/

























































































































































































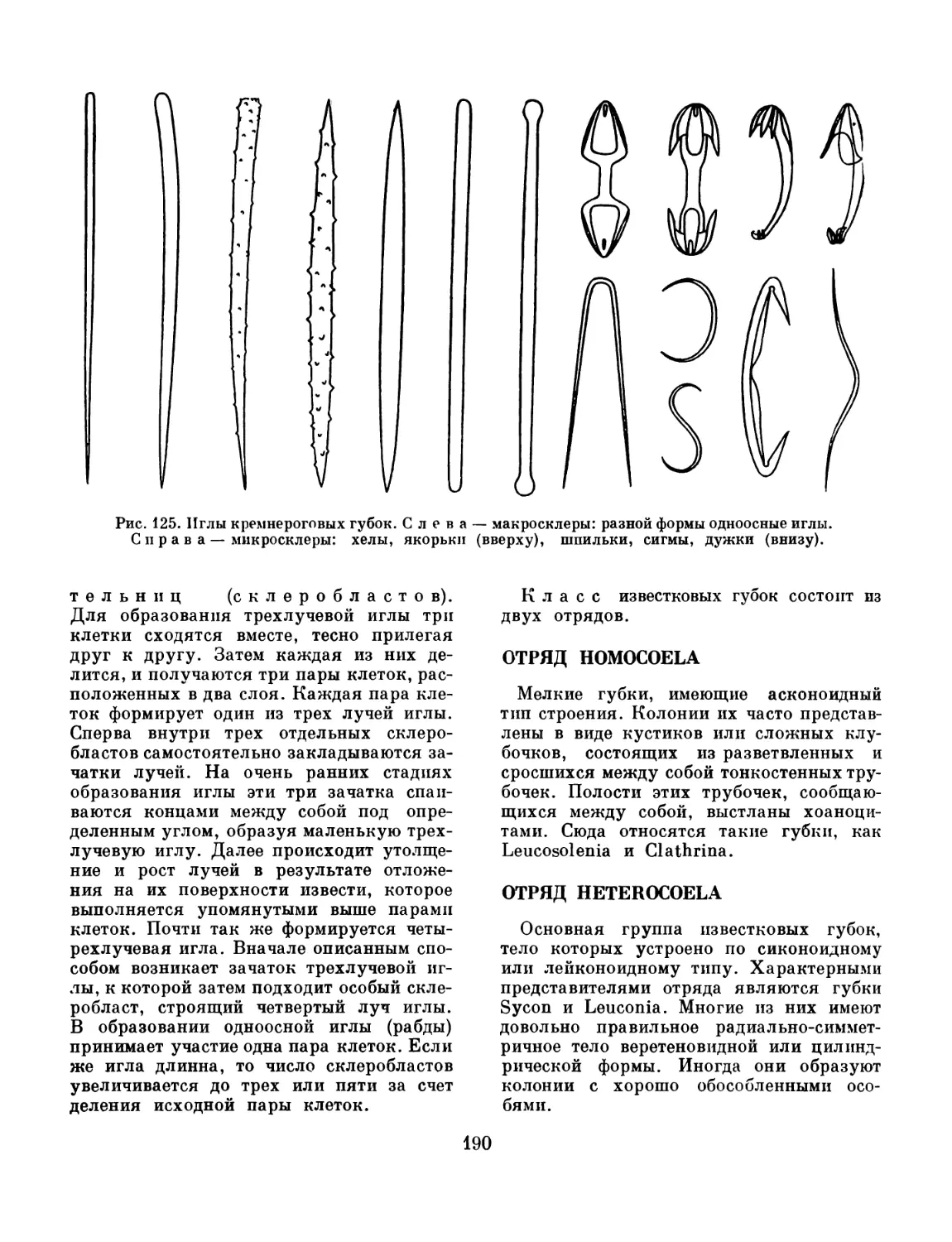





































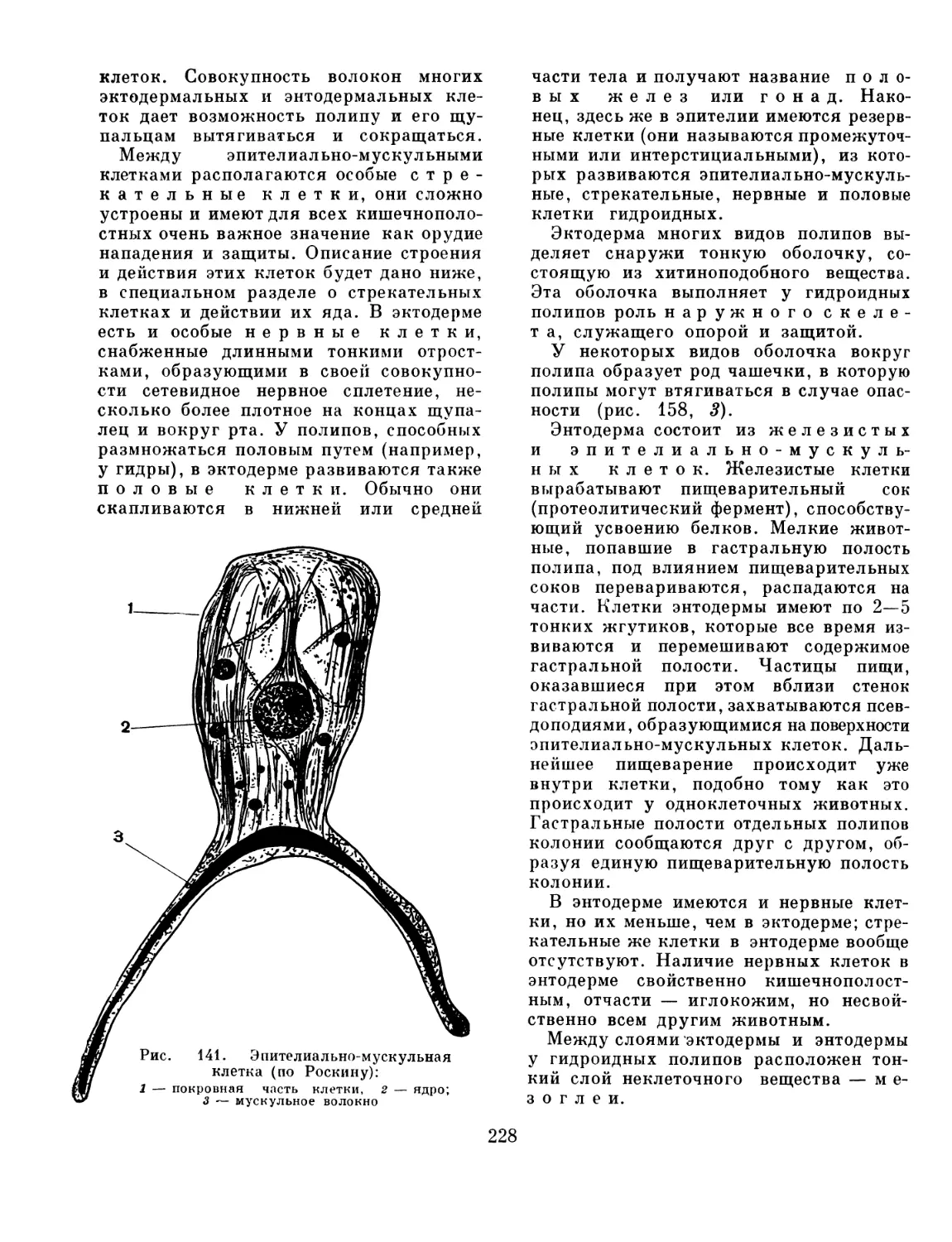



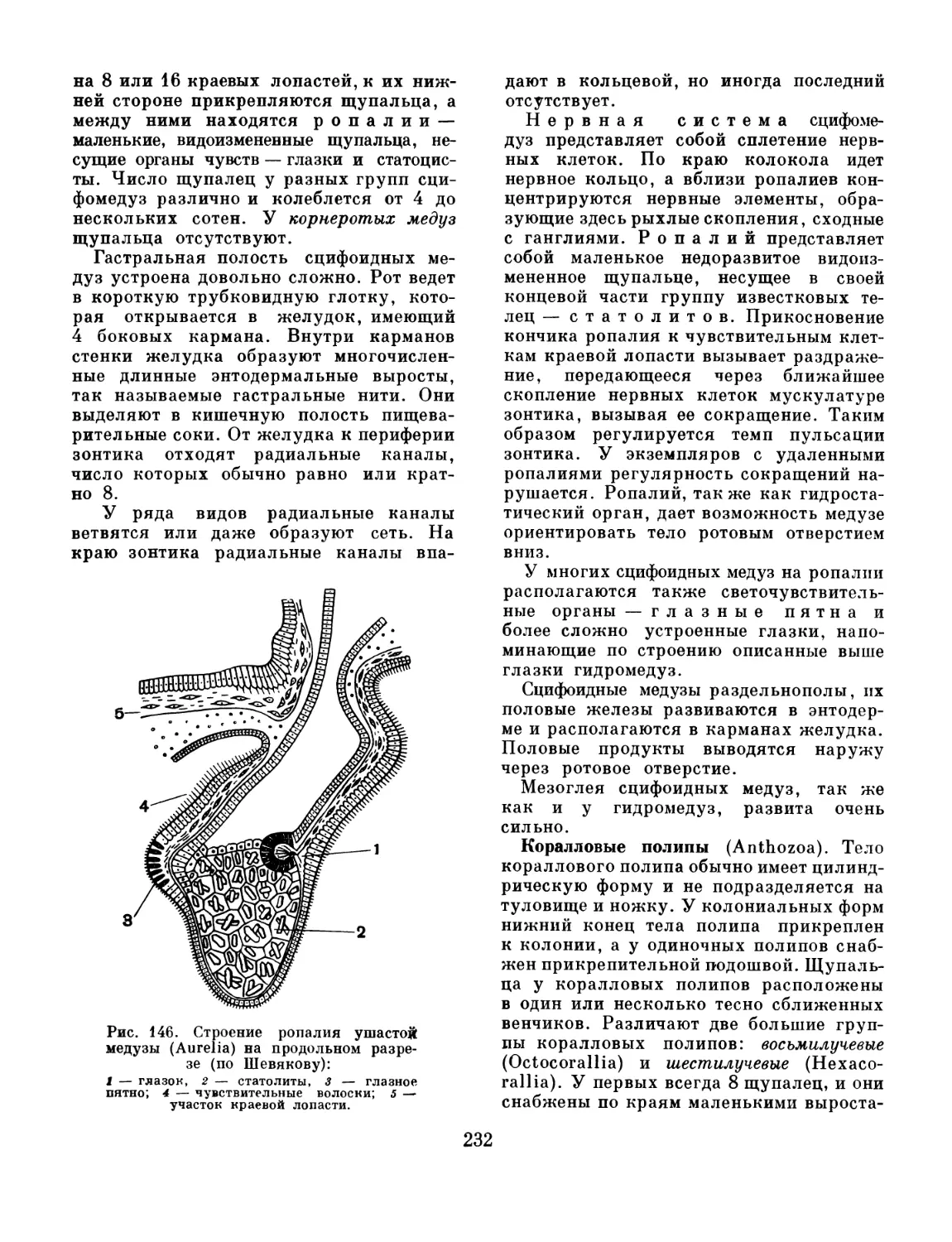


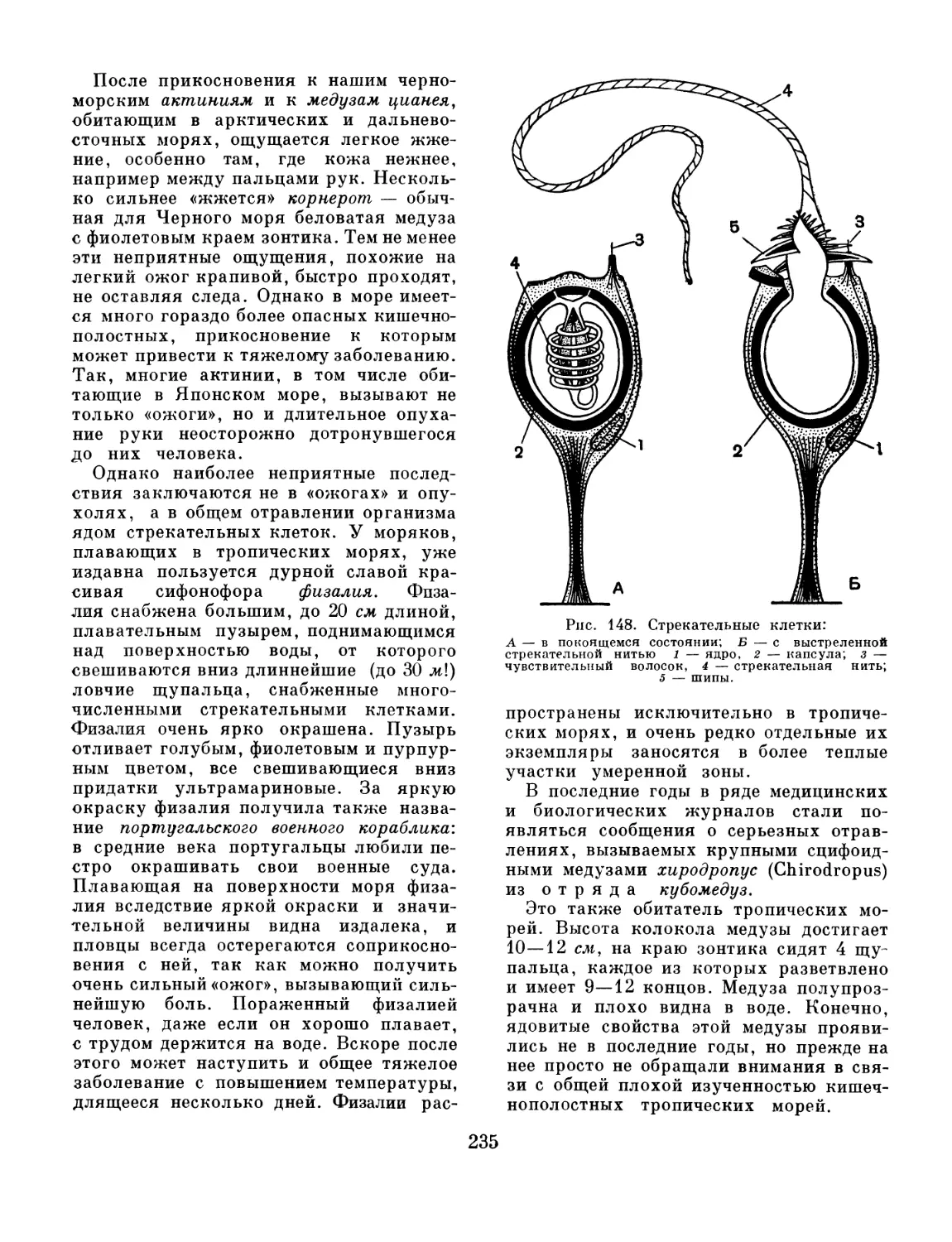










































































































































































































































































































































































Text
жизнь животных
в
6
ТОМАХ
РЕДАКЦИОННАЯ КОЛЛЕГИЯ]
проф. Л. А. ЗЕНКЕВИЧ (главный редактор)
профессора:
А. Г. БАННИКОВ, М. С. ГИЛЯРОВ,
Н. А. ГЛАДКОВ, А. П. КУЗЯКИН,
А. В. МИХЕЕВ, С. П. НАУМОВ,
Ф. Н. ПРАВДИН, Т. С. РАСС •
МОСКВА • 1968
БЕСПОЗВОНОЧНЫЕ
ЙОД РЕДАКЦИЕЙ
члена-корреспондента АН СССР
профессора Л. А. ЗЕНКЕВИЧА
1
ТОМ
ИЗДАТЕЛЬСТВО
«ПРОСВЕЩЕНИЕ»
жизнь животных
II РЕ Д И С Л О ВИ Б
«Жизнь животных» — это особый вид зоологической книги. В ней
находит отражение вся совокупность зоологических знаний, но в особом
соотношении разных разделов науки о животных. Зоология может быть
прежде всего подразделена на систематику, филогенетику, морфологию,
физиологию, экологию, зоогеографию (которая одновременно является и
разделом географии), палеонтологию, генетику, биохимию и биофизику
животных. Каждый из этих разделов зоологии может быть в свою оче-
редь разделен на ряд отдельных дисциплин; так, морфология делится на
анатомию, цитологию, гистологию, эмбриологию, а они в свою очередь
могут подразделяться на подчиненные дисциплины. В цитологии и гисто-
логии выделяют цитохимию и гистохимию, анатомию делят на сравни-
тельную, нормальную, патологическую и т. п. В физиологии в свою оче-
редь выделяют сравнительную физиологию, физиологию высшей нервной
деятельности, биохимию и т. д.
Кроме того, зоологические дисциплины могут быть разделены по
изучаемым группам животных. Так, различают протозоологию (науку
об одноклеточных животных), гельминтологию (науку о червях), энтомо-
логию (науку о насекомых), малакологию (науку о моллюсках), орнито-
логию (науку о птицах), маммалиологию (науку о млекопитающих),
герпетологию (науку о пресмыкающихся). Зоологические науки могут
выделяться также по биологическим особенностям животных объектов,
как, например, паразитология, охватывающая многие группы по их спо-
собности паразитировать на других животных или растительных орга-
низмах или быть «хозяином» паразита. Наконец, существует много смеж-
ных наук, в которые изучение животных входит как составная часть.
К ним относятся география, геология, океанология, лимнология (наука
об озерах), гидробиология, почвоведение и ряд других. Каждая из пере-
численных наук рассматривает тех или иных животных в определенном
аспекте: или как элемент внешнего мира, или по их значению для челове-
ка, или по каким-либо другим их особенностям.
План содержания книги «Жизнь животных» очень широк. Едва ли
хоть одна из перечисленных зоологических наук или смежных с ней
может оказаться полностью вне круга внимания этого сочинения,
но с использованием их достижений в разной степени.
В монографии «Жизнь животных» собраны описания животных в сис-
тематическом порядке — от простейших до млекопитающих. Для каждой
группы животных даны краткое морфологическое описание, некоторые
физиологические особенности, образ жизни, их индивидуальное раз-
витие, распространение в природе, практическое значение.
5
Сто лет назад A863—1869) появилось обширное сочинение немецкого
ученого — натуралиста и путешественника А. Б рема «Жизнь живот-
ных». За короткий срок это произведение было переведено на многие языки
и получило широкую мировую известность. Зоологические сочинения,
охватывающие все животное царство, ставят разные задачи и имеют раз-
ный характер. Мы знаем учебники, руководства и учебные пособия.
«Жизнь животных» не то, не другое и не третье. И в то же время это очень
нужная книга. Поэтому она была много раз издана в разных странах,
в том числе и у нас.
Уже давно «Жизнь животных» стала нарицательным названием,
и поэтому, когда сочинение Брема оказалось уже не на уровне современ-
ных знаний, стала выходить «Жизнь животных по Брему». Тип зооло-
гического сочинения, задуманного и осуществленного впервые Альфре-
дом Бремом, вероятно, сохранится еще на много лет.
В нашей стране сочинения Брема выходили многими изданиями: в
1866—1876 гг. под редакцией В, О. Ковалевского вышла в переводе 6-томная
«Жизнь животных». В 1892 г. начала выходить 10-томная «Жизнь живот-
ных» под редакцией Я. К. Сент-Илера, а в 1900 г. — малое трехтомное
издание под редакцией Я. Ф. Лесгафта. В начале века материал бремов-
ского сочинения стал устаревать. В Германии появилось в свет 13-томное
издание «Жизни животных» в переработке ряда специалистов под общей
редакцией Цур-Штрассена. В 1911 г. оно стало выходить в русском пе-
реводе под редакцией Я. М. Книповича. Наконец, в 1948 г. у нас вышло
5-томное издание «Жизнь животных по Брему» под редакцией А. Я. Се-
верцова. В основу его было положено также цурштрассеновское издание.
В создании первого тома настоящего издания участвовали:
доктор биол. наук Л, А. Зенкевич, доктор биол. наук Е. И. Лукин,
доктор биол. наук Д. В. Наумов, доктор биол. наук Я. Г. Ошмарин, доктор
биол. наук А. А. Парамонов, доктор биол. наук Ю. И. Полянский, доктор
биол. наук Я. М. Рыжиков, канд. биол. наук Г. Г. Абрикосов, канд.
биол. наук А. С. Богословский, канд. биол. наук В, М. Колтун, канд.
биол. наук Я. Я. Малевич, канд. биол. наук Ф. А. Пастернак, канд. биол.
наук Я. А. Свешников.
В распределении материала по томам пришлось несколько отступить
от системы. Иглокожие и низшие хордовые (из вторичноротых) включены
во второй том, который объединяет основные группы беспозвоночных, оби-
тающих в водной среде. Третий том посвящен сухопутным членистоногим.
Карты зоогеографического районирования суши и Мирового океана со-
ставлены по материалам Физико-географического атласа мира АН СССР и
ГУГК ГГК СССР издания 1964 г., с изменениями и дополнениями
Л. А. Зенкевича.
ВВЕДЕНИЕ
С животными человек был связан
в течение всей своей истории — исполь-
зовал их в качестве пищи, для изготовле-
ния украшений, а также и для других
хозяйственных надобностей и даже в каче-
стве менового товара и денег. Постепенно
различные животные становились объек-
тами охотничьего и рыбного промыслов,
а некоторые приручались человеком.
Стоянки древнего человека, как нео-
лита, так и палеолита1, служат хоро-
шей иллюстрацией. Уже палеолитиче-
ские стоянки человека сопровождаются
иногда огромными, так называемыми ку-
хонными кучами (иначе говоря, кучами
мусорных остатков). В них всегда можно
обнаружить кости рыб, птиц и зверей,
а у морских побережий — раковины
моллюсков и панцири ракообразных.
Совершенно естественно, что по мере
развития промысла зарождались и полу-
чали развитие и зоологические познания.
На пещерных и наскальных рисунках чело-
века раннего палеолита изображены сцены
охоты и объекты ее: мамонт, бизон, носо-
рог, олень, медведь, разные рыбы и дру-
гие животные. Очевидно, охотник дол-
жен был уже располагать определенными
познаниями о распространении, биологии
и внутреннем строении объектов своего
промысла. В дальнейшем определенные
зоологические познания возникли в связи
с приручением животных, развитием охот-
ничьего и рыбного промыслов.
1 Неолит и палеолит от греческих слов neos —
новый, lithos — камень, palaios — ста-
рый. Новый и древний периоды каменного
века.
Родоначальником научной зоологии
считается греческий ученый Аристотель
C84—322 гг. до н. э.). Из его замечатель-
ных зоологических сочинений наиболь-
шей известностью пользовались «История
животных», «О частях животных» и «О воз-
никновении животных». Аристотель дал
описание 454 видов животных. Ему
принадлежит и первая попытка дать
систему животных. Им же впервые вве-
дены понятия вида (eidos) ирода
(genos), конечно, совсем не в том смысле,
в котором мы принимаем эти понятия
сейчас. Аристотель делил всех живот-
ных на обладающих кровью и лишенных
ее. Главные 8 групп животных Аристо-
тель подразделял на более мелкие груп-
пировки. В античной Греции хорошо
знали головоногих моллюсков, так как
это обычная и излюбленная пища на-
родов, живших на побережье Средизем-
ного моря с древнейших времен, и Ари-
стотель подразделял их (Malakia) на деся-
тиногих и восьминогих, как и в современ-
ной зоологии. Такое подразделение живот-
ных, снабженных кровью, сохранилось
и доныне, только амфибии и рептилии
у Аристотеля объединены в одну группу.
Очень правильно большинство ракооб-
разных объединены у него в группе Mala-
kostraca и т. д. Этим объясняется, что
система животных, созданная гениальным
греческим естествоиспытателем, опере-
дила свое время на два тысячелетия и
просуществовала в малоизмененном виде
до начала XVIII столетия. Даже Лин-
ней, создатель основ современной систе-
матики животных, не внес существен-
ных изменений в систему Аристотеля.
Аристотель считал, что некоторые
ведущие прикрепленный образ жизни
животные, имеющие внешнее сходство
с растениями, имеют двойную природу—
растительную и животную. Идея о суще-
ствовании организмов смешанной при-
роды восходит к временам глубокой древ-
ности. Таким образом родилась группа
Zoophyta х. Высказывая правильную
в своем существе идею о родственной бли-
зости растений и животных, ученые,
однако, искали эту двойную природу не
среди одноклеточных организмов (о кото-
рых, правда, ничего не знали), что было
бы совершенно правильно, а среди много-
клеточных.
Крупнейшим биологом был также
римский естествоиспытатель Плиний Стар-
ший B7—79 гг. н. э.), погибший при извер-
жении Везувия, похоронившем под пеп-
лом Помпею и Геркуланум, автор «Есте-
ственной истории» в 37 книгах. В систему
животных Аристотеля Плиний, несмотря на
прошедшие 400 лет, не внес никаких изме-
нений.
СИСТЕМЫ ЖИВОТНОГО МИРА
Характерная особенность человече-
ского ума — это его стремление позна-
вать окружающий мир во всем его много-
образии, потребность систематизировать,
группировать явления по их сходству
или различию в соподчиненные катего-
рии. Если бы множество фактов не соби-
ралось в классифицированную систему,
невозможно было бы ни запомнить их,
ни тем более осмыслить. Даже самый изо-
щренный мозг систематика не может за-
помнить более нескольких тысяч назва-
ний. Однако все однородные биологиче-
ские явления природы обладают в силу
большего или меньшего родства большим
или меньшим сходством. Градации сход-
ства или различия находят свое выражение
в групповых объединениях, также свя-
1 Zoon (греч.) — животное, a phytdn —
растение. Zoophyta — организмы, объединя-
ющие в своей организации признаки животных
и растений.
занных единством происхождения. Так,
например, среди шмелей мы различаем
много видов: лесной, луговой, садовый,
каменный и т. д. Все они различаются
видовыми признаками, но все объединя-
ются родовыми— все они шмели и состав-
ляют род Bombus и подсемейство шмели-
ных (Bombinae).
В семействе пчелиных есть другие под-
семейства (Bombinae, Andreninae и др.),
объединяемые в группу Mellifera отряда
перепончатокрылых (Hymenoptera)— один
из 33 отрядов класса насекомых, а этот
последний отличается группой признаков,
выделяющих его среди типа членистоногих
(Arthropoda). Таким образом, любое жи-
вотное имеет видовое название и относится
к определенным роду, семейству, отряду,
классу и типу животных, а этот тип в
свою очередь вместе с другими типами
составляет царство животных, отличное
по ряду признаков от царства растений и
микробов.
Для живых организмов, в отличие
от неживой природы, характерна спо-
собность к питанию, обмену, росту, дви-
жению, ощущению, размножению и эво-
люционным изменениям.
Вероятно, нет ни одной области
науки, техники и искусства, в которой
не использовалась бы в большей или мень-
шей степени классификация. В опреде-
ленном аспекте она отражает все достиже-
ния в данной области человеческого зна-
ния и в значительной степени выражает
высоту достигнутого ею уровня.
Для биологии, если учесть, что на Зем-
ле существует не менее полутора миллио-
нов видов животных и полмиллиона видов
растений, систематика приобрела особое
значение и выросла наряду с морфоло-
гией, палеонтологией и физиологией
в самостоятельный раздел биологии. Зна-
чение ее чрезвычайно велико, и без даль-
нейшего ее развития и внедрения во все
разделы биологии невозможно и ее полно-
ценное развитие. Систематика нужна пре-
жде всего потому, что она сводит в систему
все многообразие живого и дает возмож-
ность легко находить в этой системе
место для нового факта. Систематика
дает наиболее точную характеристику
объекта экспериментальных и биологиче-
ских исследований, без чего само иссле-
дование теряет значительную долю, а
часто и весь смысл, так как биологиче-
ские свойства, которыми обладает опре-
деленный вид, могут быть не свойственны
другому, даже очень близкому виду.
Система дает яркую картину фило-
генетического развития животного мира,
отражая родственные связи между от-
дельными группами и предоставляя воз-
можность решать одну из наиболее важ-
ных в теоретическом и практическом
отношении проблем биологии — проблему
возникновения новых видов, а также
и других систематических категорий.
Какой бы биологический вопрос мы ни
взяли, нам прежде всего необходима
точная классификационная характери-
стика избранных нами объектов и общие
представления о происхождении и раз-
витии той группы, к которой они относятся.
Систематику справедливо называют мате-
матикой биологии. Следует при этом
отметить, что отдельным одноименным
систематическим группам может быть
свойственно различное современное ви-
довое разнообразие. Так, в класс насе-
комых включают около 1000 000 извест-
ных науке видов, в класс брюхоногих
моллюсков — около 90 тыс. видов, в боль-
шинство классов включают по нескольку
тысяч или сот живущих ныне видов,
а в классы наутилоидей и мечехвостов—
только по 4—5 видов, в класс однокрышеч-
ковых моллюсков (Monoplacophora, или
Neopilina) — два вида, а к классу кисте-
перых рыб относят только одну латиме-
рию. Вероятно, все классы с очень малым
числом видов — это вымирающие группы,
уходящие с арены жизни. Действительно,
многие из них в прежние геологические
периоды были представлены многими
десятками, сотнями, а иногда и тысячами
видов. Поэтому особенный интерес вызы-
вает их систематическая обособленность
от других ныне живущих групп.
Давно уже в биологии трактуются
понятия — искусственная и естественная
система. Большинство биологов считают,
что естественная система должна основы-
ваться на родственных связях, т. е. иметь
генеалогическую основу, и поэтому есте-
ственная система может быть только одна.
Ч.Дарвин выразил эту идею словами:
«Всякая истинная классификация есть
генеалогическая». Искусственные системы
строятся по немногим признакам, не бази-
руются на родственных связях, и поэтому
их может быть сколько угодно.
Начало построения современной на-
учной системы животных мы находим
в трудах шведского натуралиста Карла
Линнея A707—1778), и главным образом
в сочинении «Systema naturae» (первое
издание вышло в 1735 г.), выдержавшем
в течение 43 лет 13 изданий. Первой за-
слугой его явилась формулировка поня-
тия вида. По К. Линнею, вид— это сово-
купность организмов, сходных между
собою, как сходны дети одних родителей,
и способных давать плодовитое потом-
ство.
Таким образом, если плодовитого по-
томства не получается, значит, роди-
тели относятся к разным видам. Обычно
при скрещивании разных видов потом-
ства не получается. Иногда потомство
бывает, но оно не способно к дальнейшему
размножению, как, например, мулы (мать-
лошадь, отец — осел) и лошаки (мать-
осел, отец —лошадь).
Второе чрезвычайно важное поло-
жение Линнея заключалось в создании
иерархической системы соподчиненных
категорий, которых у Линнея было четыре:
вид, род, отряд и класс. Иерар-
хический характер системы Линнея за-
ключался в том, что каждая категория
включает несколько категорий низшего
порядка: класс — отрядов, отряд — ро-
дов, род — видов.
Линней предложил каждый вид имено-
вать двумя словами, первое из которых
означает название рода, а второе — соб-
ственно видовое. Например, зайца-беляка
он назвал Lepus timidus. Слово Lepus
(заяц) означает название рода. Второе
слово в названии собственно видовое:
timidus означает «трусливый». Позд-
нее П. Паллас описал другой вид — зай-
ца-русака и назвал его Lepus europaeus
(заяц европейский). По этим названиям
видно, что речь идет о двух разных видах,
но относящихся к одному роду (с одина-
ковым родовым названием). Обозначение
вида двумя латинскими словами называют
бинарной (двухсловной) номенклатурой.
За каждым видом закрепляется одно ла-
тинское название (из двух слов), которое
применяется в любой стране, независимо
от местных (национальных) названий.
Накопилось бесконечное множество слу-
чаев, когда один и тот же вид оказался
описанным разными авторами, в разное
время и под разными названиями. При
обнаружении таких случаев за видом за-
крепляют название, которое было дано
раньше.
Кроме того, общепринято к названию
добавлять имя ученого, давшего это назва-
ние, обычно в сокращенном виде. Так,
лев обозначается Felis leo L. Это значит,
что название дал Линней.
В дальнейшем система животных
усложнилась. Ж. Кювье A769—1832)
ввел еще понятие типа как высшей
систематической категории, позднее была
добавлена еще одна категория между отря-
дом и родом — семейство. Таким
образом возникло 6 основных катего-
рий: тип, класс, отряд, семей-
ство, род и вид. Однако этого ока-
залось недостаточно, и стали возникать
промежуточные категории с приставками
над- и под-, и, таким образом, добави-
лось еще около 10 категорий — под-
царство, надцарство, под-
тип, надклас с, подкласс, на д-
семейство и т. д. Но и этого оказалось
недостаточно, и появились еще категории
раздел, надраздел, триба
и т. п.
Вместе с развитием систематики живот-
ных возрастало количество научно описан-
ных видов. Аристотель дал описание 454
видов, у Линнея их уже 4208, у Гмели-
на—18 338. Наибольшее количество опи-
саний новых видов приходится на XIX в.
К началу века было описано около 50 тыс.
видов, а к концу — около 400 тыс. ви-
дов. В настоящее время известно около
миллиона видов.
В эпоху Возрождения, со второй
половины XVI в. и в течение последую-
щих трех столетий неуклонно идет накоп-
ление зоологических знаний. Если в на-
чале этого периода в обширных зоологи-
ческих сочинениях замечательные по своей
точности описания были перемешаны со
сведениями и описаниями совершенно фан-
тастическими, то уже в XVII в. появля-
ются обстоятельные зоологические сочи-
нения, удивляющие своей детальностью
и научностью.
В XVII в. появились замечательные
сочинения итальянца Мальпиги A628—
1694), главным образом по насекомым,
голландца Сваммердама A637—1680) по
моллюскам и насекомым и ряд других.
В это же время были введены в работу
оптические системы и Левенгук A652—
1723) открыл мир микроскопических су-
ществ. В конце XVII в. A693) появилось
замечательное сочинение английского
биолога Дж. Рея A628—1705) «Synopsis
methodica animalium» и целая серия бле-
стящих анатомических сочинений второй
половины XVII в. XVIII в. ознамено-
вался выходом в свет A735) основного
сочинения К. Линнея («Systema natu-
rae»).
Однако представления о системе жи-
вотных оставались в основном на уровне
аристотелевских. Только начало XIX в.
принесло новую систему животных и
эволюционное учение с определенными
представлениями в области филогении
животных. Основоположниками нового
этапа в развитии зоологии были два гени-
альных французских биолога: противник
эволюционных представлений Ж. Кювье
A769—1832) и основоположник эволю-
ционного учения Ж. Ламарк A744—1829),
причем наибольшее значение длл даль-
нейшего развития зоологии имели сочине-
ния Ламарка «Systeme des animaux
sans vertebres» A801) и «Philosophie zoolo-
gique» A809) и вышедшее несколько
позднее сочинение Кювье «Le regne animal,
distribue d'apres son organisation» A817).
Заслуга Ламарка прежде всего в том, что
он разделил всех животных на беспозво-
ночных и позвоночных. Он выделил три
основные группы червей — плоских, круг-
лых и кольчатых. В то же время в системе
Ламарка имеется группа радиальных
(Radiata), в которую включены и игло-
кожие, и кишечнополостные, и оболочни-
ковые, и еще некоторые другие группы.
Это по существу та же искусственная
группа — Zoophyta, только под другим
названием.
10
Кювье разделил животных на 4 ти-
па, которые он назвал «ветвями» (сам
термин «тип» был введен в зоологию
французским зоологом Блэнви-
л е м в 1825 г.): позвоночные, моллюски,
членистые и зоофиты (или радиаты).
Тем более удивительно, что одно-
временно с выходом в свет сочинений Ла-
марка и Кювье и еще несколько десяти-
летий позднее пышно расцветали схола-
стические натурфилософские системы жи-
вотных.
Мы не будем излагать детали после-
дующих достижений в создании современ-
ной системы животных и ознакомимся
с ней на прилагаемых таблицах.
В настоящее время зоология распола-
гает не менее чем 25 категориями в сис-
теме животных. Совершенно очевидно,
что одноименные категории в разных ча-
стях системы животных неравноценны.
В настоящем издании «Жизни живот-
ных» мы принимаем следующую систему
(табл. 1).
К изложенной системе животных сле-
дует дать некоторые пояснения. Мы ввели
5 категорий выше типа. Прежде всего это
два подцарства — одноклеточные
и многоклеточные.
Эти две группы животных очень резко
отделены одна от другой. С другой сторо-
ны, подцарство многоклеточных следует
подразделить на три отдельные группы,
которые можно назвать надразде-
л а м и. Первый из них — небольшая
группа очень примитивных организмов
(Mesozoa), состоящих по существу из
одного слоя клеток (см. дальше). Их ор-
ганизация напоминает ту предковую для
многоклеточных гипотетическую форму,
которую И. И. Мечников назвал парен-
химеллой. Однако это паразитические
формы, и, возможно, их организация
подверглась вторичному упрощению.
Замечательным строением и развитием
обладают губки. С полным основанием
они противополагаются всем остальным
многоклеточным.
Губки лишены настоящих органов и
тканей и индивидуальности (Dividualia),
и в большинстве случаев бывает неясно,
с чем мы имеем дело, с отдельным орга-
низмом или колонией.
У губок удивительным образом «извра-
щены» зародышевые листки — эктодерма
и энтодерма. (Enantiozoa — вывернутые).
Хоаноциты губок чрезвычайно схожи
с клетками колониальных жгутиковых
(Choanoflagellata), а клетки тела сохраняют
самостоятельность и переходят друг в
друга.
Кроме того, губки обладают почти не-
ограниченной способностью к восстанов-
лению: из самого маленького кусочка губ-
ки легко восстанавливается целая. Тре-
тий надраздел настоящих многоклеточных
(Eumetazoa) состоит из двух весьма
различных разделов: одному (Radiata)
свойственна радиально-осевая симметрия,
а другому (Bilateria) — двусторонняя
симметрия, возникшая из радиальной.
Кроме того, Radiata могут быть отнесены
к двуслойным, так как у них получили
развитие только два зародышевых листка,
a Bilateria — к трехслойным: у них,
кроме экто- и энтодермы, получает разви-
тие и средний зародышевый листок —
мезодерма. Наконец, Bilateria можно под-
разделить на подразделы первичноротых
(Protostomia) и вторичноротых (Deute-
rostomia).
К первичноротым можно отнести 4 над-
типа:
н а д т и п низшие черви — без сегмен-
тации, кровеносной системы и целома
(Amera s. Scolecida). Сюда относят типы
плоских червей, колючеголовых, круглых
червей, нематоморф и немертин;
н а д т и п моллюски — с одним типом
того же названия;
н а д т и п малосегментные (Oligomera),
объединяющий типы мшанок, плеченогих
и форонид;
н а д т и п полимерные (многосегмент-
ные), включающий типы аннелид, первич-
нотрахейных и членистоногих.
К вторичноротым относят семь типов,
для большинства которых свойственно
развитие, хотя бы только в личиночном
состоянии, хорды и расположение глав-
ных органов по иному плану, чем у пер-
вичноротых.
Таким образом, мы делим царство жи-
вотных на 2 подцарства, 4 надраздела,
5 разделов, 6 подразделов, 9 надтипов,
23 типа и 71 класс.
11
Таблица 1
ОСНОВНЫЕ ПОДРАЗДЕЛЕНИЯ ЖИВОТНОГО ЦАРСТВА (ДО ТИПОВ)
Подцарство Надраздел Раздел
Подраздел
Надтип
Тип
Одноклеточные
(простейшие)
животные
Protozoa
Protozoa
Protozoa
Protozoa
Protozoa
Простейшие (Protozoa)
Многоклеточ-
ные животные
(Metazoa)
Mesozoa
Mesozoa
Parazoa
Настоящие
многоклеточ-
ные животные
(Eumetazoa)
Parazoa
Радиальные
(Radiata)
Д ву сторон не-
симметричные
(Bilateria)
Mesozoa
Parazoa
Radiata
Mesozoa
Mesozoa
Parazoa
Radiata
Перви чноротые
(Protostomia)
Несегментиро-
ванные
(Amera)
Моллюски
(Mollusca)
Малосегмент-
ные, или ол и то-
ме рные (Oligo-
mera)
Многосегмент-
ные, или по-
лимерные
(Polymera)
Вторичноротые
(Deuterostomia)
Губки (Spongia)
Кишечнополостные
(Coelenterata)
Гребневики
(Ctenophora)
Плоские черви (Plat-
helminthes)
Колючего л овые
(Acanthocephala)
Круглые черви
(Nemathelminthes)
Немертины (Nemer-
tini)
Моллюски
(Mollusca)
Форониды (Phoronoidea)l
Мшанки (Bryozoa)
Плеченогие (Brachio
poda)
Кольчатые черви
(Annelides)
Первичнотрахейные
(Onychophora, Pro-
trachea ta)
Членистоногие
(Arthropoda)
Щетинкочелюстные
(Chaetognatha)
Иглокожие (Echino-
dermata)
Chordaria
Погонофоры
(Pogonophora)
Полухордовые
(Hemichordata)
Оболочниковые
(Tunicata)
Головохордовые
(Cephalochordata)
Позвоночные
(Vertebrata)
12
Таблица 2
ТИПЫ ЖИВОТНЫХ И ИХ ПОДРАЗДЕЛЕНИЯ
1 Типы
1 1. Protozoa
1 2. Mesozoa
1 3. Spongia
1 4. Coelenterata
1 5. Ctenophora
1 6. Plathelminthes
1 7. Nemathelminthes
1 8. Acanthocephala
1 9. Nemertini
110. Annelides
111. Phoronoidea
\i2, Bryozoa
113. Brachiopoda
114. Mollusca
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
18.
19.
Классы
Саркодовые (Sarcodina)
Жгутиковые (Flagellata,
Mastigophora)
Споровики (Sporozoa)
Инфузории (Infuzoria)
Книдоспоридии (Cnidospo-
ridia)
Ортонектиды (Orthonectida)
Дициемиды (Dicyemida)
Губки (Spongia)
Гидрозои (Hydrozoa)
Сцифоидные (Scyphozoa)
Кораллы (Anthozoa)
Гребневики (Ctenophora)
Турбеллярии (Turbellaria)
Сосальщики (Trematoda)
Темноцефалы (Temnocep-
hala)
Моногенетические (Mono-
genoidea) сосальщики
Гирокотилиды (Gyrocoty-
loidea)
Цестоды (Cestoidea)
Нематоды (Nematoda)
Добавочные классы к типу
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
Nemathelminthes
Ресничнобрюхие (Gastrot-
richa)
Коловратки (Rotatoria)
Киноринхи (Kinorhyncha)
Волосатики (Nemathomor-
pha, Cordiacea)
Колючеголовые (Acantho-
cephala)
Приапулиды (Priapulloidea)
Камптозои (Kamptozoa)
Немертины (Nemertini)
Многощетинковые (Poly-
chaeta)
Малощетинковые (Oligo-
chaeta)
Пиявки (Hirudinea)
Эхиуровые (Echiuroidea)
Добавочный класс
Sipunculoidea
Форониды (Phoronoidea)
Мшанки (Bryozoa)
Плеченогие (Brachiopoda)
Однокрышечковые (Mono- i
placophora)
Количество
современных
видов
10000
6000—8000
1340
6000
1000
23
5000
2700
200
6100
84
1500
3000
1000
1800
10 000
175—200
1500
100
100
250—300
5
32
300
4000
2400
300
100
300
15
2700
170
2
Основная среда обитания 1
Все среды обитания. Из cap-1
кодовых фораминиферы и ра-1
диолярии — обитатели моря. 1
Споровики исключительно па-1
разиты. Среди представителей!
остальных трех классов много!
паразитов 1
Паразиты 1
Море. Около десятка видов!
семейства Spongellidae обитает!
в пресной воде 1
Море. В пресной воде нес-1
колько видов гидр и 2—3 ви-1
да медуз 1
Море 1
Море 1
Море 1
Все среды обитания 1
Паразиты 1
Паразиты 1
Паразиты 1
Паразиты 1
Паразиты 1
Все среды обитания. Поло-1
вина паразитов 1
Все среды обитания 1
Все среды обитания 1
Море 1
Паразиты в личиночном со-1
стоянии 1
Паразиты 1
Море 1
Море 1
Море. Один вид в пресной!
воде 1
Море. В пресной воде нес-|
колько видов 1
Пресные воды и почва. В|
море несколько видов 1
Пресные воды. Несколько 1
видов в морской воде и воз-1
душной среде 1
Море 1
Море
Море 1
Море. В пресной воде нес-|
колько десятков видов отряда 1
Море 1
Море 1
13
Продолжение
1 Типы
Il5. Arthropoda
116. Onychophora s. Pro-
1 trachea ta
J17. Echinodermata
118. Chaetognatha
119. Pogonophora
120. Hemichordata
121. Tunicata
122. Cephalochordata
123. Vertebra ta (с подти-
1 памп Cyclostomata и
1 Gnathostomata)
1
Классы
37. Беспанцирные (Aplacopho-
га)
38. Панцирные (Loricata)
39. Брюхоногие (Gastropoda)
40. Лопатоногие (Scaphopoda)
41. Двустворчатые (Bivalvia),
или пластинчатожаберные
(Lamellibranhiata)
42. Головоногие (Cephalopoda)
43. Ракообразные (Crustacea)
44. Трилобиты (Trilobita)
45. Меростомовые (Xiphosura)
46. Паукообразные (Arachnida)
47. Коленчатоногие, или мор-
ские пауки (Pantopoda)
48. Многоножки (Myriapoda)
49. Насекомые (Insecta)
Добавочные классы
50. Тихоходки (Tardigrada)
51. Пятиустки (Pentastomida)
52. Онихофоры (Onychophora)
53. Морские звезды (Asteroi-
dea)
54. Змеехвостки (Ophiuroidea)
55. Морские ежи (Echinoidea)
56. Голотурии (Holothuroidea),
или морские кубышки
57. Морские лилии (Crinoidea)
58. Щетинкочелюстные (Chae-
tognatha)
59. Погонофоры (Pogonophora)
60. Кишечнодышащие (Ente-
ropneusta)
61. Крыложаберные (Ptero-
branchia)
62. Асцидии (Ascidiacea)
63. Сальпы (Thaliacea)
64. Аппендикулярии (Appen-
dicularia)
65. Бесчерепные (Acrania)
66. Круглоротые (Cyclostomata)
67. Рыбы (Pisces)
68. Земноводные (Amphibia)
69. Пресмыкающиеся (Reptilian
70. Птицы (Aves)
71. Млекопитающие (Mamma-
Количество
современных
1 видов
150
1000
90 000
300
15 000
600
25 000
4000
5
35 000
300
8100
1 000 000
300
75
65
1 6000
30
100
56
20
^ |
>
, 1600
22 1
свыше 40
20 000
1 6000
8590
lia) 4000-4500
Основная среда обитания 1
Море 1
Море 1
Главным образом море. Мно-1
го в пресных водах и в воз-1
душной среде. Есть паразиты 1
Море 1
Море. Семейство Unionidae — 1
в пресной воде 1
Море 1
Все среды обитания. Четы-|
рем отрядам низшим рако-1
образным обычен паразитизм 1
Море, пресные воды, воздушная 1
среда 1
Море 1
Воздушная среда. Среди кле-1
щей много паразитов. Предста-1
вители одного семейства кле-|
щей (Halocaridae) обитают в|
море 1
Море 1
Воздушная среда 1
Воздушная среда и пресные 1
воды. Виды одного семейства!
клопов (Halotatidae) обитают 1
в море 1
Море, пресные воды, воз-|
душная среда 1
Паразиты 1
Воздушная среда 1
Море I
Море 1
Море 1
Море 1
Море 1
Море 1
Море 1
Море и пресные воды 1
Море и пресные воды 1
Пресные воды I
Воздушная среда. Некоторые!
группы вторично возвратились!
в море: некоторые рептилии, I
а главным образом млекопи-1
тающие |
14
Представители 10 классов животных
целиком ведут паразитическое существо-
вание, а животные 60 классов в основном
свободноживущие, но 9 классов из них
частично являются паразитами. При этом
явление паразитизма свойственно только
группам, входящим в состав первичноро-
тых, и отсутствует у вторичноротых, если
не считать хищничества, переходящего в
паразитизм у круглоротых, и самцового
полупаразитизма на самках у некоторых
глубоководных рыб. Также отсутствует
паразитизм и у Oligomera, но широко
развит у Amera.
Весьма существенно, что истинными и
первичными обитателями воздушной среды,
у которых выработались воздушные орга-
ны дыхания (легочные мешки, трахеи
и легкие), являются только представители
типа Onychophora, 2 класса (из 8) типа
членистоногих и 4 класса (из 6) типа
позвоночных.
Также заслуживает внимания факт, что
эти 6 классов представляют собой высшие
группы как первично-, так и вторичноро-
тых (членистоногие и хордовые). Очевидно,
для выхода в воздушную среду были необ-
ходимы какие-то особенности организации,
отсутствующие у низших групп живот-
ных. Можно думать, что в данном случае
основная роль принадлежала свойствам
кожных покровов, препятствовавших вы-
сыханию тела в воздушной среде. У чле-
нистоногих таким образованием явилась
хитинизированная кутикула, а у рыбо-
образных — чешуя и слизь, покрывающая
тело, которая в свою очередь была образо-
ванием, обеспечивавшим быстрое (сколь-
зящее) движение в водной среде.
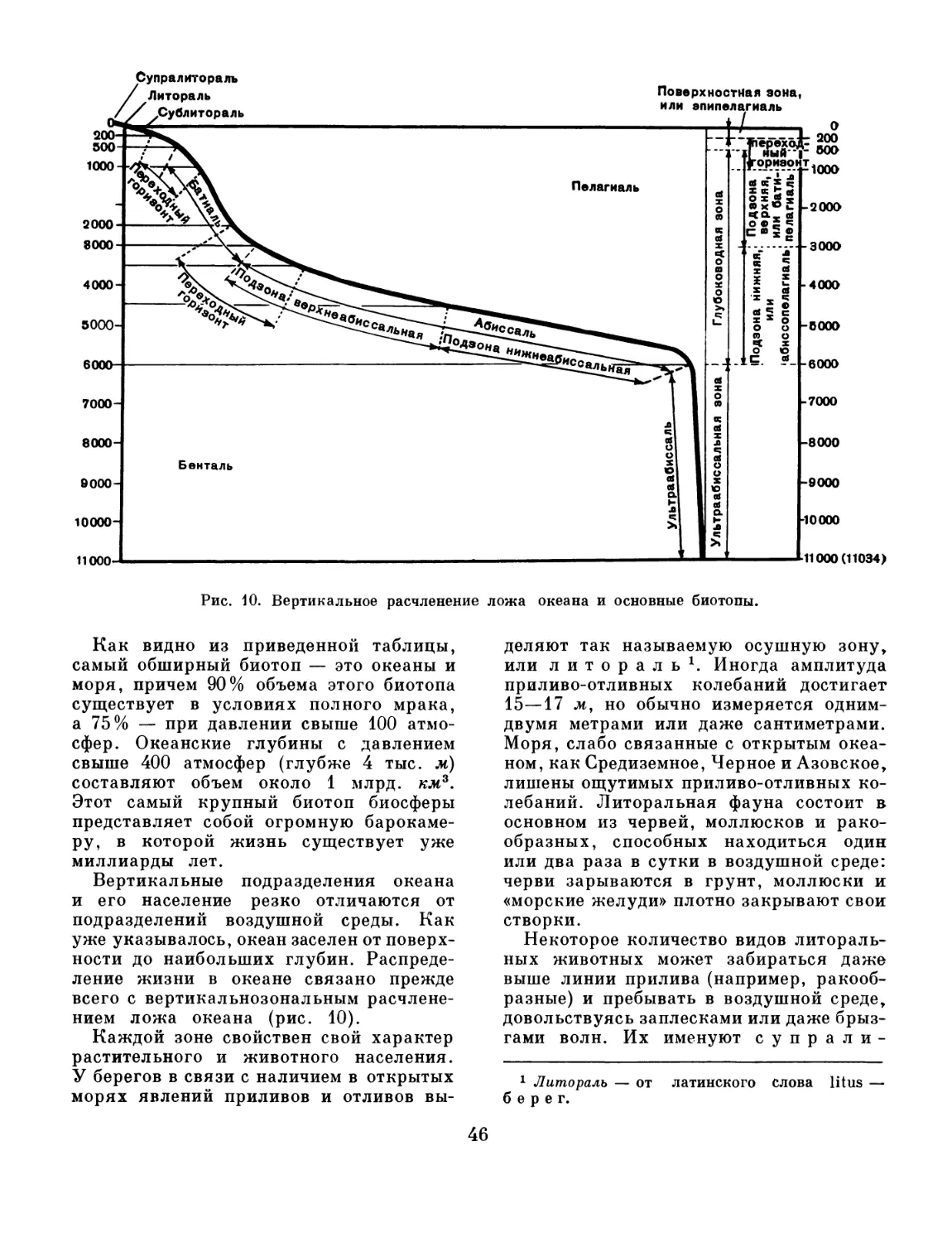
ФИЛОГЕНИЯ И ГЕОЛОГИЧЕСКОЕ
ПРОШЛОЕ ЖИВОТНОГО МИРА
Как мы уже указывали, система живых
существ неразрывно связана с их фило-
генией. Это две стороны одного и того же
явления: система — это статическое
состояние современного животного мира,
филогенетика дает представление
о процессе становления современных так-
сономических подразделений и тем самым
об их родственных связях. Естественная
система без филогении, так же как и фи-
логения без естественной системы, теряет
свой реальный смысл.
В прилагаемой схеме филогенетического
развития животных (рис. 1) отражены
современные представления. Многие части
этой схемы достаточно общепризнанны
и не вызывают сомнений. К ним можно
отнести возникновение губок и кишечно-
полостных от одноклеточных животных,
возникновение низших червей (турбелля-
рий) от кишечнополостных и немертин
от низших червей (турбеллярий). Несом-
ненно возникновение членистоногих от
кольчатых червей (морских кольчецов —
полихет).
Также общепризнанно близкое родство
групп, относящихся к вторичноротым.
В остальном филогенетические связи
более или менее спорны. Прежде всего
вызывает сомнения генезис губок. Еще
более неясны происхождение и генети-
ческие связи олигомерных групп пер-
вичноротых — форонид, мшанок и плече-
ногих, ошибочно объединявшихся иногда
термином Tentaculata (щупальцевые)
или Prosopygii (переднепорошицевые).
Развитие эмбриологии, сравнительной
анатомии, экспериментальной генетики
и сравнительной биохимии будет способ-
ствовать построению достаточно досто-
верной филогенетической схемы. Много
очень важных данных для построения
филогении дает также и палеонтология.
Так, отпечатки трилобитов и их четырех-
сегментных личинок дали возможность
сблизить их с классом хелицерат
и приблизить к ним представителей к л а с-
с а мечехвостов (Xiphosura), имеющих
трилобитообразную четырехсегм«нтную
личинку. Самые древние иглокожие кар-
поидеи (Carpoidea) были уже сидячими
организмами (вели прикрепленный образ
жизни), но сохраняли еще двусто-
роннюю симметрию. Это одно
из доказательств, что предками современ-
ных иглокожих были двусторонне-сим-
метричные организмы. Генетические взаи-
моотношения внутри типа моллюсков,
особенно головоногих, смогли быть разъ-
яснены благодаря богатейшим ископае-
мым остаткам вымерших групп. Но, ко-
45
Хронология
Периоды
ей
w L ii
0 «л
1 «О
О Четвертичный
Неогеновый
N 26-30
Палеогеновый
Pg 30 - 35
Меловой
65- 60
Юрский
25- 36
Триасовый
Т 30-35
Пермский
Р 26-30
Каменно
угольный
60-55
Девонский
D 45-60
Силурийский
S 40-45
ОрДОЙИиСКИЙ
О 70
Кембрийский
Cm 70 - 90
Ж
Растения
Рис. 1. Схема филогенетического развития животных.
16
нечно, наиболее замечательные факты
дает палеонтология для суждения о гене-
алогии современных групп позвоночных
(группа кистеперых рыб). Это касается
в равной степени и вопроса о возникно-
вении наземных позвоночных, про-
исхождении рептилий, птиц и млекопи-
тающих.
В филогенетических построениях очень
большое значение имеет принцип эволю-
ционного развития, который лежит в их
основе (принцип монофилии или полифи-
лии). Большинство зоологов придержи-
ваются дарвиновского принципа монофи-
лии, однако имеются и сторонники поли-
филии.
Принцип монофилии требует единого
корня происхождения как для всего жи-
вотного мира, так и для каждой систе-
матической группы.
Полифилитическая концепция, наобо-
рот, допускает возможность возникнове-
ния отдельных систематических групп
животных, как и всего животного мира,
из нескольких корней путем конвергент-
ного схождения. Эта концепция находит
свое применение в палеонтологии.
Борьба двух концепций идет и по
вопросу о происхождении человека. Со-
ветские антропологи, так же как и мно-
гие ученые за рубежом, считают, что все
человеческие расы произошли от одной
предковой группы. Некоторые зарубежные
антропологи склонны искать разные корни
вида Homo sapiens — азиатские, африкан-
ские, австралийские.
В истории выработки естественной сис-
темы и филогении таких примеров можно
было бы привести очень много. Всего
150 лет назад великий французский спе-
циалист по сравнительной анатомии
Ж. Кювье объединил фораминифер
и моллюсков за внешнее сходство в строе-
нии раковины, присоединяя к ним и усо-
ногих ракообразных, что уже не соответ-
ствует современным представлениям.
Вся история современной системы живот-
ных — это стремление к естественной
системе и утверждения принципа моно-
филии.
Принятая в настоящем издании фило-
гения животных построена по монофи-
литическому принципу.
Геология дает возможность располо-
жить в хронологическом порядке время
появления, наибольшего развития и выми-
рания многих групп животных, остатки
которых сохраняются в ископаемом виде
(рис. 2). Прежде чем рассмотреть эту
схему, следует сделать некоторые предва-
рительные замечания.
Более или менее достоверная пелеонтоло-
гическая летопись застает животный мир
не на первых этапах его развития, а в
довольно зрелом состоянии. Период с кем-
брия до наших дней — это, возможно,
только одна десятая всего времени разви-
тия на Земле животных. При этом следует
иметь в виду, что было бы весьма оши-
бочно переносить представления о скоро-
сти развития наземной фауны на все жи-
вотное царство, а именно на гораздо
более богатое типами и классами живот-
ное население морей и океанов, иначе
говоря, галосферы1. Темп филоге-
нетического развития подтипов наземных
членистоногих — трахеат и хелицерат
(классы многоножек, насекомых и пауко-
образных) — и подтипа наземных позво-
ночных (большинства амфибий, репти-
лий, птиц и млекопитающих), а также
некоторых других небольших групп поч-
венных и пресноводных беспозвоночных
(саркодовых, жгутиковых, инфузорий,
турбеллярий, коловраток, олигохет, мол-
люсков, ракообразных и некоторых дру-
гих) в истории развития животного мира
занимает особое место и представляет
собой бурный процесс заселения не за-
селеной до того наземной и пресноводной
среды выходцами из океана. Если мы
перенесем темпы развития с наземной
фауны на морскую, то получим зна-
чительное несоответствие. Совсем други-
ми темпами характеризуется развитие
морских животных, включающих 15 типов
фауны. Пока суша не была заселена ра-
стительностью, на ней было невозможно
и развитие жизни животных. Только
в силурийское время у морских побе-
режий стали возникать предковые для
наземной растительности формы (псило-
1 Галосфера — морские воды, от греческого
halos — соленый.
2 Жизнь животных, т. i
17
Вторичноротые
Vertebrate
CephalocKor-
data ^
Enteropneusta
Echinodermata
Chaetognatho
Tunlcata
Pogonophora
Первичноротые
Arthropoda
Phoronoidea
Bryozpa
Brachiopoda
Mollusca
Nemathelmi nthes|
lAcanthocephala
Plathelminthes
Рис. 2. Филогенетические связи некоторых групп
животных.
фитовая флора) и быстрым темпом стала
развиваться наземная растительность.
Что же касается морей и океанов, то
в них хлорофилл констатирован в отло-
жениях, отстоящих от нас на ЪУг млрд.
лет. В силурийское время у морских
побережий и в сильно опресненных при-
брежных водах уже подготовлялись
«десантные части» для захвата воздуш-
ной среды и пресных вод суши; это
были членистоногие, некоторые группы
червей и позвоночные. Морфофизиологи-
ческие особенности ряда групп морской
фауны и огромное пространство располо-
женной рядом суши, пре-
доставившей к тому вре-
мени первичные (расти-
тельные) пищевые ресур-
сы, создали условия для
бурно развернувшегося и
протекавшего быстрыми
темпами процесса видооб-
разования, на первых эта-
пах лишенного острой кон-
куренции. В этом процес-
се ярче всего сказалась
свойственная живым орга-
низмам характерная спо-
собность расширять свой
жизненный ареал и за-
хватывать новые среды
обитания в пределах воз-
можного для их морфоло-
гических и физиологиче-
ских структур.
Что касается морской
фауны, то можно с пол-
ной уверенностью конста-
тировать, что из нижнего
кембрия уже известны мно-
гочисленные представите-
ли современной морской
фауны. В этой истории мы
совсем не знаем каких-
либо катастрофических
смен фауны, о которых час-
то, со времен Ж. Кювье,
говорят специалисты по
геологической истории на-
земных рептилий и позво-
ночных.
Эволюция морской фау-
ны протекала за палеон-
тологическое время (с начала палео-
зоя) очень медленно и спокойно. Сколь
относительно медленны темпы эволюции
морской фауны, можно видеть на сле-
дующих примерах.
Близкий к замечательному моллюску
Neopilina ископаемый род Tryblidium
существовал в кембрии 400—500 миллио-
нов лет тому назад, головоногие моллю-
ски наутилиды известны с юры (свыше
100 миллионов лет) и дожили до нашего
времени, род Limulus (мечехвост)
известен с верхней перми (около 200 млн.
лет), плеченогие Lingula и Crania изве-
18
стны с девона B50—300 млн. лет). Кисте-
перые рыбы, к которым относится заме-
чательная латимерия, впервые пойман-
ная в живом виде у южных берегов
Африки, а затем у Коморских островов,
также имеют возраст 250—300 млн. лет.
Такие обычные ныне роды двустворчатых
моллюсков, как Nucula, Leda, Modiolus,
Ostrea, Lima, Avicula и др., существуют
с кембрия, т. е. 400—500 млн. лет. С и м п-
с о н в своей известной книге «Темпы
и формы эволюции» A944) определяет
длительность существования некоторых
родов двустворчатых моллюсков не менее
как в 275 млн. лет, а некоторых —более
400 млн. лет. Средний геологический
возраст для родов двустворчатых мол-
люсков он определяет в 80 млн. лет. На-
ряду с этим можно упомянуть, что прес-
новодное ракообразное щитень (Apus
cancriformis) существует с триаса, т. е.
свыше 150 млн. лет, с теми же родовыми
признаками.
Таким образом, можно считать несомнен-
ным положение, что с кембрия по наше
время морская фауна не претерпела
(по сравнению с наземной) каких-либо
очень крупных принципиально важных
изменений и находилась примерно на
том же этапе эволюционного развития,
что и ныне. Во всяком случае, все типы
и большинство классов морских животных
уже существовали. Если представить себе
длительность всего исторического разви-
тия современной морской фауны, то пе-
риод с начала кембрия до наших дней
должен рассматриваться только как по-
следний короткий этап. Если при этом
допустить, что темп эволюции в докем-
брийское время не мог существенно отли-
чаться по своему характеру и длитель-
ности от послекембрийского, то необхо-
димое время для предшествующей кемб-
рию эволюции морской фауны должно
было во много раз превышать послекем-
брийское.
Геологическая история животных вскры-
вает еще одно явление исключительного
интереса — это процесс вымирания.
Можно сказать, что почти в наше вре-
мя (в конце XVII в.) вымерла птица
дронт (или додо) на Маврикиевых остро-
вах, а у нас на Дальнем Востоке —мор-
ское млекопитающее стеллерова {морская)
корова (во второй половине XVIII в.).
В настоящее время почти полностью
могли бы быть уничтожены такие ценные
животные, как морские котики, калан
(морская выдра) и другие, если бы человек
не взял их под охрану. Исчезли дикие
лошади, коровы и т. д. Все эти случаи—
результат встречи с человеком, его про-
мыслом или бытом.
Но геология знает другие более инте-
ресные примеры вымирания, в которых
человек не играл никакой роли: трило-
биты, аммониты, наутилоидеи, белемни-
ты, некоторые классы иглокожих, пан-
цирные земноводные (стегоцефалы), пте-
родактили и др. Все это уже вымирание
не отдельных видов, а целых больших
групп и очень важных элементов фауны,
в состав которых входили сотни, а иногда
и многие тысячи видов. В современной
фауне имеются резко обособленные систе-
матически и морфологически группы, пред-
ставленные несколькими видами, являю-
щимися осколками древних богатых ви-
дами групп. Упомянутая выше Neopi-
lina, представленная ныне тремя вида-
ми в глубинах восточной стороны Ти-
хого океана,— это, вероятно, последний
осколок, может быть, многочисленного
в палеозое и в допалеозойское время
класса однокрышечковых моллюсков (Мопо-
placophora). Целый класс Priapuloidea
представлен в настоящее время всего
5 видами, но его остатки обнаружены
в палеозойских отложениях. Вероятно,
это была в прошлом обильная видами
группа животных, но отсутствие скелет-
ных частей стерло из прошлого их остат-
ки. Замечательная группа погонофор,
ранга типа, блестяще описанная А. Ива-
новым, представлена в наше время
несколькими десятками глубоководных
видов древнего облика, но в прошлом
также, вероятно, имела довольно широ-
кое распространение.
Удивительный существующий ныне
осколок древних гигантострак (из хели-
церовых членистоногих) — мечехвосты
(Limulus) существуют в количестве 5 ви-
дов, так же как остаток весьма многочис-
ленной группы Nautiloidea из голово-
ногих моллюсков, существующий ныне
2*
19
в числе 4 видов рода Nautilus. Подобных
примеров можно было бы привести не-
мало, и в каждом случае можно выска-
зать подозрение: не вымирающие ли это
организмы? Тем более удивительно, что
немногочисленные остаточные виды бога-
тых видами вымерших групп часто произ-
водят впечатление благоденствующих
ныне, форм, как, например, Nautilus,
Limulus, Priapulus.
Как объяснить причины их вымирания?
Прежде всего, конечно, конкуренцией
с другими, более молодыми и биологиче-
ски более сильными группами. Для трило-
битов это могли быть ракообразные,
для головоногих моллюсков — рыбы и ки-
ты, для птеродактилей — птицы и т. п.
Однако только конкуренцией не объяс-
нить всего явления вымирания.
Помимо конкуренции между видами
и отдельными особями, вымирание диких
животных объясняется резкой сменой
природных условий и невозможностью
для этих животных быстро приспособиться
к новым условиям среды. Мамонт вымер,
вероятно, одновременно с исчезновением
оледенения в северных областях Сибири
и Америки и в значительной степени под
воздействием человека. Возможно, что
основная причина вымирания животных
часто кроется в сочетании этих двух фак-
торов.
Геологическое прошлое Земли и геоло-
гическая хронология как бы подразде-
ляются на две части — докембрийский
период и от кембрия до наших дней.
Как хронология, так и животные после-
кембрийского периода изучены значи-
тельно лучше, чем докембрийского.
На прилагаемой схеме дана геологиче-
ская история различных групп живот-
ных, известных по ископаемым остаткам,
которая сопоставлена с историей неко-
торых групп растений, начиная с кембрий-
ского периода (рис. 1). Как видно, весь
этот период укладывается всего лишь
в 500—600 млн. лет. Точность знаний об
отдельных эпохах этого периода и его
хронологии тем меньше, чем дальше в
глубь веков уходит его история. Докем-
брийское время, которое еще недавно
исчислялось учеными сотнями миллионов
или в крайнем случае одним миллиардом
лет, в настоящее время все более и более
растягивается. Крупнейший специалист
по докембрию академик А. А. Полка-
нов дает следующую хронологическую
схему для докембрия Северной Европы.
Периоды и эпохи
Протерозой Верхний
Средний
Нижний
Архей Верхний
Нижний
Ката рх ей
Абсолютный возраст
в млн. лет
1670-715
1880—1750
2140—1900 1
2700—2200
3590-2710
Очевидно, это тот период в истории
Земли, когда земная кора уже была по
температурному режиму доступна для раз-
вития жизни. Академик А. П. Вино-
градов тоже считает, что темпера-
турные условия на поверхности Земли
приближались к современным в течение
2—3 млрд. лет, а возраст земной коры
он определяет в 5 млрд. лет. Академик
О. Ю. Шмидт допускал время сущест-
вования твердой оболочки Земли в 7 млрд.
лет.
Когда и как появилась жизнь на нашей
планете, остается неясным, но, в частно-
сти, можно допустить, что возникновение
фотосинтетической деятельности расти-
тельных организмов на Земле надо отно-
сить к периоду, отдаленному от нас на
3—4 млрд. лет. Уже давно стало очевид-
но, что остатки растений и животных из
докембрийских отложений редки, спорны
и неясны. Создалось мнение, что все эти
остатки подверглись перекристаллизации
(метаморфизации) и как документы науки
утеряны.
Существует также допущение, что отсут-
ствие ископаемых организмов в докембрии
объясняется тем, что докембрийские жи-
вотные не имели скелетов. Допускают
также, что основные группы палеозой-
ских животных сформировались на рубе-
же кембрия. Однако все эти допущения
не обоснованы в нужной мере, и вопрос
о ничтожном количестве докембрийских
20
ископаемых остатков животных пока
остается без ответа. Таблица (рис. 1) пояс-
няет то, что было сказано в предыдущем
изложении. В самом начале кембрия мы
застаем уже все основные группы совре-
менных морских животных, имеющих
скелеты, за исключением рыб: моллю-
ски, ракообразные, мшанки, плеченогие,
иглокожие. На этой схеме хорошо видно,
как растения и животные выбирались
на сушу и на сколько наземные животные
моложе морских.
НЕКОТОРЫЕ ЭТАПЫ ЭВОЛЮЦИИ
ЖИВОТНЫХ
Эволюционное развитие живых существ
следует рассматривать как целост-
ный процесс развития живого населения
биосферы от начальных, примитивных
форм до современных, наиболее совершен-
ных. Это относится в равной мере к мор-
фологическим и биохимическим структу-
рам и к физиологическим процессам.
Один из интереснейших вопросов в эво-
люции организмов — происхождение мно-
гоклеточности. Никто не подвергает сом-
нению, что многоклеточные организмы
произошли от одноклеточных, и большин-
ство ученых сходится на том, что родо-
начальниками многоклеточных были коло-
ниальные простейшие.
Биологический смысл возникающей
колониальности1 заключается
в ее защитной роли от врагов и от фак-
торов абиотической среды.
Среди высших многоклеточных коло-
ниальность очень ярко выражена у мша-
нок и оболочникових. Впрочем, следует
отметить, что колониальность отсутствует
у многих больших групп животных, на-
пример у моллюсков, членистоногих, анне-
1 Термин колониальность в зоологии
употребляется в двух смыслах: в данном случае
имеется в виду морфологическое объединение
нескольких особей общими образованиями, а иног-
да путем срастания в индивидуум высшего по-
рядка. Однако колонна л ьностью именуют и сосу-
ществование многих особей скоплениями или
колониями. В этом смысле говорят о колониях
птиц, о гнездах общественных насекомых и т. и.
лид, иглокожих, позвоночных и некоторых
других, но очень сильно выражена у про-
стейших и кишечнополостных, т. е. в тех
группах, которые стоят у истоков много-
клеточности. Особенно интересны, ко-
нечно, те колониальные организмы,
у которых наблюдается морфологическое
и функциональное разделение между
особями колонии по основным жизненным
функциям, как движение, питание, раз-
множение, защита. На низшей ступени
развития колониальности находятся такие
формы, как Pandorina или Eudorina
среди жгутиковых, между сочленами коло-
нии которых не наблюдается различий. В
данном случае колониальный организм —
индивидуум второго порядка — еще не
подчиняет себе отдельные организмы, вхо-
дящие в состав колонии. С усложнением
колониальности происходит все большее
подчинение отдельной особи целому —
индивидууму второго порядка, а «разде-
ление труда» между особями идет па
линии функций движения и размножения,
а в дальнейшем и питания. Развитию
колониальности сопутствуют явления
полиморфизма, которые наблюдаются так-
же и при формах колониального сущест-
вования насекомых. Совместное существо-
вание обычно ведет к морфологическому
п функциональному разделению между
особями.
Явление полиморфизма (мно-
гоформия), так же как и колониальность,
возникает независимо в разных группах
животных. Даже в пределах одного клас-
са (например, термиты и муравьи среди
насекомых) явления полиморфизма могуг
возникать совершенно независимо и да-
вать при этом поразительное конвергент-
ное сходство. Полиморфизм наиболее
ярко в животном царстве выражен у
кишечнополостных, хотя природа их сов-
сем другая, чем у насекомых. Эта ши-
роко развитая у простейших и кишечно-
полостных способность к образованию ко-
лоний дает основание думать, что имен-
но таким образом сформировалась много-
клеточность, а многоядерность простейших
следует рассматривать как усложнение
строения ядра, не ведущее к образованию
многоклеточности. При этом большин-
ство многоядерных простейших харак-
21
терно для паразитических форм (Sporo-
zoa), организация которых не может
быть положена в основу филогенетиче-
ских построений, а должна рассматри-
ваться как результат крайне специфиче-
ских условий существования (см. раздел
«Простейшие»).
С формированием многоклеточности
получил развитие эмбриональный про-
цесс—процесс формирования из оплодот-
воренного яйца взрослой формы.
Прежде чем перейти к некоторым момен-
там, связанным с эмбриональным про-
цессом, следует затронуть одно из наи-
более замечательных обобщений, связан-
ных с индивидуальным развитием живот-
ных, так называемый биогенети-
ческий закон, или, как его
сейчас обычно именуют, правило
рекапитуляции. Это правило
глубоко проникло в современные эмбри-
ологию и сравнительную анатомию, так
же как принципы монофилии и дивер-
гентной эволюции. Биогенетический за-
кон связывается с именами двух круп-
нейших последователей Ч. Д а р в и-
н а — Э. Геккеля и Ф. Мюл-
лера, хотя еще и до них некоторые
зоологи близко подходили к этому обоб-
щению (М е к к е л ь, Б э р), в том числе
и Дарвин. В основном это обобщение за-
ключается в том, что в онтогенетическом
развитии организмов мы обнаруживаем
в некоторой последовательности отраже-
ние этапов, пройденных в прошлом, или,
как иногда говорили в краткой форме,
«онтогенез повторяет филогенез».
Конечно, в такой категорической форме
сейчас биогенетический закон уже не
принимают. В развитии организма ча-
сто смещается последовательность по-
явления признаков эволюционного прош-
лого.
Наиболее характерная особенность
процесса совместного овладения и дру-
гими живыми существами жизненным
пространством — это переход от менее
совершенных форм к более совершенным,
от менее сложных к более сложным, от
менее дифференцированных к более диф-
ференцированным, от менее специализи-
рованных к более специализированным,
при этом от более разнообразных к менее
разнообразным, но более усложненным и
специализированным в соответствии с дар-
виновским принципом дивергентной эво-
люции. В отдельных случаях, однако,
при изменении биологии организма и его
организации, при смене среды обитания
можно наблюдать на отдельных системах
органов регрессивные изменения, приво-
дящие к значительной редукции, а иногда
и к полному исчезновению тех или иных
особенностей организации. Параллельно
с такими процессами наблюдается обычно
сильное развитие или появление других
систем органов, которые и сопутствуют
указанной редукции. Прежде всего эти
процессы сопутствовали выходу из водной
среды в воздушную, переходу к суще-
ствованию в глубинах океана, в пещерах,
переходу к паразитическому существо-
ванию, изменениям в характере питания
и т. д.
В эволюции животных изменения сход-
ного типа имеют место при увеличении
размеров тела, при повышении уровня
организации в эволюционном процессе.
Так, нематоды и членистоногие утеряли
ресничный эпителий в связи с кутикуля-
ризацией покровов, а выход многих жи-
вотных в воздушную среду привел к ре-
дукции дыхательного аппарата, свойствен-
ного водным организмам (жабры).
