аддитивные эффекты кумулятивной Feedback-ингибиции.
В отличие от Feedback-ингибиции Feedback-репрессия заклю-
чается в том, что производное (дериват) конечного продукта (ре-
прессор) подавляет образование ферментов (а не влияет на
их активность) в данном метаболическом пути:
А Ч1 ,б Ф-2 ,В—^Г-24-ffl (конечный .
\ \ I / Г продукт)
--------------J---репрессор
| аминокислоты]
Регуляторную роль в каталитической активности может играть
так называемая ковалентная модификация, когда к ферменту
присоединяется фосфатная группа (обычно у эукариот) или нук-
леотид (обычно у прокариот). Ферменты, подвергающиеся кова-
лентной модификации, сопровождаю-
щейся модуляцией их активности, на-
зываются интерконвертирующими.
Они существуют в двух активных со-
стояниях — высокой и низкой катали-
тической активности (рис. 17).
В ряде случаев конечный или про-
межуточный продукт используется в ка-
честве активатора какого-либо фермен-
та — это аллостерическая активация
или Feedback-активация.
Наряду с грубой регуляцией актив-
ности ферментов по механизму
Feedback-ингибиции известен тонкий
контроль, в котором ферменты — алло-
стерические белки имеют не только ка-
талитические центры для связывания с
субстратом, но и близко расположен-
ные другие места (одно или более), с
77
Е
НДФ
ФФн\£
НТФ
Mg h-*
НМФ-Е
Рис. 17. Регуляция фермента-
тивной активности ковалент-
ной модификацией (Е-фермент;
НТФ, НДФ и НМФ-соответст-
венно нуклеозид три-, ди- и мо-
нофосфаты, ФФн-неорганиче-
ский пирофосфат).
которыми связываются небольшие молекулы — эффекторы. При-
соединяясь к своему месту, эффектор вызывает конформационное
изменение в ферменте. При этом родство каталитического центра
фермента к субстрату либо уменьшается — наступает аллостери-
ческая ингибиция, либо, напротив, возрастает — наступает алло-
стерическая активация (рис. 18).
Рис. 18. Схематичное изображение модификации активного центра (АЦ) фермента
(Ф) с помощью эффектора (Э), взаимодействующего на аллостерическом сайте (А);
S-субстрат, (а)—аллостерическая ингибиция, (б)—аллостерическая активация.
При реализации инженерно-энзимологических процессов важ-
но знать и, следовательно, применять эффекторы на практике.
Вследствие компартментализации клеточных процессов у эукариот
остаются неизвестными концентрации эффекторов (а нередко —
и сами эффекторы) в клетках. Ярким примером здесь служит
фосфофруктокиназа - давно и хорошо изученный фермент глико-
лиза. Оказалось, что она имеет эффектор — фруктозб-2,6-дифос-
фат, открытый лишь 10 лет назад X. Г. Херсом, Л. Хью и Е. Ван
Шафтингеном.
Очевидно, что во всех рассмотренных примерах регуляции
активности ферментов важное значение имеет чувствительность
биокатализаторов к регуляции, то есть нередко усиление (ампли-
фикация) "сигнала" может оказать решающее влияние на включе-
ние—выключение каскада реакций.
78
Глава 4
ФУНДАМЕНТАЛЬНЫЕ ИССЛЕДОВАНИЯ
В ОБЛАСТИ СТРУКТУРНО-ФУНКЦИОНАЛЬНОЙ
ОРГАНИЗАЦИИ ВИРУСОВ, КЛЕТОК И ТКАНЕЙ
Чтобы с максимальной, продуктивностью использовать биообъ-
ект в биотехнологическом процессе, необходимо знать и учитывать
его структурно-функциональные особенности применительно к
конкретным условиям производства. В большинстве случаев каче-
ство и количество целевого продукта в значительной степени
находятся в прямо пропорциональной зависимости от качества и
количества продуцента. Совершенно очевидно, что подходы здесь
к акариотам, прокариотам и эукариотам должны быть различными,
поскольку первые, например, являются облигатными паразитами
и могут развиваться лишь в живых клетках (тканях); бактерии
структурно менее дифференцированы, чем эукариоты и поэтому
они в большинстве своем менее требовательны к условиям обита-
ния, чем грибные, растительные и животные организмы.
4.1. Акариоты. Ряд бактериофагов, вирусов растений и млеко-
питающих широко используют в биотехнологии, особенно — в
рДНК-биотехнологии (см. ). У них наследственный материал уст-
роен наиболеё просто и работа с ним по этой причине несколько
облегчается, хотя понятно, что она на самом деле "ювелирная".
Как уже было сказано в главе 2, вирион представляет собой
нуклеокапсид и архитектоника вирусных частиц не одинаковая
(см. рис. 1а и 226). Многие вирусы животных представляют собой
"голые" нуклеокапсиды (аденовирусы), тогда как другие (вирусы
герпеса и оспы) имеют дополнительную оболочку (суперкапсид),
нередко приобретаемую во время освобождения нуклеокапсида из
клетки организма-хозяина. "Голый" спиральный нуклеокапсид ви-
руса табачной мозаики — ВТМ (вирион) имеет молекулярную
массу (ММ) 39» 106 дальтон (Да), а его единственная молекула РНК
— 2,06*106 Да. Некоторые вирусы имеют двухнитевую РНК, на-
пример, реовирусы, ММ которой равна 9*106 Да.
У большинства вирусов, содержащих ДНК, нуклеиновая кис-
лота является двухнитевой (равно как и у многих бактериофагов).
В качестве исключения можно назвать парвовирусы и coli-фаг
<рХ 17 4, имеющих однонитевую ДНК. Молекулярные массы ДНК
различны и достигают следующих величин: у coli-фага <рХ174 —
1,7*10® Да, у четных бактериофагов Т2, Т4 и Те — 1,2*10® Да, у
герпесвирусов — 1*10® Да, у вируса оспы — 1,5*10® Да и т. д.
Капсид любого вируса построен из 5—6 белковых субъединиц
— капсомбров; это - морфологические единицы, расположенные,
79
Рис. 19. Схема строения вируса мозаики
табака: А — вид сбоку, В — вид сверху; 1
— одноцепочечная РНК, 2 — Белковые
субъединицы.
как правило, строго регуляр-
но, благодаря чему капсид
приобретает специфический
тип молекулярной
симметрии — спи-
ральный или куби-
ческий. Например, вирус
табачной мозаики имеет спи-
ральный тип симметрии. Его
спиралевидная РНК упакова-
на в винтообразно располо-
женные капсомеры и, в целом,
вирион представляется палоч-
ковидным (рис 19). Спираль-
ный тип симметрии характерен также для вирусов: бешенства,
гриппа, желтухи свеклы, кори, ложной чумы кур, парагриппа,
паротита, сендай, чумы рогатого скота, чумы собак и др.
Кубический тип симметрии является наиболее распространен-
ным среди большинства известных ныне вирусов. Из трех типов
Рис. 20. Икосаэдр с тремя осями симметрии (а, б, в).
фигур с кубической симметрией: тетраэдр (оси симметрии 2:3,
минимальное число структурных единиц 12), октаэдр (оси симмет-
рии 4:3:2, число структурных единиц 24) и икосаэдр, или правиль-
ный многогранник (оси симметрии 5:3:2, число структурных еди-
ниц 60) последний наиболее част у вирусов. Это объясняется тем,
что в случае икосаэдра минимальное число структурных единиц
оказывается наибольшим, и, следовательно, данный тип симметрии
пространственно наиболее экономичен (не требуется изготовления
крупных молекул при сборке вибриона). На рис. 20 изображен
икосаэдр с тремя осями симметрии (5:3:2). В правильном икосаэдре
имеется 6 осей симметрии пятого порядка, проходящие через
80
вершины (а), 10 осей симметрии третьего порядка, проходящие
через центры треугольных граней (б) и 15 осей симметрии второго
порядка, соединяющих середины ребер (в). Икосаэдр обладает 12
вершинами, 20 гранями и 30 ребрами.
Капсиды некоторых вирусов "осложнены" дополнительными
структурами. Так, например, у аденовируса капсид формируется
в виде икосаэдра с 6 капсомерами вдоль каждого ребра; всего в
капсиде 252 капсомера, из которых 240 имеют сферическую форму
и располагаются вдоль ребер и на гранях икосаэдра. Каждый
капсомер соседствует с 6 другими капсомерами, поэтому они
называются гексамерами, или гексонами (в
рассматриваемом примере их 240). Остальные 12 капсомеров
располагаются на 12 вершинах икосаэдра и соседствуют с 5
капсомерами (п е н т а м е р, или пентон). Эти 12 капсомеров
(пентонов) состоят из сферического основания и длинной нити,
которая может обеспечивать прикрепление вириона к клетке (рис.
21).
Рис. 21. Схематичная модель аденовируса с внутрен-
ним ДНК-протеиновым кором и внешним капсидом
— икосаэдром (П-гексон — 120000 Да; Ш-пентоновое
основание — 85000 Да; Ша, VI-, VIII и 1Х-белки,
ассоциированные с гексоном, имеющие ММ соответ-
ственно 66000 Да, 24000 Да, 13000 Да и 12000 Да;
IV-белковая "нить" — 62000 Да, V и VII-протеины кора
с ММ 48000 Да и 18000 Да).
Сложные капсиды присущи и бактери-
офагам, например, Т-четным coli-фагам
(см. рис. 1а), у которых имеется икосаэд-
рическая головка и гексагональный отро-
сток. В таблице 6 приведен перечень неко-
торых вирусов прокариотических и эукариотических организмов
с различными типами симметрии капсидов.
Необходимо подчеркнуть, что вирусы не размножаются, а
репродуцируются, или воспроизводятся, то есть правильнее гово-
рить не об их потомстве, а их "копиях" (братьях и сестрах).
Репродукция происходит только в живых клетках (тканях) и поэ-
тому биотехнологические процессы, основанные на использовании
вирусов, исключительно дорогостоящи. Достаточно сравнить, на-
пример, кишечную палочку и вирус гриппа. Первая растет на
мясопептонном бульоне, второму необходимы куриные эмбрионы;
чтобы проконтролировать рост кишечной палочки и увидеть ее в
81
среде, необходим обычный "световой” микроскоп, а чтобы увидеть
вирусные частицы, нужен электронный микроскоп, стоимость
которого во много раз превосходит стоимость светового. Подобные
сравнения можно продолжать и далее, и в этом смысле выгоды
будут складываться в пользу бактерий. Однако создание противо-
гриппозных биопрепаратов (вакцин) сегодня более насущная за-
дача, чем борьба с заболеваниями, вызванными энтеропатогенны-
ми кишечными палочками и родственными ей бактериями. Кроме
того, решение многих фундаментальных проблем в области моле-
кулярной биологии нередко возможно исключительно на вирусных
Таблица 6 Общая характеристика некоторых вирусов
Вирусы Наличие (-1-) или отсутствие (-) и ММ (Да) хЮ6 Супер- капсид Тип симмет- рии Вирион
ДНК РНК ММ (Да) хЮ6 Средний размер, нм
Бактерий (бактериофаги) 1. Колифаг <рХ174+ 2. Колифаг Л + 3. Колифаг Тг 4. Колифаг MS=2+ Растений 1. Мозаики табака (ВТМ) 2. Желтой мозаики турнепса Животных 1. Восточного энцефаломиелита лошадей+ 2. Обезьян SV-40 Человека 1. Аденовирусных инфекций 2. Гриппа+ 3. Полиомиелита4" ( + ) 1.6 ( + ) зз ( + ) 130 Н Н Н (-) ( + )3,2 ( + )23 (-) Н Н (-) (-) ( + ) 1 ( + )2,1 ( + ) 1.7 ( + )2 (-) (-) ( + )4 ( + )2,2 Отсут- ствует Име- ется Отсут- ствует Имеет- ся ке и _ 1» к с к с к 6,2 100 300 3,6 40 5 50 3 177 300 6,8 (100x50) х(80х95- 100) ' 25 15-17 хЗОО 30 50-70 40-50 70-90 80-120 28
Примечание: -I---одноцепочечная нуклеиновая кислота, К — кубический (у фа-
гов в головке), 0 — с отростком(-ами), С — спиральный
82
системах (см. раздел 5.1). Репродукцию вирусов можно подразде-
лить на семь этапов: адсорбцию, пенетрацию (проникновение
вируса в клетку), транскрипцию, трансляцию, репликацию, сборку
вирусных частиц и выход организованных вирусных частиц из
клетки. На каждом этапе совершаются многоактовые процессы,
зависимые как от вирусов, так и от клеток — их реципиентов.
На рис. 22 а, б представлены схемы строения вируса гриппа (а)
и ретровируса — ВИЧ (вирус иммунодефицита человека) — воз-
будителя СПИД (б). При репродукции вируса гриппа генетическая
информация передается по схеме РНК —► РНК —- Белок. У них
биосинтез РНК осуществляется на матрице вирионной РНК под
каталитическим действием вирионной РНК-зависимой РНК-пол-
имеразы, или вирионной транскриптазы. У ВИЧ генетическая
информация передается по схеме РНК—'ДНК—»РНК—►Белок, то
есть вначале по матрице РНК синтезируется ДНК с помощью
фермента обратной транскриптазы, а затем все протекает по
универсальной схеме ДНК—РНК—►Белок. Структурный материал
в обоих приводимых случаях используется из содержимого реци-
пиентных клеток.
Рис. 22. Схемы строения вирусов гриппа (а): 1 — гемаглютинин, 2 — нейраминидаза,
3 — липидный бислой, 4 — белковый слой, 5 — рибонуклеопротеин; и ретровируса
ВИЧ (б): 1 — гликопротеин-120, 2 — сердцевина, 3 — гликопротеин-41, 4 —
липоидная мембрана, 5 — РНК, 6 — обратная транскриптаза.
Рибонуклеиновая кислота вируса гриппа состоит из 8 сегмен-
тов, каждый из которых на З'-конце заканчивается уридиновым
остатком. Общая молекулярная масса РНК составляет 2 — 4 тыс.
83
кДа. Вследствие такой сегментированное™ геном вируса гриппа
отличается высокой частотой рекомбинации, способностью к ре-
активации и синтезировать гемагглютинин и нейраминидазу после
химического подавления вирусной инфекционности.
Частицы ВИЧ образуются из трех различных по составу и
молекулярной массе белков — протеина 25(24), гликопротеинов 41
и 120. Первый из них р25(24) входит в состав сердцевины вириона,
гликопротеины (др41 и др 120) формируют "шипы” оболочки час-
тицы. В сердцевину также входят однонитевая РНК, несущая
генетическую информацию, и фермент обратная транскриптаза,
под действием которой синтезируется провирус — ДНК-копия
вирусной РНК. Провирус встраивается в хромосомную ДНК клет-
ки — реципиента и персистирует (от лат. persistens — стойкий,
сохраняющийся, остающийся) до того времени, пока не будет
активирован. После такой активации происходит его репродукция.
Если ретровирусные РНК структурно заканчиваются прямыми
повторами нуклеотидов числом 10 — 80 пар, то свободные линей-
ные двухнитевые ДНК-копии заканчиваются длинными одинако-
выми повторами — LTR (от англ, long terminal repeat) числом 250
— 1400 нуклеотидных пар, укороченных на две пары с каждого
конца (рис. 23).
Y3’ S Y5’
LTR
9а9 ро1
Y3' S Y5'
провирусная ДНК
LTR
Рис. 23. LTR в составе линейной ДНК копии РНК вируса иммунодефицита человека
Сборка новых вирионов происходит на клеточной мембране.
В процессе агрегации вирусных белков она начинает набухать и
выпячиваться, высвобождал! в конечном итоге новоорганизован-
ные частицы вирусов.
Вирусы прокариот, или бактериофаги, в большинстве своем
имеют структурно-функциональное сходство. В частности, они
содержат лишь один какой-то тип нуклеиновой кислоты — ДНК
или РНК в качестве генетического материала (у большинства фагов
имеется двунитевая ДНК); нуклеиновая кислота упаковывается в
головку фага; преобладающее большинство фагов имеет хвостовой
отросток для прикрепления к поверхности реципиентной клетки;
наконец, бактериофаги сходны по характеру индуцируемых ими
событий, развивающихся в клетке после ее заражения — фаги
являются облигатными паразитами.
Различия бактериофагов обусловлены тонкой структурой нук-
леиновых кислот (в последовательности нуклеотидов, наличием
84
или отсутствием липких концов, в линейности или кольцевидной
замкнутости и пр.), типами молекулярной симметрии капсидов,
деталями строения хвостовой части, специфичностью действия в
отношении чувствительных клеток.
Известны три состояния, в которых могут находиться недефек-
тные фаги и три типа влияния фаговой инфекции на судьбу
зараженной клетки. К числу первых относят: свободное состояние,
вегетацию и состояние профага (для так называемых умеренных
фагов); к числу вторых — гибель зараженной клетки (фаги здесь
называют истинно вирулентными); переход клетки, несущей уме-
ренный фаг (профаг), на путь лизогенного развития, или, в случае
индуцибельности профага и воздействия индуцирующими факто-
рами (УФА, некоторые мутагены и др.) — на путь лизиса; наконец,
при третьем типе влияния фаговой инфекции не наблюдается
каких-либо заметных отклонений в характере поведения заражен-
ных клеток — гибели их не происходит; фаги при этом могут
высвобождаться из клеток или постоянно реплицироваться, нахо-
дясь внутри их и слегка замедляя скорость размножения клеток.
Учитывая сказанное, следует подчеркнуть, что бактериофаги име-
ют большое значение в биотехнологии еще и потому, что они могут
выступать ощутимыми вредителями в микробиологических произ-
водствах, базирующихся на эксплуатации прокариотических орга-
низмов.
Результаты фундаментальных исследований структуры вирои-
дов упрочили самостоятельность этой группы микробов в царстве
Vira. Ныне вироиды подразделяют на три группы согласно нукле-
отидных последовательностей в их РНК:
1) группа ВВКК, включающая вироиды веретеновидности клуб-
ней картофеля и все другие вироиды кроме ВКККП и ВСПА (см.
ниже); у них имеются значительные области гомологии последо-
вательностей (50 — 80%) и относительно протяженная полипури-
новая последователь-
ность; они содержат
высококонсерватив-
ную центральную об-
ласть (рис. 24); 2) виро-
иды болезни каданг-ка-
Рис. 24. Вироиды: 1 — линей-
ные и 2 — кольцевые формы
денатурированных молекул
вироида ВВКК (электронно-
микроскопический снимок)
(по Т. О. Динеру, 1984).
85
данг кокосовой пальмы (ВКККП), содержащий консервативную
центральную область, короткую последовательность пуринов, а за
пределами центральной области проявляет малую гомологию по-
следовательностей;
3) вироиды болезни "солнечная пятнистость авокадо" (ВСПА)
с небольшой центральной консервативной областью и малой го-
мологией последовательностей с последовательностями у любого
другого вироида.
Высказывается предположение в том, что вироиды произошли
от интронов (см.) и что недалеко то время, когда они будут
рекомендованы в качестве векторов в фитобиотехнологии реком-
бинантных ДНК.
4.2. Клетки прокариот. Почти во всех больших траксономиче-
ских группах имеются представители, с успехом использующиеся
в соответствующих биотехнологических процессах — будь это
сапрофитные или патогенные виды. Для сравнения можно указать
на лактобактерии и туберкулезные микобактерии. Первые исполь-
зуются для приготовления молочнокислых продуктов и в силосо-
вании кормов, вторые — для приготовления вакцины BCG и
туберкулина (диагностическое средство при туберкулезе). Из ар-
хеобактерий большое значение имеют метаногенные бактерии —
продуценты метана. Пневмококки используются для изготовления
полисахаридных вакцин, применяемых для профилактики и лече-
ния пневмоний, вызванных различными сероварами Streptococcus
pneumoniae.
Археобактерии заметно отличаются от эубактерий по
химическому составу клеточной стенки и клеточной мембраны, по
последовательности нуклеотидов в рибосомных нуклеиновых кис-
лотах, или рРНК (в 5S- и 16S рРНК) и по
некоторым другим признакам. Так, в кле-
точной стенке у них отсутствует типичный
пептидогликан — муреин, хотя у некото-
Н.ОН рых археобактерий имеется псевдомуре-
ин, в котором вместо ацетилмурамовой
кислоты содержится талозаминуроновая
кислота; интерпептидные мостики вклю-
талозаминуроновая кислота чают лишь L-аминокислоты, так как D-ами-
нокислоты отсутствуют в псевдомуреине.
У метанококков клеточная стенка формируется из белка, у мета-
носарцин — из нейтральных углеводов, уроновых кислот и ами-
носахаров, у метаноспирилл имеется полипептидный чехол.
Особенностями химического состава клеточной стенки архео-
бактерий можно объяснить неэффективность пенициллина, цефа-
86
соон
н н
лоспорина и D-циклосерина против этих микроорганизмов. Иначе
говоря, для данных антибиотиков отсутствуют мишени в клетках
археобакгерий. Клеточная мембрана их также существенно отли-
бифитанил-диглицерин (бифитанил-диглицерол-тетраэфир)
чается от клеточной мембраны эубактерий. В частности, в ней не
обнаружены фосфатидилглицерины, но содержатся бифитанил
(С 20)-(изопреноид)-глицерин, а у некоторых представителей архе-
обактерий — бифитанил (С40)-(изопреноид)диглицерин. Обнару-
жены и нейтральные липиды в форме свободных С15- и Сзо-изо-
преноидных углеводородов (сесквитерпены и тритерпены).
Рибосомы археобакгерий и эубактерий сходны между собой,
н2с=с-сн=сн-сн=(р-сн=сн-сн=с-сн=сн2
сн3 сн3 сн3
сесквитерпен (С]5)
тритерпен (Сзо)
по константе седиментации относятся к типу 70S, но последова-
тельность оснований в 5S- и 16S- рибосомных РНК у археобакгерий
87
рифампицин-3=(4'-метил-1 '-пиперазинилиминометил)-рифамицин SV
иная. ДНК-зависимые РНК-полимеразы, в отличие от эубакгерий,
состоят у них из более чем четырех субъединиц, и эти ферменты
не чувствительны к антибиотику рифампицину.
В геноме археобактерий выявлены интроны, что ранее рассмат-
ривалось принадлежностью генома эукариот. Поэтому в настоящее
время сформировалось два направления по трактовке места архе-
обактерий и их происхождения. Ученые — приверженцы первого
направления считают, что интроны в геноме эукариот есть резуль-
тат эндоцитоза древними эукариотическими микроорганизами
клеток прокариотических археобактерий, которые впоследствии
(в процессе эволюции) трансформировались в митохондрии. При-
верженцы второго направления считают археобактерий прароди-
телями эубактерий, то есть по их мнению эволюция шла по линии:
Археобактерий —> Эукариоты Эубактерии.
Промежуточное положение эукариот в приведенной последо-
вательности как бы разрывает одно надцарство прокариот на два,
однако принципиальная организация ядерного аппарата у архео-
бактерий и эубактерий во многом сходна , следовательно, приве-
денная схема эволюционных событий представляется как бы не-
достаточно обоснованной. С учетом современных знаний о прока-
риотах эволюцию их можно представить в двух следующих вари-
антах (см. стр. 89).
Во второй схеме отсутствует прямая связь между археобакте-
88
I) Археобактерий-------» Эубактерий
•Эукариоты
Эукариоты
Прокариоты
или Археобактерий
Эу бактерии
Древние предковые формы
риями и эубактериями. Не следует забывать и того факта, что без
бактерий жизнь всех других, более высоко организованных (эука-
риотических) существ на Земле, вообще прекратилась бы.
Из приведенных данных следует, что представления об эволю-
ции микроорганизмов (равно как и других существ) до сих пор
остаются во многом не разрешенными. Тем не менее, сегодня мы
больше знаем, чем два — три десятилетия назад, о структурно-фун-
кциональной организации клеток различных организмов и заметно
расширили арсенал промышленно полезных видов.
Эубактерий включают все прокариоты кроме археобак-
терий. Их подразделяют на две группы — фототрофные и хемо-
трофные бактерии. Этим подчеркивается их принципиальное раз-
личие по используемому источнику энергии. Фототрофы исполь-
зуют кванты солнечного света, тогда как хемотрофы — энергию
химических связей в различных химических соединениях. Среди
фототрофов различают оксигенные цианобактерии, в процессе
жизнедеятельности которых выделяется молекулярный кислород
(1), и аноксигенные пурпурные и зеленые бактерии, не выделяю-
щие кислорода (2а,б,в).
В реакциях 2а,б,в донорами электронов соответственно явля-
ются сероводород, газообразный водород и изопропанол.
Цианобактерии являются важными поставщиками кислорода
в атмосферу, а из атмосферы они поглощают (фиксируют) моле-
кулярный азот.
Хемотрофные эубактерий могут образовывать (или не образу-
ют) эндоспоры. Клетки их весьма разнообразны по морфологии:
прямые, изогнутые, палочковидные, круглые, овальные, бобовид-
ные, спиралевидные, нитевидные и др. Удельный вес микробной
клетки составляет примерно 1,038 — 1,065. Тогда, исходя из средних
89
1. 6Н7О+6СО7 ---------------------- (СН7О),+6О2
Здесь донор электронов - вода. Следует иметь в виду, что среди цианобактерий
есть виды, способные окислять сероводород и переходить на фотосинтез без
выделения кислорода.
2. 2H,S+CO7 --------------—---------------* CH7O+H7O+2S
2 I пурпурные серные и зеленые Z 2
серные бактерии, некоторые
цианобактерии
Сера затем окисляется до сульфата. Тогда суммарная реакция будет следующей:
L v
2а) H7S+2CO7+2H7O -------------—----------» (CH7O)7+H7SO4
4 4 4 пурпурные серные н зеленые 4 4 4
серные бактерии, некоторые
цианобактерии
2б) 2Н, + СО7 --------—--------* СН7О+Н7О
7 Z Z пурпурные несерные и Z Z
зеленые несерные бак-
терии
2в) 2СН3СН(ОН)СН3+СО2 nypnyphHVb,e не. ^СНгО+ДСН^СО-ьН/)
изопропанол сеРные бактерии ацетон
размеров бактерии 2 х 0,5 мкм, вес ее составит 4,12» 10'10 мг, то есть
в 1 г будет содержаться 2,42«1012 таких клеток.
Важным параметром, характеризующим большинство прока-
риотических клеток, является скорость их размножения, измеря-
емая минутами (в среднем от 8 — 10 мин до 30 — 40 мин). Это
важно в тех случаях, когда биомасса клеток является целевым
продуктом в конкретном производстве, или когда количество
образующегося метаболита (целевого продукта) находится в пря-
мопропорциональной зависимости от количества вырастающих
клеток за конкретный временной интервал.
90
Все обменные процессы развивающихся прокариотических
клеток осуществляются с участием клеточной мембраны. Через
мембрану поступают внутрь клетки питательные вещества, через
нее же секретируются определенные продукты из клетки в окру-
жающую среду; мембраны участвуют в регуляторных процессах и
энергообеспечении клетки. Вот почему, начиная с 70-х — 80-х годов
XX в., когда удалось в чистом виде изолировать и исследовать
мембранные фракции различного происхождения, интерес к мем-
бранам резко возрос. Сформировалась самостоятельная научная
дисциплина — мембранология, а специалисты, работающие
в этой области, стали называться мембранологами.
Клеточная стенка прокариот занимает свое особое место в
структуре и архитектонике клеток. Ее нельзя исключать из мета-
болических процессов, так как она занимает пограничное положе-
ние между внутренней (протопласт) и внешней средами, и через
нее должны проходить различные вещества в обоих направлениях.
Однако главные функции ее — поддержание формы клетки и
защитная, тогда как основная функция клеточной мембраны —
регуляторно-метаболическая. Клеточная стенка и клеточная мем-
брана вместе формируют оболочку.
В эволюционно-биохимическом плане клеточная стенка у про-
кариот претерпела структурные изменения в направлении: Архе-
обактерии —> Грамположительные бактерии Грамотрицатель-
ные бактерии. У археобактерий нет типичного пептидогликана,
хотя задатки к нему имеются; пептидогликан у грамположительных
бактерий многослоен, тогда как у грамотрицательных бактерий он
однослоен. Лишая клетки прокариот клеточной стенки, их относи-
тельно легко удается получать в виде протопластов или сферопла-
стов, которые могут быть использованы в клеточно-инженерных
работах.
Вместе с возможным капсульным слоем на оболочку приходит-
ся 20% и более сухой массы клетки. В оболочке имеются поры для
транспорта питательных веществ (диаметр их от 0,001 до 0,01 мкм)
и рецепторы (белки- порины) для фагов и бактериоцинов, а также
места взаимодействия с антителами и комплементом. В оболочках
грамотрицательных бактерий содержатся токсические и аллерген-
ные соединения.
Электронно-микроскопические снимки поверхностей прока-
риотических клеток отличаются большим разнообразием. Причем,
91
в противоположность сравнительно тонко очерченной поверхно-
сти у грамположительных бактерий, большинство представителей
грамотрицательных видов имеет исключительно складчатую по-
верхность.
У многих прокариот кнаружи от клеточной стенки располага-
ется капсульн...й материал, состоящий в большинстве случаев из
полисахаридов — гликанов (например, у Acinetobacter spp.), либо
— из протеинов (например, у Вас. licheniformis).
Клеточная стенка утолщается с возрастом клетки и может
достигать, например у Lactobacillus acidophilus, 0,8 мкм. Клеточная
мембрана, напротив, остается более или менее постоянной по
толщине вовсе периоды развития прокариотических клеток (0,0075
мкм) и с более или менее постоянными порами диаметром около
1 нм.
В клеточной стенке грамоположительных бактерий сосредото-
чен пептидогликан — муреин, имеются тейхоевые кислоты, белки;
у стрептококков группы А в диффузно-распределенном виде в
наружном слое клеточной стенки Находится также М протеин,
являющийся фактором вирулентности этих микробов. М протеин
может быть гидролизован с помощью трипсина без нарушения
жизнеспособности клеток.
У грамотрицательных бактерий четко обозначается трехслой-
ность клеточной стенки: липополисахаридный слой (О-антиген),
наружный слой (нередко обозначаемый как "внешняя мембрана"),
состоящий из двух фосфолипидных листков, и подлежащий липоп-
ротеиновый слой. Липополисахарид проявляет свойства эндоток-
сина, он занимает пограничное положение между внешней средой
и подлежащим фосфолипидом (преимущественно — фосфатиди-
лэтаноламином).
Липополисахариды получают в производственных условиях В
качестве средств, обладающих различными биологическими свой-
ствами: токсическими (связанными с Липидом А в липополисаха-
риде), пирогенными, митогенными (для мышиных лимфоцитов),
стимуляторами развития клеток костного мозга, активаторами
фактора VII свертывания крови (фактор Хагемана) в тромбоцитах
и комплемента; липополисахарид в концентрации 1*1012 г вызы-
вает свертывание лизата амебоцитов дальневосточного краба
Limulus poliphemus. Реакцию широко используют при выявлении
липополисахарида в каких-либо субстратах, лекарственных сред-
ствах и др. Взаимодействие между эндотоксином и лизатом аме-
боцитов протекает по следующей схеме:
92
LAL= профермент
эндотоксин
активироваиный( — е)
фермепт(ы)
активированные ферменты
коагулоген
лизат
белковые агрегаты
Фосфолипидный слой имеет внешний и внутренний "листки",
из которых внешний содержит значительную долю липополисаха-
ридных молекул. В целом этот слой имеет жидкостно-мозаичную
структуру, содержащую набор из 3 — 4 мажорных белков, погру-
женных в фосфолипидный матрикс, и на долю которых приходится
до 70% всех белков наружного слоя, а также набор из 10 — 20
минорных белков. С фосфолипидным слоем (его внутренним
листком) нековалентно связан липопротеин из третьего слоя.
Липопротеиновый слой выступает как бы посредником, неко-
валентно связывающим наружный слой с пептидогликаном. Он
включает жирные кислоты, аминокислоты, глицерин; молекуляр-
ная масса его порядка 7 кДа. Так липопротеин клеточной стенки
Escherichia coli содержит 15 аминокислот в разной повторности
(всего 58 аминокислотных остатков), среди которых не обнаруже-
ны гистидин, глицин, пролин, триптофан и фенилаланин. Он
представляется в виде блочной структуры, на конце которой
имеется глицерил-цистеин, этерифицированный остатками жир-
ных кислот. Значительная часть такого липопротеина (4,8* 105 мо-
лекул на клетку) находится в свободном состоянии, меньшая часть
(2,4* 105) ковалентно связана с пептидогликаном (с диаминопиме-
линовой кислотой в тетрапептиде муреинового "каркаса").
Белки клеточной стенки отличаются от белков клеточной мем-
браны. Ранее упомянутые белки-порины образуют трансмембран-
93
ные диффузионные каналы для прохождения неспецифических
малых гидрофильных молекул, некоторые из них выступают ре-
цепторами фагов. Выделяют мажорные, или отр-протеины (от
англ, one of major protein), и минорные, или малые протеины. По
кодирующим структурным генам мажорные белки подразделяют
на omp А, отр С, отр F, отр Н:1°; они имеют различную
молекулярную массу (от 16 кДа до 42 кДа). Первый из них (отр
А) выступает местом взаимодействия с F-пилями при конъюгации
бактерий; omp С и отр F (белки-порины) в комплексе с пептидо-
гликаном формируют поры, отр Н:1° обладает катионными свой-
ствами и причастен к ионным взаимодействиям в среде обитания
клетки. Минорные белки участвуют в транспорте специфических
малых молекул через наружный слой и в рецепции фагов, некото-
рых бактериоцинов (например, рецептор для фага Тб и колицина
"заинтересован" в транспорте нуклеозидов). Минорные белки
вовлекаются в транспорт мальтозы, комплексов Fe2+, витамина В12.
Белки внешнего слоя выполняют и некоторые другие функции.
Среди протеинов клеточной стенки грамотрицательных бакте-
рий обнаруживаются многие ферменты (аспарагиназа, фосфатазы,
эндонуклеаза и др.).
Клеточную стенку удается достаточно легко отделить от содер-
жимого клетки, окруженного клеточной мембраной (протоп-
ласт). Если часть клеточной стенки каким-то образом удержива-
ется на клеточной мембране, то в этом случае говорят о с ф е -
ропласте. Имеются разноплановые мнения о легкости
формирования протопластов у грамположительных или, напротив,
у грамотрицательных бактерий. Очевидно, и протопласты и сфе-
ропласты могут быть дериватами тех и других бактерий — все
зависит от особенностей штамма (вид, возраст, условия культиви-
рования, используемый стабилизатор — сахароза или какие-либо
соли, и т. д.), воздействующего агента (лизирующий клеточную
стенку фермент, блокатор биосинтеза компонента или компонен-
тов клеточной стенки).
Между клеточной стенкой и клеточной мембраной имеется так
называемое периплазматическое пространст-
в о, лучше выявляемое у грамотрицательных бактерий. Это оче-
видно связано с тем, что внутреннее осмотическое давление выше
у грамположительных бактерий [(8,1 - 20,2)«105 Па[, чем у грамот-
рицательных [(3,03-5,05)«105 Па]. В периплазматическом простран-
стве обнаружены ферментные и неферментные белки.
Известны прокариоты, оболочки которых существенно отли-
чаются по составу от оболочек грамположительных и грамотрица-
тельных бактерий. В качестве примера можно назвать кислото-
устойчивые микобактерии (таблица 7). Из таблицы видно, что
94
Таблица 7 Компонентный состав оболочек грамположительных, грамот-
рицательных и кислотоустойчивых бактерий
Бактерии
грамположительные кислотоустойчивые грамотрицательные
Пептидогликан (многослоен, толщина 0,02-0,6 мкм) Пептидогликан (однослоен!?, толщина 0,01 мкм) Пептидогликан (однослоен, толщина 0, 01 мкм)
Протеины Полипептиды Липопротеины
Липотейхоевые кислоты Миколовая кислота - * гликолипиды Фосфолипиды
Тейхоевые кислоты Арабиногалйктаны Липополисахарид
Тейхуроновые кислоты Воск Д Протеины
Полисахариды Корд-фактор Сульфолипиды МИКОЗЦДЫ Полисахариды
различия в компонентном составе клеток, относящихся к разным
группам, достаточно велики. Например, оценивая строение пепти-
догликанов у Е. coli и Staphylococcus aureus, можно видеть различия
между ними в характере интерпептидных мостиков. Так, у Е. coli
(равно как и у всех грамотрицательных бактерий, коринебактерий
дифтерии, нокардий, микобактерий и видов, относящихся к роду
Bacillus) пептидогликан относится к типу А (рис. 25а), у
Staphylococcus aureus (стрептококков, микрококков) — к типу В
(рис. 256); пептидогликан типа С (рис. 25в) обнаруживается редко.
Рис. 25 Схемы (а, б, в), построения трех типов пептидогликанов (А, В, С) в которых
М—N-ацетилмурамовая кислота, Г—N-ацетилглюкозамин; 1,2,3,4-тетрапептид, со-
стоящий из L-аланина (1), D-iso-глутаминовой кислоты (2), meso-DAP (или L-лизина)
(3), D-аланина (4), х и z-интерпептидные мостики, У-амидный заместитель, связанный
с а-карбоксигруппой D-iso-глутаминовой или 3-гидроксиглутаминовой кислоты.
95
Инвариантность тетрапептида обусловлена постоянным нали-
чием D-аланина, выступающего связующей единицей между цепя-
ми пептидогликана. Дисахаридные блоки МГ (см. рис. 25а, б, в)
численно варьируют от менее 10 до более 170, что зависит от вида
бактерий. С учетом сказанного можно изобразить связь пептидо-
гликана с липопротеином в клеточной стенке Е. coli (рис. 26).
Рис. 26. Фрагмент пептидогликана Е. coli, связанный с липопротеином через
meso-DAP в тетрапептиде (обозначения смотри на рис. 25).
Углеводные звенья в пептидогликанах могут быть различными.
Например, имеются О-ацетильные группы в мурамовой кислоте
или глкжозамине, в эндоспорах бацилл может присутствовать
лактам мурамовой кислоты.
н о
гликолилмурамовая кислота,
где R - гпикопип
лактам мурамовой кислоты
макнозамим мурамовой кислоты
96
Находят (около 2%) маннозамин мурамовой кислоты, например
у М. luteus. У микобактерий и некоторых нокардий (N. kirovani)
присутствует N-гликолилмурамовая кислота. В позиции С6 гидро-
ксильных групп мурамовой кислоты или глюкозами на могут нахо-
диться фосфодиэфирные остатки, которые у грамположительных
бактерий способны связывать тейхоевую, тейхуроновую кислоты,
?н
НО-Р-О-СН,-СН-СН-СН-СН,-СР-О-СН,-СН-СН-СН-СН,-О II III “ III Р-О-СН -СН-СН-СН-СН,ОН II 1 1 1
III 0 1 О OR О О OR О О ° OR О О
СО СО со 1
। । CH-NH7 CH-NH7 Iх Iх ch-nh9 1
1 1 сн3 сн3 снз
7
рибиттейхоевая кислота Вас. subtilis, R - остаток глюкозы (Р)
а также другие полисахариды. С полисахаридами клеточных стенок
у микобактерий и некоторых нокардий связаны эфирной связью
миколовые кислоты. Они же могут быть компонентами свободно
тейхуроновая кислота из
клеточных стенок бацилл
экстрагируемых гликоли-
пидов, известных как к о
рд-факторы (от англ,
cord — веревка, струна),
которые выявлены также
у некислотостойких пато-
генных для человека ко-
ринебактерий. Число уг-
леродных атомов в кори-
немиколовых кислотах
достигает 32 — 36, в но-
кардиомиколовых кисло-
тах примерно 50, в мико-
ловых кислотах микобак-
терий порядка 90. Мико-
ловые кислоты являются о-замещенными Р-гидроксижирными
кислотами с общей формулой:
R-CH(OH)-CH(Ri)-COOH
4 т. 8524
97
Различают а-, 0- и у-миколовые кислоты туберкулезных мико-
бактерий; первые содержат от 78 до 88 углеродных атомов, вторые
— от 83 до 89, третьи — 91.
СН3-(СН,) -CH-CH-(CH,)V-CH-CH-(CH7) -СН-СН-СООН
\/ 7 \/ I I
сн2 сн2 он с24н49
ct-миколовая кислота
CH3-(CH2)x-CH-CH-(CH2)y-CH-CH-(CH2)z-CH-CH-COOH
сн3осн3 сн2 он с24н49
р-миколсвая кислота
CHj-fCH^T- СН-(СН2)19-СО-СН-(СН2)]9-СН-СН-СООН
сн3 СН3 ОН С24Н49
у-миколовая кислота
Корд-факторы представляют собой 6,6'-димиколиловые эфиры
ос,ос,-1, Г- диглюкозы (трегалозы).
микояовая кислота
миколовая
кислота
Корд-фактор обладает токсичностью, которая потенцируется
сульфогликолипидами, локализующимися с ним в клеточной стен-
ке микобактерий (например, у М. tuberculosis сульфогликолипид
представляет собой 2,3,6,6'-тетраацетилтрегалозо-2'-сульфат).
В клеточной, стенке микобактерий имеются еще микозиды,
ответственные за рецепцию бактериофагов. Микозиды содержат
6-дезокситалозу и ее О-метиловые эфиры, фукозу, рамнозу. Раз-
личают микозиды А, В и С. Микозиды А и В являются фенольными
гликолипидами, микозиды С — пептидогликолипидами.
98
co—сн3
о—CH2 OSO3NH4
сульфогликолипид из М. tuberculosis
При оценке биологических свойств продуктов клеточных сте-
нок микобактерий вначале предполагали, что иммуноадьювантной
активностью обладают их уникальные липидные компоненты.
Однако на основании детального анализа установлено, что таким
действием обладают водорастворимые пептидогликаны, содержа-
щиеся в клеточных стенках большинства бактерий. Самый малый
активный фрагмент оказался мурамилдипептидом, представляю-
щим собой М-ацетил-мурамил-Е-аланил-П-изоглутамин. Мурамил-
дипептиды — высокоактивные иммуноадьюванты нашли практи-
ческое применение, в том числе — в качестве компонентов вакцин,
рекомендованных для профилактики и лечения СПИД.
сн3
сн
I
сн2он
М—о
соон
H3COCN
мурамиддипеп тид
Клеточная мембрана прокариот принципиально сходна по ар-
хитектонике с мембраной эукариотической клетки. Тем не менее,
она не является тем трехслойным сэндвичем (от англ, sandwich —
бутерброд), о котором говорили до недавних пор. В состав мембран
входят фосфолипиды с полярными "головками", обращенными
кнаружи; взаимодействующие между собой их гидрофобные хво-
сты из остатков жирных кислот обращены внутрь. Мембранные
99
белки полностью или частично погружены в липидный слой (рис.
27), в котором имеют место гидрофобные взаимодействия между
белками и липидами. Гидрофильные участки белков взаимодейст-
вие. 28. Жидкостно-мозаичная
модель клеточной мембраны.
вуют с полярными головка-
ми фосфолипидов; Мемб-
рана в целом приобретает
жидкости о-мозаичную
структуру (рис. 28), в кото-
рой имеющиеся слои пред-
ставляют собой нековален-
тно собранные термодина-
мически стабильные, но метаболически лабильные структуры.
Белковые молекулы как бы заякорены в мембране и несут специ-
фические функции. Лишь гликопротеины в мембране не сопряже-
ны с другими компонентами и могут диффундировать латерально
в плоскости мембраны; другие, напротив, интегрированы с мат-
риксом мембраны прочно. Не исключена возможность их роли в
признании и селекции функциональных молекул, в которых про-
является (или должна проявиться) заинтересованность клеточной
мембраны. Контактирующие с ней ферменты и ферментные сис-
темы подвергаются специфическим конформационным изменени-
ям, что сказывается на характере их взаимодействия с субстратами
и лигандами. Другими словами, клеточная мембрана выступает
организатором и регулятором активности связанных с нею фер-
ментов и ферментных систем. При этом на уровне мембраны
происходят химические и физические процессы.
Какие же принципиальные различия существуют между кле-
точными мембранами прокариот и эукариот? Эти различия каса-
ются химического состава и функций (таблица 8). По химическому
составу клеточные мембраны представляют собой гликопротеоли-
пиды или гликолипопротеины, или, наконец, липогликопротеины,
юо
что зависит от таксономического положения организма. В клеточ-
ной мембране золотистого стафилококка на долю углеводного
компонента приходится до 40%, столько же — на долю белкового
компонента и лишь 20% — на липидный компонент.
В усредненном варианте содержание указанных компонентов
в клеточной мембране большинства исследованных прокариот
распределяется следующим образом: белки — до 50%, липиды —
до 30%, углеводы — до 20% (из расчета на сухую массу); для
эукариот эти данные соответственно таковы: белки — до 70-80%,
липиды — До 15 — 25%, углеводы — до 5 — 15%. Однако
применительно к структуре различных мембран указанные вели-
чины могут сильно различаться и, например, на липиды может
приходиться до 50% от сухой массы мембранной фракции эндоп-
лазматического ретикулума, тесно связанного с клеточной мемб-
раной. А в миелиновых мембранах нервных волокон содержание
липидов достигает 80%.
Клеточные мембраны у всех организмов проявляют полифун-
кциональные свойства: осморегуляция, барьерные функции с се-
лективной проницаемостью за счет пор, насосов, рецепторов;
транспорт веществ (в том числе активный с затратой энергии),
участие в создании мембранного потенциала, в превращении
энергии при фотосинтезе и окислительном фосфорилировании.
Глубокое знание структуры и функции оболочки бактерий (или
клеточной мембраны у видов, лишенных клеточной стенки) облег-
чает использование биообъектов в конкретных целях, например,
для получения вторичных метаболитов (см.), когда процессы пере-
носа вещества в клетку (равно как и секреторные процессы)
выступают на первый план. В целях сравнения можно привести
пример с Leuconostoc mesenteroides и Xanthomonas compestris. В
определенных условиях первый из них секретирует фермент
декстрансахаразу, с помощью которой возможен синтез полиса-
харида декстрана без участия клеток, то есть in vitro; второй вид
синтезирует полисахарид ксантан, но уже на уровне клеточной
мембраны и в этом случае без клеток X. campestris биосинтез in
vitro невозможен. Следовательно подходы здесь к осуществлению
процессов биосинтеза полимеров различны, поскольку механизмы
функционирования соответствующих биосистем (в том числе —
оболочки бактерий) неравнозначны.
101
Таблица 8 Основные различия между клеточными мембранами и их про-
изводными у прокариот и эукариот.
Признак Прокариоты „ 1 Эукариоты
Мезосомы Тилакоиды Фикобилисомы Аэросомы Хлоросомы Карбоксисомы Гидрогеносомы Лизосомы Пероксисомы Гликосомы Митохондрии Эндоплазматичес- кий ретикулум (ЭПР) Компартмента- лизация клетки за счет ЭПР Система внутрицитопла- зматической мембраны (не ЭПР) Комплекс Гольджи Хитосомы Ломасомы Вакуоли Хлоропласты Эндоцитоз Экзоцитоз Насыщенные и мононенасы- щенные жирные кислоты Полиненасы- щенные жирные кислоты Простагландины Имеются преимущественно у грамположительных бактерий Имеются у цианобактерий Имеются у фототрофов Имеются у зеленых фототрофных бактерий Имеются у фототрофов и некоторых хемолитотро- фов Отсутствуют Отсутствует Имеется у метанокисляющих, нитрифицирующих и фототрофных бактерий Отсутствует Отсутствуют Не образуются у большинства бактерий Отсутствуют Отсутствует 11 Имеются Отсутствуют0 Отсутствуют Имеются у некоторых трихомонасов Имеются п Имеются у Protozoa Имеются Имеется Отсутствует Имеется Имеются у грибов Могут образовываться за счет ЭПР Имеются в клетках растений (хроматофоры у водорослей) Присущ Отсутствуют Имеются Имеются у некоторых грибов
102
(продолжение таблицы 8)
Признак Прокариоты Эукариоты
Кардиолипин Фосфатидилглицерин Липотейхоевые кислоты Моногалакто- зилдиглицериды Дигалактозилдиг- лицериды Сульфохиново- зилдиглицериды Ундекапренол Долихол Сфингомиелин Стерины Связь с ядерной ДНК Отсутствует Имеется Имеются у грамположительных бактерий Имеются (у зеленых бактерий и цианобактерий) Имеются у цианобактерий Имеется Отсутствует Отсутствуют+ Имеется Имеется Отсутствует или имеется в следовых количествах Отсутствуют Отсутствуют Отсутствуют Отсутствует Имеется Имеются Отсутствует
6 кроме цианобактерий, способных синтезировать полиненасыщенные жирные
кислоты.
Микоплазмы могут включать экзогенные стерины в состав мембраны.
Клеточные мембраны после выделения из клеток и тщательной
очистки остаются связанными с рибосомами и ДНК, представляя
собой мембранополирибосомо-ДНКовые агрегаты.
Рибосомы в прокариотической клетке (числом порядка 104 на
клетку) состоят приблизительно из 30% белка и 70% РНК, что в
расчете на всю клетку с оставляет до 40% белка и 90% РНК. "Мягкий"
лизис растущих клеток сопровождается выделением почти всех
рибосом в виде полирибосомомембранных агрегатов, содержащих
все компоненты белоксинтезирующей системы. Полирибосомы
представляют собой цепочки, состоящие из 70S рибосомных мо-
номеров с диаметром порядка 0,02 мкм, присоединенных к мРНК.
При низких концентрациях ионов магния — меньше 10‘4 М 70S
рибосомы диссоциируют на 30S и 50S субъединицы. Размер первых
приблизительно 0,007 — 0,016 мкм, молекулярная масса 800 кДа.
Каждая 30S субъединица включает одну молекулу 16S РНК с ММ
около 500 кДа и 21 молекулу разных белков; 50S субъединица
размером 0,014 — 0,016 мкм имеет ММ 1,8»103 кДа и содержит
юз
одну молекулу 23S РНК с ММ 1,1 х 103 кДа, 5S РНК с ММ 40 кДа
и 34 молекулы разных белков. Участие рибосом в матричном
синтезе белка см. в разделе 5.1.
Ядро прокариот (нуклеоид) представляет собой тонкую, непра-
вильную, фибриллярную сеть из ДНК, часто располагающуюся
параллельно осевой линии клетки. Во многих случаях ДНК-связы-
вающей структурой выступает мезосома. Замкнутость нити ДНК
в виде кольца доказано с помощью радиоавтографии. Это и есть
единственная хромосома в бактериальной клетке. На нее прихо-
дится 2-3% клеточной массы и 10% или более объема клетки. В
такой хромосоме у Е. coli имеется от 20 до 70 суперспирализован-
ных доменов. До недавнего времени признавали, что нуклеосомы
(см.) присущи только эукариотам. Однако нуклеосомоподобные
структуры, как оказалось, обнаруживаются и у прокариот. Из ДНК
Е. coli отделены 4 гистоноподобных белка. Они были обозначены
как HLPlla, HLPUb, HLPI и Н-протеин. У первых двух аминокис-
лотный состав подобен аминокислотному составу гистона Н2В у
эукариот, а белок Н перекрестно реагирует с антителами против
гистона Н2А из зобной железы телят. Названные гистоноподобные
белки помогают ферменту гиразе, или топоизомеразе II стабили-
зировать суперспирализацию ДНК. ДНК-гираза способна вводить
отрицательную суперспирализацию в релаксированную (от лат.
relaxatio — уменьшение напряжения, расслабление) кольцевую
молекулу ДНК (рис. 29). Одна мо-
лекула фермента вводит до 100
спиралей в 1 минуту.
Кроме гистоноподобных бел-
ков — HLP (от англ, histone-like
protein) в клетках прокариот име-
ются полиамины, связанные с ри-
босомами и мембранами. Главные
из них — путресцин и спермидин:
H2N—(СН2)з—NH2;
путресцин
H2N— (СН2)4—NH— (СН2)з—NH2
рыва с внешней стороны.
спермидин
Эти полиамины характеризуются антимутагенным эффектом,
способностью повышать устойчивость протопластов к осмотиче-
скому лизису, стабилизировать 70S рибосомы (предупреждают их
диссоциацию на субчастицы).
В цитоплазме прокариот могут накапливаться гликоген, напри-
104
Рис. 29. Инверсия положительной су-
перспирали двухцепочечной ДНК при
участии ДНК-гиразы: 1 — стабилизация
положительного витка, 2 — разрыв в
заднем сегменте, 3 — "залечивание" раз-
мер у энтеробактерий (до 40% массы клеток). Спорообразующие
бактерии и псевдомонасы накапливают до 30% и более поли-Р-гид-
роксимасляной кислоты, у большинства прокариот выявляются
полифосфаты, волютин, липиды.
Рис. 30. Эндоспора: 1 — кор. или цитоплазма
споры; 2 — стенка споры; 3 — оболочка споры;
4 — кортекс; 5 — экзоспориум.
Отдельные прокарио-
ты образуют внутрикле-
точные споры (эндоспо-
ры). Аэробные из них
представлены видами ро-
дов Bacillus и Sporosarcina
(S. ureae), микроаэрофи-
лы — видом Sporolacto-
bacillus inulinus, анаэроб-
ные — видами родов
Clostridium, Desulfo-
tomaculum. Размер, фор-
ма и расположение споры
в клетке (терминальное,
субтерминальное, цент-
ральное) достаточно по-
стоянны и поэтому ис-
пользуются для характе-
ристики видов. Содержа-
ние в спорообразующих
клетках пар нуклеотидов
ГЦ достаточно низкое, особенно — у клостридий (22-28%); у них,
аланиновая субъединица тетрапептидная субъединица гликолактамовая субъединица
фрагмент пептидогликана в кортексе спор бацилл
105
как правило, отсутствует meso-ДАП в муреине. Эндоспоры —
сложные образования и, как видно из рис. 30, эндоспора Вас. cereus
представляется сферическим многослойным телом, формирование
которого, как правило, диктуется неблагоприятными условиями. В
кортексе спор у большинства спорообразующих видов найден
специфический пептидогликан.
В цитоплазме спор содержится до
15% дипиколината кальция. В белках
оболочек споры отмечается повышен-
ное количество гидрофобных амино-
кислот и цистеина. В связи со споро-
образованием культуры ряда бацилл и
клостридий образуют белки, которые
имеют важное народнохозяйственное
значение (циклопептидные антибиоти-
дипиколинат кальция
ки, энтобактерин). С другой стороны, спорообразующие прокари-
оты могут быть вредителями биотехнологических процессов. Спо-
ры отличаются высокой устойчивостью к различным внешним
факторам, хорошо переносят их воздействия и поэтому могут
попасть в ферментационные среды или в готовый продукт при
несоблюдении правил асептики, антисептики и стерилизации.
Некоторые прокариоты (отдельные актиномицеты) могут обра-
зовывать экзоспоры, которые выступают у них репродукционными
образованиями и покоящимися формами одновременно (эндоспо-
ры — это покоящиеся формы и/или формы дифференцировки в
ходе развития и созревания клеток, например, у Вас. subtilis).
Покоящимися формами являются и цисты, которые могут
образовываться азотобактериями, миксобактериями, риккетсия-
ми, спирохетами.
4.3. Клетки эукариот. В биохимической технологии используют
эукариотические клетки различного происхождения. Источниками
их являются виды, относящиеся к царствам Mycota, Plantae и
Animalia. К настоящему времени ассортимент первых более ши-
рокий, чем ассортимент растительных и животных клеток, эксплу-
атируемых в производстве. Клетки эукариот во многом сходны
между собой, тем не менее, имеющиеся различия привносят свои
особенности, сказывающиеся на их структуре и функциях. На рис.
31 достаточно объективно отражены фактические данные, накоп-
ленные исследователями при изучении строения клеток эукариот.
Клетки грибов и растений обладают прочной клеточной стен-
106
кой, которая отсутствует у
клеток животного проис-
хождения. При этом мар-
керной структурой для
большинства грибов стано-
вится хитин, для большин-
ства растений — целлюлоза.
Молекулярные массы их
близки и достигают величи-
ны порядка 500-600 кДа.
Клеточные стенки грибов и
растений — двухфазные си-
стемы. Одной фазой явля-
ются микрофибриллярные
структуры, второй фазой —
аморфный наполнитель.
Следовательно, грибная и
растительная клеточные
стенки являются природны-
ми композитами, в химиче-
ском составе которых пре-
обладают углеводные ком-
Рис. 31. Клетка эукариот: 1 — плазматическая
мембрана, 2 — пероксисома, 3 — ядро, 4 —
ядрышко, 5 — аппарат Гольджи, 6 — шерохо-
ватый эндоплазматический ретикулум, 8 —
центриоль, 9 — цитоскелет, 10 — секреторная
гранула, 11 — экзоцитотический пузырек, 12
— эндоцитотический пузырек, 13 — эндосома,
14 — лизосома, 15 — цитозоль, 16 — митохон-
дрия, 17 — рибосомы.
звено хитина
звено целлюлозы
поненты. Из других веществ в ней обнаружены гликопротеины,
белки, в незначительных количествах — липиды и липоконъюгаты.
В связи с меньшей дифференциацией клеточных структур у грибов
в сравнении с растениями, можно привести усредненные данные
по составу компонентов в клеточных стенках грибов (таблица 9).
107
Таблица 9 Химический состав клеточных стенок некоторых грибов, %
Компонент Виды грибов
Allomyces macroginus Mucor rouxii (нитчатая форма) Saccharomyces cerevisiae
1 Азот Белок Глюкан Липиды Маннан Фосфаты Хитин Хитозан Другие углеводы Сумма углеводных компонентов 5,5 10,0 16,0 58,0 74,0 6,3 7,8 3,8 23,3 9,4 32,7 9,5 55,4 2,1 13,0 28,8 8,5 31,0 0,31 1,0 60,8
Из таблицы видно, что у некоторых видов грибов (мукоровых)
в клеточной стенке присутствуют и хитин и хитозан (деацетили-
рованная форма хитина), у других — только хитин.
Как уже было сказано, целлюлоза — маркерный компонент
клеточных стенок растений, однако в незначительных количествах
она обнаруживается, например, у акразиевых, гифохитридиевых,
сапролегниевых, пероноспоровых грибов.
Уроновые кислоты и лигнин достоверно никто не обнаруживал
в клеточной стенке грибов, но многие выявляли пигменты, в том
числе — меланиновые. Меланины, включающие в свой состав
остатки 5,5-индолхинона и пирокатехина, обычно связаны с бел-
ками (меланопротеины) или с гликопротеинами (меланогликопро-
теины), и, наряду с ферментами — супероксиддисмутазой (СОД),
каталазой и пероксидазами, обладают функцией протекторов (за-
щитников) от кислородных радикалов (О2) и синглетного кисло-
рода ( О2), являющихся сильными окислителями.
Различные грибы имеют капсулы, образующиеся в результате
гиперпродукции углеводных полимеров (Aureobasidium spp.,
Cryptococcus spp., Rhodotorula spp. и др.). Эти полимеры приобрели
или приобретают важное значение для народного хозяйства (в
нефтедобыче, здравоохранении, косметике).
Как и у грибов в клеточной стенке растений преобладают
углеводные полимеры — целлюлоза, гемицеллюлозы, пектины. К
гемицеллюлозам относятся полисахариды (гликаны), входящие в
108
состав клеточных стенок растительных тканей и способные рас-
творяться в разбавленных растворах щелочей и гидролизоваться
разбавленными растворами кислот в мягких условиях. Это —
арабинаны, галактаны, ксиланы, маннаны, фруктаны. Гемицеллю-
лозы находят широкое применение в народном хозяйстве.
Пектины — это полигалактурониды, входящие в состав клеточ-
ных стенок и сока всех высших и низших растений. Галактуроно-
вая кислота главный мономер пектинов (до 92%). Часть уроновых
кислот может этерифицироваться по карбоксигруппе метанолом;
остатки ее соединены С1----С4 гликозидными связями. Пектины
широко используются в пищевой промышленности.
Следует особо выделить
лигнин — полимер полифе-
нольной природы, образую-
щийся только у растений. Его
содержание у некоторых ви-
дов может достигать 38% и с
ним связано одревеснение
растений (лигнификация). По
распространенности среди природных биополимеров он уступает
место лишь гликанам.
Лигнин относится к нерегулярным разветвленным полимерам,
состоящим преимущественно из остатков замещенных фенолспир-
тов: n-гидроксикоричного, или п-кумарового; 3,5-диметокси-4-гид-
роксикоричного, или синапового (синапинового) и 3-метоксигид-
рОкоричного, или кониферилового.
п-ГЯДрОКенкоричный, 3,3-дим«токси-4- 3-маточсигидроксикоричный,
или кумаровый спирт гидроксчкоричный, или конифариловый спирт
или синаповый (си-
напиновый) спирт -----
Механизм биосинтеза лигнина окончательно не выяснен, хотя
доказано, что исходным продуктом является глюкоза, а непосред-
ственным предшественником его транс-кумаровый, транс-синапо-
вый и транс-конифериловый спирты.
109
В древесине он связан с гликанами, главным образом — с
гемицеллюлозами преимущественно тремя типами связей — гли-
козидными, сложноэфирными и простыми бензилэфирными.
Лигнин получают в качестве побочного продукта в целлюлоз-
ном и гидролизном производствах (более двух миллионов тонн в
год лишь в государствах бывшего СССР) и широко используют в
народном хозяйстве.
Клеточные стенки растений могут покрываться снаружи таки-
ми липидами, как воск и кутин или пропитываться суберином. Все
эти соединения выполняют главным образом защитную роль. Воска
— сложные эфиры длинноцепочечных жирных кислот — насы-
щенных или ненасыщенных (число углеродных атомов от 14 до 36)
с длинноцепочечными спиртами (число углеродных атомов от 16
до 22).
О
&н3-(сн2)^-сн=сн-сн2— (снг)—с^4о -(с^^-сн =сн-(сн2)7—сн31
остаток линолевой кислоты
остаток олеинового спирта
воск (сложный эфир линолевой кислоты и олеинового спирта)
Кутин (от лат. cutis — кожа) состоит из смеси высших жирных
кислот и их эфиров; суберин (от лат. suber — корковое дерево) —
эфир жирных кислот и суберилового спирта (циклогептанола).
Клеточные стенки растений могут утолщаться за счет утолще-
ния внутренних слоев, прилегающих к клеточной мембране —
формируется вторичная клеточная стенка (в противоположность
первичной — без внутренних от-
ICHOH ложений). Такая стенка несет,
u u / как правило, механическую фун-
2 2 2 кцию. В ней количество целлю-
лозы может достигать 50% от су-
субериповый спирт х°й массы клетки.
Животные клетки лишены
клеточных стенок. Лишь отдельные протозоа способны в опреде-
ленных условиях образовывать так называемые цисты (дизенте-
рийная амеба, кишечная балантидия и др.), которые покрываются
одно-трехслойной, преимущественно белковой природы, оболоч-
кой.
сн2—сн2—сн2\
ио
Между клетками у грибов и растений существуют сообщаю-
щиеся отверстия — п о р ы. У большинства грибов имеются простые
поры (рис. 32а). Базидиальным грибам присущи сложные поры,
или долипоры (от лат. dolium — бочонок) (рис. 326).
Предполагают, что долипоры способствуют сохранению мицелия
базидиомицетов в дикариотическом состоянии и поэтому такие
"запирающие" перегородки формиру-
ются во вторичном и третичном мице-
лии; в первичном мицелии перегород-
ки простые.
Во вторичных клеточных стенках
соседствующих клеток растений так-
же образуются поры, в которых раз-
деляющими клетки структурами явля-
ются лишь первичная оболочка и се-
рединная пластинка (рис. 33а, б).
Различают простые поры с одина-
ковым диаметром по всей длине и
окаймленные поры, расширяющиеся
на месте "вхождения" в клетку, где она
приобретает форму сводчатого окайм-
Рис. 32. Поры в перегородках
грибов — у сумчатых (а): простая
пора (1) в септе (2) между двумя
клетками; у базидиальных (б): до-
липора с ее элементами (пора —
1, парентосома — 2, долипоровая
перегородка — 3).
ления. Срединная пластинка — продукт
деления меристемных (см.) клеток. Она
состоит главным образом из аморфных
пектиновых веществ равномерно (со сто-
роны каждой клетки) "обрастающих" ге-
мицеллюлозами. Образуется первичная
клеточная стенка, в которой свободно
переплетаются целлюлозные волокна ди-
аметром порядка 10 нм и содержащие
8-12 тыс. остатков глюкозы. Центральная
часть таких волокон имеет кристалличе-
скую структуру и диаметр около 4 нм.
В состав первичной клеточной стенки
двудольных растений входят ксилоглю-
каны (в боковых цепочках которых со-
держатся ксилоза, галактоза, фукоза), а
также арабиногалактаны и рамногалак-
туронаны, ковалентно связанные друг с
другом и целлюлозными фибриллами.
Позднее формирующаяся вторичная
111
Рис. 33. Схематичное изобра-
жение простой (а) и окаймлен-
ной (б) пор у некоторых рас-
тений (продольный разрез): 1
— срединная пластинка вме-
сте с первичной клеточной
стенкой, 2 — свод окаймлен-
ной поры, 3 — окаймленная
пора в плоскостном изобра-
жении.
галакто»
клеточная стенка состоит из многочисленных слоев плотно упако-
ванных послойно разнонаправленных фибрилл. Такие фибриллы
чаще состоят из целлюлозы, но могут включать и другие полиса-
хариды, например, ксилан и маннан у некоторых водорослей.
Таким образом, углеводные полимеры и лигнин составляют
основное содержание клеточных стенок растений. В качестве
минорного компонента в них обнаруживается гликопротеин —
экстензии, содержащий в большом количестве аминокислоту 4-
гидроксипролин. Олигосахаридные остатки в экстензине состоят
из арабинозы и галактозы.
Несмотря на наличие разделительных перегородок (септ) меж-
ду клетками, они сообщаются между собой благодаря цитоплазма-
тическим нитям — плазмодесмы.
н0 н »1 Для грибов и растений типичным является об-
кн соон разование тканей. Ткань — это большая группа
4-гидроксичюпии клеток, единого (общего) происхождения, облада-
ющая сходными структурами и функциями. Дру-
гими словами такие клетки,
обладающие генетической и
структурно-функциональ-
ной общностью, формиру-
ют систему — ткань. Раз-
личают ложные и ис-
тинные ткани. Ложные
ткани образуются из групп
нитей (филаментов), кото-
рые могут переплетаться или срастаться клеточными стенками, но
сохраняют свою самостоятельность, например, при поперечном
делении. Такая ткань присуща грибам и называют ее п л е к-
тенхимой, или псевдопаренхимой (от лат. plexus
— сплетение, parenchyma — живая ткань, паренхима, от греч.
pseudes — ложь, фальшь). Она напоминает меристему растений
лишь морфологически и типична для плодовых тел базидиальных
грибов, склероциев аскомицетов. Истинные ткани у грибов выяв-
ляются редко, однако они имеются в перитециях сумчатых грибов
(на ранних стадиях их формирования), в пикнидах у несовершен-
ных грибов.
Растениям присущи истинные ткани — простые (из одной
системы клеток) и сложные (из разных систем клеток).
Ложные и истинные ткани подразделяются по функциям на 6
112
групп: образовательные (меристемы), покровные, проводящие, ме-
ханические, секреторные (выделительные) и базисные (основные).
Образовательные ткани, или меристемы (от
греч. meristos — делимый) состоят из активно метаболизирующих
клеток, способных делиться и образовывать новые клетки. Те из
них, которые служат родоначальницами всех других клеток, диф-
ференцирующихся и превращающихся в клетки постоянных тка-
ней, называются инициальными (отлат. initialis — первичный,
начальный).
У грибов настоящие меристемы не развиты, деление клеток
сосредоточено на верхушках гиф.
Топологически, или по местоположению у растений (рис.34)
различают меристемы верхушечные, или апикальные (от лат. apex,
apicis — верхушка); боковые,
боковой); промежуто
intercalaris — промежуточ-
ный, вставной). Верхушеч-
ные меристемы обеспечива-
ют рост растений в длину,
они же образуют конусы
корней; боковые обеспечи-
вают рост растений в тол-
щину (к боковым меристе-
мам относятся прокамбий,
камбий, перицикл, фелло-
ген; от лат. cambium — сме-
на, обмен; от греч. pro —
пред, вперед, ранее, реп —
вокруг, около; kiklos — круг,
цикл; fellos — пробка; genos
— род, происхождение).
В основаниях черешков
или латеральные (от лат. lateralis —
н ы е, или интеркалярные (от лат.
Рис. 34. Топология меристем в растениях. Схе-
матичный продольный разрез растительной
почки: 1 — верхушечная меристема, 2 — лис-
товые побеги, 3 — конус нарастания.
листьев, в междоузлиях локализуются промежуточные меристемы.
Клетки меристемы тотипотентны (от лат. totum — все,
. целое, potentium — способность, потенция), то есть они полностью
реализуют свой потенциал развития с образованием целого орга-
низма. В биотехнологии особое значение приобрела верхушечная
меристема (см. рис. 34), так как она всегда остается свободной
(здоровой) от фитопатогенных микроорганизмов, например, виру-
113
сов (даже в тех случаях, когда все растение поражено патогеном).
Культивирование in vitro меристемных клеток от больного расте-
ния в стерильных условиях завершается получением здорового
растения — посадочного материала.
Известны еще так называемые раневые меристе-
м ы, образующиеся в местах повреждения органов и тканей
растений. В таких поврежденных участках разрастаются однород-
ные паренхимные клетки, прикрывающие повреждения. Такие
ткани называются каллусами (от лат. callus — мозоль).
Каллусные ткани стали широко культивировать на питательных
средах в целях получения посадочного и прививочного материала,
а также ценных метаболитов.
С меристемной тканью связан рост растений. Так, у большин-
ства из них стебли и корни обладают верхушечным ростом, листья
нарастают благодаря базальному росту; в стеблях злаков преобла-
дает промежуточный рост. Для грибов также характерен верху-
шечный рост. В общем виде он протекает следующим образом. В
молодой нити гриба на всем ее протяжении достаточно легко
отдифференцировать три зоны: апикальную, субапикальную и
вакуолизированную дистальную (рис. 35). В апикальной зоне скап-
ливается большое количество везикул (пузырьков), в субапикаль-
ной - ядра, рибосомы, митохондрии, эндоплазматический ретику-
лум, микротельца, микротубули и т. д. Субапикальная зона пере-
ходит в зону вакуолизации (дистальную), которая тем выраженнее,
чем дальше расположена от верхушки. В ней возрастает также
содержание липидов. Апикальные везикулы, образующиеся в суб-
апикальной' зоне и перемещающиеся к верхушке, участвуют в
синтезе клеточной стенки. Там они секретируют свое содержимое
или сливаются с мембраной. В них так же могут быть экзофермен-
ты, способные к экструзии (от лат. extrusio — выталкивание) на
кончике гифы (рис. 36).
Как видно из рисунка везикулы, воз-
ш п I
Рис. 35. Апикальная — I, суб-
апикальная — II, вакуолизиро-
ванная — III зоны в молодой
растущей гифе одного из ни-
зших грибов (схема).
никающие из эндоплазматического рети-
кулума, сливаются и образуют цистерну
внутренней (проксимальной) части дик-
тиосомы; содержимое цистерны и мемб-
раны последовательно трансформируют-
ся, перемещаясь к наружной (дистальной)
части диктиосомы в связи с продолжаю-
114
Рис. 36. Схема формирования и перемещения апикальных визикул из субапи-
кальной зоны к верхушке гифы (КС — клеточная стенка, М — клеточная
мембрана, В — везикулы, Д — диктиосомы, Р — рибосомы, ЭР — эндоплазма-
тический ретикулум, Ц — цистерны).
щимся образованием новых цистерн. Здесь цистерны превраща-
ются в пузырьки, а затем — в секреторные везикулы, которые
мигрируют к верхушке гифы. Некоторые из везикул увеличива-
ются в размере, другие сливаются между собой и таким образом
формируются большие секреторные везикулы. Отдельные везику-
лы непосредственно достигают клеточной мембраны и сливаются
с ней. Слившиеся с мембраной визикулы освобождают свое со-
держимое в периплазматическую область апикальной части гифы,
где осуществляется синтез клеточной стенки и сбалансированный
лизис скелетных микрофибрилл.
У всех высших грибов в апикальных частях мицелиальных
нитей имеется так называемое верхушечное тельце сферической
формы (рис. 37). При остановке роста гифы оно исчезает. Поло-
жение тельца на верхушке служит направляющим вектором про-
странственного роста гифы. Например, эксцентрическая позиция
его указывает на то, что верхушка изогнется в ту же сторону.
Хотя гифы всех грибов растут за счет апикальных частей (а к-
ропетальный или базифугальный рост; от греч. acros
— самый высокий, верхушка, кончик; petalon — лист; basieos —
основание, fugas — изгоняющий), утолщение клеточных стенок
может наблюдаться и около верхушек, а иногда — ив дистальных
частях, например, при образовании гемм у Aureobasidium pullulans.
Тем не менее, если неограниченный рост мицелия в длину потен-
циально возможен, то увеличение ширины его весьма лимитиро-
вано. Верхушечный рост растений морфологически сходен с вер-
115
тельце
Рис. 37. Схематическое изображение
апикальных частей у оомицетов (А),
зигомицетов (Б) и высших грибов (В).
хушечным ростом грибных нитей,
однако у растений он характеризу-
ется более выраженной ритмично-
стью с чередованием процессов ус-
коренного и замедленного роста
при возможном переходе всего рас-
тения или его каких-либо частей в
состояние покоя (семена, клубни).
Ростовые ритмы могут быть эндо-
генного и экзогенного происхожде-
ния. Причем воздействующие
внешние факторы могут быть одни-
ми и теми же, например, свет и
температура. Так, у некоторых гри-
бов проявляются ростовые зоны
при освещении культур через опре-
деленные промежутки (чередование темноты и света). Причем на
одинаковое физическое возбуждение ответная реакция грибов
весьма различная. На этом основании грибы условно подразделяют
на три главные группы: 1) грибы, не отвечающие на свет и тепло,
их ростовые зоны, относимые к эндогенным ритмам, всецело
являются ответной функцией на условия обитания в питательной
среде. К этой группе относятся Ascochyta chrysanthemi, некоторые
мутанты Ascobolus immersus, Pestalotia annulata и Podospora
anserina; 2) грибы, ростовые зоны которых, относимые к экзоген-
ным ритмам, возникают в ответ на физические возбуждения. Сюда
относятся Alternaria tenuis и Trichoderma viride, чувствительные к
фотоциклам, Aspergillus ochraceus и A. niger, чувствительные к
фото- и термоциклам; 3) в эту группу включены грибы, которые
при слабом физическом возбуждении проявляют зональный рост
как экзогенный ритм, в то время как сильное возбуждение прово-
цирует эндогенный ритм (зональный рост), который продолжается
некоторое время после прекращения стимула. Эндогенный ритм,
наведенный разовым освещением Sclerotinia fructigena, S. laxa и
Leptospheria michotii, имеет период в 24 часа. Культуры S. fructicola,
один раз освещенные достаточно интенсивным белым светом или
светом с длиной волны меньше 500 мкм, а затем оставленные в
темноте или при очень слабом освещении, проявляют суточный
ритм в течение трех дней.
116
Ежедневное освещение интенсивностью 1000-3000 лк в течение
нескольких секунд вполне достаточно, чтобы индуцировать зональ-
ный рост у Fusarium discolorsulfureum, Trichotecium roseum и
Verticillium lateritium (экзогенные ритмы).
Фототропизм присущ и грибам (например, фикомицетам) и
растениям. Так называемые циркадные ритмы (от лат.
circus — круг) связаны со сменой дня и ночи, иногда — с
длительностью дня (фотопериодизм). Они контролируются
у всех эукариотических организмов специальным внутренним
механизмом, называемым ф и з и о л о г и ч е с к и м и, или б и-
ологическими часами.
В регуляции ростовых ритмов существенна роль фитогормонов
— ауксинов, гиббереллинов и кининов (цитокининов). Из ауксинов
(от-греч. аихо — выращиваю, увеличиваю) наиболее распростра-
ненным является гетероауксин — ИУК (3-индолилуксусная кисло-
та), из гиббереллинов (продуцент Gibberella fujikuroi) — гибберел-
ловая кислота (ГАз), из кининов (от греч. kineo — двигаю, побуждаю
— кинетин (6-фурфурилметиламинопурин).
Все названные выше ритмы отражают адаптивные (приспосо-
бительные) потенции наследственно компетентных организмов,
обитающих на разных географических широтах земного шара.
Покровные ткани имеются у грибов (на склероциях, на шляпках
плодовых тел базидиомицетов и других образованиях). Они воз-
никают из плектенхимы. У растений покровные ткани имеют
меристемное происхождение. К ним относятся: эпидерма с устьи-
цами, трихомами (от греч. trichos — волос) и эмергенцами (от англ,
emergency — крайность) на листьях и молодых побегах, нередко
покрытая кутикулой или воском, с полисахаридами, кристаллами
в клетках или кремнеземом — в качестве пропитки; перидерма (от
греч. peri — вокруг, derma — кожа) с чечевичками — вторичная
покровная ткань на корнях и стеблях преимущественно у много-
летних растений (пробка); корка на ветвях, корнях и стволах
многолетних деревьев.
Механические (опорные) ткани у грибов представлены гифами
с утолщенными клеточными стенками в плодовых телах у труто-
виков, в тяжах у домового гриба, в ножках апотециев у некоторых
сумчатых. У растений известны два основных тийа опорных тканей
— колленхима (уголковая, пластинчатая, рыхлая) и склеренхима
(от греч. kolla — клей, skleros — твердый, enchima — налитое, здесь
117
— ткань). К склеренхиме относятся волокна (лубяные, древесин-
ные) и склереиды — структурные элементы механической ткани.
В промышленности используют преимущественно лубоволокни-
стые растения (лен, липа, кенаф, конопля и др.).
Проводящие ткани в виде дифференцированных структур от-
сутствуют у грибов. Сами мицелиальные нити обеспечивают по-
ступление и проведение влаги между клетками. Тем более, что у
высших грибов септы не обладают абсолютной герметизацией, а
у низших они отсутствуют или встречаются редко по ходу гиф.
У растений имеются специализированные ткани — ксилема,
флоэма (от греч. ksilon — дерево, floos — кора, лыко) и проводящие
пучки. Ксилема состоит из трахеидов — омертвевших клеток,
суженных на концах; сосудов — полых трубок, и сердцевинных
лучей, состоящих из тонкостенных паренхимных клеток. К флоэме
относятся живые (не омертвевшие) ситовидные элементы, парен-
химные клетки (лубяная паренхима), сердцевидные лучи и меха-
нические элементы. В стеблях флоэма локализуется кнаружи от
ксилемы. Проводящие пучки представляют собой обособленные
тяжи, состоящие из элементов ксилемы и флоэмы.
Ксилема обеспечивает восходящий ток (от корня к листьям)
водных растворов солей, флоэма — нисходящий ток продуктов
фотосинтеза.
Секреторные (выделительные) ткани в виде специализирован-
ных морфологических структур отсутствуют у грибов, хотя экзо-
цитоз (см.) присущ клеточной мембране и по этой функции она
уподобляется секреторной ткани. Из клеток грибов секретируются
многие первичные и вторичные метаболиты (ферментные и не-
ферментные белки, пигменты и другие вещества). Структуры,
подобные млечникам у растений, имеются у некоторых базидио-
мицетов (скрипица, рыжик). В этих случаях между гифами и
окружающими их клетками образуются ситовидные пластинки и
гифы становятся как бы млечными трубками, содержащими млеч-
ный сок (эмульсия смол, масел, пигментов, горьких и других
веществ в воде).
У растений различают ткани наружной и внутренней секреции.
К числу первых относятся гидатоды (от греч. odos — путь),
выделяющие водно-солевые растворы; железистые трихомы (же-
лезки, нектарники, головчатые волоски). Ко вторым относятся
клетки — идиобласты (от греч. idios — своеобразный, особенный,
blastos — отросток, побег), располагающиеся среди других тканей
и накапливающие жидкие или плотные секреты — преимущест-
ве
венно продукты вторичного метаболизма (оксалат кальция, по-
лифенольные соединения, слизи, таннины, терпеноиды); смоляные
ходы, эфиромасляные каналы, млечники, пронизывающие все
растение.
Базисные ткани относятся к разряду мало специализирован-
ных, возникающих у растений из клеток апикальных меристем; у
грибов имеются немногие соответствующие органоиды (не ткани),
которые функционально сходны с базисными тканями — это
преимущественно вакуоли с запасными питательными вещества-
ми.
У растений выделяют ассимиляционную (хлоренхима), возду-
хоносную (аэренхима) и запасающие ткани, все относящиеся к
базисным. В запасающих тканях откладываются белки, вода (на-
пример, у кактусов), жиры, пигменты, углеводы и др.
Ткани высокоорганизованных животных организмов подраз-
деляют на 4 группы: эпителиальные (ведущая роль их — отграни-
чивающая), ткани внутренней среды с сильно развитым межкле-
точным веществом в виде волокнистых структур и аморфной
субстанцией (кровь, рыхлая и плотная соединительная ткань,
хрящевая и костная ткани, репродуктивная ткань), мышечные
ткани и нервная ткань. Органы животных и человека могут
состоять из разных тканей (аорта, органы пищеварительного трак-
та) или почти целиком из одной ткани (кость, печень, сухожилия
и др.). Целостный организм — как система высшего порядка
"диктует" запросы органам, структурными компонентами которой
они являются.
У человека различают двенадцать систем органов: покровная,
опорная (скелетная), мышечная, кровеносная, дыхательная, пище-
варительная, выделительная, нервная, сенсорная (от лат. sensorius
— чувствительный), эндокринная (от греч. endon — внутри, krino
— отделяю), иммунная и репродуктивная.
В настоящее время успешно культивируют in vitro клетки и
ткани (преимущественно эмбриональные) в соответствующих ус-
ловиях, что приобрело большое значение в биотехнологии.
Клетки млекопитающих существенно различаются по химиче-
скому составу, поскольку они достаточно сложны по набору орга-
нелл, а кроме того клетки специализированы в соответствии с
принадлежностью к органам и системам органов, к видам живо-
тных и т. д. Тем не менее, в усредненном варианте состав их
следующий (таблица 10).
119
Таблица 10 Усредненный химический состав клеток млекопитающих
Компонент содержание %
пикограммы (10‘12 г/клетка)
пределы размаха величин усредненно в клетке
Вода 2700-4800 2900 82,9
Белки 200-300 250 10-20
Углеводы 40-200 150 1-5
Липиды 100-200 120 1-2
РНК 20-40 25 0,7
ДНК 8-17 10 0,3
Как видно из таблицы наименьшее количество массы клетки
приходится на ДНК, то есть на тот компонент, который определяет
ее наследственные особенности. Эта закономерность присуща
всем представителям живых существ.
Некоторые свободно живущие клетки эукариот склонны к
хемотаксису (от греч. taxis — расположение), то есть
передвигаться к химическому (хемотаксическому) сигналу. Это,
например, типично для лейкоцитов крови, выходящих из сосуди-
стого русла к очагу воспаления. Хемотаксис присущ и прокарио-
там.
К 1984 г. большинство цитологов утвердилось во мнении, что
в эукариотической клетке имеющийся цитоскелет играет
важнейшую роль в различных биологических явлениях. Цитоске-
лет - это внутриплазматическая
Рис. 38. Клетка млекопитающего животно-
го (на основании иммунофунофлуорес-
ценного анализа): цитоскелет - 1, ядро-2.
поддерживающая система пере-
крещивающихся плотных воло-
кон (филаментов) и полых тру-
бочек (тубул, микротрубочек)
— "балки и канаты" — по К. Де
Дюву (1987). Используя метод
иммунофлуоресценции, удает-
ся выявлять цитоскелет клеток
во всем многообразии (рис. 38).
В 1987 г. был предложен термин
"цитоскелетология" (В. Бирх-
майер), относящийся к части на-
уки о клетке. С начала 70-х годов
стали выделять новые белки, ас-
социированные с цитоскелетом, например, белок глиальных эле-
ментов, образующий промежуточные филаменты в глиальных
нервных клетках; спектрин — основной белок цитоскелета мемб-
ран эритроцита; динеин — высокомолекулярные белки с АТФ-аз-
ной активностью; винкулин — белок, присутствующий в факаль-
ных контактах фибробластов, образует пучки актиновых филамен-
тов; и другие. Много лет тому назад были открыты актин, миозин,
сс-актинин, тубулин, тропонин, тропомиозин. Структурно-функци-
ональные особенности их изучены в достаточной степени, что
показано в таблице 11 на примере миозина, актина и тубулина.
Таблица 11. Некоторые характеристики актина, миозина и тубулина в
элементах цитоскелета животных клеток.
Элементы цитоскелета характеристика
структурная функциональная
диаметр, нм состав- ляющий белок ММ белка, кДа агенты СВЯЗЬ
диссо- циа- ции ассоци- ации
Микрофила- менты 2 миозин <150 АТФ с актином и Са+ + в сократит, акте
Микроволокна 6-7 актин 42-46 цито- холазин В АТФ с цитокине- зом, пино- и эндоци- тозом
Микротру- бочки 25-28 тубулин 120 вин- блас- тин, вин- крис- тин, колхи- цин ГТФ с митоти- ческим верете- ном Л* центри- олями, реснич- ками, жгутика- ми и экзоци- тозом
121
Миозин в виде двойной молекулы формирует стержень, рав-
ного которому по длине нет в природе аналогичных молекул
("цитомышца"). На рис. 39 представлен поперечный срез актоми-
озина, где показана ассоциация шести тонких актиновых фила-
ментов с толстым миозиновым филаментом. В зависимости от
диаметра филаменты подразделяют на тонкие (до
rtf 6-7 нм), промежуточные (8-10 нм) и толстые (15-20
нм).
Рис 3®' Схематичное строение актомиозинового пучка в попе-
СцТу'"' речном разрезе: 1 -актин, 2-миозин.
Актиновые филаменты ("цитокости"), чаще группирующиеся в
форме тонких пучков, составлены из глобулярных белков. Такие
глобулы имеют полярный и боковой сайты связывания, благодаря
которым они растут в длину в виде двойной цепи. В желобке
двухспирального филамента актина располагается тонкая белковая
нить тропонина (рис. 40), образующего вместе с миозином тропо-
миозин (от греч. tropos — поворачивать, mis — мышца).
Микроволокна актина могут разобщаться при вза-
имодействии с цитохолазином В, или фомином [7,20-
дигидрокси-16-метил- 10-фенил-24-окса- [ 14 J-цитохала
за - 6(12),13(E),21(Е)-триен,1,23-дион]. Цитохолазин В
образуется грибами Helminthosporium dematioideum и
Phoma exigua. В ряду цитохолазинов известны А, В, С,
D, Е, F, G, Н, J, родственными им явяются хетоглобо-
зины.
Со времени открытия цитохолазинов в 1964 г.
выполнено большое количество работ с ними. Показа-
Рис. 40. Тро-
помизин (1),
располагаю-
щийся вок-
руг актино-
вого фила-
мента (2).
цитохолазин В
но, что все они тормозят процессы, происходящие в
цитоплазме (не в ядре), а в относительно больших дозах
вызывают энуклеацию клеток, что особенно наглядно
проявляется на клетках млекопитающих.
122
винбластин: И=СНз
винкристин: R= СНО
Тубулин входит в состав микротрубочек. Это глобулярный
белок с диаметром порядка 4 нм, но после сборки 13 его протофи-
ламентов (от греч. protos — первый, от лат. filamentum — ниточка)
в цилиндрическую структуру, внешний диаметр ее составляет 28
нм (внутренний — 14 нм).
Тубулин диссоциируется при действии алкалоидов индольного
ряда(винбластина.винкристина) и трополона (кольцо С в колхици-
не).
Белки цитоскелета по своим функциям могут быть подразделе-
ны на гелеобразующие, фрагментирующие, кэпирующие (от англ.
cap—шапка, кепка, колпак),
адгезирующие, регулирую-
щие и другие.
С учетом новых групп
белков, выявленных в цито-
скелете, становится очевид-
ным, что само название ка-
жется не совсем адекват-
ным, поскольку цитоскелет
— это не неподвижный кар-
кас, или скелет, а сложная и
гибкая система, способная в
ряде случаев к фрагмента-
ции и самосборке, к сколь-
жению и другим функциям.
Тем не менее, определенные
элементы цитоскелета дей-
ствительно служат для со-
здания статичного остова
клетки (кератин в клетках
эпителия, десмин в мышеч-
ных клетках, и т. д.).
При выделении молекулярных компонентов клетки и тополо-
гической привязки их в фиксированных клетках необходимо
учитывать тот факт, что цитоскелетные структуры находятся в
процессе постоянных сборки и разборки в живых клетках.
В настоящее время выделяют 5 основных классов белков,
образующих так называемые промежуточные фила-
менты: виментиновые (ММ=55 кДа) в ткани мезенхимы
(зародышевая соединительная ткань большинства многоклеточных
животных и человека), глиальные (ММ=53 кДа) в клетках глии
123
колхицин
(клетки в головном и спинном мозге), десминовые (ММ = 52 кДа)
в мышцах, нейронные (три белка с ММ = 70 кДа, 150 кДа и 200
кДа) в нейронах, кератиновые (около 20 белков с ММ от 40кДа до
70 кДа). Функция их окончательно не установлена. Предполагают,
что они причастны к митозу, к развитию клеток в норме и при
опухолевой патологии, выполняют роль механического скелета.
Элементы цитоскелета поддерживают клеточную мембрану,
взаимодействуют с мембранами органелл и, как считают, участву-
ют в трансмембранной передаче сигналов в обоих направлениях,
что исключительно важно при объединении отдельных клеток в
ткани. В трансмембранной передаче сигналов участвуют мембран-
ные гликопротеины, или гликопротеиновые рецепторы, контакти-
рующие с филаментами. Эти последние индуцируют систематиче-
ское движение рецепторов. За обратимое "заякоривание" рецеп-
торов ответственны тубулы, или микротрубочки. Эти данные
представляют большой интерес не только с точки зрения раскры-
тия и познания фундаментальных биологических явлений, но и с
практической точки зрения, например, при использовании лекти-
нов (от лат. legere—читать, отбирать, выбирать) — как митогенных,
токсических и цитолитических агентов.
Цитоскелетные структуры имеют прямые связи, например,
микротрубочки с митохондриями, синаптическими пузырьками
(области контактов нервных клеток, или нейронов друг с другом
и с клетками исполнительных органов), ядром; промежуточные
филаменты — с ядром.
Эукариотические клетки многоклеточных организмов отлича-
ются друг от друга в зависимости от ткани (органа), из которой
они изолированы. Например, у млекопитающих клетки кожи
отличаются от клеток печени, у растений клетки эпидермы отли-
чаются от клеток колленхимы, у грибов рода Penicillium зароды-
шевые клетки — экзоспоры, или конидии отличаются от клеток
вегетативного мицелия.
Клетки эукариот в большинстве своем крупнее клеток прока-
риот по диаметру и объему, пространственно более организованны
и дифференцированны, чему во многом способствует цитоскелет.
Внутреннее содержимое клетки разделено на отграниченные мем-
бранами пространства — отсеки, или компартменты (от англ,
compartment — отделение, купе, отсек). Поэтому компартментали-
зация типична для эукариотической клетки и несвойственна по-
давляющему большинству прокариотических клеток.
Клеточная мембрана эукариот сходна по составу и строению с
124
клеточной мембраной прокариот — в ней содержатся белки (в том
числе — маркерный фермент — 5'-нуклеотидаза), липиды и угле-
воды. Однако компонентный состав каждого из названных пол-
имеров у эукариот и прокариот не идентичен. Например, ключе-
вым гликозиллипидом мембраны прокариот является бактопренол
(ундекапренол). Это — С55-соединение, содержащее 9 цис-двойных
связей и 2 транс-двойные связи и относящееся к изопреноидным
липидам. Бактопренол участвует в процессах биосинтеза О-анти-
генов грамотрицательных бактерий, а так же пептидогликанов в
клеточных стенках грамположительных бактерий.
бактопренол
В клеточной мембране дрожжей содержится долихол, сходный
с бактопренолом, но не тождественный ему. Долихол, будучи
гликолипидом, также является изопрениловым спиртом, способ-
ным фосфорилироваться и участвовать в синтезе маннопротеинов
клеточной стенки и неорганических полифосфатов (рис. 41); в нем
содержится 16-20 пренильных остатков и в среднем 80-100 угле-
родных атомов.
сн3
сн3
сн3
сн3
zCHj
н2 сн
СН2
СН£
с=сн —
сн2—о
он—р=о
?н3 сн
ЖЗс/
7-S
о
Man
Дрожжевая долихолфосфатманноза
125
Гидрофобная часть изопреноидов закрепляется в мембране, а
гидрофильная — выступает в цитоплазму.
Рис. 41. Схема строения клеточной стенки у Saccharomyces cerevisiae (по В.
Фаркашу, 1985).
витамины К (п-1-6)
витамин К1 (филпохинон)
Долихолы являются также участниками реакций переноса ман-
нозильных и N-ацетилглюкозильных единиц на гликопротеины в
клетках млекопитающих.
К производным полмпреноидов относятся такие БАВ, как ви-
тамины К, убихиноны, пластохиноны, токоферолы (в частности,
витамин Е), каротиноиды, фитольная группа хлорофилла, способ-
126
ОС. - токоферол (в^-токэфероле содержится Н в позиции 7;
вЯ-токофвропв содержится Н в позиции 6;
в^-тоиоферопе содержится Н в позициях 5 и 7)
каротин из зеленых листьев растений - предшественник
витамина А у животных
Н С—СН—(СН ) —СН—(СН ) —СН—(СН ) —С=СН—СН,О—СО—R
3 I -с 3 I 2 3 | 2 3 | 2
сн3 СН3 CHj CHj
остаток фитола
R - Mg-порфириновый скелет
ствующая встраиванию пигмента в липидный слой мембраны,
эфирные масла.
В состав эфирных масел входят монотерпены и сесквитерпены,
относящиеся к классу терпеноидов, то есть соединений, кратных
по составу изопрену (С5Н8).
скелетная формула
монотерпенов типа
п-ментана (ментол,
цинеол, лимонены,
И др )
скелетная формула
секвитерпенов типа
кадинана (кадинены,
калакарен и др.)
Почти все мембранные
образования внутри клетки
являются либо производны-
ми клеточной мембраны или
непосредственно связаны с
ней: комплекс Гольджи (дик-
тиосома у грибов и расте-
ний) и эндоплазматический
127
Рис. 42. Основные мембранные структу-
ры в эукариотической клетке: 1 — кле-
точная мембрана, 2 — мембрана мито-
хондрий, 3 — ядерная мембрана, 4 —
мембрана эндоплазматического ретику-
люма, 5 — мембрана диктиосом, 6 —
мембрана вакуоли.
ретикулум (ЭПР), эндосомы и ли-
зосомы, пероксисомы и др. Иск-
лючение представляют митохон-
дрии, о возникновении которых
сказано ранее (рис. 42).
Комплекс Гольджи — пред-
ставляется параллельными труб-
чатыми структурами, связанны-
ми с шероховатым ЭПР (шерохо-
ватость ЭПР обусловливается
присоединенными к его наруж-
ной поверхности рибосомами).
Принадлежностью мембраны
комплекса Гольджи являются
специфические гликозилтранс-
феразы, катализирующие реак-
ции присоединения моносахари-
дов из УДф- или ЦМФ-сахаров к
белкам с помощью гликозидных
связей — через гидроксильную
группу серина или треонина, ре-
же — через амидную группу ас-
парагина. Синтезированный гли-
копротеин может выделяться из клетки благодаря экзоцитозу или
отлагаться до времени в вакуолях комплекса Гольджи в виде гранул
или, наконец, переноситься в другие места клетки. В растительных
и грибных клетках аналогом комплекса Гольджи является диктио-
сома — немногочисленные параллельные дисковидные пластинки,
по периферии которых имеются валикообразные расширения и
множество разной величины пузырьков. Не исключается возмож-
ность участия диктиосомы в синтезе и секреции компонентов
клеточных стенок.
ЭПР — мембранная структура, локализующаяся вблизи ядра.
Считается, что секретируемые белки мембран синтезируются при
участии шероховатогоЭПР, а белки, которые используются
клеткой — на свободных рибосомах. Без присоединенных рибосом
ЭПР называют гладким. В нем локализуются различные
ферменты, включая оксидазы, участвующие в детоксикации ядо-
витых для клетки веществ. При участии гладкого ЭПР осуществ-
ляется синтез липидов и гидролитическое расщепление гликогена
(гликогенолиз).
128
Эндосома — это мембранная везикула (от лат. vesicula —
пузырек), возникающая в результате эндоцитоз а; под термином
эндоцитоз понимают проявления общего механизма захвата плот-
ных частиц (фагоцитоз, от греч. phagos — поедание), вирусов
— виропексисот лат. pexis (в конце слова) — соединение,
скрепление, коллоидов (коллоидопексис)и капелек жидкости
(пиноцитоз, от греч. pino — пью, впитываю). Однако чаще
захват водонерастворимых веществ (микробов, эритроцитов и
других частиц с диаметром более 1 мкм) с образованием фаго-
сомы относят к фагоцитозу, тогда как поглощение водораство-
римых веществ с ММ от 180 до 150000 Да и более (антитела,
гормоны, маннит, пептиды, сахароза, ферменты) с образованием
пиносомы относят к пиноцитозу. Пиноцитоз может быть
неселективным (жидкостно-фазным), когда поглощаемое
вещество предварительно не связывается с мембраной, и с е-
лективным (неспецифическим, адсорбционным
и рецепторно-опосредованным), когда вещество предварительно
связывается с определенными участками мембраны.
Следовательно, эндосома возникает при эндоцитозе благодаря
инвагинации (впячиванию) клеточной мембраны, проявляющей
свойства текучести и самозамыкания (жидкостно-мозаичная
структура). Во многих случаях эндоцитоз опосредуется рецептора-
ми, находящимися на поверхности мембраны (мембранные "пэт-
чи", от англ, patch — лоскут, обрывок, заплата). У простейших и
низших позвоночных эндоцитоз является единственным механиз-
мом питания. На поверхности клетки может располагаться множе-
ство рецепторов. Например подсчитано, что на одном нейтрофиле
размещается порядка 2» 10s рецепторов лишь для фракции компле-
мента 5а, а клетка Candida albicans связывает около 2,5-3» 10s
молекул СЗ-фракции комплемента. Рецепторами в гепатоцитах
(клетки печени) являются структуры, распознающие галактозу и
N-ацетилгалактозамин в гликопротеинах (Гал/Гал-ИАц-рецепто-
ры), в том числе — в IgG-иммунных комплексах; секреторные
компоненты, связывающие IgA-иммунные комплексы; рецепторы,
специфичные для маннозо-6-фосфата, и т. д.
Рецепторы животных организмов подразделяют на э к с т е-
рорецепторы и интерорецепторы. Первые из
них воспринимают сигналы извне: слуховые, обонятельные, вку-
совые, тактильные (от лат. tactilis — осязательный), и др.; вторые
воспринимают из внутренней среды — гормональные, медиатор-
ные. Так, наиболее чувствительные к нервным медиаторам участки
между нервными волокнами и клеточной мембраной называют
5 т. 8524 129
медиаторными рецепторами, а мембранные сайты,
чувствительные к гормонам, называют гормональными
рецепторами. Иногда используют термин химичес-
кие рецепторы, когда хотят сказать о биомолекулах клетки,
взаимодействующих с лекарственными или токсическими вещест-
вами. Так, веротоксин Е. coli 0157:Н7 взаимодействует с рецепто-
рами ряда клеточных линий, включая Hela и Vero. Вначале его
В-субъединица присоединяется к специфическому рецептору на
поверхности клетки с интернализацией (от лат. intemus — внут-
ренний) активной субъединицы А, которая прерывает функции
клетки благодаря взаимодействию со специфическими компонен-
тами субклеточной "машины". Указанный выше рецептор специ-
фичен для веротоксинов 1, 2 и Shiga-токсина. Он представляет
собой гликолипид — глоботриозилцерамид (СЬЗ).
Рецепторами также могут быть сульфгидрильные, карбониль-
ные, аминные и другие функциональные группы. Однако рецеп-
торы обладают не только химическими группировками или моле-
ОН NH-COR
I I
НзС-ССН^ 2-сн=сн-сн-сн-сн2он
церамид (R-остаток жирной кислоты)
Рис. 43. Синаптические бляш-
ки на нервной клетке в ганг-
лии сердца лягушки (1 — си-
наптическая щель, 2 — мы-
шечное волокно, 3 — синапти-
ческая бляшка, 4 — тело по-
стсинаптической клетки, 5 —
пресинаптический аксон, 6 —
постсинаптический аксон).
кулами взаимодействия с лигандами, но
и своей архитектоникой (от греч.
architectonike — строительное искусст-
во). Так, в вегетативной нервной системе
место контакта нервных элементов —
синапс включает: пресинаптическое
нервное окончание, в котором синтези-
руется медиатор; синаптическую щель,
куда поступает медиатор; постсинаптиче-
скую мембрану, в которой имеются ре-
цепторы, взаимодействующие с медиато-
рами (рис. 43). Широко известными ме-
диаторами являются адреналин, норадре-
налин, ацетилхолин.
Поэтому говорят об адренорецепто-
рах и холинорецепторах (рис. 44). Изве-
стны al-, a2-, pi-, Р2- и у-адренорецепто-
ры, а также холинорецепторы нервно-
мышечных синапсов (рис. 45). Одни хо-
линорецепторы проявляют Ьысокую чув-
130
ствительность к алкалоиду мускарину, поэтому их называют
м-холинорецепторами, другие — к алкалоиду никотину,
их называют н-холи-норецепторами. На примере
н-холинорецептора из электрического органа электрического ска-
та доказано, что рецептор существует в виде розеток в двух формах
—i легкой (95 кДа) и тяжелой (135 кДа); первая — мономер, вторая
— димер (рис. 46).
Специфическими лигандами для рецепторов в «-бунгаротокси-
не являются аминокислоты Арг-37 и Асп-31. а-Бунгаротоксин
образуется крайтом Bungarus (змея — аспид); он представляет
адреналин норадреналин
/
Н3О—СО—О—(СН2)2—Ns (CHj)3
ацетилхолин
собой полипептид из 71-74 аминокислотных остатков, стабилизи-
рованный четырьмя дисульфидными мостиками (жесткая структу-
ра).
Рис. 44. Адренорецепторы (а и р) в
стенке артерии: 1 — стенка артериаль-
ного сосуда, 2 — миофибриллы, 3 —
адренорецепторы, 4 — возбуждение, 5
— сокращение, 6 — торможение, 7 —
расслабление.
Рис. 45. Схематичное строение нервно-
мышечного синапса по С. Куфлеру и
Дж. Николсу (1979): 1 — частички, 2 —
ямки, 3 — синаптические визикулы, 4
— пресинаптическая мембрана, 5 —
постсинаптическая мембрана, 6 —
складки постсинаптической мембраны,
7 — синаптическая щель.
Из рис. 46 следует, что ионный канал в н-холинорецепторе
создается самим рецептором. Теперь доказано, что женщины,
болеющие раком молочной железы и не имеющие на раковых
клетках рецепторов эстрогена, излечиваются эндокринными пре-
паратами лишь в 10% случаев. Напротив, при наличии таких
рецепторов излечиваются подобными средствами в 50-65% случаев
и более. Заболевания типа тяжелой миастении (Myastenia gravis)
и редкой устойчивой к инсулину формы диабета обусловливаются
аутоиммунными реакциями, когда собственные антитела блокиру-
ют рецепторы, вызывают их транспозицию (перемещение) внутрь
клетки и разрушение.
Рис. 46. Н-холинорецептор электрического
ската; молекулы рецептора в составе фосфо-
липидных пузырьков (А); розетка рецептора
(Б) с двумя местами связывания a-бунгароток-
син а.
Рецепторы, соединяю-
щиеся с молекулами — ли-
гандами, группируются в
образовывающихся в мемб-
ране окаймленных ямках,
или углублениях. Диаметр
углублений около 100 нм. В
структуре их выделяют тя-
желые (ММ= 180 кДа) и лег-
кие (ММ= 35 кДа) цепи клат-
ринов (от лат. clathrum — решетчатый барьер), формирующих
ассамблею протомеров в форме корзинок, которые и покрывают
окаймленные углубления. При эндоцитозе клатриновая клетка —
ловушка втягивает внутрь участок мембраны — пэтч. При этом
углубления как бы опускаются внутрь клетки, края их сужаются,
остается маленькое отверстие, которое вскоре исчезает, и мемб-
рана становится гладкой. Мембрана таким образом лишается пэтча,
трансформировавшегося внутри клетки в свободно передвигаю-
щуюся везикулу (пузырек). В зависимости от процесса (фагоцитоз,
пиноцитоз) везикулу называют: "фагосома”, "пиносома", "фагоци-
тарная", "пиноцитарная” или "эндоцитарная вакуоль", эндосома.
Окаймленные ямки дают начало окаймленным пузырькам, теряю-
щим впоследствии клатриновое покрытие и способным сливаться
с другими неокаймленными везикулами — эндосомами, лизосома-
ми по цис- и транс-м еханизмам. Цис-механизм заключается
в слиянии поверхностей мембран, непосредственно контактирую-
щих с цитоплазмой. Транс-механизм заключается в сближении
наружных поверхностей мембраны. По цис-механизму протекает
132
экзоцитоз, слияние везикул в цитоплазме, поглощение одной ве-
зикулы другой (аутофагия), почкование; по транс-механизму про-
текают: захват мембранных везикул, разделение везикул, эндоци-
тоз.
Функция эндосомы — первичная внутриклеточная переработка
захваченных веществ при pH ниже 7,0 с последующим транспор-
том их в лизосомы или, обходя последние, в места хранения
(гранулы).
Эндосомы могут сливаться с клеточной мембраной, и тогда их
содержимое выводится из клетки — экзоцитоз. Участки
мембраны (пэтчи) могут' отделяться с частью содержимого от
эндосомы и возвращаться в одних случаях — на прежнее место
(такой процесс называется регургитацией, от франц,
regurgitation — излияние, извержение), в других случаях — на
противоположную сторону клетки, где сливаются с мембраной и
немодифицированный материал выводят наружу — д и а ц и-
т о з (от греч. dia — через). Продукты экзоцитоза (литические
ферменты, гормоны и другие вещества) поступают в околоклеточ-
ные пространства, в кровяное русло, в специализированные сек-
реторные протоки (например, слюна, секрет поджелудочной же-
лезы и т. д.).
В некоторых случаях эндосомы на время превращаются в
вакуоли, то есть когда по каким-либо причинам слияние их с
лизосомами приостанавливается. Регуляторами таких процессов
могут выступать некоторые гликопротеины (например, конканава-
лин А), микобактерии туберкулеза и другие факторы.
Весьма существенной для жизнедеятельности эукариотических
клеток является способность мембран любых компартментов клет-
ки сливаться и разъединяться. Поэтому регулируемый поток в
мембранных образованиях в направлении: клеточная мембра-
на (эндоцитоз) —> эндосома —> лизосома —> комплекс Голь-
джи -> ЭПР —> коплекс Гольджи -> секреторная гранула
(экзоцитоз) в основном экспериментально установлен и, следова-
тельно, реален.
Лизосомы (0,5 х 2-3 мкм в диаметре) происходят из эндосом.
Их число в клетке может достигать нескольких сотен. Лизосомы
характеризуются большим полиморфизмом и размерами. Они
представляют собой мембранные образования, в которых проис-
ходит "переваривание" поступающих в клетку веществ с помощью
гидролаз при pH =3,5-5,0. В лизосомах обнаружено свыше 50
133
гидролитических ферментов. Продукты гидролиза ранее поступив-
шего материала проникают через мембрану лизосом во вне — в
цитозоль (основной наполнитель клетки, состоящий из воды с
водорастворимыми компонентами), в том числе — связанные с
выработкой энергии (гликолиз).
Известны лизосомотропные вещества, которые в непротони-
рованной форме (незаряженной) проникают через мембрану ли-
зосом и могут накапливаться в них. Они, как правило, являются
лиофильными слабыми основаниями и могут быть использованы
в практических целях. К их числу относятся противомалярийный
NH—CH—(CH ) — N=(C,HJ, препарат хлорохин —про-
I I 23 z 5 2 ИзВОдное 4-аминохиноли-
на. Накапливаясь в лизо-
kJLj Л сомах, хлорохин блокиру-
ем ет Метаболизм гемоглоби-
Рис. 47. Лизосома — протонная ловуш-
ка; П —препарат. ПН — протонизиро-
ванный препарат, 1 — быстрое проник-
новение, 2 — протонный насос, 3 —
медленный выход.
на — единственного ис-
хпорохим
точника питания малярий-
ного плазмодия.
Лизосомотропизм слабых оснований базируется на механизме
протонной ловушки. Такой механизм присущ и эндосомам, неко-
торым отделам комплекса Гольд-
жи. (рис. 47).
В клетках печени и почек мле-
копитающих, растений и грибов
имеются пероксисомы —
неровные округловатые мембран-
ные образования 0,5-10 мкм в ди-
аметре, заполненные компактным
аморфным матриксом с плотной
кристаллоидной сердцевиной. В
них осуществляется окислитель-
ный метаболизм, главным
участником которого высту-
пают пероксисомальные ок-
сидазы и каталаза: Можно
О
О2 2Н2°' где
АН2 А ан2 А
полагать, что "пероксисо-
мальный тип дыхания" (не
А и А" — субсраты (А‘ — низкомолекулярный)
134
экономичный, но простой по реализации) возник задолго до того,
как были "включены" в аэробное дыхание митохондрии. Перокси-
сомы — органеллы, возникшие в ответ на появление кислорода в
атмосфере. В пероксисомах происходит р-окисление жирных кис-
лот. Субстратами окисления (кроме жирных кислот) могут быть
разные соединения, включая D- и L-аминокислоты, гидроксикис-
лоты, спирты, амины, пурин, но никогда НАД»Н, который окисля-
ется в митохондриях. Пероксисомы участвуют также в синтезе
углеводов.
У инфузорий (Tetrahymena pyriformis) пероксисомы называют
глиоксисомами, в которых содержатся лишь два фермента
глиоксилатного пути окисления веществ — изоцитрат-лиаза и
малат-синтаза.
У некоторых протозойных организмов имеются одномембран-
ные органеллы — гликосомы и гидрогеносомы. В
первых протекает гликолиз, во вторых — окисление пирувата,
сопряженное с синтезом АТФ (у Trichomonas vaginalis нет мито-
хондрий, но есть пероксисомы). При аэробных условиях высво-
бождающиеся при окислении электроны передаются на кислород
с образованием Н2О; при анаэробиозе электроны переходят к
протонам (Н+) и образуется водород (Н2). В этойпоследней реакции
участвуют гидрогеназа и ферредоксин (белок с низким окисли-
тельно-восстановительным потенциалом).
У грибов имеются хитосомы, содержащие фермент
хитинсинтазу, катализирующую реакцию синтеза микрофибрилл
хитина. Хитосомы транспортируют микрофибриллы к клеточной
стенке, где и собирается хитин. Так же у грибов известны л о-
м а с о м ы, располагающиеся между клеточной стенкой и клеточной
мембраной. Как производные клеточной мембраны они могут
возникать в результате дисбаланса пластичности и тургора клеток,
то есть когда мембрана образуется "с избытком". Функция их
окончательно не выяснена, но, очевидно, ломасомы могут быть
причастными к процессам экзоцитоза и секреции веществ, хотя
такой вывод не является окончательным.
При выделении органелл и последующей работе с ними опи-
раются на маркерные структуры, топологически привязанные или
ассоциирующиеся с ними. Чаще всего такими маркерами являются
ферменты (таблица 12).
135
Т а б л и ц a 12. Маркерные ферменты клеточных органелл
Фермент - маркер Классификационный номер фермента (КФ) Органелла 1
Аденилатпиклаза 4.6.1.1 Клеточная мембрана (базолатеральная)
Na+, К+- АТФ - аза 3.6.1.37 ”
5' - Нуклеотидаза 3.1.3.5 Клеточная мембрана (апикальная)
Лейцинаминопептидаза 3.4.11.1 -— " —
Т - Глутамилтранспептидаза 2.3.2.12 — " —
Глюкозо-6-фосфатаза 3.1.3.9 ЭПР
НАДФ»-цитохром- О-редуктаза 1.6.2.4 — —
Эпоксигидролаза 3.3.2.3 — " —
Галактозилтрансфераза 2.4.1.38 Аппарат Гольджи
Сиалилтрансфераза 2.4.99.1 — —
НАД ф-фосфатаза 3.6.1.22 — " —
Сукцинатдегидрогеназа 1.3.99.1 Митохоцдрии (внутренняя мембрана)
Цитохромокисидаза Ротенон+-нечувствительная НАД»Н-цитохром 1.9.3.1
с-редуктаза 1.6.99.1 — —
Моноаминооксидаза 1.4.3.4 Митохондрии (наружная мембрана)
Кинуренин + + -3-гидроксилаза 1.14.13.9 — •’ —
Кислая фосфатаза 3.1.3.2 Лизосомы
Р - Глюкуронидаза 3.2.1.31 — —
Каталаза 1.11.1.6 Пероксисомы
Лактатдегидрогеназа 1.1.1.22 Цитозоль
Примечание: + Ротенон является производным изофлавона, ++Кинуренин — производ-
ное триптофана
В последние годы широко используется термин г л и к о к а -
лике (от греч. glykis — сладкий, calix — оболочка) преимущест-
венно в отношении животных клеток, хотя нередко используют
его и в более обобщенном виде, то есть когда гликокаликсом
называют наружную часть клеточной мембраны эукариот, вклю-
136
чающую гликопротеины и гликолипиды, предназначенную для
узнавания однотипных и чужеродных клеток. Иногда вместо на-
звания "гликокаликс" пользуются термином "пушистая оболочка",
особенно тогда, когда идет речь о толстой оболочке, выстилающей
кишечник. Как бы то ни было, термин "гликокаликс" приложим
ко всем клеткам (в том числе — безъядерным как, например,
эритроциты), у которых мембранные олигосахаридные цепи в
гликоконъюгатах выполняют функции признания (рекогниции).
Из мембранных гликопротеинов наиболее полно изучены г л и-
кофорин и фибронектин (от греч. foros — нести, от
лат. fibra — волокно, necter — связывать, присоединять). Первый
из них содержится в эритроцитах, в нем около 50% углеводов и
130 аминокислотных остатков, ММ его 30 кДа. Фибронектин
содержится в клеточных мембранах других клеток, он обладает
выраженной адгезивностью (от лат. adhaesio — слипание,
сращение) и способствует конглютинации (от лат. conglutinatio
— склеивание) гомологичных клеток.
К мембранным белкам относятся также лектины, выпол-
няющие функции признания и защиты. Известны бактериолекти-
ны, миколектины, лихенолектины, ихтиолектины, зоолектины (все-
го порядка 1000 лектинов). Ряд из них широко используется на
практике для исследования клеточных мембран, в гистохимии, в
структурной химии углеводов и т. д.
Эукариотическая клетка содержит ядро, состоящее из ДНК,
РНК, белков и ряда низкомолекулярных веществ. Низшие грибы
полинуклеарны. Ядро окружено двухслойной липопротеиновой
мембраной, в которой имеются ядерные поры — поросомы. ДНК
вместе с гистонами (нуклеосомы) формируют хромосомы, число
которых различно у разных организмов (чаще число их находится
между 10 и 50). Ближе к центру каждой хромосомы располагаются
центромеры, связывающиеся с ахроматиновым веретеном в пери-
од деления клеток. Оформленное ядро эукариотической клетки
содержит ядрышко, в котором сосредоточены гены, кодирующие
синтез 28S, 18S и 5,8S рРНК. Ядрышко является также местом
образования рибосом. Около ядра клетки располагаются две цен-
триоли, представляющие собой белковые микротрубочки, содер-
жащие белок тубулин (в растительных и грибных клетках центри-
оли не обнаружены). Центриоли входят в состав митотического
аппарата — биполярной структуры в форме веретена.
Рибосомы эукариот по морфологии и функции напоминают
рибосомы прокариот. Однако они сами и их субчастицы характе-
ризуются другими константами седиментации — для недиссоции-
рованных рибосом 80S, для большой субчастицы 60S, для малой
40S.
137
В цитоплазме эукариотической клетки находится большое ко-
личество (до нескольких тысяч) энергообеспечивающих структур
— митохондрий, возникших эндосимбионтно. Средние прибли-
женные размеры их равны 1x7 мкм. Это — двухмембранная
Рис. 48. Эукариотические клетки: I
— грибная, II — растительная, III —
животная. (1 — клеточная стенка, 2
— клеточная мембрана, 3 — ядро, 4
— ядрышко, 5 — конденсированный
хроматин, 6 — рибосомы, 7 — пол-
ирибосомы, 8 — шероховатый эндоп-
лазматический ретикулум, 9 — аппа-
рат Гольжди (диктиосома — у грибов
и растений), 10 — митохондрии, 11
— ядерные поры, 12 — вакуоль, 13
— пиноцитозный пузырек, 14 —
окаймленный пузырек, 15 —лизосо-
ма, 16 — окаймленная ямка, 17 —
волокнистая область, 18 — тонофи-
ламенты, 19 — десмосома, 20 — плаз-
модесма, 21 — плотный контакт, 22
— жировая капля, 23 — центриоль
(триплеты микротрубочек).
отехнологического процесса,
органелла, причем внутренняя мем-
брана выраженно складчатая.
Складки ее, называемые крис-
тами (от лат. crista — гребень),
образуют митохондриальный мат-
рикс, внутри которого локализуют-
ся рибосомы и кольцевидно-замк-
нутая ДНК, кодирующая некото-
рые митохондриальные белки (ми-
тохондрии во многом сходны с бак-
териальным протопластом). Во
внутренней мембране локализует-
ся цепь переноса электронов и фер-
ментный комплекс, катализирую-
щий реакции образования макро-
эргического АТФ.
На рис 48 показано принципи-
альное сходство эукариотических
клеток млекопитающих, растений и
грибов.
4.4. Некоторые функциональ-
ные особенности клеток и клеточ-
ных систем. Биотехнология базиру-
ется на практической реализации
метаболической активности кле-
ток, выделенных из природных суб-
стратов или экспериментально со-
зданных в лабораторных условиях.
В зависимости от целевого продук-
та, получаемого при реализации би-
поддерживают соответствующий
уровень метаболической активности биообъекта. Если речь идет о
каком-либо метаболите, тогда задают такие параметры культиви-
рования, которые обеспечивают максимальный выход данного
вещества. Если же конечным продуктом является биомасса клеток
138
или тканей, то в процессе выращивания создают условия для
максимального выхода их в расчете на число одиночных клеток,
на вес массы по отношению к объему среды, либо количеству
потребленного субстрата (экономический коэффициент).
Питательные среды, рекомендуемые для культивирования
представителей акариот, прокариот и эукариот принципиально
отличаются между собой в том смысле, что для "выращивания"
акариот необходимы живые клетки или ткани. Так, вирусы гриппа
накапливают в куриных эмбрионах, вирус табачной мозаики — на
растениях табака, фаги — в клетках бактерий и т. д.
Большинство прокариот достаточно легко культивируется на
доступных питательных средах, однако среди них есть виды,
которые с трудом или совсем не растут в таких условиях — им,
как и акариотам, нужны живые ткани (некоторые спирохеты,
пневмоцисты, риккетсии, хламидии). Такие виды называют обли-
гатными паразитами, что необходимо учитывать при организации
соответствующих производств.
Эукариотические клетки, как правило, удается культивировать
в лабораторных и производственных условиях с большим или
меньшим успехом. При сравнении, например, клеток дрожжей —
сахаромицетов, используемых в производстве вин, и клеток —
бластов человека, применяемых в производстве интерферона,
культивирование последних сопряжено с большими трудностями,
чем культивирование первых. В такой же мере можно говорить о
различиях клеток растений и животных. Для клеток растений
характерна тотипотентность, то есть способность любой отдельной
растительной клетки в соответствующих условиях культивирова-
ния трансформироваться в целое растение. Клетки животных не
обладают такой способностью и выращивать их труднее, чем
клетки растений.
Питательные среды для прокариот и эукариот должны содер-
жать все необходимые ингредиенты, используемые в конструктив-
ном и энергетическом обмене (источники азота, углерода, серы,
кислорода, водорода, фосфора, витамины). В качестве примеров
можно привести питательные среды для Escherichia coli, Penicillium
chrysogenum, культур клеток табака и В-лимфобластов человека
(таблицы 13-16). Е. coli широко используют в биотехнологии, в
частности, штаммы, несущие чужеродную генетическую инфор-
мацию о синтезе человеческого гормона соматотропина, или штам-
139
мы, секретирующие энтеротоксин, и другие; отдельные виды
пенициллов являются продуцентами антибиотиков пенициллино-
вого ряда: бензил-, оксибензил-, феноксиметил-, 82-пентил-, п-геп-
тил-, п-амил-, аллилмеркаптометилпенициллинов с общей форму-
лой:
где R - один из названных радикалов
Культуру клеток N.
tabacum используют для пол-
учения алкалоида никотина,
а культуру клеток лимфобла-
стов, например, В-ряда —
для получения белка «-ин-
терферона.
Т а б л и ц а 13. Состав питательной среды для Escherichia coli
Ингредиент Миллимоли мг/л Н2О мГ%
Мясной экстракт — 1»104 1»103
Пептон — 1»104 1»103
Натрия хлорид 51 з»ю3 300
Натрия 14 2*103 200
гидро фосфат
Таблица 14. Состав питательной среды для Penicillium chrysogenum
Ингредиент МИЛЛИМОЛИ мг/л Н2О МГ%
Кукурузный — 3»103 300
экстракт Гидрол — 5»103 500
Лактоза 8,7 з»ю3 300
Аммония нитрат 15 1,25*103 125
Натрия сульфит •5НгО 4,6 1*103 100
Натрия сульфат •1ОН2О 1,6 500 50
Магния сульфат •7Н2О 1 250 25
Марганца сульфат • 5Н2О 0,008 20 2
Цинка сульфат 0,1 20 2
Калия дигидрофосфат 14 2»103 200
Кальция карбонат 30 з»ю3 300
Кислота фенилуксусная 7 1»103 100
140
Питательная среда Т. Мурасиге и Ф. Скуга (MS) для культиви-
рования клеток N. tabacum включает следующие компоненты
(таблица 15).
Таблицам Питательная среда для N. tabacum
Ингредиент Миллимоли мг/л НгО МГ%
Калия нитрат Аммония нитрат Магния сульфат «УНгО Кальция хлорид »2НгО Калия дигидрофосфат Марганца сульфат »4НгО Цинка сульфат »4НгО Кислота борная Калия иодид Меди сульфат »5H?O Натрия молибдат »2НгО Кобальта хлорид *6НгО Железа закисного сульфат «УНгО Натрия ЭДТА* «2Н2О Мезоинозит Тиамин гидрохлорид Пиридоксин гидрохлорид Ксилота никотиновая Глицин Гидролизат казеина Сахароза БАП* НУК ДФУК 19 20 1,5 2,9 1,2 0,1 0.003У 0,1 0,005 0,0001 0,001 0,0001 0,1 0,1 0,6 0,0014 0,0024 0.004 0.02У 8У,У 0,002 0,0053 0,0009 1900 1650 ЗУО 440 1У0 22,3 8,6 6,2 0,83 0,025 0,25 0,025 2У,8 ЗУ,3 100 0,4 0,5 0,5 2 10000 30000 0,5 1 0,2 190 165 ЗУ 44 1У 2,23 0,86 0,62 0.083 0,0025 0,025 0,0025 2,У8 3,УЗ 10 0,04 0,05 0,05 0.2 1000 3000 0,05 0,1 0,02
Примечание:* ЭДТА — этилендиаминтетраацетат; БАЛ (цитокинин) — 6-бензиламинопу
рин; НУК (ауксин) — а-нафтилацетат; ДФУК (ауксин) —
2,4-дихлорфеноксиацетат.
MS-среда многоцелевая, универсальная, подходит для каллусо-
образования и индуцирует морфогенез многих двудольных расте-
ний.
Среды для культивирования клеток животных являются более
сложными по сравнению с приведенными выше средами, предназ-
наченными для бактерий, грибов и клеток табака. Так, X. Игл
предложил минимальную незаменимую среду (МЕМ), вариант
которой по Р. Далбекко (ДМЕМ) содержит следующие компоненты
(таблица 16).
141
Таблица 16. Состав среды ДМЕМ для культивирования В-лимфоцитов человека
Ингредиент Миллимоли мг/л НгО мГ%
Глюкоза 5,5 1000 100
Натрия пируват 1 ПО И
L-Аргинин гидрохлорид 0.4 84 8,4
L-Валин 0,3 94 9,4
L-Гистидин гидрохлорид • НгО 0,2 42 4,2
Глицин 0,4 30 3
L-Глутамин 4 584 58,4
L-Изолейцин 0,8 104,8 10,48
L-Лейцин 0,8 104,8 10,48
L-Лизин гидрохлорид 0,86 146,2 14,62
L-Метионин 0,2 30 3
L-Серин 0,4 42 4,2
Тирозин (двунатриевая соль) 0,4 89,5 8,95
L-Треонин 0,8 95,2 9,52
L-Триптофан 0,078 16 1,6
L-Фенилаланин 0,4 66 6,6
L-Цистеин дигидрохлорид 0,2 62,6 6,26
i-Инозитол 0,044 7,2 0,72
Никотинамид 0,03 4 0,4
Кальция пантотенат 0,0096 4 0,4
Пиридоксаль (гидрохлорид) 0,02 4 0,4
Рибофлавин 0,001 0,4 0,04
Тиамин гидрохлорид 0,013 4 0,4
Кислота фолиевая 0,006 4 0,4
Холин битартрат 0,02 7,2 0,72
Натрия хлорид 109,4 6400 640
Калия хлорид 5,4 400 40
Кальция хлорид 1,8 200 20
Магния сульфат 0,8 97,7 9,77
Натрия дигидрофосфат • 2НгО 0,8 125 12,5
Натрия гидрокарбонат 21,4 1800 180
Железа закисного нитрат 0,0002 6.1 0,01
•9Н2О
Феноловый красный 0,02 5 0,5
По сравнению со средой МЕМ в среде ДМЕМ почти в два раза
увеличино содержание аминокислот и в 4 раза — витаминов. Ее
используют для культивирования различных видов клеток.
С учетом особенностей клеток в среды МЕМ и ДМЕМ можно
вносить какие-либо добавки, например, трансферрин, инсулин,
бычий сывороточный альбумин, цельную или диализованную сы-
воротку крови (5-10% по объему).
Кроме подобных сред предложены также бессывороточные
142
среды, в которых сыворотка крови замещается смесями из очи-
щенных белков, пептидов, липидов, микроэлементов и других
веществ. Однако они, в большинстве своем, не являются универ-
сальными.
Разнообразие питательных веществ в средах для культивиро-
вания клеток млекопитающих служит предпосылкой к строгому
соблюдению мер предосторожности в целях предотвращения за-
грязнения их вирусами, микоплазмами, бактериями, грибами. В
сравнительном плане клетки прокариот и эукариот заметно раз-
личаются по скорости роста. Поэтому, например, животные и
большинство растительных клеточных систем уступают в конку-
ренции микробным системам.
С биотехнологической точки зрения важными являются поня-
тия о первичных и вторичных метаболитах или о реакциях
первичного и вторичного обмена, которые сходны у всех живых
организмов. К реакциям первичного обмена относят образование
и расщепление нуклеиновых кислот и оснований, белков и их
предшественников, а также большинства углеводов, липидов, не-
которых карбоновых кислот. К реакциям вторичного обмена от-
носят те из них, которые сопровождаются образованием алкалои-
дов, антибиотиков, триспоровых кислот, гиббереллинов и некото-
рых других веществ, расцениваемых несущественными для про-
дуцента. Более того, образование вторичных метаболитов свойст-
венно не всем видам, а лишь только некоторым.
Реакции первичного и вторичного обмена трудно дифферен-
цируемы, а существенность или несущественность вторичного
метаболита для продуцента оценивается весьма субъективно и
нередко без учета роли его в естественных ассоциациях различных
организмов. Вот почему научно более обоснованным является
представление о первичных метаболитах как продуктах матрично-
го синтеза, то есть белках (преимущественно — ферментных), а
вторичные метаболиты представляют собой продукты реакций,
катализируемых ферментами (Н. П. Блинов, 1979):
।------------------► Преметаболиты <------------------
|---------► Интермедиаты, или прометаболиты
Г Информационные I
молекулы * ----
—» Первичные-метаболиты
а) ферментные белки *
б) неферментные белки Вторичные метаболиты •«-----
Преметаболиты в схеме представляют собой простые питатель-
ные вещества, поступающие извне (аммоний, ионы металлов,
углекислота, сульфаты, фосфаты, нитраты, для гетеротрофов —
моносахариды и некоторые другие).
143
К интермедиатам, или прометаболитам относятся простые са-
хара, аминокислоты, нуклеиновые основания и др. Информацион-
ные молекулы ДНК и РНК вычленены из состава других реакций,
хотя их синтез и распад (прерывистые стрелки) также катализи-
руются ферментами. Важно подчеркнуть, что, в отличие от пер-
вичных метаболитов образование вторичных метаболитов непос-
редственно не кодируется ядерной или цитоплазматической ДНК.
Согласно такому представлению все живые организмы синтези-
руют присущие им первичные и вторичные метаболиты.
/’'На основании положений, сформулированных В. Н. Шапошни-
ковым (1939), каждый продуцент в своем развитии проходит две
фазы, названных Ж. Д. Бу'Локком (1961, 1967) трофофазой и
идиофазой (от греч. trofe —- питание, idios — свой, специфический).
В период трофофазы активно протекают конструктивный и энер-
гетический обмен — в клетках преобладают синтетические про-
цессы, нарастает количество первичных метаболитов и ряда внут-
риклеточных вторичных метаболитов — липидов, гликанов, глико-
конъюгатов; скорость роста и размножения организма при этом
высокая, а продукция экзогенных вторичных метаболитов низкая,
и, напротив, в идиофазу скорость роста и размножения низкая, а
продукция экзогенных и эндогенных вторичных метаболитов вы-
сокая (рис. 49). Продуктивность культуры может быть повышена
за счет внесения предшественников метаболитов (преимуществен-
но в период времени, приходящийся на конец идиофазы).
Рис. 49. Соотношение биомассы клеток (а) и метаболитов (первичных — б,
вторичных — в) в культурах Saccharomyces cerevisiae (A), Penicillium chrysogenum
(Б), Nicotiana tabacum (В), кератоцитов (Г); М — масса высушенных клеток, М‘ —
число животных клеток, t — время в сутках, I — трофофаза (заштрихованная часть),
II — идиофаза.
144
Из рис. 49 видно, что продолжительность трофофазы более
короткая у дрожжей, чем у пеницилла и клеток табака. Накопление
этанола S. cerevisiae сопровождается возрастанием ингибирующей
активности его на продуцент и поэтому кривые, приходящиеся на
идиофазу идут почти параллельно, повторяя характер кривой для
первичных метаболитов, биосинтез которых начинается в период
трофофазы.)
Биосинтез фибринолитического фермента (первичный метабо-
лит) клетками кератиноцитов млекопитающих полностью корре-
лирует с ростом числа клеток (2) и не коррелирует с числом их в
стационарной фазе (3) или, тем более, в фазе отмирания (4).
Пенициллин, синтезируемый Р. chrysogenum, и не ингибирую-
щий продуцент, выражение накапливается в идиофазу.
Алкалоид никотин синтезируется клетками табака замедленно
и при переходе культуры в стационарную фазу выход его заметно
уменьшается.
В каждом из приведенных примеров можно отметить свои
особенности биосинтеза первичных и вторичных метаболитов, что
определяется возрастанием сложности клеточных систем при пе-
реходе в ряду —> Mycota —> Plantae —> Animalia среди
представителей эукариот. В любом случае первичные и вторичные
метаболиты образуются клетками как естественные продукты в
процессе их культивирования в соответствующих средах и под
каталитическим действием ферментов. Эти последние могут инги-
бироваться с помощью специальных веществ, называемых анти-
метаболитами, имеющими структурное сходство с метаболитами.
Такие вещества являются исключительно важными для выяснения
и познания путей обмена веществ, а также для использования
наиболее активных из них как лечебных средств. В качестве
примеров можно попарно назвать следующие метаболиты и анти-
метаболиты:
СООН
(сн2)2
I
СООН
СООН
сн2
СООН
янтарная кислота (метаболит)
малоновая кислота (антиметаболит)
145
NH=C—NH—(CH,)—СН—СООН
I 23 I
NH2 NH
аргинин (метаболит)
СООН
NH=C—NH—0—(CH )-CH-COOH
I 22 I
nh2 nh2
канаванин (антиметабопит)
никотиновая пиридин-3-сульфоновая
кислота кислота
(метаболит) (антиметаболит)
п-аминобензойная сульфаниламиды
кислота (метаболит) (антиметаболиты)
6-меркаптопурин (антиметабопит)
С учетом более медленного (примерно в 100 и более раз) роста
и размножения клеток животных и растений в сравнении с
микробными клетками трофофаза и идиофаза у них значительно
растянуты во времени. Тем не менее всегда следует стремиться к
сокращению временных интервалов трофофазы, поскольку это
поможет сократить срок проведения всего биотехнологического
процесса и удешевить конечный продукт.
При выращивании клеток в культурах следует иметь в виду их
следующие особенности:
1. дифференцированность, 2. склонность к аггломерации, 3.
изменение размеров и цитоархитектоники в процессе развития, 4.
дизруптивность, 5. способность к трансформации.
Клетки многоклеточных растений и животных, в противопо-
ложность одноклеточным видам, по мере роста и развития орга-
низма становятся все более специализированными, возникают
ткани с конкретными функциями (дифференциация). В диффе-
ренцированных клетках специализация функций обусловливается
генетической детерминированностью, проявляющейся в феноти-
пах. Иными словами — дифференциация является результатом
различной экспрессии генетической информации в разных клет-
ках.
146
С другой стороны с дифференциацией связаны биосинтез и
накопление вторичных метаболитов. В качестве примеров можно
назвать образование и отложение лигнина в сосудистых элементах
ксилемы и трахеидах растений, биосинтез андрогенных гормонов
в клетках половых желез у мужских особей животных, и т. д.
Торможение или подавление дифференцировки может сопро-
вождаться существенным изменением в характере реакций вто-
ричного обмена. Например установлено, что каллусы дифферен-
цирующихся корней растения красавки (Atropa belladonna) синте-
зируют так называемые тропановые алкалоиды (атропин; скопо-
ламин; гиосциамин — аналог атропина, содеращий L-троповую
CH.-CH-CH,
I
сн2-сн-сн2
сн-сн2
N-CH,
сн-сн2
сно-со-сн
/ I
сн2о
атропин
скополамин
кислоту):
Те же, но не дифференцированные культуры, не синтезируют
названные алкалоиды.
В других случаях не дифференцированные культуры тканей
способны синтезировать вторичные метаболиты, но в меньших
количествах, чем дифференцированные аналоги их (например,
алкалоид никотин, образуемый Nicotiana tabacum).
Применительно к бактериальным культурам можно назвать
Вас. thuringiensis, формирующей при дифференциации — фаза
спорообразования белковый ромбовидный параспоральньГй кри-
сталл, обладающий токсическим действием ‘преимущественно в
отношении насекомых из отряда чешуекрылых, и поэтому исполь-
зующийся в борьбе с вредителями, например, лесного хозяйства.
В отличие от бактерий грибы более дифференцированные
организмы, и у них также отмечена зависимость синтеза ряда
вторичных метаболитов от степени дифференцировки. Например,
биосинтез пигментов у большинства дейтеромицетов приурочен к
началу конидиеобразования или, у некоторых видов
(Aureobasidium pullulans), он связан по времени с формированием
хламидоспор.
Казалось бы, что приведенные примеры могут служить дока-
зательством существования общей закономерности, относящейся
11/
к прямо пропорциональной зависимости синтезируемого вторич-
ного метаболита от уровня дифференцировки клеток, то есть, чем
выше дифференцировка, тем больше образуется метаболита. Во
многих случаях зто так и есть. Однако, что касается растений, то
имеются многочисленные данные о высокой активности дедиффе-
ренцированных культур, например раувольфии змеевидной
(Rauwolfia serpentia), синтезирующих индольные алкалоиды. Оче-
видно, тотипотентность клеток растений является одним из важ-
нейших задатков способности их синтезировать вторичные мета-
болиты при подборе адекватных условий культивирования и сти-
мулирования, то есть воздействуя на триггерные (от англ, trigger
— спусковой крючок) или эффекторные механизмы метаболиче-
ских путей вторичного обмена.
Растительные и животные клетки примерно в 10 - 100 раз
больше по размерам, чем клетки бактерий и грибов, диаметр
многих из них варьирует в пределах 20 - 150 мкм. Однако только
животные клетки и прокариотические микоплазмы лишены ригид-
ной клеточной стенки. Тем не менее прокариотические и эукари-
отические клетки в процессе роста и развития стремятся к орга-
низационной завершенности (бактериальные — в пределах минут-
часов, грибные — в пределах часов-суток, растительные и живо-
тные — в пределах нескольких суток).
В случаях поддержания многоклеточных популяций в гистоло-
гически организованной форме, соответствующей исходным тка-
ням, и при сохранении специализированных для этих тканей
функций, говорят о культурах тканей. При диссоциации тканей
на клетки с утратой ими гистологической организации и специа-
лизированных функций при выращивании in vitro говорят о куль-
турах клеток. Разъединение клеток стимулирует их рост.
При выращивании животных клеток в культуре в целях пол-
учения специальных продуктов необходимо принимать во внима-
ние и учитывать их генетическую нестабильность и, как следствие,
вариабельность фенотипической экспрессии с последующими ста-
рением и отмиранием. Всего этого удается избегать при использо-
вании гибридомных клеток.
Если старение и гибель клетки были бы предопределены гене-
тически, то это значило бы, что позитивная дифференциация
является логическим следствием роста и развития клетки, достиг-
шей высшей стадии специализации, и она завершается смертью.
Согласно все более подтверждающейся на практике клонально-се-
лекционной (стохастической, от греч. stochasticos — случайный,
вероятностный) теории смерть клетки происходит вследствие по-
148
степенного накопления трансляционных ошибок в работе генома
под влиянием инфекций, радиационного облучения и других му-
тагенных воздействий, то есть когда наступат критическое возра-
стание энтропии.
Кроме дифференциации клеток важной особенностью их яв-
ляется склонность к аггломерации (от лат. agglomeratus — скоп-
ление). Это может происходить в различных условиях и с клетками
различного уровня организации — прокариотами и эукариотами.
Те из них, которые имеют клеточную стенку, чаще аггломерируют
за счет химических компонентов, локализованных в ней. Причем
процесс "скучивания" является физико-химическим (адсорбция,
ионное и ковалентное взаимодействия), зависящим не только от
особенностей клеток, но и от компонентов среды, используемой
для их культивирования. Поэтому аггломерация может быть след-
ствием: 1. адгезии (от лат. adhaesio — склеивание, слипание) клеток
друг к другу или к поверхности культурального сосуда за счет
веществ — адгезинов, расположенных на их поверхности, и других
причин; 2. агглютинации по схеме "антиген-антитело", когда в
качестве антигена оказываются культивируемые клетки, а в каче-
стве антитела — гомологичные или гетерологичные агглютиниру-
ющие иммунные сыворотки; 3. слияния клеток с образованием
гибридов.
Известные к настоящему времени адгезины, например, в клет-
ках дрожжей и ряде растений представляют собой преимущест-
венно гликопротеины. В поверхостных структурах гриба
Histoplasma capsulatum за адгезию к эпителиальным клеткам от-
ветственны галактоза, манноза и фукоза (в меньшей степени —
глюкоза), но не N-ацетилглюкозамин.
На поверхности ростковых трубок Candida albicans располага-
ются антигенные эпитопы, или детерминанты (от греч. epi — на,
поверх, к; topos — место; от лат. determinatio — определение) с
ММ 60-230 кДа, проявляющие адгезию к фибриногену. Такие
эпитопы называют еще фибриногеносвязывающими факторами.
На каждой ростковой трубке располагается до 4000 мест фиксации
фибриногена. На поверхности дрожжевых псевдомицелиальных
клеток того же вида располагается маннопротеин (ММ 60 кДа),
белковая часть которого может связываться с производным СЗ-
фракции комплемента (iC3b), проявляя свойства адгезина.
У бактерий и некоторых грибов функцию адгезинов выполняют
белки фимбрий. Вместе с тем у грамотрицательных бактерий могут
образовываться так называемые локальные зоны адгезии между
клеточной мембраной и клеточной стенкой, лишенной пептидог-
ликанового слоя. В этих зонах два фосфолипидных слоя входят в
149
соприкосновение друг с другом (рис. 50). Таких зон может быть
порядка 200-400 на клетку (около 5% поверхности клеточной
мембраны). Локальные зоны адгезии служат воротами для транс-
порта в обоих направлениях.
4 .4
Рис. 50. Схема образования локальной зоны адгезии в оболочке грамотрицательных
бактерий (в направлении а - в): 1 — мембранный фосфолипид, 2 — периплазмати-
ческое пространство, 3 — пептидогликан, 4 — фосфолипидный слой, 5 — белок, 6
— углевод.
Клетки животных, находясь в системах органов и тканей,
взаимодействуют между собой за счет ионов, десмосом, электо-
ронноплотных гликопротеинов.
Следует иметь в виду, что клетки прокариот и эукариот имеют
отрицательный заряд и поэтому ионные взаимодействия, сущест-
вующие между ними, присущи так же клеткам и субстратам,
несущим электрический заряд. Если этот заряд отрицательный (как
и заряд клетки), то в среде должны быть двухвалентные катионы
и адгезии, например, фибронектин клеточного или плазматическо-
го происхождения. Фибронектин — гликопротеин с ММ 200-250
кДа. Он найден на поверхности глиальных клеток и фибробластов,
в плазме, амниотической и спинно-мозговой жидкостях.
Реакция агглютинации клеток специфическими антителами
(иммуноглобулинами — 1g) с давних пор используется в иммуно-
логии. Антитела взаимодействуют с антигенными детерминантами,
локализующимися на клеточной поверхности. При этом отмечает-
ся аггломерация клеток, где связующими звеньями выступают 1g.
Вследствие различной антигенной структуры микробов в их агг-
лютинации принимают участие антитела различной специфично-
сти.
Прилипание прокариот к поверхности В-лимфомицитов про-
исходит, например тогда, когда животное, от которого взяты
лимфоциты, было предварительно иммунизировано этими же бак-
териями. Это так называемый феномен иммуноклеточного при-
липания. Вариантом его является непрямая гемадсорбция, когда
вместо клеток прокариот используют, например, гаптены, которые
сорбируются на эритроцитах in vitro. Такие эритроциты формиру-
ют розетки вокруг лимфоидных клеток животного, ранее иммуни-
зированного микроорганизмом, из которого выделен гаптен.
1.Ю
Что касается спонтанного слияния клеток прокариот или эука-
риот, то такие процессы происходят редко и преимущественно
между макрофагами или миобластами (от лат. myo — мышечный,
blastus —зародышевая' материнская-клетка) с образованием мно—
гоядерных клеток — синтиций (от лат. syncytium — многоядерная
протоплазматическая масса).
Частоту слияния можно существенно повысить, направленно
используя для этого полиэтиленгликоль, лизолецитин, ДНК-содер-
жащие вирусы герпеса или РНК-содержащие вирусы Сендай.
НО—(СН2СН2—О)п-Н Лизолецитин — это продукт, образую-
полиэтиленгликоль Щийся из лецитина под действием леци-
тиназы. В случае образования гетерока-
рионов и последующей пролиферации гибридной клеточной линии
(увеличение числа клеток), то хромосомы будут частично теряться,
в то время как оставшиеся хромосомы будут определять один или
комбинацию других признаков. Это важно для экспрессии генов
(в том числе — генов злокачественности). О гибридизации клеток
см. также 5.2.1.
В процессе роста и развития клеток происходят изменения в
размерах и архитектонике структурных компонентов клеток. Для
прокариот такие изменения трудно уловимы при их быстром
размножении простым делением. В случае спорообразования такие
изменения можно уловить с большей определенностью. Используя
цейтраферную киносъемку удается четко зафиксировать проис-
ходящие события, например, через интервалы времени, равные
нескольким секундам. Грибные, растительные и животные клетки
в этом смысле оказываются более удобными объектами для наблю-
дения. Можно проследить их рост по размерам, равно как и
формирование дифференциальных структур в течение часов и
суток.
Животные клетки образуют псевдоподии (ложноножки), с
помощью которых передвигаются по субстрату. Псевдоподии об-
ладают адгезинами. При образовании или наличии второй клетки,
с которой контактирует первая, их псевдоподии осуществляют
движения в противоположном направлении — наступает контак-
тное ингибирование. Развиваясь в виде монослоя, клетки полно-
стью перестают двигаться и приостанавливают свой рост. Плот-
ность клеток может возрасти и за счет многослойности их в
культуре. Одиночная оплодотворенная яйцеклетка служит источ-
ником всех 1014 клеток в организме взрослого человека и некоторая
часть из них делится (всего по расчетам, происходит 206 делений
в секунду). Здесь исключительно важна роль ЦНС, гормонов и
других регулирующих факторов.
151
Дизруптивность (от лат. disruptio — разрыв) клеток прокариот
и эукариот — еще одна их особенность. Причем нельзя провести
четкой границы между представителями этих надцарств по диз-
руптивности. Понятно, что клетки, имеющие клеточную стенку,
значительно устойчивее клеток, не обладающих ею (для сравнения
можно назвать микоплазмы, протопласты и сферопласты, клетки
Е. coli, конидии Aspergillus niger, меристемные клетки картофеля
— Solanum tuberosum, Т-лимфоциты человека). Даже при разру-
шении в дезинтеграторах различного типа бактериальные клетки
несколько менее устойчивы на разрыв, чем грибные.
Клетки животных без клеточных стенок относятся к разряду
высоко дизруптивных, тогда как большинство микробных клеток
— к разряду низко дизруптивных. Этот показатель имеет особое
значение для организации соответствующих производств, в кото-
рых предусматриваются перемешивание и барботаж культураль-
ной жидкости, а также при реализации конечного продукта в виде
клеток, когда, например, они должны быть разрушены и примене-
ны в качестве белковых добавок к кормам для животных. Без
дизрупции клетки пройдут через пищеварительный тракт, остава-
ясь мало измененными. В этой связи перспективны способы
ферментативного гидролиза клеточных стенок. Причем лизис кле-
ток может происходить двояко — за счет собственных ферментов
(автолиз) и под воздействием внешних ферментов (гликозидаз,
гликозаминидаз, амидаз, пептидаз) — экзолиз. Глубина лизиса
клеток зависит от ряда превходящих факторов: химического со-
става и архитектоники клеточных стенок, температуры и состава
окружающей среды, возраста клеток и других. При полном осво-
бождении от стенки клеточного содержимого, окруженного мем-
браной, образуются протопласты; при частичном — сферопласты.
Без стабилизаторов в среде время существования тех и других
весьма ограничено.
В нашей стране и за рубежом ферментная промышленность
производит отдельные литические ферменты для продажи. К числу
их относятся дрожжелитин, лизосубтилин, лизоцим, проназа и
другие.
Солюбилизирующим действием на клетки обладают некоторые
органические соединения (мочевина и ее производные, толуол,
бутанол, диметилформамид, поверхностно-активные вещества и
другие).
В ряде случаев важна синхронизация клеток, когда большин-
ство из них находятся в одинаковом фазовом состоянии. Этому
соответствует экспоненциальная фаза размножения одноклеточ-
ных видов. Синхронизации микроорганизмов добиваются следу-
152
ющими методами: сменой температурных режимов выращивания,
голоданием с последующим переносом на полные среды, фильтра-
цией клеток одинаковых размеров. В синхронизированном состо-
янии они сохраняются в течение 2-4 генераций, после чего пере-
ходят к асинхронному росту, развитию и размножению.
В случаях синхронизации клеток млекопитающих применяют
сепарацию и индукцию. В первом случае проводят дифференци-
альное центрифугирование в градиентах бычьего сывороточного
альбумина, сахарозы или фиколла (синтетический высокомолеку-
лярный сополимер сахарозы и эпихлоргидрина.
При этом отделяются, в основном, клетки
одинакового возраста и объема, перешедшие
из фазы митоза в фазу G-2 (см. раздел 5.1.),
С1-СН2-СН-СН2
эпихлоргидрин О
когда их объем возрастает примерно вдвое. Синхронизированные
клетки составляют! 65-70%. Во втором случае (при индукции)
блокируют клетки в определенной фазе жизненного цикла с
помощью ингибиторов синтеза ДНК (аметоптерина, 5-аминоура-
цилла, гидроксимочевины) или ингибиторов митоза (винобластин
— см. раздел 4.3, колхицин и его производные).
аметоптерин (метотрексат; Л-амино-N -метил фолиевая кислота)
5-амин оурацил
nh2
со
I
NHOH
гидрокси-
мочевина
Клетки затем промывают для освобождения от ингибиторов
(блокаторов) и дают им синхронно развиваться, учитывав тот факт,
что после одной-двух генераций они становятся асинхронными.
При индукции достигают меньшей синхронизации клеток (2-50%),
чем при сепарации, однако данный показатель можно повысить
при осуществлении повторных блокировок.
Степень синхронизации определяется митотическим индексом
(MI), то есть индексом частоты митозов на 1000 клеток (оценивают
в окрашенных препаратах): MI = 1000:МС, где МС — число клеток
с признаками митоза. Зависимость здесь обратно пропорциональ-
ная — чем меньше Ml, тем синхронизация больше.
153
Для некоторых представителей эукариот (не прокариот) изве-
стны фазовые состояния, сопровождающиеся изменением росто-
вых характеристик — трансформация. Так, отдельные грибы
являются диморфными, то есть фенотипически двойственными,
растущими либо в мицелиальной форме, либо в дрожжевой
(Aureobasidium spp., Histoplasma spp., Candida spp., Mucor spp. и
др.). Так, например, A pullulans — продуцент экзогликана в
глубинных условиях растет в виде одноклеточных, почкующихся
дрожжевых клеток, тогда как на поверхности агаризованных
питательных сред дрожжевые клетки трансформируются в мице-
лиальные, мало или совсем не образующие экзогликана. Подобная
морфо-физиологическая трансформация является обратимой. В
этой связи выделяют три главные группы диморфных грибов —
температурозависимые (Blastomyces dermatitidis), зависимые от
температуры и от питательных веществ (Histoplasma capsulatum),
и зависимые только от питательных веществ (Candida albicans).
У животных клеток трансформация — процесс необратимый.
Он может протекать под влиянием онкогенных РНК- или ДНК-ви-
русов и называется вирусной трансформацией. К РНК-онкоген-
ным вирусам относятся лейковирусы кошек, мышей и птиц, вирус
опухоли Биттнера, поражающий млекопитающих, вирусы сарко-
мы; к ДНК-онкогенным вирусам относятся: аденовирусы, вирусы
папилломы, полиомы и SV-40 (от лат. simian virus — вирус обезьян)
из группы папова-вирусы, герпес-вирусы и вирусы оспы. Следует
иметь в виду и тот факт, что ДНК нормальных клеток могут
содержать в себе провирусный материал и поэтому такие клетки
способны к необратимой трансформации под влиянием физиче-
ских и химических мутагенов. Провирусные сегменты ДНК назы-
вают онкогенами.
В условиях, ограничивающих рост нормальных клеток, транс-
формированные клетки как менее требовательные к факторам
роста, пролиферируют до более высокой плотности и они стано-
вятся способными культивироваться в глубинных условиях (сус-
пензионные культуры) при изменении своей формы до сфериче-
ской. Следовательно, и у грибных, и у животных, а также у ряда
растительных клеток их морфологическое сходство (сферичность,
округлость) в глубинных условиях выращивания находится под
контролем генотипа и сопровождается изменением преимущест-
венно клеточной мембраны.
Трансформированные in vitro клетки представляют собой по-
тенциально важные объекты для выбора оригинальных продуцен-
тов ценных веществ.
154
Глава 5.
ФУНДАМЕНТАЛЬНЫЕ ИССЛЕДОВАНИЯ
В ОБЛАСТИ ГЕНЕТИКИ И МОЛЕКУЛЯРНОЙ
БИОЛОГИИ ВИРУСОВ, КЛЕТОК
И КЛЕТОЧНЫХ СИСТЕМ
Генетика — наука о наследственности прошла сложный путь
своего развития; фактические данные и обоснованные гипотезы в
ней были, к сожалению, использованы даже для утверждения таких
социально-политических доктрин, которые противоречили науч-
ным истинам, этике и здравому смыслу (например, попытка обос-
нования превосходства одних рас людей над другими; полное
отрицание генов как якобы надуманных, "мифических" и несуще-
ствующих структур в зародышевых клетках любых организмов;
доминирующую зависимость наследственности от условий внеш-
ней среды обитания того или иного вида; отрицание внутривидовой
борьбы и признание межвидовой в параллели с борьбой классов
в человеческом обществе, и т. д.). Подобные грустные страницы в
истории генетики канули в вечность и этому помогло выдающееся
событие в науке, когда Дж. Уотсон и Ф. Крик в 1953 г. расшифро-
вали двойную спираль ДНК и подвели материальную базу под ранее
упомянутый "мифический” ген — материализация гена. С тех пор
прошло более 40 лет, и трудно охватить все области генетической
науки, где бы ни были сделаны открытия или которые не получили
бы мощного стимула для своего развития, включая современную
биотехнологию. Однако на фоне всех достижений в течение
последних 20 лет, нельзя забывать о том, что М. Фишер еще в 1868
г. открыл нуклеин, Ф. Гриффит в 1928 г. описал явление трансфор-
мации у бактерий, а О. Т. Эйвери, К. М. Мак-Леод и М. Мак-Карти
в 1944 г. доказали, что трансформирующим агентом является ДНК;
Ж. Ледерберг в 1947 г. открыл процесс конъюгации у Е. coli, а
позже было доказано, что спаривание клеток бактерий обусловле-
но генетически.
К времени расшифровки ДНК Э. Чаргафф (1950) уже сформу-
лировал свои правила, согласно которым: 1) число оснований с
аминогруппами в шестой позиции равно числу оснований с кетог-
руппами в той же позиции, то есть А (аденин) + Ц (цитозин) — Г
(гуанин) + Т (тимин); это единственное правило, приложимое к
ДНК и к большинству типов РНК, в которых Т заменен на У
(урацил);
2) молярное содержание аденина равно молярному содержа-
нию тимина (А=Т или А/Т= 1);
155
3) молярное содержание гуанина равно молярному содержа-
нию цитозина (Г = Ц или Г/Ц= 1);
4) сумма пиримидиновых оснований равна сумме пуриновых
оснований, то есть Ц + Т = А+Г (Пир/Пур= 1);
5) у бактерий отношения А/Т и Г/Ц близки единице, тогда как
отношение А/Г изменяется в интервале 0,4—2,7. Интервал изме-
нения А/Г для растений (1,1—1,7) и животных (1,3—2,2) меньше,
чем для ДНК бактерий. По соотношению нуклеотидов выделены
АТ-тип ДНК, когда А + Т > Г + Ц, или ГЦ-тип ДНК, когда Г + Ц >
А + Т.
Последующие успехи в области генетики нарастали исключи-
тельно быстро: в 1956 г. А Корнберг выделил ДНК-полимеразу, в
1961 г. М. Ниренберг предложил подходы и сам участвовал в
расшифровке генетического кода, в 1964 г. осуществлен первый
синтез полирибонуклеотидов (X. Г. Корана), в 1965 г. В. Арбер
открыл ферменты-рестриктазы, или рестрикционные эндонуклеа-
зы, в 1969 г. Дж. Бекуит с сотрудниками выделил из кишечной
палочки лактозный оперон, в 1970 г. Г. Темин и Д. Балтимор
открыли ревертазу, или обратную транскриптазу, в 1972 г. П. Берг
с сотрудниками выполнили первый генно-инженерный экспери-
мент — объединили ДНК R-плазмиды (плазмида множественной
устойчивости к лекарственным веществам) с ДНК мушки дрозо-
филы и размножили рекомбинант в кишечной палочке. В 1975—
1978 гг. ученые овладели методами выделения из хромосомной и
плазмидной ДНК любых генов и исследования их структуры (У.
Гилберт, Р. Дейвес, Ф. Курильский, П. Ледер, А. Максам, Т.
Манниатис, Б. Мах, Ф. Ружон, Ф. Сэнгер, С. Тонегава).
Каждый год удваивается число прочитанных генов у разных
организмов, которое собирается в банках данных. Эти банки
накопили уже порядка 2,5 • 10® пн суммарного текста, пользование
которым стало возможным лишь с привлечением ЭВМ.
В 1976 г. была основана фирма "Генентек" в г. Сан-Франциско
(США),' а в 1977 г. в этой фирме был осуществлен синтез челове-
ческого соматотропного гормона — соматостатина клетками Е. coli
в ферментаторе объемом 8 л.; в 1978 г. там же был получен гормон
инсулин при выращивании Е. coli в глубинных условиях. В начале
80-х годов так же с помощью Е. coli были получены эндорфины
(эндогенные пептиды мозга с подобным морфину действием). В
1980 г. в компании "Биоген" (США) впервые получили с помощью
биосинтеза (продуцент — кишечная палочка) интерферон.
Таким образом, расшифровка строения ДНК стала эпохальным
156
событием не только в генетике, но и в биологии в целом. Общая
генетика и общая биология обогатились фундаментальными дан-
ными по строению и составу ядерного аппарата, по углублению
подходов к пониманию эволюции жизни на Земле, по изменчиво-
сти и наследственности организмов и по другим направлениям.
Частные генетика и биология (например, микроорганизмов, рас-
тений, животных) получили почти неограниченные возможности
манипулировать с генетическим материалом, определять механиз-
мы и диапазон его контролирующих функций в процессах мета-
болизма.
Биологическая технология как пограничная научная дисципли-
на с 1972 г. перешла на новый генотехнический уровень (см. главу
1), когда методы генной инженерии были перенесены из лабора-
торных условий в производственные —получила развитие реком-
бинантная ДНК-биотехнология, или сокращенно рДНК-биотехно-
логия. По существу рДНК-биотехнология базируется на природных
возможностях наследственных структур различных представите-
лей живого мира, даже когда приходится вводить чуждую реципи-
енту, но воспринимаемую им генетическую информацию. Такая
рецепция генетического материала и определяется природными
возможностями реципиента.
5.1. Природа и передача генетической информации. Исходя их
установленных данных о том, что ДНК, в силу комплементарности
нуклеиновых оснований, представляет собой двойную спираль, что
в бактериальных клетках эта спираль кольцевидно замкнута и
составляет единственную хромосому, что в клетках любых орга-
низмов имеются механизмы синтеза и гидролиза ДНК с помощью
специфических ферментов, и, наконец, что процессы размножения
обусловлены и непосредственно связаны с функцией ДНК микро-
организмов или зародышевых клеток макроорганизмов, целесооб-
разно расширить представление о ядре и ядерном аппарате клеток,
считая первое составной частью второго. В таком случае к ядерному
аппарату должны быть отнесены все те структуры, которые обес-
печивают его запрограммированные функции — у прокариот:
нуклеоид, рапидосомы (у некоторых видов), перегородочные ме-
зосомы, рибосомы; у эукариот: ядро, ядрышко, нуклеолемма, по-
росомы, центриоли, митотический аппарат, рибосомы.
Биохимическим выражением наследственности является мат-
ричный синтез специфического белка согласно уточненному прин-
ципу Дж. Бидла и Э. Тейтама "один ген — одна белковая молекула".
157
Матричный синтез
подразделяют на ре-
пликацию, транс-
крипцию и трансля-
цию. На уровне транс-
крипции (по крайней
мере, у прокариот)
осуществляется в ос-
новном контроль экс-
прессии генов, а у
эукариот — и на уров-
не трансляции.
.При матричном
синтез^матрйцейвы-
ступает одна из нитей
ДНК, "слепок" с кото-
ти^а * рои — комплементар-
ная матричная (ин-
формационная) РНК, или мРНК, обеспечивает синтез белковой
молекулы в рибосомах с участием транспортных РНК (тРНК) и
соответствующих ферментов — полимераз. Участок мРНК, комп-
лементарный участку смысловой цепи ДНК, называется транс-
криптом.
Показанная на рис. 51 схема синтеза белковой молекулы в
действительности значительно сложнее. Вспомним, что среди по-
следовательно расположенных генов в хромосоме имеются гены
— регуляторы, операторы, структурные, терминаторы, и что каж-
дая хромосома представлена одной молекулой ДНК. Поскольку
хромосомы в клетках находятся в незамкнутом состоянии, постоль-
ку у них имеются 3' и 5' свободные концы, которые отсутствуют
в единственной кольцевидно замкнутой хромосоме прокариотиче-
ской клетки или организованной вирусной частицы (таблица 17).
Различают следующие формы ДНК по архитектонике двойной
спирали: В, А, С, Z и SBS. В форме В на шаг спирали (3,4 нм)
приходится 10 пн (пар нуклеотидов), располагающихся перпенди-
кулярно к оси спирали; форма А является трансформантом формы
В при снижении влажности ее ниже 75%; шаг спирали уменьшается
до 2,8 нм, на виток приходится уже 11 пн, располагающихся под
утлом 20° к оси спирали, а длина цепи укорачивается примерно на
25%. Форма С имеет шаг спирали, равный 3,3 нм и на один виток
158
SgT а б л и ц а 17. Характеристика молекул днк в некоторых хромосомах
¥ Организм/ вирион Хромосомы Среднее
•д форма длина, см число пар оснований в
1 ЧИСЛО (гаплоид) ДНК на хромосому, х103кЪ
1 Человек Saccharomyces 23 незамкнутая 4.1 125 000
1 cerevisiae Esctiexichia 17 — " — 0,33 1 000
coli Бактериофаг 1 замкнутая 0,14 4000
X174 1 и 0,00018 5,4
Примечание: кЬ-килооснования (от англ, kilobases).
ее приходится 9 пн. Форма Z (зигзаг) присуща углеводно-фосфат-
ному остову ДНК в участке с чередующимися последовательностя-
ми гуанина (Г) и цитозина (Ц). Это левая спираль (все предыдущие
формируются в виде правых спиралей) с 12 пн в одном витке,
форма SBS (от англ, side by side — бок о бок) лишена взаимозак-
рученности цепей в двойную спираль, что важно для биосинтеза
ДНК.
Названные выше гены в хромосомной ДНК обладают специ-
фическими функциями (средний размер гена оценивают в 1500
пн). Ген-регулятор определяет синтез белка-репрессора, способно-
го связываться с оператором (см.) на ДНК или с РНК, предотвращая
соответственно транскрипцию или трансляцию. Ген-оператор —
участок ДНК, связываясь с которым белок-репрессор предотвра-
щает инициацию (начало) транскрипции на прилежащем промо-
торе, ответственном за связывание фермента РНК-полимеразы,
инициирующей транскрипцию гена. На промоторе гена эукарио-
тической клетки имеется специфический локус (участок), в десят-
ки—сотни тысяч раз повышающий число посадок РНК-полимера-
зы на промотор ближайшего гена. Этот локус называется энхан-
сером, или усилителем (от англ, enhancer — усилитель). Энхансеры
тканеспецифичны. Они представляют собой большую разнообраз-
ную группу регуляторных элементов клетки. Другими словами это
элементы позитивного контроля. К элементам негативного конт-
роля относятся сайленсеры (от англ, silencer — глушитель), угне-
тающие транскрипцию. Энхансеры и сайленсеры обладают только
цис-действием, влияя на гены, локализующиеся на той же молекуле
159
ТАА ЦЦГЦ ГГЦГЦГ ТПТ
^ТТГАЦЛ ТАТААТ ЦАТ АТТА АТТ
В5’Ф;атл^у
г Nil; Аттгтптгпттгтгтатзтпг^ СООН
Л NHj -*ТЛЛПП ft о о а? ЛГВТЯЛПГЗгяпГ'! СООН
Рис. 52. Схематичное строение (а) и экспрессия (в-д)
гена прокариотической клетки: Р-промотор, СР-сайт
связывания, ИК-кодон инициации и ТК-кодон терми-
нации транскрипции, t —терминатор транскрипции;
б — некодирующая нить ДНК с характерными сай-
тами; координаты нуклеотидов промотора имеют от-
рицательный знак (-10 для последовательности При-
бноу, -35 для последовательности Хогнесса); транс-
крибируемые нуклеотиды — положительный знак;
в-матричная РНК, г-белок-предшественник, д-актив-
ный белок; п-палиндром, SD — последовательность
Шайна-Далгарно.
СР'ИК.
ДНК, где располагают-
ся названные элемен-
ты.
Структурные гены
ответственны за син-
тез белковых молекул
(рис. 52). Ген-термина-
тор блокирует транс-
крипцию и прерывает
синтез белка. К этому
гену примыкает тер-
минатор (стоп-сигнал),
состоящий из опреде-
ленной последователь-
ности нуклеотидов в
ДНК. У Е coli он эф-
фективен только в
присутствии р-факто-
ра белковой природы
с ММ 50 кДа, не свя-
занного с РНК-пол-
имеразой. В терминации у Е. coli участвует и другой белок,
названный к-частицей.
Оперон — это группа сцепления структурных генов, находя-
щихся под контролем гена-оператора и гена-регулятора, а также
контролирующих элементов, узнаваемых продуктами регулятор-
ного гена. На рис. 52 изображена схематичная структура гена
прокариотической клетки. В его состав входят участки, располо-
женные перед кодирующей последовательностью (лидерная 5'-об-
ласть) и после нее (концевая З'-область); синтез мРНК происходит
в направлении от 5'-конца к З'-концу. Отсчет нуклеотидов в генах
ведут от первого транскрибируемого нуклеотида, обозначаемого
как +1 (или 1); отсчет в противоположную сторону начинают с
—1. Нд и РЬ на рисунке обозначают консервативные последова-
тельности Хогнесса и Прибноу соответственно. Первая из них
представляет собою гексамер 5'ТТГАЦАЗ', локализующийся около
координаты —35. Последовательность Хогнесса необходима для
узнавания промотора РНК-полимеразой. РЬ-последовательность
является также гексамером 5'ТАТААТЗ', локализующимся около
координаты —10. Эта последовательность необходима для тесного
связывания с РНК-полимеразой. За ней закрепилось название
160
"TATA-box", или "ТАТА-блок". Ядром последовательности SD (Шай-
на—Далгарно) служит тетрамер 5'АГГАЗ’ на мРНК, являющийся
местом (сайтом) связывания рибосом. Всего в SD= последователь-
ности имеется 5—9 нуклеотидов.
В ДНК некоторых прокариот (археобактерий), в ядре и мито-
хондриях эукариот кодирующие области прерываются большими
некодирующими ДНК-последовательностями (до 5000 пар нуклео-
тидов). По предложению У. Гилберта (1978) кодирующие области
называют экзонами, или доменами, некодирующие — интронами.
Например, в генах тяжелой Н-цепи иммуноглобулинов (1g) нахо-
дится не менее 4 интронов и 5 экзонов, в гене яичного белка
(овальбумина)? интронов и 8 экзонов; из 3,5 биллионов пар нук-
леотидов в ДНК гаплоидного генома человека кодирующими явля-
ются менее 10%. Прерывистость генов у эукариот — явление
обычное, хотя известны гены, в которых подобная прерывистость
не обнаружена (в генах интерферона, гистонов).
В интронах отсутствует более или менее заметная гомология
между двумя концами их, а на границах с экзонами в разных генах
присутствует так называемая каноническая (усредненная) после-
довательность нуклеотидов (относительно короткая) — нередко
GT—слева и AG—справа:
КП КП
I I I------1
A64G73G1OOT100A62A68T63 ... 6Ру?4—87 NC65A100G10 0 N
I 11____________________ _________________________। ।
экзон
интрон
экзон
КП обозначает каноническую последовательность, цифры —
проценты случаев обнаружения
оснований на границе экзон-ин-
трон.
Экзоны во много раз меньше
(1000 пн) интронов. Интрон пред-
ставляет собой транскрибируе-
мый (но не кодирующий) участок
ДНК, который удаляется из со-
става транскрипта при сплайсин-
ге (от англ, splicing — сплетение,
сшивание). Сплайсинг протекает
в ядре, и он заканчивается объе-
динением экзонов в зрелую
Рис. 53. Сплайсинг (процессинг) РНК: 1
— транскрипция, 2 -— трансляция, а, б,
в — экзоны, г, д — интроны; I — ДНК,
II — РНК, III — мРНК, IV — Белок.
мРНК (рис. 53). Участок ДНК между правым концом интрона и
левым концом экзона называется акцепторной точкой сплайсинга.
В ДНК митохондрий интроны кодируют синтез отдельных белков,
которые сами могут участвовать в последующем вырезании инт-
ронов. Некоторые молекулы белков (ферментов) кодируются од-
новременно экзонами и интронами, например, матураза (от англ,
mature — созревать). Возможно, что для каждого интрона сущест-
вует своя матураза. Интроны митохондриальной ДНК похожи на
бактериальные транспозоны и, возможно, матуразы происходят из
транспозаз (см.).
Утверждается, что существуют белки, кодируемые ядерными
экзонами и интронами (м-белки), которые принимают участие в
Рис. 54. Выведение зрелой мРНК через пору ядерной
мембраны по П. Слонимскому (1985).
образовании и перено-
се мРНК через ядер-
ную мембрану (П. П.
Слонимский, 1980).
При этом часть белка,
кодируемая интроном,
включает преимуще-
ственно гидрофобные
аминокислоты, вслед-
ствие чего охотно ло-
кализуется в богатой
липидами ядерной
мембране и притяги-
вает мРНК. Часть м-
белка, кодируемая эк-
зоном, притягивает но-
вую полипептидную цепь, транслирующуюся с того же экзона. На
уровне ядерной мембраны находится ферментный комплекс сплай-
синга интронов. По мере прохождения сплайсинга зрелая мРНК
(без интронов)выводится через пору ядерной мембраны (рис. 54).
В процессе дифференцировки эукариотических клеток обна-
ружена неоднотипность сплайсинга, а именно то, что в одном
случае предстает интроном, в другом может оказаться экзоном, и
наоборот. В частности, доказано, что интрон цитохрома Ъ - это
экзон, кодирующий матуразу. Следовательно, существуют гены,
способные кодировать не один белок. Например, один из генов
тканевой совместимости кодирует 2 белка; один из прерывистых
генов крысы кодирует синтез парат-гормона в паращитовидной
железе и нейропептида — в гипофизе. Поэтому нельзя абсолюти-
зировать понятие "один ген — одна белковая молекула". В биохи-
мической технологии интроны привносят определенные затрудне-
ния при экспрессии (проявление, выражение свойств) эукариоти-
ческих генов в клетках многих прокариот, лишенных аппарата
сплайсинга мРНК.
162
Некоторые из нуклеотидных фрагментов входят не только в
состав интронов, или фланкирующих (от англ, flank — бок,
сторона, прикрывать с фланга) последовательностей мРНК, но и
могут выполнять функции сигнальных последовательностей,
узнаваемых белками (промоторы транскрипции, точки начала ре-
пликации ДНК, сайты скручивания хромосом и др.). Все эти
сигнальные последовательности, повторяющиеся в геноме в виде
идентичных или сходных тандемных (от англ, tandem — гуськом,
цугом) копий, имеют небольшую длину. При разделении ДНК в
градиенте плотности хлористого цезия сигнальные последователь-
ности (порядка 5%) удается отделять от основной массы ДНК в
виде сателлитной ДНК (дополнительный(е) пик(и) при центрифу-
гировании). Эти ДНК могут быть тяжелее или легче основной
фракции ДНК и могут быть метилированными. Множество тан-
демных повторов в сателлитной ДНК придает ей определенную
аномальность при центрифугировании (сходство поведения с ос-
новной ДНК в градиенте плотности хлористого цезия). В таких
случаях говорят о криптической (от лат. cryptus — скрытый)
сателлитной ДНК.
Установлено, что высокоповторяющиеся последовательности в
сателлитных ДНК обычно не транскрибируются. Следует отметить,
что сателлитные ДНК локализуются в области центромеры хромо-
сомы (выполняет структурную функцию). Предполагается, что
сателлитные ДНК произошли от мозаики последовательностей,
состоящих из 9 пн в трех повторах:
В начале 80-х годов в геноме человека были
А TGA обнаружены последовательности ДНК, обладаю-
GAAA А щие свойствами структурного полиморфизма —
Т ACT это так назЫваемые гипервариабельные области
(ГВО), обычно содержащие короткие, ГЦ-обога-
щенные и тандемно повторенные единицы. ГВО рекомендуются в
качестве маркеров-зондов при картировании генов. Варианты
кор-последовательности ГВО гена человеческого миоглобина были
названы минисателлитными.
В 1985 г. был предложен метод "генетической дактилоскопии
— ДНК" (от греч. dactilos — палец, scopein — смотреть) в целях
оценки эволюции человека по отцовской и материнской линиям
(А. Дж. Джеффрис, У. Уилсон, С. Л. Тейн). В последовательностях
ДНК отражаются происшедшие в прошлом мутации или, по Ф.
Крику, "замороженные события”. Это, в частности, поможет кар-
тировать варианты последовательностей в локусе минисателлит-
ной ДНК. Эволюцию по женской линии удобно картировать по
163
митохондриальной ДНК, так как в сперматозоидах практически
нет митохондрий, но ими "начинены" яйцеклетки. Вот почему ДНК
клеточного ядра является сочетанием материнской и отцовской
ДНК, тогда как ДНК митохондрий передается только яйцеклеткой.
Отсюда становится понятным, почему в конце 70-х годов текущего
столетия возникла новая научная дисциплина молекулярная ант-
ропология.
В ДНК различных организмов содержатся еще так называемые
палиндромы (от греч. palindrome — перевертыш) — последова-
тельности, повторяющиеся в обратном порядке:
________А__________
5* - ЦТТЦГАТГГААГ - 3’
- ' — - . - т д
3’ - ГААГЦТАЦЦТТЦ - 5’
Б
5’
дтц
ГААГ
I
3’
В суперспирализованном состоянии длинные палиндромы (10
и более пар оснований) образуют крестообразные структуры,
служащие сигналами для узнавания определенных участков ДНК
ферментами-метилазами, рестриктазами и регуляторными белка-
ми, регулирующими действия генов.
В хромосомных ДНК прокариотических и эукариотических
клеток имеются также контролирующие или так называемые
"прыгающие" подвижные гены — транспозоны (Тп), впервые
открытые Б. Мак-Клинток в 1940 г. у кукурузы. Они находятся на
значительном расстоянии от других генов, на которые оказывают
влияние. Благодаря мутациям, названным "транспозонными взры-
вами", возможно массовое и в известной мере направленное
перемещение генетических элементов. Транспозоны способны
реплицироваться и внедряться (инсерция) в виде одной из копий
в новое место генома (ДНК ядра). У бактерий преобладающая часть
транспозонов кодирует фермент транспозазу, катализирующую
реакцию встраивания транспозона в ДНК. В последнее время их
отождествляют с интронами, рассмотренными выше.
При сравнении последовательности нуклеотидов в хозяйской
ДНК до и после встраивания транспозона оказывается, что не-
сколько нуклеотидов этой ДНК после встраивания удваиваются —
дуплицируются. Эти дупликации ДНК окаймляют транспозон.
164
Причем, для каждого транспозона характерно определенное коли-
чество дуплицированных нуклеотидов. Принято считать, что при
встраивании транспозонов хозяйская ДНК перед этим фермента-
тивно расщепляется с образованием липких концов, к выступал!
которых присоединяется транспозон, а оставшиеся бреши затем
заполняются нуклеотидными последовательностями, в результате
чего и образуются небольшие дупликации.
Следует подчеркнуть, что транспозонов много и они различны
(лишь перечень Tn-ов у дрозофилы составит целую книгу). Следо-
вательно необходима точная классификция их, и, возможно, в
недалеком будущем удастся ее создать.
К настоящему времени принято считать, что механизм транс-
позиции заключается в удвоении подвижных элементов и после-
дующем встраивании одной из копий транспозона в новое место
генома, а другая копия остается в прежнем месте. Вот почему
термин "транспозиция" неточен, поскольку транспозон не покида-
ет своего первоначального места, или сайта. Более правильно
рассматривать транспозицию процессом, в результате которого
возрастает число копий транспозона. Во благо сохранения струк-
туры генома (консерватизма его) транспозиции происходят очень
редко. Так, в среднем, частота их сравнима с частотой спонтанных
мутаций, то есть 10'5—10‘7 на поколение, а частота реверсии путем
делеций, или выпадений, отмечается еще реже (10’6—1О'10).
Обращает на себя внимание тот факт, что Tn copia из мушки
дрозофилы ведет себя и как транспозон, и как ретровирус (к их
числу, кстати сказать, относится вирус иммунодефицита человека
— причина СПИД). В та-
ких вирусах интеграция
ДНК осуществляется
способом, сходным с
транспозицией. Поэтому
небеспочвенной стала
гипотеза о том, что виру-
Рис. 55. Встраивание транспозона в ДНК-мишень
(схема): I — транспозон, 2 — центральная часть,
3 — концевые повторы, 4 — дупликация ДНК-ми-
сы — это транспозоны,
приобретшие дополни-
тельные функции или,
напротив, транспозоны шеви.
— это выродившиеся вирусы. Проблема остается открытой. Тем
не менее, обнаружены самые короткие транспозоны, кодирующие
165
Рис. 56. Ori-сайты в плазмиде pSClOl E.coli
(цифры по внешнему кругу 1—6 обозначают
места действия различных ферментов-ре-
стриктаз).
белки, которые участвуют
только в транспозиции. Сле-
довательно ДНК таких
транспозонов можно отне-
сти к разряду "эгоистиче-
ской”, работающей лишь на
себя, то есть она функцио-
нирует во имя собственного
размножения.
Схематичное встраива-
ние транспозона (после ре-
пликации) в ДНК-мишень
изображено на рис. 55.
Репликация, или самоудвоение присуще ДНК и РНК, то есть в
таких случаях происходит перенос генетической информации
соответственно от ДНК к ДНК или, например у ряда вирусов, от
РНК к РНК. Репликация осуществляется полуконсервативным
способом, когда двухспиральная ДНК деспирализуется и каждая
нить индуцирует синтез комплементарной себе нити при участии
ДНК- или РНК-полимеразы.
Геномы бактерий и фагов реплицируются как единое целое, то
есть как организованные единицы репликации, или репликоны.
Каждый репликон содержит место (точку) инициации Ori (от англ,
origin — начало) — ориентированное направление репликации,
например OriC у Escherichia coli (рис. 56). В некоторых репликонах
содержится около 240—600 пн. В отдельных репликонах сущест-
вует два Ori, например, в так называемых челночных векторах,
способных реплицироваться в клетках прокариот и эукариот.
Инициация репликации осуществляется с помощью специфиче-
ских белков.
Репликация ДНК необходима функционирующей клетке для
восстановления (репарации), обмена генами между участками
хромосом (рекомбинация) и перемещения генов (транспозиции).
В течение однократного деления прокариотической или эука-
риотической клетки, независимо от числа хромосом в ней, весь
геном ее реплицируется также один раз, и только после завершения
репликации может произойти последующее деление. Удвоенный
геном подразделяется (сегрегирует) поровну в каждую дочернюю
клетку. Единицей сегрегации является хромосома, а единицей
репликации — репликон. Кроме точки Ori в репликоне имеется
166
12 3
--1-1-
Рис. 57. Две репликационцые вилки в ДНК: I и
3 — нереплицированная ДНК, 2 — репликаци-
онный глазок, 4 — стационарная точка начала,
5 — движущаяся репликационная вилка, 6 —
две репликационные вилки.
также точка (место) оста-
новки репликации ter (от
лат. terminalis — конечный,
пограничный). Единицы
сегрегации и репликации в
бактериальной хромосоме
совпадают, поскольку бак-
териальная хромосома со-
держит только один репли-
кон. В то же время каждая
плазмида, если она имеется
в бактериальной клетке,
представляет собой авто-
номную кольцевидную ге-
нетическую структуру и
является самостоятельным
репликоном.
Плазмиды, представленные в бактериальной клетке лишь в
одной копии, называются однокопийными, другие плазмиды, пред-
ставленные более чем одной копией, называются многокопийны-
ми.
В эукариотической клетке содержится большое число репли-
конов и единица сегрегации у них включает много единиц репли-
кации. Все компоненты аппарата ре-
пликации называют реплисомой. Ре-
пликация ДНК начинается в стартовой
точке, или репликационной вилке, в
одном (однонаправленная реплика-
ция) или в двух противоположных
(двунаправленная репликация) на-
правлениях. В последнем случае будет
две репликационных вилки (рис. 57).
Реплицированная часть ДНК приобре-
тает форму "глазка", представляюще-
гося 0-структурой.
В случае кольцевой матричной
структуры ДНК точка роста одной раз-
резанной цепи спирали "скользит"
вокруг второй кольцевой матричной
цепи. Возникает структура катящегося
кольца (рис. 58).
Рост клеток бактерий и репликация
ДНК в них тесно связаны между собой.
Рис. 58. "Катящееся” кольцо мат-
ричной цепи.
167
25
Точка начала 35/0 р Концевая точка
Рис. 59. Цикл деления Е. coli -—
образование хромосомы со
многими вилками в процессе
репликации (а — инициация, б
— деление, в — терминация);
цифры обозначают минуты кле-
точного цикла.
Цикл клеточного деления на примере Е. coli можно представить
двумя временными интервалами, обозначаемыми латинскими бук-
вами С и D. Первая из них обозначает фиксированное время,
необходимое для репликации всей бактериальной хромосомы (на-
пример, 15 мин.), что соответствует скорости движения отдельной
репликационной вилки с присоединени-
ем около 50000 пар оснований в 1 мин.;
D соответствует промежутку времени
(порядка 20 мин.) между завершением
репликации ДНК и делением клетки —
это как бы накопительный период. Сле-
довательно, весь так называемый корот-
кий клеточный цикл деления Е coli ук-
ладывается в среднем в 35 минут (рис.
59).
Длинный цикл клеточного деления
наблюдается тогда, когда велико время
до начала инициации. Для диплоидных
клеток эукариот характерно митотиче-
ское деление ядра с последующим обра-
зованием новых (дочерних) клеток. Этот
митотический цикл подразделют на 4 фазы: Gi, S, G? и М. Фаза
Gi обозначает время роста клетки до синтеза ДНК (от англ, дар —
пробел); в фазу S происходит синтез ДНК (от англ, synthesis); G2
— вторая фаза роста в период , после синтеза ДНК; М — фаза
митоза (от англ, mytosis). На рис 60 представлена схема цикла
эукариотических клеток с обозначением продолжительности фаз
в культивируемых линиях дрожжевых клеток Schizosaccharomyces
pombe и клеток млекопитающих. На весь цикл деления клеток
эукариот усредненно приходится следующая доля времени: на
1 г
Рис. 60. Схема цикла клеточного деления эукариотических клеток: 1
Schizosaccharomyces pombe, 2 — клетки млекопитающих.
фазу Gi — 30—40%, на фазу S — 30—50%, на фазу G2 — 10—20%,
на фазу М — 5—10%.
168
За время движения репликационных вилок в клетках эукариот
присоединяется от 1000 до 3000 пн в 1 минуту — у млекопитающих
и до 1000 пн/мин — у растений (здесь не исключается роль
пониженных температур для размножения клеток растений).
Синтез комплементарных цепей во время репликации двухни-
тевой ДНК у прокариот и эукариот катализируется ферментами,
называемыми ДНК-полимеразами. Их известно 3 (pol I, pol II и ро]
III у прокариот; а, b и у - у эукариот). Характеристика ДНК-по-
лимераз приведена в таблице 18.
Таблица 18. Характеристика прокариотических и эукариотических ДНК-
полимераз
ДНК-полиме- раза Размер, кДа Состав Ферментативная активность 1
из клеток Е. coli: 109 одна цепь а) элонгация в направлении 5' —> 3' от 3' - ОН - затравки б) 3' —> 5' - экзонуклеаза в) 5’ —» 3’ - экзонуклеаза
I
II 120 — см. для I б) 3' —> 5' - экзонуклеаза
III 250 гетеро муль- тимер а - в) см. для I 1
из клеток млекопита- ющих: 110-120 несколько субъединиц репликативный синтез | ядерной ДНК (ядерная ДНК- 1 репликаза) - 80%
а
р 45 I субъединица репарация сегментов | поврежденной ДНК
Y 60 2 репликативный синтез митохондриальной ДНК - ' 2-15% =
На раскрученной ДНК происходит непрерывный синтез веду-
щей цепи в направлении 5' —> 3', тогда как на комплементарной
цепи имеет место прерывистая репликация, то есть в направлении
169
5' —> 3' на ней синтезируется серия фрагментов ДНК (фрагменты
Оказаки), которые затем соединяются, формируя отстающую цепь
(рис. 61). Фрагменты Оказаки содержат примерно 1000—2000
Рис. 61. Фрагменты Оказаки, присоединенные к РНК — инициатору: а — матричная
ДНК, б — комплементарная ДНК, в — РНК—праймер, НТ—входящий нуклеозид-
трифосфат.
нуклеиновых оснований, и синтез фрагментов под каталитическим
действием ферментов РНК-полимераз, включая праймазу (ММ-60
кДа), инициируется РНК—затравками, каждая из которых по длине
соответствует приблизительно 10 основаниям. РНК—затравка уд-
линяется под действием ДНК-полимеразы III, катализирующей
синтез цепи ДНК. РНК удаляется под действием ферментов экзо-
нуклеаз.
В так называемую затравочную реакцию вовлекается комплекс
белков — праймосома. Например, праймосомный белок SSB свя-
зывается с одноцепочечной ДНК, стабилизируя ее; белок Dna В
образует предпраймерную РНК и участвует в передвижении прай-
мосомы; белок Dna С действует в комплексе с Dna В; белок Lig
сшивает разрывы между фрагментами Оказаки; белок п'-АТФ-аза,
белки пип" образуют предзатравочную РНК, и др.
Предполагают, что праймосома собирается в каком-то одном
участке с последующим перемещением по одноцепочечной ДНК
к сайтам,где инициируется синтез затравки. Направление движе-
ния праймосомы противоположно направлению синтеза ДНК от-
стающей цепи, но синхронно движению репликационной вилки.
По расчетным данным репликация ДНК у прокариот (вместе с
170
раскручиванием молекулы по 10 нуклеотидных пар одномоментно)
должна происходить со скоростью 400 000 оборотов в секунду, что
явно превышает реальную скорость репликации. Поэтому было
предположено, что должны существовать фиксаторы, включающи-
еся в молекулы ДНК у всех организмов. Такие фиксаторы были
обнаружены. К ним относятся ферменты — топоизомеразы. От-
дельные из них катализируют реакции объединения молекул ДНК
в зацепленные кольца — катенаны, топоизомераза II, или гираза,
катализирует суперспирализацию ДНК, топоизомераза I способна
разрезать одну из цепей суперспирализованной ДНК, при этом
цепи раскручиваются и число супервитков уменьшается, после
чего этот же фермент устраняет разрыв в нити ДНК.
Двигателем в распределении реплицированных молекул ДНК
по дочерним клеткам (по крайней мере - у прокариот) выступает
клеточная мембрана, к которой прикрепляется ДНК.
Таким образом, весь катализируемый ферментами процесс
репликации ДНК можно подразделить на 3 этапа: инициацию,
элонгацию (рост цепи) и терминацию. В процессе инициации
происходит разделение нитей ДНК с образованием репликацион-
ной вилки, формирование праймосомы и синтез затравочной РНК.
Этап роста цепи, или элонгации, реализуется в синтезе ДНК с
помощью ДНК-полимераз. Терминация, или окончание синтеза
ДНК, происходит благодаря выключению реакции с помощью
специфического "стоп-сигнала" от специального кодона (термина-
тора) в матричной цепи.
Такие же 3 этапа выделяют в процессах транскрипции и
трансляции ДНК.
Транскрипция — это процесс переписывания закодированной
в ДНК информации и перенос ее к месту синтеза белка (на
рибосомы). Этап инициации при транскрипции заключается во
взаимодействии РНК-полимеразы с ДНК-матрицей; элонгация - в
ферментативном синтезе мРНК на матрице ДНК; терминация —
в остановке синтеза мРНК благодаря "стоп-сигналу" от гена —-
терминатора.
Трансляция заключается в переводе закодированной в мРНК
информации в полипептидную цепь. Организующими центрами
процесса трансляции являются рибосомы. При трансляции на
этапе инициации происходит активация аминокислот с помощью
ферментов аминоацил-тРНК-синтетаз (АРСаз) при использовании
энергии АТФ с последующим образованием комплекса
инициации, включающего 3 фактора инициации (IF-1, 1F-2,
1F-3 — у прокариот, eIF-2, eIF-3, eIF-5 и др. — у эукариот), мРНК,
171
гуанозилтрифосфат (ГТФ) и 30S(40S) — субчастицу рибосомы.
Указанный комплекс соединяется с 50S(60S) — субчастицей рибо-
сомы и формирует функциональную 70S (80S) рибосому. На этапе
элонгации в ходе трансляции осуществляется синтез полипептид-
ной цепи функционирующей рибосомой с участием факторов
элонгации (EF-Tu, EF-Ts, EF-G — у прокариот; EF-1 и EF-2 — у.
эукариот). Указанные факторы не входят в структуру рибосом, а
присоединяются к ним на определенных этапах.
Во время синтеза белка рибосомы движутся вдоль мРНК,
последовательно считывая триплеты и поэтапно наращивая по-
липептидную цепь. У мРНК, как правило, имеется постоянная
рамка считывания — кодон AUG. При элонгации одна мРНК
связывается с несколькими рибосомами, образуя функционирую-
щий комплекс - полирибосомы, или полисомы. Из указанных трех
этапов с наибольшей скоростью протекает элонгация. Терминация
процесса трансляции осуществляется стоп-кодоном в мРНК.
Как следует из представленных выше данных РНК занимает
место посредника между ДНК и белком. Причем эта центральная
функция присуща мРНК, тогда как тРНК и рРНК являются транс-
криптами, обладающими активностью как завершенные в постро-
ении и функции молекулы. Исключение представляют некоторые
РНК — содержащие вирусные геномы, по матрице которых син-
тезируется РНК, причем реализация генетической информации
может осуществляться по схемам: для одних вирусов, у которых
отсутствует транскрипция, РНК -> Белок (вирус полиомиелита
и др.), для других, располагающих собственной вирионной РНК —
зависимой РНК-полимеразой, или вирионной транскриптазой,
РНК -» РНК -» Белок (вирусы гриппа, кори и др.), и, наконец, для
третьих — РНК -> ДНК —> РНК —> Белок (ретровирусы, в том числе
— ВИЧ, или вирус СПИД).
мРНК, рРНК и тРНК у бактерий синтезируются под каталити-
ческим действием одной и той же РНК-полимеразы (ММ-480 кДа).
В клетках эукариот обнаружены три ядерные РНК-полимеразы (I,
II и III), а также РНК-полимеразы митохондрий и хлоропластов.
Установлено, что РНК-полимераза I, находящаяся в ядрышке,
отвечает за синтез про-рРНК, из которой впоследствии образуются
28S и 18S рРНК; РНК-полимераза II катализирует реакцию синтеза
про-мРНК и только с этим ферментом связано так называемое
кэпирование РНК. А. Шаткин в 1976 г. впервые описал характер-
ную группировку, присоединяющуюся к 5'-концу фактически всех
про-мРНК и мРНК посттранскрипционно. Автор назвал ее КЭПом
172
(от англ, cap — шапка, кепка). КЭП повышает устойчивость
про-мРНК и мРНК к действию ряда рибонуклеаз и других гидро-
лизующих агентов. Пример КЭПа: 7meG5' ррр5' Nf^p ,..(7'-метил-
гуанозил-5'-трифосфорил-5'-нуклеозил-метилрибозил-фосфорил
-нуклеотид...); локализация КЭПа и предшествующие ему нуклео-
тидные последовательности в ре-
комбинантной плазмиде рТК 1
показаны на рис. 62. З'-Конец
мРНК, как правило, полиадени-
лирован.
РНК-полимераза III катали-
зирует синтез 5S РНК, всех
тРНК, ряда малых РНК, а также
часть гяРНК.
Скорость синтеза белков у
различных представителей над-
царств прокариот и эукариот
варьирует в широких пределах.
Это зависит от многих внутрен-
них и внешних факторов. Тем не
менее, у бактерий при 37°С за 1
секунду может включаться в рас-
тущую полипептидную цепь от
10 до 20 аминокислот. Подсчита-
но, например, что белковая мо-
лекула, включающая 300 амино-
Рис. 62. Локализация КЭПа в рекомби-
нантной плазмиде рТК1 (показаны нук-
леотидные последовательности, участву-
ющие в регуляции экспрессии гена ти-
мидинкиназы — tk; нумерация нуклео-
тидов проставлена относительно КЭП-
участка; буквами внутри круга показаны
места действия соответствующих фер-
ментов-рестриктаз; Amp — устойчивость
к ампициллину.
кислот, синтезируется бактериальной клеткой за 20 секунд.
Скорость синтеза белков в эукариотических клетках заметно
ниже (в среднем, за 1 секунду присоединяется 1—2 аминокислоты
к растущей цепи полипептида). Здесь следует иметь в виду и тот
факт, что для синтеза белка с прерывистого гена, включающего
экзоны и интроны, требуется дополнительное время. Ген вначале
транскрибируется целиком (со всеми экзонами и интронами) в
пре-мРНК, которая затем в ходе сплайсинга освобождается от
интронов и превращается в мРНК (см. рис. 53), в которой экзоны
соединены последовательно конец в конец. Только после этого
образованная мРНК включается в процесс синтеза белка. Следует
иметь в виду, что, например, у человека 80—90% ДНК оказывается
некодирующей.
Вырезание интронов из пре-мРНК происходит с участием
ферментов, распознающих границы интронов. Если это распозна-
ние окажется неточным, то соединившиеся экзоны будут кодиро-
вать другой белок — произойдет сдвиг рамки считы -
173
вания. Свершившуюся поломку в механизме биосинтеза
полипептида можно сравнить с типографским браком. Например,
в словах "копна сухого сена" сдвинули рамку считывания и пол-
учили бессмыслицу — "коп насухо госена" (интервалы между
словами засчитываются за одну букву). Механизмы сплайсинга
могут быть различными в зависимости от типа генов (ядерных и
внеядерных) и классов интронов (рис. 63). С распознаванием
механизмов сплайсинга связано выдающееся открытие конца 1982
г., когда Т. Чех с сотрудниками выявил авторестрикцию (самовы-
резание) интрона в гене рРНК у одного из видов Protozoa. При
этом не требовалось каких-либо ферментов и приложения энергии
Дрожжи
Не La
Tetrahymena
Рис. 63. Транскрибирование гена (сравнение механизмов сплайсинга тРНК в
дрожжах, в экстрактах клеток HeLa и самосплайсинга рРНК Tetrahymena). Символы
вокруг фосфатных групп отражают судьбу каждой из них в ходе реакции.
174
извне. Этим доказываются автокаталитические свойства нуклеи-
новых кислот, и предложенный Т. Чехом термин "рибозим" не
представляется алогичным.
Компартментализация эукариотической клетки служит веским
доказательством специального (более высокого, чем у прокариот)
разграничения функций и привязки их к определенным структу-
рам. Это относится и к белковому синтезу. Мономерные рибосомы,
в отличие от полисом, находятся в цитоплазме клетки в свободном
состоянии. Полисомы, как правило, располагаются либо цепочкой
ближе к ядерной мембране (связанные полисомы) и в тех местах,
где мРНК входит в цитоплазму, либо они (так называемые "сво-
бодные полисомы") ассоциируются через посредство мРНК с
клеточным цитоскелетом (см.). Это обусловлено качеством синте-
зируемых белков на полисомах.
В 1980 г. Трифонов и Зусман выяснили, что в геноме человека
пары адениновых нуклеотидов встречаются с периодичностью 10,5
на протяжении любой последовательности ДНК, которая взаимо-
действует с единицей гистонового ок-
тамера и образует нуклеосому. Эта
периодичность хорошо коррелирует с
периодичностью завитков в В-спирали
(см.) молекулы ДНК. Указанная пери-
одичность адениновых пар кодирует
закручивание спирали ДНК в одном
направлении вокруг комплекса гисто-
новых октамеров (рис. 64) и этот код
был назван "хроматиновым" или "код
упаковки хроматина". Высказано
предположение, что в структурах бло-
ков тандемно повторяющихся после-
довательностей потенциал скручива-
ния спирали как бы амплифицируется
(от англ, amplification — обилие). Более
того, показано, что и при альтернатив-
ных вариантах закручивания спирали
(например, левостороннем) обнару-
живаются тандемные блоки пурино-
пиримидиновых оснований. Очевидно
блоки тандемных повторов кодируют
локус-специфичную упаковку ДНК и
структуру хроматина в ядре клеток
человека.
Рис. 64. Упаковка хроматина на
примере нуклеосомы: 1 — гистон
Н1, 2 — двухнитевая ДНК, обви-
тая вокруг структуры, содержа-
щей 8 молекул гистонов — по 2
молекулы Н2А, Н2В, НЗ и Н4, 3
— спейсерный участок ДНК.
175
Локус-специфичный "код упаковки хроматина" участвует не
только в структурной организации хромосом в ядре клетки, но
может выступать как "код экспрессии гена", "код репликации”
(сайты репликации на протяжении данной повторяющейся после-
довательности) и "код рекомбинации".
Суммируя основные представления о структуре и функциях
генома акариотических, прокариотических и эукариотических ви-
дов, необходимо подчеркнуть следующие его особенности:
1) Геном акариот и прокариот, а также митохондрий и пластид
эукариот является компактной совокупностью генов с небольшим
содержанием структурных повторов, что характеризует его эко-
номичность. Он кольцевидно замкнут (непрерывен), интервалы
между генами минимальны. Например, вся генетическая инфор-
мация умеренного фага Л. размещается в кольцевой молекуле ДНК
длиной в 50 kb, где содержится порядка 40 генов; плазмидная ДНК
из 95—97 kb включает до 100 генов; замкнутая ДНК Е. coli из 400
kb может содержать до 3000 генов (примерно 1500 пн составляют
один ген);
2) Генетический аппарат в клетках эукариот организован в
форме нескольких линейных хромосом, в которых ДНК прочно
связана с белками-гистонами, обеспечивающими упаковку и упо-
рядочение ДНК в виде структурных единиц—н уклеосом
(учитывая при этом "код упаковки хроматина” и экстраполируя
его на клетки большинства эукариот). Так, в гаплоидной клетке
Saccharomyces cerevisiae содержится 17 хромосом, в каждой из
которых детектировано 1000 kb и, следовательно, число генов могло
бы достигать в такой клетке 11 000; для 23 хромосом в гаплоидной
клетке человека, где в одной хромосоме содержится 125 000 kb,
число генов должно бы возрасти до 2 млн. Предположительно
близкое число генов могло бы оказаться в гаплоидных клетках
кукурузы, где имеется 10 хромосом, в клетках кролика с 22
хромосомами, или мыши с 20 хромосомами. Однако, в хромосомах
эукариотических организмов содержится генов меньше, чем неко-
дирующих участков (спейсеров, или разделителей), и также име-
ется масса сходных между собой фрагментов ДНК, повторяющихся
десятки-сотни тысяч раз. Вот почему, например, у человека лишь
10 - 20% всей ДНК относится к разряду кодирующей. Но и в этом
случае число генов в 23 хромосомах гаплоидной клетки может
достигать порядка 200 000.
176
3) Гены эукариот, нередко располагаясь в хромосомной ДНК
рядом, образуют мультигенные семейства, состоящие
из небольшого числа родственных последовательностей. Напри-
мер, гены, кодирующие рРНК у млекопитающих, обнаруживаются
в геноме сотнями своих копий, сгруппированных в зоны, а это
свидетельствует об избыточности генетических программ у вы-
сших организмов (создание "повышенной прочности").
4) Представители микробного, растительного и животного мира
имеют в составе генетического материала одни и те же строитель-
ные блоки, то есть их "кодовый словарь” в основном однотипен,
или универсален, и функционирует преимущественно в соответ-
ствии с центральными постулатом молекулярной генетики, сфор-
мулированным Ф. Криком в 1967 г.: генетическая информация
переносится по схеме ДНК -» РНК -» Белок (у вирусов может быть
несколько видоизменена), но никогда — от белка к РНК.
5) Каждый ген проявляет себя (экспрессируется) путем обра-
зования, или биосинтеза мРНК (транскрипция гена) с последую-
щим переводом (трансляция) информации в специфическую по-
липептидую цепь, фактически являющуюся продуктом гена. Ген и
его продукт колинеарны, то есть последовательность кодонов
(триплетов) в гене точно соответствует последовательности амино-
кислот в белке.
6)’ Ген кодирует строение белковой молекулы и регулирует
процесс ее синтеза.
В таблице 19 суммированы основные представления о строении
и функциях генетического аппарата клеток разного уровня орга-
низации.
Зная строение и функцию генов, а также владея методами их
выделения и переноса в различные клетки или молекулы нуклеи-
новых кислот, можно с достаточной уверенностью подходить к
постановке генноинженерных работ.
5.2.КЛОНИРОВАНИЕ ГЕНОВ МЕТОДАМИ ГЕНЕТИЧЕСКОЙ
ИНЖЕНЕРИИ;
рДНК-БИОТЕХНОЛОГИЯ
5.2.1. Общая характеристика генетической инженерии. Гене-
тическая инженерия — это методы получения рекомбинантных
ДНК, объединяющих последовательности разного присхождения.
Некоторые ученые трактуют генетическую инженерию "как ис-
кусство использования знаний, методов и техники физико-хими-
ческой биологии и молекулярной генетики для конструирования
177
организмов с заданными наследственными свойствами” (В. Н.
Рыбчин, 1986).
Таблицам. Основные характеристики генетического аппарата у прокариот
и эукариот
Характеристика Прокариоты Эукариоты
генов по строению непрерывны прерывны (содержат интроны)
по стабильности стабильны возможны перестройки
по координации экспрессии опероны (регулоны) управляются операторами и сайтами связывания регуляторных белков оперонов нет. В индуцибельных генах имеются регуляторные элементы
про- це- ссов транскрипции по числу' РНК-полимераз одна три
по наличию энхансеров отсутствуют имеются. Энхансеры тканеспецифичны
по строению промоторов типичен единый план строения; наличие двух консервативных последованостей: ТАТААТ (-10) и ТГГАЦА (-35) промоторы генов, кодирующих белки, имеют одну консервативную последователь- ность ТАТА (А/Т) А(А/Т) (-30) и богатый ГЦ-участок (-40... - 100)
по мРНК колинеарна гену или оперону первичный транскрипт колинеарен гену. ’'Зрелая" мРНК образуется в процессе сплайсинга
1 трансляции по константе седиментации рибосом 70S (50S и 30S) 80S (60S и 40S)
по сайту связывания рибосом последователь- ность с ядром АГГА консервативной последователь- ности не выявлено
.по ко- 1 донам1 инициации АУТ, редко ГУТ АУГ
терминации УАЛ, УАГ, УГА УАА, УАГ, УГА
178
Очевидно, что под искусством автор понимает умение органи-
зовать и методически реализовать генноинженерную задачу —
получить рекомбинантную ДНК с последующим включением ее в
реципиентную клетку или осуществить перенос целых хромосом
от клеток-доноров в клетки-реципиенты.
Иногда понятия "генная инженерия" и "биотехнология" отож-
дествляются (А. А. Баев, 1984), хотя несомненно генная инженерия
представляет собой один из методов науки биотехнология. В основу
генноинженерных методов заложена способность ферментов —
рестриктаз расщеплять ДНК на отдельные нуклеотидные последо-
вательности, которые могут быть использованы для встраивания
их в геномы бактериальных плазмид и фагов с целью получения
гибридных, или химерных форм, состоящих из собственной
ДНК и дополнительных встроенных фрагментов несвойственной
им ДНК. Поэтому методами генетической инженерии добиваются
клонирования генов, когда выделяют нужный отрезок ДНК
из какого-либо биообъекта и затем получают любое количество
его, выращивая колонии генетически идентичных клеток, содер-
жащих заданный участок ДНК. Другими словами клонирование
ДНК — это получение ее генетически идентичных копий.
Генетическую инженерию подразделяют на генную инжене-
рию, геномную инженерию и хромосомную инженерию. Сущ-
ность первой состоит в целенаправленном использовании пере-
строек естественного генома, осуществляемых in vivo или in vitro,
для изменения генетических характеристик известных вирусов и
клеток. В качестве примеров генной инженерии in vivo можно
назвать: транслокацию (перемещение) в вирусные геномы некото-
рых клеточных генов, придающих вирусам свойства онкогенности;
трансдукцию (перенос) клеточных генов от клеток-доноров в клет-
ки-реципиенты с помощью фагов, и др. Примером генной инже-
нерии in vitro является создание молекулярных химер из фрагмен-
тов ДНК разного происхождения, включение их в реципиентные
клетки Е. coli, Вас. subtilis и др. с последующим культивированием
этих организмов в целях получения необходимых белковых про-
дуктов (пептидных гормонов, ферментов и т. д.).
Сущность геномной инженерии заключается в целенаправлен-
ной глубокой перестройке генома акариот, прокариот или эукари-
от, вплоть до создания новых видов. При геномной инженерии
добиваются внесения большого количества дополнительной гене-
тической информации и в результате получают гибридный орга-
низм, отличающийся от исходного по многим признакам. Гибриды
179
оказываются жизнеспособными лишь в тех случаях, когда они
содержат все безусловно необходимые (облигатные) гены, когда у
них налажены и взаимно согласованы регуляторные связи между
совмещенными генами, и, наконец, когда имеется структурное
соответствие между продуктами матричного синтеза (белками).
При геномной инженерии возможно получение половых (сли-
яние гамет) или соматических (слияние неполовых клеток) гибри-
дов. Половые гибриды получают либо в естественных, либо в
искусственных (экспериментальных) условиях. Соматические гиб-
риды способны формироваться лишь в искусственных условиях у
прокариот и эукариот, то есть у клеточных форм (клеточная
инженерия).
Примером естественной (природной) геномной инженерии
является рекомбинация геномов вирусов гриппа, относящихся к
типу А. На основании антигенных характеристик рибонуклеопро-
теинов выделяют вирусы гриппа А, В и С. Изменения антигенных
свойств постоянно происходят у вирусов типа А, меньше — у типов
В, тогда как вирусы типа С являются антигенно стабильными. К
тому же известны штаммы вируса гриппа А, изолируемые от
свиней, лошадей, уток, цыплят. Некоторые изоляты вируса от
животных антигенно подобны штаммам, циркулирующим среди
людей. Поэтому более полно изученными к настоящему времени
также оказались вирусы типа А. Их геном состоит из 8 различных
однонитевых сегментов РНК с общей молекулярной массой 2—
4-103 кДа (см. рис. 22а). Большая часть полинуклеотидов, состав-
ляющих вирусные РНК-сегменты, содержат уридин на З'-конце. С
вирусным геномом ассоциирована РНК-зависимая РНК-полимера-
за. Рибонуклеопротеин окружен белковым слоем (М), образующим
внутреннюю часть вирусной оболочки. Белок М включает малый
протеин с ММ 26 кДа, составляющим 40% от всего белка вирусной
частицы. Порядка 20% массы частицы приходится на липидный
бислой, который, по-видимому, имеет клеточное происхождение.
Гемагглютинин отвечает за агглютинацию эритроцитов вирусом
А. Он представляет из себя гликопротеин с ММ 75 кДа. Нейрами-
нидаза ответственна за рецепторцо-разрушающую активность ви-
руса, обеспечивая ему выход из эритроцита или клетки-,,хозяина”.
Нейраминидаза состоит из четырех полипептидных молекул с ММ
порядка 60 кДа, и, наряду с гемагглютинином, обладает антигенной
активностью. Их используют для регистрации и объяснения анти-
генных изменений вируса. К началу 90-х годов было известно 11
подтипов гемагглютинина (Н1—Hl 1) и 8 подтипов нейраминидазы
180
(N1—N8) у вирусов А. Поэтому система обозначения вируса гриппа
включает название штамма, номера субтипов гемагглютинина и
нейраминидазы. Например, вирус гриппа А(свиной) Тайвань
/l/70(H3N2). Это значит, что вирус изолирован от свиней на
острове Тайвань в 1970 году, содержит гемагглютинин З(НЗ) и
нейраминидазу 2 (N2). Оба антигена (Н и N) формируются и
контролируются генами (РНК-фрагментами) независимо друг от
друга. Вследствие сегментарности генома вирус проявляет реком-
бинацию с высокой частотой и, как следствие, происходят изме-
нения в антигенных свойствах вирусов (прежде всего — по
гемагглютинину и нейраминидазе).
В экспериментальных условиях возможно проведение РНК —
РНК-гибридизации с последующим определением профилей плав-
ления молекул гибридных РНК в присутствии формальдегида.
Известны некоторые вирусы бактерий, объединяющие в себе
свойства фагов и плазмид. Их назвали фазмидами. Они могут быть
природного и экспериментального (искусственного) происхожде-
ния. К природным относятся некоторые умеренные coli-фаги (N15,
Р1), к искусственным — фазмида col 106, способная при 32°С
проявлять свойства плазмиды, а при 37°С индуцирует лизис реци-
пиентных клеток.
Благодаря высокой степени генетической гомологии Е. coli,
Salmonella spp. и Shigella spp. можно получать хромосомные гиб-
риды между ними:
Доноры ДНК (хромосомы) Реципиенты ДНК (хромосомы)
Е. coli Различные штаммы Е. coli
Shigella spp.
Salmonella spp.
Salmonella typhimurium Различные виды и штаммы Salmonella spp.
Shigella flexneri Е. coli, Salmonella typhi и др.
В этих целях чаще прибегают к методу конъюгации бактерий.
Однако следует иметь в виду, что виды с небольшой генетической
гомологией (тем более — при отсутствии ее ) слабо конъюгируют
или не конъюгируют совсем и обмена генами между ними прак-
тически не наблюдается. Здесь перспективным оказывается метод
слияния протопластов, ныне широко используемый в работе с
прокариотами и эукариотами.
181
Клеточные стенки обычно у молодых клеток частично или
тотально лизируют с помощью гидролитических ферментов. Воз-
никающие протопласты затем подвергаются слиянию в среде с
полиэтиленгликолем и соответствующими стабилизаторами про-
топластов, либо используют электрическое или температурное
воздействие в целях деполяризации мембраны. Таким образом
можно создавать межвидовые и, даже, межродовые гетерокарио-
тические гибриды. Другими словами, таким путем удается совме-
щать в одной клетке геномы клеток (организмов) с половой несов-
местимостью. Например, удается слить протопласты клеток мор-
кови и ячменя, кукурузы и сои, дурмана и красавки, картофеля и
томата (pomato), некоторых бактерий и растений, мыши и моркови,
мыши и человека. Однако в большинстве случаев такие гибридные
клетки не развиваются в полноценные организмы. К тому же
стабильность (жизнеспособность) гибридов находится в обратно-
пропорциональной зависимости от эволюционной отдаленности
гибридизуемых видов, то есть эволюционно далекие виды менее
жизнеспособны в гибридном состоянии, чем близкородственные
виды. Жизнеспособные гибриды сохраняют большую часть генома
одного из слившихся партнеров. Поэтому соматическая гибриди-
зация практически реализуется пока на ограниченном числе видов
и родов. Так, используя протопласты из листьев петунии дикого
типа одного вида и "альбиносные" протопласты суспензионных
культур другого вида, удалось сконструировать амфидиплоидные
(от греч. amfi— с обеих сторон, diploos — двойной, eidos - вид),
или аллотетраплоидные соматические гибриды с 28 хромосомами,
способные трансформироваться в цветущие растения:
а) вид Petunia parodii (2n= 14)
б) вид Petunia hybrida (2n= 14) •
в) вид Petunia inf lata (2n= 14)
Соматические гибриды: 1) P.parodii x P.hybrida (4n = 28)
2) P.parodii x P.inflata (4n = 28).
К. Келер и Ц. Милыптейн в 1975 г. смогли впервые получить
соматические гибриды клеток млекопитающих — гибридо-
м ы, продуцирующие так называемые моноклональ-
ные антитела. В иммунном организме антитела образуются
плазматическими клетками, возникающими в результате диффе-
ренциации В-лимфоцитов в ответ на антигенную стимуляцию.
В-лимфоциты — высоко специализированные клетки, их в орга-
низме порядка 107 клонов и каждый клон синтезирует антитела
182
только одной специфичности — моноклональные антитела (вари-
антов антител может быть также 107). Если антиген содержит
несколько специфических (детерминантных) групп, то антитела
образуются против каждой детерминанты. В-лимфоциты не удает-
ся поддерживать в культуре.
Если В-лимфоцит перерождается и трансформируется in vivo
в злокачественную опухоль, то такая опухоль — миелоМа
синтезирует большое количество антител, специфичность которых
неизвестна. Однако возникают и такие варианты миеломных
клеток, в которых отсутствует экспрессия генов в отношении
синтеза тяжелых и легких цепей иммуноглобулинов. Миеломные
клетки могут поддерживаться в культуре неопределенно долго.
Совместив способность миелом к длительному культивированию
в условиях in vitro и способность В-клеток продуцировать моно-
клональные антитела, Келер и Милыптейн получили гибрид, сек-
ретирующий моноклональные антитела. Для этих целей они ис-
пользовали мышиные клетки миеломы и нормальные В-клетки
селезенки мыши, иммунизированной заданным антигеном (см.
специальную часть). Подобные гибридомы широко используются
теперь для получения различного рода моноклональных антител,
применяемых в диагностических, лечебных и других целях.
Необходимо иметь в виду, что, в отличие от половой гибриди-
зации, соматическая гибридизация эукариотических клеток завер-
шается объединением под одной мембраной не только ядерных
геномов двух (или более) особей, но и генов цитоплазмы (митохон-
дриальных, хлоропластных, емкостью в 1000—2000 генов), что
может отразиться на функциональной активности гибрида. У
межвидовых гибридов часть хромосом может утрачиваться за счет
элиминации, которая оказывается видоспецифичной. Так в гибри-
дах протопластов клеток "мышь х человек” и "человек х комар"
элиминируются хромосомы человека и комара соответственно.
При морфологическом различии хромосом такие гибриды удобны
для картирования генов. Напомним, что в соматических клетках
мыши содержится 20 пар хромосом, в клетках человека 23 пары
хромосом и три пары — в диплоидных клетках комара.
Таким образом, на практике стремятся осуществлять сомати-
ческую гибридизацию для заметного расширения рамок скрещи-
вания, для включения (переноса) внеядерных генов и их функций
в гибридное потомство и для локализации генов в хромосомах.
В случае переноса изолированных хромосом от клетки-донора
183
одного организма в клетку-реципиент другого организма говорят
о хромосомной инженерии. О. Мак Брайд и X. Озер
впервые в 1973 г. осуществили такой перенос хромосом (в стадии
метафазы) китайского хомячка в клетки мыши.
Хромосомная инженерия — ветвь генетической инженерии.
Объектами ее являются хромосомы клеток прокариот и эукариот.
Донорами хромосом могут быть различные суспензионные и суб-
страт-зависимые клеточные линии. Из клеток прокариот хромосо-
му (ДНК) выделяют из супернатанта после центрифугирования
дезинтеграта или лизата клеток (протопластов). Клетки эукариот
блокируют на стадии мейоза, хромосомы выделяют, применяя
"гипотонический шок" и гомогенизацию с последующей очисткой
их дифференциальным центрифугированием. Хромосомы осажда-
ют на поверхности реципиентных клеток хлоридом кальция и через
несколько часов клетки обрабатывают реагентом — "перфорато-
ром" (например, глицерином). Реципиентные клетки могут содер-
жать донорный материал в широком диапазоне (встроенным в
геном, изолированно).
Благодаря хромосомной инженерии стали возможными пол-
учение высокомолекулярных БАБ, присущих человеку, лечение
наследственных заболеваний, селекция пород домашних живо-
тных и различных видов растений.
Естественное или искусственное скрещивание растений и жи-
вотных, когда происходит оплодотворение женской гаметы муж-
ской, по сути своей касается переноса и слияния хромосом с
последующим возникновением жизнеспособных гибридов. Меж-
видовое скрещивание как правило сопровождается худшими ре-
зультатами — получаемые гибриды обладают сниженной плодови-
тостью или могут быть совершенно бесплодными.
Целые наборы хромосом от разных видов могут соединяться
вместе и в результате возникают аллополиплоидные формы (от
греч. alios — другой). Можно соединять и неполные хромосомные
наборы. Так, при скрещивании 42-хромосомной пшеницы с пше-
нично-ржаным амфидиплоидом, содержащим 56 хромосом, возни-
кает гибрид с 49 хромосомами, у которого 42 хромосомы принад-
лежат пшенице и 7 хромосом принадлежит ржи.
При искусственном скрещивании растений или животных
стремятся получать в потомстве комбинацию различных ценных
родительских признаков.
Теоретически любые клетки могут бь!ть использованы либо в
качестве донора хромосом, либо в качестве реципиента их, хотя
184
на практике стремятся иметь подходящие реципиентные линии
клеток, акцептирующие чужеродную ДНК с повышенной актив-
ностью, например, клетки карциномы мочевого пузыря человека
(линия EJ).
В процессе выделения хромосом из клеток на стадии митоза
эксперименты проводят при пониженных температурах ( + 4°С),
используя пластиковые пипетки во избежание деструкции хромо-
сом. Перед механическим разрушением клеток их суспендируют
(107 клеток/мл) в гипотоническом растворе, а затем центрифуги-
руют при 2500 g на холоду (примерно 25 мин. при 4°С). Выход
хромосом составляет около 10% от всех хромосом клеток-доноров.
Очищенные хромосомы должны быть использованы в опытах по
переносу в реципиентные клетки (трансфекция) немедленно.
Хромосомы можно выделять из клеток, блокированных в ме-
тафазе, различными способами, в том числе с помощью щадящих
поверхностно-активных веществ, например, стероидного сапони-
на-гликозида дигитонина, представляющего собой пентагликозид
дигитогенина. Олигосахаридная часть присоединяется через гид-
роксил в позиции СЗ агликона. Она включает 1 остаток ксилозы
и по 2 остатка галактозы и глюкозы.
Таким образом, соподчиненность генной, геномной и хромо-
сомной инженерий можно представить в виде схемы:
^^Генетическая инженерия^^^
генная инженерия геномная инженерия хромосомная инженерия
половая соматическая
гибриди- гибридизация
зация (клеточная
инженерия)
Такое подразделение генетической инженерии и терминология
185
достаточно условны, поскольку, в конечном итоге, результаты
манипуляций с рекомбинантной ДНК получают на клеточных
культурах. Поэтому при выделении двух уровней биотехнологии
— молекулярный и клеточный (включая клеточ-
но-инженерный) следует иметь в виду, что первый относится
к получению и клонированию рекомбинантной ДНК, второй — к
выращиванию вирусов, клеток и тканей (клеточно-инженерный —
к протопластированию и гибридизации протопластов), хотя, как
уже было сказано, все уровни можно свести к одному — клеточ-
ному.
А А А А А мРНК
ревертаза, или обратная
транскриптаза;
нуклеозидтрифосфаты;
олигодезокситимидин (dT)
А А А А А мРНК
II II II II II
_________—________—_______ Т Т Т Т Т комплементарная цепь
денатурация
ДНК=полимераза I
(фрагмент Кленова) + дезоксинуклеозидтри-
фосфат (dHT4>)
А А А А А
II II II II II -№Кс одноцепочечной
т т т т т петлей
нуклеаза S1 из Aspergillus oryzae
А А & & А двуцепочечная компле-
Т Т Т Т Т ментаРная ДНК(кДНК)
терминальная трансфераза, dL(T<t>
ЦЦЦЦЦЦЦ.
АААААЦЦЦЦЦЦЦ
II II II II II
т т т т т
фрагмент
ДНК с го-
мополи-
мерными
последо-
ватель-
ностями
ЙЦ
Рис. 65. Схема получения из мРНК фрагмента ДНК, пригодного для клонирования.
186
5.2.2. рДНК-биотехнология. Практическое использование ре-
комбинантных ДНК различного происхождения составляет основу
так называемой "рекомбинантной ДНК-биотехнологии", или со-
кращенно рДНК-биотехнологии. Теоретически все 50—200 тысяч
структурных генов человека доступны экспериментальному ана-
лизу, хотя это еще не было бы проникновением в природу человека
в целом. Тем не менее, с помощью рДНК-биотехнологии открыва-
ются невиданные доселе перспективы производства различных
веществ для народного хозяйства.
рДНК-биотехнологию подразделяют на следующие этапы: по-
лучение чужеродной ДНК, разрезание полученной ДНК на фраг-
менты и их очистка, включение фрагмента чужеродной ДНК в
векторную плазмиду и получение рекомбинантной ДНК, или
рДНК, введение рДНК в пермиссивные клетки и клонирование
генов, амплификация и экспрессия рДНК.
Получение чужеродной ДНК. Чужеродную ДНК можно по-
лучить с помощью химического или ферментативного синтеза,
либо из любого организма или из вирусов с последующим опре-
делением ее первичной структуры. Если ДНК выделяют из клеток
эукариот, то для клонирования, амплификации и экспрессии не
нужны интроны. Поэтому в таких случаях используют мРНК,
образующуюся в процессе сплайсинга (рис. 65).
Как видно из рис. 65,терминальная трансфераза катализирует
реакцию присоединения гомополимерных последовательностей
дезоксицитидина. Таким образом создаются предпосылки для по-
следующего клонирования, в частности, используя плазмидный
вектор (см.) PBR322, несущий гены устойчивости к ампициллину
и тетрациклину. После расщепления рестриктазой (см.) Pst 1 (из
Providencia stuartii) эта плазмида имеет гомополимерные последо-
вательности dT:
Pstl
—Ц—Т—Г—Ц—А—Г
—Г—А—Ц—Г—Т—Ц—
I
Pst 1
За счет этого вектор можно использовать для встраивания
полученного фрагмента ДНК с гомополимерными последователь-
ностями dL[.
Для изоляции неповрежденных генов из ДНК можно исполь-
зовать ультразвук или гидродинамическое дробление ДНК в соот-
ветствии с нижеследующей последовательностью этапов.
В соответствии с особенностями структуры полученного фраг-
187
мента его включение в pColi Е1 будет обеспечиваться связыванием
типа поли-dA—dT.
ДНК из E.coli
^гидродинамическое дробление (или ультразвук)
фрагменты ДНК
5'— рестриктаза фага X
дополнительная специфическая фрагментация
dA
терминальная трансфераза, <1АТф
~| ДНК,пригодная для включе-
ния вплазмидный вектор,
например, Coli Е1
Разрезание ДНК на фрагменты. Любая нативная ДНК может
быть "измельчена" с помощью ферментов эндонуклеаз, среди
которых особое место занимает рестриктазы (рестрикционные
эндонуклеазы). Биологическая роль их, например, в клетках про-
кариот заключается в
гидролизе чужеродной
ДНК, проникающей в
клетки извне. Собст-
венная хромосомная
ДНК при этом остается
незатронутой рестрик-
тазами, что обусловле-
но наличием фермен-
тов, называемых моди-
фикационными мети-
лазами, узнающими
одинаковые с рестрик-
тазами сайты. Назван-
ии— СН—(СН2)2-
соон
SAM
ные метилазы катализируют реакции метилирования А или Ц в
немногих специфических сайтах в хромосомах, в результате чего
метилированная ДНК оказывается нечувствительной к атаке ре-
стриктазами. Переносчиком метильных групп выступает S-адено-
зил-Б-метионин (SAM).
В настоящее время известно свыше 500 рестрикгаз и для многих
из них определены узнаваемые последовательности. Значительное
число рестриктаз производится в качестве конечных продуктов
биотехнологии, например, такими компаниями как Sigma (США),
188
Pharmacia (Швеция), Serva (Германия) и др. Эти ферменты способ-
ны узнавать специфические места (с а й т ы) и расщеплять
нуклеотидные последовательности в ДНК, индуцируя формирова-
ние тупых или липких концов.
Рестриктазы вначале подразделяли на 3 группы в зависимости
от длины узнаваемой последовательности:
I — узнают тетрануклеотиды, например Alu I из Arthrobacter
luteus,
II — узнают пентануклеотиды, например Есо RII из Е. coli,
III — узнают гексануклеотиды, например Есо RI из Е. coli;
Alu I катализирует расщепление последовательности А^Д^Ц'Г
с образованием тупых концов во фрагменте ДНК, EcoRII и EcoRl
расщепляют последовательности^^- ЦЦ(А/Т) ГГ и -ААТТЦ
соответственно с образованием липких концов во фрагментах
ДНК.
В настоящее время рестриктазы подразделяют на три класса с
учетом RMS-системы, в которой RMS — белки рестрикции, мети-
лирования (модификации) и посадки соответственно. К первому
из них относят те рестриктазы, которые расщепляют ДНК в
произвольных точках с образованием различных фрагментов ДНК
(для обеих нитей ДНК известны системы R2M2S); ко второму классу
(их известно около 500) относят ферменты, для которых сайты
рестрикции и посадки совпадают — RS и MS. Образующиеся при
этом фрагменты (рестрикты) воспроизводятся по длине.
Рестриктазы этого класса, объединяющие рестриктазы вышеука-
занных трех групп, преимущественно используются на практике.
Рестриктазы III класса (например из фага Р1, системы 2R MS)
объединяют все прочие рестрикционные эндонуклеазы. Отдель-
ные из них, например, не узнают сайтов посадки. Их редко
используют на практике.
По предложению X. Смита и Д. Натанса
(1973) номенклатура рестриктаз строится
по следующему принципу: название фер-
мента складывается из букв и цифр: первая
буква принадлежит родовому названию
источника рестриктазы, две другие буквы
относятся к названию вида источника; в
некоторых случаях дополнительно указы-
вают штамм или серовар; далее следуют 66. взаимодействие Есо
обозначения RM-системы (не всегда) и ну- ri с днк: а — вид сверху, б
мерация фермента (таблица 20). — вид сбоку.
Из таблицы видно, что число узнаваемых рестриктазами нук-
леотидных последовательностей варьирует от 4 до 13 (преимуще-
ственно 4—6). На рис. 66 показано взаимодействие Есо RI с ДНК.
189
В зависимости от источника ДНК число расщепляющихся
сайтов в ней будет неодинаковым. Например, действию рестрик-
ционных эндонуклеаз подвергаются ДНК фага X (инфицирует Е.
coli К — 12), аденовируса 2 и вируса SV — 40; в этом случае число
расщепляющихся сайтов соответственно будет следующим: для
Alul - более 50, более 50, 32; для BamHI — 5,3,1; для Ball — 15,17,0;
для Hind III — 6, И, 6; для Mbo I — 50,50,6 и т. д.
Т а б л и ц а 20. Характеристика некоторых рестрикционных эндонуклеаз
(рестриктаз)
Наименование фермента Источник получения Сайт узнавания в последовательности ДНК
Aatll Acetobacter aceti 5х - ГАЦГТ/Ц - 3х
Асе I Acinetobacter calcoaceticus 5Х - ГТ/(А, Ц) (Г, Т) - АЦ - 3х
Alu I Arthrobacter luteus 5х - АГ/ЦТ - 3х
Ava I Anabaena variabilis 5х - Ц/РуЦГРиГ - 3х
Ava II — ” — " — 5х - Г/Г(А, Т) ЦЦ - 3х
Bal I Brevibacterium albidum 5х - ТГГ/ЦЦА - 3х
Bam HI Вас. amyloliquefaciens H 5х - Г/ГАТЦЦ - 3х
Ban II Вас. aneurinolyticus 5х - ГРиГЦРу/Ц - 3х
Bel I Вас. caldolyticus 5х - Т/ГАТЦА - 3х
Bgl I Вас. globigii 5х - rL(L[NNNN/Nrn_( - 3х
Bgl II _ " — " — 5х - А/ГАТЦТ - 3х
| BstEII Вас. stearothermophilus ET 5х - Г/ГПЧАЦЦ - 3х
Cla I Caryophanon latum L 5х - АТ/ЦГАТ - 3х
Dpn I (должно быть согласно принятой номенклатуры Spn I) Diplococcus pneumoniae (Streptococcus pneumoniae) 5х - ГМА/ТЦ - 3х
Dral Deinococcus radiophilus 5х - ТТТ/ААА - 3х
Eco RI (рекомбинант) E. coli, несущая плазмиду гиперпродукции Eco RI 5х - Г/ААТТЦ - 3х
Eco RI E. coli RYI3 5х - Г/ААТТЦ - 3х
Eco RV E. coli 5х - ГАТ/АТЦ - 3х !
190
Продолжение табл. 20
Наименование фермента Источник получения Сайт узнавания в последовательности ДНК
Fsp I Fischerella species 5Z - ТПД/ГЦА - 3Z
Нае II Haemophilus aegyptius 5Z - РиГЦГЦ/Ру - 3Z
Нае III И 11 5Z - ГГ/ЦЦ - 3Z
Hhal — ” — haemolyticus 5Z - ГЦГ/Ц - 3Z
Hine II — “ — influenzae Rc 5Z - ГТРу/РиАЦ - 3Z
Hind III Из штамма E. coli, несущего плазмиду гиперчувствительности Hind III 5Z - А/АГЦТТ - 3Z
Hinf I Haemophilus influenzae серовар f 5Z - Г/АМТЦ - 3Z
Hpa I Haemophilus parain fluenzae 5Z - ГТТ/ААЦ - 3Z
Hpall 5Z - Ц/ЦГТ - 3Z
Kpn I Klebsiella pneumoniae 5Z - ГТТАЦ/Ц - 3Z
Mbo I Moraxella bovis 5Z - N/ГАЦТ - 3Z
Mbo II •• n 5Z - ГААГА (N)8/ - 3Z
Mln I Micrococcus luteus 5Z - А/ЦГЦГТ - 3Z
Msp I Moraxella species 5Z - Ц/ЦГГ - 3Z
Neo I Nocardia carollina 5Z - Ц/ЦАТГГ - 3Z
Nde I Neisseria denitrificans 5Z - ЦА/ТАТГ - 3Z
Pst I E. coli, несущая плазмиду гиперпродукции Pst I 5Z - ЦТГЦА/Г - 3Z
Pst I Providencia stuartii 5Z - ЦТГЦА/Г - 3Z
Pvu I Proteus vulgaris 5Z - ЦГАТ/ЦГ - 3Z
Pvu II 11 II 5Z - ЦАГ/ЦТГ - 3Z
Sma I Serratia marcescens Sb 5Z - ЦЦЦ/ГТГ - 3Z
Xbal Xanthomonas bardii 5Z - Т/ЦТАГА - 3Z
Примечание: 1) в скобках указаны альтернативные нуклеотиды.
2) Ри.Ру — любое пуриновое, пиримидиновое основание,
3) N — любой нуклеотид,
4) Гм — метилированный гуанин,
5) (N)e — нуклеотид, повторяющийся 8 раз.
191
Расщепление сайтов может происходить симметрично (см. Alu
I, Bal I, Dpn I и др.) с образованием тупых концов в ДНК
и асимметрично (Aat II, Асе I, Ava I, II, Bam HI и др.) с образованием
липкихконцов:
Hine II
—ЦТАГГЦАГЦТГ Г АЦГТ—
- Г АТЦЦ Г ТЦ Г АЦЦ Т Г ЦА—
Hinc II
ДНК
-ЦТАГГЦАГ ЦТГГАЦГТ—
• •••• •
рГАТЦЦГТЦ, ,ГАЦЦТГЦА- ।
фрагменты ДНК с тупыми концами
Hind III
/
- ГТЦГТААГЦТТЦЦГТ Г —
- ЦАГЦАТТЦГ^АГГЦАЦ-
Hind III
ДНК
- ГТЦГТА АГЦТТЦЦГТГ-
|-ЦАГЦАТТЦГА ! । АГГЦАЦ-
фрагменты ДНК с липкими концами
Рестриктазы расщепляют ДНК примерно через 250 нуклеоти-
дов, исходя из формулы 4П (4 — нуклеотиды — А, Г, Т, Ц; п —
число повторов сайтов в ДНК).
Фрагменты ДНК, полученные тем или иным методом, разделя-
ют с помощью электрофореза в гелях (агарозном, полиакриламид-
ном — ПААТ), из которых их экстрагируют, вырезая соответству-
ющие участки геля; эти участки геля, например агарозного, рас-
плавляют в пробирках при 68°С и ДНК экстрагируют фенолом,
затем концентрируют изобутанолом, переосаждают этанолом и
проводят анализ выделенных и очищенных фрагментов. При не-
192
обходимости можно дополнительно провести препаративный гель-
электрофорез. Из 160 мкг ДНК нематоды Caenorabiditis elegans
удается получать 2-4 мкг какого-либо одного фрагмента.
Рестрикты (особенно — с липкими концами) могут быть сшиты
с помощью ДНК-лигаз. При необходимости липкие концы отжи-
гают (отжиг—это специфическая реассоциация комплементарных
цепей с образованием двухцепочечных молекул):
-••5-Т
------>
Ц - Т • А • Г • А - 3’- + • • -З’-А • Г А Т • Ц Т - 5‘-
>
... 5'-т * ц. т А Г А - 3'-
З'-А — Г• А-Т• Ц Т —5'-
Т
реассоциант с
пробелами между Т и Ц
в обеих нитях ДНК
В пробелах между нуклеотидами отсутствуют фосфодиэфир-
ные связи, которые могут быть воссозданы с помощью ферментов
лигаз, катализирующих реакции синтеза за счет З^гидроксильной
и 5/-фосфатной групп
7 т. 8524
193
Таким путем восстанавливаются ковалентные связи в пробелах:
• ’' 5х—Т—Ц—Т—А—Г—А—3х • • •
II III IIII III II
• • ’3х—А—Г—А—Т—Ц—Т—5х' • •
Для сшивания фрагментов ДНК используют также линке-
ры и адаптеры. Линкер (от англ, link — соединять) — это
синтетический, короткий двухцепочечный олигонуклеотид, содер-
жащий сайты узнавания для ряда рестриктаз; он может быть
присоединен к концам фрагмента ДНК, полученного с помощью
какой-либо рестриктазы, в процессе реконструирования рДНК.
Например, линкер Есо RI представляет собой ds (ГГААТТЦЦ)3.
Адаптер — это линкер, содержащий больше, чем один сайт
узнавания рестриктазой, и обеспечивающий получение фрагмен-
тов от действия одной рестрикционной эндонуклеазы для включе-
ния в сайт другой рестриктазы, то есть адаптер предназначен для
объединения фрагментов с несовместимыми концами. Пример
адаптера Есо RI к Sma I: (ААТТЦЦЦГГГ). Тупые концы ДНК,
образованные после рестрикции, могут относительно легко сши-
ваться под действием ДНК-лигазы фага Т4.
Метод клонирования широко используют для выделения инди-
видуальных генов прямо из генетического материала, располагая
при этом высокоспецифичными радиоактивными РНК- и ДНК-зон-
дами. Эти последние взаимодействуют только с определенными
последовательностями и гибриды их с генами регистрируются
радиоавтографией на рентгеновских пленках. Пример анализа по
Саузерну (по англ, southern-blotting — промокание по Саузерну)
показан на рис. 67. Аналогичный подход применим и для анализа
РНК (метод Нозерн* -блотинга).
В генетической инженерии следует различать копийную
ДНК (кДНК), являющуюся копией мРНК, и векторы, несущие
клоны геномной или хромосомной ДНК. При
клонировании всего генома ("шотган-эксперимент", от англ,
shotgun — дробовик) его разделяют с помощью рестриктазы,
узнающей короткую (4 пн) последовательность нуклеотидов. Раз-
рушение проводят в таких условиях, чтобы осуществлялась час-
* — Здесь использована игра слов Southern и Northern, то есть южный и север-
ный, хотя в первом из них отражена фамилия ученого.
194
Рис. 67. Схема выделения фраг-
ментов ДНК из смеси рестриктов
ДНК по Саузерну: 1 — расщепление
геномной ДНК на случайные фраг-
менты с помощью рестриктазы (Р), 2
— электрофорез фрагментов ДНК в
агарозном геле, 3 — недифференци-
рованное пятно ДНК, 4 — тепловая
денатурация ДНК с образованием од-
ноцепочечных фрагментов и nepei юс
их на нитроцеллюлозный фильтр для
фиксации (blotting), 5 — гибридиза-
ция с радиоактивным зондом, 6 —
] 6 гибридные последовательности, вы-
явленные с помощью радиоавтогра-
фии, 7 — выделение фрагментов из
агарозного геля для исследования.
тичная рестрикция ДНК. Образующиеся при этом фрагменты
будут разной длины, но каждый из них заканчивается одной и той
же последовательностью. Наличие во фрагменте липкого конца
делает его удобным для клонирования. Разделенные фрагменты
всего генома встраивают затем в клонирующий вектор с образо-
ванием набора химер, который может храниться в течение неог-
раниченного времени. Такой набор клонированных фрагментов
ДНК называют библиотекой генома. С помощью нового
(оригинального) зонда библиотека может быть сразу же использо-
вана для поиска специфического фрагмента.
Включение фрагмента чужеродной ДНК
в векторную плазмиду
На латинском языке vector значит везущий, несущий; в мате-
матике вектор — это прямолинейный отрезок, которому придано
определенное направление. Понятие "вектор” в молекулярной
биологии объединяет сущность данных выше двух определений и
название "Вектор для клонирования" введено М. Томасом в 1974
г. для ДНК, способной переносить в клетку — реципиент чужерод-
ную ДНК любого происхождения. Вектор — самый важный ком-
понент процесса клонирования. Он должен удовлетворять следу-
ющим требованиям: 1) относительная легкость накопления и вы-
деления его в достаточном количестве,
2) он должен содержать генетические маркеры для отбора
последующих трансформантов,
3) наличие в векторных ДНК нескольких сайтов узнавания
различными рестриктазами в целях получения для клонирования
серии разных молекул ДНК.
195
Следовательно, векторы — это молекулы ДНК, обеспечиваю-
щие амплификацию фрагмента (образование дополнительных ко-
пий его) в растущей популяции клеток соответствующего вида
организма.
Для клонирования ДНК например в клетках Е. coli можно
использовать два класса векторов — плазмиды и фаги. Из числа
ДНК-плазмид для практических целей рекомендованы и имеются
на рынке следующие: Col EI из Е. coli штамм С600, Col EI Amp из
Е. coli штамм KR245, резистентный к антибиотику ампициллину;
pBR322 из Е. coli штамм RRI, резистентный к ампициллину и
тетрациклину; pBR325 из Е. coli штамм НВ101, резистентный к
ампициллину, тетрациклину и хлорамфениколу (левомицетину);
pUC-плазмиды (pUC8, pUC9, pUC18, pUC19) из E coli штамм RRI,
резистентный к ампициллину и дефектный по Р-галактозидазе;
pUBl 10 из Вас. subtilis штамм BD170, резистентный к канамицину;
рУАС-клонирующие векторы, впервые предложенные в 1987 г. (Д.
Т. Бурке, Ф. Г. Карл, М. В. Олсон) — рУАСЗ, рУАС4, рУАС5. Они
представляют собой дрожжевую (от англ, yeast), искусственную
(А, от англ, artificial) хромосомную (С, от англ, chromosome) кло-
нирующую систему для клонирования больших (50-100 кЬ в длину)
фрагментов ДНК; Ti-плазмиды (от англ. Tumor inducing — вызы-
вающие опухоль) из Agrobacterim tumefaciens (pTi — В6-806) штамм
А277, утилизирующий октопин, и pTi — С58 из A. tumefaciens
штамм С58, утилизирующий нопалин.
h2n-c=nh
I
NH-(CH2) -СН-СООН
NH-CH-COOH
I
сн3
октопин
(синтезируется из аргинина и пирувата)
HOOC-CH-NH-CH-COOH
I I
(сн2)2 (сн2)3
I I
СООН NH
I
nh2-c=nh
нопалин
1
196
Октопин и нопалин, равно как и атропин, относятся к группе
о
к
с
/ \
H2N-CO-(CH2)2-CH о
NH СН-(СНОН) -сн,он
\ / i 1
агропин
опинов — низкомолекулярных веществ, синтезируемых
клетками злокачественной опухоли двудольных растений — ко-
рончатого галла. В этой связи различают опухолевые клетки
агропинного, нопалинового и октопинового типов.
Из бактериофагов используют ДНК-содержащие колифаги Т2,
Т4, Т5, Т7, фаги X, М13тр8, М13тр9, М13тр18, М13тр19, фХ174;
из ДНК вирусов — SV40 и аденовирус 2.
Векторные системы можно конструировать из ДНК, несущих
маркер(-ы), с последующим включением их, например, в клетки
эукариот, в том числе — млекопитающих.
В качестве иллюстраций векторных систем приводим рис. 68а,
на котором изображены плазмиды Col El, pBR322, вирус SV40 и
челночный вектор.
Вирусы растений — как векторы обычно мало пригодны из-за
своей патогенности для растительных организмов и неспособности
встраиваться в хромосомы хозяйской эукариотической клетки. В
настоящее время наметились подходы к изучению и оценке трех
векторных систем: двухцепочечной ДНК вируса мозаики цветной
капусты, одноцепочечной РНК вируса погремковости табака, од-
ноцепочечной ДНК вируса золотистой мозаики фасоли. Из них
лишь первая оставляет надежды на дальнейшее продвижение этой
системы в сторону практической реализации пока в лабораторных
условиях. Не исключается возможность объединения ДНК вируса
мозаики цветной капусты с Т-ДНК Ti-плазмиды из Agrobacterium
и расширить круг растений — реципиентов такой векторной
системы.
Дж. Коллинс и Б. Хон в 1978 г. впервые ввели в практику так
называемый космидный вектор, или cos-вектор, То есть комбини-
рованную векторную систему "плазмида-ДНК фага лямбда". Дан-
ный вектор способен акцептировать до 40-50 тыс. пн. При этом
совмещаются плазмидные репликаторы и cos-сайты фага X в одном
197
геноме. Космидам присущи свойства фагов и плазмид, то есть
способности упаковывать свою ДНК в головке фагов и автономно
реплицироваться. Свойства фагов и плазмид объединяют в себе
ф а з м и д ы, к которым можно отнести и космиды.
Включение фрагмента чужеродной ДНК в вектор осуществля-
ют по следующей схеме (рис. 686). Многие плазмиды несут транс-
позоны.
Рис. 68а, б. а — плазмиды: I — Col El; II — pBR322; III — вирус SV40; IV —
челночный вектор для кишечной палочки и клеток обезьяны (1 — ori-сайт SV40; 2
— промотор SV40; 3 — вероятная сигнальная последовательность; 4 — последова-
тельность, кодирующая гамма-интерферон; 5 — фрагмент кДНК; 6 — оп-сайт
pBR322, 7 — ген устойчивости к ампициллину;
б — включение чужеродной ДНК в плазмидный вектор (схема): 1 — вектор, 2
— ген, 3 — рестрикты, образовавшиеся под влиянием рестриктазы Есо RI, 4-гиб-
ридная (химерная) ДНК, Л-ДНК-лигаза.
Клонирование рДНК
Для клонирования рДНК используют так называемые
пермиссивные клетки (от англ, permission — разрешение).
Клон — это большое число молекул или клеток, идентичных
198
’ж
исходной родительской молекуле или клетке. К числу пермиссив-
ных клеток для включения в них рДНК относятся такие, в которых:
1) не разрушаются чужеродные ДНК или РНК собственными
ферментами — нуклеазами,
2) срабатывает механизм репликации вектора,
3) проявляется выраженная активность промотора и/или тер-
минатора транскрипции рДНК,
4) отмечается полный сплайсинг мРНК,
5) наблюдается эффективная трансляция мРНК,
6) нет выраженной активности пептидогидролаз, катализиру-
ющих реакции гидролиза экспрессируемых чужеродных белков.
Излюбленными объектами в генной инженерии являются клет-
ки Е. coli, Вас. subtilis, Saccharomyces cerevisiae и некоторые другие.
Е. coli — грамотрицательные, перитрихальные, прямые палочки
(1,1-1,5x2,0-6 мкм — живые, или 0,4-0,7x1,0-3,0 мкм — высушенные
и окрашенные); факультативный анаэроб, хорошо растет на про-
стых питательных средах при 37°С, содержание Г+Ц в ДНК 50-51
мол% (по температуре плавления и плавучей плотности). Некото-
рые серовары Е. coli патогенны для человека (01, 02, 06, 08, 09, 011,
015, 026, 055, 078, 0111, 0124, 0144, 0148 и др.). Е. coli чувствительна
к включению векторов.
Вас. subtilis — почвенные, спорообразующие, грамположитель-
ные, непатогенные, прямые, перитрихальные палочки (0,7-0,8 х
2,0-3,0 мкм), хорошо растет на простых питательных средах при
разных температурах (минимальная 5-20’С, максимальная 45-55°С),
содержание Г+Ц в ДНК от 32 до 62 мол%; обладает высокой
частотой трансформации (4%), с кольцевой хромосомой, на которой
известно более 2000 локусов; содержит сайт-специфические нук-
леазы, чувствительна к включению векторов.
Saccharomyces cerevisiae — аскомицетные, одноклеточные, сап-
рофитные дрожжи (группа низших грибов), широко используемые
в микробной биотехнологии для получения разных биологически
активных веществ (БАВ). Вегетативные клетки этого вида однокле-
точные, почкующиеся, круглые или, чаще, овальные, размером
2,5-11x2,5-30 мкм, хорошо растут на средах Сабуро, с пивным
неохмеленным суслом и других при температуре 26°С; содержание
Г+Ц в ядерной ДНК колеблется в средних пределах от 48 до 63
мол%; в отличие от бактерий они генетически более стабильны, не
подвергаются фаголизису, способны гликозилировать собствен-
ные белки (продуцируемые эукариотическими клетками белки, как
199
правило, функционально активны в форме гликопротеинов), их
геном лишен оперонов, обладают тремя классами РНК-полимераз,
и т.д. Другими словами — сахаромицетные дрожжи являются
типичными эукариотами, поэтому в них лучше клонировать гены
растений и животных. У S. cerevisiae имеется 17 хромосом, на
которых картировано свыше 600 генов. Для клонирования чуже-
родных ДНК в дрожжевых клетках нередко используют челночные
векторы, имеющие ori-сайты бактериальных и дрожжевых плаз-
мид, а поэтому способные самостоятельно реплицироваться в
бактериальных и дрожжевых клетках. На рис. 68 схематично
изображен челночный вектор иммунного интерферона (INF-y),
который реплицируется в клетках обезьяны и Е. coli. В этом векторе
содержится фрагмент из 342 пн, который включает ori-сайт и
промотор из вируса SV-40; данный фрагмент связан с фрагментом
кДНК, включающим кодирующую последовательность npe-INF-y,
и с фрагментом pBR322, содержащим ген резистентности к ампи-
циллину и ori-сайт репликации pBR322.
У дрожжей выявлены 3 вида циклически замкнутых плазмид:
двух- и трехмикронная, митохондриальная (длина 24 мкм) ДНК.
Первые две относятся к разряду "эгоистических" критических,
поскольку биологическое значение их остается скрытым, неясным.
Лишенные селективных маркеров они представляют ценность в
качестве векторов только после включения в них соответствующих
хромосомных генов дрожжей, функционирующих как в дрожже-
вых, так и в бактериальных (Е. coli) клетках. К таким генам
относятся те из них, которые отвечают за биосинтез таких фер-
ментных белков, как аргининсукцинатлиаза, аспартат-транскарба-
милаза, галактокиназа, дигидрооротатдегидрогеназа, триптофан-
синтаза и др.
Известно 5 типов дрожжевых векторов, образующих У-группу
(от англ. Yeasts — дрожжи): Yip, YEp, YRp, YCp, YLp. Первые из
них (Yip) представляют собой интегративные плазмиды, в которых
только один ген принадлежит дрожжевой ДНК, остальные гены
относятся к бактериальным. Yip не способны реплицироваться в
клетках S. cerevisiae, но осуществляют их трансформацию, интег-
рируясь в хромосому через рекомбинацию.
Векторы YEp — эписомальные плазмиды, сконструированные
на основе двухмикронной плазмиды. Частота трансформации S.
cerevisiae этими векторами на три порядка выше, чем в случае
использования Yip, но, к сожалению, получаемые трансформанты
мало стабильны.
200
Векторы YRp — репликативные плазмиды, включающие хро-
мосомные репликаторные последовательности, именуемые ars-no-
следовательностями (от англ, autonomously replicating sequence —
автономно реплицирующаяся последовательность). Трансформан-
ты дрожжей, получаемые с помощью YRp, нестабильны.
Векторы YCp в виде кольцевых мини-хромосом содержат цен-
тромеры дрожжевых хромосом и репликаторы arsl и ars 2. Они
стабильны при митозе и мейозе, и наиболее перспективны для
клонирования генов.
Векторы YLp — линейные мини-хромосомы, конструируемые
на базе YCp. Для этого вводят специфические, шпилечные по
форме, нуклеотидные последовательности на концах хромосом
(теломеры) в кольцевые плазмиды YCp с последующей линеариза-
цией плазмид.
Плазмиды и фаги обеспечивают перенос чужеродной ДНК в
качестве инертной части генома, поэтому их называют еще кло-
нирующими векторами. В биологической технологии
выгодны мультикопийные плазмиды (10-20 на клетку). Если же
плазмиды находятся под ослабленным контролем репликации,
когда прекращается размножение бактерий, то они (плазмиды)
накапливаются числом до 1000 на клетку — в результате больше
образуется целевого продукта. В отличие от хромосомы репликация
плазмиды в пермиссивной клетке может происходить при останов-
ке синтеза белка, Вот почему, например, при добавлении левоми-
цетина к культуральной жидкости сопровождается заметным воз-
растанием числа копий плазмиды (увеличение степени клониро-
вания).
Для включения рДНК в пермиссивные клетки пользуются
различными методами: трансформации, инфекции, микроинъек-
ций. При естественной трансформации рДНК может проходить
через интактную клеточную стенку Вас. subtilis, Streptococcus
pneumoniae, некоторых штаммов Е. coli с ингибированной систе-
мой деградации ДНК, и др.
В 1970 г. М. Мандель и А. Хига предложили метод обработки
бактерий хлористым кальцием на холоду в целях трансформации
ДНК фага в клетки Е. coli. Этот метод используется и теперь. Этапы
его примерно следующие:
1) выращивание культуры в питательной среде при оптималь-
ной температуре в течение суток с последующим разведением
примерно в 50 раз свежим питательным бульоном, выращиванием
201
до Абоо=О,2 (показатель плотности) и охлаждение во льду;
2) центрифугирование суспензии при 3000 g 10 мин, темпера-
туре 4°С и разведение осадка клеток в 0,1 М CaCh, охлажденного
во льду, с последующим центрифугированием при тех же условиях;
3) ресуспендирование осадка в минимальном количестве (на-
пример, в 1 мл) охлажденного во льду 0,1 М растворе CaCh,
хранение до использования при 4°С. Это и будут пермессивные
компетентные клетки, готовые претерпевать трансформацию; если
такие клетки, ресуспендированные в 0,1 М CaCh, 30% глицерине
заморозить при -70°С, то их можно хранить в течение нескольких
месяцев;
4) добавление плазмидной ДНК к компетентным клеткам (при-
мерно 0,1 мкг ДНК в 0,2 мл суспензии клеток), выдерживание во
льду 1 час;
5) тепловой шок смеси при 42° С 5 мин.;
.6) разбавление пробы в 10 раз питательным бульоном, инкуба-
ция 1 час на качалке при 37°С; за это время, например, признак
резистентности к какому-либо антибиотику успевает экспресси-
роваться;
7) посев клеток в серии разведений на чашки Петри с пита-
тельным агаром, содержащим соответствующий(-ие) антиби-
отик (-и), выдерживание в течение суток при оптимальной темпе-
ратуре;
8) изоляция нужной колонии и размножение клона.
рДНК можно ввести в клетки млекопитающих путем слияния
протопластов, например Е. coli, содержащих рДНК, с культивиру-
емыми эукариотическими клетками в присутствии полиэтиленгли-
коля, или ПЭГ, способствующего слиянию протопластов.
Перенос рДНК в клетки прокариот или введение последова-
тельностей клонированной ДНК в рецепторные эукариотические
клетки (например, млекопитающих) с помощью фаговых векторов
называют трансфекцией. Альтернативный метод —
использование вирусов эукариот, то есть когда эукариотическая
клетка инфицируется — (заражается) вирусом. В качестве
векторов при этом чаще используют литические вирусы типа
вируса полиомы и SV40, а также челночные векторы, сконструи-
рованные на основе ретровирусов и папилломавирусов.
На растительных объектах можно выполнить генно-инженер-
ные эксперименты, используя методы трансформации и инфици-
рования (см. рис. 141).
202
Метод микроинъекцйи используют в случаях механического
введения ДНК (в том числе рДНК) в ядра культивируемых клеток
млекопитающих, растений, используя для этого стеклянные мик-
роиглы (рис 69). Этот метод впервые продемонстрировали Дж. Е.
Мерц и Дж. Б. Гердон в 1977 г. на ооцитах лягушек Xenopus.
Рис. 69. Микроинъекция в ядро
( чЯездЦ) 4 ооцита (1 — ооцит, 2 — пигментиро-
\ ванная половина, 3 — микропипет-
\ ка, — ядро ооцита, 5 — часть
_____ х5 нейлоновой сетки для чашки Петри).
рДНК можно переносить в пермиссивные компетентные клет-
ки с помощью липосом, приготавливаемых из смеси фосфатидил-
серина и холестерина (1:1). Например, удается ввести ген Р-лакта-
мазы из плазмиды pBR322 в отрицательно заряженные липосомы
при обработке их ультразвуком. Последующая инкубация "озву-
ченных" липосом с клетками различных культур сопровождается
приобретением р-лактамазной активности у этих клеток. На плот-
ной питательной среде клетки, несущие рДНК, размножаются и
огромное потомство их (при этом все клетки тождественны) назы-
ваются клоном. Сумму всех клонов, в которую входят почти все
кДНК, соответствующие множеству разных мРНК, образуемых
клеткой, называют "библиотекой кДНК".
Амплификация и экспрессия генов
Амплификация — это образование дополнительных копий
хромосомных последовательностей. Обычно в биотехнологии при-
меняют малые плазмиды длиной около 3 мкм в разомкнутом
состоянии. Такие плазмиды не передаются при конъюгации,
например, прокариотических клеток. Эти плазмиды называют
нетрансмиссибельными. Однако они могут быть
переданы методом трансформации. В результате амплификации
на одну клетку нередко приходится 10-30 копий нетрансмиссибель-
ных плазмид, что существенно повышает клеточную продуктив-
ность применительно к целевому продукту (белку). Схематично
амплификация представлена на рис. 70.
203
Рис. 70. Амплификация нетрансмиссибельной плазмиды в реципиентной про-
кариотической клетке (схема): 1 — клетка-донор (а — нуклеотид, б — нетрансмис-
сибельная плазмида), 2 — клетка-реципиент, 3 — лизис, 4 — обработка раствором
кальция хлорида на холоду (или протопластирование, голодание, обработка
ионами лития), 5 — клетка реципиент с “разрыхленной” клеточной стенкой, 6 -
трансформация, 7 - клетка-реципиент с переконструированной клеточной стенкой,
содержащая плазмиду, 8 - реконструкция клеточной стенки, 9 - клетка-реципиент
с реконструированной клеточной стенкой, содержащая плазмиду, 10 - амплифи-
кация плазмиды, 11 - плазмиды (25), образовавшиеся в реципиентной клетке в
результате амплификации.
Множественные (аплифицированные в миллион раз) копии
определенных фрагментов ДНК получают in vitro с помощью
широко используемой в настоящее время полимеразной
цепной реакции, или ПЦР. Ее впервые описали в 1985 г.
Р. К. Сейки, С. Шарф, Ф. Фалуна, К. Б. Муллис, Г. Т. Хорн, X. А.
Эрлих и Н. Арнхейм. ПЦР высоко специфична и чувствительна, с
ее помощью можно обнаружить, размножить и исследовать даже
единичную копию гена в исходном материале. В течение трех часов
обычно протекает 30 циклов, амплификации.
Для постановки ПЦР необходимо знать последовательность как
минимум 17 пн с обеих сторон исследуемого фрагмента ДНК.
Обычно синтезируют два дезоксинуклеотида длиной 20-30 основа-
ний, представляющие собой концевые последовательности инте-
ресующего нас фрагмента ДНК. Используя комплементарные к
этим участкам олигомеры — праймеры, запускают амплификацию.
Избыточные количества праймеров смешивают с геномной ДНК,
а затем последовательно осуществляют реакции денатурации, от-
жига (реассоциации) и наращивания цепи (удлинение праймера).
Тепловая денатурация сопровождается расплетением двойной спи-
рали ДНК. При снижении температуры имеет место отжиг олиго-
нуклеотидов, то есть происходит гибридизация олигонуклеотидных
праймеров со своими комплементарными последовательностями.
Наращивание цепи праймеров катализируется ДНК-полимеразой
в присутствии дезоксирибонуклеозидтрифосфатов, и в результате
достраивается новая комплементарная цепь ДНК.
При повторных циклах тепловой денатурации, отжига и синтеза
образовавшиеся новые нити ДНК выступают в качестве шаблонов
(матриц) для праймеров, что вызывает экспоненциальное нараста-
ние участка ДНК, ограниченного праймерами, входящими в состав
этих фрагментов.
К завершению 20-30 циклов амплификации фрагменты ДНК
представлены с избытком в сравнении с остальной геномной ДНК
со свободными концами и образовавшейся в первом цикле ампли-
фикации (рис. 71-1).
Практически эффективность каждого цикла амплификации
составляет 20-50% от теоретически возможной (2го > 106при 100%-й
эффективности). Следовательно, можно получить возрастание ко-
личества специфической последовательности кратное миллиону.
В случаях амплификации геномной ДНК из клеток (тканей)
позвоночных животных иногда происходит накопление фрагмен-
тов неясного происхождения. Тогда проводят серию дополнитель-
ных амплификаций, применяя другой набор олигонуклеотидных
праймеров — "nested primers” (от англ, nest — гнездо, гнездиться,
primer — начальный, основной). В этой связи и метод называют
nested oligo" (от греч. oligos — малый). В данном методе продукт
первичной ПЦР используется в качестве матрицы в последующих
циклах ПЦР, но уже с двумя другими олигонуклеотидами, имею-
щими гомологии с участками ДНК внутри первичного амплифи-
цированного фрагмента. При этом получается множество укоро-
ченных (по сравнению с исходным) фрагментов индивидуальной
последовательности ДНК, которые затйэм подвергаются необходи-
мому анализу.
Таким образом, под названием ПЦР понимают основной цикл
денатурации, отжига и синтеза фрагментов ДНК в присутствии
ДНК-полимеразы, дезоксирибонуклеозидтрифосфатов и прайме-
ров. Процесс по существу является цепным, так как продукты
данной реакции служат матрицей для последующих реакций.
На первых этапах использовали в ПЦР так называемые "кле-
новские фрагменты" ДНК-полимеразы I Е. coli (названы по фами-
лии ученого — X. Кленова), которые оказались термолабильными,
и после каждого цикла денатурации необходимо было добавлять
новую порцию фермента. Кроме того, при пониженной темпера-
туре отжига снижалась специфичность амплификации.
С выделением термостабильных ДНК-полимераз, в частности,
из бактерий Thermos aquaticus (Тад-полимераза)и Pyrococcus
205
t=j 1=1 = а
| Цикл 1
Цикл 2
Циклы 3-25
□ = 6
DDDDDD = в
□ODDDOD [> = г
ГГНУ1ЧП
ммм^м2
<-2
в
30 (0,5 рМ)
O.SpMDS ДНК
1 ааммавма
5-10 РМ SS ДНК
Рис. 71. Схема полимеразной цепной реакции (ПЦР): I — в классическом
исполнении (а—ДНК-мишень, б — праймер в ПЦР. в — новая ДНК, г — направление
амплификации, дс — денатурация и синтез); II — заякоренная ПЦР (1 — мРНК. 2
— синтез кДНК, 3 — наращивание полидезоксигуанозинового конца, 4 — приго-
товление заякоренного полидезоксицитидинового праймера и синтез второй нити
ДНК, 5 —добавление специфического праймера, амплификация при использовании
обоих праймеров, 6 — концевой праймер, 7,8 — якорный праймер); III — инверти-
рованная ПЦР (1 — кор с известной нуклеотидной последовательностью, 2 — место
действия рестриктазы, 3 — "переваривание", 4 — циклизация, 5 — рестрикция в
пределах кора, , 6 — добавление праймеров, дезоксинуклеозидтрифосфатов и
Taq-полимеразы, 7 — амплифицированная область, фланкирующая кор); IV —
асимметричная ПЦР (А — праймер в избытке, 50 пикмолей -рМ; В — лимитирован-
ный праймер — 0,5 рМ; ц — циклы, ds — двухнитевая ДНК, ss — однонитевая ДНК).
woesii (Pwo-ДНК-полимераза), существенно возросла эффектив-
ность ПЦР. Taq-полимераза сохраняет активность после тепловой
денатурации ДНК и не возникает необходимости заменять фер-
206
мент после каждого цикла. Температурный оптимум реакции
составляет 70-75’С, что значительно повышает специфичность,
количественный выход и возможную длину копий амплифициру-
ющихся фрагментов ДНК.
На рис. 71-1 полностью показаны лишь два первых цикла.
Начиная с третьего цикла, не отражена судьба исходной ДНК и
продуктов достройки, синтезированных на ее цепях. Длинные
! фрагменты, синтезируемые на исходной цепи, накапливаются в
/ арифметической прогрессии, тогда как короткие фрагменты, ог-
раниченные на концах праймерами и впервые появляющиеся в
X конце третьего цикла амплификации, накапливаются в геометри-
! ческой прогрессии.