При переходе животных к малоподвиж-
ному или неподвижному образу жизни
и к паразитизму происходят значительные
изменения в отдельных органах и целых
системах, причем эти изменения могут
идти в диаметрально противоположных
направлениях: у свободноживущих орга-
низмов переход к неподвижному суще-
ствованию сопровождается сильным раз-
витием механических или химических за-
щитных образований (скелеты губок, ки-
шечнополостных и мшанок, раковины
моллюсков и плеченогих, домики усоно-
гих ракообразных, стрекательный аппа-
рат кишечнополостных, ядовитые или от-
талкивающие выделения кожных желез,
слизь и т. д.)- Но наряду с этим редук-
ции подвергаются мускулатура, нервная
система и органы чувств, изменяются спо-
собы питания и захвата добычи. У пара-
зитов скелетные образования отсутствуют,
но развиваются различные органы при-
22
крепления к органам хозяина (присоски,
крючки, хоботки). В обоих случаях
имеет место сильнейшее увеличение пло-
довитости, ослабление роли особей муж-
ского пола и часто развитие гермафро-
дитизма (усоногие ракообразные). При
паразитизме часто наблюдается полная
редукция кишечника (цестоды, колюче-
головые), что до недавнего времени не
было известно у свободноживущих жи-
вотных. Совершенно парадоксальным слу-
чаем оказалось установленное А. В. И в а-
новым полное исчезновение кишечника
у глубоководной группы погонофор, что
пока не находит себе объяснения.
Весьма интересно, что часто изменения
организации, сопутствующие смене среды
обитания, даже у родственных форм про-
исходят не однообразно, а иногда идут
диаметрально противоположными путями.
Например, у одних глубоководных рыб
наблюдается темная, почти черная окра-
ска, а другие становятся бесцветными
и иногда прозрачными. У одних глубо-
ководных рыб наблюдаются гипертрофи-
рованные, иногда стебельчатые глаза,
а у других — полная редукция органов
зрения. При выходе из водной среды
в воздушную у большинства животных
выработались рычажные органы движе-
ния — ноги (членистоногие, позвоночные),
однако многоножки и змеи имеют длин-
ное тело и передвигаются изгибанием
тела, что чаще свойственно водным орга-
низмам.
Крайнюю форму частичной или полной
редукции многих органов можно наблю-
дать у паразитических усоногих рако-
образных (Sacculina, Dendrogaster и др.)
и брюхоногих моллюсков (Enteroxenos и
Parenteroxenos).
Все эти бесчисленные случаи редукции
органов нельзя рассматривать как про-
цессы регресса вообще. Вернее считать
их формами выработки узкоспециализи-
рованных черт организации как приспо-
собления к аномальным условиям суще-
ствования и видеть в них яркое выражение
огромной способности живых существ при-
спосабливаться к самым различным усло-
виям существования и к расширению сво-
его жизненного ареала. Таким образом,
все это следует рассматривать как формы
приспособления, а не явления общего
регресса.
Возвращаясь к общей тенденции эво-
люционного процесса — переходу от ме-
нее сложных, но более разнообразных
структур к немногим (иногда всего лишь
одной), но высоко и многообразно
специализированным, остановимся на ряде
примеров. Хорошим примером может
быть эволюция форм движения. Простей-
шие обладают весьма разнообразными
формами движения. Саркодовые (особенно
корненожки) имеют псевдоподиальное дви-
жение, которое осуществляется по гидра-
влическому принципу — эндоплазма уст-
ремляется в тот или иной участок перифе-
рии и «натягивает» в более плотной и
упругой эктоплазме вырост — псевдо-
подию.
Движение жгутиконосцев обеспечива-
ется винтообразным биением жгутиков,
а инфузорий — многочисленными рес-
ничками. В противоположность жгути-
кам реснички бьют в одной плоскости,
но сами они служат, помимо плавания,
разным целям: окружающие ротовое
отверстие реснички создают сложный ток
воды, загоняющий пищевые частицы
в глотку. Реснички могут слипаться
в «кисточки» — ц и р р ы и на брюшной
стороне брюхоресничных инфузорий ими-
тируют конечности. На этих циррах инфу-
зории могут быстро «бегать» по суб-
страту. Соединяясь в продольные ряды,
реснички превращаются в мембраны, спо-
собные к волнообразным колебаниям.
У трипанозом вдоль всего тела идут
также мембраны, по краю их прохо-
дит основание жгута и мембрана все
время находится в состоянии волновых
колебаний. Грегарины движутся реактив-
ным способом, отталкиваясь от стекающей
с заднего конца тела слизи. Простейшим
свойственны также многообразные формы
парения в воде при помощи радиально
расходящихся скелетных игл или псевдо-
подий.
Ресничное движение свойственно так-
же бесчисленным личинкам донных
животных (черви, моллюски, иглокожие
и др.) и самым мелким многоклеточным,
сходным по размерам с простейшими (ко-
ловратки и некоторые турбеллярии).
23
Ресничное и жгутиковое движения име-
ют еще одну особенность — двигающиеся
таким образом организмы при плавании
совершают вращательное движение вокруг
собственной оси (кроме турбеллярий)
и, кроме того, двигаются не прямолинейно,
а по винтовой линии. Таким способом
осуществляется у этих микроскопических
существ, имеющих удельный вес, близкий
к воде, двойное двигательное вбуравлива-
ние в водную среду. Этот способ движения
в воде весьма эффективен. Относительная
скорость инфузорий (отношение скоро-
сти проходимого в секунду пути к длине
тела), перенесенная на размеры тела
человека, составила бы скорость сприн-
тера.
Таким образом, у простейших мы на-
блюдаем всевозможные способы движения,
кроме полета. У кишечнополостных
способностью к свободному движению
обладают только медузы, сифонофоры и
ктенофоры. Ктенофоры движутся при
помощи ресничного эпителия, но движе-
ние их крайне медленное. Ресничное
движение эффективно только при малых
величинах тела, измеряемых долями мил-
лиметра. С увеличением размеров тела
ресничное и жгутиковое движения ста-
новятся неэффективными, так как объем
тела возрастает гораздо быстрее (в кубе),
чем его поверхность (к квадрате). В даль-
нейшем у червей сформировалась новая фор-
ма движения — изгибанием тела, а в свя-
зи с этим у них развилась мощная двига-
тельная мускулатура. У кишечнополост-
ных мускулатура была совершенно недо-
статочной для выработки новой формы
движения. 95% кишечнополостных ведут
неподвижное существование, защищаясь
от врагов мощным скелетом и стрека-
тельным аппаратом. Еще более ярко это
сказалось на губках, совсем лишенных
мускулатуры.
Движение медуз и сифонофор совер-
шается реактивным способом — колокол
медузы сжимается, выталкивает из-под
себя воду, и медуза получает реактивный
двигательный толчок верхней стороной
колокола вперед. Для такого движения
достаточно небольшой двигательной му-
скулатуры, которая составляет едва
1 — 2% от объема тела. Только у червей
начинает накапливаться двигательная
мускулатура в виде кожно-мускульного
мешка. У немертин и высших червей
она достигает наибольшего развития.
У кишечнополостных между эктодер-
мой и энтодермой уже создается мышеч-
ный слой (первоначально в очень прими-
тивной форме) в виде слоев сократитель-
ных отростков эпителиально-мышечных кле-
ток, образующих два взаимно перпенди-
кулярных волокна — клетки эктодермы
образуют систему продольных по оси тела
волокон, а клетки энтодермы — кольце-
вых. Эта система волокон как бы имити-
рует кожно-мускульный мешок червей,
но не осуществляет изгибательных дви-
жений, а осуществляет только сократи-
тельные — по основной оси тела. Кожно-
мускульный мешок червей состоит из двух
основных слоев мышц — кольцевого а
продольного. Есть еще и другие мышцы:
тело червей как бы набито мышцами, ко-
личество их достигает у некоторых червей
60—70% общего объема тела (немертины,
пиявки). Огромное количество низших и
высших червей передвигается при помощи
изгибаний тела. Таким образом, внутри
группы кишечнополостных вместе с выра-
боткой двустороннесимметричного плана
строения сформировались мускульные
формы животных, выработавшие изгиба-
тельный способ движения. Это были предки
турбеллярий, использующие более мощ-
ную мускулатуру для изгибаний тела.
Хотя изгибательное движение и домини-
рует у червей, но наряду с ним есть и не-
которые другие формы движения.
Весьма сходные процессы, которые при-
вели к тому же изгибательному движе-
нию, имели место также и в эволюции
вторичноротых.
Масса низших хордовых и родственных
им форм, ведших или малоподвижное,
или неподвижное существование, дала
начало рыбообразным с характерным для
них изгибательным движением и мощной
туловищной мускулатурой. В течение дол-
гого периода эволюции позвоночных живот-
ных основной формой их передвижения
было изгибательное.
В дальнейшем усложнение изгибатель-
ной формы движения наступило с выработ-
кой посегментных складок — пара-
24
подий, снабженных щетинками (х е-
т а м и). Это был добавочный орган дви-
жения, если можно так сказать, чрева-
тый последствиями, так как в дальней-
шем из них сформировались конечности
членистоногих. Полимерное строение коль-
чецов соответственно дало формы с боль-
шим числом ног (ракообразные, пауко-
образные),
С формированием рычажных конечно-
стей, за немногими исключениями (много-
ножки и змеи), вся двигательная функция
перешла на ножные придатки. Путь фор-
мирования конечностей у позвоночных был
несколько иным по сравнению с тако-
вым у членистоногих. У членистоногих
конечности сформировались уже в водной
среде (ракообразные), а у позвоночных —
в процессе выхода в воздушную среду.
Замечательно, что и у предков позвоноч-
ных плавники тоже сформировались из
складок по бокам тела, но только в виде
двух поясов — плечевогои та-
зового — с двумя парами конечностей.
Столь же интересно, что первоначально
плавники у предков членистоногих и позво-
ночных играли роль не основных орга-
нов движения, а только подсобных, но
важно, что в обоих случаях рычажные
конечности, при всей их чрезвычайной
функциональной усложненности (полет),—
единственный аппарат движения. Много-
образие форм движения сменилось одной
формой, однако в крайней степени услож-
ненной и дифференцированной.
Подобный же эволюционный путь про-
шло большинство систем органов и функ-
ций. Остановимся еще только на одной,
например на функции размножения. Труд-
но найти другую функцию у животных,
которая давала бы столь же исключитель-
ное разнообразие. Прежде всего, конечно,
следует указать на обычность у низших
групп животных бесполого размножения,
иногда полностью заменяющего половое,
как, например, у многих отрядов жгути-
ковых и некоторых саркодовых. При этом
формы бесполого размножения бесконечно
разнообразны: от создания огромных ко-
лоний кишечнополостных или мшанок,
при добавочных явлениях полиморфизма,
до правильного чередования г а м о г о-
н и и (полового) и агамогонии (бес-
полого) у фораминифер и споровиков*
Бесполое размножение у простейших
носит характер деления и почкованияг
простого и множественного, вплоть да
сложных форм шизогонии. Сам половой
процесс имеет двойную форму — копу-
ляции и конъюгации, а кроме
того, партеногенеза и опло-
дотворения. Столь же многообразны
формы чередования полового и бесполога
развития.
С повышением организации диапазон
и разнообразие форм бесполого размно-
жения уменьшились, среди первичноро-
тых оно встречается еще у кольчатых
червей, а из вторичноротых — только-
у оболочниковых. В конце концов, беспо-
лое размножение уступает место поло-
вому.
Партеногенез, как исключе-
ние, встречается у всех групп членисто-
ногих и позвоночных, и существуют виды
насекомых и рептилий, у которых самцы
совсем неизвестны и размножение совер-
шается только партеногенетически.
У кишечнополостных впервые появ-
ляются нервная система и органы чувств
(зрения и равновесия). Нервные клетки
генетически связаны не только с экто-
дермой, как у всех прочих животных,
кроме иглокожих, но и с энтодермой.
Расположение частей тела и органов под-
чинено у кишечнополостных радиально-
осевой симметрии *. Симметрией назы-
вается определенный геометрический поря-
док в расположении сходственных частей
тела.
Элементами симметрии являются точка
(центр), линия (ось) и плоскость. Пре-
красный пример радиально-лучевой сим-
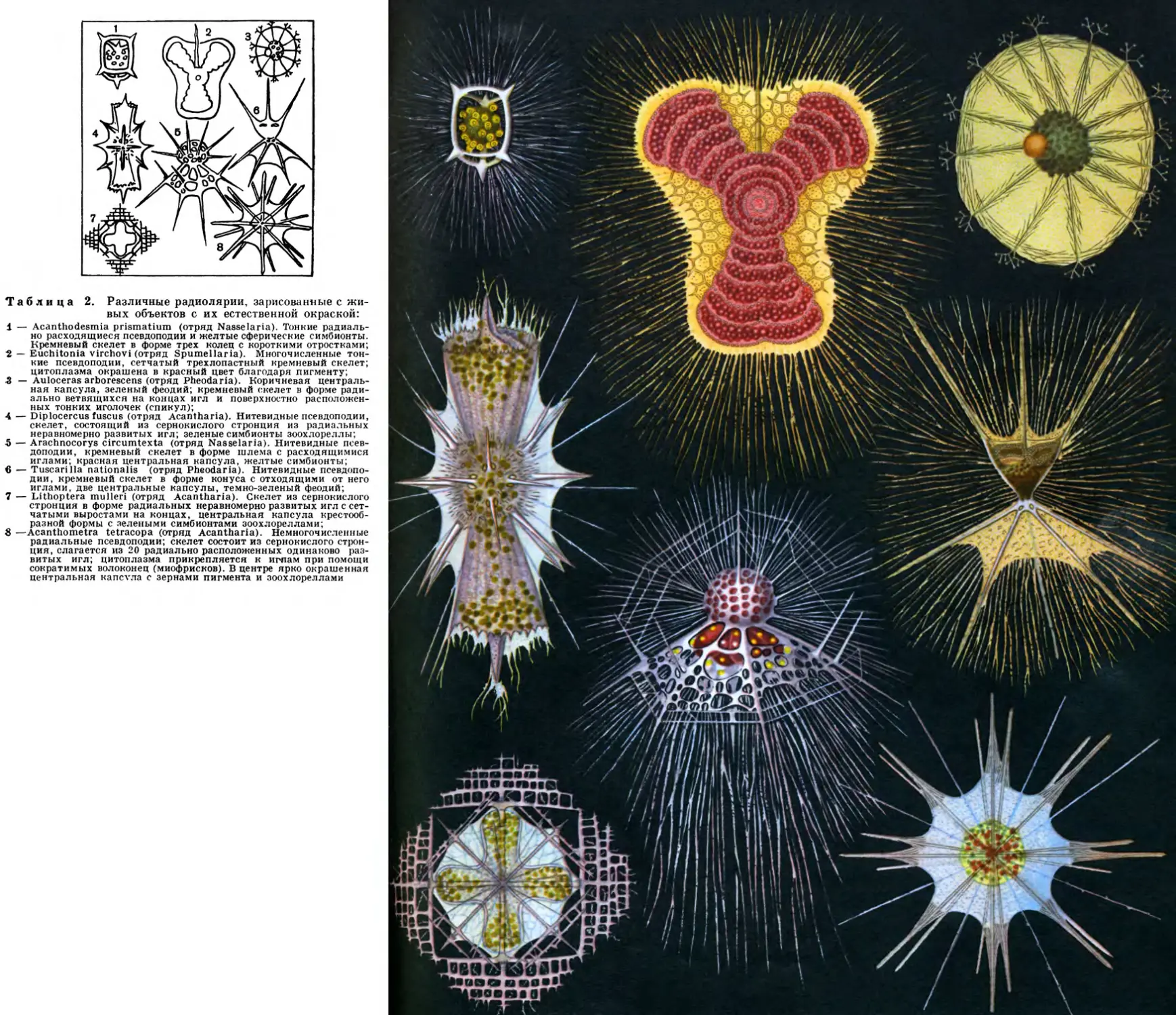
метрии дают радиолярии (рис.3). Сход-
ственные части тела расположены вокруг
центра симметрии в радиальном направ-
лении. Радиально-лучевая симметрия свой-
ственна организмам, взвешенным в воде
и имеющим со всех сторон одинаковую
среду, в силу чего и реакция организма
«одинакова во все стороны». Радиально-
лучевая симметрия наилучшим образом
1 Симметрия—от греческих слов sym — вме-
сте и metron — масса, в смысле часть
тела.
25
Рис. 3. Различные типы симметрии у животных.
А — радиально-лучевая симметрия; Б — радиально-
осевая симметрия у кишечнополостных; В — двусто-
ронняя симметрия.
соответствует биологии радиолярий.
Радиально-лучевую симметрию мы находим
также у колониальных фитомонадовых
(вольвокс, эвдорина, пандорина и др.)
и некоторых колоний многоклеточных,
например у колониальной коловратки
Conochilus.
Однако радиально-лучевая симметрия
некоторых простейших не является самой
примитивной формой строения тела. В рав-
ной мере планктонное существование так-
же нельзя считать самой примитивной
биологической формой. Самые просто
организованные формы саркодовых (Атое-
Ыпа) имеют асимметричное строение,
и, видимо, оно соответствует примитив-
ным формам организации и поведения
(псевдоподиальная форма движения и пи-
тания). Кроме того, можно думать, что
все пелагические формы существования
являются вторичными производными от
придонных. Асимметричное строение
свойственно и инфузориям, и жгутико-
вым. В частности, радиолярии обладают
необыкновенным богатством планов сим-
метрии своего скелета — радиально-осе-
вой, как гомополярной, так и гетеро-
полярной, двусторонней, двубоковой, с
обычным отклонением всех этих типов
симметрии в асимметрию. Следует отме-
тить, что в подавляющем числе случаев
при этом разные формы симметрии отно-
сятся только к скелету, что же касается
протоплазмы, то она, как правило, имеет
асимметричное расположение включе-
ний (ядро, пульсирующие и пищевари-
тельные вакуоли и иные включения).
Кишечнополостным, как сидячим, так
и пелагическим (медузы), свойственна
радиально-осевая симметрия, при которой
сходственные части расположены вокруг
оси вращения, причем эта симметрия
может быть самого различного порядка
в зависимости от того, на какой угол сле-
дует повернуть тело животного, чтобы
новое положение совпало с исходным. Та-
ким образом, может получаться 4-, 6-, 8-
лучевая симметрия и более, до симмет-
рии порядка бесконечности. У радиоля-
рий встречается радиально-осевая сим-
метрия с одинаковыми полюсами, или,
как говорят, гомополярная. У кишечно-
полостных — гетерополярная осевая сим-
26
метрия: один полюс симметрии несет рот
и щупальца (оральный), другой
(а б о р а л ь н ы й) служит для прикреп-
ления (стадия полипа), или у плавающих
форм несет орган чувств (ктенофо-
ры), или ничем не вооружен (медузы).
У некоторых медуз на этой аборальной
стороне образуется стебелек для прикреп-
ления к подводным предметам (Lucer-
nariida). Нарушение радиал ьно-осевой
симметрии возникает при уменьшении
числа шупалец или изменении формы рото-
вой щели, пищевода и разветвлений пище-
варительной системы. Количество щупа-
лец может уменьшаться до одного (Мопо-
brachium), и тогда их радиальное рас-
положение сменяется двубоковым. Глот-
ка может сплющиваться, и тогда тоже
получается двубоковая симметрия,
этому способствует и образование в глот-
ке сифоноглифов (желобок вдоль
глотки).
Наибольшее усложнение радиально-осе-
вой симметрии наблюдается у ктенофор,
где, помимо 8-лучевой симметрии, в рас-
положении отдельных частей тела и орга-
нов наблюдается 4-лучевая и двубоковая
симметрия. Это весьма существенный мо-
мент, так как большинство зоологов имен-
но от ктенофорообразных предков выво-
дит оба ствола высших животных, как
первично-, так и вторичноротых.
Гетерополярная радиально-
осевая симметрия вполне соответствует
образу жизни кишечнополостных — непо-
движному существованию в прикреплен-
ном положении или медленному плава-
нию при помощи реактивного движения.
С другой стороны, от сложного типа
радиально-осевой симметрии ктенофор
можно перейти к двусторонней симметрии,
или, как говорят, симметрии зеркального
изображения, единственного плана сим-
метрии трехслойных животных, симмет-
рии быстрого движения, с выработкой
переднего по движению конца тела,
с центральным мозговым скоплением и
основными органами чувств, спинной и
брюшной, правой и левой сторонами тела.
Однако живых или ископаемых свидете-
лей этого перехода мы не знаем. Здесь
можно пользоваться только более или
менее достоверными гипотезами.
В 1880 г. знаменитый эмбриолог А. Ко-
валевский открыл своеобразный
организм — ползающую ктенофору,—на-
званный им Coeloplana metschnikowi.
Родовым названием Ковалевский хотел
показать, что этот организм объединяет
в себе признаки целентерат и планарий,
т. е. плоских червей. В 1886 г. другой рус-
ский зоолог — А. Коротнев, рабо-
тая на о. Яве, открыл другую подобную
форму, которую назвал Ctenoplana kowa-
lewskii, также указывая в названии объе-
динение в этом организме особенностей
ктенофоры и планарий.
В настоящее время в морях Юго-Восточ-
ной Азии описан целый ряд подобных
форм, объединенных в группу Platycte-
nidae (плоских ктенофор), но изучение
их показало, что не среди них надо
искать предков плоских червей, что это
просто ползающие ктенофоры без предко-
вых черт организации плоских червей.
Вопрос этот приходится решать иным
путем. Возможны два варианта. По одному
допущению, ктенофорообразные предки
первоначально были ориентированы ро-
товым полюсом ко дну, а аборальным
вверх. Затем они испытали расплющива-
ние по основной оси тела и сближение
орального полюса с аборальным. В
дальнейшем аборальный орган чувств,
зачаток мозгового скопления, должен
был сместиться на тот участок сплюс-
нутого тела, который стал передним по
направлению движения. Таким образом
выработались спинная и брюшная по-
верхности, а ротовое отверстие, как и у
многих турбеллярий, осталось в средней
части брюшной поверхности. Однако допу-
щение такого пути формирования тела
плоских червей должно уступить место
другому. Гораздо вероятнее допустить,
что ктенофорообразные предки червей
ориентировались к дну боком, в таком
случае у них сразу формировалась перед-
няя по движению сторона тела, а ротовое
отверстие должно было несколько сме-
ститься по брюшной стороне вперед.
Такое допущение больше соответствует
расположению нервной системы турбел-
лярий.
Что касается строения тела турбелля-
рий, то они сохраняют ряд черт радиаль-
27
Рис. 4. Различные формы движения
у животных:
1 — движение трипаносомы при помощи ун-
дулирующей мембраны; 2 — изгибательное
движение полпхеты и 3 — рыбы.
ной симметрии предков, особенно в строе-
нии нервной системы. Они сохраняют
также мерцательный эпителий на поверх-
ности тела, расположение рта на брюшной
стороне и ряд других особенностей, заим-
ствованных от целентератных предков.
Пока не сформировались рычажные ко-
нечности, основным механизмом движе-
ния оставалось изгибательное движение
(рис. 4). Этот тип движения возможен
при достаточно мощной мускулатуре и
определенном ее расположении «пластами»
вдоль всего тела. Оба эти условия соче-
таются в кожно-мускульном мешке чер-
вей. При этом двигательная мускула-
тура составляет около половины общего
объема тела, а иногда (немертины, пиявки)
я значительно больше.
С образованием конечностей кожно-
мускульный мешок распадается на от-
дельные мышцы. Морфологическая основа
двигательной функции кожно-мускуль-
ного мешка — расположение сократитель-
ных волокон во взаимно перпендикуляр-
ном направлении. Это пласты кольцевых
и продольных мышц. Еще у грегарин
миофибриллы образуют систему продоль-
ных и поперечных нитей. Отростки эпи-
телиально-мышечных клеток низших
кишечнополостных также образуют слой
продольных (от эктодермы) и слой коль-
цевых сократительных волокон. Однако
у кишечнополостных количество муску-
латуры невелико, кожно-мускульный ме-
шок не формируется и движение осущест-
вляется реактивным способом — только у
ктенофор сохраняется ресничное движе-
ние, дающее, однако, при крупных раз-
мерах ктенофор очень слабый эффект, Изги-
бательное, волнообразное движение —
весьма целесообразная форма передвиже-
ния в водной среде, однако в плотной среде
грунтов, особенно морских, эта форма
движения не эффективна: у животных воз-
никает гидравлическое прямолинейное
движение. При этом формируется обшир-
ная полость тела, наполненная полостной
жидкостью. Количество мускулатуры
в кожно-мускульном мешке уменьшается,
но оно достаточно, чтобы сокращением
кольцевой мускулатуры тела и перекачкой
полостной жидкости вперед обеспечить
продавливание хода в грунте, а затем,
расширив передний конец тела и закли-
нив его в ходе, подтянуть заднюю часть
тела сокращением продольной мускула-
туры.
Принципиально тот же способ движе-
ния свойствен двустворчатым моллю-
скам, пробивающим ход в грунте клино-
видной ногой, способной к расширению
при перекачке в ее лакуны полостной
жидкости, с последующим подтягиванием
тела и одевающей его раковины. Интере-
сно отметить, что отличные пловцы — го-
ловоногие моллюски — освоили реактив-
ное движение и им не свойственно изги-
бательное движение тела. У их предков
кожно-мускульный мешок уже распался
(как и у остальных моллюсков) и была
утрачена основа для создания изгибатель-
ного движения.
В водной среде возникли и членистоно-
гие, и свойственное им движение с по-
28
ующыо рычажных конечностей, но, преж-
де чем сказать о свойственной им фор-
ме движения, следует остановиться на
метамерии (сегментации) и ее происхож-
дении.
Метамерия иногда рассматривается
как своеобразный тип симметрии. При го-
мономной сегментации, так же как и при
других типах симметрии, имеет место по-
вторение сходственных частей тела — м е-
т а м е р (или сомитов) с одинаковым
расположением половой системы, выде-
лительных органов, разветвлений нервной
(невросомит) и кровеносной (а н-
гиосомит) системы, с двумя парапо-
диями по бокам тела, с поперечными пере-
городками — диссепиментами,
отгораживающими сегменты друг от друга,
с обособленными участками продольной
и кольцевой мускулатуры (м и о с о-
м и т). Только в данном случае сходствен-
ные части — метамеры — располо-
жены не вокруг точки или линии и не
по обе стороны плоскости, а в линейном
направлении по главной оси тела. Мета-
мерное строение появилось вместе с раз-
витием вторичной полости тела и крове-
носной системы (рис. 5, 6). Низшие черви—
сколециды — лишены настоящей мета-
мерии (Amera), она появляется только
у высших червей — аннелид — и прояв-
ляется либо в малом числе сегментов
(Oligomera — мшанки, плеченогие и все
вторичноротые), либо в большом (Po-
ly mera — аннелиды и членистоногие).
Среди основных этапов развития живот-
ного мира наряду с рассмотренными вы-
ше — возникновением многоклеточности,
радиальных типов симметрии, появле-
нием двусторонней (билатеральной) сим-
метрии — развитие метамерности занима-
ет важное место. Наряду с ними можно
поставить еще только формирование ры-
чажных конечностей как основного орга-
на движения и выход из водной среды
обитания в воздушную.
Конечности членистоногих и позвоноч-
ных возникли совершенно независимо,
хотя они и обладали поразительным био-
лого-эволюционным и функциональным
сходством при резко выраженных чертах
различий как следствии различия в харак-
тере скелета тех и других. У членистоно-
гих хитинизированный трубчатый скелет
наружный и по расположению
и по происхождению (производное экто-
дермы), у позвоночных скелет костный
и внутренний (мезодермиче-
ский). Функциональное сходство обслу-
живающей конечности мускулатуры—в ан-
тагонистической системе мышц сгибателей
и разгибателей, но у членистоногих они
прикрепляются к выростам хитинового
панциря, а у позвоночных — к выростам
костей.
Что же могло вызвать развитие сег-
ментированного тела? Следует прежде
всего отметить, что подразделение тела
на сходственные части с линейным их
расположением наблюдается у разных
организмов, стоящих на более низком
уровне, чем животные с истинной мета-
мерией. Такие случаи (псевдометамерия)
наблюдаются у раковинок фораминифер
(рис. 5), турбеллярий, немертин. В этом
сказывается упорядочение расположения
множественных образований и органов,
имеющих полимерный характер.
Истинная метамерия, появляющаяся
у кольчатых червей, — гораздо более
Рис. 5. Примеры псевдометамерии:
1 —2 раковинки фораминифер; 3 — немертина.
29
общее и глубокое явление, наложившее
свою печать на большой и важный этап
эволюции полимерных животных — ан-
нелид и членистоногих.
Несегментированные, низшие черви
(Amera) немногочисленны, имеют сла-
бые способы передвижения и не имеют
в водной среде такого значения, как анне-
лиды. Наиболее существенный для всех
них фактор биотической среды — враги,
для которых они могут являться пищей.
Турбеллярии ведут придонный образ жиз-
ни, малы по размерам, очень медленно
движутся, прячутся между водорослями
и под камнями. Тело их выделяет много
слизи, вероятно, выполняющей защит-
ную функцию. Нематоды ведут скрытый
образ жизни — они наполовину паразиты,
наполовину живут в грунте и малодоступ-
ны для врагов. Среди них нет пелагических
форм, и они очень малоподвижны. Такой
же образ жизни свойствен приапулидам —
они живут в скрытых биотопах, глав-
ным образом в грунте, и либо лишены
органов движения, либо способны лишь
к самому слабому передвижению. К ним
относятся коловратки, но они имеют ми-
кроскопические размеры и распростра-
нены преимущественно в пресных водах.
Это единственная группа среди сколецид,
ведущая планктонное существование.
Единственная группа среди низших,
несегментированных червей, ведущая и
придонное и пелагическое существование,
многочисленная в морских бассейнах —
немертины— агрессивные хищники, обла-
дающие органом нападения и защиты —
хоботом. Они движутся волнообразно,
изгибая тело, но движение это очень слабо.
Кожа их выделяет огромное количество
слизи, назначение которой, возможно,
двойное: она отпугивает врагов и уско-
ряет движение в воде. Если не считать
еще нескольких немногочисленных групп
микроскопических организмов, ведущих
донную жизнь, то это и все, что мы нахо-
дим в морской воде из низших червей.
Конечно, самое большое внимание при-
влекают среди них немертины. Однако
и они не смогли занять среди населения
моря видного положения.
Для большинства перечисленных групп
сколецид характерно отсутствие способ-
ности к быстрому движению и плотных,
защищающих тело покровов. Для даль-
нейшей эволюции животных необходимо
было разрешить эти затруднения. Неко-
торые организмы, генетически связанные
с червями, находили защиту от врагов
в формировании крепких покровов (пле-
ченогие, мшанки, моллюски), но теряли
в скорости движения или даже станови-
лись неподвижными, прикрепленными
формами. И только многощетинковые
кольчецы получили дальнейшее филогене-
тическое развитие: у них выработались
уплотненные, но тонкие покровы и сегмен-
тированное строение, которое давало воз-
можность легко сгибать тело во все сто-
роны. Очевидно, первые этапы формиро-
вания метамерности были связаны с форми-
рованием только поверхностной сегмента-
ции, но постепенно она захватывала все
более и более глубоко расположенные
части тела и органы. Полимеризация
захватила некоторые системы внешних
и внутренних органов — параподии, жа-
берные выросты, половую, нервную,
кровеносную и выделительную системы.
Внутреннее упорядочение в расположе-
нии органов подчинилось метамерному
строению, и в результате развившейся
метамерии возникла гомономная х
множественность посегментного располо-
жения систем органов. Так необходимость
выработки быстрого движения и защит-
ных покровов нашла свое выражение
в метамерном строении.
Очень часто на протяжении эволюции
отдельных групп, особенно среди кишечно-
полостных и червей, наблюдается множе-
ственное повторение одноименных обра-
зований и органов. Такие структуры име-
нуют полимерными. Особенно часто
встречается полимерный тип в расположе-
нии половых желез. Их четыре у гидрозоа
и сцифозоа, много у антеозоа, 8 мужских
и 8 женских желез у гермафродитных
ктенофор, очень много у ленточных червей
(цестод), много у полихет.
Целый ряд полимерных органов, помимо
гонад с посегментным расположением,
1 Гомономный — однообразный, от гре-
ческих слов homos — одинаковый и
nomos -поделенный.
30
возник при формировании метамерных
кольчецов — параподии, жабры, выде-
лительные органы, кольцевые нервные
тяжи и кровеносные сосуды, ганглиозные
скопления в брюшной нервной системе
и многое другое. Полимерный характер
имеет расположение многих органов
у иглокожих в связи с их пятилучевои
симметрией. Подобных примеров можно
было бы привести очень много и для чле-
нистоногих —количество ног у ракообраз-
ных и особенно у многоножек, трахеаль-
ные образования у них же и у насекомых
и т. д. Однако в дальнейшем почти всегда
наблюдается процесс уменьшения коли-
чества одноименных образований, или,
как это называют, процесс о л и г о м е-
ризации, в количестве ног, сегмен-
тов тела, половых желез, органов выде-
ления, дыхания и т. д.
Гомономная сегментация стала
уступать место гетерономной1
и особенно в группе членистоногих,
а покровы становились все более проч-
ными (хитинизированными), но не уте-
рявшими своей легкости и не мешавшими
подвижности благодаря мягким и растя-
жимым сочленениям между отдельными
более плотными частями хитинового пан-
циря. В дальнейшем в эволюции обеих
основных ветвей животных — первично-
ротых и вторичноротых — проявляется
поразительное сходство в выработке ры-
чажных двигательных придатков— конеч-
ностей. У членистоногих прообразом их
были параподии полихет, у позвоночных—
плавники рыб. У членистоногих конеч-
ности появились еще в водной среде (у рако-
образных), а у позвоночных только с вы-
ходом в воздушную среду. Эволюция
параподии и конечностей у полихет и чле-
нистоногих — очень хороший пример оли-
гомеризации и перехода от гомономных
образований к гетерономным. У многих
полихет параподии имеются на всех сег-
ментах и иногда на сотнях сегментов
имеют совершенно одинаковый характер
(гомономная полимерия). У ракообраз-
ных и тем более у паукообразных и насе-
комых количество конечностей подвергает-
ся неуклонно идущей олигомеризации и
гетерономному типу изменений, т.е. умень-
шается не только количество ног, но и раз-
Рис. 6. Примеры истинной метамерии:
слева — гомономная (полихета); справа-
гетерономная (скорпион).
ные конечности отличаются друг от друга
по строению и функциям. Этот процесс
представляет собой только часть более
общих изменений организации процесса
кефализации2 (формирование го-
ловы) и тегматизации3 (форми-
рование отделов тела). Из передних
пяти-шести сегментов тела формируется
голова — передняя тегма тела, а конеч-
ности этих сегментов превращаются
в органы чувств и ротовые придатки. Сле-
дующие сегменты образуют грудной отдел
(у насекомых 3 сегмента, у паукообраз-
ных — 4, у высших ракообразных — 8
и т. д.).
Конечности грудного отдела обычно слу-
жат для передвижения. Брюшной отдел,
сформированный из различного количе-
ства сегментов, либо лишен конечностей
(насекомые, паукообразные), либо они вы-
1 Гетерономный — различно поделен-
ный, от греческого heteros — неодинако-
вый.
2 Kephalon (греч.) — г о л о в а.
3 Tegma (греч.) — отдел.
31
полняют другую функцию (половую,
дыхательную и т. д.). Грудь и брюшко
образуют вторую и третью тегмы тела.
У наземных групп конечности выпол-
няют основную двигательную функцию
(если не считать многоножек, змей и неко-
торых других групп), а сегментация как
во внешнем облике животного, так и во
внутреннем строении исчезает.
Метамерное расположение сохраняется
лишь частично и только на некоторых
системах, таких, как дыхательная, нерв-
ная и др. Крайнюю форму подавления
(исчезновения) метамерного строения
можно наблюдать у клещей, в мешкооб-
разном теле которых слиты все отделы,
не обнаруживающие сегментированного
плана строения. И у других членистоно-
гих намечается слияние трех основных
отделов тела, прежде всего головы и
груди в головогрудь (высшие ракообраз-
ные и паукообразные). При этом иногда
брюшной отдел тоже испытывает редук-
цию, и тогда, как, например, у крабов,
тело кажется состоящим из одного ком-
пактного образования, хорошо защищен-
ного крепким панцирем от врагов, а хоро-
шая подвижность его обеспечивается пя-
тью парами конечностей.
Столь же нагляден этот процесс и у
позвоночных. Две пары конечностей,
сходные между собой у низших позвоноч-
ных, особенно у рыб, у птиц совершенно
различны — передние служат для полета,
задние для передвижения по грунту.
У многих млекопитающих строение и функ-
ции передних и задних конечностей также
весьма различны. Здесь процесс олигоме-
ризации выражен очень ясно.
НЕКОТОРЫЕ БИОЛОГИЧЕСКИЕ
ОСОБЕННОСТИ ЖИВОТНЫХ
Приспособления, которые появляются
у организмов в связи с изменениями внеш-
них условий, чрезвычайно многообразны.
С одной стороны, организмы избегают
неблагоприятных и необычных для них
условий среды, а с другой — характер-
нейшая особенность организмов — рас-
ширяют свое жизненное пространство,
свой ареал, что приводит к соответствен-
ным изменениям их организации. Сочета-
ние всех этих свойств организмов создает
существующую сложную систему приспо-
соблений к условиям абиотической и био-
тической среды и характер ареалов.
Если допустить, что первоначальной
средой жизни были теплые мелкие моря
и что первичной формой существования
было дно — твердый субстрат (рис. 7),
то в дальнейшем шло завоевание живот-
ными всех глубин океана и его водной тол-
щи, проникновение их в грунт, переход
в пресную воду и воздушную среду.
Кроме того, на всех этих этапах животные
проникают в глубоко скрытые под покро-
вом земли пещеры и подземные воды и
в тела других животных (переход к пара-
зитизму). Животные, освоившие воздуш-
ную среду, нередко вновь возвращаются
в морскую среду: в меньшей степени —
коловратки, олигохеты, пиявки, насекомые
(морские клопы) и паукообразные (мор-
ские клещи), в большей степени — рыбы,
рептилии (змеи и черепахи), птицы (пин-
гвины и др.) и особенно млекопитающие
(ластоногие и китообразные). Все пере-
численные группы обладают хорошо защи-
щающими от неблагоприятного действия
соленой воды покровами, что облегчило
для них переход в соленую воду. Но среди
амфибий нет ни одного примера возвра-
щения в морскую среду. Вероятно, этому
противодействовала их тонкая и нежная,
ничем не защищенная кожа.
У многих животных вырабатывалась
способность к полету (рептилии, насеко-
мые, птицы и млекопитающие) или к его
имитации (пауки, летучие рыбы, каль-
мары). Наиболее быстрые пловцы и ле-
туны приобрели способность преодоле-
вать большие пространства в гори-
зонтальных миграциях —до
V3 окружности Земли, что помогало им
в отыскании областей, наиболее выгод-
ных для размножения и питания (гене-
ративно-алиментативные миграции). Мно-
гие киты в летний сезон передвигаются
Таблица 1. Распространение паразитизма в разных группах животных.
На генеалогическом древе синим цветом изображены классы, целиком,состоящие из паразитов, зеленым—.
классы, содержащие наряду со свободно живущими, также и паразитические формы, и красным — клас-
сы, образованные исключительно свободноживущими видами.
1 — Gregarina-; 2 — Mesozoa; 3 — Cestodes; 4— Monogenea; 5 — Digenea; fi — Acanthocephala; 7— Nematodes;
8 — Myzostomida; 9 — Pentastomida.
в северные широты, где нахо-
дят обильную пищу, а на зиму
уходят в экваториальные ши-
роты. Огромные пути проходят
в океанах тунцы. Широко из-
вестны тысячекилометровые пу-
ти личинок речных угрей из
глубин Саргассова моря в реки
Европы, Азии и Северной Аме-
рики и возвращение половозре-
лых самок для нереста на
родину. Такие же пути из мо-
ря на нерест в реки соверша-
ют лососевые рыбы, а молодые
лососи — из рек на откорм в
океан.
Сходного типа миграции яр-
ко выражены у птиц и некото-
рых наземных млекопитающих.
Использование разных участ-
ков среды у многих морских
животных выражается (главным образом
ракообразные, молодь рыб и некоторые дру-
гие) в суточных вертикальных миг-
рациях. Днем они прячутся в темных
глубинах, а ночью поднимаются в обиль-
ные пищей верхние слои.
Стремление многих морских животных
к расселению отражается и на биологии
их личинок.
Многие донные животные (черви, рако*
образные, моллюски, иглокожие, рыбы
и др.) имеют пелагических личинок,
в течение недель, а иногда месяцев ве-
дущих планктонное существование. За
этот срок их подхватывает течение и от-
носит на сотни, иногда тысячи километров
от места их рождения, и, если, опустив-
шись на дно, личинки встретят благо-
приятные условия существования, они
заселят новые районы. Таким образом,
миграции животных могут иметь различ-
ный характер.
Своеобразен способ расселения некото-
рых животных в виде пассажиров на
других животных или на судах, о чем
речь будет впереди.
Едва ли не самым интересным примером
способности осваивать новые среды оби-
тания является паразитизм, к ко-
торому приводят х ищн ичество, ком-
менсализм, симбиоз и тро-
фические связи.
•ZZ5i5^2???~xsttzl?z^zz?C
1111113
Рис. 7. Пути расселения фау-
ны из ее прародины — мелко-
го теплого моря:
1 — море; 2 — воздушная среда;
3 — пещеры; 4 — вертикальные
миграции планктона; 5 —гори-
зонтальные миграции нектонных
организмов; 6 — перелеты птиц;
7 — путь формирования глубоко-
водной фауны; 8 — путь формиро-
вания высокогорной фауны.
Симбиоз, комменсализм и паразитизм г.
Чрезвычайно многообразны взаимоотно-
шения, которые складываются между ор-
ганизмами и их группами, а иногда и це-
лыми биоценозами в длительном процес-
се эволюции приспособлений и биологиче-
ских связей.
Об одной такой системе связей между
растительным и животным царствами в
рамках всей биосферы мы уже гово-
рили.
Планктонные биоценозы дают основной
пищевой ресурс для донной фауны океа-
нов и морей, а также и для рыб. Многие
организмы моря и суши совершают гори-
зонтальные, а в море, кроме того, и
вертикальные миграции и на разных
этапах миграций вступают в биологиче-
ские связи с различными группами
организмов.
Многие паразиты часть своей жизни
проводят в свободном состоянии, и тогда
они подчиняются иным системам связей с
другими организмами и с абиотической
средой.
1 Симбиоз — совместное существо-
вание, от греческих слов sym — вместе
и bios — жизнь; комменсализм — н а х л е б-
ничество, от латинских слов cum — с
и mensa — стол; паразитизм от греческих
слов рага — о к о л о и sitos — пища.
w 3 Жизнь животных, т. 1
33
Одна из ведущих связей между орга-
низмами — это потребность в пище. В
этом отношении животные в первую оче-
редь могут быть разделены по характеру
пищевого объекта на растительноядных,
детритоядных, грунтоядных, плотоядных,
в том числе и хищников.
Понятие «хищник» употребляется в раз-
личном смысле.
Животных, питающихся моллюсками,
насекомыми или червями, обычно не на-
зывают хищниками, но совершенно оче-
видно, что треска, питающаяся сельдью,—
хищник, так же как акула, пожирающая
других рыб, или волк, гоняющийся за
зайцем. Обычно хищниками называют жи-
вотных, питающихся другими животны-
ми, близкими по систематическому поло-
жению (в рамках позвоночных, или насе-
комых, или моллюсков и т. д.), но иногда
говорят о хищных растениях, питающихся
насекомыми, как, например, росянка, или
о хищных грибах, питающихся червями,
и т. п. Более точное определение и клас-
сификация способов питания, вероятно,
затруднительны в силу того, что эти спо-
собы создаются независимо у разных
животных в совершенно разных формах,
на различной морфологической основе и
не имеют никакой генетической связи.
В силу этого создание естественной систе-
мы форм питания едва ли возможно. Си-
стема всегда будет носить более или ме-
нее искусственный характер. Она возмож-
на, пожалуй, только в рамках небольших
однородных групп.
Животных можно различать и по
способу захвата пищи — фильтрат о-
ры1 (губки, двустворчатые моллюски, усо-
ногие ракообразные, усатые киты и др.),
грунтоеды и различные группы живот-
ных, обладающих в питании избирательной
способностью,— зерноядные и на-
секомоядные птицы, хищники
и т. д.
Часто среди животных, входящих в
один морской донный биоценоз, наблю-
дается мозаичность в использовании пище-
вых ресурсов: фильтраторы могут жить
рядом с грунтоедами и собирающими, а
иногда они специализируются по отдель-
ным кормовым объектам. Обычно именно
таким образом подбираются по способу
питания животные в биоценозе. Так, в
Каспийском море осетр питается почти
целиком моллюском синдесмией, а севрю-
га почти нацело — полихетой нереис. Оба
эти организма перед второй мировой вой-
ной были перенесены в Каспий из Азов-
ского моря и получили в нем огромное раз-
витие.
По объектам питания животных можно
также разделить на монофагов и
полифагов (т. е. питаются одним
объектом или многими).
В системе биологических связей между
животными организмами можно выделить
несколько типов с постепенным усилени-
ем биологической связи — комменса-
лизм, симбиоз и парази-
тизм.
Комменсализм, или н а х л е б-
ничество,— очень обычное явление
у общественных насекомых. Особенно
обильны комменсалы в гнездах муравьев,
где они существуют за счет запасов, на-
копляемых хозяевами гнезда. Среди них
есть и несколько видов жуков семейств
Staphylinidae, Pselaphinidae pi др. Ти-
пичные комменсалы — рыбы-лоцманы и ры-
бы-прилипалы, сопутствующие акулам,
жуки в гнездах муравьев и т. п.
Чрезвычайно разнообразно явление сим-
биоза, который может быть двусторон-
не полезным (мутуализ м2), односто-
ронне полезным или двусторонне ней-
тральным. Классический пример двусто-
ронне полезного симбиоза дают рак-от-
шельник и актиния, сидящая на его ра-
ковине. Актиния пользуется остатками со
стола своего хозяина, который, кроме
того, является для нее извозчиком, а рак-
отшельник (Eunagurus) имеет защитника —
актинию, отпугивающую его врагов. Рак-
отшельник активно разыскивает актиний,
снимает их клешней с субстрата, на ко-
тором они сидят, и пересаживает к себе
на раковину, в которой живет.
Обычны различные формы симбиоза меж-
ду некоторыми морскими животными и од-
1 Фильтраторы—животные, пропускающие че-
рез особые полости большое количество воды и
отфильтровывающие при этом планктон.
2 Мутуализм—двусторонне полезный симбиозг
от латинского слова mutuus — взаимный.
34
ноклеточными водорослями — так назы-
ваемыми зоохлореллами (зелеными) и зоо-
ксантеллами (желтыми). Зооксантеллами
набита протоплазма некоторых радиоля-
рий, ими набиты ткани некоторых корал-
лов, причем если кораллы освободить от
зооксантелл, то нормальная жизнедея-
тельность кораллов нарушается. Недавно
было показано, что зооксантеллы — это
молодые стадии развития одноклеточных
водорослей перидиней.
Самый огромный моллюск Tridacna, до-
стигающий размера в 1 /4 м и веса в 200—
300 кг, дает приют огромной массе одно-
клеточных водорослей, без которых суще-
ствовать не может. Турбеллярия Convolu-
ta, развивающаяся, в огромном количестве
на литорали Англии и Нормандии, имеет
ярко-зеленую окраску из-за набивающей
ее ткани одноклеточной водоросли Carte-
ria (зоохлорелла). Это также двусторонне
аолезный симбиоз: животное защищает
водоросли, дает им углекислоту и азоти-
стые соединения, а само получает от во-
дорослей кислород и частично исполь-
зует их как пищу. Известно, что если
молодая конволюта не получит порцию
водорослей для дальнейшего развития, она
погибает.
Различного типа симбиозы, правда, не
столь резко выраженные, весьма много-
численны. Стайки молодых рыб находят
приют среди щупалец некоторых медуз,
что является примером односторонне по-
лезного симбиоза. В заднем отделе кишки
голотурии Holothuria tubulosa обитает
маленькая рыбка Teraster umberis; она
выплывает из голотурии, но при опасно-
сти мгновенно скрывается в своем убе-
жище.
Совсем особая группа биологических
связей между организмами — явление п а-
разитизма. Паразитизм очень широ-
ко распространен среди многих групп жи-
вотных. Особенно широко паразитизм рас-
пространен среди простейших, червей и
членистоногих (цв. табл. I).
Имеется немало классов животных,
ведущих исключительно паразитиче-
ское существование. К ним относятся
споровики, трематоды, цестоды, колюче-
головые и некоторые другие черви, из
ракообразных — мешкогрудые (Ascothora-
cida). С другой стороны, ни одна группа
животных не свободна от паразитов и сим-
бионтов. Существует очень много попыток
дать общее определение и классифика-
цию явлений паразитизма, но удачного ре-
шения этой задачи пока не найдено. Яв-
ления паразитизма возникали в животном
царстве много раз и независимо друг от
друга. В силу крайнего многообразия яв-
лений паразитизма и множественности пу-
тей возникновения их классификация
имеет искусственный характер.
Определение паразитизма включает два
положения: 1) организм хозяина служит
средой обитания (паразит обитает на теле
хозяина или внутри него) для паразита,
и внешняя среда по отношению к телу хо-
зяина воспринимается паразитом опо-
средствованно через тело хозяина, 2) па-
разит использует хозяина как источник
питания, он питается тканями хозяина или
переваренной им пищей.
Естественно задать вопрос: каким же
образом возникает паразитическое суще-
ствование? Что заставляет свободно живу-
щий организм переходить к паразитизму
и существованию внутри другого организ-
ма, казалось бы, в очень сложных и труд-
ных условиях? Этот вопрос аналогичен
такому же вопросу в отношении глубоко-
водной фауны, фауны пещер и почв. Най-
ти правильный ответ на этот вопрос мы
можем, только учитывая способность жи-
вых существ к расселению и освоению но-
вых биотопов и новых биологических осо-
бенностей. Это свойство настолько резко
выражено, что многие организмы путем
длительного процесса приспособления за-
селяют чуждые им аномальные биотопы,
соответственно которым изменяется их
организация. Освоение этих биотопов при
переходе к паразитическому существова-
нию идет обычно несколькими каналами.
Очень часто паразит проникает в тело хо-
зяина, внедряясь в его пищеварительную
систему в личиночном состоянии вместе
с пищей. Бесконечно повторяющаяся ин-
вазия может кончаться успехом, и внед-
ряющийся организм вырабатывает спо-
собность существовать, а затем питаться,
расти и размножаться в кишечнике дру-
гого животного. К паразитическому су-
ществованию может приводить хищниче-
3*
35
ство, если хищник очень мелкий по срав-
нению с жертвой — хозяином. Он может
сам все теснее и теснее вступать в связь
с поверхностью тела хозяина и в конце
концов проникать в него. Кровососущие
животные (мухи, комары, клопы, клещи)
могут при этом заносить в тело жертвы
паразитов. Часто на теле крупных жи-
вотных поселяются мелкие животные,
которые становятся их постоянными
спутниками, а в дальнейшем также про-
никают под покровы их тела во внутрен-
ние органы.
Существуют, конечно, и другие пути
формирования паразитизма. Вероятно, пу-
тем постепенного приспособления к хо-
зяину к паразитизму могут переходить
комменсалы и симбионты.
Явления паразитизма связаны с мас-
сой замечательных и своеобразных осо-
бенностей. Нервная система и органы
чувств, а также мускулатура и органы
движения обычно испытывают у парази-
тов явления регресса. В силу всех этих
изменений иногда взрослый паразит те-
ряет всякое сходство с родственными
группами, как, например, у некоторых
брюхоногих моллюсков (Parenteroxenos) или
ракообразных (из веслоногих и усоногих,
в том числе знаменитая Sacculina).
Сильное развитие приобретают органы
прикрепления к телу хозяина (крючки,
присоски, хоботки) и защитные от хозя-
ина образования покровов тела, но наи-
более характерное отличие — в сильном и
часто в колоссальном развитии плодови-
тости. Нередко из одного оплодотворен-
ного яйца образуются (партеногенетиче-
ски) тысячи, сотни тысяч и миллионы осо-
бей. Количество яиц, продуцируемых од-
ной самкой турбеллярий (свободноживу-
щих плоских червей), исчисляется едини-
цами или десятками, а у близкородствен-
ных им трематод — многими тысячами и
даже десятками тысяч. Интересно при
этом отметить, что у родственных между
собою трематод и цестод имеются еще
дополнительные механизмы к увеличению
числа потомства, при этом совершенно
разные у двух несомненно родственных
форм. У ленточных червей увеличивается
число половых систем, возникающих поч-
кованием, причем их количество в одном
черве может достигать нескольких тысяч.
Совершенно иначе увеличивается плодо-
витость трематод — в спороцистах и ре-
диях (см. раздел о трематодах) происходит
партеногенетическое размножение яиц
(шары дробления) на ранних стадиях
развития. Часто организм ведет парази-
тическое существование только в опреде-
ленные периоды своего жизненного цикла:
или в раннем личиночном возрасте, или
во взрослом состоянии. В остальное вре-
мя он существует во внешней среде как
свободноживущий организм (различные
нематоды). В других, довольно много-
численных случаях паразит оказывается
на свободе только в стадии яйца (трема-
тоды, цестоды и нематоды) и, наконец,
может совсем не существовать в свобод-
ном состоянии (малярийный паразит, три-
хина и др.). Усложнение биологии пара-
зита возникает в связи со сменой хозяев:
паразит проводит часть цикла в одном
хозяине, часть в другом.
При этом основным хозяином считают
организм, в котором женские гаметы, или
яйца, проходят оплодотворение, а другого
хозяина называют промежуточным. В наи-
более сложных циклах промежуточных
хозяев может быть два, например у ши-
рокого лентеца. Первый промежуточный
хозяин — пресноводный рачок циклоп, а
второй — рыба. Своеобразное усложне-
ние паразитического жизненного цикла у
споровиков заключается в множествен-
ной повторности бесполого размножения
(шизогони я1).
Очень интересные формы сверхпа-
разитизма наблюдаются у насеко-
мых (мухи и перепончатокрылые): насеко-
мое откладывает яйца в паразитическую
личинку другого насекомого, находящую-
ся внутри личинки третьего.
Не менее интересен вопрос специфич-
ности хозяина в явлениях паразитизма.
Некоторые паразиты могут существовать
в теле только одного или двух хозяев.
Так, некоторые виды споровиков рода
Plasmodium могут развиваться только в
теле комара Anopheles, где проходят по-
1 Шизогония — множественное деле-
ние; от гречевких слов schizein — расщеп-
ляться и goneia — воспроизводство.
36
ловой цикл, и в теле человека, где проте-
кает бесполое развитие (шизогония). Воз-
будитель сонной болезни — Trypanosoma
gambiense — нормально обитает в крови
антилопы, для которой, видимо, не пато-
генен, переносчиком его является муха
цеце (Glossina morsitans), а человек —
дополнительным, случайным носителем.
Свиной солитер может развиваться только
в теле свиньи (промежуточный хозяин) и
человека (основной хозяин). Наконец, ча-
сто хозяин не специфичен и паразит мо-
жет развиваться в различных животных.
Так, эхинококк может и во взрослой
стадии, и в стадии финки существовать в
различных животных, в том числе и в
человеке. Имеются паразиты с еще более
широким кругом хозяев.
Весьма замечательны, хотя еще мало
изучены, явления физиологических и био-
химических связей между хозяином и
паразитом. Разве не замечательно, что
при нормально протекающей малярии сов-
падают циклы шизогонии паразита, си-
дящего в бесчисленных эритроцитах кро-
ви хозяина (наступление пароксизма), и
что микрофилярии появляются в перифе-
рических кровеносных сосудах в ночные
часы, когда на человека нападают крово-
сосущие насекомые, переносящие микро-
филярии на другого хозяина (см. в соот-
ветствующих разделах книги)!
Явления паразитизма, симбиоза и ком-
менсализма — интереснейший раздел био-
логии, имеющий большую перспективу
дальнейшего изучения, полный глубокого
научного смысла и огромного практиче-
ского значения.
Стадии покоя, спячки и анабиоза у жи-
вотных. Животные в состоянии нормаль-
ной жизнедеятельности могут существо-
вать в различных условиях. В арктиче-
ских морях существует немало форм, вся
жизнь и размножение которых протекают
при температуре ниже нуля, а иногда 1,5—
1,8°С ниже нуля, например моллюск Рог-
tlandia arctica, офиуры —- Ophiopleura
borealis и Gorgonocephalus arcticus, звез-
ды — Asterias panopla и Pontaster tenuis-
pinus, креветка Sclerocrangon ferox, мор-
ской таракан Mesidothea sabini и др.
С другой стороны, коралловые рифы не мо-
гут развиваться при температуре ниже
21,5° С. И те и другие относятся к так на-
зываемым стенотермным1 орга-
низмам.
Множество пресноводных организмов
погибает при повышении солености, что
и происходит в течение миллионов лет
в устьях рек при изливании их вод в мо-
ря. Но вместе с тем в пересоленных
(гипергалинны хJ озерах при почти
предельном насыщении солями сущест-
вуют и развиваются, да еще в огромных
количествах, листоногий рачок Artemia
salina и служащее ему пищей жгутиковое
Dunaliella. С другой стороны, существует
фауна горячих источников — это живот-
ные, жизнедеятельность которых может
протекать при температуре до 40—50°
(нематоды, личинки мух, брюхоногие мол-
люски) .
То же самое можно сказать и о способ-
ности животных переносить пониженное
количество кислорода. Громадное боль-
шинство животных может существовать
только при достаточном насыщении среды
кислородом. Имеются и такие, которые
не выносят даже слабого понижения в
содержании кислорода. Например, тур-
беллярия Planaria alpina или ракооб-
разное Bythotrephes — стеноокси-
бионтные3 организмы. Однако имеют-
ся животные из разных групп, которые
могут существовать при очень малом ко-
личестве кислорода в окружающей среде
или совсем без него, как некоторые вод-
ные личинки двукрылых, некоторые низ-
шие ракообразные, нематоды и др.
Среди приспособлений, при помощи ко-
торых животные переживают неблагоприя-
тные для них условия существования, вид-
1 Стенотермный — существующий в усло-
виях слабых колебаний температуры, от грече-
ских слов stenos — узкий и termos — тепло.
Противоположный термин—эвритермный, eurys—
широкий.
2 Гипергалинные — существующие в условиях
повышенной солености; hiper — сверх, halos
(греч.) — соленый. Противоположный тер-
мин — олигогалинный; oligo (греч.) — мало.
3 Стенооксибионтные — требующие для сущест-
вования строго определенного снабжения кислоро-
дом, от греческих слов stenos —узкий, oxys —
кислый, bios — жизнь. Противоположный
термин — эвриоксибионтный, от eurys — широ-
кий.
37
ное место занимает временный переход в
состояние покоя (латентное состоя-
ние). Все животные в большей или меньшей
степени способны в течение очень корот-
кого срока переносить неблагоприятные
условия без специальных приспособлений.
Правда, клещи, клопы и некоторые дру-
гие кровососущие животные могут обхо-
диться без пищи в течение очень длитель-
ного времени, даже нескольких лет, но
это редкое исключение. Способность со-
хранять жизнь при наступлении неблаго-
приятных условий значительно расширяет-
ся переходом в нежизнедеятельное
состояние, в стадию покоя — зимующие
яйца, цисты, споры, спячка, анабиоз.
Формирование этих образований может
наступать или при изменении условий
среды в неблагоприятную сторону, или
связано у паразитических форм с выходом
во внешнюю среду. В северных и умерен-
ных широтах многие животные впадают
в спячку в зимнее время, а в жарких и
сухих странах — в период летней засухи.
Зимние, или покоящиеся, яйца свойст-
венны очень многим пресноводным микро-
скопическим животным — коловраткам,
ластоногим и ветвистоусым ракообразным,
которые способны переносить высыхание
окружающей среды и низкую (зимнюю)
температуру. Яйца глистов также могут
длительное время находиться во внешней
среде в покоящемся состоянии. Яйца рач-
ка Artemia salina сохраняют жизнеспособ-
ность в течение 9 лет. Сходного типа об-
разования имеются у пресноводных губок
и мшанок (геммулыи склеробла-
сты), но это не яйца, а многоклеточные
комплексы (см. соответствующие разде-
лы книги).
Иного типа образования — споры и
цисты. Первые свойственны спорови-
кам и входят в их цикл как определенная
стадия развития, связанная обычно с вы-
ходом во внешнюю среду, и поэтому у
кровяных споровиков, которые никогда
не оказываются во внешней среде, стадия
споры утрачивается.
Многие пресноводные простейшие
(корненожки и инфузории) и микроскопиче-
ские многоклеточные (тихоходки, немато-
ды, коловратки и некоторые другие) в
случае наступления неблагоприятных
условий способны выделять оболочки и
переходить на длительное время в состоя-
ние покоя. Нередко при этом можно раз-
личить две оболочки цисты — эндо-
цисту, очень тонкую и обладающую боль-
шой химической стойкостью, и э к з о -
цисту, обладающую механической про-
чностью.
При образовании цист у простейших
наблюдаются существенные изменения в
протоплазме — выделяется большое ко-
личество воды, протоплазма густеет, те-
ряются реснички и т. п. Видимо, такого
же характера изменения происходят и
при образовании любых стадий покоя.
Эти изменения дают организмам возмож-
ность переносить сильное повышение и
понижение температуры, высыхание, по-
нижение количества кислорода. Мелкие
нематоды и тихоходки (Tardigrada) в со-
стоянии цист могут переносить времен-
ный нагрев до 150° С выше нуля и охлаж-
дение до 270° С ниже нуля.
Споры многих бактерий, коловраток,
тихоходок и некоторых нематод в сильно
высушенном состоянии могут переносить
охлаждение до температуры жидкого воз-
духа и даже до—250° С. Многие насекомые,
млекопитающие и другие животные могут
переносить в состоянии переохлаждения
(но не замерзания) температуру ниже ну-
ля: у сусликов до —0,2°С, у летучих
мышей до 1—2° Сит. п., не погибая даже
при длительном пребывании в этой темпе-
ратуре. Все описанные стадии покоя имеют
и другое биологическое значение: в виде
спор и цист простейшие в высушенном
состоянии переносятся на большие рас-
стояния и таким образом расселяются.
Поэтому среди образующих цисты про-
стейших наблюдается нередко космо-
политизм1.
Явление перехода в недеятельное
состояние (спячка, анабиоз) из-
вестно и у многих выше стоящих по уров-
ню развития организмов—у высших червей,
моллюсков, насекомых и у всех классов
позвоночных — без образования спор и
1 Космополитизмом в биологии называют очень
широкое распространение в пределах своего основ-
ного биотопа, от греческих слов cosmos — мир
и polyteien — гражданин.
38
цист, причем степень перехода в покоящее-
ся состояние (спячки, как это иногда
называют) бывает различна. Для растений
и некоторых животных переход в такое
состояние дает основание говорить о пол-
ной приостановке жизненных процессов,
но мнения ученых по этому вопросу рас-
ходятся: одни считают, что полный пере-
рыв жизненного обмена невозможен,
другие считают, что возможен. Какие же
явления сопровождают переход в спячку?
Прежде всего потеря огромного количест-
ва воды иногда почти до полного ее от-
сутствия. Дождевые черви и наземные
моллюски могут при высыхании терять до
80% и более воды и сохранять способность
к «оживанию». Наиболее способные к вы-
сыханию земноводные и пресмыкающиеся
теряют до 50% воды.
Во время спячки все процессы жизне-
деятельности организма замедляются (ды-
хание, выделение) или даже приостанав-
ливаются (питание). Ранее предполагали,
что при полном промерзании организма
(лягушки, рыбы, насекомые) он может ожи-
вать, как часы с остановленным маятни-
ком, если он вновь пущен. Однако круп-
ные специалисты в этом вопросе П. Ю.
Шмидт и Н. И. Калабухов ут-
верждают, что полное замерзание жидко-
стей тела всегда убивает организм.
Переселения и акклиматизация. Каждый
организм характеризуется определенным
ареалом1, в границах которого он
находит необходимые для него условия
существования и размножения. Внутри
ареала организмы большей частью рас-
пространены неравномерно, в некоторых
его участках достигая наибольшего разви-
тия, а в других — среднего, или встре-
чаются спорадически. В связи с этим
можно различать распространение целост-
ное или разорванное. Последнее обычно
характерно для краевых зон ареала и для
реликтовых форм. Под реликтами2
подразумевают формы, основной ареал
которых расположен в другом районе, в
условиях, которые ранее существовали и
1 Ареал от греческого слова area — поле,
площадь.
2 Реликт от латинского слова relinquere —
оставаться.
в данном месте, но в силу климатических
или других изменений ему уже не свой-
ственны. Так, в Московской области мы
находим в разных местах, часто на боль-
шом расстоянии одно от другого, ледни-
ковые озера, окруженные торфяниками,
со всем типичным для торфяника набором
растительных и животных форм, мы их
называем ледниковыми реликтами. Далеко
на севере эти торфяники имеют свой ос-
новной ареал, а под Москвой сохраняются
как реликты.
Реликты могут быть связаны не только
с низкой температурой. В Белом море в
изобилии произрастает морская трава —
зостера. Эта часть ее ареала оторвана от
основного ареала, доходящего на севере
до западного Мурмана. На восточном Мур-
мане ее нет, но в литориновое более теплое
время несколько тысяч лет назад ареал
зостеры был сплошным. Последующее по-
холодание разорвало ареал зостеры, и она
сохранилась в Белом море как тепловод-
ный реликт. Под воздействием изменяю-
щихся условий основной и часто более
обширный в прошлом ареал распадается
на отдельные реликтовые местонахожде-
ния. Таким холодновидным реликтом мо-
жет считаться в Северной Европе и Си-
бири равноногое ракообразное — морской
таракан (Mesidothea etomon) (кроме того,
он еще и солоноватоводный реликт, так
как сформировался в водоемах малой соле-
ности послеледникового времени). По своему
происхождению и распространению сходно
с ним другое ракообразное — Mysis relicta.
Обе эти формы вместе с рядом других
подобных форм можно найти в наиболее
опресненных участках окраин наших се-
верных морей, а также в пресных озерах
Швеции, Северной Европы и Сибири и
даже в Каспийском море. У всех них де-
сяток тысяч лет назад был огромный
сплошной ареал, а ныне они остались
только в различных участках как аркти-
ческие солоноватоводные реликты. Ухо-
дящий на север тающий ледниковый по-
кров разбросал их в виде реликтов на
огромном пространстве Европы и Азии.
Таким образом, с понятием реликта обыч-
но связывается существование организма
в географических зонах, общие климати-
ческие условия которых претерпели зна-
39
чительные изменения. В каком-то смысле
реликтами можно считать и различные
формы, исчезнувшие, исчезающие или со-
кратившие численность под воздействием
человека. Возможно, к ним надо отнести
и еще недавно исчезнувших стеллерову
корову в морях Дальнего Востока, птицу
додо с Маврикиевых островов, лошадь
Пржевальского и др., а сейчас относятся
зубр, морской котик, морская выдра и
многие другие.
Если брать ареалы современных живот-
ных, находящихся в условиях процвета-
ния, то и среди них мы можем найти фор-
мы с очень широкими, почти всесветными
ареалами распространения. Их иногда
именуют убиквистами1, или космо-
политами. Абсолютных убиквистов мы,
пожалуй, не знаем, обычно они ограничены
или водной средой, или воздушной. Мно-
гие из них развезены по всему миру чело-
веком. Имеется немало и таких форм, ко-
торые имеют небольшие ареалы. Но по-
давляющее большинство животных имеет
ареалы, ограниченные определенными не-
преодолимыми для них преградами — гра-
ницами суши и морей, горными хребтами,
пустынями, широкими реками и т. п.
Вряд ли можно представить себе такое
животное, которое заселяло бы все те био-
топы биосферы, которые благоприятны
для его существования. Наоборот, в ог-
ромном большинстве случаев фактический
ареал — это только часть того ареала,
в котором данный организм мог бы суще-
ствовать, если бы непреодолимые для него
препятствия не ограничивали его стрем-
ления и возможностей к расселению. Надо
учитывать и то, что указанные непреодо-
лимые преграды претерпевают в геоло-
гическом масштабе существенные изме-
нения.
Английские острова были еще недавно
частью Европы, обе Америки соедини-
лись только в середине третичного перио-
да, а Азия и Америка разъединились в не-
давнее послеледниковое время. Мадага-
скар в мезозойское время был частью аф-
риканского материка.
Еще в третичное время Черное, Азов-
ское и Каспийское моря входили в один
морской бассейн и т. д. С другой стороны,
существуют и очень древние разрывы ча-
стей суши. Еще Уоллес отметил, что
глубокий морской пролив между острова-
ми Бали и Лембок разорвал пути разви-
тия австралийской и азиатской фауны на
долгие геологические времена.
Ограниченность ареалов зависит еще
и от способности организмов к расселению.
Например, некоторые простейшие, осо-
бенно пресноводные корненожки, колов-
ратки, тихоходки и некоторые другие,
обладающие способностью образовывать
цисты и стадии покоя и переносить в этом
состоянии неблагоприятные для жизнедея-
тельных форм условия, в том числе и вы-
сыхание, могут разноситься ветром, пти-
цами и разными другими способами на
огромные пространства и приобретать все-
светные ареалы. Все другие животные,
однако, ограничены в своем распростра-
нении, и для большинства из них потен-
циальные ареалы, т. е. части биосферы,
где они могут существовать, превышают
фактические ареалы, и часто в огромных
масштабах. Это подтверждается множе-
ством примеров.
За последние 40 лет наши арктические
моря претерпели значительное потепление,
и в результате вся населяющая их фауна
переместилась с запада на восток: тепло-
водная фауна — из Северной Атлантики
в Баренцево море, из него дальше —
в Карское море, а холодноводная фауна,
наоборот, отступила из западной и юго-
западной частей Баренцева моря на восток
и север. Однако гораздо показательнее те
случаи, когда животное преодолевает не-
преодолимые для него преграды, в подав-
ляющем числе случаев с сознательным
или бессознательным участием человека.
Каспийский эндемик2 гидроид Сог-
dylophora caspia, очевидно, с помощью су-
дов, днища которых он обрастает, во вто-
рой половине прошлого века по Мариин-
ской системе проник в Балтийское море, а
оттуда кораблями был развезен по всем
1 Убиквисты —от латинского ubique — и о-
всюду, то же, что космополиты.
2 Эндемик—от греческого endemos—м е с т н ы й,
живущий только в данном месте. Могут быть
эндемики узкие, например озеро Севан, и энде-
мики широкие, например озера Северной Аме-
рики или Европы.
40
крупнейшим портам мира. Китайский мох-
натоногий краб (Eriocheir sinensis) тоже
с кораблями перед первой мировой войной
проник с Дальнего Востока в устье Эльбы,
откуда широко расселился по побережьям
Северного и Балтийского моред и речным
бассейнам. В новом ареале он достиг ог-
ромного развития. Тем же путем брюхо-
ногий моллюск рапана (Rарапа besoar)
переселился около тридцати лет назад из
Японского моря в Черное, размножился
в нем и стал злостным вредителем устрицы
и мидии и излюбленным сувениром чер-
номорских туристов. Еще один краб,
Rhithropanopeus harrisi, совершил более
сложное переселение с тремя «пересадка-
ми». Сначала от побережий Северной Аме-
рики через Атлантический океан в Зюй-
дерзее, оттуда — к нам в Черное море,
а из него Волго-Донским каналом в
Каспийское море, и в каждом новом
местообитании этот маленький крабик по-
лучил массовое развитие. Крабик был за-
везен также с западного побережья Се-
верной Америки на восточное. Любопытно,
что все эти путешествия вплоть до Кас-
пийского моря крабик ритропанопеус со-
вершил с полихетой Marcierella enigma,
вместе с которой живет и в исходном ареа-
ле. И таких примеров непрошеных «нару-
шителей границ» мы знаем великое мно-
жество — американская виноградная тля
филлоксера, колорадский жук, проникший
90 лет назад из Северной Америки в Гер-
манию, а сейчас неуклонно стремящийся
на восток. Иногда эти пути идут и в об-
ратном направлении.
Развитие авиационных путей сообщения
привело к завозу из Северной Америки
малярийных комаров в Бразилию. Через
несколько лет комары распространились в
глубь страны и вызвали жестокую эпиде-
мию малярии. В начале нашего столетия
из Европы в Северную Америку был заве-
зен европейский скворец (Sturnus vulgaris),
распространившийся к середине столетия
в огромном количестве по всей стране.
Есть много и других подобного типа при-
меров.
Понимание того, что организм может
существовать не только в пределах его
фактического ареала, но и далеко за его
пределами, возникло давно, и на нем ба-
1 40
0
40
160 80
—^шш 'Л1! t*~ I _j
""""""n. vi
* V
ч \ \А
I^^Gambusia afflnis
^Ш Другие виды6атЬиа!а
0 80
•^¦К
N »
/ 1
J&
Ь
•
0 80
160
5^С_*»\
160
120
s^S^f^
Ж* \
sgjfc-l
^^-'^< ^
120
40
0
401
Рис. 8. Распространение североамериканской
рыбки гамбузии по земному шару.
Рис. 9. Распространение американского грызуна
ондатры в Европе и Азии.
зируется вся деятельность человека по
акклиматизационным мероприятиям. Чи-
сло таких мероприятий огромно. Начало
им было положено переселением в новые
места обитания домашних животных и
культурных растений. Нынешние флора
и фауна Австралии и Новой Зеландии
имеют совсем иной характер, чем сто лет
назад. В значительной степени изменились
флора и фауна Европы и Северной Амери-
ки, акклиматизационные мероприятия по-
степенно захватывают весь земной шар.
Из наиболее ярких примеров можно при-
вести рыбку гамбузию (рис. 8) и ондатру
(рис. 9). Однако известны случаи, когда
очень хорошее по первоначальным резуль-
татам акклиматизационное мероприятие
41
давало в дальнейшем и отрицательный ре-
зультат. Так, рыбка гамбузия настолько
размножилась в некоторых водоемах, на-
пример, Средней Азии, что выела все
корма для других рыб и подорвала их
запасы. С безобидным, казалось бы, кро-
ликом в Австралии пришлось вести борь-
бу, так как огромные его стаи стали угро-
зой культурным растениям.
Большое количество подобных фактов
описывается в интересной книге англий-
ского эколога Элтона «Экология наше-
ствий животных и растений», переведен-
ной и изданной в СССР в 1960 г.
Интереснейшая деталь очень многих
успешных акклиматизаций в новом ареа-
ле — это исключительная массовость раз-
вития, превышающая во много раз сте-
пень развития в исходном ареале. Такую
массовость развития вновь появившейся
в новом ареале формы можно уподобить
как бы некоему биологическому взрыву.
В 1911 г. из Японии в США был случайно
завезен майский жук Popillia japonica,
достигший в новом местообитании колос-
сального развития. Уже через 8 лет в
местах его массового развития один сбор-
щик мог собрать в течение дня до 20 000
жуков.
В течение последних 40 лет яркие при-
меры таких биологических взрывов дают
многие новые «вселенцы» в Каспийское
море. Во время гражданской войны слу-
чайно в Каспий попал из Черного моря
двустворчатый моллюск Mytilaster linea-
tus, ставший вскоре в этом море самым
массовым. В тридцатые годы такого же
массового развития достигли два вида
креветок рода Leander. Перед второй
мировой войной в Каспийское море были
перенесены из Азовского полихета Nereis
diversicolor и двустворчатый моллюск Syn-
desmya ovata для обеспечения пищей
осетровых рыб. Последующее развитие
обеих форм в Каспии также оказалось
подобным биологическому взрыву, и их
общее количество измеряется многими
миллионами центнеров. С прорытием Вол-
го-Донского канала переселение из Азов-
ского моря в Каспийское различных расте-
ний и животных приобрело стихийный ха-
рактер, и в настоящее время «вселенцы»
составляют в Каспии гораздо большую
массу, чем своя исконная фауна. Было
несколько удивительно, что балтийская
сельдь салака дала в Аральском море,
куда ее пересадили, крупную по разме-
рам, вполне жизнеспособную расу. Исклю-
чительный интерес представляет успешная
акклиматизация дальневосточных лососей
в водах Новой Зеландии и Южной Аме-
рики.
Все изложенное хорошо поясняет, ка-
кой огромной важности оружие приобре-
тает человек для своего хозяйства, ис-
пользуя способность многих растений и
животных давать массовое развитие в
новом ареале, и сколь необходима теоре-
тическая разработка проблемы акклима-
тизации для наиболее разумного исполь-
зования этой способности.
БИОСФЕРА И БИОЦЕНОЗ
Академику В. И. Вернадскому
принадлежит большая заслуга разработки
учения о биосфере. Сам термин был
введен в науку немецким геологом Э. Зюс-
с о м в 1875 г., однако понятие это не
было им разработано и не было нужным
образом воспринято в науке. Только через
50 лет это было с исключительной глу-
биной и широтой охвата осуществлено
В. И. Вернадским.
Организмы и среда находятся в нераз-
рывной связи. Невозможно представить
себе организм вне среды обитания; с дру-
гой стороны, каждый организм находится
в определенной связи с другими организ-
мами. Таким образом возникли понятия
биотопа и биоценоза1 (части
среды и части населения). Академик
В. Н. Сукачев для определения со-
вокупности населения и среды создал
понятие биогеоценоза. Биогеоце-
ноз, по В. Н. Сукачеву,— участок земной
поверхности, где на известном протяже-
нии биоценоз и отвечающие ему части
1 Биотоп — среда обитания, от гре-
ческого bios — жизнь и topos — место.
Биоценоз — совокупность организмов, населяю-
щих ту или иную часть биосферы, от греческого
bios — жизнь и koinos — вместе.
42
литосферы, гидросферы и пе-
досферы1 остаются однородными, тесно
связанными между собой также однород-
ными взаимодействиями и поэтому в сово-
купности образующими единый, внутрен-
не взаимообусловленный комплекс.
Вся Земля в целом представляет собой
биотоп высшего порядка, а все ее населе-
ние — биогеоценоз также высшего поряд-
ка. Учение В. И. Вернадского о био-
сфере — одно из крупнейших естествен-
нонаучных обобщений, к которому впол-
не применимы его же собственные слова:
«Века и тысячелетия прошли, пока чело-
веческая мысль могла отметить черты еди-
ного связного механизма в кажущейся
хаотической картине природы». Биосфера
Вернадского и есть такой «единый связан-
ный механизм». «Живое вещество биосфе-
ры,— говорит он,— единственной области
планеты, закономерно связанной с косми-
ческим пространством, есть совокупность
ее живых организмов, ее живого вещества
как планетного явления». Все живое ве-
щество в целом он рассматривает как
единую, особую область накопления сво-
бодной химической энергии в биосфере,
превращения в нее световых излучений
солнца. «Биосфера представляет собой
определенную геологическую оболочку,
резко отличную от всех других геологи-
ческих оболочек нашей планеты... И это
не только потому, что она заселена живым
веществом, которое является огромного
значения геологической силой, ее совер-
шенно перерабатывающей, меняющей ее
физические, химические и механические
свойства. Но это единственная оболочка
планеты, куда проникает заметным об-
разом космическая энергия, ее еще более
живого вещества изменяющая. Главным
ее источником является солнце... Жи-
вое вещество проникает всю биосферу и
ее в значительной степени создает».