Недостатки ПЦР: возможное загрязнение при взятии образцов
! материала для анализа, для каждой пары праймеров требуются
индивидуальные условия реакции, ПЦР опирается только на точ-
ную информацию о секвенировании последовательностей.
Однако преимущества ПЦР несомненны и практические воз-
можности ее огромны. Затравочным материалом для ПЦР можно
использовать мРНК. Тогда с помощью обратной транскриптазы
; f синтезируют кДНК, которую затем используют в качестве матрицы
[ для амплификации. В случае применения одного известного прай-
} мера, второй праймер будет искусственным, представляющим
собой якорную последовательность, присоединенную к гомополи-
| мерному "хвосту". Такой подход называют якорнойПЦР (рис.
| 71-П). Известна еще инвертированнаяПЦР, когда пара
i праймеров отжигается с известной последовательностью, но ори-
I ентируется таким образом, что синтез идет в противоположном
направлении через зону с неизвестной последовательностью (рис.
71-Ш).
В 1988 г. описана асимметрическаяПЦР, в которой
происходит амплификация однонитевой ДНК (рис. 71-IV).
р В 1987 г. фирмой Perkin-Elmer Cetus (США) разработана авто-
j матизированная приборная техника для ПЦР, которая благотворно
сказалась на генетических исследованиях. В 1991 г. этойже фирмой
предложена вторая генерация ПЦР-технологии с использованием
амплификатора 9600 (Gen Amp PCR System 9600), благодаря кото-
рой удалось достичь новых порогов чувствительности и продук-
тивности ПЦР (таблица 21).
Фирма "Ока" (г. Пущино, Московская область) предлагает
собственный оригинальный амплификатор высокого качества, по
стоимости более дешевый, чем зарубежные аналоги.
20/
Т а б л и ц а 21. Сравнительные данные ПЦР при использовании автоматизиро-
ванных приборов — амплификаторов первого (ДНК-термоцик-
лера 480) и второго (Gen Amp PGR System 9600) поколений.
ПЦР, мишень t°,°C Скорость цикла (мин.) в:
термоциклере 480 генном амплификаторе 9600
единичного полного единичного ПОЛНОГО
Фаг X 94 1 4,33 0,1 2,42
68 2 4,33 1,25 2,42
ИЛ1 а 94 1 4,8 0,25 2,05
68 1 4,8 0,25 2,05
ВИЧ-1 95 1 5,0 0,25 2,62
60 2 5,0 1,0 2,62
HLA-DQa 94 1 4,58 0,5 1,92
60 0,5 4,58 0,16 1,92
72 0,5 4,58 0,16 1,92
Функциональная активность генов проявляется в периоды
транскрипции и трансляции и называется экспрессией
генов. Степень сродства РНК-полимеразы к промотору оперона
обусловливает эффективность транскрипции, а стабильность
мРНК и ее способность связываться с рибосомами обусловливает
эффективность трансляции.
Экспрессия прокариотических генов преимущественно зави-
сит от выбора промотора в клонируемых генах тех же или близ-
кородственных видов. Чем отдаленнее геномы прокариотических
клеток друг от друга, тем система "транскрипции-трансляции"
генов срабатывает хуже или вообще не срабатывает. Вот почему
здесь важными оказались векторы.
Экспрессия эукариотических генов в прокариотических клет-
ках не реализуется или реализуется с большим трудом. В ядерных
генах эукариот имеются интроны, а у бактерий нет системы
сплайсинга, поэтому образующиеся чужеродные конечные про-
дукты в бактериальных клетках, как правило, неактивны. Однако
использование векторных систем (в том числе "челночных") и ПЦР
позволило успешно решать проблемы, связанные с созданием
рДНК их клонированием и экспрессией в различных реципиент-
ных клетках (прокариотических и эукариотических).
Экспрессия генов сопровождается, как правило, выработкой
белковых молекул. Поэтому важно использовать такие клетки,
208
несущие рДНК, которые секретировали бы белок во внешнюю
(культуральную) среду. Если этого не происходит, то для получения
целевого продукта приходится разрушать (дезинтегрировать) клет-
ки и уже из "экстракта" — дезинтеграта выделять соответствующий
белок. Е. coli, выступая на первых порах в качестве монопольной
"рабочей лошадки" биотехнологии, оказалась затем менее пригод-
ной, чем, например, Вас. subtilis, Р. aeruginosa, S. cerevisiae и другие
виды, в отличие от которых Е. coli почти совсем не секретирует
белки. Лишь недавно с помощью BRP-векторов удалось преодолеть
барьерную функцию оболочки Е. coli и трансформировать ее в
секретирующую клетку (от анг. Bacteriocin Release Protein).
В настоящее время, на основании экспрессии соответствующих
генов в рДНК удалось внедрить в производство штаммы бактерий
и дрожжей, продуцирующие инсулин, соматотропин, интерферо-
ны, интерлейкины и т. д. Получены штаммы — суперпродуценты
некоторых веществ; выведены бактерии, способные очищать ка-
менный уголь от серы и превращать его в жидкое и газообразное
топливо. Применительно к фитобиотехнологии удалось создать
растения повышенной кормовой и питательной ценности, с воз-
росшей активностью фотосинтеза и азотофиксации, резистентные
к некоторым микробам — фитопатогенам и др.
Генно-инженерные разработки в области зообиотехнологии (в
частности^ соматотропином, или гормоном роста) оказались важ-
ными для животноводства (возрастание привесов животных, лак-
тации). Используя метод микроинъекций рДНК в зародышевые
клетки млекопитающих (в пронуклеусы оплодотворенных ооцитов,
или в яйцеклетки) удается получать так называемых транс-
генных животных. Клоны реципиентных клеток, содержащие
донорный хромосомный материал, или трансгены, различаются
по количеству его в широком диапазоне, и если функциональная
активность и регуляция экспрессии клонированных генов изуча-
ются на целом организме, то и говорят о трансгенном
организме. Схема получения, например, трансгенных мышей
введением клонированной ДНК в оплодотворенное одноклеточное
яйцо приведена на рис. 72.
С получением трансгенных животных открываются перспек-
тивы развертывания заманчивых исследований на рубеже XX и
XXI вв. Уже сегодня имеется информация о том, что в Эдинбурге
(Великобритания) есть стадо овец, несущих гены человека, инду-
цирующие выработку факторов свертываемости крови (борьба с
гемофилией у человека благодаря, например, использованию в
209
линия C57BL/B6 линия PARKES
линия CBA/J
спаривание
самец
гиперовуляция *
псевдобеременная
реципиентная
самка
коллекция преимплантированных
эмбрионов
оплодотворенное
одноклеточное яйцо
мышь-кормилица
выделение и
очистка ДНК
| микроинъекции |
кесарево сечение
и вскармливание
рождение
естественным путем
выделение ДНК
из тканей хвоста *—
анализ выделенной
ДНК
идентификация трансгенных
мышеи
скрещивание
анализ экспрессии генов
Рис. 72 Схема получения трансгенных мышей.
210
пищу овечьего молока, где содержались бы такие факторы); удалось
ввести ген гормона роста крупнорогатых животных в геном клеток
поросят, заставили расти их быстрее и с меньшим количеством
жира. Однако подобные результаты, очевидно, будут сопряжены
и с неудачами. Необходимы кропотливые исследования во всех
направлениях генетической инженерии, чтобы добиться успехов
в практическом воплощении наиболее приемлемых результатов в
биологическую технологию.
В связи с экспрессией генов большое значение приобретает
"белковая инженерия”, дорожку которой проторила генетическая
инженерия, первая вытекает из второй и, следовательно, белковая
инженерия также является методом науки "Биологическая техно-
логия". Целевые установки здесь сводятся к изучению и понима-
нию главного звена в структуре ферментов (почему они функци-
онируют так, а не иначе?), к возможности научиться видоизменять
природные белки или, того больше, уметь их заново проектировать,
к выяснению причин и механизмов изменения фенотипа под
влиянием генотипа.
Природные белки имеют свою молекулярную архитектонику
— то это вытянутые, то специфически изогнутые или скрученные
структуры. Поэтому в научной литературе, посвященной белковой
инженерии, можно встретить выражение "архитектурный дизайн”,
кода говорят о конструировании молекул белков с заданной архи-
тектурой по заведомо и специально подобранным аминокислотным
текстам. Белковая инженерия в любом проявлении будет базиро-
ваться в основном на продуктах экспрессии естественных и ре-
комбинантных генов.
Таким образом, последовательность этапов клонирования и
экспрессии гена может протекать в следующей очередности:
1) сортинг хромосом (может быть автоматизированным) и
получение ДНК (или только выделение ДНК, например, из прока-
риотических клеток);
2) введение ДНК в вектор (плазмидный);
3) трансформация (трансфекция, инфекция, инъекция);
4) обнаружение и накопление клона (клонирование) клеток;
5) выделение рДНК (плазмиды);
6) определение нуклеотидной последовательности в клониро-
ванном фрагменте рДНК (может быть автоматизировано);
7) конструирование и построение плазмиды для экспрессии ее
функций;
8) трансформация;
211
9) обнаружение и накопление клона;
10) выделение плазмиды;
11) проверка нуклеотидной последовательности в рДНК (может
быть автоматизировано);
12) трансформация;
13) выращивание культуры на предмет получения экспресси-
руемого белка;
14) выделение белка.
Каковы же место и роль рДНК-биотехнологии в человеческом
обществе?
На фоне растущих народонаселения на планете Земля и эко-
логических бед место и роль рДНК-биотехнологии определяются
потребностью людей в пищевых продуктах и, прежде всего, в
белках, в различных лекарствах; в исправлении многочисленных
наследственных заболеваний. В общенаучном плане важными
являются исследования: а) процессов переноса генетической ин-
формации между видами и расшифровки механизмов функциони-
рования клеток;
б) путей и методов создания новых существ — химер и новых
организмов, возможно — монстров;
в) эволюции различных существ, включая человека (при допу-
щении альтернативной оценки всего живого как вечной материи,
подвергающейся изменчивости в пространстве и времени).
5.3. Изменчивость организмов и ее значение в биотехнологии.
С каждым годом взаимоотношения различных организмов с окру-
жающей средой все более осложняются—растет народонаселение
Земли и неуклонно истощаются ее ресурсы. Уже сегодня количе-
ство веществ, загрязняющих биосферу, трудно поддается учету
или, в ряде случаев, не поддается совсем. По имеющимся данным
первое место среди загрязнителей природы занимают пестициды
(от лат. pestis — зараза, caedo — убиваю), второе — тяжелые
металлы. С 1985 по 1990 год химическая промышленность произ-
водила ежегодно не менее 250 млн. тонн продукции, из которых
до 30% поступало в окружающую среду.
Несмотря на резкое отрицательное отношение людей к атом-
ным электростанциям, не просматривается альтернативы термо-
ядерной энергии, которая вместе с генной инженерией могут
надолго обеспечить выживание человечества на нашей планете. С
другой стороны, возрастание в обществе роли ядерных изотопов
влекут за собой увеличение числа нежелательных мутаций (от лат.
mutare — превращаться) в соматических и половых клетках. Все
212
это не может не отразиться на изменчивости организмов, включая
и те из них, которые используются в биотехнологии.
Под изменчивостью понимают возникновение организмов с
новыми свойствами (признаками) среди существующего разнооб-
разия живых организмов. Из трех типов изменчивости — моди-
фикации, длительные модификации, мутации — наибольшее зна-
чение для биотехнологии имеют мутации. Индуцированные мутан-
ты теперь широко используются в производстве антибиотиков,
аминокислот, ферментов и других продуктов.
Длительные модификации присущи только клеточным формам
из надцарств прокариот и эукариот, но не акариот. При этом
генотип клеток остается неизменным, а измененный фенотип,
проявляющийся в формировании какого-либо устойчивого мета-
болического цикла, кажущегося независимым от внешних условий,
выступает мерилом автономности внутриклеточной среды. При
многократных пересевах происходит как бы разбавление цитоп-
лазмы в ряду последующих поколений клеток и исходные метабо-
литы материнской клетки достигают ниже порогового предела,
когда указанный выше метаболический цикл исчезает. В других
случаях такой же эффект наблюдается при создании окружающих
условий, не поддерживающих один из типов метаболизма и клетки
из состояния длительной модификации возвращаются в исходное
с присущей виду комбинацией признаков (фенотип). Таким обра-
зом, при возникающих или исчезающих модификациях наблюда-
ются массовые однонаправленные изменения клеток однородного
генотипа, находящихся в среде обитания.
Особенности генотипа определяют фенотипические проявле-
ния организма в заданных условиях. Поэтому и адаптацию можно
определить как приспособление наследственно компетентной клет-
ки (культуры, организма) к конкретным условиям существования.
При мутациях изменяется генотип, или наследственная консти-
туция клетки (организма). Мутации могут быть неконтролируемы-
ми (спонтанными) и контролируемыми (индуцированными). Пер-
вые происходят с частотой 1 • 10’5 — 1 • 10'7 на одну генерацию для
разных организмов; это значит, что за одну генерацию ненаправ-
ленно изменяется одна клетка из 100 000 или из 106 клеток. Частота
индуцированных мутаций возрастает в десятки-тысячи раз.
Факторы, вызывающие мутации, относят к разряду физиче-
ских, химических или биологических, и называют мутагенами.
Мутагены по действию подразделяют на два класса — прямого (на
нуклеиновые кислоты) и непрямого, или опосредованного дейст-
213
вении из клеточной воды свободных радикалов, вступающих в
реакции с кислородом клеточных структур (образование переки-
сей), с органическими молекулами или взаимодействующих между
собой. Происходит окисление или передача энергии молекулам
ДНК (РНК). Свободно-радикальные процессы могут завершиться
дезаминированием и дегидроксилированием оснований, разрывом
N-гликозидных связей между основаниями и пентозой, деструк-
цией пиримидинов, окислением пентозы, высвобождением пиро-
фосфата. Вот почему различные излучения могут индуцировать
самые разнообразные мутации. Так, например, одновременный
разрыв сестринских хроматид у традесканции составил (по числу
разрывов на 100 клеток/г) 0,99 — для нейтронов (Li + D); 2,1 —
для ос-частиц (родон) и 3,02 — для тепловых нейтронов; 0,27; 0,26
и 0,44 — для рентгеновских лучей с X 0,015нм, 0,15нм и 0,41 нм
соответственно. Соотношения УФЛ и ионизирующих излучений
приведены в таблице 22.
Т а б л и ц а. 22. Некоторые характеристики части электромагнитного спектра
Название диапазона Д лина волны, нм. Частота, гц Энергия кванта, эв
Далекая область инфракрасного излучения 3-105 • 1012 —
Инфракрасныйдиапазон (от 770 до 4 • 10* нм) 3-104 1013 —
Инфракрасный з-ю3 1014 1
Видимый свет (от 390 до 770 нм) 3-102 ю15 10
Ультрафиолетовый диапазон (от 13,6 до 390 нм) 30 1016 ю2
Мягкие рентгеновские лучи 3 10П 103
Средние рентгеновские лучи 0,3 ю18 ю4
Жесткие рентгеновские лучи 0,03 ю19 10s
Жесткие рентгеновские и у-лучи 0.003 0,0003- 0,000003 ю20 1021- ю23 106 ю7- ю9
Применительно к ультразвуковым колебаниям (акустические
колебания частотой свыше 2 • 104 герц, гц) считают, что под их
влиянием наблюдаются последовательные изменения вначале в
пиримидинах, , а затем — в пуринах.
216
Ежегодно в мире синтезируется не менее 250 тысяч новых
химических веществ, многие из которых (особенно — при круп-
номасштабном производстве) попадают в окружающую среду.
Подсчитано, что примерно 10% человечества подвергается дейст-
вию потенциально мутагенных и токсичных химических соедине-
ний.
Становление и развитие работ по химическому мутагенезу
связано с исследованиями отечественных генетиков: В. В. Сахарова
(1933), М. Е. Лобашова (1934), И. А. Рапопорта (1938), а также
зарубежных ученых: Ш. Ауэрбах (1940), Вестергаард (1959), Мандол
и Гринберг (1960) и др. В 1966 г. И. А. Рапопорт предложил термин
"супермутагены" для веществ, обладающих исключительно высо-
кой мутагенностью и заметно не влияющих при этом на жизне-
способность клеток и организмов.
К химическим мутагенам относят: ингибиторы синтеза нукле-
иновых кислот (НК), аналоги азотистых оснований, алкилирующие
соединения, окислители, восстановители, свободные радикалы,
акридиновые красители, некоторые антибиотики (таблица 23).
Т а б л и ц а 23. Некоторые химические мутагены и супермутагены
Группа Мутаген Супер- мутаген По химическому устроению
1 2 3 4 ’
Ингибиторы синтеза предшественников НК Азасерин Азогуанидин 2-Амино-6-гидроксила- минопурин (АГАП) Бензимидазол 5-Бромурацил 2,6-Диаминопурин Гидразиноурацил N-6-Г идроксилами- нопурин (ГАП) Кофеин 6-Меркаптопурин Параксантин Тетраметилмочевая кислота Уретан 5-Фтордезоксиуридин Этил-уретан 8-Этоксикофеин II 1 1 1 1 1 Illi 1 + 1 + 1 Диазосоединение Пурин Пурин Бензимидазол Пиримидин Пурин Пиримидин Пурин Пурин Пурин Пурин Пурин Карбаминовая к-та Пиримидин Карбаминовая к-та 1 Пурин
217
(Продолжение таблицы 23)
1 2 3 4
Алкилиру- Азасерин — Диазосоединение
ющие соединения Афлатоксин Bi 1,4-Бис-диазоцетилбутан + Производное фурана
(ДАБ) + Диазоалкан
Бутилхлорэтилсульфид — Хлорэтилсульфид
Глицидол — Эпоксид
Диметилсульфат — Диалкилсульфат
Ди этилнитро зам ин + N-Нитрозозамещенное соединение
Диэпоксибутан — Эпоксид
Диэтилоксибутан + Алкан
Диэтилсульфат — Диалкилсульфат
Иприт N-Метил-бис /хлорэтил- + Хлорэтилсульфид
амин '(аналог иприта) + Хлорэтилсульфид
Метилметансульфонат ЬБМетил-М'-нитро-ЬГ- — Алкилалкансульфонат Нитрозозамещенное
нитрозогуанидин + соединение
Митомицин С — Антибиотик
1Ч-Нитрозо-1Ч-метилуретан М-Нитрозо-М-этил- + Диазосоединение
мочевина + N-Нитрозозамещенное соединение
Нитрозометилоксаимид + N-Нитрозозамещенное соединение
Пропиленоксид — Эпоксид
Р-Пропиолактон Фенол (карболовая Лактон
кислота) — Фенол
Формальдегид — Альдегид
Эпихлоргидрин — Эпоксид
Окислители Азотистая кислота — Кислота
Гидроксиламин — Амин
Диоксид водорода — Перекись
Удлинители нитей ДНК Акридиновый оранжевый — Акридиновый краситель
Бромистый этидий — Производное изохинолина
Профлавин — Акридиновый краситель
Ингибиторы синтеза РНК Актиномицины — Антибиотики
Комплексное Н, ОН — Свободные радикалы
действие на ДНК. Стрептомицин,гигро- мицин В и др. Антибиотики
218
Как видно из таблицы 23 среди ингибиторов синтеза предше-
ственников НК имеются антиметаболиты (азогуанидин, 5-аминоу-
рацил, 6-меркаптопурин, 5-фтордезоксиуридин). Нуклеозиды, со-
держащиеся в клетках или добавляемые извне, играют роль анти-
мутагенов, снижающих мутагенный эффект названных веществ.
На примере 5-аминоурацила —
аналога урацила можно показать
путь возникновения 50% потомков
после двух циклов репликации му-
тировавшей клетки под действием
мутагена. 5-Аминоурацил по хими-
ческому строению близок к тимину
и поэтому легко комплексируется
с аденином. Кето-форма 5-аминоу-
рацила более стабильна, чем его
энольная форма, однако последняя
образуется и существует какое-то
время, в течение которого он мо-
жет спариваться с гуанином — это
так называемая ошибка включе-
5-аминсурацил
(кето-форма)
5-аминоурацил
(енольная форма) гуанин
ния. Тогда очевидно, что вместо пары Г-Ц появится пара А-Т или
А-5аминоУ (А-амУ). Аналогичная ситуация мо-
жет складываться при использовании 5-брому-
рацила (5-БУ) и прибавка в числе мутантов
будет однократной. Мишень действия этого
мутагена — фермент рибонуклеотидредукта-
за. Мутации подобного рода происходят с не-
большой частотой, поскольку они всецело за-
висят от времени существования аналога нук-
леинового основания в энольной форме (а
время это, как прави-
ло, небольшое).
Смещение в комп-
лементарности пар,
или ошибка реплика-
ции может происхо-
дить по другой схеме:
219
xr>
АЛ?
Мутация здесь сохраняется до конца репликации ДНК, содер-
жащей 5амУ (или БУ) и общее число мутаций возрастает с ростом
числа циклов репликаций.
Подобный мутагенный эффект оказыва-
ют и другие аналоги НК, например 6-мер-
капто- или 6-аминопурин. Мутагенное дей-
ствие 2-аминопурина проявляется на бакте-
риях и бактериофагах, хотя в норме он
имеется в цианофагах. Будучи аналогом азо-
тистых оснований нуклеиновых кислот, 2-
аминопурин (Ап) индуцирует ошибку вклю-
Hjlv XN
U
2-амююпурин
чения. В обоих случаях произош-
ли мутации типа транзиций (см.).
Ап может вызывать также и му-
тации типа трансверзий (см.), Хо-
тя и редко. Для всех аналогов
азотистых оснований характер-
ны прямые и обратные мутации
типа простых замен, например
А=Т на Г = Ц или Ц = Г на Т=А
(обратные мутации здесь Г s Ц
ошибка включения
заменяется наА=ТиА=Т — на
Г=Ц).
Азасерин относится к двум подгруппам (см. таблицу 23). Его
мутагенный эффект связывают с нарушением биосинтеза пуринов
de novo и с радиомиметическими свойствами (от греч. mimicos
имитирующий).
Афлатоксин Bi вызывает сдвиг рамки считывания ДНК. Это
можно показать на следующем примере:
ТГГГГАТГЦ
АЦЦЦЦТАЦГ
выпадение одного Г
добавление одного Г
Супермутагены часто приводят к мутациям, возникающих на
пути 2. Кофеин в больших дозах оказывает прямой мутагенный
эффект, вызывая разрывы хромосом, например в клетках дрозо-
220
филы, человека и других эукариот. В меньших концентрациях он
действует как синергист с радиацией, ингибируя репарирующие
ферменты.
В основе алкилирования НК лежат такие реакции как депури-
низация с разрывом сахарно-фосфатной цепи в ДНК; взаимодей-
ствие алкилирующих агентов с адениловой, цитидиловой и тими-
диловой кислотами с последующей утратой способности алкили-
рованных А, Ц и Т спариваться с комплементарными основаниями;
взаимодействие с фосфатными группами НК; взаимодействие двух
функциональных групп алкилирующего агента с нуклеофильными
труппами НК. Так, например, в случае удаления .гуанина из ДНК
под влиянием какого-либо алкилирующего агента могут обнару-
житься различные варианты изменений: восстановление исходной
пары Г - Ц, ее выпадение, перекрестная замена Г = Ц на Т = А
или на Ц = Г, простая замена Г = Ц на А = Т.
Супермутагены, вызывающие до 24% летальных мутаций, в
основном относят к труппе алкилирующих агентов. Эту группу
подразделяют на следующие подгруппы:
1. Металлорганические соединения R-Me, где R— органический
радикал; например метилированная ртуть;
2. Галоидные алкилы (Алк) — (nH2n+i)R, где R — фтор, хлор,
бром, йод; например, 1,2-дихлорэтан, 1,2-дибромэтан, 1,3-дихлорп-
ропан;
3. Производные этилена (СН2 = СН-R, где R — двуокись азота,
двуокись серы и др.); например, акрилоилэтиленимин;
4. Альдегиды (R-СОН, где R=H, СН2-СН- и др.); например,
формальдегид, акролеин;
5. Уретаны (Н^МСООАлк); например, этил-уретан;
6. Диазоалканы (Алк=N + = N-); например, диазоуксусный эфир,
диазометан, 1,4-бисдиазоацетилбутан;
7. Нитрозозамещенные соединения (
*2
где Ri-H, Алк, a R2-Aak, остаток уретана, мочевины, гуанидина);
например, нитрозометилуретан, нитрозогуанидин, нитрозоалкил-
мочевины; Алк—О л°
8. Диалкилсульфаты ^8^
Алк—О О
например, диметил- и диэтилсульфаты;
221
9. Алкилалкансульфонаты (эфиры сульфоно- О
вых кислот с общей формулой, например, метил-
метансульфонат и этилмеметансульфонат);
7 л
Апк О
10. Эфиры фосфорной и фосфористой кислот
АЛК-О.
Алк—О-^Р=О
Апк— О
АПК—CL
₽—С—Алк
Z
Алк—О
11. Р -Хлорэтилсульфиды (иприты) с общей формулой и их
азотистые аналоги, где R=H, Алк и др.;
CI—Алк
CI—Алк^
К—R R=H,AnK
Cl—Алк
12. Эпоксиды,
СН2—CH —R R=H. Апк
13. Лактоны (Сн2)п—С=О
где R = H, Алк и др.,:
например, этиленок-
сид,
или окись этилена;
например,
р-пропиолактон;
н
14. Этиленимины 1 , где r.Aak> н и .
сн2—сн—R
15. Производные фурана, например аф-
латоксин В1, хлорбензфуран и др.
16. Прочие.
Указанные соединения относят к разря-
ду реактогенных, способных передавать
свои радикалы азотистым основаниям в НК
и взаимодействовать с остатками фосфор-
ной кислоты.
афлатоксин В,
По числу активных групп алкилирующие
соединения делят на моно- , би- и полифункциональные. Моно-
функциональными, например, являются диметилсульфат, этилени-
мин, этилметансульфонат и др., бифункциональными — Р-хлорэ-
тилсульфиды и их азотистые аналоги, полифункциональными —
222
дихлордиэтил-, метилдихлордиэтиламины, эфиры фосфорной и
фосфористой кислот.
Нитросоединения (нитраты, нитриты, нитрозосоединения,
окислы азота и др.) проявляют мутагенную активность на про- и
эукариотах. Нитрозосоединения могут образоваться из аминосое-
динений и нитритов в кислой среде желудка млекопитающих.
Под действием алкилирующих агентов возникают аддукты (от
лат. adductus — приведенный, притянутый): 7-алкилгуанин (наи-
более часто), 3-метиладенин, 0-6-метилгуанин, 0-4-алкилтимин и
др. Малые дозы алкилирующих агентов индуцируют фермент
метилтрансферазу, "принимающую на себя" алкильные группы.
Клетки в этих случаях выживают. Данный фермент отсутствует у
дрожжей, но есть у людей.
Еще в 50-е годы был изучен механизм мутагенного действия
азотистой кислоты, проявляющийся в окислительном дезаминиро-
вании нуклеиновых оснований, то есть в замене аминогрупп на
гидроксил или кислород. Тогда аденин трансформируется в гипо-
ксантин (Гкс), гуанин в ксантин (Кс), цитозин в урацил. Первый
спаривается с цитозином, что приводит к замене А = Т на Г = Ц,
ксантин сохраняет способность спариваться (как и гуанин) с
цитозином, что сопровождается заменой Г= Ц на А=Т.
Ошибки при спаривании оснований происходят путем транзи-
ции или трансверсии. В первом случае один пурин заменяется
другим пурином, один пиримидин — другим пиримидином. Во
втором случае пурин замещается пиримидином или наоборот —
пиримидин замещается пурином. Под влиянием М-метил-М-нитро-
N-нитрозогуанидина больше возникает транзиций, чем трансвер-
сий.
Мутагенный эффект гидроксиламина обусловлен его взаимо-
действием с цитозином, тогда как гидразин разрывает кольца
урацила и цитозина.
Метаболическая активность различных организмов может со-
провождаться образованием перекисей, которые в определенных
случаях могут быть мутагенами. Более того, перекиси выступают
как бы связующим звеном между химическим и физическим
мутагенезом (в частности, лучевым), учитывая факт их образования
во время облучения клеток или тканей. При этом перекиси оказы-
ваются мутагенами первого порядка, а например, калия цианид —
мутагеном второго порядка. Это связано с тем, что KCN блокирует
фермент каталазу (антимутаген) и, как следствие, клетка лишается
защиты против мутагенного действия Н2О2.
2Н2О2
каталаза
блокада реакции
калия цианидом
2НгО + О2
“1—I—I—I—I—Г
A T Ц А Г T
II II II III II
Т А АОТ Ц А
J__I__I_I__[_L
Т—I—I—I—I—Г
A T Ins А Г T
II II II III II
Т А АО Т Ц А
1__1_I___I_I_L
т—।—I—I—।—г
А Т Ц А Г Т
II II III И III II
Т А Г T Ц А
। ВыпадениеL|T
А Т А Г Т
II II II III-II
T А Т Ц А
J_____1_I______I—J_
“1—I—I—I—I—Г
A T Ins А Г Т
II II III II III II
T A Ins' T Ц А
J__I__J___I____I_L
Вставка Ins=Ins’
1---1
A T А Г Т
II II II III II
T А Т Ц А
J--L4J----1--1----
Выпадение Ins — АО
224
Производные акридина (акридиновый оранжевый — АО, проф-
лавин) и некоторые производные изохинолина (бромистый этидий)
вызывают удлинение нитей ДНК (растяжение) на 1-2 нуклеотида,
что приводит в итоге к выпадению или вставке мононуклеотида
(прямые мутации).
МИТОМИЦИН С
Некоторые антибиотики — как мутагены
обладают различными механизмами дейст-
вия. Так, например, митомицин С подобен
алкилирующим агентам. Он расщепляет
двойную спираль ДНК за счет образования
перекрестных связей. Антибиотики из груп-
пы актиномицинов (в частности, актиноми-
цин Д, или дактиномицин), являющиеся ди-
олигопептидилактиномицинами,
ингибируют ДНК-зависимый
синтез РНК.
Они специфически взаимо-
действуют лишь со спирализо-
ванными полидезоксирибонук-
леотидами, содержащими гуанин
(при этом существенна роль 2-
аминогруппы пуринов в образо-
вании водородной связи с хино-
идным кислородом хромофора
дактиномицин
В блеомицине А2 радикал R представляет собой - $(СНз)>
в блеомицине ДМА2 радикалом является -NH-fCHJj -S-CHj, в блеоммцюга
В2 ом следукмций:
-NH-JCHjh -NH-C-NH,. и т. д.
НН
8 т. 8524
225
актиномицина). Они располагаются в малой бороздке нативной
ДНК, мешая функционированию РНК-полимеразе.
Гликопептидные антибиотики блеомицины ингибируют синтез
ДНК и клеточный митоз.
Аминогликозидные антибиотики (гигромицин В, стрептомицин)
являются непрямыми мутагенами. Они нарушают белковый синтез
благодаря взаимодействию с 30 S субъединицами бактериальных
рибосом и предотвращению связывания отдельных тРНК. При
этом неправильно считываются пиримидиновые основания.
. Ряд алкалоцдов высших растений (из родов Heliotropium —
гелиотроповых, Senecio — крестовниковых и др.) и синтетических
лекарственных веществ (изониазид, хлорпромазид, нитрофураны
и др.) могут обладать мутагенным действием.
К биологическим мутагенам относят вирусы (фаги), живые
вакцины, некоторые биотоксины, образуемые грибами, протозоа
и гельминтами. Так, изменения в хромосомах вызывают цитоме-
галовирус, вирус простого герпеса, герпетиформный вирус, вирус
Сендай, вирус саркомы Роуса и др. Из грибных токсинов —
мутагенов ранее был назван афлатоксин Bi. Можно назвать еще
треморген, образуемый рядом аспергиллов и др.
Фагоустойчивые мутанты актиномицетов и лактобактерий пред-
ставляют интерес для соответствующих производств, например,
антибиотиков и молочнокислых продуктов.
Мутагены различных классов (физические, химические, биоло-
гические) используют в работе с биообъектами. При этом стремят-
ся получить штаммы либо с какими-то маркерными признаками,
важными для расшифровки механизмов мутагенеза, либо штаммы
— суперпродуценты БАВ, существенно увеличивающие выход
конечного продукта в сравнении с контролем.
При целенаправленном получении мутантов необходим их от-
бор, или селекция значение которой исключительно велико в
выведении различных пород животных, многих сортов злаковых,
декоративных и многих других растений, в получении микробов
— суперпродуцентОв БАВ (ряда антибиотиков, аминокислот, вита-
минов и других субстанций). Для сравнения можно указать на
успехи с селекцией мутантов пеницилла, исходный штамм которо-
го, выделенный из воздуха А. Флемингом в 1928 г., продуцировал
лишь 50 единиц антибиотика пенициллина в 1 мл культуральной
жидкости.- Тяготы второй мировой войны 1939-1945 гг. вынудили
ученых вернуться к этому виду гриба и заняться генетико-селек-
ционной работой с ним. В 1951 г. путем рассева естественной
популяции клеток, выросших в глубинных условиях культивиро-
226
вания Penicillium chrysogenum, был выделен штамм № 1951 с
ативностью 100 ЕД/мл. Его колония и стала родоначальницей всех
производственных штаммов в различных странах мира. Используя
селекцию и комбинированное воздействие различными мутагена-
ми (преимущественно — УФА и алкилирующие агенты), уже в
начале 60-х годов удалось передать в производство штамм, обра-
зующий пенициллин в количестве 5 000 ЕД и более в 1 мл
культуральной жидкости. В настоящее время используются штам-
мы, продуцирующие десятки тысяч единиц антибиотика в 1 мл
среды.
Следует различать понятия "селекция" и "интродукция" (от лат.
introductio — введение). При селекции используют методы выве-
дения новых форм организмов, отличающихся по свойствам от
исходных родительских организмов. При интродукции выделяют
из естественных (природных) и искусственных (производствен-
ных) условий существующие организмы, наиболее пригодные для
практических целей. В успехах селекции велика роль контролиру-
емого мутагенеза, тогда как успехи интродукции опираются на
генофонд, заведомо присущий организму. Селекция и интродукция
животных и растений возникли раньше генетики. Основные под-
ходы при этом концентрировались преимущественно на скрещи-
вании организмов и отборе возникающего потомства.
Применительно к микробам сформировалось четыре подхода,
используемых в селекционной работе:
1. отбор естественных штаммов, обладающих ценными призна-
ками, проявляющимися в конкретных условиях их существования
(этот подход во многом аналогичен интродукции высших эукариот);
при размножении такой культуры (с учетом частоты спонтанных
мутаций) наступает популяционное давление, когда наиболее при-
способленная форма вытесняет исходную в популяции. В анализе
таких популяций полезны флуктуационный тест (от лат. fluctuatio
— колебание) и метод отпечатков. С помощью первого доказывают
спонтанность мутаций по маркерному признаку (от англ, mark —
знак, метка), с помощью второго отбирают нужные колонии,
вырастающие на селективной среде. Этот первый подход опира-
ется на естественный ход событий у микроорганизмов в изменен-
ных условиях (естественный отбор);
2. искусственный отбор клонов микроорганизмов с полезными
для человека признаками, возникшими на основе естественной
изменчивости родительских форм. К такому отбору прибегают
тогда, когда контролируемый признак биологически малоценен для
микроба и поэтому трудно создать условия выращивания культуры,
227
в которых относительно легко удавалось бы выделять нужные
варианты. При этом необходимо проверять большое число клонов
и субклонов, и если вариации между ними по контролируемому
признаку невелики, то успех выборки оказывается мало вероятным
или небольшим;
3. ступенчатая селекция — эффективный метод искусственного
отбора форм с применением мутагенов (см. ранее приведенный в
этом разделе пример с продуцентом пенициллина). С помощью
мутагенов увеличивают изменчивость тест-культур, а среди послед-
них отбирают наиболее перспективные; такой подход, как правило,
использх ют многократно (ступенчато), добиваясь существенного
возрастания контролируемого показателя (активность, продуктив-
ность и др.);
4. гибридизация — как метод выведения полезных форм мик-
роорганизмов. Здесь имеется аналогия с методом скрещивания у
высших эукариот. О гибридизации см. в этой главе ранее.
Микроорганизмы, как и высшие организмы, способны соби-
рать и перераспределять уже имеющуюся информацию между
родственными, но генотипически неоднородными клетками. Это
происходит при трансформации, трансдукции и конъюгации у
бактерий, при половой и соматической гибридизации у растений
и животных. Яркий пример здесь с гибридомами, продуцирующи-
ми моноклональные антитела (см.).
228
Часть III.
Процессы и аппараты в биотехнологии
Глава 6.
ПРОЦЕССЫ В БИОТЕХНОЛОГИИ
Биотехнология как наука базируется на использовании биоло-
гических процессов в технике и промышленном производстве (см.
главу 1). Эти процессы (от лат. processus — продвижение) — как
совокупность последовательных действий специалистов направле-
ны на достижение соответствующих результатов при эксплуатации
биообъекта(-ов). Говоря о процессах в биологической технологии,
нельзя путать их с процессами в химической технологии. Так,
главным компонентом первых является какой-либо биообъект
(вирус, бактерия, гриб, растительные или животные клетки, био-
молекулы). Такие объекты отсутствуют в химической технологии.
Другой пример с высокими температурами, которые, как правило,
неприемлемы в биотехнологии, но часто используются в химиче-
ской технологии. Наконец, многостадийность и высокие давления
также являются атрибутами (от лат. atributio — придаю, наделяю)
химической технологии, а не биотехнологии. Однако биохимиче-
ские и химические реакции следуют принципу ле Шателье, соглас-
но которому равновесие системы смещается в направлении умень-
шения эффекта произведенного воздействия.
229
Т а б л и ц а 24. Систематизация биотехнологических процессов
230
Рис. 75. Примерная обобщенная схема процессов в биотехнологии.
Исходя из обобщенной схемы, нельзя учесть все виды и разно-
видности биотехнологических процессов (сравнить данные на рис.
75 и в таблице 24). В основу подразделения биотехнологических
процессов могут быть положены различные принципы, например,
оценка: принадлежности биообъектов к надцарствам живых су-
ществ (процессы на основе использования акариот, прокариот,
эукариот), функциональной активности биообъекта (биосинтез,
биотрансформация), возможности вычленения отдельных этапов
из биотехнологических схем производства в виде самостоятельных
процессов (подготовка питательных сред и оборудования; стери-
лизация питательных сред оборудования, воздуха, ферментация
(культивирование) биообъекта; выделение, очистка и упаковка
готового продукта и т. д.
Классификационные схемы подобного рода оправданы и ими
можно пользоваться как равноправными.
Биотехнологические процессы условно можно подразделить на
биологические, биохимические и биоаналогичные. К первым от-
носят те из них, которые основываются на использовании акариот,
прокариот и эукариот, вторые — на использовании ферментов, и
третьи (биоаналогичные) — на химическом синтезе или полусин-
тезе веществ, функционально близких или эквивалентных первич-
ным или вторичным метаболитам живых, организмов (получение
производных пенициллина и цефалоспорина, тетрациклина, нук-
леиновых оснований и др.).
231
А /снз
C H -CH —CO—NH—CH—CH С__гн R-CO-NH-CH-CH-R,
.° 5 2. I I I *-мз I I
R X.—N—CH-COOH z-c—N—SO3H
o °
бензилпенициллин монобактам
В полусинтетических ампициллине, метициллине, оксациллине
R представлены соответственно: а - D - (-) - аминобензилом
СбН5—CjH—NH2,2,6 - диметоксифенилом СбНз(ОСНз)2—, 5 - метил
- 3 - фенил - 4 - изоксиазолилом
с6н5
(СН2)3
С—N С-СК-О-СО-СН,
и 'V*
CH-NH3 0 1
I СООН
СООН
цефалоспорин
В полусинтетическом цефалоглицине радикал R представлен
аминобензилом СбН5—СН—NH2, в цепорине (цефалоридине) при-
соединены два радикал^ в позициях 3 и 7
zsx
О=С—NH—СН—СН СН2
сн2
С—N С-СН2—N
СООН
цепорин
В полусинтетических производных тетрациклина - доксицик-
лине и метациклине (рондомицине) Ri—R4 представлены соответ-
ственно: —ОН, —СНз, —Н, —Ни —OH(Ri), —СН2—(R2 и R3),
—H(R4).
232
И>«Ч*п 64>то₽урвцил
•4 R3 r2 ^снз)2 Полусинтетическим аналогом
урацила является противораковый
Г |Г ПГ | [] препарат 5 - фторурацил и т. д.
^4**^s»fz,'^zZls>>ff^4CONH2 Многие процессы биологиче-
он о он о ской технологии являются общими,
например на стадии ферментации,
поэтому можно выделять общие основы
биологической технологии. Это особен-
но показательно на аппаратурном офор-
млении отдельных стадий биопроиз-
водств и, в частности, на выборе биоре-
акторов, сепарирующего оборудования;
сушилок и т. д. (см. главу 7).
Специальные биотехнологические процессы связаны в большей
степени с особенностями биообъектов. Достаточно сравнить здесь
культивирование вирусов гриппа на куриных эмбрионах для при-
готовления вакцин и выращивание пеницилла — продуцента ан-
тибиотика бензилпенициллина в биореакторах емкостью до 100 м3
и более; другой пример с гибридомными клетками млекопитаю-
щих, продуцирующих моноклональные антитела, и с лейконосто-
ком — продуцентом полисахарида декстрана.
В каждом из этих процессов имеются свои специфические
особенности, благодаря которым выделяют биотехнологический
процесс в самостоятельный. С учетом сказанного мы группируем
все специальные биотехнологические процессы на микробиологи-
ческие, фито- и зообиотехнологические. Другими словами, мы
выделяем микробную биотехнологию, фитобиотехнологию и зоо-
биотехнологию/ Общие процессы изложены в первой части учеб-
ника, специальные - во второй.
6.1. Взаимосвязь процессов и биообъектов. Биообъекты харак-
теризуются такими показателями, как уровень структурной орга-
низации, способность к размножению (или репродукции), наличие
или отсутствие собственного метаболизма при культивировании в
подходящих условиях. Что касается характера биообъектов, то под
этим следует понимать их структурную организацию. В таком
случае биообъекты могут быть представлены молекулами (фермен-
ты, иммуномодуляторы, нуклеозиды, олиго- и полипептиды, и т.
д.), организованными частицами (вирусы, фаги, вироиды), одно-
клеточными (бактерии," дрожжи) и многоклеточными особями
(нитчатые высшие грибы, растительные каллусы, однослойные
233
культуры клеток млекопитающих), целыми организмами растений
и животных.
Молекулярные биообъекты накладывают свой отпечаток на
организацию и аппаратурное оформление соответствующих био-
технологических процессов.
Вирусы и фаги как облигатные паразиты могут культивиро-
ваться только на живых клетках и тканях, то есть фактически
биотехнологические процессы здесь основываются на использова-
нии клеток, зараженных вирусами или несущих вирус(-ы),
Одноклеточные виды прокариот и эукариот могут использо-
ваться в биотехнологических процессах в виде монокультур или в
ассоциациях. Для сравнения можно назвать производство какого-
либо антибиотика (пенициллина, рифамицина и др.) с помощью
чистой культуры соответствующего продуцента, а также производ-
ство кефира с помощью кефирных "зерен" ("грибков"), в состав
которых входят лактобактерии и дрожжи. Следовательно, в по-
следнем случае применяют природную ассоциацию микроорганиз-
мов, и кефир является продуктом смешанного брожения — молоч-
нокислого и спиртового. Пировиноградная кислота как ключевой
продукт, возникающий из моносахаридов (моноз) после гидролиза
лактозы, трансформируется лактобактериями до молочной кисло-
ты, а дрожжи "доводят" тот же пируват до этанола.
лактобактерии
н q и дрожжи
"2—-------------
лактоза
глюкоза и
галактоза
дрожжи
/3—галактозидата
-6СО
+12Н
;он)соон
6СН3СН
+12Н
6СН3СОН----->6СН3СН2ОН
6СН3СОСООН пируват
лактобактерии
лактат
В природных условиях микроорганизмы, как правило, находят-
ся в ассоциативных взаимоотношениях, часть из которых несом-
ненно может быть привнесена в производство в виде самостоя-
тельных биотехнологических процессов. Сегодня такие процессы
немногочисленны, так как природные ассоциации изучены крайне
234
недостаточно с точки зрения биологической технологии. К тому
же такие ассоциации складываются не только между микробами
— микробоценозы, но и другими организмами — микробами и
растениями, микробами и животными, между растениями, и т. д.;
в таких случаях говорят о биоценозах (от греч. bios — жизнь, coinos
— община). Если организмы выживают, но не размножаются в
данных конкретных условиях, то под этим подразумевают их
нахождение в состоянии анабиоза (от греч. ana — назад, обратное
действие, подобно, соответственно). Если же организмы растут,
развиваются и размножаются в среде обитания, то под этим
понимают их нахождение в состоянии метабиоза (от греч. meta -—
между, посредине, изменение, следование в пространстве и вре-
мени). Наконец, гибель организма(-ов) в ассоциации или вне ее
можно охарактеризовать понятием абиоз (от греч. ап — нет, bios
— жизнь). В биотехнологии приходится постоянно встречаться с
такими явлениями как метабиоз, абиоз и анабиоз. Имеются и такие
ассоциации организмов, когда один (или более) из ассоциантов
может расцениваться хищником. Например, гриб Arthrobotrys
oligospora формирует сеть из мицелиальных петель — ловушек
нематод, в частности, относящихся к числу вредителей растений
(лука, клевера и др.). Такой нематодой является Ditylenchus dipsaci
— стеблевая нематода. В сельском хозяйстве заметен вред от
картофельной нематоды — Heterodera rostochiensis. Поэтому гри-
бы-хищники оказываются союзниками человека в борьбе за уро-
жаи ряда ценных культур растений, повреждающихся нематодами.
Растением-хищником является росянка (насекомоядное расте-
ние). У нее на листьях имеются железистые волоски, выделяющие
ферментативно активную липкую жидкость, с помощью которой
схваченные насекомые прочнее адгезируются и перевариваются.
Среди микробов хищником является миксомицет Dictyostelium
discoideum, а его жертвой — кишечная палочка.
При хищничестве, присущем только эукариотическим организ-
мам, жизнь партнера-жертвы ограничена во времени, то есть здесь
ассоциация организмов кратковременная.
Таким образом, анабиоз, метабиоз, абиоз и хищничество отно-
сятся к разряду понятий об уровнях жизнедеятельности организ-
мов. При метабиозе всегда предполагается наличие организмов-
партнеров, вступающих или вступивших во взаимоотношения.
Каковы же эти взаимоотношения? — Фактически их два —
симбиоз и антибиоз (таблица 25).
235
Т а б л и ц а 25. Виды взаимоотношений при метабиозе между организмами в
природных условиях.
Взаимооотношения
Вид Подвид Краткая характеристика
Симбиоз Комменсализм (от лат. commensalis - сотрапезник) Один вид из ассоциантов живет за счет другого, не причиняя ему вреда
Мутуализм (от лат. mutualismus - взаимосвязь, сожительство) Оба ассоцианта помогают ДРУГ другу
Нейтрализм (!?) Ассоцианты не влияют друг на друга
Паразитизм Один из ассоциантов живет за счет другого, нанося ему вред
Антибиоз Собственно антибиоз и антагонизм 1) односторонний (моно- и полинаправленный) Один вид ингибирует развитие другого (других) или убивает его (их) своими метаболитами
а) спонтанный (от лат. spontaneus - самопроизвольный, без постороннего вмешательства) В природных условиях
б) направленный (насильственный) В искусственных условиях
2) двусторонний Оба ассоцианта ингибируют или убивают друг друга за счет своих метаболитов
Аутоантибиоз Один вид или ассоциация видов образует метаболиты, которые ингибируют или убивают их процудент (ы)
Симбиотические взаимоотношения типа комменсализма при-
сущи, например, рыбам прилипале и акуле, грибу пенициллу,
образующему пенициллин, и кишечной палочке, продуцирующей
фермент пенициллиназу, которая гидролизует антибиотик до пе-
нициллоевой кислоты — он становится не активным.
236
ZS. CH3 пенициллиназа
R—CO—NH—CH—^H С—CH3 _________► R—
O=C N—CH—COOH
A/"’
CO—NH—CH—<j:H c£-CH3
O=c N—CH—COOH
LI
пенициллин
пенициллоевая кислота
Оба вида могут расти на мясо-пептонном агаре, но кишечная
палочка в этом случае использует вторичный метаболит пеницилла.
Третий пример — ассоциация аэробных и анаэробных микроор-
ганизмов. Представители второй группы начинают расти на среде
лишь тогда, когда аэробы снизят концентрацию кислорода до
необходимой им величины. Следовательно, вторые живут за счет
функциональной активности первых, не принося им вреда.
Яркими природными симбиотическими ассоциациями типа
мутуализма являются лишайники, где партнерами выступают ци-
анобактерии (или зеленые водоросли) и грибы; муравьи-термиты
и некоторые грибы; микробы рубца жвачных животных; грибы-
микоризообразователи и соответствующие растения; нитрифици-
рующие бактерии-симбионты в клубеньках бобовых растений;
отдельные представители нормальной микрофлоры кишечника
человека; некоторые бактерии — эндосимбионты внутри клеток
простейших, и т. д.
При редком явлении — нейтрализме партнеры —
ассоцианты ведут себя так же, как если бы их культивировали по
отдельности — скорости роста од инаковы в обоих случаях. Напри-
мер, молочнокислые бактерии и молочнокислые стрептококки в
закваске для кисломолочного продукта (йогурта) не проявляют
отклонений в скоростях роста в сравнении с таковыми для культур
в изолированном состоянии. По другим показателям (кроме ско-
рости роста) данные не накоплены, поэтому возможно "голый"
нейтрализм в природных ассоциациях отсутствует.
В случаях паразитизма один из ассоциантов всегда
наносит вред другому. Среди микробов хорошо известно пораже-
ние прокариотических видов фагами; растений, теплокровных
животных и человека — вирусами, патогенными бактериями и
грибами; комнатных мух — энтомофторовыми грибами (например,
Empusa muscae), некоторых холоднокровных животных (лягушек,
рыб) грибами из родов Basidiobolus и Saprolegnia, и т. д. В случае
использования таких видов (паразитов) в биотехнологии требуются
особые подходы и условия работы с ними.
237
Антибиоз присущ многим видам микро- и макроорганизмов.
Ведущую роль в подавлении партнера (-ов) в ассоциации могут
играть собственно антибиоз и конкуренция
за источники питания. Собственно антибиоз обеспечивается пре-
имущественно за счет первичных (ферменты) ,или/и вторичных
метаболитов, например, токсинов, антибиотиков (или фитонцидов
растений — по Б. П. Токину); он может быть односторонним и
двусторонним. Пример одностороннего — Streptomyces
kanamyceticus образует аминогликозидный антибиотик канами-
цин, который губительно действует на партнера—Salmonella typhi;
пример второго — разные виды стрептомицетов, угнетающих друг
друга при росте на питательной среде, например взаимно антаго-
нистичны Streptomyces globivulgaris и Streptomyces griseus, S.
globivulgaris и S. levoris, и др.
В случае конкурентного антибиоза проявляется антаго-
низм (от греч. antagonisma — спор, борьба) между видами прежде
всего из-за потребностей в питательных веществах; преимущество
получает здесь наиболее приспособленный вид, менее приспособ-
ленный угнетается или погибает (при наличии в сре^е Е. coli и
Staph, aureus преимущество остается за Е. coli).
Антибиоз может быть моно- и полинаправленным, или одно- и
многонаправленным, то есть антибиотически или антагонистиче-
ски активный вид проявляет свое ингибирующее (губительное)
действие лишь в отношении одного
вида или более, чем одного вида.
Такой эффект зависит от сложив-
шейся ситуации в природных усло-
виях (спонтанно) или в искусствен-
ных условиях — производственных
или лабораторных (направленно). В
соответствующих экспериментах не-
редко определяют активность каких-
либо продуцентов антибиотиков в от-
ношении многих бактерий и грибов,
подсевая их, например, к "штрихово-
му росту" такого продуцента (рис.
76). Здесь гриб естественно или вы-
нужденно (насильственно) продуци-
рует пенициллин, который тормозит
рост чувствительных бактерий. По-
добные ситуации вынужденного (на-
238
Рис. 76. Полинаправленный ан-
тагонизм пеницилла (а) в отноше-
нии бактерий: Staphylococcus
aureus (б), Streptococcus
haemolyticus (в), Escherichia coli
(г), Neisseria meningitidis (д),
Streptococcus pneumoniae (e).
Candida albicans (ж).
сильственного) антибиоза или антагонизма могут возникать не
только в природных условиях, но и в макроорганизме (И. Г.
Шиллер, 1952).
Под аутоантибиозом следует понимать ингибирование
или гибель организма(-ов) от действия собственных метаболитов.
Например, молочнокислые кокки и бактерии, накапливающие
молочную кислоту в среде обитания, погибают в результате дей-
ствия этого метаболита. Другой пример с дрожжами — сахароми-
цетами, использующимися в производстве этилового спирта. В
1 процессе брожения лишь отдельные расы их могут накапливать до
15% этанола. Более высокие концентрации губительно действуют
| на Saccharomyces cerevisiae.
Аутоантибиоз во многом зависит от возраста культуры, и он
проявляется, как правило, в конце стационарной фазы роста и
начале фазы отмирания. Все другие виды ассоциативных взаимо-
отношений складываются преимущественно раньше.
Следует иметь в виду и адаптивные возможности видов в
ассоциациях. Если чистые культуры, используемые в соответству-
ющих производствах, не проявляют, например, какой-либо фер-
ментативной активности, то это не значит, что в ассоциациях с
другими видами они поведут себя аналогичным образом по данно-
му признаку. При наличии соответствующего субстрата в среде
может индуцироваться выработка адаптивных ферментов, не вы-
1 рабатывавшихся ранее без индуктора. И в результате этого ассо-
циант может подвергнуться, например, лизису (растворению).
По условиям проведения процесса различают нестериль-
ные (крупнотоннажное производство кормовых дрожжей) и
I стерильные производства (получение антибиотиков,
витаминов, моноклональных антител и др.); аэробные, или с
подачей воздуха и анаэробные (без подачи воздуха) —
соответственно производства лимонной кислоты и полисахарида
, декстрана.
1 Многие каллусные культуры растений выращивают поверхно-
стно, то есть используя подложку из питательных сред (чаще —
, агаризованную), тогда как микробная биотехнология рассчитана
преимущественно на глубинные процессы (погружение биообъек-
тов в жидкие питательные среды).
При реализации процессов их стремятся вести в одном из трех
режимов: периодическом, по л у непрерывном
I и непрерывном. В первом случае биотехнологический
процесс проводят от начала до конца согласно регламенту, и после
239
завершения всех операций его повторяют. Во втором случае
осуществляют так называемый отливно-доливной процесс, когда,
например, на "пике" биосинтеза какого-либо антибиотика отбира-
ют 25—75% культуральной жидкости и однократно добавляют
столько же свежей питательной среды. Непрерывные процессы
рассчитаны на непрерывный отбор культуральной жидкости и
непрерывное поступление равного количества свежей питательной
среды. Непрерывный режим используется в производстве дрож-
жей на гидролизатах древесины, в производстве пива, бутанола и
ацетона, и др.
Выход микробных клеток при непрерывном процессе в срав-
нении с периодическим будет выражаться следующими соотноше-
ниями (с учетом времени удвоения массы — в. у.); 11,9 (в. у. 0,5);
7,6 (в. у. 1,0); 5,3 (в. у. 2,0); 4,3 (в. у. 4,0). Из этого следует, что с
возрастанием времени генерации выходы биомассы в обоих про-
цессах уравниваются.
Согласно Д. Герберту, Р. Элсуорту и Р. Теллингу при непре-
рывном процессе максимальное образование продукта происходит
при
DmaxX = ЦтаХГ5о(^^-лф)2, где
•JO Оо
Dmax — максимальная скорость разбавления, % — концентрация
микроорганизмов в биореакторе при установившемся режиме,
ц max — максимальная удельная скорость роста, Y — выход клеток
в расчете на использованный субстрат, So — концентрация суб-
страта в подаваемой свежей среде, Ks — константа насыщения.
Применительно к фазовому состоянию ингредиентов в биотех-
нологических производствах различают твердофазные
процессы, например, при флуидизаций (от англ, fluid — жидкость,
жидкий) или протеинизации грубых кормов на основе соломы
злаковых растений с участием некоторых грибов, получение тка-
невых культур лекарственных растений на уплотненных средах,
из которых затем экстрагируют действующие вещества, производ-
ство сыра из белков молока и др.; газофазные процессы
основаны на использовании газа (например, метана) для получения
микробного белка с помощью ассоциаций метилотрофных бакте-
рий.
Наконец, по условиям проведения процессов выделяют од-
ноступенчатые, двухступенчатые и много-
ступенчатые. Одноступенчатые проводятся, например, при
. 240
получении 6-АПК из бензилпенициллина с помощью пеницилли-
нацилазы, или пенициллинамидазы на колоннах, содержащих им-
мобилизованный фермент.
/ место действия фермента
J ZSK ZCH3 А/СН’
CjHg СНг—СОТ-NH—CH—C^-CHj -----» HH2—CH—CH C—CHj t
O=C N—CH—COOH O=C N—CH—COOH
+ CgHs—CH2—COOH
фенилуксусная кислота
6-аминопеницилилланоеая
кислота (6-АПК)
Двухступенчатые процессы базируются на использовании кле-
ток, находящихся в разном фазовом состоянии (в трофофазе и
идиофазе, см. 4.4). Так, например, двухступенчатый процесс воз-
можен при получении полисахарида курдлана—на первой ступени
выращивают продуцент (Alcaligenes faecalis var. myxogenes) на
питательной среде, поддерживая его в трофофазе; на второй
ступени культуру переносят в другой биореактор, где нет пита-
тельной среды, но имеется глюкоза, из которой синтезируется
курдлан.
Многоступенчатые процессы присущи генетической инжене-
рии и рДНК-биотехнологии (см.).
Процессы биохимической технологии подразделяют по ста-
диям реализаци и технологической схемы производства:
подготовка оборудования и питательных сред, их стерилизация,
посев биообъекта и ферментация, выделение, очистка, сушка,
упаковка. В зависимости от целевого продукта число стадий про-
цесса может быть то больше, то меньше. Д ля сравнения можно
назвать производство кормовых дрожжей и антибиотика стрепто-
мицина. В первом случае целевым продуктом являются
дрожжевые клетки, во втором — вторичный метаболит, предназ-
наченный для парентерального введения больным людям и живо-
тным. При получении антибиотика имеется больше стадий, чем в
случае получения дрожжевых клеток.
Целевыми продуктами могут быть и первичные метаболиты —
ферментные белки. Очевидно, что, например, экзогидролазы тех-
нологически получать легче, нежели какие-нибудь эндоферменты,
локализующиеся внутри клеток, и число стадий в этом последнем
случае может возрастать.
Очевидно, что подготовка оборудования, питательных сред, и
все другие этапы регламентированной схемы производства како-
241
го-либо целевого продукта различны по многим показателям, если
зто будут, например, процессы получения экзотоксинов и органи-
ческих кислот. С биотехнологических позиций работа с токсиген-
ными культурами должна проводиться на таком уровне, чтобы
исключить возможность попадания ее за пределы биореактора в
живом состоянии. Строгий контроль осуществляется также за
внутрипроизводственной обработкой и транспортировкой культу-
ральной жидкости, содержащей экзотоксин. Эксплуатация воз-
можностей, например Aspergillus niger продуцировать лимонную
кислоту не сопряжена с подобной опасностью. Тем не менее, во
избежание рассеивания грибных спор во внешней среде желатель-
но и в этом случае ферментацию осуществлять в герметизирован-
ных биореакторах.
После завершения ферментации отделяют либо клетки (кле-
точную массу), содержащие необходимое нам вещество, либо
жидкость, в которой накопился конечный продукт. В первом случае
отходом является жидкая часть культурального "бульона", во вто-
ром — плотная часть (клетки). Культуральная жидкость содержит
биообъект, недоиспользованные компоненты питательной среды,
продукты метаболизма, включая ожидаемый , конечный (целевой)
продукт. От качества целевого продукта зависит выбираемый
метод сепарирования клеток. Например, при производстве экзо-
полисахаридов, или экзогликанов культуральные жидкости стано-
вятся вязкими и, даже, гелеобразными, из которых выделить
клетки продуцента удается лишь после разбавления их водой в
10—20 раз с последующим нагреванием, например, до 80°С, и
сепарированием в подогретом виде на мощных сепараторах типа
"Альфа Лаваль" (Швеция). Полученный разбавленный раствор
полисахарида подвергается концентрированию путем мембранной
фильтрации и вакуум-упаривания. Если минорные компоненты в
полидисперсном полисахариде количественно малы, то такой поли-
сахарид может быть использован в качестве готового продукта,
например, как вспомогательное вещество при изготовлении раз-
личных лекарственных средств.
В других случаях необходимо спиртовое осаждение гликана с
последующим удалением надосадочной жидкости и высушивание
полисахарида, поступающего на хранение и на реализацию.
В производстве некоторых антибиотиков используют ротаци-
онные вакуум-барабанные фильтры непрерывного действия. Если
биообъектами оказываются бактерии или дрожжи, то можно
применить центрифугирование в целях концентрирования суспен-
242
зий и отделения клеточной биомассы. Применяют центрифуги
непрерывного или осадительного действия. В первом случае клетки
выгружаются через сопла, во втором — с помощью шнеков.
Клеточную массу можно отделять методом осаждения (седи-
ментации), используя коагулянты (например, поликатионные ве-
щества). Некоторые штаммы дрожжей, использующиеся в пиво-
варении, обладают выраженной флокулирующей способностью, и
поэтому они легко могут быть отделены таким способом.
В целях концентрирования клеток может быть полезной фло-
тация (от франц, flottation — плавание), когда на границе раздела
фаз "воздух — жидкость” собираются, например, клетки бактерий
(Acinetobacter sp.), рекомендованные в качестве продуцентов бел-
ка.
В целях выделения конечных продуктов из культуральных
жидкостей после сепарирования клеток прибегают к осаждению,
экстракции, сорбции, ультрафильтрации (см. 7.4) или их сочета-
нию.
инозин (рибоксин)
Применительно к механизму образования конечных продуктов
можно говорить о процессах биосинтеза и биотран-
сформации. При биосинтезе имеет место конститутивное или
адаптивное образование метаболи-
тов, относящихся к числу преметабо-
литов, первичных или вторичных ме-
таболитов. Так, нуклеозид инозин
синтезируется некоторыми бактери-
ями (Вас. subtilis, Вас. pumilis,
Brevibacterium ammoniagenes и др.).
Пентоза в структуре инозина образу-
ется в гексозомонофосфатном шунте
(путь окисления
глюкозы по Вар-
бургу- Дикенсу
-Хорекеру), а пу-
риновое ядро
(гипоксантин,
или 6-оксипурин) в общем виде синтезируется
из глицина (атомы в позициях 4, 5, 7), форми-
ата (позиции 2 и 8), диоксида углерода (пози-
ция 6), аспарагиновой кислоты (позиция 1) и
глутамина (позиции 3 и 9).
пуриновое ядро
243
тетрациклин(ы)
Из первичных метаболитов можно назвать производство фер-
ментов амилаз для хлебопекарной и текстильной промышленности,
L-аспарагиназы — для здравоохранения и др. Из вторичных мета-
болитов хорошо известны поликетидные антибиотики — тетрацик-
лины, продуцируемые некоторыми стрептомицетами. Они образу-
ются путем конденсации молекул малонилКоА. Донором метиль-
ной группы для некоторых
тетрациклинов является
метионин, донором ами-
ногруппы — глутамин.
В процессах биотранс-
формации имеет место ви-
доизменение какой-либо
молекулы — предшествен-
ника конечного продукта
или превращение одного
продукта в другой. Напри-
мер, почвенные грибы и некоторые другие микроорганизмы пре-
вращают фенолкарбоновые кислоты в другие формы фенолов с
помощью реакций окисления, гидроксилирования и декарбокси-
лирования.
СООН соон
галловая кислота
шикимовая кислота пирогаллол
Биотрансформация имеет место при компостировании отходов,
при получении биогаза, силосовании кормов и др.
Тем не менее, подразделение процессов в биохимической
технологии на биосинтез и биотрансформацию связано в основном
с результирующим эффектом, а не с механизмом протекающих
при этом биохимических реакций.
Подразделение биотехнологических процессов на управ-
244
I
ляемые и неуправляемые опирается на глубину и
масштабность контроля, осуществляемого с применением средств
автоматики и ЭВМ. К числу неуправляемых процессов можно
отнести спонтанно протекающие компостирование плотных отхо-
дов в животноводческих комплексах и фермах. К управляемым
относятся все производственные процессы, основанные на исполь-
зовании микробных, растительных и животных клеток. При этом
уровень управления может быть двояким — операторским и
автоматическим. В первом случае поддержание заданных режимов
осуществляет оператор, во втором — соответствующие контроль-
но-измерительные приборы, автоматы, ЭВМ.
Некоторые авторы (Ф. X. Дейндорфер, 1960) подразделяют
биотехнологические процессы по типу на простые, совместные,
последовательные и ступенчатые. Под простыми понимается
трансформация питательных веществ в продукты метаболизма без
накопления интермедиатов при наличии строго определенной
стехиометрии в утилизации субстрата (или клеточном росте) и
образовании метаболита. Скорость образования метаболита можно
выразить следующим уравнением: г/р= - Yp/Sr, где г — скорость,
fp — контролируемый продукт метаболизма, YP/ST — коэффициент
отношения метаболита (р) к субстрату (s). В молярном выражении
коэффициент Y составляет: Yp/s = где p/s — число молей
метаболита, образующегося на 1 моль использованного субстрата,
f— метаболит, ns — число атомов углерода в молекуле субстрата,
с-число молекул субстрата, пр-число атомов углерода в молекуле
продукта. Кинетика простого процесса сопряжена с ростом клеток.
Примером такого процесса является получение этанола из глюкозы
при спиртовом брожении.
Совместными процессами считаются те из них, в которых
имеет место превращение питательных веществ в продукты мета-
болизма без накопления интермедиатов, но с возможным измене-
нием стехиометрии процессов. Следовательно в таких случаях
стехиометрию можно выразить только с помощью уравнения
реакции конечного продукта. Нередко скорость образования ме-
таболита принимается пропорциональной концентрации клеток, а
не скорости их размножения. В качестве примера такого типа
процессов можно назвать молочнокислое брожение.
245
Последовательные процессы биохимической технологии
характеризуются накоплением интермедиатов в ходе трансформа-
ции питательных веществ в целевые продукты. Это имеет место,
например, при ацетоно-бутиловом брожении, когда вначале из
глюкозы образуются интермедиаты — ацетат и бутират, которые
затем (наряду с глюкозой) превращаются в конечные продукты —
ацетон и бутанол.
При ступенчатых процессах биологической технологии
питательные вещества вначале полностью трансформируются в
интермедиат (и эта трансформация протекает избирательно в
какой-либо определенной последовательности) и лишь после этого
образуется конечный продукт. Пример — биосинтез лимонной
кислоты.
6.2. Значение асептики в биотехнологических процессах. Би-
отехнологические процессы, как правило, проводят в асептичных
условиях, хотя могут быть исключения для некоторых из них.
Например, при культивировании отдельных эукариот (дрожжи) в
негерметизированных ферментаторах (нестерильный процесс)
происходит заметное снижение pH среды, где доминирующее
положение дрожжей не изменяется при попадании контаминиру-
ющих бактерий (от лат. contaminatio — загрязнение, заражение,
смешение) — они не могут составить конкуренции основному виду.
Асептика (от греч. а—не, нет, sepsis—гниение) — это комплекс
мероприятий, направленных на предотвращение попадания в среду
(объект) посторонних микроорганизмов, включая болезнетворные.
Следовательно, асептика в биологической технологии и, например,
в хирургии — это не одно и то же понятие. В первом случае
предполагают использование какого-либо биообъекта (в том числе
— микроба) и полное исключение попадания других микроорга-
низмов, являющихся загрязнителями. Во втором случае стремятся
исключить любук) возможность попадания патогенных микробов
и микробов — контаминантов на операционное поле или в рану.
Каждый из материальных потоков в биотехнологических про-
цессах — потенциальный источник микробов — контаминантов.
Асептика может включать влажную уборку помещений, обра-
ботку их ультрафиолетовыми лучами, антисептическими средства-
ми, использование стерильных инструментов, сред, технологичес-
246
кой одежды, подачу стерильного воздуха (столы с ламинарным
потоком стерильного воздуха в боксированных помещениях, по-
ступление в ферментатор стерильного воздуха через барботер —
от франц, barbotage — перемешивание) и пр. Следовательно
комплекс мер, обеспечивающих асептику биотехнологических
процессов, включает: механическую, физическую и химическую
защиту биообъекта и среды его обитания, а при необходимости —
и конечный продукт. К механической защите относятся: удаление
механических примесей, например, из воздуха, культиваторов,
герметизация оборудования, изоляция узлов и соединений; к фи-
зической — обработка воздуха и поверхностей приборов и аппа-
ратов ультрафиолетовыми лучами, кипячение, стерилизация паром
под давлением, обработка ультразвуком; к химической — обработ-
ка поверхностей химическими антисептиками.
В производственных условиях источниками микробов-конта-
минантов могут быть почва, вода, окружающий воздух, люди. Из
почвы в сферу биотехнологических процессов попадают спорооб-
разующие палочки-бациллы, конидии грибов, актиномицеты; эти
же микроорганизмы с пылью могут попасть в воздух, через по-
средство которого они способны проникнуть в среду выращивания
биообъекта или в конечный продукт производства.
Качественный состав и размеры частиц в воздушной пыли
колеблются в широких пределах. В производственных помещениях
это зависит от конструкционных особенностей здания, розы вет-
ров, географической зоны расположения города и предприятия,
наличия или отсутствия потоков автомобильного и другого транс-
порта, количества непосредственно занятых в технологическом
процессе людей, характера и локализации складских помещений
и т. д.
Образующиеся пыль или/и капельки влаги в воздухе, как
правило, содержат на своей поверхности слой адсорбированного
воздуха и большее или меньшее количество микроорганизмов.
Газовая оболочка предохраняет частицы от смачивания. Такие
частицы представляют собой дисперсную фазу аэрозоля, устойчи-
вость которой зависит от размеров (величины) частиц, их элект-
рического заряда и поверхностной энергии. Необходимо помнить,
что в случае нахождения на частицах аэрозоля микробных клеток,
то их отрицательный электрический заряд будет привносить свою
247
долю в общий заряд частицы. Опираясь лишь на величину аэрозоля,
содержащего микроорганизмы, можно выделить три фазы его:
крупноядерную (диаметр частиц более 100 мкм), мелкоядерную
(диаметр частиц менее 100 мкм) и фазу бактериальной пыли
(диаметр частиц от 1 мкм до 100 мкм). Частицы крупноядерной
фазы в течение нескольких секунд оседают из воздуха, тогда как
частицы двух других фаз могут длительно находиться в воздухе,
образуя устойчивую коллоидную систему. В таблице 26 представ-
лены данные о скорости седиментации частиц в воздухе и, для
сравнения, в воде.
Т а б л и ц а 26. Седиментационная скорость (мкм/с) для сферических частиц в
воздухе при нормальных температуре и давлении
(18°с, 1,01* 105Па) и в воде при 2ГС
Диаметр частиц, мкм В воздухе В.воде
0,1 1.0 0,001
1.0 42,0 0,11
5,0 960,0 2.8
10,0 3650,0 11.1
Бактериальная пыль может формироваться из первых двух фаз
после их высыхания и повторного попадания в воздух. В разряд
частиц с диаметром от 0,001 мкм до 1 мкм подпадают вирусы и
некоторые бактерии. Аэрозоли могут быть вредными и для чело-
века не только из-за микробов, наход ящихся на частицах пыли или
капельках жид кости, но и сами по себе вследствие проникновения
в альвеолы дыхательной системы с последующим расстройством
ее функций. В таком понимании вредными являются следующие
аэрозольные частицы: асбеста, алебастра, абразивного порошка,
графита, гипса, д иоксида титана, дорожной пыли, извести, каолина,
корунда, карбида кремния, мрамора, оксида олова, стекловолокна
и др. В альвеолы проникают частицы размером менее 3 мкм при
скорости потока вдыхаемого воздуха уже около 1 см/с.
По степени загрязненности воздуха микробами и механически-
ми частицами в расчете на 1м3 производственные помещения, в
которых асептично изготавливают лекарственные средства, клас-
сифицируют по классности следующим образом:
1-й класс чистоты с ламинарным потоком стерильного воздуха
— для изготовления стерильных лекарств — не должно быть
микробов, а механических частиц размером до 0,5 мкм — не более
3500;
248
2-й класс чистоты — до 50 микробных клеток, до 2500 частиц
размером 5 мкм и до 350000 частиц размером 0,5 мкм;
3-й класс чистоты — до 100 микробных клеток (ЗА класс — до
200 и ЗБ класс — до 500 клеток), до 25000 частиц размером 5 мкм
и до 3500000 частиц размером 0,5 мкм;
4-й класс чистоты — по ГОСТ 12.1.005 — 86.
Для помещений 2-го и 3-го классов чистоты, в которых изго-
тавливают нестерильные лекарственные средства, нормирование
воздуха по содержанию механических частиц не предусматрива-
ется.
Лекарственные средства по "микробной чистоте” разделяют на
4 категории: 1) стерильные препараты для инъекций, приготовлен-
ные на апирогенной воде;
2) глазные лекарства, препараты для введения в закрытые
полости тела, средства для лечения обширных ожогов и открытых
ран не должны содержать живых микроорганизмов — они также
относятся к разряду стерильных;
3) лекарственные средства для наружного применения и для
введения в открытые полости не должны содержать более 100
живых микробных клеток в 1 г(мл) препарата и при безусловном
отсутствии в них бактерий, относящихся к семейству
Enterobacteriaceae, а также Pseudomonas aeruginosa и
Staphylococcus aureus;
4) все прочие лекарственные средства должны содержать в 1
г(1мл) не более 1000 жизнеспособных бактериальных клеток сап-
рофитов и не более 100 клеток непатогенных грибов при отсутст-
вии болезнетворных и условно патогенных микроорганизмов,
включая энтеробактерии, Pseudomonas aeruginosa, Staphylococcus
aureus, аспорогенные дрожжи рода Candida.
Работающие в помещениях различной степени чистоты должны
одевать рекомендуемую и пригодную для таких целей технологи-
ческую одежду (согласно требованиям системы GMP — см.). Так,
в помещениях первого класса, где кратность обмена воздуха в час
600—200, одевают стерильный костюм, головной убор должен
полностью закрывать волосы, включая бороду, и Заворачиваться
под ворот костюма; на лицо одевается маска во избежание попа-
дания частиц и капель в окружающую среду; на руки одевают
стерилизованные без сыпучих материалов перчатки из каучука
или пластичных материалов, на ступни - стерилизованную или
продезинфицированную обувь, включая бахилы. Низ брюк подво-
249
рачивают в обувь (как и рукава костюма — в перчатки). От
защитной одежды не должны попадать в воздух частицы и волокна,
а сама она должна задерживать частицы, исходящие с тела опера-
тора. Указанная одежда должна быть разового использования или
использоваться в течение одного дня, если результаты проверки
подтверждают такую возможность. Перчатки рекомендуется по-
стоянно дезинфицировать во время операций, маски и перчатки
необходимо менять перед каждой рабочей процедурой. Стериль-
ную зону желательно проектировать таким образом, чтобы все
операции можно было наблюдать извне. В рабочих зонах таких
помещений все открытые поверхности должны быть гладкими,
непроницаемыми, неразбитыми, удобными для очистки и дезин-
фекции, где и когда это необходимо, без труднодоступных высту-
пов и углублений, полок, шкафов, излишнего оборудования, раз-
движные двери здесь нежелательны из-за возможности скопления
пыли в пазах; сточные и канализационные трубы не должны
проходить в стерильных зонах. Комнаты для смены одежды необ-
ходимо проектировать и строить с воздушными шлюзами, снабжа-
ющимися стерильным воздухом. Двери с воздушными шлюзами
не должны открываться одновременно; между шлюзами должна
быть система для визуального или аудиоконтроля (от лат. visus —
зрение, auditus — слух, слышание). Мытье рук и средства для этого
должны быть только в комнате для' смены одежды.
В рабочие помещения должен подаваться стерильный воздух
под положительным давлением.
В помещениях второго класса чистоты с кратностью обмена
воздуха 20—60 следует одевать гладкий (без складок), не отделяю-
щий ворса, комбинезон, стянутый на поясе, с манжетами, плотно
облегающими щиколотки ног; на голову необходимо одевать шлем-
капюшон, полностью закрывающий волосы, нос й подбородок; на
лицо — маску, не отделяющую ворса; на руки — резиновые (или
из эластичных полимеров) перчатки, на ноги — стерильную или
продезинфицированную обувь, поверх которой рекомендуется
одевать бахилы, полностью закрывающие ступню. Нижняя часть
брюк должна заправляться в бахилы, а рукава комбинезона — в
перчатки. Ни одна часть тела или разрешенного для использования
нижнего белья не должна быть открыта.
В помещениях третьего класса чистоты с кратностью обмена
воздуха 1—15 рекомендуется одевать не отделяющий ворса ком-
бинезон или куртку с собранными рукавами на запястьях и
250
воротником-стойкой, шапочку или косынку, брюки, бахилы и
маску.
В помещениях четвертого класса чистоты рекомендуется оде-
вать комбинезон, или куртку и брюки, или халат, шапочку или
косынку из хлопчатобумажных или льняных тканей.
Вот почему важно глубоко продумывать размещение помеще-
ний в производственных зданиях (особенно — для стерильных
лекарств).
С водой в сферу технологического процесса могут попасть
грамотрицательные бактерии из групп энтеробактерий, псевдомо-
насов и некоторых других. В природных открытых водоемах
обнаруживаются целлюлозоразрушающие, нитрифицирующие и
денитрифицирующие бактерии, цианобактерии, аммонификато-
ры, железобактерии и многие другие. Лишь вода артезианских
колодцев, глубоких скважин и родников отличается высокой чис-
тотой. Следует помнить, что чем больше вода загрязнена органи-
ческими веществами, тем больше в ней содержится микробов.
По степени загрязненности открытых водоемов различают 3
зоны сапробности (от лат. saprotes — гниль, гниение): полиса-
про.бная — сильно загрязненная, содержащая в 1 мл несколько
миллионов микробных клеток, включая гнилостные и кишечные
бактерии; мезосапробная — умеренно загрязненная,
содержащая в 1 мл до 100000 микробных клеток с преобладанием
аэробных видов; олигосапробная — зона чистой воды,
содержащей в 1 мл не более 1000 микробных клеток из предста-
вителей железо-, серобактерий и некоторых других видов. Поли-
сапробные зоны характерны д ля рек, протекающих по населенным
пунктам или вблизи них, и где в воду попадают жидкие отходы из
крупных свиноводческих ферм, канализационные стоки и стоки
промышленных предприятий. В такой воде могут быть санитарно-
показательные (Е. coli, Streptococcus faecalis), условно патогенные
псевдомонасы, протей и другие виды, а также болезнетворные
микроорганизмы из группы энтеробактерий.
Седиментационная устойчивость бактерий в воде достаточно
выраженная (см. таблицу 26). Следует также иметь в виду и
минимальные значения активной воды (aw) для различных микро-
организмов, под которой понимают отношение давления паров
раствора (Vs) к давлению паров чистой воды (Vpw):
(таблица 27).
Vs
aW~ -Tr
Vpw
251
Таблица??. Минимальные значения aw для микроорганизмов
Группа микробов-сапрофитов Минимальные значения а«
Мезофильные бактерии 0,91
Галофильные бактерии 0,75
Психрофильные бактерии 0,75
Дрожжи 0,88
Нитчатые грибы 0,62 - 0,93
Осмофильные дрожжи 0,60
Нитчатые грибы — ксерофиты 0,65
Для сырья опасным уровнем влажности при длительном хра-
нении является показатель aw, превышающий 0,6, реже — 0,7.
С учетом всех характеристик необходимо осуществлять подго-
товку воды для использования ее в биологической технологии.
Люди, занятые в биотехнологическом производстве, также
могут быть источником контаминирующей микрофлоры — гра-
мотрицательных бактерий, кокков, микоплазм, вирусов и др. Толь-
ко на поверхности кожи может сосредоточиваться до 1О10 микро-
бных клеток. Наиболее загрязненными являются кисти рук, ступ-
ни, локти, шея, грудь, промежность, паховые области. Разнообраз-
на и многочисленна микрофлора ротовой полости: бактериальные
и кокковые формы, вибрионы, спириллы и спирохеты, нокардии,
дифтероцды, протозойные организмы, аспорогенные дрожжи рода
Candida, микоплазмы, вирусы и др. При разговоре, кашле, чихании
микробы в большом числе попадают в воздух. Установлено, что
здоровый человек за одно чихание выделяет до 20000 микробных
клеток, способных распространяться по горизонтали, в среднем,
до 1,5 м. Капельки носовой слизи, слюны и мокроты, подсыхая,
образуют частицы, покрытые белковой или гликопротеиновой
оболочкой, и содержащие микробные клетки. В таких частицах
микроорганизмы длительно сохраняются и могут являться причи-
ной нестерильности материалов (объектов) на каких-либо техно-
логических операциях.
Источником микробов-загрязнителей могут быть некоторые
компоненты питательных сред, например, кукурузный экстракт
(фаги, дрожжи и др.).
Растительные вирусы часто выявляются в культурах каллусных
тканей; высокую озабоченность в этом смысле вызывают также
культуры клеток животных тканей и клеток человека, будучи
исключительно благоприятными средами для контаминирующей
микрофлоры.
252
Микробы-контаминанты не только могут подавить развитие и
функции биообъекта в силу конкуренции и антибиоза, но и
дезорганизовать какую-либо ткань — среду выращивания; более
того, некоторые из них способны продуцировать токсические
вещества, которые могут попасть в целевой продукт (равно как и
сами микробы-загрязнители).
6.3. Борьба с микробами-контаминантами в биотехнологиче-
ских производствах. Защита биотехнологических процессов от
микробов-контаминантов эффективно осуществляется с помощью
различных фильтров. В последнее десятилетие широкое распрост-
ранение приобрела мембранная фильтрация в целях получения
стерильных воздуха и различных жидкостей (разновидность хо-
лодной стерилизации). Более того, мембраны нашли применение
в рДНК-биотехнологиии, в дисперсионном и других анализах
* биомолекул. Многие термолабильные вещества стерилизуют таки-
ми же способами. Следовательно, мембранная фильтрация может
рассматриваться как самостоятельный (лабораторный или про-
мышленный) процесс.
В биологической технологии, независимо от условий проведе-
ния процессов (поверхностно и глубинно, в периодическом, полу-
непрерывном и непрерывном режимах, в виде твердофазных или
газофазных процессов, одно-, двух- и полиступенчато), широко
используют стерильный воздух, подаваемый: в ферментаторы для
аэробных организмов (клеток, клеточных систем); в специальные
помещения в виде ламинарных потоков для асептичного приготов-
ления лекарственных средств, в боксы и операционные виварии,
в распылительные сушилки для высушивания некоторых веществ,
в шлюзы между (перед) асептическими блоками. Стерилизующая
мембранная фильтрация здесь оказывается наиболее приемлемой.
Промышленный выпуск мембранных фильтров был начат с
конца 40-х годов текущего столетия. Согласно Р. Е. Кестингу (1971)
наиболее распространенными процессами фильтрации являются
обычная фильтрация (ее можно назвать макрофильтрацией, Н. Е.),
микрофильтрация, ультрафильтрация, диализ (и обратный осмос).
Для макрофильтрации применяют обычно бумажные или стеклян-
ные фильтры. При этом отделяют частицы, размеры которых
находятся в пределах от 1 до 103 мкм. Для других типов фильтрации
используют нитроцеллюлозные, ацетилцеллюлозные, поливиниль-
ные, полиамидные, фторуглеводородные мембраны толщиной ме-
нее 0,1 мкм с высокой степенью пористости и с диаметром пор в
253
пределах от 10'4 до 10 мкм. В случаях микрофильтрации отделяют
частицы размерами 2* 10'2—10 мкм, в случаях ультрафильтрации
размеры отделяющихся при этом макромолекул находятся в при-
мерных пределах от 0,001 до 0,02 мкм, а их молекулярные массы
соответствуют 1—1000 кДа.
При диализе через мембраны отделяют небольшие молекулы,
сопоставимые по размеру с молекулами растворителя (10 3 мкм и
менее, или до 1 нм). Здесь вещества разделяются вследствие
различных скоростей диффузии через мембрану.
Осмос относится к разряду молекулярно-диффузионных про-
цессов, когда через мембрану переносится не вещество (из области
высокой концентрации в область низкой концентрации), а раство-
ритель. Если заставить двигаться растворитель в обратном направ-
лении через мембрану, то это и будет обратный осмос. Превысить
осмотическое давление в подобных случаях удается с помощью
внешнего повышенного давления. Обратный осмос рекомендуют
обычно для понижения концентрации молекул или ионов в водных
растворах (таблица 28).
В случаях стерилизации растворов фильтрованием они должны
содержаться перед розливом и при последующем розливе в асеп-
тичных условиях. При этом необходимо помнить и следить за тем,
чтобы время между началом приготовления раствора и его стери-
лизацией фильтрованием должно быть возможно кратким.
Для очистки растворов от бактерий-контаминантов в США,
например, выпускают в продажу так называемый Criti Сар-капсу-
лы, содержащие двойной слой гидрофильной полисульфоновой
микропористой (диаметр пор 0,2 мкм) мембраны, обеспечивающей
задерживание бактерий при высоком потоке жидкости.
Если применяют какой-либо газ для очистки раствора, то он
перед использованием должен подвергаться фильтрующей стери-
лизации.
В случаях, когда целевой продукт в виде раствора или жидкости
нельзя стерилизовать упакованным в контейнеры, то его фильтру-
ют через стерильный фильтр с диаметром пор не более 0,22 мкм.
При обнаружении вирусов и/или микоплазм в растворе (жидко-
сти), стерилизуемом (-ой) фильтрованием необходимо предусмот-
реть возможность использования дополнительного метода стери-
лизации (физического или химического).
Нельзя применять асбестовые фильтры для стерилизации инъ-
екционных лекарственных форм.
254
Т а б л и ц a 28. Некоторые области практического применения мембранной
фильтрации
Процесс рабочее давление, МПа Проходящие через мембрану вещества Частицы и вещества, задерживающиеся мембраной Области использования
Микрофильтрация, 0,05 Вода, растворенные вещества Суспендированные и коллоидные, частицы, бактерии и другие микробы размером более 0,1 мкм Стерилизация растворов в биотехнологичес ких и химико- фармацевти- ческих производствах, в бактериологии, анализах, при разделении веществ
Ультрафильтрация 0,5 Вода, растворенные неорганические вещества Бактерии, коллоидные и суспендированные частицы размером более 1O'J мкм, растворенные органич. полимеры с ММ 1 кДа Очистка растворов биополимеров, концентриро- вание, фракциони- рование, освобождение от пирогенов (депироге- низация) 1
Обратный осмос 5 • Вода, растворенные высоко полярные ни зкомолеку- лярные вещества Бактерии, вирусы, суспендированные и коллоидные частицы, растворенные неорганические вещества, органические вещества с ММ 0,2 кДа Концентриро- вание растворов в биотехнологии, глубокое обессоливание воды и т. д. I
Любой фильтр, используемый для стерилизующей фильтрации,
не должен качественно и количественно изменять конечный про-
дукт (в том числе и антибиотическую активность культуральных
жидкостей после их стерилизующей фильтрации через батареи
фарфоровых свечей).
255
Качество мембран проверяют обычно биологическим методом,
используя для этого виды из родов Pseudomonas и Serratia.
При всех положительных качествах стерилизующей фильтра-
ции через мембраны нельзя не отметить и недостатки этого
способа, к которым относятся: адгезия частиц к мембранам, нео-
днородность пор по диаметру ("абсолютных" мембран по стерили-
зующей эффективности не существует, но стерильность может
быть достигнута и достигается вследствие наложения других при-
чин, например, адсорбции частиц на мембране), удержание части
стерилизуемой дорогостоящей жидкости на мембране при фильт-
рации малых объемов ее, а также возможная селективная адсор-
бция ионов (чаще — катионов) из небольших объемов растворов,
недостаточная или плохая смачиваемость мембран водой и др. К
тому же по-прежнему актуальной остается проблема вирусного
загрязнения БАВ и очистки БАВ от вирусов. Ситуация, связанная
с очисткой биопродуктов от вирусов, обострилась еще и потому,
что появилось сообщение о контаминации гормона роста человека,
получаемого из гипофиза, "медленным” вирусом болезни Крейтц-
фельда-Якоба, и это на фоне возрастающей роли ретровирусов
(включая ВИЧ). Как следствие — усилилась настороженность к
препаратам из крови, гормонам, экстрагированным из тканей
млекопитающих; рекомбинантным белкам, образуемым культиви-
руемыми клетками животных. Более того, ряд вирусов животных
являются патогенными для человека (зоонозные вирусные инфек-
ции).
Потенциальная контаминация названных выше продуктов воз-
можна из различных источников: от лиц, чьи клетки используются
в биотехнологических процессах; от миеломных клеток, применя-
емых для получения гибридов; из питательных сред для культиви-
рования клеток млекопитающих, из реагентов, используемых для
очистки белковых продуктов, например, моноклональные антите-
ла; наконец, несоблюдение требований GMP.
Принятая проверка инактивации вирусов-контаминантов в био-
продуктах заключается в их температурной или химической обра-
ботке. Однако эти методы не универсальны. К тому же выявление
вирусов базируется не только на дорогостоящих, но и мало чувст-
вительных процедурах. Корпорация "MiHipore" разработала тех-
нологию так называемой наноселективной фильт-
рации с включением вирусудаляющего модуля (Viresolve/70),
снабженного уникальной запатентованной мембраной. Эта техно-
256
логия значительно отличается от технологий с использованием
ультрафильтрационных мембран, хотя бы, например, тем, что в
модуле не предусматривается применение открытой матриксной
поддерживающей структуры, а поры уникальной мембраны иск-
лючительно малы, благодаря чему проявляются отличные качества
ее по задержке вирусов размерами от 23 нм и выше. За этими
пределами остаются тогавирусы (18 нм) и вироиды (15 нм). К тому
же однообразный матрикс мембраны обеспечивает эффективный
поток жидкости и ее физическую стабильность. Модуль предназ-
начен для фильтрации белковых растворов. Они (модули) выпу-
скаются с мембранами разных размеров, в зависимости от объемов
фильтруемых растворов. В промышленном модуле применена мем-
брана с площадью 0,92 м2 для фильтрации нескольких сотен литров.
Воздух, подаваемый в ферментаторы, должен быть чистым и
стерильным. В этих случаях еще используют и другие фильтрую-
щие материалы, которыми заполняются общие и индивидуальные
(для каждого ферментатора) фильтры, например, из перхлорвини-
ла (в фильтрах Петрянова — ФП), стекловаты и др. Профильтро-
ванный воздух сжимается в компрессорах, охлаждается в тепло-
обменнике, поступает в ресивер (от англ, reciever — емкость),
устраняющий пульсации воздуха, из которого через брызгоулав-
ливатель проходит общий и индивидуальный фильтры, и только
после этого поступает через барботер в культуральную жидкость.
Питательная среда перед засевом каким-либо биообъектом
также должна быть стерильной. В таких случаях прибегают к
тепловой стерилизации, заботясь при этом о сохранении стабиль-
ности ингредиентов среды. В микробной биотехнологии обычно
используют методы периодической и непрерывной стерилизации.
Первую из них осуществляют в аппаратах малой емкости непос-
редственно в ферментаторах глухим или острым паром под давле-
нием в течение 30—40 мин. при температуре порядка 134°С
(2,02650x10s Па) после удаления воздуха из аппарата при нагреве
до 100°С. Затем среду охлаждают водой через змеевик или рубашку
аппарата и засевают тем или иным биообъектом.
Метод непрерывной стерилизации основан на том, что концен-
трат питательной среды подают насосом через систему конструк-
ций, включающую нагреватель, выдерживатель (собственно сте-
рилизатор) и теплообменник (охладитель, в котором охлаждение
среды происходит до температуры, оптимальной для культивиро-
вания клеток).
9 т. 8524 257
В лабораторных условиях стерилизацию питательных сред и
некоторых других объектов осуществляют в автоклавах. Стерили-
зацию проводят паром под давлением, соблюдая необходимые
режимы (табл. 29). В отдельных случаях прибегают к сухожаровой
стерилизаций.
Таблица 29. Режимы стерилизации
t°, °C Паром под давлением Сухим жаром, I время, мин. [
время, мин. давление, Па (атм.)
121 15 1,01»105/1) —
126 10 l,414»10S (1,4) —
134 3 2.02.105 (2) —
140 — — 180
150 — — 150
160 -г- — 120
170 — — 60
Различные материалы по-разному влияют на рост и развитие
биообъектов, к тому же — не все из них можно подвергать
автоклавированию (таблица 30).
Таблица 30. Влияние некоторых материалов на рост клеток в культурах и
возможность их стерилизации паром под давлением
(выборочно)
Материал Рост клеток, % к контролю Возмо- жность авток- лавиро- вания мате- риала J
бактерий микро- водо- рослей в культуре тканей
метано- окисля- ющих- других разных расти- тель- ных живот- ных
Стекло (контроль) 100 100 100 100 100 Да
Дюралюминий Д1АТ — — 85,9 — — Да
Оргстекло 40ПБ 100 — 85 97 60-80 Нет
Полиэтиленовая трубка 105 133,8 91,4 43 25-30 Нет
Резина белая (пищевая) Резина красная 102 5,7 — 184 100 0 100 0 Да Да
Резина черная 6,2 0 0
(вакуумная) 110 76,4 92 50 Да
Сталь Х18Н10Т 102 — 112,1 93 60-70 Да
Фторопласт-4 Хлорвиниловая трубка 132 — 175 0 0 Нет
Эпоксидная смола полимеризованная 120 — — 100 — Да
Примечание: (-) — данные отсутствуют, (0) — клетки не растут.
258
Из приведенных в таблице 30 данных видно, что лишь немногие
перечисленные материалы могут быть использованы в биотехно-
логических процессах-, оставаясь нейтральными в отношении био-
объектов (резина пищевая, с оговорками — фторопласт).
Названные выше методы обеспложивания относят к числу
наиболее распространенных, однако, учитывая необходимость сте-
рилизации весьма широкого ассортимента различных материалов,
приходится прибегать и к другим способам. Этим подтверждается
то положение, что универсальных методов стерилизации не суще-
ствует.
- Термолабильные Материалы, например, не могут подвергаться
тепловой стерилизации и тогда можно использовать холодную
химическую стерилизацию, или газовую стерилизацию этиленок-
сидом (температура его кипения + 12,5°С)
Этиленоксид (окись этилена) относится к раз- СН
ряду химических веществ, неспецифически губи- X /
тельно действующих на различные микроорганиз- О
мы — вегетативные клетки бактерий и грибов,
бактериальные споры и грибные конидии, вирусы. Стерилизуемые
при этом объекты не должны вступать во взаимодействие с
этиленоксидом и должны быть нейтральными, хорошо очищенны-
ми, при необходимости — продезинфицированными, соответству-
юще упакованными. Не рекомендуется использовать для упаковки
пергаментную бумагу, стеклянную и металлическую тару, шелко-
вые ткани и некоторые другие, не пропускающие этиленоксид.
Упаковка должна обеспечивать диффузию газа и водяных паров к
стерилизуемому объекту. В упаковку вкладывают индикатор (на-
пример, лакмусовую бумагу, изменяющую свой цвет из желтого в
синий при достижении стерильности; это — индикатор типа 1—21
производства "Медикор", Венгрия) и сертификат с указанием
номера серии стерилизации и гарантийный срок хранения (обычно
— в течение Года) в стерильном состоянии, после чего ее помещают
в полиэтиленовый пакет.
После стерилизации этиленоксидом упаковки выдерживают на
предмет испарения газа. Для этого используют проветривание при
температуре не ниже 20°С в течение 4—5 суток, удаление газа в
рабочей камере циркуляционного парового стерилизатора при
52—55°С в течение 3—4 суток, удаление газа с помощью проточного
вакуума (остаточное давление 101—101,2 кПа) при 52—55°С в
течение 2-3 суток.
Этиленоксид - токсичный газ, поэтому простерилизованные с
259
его помощью объекты и надлежащим образом проветренные хра-
нят в специальном помещении.
Эффективность такой стерилизации определяется следующи-
ми параметрами: концентрацией окиси этилена, давлением смеси
газов, влажностью, температурой и временем стерилизации. Сте-
рилизацию проводят в специальных газовых стерилизаторах раз-
личной емкости (например, 20—25 л) с длительностью разовой
стерилизации при этом до 5 часов, поддерживая температуру
50 ± 8°С (323± 8°К) и концентрацию этиленоксида около 750 мг/л
(17,04 моль/м3). Запрещается выводить газовую смесь после стери-
лизации в вентиляционную шахту; она не должна попадать в другие
помещения; ее следует собирать в виде конденсата.
В некоторых случаях применяют для стерили-
СН2---СН2 зации p-пропиолактон, который отличается высо-
I I кой реакционной способностью и, например, лег-
q_____ко взаимодействует с аминами и многими другими
веществами. В малых концентрациях он губитель-
но действует на вирусы, бактерии и грибы, особен-
но — при повышении температуры, однако водные растворы его
быстро инактивируются при комнатной температуре. Р-Пропио-
лактон токсичен для кожи, агрессивен в отношении некоторых
металлов и многих пластмасс, в повышенных концентрациях про-
являет свойства мутагена. Поэтому в биологической технологии
его не следует применять в качестве стерилизующего агента.
В лабораторных и производственных условиях иногда приме-
няют ультразвуковые установки, например, для мойки посуды
(частота звука здесь выше 20000 гц). В случае распространения
звука в воздухе за нулевую отметку принимают порог нормальной
слышимости 1000 гц. Более высокие величины оценивают в деци-
беллах — дц [от франц, decibel — единица акустической (электри-
ческой) мощности, равная 0,1 бела и принятая в электроакустике],
и если этот показатель выше 100дц, то звук оценивается вредным
для людей и от такого звука следует экранировать занятых в
соответствующих производствах операторов (особенно — при
длительном воздействии). Ультразвук можно использовать для
выделения некоторых действующих веществ из растений (напри-
мер, алкалоиды), антигенов из некоторых бактерий.
Тем не менее, ультразвуковые генераторы не нашли широкого;
распространения в биотехнологии, хотя в отдельных исследовани-
ях было показано, что с помощью ультразвука можно стимулиро-
260
вать рост клеток, гасить ленообразование в биореакторах при
ферментациях и т. д.
Для обработки стен некоторых помещений, мебели — можно
использоватькатионные поверхностно-активные вещества—ПАВ,
проявляющие губительное действие в отношении широкого спек-
тра микроорганизмов. Растворы ПАВ можно применять не только
для протирания стен и мебели, но, например, в боксированных
помещениях заблаговременно распылять в виде аэрозолей (до
прихода оператора).
Во избежание микробного загрязнения биотехнологических
производств используют и специальные меры, как то медицинский
осмотр персонала работниками медицинской службы предприятия
и СЭС в целях выявления заболевших и носителей патогенной и
условно патогенной микрофлоры и для исключения микробного
загрязнения ими целевых продуктов; контроль за водоснабжением,
особенно — на тех производствах, в которых воду забирают из
открытых водоемов; предупреждение попадания микробов в скла-
ды готовой продукции — здесь наиболее эффективен способ
подачи воздуха через приточные вентиляционные устройства, в
которых предусмотрена фильтрация поступающего воздуха в
складские помещения.
6.4. Биотехнологические процессы в связи с массообменом.
Принято считать, что диффузионные процессы, протекающие в
биореакторах, не накладывают заметных ограничений на макси-
мально проявляющуюся функциональную активность клеток и
клеточных структур; главное при этом — поддержание массооб-
мена (и прежде всего — кислорода в системе "газ-жидкость”
применительно к аэробным организмам) на оптимальном уровне.
При выращивании биообъекта в реакторе образуется сложная
система массопереноса О2, а именно - "газ-жидкость-твердое тело",
понимая под твердым телом биообъект. Названная тройная система
легко расчленяется на три самостоятельные системы: "газ-жид-
кость", "жидкость-жидкость" и "жидкость-твердое тело". Это мож-
но продемонстрировать следующим образом (рис. 77).
Перемещение О2 в этих системах неравноценно по интенсив-
ности и зависит от растворимости газа в жидкой фазе, от мощности
барботажа, размера пузырьков, скорости вращения вала и формы
мешалки, химического состава питательной среды, температуры,
толщины невозмущаемых слоев жидкости вокруг газового пузырь-
ка и клетки, от наличия и химического состава капсул, толщины
и особенностей клеточной стенки и клеточной мембраны биообъ-
екта и некоторых других причин.
261
Рис. 77. Системы, образующиеся в биореакторе (8) при массопереносе кислорода
(1) из среды (4) в клетку-продуцент (5); 2 — невозмущаемый слой жидкости, 3 —
пограничная область раздела фаз "газ-жидкость", 6 — пограничная область раздела
фаз "Жидкость-твердое тело", 7 — условный путь перемещения О2 в системе
"жидкость-жидкость" от (1) до (5), где он расходуется в биохимических реакциях.
Из рис. 77 можно сделать вывод, что зона 2 вокруг клетки может
не формироваться в случаях образования ее вокруг пузырька и
клетки одновременно. Тогда и путь перемещения О2 от газового
пузырька в клетку сокращается, а его содержание в жидкой фазе
(4) не будет коррелировать со скоростью поступления его в клетку
для поддержания реакций биологического окисления (в ряде слу-
чаев О2 выступает химическим реагентом, обеспечивающим био-
трансформацию, например, глюкозы в 5-кетоглюконовую кислоту
с помощью уксуснокислых бактерий). Следует иметь в виду, что
при избытке растворенного кислорода проявляется его токсиче-
ское действие на биообъект. Вот почему желателен автоматический
контроль за поддержанием оптимальных концентраций О2 в среде
для соответствующих культур. Теперь известны РО-статы (прибо-
ры для поддержания необходимого уровня растворимого О2),
обеспечивающие подобный контроль.
Если молекула кислорода приобретает дополнительный (экс-
тра-) электрон, то образуются свободные кислородные радикалы:
супероксидный (Ог), гидроксильный (НО ) и синглетный кислород
('Оз). Эти радикалы — потенциальные деструкторы липидов, бел-
ков, нуклеиновых кислот. В частности, к ним чувствительны кле-
точные мембраны, в которых первичной мишенью выступают
липиды, протоны которых взаимодействуют с .радикалами и насту-
пает так называемая "липидная пероксидация" с образованием
пероксидов. Свободнорадикальное повреждение мембраны схема-
тично представлено на рис. 78.
Рис. 78. Поперечный срез клеточной
мембраны, повреждающейся от свобод-
но-радикальных процессов, индуцируе-
мых супероксидным и гидроксильными
радикалами, синглетным кислородом: 1 —
ядро, 2 — нарушенные белковые нити, 3
— перекрестное связывание липидов, 4
— липидно-протеиновое перекрестное
связывание, 5 — поврежденный мемб-
ранный липид.
Известно, что с возрастанием температуры окружающей среды
растворимость кислорода в воде уменьшается. Так, например, при
35°С и давлении кислорода в воде 1,01 • 105 Па его растворимость
составляет 1,09 ммоля/л или в соотношении 1:28600; при 20°С
растворимость возрастает до 1,38 ммоля/л или в соотношении
1:22640; при 14°С в воде растворяется 1,5 ммоля/л О? или в
соотношении 1:21000. В одномолярном растворе натрия хлорида
при 25°С растворяется 0,89 ммоля О?/л или в соотношении порядка
1:35000.
Таким образом, для переноса кислорода из одной фазы в другую
(см. рис. 77) необходимо добиться разницы его концентрации в
разных фазах с преобладанием в направлении слева направо, то
есть от газового пузырька к клетке. Но поскольку растворимость
О2 в воде и водных растворах мала, постольку необходимо подавать
его в биореактор в несколько повышенных количествах, добиваясь
указанной разницы в фазовых системах. При этом достигается
некая критическая концентрация кислорода в культуральной сре-
де, когда все аэробные клетки насыщаются Ог- Для клеток микро-
организмов показатели таких концентраций приходятся на диапа-
зон 0,003—0,05 ммоля/л (примерно 0,1—10% наибольшей раство-
римости О2 в воде) или, применительно к воздуху, от 0,5 до 50%-го
насыщения. Так, для клеток кишечной палочки критическая кон-
центрация растворенного кислорода составляет 0,0082 ммоля/л при
38°С, для пеницилла — продуцента пенициллина при 24°С — около
0,022 ммоля/л, для Saccharomyces cerevisiae при 20°С - 0,0037
ммоля/л.
Для некоторых нитчатых грибов, например, Myrothecium
verrucaria, удельная потребность в кислороде становится наивыс-
шей в начале Log-фазы размножения клеток, хотя плотность
263
I
клеточной биомассы продолжает нарастать. В то же время общая
потребность в О2 достигает максимума в конце Log-фазы и начале
const-фазы, то есть удельная скорость размножения клеток дости-
гает своего пика раньше, чем скорость использования О2.
Скорость переноса кислорода (QoJ рассчитывают на единицу
объема биореатора, исходя из отношения произведения плотности
потока и площади поверхности раздела фаз (газ-жидкость) к
объему жидкой фазы в реакторе, то есть Q02 =-у----. где К?
— коэффициент массопередачи, С'1 — концентрация растворен-
ного вещества в жидкой фазе, находящейся в равновесии с газовой
фазой, А — площадь поверхности раздела фаз "газ-жидкость”, V
— объем жидкой фазы. Если весь кислород, поступающий в
жидкую среду, быстро усваивается клетками, тогда С/=О и мас-
сообмен кислорода здесь максимален.
Средняя объемная скорость поглощения О2 (Q02 моль*с/л) в
объеме V выражается уравнением:
V
Qo2 = 4jQo2dv
о
Когда гидродинамические условия в биореакторе, концентра-
ция О2 и отношение площади поверхности раздела фаз к объему
одинаковы, тогда Оо2 равна Оог-
Обычно при вполне благоприятном аэрировании среды кон-
центрация клеток в биореакторе может достигать величины 108
/мл. Исходя из усредненцых размеров клеток бактерий по диамет-
ру 1 мкм, дрожжей — 7 мкм, нитчатых грибов = 20 мкм объемы
их в процентах к объему среды составят около 0,005, 1,8 и 2
соответственно, то есть бактериальная масса будет примерно в 400
раз меньше грибной массы. Удельная поверхность раздела всех
клеток будет 3,1 см2/см3 - для бактерий, 153 см2/см3 - для дрожжей
и 40 см2/см3 — для нитчатых грибов. Следовательно, эффективная
поверхность в данных примерах будет больше у дрожжей. Поэтому
адекватная доставка кислорода — как лимитирующего фактора
зависит от морфофункциональных особенностей культивируемого
биообъекта и условий его выращивания.
На скорость потребления кислорода биообъектом влияют:
1) возраст культуры (делящиеся, или размножающиеся клетки
потребляют кислорода больше, чем не делящиеся);
2) межклеточная адгезия, когда при выраженной адгезии обра-
264
i
зуются аггломераты, или комочки клеток, и, напротив, при малой
или совсем не проявляющейся адгезии клетки находятся в изоли-
рованном состоянии друг от друга; в первом случае потребление
кислорода уменьшается в сравнении с вторым, когда клетки обла-
дают большей площадью своей поверхности;
3) от быстроты динамических изменений в среде выращивания
биообъекта (например, в производстве экзополисахаридов, проду-
цируемых аэробными микроорганизмами, заметно возрастающая
вязкость среды снижает поступление О2 к клеткам);
4) скорость накопления биомассы клеток — чем больше био-
масса, тем скорость поглощения кислорода быстрее снижается, то
есть зависимость здесь обратно пропорциональная;
5) качество источников питания, подлежащих окислению. Для
сравнения можно назвать такие источники углерода, как глюкоза
и предельные углеводороды из нефти. Молекулы глюкозы в пита-
тельной среде и в клетках одинаковы, так как степень восстанов-
лены ости атомов углерода в данных молекулах одинаковы. Поэтому
и отношение потребленного кислорода к количеству превращен-
ной глюкозы меньше, чем это имеет место в случае использования
углеводородов;
6) пеногасители, добавляемые к вспенивающимся культураль-
ным жидкостям в процессе выращивания некоторых биообъектов.
Так, например, натрия лаурилсульфат в концентрации 10 млн '*
снижает коэффициент массопередачи О2 на 56% (в сравнении с
водой);
7) продукты метаболизма, например, секретируемые белки,
также снижают массопередачу кислорода.
Действие всех перечисленных факторов проявляется во време-
ни и сказывается не только на показателях массообмена, но и
теплообмена — ферментации протекают в растворах неньютонов-
ских псевдопластических жидкостей. В биотехнологических про-
цессах теплообмен — постоянно учитываемый и контролируемый
фактор — либо требуется подача тепла, например, в случаях
стерилизации питательных сред или при культивировании анаэро-
бов при 55—56°С (биологическая обработка отходов в бескисло-
родных условиях), либо, напротив, необходим отвод тепла, обра-
зующегося в биореакторах, в которых выращивают аэробные
клетки. В этих целях пользуются рубашками аппаратов, в которые
подается горячая или холодная вода, змеевиками, вмонтированны-
ми внутри аппаратов, "выносными" теплообменниками и др. Вза-
имосвязь скоростей отвода и образования теплоты описывается
265
уравнением Q = UAAT, где Q—теплота, U—общий коэффициент
теплообмена, А—площадь поверхности теплообмена, ДТ—харак-
терная межфазная разность температур между культуральной
жидкостью и нагревающей (охлаждающей) жидкостью. Под общим
коэффициентом теплообмена (U) понимают количество теплоты,
передаваемое в единицу времени через единичную поверхность
при разности температур в 1°С. Поэтому и теплообмен в общем
виде можно определить как перераспределение тепловой энергии
между взаимодействующими фазами в биореакторе. Теплообмен
зависит от ряда факторов:
1) ламинарного или турбулентного движения теплоносителя,
2) толщины и качества материала стенок биореактора,
3) вязкости среды,
4) скорости потока при полунепрерывном и непрерывном
способах культивирования биообъектов,
5) характера охлаждения биореактора.
Познание этих факторов является необходимым для констру-
ирования оптимальных биореакторов и наиболее выгодного про-
ведения биотехнологических процессов.
Массообмен в биологических системах связан не только с
доставкой кислорода к клеткам аэробных видов и теплообменом,
но и с выделением диоксида углерода как конечного продукта
различных катализируемых реакций, транспортом других веществ
через клеточные мембраны (в том числе — анаэробных организ-
мов).
Диоксид углерода может находиться в четырех различных
формах, с различной степенью растворяясь в воде: СО5", НСОз,
Н2СО3 и СО2. Растворимость СО2 зависит от pH жидкости и он j
переносится через границу раздела фаз "газ-жидкость" лишь в
растворенном виде.
Для оценки газообмена при культивировании отдельных био-
объектов, а также в целях контроля за микробами — загрязните-
лями зерна, пищевых продуктов, других жидких и плотных мате- -
риалов используют специальное оборудование типа "О2/СО2 ре-
спирометров", в том числе — с компьютерным контролем (напри- J
мер, "Микро-цксимакс О2/СО2 респирометр" фирмы Columbus ]
Instruments, США). ‘
Транспорт питательных веществ через мембраны клетки осу-
ществляется различными путями в системах "жидкость-твердое |
тело”: простой (пассивной) диффузией, облег- |
266 ’
ч е н н о й диффузией и благодаря активному
транспорту, ау ряда эукариотических организмов — с
помощью эндоцитоза. В нормальном состоянии мембрана непро-
ницаема для большинства полярных молекул, но некоторые газы
(СО2, N2) и водные растворы многих веществ свободно диффун-
дируют через мембрану благодаря соответствующим концент-
рационному и электохимическому гради-
ентам. Градиент концентраций - это разность концентраций, а
диффузия — суммарное перемещение вещества, зависимое от
градиента концентраций. Электрохимический потенциал склады-
вается из суммы зарядов диффундируемых веществ. Электроны
медленно проходят через мембрану, и чем больше их заряд, тем
меньше транспорт.
При пассивной диффузии молекула растворенного вещества
не изменяет свою первоначальную молекулярную форму. Значе-
ние свободной энергии (ДС7) при этом всегда отрицательно.
dtt Ci _
AG = KTLn-gr , где Ci — область пониженной концентрации
вещества, С2 — область повышенной концентрации вещества.
Ионизированные и полярные молекулы, плохо растворимые в
полярных растворителях, обычно не транспортируются через мем-
брану пассивной диффузией. Для этого существует механизм
облегченной диффузии, или сопряженного транспорта, когда ве-
щество связывается с молекулой-переносчиком, локализованной
на наружной поверхности мембраны. Образующийся комплекс
достигает внутренней поверхности мембраны, где и разъединяется,
перенеся молекулу вовнутрь клетки.
Облегченная диффузия осуществляется при участии мембран-
ных пор, генерирующих ион-проводящие пути; ионофоров-пепти-
дов, формирующих ионные каналы (например, декапептиды—
антаманид у базидиомицетов, грамицидин С у некоторых бацилл
и др.); интегральных и периферических мембранных белков, дей-
ствующих взаимосвязанно (например, пермеазы и др.).
Транспорт веществ при облегченной диффузии протекает не
линейно (как при пассивной диффузии), ас определенным уровнем
насыщения (maximum), после которого возрастание разности кон-
центраций вещества не сказывается на скорости его переноса.
Этот вид транспорта кинетически сходен с ферментативным ката-
лизом.
Система активного транспорта через мембрану является энер-
гозависимой. Она включает фермент АТФ-азу, белки-переносчики
267
различных веществ, фосфолипиды. АТФ-аза проявляет активность
в присутствии фосфолипидов. Активный транспорт подразделяют
на первичный и вторичный. Энергия для первого поставляется
одним их двух способов:
1) когда транспорт сопряжен с одновременным прохождением
через мембрану второго вещества, движущегося по градиенту
концентраций и в том же направлении, что и первое вещество —
симпорт, или в обратном направлении — антипорт; свободная
3Na+ Na^~ К1— АТФ — аза
4
АТФ
АДФ + Фн
6
Рис. 79. Мембранная Na+ —
КН---АТФ-аза (Nad---К+—на-
сос), участвующая в активном
транспорте катионов (1—олиго-
сахариды, 2—периферические
белки, 3—трансмембранные бел-
ки, 4—периплазматическое про-
странство или внеклеточная сре-
да, 5—мембрана, 6—цитоплазма).
энергия (AG) этого процесса большая и отрицательная величина;
2) когда энергия поставляется за счет сопряженного гидролиза
АТФ или какого-либо другого высокоэнергетического соединения
(фосфоенолпируват, карбомоилфосфат, 1,3-дифосфоглицерат и
др.). Этот способ называют насосом (AG) здесь положительная
величина) (рис. 79).
За счет работы систем первичного активного транспорта на
мембране создается градиент ионов, прежде всего — протонный
потенциал АДЕГ, который индуцирует
вторичный активный транспорт — пе-
ренос с помощью белков-переносчи-
ков (в частности, пермеаз) аминов,
аминокислот, сахаров и других ве-
ществ (например, М-белок у Е. coli
является компонентом трансфераз-
ной системы, и он переносит 0-галак-
тозиды, и т. д.).
Функции системы активного
транспорта: поддержание постоянст-
ва концентраций метаболитов незави-
симо от колебаний содержания их во
внешней среде; стабильное поддержа-
ние оптимальных концентраций неор-
ганических ионов как кофакторов
ферментативных реакций и для акти-
вирования других процессов; извлечение из окружающей среды
необходимых веществ даже при низкой их концентрации; регуля-
ция метаболизма.
Массообмен для многих веществ в различных системах еще
далеко не изучен полностью и здесь биологическая технология
имеет резерв совершенствования биотехнологических процессов
в целях достижения более высокого выхода конечных продуктов.
268
6.5. Биотехнологические процессы в связи с особенностями
метаболизма клеток. Процессы в биохимической технологии в
большинстве своем базируются на использовании продуктов вто-
ричного метаболизма. Даже в тех случаях, когда преследуют цель
промышленного производства биомасса клеток или тканей, опти-
мизация условий ее выращивания также основывается на знаниях
особенностей метаболизма тест-культур. Эффективность накопле-
ния такой биомассы по-прежнему оценивается экономическим
коэффициентом (ЭК), то есть отношением веса сухой массы клеток
(ткани)-У к весу потребленного углевода (Сп). Величину экономи-
ческого коэффициента выражают в процентах, и она, как правило,
обратно пропорциональна концентрации сахара: ЭК%=У/Сп-100.
Известно, что метаболизм различных организмов включает
реакции анаболизма, амфиболизма и катаболизма. , Анабо-
лизм (от греч. anabole — подъем) — это синтез веществ клетки;
амфиболизм (от греч. amfi—оба) объединяет реакции
промежуточного обмена; катаболизм (от греч. catabole
—сбрасывание, разрушение) — расщепление веществ. В связи с
энергетикой процессов первый и второй из них протекают с
потреблением энергии, третий — с выделением энергии. Поэтому
метаболизм в целом можно подразделить на энергетический и
конструктивный. Энергия выделяется при брожении и дыхании,
а потребляется в реакциях синтеза нуклеиновых кислот, белков,
углеводов, липидов, их конъюгатов, при активном транспорте
различных веществ через клеточные мембраны, и т. д.
Все организмы, использующие энергию солнечного света, на-
зывают фототрофными; организмы, использующие энергию
при окислении химических веществ, называют хемотроф-
н ы м и.
Реакции обмена веществ у прокариот и эукариот (акариоты не
обладают собственным обменом веществ) катализируются фер-
ментами, число которых в клетке может достигать 1,5—2 тысяч.
Скоординированные биохимические и биофизические процес-
сы размножения, роста и развития клеток в конкретных условиях
среды обитания и составляют сущность обмена веществ, или
метаболизма. Наличие большого количества ферментов в клетках
различных организмов обеспечивает им широкий диапазон адап-
тивных, или приспособительных возможностей (хотя и не безгра-
ничный). Среди различных групп организмов наиболее лабильны-
ми являются одноклеточные виды и, прежде всего, бактерии и
дрожжи. Высшие животные и растения имеют собственные меха-
269
низмы для поддержания своего, гомеостаза (от греч. omoios—подо-
бный, одинаковый, stasis—неподвижность, состояние), то есть
динамического постоянства состава и свойств внутренней среды,
а это значит, что при заметных отклонениях в среде обитания
наступают отклонения от нормы либо в их структуре, либо в
функциях (признаки патологии, или болезни).
Следовательно, кроме обеспечения тест-организмов питатель-
ными веществами им необходимы более или менее постоянные
условия функционирования в заданном направлении (накопление
биомассы или метаболитов), в частности, стабильность температу-
ры, снабжение или, напротив, исключение Ог, поддержание pH на
определенном уровне.
Все биохимические реакции, катализируемые ферментами, не
выходят за пределы классических законов термодинамики, и к ним
(как и к чисто химическим системам) приложим принцип ле
Шателье — равновесная точка системы смещается в направлении
. уменьшения эффекта произведенного воздействия. Очевидно, что
"метаболическое разнообразие" определяется не столько механиз-
мами реакций, а сколько ферментным набором в клетках различ-
’ ных организмов и подходящими условиями для их проявления, то
есть первичными здесь являются ферменты, а вторичными —
катализируемые ими реакции. .
Метаболизирующие клетки ограничены сроком выживания в
конкретной среде обитания, а после отмирания в природе они
подвергаются разложению и минерализации. Если проследить за
круговоротом веществ, входящих в состав клеток, то можно выде-
лить определенную цикличность процессов для азота, углерода и
других биоэлементов. Метаболизм детально рассматривается в
курсах биологической химии и химической микробиологии.
Микроорганизмы, например, из рода Bacillus, продуцирующие
активную фосфатазу, превращают мало растворимые фосфаты в
растворимые и легко доступные другим организмам, у которых он
включается в состав нуклеиновых кислот, некоторых макроэрги-
ческих соединений (например, АТФ) и др.
Выдающаяся роль серы в живых организмах обусловлена ти-
ольными группами, которые в составе серусодержащих аминокис-
лот участвуют в формировании и стабилизации третичных струк-
тур белковых молекул.
Зная пути и механизмы превращения различных веществ, мож-
но использовать отдельные процессы в практических целях либо
для получения метаболитов, либо клеточной массы.
Трансформация различных веществ в природных или искусст-
270
венных условиях сопряжена с расходованием и накоплением
энергии. Основными источниками энергии для живых существ
является солнечная радиация, углеводы; только для прокариот —
Нг, СОг, NHs, Nz и его неорганические производные (H2S,
Na2S2O3, SOl~), а также Fe2+. К первичным генераторам энергии, в
которых происходит преобразование световой и химической энер-
гии в доступную клеткам форму, относятся бактериородопсин,
редокс-цепь фототрофов и дыхательная цепь. Все эти генераторы
локализованы в мембранах (в пурпурных мембранах у галобакте-
рий, в хроматофорах фотосинтезирующих бактерий, в тилакоидах
у цианобактерий, в хлоропластах у водорослей и растений, цитоп-
лазматической мембране у прокариот, и внутренней мембране
митохондрий у эукариот). Под "редокс-цепью" понимают последо-
вательность переносчиков восстановительных эквивалентов с лю-
бым акцептором этих эквивалентов, кроме О2; "дыхательная цепь"
— это последовательность переносчиков восстановительных экви-
валентов с кислородом в качестве их конечного акцептора. Функ-
ционирование "дыхательной цепи" всегда сопряжено с генерацией
энергии, доступной для клетки, тогда как у "редокс-цепи” эта
функция может отсутствовать. У фототрофных, или фотосинтези-
рующих организмов возможны следующие окислительно-восста-
новительные системы (рис. 80 а, б)
свет (hv)
_____4_____
(-) хлорофилл (р)
,е
свет (hv)
'I______
(^j хлорофилл
растения,
водоросли, цитохром
цианобак- 7^
терии &/
е Н;А^-А+2Н*
•АДФ+Р+Э
кофактор
2е (Н* z
\ лдф+р-Хлтф)
НАДФ*-»НАДФ
восст.
цитохром
2е
(а)
(6)
растения,
водоросли,
бактерии
Рис 80. Схемы окислительно-восстановительных процессов при цикличном (а) и
нецикличном (б) фосфорилировании; НаА соответствует НаО — у растений и водорослей,
На, Н aS или разные органические вещества — у других фотосинтезирующих аноксиген-
ных бактерий.
У хемотрофных, или хемосинтезирующих видов набор окисли-
тельно-восстановительных систем шире:
1) доноры водорода — неорганические вещества (литотрофия);
акцепторы водорода:
а) кислород (аэробное дыхание)
271
H2S ------► S
+ 40
SOl' (Beggiatoa, Thiobacillus)
NH3 --------—► NO2 (Nitrosomonas)
NO? --------—► NO3 (Nitrobacter)
, +0
Fe2+ ---------► Fe3+ (Leptothrix)
+0
H2 ----------► H2O (Hydrogenomonas)
CO -------► CO2 (Carboxydomonas)
б) -неорганические вещества (анаэробное дыхание):
NO3, SO1', со§-
Н2 ------► НгО
} некоторые водородные бактерии
NO3------► N2
Н2 —------► Н2О
} некоторые виды Desulfovibrio
SO1'______, Н2О
Н2 _______► Н2О
} некоторые метайогенные бактерии
СОЗ’________►СН4
4Н2.+ СО2
СНз СООН + 2НгО Clostridium aceticum
S------► SOl'
} Thiobacillus nitrificans
NO3 — N2
2) доноры водорода — органические вещества (органотрофия);
акцепторы водорода:
а) кислород (аэробное дыхание); в эту группу относят все
гетеротрофные организмы, которые могут использовать органиче-
ские вещества в качестве доноров водорода;
б) неорганические’ вещества (анаэробное дыхание):
NO3, SOi, СОЗ’
272
NO3 --------► N2 денитрификаторы
NO3 --------► NH3 нитратредуцирующие бактерии
SOl- ------- H2S Desulfovibrio
СОЗ'--------»СН4 метаногенные бактерии
в) органические вещества (брожение); сюда относят все виды
бродильных процессов (молочнокислое, маслянокислое, спиртовое
и др.).
Уравнение, с помощью которого описывают взаимоотношение
энергетических компонентов, включает общую энергию системы,
или энталиию (Н, в Джоулях), меру разупорядоченности системы,
или энтропию (S, в Джоулях/градусы) и свободную энергию
системы (G, в кДж/моль), представляется в следующем виде:
G = AH — TAS (Т— абсолютная температура),
G АТФ= —30,564 кДж/моль.
В большинстве случаев изучение энергетического метаболизма
сводится к оценке продукции АТФ и трансмембранного потенциала
(АрТГ), пороговая величина которого, необходимая для окислитель-
ного фосфорилирования, составляет 180—270 мВ; ДрТГ формиру-
ется на клеточных мембранах у различных организмов (кроме
облигатных анаэробов).
Клетки, как правило, метаболизируют при изотермических
условиях и значительная часть энергии ими не используется (в
противном случае мог бы возникнуть перегрев системы) — она
теряется. При этом возрастает энтропия, и клетки могут погибнуть.
Смерть — это большая положительная величина энтропии, тогда
как негативная величина ее и упорядоченность — это жизнь.
Суммируя дыхательные процессы, сопряженные с получением
энергии, приводим таблицу 31, в которой отражены основные типы
дыхания и указаны представители, "ведущие" эти процессы, а на
рис. 81 представлена схема путей образования и расходования
унифицированных форм энергии [по В. П. Скулачеву (1984) в
некотором видоизменении В. К. Акименко (1989)]. На рис. 81
обобщена сущность хемиосмотического принципа П. Митчелла о
сопряжении процессов генерации и потребления энергии, доступ-
ной для кле,тки. Здесь речь идет’о трансмембранном потенциале
hqhob водорода — А pH* , который представляет собой унифици-
рованную форму энергии, обеспечивающей (наряду с АТФ при
гликолизе у анаэробов, и у некоторых видов - ApNa+) все энерго-
потребности клеток.
273
Название дыхательной цепи с учетом акцепторов восстановительных эквивалентов (е‘, ЬГ) Название дыхательного процесса с учетом потребностей в кислороде 1 Тривиальное название I дыхания Организмы, участвующие в дыхательном процессе Название типа дыхания Образующийся конечный продукт Акцепторы водорода (электронов) Доноры водорода (электронов)
р е д о к с (О2 здесь н< & X сь и и 0 а н а э р о । факульта- тивные анаэроб- ные и аэробные бактерии нитратное NO1 N2O n2 9 Неорг аниче
г. 4? % СА X £ о 3 "О б н о е д ы облигат- ные анаэробы сульфат- ное % О кжие или орг
1 I i 0 х: о Г) £ х а н и е облигат- ные и факуль- тативные анаэробы серное % анические ве
3 • анаэроб- ные аце- тогенные бактерии карбонатное д а: о о £> 5 щества (Hi, 1
анаэроб- ные ме- танобра- зующне бактерии § 1гО, HjS, гик
анаэроб- ные желе- зобакте- рии "железное' ¥ 5 ¥ ►коза и др.)
анаэроб- ные сукцино- генные бактерии 3$ н сукцинат фумарат органическ
брожение анаэроб- ные дрожжи, лакто- бактерии брожение этанол, пактат и др. продукты брожения ацеталь- Дегид, пируват и др. ие вещества
дыха- тельная цепь (О? - акцептор е‘, И) аэробные процессы аэробное 1 дыхание В аэробные прокариоты и эукариоты 9? Н2О s>
Таблица 31. Механизм дыхательных процессов у различных организмов
274
Рис. 81. Схема путей образования и расходования унифицированных форм
энергии^
Среди названных в таблице 31 представителей многие нашли
применение в биотехнологии (аэробные и Анаэробные вцды), а ряд
из них относится к числу перспективных для практического ис-
пользования (метанобразующие бактерии, клеточные культуры
лекарственных растений и животных).
Следует подчеркнуть, что в сравнительном плане экономически
более выгодными являются анаэробные биотехнологические про-
цессы, когда резко сокращаются затраты, связанные с подачей
стерильного воздуха в биореакторы (он здесь не нужен), с работой
мешалок и барботеров и др. А с этим упрощаются конструкции
аппаратов и происходит их удешевление.
6.6. Управление биотехнологическими процессами. Все био-
технологические процессы относят к числу управляемых, хотя
уровень управления различен, и он изменяется со временем.
Поэтому неуправляемые процессы могут быть в природных усло-
виях (тогда они не относятся к разряду биотехнологических) или
они осуществляются с расчетом на саморегулируемость биообъек-
та. Абсолютно неуправляемых биотехнологических процессов не
существует. Например, микробиологические процессы характери-
зуются рядом особенностей, которые необходимо учитывать при
конструировании оборудования. К таким особенностям относятся:
275
1) необходимость обеспечения стерильности (см. раздел 6.3) в
биореакторе, включая коммуникации, датчики, все продуктовые
потоки, идущие в реактор;
2) проведение культивирования аэробов и анаэробов в системах
"газ-жидкость-твердое тело" или анаэробов в системах "жидкость-
твердое тело" при низких скоростях биохимических процессов и
многокомпонентности состава питательных сред для выращивания
биообъектов;
3) исключительная сложность механизмов регуляции биосинте-
за БАВ (или выращивания биомассы клеток) при заметных изме-
нениях физико-химических показателей культуральных жидко-
стей;
4) заметная чувствительность биообъектов к воздействию фи-
зико-механических факторов при перемешивании и других опе-
рациях.
Чистота и стерильность — важнейшие составляющие благопо-
лучно проведенного биотехнологического процесса. Это достига-
ется при осуществлении подготовительных операций перед куль-
тивированием биообъекта, в том числе — стерилизации питатель-
ных сред, пеногасителей, различных жидких добавок, оборудова-
ния и коммуникаций, фильтров для очистки воздуха, использова-
ния высококачественного посевного материала.
Нарушение гарантированного "чистого" культивирования био-
объекта согласно регламентным условиям сопровождается, как
правило, его загрязнением контаминирующей микрофлорой. При
этом выявление микробов-загрязнителей может быть трудным
из-за присутствия единичных клеток, например, бактерий — кон-
таминантов в бактериальных вакцинах, или грибов-контаминантов
при культивировании грибов-продуцентов ферментов. В равной
мере можно ожидать загрязнения на промежуточных стадиях или
в конечном продукте бактериальных или грибных токсинов. К тому
же, обнаружение микробов-контаминантов или их токсинов, как
правило, возможно после завершения какой-то стадии биотехно-
логического процесса, либо в готовом продукте. Следует еще
подчеркнуть и тот факт, что методы выявления посторонней
микрофлоры, к сожалению, в большинстве случаев низко чувст-
вительны.