Состав и структура живого населения
биосферы и его роль в изменениях био-
сферы в настоящем и геологическом прош-
лом составляет основное содержание нау-
ки геобиологии, которая вместе с гео-
логией, геофизикой и геохимией должна
осветить историю нашей планеты Земли.
Миллиарды лет назад на Земле появи-
лась первая жизнь. Вероятно, это про-
изошло на дне теплого мелкого моря в
форме прототрофных2 организ-
мов. После долгого пути развития сло-
жился современный «трехликий» круго-
ворот органического вещества в биосфере:
аутотрофные3 организмы создают
органическое вещество и при этом обо-
гащают окружающую их среду кислоро-
дом, обеспечивая тем возможность суще-
ствования гетеротрофных4 ор-
ганизмов, в свою очередь обогащающих
среду углекислым газом, необходимым
растительным организмам для сущест-
вования.
Растительный мир как продуцент мо-
жет быть противопоставлен животным
организмам как консументам5. На-
конец, третий компонент этого круговоро-
та вещества — микроорганизмы-р е д у -
центы6. Одни из них разлагают ор-
ганические вещества, минерализуют их,
другие ведут обратные процессы, создают
более сложные соединения, пригодные
для жизнедеятельности растений. Сотни
миллионов лет назад из моря на сушу
вышли первые растения, а до того, ве-
роятно, поверхность суши была лишена
жизни. Но к моменту выхода растений
на сушу поверхность океана снабдила ат-
мосферу кислородом и углекислотой и
создались условия, пригодные для раз-
вития жизни и на суше. Поверхность
суши и пресные водоемы стали быстро
заселяться растительными и животными
организмами.
И растительный и животный мир су-
ши — это очень молодое образование по
сравнению с населением океана, с ис-
торией развития, по крайней мере в
десять раз более короткой.
1 Педосфера — почва.
2 Прототрофные организмы — микроорга-
низмы, развитие которых происходит за счет
перевода одних неорганических соединений в дру-
гие. К ним относятся серобактерии, железобакте-
рии и др.
3 Аутотрофный — сам себя питающий, от
греческих слов autos—с а м и trophon — пита-
ние.
4 Гетеротрофный—от греческого слова heteros—
другой.
5 Консумент — потребитель, от латин-
ского слова consumere — потреблять.
6 Редуценты—р азлагатели, от латинского
слова reducere—р азлагать.
43
Жизнь в океане получила уже свое
полное развитие, когда первые пере-
селенцы только выходили на сушу.
Завоевание суши и материковых водое-
мов шло двумя путями — прямо из моря
в воздушную среду через прибреж-
ную, осушную зону (см. дальше) и вод-
ным путем — из морской воды в прес-
ную. Таким образом создавались основ-
ные части биосферы: древняя пра-
матерь жизни — океан, воздушная среда,
пресные воды и, наконец, наземные поч-
вы. Эти четыре подразделения биосферы
весьма различны по объему. Объем морей
и океанов составляет 1370 млн. км3,
и он заселен, за очень немногими исклю-
чениями, весь от поверхности до наиболь-
шей глубины (И 034 м в Марианской
впадине).
Объем атмосферы, заселенной жизнью,
над сушей, если принять ее мощность в
50 м (средняя высота древесного покро-
ва), составляет не более 7 444 600 км3,
или 1/183 часть объема океана. Объем
почвенного слоя суши, пригодного для
жизни, составляет около 100 тыс. км3,
т. е. 1/137оо часть объема океана.
Материковые воды суши (реки, озера,
водохранилища, подземные воды) имеют
объем около 8 млн. км3, или 1/110 часть
объема океана. Таким образом, моря и
океаны составляют по объему около 98%
всей биосферы.
Мы уже указывали, что большинство
типов и классов животных, к которым
относится 150 тыс. видов,— обитатели
морей и океанов. Если не считать насе-
комых, создавших огромное видовое разно-
образие (не менее миллиона видов), то
воздушной среде и пресным водам свой-
ственно всего около 80 тыс. видов живот-
ных, а пресным водам — около четверти
этого числа. В биосфере имеются две
природные среды, обладающие свойством
плодородия,— вода и почва. В обоих
средах минеральная основа насыщена
продуктами жизнедеятельности бесчис-
ленных предыдущих поколений расти-
тельных и животных организмов, нахо-
дящихся на всех стадиях распада, ми-
нерализации и восстановления. Морские
растения, даже прикрепленные ко дну,
поглощают все нужные им вещества из
окружающей их воды, а наземные —
из почвы (за исключением углекислого га-
за). Однако это различие только кажу-
щееся. Дело в том, что вся водная толща
является аналогом почвы. Растение, це-
ликом погруженное в водную толщу, не
лишено света. В почву же погружена
только корневая система растений, так
как иначе они были бы лишены света.
У наземных растений развиваются стеб-
левая и древесинная части, выносящие
листья к свету.
Крупные биотопы делятся на более
мелкие, второго, третьего, четвертого,
пятого и т. д. порядка вплоть до самых
мелких. Верхняя и нижняя листовая
поверхность погруженного в воду расте-
ния, его стебель — это также мелкие био-
топы, но каждому из них свойствен свой
набор животных, свой биоценоз. Так же
как и биотоп, биоценозы могут иметь
все более и более дробный характер. Одну
из самых простых форм биоценоза пред-
ставляет популяция1 — поселение,
состоящее всего лишь из одного вида.
Иногда биоценоз состоит из двух-трех
видов. В качестве примера такого о л и-
гомикстного2 биоценоза можно при-
вести население пересоленных озер
(Баскунчак, Кара-Богаз-Гол и др.).
В этот биоценоз входят только два орга-
низма — жгутиковая водоросль Duna-
liella, продуцирующая органическое ве-
щество, и питающийся им рачок Artemia.
Олигомикстные биоценозы соответству-
ют обычно уклоняющимся от нормальных
условий (аномальным) биотопам либо по
температуре, либо по солености, либо по
другому фактору среды. Олигомикстные
биоценозы мы встречаем в пещерах, в
глубинах океана, на вершинах гор. Обыч-
но в нормальных условиях биоценозы
состоят из нескольких десятков или даже
сотен видов. Их называют полимик-
ст н ы м и 3.
1 Популяция — одновидовое поселение, от ла-
тинского слова populus — народ.
2 Олигомикстный — составленный из малого
числа компонентов, от греческого слова oligos —
малый и латинского mixtus — состав-
ленный.
3 Полимикстний — составленный из многих
компонентов, от греческого poly — много.
44
Организмы, входящие в состав биогео-
ценоза, связаны друг с другом (био-
тические факторы) и со средой
обитания (абиотические фак-
торы) сложной системой связей: хищ-
ник — жертва, паразит — хозяин, кон-
куренция за пространство, пищу, кисло-
род и т. д. Единым местообитанием не-
редко бывают связаны организмы, наи-
менее зависимые друг от друга.
Биоценоз можно определить как со-
вокупность организмов, населяющих
определенную часть биосферы и исполь-
зующих ее для питания, роста и размно-
жения.
Изучением сложной системы отноше-
ний организмов друг с другом и с внеш-
ней средой занимаются науки эколо-
гия1 и биоценология. Принято
различать два ее раздела — аутэколо-
гию и синэкологию, иначе говоря, систе-
му связей организма с абиотической сре-
дой и систему связей организмов между
собой. Однако нельзя разделять эти две
системы связей, поскольку отдельный
1 Экология — наука о связи организма со сре-
дой, от греческого слова oikos —дом.
организм представляет собой только эле-
мент биотической среды, от которой во
многом зависит и вместе с которой пре-
терпевает изменения. Это, однако, не
значит, что отдельный организм не вос-
принимает непосредственно факторы аби-
отической среды — температуру, соле-
ность, влажность и т. д. Но биотиче-
ская система в определенной степени
контролирует, а часто определяет реак-
ции организма на эти факторы.
К четырем указанным подразделениям
биосферы можно добавить еще несколько
крупных и характерных биотопов с осо-
быми более или менее резко отличными
условиями существования. Среди морских
вод можно выделить еще воды промежу-
точной солености, или солоноватые, а
среди пресных вод — подземные воды.
Среди биотопов воздушной среды можно
также выделить почву и пещеры. Совсем
особняком надо поставить существование
внутри тела другого животного, или
паразитическое существование. Таким
образом, можно выделить 10 биотопов,
каждый из которых в свою очередь может
быть подразделен на биотопы второго и
третьего порядка (табл. 3).
Группа биотопов
Свободное существо-
1 вание
1 Паразитическое су-
I ществование
Основные биотопы
Водная среда
Воздушная среда
Тело других орга-
низмов
Таблица 3
Их подразделения и объем в клс3
Морские воды 1 370 000 000
Пресные воды (не считая
материковых льдов)
8 300 000
Открытые материковые воз-
душные биотопы 7 440 600
Скрытые воздушные био-
топы
Полносоленая морская вода
Солоноватые воды
Морские грунты
Надземные воды
суши 4 000 000
Подземные воды 4 000 000
Лес, степь, поле, пустыня
и т. д.
Наземные почвы 149 000
Пещеры
Тело животных 1
Тело растений
45
Супралиторалъ
Литораль
.Сублитораль
Поверхностная зона,
или эпипелагиаль
Рис. 10. Вертикальное расчленение ложа океана и основные биотопы.
Как видно из приведенной таблицы,
самый обширный биотоп — это океаны и
моря, причем 90% объема этого биотопа
существует в условиях полного мрака,
а 75% — при давлении свыше 100 атмо-
сфер. Океанские глубины с давлением
свыше 400 атмосфер (глубже 4 тыс. м)
составляют объем около 1 млрд. км3.
Этот самый крупный биотоп биосферы
представляет собой огромную барокаме-
ру, в которой жизнь существует уже
миллиарды лет.
Вертикальные подразделения океана
и его население резко отличаются от
подразделений воздушной среды. Как
уже указывалось, океан заселен от поверх-
ности до наибольших глубин. Распреде-
ление жизни в океане связано прежде
всего с вертикальнозональным расчлене-
нием ложа океана (рис. 10).
Каждой зоне свойствен свой характер
растительного и животного населения.
У берегов в связи с наличием в открытых
морях явлений приливов и отливов вы-
деляют так называемую осушную зону,
или литораль1. Иногда амплитуда
приливо-отливных колебаний достигает
15—17 м, но обычно измеряется одним-
двумя метрами или даже сантиметрами.
Моря, слабо связанные с открытым океа-
ном, как Средиземное, Черное и Азовское,
лишены ощутимых приливо-отливных ко-
лебаний. Литоральная фауна состоит в
основном из червей, моллюсков и рако-
образных, способных находиться один
или два раза в сутки в воздушной среде:
черви зарываются в грунт, моллюски и
«морские желуди» плотно закрывают свои
створки.
Некоторое количество видов литораль-
ных животных может забираться даже
выше линии прилива (например, ракооб-
разные) и пребывать в воздушной среде,
довольствуясь заплесками или даже брыз-
гами волн. Их именуют супрали-
1 Литораль — от латинского слова litus —
берег.
46
торальными1. От самого низкого
уровня до 200—400 м (для разных морей)
на так называемом континентальном
склоне располагается сублитораль,
самая густо населенная наряду с лито-
ралью' растениями и животными морская
зона. В этой зоне расположены и самые
обильные рыбопромысловые угодья.
Глубже, до 2500—3000 м, идет ба-
тиаль2, или материковый
с к л о н, а еще ниже расположена самая
обширная часть океанского дна, или
ложе океана — абиссаль3. Она за-
нимает около 4/б всего дна океана. С пе-
реходом в батиаль и абиссаль фауна
беднеет и качественно и количественно.
Вместо 150 тыс. видов животных сохра-
няется всего несколько тысяч, а плотность
населения с сотен и тысяч граммов на
квадратный метр дна падает до несколь-
ких сот или даже десятков миллиграм-
мов. Наконец, глубоководные впадины,
которые занимают едва 1 % площади оке-
анского дна и названы ультра-
абиссалью4, имеют самый бедный
по количеству видов состав фауны (две-
три сотни видов).
Соответственным образом подразделяет-
ся и толща океанских вод (пелагиаль5).
Самый верхний слой, или, как его назы-
вают, эпипелагиаль6, или э в -
фотическая7 зона,— зона разви-
тия водорослей, продуцирующая органи-
ческое вещество для всей толщи океана.
Батиали в пелагической зоне соответству-
ет батипелагиаль, а абиссали —
абиссопелагиаль. Каждой вер-
тикальной зоне соответствуют особый
состав населения и особый набор свой-
ственных ему биологических и морфоло-
гических особенностей (окраска, органы
чувств, способы размножения и т. п.).
1 Супралиторалъный—от латинского super—
вверху.
2 Батиаль—от греческого bathos —глуби-
н а.
3 Абиссаль — от греческого abyssos — мор-
ская пучина.
4 Ультра —от латинского ultra — дальше.
5 Пелагиаль —от греческого pelagos — от-
крытое море.
6 Эпипелагиалъ — от греческого epi — над.
7 Эвфотический—от греческих слов eu—x оро-
шо, по-настоящему и photos — свет.
Многие считают, что абиссальной фауне
свойственна большая геологическая
древность и определяемая этим прими-
тивность строения.
Распределение жизни во всех указан-
ных выше средах зависит от факторов
среды, причем они различны для разных
биотопов. В воздушной среде огромное
определяющее значение имеет фактор
влажности, а в водной среде он отсут-
ствует, но зато в воздушной среде, за
очень редкими исключениями, не имеет
места дефицит кислорода и наличие не-
которых других газов, а в водной среде
газовый состав приобретает часто перво-
степенное значение (например, в глубинах
Черного моря, зараженных сероводоро-
дом). В наземной среде соленость редко
приобретает определяющее значение (под-
соленные почвы), и то только для скрытых
биотопов (почвы, пещеры), а в водной
среде это один из наиболее значимых
факторов. Фактор давления проявляется
по-разному в воздушной среде и в мор-
ской. В первом случае давление может
быть ниже одной атмосферы, а в морской
среде значительно превышать ее (до 1100
атмосфер).
Распределение температуры и света
имеет и в водной и в воздушной среде
первостепенное значение. Колебания тем-
пературы в водной среде имеют гораздо
меньшую амплитуду, чем в воздушной.
Морская среда вообще характеризуется
устойчивостью режима и слабыми коле-
баниями (сезонными, суточными и локаль-
ными) факторов среды, а воздушная сре-
да — резкими и сильными. Поэтому ино-
гда моря называют царством однообра-
зия, противопоставляя их воздушной
среде как царству разнообразия.
Сезонные и суточные колебания в све-
товом режиме одинаковы для открытых
биотопов суши и моря, с тем различием,
что в океанах 90% водной массы сущест-
вует в полном мраке, а в воздушной
среде это имеет место только в скрытых
биотопах (почвы, пещеры и подземные
воды), составляющих малую долю био-
топов воздушной среды.
Между столь удаленными друг от друга
биотопами, как глубины океана, пещеры
и подземные воды и почвы (как морские,
47
Рис. 11. Планктонные веслоногие ракообразные.
так и наземные), существует сходство в
некоторых факторах среды. Их сближает
отсутствие света, очень слабые колеба-
ния температуры (а иногда и отсутствие
их) и, кроме того, отсутствие дефицита
влажности. Эти черты сходства обуслов-
ливают и сходство в ряде признаков,
свойственных населяющим их животным:
ослабление или утеря органов зрения,
развитие органов осязания, депигмен-
тация, замедленные темпы филогенети-
ческого развития и т. п.
Население водной среды по характеру
поведения и распределения в его вод-
ной массе можно разделить на ряд био-
логических групп, а именно п л а н к -
т о н1, бентос2 и нектон3.
Планктоном называют всю совокупность
мелких организмов, лишенных активных
органов передвижения и как бы парящих
в воде. Они перемещаются вместе с вод-
ными массами (рис. 11). Это, впрочем,
правильно в отношении только части
планктонных организмов, так как мно-
гие из них способны совершать суточ-
ные вертикальные миграции в сотни
метров.
Планктон включает как растительные
организмы (фитопланктон), так
и животные (зоопланктон). В мор-
ском фитопланктоне особенно видное
место занимают одноклеточные водорос-
ли — диатомовые, перидиниевые, сине-зе~
леные, жгутиковые, а в зоопланктоне —
фораминиферы и радиолярии из однокле-
точных, а из многоклеточных — кишеч-
нополостные (медузы, ктенофоры, сифоно-
форы), некоторые группы червей и обо-
лочниковых, ракообразные и разнообраз-
ные личинки беспозвоночных животных
и рыб. Кроме того, среди населения
моря выделяют еще несколько биоло-
гических групп — плейстон, нейстон, ги-
понейстон.
Плейстон — это группа организ-
мов, держащаяся полностью или ча-
стично на поверхности воды, использую-
1 Планктон—от греческого слова planktos —
парящий, носящийся туда-сюда.
2 Бентос—от греческого слова benthos —
придонный.
3 Нектон— от греческого слова nektos — пла-
ва ющий.
щая в качестве опоры поверхностную
пленку натяжения. Замечательными при-
мерами плейстонных организмов являют-
ся некоторые сифонофоры, как парусница
(Velella) или физалия (Physalia), так
называемый «португальский военный ко-
раблик». Они поднимают над поверхно-
стью воды гребни своих пневматофоров,
выполняющих роль паруса. Н е й с т о н—
это обитатели поверхностной пленки, а
гипонейстон — совокупность организ-
мов, скопляющихся под пленкой поверх-
ностного натяжения в нескольких верх-
них сантиметрах воды. Здесь собирают-
ся часто в огромном количестве икра рыб,
различные личинки и некоторые другие
планктонные организмы (рис. 12).
Бентос — это совокупность орга-
низмов, ведущих придонный образ жиз-
ни. Как и для планктона, различают
фитобентос {водоросли) и з о о -
бентос, в состав которого входят
фораминиферы, разнообразные губки,
кишечнополостные, черви, моллюски, ра-
кообразные, мшанки, плеченогие, иглоко-
жие, асцидии, некоторые рыбы и ряд
других.
Но в морях и пресных водоемах живет
еще одна группа более крупных организ-
мов, хороших пловцов. Их называют
нектоном и относят к ним рыб, мор-
ских млекопитающих и кальмаров.
Океаническая фауна существует при
солености, близкой к 35°/00 C,5°/0). В
экваториальной зоне на поверхности она
повышается до 36°/00, в Северном Ледо-
витом океане понижается до 33—34°/0о
(также в поверхностном слое). В Среди-
земном море и особенно в Красном
море и в Персидском заливе вследствие
сильного испарения и малого речного
стока соленость повышается до 37—38°/00.
Все указанные колебания солености не
сказываются заметным образом на соста-
ве фауны, который определяется в ос-
новном температурой. Это не относится,
однако, к некоторым окраинным полузамк-
нутым морям северного полушария, обиль-
но снабжаемым речной водой и в силу
этого имеющим пониженную соленость.
Такие моря характерны для окраин Ев-
разии — Балтийское, Белое, южные ча-
сти сибирских морей — Карского, Лап-
тевых и Восточно-Сибирского, а также
Черное, Азовское и Каспийское. Все
эти моря имеют пониженную соленость— в
Белом море 24—30°/00, в Балтийском около
8°/00, в Черном, в верхних слоях, на-
селенных жизнью, около 18°/00, в Азов-
ском 10—12°/0о> в Каспийском 5—13°/00.
Соленость Белого и Черного морей еще
не меняет характера фауны, но только
обедняет ее.
В Средиземном море обитает 7—8 тыс.
видов животных, а в Черном — только
около 1500. В Баренцевом море фауна
состоит из 2500 видов, а в Белом — всего
около 1000. Основная часть Азовского
моря тоже заселена фауной средизем-
номорского типа, но в самых опреснен-
ных уголках (заливы, лиманы, устья
рек) Черного, Азовского, Белого, си-
бирских морей к наиболее эвригалинной
фауне морского типа примешивается осо-
бая солоноватоводная фауна (рис. 13).
В Балтийском и Каспийском морях
солоноватоводная фауна имеет основные
ареалы своего распространения. Харак-
терная ее особенность в том, что она не
распространяется в воды морской солено-
сти, распространена в малой степени в
пресных водах и имеет реликтовый ха-
рактер. Мы уже указывали, что морская
и пресноводная фауна в основном имеют
разный состав и существуют раздельно,
подавляющее количество групп совсем
лишено способности проникать в чуждую
им среду, даже за геологически длитель-
ные периоды. Только рыбы и некоторые
крабы обладают способностью существо-
вать и в морской и в пресной воде, правда,
в разные периоды своей жизни. На Севере
и Дальнем Востоке — это лососевые, си-
говые и корюшковые рыбы, а в наших
южных морях — сельди, осетровые, судак
и многие другие. Однако такое проникно-
вение совершается и у других групп жи-
вотных — известны преоноводные губки,
кишечнополостные, мшанки, полихеты,
крабы, морские коловратки, олигохеты,
пиявки и др. Но когда и каким образом
произошло проникновение этих живот-
ных в пресные воды, мы не знаем.
Гораздо яснее вопрос о происхожде-
нии солоноватоводной фауны. Ее генезис
связан с палеогеографическими измене-
W 4 Жизнь животных, т. i
49
Рис. 12. Организмы гипонейстона:
1 — личинка хамсы; 2 — личинка Blennius; з — личинка Trachinus; 4 — молодой крабик Portunus; 5 —ли-
чинка кефали (остроноса); 6 — Ponlella raediterranea; 7 — Anomalocera patersoni', 8 — личинна саргана;
9 — малек Gallionymus, 10 — малек Solea; 11— малек саргана; 11 а—обрывок Zostera marina; 12 — равно-
ногий рачок Idothea stephenseni на кусочке плавника; 12 а — обрывок бурной водоросли Cystoseira barbata.
ниями в судьбе тех водоемов, где она
зародилась. Вообще солоноватоводная
фауна имеет морской генезис, а форми-
ровалась она тогда, когда морские водое-
мы вместе с населявшей их фауной под-
вергались медленному геологически дли-
тельному процессу опреснения. Большин-
ство видов морской фауны при этом про-
цессе вымирало, но некоторая часть, глав-
ным образом из рыб и ракообразных, при-
спосабливалась к существованию в воде
малой солености C—4—5—8°/00), и, что
замечательно, животные теряли при этом
способность к возврату в морские воды
полной солености.
Обширные полузамкнутые и замкнутые
водоемы малой солености в третичное
время создавались на месте современных
Черного и Каспийского морей, а на севе-
ре в области Полярного бассейна — в
ледниковое и послеледниковое время.
Эти бассейны и оставили реликтовую
солоноватоводную фауну. У морских по-
бережий в мелком песке обнаружена весь-
ма разнообразная псаммофил ьная1
фауна, иначе называемая интерсти-
циальной2. Эта микроскопическая
фауна, обитающая в пустотах между
песчинками, включает инфузорий, тур-
беллярий, нематод, коловраток, ракооб-
разных.
Пресноводная фауна по составу суще-
ственно отличается от морской, в ней
отсутствуют наиболее характерные мор-
ские группы животных и преобладают в
планктоне низшие ракообразные и коло-
вратки, а в бентосе — личинки насеко-
мых, брюхоногие моллюски (главным
образом из подкласса легочных), дву-
створчатые моллюски (из семейства унио-
нид), олигохеты, пиявки, некоторые ра-
кообразные (ракушковые, бокоплавы, во-
дяной ослик), амфибии и некоторые дру-
гие. Обычны в пресных водоемах также
пресноводные губки семейства Spongil-
lidae (бадяги), гидры, мшанки. Имеются
и такие редкие формы, как пресноводные
медузы и полихеты.
1 Псаммофильная от греческого psammos —
песок.
2 Интерстициальный от латинского interes-
titium —промежуток.
О б 10 15 20 26 30 36
Рис. 13. Положение солоноватоводной
фауны (косая штриховка) между морской
и пресноводной.
Рыбное население пресных водоемов не
столь разнообразно, как морских, одна-
ко достаточно богато. Особенно выделяют-
ся разнообразием карповые рыбы. Особый
тип пресных водоемов представляют под-
земные и пещерные воды с наиболее ха-
рактерными для них некоторыми ракооб-
разными.
Встречаются в подземных водах и по-
звоночные: в балканских пещерах изве-
стна хвостатая слепая амфибия — про-
тей, а в пещерах Северной Америки—
слепые рыбы.
Переходя к основным биотопам воз-
душной среды, можно прежде всего вы-
делить население почвы, или э д а ф о н.
Состав эдафона весьма разнообразен, в
него входят корненожки, инфузории, не-
матоды, олигохеты, огромное количест-
во насекомых, клещей и мелких млекопи-
тающих, а также растения. Свободная
воздушная среда также может быть разде-
лена на ряд основных биотопов, каждому
из которых свойствен свой набор жи-
вотных,— пустыня, степь, лес (болото,
тундра, высокие горы).
i*
51
ФАУНИСТИЧЕСКИЕ ОБЛАСТИ
ОКЕАНА И СУШИ
Фаунистическое районирование океана
н материков имеет сходные черты, но
также и резкие различия. Сходство обу-
словливается общей температурной гео-
графической зональностью,одинаково воз-
действующей на распределение и мор-
ских и наземных животных. Различия
являются следствием целостности Миро-
вого океана и разобщенности материков.
Материковые массивы — американский,
европейско-африканский и азиатско-ав-
стралийский — имеют меридиональную
протяженность и в южном полушарии
разделены огромными океаническими
просторами. Океаны также разобщены
материками, но не полностью. Полно-
стью (если не считать Панамского и Суэц-
кого каналов) разобщены только фауны се-
верного умеренного пояса и тропической
зоны Атлантического и Тихого океанов:
яа севере — холодными арктическими мо-
рями, на юге — слишком теплым режи-
мом тропического пояса, а для тропиче-
ской фауны — холодными водами уме-
ренной зоны южного полушария. Тем
не менее океанические фауны, атланти-
ческая и индо-тихоокеанская, обладают
сходством в гораздо большей степени,
чем материковые (Южной Америки, Аф-
рики и Австралии).
Для океана обычно различают 8 основ-
ных областей (карта 1): Кругопо-
л я р н у ю (Арктическую), Б о р е а л ь-
н у ю (Умеренную)— атлантичес-
кую и тихоокеанскую, Тро-
пическую Индийско-Запад-
но-Тихоокеанскую, Тропи-
ческую Западно-Африканскую,
Тропическую Американскую,
Нотальную (или Южную Умерен-
ную) и Антарктическую кру-
гополярную. Кроме этого, иногда выде-
ляют Субарктическую, Суб-
тропическую и Субантарк-
тическую переходные области. Арк-
тическую область делят обычно на две
подобласти: Высоко-Арктическую и Ниж-
не-Арктическую (Баренцево и Белое моря
и некоторые районы, прилежащие к Грен-
ландии и к северо-восточной части Север-
ной Америки). И области и подобласти
делятся на более мелкие подразделе-
ния — провинции, округа и т. д.— по
степени сходства и различия населяющих
их фаун.
Вряд ли возможно для океанической и
наземной фауны создать единые зоогеогра-
фические подразделения, в силу того что
факторы среды, обусловливающие рас-
пределение морских и наземных живот-
ных, в значительной части различны, о
чем уже говорилось выше. Различно
также палеогеографическое и палеокли-
матологическое прошлое океанов и суши.
Так, Северная и Южная Америка до се-
редины третичного времени были разоб-
щены, а Атлантический и Тихий океаны
соединены широким проливом. В тре-
тичное время в широтном направлении в
тропическом поясе через Европу и Азию
тянулось огромное море Тетис, перекры-
вавшее и наши южнорусские моря. Иная
картина наблюдалась в северной части
Тихого океана. Еще недавно северовос-
точное окончание Азии было соедине-
но широкой связью с северо-западной
частью Северной Америки. Австралий-
ская фауна резко отлична от фауны Ев-
разии, и причиной этого был глубокий
и геологически длительно существовав-
ший, хотя и неширокий, пролив между
двумя островами Малого Зондского ар-
хипелага — Бали и Лембоком, кото-
рый оказался непреодолимым в течение
долгого геологического периода для выс-
ших млекопитающих Азии. Поэтому мле-
копитающие и остались в Австралии на
уровне развития сумчатых.
Зоогеографическое районирование оке-
ана усложняется двумя обстоятельства-
ми. Для пелагической и донной фауны зоо-
географические подразделения имеют
различный характер, так же как и для
различных вертикальных зон. Законо-
мерности горизонтального распределения
абиссальной фауны значительно отлича-
ются от закономерностей распределения
литоральной, сублиторальной и абис-
сальной фауны.
Для наземной фауны также различают
от 6 до 9 областей (карта 2): Палеаркти-
ческую (Европа, Средняя и Северная
Азия и северная часть Африки), соответ-
52
ствующую ей Неоарктическую (Северная
Америка). Палеарктическая и Неоаркти-
ческая области (объединяемые в Голарк-
тическую) простираются далеко на юг, до
20° с. ш., и делятся на 7 подобластей.
Экваториальный материковый пояс за-
нят Неотропической (Южная Америка),
Эфиопской (Средняя и Южная Африка),
Индо-Малайской (Юго-Западная Азия)
и Австралийской областями, в состав
которых входят 14 подобластей. Антар-
ктика занята единой Кругоантарктиче-
ской областью.
Одна из наиболее крупных закономер-
ностей в распределении современной фа-
уны, определяемой палеоклиматологи-
ческим прошлым,— это явление биполяр-
ности. В высоких широтах умеренных
зон обоих полушарий, в северобореальной
и южнонотальной частях этих областей,
наблюдается частичное сходство система-
тического состава и ряда биологических
явлений для наземной фауны и флоры и
особенно для морской. Очень хорошим
примером может быть географическое рас-
пространение сардин (рис. 14). Биполяр-
ные ареалы обычно имеют несколько со-
тен видов различных групп фауны и
флоры.
В настоящее время соединению ареа-
лов биполярных форм препятствует вы-
сокая температура тропической зоны, но
если допустить, что в ледниковое время
температура тропической зоны была не-
сколько ниже современной, то северные
формы могли проникать на юг, а южные —
на север. Последующее потепление разор-
вало их первоначально сплошные ареа-
лы. Такое объяснение и было дано акад.
Л. С. Бергом. Он же указывает, что,
помимо этого недавнего периода воз-
никновения биполярности, имеются и бо-
лее древние источники биполярности —
остатки ледниковых периодов более древ-
них геологических периодов.
Биполярность сказывается не только
на систематическом составе, но и на
биологических и экологических особен-
ностях: размерах тела, темпе роста,
жирности, явлениях живорождения, био-
ценотических связях и многом другом.
Рис. 14. Биполярное распространение сардин.
К явлениям биполярности тесно примы-
кают и поширотные различия в качест-
венном разнообразии. В тропической зо-
не видовое обилие значительно больше,
чем в высоких широтах: море Лаптевых
населяют 400 видов животных, Карское
море — 1200, Баренцево — 2500, Север-
ное море — 3—4 тыс., Средиземное мо-
ре— 6—7 тыс., а Малайский архипелаг—
около 40 тыс. видов. В тропической зоне
одних крабов насчитывается свыше 700
видов, а в арктической области — всего
несколько видов. Мадрепорових кораллов
в тропической зоне свыше 2500 видов, а в
Арктике — один вид. Моллюсков соответ-
ственно — 6 тыс. и 250 видов (для Ба-
ренцева моря). То же самое наблюдается
и в наземной фауне и флоре.
До соединения Южной и Северной Аме-
рики, в середине третичного периода, тро-
пическая фауна прилежащих частей Ат-
лантического и Тпхого океанов была еди-
ной.
После образования Панамского перешей-
ка, несмотря на значительное сходство ус-
ловий существования по обе стороны от
него, изолированные друг от друга фауны
пошли по самостоятельному пути разви-
тия, и за прошедшие 5—8 млн. лет в них
сформировалось около половины новых под-
водов и видов.
53
КЛЕТКА
Начиная знакомство с животным миром,
необходимо сначала в самых общих чер-
тах остановиться на строении и отправ-
лениях клетки.
Клетка представляет собой струк-
турную и функциональную единицу, ле-
жащую в основе строения и развития ор-
ганизмов х. В многоклеточном организме
форма строения клеток в соответствии с
выполняемыми ими функциями очень раз-
лична. Однако основные черты их организа-
ции свойственны как простейшим, так
и многоклеточным животным и растениям.
Каждая клетка состоит из цитоплаз-
мы и ядра. Оба эти компонента
представляют собой единую и неделимую
структурную и функциональную систему,
части которой не могут существовать раз-
дельно.
Цитоплазму часто обозначают другим
термином —п ротоплазмой. Однако
многие ученые в слово «протоплазма» вкла-
дывают иное содержание, обозначая им
все живые части клетки, в том числе и ядро.
Ввиду некоторой неопределенности терми-
на «протоплазма» в дальнейшем изложе-
нии д:ы не будем им пользоваться.
Наука, изучающая строение и отправ-
ление клеток, называется цитологией.
За последнее десятилетие она достигла
больших успехов, что в значительной мере
связано с разработкой новых методов ис-
следования клетки.
Основным «орудием» цитологии служит
микроскоп, позволяющий изучать строе-
ние клетки при увеличении в 2400—2500
раз. Клетки изучают в живом виде, а так-
же после специальной обработки. Послед-
няя сводится к двум основным этапам.
Сначала клетки фиксируют, т. е. убивают
их быстродействующими ядовитыми для
клеток веществами, не разрушающими
их структуры. Вторым этапом является
окраска препарата. Она основана на
том, что разные части клетки с разной
степенью интенсивности воспринимают
1 Не все организмы имеют клеточное строение.
Клеточная организация явилась результатом
длительной эволюции, которой предшествовали
неклеточные (доклеточные) формы жизни.
некоторые красители. Благодаря этому
удается отчетливо выявить различные
структурные компоненты клетки, которые
без окраски благодаря сходному коэф-
фициенту преломления не видны. Очень
часто применяют метод изготовления сре-
зов. Для этого ткани или отдельные клетки
после специальной обработки заключают
в твердую среду (парафин, целлоидин),
после чего при помощи особого прибора —
микротома, снабженного острой брит-
вой, раскладывают на тонкие срезы толщи-
ной от 3 микрон (микрон = 0,001 мм).
Фиксированные и окрашенные препараты
перед изучением заключают в среду с вы-
соким коэффициентом преломления (глице-
рин, канадский бальзам и др.). Благодаря
этому они становятся прозрачными, что
облегчает исследование препарата.
В современной цитологии разработан
ряд новых методов и приемов, примене-
ние которых чрезвычайно углубило зна-
ния о строении и физиологии клетки.
Очень большое значение для изучения
клетки имеет применение биохими-
ческих и цитохимических
методов.В настоящее время мы можем
не только изучать строение клетки, но и
определять ее химический состав и изме-
нения его в процессе жизнедеятельности
клетки. Многие из этих методов основаны
на применении цветных реакций, позво-
ляющих различать определенные химиче-
ские вещества или группы веществ. Изуче-
ние распределения разных по своему хи-
мическому составу веществ в клетке путем
цветных реакций представляет собой ци-
тохимический метод. Он имеет большое
значение для исследования обмена веществ
и других сторон физиологии клетки.
В современной цитологии широко при-
меняют ультрафиолетовую ми-
кроскопию. Ультрафиолетовые лу-
чи невидимы для человеческого глаза, но
воспринимаются фотографической пластин-
кой. Некоторые играющие особо важную
роль в жизни клетки органические веще-
ства (нуклеиновые кислоты) избирательно
поглощают ультрафиолетовые лучи. По-
этому по снимкам, изготовленным в ульт-
рафиолетовых лучах, можно судить о
распределении нуклеиновых веществ
в клетке.
54
Разработан ряд тонких методов, позво-
ляющих изучать проникновение разных
веществ в клетку из окружающей среды.
Для этого, в частности, применяют при-
жизненные (витальные) красители. Это
такие красящие вещества (например, ней-
тральный красный), которые проникают
в клетку, не убивая ее. Наблюдая за жи-
вой витально окрашенной клеткой, можно
судить о путях проникновения и накоп-
ления веществ в клетке.
Особенно большую роль в развитии ци-
тологии, а также в изучении тонкого строе-
ния простейших сыграла электронная
микроскопия.
Электронный микроскоп
основан на ином принципе, чем световой
оптический микроскоп. Объект изучают
в пучке быстро летящих электронов. Дли-
на волны электронных лучей во много ты-
сяч раз меньше длины волны световых лу-
чей. Это позволяет получить значительно
большую разрешающую способность, т. е.
гораздо большее увеличение, чем в све-
товом микроскопе. Пучок электронов про-
ходит сквозь изучаемый объект и затем
падает на флуоресцирующий экран, на
котором и проецируется изображение
объекта. Чтобы объект был проницаемым
для электронного пучка, он должен
быть очень тонким. Обычные микротом-
ные срезы толщиной в 3—5 мк для это-
го совершенно непригодны. Они пол-
ностью поглотят пучок электронов. Были
созданы особые приборы — ультрамикро-
томы, которые позволяют получать срезы
ничтожной толщины, порядка 100—300 анг-
стрем (ангстрем — единица длины, рав-
ная одной десятитысячной микрона). Раз-
личия в поглощении электронов разными
частями клетки настолько малы, что без
специальной обработки на экране элект-
ронного микроскопа они не могут быть
обнаружены. Поэтому изучаемые объекты
предварительно обрабатываются вещества-
ми, непроницаемыми или труднопроницае-
мыми для электронов. Таким веществом
является четырехокись осмия (Os04). Она
в различной степени поглощается разными
частями клетки, которые благодаря это-
му по-разному задерживают электроны.
Применяя электронный микроскоп, мож-
но получить увеличения порядка 100000.
Электронная микроскопия открывает но-
вые перспективы в изучении организации
клетки.
На рис. 15 и рис. 16 сопоставлена схема
строения клетки, как она представлялась
в двадцатых годах этого столетия и как
она представляется в настоящее время.
Снаружи клетка отграничена от окру-
жающей среды тонкой клеточной
мембраной, которая играет важную
роль в регуляции поступления веществ в
цитоплазму. Основное вещество цитоплаз-
мы имеет сложный химический состав.
Основу его составляют белки, которые
находятся в состоянии коллоидного рас-
твора. Белки — это сложные органи-
ческие вещества, обладающие крупными
молекулами (молекулярный вес их очень
высок, измеряется десятками тысяч по
отношению к атому водорода) и большой
химической подвижностью. Кроме белков,
в цитоплазме присутствуют и многие дру-
гие органические соединения (углево-
ды, жиры), среди которых особенно
55
большое значение в жизни клетки играют
сложные органические вещества — ну-
клеиновые кислоты. Из неорга-
нических составных частей цитоплазмы
следует прежде всего назвать воду, кото-
рая по весу составляет значительно боль-
ше половины всех веществ, входящих в
состав клетки. Вода важна как раство-
ритель, так как реакции обмена веществ
протекают в жидкой среде. Кроме того, в
клетке присутствуют ионы солей (Са2+,
К+, Na+, Fe2+, Fe3+ и др.).
В основном веществе цитоплазмы рас-
полагаются органоиды — постоян-
но присутствующие структуры, выполняю-
щие определенные функции в жизни клет-
ки. Среди них важную роль в обмене ве-
ществ играют митохондрии. В све-
товом микроскопе они видны в форме
небольших палочек, нитей, иногда гранул.
Электронный микроскоп показал, что
структура митохондрий очень сложна.Каж-
дая митохондрия имеет оболочку, состоя-
щую из трех слоев, и внутреннюю полость.
От оболочки в эту полость, заполненную
жидким содержимым, вдаются многочис-
ленные перегородки, не доходящие до про-
тивоположной стенки, называемые к р ис-
та м и. Цитофизиологические исследования
показали, что митохондрии являются орга-
ноидами, с которыми связаны дыхательные
процессы клетки (окислительные). Во внут-
ренней полости, на оболочке и кристах
локализуются дыхательные ферменты (ор-
ганические катализаторы), обеспечиваю-
щие сложные химические превращения,
из которых слагается процесс дыхания.
В цитоплазме,кроме митохондрий,имеет-
ся сложная система мембран,
образующая в совокупности э н д о -
плазматическую сеть (рис. 16).
Как показали электронномикроскопиче-
ские исследования, мембраны эндоплаз-
матической сети двойные. Со стороны, об-
ращенной к основному веществу цитоплаз-
мы, на каждой мембране расположены
многочисленные гранулы (называе-
мые «тельцами Паллада» по имени открыв-
шего их ученого). В состав этих гранул
входят нуклеиновые кислоты (а именно
рибонуклеиновая кислота), бла-
годаря чему их называют также рибо-
сомами. На эндоплазматической сети
при участии рибосом осуществляется один
из основных процессов жизнедеятельно-
сти клетки — синтез белков.
Часть цитоплазматических мембран ли-
шена рибосом и образует особую систему,
называемую аппаратом Гольджи.
Это образование обнаружено в клетках уже
довольно давно, ибо его удается выявить
особыми методами при исследовании в
световом микроскопе. Однако тонкая
структура аппарата Гольджи стала из-
вестна лишь в результате электронно-
микроскопических исследований. Функ-
циональное значение этого органоида сво-
дится к тому, что в области аппарата кон-
центрируются различные синтезируемые в
клетке вещества, например зерна секрета
в железистых клетках и т. п. Мембраны
аппарата Гольджи находятся в связи с
эндоплазматической сетью. Возможно, что
на мембранах аппарата Гольджи протекает
ряд синтетических процессов.
Эндоплазматическая сеть связана с на-
ружной оболочкой ядра. Эта связь играет,
по-видимому, существенную роль во взаи-
модействии ядра и цитоплазмы. Эндоплаз-
матическая сеть имеет также связь с на-
ружной мембраной клетки и местами не-
посредственно переходит в нее.
При помощи электронного микроскопа
в клетках был обнаружен еще один тип
органоидов — лизосомы (рис. 16).
По размерам и форме они напоминают
митохондрии, но легко отличаются от них
по отсутствию тонкой внутренней струк-
туры, столь характерной и типичной для
митохондрий. По представлениям боль-
шинства современных цитологов, в лизо-
сомах содержатся переваривающие фер-
менты, связанные с расщеплением круп-
ных молекул органических веществ, по-
ступающих в клетку. Это как бы резерву-
ары ферментов, постепенно используемых
в процессе жизнедеятельности клетки.
В цитоплазме животных клеток обычно
по соседству с ядром располагается цен-
тросома. Этот органоид имеет постоянную
структуру. Он слагается из девяти
ультрамикроскопических палочковидных
образований, заключенных в особо диф-
ференцированную уплотненную цитоплаз-
му. Центросома — органоид, связан-
ный с делением клетки (см. стр. 60).
56
Рис. 16. Схема строения клетки, по современным данным, с учетом электронномикроскопических
исследований:
1 — цитоплазма; 2 — аппарат Гольджи, з— центросома; 4 — митохондрии; 5 — эндоплазматическая сеть;
6 — ядро; 7 — ядрышко; 8 — лизосомы.
К роме перечисленных цитоплазматиче-
ских органоидов клетки, в ней могут при-
сутствовать различные специальные струк-
туры и включения, связанные с обменом ве-
ществ и выполнением различных специаль-
ных, свойственных данной клетке функ-
ций. В животных клетках обычно при-
сутствует гликоген, или живот-
ный крахмал. Это резервное веще-
ство, потребляемое в процессе обмена
веществ как основной материал для оки-
слительных процессов. Часто имеются жи-
ровые включения в форме мелких капель.
В специализированных клетках, таких,
как мышечные клетки, имеются особые
сократимые волоконца, связанные с со-
кратительной функцией этих клеток. Ряд
специальных органоидов и включений
имеется в растительных клетках. В зеле-
ных частях растений всегда присутствуют
хлоропласты — белковые тела, со-
держащие зеленый пигмент хлоро-
филл, при участии которого осуществ-
ляется фотосинтез — процесс воз-
душного питания растения. В качестве
резервного вещества здесь обычно находят-
ся крахмальные зерна, отсут-
ствующие у животных. В отличие от жи-
вотных, растительные клетки обладают,
кроме наружной мембраны, прочными о б о-
57
лочками из клетчат-
к и, что обусловливает особую
прочность растительных тка-
ней.
Клеточное ядро — вто-
рой основной компонент клет-
ки, образующий вместе с ци-
топлазмой единую структурную
и функциональную систему. Яд-
ро отграничено от цитоплазмы
оболочкой. Оболочка сос-
тоит из двух мембран, из кото-
рых наружная связана с эндо-
плазматической сетью цитоплаз-
мы. Электронномикроскопиче-
ские исследования показали, что
ядерная оболочка пронизана
многочисленными мельчайшими
порами диаметром около
600—700 ангстрем. Эти поры
играют, вероятно, важную роль
в обмене веществ между цито-
плазмой и ядром. Внутри ядра
имеется жидкий ядерный сок —
к а р иолимфа,в которой рас-
положены структурные компо-
ненты ядра. Количество карио-
лимфы в разных ядрах различ-
но. Иногда ее может быть очень
много, и ядро приобретает пу-
зырьковидное строение. В дру-
гих случаях кариолимфы мало
(пример — макронуклеусы ин-
фузорий, см. стр. 138), и ядро становит-
ся массивным и плотным. В кариолим-
фе растворены различные органические,
в том числе белковые, вещества. Основ-
ной структурный компонент ядра — хро-
матин (хромосом ы), который
представлен обычно беспорядочно рас-
положенными нитями и тяжами, не-
редко образующими скопления в виде
гранул. С биохимической стороны хрома-
тин состоит из особого содержащего
фосфор вещества — нуклеопротеи-
дов дезоксирибонуклеино-
вой кислоты (сокращенно ДНК).
Нуклеопротеид ы—это белки (про-
теиды), химически связанные с нуклеи-
новой кислотой ДНК. Роль хромосом
в жизни клетки чрезвычайно велика.
Об этом будет сказано ниже. Наконец,
в ядре всегда присутствует одно или
Рис. 17. Схема
строения части мо-
лекулы дезоксири-
бону клеи новой ки-
слоты по Ватсон-
Крику. Цифрами
указаны расстоя-
ния в ангстремах
между отдельны-
ми частями моле-
кулы.
несколько ядрышек ?нук-
леол), которые тоже в основном
состоят из нуклеопротеидов, но
уже другой, а именно ри-
бонуклеиновой кис-
лоты (РНК).
В настоящее время доказано,
что ДНК обладает замечатель-
ной способностью к самовос-
произведению (ауторепро-
дукции, репликации).
Строение молекулы ДНК очень
сложно. Она слагается из двух
закрученных спирально одна во-
круг другой нитей, в свою оче-
редь состоящих из повторяю-
щихся по продольной оси групп
молекул (нуклеотидов,
рис. 17). Каждый нуклеотид
включает в свой состав орга-
ническое основание (из групп
пуриновых и пиримидиновых
оснований). Связь между дву-
мя цепочками в молекуле ДНК
осуществляется при помощи
этих оснований, которые обра-
щены друг к другу. Обе нити
(цепи нуклеотид) относительно
слабо связаны между собой.
Эти связи могут нарушаться,
в результате чего нити раскру-
чиваются своими внутренними
сторонами наружу. Из окружаю-
щей кариолимфы к ним на освободившиеся
химические связи органических оснований
присоединяются нуклеотиды, точно такие же,
какие входили в состав второй цепи.
В результате этого замечательного про-
цесса из одной молекулы получается две,
точно такие же.
Одиночная нить ДНК является как
бы «матрицей» (формой), воспроизво-
дящей вторую нить. Этот процесс яв-
ляется молекулярной основой размно-
жения клетки, который мы рассмотрим
ниже.
Молекулы ДНК способны не только
воспроизводить себе подобные, но и уча-
ствовать через посредство РНК (в рибо-
сомах) в синтезе белков. За последние
годы этот сложный процесс в основных
своих чертах изучен. Сущность его сво-
дится к следующему.
58
На «матрице» ДНК из нуклеотидов ка-
риолимфы синтезируются молекулы РНК,
отличающиеся от ДНК по химическому
составу некоторых нуклеотидов, а именно
органических оснований.
Синтезированная в ядре РНК выхо-
дит в цитоплазму и локализуется на
рибосомах. Эта РНК в свою очередь слу-
жит «матрицей» для синтеза белков. Как
известно, в основе строения молекулы
белка лежат аминокислоты — органиче-
ские соединения, обладающие одновремен-
но кислотными и щелочными свойствами.
Основу белковой молекулы составляют
цепочки аминокислот, соединенные в стро-
го определенном порядке. Число входя-
щих в состав белков аминокислот неве-
лико, оно равняется всего 20. Количество
же различных белков поистине колос-
сально, ибо не только в каждом организме
имеется много разных белков, но и раз-
ные виды организмов обладают различ-
ными белками. Каждому виду животных
и растений свойственны свои специфиче-
ские для данного вида белки. В этом за-
ключается одно из замечательных биоло-
гических свойств организмов. Все это
разнообразие белков определяется после-
довательностью аминокислот в цепочке
белковой молекулы. Достаточно изменить
эту последовательность или на место од-
ной аминокислоты поставить другую, как
свойства белков меняются.
Вернемся к вопросу о синтезе белка
на «матрицах» РНК — на рибосомах.
Аминокислоты, находящиеся в цитоплаз-
ме, переносятся на цепочки РНК, где
и располагаются в строго определенном
порядке. Этот порядок определяется после-
довательностью нуклеотидов, вернее, орга-
нических оснований (известных в органи-
ческой химии под именем пуриновых и
пиримидиновых оснований). Каждая ами-
нокислота связывается тремя основания-
ми. Таким образом создается «триплет-
ный» (состоящий из трех расположенных
рядом оснований) код для каждой ами-
нокислоты. Последовательно «триплеты»
цепочки РНК определяют последователь-
ность аминокислот, а следовательно, и
первичную структуру синтезирующих бел-
ков (последовательность аминокислот).
Поскольку структура РНК, в которой
«закодирована» структура белка, в свою
очередь определяется химической струк-
турой ДНК хромосом ядра, то очевидно,
что именно в хромосомах «закодированы»
все свойства белков организма.
Разгадка механизма синтеза белка в
клетке представляет собой величайшее
открытие биологии за последнее десяти-
летие. Удалось поднять завесу над одной
из основных проблем науки о жизни —
проблемой образования белка. Ее реше-
ние осуществлено на основе тесного со-
трудничества двух биологических наук —
цитологии и биохимии.
Наряду с проблемой синтеза белка ос-
новным вопросом физиологии клетки яв-
ляется вопрос об источниках энергии.
Все те синтетические процессы, которые
мы только что рассмотрели, протекают
с затратой энергия. Энергия необходима
и для ряда других жизненных процессов,
в частности движения. В отношении клет-
ки справедливо будет утверждение, что
самосохранение ее сложной структуры
и основные жизненные функции осуществ-
ляются лишь благодаря непрерывному
потреблению энергии. Где же источники
этой энергии? В каких формах происхо-
дит превращение энергии в клетке? Основ-
ным источником энергии в клетке явля-
ются процессы, связанные в конечном
счете с окислением углеводов и других
органических веществ вплоть до образо-
вания углекислого газа и воды. Наряду
с окислением источником энергии могут
являться и расщепительные анаэробные
процессы, ведущие к образованию из бо-
лее сложных органических соединений
(например, глюкозы) менее сложных (на-
пример, молочной кислоты). Материаль-
ным субстратом, на котором протекают
окислительные процессы, служат мито-
хондрии. Однако нужно подчеркнуть, что
окислительные процессы в клетке с хими-
ческой стороны протекают совсем не так,
как горение органических тел на воздухе,
хотя и в том и в другом случае конечные
продукты одинаковы — углекислый газ и
вода. Окисление в клетке протекает при
температурах относительно низких, при
которых вне организма органические ве-
щества не окисляются. Процессы, проте-
кающие в живой клетке, обусловлены
59
наличием сложной системы ферментов
(органических катализаторов), каждый из
которых обусловливает строго определен-
ную реакцию. Процесс этот очень сложен
и состоит из ряда этапов. Во время пер-
вого этапа углевод (глюкоза), обладаю-
щий шестью атомами углерода, дает нача-
ло двум трехуглеродным молекулам мо-
лочной кислоты. На втором этапе, сла-
гающемся из множества промежуточных
звеньев, рассматривать которые мы не
имеем здесь возможности (эта серия по-
следовательных превращений известна под
названием цикла Кребса), часть вещества
дает начало углекислому газу и воде.
Все эти превращения связаны с освобож-
дением энергии. Одной из самых замеча-
тельных сторон этого процесса является
связь окисления с фосфорилированием.
Под фосфорилированием по-
нимают процесс образования молекул осо-
бого органического вещества — а д е н о-
зинтрифосфата (сокращенно —
АТФ), в состав молекулы которого вхо-
дят три фосфатные группы (фосфатная
группа — это атом фосфора, соединенный
с атомами кислорода и гидроксильной
группой). Две фосфатные группы АТФ
соединены с остальной частью молекулы
особой химической связью, носящей наз-
вание макроэргической, что
означает «богатая энергией». Один из фос-
фатных остатков может легко отделяться
от АТФ, и, высвобождая энергию, АТФ
при этом переходит в аденозиндифосфат
(АДФ), который гораздо беднее химиче-
ской энергией, чем АТФ. За счет энергии,
освобождаемой при окислении, АДФ
вновь может подвергаться фосфорилиро-
ванию, в результате чего происходит при-
соединение фосфатной группы и восста-
навливается макроэргическая связь.
АТФ, таким образом, можно рассматри-
вать как богатую энергией «заряженную»
форму, тогда как АДФ — как «разряжен-
ную», относительно более бедную энерги-
ей форму аденозинфосфата. АТФ в клетке
является своеобразным химическим акку-
мулятором энергии, за счет которого
выполняется самая различная работа:
механическая, химическая (в том числе
синтез белков), осмотическая, электриче-
ская.
Мы видим, таким образом, что в живот-
ных клетках в ходе сложной цепи превра-
щений, из которых слагается дыхание,
энергия, заключенная в питательных ве-
ществах, в результате окисления расхо-
дуется на построение АТФ из АДФ. Если
этим процессам дать количественное выра-
жение, то оказывается, что немного бо-
лее половины высвобождаемой при окис-
лении энергии идет на фосфорилирование,
на «зарядку» аденозинфосфатов — этих
замечательных органических аккумуля-
торов энергии.
Хорошо известно, что животный орга-
низм не способен создавать органические
вещества из неорганических. В качестве
пищи животное получает органические
вещества, о судьбе которых в клетке мы
только что говорили. Лишь зеленое расте-
ние при помощи хлорофилла создает орга-
нические вещества из неорганических в
результате как воздушного, так и мине-
рального питания из почвы. Эти процессы
требуют затраты энергии. Источник ее —
солнечный луч. Энергия солнца перехо-
дит в потенциальное состояние в форме
органических соединений, создаваемых
растением. Эта энергия с пищей передает-
ся животному, где она претерпевает ряд
превращений, в которых особо важная
роль принадлежит АТФ — органическо-
му аккумулятору энергии в клетке.
Одним из основных биологических про-
цессов, обеспечивающих преемственность
форм жизни и лежащих в основе всех
форм размножения, является процесс де-
ления клетки. Этот процесс, известный
под названием кариокинеза, или
митоза, с удивительным постоянством,
лишь с некоторыми вариациями в дета-
лях, осуществляется в клетках всех расте-
ний и животных, в том числе и простей-
ших. При митозе происходит равномер-
ное распределение хромосом, претерпева-
ющих удвоение между дочерними клет-
ками. От любого участка каждой хромо-
сомы дочерние клетки получают полови-
ну. Не вдаваясь в детальное описание
митоза, отметим лишь его основные мо-
менты (рис. 18).
В первой стадии митоза, называемой
профазой, в ядре становятся отчет-
ливо видимыми хромосомы в форме нитей
60
:::$&*^
':%!~*.
А
W
• "..•**/ i \ "-¦
10
11
12
Рис. 18. Схема митотического деления клетки:
1 — неделящееся ядро; 2—6 — последовательные этапы изменения ядра в профазе; 7—9 — метафаза;
Ю — анафаза; 11—13 — телофаза.
разной длины. В неделящемся ядре, как
мы видели, хромосомы имеют вид тонких,
неправильно расположенных нитей, пере-
плетающихся друг с другом. В профазе
происходит их укорачивание и утолще-
ние. Вместе с тем каждая хромосома ока-
зывается двойной. По длине ее проходит
щель, разделяющая хромосому на две
рядом лежащие и совершенно подобные
друг другу половины.
На следующей стадии митоза — мета-
фазе — оболочка ядра разрушается,
ядрышки растворяются и хромосомы ока-
зываются лежащими в цитоплазме. Все
хромосомы располагаются при этом в один
ряд, образуя так называемую эквато-
риальную пластинку. Суще-
ственные изменения претерпевает цент-
росома. Она делится на две части,
которые расходятся, и между ними обра-
зуются нити, формирующие а х р о м а-
тиновое веретено. Экваториаль-
ная пластинка хромосом располагается
по экватору этого веретена.
На стадии анафазы происходит
процесс расхождения к противоположным
61
полюсам дочерних хромосом, образовав-
шихся, как мы видели, в результате про-
дольного расщепления материнских хро-
мосом. Расходящиеся в анафазе хромо-
сомы скользят по нитям ахроматинового
веретена и в конце концов собираются
двумя группами в области центросом.
Во время последней стадии митоза —
телофазы — происходит восстанов-
ление структуры неделящегося ядра. Во-
круг каждой группы хромосом образует-
ся ядерная оболочка. Хромосомы вытя-
гиваются и утончаются, превращаясь в
длинные, беспорядочно расположенные
тонкие нити. Выделяется ядерный сок,
в котором появляется ядрышко.
Одновременно со стадиями анафазы и
телофазы происходит разделение на две
половины цитоплазмы клетки, которое
осуществляется обычно путем простой
перетяжки.
Как видно из нашего краткого описа-
ния, процесс митоза сводится в первую
очередь к правильному распределению
хромосом между дочерними ядрами. Хро-
мосомы состоят из пучков нитевидных
молекул ДНК, расположенных по про-
дольной оси хромосомы. Видимому нача-
лу митоза предшествует, как это теперь
установлено точными количественными из-
мерениями, удвоение ДНК, молекуляр-
ный механизм которого мы уже рассмот-
рели выше.
Таким образом, митоз и расщепление
хромосом во время него является лишь
видимым выражением процессов удвое-
ния (ауторепродукции) молекул
ДНК, осуществляемого на уровне моле-
кул. ДНК определяет через посредст-
во РНК белковый синтез. Качествен-
ные особенности белков «закодированы»
в структуре ДНК. Поэтому очевидно,
что точное разделение хромосом в митозе,
базирующееся на редупликации (ауто-
репродукции) молекул ДНК, лежит в ос-
нове «наследственной информации» в ряде
следующих друг за другом поколений кле-
ток и организмов.
Число хромосом, так же как их форма,
размеры и т. п., является характерным
признаком каждого вида организмов.
У человека, например, имеется 46 хромо-
сом, у окуня — 28, у мягких пшениц —
42 и т. п.
Простейшие
Protozoa
ТИП ПРОСТЕЙШИЕ (PROTOZOA)
ОБЩАЯ ХАРАКТЕРИСТИКА
Первый тип, с которого мы начинаем
знакомство с миром животных,— это
тип простейших (Protozoa). Он состоит
из многих классов, отрядов, семейств и
включает примерно 20—25 тыс. видов.
Простейшие распространены на всей
поверхности нашей планеты и живут в са-
мых различных средах. В большом коли-
честве мы найдем их в морях и океанах,
как непосредственно в толще морской
воды, так и на дне. Обильны простейшие
в пресных водах. Некоторые виды живут
в почве.
Многие простейшие в качестве среды
обитания используют другие организмы,
ведут паразитический образ жизни.
Среди паразитических простейших име-
ются возбудители тяжелых заболева-
ний человека, домашних и промысловых
животных.
По своему строению простейшие чрез-
вычайно разнообразны. Подавляющее
большинство их обладает микроскопиче-
ски малыми размерами, для изучения их
приходится пользоваться микроскопом.
Каковы же общие признаки типа про-
стейших? На основании каких особенно-
стей строения и физиологии мы причис-
ляем животных к этому типу? Основной
и самой характерной чертой простей-
ших является их одноклеточность. Про-
стейшие являются организмами, тело
которых по строению соответствует одной
клетке.
Все другие животные (а также и расте-
ния) тоже состоят из клеток и их производ-
ных. Однако, в отличие от простейших,
в состав тела их входит большое количе-
ство клеток, различных по строению и
выполняющих в сложном организме раз-
ные функции. По этому признаку все
остальные животные могут быть противо-
поставлены простейшим и отнесены к
многоклеточным (Metazoa).
Сходные по строению и функции клетки
их слагаются в комплексы, называемые
тканями. Органы многоклеточных со-
стоят из тканей. Различают, например,
покровную (эпителиальную)
ткань, мышечную ткань, нерв-
ную ткань и др.
Если по строению своему простейшие
соответствуют клеткам многоклеточных
организмов, то в функциональном отно-
шении они несравнимы с ними. Клетка
в теле многоклеточного всегда представ-
ляет собой только часть организма, ее
отправления подчинены функциям мно-
гоклеточного организма как целого. На-
против, простейшее — это самостоятель-
ный организм, которому свойственны все
жизненные функции: обмен веществ, раз-
дражимость, движение, размножение.
К окружающим условиям внешней сре-
ды простейшее приспосабливается как
^ 5 Жизнь животных, т. i
65
целый организм. Следовательно, можно
сказать, что простейшее — это самостоя-
тельный организм на клеточном уровне
организации.
Размеры тела подавляющего большин-
ства простейших микроскопические. Наи-
более мелкие представители типа имеют
2—4 микрона * в поперечнике (например,
паразитические жгутиконосцы из рода
лейшманий — Leichmania, разные виды
семейства пироплазмид, паразити-
рующие в красных кровяных клетках
млекопитающих).
Наиболее обычные размеры простей-
ших — в пределах 50—150 мк. Но среди
них имеются и гораздо более крупные
организмы.
Инфузории Bursaria, Spirostomum до-
стигают 1,5 мм длины—их хорошо видно
простым глазом, грегарины Porospora
gigantea — длины до 1 см.
У некоторых корненожек фораминифер
раковина достигает 5—6 см в диаметре
(например, виды рода Psammonix, ис-
копаемые нуммулиты и др.).
Низшие представители простейших (на-
пример, амебы) не обладают постоянной
формой тела. Их полужидкая цитоплазма
постоянно меняет свои очертания благо-
даря образованию разнообразных выро-
стов — ложных ножек (рис. 24),
служащих для движения и захвата пищи.
Большинство же простейших обладает
относительно постоянной формой тела,
которая обусловлена наличием опорных
структур. Среди них наиболее обычной
является плотная эластичная м е м-
брана (оболочка), образуемая
периферическим слоем цитоплазмы
(эктоплазмой) и носящая назва-
ние пелликулы.
В одних случаях пелликула относи-
тельно тонка и не препятствует некото-
рому изменению формы тела простейшего,
как это имеет место, например, у способ-
ных сокращаться инфузорий (см. стр. 141).
У других простейших она образует проч-
ный и не меняющий своей формы наруж-
ный панцирь.
У многих жгутиконосцев, окрашенных
в зеленый цвет благодаря наличию хлоро-
Микрон (мк) — ©дна тысячная миллиметра.
филла, имеется наружная оболочка из
клетчатки — признак, характерный
для растительных клеток.
Что касается общего плана строения
и элементов симметрии, то простейшие
обнаруживают большое разнообразие. Та-
кие животные, как амебы, не обладающие
постоянной формой тела, не имеют по-
стоянных элементов симметрии.
Широко распространены среди Proto-
zoa разные формы радиальной
симметрии, свойственной главным обра-
зом планктонным формам (многие радио-
лярии, солнечники, стр. 88—94).* При
этом имеется один центр симметрии, от
которого отходит различное число пере-
секающихся в центре осей симметрии,
определяющих расположение частей тела
простейшего.
У многих радиально построенных форм
можно выделить одну главную ось, опре-
деляющую передний и задний концы
тела, вокруг которой радиально распо-
лагаются части тела простейшего (неко-
торые радиолярии, табл. 2, 3, инфузории
Didinium, стр. 147).
Относительно редко встречается у про-
стейших двубоковая (билате-
ральная) симметрия, при ко-
торой можно провести одну-единственную
плоскость симметрии, делящую тело жи-
вотного на две равные зеркальные поло-
вины (раковины некоторых фораминифер,
рис. 32, 33, радиолярии, табл. 2 и 3,
некоторые виды жгутиконосцев, например
лямблия, рис. 57). Большинство про-
стейших из разных классов являются
асимметричными.
У сложно организованных простейших
из класса инфузорий и у некоторых жгу-
тиконосцев, кроме пелликулы, имеются
еще и другие опорные структуры, поддер-
живающие и определяющие форму тела.
К ним относятся тончайшие волоконца
(фибрилл ы), проходящие в различ-
ных направлениях. Примером могут слу-
жить опорные волоконца одной из инфу-
зорий.
На рисунке 19 видно, какой боль-
шой сложности может достигать эта систе-
ма, образующая прочный и эластичный
каркас, поддерживающий полужидкую
цитоплазму простейшего.
66
К числу опорных и вместе с тем защит-
ных образований у простейших относятся
различные формы минерального
скелета, свойственного преимущест-
венно многим представителям класса
саркодовых. Эти скелетные образования
чаще всего имеют форму раковинок, иног-
да очень сложно устроенных (в отряде
фораминифер). В других случаях основу
скелета составляют отдельные иглы
(с п и к у л ы), обычно соединяющиеся
между собой, По химическому составу
минеральный скелет простейших разли-
чен. Наиболее обычными компонентами
его являются углекислый кальций
(СаС03) или окись кремния (Si02). Более
подробно строение скелета будет рассмот-
рено при знакомстве с отдельными клас-
сами простейших.
Большинству простейших, за исключе-
нием некоторых паразитических форм,
свойственна способность к движению —
перемещению в пространстве. Формы дви-
жения простейших разнообразны. Наибо-
лее простой и, вероятно, исходной фор-
мой является амебоидное дви-
жение. Оно выражается в образова-
нии ложных ножек (псевдо-
подий) — выростов цитоплазмы разной
формы. Все содержимое клетки как бы
медленно перетекает в направлении обра-
зующейся псевдоподии, и таким путем
осуществляется перемещение простейшего
в пространстве. Эта форма движения пре-
имущественно свойственна представите-
лям класса саркодовых. Разным видам
свойственна различная форма псевдопо-
дий (рис. 26).
Более сложной формой является дви-
жение, осуществляемое при помощи жгу-
тиков и ресничек. Жгутиковая
форма движения характерна для к л а с-
с а жгутиконосцев.
Жгутики представляют собой тон-
чайшие выросты тела. Количество их
у разных видов различно — от одного до
многих десятков и даже сотен (рис. 40, 63).
Каждый жгутик берет начало от не-
большого базального зернышка, называемо-
го блефаропластом и расположен-
ного в цитоплазме. Таким образом, непо-
средственно граничащая с базальным зер-
ном часть жгутика проходит внутри цито-
Рис. 19. Опорные (скелетные) фиб-
риллы и ресничный аппарат круго-
ресничной инфузории Trichodina.
плазмы (она носит название корневой
нити), а затем проходит через пелли-
кулу наружу. Механизм жгутикового дви-
жения у разных видов различен. В боль-
шинстве случаев он сводится к вращатель-
ному движению. Жгутик описывает фигу-
ру конуса, вершиной обращенного к ме-
сту его прикрепления. Наибольший меха-
нический эффект достигается, когда угол,
образуемый вершиной конуса, составляет
40—46°. Быстрота движения различна,
она колеблется у разных видов между 10
и 40 оборотами в секунду. Простейшее
как бы «ввинчивается» в окружающую его
жидкую среду.
Нередко вращательное движение жгу-
тика сочетается с его волнообразным дви-
жением. Обычно при поступательном дви-
жении само тело простейшего вращается
вокруг продольной оси.
Изложенная схема справедлива для
большинства одножгутиковых форм.
У многожгутиковых движение жгутиков
может носить иной характер, в частности
жгутики могут находиться в одной плос-
кости, не образуя конуса вращения.
&
67
Рис. 20. Поперечный разрез жгутиков Pseu-
dotrichonympha (рисунок сделан с электронно-
микроскопической фотографии). Видны в каж-
дом жгутике девять периферических двойных
фибрилл и центральная пара фибрилл.
Электронномикроскопические исследо-
вания последних лет показали, что внут-
ренняя ультрамикроскопическая струк-
тура жгутиков весьма сложна. Снаружи
жгутик окружен тонкой мембраной, кото-
рая является непосредственным продол-
жением самого поверхностного слоя экто-
плазмы — пелликулы. Внутренняя полость
жгутика заполнена цитоплазматическим
содержимым. По продольной оси жгутика
проходит одиннадцать тончайших нитей
(фибр и л л), которые нередко являются
двойными (рис. 20). Эти фибриллы рас-
полагаются всегда закономерно. Девять
из них (простых или двойных) лежат по
периферии, образуя в совокупности как
бы цилиндр. Две фибриллы занимают
центральное положение. Чтобы составить
себе представление о размерах всех этих
образований, достаточно сказать, что диа-
метр периферических фибрилл составляет
около 350А (ангстрем). Ангстрем —
единица длины, равная 0,0001 мк, а мик-
рон равняется 0,001 мм. Вот какие нич-
тожные по своим размерам структуры ста-
ли доступными для изучения благодаря
внедрению в микроскопическую технику
электронного микроскопа.
Функциональное значение фибрилл жгу-
тиков не может считаться окончательно
выясненным. По-видимому, часть их (ве-
роятно, периферические) играет активную
роль в двигательной функции жгутика
и содержит особые белковые молекулы,
способные сокращаться, другие же явля-
ются опорными эластическими структу-
рами, имеющими поддерживающее зна-
чение.
Реснички служат органоидами
движения инфузорий. Обычно число их
у каждой особи очень велико и измеря-
ется несколькими сотнями, тысячами и да-
же десятками тысяч. Механизм движения
ресничек несколько иной, чем жгутиков.
Каждая ресничка совершает гребные дви-
жения. Она быстро и с силой сгибается
в одну сторону, а затем медленно выпрям-
ляется.
Совместное действие большого числа
ресничек, биение которых координиро-
вано, вызывает быстрое поступательное
движение простейшего.
Каждая ресничка инфузории, как пока-
зали новейшие исследования, является
сложным образованием, по своему строе-
нию соответствующим жгутику. У осно-
вания каждой реснички всегда распо-
лагается так называемое базальное
зерно (иначе, кинетозома)-
важная часть ресничного аппарата
(см. стр. 135).
У многих инфузорий отдельные реснич-
ки соединяются друг с другом, образуя
структуры более сложного строения
(мембранеллы, цирры и др.)
и более эффективного механического дей-
ствия.
Некоторым высокоорганизованным про-
стейшим (инфузориям, радиоляриям) свой-
ственна еще одна форма движения —
сокращение. Тело таких простей-
ших способно быстро менять свою форму,
а затем вновь возвращаться к исходному
состоянию (стр. 142).
Способность к быстрому сокращению
обусловлена наличием в теле простейшего
особых волоконец — мионем — обра-
зований, аналогичных мышцам многокле-
точных животных.
У некоторых простейших существуют
еще и другие формы движения (см. стр. 118).
По способам и характеру питания, по
типу обмена веществ простейшие обнару-
живают большое разнообразие.
В классе жгутиконосцев имеются
организмы, способные подобно зеленым
растениям при участии зеленого пигмента
68
хлорофилла усваивать неорганические
вещества — углекислый газ и воду, пре-
вращая их в органические соединения
(аутотрофный тип обмена). Этот
процесс фотосинтеза протекает с поглоще-
нием энергии. Источником последней яв-
ляется лучистая энергия — солнечный
луч.
Таким образом, эти простейшие орга-
низмы правильнее всего рассматривать
как одноклеточные водоросли. Но наряду
сними в пределах того же класса жгутико-
носцев имеются бесцветные (лишенные
хлорофилла) организмы, неспособные к фо-
тосинтезу и обладающие гетеротроф-
ным (животным) типом обмена веществ,
т. е. питающиеся за счет готовых органи-
ческих веществ. Способы животного пита-
ния простейших, так же как и характер
пищи их, очень разнообразны. Наиболее
просто устроенные простейшие не обла-
дают специальными органоидами захвата
пищи. У амеб, например, псевдоподии
служат не только для движения, но вме-
сте с тем и для захвата оформленных ча-
стиц пищи (стр. 75). У инфузорий для
захвата пищи служит ротовое отверстие.
С последним обычно связаны разнообраз-
ные структуры — околоротовые мерца-
тельные перепонки (мембранеллы), спо-
собствующие направлению пищевых ча-
стиц к ротовому отверстию и далее в осо-
бую трубку, ведущую в эндоплазму —
клеточную глотку.
Пища простейших очень разнообразна.
Одни питаются мельчайшими организма-
ми, например бактериями, другие —
одноклеточными водорослями, некоторые
являются хищниками, пожирающими
других простейших, и т. п. Неперева-
ренные остатки пищи выбрасываются на-
ружу — у саркодовых на любом участке
тела, у инфузорий через особое отверстие
в пелликуле.
У пресноводных и у части морских
простейших имеется особый органоид, свя-
занный с регуляцией осмотического дав-
ления в клетке простейшего и с выделе-
нием. Это сократительная ва-
куоля. Она представляет собой перио-
дически появляющийся в цитоплазме пу-
зырек, наполняющийся жидкостью и опо-
рожняющийся наружу. Сократительная
вакуоля регулирует количество воды в ци-
топлазме, поступающей из окружающей
среды благодаря разности осмотических
давлений. У паразитических простейших
и у многих морских форм, живущих в сре-
де с повышенным осмотическим давлением,
сократительные вакуоли отсутствуют.
Особых органоидов дыхания у простей-
ших нет, они поглощают кислород и выде-
ляют углекислоту всей поверхностью
тела.
Как и все живые существа, простейшие
обладают раздражимостью, т. е.
способностью отвечать той или иной реак-
цией на факторы, действующие извне.
Простейшие реагируют на механические,
химические, термические, световые, элект-
рические и иные раздражения. Реакции
простейших на внешние раздражения ча-
сто выражаются в изменении направления
движения и носят название таксис.
Таксисы могут быть положительными,
если движение осуществляется в направ-
лении раздражителя, и отрицательными,
если оно осуществляется в противополож-
ную сторону.
Реакции многоклеточных животных на
раздражения осуществляются под воз-
действием нервной системы. Многие иссле-
дователи пытались обнаружить и у прос-
тейших (т. е. в пределах клетки) аналоги
нервной системы. Американские ученые,
например, описывали у многих инфузо-
рий наличие особого нервного цен-
тра (так называемого моториума),
представляющего собой особый уплотнен-
ный участок цитоплазмы. От этого центра
к различным участкам тела пнфузории
отходит система тонких волоконец, кото-
рые рассматривались как проводники нер-
вных импульсов. Другие исследователи,
применяя особые методы серебрения пре-
паратов (обработка азотнокислым сереб-
ром с последующим восстановлением ме-
таллического серебра), обнаружили в эк-
топлазме инфузорий сеть тончайших воло-
конец. Эти структуры (рис. 21) также рас-
сматривались как нервные элементы, по
которым распространяется волна возбуж-
дения. В настоящее время, однако, боль-
шинство ученых, изучающих тонкие фиб-
риллярные структуры, придерживаются
иного мнения об их функциональной роли
69
в клетке простейшего. Эксперименталь-
ных доказательств нервной роли фибрил-
лярных структур не получено. Напротив,
имеются опытные данные, которые дают
возможность предполагать, что у простей-
ших волна возбуждения распространяется
непосредственно по наружному слою ци-
топлазмы — эктоплазме. Что же касается
различного рода фибриллярных структур,
которые еще недавно рассматривались как
«нервная система» простейших, то они
имеют скорее всего опорное (скелетное)
значение и способствуют сохранению фор-
мы телл простейшего.
Как и всякая клетка, простейшие имеют
ядро. Выше, при рассмотрении строе-
ния клетки, мы уже ознакомились с основ-
ными структурными компонентами ядра.
В ядрах простейших, так же как и в яд-
рах многоклеточных, имеется оболоч-
ка, ядерный сок (кариолимфа),
хроматин (хромосомы) и я д р ы ш-
к и. Однако по размерам и строению ядра
разные простейшие весьма разнообразны
(рис. 22). Эти различия обусловлены соот-
ношением структурных компонентов ядра:
количеством ядерного сока, количеством
Рис. 21. Инфузория Urocentrum turbo. Базаль-
ные зерна ресничек и система эктоплазматиче-
ских волоконец (фибрилл), выявляемые методом
импрегнации серебром.
и размерами ядрышек (н у к л е о л), сте-
пенью сохранения строения хромосом
в интерфазном ядре и т. п.
У большинства простейших имеется од-
но ядро. Однако встречаются и много-
ядерные виды простейших.
У некоторых простейших, а именно
у инфузорий и немногих корненожек —
фораминифер, наблюдается интересное яв-
ление дуализма (двойственности)
ядерного аппарата. Оно сводится к тому,
что в теле простейшего имеются два ядра
двух категорий, различающиеся как по
своему строению, так и по физиологичес-
кой роли в клетке. У инфузорий, напри-
мер, имеется два типа ядер: большое, бо-
гатое хроматином ядро — макронук-
леус и маленькое ядро — микро-
нуклеус. Первое связано с выполне-
нием вегетативных функций в клетке,
второе—с половым процессом (см. стр. 138).
Простейшим, как и всем организмам,
свойственно размножение. Суще-
ствуют две основные формы размножения
простейших: бесполое и поло-
вое. В основе того и другого лежит
процесс деления клетки (стр. 60).
При бесполом размноже-
нии число особей возрастает в резуль-
тате деления. Например, амеба при беспо-
лом размножении делится на две амебы
путем перетяжки тела. Процесс этот начи-
нается с ядра, а затем захватывает цито-
плазму. Иногда бесполое размножение
приобретает характер множественного де-
ления. При этом ядро предварительно
делится несколько раз и простейшее ста-
новится многоядерным. Вслед за этим ци-
топлазма распадается на число отдельно-
стей, соответствующих количеству ядер.
В результате организм простейшего сра-
зу дает начало значительному количеству
мелких особей. Так происходит, напри-
мер, бесполое размножение малярийного
плазмодия — возбудителя малярии чело-
века (стр. 127).
Половое размножение про-
стейших характеризуется тем, что собст-
венно размножению (увеличению числа
особей) предшествует половой процесс,
характерным признаком которого явля-
ется слияние двух половых клеток (г а-
м е т) или двух половых ядер, ведущее
70
к образованию одной клетки — з и г о-
т ы, дающей начало новому поколению.
Формы полового процесса и полового раз-
множения у простейших в высшей степени
разнообразны. Основные формы его будут
рассмотрены при изучении отдельных
классов.
Многие виды простейших имеют не од-
ну, а несколько форм размножения, кото-
рые могут закономерно чередоваться друг
с другом. В результате получается слож-
ный цикл развития, отдельные этапы кото-
рого могут протекать в разных условиях
среды. Особенной сложностью отличаются
циклы развития у паразитических про-
стейших в классе споровиков (стр. 116).
Простейшие обитают в самых раз-
личных условиях среды. Большинство
их — водные организмы, широко распро-
страненные как в пресных, так и в мор-
ских водоемах. Многие виды их живут
в придонных слоях и входят в состав бен-
тоса. Большой интерес представляет при-
способление простейших к жизни в толще
песка, в толще воды (планктон).
Небольшое число видов Protozoa при-
способилось к жизни в почве. Их средой
обитания являются тончайшие пленки
воды, окружающие почвенные частицы и
заполняющие капиллярные просветы в
почве. Интересно отметить, что даже в пе-
сках пустыни Каракум живут простей-
шие. Дело в том, что под самым верхним
слоем песка здесь расположен влажный
слой, пропитанный водой, приближаю-
щейся по своему составу к морской воде.
В этом влажном слое и были обнаружены
живые простейшие из отряда форамини-
фер (стр. 81), являющиеся, по-видимому,
остатками морской фауны, населявшей
моря, ранее находившиеся на месте
современной пустыни. Эта своеобразная
реликтовая фауна в песках Каракумов
впервые была обнаружена проф.
Л. Л. Бродским при изучении воды,
взятой из колодцев пустыни.
Очень многие простейшие, относящиеся
к различным классам этого типа, ведут
паразитический образ жизни. Их среда
обитания и источники пищи — другие
живые организмы. Сыше 3,5 тыс. видов
простейших — паразиты. Хозяевами их
являются самые различные животные и ра-
Рис. 22. Ядра различных простейших:
А — микронуклеус инфузории Paramecium aurelia;
Б — жгутиконосец Trypanosoma brucei, В — амеба
Amoeba sphaeronuclei; Г — панцирный жгутиконосец
Geratium fuscus; Д — радиолярия Aulacantha scoly-
mantha; E — макрогамета кокцидий Aggregata eberthi;
Ж — макронуклеус инфузории Epidinium ecaudatum.
стения. Многие паразитические простей-
шие живут в организме человека, домаш-
них и промысловых животных, нанося
им большой вред. Один из классов типа
простейших — споровики — целиком со-
стоит из паразитов.
Практическое значение простейших для
человека велико. В особенности это отно-
сится к паразитам. До настоящего времени
в тропических зонах земного шара широ-
ко распространена малярия — тяжелое за-
болевание, поражающее ежегодно десятки
миллионов человек в Индии и других
тропических частях Азии, в Африке и дру-
гих странах. Возбудитель этого заболе-
вания относится к классу спорови-
ков типа простейших (стр. 124). Тяже-
лым заболеванием человека в Централь-
ной Африке является сонная болезнь,
вызываемая паразитом из к л а с с а жгу-
тиконосцев (стр. 107). Большой ущерб
наносят животноводству заболевания до-
71
машних животных, вызываемые простей-
шими. Сюда относятся различные пиро-
плазмозы, кокцидиозы, трипанозомозы и
многие другие. Важнейшие из них в прак-
тическом отношении мы рассмотрим ниже.
Имеется ряд простейших паразитов из
отряда кощидий (стр. 120), поражаю-
щих домашнюю птицу, в особенности кур.
Борьба с этими многочисленными и опас-
ными протозойными заболеваниями тре-
бует детального изучения биологии воз-
будителей, их циклов развития.
Некоторый практический интерес пред-
ставляют и свободноживущие простей-
шие. Разные виды их приурочены к опре-
деленному комплексу внешних условий,
в частности к различному химическому
составу воды.
Определенные виды простейших живут
при разной степени загрязненности пре-
сных вод органическими веществами. По-
этому по видовому составу простейших
можно судить о свойствах воды водое-
ма. Эти особенности простейших исполь-
зуют для санитарно-гигиенических целей
при так называемом биологическом ана-
лизе воды.
В общем круговороте веществ в природе
простейшие играют заметвую роль. В во-
доемах многие из них являются энергич-
ными пожирателями бактерий и других
микроорганизмов. Вместе с тем сами они
служат пищей для более крупных живот-
ных организмов. В частности, выклевы-
вающиеся из икринок мальки многих
видов рыб на самых начальных этапах
своей жизни питаются преимущественно
простейшими.
Тип простейших в геологическом от-
ношении является весьма древним. В ис-
копаемом состоянии хорошо сохранились
те виды простейших, которые обладали
минеральным скелетом (фораминиферы,
радиолярии). Ископаемые остатки их из-
вестны начиная с самых древних нижне-
кембрийских отложений.
Морские простейшие — корненожки и
радиолярии — играли и играют весьма
существенную роль в образовании мор-
ских осадочных пород. В течение многих
миллионов и десятков миллионов лет мик-
роскопически мелкие минеральные ске-
леты простейших после отмирания живот-
ных опускались на дно, образуя здесь
мощные морские отложения. При измене-
нии рельефа земной коры, при горнообра-
зовательных процессах в прошлые геоло-
гические эпохи, морское дно становилось
сушей. Морские осадки превращались
в осадочные горные породы. Многие из
них, как, например, некоторые извест-
няки, меловые отложения и др., в значи-
тельной своей части состоят из остатков
скелетов морских простейших. В силу
этого изучение палеонтологических остат-
ков простейших играет большую роль
в определении возраста разных слоев
земной коры и, следовательно, имеет су-
щественное значение при геологической
разведке, в частности при разведке по-
лезных ископаемых.
ИСТОРИЯ ИЗУЧЕНИЯ
ПРОСТЕЙШИХ
Изучение простейших началось значи-
тельно позже, чем изучение большинст-
ва других групп животного мира. Оно ста-
ло возможным лишь после изобретения
микроскопа, что произошло в начале
XVII в.
В 1675 г. голландец Антон Л е в е н-
г у к, рассматривая под микроскопом
каплю воды, впервые открыл в ней мно-
жество микроскопических, ранее неведо-
мых организмов, среди которых были и
простейшие. Наблюдения Левенгука воз-
будили большой интерес к этому новому
миру живых существ. В конце XVII и пер-
вой половине XVIII в. появляется боль-
шое число сочинений, посвященных изу-
чению микроскопических организмов. Од-
нако современного представления о про-
стейших как одноклеточных организмах
тогда не существовало, ибо само понятие
о клетке было сформулировано лишь в
конце первой половины XIX в. К этому
вновь открытому миру микроскопических
живых существ, которых чаще всего назы-
вали «мелкими наливочными животными»
(Animalcula infusoria), относили самые
различные организмы (простейших, круг-
лых и ресничных червей, коловраток, одно-
клеточные водоросли ит. п.) по признаку их
микроскопических размеров. Термин «ин-
72
фузории», который в настоящее время обо-
значает один из классов простейших, в
XVII—XVIII вв. имел совсем другое зна-
чение. Микроскопические организмы обиль-
но развиваются в разных растительных
настойках — infusum. Отсюда и произо-
шло название, которое сначала не было
связано с систематическим положением
организмов, а означало «наливочные» или
«настоечные» животные, т. е. разви-
вающиеся в настойках.
Представления о строении и жизни микро-
скопических существ в XVII—XVIII вв.,
несмотря на большое количество посвя-
щенных им сочинений, были крайне неоп-
ределенными и хаотичными. Это дало
знаменитому систематику Карлу Лин-
нею основание для объединения в своей
«Системе природы» (издание 1759 г.) всех
известных ему простейших в один род,
который он назвал весьма выразительно —
Chaos infusorium.
Большое значение для познания микро-
скопических существ имело сочинение
О. Ф. Мюллера «Animalcula infusoria»
A770), в котором описано 377 видов мик-
роскопических организмов, главным обра-
зом простейших. Многие предложенные
им родовые и видовые названия сохрани-
лись и в современной системе простейших.
Мюллера нередко называют «Линнеем про-
тистов», подчеркивая этим то большое зна-
чение, которое имели его работы для
изучения мира микроскопических орга-
низмов.
Воззрения ученых на простейших в
XVIII и начале XIX в. носили еще край-
не противоречивый и подчас даже диамет-
рально противоположный характер. Так,
например, Эренберг в своем извест-
ном сочинении «Наливочные животные
как совершенные организмы» A838) опи-
сывает простейших как сложно организо-
ванных существ, обладающих различными
системами органов и отличающихся от
других животных лишь своими размерами.
В противоположность Эренбергу дру-
гой крупный ученый этого периода, Дю-
жардэн, в ряде сочинений утверждает,
что простейшие не обладают никакой
внутренней организацией и построены из
бесструктурного полужидкого живого ве-
щества — саркоды.
Название типа Protozoa впервые было
введено в науку Гольдфусом в 1820 г.
Однако наряду с простейшими в совре-
менном понимании к Protozoa он отно-
сил коловраток, мшанок, гидроидных по-
липов.
Понадобились еще многие годы работы,
прежде чем удалось выяснить истинную
природу простейших. Это стало возмож-
ным лишь после того, как в конце 30-х
годов XIX в. трудами Шлейдена, Шванна
и ряда других ученых было разработано
учение о клетке.
Впервые в 1845 г. Зибольдом и
Кёлликером было сформулировано
представление о простейших как об одно-
клеточных организмах. Таким образом,
тип Protozoa был четко отграничен от
других типов микроскопических животных.
Для того чтобы определить границы
типа и природу Protozoa, понадобилось
200 лет (со времен Левенгука) напряжен-
ных исследований.
Во второй половине XIX в. в изучении
простейших особенно большую роль сыг-
рали исследования немецкого биолога
Б ю ч л и и его многочисленных учени-
ков. Они изучили с позиций клеточной
теории основные черты строения простей-
ших и положили начало исследованию
форм их размножения. Особенно большую
роль для изучения половых процессов
в размножении инфузорий сыграли рабо-
ты М о п а.
В конце XIX и в первой четверти XX в.
внимание исследователей привлекает изу-
чение паразитических простейших и их
циклов развития, что связано с большим
практическим значением Protozoa как воз-
будителей опасных заболеваний человека
и животных. В самом начале XX в. Ш а у-
д и н о м (Германия) был впервые опи-
сан жизненный цикл кокцидий. Трудами
Г р а с с и (Италия), Данилевско-
г о (Россия) и других раскрыт жизненный
цикл кровяных споровиков, что имело
значение для борьбы с малярией.
В XX в. изучение простейших разви-
вается очень быстро, что, в частности,
связано с разработкой новых методов ис-
следования их строения и физиологии:
изучается размножение простейших из
разных групп, физиологическая роль по-
73
ловых процессов (К а л к и н с, В у д-
руф, Дженнингс — США; Герт-
в и г — Германия; Метальнико в—
Россия); исследуется изменчивость и на-
следственность; разрабатываются проб-
лемы экологии и т. п. Изучение Protozoa
все более и более тесно переплетается
с проблемами исследования клетки (цито-
логии) и общей биологии.
За последние годы широкое применение
в изучении простейших нашли методы
электронной микроскопии, цитохимии,
ультрафиолетовой микроскопии и др.,
о которых уже говорилось выше (стр. 54).
Русские и советские ученые внесли зна-
чительный вклад в изучение простейших.
В конце XIX и начале XX в. профессор
Петербургского университета Шевя-
к о в опубликовал ряд крупных исследо-
ваний по инфузориям и радиоляриям. Осо-
бенно большой вклад в изучение система-
тики, строения, размножения и жизнен-
ных циклов простейших в течение второй
четверти XX в. был внесен В.А. До ге-
ле м и его многочисленными ученика-
ми — протозоологами.
К классу саркодовых относятся наи-
более просто устроенные среди простей-
ших организмы. Как уже было сказано
в общем очерке организации типа простей-
ших, в подразделении его на классы суще-
ственную роль играют органоиды движе-
ния. Для саркодовых характерной фор-
мой их являются ложноножки, или псев-
доподии, которые представляют собой вре-
менно образующиеся выросты цитоплаз-
мы. Псевдоподии служат вместе с тем для
захвата пищи.
Строение саркодовых, несмотря на отно-
сительную простоту их организации, от-
личается большим разнообразием. Глав-
ным образом это касается скелетных обра-
зований, которые, как увидим дальше,
достигают у саркодовых большой слож-
ности и совершенства.
Общее количество видов живущих в на-
стоящее время саркодовых измеряется
В области медицинской протозоологии
(протозоология — область зоологии, изу-
чающая простейших) большое значение
имеют труды Данилевского,
Марциновского, Эпштейна,
Филипченко; в области ветеринар-
ной протозоологии— Якимова, Мар-
кова и многих других.
В настоящее время существует несколь-
ко международных научных журналов, где
печатаются работы, посвященные изуче-
нию простейших. В ряде стран, в том числе
и в Советском Союзе, изданы большие
руководства, освещающие разные стороны
протозоологии.
В 1961 г. в Праге состоялся первый
международный конгресс протозоологов,
на котором из всех стран мира собрались
ученые, изучающие простейших. Второй
международный конгресс протозоологов
проходил в Лондоне в 1965 г.
Тип простейших (Protozoa) состоит
из 5 классов: Саркодовые (Sarco-
dina), Жгутиконосцы (Mastigophora), Спо-
ровики (Sporozoa), Книдоспоридии (Cni-
dosporidia) и Инфузории (Infusoria).
чпслом 8—10 тыс. Очень большое количе-
ство видов известно в ископаемом состо-
янии благодаря хорошей сохранности
скелетов многих групп саркодовых.
Свыше 80% всех современных саркодо-
вых являются обитателями моря. Некото-
рые виды живут в пресной воде, немно-
гие приспособились к жизни в почве.Име-
ются также паразитические виды.
Класс саркодовых состоит из трех
подклассов, весьма неравноценных
по количеству относящихся к ним видов:
1. Корненожки (Rhizopoda) — около
2 тыс. видов;
2. Солнечники (Helizoa) —всего несколь-
ко десятков видов;
3. Лучевики (Radiolaria) — 7—8 тыс.
видов.
Рассмотрим наиболее интересных и ха-
рактерных представителей этих трех под-
классов.
КЛАСС САРКОДОВЫЕ (SARCODINA)
74
ПОДКЛАСС КОРНЕНОЖКИ (RHIZOPODA)
Наиболее просто устроенными организ-
мами среди корненожек являются голые
амебы (Amoebina), образующие первый
отряд подкласса корненожек.
Чтобы познакомиться со строением и об-
разом жизни голых амеб, рассмотрим
сначала какого-нибудь одного характер-
ного и часто встречающегося представи-
теля.
Амеба протей (Amoeba proteus). В прес-
ных водах, в небольших прудах и кана-
вах с илистым дном, нередко удается обна-
ружить амебу протея (Amoeba proteus).
Культуру этого вида легко развести в ла-
бораторных условиях. Амеба протей —
одна из крупных свободноживущих амеб.
В активном состоянии она достигает раз-
мера 0,5 мм, ее видно простым глазом.
Если наблюдать под микроскопом за жи-
вой амебой (рис. 23, 24), видно, что она
образует несколько довольно длинных
лопастных, тупо заканчивающихся псев-
доподий. Псевдоподии все время
меняют свою форму, часть их втягивается
внутрь, часть, напротив, удлиняется,
иногда разветвляется. Тело амебы как бы
переливается в псевдоподии, которые в не-
скольких точках прикрепляются к суб-
страту, и благодаря этому образование
ложных ножек приводит к поступатель-
ному движению всей амебы. Псевдоподии
служат не только для движения, но и для
заглатывания пищи. Если псевдоподия
в процессе своего образования наталки-
вается на какую-либо органическую ча-
стицу (водоросль, мелкое простейшее и
т. п.), она «обтекает» ее со всех сторон
(рис. 25) и включает внутрь цитоплазмы
вместе с небольшим количеством жидко-
сти.
Таким образом в цитоплазме обра-
зуются пузырьки с пищевыми включени-
ями, которые называют пищевари-
тельными вакуолями. В них
происходит переваривание пищи (внутри-
клеточное пищеварение).
Непереваренные остатки пищи через
некоторое время выбрасываются наружу
<рис. 24).
Вся цитоплазма амебы ясно подразде-
лена на два слоя. Наружный, светлый,
вязкий, всегда лишенный пищеваритель-
ных вакуолей, носит название экто-
плазмы. Внутренний, зернистый, го-
раздо более жидкий, несущий многочис-
ленные пищевые включения, называют эн-
доплазмой. В состав псевдоподий
входят оба слоя цитоплазмы. Эктоплазма
и эндоплазма не представляют собой резко
разграниченных частей тела амебы. Они
могут превращаться друг в друга. В обла-
сти образования и нарастания псевдопо-
дии, куда устремляется жидкая эндоплаз-
ма, периферические части ее желатинизи-
руются (уплотняются) и превращаются
в эктоплазму.
Напротив, на противоположном конце
тела протекает обратный процесс — раз-
жижение эктоплазмы и частичное пре-
вращение ее в эндоплазму. Это явление
обратимого превращения эндоплазмы в
эктоплазму и обратно лежит в основе
образования псевдоподий.
Кроме пищевых включений (часто сосре-
доточенных в пищеварительных
вакуолях), в цитоплазме амебы про-
тея обычно отчетливо бывает виден свет-
лый пузырек, который периодически -то
появляется, то исчезает. Это сократи-
тельная вакуоля, играющая
очень важную роль в жизненных отправ-
лениях амебы. Сократительная вакуоля
заполняется жидкостью (в основном во-
дой), которая поступает в нее из окружаю-
щей цитоплазмы. Достигнув определен-
ного, характерного для данного вида амеб
размера, сократительная вакуоля сокра-
щается. Ее содержимое при этом изливает-
ся наружу через пору. Весь период на-
полнения и сокращения вакуоли при ком-
натной температуре длится у амебы протея
обычно 5—8 минут.
Концентрация различных растворенных
органических и неорганических веществ
в теле амебы выше, чем в окружающей
пресной воде. Поэтому в силу законов
осмоса вода проникает в протоплазму
амебы. Если бы избыток ее не выводился
75
Рис. 23. Амебы в капле воды
(Amoeba proteus).
наружу, то через короткий промежуток
времени амеба «расползлась» бы и раство-
рилась в окружающей воде. Благодаря
деятельности сократительной вакуоли это-
го не происходит. Таким образом, сокра-
тительная вакуоля — это прежде всего
органоид осморегуляции, регулирующий
постоянно осуществляемый ток воды через
тело простейшего. Однако наряду с этим
она связана и с другими жизненными
функциями. Вместе с выводимой из тела
амебы жидкостью выводятся и продукты
обмена веществ. Следовательно, сократи-
тельная вакуоля участвует в функции вы-
деления.
Постоянно поступающая в цитоплазму
вода содержит кислород. Поэтому сокра-
тительная вакуоля косвенно участвует и
в функции дыхания.
Как и во всякой клетке, в теле амебы
есть ядро. На живом объекте оно почти
не видно. Для выявления ядра приме-
няют некоторые красители, избирательно
окрашивающие нуклеиновые вещества яд-
ра. У амебы протея ядро довольно круп-
ное, расположено в эндоплазме, примерно
в центре тела.
Как размножаются амебы? Единствен-
ной известной у них формой размножения
является деление надвое в свободнопод-
вижном состоянии. Процесс этот начи-
нается с кариокинетического
деления ядра (стр. 60). Вслед за тем на
теле амебы появляется перетяжка, кото-
рая в конце концов перешнуровывает
тело ее на две равные половинки, в каж-
дую из которых отходит по одному ядру.
Темп размножения амебы протея зависит
от условий, и прежде всего от питания
и температуры. При обильном питании
и температуре 20—25° С амеба делится
один раз в течение 1 — 2 суток.
Рис. 24. Амеба протей (Amoeba proteus):
I — эктоплазма; 2 — эндоплазма; з — непереваренные
частицы пищи, выбрасываемые наружу; 4 — сократи-
тельная вакуоля; 5 — ядро; 6 — пищеварительная,
вакуоля.
76
В пресной и морской воде живет не-
сколько десятков видов амеб. Они разли-
чаются размерами, формой псевдоподий
(рис. 26). Ложные ножки могут сильно
отличаться по форме и размерам. Есть
виды амеб (рис. 26), у которых образуется
всего одна толстая короткая псевдоподия,
у других — несколько длинных заострен-
ных, у третьих — много коротких тупых
и т. п. Следует отметить, что даже в пре-
делах одного вида амеб форма псевдопо-
дий может довольно широко варьировать
в зависимости от условий окружающей
среды (солевой состав, кислотность среды
и т. п.).
Паразитические амебы. Некоторые виды
амеб приспособились к паразитическому
образу жизни в кишечнике позвоночных
и беспозвоночных животных. В толстых
кишках человека живет пять видов пара-
зитических амеб. Четыре вида их явля-
ются безобидными «квартирантами». Они
питаются бактериями, которые в огром-
ном количестве населяют толстую и сле-
пую кишку человека (так же как и всех
позвоночных животных), и не оказывают
никакого влияния на хозяина. Но один
из видов паразитирующих в кишечнике
человека амеб — дизентерийная амеба
(Entamoeba histolytica) — при опреде-
ленных условиях может вызвать у челове-
ка тяжелое заболевание — особую фор-
му кровавого поноса (колита), болезни,
носящей название амебиаза.
Что же представляет собой дизентерий-
ная амеба человека, почему она вызывает
заболевание, как она проникает в кишеч-
ник?
Дизентерийные амебы живут в толстом
кишечнике человека. Это очень мелкие
(по сравнению, например, с только что
описанной амебой протеем) простей-
шие. Размеры их — 20—30 мк. При изу-
чении живой амебы под микроскопом ясно
видно, что у нее резко разграничены экто-
и эндоплазма, причем зона эктоплазмы
относительно широка (рис. 27). Дизенте-
рийная амеба характеризуется очень ак-
тивной подвижностью. Она образует не-
многочисленные короткие широкие псевдо-
подии, в формировании которых прини-
мает участие почти исключительно экто-
плазма.
Дизентерийная амеба широко распро-
странена по всему земному шару. В зави-
симости от географического положения
процент зараженности людей этим пара-
зитом варьирует в среднем от 10 до 30.
Но заболевание амебпазом встречается
очень редко и приурочено преимущест-
венно к субтропическим и тропическим
районам земного шара.
В умеренных и северных широтах в по-
давляющем большинстве случаев дело ог-
раничивается носительством и клиничес-
ки выраженные формы амебиаза представ-
ляют большую редкость.
Почему же имеется такое расхождение
между частотой встречаемости паразита
и частотой вызываемого им заболевания?
Оказывается, дело в том, что далеко не
всегда наличие дизентерийной амебы
в кишечнике человека сопровождается
болезненными явлениями. В большинстве
случаев амеба не приносит своему хозяи-
ну-человеку никакого вреда. Она жи-
вет в просвете кишечника, активно дви-
гается и питается бактериями. Это явле-
ние, когда возбудитель какого-либо забо-
Рис. 25. Последовательные стадии заглатывания
пищи амебой (Amoeba terricola).
77
Рис. 27. Дизентерийная амеба (Entamoeba his-
tolytica), рисунки с живого объекта в различных
стадиях движения:
1 — эктоплазма, 2 — эндоплазма; 3 — ядро.
Рис. 26. Разные виды амеб с различной формой
псевдоподий:
1 — Amoeba limax; 2 — Pelomyxa binucieata, з —
Amoeba proteus; 4 — Amoeba radiosa; 5 —Amoeba
verrucosa; 6 — Amoeba polypodia.
левания присутствует в организме хозяи-
на, но не вызывает патологических явле-
ний, носит название носительства. В
отношении дизентерийной амебы имеет
место именно носительство.
Иногда амеба меняет свое поведение.
Она активно внедряется в стенки кишеч-
ника, разрушает эпителий, выстилающий
кишку, и проникает в соединительную
ткань. Происходит изъязвление стенки
кишечника, которое приводит к тяжелой
форме кровавого поноса. Амебы, проник-
шие в ткани, меняют и характер своего
питания. Вместо бактерий они начинают
активно пожирать красные кровяные клет-
ки (эритроциты). В цитоплазме амеб скап-
ливается большое количество эритроци-
тов на разных стадиях переваривания
(рис. 28). Медицине в настоящее время
известны некоторые специфические ле-
карственные вещества, применение кото-
рых убивает амеб, что приводит к выздо-
ровлению. Если не прибегать к лечению,
то амебиаз переходит в хроническую фор-
му и, вызывая тяжелое истощение орга-
низма человека, иногда приводит к смер-
тельному исходу.
До сих пор остаются неизвестными при-
чины, которые превращают безобидного
«квартиранта» кишечника в «агрессив-
ного» пожирателя тканей. Высказывалось
предположение, что существуют разные
формы дизентерийной амебы, не отли-
чающиеся друг от друга по своему строе-
нию.
Одни из них, распространенные в уме-
ренном и северном поясе, редко переходят
к паразитизму в тканях и почти всегда
питаются бактериями. Другие — юж-
ные — относительно легко становятся «аг-
рессивными» пожирателями тканей.
Рис. 28. Дизентерийная амеба (Entamoeba histo
lytica):
А — амеба с заглоченными красными кровяными
клетками; Б — амеба без эритроцитов. 1 — ядро;
2 — эритроциты.
78
Каким образом дизентерийная и другие
амебы, паразитирующие в кишечнике че-
ловека, попадают в организм хозяина?
Активно подвижные формы амеб могут
жить только в кишечнике человека. Бу-
дучи выведены из него, например в воду,
в почву, они погибают очень быстро и не
могут служить источником заражения.
Заражение осуществляется особыми фор-
мами существования амеб — цистами.
Посмотрим, как происходит у дизентерий-
ной амебы процесс формирования цист.
Попадая вместе с содержимым толстого
кишечника в его нижние отделы и в пря-
мую кишку, амебы претерпевают значи-
тельные изменения. Они втягивают псев-
доподии, выбрасывают пищевые частицы,
округляются. Затем эктоплазма выделяет
тонкую, но весьма прочную оболочку.
Этот процесс представляет собой инци-
стирование.
Одновременно с выделением оболочки
цисты претерпевает изменение и ядро.
Оно дважды последовательно делится,
причем деление ядра не сопровождается
делением цитоплазмы. Таким образом об-
разуются столь характерные для дизен-
терийной амебы четырехъядерные цисты
(рис. 29).
В таком виде вместе с фекальными мас-
сами цисты выводятся наружу. В отличие
от активно подвижных вегетативных форм
цисты обладают большой стойкостью. По-
падая в воду или в почву, они долгое
время сохраняют жизнеспособность (до
2—3 месяцев).
Подсыхание и нагревание гибельны для
цист. Доказано, что цисты могут распро-
страняться мухами, сохраняя при этом
жизнеспособность.
Попадая в кишечник человека с пищей,
водой и т. п., амеба эксцистиру-
е т с я. Ее наружная оболочка раство-
ряется, после чего следуют два деления,
не сопровождающиеся делением ядра (ци-
сты, как мы видим, четырехъядерны).
В результате получаются четыре одноядер-
ные амебы, которые переходят к активной
жизни.
Другие, не патогенные виды амеб ки-
шечника человека распространяются та-
ким же путем при помощи цист. По своему
строению (размеры, число ядер) цисты
Рис. 30. Разные виды раковинных корненожек:
Л — Arcella; Б — Difflugia; В — Euglypha — рако-
винка; Г — Euglypha с псевдоподиями. J — псевдо-
подии; 2 — ядро.
79
разных видов несколько отличаются друг
от друга. На этом основывается их диаг-
ностика.
Раковинные корненожки. Кроме амеб,
в пресных водах встречаются представи-
тели и другого отряда корненожек —
раковинные корненожки (Testacea). В море
они не встречаются.
По своему строению раковинные корне-
ножки напоминают амеб. В отличие от
них часть протоплазматического тела кор-
неножек заключена внутри раковинки,
играющей роль защитного образования.
В раковинке есть отверстие (устье), через
которое наружу выдаются псевдоподии.
У арцеллы (Arcella, рис. 30) раковинка
имеет форму блюдечка. Устье ее располо-
жено в центре. Раковинка, часто коричне-
вой окраски, состоит из органического
вещества, напоминающего по консистен-
ции рог. Выделяется она веществом цито-
плазмы подобно тому, как выделяется
оболочка цисты. У диффлюгии (Difflugia,
рис. 30) раковинка грушевидная. Она
состоит из песчинок — мелких посторон-
них частичек, заглоченных, а затем от-
ложенных на поверхности тела. У эугли-
фы (Euglypha) раковинка башневидная
(рис. 30), но, в отличие от диффлюгии, она
слагается из кремневых пластиночек пра-
вильной овальной формы. Эти пластиночки
образуются в толще цитоплазмы корне-
ножек, а затем выделяются на поверхность.
Размеры раковинных корненожек невели-
ки. Обычно они варьируют в пределах
50—150 мк.
Выдающиеся из устья наружу псевдопо-
дии выполняют двоякую функцию. Они
служат органоидами движения и захвата
пищи. Последнее осуществляется по тому
же типу, как и у голых амеб.
В связи с наличием раковины несколь-
ко видоизменяется, по сравнению с аме-
бами, способ бесполого размножения —
деления. Раковинка служит прочным ске-
летным образованием, и понятно, что она
не может перешнуроваться пополам. По-
этому процесс деления раковинных кор-
неножек связан с развитием новой рако-
вины. Обычно он осуществляется следую-
щим образом. Сначала примерно поло-
вина цитоплазмы выступает из устья.
Вокруг этой части образуется новая рако-
винка. Одновременно с этим процессом
делится ядро и одно из ядер переходит
в дочернюю особь (рис. 31). На этой ста-
Таблица 2. Различные радиолярии, зарисованные с жи-
вых объектов с их естественной окраской:
1 — Acanthodesmia prismatium (отряд Nasselaria). Тонкие радиаль-
но расходящиеся псевдоподии и желтые сферические симбионты.
Кремневый скелет в форме трех колец с короткими отростками;
¦2 — Euchitonia virchovi (отряд Spumellaria). Многочисленные тон-
кие псевдоподии, сетчатый трехлопастный кремневый скелет;
цитоплазма окрашена в красный цвет благодаря пигменту;
.3 — Auloceras arborescens (отряд Pheodaria). Коричневая централь-
ная капсула, зеленый феодий; кремневый скелет в форме ради-
ально ветвящихся на концах игл и поверхностно расположен-
ных тонких иголочек (спикул);
4 — Diplocercus fuscus (отряд Acantharia). Нитевидные псевдоподии,
скелет, состоящий из сернокислого стронция из радиальных
неравномерно развитых игл; зеленые симбионты зоохлореллы;
5 — Arachnocorys circumtexta (отряд Nasselaria). Нитевидные псев-
доподии, кремневый скелет в форме шлема с расходящимися
иглами; красная центральная капсула, желтые симбионты;
6 — Tuscan 11a nationalis (отряд Pheodaria). Нитевидные псевдопо-
дии, кремневый скелет в форме конуса с отходящими от него
иглами, две центральные капсулы, темно-зеленый феодий;
7 — Lithoptera mulleri (отряд Acantharia). Скелет из сернокислого
стронция в форме радиальных неравномерно развитых игл с сет-
чатыми выростами на концах, центральная капсула крестооб-
разной формы с зелеными симбионтами зоохлореллами;
$ —Acanthometra tetracopa (отряд Acantharia). Немногочисленные
радиальные псевдоподии; скелет состоит из сернокислого строн-
ция, слагается из 20 радиально расположенных одинаково раз-
витых игл; цитоплазма прикрепляется к игпам при помощи
сократимых волоконец (миофрисков). В центре ярко окрашенная
центральная капсула с зернами пигмента и зоохлореллами
дии обе особи оказываются еще
связанными друг с другом мо-
стиком цитоплазмы и обе рако-
винки (старая и вновь образо-
вавшаяся) направлены одна к
другой устьями. Вскоре после
этого цитоплазматический мостик
между особями утончается и пе-
решнуровывается и обе корне-
ножки переходят к самостоя-
тельному существованию. По
существу этот процесс мало чем
отличается от деления амеб,
он несколько усложнен лишь
процессом образования новой
раковинки.
Как уже говорилось выше,
раковинные корненожки — оби-
татели пресных вод. Они входят
в состав донного населения,
причем большая часть видов
приурочена к прибрежной зоне.
Преимущественно это обита-
тели мелких стоячих водоемов — прудов,
канав, богатых органическими вещест-
вами.
Довольно богатая фауна корнено-
жек (несколько десятков видов) встре-
чается в сфагновых болотах, в самом
сфагновом мху. Этот мох очень гигроско-
пичен и всегда впитывает большое коли-
чество воды. В прослойках воды, между
стебельками и листочками мха, живут
многочисленные раковинные корненожки.
Здесь же встречаются и некоторые виды
инфузорий.
Таким путем создается чрезвычайно ха-
рактерный биоценоз обитателей сфагно-
вых мхов.
ОТРЯД ФОРАМИНИФЕРЫ
(FORAMINIFERA)
Самым обширным отрядом среди корне-
ножек являются обитатели моря — фо-
раминиферы (Foraminifera). В составе со-
временной морской фауны известно свыше
1000 видов фораминифер. Небольшое
число видов, представляющих, вероятно,
остаток морской фауны, обитает в подпоч-
венных соленых водах и солоноватых
колодцах Средней Азии (стр. 71).
Рис.31. Бесполое размножение
путем деления раковинной кор-
неножки Euglypha alveolata:
А — корненожка перед делением; Б—образование цитоплазм этиче-
ской почки, на поверхности которой располагаются скелетные
пластинки; В — деление ядра, скелетные пластинки образуют новую
раковину; Г—конец деления, одно из ядер переместилось в дочернюю
особь. 1 — ядро; 2 — псевдоподии.
Подобно раковинным корненожкам, все
фораминиферы имеют раковину. Строе-
ние скелета достигает здесь большой слож-
ности и огромного разнообразия.
В океанах и морях фораминиферы рас-
пространены повсеместно. Их находят во
всех широтах и на всех глубинах, начи-
ная от прибрежной литоральной зоны
и кончая самыми глубокими абиссальными
впадинами. Все же наибольшее разнооб-
разие видов фораминифер встречается на
глубинах до 200—300 м. Подавляющее
большинство видов фораминифер являет-
ся обитателями придонных слоев, вхо-
дят в состав бентоса. Лишь очень немногие
виды живут в толще морской воды, являют-
ся планктонными организмами.
Познакомимся с некоторыми наиболее
характерными формами скелета форами-
нифер (рис. 32).
Среди огромного разнообразия строе-
ния раковин фораминифер можно разли-
чить по составу их два типа. Одни из них
состоят из посторонних телу корненожки
частиц — песчинок. Подобно тому как
мы это видели у диффлюгии (стр. 80,
рис. 30), фораминиферы, обладающие та-
кими агглютинированными
раковинами, заглатывают эти посторон-
ние частицы, а затем выделяют их на по-
W 6 Жизнь животных, т. 1
81
Рис. 32. Раковинки различных фораминифер:
! — Saccamina sphaerica; 2 — Lagena plurigera; з — Hyperammina elongata; 4 — то же в разрезе; 5 — Rhabdam-
mina linearis; 6 — то же в разрезе, 7 — Astrorhiza limicola; 8 — Ammodiscus incertus, вид сбоку, 9 — то же со сто-
роны устья; 10 — Cornuspira involvens; 11 — Rheopax nodulosus; 12 — Nodosaria hispida; 13 — Haplophragmoides
canariensis, вид сбоку; 14 — то же со стороны устья; 15 — Nonion umbilicatulus; 16 — то же со стороны устья,
17 — Discorbis vesicularis; 18 — тоже, вид со стороны основания; 19 — Quinqueloculina seminulum (вид сбоку);
20 — то же со стороны устья; 21 — Spiroloculina depressa; 22— Textularia sagittula; 23— Globigerina sp.
верхности тела, где они закрепляются
в тонком наружном кожистом слое цито-
плазмы. Такой тип строения раковины
имеют часто встречающиеся представители
родов Hyperammina, Astrorhiza (рис. 32,
3—7) и др. Например, в некоторых районах
наших северных морей (море Лаптевых,
Восточно-Сибирское море) эти крупные
фораминиферы, достигающие 2—3 см дли-
ны, почти сплошным слоем покрывают
дно.
Число видов фораминифер с агглю-
тинированной раковиной относительно не-
велико (хотя число особей этих видов
может быть огромным). Большая часть
обладает известковыми раковинами, со-
стоящими из углекислого кальция (СаС03).
Эти раковины выделяются цитоплазмой
корненожек, которые обладают замечатель-
ной особенностью концентрировать в сво-
ем теле кальций, содержащийся в морской
воде в небольших количествах (соли каль-
ция в морской воде составляют немногим
более 0,1%). Размеры известковых рако-
вин разных видов фораминифер могут
быть очень различны. Они варьируют
в пределах от 20 мк до 5—6 см. Это при-
мерно такое же соотношение размеров,
как между слоном и тараканом. Наиболее
крупных из фораминифер, раковина кото-
рых имеет 5—6 см в диаметре, уже нельзя
назвать микроскопическими организма-
ми. Наиболее крупные (роды Cornus-
pira и др.) живут на больших глубинах»
82
Среди известковых раковинок форами-
нифер в свою очередь можно различить
две группы.
Однокамерные фораминиферы
имеют одну-единственную полость внутри
раковины, которая сообщается с наружной
средой устьем. Форма однокамерных рако-
вин разнообразна. У одних (например,
Lagena) раковинка напоминает бутылочку
с длинным горлышком, иногда снабжен-
ную ребрами (рис. 32, 2).
Очень часто происходит спиральное
закручивание раковины, и тогда внутрен-
няя полость ее становится длинным и
тонким каналом (например, Ammodiscus,
рис. 32, 8, 9).
Большинство известковых раковин кор-
неножек является не однокамерными, а
многокамерными. Внутренняя
полость раковины поделена перегород-
ками на ряд камер, число которых может
достигать нескольких десятков и сотен.
Перегородки между камерами не сплош-
ные, в них имеются отверстия, благодаря
чему протоплазматическое тело корненож-
ки не расчленено на части, а представляет
собой единое целое. Стенки раковинок
не у всех, но у многих фораминифер про-
низаны мельчайшими порами, которые
служат для выхода наружу псевдоподий.
Об этом подробнее будет сказано ниже.
Число, форма и взаимное расположе-
ние камер в раковинке может быть очень
различным, что и создает огромное раз-
нообразие фораминифер (рис. 32). У неко-
торых видов камеры расположены в один
прямой ряд (например, Nodosaria, рис.
32, 12), иногда же их расположение дву-
рядное (Textularia, рис. 32, 22). Широко
распространена спиральная форма рако-
вины, когда отдельные камеры располо-
жены по спирали, причем по мере при-
ближения к камере, несущей устье, раз-
меры их возрастают. Причины этого по-
степенного нарастания размеров камер
станут понятны, когда мы рассмотрим ход
их развития.
В спиральных раковинках форамини-
фер имеется несколько оборотов спира-
ли. Наружные (более крупные) обороты
могут быть расположены рядом с внутрен-
ними оборотами (рис. 32,17,18) так, что все
камеры видны снаружи. Это эволют-
н ы и тип раковины. У других форм наруж-
ные (более крупные) камеры целиком или
частично охватывают внутренние камеры
(рис. 33, 1). Это инволютный тип ра-
ковины. Особую форму строения раковины
находим у фораминифер милиолид (се-
мейство Miliolidae, рис. 32, 19). Здесь
камеры сильно вытянуты параллельно
продольной оси раковины и расположены
в нескольких пересекающихся плоскос-
тях. Вся раковина в целом оказывается
продолговатой и по форме несколько
напоминает тыквенное зерно. Устье рас-
положено на одном из полюсов и обычно
снабжено зубцом.
Большой сложностью строения отли-
чаются раковинки, относящиеся к цик-
Рис. 33. Раковины фораминифер:
i — Elphidium strigilata; 2 — Archiacina
verworni
6*
83
Рис. 34. Фораминифера Orbitolites complanatus
с ризоподнями.
лическому типу (роды Archiacina,
Orbitolites и др., рис. 33, 2\ 34). Число
камер здесь очень велико, причем внутрен-
ние камеры располагаются по спирали, на-
ружные же—концентрическими кольцами.
В чем заключается биологическое зна-
чение столь сложного строения многока-
мерных раковинок корненожек? Специ-
альное исследование этого вопроса пока-
зало, что многокамерные раковинки по
сравнению с однокамерными обладают
гораздо большей прочностью. Основное
биологическое значение раковинки — это
защита мягкого протоплазматического те-
ла корненожки. При многокамерном
строении раковинки эта функция осуще-
ствляется весьма совершенно.
Как устроено мягкое протоплазматиче-
ское тело фораминифер?
Внутренняя полость раковины за-
полнена цитоплазмой. Внутри ра-
ковинки помещается иядерный ап-
парат. В зависимости от стадии раз-
множения (о чем будет сказано несколько
ниже) ядро может быть одно или их несколь-
ко. Из раковинки через устье наружу вы-
даются многочисленные очень длинные
и тонкие псевдоподии, ветвящиеся
и анастомозирующие между собой. Эти
особые свойственные фораминиферам лож-
ные ножки называют ризоподия-
м и. Последние образуют вокруг рако-
винки тончайшую сеточку, общий диаметр
которой обычно значительно превосходит
диаметр раковинки (рис. 34). У тех видов
фораминифер, у которых имеются поры,
ризоподии выдаются наружу через поры.
Функция ризоподии двоя-
к а. Они являются органоидами движе-
ния и захвата пищи. К ризоподиям «при-
липают» различные мелкие пищевые ча-
стицы, очень часто это бывают одноклеточ-
ные водоросли. Переваривание их может
происходить двояко. Если частичка ма-
ла, она постепенно как бы «скользит» по
поверхности ризоподии и через устье втя-
гивается внутрь раковинки, где и проис-
ходит переваривание. Если пищевая ча-
стица велика и не может быть втянута
внутрь раковины через узкое устье, то
переваривание происходит вне раковин-
ки. Вокруг пищи при этом собирается
цитоплазма и образуется местное, иногда
довольно значительное утолщение ризо-
подии, где и осуществляются процессы
пищеварения.
Выполненные за последние годы с при-
менением цейтраферной киносъемки иссле-
дования показали, что цитоплазма, вхо-
дящая в состав ризоподии, находится
в непрерывном движении. Вдоль ризопо-
дии в центростремительном (к раковине)
и центробежном (от раковины) направ-
лениях довольно быстро текут токи цито-
плазмы. По двум сторонам тонкой ризо-
подии цитоплазма как бы струится в про-
тивоположных направлениях. Механизм
этого движения до сих пор остается не
выясненным.
Размножение фораминифер
происходит довольно сложно и у боль-
шинства видов связано с чередованием
84
двух разных форм размножения и двух
поколений. Одно из них бесполое,
второе — половое. В настоящее время
процессы эти изучены у многих видов
фораминифер. Не вдаваясь в детали, рас-
смотрим их на каком-нибудь конкретном
примере.
На рисунке 35 изображен жизненный
цикл фораминиферы Elphidium crispa.
Этот вид представляет собой типичную
многокамерную фораминиферу со спи-
рально закрученной раковинкой. Начнем
рассмотрение цикла с многокамерной кор-
неножки, обладающей маленькой зароды-
шевой камерой в центре спирали (м и-
кросферическое поколение).
В цитоплазме корненожки первоначаль-
но имеется одно ядро. Бесполое
размножение начинается с того,
что ядро последовательно несколько раз
делится, в результате чего образуется мно-
жество небольшого размера ядер (обычно
несколько десятков, иногда свыше сотни).
Затем вокруг каждого ядра обособляется
участок цитоплазмы и все протоплазма-
тическое тело корненожки распадается
на множество (по числу ядер) одноядер-
ных амебообразных зародышей, которые
выходят через устье наружу. Сразу же
вокруг амебовидного зародыша выделяет-
ся тонкая известковая раковинка, кото-
рая и явится первой (эмбриональ-
ной) камерой будущей многокамерной
раковины. Таким образом, при бесполом
размножении на первых стадиях своего
развития корненожка является однока-
мерной. Однако очень скоро к этой пер-
вой камере начинают добавляться следую-
щие. Происходит это так: из устья сразу
выступает наружу некоторое количество
цитоплазмы, которая тотчас же выделяет
раковинку. Затем наступает пауза, в те-
чение которой простейшее усиленно пи-
тается и масса протоплазмы его увеличи-
вается внутри раковины. Затем вновь
часть цитоплазмы выступает из устья
и вокруг нее образуется очередная извест-
ковая камера. Этот процесс повторяется
несколько раз: возникают все новые и но-
вые камеры, пока раковина не достигнет
характерных для данного вида размеров.
Таким образом, развитие и рост раковины
носит ступенчатый характер. Размеры и
Рис. 35. Жизненный цикл фораминиферы
Elphidium crispa:
слева внизу — выход зародышей, образовав-
шихся в результате бесполого размножения; свер-
ху—выход гамет и их копуляция.
взаимное положение камер определяется
тем, какое количество протоплазмы вы-
ступает из устья и как эта протоплазма
располагается по отношению к предшест-
вующим камерам.
Мы начали рассмотрение жизненного
цикла Elphidium с раковинки, обладав-
шей очень маленькой эмбриональной ка-
мерой. В результате бесполого размноже-
ния получается раковинка, эмбриональ-
ная камера которой значительно крупнее,
чем у той особи, которая приступала к бес-
полому размножению. В результате беспо-
лого размножения получаются особи
макросферического поколе-
ния, которые существенно отличаются от
дающего им начало микросфериче-
85
ского поколения. В данном случае
потомство оказывается не вполне похожим
на родителей.
Каким же путем возникают особи ми-
кросферического поколения?
Они развиваются в результате поло-
вого размножения макросферического по-
коления. Происходит это следующим об-
разом. Как и при бесполом размножении,
половой процесс начинается с деления
ядра. Количество образующихся при этом
ядер значительно больше,чем при бесполом
размножении. Вокруг каждого ядра обо-
собляется небольшой участок цитоплаз-
мы, и таким путем формируется огромное
количество (тысячи) одноядерных кле-
ток х. Каждая из них снабжена двумя
жгутиками, благодаря движению которых
клетки активно и быстро плавают. Эти
клетки являются половыми клетками (г а-
метами). Они сливаются друг с дру-
гом попарно, причем слияние затрагивает
не только цитоплазму, но и ядра. Этот про-
цесс слияния гамет и есть половой про-
цесс. Образующаяся в результате слияния
гамет (оплодотворения) клетка
носит название зиготы. Она дает начало
новому микросферическому поколению фо-
раминиферы. Вокруг зиготы сразу же по
ее образовании выделяется известковая
раковина — первая (эмбриональ-
ная) камера. Затем процесс развития
и роста раковины, сопровождающийся
увеличением числа камер, осуществляется
по тому же типу, как и при бесполом раз-
множении. Раковина получается микро-
сферической потому, что размер зиготы,
выделяющей эмбриональную камеру, во
много раз меньше одноядерных амебовид-
ных зародышей, образующихся при бес-
полом размножении. В дальнейшем микро-
сферическое поколение приступит к бес-
полому размножению и вновь даст начало
макросферическим формам.
На примере жизненного цикла форами-
нифер мы встречаемся с интересным био-
логическим явлением закономерного ч е-
1 У некоторых видов фораминифер не все
ядра участвуют в формировании гамет. Часть
их остается в виде вегетативных ядер, не уча-
ствующих в процессах размножения. Этим веге-
тативные ядра напоминают макронуклеусы инфу-
зорий.
редования двух форм раз-
множения — бесполого и полово-
го, сопровождающегося чередованием двух
поколений — микросферического (разви-
вается из зиготы в результате оплодотво-
рения) и макросферического (развивается
из одноядерных амебоидных зародышей
в результате бесполого размножения).
Отметим еще одну интересную особен-
ность полового процесса фораминифер.
Известно, что у большинства животных
организмов половые клетки (гаметы) бы-
вают двух категорий. С одной стороны,
это крупные, богатые протоплазмой и за-
пасными питательными веществами непод-
вижные яйцевые (женские) клетки,
а с другой — мелкие подвижные спер-
матозоиды (мужские половые клет-
ки). Подвижность сперматозоидов обычно
связана с наличием у них активно двигаю-
щегося нитевидного хвостового отдела.
У фораминифер, как мы видели, морфоло-
гических (структурных) различий между
половыми клетками нет. По строению
своему все они одинаковы и благодаря
наличию жгутиков обладают подвижно-
стью. Здесь нет еще структурных разли-
чий, которые позволили бы различать
мужские и женские гаметы. Такая форма
полового процесса является исходной,
примитивной.
Как уже было сказано (стр. 81), огром-
ное большинство современных видов фо-
раминифер являются придонными (бенти-
ческими) организмами, встречающимися
в морях всех широт от прибрежной зоны
вплоть до самых больших глубин мирового
океана. Изучение распределения корне-
ножек в океане показало, что оно зависит
от ряда факторов внешней среды — от
температуры, глубины, солености. Для
каждой из зон типичны свои виды форами-
нифер. Видовой состав фораминифер мо-
жет служить хорошим показателем усло-
вий среды обитания.
Среди фораминифер имеются немного-
численные виды, ведущие планктонный
образ жизни. Они постоянно «парят»
в толще водяной массы. Типичный пример
планктонных фораминифер — разные виды
глобигерин (Globigerina, рис. 36). Строение
их раковинок резко отличается от строе-
ния раковинок донных корненожек. Ра-
86
новинки глобигерин более тонкостенные, а
главное, несут многочисленные расходя-
щиеся во все стороны придатки — тончай-
шие длинные иглы. Это одно из приспо-
соблений к жизни в планктоне. Благо-
даря наличию игл поверхность тела, а
именно отношение поверхности к массе —
величина, называемая удельной по-
верхностью, возрастает. Это уве-
личивает трение при погружении в воду
и способствует «парению» в воде.
Широко распространенные в современ-
ных морях и океанах фораминиферы были
богато представлены и в прежние геоло-
гические периоды начиная с самых древ-
них кембрийских отложений. Известко-
вые раковинки после размножения или
смерти корненожки опускаются на дно
водоема, где входят в состав отлагающе-
гося на дне ила. Процесс этот совершается
десятки и сотни миллионов лет; в резуль-
тате на дне океана образуются мощные
отложения, в состав которых входит не-
сметное количество раковинок корнено-
жек. При горообразовательных процес-
сах, которые совершались и совершаются
в земной коре, как известно, некоторые
области дна океана поднимаются и стано-
вятся сушей, суша опускается и стано-
вится дном океана. Большая часть совре-
менной суши в различные геологические
периоды была дном океана. Это отно-
сится в полной мере и к территории
Советского Союза (за исключением немно-
гих северных районов нашей страны:
Кольский полуостров, большая часть Ка-
релии и некоторые другие). Морские дон-
ные отложения на суше превращаются
в горные осадочные породы. Во всех мор-
ских осадочных породах присутствуют ра-
ковинки корненожек. Некоторые же отло-
жения, как например меловые, в основ-
ной своей массе состоят из раковин корне-
ножек. Столь широкое распространение
фораминифер в морских осадочных поро-
дах имеет большое значение для геологи-
ческих работ, и в частности для геологи-
ческой разведки. Фораминиферы, как и
все организмы, не оставались неизмен-
ными. В течение геологической истории
нашей планеты происходила эволюция
органического мира. Изменялись и фора-
миниферы. Для разных геологических пе-
Рис. 36. Планктонная фораминифера
Globigerina bulloides.
риодов истории Земли характерны свои
виды, роды и семейства фораминифер.
Известно, что по остаткам организмов
в горных породах (окаменелостям, отпе-
чаткам и т. п.) можно определить геоло-
гический возраст этих пород. Для этой
цели могут быть использованы и форами-
ниферы. Как ископаемые они благодаря
своим микроскопическим размерам пред-
ставляют очень большие преимущества,
так как могут быть обнаружены в очень
небольших количествах горной породы.
При геологической разведке полез-
ных ископаемых (в особенности при раз-
ведке нефти) широко используется ме-
тод бурения. При этом получается ко-
лонка породы небольшого диаметра, охва-
тывающая все слои, через которые про-
шел бур. Если эти слои представляют
собой морские осадочные породы, то в них
при микроскопическом анализе всегда об-
наруживаются фораминиферы. Ввиду
большой практической важности вопрос
о приуроченности определенных видов
фораминифер к тем или иным осадочным
породам известкового возраста разработан
с большой степенью точности.
87
ПОДКЛАСС ЛУЧЕВИКИ, ИЛИ РАДИОЛЯРИИ (RADIOLARIA)
Еще более обширную по числу видов
группу морских саркодовых, чем форами-
ниферы, образуют лучевики, или радиоля-
рии (Radiolaria).
Это отдельный подкласс в классе сар-
кодовых, насчитывающий не менее 7—
8 тыс. видов. Кроме современных видов,
радиолярии богато представлены и в ис-
копаемом состоянии. Это обусловлено тем,
что у большинства их, так же как и у фо-
раминифер, имеется минеральный скелет.
Радиолярии, так же как и форамини-
феры,— исключительно обитатели моря.
Все радиолярии — планктонные орга-
низмы. Жизнь их протекает в состоянии
«парения» в морской воде. Наибольшее
число видов радиолярий приурочено к тро-
пическим и субтропическим водам. В хо-
лодных морях число видов их невелико.
Строение радиолярий сложно и
разнообразно. Вся их организация несет
ясно выраженные черты приспособления
к планктонному образу жизни, которые
весьма совершенны и затрагивают разные
стороны строения.
Размеры радиолярий варьируют в до-
вольно широких пределах — от 40 —
50 мк до 1 мм и более. Имеются немного-
численные колониальные формы радиоля-
рий, размеры которых достигают величины
нескольких сантиметров.
Обратимся прежде всего к рассмотре-
нию протоплазматических частей тела ра-
диолярий. Большинство их имеет более
или менее ясно выраженную сферическую
форму. Характерная особенность строе-
ния радиолярий — это наличие цент-
ральной капсулы (рис. 37).
Центральная капсула пред-
ставляет собой мембрану, состоящую из
органического вещества и окружающую
центральные части цитоплазмы с ядром.
Стенки центральной капсулы обычно про-
низаны многочисленными мелкими по-
рами, через которые внутрикапсу-
л я р н а я цитоплазма сообщается с э к-
стракапсулярной. Централь-
ную капсулу следует рассматривать как
скелетное образование, защищающее внут-
ренние части цитоплазмы и ядерный
аппарат. У некоторых радиолярий внутри-
капсулярная полость сообщается с экст-
ракапсулярным пространством не много-
численными мелкими отверстиями, а ши-
роким отверстием, напоминающим устье
раковинок корненожек.
Наружный слой цитоплазмы — экто-
плазма — образует у радиолярий широ-
кую зону. В этой зоне располагаются раз-
нообразные включения, составляющие
главную массу наружного слоя тела ра-
диолярии. Сама цитоплазма представлена
лишь тонкими прослойками между вклю-
чениями. Основная масса этих включе-
ний — слизь, образующая в совокуп-
ности мощный слой, называемый калим-
м о й. Кроме слизи, в цитоплазме радио-
лярий имеются и другие включения, в
частности очень часто капли жира.
Все эти разнообразные включения умень-
шают удельный вес животного и могут
рассматриваться как одна из форм при-
способления к «парению» в толще воды.
У многих радиолярий в цитоплазме
имеются иногда в значительных количе-
ствах зеленые (зоохлореллы) и желтые
(зооксантеллы) включения. Это однокле-
точные водоросли. Некоторые из этих
водорослей относятся к отряду пан-
цирных жгутиковых — Dinoflagellata
(стр. 100). Перед нами типичный пример
симбиоза простейшего животного ор-
ганизма с растительным. Это сожительст-
во полезно для обоих компонентов. Водо-
росли получают в теле радиолярии защи-
ту и, вероятно, некоторые питательные
вещества, а также углекислоту, образую-
щуюся при дыхании. Углекислота необ-
ходима для фотосинтеза зеленого расте-
ния. Водоросли в результате фотосинтеза
выделяют свободный кислород, использу-
емый радиолярией для дыхания. Кроме
того, часть водорослей может перевари-
ваться радиолярией, т. е. служит источ-
ником пищи. Водоросли встречаются лишь
у радиолярий, живущих на небольших
глубинах, куда проникает свет. У глубо-
ководных форм они отсутствуют.
От тела радиолярии наружу отходят
многочисленные тончайшие псевдоподии
88
Рис. 37. Радиолярия Thalassicola nucleata. В центре темно окрашенная центральная
капсула, две зоны сильно вакуолизированной цитоплазмы (мелкоячеистая и крупно-
ячеистая), наружу выдаются радиально расположенные тончайшие псевдоподии.
(рис. 37, табл. 2 и 3), у некоторых видов
анастомозирующие между собой. Они слу-
жат для улавливания пищи.
Лишь очень немногие виды радиоля-
рий лишены скелета. У огромного боль-
шинства их имеется скелет, выпол-
няющий двойную функцию — защитную
и способствующую «парению» в толще
воды. Разнообразие форм их скелетов
очень велико. Многие скелеты радиоля-
рий, имеющие часто правильную геомет-
рическую форму, необычайно привлека-
тельны. В скелетах радиолярий сочетает-
ся большая легкость (у планктонных ор-
ганизмов скелет не может быть тяжелым)
с прочностью и часто с наличием разнооб-
разных выростов, увеличивающих поверх-
ность животного. Радиолярии — это одни
из наиболее красивых и изящных орга-
низмов. Известный немецкий зоолог и дар-
винист второй половины XIX в. Эрнст
Г е к к е л ь, бывший вместе с тем незау-
рядным художником, издал интересный
атлас рисунков, который он назвал «Кра-
89
сота форм в природе». В этом обширном
атласе собраны изображения множества
беспозвоночных животных, которые Гек-
келю, как художнику, казались наиболее
привлекательными с эстетической точки
зрения. В атласе (имеется его русское из-
дание) значительное место отведено ради-
оляриям. Геккель много занимался их
изучением. Он является автором больших
монографических исследований в этой об-
ласти зоологии.
Мы не имеем возможности в этой книге
дать подробное описание различных форм
радиолярий и ограничимся лишь некото-
рыми наиболее интересными представи-
телями этой обширной группы про-
стейших (см. табл. 2 и 3).
Радиолярии обладают минеральным ске-
летом. У большинства он слагается из
кремнезема (Si02). В одном из отрядов
радиолярий скелет состоит из сернокис-
лого стронция (Sr2S04).
Подкласс радиолярий состоит из
4 отрядов. Для каждого из отрядов
характерны свои типичные формы скеле-
та. Остановимся кратко на рассмотрении
этих отрядов (табл. 2—3).
В отряде Spumellaria встречаются
единичные виды, лишенные скелета
(рис. 37), но у большинства имеется крем-
невый скелет. Исходная и наиболее при-
митивная форма его — это отдельные раз-
бросанные в эктоплазме одноостные или
трех- и четырехостные микроскопические
иглы. У многих Spumellaria эти иглы спа-
иваются друг с другом, в результате чего
получаются ажурные скелетные шары
(табл. 3, 4,5).
Очень часто от шаров отходят радиаль-
ные иглы. У некоторых видов образуется
не один, а несколько шаров, вложенных
друг в друга и соединенных радиальными
иглами. Вероятно, эти вложенные друг
в друга шары образуются последователь-
но, по мере роста простейшего (процесс
этот остается неизученным). Очень раз-
нообразны кремневые скелеты в отряде
Nasselaria. Исходным форматом здесь, по-
видимому, является четырехлучевая спи-
кула. Три луча ее образуют треножник,
поддерживающий центральную капсулу,
четвертый же направлен вверх, образуя
апикальную иглу. К этой основной спи-
куле присоединяются кольца, соединяю-
щие иглы спикулы (табл. 3, 3). Эти коль-
ца, разрастаясь, образуют очень разнооб-
разные и причудливые формы скелета
в виде ажурных шапочек, шлемов, шаров
и т. п. (табл. 3, 2, 3, 6). Весьма характерен
и типичен скелет радиолярий, относящих-
ся к отряду Acantharia. В химичес-
ком отношении он имеет иной состав, чем
Spumellaria и Nasselaria, а именно состоит
из сернокислого стронция, который до-
вольно легко растворим в морской воде.
Поэтому после отмирания животного ске-
лет акантарий растворяется, тогда как крем-
невые скелеты Spumellaria и Nasselaria
опускаются на дно и входят в состав ила.
Основу скелета акантарий составляют
20 радиально расположенных игл, сходя-
щихся в центре животного (табл. 3, 8).
Эти иглы образуют пять поясов, по 4 иглы
в каждом. Их свободные концы торчат
из тела радиолярии наружу. Эта исходная
для Acantharia форма скелета у разных
видов претерпевает разнообразные видо-
изменения. Иглы могут быть развиты
в различной степени. Например, у Diploce-
rcus (табл. 2, 4) преобладают две иглы. Очень
красив скелет Lithoptera (табл. 2, 7).
Здесь преимущественно развиты 4 иглы,
на которых развиваются ответвления, об-
разующие решетки. Весь скелет приоб-
ретает характер ажурной пластинки. Ске-
летные иглы акантарий прикрепляются
к наружному слою цитоплазмы при
помощи особых волоконец, расположен-
ных вокруг игл. Эти волоконца способны
сокращаться. При их сокращении или,
наоборот, удлинении меняется общий объ-
ем протоплазматического тела радиоля-
рии. Эти изменения представляют собой
очень тонкую «настройку» для поддержа-
ния животного в состоянии «парения»
в толще воды. При колебании температу-
ры или солености плотность морской воды
не остается постоянной. Увеличение или
уменьшение объема тела радиолярии ме-
няет удельный вес животного в соответ-
ствии с физическими свойствами воды.
В высшей степени разнообразны и труд-
но сводимы к какой-либо общей схеме
кремнеземные скелеты четвертого отря-
д а радиолярий — Pheodaria (табл. 2,
3, 6), куда относятся наиболее глубоко-
90
водные виды. Скелет одних, аналогично
Spumellaria, представляет вложенные
друг в друга шары. У других имеются
2 створки, окружающие центральную
капсулу. К этим элементам скелета до-
бавляются разнообразные, иногда ветвя-
щиеся радиальные отростки — иглы.
Нередко имеется периферически лежащая
зона тонких полых игл, расположен-
ных в несколько слоев. Многообразие
в строении скелета у Pheodaria порази-
тельно!
От представителей других отрядов ра-
диолярий представители Pheodaria отли-
чаются тем, что центральная капсула
их имеет одно или три широких отверстия,
сообщающих внутрикапсулярную полость
с экстракапсулярным пространством (вме-
сто многочисленных пор, имеющихся
у других радиолярий). Кроме того, в экс-
тракапсулярной цитоплазме в области от-
верстия, ведущего в полость центральной
капсулы, у Pheodaria имеется особое,
обычно окрашенное в коричневый цвет
скопление пигмента, выделительных те-
лец и нередко пищевых включений. Эти
резко выделяющиеся благодаря своему
яркому цвету на фоне бесцветной цито-
плазмы образования носят название ф е о-
д и у м а. Вопрос о физиологическом зна-
чении феодиума остается неясным.
Процессы размножения
радиолярий до сих пор изучены недоста-
точно, несмотря на то что многие ученые
занимались исследованием этих интерес-
ных животных. Объясняется это в значи-
тельной мере тем, что никому еще не уда-
лось длительное время содержать культу-
ру радиолярий в аквариумах. Эти подлин-
ные «дети моря» не выносят лабораторных
условий существования. У некоторых
крупных видов, которые имеют скелет,
состоящий из отдельных игл, наблюда-
лось размножение путем деления надвое.
У видов, обладающих сложным монолит-
ным скелетом, такой способ размножения
невозможен, так как прочный минераль-
ный скелет не может разделиться на две
половинки. По-видимому, у таких видов
происходит формирование одноядерных
зародышей (бродяжек), подобно тому
как это происходит при бесполом размно-
жении фораминифер (стр. 85).
Свойствен ли половой процесс всем ра-
диоляриям, остается неясным. Выяснение
этого вопроса требует дальнейших иссле-
дований. Дело чрезвычайно усложняется
еще и тем обстоятельством, что в теле ра-
диолярий часто живут (стр. 88) симбио-
тические, а иногда и паразитические водо-
росли и жгутиконосцы. Эти организмы
в свою очередь при размножении обра-
зуют снабженные жгутиками бродяжки,
которые выходят из тела радиолярии.
Не всегда легко бывает решить вопрос,
имеем ли мы дело с бродяжками, принад-
лежащими самой радиолярии, или же
с бродяжками живущих в их теле расти-
тельных организмов.
Как уже указывалось, радиолярии яв-
ляются преимущественно обитателями теп-
лых морей. Для Атлантического океана,
например, установлено, что в эквато-
риальной области число видов радиоля-
рий из отряда Acantharia в 10 раз
превышает таковое в северных районах.
Такие же примерно соотношения наблю-
даются и в Тихом океане. В арктических
морях радиолярий мало. Например, в Кар-
ском море их найдено только 15 видов.
Указанные закономерности в географи-
ческом распределении радиолярий спра-
ведливы для поверхностных слоев океана,
температура которых определяется ши-
ротным фактором. По мере углубления
в толщу водной массы различия в темпе-
ратурах между южными и северными ши-
ротами постепенно стираются, в связи
с чем уменьшаются и различия в фауне
радиолярий.
Кроме отчетливо выраженной зависи-
мости распределения радиолярий от ши-
ротного фактора, что связано в первую
очередь с температурой, у них отчетливо
выражена и вертикальная зональность.
Этот вопрос был изучен, например, до-
вольно подробно в области Курило-Кам-
чатской впадины Тихого океана, где глу-
бины достигают 10 тыс. м. Среди радио-
лярий можно различить 2 группы видов.
Одна не приурочена или слабо приуро-
чена к какой-либо определенной глубине
и встречается в разных глубинных зонах.
Такие виды называют эврибатными. Дру-
гие, напротив, более или менее характер-
ны для определенной глубины — это сте-
91
нобатные формы. В области Курило-Са-
халинской впадины было обнаружено до-
вольно много таких стенобатных видов,
причем некоторые из них были найдены
лишь на глубинах, превышающих 4000 м.
Для стенобатных видов радиолярий Ку-
рило-Камчатской впадины Тихого океа-
на их приуроченность к определенным
глубинам совпадает с приуроченностью
к определенным температурам. Наиболее
глубоководные (абиссальные) ви-
ды живут при постоянной температуре
1,5—2,0° С. Среднеглубинные виды рас-
пространены в относительно теплой водной
массе с температурой 3,5° С. Виды, приуро-
ченные к небольшим глубинам, заселяют
характерный для этой области океана хо-
лодный слой воды с температурами около
0° С. Наконец, виды, живущие в поверх-
ностных слоях, подвергаются сезонным
колебаниям температуры (средняя годо-
вая температура этого слоя 2,6° С).
В Неаполитанском заливе Средизем-
ного моря русский ученый проф.
В. Т. Шевяков проводил в течение
нескольких лет наблюдения за распреде-
лением радиолярий. В частности, оказа-
лось, что радиолярии отряда Acant-
haria чрезвычайно чувствительны к ма-
лейшим изменениям условий среды, в том
числе, например, к опреснению. Обычно
акантарии распределены преимуществен-
но в поверхностных слоях моря. Однако
после сильных дождей они «спасаются»
от опреснения и опускаются на глубины
100—200 м. Через 1—2 суток акантарии
вновь поднимаются в поверхностные слои.
Acantharia оказались очень чувстви-
тельными и к волнению. При сильной
волне они уходят на глубину 5—10 м.
В зимние месяцы в связи с похолоданием
поверхностных слоев моря акантарии так-
же опускаются на глубины 50—200 м,
где температура воды выше.
Среди четырех рассмотренных выше
(стр. 90) отрядов радиолярий Acantharia
являются по преимуществу обитателями
поверхностных слоев моря. Три остальных
отряда (Spumellaria, Nasselaria, Pheoda-
ria) в большей своей части приурочены
к глубинным частям.
Из морей, омывающих берега Совет-
ского Союза, наиболее богаты радиоля-
риями дальневосточные моря. Внутрен-
ние моря (Каспийское, Азовское) совер-
шенно лишены радиолярий. Это связано,
очевидно, с их опреснением по сравнению
с Мировым океаном. В северных морях
как европейской, так и азиатской части
СССР радиолярии очень немногочислен-
ны, что связано с преобладающими здесь
низкими температурами.
Радиолярий нередко находят и в иско-
паемом состоянии в осадочных морских
породах. В ископаемом состоянии извест-
ны представители двух отрядов —
Spumellaria и Nasselaria. Скелет Acant-
haria, состоящий из сернокислого строн-
ция (SrSOJ, в ископаемом состоянии не
сохраняется, так как довольно легко рас-
творяется в воде. Ископаемые Pheodaria
не описаны.
Ископаемые радиолярии встречаются
в осадочных породах различного геологи-
ческого возраста, начиная с кембрийских
отложений. Это говорит о том, что радио-
лярии представляют собой очень древнюю
группу животного мира. Существуют не-
которые древние осадочные породы, в ко-
торых среди прочих органических остат-
ков скелеты радиолярий преобладают,
составляя основную массу их (такие поро-
ды получили название радиоляри-
т ы). На территории Советского Союза
радиоляриты известны в силурийских и
девонских отложениях Урала, в Западной
Сибири, на Дальнем Востоке (в пермских
отложениях Сихотэ-Алиня). Остров Бар-
бадос (Карибское море), входящий в груп-
пу Малых Антильских островов, в основ-
ном слагается из трепела — породы,
состоящей из скелетов радиолярий. На ост-
рове есть гора высотой 360 м, построен-
ная из трепела. Здесь найдено свыше 200
видов радиолярий, причем скелеты их
очень хорошо сохранились.
Радиолярии, кроме пород, состоящих
преимущественно из их скелетов, присут-
ствуют в различных количествах во мно-
гих других осадочных морских породах
(известняки, мел, сланцы и т. п.).
В связи с таким широким распростране-
нием в морских отложениях радиолярии
наряду с фораминиферами (стр. 87) иг-
рают важную роль при определении воз-
раста горных пород.
92
ПОДКЛАСС СОЛНЕЧНИКИ (HELIOZOA)
Третий подкласс класса саркодовых —
это солнечники (Heliozoa). Сюда относится
всего несколько десятков видов простей-
ших, часть которых обитает в пресной,
часть в морской воде. Большинство сол-
нечников — свободноплавающие в толще
воды организмы, лишенные минерального
скелета. Некоторые виды ведут прикреп-
ленный к субстрату образ жизни.
В небольших пресноводных водоемах —
прудах, канавах — в летнее время неред-
ко можно встретить солнечника Actino-
sphaerium eichhorni (рис. 38).
Тело Actinosphaerium шарообразно, до-
стигает в диаметре 1 мм, так что его хоро-
шо видно простым глазом. Многочислен-
ные тонкие совершенно прямые псевдо-
подии расходятся по радиусам во все сто-
роны. Постоянство формы псевдоподий
обусловлено тем, что внутри них прохо-
дит прочная эластичная осевая нить.
Такой тип строения псевдоподий носит
название а к с о п о д и й.
Цитоплазма солнечника имеет ячеистую
структуру и распадается на два слоя:
наружный, более светлый, — корковый
(эктоплазма) и внутренний — сердцевин-
ный (эндоплазма). Эктоплазма занимает
примерно 1/6—1/10 всего диаметра тела
животного. В этом наружном слое вакуо-
ли особенно крупны и заполнены прозрач-
ным содержимым. Среди них выделяются
но своим размерам две (а иногда и боль-
шее количество) особенно крупные вакуо-
ли, которые периодически сокращаются.
Это сократительные вакуоли. Частота их
сокращения при комнатной температуре
около минуты.
Внутренний слой цитоплазмы (эндо-
плазма) обладает гораздо более мелкой
ячеистостью, чем эктоплазма, и заполнен
обычно многочисленными пищевыми вклю-
чениями. Здесь происходит внутриклеточ-
ное пищеварение. Непереваренные остат-
ки пищи выбрасываются наружу через
слой эктоплазмы. Существенным отли-
чием солнечников от радиолярий является
отсутствие у них центральной капсулы.
Каким образом солнечник ловит добычу
и чем он питается? Добыча улавливается
псевдоподиями. Если какое-либо мелкое
животное или растение прикоснется к
псевдоподии, то оно тотчас же прилипает
к ней и очень скоро теряет способность
двигаться. Вероятно, животных парали-
зует какое-то ядовитое, выделяемое цито-
Рис. 38. Солнечник Actinosphaerium eichhorni:
А — общий вид; В — участок тела при большем уве-
личении; В — строение ядра при большом увеличе-
нии. I — эктоплазма; 2 — эндоплазма; 3 — пища;
4 — аксоподии; 5 — ядро.
93
Рис. 39. Солнечник Clathrulina elegans:
I — аксоподии; 2 — скелет, 3 — дочерние особи
внутри скелета материнской особи.
плазмой солнечника вещество. К захва-
ченной добыче склоняются соседние псев-
доподии (аксоподии), и она их совместны-
ми движениями подводится к поверхности
коркового слоя цитоплазмы. Нередко на-
встречу пище со стороны эктоплазмы обра-
зуется временная короткая тупая псев-
доподия, способствующая захвату ее.
В эктоплазме пища не задерживается,
а тотчас же переходит в эндоплазму, где
и переваривается.
Пищевые объекты солнечников разнооб-
разны. Это могут быть различные простей-
шие (инфузории, жгутиконосцы), но так-
же и мелкие многоклеточные животные
(например, коловратки, мелкие реснич-
ные черви).
Ядерный аппарат солнечника Actino-
sphaerium состоит из множества одина-
ковых ядер (у крупных особей число их
может достигать 200 и даже более), всегда
расположенных только в эндоплазме. Acti-
nosphaerium — пример многоядерного про-
стейшего.
Солнечники могут вести себя в водоеме
различным образом. Часто они «парят»
в придонном слое толщи воды. Кроме
того, они могут передвигаться и по суб-
страту, как бы катиться по дну, медлен-
но изгибая при этом аксоподии.
Размножаются солнечникп Acti-
nosphaerium делением. Тело солнечника
при этом перетягивается на две более или
менее одинаковые половинки. Этому пред-
шествует деление ядер, которое, однако,
не протекает одномоментно, а растяги-
вается на весь промежуток времени меж-
ду двумя делениями солнечника.
Кроме бесполого, у солнечни-
ков имеется и половое размножение,
протекающее довольно сложно.
Кроме крупного Actinosphaerium, в пре-
сных водах встречаются и другие, более
мелкие виды солнечников, близких по
строению к Actinosphaerium. Укажем,
например, на часто встречающихся мел-
ких Actinophrys sol, обладающих, в отли-
чие от Actinosphaerium, лишь одним ядром.
В море тоже встречаются солнечники, по
своему строению напоминающие Actino-
sphaerium, например многоядерные Сатр-
tonema nutans.
Среди солнечников, как пресноводных,
так и морских, имеются некоторые виды
с минеральным скелетом из кремнезема,
подобно радиоляриям. Примером может
служить пресноводная Clathrulina ele-
gans (рис. 39). Это небольшой солнечник
с типичными радиальными аксоподиями,
тело которого окружено тонким скеле-
том в форме ажурного шарика. При по-
мощи тонкой ножки организм этот при-
крепляется к субстрату. В эндоплазме
имеется одно ядро. При размножении
Clathrulina протоплазма делится внутри
скелета на две дочерние особи, которые
выходят наружу через отверстие в ске-
лете, а затем образуют свой новый скелет
и стебелек.
В процессе эволюции саркодовые, при-
спосабливаясь к различным средам оби-
тания, дали большое разнообразие форм.
Организация саркодовых еще очень при-
митивна. На примере амебы мы видим од-
ного из наиболее просто устроенных жи-
вотных организмов. Но и в пределах при-
митивной организации саркодовые проде-
лали большой эволюционный путь от
амебообразных организмов к сложно уст-
роенным фораминиферам и радиоляриям.
Однако усложнение строения в пределах
94
класса саркодовых лишь в относительно
малой степени затронуло живые прото-
плазматические части клетки. Органоида-
ми движения и захвата пищи на всех эта-
пах эволюции саркодовых остаются псев-
доподии. Зато большого прогрессивного
развития достигают в пределах саркодо-
вых скелетные элементы, выполняющие,
с одной стороны, защитную и опорную
функцию, а с другой — способствующие
«парению» в воде (увеличение удельной
поверхности), что имеет важное биологиче-
ское значение, так как многие морские
саркодовые — планктонные организмы.
Развитие сложных скелетных образо-
ваний сыграло большую роль и в эволю-
ции донных (бентических) саркодо-
вых — фораминифер. Развитие сложного
и разнообразно устроенного скелета обес-
печило фораминиферам биологическое про-
цветание как в прежние геологические
эпохи, так и в современной морской фауне.
Наконец, относительно небольшое чис-
ло видов саркодовых избрало в качестве
среды обитания другие организмы и стало
паразитами. В отношении сложности
строения паразитические саркодовые оста-
лись на уровне голых амеб.