Биообъект (клетки, ткани), будучи сложной системой, не под-
дается охвату единой математической моделью, а это исключает
возможность описания процесса культивирования конечным чис-
лом уравнений химической кинетики. Даже при гомогенно-непре-
рывном процессе культивирования в околоклеточных областях
276
биосистема гетерогенна (различие в массообмене в многофазной
среде — "газ-жидкость-твердое тело", "жидкость-твердое тело” при
различии структур оболочек, клеточных стенок, мембран, их адге-
зивной способности и пр.).
Биологические системы являются саморегулирующимися
(включая способность к мутациям), что может привести к непред-
виденному изменению биотехнологического процесса.
Утрата качеств биообъекта равноценна катастрофе. Здесь мож-
но указать на автоселекцию биообъекта в биореакторе, например,
какого-либо микроба-продуцента вторичного метаболита. Если в
ферментаторе объемом 50 м3 при коэффициенте заполнения 0,6 и
длительности культивирования 4—5 суток будет содержаться 1012'15
и более клеток, то, учитывая частоту спонтанных мутаций 1 • 105
— 1 • 10'7, число мутантов может достичь величин 105—107 за
генерацию. Поэтому не исключено, что процесс ферментации
может протекать вопреки регламентным требованиям.
Иногда автоселекция приобретает положительное значение в
том смысле, что, например, возврат к "дикому типу" в результате
спонтанных мутаций сопровождается более высокой скоростью
размножения культуры и тогда, стремясь к накоплению клеточной
белковой массы, мы получаем желанный результат.
Квалифицированное управление любым биотехнологическим
процессом основывается на глубине познания "поведенческих
реакций" биообъекта в конкретных условиях культивирования при
получении целевого продукта, когда с помощью соответствующего
приборного оснащения необходимо непрерывно или периодически
фиксировать контролируемые показатели (t°, pH, количество био-
массы клеток, скорость потребления источников питания, количе-
ство растворенного кислорода, количество образующегося мета-
болита и др.). При размножении и развитии клеток происходит
постоянное изменение отдельных параметров биотехнологическо-
го процесса, и в каждый данный момент эти клетки функциони-
руют в иных условиях.
Сложность биологических систем сказывается на необходимо-
сти многопрофильного контроля за ними, например, в биореакто-
рах (таблица 32). Указанные в таблице виды контроля в основном
характерны для культивирования биообъектов в реакторах боль-
шой емкости. На небольших биореакторах число контролируемых
параметров заметно сокращается (например, чаще регистрируют
лишь температуру, pH, подачу стерильного воздуха и количество
клеток и/или метаболита).
277
Таблица 32. Примерный перечень контролируемых параметров при
выращивании клеток прокариот и эукариот в биореакторах
№ п/п Параметр Вид контроля № п/п Параметр. Вид контроля
1 Коэффициент заполнения Физический (светоопти- ческий) ' и Количество биомассы физический (спектрофото- метрия, нефелометрия)
2 Мощность мешалки Механический (внешний- торсионная динамометрия, вйутренний- тензодатчик) 12 Температура Физический. (термометрия) •
13 Скорость потока газа - Механический (ротаметры, расходомеры).
3 Скорость вращения мешалки Механический (тахометрия) 14 Скорость добавления питательных - веществ в растворах Механический / расходомеры (счетчик капель, . динамомет- рические дат- чики и др.)/
4 Redox- потенциал Физический / электропро- водность (определение окислительно- восстановит. потенциала)/ 15 Давление Механический . (манометрия)
16 Вязкость Физический (вискози- метрия)
17 рн Химический (рН-метрия)
5 Количество растворенного кислорода Физико- химический (электрохими- ческий и др.) 18 Отбор (слив) культуральной жидкости Механический (регулиро- вание уровня, динамо- метрический датчик)
6 Количество растворенного СО2 Физико- химический (ИК- спектрометрия и др.) 19 Фермента- тивная активность Биохими- ческий, физико- ; химический
7 Обнаружение пены Физический (электро- проводность) 20 Антибиотичес- кая активность Биологический, химический, физико- химический
278
Продолжение табл. 32
8 Регулирование пенообразо- вания Механический (механич. устройство), физико- химический (например, силиконовый эмульсионный пеногаситель) 21 Определение ДНК, РНК Химический, физико- химический
22 Определение АТФ Химический, физико- химический
9 Потребление глюкозы Химический физико- химический
10 Потребление азота Химический, физико- химический
В случаях накопления биомассы клеток в качестве антигенного
вещества можно воспользоваться серологическими методами,
включая иммуноферментные. Меченые радиоактивными изотопа-
ми антитела рекомендуют применять для определения антигенных
детерминант на поверхности клеток.
Контроль и управление биотехнологическими процессами, ре-
ализуемыми в периодическом или непрерывном режимах (даже
при использовании одних и тех же биообъектов), будут не одина-
ковыми. Например, одноклеточные культуры в глубинных услови-
ях в периодическом режиме проходят все фазы своего развития
(скрытую — lag, экспоненциальную — log, стационарную — const.,
отмирания— let.). Поэтому продукция какого-либо вторичного
метаболита будет далеко неравнозначной в ту или иную фазу
развития продуцента. В непрерывном режиме клетки находятся,
как правило, в одной определенной фазе.
Нитчатые грибы растут верхушками гиф, поэтому у них со
временем изменяется не только масса грибницы, но и морфология
клеток. Если у прокариот количество клеточной массы возрастает
в течение log-фазы экспоненциально (в геометрической прогрес-
сии), то грибы растут медленнее и не экспоненциально, биомасса
их клеток приблизительно пропорциональна третьей степени вре-
мени. Так, в глубинных условиях продуцент антибиотика цефалос-
порина С — Cephalosporium acremonium имеет три морфологиче-
ские формы: артроспоральную, фрагментарно-гифальную (набух-
шую) и гифальную, способных превращаться друг в друга при
изменении среды культивирования. Однако, антибиотик образу-
ется главным образом второй формой. Поэтому контроль и управ-
ление таким биотехнологическим процессом должен проводиться
с учетом указанных морфо-функциональных изменений продуцен-
та.
Уровень управления различается также в зависимости от ре-
жима проведения биотехнологического процесса: периодический,
полунепрерывный и непрерывный. Периодические процессы чаще
используются в случаях получения метаболитов (аминокислоты,
этанол, уксусная кислота и др.), полунепрерывные и непрерывные
— при наработке клеточной биомассы, например, дрожжей. Пол-
унепрерывные процессы отличаются от непрерывных тем, что при
их реализации периодически отбирают 25—75% культуральной
жидкости и добавляют равный объем свежей питательной среды.
Непрерывные процессы ведут в проточных реакторах-хемостатах
или турбидостатах (по лат. turbidus — мутный), в которые непре-
рывно дозированно поступает свежая питательная среда и непре-
рывно в таких же количествах отводится культуральная жидкость.
Таким образом, содержимое биореактора тождественно содержи-
мому отбираемой культуральной жидкости в каждый данный
момент. Такие хемостаты или турбидостаты используют в лабора-
торных условиях для изучения кинетики клеточного роста и мета-
болизма с последующей экстраполяцией результатов при масшта-
бировании процессов в крупногабаритных реакторах.
Различие хемостатов и турбидостатов заключается лишь в том,
что контроль за процессом включает фотоэлектрические элементы
за регуляцией подачи свежей питательной среды и отвода культу-
ральной жидкости.
Если бы рост (Н) ограничился вследствие истощения питатель-
ного вещества, то Н = КС, где К — константа, а С — начальная
концентрация лимитирующегося питательного вещества. Следова-
тельно, между С и Н существует прямая зависимость. Если какое-то
питательное вещество изначально содержится в довольно низкой
концентрации, то промежуточные метаболиты будут образовы-
ваться с ограниченной скоростью, и общая скорость роста будет
функцией концентрации названного выше вещества. Эксперимен-
тально установлено, что в соответствии с уравнением
Hv=Hvk
С
С1+С
получается кривая типа гиперболы. В этом уравнении
Hv—скорость роста, HVk—максимальная скорость роста, достигае-
мая при повышении концентрации питательного вещества; С—
концентрация лимитируемого питательного вещества; Ci—величи-
280
на С, при которой Hv=l/2Hvk. Общий рост (Н) и скорость роста
(Hv) являются существенными критериями во многих биотехноло-
гических процессах. Ими часто пользуются при выборе контроля,
например, за ходом накопления клеточной биомассы в качестве
целевого продукта. При периодических процессах с помощью
контрольно-измерительных приборов и автоматики (таймеры, ре-
ле, датчики и др.) управляли биотехнологическими процессами.
Теперь пытаются заменять их ЭВМ, или компьютерами.
При непрерывных биотехнологических процессах управление
ими не усложняется, так как биосистемы здесь работают в опре-
деленных стационарных состояниях и на определенном этапе
сходны с системами, функционирующими в периодических режи-
мах. Однако биосистема изменяется на пути от начального (стар-
тового) периода к стационарному, и чем этот путь, короче, тем
лучше. Таким образом, в подобных случаях цель управления
фокусируется на поддержании заданного стационарного состоя-
ния, используя, например, микро-ЭВМ. На рис. 82 показана прин-
ципиальная схема работы ЭВМ.
Рис. 82. Архитектура и
принцип работы ЭВМ (по Л.
В. Зудину, В. М. Кантере, Г. А.
Угодчикову, 1987): 1 — уст-
ройство управления, 2 —
ввод, 3 — вывод, 4 — арифме-
тическое устройство, 5 — ис-
ходные числа, 6 — промежу-
точный результат, 7 — очеред-
ная команда, 8 — адресная
команда, 9 — код операции,
а—программа, в—запомина-
ющее устройство, б—данные,
г—результаты.
Некоторые микро-ЭВМ с количеством операций от 1000 до
500000 операций в секунду используются главным образом как
вычислительные (управляющие) блоки в системе управления и
регистрации.
Упомянутые ранее РО-статы (см. раздел 6.4), применяемые для
контроля и поддержания необходимого уровня растворенного
кислорода в питательных средах, обычно включают микрокомпь-
ютер, интерфейс, расходомеры для воздуха и кислорода, и сенсоры
для растворенного Ог.
Микрокомпьютеры используют также в комплексном контроле
за ферментацией при получении некоторых вторичных метаболи-
тов (например, этанола).
ЭВМ удобны в случаях одновременного контроля и учета
многих параметров. Функционально-законченный программно-уп-
равляемый малоразрядный блок, выполненный в виде одной или
нескольких больших интегральных схем, называют микро-
процессором (выполняет до нескольких миллионов операций
в секунду). Его используют для управления ферментационными
процессами. Микропроцессоры можно устанавливать непосредст-
венно на обслуживаемых аппаратах.
Оптимизацию процессов биологической технологии можно
расчленить на три уровня сложности. Первый — низший, решае-
мый обычно с помощью микро-ЭВМ, связан с регулированием
параметров биотехнологического процесса с помощью системы
Рис. 83. Главные элементы системы управления с обратной связью: (1 —
регулятор, 2 — исполнительный механизм, 3 — процесс, 4 — измерительный
прибор).
Второй (средний) уровень рассчитан на более подробный ана-
лиз данных с последующим диалогом оператора с ЭВМ через
терминал. Здесь допустима работа с мини-ЭВМ с числом операций
от 100—300 тыс. до 1 млн. в секунду.
На третьем (наивысшем) уровне сложности применяют боль-
шие и супер-ЭВМ с числом операций от десятков-сотен тысяч до
12 млн. операций в секунду. Информация, поступающая от первого
и второго уровней, используется для решения сложных модельных
уравнений в целях совершенствования режимов биотехнологиче-
ского процесса в целом. Указанные три уровня сложности расчетов
оптимизации условно представлены на рис. 84.
282
Рис. 84. Три уровня сложности расчетов оптимизации микробиологического
процесса в биотехнологии (Л. Валентини, Ф. Андреони, М. Буволи, П. Пенелла,
1982).
В последнее десятилетие все больше расширяется применение
различных ЭВМ и микропроцессоров для управления биотехноло-
гическими процессами. Их конструкции постоянно совершенству-
ются, цены снижаются, а программное обеспечение заметно убы-
стряется. Тем не менее, многие существующие биотехнологиче-
ские производства частично или полностью еще нельзя отнести к
автоматизированным системам управления (АСУТП).
6. 7. Системы GLP и GMP в связи с качеством биотехнологи-
ческих продуктов. В целях организации качественного проведения
283
доклинических испытаний лекарственных и других биологически
активных веществ (пищевых добавок, агрохимикатов и др.) в
промышленно развитых странах (Англия, Германия, США, Фран-
ция, Япония и др.) утверждены единые правила системы GLP (Good
Laboratory Practice). Существует группа GLP в Европейском Центре
по экологии и токсикологии химической промышленности; в США
система GLP действует с июня 1979 г. Главными в такой системе
являются следующие основные действия:
1) заблаговременная разработка стандартной методики прове-
дения испытаний, или SOP (Standard Operating Procedure) приме-
нительно ко всем ее этапам;
2) назначение руководителя и ответственных за каждый вид
испытаний;
. 3) каждому ответственному исполнителю поручается строго
выполнять все операции в отведенных ему пределах;
4) результаты выполнения операций должны быть внесены в
специальный протокол, датированы и подписаны;
5) в. случае выполнения сложных операций, во избежание
ошибок, рекомендуется прибегать к двойной проверке;
6) в установленном порядке исполнитель докладывает руково-
дителю о ходе испытаний. Руководитель должен быть компетент-
ным во всех делах, связанных с испытанием;
7) фактические данные) записи и препараты (вещества) должны
храниться в полном порядке таким образом, чтобы всегда можно
было отыскать требуемое (необходимое);
8) окончательный отчет по своему содержанию должен отра-
жать свежие и еще не обработанные данные, а также сопровож-
даться обсуждением, составленным ответственным исполнителем;
на отчете проставляются дата и подписи (ответственного исполни-
тсля^ лйц. полтверждающих содержание отчета);
9) должна быть служба качественной оценки испытаний —QAU
(Quality Assuarance Unit). Лица, занятые в этой службе, обязаны
стремиться к тому, чтобы свою внутреннюю инспекцию проводить
в установленном порядке и по необходимости выдавать рекомен-
дации. Направленные на совершенствование процессов проведе-
ния испытаний.
На систему GLP опираются в случаях испытания веществ: на
микробную обсемененность, на пирогенность; острую, подострую
и хроническую токсичность, на Специфическую токсичность (кан-
церогенность, антигенность, лекарственную зависимость, повреж-
284
дение зародышевых клеток; раздражение слизистых оболочек,
кожи и в месте введения вещества; мутагенность, тератогенность
— от греч. teratos — чудовище, урод; цитотоксичность), на безопас-
ность для макроорганизма при введении in vivo (абсорбция, рас-
пределение, скорость выведения, метаболизм); проводят фармако-
логические испытания с оценкой фармакокинетики (действие
изучаемого лекарственного вещества на организм) и фармакоди-
намики (изучение силы действия лекарственного вещества).
В связи с необходимостью проведения названных испытаний
создают специальные группы: общую (в том числе для контроля за
гигиеной и санитарией личного состава), микробиологическую,
метаболизма, общефармакологических испытаний, общих клини-
ческих исследований, патологоанатомическую, проведения экспе-
риментов на животных, обработки данных (с включением управ-
ления ЭВМ), по приготовлению проб, аналитическую, по управле-
нию исследованием и, при необходимости, другие. Во главе каждой
группы утверждается руководитель, который не должен совмещать
свои прямые обязанности с работой в группе инспекций.
Соблюдение требований системы GLP должно быть подкреп-
лено совершенством организации всех вспомогательных служб и
достаточным материальным обеспечением. Желательно иметь от-
дельное здание для проведения биологических испытаний, где
экспериментальные животные размещались бы в помещениях
соответствующих классов: для гнотобионтов, зараженных, конт-
рольных, предназначенных для работы с радиоизотопами, для
карантинизации и т. д.
В работе с животными должны учитываться все инфекционные
заболевания, которые могут сказаться на результатах эксперимен-
тов. При этом необходимо иметь в виду и тот факт, что отдельные
возбудители инфекционных заболеваний могут передаваться от
человека к животным и наоборот. К их числу относятся вирусы
бешенства, лимфоцитарного хориоменингита, шигеллы, некоторые
бруцеллы (Brucella canis), сальмонеллы, микобактерии туберкулеза,
токсоплазмы (Toxoplazma gondii), дизентерийная амеба.
Одобренный препарат (вещество) после лабораторных пред-
клинических испытаний по системе GLP и последующей клиниче-
ской проверки разрешается к выпуску в условиях промышленного
производства. Для обеспечения изготовления высокого качества
продукта Всемирная Организация Здравоохранения (ВОЗ) еще в
1968 г. утвердила "Требования для практики качественного произ-
285
водства при изготовлении и контроле качества лекарств и к
специалистам в области фармации". Годом позже эти требования,
вошедшие (с небольшими уточнениями и изменениями) в правила
системы GMP (Good Manufacturing Practice) были рекомендованы
Ассамблеей ВОЗ для международной торговли, а в 1971 г. они были
изданы в качестве приложения ко второму изданию Международ-
ной Фармакопеи. -
GMP — это единая система требований rib контролю качества
лекарственных средств с начала переработки сырья до производ-
ства готовых прейаратов, включая общие требования к помещени-
ям, оборудованию и персоналу. С 1975 г. правила GMP расширены,
и они касаются различных химических и биологических веществ
в индивидуальном виде, ветеринарных препаратов, применяемых
в животноводстве; исходных материалов для использования в
дозированных формах, если они включены в законодательства
стран-экспортеров и стран-импортеров; и, наконец, информации
о безопасности и эффективности перечисленных веществ, мате-
риалов и препаратов.
С учетом издания в 1987 г. руководств Международной Орга-
низации Стандартизации (ISO) серии ISO 9000—9004 по системам
качества возникла необходимость пересмотреть существовавшие
требования GMP. В сентябре 1991 г. на специальной конференции
по GMP в г. Москве представлен пересмотренный проект требо-
ваний GMP, включающий три части:
1) "Управление качеством в промышленном производстве ле-
карственных средств: философия и основные составляющие";
2) "Практика качественного производства и контроль качества";
3) "Дополнительные и вспомогательные направления".
Первая часть содержит 12 разделов, касающихся организации
контроля за качеством производства, санитарии и гигиены, заклю-
чения контрактов, стандартных рабочих методик, оформления
необходимой документации и др.
Вторая часть содержит два раздела — производство и контроль
качества. Применительно к производству лекарственных средств
указано, что оно должно опираться на принцип четкого соблюдения
методов ведения технологического процесса согласно норматив-
но-технической документации с целью получения продукта требу-
емого качества и в соответствии с разрешением на его изготовле-
ние и продажу. По возможности избегать любых отклонений от
методик или инструкций. При наличии таких отклонений необхо-
286
дим о согласование, разрешение, утверждение и подпись назначен-
ного ответственного лица, а при необходимости — привлечение
службы отдела контроля качества.
Операции с различными продуктами не должны выполняться
одновременно и последовательно в одном и том же помещении
пока не устранен риск перемешивания или перекрестного загряз-
нения.
Доступ в производственные помещения должен быть ограничен
лишь определенным кругом лиц, занятых в производстве. Избегать
изготовления немедицинской продукции в зонах и на оборудова-
нии, предназначенных для изготовления фармацевтической про-
дукции. При работе с сухими материалами и продуктами необхо-
димы меры предосторожности для предупреждения возникнове-
ния, накопления и распространения пыли, что может привести к
перекрестному загрязнению изготавливаемых продуктов или к их
микробному загрязнению. Микробы могут попадать в воздух и на
частицы пыли из обсемененных ими материалов и продуктов при
изготовлении, с загрязненных оборудования и одежды, кожи
работающих людей. Перекрестное загрязнение может быть пред-
отвращено изготовлением каждого целевого продукта в раздель-
ных зонах (пенициллины, живые вакцины и другие БАВ) или, по
крайней мере, разделением изготовления их по времени; обеспе-
чением соответствующих воздушных шлюзов; ношением защит-
ной технологической одежды; использованием средств эффектив-
ной деконтаминации оборудования, стен, и пр.; использованием
"закрытых систем" производства и т. д.
Необходимо проверять правильность и надежность сочленения
трубопроводов и другое оборудование, используемое для транс-
портировки продуктов (материалов) из одной зоны в другую.
Дистиллированная или деионизированная вода, поступающая по
трубам, должна соответствовать санитарно-микробиологическим
нормативам. Операции по техническому обслуживанию или ре-
монту не должны сказываться на качестве продукции.
Контроль качества продукции касается процесса забора проб,
проведения исследований, документации и пр. Все исследования
должны проводиться согласно утвержденным инструкциям для
каждого материала или продукта.
Забор проб осуществляют таким образом, чтобы не загрязнить
их или не подвергнуть нежелательному воздействию, сказываю-
щемуся на качестве продукта или, напротив, чтобы отбираемый
287
материал не был токсичным (вредным) для здоровья оператора.
• Для каждой партии продукта до выпуска должна иметься
лабораторная документация с подтверждением соответствия ко-
нечного продукта спецификациям.
Из каждой партии целевого продукта оставляют пробы на
хранение при рекомендуемых условиях сроком не менее года,
превышающего срок годности. Пробы должны храниться в таком
количестве, чтобы можно было при необходимости провести как
минимум два повторных исследования.
Третья часть требований GMP включает разделы о стерильных
фармацевтических продуктах и практике качественного производ-
ства основной массы лекарственных субстанций.
Необходимо помнить о том, что лица, обладающие повышенной
чувствительностью к конкретному веществу — действующему или
вспомогательному, не должны включаться в группу исполнителей.
Для них допустима работа в отделении или цехе упаковки, где
исключен контакт с аллергеном.
В 1991 г. правила GMP утверждены.в нашей стране примени-
тельно к производству и контролю качества лекарственных
средств. Эти правила соответствуют Международной Системе
GMP и включают следующие разделы: введение, терминология,
персонал, здания и помещения, оборудование, процесс производ-
ства, отдел технического контроля, аттестация и контроль произ-
водства; выделены требования к стерильным лекарственным сред-
ствам и описаны особенности их производства.
Соблюдение правил GMP обеспечивает выпуск качественных
продуктов и гарантирует благополучие потребителей. В 1995 г. ВОЗ
планирует утвердить GPP (Good Pharmacy Practice) по предложе-
нию Международной Фармацевтической Федерации (FIP).
Глава 7.
ТЕХНИЧЕСКАЯ ВООРУЖЕННОСТЬ
БИОТЕХНОЛОГИЧЕСКИХ ПРОИЗВОДСТВ
В большинстве случаев биотехнологические процессы
рассчитаны на использование живых клеток и тканей различного
происхождения, и даже в инженерной энзимологии применение
молеклул ферментов сопряжено с рядом ограничений, присущих
288
живым клеткам или тканям. К их числу относят поддержание
оптимальных температуры (достаточно низкой), pH, стерильности,
концентрации растворенного кислорода (в случаях эксплуатации
биообъектов — аэрофилов). Микробные клетки, за редким исклю-
чением (например, микоплазмы), и клетки растений имеют кле-
точную стенку , которой не обладают клетки животных организ-
мов; вирусы как организованные частицы реплицируются и пара-
зитируют на живых культурах клеток или в тканях, поэтому
отмеченные выше ограничения относятся больше к этим послед-
ним, а не к вирусным частицам. Наличие клеточной стенки у
биообъекта заведомо обеспечивает ему более высокую устойчи-
вость в различных условиях существования, чем биообъектам,
лишенным клеточной стенки. Даже при большой густоте, напри-
мер, бактерий, они активно размножаются, если им обеспечива-
ются достаточные пищевые потребности.
Вирусам — как облигатным паразитам — присуща еще и весьма
высокая специфичность применительно к организмам-хозяевам
(вирусы бактерий, грибов, растений, животных) и, даже тканям
(вирусы бешенства и полиомиелита—в отношении нервной ткани,
вирус оспы — в отношении кожи, аденовирусы — в отношении
железистой ткани).
Таким образом, особенности биообъектов обусловливают сле-
дующие принципы технического оснащения биопроизводств:
1. конструкционное совершенство и относительная универсаль-
ность биореакторов;
2. инертность, или коррозионная стойкость материалов биоре-
акторов и другого технологического оборудования, вмещающих
биообъект или контактирующих с ним или продуктами его мета-
болизма;
3. эксплуатационная надежность технологического оборудова-
ния;
4. доступность, эстетичность и легкость обслуживания, замены,
смазки, чистки, обработки антисептиками или дезинфектантами
узлов и соответствующих частей оборудования.
Эти принццпы рассматриваются более подробно в самостоя-
тельной научной дисциплине "Процессы и аппараты биотехноло-
гий", или "Биоинженерия". В настоящем учебнике они изложены
в объеме, необходимом для целостного восприятия общих и спе-
циальных разделов биотехнологии, и данная глава, в основном,
посвящена биореакторам.
10 т. 8524 289
Согласно первому принципу желательно конструировать такие
биореакторы, которые можно использовать для реализации био-
технологических процессов, основанных на использовании разных
биообъектов (бактерий, грибов, клеток растений и млекопитаю-
щих). Под совершенством и относительной универсальностью
следует понимать возможность создания оптимальных условий
культивирования разных биообъектов в одном и том же биореак-
торе с автоматическим регулированием заданных параметров.
Многоцелевые реакторы высоких стандартов изготавливает нидер-
ладская фирма ADI (Applikon Dependable Instruments). Они при-
годны для флокулирующих организмов, иммобилизованных клеток
(микробных, растительных, животных) в целях повышения выхода
биомассы и достижения их выраженной продуктивности.
Чтобы обеспечить биореакторы компьютерным контролем,
фирма ADI выпускает биопроцессор, с помощью которого обеспе-
чивается полное регулирование заданных параметров в течение
роста биообъекта (рис. 85).
Рис. 85. Многоцелевой биопроцессор ADI 1020 Рис. 86. Циклонный колон-
с компьютерным контролем различных параметров ный ферментатор (схема),
во время культивирования биообъектов.
Заманчивой представляется конструкция ферментатора в виде
циклонной колонны (рис. 86), создающей возможность проведения
периодических, непрерывных и фазовых ферментаций, и обеспе-
290
чивающей новые подходы к оценке клеточных циклов и поведению
самих клеток.
Поддержание заданных параметров — дело не простое, когда
непрерывно изменяются условия культивирования биообъекта
вследствие многокомпонентное™ исходной питательной среды и
культуральной жидкости на заключительной стадии ферментации,
изменения pH в процессе роста, размножения и развития биообъ-
екта, сложности регуляторных механизмов при биосинтезе целе-
вых продуктов, к тому же возможно — нестабильных, различный
уровень структурно-функциональной дифференцировки акариот,
прокариот и эукариот, и пр. Лишь при эксплуатации в колоннах
иммобилизованных ферментов аппаратурное оформление техно-
логических процессов существенно отличается от оформления
процессов, когда используют, прежде всего, специальные биоре-
акторы.
Второй принцип технического оснащения касается инертности
материалов, используемых для изготовления биореакторов и дру-
гого технологического оборудования. Это означает, что в процессе
культивирования биообъекта как будто не будет происходить его
взаимодействия (равно как и продуктов метаболизма) с металли-
ческими и неметаллическими конструкциями, постоянно или вре-
менно находящимися в контакте с ним.
Реально при выборе конструкционных материалов фермента-
торов необходимо учитывать тот факт, что эти аппараты в процессе
работы подвергаются механическим, химическим и физическим
(тепловым) воздействиям. Стерилизацию ферментаторов осущест-
вляют острым паром при 127—130’С с последующим охлаждением
до 24—28°С. В результате непрерывной аэрации культуральной
жидкости внутренние поверхности аппаратов испытывают посто-
янное окисляющее действие кислорода; кроме того, развитие
продуцента сопровождается выделением в среду агрессивных ме-
таболитов (например, органических кислот).
Весьма существенно также влияние на стенки аппарата интен-
сивных тазожидкостных потоков, кавитации, вибрации; содержа-
ние в среде абразивных примесей—частиц масла и муки — создает
дополнительный истирающий эффект.
Для обеспечения стерильности необходима систематическая
обработка аппарата моющими средствами и антисептиками (рас-
творами триполифосфата, хлорамина, формалина).
Материалы, идущие на изготовление ферментационного обо-
291
рудования, должны быть химически стойкими и не подвергаться
коррозии в течение длительного времени эксплуатации. Корро-
зия (от лат. corrosion — разъедание) — это процесс молекулярного
разрушения материала (ограниченного — локального или распро-
страненного) под воздействием внешних факторов. Коррозионные
процессы можно подразделить на 3 большие группы: химическая,
физическая (электрохимическая) и биологическая коррозии. Хи-
мическая коррозия индуцируется сухими газами и жидкостями,
неспособными проводить электрический ток. Физическая— (элек-
трохимическая) коррозия имеет место в случаях возникновения
местных электрических токов — гальванокоррозия, например, в
конструкциях, где контактируют несколько металлов с разными
электродными потенциалами (электролиты здесь особенно благо-
приятны для протекания электрохимической коррозии). Возник-
новение такой коррозии возможно даже при неоднородности
одного и того же металла в случае его неравноценной обработки
на отдельных участках.
Биологическая коррозия обусловливается биологическими
объектами (бактериями, микромицетами и другими организмами).
По механизму развития биокоррозию можно отнести к химиче-
ской. Однако наличие биообъекта в среде и на объекте коррозии
привносит дополнительный фактор — клеточную биомассу (фак-
тор обрастаниянли/и зарастания). Поэтому биокоррозия нередко
сопрягается с биоповреждением. Биоповреждение — понятие
более широкое, чем биокоррозия;
биоповреждение
биокоррозия механическое
(химическая (физическое)
молекулярная повреждение
деструкция конструкции (аппарата,
материала) прибора, сооружения)
биообъектом
Коррозии подразделяют еще по средам, где они протекают:
атмосферная,почвенная, водная; по распространенности
— ограниченная и распространенная
(поверхностные, сквозные); по локализации: ножевая (по
сварным швам), межкристаллитная, послойная.
Интенсивность коррозионных процессов оценивают в так на-
зываемых массовом (Кщ) и глубинном (D) показателях. Первый из
292
них рассчитывают в случаях неравномерной коррозии, исходя из
уменьшения массы материала, в кг на 1 м2 поверхности за 1 час:
„ Mj-M2
где М — масса материала до (Mi) и после (М2) коррозии, S —
поверхность материала (м2), t — длительность коррозии в часах.
Глубинный показатель оценивают при равномерной коррозии
по уменьшению толщины металл в мм за год:
„ Кт-24-365 1000 , , А „
D =-----*------- ( р плотность металла, кг/м ). Исходя из
Р
оценочных показателей Кт и D, выделяют группы стойкости
металлов. Например, D более 10, мёталл оценивают баллом 9 и
относят к разряду нестойких (группа VI); D менее 0,001, металл
оценивают баллом 0 и относят к совершенно стойким (группа 1).
Весьма стойкие и просто стойкие металлы характеризуются соот-
ветственно следующими показателями: D=0,001 —0,005 (1 балл)
и 0,005 — 0,01 (2 балла) — II группа; D=0,01—0,05 (3 балла) и
0,05—0,1 (4 балла) — III группа. Очевидно, что в различных средах
стойкость одного и того же материала к коррозии различна.
В биотехнологии, где среды чаще не агрессивны, применяют
обычные углеродистые стали с повышенной прочностью и выра-
женной упругостью. Нержавеющая сталь сохраняет большую стой-
кость в условиях атмосферной коррозии.
Крупнейшие отечественные заводы антибиотиков были осна-
щены ферментаторами из углеродистой стали марки Ст-3. Однако
по отношению к углеродистой стали культуральные жидкости
оказались агрессивными. По данным ВНИИА скорость коррозии
стали марки Ст-3 достигает 0,2—0,4 мм/год; по стандартной клас-
сификации такой показатель соответствует 6 баллам, т. е. харак-
теризует материал пониженной стойкости. При этом происходит
не только уменьшение толщины стенок аппаратов, но возможно
также появление раковин и каверн. Железо, попадая в среду,
подавляет активность ферментов продуцента, угнетает биосинтез,
инактивирует некоторые антибиотики.
Несмотря на коррозию, возможность использовать стальные
ферментаторы обусловлена образованием на внутренних поверх-
ностях аппаратов пассивирующей пленки, которая состоит из
окислов железа и веществ, выделившихся из среды. Эта пленка
замедляет коррозию.
Хорошо удовлетворяет всем требованиям, предъявляемым к
293
материалам для ферментационной аппаратуры, хромоникелевая
нержавеющая сталь марки Х18Н10Т, которая содержит 0,01%
углерода, 18% хрома, 10% никеля и менее 1% титана. Аппараты из
этой стали долговечны, удобны в эксплуатации, "универсальны”,
то есть пригодны для производства любых антибиотиков, витами-
нов, ферментов и других БАВ. Однако высоколегированная сталь
очень дорога, поэтому ферментаторы вместимостью 32 м3 и более
изготовляют из двухслойной стали: корпус — из Ст-3, внутренний
тонкий слой — из хромо-никелевой стали.
Обычно кислотоустойчивые и нержавеющие стали — это спла-
вы железа с хромом и легированные в целях улучшения их
сопротивляемости молибденом, никелем, титаном, марганцем и
другими элементами. Содержание углерода в них порядка 0,15%.
Жаропрочные стали включают железо, хром, никель; их исполь-
зуют для изготовления арматуры печей, муфелей, воздухоподогре-
вателей. Вольфрам и молибден используют в качестве легирующих
веществ.
В биотехнологии также широко применяют чугун, из которого
делают компрессоры, поршневые кольца, рамы фильтрпрессов и
др.; некоторые чугунные аппараты покрывают эмалью. Хром,
никель, молибден — как легирующие элементы повышают жаро-
стойкость и химическую стойкость чугуна. Такой чугун полезен,
например, для изготовления отдельных частей барабанных суши-
лок, работающих при повышенных температурах.
Различные конструкции, используемые во многих аппаратах,
состоят из алюминия (или включают его). Этот металл быстро
подвергается электрохимической коррозии, но вследствие легко
образующейся окисной защитной пленки, дальнейшее разрушение
алюминия прекращается. Продукты его коррозии неядовиты.
Стекло, полиэтилен, "пищевая” резина и другие полимерные
материалы также широко используют в биотехнологических про-
цессах.
Защиту материалов от коррозионных процессов осуществляют
различными способами: применяют специальные ингибиторы хи-
мической и биологической коррозии, катодную и протекторную
защиту- от электрохимической коррозии.
Третий принцип технического оснащения биопроизводств ка-
сается эксплуатационной надежности технологического оборудо-
вания. Эта надежность обеспечивается соответствием аппаратов,
приборов и другого оборудования целевому назначению, в част-
ности, по конструкционному совершенству, полностью обеспечи-
294
вающему оптимальные условия для протекания технологического
процесса и контроля за ним (в том числе, с учетом требований по
технике безопасности). Лишь в подобных случаях достигают мак-
симально возможной интенсивности работы оборудования, то есть
его производительности, отнесенной к выбранной основной еди-
нице, например, количество упаренной воды в расчете на 1 час и
на 1 м2 поверхности нагрева:
Qw —
V1-V2
St
где Qw — масса упаренной воды, V — масса воды
^о упаривания (Vi) и после упаривания (V2), S — поверхность
нагрева (м2), t — время в часах.
Критерием оптимальности работы аппарата выступает себесто-
имость продукции.
Интенсивность работы оборудования может быть повышена за
счет автоматизации и замены периодических процессов непрерыв-
ными, снижения энергоемкости аппаратов благодаря уменьшению
расхода энергии на единицу сырья или конечного продукта.
Эксплуатационная надежность технологического оборудова-
ния обеспечивается также стабильностью его работы в заданном
режиме и временном интервале, при максимальном соответствии
условий труда физическим и психическим возможностям людей,
занятых в конкретном производстве (эргономика).
Рассмотренный третий принцип напрямую связан с экономи-
кой биотехнологического процесса. Оборудование с высокими
конструктивными и эксплуатационными показателями обеспечи-
вает, как правило, и высокие экономические показатели.
Четвертый принцип технического оснащения биопроизводств
касается эстетичности, легкости и удобства обслуживания, замены,
смазки, чистки, обработки антисептиками или дезинфектантами
узлов и соответствующих частей оборудования.
Любой прибор или аппарат, легко доступный для сборки и
разборки, загрузки материалами, питательными средами и выгруз-
ки, для чистки, мойки, смазки, ремонта и пр.; оценивается выше,
чем оборудование с усложненным доступом к его частям.
Масса аппаратов, используемых, например, в микробной био-
технологии, различна, и требования здесь определяются большей
частью экономическими соображениями. Применительно к фер-
ментаторам различают следующие типы их: лабораторные емко-
стью 0,5—100 л, пилотные емкостью 100л—10 м3, промышленные
емкостью 10—100 м3 и более.
295
При масштабировании добиваются соответствия важнейших
характеристик процесса, а не сохранения принципа конструкции.
Применяемое в биотехнологии оборудование должно вносить
определенную долю эстетичности в интерьер цеха или отделения
("ласкать глаз"). В ходе его эксплуатации и вне ее оборудование
должно быть легко доступным, содержащимся и функционирую-
щим в определенных рамках требований гигиены и санитарии.
В случае замены каких-либо частей или деталей в аппарате,
смазки и чистки узлов при текущем ремонте, и т._ д., загрязнения
не должны попадать внутрь биореакторов, в материальные поточ-
ные коммуникационные линии, в конечные продукты.
Техническую вооруженность биотехнологических процессов
целесообразно условно ограничить аппаратурным оформлением
производств, базирующихся на культивировании: 1) бактерий и
грибов, 2) клеток и тканей растений, 3) клеток и тканей животных
организмов и человека. Такое подразделение обусловлено тем, что
бактерии и грибы в большинстве своем выращивают в однотипных
биореакторах, имеющих почти однотипную обвязку, в которую
входят: ферментатор, многокорпусный вентиль стерильный (для
подачи питательной среды, посевного материала, подпитки и пр.),
системы регулирования pH, t°, подачи пеногасителя, система кон-
троля расхода воздуха, пробоотборник, электродвигатель.
Растительные клетки, имеющие клеточную стенку (также как
бактерии и грибы) растут, размножаются и развиваются значи-
тельно дольше, чем большинство бактерий и грибов, а это вносит
определенные коррективы в аппаратурное оформление соответст-
вующих биотехнологических процессов.
Культуры клеток животных и человека, не имеющие клеточных
стенок, являются более ранимыми и требовательными к условиям
своего существования, чем клетки других эукариот и прокариот.
Поэтому оборудование для них можно отнести к разряду "тихо-
ходного", обеспечивающего нежное обращение с биообъектами.
Несомненно, в отдельных случаях допустимы исключения,
например, когда возможно культивирование в глубинных условиях
некоторых растительных клеток (суспензионная культура жень-
шеня), используя ферментационное оборудование, рассчитанное
на выращивание, например, бактерий или грибов.
К. Шюгерль в 1982 г. предложил подразделить биореакторы на
3 основные группы согласно способу потребления энергии для
перемешивания и диспергирования стерильного воздуха (газа):
296
— в биореакторах I типа энергия расходуется на механическое
движение внутренних устройств;
— в биореакторах II типа' энергия расходуется на работу
внешнего насоса, обеспечивающего рециркуляцию жидкости
и/или газа;
— в биореакторах III типа энергия расходуется на сжатие и
подачу газа в культуральную жидкость.
На рис. 87 схематично изображены некоторые из таких био-
реакторов.
Рис. 87. Биореакторы для аэробных процессов: с расходом энергии на механи-
ческое движение внутренних устройств а — 1, 2, 3; с расходом энергии на работу
насоса, обеспечивающего рециркуляцию культуральной жидкости б — 4; с расхо-
дом энергии на сжатие и подачу газовой фазы в — 5 (г — газ, ж — жидкая фаза,
Д — двигатель):
7.L Аппаратурное оснащение микробиологических произ-
водств. Человек с древнейших времен эмпирически применял
.дрожжевое организмы в примитивных по аппаратурному оформ-
лению биотехнологических процессах (хлебопечение, виноделие
и пр.). Развитие промышленности антибиотиков продвинуло дале-
ко вперед проблему создания специальной аппаратуры для куль-
тивирования микробов — продуцентов БАВ (аминокислот, анти-
биотиков, полисахаридов, витаминов, ферментов и других соеди-
нений). Были предложены различного типа биореакторы для вы-
ращивания микроорганизмов, однако все конструкции фермента-
торов (ферментеров) оставались в основном сходными по боль-
шинству параметров и, усредненно, их можно подразделить на 2
типа: без подводки стерильного воздуха (для анаэробов) и с под-
водкой его (для аэробов). Аэрируемые биореакторы могут быть с
мешалками и без них (рис. 88).
297
Рис. 88. Ферментатор периодического действия (1 — турбинная трехярусная
мешалка, 2 — охлаждающий змеевик, 3 — секционная рубашка, 4 — отражательная
перегородка, 5 — барботер, П-пар); I—XI — материальные и вспомогательные
трубопроводы с запорно-регулирующими устройствами (I — посевная линия, II
—подача стерильного сжатого воздуха, III — подача пара, IV — удаление отрабо-
танного воздуха, V — загрузочная линия, VI — линия введения добавок, VII —
подача пеногасителя, VIII — подача моющего раствора, IX — пробоотборник, X —
выдача продукта, XI — выдача в канализацию через нижний спуск).
В последние годы апробированы мембранные биореакторы,
биореакторы с полыми волокнами и некоторые другие.
При расчете и конструировании биореакторов необходимо
учитывать время протекания различных биологических процессов
у представителей различных групп организмов (рис. 89).
298
, 4 } 6 Рис. 89. Ориентировочное усреднен-
I I ное время протекания биологических
|||| процессов у различных групп организ-
мов (по Дж. Роуилсу, 1982): 1 — репли-
кация хромосомы, 2 — продолжитель-
2 пость клеточного цикла, 3 — бактерии,
4 — дрожжи, 5 — плесени, 6 — расти-
JL тельные и животные клетки, 7 — эле-
, iff5, 10л, 10л| । io t io ! 10*11)' с г ментарные химические реакции, 8 —
7 ю’ _ ? регуляция транскрипции, 9 — аллосте-
~*~ о____ рическая регуляция белков, 10 — изме-
"* нение концентрации ферментов, 11 —
“ возникновение мутантов.
Некоторые технические характеристики промышленного би-
ореактора в сравнении с пилотным и лабораторным приведены в
таблице 33.
Т а б л и ц а 33. Технические характеристики биореакторов
Характеристика Показатели для аппаратов
промышленного на 100 м3 пилотного на 150 д лабораторного на 10 л
Внутренний диаметр, мм Высота, мм Рабочий объем, л Диаметр турбин, мм Число турбин Число отбойников Частота вращения вала мешалки, об/мин Мощность электродвигателя мешалки, кВт Мощность электродвигателя пеногасителя, кВт Максимальное количество отработанного пеногасителем газа, м3/мин Частота вращения вала пеногасителя, об/мин 3600 15715 1* 900 1-2 (диаметр рабочего колеса 960 мм) 4 173 160 4 100-110 725 420 1140 100 140 3 4 125-990 2,2 0,73 0,3 3000 2-6 2*. ± 200-15003’ Не более 2
Примечание: 1 — зависит от коэффициента заполнения; 2 — имеется одна мешалка; 3 —
диапазон регулируемой частоты вращения мешалки.
299
Размеры ферментаторов определяются соотношением внешне-
го диаметра к высоте, который варьирует обычно в пределах ot
1:2 до 1:6. Почти универсальными и чаще используемыми являются
ферментаторы для анаэробных и аэробных процессов. Эти фер-
ментаторы в свою очередь классифицируют по способу ввода в
аппарат энергии для перемешивания (таблица 34): газовой фазой
(ФГ), жидкой фазой (ФЖ), газовой и жидкой фазами (ФЖГ).
С использованием указанных выше классификаций удается
разработать единые методы инженерных расчетов основных кон-
структивных элементов и режимов работы ферментаторов.
Таблица 34. Классификация ферментаторов по способу ввода энергии для
перемешивания
Ферментаторы Характеристика конструкции аппарата Тип аппарата
ФГ с подводом энергии газовой фазой Простота конструктивного оформления и высокая надежность в связи с отсутствием движущихся узлов и деталей Барботажный, барботажно- эрлифтный, колоночный (колонный), форсуночный
ФЖ с подводом энергии жидкой фазой Обычно энергия передается жидкой фазе самовсасывающей мешалкой или насосом Эжекционный, с циркуляционным контуром, с всасывающей мешалкой
ФЖГ (комбинированные) Основным конструктивным элементом является перемешивающее устройство, обеспечивающее высокую интенсивность растворения кислорода и высокую степень диспергирования газа. В то же время энергия газовой фазой выводится обычным способом Барботажный с механическим перемешиванием
Ферментаторы указанных трех групп имеют большое количе-
ство общих элементов. Различие же состоит в конструкциях
аэрирующих и перемешивающих устройств. Примером конструк-
тивного оформления ферментатора группы ФГ может быть аппарат
с эрлифтом вместимостью 63 м3 (рис. 90). В аппарате отсутствует
механическое перемешивание, поэтому проще поддерживать асеп-
тические условия. Воздух для аэрации среды подается по трубе,
зоо
расположенной верти-
кально в ферментаторе.
Аэратор, конструкция
которого обеспечивает
вихревое движение вы-
ходящего воздуха, распо-
ложен в нижней части
диффузора и насыщает
питательную среду воз-
духом. Газожидкостная
смесь поднимается по
диффузору и перемеши-
вается через его верхние
края. В этой же зоне
часть воздуха уходит из
аппарата, и более плот-
ная среда опускается
вниз в кольцевом про-
странстве между корпу-
сом ферментатора и
диффузором. Так проис-
ходит многократная цир-
куляция среды в фермен-
таторе. Для отвода био-
логического тепла внут-
ри ферментатора уста-
новлен змеевик, а также
аппарат снабжен секци-
Рис. 90. Ферментатор с эрлифтом: 1 — штуцер
для слива, 2 — аэратор, 3 — змеевик, 4 — штуцер
для загрузки, 5 — люк, 6 — корпус аппарата, 7
— диффузор, 8 — рубашка, 9 — труба передав-
ливания.
онной рубашкой. Недостатком этих аппаратов является низкая
интенсивность массообмена по кислороду. Известны ферментато-
ры этого типа объемом 25, 49, 63 и 200м3.
Широкое распространение в производстве кормового белка
получили ферментаторы с самовсасывающими мешалками (рис.
91). Это ферментаторы из группы ФЖ. Для выращивания чистой
культуры дрожжей созданы ферментаторы вместимостью 0.32, 3.2
и 50 м3. Ферментатор представляет собой вертикальный цилинд-
рический аппарат, снабженный циркуляционными, теплообмен-
ными и аэрирующими устройствами. В качестве циркуляционных
устройств использованы системы направляющих диффузоров, раз-
граничивающих восходящие и нисходящие потоки. Теплообмен-
301
Рис. 91. Ферментатор с самовсасывающей
мешалкой непрерывного действия: 1 — кор-
пус, 2 —- диффузор, 3 — самовсасывающая
мешалка, 4 — теплообменник, 5 — фильтр.
ные устройства выполнены
в виде трубок, установлен-
ных в трубных решетках
диффузоров.
На предприятиях микро-
биологической промышлен-
ности при выращивании
дрожжей в средах с жидки-
ми парафинами также при-
меняют ферментаторы с са-
мовсасывающими мешалка-
ми непрерывного действия.
Аппарат такого типа пред-
ставлен на рис. 91. Емкость
его 800 м3 (рабочий объем
320 м3) разделена на 12 сек-
ций. Ферментационная сре-
да последовательно прохо-
дит все секции, и из послед-
ней выходит культуральная
жидкость с минимальным
содержанием н-парафинов
и максимальной концентра-
цией биомассы. В каждой
секции установлено переме-
шивающее и аэрирующее
устройство и змеевики для
отвода тепла.
Ферментаторы периоди-
ческого действия из групп
ФЖГ применяют с 1944 г. в
промышленности для получения антибиотиков, витаминов и
других биологически активных веществ (см. рис. 88). Его конст-
рукция обеспечивает стерильность ферментации в течение дли-
тельного времени (нескольких суток) при оптимальных условиях
для роста и жизнедеятельности продуцента. Ферментаторы такой
конструкции изготавливают на 1,25; 2,0; 2,5; 3,2; 4,0; 5,0; 6,3; 10,0;
16,0; 20,0; 32,0; 50,0; 63,0; 100,0 и 160,0 м3. Как видно из рисунка,
это цилиндрический вертикальный аппарат со сферическим дни-
щем, снабженный аэрирующим, перемешивающим и теплопере-
дающим устройствами. Воздух для аэрации поступает в фермен-
302
татор через барботер, установленный под нижним ярусом мешал-
ки. С точки зрения эффективности диспергирования воздуха
конструкция барботера принципиальной роли не играет при нали-
чии мешалки, однако, с точки зрения эксплуатации, наиболее
удобным является квадратный барботер, который получил наи-
большее распространение. Отверстия в барботере направлены
вниз, во избежание засорения биообъектами. Общая площадь
отверстий должна быть на 25% больше площади поперечного
сечения трубопровода, подводящего воздух. Барботер по своим
размерам должен соответствовать диаметру мешалки, чтобы вы-
ходящий из него воздух попадал в зону ее действия.
Эффективность работы ферментатора определяется прежде
всего необходимой интенсивностью перемешивания. Перемеши-
вающие устройства служат для сохранения равномерного темпе-
ратурного поля по всему объему аппарата, своевременного подвода
продуктов питания к клеткам и отвода от них продуктов метабо-
лизма, а также интенсификации массопередачи кислорода.
Для культуральных жидкостей с высокой структурной вязко-
стью наиболее эффективными являются открытые турбинные
мешалки с шестью лопастями. Выбраны более или менее оптималь-
ные соотношения размеров мешалки (бм/Д=0,3—0,4, где бм —
диаметр мешалки, Д — диаметр аппарата). Количество ярусов
мешалки определяется высотой столба жидкости. Оптимальное
межярусное расстояние (Ья) = (1,5—1,8) бм. Расстояния от верх-
него яруса мешалки до уровня жидкости в ферментаторе (Нж1)
должно быть > бм и от нижнего яруса мешалки до дна аппарата
(0,8—1,1) бм.
Для создания в ферментаторе условий "полного отражения", во
избежание образования вращательного контура, который резко
снижает интенсивность перемешивания, в аппарате устанавлива-
ют-отражательные перегородки (отбойники). Ширина их состав-
ляет (0,1—0,12) бм. Обычно рекомендуют устанавливать 4 отража-
тельных перегородки, несколько отступая от стенок ферментатора.
Важным элементом в конструкции ферментатора являются
теплообменные устройства. Применение высокопродуктивных
штаммов биообъектов, концентрированных питательных сред, вы-
сокий удельный расход мощности на перемешивание — все эти
факторы сказываются на существенном возрастании тепловыде-
лений, и для отвода тепла в ферментаторе устанавливают наруж-
ные и внутренние теплообменные устройства. Промышленные
ферментаторы, как правило, имеют секционные рубашки, а внутри
аппарата — четыре змеевика.
зоз
Рис. 92. Ферментатор емкостью
200 mj (внутренний вид).
Разработчики аппаратуры в нашей стране и за рубежом посто-
янно совершенствуют конструкции биореакторов. Так, например,
фирма New Brunswick Scientific Со., Inc. (США) предложила следу-
ющие типы ферментаторов: Био-Фло III — для периодического и
непрерывного культивирования микробных, животных и расти-
тельных клеток, совмещенный с микропроцессором и персональ-
ным компьютером; Микрос I —
для культивирования микроорга-
низмов (совмещен с микропро-
цессором) и промышленные фер-
ментаторы емкостью от 40 до 4000
литров и более (совмёщены с мик-
ропроцессорами). В Датской
мультинациональной компании
Gist-Brocades в 1987 г. сконструи-
рован и изготовлен самый боль-
шой промышленный фермента-
тор для производства пеницил-
лина (200 м3) (рис. 92);
Общая продуктивность процесса (Рар) в биореакторе опреде-
ляется количеством целевого продукта в ЕД активности или в кг,
получаемого с 1 м3 ферментационной емкости в час. Расчет ведут
по формулам, отдельно — для периодического и непрерывного
процессов:
периодический процесс
_ УсМсгЮ6. ЕД ,
ар VfTc м3-час
непрерывный процесс
_ _ WcrAct 106 ЕД .
Рор- у{
П vcrC кг WcfC
Рар = —— [ 3---1 Рар ~ • тл - где Vcf (мл) — объем
р VfTc м-час р Vf
культуральной жидкости за весь процесс ферментации; Acf (ЕД/мл)
— активность культуральной жидкости; С (кг/м3) — концентрация
целевого продукта в культуральной жидкости; Wcf (м3/час) —
скорость слива культуральной жидкости из ферментатора; Vf (м3)
— вместимость ферментатора; Тс (час) — время цикла работы
ферментатора.
Общую продуктивность для непрерывных процессов (рис. 93)
определяют в установившемся режиме, а для периодических про-
цессов и полунепрерывных — с учетом времени на подготовку
ферментатора к работе.
304
Объемная продуктивность про-
цесса (Рср) — это количество целе-
вого продукта в ЕД активности или
в кг, получаемое с 1 м3 питательной
среды в час. Расчет ведут следую-
щим образом:
периодический процесс
lbw*
р VcMcf Ю6 . ЕД .
Ср Vnm-tc м3-час
р = V<*C Г •
СР Vnm-Tc л^час
Рис. 93. Ферментатор непрерывного
действия с эжекционной системой аэра-,
ции (Б-50): 1 — кольцевой Канал, 2 —
воздуховод, 3 — пеногаситель, 4 — се-
паратор, 5 — цилиндр, 6 — привод, 7 —
теплообменник. 8 — диффузор, 9 —
цилиндрический стакан, 10 — эжекци-
онное устройство.
непрерывный процесс
_ WcMcflO6 . ЕД
ср Vrun мз-час
_ Wcf-C .кг •
Рср = -J7— (-5--1, где
Vnm м-час
Vnm(M3) — объем питательной среды.
Выход продукта от субстрата (cts) — это количество целевого
продукта в ЕД активности или в кг, полученное из 1 кг компонента
ферментационной среды, являющегося энергоносителем. Тогда для
периодического процесса:
VcMcf Ю6 ГЕД, Vcf-C ,кг, , ,
as =--------I-—) или as -------[—], где ms (кг) —
ms кг‘ ms кг1
исходное содержание энергоносителя в субстрате; для непрерыв-
ного процесса:
Acf-106 ЕД Ccf .кг. с . . 3.
as = —е— [-°] или as = -ZT- [—J, где So (кг/м ) — исходная
О0 кг* оо кг*
концентрация энергоносителя в субстрате. Степень использования
субстрата (U) выражается формулой:
[7 = ^7^ , где So (кг/м3) — исходная концентрация энергоно-
•-Х)
сителя в субстрате, Sk (кг/м3) — конечная крнцентрация энерго-
емкого компонента в субстрате. *
305
7.2. Некоторые особенности культивирования биообъектов.
При периодическом культивировании целесообразно создать ис-
кусственно такое установившееся состояние, при котором концен-
трация клеток, удельная скорость роста и окружающая клетки
среда не изменялись бы со временем. Такие условия возможны
при непрерывном культивировании, когда клетки продуцента раз-
множаются со скоростью, зависящей от притока питательных
веществ и некоторых других условий. Часть объема культуральной
жидкости постоянно вытекает с той же скоростью, с какой подается
среда в аппарат. Метод проточного культивирования может быть
организован как процесс полного вытеснения и как процесс
полного смешения. Осуществление первого возможно для культи-
вирования анаэробных микроорганизмов в ферментаторе, пред-
ставляющем собой трубу, в которую с одного конца непрерывно
подают питательную среду и посевной материал, а из другого конца
отбирают культуральную жидкость. Процесс происходит без пе-
ремешивания и аэрации. Когда среда и посевной материал попа-
дают в ферментатор, популяция находится в лаг-фазе, а на выходе
из ферментатора культура может находиться в любой фазе в
зависимости от скорости подачи среды. В ферментаторе воспро-
изводится полная кривая размножения, но не во времени, а в
пространстве.
В процессе полного смешения размножение культуры происхо-
дит в ферментаторе при интенсивном перемешивании и аэрации.
Во всем объеме культуральной жидкости условия должны быть
одинаковыми. При этом в ферментаторе могут быть созданы
условия, соответствующие любой точке кривой размножения куль-
туры, выращиваемой периодическим способом. Процесс полного
смешения может быть организован по типу системы "турбидостат"
и "хемостат”.
В системе "турбидостат" в ферментаторе поддерживают плот-
ность популяции постоянной, и при быстром протоке среды созда-
ются условия, близкие к тем, которые соответствуют логарифми-
ческой фазе, при медленном — приближаются к условиям, соот-
ветствующим стационарной фазе. В установившемся режиме ра-
боты ферментатора удельная скорость протока среды равна удель-
ной скорости размножения культуры. Повышение скорости про-
тока или воздействие, замедляющее рост, приводят к тому, что
скорость размножения оказывается меньше скорости протока и
клетки культуры будут "вымыты" из ферментатора.
В системе "хемостат" величину клеточной популяции контроли-
руют с помощью отдельных компонентов питательной среды.
306
Среду составляют так, чтобы один из компонентов, необходимый
для роста биомассы клеток, был в недостатке или лимитировал
рост, поддерживая тем самым культуру в нужном состоянии.
Используя батарею ферментаторов, можно в каждом аппарате
постоянно поддерживать продуцент в определенной фазе размно-
жения.
В настоящее время непрерывное культивирование применяют
для получения белково-витаминных концентратов (БВК), кормовых
дрожжей, лимонной кислоты, лизина.
Культивирование макромицетов в специально подобранных
условиях характеризуется некоторыми особенностями. Во-первых,
в стационарных условиях базидиомицеты формируют плотные
пленки на поверхности жидких питательных сред; во-вторых, в
глубинных культурах они образуют агломераты. Поэтому, напри-
мер, при выращивании вешенки обыкновенной (Pleurotus ostreatus)
соблюдают следующую очередность этапов: культивирование
грибного мицелия в колбах на качалке, дезинтеграция мицелиаль-
ных агрегатов да частиц размером 100—200 мкм (инокулят), засев
ферментационной среды (0,4—0,5 г/л'по сухой массе гриба) и
выращивание гриба в течение 5 суток на специальной синтетиче-
ской питательной среде до получения примерно 6 г/л биомассы
клеток по. сухой массе (рис. 94).
Рис. 94. Схема получения мицелия вешенки обыкновенной для пищевых целей
(периодическое культивирование): 1 — семисуточная культура P.ostreatus в пробир-
ке на сусло-агаре, 2 — шестисуточная стационарная культура на жидкой среде в
колбе с отбойниками и периодическим встряхиванием в течение 5 мин., 3 —
четырехсуточная культура в колбах на качалке, 4 — дезинтеграция грибных
агломератов до частиц размером 100—200 мкм, 5 — инокулят 0,4—0,5 г/л, 6 —
культивирование в ферментаторе в течение 5сут., 7—отделение конечного продукта
(мицелия).
В производстве антибиотиков, наряду с периодическим куль-
тивированием, все чаще начинают использовать методы, занима-
ющие промежуточное положение между периодическим и непре-
рывным культивированием, то есть полунепрерывный отьемно-до-
ливной метод. Таким способом удается в 2-3 раза увеличить время
307
пребывания продуцента в активной фазе. Недостаток состоит в
том, что доливы осуществляют питательной средой полного соста-
ва. Этого недостатка лишен метод полунепрерывной регулируемой
ферментации. Суть этого метода состоит в том, что в определенное
время, начиная с логарифмической фазы размножения, по специ-
ально разработанной программе, в культуральную жидкость до-
бавляют отдельные компоненты питательной среды (раствор саха-
ров, аммония сульфат, жир и т. д.), поддерживая их концентрацию
на постоянном благоприятном уровне — вначале для роста био-
массы клеток, а затем — для синтеза целевого продукта.
Периодически из ферментатора отбирают определенные объ-
емы культуральной жидкости со все возрастающей концентрацией
целевого продукта. Ферментацию прекращают после того как
активность продукта достигла максимума. Этим способом удается
не только прод лить активную фазу, в которой наход ится продуцент,
но и повысить степень использования им субстрата, а в конечном
итоге — продуктивность процесса, то есть увеличить выход конеч-
ного продукта в расчете на потребленный субстрат.
Из всего разнообразия способов проведения процессов био-
технологии наиболее сложными являются регулируемые фермен-
тации с соблюдением условий асептики. Реализация таких процес-
сов рассчитана на использование ЭВМ при соответствующем
программном обеспечении. Кроме того для проведения процесса
в асептичных условиях необходимо введение дополнительных
стадий, обеспечивающих стерилизацию питательных сред и пода-
ваемого в ферментаторы воздуха.
Стерильную питательную среду в инокуляторе — посевном
аппарате первой ступени (рис. 95) засевают с соблюдением правил
асептики через специальное устройство посевным материалом,
который предварительно был выращен в колбах (в лаборатории)..
Технологи создают и поддерживают необходимые режимы в
аппарате для размножения клеток продуцента (температуру, аэра-
цию и перемешивание), а микробиологи контролируют и оцени-
вают развитие культуры. При достижении требуемых стадий раз-
вития и количества биомассы посевной материал передавливают
стерильным сжатым воздухом по посевному коллектору в посевной
аппарат большей вместимости. На этой второй ступени выращи-
вания посевного материала стремятся получить больше биомассы
клеток, чтобы в ферментаторе можно было создать необходимую
для данного штамма продуцента исходную плотность популяции.
308
Рис. 95. Инокулятор: 1 — кор-
пус, 2.— лаз, 3 — смотровое окно,
4 — аэратор, 5 — труба для пере-
пуска посевной культуры в фер-
ментатор, 6 — диффузор, 7 — ру-
башка, 8 —.гильза для термометра,
9 — розетка аэратора, 10—12 —
штуцера для манометра, гильзы
• термометра и трубы для передав-
ливания, 13—17 — штуцера для
загрузки среды, аэратора, выхода
воздуха, отбора проб и выхода во-
ды, 18 — штуцер для спуска жид-
кости.
Если это требование выполнимо без второй ступени, то фермен-
тационную среду засевают непосредственно из инокулятора.
Биосинтез целевого продукта в ферментаторе происходит при
заданных температурном режиме, аэрации, перемешивании и pH
культуральной жидкости; величину pH обычно регулируют пери-
одической подачей аммиачной воды через барботер ферментатора.
В случае пенообразования подают стерильный пеногаситель из
специального аппарата по сигналу датчика уровня йены.
Для поддержания постоянства концентраций отдельных ком-
309
понентов питательной среды их подают в виде стерильных раство-
ров по команде ЭВМ с определенной скоростью и периодичностью
в ферментатор из специальных аппаратов.
Все ферментаторы цеха соединены между собой несколькими
коллекторами: 1 — посевным, благодаря которому можно засевать
среду в любом ферментаторе из любого посевного аппарата; 2 —
коллектором подачи стерильной питательной среды; 3 — коллек-
тором подвода стерильного сжатого воздуха к индивидуальным
фильтрам; 4 — коллектором отработанного воздуха, выходящего
из ферментатора; 5 — коллектором перекачивания культуральной
жидкости из ферментатора (рис. 96).
Рис. 96. Аппаратурно-технологическое оформление процесса ферментации в
производстве пенициллина: 1 — сборник нативного раствора, 2, 3, 7, 11, 13, 15, 16
— эмульгаторы, 4 — сборник серной кислоты, 5 — сборник бутилацетата, 6, 9, 12,'
14,17 — экстракторы-сепараторы, 8 •— сборник обессоленной воды, 10 — сборник
I бутилацетатного экстракта, 18—сборник бикарбонатного раствора, 19—аппарат
для вымораживания и обработки углем, 20, 24, 25, 28 — фильтры, 21 — реактор
для получения калиевой соли бензилпенициллина, 22 — сборник раствора калия
карбоната, 23 — сборник раствора калия ацетата, 26 — вакуум-выпарной аппарат,
27 — теплообменник, 29 — сборник бутанола.
В случаях проведения ферментаций заведомо в нестерильных
условиях питательную среду и воздух для аэрации не стерилизуют,
но посевной материал всегда выращивают на стерильных пита-
тельных средах в асептических условиях.
7.2.1. Приготовление и стерилизаций питательных сред. В
статьях расходов в производстве продуктов микробного синтеза
одно из первых мест занимает сырье. При производстве 1 • 109. ЕД
310
пенициллина, например, необходимо израсходовать 6,7 кг сырья.
Для проведения ферментационных процессов, наряду с очень
дорогим пищевым сырьем, таким как мука кукурузная, соевая,
пшеничная, крахмал, пищевой сахар, глюкоза, лактоза, раститель-
ные масла, лярд, применяют и значительно более дешевые, явля-
ющиеся отходами пищевой промышленности — гидрол, меласса,
зеленая патока, молочная сыворотка, кукурузный экстракт, рыб-
но-костная мука, гидролизаты кукурузных кочерыжек, соломы,
подсолнечной мезги, а также специально получаемые продукты —
гидролизаты древесины и торфа, парафины нефти, метан, этанол
и др.
Жидкие компоненты питательных сред (кукурузный экстракт,
зеленую патоку, мелассу, гидрол, растительные масла, рыбий жир)
доставляют в железнодорожных цистернах и хранят в специальных
сборниках на складах заводов и транспортируют по коммуника-
циям с помощью вакуума, сжатого воздуха или перекачивают
насосами. Дозировку жидких компонентов осуществляют по массе
или по объему в соответствии с прописью среды и контрольными
показателями каждой партии этого нестандартного вида сырья.
Сыпучие компоненты сред из транспортной тары забирают или
в специальные бункеры или хранят на складах в исходной упаков-
ке. Для транспортировки сыпучих компонентов используют лен-
точные и винтовые конвейры, элеваторы, пневматический транс-
порт.
Процесс приготовления питательных сред независимо от того,
сложен он технологически или прост, всегда очень ответственен,
так как от качества питательной среды во многом зависит результат
проведения ферментации.
Жидкие питательные среды приготовляют в аппаратах-смеси-
телях с мешалкой, куда загружают отдельные компоненты в опре-
деленной последовательности, установленной по регламенту
(рис. 97).
Приготовление сложных комплексных сред в состав которых,
кроме минеральных компонентов и сахаров, входит мука, крахмал,
кукурузный экстракт, проводят в нескольких смесителях. Куку-
рузный экстракт обычно кипятят с мелом для нейтрализации
содержащихся в нем аминокислот и органических кислот. Муку,
крахмал предварительно заваривают и тщательно перемешивают,
чтобы не допустить образования крупных комков, которые могут
быть причиной нестерильных операций, поэтому реакторы должны
быть снабжены барботерами для подачи пара.
311
Рис. 97. Принципиальная схема процесса приготовления и тепловой
стерилизации питательной среды.
312
Часто для снижения вязкости питательной среды, содержащей
достаточно большую концентрацию кукурузной муки или крахма-
ла, проводят их частичный гидролиз амилолитическим ферментом
— оризином (продуцент — Aspergillus oryzae) с последующей его
инактивацией нагреванием. Иногда необходимость проведения
такой операции обусловлена физиологией продуцента, для кото-
рого предназначается среда.
В связи с тем, что в большинстве своем питательные среды
имеют жидкую и твердую фазы, возникает необходимость тонкого
измельчения твердых компонентов — отрубей, муки грубого по-
мола, рыбно-костной муки, соевого жмыха. В этих целях с большой
эффективностью используют роторно-пульсационный аппарат,
или РПА, через который пропускают суспензию компонента перед
завариванием или после него. Благодаря этой процедуре не только
избавляются от комков, образовавшихся при заваривании, но и
повышают степень использования сырья, равно как и получают
возможность применять отдельные виды сырья (например, среды
с рыбно-костной мукой, плохо поедающиеся стерилизации из-за
большого количества рыбьих глаз).
Растворы сахаров, нуждающиеся в более щадящих режимах
стерилизации, рекомендуют готовить и стерилизовать отдельно,
смешивая с основной средой только в ферментаторе.
Некоторые виды сырья, например, соевая мука, вызывают
повышенное вспенивание среды, поэтому для снижения пенооб-
разования при стерилизации в такие среды добавляют жир в
качестве пеногасителя. Подобная мера вызвана технологической
необходимостью, в принципе, добавление жира в среду повышает
устойчивость спор к тепловому воздействию и поэтому крайне
нежелательно. Все жировые компоненты сред необходимо стери-
лизовать отдельно. Для этого, как правило, предварительно готовят
водно-масляную или водно-жировую эмульсию с хозяйственным
мылом, повышающим ее стойкость.
Качество используемой воды зависит от назначения питатель-
ной среды. Чаще всего применяют артезианскую, реже — водо-
проводную воду. В крупнотоннажных производствах кормовых
дрожжей и белкововитаминных концентратов (ЕВК) используют
воду, полученную по замкнутому циклу этого производства, то есть
прошедшую очистные сооружения. В производстве кровезамени-
телей используют только апирогенную воду (бидистиллят).
Под стерилизацией сред обычно понимают любой метод воз-
действия, обеспечивающий удаление из них микробов — конта-
313
минантов или разрушение последних. Наиболее распространен-
ным и универсальным среди возможных методов, вызывающих
деструкцию микроорганизмов, является метод, основанный на
использовании влажного тепла. Характерной особенностью, отли-
чающей действие влажного тепла на микроорганизмы и химиче-
ские вещества, является высокая энергия активации р, которая
характерна для микробов (таблица 35).
Таблица 35. Значение энергии активации бактериальных спор и некоторых
витаминов
Объект Энергия активизации ц, Дж/моль
Baccillus subtilis 318440
-" - stearothermophilus 266903
Clostridium botulinum 343580
Тиамин гидрохлорид 92180
Фолиевая кислота 70392
По разнице значений энергии активации можно говорить о
том, что одинаковое приращение температуры оказывает разное
влияние на гибель спор и термическую деструкцию химических
соединений. Как видно из таблицы 35, споры палочки ботулизма
наиболее чувствительны к воздействию влажного тепла, а фолие-
вая кислота меньше подвержена деструкции по сравнению с
тиамином. На практике главная цель стерилизации — достижение
стерильности, но сохранение качества питательной среды также
имеет важное значение, так как непосредственно от этого будет
зависеть результат процесса ферментации. Благодаря различиям
в энергии активации отмирание термостойких спор при высоких
температурах происходит значительно быстрее, чем скорость хи-
мической реакции. Длительность экспозиции, или время вы-
держки— это тот временной интервал, в пределах которого
погибают микроорганизмы. Гибель последних спор в среде явля-
ется случайным процессом, поэтому введено понятие "критерий
стерильности" (N) — отношение числа операций стерилизации, в
результате которых выжили по одной термостойкой споре, к
общему числу проведенных операций. Для стерилизации сред
принимают критерий стерильности, равный 0,01 ± 0,001. Если
исходное количество спор в среде принять No, то получим соотно-
шение N/No — уровень стерильности или коэффициент выжива-
ния, который означает, что для достижения заданного критерия
314
стерильности (например 0,01) среда должна выдерживаться при
температуре стерилизации строго определенное время, чтобы "по-
пуляция" спор снизилась от исходного значения No до N/No
(например, до 10’16).
Тепловую стерилизацию сред (по способу ее проведения) под-
разделяют на периодическую и непрерывную. При периоди-
ческом способе стерилизации процессы: нагрев, выдержка и
охлаждение среды (рис. 98) протекают последовательно во време-
Рис. 98. Установка непрерывной стерилизации питательной среды: 1 — приемник
концентрата питательной среды, 2 — фильтр для отделения комков среды, 3 —
насос, 4 — паровой инжектор, 5 — выдерживатель трубчатого типа, свернутый в
виде плоской спирали, 6 — теплообменник.
ни в одном аппарате. Это может быть ферментатор, посевной
аппарат или специальный стерилизатор. Весь объем среды нагре-
вают в аппарате до заранее выбранной температуры, выдерживают
при этой температуре строго определенное время и охлаждают
водой, подаваемой в рубашку аппарата или змеевик. Метод отли-
чается простотой и надежностью, однако имеет и свои недостатки.
В частности, ухудшается качество питательной среды из-за дли-
тельного воздействия высокой температуры, при этом происходит
карамелизация сахаров (образование ангидридов сахаров), де-
струкция витаминов; излишне длительный нагрев приводит не
только к разрушению питательных веществ, но и к образованию
в среде потенциальных ингибиторов процесса ферментации, таких
как аминосахара. Второй недостаток связан с тем, что требуется
повышенный расход пара за короткий период нагрева. Третий
недостаток обусловлен невозможностью регенерировать тепло.
Последний, четвертый, недостаток касается трудности автомати-
зации процесса периодической стерилизации по сравнению с
непрерывным.
315
При непрерывном способе стерилизации каждый
элементарный процесс — нагрев, выдержка, охлаждение (рис. 99)
осуществляется в специально предназначенных для этого аппара-
тах: нагревателе, выдерживателе, теплообменнике, которые со-
Рис. 99. Установка непрерывной стерилизации питательной среды с рекупера-
цией тепла: 1 — приемник концентрата среды, 2 — фильтр. 3 — насос, 4 —
пластинчатый теплообменник-рекуператор, 5 — теплообменник-доохладитель, 6 —
паровой эжектор, 7 — выдерживатель трубчатого типа, разделенный на две секции.
ставляют систему аппаратов для непрерывной стерилизации —
установку непрерывной стерилизации (УНС). Непрерывная стери-
лизация имеет следующие преимущества по сравнению с перио-
дической:
1) при непрерывном методе стерилизации каждый элементар-
ный объем среды (бесконечно малый объем, содержащий одну
спору) находится при высокой температуре короткое время;
2) благодаря более высоким температурам стерилизации и
короткой экспозиции деструкция компонентов питательной среды
минимальна;
3) процесс стерилизации всего объема питательной среды
растянут во времени, этим обеспечивается более равномерная
загрузка котельной;
4) процесс легко контролируем и управляем;
5) возможна частичная регенерация тепла.
Непрерывное нагревание среды может быть осуществлено без
прямого контакта с теплоносителем в трубчатом, пластинчатом или
спиральном теплообменнике. Но чаще всего среда нагревается до
нужной температуры в течение нескольких секунд прямым инжек-
316
тированием пара в паровых контактных нагревателях. Наиболее
распространены те конструкции теплового инжектора, в который
подают вихревым насосом нестерильную питательную среду и пар
под давлением — 0,3—0,6 МПа. Производительность нагревателей
5, 10, 15, 20, 25, 50 м3/ч стерильной среды. Подбирают нагреватель
в зависимости от объема стерилизуемой среды так, чтобы время
операции стерилизации всего объема среды составляло 40—120
минут. После этого среду подают в выдерживатель — проточный
аппарат непрерывного действия, работающий по принципу вытес-
нения вновь поступающими объемами среды предыдущих. За
время прохождения через выдерживатель элементарного объема
питательной среды содержащаяся в этом объеме спора погибает.
Следовательно принцип работы выдерживателя может быть запи-
сан в следующем виде: т выдержки = т отмирания термостойких
спор = т заполнения. По конструктивным особенностям выдер-
живатели подразделяют на два типа — емкостные и трубчатые.
Чтобы приблизить течение жидкости в емкостном выдерживателе
к поршневому, снизить скорость потока по центральной оси и
уменьшить застойные зоны у корпуса, необходимо предусматри-
вать специальные устройства для ввода среды, обеспечивающие
равномерное распределение потока, и
для вывода среды, где надо равномерно
собрать поток со всего сечения.
Наиболее простым по конструкции и
надежным является емкостной выдержи-
ватель, вместимостью не менее 2 м3 с
подачей среды сверху по трубе, опущен-
ной до днища аппарата; днище при этом
играет роль отбойника, разбивающего
струю жидкости, а сбор стерильной сре-
ды осуществляется через перфорирован-
ное кольцо, расположенное в верхней
части выдерживателя (рис. 100). Но в
процессе работы емкостного выдержива-
теля возможно возникновение пульса-
ций, вызывающих выбросы нестериль-
ных объемов среды.
Практически идеальный поршневой
поток осуществить невозможно. При ла-
минарном и турбулентном потоке сред-
няя скорость жидкости является функ-
317
Рис. 100. Выдерживатель с
перфорированным кольцом в
верхней части: 1 — труба вво-
да нестерильной среды, 2 —
кольцевой коллектор вывода
стерильной среды, 3 — шту-
цер слива конденсата после
стерилизации системы, 4 —
корпус аппарата.
цией распределения скоростей отдельных объемов жидкости по
сечению потока. Средняя скорость ламинарного потока вязкой
жидкости в трубе составляет 50% максимальной скорости по оси
трубы, а турбулентного — 82%. Степень отклонения от поршневого
течения может быть выражена безразмерным критерием Пекле
(Ре):
_ Wcpl ...
Ре = ~где Wcp
— средняя скорость потока, 1 - длина
трубы, Dl — коэффициент продольного перемешивания. Для иде-
ального течения в поршневом режиме критерий Ре равен беско-
нечности, а при полном смешении — нулю.
Расчетными данными показано, что из различных по длине и
диаметру трубчатых выдерживателей, дающих один и тот же
уровень стерильности, экономически наивыгоднейшим является
тот, который характеризуется потоком с критерием Ре = 50. Такие
трубчатые выдерживатели получили название оптимальных.
Трубчатые выдерживатели большого диаметра и длины разде-
ляют на секции и укрепляют вертикально как колонные аппараты.
Выдерживатели с диаметром трубы 150—200 мм сворачивают в
виде объемной или плоской спирали и устанавливают над нагре-
вателем. Все выдерживатели должны быть покрыты слоем изоля-
ции для предотвращения теплопотерь.
При высокой температуре среда находится в выдерживателе
строго определенное расчетное время, а затем ее необходимо очень
быстро охладить до температуры ферментации, иначе в среде
пойдут деструктивные процессы. Теплообменник, в котором воз-
можен подобный процесс, должен обладать не только высокими
теплообменными характеристиками, но и конструктивным совер-
шенством, обеспечивающим поддержание среды в стерильном
виде, следовательно, он должен быть герметичен.
Наиболее широкое распространение в нашей стране получили
теплообменники типа "труба в трубе", тогда как за рубежом
применяют более эффективные пластинчатые и спиральные теп-
лообменники. Использование пластинчатых теплообменников ог-
раничивается невозможностью их применения для охлаждения
вязких сред, содержащих твердую фазу.
В настоящее время наиболее часто используют УНС, схема
которой представлена на рис. 98. Но в такой схеме не реализуется
одно из преимуществ непрерывного способа стерилизации —
возможность рекуперации тепла.
До 70% тепла можно вернуть, если дополнительно установить
318
теплообменник —рекуператор, в котором стерильная среда, про-
шедшая через выдерживатель и имеющая температуру 130—140°С,
нагревает нестерильную питательную среду до 115—125°С, а затем
доохлаждается в теплообменнике-доохладителе до температуры
ферментации (см. рис. 99). При этом сокращается расход пара на
нагрев нестерильной среды до температуры стерилизации и расход
воды на охлаждение стерильной среды.
Все аппараты УНС выполняют из нержавеющей стали, а наи-
более надежной арматурой является сильфонная нержавеющая.
Перед началом операции стерилизации всю систему УНС стери-
лизуют водяным паром, после чего стерилизуют концентрат пита-
тельной среды. Для удаления остатков среды из системы и для
разбавления концентрата через УНС пропускают воду. Перед
отключением УНС ее вновь стерилизуют.
Сыпучие среды, используемые в настоящее время для получе-
ния ферментов поверхностным методом и кормовых добавок,
стерилизуют паром, инфракрасными лучами, а также используют
другие губительные факторы. Компонентами таких сред являются
отруби, биошрот, свекловичный жом, древесные опилки, солома.
Компоненты предварительно тщательно измельчают и смешивают
в оптимальных соотношениях.
В промышленности наиболее широко применяют обработку
сред глухим и острым паром, которую условно можно назвать
стерилизацией. В действительности деструкция спор происходит
только на поверхности твердых комков среды. Споры, находящи-
еся в глубине комка, могут выжить. Уровень стерильности таких
сред относительно невысок, но достаточен для проведения фер-
ментации на твердой сыпучей среде.
7.2.2. Подготовка стерильного сжатого воздуха и очистка
отработанного воздуха. Одной из важных задач биотехнологии
является получение большого количества стерильного воздуха. В
наибольших масштабах стерильный воздух применяется в процес-
сах ферментации для аэрации. Его также используют для венти-
ляции участков цехов так называемой стерильной зоны, где в
асептических условиях проводят, например, последние стадии
очистки готового продукта. В атмосферном воздухе, наряду с
инертными газами, азотом, кислородом, диоксидом углерода, со-
держатся пары воды и мелкодисперсные частицы. Более 30% массы
частицы имеют размер 1—2 мкм и около 50% — меньше 0,5 мкм.
В состав дисперсных частиц, наряду с частицами пыли, копоти,
входят клетки и споры микроорганизмов как в свободном, так и в
319
сорбированном на пылевых частицах виде. Результаты анализа
технологического воздуха на содержание микробов-контаминан-
тов представлены в таблице 36.
Таблица 36. Типы микробов и частота их обнаружения в технологическом
воздухе
Микробы Размеры клеток микробов, мкм Частота обнаружения, %
Актиномицеты Кокки Палочки Плесневые грибы Споры бацилл 0,8 - 1,2 (диаметр клеток) 0,47 - 1,7 0,77 - 0,98 х 2,0 - 5,8 0,8 - 12,0 (диаметр нитей) 0,55- 1,2 х 2,3-7,1 4,5 33,5 22,5 8,1 18,7
Температура и влажность наружного воздуха, количество в нем
пылинок и микроорганизмов непостоянны и зависят от времени
года (микроорганизмов летом в 10 раз больше, чем зимой), погод-
ных условий — наибольшее количество пыли и, соответственно,
микроорганизмов приходится на сухую ветреную погоду, геогра-
фического расположения предприятия, высоты забора воздуха и
т. д. Особенно много микробов у поверхности земли, с высотой
концентрация их убывает и становится постоянной на уровне
около 30 м над землей.
7.2.2 Л. Способы очистки воздуха. Очистку воздуха можно
осуществить принципиально разными методами, основанными
либо на уничтожении микроорганизмов, либо на удалении их.
Одним из самых эффективных способов стерилизации воздуха
является облучение ультрафиолетовыми лучами. Этот метод ис-
пользуется для обеззараживания воздуха в боксах.
Отечественным и зарубежным опытом показано, что техноло-
гически и экономически оправданным в промышленности является
способ очистки воздуха с помощью волокнистых, и пористых
материалов. Таким путем удается получить воздух со степенью
чистоты 99,9999%. Взвешенные в воздухе частицы задерживаются
волокнистым материалом благодаря инерционному и диффузион-
ному механизмам осаждения. В общих чертах механизм инерци-
онного осаждения основан на том, что когда воздушный поток
начинает обтекать нить волокна на своем пути, взвешенные в этом
потоке частицы движутся по инерции, отклоняются от потока
воздуха и осаждаются на волокне. Эффект инерционного осажде-
ния высок на сравнительно грубых волокнах для относительно
крупных частиц и высоких скоростей воздуха. Малые частицы
320
способны к броуновскому движению. Движущиеся вблизи волокна
частицы диффундируют в случайных направлениях и могут задер-
живаться на поверхности волокон. Этот эффект осаждения увели-
чивается с уменьшением диаметра волокна фильтрующего мате-
риала, диаметра частицы и скорости воздуха.
Качество фильтрующего материала характеризуется коэффи-
циентом осаждения (h):
Cn0-Cn
п = —, где Ск0 — начальная концен-
f-No
трация микробов в воздухе до фильтра, может быть принята из
расчета максимального содержания 5000 клеток в 1 м3; Cn = 0,001
— конечная концентрация микробов в воздухе после фильтра,
выбранная, исходя из вероятности проскока одной клетки на 1000
операций. Фильтрующий материал тем лучше, чем выше коэффи-
циент осаждения.
Качество аэрозольных фильтров для очистки воздуха характе-
ризуется коэффициентом проскока (Кпр):
Кпр~с^0
Чем меньше Кпр, тем качество фильтра выше.
7.2.2.2. Характеристика фильтрующих материалов, применяе-
мых в промышленности для стерилизации воздуха. Поскольку
отдельные механизмы осаждения имеют различную природу и
обычно один из них преобладает над другим, необходимо приме-
нять фильтрующий материал различной структуры для полноцен-
ной очистки воздуха. Так называемые головные фильтры заполня-
ют более грубым волокном, на этих фильтрах удаляется около 98%
микробов-контаминантов, а следующую ступень — индивидуаль-
ные фильтры заправляют супертонким волокном или мембранами.
На этой последней ступени удаляют остальные 2% контаминантов.
Кроме высокой пылеемкости фильтрующие волокна должны
обладать еще одним свойством — работать при малых перепадах
давления так, чтобы их разность до и после фильтра были как
можно меньше.
В головных фильтрах для грубой очистки используют следую-
щие фильтрующие материалы:
а) грубое базальтовое волокно ВРВ (вертикальный раздув воз-
духом) с диаметром волокна 16 мкм и 26 мкм, толщина слоя 1000 мм;
волокно обладает высокой паростойкостью, не теряет своих
свойств при нагревании до 1100°С, отличается механической проч-
ностью и высокой пылеемкостью;
Пт. 8524 321
б) высокообъемный нетканый фильтрующий материал
(ВНФМ), или ткань Каминской с диаметром волокна 16,9 мкм,
выпускают в виде слоев (матов) толщиной 20—23 мм; особенностью
этого материала является то, что в прядильный раствор при его
изготовлении вводят антисептик гексахлорофен, поэтому фильтры,
заполненные такой тканью, не подвергают стерилизации;
в) стеклосрезы с диаметром волокна 6 мкм, толщиной слоя 600
мм, термостойкость 300°С.
В индивидуальных фильтрах для тонкой очистки воздуха ис-
пользуют фильтры Петрянова (ФП) с тканью Петрянова, состоя-
щую из ультратонких полимерных волокон, сформированных в
виде ткани на. марлевой подложке. В настоящее время изготавли-
вают много видов ФП. Так ФПП из перхлорвинила и фторполиме-
ров обладают устойчивостью к кислотам и щелочам. ФПП из
полиакрилонитрила стойкие к органическим растворителям. Но
эти ФПП выдерживают температуру только до 60°С и поэтому их
стерилизуют формалином с последующей нейтрализацией амми-
аком. В последние годы разработаны ФП, стойкие к высоким
температурам. ФПАР (полиакрилатные) и ФПФС (полифторстиро-
ловые) выдерживают температуру до 250—270°С; их стерилизуют
острым паром.
Базальтовое суперволокно (БСТВ) диаметром 1,0—1,5 мкм вы-
пускают в виде матов толщиной 30—40 мм. Недостатком этих
волокон является небольшая механическая прочность. Этого недо-
статка лишены базальтовые бумаги и картон, разработанные Ук-
раинским НИИ бумаги. Базальтовые картон и бумаги имеют, кроме
того, высокие коэффициенты осаждения, что позволяет уменьшить
габариты фильтров. В 1989 г. Украинский НИИ химического
машиностроения (НИИХИММАШ) и Пензенский филиал Всерос-
сийского НИИ антибиотиков (ВНИИА) разработали конструкции
фильтров с гофрированными элементами из базальтовой бумаги
ФГ-5 и ФГ-160 производительностью 5 и 160 м3/мин.
В последние годы для тонкой очистки воздуха используют
жесткие пористые перегородки из пластмасс и металлокерамики.
Преимуществом этих материалов по сравнению с волокнистыми
является стабильность структуры, устойчивость к повышенным
температурам, простота конструктивного оформения, легкость
обслуживания фильтров. Существенный недостаток — высокий
перепад давления.
322
Наибольшее применение находят фильтрующие материалы из
фторопласта и металлокерамики в виде втулок ФЭП — фильтру-
ющий элемент патронного типа (таблица 37).
Т а б л и ц а 37. Характеристика фильтрующих элементов, применяемых в про-
мышленности
Фторопластовый элемент Размеры мм Площадь, Производи- тельность, М3/ч Срок службы, мес.
ФЭП-46 46x38x110 0,015 60 10-11
ФЭП-120 120x96x170 0,06 230 10-11
Для стерилизации воздуха рекомендуют также мембраны с
диаметром пор 0,45 мкм. • Пористость мембран достигает 80%.
Удаление микроорганизмов с помощью мембраны основано на
ситовом эффекте. В основном фильтрующие мембраны поставляет
фирма "Миллипор” (США), в нашей стране (г. Владимир) налажен
выпуск мембраны "Владипор". Мембранам не требуются высокие
перепады давления, но для их надежной работы необходимо точное
выполнение условий стерилизации. Стерилизовать мембраны
можно только насыщенным водяным паром, от перегретого пара
в мембранах появляются трещины, и они выходят из строя.
Высокоэффективную фильтрацию воздуха (и жидкостей) обес-
печивают также фильтры, изготавливаемые фирмами Domnick
Hunter (Англия), DDS-De Danske Suckerfabriker (Дания) и др. (рис.
101а).
7.2.3. Технологическая схема сжатия и очистки воздуха. Сис-
тема производства. сжатого, очищенного от микроорганизмов,
воздуха, имеющего определенную температуру, является сложной
технологической системой (рис. 1016).
Она состоит их трех последовательно соединенных подсистем:
очистки от пыли и сжатия; приведения воздуха к термодинамиче-
скому состоянию, благоприятному по влажности и температуре
для разделения аэрозоля; разделение аэрозоля в фильтрах грубой
и тонкой очистки.
Первая подсистема. Атмосферный воздух забирают турбоком-
прессором через заборную шахту высотой 20—30 м, где концент-
рация микроорганизмов стабилизирована. Прежде всего воздух
попадает в предфильтры, где он освобождается от грубого аэрозоля
— пыли. Предфильтры не только предохраняют компрессоры от
323
Рис. 101а, рис. 1016. Фильтр тонкой очистки воздуха в разрезе (а), схема очистки
воздуха в масляном циклофильтре (б): 1 - циклофильтр, 2 - ванна с маслом, 3 -
отбойный козырек, 4 - направляющий короб.
загрязнения, но и существенно снижают количество контаминан-
тов, которые могли бы попасть во 2-ю подсистему.
За рубежом в настоящее время в предфильтрах применяют
рулонные пористые материалы. В нашей стране успешно испытан
пенополиуретан, который обеспыливается пылесосом или теплой
водой с мылом. Срок службы материала 1,5—2 года. Однако до сих
пор на большинстве заводов используют масляные фильтры (см.
рис. 1016). После этого воздух сжимают в турбокомпрессоре до
0,35—0,5 МПа. Давление сжатия воздуха в компрессоре определя-
ют из расчета давления на преодоление сопротивления в системе
воздухоподготовки, давления столба жидкости в ферментаторе и
создания в нем давления 0,13—0,14 МПа. Сжатие воздуха в комп-
рессоре приводит к повышению его температуры до 120—250°С и
увеличению влагос одержания в единице объема.
Вторая подсистема. В случае высокого содержания влаги в
исходном атмосферном воздухе конденсируется еще большее ко-
личество влаги при его охлаждении. Выпадение влаги на фильтрах
324
недопустимо, так как это приводит к слипанию волокон и образо-
ванию каналов, и тогда эффекты, осаждения частиц на волокне не
проявляются. Кроме того, на увлажненных волокнах фильтров
возможно размножение осевших микроорганизмов, что приводит
к дополнительному обсеменению воздуха
Чтобы обеспечить выпадение влаги в каплеуловителе, воздух
“переохлаждают" до температуры 25-40°С в теплообменном аппа-
рате. Затем, для обеспечения надежной работы фильтров 2-й и 3-й
ступеней, воздух нагревают до температуры 70—90°С. При таких
температурах исключается конденсация паров воды на волокнах
фильтра. С этой целью воздух после брызгоуловителя подогревают
в теплообменнике, при этом допускается частичное подмешивание
горячего воздуха после компрессора Количество подмешиваемого
воздуха определяется условиями относительной влажности, кото-
рая не должна быть больше 40%.
Третья подсистема состоит из двух фильтров второй и третьей
ступеней очистки. Фильтр второй ступени, или головной фильтр
обычно расположен на территории завода рядом с цехом. На
головном фильтре очищают воздух для всех ферментаторов цеха
Из головного фильтра воздух по коллектору подается в индивиду-
альные фильтры третьей ступени, установленные у каждого фер-
ментатора, независимо от его вместимости.
Конструкция индивидуального фильтра зависит от типа исполь-
зуемого фильтрующего материала. Для ткани Петрянова применя-
ют конструкцию, представленную на рис. 102а. В цилиндрический
корпус монтируют прямоугольный пакет, собранный из П-образ-
ных алюминиевых рамок, между которыми зажимается лента из
ткани Петрянова.
Для фильтрующего материала, уложенного в виде матов, ис-
пользуют конструкцию, представленную на рис. 102(6), а на рис.
102(b) изображена конструкция, предназначенная для установки
фторопластовых фильтрующих элементов.
При эксплуатации фильтров необходима их стерилизация.
Наиболее эффективным методом является нагревание влажным
паром и выдержка в течение определенного времени при темпе-
ратуре 125—130°С. Применение более высокой температуры вы-
зывает деструкцию герметизирующих прокладок в фильтрах. По-
сле стерилизации фильтрующий материал высушивают горячим
воздухом.
325
Рис. 102. Конструкция фильтров с тканью Петрянова (а), аэрозольного фильтра
фланцевого типа (б): 1 — сетка для равномерного распределения воздуха, 2 —
крышка фильтра, 3 — корпус, 4 — паронитовая прокладка, 5 — решетка, 6 — слой
фильтрующего материала; фильтра с фторопластовыми фильтрующими элементами
(в): 1 — корпус, 2 — крышка, 3 — фторопластовые фильтрующие элементы.
В зарубежных системах очистки воздуха значительно повыше-
на надежность работы за счет установки дополнительной ступени
очистки и дублирования основного оборудования. Кроме того, для
крупных ферментаторов применяют автономные системы очистки
воздуха, что облегчает задачу поддержания термодинамического
режима, так как не требуется большой протяженности трубопро-
водов; наконец, в фильтрах используют стандартные фильтрующие
элементы, изготовленные промышленным способом.
7.2.4. Очистка отработанного воздуха. В процессе фермента-
ции в качестве отхода производства образуется большое количе-
ство отработанного воздуха, выбрасываемого в атмосферу. Уста-
новлено, что такой воздух на заводах антибиотиков содержит от 2
до 4 мг/м3 веществ с неприятным запахом.
С отработанным воздухом выбрасывается в атмосферу несколь-
ко десятков органических соединений: амины, альдегиды, жирные
326
кислоты, кетоны, спирты, эфиры и т. д . Относительная влажность
воздуха, выходящего из ферментатора, приближается к 100%;
кроме того он включает культуральную жидкость в виде мелких
брызг, а содержание клеток продуцента зависит от вида его,
времени ферментации, и составляет от 1 • 103 до 1 • 105 клеток
в 1 м3.
В настоящее время применяют несколько принципиально раз-
личающихся методов очистки отработанного воздуха. Метод ката-
литического дожигания относится к разряду энергоемких. Суть его
состоит в том, что отработанный воздух прокачивают при темпе-
ратуре 320-350°С через комбинированный катализатор, состоящий
из слоя пиролюзита и слоя палладиевого катализатора. Степень
обезвреживания воздуха 87—98,5%.
Менее энергоемким является метод жидкофазного окисления
с применением в качестве окислителей перманганата калия или
гипохлорита натрия. Отработанный воздух по мере прохождения
скруббера орошается раствором натрия гипохлорита, 20% раство-
ром едкого натра, водой и выбрасывается в атмосферу. Орошаю-
щие растворы обращаются в замкнутом цикле. Смену отработан-
ных растворов проводят 1 раз в неделю. Эффективность очистки
90—95%. Недостатком метода является то, что при его реализации
накапливаются в небольшом количестве сточные воды, которые
нужно утилизировать. Такие установки успешно работают, напри-
мер, в Италии в производстве пенициллина, цефалоспорина С и
других антибиотиков.
Известен также метод с применением сетчатых фильтров (ФС);
конструкция ФС разработана во ВНИ проектно-конструкторском
институте прикладной биохимии. Фильтр состоит из цилиндриче-
ского корпуса с крышкой и днищем, внутри помещен фильтрую-
щий элемент, изготовленный из металлических сеток трикотажно-
го плетения с диаметром проволоки 0,28 мм из нержавеющей стали
(рис. 103).
Воздух, проходя через фильтр, освобождается от капелек куль-
туральной жидкости с микроорганизмами. Эффективность очист-
ки 99,6%. Для повышения эффективности очистки на ряде заводов
осуществлена схема, состоящая из циклона и сетчатого фильтра
"Ц-ФС". Эффективность этой системы составляет 99,97%.
327
Рис. 103. Схема сетчатого фильтра для очистки отработанного
воздуха: 1 — крышка, 2 — корпус, 3 — фильтрующий элемент.
7.3. Тепловые процессы в ферментаторах. Интерес к проблемам
выделений тепла в процессе ферментаций связан, прежде всего, с
необходимостью его отведения охлаждающей водой, а также с
возможностью использования интенсивности тепловыделения в
качестве индикатора метаболической деятельности продуцента.
Около 40—50% всего тепла, выделяющегося в процессе фермента-
ции, приходится на тепло, образующееся в результате жизнедея-
тельности продуцента и, прежде всего, за счет энергетического
обмена Накопленные макроэргические структуры (преимущест-
венно — АТФ) расходуются в реакциях биосинтеза. Так, напри-
мер, на образование одной эфирной связи в молекуле какого-
либо метаболита требуется 12570 Дж. При этом в реакции:
АТФ—>АДФ + 50280Дж из одной молекулы АТФ высвобождается
столько энергии, что большая часть ее (37710 Дж) превращается в
тепло. Это тепло выделяется неравномерно — с максимумом,
приходящимся на период максимальной интенсивности дыхания
и наивысшей скорости потребления углеводов в период активного
размножения клеток (рис. 104).
328
33000
2jooo Рис. 104. Соотношение скорости
потребления углевода, интенсивно-
17000 сти дыхания и тепловыделения: 1 —
накопление антибиотика, 2 — выде-
sooo ление диоксида углерода, 3 — тепло
биосинтеза, 4 — потребление углево-
дов.
Тепло также образуется в результате работы мешалки, переме-
шивающей культуральную жидкость — "тепло перемешивания"
(Оперем-)
Оперем. = Nr’Тф* 3600, (Дж), где Nr(BT) — мощность, затрачи-
ваемая на перемешивание газожидкостной смеси, т<р (час) — время
ферментации.
Тепловой эффект перемешивания при одном и том же числе
оборотов мешалки будет тем выше, чем больше вязкость культу-
ральной жидкости. Поэтому в процессе ферментации, по мере
накопления биомассы, тепловой эффект перемешивания возраста-
ет, если не уменьшать интенсивность перемешивания, снижая
число оборотов мешалки.
Третий источник тепла —горячий стерильный воздух, .подава-
емый в ферментатор для аэробных организмов; при барботаже он
мгновенно охлаждается культуральной жидкостью и отдает при-
внесенное тепло (ОвоЗД-)- ОвОЗД- = Vno3A- • Рвозд. • Свозд. (tflO3A- 'ф). ГДе
Vbosa., рвозд., Свозд-, ^возд- — соответственно объем, плотность,
теплоемкость и температура подаваемого воздуха, 1ф — темпера-
тура, при которой проводят ферментацию.
Чаще всего воздух расходуют в процессе ферментации ступен-
чато, с максимумом, приходящимся на период лагарифмической
фазы размножения, например одноклеточных микроорганизмов;
к концу ферментации расход воздуха снова снижают. Следова-
тельно, количество тепла, привносимое с воздухом, различно в
разные периоды ферментации.
В процессе ферментации происходит унос влаги из аппарата с
отработанным воздухом за счет испарения, а, следовательно, и
части тепла, затраченного на этот процесс (Оисп.):
329
Оисп- = пхвлаги. • г, где гпВлаги — масса испарении влаги,
г (Дж/кг) — удельная теплота парообразования.
По абсолютному значению Оисп. — величина небольшая,
поэтому ее колебания, пропорциональные количеству подаваемого
воздуха, не оказывают заметного влияния на общее количество
тепла, образующегося в процессе ферментации.
Рис. 105. Теплотрубный теплооб-
менник Херсонского индустриального
университета: а — общий вид в сборке,
б — устройство тепловой трубы (1 —
подвод тепла, 2 —- фитиль, 3 — жид-
кость, 4 — пар, 5 — отвод тепла, 6 —
зона испарения, 7 — адиабатическая
зона, 8 — зона конденсации).
Как правило, тепловой режим
в процессе ферментации поддер-
живается на определенном уров-
не и строго контролируется. Все
образующееся тепло должно быть
отведено охлаждающей водой.
В общем виде тепловой ба-
ланс процесса ферментации мо-
жет быть представлен следую-
щим уравнением:
Ож + Оп + Овозд.—Оисп.= Оохл-,
где Оохл. — тепло, отводимое ох-
лаждающей водой, подаваемой в
теплообменные устройства;
Оохл- = ШвОДЫ • Своды (t?—11), где
тпводы — масса охлаждающей во-
ды, Своды — теплоемкость ох-
лаждающей воды, ti и t? — тем-
пература воды соответственно на
входе и выходе из теплоотводя-
щих устройств.
Для поддержания теплового ре-
жима в ферментаторе в настоя-
щее время чаще всего используют
артезианскую воду 12—15°С или
смесь артезианской и оборотной
воды 15—16°С; отработанная вода,
выходящая из теплообменных ус-
тройств с температурой 18—20<>С,
в этом случае поступает в линию
оборотной воды и может быть ис-
пользована в качестве охлаждаю-
щего агента в тепловых процессах
с высокими температурными ре-
жимами (рис. 105).
ззо
Наиболее эффективно использование так называемой "захоло-
женной" воды (10°С), получаемой для цеха ферментации на спе-
циальных установках. В теплоотводящих устройствах ферментато-
ра она нагревается до 15—16°С и по коллектору возвращается на
станцию получения "захоложенной” воды.
Таким образом, применительно к тепловым процессам, проис-
ходящим в ферментаторе, можно сказать, что все они (тепловой
эффект жизнедеятельности, перемешивания, тепло, привносимое
воздухом, тепло испарения, теплопередача при охлаждении водой)
протекают одновременно с разной интенсивностью и динамично-
стью, отсюда понятны высокие требования к конструктивным
характеристикам теплообменных устройств.
7.4. Аппаратурное оформление процессов выделения и очист-
ки некоторых продуктов микробного синтеза. Культуральная
жидкость, образующаяся в процессе ферментации, представляет
собой сложную многофазную систему: в водной фазе содержатся
клетки продуцента, продукты их жизнедеятельности, непотреблен-
ные компоненты питательной среды, мельчайшие капельки жира,
пузырьки воздуха, как правило, мел. В свою очередь водная фаза
культуральной жидкости (нативный раствор) включает большое
число органических и неорганических веществ, коллоидных фрак-
ций белков, сухой остаток культуральной жидкости — до 17% и
более; содержание биомассы в культуральной жидкости достигает
8—10%. Концентрация целевого продукта чаще всего не превышает
1,5%, что составляет менее 10% сухого остатка.
В зависимости от целевого назначения конечного продукта (для
здравоохранения, технических целей, сельского хозяйства и т. д.)
реализуют различной степени сложности схемы производства, при
этом учитывают и место накопления целевого продукта — внут-
риклеточно или внеклеточно.
Если способы выделения целевых продуктов можно разделить
на несколько типовых групп (рис. 106), то очистка каждого из них
практически индивидуальна, поскольку зависит от физико-хими-
ческих свойств конкретного вещества.
Примером наиболее простой схемы может служить производ-
ство биовита, базирующегося на использовании продуцента анти-
биотика хлортетрациклина. Кальциевый комплекс хлортетрацик-
лина осажадают из культуральной жидкости при pH выше 7,0.
Формирующийся при этом осадок увлекает с собой и клетки
продуцента. Так в осадке вместе с биомассой оказываются внекле-
точный антибиотик и внутриклеточный витамин В12. Осадок филь-
331
труют, высушивают, размельчают, добавляют отруби в качестве
наполнителя и фасуют. Этот комплекс используют в качестве
добавки к корму для скота.
Рис. 106. Возможные способы выделения целевого продукта.
Если целевым продуктом является биомасса клеток, например,
кормовые или пекарские дрожжи, необходимо их концентрирова-
ние (сгущение). При этом широко применяют флотацию, с по-
мощью которой увеличивают концентрацию дрожжей в 4—6 раз.
Основной движущей силой флотации являются всплывающие
пузырьки газа, которые захватывают клетки и выносят их на
поверхность. В результате над слоем отработанной культуральной
жидкости образуется пенный слой, в котором сконцентрированы
дрожжи. Процесс проводят в специально предназначенных для
этой цели .емкостных аппаратах — флотаторах, в которые нагне-
тают роздух и, в зависимости от их конструкции, диспергируют
его через барботер или эжектор, пену гасят в специальной части
аппарата с помощью пеногасителя и выводят сконцентрированную
суспензию дрожжей.
Для последующего концентрирования дрожжей широко приме-
няют сепарирование, например, с помощью сепаратора—сгусти-
теля, непрерывного действия тарельчатого типа СОС-501 К-3. Для
сгущения суспензии дрожжей от 20—30 г/л до 550—600 г/л
необходимо последовательное сепарирование в 2—3 ступени.
Методы извлечения целевых продуктов из клеток зависят от
свойств изолируемых веществ. Полиеновые антибиотики, напри-
мер, экстрагируют органическими растворителями из нативных
клеток, тогда как эндоферменты получают из разрушенных (дез-
332
интегрированных) клеток. На рис. 107 представлена схема процес-
са выделения L-аспарагиназы. Клетки отделяют на дисковой цен-
трифуге (позиция 4), позволяющей концентрировать от 6 г/л до
120 г/л по массе сухих веществ. Разрушение клеток проводят в.
гомогенизаторе (позиция 5) при двукратном пропускании высво-
бождается до 97% фермента. От остатков клеток освобождаются
после добавления этанола (позиция 9) до 30% концентрации с
последующей фильтрацией на ротационном фильтре (позиция 10).
Затем фермент осаждают метанолом при его концентрации в.
маточнике 50% (позиция И), фильтруют (позиция 12),-.растворяют
в воде и переосаждают (позиция 13) при концентрации метанола
60%. Осадок центрифугируют (позиция 12), снова растворяют в
воде (позиция ,14) и концентрируют ультрафильтрацией (позиция
16), затем выделяют с помощью*еефадексаДЕАЕ-А50 (позиция 19),
осаж'адют этанолом (0,6 объема) прирН 5,0 (позиция 27) и собирают
в корзинчатой центрифуге (позиция 28). Осадок еще раз раство-
ряют.в воде (позиция 29), стерилизуют фильтрацией через мемб-
рану [позиция 30), разливают (позиция 32) й подвергают сублима-
ционной сушке (позиция 34).
Рис. 107. Схема процесса выделения L-аспарагиназы: 1,5,17 — промежуточные емкости,
2, 6, 15, 18, 26 — насосы, 3 — теплообменник, 4 — дисковая центрифуга, 7 — аппарат для
получения суспензии микробных клеток, 8 — гомогенизатор, 9 — аппарат для спиртового
осаждения остатков клеток и белка, 10—ротационный фильтр, 11—аппарат для осаждения
фермента метанолом, 12 — центрифуга, 13 — аппарат для переосаждения фермента, 14
— аппарат для растворения фермента, 16 — аппарат для ультрафильтрации, 19 — колонка
с сефадексом, 20, 21 — емкости для буферных растворов, 22, 23 — насосы, для подачи
буферных растворов, 24, 25 —- емкости для сбора фракций, 31 — аппарат для осаждения
фермента, 28 — центрифуга, 29 — аппарат для переосаждения, 30 — стерилизующая
мембрана, 31 — осадитель, 32 - кювета, 33 - фасовочное устройство, 34 — сублимационная
сушилка.
333
Выделение большей части продуктов микробного синтеза из
культуральной жидкости начинают при их содержании около 1,5%.
Чтобы иметь возможность выделить вещество, используя осажде-
ние, кристаллизацию или высушивание, необходимо оперировать
с растворами, содержащими 15—20% целевого продукта. Вот по-
чему приходится применять разномасштабное оборудование по
ходу процесса: вначале необходимы емкости в несколько десятков
м3, высокопроизводительное оборудование непрерывного или по-
лунепрерывного действия, а завершают процесс небольшими ап-
паратами вместимостью порядка нескольких сотен и даже десятков
литров с периодической загрузкой и выгрузкой, или лабораторное
оборудование.
Технологические показатели фильтрации культуральных жид-
костей определяются свойствами их как дисперсных систем. Клет-
ки микробов-продуцентов, например, грибов чаще всего образуют
микроколонии различной плотности размером 16—600 мкм. Филь-
трационные свойства жидкости обусловлены размерами отдель-
ных гиф, типом микроколоний, характером роста мицелия. Эти
факторы зависят от вида и штамма биообъекта, условий его
культивирования.
Пенициллы и аспергиллы образуют легко фильтруемую био-
массу, поэтому для получения нативного раствора в этом случае
применяют барабанные вакуум-фильтры — аппарат непрерывного
действия с фильтрующим основанием, расположенным на наруж-
ной цилиндрической поверхности горизонтального вращающегося
барабана, частично погруженного в суспензию.
Культуральные жидкости после ферментации проактиномице-
тов представляют собой густые суспензии с относительной вязко-
стью 50—1000 и с высоким содержанием твердой фазы. Они
практически не расслаиваются при стоянии. Характерной особен-
ностью и других бактериальных суспензий является изменчивость
их фильтрационных свойств как во времени, так и от одной
операции до другой.
Необходимо также отметить, что образуемые осадки заметно
сжимаемы (показатель сжимаемости 0,9), значит процесс фильт-
рации не может быть интенсифицирован повышением разности
давления из-за соответственно возрастающего удельного сопро-
тивления осадка. Такие суспензии без предварительной обработки
культуральной жидкости отфильтровать практически невозможно.
Предварительную обработку культуральной жидкости проводят с
целью максимального перевода конечного продукта в ту фазу, из
334
которой предполагают его выделить, а также для коагуляции
коллойдных примесей и, следовательно, улучшения процесса филь-
трации, для удаления или связывания в растворимые комплексы
тех побочных веществ, которые затрудняют последующий процесс
химической очистки. К тому же при коагуляции изменяется струк-
тура твердой фазы культуральной жидкости.
Применяют несколько видов коагуляции: неорганическими
солевыми электролитами, органическими и неорганическими кис-
лотами, термической обработкой, добавлением веществ, образую-
щих наполнители. Например, при добавлении к культуральной
жидкости раствора жидкого стекла (Na2SiOs).H натрия моногидро-
фосфата, в результате взаимодействия с ионами кальция, находя-
щимися в культуральной жидкости, образуется осадок наполните-
лей:
Са++ + Na2SiO3—>CaSiO3J< + 2Na+
Са++ + Na2HPO4-»CaHPO4>L + 2Na+
Процесс образования хлопьевидного осадка наполнителя про-
текает с большой скоростью, осадок захватывает гифы мицелия и
молекулы белка.
Процесс коагуляции улучшается при использовании комбини-
рованных методов таких, как кислотная и тепловая, электролитная
и тепловая коагуляции.
Эффективным методом коагуляции дисперсных систем явля-
ется обработка их высокомолекулярными полиэлектролитами —
флоккулянтами. При этом в отличие от обычной коагуляции,
образуются флоккулы или осадки рыхлой структуры, что значи-
тельно улучшает процесс фильтрации.
Для получения нативного раствора из труднофильтруемых
культуральных жидкостей после коагуляции применяют вакуум-
барабанный фильтр с намывным слоем — аппарат полунепрерыв-
ного действия. Перед работой на фильтрующую поверхность ба-
рабана намывают дренажный слой наполнителя. В качестве напол-
нителя используют суспензию порошков целлюлозы, перлита,
древесной муки. Особенность работы такого фильтра (в отличие
от вакуум-барабанного) состоит в том, что нож, предназначенный
для съема осадка с барабана, имеет специальную микрометриче-
скую подачу. С каждым оборотом барабана нож подается к центру,
срезая нефильтрованный осадок вместе с тонким слоем дренажа,
обновляя таким образом фильтрующую поверхность; поэтому
скорость фильтрации не снижается. На рис. 108 представлена
схема аппаратурного оформления процесса фильтрации культу-
335
ральной жидкости на вакуум-барабанном фильтре с намывным
слоем.
Рис. 108. Схема аппаратурного оформления процесса фильтрации на вакуум-
барабайном фильтре с намывным слоем: 1 — аппарат для суспензии вспомогатель-
ного материала, 2, 4, 7 — насосы, 3 — коагулятор, 5 — вакуум-барабанный фильтр
с намывным слоем, 6 — ресивер. . • . , .
Названный выше перлит представляет собой стекловидную
горную породу вулканического происхождения, в состав его входят
окислы кремния, калия и кальция. Для получения вспомогательного
вещества природный перлит нагревают до начала плавления
(1000°С). Образуются небольшие бусинки неправильной формы,
состоящие из очень большого числа полых ячеек. Для получения
вспомогательного фильтрующего материала бусинки измельчают-
ся. Получают порошок с пористостью 85—90% и объемной массой
500—1000 кг/м3.
Экстракционный метод выделения целевого вещества из на-
тивного раствора основан на различной растворимости разных
химических форм этого вещества в воде и органическом раство-
рителе, не смешивающимся с водой. Основной закон экстракции
— закон распределения Нернста:
Ср
Со
где К — коэффициент
К =
распределения, Со — концентрация компонента в экстрагируе-
мой жидкости, Со —концентрация компонента в экстракте. Коэф-
336
фициент распределения — величина, определяемая опытным пу-
тем, показывает во сколько раз равновесная концентрация веще-
ства в экстракте больше, чем в обработанном растворе. Например,
в случае экстракции пенициллина из нативного раствора бутила-
цетатом при температуре 0°С и pH 2,5 коэффициент распределения
К=35. Это значит, что при остаточном содержании пенициллина
в нативном растворе 200 ЕД/мл, равновесная активность бутила-
цетатного экстракта составит 7000 ЕД/мл. Это позволяет одновре-
менно и концентрировать целевой продукт и очищать его от
сопутствующих примесей.
Классическим примером применения экстракционного метода
в системе "жидкость-жидкость" для выделения й очистки продук-
тов микробного синеза является схема получения пенициллина,
представленная на рис. 96.
Нативный раствор, содержащий бензилпенициллин, из сборни-
ка (1) поступает на первую экстракцию бутилацетатом. Экстракция
—массообменный процесс, и он протекает тем быстрее, чем ин-
тенсивнее входят в соприкосновение друг с другом несмешиваю-
щиеся жидкости. Поэтому очень важно провести эмульгирование
(3), нб при этом эмульсия должна хорошо разделятьсяв сепараторе
(6). В связи с тем, что нативный раствор содержит большое
количество поверхностно-активных веществ белковой природы, в
процессе экстракции образуются весьма стойкие, трудноразделя-
емые эмульсии. Это требует применения специальных дезэмуль-
гаторов, например, поверхностно-активное вещество — авироль.
Действие'авироля основано на том, что он вытесняет белковые
вещества из межфазовой поверхности, образуя пленку на границе
раздела фаз (между Водой и бутилацетатом). Пленки, образованные-
ПАВ, обладают незначительной прочностью по сравнению с плен-
ками из белка, поэтому эмульсии легко разрушаются под влиянием
центробежных сил, развиваемых в сепараторах. Для разделения
эмульсий достаточно добавить к нативному раствору 0,05—0,1%
ПАВ.
Экстракцию проводят при pH 2,8±3,0 и температуре 10°С.
Подкислением нативного раствора серной кислотой (2) подавляют
диссоциацию карбоксильной группы молекулы пенициллина, в
недиссоциированном состоянии он переходит н органическую
фазу (бутилацетат). Высокая селективность экстракции достигает-
ся благодаря тому, что в области pH 2,8± 3,0 более Сильные кислоты,
чем пенициллин, находятся в диссоциированном состоянии и
поэтому не растворяются в бутилацетате; в водной фазе остаются
337 • '
минеральные компоненты (загрязнения), большая часть азотистых
соединений и других органических веществ, так что в результате
экстракции чистота продукта увеличивается в 4—5 раз.
Первый бутилацетатный экстракт пенициллина промывают
обессоленной водой в эмульгаторе (7) и сепараторе (9), затем
подают на бикарбонатную экстракцию-двухступенчатую. На пер-
вой ступени экстракцию проводят в эмульгаторе (11) и экстракторе
— сепараторе (12) обедненным бикарбонатным экстрактом со
второй ступени бикарбонатной экстракции. Обедненный бикар-
бонатный экстракт пенициллина поступает на новую ступень
бикарбонатной экстракции в эмульгатор (13) и экстрактор-сепара-
тор (14). Экстракцию бикарбонатом натрия проводят при pH
7,3-5-7,8. При этих условиях пенициллин находится в диссоцииро-
ванном состоянии и поэтому хорошо растворяется в водной фазе,
но не растворяется в органической фазе. На данной стадии удается
освободиться от примесей, являющихся более слабыми кислотами,
чем бензилпенициллин, и сконцентрировать раствор в 2 раза.
Из бикарбонатного экстракта антибиотик вторично экстраги-
руют бутилацетатом при pH 2,2 - 2,5 в эмульгаторах (15,16) и
экстракторе-сепараторе (17).
Второй бутилацетатный экстракт бензилпенициллина поступает
на осветление углем и на вымораживание (19). С помощью угля
освобождаются от оставшихся пигментов. При вымораживании
(—5°С) вместе с замерзающей водой удаляются сульфаты.
Освобождение от угля и льда осуществляют с помощью филь-
трации (20), затем второй бутилацетатный экстракт поступает в
реактор (21) для получения калиевой соли бензилпенициллина. В
реактор добавляют насыщенный раствор калия уксуснокислого и
раствор калия углекислого (донор ионов калия).
S S-
C,H.-CH,-CO-NH-CH-CH C(CHJ,
6 5 2 I | I 32 + CH3COOK -»
ОС---N-----CH-COOK
—> C.H,-CH,-CO-NH-CH-CH C(CH,),
6 5 2 || I 2 + CH3COOH
ОС N------CH-COOK
2CH3COOH+K2CO^2CH3COOK +H2O+CO2
338
Образовавшаяся калиевая соль бензилпенициллина переходит
снова в водную фазу. Водный слой и бутилацетат разделяют при
отстаивании (контроль за разделением проводят визуально через
смотровое стекло, установленное на трубопроводе).
Водный концентрат калиевой соли бензилпенициллина после
фильтрации (24) передают на вакуум-упаривание (26) и кристал-
лизацию. Этот процесс является процессом кристаллизации со
сменой растворителя. Калиевая соль бензилпенициллина кристал-
лизуется в бутаноле после выпаривания воды. В случае проведения
процесса при пониженной температуре к водному раствору добав-
ляют бутанол. Образовавшуюся азеотропную смесь "бутанол-вода"
отгоняют при 20°С. Кристаллы отфильтровывают от маточника (28),
промывают безводным бутанолом и высушивают.
Выделение и очистка канамицина из нативных растворов ос-
нована на ионообменных методах. Канамицин — антибиотик
основного характера, его выделяют из нативных растворов сорб-
ционным методом. Для удаления ионов кальция культуральную
жидкость обрабатывают щавелевой кислотой. После отделения
осадка нативный раствор . (pH 6,8+7,2), содержащий канамицин
поступает на ионообменные колонны (рис. 109). При этих условиях
канамицин представляет собой шестивалентный катион.
Антибиотик сорбируется на карбоксильном катионите КБ-2 в
натриевой форме, так как активные центры этого сорбента заняты
ионом натрия. Процесс сорбции идет по схеме:
6RCOONa+KaH.+6 -+ (RCOO)6 Кан.+ 6Na+
Нативный раствор канамицина проходит через батарею после-
довательно соединенных колонн (5—7). Раствор подают снизу, так
как сорбция протекает ₽о взвешенном слое сорбента. Здесь важ-
ным является выбор рабочей скорости (Wpa6.), которая должна
быть больше критической скорости (Wkpht.), определяющей про-
цесс псевдоожижения, и меньше скорости уноса (WyH-) — скорость,
при которой сорбент уносится с раствором из колонны,
Сорбцию ведут до тех пор, пока в отработанном растворе не
останется порядка 5—6% концентрации антибиотика от исходной.
Каждую колонну в батарее отключают после полного насыще-
ния, промывают обессоленной водой (1) и подвергают десорбции.
Десорбцию канамицина проводят 1N раствором аммиака (2) или
элюатом третьей фракции (10), укрепленным 25% раствором ам-
миака. Процесс проводят в соответствии с реакцией (при подаче
раствора сверху):
339
Рис. 109-Принципиальная схема получения канамйциИЧ: 1 сббрник обессо-
ленной воды, 2—сборник Гн. р-ра NH4OH , 3 — сборник 1 н. р-наHCL, 4 —
сборник 1 н. р-на NaOH, 5, 6, 7 — колонны с катионитом КБ-2, 8, 9, 10 — сборник
элюата, I, II и III фракций соответственно, 11,12 — кОлонны-с анионитом АВ-1-7-2П.
13.— сборник осветленного элюата, 14'— вЗгкуум-выйарной аппарат, 15 — реактор
для подкисления и обработки углем, 16,21 — фильтры, 17 — сборник осветленного
концентрата, 18, 19 — колонны с катионитом КУ-2-"20, 20 — реактор для подщела-
чивания,-22 — аппарат для кристаллизации, 23 •— центрифуга.
(RCOO)6 Кан + 6NH4-> 6RCOONH4 + Кан+6
ж - . ’ •
Ионы NH4 в растворе аммиака не вытесняют катионы со
смолы, поэтому элюаты содержат незначительные количества при-
месей минеральных солей. При этом образуются 3 фракции, из
которых первая с небольшим содержанием канамицина (8) посту-
пает на очистку стоков; вторую фракцию — основную собирают
в сборнике (9); третью фракцию из сборника (10) возвращают для
десорбции канамицина с сорбента.
Сорбент КБ-2 в колонне после трехкратного использования
подвергают регенерации, которую последовательно проводят 1N
340
раствором HCL (3), обессоленной водой, IN раствором NaOH(4) и
повторно — обессоленной водой. Все отработанные растворы
сливают в специальную "кислотно-щелочную канализацию".
Таким образом, на каждой колонне с сорбентом КБ-2 много-
кратно повторяются процессы сорбции, десорбции, регенерации.
Дальнейшей очистке подвергают вторую фракцию элюата кана-
мицина. Эта фракция поступает на осветление на колонны с
анионитом АВ—-17—2П в ОН" форме (И, 12). Данный анионит
избирательно поглощает пигментные вещества и не сорбирует
антибиотик. Раствор подают сверху; он проходит через две после-
довательно соединенные колонны. Осветленный раствор канами-
цина собирают в сборнике (13), а сорбент регенерируют последо-
вательным пропусканием растворов -HCL, обессоленной воды,
NaOH и вновь — обессоленной воды. ИЗ сборника (13) раствор
подают для концентрирования в вакуум-выпарнЬй апйарйт (14),
представляющий собой вертикальный кожухотрубчатый испари-
тель пленочного типа. Испарение протекает При 80°С под вакуумом.
Концентрат направляют в реактор (15) для обработки углем. Про-
цесс осуществляют при pH 4,2—5,2. В качестве подкисляющего
агента используют 25% раствор H2SO4. После фильтрации (16)
прозрачный, осветленный концентрат канамицина (17) поступает
. на обеззоливание, которое проводят на колоннах с катионитом
КУ-2-20 в Н+ форме. ’
После пропускания раствора канамицина сорбент регенериру--
ют с- помощью IN .раствора HCL g последующей промывкой
обессоленной водой. - - ,
Обессоленный концентрат канамицина (20) После фильтрации
(21) поступает на кристаллизацию (22). Кристаллизацию моносуль-
фата канамицина проводят при добавлении двойного объема 98%
метанола при pH 9,3 — 9,7. Кристаллы отделяют на центрифуге (23)
и передают' на высушивание. * ' \
7.5. Аппаратурное оснащение фитобиотехнологйческих про-
изводств. Особенностями растительных клеток . являются; их
склонность к объединению в тканевые' структуры, которые .в
целостном организме "омываются" жидкостями определенного
состава, содержащими многие регуляторные вефества, оказываю-
щие заметное влияние на функцию тканей; малые скорости роста
и развития растительных клеток, а некоторые из них вообще не
культивируются в глубинных условиях на жидких питательных
средах; хемогетеротрофизм (а не фотоавтотрофизм) растущих
клеток в культуре; синтез целевого продукта лишь агломератами
341
клеток в одних случаях (не отдельными клетками) — здесь сказы-
вается позитивная роль дифференциации в клеточных агломера-
тах, и, напротив, недифференцирован-
ное состояние клеток в культуре может
быть более предпочтительным — в дру-
гих случаях.
В зависимости от вида и потребно-
стей клеток их культивируют в разных
биореакторах (с механическим переме-
шиванием,. колонного типа с переме-
шиванием (рис. 110)). Фирма "Sigma"
(США) выпускает мембранные наборы
(Rafts) для растительных культур тка-
ней, что помогает исключить ингибито-
ры, нередко присутствующие в агар-
агаре и других желирующих агентах.
Мембранные наборы (Rafts) предназна-
чены для выращивания протопласти-
рованных и других культур, которые
могут поддерживаться длительное
время без смены культурального кон-
тейнера.
Мембранные наборы (Rafts) име-
ют квадратную (60x60 мм и др.) или
круглую (73 мм в диаметре и др.)
форму. Их изготавливают из поли-
пропилена с размером пор, в среднем,
0,075x0,25 мкм (площадь всех пор за-
нимает порядка 45% мембраны), тол-
Рис. 111. Мембрана, используе-
мая при выращивании раститель-
ных клеток, х 10000.
щиной 25 мкм; они пригодны для про-
тока воды 288 л/м2/час. Мембраны ус-
тойчивы к действию сильных кислот,
оснований и к органическим раствори-
телям (за исключением галогенизиро-
ванных углеводородов). На рис.111,112
представлены мембрана и мембранный
контейнер.
Та же фирма "Sigma" предлагает так
называемый фитатрей (от англ, tray —
поднос) для растительных клеточных
культур (рис. ИЗ); размеры его
342
Рис. 110. Биореакторы для вы-
ращивания растительных клеток:
а — ферментатор с подачей сте-
рильного воздуха снизу, б — фер-
ментатор с продувкой стериль-
ным воздухом и отводной трубой.
Рис. 112. Мембранный кон-
тейнер для выращивания расти-
тельных культур тканей.
92x114x89 мм. Каждый фитатрей упако-
Звывают в ящики по 100 кювет и крышек
Дна ящик. Составные части фитатрея сте-
Дрилизуют гамма-облучением, после чего
они готовы к применению по назначе-
Днию.
Для культивирования растительных
jar клеток разработаны специальные био-
реакторы различной емкости. Принци-
пиальная схема одного из них показана
на рис. 85. Здесь также можно исполь-
Рис. из. Фитатреи. зовать ферментатор Био-Фло III (см. раз-
дел 7.1.).
7.6. Аппаратурное оснащение зообиотехнологических произ-
водств. К зообиотехнологическим относятся производства: вакцин,
моноклональных антител, лимфокинов (медиаторов иммунокомпе-
тентных клеток), других белков (в том числе — ферментов),
гормонов и пр. Ряд биологически активных веществ образуется
эукариотическими клетками лишь в результате посттрансляцион-
ной модификации. К этому неспособны прокариотические клетки.
Клетки млекопитающих не имеют клеточной стенки и их
протоплазматическое содержимое защищено лишь тонкой клеточ-
ной мембраной. Они всегда представляют собой часть какой-то
специализированной ткани и, следовательно, зависят от жизнеде-
ятельности других клеток. Поэтому существование животных кле-
ток в изолированном состоянии весьма ограничено и, порой,
исключительно трудно воспроизводимо в ла-
бораторных и, тем более, в производствен-
ных условиях. К тому же и растут они срав-
нительно медленно (см. рис. 89). Тем не
менее, успехи в зообиотехнологии весьма
ощутимы и этому способствовали приемле-
мые методы культивирования их в подходя-
щих условиях (рис. 114).
Выращивание животных клеток условно
можно подразделить на следующие этапы: I
— механическая и ферментативная дезин-
теграция ткани животного до получения сме-
си клеток или их агломератов;
II — перенос клеток или их агломератов
в питательную среду во флаконах;
343
Рис. 114. Раковые клет-
ки человека (HeLa) в пла-
стиковом культуральном
сосуде (увеличение х40).
Ill — отделение клеток от дна флакона с помощью фермента и
перенос их в другой флакон со свежей питательной средой для
размножения;
IV — перенос клеток в биореактор и накопление целевого
продукта;
V — выделение целевого продукта.
Первый этап исключают в случае использования клеток крови,
лимфы, опухолей и некоторых других трансформированных кле-
ток, поскольку они растут в виде суспензионных культур. Все
другие животные клетки прикрепляются к какой-то поверхности.
Поэтому ткани, из которых получают такие клетки, дезинтегриру-
ют в тканевых ^гомогенизаторах и обрабатывают протеазой, цент-
рифугируют при небольших скоростях ротора во избежание трав-
мирования клеток.
На втором и третьем этапах осуществляют выращивание и
наращивание клеток во флаконах или в горизонтально располо-
женных и медленно вращающихся (порядка одного оборота в мин.)
бутылях. Самые большие бутыли имеют пло-
щадь около 1600 см-2. Отделение клеток от
доньев флаконов после П-го этапа также
осуществляют с помощью протеолитиче-
ских ферментов.
Используемые при этом питательные
среды весьма сложные и дорогие (см. спе-
циальную часть книги), обычно содержащие
антибиотик и сыворотку животных (от 5 до
20%). К сожалению, сыворотки могут высту-
пать источником заражения (загрязнения)
культуры животных клеток бактериями, ви-
русами, микоплазмами, или какими-либо
ингибиторами. Поэтому синтетические сре-
ды представляются более удобными и вы-
годными с экономической точки зрения.
На IV этапе клетки переносят в биоре-
актор большого объема. Биореакторы для
культур животных клеток считают больши-
ми, начиная с объема 10 литров, а если
имеются 100—200-литровые биореакторы,
то это исключительно большие аппараты.
Они отчасти напоминают биореакторы,
применяемые в промышленной микробио-
344
Рис. 115. Мешалки в би-
ореакторах для выращива-
ния клеток: микроорганиз-
мов (1 — турбинная) и жи-
вотных- (2 — пропеллер-
ная, 3 — вибромешалка).
логии, так как в обоих случаях принципиальные подходы к выра-
щиванию клеток во многом сходны (хотя животные клетки нежнее
Рис. 116. Роллерный био-
реактор: 1 — вращающийся
цилиндр, 2 — двигатель, 3 —
вход и 4 — выход газа-возду-
ха.