КЛАСС ЖГУТИКОНОСЦЫ
(MASTIGOPHORA, ИЛИ FLAGELLATA)
Жгутиконосцы представляют со-
бой очень обширную и разнооб-
разную группу простейших орга-
низмов, широко распространенную
в природе. Характерная черта
их — наличие жгутиков — орга-
ноидов движения (рис. 40). Жгу-
тики представляют собой тончай-
шие волосовидные выросты цитоп-
лазмы, длина которых иногда зна-
чительно превосходит длину тела
(см. стр. 68). Активное и быстрое
движение жгутиков обусловливает
поступательное движение простей-
шего. Число их может быть раз-
личным. Чаще всего имеется лишь
один жгутик (рис. 40), нередко их
бывает два (рис. 41), иногда 8.
Имеются некоторые виды жгути-
коносцев, число жгутиков у ко-
торых может достигать несколь-
ких десятков и даже сотен (рис.
63, 64).
В отличие от саркодовых, боль-
шинство жгутиконосцев обладает
более или менее постоянной фор-
Рпс. 40. Жгутиконосец Euglena viridis:
J — жгутик; 2 — глазное пятнышко (стиг-
ма); 3 — сократительная вакуоля; 4 — хро-
матофоры; 5 — ядро.
•*i
'jr.:-*
мой тела. Это обусловливается
тем, что наружный слой эктоплаз-
мы образует плотную эластич-
ную пелликулу. Однако у неко-
торых жгутиконосцев пелликула
очень тонка, и они способны в до-
вольно широких пределах менять
форму тела, а немногие виды со-
храняют даже способность образо-
вывать ложные ножки (псевдопо-
дии).
Как и все простейшие, жгути-
коносцы имеют одно или несколь-
ко ядер.
Среди жгутиконосцев мы встре-
чаем поразительное разнообразие
типов обмена веществ. По этому
важному признаку жгутиконосцы
занимают как бы промежуточное
положение между растительным
и животным миром: у них можно
наблюдать все переходы от типич-
но растительного к животному ти-
пу питания.
Среди жгутиконосцев имеются
организмы как с аутотроф-
н ы м, так и с гетеротроф-
ным типом питания, а также
виды, которые сочетают в себе
черты обоих этих типов обмена.
Таким образом, некоторые (ауто-
95
трофные) жгутиконосцы представляют собой
объект ботаники, тогда как другие (гетерот-
рофные) — объект изучения зоологии. Мы
рассмотрим в дальнейшем изложении и тех
и других, так как среди жгутиконосцев
иногда даже относительно близкие виды
могут обладать разными типами обмена
веществ: одни — растительным, другие —
животным.
Жгутиконосцы как по строению, так и по
образу жизни чрезвычайно разнообразны.
Число видов их велико, оно достигает
6—8 тыс. Многие из них являются обита-
телями моря. Значительная часть морских
видов жгутиконосцев входит в состав
планктона, где они развиваются иногда
в огромных количествах. Пресные воды
тоже богаты жгутиконосцами. Видовой
состав их в пресноводных водоемах в боль-
шой степени зависит от степени загряз-
ненности вод органическими веществами.
Для разных типов озер, прудов и других
водоемов характерны определенные виды
жгутиковых. Такая довольно строгая при-
уроченность их к определенным условиям
жизни позволяет использовать жгутико-
носцев (наряду с другими группами орга-
низмов) как своеобразные «биологические
индикаторы» при санитарной оценке во-
доема.
Многие жгутиконосцы паразитируют в
организмах различных животных. Хозя-
ева паразитических форм жгутиковых
разнообразны. Среди них встречаются бес-
позвоночные животные (чаще членисто-
ногие, в том числе насекомые); особенно
часто жгутиконосцы паразитируют в раз-
ных классах позвоночных (в том числе
домашних животных и птицах).
Средой обитания паразитических форм
жгутиконосцев могут быть различные ор-
ганы: кишечный канал, кровяное русло,
кожа, половые пути. Человек также яв-
ляется хозяином некоторых видов жгу-
тиконосцев; среди них имеются весьма
патогенные виды (трипанозомы, лейшма-
нии, лямблии и др.), вызывающие тяже-
лые заболевания.
В отличие от фораминифер и радиоля-
рий, в ископаемом виде жгутиконосцы
почти неизвестны. Это объясняется отсут-
ствием у подавляющего большинства их
минерального скелета. Существует толь-
ко одна небольшая группа планктонных
жгутиконосцев (сем. Silicoflagellidae), об-
ладающих очень нежным кремневым ске-
летом. Представители этой группы жгути-
коносцев найдены в ископаемом состоя-
нии в морских отложениях.
По характеру питания и обмена ве-
ществ класс жгутиконосцев, естественно, де-
лят на два подкласса: растительных жгу-
тиконосцев, или фитомастигин (Phyto-
mastigina), и животных жгутиконосцев,
или зоомастигин (Zoomastigina). Мы рас-
смотрим некоторых наиболее интересных
представителей, относящихся к тому и к
другому подклассу.
Таблица 3. Различные радиолярии (скелеты):
1 — Dorcadospyris dinoceras (отряд Nasselaria). Кремневый скелет
в виде центрального шара с двумя дугами;
2 — Calocyclus monumentum (отряд Nasselaria). Кремневый скелет
в форме решетчатого колокола с перетяжками и радиальными
иглами; многочисленные нитевидные псевдоподии;
3— Coronidium acacia (отряд Nasselaria). Кремневый скелет из
трех взаимно пересекающихся колец с радиальными выростами;
4 — Perypanicium amphocorona (отряд Spumellaria). Кремневый
скелет из вложенных друг в друга двойных сфер, соединенных
радиальными иглами;
5 — Hexancistra quadricuspis (отряд Spumellaria). Кремневый ске-
лет из вложенных друг в друга решетчатых шаров, соединенных
радиальными иглами с разветвлениями на концах;
6 — Tympaniscus tropodiscus (отряд Nasoelaria). Кремневый скелет
из трех взаимно пересекающихся колец с радиально отходящи-
ми вторично ветвящимися иглами.
ПОДКЛАСС РАСТИТЕЛЬНЫЕ ЖГУТИКОНОСЦЫ,
ИЛИ ФИТОМАСТИГИНЫ (PHYTOMASTIGINA)
Многим, вероятно, приходилось видеть
в природе своеобразное явление «цветения
воды», когда вода в луже, пруду или озере
становится изумрудно-зеленой. Причиной
«цветения» являются различные микроско-
пические растительные организхмы, раз-
вивающиеся в несметных количествах.
Очень часто это бывают разные представи-
тели фитомастигин.
Хламидомонады. Эти одноклеточные ор-
ганизмы часто являются причиной «цве-
тения». К роду хламидомонада (СЫа-
mydomonas) относится свыше 150 видов.
Тело большинства их яйцевидное и шаро-
видное (рис. 41), очень небольших разме-
ров A0—30 мк). Все тело покрыто доволь-
но прочной оболочкой из целлюлозы —
вещества, из которого состоят оболочки
растительных клеток. На переднем конце
расположены два жгутика одинаковой
длины. Ярко-зеленый цвет жгутиконосца
обусловлен наличием в цитоплазме его
чашевидного хроматофора, несущего зе-
леный пигмент — хлорофилл. Весь
хлорофилл сосредоточен в хроматофоре
и вне его никогда не встречается. В обла-
сти прикрепления жгутиков имеется одна
или две сократительные вакуоли — это
органоид осморегуляции и выделения (см.
стр. 69). У некоторых видов хламидомо-
над на переднем конце тела имеется еще
одно образование в виде не-
большого пятна яркого крас-
но-коричневого цвета. Это \ А
стигма, или глазок. \
Его окраска обусловлена на- ' \ ,
личием особого пигмента, на- \ /
зываемого гематохро- \
м о м. Хламидомонады обла- % \
дают резко выраженным по- \Аа
ложительным фототропиз- /д&**Ш
мом: они всегда плывут в з—/йрЧ&гё
сторону источника света. Не- /НоС^У*
которые наблюдения позво- 1Ш®*в•**
ляют считать, что стигма \w|gjggj
представляет собой органоид, ушШН
связанный с восприятием све- N§jggj$3
тового раздражения, и явля-
ется по своей функции действительно кле-
точным глазком. В цитоплазме обычно
присутствуют зерна крахмала, являющие-
ся продуктом фотосинтеза.
Клеточное ядро хламидомонад сфе-
рической формы и расположено примерно
в середине клетки.
Размножение большинства хла-
мидомонад происходит в неподвижном со-
стоянии (рис. 41): жгутиконосец теряет
жгутики, опускается на дно. Внутри обо-
лочки происходит деление протоплазмати-
ческой части клетки (протопласта) сна-
чала на две, затем на четыре части. Через
разрыв оболочки материнской клетки об-
разовавшиеся в результате деления до-
черние клетки выходят наружу, приобре-
тают жгутики, дорастают до величины
материнской клетки и выделяют оболочку.
Рассмотренный только что процесс пред-
ставляет собой бесполое размно-
жение.
Кроме бесполого размножения, у боль-
шинства хламидомонад известен поло-
вой процесс. Он сводится к тому, что
вегетативные клетки становятся гамета-
ми, которые попарно сливаются. Слияние
гамет сопровождается слиянием ядер. Об-
разовавшаяся в результате оплодотворе-
ния зигота теряет жгутики и покры-
вается прочной оболочкой. Внутри нее
• 7 Жизнь животных, т. 1
97
происходит деление протопласта на четы-
ре клетки, у которых развиваются жгу-
тики. Клетки превращаются в четыре
особи хламидомонад, переходящих к сво-
бодноподвижному образу жизни.
Колониальные фитомастигины. Среди
фитомастигин есть немало колониальных
видов. Колонии образуются в результате
бесполого размножения путем деления,
при котором продукты деления остаются
связанными друг с другом. Таким образом
возникает как бы особь высшего порядка,
включающая в себя некоторое (иногда
очень значительное) число отдельных кле-
ток, каждая из которых соответствует
одной хламидомонаде. Рассмотрим неко-
торые наиболее широко распространенные
виды колониальных фитомастигин.
В пресных водах часто встречаются
маленькие изящные колонии гониумов (Go-
nium pectorale), представляющие собой
зеленые пластиночки, каждая из которых
состоит из 16 клеток (рис. 42). Клетки
колонии по своему строению полностью
соответствуют хламидомонадам. Отдел ь-
Рис. 42. Колониальная фитомастпгина Goni-
um pectorale:
2 — жгутики; 2 — глазок (стигма); з — сократи-
тельная вакуоля; 4 — ядро.
ные клетки связаны друг с другом бесцвет-
ной прозрачной студенистой массой. Эта
масса образуется в результате ослизне-
ния наружного слоя оболочек клеток.
Благодаря биению жгутиков вся колония
активно плавает в воде.
Другой часто встречающейся пресно-
водной колониальной формой фитомасти-
гин является эвдорина (Eudorina elegans).
Колонии этого вида, в отличие от гониу-
мов, шаровидны (рис. 43) и в большинстве
случаев слагаются из 32 клеток (изредка
число клеток колонии может быть и мень-
ше, а именно 16 и 8). Как и у гониумов,
отдельные особи колонии эвдорин связаны
между собой студенистым прозрачным ве-
ществом. Диаметр колоний этого вида
варьирует в довольно широких пределах
(от 50 до 200 мк).
Каждая клетка колоний гониумов и
эвдорин сохраняет способность к беспо-
лому размножению, которое удается
наблюдать довольно часто. К делению при-
ступают одновременно все клетки коло-
нии (рис. 43, 2). Каждая клетка у гониу-
ма претерпевает 3, а у эвдорин — 4 после-
довательных, быстро следующих друг за
другом деления. В результате внутри об-
щей слизистой оболочки колонии обра-
зуются новые, дочерние колонии, число
которых соответствует числу приступив-
ших к размножению клеток. В дальней-
шем слизистый чехол материнской коло-
нии распадается, и дочерние колонии при-
ступают к самостоятельному существова-
нию.
Очень большой сложности достигает
строение колоний у вольвокса (Volvox).
Несколько видов этого рода — частые
обитатели небольших пресноводных водое-
мов (рис. 44).
Вольвоксы образуют крупные шаровид-
ные колонии. У наиболее часто встречаю-
щегося вида Vol vox aureus диаметр коло-
ний равен 500—850 мк, а размеры колонии
Volvox globator достигают 2 мм. У Volvox
aureus в состав колонии входит 500—1000
отдельных клеток (рис. 44), а у Volvox
globator — до 20 тыс. Основная масса
колонии состоит из студенистого вещества,
образующегося в результате ослизнения
клеточных оболочек. Самый наружный
слой его представляет собой особый плот-
98
1
2
Рис. 43. Шаровидная колония Eudo-
rina elegans:
1 — колония из 32 клеток; 2 — бесполое
размножение (каждая особь колонии в
результате последовательных делений
дает начало новой, дочерней колонии).
ныи кожистый слои, придающий всей ко-
лонии значительную прочность. Централь-
ные части студенистого вещества гораздо
менее плотной консистенции — они полу-
жидкие. Отдельные клетки колонии распо-
лагаются в самом периферическом слое.
Каждая клетка имеет в основных чертах
такое же строение, как и одиночные хла-
мидомонады. В колонии вольвокса отдель-
ные особи не вполне изолированы друг
от друга, они связаны между собой тон-
чайшими цитоплазматическими мости-
ками (рис. 45).
Движение вольвокса осуществляется бла-
годаря совместному действию жгутиков.
Несмотря на то что колония шаровидна,
она обладает некоторой полярной диффе-
ренцировкой. Последняя выражается в
том, что на одном полюсе стигмы (глазки)
развиты сильнее, чем на другом. Движе-
ние вольвокса тоже оказывается ориен-
тированным: он плывет вперед тем полю-
сом, на котором глазки сильнее развиты.
Мы видели, что у относительно просто
устроенных колоний, таких, как гониум
и эвдорина, все клетки способны, путем
бесполого размножения давать начало но-
вой колонии. У вольвокса дело обстоит
иначе. Огромное большинство клеток ко-
лонии не способно делиться. Этим свойст-
вом обладают лишь очень немногие особи.
Число их в колонии относительно неве-
лико: чаще всего 4—10. Они располага-
ются в большинстве своем в нижней части
колонии (в той части ее, где глазки отно-
сительно слабо развиты). Эти клетки, на-
зываемые «вегетативными клетками раз-
множения», удается рассмотреть лишь в
Рис. 44. Колония Volvox aureus с дочер-
ними колониями внутри материнской.
Рис. 45. Небольшой участок колонии
Volvox aureus (схематизированно):
1 — вегетативная особь колонии; 2 — цитоплаз-
матический мостик; 3 — более крупная особь, в
результате развития которой получаются новые,
дочерние колонии.
7»
99
относительно молодых колониях. В более
старых они находятся на различных ста-
диях развития в дочерние колонии. Не
вдаваясь в детали этого довольно слож-
ного процесса, укажем, что дочерние
колонии развиваются внутри материнской
в результате многократного деления «веге-
тативных клеток размножения» (рис. 46).
Когда размеры дочерних колоний увели-
чатся настолько, что они уже не смогут
поместиться внутри старой, последняя ло-
пается и погибает, а дочерние колонии
выходят наружу. Летом, в период энер-
гичного размножения, нередко можно на-
блюдать, как внутри дочерних колоний,
находящихся еще внутри материнской,
начинают развиваться внучатые колонии.
Кроме бесполого размножения, у коло-
ниальных фитомастигин наблюдается и
половой процесс, который протекает у раз-
личных видов по-разному. Укажем
лишь, что при половом процессе в коло-
нии образуются гаметы, в результате слия-
ния (оплодотворения) которых получается
зигота. Из зиготы путем ряда последова-
тельных делений развивается новое поко-
ление колоний.
Рис. 46. Разрез через колонию вольвокса с тре-
мя дочерними колониями внутри материнской
(схематизированно).
ОТРЯД ПАНЦИРНЫЕ
ЖГУТИКОНОСЦЫ, ИЛИ ПЕРИДИНЕИ
(DINOFLAGELLATA, ИЛИ PERIDINEA)
Панцирные жгутиконосцы — обширная
группа простейших, широко распростра-
ненная в морях и пресных водах. Подав-
ляющее большинство видов этого отряда
относится к планктонным формам.
Характерной особенностью панцирных
жгутиконосцев является своеобразное
строение жгутикового аппара-
т а (рис. 47). Имеющиеся у них два жгу-
та берут начало рядом друг с другом на
одной стороне тела простейшего, которую
условно называют брюшной. Один жгу-
тик направлен назад и свободно выдается
в окружающую среду. Начальная часть
его лежит в небольшом желобке, идущем
вдоль тела. Второй жгутик, называемый
поперечным, опоясывает все тело в эква-
ториальной плоскости и расположен в до-
вольно глубоком желобе, называемом поя-
сом (cingulum), который опоясывает все
тело. Поперечный жгут производит ха-
рактерные волнообразные движения, что
создает ложное впечатление, будто в по-
перечной борозде расположен ряд рес-
ничек.
Вторая характерная особенность боль-
шинства (но не всех) панцирных жгутико-
носцев — наличие у них оболочки,
состоящей из клетчатки. Эта обо-
лочка складывается из строго определен-
ного числа закономерно расположенных
отдельных пластинок, соединенных друг
с другом швами.
Цитоплазма жгутиконосцев обычно бы-
вает окрашена в буроватый цвет, который
обусловлен наличием в ней большого ко-
личества мелких дисковидных хроматофо-
ров, содержащих хлорофилл и некоторые
дополнительные пигменты. Продуктом ас-
симиляции является крахмал, зерна кото-
рого рассеяны в цитоплазме.
Рассмотрим представителей двух наибо-
лее широко распространенных родов
динофлагеллят.
На рисунке изображен часто встречаю-
щийся в пресных водах перидиний (Peri-
dinium tabulatum). У него отчетливо
выражены упомянутые выше пластин-
100
ки. Форма тела более или менее правильно
яйцевидная. Иногда бурное развитие пе-
ридиний вызывает «цветение» воды.
Широко распространены как в пресной,
так и в морской воде многочисленные виды
рода церациум (Ceratium). Характер-
ной особенностью представителей этого
рода являются длинные выросты. У прес-
новодного Ceratium hirudinella (рис. 48)
один вырост (апикальный) направлен впе-
ред и три (антапикальных) — назад. Осо-
бенно сильно эти выросты выражены
у морских видов Ceratium, у которых
они вторично ветвятся (рис. 49). Для
морских же видов характерно образование
временных колоний-цепочек (рис. 50), ко-
торые возникают в результате того, что
разделившиеся особи остаются некоторое
время связанными друг с другом. Длин-
ные отростки у морских видов Ceratium
представляют собой приспособление
к планктонному образу жизни. Отростки
увеличивают поверхность тела, что способ-
ствует «парению» в воде. Аналогичные
приспособления мы уже видели у радио-
лярий.
Особенно причудливую форму приобре-
тает панцирь у представителей рода
Ornithocercus, где по краям борозд обра-
зуются длинные крыловидные выросты
(рис. 51). Это тоже приспособление к «па-
рению» в воде.
В планктоне теплых и тропических
морей очень часто, и нередко в огромных
количествах, встречается жгутиконосец
ночесветка (Noctiluca miliaris). В летнее
время это простейшее обычно развивается
и в Черном море.
Ночесветка имеет шаровидное тело до
2 мм в диаметре. В отличие от других
панцирных жгутиконосцев, она не имеет
оболочки из клетчатки и лишена хромато-
форов (рис. 52). Поэтому ночесветка не
способна к фотосинтезу. В отличие от
большинства Dinoflagellata, ночесветке
свойственно животное (анимальное) пита-
ние. Она заглатывает и переваривает раз-
личные оформленные пищевые частицы:
мелкие водоросли, мелкие виды простей-
ших.
На одной стороне шаровидного тела но-
чесветки имеется углубление, соответст-
вующее продольному желобку типичных
Рис. 47. Панцирный жгутиконосец Peredinium
tabulatum:
А — вид с брюшной стороны; Б — вид со спинной
стороны. 1 — поперечный жгутик в пояске, 2 — сво-
бодный жгутик.
Рис. 48. Ceratium hirudinella:
А — неделящаяся особь; Б — деление; В — только
что разделившаяся особь, часть пластинок на заднем
конце недоразвита. 1 — апикальный вырост; 2 — ан-
тапикальный вырост; 3 — поясок, 4 — задний жгу-
тик; 5 — ядро.
101
• ¦ Рис. 50. Морской
панцирный жгутико-
Рис. 49. Морской носец Ceratium voltur.
панцирный жгутико- Цепочка, образовав-
носец Ceratium pal- шаяся в результате
ma turn. делений.
панцирных жгутиконосцев. В глубине его
помещается отверстие, выполняющее
функцию рта. Через него и происходит
поглощение пищи. В области этого же
углубления помещаются два жгута. Один
из них короткий и толстый. Он скорее
напоминает щупальце. Этот жгут совер-
шает относительно очень медленные коле-
бательные движения (не более 10 в ми-
нуту), способствующие направлению пи-
щевых частиц к ротовому отверстию.
Второй, более короткий и тонкий жгутик
также расположен в области ротового
углубления (рис. 52, 4).
Очень своеобразно строение эндоплаз-
мы ночесветки. Непосредственно под ро-
товым отверстием располагается централь-
ное скопление цитоплазмы, от которого
к периферии во всех направлениях отхо-
дят тонкие тяжи. Вся же остальная масса
тела заполнена жидкостью и представляет
собой систему вакуолей. Ядро располо-
жено в центральном скоплении цитоплаз-
мы. В цитоплазматических тяжах имеются
многочисленные жировые включения.
Описанное выше своеобразное строение
цитоплазмы следует рассматривать как
одну из форм приспособления к планктон-
ному образу жизни. Сильная вакуолиза-
ция цитоплазмы и наличие жировых вклю-
Рис. 51. Панцирный жгутиконосец
Omithocercus magnificus-
Рис. 52. Ночесветка Noctiluca miliaris:
I — центральное скопление цитоплазмы; 2 — ядро;
3 — пищеварительные вакуоли; 4 — жгутик, рас-
положенный в области ротового углубления; 5 —
жгутик «щупальце».
чений ведут к уменьшению удельного веса
и, следовательно, способствуют «парению»
в толще воды.
У ночесветки есть еще одна интерес-
ная особенность, за которую она и по-
лучила свое название. При раздраже-
нии (механическом, химическом и т. п.)
ночесветка ярко «вспыхивает».
Это явление незаметно днем, при
ярком свете, но очень отчетливо
выражено в темноте, ночью. Но-
чесветка — это один из организ-
мов, вызывающих явление све-
чения, столь характерное для
теплых морей. Всякий, кому при-
ходилось в теплую летнюю ночь
плыть по Черному морю в лодке
или на пароходе, вероятно, имел
возможность наблюдать это эф-
фектное явление. При ударе ве-
сел по воде, при падении капель
воды с весел в море, при враще-
нии пароходного винта и т. п.
вода начинает светиться слабым
фосфорическим, но вполне отчет-
ливым светом. При этом видны бы-
вают отдельные вспышки, напо-
минающие искорки.
Свечение наступает в результате
окисления жировых включений, о
наличии которых уже говорилось
выше.
Свечение имеет, вероятно, за-
щитное значение (отпугивает хищ-
ников).
Некоторые виды панцирных
жгутиконосцев являются симбион-
тами радиолярий (стр. 88). Недав
плазме содержатся зеленые х р о-
.1 матофоры, несущие хлоро-
филл. Форма хроматофоров у раз-
ных видов различна. Наружный
слой цитоплазмы образует пел-
ликулу. У многих видов она
бывает структурирована: несет
расположенные рядами утолще-
ния (рис. 53). Тело некоторых эвг-
лен покрыто очень тонкой эласти-
чной пелликулой. Эти организмы
способны сокращаться и вытяги-
ваться по продольной оси. У эвг-
леновых имеется одно ядро с круп-
ным ядрышком.
Замечательной особенностью
многих видов эвглен является спо-
собность менять характер пита-
ния и обмена веществ в зависи-
мости от условий среды. На свету
при наличии в окружающей сре-
де минеральных солей, содержа-
щих необходимые химические эле-
менты, эвгленам свойствен типич-
ный аутотрофный обмен. В их
теле протекает фотосинтез и ус-
воение неорганических солей. Ес-
ли тех же эвглен поместить в тем-
ноту, в раствор, содержащий ор-
Рис. 53. Euglena spiro- ганические вещества, то они теря-
дУГ%™оКвУанаСТРУ" ют хлорофилл, становятся бес-
но выяснено, что они в большом 2 __ жгуТик; 2 — глазок цветными и начинают усваивать из
количестве поселяются и в мяг-
ких тканях коралловых полипов.
(стигма); з — резервуар окружающей среды готовые орга
сократительной вакуо- rj ' г ^ ,_ r ^
ли; 4 — ядро. нические вещества. Гаким об
ОТРЯД ЭВГЛЕНОВЫЕ (EUGLENOIDEA)
Значительный интерес представляет о т-
р я д эвгленовых (Euglenoidea), так как
в пределах этой группы встречаются все
типы питания — от типично растительного
(аутотрофного) до типично животного
(анимального). Многочисленные виды р о-
д а эвглена (Euglena) широко распростра-
нены в пресноводных водоемах, особенно
часто они встречаются в загрязненных
водах. Строение эвглен представлено на
рисунках 40 и 53. Эвглены снабжены од-
ним жгутиком. В передней части тела их
расположена сократительная вакуоля и
ярко-красный глазок (стигма). В цито-
вещества.
разом эвглены переходят от ау-
тотрофного к гетеротрофному пи-
танию. Гетеротрофное питание осуществ-
ляется у них не путем заглатывания
оформленных частиц пищи, а путем ос-
моса: поглощения растворенных в ок-
ружающей среде органических питатель-
ных веществ через пелликулу. Такой
способ питания называют сапрофит-
ным. Часто, развиваясь в загрязнен-
ных водах, где имеется большое коли-
чество растворенных органических веществ,
эвглены сочетают оба типа питания —
аутотрофный и сапрофитный. Если обес-
цвеченных, культивируемых в темноте
эвглен вновь перенести на свет, то через
короткий промежуток времени они вновь
зеленеют и переходят к аутотрофному пи-
танию. Таким образом, мы видим, что на
103
низших ступенях развития органического
мира животный и растительный тип
обмена нерезко отграничены друг от
друга.
В отряде эвгленовых имеются роды и ви-
ды, которые окончательно утеряли спо-
собность к аутотрофному типу обмена
Среди зоомастигин имеется значитель-
ное количество видов, живущих в мор-
ской и пресной воде и питающихся либо
сапрофитно (осмотическим путем), либо
путем заглатывания оформленных частиц
пищи (анимальное питание). Однако боль-
шая часть зоомастигин перешла к парази-
тическому образу жизни в различных рас-
тительных и животных организмах. Рас-
Рис. 54. Bodo edax.
А — неделящаяся особь; Б—К — последовательные
стадии деления. 1 — жгутики; 2 — сократительная ва-
куоля; 8 — парабавальное тело; 4 — ядро.
веществ и при всех условиях не синтези-
руют хлорофилл. Некоторые из них,
способные заглатывать оформленные ча-
стицы пищи, перешли уже к настоящему
животному типу питания и обмена ве-
ществ (например, роды Peranema, Ur-
ceolus).
смотрим сначала некоторых наиболее ха-
рактерных и широко распространенных
представителей свободноживущих зоо-
мастигин.
В сильно загрязненных органическими
веществами водах почти всегда в больших
количествах можно встретить разные виды
рода Bodo, относящегося к отряду
протомонад (Protomonadina). Это очень
мелкие жгутиконосцы (рис. 54) длиной
10—25 мк, с двумя жгутиками. Один из
них направлен вперед и активно движется,
вызывая поступательное движение про-
стейшего. Второй жгутик направлен на-
зад и более или менее пассивно волочится
в воде. Он выполняет, вероятно, функцию
руля. Форма тела яйцевидная, довольно
сильно варьирующая. Ядро одно, распо-
ложено в передней трети тела. У основа-
ния жгутиков имеется довольно крупное
сферической формы образование, называе-
мое парабазальным телом. Предполагают,
что в нем сосредоточены вещества, потреб-
ляемые жгутиком в процессе его дви-
жения.
Bodo свойствен анимальный способ пи-
тания. У основания жгутиков располо-
жен небольшой участок цитоплазмы, ли-
шенный пелликулы. В этой области и про-
исходит заглатывание пищи, каковой
обычно являются бактерии. В передней
трети тела (на уровне парабазального
тела) расположена сократительная ва-
куоля.
Большой интерес для понимания свя-
зей между саркодовыми и жгутиконос-
цами представляют простейшие, объеди-
няемые в группу ризомастигин (Rhizomas-
tigina), которая некоторыми зоологами
ПОДКЛАСС ЖИВОТНЫЕ ЖГУТИКОНОСЦЫ,
ИЛИ ЗООМАСТИГИНЫ (ZOOMASTIGINA)
104
рассматривается как самостоятельный о т-
р я д или подотряд жгутиконосцев,
а другими (например, известным фран-
цузским зоологом Г р а с с е) включает-
ся в к л а с с саркодовых. Это расхож-
дение во мнениях не случайно. Оно объ-
ясняется тем, что ризомастигины соче-
тают в своем строении признаки обоих
классов: у них имеются и псевдоподии
и жгутик (рис. 55). На прилагаемом ри-
сунке изображено строение одного из
представителей этой группы — Mastiga-
moeba aspersa. На переднем конце тела
расположен жгутик, а на поверхности
всего тела — хорошо развитые многочис-
ленные псевдоподии, несущие, особенно
на заднем конце, щетинкоподобные вы-
росты. Вся цитоплазма отчетливо разде-
лена на два слоя: наружную (светлую)
эктоплазму и внутреннюю (зернистую)
эндоплазму. Ядро помещается на переднем
конце тела, сократительная вакуоля —
на заднем. Размеры Mastigamoeba aspersa
достигают 100 мк. Встречаются эти орга-
низмы не очень часто, преимущественно
в болотных, богатых гумусовыми веще-
ствами водах.
Наличие таких форм, как различные
ризомастигины, показывает, что между
саркодовыми и жгутиконосцами сущест-
вуют тесные связи, что эти два класса
связаны филогенетически.
Паразитические жгутиконосцы, как уже
говорилось выше, многочисленны и разно-
образны. Мы остановимся лишь на не-
многих представителях, наиболее интерес-
ных в практическом или биологическом
отношении. Рассматривать мы их будем
не в порядке зоологической системы,
а по месту и характеру паразитирова-
ния.
Жгутиконосцы, паразитирующие в ки-
шечном тракте человека и позвоночных
животных. Кишечник позвоночных, а так-
же и человека является местом обитания
довольно многих видов жгутиконосцев.
Рассмотрим некоторых из них. Широко
распространен род трихомонад (Tricho-
monas), обладающий многочисленными ви-
дами. В кишечнике человека паразити-
рует Trichomonas hominis (рис. 56). Ме-
стом локализации являются тонкие и тол-
стые кишки. Строение его, несмотря на
небольшие размеры G—10 мк в длину,
4—5 мк в ширину), довольно сложно.
Тело почти округлое. На переднем конце
расположены 4 жгутика, каждый из кото-
рых берет начало от базального зерна
(блефаропласта). Пятый жгутик завора-
чивает назад и идет вдоль края тела,
прирастая к нему при помощи тонкой
цитоплазматической перепонки. Лишь са-
мый задний конец его торчит свободно.
Этот пятый жгутик вместе с перепонкой
образует так называемую у н д у л и-
рующую мембрану, которая на-
ходится в постоянном волнообразном дви-
жении и играет существенную роль в дви-
жении самого жгутиконосца. Через всю
цитоплазму от области расположения ба-
зальных зерен до самого заднего конца
проходит аксостиль. Он представляет
собой опорную эластичную нить, которая
Рис. 55. Жгутиконосец Mastigamoeba
aspersa, обладающий жгутиком и псев-
доподиями:
1 — жгутик; 2 — ядро; 3 — псевдоподии:
4 — эктоплазма; 5 — эндоплавма; 6 — со-
кратительная вакуоля.
105
Рис. 56. Жгутиконосцы рода Trichomonas:
А — схема строения, Б — Trichomonas hominis из ки-
шечника человека. J — жгутики; 2 — ундулирующая
мембрана, з — базальные зерна жгутиков; 5 — ак-
состиль.
является внутренним скелетом простей-
шего. Наличие его свойственно и мно-
гим другим паразитическим жгутиконос-
цам. Трихомонады живут в содержимом
кишечника, обладающем довольно густой
консистенцией, им приходится передви-
гаться в этой среде. Одно ядро помещает-
ся на переднем конце тела.
У трпхомонад имеется ротовое отвер-
стие, расположенное на переднем конце,
вблизи основания жгутиков, на стороне,
противоположной той, по которой прохо-
дит ундулирующая мембрана. Размно-
жаются трихомонады продольным деле-
нием.
В кишечнике человека трихомонады
встречаются довольно редко. При массо-
вом размножении могут, по-видимому,
вызывать понос.
Трихомонады встречаются, кроме ки-
шечника человека, в кишечнике многих
позвоночных животных, в частности кро-
ликов, мышей, ящериц.
В верхних отделах тонкого кишечника
человека довольно часто встречается еще
один вид паразитических жгутиконосцев—
лямблии (Lamblia intestinalis, рис. 57).
Этот вид обладает весьма сложным строе-
нием. Форма тела грушевидная, заостряю-
щаяся к заднему концу. Длина около
15 мк, ширина в передней части 7—8 мк.
Все строение лямблип строго двусторонне-
симметрично. По продольной длинной оси
ее можно рассечь на две зеркально рав-
ные половинки. Все тело довольно сильно
сплющено в спинно-брюшном направле-
нии. Спинная сторона выпуклая, брюш-
ная плоская. В передней части брюш-
ной поверхности лямблий имеется углуб-
ление, представляющее собой присоску,
при помощи которой жгутиконосец плот-
но присасывается к эпителиальным клет-
кам кишечника (рис. 57). Посередине тела
до самого заднего конца проходит двой-
ной аксостиль, состоящий из двух эластич-
ных нитей. Имеется четыре пары жгу-
тиков, расположение которых ясно видно
на рисунках. Задняя пара их является
продолжением аксостилей. Ядер два, они
лежат справа и слева от аксостилей
в области присоски.
Лямблии — очень нежелательные «го-
сти» кишечника человека. Нередко зара-
жение ими проходит без каких-либо
болезненных симптомов. Вместе с тем изве-
стны многочисленные случаи, когда за-
ражение лямблиями сопровождалось ки-
шечными расстройствами. Через желч-
ный проток из двенадцатиперстной кишки
лямблии могут проникать в желчный
пузырь, вызывая заболевания типа холе-
цистита. У человека лямблии гораз-
до чаще встречаются у детей, чем у взро-
слых.
Попадая в нижние отделы кишечника,
лямблии могут инцистироваться: они те-
ряют жгутики и одеваются оболочкой.
В состоянии цист они и выходят из кишеч-
ника наружу. Человек заражается, про-
глатывая цисты лямблий.
Лямблии встречаются и у многих мле-
копитающих, в частности у кроликов
и мышей. Однако у каждого вида хозяина
существует свой, свойственный только ему
106
вид лямблий. Лямблии кролика или мы-
ши не могут заразить человека. В этом
выражается явление узкой специфичности
этих видов паразитов.
Паразиты крови. Значительное число
видов жгутиконосцев являются парази-
тами крови. Их среда обитания — жидкая
часть (плазма) крови. Кровяные жгути-
коносцы относятся к р о д у трипаносом
(Trypanosoma). У человека паразитирует
Trypanosoma gambiense, возбудитель тя-
желого заболевания — сонной бо-
лезни. Это заболевание широко рас-
пространено в Экваториальной Африке.
На ранних стадиях заболевания трипано-
сома живет в плазме крови, на поздних
стадиях она проникает в спинномозговую
жидкость.
Строение трипаносом чрезвычайно свое-
образно и характерно (рис. 58). Тело их
продолговатое, суженное на обоих концах
и сплющенное в одной плоскости. Длина
равняется 15—30 мк. От переднего конца
вперед выдается короткий жгутик. Он про-
должается вдоль почти всего тела и соеди-
няется с его краем тонкой протоплазмати-
ческой перепонкой, образуя, как и у
трихомонад, ундулирующую мем-
брану. Несколько не доходя до зад-
него конца тела, базальная часть жгутика
входит внутрь цитоплазмы. У основа-
ния его расположено базальное зерно
и особое, резко окрашивающееся различ-
ными красками довольно крупное тельце,
которое носит название кинетопла-
с т а. Наблюдения за живыми трипано-
сомами в крови показывают, что движение
их осуществляется благодаря энергичным
волнообразным движениям ундулирую-
щей мембраны. Ядро одно, несколько
сдвинуто с середины тела кзади. В цито-
плазме содержатся резервные питательные
вещества в виде разной величины гра-
нул. Питание чисто осмотическое
(сапрофитное). Размножение обыч-
но осуществляется путем продольного
деления.
Заболевание человека сонной болезнью
при отсутствии лечения протекает очень
тяжело. Первый период заболевания ха-
рактеризуется явлениями лихорадки. Во
втором периоде (когда трипаносомы про-
никают в спинномозговую жидкость) на-
Рис. 57. Паразитирующий в кишечнике человека
жгутиконосец Lamblia intestinalis:
А — вид сбоку; Б — вид с брюшной стороны; В —
лямблия, присосавшаяся к эпителиальной клетке.
1 — жгутики, 2 — базальные зерна, з — присоска;
4 — ядро, 5 — парабазальное тело; 6 — аксостиль.
блюдаются нервно-психические явления,
сонливость, резкое истощение, которые
приводят к смерти. В настоящее время
существуют специфические лекарствен-
ные вещества (например, германии), кото-
рые излечивают это тяжелое заболевание.
В XIX в., когда терапия сонной болезни
еще не была разработана и неизвестны
были специфические лекарственные веще-
ства, сонная болезнь представляла страш-
ный бич Африки, от нее погибало огром-
ное количество людей.
Какими путями происходит заражение
человека трипаносомами, как проникают
они в кровяное русло? Этому вопросу были
посвящены многочисленные исследования,
в результате которых удалось выяснить
весь жизненный цикл возбудителя сонной
болезни. Оказалось, что переносчиком
трипаносомы является кровососущая му-
ха цеце (Glossina palpalis), широко рас-
пространенная в Африке (рис. 59).
107
Рис. 58. Различные формы жгутиконосца Trypa-
nosoma gambiense — возбудителя сонной боле-
зни человека.
Когда муха насосется крови от боль-
ного сонной болезнью, то в кишечнике
ее трипаносомы не погибают. Напротив,
они энергично размножаются, несколько
меняя при этом свою форму. Образуется
так называемая критидиальная
стадия, которая отличается от трипаносом
крови тем, что кинетопласт у нее сдвигает-
ся несколько вперед и оказывается рас-
положенным на уровне ядра. После пе-
риода размножения трипаносомы вновь
приобретают типичную форму и активно
проникают в слюнные железы и хоботок
мухи. Эти «метациклические формы» яв-
ляются инвазионными, т. е. способными
заражать. Если цеце с метациклическими
трипаносомами укусит человека, то три-
паносомы проникают в кровяное русло,
где и размножаются, вызывая заболева-
ние сонной болезнью.
Trypanosoma gambiense паразитирует,
кроме человека, в крови антилоп. Однако,
в отличие от человека, антилопы не стра-
дают от трипаносом, не обнаруживают
никаких симптомов заболевания. В при-
родных условиях антилопы являются по-
стоянным источником заражения челове-
ка, так как мухи цеце в одинаковой сте-
пени нападают на человека и на антилоп.
Сонная болезнь — это типичный
пример трансмиссивного (передающегося
через переносчика) заболевания с природ-
ной очаговостью (по терминологии акад.
Е. Н. Павловского). Борьба с та-
кими заболеваниями особенно затрудни-
тельна, так как не только больной человек
является источником распространения
заболевания, но, кроме того, сущест-
вует природный очаг возбудителя, пред-
ставляющий постоянную угрозу как пето1
чник инфекции.
Трипаносомы паразитируют, кроме че-
ловека, в самых разнообразных позвоноч-
ных животных (в рыбах, амфибиях, пре-
смыкающихся, птицах и млекопитающих).
Ряд видов трипаносом паразитирует и в
домашних животных, вызывая подчас
очень тяжелые заболевания.
Для крупного рогатого скота в Африке
одним из наиболее опасных и широко рас-
пространенных заболеваний является бо-
лезнь, называемая нагана. Она вызы-
вается одним из видов трипаносом (Trypa-
nosoma brucei), которая также передается
кровососущими мухами из рода Glossina.
В Южной Азии не менее распространена
другая болезнь — с у р р а, вызываемая
трипаносомой Trypanosoma brucei. Пере-
носчики ее — насекомые из отряда дву-
крылых—слепни. Однако в слепнях, в от-
личие от Glossina, трипаносомы не раз-
множаются и никаких морфологических
изменений не претерпевают. Слепни яв-
ляются чисто механическими передатчи-
ками инвазионного начала, так как трипа-
носомы, находясь у них в хоботке, сохра-
няют жизнеспособность несколько часов.
В Советском Союзе трипаносомные за-
болевания рогатого скота не играют сколь-
ко-нибудь заметной роли. Гораздо более
распространены они в южных районах
СССР у верблюдов и лошадей. Возбуди-
тель трипаносомозов верблюдов, по-ви-
димому, тождествен Trypanosoma evan-
si и передается слепнями. У лошадей
трипаносомы вызывают случную бо-
лезнь (возбудитель — Trypanosoma
108
equiperdum). Возбудитель этого заболе-
вания не передается переносчиками, а пе-
реходит от одного животного к другому
через слизистые половых путей в момент
случки.
К семейству трипаносомид от-
носятся также представители рода лей-
шманий (Leishmania), два вида которых
являются возбудителями тяжелых забо-
леваний человека. Leishmania tropica вы-
зывает кожное заболевание, известное под
именем л е й ш м а н и о з а или пен-
динской язвы. Эта болезнь широко
распространена в Южной Азии, Северной
Африке, в некоторых районах Южной
Европы (Италия). В дореволюционное вре-
мя пендинская язва часто наблюдалась
в России в Средней Азии (Туркестан, Уз-
бекистан). В настоящее время благодаря
большой лечебной и профилактической
работе, проведенной советскими органами
здравоохранения, заболевание это сведено
до минимума и встречается в виде редких
единичных случаев.
Заболевание начинается с того, что на
коже (чаще всего на открытых частях те-
ла — на руках, лице) появляется узелок,
увеличивающийся до размеров ореха, а за-
тем на этом месте возникает открытая
язва. Она очень долго не заживает A —
2 года), затем зарубцовывается. Возбуди-
тель заболевания — Leishmania tropica —
локализуется в подкожной клетчатке, пре-
имущественно в соединительнотканых клет-
ках (рис. 60), частью — в межклеточных
промежутках. Возбудитель представляет
собой мелкие овальные тельца длиной
4—7 мк, шириной 3—4 мк. Они в боль-
шом количестве заполняют клетки хозяи-
на. Каждая клетка паразита имеет одно
ядро и окрашивающееся тельце, которое
является не чем иным, как кинетонуклеу-
сом, столь характерным для трипаносом.
От кинетопласта отходит базальная часть
жгута. Однако ундулирующая мембрана
и свободная часть жгутика не выражены.
Это вполне понятно, так как при внутри-
клеточном паразитизме лейшмании, в
отличие от трипаносом, неподвижны. Одна-
ко если их выделить в культуру (они легко
культивируются на питательных средах),
то у лейшмании развиваются жгутики и
они активно плавают (рис. 61).
Каким же путем происходит заражение
лейшманиями? В настоящее время можно
считать доказанным, что переносчиками
лейшмании являются москиты — очень
мелкие кровососущие насекомые из отряда
двукрылых (рис. 62), нападающие на че-
ловека преимущественно в ночное время.
Москиты широко распространены в теп-
лых странах и многочисленны у нас
в Крыму, на Кавказе, в Средней Азии.
Личинки москитов развиваются во влаж-
ной почве. Экспериментально было выяс-
нено, что москиты заражаются лейшма-
ниями, которые в кишечнике у них пере-
Рис. 59. Переносчик сонной болезни кровососу-
щая муха Glossina palpalis (цеце):
1 —.со спинной стороны; 2 — сбоку, насосавшаяся
крови.
Рис. 60. Возбудитель кожного лейшманиоза
(пендинской язвы) Leishmania tropica:
1 — группа паразитов, выпавшая из клеток хозяи-
на; 2— клетка с многочисленными паразитами.
109
Рис. 61. Возбудитель висцерального лейшма-
ниоза (кала-азара) Leishmania donovani в куль-
туре. У жгутиконосца развиты жгутики, отсут-
ствующие при паразитировании его в тканях
хозяина.
Рис. 62. Переносчик лейшманиозов москит РЫе-
botomus pappatasii. Самка, увеличенная в 13 раз.
ходят в жгутиковую стадию (так называе-
мая «лептомонадная форма» — название,
обязанное сходству со жгутиконосцами
рода Leptomonas). Здесь же происходит
размножение лейшманий. Затем часть их
перемещается в самый передний отдел
кишечника и при укусе вводится в кожу
жертвы.
Многолетние исследования советских
ученых (главным образом проф. Н.Ла-
тышева и его сотрудников) показали,
что пендинская язва также является
трансмиссивным заболеванием с природ-
ной очаговостью. Лейшманиями могут за-
ражаться грызуны, а также собаки. Есте-
ственный цикл паразита обычно завер-
шается грызунами и обитающими в их же
норах москитами, минуя человека. До-
статочно человеку попасть в такой есте-
ственный очаг лейшманиоза, чтобы он за-
разился пендинской язвой вне зависимо-
сти от наличия таковой у окружающих
его людей.
Вторая болезнь человека, вызываемая
лейшманиями, — висцеральный
(внутренностный) лейшманиоз, или
кал а-а з а р. Возбудитель его Leish-
mania donovani морфологически неотли-
чим от описанной выше Leishmania tro-
pica. Переносчиками и здесь являются
москиты. Кала-азар также распростра-
нен в Южной Азии, Северной Африке,
Малой Азии. В Советском Союзе известен
в Средней Азии (Туркменской ССР). В Ев-
ропе встречается в Южной Италии, Гре-
ции. Это очень тяжелое заболевание, про-
являющееся в лихорадке, малокровии,
общем истощении и гипертрофии печени
и селезенки, при отсутствии лечения обыч-
но приводящее к смертельному исходу.
Возбудитель поражает различные внут-
ренние органы, особенно часто встречает-
ся в эндотелии кровеносных срсудов,
в селезенке, печени, костном мозгу, лим-
фатических железах. Это заболевание ча-
ще поражает детей, чем взрослых.
Кроме человека, носителями лейшма-
ний, вызывающих кала-азар, могут быть
собаки, которые в распространении этого
заболевания играют, по-видимому, нема-
ловажную роль.
Паразитические жгутиконосцы земно-
водных. В заднем отделе кишечника зем-
новодных обитают многочисленные виды
паразитических простейших — жгутико-
носцев и инфузорий, легко доступных для
наблюдения. Из жгутиконосцев особенно
богато представлены разные виды семей-
ства опалинид (Opalinidae). В заднем
отделе кишечника лягушек в огромных
количествах встречаются разные виды р о-
110
д а опалин (Opalina). Это очень крупные
простейшие, достигающие 1 мм. У наи-
более обычной в наших травяных лягуш-
ках О. ranarum, как и у других опалин,
тело плоское, листовидное. Оно не вполне
симметрично: косо срезано спереди и за-
круглено сзади (рис. 63). Поверхность
тела покрыта многочисленными жгутика-
ми, количество которых измеряется мно-
гими тысячами. Они расположены про-
дольными и несколько S-образно изгибаю-
щимися рядами. Хорошо разграничены
эктоплазма и эндоплазма. В эндоплазме
разбросано несколько десятков неболь-
ших ядер. Здесь мы встречаемся с при-
мером многоядерного простейшего. Со-
кратительных вакуолей у опалин нет.
Среди опалин нередко встречаются деля-
щиеся особи. Борозда деления при этом
проходит косо (рис. 63), что не позволяет
это деление рассматривать ни как продоль-
ное, ни как поперечное.
До недавнего времени опалин причис-
ляли к инфузориям в качестве самостоя-
тельного подкласса. Основанием для это-
го служило большое количество ресни-
чек (жгутиков), которые целиком покры-
вают все тело сплошными рядами (см.
ниже об инфузориях, стр. 134). Однако
у опалин отсутствуют основные черты,
характерные для инфузорий: ядерный ду-
ализм и половой процесс типа конъюга-
ции (см. стр. 139). Это заставило выделить
их из инфузорий и рассматривать как
своеобразное многожгутиковое простей-
шее из класса жгутиконосцев.
У опалин нет ротового отверстия и пи-
тание совершается осмотическим путем.
Как осуществляется заражение новых
особей хозяина, как заражаются ими
головастики? Основные этапы жизненного
цикла хозяина (лягушки) и паразита
(опалин) очень точно «синхронизированы»,
что обеспечивает заражение.
В лягушках, пойманных ранней весной
в момент икрометания, встречаются очень
мелкие особи опалин, образующиеся в ре-
зультате многократных делений. Это пред-
цистные формы. Они инцистируются и вы-
ходят из лягушки, падая на дно водоема.
Там они лежат до тех пор, пока не будут
проглочены головастиками. В их кишеч-
нике из цист вылупляются мелкие опали*
ны, которые путем деления дают начало
еще более мелким одноядерным клеткам,
являющимся половыми (гаметами).
Эти клетки попарно сливаются (половой
процесс — копуляция), причем про-
исходит слияние не только цитоплазмы,
но и ядер. Из образовавшихся зигот
в дальнейшем развиваются многоядерные
опалины. Интересно отметить, что поло-
вой процесс опалин совершается только
на одном этапе цикла — в головастиках.
Полового процесса у опалин в теле ля-
гушек никогда не происходит, и размно-
жаются они бесполым путем.
Жгутиконосцы, обитающие в кишечни-
ке термитов и тараканов. Термиты — это
большая группа (отряд) преимущественно
тропических насекомых, характерных сво-
им «общественным» образом жизни. На
территории Советского Союза термиты
встречаются в Средней Азии и в окрест-
ностях Одессы. Эти насекомые, образую-
щие большие колонии и строящие сложные
гнезда — термитники, питаются преи-
мущественно растительной пищей, в том
числе древесиной, поглощая ее в больших
количествах. В кишечнике термитов, а
именно в задней кишке, образующей боль-
шое расширение, живет огромное количе-
ство жгутиконосцев. Видовой состав их
исключительно разнообразен. В настоя-
Рис. 63. Паразитический жгутиконосец из задней
кишки лягушки Opalina ranarum. Видно большое
количество жгутиков и ядер.
1 — неделящаяся особь; 2 — деление, бороэда про-
ходит косо по отношению н длинной оси простейшего.
in
щее время описано свыше 200 видов жгу-
тиконосцев, обитающих в кишечнике тер-
митов. При этом нужно иметь в виду, что
в паразитологическом отношении изучено
не более 1/3 общего числа видов термитов.
Учитывая, что некоторые паразитические
простейшие обладают довольно узкой спе-
цифичностью, можно думать, что число
видов их значительно больше известного
в настоящее время.
Жгутиконосцы из кишечника термитов
относятся к двум отрядам: Polymas-
tigina и Hypermastigina. Это наиболее
сложно устроенные среди жгутиконосцев
формы. Их паразитический образ жизни
не только не привел к упрощению органи-
зации, но, напротив, связан с ее усложне-
нием. В этом сходство их с инфузориями,
населяющими кишечный тракт копытных
(рис. 64). Количество простейших в кишеч-
нике термитов бывает настолько велико,
что оно составляет иногда до */з веса
тела насекомого!
На рисунке 64 изображены некоторые
представители жгутиконосцев, обитаю-
щих в кишечнике термитов. Для многих
из них характерно большое количество
жгутиков, которые или сосредоточены на
переднем конце тела, или — в других
случаях — занимают значительную часть
его поверхности. Жгутики могут быть
расположены продольными рядами, по
спирали (относительно продольной оси
тела) или поперечными рядами. Иногда
жгутики собраны пучками на переднем
конце (рис. 64). Размеры жгутиконос-
цев различны, они варьируют от 30—40 мк
до 0,5 мм и более.
Ядерный аппарат у разных представи-
телей устроен неодинаково. Некоторые
виды (рис. 64,Д) имеют одно крупное ядро.
Другие являются многоядерными. Иногда
число ядер невелико (рис. 64,Б), и они рас-
полагаются венчиком на переднем конце
тела. Иногда же число ядер достигает не-
скольких десятков (рис. 64,Г), и они сосре-
доточены в передней трети тела.
Характерной особенностью строения не-
которых жгутиконосцев, обитающих в ки-
шечнике термитов, является наличие у них
опорных образований — аксостилей.
Пример таких эластических скелетных
структур мы уже видели у трихомонад
и лямблий (стр. 106). Часто имеется не
один, а целый пучок аксостилей (рис. 64,Г).
В цитоплазме жгутиконосцев, обитаю-
щих в кишечнике термитов, обычно имеет-
ся большое количество пищевых вклю-
чений. Это кусочки растительных тканей
разных размеров. Заглатывание пищи про-
исходит различными способами. Ротовое
отверстие у большинства видов отсут-
Таблица 4. Разные стадии жизненного цикла возбудите-
лей человеческой малярии в эритроцитах крови человека
A—15) и в слюнных железах комара-переносчика Anopheles
A6, 17) с препаратов, окрашенных по Гимза-Романовскому:
1—5 — возбудитель тропической малярии Plasmodium falciparum:
1—начало роста шизонта в эритроците, в одном эритроците два мел-
ких «кольца», в каждом из них по одному окрашенному в крас-
ный цвет ядру;
2, 3—более крупные «кольца», одно B) с двумя, другое C) с одним
ядром;
4—бобовидная макрогамета; 5—микрогаметоцит.
6—11 — возбудитель трехдневной малярии P. vivax:
6—молодой шизонт в начале роста в форме кольца;
7—более крупный одноядерйый шизонт амебоидной формы;
8—растущий шизонт с четырьмя ядрами;
9—выросший шизонт, распадающийся на одноядерные мерозоиты;
10—макрогамета; 11—микрогаметоцит.
12—15 — возбудитель четырехдневной малярии P. malariae:
12—растущий шизонт в форме пояска;
13—выросший шизонт, распадающийся на одноядерные мерозоиты;
14—макрогамета; 15—микрогаметоцит.
16— продольный разрез через слюнную железу комара Anopheles
maculipennis; в клетках железы, обладающих крупными ядрами,
расположены многочисленные веретеновидные спорозоиты, часть
их вышла в проток железы.
17—мазок слюнной железы комара Anopheles superpictus, заражен-
ного малярийным плазмодием; в центре крупная клетка железы
с ядром, вокруг нее многочисленные веретеновидные одноядер-
ные спорозоиты.
Рис. 64. Различные жгутиконосцы из кишечника термитов.
А — Teratonympha mirabilis; Б — Spirotrichonympha flagellata; В — Coronympha octonaria, Г — Calonympha
graesi; Д — Trichonympha turkestanica; E — Rhynchonympha tarda, l — ядра, 2 — аксостили.
ствует. Часть их цитоплазмы не имеет резко
очерченной пелликулы, а сохраняет спо-
собность образовывать выросты, напоми-
нающие псевдоподии. При помощи таких
выростов жгутиконосец заглатывает ча-
стицы растительных тканей, присутствую-
щие всегда в большом количестве в кишеч-
нике термита. На рисунке 65 изображены
последовательные стадии заглатывания
жгутиконосцем из рода трихонимфа
(Trichonympha) довольно крупного ку-
сочка древесины. Захват пищи осуще-
ствляется и другим путем. Для за-
глатывания мелких частиц на заднем
конце жгутиконосца образуется времен-
ное впячивание, в которое втягиваются
кусочки пищи. Впячивание замыкается
сзади, и пищевые частицы оказываются
лежащими в цитоплазме простейшего.
Установлено, что взаимоотношения меж-
ду термитами и населяющими их кишеч-
ник жгутиконосцами — это явление тес-
ного симбиоза: жгутиконосцы мо-
гут существовать только в кишечнике тер-
митов, содержимое которого является для
них средой обитания и источником пищи.
Но и термиты не могут существовать без
жгутиконосцев. Пищей для термитов слу-
жит почти исключительно клетчатка (дре-
весина). Переваривание заглоченной пи-
щи происходит в основном в заднем отделе
кишечника, заселенном жгутиконосцами.
В кишечнике самих термитов не выделяет-
ся ферментов, расщепляющих клетчат-
8 Жизнь животных, т. 1
113
Рис. 65. Жгутиконосец из кишечника термитов
Trichonympha sp., заглатывающий кусочек дре-
весины.
ку. Напротив, в теле симбиотических жгу-
тиконосцев исследованиями Хангейта об-
наружен фермент целлюлаза, расщепляю-
щий клетчатку. Жгутиконосцы переводят
клетчатку в растворимое состояние, ис-
пользуя при этом лишь часть переработан-
ного вещества. Таким образом, питание
термитов в значительной мере осуществля-
ется при участии симбиотических простей-
ших. Это доказывается прямыми опытами,
проведенными Кливлендом и под-
твержденными многими исследователями.
Оказывается, довольно простым методом
можно освободить термитов от жгутико-
носцев, не повреждая самих насекомых.
Если их поместить в чистый кислород при
давлении в 3 атмосферы, все простейшие
вскоре погибают, тогда как термиты оста-
ются живыми. После освобождения насе-
комых от простейших Кливленд помещал
их в нормальные условия с обильным ко-
личеством древесной пищи. В таких же
условиях жили и нормальные термиты,
т. е. содержащие простейших в кишечни-
ке. Термиты со жгутиконосцами жили
в условиях опыта в стеклянных сосудах
долгое время, до 1 — 1V2 лет, пока не на-
ступала естественная смерть их от старо-
сти. Напротив, насекомые, лишенные жгу-
тиконосцев, выживали не более 10—
14 дней и потом неизбежно погибали. При
этом они энергично поглощали пищу, но
она не могла у них перевариваться. Стои-
ло искусственно заразить таких «стериль-
ных» термитов жгутиконосцами, как они
приобретали способность переваривать пи-
щу и жить продолжительное время.
Мы уже говорили, что каждая особь
термита имеет в кишечнике жгутиконос-
цев. Возникает вопрос: какими путями
происходит проникновение в кишечник
молодых личинок их симбионтов — жгу-
тиконосцев? У термитов существует осо-
бый инстинкт слизывания друг у друга
жидких экскрементов, выступающих в
форме капельки из анального отверстия.
Термит, проглатывающий содержимое зад-
ней кишки особи того же вида, получает
некоторое количество усвояемой пищи,
переведенной в растворенное состояние
благодаря деятельности жгутиконосцев.
С другой стороны, проглатывая жгутико-
носцев, он вводит их в свой кишечник, где
они продолжают жить и размножаться.
Так же заражаются и молодые личинки.
В колонии термитов лишены жгутико-
носцев лишь половозрелые взрослые осо-
би. В термитнике обычно имеется одна
самка («царица»), непрерывно отклады-
вающая яйца. Эта самка не питается дре-
весиной. Ее кормят «рабочие», которые
представляют собой недоразвитых в поло-
вом отношении особей. Пищей служит
отрыжка из переднего отдела кишечника,
содержащая легкоусвояемые вещества.
Фауна жгутиконосцев развивается, кро-
ме термитов, в кишечнике растительнояд-
ных тараканов рода Cryptocercus.
Рост и развитие термитов, как и других
насекомых, сопровождается периодиче-
ской линькой, в процессе которой сбрасы-
вается тонкая хитиновая выстилка зад-
него отдела кишечника. При этом у боль-
шинства видов термитов выходят наружу
и жгутиконосцы, и насекомое после линь-
ки оказывается лишенным простейших.
Очень скоро они вновь появляются,
так как слинявшее насекомое начинает
энергично слизывать жидкие экскременты
окружающих особей. У тараканов Cryp-
tocercus во время линьки жгутиконосцы
заползают в просвет между отделяемой
внутренней выстилкой и эпителиальной
114
стенкой кишки, откуда по окончании
линьки вновь выходят и заполняют про-
свет кишечника.
У жгутиконосцев, обитающих в кишеч-
нике термитов, известно только беспо-
лое размножение путем деления.
Половой процесс у них отсутствует. Иное
дело жгутиконосцы из кишечника тара-
кана Cryptocercus. Работами Кливлен-
д а установлено наличие у них полового
процесса по типу копуляции, который про-
текает строго периодически. Оказалось,
что половые процессы жгутиконосцев на-
ходятся в строгом соответствии с линькой
личинок. Как известно, рост насекомых
на личиночных стадиях сопровождается
периодическим сбрасыванием хитинового
покрова, так как последний может рас-
тягиваться лишь до определенного пре-
дела, а затем, после линьки, заменяется
новым хитиновым покровом. Процесс
линьки — очень сложное явление. На-
ступление его связано с действием гормо-
нов, которые выделяются некоторыми эн-
докринными железами насекомого и обу-
словливают отслаивание старой хитино-
вой кутикулы. У разных видов жгутико-
носцев половой процесс происходит строго
закономерно в разные сроки подготови-
тельного к линьке периода или же вскоре
(через 1—2 дня) после завершения линьки.
Для каждого вида жгутиконосцев соотно-
шение срока наступления полового про-
цесса и линьки является строго опреде-
ленным. Очевидно, выделяемые хозяином
гормоны линьки воздействуют не только
на насекомое, но и на обитателей его кишеч-
ника, стимулируя половое размножение.
У взрослых тараканов, которые не растут
и не линяют, половых процессов у насе-
ляющих кишечник жгутиконосцев не про-
исходит.
Жгутиконосцы — паразиты растений.
Некоторые жгутиконосцы приспособились
к паразитированию в растительных тка-
нях. Хозяевами их являются растения
с млечным соком. Это виды семейств моло-
чайных, сложноцветных, маковых, ластов-
никовых и др. Жгутиконосцы—паразиты
растений относятся к р о д у Leptomo-
nas, принадлежащему к семейству
тршганосомид, с некоторыми представи-
телями которых мы познакомились выше
8*
Рис. 66. Жгутиконосец Leptomonas davidi
из млечного сока молочая.
(стр. 107). Виды этого рода имеют одно
ядро, один жгутик и кинетопласт, рас-
положенный впереди ядра (рис. 66).
Живут Leptomonas в сосудах, напол-
ненных млечным соком, в межклетниках,
в вакуолях отдельных клеток. Нередко
эти простейшие развиваются в раститель-
ных тканях в огромных количествах.
Массовое развитие жгутиконосцев на-
носит растениям большой вред, вызывает
разрушение хлорофилла, опадение листь-
ев и в конечном счете гибель растения.
Жгутиконосцы—паразиты растений рас-
пространены преимущественно в тропиче-
ских и субтропических странах, в умерен-
ной зоне встречаются реже. Очень боль-
шой вред паразитические жгутиконосцы
наносят в некоторых странах плантациям
кофе. Особенно широко это заболевание
кофе распространено в Гвиане (Южная
Америка). Оно носит название некроз фло-
эмы, сопровождается пожелтением и опа-
дением листвы. При сильном заражении
кофейные деревца погибают в течение
3—12 месяцев.
Перенос возбудителя и заражение рас-
тений осуществляется насекомыми. Обыч-
но переносчиками являются растительно-
ядные клопы, сосущие соки растений.
115
Обзор важнейших представителей
класса жгутиконосцев показал, на-
сколько разнообразны эти организмы.
Этот класс стоит как бы на границе рас-
тительного и животного мира, и многие
представители его совмещают столь раз-
личные типы обмена веществ, как ауто-
трофный и гетеротрофный.
Эволюция жгутиконосцев
шла в различных направлениях. Многие
ветви их развития привели к образованию
планктонных форм, примером которых яв-
ляются панцирные жгутиконосцы. Осо-
бенно широкое развитие получил у жгу-
тиконосцев паразитизм. Многократный пе-
реход к паразитизму осуществлялся в
разных линиях их филогенетического
развития. В ряде случаев на основе парази-
тизма возникли очень тесные симбиоти-
Класс споровиков представляет со-
бой обширную группу простейших орга-
низмов, ведущих исключительно пара-
зитический образ жизни. Приспособление
к паразитизму у них очень глубокое и со-
вершенное. У всех споровиков отсутству-
ют специальные органоиды захвата пищи,
и они питаются осмотическим путем. Хо-
зяевами их являются самые различные
беспозвоночные и позвоночные животные.
Несколько видов споровиков паразитирует
в человеке.
В процессе эволюции они приспосо-
бились к паразитированию в самых
различных органах и тканях. Многие спо-
ровики — паразиты кишечника и различ-
ных органов, связанных с пищеваритель-
ной системой (в том числе печени). Имеют-
ся виды, паразитирующие в органах выде-
лительной системы — почках. Органы
кровеносной системы и кровь также слу-
жат средой обитания некоторых споро-
виков.
В этой группе простейших есть немало
видов, приспособившихся к внутриклеточ-
ному паразитизму. Паразиты проникают
внутрь клеток различных тканей хозяи-
ческие взаимоотношения, примером кото-
рых могут служить жгутиконосцы кишеч-
ника термитов и некоторых тарака-
нов.
Интересным путем эволюции жгутико-
носцев является переход к колониальным
формам организации. Колонии их очень
разнообразны и достигают у высших пред-
ставителей (например, Vol vox) очень слож-
ной дифференцировки, выходящей уже
за пределы одноклеточности. Колонию
вольвокса, может быть, правильнее рас-
сматривать как многоклеточный организм,
поскольку, как мы видели, не все клетки
колонии равноценны (стр. 99). Возмож-
но, что колонна л ьность имела большое
значение в эволюции органического мира
и явилась переходным этапом к многокле-
точным животным.
на, питаются, растут и развиваются за
их счет.
Одной из форм глубокого приспособ-
ления споровиков к паразитизму явилась
выработка сложных и разнообразных жиз-
ненных циклов, обеспечивающих зараже-
ние хозяина. По ходу циклов происходит
смена разных форм размножения, веду-
щих, с одной стороны, к увеличению
числа паразитов в данной особи хозяина,
а с другой — к образованию таких ста-
дий, которые служат для заражения но-
вых особей хозяина.
У некоторых споровиков выработалось
не только чередование различных форм
размножения, но и смена хозяев, относя-
щихся к разным видам и группам живот-
ного мира.
Многие виды споровиков приносят боль-
шой вред как возбудители заболеваний
человека, домашних и промысловых жи-
вотных.
В данном разделе мы ограничимся
рассмотрением лишь немногих споро-
виков, представляющих наибольший био-
логический интерес или особенно большое
практическое значение.
ЕЛАСС СПОРОВИКИ (SPOROZOA)
не
ОТРЯД ГРЕГАРИНЫ (GREGARINIDA)
Грегарины — большая группа споро-
виков, включающая около 500 видов.
Все без исключения грегарины — пара-
зиты беспозвоночных. Наибольшее число
видов их обитает в кишечнике членисто-
ногих, в особенности часто у насекомых.
Некоторые грегарины паразитируют и в
других группах беспозвоночных — в коль-
чатых червях, иглокожих, оболочниках.
Кроме кишечных грегарин, каковых боль-
шинство, имеются виды, живущие в по-
лости тела, в органах размножения и др.
Наиболее сложно устроены грегарины,
живущие в кишечнике членистоногих. На
их примере мы и ознакомимся с организа-
цией представителей этого отряда.
На рисунке 67 показано строение взрос-
лой формы Corycella armata — грегарины
из кишечника личинки жука-вертячки
Gyrinus natator. Тело ее, достигающее
довольно значительной длины, разделено
на 3 отдела. В наиболее крупном заднем
отделе помещается ядро. Этот отдел называет-
ся дейтомеритом. Впереди от него рас-
Рис. 67. Грегарина Corycella ar-
mata из кишечника насекомых:
А — взрослая грегарина; Б — эпиме-
рит грегарины, внедрившийся в эпи-
телиальную клетку кишечника; В —
грегарина, сбросившая эпимерит.
1 — эпимерит; 2 — протомерит; 3 —
дейтомерит; 4 — ядро.
положен протомерит. Он отделен от
дейтомерита тонкой прослойкой светлой
цитоплазмы, представляющей собой про-
должение слоя эктоплазмы. На самом пе-
реднем конце грегарины расположен эпи-
мерит. Этот участок тела служит для при-
крепления грегарины к стенке кишечника.
Часто он бывает снабжен крючьями, иног-
да нитевидными выростами и может при-
нимать весьма причудливую форму. На
рисунке 68 изображены некоторые харак-
терные формы эпимеритов разных видов
грегарин. По достижении предельных раз-
меров, когда у грегарины наступают про-
цессы полового размножения, эпимерит обыч-
но отбрасывается. Он целиком состоит из
эктоплазмы.
Разделение цитоплазмы грегарин на
два слоя — эктоплазму и эндоплазму —
выражено всегда очень отчетливо. На-
ружный слой эктоплазмы образует проч-
ную пелликулу, определяющую постоян-
ство формы тела грегарин. У некоторых
грегарин в эктоплазме найдены тонкие
скелетно-опорные волоконца, еще более
увеличивающие прочность наружных сло-
ев тела. Немногие виды грегарин способ-
ны несколько менять форму своего тела,
сокращаться вдоль продольной оси. У та-
ких видов в эктоплазме, на границе с эндо-
плазмой, найдены сократимые волокон-
ца — миофибриллы.
Эндоплазма грегарины, резко отграни-
ченная от эктоплазмы, при рассматрива-
нии под микроскопом живых грегарин
выглядит темно-серой. Это обусловлено
наличием в эндоплазме большого количе-
ства гликогена, нередко сконцентриро-
ванного в виде мелких гранул. В этом
проявляется одна из характерных особен-
ностей многих внутренностных паразитов,
тело которых богато гликогеном. Кишеч-
ные паразиты живут в среде, в которой
свободный кислород присутствует в нич
тожяых количествах. Это затрудняет или
делает невозможными дыхательные про
цессы, сопровождающиеся поглощением
кислорода. В этих условиях паразитиче-
ские организмы вынуждены искать вза-
мен дыхания другие источники энергии.
Такими источниками оказываются про-
цессы анаэробного (без участия свобод-
ного кислорода) расщепления гликогена,
117
Рис. 08. Эпимериты различных
видов грегарин:
J — Gregarina longa; 2 — Sycia ini-
pitata; 3 — Pileocephalus heeri; 4 —
Stylocephalus longicollis; 5 — Beloides
firnus; 6 — Gometoides crinitus; 7 —
Geniorhynchus monnieri; 8 — Echi-
погаегч hispida.
процессы гликолиза. Не вдаваясь в хими-
ческую характеристику этих процессов,
отметим лишь, что при этом сложная мо-
лекула гликогена распадается на более
простые органические соединения (в ча-
стности, некоторые органические кисло-
ты). Этот процесс сопровождается выделе-
нием энергии. Однако в отношении коли-
чества освобождаемой энергии на единицу
веса (на грамм-молекулу) гликогена при
окислении выделяется во много раз боль-
ше энергии, чем при гликолизе. Окисле-
ние — гораздо более «экономичный» про-
цесс, чем гликолиз. Поэтому обычно у ор-
ганизмов, основным источником энергии
которых являются анаэробные гликолити-
ческие процессы, в теле имеются значи-
тельные скопления гликогена. Грегари-
ны — один из ярких примеров, иллюстри-
рующих это правило.
Ядра, расположенные всегда в дей-
томерите, имеют обычно вид пузырька,
богатого ядерным соком и снабженного
одним или несколькими крупными ядрыш-
ками.
Рассмотренное выше строение грегари-
ны свойственно преимущественно кишеч-
ным паразитам. Оно характерно для груп-
пы видов, объединяемых в семейство
Polycystidae. У представителей другого
семейства — Monocystidae, которые
паразитируют главным образом в поло-
сти тела, половых железах и других орга-
нах, строение более простое. У Monocys-
tidae нет разделения на эпи-, прото- и дей-
томерит, и тело их часто имеет червеобраз-
ную форму.
Наблюдая под микроскопом за живыми
грегаринами, легко обнаружить, что они
способны двигаться, медленно перемеща-
ясь в содержимом кишечника. Это движе-
ние носит характер плавного скольже-
ния, не сопровождающегося изменением
формы тела.
Механизм этого процесса до настоящего
времени не вполне ясен, поскольку у гре-
гарин нет никаких специальных органои-
дов движения. Исследования Ш е в я к о-
в а и других авторов показали, что на
заднем конце движущейся грегарины вы-
деляется слизь. По-видимому, она выбра-
сывается наружу с известной силой, бла-
годаря чему и происходит — по принципу
ракетного двигателя — поступательное
движение грегарины.
У настоящих грегарин х отсутствует
бесполое размножение, они размножаются
только половым путем. Рассмотрим типич-
ный цикл развития грегарин (рис. 69).
Живущие в кишечнике насекомого гре-
гарины, достигнув предельного роста, те-
ряют эпи мерит и соединяются по-
парно. Вокруг такой парочки (сизи-
гия) выделяется общая оболочка (ц и-
с т а). Вскоре после инцистирования в
1 Кроме настоящих грегарин (отряд Eugrega-
rinida), имеется небольшой отряд шизогрегарин
(Schizogregarinida), размножающихся бесполым
путем.
118
каждой из соединившихся в сизи-
гий особи начинается многократ-
ное последовательное деление яд-
ра. В каждой особи образуется
множество ядер. К концу этого
процесса все ядра располагаются
по периферии грегарины и вокруг
каждого из них обособляется не-
большой участок цитоплазмы. Так
формируется множество мелких
одноядерных клеток. Большая
часть цитоплазмы при этом оста-
ется неиспользованной. Это так
называемое остаточное тело, кото-
рое в дальнейшем процессе раз-
множения участия не принимает.
Мелкие одноядерные клетки пред-
ставляют собой половые клетки —
гаметы. Эти клетки сближаются
попарно и сливаются.
У некоторых видов грегарин
гаметы, образуемые обоими соеди-
нившимися в сизигий особями, по
размерам и строению неотличимы
друг от друга. У других видов
они различны. Одна особь дает на-
чало округлым и неподвижным
гаметам. Это женские гаметы.
Другая особь образует удлинен-
ные гаметы, снабженные жгутико-
видным выростом. Это мужские
подвижные гаметы.
Образовавшиеся в результате
оплодотворения зиготы сразу же
покрываются оболочкой, принимая
при этом веретенообразную, а не-
редко бочонковидную форму. По-
крывшаяся оболочкой зигота но-
сит название о о ц и с т ы. Обо-
лочка цисты в этот период цикла
развития еще не разрушается. Та-
ким образом, многочисленные образовав-
шиеся после копуляции гамет ооцисты
лежат внутри оболочки цисты. На этой ста-
дии, а иногда и несколько позже, цисты с
ооцистами выводятся из кишечника нару-
жу. Последним этапом цикла развития гре-
гарины является процесс формирования
ооцисты, получивший название спорого-
нии. Он сводится к следующему. Внутри
ооцисты ядро (образовавшееся в резуль-
тате слияния ядер гамет) трижды делится.
Вокруг каждого из возникших таким пу-
Рис. 69. Цикл развития грегарины Stylocephalus longi-
collis:
1—4 — в кишечнике жука Blaps mortisaga; 1 — спорозоит;
2 — гамонт; 3 — соединение двух особей, образование сизи-
гия; 4 — циста с двумя округлившимися гамонтами; 5—15 —
во внешней среде; 5, 6 — деление ядер гамонтов, ведущее к
образованию половых клеток (гамет); 7 — сформированные га-
меты; 8 — копуляция гамет; 9 — зигота, образовавшая оболочку
(ооциста); 10—14 — развитие спорозоитов в ооцисте (спорого-
ния); 15 — спорозоиты, выходящие из споры.
тем ядер обособляется участок цитоплаз-
мы. В каждой ооцисте получается восемь
очень маленьких червеобразной формы кле-
ток, называемых сп о р о з о итам и. На
этой стадии ооцисты способны к зараже-
нию новых особей хозяина. Если циста
с ооцистами или даже отдельная ооциста
будет проглочена насекомым — хозяином
данного вида грегарин, то под воздейст-
вием кишечного сока спорозоиты выходят
из ооцисты. Они подвижны. В дальнейшем
из каждого спорозоита развивается гре-
119
гарина. Достигнув типичного для данного
вида размера, грегарины вновь соеди-
няются попарно, инцистируются, и цикл
начинается снова.
Таким образом, рассмотренный выше
цикл развития грегарин можно отчетливо
разделить на три этапа. Первый — это
рост и развитие взрослой грегарины.
Второй — образование половых клеток
(гамет) и оплодотворение. Третьим эта-
пом цикла является спорогония — раз-
витие зиготы (ооцисты) и формирование
спорозоитов.
ОТРЯД КОКЦИДИИ (COCCIDIA)
Так же как и грегарины, кокцидии пред-
ставляют собой обширную группу парази-
тических простейших, включающую около
400 видов. Паразитируют они в самых раз-
личных группах беспозвоночных и позво-
ночных животных: кольчатых червях, мол-
люсках, членистоногих, позвоночных. Для
кокцидии характерен внутриклеточный
паразитизм в различных органах и тка-
нях. Их жизненный цикл слагается (за
единичными исключениями) из законо-
мерного чередования бесполого размно-
жения, полового процесса и спорогонии.
У одних видов спорогония совершается
вне организма хозяина, в наружной сре-
де; у других, при наличии смены хозяев,
бесполое размножение и спорогония про-
исходят в организме разных хозяев.
Некоторые виды кокцидии являются
опасными паразитами домашних живот-
ных, домашней птицы и некоторых разво-
димых в рыбоводческих хозяйствах рыб.
Один очень редкий вид кокцидии парази-
тирует в человеке. Рассмотрим один при-
мер из числа наиболее обычных и вместе
с тем патогенных видов.
В кроликах паразитирует несколько
видов рода эймерия (Eimeria). Большая
часть их — паразиты кишечника, один
вид локализуется в печени. Эймерии —
кокцидии, развитие которых не сопровож-
дается сменой хозяев. В их жизненном
цикле имеется два основных этапа. Пер-
вый этап цикла протекает в кишечнике
кролика. Это эндогенная часть цикла.
Второй этап проходит вне тела хозяина,
во внешней среде—экзогенная часть цикла.
Eimeria magna — один из наиболее ши-
роко распространенных видов кокцидии
кролика. На рисунке 70 схематически
представлены основные этапы жизненно-
го цикла этого паразита.
Начнем рассмотрение жизненного цикла
Eimeria magna с момента, когда паразит
проникает в кишечник хозяина. Первой
стадией эндогенной части цикла является
спорозоит, выходящий из заглоченной
кроликом ооцисты (строение и развитие
ооцист будет рассмотрено ниже). Споро-
зоиты — это очень маленькие клетки (дли-
ной около 8 мк, шириной 2—3 мк), верете-
новидной формы с одним ядром. Они ак-
тивно внедряются в клетки эпителия ки-
шечника, где тотчас же округляются,
становятся шаровидными. За внедрением
кокцидии в клетку хозяина следует ак-
тивный рост: паразит увеличивается в раз-
мерах. Питание его осуществляется осмо-
тическим путем. На этой стадии развития
паразит носит название шизонта в соот-
ветствии с тем способом размножения,
к которому он приступает.
По мере роста паразита начинается деле-
ние ядра, благодаря чему шизонт стано-
вится многоядерным. Число ядер варьиру-
ет в довольно широких пределах — от 8
до 60. Растущий шизонт разрушает эпите-
лиальную клетку и постепенно переме-
щается в субэпителиальный слой соеди-
нительной ткани. Развитие шизонта завер-
шается бесполым размножением. Вокруг
каждого из ядер обособляется участок
цитоплазмы, в результате чего весь шизонт
распадается на одноядерные мелкие клет-
ки, каждая из которых имеет веретено-
видную форму (рис. 71). На этом закан-
чивается процесс бесполого размножения
шизонта. Такое размножение носит на-
звание множественного деления
или шизогонии. Весь процесс от момента
внедрения спорозоита в эпителиальную
клетку до завершения шизогонии занимает
80—96 часов. Одноядерные веретеновидные
клетки, образовавшиеся в результате ши-
зогонии, называют мерозоитами. Какова
их дальнейшая судьба?
Мерозоиты вновь внедряются в эпите-
лиальные клетки кишечника и дают на-
120
чало второму поколению шизонтов. Про-
цесс развития шизонтов второго поколе-
ния занимает несколько больше времени,
чем первого,— 110—132 часа.
Часть мерозоитов от второго поколения
дает начало третьему поколению шизон-
тов. Образующиеся в результате его меро-
зоиты, так же как и часть мерозоитов вто-
рого поколения, не образуют шизонтов.
Внедряясь в эпителиальные клетки, они
дают начало гаметам (половым клеткам).
Гаметы кокцидий резко дифференциро-
ваны в половом отношении на мужские
микрогаметы и женские макрогаметы. Об-
разование тех и других из мерозоитов
протекает различно.
Стадии развития кокцидий, дающие на-
чало микрогаметам, называют
микрогаметоцитами. При раз-
витии их, сопровождающемся энергичным
ростом, деление ядер начинается на самых
ранних стадиях роста, оно совершается
гораздо чаще, чем при шизогонии (рис.
72). В результате в цитоплазме распола-
гается несколько сот мелких ядер. Микро-
гаметы образуются путем вытягивания
отдельных ядер. Тело их почти целиком
состоит из веретеновидно вытянутого яд-
ра с очень тонким периферическим слоем
цитоплазмы. На переднем конце каждой
зрелой гаметы имеются два жгута. Длина
их очень мала — 4—3 мк. Гаметы подвиж-
ны. Большая часть цитоплазмы микрога-
метоцита не идет на образование гамет,
а остается неиспользованной в виде боль-
шого так называемого остаточного тела.
Совсем иначе происходит развитие жен-
ских половых клеток — макрогамет
(рис. 73). Происходит рост цитоплазмы, не
сопровождающийся делением ядра, раз-
меры которого увеличиваются. Ядро при-
обретает пузыревидное строение, в центре
его расположено крупное ядрышко. В ци-
топлазме по мере роста происходит накоп-
ление большого количества гранул. Мак-
рогаметы сначала имеют сферическую фор-
му, к концу развития они становятся
овальными. Часть гранул цитоплазмы
приближается к поверхности и распола-
гается периферически в один слой. За
счет этих гранул в конце развития макро-
гаметы образуется оболочка. На одном из
полюсов макрогаметы в оболочке остается
Рис. 70. Цикл развития кокцидий Eimeria magna:
J—первое поколение шизонтов; //—2-е поколение ши-
вонтов; 111 — 3-е поколение шизонтов; IV — гаметого-
ния; V — спорогония. 1 — спорозоиты; 2 — молодой
шизонт; 3 — растущий шизонт с многими ядрами;
4 — шизонт, распавшийся на мерозоиты; 5 — меро-
воиты; 6 — развитие макрогаметы; 6а — развитие ми-
крогамет; 7 — микрогамета; 8 — ооциста; 9 — ооциста,
вышедшая из кишечника кролика; 10 — ооциста с че-
тырьмя споробластами и остаточным телом; 11 — раз-
витие спор; 12 — ооциста с 4 зрелыми спорами
(в каждой споре по 2 спорозоита).
отверстие (микропиле), через которое про-
никают микрогаметы.
Следующий важный этап развития кок-
цидий — это процесс оплодотворения. По-
движные микрогаметы приближаются
вплотную к макрогаметам. Одна микрога-
мета проникает через микропиле в макро-
гамету, причем ядра гамет сливаются, об-
разуя характерную веретеновидную фи-
гуру. В момент оплодотворения форми-
руется вторая (внутренняя) оболочка,
микропиле закрывается особой слизистой
121
Рис. 71. Рост шизонта и шизогония кокцидии
Eimeria magna:
1 — молодой шизонт, ядро еще не приступило к
делению; 2—4 — увеличение числа ядер путем де-
ления и рост шизонта; 5 — распад шизонта на
мерозоиты •
Рис. 72. Развитие микрогамет Eimeria magna:
1 — молодой микрогаметоцит с неразделившимся
ядром; 2—4 — стадии роста микрогаметоцита, сопро-
вождающиеся делением ядер, 5 — формирование мик-
рогамет; 6 — микрогаметы, каждая снаб :,енная двумя
жгутиками.
пробкой и зигота попадает в просвет ки-
шечника. Обладающая двумя оболочками
зигота получает название ооцисты.
На этом заканчивается эндогенная часть
цикла, так как в кишечнике в отсутствие
кислорода ооцисты эймерий кролика даль-
ше развиваться не могут.
Экзогенная часть цикла сводится к про-
цессу спорогонии и осуществляется вне
тела хозяина. Из кишечника наружу ооци-
сты выходят вместе с фекальными масса-
ми. Только что вышедшие из кишечника
ооцисты имеют овальную форму (рис.
70). Все протоплазматическое содержи-
мое отделено от оболочки и занимает
центральную часть ооцисты. Споруляция
сводится к тому, что после двукратного
деления ядра протоплазматическое содер-
жимое ооцисты распадается на четыре од-
ноядерных споробласта. Каждый из них
выделяет свою оболочку, превращаясь
в спору. Обычно не все протоплазматиче-
ское содержимое ооцисты идет на обра-
зование споробластов, часть его остается
неиспользованной в виде остаточного тела.
В каждой споре ядро делится еще один
раз, и образуются два червеобразных од-
ноядерных спорозоита. На этом процесс
спорообразования заканчивается. Размеры
зрелых ооцист Eimeria magna: длина
25—40 мк, ширина 18—30 мк.
Таким образом, зрелая, способная к за-
ражению ооциста содержит четыре споры,
с двумя спорозоитами каждая.
При попадании зрелой ооцисты в кишеч-
ник кролика под воздействием кишечного
сока пробка ооцисты разрушается, споро-
зоиты выходят из оболочки спор и про-
никают в клетки эпителия кишечника,
начиная новую, эндогенную часть цикла.
В рассмотренном выше жизненном цик-
ле кокцидии эймерий следует различать
три основных этапа, из которых два пер-
вых относятся к эндогенной, а третий —
к экзогенной части цикла.
Цикл начинается с нескольких по-
колений бесполого размножения — ши-
зогонии. Это первый этап. Биологи-
ческое значение его очевидно: это увеличе-
ние числа особей паразита в данном экземп-
ляре хозяина. Благодаря шизогонии коли-
чество паразитов возрастает в огромной
степени. У кролика, проглотившего даже
небольшое количество ооцист, благодаря ши-
зогонии через короткий срок в кишечнике
окажется огромное количество паразитов.
Второй этап — это образование
половых клеток и оплодотворение. Он
завершается формированием тех стадий
жизненного цикла паразита (в данном
случае ооцист), при помощи которых про-
исходит распространение паразита на но-
вые особи хозяина, т. е. расселение вида.
Третий этап — спорогония.
Он приводит к тому, что «расселительные»
стадии жизненного цикла становятся спо-
собными к инвазированию (заражению) хо-
зяина. Зрелые ооцисты кокцидий обладают
большой стойкостью к действию различ-
ных неблагоприятных факторов среды.
Наличие двух оболочек является не толь-
ко механической, но и химической защи-
той протоплазмы паразита. Это чрезвы-
чайно затрудняет борьбу с кокцидиями,
так как применение большинства распро-
страненных дезинфицирующих средств
оказывается неэффективным или мало-
эффективным. Ооцисты очень долго сохра-
няют жизнеспособность: в течение многих
месяцев, а в некоторых случаях до года
и более.
Кокцидий — паразиты с очень узкой
специфичностью. Почти все виды их строго
приурочены к одному виду хозяина и не
могут паразитировать даже в близких
видах. Например, кокцидий кролика не
могут заражать зайцев, а кокцидий зай-
цев — развиваться в кроликах. В преде-
лах организма хозяина кокцидий локали-
зуются в строго определенных органах
и даже частях органов. Например, Е. М.
Хейсиным было доказано, что у кро-
лика разные виды кокцидий локализу-
ются в строго определенных отделах ки-
шечника (рис. 74).
Как уже отмечалось, кокцидий прино-
сят большой вред. Следует отметить, что
большинство кокцидиозов (заболева-
ний, вызываемых кокцидиями) поражает
молодых птиц и млекопитающих, это глав-
ным образом «детские болезни», хотя и
взрослые животные могут заражаться
кокцидиями.
Ряд видов кокцидий (Eimeria tenella,
Е. praecox и др.) поражает кур (преиму-
щественно цыплят), вызывая нередко ги-
бель молодняка.
Крупный рогатый скот также страдает
от кокцидий, причем заболевает преиму-
щественно молодняк (Eimeria zurni,
Е. smithi, Isospora aksaica).
В рыбоводных хозяйствах (особенно на
Украине) кокцидиозы неоднократно пора-
Рис. 73. Развитие макрогамет Eimeria magna:
J—5 — последовательные стадии роста и формирова-
ния макрогаметы.
Рис. 74. Локализация разных видов кокцидий
рода Eimeria в кишечнике кролика:
1 — Eimeria media; 2 — Е. irresidua, 3 — Е. рег-
forans; 4 — Е. magna; 5 — Е. intestinalis; 6 — Е.
coecicola; 7 — Е. piriformis.
жали карпов (Eimeria cyprini), что также
часто приводило к гибели рыб.
Борьба с кокцидиозами — важная зада-
ча животноводства и ветеринарии. Она
строится главным образом на профилак-
тических мероприятиях, направленных на
предохранение животных от заражения.
ОТРЯД КРОВЯНЫЕ СПОРОВИКИ
(HAEMOSPORIDIA)
Кровяные споровики, как показывает и
само название отряда, приспособились к
паразитированию в крови позвоночных
животных, главным образом млекопитаю-
щих и птиц. Они являются внутриклеточ-
ными паразитами. Место локализации па-
разитов — кровяные клетки. Число видов
кровяных споровиков превосходит сотню.
Жизненные циклы кокцидий и кровя-
ных споровиков очень сходны. Они сла-
гаются из тех же основных этапов: зако-
номерной смены шизогонии, развития га-
мет, оплодотворения и спорогонии. В от-
личие от кокцидий, здесь совершенно от-
сутствуют экзогенные стадии, половая и
бесполая части цикла распределяются
между двумя хозяевами, из которых один
является позвоночным (бесполая часть
цикла), а другой — беспозвоночным, обыч-
но насекомым (половая часть цикла и
спорогония).
В этих условиях защитные оболочки
ооцист и спор, столь характерные для
кокцидий, лишены биологического зна-
чения и в процессе эволюции исчезают.
Кровяные споровики имеют очень боль-
шое практическое значение для медици-
ны, так как к этому отряду относится
возбудитель тяжелого заболевания чело-
века — малярии. Существует четыре
вида рода Plasmodium, вызывающие ма-
лярию (P. vivax, P. malaria, P. falcipa-
rum, P. ovaleI. Клиническая картина
болезни, вызываемой разными видами па-
разита, несколько различна. О некоторых
из этих различий будет сказано ниже.
Жизненный цикл всех четырех видов про-
текает сходно, что позволяет рассмотреть
его для всего рода Plasmodium.
Человек является для Plasmodium хо-
зяином, в котором протекают стадии бес-
полого размножения (шизогония). Поло-
вой процесс и спорогония осуществля-
ются в переносчике, каким для возбу-
дителей человеческой малярии служат
1 Первые три вида распространены широко,
Plasmodium ovale встречается редко—в тропиче-
ском поясе Африки и Азии.
виды комаров, относящиеся к роду Ano-
pheles.
Обратимся к рассмотрению жизненного
цикла Plasmodium и начнем его с той же
стадии спорозоита, как мы это делали
выше для кокцидий (рис. 75). Спорозоит
проникает в кровь человека при укусе
зараженным комаром. Он представляет
собой очень маленькую одноядерную вере-
теновидно вытянутую клетку длиной 10—
15 мк при ширине всего в 1 мк. С током
крови спорозоиты разносятся по телу
человека и попадают в печень. Здесь они
активно внедряются в клетки печени и
превращаются в шизонтов, достигающих
значительной величины. Каждый шизонт
распадается в процессе шизогонии на
большое количество мерозоитов, которые
могут вновь внедряться в клетки печени.
После одного или большего числа бес-
полых поколений в печени шизонты дают
мерозоитов, судьба которых оказывается
иной. Они внедряются не в печеночные
клетки, а в эритроциты крови, где дают
начало шизонтам гораздо меньших раз-
меров, чем шизонты, развивающиеся в
печени. Начинается период шизогонии в
кровяном русле. Число мерозоитов, на
которые распадаются шизонты крови, от-
носительно невелико, обычно 8—16.
Шизонты на ранних стадиях развития
имеют характерную форму колечка благо-
даря тому, что центр их тела занят боль-
шой вакуолей (табл. 4). По мере роста
шизонта вакуоля исчезает и он приобре-
тает форму маленькой амебы. Наблюде-
ния за живыми паразитами показал pi, что
шизонтам свойственно довольно активное
амебоидное движение внутри красного
кровяного тельца. Мерозоиты, получаю-
щиеся в результате размножения шизон-
та, представляют собой мелкие клетки
овальной формы диаметром около 2 мк.
По завершении шизогонии оболочка эрит-
роцита лопается и мерозоиты оказываются
плавающими в плазме крови. По мере
роста шизонт поглощает содержимое
эритроцита. Красный пигмент крови (ге-
моглобин) при этом изменяется химически
и превращается в темно-коричневый, поч-
ти черный меланин, откладывающийся в
цитоплазме паразита в виде мелких зерен.
В момент шизогонии пигмент выбрасы-
124
вается из тела паразита и остается в крови
в виде остаточного тела.
Шизогония в крови осуществляется с
большой правильностью в отношении вре-
мени роста и размножения шизонта. У ви-
дов Plasmodium vivax, P. falciparum,
P. ovale шизогония происходит каждые
48 часов, у P. malaria — каждые 72 часа.
С моментом завершения шизогонии свя-
заны характерные клинические явления:
повышение температуры до 40° С и выше,
озноб. Эти явления обусловлены тем, что
при разрушении эритроцитов в кровь
поступают токсические вещества — ре-
зультат жизнедеятельности паразита, ко-
торые в период роста шизонта были изо-
лированы оболочкой эритроцита.
Мерозоиты вновь внедряются в эритро-
циты, и весь цикл шизогонии начинается
сначала. После каждой шизогонии число
паразитов в крови все более и более воз-
растает.
Шизогония, однако, идет не беспре-
дельно. Через несколько циклов бесполого
размножения часть внедрившихся в эрит-
роциты мерозоитов превращается в ста-
дии развития, подготовительные к обра-
зованию половых клеток, которые полу-
чают название гаметоцитов. Гаметоциты
образуются двух категорий: микро- и мак-
рогаметоциты. Первые в дальнейшем дают
начало микрогаметам (мужским половым
клеткам), вторые — макрогаметам (жен-
ским половым клеткам). Между этими
двумя категориями гаметоцитов имеются
некоторые небольшие различия в строе-
нии, которые в основном сводятся к тому,
что макрогаметоциты богаче снабжены ре-
зервными запасными включениями.
Для дальнейшего развития гаметоциты
должны попасть в кишечник комара ано-
фелес, что происходит при сосании им
крови больного малярией. Судьба микро-
и макрогаметоцитов различна. Каждый
микрогаметоцит в желудке комара дает
начало 4—8 нитевидным подвижным мик-
рогаметам. Макрогаметоциты без деления
преобразуются в макрогаметы. В просвете
желудка комара происходит слияние муж-
ских и женских гамет (оплодотворение),
и образуется овальной формы зигота.
Зигота (которая здесь благодаря своей
подвижности получает название оокине-
ты) прободает желудок и закрепляется
на его стенке, обращенной в сторону
полости тела, превращаясь в ооцисту.
Здесь она одевается оболочкой (эта обо-
лочка несравнима с оболочкой ооцист
кокцидий: у плазмодия она образуется
за счет тканей комара), после чего начи-
нается очень быстрый рост, в результате
которого объем ооцисты увеличивается
во много сот раз. По мере роста протоплаз-
мы ооцисты происходит многократное де-
ление ядра. Процесс развития ооцисты
на кишечнике завершается образованием
длинных (до 14 мк) спорозоитов, очень
тонких A мк) одноядерных клеток (но
спор, как у кокцидий, здесь не образу-
ется). В каждой ооцисте формируется ог-
ромное количество (до 10 тыс. в одной
ооцисте) спорозоитов. Зрелая ооциста ло-
пается, и спорозоиты попадают в запол-
ненную гемолимфой (кровью) полость тела
комара. Интересна их дальнейшая судь-
ба. Спорозоиты активно перемещаются,
совершая червеобразные движения. Пере-
мещение их не беспорядочно. Они направ-
ляются в слюнные железы комара, в кото-
рых собираются в огромных количествах
(табл. 4).
Анофелесы, у которых спорозоиты запол-
няют слюнные железы, являются источ-
ником заражения человека. Спорозоиты
при укусе проникают в кровяное русло
человека, и цикл развития плазмодия на-
чинается вновь.
Сколько времени требуется для завер-
шения цикла развития малярийного плаз-
модия в комаре и при каких условиях
среды он протекает?
Быстрота развития плазмодия в комаре
в большой степени зависит от темпера-
туры. Цикл развития Plasmodium vivax
при 17—18° С завершается в комаре в те*
чение трех недель. При более высоких
температурах среды он протекает значи-
тельно быстрее. Например, при 25—27° С
для развития плазмодия требуется лишь
одна неделя. Если температура окружаю-
щей среды ниже 15—17° С, то развитие
его начинается нормально, происходит
оплодотворение и образование оокинеты,
но спорозоиты не формируются.
Указанные закономерности зависимости
развития человеческих видов Plasmodium
125
в переносчике от температуры объясняют
основные факты географического распро-
странения малярии. Ареал комаров рода
Anopheles значительно шире, чем ареал
возбудителя малярии. Комары могут жить
и развиваться при температурах более
низких, чем это необходимо для развития
Plasmodium. Поэтому наличие Anopheles
далеко не всегда означает возможность
существования в данной местности маля-
рии.
Малярия — болезнь преимущест-
венно теплых стран, обладающих необхо-
димыми условиями для развития кома-
ров анофелесов. Личинки этих насеко-
мых, развивающихся с полным превра-
щением, живут в воде, в мелких стоячих
водоемах (рис. 76), сохраняя, однако, воз-
душное дыхание. На заднем конце тела
личинки расположены дыхальца, через
которые воздух проникает в их дыхатель-
ную (трахейную) систему. Личинки перио-
дически поднимаются на поверхность воды
для дыхания. Куколки комаров тоже
живут в воде и дышат атмосферным воз-
духом. Лишь взрослый комар является
воздушным насекомым. Яйца анофелесы
откладывают в воду.
Борьба с малярией осуществляется дву-
мя основными путями. Первый — это ле-
чение маляриков специальными лекарст-
венными средствами (хинин, акрихин,
плазмоцид и др.)- Отсутствие возбудителя
в крови людей делает невозможным зара-
жение переносчиков и, следовательно, рас-
пространение малярии.
При этом необходимо отметить одну
важную особенность, имеющую сущест-
венное значение для борьбы с малярией.
Дело в том, что большинство специфич-
ных лекарственных средств убивает ши-
зонтов в крови. Гаметоциты же являются
гораздо более стойкими и могут сохра-
няться в эритроцитах продолжительное
время в отсутствие шизонтов. Больной
малярией после лечения становится впол-
не здоровым и часто не подозревает, что
является носителем гаметоцитов. Он мо-
жет явиться источником заражения дру-
гих людей, если комар насосется его кро-
ви с гаметоцитами. Поэтому в малярий-
ных местностях необходим систематиче-
ский контроль за всеми переболевшими
малярией лицами. У них периодически
берутся мазки крови и исследуются на
носительство гаметоцитов.
В случае обнаружения гаметоноситель-
ства проводится специальный курс лече-
ния лекарственными препаратами, разру-
шающими гаметоцитов.
Второй путь борьбы с малярией — это
уничтожение переносчиков. Методы этой
борьбы разнообразны. В основном это
уничтожение личинок путем заливки во-
доемов керосином или нефтью. Личинки,
поднимающиеся на поверхность для дыха-
ния, наталкиваются на пленку нефти,
которая, заполняя трахеи, вызывает их
гибель.
Существуют и другие методы борьбы
с личинками. Особенно интересен биоло-
гический метод борьбы с использованием
небольших живородящих рыбок — гам-
бузий (Gambusia affinis, рис. 77). Эти
теплолюбивые рыбки (родина их Америка)
особенно охотно поедают личинок кома-
ров. Водоемы, являющиеся местами вы-
плода личинок Anopheles, заселяют гам-
бузиями, которые быстро уничтожают всех
личинок. Этот способ применяют в теплых
странах, так как гамбузии не могут жить
в холодных водоемах.
Борьба со взрослыми летающими кома-
рами представляет большие трудности. Ее
Рис. 75. Цикл развития кровяного споровика Plasmodium vivax (возбудителя малярии):
1 — спорозоиты; 2—4 — бесполое размножение (шизогония) паразита в клетках печени; 2 — спорозоит,
внедрившийся в печеночную клетку; 3 — растущий шизонт с многочисленными ядрами; 4 — шизонт, рас-
падающийся на мерозоиты; 5—ю — бесполое раэмножение (шиэогония) в красных кровяных клетках
(эритроцитах); 5 — молодой шизонт в форме кольца; 6 — растущий шизонт с псевдоподиями; 7, 8 — даль-
нейшие стадии роста шизонта, деление ядер; 9 — распад шиэонта на мерозоиты; 10 — выход мерозоитов из
эритроцита; 11 — молодой макрогаметоцит; 11а — молодой микрогаметоцит; 12 — зрелая макрогамета;
12а — зрелый микрогаметоцит, 13 — макрогамета; 14 — микрогаметоцит; 15 — образование микрогамет,
16 — копуляция микро- и макрогаметы; 17 — зиготы; 18 — подвижная зигота (оокинета); 19 — оокинета,
проникающая через стенку кишечника комара; 20 — оокинета, прикрепившаяся к наружной стенке ки-
шечника комара и превращающаяся в ооцисту; 21, 22 — растущая ооциста с делящимися ядрами; 23 —
зрелая ооциста со спорозоитами; 24 — спорозоиты, покидающие оболочку ооцисты; 25 — спорозоиты в
слюнной железе комара.
Рис. 76. Комар Anopheles maculipennis, пе-
реносчик возбудителя малярии:
1 — личинка в момент захвата атмосферного возду-
ха через дыхальца у поверхности воды; 2 — кукол-
ка; 3 — взрослая самка.
проводят в зимнее время на местах зимо-
вок. Анофелесы скопляются в значитель-
ных количествах в хлевах, так как, кроме
человека, они охотно нападают на рога-
тый скот. Здесь их и уничтожают, при-
меняя для этого различные инсектициды.
В дореволюционной России малярия
была широко распространена и для мно-
гих областей страны была массовым забо-
леванием. Десятки тысяч людей болели
малярией. Особенно неблагополучными
были некоторые районы Кавказа (окрест-
ности Батуми, Сухуми, Сочи), Средней
Азии, Поволжья. Большие эпидемические
вспышки возникали и в первые годы Со-
ветской власти, особенно в период граж-
данской войны. Органы здравоохранения
Советского Союза провели огромную ра-
боту по борьбе с малярией. Этим в первую
очередь занималась широкая сеть спе-
циальных учреждений — малярийных
Рис. 77. Рыбка Gambusia affinis. Слева самец,
справа самка. Естественная величина.
станций, развернутых во всех централь-
ных и южных районах страны. Эта борь-
ба, шедшая по разным направлениям,
увенчалась полным успехом. В настоя-
щее время малярия как массовое заболе-
вание на территории нашей Родины не
существует.
В других странах Европы малярия как
массовое заболевание тоже ликвидиро-
вана. Но в азиатских и африканских
странах (в особенности в Индии с ее влаж-
ным и теплым климатом) малярия —
страшный бич населения, от которого
страдают десятки миллионов людей. По-
надобятся еще большие усилия, чтобы
окончательно освободить человечество от
этого тяжелого заболевания.
Кровяные споровики, так же как и кок-
цидии,— паразиты с узкой специфич-
ностью. Виды рода Plasmodium, пара-
зитирующие в человеке, в других млеко-
питающих развиваться не могут. Благо-
даря этому не существует природных
очагов малярии, не связанных с челове-
ком, какие мы видим, например, у вызы-
ваемой трипаносомами сонной болезни
(стр. 108).
Среди многочисленных видов гемоспо-
ридий известное практическое значение
имеет Plasmodium gallinaceum, паразити-
рующий в крови домашней курицы. Этот
паразит встречается в тропических стра-
нах, где он может вызывать тяжелые
эпизоотии среди кур. Особенно тяжело
протекает заболевание цыплят, обычно
приводящее к гибели. Переносчиками
P. gallinaceum являются комары рода
Aedes.
P. gallinaceum широко используют в
лабораторной практике для изучения жиз-
ненного цикла, так как исследование чело-
веческих видов (особенно шизогонии, про-
текающей во внутренних органах) пред-
ставляет большие трудности.
Приведенный выше обзор основных
групп споровиков показывает их глубо-
кую приспособленность к паразитиче-
скому образу жизни. Приспособления эти
разнообразны. Характерный для боль-
шинства споровиков способ бесполого раз-
множения — множественное деление (ши-
зогония) обеспечивает быстрое увеличение
числа паразитов в хозяине. При наличии
128
в жизненном цикле экзогенных стадий
вырабатываются специальные приспособ-
ления защитного характера, допускаю-
щие длительное существование паразита
в неблагоприятных условиях внешней сре-
ды. Примером могут служить ооцисты
кокцидий. Наконец, наиболее сложным
и биологически совершенным приспособ-
лением к паразитизму является полное
выключение экзогенных стадий и появ-
ление двух хозяев, в одном из которых
осуществляется шизогония, а в другом —
половой процесс и спорогония. По этому
пути пошла эволюция кровяных споро-
виков.
Среди какой группы свободноживущих
простейших следует искать корни про-
исхождения споровиков? Весьма вероят-
Этот класс, так же как и споровики,
состоит исключительно из паразитических
организмов. Жизненный цикл книдоспо-
ридий начинается с амебоидного одноядер-
ного зародыша. Характерной особен-
ностью книдоспоридий является наличие
у них особых спор, обладающих стрека-
тельными капсулами. Из этих капсул в
момент проникновения книдоспоридий в
хозяина выбрасывается длинная тонкая
нить, способствующая закреплению споры
в тканях.
Рассмотрим наиболее характерных пред-
ставителей двух отрядов книдоспо-
ридий — миксоспоридий (Myxosporidia)
и микроспоридий (Microsporidia).
ОТРЯД МИКСОСПОРИДИЙ
(MYXOSPORIDIA)
Миксоспоридий (их называют также
слизистыми споровиками)—почти исключи-
тельно паразиты морских и пресновод-
ных рыб. Некоторые виды миксоспоридий
вызывают заболевание и нередко гибель
но, что они взяли начало от жгутиконос-
цев. Серьезным доказательством в пользу
этой точки зрения является наличие в
жизненном цикле большинства спорови-
ков жгутиковых стадий. Например, мик-
рогаметы кокцидий по своему строению
являются как бы небольшими жгутико-
носцами, снабженными двумя жгутиками.
Характер полового процесса споровиков—
копуляция — весьма близок к тому, что
наблюдается у жгутиконосцев. Наконец,
как мы видели выше, представители клас-
са жгутиконосцев довольно легко пере-
ходят к паразитизму.
Глубокая приспособленность к парази-
тическому образу жизни обеспечивает спо-
ровикам широкое распространение и боль-
шое видовое разнообразие.
хозяина (в особенности мальков), чем на-
носят огромный ущерб рыбному хозяй-
ству. В природе миксоспоридий распро-
странены очень широко во всех широтах
и во всех типах водоемов. Число их видов
равно приблизительно 700.
Локализация миксоспоридий в теле хо-
зяина бывает самой разнообразной. Часть
видов поражает ткани различных орга-
нов — соединительную (в том числе и
хрящ), мускульную, нервную, ткани по-
чек. Некоторые виды живут в полостях
органов, чаще всего в желчном и мочевом
пузыре. Миксоспоридий никогда не пара-
зитируют в полости кишечника.
Полостные виды миксоспоридий имеют
амебоидное тело с несколькими ядрами.
Они образуют псевдоподии и способны
передвигаться (рис. 78). Размеры этих
миксоспоридий невелики, они измеряются
обычно десятками микрон. Цитоплазма
разделена на два слоя: наружную, более
светлую эктоплазму и внутреннюю —
эндоплазму. В эндоплазме у зрелых осо-
бей лежат споры.
Тканевые миксоспоридий представляют
собой плазматические массы, лежащие в
КЛАСС КНИДОСПОРИДИИ (CNIDOSPORIDIA)
Ф 9 ЖИ8НЬ ЖИВОТНЫХ, Т. 1
129
Рис. 78. Плазмодии полостных миксоспоридий
со спорами:
А — Ceratomyxa appendiculata; Б — Leptotheca agi-
lis; В — Chloromyxum leidigi. 1 — псевдоподии; 2 —
споры со стрекательными капсулами.
Рис. 79. Плотва с опухолями, образованными
плазмодиями миксоспоридий.
толще тканей и окруженные оболочкой.
В плазматическом теле их обычно имеется
большое число ядер (измеряемое иногда
сотнями и тысячами) и спор. Размеры
многоядерных плазмодиев тканевых мик-
соспоридий могут быть очень различны.
Некоторые из них достигают гороши-
ны или даже ореха. В этих случаях на
теле рыб образуются опухоли (рис. 79),
которые затем прорываются и превраща-
ются в язвы.
Как же осуществляется цикл развития
миксоспоридий, как паразит проникает
в тело рыбы?
Для распространения миксоспоридий
служат споры, с рассмотрения строения
и развития которых мы и начнем изложе-
ние хода жизненного цикла.
Споры формируются в эндоплазме мик-
соспоридий. В плазмодиях многих мелких
полостных видов образуются всего две
споры. В крупных плазмодиях тканевых
видов число спор может достигать сотен
и тысяч. Зрелая спора миксоспоридий
обладает характерной и сложной струк-
турой (рис. 80). Снаружи спора имеет
двустворчатую прочную оболочку. Створ-
ки ее соединены друг с другом по линии,
называемой швом. В этой области они
соприкасаются друг с другом, как два
часовых стекла, сложенных краями. Как
видно на рисунке 80, форма створок у раз-
ных видов различна, кроме того, они
могут нести снаружи отростки разной
формы и длины.
Внутри створок в задней части споры
помещается маленький амебовидный заро-
дыш, обычно двухъядерный. На переднем
конце споры (а у сильно вытянутых
спор — у линии шва или на противопо-
ложных концах) помещаются две стрека-
тельные капсулы (у немногих видов их
бывает четыре). Стрекательная капсула
представляет собой пузырек с жидкостью,
имеющий собственную оболочку, внутри
которого расположена скрученная спи-
ралью длинная тонкая нить. Длина нити
в несколько раз превосходит длину спо-
ры. Эта нить играет важную роль при
заражении хозяина миксоспоридиями.
Развитие споры протекает сложно. На
деталях его мы останавливаться не будем,
укажем лишь, что в формировании каж-
дой споры принимают участие 6 ядер,
из них два переходят в амебоиды, за счет
двух формируются створки, два осталь-
ных участвуют в образовании стрекатель-
ных капсул.
Зрелые споры попадают в воду. Это
совершается разными путями: у полост-
ных миксоспоридий — через кишечник
130
или почки, у тканевых — прп разрыве
цисты и образовании язв. У некоторых
видов споры освобождаются лишь после
смерти хозяина, как, например, у миксо-
споридий, паразитирующих в хрящевой
и нервной ткани. Если спора будет загло-
чена хозяином (рыбой), то под воздейст-
вием пищеварительных соков стрекатель-
ные нити с силой выбрасываются и внед-
ряются в стенку кишечника. Таким путем
спора прочно закрепляется в теле хозяи-
на. После этого створки расходятся по
линии шва — спора как бы раскрывается.
Амебоидный зародыш выходит из споры
и активно внедряется в ткани. Далее
через ткани хозяина он направляется в
тот орган, в котором паразитирует дан-
ный вид миксоспоридий.
Рассмотренный выше жизненный цикл
миксоспоридий не имеет ничего сходного
с циклами споровиков (грегарин, кокци-
дий, гемоспоридий). Здесь отсутствуют
шизогония, гаметогенез, оплодотворение
и спорогония, закономерное чередование
которых столь характерно для класса
споровиков. Размножение и распростра-
нение связано у миксоспоридий с образо-
ванием спор. Строение спор и их развитие
совершенно иное, чем у споровиков. Оче-
видно, что развитие спор миксоспоридий
следует рассматривать как своеобразный
процесс бесполого размножения, несколь-
ко напоминающий внутреннее почкование.
Имеется ли у миксоспоридий в какой-либо
форме половой процесс? Вопрос этот до
настоящего времени не может считаться
окончательно решенным. Мы видели вы-
ше, что амебоид, заключенный в споре,
обладает двумя ядрами. По наблюдениям
многих авторов, по выходе амебоида из
споры ядра сливаются. В этом усматри-
вают вторично упрощенную форму поло-
вого процесса, поскольку основным момен-
том в оплодотворении является слияние
ядер.
Как уже отмечалось, миксоспоридий
являются возбудителями заболеваний рыб.
Перечислим некоторые наиболее опасные
из них.
У карпа, являющегося одним из основ-
ных объектов разведения в прудовых ры-
боводческих хозяйствах, известно несколь-
ко заболеваний, вызываемых миксоспо-
ридиями. Наиболее часто встречается
Myxobolus cyprini, вызывающий злока-
чественную анемию. Вегетативные стадии
паразита и цисты со спорами поражают
ткани различных органов карпа: жабры,
мускулатуру, почки, печень, селезенку.
Имеется много видов миксоспоридий
(преимущественно виды родов Myxo-
bolus и Henneguya), которые паразити-
руют на жабрах. Они поражают карпо-
вых рыб, судаков, щуку и других промыс-
ловых рыб (рис. 81). При массовом пора-
жении жабр нарушается нормальный про-
цесс дыхания, что приводит к заболева-
ниям.
Тяжелое заболевание особенно ценных
лососевых рыб (форель, лосось) вызывает
миксоспоридия Myxosoma cerebral is. Этот
паразит поражает преимущественно маль-
ков, вызывая их массовую гибель.
Болезнь получила название «вертеж форе-
лей» по характерному признаку заболе-
А Б
Л
Рис. 80. Споры различных миксоспоридий:
А — Henneguya acerina; Б — Ceratomyxa truncata;
В — Sinuolinea dimorpha; Г — Sphaerospora divergens;
Д — Mitraspora cyprini; E — Myxobolus sp. 1 — стре-
кательные капсулы; 2 — выброшенная стрекательная
нить; 3 — амебоид с двумя ядрами; 4 — отростки
оболочки споры.
9*
131
вания. Myxosoma cerebralis лока-
лизуется в хрящевой ткани. При
массовом заражении споры встре-
чаются и в других органах: го-
ловном мозгу, мышцах. Особенно
часто миксоспоридия локализуется
в полукружных каналах, вызывая
разрушение органа равновесия.
Это нарушает нормальное движе-
ние. Рыбки начинают кружиться
с большой быстротой, после чего
наступает период утомления. Вто-
рой характерный симптом заболе-
вания — искривление позвоноч-
ника (рис. 82), связанное с нару-
шением целостности хряща позво-
ночника. Кроме искривления хво-
стового отдела, наблюдается не-
нормальная темная пигментация
хвоста. «Вертеж форелей» часто
приводит к массовой гибели мо-
лодых рыбок. После гибели рыбки
при разложении трупа споры по-
падают в воду.
Как осуществляется в рыбовод-
ных хозяйствах борьба с миксо-
споридиозами? Мероприятия эти
носят в основном профилактиче-
ский характер. Следует избегать
контакта больных и здоровых рыб.
Зараженные водоемы рекомендуют
вреживать, спуская их на зиму и дезин-
фицируя ложе водоема негашеной из-
вестью и другими химикатами.
ОТРЯД МИКРОСПОРИДИИ
(MICRQSPORIDIA)
В отличие от миксоспоридий, микроспо-
ридии паразитируют как в позвоночных
животных (преимущественно в рыбах),
так и в беспозвоночных. Это внутрикле-
точные паразиты. Число известных видов
их около 200. Наибольший вред причи-
няют паразиты насекомых, так как неко-
торые виды микроспоридий вызывают тя-
желое заболевание полезных для чело-
века пчелы и тутового шелкопряда. У медо-
носной пчелы болезнь, вызываемая микро-
споридиями (Nosema apis), называется
белым поносом. Она сильно исто-
щает пчелиную семью и нередко
вызывает ее гибель. Микроспори-
диоз тутового шелкопряда носит
название п е б р и н ы (Nosema
bombycis). Это заболевание в боль-
шинстве случаев смертельно. Не-
которые микроспоридии поража-
ют насекомых — вредителей сель-
скохозяйственных растений и этим
приносят пользу.
Число видов микроопоридий, па-
разитирующих в рыбах, невелико.
Они могут поражать разные ор-
ганы, локализуясь чаще всего в
соединительной ткани и мышцах.
Из рыб, имеющих промысловое
значение, микроспоридиями пора-
жаются снетки, корюшка. Особенно
сильно страдает от микроспоридии
Glugea hertwigi снеток, имеющий
немаловажное промысловое значение
в северных пресноводных водоемах
СССР (Псковско-Чудское озеро, оз.
Белое п другие). Сильное заражение
снетков микроспоридиями вызывает
у них воспаление и нарушение
функций большинства внутренних
органов и, как результат, массовую
гибель рыбок. В некоторые годы
зарегистрирована гибель 25—50%
снетков от микроспоридиоза.
Один пз видов микроспоридий (Соссопе-
ma sulci) паразитирует в икринках осетро-
вых рыб бассейнов Черного и Каспийского
морей, вызывая гибель икринок. Однако
массового заражения осетровых этим парази-
том пока не отмечалось. Нередко микроспо-
ридии рода Glugea паразитируют в ко-
люшках, вызывая образование крупных
белых цист в коже (рис. 83).
Познакомимся с жизненным циклом
микроспоридий на примере Nosema apis
(из медоносной пчелы) и N- bombycis
(из тутового шелкопряда). Он сходен
с циклом миксоспоридий. У пчелы пара-
зит развивается в клетках эпителия ки-
шечника, у тутового шелкопряда —
в клетках самых различных органов. Спо-
ры (рис. 84) очень мелкие (у Nosema apis
длина 4,5—6,5 мк, ширина 2,5—3,5 мк).
Имеется прочная и относительно тонкая
оболочка, которая, в отличие от таковой
у миксоспоридий, не является двуствор-
Рис. 81. Жаберный
лепесток щуки,
пораженный ци-
стами миксоспори-
дий Henneguya
lobosa.
l — цисты миксо-
споридий.
132
чатой. Значительная часть пространства
внутри оболочки споры занята одной стре-
кательной капсулой с расположенной
в ней спирально закрученной длинной
тонкой нитью. Сзади к капсуле примы-
кает очень маленький, вероятно двухъ-
ядерный, амебоидный зародыш.
При проникновении споры в кишечник
хозяина нить стрекательной капсулы с си-
лой выбрасывается и внедряется в клетку
эпителия. Длина ее в выброшенном состоя-
нии достигает 70 мк, что превосдодит
длину споры более чем в 10 раз.
Вместе с нитью (по-видимому, на конце
нити) из споры выносится и амебоидный
зародыш, который таким путем проникает
в клетку хозяина. Здесь амебоид начи-
нает энергично делиться, образуя харак-
терные цепочки, состоящие из отдельных
клеток паразита. Многоядерных плазмо-
диев не образуется. Вскоре начинается
формирование спор, строение которых
уже рассмотрено выше. У Nosema bomby-
cis (возбудителя пебрины шелкопряда)
паразит при сильном заражении прони-
кает в цитоплазму яйцевых клеток, и в
этом случае, если родительская особь не
погибает, паразит передается следующему
поколению. Получается передача возбу-
дителя «по наследству».
Борьба с микроспоридиозами пчелы и
тутового шелкопряда представляет собой
сложную задачу. В основном дело сво-
дится к профилактическим мероприя-
тиям. Зараженные ульи дезинфицируют.
Сильно зараженные семьи приходится
уничтожать. Важной мерой борьбы с пеб-
риной шелковичного червя является мик-
роскопический контроль отложенных яиц.
Часть яиц изучают под микроскопом. Если
в них найдут споры микроспоридий, то
всю партию яиц уничтожают.
* * *
Книдоспоридии представляют собой
группу простейших, глубоко измененную
паразитизмом. В их происхождении и эво-
люции важную роль сыграло замечатель-
ное приспособление — образование слож-
ных спор со стрекательными нитями. Эта
особенность их строения обеспечивает ши-
рокое распространение паразитов. С ка-
кой группой свободноживущих простей-
ших вероятнее всего связаны книдоспо-
ридии в своем происхождении? Наличие
в их жизненном цикле амебоидной стадии
и полное отсутствие жгутиковых стадий
позволяет предположить, что книдоспо-
ридии происходят от саркодовых в резуль-
тате их глубокого приспособления к пара-
зитизму, выразившегося в появлении осо-
бо устроенных спор.
Рис. 82. Молодая форель, больная «вертежом»
(возбудитель: Myxosomacerebralis). Позвоночник
искривлен, задняя треть тела сильно пигмен-
тирована.
Рис. 83. Трехиглая колюшка с цистами мпксо-
споридии Glugea anomala в коже.
Рис. 84. Микроспоридии рода Nosema:
А — строение споры Nosema bombycis; Б — срез че-
рез эпителий кишечника шелковичного червя, наби-
того шизонтами и спорами. 1 — стрекательная кап-
сула; 2 — нить стрекательной капсулы; 3 — амебоид
с ядрами.
133
КЛАСС ИНФУЗОРИИ (INFUSORIA, или CILIATA)
Простейшие этого обширного по коли-
честву видов —около 6 тыс.— класса широ-
ко распространены в природе. (Эта цифра
приводится в сводке Корлисса, 1961 г.). К ним
относятся многочисленные обитатели мор-
ских и пресных вод. Некоторые виды при-
способились к жизни во влажной почве.
Немалое количество видов инфузорий ведет
паразитический образ жизни. Хозяевами
для паразитических инфузорий являются
самые разнообразные беспозвоночные и по-
звоночные животные до высших обезьян
и человека включительно.
По сравнению с другими группами про-
стейших инфузории имеют наиболее слож-
ное строение, что связано с разнообразием
и сложностью их функций.
Откуда взялось название «инфузория
туфелька»? Вы не будете удивлены, если
взглянете под микроскопом на живую
инфузорию или даже на ее изображение
(рис. 85).
Действительно, форма тела этой инфу-
зории напоминает изящную дамскую ту-
фельку.
Инфузория туфелька находится в непре-
рывном довольно быстром движении. Ско-
рость его (при комнатной температуре)
около 2,0—2,5 мм/сек. Для такого малень-
кого животного это большая скорость!
Ведь это означает, что за секунду туфель-
ка пробегает расстояние, превышающее
длину ее тела в 10—15 раз. Траектория
движения туфелькд довольно сложна. Она
движется передним концом прямо вперед
ИНФУЗОРИЯ ТУФЕЛЬКА
(PARAMECIUM CAUDATUM)
Чтобы ознакомиться со строением и об-
разом жизни этих интересных одноклеточ-
ных организмов, обратимся сначала к од-
ному характерному примеру. Возьмем
широко распространенных в мелких прес-
новодных водоемах инфузорий туфелек
(виды рода Paramecium). Этих инфузорий
очень легко развести в небольших аква-
риумах, если залить прудовой водой обыч-
ное луговое сено. В таких настойках раз-
вивается множество различных видов про-
стейших и почти всегда развиваются инфу-
зории туфельки. При помощи обычного
учебного микроскопа можно рассмотреть
многое из того, о чем будет дальше рас-
сказано.
Среди простейших инфузории туфельки
являются довольно крупными организ-
мами. Длина тела их около 1/в—1/ъ мм.
Рис. 85. Инфузория туфелька (Paramecium cau-
datum)
1 — реснички; 2 — пищеварительные вакуоли; з —
большое ядро (макронуклеус); 4 — малое ядро (ми-
кронуклеус); 5 — ротовое отверстие и глотка; 6 —
непереваренные остатки пищи, выбрасываемые нару-
жу; 7 — трихоцисты; 8 — сократительная вакуоля.
134
и при этом вращается вправо вдоль про-
дольной оси тела.
Столь активное движение туфельки за-
висит от работы большого количества
тончайших волосковидных придатков —
ресничек, которые покрывают все тело
инфузории. Количество ресничек у одной
особи инфузории туфельки равняется
10—15 тыс.!
Каждая ресничка совершает очень час-
тые веслообразные движения — при ком-
натной температуре до 30 биений в се-
кунду. Во время удара назад ресничка
держится в выпрямленном положении.
При возвращении же ее в исходную пози-
цию (при движении вниз) она движется
в 3—5 раз медленнее и описывает полу-
круг.
При плавании туфельки движения мно-
гочисленных покрывающих ее тело рес-
ничек суммируются. Действия отдельных
ресничек оказываются согласованными,
в результате чего получаются правильные
волнообразные колебания всех ресничек.
Волна колебания начинается у переднего
конца тела и распространяется назад.
Одновременно вдоль тела туфельки про-
ходят 2—3 волны сокращения. Таким об-
разом, весь ресничный аппарат инфузории
представляет собой как бы единое функ-
циональное физиологическое целое, дей-
ствия отдельных структурных единиц ко-
торого (ресничек) тесно связаны (коорди-
нированы) между собой.
Строение каждой отдельной реснички
туфельки, как показали электронномик-
роскопические исследования, является
весьма сложным.
Направление и быстрота движения ту-
фельки не являются величинами постоян-
ными и неизменными. Туфелька, как и все
живые организмы (мы видели это уже на
примере амебы), реагирует на изменение
внешней среды изменением направления
движения.
Изменение направления движения про-
стейших под влиянием различных раздра-
жителей называют таксисами. У ин-
фузорий легко наблюдать различные
таксисы. Если в каплю, где плавают ту-
фельки, поместить какое-либо неблаго-
приятно действующее на них вещество
(например, кристаллик* поваренной соли),
.••а
О!
о
Рис. 86. Хемотаксис инфузорий туфелек:
1 — положительный хемотаксис, инфузории концент-
рируются в области введенного привлекающего их ве-
щества; 2 — то же при введении повышенной кон-
центрации того же вещества, вредно действующей на
инфузорий; з — введен пузырек углекислого газа,
инфузории обнаруживают к нему положительный хе-
мотаксис; 4 — тот же препарат через несколько ми-
нут; углекислый газ диффундирует в воду, высокая
концентрация его вызывает отрицательный хемотаксис
туфелек, которые собираются в зону, где концентра-
ция углекислоты является для них оптимальной.
то туфельки уплывают (как бы убегают)
от этого неблагоприятного для них фак-
тора (рис. 86).
Перед нами пример отрицательного
таксиса на химическое воздействие (от-
рицательный хемотаксис). Можно наблю-
дать у туфельки и положительный х е-
мотаксис. Если, например, каплю
воды, в которой плавают инфузории,
прикрыть покровным стеклышком и под-
пустить под него пузырек углекислого
газа (С02), то большая часть инфузорий
направится к этому пузырьку и располо-
жится вокруг него кольцом.
Очень наглядно явление таксиса прояв-
ляется у туфелек под влиянием электри-
ческого тока. Если через жидкость, в ко-
торой плавают туфельки, пропустить сла-
бый электрический ток, то можно наблю-
дать следующую картину: все инфузории
ориентируют свою продольную ось парал-
лельно линии тока, а затем, как по
команде, двинутся в направлении катода,
в области которого и образуют густое
скопление. Движение инфузорий, опреде-
ляемое направлением электрического то-
ка, носит название гальванотак-
сиса. Различные таксисы у инфузорий
135
могут быть обнаружены под влия-
нием самых разнообразных фак-
торов внешней среды.
Все цитоплазматическое тело
инфузории отчетливо распадается
на 2 слоя: наружный — более
светлый (эктоплазма) и внутрен-
ний — более темный и зернистый
(эндоплазма). Самый поверхност-
ный слой эктоплазмы образует на-
ружную очень тонкую и вместе с
тем прочную и эластичную оболоч-
ку — пелликулу, которая играет
важную роль в сохранении посто-
янства формы тела инфузории.
В наружном слое (в эктоплаз-
ме) тела живой туфельки хорошо
видны многочисленные коротень-
кие палочки, расположенные пер-
пендикулярно к поверхности
(рис. 85,7). Эти образования носят
название трихоцисты. Функ-
ция их очень интересна и связана с
защитой простейшего. При механи-
ческом, химическом или каком-
либо ином сильном раздражении
трихоцисты с силой выбрасывают-
ся наружу, превращаясь в тонкие
длинные нити, которые поражают
хищника, нападающего на ту-
фельку. Трихоцисты представля-
ют собой мощную защиту. Они
располагаются закономерно между
ресничками, так что число трихо-
цист приблизительно соответст-
вует числу ресничек. На месте ис-
пользованных («выстреленных»)
трихоцист в эктоплазме туфельки
развиваются новые.
На одной стороне, приблизитель-
но по середине тела (рис. 85,5), у
туфельки имеется довольно глу-
бокая впадина. Это ротовая
впадина, или перистом.
По стенкам перистома, так же как и
по поверхности тела, расположены рес-
нички. Они развиты здесь гораздо бо-
лее мощно, чем на всей остальной по-
верхности тела. Эти тесно расположенные
реснички собраны в две группы. Функ-
ция этих особо дифференцированных рес-
ничек связана не с движением, а с пита-
нием (рис. 87).
Как и чем питаются туфельки,
как осуществляется у них пище-
варение?
Туфельки относятся к числу
инфузорий, основную пищу кото-
рых составляют бактерии. Наряду
с бактериями они могут заглаты-
вать и любые другие взвешенные
в воде частицы независимо от их
питательности. Околоротовые ре-
снички создают непрерывный
ток воды со взвешенными в ней
частицами в направлении ротово-
го отверстия, которое расположе-
но в глубине перистома. Мелкие
пищевые частицы (чаще всего
бактерии) проникают через рот
в небольшую трубковидную глот-
ку и скапливаются на дне ее,
на границе с эндоплазмой. Рото-
вое отверстие всегда открыто. По-
жалуй, не будет ошибкой сказать,
что инфузория туфелька — одно
из самых прожорливых живот-
ных: она непрерывно питается.
Этот процесс прерывается только
в определенные моменты жизни,
связанные с размножением и по-
ловым процессом.
Скопившийся на дне глотки
пищевой комочек в дальнейшем
отрывается от дна глотки и вме-
сте с небольшим количеством жид-
кости поступает в эндоплазму,
образуя пищеваритель-
ную вакуолю. Последняя
не остается на месте своего обра-
зования, а, попадая в токи эн-
доплазмы, проделывает в теле
туфельки довольно сложный и
закономерный путь, называемый
циклозом пищеварительной ваку-
оли (рис. 88). Во время этого до-
вольно длительного (при комнат-
ной температуре занимающего около часа)
путешествия пищеварительной вакуоли
внутри ее происходит ряд изменений,
связанных с перевариванием находящей-
ся в ней пищи.
Здесь, так же как у амеб (стр. 75)
и некоторых жгутиконосцев, происходит
типичное внутриклеточное пищеварение.
Из окружающей пищеварительную вакуо-
Рис. 87. Продоль-
ный разрез инфу-
зории туфельки в
области глотки:
1 — густо располо-
женный ряд ресни-
чек глотки; 2 — об-
разующаяся пище-
варительная вакуо-
ля; 3 — пищевари-
тельная вакуоля;
4 — макронуклеус;
5 — трихоцисты.
Рис. 88. Путь пи-
щеварительной ва-
куоли (показан
стрелками) в теле
инфузории ту-
фельки.
136
лю эндоплазмы в нее поступают пищева-
рительные ферменты, которые воздейст-
вуют на пищевые частицы. Продукты
переваривания пищи всасываются через
стгнку пищеварительной вакуоли в эндо-
плазму.
По ходу циклоза пищеварительной ва-
куоли в ней сменяется несколько фаз пи-
щеварения. В первые моменты после обра-
зования вакуоли заполняющая ее жид-
кость мало отличается от жидкости окру-
жающей среды. Вскоре начинается по-
ступление из эндоплазмы в вакуолю пище-
варительных ферментов и реакция среды
внутри нее становится резко кислой. Это
легко обнаружить, добавляя к пище ка-
кой-либо индикатор, цвет которого ме-
няется в зависимости от реакции (кислой,
нейтральной или щелочной) среды. В этой
кислой среде проходят первые фазы пище-
варения. Затем картина меняется и реак-
ция внутри пищеварительных вакуолей
становится слабощелочной. В этих усло-
виях и протекают дальнейшие этапы
внутриклеточного пищеварения. Кислая
фаза обычно более короткая, чем щелоч-
ная; она длится примерно х/6—х/4 часть
всего срока пребывания пищеваритель-
ной вакуоли в теле инфузории. Однако
соотношение кислой и щелочной фаз
может варьировать в довольно широких
пределах в зависимости от характера
пищи.
Путь пищеварительной вакуоли в эндо-
плазме заканчивается тем, что она при-
ближается к поверхности тела и через
пелликулу содержимое ее, состоящее из
жидкости и непереваренных остатков пи-
щи, выбрасывается наружу — происхо-
дит дефекация. Этот процесс, в отличие
от амеб, у которых дефекация может
происходить в любом месте, у туфелек, как
и у других инфузорий, строго приурочен к
определенному участку тела, расположенному
на брюшной стороне (брюшной условно на-
зывают ту поверхность животного, на кото-
рой помещается околоротовое углубление),
примерно посередине между перистомом и
задним концом тела.
Таким образом, внутриклеточное пище-
варение представляет собой сложный процесс,
слагающийся из нескольких последовательно
сменяющих друг друга фаз.
Рис. 89. Последовательные стадии работы сокра-
тительной вакуоли инфузории туфельки.
Кроме пищеварительных вакуолей, в теле
инфузории туфельки имеются выполняющие
также очень важные жизненные функции
сократительные вакуоли. У ту-
фельки их две, они расположены в передней
и задней трети тела. Каждая из вакуолей
состоит из центрального резервуа-
ра и приводящих каналов E—7),
которые расположены радиально вокруг цент-
рального резервуара. Цикл работы сократи-
тельной вакуоли начинается с того, что
приводящие каналы заполняются жидкостью
и становятся хорошо видимыми (рис. 89).
Затем жидкое содержимое их изливается
в центральный резервуар, сами каналы
после опорожнения становятся на некото-
рое время невидимыми. Последний этап
цикла работы сократительной вакуоли
заключается в том, что жидкость из
центрального резервуара изливается че-
рез особую пору в пелликуле наружу.
После этого центральный резервуар на
короткий срок перестает быть видимым.
В это время приводящие каналы вновь
начинают заполняться жидкостью и весь
цикл начинается сначала. Обычно перед-
няя и задняя сократительные вакуоли
работают последовательно, как бы по
очереди. Каков темп пульсации вакуо-
лей? Какое количество жидкости выво-
дится ими наружу? Частота сокращения
сократительной вакуоли в большой сте-
пени зависит от условий внешней среды,
и особенно от температуры и осмотического
давления. При комнатной температуре
у туфельки сократительная вакуоля про-
делывает весь цикл пульсации за 10—15
секунд. У морских и паразитических
инфузорий темп пульсации сократитель-
ных вакуолей обычно значительно ниже,
чем у пресноводных.
Подсчеты показывают, что примерно
за 30—45 минут у туфельки через сокра-
тительные вакуоли выводится объем жид-
137
кости, равный объему тела инфузории.
Таким образом, благодаря деятельности
сократительных вакуолей через тело ин-
фузории осуществляется непрерывный ток
воды, поступающей снаружи через рото-
вое отверстие (вместе с пищеварительными
вакуолями), а также осмотически непо-
средственно через пелликулу. Сократи-
тельные вакуоли играют важную роль
в регулировании тока воды, проходящего
через тело инфузории, в регулировании
осмотического давления (стр. 76). Этот
процесс здесь протекает в принципе так же,
как у амеб, только структура сократитель-
ной вакуоли намного сложнее.
В течение долгих лет среди ученых, за-
нимающихся изучением простейших, шел
спор по вопросу о том, имеются ли в ци-
топлазме какие-нибудь структуры, свя-
занные с появлением сократительной ва-
куоли, или же она образуется всякий раз
заново. На живой инфузории никаких
особых структур, которые предшествовали
бы ее образованию, наблюдать не удается.
После того как произойдет сокращение
вакуоли — систола, в цитоплазме на ме-
сте бывшей вакуоли не видно решительно
никаких структур. Затем заново появля-
ются прозрачный пузырек или приводя-
щие каналы, которые начинают увеличи-
ваться в размерах. Однако никакой связи
вновь возникающей вакуоли с сущест-
вовавшей ранее не обнаруживается. Со-
здается впечатление, что преемственности
между следующими друг за другом цик-
лами сократительной вакуоли нет и вся-
кая новая сократительная вакуоля обра-
зуется в цитоплазме заново. Однако спе-
циальные методы исследования показали,
что на самом деле это не так. Применение
электронной микроскопии, дающей очень
большое увеличение (до 100 тыс. раз),
убедительно показало, что у инфузории
на том участке, где формируются сокра-
тительные вакуоли, имеется особо диффе-
ренцированная цитоплазма, состоящая из
переплетения тончайших трубочек. Та-
ким образом, оказалось, что сократитель-
ная вакуоля возникает в цитоплазме не на
«пустом месте», а на основе предшествую-
щего особого органоида клетки, функцией
которого является формирование сокра-
тительной вакуоли.
Как и у всех простейших, у инфузорий
имеется клеточное ядро. Однако по строе-
нию ядерного аппарата инфузории резко
отличаются от всех других групп простей-
ших.
Ядерный аппарат инфузо-
рий характеризуется своим дуализ-
мом. Это означает, что у инфузорий
имеется два разных типа ядер — боль-
шие ядра, или макронуклеусы,
и м а л ы е ядра, или микронукле-
усы. Посмотрим, какое строение имеет
ядерный аппарат у инфузории туфельки
(рис. 85).
В центре тела инфузории (на уровне пе-
ристома) помещается большое массивное
ядро яйцевидной или бобовидной формы.
Это макронуклеус. В тесном соседстве
с ним расположено второе ядро во много
раз меньших размеров, обычно довольно
тесно прилежащее к макронуклеусу. Это
микронуклеус. Различие между этими
двумя ядрами не сводится только к раз-
мерам, оно более значительно, глубоко
затрагивает их структуру.
Макронуклеус по сравнению с микро-
нуклеусом гораздо богаче особым ядерным
веществом (хроматином, или, точнее, дезо-
ксирибонуклеиновой кислотой, сокращен-
но ДНК), входящим в состав хромосом
(см. стр. 58).
Исследования последних лет показали,
что макронуклеус обладает в несколько
десятков (а у некоторых инфузорий и со-
тен) раз большим числом хромосом, чем
микронуклеусы. Макронуклеус является
совсем своеобразным типом многохромо-
сомных (полиплоидных) ядер. Таким об-
разом, различие между микро- и макро-
нуклеусами затрагивает их хромосомный
состав, отчего и зависит большее или
меньшее богатство их ядерным вещест-
вом — хроматином.
У одного из наиболее обычных видов
инфузории — туфельки (Paramecium cau-
datum) — имеется один макронуклеус (со-
кращенно Ма) и один микронуклеус (со-
кращенно Mi). Такая структура ядер-
ного аппарата свойственна многим инфу-
зориям. У других может быть по несколь-
ку Ма и Mi. Но характерная особенность
всех инфузорий — дифференцировка ядер
на две качественно различные группы, на
138
Ma и Mi, или, другими сло-
вами, явление ядерного дуа-
лизма *.
Как размножаются
инфузории? Обратимся в каче-
стве примера опять к инфузо-
рии туфельке. Если отсадить
в небольшой сосуд (микро-
аквариум) один-единственный
экземпляр туфельки, то уже
через сутки там окажется две,
а нередко и четыре инфузории.
Как это происходит? После не-
которого периода активного
плавания и питания инфузо-
рия несколько вытягивается в
длину. Затем точно по сере-
дине тела появляется все уг-
лубляющаяся поперечная пе-
ретяжка (рис. 90). В конце
концов инфузория как бы пе-
решнуровывается пополам и из
одной особи получаются две,
первоначально несколько мень-
ших размеров, чем материн-
ская особь. Весь процесс де-
ления занимает при комнат-
ной температуре около часа.
Изучение внутренних процес-
сов показывает, что еще до то-
го, как появляется поперечная пере-
тяжка, начинается процесс деления ядер-
ного аппарата. Первым делится Mi и
лишь после него — Ма. Мы не будем
здесь останавливаться на детальном
рассмотрении процессов ядерного де-
ления и лишь отметим, что Mi делится
путем митоза (стр. 60), тогда как деле-
ние Ма по внешности напоминает прямое
деление ядра — амитоз. Этот бесполый
процесс размножения инфузории туфель-
ки, как мы видим, сходен с бесполым раз-
множением амеб (стр. 77) и жгутико-
носцев (стр. 104). В отличие от них
инфузории в процессе бесполого размно-
жения делятся всегда поперек, тогда как
у жгутиконосцев плоскость деления па-
раллельна продольной оси тела.
1 Из этого правила существует, может быть,
одно исключение — у морских инфузорий рода
Stephanopogon ядерный дуализм не обнаружен.
Этот случай требует дальнейших исследований.
Рис. 90. Бесполое раз-
множение путем по-
перечного деления
инфузории туфельки
(схематизированно):
2 — мякронуклеус; 2 —
макронуклеус.
Во время деления происхо-
дит глубокая внутренняя пере-
стройка тела инфузории. Обра-
зуется два новых перистома,
две глотки и два ротовых от-
верстия. К этому же времени
приурочено деление базальных
ядер ресничек, за счет которых
образуются новые реснички.
Если бы при размножении чис-
ло ресничек не возрастало, то
в результате каждого деления
дочерние особи получили бы
примерно половину числа рес-
ничек материнской особи, что
привело бы к полному «облы-
сению» инфузорий. На самом
деле этого не происходит.
Время от времени у большин-
ства инфузорий, в том числе и у
туфелек, наблюдается особая и
чрезвычайно своеобразная фор-
ма полового процес-
с а, которая получила назва-
ние конъюгации. Мы
не будем здесь детально разби-
рать все те сложные ядерные
изменения, которые сопровож-
дают этот процесс, а отметим
только самое главное. Конъюга-
ция протекает следующим образом (рис. 91),
Две инфузории сближаются, тесно прикла-
дываются друг к другу брюшными сторона-
ми и в таком виде плавают довольно дли-
тельное время вместе (у туфельки пример-
но в течение 12 часов при комнатной тем-
пературе). Затем конъюганты расходятся.
Что же происходит в теле инфузории во
время конъюгации? Сущность этих про-
цессов сводится к следующему (рис. 91).
Большое ядро (макронуклеус) разрушает-
ся и постепенно растворяется в цитоплаз-
ме. Микронуклеусы сначала делятся, часть
ядер, образующихся в результате деле-
ния, разрушается (см. рис. 91). В каждом
из конъюгантов сохраняется по два ядра.
Одно из этих ядер остается на месте в той
особи, в которой оно образовалось (ста-
ционарное ядро), другое же активно
перемещается в партнера по конъюгации
(мигрирующее ядро) и сливается
с его стационарным ядром. Таким образом,
в каждом из конъюгантов на этой стадии
139
5 6
7 8 g ю
Рис. 91. Конъюгация инфузорий (схематизи-
рованно):
J — две особи соединились брюшными сторонами, в
каждой имеется один Mi и один Ма', 2 — Mi проделы-
вает первое деление; 3 — в левой особи Mi заканчивает
первое деление, в правой оба ядра, образовавшиеся в
результате деления, приступили ко второму делению,
Ма распался на фрагменты; 4 —в каждой особи вто-
рое деление Мг; 5 — из образовавшихся в каждой ин-
фузории в результате двух делений Mi четырех ядер
три дегенерируют, по одному приступают к третьему
делению; 6 — из двух ядер, образовавшихся после 3-го
деления Мг, одно переходит в партнера (мигрирующее
ядро), другое остается на месте (стационарное ядро);
7 — мигрирующее и стационарное ядра сливаются (оп-
лодотворение), образуя синкарион, на этой стадии конъ-
югирующие особи расходятся; 8 — синкарион делится;
9 — из двух ядер, образовавшихся в результате деле-
ния синкариона, одно ядро начинает расти, превра-
щаясь в Ма, другое дает Mi, старый Ма окончательно
растворяется в цитоплазме; реконструкция ядерного
аппарата закончилась, имеется один Ма и один Мг.
имеется по одному ядру, образовавшемуся
в результате слияния стационарного и
мигрирующего ядер. Это сложное ядро
получает название синкарион. Об-
разование синкариона представляет со-
бой не что иное, как процесс оплодотво-
рения. И у многоклеточных существенным
моментом оплодотворения является слия-
ние ядер половых клеток. У инфузорий
половые клетки не образуются, имеются
лишь половые ядра, которые и сливаются
между собой. Таким образом происходит
взаимное перекрестное оплодотворение.
Вскоре после образования синкариона
конъюганты расходятся. По строению сво-
его ядерного аппарата они на этой стадии
еще очень существенно отличаются от
обычных так называемых нейтральных (не
конъюгирующих) инфузорий, так как
у них имеется лишь по одному ядру.
В дальнейшем за счет синкариона проис-
ходит восстановление нормального ядер-
ного аппарата. Синкарион делится (один
или несколько раз). Часть продуктов
этого деления путем сложных преобразо-
ваний, связанных с увеличением числа
хромосом и обогащением хроматином, пре-
вращается в макронуклеусы. Другие со-
храняют структуру, характерную для ми-
кронуклеусов. Таким путем восстанав-
ливается характерный и типичный для
инфузорий ядерный аппарат, после чего
инфузории приступают к бесполому раз-
множению путем деления.
Таким образом, процесс конъюгации
включает в себя два существенно важных
биологических момента: оплодотворение
и восстановление нового макронуклеуса
за счет синкариона.
В чем заключается биологическое зна-
чение конъюгации, какую роль играет
она в жизни инфузорий? Мы не можем
назвать ее размножением, ибо при этом
увеличения числа особей не происходит.
Поставленные выше вопросы послужили
материалом для многочисленных экспе-
риментальных исследований, осущест-
влявшихся во многих странах. Основной
результат этих исследований сводится
к следующему. Во-первых, конъюгация,
как и всякий другой половой процесс,
при котором происходит объединение в од-
ном организме двух наследственных на-
чал (отцовского и материнского), ведет
к повышению наследственной изменчиво-
сти, наследственного многообразия. По-
вышение наследственной изменчивости
увеличивает приспособительные возмож-
ности организма к условиям окружающей
среды. Второй биологически важной сто-
роной конъюгации является развитие
нового макронуклеуса за счет продуктов
деления синкариона и одновременно с этим
разрушение старого. Экспериментальные
данные показывают, что именно макро-
нуклеус играет исключительно важную
роль в жизни инфузорий. Им контроли-
руются все основные жизненные процессы
140
и определяется важнейший из них —
образование (синтез) белка, составляю-
щего основную часть протоплазмы жи-
вой клетки. При длительном бесполом
размножении путем деления происходит
как бы своеобразный процесс «старения»
макронуклеуса, а вместе с тем и всей
клетки: снижается активность процесса
обмена веществ, снижается темп деления.
После конъюгации (в процессе которой,
как мы видели, старый макронуклеус раз-
рушается) происходит восстановление
уровня обмена веществ и темпа деления.
Поскольку при конъюгации происходит
процесс оплодотворения, который у боль-
шинства других организмов связан с раз-
множением и появлением нового поколе-
ния, у инфузорий особь, образовавшуюся
после конъюгации, тоже можно рассмат-
ривать как новое половое поколение, ко-
торое возникает здесь как бы за счет «омо-
лаживания» старого.
На примере инфузории туфельки мы по-
знакомились с типичным представителем
обширного класса инфузорий. Однако
класс этот характерен чрезвычайным раз-
нообразием видов как по строению, так
и по образу жизни. Познакомимся ближе
с некоторыми наиболее характерными и
интересными формами.
У инфузории туфельки реснички рав-
номерно покрывают всю поверхность те-
ла. Это характерная черта строения о т-
р я д а равноресничных инфузорий (Holot-
richa). Многим инфузориям свойствен иной
характер развития ресничного покрова.
Дело в том, что реснички инфузорий спо-
собны, соединяясь вместе, образовывать
более сложные комплексы. Например, не-
редко наблюдается, что расположенные
в один или два ряда близко друг к другу
реснички соединяются (слипаются) вме-
сте, образуя пластинку, которая, так же
как и реснички, способна к биению.
Такие пластинчатые сократимые образо-
вания получили название м е м б р а-
не л л (если они короткие) или мемб-
ран (если они длинные). В других
случаях соединяются вместе реснички,
расположенные тесным пучком. Эти образо-
вания - ц и р р ы — напоминают кисточ-
ку, отдельные волоски которой слеплены
друг с другом. Разного рода сложные
ресничные образования характерны для
многих инфузорий. Очень часто реснич-
ный покров развивается не равномерно,
а лишь на некоторых участках тела.
ИНФУЗОРИЯ ТРУБАЧ
(STENTOR POLYMORPHIC)
В пресных водах очень часто встреча-
ются виды крупных красивых инфузорий,
относящихся к р о д у трубачей (Stentor).
Это название вполне соответствует форме
тела этих животных, которая действитель-
но напоминает трубу (рис. 92), широко
раскрывшуюся на одном конце. При пер-
Рис. 92. Инфузория трубач (Stentor coeruleus)
в расправленном состоянии:
J — околоротовая (адоральная) зона мембранелл; 2 —
сократительная вакуоля, состоящая из резервуара и
двух приводящих каналов; 3 — четковидный макро-
нуклеус; 4 — микройукяеусы.
141
вом же знакомстве с живыми трубачами
можно заметить одну особенность, кото-
рая не свойственна туфельке. При малей-
шем раздражении, в том числе механиче-
ском (например, постукивание каранда-
шом по стеклу, где имеется капля воды
с трубачами), тело их резко и очень быст-
ро (в долю секунды) сокращается, прини-
мая почти правильную шарообразную фор-
му. Затем довольно медленно (время изме-
ряется секундами) трубач расправляется,
принимая характерную для него форму.
Эта способность трубача быстро сокра-
щаться обусловлена наличием особых мус-
кульных волоконец, расположенных вдоль
тела и в эктоплазме. Таким образом, и в
одноклеточном организме может развиться
мышечная система.
В роде трубачей имеются виды,
некоторые из которых характеризуются
довольно яркой окраской. Очень обычен
в пресных водах голубой трубач (Stentor
coeruleus), который окрашен в ярко-го-
лубой цвет. Эта окраска трубача обуслов-
лена тем, что в эктоплазме его располо-
жены мельчайшие зерна синего пигмента.
Другой вид трубача (Stentor polymor-
phus) нередко бывает окрашен в зеленый
цвет. Причина этой окраски совсем иная.
Зеленый цвет обусловлен тем, что в эндо-
плазме инфузории живут и размножаются
мелкие одноклеточные зеленые водорос-
ли, которые и придают телу трубача ха-
рактерную окраску. Stentor polymor-
phus — типичный пример взаимно полез-
ного сожительства — симбиоза. Трубач и
водоросли находятся во взаимно симбио-
тических отношениях: трубач защищает
водоросли, живущие в его теле, и снаб-
жает их углекислотой, образующейся в
результате дыхания; со своей стороны
водоросли дают трубачу кислород, освобож-
дающийся в процессе фотосинтеза. По-види-
мому, часть водорослей переваривается
инфузорией, являясь пищей для трубача.
Трубачи медленно плавают в воде ши-
роким концом вперед. Но они могут также
временно прикрепляться к субстрату зад-
ним узким концом тела, на котором при
этом образуется небольшая присоска.
В теле трубача можно различить рас-
ширяющийся сзади наперед туловищный
отдел и почти перпендикулярно к нему
расположенное широкое околоротовое
(перистомальное) поле. Это поле
напоминает асимметричную плоскую во-
ронку, с одного края которой имеется
углубление — глотка, ведущая в эн-
доплазму инфузории. Тело трубача по-
крыто продольными рядами коротких рес-
ничек. По краю перистома л ьного поля
по кругу располагается мощно развитая
околоротовая (адоральная) зона
мембранелл (рис. 92). Эта зона состоит
из большого числа отдельных мерцатель-
ных пластинок, каждая из которых в свою
очередь слагается из множества слип-
шихся друг с другом, расположенных
двумя тесно сближенными рядами ресни-
чек.
В области ротового отверстия околоро-
товые мембранеллы заворачивают в сто-
рону глотки, образуя левозакрученную
спираль. Ток воды, вызываемый колеба-
нием околоротовых мембранелл, направ-
лен в сторону ротового отверстия (в глу-
бину воронки, образуемой передним кон-
цом тела). Вместе с водой в глотку попа-
дают и взвешенные в воде пищевые ча-
стицы. Пищевые объекты у трубача раз-
нообразнее, чем у туфельки. Наряду с бак-
териями он поедает мелких простейших
(например, жгутиковых), одноклеточные
водоросли и т. п.
У трубача хорошо развита сократи-
тельная вакуоля, имеющая своеобразное
строение. Центральный резервуар распо-
ложен в передней трети тела, несколько
ниже ротового отверстия. От него отхо-
дят два длинных приводящих канала.
Один из них идет от резервуара к заднему
концу тела, второй расположен в области
перистомального поля параллельно около-
ротовой зоне мембранелл.
Очень своеобразно устроен ядерный ап-
парат трубача. Макронуклеус здесь поде-
лен на четки (их около 10), связанные
друг с другом тонкими перемычками. Мик-
ронуклеусов несколько. Они очень мел-
кие и обычно тесно прилегают к четкам
макронуклеуса.
Инфузория трубач — излюбленный
объект для экспериментальных исследо-
ваний по регенерации. Многочисленными
опытами была доказана высокая регене-
ративная способность трубачей. Инфузо-
142
рию тонким скальпелем можно разрезать
на множество частей, и каждая из них че-
рез короткое время (несколько часов,
иногда сутки и более) превратится в про-
порционально построенного, но малень-
кого трубача, который затем в результате
энергичного питания достигает типичного
для данного вида размера. Для заверше-
ния восстановительных процессов в реге-
нерирующем кусочке должен быть хотя
бы один сегмент четковидного макронук-
леуса.
Трубач, как мы видели, обладает раз-
ными ресничками: с одной стороны корот-
кими, покрывающими все тело, а с дру-
гой — околоротовой зоной мембранелл.
В соответствии с этой характерной чер-
той строения отряд инфузорий, к кото-
рому относится трубач, получил назва-
ние разноресничних инфузорий (Hetero-
tricha).
ИНФУЗОРИЯ БУРСАРИЯ
(BURSARIA TRUNCATELLA)
Второй интересный представитель раз-
норесничных инфузорий — часто встре-
чающаяся в пресных водах бурсария (Виг-
saria truncatella, рис. 93). Это гигант сре-
ди инфузорий: ее размеры могут дости-
гать 2 мм, наиболее обычные величины —
0,5—1,0 мм. Бурсарию хорошо видно не-
вооруженным глазом. В соответствии со
своим названием бурсария имеет форму
мешка, широко открытого на переднем
конце (bursa — латинское слово, в пере-
воде на русский язык означает «кошель»,
«мешок») и несколько расширенного на
заднем конце. Все тело инфузории покры-
то продольно идущими рядами коротких
ресничек. Биение их обусловливает до-
вольно медленное поступательное движе-
ние животного. Бурсария плывет как бы
«переваливаясь» с бока на бок.
От переднего конца в глубь тела (при-
мерно на 2/3 его длины) вдается околоро-
товое углубление — перистом. С брюш-
ной стороны оно сообщается с наружной
средой узкой щелью, на спинной стороне
полость перистома с наружной средой не
сообщается. Если посмотреть на попереч-
Рис. 93. Разно ресничная инфузория Bursaria
truncatella:
А — общий вид инфузории; Б — поперечный разрез
через середину тела. 1 — отверстие, ведущее в перис-
томальное углубление; 2 — реснички; 3 — большое
ядро (макронуклеус); 4 — околоротовые (адоральные)
мембранеллы; 5 — пищевые включения; 6 — скелет,
поддерживающий перистомальную впадину; 7 — ро-
товая щель; 8 — эктоплазма; 9 — эндоплазма; 10 —
ряды ресничек, спускающиеся в перистомальную
впадину; 11 — полость перистомальной впадины.