и во много раз крупнее бактериальных или многих других микро-
бных клеток).
Чтобы меньше повреждать клетки,
турбинные мешалки заменяют на пропел-
лерные (винтовые) или на вибромешалки
(рис. 115). При использовании мешалок
среда перемешивается, а биореактор ос-
тается неподвижным. В других случаях
среда остается неподвижной или мало по-
движной, а перемещению подвергается
биореактор (рис. 116).
Увеличение отношения площади по-
верхности биореактора (S) к его объему (V) (во благо возрастания
объемной плотности растущих адгезированных клеток) возможно
либо за счет увеличения числа реакторов, или за счет его конст-
рукционных модификаций. Последнее оказалось предпочтитель-
нее со всех точек зрения. Были предложены способы выращивания
клеток на гранулированных микроносителях, на губчатых полиме-
рах, на стопках тонких пластинок, на тонких нитях и пористых
трубочках. В подобных реакторах удается в 10 и более раз повысить
плотность клеток на 1 см2 поверхности (рис. 117).
Впервые культивирование клеток животных на микроносите-
лях осуществил А. Ван-Везель (Голландия) в 1967 г. Гранулирован-
ные микроносители изготавливают из альгината, декстрана, син-
тетических полимеров размером, в среднем, 50—200 мкм в диамет-
ре. Клетки прикрепляются к поверхности гранул и образующаяся
Рис. 117. Проточный реактор с подушкой из пластмассовых трубочек для роста
животных клеток (а); клетки HeLa, растущие на полой нити; сканирующий элект-
ронный микроскоп; ув. х250; (б).
345
система представляет собой монослойную и суспензионную куль-
туру одновременно. В проточном реакторе с мешалкой, несущей
гибкие лопасти (перемешивание по типу движения рыбьего хвоста)
удается создавать высокую концентрацию гранул (12 г/л) без
заметного повреждения растущих на них клеток.
В компании Monsanto (США) созданы биореакторы для массо-
вого культивирования клеток млекопитающих, в том числе —
проточные. Впервые проточная система была разработана Ф.
Химмельфарбом и Ф. Тейером в 1969 г. Она и была положена в
основу биореактора, предложенного Дж. Фидером и У. Р. Толбер-
том в начале 80-х годов (рис. 118). В таком биореакторе осущест-
вляется комбинация выращивания культур с высокой плотностью
клеток и переноса их в реактор большего объема. Здесь демонст-
Рис. 118. Проточный реактор с гибкими лопастными мешалками (по Дж. Фидеру
и У. Толберту, 1983).
ративен следующий пример: в 4-литровом проточном реакторе
получено 40 млрд, фибробластов человека (если их выращивать в
роллерных сосудах); после отделения фибробластов от гранул
микроносителя и переноса их в 44-литровый биореактор с 400 г
новых гранул с общей площадью их поверхности 188 м2 удается
получить 340 млрд, клеток, то есть в 8,5 раза больше (для накопления
такого числа клеток в роллерных сосудах потребовалось бы 11000
вращающихся бутылей).
Известны и другие типы биореакторов, принципы работы
которых во многом сходны. К их числу можно отнести биореактор
CellGen, предложенный фирмой New Brunswick Scientific Со., Inc.
(США). Его особенности: 1) многоцелевое назначение (для суспен-
зионного культивирования, в том числе, на микроносителях, для
проточного культивирования); 2) при малых скоростях перемеши-
346
вания достигается эффективная массопе-
редача О2; 3) надежность контроля темпе-
ратуры с исключением возможности ло-
кального перегрева; 4) обеспечение на-
дежного контроля за pH и растворенным
кислородом (рис. 119); 5) взаимозаменяе-
мость реакторов емкостью 1,5; 2,5 и 5,0
литров; 6) оснащенность биореактора мик-
ропроцессором. В таком биореакторе за-
метно повышается выход конечных про-
дуктов, а жизнеспособность клеток пре-
вышает 90%. Применительно к гибридом-
ным клеткам показана возможность их
выращивания в так называемой проточ-
ной или перфузионной системе, состоя-
щей из биореактора и фильтрационного
устройства с тангенциальным потоком
жидкости (рис. 120). Клеточную культуру удается поддерживать до
1,5 месяцев и более (с числовым показателем, достигающим 1,8 • 107
клеток/мл и максимальной концентрацией моноклональных анти-
тел до 800 мкг/мл, обеспечивая их продуктивность 504 мг/день).
В целях реализации крупномасштабного культивирования кле-
ток млекопитающих предложена так называемая система Micro-
Lift, функционирующая совмещенно с многоканальным процессо-
ром ML-4100.
Рис. 120. Схема включения биореактора Cell Gen в проточную систему с
тангенциальным потоком культуральной среды (1 — свежеприготовленная среда, 2
— контроль уровня, 3 — культуральная система Cell Gen, 4 — перфузионная ячейка
с тангенциальным потоком жидкости, 5 — культуральная среда, 6 — клетки, 7 —
освобожденная от клеток среда, содержащая МкАТ, 8 — насос.
347
Во всех случаях культивйрования животных клеток в больших
реакторах технологический процесс ведут- ступенчато, начиная с
сосудов малого объема:
поверхностная культура
L чистая культура клеток
в жидком азоте
2. культура во флаконах
(3 • 105 клеток 7см 2)
3. культура в сосудах
(8 • 10° клеток/см2)
4. культура в роллерных
сосудах (5 • 109 клеток/см2)
5. культура в аппаратах
с пластинами
(2 • 10'°клеток/см2)
6. культура в аппарате
суспензионная культура
1. чистая культура клеток
в жидком азоте
2. культура во флаконах
/(5—10) • 10е клеток/мл/
3. культура в роллерных сосудах
(7 • 109 клеток/см2)
4. культура в биореакторе
на 20 л (7,5 • 10" клеток/см2)
5. культура в биореакторе
на 500 л
(3,7 • 1012 клеток/см2)
с микроносителем
(3,7 • 10" /см2).
Из-за повышенной чувствительности животных клеток к раз-
личным внешним воздействиям важно исключить не только по-
вреждающее действие перемешивающих систем в биореакторах,
но и заметные колебания температуры, поскольку ниже 37°С
клетки растут очень медленно, а выше 37°С они теряют жизнеспо-
собность Температурный контроль при этом осложняется тем, что
скорость перемешивания среды с микроносителем малая и, следо-
вательно, эффективность теплопередачи заметно снижается. Од-
нако в отличие от микробных ферментаций, здесь нет проблемы
с отводом тепла, образующегося в процессе метаболизма клеточной
культуры. Определено, что теплоемкость должна быть
712*103 Дж/час/л культуры, содержащей 109 клеток.
К настоящему времени сделаны базисные разработки для
выращивания клеток млекопитающих и есть все основания наде-
яться на то, что аппаратурное оформление биотехнологических
процессов будет постоянно совершенствоваться. Как пример ска-
занному можно назвать геновый ассемблер, с помощью которого
синтезируют олигонуклеотидные последовательности ДНК длиной
около 200 оснований. Олигонуклеотиды "растут" на твердых бу-
синках, помещенных в специальные кассеты, имеющие цветовой
код (A/О Фармация, АКБ-приборы, Швеция). С применением
уникальной кассетной системы (объемом 0, 2, 1, 3 или 10 микро-
молей) исключается необходимость взвешивания матрицы перед
348
ее применением, равно как и работа с незакрепленными бусинка-
ми.
Перед началом синтеза соответствующую кассету вставляют в
один из реакторов колонки, а после окончания синтеза ее выни-
мают й выделяют олигонуклеотиды. При этом колон очный .реактор
пригоден для повторного использования. Работа оператора состоит
лишь в передаче геновому ассемблеру желаемой информации о
синтезе олигонуклеотида с заданной последовательностью. Подо-
бные приборы обеспечивают потребности в генно-инженерных
разработках для биотехнологии в целом.
Глава 8.
ОТХОДЫ БИОТЕХНОЛОГИЧЕСКИХ ПРОИЗВОДСТВ,
ИХ ОБЕЗВРЕЖИВАНИЕ И УТИЛИЗАЦИЯ
Общая принципиальная схема любого биотехнологического
производства включает какой-либо биообъект (или ассоциацию
их) и питательную среду (культуральную жидкость, растворы,
подлежащие обработке). Целевым продуктом оказывается либо
биомасса клеток (тканей), либо метаболит(-ы). Следовательно, в
каждом производстве отходом могут быть эти же компоненты —
клетки (ткани) и культуральные жидкости после извлечения из них
нужных метаболитов (рис. 121).
Рис. 121. Биотехнологический процесс, в
котором клетки и супернатант являются ли-
бо целевыми продуктами, либо отходами: 1
— биообъект, 2 — питательная среда, 3 —
культивирование в соответствующих усло-
виях, 4 — диоксид углерода, отработанный
воздух, 5 — сепарирование клеток от куль-
туральной среды, 6 — клетки, 6а — отход,
66 — целевой продукт, 7 — супернатант, 7а
— целевой продукт, 76 — отход.
Если условно принять скорость удвоения каждой генерации,
например, микробной клетки весом 4х1О'10 мг, в течение 20 минут,
то за двое суток возникло бы 2144 клеток или, примерно, 8 • 10131 г,
или 8 • 10,и т. Такая величина примерно в несколько тысяч раз
превышает вес Земного шара. Ясно, что в процессах биологической
технологии на размножение биообъектов влияют многие факторы,
ограничивающие такой безудержный прирост биомассы клеток.
349
Однако в случаях с производством пенициллина расчеты подтвер-
ждают возможность получения 2 т сухого мицелия через ряд
генераций пеницилла в течение 10—12 дней выращивания при
первоначальном засеве 10‘5 г спор гриба с доведением объема
влажной культуры до 100 м 3.
Даже при использовании биомолекул (например, ферментов)
в иммобилизованном состоянии своеобразная двухкомпонент-
ность системы сохраняется и здесь. Метаболит(-ы) будут в раство-
ре, носитель с биомолекулами остается в твердом состоянии.
При наличии крупномасштабных биотехнологических произ-
водств возникают проблемы общего и частного характера. К ним
можно отнести: 1) необходимость решения задач по экологическо-
му выравниванию нагрузок, оказываемых производством на окру-
жающую среду: а) вследствие непомерного потребления природ-
ной воды и столь же непомерного количества выбросов во внеш-
нюю среду; только в промышленности расходуется примерно
следующее количество воды (в литрах): на 1 т нефти — 10, на одну
банку овощных консервов — 40, на 1 кг бумаги — 100, на 1 кг
шерстяной ткани — 600, на 1 т сухого цемента — 3500, на 1 т стали
— 20000, на 1 т сухих дрожжей — более 100000 литров воды и
более 10 т пара; в США ежегодно генерируется более 145 млн тонн
городских сточных вод (данные на 1990 год), из них порядка 94
млн. т (65%) легко очищаемых биодеградацией; 23,2 млн. т (16%) —
потенциально биодеградируемых и 27,6 млн. т (19%) — не подда-
ющихся очистке с помощью биологических процессов (стекло,
металлы, пр.); б) вследствие подавления и, даже, гибели естествен-
ных экосистем вокруг биотехнологических предприятий или вы-
ражение неадекватное популяционное давление одних видов жи-
вых существ на другие (например, разрастание цианобактерий в
водохранилищах); в) вследствие стрессовых нагрузок на людей,
проживающих вблизи крупных биотехнологических предприятий
(выхлопные газы, шум, испарения, корпускулярные аллергены в
атмосфере и пр.).
2) необходимость сохранения воды, памятуя о том, что ее гибель
— наша гибель! К сожалению, даже океаны превращаются в
крупнейшие сточные резервуары Земли. Для сравнения можно
назвать прежнее состояние вод и теперь в озерах Ладожском,
Байкале, в крупнейших реках нашей страны; сбросы с рыболовных
кораблей, загрязненные кишечной палочкой, достигают районов
Антарктиды;
3) борьбу с мусором и грязью, не забывая о .том, что при их
350
наличии восстанавливаются прерванные цепочки: мусор
(грязь)—>переносчики (крысы, мухи и др.) -^инфекционные болез-
ни (чума, кишечные заболевания и др.);
4) борьбу с загрязнением воздуха, в который от предприятий
попадает тепло (равно как и в водоемы), жидкие, газообразные,
твердые (пыль) отходы — изменяется местный климат в селитеб-
ных (от слова "селитьба" — поселение, водворение) зонах, а
накопление СО? в атмосфере от биотехнологических производств
вносит свой вклад в "тепловую копилку" в глобальном масштабе;
5) регулирование народонаселения в различных регионах пла-
неты Земля. Биотехнологические производства прямо или косвен-
но нацелены на обеспечение здоровья людей. Тем не менее, ни с
помощью биотехнологии, ни с помощью сельскохозяйственных
наук до сих пор не удалось полностью решить проблему недоста-
точности питания (или даже голода) во многих странах мира. В то
же время на Земле каждые две секунды рождается человек и через
20 лет ожидается возрастание народонаселения до 6 млрд, человек.
Следовательно, эффективность фитобиотехнологических произ-
водств, включая другие предприятия агропромышленных комплек-
сов, должна существенно возрасти. При этом уже невозможно
увеличить нагрузку на почву пестицидами в целях борьбы за
урожайность сельскохозяйственных культур. По общему обороту
химической промышленности все государства бывшего СССР
занимали второе место после США. В настоящее время в мире
вырабатывается около 300 млн. т органических веществ, в том
числе 1,2 млн. т. пестицидов, которые вместе с другими реактоген-
ными химическими веществами, попадающими в среду обитания
человека, могут проявлять канцерогенное, мутагенное и токсиче-
ское действие.
Так называемые "кислотные дожди", выпадающие в различных
географических регионах, пагубно сказываются на всем живом,
включая и "микробный пейзаж".
Качество и количество отходов биотехнологических произ-
водств зависит от ряда причин, среди которых можно назвать:
характер производства по номенклатуре выпускаемой продукции
(например, производство микробного белка, антибиотиков, вита-
минов, аминокислот, полисахаридов, ферментов и др.); особенно-
сти технологии производства — аэробное или анаэробное культи-
вирование биообъекта, в герметизированных или негерметизиро-
ванных биореакторах, в периодическом, полунепрерывном или
непрерывном режимах; объемы производства — малотоннажные
351
(некоторые ферменты) и крупнотонажные (кормовые дрожжи);
уровень профессиональной подготовки кадров, занятых в биотех-
нологических производствах; культуру производства.
К сожалению, нет информации о количестве отходов всех
биопроизводств в мире, но, несомненно, они ошеломляюще огром-
ны. Подсчитано, например, что на одну тонну лимонной кислоты
образуется 150—200 кг сухого мицелия Aspergillus niger и 7 м3
фильтрата. При выработке в мире порядка 30000 тонн в год
антибиотиков плотные и жидкие отходы составляют исключитель-
но большие величины. Так, с одного пятидесятикубового аппарата
при производстве пенициллина может быть получено около 1 т
мицелия в расчете на сухую массу. Применительно к комплексной
биотехнологии (микробной на базе зоопредприятий), в связи с
микробной переработкой стоков свинооткормочных комплексов
показано, что каждый животноводческий комплекс (54—108—216
тыс. голов скота) ежегодно производит соответственно 0,5—1—2
млн. м3 навозосодержащих стоков. Из 1,5 млрд, т отходов живо-
тноводства, получаемых каждый год в нашей стране, большая часть
попадает в открытые водоемы, загрязняя окружающую среду (И. А.
Архипченко, 1991). Вот почему бесспорным является утверждение
о том, что от состояния окружающей среды во многом зависит
выживание человечества. Наука о биологии окружающей среды
называется экологией (от греч. oikos — дом, жилище, logos —
учение). Она посвящается изучению биологических систем надор-
ганизменного уровня; другими словами, экология — это совокуп-
ность или структура связей между организмами и их средой. Ее
подразделяют на аутоэкологию и синэкологию. Первая изучает
отдельные виды или организмы и их среду обитания, вторая —
группы организмов в ассоциациях, составляющих определенные
единства во внешней среде.
При общей взаимосвязи всего живого на земле биотехнолог
обязан обеспечивать безотходность биотехнологических произ-
водств во имя поддержания нормальной экологической ситуации
(в месте производства и на расстоянии). Неограниченное истоще-
ние ресурсов земли и ее непрекращающееся загрязнение могут
привести человечество к трагическому концу. Поэтому в любом
производстве, в том числе — биотехнологическом, должны исполь-
зоваться экосистемные (целостные) подходы, девиз которых: "По-
лучай, бери, но не вреди Природе, частью которой ты являешься!".
Неоправданно большие количества отходов в производстве —
это значит большая потеря ресурсов, так как отходы нередко
352
Рис. 122. Примерные рас-
ходы на обработку различных
отходов (1 — сточных вод, 2
— твердых отходов, 3 — отхо-
дов от автотранспорта, 4 —
отходов от всех видов про-
мышленности, 5 — отходов от
производства электроэнер-
гии); $ — затраты в млн. дол-
ларов, t — время в годах.
представляют собой ресурсы, оказавшиеся не на своем месте; это
значит возрастание стоимости ликвидации отходов и.контроля за
ними; это значит ухудшение состояния здоровья людей (особенно
— в крупных промышленных центрах).
На рис. 122 представлены ориентиро-
вочные расходы на отработку различных
отходов к 2000 году. Из рисунка видно,
что в структуре расходов наибольшие
затраты ожидаются на обезвреживание
отходов от автотранспорта; от 600 млн.
до 1 млрд, долларов будут расходоваться
на обработку всех прочих отходов, вклю-
чая жидкие и твердые от биотехнологи-
ческих производств.
Что касается инфекционных заболе-
ваний, связанных с экологической обста-
новкой, то их распределение неравноз-
начно в различных регионах земного
шара и, даже, на территории одной стра-
ны. Если общая тенденция к снижению
туберкулеза и кишечных заболеваний
прослеживается в странах Западной Ев-
ропы, Северной Америки и некоторых других, то применительно
к независимым государствам бывшего СССР этого сказать нельзя
— ежегодные вспышки сальмонеллезов, дизентерии, диарреи ста-
новятся все более беспокойнее на фоне острой нехватки химио-
терапевтических средств; растет заболеваемость туберкулезом ма-
териально плохо обеспеченных слоев населения.
8.1. Обезвреживание отходов биотехнологических произ-
водств. Отходы биотехнологических производств относятся, как
правило, к типу разлагающихся в природных условиях под дейст-
вием различных факторов (биологических — минерализация с
участием микроорганизмов, химических — окисление, физико-хи-
мических благодаря комплексному воздействию, например, лучи-
стой энергии и химических веществ).
Плотные отходыв биотехнологических производствах
представляют собой: микробную массу, отделяемую от культураль-
ного фильтрата, поступающего на последующие стадии выделения
целевого продукта; шламы (от нем. Schlamm — грязь); раститель-
ную биомассу после экстракции из нее действующих веществ (а
в случае суспензионной культуры, продуцирующей метаболит в
12 т. 8524 353
питательную среду, отходом являются клетки); остатки куриных
эмбрионов при культивировании, например, вируса гриппа; неко-
торые тканевые культуры млекопитающих; осадки из сточных вод
(ил). Подсчитано, что в коммунальных очистных сооружениях
сточные воды от одного горожанина образуют за год около 500
литров ила со средней влажностью 5%. Если городское население
в стране составляет 100 млн. человек, то за год накопится 47,5 млн.
м3 такого ила. Если сюда приплюсовать почти такое же количество
промышленных осадков, включая плотные отходы биотехнологи-
ческих производств, то необходимо приложить большие усилия и
средства для обезвреживания их или утилизации.
Давно освоенными биотехнологическими производствами во
многих странах мира являются промышленные способы получения
пива, дрожжей, вин и др. (см. специальную часть). На примере
лишь пивоварения можно указать, что плотными отходами здесь
являются дрожжевые клетки (0,25—0,40 кг на 1 гл. пива), солодовая
и хмелевая дробины, белковый осадок из сепараторов. Остатки
хмеля (хмелевая дробина) и белка содержат горечи, из-за которых
они не употребляются в качестве добавок к рационам кормов для
животных. Поэтому такие остатки либо сжигаются (что нерента-
бельно), либо передаются на биологическое обезвреживание.
При оптимальных средах и аэрации биомасса клеток нитчатых
грибов и дрожжей может составить 2,5% в пересчете на сухую
массу, причем, около 50% в ней приходится на белки.
В спиртовом производстве отходом является барда, состав
которой зависит от качества используемого сырья (зерно, карто-
фель). Сугубо усредненные данные по основному составу зерно-
картофельной барды представляются следующими: вода — 91—
93%, сухой остаток — 7—9%, в составе которого зольность состав-
ляет от 6 до 12%, общий азот 21—23%, липиды — 2—8%, целлюлоза
— 9—10%, безазотистые экстрактивные вещества — 50—59%.
Отжатая или высушенная барда используется в качестве добавок
к корму для сельскохозяйственных животных.
В производстве этанола, пива, хлебного кваса используют солод
— пророщенное зерно (ячмень, овес, просо, пшеница, рожь — на
спиртовых заводах; ячмень — на пивоваренных заводах; рожь и
ячмень — в производстве кваса). В процессах получения солода
образуются отходы в форме очисток, сплава, солодовых ростков,
которые с успехом используются в животноводстве, а также в
целях получения ряда биологически активных веществ (прежде
всего — ферментов из солодовых ростков).
354
Качество плотных отходов в определенной мере диктует выбор-
метода их обеззараживания. Так, патогенные микробы — проду-
центы сильных ядов (токсинов) должны быть обезврежены полно-
стью,. и, очевидно, наиболее эффективный способ для этого —
сжигание. Если отходом является биомасса клеток стрептомицетов,
то их достаточно убить нагреванием с последующим вывозом на
фермы, где она может добавляться в корм скоту (например,
уплотненный отход в производстве тетрациклиновых антибиоти-
ков, содержащий белки и витамин Bi?), вноситься в почву в
качестве органического удобрения; можно передавать на общего-
родские очистные сооружения, а также на метановое брожение.
Если по технологической схеме плотные и жидкие отходы
подаются в виде смешанного стока, то вначале осуществляют
грубое разделение первых от вторых, затем производят отжим
влаги с последующей передачей уплотненной биомассы клеток на
обезвреживание вышеуказанными путями.
Аналогичным образом подходят к плотным отходам раститель-
ного или животного происхождения — токсичные из них сжигают,
не токсичные, по возможности, отправляют на утилизацию.
При обезвреживании плотных отходов в микробиологических
производствах лишь убиванием необходимо иметь в виду антиген-
ные особенности такой микробной биомассы (способность вызы-
вать образование антител in vivo) — в любом случае необходимо
исключить сенсибилизирующее (от лат. sensibilis — чувствитель-
ный) действие ее на макроорганизм во избежание возникновения
аллергических заболеваний.
В аэротенках очистных сооружений, где происходит обезвре-
живание отходов, лимитирующими факторами выступают главным
образом качество и площадь биологической пленки, состоящей из
микро- и макрофлоры, микро- и макрофауны. В этой связи необ-
ходимо быть убежденным, что привносимые плотные отходы,
богатые органическими веществами, не приведут к ухудшению
работы аэротенков.
При анаэробном метановом брожении практически любые
органические вещества (за исключением лигнина) могут выступать
субстратами, трансформирующимися до метана и диоксида угле-
рода. Метан используют в качестве топлива, углекислоту — в
пищевой промышленности в виде "сухого льда". Остающийся
плотный остаток после метанового брожения (примерно 40% от
первоначального количества) представляет собой гумус, который
используют в качестве удобрения при возделывании сельскохозяй-
355
ственных культур растений. По ориентировочным расчетам (Е. С.
Панцхава, 1987; А. X. Авизов, Ю. В. Синяк, 1987), переработка
органических отходов в нашей стране могла бы дать 37 млн. т
условного топлива в год. На крупнейшей в стране Московской
очистной станции ежесуточно перерабатывается 28 тыс. м3 актив-
ного ила с получением 700 тыс. м3 биогаза.
Жидкие отходыв биотехнологических производствах
достаточно разнообразны по своему составу. Это объясняется
неполным использованием биообъектами компонентов, входящих
в состав питательных сред; наличием веществ (кроме целевых
метаболитов), секретируемых клетками; присутствием раствори-
телей, используемых, например, для экстракции конечных продук-
тов, и т. д. Рассмотрим, к примеру, сульфитные щелока, образую-
щиеся на предприятиях целлюлозно-бумажной промышленности
в результате гидролиза древесины и используемые для выращива-
ния кормовых дрожжей. Они содержат в среднем 50—60% суль-
фоната лигнина, 7—8% сахарных сульфокислот, около 18% различ-
ных сахаров, 10% диоксида серы, 8% солей кальция. После значи-
тельного удаления сульфита и подготовки щелока (разбавление,
внесение некоторых питательных ингредиентов) его используют
для выращивания адаптированной расы дрожжевых организмов.
Образующаяся клеточная масса здесь является целевым продук-
том, а отходом — культуральная среда после отделения дрожжей.
Жидкие отходы дрожжевых заводов, где производят дрожжи
на мелассном сусле, содержат органические и минеральные веще-
ства (мг/л в среднем): этанол — 0,45, углеводы (в том числе —
сбраживаемые) — 1,0, общий азот — 0,8, азот неорганический —
0,13, зольные элементы — 5,4. ВПК таких отходов составляет около
20000 частей Ог на 1 млн., то есть примерно столько, сколько и
ВПК для канализационных вод. Отходы, образующиеся от 1000 т
мелассы, соответствуют бытовым стокам города с населением
около 0,5 млн. жителей. Подобные жидкие отходы подвергают
микробиологической обработке (анаэробной или аэробной).
Сточные воды бродильных предприятий неравноценны. Так,
одни из них могут быть названы условно чистыми, поскольку они
почти не отличаются от потребляемой в производстве природной
воды (конденсаты, вода из теплообменников). Другие воды явля-
ются загрязненными неорганическими и органическими примеся-
ми, попадающими 1) от сырья (загрязнения при транспортировке,
мойке картофеля, свеклы), 2) от оборудования (мойка технологи-
ческой аппаратуры). Чистые воды могут быть использованы по-
356
вторно в технологических процессах, либо направлены в чистые
водоемы; загрязненные воды освобождают от механических при-
месей, а затем направляют на обезвреживание.
В производстве антибиотиков, кроме воды, используют углево-
ды и углеводистые продукты, масла, соевую муку, кукурузный
экстракт, нитраты, соли аммония, серо- и фосфорсодержащие
соединения, возможные предшественники антибиотиков (напри-
мер, амид фенилуксусной кислоты в качестве предшественника
пенициллина; н-пропанол в качестве предшественника эритроми-
цина и т. д.), неорганические кислоты и щелочи, органические
экстрагенты и пр.
Отличительной особенностью биотехнологических процессов,
основанных на выделении метаболитов из культуральных жидко-
стей, является неравновесное соотношение целевого продукта и
жидкости (жидкой среды), в которой он содержится (чаще — 1:100,
1:200, то есть 1%, 0,5% и менее). В подобных производствах
количество жидких отходов заметно больше, чем плотных. Если
последние содержатся в ощутимых количествах в сточной жидко-
сти, то их отделяют, отжимают и обезвреживают (см. плотные
отходы).
Сточные воды на зоопредприятиях содержат 1—2% органиче-
ских веществ и 98—99% воды. При использовании микробной
Очищенные .
воды
Рис. 123. Схема биологической очистки сточных вод.
357
обработки таких стоков в аэротенках (например, с участием сап-
рофитных бактерий из рода Afthrobacter, Corynebacterium,
Mycobacterium, Nocardia и некоторых других) можно получать
ценные микробные удобрения.
В зависимости от качества сточных вод возможна также их
очистка до целесообразного уровня (например, получение оборот-
ной воды, реализуемой повторно в том же биотехнологическом
производстве). На рис. 123 представлена схема биологической
очистки сточных вод.
Растворенные органические вещества можно удалять с по-
мощью активного ила в аэротенках или при аэробной обработке,
на биологических капельных фильтрах; нитраты обезвреживают с
помощью микробов-денитрификаторов (Pseudomonas spp., Вас.
licheniformis, Paracoccus denitrificans, Thiobacillus denitrificans), соли
фосфорной кислоты коагулируют и осаждают. Вновь образующи-
еся твердые (плотные) осадки концентрируют, обезвоживают
(фильтрованием, центрифугированием, отстоем на песчаном слое),
а затем сжигают, либо используют в качестве удобрения.
Таким образом, при обработке сточных вод до уровня чис-
той воды можно выделить следующие фазы: отделение крупных,
легко осаждающихся частиц и масляных пленок (грубая очистка),
отделение суспендированных частиц и растворенных органиче-
ских веществ (умеренно тонкая очистка), и, наконец, отделение
всех других примесей (тонкая очистка). При грубой очистке отде-
ляются частицы размером 100 мкм и более, при умеренно тонкой
— от 1 мкм до 100 мкм, при тонкой - от 0,1 нм до 1 мкм.
Тонкой очистки сточных вод последовательно достигают с
помощью фильтрации через песчаные слои, хлорирования, филь-
трации через активированный уголь, упаривания (жидкостной,
экстракции, вымораживания, обратного осмоса), ионного обмена.
Если в эту фазу образуются осадки (плотные вещества), то их
присоединяют к другим осадкам и обрабатывают как сказано выше.
Во всех случаях организации биотехнологических производств
необходимо предусматривать раздельные системы стоков — тех-
нологических и коммунальных.
На данном этапе развития биотехнологических производств
далеко не всегда (и не везде) применяют не только тонкую очистку
сточных вод, но, более того, известны случаи, когда необработан-
ные стоки от промышленных предприятий поступают непосредст-
венно в водоемы или в почву. В подобных случаях трудно сохранять
биологическое равновесие в водоеме — вместо чистой и красивой
358
реки, например, может сформироваться безжизненный, грязный
поток жидкости, попадающий в еще больший водоем (озеро, море),
где так же могут происходить пагубные изменения. Биологическое
равновесие в природном ("живом") водоеме нарушается за счет
следующих факторов:
1) изменения степени аэрации воды,
2) возрастания уровня органических субстратов,
3) изменения количества неорганических веществ (фосфор,
сера),
4) изменения pH,
5) изменения температуры.
Степень аэрации воды заметно уменьшается при попадании в
нее стоков, содержащих различные вещества, способные "погло-
щать" растворенный в воде кислород, иначе говоря, способные
окисляться. Маслянистые отходы могут физически мешать про-
никновению кислорода в водоем. Органические вещества также
нуждаются в повышенных количествах растворенного кислорода,
идущего на их минерализацию. В такой ситуации органические
структуры конкурируют с рыбами и другими водными обитателями
за такой кислород, но поскольку поступление загрязненных сточ-
ных вод может быть почти непрерывным, то живые существа не
выдерживают этой "конкурентной борьбы" и погибают.
Поступление в водоемы больших количеств фосфора и серы
приводит в конечном результате к такому же плачевному итогу —
образуются плохо растворимые или нерастворимые осадки фос-
форных солей и сульфидов [Са2(НРО4)2, Саз(РС>4)2; MeSJ, благодаря
чему выводятся из круговорота такие элементы-органогены как
фосфор и сера.
Физиологические значения pH и температуры хорошо извест-
ны для различных групп организмов. Например, большинство
бактерий лучше обитает в средах с pH 7,3—7,5; большинство грибов
— при pH 5,6—6,5; оптимальная температура для роста мезофиль-
ных микроорганизмов соответствует 20—40°С с колебаниями в
пределах от 10° до 50°С, и т. д. Холоднокровные животные относятся
к пойкилотермным организмам (от греч. poikilos — различный,
terme — тепло), тогда, как теплокровные животные — к гомойо-
термным (от греч. omoios — подобный, одинаковый) организмам.
Исходя из приведенных данных, можно представить реакцию
микро- и макрофлоры, микро- и макрофауны на непрерывные
температурные изменения или изменения pH среды обитания. В
таких ситуациях нормальные обитатели водоема погибают либо
359
из-за популяционного довления одних видов над другими, либо
вследствие невыносимости экстремальных (от лат. extremus —
крайний) условий жизни.
Растворимость чистого кислорода в воде составляет 48 частей
О? на 1 млн. частей Н2О при 14°С. При такой же температуре и
насыщении воды воздухом (содержание О2 в воздухе 20,9%) рас-
творимость кислорода’ составляет бкрЛо 10 частей на 1 млн. В
естественных водоемах растворимость оказывается еще меньше.
Например, в морской воде с соленостью 3,4% растворяется 80% О2
от растворенного в чистой воде, то есть 38,4 части на 1 млн.
Экстраполируя эти данные в пересчетах на моли других веществ,
можно прогнозировать потери растворенного кислорода в естест-
венных водоемах, куда сбрасываются стоки от биопроизводств,
содержащие органические и неорганические примеси. Все это
отрицательно сказывается на водных экосистемах. К тому же из-за
многокомпонентное™ стоков, трудностей определения каждого
компонента прибегают к анализу плотных остатков, общего азота,
органического углерода и биохимической потребности кислорода
(ВПК). Опираясь на фактические данные, полученные в результате
проведенных анализов, выдают рекомендации по обработке жид-
ких стоков. ВПК означает количество потребляемого растворен-
ного кислорода при инкубации стоков в течение 5 дней и темпе-
ратуре 20°С. Растворенный кислород определяют различными
методами—химическим, биологическим или физико-химическим.
ВПК можно выразить в мг О2 на 100 мл или на 1 л пробы, в частях
на 1 млн в мл О2 на 1л пробы при О°С и 1,01 • 10s Па. Если,
например, ВПК воды больше 10 частей на 1 млн., то она непригодна
для использования человеком. ВПК для неочищенных стоков в
производстве пенициллина 32000 частей на 1 млн.
Загрязненные воды и промышленные стоки должны проходить
обработку на предмет обезвреживания и очистки перед поступле-
нием их в природные водные резервуары. Наличие в стоках (воде)
ингибиторов или токсических веществ для микроорганизмов от-
рицательно сказывается на правильности измерения ВПК, а в
случае попадания таких стоков в аэротенки или природные нако-
пители, содержащие микрофлору и микрофауну, происходит на-
рушение процесса их обезвреживания. Из различных токсических
веществ и ингибиторов можно назвать соли тяжелых металлов,
поверхностно-активные вещества (ПАВ), фенольные соединения
и др. Тяжелые металлы необратимо ингибируют ферменты вслед-
ствие блокады тиольных групп:
360
/SH 2+ A
ферментный белок +Hg ---------> ферментный белок rig
XSH V
нативный денатурированный
В порядке снижения активности, например, в отношении мик-
ромицетов металлы можно расположить в следующем порядке:
Ag > Нд > Си > Cd > Сг > Ni > Pb > Со > Zn > Fe. '
Начиная с 1949 г.'(в странах Западного полушария) и несколько
позже (в нашей стране и некоторых государстах Востока), про-
мышленное производство ПАВ развивалось исключительно быстро
— многие из.них были включены в состав моющих средств. По
заряду их подразделяют на катионные, анионные, амфотерные и
неиодогрнные. Катионные ПАВ содержат гидрофобный радикал
типа алифатической цепочки предельных углеводородов, бензо-
льного или нафталинового кольца с алкильным остатком, ‘а также
положительно заряженную гидрофильную группу, например, чет-
вертичного аммония, сульфония,-фосфония, арсония, йодония. Из
них широко известны на практике хлоргексидин биглюконат,
цетавлон и др.
NH NH
। ।
C|-C6H4-NH-C-NH-C-NH-(CH2)6-NH-C-NH-C-NH-C6H4-CL
NH NH
2C6Hi/>^
хлоргексидин биглюконат (гексам тилем-1,6-бис-р-хлорфенилбигуанидина диглюконат)
[НзС—(CH2)i5—N = (СНз)зГВг-
Цетавлон
Катионными ПАВ являются
циклопептидные антибиотики
(грамицидин С, полимиксин В,
циклоспорин А и др.).
Потммксмн В
(ДАМ - с, у диаминобутираг.б - МОК - м«т-
поктаиат)
361
L-Ane------► L-Ал»-------► Me-L-Лвй--------► Me-L-Лей
t
Me-L-ЛвА Y
Ma-L-Лвй
L-Ban
t ______2
Me-L-Лей --------Kte-Гли-^--------AMK^---------Me-Fen
циклоспоринA
AMK - «миноыасляная кислота
Ме-Геп - г-Ычиетмл-З-оиси-гвлтеновая (5,6 ей) кислот»
К анионным ПАВ относятся структуры, сходные с катионными
ПАВ в гидрофобной части, но имеющие отрицательно заряженную
группу — карбоксильную, сульфатную, сульфонатную, фосфат-
ную:
О
II
R—COO- R—OSO3- R—SO3 R — О— ₽—О
о-
По объему производства они занимали (и пока еще занимают)
ведущее место среди всех ПАВ. Хорошо известны изомерные смеси
алкилбензолсульфонатов:
__ СН, сн,
Л-\ J 3 j 3
NaO—SO2~^ ff- С-СН2-СН2-С-СН2-СН-СН
сн3 сн3 сн3
2-метил-4.7-диметил-гептил-п-бензолсульфонат
2
NaO-SOy
СН3 СН3 СН,
al j 61 3 5 41 3 3
- С-СН,-СН-СН,~СН-СН,-СН,-СН,
СН 2 2 3
4,6-диметил-8-диметил-октил-п-бензолсульфонат
362
сн,
I 3
СН3—(СН2)9—СН—С6Н4—SO3Na
11-метил-ундецил-п-беизол-сульфонат
VH=
CHj—(СН2)8— с—С6Нд—SO3Na
сн3
10-диметил-децил-п-бензол-сульфонат
Доступность таких структур для деградации микроорганизмами
определяется природой алкила и а-углеродным атомом. Так, если
алифатическая (гидрофобная) часть будет в виде неразветвленной
цепи, то такое анионное ПАВ подвержено более глубокой дегра-
дации, чем такое же ПАВ, но с разветвленной цепью. В зависимости
от характера гидрофильной группы в замещенных бензолах мик-
робы активного ила по-разному разрушают эти структуры. "Био-
логическую податливость” таких веществ можно расположить в
следующем порядке:
С6Н5ОН > СбНзСООН > C6H5NH2 > C6H5SO3H.
I
Из вышеприведенных структур алкилбензолсульфонатов две
последние легче подвергаются биологическому окислению.
К числу амфотерных ПАВ относятся такие структуры, у кото-
рых гидрофильная часть представлена катионной и анионной
группами одновременно. Таковым является, например, амфолан
(ТЕГО-103).
[сн3-(сн2)1 _nh-(ch2)2-nh-(ch2)2-nh-ch2-cooh] HCI
Амфолан (хлоргидрат алкилдиаминоэтилглицина)
Они меньше других ПАВ используются на практике, и поэтому
не возникает особых проблем с их обезвреживанием.
Неионогенные ПАВ типа спанов и твинов изменяют поверхно-
стное натяжение на границе раздела фаз "жидкость-твердое тело"
(применительно к сточным водам), однако они не обладают выра-
женным губительным действием в отношении различных организ-
мов, но сильно изменяют водные среды по качеству (пенообразо-
вание) и по БПК. Спаны — это сложные эфиры жирных кислот и
спирта сорбита: НОН2С—(СНОН) 4—CH2OCOR, -где R — остаток
363
жирной кислоты. Твины являются сложными эфирами ангидро-
сорбита и жирных кислот, алкилированные окисью этилена; дру-
гими словами — это эфиры циклизованных полиэтиленоксидов и
жирных кислот
<^\h-ch2-o-r
HOH2C-f-CH2—О—СН2-^-СН2—О—С^^СН—О—СН2-£-СН2—О—CHj-JjCHjOH
6-СН2-[-СН2-О-СН2-}-СН2ОН
Фенольные соединения типа карболовой кислоты и ее аналогов
также являются токсическими структурами и уже в концентрации
1-2% коагулируют белки, а в более высоких проявляют гидролити-
ческий эффект.
ОН ’
фенол, или
оксибензол
(карболовая
кислота)
ОН
диоксибензол
(резорцин)
крезолы
Следовательно, токсические вещества и ПАВ в сточных водах
должны быть исключены или многократно уменьшены по своему
количественному содержанию усилиями заводских коллективов,
то есть за счет внедрения новых технологий или совершенствова-
ния существующих, благодаря которым отходы производства не
будут поступать в общегородские и другие стоки и пагубно влиять
на природные экосистемы.
Таким образом, обработка отходов может быть условно под-
разделена на 4 стад ии: 1) разрушение сложных белковых комплек-
сов (главным образом — их конъюгатов) до простых растворимых
веществ и отделение их от нерастворимых субстанций,
2) разжижение и анаэробная обработка нерастворимого остат-
ка с помощью соответствующих микроорганизмов,
364
3) трансформация органического азота до NH4 (аммонифика-
ция) с последующим окислением аммония до нитратов,
4) превращение органического углерода до СО?.
В этих стадиях (преимущественно — в первой, третьей и
четвертой) подчеркнута биохимическая сущность происходящих
явлений, а процессуальная технологическая схема обработки от-
ходов в обобщенном виде приведена на рис. 123. Нерастворимый
остаток (стадия 2) может быть использован для получения метана.
Газообразные отходыв процессах биологической
технологии немногочисленны в ассортименте. Это определяется
биохимической сущностью реакций, катализируемых фермента-
ми. Как правило, энергетическим субстратом для биообъектов
являются углеводы. В аэробных и анаэробных условиях из них
образуется диоксид углерода. Так, при брожении (гликолитический
процесс) из глюкозы образуётся 2 моля СОг:
СбН12О6 ----------- 2С2Н5ОН + 2СО2
При аэробном дыхании (окислительный процесс) — 6 молей
СО2:
+ 6O2
СбН12Об-----------► 6СО2 + 6Н2О
При выращивании метанокислящих бактерий из СН4 также
образуется СО2:
СН4 + 202---------► СО2 + 2Н2О
Выделяющийся диоксид углерода улавливается и утилизирует-
ся в пищевой промышленности в качестве хладагента.
Газообразным отходом биотехнологических производств, бази-
рующихся на использовании аэробных микроорганизмов, является
"отработанный воздух". Он не должен поступать в атмосферу без
очистки и обезвреживания. Отработанный воздух при этом чаще
всего представляет собой высокодисперсный аэрозоль, в котором
дисперсной фазой оказываются капельки жидкости и/или микро-
организмы. Скорости падения аэрозольных частиц зависят от их
размеров и смещения воздушных потоков. Для некоторых из них
скорости падения приведены в таблице 38.
365
Таблица 38. Скорости седиментации частиц (плотность, г/см3) из высокодис-
персных аэрозолей (вязкость воздуха — 1,82* 10'4 пуаз,
t° -293°k)
Радиус частиц, мкм Критерий Рейнольдса Скорость седиментации, м/сек. Броуновское движение, м/сек.
3 0,366 0,11 • 10'2 2.84 • 10'4
1 1,43 • 10'2 1,3 • 10'4 5,07 • 10’6
0,3 4,62 • 10'2 1,39 • 10'5 1,0 • 10'5
0,1 2,45 • 10 s 2,23 • 10‘6 2,1 • 10’5
0,03 1,37 • 10‘6 4,16 • 10’7 5,5 • 10’5
U,U1 1,26 • 10’7 1,14 • IO’7 1,06 • 10’4
Из таблицы видно, что с уменьшением размера частиц их
седиментация заметно уменьшается, но возрастает броуновское
смещение. Вот почему высокодисперсные аэрозоли легко могут
переноситься воздушными потоками на большие расстояния, и
поэтому не исключено неблагоприятное воздействие их на чувст-
вительные контингенты людей, вдыхающих микробные аэрозоли.
Отработанный воздух, содержащий микроорганизмы (в том
числе, например, болезнетворные продуценты экзотоксинов) дол-
жен быть термически обработан и только после этого подвергаться
фильтрационной очистке.
При реализации биотехнологических процессов необходимо
осуществлять постоянный (непрерывный) контроль за качеством
отработанного воздуха, выбрасываемого в атмосферу.
Многие биотехнологические процессы протекают с выделени-
ем тепла, которое оказывается в ряду побочных продуктов биохи-
мических реакций. Нередко такое тепло теряется и практически
не используется. Лишь в отдельных производствах оно реутилизи-
руется (см. раздел 7.2).
8.2. Утилизация отходов биотехнологических производств. От
качества плотных и жидких отходов, образующихся в биотехноло-
гических производствах, зависит выбор путей использования их
на практике. Так, в производстве пива из ячменя отходами явля-
ются дрожжевые клетки, солодовая дробина и некоторые другие
вещества. Можно предположить, что все отходы такого производ-
ства равно используемы, например, для откорма сельскохозяйст-
венных животных (таблица 39). Из таблицы видно, что по пита-
тельной ценности и усвояемости все компоненты плотных отходов
могли бы быть рекомендованы к употреблению на животноводче-
ских фермах. Однако горечи, имеющиеся в женских соцветиях
366
хмеля, и продукты их превращения при кипячении солодового
сусла с хмелем, являются главным препятствием к использованию
их в животноводстве. Такими веществами являются гумулон (а-
горькая кислота), лупулон (P-горькая кислота) и продукты их
окисления — смолы А и В, дигидрогумулиновая кислота и др.
Т а б л и ц а 39. Состав пивных дрожжей, белкового остатка, солодовой и хме-
левой дробин (выборочно)
Вид продукта Содержание, % Усвояемость % Эквива- лент- ность (в кг) 100 кг крахмала
во- ды Л Пр Ц БЭВ 3 У сви- ней У жва- чных
Влажные дрожжи 86 0,2 6,9 0,4 5,9 0,9 85 85
Высушенные дрожжи 8-12 2-3 43-55 0,4- 1,7 25-35 6-8 85 85 76,6
Белковый остаток+ 8,5 3,9 43,6 6,8 32,9 4,3
Солодовая дробина 77 1,4 5,3 4 11 1,3 52,25 50 12,7
Хмелевая дробина+ 62 3.21 4,46 5,4 26,15
Примечание: Л — липиды неочищенные. Пр — протеин сырой, Ц — клетчатка, БЭВ —
безазотистые экстрактивные вещества, 3 — зола, + — животные не употреб-
ляют из-за горечи продукта, ‘ — ароматические экстрактивные вещества.
Из 1 кг хмеля, примененного для варки пива, получается около
7 кг хмелевой дробины. Она может быть использована либо в
со
(СН ) =С=СН-СН2-СН С-СО-СН2-СН=(СН3)2
I II
со с-он
он—С----СН2—CH=C=(CHj)2
гумулон
СО
(СН,)=С=СН-СН.-СН С-СО-СН,-СН=(СНД
1/ОН И
(СН3)2=С=СН-СН?_СО—с — с-он
изогумулон (мягкая смола А)
367
СОЧ
(CH3)2=C=CH-CH2—СН с—СО—СН2-СН=(СН3)2
- • • I II
сн3—СО—-С—С—ОН
он "
гумулиновая кислота (мяпвя смола В)
компостах, либо в качестве подстилки на скотных дворах. В других
случаях ее сжигают' (например, добавляя к каменному углю). К
сожалению, названные горечи переходят и в белковый отстой. Цо
этой причине он не используется в качестве добавок к корму
животных. Имеются данные о том, что такой белок пригоден для
подкормки рыбы, разводимой, в прудовых хозяйствах.
• zCO\ *
(СН3)2==СН-СН£-СН2—СН с—со—сн2—СН----(СН3)2
. ТСН—с—ОН
он
дигидрогумулиновая кислота
СО
(СН3)2=С=СН-СН2—СН хс—со—сн2—СН=(СН3)2
СО он
(СН3)2=С=СН-СНг—С— СН2-СН=С=(СН3)2
лупулон
Дрожжевые клетки и солодовая дробина — хороший корм для
животных. В высушенном виде их можно сохранять впрок. Отмы-
тые от горечей клетки дрожжей соответствуют по калорийности
говяжьему мясу при соотношении 1:2,5, то есть 1 кг сухих пивных
дрожжей равноценен 2,5 кг говядины. В них содержатся также
витамины 1руппы В, в частности, тиамин и рибофлавин (50—100
мг/кг), эргостерин (0,56%), микроэлементы. Следовательно, пивные
дрожжи полезны и человеку, и не, только в качестве пищевого
продукта, но и лечебного. Применение пивных дрожжей норма-
лизует некоторые обменные процессы, восполняет дефицит от-
дельных коферментов.
Солодовая дробина состоит из солода, оболочек зерен ячменя,
368
липидов, белков и других веществ (см. таблицу 39). При скармли-
вании солодовой дробины, например, коровам, возрастает их
удойность. Если в день давать 2,5 кг сухого солода на голову скота,
удой молока возрастает примерно на 5 литров.
Таким образом, пивоваренное производство можно отнести к
разряду малоотходных и, следовательно, выгодных с
. экономической и экологической точек зрения.
Производство многих антибиотиков, базирующееся на исполь-
зовании бактерий —.стрептомицетов, также приближается (или
может быть приближено) к малоотходным. Плотные отходы после
термообезвреживания могут быть, использованы в качестве корма
для сельскохозяйственных животных, так как белковое, минераль-
ное и, вреде случаев, витаминное содержание (Вщ) клеток является
важной добавкой к их дневному рациону. Однако по некоторым
данным, в клеточной биомассе не должно содержаться ощутимых
количеств остаточных антибиотических веществ, что, якобы, мб-
жет сказаться на селекционировании резистентной микрофлоры,
включая патогенную и условно паторенную (в частности, из семей-
ства Enterobacteriaceae). По другим данным антибиотики — не
только профилактические и лечебные средства, но и стимулируют’
прибавление веса, увеличивают эффективность, использования
кормов. Однако учитывая высокую биологическую активность
антибиотиков, целесообразно придерживаться первых рекоменда-
ций. . ' Г ' \
Жидкие отходы в производстве антибиотиков передаются на
обезвреживание.
Применительно к микробной биотехнологии имеются данные
об использовании плотных отходов в производстве антибиотиков
и ферментов (биомассы клеток) в качестве армирующего накопи-
теля в бетонных изделиях.
В призводстве декстрана возможна организация безотхо-
дной технологии. Исходя из механизма биосинтеза полисахарида
видно, что отходом производства является фруктоза. К сожалению,
декстрансахароза
С12Н22°11 <С6Н12°5>П ♦ "С6Н12О6
сахароза декстран фруктоза
на предприятиях, изготавливающих декстран, фруктоза чаще вме-
сте с маточными растворами спускается в трап. Между тем фрук-
тоза может быть очищена и использована для пищевых целей,
например, в виде сиропа.
369
Полисахарид аубазидан продуцируется штаммом № 8
Aureobasidium pullulans. Биомасса клеток как отход производства
составляет около 50% от так называемого "нативного полисахарида"
(клетки + гликан). Они содержат порядка 31% белков, включаю-
щих ряд незаменимых аминокислот в количествах, превышающих
их концентрацию в кормовых дрожжах (аргинин, гистидин, трип-
тофан, фенилаланин). Биологическая ценность белка составляет
83,4% и она выше, чем у белков шелушоного риса (71,2%), рисовой
крупы (65,23%) и кормовых дрожжей (40—60%). В клетках
A.pullulans содержится 35% углеводов и 14% липидов. По составу
жирных кислот эти липиды близки пальмовому маслу.
Зольные вещества клеток представлены (по убыванию содер-
жания) железом, алюминием, кремнием, марганцем, магнием,
медью, барием, натрием, калием, кальцием, серой, хлором; из числа
витаминов в нативных клетках найдены витамины Bi, Вб, РР, В?,
Вв- Таким образом, биомасса клеток Apullulans является тем
отходом, который с успехом может быть использован для вскарм-
ливания сельскохозяйственных животных.
При винокурении в качестве сырья используют различные
источники (зерно, картофель, мелассу и пр.). Плотные отходы
(барда) утилизируются в качестве кормовых добавок; выделяюща-
яся при спиртовом брожении углекислота улавливается и очища-
ется (обычно — на водяных скрубберах, затем кислотой), дезодо-
рируется, сжижается и поступает в продажу в виде "сухого льда".
Из 1 т патоки, содержащей 45% сахара (при фактическом исполь-
зовании, равном 65%), выход углекислоты (жидкой) составляет 135
кг. Жидкая углекислота широко применяется при изготовлении
безалкогольных напитков и в технике.
Жидкие отходы после извлечения этанола могут быть упарены
до сиропообразного состояния, высушены (высушенный фильтрат
барды) и так же применены в качестве корма для животных.
При виноделии получают такие отходы как виноградные вы-
жимки (до 20% и более от веса винограда), дрожжевые клетки, а
также отстойные осадки. При усредненной оценке отходов (преж-
де всего — выжимок) в них находят этанол, соли винной кислоты,
масло (в косточках), реже — сахаристые вещества. Виноградный
спирт используют при производстве крепленых вин, виноградное
масло рафинированное — ценный пищевой продукт, а также ценен
в изготовлении олифы. Винную кислоту применяют в медицинской
практике, в кондитерской промышленности.
370
После выделения этанола из виноматериалов остается винная
барда (винасса), содержащая органические кислоты, белки, мине-
ральные соли, красящие вещества. Из барды выделяют винную
кислоту. Из отходов виноделия получают дубильное вещество —
танин, пищевой пектин, красящие вещества. Не подвергающиеся
дальнейшей обработке осадки передают на аэротенки или на
сбраживание (см. плотные отходы).
При изготовлении вин из плодов и ягод отходами являются
дрожжи, клеевые осадки и отпрессованная мезга. Нерастворимые
вещества отходов представляют собой клетчатку, крахмал, прото-
пектин, белки, жиры, остатки органических кислот и сахаров.
Протопектин выделяют в виде пектина, широко использующегося
в пищевой и кондитерской промышленности.
Другие составные части плотных отходов являются ценным
кормом для сельскохозяйственных животных. Косточки и семена
плодов можно отделять от выжимок и затем использовать для
получения масла.
В фитрбиотехнологии обезвреживание отходов и их утилиза-
ция сходны с таковыми для микробной технологии.
Утилизация отходов зообиотехнологических производств про-
исходит с учетом того, что плотные и жидкие отходы содержат
большое разнообразие веществ, пригодных для гетеротрофных
видов микроорганизмов. Это, прежде всего, создание питательных
сред для микробов в экспериментальных (лабораторных) и приро-
дых условиях (удобрения).
В связи с обезвреживанием и утилизацией отходов биотехно-
логических производств во имя поддержания природного экологи-
ческого баланса находится проблема организации и развития
ресурсосберегающих безотходных процессов биологической тех-
нологии. Это решаемо и может быть решено в двух приоритетных
направлениях: 1) полное использование сырьевых источников
путем применения биообъектов в ассоциациях, когда имеет место
более глубокая трансформация сырья и получаются различные
конечные продукты (примеры — обработка жидких отходов жи-
вотноводческих комплексов с участием разных организмов; пол-
учение декстрана и аубазидана с использованием соответствую-
щих штаммов Leuconostoc sp. и Aureobasidium pullulans, из которых
первый использует глюкозу из сахарозы, второй — оставшуюся в
отходах фруктозу; иммобилизация клеток как полиферментных
систем, например, микроб, несущий гены сс-амилазы, глюкоамила-
371
зы и глюкозоизомеразы, способен в одну стадию трансформиро-
вать крахмал в глюкозу и фруктозу; в этом случае глюкозо-фрук-
тозный сироп (заменитель сахара) является конечным продуктом;
и т. д.,
2) экономия сырья, вспомогательных материалов и энергии. В
этом случае, например, в качестве источников углерода в микро-
бной биотехнологии могут быть использованы отходы деревопе-
рерабатывающей промышленности (стружка, тара), сельского хо-
зяйства (солома, подсолнечная лузга); источников ряда витаминов
— гидролизаты (или экстракты) дрожжей, являющихся отходом
производств,-основанных на спиртовом брожении; энергетически
целесообразны рециркуляционные схемы использования тепла,
получаемого с помощью теплообменников при стерилизации и
других термопроцессах и т. д.
Важно помнить и иметь в виду необходимость глубокой про-
работки проектно-сметной документации на предмет создания
ресурсосберегающих и экологически чистых безотходных или
мало отходных биотехнологических процессов.
372
Часть IV.
Специальные биотехнологии
' Во второй части учебника изложены основные представления
о биотехнологии, базирующейся на использовании биообъектов
микробного, растительного и животного происхождения. В срав-
нительном плане можно подчеркнуть следующие особенности
таких биотехнологий: во-первых, имеется много аналогий при
реализации биотехнологических процессов, в которых биообъекты
используются на молекулярном и клеточном уровнях; во-вторых,
только некоторые виды биотехнологий, осуществляемые с приме-
нением клеток, могут быть подведены под рубрику "технологий на
организменном уровне" (микробная биотехнология); в-третьих,
лишь при использовании в биотехнологии микроскопических ор-
ганизмов необходимо специальное аппаратурное оформление про-
цессов, тогда как в случаях выращивания макроорганизмов (рас-
тений и животных) в естественных, или природных условиях этого
делать не требуется, и, следовательно, посевы и сборы урожаев,
выращивание сельскохозяйственных животных в целях получения
373
молока, мяса, кож, шерсти и пр. не относится к новым и новейшим
биотехнологическим процессам в таком понимании предмета, по-
скольку здесь выпадает ведущее звено, связанное с управляемым
культивированием биообъекта, например, в ферментаторах или в
других аппаратах. Однако третья особенность не является универ-
сальной. В частности, к истинной биотехнологии тесно примыкает
технология выращивания некоторых растений на основе метода
гидропоники. Тем не менее, особенности каждой группы биотех-
нологических процессов сохраняются и поэтому есть все основа-
ния рассматривать их в качестве самостоятельных. К ним относят
микробную биотехнологию, фитобиотехнологию и зообиотехно-
логию, то есть главным критерием подразделения биотехнологий
являются биообъекты. Микроорганизмы в этом смысле занимают
первенствующее положение (сравнить число реализованных на
практике микробиологических и других производств по объему
выпускаемой продукции).
Существующие ветви биотехнологических субдисциплин (им-
мунобиотехнология, инженерная энзимология), применительно к
конкретным производствам, рассмотрены в соответствующих гла-
вах второй части учебника. Например, генно-инженерные проти-
вовирусные вакцины включены в главу "Микробная биотехноло-
гия", тогда как вирусные вакцины, получаемые на культивируемых
клетках и тканях животных организмов, приведены в главе "Зоо-
биотехнология". Биохимические процессы на основе инженерной
энзимологии рассмотрены на примерах ферментов преимущест-
венно микробного происхождения, поэтому и этот раздел включен
в главу 9.
Глава 9.
МИКРОБИОТЕХНОЛОГИЯ
Микробиотехнология, или микробная биотехнология базиру-
ется на интегрированном использовании микробиологии, биохи-
мии и инженерных наук. с целью реализации потенциальных
способностей микроорганизмов в технике и промышленном про-
изводстве. По сути своей микробиотехнология тождественна про-
мышленной (технической) микробиологии. Ее объектами являются
микробы-вирусы (включая вироиды и фаги), бактерии, грибы,
лишайники, протозоа (см. главу 2). В ряде случаев биообъектами
являются первичные метаболиты микробного происхождения —
ферменты, каталитическая активность которых лежит в основе
инженерной энзимологии.
374
В сравнении с растительными и животными клетками микробы
размножаются, как правило, быстрее и, следовательно, у них
быстрее протекают все метаболические (обменные) процессы.
Относительные преимущества большинства микробов как биообъ-
ектов следующие:
1) большая "простота" организации генома,
2) достаточно легкая приспособляемость (лабильность) к среде
обитания в естественных и искусственных условиях,
3) выраженные скорости протекания ферментативных реакций
и нарастания клеточной массы в единицу времени.
Первое преимущество обеспечивает микробным клеткам луч-
шие возможности для изменения и перестроек наследственного
материала, например, включение в него чужеродной генетической
информации, привнесение в клетки или, напротив, элиминация из
них плазмид, и пр.
Второе преимущество, связанное с лабильностью микробов,
можно показать на примере бактерий и грибов. Так, применитель-
но к температуре микробы подразделяют на психрофилы, мезо-
филы и термофилы (от греч. psichria — холод, mesos — средний,
terme — теплота, fileo — люблю) со следующими оптимумами
показателей для них: ниже 20°С, 20—45°С и выше 45°С соответст-
венно. Однако указанные интервалы, как правило, перекрываются.
Например, психрофильный вид Xanthomonas pharmiicola может
расти при температуре от 0°С до + 40°С, мезофильный Lactobacillus
lactis — от + 20°С до + 50°С, термофильный Вас. coagulans — от
+ 20°С до + 65°С. Поэтому выделили дополнительные подгруппы:
облигатные психрофилы, факультативные психрофилы, стено-
термофилы и эвритермофилы. Первые из них не растут при
температуре выше +20°С, для вторых верхний температурный
предел превышает + 20°С, третьи не растут при температуре ниже
+ 37°С, четвертые растут при температуре ниже + 37°С.
Кроме стенотермофилов и эвритермофилов целесообразно вы-
делить еще одну подгруппу термофильных микроорганизмов под
названием супертермофилов, для которых температурный опти-
мум составляет 105°С, то есть выше точки кипения воды (100°С).
К числу таких микробов относят бактерии родов Pyrodictium и
Pyrococcus. Так для Pyrococcus furiosus, изолированного из подвод-
ного гидротермального излияния около средиземноморского ост-
рова Вулкано, кардинальные точки температуры (minimum,
optimum, maximum) составляют 60°С, 105°С, 110°С. Среди супер-
термофилов отдельные представители являются или могут быть
375
источником термостабильной ДНК-полимеразы, активной к тому
же при высоких давлениях (рис. 124).
Рис.* 124. Скорость роста
или другой физиологиче-
ской функции микроорга-
низмов (V max) в процентах
от максимального значения
для данных условий: 1 —
психрофилы, 2 — мезофи-
лы, 3 — термофилы, 4 —
супертермофилы.
Среди грибов имеются психрофилы и мезофилы, реже — .
эвритермофилы (от греч. euris —. широкий), но до сих пор не
описаны грибы — стено- и супертермофилы. Например, аспоро-
генные дрожжи Candida включают виды, способные расти при
температуре* в диапазоне от —10°С до +37вС; диморфный гриб
Aureobasidium pullulans растет при температуре от 16°С до 30°С.
Грибы-сапрофиты — продуценты БАВ выращивают в производст-
венных условиях обычно при 24—26°С.
Клетки микроорганизмов проявляют также лабильность к из-
менению в среде концентрации водородных ионов. Давно известна
следующая закономерность — бактерии более выражение прояв-
ляют свою физиологическую активность в средах при значениях
pH выше 7,0, тогда как грибы — при pH ниже 7,0. Однако это не
значит, что" отдельные микробы не будут расти и размножаться
при существенных отклонениях pH. Так, молочнокислые бактерии
в процессе размножения закисляют среду до pH 3,0 и ниже;
некоторые представители дрожжей рода Candida на питательных
средах с pH 8,0—10,0 способны накапливать биомассу клеток
достаточно эффективно после заметной пролонгации lag- и log-
фаз. В это время происходит не только адаптация к среде обитания,
но и постепенное закисление.
Диморфный дрожжеподобный гриб Aureobasidium pullulans
хорошо развивается при pH 6,0—6,5, но продуцирует экзогликан
— аубазидан более выраженно при pH 3,0—3,5.
Образование ряда вторичных метаболитов нитчатыми грибами
имеет место на грани между стационарной и летальной фазами,
когда происходит защелачивание сред. Так, Penicillium notatum
лучше растет при pH 4,5, тогда как продукция пенициллина более
выражена при pH 7,5.
376
Третье преимущество микробов, связанное с большими скоро-
стями ферментативных реакций, можно показать на примерах
размножения отдельных видов различных микроорганизмов. Так,
удвоение числа клеток E.coli и Вас. subtilis на благоприятных
питательных средах наблюдается, в среднем, через 20 минут;
почкование Candida albicans на жидком сусле —’ через 30 минут;
после проникновения Т-четной фаговой частицы в клетку Е. coli-
через 20 минут появляется от 20 до 200 новых фаговых частиц, и
т. п. Конечно, в мире микробов имеются представители, скорость
размножения которых определяется сутками, например, микобак-
терии туберкулеза. С учетом условий среды культивирования и
активности ферментов (чаще — иммобилизованных), скорость
реакции является, в основном, результатом скорости реакции в
непосредственном микроокружении активных центров ферментов
и диффузии субстратов и веществ в направлении к этим центрам
и от них — в противоположную сторону.
Микробы относятся к живым саморегулирующимся системам,
восстанавливающим равновесие после какого-либо воздействия,
если оно не было чрезмерно сильным и продолжительным. Если
же воздействие действительно было сильным и продолжительным,
то организм погибает или переходит в новое устойчивое (равно-
весное) состояние. Это может произойти, например, после воздей-
ствия мутагенных факторов и образования мутантов (см. выше
первое преимущество микробных клеток). В этой связи необходи-
мо отметить, что микроорганизмы легче приспосабливаются, или
адаптируются к изменившимся условиям существования, чем рас-
тения и животные, хотя и среди микробов адаптивные возможно-
сти у видов-сапрофитов более выраженные, чем у видов-паразитов;
это связано с более широким набором ферментов у первых, нежели
у вторых. Поскольку ферменты — это первичные метаболиты, то,
в конечном итоге, их арсенал определяется генотипом.
Микробиотехнология на клеточном уровне практически реа-
лизована значительно раньше, чем фито- и зообиотехнологии. Для
примера можно назвать приготовление ряда пищевых продуктов
(кисломолочных, хлеба, сыров), производство вин, пива, спиртов,
органических кислот, аминокислот, антибиотиков, ферментов, от-
дельных витаминов, белковых и других кормов для сельскохозяй-
ственных животных, а также веществ, производство которых
основано на методах генетической инженерии, и пр.
В данной главе не приведены материалы о биотехнологии
вирусных препаратов и о процессах, в которых применяют прото-
377
зойные организмы. Вирусы — как облигатные, или безусловные
паразиты выращивают в живых тканях животных организмов, а
протозоа пока не нашли широкого практического воплощения в
производстве. Поэтому определенные сведения о тех и других даны
в главе 11.
Микроскопические водоросли — как биообъекты рассмотрены
в главе 10. Настоящая глава посвящена биотехнологическим про-
цессам, основанным на использовании биообъектов — представи-
телей двух царств — бактерий и грибов.
9.1. Принципы культивирования микроорганизмов. С момента
внесения микробов (засева) в питательную среду имеет место
индукция их физиологической активности, особенно — в логариф-
мическую и/или стационарную фазы размножения. При этом
одновременно сопряженно протекают многие реакции, катализи-
руемые иммобилизованными или свободными ферментами. В ре-
акции, особенно — на первых этапах, нередко вовлекаются высо-
комолекулярные вещества с определенной конфигурацией моле-
кул (сравнить такие источники углерода как глюкоза и крахмал
или источники азота—аммония сульфат, какая-либо аминокислота
и нативный белок). Поэтому следует учитывать специфику выра-
щивания микроорганизмов.
Главные особенности культивирования микробов в целях пол-
учения большинства первичных и вторичных метаболитов следу-
ющие:
1) необходимость применения специальных биореакторов вме-
стимостью 63, 200, 1000 и более м3, в которых возможно поддер-
жание асептических условий в течение сравнительно длительного
времени;
2) видовые различия биообъектов, с которыми связаны специ-
фические характеристики питательных сред, кардинальные точки
(минимальные, оптимальные и максимальные) температуры и pH;
3) невозможность одновременного поддержания постоянства
критериев химического, теплового, диффузионного и гидродина-
мического подобия, с чем связаны трудности масштабирования
биотехнологических процессов;
4) различия в массообменных процессах у аэробов и анаэробов
— культивирование аэробов осуществляют в трехфазных системах
["твердое тело (клетки)-жидкость-газ"], анаэробы выращивают в
виде двухфазных систем ["твердое тело (клетки)-жидкость"];
5) необходимость перемешивания культуральных жидкостей в
целях улучшения массообмена (кислорода воздуха для аэробов —
378
прежде всего), а это, в свою очередь, индуцирует новообразование
и, как следствие, диктует необходимость прибегать к пеногашению;
6) микроорганизмы чувствительны к воздействию механиче-
ских, физических и химических факторов;
7) при микробном синтезе целевых продуктов имеют место
индукция, активация, ингибирование, репрессия и некоторые дру-
гие регуляторные процессы, усложняющие в целом регуляцию
размножения продуцента и биосинтез им конечного продукта;
8) скорость биосинтеза целевых продуктов более медленная по
сравнению со скоростями химического синтеза;
9) отдельные виды микроорганизмов, используемых в биотех-
нологических процессах, являются болезнетворными и работа с
ними должна проводиться с особой тщательностью (дифтерийные
и столбнячные палочки, микобактерии туберкулеза, холерный
вибрион и др.);
10) некоторые представители микробного мира должны куль-
тивироваться только на (в) живых тканях/клетках (куриные эмб-
рионы, клетки человека и животных и т. д.); к таким представителям
можно отнести вирусы, риккетсии.
Любой биотехнологический процесс, реализуют условно в два
этапа. Первый из них — предферментация, когда необходимо
выполнить все подготовительные работы для реализации второго
этапа — ферментации, то есть накопить и выделить целевой
продукт.
Предферментация
Предферментационный этап включает подготовку питательных
сред, биообъекта, воздуха для аэробов и биореактора. Компоненты
питательных сред подбирают на основании расчета материального
баланса, связанного с трансформацией того или иного источника
питания в клеточную биомассу и/или метаболит при учете расхо-
дуемой (выделяемой) энергии. Обычно качественный и количест-
венный составы питательных сред указаны в регламентной доку-
ментации.
Питательные среды, используемые на подготовительном этапе,
могут несколько отличаться от среды, применяемой на втором
этапе. Так, например, при пересеве лиофильно высушенной ма-
точной культуры обычно рекомендуют обогащенные питательны-
ми ингредиентами жидкие среды. Последующие пересевы осуще-
ствляют вначале на агаризованную ферментационную среду, а
затем — на жидкую.
На всех этапах подготовки биообъекта питательные среды
перед их засевом должны быть стерильными (см. главу 7). Биообъ-
379
ект, или промышленный штамм в идеале должен удовлетворять
следующим основным требованиям:
1) стабильность структурно-морфологических признаков и фи-
зиологической активности при длительных хранении и эксплуата-
ции в производстве;
2) повышенные скорости роста и биосинтеза целевого (-ых)
продукта(-ов) в лабораторных и производственных условиях;
3) достаточно широкий диапазон устойчивости к воздействию
неблагоприятных внешних факторов (колебания температуры, pH,
аэрация, перемешивание, вязкость среды);
4) умеренная требовательность к ограниченному числу источ-
ников питания; чем более широкий набор 'источников углерода,
азота и других элементов может использовать производственный
штамм, тем легче его культивировать и с большей экономической
выгодой.
В действительности каждый штамм имеет свои особенности и
не по всем показателям отвечает вышеперечисленным требовани-
ям. Как правило, чем богаче усвояемыми ингредиентами питатель-
ная среда, тем лучше растет и метаболизирует на(в) ней микроор-
ганизм. Уже многие годы используют кукурузный экстракт в
качестве добавки к питательным средам, поскольку он богат не
только источниками углерода и азота, но также микроэлементами
и витаминами. Сухие вещества в нем составляют 45—55%, в их
состав входят зольные вещества —. 1,5—4,5% (таблица 40).
Таблица 40. Химические ингредиенты в составе сухих веществ (СВ)
и золы кукурузного экстракта
Ингредиент Содержание, % Ингредиент Содержание, %
Аминокислоты (в СВ) Аланин Аргинин Валин Гистидин Изолейцин Кислота аспарагиновая Кислота глутаминовая 2,4-5,9 1.0-2,4 0,8-1,8 0,2-0,4 3,5-4,2 1,0-2,7 3,5-8,8 Зольные элементы Алюминий Железо Калий Кальций Магний Марганец Медь Натрий Фосфор (Р2О5) 0,007-0,02 0,023-0,068 0,450-1,350 0,225-0,675 0,188-0,563 0,006-0,018 0,001-0,002 0,075-0,225 0,006-0,018
380
Продолжение табл. 40
Ингредиент Содержание, % Ингредиент Содержание, %
Лейцин 2,7-4,2 Цинк 0.005-0,016
Лизин 1,6-3,7 Витамины (в СВ):
Метионин 0,2-0,6 Биотин (1,5-5,5) • IO 3
Пролин 1.6-2,0 Кислота
Серин 1,2-2,0 никотиновая Кислота (1,2-1,8) • ю'2
Тирозин 0,5-1,0 пантотеновая Кислоты (0,8-1,4) • 10'2
Треонин 0,4-1,1 органические (летучие,
молочная) в СВ 5,1-12,0
Триптофан 0,5-1,0 Сахароза в СВ 0,1-11,0
Фенилаланин Цистин 0,8-1,3 0,2-0,4 -
Не менее часто применяют дрожжевой экстракт из клеток
Saccharomyces cerevisiae, богатый различными веществами —ами-
нокислотами (аргинином — 5%, валином — 5,5%, гистидином —
4%, изолейцином — 5,5%, лейцином — 7,9%, лизином — 8,2%,
метионином — 2,5%, тирозином — 5%, треонином — 4,8%, трипто-
фаном — 1,2%, фенилаланином — 4,5%, цистином — 1,5%) и
витаминами (биотином — 0,06%, инозитом — 0,3%, кальция
пантотенатом — 0,01%, кислотой р-аминобензойной — 0,016%,
кислотой никотиновой — 0,059%, кислотой фолиевой — 0,001%,
пиридоксина монохлоридом — 0,002%, рибофлавином — 0,01%,
тиамина монохлоридом - 0,017%, холинхлоридом — 0,27%) в расчете
на сухое вещество. К тому же в биомассе клеток дрожжей содер-
жится до 50% белков.
Вместо экстракта можно добавлять автолизат или гидролизат
дрожжей.
Объемное и дозирующее оборудование для измерения, транс-
портировки и загрузки биореакторов аналогично оборудованию,
применяемому, например, в пищевой (или в химической) промыш-
ленности: различных типов весы, насосы (например, вакуумные),
транспортеры (ленточные, шнековые), элеваторы, контейнеры и
др. Газообразные и жидкие продукты обычно подают в биореак-
торы по системам стерильных трубопроводов. В крупномасштаб-
ном производстве питательные среды и некоторые их компоненты
стерилизуют нагреванием и/или фильтрованием через пористые
381
мембраны. Тепловая стерилизация может быть периодической и
непрерывной; в биотехнологии применяют оба вида стерилизации.
В случае получения лекарственных средств, например, антиби-
отиков для парентерального введения, необходима исключительно
высокая степень стерильности питательных сред и целевых про-
дуктов. При этом необходимо стремиться снизить, например,
вероятность выживания бактериальных спор до величины менее
10'12, исходя из уравнения:
1—Po(t) = l — e-N', где Р — популяция микроорганизмов, е —
логарифм натуральный, t-время, Nt=No е~к"‘, No — численность
жизнеспособных микроорганизмов в среде до начала стерилиза-
ции, kd — величина, обратная средней продолжительности жизни
микроорганизма, 1 — Po(t) - вероятность выживания хотя бы одной
микробной особи при вероятности гибели
Po(t) = l—(1—| е-*-‘ | ) No
Понятно, что перед засевом биообъекта стерильными должны
быть и питательная среда и биореакторы. Стерилизацию биореак-
торов часто проводят одновременно со стерилизацией питательной
среды в них.
Подготовку биообъектов проводят согласно прилагаемым к
регламентам инструкциям. В заводской или цеховой лаборатории
должна быть подготовлена культура для последующей наработки
инокулюма (инокулята), или посевного материала. В этих целях
исходный штамм микроорганизма, сохраняемый в условиях, близ-
ких к анабиозу или анабиоза (высушенным в стерильной почве,
песке, на пшене, путем лиофилизации, или сублимационной суш-
ки), оживляют после добавления стерильной жидкой питательной
среды с последующим высевом на уплотненную питательную
среду. Убедившись в подлинности и чистоте культуры (культура
называется чистой, если родительские и дочерние клетки в ней
практически неразличимы и между ними нельзя установить род-
ственные связи), операции по пересеву штамма на среду возраста-
ющих объемов (площади) повторяют несколько раз и проводят в
асептических условиях, переходя от пробирок к колбам, помеща-
емым на качалочные устройства (шюттель-аппараты).
Последующую подготовку биообъекта осуществляют в цехе,
используя небольшие ферментаторы-инокуляторы, в которых на-
ращивают посевной материал для промышленных ферментаций.
При этом одноклеточные культуры чаще доводят до середины —
окончания Log-фазы, то есть когда клетки делятся синхронно.
Известно понятие степень синхронизации, то есть степень участия
382
/4^-1
клеток популяции в синхронном делении (О. Шербаум, 1959—1960),
выражающаяся в индексе синхронизации (Is):
1 И
. 5J'
No — число клеток перед началом их синхронного деления, Ni
— число клеток после синхронного деления, Т — время, приходя-
щееся на Log — фазу, g — продолжительность одной генерации.
Индекс синхронизации характеризует степень однородности по-
пуляции.
При необходимости синхронизацию деления можно индуциро-
вать, например, метаболическим шоком (предварительный посев
культуры на голодные среды), температурным шоком (смена тем-
ператур, в частности, пониженных в начале на оптимальные в
последующем), или используя одинаковые по размеру клетки,
механически разделенные, например, фильтрованием (селектив-
ные методы) и т. п.
В зависимости от плотности суспензии ее необходимое коли-
чество может достигать 1—20% объема производственного фер-
ментатора. Для аэробных микроорганизмов в инокулятор достав-
ляют очищенный стерильный воздух.
В целом, предферментация схематично представлена на рис.
125. В инокуляторах, как и в промышленных ферментаторах,
целесообразно поддерживать незначительное избыточное давле-
ние воздуха, когда случайные утечки будут происходить только в
направлении из системы, а не наоборот. Этим дополнительно
обеспечивается асептичность биотехнологического процесса.
В условиях
цеховое
ЗЛЯОДСХйв
лаборжторп
В условиях
рокзвод-
спеожого
цеха
(участка|
Рис. 125. Схема получения посевного материала (1 — инокулятор, 2 — мешалка,
3 — подача очищенного стерильного воздуха для микроорганизма-аэроба, 4 —
регулирование процесса, 5 — сливной штуцер).
383
Инокуляторы должны отвечать следующим основным требова-
ниям: конструктивные простота, удобство и надежность эксплуа-
тации. Посевные аппараты отечественного производства имеют
следующие объемы: 10, 5, 2 и 0,63 м3, диаметром от 0,9 до 2 м и с
частотой вращения мешалки от 180 до 270 оборотов в минуту.
Ферментация
Второй этап биотехнологического процесса, называемый фер-
ментацией, проводят в производственных биореакторах. По био-
химической сущности он во многом имитирует предферментацию
и поэтому названный термин является условным. Тем не менее,
он принят на практике и в специальной литературе и не нуждается
в каких-либо дополнительных пояснениях.
В процессе ферментации также необходимо использование
стерильных питательных сред, воздуха и биореакторов, выбор
которых обусловлен особенностями культивируемых микроорга-
низмов. На рис. 126 представлены подходы к такому выбору (см.
также главу 7).
Одноклеточные
(бактерии, дрожжи)
Одноклеточные (бактерии,
дрожжи), нитчатые грибы
микроорганизмы
Рис. 126. Подход к выбору биореакторов для аэробных микроорганизмов.
384
Микроорганизм в виде суспензии определенной плотности
подают из инокулятора(-ов) в промышленный биореактор, или
ферментатор, в котором содержится стерильная жидкая питатель-
ная среда. При этом не должно произойти попадания каких-либо
посторонних микробов в питательную среду вместе с продуцентом
— все соединения системы должны быть герметично закрытыми.
Схема запорно-регулирующих устройств представлена на рис. 127.
Рис. 127. Запорно-регулирующее устройство
в системе трубопроводов для засева промыш-
ленного ферментатора (2) из инокулятора (1);
3—10 — клапаны, 11 — ловушки конденсата;
АБ — отрезок трубопровода.
Исходя из этой схемы, последовательность операций заключа-
ется в следующем: открывают клапаны 4 и 7 (клапан 3 закрыт) и
стерилизуют участок трубпровода АБ паром под давлением 1,055
кг/см2 20 минут; конденсат собирается в ловушках (11); закрывают
клапаны 7, 8, 9, 10 и открывают клапаны 4, 5, 6; ферментатор
охлаждают под давлением очищенного стерильного воздуха, сте-
рильная среда заполняет соединительный трубопровод; повышают
давление в посевном аппарате до 0,7 кг/см2 при его снижении в
ферментаторе до 0,14 кг/см2; открывают клапан 3 и посевной
материал переводят в ферментатор, после чего отключают иноку-
лятор и ферментатор от системы подачи пара, закрыв клапаны 3
и 6; открывают клапаны 7 и 8, спускают пар и конденсат при
частично открытых клапанах 4 и 5.
Общий объем ферментатора заполняют инокулированной сре-
дой на 70—80%, 20—30% объема заполняют газами (инертным —
для анаэробов, воздухом — для аэробов).
Аэрация жидкости способствует пенообразованию, снижаю-
щему качество ферментаций, поэтому используют пеногашение
либо механическое (установка в верхней части ферментатора
специальной дополнительной мешалки), либо физико-химическое
(использование ПАВ для снижения поверхностного натяжения на
границе раздела фаз "газ-жидкость").
13 т. 8524 385
Длительность ферментаций колеблется в пределах от 4—5 до
14 суток и дольше, что зависит от особенностей физиологической
активности биообъектов. Применительно к биосинтезу антибио-
тиков и экзогликанов периодические ферментации проводят обыч-
но в течение 4—5 суток.
9.2. Выделение конечных продуктов ферментации. В зависи-
мости от целей проведения ферментаций конечным продуктом
может быть биомасса клеток или какой-либо внеклеточный мета-
болит. Тогда в первом случае отходом будет жидкая часть культу-
ральной среды, во втором — клетки (см. также главу 8). Культу-
ральная среда представляет собой смесь клеток биообъекта, его
растворимых продуктов метаболизма, нерастворимых компонен-
тов, выделившихся после автолиза части клеток или вследствие
нарушения клеточных барьеров проницаемости, а также полно-
стью неиспользованных компонентов питательной среды. Исход-
ные характеристики культуральной среды (концентрация клеток
и продуктов метаболизма, вязкость, морфология клеток и клеточ-
ных элементов и др.) накладывают отпечаток на выбор способа
отделения биомассы клеток от жидкой фазы.
В зависимости от целевого продукта используют наиболее
приемлемый подход для его выделения:
Клетки Растворимый метаболит
1. Седиментация 1. Экстракция
и декантация 2. Сорбция
2. Фильтрование 3. Осаждение
3. Центрифугирование 4. Хроматография
4. Отстаивание 5. Выделение с помощью
5. Флотация мембран
Приближенная градация нерастворимых веществ и частиц
(включая микробные клетки) по размерам, сказывающаяся на
выборе методов их отделения и разделения, может быть представ-
лена следующим образом:
1. Крупные частицы — от 0,1 до 1 мм
2. Мелкие частицы — от 0,01 до 0,1 мм
3. Инфрамелкие частицы — от 0,001 до 0,01 мм
4. Высокомолекулярные вещества — от 10 до 1000 нм
(10 5—10 3 4 5 мм)
5. Низкомолекулярные вещества — от 0,1 до 10 нм
(10 -7—10 5 мм).
Для отделения крупных и мелких частиц возможно использо-
вание: седиментации, тканевых и волокнистых фильтров, сит и
386
сетчатых фильтров, фракционирование в пене и пузырьках; для
отделения низкомолекулярных нерастворимых веществ—диализа
и электродиализа, ионного обмена, экстракции растворителями,
обратного осмоса.
Некоторые методы применимы к отделению частиц с более
широким диапазоном размеров, например, ультрафильтрация (для
частиц 2—5 групп), ионный обмен (для частиц 3—5 групп), гель-
хроматография (для частиц 2—4 групп), фракционирование в пене
и пузырьках (для частиц 1—4 групп), чем другие методы: ультра-
центрифугирование (длячастиц, в основном, 5 группы), экстракция
растворителями (для частиц 4—5 групп), жидкостные и циклонные
сепараторы (для частиц 2—3 групп).
Седиментация происходит в поле гравитационных сил. Ею
пользуются для отделения конгломератов типа "кефирных зерен"
при некоторых видах молочнокислого или смешанного брожений
(молочнокислого и спиртового), а также при биологической обра-
ботке отходов с помощью активного ила.
Микробные клетки легко коагулируют под влиянием полика-
тионов или привносимых извне полимеров. Возникающие при этом
хлопья вместе с клетками достаточно легко отделяются седимен-
тацией. Известны легко флокулирующие производственные штам-
мы дрожжей, формирующие осадки.
Декантацию, или слив надосадочной жидкости (декантат, су-
пернатант) можно заменить вакуум-отсасыванием.
Фильтрование малых объемов культуральной жидкости можно
проводить на рамных фильтрах, при больших объемах — на
барабанных вакуум-фильрах. Процесс фильтрации можно суще-
ственно ускорить (в 10—100 раз), если культуральную жидкость
подвергнуть предварительной обработке — тепловой, добавление
флокулянтов (глинозема, кальция хлорида, полиэлектролитов, и т.
д.). Если биомасса клеток оказывает выраженное сопротивление
фильтрованию, то прибегают к использованию фильтрующего слоя
на подкладке. Осадок микробных клеток относится к разряду
сжимаемых, то есть уплотняющихся, поэтому со временем скоро-
сть фильтрации будет заметно уменьшаться. Чтобы исключить
перепады в скоростях фильтрации, слой клеточной массы (напри-
мер, мицелия) срезают с помощью ножа.
Центрифугирование — принудительное осаждение частиц (ве-
ществ) за счет скорости возрастания центробежных сил. В микро-
биотехнологии применяют различные типы центрифуг: осадитель-
ные шнековые, непрерывного действия с выгрузкой осадка через
387
сопла, саморазгружающиеся сепараторы и др. С помощью цент-
рифугирования удается отделять из культуральных жидкостей
бактерии, дрожжи, мицелиальные грибы. Однако например, в
случаях высоковязких культуральных жидкостей прибегают к их
предварительному разбавлению водой (иногда — теплой или горя-
чей) во много раз, после чего приступают к центрифугированию
— сепарированию. Это характерно для отделения продуцентов
высоковязких экзополисахаридов, например, некоторых грибных
гликанов типа аубазидана, пуллулана и др.
Отстаивание реализуют в традиционных процессах брожения,
а также в крупномасштабных процессах переработки отходов. Это
как бы продолжение процесса седиментации.
Флотацию (от англ, floatation — всплывание) используют,
например, для концентрирования микобактерий туберкулеза из
патологического материала в диагностических целях. В микроби-
отехнологии флотацию применяют в производстве белка некото-
рых одноклеточных микроорганизмов, в пивоварении. Пенная
флотация ответственна за образование устойчивой пены на пиве
за счет концентрирования растворенных белков на границе раз-
дела "воздух-жидкость”.
Если целевой продукт представляет собой растворимый мета-
болит или он синтезируется внутри клетки и не секретируется
вовне, то прибегают к другим методам выделения: экстракции,
сорбции, осаждению, хроматографии, выделению с помощью мем-
бран. Экстракцию проводят органическими растворителями из
клеток (твердая фаза), например, антибиотика гризеофульвина
ацетоном, или бензилпенициллина при pH 2,0—3,0 — бутил-аце-
татом из культуральной жидкости после отделения клеток проду-
цента (система "жидкость-жидкость”), или, наконец, экстракция
ферментов (в частности, пуллуланазы) в двуфазных водных систе-
мах, например, глюкана-декстрана и несовместимого с ним поли-
этиленгликоля (ПЭГ-6000).
Сорбция — это. поглощение каким-либо телом газов, паров или
растворенных веществ из окружающей среды. Различают адсо-
рбцию, когда в сорбции принимает участие поверхность твердого
тела; абсорбцию — поглощение вещества всем объемом
жидкости поглотителя; хемосорбцию — поглощение газа,
при котором имеет место химическое взаимодействие между
поглотителем и газом; десорбция — процесс обратный сорбции,
то есть это выделение вещества, поглощенного твердым телом или
жидкостью. Из адсорбентов наиболее известны активированный
388
древесный уголь, кизельгур, силикагель, целлюлоза. Все адсорбен-
ты должны обладать большой удельной поверхностью. Так, 1 г
активированного угля имеет поверхность от 600 до 1700 м2, благо-
даря чему он обладает высокой поглотительной способностью.
Как осветлители адсорбенты широко применяют в микробио-
технологии.
Хорошо известна ионообменная сорбция, используемая, в ча-
стности, при выделении аминогликозидного антибиотика стрепто-
мицина н^ катионообменной смоле, содержащей карбоксильные
группы:
стрептомицин п+ +п (смола)‘Н+<=> п (смола)’ стрептомицин п+
+ Н+. Стрептомицин элюируют (десорбируют) подкисленной во-
дой, смола при этом регенерируется.
В ряде случаев ионообменники можно добавлять непосредст-
венно в культуральную жидкость для избирательной сорбции
несущего заряд действующего вещества.
Осаждение широко используют в микробиотехнологии при
получении белков (например, ферментов), полисахаридов, ряда
антибиотиков и других веществ. Белки выделяют при высаливании,
изменении pH до изоэлектрической точки, снижении диэлектри-
ческой проницаемости раствора, снижении степени сальватации
белковых молекул, и другими процедурами.
Наиболее часто используют метод высаливания белков концен-
трированными растворами солей. Еще в конце XIX в. было уста-
новлено, что различные ионы можно расположить по уменьшению
катионы: Th , Al , Н , Ba , Sr , Са ,Mg ,Cs , Rb ,
NHj Kt Nat Lit
анионыГООС - CH2- C(OH) - CH2~ COO 7
COO-
-OOC-CH(OH)-COO7F_,IO2, НзРОф CH3COO ,
Br^CltCOt Br“, NO3,C1O4, I , CNS .
высаливающей активности в виде лиотропных рядов, или рядов
Гофмейстера:
Несмотря на то, что аммония сульфат наиболее часто приме-
няют в качестве высаливающего агента, его с успехом можно
заменить другими солями.
Высоковязкие экзополисахариды (аубазидан, декстран, ксан-
тан, пуллулан и др.) выделяют осаждением этанолом или ацетоном,
то есть смешивающимися с водой органическими растворителями.
389
Альгинаты—полианионные биополимерные углеводы осаждаются
раствором кальция хлорида в виде геля.
Некоторые осадители образуют с растворенным действующим
веществом нерастворимые соли, выпадающие в осадок в присут-
ствии органического растворителя:
стрептомицин + H2SO4+ органический растворитель —>
—> сульфат стрептомицина
Хроматографическое разделение БАВ используют в различных
вариантах: гель-фильтрация, или гель,-проникающая хроматогра-
фия — хроматография на молекулярных ситах, ионообменная
хроматография.
Гель-фильтрацию успешно применяют для разделения смеси
веществ, различающихся по молекулярным массам. Это могут быть
различные метаболиты микроорганизмов, в том числе — полидис-
персные белки и полисахариды. Причем небольшие по размеру
молекулы разделяемых веществ способны проникать в гель-насад-
ку, и поэтому они движутся в хроматографической колонне более
медленно, тогда как большие молекулы не проникают в частицы
геля и движутся быстрее — они выходят из колонны в первую
очередь.
При ионообменной хроматографии ионообменная смола пред-
ставляет собой неподвижный слой, тогда как, например, раствор
смеси белков является подвижным слоем. Так, белки в катионной
форме связываются с катионобменной карбоксиметилцеллюлозой
(КМЦ), несущей отрицательные заряды на целлюлозной матрице.
Последующую элюцию белков проводят с помощью буферных
растворов возрастающей ионной силы. Первыми элюируют белки,
более слабо связанные с КМЦ.
Подобный процесс проводят и на анионобменниках. Наиболее
часто применяемым анионитом является диэтиламиноэтилЦеллю-
лоза.
В случае афинной хроматографии используют высокую специ-
фичность таких природных веществ, как ферменты, антитела и
лектины. Ферменты образуют комплексы с ингибиторами, анти-
тела —- с соответствующими антигенами (иммуносорбционная
хроматография), лектины — со специальными рецепторами кле-
точных стенок.
Метод выделения различных веществ или клеток с помощью
мембран все шире внедряется в биотехнологию. Диаметр молекул
или частиц является определяющим фактором в выборе обратного
осмоса или ультрафильтрации. Для примера можно привести
390
приближенные размеры (диаметр) некоторых молекул и клеток в
мкм:
вода (ММ 18 Да) — 0,0002, органические кислоты с ММ от 100
до 500 Да — 0,0004—0,0008, монозы и биозы с ММ от 180 до 400
Да — 0,0008—0,001, некоторые антибиотики с ММ от 300 до 100
Да — 0,0006—0,0012, протеины и гликаны с ММ от 10000 до 1 млн.
Да—0,002—0,01, клетки ряда бактерий — 0,3— 1, клетки некоторых
дрожжей и мицелиальных грибов — 1—10.
Явление уравновешивания химических потенциалов какого-ли-
бо вещества в растворе и в чистом растворителе, разделенных
непроницаемой для вещества мембраной, называют осмосом.
Избыточное давление, приложенное к раствору и превышаю-
щее его осмотическое давление, побуждает перенос растворителя
против градиента концентрации (из разбавленного раствора в
чистый растворитель). При этом происходит возрастание концен-
трации растворенного вещества, то есть имеет место обрат-
ныйосмос.
В ряду с осмотическими процессами находится диализ, когда
происходит непрерывная диффузия малых молекул в циркулиру-
ющий растворитель с удерживанием больших молекул. При этом
отсутствует выраженный перенос растворителя против градиента
концентрации. Диализ применяют, например, при очистке анти-
генных препаратов, представляющих собой протеины, гликопро-
теины или более сложные комплексы. С помощью диализа через
раличные мембраны освобождаются от неорганических солей.
Ультрафильтрация (см.) применима для концентрирования и
очистки молекуле диаметром от 1 до 100 нм (отдельных ферментов)
от низкомолекулярных примесей. Растворитель при этом частично
удаляется.
На практике нередко используют комбинацию методов для
разделения клеток и молекул биологически активных веществ.
Процесс ферментации можно оценивать по различным пока-
зателям, используя для этого соответствующие расчетные форму-
лы:
1) продуктивность по биомассе (Qx, г/л*ч):
а) для периодического процесса Qx =
ч— to
б) для непрерывного процесса Qx — DX, где Хо — концент-
рация биомассы (г/л) на период времени to (ч), Xi — концентрация
биомассы (г/л) на период времени tt (ч), D — коэффициент разбав-
ления или скорости протока, (1/ч), равный удельной скорости (ц)
391
для непрерывного процесса; D можно нарушить, если изменить
скорость протока среды или концентрацию субстрата в ней;
2) удельная скорость роста (1/ч):
Xi-Xo
3) концентрация биомассы: Xi=Xoe ц v,~ w
для log-фазы размножения е=2,718;
4) продуктивность по целевому продукту (Ор, г/л«ч):
v л Pl— Рр
а) для периодического процесса: Qp = —-
tr~ to
б) для непрерывного процесса: Op=DP, где Р — концентра-
ция продукта (г/л • ч);
5) удельная скорость образования целевого продукта (qp) в
г/г« ч:
Л-Ро
Чр Х^- to)
6) удельная скорость потребления субстрата (qs, г/г*ч):
•Sfl— St а /
~ —77 . где S — концентрация субстрата в г/л;
Хцц— to)
7) выход биомассы из субстрата, или экономический коэффи-
циент (Yx/S) в г/г:
v — Р _
x/s" 9s " So-S. :
8) выход целевого продукта (Ye/S) в г/г:
V — 4р _ Pi~ Pq
P,s“ 9s" So- S. •
9.3. МИКРОБИОТЕХНОЛОГИЧЕСКИЕ ПРОЦЕССЫ
9.3.1. Получение продуктов брожения. Брожение — это одна
из разновид ностей биологического окисления субстрата у гетерот-
рофных микробов в целях получения энергии, когда акцептором
электронов или атомов водорода является органическое вещество.
Биотехнологические бродильные процессы изучены давно в срав-
нении, например, с биотехнологией антибиотиков, аминокислот и
других продуктов. Однако некоторые брожения реализованы на
практике относительно недавно, например, брожения с участием
Zymomonas spp.
В основе многих бродильных процессов лежит универсальная
реакция превращения глюкозы (источник углерода) в ключевой
392
промежуточный продукт (интермедиат) — пировиноградную кис-
лоту, или пируват, из которого синтезируются различные конечные
продукты. По метаболиту, образующемуся в наибольшем количе-
стве, называют соответствующее брожение: спиртовое, масляно-
кислое, молочнокислое, и т. д.
сн3сосн3
ацетои i i
СООН
уксусная
КИСЛОТ» ( f
СН3(СН2)3ОН
бутанол
пропионов»
кислота
9.З.1.1. Спиртовое брожение. Оно лежит в основе получения
этилового спирта, кормовых и пищевых дрожжей, пивоварения и
виноделия. Возбуд ителями спиртового брожения могут быть дрож-
жи — сахаромицеты, некоторые мицелиальные грибы (Aspergillus
oryzae) и бактерии (Erwinia amylovora, Sarcina ventricula, Zymomonas
mobilis, Z. anaerobia). Среди названных организмов дрожжи зани-
мают ведущее место, а получение с их помощью этанола относят
к разряду наикрупнейших в мире. Этанол используют в различных
отраслях народного хозяйства: то как растворитель, то как сырье
для химического синтеза, широко используют в медицине, и т. д.
Специальные штаммы Saccharomyces cerevisiae рекомендованы
как биообъекты в производстве этанола (на африканском конти-
нете чаще применяют Schizosaccharomyces pombe и S. octosporus).
Штаммы подразделяют в свою очередь на расы верхового и
низового брожений, а по способности к флокуляции — на хлопь-
евидные и пылевидные. Расами верхового брожения являются
спиртовые, хлебопекарные и некоторые пивные дрожжи, расами
низового брожения — большинство винных и пивных дрожжей.
Клетки обеих рас могут быть подвержены флокуляции. При этом
следует помнить, что пылевидные дрожжи находятся в дисперги-
рованном состоянии в течение бродильного процесса. Они менее
стойки к автолизу, но более полно сбраживают сусло; хлопьевид-
ные — оседают на дно или всплывают на поверхность, они более
выраженные ароматизаторы.
393
Особенности некоторых продуцентов этанола приведены в
таблице 41.
В отличие от S. cerevisiae аэротолерантные бактерии Z. mobilis
меньше чувствительны к этанолу, у них отсутствует катаболитная
репрессия, а удельная скорость потребления глюкозы и образова-
ния этанола в 2—3 раза выше (qC2H5OH=:: 1,87 г/г *ч). Катаболизм
глюкозы протекает по Энтнеру-Дудорову. Однако скорость раз-
множения этой бактерии низка и продуктивность по спирту не
столь высока. Здесь остается резерв надежд на позитивные резуль-
таты генетико-селекционной работы с продуцентом.
Значительный интерес представляет S. rosei. Эти дрожжи спо-
собны образовывать этанол при использовании топинамбура (зем-
ляная груша), в котором из углеводов содержится преимуществен-
но инулин, сбраживаемый после гидролиза до этанола. Топинамбур
хорошо растет даже в северных регионах (Архангельская, Ленин-
градская и другие области) на бедных супесчаных почвах.
Таблица 41. Характеристика микроорганизмов — продуцентов этанола
Микроорганизм Оптимальные параметры культивирования Выход этанола, % от макси- маль- ного Макси- маль- ная кон- цент- рация этанола, г/л Примечание
pH t°. °C
Saccharomyces cerevisiae 3-4 30 100 130 Не использует пентозы
Saccharomyces rosei 4,6 35 88 42,5 Конвертирует инсулин в этанол
Kluyveromyces marxianus 4,4 35 88 44 и
Pachysolen tarmophilus 4,2 25 40-60 20 Растет в смешанной культуре. Использует ксилозу
Zymomonas mobilis 5.5 30 95 130 Аэротолерантен
Zymomonas anaerobia 5,6 35 90-95 96 Термоустойчив
394
(Продолжение таблицы 41)
Микроорганизм Оптимальные параметры культивирования Выход этанола, % от макси- маль- ного Макси- маль- ная кон- цент- рация этанола, г/л Примечание
pH t°, °C
Clostridium thermocellum 7 62 50 1,5 Образует активный целлюлазный комплекс, не утилизирует ксиланы, ксилозу. Соотношение этанола и ацетата «1:1, устойчив в ассоциациях
Clostridium thermohyd- rosulfuricum 6,9-7,5 69-70 80 — Аэротолерантен, время генерации 70 мин.
Clostridium thermosac- charolyticum — 55-60 70 27 на кси- лозе Образует бутират
Thermoanaerobacter ethanolicus 5,8-8,5 69 90 — Широкий оптимум pH; время генерации 90 мин.
В качестве сырья для производства этанола в различных странах
используют доступные растительные источники: зерновые, карто-
фель и свекловичная меласса — в России, Украине, Беларуси;
сахарозу и тростниковую мелассу — в США, рис — в Японии и т.
д. В принципе любой источник гексозанов может быть использован
в качестве сырья для получения этилового спирта, например,
целлюлоза в древесине хвойных, соломе, торфе и пр. Поэтому
сульфитные щелока — отходы целлюлозно-бумажной промышлен-
ности нашли широкое применение в производстве этилового спир-
та.
Ниже представлена технологическая схема получения этанола,
из которой видно, что первым делом необходимо трансформиро-
вать крахмал в глюкозу, чего добиваются при обработке сырья
395
амилолитическими ферментами. На практике обычно применяют
грибную амилазу (Aspergillus niger, A.oryzae и др.) или пророщен-
ное зерно (солод).
Крахмал
Клейстеризация
+ ферменты солода (преимущественно ячменного)
или грибов
Осахаренная масса (затор)
1 + дрожжи
Бражка
1 дистилляция
Этанол
Крахмал для получения этилового спирта может быть различ-
ного происхождения (картофельный, кукурузный, пшеничный,
рисовый). Крахмалистое сырье предварительно дробят (измельча-
ют), применяя для этого вальцовые, молотковые или другие дро-
билки. Крахмал необходимо клейстеризовать при разваривании.
Например, пшеница и пшеничная крупчатка способны полностью
клейстеризоваться при 68°С в течение 30 мин. Крахмал затем
должен быть гидролизован до низших сахаров (моноз, биоз),
поскольку более высоко полимеризованные углеводы не сбражи-
ваются дрожжевыми организмами. В качестве гидролизующих
агентов применяют соответствующие ферментные препараты из
нитчатых грибов или солода. Крахмал состоит из амилозы и
амилопектина. Неразветвленная амилоза почти полностью гидро-
лизуется до биозы — мальтозы, тогда как разветвленный амило-
пектин гидролизуется лишь частично — до декстринов, медленно
разлагаемых дрожжами до мальтозы в процессе брожения.
Кроме сахаров и декстринов в заторах обычно содержатся
аминокислоты, пептиды, макро- и микроэлементы в виде неорга-
нических солей (фосфор — еще и в виде фосфорорганических
соединений).
Сбраживание затора .осуществляется с помощью чистых куль-
тур либо периодическим, либо непрерывным способом. Относи-
тельно высокие начальные концентрации сахаров и декстринов в
заторах неблагоприятны для бактерий — контаминантов из-за
повышенного осмотического давления. Позже, когда осмотическое
давление снижается, образующийся этанол и естественно возра-
стающая или искусственно создаваемая подкислением серной
кислотой) кислотность среды (pH 3,8—4,0) выступают основными
396
факторами, предотвращающими развитие контаминирующих бак-
терий.
В период брожения поддерживают темпрературу от 30°С до
38°С, (в зависимости от расы дрожжей).
На сбраживание затора влияют не только вид и раса дрожжей,
температура и pH, но и конструктивные особенности фермента-
ционных аппаратов (система охлаждения/нагрева, способ и интен-
сивность перемешивания). Длительность сбраживания составляет
в среднем от 1,5 до 3 суток.
В бражке накапливается от 1—1,5% до 6,5—8,5% этанола; его
перегоняют и ректифицируют до 96%. Кроме того, в бражке
содержатся так называемые "сивушные масла" (высококипящая
фракция;— 90°—150°С) и 5—10% альдегидов с эфирами. Сивушные
масла представляют собой смесь изопропилового и н-пропилового,
изобутилового и н-бутилового, изоамиловых (2-метил- и 3-метил-
бутанолы) спиртов. Доля последних двух обычно составляет 50%;
в сивушных маслах находят также [3 -фенил- и р-оксифенилэтило-
вые спирты.
Исходя из расчетов по содержанию крахмала на сухое веще-
ство, различные сорта кукурузы, пшеницы, риса, сорго накапли-
вают в среднем 65—75% крахмала, из которого можно получить до
45 дкл этанола.
Отходами производства являются барда и диоксид углерода
Барду используют для откорма скота и птиц, диоксид углерода —
в пищевой промышленности, например, в виде "сухого льда".
Этанол можно получать также при сбраживании гидролизатов
древесных и травянистых растений, содержащих целлюлозу. В
таких гидролизатах обычно содержится 2—3,5% редуцирующих
сахаров (преимущественно — гексозы и, меньше, пентозы, в
большем количестве присутствующие в гидролизатах древесины
лиственных растений).
Применив методы генетической инженерии, удалось включить
в дрожжи Schizosaccharomyces pombe ген, кодирующий биосинтез
фермента ксилозоизомеразы. Этот фермент катализирует реакцию
превращения D-ксилозы в D-ксилулезу. Векторной системой при
этом была "ксилозоизомеразная" плазмида Escherichia coli.
Как известно, D-ксилулоза может включаться в пентозный
цикл, способный переключаться на гликолиз с помощью образую-
щихся ключевых веществ — фруктозо-6-фосфата и 3-фосфогли-
церинового альдегида (интермедиаты в пентозном цикле). В итоге
удалось добиться прямой конверсии ксилозы в этанол. Следова-
тельно, клетки S.pombe, несущие ген ксилозоизомеразы, стали
397
способными одновременно сбраживать глюкозу и ксилозу в гид-
ролизатах разных пород древесины.
Известны природные виды бактерий, сбраживающие глюкозу
и ксилозу, например, Thermoanaerobacter ethanolicus. Однако не-
обходимо иметь в виду рентабельность бактериальных анаэробных
ферментаций гидролизатов растительных отходов — она реальна
лишь в тех случаях, когда концентрация этанола в бражке будет
не ниже 4,5—5%.
Так называемый "гидролизный спирт" после ректификации
содержит до 0,05—0,1% метанола и относительно повышенные
количества альдегидов, органических кислот и эфиров в сравнении
с этанолом — ректификатом из картофеля или зерен.
Отходами целлюлозного производства являются сульфитные
щелока, содержащие около 3% редуцирующих веществ (таблица
42).
Т а б л и ц а 42. Примерный состав сульфитных щелоков
Ингредиенты %
Сухие вещества 11,5-12,2
Редуцирующие вещества в пересчете на глюкозу 2,0-2,7
Сбраживаемые углеводы в пересчете на глюкозу 1,3-1,9
Метоксильные группы 0,78-0,8
Зола 1,2-2,1
Диоксид серы 0,4-1,4
Оксид кальция 0,6-1,0
Общая сера 0,9-1,6
Сульфаты 0,1
Сахара в сульфитных щелоках, например, из ели, обычно
включают глюкозу—около 29%, галактозу — 4,2%, маннозу — 43%,
пентозы — 17%, уроновые кислоты — 3%, фруктозу — 4%.
Сорта и происхождение древесины, условия сульфитной варки
сказываются на качественных и количественных показателях для
сульфитных щелоков.
398
Из щелоков извлекают прежде всего целлюлозу, после гидро-
лиза которой сбраживание гексоз (глюкозы) осуществляют с по-
мощью специальных рас Saccharomyces cerevisiae. "Сульфитный"
спирт — один из наиболее дешевых, он содержит до 2—8%
метанола, а также другие примеси. Его используют в технических
целях.
Применяя древесину, обычно стремятся совместить получение
этанола и дрожжей, хотя оба этих процесса можно вести по
отдельности. Ниже приведена схема комбинированного использо-
вания целлюлозы.
Древесина
пар ->j+ H2SO3«----------------SO2---------
Варка целлюлозы
Гидро- Гсепарирование '^Сульфитный щелок
™з | i
Целлюлоза <-------------Сепарирование
|<- пар
Продувание щелока -------
1 + известковое молоко
(аммиачная вода)
----------<----------------> Нейтрализация
Осадок (отход) Ч-
Осветленный раствор
„ 'I'
Охлаждение
Спиртовое брожение
|пар
Дистилляция и ректификация
Этанол, метанол и пр. Ч------
Охлаждение
аэрация --->|+ питательные соли и
Iдрожжи
Флотация, промывание, сепариро-
вание, высушивание
Кормовые дрожжи
399
Расчетами показано, что при совмещенном производстве
спирта и кормовых дрожжей "выходные" показатели на 1 т
абсолютно сухой древесины выглядят следующим образом:
этанол (абс.) — 175—182 л, метанол — 2 кг, сивушные масла —
0,3 кг, фурфурол (94%-й) — 5,6 кг, диоксид углерода (жидкий)
— 70 кг, дрожжи с остаточной влажностью 10% — 32 кг, лигнин
(абсолютно сухой) — 380 кг, гипс — 225 кг.
На сульфитных щелоках можно получать грибную кормо-
вую массу, используя, например, сапрофитный Paecilomyces
varioti, который утилизирует гексозы, пентозы и ацетат. Подо-
бная технология разработана в Финляндии.
При спиртовом брожении часто используют мелассу —
отход производства свекловичного или тростникового сахара
(патока). Она содержит в среднем 80% сухих веществ и 20%
воды. Примерно от 30—40% до 45—50% сухих веществ состав-
ляет биоза-сахароза, 0,5—2% — раффиноза и до 12—18% —
инвертный сахар (смесь глюкозы и фруктозы), прочие вещества
представлены аминокислотами, бетаином (органическое осно-
вание), некоторыми витаминами группы В, неорганическими
солями, пигментами; pH мелассы находится в диапазоне 7,2—
8,9.
Мелассу применяют одновременно для получения этанола
и -кормовых дрожжей (Рис. 128а). Сравнивая "паточные" и
"гидролизные” дрожжи, можно привести следующие данные по
основным ингредиентам клеток (таблица 43).
В паточных дрожжах сравнительно меньше липидов, неко-
торых витаминов (биотина, инозита, пиридоксина, рибофлави-
на), углеводов, но несколько больше кислоты пантотеновой,
тиамина, холина и зольных элементов; паточные и гидролизные
дрожжи примерно одинаковы по содержанию белков и по
коэффициенту отягощения белка азотом нуклеиновых кислот,
или КОБА-коэффициенту, равному 20% (Nh^/N^. • 100). Для
сравнения можно указать на КОБА-коэффициент для белка
нитчатых грибов — он равен 2—5%, что выгодно отличает его
от белка дрожжей и, тем более, от белка бактерий (КОБА-ко-
эффициент 30%).
400
Т а б л и ц a 43. Химический состав "паточных” (ПД) и "гидролизных" дрожжей
(ГД). % от сухих веществ
Ингредиенты ПД ГД
Белки 46—52 43—58
Биотин 3-105—1,6’КГ4 6-Ю5—2,3-Ю'4
Зольные элементы 12—14 5—11
Инозит 0,045—0,35 0,12—0,48
Кислота никотиновая 0,036—0,045 0,04—0,06
Кислота пантотеновая 0,009—0,011 0,006—0,01
Липиды 0,5—2 3—4
Пиридоксин 7 • 10'4—1,2’Ю3 1-10'3—2-Ю3 1
Рибофлавин 0,004—0,007 0,004—0,013
Углеводы 10—18 11—23
Холин 0,38—0,75 0,25—0,45
Одна из схем получения этанола и хлебопекарных дрожжей на
мелассе приведена на рис. 128(6). Перед перегонкой бражки
(позиция 7) хлебопекарные дрожжи сепарируют. Барду в данном
производстве можно использовать для выращивания кормовых
дрожжей.
Рис. 128. Технологическая схема получения: кормовых дрожжей (а): 1 —
ферментатор, 2 — сборник, 3,5 — сепараторы, 4 — сборник, 6 — термонагреватель,
7 — сушилка, 8—упаковка, фасовка; этанола и хлебопекарных дрожжей на мелассе
(б): 1 — рассиропники, 2—4 — аппараты с чистой культурой, 5— стерилизатор, 6
—дрожжегенератор, 7—насос, 8 — бродильный аппарат, 9—головной брод ильный
аппарат.
401
Дрожжевые организмы, применяемые для получения этанола
из патоки, должны быстро и эффективно сбраживать высокие
концентрации сахаров (включая рафинозу) и быть устойчивыми к
повышенной температуре (до 35°С).
Чтобы приготовить затор, патоку разбавляют водой до получе-
ния 14—18%-й концентрации сахаров, подкисляют серной кисло-
той (можно соляной или молочной) до pH 4—5, при необходимости
добавляют 0,1% аммонийных и 0,01% фосфорных солей, а затем
2—4% по объему, суспензии активных дрожжей Saccharomyces
cerevisiae [в России чаще используют расы V-30 (рис. 129), а также
диплоидные гибриды 67 и 73, полученные в результате соматиче-
ской гибридизации расы Я и
а-галактозидазу].
пивных дрожжей, образующих
Брожение ведут при темпе-
ратуре 21—27°С в начале про-
цесса, при 32—33°С — в после-
дующее время. Длительность
брожения зависит от качества
мелассы, в среднем продолжи-
тельность составляет 36—72 ча-
са, содержание этанола в браж-
ке составляет 6—9%.
Бактериальная контамина-
ция заторов, как правило, иск-
лючена благодаря их низким
значениям pH и высокому со-
держанию сахаров (оба фактора
Рис. 129. Дрожжи Saccharomyces
cerevisiae раса V-30 на мелассной среде.
Х7000.
являются основными, определяющими ингибирование роста бак-
терий).
. Процесс брожения можно осуществлять в различных вариан-
тах — непрерывном, двухстадийном. На рис. 128 показан двухста-
дийный процесс, когда в дрожжегенераторах (позиция 6) среду
аэрируют 3—4 м3/м3*ч при 28—30°С и pH 4,2—4,5; дрожжи
наращивают до 2,5—6,5% на сухие вещества, затем передают в
бродильные аппараты (позиции 8,9), где в анаэробных условиях
протекает спиртовое брожение. Перегонку или дистилляцию спир-
та проводят с целью его отделения от примесей и последующей
ректификации до получения 96% этанола или абсолютного (100%)
спирта.
Спиртовое брожение можно интенсифицировать различными
путями:
402
1) использованием непрерывной ферментации вместо перио-
дической, что повышает продуктивность системы по этанолу более
чем в 2 раза; при этом желательно применение флокулирующих
рас активных дрожжей, осуществление рециркуляции биомассы,
иммобилизации клеток;
2) проведением вакуумной ферментации при разрежении
4265,6—4665,5 Па с целью удаления этанола и снижения его
ингибирующего действия (не забывать при этом о возможной
контаминации);
3) осуществлением флеш-ферментации, то есть, когда часть
культуральной жидкости периодически подают в вакуумную ка-
меру для удаления этанола;
4) благодаря селекции этанол-толерантных штаммов микроор-
ганизмов, способных в ходе брожения образовывать сравнительно
высокие концентрации спирта.
В различных странах мира названные пути интенсификации
спиртового брожения уже применяют на практике.
В последние годы реализованы усилия исследователей по не-
прерывному бессепарационному сбраживанию углеводсодержаще-
го сырья с использованием дрожжей, иммобилизованных на по-
верхности раздела фаз "газ-жидкость", то есть на пузырьках газа.
С этой целью используют определенные штаммы дрожжей, напри-
мер, Candida tropicalis, выращиваемых в анаэробных условиях.
Коэффициент иммобилизации (Kta) рассчитывают по формуле
.. X] - х2 _ _
Ли =------, где Xi — концентрация в биореакторе абсолютно
сухих дрожжей в г/л, х2 — концентрация таких же дрожжей (г/л)
в оттоке. Для различных дрожжевых организмов К1Н1 неравнозна-
чен. Для Saccharomyces cerevisiae и S.uvarum он равен 0,1; для S.vini
— 0,22, для Schizosaccharomyces sp. штамм ЛГС-1 — 0,07; для
С.tropicalis — от 0,62 до 1,0.
Основные условия реализации данного процесса: способность
клеток к флотации, работа ферментатора в режиме газожидкост-
ной эмульсии, осуществление отбора культуральной жидкости в
зоне, лишенной подвода энергии на перемешивание.
Приемлемый газо-эмульсионный режим возникает при интен-
сивном барботаже пенящихся субстратов. Для этой цели пригодны
ферментаторы с ярусными турбинными мешалками (рис. 130):
403
Рис. 130. Ферментатор для не-
прерывного бессепарационного
сбраживания углеводсодержаще-
го сырья дрожжевыми организма-
ми, иммобилизованными на по-
верхности раздела фаз в системе
"газ-жидкость": 1 — ферметатор,
2 — слой пены с адгезированными
клетками, 3—зона, лишенная под-
вода энергии на перемешивание,
4 — ложное дно (карман), 5 —
мешалка, 6 — подвод питательной
среды, 7—насос, 8 — компрессор,
9 — газ (подведение), 10 — газ
(отведение).
Описанный способ имеет следующие преимущества: отсутст-
вие диффузионного барьера между клетками и средой культиви-
рования, прочная адгезия клеток, нет необходимости регенериро-
вать носитель, возможность проведения процессов накопления
клеток, их иммобилизации и осуществление основного биотехно-
логического процесса получения спирта на одном и том же обору-
довании.
На примере спиртового брожения показано, что выход этанола
составляет 95% от используемого сахара.
Целенаправленное получение хлебопекарных дрожжей (расы
верхового брожения) реализуют на мелассной среде при аэрации
(pH 4,4—4,5) по так называемому приточному методу, когда среду
подают в биореактор преимущественно непрерывным, умеренно
возрастающим (по объему) потоком. На первом и последнем часе
ферментации аэрация равна 1:1, в период интенсивного размно-
жения дрожжей — 1,5—2 об/об*мин. Клетки при этом проходят
все фазы размножения. Выход прессованных дрожжей составляет
около 38% сухой, биомассы (порядка 150% от использованного
сахара).
Отсепарированные, промытые водой и спрессованные хлебо-
пекарные дрожжи хранят при 4—5°С в рефрижераторах. Требо-
вания к таким дрожжам следующие: влажность не более 75%,
быстрая реактивация в замесах муки, активное сбраживание са-
харов (высокие подъемная сила и мальтазная активность), солето-
лерантность и устойчивость к примесям в мелассе.
Изготавливают также сухие хлебопекарные дрожжи, высуши-
вая прессованные до влажности 7—10%, и так называемые "жидкие
дрожжи”, представляющие собой полуфабрикат, содержащий хле-
404
бопекарные дрожжи, выращенные на осахаренной мучной завар-
ке, которая предварительно была заквашена молочнокислыми
бактериями — Lactobacillus delbrueckii.
Спиртовое брожение находится также в основе пивоварения.
Пиво относят к так называемым солодовым слабо алкогольным
напиткам, получаемым в результате сбраживания дрожжами экс-
трактов из проросших семян хлебных злаков (солода). В подобных
экстрактах содержатся сбраживаемые углеводы.
В различных сортах пива находятся этанол, углеводы (глюкоза,
мальтоза, мальтотриаоза, мальтотетраоза, декстрины), азотистые
вещества (амиды, аминокислоты, пептоны), диоксид углерода ‘—
продукты ферментативного гидролиза осоложенного зерна; горе-
чи, смолы, танин, эфирные масла — из соцветий женских особей
хмеля, следы неорганических солей и жира. Окраска, аромат и
крепость пива в значительной степени зависят от штамма дрожжей
(Saccharomyces cerevisiae, S. carlsbergensis и др.). В России наиболее
широко используют S. cerevisiae низового брожения, расы 776, 41,
44, И, 8а(М), РиЕ
На практике чаще всего применяют ячменный солод. Для его
приготовления отсортированные зерна ячменя увлажняют, прора-
щивают при 15—25°С до тех пор, пока зародышевый листок
становится в 3—4 раза длиннее зерна, затем проросший ячмень
высушивают до конечной влажности солода 5%. В таком виде солод
может хорошо сохраняться, при этом он имеет специфические
окраску и аромат. Солод светлого цвета получается при более
низкой температуре высушивания, более темного цвета — при
повышенной температуре (светлые сорта пива содержат меньше
углеводов, чем темные сорта). Отделенный от проростков сухой
солод может быть использован не только в пивоварении, но и в
винокурении (получении спирта), в кондитерском производстве.
Если пиво изготавливают из солода, хмеля и воды (без каких-
либо "дополнителей"), то следующим этапом является затирание,
когда стремятся перевести в раствор наибольшую часть содержи-
мого солода. Затирание чаще осуществляют либо настаиванием,
либо вывариванием. По первому методу солод дробят, размешива-
ют в воде при температуре 38—50°С (выдерживают 1 час), когда
активизируются протеазы, затем температуру повышают до 65—
70°С и оставляют затор на несколько минут для гидролиза крахмала.
После этого температуру повышают до 75—77°С для денатурации
ферментов и затор фильтруют.
405
По второму методу (вываривание) размолотый солод вносят в
теплую (40°С) воду, размешивают и постепенно повышают темпе-
ратуру затора до 75°С; около 1/3 такого затора отбирают и непро-
должительно кипятят, после чего его возвращают в основной затор.
При этом ферменты разрушаются, клеточные стенки набухают,
крахмал "распускается" (разжижается), чем облегчается его гидро-
лиз в основном заторе. Кипячение и возврат части затора можно
повторять 2—3 раза.
Пиво, полученное на настоенном заторе, более ароматное,
поскольку в него переходит меньше горьких веществ.
Последующие этапы технологического процесса пивоварения
заключаются в следующем:
Затор
фильтрация
Дробина <-
(отход)
4 Сусло неохмеленное
Осадок «------
(альбумины,
Хмелевы смолы,
неорганические
соли)
кипячение
+ хмель
— Фильтрация
охлаждение
Сусло охмеленное
+ дрожжи
Сусло сброженное
(пиво неосветленное)
выдержка в лагер-
ных емкостях в
Осадок <------------
(клетки дрожжей,
нерастворимые
фосфаты)
течение нескольких
недель
Пиво осветленное
фильтрация
Пиво на розлив
(пастеризация)
Пивные дрожжи относят к разряду флокулирующих, оседаю-
щих при осветлении молодого пива и в конце дображивания; они
не сбраживают декстрины (эти полимерные углеводы вносят оп-
ределенный вклад в создание вкуса пива). Отсепарированные
пивные дрожжи иногда рекомендуют в качестве лекарственного
средства.
В последние годы удалось перенести ген Bac.subtilis, детерми-
нирующий Р-глюканазу в пивные дрожжи S.cerevisiae. Этот реком-
406
бинантный штамм оказался способным перерабатывать крахмал
непосредственно в этанол (исключается стадия солодования ячме-
ня). В этом направлении также перспективно использование бро-
дильных штаммов дрожжей S.cerevisiae, несущих киллер-плазми-
ды, благодаря которым они могут быть устойчивы к "диким"
дрожжам.
Из алкогольных напитков можно упомянуть русский хлебный
квас, содержащий менее 0,5% этанола, популярный в Японии
алкогольный продукт Саке (12—24% этанола), таэте — алкогольный
напиток, приготовляемый из молока и с давних пор применяемый
в Скандинавских странах — содержит менее 2% этанола, и другие.
Спиртовое брожение находится также в основе виноделия.
Вина обычно получают из сока спелого неиспорченного винограда,
отделенного или неотделенного от мезги (например, при изготов-
лении красных вин). Индукторами брожения являются различные
расы Saccharomyces cerevisiae. В винах, кроме этанола, содержатся:
белки, пигменты, неорганические соли, летучие и нелетучие орга-
нические кислоты, танин, в некоторых сортах — углеводы, глице-
рин.
Изготовление вин отрабатывалось веками, поэтому в техноло-
гии каждого сорта существуют особые подходы, хотя принципи-
альные стадии укладываются в следующую схему:
Виноградный сок, или сусло
+ дрожжи (температура брожения
21-32,2°С)
Молодое вино
Л слив
(дрожжи) Дображ> 1°=21,1-; евание 7-11 дней, !9,4°С переливание
[КС4Н5О6(винный камень), дрожжи] Хранение в деревя Готовое ви> и созревание нных герметичных сборниках осветление (при необходимости - крепление виноградным спиртом), охлаждение но. розлив
407
Вина классифицируют по-разному. Так различают: сортовые
— по сорту винограда, купажные — из смеси сортов; сладкие и
сухие — по содержанию сахара; натуральные и крепленые, столо-
вые и десертные — по содержанию спирта; игристые и неигристые
— по содержанию углекислоты; белые и красные — по цвету;
ординарные и марочные — по срокам выдержки.
Как пояснение к классификации можно отметить, что в сухом
вине сахар фактически полностью сброжен, а если он имеется, то
в таком количестве, что не ощущается на вкус. В сладких винах
сахар выражение ощущается на вкус. Натуральные вина содержат,
как правило, 9—11% этанола, реже — 13%. В крепленые сухие вина
добавляют коньяк или винный спирт. Столовые вина содержат
менее 14% спирта, десертные — более 14% (в среднем около 20%)
и некоторое количество сахара. Игристые вина содержат значи-
тельное количество диоксида углерода, образующегося при добра-
живании вина в толстостенных сосудах или добавляемого к нату-
ральным винам; к игристым относят шампанское — продукт
вторичного брожения вина, когда к недобродившему вину перед
розливом в герметизированные бутылки добавляют ликер до со-
держания сахара 2,2%. В России разработана технология производ-
ства шампанского непрерывным методом. В шампанском содер-
жится не только повышенное количество углекислоты, но и ряд
ценных метаболитов, сказывающихся на специфическом вкусе
этого вина.
Вина, выпускаемые в продажу на первом году после изготов-
ления, называют ординарными, а выдержанные не менее 1,5 лет и
сохраняющие свои высокие качества — марочными.
Известны так называемые плодовые вина (кроме виноградных),
получаемых при спиртовом брожении соков зрелых плодов: ягод-
ное, яблочное и др.
На виноградных ягодах поселяются различные микроорганиз-
мы (дрожжи, нитчатые грибы, бактерии), которые необходимо
подавить, так как в противном случае будет трудно гарантировать
получение вина высокого качества. Как ингибитор микробов —
контаминантов давно и эффективно используют сернистый газ или
сульфит, например, в виде метабисульфита калия (примерно от 0,1
до 0,2% SO2), не подавляющих производственный штамм дрожжей
в его активную фазу. Пастеризация здесь оказывается менее
благоприятной.
Концентрация сахара в винограде — важный фактор для
ферментации (концентрация его в сусле выше 28% будет тормозить
408
брожение). Определенную роль играют исходное значение pH и
температура. Чтобы избежать повышенной кислотности готового
вина, было предложено устанавливать pH сусла ниже 3,6; опти-
мальная температура для большинства рас дрожжей 27—29°С, но
есть и психрофильные виды, сбраживающие виноградное сусло
при 10°С. При низкой температуре и медленном брожении фор-
мируется более яркий букет вина, чем при кратковременном
брожении и повышенной температуре.
Аэрирование сусла возможно и целесообразно в самом начале
процесса, чтобы быстрее наросла биомасса клеток для ведения
последующего анаэробного процесса. Количество привносимой в
сусло суспензии дрожжей обычно составлет 1% по объему.
В случае применения биореакторов больших емкостей для
производства столовых вин бродящий сок принудительно охлаж-
дают, используя теплообменники, змеевики или другие устройства.
Мезга (оболочки виноградных ягод, семена, частички стеблей
и т. п.) привносит определенные сложности в связи с теплообменом
при брожении — образование "шапки".
Очистка вин при естественном хранении и созревании не
всегда завершается его полным осветлением. В этих случаях при-
бегаются к очистке путем осветления, старения и созревания до
розлива в бутылки. Дополнением к осветлению являются фильт-
рация (в том числе — стерилизующая), пастеризация, охлаждение
— для удаления винного камня и коллоидов.
9.3.1.2. Ацетонобутиловое брожение. Ранее упоминаемые угле-
водные источники (крахмал, патока, гидролизаты целлюлозы раз-
личного происхождения и др.), применяемые для дрожжей при
спиртовом брожении, могут быть использованы при ацетонобути-
ловом брожении, протекающем в анаэробных условиях в процессе
жизнедеятельности спорообразующих бактерий — Clostridium
acetobutylicum. В результате образуются такие нейтральные про-
дукты, как ацетон и бутанол, а также уксусная и масляные кислоты,
диоксид углерода, водород. Нейтральные продукты представляют
большой интерес для промышленного органического синтеза (эфи-
ров для лакокрасочной промышленности), как экстрагенты и рас-
творители, и т. д. Несмотря на то, что синтетические и биотехно-
логические процессы получения ацетона и бутанола являются
конкурирующими, на практике реализуют оба вида производств,
так как все зависит, главным образом, от стоимости сырья (эконо-
мические проблемы). Очевидно "сосуществование" синтетических
409
и биологических технологий получения одних и тех же продуктов
оправдано временем и жизнью общества.
Биосинтез ацетона и бутанола (в ФДФ-пути по Эмбдену-Мей-
ергофу-Парнасу) существенно зависит от pH сред — при низких
значениях активируются ферменты, участвующие в трансформа-
ции ацето-АцКоА в ацетон и, соответственно, больше расходуется
НАД«Н для восстановления бутирил-КоА в бутанол Уксусная и
масляная кислоты выступают здесь в роли минорных компонентов,
причем, первая из них может подвергаться конденсации до бути-
рата, который, в свою очередь, через бутирил-КоА превращается
в бутанол (в незначительном количестве может образовываться
этанол). Таким образом, бутанол и ацетон являются основными
продуктами ацетонобутилового брожения.
С6В12О6 ►„СН3СОСООН ► АдКоА * Адепфсфп ► СН^СООН
глав» I jeyannum
(CHgtgCOOIH-- БячиЛоА -<---1----► Адетодепт-►CHjCOCHj
СН3 I цетя
шап у
num «ущимачд
CH3(CHg)2CH2OH
fyau
Известно, что первичные алифатические спирты обладают
антимикробным действием, поэтому в питательных средах концен-
трация сбраживаемых углеводов не должна превышать 6%, так как
Cl. acetobutylicum не будет их утилизировать, если концентрация
бутанола приближается к ингибирующей — 1,5%.
Сырьем для производства целевых продуктов обычно является
меласса или сульфитные щелока, смешиваемые с кукурузным и
ржаным мучным затором. Инокулят готовят из свежих спор (тем-
пература выращивания 37°С), активных по образованию бутанола,
на такой же среде. Основные ферментации проводят в периоди-
ческом, полунепрерывном и непрерывном режимах. Исходное
значение pH среды примерно 6,0. Уже через 12 часов pH снижается
до 4,1—4,2 и на этом уровне остается до конца ферментации.