Рис. 94. Деление и циста Bursaria truncatella:
А — начало деления, старый перистом редуцирован, заложены два зачатка новых перистомов и адо-
ральных мембранелл дочерних особей; Б — деление, между дочерними особями образовалась пере-
тяжка, В — только что разделившаяся особь, продолжается развитие перистома; Г — циста, хорошо
выражены две оболочки. 1 — зачаток перистома, 2 — макронуклеус.
ный разрез верхней трети тела бурсарии
{рис. 93, Б), то видно, что полость перисто-
ма занимает большую часть тела, тогда
как цитоплазма окружает ее в виде ободка.
На переднем конце тела, слева берет
начало очень мощно развитая у бурсарии
зона околоротовых (адоральных)
мембранелл (рис. 93, 4). Она спускается
в глубину полости перистома, заворачи-
вая налево. Адоральная зона доходит до
самой глубокой части перистома. Кроме
околоротовых мембранелл, других рес-
ничных образований в полости перистома
нет, за исключением ресничной полоски,
идущей по брюшной стороне полости пери-
стома (рис. 93, 10). На внутренней задней
стенке перистомальной полости почти
но всей ее длине проходит узкая щель
(рис. 93, 7), края которой обычно тесно
прилегают друг к другу. Это ротовая
щель. Ее края раздвигаются только в
момент приема пищи.
Бурсарии не обладают узкой пищевой
специализацией, но в основном это хищ-
ники. При поступательном движении они
наталкиваются на различных мелких жи-
вотных. Благодаря работе мембранелл
околоротовой зоны добыча с силой втя-
гивается в обширную перистомальную
полость, откуда уже не может выплыть
наружу. Пищевые объекты прижимаются
к спинной стенке перистомальной полости
и через раздвигающуюся ротовую щель
проникают в эндоплазму. Бурсарии очень
прожорливы, они могут проглатывать до-
вольно крупные объекты: например, из-
любленной пищей их являются инфузо-
рии туфельки. Бурсария способна под-
ряд проглотить 6—7 туфелек. В резуль-
тате в эндоплазме бурсарии образуются
очень крупные пищеварительные ваку-
оли.
Ядерный аппарат бурсарии
устроен довольно сложно. Они имеют
один длинный колбасовидный макронук-
леус и большое (примерно до 30) количе-
ство мелких микронуклеусов, беспоря-
дочно разбросанных в эндоплазме инфу-
зории.
Бурсарии относятся к числу немногих
видов пресноводных инфузорий, у кото-
рых отсутствует сократительная вакуоля.
Как осуществляется у этой крупной инфу-
зории осморегуляция, до сих пор остается
не вполне ясным. Под эктоплазмой бур-
сарии в разных участках тела можно на-
блюдать различной формы и размеров
пузырьки жидкости— вакуоли, кото-
рые меняют свой объем. По-видимому,
эти неправильной формы вакуоли и соот-
ветствуют по своей функции сократитель-
ным вакуолям других инфузорий.
Интересно наблюдать за последователь-
ными стадиями бесполого раз-
множения бурсарии. На начальных
стадиях деления происходит полная ре-
дукция всей полости перистома и околоро-
товой зоны мембранелл (рис. 94). Сохра-
няется лишь наружный ресничный пок-
ров. Инфузория приобретает форму яйца.
После этого тело перешнуровывается по-
перечной бороздой на две половины.
В каждой из образовавшихся дочерних
инфузорий путем довольно сложных пре-
образований развивается типичный пери-
стом и околоротовая зона мембранелл.
Весь процесс деления бурсарии протекает
быстро и занимает немногим больше
часа.
Очень легко наблюдать у бурсарии еще
один важный жизненный процесс, наступ-
ление которого связано с неблагоприят-
ными для инфузории условиями, — про-
цесс образования цист (и н ц и с т и р о-
в а н и е). Это явление свойственно, на-
пример, амебам (стр. 79). Но оказы-
вается, что и такие сложно организован-
ные простейшие, как инфузории, способ-
ны переходить в недеятельное состояние.
Если культуру, где живут бурсарии, во-
время не покормить или сильно охла-
дить, то уже через несколько часов нач-
нется массовое инцистирование. Проте-
кает этот процесс следующим образом.
Бурсарид, так же как перед делением,
теряют перистом и околоротовую зону
мембранелл. Затем они становятся совер-
шенно шаровидными, после чего выделяют
характерной формы двойную оболочку
(рис. 94, Г).
В состоянии цист бурсарии могут на-
ходиться месяцами. При наступлении
благоприятных условий оболочка цисты
лопается, бурсарии выходят из нее, раз-
вивают перистом и переходят к активной
жизни.
фЮ Жизнь животных, т. 1
145
стилонихия
(STYLONICHIA MYTILUS)
Очень сложно и разнообразно диффе-
ренцированный ресничный аппарат имеют
инфузории, относящиеся к отряду
брюхоресничных (Hypotricha), многочис-
ленные виды которых обитают как в пре-
сной, так и в морской воде. Одним из наи-
более обычных, часто встречающихся
представителей этой интересной группы
можно назвать стилонихию (Stylonichia
my til us). Это довольно крупная инфузо-
рия (длина до 0,3 мм), живущая на дне
пресноводных водоемов, на водной рас-
тительности (рис. 95). В отличие от ту-
фельки, трубача и бурсарии у стилони-
хии отсутствует сплошной ресничный по-
кров, а весь ресничный аппарат пред-
Рис. 95. Брюхоресничная инфузория Stylonichia
mytilus:
А — с брюшной стороны; Б — сбоку 1 — брюшные
цирры; 2 — мембранеллы адоральной зоны; 3 — пери-
стом с околоротовыми ресничками и мембраной; 4 —
хвостовые (каудальные) цирры; 5 — пищевые вакуоли;
6 — макронунлеус; 7 — микронуклеус; 8 — спинные
щетинки; 9 — сократительная вакуоля.
ставлен ограниченным числом строго опре-
деленно расположенных ресничных обра-
зований.
Тело стилонихии (как и большинства
других брюхоресничных инфузорий) силь-
но сплющено в спинно-брюшном направле-
нии, и у нее ясно различимы спинная
и брюшная стороны, передний и задний
концы. Спереди тело несколько расширено,
сзади — сужено. При рассматривании
животного с брюшной стороны ясно видно,
что в передней трети слева расположен
сложно устроенный перистом и ротовое
отверстие.
На спинной стороне довольно редко
расположены реснички, которые не спо-
собны к биению. Их скорее можно назвать
тонкими эластичными щетинками. Они не-
подвижны и не имеют отношения к функ-
ции движения. Этим ресничкам приписы-
вают обычно осязательную, чувствитель-
ную, функцию.
Все ресничные образования, связанные
с движением и захватом пищи, сосредо-
точены на брюшной стороне животного
(рис. 95). Здесь имеется небольшое коли-
чество толстых пальцевидных образова-
ний, расположенных несколькими груп-
пами. Это брюшные цирры. Каж-
дая из них — сложное ресничное обра-
зование, результат тесного соединения
(слипания) многих десятков отдельных ре-
сничек. Таким образом, цирры представ-
ляют собой как бы кисточки, отдельные
волоски которых тесно сближены и соеди-
нены между собой.
При помощи цирр животное довольно
быстро передвигается, «бегает» по субст-
рату. Кроме «ползания» и «бегания» по
субстрату стилонихия способна производить
довольно резкие и сильные скачки, сразу
отрываясь при этом от субстрата. Эти рез-
кие движения осуществляются при по-
мощи двух мощных хвостовых цирр (рис.
95), которые в обычном «ползании» уча-
стия не принимают.
По краю тела справа и слева располо-
жены два ряда краевых (маргиналь-
ных) цирр. С правого края животного
они проходят вдоль всего тела, с левого
же края доходят лишь до области пери-
стома. Эти ресничные образования служат
для поступательного движения животного,
146
когда оно оторвано от субстрата и свобод-
но плавает в воде.
Moi видим, таким образом, что разнооб-
разный и специализированный реснич-
ный аппарат стилонихии позволяет ей
делать весьма разнообразные движения
в отличие, например, от простого сколь-
жения в воде, как у туфельки или тру-
бача.
Сложно устроен также ресничный ап-
парат, связанный с функцией питания.
Мы видели уже, что околоротовое углуб-
ление (перистом), на дне которого
помещается ротовое отверстие, ведущее
в глотку, расположен в передней поло-
вине животного слева. По левому краю
начиная с самого переднего конца тела
проходит сильно развитая зона околоро-
товых (адоральных) мембранелл.
Своим биением они направляют ток воды
в сторону ротового отверстия. Кроме
того, в области перистомального углубле-
ния расположены еще три сократимые
перепонки (мембраны), своими внутрен-
ними концами заходящие в глотку, и ряд
особых околоротовых ресничек (рис. 95).
Весь этот сложный аппарат служит для
улавливания и направления пищи в
ротовое отверстие.
Стилонихия относится к числу простей-
ших с очень широким диапазоном пище-
вых объектов. Ее с полным правом мож-
но назвать всеядным животным. Она мо-
жет питаться, как и туфелька, бактери-
ями. В число ее пищевых объектов вхо-
дят жгутиконосцы, одноклеточные водо-
росли (нередко диатомовые). Наконец,
стилонихия может быть и хищником, на-
падая на другие, более мелкие виды ин-
фузорий и поглощая их.
У стилонихии имеется сократительная
вакуоля. Она состоит из центрального
резервуара, расположенного у левого зад-
него конца перистома, и одного направ-
ленного назад приводящего канала.
Ядерный аппарат, как всегда у инфузо-
рий, состоит из макронуклеуса и микро-
нуклеуса.
Макронуклеус слагается из двух поло-
винок, соединенных тонкой перетяжкой;
микронуклеусов два, они расположены
непосредственно около обеих половинок
Ма.
Рис. 96. Инфузории дидинии
(Didinium nasutum), пожираю-
щие инфузорию туфельку.
Стилонихия, отчасти бурсария, тру-
бач — это все инфузории с широким диа-
пазоном пищевых объектов. Способность
поглощать различную пищу свойственна
большинству инфузорий. Однако среди
них можно найти и такие виды, которые
в отношении характера пищи строго спе-
циализированы.
ИНФУЗОРИИ-ХИЩНИКИ
Среди инфузорий есть хищники, кото-
рые очень «разборчивы» в отношении своей
жертвы. Хорошим примером может слу-
жить инфузория дидинии (Didinium nasu-
tum). Дидиний — относительно неболь-
шая инфузория, длиной в среднем около
0,1—0,15 мм. Передний конец вытянут
в виде хоботка, на конце которого поме-
щается ротовое отверстие. Ресничный ап-
парат представлен двумя венчиками рес-
ничек (рис. 96). Дидиний быстро плавает
в воде, часто меняя направление движе-
ния. Предпочитаемая пища дидиниев —
инфузории туфельки. В данном случае
хищник оказывается меньше своей жерт-
вы. Дидиний внедряется в добычу хобо-
том, а затем, постепенно все более и более
расширяя ротовое отверстие, проглатывает
туфельку целиком! В хоботке имеется
особый, так называемый палочковый, ап-
парат. Он состоит из ряда эластичных проч-
10*
147
Рис. 97. Последовательные стадии за-
глатывания инфузорией Nassula нити
сине-зеленой водоросли.
Рис. 98. Кругоресничная инфузория _*.
Vorticella:
1 — мерцательные (ундулирующие) мембра-
ны; 2 — перистомальный валик; 3 — углуб-
ление ротовой воронки; 4 — сократительная
вакуоля; 5 — стебелек.
ных палочек, располагающихся в цито-
плазме по периферии хоботка. Предпо-
лагают, что этот аппарат увеличивает
прочность стенок хоботка, который не
разрывается при проглатывании такой
огромной по сравнению с дидинием добы-
чи, как туфелька. Дидиний — пример
крайнего случая хищничества среди про-
стейших. Если сравнить заглатывание
дидинием своей добычи — туфельки —с
хищничеством у высших животных, то
аналогичные примеры найти трудно.
Дидиний, проглотивший парамецию, ра-
зумеется, очень сильно раздувается. Про-
цесс переваривания протекает очень бы-
стро, при комнатной температуре он за-
нимает всего около двух часов. Затем
непереваренные остатки выбрасываются
наружу и дидиний начинает охотиться
за очередной жертвой. Специальными ис-
следованиями было выяснено, что суточ-
ный «рацион» дидиния со-
ставляет 12 туфелек — по-
истине колоссальный ап-
петит! Нужно иметь в ви-
ду, что в промежутках
между очередными «охота-
ми» дидинии иногда делят-
ся. При недостатке пищи
дидинии очень легко ин-
цистируются и так же лег-
ко вновь выходят из цист.
РАСТИТЕЛЬНОЯДНЫЕ
ИНФУЗОРИИ
Гораздо реже, чем хищ-
ничество, встречается сре-
ди инфузорий «чистое ве-
гетарианство» — питание
исключительно раститель-
ной пищей. Одним из не-
многих примеров инфузо-
рий-«вегетарианцев» могут
служить представители
рода нассула (Nassula).
Объектом питания их яв-
ляются нитчатые сине-зе-
леные водоросли (рис. 97).
Они проникают в эндо-
плазму через рот, распо-
ложенный сбоку, а затем закручиваются
инфузорией в плотную спираль, которая
постепенно переваривается. Пигменты во-
дорослей частично поступают в цитоплаз-
му инфузории и окрашивают ее в яркий
темно-зеленый цвет.
СУВОЙКА (VORTICELLA NEBULIFERA)
Интересную и довольно большую по
числу видов группу инфузорий состав-
ляют сидячие, прикрепленные к субстра-
ту формы, образующие отряд кругоре-
сничных (Peritricha). Широко распростра-
ненными представителями этой группы
являются сувойки (виды рода Vorticella).
Сувойки напоминают изящный цветок
вроде колокольчика или ландыша, сидя-
щий на длинном стебельке, который своим
концом прикреплен к субстрату. Боль-
1
2
3
4
шую часть жизни сувойка проводит в при-
крепленном к субстрату состоянии.
Рассмотрим строение тела инфузории.
У разных видов их размеры варьируют
в довольно широких пределах (примерно
до 150 мк). Ротовой диск (рис. 98) располо-
жен на расширенном переднем участке
тела, которое совершенно лишено ресни-
чек. Ресничный аппарат расположен лишь
по краю ротового (перистомального) ди-
ска (рис. 98) в особой бороздке, снаружи
от которой образуется валик (п е р и-
стомальная губа). По краю вали-
ка идут три мерцательные мембраны, из
которых две расположены вертикально,
одна (наружная) — горизонтально. Они
образуют несколько больше одного пол-
ного оборота спирали. Эти мембраны на-
ходятся в постоянном мерцательном дви-
жении, направляя ток воды к ротовому
отверстию. Ротовой аппарат начинается
довольно глубоко воронкой у края пери-
стомального поля (рис. 98), в глубине
которой располагается ротовое отверстие,
ведущее в короткую глотку. Сувойки,
так же как и туфельки, питаются бакте-
риями. Их ротовое отверстие постоянно
открыто, и возникает непрерывный ток
воды в направлении рта.
Одна сократительная вакуоля без при-
водящих каналов расположена недалеко
от ротового отверстия. Макронуклеус име-
ет лентовидную или колбасовидную фор-
му, к нему тесно примыкает единственный
маленький микронуклеус.
Сувойка способна резко сокращать сте-
белек, который в долю секунды закручи-
вается штопором. Одновременно с этим
сокращается и тело инфузории: перисто-
мальный диск и мембраны втягиваются
внутрь и весь передний конец замыкается.
Естественно возникает вопрос: посколь-
ку сувойки прикреплены к субстрату,
каким путем осуществляется их расселе-
ние по водоему? Это происходит путем
образования свободноплавающей стадии—
бродяжки. На заднем конце тела инфу-
зории возникает венчик ресничек (рис.
99). Одновременно перистомальный диск
втягивается внутрь и инфузория отде-
ляется от стебелька. Образовавшаяся бро-
дяжка способна плавать в течение не-
скольких часов. Затем события разыгры-
Рис. 99. Кругоресничная инфузория Vorticella
microstoma:
J — инфузория с расправленным передним концом,
несущим ротовой аппарат; 2 — инфузория с сокра-
тившимся стебельком и втянутым передним концом;
3 — 6 — последовательные стадии образования бро-
дяжки.
Рис. 100. Разные представители кругоресничных
инфузорий:
1 — Pyxidium ventriosa, не образующие колоний,.
2 — колония Carchesium polypinum, 3 — участок ко-
лонии Campanella umbellaria
149
I
V
Рис. 101 Общий вид колонии Zoothamnium
arbuscula.
На ветвях колонии множество мелких особей
и крупные особи-расселительницы (макрозоиды).
ваются в обратном порядке: инфузория
прикрепляется к субстрату задним кон-
цом, вырастает стебелек, редуцируется
задний венчик ресничек, на переднем конце
расправляется перистомальный диск, на-
чинают работать адоральные мембраны.
Образование бродяжек у сувойки нередко
связано с процессом бесполого размноже-
ния. Инфузория на стебельке делится,
причем одна из дочерних особей (а иногда
и обе) становится бродяжкой и уплывает.
Многие виды сувоек при неблагоприят-
ных условиях способны инцистироваться.
Среди сидячих инфузорий, относящих-
ся к отряду кругоресничных, лишь отно-
сительно немногие виды вроде рассмот-
ренных выше сувоек являются одиночно
живущими формами. Большая часть отно-
сящихся сюда видов — организмы коло-
ниальные.
Обычно колониальность воз-
никает в результате не вполне завершен-
ного бесполого или вегетативного размно-
жения. Образующиеся в результате раз-
множения особи в большей или меньшей
степени сохраняют связь друг с другом и
все вместе образуют органическую индиви-
дуальность высшего порядка, объединя-
ющую большие количества отдельных
особей, которая и получает название ко-
лонии (с примерами колониальных орга-
низмов мы уже познакомились в к л а с-
с е жгутиконосцев, стр. 98).
Колонии кругоресничных инфузорий об-
разуются в результате того, что разде-
лившиеся особи не превращаются в бро-
дяжек, а сохраняют связь друг с другом
при помощи стебельков (рис. 100). При
этом основной стебелек колонии, так же
как и его первые разветвления, не может
быть отнесен ни к одной из особей, а при-
надлежит всей колонии в целом. Иногда
колония состоит лишь из небольшого
числа особей, у других же видов инфузо-
рий число отдельных особей колонии мо-
жет достигать нескольких сотен. Однако
рост любой колонии не беспределен. По
достижении характерных для данного ви-
да размеров колония перестает увеличи-
ваться и образующиеся в результате де-
ления особи развивают венчик ресничек,
становятся бродяжками и уплывают, да-
вая начало новым колониям.
Колонии кругоресничных инфузорий
бывают двух типов. У одних стебелек ко-
лонии несократим: при раздражении со-
кращаются лишь отдельные особи коло-
нии, втягивая перистом, вся же колония
в целом не претерпевает изменений (к та-
кому типу колоний относятся, например,
роды Epistylis, Opercularia). У других
(например, род Carchesium) стебелек
всей колонии способен сокращаться, так
как цитоплазма проходит внутри всех
веточек и связывает, таким образом, меж-
ду собой всех особей колонии. При раздра-
жении таких колоний они сокращаются
целиком. Вся колония в данном случае
реагирует как единое целое, как органиче-
ская индивидуальность.
Среди всех колониальных кругореснич-
ных инфузорий особый интерес представ-
ляет, пожалуй, зоотамний (Zoothamnium
«arbuscula). Колонии этой инфузории отли-
чаются особенной правильностью строе-
ния. Кроме того, в пределах колонии здесь
намечается интересное биологическое яв-
ление полиморфизма.
Колония зоотамния имеет вид зонтика.
На одном, главном, стебельке колонии
располагаются вторичные ветви (рис. 101).
Размер взрослой колонии 2—3 мм, так
что они хорошо видны простым гла-
зом. Живут зоотамний в небольших пру-
дах с чистой водой. Колонии их обычно
находят на подводных растениях, чаще
всего на элодее (водяной чуме).
Стебельки колонии зоотамния обладают
сократимостью, так как сократимая ци-
топлазма проходит через все ветви коло-
нии, за исключением базальной части
главного стебелька. При сокращении, ко-
торое происходит очень быстро и резко,
вся колония собирается в комочек.
Для зоотамния характерно строго зако-
номерное расположение ветвей. К суб-
страту прикрепляется один главный сте-
белек. От верхней части его в плоскости,
перпендикулярной к стебельку, отходит
девять главных ветвей колонии, строго
закономерно расположенных друг относи-
тельно друга (рис. 102, 6). От этих ветвей
отходят вторичные веточки, на которых
сидят отдельные особи колонии. На каж-
дой вторичной веточке может быть до
50 инфузорий. Общее количество особей
в составе колонии достигает 2—3 тыс.
Большая часть особей колонии по свое-
му строению напоминает небольших оди-
ночных сувоек, размером 40—60 мк. Но
кроме мелких особей, которые называются
микрозоидами, на взрослых колониях,
примерно на середине главных ветвей,
развиваются особи совершенно другого
вида и размера (рис. 102, 5). Это крупные
шаровидные особи диаметром 200—250 мк,
превосходящие по своей массе объем мик-
розоида в сотню и более раз. Крупные
особи получили название макрозоидов.
По своему строению они существенно от-
личаются от мелких особей колонии. Пе-
ристом у них не выражен: он втягивается
внутрь и не функционирует. Макрозоид
с самого начала своего развития из микро-
зоида перестает самостоятельно принимать
пищу. У него отсутствуют пищеваритель-
ные вакуоли. Рост макрозоида осуществ-
ляется, очевидно, за счет веществ, посту-
пающих через цитоплазматические мости-
ки, соединяющие между собой всех осо-
бей колонии. В участке тела макрозоида,
которым он прикрепляется к стебельку,
имеется скопление особых зернышек (гра-
нул), которые, как увидим, в его дальней-
шей судьбе играют существенную роль.
Что же представляют собой эти крупные
шаровидные макрозоиды, какова их био-
Рис. 102. Развитие и строение колонии Zooth-
amnium arbuscula:
1—4 — последовательные стадии развития B-, 4-, 8- и
16-клеточные стадии); 5 — небольшой участок колонии
с микрозоидами и двумя макрозоидами; один макро-
зоид в начале роста, второй выросший, готовый к отде-
лению от колонии; 6 — схема расположения девяти
ветвей колонии, видны места образования макрозоидов.
151
логическая роль в жизни колонии зоотам-
ния? Наблюдение показывает, что макро-
зоиды — это будущие бродяжки, из кото-
рых развиваются новые колонии. Достиг-
нув предельного размера, макрозоид раз-
вивает венчик ресничек, отделяется от коло-
нии и уплывает. Его форма при этом не-
сколько меняется, из шаровидного он
становится коническим. Через некоторое
время бродяжка прикрепляется к субст-
рату всегда той стороной, на которой рас-
положена зернистость. Сразу же начи-
нается образование и рост стебелька, при-
чем на построение стебелька затрачива-
ются гранулы, которые локализованы на
заднем конце бродяжки. По мере роста
Рис. 103. Разные виды сосущих инфузорий:
1 — Tokophrya qua (tripartita, щупальца расположены
четырьмя пучками; 2 — Ephelota gemmipara, много ко-
ротких щупалец; 3 — Dendrosoma radians, инфузория
образует древовидно ветвящуюся колонию со многими
группами щупалец.
стебелька зернистость исчезает. После
того как стебелек достигнет окончатель-
ной, характерной для зоотамния длины,
начинается ряд быстро следующих друг
за другом делений, ведущих к образова-
нию колонии. Деления эти совершаются
в строго определенной последовательно-
сти (рис. 102). На рассмотрении деталей
этого процесса мы останавливаться не
будем. Обратим лишь внимание на следую-
щее интересное явление. Во время первых
делений бродяжки зоотамния при развитии
колонии у образующих особей перистом и
рот не функционируют. Питание начина-
ется позже, когда молодая колония состоит
уже из 12—16 особей. Таким образом, все
первые стадии развития колонии осуще-
ствляются исключительно за счет тех за-
пасов, которые образовались в теле мак-
розоида в период его роста и развития на
материнской колонии. Существует бес-
спорное сходство между развитием бро-
дяжки зоотамния и развитием яйца у мно-
гоклеточных животных. Это сходство вы-
ражается в том, что развитие в обоих слу-
чаях осуществляется за счет ранее на-
копленных запасов, без восприятия пищи
из внешней среды.
При изучении сидячих кругоресничных
инфузорий возникает вопрос: а как же осу-
ществляется у них характерная для инфу-
зорий форма полового процесса — конъ-
югация (стр. 139)? Оказывается, что
в связи с сидячим образом жизни она пре-
терпевает некоторые существенные изме-
нения. К началу полового процесса на
колонии образуются особые, очень мел-
кие бродяжки. Активно двигаясь при по-
мощи венчика ресничек, они ползают в те-
чение некоторого времени по колонии, а
затем вступают в конъюгацию с крупными
нормальными сидячими особями колонии.
Таким образом, здесь происходит диффе-
ренцировка конъюгантов на две группы
особей: мелкие, подвижные (микроконъ-
юганты) и более крупные, неподвижные
(макроконъюганты). Эта дифференцировка
конъюгантов на две категории, из которых
одна (микроконъюганты) подвижная, яви-
лась необходимым приспособлением к си-
дячему образу жизни. Без этого нормаль-
ный ход полового процесса (конъюгации)
не мог быть, очевидно, обеспечен.
152
СОСУЩИЕ ИНФУЗОРИИ (SUCTORIA)
Весьма своеобразную в отношении спо-
соба питания группу представляют сосу-
щие инфузории (Suctoria). Эти организмы,
подобно сувойкам и другим кругореснич-
ным инфузориям, сидячие. Количество
относящихся к этому отряду видов изме-
ряется несколькими десятками. Форма
тела сосущих инфузорий весьма разнооб-
разна. Некоторые характерные виды их
изображены на рисунке 103. Одни сидят
на субстрате на более или менее длинных
стебельках, другие не имеют стебельков,
у некоторых тело довольно сильно вет-
вится и т. п. Однако, несмотря на разнооб-
разие формы, все сосущие инфузории ха-
рактеризуются следующими двумя при-
знаками: 1) полным отсутствием (у взрос-
лых форм) ресничного аппарата, 2) на-
личием особых придатков — щупалец,
служащих для высасывания добычи.
У разных видов сосущих инфузорий
количество щупалец неодинаково. Не-
редко они бывают собраны группами. При
большом увеличении микроскопа можно
рассмотреть, что на конце щупальце снаб-
жено небольшим булавовидным утолще-
нием.
Как функционируют щупальца? На
этот вопрос нетрудно ответить, наблюдая
в течение некоторого времени за сосущи-
ми инфузориями. Если какое-нибудь не-
большое простейшее (жгутиконосец, ин-
фузория) прикоснется к щупальцу сук-
тории, то оно к нему мгновенно прилип-
нет. Все попытки жертвы оторваться бы-
вают обычно тщетными. Если продолжить
наблюдение за прилипшей к щупальцам
жертвой, то можно видеть, что она посте-
пенно начинает уменьшаться в размерах.
Ее содержимое через щупальца постепен-
но «перекачивается» внутрь эндоплазмы
сосущей инфузории до тех пор, пока от
жертвы не останется одна пелликула, ко-
торая отбрасывается. Таким образом, щу-
пальца сосущих инфузорий — это совер-
шенно своеобразные, нигде больше в жи-
вотном мире не встречающиеся органы
улавливания и вместе с тем высасывания
пищи (рис. 103).
Сосущие инфузории — это неподвижные
хищники, которые не гоняются за добы-
чей, но мгновенно улавливают ее, еслет
только неосторожная добыча к ним при-
коснется сама.
Почему же эти своеобразные организмы
мы относим к классу инфузорий? На пер-
вый взгляд они не имеют с ними ничего
общего. О принадлежности сукторий к ин-
фузориям говорят следующие факты. Во-
первых, они обладают типичным для инфу-
зорий ядерным аппаратом, состоящим из
макронуклеуса и микронуклеуса. Во-вто-
рых, во время размножения у них появ-
ляются реснички, отсутствующие «у взро-
Рис. 104. Почкование у разных видов сосущих
инфузорий:
А — Paracineta patula, образующая одну почку;
Б — отделившаяся бродяжка Paracineta patula; В —
множественное наружное почкование Ephelota gem-
mipara; Г — внутреннее почкование Tocophrya cyclo-
pum, отделение почки в выводковую камеру еще не
завершено; Д — то же, в выводковой камере находит-
ся вполне сформированная бродяжка с ресничками.
1 — почка; 2 — макронуклеус; 3 — выводковая камерг;
4 — отверстие выводковой камеры.
153
слых» особей. Бесполое размножение и вме-
сте с тем расселение сосущих инфузорий
осуществляется путем образования бро-
дяжек, снабженных несколькими кольце-
выми венчиками ресничек. Образование
бродяжек у сукторий может происходить
по-разному. Иногда они образуются в ре-
зультате не вполне равномерного деления
(почкованием), при котором каждая отде-
ляющаяся наружу почка получает участок
макронуклеуса и один микронуклеус (рис.
104,Л). На одной материнской особи мо-
жет образоваться сразу несколько дочер-
них почек (рис. 104,5). У других видов
{рис. 104,Г,Д) наблюдается очень своеоб-
разный способ «внутреннего почкования».
При этом внутри тела суктории-матери
образуется полость, в которой и форми-
руется почка-бродяжка. Наружу она вы-
ходит через специальные отверстия, сквозь
которые с известным трудом «протиски-
вается».
Такое развитие зародыша внутри тела
матери, а затем акт деторождения —
интересная аналогия простейшего с тем,
что происходит у вышестоящих много-
клеточных организмов.
На предыдущих страницах было рас-
смотрено несколько типичных свободно-
живущих представителей класса инфузо-
рий, по-разному приспособленных к
различным условиям среды. Интересно
подойти к вопросу приспособления инфузо-
рий к условиям жизни и, с другой сторо-
ны, посмотреть, каковы характерные
общие черты инфузорий, живущих в опре-
деленных, резко очерченных условиях
среды.
Рис. 105. Планктонные инфузории:
Liliomorpha viridis; 2 — Marituja pelagica; з — Tintinnopsis beroidea; 4 — Mucophrya pelagica (Suctoria).
I, 2 и 4 — планктонные инфузории озера Байкал; 3 — морской вид.
154
В качестве примера возьмем две очень
резко различающиеся среды обитания:
жизнь в составе планктона и жизнь на дне
в толще песка.
ПЛАНКТОННЫЕ ИНФУЗОРИИ
В составе как морского, так и пресно-
водного планктона встречается довольно
большое число видов инфузорий.
Особенно ярко выражены черты приспо-
соблений к жизни в толще воды у радио-
лярий (стр. 88). Основная линия при-
способления к планктонному образу жиз-
ни сводится к выработке таких черт строе-
ния, которые способствуют парению орга-
низма в толще воды.
Типичным планктонным, к тому же поч-
ти исключительно морским семейством
инфузорий является тинтинниды (Tin-
tinnidae, рис. 105,5). Общее число извест
ных до сих пор видов тинтиннид около
300. Это мелкие формы, характерные тем,
что протоплазматическое тело инфузории
помещается в прозрачном, легком и вместе
с тем прочном домике, состоящем из орга-
нического вещества. Из домика выдает-
ся наружу диск, несущий венчик ресничек,
находящихся в постоянном мерцательном
движении. В состоянии парения инфузо-
рий в толще воды поддерживает здесь
главным образом постоянная активная
работа ресничного аппарата. Домик, оче-
видно, выполняет функцию защиты ниж-
ней части тела инфузории. В пресной воде
обитает всего 2 вида тинтиннид (не считая
7 видов, характерных только для озера
Байкал).
У пресноводных инфузорий наблюда-
ются некоторые другие приспособления
к жизни в планктоне. У многих из них
цитоплазма очень сильно вакуолизиро-
вана (Loxodes, Condylostoma, Trachelitis),
так что напоминает пену. Это ведет к зна-
чительному уменьшению удельного веса.
Все перечисленные инфузории обладают,
кроме того, ресничным покровом, благо-
даря работе которого тело инфузории, по
удельному весу лишь немногим превы-
шающее удельный вес воды, легко под-
держивается в состоянии «парения». У не-
которых видов форма тела способствует
увеличению удельной поверхности и облег-
чает парение в воде. Например, некото-
рые планктонные инфузории озера Бай-
кал напоминают по форме зонтик или па-
рашют (Liliomorpha, рис. 105,2). Сущест-
вует в озере Байкал одна планктонная
сосущая инфузория (Mucophrya pelagica,
рис. 105,4), которая резко отличается от
своих сидячих родичей (стр. 153). Этот
вид лишен стебелька. Его протоплазмати-
ческое тело окружено широким слизистым
футляром — приспособлением, ведущим к
уменьшению веса. Наружу торчат длин-
ные тонкие щупальца, которые наряду со
своей прямой функцией, вероятно, вы-
полняют и другую — увеличение удельной
поверхности, способствующее парению в
воде.
Нужно, наконец, упомянуть еще об од-
ной, так сказать, косвенной форме при-
способления инфузорий к жизни в планк-
тоне. Это прикрепление мелких инфузо-
рий к другим организмам, ведущим планк-
тонный образ жизни. Так, среди кругоре-
сничных инфузорий (Peritricha) имеются
довольно многочисленные виды, которые
прикрепляются к планктонным веслоно-
гим рачкам. Это обычный и нормальный
для данных видов инфузорий образ жизни.
Наряду с кругоресничными инфузо-
риями и среди сосущих (Suctoria) имеются
виды, поселяющиеся на планктонных ор-
ганизмах.
ИНФУЗОРИИ, ЖИВУЩИЕ В ПЕСКЕ
Чрезвычайно своеобразную среду оби-
тания представляют песчаные пляжи и от-
мели. По побережью морей они занимают
огромные пространства и характеризуются
своеобразной фауной.
Проведенные за последние годы в раз-
личных странах многочисленные исследо-
вания показали, что толща многих мор-
ских песков очень богата разнообразной
микроскопической или приближающейся
по своим размерам к микроскопической
фауной. Между частичками песка имеются
многочисленные заполненные водой мел-
кие и мельчайшие пространства. Оказы-
вается, что эти пространства богато засе-
лены организмами, относящимися к са-
155
мым различным группам животного мира.
Здесь живут десятки видов ракообраз-
ных, кольчатые черви, круглые черви,
особенно многочисленные плоские черви,
некоторые моллюски, кишечнополостные.
В большом количестве здесь встречаются
и простейшие, главным образом инфузории.
По современным данным, в состав фауны
инфузорий, населяющих толщу морских
песков, входит примерно 250—300 видов.
Если иметь в виду не только инфузорий,
а и другие группы населяющих толщу
песка организмов, то общее число видов
их будет очень велико. Всю совокупность
животных, населяющих толщу песка, жи-
вущих в мельчайших просветах между
песчинками, называют псаммофиль-
ной фауной.
Богатство и видовой состав псаммо-
фильной фауны определяется многими
факторами. Среди них особенно важное
значение имеет размер частиц песка.
Крупнозернистые пески имеют бедную
фауну. Бедна также фауна очень мелко-
зернистых заиленных песков (с диамет-
ром частиц менее 0,1 мм), где, очевидно,
просветы между частицами слишком мел-
ки для обитания в них животных. Наибо-
лее богаты жизнью пески средне- и мел-
козернистые.
Второй фактор, играющий важную роль
в развитии псаммофильной фауны,— бо-
гатство песка органическими остатками,
разлагающимися органическими вещест-
вами (так называемая степень сапроб-
ности). Пески, лишенные органических ве-
ществ, бедны жизнью. С другой стороны,
почти безжизненны и пески, очень бога-
тые органическими веществами, так как
распад органических веществ ведет к обед-
нению кислородом. Нередко к этому до-
бавляется анаэробное сероводородное бро-
жение.
Наличие свободного сероводорода
является крайне отрицательно действую-
щим на развитие фауны фактором.
В поверхностных слоях песка иногда
развивается довольно богатая флора одно-
клеточных водорослей (диатомовые, пери-
диниевые). Это фактор, благоприятствую-
щий развитию псаммофильной фауны, так
как многие мелкие животные (в том числе
и инфузории) питаются водорослями.
Наконец, фактор, очень отрицательно
действующий на псаммофильную фау-
ну,— прибой. Это вполне понятно, так
как прибой, перемывающий верхние слои
песка, убивает здесь все живое. Наиболее
богата псаммофильная фауна в защищен-
ных, хорошо прогреваемых бухточках.
Приливы и отливы не препятствуют раз-
витию псаммофильной фауны. Когда в от-
лив вода временно уходит, обнажая пе-
сок, то в толще песка, в промежутках меж-
ду песчинками, она сохраняется, и это
не препятствует существованию живот-
ных.
У инфузорий, входящих в состав псам-
мофильной фауны и относящихся к раз-
личным систематическим группам (отря-
дам, семействам), вырабатываются в про-
цессе эволюции многие общие черты, яв-
ляющиеся приспособлениями к своеоб-
разным условиям существования между
частицами песка.
На рисунке 106 изображены некоторые
виды псаммофильной фауны инфузорий,
относящиеся к разным отрядам и семейст-
вам. Между ними много сходства. Тело
большинства их более или менее сильно
вытянуто в длину, червеобразно. Это
дает возможность легко «протискиваться»
в мельчайшие отверстия между песчин-
ками. У очень многих видов (рис. 106)
удлинение тела сочетается с его уплоще-
нием. Ресничный аппарат всегда хорошо
развит, что позволяет активно, с извест-
ной силой, двигаться в узких просветах.
Нередко реснички развиваются на одной
стороне червеобразного уплощенного те-
ла, противоположная сторона оказывается
голой. Эта особенность связана, вероят-
но, с резко выраженной у большинства
псаммофильных в»идов способностью очень
тесно и очень прочно при посредстве рес-
ничного аппарата приставать (прикреп-
ляться) к субстрату (явление, называемое
тигмотаксисом). Это свойство позволяет
животным оставаться на месте в тех слу-
чаях, когда в узких просветах, где они
живут, возникают токи воды. При этом,
вероятно, выгоднее, чтобы сторона, про-
тивоположная той, которой животное при-
крепилось к субстрату, была гладкой.
Чем питаются псаммофильные инфу-
зории? Значительную часть «рациона» у
156
wan i
m
Ш
1
I
m
Ш
Ш
1
f
^
ж
Рис. 106. Разные виды морских инфузорий, входящие в состав псаммофильной фауны (жи-
вущие в толще песка):
1, 2 — Trahelonema grassei; 3,4 — Centrophorella grandis; 5 — Centrophorella fasciolata, 6 —
Helicoprorodon multinucleatum; 7 — Pseudoprorodon arenicola; 8 — Remanella caudata; 9 — Condy-
lostoma remanei; 10 — Uroleptus rattulus, 11 — Strongilidium arenicolus.
Рис. 107. Схема жизненного цикла Spirophrya
subparasitica:
1 — инцистированная инфузория на рачке (стадия
форонта); 2—молодой трофонт, проглоченный полипом;
3 — выросший трофонт в теле полипа; 4—6 — выход
трофонта из пищеварительной полости полипа и инци-
стирование на полипе; 7 — образование в цисте томи-
тов (расселительниц), выход их в воду и оседание на
рачке.
многих видов составляют водоросли, в осо-
бенности диатомовые. Бактерии в меньшей
степени служат им пищей. Это зависит
в значительной мере и от того, что в пес-
ках, не сильно загрязненных, бактерий
мало. Наконец, особенно среди наиболее
крупных псаммофильных инфузорий име-
ется немалое количество хищных форм,
которые поедают других инфузорий, от-
носящихся к более мелким видам. Псаммо-
фильные инфузории распространены, по-
видимому, повсеместно.
ИНФУЗОРИИ АПОСТОМАТЫ
В процессе приспособления к условиям
жизни очень сложные жизненные циклы,
сопровождающиеся сменой нескольких
форм строения, выработались у своеобраз-
ной морской группы инфузорий — апосто-
мат (Apostomata). Сложность и своеоб-
разие жизненного пути этих инфузорий за-
висит от того, что часть стадий цикла
они проводят как свободноживущие орга-
низмы, другие же стадии являются пара-
зитическими или полупаразитическими.
Рассмотрим на конкретном примере жиз-
ненный цикл апостомат.
Инфузория спирофрия (Spirophrya sub-
parasitica) в инцистированном состоянии
часто может быть обнаружена сидящей
на небольшой ножке на мелких планк-
тонных морских ракообразных (особенно
часто на рачках рода Idia). Пока рачок
активно плавает в морской воде, сидящие
на нем спирофрии не претерпевают ника-
ких изменений. Для дальнейшего разви-
тия инфузорий необходимо, чтобы рачок
был съеден морским гидроидным поли-
пом, что случается нередко (рис. 107).
Как только цисты спирофрии вместе с рач-
ком проникнут в пищеварительную по-
лость, из них сразу же выходят мелкие
инфузории, которые начинают энергично
питаться пищевой кашицей, образующей-
ся в результате переваривания заглочен-
ного рачка. В течение часа размеры инфу-
зории увеличиваются в 3—4 раза. Однако
размножения на этой стадии не происхо-
дит. Перед нами типичная стадия роста
инфузории, которую называют трофонтом.
Через некоторое время вместе с неперева-
ренными остатками пищи трофонт выбра-
сывается полипом наружу в морскую во-
ду. Здесь он, активно плавая, спуска-
ется по телу полипа к его подошве, где
и прикрепляется, окружаясь при этом
цистой. Эта стадия инцистированной, си-
дящей на полипе крупной инфузории
получила название томонта. Это фаза раз-
множения. Томонт не питается, но быстро
последовательно несколько раз делится
(рис. 107,7). В результате получается це-
лая группа очень мелких инфузорий. Ко-
личество их зависит от размеров томонта,
который определяется в свою очередь ве-
личиной трофонта, давшего ему начало.
Мелкие инфузории, образовавшиеся в ре-
зультате деления томонта (они называются
томитами или бродяжками),
представляют собой стадию расселения.
158
Они выходят из цисты, быстро плавают
(не питаясь при этом, а используя имею-
щиеся у них в цитоплазме запасы). Бели
им «посчастливится» натолкнуться на вес-
лоногого рачка, то они тотчас же прикреп-
ляются к нему и инцистируются. Это ста-
дия, с которой мы начали рассмотрение
цикла.
В рассмотренном нами жизненном цикле
спирофрии обращает на себя внимание
резкая разграниченность стадий, имею-
щих разное биологическое значение.
Трофонт — это стадия роста. Он только
растет, накапливает большое количество
цитоплазмы и всякого рода резервных
веществ за счет энергичного и быстрого
питания. К размножению трофонт не спо-
собен. Обратное явление наблюдается у
томонта — неспособность питаться и
энергичное быстрое размножение. После
каждого деления не происходит роста, и
поэтому размножение томонта сводится
к быстрому распаду на множество бро-
дяжек. Наконец, бродяжки выполняют
свою специальную и только им свойст-
венную функцию: это особи — рассели-
тельницы и распространительницы вида.
Они не способны ни питаться, ни раз-
множаться.
ЖИЗНЕННЫЙ ЦИКЛ
ИХТИОФТИРИУСА
Среди инфузорий, часть жизненного
цикла которых протекает в паразитиче-
ском, часть в свободноживущем со-
стоянии, большой интерес, особенно
в практическом отношении, представляет
ихтиофтириус (Ichthyophthirius multifi-
liis). Эта инфузория в течение большей
части своей жизни является паразитом
рыб. Она может вызвать тяжелое заболе-
вание, в особенности поражающее маль-
ков, часто приводящее к гибели хозяина
(рис. 108).
Паразитарные стадии развития ихтиоф-
тириуса протекают непосредственно в по-
кровах рыбы: в коже, на плавниках,
очень часто на жабрах рыбы.
Рыба заражается очень мелкими сво-
бодноплавающими бродяжками B0—30 мк
диаметром), плавающими в воде. Бродяж-
ки при соприкосновении с рыбой немед-
ленно прикрепляются к ее поверхности
и сразу же активными движениями внед-
ряются в ткани. Проникнув в живую
ткань, инфузории начинают активно пи-
таться и расти. Питание осуществляется
путем заглатывания при помощи помещаю-
щегося на переднем конце тела ротового
отверстия клеточных элементов хозяина.
Растущие в тканях рыбы ихтиофтириусы
вскоре становятся видны простым глазом.
Рыба как бы обсыпана манной крупой
(рис. 108). Продолжительность стадии ро-
ста и паразитирования ихтиофтириуса
в субэпителиальном слое рыбы в высокой
степени зависит от температуры. По иссле-
дованиям Бауера, при 28—30°С этот срок
составляет 4 суток, при 19—21°С—7Г
при 13—15°С—10—И, а при 8— 9°С пе-
риод этот затягивается до 40 дней.
К концу периода роста ихтиофтириус
по сравнению с бродяжками достигает
очень большой величины: 0,5—1 мм в ди-
аметре. По достижении предельной вели-
чины инфузории активными движениями
выходят из тканей рыбы в воду и неко-
торое время медленно плавают при помо-
щи покрывающего все их тело ресничного
аппарата. Вскоре крупные ихтиофтири-
усы оседают на каком-нибудь подводном
предмете и выделяют цисту. Тотчас же
вслед за инцистированием начинаются
последовательные деления инфузории: сна-
чала пополам, затем каждая дочерняя особь
делится опять на две и т. д. до 10—11 раз.
В результате внутри цисты образуется
до 2000 мелких почти округлых особей,
покрытых ресничками. Внутри цисты бро-
дяжки активно двигаются. Они пробо-
дают оболочку и выходят наружу. Актив-
но плавающие бродяжки заражают новых
особей рыб.
Быстрота деления ихтиофтириуса в ци-
стах, так же как и темп его роста в тка-
нях рыб, в большой степени зависит от
температуры. По исследованиям различ-
ных авторов, приводятся следующие циф-
ры: при 26— 27°С процесс развития бро-
дяжек в цисте занимает 10—12 часов, при
15—16°С требуется 28—30 часов, при
4—5°С он тянется уже в течение 6—7 су-
ток.
159
Ихтиофтириус — широко распростра-
ненный и очень опасный паразит рыб. Осо-
бенно большой вред он приносит в рыбо-
водческих хозяйствах, где происходит не-
рест и выращивание мальков. Ихтиофти-
риус, нападая на мальков, в короткий
срок приводит их к гибели. Известны
многочисленные случаи поголовного унич-
тожения мальков как карповых, так
и лососевых рыб в прудовых хозяйст-
вах.
Борьба с ихтиофтириусом представляет
значительные трудности. Главное значе-
ние здесь имеют профилактические меро-
приятия, направленные на то, чтобы не
допустить свободно плавающих в воде бро-
дяжек проникнуть в ткани рыбы. Для это-
Рис. 108. Жизненный цикл Ichthyophthirius multifiliis:
1 — рыба, пораженная ихтиофтириазитом; 2 — зрелый паразит из
кожи рыбы; 3 — паразит, покинувший рыбу; 4 — циста размноже-
ния, 5 — выход бродяжек из цисты.
го полезно осуществлять частую пере-
садку больных рыб в новые водоемы или
аквариумы, создавать условия проточ-
ности, что особенно эффективно в борьбе
с ихтиофтириусом.
ИНФУЗОРИИ ТРИХОДИНЫ
Очень своеобразны паразитирующие
главным образом на рыбах инфузории
семейства урцелариид (Urcelarii-
dae) отряда кругоресничных.
Особенно характерны многочисленные
представители рода триходин (Tricho-
dina) и некоторых других близких к нему
родов (рис. 109). За немногими
исключениями, все триходины
и близкие к ним формы явля-
ются эктопаразитами, т. е. они
живут на поверхности (а не в
толще тканей и не внутри орга-
нов) своих хозяев (рыб). Место
локализации триходин — кож-
ные покровы, плавники, очень
часто жабры рыб. Они встреча-
ются как на морских, так и на
пресноводных рыбах. Количе-
ство видов триходин измеряет-
ся несколькими десятками. Три-
ходины не являются прикреп-
ленными животными, а, напро-
тив, активно передвигаются по
поверхности своих хозяев. Вне
организма хозяина, в воде, они
долго жить не могут и, будучи
отделены от рыбы, довольно
быстро погибают.
Вся система приспособлений
триходин к жизни на поверхно-
сти хозяина направлена на то,
чтобы не оторваться от тела хо-
зяина (что почти всегда равно-
сильно гибели), сохраняя при
этом подвижность. Приспособ-
ления эти весьма совершенны.
Тело большинства триходин
имеет форму довольно плоско-
го диска, иногда шапочки. Сто-
рона, обращенная к телу хо-
зяина, слегка вогнутая, она
образует прикрепительную при-
160
Рис. 109. Инфузория триходина:
А — Trichodina pediculus сбоку; Б — прикрепитель-
ный аппарат Trichodina domerguei. l — нижний вен-
чик ресничек, 2 — околоротовые (адоральные) мем-
браны; 3 — сократительная вакуоля, 4 — ядро.
соску. По наружному краю присоски
расположен венчик хорошо развитых рес-
ничек, при помощи которых главным
образом и происходит передвижение
(ползание) инфузории по поверхности
тела рыбы. Этот венчик соответствует
венчику, имеющемуся у бродяжек си-
дячих кругоресничных инфузорий, рас-
смотренных выше (стр. 149). Таким обра-
зом, триходину можно сравнить с бродяж-
кой. На брюшной поверхности (на при-
соске) у триходин имеется очень сложный
опорный и прикрепительный аппарат, спо-
собствующий удержанию инфузории на
хозяине. Не вдаваясь в детали его строе-
ния, отметим, что основу его составляет
сложной конфигурации кольцо, слагаю-
щееся из отдельных сегментов, несущих
наружный и внутренний зубцы (рис. 109,
Б). Это кольцо образует эластичную и вме-
сте с тем прочную основу брюшной поверх-
ности, выполняющей функцию присоски.
Разные виды триходин отличаются друг
от друга по количеству сегментов, обра-
зующих кольцо, и по конфигурации на-
ружного и внутреннего крючьев.
На противоположной от диска стороне
тела триходины расположен перистом и
ротовой аппарат. Строение его более или
менее типично для кругоресничных инфу-
зорий (стр. 148). Закрученные по ча-
совой стрелке адоральные мембраны ве-
дут в углубление, на дне которого рас-
положен рот. Ядерный аппарат триходин
устроен типично для инфузорий: один
лентовидный макронуклеус и один распо-
ложенный рядом с ним микронуклеус.
Имеется одна сократительная вакуоля.
Триходины широко распространены
в водоемах всех типов. Особенно часто
их находят на мальках разных видов рыб.
При массовом размножении триходины
наносят большой вред рыбе, в особенно-
сти если массами покрывают жабры. Это
нарушает нормальное дыхание рыбы.
Для того чтобы очистить рыбу от три-
ходин, рекомендуют делать лечебные ван-
ны из 2-процентного раствора поваренной
соли или 0,01-процентного раствора мар-
ганцовокислого калия (для мальков —
в течение 10—20 минут).
ИНФУЗОРИИ КИШЕЧНОГО ТРАКТА
КОПЫТНЫХ
Среди инфузорий имеется множество
видов, ведущих эндопаразитический образ
жизни, т. е. живущих внутри (в различных
органах и тканях) животных-хозяев. Осо-
бенно многочисленны и разнообразны ин-
фузории, населяющие кишечный тракт
копытных животных. Большинство их об-
разует самостоятельный отряд инфузо-
рий, называемый энтодиниоморфами (Еп-
todiniomorpha). У жвачных (рогатый скот,
овцы, козы, антилопы, олени, лоси) эти
инфузории в огромных количествах насе-
ляют передние отделы желудка. Желудок
жвачных состоит из чэтырех отделов —
рубца, сетки, книжки и сычуга. Из них
только сычуг соответствует желудку дру-
гих млекопитающих: он имеет пищевари-
тельные железы, в него выделяются
пищеварительные ферменты и соляная
кислота. Реакция среды в сычуге резкокис-
лая. Первые три отдела не имеют пище-
варительных желез. Их внутренняя стен-
ка частично ороговевает. Эти три отдела
по происхождению следует рассматривать
как нижние расширенные части пищевода.
Названия сетки и книжки даны по форме
и характеру складок их внутренней стен-
ки. Самый вместительный отдел — рубец.
Заглоченная и почти не пережеванная
растительная пища сначала проходит в ру-
бец. Сюда же попадает большое количество
ф 11 ЖИЗНЬ ЖИВОТНЫХ, Т. 1
161
Рис. 110. Инфузории семейства Ophryoscolecidae из рубца жвачных:
А — Entodinium simplex; Б — Anoplodinium denticulatum, В — Eudiplodinium neglectum; Г — Poly-
plastron multivesicu latum с левой стороны; Д — то же с правой стороны; Е — Ophryoscolex caudatus; Ж —
Epidinium ecaudatum. J — околоротовая ресничная зона; 2 — большое ядро (макронуклеус); 3 —
малое ядро (микронуклеус), 4 — сократительная вакуоля; 5 — спинная ресничная зона; 6 — скелетные
пластинки.
слюны. В рубце пища подвергается слож-
пым процессам брожения, которые увели-
чивают ее усвояемость. В этом отделе в
огромных количествах живут разнооб-
разные микроорганизмы, вызывающие
процессы брожения, которые протекают
в условиях нейтральной или слабощелоч-
ной реакции. В рубце наряду с бактерия-
ми обитает масса простейших — очень мел-
ких жгутиконосцев и инфузорий, так
как здесь создаются весьма благоприят-
ные условия для их развития.
Из рубца через сетку пища отрыги-
вается в ротовую полость, где дополни-
тельно пережевывается (жвачка). Прогла-
тываемая вновь пищевая пережеванная
масса по особой трубке, образуемой склад-
ками пищевода, идет уже не в рубец, а в
книжку и оттуда в сычуг, где подверга-
ется действию пищеварительных соков
жвачного. В сычуге в условиях кислой
реакции и наличия пищеварительных фер-
ментов инфузории погибают. Попадая ту-
да со жвачкой, они перевариваются.
Количество простейших в рубце (а так-
же в сетке) может достигать колоссаль-
ных величин. Если взять каплю содержимо-
го рубца и рассмотреть ее под микроскопом
(при нагревании, так как при комнат-
ной температуре инфузории останавли-
ваются), то в поле зрения инфузории бук-
вально кишат. Трудно даже в условиях
культуры получить такую массу инфузо-
рий. Количество инфузорий в 1 см3 содер-
жимого рубца достигает миллиона, а не-
редко и более. В пересчете на весь объем
рубца это дает поистине астрономичес-
кие цифры! Богатство содержимого рубца
инфузориями в большой степени зависит
от характера пищи жвачного. Если пища
богата клетчаткой и бедна углеводами
и белками (трава, солома), то инфузорий
в рубце относительно немного. При добав-
лении в пищевой рацион углеводов и бел-
ков (отруби) количество инфузорий резко
увеличивается и достигает огромных цифр.
Нужно иметь в виду, что существует по-
стоянный отток инфузорий. Попадая вме-
сте с жвачкой в сычуг, они погибают. Вы-
сокий уровень количества инфузорий под-
держивается энергичным размножением их.
У непарнокопытных (лошадь, осел, зеб-
ра) в пищевом тракте тоже имеется боль-
шое количество инфузорий, однако лока-
лизация их в хозяине иная. У непарно-
копытных нет сложного желудка, благо-
даря чему возможность развития простей-
ших в передних отделах пищевого тракта
отсутствует. Зато у непарнокопытных
очень сильно развиты толстая и слепая
кишка, которые обычно забиты пищевыми
массами и играют существенную роль
в пищеварении. В этом отделе кишечника,
подобно тому как в рубце и сетке жвач-
ных, развивается очень богатая фауна
простейших, преимущественно инфузо-
рий, большинство которых также отно-
сится к отряду энтодиниоморф. Однако
по видовому составу фауна рубца жвач-
ных и фауна толстого кишечника непар-
нокопытных не совпадают.
ИНФУЗОРИИ КИШЕЧНОГО ТРАКТА
ЖВАЧНЫХ
Наибольший интерес представляют ин-
фузории семейства офриосколецид
(Ophryoscolecidae), относящегося кот-
ряду энтодиниоморф. Характерный
признак этого отряда — отсутствие сплош-
ного ресничного покрова. Сложные рес-
ничные образования — цирры — располо-
жены на переднем конце тела инфузорий
в области ротового отверстия. К этим ос-
новным элементам ресничного аппарата
могут прибавиться еще дополнительные
группы цирр, расположенные либо на
переднем, либо на заднем конце тела. Об-
щее количество видов инфузорий семей-
ства офриосколецид — около 120.
На рисунке 110 изображены некоторые
наиболее типичные представители офрио-
сколецид из рубца жвачных. Наиболее
просто устроены инфузории рода энто-
диний (Entodinium, рис. 110,Л). На пе-
реднем конце тела их имеется одна около-
ротовая зона цирр. Передний конец тела,
на котором расположено ротовое отвер-
стие, может втягиваться внутрь. Резко
разграничены эктоплазма и эндоплазма.
Хорошо видна на заднем конце анальная
трубка, служащая для выведения наружу
непереваренных остатков пищи. Несколь-
ко сложнее строение аноплодиния (Anoplo-
ll*
163
dinium, рис. 110, Б). У них имеются две
80ны ресничного аппарата — околорото-
вые цирры и спинные цирры. Обе они рас-
положены на переднем конце. На заднем
конце тела изображенного на рисунке вида
имеются длинные острые выросты — это
довольно типично для многих видов оф-
риосколецид. Высказывалось предполо-
жение, что эти выросты способствуют
«проталкиванию» инфузорий между рас-
тительными частицами, заполняющими ру-
бец.
Виды рода эудиплодиний (Eudip-
lodinium, рис. 110,Б) сходны с аноплоди-
нием, но, в отличие от них, имеют скелет-
ную опорную пластинку, расположенную
по правому краю вдоль глотки. Эта скелет-
ная пластинка состоит из вещества, близ-
кого по химической природе к клетчатке,
т. е. к веществу, из которого состоят обо-
лочки растительных клеток.
Урода полипластрон (Polyplastron,
рис. 110, Г, Д) наблюдается дальнейшее
усложнение скелета. Строение этих инфу-
зорий близко к эудиплодиниям. Отли-
чия сводятся прежде всего к тому, что
вместо одной скелетной пластинки эти
инфузории имеют пять. Две из них, наи-
более крупные, расположены по правой
стороне, а три, более мелкие,— по ле-
вой стороне инфузории. Вторая особен-
ность полипластрона — увеличение коли-
чества сократительных вакуолей. У энто-
диниев одна сократительная вакуоля, у
аноплодиниев и эудиплодиниев две сокра-
тительные вакуоли, у полипластрона их
около десятка.
У эпидиниев (Epidinium, рис. 110), об-
ладающих хорошо развитым углеводным
скелетом, расположенным по правой сто-
роне тела, спинная зона цирр смещается
с переднего конца на спинную сторону.
На заднем конце инфузорий этого рода
часто развиваются шипы.
Наиболее сложное строение обнаружи-
вает род офриосколекс (Ophryoscolex),
по которому названо и все семейство инфу-
зорий (рис. 110, Е). У них хорошо развита
спинная зона цирр, охватывающая около
2/3 окружности тела и скелетные пластин-
ки. На заднем конце образуются много-
численные шипы, из которых один бывает
обычно особенно длинным.
Знакомство с некоторыми типичными
представителями офриосколецид показы-
вает, что в пределах этого семейства про-
изошло значительное усложнение орга-
низации (от энтодиниев до офриосколек-
са).
Очень часто, как мы это увидим, напри-
мер, при знакомстве с червями, парази-
тизм, особенно эндопаразитизм, приво-
дит к значительному упрощению орга-
низации, к регрессивным изменениям
многих систем органов. У инфузорий, живу-
щих в кишечнике копытных, этого не про-
изошло. Напротив, здесь имело место про-
грессивное развитие, не упрощение, а
усложнение организации. Это связано,
вероятно, с особенностями среды обита-
ния инфузорий рубца. Они живут как бы
в своеобразном водоеме с очень сложными
условиями (наличие многочисленных ме-
ханических препятствий, сложный бак-
териальный состав среды и многое дру-
гое), приспособление к которым было
связано с усложнением строения инфу-
зорий.
Кроме инфузорий семейства оф-
риосколецид, в рубце жвачных встречают-
ся в небольших количествах представи-
тели уже известного нам отряда рае-
поресничных инфузорий. Они представле-
ны небольшим числом видов. Их тело рав-
номерно покрыто продольными рядами
ресничек, скелетные элементы отсутст-
вуют. В общей массе инфузорного населе-
ния рубца они не играют заметной роли,
и поэтому мы не будем здесь останавли-
ваться на их рассмотрении.
Чем и как питаются инфузории офриос-
колециды? Этот вопрос подробно изу-
чался многими учеными, особенно деталь-
но профессором В. А. Догелем.
Пища офриосколецид довольно разнооб-
разна, и у разных видов их наблюдается
известная специализация. Самые мелкие
виды рода энтодиниев питаются бакте-
риями, зернами крахмала, грибками и
другими небольшими частицами. Очень
многие средние и крупные офриосколе-
циды поглощают частицы растительных
тканей, которые составляют главную мас-
су содержимого рубца. Эндоплазма неко-
торых видов бывает буквально забита
растительными частицами. Можно видеть,
164
Рис.111. Инфузории семейства Ophryoscolecidae,
пожирающие растительные ткани:
1 — Opisthotrichum janus; 2 — Ostracodinium sp ;
3—Anoplodinium bubalidis, 4 — Anoplodinium gracile.
как инфузории набрасываются на об-
рывки растительных тканей, буквально
разрывают их на кусочки и затем загла-
тывают, нередко закручивая их в своем
теле спиралью (рис. 111,4). Иногда при-
ходится наблюдать такие картины (рис.
111,2), когда само тело инфузории оказы-
вается деформированным благодаря загло-
ченным крупным частицам.
У офриосколецид наблюдается иног-
да хищничество. Более крупные виды по-
жирают более мелкие. Хищничество (рис.
112) сочетается со способностью тех же
видов питаться растительными частицами.
Каким путем инфузории проникают
в рубец жвачного? Каковы пути зараже-
ния офриосколецидами? Оказывается, что
новорожденные жвачные не имеют еще
в рубце инфузорий. Они отсутствуют
также, пока животное питается молоком.
Но как только жвачное переходит на рас-
тительную пищу, сразу же в рубце и сет-
ке появляются инфузории, количество
которых быстро нарастает. Откуда они
берутся? Долгое время предполагали, что
инфузории рубца образуют какие-то поко-
ящиеся стадии (вероятнее всего, цисты),
которые широко рассеиваются в природе
и, будучи проглоченными, дают начало
активным стадиям инфузорий. Дальней-
шие исследования показали, что у инфу-
зорий рубца жвачных никаких покоящих-
ся стадий нет. Удалось доказать, что за-
ражение происходит активными подвиж-
ными инфузориями, которые проникают
в ротовую полость при отрыгивании жвач-
ки. Если исследовать под микроскопом
взятую из ротовой полости жвачку, то
в ней всегда имеется большое количество
активно плавающих инфузорий. Эти ак-
тивные формы легко могут проникнуть
в рот и далее в рубец других особей жвач-
ного из общего сосуда для пойла, вместе
с травой, сеном (на которых может ока-
заться слюна с инфузориями) и т. п. Этот
путь заражения был доказан эксперимен-
тально.
Если покоящиеся стадии у офриосколе-
цид отсутствуют, то, очевидно, легко
получить «безинфузорных» животных, изо-
лировав их, когда они еще питаются мо-
локом. Если не допускать прямого кон-
такта между растущим молодняком и
165
Рис. 112. Хищные инфузории семейства Ophry-
oscolecidae:
1 — Anoplodinium costatum, заглотивший несколько
инфузорий рода Entodinium; 2 — Entodinium vorax,
проглотивший Entodinium simplex.
имеющими инфузорий жвачными, то моло-
дые животные могут остаться без инфу-
зорий в рубце. Такие опыты были прове-
дены несколькими учеными в разных стра-
нах. Результат был однозначен. При от-
сутствии контакта между молодняком (от-
нятым от матери еще в период кормления
молоком) и имеющими в рубце инфузо-
рий жвачными, животные вырастают сте-
рильными в отношении инфузорий. Одна-
ко достаточно даже кратковременного кон-
такта с животными, имеющими инфузорий
(общая кормушка, общее ведро для питья,
общее пастбище), чтобы в рубце стериль-
ных животных появилась фауна инфу-
зорий.
Массовое развитие фауны простейших
в рубце и сетке, естественно, ставит во-
прос о характере взаимоотношений между
простейшими и их хозяевами — жвач-
ными. Мы уже видели (стр. 111) на при-
мере жгутиконосцев, паразитирующих в
кишечнике термитов, какие тесные симбио-
тические взаимоотношения могут возни-
кать между простейшими, населяющими
кишечный тракт, и их хозяевами (стр.
113). Бе существует ли между инфузо-
риями, населяющими рубец и сетку, ана-
логичных симбиотических взаимоотноше-
ний? Этот вопрос ставился многими иссле-
дователями. Однако до сих пор оконча-
тельный ответ на него не получен, и тре-
буются дальнейшие исследования. Ввиду
большого биологического и практического
интереса этого вопроса рассмотрим его
несколько подробнее.
Выше (стр. 165) были приведены ре-
зультаты опытов по содержанию жвач-
ных, совершенно лишенных инфузорий
в рубце и сетке. Это достигается, как
мы видели, путем ранней изоляции молод-
няка. Опыты проводились на овцах и на
козах.
Таким путем удавалось проводить на-
блюдения за «безинфузорными» животны-
ми в течение значительного промежутка
времени (свыше года). Как отражается
отсутствие инфузорий в рубце на жизни
хозяина? Отрицательно или положитель-
но влияет отсутствие инфузорий на хо-
зяина? Для ответа на этот вопрос на козах
провели следующие опыты. Были взяты
козлята-близнецы (одного помета и одного
пола), чтобы иметь более сходный матери-
ал. Затем один из близнецов данной пары
воспитывался без инфузорий в рубце
(ранняя изоляция), другой же с самого на-
чала питания растительной пищей обиль-
но заражался множеством видов инфузо-
рий. Оба получали совершенно одинако-
вый рацион и воспитывались в одинако-
вых условиях. Единственное различие
между ними сводилось к наличию или от-
сутствию инфузорий. На нескольких изу-
ченных таким образом парах козлят не
было обнаружено каких-либо различий
в ходе развития обоих членов каждой па-
ры («инфузорного» и «безинфузорного»).
Таким образом, можно утверждать, что
сколько-нибудь резкого влияния на жиз-
ненные функции животного-хозяина инфу-
зории, живущие в рубце и сетке, не оказы-
вают.
Приведенные выше результаты опытов
не позволяют, однако, утверждать, что
инфузории рубца совершенно безразлич-
ны для хозяина. Эти опыты проводились
при нормальном пищевом рационе хозяи-
на. Не исключена возможность, что при
других условиях, при ином пищевом ре-
жиме (например, при недостаточном корм-
лении) удастся выявить влияние на хо-
зяина фауны простейших, населяющих
рубец.
В литературе высказывались различные
предположения о возможном положитель-
ном влиянии протозойной фауны рубца
166
на пищеварительные процессы хозяина.
Указывалось, что многие миллионы инфу-
зорий, активно плавающие в рубце и раз-
мельчающие растительные ткани (см. вы-
ше, стр. 161 —162), способствуют брожению
и перевариванию пищевых масс, находя-
щихся в передних отделах пищеваритель-
ного тракта. Значительное количество ин-
фузорий, попадающее вместе с жвачкой
в сычуг, переваривается, и белок, состав-
ляющий значительную часть тела инфузо-
рий, усваивается. Инфузории, таким об-
разом, могут быть дополнительным источ-
ником белка для хозяина. Высказывались
также предположения, что инфузории спо-
собствуют перевариванию клетчатки, со-
ставляющей основную массу пищи жвач-
ных, переводу ее в более усвояемое состо-
яние.
Все эти предположения не являются
доказанными, и против некоторых из них
выдвигались возражения. Указывалось,
например, что инфузории строят прото-
плазму своего тела из белков, которые
поступают в рубец с ппщей хозяина. Погло-
щая белок растительный, они перево-
дят его в животный белок своего тела,
который затем переваривается в сычуге.
Дает ли это какие-нибудь преимущества
хозяину, остается неясно. Все эти вопро-
сы имеют большой практический инте-
рес, так как речь идет о пищеварении
жвачных — основных объектов животно-
водства. Дальнейшие исследования о ро-
ли инфузорий рубца в пищеварении жвач-
ных очень желательны.
Насколько строго приурочены опреде-
ленные виды офриосколецид к определен-
ным видам хозяев? Имеются ли, напри-
мер, в рогатом скоте свои виды инфузо-
рий, в овцах — свои и т. д.? Приспо-
собленность паразита к определенному
кругу хозяев называют его специфично-
стью. Если паразит строго приурочен к од-
ному виду хозяев, это говорит об узкой
специфичности. Если данный вид парази-
та может жить в разных видах хозяев,
то это проявление широкой специфич-
ности.
Офриосколециды жвачных обладают,
как правило, широкой специфичностью.
В видовом отношении население рубца
и сетки рогатого скота, овец, коз очень
близко между собой. Если сравнить видо-
вой состав рубца африканских антилоп
с рогатым скотом, то и здесь около 40%
от общего числа видов окажется общим.
Однако существует немало видов офрио-
сколецид, которые встречаются только
в антилопах или только в оленях. Таким
образом, на фоне общей широкой специ-
фичности офриосколецид можно говорить
об отдельных более узко специфичных ви-
дах их.
ИНФУЗОРИИ КИШЕЧНИКА
НЕПАРНОКОПЫТНЫХ
Обратимся теперь к краткому знаком-
ству с инфузориями, населяющими тол-
стую и слепую кишку непарнокопытных.
В видовом отношении эта фауна, как и
фауна рубца жвачных, тоже весьма раз-
нообразна. В настоящее время описано
около 100 видов инфузорий, обитающих
в толстом кишечнике животных семей-
ства лошадиных. Встречающиеся здесь
инфузории в смысле их принадлежности
к разным систематическим группам более
разнообразны, чем инфузории рубца жвач-
ных.
В кишечнике лошадиных обитает до-
вольно много видов инфузорий, относя-
щихся к отряду равноресничных (стр. 141),
т. е. инфузорий, у которых ресничный
аппарат не образует около ротовой зоны
мембранелл или цирр (рис. 113,7).
Отряд энтодиниоморф (Entodinio-
morpha) тоже богато представлен в ки-
шечнике лошади. В то время как в рубце
жвачных встречается лишь одно семей-
ство энтодиниоморф (семейство офриоско-
лецид), в кишечнике лошади живут пред-
ставители трех семейств, на характери-
стике которых мы, однако, останавливать-
ся здесь не будем, ограничившись лишь
несколькими рисунками типичных для
лошади видов (рис. ИЗ).
Детальными исследованиями А.Стрел-
к о в а было показано, что разные виды
инфузорий далеко не равномерно распре-
деляются по ходу толстого кишечника ло-
шади. Имеются две разные группы видов,
как бы две фауны. Одна из них населяет
167
Рис. ИЗ. Инфузории из кишечника лошади:
1 — Blepharoprosthium pireum (отряд Holotricha); 2 — Spirodinium equi (отряд Entodiniomorpha);
3 — Triadinium galea (отряд Entodiniomorpha), 4 — Cycloposthium bipalmatum (отряд Ento-
diniomorpha).
слепую кишку и брюшной отдел большой
ободочной кишки (начальные отделы толсто-
го кишечника), а другая—спинной отдел
большой ободочной и малую ободочную.
Эти два комплекса видов довольно резко
разграничены. Видов, общих для этих
двух разделов, немного — менее десятка.
Интересно отметить, что среди много-
численных видов инфузорий, населяющих
толстый кишечник непарнокопытных, име-
ются представители одного рода, относя-
щегося к сосущим инфузориям. Как мы ви-
дели выше, сосущие инфузории (Suctoria) —
это типичные свободноживущие сидячие
организмы с совершенно особым способом
питания при помощи щупалец (стр. 153,
рис. 103). Один из родов сукторий при-
способился к такой, казалось бы, необыч-
ной среде обитания, как толстый кишеч-
ник лошади, например несколько видов
аллантозом (Allantosoma). Эти очень свое-
образные животные (рис. 114) не имеют
стебелька, реснички отсутствуют, булаво-
видные, утолщенные на концах щупальца
хорошо развиты.
При помощи щупалец аллантозомы
присасываются к разным видам инфузо-
рий и высасывают их. Нередко при этом
добыча во много раз превосходит хищ-
ника.
Неясен еще вопрос о характере взаи-
моотношений инфузорий толстого кишеч-
ника непарнокопытных с хозяевами. Ко-
личество инфузорий может быть столь
же велико, а иногда даже больше, чем
в рубце жвачных. Имеются данные, по-
казывающие, что количество инфузорий
в толстых кишках лошади может дости-
гать 3 млн. в 1 см3. Симбиотическое зна-
чение, предполагавшееся некоторыми уче-
ными, еще менее вероятно, чем для инфу-
зорий рубца.
Наиболее вероятно мнение, что они на-
носят некоторый вред хозяину, поглощая
значительное количество пищи. Часть
инфузорий выносится наружу с фекаль-
ными массами, и таким образом органиче-
ские вещества (в том числе и белок), вхо-
дящие в состав их тела, остаются не ис-
пользованными хозяином.
Вопрос о путях заражения непарноко-
пытных инфузориями, населяющими тол-
стый кишечник, до сих пор не разрешен.
ВАЛАНТИДИЙ-ПАРАЗИТ
КИШЕЧНИКА ЧЕЛОВЕКА
Один вид паразитических инфузорий
из отряда равноресничных — балантидий
коли (Balantidium coli) — паразитирует
в толстом кишечнике человека. Случаи
заражения человека балантидиями очень
редки. В этом отношении балантидий не-
сравним с кишечными амебами, которые,
как мы видели, встречаются часто (стр.
77—78). Паразитирование балантидия в
кишечнике человека обычно вызывает тя-
жело протекающий колит. Поэтому па-
разит этот требует к себе пристального
внимания, несмотря на то что встречает-
ся он относительно редко.
Тело балантидия, паразитирующего в
кишечнике человека, имеет яйцевидную
форму (рис. 115). Размеры варьируют
в очень широких пределах: длина 30—150,
ширина 20—НО мк. Эти вариации величи-
ны в значительной степени зависят от
того, в какой мере инфузория набита пи-
щей. Все тело густо покрыто короткими
ресничками, расположенными продоль-
ными рядами от переднего конца к задне-
му. На переднем конце имеется углубле-
ние в виде неглубокой щели. Это около-
ротовое углубление (перистом), на дне
которого помещается ротовое отверстие.
По краю перистома расположены особые
сильно развитые реснички, направляю-
щие пищу в глубину его. Имеются две
сократительные вакуоли (одна в перед-
ней трети тела, вторая на заднем конце).
Ядерный аппарат типичный для инфузо-
рий: бобовидный, расположенный пример-
но в центре тела макронуклеус и в непо-
средственном соседстве с ним один малень-
кий микронуклеус.
Балантидий захватывает разнообразные
пищевые частицы из содержимого тол-
стой кишки. Особенно охотно он питается
крахмальными зернами. Если балантидий
живет в просвете толстой кишки челове-
ка, то он питается содержимым кишеч-
ника и не оказывает никакого вредного
влияния. Это типичное «носительство»,
с которым мы познакомились уже при рас-
смотрении дизентерийной амебы. Однако
балантидий реже, чем дизентерийная аме-
169
Рис. 114. Сосущие инфузории (Suctoria), живу-
щие в кишечнике лошади:
1 — Allantosoma intestinalis, 2 — Allantosoma cucu-
mis, присосавшиеся к ресничным пучкам Cyclopos-
thium.
Рис. 115. Balantidium coli — инфузория, парази-
тирующая в кишечнике человека:
А — активноплавающая форма; Б — циста неокра-
шенная; В — циста — окрашенный препарат. 1 —
ротовое отверстие; 2 — ядро (макронуклеус); 3 — обо-
лочка цисты.
ба, остается таким «безобидным кварти-
рантом».
Он активно внедряется в слизистую ки-
шечника и проникает в толщу его стенки.
Здесь балантидий переключается на пи-
тание клетками хозяина и особенно «охот-
но» заглатывает красные кровяные тель-
ца (эритроциты). В результате на-
рушения целостности внутренней стенки
кишечник изъязвляется, и это сопровож-
дается тяжелой формой кровавого поно-
са (колита). Поскольку паразит внед-
ряется в самую толщу тканей кишечника,
лечение балантидиоза очень затрудни-
тельно.
В настоящее время в медицине изве-
стны некоторые специфические лекарства,
убивающие балантидия. Однако успех ле-
чения не всегда бывает положительным
именно в силу внутритканевой локализа-
ции паразита.
Может ли человеческий балантидий па-
разитировать в других хозяевах, кроме
человека? Оказывается, что эта паразити-
ческая инфузория часто встречается в ки-
шечнике свиней. Но, в отличие от челове-
ческого, этот балантидий обычно не вы-
зывает у свиней болезненных явлений
и живет в кишечнике их как «безобидный
квартирант». Основным хозяином балан-
тидия следует считать свинью, тогда как
человек заражается редко, причем почти
всегда источником заражения являются
свиньи.
Каким же путем осуществляется зара-
жение балантидием? У этой инфузории
имеются покоящиеся стадии — цисты.
Инцистирование происходит при проник-
новении паразита в заднюю кишку. Ци-
сты выводятся наружу с фекальными мас-
сами. Они долгое время (свыше двух меся-
цев при комнатной температуре) сохраняют
жизнеспособность. Если циста будет слу-
чайно проглочена свиньей или человеком,
то она проходит в неизменном виде через
желудок и тонкие кишки. Когда циста до-
стигнет толстой кишки, оболочка ее раз-
рушается и из нее выходит активная
стадия балантидия.