После окончания процесса сепарируют ацетонобутиловую бар-
ду, а дистиллят упаривают, примерно наполовину и ацетон отде-
ляют от этанола и бутанола перегонкой при различных темпера-
410
турах — ацетон кипит при 56,2°С, этанол — при 78,4°С, азеотроп
бутанола с водой — при 93,4°С, чистый бутанол — при 117,7°С.
Отходами производства являются газообразные водород и ди-
оксид углерода (порядка 30 м3 на 100 кг сахарозы, из которых около
70% составляет СО2) и плотная ацетонобутиловая барда. Газы
можно улавливать и применять для синтеза аммиака и метанола,
или применять как таковые. Барда — ценный продукт, содержащий
заметные количества рибофлавина; сухие вещества (преимущест-
венно — азотистые) в ней составляют 3—5%. Ранее барду самосто-
ятельно использовали в высушенном виде для кормления скота, в
настоящее время ее применяют для выращивания кормовых дрож-
жей.
9.3.2. Получение органических кислот. Прежде чем рассмот-
реть конкретные биотехнологические процессы получения орга-
нических кислот, необходимо оговориться, что под рубрику "бро-
жения" должно быть отнесено образование в анаэробных условиях
только молочной и пропионовой кислот с помощью соответствую-
щих бактерий, тогда как биосинтез лимонной, глюконовой, итако-
новой и некоторых других органических кислот определенными
микромицетами представляет собой разновидность того или иного
окислительного (аэробного) процесса и поэтому отнесение их к
брожениям является условным.
9.3.2.1. Бродильные процессы получения органических кислот.
Получение молочной кислоты. Образование молочной кислоты
(СН3СНОНСООН) лактобактриями происходит в естественных
условиях при скисании молока и молочных продуктов, а также при
ее целенаправленном получении в производственных условиях.
Молочнокислые бактерии относят к 4 родам: Lactobacillus,
Leuconostoc, Streptococcus и Pedicoccus. Род Lactobacillus включает
3 подрода — Thermobacterium, Streptobacterium и Betabacterium.
Представители первого из них не растут при 15°С, но могут
выдерживать температуры выше 50°С. Стрептобактерии не явля-
ются термофилами. Бетабактерии образуют DL-молочную кислоту
из глюкозы. Одни из них (термобактерии, стрептобактерии, стреп-
тококки и педикокки) являются гомоферментативными, образую-
щими при сбраживании гексоз преимущественно молочную кис-
лоту, другие (бетабактерии и лейконостоки) — гетерофермента-
тивными, образующими молочную и уксусную кислоты, диоксид
углерода, возможно — этанол; молочнокислые бактерии могут
использовать мальтозу, глюкозу, лактозу, осахаренный крахмал и
пр. В целом, лактобактерии — требовательны к питательным
411
средам — многие из них нуждаются в ряде витаминов из группы
В, некоторых аминокислотах, пуринах и пиримидинах, отдельных
органических кислотах алифатического ряда (уксусной, лимонной,
олеиновой). Для сбраживания глюкозы и гидролизатов крахмала
на практике применяют обычно Lactobacillus delbrueckii, L.
bulgaricus, L. leichmanii (одни или в смеси между собой или со
Streptococcus lactis), для сбраживания мальтозы иногда используют
L. casei.
В промышленном производстве молочной кислоты обычно
используют термофильные гомоферментативные виды, активно
синтезирующие целевой продукт, например, при 50°С. Таким
видом яляется L. delbrueckii штамм Л-3, отличающийся высокими
стабильностью й активностью кислотообразования (выход молоч-
ной кислоты составляет 95—98% от потребленной сахарозы). Этот
вид внедрен в промышленность еще в 1923 г. под руководством
В. Н. Шапошникова.
Принципиальная технологическая схема получения L( + )-мо-
лочной кислоты состоит в следующем: мелассную среду, содержа-
щую 5-—20% сахара, вытяжку солодовых ростков, дрожжевой
экстракт, витамины, аммония фосфат, засевают L. delbrueckii.
Брожение протекает при 49—50°С при исходном pH 6,3—6,5. По
мере образования молочной кислоты ее периодически нейтрали-
зуют мелом. Весь цикл ферментации завершается за 5—10 дней;
при этом в культуральной жидкости содержатся И—14% лактата
кальция и 0,1—0,5% сахарозы (80—90 г лактата образуются из 100
г сахарозы). Клетки бактерий и мел отделяют фильтрованием
(отход), фильтрат упаривают до концентрации 30%, охлаждают до
25°С и подают на кристаллизацию, которая длится 1,5—2 суток.
Кристаллы лактата кальция обрабатывают серной кислотой при
60—70°С, гипс выпадает в осадок, а к надосадочной жидкости
добавляют желтую кровяную соль при 65°С для удаления ионов
железа, затем натрия сульфат для освобождения от тяжелых
металлов. Красящие вещества удаляют с помощью активирован-
ного угля. После этого раствор молочной кислоты подвергают
вакуум-упариванию (при остаточном давлении 800—920 кПа) до
50% или 80%. Оставшийся не до конца очищенный раствор молоч-
ной кислоты используют для технических целей. Более очищенную
кислоту можно получать при перегонке ее сложных метиловых
эфиров, при экстракции простым изопропиловым эфиром в про-
тивоточных насадочных колоннах.
412
С помощью L. bulgaricus получают молочную кислоту из мо-
лочной сыворотки; гидролизаты пентозанов в кукурузных коче-
рыжках, соломе и других видах пентозного сырья можно исполь-
зовать для сбраживания клетками L. brevis.
В конце 80-х годов разработана технология получения молочной
кислоты с помощью клеток Streptococcus thermophilus, адгезиро-
ванными на микросферах из активированного угля и помещенны-
ми в биореактор, работающий по принципу "кипящего" или псев-
доожиженного слоя, через который перемещаются микросферы.
В нижней части они сорбируют субстрат, в верхней — молочную
кислоту, благодаря чему нет надобности в регуляции pH в процессе
ферментации. Продуктивность системы — 12 г/л’Ч'1 молочной
кислоты.
Необходимо помнить о том, что молочная кислота является
выраженным коррозирующим агентом. В то же время она быстро
полимеризуется, а большинство ее солей хорошо растворимо в
воде, благодаря чему молочную кислоту широко используют в
пищевой; текстильной и фармацевтической промышленности, в
изготовлении растворителей и пластификаторов в лаках, олифах
и т. п.
Гомо- и гетероферментативные молочнокислые бактерии давно
используются в хлебопечении. Их ассоциации с дрожжами, благо-
приятные для создания аромата, вкуса, пористости, окраски и
свежести, называют заквасками.
В России, особенно — в сельской местности, где хлеба выпе-
кают в домашних условиях, закваски поддерживаются длительное
время более или менее стабильными. Это обусловлено антагони-
стическим влиянием лактобактерий на гнилостные, уксуснокислые
и маслянокислые бактерии, энтеробактерии и др., но не на дрожжи,
содержащиеся в заквасках. Тем не менее, специально приготов-
ленные закваски из чистых культур предпочтительнее спонтанных
(не контролируемых) ассоциаций, так как первые предсказуемы в
своем "поведении" и легче управляемы.
Молочнокислое брожение находится в основе силосования
кормов и квашения овощей (капусты, огурцов), плодов, ягод (маслин,
яблбк) и т. д. Эти процессы протекают за счет естественных
микроорганизмов, находящихся на заквашиваемом объекте. В
последнее время используют специальные закваски, обеспечива-
ющие проведение процессов в прогнозируемых режимах и с
ожидаемыми результатами.
413
Молочнокислыми продуктами являются различные сыры, пол-
учаемые из обезжиренного или цельного молока. Такое молоко
под влиянием лактобактерий и молочной кислоты створаживается
— творог отделяют от сыворотки и засевают специальными мик-
роорганизмами (в соответствии с сортом изготавливаемого сыра)
с последующим выдерживанием его от нескольких недель до 8
месяцев (например, сыр "Чеддер") для созревания.
Створаживание молока проводят также с помощью реннина —
сычужного фермента из желудка (сычуга) молодых телят, или
реннина микробного происхождения.
Молочнокислые бактерии включают в разные композиции
профилактических и лечебных препаратов: бифидумбактерин, би-
фикол, колибактерин, лактобактерии. Первый из них состоит из
живых высушенных бифидобактерий определенного штамма, вто-
рой — из живых бифидобактерий (штамм 1) и кишечной палочки
(штамм М-17), третий содержит живые кишечные палочки штамма
М-17, четвертый представляет собой сочетание лиофильно высу-
шенных лактобацилл (L.fermenti и Lplantarum).
За рубежом изготавливают так называемый пробиотик "Фер-
лак-5", содержащий молочнокислые бактерии с добавками вита-
минов A, D3 и Е. Его смешивают с кормом из расчета 1 млн. клеток
бактерий на 1 г корма. Этот пробиотик рекомендуют поросятам,
телятам и птицам.
Получение пропионовой кислоты. Пропионовокислое броже-
ние характерно для пропионовых бактерий, культивируемых в
средах, где глюкоза является источником углерода. Из трех молекул
глюкозы образуется 4 молекулы пропионовой кислоты, 2 молекулы
уксусной кислоты, 2 молекулы диоксида углерода и 2 молекулы
воды:
ЗС6Н12О6 —> 4СНзСН2СООН + 2СН3СООН + 2СО2 + 2Н2О.
Фактически механизм образования пропионовой кислоты значи-
тельно сложнее; ключевыми интермедиатами здесь являются пи-
руват, метил-малонил-КоА, пропионил-КоА, сукцинил-КоА, окса-
лоацетат.
Пропионовые бактерии представляют собой грамположитель-
ные, бесспоровые, неподвижные палочки, подразделямые на "кож-
ные” (P.acnes, P.avidum, P.granulosum) и "классические"
(P.freudenreichii, P.thoenii, P.jensenii, P.acidipropionici) в едином
семействе Propiornbacteriacae. "Кожные" обитают на кожных по-
кровах человека и в желудке жвачных животных — они могут
414
быть причиной определенных патологических процессов (опти-
мальная температура для их роста 37°С); "классические" пропио-
новые бактерии обитают, как правило, в молоке и молочных
продуктах. Температурный оптимум для них 37°С, анаэробы, но
образуют каталазу, пероксидазу и супероксиддисмутазу (СОД),
фиксируют СО2, некоторые виды фиксируют молекулярный азот,
утилизируют элементарную серу, нуждаются в витаминах группы
В — биотине, пантотенате и тиамине.
При контакте с О2 у пропионовых бактерий образуются супер-
оксидные радикалы, превращающиеся благодаря СОД в пероксид
водорода, а этот последний разлагается под действием каталазы и
пероксидазы.
Перспективными для производства пропионовой кислоты ока-
зались виды P.freudenreichii и P.acidipropionici, а для получения
витамина Bi2 — P.freudenreichii и Р.аспе.
Биосинтез кислоты проводят на достаточно простых средах,
например, такого состава (в %): углевод— 1—2; аммония сульфат
— 0,3; гидрофосфат калия — 0,2; кобальта хлорид — 0,0001; биотин
— 0,00001; пантотенат — 0,1; тиамин — 0,01.
Заметный прогресс наблюдается в совершенствовании процес-
сов биосинтеза пропионовой кислоты иммобилизованными клет-
ками в фиксированном слое, например, в ПААТ в колоннах. Так,
в случае с P.acidipropionici при D = 0,05 ч '1 удается получать до 15
г/л целевого продукта.
Биосинтетическая пропионовая кислота может конкурировать
с пропионатом "нефтяного происхождения", особенно в случае ее
применения в пищевой и фармацевтической промышленности в
качестве консерванта.
Конечные продукты ферментации (пропионат и ацетат) можно
не разделять, поскольку обе кислоты обладают консервирующими
свойствами. Клетки, отсепарированные от культуральной жидко-
сти, могут быть экстрагированы соответствующими растворителя-
ми в целях извлечения СОД, каталазы, пероксидазы и витамина
В12, высушенный экстракт в виде порошка используют в пищевой
промышленности как антиоксидант и витаминизированный пре-
парат.
9.3.2.2.Окислительные процессы. Получение уксуса. Известны
различные виды и подвиды уксуснокислых бактерий, включенных
в роды Acetobacter и Gluconobacter. Форма их непостоянна (пол-
иморфизм) — от эллиптических до палочковидных, молодые клетки
не окрашиваются по Граму (грамотрицательны). Одни из них
415
подвижны — имеют жгутики, другие непод вижны. Ацетобактерии
окисляют этанол в уксусную кислоту по схеме:
С2Н5ОН + 1/2О2 —> СН3СОН + Н2О
СНзСОН + 1/2О2 —> СНзСООН
Некоторые штаммы могут содержать порфирины (биомасса
клеток приобретает розовую окраску) или продуцируют водораст-
воримый коричневый пигмент. Большинство ацетобактерий обхо-
дится без витаминов в питательных средах. В естественных услових
их находят на овощах, фруктах и скисших фруктовых соках, в
уксусе и в некоторых слабоалкогольных напитках. Типовой вид
—Acetobacter aceti.
Уксус (4—9%-й) непосредственно используют как приправу к
различным блюдам, а также при изготовлении майонеза, горчицы,
маринадов, соусов, хрена. Поэтому уксус, ранее получаемый из
сахарных и фруктовых сиропов, вин, ягод и других аналогичных
продуктов, предварительно сбраживаемых дрожжами до этанола,
отличается высокими вкусовыми качествами. Однако в настоящее
время основное количество уксусной кислоты получают синтезом
ее из ацетилена:
СН=СН + НОН-—> СН^СНОН------>СН3СОН±^г>СН3СООН
или выделением из продуктов сухой перегонки древесины (свя-
зывание известью с последующим разложением серной кислотой
до уксусной кислоты и гипса); полученная таким способом кислота
называется "ледяной" (77—80%-й концентрации). Разбавляя "ледя-
ную" уксусную кислоту, в 10—20 раз, можно получить близкий к
столовому уксус. Качество уксуса не изменяется и в случае при-
готовления его из чистого эталона. В то же время уксус, полученный
из фруктовых сиропов, вин, ягод и т. п., способен созревать,
улучшая свои потребительские качества при хранении (прежде
всего — за счет образования эфиров уксусной кислоты).
Поскольку уксуснокислые бактерии не превращают углеводы
непосредственно в уксусную кислоту, постольку исходное сырье
должно подвергаться спиртовому брожению (рис. 131). Самой
старой технологией получения столового уксуса лучшего качества
был так называемый медленный, или орлеанский (французский)
процесс, в котором исходным сырьем было легкое виноградное
вино, которым на 3/5 заполняли плоские деревянные чаны и туда
же наливали столовый уксус, заполняя оставшиеся 2/5 объема (в
416
уксусе содержатся уксуснокислые бактерии). На поверхности
смеси развивалась пленка ацётобактерий.
Патока Виноградный сок Яблочный сок Зерновой затор Этанол
Разбав-
ление до *
содержа-
ния са-
хара не
менее 8%
' I___________________________________________' ________
(+ дрожжи - сахаромицеты) Спиртовое брожение (3-7 суток)
| + СН^СООН (в соотношении 1:100)
Выдерживание в анаэробных
условиях нескольких недель
Сепарирование ----»— Осадок дрожжевых
| клеток (отход)
Жидкость после брожения ------------
(уксусная смесь)
Получение уксуса
Получение уксуса
по орлеанскому J J
\ по немецкому, или
(французскому), или
, быстрому способу
медленному способу г
Рис. 131. Технологическая схема получения уксуса из различного сырья.
После завершения окисления этанола отбирали из чана 10%
жидкости, а взамен добавляли столько же вина. Вместо плоских
чанов можно использовать деревянные бочки, например, дубовые,
емкостью до 20 дкл. В них должны быть отверстия для поступления
воздуха и сливной кран. Процесс можно вести полунепрерывно,
отрегулировав подачу свежих порций вина и слив готового уксуса.
В настоящее время почти везде "пищевой" уксус получают
согласно так называемому "генераторному", или немецкому (быс-
трому) процессу, впервые реализованному в 1832 г. Особенность
его — создание максимальной поверхности для снабжения возду-
14 т. 8524
417
хом уксуснокислых бактерий и, как следствие, быстрое окисление
спирта в уксусную кислоту. Названную технологию осуществляют
так же в деревянных емкостях — трехсекционных генераторах (из
дуба, пихты, красного дерева и пр.) В верхней секции размещено
распылительное устройство для разбрызгивания 3—10% водного
раствора этанола (например, сегнерово колесо), в средней —
наполнитель для создания хорошо аэрируемой поверхности
(стружки — буковые, березовые; стержни кукурузных початков;
виноградные выжимки; керамические материалы и др.), в нижней
— собирается готовый уксус. Секции в генераторах разделены
дырчатыми перегородками. Воздух подается снизу и выходит через
негерметично закрытый верх.
Генераторы могут быть рецеркуляционного типа на 30—60 м3,
в которых воздух перекачивается с относительно постоянной
скоростью и регулирование температуры (27—29°С) осуществляют
путем охлаждения уксусной смеси.
При получении уксуса необходимо хорошо контролировать
температуру и подачу воздуха, чтобы избежать заметной потери
спирта и уксусной кислоты за счет их сгорания (полного окисления)
до СО2 и Н2О. Питательные вещества (кроме спирта) в уксусной
смеси будут влиять на жизнеспособность ацетобактерий.
Необходимо иметь в виду, что качество используемой воды для
разбавления этанола могут заметно сказываться на снижении
активности микробов в этом процессе (медь, железо, олово, цинк).
Уксус, получаемый из разведенного этанола, обычно не под-
вергают "старению", так как оно не прибавляет каких-либо прият-
ных вкусовых и обонятельных ощущений. Готовый уксус осветля-
ют фильтрованием, разливают в стеклянные емкости и пастеризу-
ют.
Выход уксуса оценивают по массе уксусной кислоты, содержа-
щейся в целевом продукте, наработанном за сутки (кг/м3/сут.).
Обычно эта величина довольно постоянна и равна 1,4 кг/м3/ сут.
для обычных генераторов, 5—12 кг/м3/ сут. — для рециркуляци-
онных генераторов.
Уже в течение четверти века совершенствуют метод глубинного
культивирования селекционированных штаммов ацетобактерий с
целью получения большего количества уксусной кислоты. Так, при
периодическом выращивании A.aceti в глубинных условиях при
25—30°С на среде, содержащей 10—11% этанола, 1% уксусной
кислоты и минеральные соли выход уксусной кислоты повышался
до 18—23 кг/м3/сут. Более производительным стал непрерывный
418
процесс, реализуемый в батарее ферментаторов (например, изпяти
емкостью 6 м3 каждый и соединенных последовательно). Исходная
среда с 4% этанола, 1,5% уксусной кислоты и минеральными солями
(моногидрофосфат аммония, дигидрофосфат калия, магния суль-
фат), непрерывно поступает в первый ферментатор, обогащается
спиртом в последующих ферментаторах. Таким образом происхо-
дит обогащение среды уксусной кислотой при снижении концен-
трации этанола (таблица 44).
Т а б л и ц а 44. Содержание уксуса при непрерывной глубинной ферментации
Acetobacter acetl в батарее из пяти ферментаторов
Концентрация, % № ферментатора
1 2 3 4 5
Этанола 2,6 1.8 1.6 0,9 0,2—0,15
Уксусной кислоты 3,0 5,9 8,0 9,3 10,0
Из последнего ферментатора непрерывно вытекает уксус. Вы-
ход уксусной кислоты достигает 30 кг/м3/ сут. и более.
Получение лимонной кислоты. Около 60 лет назад лимонную
КйСАОТу выделяли преимущественно из плодов цитрусовых расте-
ний. Теперь же основную массу ёё производят с помощью опре-
деленных штаммов плесневого гриба Aspergillus niger. В настоящее
время ведущими производителями лимонной кислоты являются
КНР, США, Франция, Россия и некоторые другие страны. Ранее,
начиная с 1917 г., производство было основано на поверхностном
культивировании микроба-продуцента; в 1938—1942 гг. освоено
также глубинное культивирование в герметичных ферментаторах
(см. главу 7). Благодаря этому удалось механизировать и автома-
тизировать процесс, эффективнееиспользовать производственные
площади и снизить себестоимость целевого продукта, сократить
общую продолжительность технологического цикла, облегчитьпод-
держание асептичности в производственных условиях.
Ныне в производстве применяют селекционированные штам-
мы A.niger (например Р-3), дающие выход лимонной кислоты
98—99% в расчете на потребленную сахарозу и обладающие повы-
шенной осмотолерантностью' (при начальных концентрациях са-
хара в питательной среде порядка 12%).
419
Лимонная кислота, как трехосновная оксикарбоновая кислота,
наряду с глюконовой, фумаровой и другими, является интермеди- •
атом метаболизма в цикле трикар-
ОН боновых кислот, когда имеет место
НООС___СН__С__СН___СООН неполное окисление соединений
2 I 2 углерода в аэробных условиях. Ее
сос>н сверхсинтез возможен при лимити-
ровании гриба — продуцента по
железу и фосфору, при одновременном избытке в среде источника
углерода и при низких значениях pH.
Лимонная кислота накапливается вначале в клетках продуцен-
та, а затем выделяется в культуральную среду.
Вышеперечисленные факторы ингибируют такие ферменты,
как аконитат-гидратазу, изоцитратдегидрогеназу и, возможно, а-
кетоглутаратдегидрогеназу. Поэтому не происходит полного мета-
болизма лимонной кислоты в ЦТК и ее можно получать в доста-
точно больших количествах с коммерческими целями.
Поскольку основным сырьем для производства лимонной кис-
лоты является меласса, в которой содержится много железа, то на
стадии предферментации необходимо его осадить с помощью
желтой кровяной соли — К, [Fe(CN6)]. К тому же доказано, что
эта соль и лимонная кислота в клетках выступают ингибиторами
изоцитратдегидрогеназы.
Известны два способа ферментации A.niger — поверхностный
и глубинный. Первый из них реализуют на предприятиях малой и
средней мощности в виде жидкофазной ферментации на жидкой
среде (например, в ряде стран Европы и Америки) и в виде
твердофазной ферментации (например, в Японии) на уплотненной
среде. Технологическая схема жидкофазной ферментации пред-
ставлена по Р. Я. Карклиньшу и А. К. Пробоку (1972) на рис. 132.
В отдельном цехе осуществляют наработку спор (конидий)
гриба в виде трехстадийной схемы. В первую стадию A.niger
выращивают на скошенной агаризованной среде (например, на
сусло-агаре) в пробирках, во вторую и третью стадии его размно-
жают на плотной или жидкой среде соответственно в колбах
Эрленмейера или в алюминиевых кюветах площадью 8,5—12 дм2
и с высотой бортиков от 7 до 20 см. Продолжительность каждой
стадии — от 2 до 4 суток при температуре 32°С. При образовании
и созревании конидий вначале бесцветный мицелий становится
затем черным; конидии собирают по принципу аспирации (по лат.
aspiratio — вдыхание, надувание) специальным вакуумным насо-
420
сом, подсушивают в термокамере при 28-30°С, смешивают со
стерильным активированным углем (1:2), фасуют в стерильные
флаконы (колбы) и хранят в течение от полутора до двух лет. С 10
дм2 питательной среды в кюветах можно получить до 4—5 г сухих
конидий. Подобный посевной материал может быть самостоятель-
ным коммерческим продуктом, поставляемым на заводы лимонной
кислоты.
Поверхностный способ жидкофазной ферментации A.niger для
промышленного производства лимонной кислоты реализуют в
"бродильных камерах", где размещают на стеллажах названные
выше кюветы (8—10 штук на один стеллаж) одну над другой. На
дне каждой кюветы имеется сливной штуцер. "Бродильные каме-
ры" оборудованы приточно-вытяжной вентиляцией, обеспечиваю-
щей равномерный приток стерильного воздуха заданной темпера-
туры и влажности (3—4 м3/м2 мицелия • ч'1). Температура в камерах
поддерживается на уровне 34—36°С, высота питающего слоя жид-
кой мелассной среды 6—12 см. Максимальное тепловыделение
(500—550 кДж/м2 • ч) имеет место к 5 суткам; исходная концентра-
ция сахаров в питательной среде в среднем порядка 12%; начальное
значение pH 6,8—7,0 снижается до 4,5 в течение первых трех суток
и до 3,0 — к концу процесса (8—9 сутки). Максимальное кислото-
образование в таких условиях происходит на 5—6 сутки (100—105
г/м2 пленки гриба’Ч'1, а затем стабильно удерживается на уровне
50-60 г/м2 «ч *.
Из трех вариантов проведения технологического процесса
(периодический, или бессменный; сменный и доливной) наилуч-
шим оценивают доливной, когда через 6—7 суток от начала
процесса ферментации (концентрация сахара снижается до 3—4%)
подливают стерильный раствор мелассы без питательных солей —
30—35% начального объема (не забывать рационально использо-
вать объем кювет при первоначальном заполнении питательной
средой с учетом ее испарения). Таким путем добиваются продления
цикла ферментации до 12 суток, а с этим на 30—35% возрастает
количество перерабатываемой среды для получения целевого про-
дукта. При сменном (одно- и многосменном) методе культуральную
жидкость в конце ферментации сливают из-под пленки, пленку
снизу промывают стерильной водой и под нее же заливают свежую
стерильную питательную среду, содержащую только углевод и
лишенную минеральных солей. Ферментацию продолжают еще
4—6 суток.
421
В собранной культуральной жидкости содержится смесь орга-
нических кислот — лимонная, глюконовая, щавелевая и неисполь-
зованный сахар в примерном соотношении 45—50:3:1:7, то есть
лимонная кислота составляет от 80 до 90%. Ее выделяют химиче-
ским путем — добавляют к нагретой до 100°С культуральной
жидкости известковое молоко — Са(ОН)2 или мел—СаСО3, доводя
pH до 6,8—7,0; это количество составляет примерно 2,5—3%;
трехзамещенный кальция цитрат, хуже растворимый в горячей
воде, чем в холодной, выпадает в осадок вместе с кальция оксалатом
(кальция глюконат остается в растворе); осадок отфильтровывают,
промывают горячей водой и гидролизуют серной кислотой. Сво-
бодная лимонная кислота остается в растворе, а негидролизован-
ный кальция оксалат и образовавшийся гипс — CaSO4 остаются в
осадке. Раствор лимонной кислоты очищают, подвергают вакуум-
упариванию и кристаллизуют. Кристаллы кислоты высушивают и
Рис. 132. Технологическая схема
получения лимонной кислоты из ме-
лассы поверхностным способом
(жидкофазная ферментация): 1 —
цистерна для мелассы, 2 — центро-
бежные насосы, 3 — реактор для
разбавления мелассы, 4 — стерили-
затор, 5 — бродильная камера, 6 —
сборник сбраживаемых растворов, 7
— нейтрализатор, 8,10 — нутч-филь-
тры, 9 — расщепитель, 11 — сбор-
ник-монтежю, 12—вакуум-аппарат,
13 — дисольвер, 14 — фильтр-пресс,
15 — кристаллизатор, 16 — прием-
ник, 17 — сушилка, 18 — готовая
продукция, 19 — сборник фильтрата.
Мицелий продуцента либо используют для выделения фермен-
та пектиназы, либо высушивают и поставляют на корм скоту и
домашней птице (желательно — в обезвреженном — убитом виде);
наконец, он может быть использован, как источник флавинов.
Твердофазная ферментация на уплотненных средах для пол-
учения лимонной кислоты — наиболее простой способ из всех
известных. Ферментацию определенного штамма A.niger, рези-
стентного к высоким концентрациям металлов (особенно — желе-
за), содержащихся в семенах злаковых растений, проводят на
увлажненных отрубях риса или пшеницы, находящихся в кюветах.
Условия биосинтеза кислот при этом аналогичны условиям на
422
агаризованных или в жидких питательных средах. После оконча-
ния процесса отруби экстрагируют водой, куда переходят кислоты,
а затем выделяют цитрат кальция и чистую лимонную кислоту
согласно схеме, изложенной выше.
Глубинный способ производства базируется на использовании
специальных культур A.niger ( в России применяют селекциони-
рованный штамм № 288/9). Способ подготовки посевного матери-
ала в виде конидий описан ранее. Инокулят подращивают сначала
в инокуляторе, затем — в посевном аппарате (примерно в 1/10
объема основного ферментатора) на среде с 3—4% сахара. Спустя
1—1,5 суток инокулюм передают из посевного в основной фер-
ментатор, где процесс ведут в течение 5—7—10 суток на аналогич-
ной среде с трехразовым доливом 25-28% (по сахару) раствора
мелассы с целью доведения конечной концентрации сахара в
культуральной жидкости до 12—15%. После окончания фермента-
ции (контроль — снижение кислотообразования) мицелий гриба
отфильтровывают, а культуральную жидкость подвергают обра-
ботке, как это описано выше.
Общая технологическая схема получения лимонной кислоты
при глубинной ферментации A.niger приведена на рис. 133.
Рис. 133. Технологическая схема
получения лимонной кислоты при
глубинной ферментации продуцента:
1 — емкость с мелассой, 2 — прием-
ник мелассы, 3 — весы, 4 — варочный
котел, 5 — центробежный насос. 6 —
промежуточная емкость, 7 — стери-
лизующая колонка, 8 — выдержива-
тель, 9—холодильник, 10 — посевной
аппарат, 11 — головной ферментатор,
12 — стерилизующие фильтры, 13 —
емкость для хранения мелассы, 14 —
промежуточный сборник, 15 — бара-
банный вакуум-фильтр, 16 — прием-
ник для мицелия, 17 — вакуум-сбор-
ник для мицелия, 18 — вакуум-сбор-
ник фильтрата культуральной жидко-
сти.
Согласно подсчетам глубинный метод экономически выгоден
в тех случаях, когда мощность завода превышает 2,5 тыс. тонн
лимонной кислоты в год, в противном случае поверхностный метод
оказывается предпочтительнее из-за меньших энергозатрат и се-
бестоимости продукции.
423
Лимонную кислоту широко используют в пищевой, медицин-
ской, фармацевтической, лакокрасочной промышленности и в
некоторых других отраслях народного хозяйства.
Лимонную кислоту можно получать из н-парафинов с помощью
дрожжевых организмов рода Candida — C.lipolytica, C.tropicalis,
C.parapsilosis, C.oleophila, C.guilliermondii, Czeylanoides. Наиболее
активными из них являются дрожжи C.lipolytica. К тому же другие
виды (кроме C^oleophila) относят к разряду условно патогенных.
Кроме лимонной дрожжи образуют на н-алканах Tpeo-Ds-изо-
лимонную, представляющую собою геометрический изомер ли-
монной кислоты.
Н
СН,—СООН ОН—С—СООН
I J
НО—С—СООН НС—СООН
I I
сн2—СООН н2с—СООН
лимонная кислота Tpeo-Ds-изолимонная
кислота
При лимитирований продуцента в азоте, фосфоре, сере и
магнии, но при избытке в среде парафина, можно наблюдать
сверхсинтез равных количеств обоих изомеров. Методами генети-
ки и селекции получены мутанты, синтезирующие лишь один
какой-либо изомер лимонной кислоты. Накопление целевого про-
дукта может достигать 200 г/л и выше при выходе кислоты от
использованного парафина более чем 140%.
Культивирование C.lipolytica проводят в ферментаторах при
интенсивных перемешивании и аэрации среды. В нашей стране
отработаны методы получения лимонной и изолимонной кислот
на н-алканах. Большая заслуга в этом принадлежит Т. В. Финоге-
новой. Получаемые технические соли цитраты и изоцитраты имеют
важное значение при изготовлении, например, моющих и других
веществ.
Получение глюконовой кислоты. Используют селекционирован-
ные штаммы Aspergillus niger, культивируемые в ферментаторах
при интенсивных аэрации и перемешивании, поддерживая посто-
янными температуру (30°С) и pH среды 6,0-7,0. Технологический
процесс можно проводить в одном из двух вариантов нейтрализа-
ции среды при ее закислении в ходе биосинтеза целевого продукта
— мелом или раствором едкого натра. В первом случае исходная
питательная среда должна содержать 10—15% глюкозы, поскольку
кальция глюконат ограниченно растворим в воде при 30°С (порядка
424
40 г/л); во втором случае образуется натрия глюконат, который
хорошо растворим в воде (до 400 г/л при 30°С и 569 г/л — при
80°С) и поэтому концентрацию глюкозы в исходной среде можно
повысить до 35%.
При использовании мела ориентировочный состава питатель-
ной среды может быть следующим (%): 10—15 глюкозы; 0,016 —
магния сульфата; 0,02 — калия дигидрофосфата; 0,04 — аммония
гидрофосфата и 2,6 — кальция карбоната. Вместо глюкозы можно
применять гидролизаты крахмала.
Если культуральную среду подтитровать раствором едкого на-
тра (вместо нейтрализации мелом), то содержание глюкозы в среде
доводят до 350 г/л; выход натрия глюконата тогда составит 95% от
внесенного углевода.
Технологический процесс реализуют либо в периодическом,
либо в непрерывном режиме. Второй из них более эффективен по
скорости и по выходу целевого продукта (свыше 95% от использо-
ванной глюкозы).
Механизм образования глюконовой кислоты связан с окисле-
нием глюкозы кислородом при участии фермента глюкозооксида-
зы (Е):
НОНгС-СН-(СНОЮ3—СНОВКЕ». НОН2С-СН—(СНОН)ргС=О
''"'''"(Г
р-Втииию-^ипм
---► HOHjC—(СВ0Н)4—СООН
ггюкомовая кислота
При этом вначале образуется P-D-глюконо-б-лактон, который
затем гидролизуется до глюконовой кислоты.
Учитывая тот факт, что селекционированный штамм Aniger
продуцирует почти одну глюконовую кислоту, то для ее выделения
в виде натриевой соли культуральную жидкость просто отфильт-
ровывают от мицелия, упаривают и высушивают. В случае необ-
ходимости получения кислоты, технический натрия глюконат пе-
рекристаллизовывают и химически переводят в свободную кисло-
ту.
Аппаратурное оформление технологического процесса получе-
ния глюконовой кислоты во многом напоминает производство
лимонной кислоты глубинным методом.
Отделенный мицелий гриба используют для выделения фер-
мента глюкозооксидазы, полезной в пищевой и фармацевтической
промышленности. Глюконовую кислоту и ее соли широко приме-
425
ня ют на практике. Так, кальция глюконат является лекарственным
препаратом, ускоряющим свертывание крови (его назначают при
острых и хронических кровотечениях); натрия глюконат исполь-
зуют для изготовления моющих средств, глюконовую кислоту
применяют в фотографии, литографии, в изготовлении красок, для
очистки металлов и т. д.
Получение шпаконовой кислоты. Итаконовая, или метиленян-
ноос—HjC—синтезируется плесне-
тарная кислота выми rpJ6aMIi из рода
Aspergillus и, в частности, A.itaconicus и A.terreus. Промышленное
производство базируется на использовании A.terreus. Образование
итаконовой кислоты из углеводных источников сопряжено с ЦТК
и непосредственным ее предшественником является цис-аконито-
вая кислота:
ОН
| -Н?0 —СОо
HOOC-C^-C-CHg-COOH НООС—СНг—С=СН—СООН —
СООН СООН
лимонная кислота
цис-аконитовая кислота
----► НООС-сн?—с=сн2
СООН
итаконовая кислота
Реакцию дегидратации лимонной кислоты катализирует фер-
мент аконитат-гидратаза, а декарбоксилирование происходит при
участии аконитатдекарбоксилазы, обнаруженной у A.terreus.
Технологический процесс получения итаконовой кислоты схо-
ден с процессом получения лимонной кислоты, то есть фермента-
цию продуцента осуществляют либо поверхностным, либо глубин-
ным методом, используя в качестве посевного материала конидии
гриба — отселекционированного высокопродуктивного штамма
A.terreus (например, ЭУУ-417).
Поверхностное выращивание продуцента проводят в кюветах,
глубинное — в ферментаторах, оборудованных мешалками и бар-
ботерами.
Итаконовая кислота в концентрации 7% ингибирует рост про-
дуцента, поэтому в процессе ее биосинтеза необходима нейтрали-
426
зация кислоты каким-либо основанием. Для этой цели чаще ис-
пользуют аммония гидроксид, с помощью которого поддерживают
pH среды равным 3,8 и добиваются образования 150—200 г/л
целевого продукта. Активаторами биосинтеза итаконовой кислоты
выступают катионы магния, меди и цинка. Источником углерода,
как правило, служит меласса, разбавляемая водой до 8%-го содер-
жания углеводов. Туда же добавляют (г/л) магния сульфат — 4,5,
аммония нитрат — 2,5, цинка сульфат — 0,0044, кукурузный
экстракт —4 мл, желтую кровяную соль — 0,25. Эта среда или
близкие к ней другие варианты сред применяют для поверхност-
ного и глубинного (при уменьшении сахара на 1—2%) культивиро-
вания продуцента. Уровень pH среды, равный 1,8—2,3, является
оптимальным для биосинтеза итаконовой кислоты. Длительность
культивирования в кюветах обычно составляет 10—12 дней при
35°С; в случае глубинного культивирования длительность процесса
сокращается до 2—3 суток при аэрировании со скоростью 1/8
объема воздуха в минуту на один объем среды. >
Выход итаконовой кислоты в первом случае достигает 36—38%
в расчете на 100 г потребленной глюкозы, во втором — 65% и выше.
После окончания ферментации мицелий сепарируют, промыд_
вают, отжимают и передают в сборники отходов; культуральн'уЮ
жидкость осветляют активированным углем, упаривают менее.. чем
до 1/10 первоначального объема и передают на кристалли'зацик
итаконовой кислоты. После этого ее центрифугируют, высуши
ют и фасуют. Дополнительную очистку кислоты можно пров-'
перекристаллизацией ее из 25% водных растворов.
Итаконовую кислоту применяют в химическом сии
кокачественных смол, волокон типа "нитрон", детерг'
ственных веществ, красителей и других органическ-
Из других органических кислот, синтезируй
целиальными грибами (например, Rhizopus г‘
время привлекала внимание ученых ф у ’
т а, являющаяся транс-изомером цис-ма •
Н—С—СООН
II
СООН—С—н
фумаровая кислоте
Фумаровая кислота способна трансформироваться в малеино-
вую, широко используемую для химического синтеза некоторых
лекарственных веществ, смол, лаков и красок. Однако конкурен-
тоспособность синтетического малеинового ангидрида (из бензи-
на) для тех же целей служит веским ограничителем развития
производства фумаровой кислоты путем микробного синтеза. И,
очевидно, это будет продолжаться до тех пор, пока нефтяные
запасы в мире значительно не оскудеют.
9.3.3. Получение антимикробных веществ. К антимикробным
средствам относят антисептики, дезинфектанты и химиотерапев-
тические вещества. Антисептики — это химические вещества,
неизбирательно и губительно действующие на микроорганизмы (в
том числе — патогенные), находящиеся на поверхности тела и
слизистых оболочек открытых полостей. Дезинфектанты — это,
в основном, антисептики, действие которых направлено на пато-
генные микроорганизмы, находящиеся на (в) объектах внешней
среды. Химиотерапевтические средства — это вещества синтети-
ческого и природного происхождения, избирательно и губительно
действующие на патогенные микробы, клетки злокачественных
опухолей и гельминты.
Антисептики, дезинфектанты, неорганические и синтетиче-
ские химиотерапевтические средства не имеют непосредственного
отношения к биотехнологии. В то же время подавляющее боль-
шинство антибиотиков получают при ферментации микробов-про-
дуцентов, или, в других случаях, применяют "антибиотическое
ядро" для синтеза целевого продукта — полусинтетического анти-
биотика. Известны единичные антибиотики (ранее получаемые с
помощью биосинтеза или получаемые таким путем и ныне), кото-
рые в настоящее время производят на предприятиях органического
синтеза (левомицетин и некоторые другие).
Таким образом, определение понятия "антибиотики" теперь
относят к веществам природного, полусинтетического и, реже,
синтетического происхождения, которые в малых концентрациях
ингибируют рост (размножение) каких-либо микроорганизмов.
Продуцентами одних антибиотиков (например, бацитрацина, по-
лимиксина и др.), являются бациллы, других — актиномицеты
(например, актиномицинов, гентамицинов, канамицинов), третьих
— нитчатые грибы (например, гризеофульвина, пенициллинов,
цефалоспоринов), четвертых—неспорообразующие бактерии (на-
пример, монобактамов).
428
Антибиотики — р-лактамы
Пенициллины. Пенициллин впервые получен из
культуральной жидкости в аморфном, а затем—в кристаллическом
виде А. Флемингом, X. Флори и Е. Чейном в 1940 г. До сих пор
пенициллиновые антибиотики составляют наиважнейшую группу
химиотерапевтических средств. Ядром пенициллинов является 6-
аминопенициллановая кислота, или 6-АПК, которую используют
для получения полусинтетических пенициллинов.
Один водород в аминогруппе 6-АПК
/S\
Н^-СН-СН С=(СНз)2
-С— N —CH— СОО
о
6-АПК
может быть заменен на какой-либо ра-
дикал (R), усиливающий и/или расширя-
ющий (изменяющий) антимикробный
спектр полусинтетического производно-
го. Обычно 6-АПК получают из бензил-
пенициллина (пенициллина G).
В настоящее время в различных странах производят биосинте-
тические (с помощью Penicillium notatum или P.chrysogenum) ан-
тибиотики и на их основе — полусинтетические антибиотики —
Р-лактамы, перечисленные в таблице 45.
Цефалоспорины — другая обширная группа
экстрацеллюлярных Р-лактамных антибиотиков, в которых шести-
членное дигидротиазиновое кольцо соединено с Р-лактамным коль-
цом. Благодаря этому ядром цефалоспоринов является 7-аминоце-
фалоспорановая кислота, или 7-АЦК (цефемовое ядро], впервые
R,—NH-CH—СЙ 2СН7
к I Г
°=с— N5 зс-CHj-R,
COO”
7-АКЦ
R^’OOC—CH—(CH2)3— CO—;
NH2
R2=—О—CO—CH3
выделенная в ходе очистки
цефалоспорина С (проду-
цент Cephalosporium
acremonium), в котором
Известно около 20 це-
фалоспоринов, созданных
методом полусинтеза из
АЦК: цефалоридин (цепо-
рин), цефуроксим, цефок-
ситин, цефалотин, цефа-
цетрил, цефалексин, цефа-
золин, цефаридин, цефо-
таксим, цефтриаксон, цеф-
тазидим и др.
429
Т а б ли ц a 45. Природные и полусинтетические пенициллины
Название Радикал, R Эффек- тив- ность per os Устойчивость к р-лактамазам Гидро- лизует- ся после всасы- вания
Staph, aureus 1рам- отриц. бакте- рий
Бензилпенициллин * Бензил — — — —
Феноксиметил- пенициллин Феноксибензил + — —
Метициллин 2, 6- Диметоксифенил — + + — 1
Оксациллин 5~Метил-3'фенил- 4-изоксазолил + + + —
Клоксаццллин З-о-Хлорфенил- 5-мегил- 4-изооксазолил + + + —
Флуклоксациллин 2-Хлор-6-фтор- фенил-5-метил- 4-изоксазолил + + + —
Ампициллин а-В-(-)-аминобензил + — — —
Амоксициллин р-Окси-а-В-(-)- аминобензил + — —
Карбенициллин Натриевая соль a-D- (-)-карбоксибензил — — +
Тикарциллин Натриевая соль карбокси (тиен- 3-ил) ацетил — — +
Карфециллин" Фенил (феноксикарбонил) -ацетил + — + +
Кариндациллин* Инден-5-ил- оксикарбонилфенил- ацегил + — + +
Пивампициллин* Аминофенилацегил + — — +
430
Продолжение табл. 45
Название Радикал, R Эффек- тив- ность peros Устойчивость к р-лактамазам Гидро- лизует- ся после всасы- вания
Staph, aureus грам- отриц. бакте- рий
Таламщщиллин* См. ампициллин00 + — — +
Бакампициллин* См. ампициллин* + — — +
Пиперациллин** 2, 3- Диоксопиперазин-1- ил-карбонил-амино (фенил) ацетил — — — —
Азлоциллин** 4-Оксопиразолидин- 1 -ил-карбонилами- но-(фенил) ацетил — — — —
Мезлоциллин** З-Метилсульфонил- 2- оксоимидазолидин- 1 -ил-карбонилами- но-(фенил) ацетил — — — —
Мециллинам** Пергидроазепин- 1-ил-метилен — о ?
Пивмёциллинам** См. мециллинам0 + о ? +
Примечание: 'эфиры карбенициллина; *эфиры ампициллина; **замещенные ампицил-
лины; **6-р-амцдинозамещенные пенициллины; в позиции 3 тиазолидино-
вого цикла (в форме сложного эфира): “-пивалоилоксиметил, °°3-окСо- 1,
З-дигидроизобензофуран-1-ил, + 2-этоксикарбонилоксиэтил или 4-эток-
сикарбонилоксибугил; (+) — ответ положительный; (—) — ответ отрица-
тельный; (о) — не действует на грамположительные бактерии; (?) —
информация отсутствует.
Боковые цепочки, содержащие 2-аминотиазолил как Rj в цефо-
таксиме, цефтриаксоне, цефтазидиме и некоторых других, усили-
вают активность цефалоспоринов против стрептококков и грамот-
рицательных бактерий; иминометоксигруппа в той же позиции,
например, в цефуроксиме, обеспечивает стабильность антибиоти-
ка против плазмидо-опосредованных Р-лактамаз, и т. д.
1-Оксацефемы — сходны с цефалоспоринами, у которых
атом серы в дигидротиазиновом (цефемовом) кольце замещен на
431
R1
антибиотик
*2
—о—со— сн3
Оацатил
цефотаксим
2-ами№Тиаэол-4-ип-
(метоисиимино)ацетил
радикал тот же
цефтриаксон
Б-гцдроеси-2-метил-
5-<жсо2,6-дегмдро-
-1,2,4-триавин-З-ип
-пю...натриевая соле
пиридил
цефтазидим
2-амино-0-С1-шрбоаси
1,1-ди*1втил)-4-т>вэал-
карбогмдропамоил...
натриевая соль
кислород. Представителем такой группы антибиотиков является
широкоспектральный латамоксеф (мексалакта м), в
котором 7-а-метоксигруппа стабилизирует молекулу, как таковую,
равно как и придает устойчивость и к большинству Р-лактамаз.
ОН
СН СН2
COONa
,с—сн2—-
COONa сн3
432
К л а в а м ы — Р-лактамные антибиотики, отличающиеся от
пенициллинов тем, что в тиазолидиновом кольце последних сера
заменена на кислород (клавемовое оксазолидиновое кольцо) и в
позиции 6 нет боковой цепи. Та-
кой антибиотик — клавулановая
кислота изолирована из
Streptomyces clavuligerus. Она
слабо активна против бактерий,
но является потенциальным ин-
гибитором стафилококковой Р-
.0'
А* Ч
СН2—СН5 с=сн—сн2он
г I I
I? '¥ 3’
—Е—N сн..,
‘‘•соон
клавулановая кислота
лактамазы и Р-лактамазы, обра-
зуемой грамотрицательными
бактериями. Сочетание широкоспектральной клавулановой кисло-
ты с Р-лактамазочувствитёльными ампициллином и амоксицилли-
ном является основой комбинированного препарата аугментин.
1 — Карбапенемы — это новое семейство Р-лактамных
антибиотиков, представляющих собой аналоги пенициллинов или
клавамов, в которых сера у первых и кислород у вторых замещены
на углерод:
1-£Г“агС1Н zCH
I I II
О=с?------N4—чсн
1-карбапенемовое кольцо
К этому семейству относятся оливановые кислоты и тиенами-
цин. Их продуценты — стрептомицеты. Эти антибиотики обладают
широким - антимикробным спектром и являются выраженными
ингибиторами Р-лактамаз.
общее строение оливановых кислот (продуцент - Streplornyccs olivaccus)
тиенаыицин (продуцент - Streptomyces otvaceus)
433
Нокардицины — новая группа.Р-лактамных антибиотиков,
продуцируемых бактериями из рода Nocardia. Выделены нокарди-
цины А, В, D, Е, F, G. Наиболее активным из них против грамот-
рицательных бактерий (но не грамположительных) оказался но-
кардицин А.
нокардицим А (продуцент - Nokardia sp.)
Монобактамы — это моноциклические монобактамные
антибиотики, продуцируемые штаммами бактерий, например
Pseudomonas acidophila и Gluconobacter species. Их ядром является
3-аминомонобактамовая кислота (3-АМК). Ее можно получить из
природных монобактамов или из 6-АПК.
Природным высокоактивным против грамотрицательных бак-
терий и устойчивым к действию Р*лактамаз оказался азтреонам.
Однако он не активен против грамотрицательного анаэроба
Bacteroides fragilis, способного продуцировать Р-лактамазу, актив-
ную против данного антибиотика. Комбинация клавулановой кис-
лоты с монобактамами также неэффективна против этого микроба.
Ън—сн2
,С----
// so3-
З^АЫК
СООН
азтреонам (продуцент - Pseudomonas acidophfla)
Антибиотики-тетрациклины — это
широкоспектральные антибиотики, активные против бактерий; их
продуцентами являются стрептомицеты. Некоторые тетрациклины
относятся к ряду полусинтетических препаратов.
Антибиотики-гликозиды. Среди них можно
выделить структуры, содержащие О-, S- и N-гликозидные связи.
К числу первых относят аминогликозидные антибиотики и ново-
434
биоцин, ко вторым — клиндамицин и линкомицин, к третьим —
некоторые нуклеозидные антибиотики.
Антибиотики — О-гликозиды. Аминогликозиды
— содержат аминосахара в своей структуре. Большинство из них
(канамицины, гентамицины и др.) продуцируется актиномицетами;
тобрамицин — аналог канамицина В (З'-дезоксиканамицин В);
нетилмицин является полусинтетическим производным сизомици-
на (N-этилсизомицин).
Все антибиотики-аминогликозиды обладают широким спект-
ром действия на различные патогенные бактерии.
ниацин
(пслусйнптачюм! аммдпмимид -
производное шамяцинв)
Продуцентом ген-
тамицинов является
Micromonospora
purpurea, а сагамици-
на — M.sagamiesis.
Сизомицин по строе-
нию близок гентами-
цинам и отличается от
них лишь одним цик-
лом
Его продуцентом является мутант
Micromonospora sagamiensis KYI 1535.
Антибиотики — макроли-
ды — имеют О-гликозидные связи; они
продуцируются стрептомицетами и прояв-
ляют активность против большинства грам-
1Х>
положительных и некоторых грамотрицательных бактерий
(Neisseria spp., Haemophilus influenzae и Legionella pneumophila).
На практике чаще применяют эритромицин, олеандомицин, три-
ацетилолеандомицин и спирамицин. Они содержат в своей струк-
туре макроциклическое лактонное кольцо, связанное с углеводны-
ми остатками, из которых аминосахар (если имеется) всегда рас-
полагается ближе к макролактону, чем нейтральный углевод.
спирамицин J (продуцент - Streptomyces ambotaciens)
Олеандомицин, его эфир — триацетилолеандомицин и спирамицин
сходны по спектру антибактериального действия с эритромицином,
но несколько слабее его по активности.
Новобиоцин продуцируется культурой Streptomyces spheroides
и другими видами. Антибиотик активен против возбудителей
пневмонии, некоторых кишечных и раневых инфекций, ангин и
флегмон. По химической структуре — это О-глйкозид (амидогли-
козид), агликон которого существенно массивнее углеводной час-
ти.
Оливомицин — О-гликозидный антибиотик, в котором сахари-
стая часть представлена триаозой и биозой, между которыми как
бы "вставлен" агликон-оливин (хромомицинон).
Оливомицин является представителем большой группы анти-
биотиков — аналогов ауреоловой кислоты (митрамицина), актив-
ных против грамположительных бактерий и отдельных опухолей.
Оливомицин продуцируется Streptomyces olivoreticuli.
Дауномицин (рубомицин) — O-гликозидный антибиотик из
группы, антрациклинов, образуемый Streptomyces species. Углевод-
ный компонент в нем — даунозамин, или 2, 3, 6-тридезокси-З-ами-
ноликсоза. Антибиотик обладает противомикробным и противо-
опухолевым действием.
436
, красного цвета
дауномицин
Иммуносупрессивным и противоопухолевым действием обла-
дает О-гликозидный антибиотик блеомицин, содержащий также и
пептидные связи. Он образуется стрептомицетом; углеводный
компонент в нем представлен амидобиозой.
Ристомицин А (ристоцетин) — О-гликозидный антибиотик, в
составе которого имеется циклизованная агликоновая часть (цик-
лопентит) и углеводный компонент, включающий ристозамин,
маннозу, арабинозу, глюкозу и рамнозу. Ристомицин активен
против грамположительных бактерий.
Антибиотики — S-гликозиды
Из этой группы антибиотиков наиболее известны линкомицин
и клиндамицин, агликоном которых является метил. Другими сло-
вами — это метил-S-гликозиды. Из них клиндамицин является
производным линкомицина, у которого гидроксил в позиции 7
заменен на хлор.
(продуцент - Staptomycas iinkolneasB)
Углеводная часть обоих антибиотиков представлена 6, 8-дидё-
зокси- 1-тио-0-эритро-0-галакто-октапиранозой, в которой С6 име-
ет радикал— 1-метил-4-н-пропил-2-пирролидин-карбоксамид.
437
Клиндамицин лучше всасывается в кровь из кишечника. Оба
антибиотика активны преимущественно против грамположитель-
ных кокков.
Антибиотики — N-r ликозиды
Гужеротин — антибиотик широкого спектра антимикробного
действия. Он продуцируется культурой Streptomyces gougerotii. По
химическому строению представляет собой 1 — (цитозинил)-4-сар-
козил-В-сериламино-1,4-дидезокси-Р-В-галактопиран-уронамид
Пуромицин — N-гликозидный антибиотик, имеющий важ-
ное значение в расшифровке синтеза белка на рибосомах. Он
подобен З'-концу аминоацил-тРНК, связывающемуся с опреде-
ленным участком рибосомы, и действует как аналог аминоацил-
тРНК, прерывая удлинение полипептидной цепи.
Продуцентом пуромицина является Streptomyces alboniger. Хо-
тя антибиотик ингибирует
многие микроорганизмы,
включая трипаносомы, те-
рапевтическое значение
его невелико.
Антибиотики-
депсипептиды
Все они продуцируют-
ся, как правило, бациллами
и проявляют активность
против грамположитель-
ных и/или грамотрицатель-
ных патогенных бактерий.
К числу циклопептидных
антибиотиков относят ба-
цитрацины, грамицидины,
полимиксины и др.:
438
продуцент антибиотик активен против
Вас. lichenitormis бацитрацины (10 форм) грамположи- тельных бактерий и грам- отрицательных кокков
Вас. brevis var.G.-B. Вас. polymixa грамицидины (5 форм) полимиксины, II
Вас. circulans колистины, циркулины (22 формы) грамотри- цательных бактерий
Полимиксин В|, циркулин А и колистин различаются по еди-
ничным аминокислотам в циклах.
К циклопептидным антибиотикам относится также циклоспо-
рин А, продуцируемый мицелиальными грибами Cylindrocarpum
lucidum и Tolypocladium inflatum-Bauveria nives (рис. 134). Приме-
чательным качеством антибиотика является иммуносупрессивное
действие, в связи с чем его преимущественно используют в
трансплантологии, хотя он также проявляет антифунгальную ак-
тивность.
Рис. 134. Продуцент циклоспорина
А — Tolypocladium inflatum (сканирую-
щий электронномикроскопический сни-
мок, х20000).
Антибиотики-акт и-
номицины составляют
большую группу сходных ве-
ществ, образуемых стрептоми-
цетами. Некоторые из них обла-
дают противоопухолевым и им-
муносупрессивным действием.
По химическому строению их
относят к хромопептидам, со-
держащим феноксазиновую
хромофорную группу и два пен-
тапептида в форме циклических
лактонов. На практике хорошо
известен актиномицин (дакти-
номицин), ингибирующий ДНК-зависимый синтез РНК.
А н т и б и о т и к и - п о л и е н ы
Продуцентами антибиотиков-полиенов являются стрептомице-
ты и некоторые мицелиальные грибы, образующие структуры с
439
четырьмя-семью сопряженными двойными связями (тетра-, пента-,
гекса- и гептаены). Из группы полиеновых антибиотиков в меди-
цинской практике известны нистатин, фумагиллин (тетраены),
амфотерицин В (гептаен) и полусинтетический амфоглюкамин,
обладающие противогрибковым действием (преимущественно —
в отношении патогенных дрожжевых организмов и некоторых
других диморфных грибов); фумагиллин ингибирует стафилокок-
ки, дизентерийную амебу, бактериофаги. Нистатин (продуцент
Streptomyces noursei) — аналог амфотерицина В, но в его цепочке
(позиции 20—27) содержится 4 конъюгированных двойных связей.
Антибиотики -а нзамицины (от лат. ansa — ручка).
В их молекулах к ароматическому ядру присоединяется алифати-
ческая анза-цепь. Анзамицины образуются актиномицетами и
некоторыми растениями. Из природных и полусинтетических ри-
фамицинов наиболее известными являются: рифоцин (рифампи-
цин SV), рифампицин и рифамид. Анзамицины — щирокоспект-
ральные антибиотики, ингибирующие бактерии, отдельные вирусы
и некоторые эукариотические клетки.
• Рифампицин (продуцент — Streptomyces mediterranei) расцени-
вают одним из лучших химиотерапевтических средств при тубер-
кулезе, проявляющим бактерицидное действие против
Mycobacterium tuberculosis. Он проникает в спинномозговую жид-
кость и поэтому эффективен при туберкулезном менингите.
Антибиотики-стероиды — к ним относят фузидиевую
кислоту, изготавливаемую в виде натриевой соли. Она образуется
мицелиальным грибом Fusarium coccineum. Антибиотик активен
против многих типов грамположительных бактерий, особенно —
стафилококков.
Пр очие антибиотики
Гризеофульвин — продуцируется нитчатым грибом Penicillium
griseofulvum и другими, активен против болезнетворных грибов-
дерматофитов. Он состоит из конденсированных колец А В, С.
Трихотецин — образуется нитча-
тым грибом Trichothecium roseum и
некоторыми другими несовершен-
ными грибами. Это — антифунгаль-
ный антибиотик, полезный, прежде
всего, для ветеринарии и растение-
водства. В своем составе содержит
гетероциклы.
440
трихотацин
Ферментационные процессы получения антибиотиков еще не
достигли того совершенства, которое могло бы быть обеспечено
благодаря компьютерной технике и пониманию физиологии про-
дуцентов. Своеобразным тормозом совершенствования техноло-
гии выступают сами клеточные популяции микроорганизмов, ко-
торые изменяются качественно и количественно в течение всего
продуктивного цикла. Поэтому не удается получать полностью
идентичных результатов даже в двух смежных ферментациях,
равно как и весь технологический процесс удается сравнительно
хорошо проводить лишь в периодическом режиме.
В главе 7 приведены технологические схемы производства
пенициллина и канамицина, поскольку предферментационные и
ферментационные стадии во многом подобны, то нет необходимо-
сти рассматривать технологию производства каждого антибиотика
в отдельности.
Так как антибиотические вещества являются вторичными ме-
таболитами, то биосинтез их сопряжен с переходом культуры
продуцента в идиофазу. Следовательно, здесь целесообразно ли-
митирование роста продуцента. Таким лимитирующим ингредиен-
том (фактором) при биосинтезе пенициллина выступает глюкоза,
тогда как при биосинтезе антибиотиков стрептомицетами — фос-
фаты. Все это важно при "компановке" питательных сред, подкор-
мке штамма - продуцента в процессе биосинтеза антибиотика. Так,
среда для продукции пенициллина (она же пригодна для накопле-
ния инокулюма) включает глюкозу — 1,5%, лактозу — 5% (лактоза
снимает катаболитную репрессию глюкозы), аммония сульфат и
фосфаты — 0,5—1%, кукурузный экстракт — 2—3%, предшествен-
ники антибиотика — фенокси- или фенилуксусная кислота —
0,3—0,6%, мел — 0,5-1%, пеногаситель — 0,5—1%; температуру
ферментации поддерживают на уровне 22—26°С при pH от 5,0 до
7,5 и аэрации 1 м3 воздуха на 1 м3 среды в 1 минуту; продолжи-
тельность ферментации — 4 суток.
441
Так называемая "всеядность” актиномицетов обеспечивает им
способность расти и продуцировать антибиотики на средах, содер-
жащих белки (соевая мука, рыбная мука, белок клейковины пше-
ницы и пр.), крахмал. Тем не менее, применительно к каждому
продуценту имеются свои особенности, обычно отмечаемые в
регламентной документации. Например, посевной материал
Streptomyces kanamyceticus получают на соево-крахмальной среде
и на ней же проводят основную ферментацию при 27—28°С в
течение 4—5 суток при поддержании pH на уровне 7,1-7,6.
При ферментации Str. floridae (продуцент виомицина, или
флоримицина) рекомендуют среду, содержащую глюкозу или гид-
рол, соевую муку, кукурузный экстракт, нитраты, мел; температуру
поддерживают в пределах 27—29°С, pH на уровне 7,0—7,3.
В случае ферментации Str. erythreus в питательную среду
добавляют пропиловый спирт, как предшественник антибиотика
эритромицина.
Продуцент нистатина Str. noursei интенсивно потребляет аммо-
нийный азот (не нитратный), и т. д.
Перед выделением антибиотика из культуральной жидкости
необходимо отделить твердую, или плотную фазу от жидкой. В
этих случаях, как правило, используют фильтрацию. Качество
твердой фазы заметно сказывается на эффективности фильтрации.
Здесь можно отметить следующую закономерность — нативная
бактериальная масса фильтруется хуже мицелиальной. С учетом
того факта, что высшие актиномицеты способны формировать
нитчатые структуры, то их отделение при фильтрации происходит
несколько легче, чем других бактерий.
Пути улучшения фильтрации: обработка культуральных жид-
костей электролитами, тепловая коагуляция, добавление фильтру-
ющих наполнителей, кислотная коагуляция и некоторые другие.
Трудности выделения антибиотиков обусловлены многокомпо-
нентностью культуральных жидкостей и малой концентрацией в
них целевых продуктов. Так, пенициллин может накапливаться
примерно до 30 г/л при 8%-м выходе от субстрата. При этом сухие
выщества после отделения мицелия обычно составляют порядка
3—6%, из которых лишь 15—30% приходятся на антибиотик. Поэ-
тому любую культуральную среду необходимо обрабатывать так,
чтобы антибиотическое вещество переходило в ту фазу, из которой
затем он будет наиболее полно выделен. В ряде случаев этого можно
добиться подкислением (тетрациклины) или, напротив, подщела-
чиванием культуральной жидкости (новобиоцин), добавлением
442
солей, щавелевой кислоты (эритромицин) и т. д., до или после
коагуляции и осаждения белков из нативных растворов.
Если для выделения антибиотика применяют ионный обмен, то
нативный раствор стремятся освободить от ионов-конкурентов
(растворимые оксалаты — для удаления ионов кальция; натрия
триполифосфат — для связывания ионов магния; желтая кровяная
соль — для комплексования ионов железа).
Антибиотические вещества, накапливающиеся в клеточной
биомассе в процессе ферментации микробов-продуцентов, выде-
ляют другими методами, нежели антибиотики, содержащиеся в
культуральной жидкости. В первом случае, как правило, необхо-
дима экстракция каким-либо растворителем из нативных или
дезинтегрированных клеток (система "твердое тело-жидкость").
Примерная схема выделения целевого продукта (антибиотика)
из культуральной жидкости может быть представлена в следующем
виде (рис. 135). В приведенную схему должны быть внесены
соответствующие коррективы в зависимости от физико-химиче-
ских характеристик целевого продукта и возможностей аппара-
турного оформления процесса. В настоящее время все большее
распространение приобретают мембранные методы концентриро-
вания и выделения различных веществ, хотя до сих пор в ряде
производств БАВ (включая антибиотики, например, пенициллин)
не удалось отказаться от традиционных способов выделения и
очистки целевых продуктов (экстракция в системе "жидкость-жид-
кость", адсорбция на активированных углях, диализ).
Рис. 135. Примерная тех-
нологическая схема выделе-
ния антибиотика X из культу-
ральной жидкости (КЖ): 1 —
предварительная обработка
КЖ, 2 — подготовка раство-
ра, 3 — первый фильтр, 4 —
сборник первого фильтра, 5
— подготовка раствора для
фильтрации, 6 — дополни-
тельная фильтрация, 7 — сте-
рилизующая фильтрация, 8—
коагуляция растворителем, 9
— предварительное осажде-
ние, 10—промывание осадка,
11 — вторичное осаждение,
12 — высушивание.
443
В сравнительном плане следует иметь в виду, что механическое
обезвоживание заметно дешевле термического и это сказывается
на себестоимости производимого продукта.
Широко используют методы хроматографии, например, при
выделении новобиоцина, стрептомицина и других антибиотиков.
В подобных случаях технологическая схема аппаратурного офор-
мления процессов заметно упрощается.
9.3.4. Получение аминокислот. Аминокислоты играют большую
роль в здравоохранении, животноводстве и легкой промышленно-
сти. По значению для макроорганизма аминокислоты подразделя-
ют на заменимые и незаменимые. К незаменимым относят те
аминокислоты, которые не синтезируются в человеческом и/или
животном организме, они должны быть привнесены с пищей или
кормом для животных (таблица 46).
Т а б л и ц а 46. Незаменимые и заменимые аминокислоты
Незаменимые Заменимые
Аргинин (только для молодых растущих животных) Аланин
Валин Аспарагин
Гистидин Аспарагиновая кислота
Изолейцин Глицин
Лейцин Глутамин
Лизин Глутаминовая кислота
Метионин Пролин
Треонин Серин
Триптофан Тирозин
Фенилаланин Цистеин
Заменимые синтезируются in vivo из аммиака и различных
источников углерода. Микроорганизмы сами синтезируют все
необходимые им аминокислоты из аммиака и нитратов, а углерод-
ные "скелеты" — из соответствующих интермедиатов.
Исходя из такой оценки аминокислот, ученые давно стремятся
использовать способности микроорганизмов продуцировать заме-
нимые и незаменимые аминокислоты в ощутимых количествах.
444
Потребность людей в аминокислотах достаточно велика и этим
определяется уровень их производства в мире (порядка 500 тыс.
тонн в год).
Специфические ферменты, регулирующие биосинтез амино-
кислот, широко распространены у бактерий; они с определенной
глубиной изучены у Escherichia coli, Salmonella typhimurium,
Bacillus subtilis и пр. У грибов, в ответ на аминокислотное лимити-
рование, отмечается некоординированное, параллельное возраста-
ние уровня ферментов, катализирующих реакции биосинтеза раз-
личных аминокислот. Этот "общий контроль биосинтеза амино-
кислот" был также назван "метаболическим интерблоком", или
"перекрестнопутевой регуляцией”, впервые выявленной у
Neurospora crassa в 1965 г. М. Карсиотисом и сотрудниками, а
позднее — у Saccharomyces cerevisiae, Aspergillus nidulans и других
грибов.
В гиперпродукции отдельных аминокислот культурами E.coli,
Serratia marcescens и др. важную роль играют Feedback-ингибиция
и Feedback-репрессия, например, при биосинтезе ароматических
аминокислот на последних стадиях. Штаммы — суперпродуценты,
эксплуатируемые в производстве, как правило, получены с исполь-
зованием методов генетики и селекции.
В любом живом организме аминокислоты расходуются прежде
всего на биосинтез первичных метаболитов — ферментных и
неферментных белков. Следовательно, кроме биосинтеза амино-
кислот de novo,возможен другой путь их получения, а именно —
из гидролизатов соответствующих белков (триптофан разрушается
при кислотном гидролизе), в том числе — из нативной биомассы
микробных клеток.
Природные аминокислоты являются, как правило, оптически
активными L-формами, тогда как аминокислоты, получаемые хи-
мическим синтезом, являются рацемическими смесями L- и D-
форм, которые трудно разделить. Вот почему микробный синтез с
помощью коринебактерий и некоторых других микробов является
ныне основным и экономически выгодным. Первое место здесь по
праву занимает Япония, где лишь глутаминовой кислоты изготав-
ливается свыше 100 тыс. тонн в год; большинство природных
незаменимых аминокислот производит фирма "Такеда". С. Кино-
шита, впервые в 50-е годы открывший и доказавший перспектив-
ность микробного синтеза, уже в 1963 г. прогнозировал: "Мало
сомнения в том, что недалеко то время, когда с помощью микро-
организмов будет возможно производить все известные аминокис-
лоты", Это время наступило уже к 70-м годам. Получены микробы
— суперпродуценты из родов Brevibacterium, Corynebacterium,
Micrococcus и др., с помощью которых освоено крупнотоннажное
445
производство не только глутамата, но и L-лизина, L-валина, L-гис-
тидина и других.
При суперпродукции уровень экспрессии клонированного гена
выражается в синтезе специфического белка в количестве не менее
2% от всех растворимых белков клетки-хозяина. В настоящее время
имеются суперпродуценты, у которых количество синтезируемого
специфического белка достигает 10—50% (здесь важнейшую роль
играют многокопийные плазмиды, несущие встроенные гены).
Генно-инженерными методами во ВНИИ генетики и селекции
промышленных микроорганизмов (Москва) был получен штамм
E.coli, обладающий сверхпродукцией L-треонина (30 г/л за 40 часов
ферментации).
с любым штаммом — продуцентом какой-либо аминокислоты
необходимо внимательное и бережное обращение в целях поддер-
жания ее в активном состоянии в течение длительного времени.
Технология получения аминокислот базируется на принципах
ферментации продуцентов и выделении вторичных метаболитов,
то есть размножают маточную культуру вначале на агаризованной
среде в пробирках, затем — на жидкой среде в колбах, инокуля-
торах и посевных аппаратах, а затем в головных (основных)
ферментаторах. Отработку культуральных жидкостей и выделение
аминокислот проводят по схеме, аналогичной схеме получения
антибиотиков (см.). Изолированные чистые кристаллы целевого
продукта обычно высушивают под вакуумом и упаковывают.
Если аминокислота предусмотрена в качестве добавки к кор-
мам, то биотехнологический процесс кормового продукта включает
следующие стадии: ферментацию, стабилизацию аминокислоты в
культуральной жидкости перед упариванием, вакуум-упаривание,
стандартизацию упаренного раствора при добавлении наполните-
ля, высушивание и упаковку готового продукта, в котором должно
содержаться не более 10% основного вещества. Например, в про-
мышленности изготавливают сухой кормовой и жидкий кормовой
концентраты лизина наряду с кристаллическим лизином (рис. 136).
Рис. 136. Технологическая схема пол-
учения лизина: 1 — емкость для культу-
ральной жидкости (КЖ), 2 — ионообмен-
ные колонны, 3 — сборник элюата, 4 —
сборник фильтрата, 5 — емкость для элю-
ата, 6 — насос, 7 — вакуум-выпарной
аппарат, 8 — циклон, 9 — сушилка кор-
мового концентрата, 10 — сборник, 11 —
реактор-кристаллизатор, 12 — центрифу-
га, 13 — сушилка.
446
Если концентрат содержит 70—80%. сухих веществ, то он
достаточно устойчив против микробной порчи за счет повышенной
осмотической концентрации ингредиентов.
Известны два способа получения аминокислот: одноступенча-
тый и двухступенчатый. Согласно первому способу, например,
мутантный полиауксотрофный штамм — продуцент аминокислоты
культивируют на оптимальной для биосинтеза среде. Целевой
продукт накапливается в культуральной жидкости, из которой его
выделяют согласно схеме на рис. 137.
Рис. 137. При-
мерная технологи-
ческая схема пол-
учения ферментов:
1 — ферментатор, 2
— охладитель, 3, 9
— рефрижерато-
ры, 4 —емкость для
предварительной
обработки, 5 —
центрифуга, 6 —
вакуум - упарива-
тель, 7 — аппарат
прямой предобра-
ботки фермента, 8
— барабанный
фильтр, А, Б—пути
(при необходимо-
сти смыкающиеся),
10 — аппарат для
ультрафильтрации,
11 — емкость для
консервации рас-
твора фермента, 12
— мембранный
фильтр, 13 — нако-
питель жидкого
концентрата, 14 —
емкость для осаж-
дения фермента, 15
— фильтр-пресс, 16
— распылительная
сушилка, 17,— на-
копитель сухого
концентрата.
447
В двухступенчатом способе микроб-продуцент культивируют в
среде, где он получает и синтезирует все необходимые ингредиен-
ты для последующего синтеза (в идиофазу) целевого продукта.
Схема двухступенчатого процесса может быть представлена в
следующем виде:
Микроб — продуцент аминокислоты
л * I ступень культивирование в жидкой питательной среде
Биосинтез предшественников аминокислоты и ферментов, катализирующих биосинтез целевого продукта
биосинтез целевого продукта с
II ступень участием ферментов ' Аминокислота — целевой продукт
Если ферменты биосинтеза аминокислоты накапливаются
внутриклеточно, то после I-й ступени клетки сепарируют, дезин-
тегрируют и применяют клеточный сок. В других случаях для целей
биосинтеза целевых продуктов применяют непосредственно клет-
ки.
Экономически целесообразными являются способы получения
аминокислот с помощью иммобилизованных ферментов и клеток.
Сравнительно давно реализован процесс получения L-аспарагино-
вой кислоты из фумаровой и аммиака в одну стадию с помощью
иммобилизованных клеток E.coli или Pseudomonas aeruginosa, об-
ладающих аспартазной активностью (Е):
nh2
нс—соон _ |
й ф нн-.——► ноос—сн2—сн—соон
ноос—сн
фумаровая кислота юмрммновая кислота
Аспартаза катализирует реакцию присоединения аммиака к
фумаровой кислоте. Фермент в иммобилизованном состоянии
сохраняет активность на исходном уровне до 2—2,5 недель и более.
L-Аспарагиновую кислоту можно получить и с помощью иммо-
билизованных клеток, что существенно повышает длительность
функционирования системы, производительность которой по це-
левому продукту составляет около 2000 кг с 1 м3 биореактора.
Периодические ферментации используют при получении дру-
гих L-аминокислот (глутаминовой, фенилаланина, лизина, трипто-
448
фана и др.). При этом культивируют обычно специальные мутан-
тные штаммы, метаболизм которых по целевому продукту изучен
достаточно полно. Так, например, установлено, что лимитирующим
агентом коринебактерий, образующих глутаминовую кислоту, яв-
ляется биотин (витамин В?, витамин Н) в дозе 1—5 мкг/л. Биотин
индуцирует структурно-функциональные изменения в клеточной
мембране, благодаря чему увеличивается ее проницаемость для
глутаминовой кислоты, выходящей из клетки в культуральную
жидкость. Отдельные штаммы продуцентов способны накапливать
ее более 50 г/л на мелассных средах.
Роль биотина аналогична в случае получения L-пролина, явля-
ющегося производным L-глутаминовой кислоты.
9.3.5. Получение витаминов. Витамины поставляются в орга-
низм с пищей или их назначают в форме лекарственных препара-
тов при определенных патологических процессах. Среди липидо-
и водорастворимых витаминов известны реализованные биотех-
нологические процессы производства витаминов А] и D, рибофла-
вина, аскорбиновой кислоты, цианкобаламина (В12).
Каротиноиды — это изопреноидные соединения, синтезирую-
щиеся многими пигментными микроорганизмами из рода Aleuria,
Blakeslea, Corynebacterium, Flexibacter, Fusarium, Halobacterium,
Phycomyces, Pseudomonas, Rhodotorula, Sarcina, Sporobolomyces и
др. Всего описано около 500 каротиноидов.
Из одной молекулы Р-каротина при гидролизе образуются две
молекулы витамина At. Это имеет место, например, в кишечнике
человека.
("V"’ сиз СИ,
сн=сн—с=сн—снмн—с=сн—ск2он
СНз^СНз
•ммымнАП
Каротиноиды локализуются в виде сложных эфиров и глико-
зидов в клеточной мембране микроорганизмов, либо в свободном
состоянии — в липидных гранулах в цитоплазме. Каротиноид
"ретиналь”, например, у галофильного вида — Halobacterium
halobium — соединен с белком через остаток лизина (опсинопо-
добный белок); он участвует в синтезе АТФ благодаря генерации
трансмембранного потенциала. В целом, основная функция каро-
тиноидов — защитная. Их биосинтезу в клетках способствует свет.
15 т. 8524 449
В качестве продуцентов каротиноидов можно использовать
бактерии, дрожжи, мицелиальные грибы. Более часто применяют
зигомицеты Blakeslea trispora и Choanephora conjuncta. Спариваю-
щиеся (+) и (—) особи этих видов при совместном культивирова-
нии могут образовать 3—4 г каротина на 1 л среды.
Питательные среды для них достаточно сложные и включают
источники углерода, азота, витаминов, микроэлементов, специаль-
ных стимуляторов (гидрол, кукурузно-соевая мука, растительные
масла, керосин, Р-ионон или изопреновые димеры, и пр.). Стиму-
ляторы целесообразно вносить в культуральные среды в конце
трофофазы, то есть когда продуцент переходит в продуктивную
фазу (идиофазу).
Вначале штаммы выращивают раздельно, а затем — совместно
при 26°С и усиленной аэрации с последующим переносом в
основной ферментатор. Условия культивирования сохраняют
прежними. Длительность ферментации — 6—7 дней. Каротиноиды
извлекают ацетоном (можно каким-либо другим полярным раство-
рителем), переводят в неполярный растворитель. В случаях извле-
чения белково-каротиноидных комплексов, то применяют повер-
хностно-активные вещества в концентрации 1—2%. В целях очи-
стки и более тонкого разделения гомологов можно прибегать к
методам хроматографии или к смене растворителей. Витамин At
из Р-каротина сравнительно легко можно получить при гидролизе.
В случае изготовления каротинсодержащей биомассы для скар-
мливания животным и птицам возможно ее сочетанное примене-
ние с витамином А или без него. В медицинских целях витамин А
изготавливают в капсулах для приема через рот.
Витамин D — это группа родственных соединений, в основе
которых находится эргостерин, который обнаружен в клеточных
мембранах эукариот. Поэтому, например, пекарские или пивные
дрожжи применяют для получения эргостерина, как провитамина,
обладающего антирахитическим действием. Содержание эргосте-
рина в дрожжевых клетках колеблется в пределах 0,2—11%.
При недостатке в организме гормона 1,25-дигидроксихолекаль-
циферола, предшественником которого является витамин D3, у
детей развивается рахит (аналог рахита у взрослых — остеомаля-
ция).
Трансформация эргостерина в витамин D2 (кальциферол) про-
исходит под влиянием ультрафиолетового облучения. При этом
разрывается связь в кольце (позиции 9,10) и образуется двойная
связь в боковой цепочке (позиции 22, 23). Эта последняя гидриро-
450
вана в витамине D3 (холекальциферол). Физиологическая актив-
ность обоих витаминов (D2 и D3) равноценна.
Кроме дрожжей продуцентами эрогостерина могут быть мице-
лиальные грибы — аспергиллы и пенициллы, в которых содержится
1,2—2,2% эргостерина. Замечено, что полиеновые антибиотики,
действующие на клеточную мембрану дрожжей, заметно стимули-
руют их содержание в биомассе.
Получение эргостерина в производственных условиях можно
подразделить на следующие этапы: размножения исходной куль-
туры и накопление инокулюма, ферментация, сепарирование кле-
ток, облучение клеток ультрафиолетовыми лучами, высушивание
и упаковка целевого продукта.
Так, применительно к дрожжам, инокулюм получают на средах,
обеспечивающих полноценное развитие клеток, после чего основ-
ную среду с ацетатом (активатором биосинтеза стеринов), обога-
щенную источником углерода и содержащую пониженное коли-
чество азота (высокое значение С/N), засевают сравнительно
большим объемом инокулята.' Культивирование дрожжей (фермен-
тацию) проводят при температуре, близкой к максимальной для
конкретного штамма, и выраженной аэрации (2% О2 в газовой
фазе). Спустя 3—4 суток, в зависимости от ростовых характеристик
и биосинтетической активности культуры, клетки сепарируют и
подвергают вакуум-высушиванию. Затем сухие дрожжи облучают
ультрафиолетовыми лучами — УФЛ (длина волны 280—300 нм) в
течение оптимального по продолжительности времени, при требу-
емой температуре и с учетом примесных веществ. Эти контроли-
руемые показатели, установленные опытным путем, указываются
в регламентной документации. Облучение дрожжей можно прово-
дить до сепарирования клеток в тонком слое 5% суспензии, учи-
тывая малую проникающую способность УФЛ.
Облученные сухие дрожжи применяют в животноводстве; в
промышленности их выпускают под названием "кормовые гидро-
лизные дрожжи, обогащенные витамином D2". В таком препарате
содержится не менее 46% сырого белка, незаменимые аминокис-
лоты (лизин, метионин, триптофан) и 5000 ME витамина D2 /г.
В случае получения кристаллического витамина D2 клетки
продуцента гидролизуют соляной кислотой при 110°С, затем тем-
пературу снижают до 75—78°С и добавляют этанол. Смесь фильт-
руют при 10—15°С, оставшуюся после фильтрации массу промы-
вают водой, высушивают, измельчают, нагревают до 78°С и дважды
обрабатывают тройным объемом этанола. Спиртовые экстракты
, 451
объединяют и упаривают до 70%-го содержания сухих веществ.
Полученный "липидный концентрат" обрабатывают раствором ед-
кого натра. Эргостерин кристаллизуется из неомыленнной фрак-
ции концентрата при 0°С. Его можно очистить повторными пере-
кристаллизациями. Кристаллы высушивают, растворяют в серном
эфире, облучают УФЛ, эфир отгоняют, раствор витамина D2 кон-
центрируют и кристаллизуют.
"Кислотный фильтрат" обычно упаривают до 50%-го содержа-
ния сухих веществ и применяют как концентрат В-витаминов.
Производят также масляный концентрат витамина D2.
Рибофлавин, или витамин В2 — содержится в клетках различ-
ных микроорганизмов, будучи коферментом в составе флавопро-
теинов (прежде всего — соответствующих ферментов из класса
оксидоредуктаз — ФМН, ФАД). Поэтому в качестве продуцентов
рибофлавина (флавопротеинов) могут быть бактерии, дрожжи и
нитчатые грибы. Однако наиболее заманчивыми являются те штам-
мы, которые образуют на жидких средах 0,5 г и более рибофлавина
в 1 л среды. К подобным организмам относятся Ashbyii gossypii,
Eremotheciurii ashbyii и Candida guilliennondii. Учитывая изменчи-
вость активных продуцентов названных видов по способности
синтезировать витамин В2, необходим систематический поддержи-
вающий отбор культур в процессе их эксплуатации на производ-
стве. Обычно активные продуценты первых двух видов формируют
яркооранжевые колонии на агаризованных средах. Методами ген-
ной инженерии удалось получить штамм сенной палочки, образу-
ющий около 6 г рибофлавина в 1 л среды, включающей мелассу,
белково-витаминный концентрат и его гидролизат.
Высокий выход рибофлавина у E.ashbyii коррелирует с азотом
пуринов и другими азотистыми источниками, содержание которых
должно быть достаточным. В качестве источников углерода при-
меняют глюкозу или сахарозу, практикуют использование дрож-
жевого и кукурузного экстрактов, соевой муки, масла (жира).
Жидкие питательные среды для получения инокулюма и для
основной ферментации могут несколько различаться между собой.
Например, для получения посевного материала известна среда,
содержащая сахарозу, пептон, кукурузный экстракт, калия дигид-
рофосфат, магния сульфат, подсолнечное масло, время выращива-
ния продуцента на этой среде — 2 суток при 27—30°С (в зависи-
мости от штамма). Ферментационная среда обычно включает ку-
курузную и соевую муку, сахарозу, кукурузный экстракт, калия
452
дигидрофосфат, кальция карбонат, натрия хлорид и ненасыщен-
ный жир.
Обычно ферментацию проводят в течение 5 суток при pH 5,5—
7,7. После использования сахарозы (примерно через 30 часов)
начинает заметно накапливаться витамин Вг, вначале — в мицелии,
а затем — в культуральной жидкости. Всю биомассу можно
подвергнуть высушиванию и полученный сухой продукт с остаточ-
ной влажностью 8%, содержащий 1,5—2,5% рибофлавина, 20%
белка, тиамин, никотиновую кислоту, пиридоксин, цианкобаламин,
микроэлементы и другие вещества, рекомендуют для кормления
животных.
В случае высоких выходных показателей по рибофлавину,
витамин можно выделять в индивидуальном состоянии и, наряду
с синтетическим рибофлавином, использовать в медицине.
Для Candida guilliennondii важно регулировать содержание
железа в питательной среде; оптимальные концентрации колеб-
лются, в среднем, от 0,005 до 0,05 мкг/мл. При этом определенные
штаммы дрожжей могут образовывать за 5—7 дней до 0,5 г/л и
более витамина. Однако для целей промышленного производства
рибофлавина предпочитают использовать более продуктивные ви-
ды и штаммы грибов — E.ashbyii и Ashbyii gossypii.
Аскорбиновая кислота, или витамин С — это противоцингот-
ный витамин, имеющийся у.всех высших растений и животных;
толькс человек и микробы не синтезируют ее, но людям она
неотложно необходима, а микробы не нуждаются в ней. И, тем не
менее, определенные виды уксуснокислых бактерий причастны к
биосинтезу полупродукта этой кислоты — L-сорбозы. Таким обра-
зом, весь процесс получения аскорбиновой кислоты является
смешанным, то есть химико-ферментативным.
сн2он
- ( о
/ пярирсванкБ Gluconobacter cxydos
КI 'Л 1 н он ?С( снон UCH 2ОН в*
' " ' | D-сорбиг биологическое окисление,
| pH-5,0
D-пкмшаа
лммжескми
процесс
ном:со(снон)3сн?он в
* (трансформация)
L-сорбоеа
. L-аскорбиноаая кислота
453
Биологическая стадия процесса катализируется мембраносвя-
занной полиолдегидрогеназой, а последняя (химическая) включает
последовательно следующие этапы: конденсация сорбозы с диаце-
тоном и получение диацетон — L-сорбозы, окисление диацетон —
L-сорбозы до диацетон-2-кето-Е-гулоновой кислоты, подвергаемой
затем гидролизу с получением 2-кето-Е-гулоновой кислоты; послед-
нюю подвергают энолизации с последующей трасформацией в
L-аскорбиновую кислоту.
Ферментацию G.oxydans проводят на средах, содержащих сор-
бит (20%), кукурузный или дрожжевой экстракт, при интенсивной
аэрации (8—10 г О2/л/ч). Выход L-сорбозы может достичь 98% за
одни-двое суток. При достижении культурой log-фазы можно
дополнительно внести в среду сорбит, доводя его концентрацию
до 25%. Также установлено, что G.oxydans может окислять и более
высокие концентрации полиспирта (30—50%), создаваемые на
последних стадиях процесса. Это происходит благодаря полиолде-
гидрогеназы, содержащейся в клеточной биомассе. Ферментацию
бактерий проводят в периодическом или непрерывном режиме.
Принципиально доказана возможность получения L-сорбозы из
сорбита с помощью иммобилизованных клеток в ПААТ.
Аскорбиновую кислоту используют как антиоксидант в здра-
воохранении и пищевой промышленности.
Цианко балам ин, или витамин В12— получают только микроби-
ологическим синтезом. Его продуцентами являются прокариоты и,
прежде всего, пропионовые бактерии, которые и в естественных
условиях образуют этот витамин. Мутанты Propionibacterium
shermanii М-82 и Pseudomonas denitrificans М-2436 продуцируют на
жидкой среде до 58—59 мг/л цианкобаламина.
Учитывая важную функцию витамина в организме человека
(он является противоанемическим фактором), его мировое произ-
водство достигло 10 т в год из которых 6,5 т расходуют на
медицинские нужды, а 3,5 т — в животноводстве.
Отечественное производство цианкобаламина базируется на
использовании культуры P.freudenreichii var. shermanii, культиви-
руемой в периодическом режиме без доступа кислорода. Фермен-
тационная среда обычно содержит глюкозу, кукурузный экстракт,
соли аммония и кобальта, pH около 7,0 поддерживают добавлением
NHjOH; продолжительность ферментации 6 суток; через 3 суток в
среду добавляют 5,6-диметилбензимидазол — предшественник ви-
тамина В12 и продолжают ферментацию еще 3 суток. Цианкобала-
мин накапливается в клетках бактерий, поэтому операции по
454
выделению витамина заключаются в следующем: сепарирование
клеток, экстрагирование водой при pH 4,5—5,0 и температуре
85—90°С, в присутствии стабилизатора (0,25% раствор натрия
нитрита). Экстракция протекает в течение часа, после чего водный
раствор охлаждают, нейтрализуют раствором едкого натра, добав-
ляют коагулянты белка — хлорид железа трехвалентного и алю-
миния сульфат с последующим фильтрованием, фильтрат упари-
вают и дополнительно очищают, используя методы ионного обмена
и хроматографии, после чего проводят кристаллизацию витамина
при 3—4°С из водно-ацетонового раствора.
Кристаллический цианкобаламин можно получать с помощью
резорцина или фенола, образующих с ним аддукты, которые
сравнительно легко разлагаются на составляющие компоненты.
При реализации данного биотехнологического процесса не
забывать о высокой светочувствительности витамина В12, поэтому
все операции необходимо проводить в затемненных условиях (или
при красном свете).
На ацетонобутиловой и спиртовой бардах с добавлением солей
кобальта и метанола в нашей стране получают кормовой препарат
КМБ и — концентрат, содержащий витамин В12 и другие ростовые
вещества. Биообъектом здесь является смешанная культура мета-
ногенных бактерий.
9.3.6. Получение микробных препаратов — удобрителей почв,
стимуляторов и регуляторов роста растений. На основании более
чем векового изучения азотфиксирующих микроорганизмов ут-
вердилось представление о целесообразности производства в про-
мышленных масштабах лишь клубеньковых бактерий, поскольку
эффективность от внесения в почву препаратов из свободноживу-
щих азотфиксаторов по разным причинам является незначитель-
ной. Однако ясно, что в условиях интенсивного земледелия и
бурного роста населения земли плодородие почвы — одна из
главнейших проблем, на решение которой должны быть направ-
лены усилия различных специалистов, включая биотехнологов. И
нельзя считать, что свободноживущие азотфиксаторы не будут
вновь объектами биотехнологии. Очевидно не все изучено приме-
нительно к их ассоциативным взаимоотношениям в среде обитания
и значение самой среды, заметно изменяющейся при энергичном
воздействии на нее человека.
Микроорганизмы, фиксирующие азот из воздуха, подразделя-
ют на свободноживущие в природных условиях и на симбиотиче-
ские с растениями или другими микробами. К свободноживутцим
455
относят азотобактеры, актиномицеты, определенные цианобакте-
рии и др., к микробам-симбионтам: азотоспириллы — симбионты
папоротника азолла, ризобии — симбионты бобовых растений,
отдельные виды цианобактерий в составе лишайников (лишайники
представляют собой пример устойчивого симбиоза некоторых
грибов с цианобактериями или водорослями), и др.
Микробы-азотофиксаторы ежегодно фиксируют примерно
17,5* 107 т молекулярного азота из воздуха, из которых 10—15%
приходится на цианобактерии. Очевидно, что до тех пор, пока не
будут созданы "молекулярные гибриды" микробов — азотофикса-
торов с широким набором полезных растений методами генной
инженерии, использование соответствующих "препаратов-удобри-
телей" будет целесообразным и экономически оправданным.
В таком же ряду находятся микробные стимуляторы и регуля-
торы роста растений — гиббереллины, фузикокцин и др.
Ризотрофин — своеобразный "потомок" коммерческого пре-
парата "нитрагин", впервые созданного в 1896 г. в Германии. Он
представляет собой торфяную основу, смешанную с ризобактери-
ями (5—8*10® клеток в 1 г торфа). Производство ризотрофина
разработано во ВНИИ сельскохозяйственной микробиологии (г.
Санкт-Петербург).
Технологический процесс производства ризотрофина включает
подготовку клубеньковых бактерий и получение инокулята, подго-
товку торфа, "обсеменение" торфа ризобактериями (отбор вцдов*
с учетом целесообразных рекомендаций по выращиванию соответ-
ствующих растений-"хозяев").
В производстве используют клубеньковые бактерии, быстро
или сравнительно медленно размножающиеся на простых средах,
содержащих растительные экстракты или отвары, сахарозу (pH
6,8—7,0) — для первых, почти то же самое — для вторых, но
добавляют глюкозу, маннит, сахарозу.
Инокулюм готовят в условиях умеренной аэрации на подобных
средах, но дополнительно вводят неорганические соли: фосфаты,
сульфаты, карбонаты (температура 28—ЗО’С); необходимо пенога-
шение. Накапливают примерно 5 млрд, клеток в 1 мл. Такую
суспензию вносят в "кислый" торф (pH 3,0—6,0) при 10-15’С.
Предпочтительнее использовать древесно-осоковые или осоковые
торфа с удельной площадью поверхности порядка 300 м2/г.
Торф до "обсеменения" бактериями должен быть специально
подготовленным —гомогенным, подсушенным, размолотым, ув-
лажненным, смешанным с мелом, расфасованным в полиэтилено-
> 456
вые пакеты по 150—160 г и простерилизованным методом радиа-
ционного облучения — 2,5 Мрад. В подготовленный торф "инъе-
цируют” 50-60 мл суспензии бактерий (в торфе должно содержать-
ся 0,5—1 млрд, клеток/г). После этого содержимое пакетов пере-
мешивают во вращающемся барабане и продукт сохраняют до
полугода при 5— 10°С — применительно к быстро растущим ризо-
бактериям и при 12—15°С — применительно к медленно растущим
ризобактериям. Расход ризотрофина — 200 г на гектарную норму
семян любых бобовых растений.
В странах Юго-Восточной Азии используют цианобактерии
Anaboena azollae, обитающие в полостях листьев папоротника
Azolla. Азоллу выращивают в специальных прудах, откуда ее
вывозят по назначению. Такая симбиотическая ассоциация —
хороший "удобритель" азотом рисовых полей.
В последние годы много внимания уделяют бактериям рода
Asospirillum — активному азотофиксатору, обитающему в почве
(реже — на надземной части растений) в ассоциации с кукурузой,
просом, сахарным тростником и др.
Гибберелловая кислота — синтезируется микромицетом
Gibberella fujikuroi (ее несовершенная форма Fusarium
monilifonne); она в качестве равноправного члена находится в ряду
гиббереллинов А1..., А — группы растительных гормонов сложного
химического строения, ядром которых является тетракарбоциклич-
ный гиббан. Его образование представляет собой частный случай
биосинтеза терпенов, как и синтез каротинов и витамина D.
nh4oh н
гибберелловая кислота
Все известные гиббереллины во многом сходны с гибберелло-
вой кислотой и в зависимости от условий культивирования проду-
цента могут варьировать в той или иной степени.
Биосинтез гибберелловой кислоты происходит, начиная с аце-
тата и двух молекул АцКоА через ряд промежуточных продуктов,
в том числе — через мевалоновую кислоту, изопентенилпирофос-
фат и циклизованные структуры. Образование и накопление гиб-
береллинов — процесс длительный, он занимает до 15 суток и более
457
при поверхностном выращивании продуцента при 25°С, до 9—10
суток — при глубинных ферментациях. Питательные среды для
G.fujikuroi обычно содержат 4—6% источника углерода (глюкозу,
сахарозу или глицерин), 0,7% источника азота (соли аммония,
нитрат или пептон), 0,3% неорганического фосфата, 0,05% магния
сульфата, микроэлементы, особенно — железо, медь, марганец,
молибден, серу, цинк; pH среды устанавливают на урЬвне 3,0—4,0
соляной кислотой. Выход гиббереллинов при этом достигает 200
мг/л культуральной жидкости (поверхностный способ) или 1 г/л
(глубинное культивирование в ферментаторах).
С учетом близости структур гиббереллинов гибберелловую
кислоту получают в виде суммарного препарата из вакуум-упарен-
ной культуральнойжидкости после отделения мицелия продуцента.
Гибберелловую кислоту применяют в сельском хозяйстве в
качестве стимулятора роста различных растений с максимальным
эффектом в дозе 1 мкг на растение.
Фузикокцин — относят к регуляторам роста растений гормо-
нального типа. Он индуцирует корнеобразование у многих древес-
ных и плодовоягодных культур, стимулирует прорастание семян
моркови, томатов, сахарной свеклы и других сельскохозяйствен-
ных растений (Г. С. Муромцев, 1986).
По Химическому строению фузикокцин относят к карботри-
циклическим дитерпенам, в которых тройной цикл является агли-
коном в гликозидной структуре.
Фузикокцин образуется грибом Fusicoccus amygdali. Продукт
получают при глубинной ферментации в периодическом режиме
на средах с глюкозой или сахарозой и соевой мукой; фузикокцин
затем экстрагируют бутилацетатом или хлороформом из культу-
ральной жидкости, сорбируют активированным углем, повторно
очищают и кристаллизуют из этилацетата.
В настоящее время активно изучают трибы Botrytis cinerea и
Cercospora rosicola — продуценты фитогормона — абсцизовой
458
кислоты, а также некоторые виды рода Cladosporium — продуцен-
ты фитогормона котиленина, которые, очевидно, в скором времени
будут производить в виде коммерческих препаратов.
абсцизовая кислота
котиленин
9.3.7. Получение микробных полимеров. Биотехнологические
процессы получения микробных полимеров освоены в различных
масштабах применительно к белкам, углеводам, липидам и неко-
торым полимерным конъюгатам.
Получение белков. Белки микробного происхождения находят
применение в здравоохранении, пищевой промышленности, жи-
вотноводстве, реже — в других отраслях производства. Разнонап-
равленность практического применения зависит от химической
структуры, молекулярной массы и физико-химических свойств
белков.
Ферментные белки, ферменты, будучи биокатализа-
торами реакций в любом организме, давно приобрели важное
практическое значение. Первый патент на грибную амилазу (та-
кадиастазу) зарегистрировал И. Такамине в 1894 г. К настоящему
времени описано свыше 3000 ферментов, из которых лишь немно-
гие изготавливают в промышленности, то есть резерв здесь для
биотехнологии поистине огромен.
В качестве лекарственных средств применяют гидролазы: гиа-
луронидазу (она может быть микробного и животного происхож-
дения, например, лидаза), протеазы, липазы, амилазу, из других
классов — L-аспарагиназу, стрептазу в составе препарата стрепто-
деказы, стрептокиназу и др.
В хлебопекарной и текстильной промышленности применяют
амилазы, в мясо-молочной и кожевенной промышленности —•
пептидазы; в сельском хозяйстве используют такие полифермент-
459
ные препараты, как глюкаваморин, пектаваморин и др. Большое
количество ферментов приходится на моющие средства.
Рестрикционные эндонуклеазы — важнейший "инструмент"
для выполнения генно-инженерных работ. Часть производимых
рестриктаз приведена в таблице 20.
К началу 90-х годов текущего столетия объем продажи фермен-
тов на мировом рынке оценивали в сумме около 200 млн. долларов
в год с ежегодным возрастанием на 10—15%. Лишь зарубежные
фирмы производили свыше 200 наименований ферментных пре-
паратов, в странах СНГ — около 40; примерно 40—50% мировой
потребности в ферментах обеспечивается датской фирмой "Ново
индастра".
В клетках микроорганизмов найдено порядка 1500 различных
ферментов, причем в каждой клетке, по расчетам, содержится в
среднем 100 тыс. молекул ферментов; на единичный энзим прихо-
дится 0,1—5% от общего количества белка.
В сравнительном плане получение экзоферментов осуществля-
ется с меньшими затратами усилий и средств, чем получение
эндоферментов. В то же время успехи в области генетической
инженерии явились мотивом к расширению ассортимента произ-
водимых ферментов, в том числе :— для научных целей (пример с
рестриктазами).
За последние 40 лет почти все новые ферментные препараты
получены с помощью микроорганизмов, что объясняется быстро-
той размножения большинства микробов на доступных и относи-
тельно дешевых питательных средах, а также возможностью их
изменения в желаемом направлении с помощью методов генети-
ческой инженерии — созданы предпосылки трансформировать
любой клеточный белок во внеклеточный.
Ферменты и ферментные системы могут функционировать in
vitro, однако их источником являются живые организмы, содержа-
щие генетическую программу о синтезе первичных метаболитов,
или ферментов. Поэтому вид микроба накладывает отпечаток на
характеристику биокатализатора. Так, например, бактерии-термо-
филы синтезируют ферменты, которые будут эффективны при
повышенных температурах скорее, чем ферменты психрофилов.
В любом случае для производства важны продуцент и образуемый
им фермент (целевой продукт), о которых следует иметь представ-
ление в возможно полном объеме.
Промышленный штамм должен быть апатогенным (кроме спе-
циальных случаев), образовывать в больших количествах преиму-
460
щественно один фермент и минимум—других, если не преследуют
цель получать ферментный комплекс; желательно, чтобы такой
штамм не продуцировал аллергенные и токсические субстанции,
и, наконец, хорошо развивался в крупномасштабных ферментато-
рах.
На каждой стадии процесса ферментации необходимо контро-
лировать чистоту культуры — продуцента и скорость его размно-
жения, pH среды, отсутствие фаголизиса и выход фермента.
Установлено, что одна жизнеспособная спора микроба-контами-
нанта способна "заразить" объем среды, равный 10—50 м3. Поэтому
важно убедиться в "монолитности" инокулята и в полной стериль-
ности подготовленного к засеву ферментатора. К сожалению, в
промышленных биореакторах трудно добиться абсолютно полной
стерильности, поэтому целесообразно засевать питательную среду
в них большим по объему и физиологически активным посевным
материалом. Таким приемом обеспечивают конкурентоспособ-
ность продуцента при возможном попадании в среду его обитания
любого постороннего микроба. Этому же способствует уменьше-
ние числа фаз размножения продуцента, если основной фермен-
татор засевают культурой, находящейся в продуктивной фазе.
Так же как и в других биотехнологических процессах здесь
предусматривают выращивание продуцента по схеме: лиофилизи-
рованная стандартная культура рост в колбах на качалке до
log-фазы перенос инокулята в ферментаторы малой емкости
(300—500 л) — инокуляторы —> инокулят через 10—80 часов (в
зависимости от процесса) в const-фазе подают в промежуточный
ферментатор — посевной аппарат емкостью от 3 до 5 м3 —>
посевным материалом в стационарной фазе засевают головной
(основной) ферментатор емкостью 50 м3. Среда в инокуляторе,
посевном аппарате и головном ферментаторе, как правило, одно-
типна. Если основной биореактор имеет объем 3—5 м3, то исклю-
чают промежуточную фазу подготовки инокулята (посевного ма-
териала) в посевном аппарате, то есть в таком случае этот аппарат
является головным.
Микроорганизм-загрязнитель обычно выявляют при микроско-
пии или культурально при высеве на агаризованные питательные
среды; бактериофаги обнаруживаются на основании замедленного
роста продуцента и, как следствие, по снижению его продуктив-
ности; pH среды контролируют с помощью рН-метров.
461
В производственных условиях ферменты обычно получают в
периодических режимах культивирования продуцентов. Основная
ферментация может продолжаться от одних до семи суток в
зависимости от микроба и фермента. Примерная биотехнологиче-
ская схема получения ферментов показана на рис. 137.
Перед завершением ферментации культуральную жидкость
быстро охлаждают до +5°С, при необходимости доводят pH до
требуемого показателя; предварительная обработка жидкости мо-
жет заключаться в добавлении флокулянтов при последующих
центрифугировании и вакуум-упаривании (при температуре ниже
35°С). После этого к упаренному центрифугату добавляют раствор
электролита, чтобы освободиться от возможных загрязнителей
(часть клеток продуцента, крупные частицы из среды, не отделив-
шиеся при центрифугировании). Раствор затем фильтруют на
вакуум-барабанном фильтре с намывным слоем, фильтрат посту-
пает в рефрижератор, из которого в одном случае (для получения
жидкого концентрата фермента) его подвергают ультрафильтра-
ции, консервируют, пропускают через мембранный фильтр и
собирают в накопителе, откуда он поступает на фасовку (на схеме
— путь А). В другом случае (на схеме — путь Б) из рефрижератора
(позиция 9) раствор фермента пропускают через мембранный
фильтр, добавляют к нему электролит, например, натрия сульфат,
для высаливания ферментного белка, пропускают через фильтро-
пресс. Фермент- собирают и смешивают с встречным потоком
раствора концентрата того же фермента (путь А) с последующим
высушиванием в вакуум-распылительной сушилке. Сухой экстракт
собирают в накопителе, стандартизуют по активности и подают на
расфасовку и упаковку.
Жидкий ферментный концентрат, при необходимости, разбав-
ляют дистиллированной водой до требуемой активности, добавля-
ют к нему стабилизаторы (натрия хлорид, бензоаты, сорбинаты) и
передают на фасовку и упаковку. Из жидкого концентрата фермент
может быть осажден спиртом или ацетоном (равно как и высолен
электролитом), высушен, измельчен, смешан с наполнителем (на-
трия хлоридом, декстринами, крахмалом) до требуемой активности
и упакован.
Когда необходимо выделить фермент в более очищенном виде
и без большинства примесных (ферментных и неферментных)
полидисперсных белков, то в схему можно включить стадии диа-
лиза и гель-хроматографии.
462
Фактически целевое назначение фермента предопределяет сте-
пень его очистки — она должна быть более высокой в случае
получения фермента как медицинского препарата.
Микробные эндоферменты получают по следующей схеме:
Охлажденная до 5°С kj гльтуральная жидкость
Жидкость (отход)« Кле Цитс "Обрывки" клеток « (отход) центрифугирование и промывание клеток тки дезинтеграция клеток золь центрифугирование
Ферментный Раствор с супернатант различные методы очистки эермента
Дезинтеграцию клеток можно проводить различными способа-
ми: обработкой соответствующими ферментами-гидролазами, ме-
ханическим разрушением и пр.
Обработку супернатанта (надосадочной жидкости) проводят с
целью освобождения от нуклеиновых кислот (действие нуклеаза-
ми, высокомолекулярными катионами-полиэтилениминами и др.),
от других белков (хроматография).
Получение очищенных эндоферментов — дело дорогостоящее
и поэтому стремятся их не выделять из клеток, а использовать в
иммобилизованном виде нативные клетки, способные осуществ-
лять желаемые каталитические функции.
В ближайшем будущем получение ряда ферментных белков
потенциально возможно с помощью рДНК-биотехнологии, которая
ныне затрагивает практически все сферы биотехнологии.
Важнейшим направлением в реализации каталитических фун-
кций биокатализаторов является их иммобилизация, что обеспечи-
вает многократное использование фермента, пролонгацию его
стабильности в активной форме, возможность применения в не-
463
прерывных биотехнологических процессах, исключение, попада-
ния фермента в целевой продукт.
Иммобилизованные ферменты — это основа "инженерной
энзимологии" — самостоятельной ветви биотехнологии, впервые
оформившейся в 1971 г. Иммобилизация фермента — "это любое
ограничение свободы движения белковых молекул (или их фраг-
ментов!) в пространстве” (И. В. Березин, 1987).
В 3-й части книги частично представлен материал по Иммоби-
лизации ферментов. Здесь лишь перечислим отдельные примеры
для промышленнозначимых биокатализаторов. Наипростейшим
методом закрепления бактериальной глюкозоизомеразы было вза-
имодействие фермента с ионообменным носителем, в частности,
с ДЭАЭ-целлюлозой — удерживание фермента за счет ионного
взаимодействия. Такой же способ иммобилизации был реализован
в отношении грибной аминоацилазы (Ф), использующейся в про-
мышленности для получения L-аминокислот из синтетических
рацемических ацил — DL-аминокислот:
Ацил - DL — аминокислота » L — аминокислота + ацил — D — амино —
кислота
Глюкозоизомеразу с определенным успехом иммобилизуют на
окиси алюминия — адсорбционное взаимодействие. При этом
могут быть иммобилизованы ощутимые количества ферментов.
Ковалентное взаимодействие ферментного белка возможно на
алкиламинированном стекле, на керамике и пр. Однако такая
иммобилизация характеризуется снижением активности фермен-
тов и их эффективности в работе.
Наипростейшим способом иммобилизации оказалось сохране-
ние фермента внутри клеток продуцента. Так, прогревание стреп-
томицета, образующего глюкозоизомеразу, при 60—80°С сопро-
вождается отмиранием культуры наряду с избирательным разру-
шением автолизинов. Глюкозоизомераза остается активной, а ста-
билизированные таким образом клетки становятся проницаемыми
для малых молекул.
Внутриклеточный фермент может быть ковалентно сшит и,
следовательно, фиксирован с помощью ковалентного связывания
глутаровым альдегидом.
Микробные клетки—продуценты ферментов и сами ферменты
могут быть заключены в различные гелевые и волокнистые струк-
туры, в микрокапсулы, липосомы.
464
В производственных условиях основной технологический про-
цесс с участием иммобилизованных ферментов проводят, как
правило, в проточных биореакторах (колоннах), в которых биока-
тализатор может быть в виде неподвижного или перемешиваемого
слоя; в биореакторах со взвешенным слоем объединены принципы
двух предыдущих аппаратов (рис. 138).
Рис. 138. Биореакторы
I—V типов, используемых в
инженерной энзимологии: I
— периодического действия
(1 — биореактор, 2 — мешал-
ка, 3 — иммобилизованный
фермент), II — проточный с
перемешиванием (1 — биоре-
актор, 2 — мешалка, 3 — им-
мобилизованный фермент, 4
вход субстрата, 5 — выход
целевого продукта), III — с
подвижным ферментным
слоем, IV — со взвешенным
слоем, V — с рециркуляцией
(для III—V: 1 — колонна, 2 —
иммобилизованный фер-
мент, 3 — вход субстрата, 4
— выход целевого продукта,
5 — рециркуляция).
Если профиль водного раствора субстрата распространяется
(течет) равномерно по всему сечению аппарата, то такой реактор
функционирует по "поршневому типу”. При этом концентрация
субстрата максимальна на входе в аппарат, а концентрация целе-
вого продукта — на выходе из аппарата. Реактор IV типа напоми-
нает хемостат для культивирования микробов (удобен для контроля
pH, температуры). Если конечный продукт может выступать инги-
битором фермента, то в подобных случаях удобна конструкция III,
если же при определенных условиях ингибитором оказывается
субстрат, то лучше использовать конструкцию I. В IV реакторе
поток раствора субстрата должен быть таким, чтобы постоянно
удерживать фермент во взвешенном состоянии и не уносить его
с выходящим потоком раствора целевого продукта.
Крупномасштабное производство приходится в основном на
такие гидролазы, как протеазы и гликозидазы. Различают три
категории протеаз — сериновые, кислые и металлопротеазы. Пер-
вые термостабильны, оптимум pH для них выше 8,0. Поэтому
сериновые протеазы широко используют в моющих средствах.
Основными продуцентами этих ферментов являются бациллы.
465
Кислые протеазы имеют рН-оптимум 1,5—3,7. Они полезны в
изготовлении сыров. Их продуцируют, например, различные виды
низших грибов (Mucor, Endothea и др.). Грибной реннин с успехом
заменяет сычужный фермент из желудков телят. Однако необхо-
димо подчеркнуть, что рекомендации использовать термофильный
штамм вида Mucor pusillus, образующего реннин, должны быть
восприняты с большой осторожностью, поскольку из научной
литературы и медицинской практики известно, что этот вид может
вызывать заболевания у людей (мукороз, или мукоромикоз).
Металлопротеазы с оптимумом pH в нейтральной зоне (6,5—7,5)
применяют в пищевой промышленности. Их основными продуцен-
тами являются бациллы.
Из гликозидаз большое значение приобрели грибные и бакте-
риальные амилазы, катализирующие реакции гидролиза крахмала.
Сегодня в мире организовано крупнотоннажное производство
глюкозо-фруктозных сиропов на основе эксплуатации иммобили-
зованной глюкозоизомеразы, обеспечивающей превращение глю-
козы во фруктозу (рис. 139). Фруктоза (левулеза) слаще глюкозы
и такими сиропами заменяют сахарозу в соответствующих видах
подслащенных или сладких изделий пищевых продуктов. Следует
иметь в виду возможность получения фруктозных сиропов из
жидких отходов в производстве декстрана, где до сих пор на
заводах в странах СНГ фруктоза "уходит в трапы”.
Пектиназы успешно применяют при изготовлении фруктовых
соков. Эти ферменты включают комплекс индивидуальных биока-
тализаторов: пектинлиаз, пектатлиаз и полигалактуронидаз (пек-
тинэстераз). С их помощью осветляют и снижают вязкость соков
Лгавм Сот
Рис. 139. Технологическая схема
трансформации глюкозы во фруктозу с
помощью иммобилизованной глюкозоизо-
меразы: 1 — емкость для приготовления
раствора глюкозы, 2 — теплообменники, 3
— биореакторы с иммобилизованной глю-
козоизомеразой, 4 — реактор для глюко-
зо-фруктозного раствора с системой регу-
лирования pH, 5 — установка для обесцве-
чивания раствора углем, 6, 8 — сборники
раствора, 7 — фильтр, 9 — ионообменные
колонны, 10 — вакуум-выпарная установ-
ка, 11 — сборник глюкозо-фруктозного
сиропа (по М. Е. Бекеру, Г. К. Лиепиньшу,
Е. П. Райпулису, 1990).
466
благодаря гидролизу пектина, что весьма важно при концентриро-
вании соков. Основным продуцентом пектиназ является Aspergillus
niger.
В промышленных условиях производят и другие ферменты,
названные в таблице 47.
Т а б л и ц а 47. Промышленноважные микробные ферменты и их продуценты
Фермент Продуцент |
Гидролазы
Гликозидазы:
а-Амилаза Asp.niger, Asp. oryzae, Bac.amyloliquefaciens, Вас. 1 icheniformis
Р-Глюканаза Asp.niger, Bac.amyloliquefaciens
Глюкоамилаза Asp.niger, Rhizopus niveus, Endomycopsis sp.
Глюкоизомераза Astinoplanes missouriensis, Arthrobacter sp., Вас. coagulans, Streptomyces sp.
Декстраназа Penicillium sp.
Инвертаза Asp. sp., Saccharomyces cerevisiae
Лактаза Asp. niger, Kluyveromyces marxianus
Пектиназы Asp. awamori, Asp. niger, Klebsiella pneumoniae
Протеазы Asp. niger, Asp. oryzae, Алкалифильные бациллы, 1 Вас. amyloliquefaciens, Вас. licheniformis, Вас. stearothermophilus
Липазы Asp. awamori, Asp. oryzae, Candida cylindrica, Mucor miehei, Rhizopus sp.
Пенициллиназа (пенициллинацилаза, пенициллинамидаза) Escherichia coli
467
Другие белки и белковые продукты.
Токсины и анатоксины. Отдельные виды болезнетворных мик-
роорганизмов образуют экзотоксины, которые могут быть отнесе-
ны к "факторам агрессии”. Они представляют собою высокополи-
мерные термолабильные белки — продукты матричного синтеза,
секретирующиеся в окружающую среду. При попадании в орга-
низм человека экзотоксины вызывают серьезные повреждения
функций определенных тканей или систем. Например, столбняч-
ный токсин относят к числу нейротоксинов, нарушающих функ-
цию нервно-мышечного аппарата; гангренозные токсины являются
некротоксинами, индуцирующими повреждение тканей; экзоток-
сины определенных штаммов кишечной палочки повреждают ки-
шечник и т. д. По механизму действия на ткани они сходны с
ферментами. Некоторые токсины применяют для диагностики
соответствующих заболеваний. Например, токсин дифтерийный
рекомендуют для постановки внутрикожной реакции Шика. Ток-
син изготавливают по обычной схеме выделения экзобелков из
жидких питательных сред после выращивания определенных
штаммов дифтерийных бактерий. Препарат для реакции Шика
готовят из очищенного дифтерийного токсина, разводя его глице-
рино-желатиновой смесью до необходимой концентрации (1/40
часть одной смертельной дозы для морских свинок — одна Шик-
доза). Выпускаемый препарат — бесцветная прозрачная жидкость
в ампулах по 1 мл. Срок годности — 2 года, хранят при 3—10°С.
Обработка экзотоксинов формалином сопровождается их пол-
ным обезвреживанием при сохранении антигенных свойств. Обез-
вреженные токсины называют анатоксинами, или токсоидами. Их
используют для получения антитоксических сывороток (глобули-
нов) . В практике здравоохранения находят применение следующие
анатоксины: ботулинический, гангренозный, дифтерийный, стафи-
лококковый, столбнячный.
Технология получения экзотоксинов включает следующие ста-
дии: культивирование соответствующего штамма патогенного мик-
роба-продуцента на определенной питательной среде и при опти-
мальных режимах (pH, температура, аэрация или анаэробиоз,
продолжительность выращивания), обезвреживание формалином
при 37—40°С, сепарирование клеток (отход) от культуральной
жидкости, содержащей анатоксин; очистка, концентрирование,
добавление адсорбента, фасовка, упаковка.
В качестве адсорбентов обычно используют неорганические
вещества — алюминия гидроксид, алюминия фосфат, кальция •
468
фосфат и т. п. (в России применяют гидроксид алюминия), которые
создают депо в организме при введении анатоксинов. Таким
образом, удается повысить эффективность иммунизации.
Очищенные адсорбированные анатоксины — это жидкие пре-
параты-суспензии, расслаивающиеся на белый, светло-коричне-
вый или желтоватый осадок и прозрачную надосадочную жид-
кость.
Ботулинические и гангренозные анатоксины применяют для
иммунизации лошадей в целях получения специфических лечеб-
ных иммунопрепаратов.
Дифтерийный анатоксин рекомендуют в качестве профилак-
тического антидифтерийного средства для активной иммунизации.
Его выпускают в виде монопрепарата и в составе ассоциированных
вакцин и в каждом из них токсин адсорбирован на гидроксиде
алюминия. Монопрепарат— адсорбированный дифтерийный, или
АД-анатоксин — представляет собой очищенный концентрирован-
ный продукт, содержащий в 1 мл 60 антигенных (флоккулирующих)
единиц анатоксина и не свыше 2 мг адсорбента. Флоккуляция —
это реакция взаимодействия дифтерийного анатоксина или токси-
на с антитоксином (иммунной сывороткой). Флоккулирующая
единица — это минимальное количество антигена (токсина), обра-
зующего флоккулят (от лат. flocculi — клочок).
АДС-анатоксин — это ассоциированный препарат из очищен-
ных концентрированных дифтерийного и столбнячного анатокси-
нов, адсорбированных на А1(ОН)3 и содержащих в 1 мл 60 анти-
генных единиц дифтерийного анатоксина и 20 единиц (антиток-
синсвязывающих) столбнячного анатоксина и не более 2 мг адсор-
бента. Известен и другой вариант препарата с содержанием в 1 мл
тех же ингредиентов в соотношении 1:1, адсорбент, как и в первом
варианте.
АКДС-вакцина - ассоциированный препарат, включающий те
же вещества, что и АДС-анатоксин, а также коклюшную вакцину.
В 1 мл такой вакцины содержатся дифтерийный и столбнячный
анатоксины по 30 и 10 антигенных единиц соответственно, не более
2 мг алюминия гидроксида и 20 млрд, клеток коклюшных бактерий,
убитых формалином или мертиолатом. Мертиолат используют как
консервант в концентрации 0,01%; в 0,5 мл вышеназванных пре-
паратов содержится одна прививочная доза. Перед употреблением
их необходимо взбалтывать. Форма выпуска— ампулы с 1 мл
препарата. Хранят ампулы ’с АД- и АДС-анатоксинами при 3—10°С
до 3 лет (срок повторного контроля и определения возможности
469
дальнейшего хранения еще в течение 1 года). Что касается АКДС-
вакцины, то ее срок хранения в тех же условиях сокращается в 2
раза, т. е. до 1,5 лет с вероятностью продления его на 6 месяцев
после переконтроля. Прививки делают согласно схеме, утвержден-
ной М3 России.
В 1994 г. в Канаде введена в практику пентавалентная вакцина
против коклюша, дифтерии, столбняка, полиомиелита и заболева-
ний, вызванных гемофильной палочкой серотипа b(Hib).
Дифтерийный анатоксин, как монопрепарат используется так-
же для гипериммунизации лошадей в целях получения лечебной
противодифтерийной сыворотки, содержащей антитоксический
иммуноглобулин.
Профилактику столбняка осуществляют либо одним столбняч-
ным анатоксином (20 ЕС в 1 мл с адсорбентом), либо в ассоциации
с другими препаратами, в частности, его применяют в составе
АДС-анатоксина и АКДС-вакцины (см. выше), а также в составе
ТАВТе-вакцины — "химической" сорбированной тифо-паратифоз-
но-столбнячной вакцины, содержащей по 0,2 мг брюшнотифозного
и А-паратифозного, 0,25 мг В-паратифозного антигенов, 10 ЕС
столбнячного анатоксина и 1,5—2 мг А1(ОН)3. Выпускают во фла-
конах, содержащих 8 мл препарата для инъекций, и консервант
мертиолат. Срок хранения — 3 года при 8—10°С (срок переконт-
роля с возможностью хранения еще в течение 1 года). Иммуниза-
цию людей проводят по схеме, утвержденной М3 России.
Столбнячный анатоксин, реже — токсин используют для ги-
периммунизации лошадей в целях изготовления противостолбняч-
ной сыворотки, содержащей антитоксический иммуноглобулин.
Иммунопрепарат используют в качестве лечебного и экстренно-
профилактического средства.
Стафилококковый анатоксин представляет собой стерильный
фильтрат бульонной культуры золотистого стафилококка, содер-
жащий экзотоксин, обезвреженный формалином. Количество ток-
сина в такой жидкости должно быть не менее 5 ЕС в 1 мл препарата;
консервант не добавляют. В готовом виде нативный стафилокок-
ковый анатоксин — прозрачная желтоватая жидкость для подкож-
ного введения. Форма выпуска — ампулы по 2 мл. Из этого
анатоксина получают очищенный адсорбированный анатоксин
осаждением трихлоруксусной кислотой с последующей очисткой
этанолом и адсорбцией на гидроксиде алюминия. В 1 мл препара-
та-суспензии содержится 10 ЕС.
470
Токсические белки энтомопатогенных
бацилл.
Некоторые энтомопатогенные бациллы применяют в качестве
биологических средств защиты и борьбы с вред ными насекомыми
(от греч. entomon-насекомое). К их числу относят Вас. popilliae и
Вас. thuringiensis, образующих в процессе спорообразования так
называемые параспоральные белковые кристаллы. Синтез такого
белка находится под контролем промотора гена, содержащегося в
плазмиде.
Вас. popilliae способна вызывать хроническое инфекционное
заболевание у насекомого, передающееся другим особям и распро-
страняющееся среди популяции.
Вас. thuringiensis токсический параспоральный белковый кри-
сталл действует на вредных насекомых как пестицид. К сожалению,
во внешней среде этот токсин длительно не сохраняется, поэтому
требуются повторные обработки данной культурой территорий с
вредными насекомыми.
Вас. popilliae и родственные ей микробы (Вас. lentimorbus, Вас.
euloomarhae и др.) индуцируют так называемую молочную болезнь
у жуков-скарабеев. Эти виды бацилл трудно культивировать на
искусственных питательных средах, но все же удается, если под-
держивать анаэробные условия. Собирают спорулирующую массу,
споры сепарируют, смешивают с наполнителем, фасуют и упако-
вывают. Применяют в виде порошка - дуста. Споры Вас. popilliae
заглатываются личинками насекомых, прорастают в кишечнике и
делятся, давая потомство. Бактерии попадают в гемолимфу и через
10 дней у насекомых возникают признаки молочной болезни.
Клетки затем спорулируют, образуя к моменту гибели одной
личинки порядка 5 » 109 спор.
Препарат на основе Bacpopilliae длительное время изготавли-
вают в США в целях контроля численности японского жука—
Popillia japonica.
В России изготавливают препараты (дендробациллин, инсек-
тин, токсобактерин, энтобактерин-3 и др.) на основе Вас.
thuringiensis. Данный вид микроба образует два токсина — р и 8,
из которых Р-экзотоксин обладает широким спектром действия на
насекомых; он представляет собой адениннуклеотид, конкурентно
ингибирующий ферменты, которые катализируют реакции гидро-
лиза АТФ и пирофосфата.
471
При введении [3-токсина млекопитающим животным он оказы-
вается для них губительным. Поэтому в производстве используют
штаммы, не продуцирующие 0-экзотоксин, но образующие 6-ток-
син, который и представляет собой белковое кристаллическое
вещество - "параспоральное тело" - правильный восьмигранник.
Чтобы проявить токсичность, 6-токсин должен попасть в кишечник
личинок насекомых прежде всего - листогрызущих из отряда
чешуекрылых Lopidoptera), где растворяется в щелочной среде и
частично гидролизуется протеазами. Такой модифицированный
белок взаимодействует со стенкой кишки и изменяет ее так, что
содержимое кишечного тракта попадает в кровоток, вызывая
общий паралич. Смерть личинок наступает от септицемии.
6-Токсин безвреден для млекопитающих животных, человека,
птиц. В литературе о Вас. thuringiensis описано более 20 серотипов
(сероваров) микроба и 15 вариантов 6-токсина, часть из которых
имеет промышленное значение, например, thuringiensis или
berliner(I), alesti(III), dendrolimus(IV), galleria(V) и др.
Коммерческие препараты представляют собой сумму спор и
белковых кристаллов в клетках микроба-продуцента.
Технология получения энтомопатогенных препаратов включает
следующие стадии: 1) проверка маточной культуры на отсутствие
свободного фага, продуктивность спор и кристаллов, на вирулен-
тность; 2) выращивание культуры в колбах до титра спор не менее
1,7* 109 в 1 мл; 3) пересев культуры из колб в посевной аппарат
(0,05% на объем среды); 4) засев посевным материалом основного
ферментатора (вносят 0,0012% инокулюма от объема среды в
биореакторе). Продолжительность выращивания в посевных и
головных ферментаторах - 35-40 часов при 28-30°С (исходное
значение pH 6,3) до получения указанной выше плотности спор в
1мл среды. В процессе культивирования среда защелачивается до
pH 8, 0-8, 5, поэтому перед тем, как передать ее на сепарирование,
pH доводят до 6,0-6,2 подкислением. После сепарирования получа-
ют пасту (в среднем 100 кг из 1м3 культуральной жидкости) с
влажностью 85% и содержанием спор 2 • 1О10 в 1 г.
Готовый продукт может быть в виде стойкой вязкой жидкости
кремового или светло-серого цвета, без запаха - она содержит
наполнитель—кристаллическую микроцеллюлозу; другой вариант
- смачивающий порошок, приготовленный после высушивания на
распылительной сушилке (остаточная влажность 10%) и смешения
с каолином. Концентрация спор в препаратах 3 • 109 в 1г.
472
Белки одноклеточных и многоклеточ-
ных микроорганизмов
На белки приходится часть сухих веществ микробной клетки
от 19 до 90% (таблица 48).
Таблица 48. Содержание белка (% от сухой массы) в клетках
некоторых бактерий и грибов
Микроорганизм Бе- лок, % Микроогранизм Бе- лок, %
Aspergillus flavus 19 Rhodotorula rubra 56
Aspergillus niger 33 Saccharomyces cerevisiae 56
Rhizopus nigricans 36 Streptomyces griseus 57
Penicillium notatum 38 Bacillus subtilis 63
Bacillus megaterium 39 Staphylococcus aureus 65
Candida arborea 46 Escherichia coli 82
Lactobacillus casei 47 Eactobacillus fermentans 87
Candida utilis 53 Methanobacterium sp. 90
Hansenula suaweolens 53
Следовательно необходимо иметь в виду, что в случае исполь-
зования микробной биомассы как белкового продукта, фактически
имеют дело с суммой различных веществ (в том числе — небелко-
вых, содержащихся в клетках). Не существует природных белков
с эквимолярным количеством составляющих его аминокислот, но
их (аминокислот) относительное содержание является строго оп-
ределенным для каждого белка.
На практике чаще прибегают к использованию убитых клеток
в качестве белкового продукта, так как любые методы выделения
протеинов из клеточной биомассы сопровождаются незначитель-
ным выходом целевого продукта
473
В разные годы и в разных странах применяли следующие
микроорганизмы как источники белка: Saccharomyces cerevisiae,
Candida utilis, Fusarium graminearum, Methylomonas clara, Candida
tropicalis (апатогенные штаммы), Candida maltosa, Hansenula sp. и
др. Выращивание производили и производят на различных средах,
нередко используя отходы или конечные продукты в других про-
изводствах (деревообрабатывающей промышленности—сульфит-
ные щелока, сельского хозяйства — меласса, гидролизаты соломы,
кукурузных кочерыжек и пр., нефтеперерабатывающей промыш-
ленности — н-алканы, Си—Cf8; спиртовой промышленности —
этанол; газовой промышленности — метан).
Пивные дрожжи с разной интенсивностью стали использовать
в пищу с конца XIX в., особенно — в периоды разрухи и недостатка
пищевых продуктов, например, во времена первой и второй миро-
вых войн. В 1980 г. в Англии был разрешен к использованию в
пищу микопротеин — мицелий гриба Fusarium graminearum, тек-
стура которого удобна для включения различных наполнителей и
формования, например, мясоподобных изделий.
Технология получения кормового или пищевого белка однокле-
точных и многоклеточных микроорганизмов сравнительно неслож-
ная и заключается в наращивании по возможности наибольшего
количества биомассы клеток, ее денуклеинизации, сепарировании
и приготовлении целевого продукта. Культивирование того или
иного микроорганизма проводят в оптимальных условиях (до
получения десятков-сотен граммов дрожжей в 1 л) в периодическом
или непрерывном режиме, в стерильных или нестерильных усло-
виях; денуклеинизацию клеточной биомассы можно проводить
различными способами — экстракцией метанолом или щелочами,
обработкой нуклеазами, температурным шоком для активизации
эндонуклеаз (в частности — РНКазы); отделением белка от нукле-
иновых кислот из дезинтеграта клеток. Повышенное количество
нуклеиновых кислот в белке нежелательно из-за возможного
повышения мочевой кислоты in vivo и, как следствие, потенциаль-
ное возрастание опасности заболеть подагрой и почечно-каменной
болезнью (содержание нуклеиновых кислот в микробном протеине
не должно превышать 2 г в сутки).
Известны различные коммерческие белковые продукты, изго-
тавливаемые в промышленности тех или иных стран мира
(таблица 49).
474
Та б л и ц a 49. Белковые продукты микробного происхождения
Название и назначение Содер- жание белка, % Продуцент Источник углерода
Пищевые дрожжи 52 Sacch. cerevisiae углеводы, этанол
Лечебные (пивные) дрожжи 52 II углеводы
Паприн (дрожжевой кормовой белок) 52 Candida maltosa твердые парафины
Г{1прин (бактериальный кормовой белок) 74 разные бактерии метан
Пищевой микопротеин 47 Fusarium graminearum углеводы
Торутин 52 Candida utilis этанол
Дигитатин 53 PeniciUium digitatum крахмал картофельный
Микробный белок заманчив в перспективе, поскольку его
экономическая целесообразность представляется несомненной хо-
тя бы из следующего сравнения: на 1 кг корма для крупного
рогатого скота можно получить около 70 г говядины с содержанием
14 г белка, тогда как на 1 кг углеводов с добавкой неорганического
азота для F. graminearum в Англии получают около 1100 г. сырой
мицелиальной массы, содержащей 136 г белка.
Сепарирование клеток из культуральной жидкости проводят
одним из следующих способов (в зависимости от биообъекта):
фильтрованием через ротационные вакуум-фильтры, фильтр-прес-
сы (например для отделения мицелиальной массы); коагуляцией
или электрокоагуляцией с последующим центрифугированием,
исключая фильтрацию (например, для отделения бактериальной
массы); центрифугированием, используя центрифуги непрерывно-
го действия, например, для дрожжей, или корзинчатые — для
нитчатых грибов.
На последних стадиях отсепарированную клеточную массу
можно подвергать либо высушиванию (не чрезмерному) различ-
475
ними методами, либо подсушиванию, либо эффективному отжиму.
В конечном продукте все клетки микроорганизмов должны быть
погибшими. На рис. 128а показана принципиальная схема получе-
ния кормовых дрожжей.
Получение микробных гликанов и гликоконъюгатов
Микробные полисахариды, или гликаны можно подразделить
на внеклеточные и внутриклеточные, или экзо- и эндогликаны.
Внутриклеточные могут быть структурными в клеточных стенках
(нерастворимые хитин, глюканы) и структурно-метаболическими
(относительно растворимые маннаны, гликоманнаны, определяю-
щие антигенную специфичность видов), способными переходить
в культуральную жидкость при гиперпродукции и оказываться в
разряде внеклеточных. К эндогликанам относятся запасные угево-
ды и различные гликоконъюгаты (нуклеозиды и полинуклеотиды,
отдельные ферменты, гликолипиды, пептидогликаны и др.).
Выраженными продуцентами экзогликанов являются молочно-
кислые и уксуснокислые бактерии, ксантомонасы и псевдомонасы,
отдельные виды дрожжевых и нитчатых грибов. В разных странах
производят альгиновую кислоту с помощью Azotobacter vinelandii,
Pseudomonas aeruginosa, аубазидан — с помощью Aureobasidium
pullulans, декстран (продуценты — Leuconostoc mesenteroides,
L. dextranicum), курдлан (продуцент — Alcaligenes faecalis var.
mixogenes), маннаны (продуценты — разные дрожжи из родов
Hansenula, Rhodotorula и др.), пуллулан (продуцент — Aure-
obasidium pullulans) и др.
Из эндогликанов (в виде гликоконъюгатов) на практике изве-
стны: пирогенал, продигиозан, сальмозан, выделяемые из биомассы
клеток следующих микроорганизмов соответственно: Salmonella
spp., Serratia marcescens и Salmonella typhi; мурамилпептиды из
клеточных стенок грамположительных бактерий; некоторые гли-
копротеины или пептидогликаны (их включают в состав иммуно-
генных препаратов, например, пептидогликан из Klebsiella
pneumoniae входит в состав рибосомальной вакцины "Рибомунил",
выпускаемой во Франции).
Технологические процессы получения экзогликанов заметно
различаются на стадии их выделения, очистки и высушивания. Это
обусловлено неодинаковыми структурой и физико-химическими
свойствами экзополисахаридов (молекулярная масса, раствори-
мость, конфигурация гликозидных связей ц пр.). Ферментацию
продуцентов проводят в аэробных или анаэробных условиях, асеп-
тично, в периодическом режиме, на средах с подходящими источ-
476
никами углерода и азота. В большинстве случаев стремятся под-
держивать такие условия, когда соотношение С/N в среде больше
1, так как в противном случае микроорганизм будет находиться
преимущественно в трофофазе без выраженной продукции экзог-
ликана.
В конце ферментации, при .заметном накоплении полисахарида,
культуральная среда может приобретать консистенцию геля. Если
необходимо отделять клетки от гелеобразной культуральной жид-
кости, то нередко прибегают к многократному ее разбавлению
водой (иногда в 10—15 раз) в потоке перед подачей в сепаратор
или в специальные емкости. При необходимости клетки продуцен-
та убивают прогреванием до сепарирования. В некоторых случаях,
например, при изготовлении полисахаридов медицинского назна-
чения, сепарирование повторяют 2—3 раза.
Если экзополисахарид резко изменяет вязкостные характери-
стики в водном растворе при подкислении и подщелачивании (без
разрушения своей первичной структуры), то перед сепарировани-
ем экзогликан разжижают добавлением соответствующего реаген-
та (например, щелочи), сепарируют клетки продуцента без разбав-
ления культуральной жидкости, а затем подкисляют раствор по-
лисахарида (например, соляной кислотой), восстанавливающего
при этом свои первоначальные свойства (курдлан).
Последующая стадия заключается в вакуум-упаривании или
"мембранном" концентрировании, используя соответствующие
фильтры типа "Владипор" или "Миллипор", с последующим осаж-
дением экзогликана каким-либо полярным растворителем.
На последней стадии технологического процесса полученный
экзогликан подвергают высушиванию, измельчению с последую-
щей упаковкой в соответствующую тару, или используют его для
приготовления коммерческого продукта (раствор для инъекций,
гранулы для приема внутрь, гели косметические, пасты, кремы и
пр.). В таких случаях возможен гидролиз полисахарида на какой-
либо стадии технологического процесса в целях его деполимери-
зации (декстран).
Эндогликаны выделяют из клеточной массы либо экстракцией
соответствующими экстрагентами в водных или водно-спиртовых
растворах, либо используют предварительный гидролиз клеток
(ферментный, щелочной, кислотный) или дезинтеграцию в каких-
либо дезинтеграторах, либо протопластирование (в случае выделе-
ния небольших количеств целевого продукта).
477
Полученный тем или иным способом водный раствор эндогли-
кана (гликоконъюгата) подвергается выделению и очистке таким
же образом, как и экзогликаны.
Гликаны, выполняющие общебиологические защитные функ-
ции в живых организмах на молекулярном и клеточном уровнях,
находят широкое практическое применение в качестве основных
и вспомогательных веществ: плазмозамещающие агенты (6% вод-
ный раствор декстрана—полиглюкин), лекарственные (пирогенал,
родэксман, продигиозан) и диагностические средства (см. раздел
об антигенах и аллергенах), иммуноадьюванты (грибные полиса-
хариды, сальмозан), гидрофильные основы для мазей, суспензий,
кремов, паст и др. (аубазидан, декстран, пуллулан).
Ассортимент углеводных полимеров может быть расширен за
счет изготовления на их основе полусинтетических производных,
например, сульфатированных декстранов, маннанов и других гли-
канов, относящихся к группе гепариноподобных веществ — гепа-
риноидов.
Микробные липиды пока не внедрены в промышленное про-
изводство, хотя некоторые продуценты являются заманчивыми для
этих целей, поскольку могут накапливать липиды до 40% и более
в расчете на сухие вещества клеток. К тому же в составе этих
липидов содержатся биологически ценные ненасыщенные жирные
кислоты. К таким продуцентам могут быть отнесены: Cryptococcus
terricolus, Lipomyces lipoferus, Rhodotorula gracilis, Sporobolomyces
roseus, Trichosporon pullulans и др. На первых порах перспективно
использование "липидных дрожжей" в качестве кормовых добавок,
особенно -— тех их них, которые синтезируют также заметные
количества и витаминов (например, каротина).
Получение полиоксибутирата
Поли П(-)0-оксимасляная кислота — это запасной энергетиче-
ский материал у прокариот с ММ от 60 кДа до 250 кДа. В отсутствии
экзогенного источника энергии он деполимеризуется и участвует
в снабжении клетки АТФ.
Полиоксибутират накапливается в клетках (иногда до 70% от
сухих веществ) в форме гранул диаметром 0,1—0,7 нм, окруженных
мембраной. По своим физико-химическим свойствам он перспек-
тивен в радиоэлектронике (обладает хорошими пьезоэлектриче-
скими свойствами), в медицине (хирургический материал), фарма-
ции (вспомогательное средство при создании некоторых лекарст-
венных форм), в органическом синтезе (сходен с полиизопропи-
478
леном по термопластическим характеристикам, t° плавления 166
188°С).
НО—СН—CHj—СО---О—СН—СН2—СО О—СН—сн2—СООН
СН3 СН3 СН3
п
попиоксибутират (п^300-1300)
Заметные количества полиоксибутирата накапливают бактерии
ив родов: Alcaligenes, Chromatium, Hyphomicrobium, Methyl-
obacterium, Nocardia, Pseudomonas, Rhizobium, Spirillum,
Streptomyces, Vibrio и др. Однако лишь три рода являются перс-
пективными для промышленного биосинтеза биополимера:
Alcaligenes, Azotobacter и Methylobacterium. Они способны накап-
ливать полиоксибутират на относительно недорогих субстратах
(ацетат, водород меласса, метанод сахароза, этанол). Контроль за
накоплением полимера в клетках относительно легко проводить с
помощью ИК-спектрофотометрии.
При лимитировании роста продуцента по какому-либо источ-
нику питания (углерод азот, сера, фосфор) или по кислороду
происходит накопление полиоксибутирата, синтезирующегося че-
рез АцКоА. Процесс ферментации может быть одностадийным или
двухстадийным, когда вначале накапливают биомассу клеток на
сбалансированной среде по источникам питания и дыхания, а затем
ее переводят в условия лимитированного роста и развития.
Для выделения полимера отсепарированные клетки экстраги-
руют, например, 1,2-дихлорэтаном, или дезинтегрируют с после-
дующей обработкой, исходя из целевых установок на получение
целевого продукта.
Получение микробных иммунобиологических препаратов
К микробным иммунобиологическим препаратам относят вак-
цины, диагностикумы, аллергены.
Вакцины - по праву занимают ведущее место в профилактике
инфекционных заболеваний. Миллионы детей и взрослых ежегод-
но "прививают" во всем мире вакцинами из бактерий и вирусов.
Вакцины давно подразделяют на корпускулярные и растворимые.
К числу первых относят, как правило, вакцины из убитых или
479
ослабленных живых клеток патогенов. К вторым — так называе-
мые химические вакцины. Однако, такое деление теперь оказыва-
ется недостаточным и наиболее адекватной классификацией вак-
цин является следующая:
из клеток
прокариот и
эукариот
1. живые
2. убитые
I
из клеточных
компонентов
— Вакцины -
♦
из продуктов
обмена клеток
Т”
Вирусные
I
Генноинже —
верные
1. полисаха-
ридные
2. рибосо-
мальные
1. анатоксины
(токсоиды)
А. из вирионов
1. живые
2. инактиви-
рованные
Б. из компо-
нентов вири-
онов
1. субъеди-
ничные
Подбор или создание вакцинных штаммов—дело чрезвычайно
важное, кропотливое и ответственное. В настоящее время под
эгидой Всемирной организации здравоохранения (ВОЗ) и под
контролем соответствующих служб здравоохранения стремятся
повсеместно (в том числе — в России) проводить мероприятия по
профилактике эпидемических инфекционных заболеваний с по-
мощью стандартных вакцинных препаратов. Основные требования
к вакцинам — высокая иммуногенность и безвредность для людей
и животных.
Вакцины могут быть в виде монопрепаратов, предназначенных
для профилактики специфических инфекций, а также в ассоции-
рованной форме для создания иммунитета против нескольких
инфекций.
Вакцины из клеток патогенных микробов
Вакцины живые. Это взвеси клеток вакцинных штаммов
микроорганизмов, стойко утратившие способность вызывать ин-
фекционное заболевание у человека, животного, но приживающи-
еся в нем. Такие штаммы, называемые аттенуированными (от лат.
attenuatus — ослабленный, тонкий, уменьшенный), могут быть
природными (спонтанные мутанты) или полученными искусствен-
но в лабораторных условиях (созданные мутанты).
Технология получения живых вакцин включает следующие
этапы:
480
1) размножение вакцинного штамма в несколько пассажей от
культуры на средах в пробирках и до ферментатора в оптимальных
условиях. Продолжительность этапа зависит от скорости размно-
жения микроорганизма (для сравнения можно назвать сальмонел-
лы и микобактерии туберкулеза). Режим культивирования - как
правило периодический;
2) сепарирование клеток от культуральной жидкости, напри-
мер, центрифугированием;
3) ресуспендирование клеток в подходящем растворителе
(смесь сахарозы и желатина - для вакцины БЦЖ, вода - для
туляремийной вакцины и т. д.);
4) розлив суспензии по ампулам или флаконам;
5) лиофильное высушивание, запаивание ампул или укупори-
вание флаконов.
Живые вакцины не должны содержать консервантов или ка-
ких-либо других ингибиторов роста и развития вакцинных штам-
мов. Если живые вакцины выпускают в живом виде, то в качестве
суспензионной среды можно использовать какие-либо стабилиза-
торы или забуференный изотонический раствор натрия хлорида
В России изготавливают следующие живые вакцины: М-44
против лихорадки Ку, бруцеллезную накожную, БЦЖ для внутри-
кожного применения, сибиреязвенную (для людей) - вакцина СТИ
для накожного или подкожного применения, сыпнотифозную ком-
бинированную (ЖКСВ-Е), туляремийную накожную, чумную.
Живые вакцины вводят обычно однократно.
Вакцины убитые из клеток патогенов
представляют собой взвеси клеток болезнетворных бактерий или
грибов, обладающих выраженной иммуногенностью, но лишенные
патогенности. Технология изготовления таких вакцин принципи-
ально сводится к следующему: выращивание стандартного произ-
водственного штамма на подходящей питательной среде; обезвре-
живание (инактивирование) клеток одним из нижеуказанных ме-
тодов; сепарирование клеток (чаще — с помощью центрифугиро-
вания); ресуспендирование клеток в изотоническом растворе на-
трия хлорида до определенной концентрации; контроль на отсут-
ствие живых клеток патогена, на иммуногенность и по другим
показателям, предусмотренным в научно-технической документа-
ции.
Обезвреживание клеток проводят одним из следующих мето-
дов: прогреванием, обработкой формалином, ацетоном, этанолом.
Инактивированные разведенные взвеси микробов разливают в
16 т. 8524 . 481
ампулы или флаконы и хранят при 2—10°С. Основной способ
применения убитых вакцин — подкожные инъекции.
Убитыми являются вакцины против бруцеллеза (лечебная),
брюшного тифа, гонореи, дизентерии Флекснера-Зонне, коклюша,
лептоспироза, паратифов, холеры.
Крупномасштабного производства убитых вакцин против гриб-
ковых заболеваний пока нет, но в некоторых случаях в лаборатор-
ных условиях изготавливают аутовакцины для лиц с гипоэргией
при микозах или для пациентов, состояние которых оценивают
близким к анергии (например, при кандидозе).
Вакцину против брюшного тифа, инактивированную ацетоном,
выпускают в сухом виде.
Вакцины из клеточных компонентов патогенных микробов
Вакцины полисахаридные. Полисахариды, или
гликаны определяют антигенную специфичность ряда возбудите-
лей инфекционных заболеваний, поэтому антигенноактивные из
них выделяют в индивидуальном состоянии. Такими вакцинами
являются менингококковая и пневмококковая. Вакцинные штаммы
выращивают на жидких питательных средах, клетки сепарируют,
промывают водой и капсульный материал экстрагируют каким-ли-
бо методом, переводя полисахарид в водный раствор. Если гликан
также накапливается в культуральной среде, то его можно выде-
лить в чистом виде.
Из водных растворов менингококковые полисахариды осажда-
ют катионным ПАВ — гексадецилтриметиламмоний бромидом,
пневмококковые полисахариды — этанолом. Дополнительную очи-
стку таких препаратов можно проводить с помощью этих же
реагентов из водных растворов.
Менингококковые и пневмококковые полисахаридные вакци-
ны, как правило, являются поливалентными. Из них первые вклю-
чают обычно 4 типа гликана, вторые — 23 типа (всего типировано
свыше 80 сероваров пневмококков).
В России (на предприятии НИИЭМ им. Г. Н. Габричевского) к
1982 г. освоены менингококковые полисахаридные вакцины —
моновалентная типа А и бивалентная типов А+С. Культуры вак-
цинных штаммов менингококков выращивают 8—10 часов в опти-
мальных условиях при 37°С до плотности 6—7 • 109 клеток в 1 мл
(для менингококков типа А) и 8—10 • 109 клеток в 1 мл (для
менингококков типа С). Затем клетки сепарируют и подвергают
обработке по следующей схеме:
482
Бактериальная культура
| +цетавлон
Осадок комплекса "цетавлон—полисахарид” — ЦП
| экстрация 1М раствором СаС12
ЦП в экстракте
+этанол до концентрации 25%
Осадок — отход «-
(нуклеиновые кислоты
+ ЛПС)
Надосадок
+этанол до концентрации 80%
Осадок полисахарида—сырца
+0,ЗМ раствор натрия ацетата
(pH 7,2)
Полисахарид—сырец в растворе
+фенол
Депротеинизированный раствор полисахарида
| диализ против 0,1М раствора СаС12
Диализованный раствор полисахарида—।
А—полисахарид С—полисахарид
ультрацентри—
фугирование
Надосадок полисахарида С
+ этанол (4 объема) + |
Осадок полисахарида А Осадок полисахаридаС
промывание этанолом, ацетоном;
высушивание над СаС12
Очищенный полисахарид А Очищенный полисахарид С
Очищенные полисахариды с лактозой — наполнителем реко-
мендованы в качестве вакцинирующих препаратов.
Полисахаридный Vi-антиген из Salmonella typhi (штамм Ту2)
входит в состав брюшнотифозной спиртовой вакцины. В 1 мл
такого препарата содержится 5 • 108 клеток $. typhi и 400 мкг
Vi-антигена. Вакцину вводят подкожно.
483
В США изготавливают полисахаридные вакцины из некоторых
видов Pseudomonas (в частности, Р. fluorescens).
Рибосомальные вакцины. У прокариот рибосомы
содержат примерно 60% РНК и 40% белка, у эукариот — 55% и 45%
соответственно. В стационарной фазе размножения бактериальная
клетка содержит 104 рибосом; это число возрастает в период
log-фазы. Впервые рибосомальный препарат из авирулентного
штамма Mycobacterium tuberculosis приготовили Юманс А. С. и
Юманс Г. П. (1965). Рибосомы в чистом виде не применяют в
качестве вакцин, но обогащенные ими полисахаридные и другие
антигенные препараты используют на практике. Наиболее извест-
ный из них "Рибомунил D-53", выпускаемый во Франции. Он
включает рибосомы из клеток Klebsiella pneumoniae, Streptococcus
pneumoniae, S. pyogenes и Haemophilus influenzae с добавлением
протеогликана из К. pneumoniae; Применяют рибомунил интрана-
зально. Его рекомендуют для профилактики заболеваний дыхатель-
ной системы и для лечения ринитов, риносинусйтов и ринофарин-
гитов — в особенности.
Известен препарат в виде экстракта из клеток Streptococcus
mutans, обогащенный рибосомами. Схема его приготовления сле-
дующая:
Streptococcus mutans серотип g, нативные клетки — 100 г
Дезинтеграция клеток с помощью стеклянных бус — баллотини
(100 г) в 100 мл 10 2 М фосфатного буфера (pH 7,4) с 10'2 М МдС12
(РМВ) и 3 мкг ДНК-азы/мл.
Неразрушенные клетки и остатки клеток отделяют низкоско-
ростным центрифугированием (дважды по 10 мин. при 27000 g и
47000 д).
Рибосомы отмывают 5 раз в РМВ в течение 2,5 часов при 250000
д с последующим двухкратным центрифугированием в течение 20
мин. при 47000 д, а затем фильтруют через стерильный мембран-
ный фильтр (0,45 мкм).
Препарат-экстракт S.mutans, обогащенный рибосомами.
В Японии для предупреждения коклюша у детей используют
вакцину из филаментозного гемагглютинина и лимфоцитстимули-
рующего фактора из Bordetella pertussis.
484
Вакцины из продуктов метаболизма патогенных микроорга-
низмов
Анатоксины (токсоиды) — см. раздел "Токсины и
анатоксины" в данной главе.
Вакцины вирусные.
По аналогии с бактериальными вакцинами вирусные также
подразделяют на живые и инактивированные. Для
приготовления обоих типов вирусных вакцин необходимо нако-
пить вирусный материал (вирионы), используя либо куриные
эмбрионы, либо культуры тканей из почек обезьян, куриного
эмбриона, диплоидных клеток человека. Так вакцинный вирус
гриппа (авирулентный) накапливают в аллантоисной жидкости
эмбриона, которую затем отсасывают и центрифугируют. Если
предполагают готовить живую вакцину, то вирус суспендируют и
разводят до нужной концентрации и подвергают лиофильному
высушиванию.
Для получения вакцины против желтой лихорадки также зара-
жают куриный эмбрион, в нервных тканях которого накапливается
патоген (аттенуированный штамм 17 D). Последующие этапы
технологии получения вакцины заключаются в гомогенизации
эмбриона в стерильной воде, получают "пюре", которое центрифу-
гируют, осадок представляет отход, а надосадочную жидкость,
содержащую вирус, лиофильно высушивают.
Культуры тканей, зараженные вирусами, выращивают в жид-
ких средах, поэтому вирусный материал можно получить после
фильтрации культуральной жидкости. Учитывая тот факт, что
вакцинные штаммы вирусов являются, как правило, аттенуирован-
ными, то стадия инактивации при их изготовлении отпадает сама
собой. Однако, имеются исключения из этого правила — вакцины
против бешенства и полиомиелита; первую инактивируют 0-про-
пиолактоном, вторую — либо Р-пропиолактоном, либо формали-
ном.
Очищенный вирусный материал, готовый для приготовления
вакцин, или готовые вирусные вакцины сохраняют при -70°С.
Вирусные вакцины не сочетают в себе разные виды, но,
например, инактивированная и живая полиомиелитные вакцины
или убитая гриппозная вакцина почти всегда содержат разные
серотипы вирусов.
Живые ослабленные вирусные вакцины в виде суспензий
теряют свою потенциальную активность достаточно быстро, поэ-
тому их сохраняют либо в замороженном состоянии, или добавля-
ют стабилизаторы — сахарозу, магния хлорид.
Живыми вирусными вакцинами являются: гриппозные интра-
назальные для детей и взрослых (профилактические), гриппозная
пероральная для детей и взрослых (лечебно-профилактическая),
485
против желтой лихорадки, коревая, против краснухи, полиомие-
литная пероральная, против свинки.
К инактивированным вирусным вакцинам относятся: антира-
бические сухая МИВП и типа Ферми, против клещевого энцефа-
лита.
Субъединичные, или расщеп л.е иные вак-
цины. Такие вакцины можно получать из оболочечных вирусных
конъюгированных или неконъюгированных белков. Пока внедре-
на в производство лишь субъединичная гриппозная вакцина, вклю-
чающая гликопротеин-гемагглютинин из одного или нескольких
штаммов вируса. Клиническим, испытаниям подвергаются вариан-
ты субъединичных вакцин вируса иммунодефицита человека, уси-
ленные мурамилди- и мурамилтрипептидами.
Технология получения гемагглютинина из гриппозных вирио-
нов заключается в следующем: накопленный вирусный материал
в аллантоисной жидкости куриного эмбриона сепарируют, подвер-
гают микрофильтрации и фильтрационному концентрированию
примерно в 50 раз, при необходимости дополнительно очищают
ультрацентрифугированием в градиенте плотности сахарозы; кон-
центрированный вирионный материал обрабатывают катионным
ПАВ и отделяют гемагглютинины ультрафильтрацией с последую-
щими диализом и стерилизующей фильтрацией. На последней
стадии стандартизируют и контролируют субъединичную вакцину
(особенно — на отсутствие живых гриппозных вирусов).
Генно-инженерные вакцины
Биотехнология таких вакцин базируется на возможности пере-
носа фрагментов хромосомной ДНК или плазмид из бактерий или
вирусов в клетки других видов бактерий или дрожжей, то есть не
в природные для них реципиентные клетки. Таким путем создана
возможность продукции поверхностного антигена вируса гепатита
В (HBsAg) дрожжевыми клетками. Этот антиген отделяют от
дрожжей-продуцентов и используют для приготовления вакцины.
рДНК-биотехнология в этом направлении "становится на ноги"
и за ней большое будущее на пути создания генно-инженерных
вакцин, а также белковых иммуногенов и других белков, включая
стафилококковый белок А и стрептококковый белок М.
На подходе к широкому внедрению вакцины против некоторых
протозойных инфекционных заболеваний (например, малярии).
Диагностикумы
Диагностические приемы при инфекционных заболеваниях
могут быть сведены к серодиагностике, аллергодиагностике и
фагодиагностике, если иметь в виду лишь использование биопре-
паратов. Серодиагностику выполняют с сыворотками крови, при-
меняя соответствующие антигенные препараты, называемые
диагностикумами; аллергодиагностику проводят в форме
486
аллергических проб с аллергенами на людях (на животных — в
эксперименте или при наличии у них инфекционных заболеваний);
фаги оценивают по литическому действию на чувствительные
клетки соответствующих видов бактерий. Аллергены и бактерио-
фаги выделены в самостоятельные подразделы (см.).
Диагностикумы могут представлять из себя убитые клетки с
выраженной чувствительностью к специфическим антителам, а
также отдельные, хорошо изученные антигенные компоненты. Так
у энтеробактерий известны Н-, О- и\ Vi-антигены, из которых
первый термолабилен, второй и третий — термостабильны. На
этом основано их выделение из клеток. Жгутиковый Н-антиген
изолируют с помощью 0,2% раствора формалина спустя сутки после
его добавления и выдержки при 37"С; О- и Vi-антигены экстраги-
руют при повышенных температурах водой или другими раство-
рителями. Такие антигены используют как таковые или в адсор-
бированном виде на эритроцитах, предварительно обработанных
таннином или формалином.
Бактериальные и эритроцитарные диагностикумы применяют
соответственно в реакциях агглютинации и гемагглютинации.
Для диагностики сифилиса выпускают кардиолипиновый анти-
ген для реакции микропреципитации. Этот антиген включает 3
очищенных липида: 0,03% 1,3-дифосфатглицерина (кардиолипин),
0,27% фосфатидилхолина (лецитин) и 0,9% холестерина в абсолют-
ном этаноле. Препарат выпускают в ампулах по 2 мл. Срок
хранения — 4 год. Инактивированные вирусные диагностикумы
используют в реакциях связывания комплемента, нейтрализации
и торможения гемагглютинации.
Кроме вышеназванных диагностикумов производят: единый
бруцеллезный диагностикум из убитых фенолом бруцелл и под-
крашенный метиленовым синим; гриппозный диагностикум в виде
взвеси инактивированных формалином или мертиолатом вирионов
в аллантоисной жидкости куриных эмбрионов; диагностикум инак-
тивированного вируса клещевого энцефалита (суспензия мозга
белых мышей, зараженных этим вирусом); эритроцитарный Vi-ди-
агностикум, содержащий Vi-антиген Salmonella typhi, и О-диагно-
стикум сальмонеллезный, включающий О-антиген из разных групп
сальмонелл.
Аллергены
Аллергены по своему происхождению подразделяют на микро-
бные, растительные и животные. Их используют для диагностики
патологических, процессов, в развитии которых заметную роль
играет сенсибилизация макроорганизма соответствующим антиге-
ном-аллергеном.
Микробные аллергены изготавливают разными способами. На-
тивные аллергены — это взвесь убитых бактерий и растворимых
487
продуктов их метаболизма; очищенные аллергены — это осажден-
ные и лиофильно высушенные термостабильные фракции фильт-
ратов 5—6 суточных бульонных культур бактерий, в которых
содержатся свыше 80% белков, около 7% углеводов и до 10%
нуклеиновых кислот.
К разряду бактериальных относятся аллергены: антраксин,
бруцеллин, дизентерии, дифтероида, листерийный, малеин, ката-
ральной нейссерии, лепромин, орнитозный, палочки дифтерийной
нетоксигенной, кишечной палочки, ложнодифтерийной палочки,
синегнойной палочки, пестин, стафилококковые видовые, стреп-
тококковые видовые, токсоплазмин, энтерококковый, альттубер-
кулин Коха, туберкулин очищенный (PPD-Л), туберкулин очищен-
ный сухой (PPD), тулярин и др.
Грибковые аллергены обычно выделяют из клеток зрелых
культур, реже — из культуральных жидкостей. Они представляют
собой белково-углеводные комплексы. Выпускают их в ампулах по
1 мл.
Из грибковых аллергенов известны: бластомицин, гистоплаз- ’
мин, кандидин, кокцидиоидин, а также аллергены из некоторых
плесневых грибов (аспергиллов, пенициллов) и дерматофитов. ~ ;
Бактериофаги
Бактериофаги используют для фаготипирования бактерий и с
лечебно-профилактическими целями.
Для индикации бактерий изготавливают так называемые видо-
вые и типовые фаги: брюшно-тифозные Vi-типовые, дизентерий-
ные индикаторные, паратифозные В-типовые, сальмонеллезный
идикаторный, стафилококковые типовые.
Фаги для профилактики и лечения инфекционных заболеваний
выпускают моно- и поливалентными, их нельзя подменять диагно-
стическими фагами, равно как и наоборот—диагностические фаги
нельзя использовать вместо лечебно-профилактических фагов. Из
числа последних отечественные предприятия выпускают жидкие
моновалентные стафилофаги, стрептококковый фаг, фаг пиоциа-
неус, фаги протейный и коли-протейный, коли-фаг, поливалентный
дизентерийный (жидкий, сухой и в суппозиториях), поливалентные
сальмонеллезные (жидкий и сухой).
Производство фагов основано на заражении бактериофагами
чувствительных бульонных культур определенных видов и штам-
мов. "Вегетация" фагов сопровождается при этом лизисом бакте-
риальных клеток. От части нелизировавшихся клеток и клеточного
детрита освобождаются стерилизующей фильтрацией. Фаги про-
ходят через фильтр, фильтраты тестируют по эталонным культурам
чувствительных бактерий.
В жидкие препараты лечебно-профилактических фагов можно
добавлять консерванты.
488
Глава 10.
ФИТОБИОТЕХНОЛОГИЯ
Фитобиотехнология — составная часть биотехнологии. Объек-
тами ее являются клетки и ткани растений — фотоавтотрофных
эукариот, а также биоактивные молекулы растительного проис-
хождения (ферменты, нуклеиновые кислоты, стероиды и некото-
рые другие сложные органические вещества). Следует обратить
внимание на тот факт, что растительные клетки в культуре обычно
являются хемогетеротрофамй.
Название фитобиотехнология слагается из четырех слов гре-
ческого происхождения: phyton — растение, bios — жизнь, teken
— искусство, logos — наука, слово. Следовательно, это наука об
использовании растительных объектов в технике и промышленном
производстве. На первый взгляд выращивание растений (злаковых,
лекарственных, декоративных и т. д.) в полевых условиях подпадает
под рубрику “фитобиотехнология", однако, как и с животными,
здесь отсутствуют специфические методы промышленного куль-
тивирования цельных растений в биореакторах. Таким образом, к
фитобиотехнологическим процессам относят те из них, которые
базируются на клеточном уровне, если даже клетки несут генети-
ческую информацию, воспринятую ими в результате генно-инже-
нерного эксперимента.
Клетки и ткани высших растений, выращиваемые на питатель-
ных средах in vitro в строго контролируемых условиях, все шире
используют в фитобиотехнологии. Общирное царство растений —
потенциально неограниченный источник клеток и тканей, которые
могут быть введены в культуру и, как следствие, существуют
неограниченные возможности для создания новых фитобиотехно-
логических процессов, в которых нуждается человечество.
В настоящее время царство растений на земном шаре включает
порядка 300 000 видов. От рационального и бережного отношения
к растениям зависит благополучное выживание человечества на
планете Земля.
Представители царства растений (Plantae)
Надцарство — Eucaryota
Царство — Plantae
Отделы: 1. Водоросли — Algae
Типы: а) Красные — Rhodophyta .
б) Золотистые — Chrysophyta.
489
в) Желтозеленые — Xanthophyta
г) Диатомовые — Bacillariophyta
д) Динофлагелляты и криптомонады — Dinophyta
е) Бурые — Phaoephyta
ж) Зеленые — Chlorophyta
з) Эвгленовые — Euglenophyta
Некоторые биологи выделяют дополнительный
(девятый) тип пиррофитовых водорослей —
Pyrrophyta.
2. Печеночники и мхи — Bryophyta
3. Папоротники, плауны и хвощи — Pteridophyta
4. Семенные растения — Spennatophyta
Типы: а) Голосеменные — Gymnospermae
б) Покрытосеменные (цветковые) —
Angiospermae.
В таком смысле фитобиотехнология является "палочкой-выру-
чалочкой”, способной освободить отдельные регионы и страны от
импорта и интродукции растений, им не свойственных или совсем
неспособных культивироваться в неподходящих климатических
зонах. Это тем более резонно, когда речь идет о растениях —
продуцентах БАВ.
Хотя растительные клетки и ткани принадлежат к более диф-
ференцированным организмам в сравнении, например, с бактери-
ями, тем не менее они способны культивироваться в форме
неорганизованной клеточной массы (каллус). Каллусную ткань
можно "заставить" формировать зародышеподобные структуры,
почки, побеги, а на их основе — растения-регенеранты. Все это
происходит благодаря тотипотентности растительных
клеток (от лат. totus — все, целый, potentia — сила, потенция).
Понятие "тотипотентность" является клеточной характеристикой;
в нем отражен потенциал клетки воспроизводить все типы клеток,
присущих взрослому организму. Другими словами клетка обладает
способностью воспроизводить целый организм.
Если в качестве биообъекта применяют изолированный заро-
дыш, меристему верхушечных или пазушных почек, то есть интег-
рированную систему, то все получаемые растения-регенеранты
будут полностью соответствовать исходному растению, из которо-
го была взята какая-либо интегрированная система из числа вы-
шеназванных. Этим добиваются клонирования интересующих нас
растений (рис. 140).
490
Рис. 140. Клетки, органы и ткани, используемые в фитобиотехнологии.
В случаях применения изолированных клеток и протопластов
удается получать измененные варианты исходной формы, которые
передают полученные новые признаки потомству. Этим добивают-
ся возможностей искусственного создания новых форм растений,
пригодных для выборки, или селекции (рис. 141).
ядро
Рис. 141. Пути искус-
ственного создания но-
вых форм растений (1 —
трансгенез, 2 — хромо-
сомная инженерия, 3 —
генная инженерия. 4,8 —
клеточная (геномная) ин-
женерия, 5—7 — цибри-
дизация).
В течение последних трех десятилетий удалось массовое раз-
множение культивируемых растений с уникальными характери-
стиками, а выращивание растительных клеток стало основой для
получения многих природных веществ, образуемых различными
растениями в качестве продуктов вторичного обмена (гликозиды,
алкалоиды и другие вещества).
Таким образом, клеточная и молекулярная биология растений
составляет основу фитобиотехнологии, а главными направлениями
стали создание новых форм полезных растений для сельского и
лесного хозяйства, а также разработка промышленного производ-
ства химических веществ из растений.
В сравнении с микробной биотехнологией фитобиотехнология
является совсем молодой научной дисциплиной, становление ко-
торой на промышленные рельсы происходит в настоящее время.
491
Для нее характерна и своя терминология, совпадающая или не
совпадающая с общепринятой в биотехнологии в целом. Отдельные
термины типичны для фитобиотехнологии. Примеры некоторых
из них приведены ниже.
ЮЛ Термины, используемые в фитобиотехнологии
Адаптация — приспособление наследственно компетентной
клетки (организма) к изменившимся условиям существования.
Адвентивный — развитие из необычных точек происхождения,
например почечные или корневые ткани, возникающие из каллуса,
или зародыши, развивающиеся из других источников, а не из зигот.
Этот термин также может быть использован для описания агентов,
загрязняющих клеточные культуры.
Аксеническая культура — культура без чужеродных или не-
желательных форм жизни; аксеническая культура может включать
запланированное сокультивирование различных типов клеток, тка-
ней или организмов.
Андрогенез — процесс развития растения из микроспоры, или
пыльцевого зерна, либо через гаметический эмбриогенез, либо из
каллуса.
Анеуплоид — организм, клетка, ядро с числом хромосом,
отклоняющимся от х и от чисел, кратных х; хромосомы могут/не
могут перестраиваться.
Апекс — верхушка.
Апикальное доминирование — подавление роста боковых
почек растительного побега или наличие терминальной почки.
Апикальная (верхушечная) культура почечная — структура,
состоящая из почечной апикальной меристемы с одним — до
нескольких примордиальных листьев и обычными размерами от
0,1 до 1 мм в длину; если включаются более зрелые листья, то такая
структура может достигать в длину нескольких сантиметров.
Апикальная меристема почечная — недифференцированная
ткань, локализованная в пределах верхушечной почки, представ-
ляющаяся обычно в виде блестящей куполоподобной структуры,
дистальной к самому молодому листовому примордию и размером
менее, чем 0,1 мм в длину при вырезании.
Асептические процедуры (методы, техника работы) — проце-
дуры, обеспечивающие предотвращение попадания микробов в
клетки, ткани и органы растительных культур, или перекрестное
загрязнение одной клеточной культуры другой. Эти процедуры
могут/не могут исключить занесение инфицирующих молекул.
492
Асептическое состояние — состояние без инфекции или мик-
робов-контаминантов.
Бессмертие—приобретение ограниченной во времени клеточ-
ной культурой признаков непрерывной клеточной линии. Бессмер-
тная клетка не обязательно та, которая неопластически, или зло-
качественно трансформируется.
Вариант — культура, проявляющая стабильное фенотипиче-
ское изменение — генетическое или эпигенетическое по природе.
Вегетативный рост — воспроизведение растений, используя
бесполый процесс (части стебля или листьев).
Время генерации клетки — интервал времени между двумя
последовательными делениями клетки. Этот интервал может быть
лучше определен с помощью кинофотомикрографии (цейтрафер-
ная съемка); этот термин не является синонимом "времени удвое-
ния популяции".
Время удвоения популяции — интервал времени, в течение
которого число клеток в популяции возрастает вдвое; другими
словами — это интервал, соответствующий логарифмической фазе
роста, за который, например, число клеток 1 • 106 увеличивается до
2 • 106 клеток. Этот термин не является синонимом термина "время
генерации клетки".
Габитуация — приобретенная способность популяции клеток
расти и делиться независимо от экзогенно добавляемых ростовых
факторов.
Гаметоклон — растение, регенерированное из клеточной куль-
туры, произошедшей из мейоспоры, гаметы или гаметофита.
Гаметоклональная вариация — вариация в фенотипе, либо
генетическая или эпигенетическая по природе, выраженная гаме-
токлонами.
Гаплоид — ядро, клетка, организм с половинным набором
хромосом (п или In).
Гетерокарион—клетка, содержащая два или более генетически
различных ядер в общей цитоплазме (обычно является результатом
слияния клеток).
Гетероплоид — термин относится к клеточной культуре, в
которой клетки обладают ядрами, содержащими иное число хро-
мосом в сравнении с диплоидным числом (2п). Данный термин
используют только для описания культуры, а не отдельных клеток.
Следовательно, гетероплоидная культура должна быть той культу-
рой, которая содержит анеуплоидные клетки.
Гибридная клетка — термин используют для описания моно-
493
ядерной клетки, произошедшей из слившихся двух различных
клеток с образованием синкариона.
Гиногенез — процесс возникновения растения из клеток заро-
дышевого мешка.
Гомокарион — клетка, содержащая два или более генетически
идентичных ядер в общей цитоплазме (результат слияния клеток).
Дедифференциация — переход специализированных, неделя-
щихся клеток к пролиферации.
Диплоид — состояние клетки, в которой все хромосомы (за
исключением половых) находятся в удвоенном числе (2п) и струк-
турно идентичны хромосомам видов, от которых произошла куль-
тура.
Дифференциация—комплекс процессов, в результате которых
наступают различия между дочерними клетками, а также между
материнскими и дочерними клетками.
Дифференцированные клетки — клетки в культуре, в которых
поддерживаются все или многие из специализированных структур
и функций, типичные для этих клеток in vivo; другими словами
дифференцировка — состояние специализации клеток, отличаю-
щее их от других.
In vitro — опыты, выполненные в пробирке, колбе и т. д., то
есть вне живого организма.
In vivo — опыты, проведенные на живом организме.
Индукция — инициация структуры, органа или процесса in
vitro.
Инокулюм (трансплант) —часть суспензионной или каллусной
культуры, переносимой в свежую питательную среду.
Каллус -— неорганизованная, пролиферирующая масса диффе-
ренцированных растительных клеток.
Кариопласт — клеточное ядро, полученное из клетки посред-
ством энуклеации, окруженное узким ободком цитоплазмы и
клеточной мембраны.
Кариотип — набор хромосом, характерный для данного вида.
Клеточная линия — возникает из первичной культуры при
первом удачном субкультивировании; название "клеточная линия"
относится к таким культурам, которые включают линии клеток,
изначально присутствующих в первичной культуре. Если известен
статус культуры, то используют термины "прерывная" или "непре-
рывная" культура. Если же статус неизвестен, то достаточен термин
"линия".
Клон — популяция клеток, вози икших из одной клетки посред-
ством митоза или группа растений, развившихся вегетативным или
бесполым Путем, все члены которой произошли из одной повторно
культивируемой клетки.
Клональное микроразмножение — получение in vitro неполо-
вым путем растений, генетически идентичных исходному.
Криоконсервация — сохранение клеток, тканей, зародышей
или семян при ультранизкой температуре (ниже -100°С).
Культура зародышей — стерильное выращивание in vitro на/в
питательной среде незрелых или зрелых изолированных зароды-
шей.
Культура изолированных протопластов — выращивание про-
топластов на/в среде in vitro в присутствии стабилизатора. При
регенерации клеточных стенок культура протопластов трансфор-
мируется в клеточную культуру.
Культура каллусных тканей — длительно выращиваемая пе-
ресадочная культура тканей, возникших вследствие пролиферации
клеток изолированных фрагментов органов или самих органов
растений (пыльники, семяпочки и т. д.).
Культура корней — выращивание in vitro на питательной среде
изолированных корней в пересадочном режиме.
Культура меристем побега — выращивание in vitro на пита-
тельной среде изолированного из верхушки или пазушной почки
побега конуса нарастания с одним или двумя листовыми примор-
диями, с первоначальным размером менее 0,1 мм в длину.
Культура-нянька — рост клетки или клеток на подлежащей
культуре различного происхождения, которая в свою очередь
находится в контакте с культуральной средой. Культивируемая
клетка или ткань может быть отделена от питательного слоя
пористым материалом — фильтрованной бумагой или мембранны-
ми фильтрами.
Культура опухолевых тканей — длительная культура фрагмен-
тов, изолированных из опухолей растений разного происхождения
и освобожденных от патогенов, индуцировавших развитие опухо-
ли.
Культуры тканевые — общий термин, который относят к
изучению клеток, тканей и органов, поддерживаемых или выра-
щиваемых in vitro в течение свыше 24 часов. Обычно используют
следующие варианты термина: клеточная культура — растущие
in vitro клетки, включая культуру из одиночных клеток; в клеточных
культурах клетки не организуются в ткани; тканевая культура —
поддержание роста тканей in vitro в состоянии, при котором может
495
происходить дифференциация и сохранение их строения и/или
функции; органная культура — поддержание или рост органного
примордия или целого (или части) органа in vitro в состоянии, при
котором может происходить дифференциация и сохранение стро-
ения и/или функции; культура растительной ткани — рост или
поддержание растительных клеток, тканей, органов или целых
растений in vitro.
Линия — культура, возникшая из штамма путем селекции или
клонирования, имеющая маркерные признаки (см. также "клеточ-
ная линия").
Липосома —. закрытый липидный пузырек, содержащий вод-
ную фазу; используют для включения экзогенных веществ, достав-
ляемых в клетки посредством слияния липосом с клетками.
Микроклетка — клеточный фрагмент, содержащий одну —
несколько хромосом и который образуется при энуклеации или
разобщении микроядерной клетки.
Микрокультивирование — in vitro клональное культивирова-
ние растений из почечных верхушек или узловатых эксплантов,
обычно .с ускоренной пролиферацией почек в течение субкульти-
вирования.
Микроядерная клетка — клетка с блокированным митозом и
в которой малые группы хромосом функционируют как фокусы
для переустройства ядерной мембраны, формируя таким образом
микроядра, максимум которых должен быть равен общему числу
хромосом.
Морфогенез: а) развитие структуры от недифференцирован-
ного состояния к дифференцированному; б) процесс роста и
развития д ифференцированных структур.
Мутаген — фактор, обусловливающий мутацию.
Мутант — фенотипический вариант, возникший в результате
действия измененного или нового гена.
Мутация — генотипическое и необратимое (наследуемое) из-
менение нормы реакции клетки (организма).
Непрерывная клеточная культура — культура, способная к
неограниченному числу удвоения популяции; часто ее называют
"бессмертной культурой” клеток. Такие клетки могут или не могут
проявлять in vitro свойства неопластической или злокачественной
трансформации.
Норма реакции организма — проявление фенотипа в разных
условиях существования вида.
Омнипотентность — всесильность.
496
Органогенез — процесс дифференциации в каллусных клетках,
сопровождающийся образованием органов (корней, побегов) de
novo или из предсуществующих структур.
Пассаж — перенос или пересадка клеток (с разведением или
без разведения) из одной культуральной емкости в другую. Этот
термин является синонимом термина "субкультура", а число пере-
носа клеток или субкультур равнозначно числу пассажей.
Первичная культура — культура, берущая начало от клеток,
тканей или органов, взятых непосредственно от организмов. Пер-
вичная культура может расцениваться таковой до тех пор, пока не
удаются субкультуры в первое время. Затем она становится "кле-
точной линией".
Плотность насыщения — максимальное число клеток в куль-
туральном сосуде при специальных условиях выращивания. Плот-
ность насыщения выражается числом клеток на 1 см2 в монослой-
ной культуре или числом клеток на 1 см3 в суспензионной культуре.
Плотность популяции — число клеток на единицу поверхности
или объема среды. Общее число удвоений популяции (уровень
удвоения популяции) клеточной линии или штамма за один пассаж
in vitro определяют со времени посева в расчете на одно удвоение:
Nd = in (N/No х 3.33), где N — число клеток в культуральном сосуде
на конечный период роста, No — число клеток, засеянных в
культуральный сосуд, Nd — число популяционных удвоений. Луч-
ше всего использовать число прикрепившихся клеток.
Полиплоид — ядро клетки, организм с умноженным основным
числом хромосом (Зх, 4х и т. д.).
Популяция клеток — совокупность клеток в культуре.
Примордий — первый, первичный.
Пролиферация — разрастание клеток и тканей путем размно-
жения.
Пропагула — общий термин для обозначения органов и тканей,
служащих для вегетативного размножения (синоним термину "ди-
аспора") .
Протопласт — клетка с удаленной клеточной стенкой.
Псевдодиплоид — клетка, в которой число хромосом является
диплоидным, но в результате хромосомных перестроек нарушают-
ся нормальный кариотип и родственные связи.
Регенерация — морфогенетический ответ на стимул, который
приводит к образованию органов, зародышей или целых растений.
Редифференциация — переход специализированных клеток из
одного состояния дифференцировки в другое с предшествующими
497
делениями или непосредственно.
. Рекон — жизнеспособная клетка, реконструированная слияни-
ем кариопласта с цитопластом.
Реконструированная клетка — синоним термина "рекон".
Ростовый цикл — рост популяции клеток в цикле периодиче-
ского выращивания, характеризуется S-образной кривой. Фазы
ростового цикла: латентная, экспоненциальная, замедления роста,
стационарная, деградации.
Синкарион — гибридная клетка со слившимися ядрами.
Сомаклональная вариация — фенотипическая вариация среди
соматических клонов генетической или эпигенетической природы.
Соматическая (парасексуальная) гибридизация — слияние in
vitro растительных протопластов соматических клеток, которые
различаются генетически.
Соматический клеточный гибрид — клетка или растение,
возникшее от слияния протопластов соматических клеток, разли-
чающихся генетически.
Соматический клон, или соматоклон — растение, возникшее
из любой формы клеточной культуры растительных соматических
клеток.
Соматический эмбриогенез — образование эмбриоидов (заро-
дышевых структур) в культурах клеток и тканей способом, напо-
минающим формальный зиготический эмбриогенез; другими сло-
вами соматический эмбриогенез — это процесс эмбриоинициации
и развития из вегетативных или не гаметических клеток.
Стадии по Мурасиге: I — культивирование in vitro, характери-
зующееся посадкой растительной тканевой культуры в асептиче-
ских условиях; II — ступень в культивировании in vitro, характе-
ризующаяся быстрым численным возрастанием органов или дру-
гих структур; III — ступень культивирования in vitro, характери-
зующаяся приготовлением пропагулы для успешного переноса в
почву; при этом происходит процесс укоренения и отвердения
растений, начинается переход от гетеротрофного к аутотрофному
состоянию; IV — ступень в культивировании in vitro, характеризу-
ющаяся посадкой в почву растительной тканевой культуры, или
после претрансплантной обработки материала (пропагул) в стадии
III, либо (для некоторых видов) после прямого переноса растений
из стадии II в почву.
Субкультивирование — перенос транспланта (инокулюма) на
свежую питательную среду.
Субкультура — см. термин "пассаж".
498
Субпротопласт — протопласт, потерявший часть цитоплазмы.
Субштамм — происходит из штамма при изоляции одиночной
клетки или группы клеток, обладающих свойствами или маркера-
ми, которыми не обладают все клетки родительского штамма.
Суспензионная культура — тип культуры, в которой клетки
или их агрегаты размножаются, будучи суспендированными в
жидкой питательной среде.
Сферопласт — клетка с неполностью удаленной клеточной
стенкой.
Тотипотентность — клеточная характеристика способности к
формированию всех клеточных типов взрослого организма, или
это — свойство соматических клеток растений полностью реали-
зовать свой потенциал развития, то есть реализовать омнипотент-
ность ядра с образованием целого организма.
Трансгенез — перенос любым способом чужеродных генов в
клетки растений.
Трансформация—введение и устойчивая геномная интеграция
чужеродной ДНК в растительной клетке, сопровождающаяся ге-
номной модификацией.
Фенотип — определенная сумма признаков организма в конк-
ретных внешних условиях.
Цибрид — жизнеспособная клетка, возникшая в результате
слияния цитопласта с клеткой — протопластом или цитопластом;
при этом образуется цитоплазматический гибрид
Цибридизация — введение цитоплазматических (внеядерных)
генов в изолированный протопласт.
Цитоплазматическая наследственность — наследственность,
обусловленная внеядерными генами или хлоропластными, плаз-
мидными и т. д.
Цитопласт — интактная цитоплазма, остающаяся после энук-
леации (удаление ядра) клетки.
Штамм — происходит из первичной культуры или клеточной
.линии при селекции или клонировании клеток, имеющих специ-
фические свойства или маркеры.
Эксплант — ткань, взятая из своего оригинального места и
перенесенная в искусственную среду для роста и поддержания.
Эмбриоид — зародышеподобная структура, возникшая путем
соматического эмбриогенеза.
Эмбриокультура — in vitro развитие и поддержание изолиро-
ванных зрелых или незрелых зародышей.
Эпигенетические вариации — фенотипическое выражение
499
дифференциальной активности генов, определяющих независи-
мость клетки от экзогенных влияний. В отличие от сомаклональных
вариаций и мутаций не сохраняются в цикле "клетка -> растение
-> клетка".
Эуплоид — ядро, клетка, организм с числом хромосом, крат-
ным X.
Ювенильный (-ая, -ое) — фаза в половом цикле растения,
характеризующаяся различиями (в проявлении) от взрослого ор-
ганизма, и которая теряет способность отвечать на индуцирующие
цветение стимулы. Это фаза роста после образования проростков.
10.2 Вегетативное размножение растений методом культур
тканей. Метод культур тканей, или микроразмножение in vitro
исключительно удобен для быстрого размножения и сохранения
здоровых растений. Для этого производят отбор соответствующих
эксплантов, подвергают стерилизации с последующим переносом
на подходящую питательную среду. Качество эксплантов зависит
от вида и фазы развития растения; органа, из которого взят
эксплант; сезона года. Установлено, что меристемные ткани, сер-
дцевина луковиц, клубнелуковиц и корневищ, надежно защищен-
ные листьями и чешуйками, являются стерильными. Перед удале-
нием покрывающих структур их каждый раз протирают 70%
этанолом. Открытые органы — источники эксплантов подвергают
стерилизации ртуть- или хлорсодержащими агентами (таблица 50).
Таблица 50. Стерилизация растительных биообъектов
Объект Время стерилизации, мин.
гипохлоритами Na, Са, 5-10% диацидом 0,1% пероксидом водорода 3-12% сулемой 0,1 %
Верхушки стеблей (апексы) 3—15 1—10 2—7 0,5—7
Листья 3—6 1—3 3—5 0,5—3
Семена набухшие 10—15 6—10 6—8 2—8
Семена сухие 15—20 15—20 12—15 10—15
Ткани клубней 15—20 20—30 — 15—25
Ткани корневищ 15—20 20—30 — 15—25
Ткани стебля 20—25 ' 20—40 — 20—25
500
Как видно из таблицы, самая длительная стерилизация по
времени не превышает 40 минут, в других случаях время заметно
меньше. В усредненном варианте и применительно к различным
культурам растительных тканей, наряду с этанолом (70-^95%),
рекомендуют еще серебра нитрат (1 %), бензалконий хлорид (0,01— '
0,1%) с длительностью экспозиции 0,1—5; 5—30 и 5—20 минут
соответственно.
Экспланты (за исключением материала из надземных частей
растений) целесообразно промывать в растворах неионогенных
ПАВ перед обработкой дезинфектантом. После этого ткань тща-
тельно промывают водопроводной водой в течение 10—30 минут,
а затем погружают в раствор дезинфектанта и нерезко взбалты-
вают в течение отведенного времени. После стерилизации расти-
тельный биообъект трижды промывают свежими порциями сте-
рильной дистиллированной воды.
Стерилизующий эффект можно повысить следующими проце-
дурами: проведением стерилизации - под вакуумом для удаления
пузырьков воздуха; помещением материала в 70% этанол до того,
как обработать раствором другого дезинфектанта; использованием
таких увлажняющих агентов как твин 80 или твин 20 в виде добавок
к растворам дезинфектантов для снижения поверхностного натя-
жения и обеспечения более полного контакта дезинфектанта с
материалом.
В последние годы все чаще применяют специальные емкости
(сосуды) для изоляции в асептических условиях органов из
молодых растений. Фирма Sigma (QUIA) к 1990 г. ввела новые
мембранные наборы для культур растительных тканей. Их изго-
тавливают из микропористой полипропиленовой мембраны, обра-
ботанной специальным ПАВ для улучшения прохождения пита-
тельных веществ. Мембранные наборы могут быть использованы
при культивировании протопластов, в соматическом эмбриогенезе,
при получении культур цветов и в других направлениях.
С применением мембран существенно ускоряется рост в срав-
нении с агаризованной средой, лучше удается освобождать куль-
туры от ингибирующего влияния фенольных веществ, выделяю-
щихся в среду, а также предохранять клеточную линию от суще-
ственных повреждений при изучении метаболитов, продуцируе-
мых культурами, и др. Таким путем удается уменьшить расход
материалов и, как следствие, снизить цены, поскольку культуры
могут поддерживаться стечение длительного времени без замены
культуральных контейнеров. Ингредиенты (продукты) для культи-
501
вирования растительных тканей должны производиться в соответ-
ствии с требованиями GMP. Этим гарантируется их высокое
качество. Пригодность ингредиентов сред, обеспечивающих рост
тканевых культур, оценивают, как минимум, на трех клеточных
линиях в 1—3-х пассажах. Избранные основные среды дополняют
углеводами, витаминами, регуляторами роста (в зависимости от
используемой клеточной линии).
Применяют следующие критерии оценки роста на рекоменду-
емой питательной среде (таблица 51).
Т а б л и ц а 51. Критерии оценки роста клеточных культур
на питательных средах
Культуры тканей
Каллусная Почечная пролиферирующая Другие клеточные линии
Возрастание веса каллуса Активное образование почек или растеньиц Активно растущий • каллус
Отсутствие некротической ткани Образование сильных здоровых почек .без морфологических аберраций Увеличение плотности клеток
Внешне — активный рост — Выраженный рост почек и корней
Питательные среды стерилизуют обычно в автоклавах (таблица
52). В принципе желательна стерилизация автоклавированием
(12ГС) небольших объемов питательных сред, поскольку многие
ингредиенты разрушаются при более продолжительном нагрева-
нии и давлении. Установлено, что температура выше 12 ГС может
быть причиной нарушения качества сред, и, как следствие, плохого
роста культур.
Таблица 52. Стерилизация сред, рекомендуемых для выращивания
растительных культур тканей (t=121°C)
Объем среды, мл Минимальное время стерилизации, мин
25 20
50 25
100 28
250 31
502
Продолжение табл. 52
Объем среды, мл Минимальное время стерилизации, мин
500 35
1000 40
2000 48
4000 63
Некоторые ингредиенты в растворенном виде стерилизуют
фильтрованием (через фильтры с порами 0,22 мкм в диаметре) и
в виде стерильных растворов вносят в приготовленные среды перед
их использованием. Питательные среды по своему составу доста-
точно разнообразны, однако в какой-то мере они и однообразны.
В частности, их компоненты можно подразделить на 3 группы:
источники органического углерода (чаще — сахароза), неоргани-
ческие соли (включая источники азота) и стимуляторы роста. К
числу последних относятся некоторые витамины комплекса В и
растительные гормоны — цитокинины и ауксины. Почти универ-
сальной средой для клеток различных видов является среда MS
Т. Мурасиге и Ф. Скуга (см. главу 4). Тем не менее, при работе с
разными растительными объектами необходим специальный под-
бор ингредиентов сред для достижения поставленных целей. Пи-
тательные среды могут быть плотными за счет внесения 0,7—1%
агар-агара и жидкими. Показано, например, что для микрокультуры
ананасов предпочтительнее жидкие среды.
Микроразмножение in vitro можно осуществлять из существу-
ющих в растении тканей меристемного типа (зародыш, верхушка
основного побега) или позже образующиеся пазушные побеги,
равно как и из дополнительных меристематических тканей (точки
роста, эмбриоиды) изолированных органов растений, где они
формируются непосредственно в родительской ткани; из проме-
жуточного каллуса или клеток суспензионных культур (см. рис.
140).
Если в качестве источника углерода используют в питательных
средах сахарозу, то фотосинтез не нужен культуре, однако для
морфогенеза и биосинтеза хлорофилла применяют люминесцент-
ные лампы с интенсивностью освещения 1000—5000 люкс.
Важную роль играют регуляторы роста растений — аукси-
ны и цитокинины (растительные гормоны, или
фитогормоны). Первые усиливают или поддерживают рост каллу-
503
сов и/или корнеобразование in vitro; вторые стимулируют образо-
вание почек. Их используют в малых концентрациях, добавляя в
стерильном виде к готовым питательным средам.
Ауксинами являются: 2,4-дихлорфеноксиуксусная кислота, р-
хлорфеноксиуксусная кислота, индол-3-уксусная кислота, индол-
З-ацетил-Ь-аланин, индол-3-ацетил-аспарагиновая кислота, индол-
З-ацетил-Ь-фенилаланин, индол-3-ацетилглицин, индол-3-масляная
кислота и ее калиевая соль, а-нафталинуксусная кислота. Среди
цитокининов известны: аденин, 6-бензиламинопурин, N-бензил-З-
(2-тетрагидропиранил)-аденин, 6-у-у-диметилаллиламинопурин,
1,3-дифенилмочевина, кинетин, зеатин и некоторые другие. Сме-
шанными функциями обладают абсцизовая, коричная и гибберел-
ловая кислоты, флороглюцинол; 2,2-диметилгидразид янтарной
кислоты?
В целях предотвращения возможного бактериального и гриб-
ного загрязнения к ним добавляют такие антибиотики, как полиены
(амфотерицин В, нистатин), карбенициллин, цефалоспорин и его
производные, левомицетин, аминогликозиды (гентамицин сульфат,
канамицин моносульфат), рифампицин и др. Большинство антиби-
отиков неустойчиво при нагревании, поэтому их растворы стери-
лизуют фильтрацией через мембраны. .
Для растений характерно так называемое апикальное домини-
рование, при котором за счет регулирующего действия фитогор-
монов верхушечная почка развивается быстрее, чем пазушные
почки. Если культивировать кончик побега без фитогормонов, то
развивается одиночный проросток со строго апикальным домини-
рованием. Если же в среду внести цитокинин и засеять пазушные
побеги, то они вырастают недоразвитыми, но с появлением пучка
ростков, состоящего из второго, третьего и более порядков, кото-
рые можно разделить на составляющие и вновь выращивать на
свежей среде до получения пучков таких побегов. Этот процесс
можно вести до бесконечности, получая быстро или относительно
быстро размножающиеся побеги желаемых растений.
В большинстве случаев верхушечные (длиной 1—5 мм) и па-
зушные побеги заражены растительными вирусами, поэтому при-
бегают к использованию так называемой культуры верхушечных
меристематических тканей, когда отрезают лишь 0,3—0,5 мм
верхушки, содержащей меристему и 1—2 листовых примордия с
последующей выдержкой при 30—40°С в течение 1,5—3 меся-
цев. Этот метод удачно использован и используется для оздоров-
ления хозяйственно ценных растений.
504
С помощью метода культур тканей удалось значительно повы-
сить коэффициент размножения многих садовых древесных (яб-
лони) и травянистых декоративных растений (ирис, нарцисс, пе-
туния, фиалка и др.), пользуясь придаточными побегами, а также
каллусами, индуцированными фитогормонами и дающими побеги
или эмбриоиды. Доступными для производства стали клетки бар-
бариса — продуцента спазмолитика ятроризина, табака — проду-
цента убихинона-10.
Некоторые каллусы стабильно продуцируют растения — реге-
неранты в течение 10 лет (табак, хризантемы и др.), оставаясь при
этом смесью нерегулярно дифференцированных клеток, среди
которых постоянно содержатся диплоидные меристематические
клетки, дающие стеблевые апексы или зародыши.
Перед высадкой проростков в грунт прибегают к стимуляции
корнеобразования с помощью индолилмасляной кислоты (0,5—г 10
мг/л).
Фитобиотехнологи, занимающиеся микроклональным размно-
жением растений, должны быть хорошо подготовленными в обла-
сти биологии растений, с которыми приходится иметь дело, а также
в области микробиологической техники работы. Лишь благодаря
заинтересованности и профессиональному подходу к выполнению
задания обеспечивается высокое качество конечного продукта
(материала). Тем не менее необходимо помнить о трудностях,
возникающих при микрокультивировании. К ним относятся: ин-
фицирование растительных тканей вирусами и фитопатогеннымй
бактериями (например, Erwinia carotowora), бактериальные и гриб-
ковые контаминации, ингибирование тканей фенольными и дру-
гими, например, летучими, соединениями, возрастание числа му-
таций, и пр. И все же микрокультивирование исключительно
заманчиво не только на пути создания большого количества без-
вирусного материала, но и массовости получения образцов в
относительно короткие сроки (несколько тысяч в течение месяцев).
Микрокультивирование in vitro обеспечивает быстрое вегета-
тивное размножение не только травянистых, но и древесных
растений, также прибегая при этом к культуре побегов (например,
для декоративных, лесных, плодовых и других деревьев) или
каллусным культурам (гвинейская масличная пальма, цитрусовые
и др.). Здесь важное значение имеет состояние ювенильности (от
англ, juvenile — юношеский, юный), которое обычно растягивается
на несколько лет и характеризуется сравнительной легкостью
вегетативного размножения. Для древесных растений известно
505
также явление реювенильности, когда некоторые ткани взрослого
растения проявляют свойства проростков, например, формируют
придаточные корни. Реювенилизированные ткани все больше ис-
пользуются на практике для размножения отдельных покрытосе-
менных (например, эвкалиптов) и голосеменных деревьев (разные
виды сосны). Ускорение реювенилизации может происходить под
влиянием отдельных фенольных соединений (флороглюцин, квер-
цетин, рутин), памятуя о том, что изменение уровня эндогенных
фенольных соединений является составной частью регуляторных
процессов, индуцируемых цитокининами.
Каллусные культуры древесных растений также перспективны
как и каллусные культуры травянистых растений. Показателен
остаток G-p- L-рамнозидо-D-глюкозы
рутин
пример с гвинейской масличной пальмой (Elaeis guineensis) —
источником получения масла для населения Юго-Восточной Азии,
Америки, Африки. К тому же эта пальма не размножается вегета-
тивно. Генетико-селекционная работа по выведению более уро-
жайных сортов на основании полового размножения занимает
15—20 лет. Вот почему получение каллусов, а затем — эмбриоидов
и регенерантов масличной пальмы оказалось экономически выгод-
506
ным делом. Обычно за 3 месяца удается выращивать из эмбриоидов
побеги длиной 12 см (рис. 142).
Рис. 142. Процесс клонирования гвинейской масличной пальмы: 1 — верху-
шечные молодые листья (посевной материал), 2—5 — сменяемые питательные
среды, в которых развиваются каллусы, эволюционирующие в эмбриоиды (4) и
затем — в молодые растеньица (5), 6 — образование корней при очередном переносе
растения на свежую питательную среду, из которой их пересаживают в грунт.
Учитывая тот факт, что в 1 см3 каллусной культуры содержится
до 1 млн. клеток, каждая из которых способна трансформироваться
в новое растение, становятся понятными все преимущества метода
стерильного вегетативного размножения растений в лабораторных
условиях -на средах с фитогормонами при последующей пересадке
их в почву. Отбирая нужные клетки, можно вывести новые сорта
растений, удовлетворяющие запросы по одному или ряду показа-
телей (устойчивость к микробным заболеваниям, урожайность,
особая окраска цветов, повышенная продуктивность вторичных
метаболитов и т. д.).
Для развития и формирования каллусных культур значительна
роль не только фитогормонов, но и так называемых элисито-
ров (от англ, elect — выбирать) — БАВ, дерепрессирующих гены,
и, как следствие, активирующих клеточные деления, сравнительно
быструю дедифференцировку специализированных клеток, сопро-
вождающуюся активацией белкового синтеза. Полагают, что эли-
ситорами являются углеводные молекулы, происходящие из гид-
ролизованнйх гликанов клеточных стенок. Не исключена возмож-
ность экзогенного происхождения элиситоров, например, от мик-
роорганизмов.
10.3. Культивирование клеток растений в глубинных услови-
ях. В случае выращивания клеток растений в глубинных (погру-
женных) условиях необходимо иметь такие их линии, которые
отвечали бы следующим требованиям: склонность
к размножению в дезагрегированном со-
стоянии, морфологическая "выравнен-
ное т ь", удовлетворительная скорость раз-
множения, сохранность путей метаболиз-
507
м а. Такие клеточные линии уподобляются одноклеточным мик-
роорганизмам и могут называться "суспензионными культурами"
в случае их выращивания в жидких питательных средах. Состав и
реологические характеристики сред являются основными факто-
рами, определяющими нахождение одиночных или аггломериру-
ющихся (агрегирующихся) клеток во взвешенном состоянии. Если
число клеток в аггломератах превышает 50, то они могут выпасть
(или выпадают) в осадок. Желательно, чтобы в процессе размно-
жения клетки дольше оставались в разобщенном состоянии, тогда
поддержание суспензионной культуры в течение необходимого
времени оказывается вполне достижимым.
Что касается морфологической "выравненности" клеток, то при
этом имеют в виду их округлую (или сферическую) форму, уплот-
ненную цитоплазму, сравнительно небольшие размеры (в среднем,
от 10 до 50 мкм) и отсутствие трахеидоподобных элементов.
Одиночные растительные клетки в таких случаях напоминают
одноклеточные эукариотические микроорганизмы; их ростовые
характеристики (фазы размножения) в-некотором приближении
также оказываются совпадающими (lag-фаза, log-фаза, const-фаза,
let-фаза), хотя временные интервалы между фазами более растя-
нуты для клеток растений.
Скорость размножения растительных клеток невелика—время
удвоенна их числа равно 1—3 суткам и поэтому по данному
показателю они существенно уступают, например, бактериальным
и дрожжевым клеткам, время удвоения которых в глубинных
условиях укладывается для большинства видов в 20—30 минут.
Однако, переводя культуру растительных клеток на хемостатноё
(непрерывное) выращивание, можно добиться их высокой продук-
тивности по биомассе или по вторичным метаболитам (пример с
Nicotiana tabacum).
Исходные клеточные суспензии обычно получают из рыхлых,
оводненных каллусных тканей (2—3 г на 60—100 мл питательной
среды), подвергнутых обработке пектиназой и полигалактурона-
зой, и помещаемых в жид кую питательную среду, лишенную ионов
кальция, но содержащую ауксин — 2,4-дихлорфеноксиуксусную
кислоту. Условия выращивания поддерживают либо в периодиче-
ском, либо в непрерывном режиме, специально подобранном для
конкретного биообъекта. В других случаях источниками суспензи-
онных культур могут быть протопласты с реконструированными
клеточными стенками. Стимулировать деления отдельных клеток
изолированных протопластов можно с помощью обогащенных
508
питательных сред или использованием гомологичной ткани —
"няньки" ("кормящий слой”), находящейся в состоянии активного
роста (рис. 143). Колонии из отдельных клеток (клоны) могут быть
субкультивированы один или более раз (при необходимости), а
затем переведены в суспензионные культуры.
Рис. 143. Использование тка-
ни-"няньки" (суспензионная куль-
тура) при выращивании колоний
из одиночных клеток кукурузы или
протопластов (схема): 1 —качалоч-
ная колба, 2 — колония, возникшая
из отдельной клетки, 3 — фильтро-
вальная бумага, 4 — металлическая
сетка, 5 — суспензия клеток "кор-
мящего слоя", 6 — подложка (пе-
нополиуретан), 7 — питательная
среда.
Важно также сохранить
присущие клеткам метаболические пути при их выращивании в
суспензионных культурах. Более того, регулируя обмен, можно
добиваться заметного повышения выхода целевых продуктов. При
этом всегда необходимо учитывать тип дифференцировки, или
состояния специализации исходных клеток, так как от него зависит
видоспецифичность первичного и вторичного метаболизма.
Суспензионные культуры обычно выращивают в подходящих
емкостях роллерного типа после фильтрации первичной суспензии
через стерильные металлические, нейлоновые или марлевые сита
для освобождения от крупных аггломератов клеток.
К сожалению, выращивание растительных клеток в "погружен-
ных" условиях пока удается в редких случаях, что обусловлено
недостаточной глубиной познания всех особенностей обмена ве-
ществ у различных видов и их клеточных культур; определенные
помехи здесь связаны и с медленным ростом клеток в строго
асептических условиях, их чувствительностью к механическим
повреждениям и другими причинами.
В начале 80-х годов в компании Mitsui-Petrocherpical Industries
осуществлен первый крупномасштабный процесс по выращива-
нию воробейника (Lithospermum erythrorhizon) в погруженных
условиях в целях получения вторичного метаболита — шиконина,
509
являющегося ценным фармацевтическим препаратом и красите-
лем (рис. 144). За один периодический процесс удается получать
порядка 5 кг конечного продукта, накапливающегося в клетках. На
первой стадии клетки воробейника выращивают на среде (с пода-
чей стерильного воздуха) в 200-литровом биореакторе в течение 9
суток; затем культуру переносят в биореактор меньшего размера
со средой (М-9), стимулирующей продукцию шиконина, и, нако-
нец, в третьем реакторе на 750 л ферментацию ведут в течение 2
недель. Клетки на первой стадии белые, на последней — красные
(накоплен шиконин). Стоимость красителя в 1983 г. составляла 4000
долларов за килограмм.
2 2
Рис. 144. Схема культи-
вирования воробейника в це-
лях получения шиконина (1
— биореактор, 2 — подача
стерильного воздуха, 3 — ме-
шалка, 4 — питательная сре-
да, 5 — выпуск отработанно-
го воздуха, 6 — перенос сре-
ды в малый реактор, 7 —
фильтрат, 8 — красные клет-
ки, содержащие шиконин).
10.4. Использование методов генетической инженерии, или
рДНК в фитобиотехнологии. Растения — как многоклеточные
организмы с огромной емкостью геномов, с половым путем раз-
множения и многоступенчатыми программами развития — явля-
ются более сложными объектами для генноинженерных экспери-
ментов, чем, например, вирусы, бактерии и дрожжи. Тем не менее,
уже теперь достигнуты определенные успехи с растительными
объектами и по прогнозам ученых США к 2000 году ожидается
прирост урожайности сельскохозяйственных культур примерно на
60% по сравнению с началом 80-х годов текущего столетия в
основном на основе использования методов генетической инже-
нерии, когда стал возможным перенос отдельных генов от одного
растения другому (в противоположность естественному половому
процессу, при котором происходит замена целых блоков сцеплен-
ных генов).
10.4.1. Генная инженерия. Плазмиды, органеллы, транспозоны
и вирусы как векторные системы. В генноинженерном экспери-
менте необходимо изолировать конкретный ген, включить его в
наследственный аппарат растительной клетки и регенерировать
фертильное растение (способное к размножению) с измененным
510
наследственным признаком. В народнохозяйственном аспекте важ-
ными признаками являются: высокая урожайность (или продук-
тивность), скороспелость, устойчивость к различного рода заболе-
ваниям, засухе, к гербицидам и пестицидам, неполегаемость и др.
К сожалению каждый из названных признаков контролируется,
как правило, группой ‘ генов, что заметно усложняет проблему
клонирования соответствующих генов. Однако, Црирода с давних
пор "доказывает" нам, что генетическая инженерия также подвла-
стна и ей. Примером тому служит инфекционная злокачественная
болезнь (корончатые галлы) многих двудольных растений, вызы-
ваемая почвенной агробактерией — Agrobacterium tumefaciens.Ta-
кие однодольные растения, как пшеница, кукуруза, овес, рис, рожь,
сахарный тростник, естественно устойчивы к заражению агробак-
териями.
В 1971 г. Р. Гамильтон и М. Фолл (США) пришли к выводу о
том, что опухоль у растений индуцируется нуклеиновой кислотой.
В 1974 г. Н. Ван Ларебеке, П. Энглер и др. (Бельгия) установили,
что способность Atumefaciens вызывать образование корончатого
галла с большими по размерам плазмидами (в длину 50—80 мкм в
разомкнутом состоянии и с ММ 112—156 МДа — мегадальтон),
названными Ti- плазмидами (от англ, tumor inducing —
вызывающий опухоль). Обычно они в клетке содержатся в 1—2
копиях и составляют примерно 4% ДНК от бактериального генома.
Однажды наведенный Ti-плазмидой опухолевый рост у расте-
ния не нуждается в ее дальнейшем присутствии для поддержания
злокачественной пролиферации. Следовательно, выращивание и
поддержание опухолевых клеток возможно в стерильной культуре
(без агробактерий и без Ti-плазмиды), и, как установлено, без
ауксинов и цитокининов, то есть без фитогормонов. При этом они
образуют опины (см.), отсутствующие у растений в нормальных
условиях, и по которым можно определять опухолевую клетку. В
зависимости от клеток корончатого галла могут образовываться
атропин (см.), нопалин (см.) или октопин (см.). Поэтому и говорят
соответственно: клетки атропинового, нопалинового или октопи-
нового типа.
Фактическая ненужность Ti-плазмиды после индукции опухоли
у растения связана с тем, что стерильные опухолевые клетки в
своих хромосомах содержат ковалентно связанную часть этой
плазмиды (около 10%), получившей название Т-ДНК (от англ,
transferred — перенесенный), а применительно к клеткам
511
Atumefaciens с Ti-плазмидой — Т-участок бактерии. Т-ДНК содер-
жится исключительно в ядре клеток корончатого галла.
Таким образом, на примере Ti-плазмиды виден перенос генов
от прокариот к эукариотическим растениям в природных условиях
(рис. 145). Транскрипты интегрированной Т-ДНК в виде мРНК
поступают из ядра в цитоплазму клетки, где включаются в реакции
синтеза специфических белков на полирибосомах: ядерная ДНК-
Т-ДНК -» транскрипты —> мРНК -э полирибосома (трансляция) —>
белки —> синтез опинов (гормон-независимый рост)
Agrobacterium
tumefaciens
Растительная клетка
с встроенной Т-ДНК
в ядерную ДНК
Растение с корон-
чатым галлом
Б4 . a z. ~ ... талт-днк
___________ _____________________________транскриты
Рис. 145. А. Схема пере-
носа Т-ДНК из агробактерии
в растение с последующим
формированием корончатого
галла (1 — бактериальная
хромосома, 2 — Т-ДНК, 3 —
Ti-плазмида, 4 — ядро расти-
тельной клетки, 5 -—встроен-
ная Т-ДНК, 6 — корончатый
галл); Б. Т-ДНК в Ti-плазмиде
октопинового типа (продук-
ты генов 1 и 2 ингибируют
побеги, гена 3 — ингибируют
корни).
Каждый транскрипт определяется специфическим промотор-
ным участком на Ti-плазмиде, узнаваемым полимеразой II растения
— хозяина. Из рисунка 145 Б следует, что функция генов 1—3
проявляется в неорганизованном росте клеток, поскольку тормо-
зится формирование побегов и корней.
Фитопатогенные агробактерии способны вызывать 3 вида ра-
ковых опухолей у растений: корончатый галл, бородатый корень
и стеблевой галл. Возбудителем каждого заболевания являются
A.tumefaciens, Arhizogenes и A.rubi соответственно. Проникая в
поврежденное растение, патогенный микроорганизм с помощью
Ti-плазмиды создает свою экологическую нишу и реализует "гене-
тическую колонизацию” объекта — происходит гормон-независи-
мый рост опухолевых клеток.
Ti-плазмиды оказались хорошими векторными системами для
осуществления генноинженерных экспериментов с растениями,
поскольку Т-ДНК этих плазмид обладает достаточно высокой
эффективной емкостью (порядка 50 000 пн чужеродной ДНК могут
быть перенесены и встроены с ее помощью в хромосомную ДНК
512
растений), широким спектром хозяев, инфекционностыо и ста-
бильностью.
Перенос гипотетического гена с помощью Ti-плазмиды пред-
ставлен на рис. 146.
1 2 3 4 5
Рис. 146. Перенос гена (а) из хромосомы (б) Escherichia coli в растение (5) с
помощью Т-ДНК (в) Ti-плазмиды (г) A.tumefaciens (2,3) в ядерную ДНК (д) расти-
тельной клетки (4).
Разработаны векторные системы на основе растительных хло-
ропластов и митохондрий, ДНК которых имеют гомологичные
области с ядерной ДНК, что облегчает обмен участками между
ними при выполнении генноинженерных экспериментов.
Заманчивы в качестве векторных систем й транспозоны (см).
Их множество сулит многообещающие результаты в опытах с
однодольными растениями, устойчивыми к заражению агробакте-
риями и, следовательно, к восприятию Т-ДНК Ti-плазмиды.
В качестве векторов могут также использоваться вирусы рас-
тений. Их нуклеиновые кислоты реплицируются и проявляют свои
функциональные свойства (экспрессируют) в клетках растений-
хозяев, где потенциал вирусов и воспринимающих клеток объеди-
няется и реализуется в приумножении организованных частиц
патогена (например, для вируса мозаики табака в среднем 107
частиц на клетку, для вируса мозаики цветной капусты — порядка
104 частиц на клетку).
Однако вирусы как векторы обладают четырьмя существенны-
ми недостатками: они патогенны, их емкость небольшая, вирусы
не способны встраиваться в хромосомы растительных клеток, они,
в основном, видоспецифичны, то есть поражают определенный
круг растений — хозяев.
В качестве векторов могут быть использованы также и вироиды
(см.), представляющие собой однонитевые цепи ковалентно свя-
занных кольцевых РНК, включающих 270—300 нуклеотидов, ли-
шенных оболочки. Вироиды поражают виноград, картофель, коко-
совую пальму, томаты и другие растения как по вертикали (через
17 т. 8524 513
зародышевые клетки), так и по горизонтали (распространяются по
растительному организму через клеточный сок или переносятся
механически).
Чтобы использовать вирусную (вироидную) РНК в качестве
вектора, необходимо выполнить следующие последовательные опе-
рации (рис. 147).
РНК вируса (внроида)
кДНК «диоииттлая
Рис. 147. Последователь-
ность событий при использо-
вании вирусной (вироодной)
РНК в качестве вектора: 1 —
обратная транскриптаза, 2 —-
ДНК-полимераза, 3 — встра-
ивание в плазмиду (клониро-
вание), 4 — включение чуже-
родного гена, 5 — инфициро-
вание растений, 6 — транс-
крибирование с образовани-
ем инфекционной РНК и по-
следующее заражение расте-
ния-хозяина.
Клон кДНК с встроенным геном может обладать инфекцион-
ностью и, как следствие, им заражают хозяйское растение. Если
же кДНК с встроенным геном не обладает инфекционностью, то
вирусный вектор может быть транскрибирован с образованием
РНК, которая затем используется для заражения растения.
До сих пор признают, что векторная система на основе цитоп-
лазматического вируса мозаики цветной капусты (содержит двух-
цепочечную ДНК) оказывается наиболее приемлемой в генноин-
женерных экспериментах с растениями, хотя многое здесь остается
неизученным (например, молекулярные механизмы проявления
симптомов заболевания растения-хозяина от поражения вирусом,
неопределенность диапазона растений-хозяев’ и возможность его
расширения, неспособность векторной системы встраиваться в
геном хозяина, и др.).
Если удается объединить возможности ДНК из вируса мозаики
цветной капусты с Т-ДНК Ti-плазмиды, то такую векторную сис-
тему можно будет использовать на более широком круге растений-
хозяев.
514
10.4.2. Использование методов геномной и хромосомной ин-
женерии в фитобиотехнологии. Объединение геномов клеток
разных особей можно осуществлять при половой и соматической
гибридизации. Первая из них способна реализоваться в природных
и искусственных условиях, тогда как вторая — только в искусст-
венных. Половая гибридизация давно известна человеку, и он с
тех пор использовал ее для выявления новых сортов растений и
для повышения их продуктивности.
Соматическая гибридизация базируется в основном на приме-
нении растительных протопластов, впервые полученных Дж. Клер-
кером в 1892 г. из листьев Stratiotes aloides (водное растение,
именуемое телорезом).
Соматические клетки растений, как и интактные клетки бак-
терий или грибов (по лат. intactus — нетронутый), в обычных
условиях не сливаются друг с другом. Все они предварительно
должны быть лишены клеточной стенки и трансформированы в
протопласты.
Протопласты (от греч. protos—первый, plastos—вылепленный,
образованный) — это клетки, лишенные клеточных стенок, спо-
собные сохраняться и метаболизировать, равно как и реконстру-
ировать клеточную стенку в подходящих условиях. Протопласты
можно получать с помощью механических и биохимических (эн-
зиматических) методов. В первых случаях добиваются плазмолиза
растительных тканей, например, с помощью 0,1% раствора саха-
розы с последующим разрезанием эпидермиса и освобождением
протопластов в окружающую питательную среду. Энзиматические
методы по-разному эффективны в получении протопластов. Здесь
необходимо подбирать ферменты для каждого вида растений,
однако наиболее часто используют целлюлазы, пектиназы, геми-
целлюлазы как индивидуально, так и в комбинациях. Например,
для получения протопластов из листьев Nicotiana tabacum листовую
ткань без эпидермиса погружают в смесь ферментов, включающую
0,5% пектиназы и 2% целлюлазы в 0,7 М растворе маннита или
сорбита; для получения протопластов из листьев клевера и люцер-
ны используют 2—5% смесь гликаназ с пептидазами, и т. д.
В сравнительном плане более подходящим является энзимати-
ческий метод получения протопластов как более щадящий. Однако
при любом способе важен подбор осмотического стабилизатора,
515
без которого может наступить быстрый плазмоптиз* протопластов
(по лат. plasma — плазма, option — альтернатива). В качестве таких
стабилизаторов могут быть применены растворы неорганических
солей (СаС12, КС1, Na2HPO4), органических гликолей (маннита,
сорбита), моно- и дисахаридов (глюкозы, ксилозы, сахарозы) в
ориентировочных концентрациях 0,3—0,8 М/л. Однако примени-
тельно к виду растения, из которого получают протопласты, необ-
ходимо подбирать индивидуальные условия д ля "протопластирова-
ния" (осмотический стабилизатор, pH среды, ГС). Эти требования
сохраняются и в случаях получения протопластов из каллусных
культур и клеточных суспензий.
В жизнеспособности выделенных протопластов большую роль
играет физиологическая полноценность исходного растительного
материала. Суспензионные культуры наиболее приемлемы для
этих целей, будучи в конце логарифмической фазы роста (размно-
жения).
Полученные тем или иным путем протопласты либо используют
для регенерации растения (см. тотипотентность), либо их можно
подвергнуть слиянию с образованием гетерокариотических гибри-
дов.
Из растений, относительно легко регенерирующих из протоп-
ластов, можно назвать картофель, люцерну, маниок, рапс, табак.
Для получения растений — регенерантов, высаженных в грунт,
требуется, как минимум, 16—18 недель.
Растения, полученные из протопластов, могут отличаться до-
статочно выраженной йзменчивостью по ряду интересующих нас
признаков: урожайности, величине (размеру), морфологии отдель-
ных частей, фотопериодичности и др. Как правило, это связывают
с изменением числа и перестройками хромосом. Поэтому протоп-
ласты растений представляют собой интересные объекты и для
изучения процессов мутагенеза.
Гибридизация протопластов удается не только в тех случаях,
когда исходные растения (источники клеток для протопластирова-
ния) способны гибридизоваться половым путем, но и тогда, когда
заведомо известно, что клетки, подлежащие слиянию, относятся к
организмам с половой несовместимостью. Тем не менее, во всех
случаях соматической гибридизации один из партнеров должен
нести какой-либо маркерный признак (биохимический или дру-
* Явление, обратное плазмолизу.
516
гой), по которому возможно подтвердить достоверность получения
гибрида. В этом смысле удобны ауксотрофные мутанты.
Гетерокариотические соматические гибриды выделяют либо
вручную с помощью микроманипулятора, капиллярной пипетки и
шприца, либо автоматически на основе применения флуоресцен-
тных систем.
Для биотехнологии также важна цибридизация, когда возника-
ют гетерозиготы по внеядерным (цитоплазматическим) генам,
определяющим, например, устойчивость к гербицидам или признак
цитоплазматической мужской стерильности (рис. 148).
Рис. 148. Слияние
растительных протопла-
стов и возможная сегре-
гация хлоропластов: 1 и
2 — протопласты разной
видовой принадлежно-
сти, 3 — гетерокарион, 4
— гибриды, Г и 2' —
цибриды.
К настоящему времени из ряда растительных протопластов
получены их безъядерные фрагменты — микропласты, окружен-
ные тонопластной мембраной и включающие часть внеядерного
генетического материала. Используя приемы соматической гибри-
дизации, микропласты можно сливать с реципиентными протоп-
ластами. Таким образом, микропласты являются резервным мате-
риалом для расширения возможностей клеточной инженерии в
фитобиотехнологии. Более того, теперь удается гибридизация рас-
тительных протопластов с протопластами микроорганизмов и жи-
вотными клетками.
Применительно к хромосомной инженерии
удалось доказать проникновение изолированных метафазных хро-
мосом в обработанные полиэтиленгликолем реципиентные протоп-
ласты таких однодольных растений, как пшеница и кукуруза.
517
Открываются пути привнесения дополнительной генетической
информации по желанию экспериментатора в целях создания
трансгенных растений. Прямой перенос чужеродной ДНК в про-
топласты возможен также с помощью микроинъекции, трансфор-
мации, упаковки в липосомы с последующим их эндоцитозом
растительными протопластами, и электропорации (высоковольт-
ных электрических импульсов).
10.4.3. Генная инженерия и биологическая фиксация азота.
Азотистые удобрения, вносимые в почву (наряду с другими пита-
тельными веществами) стимулируют рост и развитие растений.
Обогащению почвы азотом помогают особые микроорганизмы,
фиксирующие молекулярный азот из воздуха. Такие микробы
подразделяют на две группы — свободноживущие в почве (напри-
мер, Azotobacter chroococcum) и микробы - симбионты с корневой
системой растений (например, Rhizobium meliloti). Те и другие
осуществляют биологическую азотофиксацию. Число азотофикса-
торов невелико, но от них в буквальном смысле слова зависит
жизнь на нашей планете. С помощью микробов - азотофиксаторов
ежегодно фиксируется около 17,5 • 107 т N2 из воздуха. Основные
компоненты системы азотофиксации являются сильными восста-
новителями: фермёнтный комплекс — нитрогеназа, ферредоксин
или флаводоксин и АТФ. Микробы — азотофиксаторы трансфор-
мируют молекулярный азот в аммиак по следующей схеме:
N2
+2Н
+2Н
NH ------► NH2—NH2 +2H
II . 2 NH3
NH t2-*j2.9^2NH2OH
Ассоциация клубеньковых бактерий из рода Rhizobium с бобо-
выми растениями является наиболее эффективной по фиксации
N2 из воздуха. Ответственными за это являются nif-гены. Соответ-
ствующие биопрепараты бактериальных удобрений (см. главу 9)
выпускают в различных странах мира.
К настоящему времени практически решена проблема увели-
чения nif-генов в микробах — азотофиксаторах, являющихся сим-
бионтами донника (Rh. meliloti). За счет этого заметно усиливается
азотофиксация и, как следствие, повышается урожайность расте-
ния — хозяина (рис. 149).
518
Рис. 149. Схема получения клеток
Rhizobium meliloti, несущих дополнитель-
ные гены азотфиксации: 1 — плазмидная
ДНК, 2 — рестриктаза, 3 — клетка
Rhaneliloti, 4 — хромосома клетки, 5 —
nif-гены, 6 — лигаза, 7 — рестриктирован-
ная плазмидная ДНК, 8 — плазмидная
ДНК с nif-геном, 9 — трансформация, 10
— клетка Rh.meliloti с nif-генами.
Имеются также предпосылки к созданию методами генной
инженерии злаковых растений — азотофиксаторов. Это сулит
большие выгоды агрохозяйствам, занимающимся выращиванием,
например, зерновых культур.
Прибегают и к простым искусственным ассоциациям азото-
фиксирующих бактерий с растениями, дающими ощутимые ре-
зультаты по урожайности (например, цианобактерии Anaboena
variabilis и табака).
10.4.4. Другие прикладные аспекты фитобиотехнологии. В
начале 80-х годов текущего столетия удалось создать подсолнечник,
несущий гены бобов фасоли, кодирующих белок фазеолин. Такое
химерное растение получило название "Санбин" (от англ, sun —
солнце, bean — боб), оно способно синтезировать во многом
полноценные белки. Каллусная культура санбина также обладает
этой способностью.
Кроме генов фазеолина локализованы и выделены гены таких
запасаемых белков, как зеин — белок кукурузы и легумин — белок
гороха. Эти гены удается переносить в отдельные негомологичные
растения.
Создан трансгенный рапс, содержащий гены отдельных ней-
ропетидов человека, например, лей-энкефалина, связанного с геном
альбумина семян рапса. С одного гектара земли, засеянного таким
рапсом, можно получать до 3 кг нейропептида.
Важное значение имеют работы по созданию гербицидоустой-
чивых растений, не боящихся сорняков. Удалось перенести из
представителей актиномицетов ген устойчивости к фосфинотри-
ацину в клетки сахарной свеклы; получены также устойчивые к
гербицидам определенные сорта табака.
Токсические белковые кристаллы, образующиеся в клетках Вас.
thuringiensis на основе матричного синтеза, убивают личинок
листогрызущих насекомых. Поэтому генноинженерная разработка
по переносу гена бактериального токсина в геном табака увен-
чалась успехом в 1987 г. Экспрессия такого гена сопровождалась
519
биосинтезом токсина, и личинки насекомых, поедающие листья,
погибали в сравнительно короткий промежуток времени. По тако-
му же принципу был создан инсектицидный трансгенный хлоп-
чатник, содержащий ген ингибитора трипсина гороха коровьего;
этот ген обусловливает синтез белка, блокирующего протеолити-
ческую активность в пищеварительной системе у насекомых.
Создан новый сорт помидоров, длительно сохраняющийся без
размягчения вследствие подавления активности фермента полига-
лактуронидазы. Методом генной инженерии получен ген, опреде-
ляющий биосинтез анти-мРНК, которая ингибирует природную
мРНК.
Благодаря удачному переносу генов фермента хальконсинте-
тазы (в антисмысловой ориентации) в петунию удалось получить
растение с белыми цветами.
Сконструированы такие генноинженерные сорта сои, которые
проявляют устойчивость к насекомым, гербицидам, вирусам и
образуют больше запасных белков, обогащенных метионином.
На основе методов геномной инженерии получены межвидо-
вые гибриды капусты, картофеля и табака с турнепсом, картофеля
с помидором ("Помат"), помидора дикого вида, устойчивого к
некоторым вирусам, и культурного сорта.
Успешными оказались работы по созданию морозоустойчивых
растений на основе их обработки культурой клеток Pseudomonas
syringae, из которой элиминирован ген, ответственный за синтез
специального белка, ускоряющего кристаллизацию воды с образо-
ванием льда. Обработка клубней картофеля таким "лед-минус"
микробом приводит к возрастанию их морозоустойчивости.
10.5. Получение биогаза и удобрений на основе использования
растений. Традиционные методы гибридизации растений помогли
достигнуть резкого повышения их урожайности (особенно —
зерновых культур); наступила так называемая "зеленая револю-
ция", с которой, например, в странах Юго-Восточной Азии связы-
вают полное самообеспечение продуктами питания.
Возрастание урожайности культур растений сопряжено с уве-
личением их отходов. Одна часть таких отходов может быть
использована в качестве корма для сельскохозяйственных живо-
тных, другая — для биотрансформации в какие-либо полезные
продукты (биогаз, удобрения и пр.). Рациональное использование
растительных отходов помогает решать отдельные экологические
и энергетические проблемы.
520
10.5.1. Получение метана из растительных отходов. Метан
(СН4) входит в состав природных газов (на него приходится 97—98%
в природных газовых месторождениях). Так называемые "попут-
ные нефтяные газы" из буровых скважин содержат 39—85% мета-
на, -остальные газы представлены, в основном, этаном, пропаном,
бутанами, пентанами.
В воздухе каменноугольных шахт всегда присутствует метан в
примерных количествах 3,5—7,5%. Смесь метана с воздухом сильно
взрывается от пламени.
Метан образуется также в природе при бактериальном разло-
жении клетчатки или в рубце жвачных животных (метановое
брожение), при окислении молекулярного водорода до СН4 (анаэ-
робное дыхание).
Метановое брожение происходит в анаэробных условиях под
влиянием смешанной микрофлоры, включая метанобразующие
бактерии, или бактерии — метаногены (археобактерии, см. главу
2)-.
В настоящее время миллионы биогазовых установок в Индии,
Китайской Народной Республике, в странах ЕЭС рассчитаны на
проведение метанового брожения в одном биореакторе (метантен-
ке) емкостью от одного до нескольких тысяч кубометров.
10.5,2. Получение кормового белка на основе биоконверсии
целлюлозосодержащих растительных материалов. Давно изве-
стен биотехнологический процесс получения дрожжей на гидро-
лизатах древесины хвойных растений. В такой древесине содер-
жатся гексозаны, тогда как в древесине лиственных растений —
преимущественно пентозаны, почти совсем не сбраживаемые по-
сле гидролиза дрожжевыми организмами.
В качестве гидролизующих агентов используют кислоты или,
реже, ферментные комплексы. В любом случае матричный лигнин
остается негидролизованным и от него стремятся избавиться пред-
варительной делигнификацией целлюлозных материалов различ-
ными способами, в том числе — паракрекингом, или обработкой
водяным паром под давлением 7—8 ати. Можно использовать
биологический способ делигнификации, когда на целлюлозосодер-
жащем материале выращивают дереворазрушающие грибы из
класса базидиомицетов. Эти последние делают целлюлозный ма-
териал доступным в качестве корма для жвачных животных (со-
держание лигнина в нем не должно превышать 18—20%).
Делигнифицированная целлюлоза становится более доступной,
например, для целлюлазного комплекса гриба Trichoderma viride,
521
обогащающей белком этот продукт при твердофазной фермента-
ции.
Получение кормовых дрожжей на гидролизатах древесины и
сульфитных щелоках (отходах целлюлознобумажной промышлен-
ности), приведено в главе 9. Здесь рассмотрены биотехнологиче-
ские процессы использования растительной массы.
Технология получения ферментированного сока и белкового
коагулята из зеленой массы растений.
Названная технология может быть реализована по так называ-
емой схеме ТПФ-1, предложенной М. Е. Бекером (Латвия). Для
этого используют свежескошенную зеленую массу клевера и
люцерны, которую измельчают лацератором, прессуют на прессе
(например,Tl-ВПО-ЗО), жом передают на приготовление травяной
муки с добавлением антиоксидантов, либо — на силосование.
Натуральный травяной сок подают в биореактор для непрерывной
анаэробной ферментации, где он обогащается белком и за счет
органических кислот консервируется естественным путем.
Сухие вещества в ферментированном соке составляют 5—10%,
сырой протеин — 1—3%, общее количество органических кислот
— 1—2%, pH 3,9—5,4.
Кислотообразующие бактерии дополняют сок метионином.
Вместе с тем в соке снижается количество сапонинов и ингибитора
трипсина.
Кроме ферментированного сока по этой биотехнологии пол-
учают белковый коагулят, содержащий 10—20% сухого остатка (в
сухом остатке содержится 25% сырого протеина).
Из нижнего штуцера биореактора коагулят отбирают периодиг
чески (1 раз в 1—2 суток) в количестве 20—30% от поданного
натурального растительного сока. На сброженный сок приходится
70—80% объема от натурального сока. Его подают по трубопрово-
дам либо в кормушки животным (прежде всего имеющим однока-
мерный желудок), либо в другой биореактор, где на нем выращи-
вают специальные расы кормовых дрожжей из родов Hansenula и
Trichosporon, с помощью которых конечный продукт вдвое обога-
щается истинным белком.
Способ ТПФ-1 пригоден для мелких и крупных агрохозяйств,
располагающих электроэнергией; он нетрудоемок и экономически
выгоден, а получаемый сброженный сок представляет собой вы-
сококачественный кормовой продукт (таблица 53).
522
Т а б л и ц a 53, Состав кислого коричневого сока люцерны до и после
выращивания на нем Trlchosporon cutaneum A52/S1, %
(по У. Э. Виестуру и др., 1987)
Компонент До (pH 5,4) После (pH 8,8)
Сухие вещества 4,57 3,99
в том числе: растворимые 4,22 2,78
нерастворимые 0,35 1,21
Сырой протеин (% от сухих веществ) 3.5 45,2
Истинный белок (% от сухих веществ) 0,47 11,82
Кислоты: молочная 0.84 0,39
I уксусная 0,37 0
Силосование кормов давно используют на практике и силос
занимает важное место в рационе сельскохозяйственных живо-
тных. На первых порах использовали примитивные силосные ямы,
куда вносили свежескошенную траву (нередко грубую, плохо
поедаемую в естественном виде жвачными животными). Ямы
герметизировали и за счет природной эпифитной микрофлоры
происходила твердофазная ферментации травы. Нередко такие
ферментации были неудачными из-за нерегулируемости желатель-
ной микрофлоры (молочнокислых бактерий) и преобладания гни-
лостных процессов.
В последующем были сконструированы и внедрены в сельско-
хозяйственную практику силосные башни или процесс силосования
начали вести в специальных траншеях, в которых стало возможным
осуществление направленных процессов с помощью специальных
бактериальных заквасок. Используемые закваски различаются по
содержащимся в них видам. Например, препарат "Казахсил ПБК"
включает только пропионовые бактерии, латвийская закваска —
чистую культуру Lactobacillus plantarum и т. д. По консистенции
закваски подразделяют на три группы: сухие, уплотненные, жид-
кие. Содержание сухих веществ в них составляет для первых —
523
I
более 90%, для уплотненных — 45—65% и для жидких — 5—10%.
Наилучшими из них признают жидкие закваски, в которых кон-
центрация живых клеток равна 1 • 109/мл. Тогда для траншейного '
силосования растительной массы в количестве 500 т необходимо
от 500 до 1000 л закваски.
Недостатки жидких заквасок: температура хранения должна
быть в пределах 4—8°С; малый срок хранения при указанной
температуре — не более двух недель; необходимость больших
емкостей в случае силосования большого количества растительной
массы, и, в этой связи, заметные транспортные расходы для
перевозки жидких заквасок (особенно — на дальние расстояния). '
Поэтому сухие и уплотненные закваски выгоднее жидких, если ;
реактивация микробных клеток при рекомендуемых температурах
(чаще при 20—25°С) происходит в короткие сроки и полностью 1
(т. е; почти 100% клеток).
Молочнокислые бактерии индуцируют соответствующее бро-
жение с накоплением молочной кислоты при снижении pH до 4,3
и ниже; это предотвращает развитие нежелательной микрофлоры
(в том числе — эпифитной) и силос обогащается "природными"
органическими кислотами — молочной и уксусной. С этим улуч-
шается качество силоса, хотя и происходит определенная утрата
сахаров, расходуемых в течение первой (спонтанной аэробной) и
второй (индуцируемой анаэробной) фаз силосования. -
До герметизации башни (траншей) трансформация углеводов
протекает под действием не только вносимой закваски, но и •
естественной микрофлоры растений, закладываемых на силосова-
ние. В первую фазу имеют место гомо- и гетероферментативное
брожение, во вторую — преимущественно гомоферментативное,
когда меньше образуется "побочных" продуктов (диоксида углеро-
да, маннита).
В приготовлении силоса важное значение приобретает также
равномерное распределение молочнокислых бактерий закваски в
толще закладываемой растительной массы, а также применение
таких химических консервантов, как муравьиная и бензойная
кислоты, другие вещества. Консерванты предотвращают потери
углеводов, (особенно — в первую фазу), а во вторую при pH 4,0
ингибируются нежелательные ферментативные реакции (в том
числе — гниение). Более того, подобные консерванты улучшают
качество конечного продукта — возрастает усвояемость корма. Из
химических консервантов известны препараты АИВ-2, Фарми-ли-
уос и др. АИВ-2 содержит 80% НСООН, 2% — Н3РО4 и 18% Н2О;
524
Фарми-лиуос включает 25% НСООН, 25% СН3 СООН, 50% добавки,
содержащей 35% "Финфермекса" (лигносульфонаты, пентаозы,
гексаозы, соли кальция, магния, серы, железа, другие органические
кислоты) и 15% других веществ.
10.5.3. Получение удобрений на основе ассоциации растений
и микроорганизмов. Круговорот веществ в природе в значительной
степени обусловлен и поддерживается за счет деятельности мик-
роорганизмов, обеспечивающих доступность для растений азота,
фосфора и других элементов. Запасы азота в природе практически
безграничны и возобновляются благодаря микробам-азотофикса-
торам. В то же время запасы фосфора менее богаты и легче
истощаются, а для растений (как и для любого живого организма)
данный элемент относится к разряду органогенов. Установлено,
что фосфор в виде ортофосфата легче освобождается из трудно-
доступных для растений соединений при действии микроорганиз-
мов, потребляющих органические вещества. Здесь важны свобод-
ноживущие азотофиксаторы, клубеньковые бактерии и грибы-ми-
коризообразователи. При их рациональном использовании удается
существенно повысить плодородие почвы для культивирования
важных растений. В регионах земного шара с теплым климатом
удается получать ценные удобрения на основе водяного папорот-
ника (Azolla pinnata) и цианобактерий (Anabaena spp.). Подобная
симбиотическая ассоциация характеризуется тем, что она фикси-
рует молекулярный азот из воздуха и обогащает почву раствори-
мыми соединениями.
Показано, что подобное ’’зеленое удобрение” повышает уро-
жайность риса на 50%, если его вносят в почву перед следующим
посевом. Такой эффект, аналогичный использованию 60 кг азот-
ного удобрения на 1 га, пролонгируется и в течение второго года.
Из представителей царства Mycota базидиальные грибы-мак-
ромицеты связаны трофическими связями с различными растени-
ями) прежде всего — древесными. Поэтому возможна и полезна
искусственная микоризация (от греч. mykes — гриб, rhiza —
корень) древесных культур макромицетами, например, при посад-
ках. Около 40% всех видов макромицетов (порядка 5000) образуют
эктомикоризу.
Поглощаемые корнями растения питательные вещества посту-
пают также и в гифы грибов, где фосфор переходит в связанное
состояние и в таком виде поступает в ткани деревьев. Фосфор
возвращается в почву после отмирания "микоризных корней".
Динамика содержания, фосфора в подстилке ( по лат.
525
stramentum — солома; в лесу она состоит из опада и чаще —
многолетнего) и дерново-подзолистом горизонте почв коррелирует
в фенологическом смысле (фенология — наука о сезонных явле-
ниях в живой природе) с динамикой плодоношения макромице-
тов-микоризообразователей. Наибольшее количество плодовых тел
и "подвижного" фосфора совпадают во времени и пространстве.
Макромицеты-симбиотрофы имеют большое экономическое и
экологическое значение. Некоторые из них, например, из родов
Laccaria, Paxillus, Suillus и др. используют для восстановления
природных ценозов.
О симбиогрофии ризобактерий и растений (в частности, бобо-
вых), об изготовлении ряда препаратов для удобрения почв см.
главу 9.
10.6. Коллекционные центры сохранения генофонда растений.
Обычно для селекции использовали древние культурные растения
и их сородичей. С ростом народонаселения на нашей планете и
существенно возросшими потребностями в продуктах питания
происходили сдвиги в сторону преобладания в севооборотах вы-
сокопродуктивных современных сортов, например, злаковых рас-
тений. К тому же включение все новых и новых площадей под
посевы (особенно — на территории России) подвинуло нас ко
времени, когда популяции диких предков, адаптировавшихся к
конкретным условиям обитания, стали исчезать с "лица Земли". А
это привело к оскудению генофонда растительного мира. Вот
почему сегодня проблема его сохранения стала наиважнейшей.
В 1974 г. был образован Международный Совет по генофонду
растений (IBPGR), поддерживаемый FAO, Международным банком
и за счет программы развития при ООН (UNDP). Цель этого совета
— создавать и координировать международное сотрудничество по
сохранению генофондов, и организовывать финансовую поддер-
жку такого рода программ. В этой связи IBPGR координирует
работу по сбору, сохранению, документированию, оценке и ис-
пользованию генофонда.
Генофонд растений можно сохранять в естественных условиях,
то есть в местах их произрастания. Здесь важную роль играют
природоохранные мероприятия, опирающиеся на соответствую-
щие законодательства и законодательные акты, принятые в меж-
дународном и региональном масштабах.
Другой путь сохранения генофонда — поддержание материала
в так называемых банках генов. Такие банки могут быть рабочими
и законсервированными. Первые используются генетиками и се-
526
лекционерами для своих непосредственных нужд и поэтому в них
\ не всегда предусмотрено длительное хранение материала; они, как
правило, небольшие по ассортименту растительных видов. Вторые
относят к разряду крупных банков генов — в них сохраняют
большой набор генотипов, включая и такие, ценность которых еще
не определена, но над которыми нависла угроза исчезновения. Из
законсервированных генофондов могут пополняться рабочие кол-
лекции.
Особенно ст и банков генов: их целесообразность
и широкий видовой набор сохраняемого материала, надежность
способов хранения, своевременность обновления материала, фун-
кциональный динамизм.
Что касается широты видового набора сохраняемого материала,
то под этим понимают спектр сохраняемых генотипов, и чем он
больше, тем богаче возможность человечества жить в прекрасном
мире различных растительных сообществ, чем и определяется
целесообразность создания банков генов. В этой связи можно
отметить, что в генобанке (коллекции) в ВИРе (Всероссийский
институт растениеводства) хранится 380000 генотипов растений.
Надежность способов хранения генофонда зависит от вида
растения и сохраняемости материала (семена, вегетативная форма
ткани, искусственные посадки), а также методов обработки и/или
подготовки материала для хранения. В отношении большинства
видов сельскохозяйственных культур приемлемым оказалось кри-
осохранение семян в течение длительного времени. Если растения
не образуют жизнеспособных семян или семян, не выдерживаю-
щих длительного хранения (многолетние древесные растения), то
стремятся сохранять относительно долго живые ткани в вегетатив-
ной форме. При этом необходимо помнить о возможности возник-
новения микробных болезней, обусловленных вирусами, бактери-
ями, грибами. При ежегодном хранении вегетативных отводков
травянистых растений существенная выбраковка происходит из-за
легкости их повреждений.
Хранение в виде искусственных посадок, например, древесных
растений, необходимы большие площади пригодных для этого
земель и достаточно внимательный уход за посадками.
Из сказанного вытекает, что универсального способа сохране-
ния генофонда не существует и, очевидно, его не может быть,
учитывая разнокачественность видов в царстве растений. Наибо-
лее надежным для семян признают криосохранение (от
греч. krios — мороз). Изучение влияния низких температур на
527
живые организмы составляет предмет науки "Криобиология". Кри-
осохранение в буквальном смысле Означает хранение в заморо-
женном состоянии. Точка замерзания чистой воды соответствует
0°С, точка переохлаждения, или образования однородных центров
кристаллизации соответствует -38’С. На практике используют
температуры значительно ниже, чем температура замерзания во-
ды, например, -79°С (температура сухого льда), -80°С и ниже
(температура в специальных морозильниках), -150°С (температура
паров жидкого азота), -196°С (температура жидкого азота). Сохра-
няемый биообъект переходит в анабиотическое состояние, когда
метаболизм замедляется или приостанавливается на время хране-
ния. При этом также не происходят какие-либо генетические
изменения за исключением повреждающего действия ионизиру-
ющей радиации.
В практике работы коллекций может наблюдаться так называ-
емый холодовый шок, или повреждение от охлаждения,
когда сохраняемый материал проявляет чувствительность к пони-
женным температурам. При этом жидкая часть цитоплазмы выхо-
дит из клеток из-за структурных нарушений в клеточной мембране.
К счастью, холодовый шок не проявляется при криосохранении
большинства типов растительных клеток.
При охлаждении клеток важную роль играет кристаллообра-
зование льда вокруг внутриклеточных гетерогенных центров кри-
сталлизации, когда образующиеся кристаллы могут физически
повреждать, например, клеточные мембраны.
Методический подход к выбору способа замораживания зави-
сит от качества материала (семена, вегетативная ткань, уровень их
обезвоживания и пр.) На рис. 150 представлены данные Дж.
Фарранта (1980) по выживаемости клеток при охлаждении.
Рис. 150. Выживаемость
клеток при быстром и мед-
ленном охлаждении: 1 — не-
замерзшая клетка, 2 — точка
замерзания, 3 — выход внут-
риклеточной воды, 4 — быс-
трое охлаждение — клетка не
сжата, внутриклеточный лед,
клетка не выживает, 5 - мед-
ленное охлаждение — клетка
сжата, нет внутриклеточного
льда, клетка выживает.
528
Чтобы уменьшить образование внутриклеточного льда в про-
цессе охлаждения клеток, используют криопротекторы
(в концентрации около 2 М) с разным механизмом действия. Они
снижают точки замерзания (точку плавления) и переохлаждения
воды (температура, при которой происходит гомогенная кристал-
лизация льда), поднимают точку девитрификации раствора (тем-
пература, при которой происходит исключение аморфности, стек-
ловидности и когда начинается внезапная кристаллизация). Рас-
твор остается аморфным, когда он охлаждается быстро, то есть
время, необходимое для роста кристаллов, ограничено. В этом
случае говорят, что раствор витрифицируется. При
медленном нагревании такого раствора происходит выделение
тепла, совпадающее по времени и внезапной кристаллизацией или
девитрификацией.
К числу криопротекторов относят глицерин, гликаны, гликоп-
ротеины, ДМСО; сахарозу, ПВС, сыворотку крови и др. Их можно
использовать по отдельности или в смесях. Лучший способ стери-
лизации растворов криопротекторов — мембранная фильтрация.
При получении культуры после криосохранения в парах азота
(-150°С) или в жидком азоте (-196°С) их оттаивание проводят
достаточно быстро в водяной бане при 35—40°С. После оттаивания
необходим тщательный уход за каждым образцом.
Установлено, что при криосохранении протопластов из многих
видов растений достигают 70—90%-й их выживаемости. При соче-
тании медленного и ступенчатого замораживания и быстрого
оттаивания в качестве криопротектора рекомендуют 10% ДМСО.
Для криосохранения каллусов обычно применяют двух- или
трехкомпонентную смесь криопротекторов: 8% глюкоза, 10%
ДМСО и 10% ПЭГ (ММ 6 кДа).
В таблице 54 приведены различные типы культур растений и
применяемые условия их хранения.
Среди факторов, лимитирующих рост отдельных растений,
могут быть и ретарданты (от лат. retardatio — замедление), что
отмечено, например, для меристематических тканей паслена.
529
Т а б л и ц а 54. Способы храпения некоторых культур растительных тканей
Растение Тип культуры Криосохранение (° С) 1 Условия хранения, лимитирующие рост
Береза обыкновенная (Betula vulgaris) Кончики побегов — Хранение при низких температурах
Дурман (Datura stramonium) Суспензия клеток —196
Красавка (Atropa belladonna) Зародыш из пыльцы —196 —
Пыльник —196 —
Суспензия клеток —196
Кукуруза(Zea mays) Зародыш из зиготы —196
Каллус —196
Суспензия клеток —196 —
Морковь (Daucus carota) Зародыш соматический — Высушивание ткани перед хранением + хранение в условиях недостатка сахарозы
Зародыш соматический/ клональный регенерант —196
Каллус — Хранение под маслом минеральным
Паслен (Solanum spp.) Кончики побегов Хранение при низкой температуре или (и) в присутствии или после применения ретардантов
530
Продолжение табл. 54
Растение Тип культуры Криосохранение Г С) Условия хранения, лимитирующие рост
Перец горький (Capsicum annuum) Каллус — Хранение при . низких температурах
Суспензия клеток —196 —
Рис (Oryza sativa) Каллус —196 —
Суспензия клеток —196 —
Табак (Nicotiana tabacum) Зародыш из пыльцы —196 —
Каллус — Хранение в условиях недостатка снабжения Ог
Кончики побегов — —
Пыльник —196 —
Суспензия клеток —196 —
Из таблицы 54 видно, что криосохранение в жидком азоте
приемлемо для преобладающего числа указанных видов культур
тканей растений.
Для суспензионных культур клеток нередко рекомендуют трех-
компонентную двухмолярную смесь криопротекторов, включаю-
щую глицерин (0,5 М), ДМСО (0,5 М) и сахарозу (1 М).
Необходимо иметь в виду, что методы криосохранения до сих
пор остаются трудоемкими и дорогостоящими, однако "цели оп-
равдывают средства".
Охлаждение суспензионных, каллусных и различных органи-
зованных культур проводят обычно медленно (1—2°С в минуту),
меристематические ткани, напротив, охлаждают очень быстро (от
50° до более чем 1000°С в минуту); также быстро проводят завер-
шающее охлаждение изначально медленно охлаждаемых образцов.
Материалом для первичного хранения образцов выбирают
стекло, полипропилен, фольгу. К сожалению, стекло разных сортов
нередко растрескивается, и поэтому оно уступает полипропилену
53J
и фольге. Полипропиленовые ампулы с завинчивающимися крыш-
ками стерилизуют заранее отдельно или их поставляют в стериль-
ном виде. Из фольги готовят пакеты, поддающиеся стерилизации
в автоклаве. К сожалению, абсолютно надежный способ сохране-
ния любых образцов еще не разработан.
Глава 11.
ЗООБИОТЕХНОЛОГИЯ
Зообиотехнология является ветвью биотехнологии. Принципи-
альное отличие ее от микробной биотехнологии и фитобиотехно-
логии состоит в том, что объектами зообиотехнологии являются
клетки животных и человека. Наипростейшими животными суще-
ствами клеточной организации являются протозойные организмы,
или Protozoa (от греч. protos — первый, zoon — животное, живое
существо), составляющие самостоятельный отдел в царстве
Animalia (таблица 55).
Таблица 55. Представители царства Animalia
№№ п/п Отдел Типы или классы
1 Protozoa (простейшие) Типы: Acanthariae — акантарии, Ascetospora — асцетоспоровые, Ciliophora — реснитчатые, ' Cnidosporida — книдоспоридевые, Mastigophora — жгутиконосцы, Microsporida — микроспоридевые, Opalinidomorpha — опалинидоформные, Sarcodina — ложноножки, Sporozoa — споровики.
2 Parazoa (губки) Классы: Calcarea — известковые губки, Demospongiae — обыкновенные губки, Hexactinellida — кремниевые шестилучевые губки.
3 Metazoa (многоклеточные гетеротрофные животные) Типы: Annelida — кольчатые черви, Arthropoda — членистоногие. Chordata — хордовые. Coelenterata — кишечнополостные, Echinodermata—иглокожие, MoIIusca — моллюски, или мягкотелые, Nematoda — круглые черви, Platyhelminthes •— плоские черви.
532
Из 65 тысяч видов Protozoa 55 тысяч являются свободно живу-
щими в природе, а 10 тысяч ведут паразитический образ жизни.
Среди выделяемых ныне 9 типов простейших организмов
жгутиковым отводят место пограничных существ, являющихся
ближайшими предками многоклеточных животных и растений и
образующих мост между ними.
Поддержание Protozoa в культуре - дело достаточно сложное,
однако многие виды удается выращивать на относительно простых
средах или на более сложных, рекомендованных для культивиро-
вания клеток и тканей животных (например, среда № 199 и др.).
Для некоторых протозойных видов целесообразно добавлять в
среды споровые бактерии (Вас. subtilis) и/или дрожжи
(Saccharomyces cerevisiae и др.). Это обусловлено их фагоцитарной
активностью.
С биотехнологических позиций попеременный интерес был
проявлен к препаратам круцин (Н. Г. Клюева, Г. И. Роскин) и его
аналогу — трипанозе (Ж. Кудер, Ж. Мишель-Брэн и др.), получа-
емым с помощью Trypanosoma (Schizotrypanum) cruzi, астазилиду
(Н. Н. Сухарева-Немакова) — полусинтётическому продукту, вклю-
чающему комплекс эфиров сахарозы и жирных кислот, выделен-
ных из жгутиконосца Astasia longa, полисахаридам. В доклиниче-
ских и клинических испытаниях они проявляли больший или
меньший противоопухолевый эффект. Однако из-за нестабильно-
сти клинических результатов, неосвоенное™ крупномасштабного
культивирования Protozoa и некоторых других причин зообиотех-
нология в этом разделе мало продвинулась вперед.
Из всех других представителей царства животных наибольший
интерес представляют хордовые. Их подразделяют на два подтипа
— бесчерепные (Protochordata, или Acrania) и черепные, или
позвоночные (Craniata, или Vertebrata), Всех черепных объединяют
в следующие классы: Cyclostomata (круглоротые), Pisces (рыбы),
Amphibia (земноводные), Reptilia (пресмыкающиеся), Aves (птицы)
и Mammalia (млекопитающие).
Значимость представителей царства животных в зообиотехно-
логии далеко неравноценная. Это связано с теми обстоятельствами,
что во многих случаях еще не разработаны способы культивиро-
вания клеток не только в производственных, но и в лабораторных
условиях, или, как говорят, "еще не дошли руки" до конкретных
биообъектов животного происхождения, хотя техника однослой-
ной культуры клеток впервые разработана еще в 1950 г. Ф. К.
Роббинсом и Дж. Ф. -Эндерсом.
533
В настоящее время в промышленном масштабе освоены лим-
фобласты человека, продуцирующие интерферон, клетки-гибри-
домы, образующие моноклональные антитела; монослойные кле-
точные культуры различных тканей, использующиеся, например,
для репродукции вирусов и т. д. Даже из этих немногочисленных
примеров можно заключить, что первостепенной задачей для
зообиотехнологических процессов является получение клеток и
клеточных культур.
11.1. Способы выращивания клеток животных. Любые клетки
животного организма происходят из оплодотворенной яйцеклетки,
которая со временем развивается и дифференцируется (рис. 151).
Зародышевый диск вклю-
чает эктодерму, эндодерму
и мезодерму, из которых
впоследствии образуются
все ткани макроорганизма.
Но как только какую-либо
ткань перевести в культу-
ру, то наступает дедиффе-
ренциация клеток и разли-
чить их становится делом
трудным или невозмож-
Рис. 151. Эмбрион млекопитающего на ран- п ПОЭТОМУ рОГИСТрЗ.-
ней стадии развития.’ 1 зародышевый диск, 2 pjx Ведут ПОЧТИ ИСКЛЮ~
эктодерма, 3 эндодерма, 4 первичнвя чительНО по происхожде-
мезодерма, 5 — трофобласт. нию. Например, для иссле-
дования в области онкологии важны культуры раковых клеток, о
которых имеются сообщения в научной литературе. Такие клетки
получены от больных раком легкого и щитовидной железы. В 1986
г. в Японии впервые получен в культуре штамм клеток рака
желчного пузыря. Он был стабилен после 103 пассажей in vitro,
сохранил прежнее число хромосом в пределах 76—101. Индуциру-
емый им опухолевый процесс и первичный рак желчного пузыря
оказались тождественными по гистограмме. Подобный штамм
оказался полезным при оценке противоопухолевых средств.
Любой штамм, представляющий определенную ценность для
научно-практического использования, должен иметь следующие
характеристики:
1) качество исходной ткани - нормальная или опухолевая (для
опухолевой необходим показатель доброкачественности или злгР
качественности),
534
2) тип ткани — эмбриональная или зрелая,
3) принадлежность ткани — вид животного,
4) источник ткани — орган,
5) тип клетки (если известно),
6) наименование штамма (буквенное — не более 4 букв и серия
цифр нумерации),
7) номер клона, если штамм клонировался,
8) источник информации (ссылка на оригинальную публика-
цию).
Все способы выращивания клеток животных могут быть соот-
несены либо к интактным (нетрансформированным с помощью
вирусов), либо к вирусотрансформированным клеткам. Успех куль-
тивирования первых во многом обусловлен наличием и плотностью
адгезинов, благодаря которым они проявляют эффект "заякорива-
ния" на подлежащей поверхности стекла, металла или пластика.
Однако такие клетки не растут в суспензионных культурах.
Вирусотрансформированные клетки животных, напротив, хо-
рошо растут в виде суспензионных культур и хуже или совсем не
растут в адгезированном состоянии.
Имеются такие штаммы животных клеток, которые могут расти
и в прикрепленном состоянии, и в виде суспензии. Прикрепивши-
еся клетки размножаются, растут и развиваются до тех пор, пока
не сольются в монослой, то есть плотность клеток выступает
тормозным сигналом. Однако при смене питательной среды на
новые порции наблюдается дальнейшее разрастание клеток в
форме нескольких слившихся слоев. Подобного результата можно
добиться при использовании других факторов воздействия на
клеточные культуры (гормоны, ферменты, pH и т. д.). Тем не менее,
на практике широко пользуются монослойными культурами.
11.1.1 .Глубинное выращивание клеток в монослое. Рост клеток
в виде монослоя зависит от адгезивных белков — фибронек-
тинов (от лат. fibra — нить, nectere — связывать или соединять),
обеспечивающих межклеточную адгезию, прикрепление клеток к
подложке и направляющих их перемещения. В животном организ-
ме клетки, способные к перемещениям (особенно в период эмбри-
онального развития, при заживлении ран), и связанные с базаль-
ными мембранами, содержат на своей поверхности крупномоле-
кулярные гликопротеиновые молекулы фибронектина, открытого
535
в 1973 г. Р.О.Хайнсом (Англия), К. Гамбергом и С.-И. Хакомори
(США) при изучении нормальных и опухолевых клеток. Фибро-
нектин, полимеризуясь, образует длинные нити вокруг клетки,
которые контактируют с клеточной мембраной и связываются с
цитоскелетным белком-актином (см.). Топология фибронектина в
экстрацеллюлярном матриксе схематично представлена на рис.
158.
Молекула фибронектина состоит из двух субъединиц с ММ
около 250 кДа, содержащих небольшие по размеру домены и
соединенных на одном конце двумя S—S-мостиками. Размеры
одной субъединицы 60—70x2—3 нм; на эту длину приходится от
2145 до 2445 остатков аминокислот. Каждый домен отвечает за одну
функцию фибронектина, например, за соединение с фибрином,
другой домен — за прикрепление к пластику и т. д. (рис. 152).
Доказано, что
фибронектин имеет-
ся у всех представи-
телей царства
Animalia. Амфотер-
ный гликопротеин-
фибронектин в изо-
лированном виде вы-
ражение стимулиру-
ет адгезию, если до-
Рис. 152. Схематичное изображение молекулы фиб- бавлять его 'К пита-
ронектина с доменами, обеспечивающими связь с фиб- тельной среде В КОН-
рином (1), с коллагеном (2), с клеткой (3).
центрации порядка
1—5 мкг/мл. Амфолит в данном случае выступает мостиком между
отрицательно заряженной поверхностью животной клетки и суб-
стратом, несущим отрицательный или положительный заряд. Сво-
бодная энергия поверхности твердого носителя может быть высо-
кой (чистые поверхности стекол, металлов, металлических окислов)
или низкой (поверхности из органических полимеров), хотя понят-
но, что при соответствующих обработках низкоэнергетические
поверхности трансформируются в высокоэнергетические.
В качестве связующих мостиков можно применять ионы каль-
ция или магния (рис. 153).
536
клетка
клетка
а б
Рис. 153. Механизм адгезии животной клетки к субстрату (например, к
подложке), несущему положительные (а) и отрицательные (б) заряды (А — гликоп-
ротеиновый адгезии, стрелки — электростатические взаимодействия, пунктиры —
связывание при участии двухвалентных катионов; АС — адсорбционный слой,
равный приблизительно 5 нм).
Наряду с плотностью заряда в прикрепляемости клеток боль-
шое значение имеют характеристики субстрата: гидрофильность
его поверхности (смачиваемость), протяженность и горизонталь-
ность. Распластываемость клеток на подложке будет выше, если
число отрицательных зарядов оказывается не ниже 5 на площади
10 нм2. Адгезивные свойства более выражены для смачиваемых
поверхностей в сравнении с гидрофобными, по протяженности
они должны быть больше нормальной длины клеток; субстраты с
искривленной поверхностью хуже гладких — растущие клетки
распространяются в направлении наименьшей кривизны субстра-
та (здесь существен вклад цитоскелета, см.). Если, например,
прикрепление клеток к агар-агару принять за ед иницу, то к тефлону
они прикрепляются в 5 раз лучше, к полиэтилену — в 8 раз, к
полипропилену — в 9 раз, к резине — в 12 раз, к алюминию — в
15 раз, к стеклу — в 16 раз, а к стали и полистирену — в 20 раз.
В 1993 г. поступил на рынок пронектин F — продукт белковой
инженерии, созданный в Великобритании. Он напоминает ранее
названный фибронектин, но пронектин F включает 13 мест связы-
вания на молекулу (в сравнении с нормальным комплементом). Его
потенциал к клеточной адгезии в 13 раз выше потенциала фибро-
нектина, полученного от млекопитающих. Пронектин F рекомен-
дуют для выращивания клеток в бессыворочных средах.
Крупномасштабное культивирование животных клеток в мо-
нослое нацелено на получение наибольшей концентрации клеток
в наименьшем объеме газовой фазы.
537
Выбор клеток не столь велик, но некоторые из них вполне
адаптированы к условиям выращивания in vitro. В 1964 г. впервые
были получены линии соединительнотканных клеток — фиброб-
ластов ВНК 21 от сирийского хомяка. Эти клетки пассируются
неограниченно долго и первоначально использовались для репро-
дукции вируса ящура д ля приготовления соответствующих вакцин.
Следует иметь в виду, что нормальные диплоидные клетки челове-
ка, например, линии WI-38, могут делиться ограниченное число раз
(50 ± 10) и проявляют феномен старения (см. раздел 3.2.4.). В то
же время такие клетки , как HeLa, выделенные из раковой опухоли
шейки матки человека, оказались бессмертными при пассирова-
нии.
При доступности донорской ткани используют клетки почек
кроликов, обезьян, собак (линия МДСР), 10—14-дневных куриных
эмбрионов, клетки из миндалин, амниона, легких эмбриона чело-
века 12—16-недельного возраста, клетки эпидермиса взрослого
человека, мышиные фибробласты (линия LS), диплоидные фиброб-
ласты человека (линия MRC-5) и другие.
Выбранные клетки выращивают в строго, асептичных условиях
в специальном оборудовании (см. главу 7) при медленном вращении
роллерной системы или покачивании для омывания большей пло-
щади культуральной среды. Клетки то погружаются в жидкую
фазу, то в газообразную. При этом с избытком обеспечиваются их
дыхательные потребности в кислороде.
Во время цикла культивирования клеток должны учитываться
разные параметры. Одни из них относятся к числу константных,
другие — к числу вариабельных. Константными являются качество
материала, форма и объем культиватора, вариабельными — ско-
рость вращения или покачивания, качество и объем среды, тип
клеток и размер (величина) посевного материала, pH и температура
среды, снабжение кислородом и содержание СО2 в культиваторе,
окислительно-восстановительный потенциал и концентрация ос-
новных источников питания. Первые три вариабельных показателя
можно поддерживать постоянными, переводя их в разряд констан-
тных.
Естественно ожидать, что среды для клеток животных должны
отличаться от сред, используемых, например, для прокариот. На
начальных этапах развития зообиотехнологии обязательным было
добавление сывороток к средам, применяемым в лабораторных и
производственных условиях. Сыворотки обеспечивали клетки не-
обходимыми гормонами (глюкокортикоиды, инсулин, половые гор-
538
моны, простагландины и др.), ростовыми факторами (эпидермаль-
ным, фибробластным, Т-клеточным, тромбоцитарным, опухоленек-
ротическим и др.), факторами адгезии (коллаген, фибронектин,
ламинин*), белками (альбумин, трансферрин, фетуин*’), микроэле-
ментами и липидами, для выживания in vitro. Обычно используют
фетальную бычью сыворотку (ФБС). Высоким качеством отлича-
ются сыворотки, рекомендуемые фирмой Sigma (США). Некоторые
из них по качеству (для отдельных линий клеток) превосходят ФБС,
например, сыворотка крови новорожденных телят в возрасте до
10 дней и менее, сыворотка от телят возраста менее 10-месячного,
лошадиная сыворотка от жеребцов, кастрированных за год до
взятия крови.
Вместо натуральных сывороток фирма IBF biotechnics (Герма-
ния) предлагает высококачественные заменители, в частности,
ultroser G — для иммобилизованных клеток. Эта среда содержит
мало белка, благодаря чему упрощается очистка биопродукта и
достигается стабильность компонентного состава.
Оптимальные показатели pH и температуры зависят от проис-
хождения и качества клеточных линий. Так, для клеток млекопи-
тающих концентрацию водородных ионов поддерживают в преде-
лах pH от 7,2 до 7,4, а температуру — в пределах + 36— + 37,5°С.
Поддержание pH осуществляют обычно за счет буферных систем
[(СО2—NaHCO3), трис- и др.), а температуру — с помощью термо-
регуляторов.
Контроль за развитием клеток проводят с помощью прямых
(подсчет) и непрямых методов (по мутности, по подсчету ядер, по
определению общего белка).
Посевной материал (инокулят, от лат. inoculatio — прививание)
заметно сказывается на скорости роста клеток в монослойной
культуре. В случае применения первичных клеток необходимо
иметь их в более высокой плотности на 1 см2 внутренней (рабочей)
поверхности культиватора.
Если же используют клеточные линии (а не первичные клетки),
то плотность инокулята может быть несколько меньшей, так как
они почти все 100% адгезируются на поверхности культиватора.
* ламинин получают из базальной мембраны мышиной саркомы Englebreth-
Holm-Swarm;
" трансферрин — плазменный белок теплокровных животных и человека,
содержащий железо. Фетуин — белок из сыворотки плода теленка, содержащий
около 0,2% свободной N-ацетилнейраминовой кислоты.
539
Покачивание или вращение омываемых средой клеток несомненно
сказывается на их прикреплении.
11.1.2. Глубинное выращивание клеток в суспензионных куль- .
турах. Площадь выращиваемых клеток можно существенно уве-
личить при использовании микроносителей, суспендируемых в
питательной среде и на поверхности которых клетки закрепляются,
а затем разрастаются в виде монослоя. Такие суспендированные
микроносители с клетками моделируют глубинные культуры, на-
пример, микроорганизмов. Следовательно, в таких системах со-
вмещаются монослойные и суспензионные культуры животных
клеток.
В качестве микроносителей применяют положительно заря-
женные ДЕАЕ-сефадексы, сефадексы с коллагеновым покрытием,
отрицательно заряженный полистирол, полые стеклянные сферы
и др. Так, например, отдельные фирмы предлагают микросферы
из пористого шлачного стекла (рис. 154), которые могут быть
использованы для иммобилизации клеток млекопитающих. Поры
их доступны для пенетрации (от лат. penetratio — проникновение)
клеток внутрь матрикса, облегчая трехмерную колонизацию внут-
ренней поверхности носителя, что способствует взаимодействию
метаболизирующих соседствующих клеток и поддержанию их
возросших жизнеспособности и продуктивности. Пористые сферы
пригодны для иммобилизации прилипающих и суспензионных
клеток (например, гибридом).
Рис. 154. Микросферы для иммобилизации клеток млекопитающих из микро-
пористых, биологически инертных, стекла (а) и стеклошлака (так называемого
Cultispher-G) (б) увеличение xlOOOO.
540
Рекомендуемый размер сферических микроносителей порядка
200± 25 мкм оценивают оптимальным с удельной плотностью 1*,03.
Требуемое количество их примерно равно 104 мл, а для инокуляции
необходимо порядка 105 клеток. При увеличении количества мик-
роносителя возрастает доза клеток в инокуляте.
Подход к выбору сред и контролируемого параметра культи-
вирования остается прежним. Клетки после выращивания отделя-
ют от микроносителей центрифугированием (в том числе — в
градиенте плотности), фильтрованием, или, чаще, обработкой трип-
сином с последующими промыванием и сепарированием.
Суспензионные клетки на микроносителях применяют в целях
получения вирусных вакцин (против бешенства, полиомиелита,
ящура).
Возможности переноса генетической информации из эукари-
отических клеток в прокариотические сняло проблему массового
культивирования животных клеток для получения таких продук-
тов, как интерферон, гормоны, лимфокины и другие. Здесь успеш-
но развивается рДНК-биотехнология.
Клеткам животных в глубинных культурах .почти во всех
случаях требуется защитная матрица, функцию которой берут на
себя белки сыворотки крови, добавляемой в среду культивирова-
ния. Этим еще раз подтверждается более высокая ранимость
животных клеток по сравнению с микробными и растительными
клетками. Тем не менее, основные подходы к выбору оборудования
и к реализации биотехнологических процессов с использованием
животных клеток во многом аналогичны с таковыми в микробной
биотехнологии. Например, системы культивирования в обоих слу-
чаях могут быть хемо- или турбидостатными, циклическими, не-
прерывными или полунепрерывными.
В противоположность суспензионным культурам клеток на
микроносителях разработаны способы инкапсулирования их в
полимерные сферы (полилизиновые, агарозные, из полимерных
смол), когда клетки, по-существу, не испытывают того механиче-
ского напряжения, которое они должны проявлять в свободном
виде. Эти способы оказались выгодными для культивирования
гибридом в целях получения моноклональных антител. Диаметр
микрокапсул, размер пор в них и их толщина могут быть незави-
симо проконтролированы в процессе изготовления и оптимизиро-
ваны для конкретного типа гибридом. Например, согласно данным
фирмы Damon Biotech (Германия) гибридомные клетки человека
или животных вначале суспендируют в биосовместимом растворе
541
натрия альгината. Образовавшиеся капельки затем пропускают в
раствор кальция хлорида, где они становятся желированными
микросферами. На поверхности таких сфер затем формируют
полилизиновую мембрану. После этого гель разжижают, оставляя
гибридомные клетки в нормальном физиологическом состоянии
внутри "дырчатых" микросфер. В процессе культивирования таких
инкапсулированных гибридом продукция моноклональных анти-
тел возрастает в сотни-тысячи раз по сравнению с обычным
культивированием гибридомных клеток. После завершения био-
синтеза антител микрокапсулы подвергают диализу для удаления
примесных веществ, а затем физически их "раскрывают", гибри-
домные клетки и фрагменты капсул легко сепарируются от иско-
мых моноклональных антител, которые могут быть подвергнуты
дальнейшей очистке или использованы по назначению. Схема
описанных процедур представлена на рис. 155.
Полилизиновая мембрана
обеспечивает диффузию неболь-
ших молекул питательных ве-
ществ внутрь микросфер, но не
позволяет выходить иммунным
глобулинам. Кроме физического
удаления полилизиновой мембра-
ны можно воспользоваться фер-
ментативным гидролизом.
В описанные микросферы
возможно инкапсулировать не
только клетки, но и ферменты,
индукторы каких-либо процессов,
а также мультисистемы.
Таким образом, глубинное вы-
ращивание животных клеток реализуют в трех вариантах:
1) монослой клеток статичен, культуральная среда подвижна,
2) среда культивирования статична, монослой клеток подвижен,
3) клетки (например, на микроносителях, в мйкросферах) и
среда культивирования подвижны. Эти варианты выращивания
клеток должны приниматься в расчет при выборе или проектиро-
вании соответствующего оборудования.
11.1.3. Другие способы выращивания клеток. В настоящее
время в опытно-промышленных условиях получают интерферон,
542
Рис. 155. Получение моноклональ-
ных антител с помощью инкапсулиро-
ванных клеток гибридомы (схема): 1 —
клетки-гибридомы, 2 — клетки-гибридо-
мы в растворе натрия альгината, 3 —
клетки-гибридомы в желированном рас-
творе натрия альгината, покрытые пол-
илизиновой мембраной, 4 — клетки-гиб-
ридомы, в "дырчатых" микросферах, 5
— антитела и клетки-гибридомы после
разрушения пблилизиновой мембраны.
выращивая лимфобласты человека в виде отъемно-доливных куль-
тур, то есть когда определенная часть клеточной суспензии заме-
щается свежей средой, например, ultroser HY (Германия), а оста-
ющаяся часть "старой" культуры выполняет роль инокулята.
Интерфероны можно получать в периодической культуре, ког-
да часть ее отбирают через определенные интервалы времени при
непрерывной подпитке свежей питательной средой и при посто-
янной скорости ее подачи. Тогда объем культуры прогрессивно
возрастает, равно как прогрессивно снижаются скорости ее роста
и разбавления.
Как бы долго не удавалось поддерживать клетки в циклических
культурах с подпиткой, в конечном итоге они погибают и весь цикл
приходится начинать сначала.
Неопределенно длительное время можно выращивать клетки
млекопитающих в непрерывных хемостатных культурах, когда
удается добиваться постоянства концентрации лимитирующего
субстрата и плотности клеток (см. главу 7). Теория и практика
непрерывного культивирования впервые сформулированы в 1950
г. Ж. Моно, и, независимо от него, А. Новиком и Л. Сцилардом,
предложившими термин "хемостат". В хемостатах скорость подачи
свежей среды и отбора культуры равны (как и объем их). Скорость
роста, развития и размножения клеток контролируется скоростью
под вода лимитирующего компонента, а численность — его концен-
трацией. В качестве лимитирующего рост агента чаще всего ис-
пользуют глюкозу, реже — фосфат и другие вещества. При
правильном подборе условий выращивания в хемостатах удается
на порядок увеличить выход клеток по сравнению с период ическим
культивированием. Причем, хемостатные культуры отличаются
накоплением физиологически однотипных клеток. Это можно
показать на примере с клетками лейкемии мышей — L 1210
(таблица 55), которые засевали (инокулят) в концентрации 2* 10s
клеток/мл для периодического культивирования, длившегося 3
суток до получения максимальной плотности 2,5 «106 клеток/мл
(суспензионные культуры). При хемостатном культивировании
скорость подачи среды и отъема культуры составляла 0,3 сут *.
543
T a 6 л и ц a 55. Накопление клеток L 1210 при периодическом и непрерывном
культивировании (по М. Ж. Тови, 1985)
Способ выращивания Объем среды, л Урожайность клеток
на 1 мг глюкозы на 1 мл среды в неделю 1
х 106 МКГ х 106
Периодический 0.3 3,5 524 2,3 1,38-109 |
Непрерывный (хемостатный) 0,3 6.2 601 6,2 1,12-10'°
Естественно, что время, затрачиваемое на подготовку смежных
циклов и их реализацию при периодическом культивировании,
является непроизводительным.
11.2. Эмбриональные и другие ткани для репродукции вирусов
и получения вирусных вакцин. Из эмбриональных тканей наибо-
лее широко используемыми являются эмбриональные ткани цып-
ленка, мыши и человека. Особенной выгодой отличаются куриные
эмбрионы (по доступности) десяти-двенадцатисуточного возраста,
используемые преимущественно для репродукции вирусов и по-
следующего изготовления вирусных вакцин. Куриные эмбрионы
введены в вирусологическую практику в 1931 г. Г. М. Вудруфом и
Е. У. Гудпасчером. Такие эмбрионы рекомендуют также для выяв-
ления, идентификации и определения инфицирующей дозы виру-
сов, для получения антигенных препаратов, применяемых в серо-
логических реакциях.
Инкубированные при 38°С куриные яйца овоскопируют (про-
свечивают), отбраковывают, "прозрачные" неоплодотворенные эк-
земпляры и сохраняют оплодотворенные, в которых хорошо видны
наполненные кровеносные сосуды хорионаллантоисной оболочки
и движения эмбрионов.
Заражение эмбрионов можно проводить вручную и автомати-
зированно. Последний способ применяют в крупномасштабном
производстве, например, противогриппозных вак-
цин. Материал, содержащий вирусы, вводят с помощью
шприца (батареи шприцов) в различные части эмбриона (эмбрио-
нов) (рис. 156).
544
Рис. 156. Структурные части деся-
тидневного куриного эмбриона: 1 - плод,
2 - аллантоисная полость, 3 - хорионал-
Все этапы работы с куриными
эмбрионами после овоскопии
проводят в асептичных условиях
(см. главу 6). Материалом для за-
ражения могут быть суспензия
растертой мозговой ткани (при-
менительно к вирусу бешенства),
печени, селезенки, почек (приме-
нительно к хламидиям орнитоза)
и т. д. В целях деконтаминации
вирусного материала от бактерий
или в целях предотвращения его
бактериального загрязнения
лантоисная оболочка, 4 - полость амни-
она, 5 - оболочка амниона, 6 - желточный
мешок, 7 - белочный мешок, 8 - воздуш-
ное пространство, 9 - скорлупа, 10 -
скорлупная оболочка, 11 - введение ви-
русного материала в аллантоисную по-
лость.
можно использовать соответству-
ющие антибиотики, например,
пенициллин с каким-либо ами-
ногликозидом порядка 150 ME
каждого на 1 мл суспензии виру-
сосодержащего материала. Для
борьбы с грибковым заражением
эмбрионов целесообразно воспользоваться некоторыми антибио-
тиками-полиенами (нистатин, амфотерицин В) или отдельными
производными бензимидазола (например дактарин и др.).
Чаще всего суспензию вирусного материала вводят в алланто-
исную полость или, реже, на хорионаллантоисную оболочку в
количестве 0,05-0,1 мл, прокалывая продезинфицированную скор-
лупу (например, иодированным этанолом) на расчетную глубину.
После этого отверстие закрывают расплавленным парафином и
эмбрионы помещают в термостат, в котором поддерживается
оптимальная температура для репродукции вируса, например 36-
37,5°С. Продолжительность инкубации зависит от типа и активно-
сти вируса. Обычно через 2-4 суток можно наблюдать изменение
оболочек с последующей гибелью эмбрионов. Зараженные эмбри-
оны контролируют ежедневно 1-2 раза (овоскопируют, поворачи-
вают другой стороной). Погибшие эмбрионы затем передают в
отделение сбора вирусного материала. Там их дезинфицируют,
аллантоисную жидкость с вирусом отсасывают и переносят в
стерильные емкости. Инактивацию вирусов при определенной
температуре проводят обычно с помощью формалина, фенола или
других веществ. Применяя высокоскоростное центрифугирование
18 т. 8524 545
или афинную хроматографию (см.), удается получать высокоочи-
щенные вирусные частицы.
Собранный вирусный материал, прошедший соответствующий
контроль, подвергают лиофильной сушке. Контролю подлежат
следующие показатели: стерильность, безвредность и специфиче-
ская активность. Применительно к стерильности имеют в виду
отсутствие: живого гомологичного вируса в убитой вакцине, бак-
терий и грибов. Безвредность и специфическую активность оце-
нивают на животных и только после этого вакцину разрешают
испытывать на волонтерах или добровольцах; после успешного
проведения клинической апробации вакцину разрешают приме-
нять в широкой медицинской практике.
На куриных эмбрионах получают, например, живую противо-
гриппозную вакцину. Она предназначается для интраназального
введения (лицам старше 16 лет и детям от 3 до 15 лет). Вакцина
представляет собой высушенную аллантоисную жидкость, взятую
от зараженных вирусом куриных эмбрионов. Тип вируса подби-
рают согласно эпидемиологической ситуации и прогнозам. Поэто-
му препараты могут выпускаться в виде моновакцины или дивак-
цины (напрмер, включающая вирусы А2 и В) в ампулах с 20 й 8
прививочными дозами для соответствующих групп населения.
Высушенная масса в ампулах обычно имеет светло-желтый цвет,
который сохраняется и после растворения содержимого ампулы в
прокипяченой остуженной воде.
Живые противогриппозные вакцины для взрослых и детей
готовят и для приема через рот. Такие вакцины представляют собой
специальные вакцинные штаммы, репродукция которых происхо-
дила в течение 5-15 пассажей (не менее и не более) на культуре
почечной ткани куриных эмбрионов. Их выпускают в сухом виде
во флаконах. При растворении в воде цвет из светло-желтого
переходит в красноватый.
Из других вирусных вакцин, получаемых на куриных эмбрио-
нах, можно назвать противопаротитную, против желтой лихорад-
ки.
Из прочих эмбриональных тканей используют эмбрионы мы-
шей или других млекопитающих животных, а также абортирован-
ные плоды человека.
Эмбриональные перевиваемые ткани доступны после обработ-
ки трипсином, поскольку в таких тканях еще не формируется
большого количества межклеточных веществ (в том числе небел-
546
ковой природы). Клетки разделяются и после необходимых обра-
боток их культивируют в специальных средах в монослое или в
суспендированном состоянии.
Ткани, изолируемые от животных после рождения, относятся
к разряду зрелых. Чем их возраст больше, тем с большим трудом
они культивируются. Однако после успешного выращивания они
затем "выравниваются" и мало чем отличаются от эмбриональных
клеток.
В целях репродукции вирусов особую роль сыграли почечные
и амниотические зрелые ткани. Из них первые получали от обезъ-
ян, вторые — из амниотических оболочек от женщин-рожениц при
нормальных родах. Свежие почки от здоровых обезьян, например
макаки резус или африканских зеленых мартышек (сразу же после
экстирпации, от лат. extirpatio — полное вырезание, вылущение,
искоренение) декапсулирутот, рассекают пополам, удаляют лохан-
ки, нарезают паренхиматозную ткань на мелкие кусочки, трипси-
низируют и клетки (порядка 3 - 103/мл) помещают в специальную
среду, термостатируют при 45°С в качающихся аппаратах. Спустя
сутки, культуру заражают, например, вирусом полиомиелита и
уже через 4-7 дней в надосадочной жидкости накапливается вирус.
Эту жидкость сливают, к ней добавляют формалин до получения
0,025%-й концентрации и смесь оставляют при 37°С на 12 дней для
полного обезвреживания вируса. После этого препарат подвергают
диализу с последующим контролем на отсутствие живых вирусных
частиц, токсичности, безвредности и на активность. Таков принцип
изготовления полиовакцины Дж. Солка. В случае приготовления
живых вакцин из ослабленных (аттенуированных) штаммов виру-
сов исключают стадию обработки формалином. Так получают,
например, живую полиовакцину А. Себина (иммунологических
типов I, II,III). В целях термостабилизации вакцины к ней добавляют
химически чистый магния хлорид. Вакцину выпускают в жидком
виде (по 5 мл) и в форме конфет-драже. Жидкая форма имеет
красновато-оранжевый цвет; цвет драже различен: розовый — для
первого иммунологического типа вируса, сиреневый—для второго
типа, голубой — для третьего, типа и белый — для смеси всех трех
типов.
Кроме полиомиелита специфическую профилактику живыми
вакцинами проводят при кори. Противокоревую живую сухую
вакцину изготавливают из вакцинного штамма, репродукция ко-
торого осуществлялась на клеточных культурах почек морских
свинок или фибробластах японских перепелок. Сухая вакцина
547
имеет желтовато-розовый цвет, который может сохраняться после
растворения ее в прилагаемом к упаковке растворителе. Выпуска-
ют во флаконах по 1,2,5 и 10 прививочных доз. После растворения
вакцину вводят в количестве 0,5 мл подкожно или по 0,1 мл
внутрикожно. Она предназначается для не болевших корью детей
в возрасте от 10 месяцев до 8 лет (по показаниям — до 14 лет).
Вакцина против клещевого энцефалита является формолвак-
циной, полученной из тканевой культуры фибробластов куриного
эмбриона и выпускаемом в жидком виде во-флаконах (по 10 мл)
или ампулах (по 5 мл) с гидроокисью алюминия (до 2 мг) в качестве
адсорбента или в лиофильно высушенном виде с желатином (до
1.5%), и сахарозой (до 8%). Вакцину вводят подкожно (жидкую или
после разведения в дистиллированной воде сухого препарата).
По способам культивирования особое место занимает вирус
бешенства. Его репродукцию осуществляют на мозговой ткани
целостных организмов, используя для этого кроликов, крыс, овец.
Л. Пастер в 80-е годы XIX века выделил от больной собаки вирус
бешенства, и после ряда его пассажей через мозг кроликов он
добился того, что вирус приобрел свойства вызывать заболевание
у данных животных после определенного и постоянного инкуба-
ционного периода (4-6 дней). Такой вирус, в отличие от вируса
"уличного" бешенства, получил название Virus fixe (фиксирован-
ный вирус). Его-то и применяют для получения вакцин.
В целях профилактики бешенства используют сухие антира-
бические вакцины Ферми и МИВП. Первую из них готовят из
мозга овец возраста до 1 года, вторую — из мозга сосунков белых
крыс (возраст 4-8 суток) после заражения тех и других фиксиро-
ванным вирусом бешенства. Подобные вакцины представляют
собой суспензии мозговых тканей, содержащих указанный вирус.
В сухом виде вакцины выпускают в форме таблеток беловато-се-
рого цвета, которые суспендируют в прилагаемом растворителе
(дистиллированная вода) и сразу же вводят подкожно. Хранение
разведенных вакцин впрок запрещается.
Вакцина против краснухи содержит живой ослабленный (ат-
тенуированный, от лат. attenuare — уменьшать) вирус, репродуци-
ровавшийся на диплоидных клетках человека или на клетках почек
кролика. Используют для профилактики краснухи.
Клетки амниона человека должны быть получены не позднее
12 часов после родов. Для этого послед помещают в стерильный
контейнер без применения антисептиков и в ближайшие после
этого часы отделяют амнион от хориона, промывают специальным
548
солевым раствором и дважды последовательно трипсинизируют.
После этого клетки центрифугируют — примерно при 100g в
течение 10 минут, ресуспендируют осадок в среде и подсчитывают
образовавшиеся изолированные клетки (необходимо получить по-
рядка 106 клеток в 1 мл). Их суспендируют и переносят в большие
емкости до получения 106 клеток в 1 мл питательной среды, сменяя
ее каждые 5-7 дней. Полученные клетки можно использовать для
культивирования вирусов.
Перевиваемые опухоли животных, в частности, асцитные,
нашли широкое применение на практике. Они хорошо растут в
форме суспензии изолированных клеток в экссудате брюшной
полости, например, мышей и крыс. Опухолевые клетки размножа-
ются в асцитической жидкости, которую отсасывают из брюшной
полости с помощью шприца (или с помощью пипетки после
оперативного вмешательства). Такие клетки можно также выра-
щивать in vitro.
Перевиваемые опухоли животных представляют определенную
ценность при изучении противоопухолевых химиотерапевтиче-
ских веществ. Для выращивания опухолевых клеток могут оказать-
ся полезными так называемые кокарциногены — вещества, инду-
цирующие формирование опухолевой ткани, хотя сами не явля-
ются карциногенами. Среди известных кокарциногенов наиболее
сильным триггерным действием (от англ, trigger — спусковой
крючок, "собачка") обладает тиглиан — природный вторичный
метаболит у растений, относящихся к семействам Euphorbiaceae
(молочайные) и Thymelaeaceae (тимиановые). Он является природ-
ным Лоооболом из rovnnbi тетрациклических дитерпенов с пер-
гидро-циклопропа-бензазуленовым
скелетом. В этом скелете содержится
7 хиральных атомов углерода, и, сле-
довательно, возможно существование
большого числа изомеров. Из всех
фороболов только производные 4₽-
форобола являются активными, но не
производные 4а стереоизомеров.
Предполагают, что их деградация про-
исходит замедленно и поэтому они
могут непрерывно активировать фермент протеинкиназу С. Это,
в свою очередь, стимулирует специфические протеазы, катализи-
рующие реакцию распада фермента с последующим серьезным
нарушением клеточной регуляции, сопровождающейся индукцией
549
опухолевого роста.
Необходимо также помнить, что фороболы еще стимулируют
клеточную пролиферацию, митогенез лимфоцитов, продукцию
простагландинов, дегрануляцию нейтрофилов, синтез ДНК, изме-
нения в свойствах мембран и др. Однако в связи с высокой
биологической активностью, эфиры форобола представляют опре-
деленную опасность для здоровья работающих с ними лиц. Поэтому
операторы должны надевать перчатки, защитные очки и прочую
одежду, чтобы защитить кожу и глаза от контакта с этими веще-
ствами.
Для инактивации эфиров форобола их необходимо растворить
в органическом растворителе, смешивающимся с водой, в равном
объеме 10% водного раствора натрия гидроксида. Гидролиз эфиров
завершается в срок до одного часа выдержки, после чего щелочной
раствор становится безопасным.
Водные растворы эфиров форбола могут быть обезврежены
добавлением около 10 % по объему натрия гипохлорита.
Способствует росту опухолевых клеток (-)-индолактам, являю-
щийся производным индола. Его эффективность в других биоло-
гических системах различна, что может быть объяснено различным
аффинитетом к С-подтипам протеинкиназы; стереоизомер (+)-ин-
долактам не обладает стимулирующим действием, но его можно
использовать в качестве субстанции для негативного контроля.
В таком же плане могут быть использованы новые, человече-
ского происхождения, гормоноподобные ростовые факторы: инсу-
лино-подобные (IGF 1 и 2), тромбоцитарные (PDGF, от англ. Platelet
Derived-Growth-Factors), трансформирующие ростовые (TGFan £),
некроза опухолей (TNFa).
IGF 1 — полипептид, состоящий из 70 аминокислотных остат-
ков, спирально закрученный и удерживаемый в таком состоянии
благодаря трем S-S — мостикам между остатками цистеина (рис.
157). Первый из них соединяет дисульфидной связью 6-й и 48-й
остатки, второй — 18-й и 61-й остатки, третий—47-й и 52-й остатки.
IGF 1 является пептидным гормоном, влияющим на рост и обмен
веществ организма после рождения. Его секреция находится под
влиянием уровня гормонов роста в сыворотке крови и характера
питания ребенка. IGF 1 влияет на рост и дифференциацию многих
типов животных клеток, включая мышиные клетки — предшест-
венники эритроцитов последней стадии и эпителиальные клетки
млекопитающих, крысиные миобласты, олигодендроциты, клетки
черепа и клетки зобной железы; человеческие хондроциты и
550
Рис. 157. Полипептид IGF-
1 (схема).
адренергические нейробластные клетки.
Вариантом IGF 1 является укорочен-
ный на N-терминальный трипептид-моз-
говой IGF, изолированный из мозга плода
и взрослого человека. Этот пептидный
гормон важен для роста и метаболизма
центральной нервной системы. Он в 4
раза сильнее IGF 1 по стимуляции синте-
за ДНК в фетальных мозговых клетках
крыс и более выраженно реагирует в
перекрестных реакциях с рецепторами
для IGF 1 на мембранах мозга у плода и взрослого человека.
IGF 2 — полипептид, состоящий из 67 аминокислот (ММ-7470).
Как и IGF 1 является сильным митогеном для многих клеток. Как
полагают, он — важный регулятор роста соматических клеток,
стимулирующий их размножение.
Тромбоцитарный ростовый фактор PDGF — гомодимерный,
мультифункциональный полипептид, который действует как силь-
ный митоген в отношении широкого набора эндотелиальных кле-
ток фибробластов. Его рассматрвают как вещество, играющее
важную роль в восстановлении тканей и воспалении. PDGF —
хемоаттрактант для фибробластов, гладкомышечных клеток, моно-
цитов и нейтрофилов, что играет немаловажную роль в заживлении
ран.
В отвечающих на PDGF клетках индуцируются оборот фосфа-
тидилинозита, метаболизм простагландинов и активируются фос-
фолипазы. В человеческих фибробластах кожи, стимулированных
PDGF, возрастает в три раза коллагеназная активность. Полагают,
что он принимает участие в трансформации клеток и вовлекается
в процесс атеросклероза.
Следует отличать PDGF от фактора активации тромбоцитов
(PAF, от англ, platelet activating factor) — это фосфолипид, образу-
емый тромбоцитами, макрофагами, нейтрофилами и эозинофила-
ми под влиянием воспалительного стимула.
Он синтезируется из клеточ-
ных мембран под влиянием фос-
фолипазы А 2 вначале в форме
предшественника — lyso-PAF,
который затем превращается в
PAF под влиянием ацетил-транс-
феразы (этот же энзим катали-
зирует реакцию обратной транс-
формации PAF в lyso-PAF).
О
СН3
О— (СН2)п,—СН3
сн3
;СН3
сн3
RAF; п может быть 1S или 17
551
PAF — исключительно потентная субстанция. Она индуцирует
агрегацию тромбоцитов, будучи в фемтомолярных концентрациях
(1015). После парентерального введения или вдыхания PAF вызы-
вает бронхоспазм и повышенную проницаемость микрокапилля-
ров; количество кровяных пластинок и нейтрофилов в крови падает
при одновременном клеточном скоплении в легких. Введенный
внутрикожно он вызывает формирование пустулы и повышение
температуры при устойчивой клеточной инфильтрации. PAF обла-
дает сильным хемотаксическим эффектом, вызывая накопление
нейтрофилов й эозинофилов, активирует их в плане образования
других воспалительных компонентов (супероксидных анионов Oi
и эйкозаноидов).
СзоНцСООН; СН^СН^- сн—сн— сн2- сн» сн - сн2- сн= сн-сн2
эйкозанкарбоновая НООС - (СН,),— CH = СН
кислота _ z
эикозантетраен-5,8,11,14-овая кислота
(арахидоновая кислота)
Ингалированный PAF повышает экскрецию лейкотриенов с
мочой, возможно потому, что он стимулирует повышенное накоп-
ление их в легких как медиаторов воспаления. Лейкотриены ранее
называли "медленно реагирующими субстанциями" при анафилак-
сии (SPS-A).
TGF — гормоноподобные вещества, которые могут индуциро-
вать временный неопластический рост и дифференциацию нор-
мальных клеток. Фибробласты в среде с TGF не нуждаются затем
в субстрате для роста и не отвечают на межклеточные контакты.
После удаления TGF из культуральной среды клетки возвращаются
к нормальному росту.
TGFa близко родственен эпидермальному ростовому фактору
(EGF) благодаря обладанию тем же клеточным рецептором.
552
TGFa — одиночный полипептид, содержащий 50 аминокислотных
остатков; он взаимодействует с тем же рецептором, что и EGF и
проявляет такую же потенцию в индуцировании тирозин-киназной
активности, связанной с EGF-рецептором. TGFa, в отличие от EGF,
"запускает" эмбриогенез и активацию протоонкогенеза в течение
регенерации поврежденных тканей.
TGF0 — кислотостойкий полипептид, состоящий из двух иден-
тичных субъединиц со 112 аминокислотами в каждой. Его ММ-25
кДа; изолированы TGFP — 1 и 2 с 71% гомологией между ними и
сходной функциональной активностью. В частности, TGFP играет
важную роль в генезе опухолей, развитии эмбриона, тканевой
репарации и иммунорегуляции. TGFP — ингибитор роста для
большинства типов клеток, но может стимулировать образование
внеклеточных матриксных белков типа фибронектина и коллагена.
TGFa и Р выступают синергистами при трансформации различных
клеточных линий.
В регуляции поведения клеток важную роль отводят семейству
из четырех синдеканов, являющихся поверхностными протеогли-
канами клеток. Из них более детально изучен синдекан - 1.В
его так называемом внешнем домене, или эктодомене содержатся
ковалентно связанные цепи гликозаминогликанов, состоящие пре-
имущественно из гепарансульфата, а также из хондроитинсульфа-
та. Синдекан - 1 селективно связывает различные внеклеточные
матриксные молекулы через его гепарансульфатные цепи. Одно-
временно он может связывать ростовые факторы и выступать
вектором сайта стимуляции роста; здесь (в месте взаимодействия
синдекана-1 с матриксом) иммобилизуется рецепторный комплекс
для тирозинкиназы (рис. 158).
Рис. 158. Внеклеточный матрикс (а): ЭЦМ — экстрацеллюлярный матрикс. БМ
— базальная мембрана, содержащая коллаген типа IV и ламинин, КН — коллагеновые
нити 1 типа, ФН — фибронйктин, ФБ — фибробласт; селективное связывание
внеклеточных матриксных молекул синдеканом-1 (б): 1 - фибронектин, 2 - фиброб-
ластный ростовой фактор - ФРБ, 3 - интегрин, 4 - протеогликан - синдекан, 5 -
рецептор ФРФ на тирозин-киназном комплексе, 6 - гепарансульфатные цепи, 7 -
димеризация рецепторов ФРФ. 8 - внутриклеточное фосфорилирование с последу-
ющим индуцированием сигнала, 9 - эффектор, 10 - клеточная мембрана, 11 -
структуры цитоскелета (по М. Ялканену, К. Элениусу и А. Раприджеру, 1993).
553
Плазматические клетки и ряд человеческих В-клеточных опу-
холевых линий экспрессируют синдекан-1 преимущественно в
виде гепарансульфат-протеогликана. Способность В-клеточных ли-
ний связывать фибронектин и фибриллярные коллагеновые мат-
рицы четко коррелирует с их экспрессией синдекана-1 на клеточ-
ной поверхности. Экспрессия этого протеогликана также сопря-
жена с пролиферацией клеток и, как предполагают, с малигниза-
цией их (от лат. malignus — злокачественный).
Кроме стимуляции роста синдекан-1 также участвует в регуля-
ции морфологии клетки благодаря содействию организации эле-
ментов цитоскелета, прежде всего, эпителиальных клеток.
Менее изученными гепарансульфат-протеогликанами являют-
ся синдекан-2 (фиброгликан), синдекан-3 (N-синдекан) и синдекан-
4 (риудокан, амфигликан). Последнему приписывают внутрикле-
точную функцию возможного связывания других мембранных
белков или цитоскелета.
Более "многоликим" представляется TNPa — полипептид с ММ
17 к Да. Он — медиатор воспаления и играет выдающуюся роль в
патогенезе эндотоксического шока, вызванного грамотрицатель-
ными бактериями. TNPa стимулирует активность коллагеназы и
продукцию простагландина Е2 синовиальными клетками, актив-
ность остеокластов и резорбцию костной ткани, способствует
ангиогенезу, стимулирует пролиферацию фибробластов, индуци-
рует высвобождение ИЛ 1 и 6, PDGF. Вместе с тем, он — цитоток-
сический и цитостатический ингибитор опухолевого роста, вовле-
кается в патологические процессы типа раковой кахексии, или
истощения (от греч. kakos — плохо, oexis — состояние) и цереб-
ральной малярии (от лат. cerebrum — головной мозг). Установлено,
что убитые клетки Staphylococcus aureus стимулируют продукцию
TNFa в культурах мононуклеарных фагоцитов или моноцитов
крови человека. Предварительная обработка клеток бисбензили-
зохинолиновым алкалоидом тетрандрином подавляет эту продук-
цию. Поэтому высказано предположение о том, что тетрандрин
можно использовать для лечения воспал ительных заболеваний, при
которых TNFa играет основную роль.
С добавлением или без добавления фороболов и ростовых
факторов клетки опухолей теряют присущий нормальным клеткам
фибронектин, округляются и в таком виде продолжают размно-
жаться и существовать в ряду поколений. Привнесение к ним извне
фибронектина сопровождается восстановлением их способности
554
к адгезии, но без реверсии в нормальные клетки по физиолого-би-
охимической сущности, то есть они все равно остаются опухоле-
выми.
Следует отметить, что в культуру давно введены клетки крови,
соединительной ткани и другие (см., например, раздел 11.4). Сое-
динительнотканные клетки — фибробласты (от лат. fibra — нить,
волокно; от греч. blastos — почка, росток, отпрыск) характеризу-
ются выраженной адгезивностью благодаря обильной продукции
ими фибронектина (см. рис. 152), поэтому их монослойные куль-
туры всегда удаются с определенным успехом.
В искусственных условиях удается культивировать клетки эпи-
дермиса кожи, то есть диплоидные и "продвинутые" в дифферен-
циации клетки. Направленность такой работы, главным образом,
практическая и результаты ее бесценны для ожоговых клиник. По
сообщению немецких ученых (1988) за 6—7 недель из кусочка
кожи размером 2 см2 можно получить "кожу" размером 1—2 м2.
Для этого у пациента, которому предстоит пересадка кожи, берут
кожный лоскут и помещают в раствор специальных ферментов,
эпидермис отделяется от подлежащих слоев кожи, его промывают
и переносят в питательную жидкость. Клетки делятся, а в присут-
ствии стимуляторов их рост еще более ускоряется.
11.3. Получение инсектопатогенных вирусов в клеточных
культурах. Культуры клеток и тканей насекомых приобрели боль-
шое значение в различных биологических науках (эндокринология,
биохимия, вирусология и др.) и в биотехнологии. Это стало воз-
можным благодаря разработкам соответствующих сред, на кото-
рых хорошо развиваются указанные биообъекты, а на объектах, в
свою очередь, можно с успехом поддерживать рост соответствую-
щих патогенов, поражающих растения, животных и человека
(членистоногие паразиты и вирусы). Более того, на культурах
клеток и тканей насекомых можно обеспечить продукцию виру-
лентных инсектовирусов и рекомбинантных белков, используя
вирусы насекомых в качестве векторов. В этой связи велика роль
таких культур в контроле за чумой, используя биотехнологию.
Прогресс в получении клеточных линий насекомых был после-
довательно обеспечен трудами В. Трагера в конце 30-х годов
текущего столетия, С. Вьятта (1956), Т. Д. С. Грейса (1962). Грейс
модифицировал среду Вьятта, включавшую 21 аминокислоту, пять
солей, три сахара, три органические кислоты (pH=6,3—6,5), доба-
вив в нее 9 витаминов комплекса В. В результате ему удалось
изолировать 4 культуры, ставшие стабильными линиями клеток
555
насекомых. В последующие годы были получены самые различные
клеточные линии от насекомых, относящихся к различным поряд-
кам, но с преобладанием Diptera (мухи, комарц) и Lepidoptera —
бабочки (от греч. di(s) — два, дважды, двух; pteron — крылышко;
lepis — чешуя). Для них наиболее часто используют среды Дж.
Митсухаси и К. Марамороша (среда ММ), Шилда и Санга (среда
М3), Шнейдера; Эхальера и Оганесяна (среда D-22), IPL-41 и другие,
фирма Sigma (США) выпускает указанные среды в сухом виде.
Технология выращивания вирусов на культурах клеток насе-
комых принципиально'мало чем отличается от выращивания дру-
гих вирусов на клетках животных и человека (см. раздел 11.1).
11.4. Интерфероны. Интерферон открыт А: Айзексом и Дж.
Линдеманом в 1957 г. в клетках цыпленка, зараженных вирусом
гриппа. Это видоспецифическое белковое вещество, синтезируе-
мое лейкоцитами в ответ на воздействие интерфероногенов. При-
менительно к млекопитающим и, прежде всего, человеку под
названием "лейкоциты" объединяют все белые клетки крови (от
греч. leikos —. белый, kytos — ячейка, клетка) — сегментоядерные
лейкоциты (нейтрофильные, эозинофильные, базофильные), или
микрофаги, лимфоциты (Т и В) и моноциты. Все эти клетки
являются ядерными и участвуют в обеспечении постоянства внут-
ренней среды макроорганизма (гомеостаза), включая неспецифи-
ческую и специфическую защиту, иди Иммунитет. Однако сегмеи-
тоядерные лейкоциты относят к разряду полинуклеарных, тогда
как моноциты — к разряду мононуклеарных. Мононуклеарными
фагоцитами (наравне с моноцитами) являются также гистиоциты,
или макрофаги. К ним относят макрофаги соединительной ткани,
звездчатые клетки Купфера в печени, альвеолярные макрофаги в
легких, свободные- и фиксированные "макрофаги в лимфоузлах й
селезенке, гистиоциты в коже, остеокласты в костной ткани,
фиксированные макрофаги в костном мозге, макрофаги в серо-
зных полостях (плевральной и брюшной); макрофаги в нервной
ткани (клетки микроглии).
С учетом топологии и/или функции макрофаги подразделяют
еще на резидентные, эксудативные (макрофаги воспалительного
эксудата), активированные, индуцированные.
Моноциты, макрофаги и их предшественники объединены в
так называемую систему мононуклеарных фагоцитов (СМФ). Ге-
неалогия (от греч. genea — рождение, происхождение, logos —
обсуждение) клеток крови изучена достаточно глубоко.
Лейкоциты и другие клетки млекопитающих, в частности, при
556
заражении вирусами продуцируют не один интерферон, а больше,
объединяемых в семейство интерферонов, ингибирующих продук-
тивный цикл репликации вирусов. Вот почему они являются
оружием первой линии защиты против вирусных инфекций.
Однако в обычных (неиндуцированных) клетках интерфероны
не выявляются. Некоторые характеристики отдельных интерфе-
ронов приведены в таблице 56.
Таблица 56 Некоторые характеристики интерферонов человека
Тип интер- ферона Характеристики
клетки - проду- центы индук- тор число генов интер- фв- рона локали- зация в хро- мосо- мах нали- чие интро- нов в генах стабиль- ность при рНг 'актив- ность в присут- ствии ддс длина сигналь- ной после- да ва- тельно- стти (ч.амк-т) ч. амк-т ММ. Ла
в пер- вичной моле- куле в окон- чатель- ной моле- куле
(X лейко- циты пери- фери- ческой крови вирус- ная индук- ция, двух- нитевая РНК 14 „9 да v -да 23 166 143 17
₽ фиб- робла- сты 2 (54) 9,2,5 — да «а 21 166 145 17 ’
г лимфо- циты митоге- ны (для несен- сибили- з и ро ван- ных ливр фоци- , Т9В|. We$n- фичес. АГ (сенси- билизи- рован- ные лим- фоциты) 1 12 + нет нет 20 166 « * «•« 146 17
Примечание: Л АС — натрия додецилсульфат; ч. амк-т — число аминокислот, ММ —
молекулярная масса в килоддльтонах; АГ — антигены.
Как следует из таблицы размеры молекул интерферонов близки
по числу аминокислот и молекулярной массе, хотя по другим
признакам они различны; интерфероны Р и у являются гликопро-
теинами, тогда как интерферон а — протеином.
557
Интерфероны — клеточные белки и поэтому они видоспеци-
фичны, то есть каждому виду животного свойственен свой интер-
ферон, но не являются вирусоспецифическими. При смешанной
вирусной инфекции один вирус подавляет другой за счет интер-
фероногенности первого — феномен вирусной интефе-
р е н ц и и. Иногда эта видоспецифичность очень узкая, например,
для курицы, утки, мыщи и крысы, но не перекрестно в группах
птиц и грызунов или между труппами. Однако есть исключения —
человеческий интерферон защищает клетки крупного рогатого
скота лучше, чем коровий интерферон.
Человеческие интерфероны а и Р продуцируются преимуще-
ственно лейкоцитами, В-лимфобластами и соединительнотканны-
ми клетками мезенхимного происхождения — фибробластами в
ответ на вцрусную инфекцию. Интерферон у прежде называли
иммунным, или тип 2; он образуется несенсибилизированными
лимфоидными клетками Т-лимфобластами в ответ на митогены, и
сенсибилизированными лимфоцитами при стимуляции специфи-
ческими антигенами (таблица 57).
Таблица 57. Виды индуцируемых интерферонов
Интерфероны Система
индуктор клетки-продуценты
лейкоцитарный-сс вирус лейкоциты периферической крови
лимфобластный-а и_ В-лимфобласты
фибробластный-{! п фибробласты
имунный-у митоген лейкоциты периферической крови или Т-лимфобласты
специфический антиген лимфоциты
Установлено, что индукторами интерферонов могут быть: инак-
тивированные температурой или УФА вирусы гриппа, рео-, рота-
и другие вирусы; двухнитевые РНК (например, реовирусная);
некоторые полианионные вещества (бактериальные эндотоксины,
поликарбоновые кислоты, сополимеры пирана); синтетические
двухнитевые полирибонуклеотиды, например, полиИ: полиЦ —
558
можно достичь супериндукции интерферона, если обрабатывать
клетки полиИ: полиЦ вместе с циклогексимидом (ингибитором
синтеза белка), а спустя 5 часов — актиномицином Д. Механизмы
индукции интерферона до конца еще не изучены и трудно объяс-
нить почему, например, двухнитевые РНК стимулируют образова-
ние интерферона, а двухнитевая ДНК не обладает аналогичным
действием.
На практике интерферон-а выделяют из лейкоцитов при низ-
коскоростном центрифугировании свежевыделенной крови чело-
века. Лейкоциты переносят в культуральную среду, содержащую
либо сыворотку крови человека или казеин молока, в среду вносят
вирус — интерфероноген (вирус Сендай или вирус ньюкаслской
болезни), выдерживают в течение ночи, после чего лейкоциты
отделяют центрифугированием, вирус — интерфероноген инакти-
вируют любым из приемлемых способов. Супернатант (от лат.
supematans — плавающий на поверхности), или надосадок пред-
ставляет собой нативный интерферон. Его лиофильно высушивают
и выпускают в ампулах. Это — пористый, серовато-коричневый
порошок, легко растворимый в воде. Растворенный препарат имеет
розовато-красноватый цвет и слегка опалесцирует. Из нативного
интерферона можно получить концентрированный интерферон
путем очистки колоночной хроматографией на сефадексах. Пол-
ученный препарат после высушивания имеет вид пористого по-
рошка серовато-белого цвета, хорошо растворимый в воде. Тот и
другой интерфероны должны быть стерильными.
Активность препаратов определяют титрованием на первичных
культурах клеток, например, кожно-мышечной ткани эмбриона
человека с вирусом везикулярного стоматита. Противовирусная
активность (так называемая удельная активность) нативного ин-
терферона должна быть не менее 32 единиц, концентрированного
— 100 единиц. Для очистки интерферона можно прибегнуть и к
высоко эффективной жидкостной хроматографии.
Интерферон Р получают из фибробластов, выращенных в
монослойной культуре, индуцированной полиИ: полиЦ в присут-
ствии циклогексимида и актиномицина Д. Обычно интерфероны
продуцируются в малых количествах (около 1 мг на 10 л тканевой
культуральной жидкости) и, к тому же, после 48—72 часов клет-
ки-продуценты отмирают. Вот почему производство лейкоцитар-
ных интерферонов относят к разряду дорогостоящих и экономи-
чески— мало выгодных. Поэтому успешно завершившиеся работы
559
по генно-инженерному интерферону помогли решить и эти про-
блемы.
В начале 80-х годов XX в. британская фирма The Wellcome
Foundation LTD передала на клинические испытания природную
смесь а-интерферонов, полученную в глубинной суспензионной
культуре вирусоиндуцированных человеческих лимфобластов.
Препарат был назван "велфероном”. Выпускается в виде стериль-
ной, прозрачной, бесцветной жидкости по 1 мл (3 мегаединицы-
МЕ). К 1988 году уже три фирмы выпускали а-интерфероны:
1) выше названный велферон — фирма Wellcome Found.,
2) интрон А — фирма Kirby-Warrick,
3) роферон А — фирма Roche.
Интрон А и роферон А являются генно-инженерными препа-
ратами, содержащими лишь один субтип а-интерферона и разли-
чающиеся по одной аминокислоте. Так, роферон А обозначается
как интерферон а2а; он содержит в полипептидной цепи лизин в
позиции 23. Интрон А - это а2Ь-интерферон, содержащий аргинин
вместо лизина. Как оказалось, роферон А индуцирует антителооб-
разование примерно у 25% больных, у которых благодаря этому
нейтрализуется введенный интерферон. Технологическая схема
получения генно-инженерных интерферонов (как одна из возмож-
ных) принципиально сводится к следующему:
1. индукция синтеза и выделение интерфероновой мРНК из
клеток,
2. получение кДНК, комплементарной интерфероновой мРНК
из лейкоцитов,
3. встраивание кДНК в плазмиду,
4. введение реконструированной плазмиды в клетки Е. coli,
5. размножение бактерий, содержащих реконструированную
плазмиду, в культуральной среде,
6. сепарирование клеток Е. coli,
7. дезинтеграция и экстракция клеток Е. coli,
8. осаждение (например, полиэтиленамином) с последующим
центрифугированием,
9. высаливание интерферона из супернатанта аммония сульфа-
том,
10. диализ осадка интерферона,
И. растворение интерферона, пропускание раствора через
колонку с иммуносорбентом (пришитыми моноклональными анти-
телами, рис. 159).
560
12. элюция интерферона с последующей хроматографией на
целлюлозном катионообменнике.
Рис. 159. Схема очистки
пых антител (МкАТ): 1 - диа-
лизат с интерфероном и дру-
гими бактериальными белка-
ми, 2 - интерферон, 3 - балла-
2 стные белки, 4 - МкАТ, 5 -
интерферон, связывавший с
МкАТ, 6 - слабая кислота.
Из указанных 12 стадий только 8 последних фактически реа-
лизуются в производственных условиях, тогда как первые 4 стадии
выполняются в лабораторных условиях. Кстати сказать, эти 4
стадии наиболее трудные и сложные (рис. 160).
Рис. 160. Этапы клониро-
вания гена интерферона:
мРНК - интерфероновая
мРНК из индуцированных
клеток, кДНК1 - комплемен-
тарная ДНК однонитевая,
кДНК2 - комплементарная
ДНК двухнитевая, 3 - обрат-
ная транскриптаза, 4 - синтез
двуспиральной кДНК, 5 - кон-
цевая трансфераза, 6 - ре-
стрикция, 7 - кДНК с липкими
концами, 8 - отжиг, 9 - интер-
фероновая ДНК, 10-включе-
ние в клетки E.coli плазмиды-
химеры, КО - клонирование
и отбор колоний, содержа-
щих клетки с заданными
плазмидами, БРП - "библио-
тека" рекомбинантых плаз-
мид.
Бактерии с генами интерферона человека продуцируют специ-
фический белок в количестве 200—250 мкг/л бактериальной сус-
пензии. Кроме Е. coli удается получать человеческий интерферон
с помощью Вас. subtilis, способную секретировать синтезируемые
белки в окружающую среду (у Е. coli они накапливаются в
561
периплазматическом пространстве), а также с помощью
Saccharomyces cerevisiae, растущих на средах с более дешевыми
субстратами, на них не действуют фаги, они крупнее бактерий и
легче сепарируются, в их клетках осуществляется процессинг
преинтерферонов. Применяют также культуры бактерий из родов
Methylomonas, Pseudomonas, Salmonella. Все названные микроор-
ганизмы конститутивно (не адаптивно) образуют интерфероны, не
отличающиеся от природных. Против всех классов интерферонов
получены моноклональные антитела, с помощью которых проводят
быструю и высококачественную очистку продуктов, применяя
иммуноафинную хроматографию.
В молекуле интерферона имеются два консервативных домена
— один локализуется на NH 2-конце, а другой — на СООН-конце.
Первый, очевидно, помогает связыванию с рецептором на повер-
хности клетки, а второй—моделирует это связывание и опосредует
другие биологические функции.
Интерфероны а, р, и у иммунологически различны и, напри-
мер, а-антисыворотка не инактивирует гетерологичные интерфе-
роны.
Интерфероны обладают двумя типами биологической активно-
сти — противовирусной и противоклеточной. В отношении виру-
сов действие трех интерферонов сравнимое по эффективности, но
в отношении клеток более активен интерферон у, причем против
опухолевых клеток он активнее, нежели против нормальных кле-
ток.
На практике интерфероны применяют при вирусных инфек-
циях, ревматоидном артрите (интерферон')), при иммунопатологии
и в онкологии.
11.5. Получение и использование гомо-, гетеро- и синкариоти-
ческих гибридов. Материалы этого раздела базируются на данных
по геномной инженерии и, в частности, на клеточной инженерии,
или соматической гибридизации.
Понятия о гомо- и гетерокарионах чаще относили к дейтеро-
мицетам и некоторым другим грибам, которым присущ так назы-
ваемый парасексу а льный процесс размножения. При
этом наблюдается слияние (копуляция) нитей одного и того же или
562
различных типов спаривания, происходит объединение в одной
клетке содержимого протопластов, включая ядра. Так возникают
гомокарионы и гетерокарионы, функционирующие самостоятель-
но, не сливаясь. Образующиеся в результате деления "дочерние"
ядра распределяются по мицелию. Время от времени такие ядра
сливаются, затем делятся при помощи меойза. Слившиеся ядра
называют синкарионами.
Владея техникой клеточно-инженерного эксперимента, можно
искусственно получать гомо-, гетеро- и синкариотические особи.
Это нашло воплощение, например, в иммунобиотехнологии. Им-
мунобиотехнология является составной частью биотехнологии и
связана с получением и производством иммунопрепаратов, обла-
дающих свойствами антигенов или антител. Теоретической базой
ее является учение об иммунитете, практической базой — иммун-
ная система макроорганизма.
11.5.1. Основные представления об иммунной системе. Иммун-
ная система — это система органов и клеток, реагирующая на
генетически чужеродную информацию и участвующая в защите
макроорганизма от такой информации. Иммунная система во
взаимодействии с другими системами (нервной, эндокринной,
сердечно-сосудистой, пищеварительной и др.) обеспечивает защи-
ту организма от внедрения и развития в нем патогенных микробов
(вирусов, бактерий, грибов, протозоа), а также от воздействия
антигенов разной другой природы и специфичности (тканевые
трансплантаты; растительные и животные яды; аутоантигены, опу-
холи). Она выполняет функцию иммунологического надзора во
благо поддержания постоянства внутренней среды (гомеостаза)
макроорганизма, обеспечивая его целостность и индивидуальность.
Иммунная система состоит из трех главных компонентов:
клеточного, молекулярного и генетического. Первый из них вклю-
чает макрофаги, Т- и В-лимфоциты, второй — пять классов иммун-
ных глобулинов (Ig): IgM, IgG, IgA, IgE, IgD, третий — множество
генов (таблица 58).
563
Таблица 58. Генетический компонент иммунной системы
Название функция » |
Иммуноглобулиновые гены Определяют» структуру вариабельных (v) и | константных (С) областей, Н (ц, у, а, е, 5).- и L (к, X - цепей иммуноглобулиновых генов на В- лимфоцитах; v - область также образует часть рецепторов для антигенов на Т-лимфоцитах .,
Гены главного комп- лекса гито- совме- стимо- сти (МНС), или HLA+- гены а- « области I (HLA-D) Ответственны за взаимодействия макрофагов, Т- и В-лимфоцитов (CI; от англ, cellinteraction)
1г-гены Определяют индивидуальную чувствительность организма к заболеванию благодаря регулированию силы ответа- на соответствующие антигены (от англ, immune response)
гены, кодирующие 1а-антигены Ответственны главным образом за смешанную реактивность лимфоцйтов и за реакции типа "трансплантат против хозяина" (от англ. Immune antigen)
гены иммуно- медиаторов вовлечены в синтез таких иммуномедиаторов, как цитцкины (лимфокины, включая интерлейкины • (IL); у-и'нтерферон, нейротрансмиттеры,' фактор задержки миграции макрофагов (MIF) и др.
другие области определяющие: большой и малый трансплантационные антигены, связанные с отторжением трансплантантов и опухолей; взаимодействия с патогенными вирусами; синтез компонентов сйстемы комплимента
Гены дифференциации клеточных антигенов и рецепторов определяющие лимфоцитарные антигены (Lyt-1, 2,3 для различных субпопуляций Т-лимфоцитов; 1 Lyb-3, 5 и др., дифференциально экспрессирующиеся на определенных В- лимфоцитах); Fc-рецепторы (FcRy - на В- лимфоцитах, макрофагах и Т-супрессорах; FcRp на макрофагах и Т-хелперах), рецепторы комплемента на субпопуляциях В- и Т-клеток
Примечание: +HLA — от англ. Human Leucocyte Antigen. МНС состоит из to генов,
расположенных в 6-й паре хромосом.
Иммунная система — одна из самых гетерогенных и сложных
систем организма. У человека она включает примерно 2 • 1012
лимфоцитов и 1О20 молекул антител, обладающих с одной стороны
564
общностью организации, а с другой -— различной специфичностью.
В характере иммунного ответа исключительная роль принадле-
жит кооперативным взаимрдёйствиям иммунокомпетентных кле-
ток (ИКК) — макрофагов, лимфоцитов и вспомогательных дендрит-
ных клеток. ИКК содержатся в периферической крови, селезенке,
лимфоузлах, зобной железе и, следовательно, кооперативные вза-
имодействия ИКК могут иметь место в различных частях тела. В
таких взаимодействиях ведущую роль играют поверхностные ан-
тигены, кодируемые HLA; в них принимают также участие различ-
ные медиаторы (посредники), или цитокины (рис. 161). К цитоки-
нам относят разнородные вещества, участвующие в физиологиче-
ских и патофизиологических реакциях лейкоцитов^и других кле-
ток, и называемые клеточными регуляторами.
Рис. 161. Кооперативные взаимодействия клеток иммунной системы с участием
медиаторов, или цитокинов: МФ — макрофаг, СК — стволовая клетка, ЭБ —
эквивалент бурсы, В —• лимфоцит, ТИ — тимус, Кп — клетка памяти, ПК —
плазматическая клетка, КомК — коммитированные клетки, Тс — Т-супрессоры, Тх
— Т-хелперы, АГ — антиген, НК — натуральные (естественные) киллеры, КК —
цитотоксические клетки-киллеры, ИЛ — интерлейкины, ФР — факторы роста, Ig-ы
— продукция циркулирующих антител: IgG, IgM, IgA, IgD, IgE, ВИЧ-вирус иммуно-
дефицита человека.
565
Цитокинам присущ ряд общих признаков: они низкомолекулярны
(ММ менее 80 кДа) и гликозилированы, действуют в пикомольных
(пМ) концентрациях, выделяются местно и непостоянно, выступа-
ют как паракринные или аутокринные (но не эндокринные) фак-
торы (от греч. krino — отделяю; para — возле, при; autos — сам;
endon — внутри), взаимодействуют с высокоафинными рецепто-
рами на клетках, изменяют спектр клеточных РНК и функциональ-
ную активность клетки после взаимодействия с рецепторами,
регулируют силу и продолжительность реакций воспаления и
иммунитета. Отдельные цитокины проявляют множественную ак-
тивность; ответ на цитокины зависит от их концентраций и типов
клеток. К тому же они взаимно индуцируют секрецию друг друга,
проявляя то синергизм, то антагонизм. Так, например, биологиче-
ская эффективность интерлейкинов (ИЛ) 1 и 6 опосредуется друг
другом.
К настоящему времени описано более десятка цитокинов,
включая лимфокины, монокины, интерлейкины, интерфероны и
др. Только интерлейкинов известно 13, первый из них ИЛ-1 открыт
в 1972 г., образуется макрофагами; ИЛ-2, ИЛ-3, ИЛ-5, ИЛ-7, ИЛ-8
и ИЛ-9 — Т-клетками; ИЛ-4 — активированными Т-клетками,
некоторыми линиями В-клеток и тучными клетками; ИЛ-6 —
различными клетками и является интерфероном Р-2. Не все ин-
терлейкины изучены с одинаковой глубиной, поэтому приводимая
характеристика их неравноценна.
ИЛ-1 стимулирует пролиферацию тимоцитов, рост и диффе-
ренцировку стволовых клеток, активирует Т-хелперы и В-клетки,
индуцирует миграцию лимфоцитов; его ММ 12—18 кДа.
ИЛ-2 называют еще Т-клеточным ростовым фактором. Он
активирует Т-клетки, стимулирует дифференциацию и пролифе-
рацию В- и Т-клеток, повышает функцию клеток-киллеров, акти-
вированных лимфокинами.
ИЛ-3 повышает колониеобразование костно-мозговых клеток,
стимулируя гемопоэз, функцию эозинофилов и клеток-киллеров,
активированных лимфокинами.
ИЛ-4, или фактор 1, стимулирующий В-клетки. Он модулирует
переключение синтеза классов 1g. ММ его 15 кДа в негликозили-
рованной форме и 20 кДа — в гликозилированной; индуцирует
синтез IgE, выполняя при этом хелперную функцию, экспрессию
Fc-рецепторов.
ИЛ-5 — гликопротеин с ММ 45—60 кДа, усиливает дифферен-
циацию В-клеток в плазматические клетки на последней стадии,
566
секрецию IgA IgA-несущими клетками, пролиферацию эозинофи-
лов и активацию их зрелых форм.
ИЛ-6 (В-клеточныйх фактор дифференциации, гибридомный
ростовый фактор, гепатоцитарный стимулирующий фактор: ин-
терферон Р-2) стимулирует В-клеточную дифференциацию. Он
включает 184 аминокислоты. ММ его 26 кДа, индуцирует секрецию
Ig В-клетками, рост гибридом, плазмацитом, проявляет синергизм
с ИЛ-1, ИЛ-3 и фактором некроза опухоли (TNF). Предполагают,
что ИЛ-6 является ключевым веществом в семействе цитокинов.
ИЛ-7, содержащий 154 аминокислотных остатка, индуцирует
селективную дифференцировку пре-В- в В-лимфоциты. Ген ИЛ-7
включает 6 экзонов и 5 интронов.
ИЛ-8, или фактор хемотаксиса нейтрофилов, активирует ней-
трофильные сегментоядерные лейкоциты.
ИЛ-9, или цитокин Р40 индуцирует миграцию лимфоцитов. Ген
ИЛ-9 содержит 5 экзонов.
Таким образом, лишь на примере 9 интерлейкинов видна
сложность регуляторных механизмов, включающихся в управле-
ние взаимодействиями клеток иммунной системы. Очевидно, что
in vivo имеется много других регуляторов (цитокинов), которые
будут открыты в ближайшие годы.
Познание и использование функций компонентов иммунной
системы на молекулярном уровне называется молекулярной им-
мунологией. На этом уровне теперь исследуют антигены и анти-
тела, их взаимодействия; рецепторы различных клеток иммунной
системы; гены и генетическое обеспечение этой системы.
В течение 1982—1986 гг. было проведено три Международных
рабочих совещания, посвященных поверхностным лейкоцитарным
дифференцирующим антигенам, обозначенным CD-антигенами
(от первых букв в английских словах Cluster Differentiation —
групповая дифференциация). Некоторые авторы используют обоз-
начения CD в качестве ссылки на антиген, например CD-2 —
синоним Т11 антигена и т. д. Другие используют CD-номер для
того, чтобы указать на антитела, определяющие антиген.
В целях определения CD-антигенов используют 3 главных
метода с моноклональными антителами:
иммунофлуоресцентный, иммуноосадочный (иммунопреципита-
ция) и иммуногистологический на тканевых срезах. К началу 90-х
годов было определено 78 CD-антигенов (CD1—CD78), часть из
которых представлена в таблице 59.
567
Т а б л и ц a 59. Характеристика некоторых лейкоцитарных CD-антигенов
Обозначе- ние антигена Характеристика |
CD1 Кортикальный маркер тимоцитов, исчезающий на последней стадии созревания Т-клетки. Его находят также на клетках Лангерганса+. Молекула CD 1 состоит из трех полипептидных цепей с ММ 49 кДа (CDla), 45 кДа (CDIb) и 43 кДа (СД1с) и каждая из них закодирована соответствующим геном на первой хромосоме. Названные цепи ассоциированы с |3-2-микроглобулином и, следовательно, антиген аналогичен классическим антигенам гистосовместимости, хотя и закодированным на другой хромосоме.
CD2 Т-клеточный антиген, являющийся рецепторной молекулой для эритроцитов барана (ЭБ), называемый еще Т11-антиген и LFA-2. Молекула его имеет ММ 46-50 кДа, и важно подчеркнуть его значимость в Т-клеточном созревании, поскольку антитела против одного эпитопа (см) могут индуцировать клеточную пролиферацию.
CD4 Молекула антигена с ММ 55 кДа представлена приблизительно на 2/3 цикрулирующих Т-клеток у человека, включая большинство Т-хелперов. CD4 называют еще Т-4 антигеном. Его находят также на моноцитах и макрофагах. Он выступает рецептором ВИЧ (вирус СПИД, см. рис. 226).
CD5 Его называют еще Т-1 антигеном. Он имеет ММ 67 кДа; представлен на большинстве Т-клеток у человека, на незрелых В-клетках плода и, в небольшом числе, на покровной зоне В-клеток в лимфоидной ткани взрослого человека.
CD7 Антиген имеет ММ 40 кДа, представлен на многих Т-клетках; он полезен в качестве маркера Т-клеточных неоплазм, когда другие Т-клеточные антигены отсутствуют. CD7 антиген, по-видимому, является Fc-рецептором для IgM.
CD8 Антиген, называемый еще Т-8 антигеном и аналогичный Lyt 2(3) антигену у мышей, имеет ММ 32-33 кДа. Широко используют в качестве маркера для субпопуляции Т-клеток — суппрессов и цитотоксических клеток. Антиген также имеется на синусоидальных клетках селезенки.
CD10 Этот антиген называют еще как антиген общей острой лимфобластной лейкемии, или CALLA-антиген (от ангд Common Acute Lympoblastic Leukaemia Antigen). Его ММ 100 кДа. Он представлен на многих типах клеток (стволовые, ростковые клеточные центры, гранулоциты++, макрофаги, почечный эпителий, фибробласты, желчные канальцы)
CD 16 Молекула антигена имеет ММ 50-60 кДа, она представлена преимущественно на гранулоцитах. Он также может быть найден на НК- и К-клетках, на некоторых микрофагах. Предположительно молекула функционирует как низкоафинный рецептор для Fc- области IgG.
568
Продолжение таблицы 59.
Обозначе- ние антигена Характеристика
CD26 Состоит из двух полипептидных цепей с ММ 120 и 200 кДа соответственно: представлен на активированных Т-клетках.
CD43 Молекула с ММ 95 кДа представлена на многих клетках, включая Т-клетки, гранулоциты, эритоциты, эпителиальные клеточные 1 линии и мозг. |
Примечание: + Клетки Лангерганса локализуются в коже.
+ +К гранулоцитам относят сегментоядерные нейтрофильные, эозино-
фильные и базофильные лейкоциты (микрофаги, полинуклеары)
Благодаря анализу рецепторов обнаружены так называемые
"двойные отрицательные (DN) Т-клетки”, содержащие у/5-рецеп-
торы, контактирующие с CD3 рецепторами на а- и р- Т-клетках,
но не имеющие CD4 и CD8 рецепторов. Таких лимфоцитов мало
в тимусе и еще меньше в других лимфоидных тканях (не более
1%). Считают, что эти Т-клетки выполняют контролирующие фун-
кции на поверхности эпителия (в частности, кишечника) и поэтому
их называют эпителиальными лимфоцитами.
Молекулярный компонент иммунной системы, представлен-
ный пятью классами иммуноглобулинов, формируется за счет
клеток В-ряда (в частности, плазматических), кооперативно взаи-
модействующих с некоторыми другими ИКК (см. рис. 161).
Молекулы антител построены, в основном, по единому плану
(рис. 162). Несмотря на огромное разнообразие антиген-связыва-
ющих мест, вариабельная часть молекул антител представлена 5—6
каноническими вариантами пространственной укладки и, как по-
лагает К. Милстейн (1990), мно-
гообразие их репертуара обус-
1_ ловливается комбинаторикой
канонических структур в соче-
тании с точковыми заменами
аминокислотных остатков в ан-
тиген-связывающих центрах.
Рис. 162. Схематичное строение мо-
лекулы антитела IgGl человека: 1 — иди-
отип, 2 — изотип, 3 — аллотип, Па —
расщепление папаином, Fc — фрагмент,
Fd —. фрагмент, С — константная об-
ласть, У — углевод, V — вариабельная
область, Н — тяжелая цепь, L — легкая
569 цопь-
Являясь белками сыворотки крови, строго специфичные анти-
тела образуются после стимулирования иммунной системы соот-
ветствующим антигеном. Благодаря своей исключительной специ-
фичности (за счет v областей), антитела находят все более широкое
применёние не только в фундаментальных исследованиях, но и в
практике выделения и очистки различных антигенных веществ, в
профилактике, диагностике и лечении многих заболеваний.
11.5.2. Получение моноклональных антител (МкАТ). Вводимый
или попадающий в организм антиген признается и перерабатыва-
ется макрофагами, затем антигенная информация представляется
лимфоцитам, содержащим рецепторы. Так, в частности, стимули-
руется формирование клонов плазматических клеток, продуциру-
ющих специфичные антитела. Качество и количество антител
зависят от качества и дозы антигена, способа его введения в
организм, от вида животного — реципиента. Однако при традици-
онной технологии удается получать преимущественно гетероген-
ные антитела. В то же время "жизнь диктовала" необходимость
получения моноклональных антител — продуктов единого клона
клеток. Важным шагом к этому было открытие Портера в 1972 г.,
связанное с установлением "антительной природы" парапротеинов
в сыворотке крови людей и животных с опухолевым поражением
лимфатической системы (миелома). Миеломные клетки представ-
ляют собой трансформантов плазматической клетки, продуциру-
ющих моноклональные антитела. К сожалению, антиген при этом
до сих пор остается неустановленным. Тем не менее, факт обра-
зования моноклональных антител миеломными клетками был ре-
шающим в создании гибридом, с помощью которых сделан огром-
ный скачок в иммунобиотехнологии. Выдающийся вклад в это
внесли Г. Келер и К. Милстейн (1975), разработавшие метод
получения МкАТ желаемой специфичности и в большом количе-
стве.
Теперь известно, что каждой антигенной детерминанте соот-
ветствует свой клон В-клеток — антителопродуцентов (в организме
человека имеется в среднем 107 клонов В-клеток). Отсюда и
происходит понятие о МкАТ.
Келер и Милстейн слили мышиные миеломные клетки P3-X63-
Ад8, культивируемые in vitro, с лимфоцитами селезенки иммуни-
зированной овечьими эритроцитами мыши. При слиянии образо-
вывались гомо (ди-, поли)кариотические гибридные клетки — гиб-
ридомы, сохраняющие признаки обеих родительских особей, а
именно — способность к неограниченному росту in vitro и in vivo
(признак миеломной клетки) и способность продуцировать специ-
570
фичное антитело (признак В-лимфоцита). Названием "гомо(ди-,
поли)кариотические гибридомы" подчеркивается факт происхож-
дения клеток от одного вида животного. Принципиально этот метод
пригоден для получения антител против любого антигена.
Таким образом, при традиционных методах получения антител
после иммунизации животных имунные глобулины оказываются,
как правило, поликлональными, тогда как с помощью
гибридом получают МкАТ.
Однако следует помнить, что не все клетки подвергаются
соматической гибридизации — специализированные функции ро-
дительских партнеров сохраняются лишь в том случае, если эти
клетки находятся на одной и той же стадии деления. Другие клетки
остаются неслившимися или сливаются с гомологичными особями.
Этапы получения гомо-, гетеро- или синкариотической гибридомы
являются следующими:
1) иммунизация антигеном (например, мыши),
2) взятие клеток селезенки (лимфоциты — плазматические
клетки) от иммунной мыши,
3) отбор миеломных клеток,
4) слияние клеток — получение гетерокариотических гибридом,
5) селекция гибридов,
6) клонирование гибридов,
7) накопление in vivo или in vitro антител и их выделение.
Клетки — гибридомы можно сохранять в замороженном виде,
например в 1% диметилсульфоксиде, или вначале их суспендируют
в смеси 5% диметилсульфоксида и 95% сыворотки в концентрации
106 — 107 клеток/мл. Замораживают при — 70°С и хранят в жидком
азоте.
Теперь имеются линии клеток, которые сохраняются в течение
ряда лет без потери способности выделять 1g. Но есть и коротко-
живущие линии; у таких мышиных линий обычно вначале исчезает
тяжелая цепь 1g (утрата 12-й хромосомы), а затем — легкая цепь
Ig (потеря 6-й хромосомы).
В целях получения гибридом используют разные линии мие-
ломных клеток (таблица 60), из которой видно, что более прием-
лемыми линиями являются 8226AR/NI Р4-1 и Sp 2/0, не продуци-
рующие Ig. Следует иметь в виду и тот факт, что гибридомы легче
получаются при использовании клеток от одного вида животного
(гомокариотические гибриды). Межвидовые гибридизации (гете-
рокариотические гибриды) всегда мало удачны, так как один из
571
партнеров в гибридоме теряет ряд хромосом, включая и те из них, ~
которые контролируют образование 1g. И все же стабильные
гетерокариотические межвидовые гибриды удается получать, на-
пример, при слиянии миеломных клеток мышей с крысиными
лимфоцитами.
Таблица 60. Линии некоторых миеломных клеток, рекомендованные
для получения гибридом
Происхождение Название Образуемые цепи Ig
полное сокращенное легкие тяжелые
человеческая SKO-007 (u = 266ARI — X 3
«1 GM15006TG- А12 — к ¥2
»» PPMJ-8226 8226AR/ NIP4-1 —
крысиная LOU 210.RCY3. .Agl.2.3 Y3 к
мышиная BALB/c P3-X63-Ag8 X63 к у1
II P3-NSI/1- Ag4-1 NS-1 к + —
и Sp2/O-Agl4 Sp2/O —
>11 MPC-11-X45- -6TG1.7 X45 к У2Ь
Примечание: +легкая цепь только образуется, но не выделяется.
Образование гомо(ди-, поли)кариотической клетки является
лишь первой ступенью к возникновению гибридомы. В последую-
щем, после первого митоза, ядра сливаются и образуются синка-
риотические гибридные клетки. Однако замечено, что лишь гиб-
ридомы с неслившимися ядрами могут развиваться дальше. Если
родительские партнеры при этом различались по фенотипу, то
соматический гибрид имеет смешанный фенотип.
В 1980 г. Л. Олесон и X. С. Каплан; К. М. Кроус, А. Линенбах
572
и др. сообщили о получении миеломных линий от людей, пригодных
для накопления и выделения человеческих МкАТ (см. таблицу 60).
В целях получения лимфоидных клеток животному (мыши)
вводят внутримышечно или внутрибрюшинно антиген в дозе при-
мерно 5—l200 мг в расчете на чистый белок. В ответ на сильный
антиген в селезенке 0,05% клеток секретируют IgM и IgG в
соотношении, близком 10:1 из примерно 1% всех секретирующих
клеток. Через 3—4 недели повторно вводят 10—100 мкг антигена.
В течение последующих 2—4 недель проверяют титры антител и
затем перед экстирпацией селезенки в течение нескольких дней
вводят внутривенно небольшие дозы антигена для активации
В-лимфоцитов. Могут быть приняты и другие схемы иммунизации.
Для целей соматической гибридизации необходимы также
миеломные клетки мышей. Обычно используют линию BALB/c, от
которой была получена перевиваемая миелома МОРС-21 (от англ.
Mineral Oil—induced Plasmocytome 21). При этом необходимо:
1) наличие у клеток селективного маркера устойчивости к
определенному ингибитору, например, к 8-азагуанину, к бромде-
зоксиуридину и др.; клетки, устойчивые, к 8-азагуанину, не содер-
жат фермента гипо-
ксантингуанинфос
форибозилтранс-
феразы (ГГФРТ) и
поэтому не способ-
ны использовать
экзогенный гипо-
ксантин для синте-
за пуринов. ГГФРТ
катализирует реак-
цию взаимодейст-
вия гипоксантина и
гуанина с 5'-фосфо-
рибозил-1-пирофосфатом с образованием соответствующих нук-
леотидов — инозин-5'-фосфата и гуанозин-5'-фосфата. Клетки,
устойчивые к бромдезоксиуридину, не обладают ферментом тими-
динкиназой (ТК); ТК катализирует реакцию превращения тимиди-
на в тимидин-5'-фосфат;
2) отсутствие образования нежелательных легких или тяжелых
цепей 1g (происхождение их может быть "миеломное'иЛи "селезе-
ночное").
573
о
он он
гуанозин-5’-фосфат
тимцдин-5'-фосфат
Последний этап на пути к собственно технологическому про-
цессу накопления МкАТ заключается в экстирпации селезенки из
организма мыши и получение В-клеток. Это осуществляют через
3—4 дня после последнего введен г и антигена.
Изолированную селезенку помещают в чашку Петри со средой,
содержащей сыворотку крови и ее промывают (перфузируют).
Клетки центрифугируют, эритроциты лизируют, лимфоциты сус-
пендируют в среде Дальбекко без сыворотки и проверяют их на
жизнеспособность.
На следующем этапе смешивают примерно 107 миеломных
клеток с 10в клеток селезенки. Клетки в среде без сыворотки
выдерживают 45 мин., центрифугируют в конических пробирках.
Супернатант удаляют, осадок встряхивают и к нему добавляют 2
мл. 40% полиэтиленгликоля (ПЭГ) с ММ примерно 1,5—4,0 кДа,
поддерживают температуру 37°С, pH среды 7,6—7,8, выдерживают
30 мин., затем в течение последних 7—8 мин. добавляют среду
Дальбекко с фетальной телячьей сывороткой (5—15%). Клетки в
разведенном ПЭГ осаждают, ресуспендируют в 30 мл. гибридомной
среды с сывороткой и распределяют в лунки объемом 2 мл для
микротитрования. При необходимости среду заменяют на свежую
(среда с гипоксантином и тимидином). В среду добавляют клетки-
фидеры, или "кормилки” (от англ, feeder — приток, питатель) —
преимущественно перитонеальные макрофаги, индуцированные
минеральными маслами) и 1% карбоксиметилцеллюлозу. В качест-
ве индукторов слияния клеток можно использовать вирус Сендай,
температурный шок, электроразряд и др.
В смешанной популяции из миеломных и селезеночных клеток
возможно возникновение нескольких типов гетерокарионов:
1) миеломные клетки — миеломные клетки,
2) миеломные клетки — В-лимфоциты,
3) миеломные клетки — любые клетки селезенки,
4) клетки селезенки — клетки селезенки.
574
На селективной среде погибают партнеры первой системы;
четвертая пара клеток не растет в культуре, поскольку оба партнера
не пролиферирующие. Гибридные клетки третьей пары немного-
численны и слабо растут, они вскоре погибают (к тому же они
отличаются по морфологии вырастающих колоний от второй пары
клеток). Гибридные клетки второй системы быстро растут при
потере хромосом у непролиферирующего партнера (В-лимфоци-
ты). Поэтому формируются две группы гибридом — секретирую-
щие и несекретирующие 1g. Первые медленно растут и поэтому
возникает проблема — не потерять их в последующих операциях.
Таким образом, особенности гибридизации заключаются в том,
что неслившиеся клетки селезенки лишены способности к длитель-
ному культивированию (они исчезают), тогда как миеломные
клетки, напротив, за счет своих потенций к росту могут вытеснить
гибридные клетки. Чтобы этого не произошло, используют селек-
тивные среды. Так, отбирают миеломного родителя с генетическим
дефектом по ГГФРТ, не использующего экзогенный гипоксантин
аминоптерин (4-аминофопиевая кислота)
для синтеза пуринов. Клетки селезенки не обладают таким гене-
тическим дефектом и поэтому миеломные клетки, не синтезирую-
щие нуклеотиды, через определенное время исчезают из культуры.
Гибридомы остаются единственными делящимися клетками. Тогда
через сутки инкубации 50% объема среды заменяют на среду,
содержащую 10‘4 М гипоксантина, 10’5 М аминоптерина и 4 • 10"5
М тимидина (среда ГАТ). Среду сменяют каждые сутки в течение
двух дней, а затем — один раз через 48 часов. Примерно через 2
недели гибридомы начинают активно расти. Необходимо следить,
чтобы не было слишком густого роста — признак развития поли-
клональной популяции; негустая суспензия клеток обеспечивает
рост моноклональных гибридом (выживаемость клеток при этом
невысокая).
В течение 10—14 дней дефектные клетки элиминируются.
Миеломные клетки, устойчивые к бромдезоксиуридину, дефектны
по тимидинкиназе (ТК), а устойчивые к 8-азогуанину, не содержат
ГГФРТ (ГГФРТ). Эти клетки не располагают обходными путями
синтеза пуринов или пиримидинов, но способны синтезировать
575
основания для ДНК и РНК, используя путь, блокируемый аминоп-
терином (а, б).
Углеводы + Аминокислоты
Аминопте—
риновый
блок
Основной путь
Нуклеотиды----» ДНК
ГГФРТ, ТК
Вспомогательный
путь
Основания + Нуклеозиды
8=Азогуанин
[~ т, 1 ГГФРТ
Исходная Г
опухолевая ---S---» Мутантные
клетка
(ГГФРТ+,ТК+)
Мутанты
0ТФРТ’,ТК’2
5=Бромдезокси—
уридин
(б)
опухолевые
клетки
(ГГФРТ“, ТК“)
Основной и вспомогательный
пути синтеза нуклеотидов у
гибридных клеток
Схема получения мутантных
опухолевых клеток
На селективной среде ГАТ клетки ТК- и ГГФРТ- не растут и
погибают; на ней растут природные клетки миеломы, но они
выживают недолго (несколько суток). Лишь гибридные клетки
выживают на среде ГАТ при условии наличия у них генов, коди-
рующих ТК и ГГФРТ (в Х-хромосоме). Схема получения гибридомы
показана на странице 577.
При наличии гибридов производят смену среды Дальбекко (без
аминоптерина) в течение последующих 14 дней и, наконец, добав-
ляют минимальную модифицированную среду Дальбекко. После
этого следует скрининг (от англ, screen — решето, просеивать)
гибридных клеток по образованию антител, используя серологи-
ческие реакции, радиоиммунный, иммунофлуоресцентный или
иммуноферментный анализ. Более чувствительный из них радио-
иммунный метод.
Только часть клеток оказывается продуцентами Ig. Положи-
тельные гибриды сразу же клонируют и реклонируют во избежа-
ние реверсии, или возврата к неактивному состоянию.
Из разных способов клонирования наиболее широкое распро-
странение получил метод предельных разведений, когда стремятся
получить одну гибридомную клетку на ячейку платы с вероятно-
стью в 63%, что следует из распределения Пуассона: f(О) = е'\ где
f (О) — число ячеек без роста, Л, — среднее число клонов на ячейку,
е" — основание натурального логарифма; при Х-1 значение f(0)
576
равно 0,37, то есть гибридомные клетки, посеянные с эффективной
концентрацией одна клетка на ячейку, не вырастут в 37 ячейках
из 100. Фактически вероятностный процент роста гибридомных
клеток будет ниже, поскольку не все из них дают растущие клоны.
Вот почему на практике высевают их с разными концентрациями.
Оценку растущего клона в ячейке проводят визуально.
Миеломные клетки Х63—Ад8 Приготовление суспензии
(селезеночных клеток
подсчет | подсчет
центрифугирование в растворе Эрла (200 д) 10 минут
7 В
10 клеток I Ю клеток
Объединение клеток, центрифугирование (200 д) 10 минут
+ к 1 мл осадка ГАЕ=гемагглютинирующая
1000 ГАЕ вируса Сендай I единица
ресуспендирование
I
20 минут на льду с периодическим встряхиванием
I
30 минут при 37*С с периодическим встряхиванием
Двухкратное отмывание в среде Дальбекко
Ресуспендирование в 50 мл той же среды
Распределение суспензии в лунки планшет (по 1 мл в 48 лунок)
На 'дующий день добавление среды Дальбекко, содержащей ГАТ
I
Смена питательной среды в течение 14 дней (каждый второй или третий
день), удаление 0,5 объема + 0,5 объема свежей среды
. Да «--------гибриды----------* нет
19 т. 8524
577
В последние годы отбор гибридом для клонирования проводят
с использованием автоматизированного оборудования. На после-
дующем этапе получают массовую культуру гибридомных клеток:
1) в культуре ткани — предпочтительный способ, так как 1g
выделяют с большей чистотой;
2) в ферментаторах — суспензионная культура;
3) в организме сингенных животных (мыши, крысы). Накоп-
ленные МкАТ (10—200 мгк/мл среды) выделяют с помощью выса-
ливания аммония сульфатом до 95% чистоты, а, при необходимости,
1g
/о
А
стабилизация
12
Рис. 163. Схема получе-
ния моноклональных анти-
тел с помощью гибридом-
ной техники: АС — агент
слияния клеток, 1 —В-клет-
ки из селезенки иммунной
мыши, 2 — культура мие-
ломных клеток мыши, 3 —
гибридизация, 4 — гибри-
домы. 5 — селекция и кло-
нирование гибридом, 6 —
гибридомы, образующие
моноклональные антитела,
7 —- введение гибридом, об-
разующих моноклональ-
ные антитела, в организм
сингенной мыши, 8 — клет-
ки гибридомы, образую-
щие моноклональные анти-
тела, культивируемые в ви-
де культуры ткани, 9 — вы-
ращивание гибридомных
клеток, образующих моно-
клональные антитела, в
ферментаторе, 1дкж — им-
муноглобулины в культу-
ральной жидкости, Igac —
иммуноглобулины в асци-
тической жидкости. 10 —
высаливание Ig-ов аммония
сульфатом, 11 —стандарти-
зация Ig-ов, 12 — лиофили-
зация Ig-ов.
578
Рис. 164. Культиватор гибридомных
клеток (общий вид) фирмы IBF
biotechnics.
Перевод гибридных клеток в большие объемы среды при
крупномасштабном культивировании всегда сопряжен с большими
трудностями, так как не каждый их клон выживает в таких
условиях. Поэтому на практике стремятся делать разбавление
"маточных" клеток постепенно, вначале 1:2—1:3, а затем и более,
используя специальные культиваторы (рис. 164).
Для выращивания гибридом в
суспензионных культурах важны
бессывороточные среды, посколь-
ку это улучшает процесс культи-
вирования в больших емкостях,
снижает примерно в 500 раз бел-
ковые примеси и заметно сказы-
вается на снижении стоимости це-
левого продукта. Фирма Sigma
(США) выпустила в продажу та-
кую среду (в сухом/жидком виде)
под названием Hybry-Мах, не со-
держащую сыворотки и белков
(SFPF-Serum Free, Protein Free).
SFPF-среда включает аминокис-
лоты, витамины, нуклеотидные
предшественники, глюкозу, липи-
ды, прогестерон, неорганические
соли и микроэлементы. Подобная
среда рекомендована для слияния,
клонирования, выращивания гиб-
ридомных и миеломных клеток, а
также для поддержания на соот-
ветствующем уровне продукции
антител. Всего в среде содержится
82 ингредиента (22 аминокисло-
ты, 12 витаминов, 27 неорганиче-
ских веществ й 21 другое соеди-
нение).
11.5.3. Применение моноклональных антител. Практическое
использование МкАТ исключительно многообразное—в медицине
и ветеринарии (в целях диагностики, терапии), в биотехнологии
для получения, выделения и очистки специфических продуктов, в
исследованиях фундаментальных и прикладных проблем. Так,
например, стволовые клетки костного мозга не обладают какими-
579
либо характерными особенностями, По которым можно было бы
их отличить от других клеток. К тому же в костном мозге на них
приходится где-то порядка 0,05% от всех клеток.
Стволовая клетка и сывороточйый белок ос-фетопротеин, от-
крытый Г. И. Абелевым, впервые появляются в желточном мешке
эмбриона. Спустя некоторое время стволовые клетки переселяют-
ся в печень эмбриона и там же интенсивно синтезируется сыво-
роточный а-фетопротеин. В желточном мешке и печени протекает
активное кроветворение. Позже синтез этого белка прекращается,
а стволовая клетка перемещается в костный мозг, где и происходит
кроветворение в течение всей последующей жизни'организма.
Используя МкАТ против поверхностного CD-белка стволовых
клеток, ученые из США и Германии близко подошли к выделению
таких клеток, массовое получение которых могло бы быть реша-
ющим в терапии лучевой болезни, талассемии, тяжелых осложне-
ний при иммунодефицитах, лейкемии и других заболеваний. На
способности стволовых клеток к переселению основывается спо-
соб лечения лучевой болезни внутривенным введением стериль-
ного костного мозга.
Au
Рис. 165. Схема взаимодействия конъюгата
"анти-1д-золото“ с моноклональным антителом
’’антитироксин1' (2), заякоренным на срезе ткани
щитовидной железы (1).
Неоценимый вклад внесен в оценку CD-рецепторов клеток с
помощью МкАТ. Это позволяет глубже понять механизмы клеточ-
ных взаимодействий в целях направленного вмешательства для их
коррекции при патологических состояниях.
Непреходяще значение МкАТ в диагностике, в том числе —
при использовании иммуноферментного анализа в различных его
вариантах, включая метку коллоидного золота. Например, чтобы
локализовать тироксин-секретирующие клетки в образце ткани
щитовидной железы, эту
ткань инкубируют с мыши-
ным моноклональным анти-
тироксином. Затем на обра-
зец наносят конъюгат "анти-
Ig-золото", специфичный к
Ig-антитироксину. Фиксиру-
ясь на моноклональном ан-
тителе, конъюгат становится
прекрасной меткой, выявля-
емой в световом биологиче-
ском микроскопе по крас-
ной окраске золота (рис.
165).
580
Очень важно использование МкАТ для прицельной доставки
например цитостатиков в злокачественные клетки на предмет их
деструкции и, как следствие, для лечения соответствующего боль-
ного с опухолевым процессом.
При диагностике, например СПИД, на покрытой моноклональ-
ными антителами ячейке выявляется концентрация антигенного
белка р24 вируса иммунодефицита человека (ВИЧ-1) порядка
1,0—1,56 пикограмма в течение 1,5—3 часов; при цитомегалови-
русной инфекции МкАТ типа Е-13 используют для выявления
позднего антигена спустя 72 часа после заражения; важен вклад
МкАТ в диагностику герпеса, краснухи, токсоплазмоза, различных
бактериальных и грибковых инфекций, и т. д. В настоящее время
около 30 компаний в мире производят МкАТ в виде диагностиче-
ских наборов, а 13 компаний разработали оборудование или тех-
нологические процессы, относящиеся к МкАТ; теперь на рынке
имеются такие антитела против классов и подклассов иммуногло-
булинов человека, отдельных белков человека и животных, гормо-
нов и лекарств, и многие другие. Например, в 1991 г. американская
компания "Centocor” выпустила в продажу новое моноклональное
лекарство "Центоксин" для лечения сепсиса, вызванного грамот-
рицательными бактериями.
Ныне издают книги и журналы, посвященные МкАТ, и уже это
одно является веским доводом в пользу огромной роли их в
фундаментальных и прикладных исследованиях самого широкого
профиля.
11.6. Трансгенные животные. Включение чужеродных после-
довательностей ДНК или трансгенов в геном макроорганизма —
реципиента с последующей устойчивой их наследуемостью в ряду
поколений обеспечивает получение так называемых трансгенных
животных. Следовательно, процесс трансгенеза по своему меха-
низму относится к генетической инженерии и, в частности, к
генной и клеточной инженерии.
Доказано, что трансгены нередко экспрессируются, вызывая
те или иные структуры и функциональные изменения вплоть до
изменения "программы” развития организма. Подобного рода экс-
перименты сулят заметный прогресс в области фундаментальных
и прикладных разработок. К разряду фундаментальных относят:
анализ различных отклонений в экспрессии нормальных и мути-
ровавших генов, включая онкогены; осуществление направленного
мутагенеза за счет введения в макроорганизм экспрессирующих
581
специальных "конструкций", предназначенных определенным ге-
нам и т. д.
К разряду прикладных исследований можно отнести, например,
получение быстро растущих трансгенных свиней, выведение
трансгенных овец, образующих и накапливающих в молочных
железах некоторые факторы свертываемости крови человека и др.
Для получения трансгенных животных, в частности — мышей,
можно воспользоваться такими методами, как микроинъекции
ДНК в оплодотворенное одноклеточное яйцо, заражение предим-
плантированных эмбрионов рекомбинантными ретровирусами, и,
наконец, применение эмбриональных стволовых ES- или ЕК-кле-
ток.
Быстрота выполнения и надежность метода микроинъекций
сделали его наиболее популярным среди специалистов-экспери-
ментаторов. Используют различных линейных мышей, обеспечен-
ных всем необходимым при хорошем их содержании в условиях
вивария (животника). Обычно в опыт по микроинъекции берут не
менее 20 гибридных самцов-производителей первого поколения
(FI) в возрасте 8—12 месяцев, когда они характеризуются хорошей
производительностью. При слабой или низкой активности самцов
к подсаживаемым самкам их заменяют. Донорами оплодотворен-
ных яйцеклеток выступают не менее 10 неполовозрелых (12—14
г) гибридных самок первого поколения (FI), подвергшихся супер-
овуляции и спариванию с гибридными самцами FI. Оплодотворен-
ные одноклеточные яйца выделяют из операционно вскрытых
яйцеводов самок-доноров спустя 12 часов после их спаривания с
самцами-производителями. Выделенные яйца помещают в культу-
ру на срок от 3 до 36 часов. От 10 самок можно получить 250 яиц,
' пригодных для микроинъекций.
Оплодотворенные яйцеклетки поддерживают на среде М16 в
микрокапельной культуре в СО2-инкубаторе тканей при 37®С и с
5% диоксида углерода. Все другие манипуляции с яйцеклетками
вне инкубатора проводят в HEPES-забуференной среде М2 (HEPES
— это N-2-гидроксиэтилпиперазин — N’-2-этансульфоновая кис-
лота). Компонентный состав сред (кроме HEPES) почти тождест-
венен. Различие касается натрия бикарбоната, фенолового крас-
ного, натрия пирувата и кальция хлорида дигидрата, доля которых
в среде М16 больше, чем в среде М2. Остальные ингредиенты
представлены KCL, NaCL, КН2РО4, MgSO 4 • 7Н2О, натрия лактатом,
Глюкозой, пенициллином, стрептомицином, бычьим сывороточным
альбумином (БСА), внесенными в бидистиллированную воду.
582
Изолированные оперативным вмешательством яйцеклетки из
яйцеводов выдерживают в атмосфере 5% СО2 при 37°С до микро-
инъекции в них клонированных ДНК любого размера в количестве
1—2 нл 0,0001—0,0005% раствора ДНК с помощью микроманипу-
лятора.
Для трансплантации оплодотворенных яйцеклеток, в которые
была инъецирована экзогенная ДНК, используют "суррогатных"
матерей — псевдобеременных реципиентных самок (12 часов post
coitum), вынашивающих такие яйцеклетки.
В целях получения псевдобеременных мышей половозрелые
самки F-1 (19—20 г) в стадии эструса (течки, овуляции) спаривают
со стерильными самцами, у которых операционно блокированы
семявыносящие протоки. Наилучшие результаты получают с сам-
цами линии Parkes, обладающих высокой половой активностью.
Обычно используют 20—30 таких самцов, которых спаривают
через день с названными выше самками (получают в среднем 5
псевдобеременных самок в день). Псевдобеременные самки при-
годны для пересадки донорских оплодотворенных яйцеклеток.
Отбирают яйцеклетки, переживших микроинъекцию ДНК, и
вводят их в специальную микропипетку, с помощью которой
привносят в яйцевод псевдобеременной мыши оплодотворенную
яйцеклетку с включенной в нее клонированной ДНК. Часть пере-
саженных яиц погибает, а часть продолжает нормально развивать-
ся. После этого естественным путем (или с помощью кесарева
сечения) получают трансгенное потомство, идентифицируемое с
помощью гибридизационного анализа высокомолекулярной геном-
ной ДНК, изолированной из хвоста (отрезают кончик хвоста
длиной примерно 1 см).
При гомологии трансгена с каким-либо участком генома мыши
используют блот-гибризацию, если же гомология незначительна
или она отсутствует совсем, то ограничиваются дот-блот- или
слот-блот-анализом (от анг. blot — пятно, blotting — промокание,
dot — точка, slot — прорез, паз). В первом случае, когда трансген
идентичен эндогенному гену, он все равно окружен другими
нуклеотидными последовательностями. Поэтому, используя ре-
стриктазы, удается доказать локализацию трансгена в иных ре-
стрикционных фрагментах. Во втором случае, когда трансген
негомологичен геному мыши, то он ограничен другими сайтами
рестрикции, и здесь удобен дот-блот- или слот-блот-анализ в
качестве экпсресс-метода. Фрагменты анализируемой геномной
ДНК на нитроцеллюлозных фильтрах выявляют при экспозиции с
рентгеновской пленкой.
583
Трансгенные линии животных получают после скрещивания
первичных трансгенных экземпляров.
Изложенные события по получению трансгенных мышей схе-
матично представлены на рис. 72.
Как уже было сказано в начале этого раздела, другим методом
получения трансгенных животных (в частности, мышей) является
заражение предимплантированных эмбрионов рекомбинантными
ретровирусами, которые оказались удобными объектами для этих
целей. Геном ретровирусов сравнительно небольшой и с ним
относительно легко манипулировать, вводя в него чужеродные
гены; ретровирусы достаточно инфекционны в отношении пермис-
сивных клеток (почти 100% заражение их), а единственная копия
ретровирусного генома ДНК прочно и строго определённым обра-
зом интегрируется с ДНК клетки-мишени и эти последние способ-
ны экспрессировать привнесенные вирусом гены. Уровень экс-
прессии клонированных генов заметно повышается вследствие
того, что в ретровирусах имеются весьма активные транскрипци-
онные энхасеры, или усилители (см.). Известные ретровирусные
векторы ведут свое происхождение от вирусов лейкоза мышей
(MLV) или от вирусов птиц (ALV).
Геном ретровирусов функционирует по схеме:
РНК обРатная транскриптаза^ > РНК-----> Белок
Каждая вирусная частица содержит две копии одноцепочечного
РНК-генома, а после проникновения в пермиссивную клетку этот
геном переводится в линейную двухнитевую ДНК под влиянием
вирусного фермента — обратной транскриптазы. Чтобы интегри-
роваться в клеточный геном клетки-мишени, линейная ДНК про-
никает в ядро, где приобретает кольцевидную форму. Интегриро-
ванная линейная ДНК-копия ретровирусного генома (провирус)
имеет на обоих концах длинные нуклеотидные повторы — LTR (от
англ, long termine repeats). 5'LTR несет промотор, с которого
начинается транскрипция генов интегрированного провйруса; 3'
LTR-сайт полиаденилирования, где происходит терминация РНК-
транскриптов (см. рис. 23).
Существует ряд векторов на основе ретровирусного генома. В
простейших случаях удаляют структурные гены и на их место
встраивают один или больше рестрикционных сайтов в целях
584
клонирования. Из семейства векторов MLV известны: рМХ1112,
рМХ1122, рМХ262 и др.
Ретровирусный вектор может включить лишь около 10000 пн.
Имея в руках готовый вирусный вектор, им можно инфицировать
подходящие клетки-мишени, например, фибробласты, экспресси-
рующие соответствующие поверхностные рецепторы.
Чтобы заразить рекомбинантным ретровирусом эмбриональ-
ные клетки, в культуральную емкость с инфицированными фиб-
робластами, продуцирующими рекомбинантный вирус, помещают
восьмиклеточную морулу (группа бластомер, возникшая после
равномерного дробления оплодотворенного яйца), освобожденную
от яйцевой оболочки. Морула инфицируется и после достижения
стадии бластоцисты ее вводят в матку псевдобеременной матери.
Часть бластоцист может погибнуть, а часть нормально развивается
и в соответствующие сроки трансформируется в трансгенное
потомство, которое затем подвергается тщательному генетическо-
му анализу и может использоваться для выведения трансгенных
линий.
Бластоцисты стали родоначальницами плюрипотентных эмбри-
ональных стволовых клеток типов ES и ЕК. Показано, что, напри-
мер, ES-клетки доступны культивированию in vitro и после каких-
либо манипуляций (например, после введения клонированных
генов путем инфекции или трансфекции) можно инъецировать их
в бластоцисту и вернуть в живой макроорганизм. ES-клетки коло-
низируют эмбрион и составляют с ним единое целое, хотя коло-
низация ими зародышевого пути по разным причинам удается не
всегда. Последующие этапы работы с трансгенными животными
во многом сходны с. ранее описанными. Таким образом, все три
метода получения трансгенных животных можно представить в
виде следующей схемы по Д. Мерфи и Дж. Хенсону 1987 г. (см.
рис. 166).
С конца XIX века и до середины текущего столетия были
разработаны методы пересадки эмбрионов от различных живо-
тных с последующим рождением полноценного потомства (клеточ-
ная инженерия). Следовательно, получение популяции, состоящей
из генетически идентичных особей (клонирование), возможно
не только с помощью рассмотренных выше трех способов (микро-
инъекций ДНК, вирусная инфекция и пересадка ES- и ЕК-клеток),
но и с помощью пересадки эмбрионов, которая может быть
585
осуществлена на раз-
личных представителях
царства Animalia, напри-
мер, на кроликах, козах,
коровах, мышах, овцах,
свиньях и др. Так, для
получения элитного ста-
да коров в сравнительно
короткий временной ин-
тервал (с учетом естест-
венного процесса от за-
чатия до отела) отобран-
ной родоначальнице
вводят фолликулостиму-
лирующий гормон
(ФСГ). В ответ на ФСГ
Рис. 166. Схема получения трансгенных живо- ее °Рганизм ~вы Расы
тных методом микроинъекции ДНК в оплодотворен- вает 10 15 яйцеклеток
ную одноклеточную яйцеклетку: 1 - яйцеклетка, 2 - вместо ОДНОЙ в норме,
микроинъекция ДНК, 3 - трансфекция ДНК, 4 - gce эти яйцеклетки оп-
пересадка ядра, 5 - пересадка клетки в бластоцисту. , т
лодотворяются. Через
4—5 дней, когда сформировавшиеся 32-клеточные эмбрионы (око-
ло 0,2 мм в диаметре) еще не прикрепились к стенкам матки и
свободно плавают в ней, их вымывают, определяют полноценность
и вводят микрошприцем непородистым коровам, находящимся в
воспроизводительном цикле благодаря предварительному введе-
нию им простагландинов. Коровы-реципиенты эмбрионов вына-
шивают и рожают элитных телят в нормальные сроки.
Таким путем удается получить от одной элитной коровы в
среднем 5 элитных телят за 15 месяцев. В этот срок с интервалом
в 2—3 месяца у нее дважды забирают яйцеклетки и один раз она
разрешается собственным теленком. На этом пути открываются
заманчивые перспективы, связанные с возможностями оплодотво-
рения яйцеклеток in vitro, а также длительным хранением эмбри-
онов в замороженном состоянии. Тогда в целях экономии можно
продавать такие эмбрионы в соответствующие агрохозяйства.
С учетом религиозных воззрений, гражданских законода-
тельств и этики в ряде стран решаются проблемы искусственного
осеменения женских яйцеклеток зародышевыми клетками от муж-
586
чин. Лишь только во Франции на начало 1992 г. родилось свыше
10000 детей от женщин-доноров.
11.7. Иммуномодуляторы. Иммуномодуляция (от лат. immunitas
— избавление, освобождение, modulatio — мерность, размерен-
ность) — термин интегративный, объединяющий представления о
каком-либо целенаправленном вмешательстве в работу иммунной
системы, функционирующей в условиях нормы и патологии. Со-
ставными частями иммуномодуляции являются: иммуностимуля-
ция и иммунодепрессия (иммуносупрессия), т. е. иммунокоррек-
ция. При иммуностимуляции происходит активизация иммунной
системы, при иммунодепрессии (иммуносупрессии) — ее частич-
ное угнетение; следовательно иммунокоррекция сопровождается
выравниванием деятельности иммунной системы. Индивидуаль-
ные вещества или средства, применяемые в качестве иммуномо-
дуляторов, соответственно подразделяют на иммуностимуляторы
и иммуносупрессоры (иммунодепрессанты).
Все иммуномодуляторы условно можно еще подразделить на
специфические и неспецифические. Первые из них выступают
преимущественно специфическими антигенами, индуцирующими
антителообразование, вторые, влияя на иммунную систему, выра-
жение или совсем не являются антигенноактивными. Неспецифи-
ческие иммуномодуляторы бывают двух типов: .
а) действующих при нормально функционирующей иммунной
системе (вакцина BCG, Corynebacterium parvum),
б) максимально действующих на супрессированную иммунную
систему (левамизол, препараты тимуса, интерфероны); они мало
активны или совсем неактивны, когда их вводят пациентам с
нормальной иммуной системой.
Все иммуномодуляторы используют в качестве средств лечения
соответствующих заболеваний, связанных с дефектами иммуной
системы, или иммунодефицитами.
11.7.1. Иммуностимуляторы. Иммуностимуляторы повышают
устойчивость организма к инфекционным заболеваниям, активи-
зируют иммунитет у онкологических больных, получающих цито-
статики, радиоактивное облучение. Их подразделяют на природные
и синтетические. К природным относят иммуноглобулин-у (Ig-y),
являющийся также средством заместительной терапии; препараты
из зобной железы крупного рогатого скота (тактивин, тималин,
тимопоэтин и др.), интерфероны (см.), фактор переноса, или
лейкоцитарный трансфер-фактор (АТФ), специфические опухоле-
вые вакцины, иммунную РНК (И-РНК), натрия нуклеинат, проди-
587
гиозан, сальмозан, грибные гликаны, апатогеннЫй штамм
Mycooacterium tuberculosis (вакцина BCG) и другие вакцинные
штаммы различных микроорганизмов, анатоксины.
Синтетическими иммуностимуляторами неспецифического *
действия являются батилол, йзопринозин, левами^рл, хлоридин и
некоторые другие. Поскольку эти вещества представляют собой
продукты органического синтеза, то они не рассматриваются в
настоящей книге.
•’ Иммунный гамма-глобулин (IgG) -ч- продукт матричного син-
теза, являющийся гликопротеином и способный проникать через
плаценту. Его получают в очищенном и концентрированном видах
из донорской, плацентарной и абортной крови человека в виде 10%
или 16%" раствора. Препарат изготавливают из смеси большого
числа сывороток крови взрослых людей, ранее болевших, напри-
мер, гриппом, корью и другими инфекционными заболеваниями,
или получавших вакцины в качестве профилактических среДс^гв.
Поэтому так называемый нормальный глобулин может содержать
Ig-ы против возбудителей дифтерии, кори, оспы и других заболе-
ваний.
Специфические гипер-1д-ы изготавливают из крови специально .
иммунизированных людей, например, против бешенства, клеще-
вого энцефалита, стафилококковых инфекций, столбняка и др. -.
Основной метод выделения иммуноглобулинов является их
фракционированное осаждение этанолом (по Е. Дж. Кону, 1945—
1946) на холоду при строгом контроле pH и ионной силы раствора.
На процесс разделения белков сыворотки крови влияют следую-
щие основные факторы: концентрация белка, диэлектрическая
постоянная раствора, концентрация этанола, изоэлектрическая
. точка, pH, ионная сила раствора, ^температура. ‘
Необходимо иметь-в виду тот факт, что в сыворотке крови
' содержатся различные белки (альбумины — 57—65%, сх-глобулины
— 13%, Р-глобулины — 12,5%, у-глобулины.— 11.—19%). В. случае
сильного разбавления растворов белков_увеличивается диссоциа-
ция молекул (закон Действующих масс) преимущественно благо-
даря внутреннему электростатическому отталкиванию, и -тогда
белки труднее осаждаются; при высокой концентрации бел^цв, в
растворе больше возможностей для пенообразования с последую-
щей денатурацией белковых молекул. Денатурация-также возра-
стает, если создаются реальные условия для формирования моно-
молекулярной белковой пленки на границе раздела фаз, например,
"жидкость—воздух" (учитывать свойства ПАВ у белков).
588
Способность белковых растворов увеличивать или уменьшать
диссоциацию находящихся в них солей, характеризуется диэлек-
трической постоянной таких растворов (D); чем выше полярность
молекул, тем больше D; для растворов неполярных соединений
величины D низкие. Существует обратнопропорциональная зави-
симость между D и силами электростатического протяжения и
отталкивания молекул — возрастание D сопровождается стабили-
зацией молекул, снижение D — их притяжением и агрегацией.
Напомним, что D для воды при 20°С равна — 80—82, "для етанола
— 24, то есть в 3,5 раза меньше,.поэтому этиловый спирт исполь-
зуют в качестве фракционирующего агента белковых молекул. К
тому же он дегидратирует эти молекулы.
В изоэлектрической точке (pl) при данном pH, когда число
положительных зарядов белковых молекул равно общему числу их
отрицательных зарядов, белок становится наименее растворимым,
и он легче выпадает в осадок. D белковых растворов изменяется в
зависимости от их pH и достигает максимума в изоэлектрической
точке.
Ионная сила раствора (ц) выступает мерой интенсивности
электрического поля, создаваемого в растворе каждым ионом; ц
равна полусумме произведений молярности каждого иона (ш) на
квадрат его валентности (z2):
' ц *i У mtz 2 . Изменяя ц, достигают более эффективного
осаждения белков. - .
Температура — важный фактор для сохранения белков в
нативном состоянии. При их фракционировании стремятся под-
держивать температуру ниже 0°С, при которой снижаются раство-
римость белковых молекул и рост случайно попавших в раствор
микробов-контаминантов.’ "• . • ‘
Известны дваЪпособа производства4! gG кам£рный.и внека-
мерный (последний применяют при крупносерийном производстве
препарата; он предложен Г. Я. Розенбергом в 1959 г.). Общая схема
внекамерногб способа фракционирования IgG представлен ниже.
Фракционирование, например, донорской крови включает се-
парировайие'формбнйых эЛембнтой^ отделенийфибрина;'получен1®’
ную сыворотку очищают от остатков фибриногёйа, балластных
белков, а затем ее передают на последующие ступени выделения
IgG. Полученный иммуноглобулин растворяют и подвергают сте-
рилизующей фильтрации.
589
В целях улучшения качества продукта указанный выше метод
неоднократно модифицировали.
П—стадия первого
осаждения
Сыворотка ---------•
I—стадия предвари —
тельного осаждения
III —стадия второго
осаждения
IV— стадия третьего
осаждения
К иммуноглобулинам, получаемым из крови человека, относят-
ся Ig-ы: антирабический, нормальный, противогриппозный, проти-
востафилококковый, противостолбнячный, противоэнцефалитный
(титрованный на антитела к вирусу клещевого энцефалита); из
590
крови животных получают Ig-ы, или сыворотки, содержащие Ig-ы:
антирабический, противоботулиновые, противогангренозные, про-
тиводифтерийный, противолептоспирозный, против клещевого эн-
цефалита, противосибиреязвенный, против столбняка.
Антитоксины — обычно это препараты лечебно-профилакти-
ческого действия, изготавливаемые из сывороток крови лошадей,
иммунизированных соответствующими анатоксинами (см. также
главу 9). Схема получения антитоксинов в общем виде может быть
сведена к следующим этапам:
Обезвреживание токсина возможно с помощью других деток-
сикаторов (Р-пропиолактон, протеазы или окислители и пр.).
В некоторых случаях отделение форменных элементов крови
осуществляют одноактно с дефибринированием крови. При этом
фибриновый сгусток включает в себя эритроциты, лейкоциты и
тромбоциты. Однако при работе с большими объемами крови
названный одноактный процесс сопровождается частичным гемо-
лизом эритроцитов, что осложняет последующую очистку антиток-
сина.
591
На практике получают от гипериммунизированных соответст-
вующими анатоксинами лошадей лечебные или лечебно-профи-
лактические антитоксические сыворотки: противоботулиновые ти-
пов А и В (в жидком виде), концентрированные жидкие противо-
гангренозные очищенные (моно- и поливалентные), противодиф-
терийную и противостолбнячную.
Очищенный сорбированный на алюминия гидроксиде столб-
нячный анатоксин применяют для ревакцинации людей, являю-
щихся затем донорами специфического 1g. Схема его выделения
аналогична той, которая описана ранее для IgG. Выпускают про-
тивостолбнячный 1g в жидком виде в ампулах.
Кроме Ig-ов другие белковые фракции сыворотки крови обла-
дают важными биологическими функциями и поэтому могут быть
выделены в очищенном виде для последующего использования в
практическом здравоохранении и медицине в целом.
Препараты из зобной железы. К ним относят тактйвин, тима-
лин, тимопоэтин и некоторые другие, получаемые методами экст-
ракции и последующей очистки из зобной железы (Glandus thymus)
крупного рогатого скота, поступающего на мясокомбинаты для
убоя и последующей комплексной переработки. Зобную железу
относят к центральным органам иммуной системы. В ней проходят
специальную обработку будущие тимоциты, приобретающие соот-
ветствующие функции. (Т-хелперы, Т-супрессоры, Т-киллеры). В
этой железе содержатся гормоноподобные субстанции, которые и
составляют основу иммуностимулирующих веществ. Одна из схем
получения иммуностимуляторов из Gl. thymus представлена на
странице.
Фактор переноса, или лейкоцитарный трансфер-фактор (ЛТФ)
— это составная часть диализата экстракта лейкоцитов, ответст-
венная за пассивный перенос реакции гиперчувствительности
замедленного типа (ГЗТ) от сенсибилизированных лиц-доноров к
несенсибилизированным реципиентам. Впервые открыт К. Ланд-
штейнером и М. Чейзом в 1942 г. в лейкоцитах морской свинки.
ЛТФ является медиатором клеточного компонента иммунной сис-
темы и его можно причислить к лимфокинам; вырабатывается
нормальными лимфоидными клетками. Кроме индукции кожных
реакций (ГЗТ) он тормозит миграцию клеток и вызывает транс-
формацию лимфоцитов. ЛТФ не является антителом, не обладает
иммуногенностью и не проявляет свойств трансплантационного
антигена. Он устойчив к ДНК-азе, РНК-азе и трипсину, но термо-
лабилен (инактивируется при 56°С в течение 30 мин.). В составе
592
ЛТФ обнаружены полипептиды и олигонуклеотиды, ММ менее 10
кДа, хорошо растворим в воде.
Выделение иммуностимулирующих средств ("суммарный препарат") из ткани
зобной железы крупного рогатого скота (ФСБ — фосфатно-солевой буфер)
Под влиянием ЛТФ отрицательная ГЗТ переходит в положи-
тельную. Такой эффект одной дозы фактора переноса продолжа-
ется до 6 месяцев. В настоящее время за одну дозу ЛТФ принято
его количество, получаемое из 10®'9 лимфоцитов. ЛТФ можно
применять в больших дозах в течение нескольких месяцев при
иммунодефицитах.
Схема получения фактора переноса заключается в следующем.
От здоровых доноров с выраженной реакцией ГЗТ на соответст-
вующйй антиген отбирают кровь в асептических условиях. Кровь
собирают в стерильные емкости, куда добавляют антикоагулянт
гепарин и 6% раствор декстрана 250 в 0,15 М натрия хлорида (из
расчета 500 ME гепарина и 10 мл декстрана на каждые 100 мл
крови). Емкость закрывают, несколько раз встряхивают и термо-
593
статируют при 37°С. Осаждение эритроцитов по времени не
должно продолжаться более часа. Надосадочную жидкость цент-
рифугируют в асептичных условиях (1500 g 10 минут), плазму
осторожно сливают (декантируют). Остающийся осадок включает
эритроциты, на поверхности которых располагается слой лейко-
цитов (1 мл осевших лейкоцитов соответствует примерно 109
клеток, что достаточно для получения 1—10 доз АТФ). Из числа
белых клеток этого слоя 70% приходится на лимфоциты. На осадок
осторожно наслаивают 0,15 М стерильный раствор натрия хлорида,
приготовленный на апирогенной воде (0,5 мл на 100 мл крови), и
осторожно отсасывают лейкоциты в виде суспензии в изотониче-
ском растворе NaCl. Полученную суспензию замораживают
смесью сухого льда с этанолом и размораживают при 37°С деся-
тикратно (до полного разрушения лейкоцитов). В полученный
вязкий раствор добавляют ДНК-азу и катионы магния для гидро-
лиза ДНК при 37°С (эта процедура возможна в процессе замора-
живания и оттаивания, поскольку ДНК-аза стабильна в таких
условиях). Деполимеризованный экстракт лейкоцитов диализуют
против дистиллированной воды (на 1 л экстракта берут 50 мл
дистиллированной воды) в течение суток при + 4°С, после чего его
подвергают фильтрующей стерилизации через мембранный
фильтр и лиофильно высушивают.
Полученный диализат экстракта лейкоцитов растворяют в сте-
рильном фосфатно-солевом буфере с pH 7,4 (0,01 М фосфатный
буфер +8,5 г/л NaCL) в соотношении 1:20 и асептично подают на
хроматографическую колонку, заполненную сефадексом G-25, ра-
нее законсервированную 0,2% натрия азидом и промытую тем же
ФСБ, разведенным в 20 раз.
Хроматографию проводят в указанном выше стерильном бу-
фере со скоростью до 90 мл/час. Собирают активные фракции
(восстановление способности лимфоцитов к розеткообразованию
трипсинизированных эритроцитов барана в присутствии фактора
переноса) — это и есть АТФ.
АТФ не рекомендуют применять при кандидозах кожи и сли-
зистых оболочек, кокцидиоидозе, лепре, туберкулезе и остеоген-
ной саркоме.
Специфические опухолевые вакцины, а таже иммунная РНК
(И = РНК) еще не вышли за пределы лабораторий, поэтому их
крупномасштабное производство пока не освоено.
594
Продигиозан, сальмозан, вакцинный штамм микобактерий и
грибные иммуностимуляторы (гликаны, нуклеинат натрия) рас-
смотрены в главе 9.
11.7.2. Иммунодепрессоры (иммуносупрессоры). Среди имму-
нодепрессоров известны синтетические вещества (азатиоприн,
меркаптопурин, преднизон, циклофосфамид и пр.), отдельные
антибиотики (блеомицин, циклоспорин А, см. главу 9), а также
зообиотехнологические продукты — антилимфоцитарный имму-
ноглобулин, aHTH-Rh0(D)Ig человека и лимфоцитарные кейлоны.
Антилимфоцитарный IG получают из сыворотки крови животных
по схеме, близкой к описанной выше для выделения IgG (см. 11.7.1).
При этом иммунизирующим агентом (антигеном) являются чело-
веческие лейкоциты.
При выделении суммарных иммуноглобулинов можно исполь-
зовать два подхода. Один из них базируется на высаливании Ig-ов
насыщенным раствором аммония сульфата (3,9 М при 0°С или 4,06
М при 20°С) с последующей хроматографией растворенного осадка
антилимфоцитарного Ig в подходящем буфере (например, фосфат-
ном — 0,01 М с pH 6,5). Хроматографию проводят на ионообмен-
никах — ДЭАЭ-сефадексе или ДЭАЭ-целлюлозе.
Поскольку антилимфоцитарный Ig содержит разные Ig-ы со
сходными характеристиками, то их можно разделить, изменяя
ионную силу элюента.
В производственных условиях получают в среднем 5,5—6,0 г
IgG из 1 л иммунной сыворотки в пересчете на сухое вещество.
Более высокого выхода Ig удается достигнуть согласно другому
методу — с помощью 2-этокси-6,9-диаминоакридинлактата, или
риванола (Дж. Хорейси, 1952). Выход IgG при этом составляет в
среднем 60—65%.
Реакция взаимодействия риванола(катион) с белками протекает
при незначительном сдвиге pH от изоэлектрической точки, то есть
при малом отрицательном заряде белковой молекулы, а образую-
щийся риванольно-белковый комплекс слабо растворим в воде —
он растворим при pH 5,0.
В слабо щелочной среде риванол не образует комплексов с IgG,
на чем и основана очистка IgG:
IgG можно выделять прямо из иммунной сыворотки, добавляя
к ней 0.75% раствор риванола в соотношении примерно 1:2.
Анти-КЬ0(В)-иммуноглобулин человека приобрел важное зна-
чение в профилактике гемолитической болезни у новорожденных.
595
IgG — "сырец" (см. стадию первого
осаждения на с. 590) разводят в 10 раз
изотоническим раствором NaCl, дово-
дят pH до 6,6—6,9
+ 0.2% раствор
риванола --------» осадок в
f = -10 - 15’СJ отходы.
-Центрифугирование ' '
к супернатанту + 26% этанол,
pH 6,9-7,5 Г=-10.-1 ГС
осадок IgG (растворение, стерили-
зующая фильтрация, лиофильное ’
высушивание)
С его помощью удается предотвращать развитие так называемой
резус-сенсибилизации примерно у 1% первородящих резус-отри-
цательных /Rh(—)/ женщин, если вводить им в послеродовом
периоде этот 1g.
Иммуносупрессивное действие анти-RhoD-Ig проявляется после
внутримышечного введения препарата в течение первый 2—3 суток
после родов первородящим нёсенсибилйзированным к Rh0(D) изо-
антигену резусотрицаТельным женщинам, которые родили Rh( +)
ребенка, совместимого с матерью по группе крови системы АВО.
Доза вводимого анти-ИЬо(1>)-1д составляет 200—250 мкг.
• Исходным материалом для получения aHTH-Rh0(E))-Ig является
. сыворотка или плазма крови, содержащая неполные анти-RhJD)- -
антитела в высоком-титре; сыворотка ИЪ(-)лиЦ, сенсибилизирован-
ных К D-изоантигену во время предшествующих "беременностей.
Rh( + ) плодом или в результате переливания Rh( +) крови. Однако
' более стабильные и высококачественные препараты получают в
случае использования сыворотки или плазмы крови от специально
иммунизированных доноров (например, Rh-отрицательных жен-
щин нёдеторрдноГр вбзраста)." *’ ***’ ”
ОчИщенный aHTH-Rh0(D)-Ig“выделяют ^принципиально тём же'
методом, который описан для донорского 1g.
Все препараты Ig-ов контролируют на стерильность, безвред-
ность, апирогенность, содержание в них IgG должно быть не менее
90% общего ^белка. ,
596
11.8. Коллекционные центры клеточных культур, их роль в
сохранении генофонда животных организмов. По данным, опуб-
ликованным в научной литературе, за последние 400 лет с лица
Земли исчезли 130 видов птиц и млекопитающих, из них 100 видов
— за последнее столетие. В Красную Книгу бывшего СССР внесено
92 вида и подвиде млекопитающих, 80 — птиц, 35— рептилий, 9
— амфибий, 9 — рыб, 209 — насекомых, 2 — ракообразных, 11 —
червей, всего — 1Ц6 видов. £ уменьшением численности видов
уменьшается генетическое разнообразие. Генофонд вида (уникаль-
ный набор его генов) во всей полноте представлен лишь в некото-
рой совокупности особей, которую надо беречь и сохранять. Для
сохранения естественных экосистем с их генофондом необходимо
заповедать не менее 30% поверхности земли (сегодня заповедники
составляют всего около 2%).
Сохранение генофондов различных видов животных возможно
в зоопарках, питомниках, на фермах, где они содержатся и раз-
множаются в неволе. Другой путь — это консервация геномов
половых или соматических клеток тех видов, численность которых
упала ниже критической, необходимой для их выживания (ниже
500 особей для позвоночных и 50000 — для беспозвоночных).
Следует иметь в виду, что с безвозвратной утратой не только вида
животных (как и-других организмов), но даже отдельной особи -
может быть навсегда потёрянй какая-то часть генов, вносящая
разнообразие в генетические характеристики представителей это-
го вида. Таким образом, благодаря консервации геномов можно
решать следующие задачи по сохранению:
1. разнообразия видов животных,
2. генетического стандарта видов, для которых отмечен выра-
женный дрейф^ёнов, ... „ , . - *
3. , лабораторных жиротнЫх с наследственными аномалиями,
экстраполируя результаты, например, на других животных и людей
• с-собтветствующими наследственными заболеваниям (от лат. extra
— сверх, вне, дополнительный; polaris -Шиолярный),
4. сниженного числа животных, содержащихся в неволе, не
опасаясь инбридинга ("От англ.1пЬгеес1фр'— врря$денный, природ-
'ный|,'и прибегну^ при Этом к искусственному осеменению ранее^,-
законсервированной спермой или к обеспечению развития ранее*
замороженных зародышей,
5. особо ценных пород сельскохозяйственных животных, а
также аборигенных пород* (от лат. ab origine — от начала, т„ е.
коренные), важных для селекционной работы.
597
Обычно для консервации используют зародышевые клетки
(сперматозоиды и яйцеклетки), а также сами зародыши; в редких
случаях прибегают к консервации соматических клеток (например,
в целях сохранения последнего представителя исчезающего на
земле вида).
Известны 3 типа консервации: физиологическая, криоконсер-
вация и создание банка генов. К физиологической консервации
относят анабиоз в естественных условиях, диапаузу
— временную остановку развития зародышей в половых путях
самок у представителей копытных, сумчатых, хищных и других
млекопитающих животных (до нескольких месяцев), и, наконец,
сохранение спермиев в половых путях самок, например у
муравьев — до 15 лет, у летучих мышей — 5—7 месяцев.
Криоконсервация основана на глубоком замораживании био-
объектов (клеток, тканей и органов). Организмы, выдерживающие
сильное обезвоживание, оказались наиболее устойчивыми при
^консервации (некоторые протозоа в стадии цист, нематоды и др.).
О сущности этого метода, в том числе — об использовании
криопротекторов см. главу 10.
В настоящее время разработаны методы криоконсервации
сперматозоидов, гонад, лимфоцитов и других соматических клеток,
зародышей. Полагают, что сохранение клеток при — 196’С обес-
печит их жизнеспособность в течение многих десятков лет. Этот
срок можно было бы растянуть до вечности, если бы не спонтанные
мутации от воздействия естественной радиоактивности.
Лиофилизация — как вариант криоконсервации не имеет су-
щественного значения в сохранении животных клеток. Он более
значим ддя микроорганизмов.
В настоящее время, благодаря достижениям в области генети-
ческой инженерии, в различных странах созданы и создаются
банки генов, приобретающие непреходящее значение для поддер-
жания, изучения строения и функциональной активности генома
животных организмов. В России большое внимание этой проблеме
уделяется в учреждениях РАН и, в том числе, в институтах
цитологии (г. Санкт-Петербург), молекулярной биологии (г. Мос-
ква), цитологии и генетики Сибирского отделения РАН (г. Ново-
сибирск), МГУ им. М. В. Ломоносова и др.
598
ЛИТЕРАТУРА
Акименко В. К. Альтернативные оксидазы микроорганизмов.
М., Наука, 1989.
Бабьева И. IL, Зенова Г. М. Биология почв. Под ред. Д. Г.
Звягинцева. МГУ, 1983.
Бациллы. Генетика и биотехнология. Под ред. К. Харвуда. М.,
Мир, 1992.
Бейли Дж., Оллис Д. Основы биохимической инженерии, в 2-х
частях. М., Мир, 1989.
Бекер М. Е., Лиепиньш Г. К., Райпулис Е. П. Биотехнология,
М., ВО Агропромиздат, 1990.
Биологические мембраны. Методы. Под ред. Дж. Финдлея, У.
Эванза, М., Мир., 1990.
Биотехнология в 8-ми томах. Под ред. Н. С. Егорова, В. Д.
Самуилова Уч. пособие для вузов. М., Высшая школа, 1987—1988.
Биотехнология клеток животных. В 2-х томах. Под ред. Р. Е.
Спиера и Дж. Гриффитса. М., ВО Агропромиздат, 1989.
Биотехнология: принципы и применение. Под ред. И. Хиггинса,
Д. Беста, Дж. Джонса. М., Мир, 1988.
Биотехнология растений. Под ред. С. X. Мантелла и X. Смита.
М., ВО Агропромиздат, 1987.
Варфоломеев С. Д., Калюжный С. В. Биотехнология. Кинети-
ческие основы микробиологических процессов. М., Высшая школа,
1990.
Виестур У. Э., Шмите И. А, Жилевич А В. Биотехнология.
Биотехнологические агенты, технология, аппаратура. Рига, Зинат-
не, 1987.
Воробьева Л. И. Промышленная микробиология. М., МГУ, 1989.
Егоров А М., Осипов А П., Дзантиев Б. Б., Гаврилова Е. М.
Теория и практика иммуноферментного анализа. М., Высшая
школа, 1991.
Блинов Н. П. Химическая микробиология. М., Высшая школа,
1989.
Иммобилизованные клетки и ферменты. Методы. Под ред. Дж.
Вудворда. М., Мир, 1988.
Иммунологические методы. Под. ред. Г. Фримеля. М., Медици-
на, 1987.
Иммунология. Под ред. У. Пола в 3-х томах. М., Мир, 1989.
Каратыгин И. В. Коэволюция грибов и растений. Санкт-Петер-
бургский Гидрометеоиздат. С.-Пбг. 1993.
599
Коваль Э. 3., Сидоренко Л. П. Микодеструкторы промышлен-
ных материалов. Киев, Наукова Думка, 1989.
Кузник Э. 3., Васильев Н. В., Цыбиков Н. Н. Иммуногенез,
гемостаз и неспецифическая резистентность организма. М., Ме-
дицина, 1989.
Ленинджер А. Основы биохимии в 3-х томах. М., Мир, 1985.
Лиепиньш Г. К., Дунцэ М. Э. Сырье и питательные субстрат!!
для промышленной биотехнологии. Рига, Зинатне, 1986.
Льюин Б. Гены. М., Мир, 1988.
Молекулярные и клеточные аспекты биотехнологии. Сб. научн.
трудов под ред. С. Г. Инге-Вечтомова. Л., Наука, 1986.
Муромцев Г. С., Бутенко Р. Г., Тихоненко Т. И., Прокофьев М.
И. Основы сельскохозяйственной биотехнологии. М., ВО Агропро-
миздат, 1990.
Перспективы биохимических исследований. Под ред. Дж. Туза
и С. Прентиса, М., Мир, 1987.
Петров Р. В. Иммунология. М., Медицина, 1987.
Промышленная микробиология. Под общей ред. Н. С. Егорова.
М., Высшая школа, 1989.
Рыбчин В. Н. Основы генетической инженерии. Минск, Вы-
шэйшая школа, 1986.
Сассон А. Биотехнология. Свершения и надежды. М., Мир.,
1987.
Седых Н. В., Кристапсонс М. Ш. Контроль качества в биотех-
нологии. Рига, Зинатне, 1990.
Щелкунов С. Н. Клонирование генов. Новосибирск, Наука,
1986.
БЛИНОВ Николай Петрович
Основы биотехнологии
Зав. редакцией Г. Н. Левина, Технический редактор В. С. Просекова, Корректор С. С. Кухарева, лицензия
ЛР №020297 от 27. 11.91.
Сдано в набор 10.02.94 г. Подписано к печати 10.09.95 г. Формат 60x84 1/16, бумага офсетная. Гарнитура Бал-
тика. Печать офсетная. Уел. печ. лист 34.88 уч. изд. лист 35.53. Тираж 10000. Заказ 8254
Издательская фирма "Наука" 119034, Санкт-Петербург, Менделеевская линия, 1.
Белгород, облтийография. ул. Б, Хмельницкого, 111а.
Перечень исправлений,
внесенных в книгу-учебник "Основы биотехнологии"
Стр. Строка Напечатано Следует читать
27 подпись к рис. 1 мендсикуты мендосикуты
37 подпись к рис. 5 боковые-2, боковые-З,
промежуточные-З промежуточиые-З
58 рис. 10, С сд СП
58 4 снизу аикислот аминокислот
65 формула витамина В12 5,6-диметилбензимида- 5,6-диметилбензимида-
золрибокуклеотид золрибонуклеотид
73 9 снизу И или
80 на рис. 19 АБ в
80 3 снизу вибриона вириона
86 13 сверху траксономических таксономических
107 подпись к рис. 31 — 7—гладкий
эндоплазматический
ретикулум
115 подпись к рис. 36 визикул везикул
115 7 сверху БИЗИКуЛЫ везикулы
141 табл. 15, 8 снизу ксилота кислота
159 табл. 17, 4 снизу Eschexichia Escherichia
168 подпись к рис. 59 Цикл деления Е Цикл подразделяют
190-
191 3 кол. табл. 20 /между проп. буквами
204 подпись к рис. 70 а-нуклеотид а-нуклеоид
215 7 сверху фотолиаза+hT] фотолиаза+hu
234 схема лактобактерии лактобактерии
234 схема Р-галактозидата Р-галактозидаза
268 3 абз. 2 снизу (AG) здесь (AG здесь
369 схема декстрансахароза декстрансахараза
389 7 и 8 снизу читать выше схемы
392 14 снизу (Y e/s) (Y p/s)
434 подпись к формуле Nokardia Nocardia
506 2 абз. 1 и 2 сверху читать ниже схемы
511 2 абз. 5 сверху галла с большими галла присуща клеткам
с большими
549 3 абз. и подпись
к формуле форобол форбол
550 1 и 2 абз. форобол форбол
551 подпись к формуле RAF PAF
553 подпись к рис. 158 ФРБ ФРФ
554 4 абз. 1 сверху TNPa TNFa
554 5 снизу фороболов форболов
557 табл. 56 рНг рН2
563 6 сверху меойза мейоза
564 11 снизу в табл. 58 трансплантантов трансплантатов
569 на рис. 162 Fa Fd
569 подпись к рис. 162 У Y
578 на рис. 163 Ig Igac
589 7 снизу IgG представлен IgG представлена
592 4 абз. 1 снизу странице странице 593