Из сказанного выше следует, что персо-
нал, обслуживающий свиней, должен со-
блюдать элементарные гигиенические пра-
вила, а главное не приступать к еде, не
170
вымыв предварительно руки. Помимо ки-
шечника человека и свиней, балантидий
коли может паразитировать (при непо-
средственном заражении) в кишечнике
крыс и человекообразных обезьян. У обе-
зьян, так же как и у человека, балантидий
вызывают тяжелую форму колита.
В настоящее время специалистами хоро-
шо разработаны различные методы, позво-
ляющие культивировать балантидиев в
искусственной среде — вне организма хо-
зяина.
ИНФУЗОРИИ-ПАРАЗИТЫ
ЧЕЛОВЕКООБРАЗНЫХ ОБЕЗЬЯН
Мы видели, что человеческий баланти-
дий может паразитировать в обезьянах,
вызывая колиты. Однако в человекообраз-
ных обезьянах, кроме того, встречаются
очень интересные и сложно устроенные
инфузории, которые в организме челове-
ка не паразитируют. Они относятся кот-
ряду энтодиниоморф (Entodiniomor-
pha), т. е. к тому же отряду, который,
как мы видели, богато представлен в ки-
шечном тракте копытных млекопитаю-
щих.
В человекообразных обезьянах (горил-
ле и шимпанзе) встречаются виды рода
троглодителла (Troglodytella), парази-
тирующие в толстой и слепой кишке (рис.
116). В природной обстановке в Африке
как гориллы, так и шимпанзе заражены,
по-видимому, поголовно. В условиях не-
воли, в зоологических садах, инфузории
постепенно исчезают из организма обе-
зьян.
Как видно из рисунка, троглодителла
относится к числу сложно устроенных
энтодиниоморф. У нее, кроме около-
ротовой зоны цирр (на переднем конце
тела), имеются еще три зоны хорошо раз-
витых цирр, кольцеобразно охватываю-
щих тело инфузории. Троглодителлы об-
ладают хорошо развитым, охватывающим
почти весь передний конец тела скелет-
ным аппаратом, состоящим из углеводов.
Размеры этих своеобразных инфузорий
довольно значительны. В длину они до-
стигают 200—280 мк.
БЕЗРОТЫЕ ИНФУЗОРИИ АСТОМАТЫ
На предыдущих страницах было рас-
сказано о различных паразитических ин-
фузориях, хозяевами которых являются
позвоночные животные. Наряду с этим
большое количество видов паразитичес-
ких инфузорий живет в самых различных
беспозвоночных животных. Рассмотрим
кратко наиболее интересных представите-
лей их.
Глубоко приспособленной к паразитиз-
му и выработавшей ряд интересных при-
способлений является обширная группа
безротых инфузорий — астомат (Astoma-
ta). Самая характерная особенность этого
отряда отражена в его названии — инфу-
зории астоматы лишены ротового отвер-
стия. Их питание осуществляется осмоти-
ческим путем — всасыванием питательных
веществ всей поверхностью тела. По-
скольку оформленной пищи астоматы не
поглощают, пищеварительные вакуоли
у них не образуются.
Общее число видов астомат превышает
сотню. Встречаются они в беспозвоноч-
ных, относящихся к самым различным
группам (типам и классам) животного
мира: в ресничных червях, кольчатых
червях, моллюсках, иглокожих. Небольшое
число видов астомат паразитирует и в
позвоночных животных, а именно в зем-
новодных. Однако астоматы распределе-
ны по разным группам хозяев совсем не-
равномерно. Существует одна группа «из-
любленных» хозяев астомат, в которой
сосредоточено свыше 75% их видов. Такой
группой являются малощетинковые коль-
чатые черви (Oligochaeta), класс живот-
ных, к которому относится всем извест-
ный дождевой червь. Однако лишь часть
видов малощетинковых кольчецов живет
в почве. Большая часть их — обитатели
пресных вод, где они образуют заметную
(а иногда и преобладающую) часть дон-
ного населения. Эти пресноводные мало-
щетинковые кольчецы и являются «излюб-
ленными» хозяевами астомат. Небольшое
число видов олигохет встречается ив море,
почти исключительно в прибрежной, лито-
ральной (осушаемой во время отлива)
зоне, но в морской фауне они не играют
сколько-нибудь заметной роли.
171
Органом, в котором локализуются
в олигохетах инфузории астоматы, явля-
ется кишечник. Поэтому многие черты
строения астомат связаны с приспособ-
лением к жизни в кишечнике. Через ки-
шечник постоянно осуществляется движе-
ние пищи, иногда довольно активное. Па-
разиты кишечника, чтобы не быть выне-
сенными наружу (что равнозначно гибе-
ли), должны обладать приспособлениями,
направленными на то, чтобы удержаться
в кишечнике. Эти приспособления выра-
жены не только у инфузорий, но и у всех
других групп кишечных паразитов, к ка-
кой бы систематической группе они ни от-
носились и в каких бы хозяевах ни пара-
зитировали. Мы увидим это в дальнейшем,
при знакомстве с разными группами пара-
зитических червей.
Тело всех паразитирующих в кишеч-
нике олигохет астомат вытянуто в длину,
что способствует их движению в просвете
кишечника. Ресничный покров всегда хо-
Рис. 116. Troglodytella gorillae — ин-
фузория, паразитирующая в кишеч-
нике гориллы.
рошо развит, реснички покрывают все
тело инфузорий и расположены тесными
продольными рядами.
В простейшем случае, например у пред-
ставителей рода аноплофрий (Anoploph-
гуа, рис. 117, Б), никаких особых прикре-
пительных и опорных образований не раз-
вивается. Но у большинства видов дру-
гих родов имеются разнообразные и под-
час довольно сложные структуры двух
категорий — опорные и прикре-
пительные, являющиеся формами
приспособления к кишечному паразитиз-
му.
Опорные скелетные образования раз-
виваются преимущественно на переднем
конце тела, которому приходится испыты-
вать механические воздействия и прео-
долевать препятствия, проталкиваясь в
просвете кишечника между частицами пи-
щи. У видов рода радиофрий (Radio-
phrya) на переднем конце с одной стороны
тела (которую условно считают брюшной)
располагаются очень прочные эластич-
ные ребра (спикулы), лежащие в поверх-
ностном слое эктоплазмы (рис. 117,В,Г,Д).
У видов рода менилелла (Mesnilella)
тоже имеются опорные лучи (спикулы),
которые на большей части своего протя-
жения лежат в более глубоких слоях
цитоплазмы (в эндоплазме, рис. 117, А).
Аналогично устроенные опорные образо-
вания развиты и у видов некоторых дру-
гих родов астомат.
У многих представителей этой интерес-
ной группы инфузорий имеются специ-
альные структуры, позволяющие паразиту
прочно прикрепляться к стенке кишеч-
ника. У известной уже нам радиофрий
на самом переднем конце торчит наружу
острый зубец, подвижно сочленяющийся
с особыми опорными образованиями, имею-
щими форму наконечника стрелы. При по-
мощи этого зубца радиофрия может
прочно прикрепляться (зацепляться) к
стенке кишечника.
Другим типом прикрепительных образо-
ваний астомат, но менее распространен-
ным, являются присоски на переднем
конце тела. Такого рода прикрепитель-
ный аппарат развит у видов рода гап-
тофрий (Haptophrya). Разные виды это-
го рода паразитируют в очень несхожих
172
друг с другом хозяевах: одни — в реснич-
ных червях (турбелляриях), другие — в
земноводных.
При изучении прикрепительных аппа-
ратов паразитирующих в кишечнике раз-
личных хозяев инфузорий астомат обра-
щает на себя внимание их большое сход-
ство с аналогичными органами у других
групп паразитов. Например, у ленточных
червей (см. стр. 382) основными форма-
ми органов прикрепления являются при-
соски и крючья. У инфузорий астомат —
одноклеточных организмов — возникают
структуры, весьма напоминающие органы
прикрепления сложных многоклеточных
животных (ленточные черви). Возникнове-
ние сходных структур,выполняющих ана-
логичную функцию у животных, относя-
щихся к совершенно различным группам
животного мира, связано с приспособле-
нием к сходным условиям существова-
ния, в данном случае с приспособлением
паразитов к жизни в кишечнике хозяев.
Это явление носит название конвер-
генции.
Бесполое размножение у
некоторых инфузорий астомат протекает
своеобразно. Вместо поперечного деле-
ния надвое, свойственного большинству
инфузорий (стр. 139), у многих астомат
происходит неравномерное деление (п о-
чкование). При этом почки, отде-
ляющиеся на заднем конце, некоторое
время остаются связанными с материнской
особью (рис. 117, В). В результате полу-
чаются цепочки, состоящие из передней
крупной и задних более мелких особей
(почек). В дальнейшем почки постепен-
но отделяются от цепочки и переходят
к самостоятельному существованию. Эта
своеобразная форма размножения широко
распространена, например, у известной
уже нам радиофрии. Возникающие в ре-
зультате почкования цепочки некоторых
астомат напоминают по внешнему виду
цепочки ленточных червей (стр. 383).
Здесь мы вновь встречаемся с явлением
конвергенции.
Ядерный аппарат астомат имеет харак-
терную для инфузорий структуру: мак-
ронуклеус, чаще всего лентовидной фор-
мы (рис. 117), и один микронуклеус. Со-
кратительные вакуоли обычно хорошо раз-
Рис. 117. Безротые инфузории отряда Asto-
mata:
А — Mesnilella maritui; Б—Anoplophrya teleuscoli-
cis; В —Radiophrya hoplites с почками на заднем ко-
нце; Г — то же, передний конец с прикрепительным
аппаратом, Д — изолированный прикрепительный
аппарат. 1 — макронуклеус; 2 — микронуклеус; 3 —
скелетные лучи; 4 — сократительные вакуоли.
виты. У большинства видов имеется не-
сколько (иногда свыше десятка) сократи-
тельных вакуолей, расположенных в один
продольный ряд.
Изучение распределения видов астомат
по разным видам хозяев показывает, что
большая часть астомат приурочена к стро-
го определенным видам хозяев. Боль-
шинству астомат свойственна узкая спе-
цифичность: хозяином для них может
служить лишь один вид животного.
Несмотря на большое количество иссле-
дований, посвященных изучению инфузо-
рий астомат, одна очень важная сторона
их биологии остается совершенно неясной:
каким образом происходит передача этих
инфузорий от одной особи хозяина к дру-
гой? Никогда еще не удавалось наблю-
дать образование цист у этих инфузорий.
173
Поэтому высказывают предположение, что
заражение происходит активно — подвиж-
ными стадиями.
Инфузории астоматы представляют со-
бой яркий пример глубокого приспособ-
ления инфузорий к паразитическому об-
разу жизни. Это приспособление отрази-
лось на различных сторонах их организа-
ции (скелет, органоиды прикрепления) и
жизненных отправлений. Они утеряли ха-
рактерный для инфузорий способ питания
путем заглатывания оформленных частиц
пищи и перешли к осмотическому пита-
нию,
ИНФУЗОРИИ КИШЕЧНИКА
МОРСКИХ ЕЖЕЙ
Разнообразный комплекс паразитиче-
ских инфузорий обитает в кишечнике мор-
ских ежей. Морские ежи—богатый видами
класс морских беспозвоночных животных,
относящихся к т и п у иглокожих. Разме-
ры морских ежей различны, но у боль-
шинства встречающихся в наших водах
видов поперечник их почти шаровидного
тела составляет 5—10 см.
Очень многочисленны морские ежи в
прибрежной зоне наших северных (Барен-
цево) и дальневосточных морей (Японско-
го моря, тихоокеанского побережья Ку-
рильских островов). Большинство мор-
ских ежей питается растительной пищей,
главным образом водорослями, которые
они соскабливают с подводных предметов
особыми острыми «зубами», окружающими
ротовое отверстие. В кишечнике этих ра-
стительноядных ежей имеется богатая фа-
уна инфузорий. Нередко они развиваются
здесь в массовом количестве, и содержимое
кишечника морского ежа под микроско-
пом почти так же «кишит» инфузориями,
как содержимое рубца жвачных. Нужно
сказать, что, кроме глубоких различий
в условиях жизни инфузорий кишечника
морского ежа и рубца жвачного, имеются
и некоторые черты сходства. Они заклю-
чаются в том, что и там и здесь инфузо-
рии живут в среде, очень богатой расти-
тельными остатками. В настоящее время
известно свыше 50 видов инфузорий, оби-
тающих в кишечнике морских ежей, кото-
рые встречаются только в прибрежной
зоне, где ежи питаются водорослями.
На больших глубинах, где водоросли уже
не растут, инфузорий в морских ежах
нет.
Что же представляет собой фауна пара-
зитических инфузорий кишечника мор-
ских ежей в отношении своего системати-
ческого состава (принадлежности их к оп-
ределенным систематическим группам)?
Оказывается, что в кишечнике морских
ежей встречаются представители разных
отрядов и семейств инфузорий. На рисун-
ке 118 изображены некоторые характер-
ные представители этой фауны из кишеч-
ника морских ежей, обитающих в при-
брежных водах Японского моря в окрест-
ностях Владивостока. Здесь имеются раз-
личные равноресничные инфузории (Holot-
richa), брюхоресничные (Hypotrycha), ма-
лоресничные (Oligotricha). Интересно от-
метить, что некоторые из видов инфузорий
из кишечника морских ежей относятся
к родам, имеющим свободноживущие ви-
ды. У них нет тех глубоких приспособле-
ний, которые можпо видеть у энтодини-
оморф (Entodiniomorpha) к жизни в ки-
шечнике копытных или у астомат (Asto-
mata) к жизни в кишечнике малощетин-
ковых кольчецов. Лишь у некоторых ви-
дов, например у энторипидиум (Entorhipi-
dium), развиваются скелетные лучи (по-
добно тому как это мы видели у радиоф-
рий, рис. 117). Другие же виды мало чем
отличаются от свободноживущих инфу-
зорий.
По образу жизни и характеру питания
большинство инфузорий кишечника мор-
ских ежей растительноядны. Они пита-
ются водорослями, которые в большом
количестве наполняют кишечник хозяина.
Некоторые виды довольно «привередли-
вы» в выборе пищи. Например, стробили-
дий (Strobilidium, рис. 118, А) питается
почти исключительно крупными диатомо-
выми водорослями. Имеются здесь и хищ-
ники, поедающие представителей других,
более мелких видов инфузорий.
У инфузорий из кишечника морских
ежей, в отличие от астомат, не наблюда-
ется какой-либо строгой приуроченности
к определенным видам хозяев. Они живут
174
в самых разных видах морских ежей, пи-
тающихся водорослями.
Как уже говорилось, простейшие из ки-
шечника морских ежей хотя и ведут эндо-
паразитический образ жизни, но почти
не отличаются от свободноживущих роди-
чей. Вероятно, эти инфузории относитель-
но недавно перешли к парази-
тизму. Этот переход осуществ-
лялся независимо для разных
видов. В новых условиях пара-
зитической жизни они нашли
среду, не очень отличающуюся
от морской воды. В кишечнике
морских ежей всегда много заг-
лоченной вместе с пищей воды
и много водорослей на разных
стадиях переваривания. Эти ус-
ловия оказались весьма благо-
приятными для жизни инфузо-
рий, которые и размножаются
здесь в больших количествах.
Пути заражения морских
ежей инфузориями не изучены.
Однако здесь с большой долей
вероятности можно допустить,
что оно происходит активными
свободноплавающими формами.
Дело в том, что инфузории из
кишечника морских ежей могут
длительное время (многие часы)
жить в морской воде. Однако
они уже настолько приспособи-
лись к жизни в кишечнике ежей,
что вне тела их, в морской воде,
рано или поздно погибают.
Заканчивая знакомство с ин-
фузориями, следует еще раз под-
черкнуть, что они представляют
собой богатую видами, обшир-
ную и процветающую группу
(класс) животного мира. Оста-
ваясь на уровне клеточной ор-
ганизации, инфузории достигли
по сравнению с другими клас-
сами простейших наибольшей
сложности строения и функций.
Рис. 118. Инфузории, живущие
в кишечнике морских ежей:
А — Strobilidium rapulum; Б — Entorhi-
pidium tenue; В — Entorhipidium triangu-
laris; Г — Entodiscus indomitus. l — мак-
ронуклеус; 2 — микронуклеус, 3 — ске-
летные лучи.
Особенно существенную роль в этом про-
грессивном развитии (эволюции) сыграло,
вероятно, преобразование ядерного аппа-
рата и возникновение ядерного дуализма
(качественной неравноценности ядер). Бо-
гатство макронуклеуса нуклеиновыми ве-
ществами связано с активными процес-
сами обмена веществ, с энергично проте-
кающими процессами синтеза белков ци-
топлазмы и ядер.
Организация инфузорий оказалась
очень пластичной, что позволило им при-
способиться к различным средам обита-
ния в морской и пресной воде и дать раз-
нообразные паразитические формы.
Заключение
Мы подошли к концу нашего обзора
строения и образа жизни обширного типа
животного мира — простейших. Харак-
терной особенностью их, как уже неодно-
кратно подчеркивалось выше, является
одноклеточность. В отношении своего
строения простейшие — клетки. Однако
они несравнимы с клетками, входящими
в состав тела многоклеточных организмов,
потому что сами являются организмами.
Таким образом, простейшие — это орга-
низмы на клеточном уровне организации.
Некоторые высокоорганизованные про-
стейшие, обладающие многими ядрами,
уже как бы выходят за морфологические
пределы строения клетки, что дает осно-
вание некоторым ученым называть таких
простейших «надклеточными». Это мало
меняет существо дела, ибо типичной для
Protozoa является все же одноклеточная
организация.
В пределах одноклеточности простей-
шие прошли длинный путь эволюционного
развития и дали огромное разнообразие
форм, приспособленных к самым различ-
ным условиям жизни. В основе родослов-
ного ствола простейших стоят два класса:
саркодовые и жгутиконосцы. До сих пор
в науке дебатируется вопрос, какой из
этих классов более примитивный. С одной
стороны, низшие представители саркодо-
вых (амебы) обладают самым примитив-
ным строением. Но жгутиконосцы обна-
руживают наибольшую пластичность типа
обмена веществ и стоят как бы на границе
между животным и растительным миром.
В жизненном цикле некоторых саркодо-
вых (например, фораминифер) имеются
жгутиковые стадии (гаметы), что указы-
вает на их связь со жгутиконосцами. Оче-
видно, что ни современные саркодовые,
ни современные жгутиконосцы не могут
быть исходной группой эволюции живот-
ного мира, ибо они сами прошли большой
путь исторического развития и вырабо-
тали многочисленные приспособления
к современным условиям жизни на Земле.
Вероятно, оба этих класса современных
простейших следует рассматривать как
два ствола в эволюции, берущих свое на-
чало от древних, не сохранившихся до
наших дней форм, которые жили на заре
развития жизни на нашей планете.
В дальнейшей эволюции простейших
произошли изменения различного харак-
тера. Некоторые из них привели к общему
повышению уровня организации, повы-
шению активности, интенсивности жиз-
ненных процессов. К числу таких филоге-
нетических (эволюционных) преобразова-
ний следует, например, отнести развитие
органоидов движения и захвата пищи, ко-
торое достигло высокого совершенства
в классе инфузорий. Бесспорно, что реснич-
ки представляют собой органоиды, со-
ответствующие (гомологичные) жгути-
кам. В то время как у жгутиконосцев,
за немногими исключениями, число жгу-
тиков невелико, у инфузорий число рес-
ничек достигло многих тысяч. Развитие
ресничного аппарата резко повысило ак-
тивность простейших, сделало более мно-
гообразными и сложными формы их взаи-
моотношений со средой, формы реакций
на внешние раздражения. Наличие диф-
ференцированного ресничного аппарата,
бесспорно, явилось одной из основных
причин прогрессивной эволюции в классе
инфузорий, где возникло большое мно-
гообразие форм, приспособленных к раз-
ным средам обитания.
Развитие ресничного аппарата инфу-
зорий — пример такого рода эволюцион-
ных изменений, которые были названы
акад. Северцовым ароморфозами. Аромор-
фозы характеризуются общим повышением
организации, развитием приспособлений
широкого значения. Под повышением ор-
ганизации понимают изменения, которые
вызывают усиление жизнедеятельности ор-
ганизма; они связаны с функциональной
дифференцировкой его частей, ведут к бо-
лее многообразным формам связи организ-
ма с окружающей средой. Развитие рес-
176
ничного аппарата инфузорий как раз и от-
носится к такого рода преобразованиям
строения в процессе эволюции. Это ти-
пичный ароморфоз.
У простейших, как это было подчерк-
нуто В. А. Догелем, изменения типа
ароморфозов обычно бывают связаны
с увеличением числа органоидов. Проис-
ходит полимеризация органоидов. Разви-
тие ресничного аппарата у инфузорий—
типичный пример такого рода изменений.
Вторым примером ароморфоза в эво-
люции инфузорий может служить их ядер-
ный аппарат. Мы рассмотрели выше осо-
бенности строения ядра инфузорий (стр.
138). Ядерный дуализм инфузорий (на-
личие микронуклеуса и макронуклеуса)
сопровождался увеличением в составе
макронуклеуса числа хромосом (явление
полиплоидии). Поскольку хромосомы свя-
заны с основными синтетическими процес-
сами в клетке, в первую очередь с синте-
зом белков, этот процесс привел к обще-
му повышению интенсивности основных
жизненных функций. И здесь имела место
полимеризация, затронувшая хромосом-
ные комплексы ядра.
Инфузории — одна из наиболее много-
численных и прогрессивных групп про-
стейших, происходит от жгутиконосцев.
Об этом говорит полное морфологическое
сходство их органоидов движения. Этот
этап эволюции был связан с двумя боль-
шими ароморфозами: один из них затра-
гивал органоиды движения, второй —
ядерный аппарат. Оба эти типа изменений
связаны между собой, так как оба ведут
к повышению жизнедеятельности и услож-
нению форм взаимосвязей с внешней сре-
дой.
Наряду с ароморфозами существует и
другой тип эволюционных изменений, вы-
ражающихся в выработке приспособлений
(адаптации) к определенным, резко очер-
ченным условиям существования. Этот
тип эволюционных изменений Север-
ц о в ы м был назван идиоадапта-
ц и я м и. В эволюции простейших этот
тип изменений играл очень большую роль.
Выше, при рассмотрении разных классов
простейших, приведепы многочисленные
примеры идиоадаптивных изменений. При-
способления к планктонному образу жиз-
ни у разных групп простейших, приспо-
собления к жизни в песке у инфузорий
(стр. 155), образование защитных обо-
лочек ооцист у кокцидий и многое дру-
гое — все это идиоадаптации, игравшие
большую роль в возникновении и развитии
отдельных групп, но не связанные с об-
щими прогрессивными изменениями орга-
низации.
Приспособления к различным конкрет-
ным средам обитания у простейших носят
весьма разнообразный характер. Они обес-
печили широкое распространение этого
типа в самых разных средах обитания, что
подробно рассматривалось выше, при опи-
сании отдельных классов.
Следует еще остановиться на вопросе о
паразитизме у простейших. В типе про-
стейших переход от свободного образа
жизни к паразитизму распространен очень
широко. Паразитические формы мы встре-
чаем среди всех классов Protozoa. Кроме
того, два класса — споровики и книдоспо-
ридии — целиком состоят из паразитов.
Один из них (споровики), как вы видели
(стр. 129), взял свое начало от жгу-
тиконосцев, второй (книдоспоридии) —от
саркодовых (стр. 133). По ходу эволю-
ции паразитических групп простейших
у них выработались весьма сложные жиз-
ненные циклы, обеспечивающие как раз-
множение паразитов, так и их широкое
распространение в природе. Многие пара-
зитические простейшие имеют большое
практическое значение для человека как
возбудители тяжелых заболеваний самого
человека, домашних и промысловых мле-
копитающих, птиц, рыб, а также полез-
ных насекомых. Поэтому борьба с прото-
зойными паразитарными заболеваниями,
профилактика и лечение их играют боль-
шую роль в медицинской и ветеринарной
практике.
W 12 Жизнь животных, т. 1
Мезозой
Mesozoa
Губки
Spongia
ТИП МЕЗОЗОЙ (MESOZOA)
Мезозой — мелкие (не свыше 1 см, обыч-
но меньше 1 мм), несколько сходные с
плоскими червями организмы, паразити-
рующие внутри некоторых морских чер-
вей, моллюсков и офиур. Мезозой покры-
ты ресничками, обычно раздельнополые,
редко гермафродитные, обладают сложным
циклом развития с чередованием партено-
генетических поколений с самцами и сам-
ками (гетерогония).
Известно несколько десятков видов ме-
зозоев.
Для них характерно очень упрощенное
строение. Снаружи их тело покрыто эпи-
телием, внутреннее пространство занято
только генеративными клетками, которые
могут рассматриваться как энтодерма.
В состав Mesozoa входят два клас-
са — Orthonectida и Dicyemida.
Систематическое положение и филогенети-
ческое значение группы мезозоев остается
неясным. Некоторые зоологи в попытке най-
ти переходные группы между одноклеточ-
ными и настоящими многоклеточными склон-
ны в организации мезозоев усматривать чер-
ты самых примитивных многоклеточных.
По другой концепции, мезозой представ-
ляют собой упростившихся в результате па-
разитизма потомков паразитических червей,
т. е. трехслойных животных, скопление ге-
неративных клеток внутри поверхностного
слоя клеток как крайне упростившуюся эн-
тодерму, а своеобразные зародышевые клет-
ки как сильно измененные стадии партено-
генетического развития. Так как в жизнен-
ном цикле мезозоев формируются и половые
клетки с последующим оплодотворением, то,
возможно, мы имеем в данном случае тот
тип развития, который называют гетерого-
нией и который довольно обычен у беспозво-
ночных. Бесполое же развитие из отдельных
клеток известно только у одноклеточных жи-
вотных, а у многоклеточных не наблюдается.
Рис. 119. Представители Mesozoa.
На правом рисунке внутри видны развивающиеся
молодые особи. Р — половая пора
181
ТИП ГУБКИ (PORIFERA, или SPONGIA)
Губки — исключительно своеобразные
животные. Их внешний вид и строение те-
ла столь необычны, что долгое время не
знали, куда отнести эти организмы, к ра-
стениям или животным. В средние века,
например, и даже значительно позднее
губки вместе с другими подобными же
«сомнительными» животными (мшанками,
некоторыми кишечнополостными и др.)
помещали среди так называемых зоофи-
тов, т. е. существ, как бы промежуточ-
ных между растениями и животными.
В дальнейшем на губок смотрели то как
на растения, то как на животных. Лишь
в середине XVIII в., когда подробнее по-
знакомились с жизнедеятельностью гу-
бок, окончательно была доказана их
животная природа. Долгое время оста-
вался нерешенным вопрос о месте губок
в системе животного царства. Вначале
ряд исследователей рассматривал эти
организмы как колонии простейших, или
одноклеточных, животных. И такой взгляд,
казалось, нашел свое подтверждение в от-
крытии Д. Кларком в 1867 г. хоано-
флагеллат, жгутиконосцев с плазмати-
ческим воротничком, которые обнаружи-
вают удивительное сходство с особыми
клетками — хоаноцитами, имею-
щимися у всех губок. Однако вскоре после
этого, в 1874—1879 гг., благодаря исследо-
ваниям И. М е ч н и к о в а, Ф. III у л ь-
це и О.Шмидта, изучавших строе-
ние и развитие губок, неопровержимо
была доказана принадлежность их к мно-
гоклеточным животным.
В отличие от колонии простейших, со-
стоящих из более или менее однообразных
и независимых клеток, в теле многокле-
точных животных клетки всегда дифферен-
цированы как в отношении строения, так
и по выполняемой ими функции. Клетки
здесь утрачивают свою самостоятельность
и являются лишь частями единого слож-
ного организма. Они образуют различные
ткани и органы, выполняющие какую-
либо определенную функцию. Одни из
них служат для дыхания, другие выпол-
няют функцию пищеварения, третьи обес-
печивают выделение и т. д. Поэтому мно-
гоклеточных животных называют иногда
еще тканевыми животными. У губок клет-
ки тела также дифференцированы и имеют
тенденцию к образованию тканей, правда,
очень примитивных и слабо выраженных.
Еще более убеждает в принадлежности
губок к многоклеточным животным нали-
чие у них в жизненном цикле сложного
индивидуального развития. Подобно всем
многоклеточным, губки развиваются из
яйца. Оплодотворенное яйцо многократно
делится, в результате чего возникает за-
родыш, клетки которого группируются
таким образом, что формируются два раз-
личных слоя: наружный (эктодерма) и
внутренний (энтодерма). Эти два слоя кле-
ток, называемые зародышевыми пластами
или листками, при дальнейшем развитии
образуют строго определенные части тела
взрослого животного.
После признания губок многоклеточ-
ными организмами прошло еще несколько
182
десятилетий, прежде чем они заняли свое
настоящее место в системе животных. До-
вольно долгое время губок относили к ти-
пу кишечнополостных животных. И хотя
искусственность подобного их объедине-
ния с кишечнополостными была очевид-
ной, лишь с конца прошлого века взгляд
на губок как на самостоятельный тип
животного царства стал постепенно завое-
вывать всеобщее признание. Этому в зна-
чительной степени способствовало от-
крытие И. Деляжем в 1892 г. так на-
зываемого «извращения зародышевых пла-
стов» при развитии губок — явления, ко-
торое резко отличает их не только от
кишечнополостных, но и от остальных
многоклеточных животных. Поэтому в на-
стоящее время многие зоологи склонны
подразделять всех многоклеточных (Meta-
zoa) на два надраздела: на Parazoa, к ко-
торому из современных животных принад-
лежит только один тип губок, и на Eume-
tazoa, охватывающий все остальные типы.
Согласно этому представлению к Parazoa
относят таких примитивных многоклеточ-
ных животных, тело которых еще не имеет
настоящих тканей и органов; кроме того,
у этих животных зародышевые пласты
в процессе индивидуального развития ме-
няются местами, н так или иначе анало-
гичные части тела взрослого организма
по сравнению с Eumetazoa возникают
у них из диаметрально противополож-
ных зачатков.
Таким образом, губка являются самыми
примитивными многоклеточными живот-
ными, о чем свидетельствует простота
строения их тела и образ жизни. Это вод-
ные, преимущественно морские, непод-
вижные животные, обычно прикреплен-
ные ко дну или различным подводным
предметам.
ВНЕШНИЙ ВИД ГУБОК
И СТРОЕНИЕ ИХ ТЕЛА
Форма тела губок чрезвычайно разнооб-
разна. Часто они имеют вид корковых,
подушковидных, ковриговидных или ком-
кообразных обрастаний и наростов на
камнях, раковинах моллюсков или на ка-
ком-нибудь другом субстрате. Нередко
среди них встречаются также более или
менее правильные шаровидные, бокало-
видные, воронковпдные, цилиндрические,
стебельчатые, кустистые и иные формы.
Поверхность тела обычно неровная,
в различной степени игольчатая или даже
щетинистая. Лишь иногда она бывает
относительно гладкой и ровной. Многие
губки имеют мягкое и эластичное тело,
некоторые более жесткие или даже твер-
дые. Тело губок отличается тем, что лег-
ко рвется, ломается или крошится. Раз-
ломив губку, можно видеть, что она со-
стоит из неровной, ноздреватой массы,
пронизанной идущими в разных направ-
лениях полостями и каналами; достаточно
хорошо различимы также элементы ске-
лета — иглы или волокна.
Размеры губок варьируют в широких
пределах: от карликовых форм, измеряе-
мых миллиметрами, до очень крупных
губок, достигающих одного метра в высо-
ту и более.
Многие губки ярко окрашены: чаще все-
го в желтый, коричневый, оранжевый,
красный, зеленый, фиолетовый цвета. При
отсутствии пигментов губки имеют белую
или серую окраску.
Поверхность тела губок пронизана мно-
гочисленными мелкими отверстиями, по-
рами, откуда и происходит латинское
название этой группы животных — Pori-
fera, т. е. пористые животные.
При всем многообразии внешнего вида
губок строение их тела может быть сведе-
но к следующим трем основным типам,
получившим специальные названия: а с-
кон, сикон и лейкон.
Аскон. В наиболее простом случае тело
губки имеет вид небольшого тонкостен-
ного бокала или мешочка, основанием
прикрепленного к субстрату, а отверстием,
которое называется устьем или
оскулумом, обращенного кверху.
Поры, пронизывающие стенки тела, ведут
в обширную внутреннюю, а т р и а л ь-
н у ю, или парагастральную,
полость. Стенки тела состоят из двух сло-
ев клеток — наружного и внутреннего.
Между ними располагается особое бес-
структурное (студенистое) вещество —
мезоглея, в котором содержатся раз-
ного рода клетки. Наружный слой тела
183
Рнс. 120. Строение губок:
^ — устройство простейшей губки (по Бриену): 1 —
устье, 2 — поры; 3 — иглы; 4 — хоаноциты; 5 — аме-
боциты; 6 — колленциты; 7 — пннакоциты; 8 — поро-
циты. Б — хоаноциты: 1 — Halichondria; 2 — Spon-
gilla (по Минчину).
состоит из плоских клеток— пинакоци-
т о в, образующих кроющий эпителий,
который отделяет мезоглею от окружаю-
щей губку воды. Отдельные более крупные
клетки кроющего эпителия, так называе-
мые п ороциты, имеют внутриклеточ-
ный канал, открывающийся наружу по-
ровым отверстием и обеспечивающий связь
внутренних частей губки с наружной
средой. Внутренний слой стенки тела со-
стоит из характерных воротничко-
вых клеток, или хоаноцитов.
Они имеют вытянутую форму, снабжены
жгутом, основание которого окружено
плазматическим воротничком в виде от-
крытой воронки, обращенной в сторону ат-
риальной полости. В мезоглее содержатся
неподвижные звездчатые клетки (к о л-
л е н ц и т ы), являющиеся соединитель-
ноткаными опорными элементами, клет-
ки-скелетообразовательницы (с к л е р о-
б л а с т ы), образующие скелетные эле-
менты губок, разного рода подвижные
амебоциты, а также а р х е о ц и-
т ы — недифференцированные клетки, ко-
торые могут превращаться во все прочие
клетки, и в том числе в половые. Так уст-
роены губки простейшего асконоидного
184
типа. Хоаноциты здесь выстилают атри-
альную полость, которая сообщается
с внешней средой посредством пор и устья.
Сикон. Дальнейшее усложнение в строе-
нии губок связано с разрастанием мезо-
глеи и впячиванием в нее участков атри-
альной полости, образующих радиальные
трубки. Хоаноциты теперь сосредоточены
только в этих впячиваниях, или жгу-
тиковых трубках, и исчезают
с остальных участков атриальной поло-
сти. Стенки тела губки становятся более
толстыми, и тогда между поверхностью
тела и жгутиковыми трубками образуются
особые ходы, получившие название при-
водящих каналов. Таким образом, при
сиконоидном типе строения губок хоано-
циты выстилают жгутиковые трубки, кото-
рые сообщаются с внешней средой, с од-
ной стороны, посредством наружных пор
или системы приводящих каналов, а с дру-
гой— через атриальную полость и устье.
Лейкон. При еще большем разрастании
мезоглеи и погружении в нее хоаноцитов
образуется самый развитый, лейконоид-
ный тип строения губок. Хоаноциты сосре-
доточены здесь в небольших жгутиковых
камерах, которые, в отличие от жгутиковых
трубок типа сикон, не открываются непо-
средственно в атриальную полость, а свя-
заны с ней особой системой отводящих ка-
налов. Следовательно, при лейконоидном
типе строения губок хоаноциты выстилают
жгутиковые камеры, которые сообщаются
с внешней средой, с одной стороны, по-
средством наружных пор и приводящих
каналов, а с другой — через систему отво-
дящих каналов, атриальную полость и
устье. Большинство губок во взрослом
состоянии имеет лейконоидный тип строе-
ния тела. У лейкона, так же как pi у сико-
на, кроющий эпителий (п и н а к о ц и-
т ы) выстилает не только наружную по-
верхность губки, но и атриальную по-
лость и систему каналов.
Следует иметь в виду, однако, что губки
в процессе роста нередко испытывают
разного рода усложнения в строении тела.
Кроющий эпителий при участии элемен-
тов мезоглеи часто утолщается, превра-
щаясь вдермальную мембра-
ну, а иногда и в корковый слои разной
толщины. Под дермальной мембраной
местами образуются обширные полости,
откуда берут начало приводящие кана-
лы. Такие же полости могут образо-
ваться и под гастральной мембраной,
выстилающей атриальную полость. Исклю-
чительное развитие тела губки, его мезо-
глеи, приводит к тому, что атриальная
полость превращается в узкий канал и ча-
сто вообще не отличима от отводящих ка-
налов. Особенно сложной и запутанной
становится система жгутиковых камер,
каналов и добавочных полостей в тех
случаях, когда губки образуют колонии.
Одновременно с этим могут наблюдаться
некоторые упрощения, связанные с почти
полным исчезновением в теле губок ме-
зоглеи и появлением синцитиев — много-
ядерных образований, получающихся в
результате слияния клеток. Кроющий
эпителий также может отсутствовать или
замещаться синцитием.
\-j
Рис. 121. Различные типы строения губок
и их канальной системы (по Гессе):
1 — аскон; 2 — сикон; 3 — лейкон. Стрелки по-
казывают направление тока воды в теле губок.
185
Рис. 122. Иглы известковых губок. Различные
формы трехлучевых, четырехлучевых и одно-
осных игл.
Кроме отмеченных выше клеток, в теле
губок, особенно около многочисленных
отверстий, полостей и каналов, встре-
чаются еще особые веретеновидные клет-
ки—м и о ц и т ы, способные к сокраще-
нию. У некоторых губок в мезоглее обна-
ружены звездчатые клетки, соединяющие-
ся между собой отростками и посылающие
отростки к хоаноцитам и клеткам крою-
щего эпителия. Эти звездчатые клетки
рассматриваются некоторыми исследова-
телями как нервные элементы, способные
к передаче раздражений. Вполне возмож-
но, что подобные клетки играют какую-то
связующую роль в теле губок, однако о пе-
редаче импульсов, которыми отличаются
нервные клетки, говорить здесь не при-
ходится. Губки очень слабо реагируют да-
же на самые сильные внешние раздраже-
ния, а передача раздражений от одного
участка тела к другому почти незаметна.
Это свидетельствует об отсутствии у губок
нервной системы.
Губкп — настолько примитивные мно-
гоклеточные животные, что образование
тканей и органов у них находится в самом
зачаточном состоянии. В большинстве сво-
ем клетки губок обладают значительной
самостоятельностью и выполняют опреде-
ленные функции независимо одна от дру-
гой, не соединяясь между собой в какие-
либо тканеподобные образования. Толь-
ко слой хоаноцитов и кроющий эпителий
образуют нечто вроде тканей, но и здесь
связь между клетками крайне незначи-
тельна и неустойчива. Хоаноциты могут
терять жгутики и уходить в мезоглею,
превращаясь в амебоидные клетки; в свою
очередь амебоциты, перестраиваясь, да-
ют начало хоаноцитам. Клетки кроющего
эпителия также, погружаясь в мезоглею,
могут превращаться в амебоидные клетки.
ОСНОВНЫЕ ЖИЗНЕННЫЕ ОТПРАВЛЕНИЯ
ГУБОК.
Как уже отмечалось, губки — непод-
вижные животные и не способны к ка-
ким-либо изменениям формы тела. Лишь
при довольно сильном раздражении у не-
которых губок наблюдается очень мед-
ленное сужение отверстий (устий и пор)
и просветов каналов, которое достигает-
ся сокращением миоцитов или протоплаз-
мы других клеток. Наблюдения над мел-
ководными губками, обитающими в при-
ливо-отливной зоне, показали, напри-
мер, что их устья замыкаются за 3 минуты
и полностью открываются за 7 —10 минут.
Большинство клеток в теле губок спо-
собно выпускать и втягивать ложно-
ножки, или псевдоподии, или
даже с их помощью передвигаться в толще
мезоглеи. Особенной подвижностью отли-
чаются амебоциты, перемещающиеся иног-
да со скоростью до 20 мк в минуту. Но
самыми деятельными клетками губок яв-
ляются хоаноциты. Их жгутики находятся
в постоянном движении. Благодаря со-
гласованным винтообразным колебаниям
жгутиков множества хоаноцитов внутри
губки создается ток воды. Вода поступает
через поры и систему приводящих каналов
в жгутиковые камеры, откуда направля-
ется по отводящей системе каналов в ат-
риальную полость и через устье выводится
наружу. Естественно, что у губок сиконо-
идного, а в особенности асконоидного
типа строения путь воды значительно со-
кращается. Очень хорошо этот ток воды
наблюдать в аквариуме, если около жи-
вущей там губки выпустить небольшое
количество мелко растертой туши. Можно
заметить, как частицы краски через поры
увлекаются внутрь губки, а спустя некото-
рое время выходят наружу. Еще более
яркая картина наблюдается, если в тело
186
губки ввести шприцем некоторое количе-
ство кармина. Очень скоро из ближайших
устьевых отверстий начинают бить фон-
танчики красной жидкости.
Наличие в канальной системе губок по-
стоянного движения воды играет важней*
шую роль в их жизни.
Дыхание. Как и большинство жи-
вотных, обитающих в водной среде, губ-
ки используют для дыхания растворен-
ный в воде кислород. Ток воды, про-
никающий во все полости и каналы губки,
снабжает близлежащие клетки и мезо-
глею кислородом и уносит выделяемую
ими углекислоту. Таким образом, газо-
вый обмен с наружной средой осуществ-
ляется у губок непосредственно каждой
клеткой или через мезоглею.
Питание. Губки питаются главным
образом взвешенными в воде остатками
отмерших животных и растений, а так-
же мелкими одноклеточными организма-
ми. Размер пищевых частиц обычно не
превышает 10 мк. Частицы пищи прино-
сятся с током воды к жгутиковым каме-
рам, где они захватываются хоаноцита-
ми и затем поступают в мезоглею. Здесь
пища попадает к амебоцитам, которые
разносят ее по всем частям тела губки.
Внутри этих клеток, в пищеварительных
вакуолях, образующихся вокруг захва-
ченных частиц, происходит переварива-
ние пищи. Этот процесс пищеварения у гу-
бок можно наблюдать непосредственно под
микроскопом. Видно, как амебоцит обра-
зует вырост тела — ложноножку, направ-
ленную в сторону частицы пищи, посту-
пающей в мезоглею. Постепенно ложно-
ножка охватывает эту частицу и втяги-
вает ее внутрь клетки. Уже в вытянутой
ложноножке появляется пищеваритель-
ная вакуоля — пузырек, наполненный
жидким содержимым, имеющим вначале
кислую, а затем щелочную реакцию, при
которой и происходит переваривание пи-
щи. Захваченная частица растворяется,
а на поверхности вакуоли появляются
зерна жироподобного вещества. Так про-
исходит переваривание и усвоение пище-
вого материала клетками губок. Более
крупные частицы, застревающие в при-
водящих каналах, захватываются высти-
W
Рис. 123. Иглы стеклянных губок. Слева — макросклеры: шестилучевые, пятилучевые и одноосные
иглы. Справа — микросклеры: разные формы гексастр и амфидиски (внизу).
187
w
Рис. 124. Иглы четырехлучевых губок. Слева
и одноосная головчатая игла. Справа
макросклеры: различные четырехлучевые пглы
микросклеры: звездчатые иглы и сигмы.
лающими их клетками и также попадают
в мезоглею. Если такая частица слишком
крупна и не помещается внутри амебоид-
ной клетки, она окружается несколькими
амебоцитами, и переваривание пищи про-
исходит внутри такой клеточной массы.
У некоторых губок переваривание пищи
происходит также и в хоаноцитах.
Выделение. Непереваренные ос-
татки пищи выбрасываются в мезоглею
и постепенно скапливаются около отводя-
щих каналов, а затем поступают в просве-
ты каналов и выводятся наружу. Иногда
сами амебоциты, приближаясь к отводя-
щим каналам, выделяют туда зернистое
содержимое своих вакуолей.
Губки не обладают избирательной спо-
собностью к захвату только пищевых
частиц. Они поглощают все взвешенное
в воде. Поэтому в тело губки постоянно
попадает большое количество мелких не-
органических частиц. О дальнейшей судь-
бе их достаточно красноречиво свидетель-
ствует опыт по окраске воды аквариума
кармином. Очень скоро красные частицы
кармина попадают внутрь хоаноцитов,
а затем в мезоглею, где подхватываются
амебоцитами. Постепенно вся губка окра-
шивается в красный цвет, а ее клетки пе-
реполняются частицами кармина. Через
несколько дней клетки губки, и в первую
очередь хоаноциты, освобождаются от этих
неорганических частиц и губка приобре-
тает нормальный цвет.
Следовательно, основные жизненные от-
правления губок осуществляются крайне
примитивным способом. При отсутствии
специальных органов процессы дыхания,
питания и выделения протекают у них
внутриклеточно, за счет деятельности от-
дельных клеток. Можно сказать,что уро-
вень физиологии губок в этом отношении
лишь немногим выше уровня физиологии
одноклеточных животных.
СКЕЛЕТ И КЛАССИФИКАЦИЯ ГУБОК
Губки почти всегда имеют внутрен-
ний скелет, служащий опорой всего
тела и стенок многочисленных каналов
и полостей. Скелет может быть известко-
188
вым, кремниевым или роговым. Мине-
ральный скелет состоит из множества
игл, или с п и к у л, имеющих разнооб-
разную форму и различным образом рас-
положенных в теле губок. Среди игл обыч-
но различают макросклеры, со-
ставляющие основную массу скелета, и бо-
лее мелкие и иначе устроенные мик-
росклеры. Макросклеры в основном
представлены простыми, или одноосными,
трехлучевыми, четырехлучевыми и шести-
лучевыми иглами. В образовании скеле-
та, кроме игл, нередко принимает участие
особое органическое вещество спон-
гин, при помощи которого отдельные
иглы склеиваются друг с другом. Иногда
соседние иглы спаиваются концами, со-
ставляя разной прочности решетчатый ске-
летный каркас губок. При отсутствии
минеральных образований скелет может
быть образован одними роговыми
(спонгиновыми) волокнами.
Классификация губок в значительной сте-
пени основана на строении скелета. Прини-
мается во внимание вещество, из которого
Исключительно морские губки, живу-
щие обычно на небольшой глубине. Это
довольно нежные организмы, одиночные
или колониальные, редко превышающие
7 см в высоту. Тело их часто имеет труб-
чатую, бочонковидную или цилиндриче-
скую форму. Иногда образуется ножка,
при помощи которой губка прикрепля-
ется к субстрату. Поверхность известко-
вых губок может быть гладкой или покры-
той торчащими из нее пучками игл. Усть-
евое отверстие бывает окружено венчиком
из длинных одноосных игл. Известковые
губки чаще всего лишены пигмента и име-
ют белый или серый цвет. Строение их те-
ла может быть асконоидного, сиконоидно-
го или лейконоидного типа. Это единствен-
ная группа губок, у которых во взрослом
состоянии наблюдаются все три типа
строения, представленные в наиболее от-
образованы иглы, их форма и общий план
строения скелета. Каждый вид губок со-
держит характерные для него иглы одного
или чаще нескольких сортов, отличающие-
ся по форме и величине.
Тип губок делят на три класса: из-
вестковые (Calcispongia), стеклянные, или
шестилучевые (Hyalospongia), и обыкновен-
ные губки (Demospongia). К первым отно-
сятся губки с известковым скелетом, ко
вторым — содержащие кремниевые шес-
тилучевые иглы, и к последним — все
остальные, т. е. губки, имеющие кремние-
вые четырехлучевые и одноосные иглы,
а также роговые губки и очень немногие
губки, совершенно лишенные скелета.
Тип PORIFERA
Класс Calcispongia, или Calcarea
Отряд Homocoela
Отряд Heterocoela
Класс Hyalospongia, или Hexactinellida
Отряд Hexasterophora
Отряд Amphidiscophora
Класс Demospongia
Отряд Tetraxonida
Отряд Cornacuspongida
четливом виде. Поэтому известковые губ-
ки являются излюбленным объектом для
изучения строения, развития и жизнедея-
тельности животных этого типа.
Скелет известковых губок состоит из
трехлучевых, четырех лучевых и одноос-
ных игл, построенных из извести. Обычно
иглы в теле этих губок лежат свободно
и не связаны друг с другом. Лишь у так
называемых фаретронных губок иглы
склеиваются известковым цементом, об-
разуя решетчатый скелет. Иглы известко-
вых губок очень мелкие и не разделяются
на макросклеры и микросклеры. Длина
лучей трех- и четырехлучевых игл редко
превышает 0,3 мм; длина одноосных игл
(они называются здесь р а б д а м и) мо-
жет достигать Зеле. Скелетные элементы
известковых губок формируются внут-
ри клеток - скелет о образова-
КЛАСС ИЗВЕСТКОВЫЕ ГУБКИ (CALCISPONGIA)
189
ft
I?
I!
Рис. 125. Иглы кремнероговых губок. Слева — макросклеры: разной формы одноосные иглы.
Справа—микросклеры: хелы, якорьки (вверху), шпильки, сигмы, дужки (внизу).
тельниц (склеробластов).
Для образования трехлучевой иглы три
клетки сходятся вместе, тесно прилегая
друг к другу. Затем каждая из них де-
лится, и получаются три пары клеток, рас-
положенных в два слоя. Каждая пара кле-
ток формирует один из трех лучей иглы.
Сперва внутри трех отдельных склеро-
бластов самостоятельно закладываются за-
чатки лучей. На очень ранних стадиях
образования иглы эти три зачатка спаи-
ваются концами между собой под опре-
деленным углом, образуя маленькую трех-
лучевую иглу. Далее происходит утолще-
ние и рост лучей в результате отложе-
ния на их поверхности извести, которое
выполняется упомянутыми выше парами
клеток. Почти так же формируется четы-
рехлучевая игла. Вначале описанным спо-
собом возникает зачаток трехлучевой иг-
лы, к которой затем подходит особый скле-
робласт, строящий четвертый луч иглы.
В образовании одноосной иглы (рабды)
принимает участие одна пара клеток. Если
же игла длинна, то число склеробластов
увеличивается до трех или пяти за счет
деления исходной пары клеток.
Класс известковых губок состоит из
двух отрядов.
ОТРЯД HOMOCOELA
Мелкие губки, имеющие асконоидный
тип строения. Колонии их часто представ-
лены в виде кустиков или сложных клу-
бочков, состоящих из разветвленных и
сросшихся между собой тонкостенных тру-
бочек. Полости этих трубочек, сообщаю-
щихся между собой, выстланы хоаноци-
тами. Сюда относятся такие губки, как
Leucosolenia и Clathrina.
ОТРЯД HETEROCOELA
Основная группа известковых губок,
тело которых устроено по сиконоидному
или лейконоидному типу. Характерными
представителями отряда являются губки
Sycon и Leuconia. Многие из них имеют
довольно правильное радиально-симмет-
ричное тело веретеновидной или цилинд-
рической формы. Иногда они образуют
колонии с хорошо обособленными осо-
бями.
190
КЛАСС СТЕКЛЯННЫЕ ГУБКИ (HYALOSPONGIA)
Стеклянные губки — своеобразные мор-
ские, преимущественно глубоководные,
губки, достигающие 50 см в высоту и бо-
лее. Тело их чаще всего бокаловидное,
мешковидное или трубчатое, мягкое и лег-
ко рвущееся наподобие непрочного вой-
лока, или при более значительном разви-
тии скелета — довольно твердое и ломкое.
Цвет серый, коричневый, белый или жел-
товатый. Обычно одиночные радиально-
симметричные, реже колониальные орга-
низмы. Это губки упрощенного ленконоид-
ного типа с большими трубчатыми жгути-
ковыми камерами. Мезоглея у них почти
совершенно отсутствует, и живая ткань
в значительной степени представлена син-
цитиями, которые в виде тонких мембран
и рыхлой сети перемычек связывают силь-
Рис. 126. Разрез через стенку тела стеклянной губки Euplectella aspergillum (по Шульце):
I — дермальная мембрана; 2 — субдермальные синцитиальные перемычки; 3 — жгутиковые камеры; 4 — суб-
атриальные синцитиальные перемычки; 5 — гастральная мембрана; 6 — макросклеры; 7 — микросклеры.
191
но развитый скелет стеклянных губок.
Жгутиковые камеры также представляют
собой синцитальные образования; отдель-
ные хоаноциты сливаются друг с другом
основаниями. Кроющий эпителий часто
отсутствует или замещен подобными же
синцитиями. Кроме ник, в теле губок об-
наруживают лишь амебоидные клетки.
Скелет стеклянных губок построен из
кремнезема в виде аморфной водной крем-
некислоты и состоит из множества разно-
образных шестилучевых игл и их произ-
водных. Лучи игл направлены по трем
взаимно перпендикулярным осям, поче-
му этих губок называют еще трехосными
или шестилучевыми. Часто один или не-
сколько лучей редуцируются, и тогда
образуются пятилучевые, четырехлуче-
вые и даже одноосные иглы, а от исчез-
нувших лучей остаются лишь небольшие
бугорки. Лучи игл стеклянных губок мо-
гут быть гладкими или шиповатыми, пря-
мыми или изогнутыми, а иногда несут на
концах вздутия. У некоторых губок иглы
срастаются своими концами так, что обра-
зуется правильный решетчатый каркас
с прямоугольными ячейками, либо срос-
шийся скелет представляет собой очень
неправильную запутанную сеть из крем-
ниевых перекладин. Микросклеры у стек-
лянных губок еще более разнообразны и
имеют довольно причудливую форму. Сре-
ди них различают гексастры и амфидиски.
Гексастры — это в основе своей ма-
ленькие шестилучевые иголочки, лучи ко-
торых на концах часто снабжены много-
численными и разнообразными придатка-
ми. Амфидиски же подобны якорь-
кам, состоящим из тонкого стерженька,
на обоих концах которого венчиком рас-
положены лопастные выросты. Скелет
стеклянных губок очень дифференциро-
ван, и иглы каждой формы занимают
строго определенное место в их теле. Вели-
чина игл этих губок значительно варьи-
рует. Макросклеры обычно измеряются
сотнями микрон, но могут достигать не-
скольких десятков сантиметров, а в ис-
ключительных случаях даже 3 метров дли-
ны. Микросклеры значительно мельче,
их размеры колеблются в среднем от деся-
ти до ста микрон.
Иглы стеклянных губок формируются
в синцитиях и имеют сложную микроско-
пическую структуру. Каждая игла содер-
жит внутри органическую осевую нить,
вокруг которой слоями расположен крем-
незем. Слои отделены друг от друга про-
слойками из органического вещества.
Класс стеклянных губок состоит из
двух отрядов: Hexasterophora и Amphidis-
cophora.
Таблица 5. Поселение губок на дне Средиземного
моря:
1 — клиона (Cliona);
2 — Calyx;
3 — Siphonochalina;
4 — Acanthella;
5 — Chondrilla;
6 — геодия (Geodia) с сидящей на ней змеехвосткой;
7 — Raspailia;
8 — Halichondria;
9 — Siphonochalina;
10 — пробковая губка (Suberites domuncula) и рак-отшельник;
11 — морской апельсин (Tethya aurantium);
12 — Hymedesmia;
13 — Axinella в сожительстве с коралловыми полипами (Parazoan-
thus axinellae);
14 — Haliclona;
15 — Axinella.
ОТРЯД HEXASTEROPHORA
ОТРЯД AMPHIDISCOPHORA
К этому отряду относятся стеклянные
губки, у которых микросклеры представ-
лены различными гексастрами. Часто
большие иглы этих губок, соединяясь друг
с другом, образуют скелет в виде прост-
ранственной решетки. Характерные пред-
ставители отряда: корзинка Венеры (сем.
Euplectellidae), имеющая цилиндрическое
тело, крупные бокаловидные или мешко-
видные губки Росса (сем. Rossellidae) и
колониальные губки из сем. Euretidae,
тело которых образовано разветвленными
и неравномерно сросшимися тонкостен-
ными трубками.
К этому классу принадлежит большин-
ство живущих в настоящее время губок.
Именно эти губки поражают наблюдателя
разнообразием форм, размеров и красок.
Подобно стеклянным губкам, они дости-
гают значительной величины. Но в то
же время среди них встречается много
представителей карликового роста. Обык-
новенные губки — это колониальные или
реже одиночные организмы, имеющие лей-
коноидный тип строения тела. Жгутико-
вые камеры у них мелкие, округлой, гру-
шевидной или слегка вытянутой формы.
Скелет образован одноосными и четырех-
лучевыми кремниевыми иглами, нередко
в сочетании с органическим веществом
спонгином. Иногда иглы отсутствуют и
скелет представлен одними роговыми во-
локнами.
Макросклеры обычно закладываются
внутри склеробластов, носящих здесь на-
звание силикобластов. Вначале образует-
ся тонкая осевая нить из органического
вещества, вокруг которой откладывается
слой аморфного кремнезема. Затем мо-
лодая иголочка освобождается из мате-
ринской клетки, и ее формирование за-
вершается другими силикобластами, от-
кладывающими на поверхности иглы кон-
центрические слои плотного аморфного
Губки этого отряда содержат микроскле-
ры в виде амфидисков, тогда как гекса-
стры в их скелете совершенно отсутству-
ют. Типичный представитель отряда —
губка гиалонема (сем. Hyalonematidae),
имеет бокаловидное или овальное тело,
сидящее на длинном пучке игл, при по-
мощи которого губка укореняется в грун-
те. Сюда же относится гигантская губка
монорафис (Monorhaphis chuni), имеющая
сильно ноздреватое цилиндрическое тело
около 1 м в высоту, которое пронизано
в виде оси иглой, достигающей в длину
3 м при толщине 8,5 мм.
кремнезема. Образование микросклер пол-
ностью завершается внутри материнской
клетки. Очень немногие, так называемые
мясистые губки (Oscarella, Halisarca), от-
носящиеся к этому классу, совсем ли-
шены скелета.
Обыкновенные губки подразделяются на
два больших отряда: Tetraxonida и Сог-
nacuspongida.
ОТРЯД ЧЕТЫРЕХЛУЧЕНЫЕ ГУБКИ
(TETRAXONIDA)
В типичном случае скелет образован че-
тырехлучевыми иглами с примесью одно-
осных игл, заостренных на обоих концах.
Наиболее характерные микросклеры —
звездчатые иголочки. Гораздо реже ске-
лет представлен только одноосными, ча-
ще всего головчатыми иглами. Спонгин,
как правило, отсутствует. Четырехлуче-
вые иглы довольно разнообразны по внеш-
нему виду. Обычно один из лучей, значи-
тельно более длинный, служит основным
стержнем, на конце которого расходятся
более или менее одинаковые остальные
три луча. Эти лучи у разных игл различ-
ным образом изогнуты, а иногда дихото-
мически разветвлены. В редких случаях
КЛАСС ОБЫКНОВЕННЫЕ ГУБКИ (DEMOSPONGIA)
V 13 Жизнь животных, т. 1
193
Рис. 127. Строение скелета четырех лучевых гу-
бок (по Шульце). Разрез через поверхностный
слой тела геодии (Geodia barretti): видны ради-
алъно расположенные большие иглы и шаровид-
ные микросклеры (стеррастры), образующие кор-
ковый слой губки.
все четыре луча могут быть одинако-
выми и, расходясь во все стороны, обра-
зуют правильную пространственную фи-
гуру. Иглы четырехлучевых губок обычно
расположены радиально, т. е. направлены
от основания или из центра тела к его по-
верхности. Сами губки имеют нередко до-
вольно правильную лучистую, или ради-
альную, симметрию тела. На поверхности
губки часто развит корковый слой, иног-
да даже в виде прочного панциря. У мно-
гих четырехлучевых губок тело жест-
кое, игольчатое; длинные иглы могут
торчать на поверхности в виде густой
щетины.
Среди четырехлучевых губок пользу-
ются известностью крупные своеобразные
губки геодии (сем. Geodiidae). Они имеют
вид шаров, неглубоких бокалов или не-
равномерно разросшихся комков величи-
ной до 50 см. Тело геодии отличается
большой прочностью, так как снаружи
оно обычно покрыто жестким панцирем
толщиной в несколько миллиметров. Устья
и поры рассеяны по всей поверхности губ-
ки или сосредоточены в особых углубле-
ниях. Так или иначе они хорошо заметны
простым глазом. Под панцирем находятся
мягкие части губки, скелет которых со-
стоит из радиально расположенных четы-
рехлучевых и одноосных крупных игл.
Сам панцирь образован исключительно
мелкими шарообразными звездчатыми
микросклерами (около 0,1 мм в диа-
метре), тесно лежащими в несколько
слоев.
Очень многочисленны и разнообразны
по внешнему виду представители с е-
м е й с т в a Polymastiidae — полимастии,
имеющие обычно подушковидное, диско-
видное или чашевидное тело величиной
до 10 см. Скелет их состоит из одноосных
головчатых игл. На поверхности губок
образуются характерные конические пли
цилиндрические выросты тела — папил-
л ы, на которых помещаются устья и
поры. Иногда папиллы по длине пре-
восходят само тело губки. К этому се-
мейству относится небольшая губка мор-
ской гриб (Tentorium semisuberites); no
форме тела она напоминает молодой
шампиньон.
Не менее интересны так называемые
пробковые губки (сем. Suberitidae). Скелет
их образован относительно мелкими го-
ловчатыми иглами, благодаря чему ком-
кообразное тело пробковых губок отли-
чается некоторой эластичностью. Многие
из них имеют очень яркую окраску, часто
красную или оранжевую.
Небольшие шарообразные или яйцевид-
ные губки из семейства Tetillidae
характерны тем, что основание их часто
окружено густым войлоком из длинных
игл, при помощи которых губка прикреп-
ляется к илистому субстрату. На поверх-
ности этих губок имеются низкие кони-
ческие выросты тела — к о н у л и, обра-
зованные пучками четырехлучевых и одно-
осных игл.
Широко распространенная губка мор-
ской апельсин (Tethya aurantium), полу-
чившая такое название за внешнее сход-
ство с апельсином, имеет шарообразное
тело, слегка бугорчатую поверхностьи час-
то яркую окраску. Радиальный или спи-
ральный скелет этой губки образован од-
ноосными крупными иглами, а также звезд-
194
чатыми микросклерами. Среди четырех-
лучевых губок булавовидные или стебель-
чатые формы почти не встречаются. Ред-
кое исключение представляет губка Sty-
locordyla, имеющая очень характерный
внешний вид: небольшое округлое или
несколько вытянутое тело ее снабжено
длинным прочным стебельком, или нож-
кой, при помощи которого губка при-
крепляется ко дну. Стебелек образован
массой вертикальных и тесно лежащих
друг подле друга одноосных игл. Близки
к геодиям представители обширного с е-
м е й с т в a Stellettidae. Они не имеют жест-
кого панциря, но их корковый слой
отличается большой прочностью. Да и все
тело этих губок нередко бывает довольно
жестким и очень колючим в силу зна-
чительного развития минерального ске-
лета.
К этому же отряду относятся оригиналь-
ные «сверлящие» губки, или клионы (сем.
Clionidae), обладающие способностью про-
делывать ходы в каком-либо известковом
субстрате, а также характерная бокаловид-
ная губка кубок Нептуна (Poterion neptu-
ni), обитающая в тропических водах.
ОТРЯД КРЕМНЕРОГОВЫЕ ГУБКИ
(CORNACUSPONGIDA)
Самая многочисленная группа губок.
Это преимущественно мягкие эластичные
формы. Скелет их образован одноосными
иглами. Всегда имеется в том или ином
количестве спонгин, с помощью которого
иглы склеиваются между собой в пучки
или волокна. Одноосные иглы кремнерого-
вых губок отличаются большим разно-
образием: они могут быть гладкими или
шиповатыми, прямыми, цилиндрическими,
веретеновидными, изогнутыми или змее-
видно искривленными, головчатыми или
булавовидными, с заостренными, тупыми
или утолщенными концами. Кроме одно-
осных больших игл (макросклер), в обра-
зовании скелета кремнероговых губок
принимают участие особые микросклеры.
Для отряда характерны микросклеры
в виде маленьких якорьков, шпилек
или сигм.
В теле кремнероговых губок иглы рас-
полагаются таким образом, что образуют
различного вида сетчатый или сетчато-
волокнистый скелет. При увеличении ко-
личества спонгина в теле губки появляются
спонгиновые (роговые) волокна, внутри
которых содержатся иглы. Иногда у на-
стоящих роговых губок иглы совершенно
отсутствуют и скелет тела представлен
одними роговыми волокнами, образую-
щими также сетчатую или древовидно раз-
ветвленную опору тела. Некоторые рого-
вые губки используют для построения ске-
лета различные инородные минеральные
частицы (песчинки, обломки игл, рако-
вины простейших), которые заключают-
ся внутри волокон этих губок.
Спонгин выделяется особыми клетка-
ми—с понгобластами. Роговые во-
локна при формировании бывают окру-
жены футляром из подобного типа клеток,
отлагающих на их поверхности концент-
рические слои спонгина. Таким образом,
в отличие от игл, роговые волокна форми-
руются межклеточно. Роговое вещество
губок по химическому составу близко
к шелку и отличается значительным содер-
жанием иода. В скелете некоторых рого-
вых губок содержится до И —14% иода.
Кремнероговые губки преимущественно
колониальные организмы. Форма тела их
отличается исключительным разнообра-
зием. Характер поверхности, очертания
тела и структура губки очень изменчивы
даже у представителей одного и того же
вида. Лишь скелет и в особенности иглы,
его образующие, дают возможность судить
о видовой принадлежности губок.
Тело кремнероговых губок имеет вид кор-
ковых или подушковидных обрастаний,
бесформенных, неравномерно разросшихся
комков с разной величины выростами
и лопастями, неравномерно утолщенных
пластин, разного вида воронок, трубок,
стебельчатых, кустистых и других обра-
зований. Невозможно перечислить все
многообразие форм, которое наблюдается
у кремнероговых губок, настолько плас-
тично их тело.
Наиболее часто среди них встречаются
различные комкообразные губки. Тако-
вы широко распространенные миксиллы
(с е м. Myxillidae), морской каравай (сем.
13*
195
Halichondriidae), некоторые роговые губ-
ка (с е м. Spongiidae), пресноводные
губки бадяги (с е м. Spongillidae) и др.
Стебельчатые и воронковидные формы
значительных размеров наблюдаются у ря-
да представителей роговых губок и различ-
ных факеллий (с е м. Axinellidae); бай-
кальская губка (с е м. Lubomirskiidae)
может иметь пальцевидное или кусти-
стое тело до 1 ж высоты. Очень своеоб-
разны карликовые формы губок в виде не-
больших тонких корочек или нежных пле-
нок различного цвета, встречающиеся
на поверхности камней, на раковинах
Лучше всего изучено размножение из-
вестковых, кремнероговых и отчасти четы-
рехлучевых губок. Относительно стеклян-
ных губок вполне достоверные сведения
имеются лишь об их бесполом размно-
жении.
Половое размножение. Сре-
ди губок встречаются как раздельнополые,
так и гермафродитные формы. Какого-либо
внешнего различия мужских и женских
особей в случае раздельнополости не на-
блюдается. Половые клетки образуются из
археоцитов в мезоглее губки. Там же про-
исходит рост и созревание яиц и формиро-
вание сперматозоидов. Зрелые спермато-
зоиды выходят из губки наружу и с током
воды по системе приводящих каналов по-
падают в жгутиковые камеры других гу-
бок, имеющих зрелые яйца. Здесь они за-
хватываются хоаноцитами и передаются
в мезоглею амебоцитам, которые транс-
портируют их к яйцам. Иногда сами хоа-
ноциты, теряя жгутики, подобно амебо-
цитам, переносят сперматозоиды к яйцам,
обычно расположенным вблизи жгутико-
вых камер.
Дробление яйца и формирование личин-
ки у большинства губок протекают внутри
материнского организма. Лишь у пред-
ставителей некоторых родов четырех-
лучевых губок (Cliona, Tethya) яйца
выходят наружу, где и развиваются.
Личинка губки, как правило, имеет
овальную или округлую форму тела вели-
моллюсков или на каком-либо дру-
гом субстрате (сем. Hymedesmiidae). Осо-
бый интерес представляют морские ер-
шики — губки, имеющие довольно пра-
вильную и симметричную форму тела (As-
bestopluma, Cladorhiza, Chondrocladia).
В основном это небольшие губки, стебле-
видное или булавовидное тело которых
несет тонкие отростки, придающие губке
вид лампового ершика, птичьего пера
или зонтика. Великаном среди морских
ершиков является гигантская хондрокла-
дия (Chondrocladia gigantea) величиной
свыше 40 см.
чиной до 1 мм. Поверхность ее покрыта
жгутиками, благодаря движению которых
личинка энергично плавает в толще воды.
Длительность свободного плавания ли-
чинки до момента ее прикрепления к суб-
страту колеблется от нескольких часов
до трех суток. У большинства губок
плавающая личинка состоит из внутрен-
ней (мезоглеальной) массы рыхло располо-
женных крупных зернистых клеток, по-
крытых снаружи слоем более мелких ци-
линдрических жгутиковых клеток. Такая
двуслойная личинка называется парен-
химулой и возникает в результате не-
равномерного и неправильного дробления
яйца. Уже на первых стадиях дробления
образуются клетки различной величины:
макро- и микромеры. Быстро делящиеся
микромеры постепенно обрастают компакт-
ную массу более крупных макромеров,
и таким образом получается двуслойная
личинка паренхимула. У известковых гу-
бок (Homocoela) и у некоторых наиболее
примитивных четырехлучевых губок (Р1а-
kina, Oscarella) личинка вначале имеет вид
пузырька, оболочка которого состоит из
одного слоя однородных призматических
клеток, снабженных жгутиками. Эта ли-
чинка называется ц е л о б л а с т у л о и.
По выходе из материнского организма она
претерпевает некоторый метаморфоз, за-
ключающийся в том, что часть клеток, те-
ряя жгутики, погружается внутрь личин-
ки, постепенно заполняя имеющуюся там
РАЗМНОЖЕНИЕ И РАЗВИТИЕ ГУБОК
196
Рис. 128. Строение скелета некоторых кремнероговых губок:
I — Haliclona gracilis; 2 — Haliclona schmidti; 3 — Dysidea palescens; 4 — Spongia officinalis.
полость. В результате личин-
ка целобластула превращается
в уже известную нам паренхи-
мулу. У другой части извест-
ковых губок (Heterocoela) личин-
ка также имеет вид однослой-
ного пузырька, но отличается
тем, что верхняя ее половина
(передняя часть) образована
мелкими цилиндрическими
клетками, снабженными жгути-
ками, а нижняя (задняя) состоит
из крупных округлых зернис-
тых клеток. Подобная однослой-
ная личинка, состоящая из
двух сортов клеток, называется
амфибластулой. Она
сохраняет такой вид до ее при-
крепления к субстрату.
Таким образом, у губок име-
ются две основные личиночные
формы: паренхимула и амфи-
бластула. Проплавав некоторое
время, личинка оседает на под-
ходящий субстрат, прикрепля-
ясь своим передним концом, и
постепенно из нее формируется
молодая губка. При этом у па-
ренхимулы наблюдается очень
интересный и свойственный
только губкам процесс пере-
мещения зародышевых пластов,
которые меняются своими мес-
тами. Жгутиковые клетки на-
ружного эктодермального слоя
личинки мигрируют во внутрен-
нюю клеточную массу, превра-
щаясь в хоаноциты, которые
выстилают образующиеся жгу-
тиковые камеры. Клетки энто-
дермы, лежащие под наружным
слоем личинки, напротив, ока-
зываются на поверхности и дают
покровный слой и мезоглею гу-
бок. Это и есть так называе-
мое «извращение зародышевых
У губок.
Ничего подобного не наблюдается у
других многоклеточных животных: из эк-
тодермы и энтодермы их личинок фор-
мируются соответственно эктодермальные
и энтодермальные образования взрослого
организма.
гу-
Рис. 129. Личинки
бок:
1 — целобластула известко-
вой губки Clathrina retic-
ulum (по Минчину); видны
клетки энтодермы, мигри-
рующие в полость личин-
ки; 2 — амфибластула Sy-
con raphanus (по Шуль-
це); 3 — паренхимула кре-
мнероговой губки Ргопах
piumosa (по Леви), внутри
имеются личиночные иглы
и спонгин.
пластов»
Несколько иначе протекает
развитие губок, которые име-
ют плавающую личинку амфи-
бластулу. Перед прикреплени-
ем такой личинки к субстрату
переднее ее полушарие с мелки-
ми эктодермальными жгутико-
выми клетками впячивается
внутрь, и зародыш становится
двуслойным. Более крупные
энтодермальные клетки амфи-
бластулы образуют наружный
слой губки, а за счет жгутико-
вых клеток формируются хоа-
ноциты жгутиковых камер. Как
можно видеть, и в данном слу-
чае имеется извращение заро-
дышевых листков. У других
многоклеточных животных, у
которых в развитии есть пузы-
ревидная личиночная стадия
(бластула), состоящая из кле-
ток различной величины, более
крупные клетки дают начало
энтодерме взрослого животно-
го, а мелкие клетки (переднее
полушарие) — эктодерме. У гу-
бок же мы наблюдаем как раз
обратные отношения. В резуль-
тате личиночного метаморфоза
губок, сопровождающегося об-
разованием атриальной полос-
ти, устьевого отверстия и ске-
летных элементов, получаются
послеличиночные стадии —
о л и н т у с или р а г о н.
Олинтус представляет собой не-
большую мешковидную губку
асконоидного типа строения.
При дальнейшем его росте фор-
мируются одиночные или ко-
лониальные известковые Но-
mocoela, а при соответствую-
щем усложнении строения —
прочие известковые губки (He-
terocoela). Рагон характерен для обык-
новенных губок. Он имеет вид сильно
сплюснутой в вертикальном направле-
нии губки сиконоидного строения с
обширной атриальной полостью и с
устьем на вершине. Через некоторое
время рагон преобразуется в молодую
губку лейконоидного типа. Интерес-
198
но, что отдельные представители обыкно-
венных губок (Halisarca), подобно извест-
ковым губкам, в своем развитии проходят
вначале стадию, имеющую наиболее при-
митивный, асконоидный тип строения.
В этом нельзя не видеть проявления био-
генетического закона, согласно которому
животные в своем индивидуальном раз-
витии последовательно проходят опре-
деленные стадии, соответствующие основ-
ным чертам строения их предков.
Бесполое размножение.
У губок очень широко распространены
разные формы бесполого размножения.
К их числу относятся наружное почкова-
ние, образование соритов, геммул и др.
При почковании отделяющаяся дочер-
няя особь может содержать все ткани
материнского организма и представлять
собой просто его обособившийся участок.
Подобное почкование наблюдается у изве-
стковых, а также у некоторых стеклянных
и примитивных четырехлучевых губок.
В других случаях почка возникает из
скопления археоцитов. Наиболее извест-
ный пример такого образования почек —
морской апельсин (Tethya aurantium). Груп-
па археоцитов скапливается на поверх-
ности губки; из них постепенно форми-
руется маленькая губка, которая через
некоторое время отшнуровывается от ма-
теринского организма, отваливается и
приступает к самостоятельному образу
жизни. Иногда почки формируются на
концах игл, выступающих из тела губок,
либо образуется четковидная серия из
последовательно соединенных друг с дру-
гом мелких почек, имеющих слабую связь
с материнским организмом. Как крайний
случай такого почкования может рассмат-
риваться особый способ бесполого размно-
жения, наблюдаемый у некоторых геодий
(Geodia barretti). Археоциты здесь выхо-
дят за пределы материнской губки; одно-
временно происходит выталкивание из
нее некоторых длинных игл, оседающих
на дно. На этих иглах как на субстрате
скапливается воспроизводящая масса ар-
хеоцитов, и совершенно независимо от
материнского организма постепенно фор-
мируется маленькая губка геодия. Обра-
зование наружных почек из скопления
археоцитов широко распространено у
многих четырехлучевых губок (Tethya,
Polymastia, Tetilla и др.), а также встре-
чается иногда у кремнероговых и других
губок.
Значительно реже наблюдается у губок
бесполое размножение, выражающееся в
отделении от материнского организма
разной величины участков, которые
далее развиваются во взрослый организм.
Очень близко примыкает к такому способу
размножения образование у губок при
неблагоприятных условиях существования
так называемых редукционных телец.
Этот процесс неизменно сопровождается
распадом значительной части тела губ-
ки. Сохранившаяся часть обособляется
в виде нескольких клеточных скоплений,
или редукционных телец, состоящих из
группы амебоцитов, одетой снаружи клет-
ками кроющего эпителия. С наступле-
нием благоприятных условий из этих
редукционных телец развиваются новые
губки. Образование редукционных телец
встречается у морских губок, в особен-
ности живущих в приливо-отливной зоне,
а также у пресноводных губок, не обла-
дающих способностью к образованию
геммул.
Характерно для губок также размно-
жение с помощью соритов и геммул. Этот
способ размножения называют иногда
внутренним почкованием. Сориты
представляют собой образования округлой
формы, значительно менее 1 мм в диаметре.
Они возникают внутри губок из скопле-
ния археоцитов. При развитии сорита
зародыш обычно формируется из одной
клетки, питающейся за счет остальных
клеток сорита, слившихся в синцити-
альную массу. Из соритов могут полу-
чаться свободноплавающие личинки, ни-
чем по существу не отличающиеся от
образующихся половым путем. Такая ли-
чинка в дальнейшем претерпевает мета-
морфоз и превращается в молодую губку.
Сориты известны у многих обыкновенных
губок, в том числе у пресноводной
байкальской губки. Нетрудно заметить,
что бесполое размножение при помощи
соритов чрез-вычайно близко к полово-
му партеногенетическому размножению,
наблюдаемому у некоторых многоклеточ-
ных животных. У губок, следовательно,
199
Рис. 130. Бесполое размножение губок наружным почкованием:
1—5 — образование почек за счет скопления воспроизводящей массы археоцитов (у стеклянных губок — Апохуса-
lyx ijimae, Polylophus philippiensis и у четырехлучевых губок — Tethya aurantium, Polymastia mammillaris,
Polymastia kurilensis).
происходит крайнее сближение явлений
бесполого и полового размножения.
Это связано с тем, что недифференциро-
ванные амебоидные клетки, или архео-
циты, не только дают начало половым
клеткам, но п принимают участие в
разных формах бесполого размножения.
Геммулы, подобно соритам, образуются
внутри губок из скопления археоцитов.
Они очень характерны для пресноводных
губок и имеют нередко довольно сложное
устройство. При образовании геммул
группа археоцитов, богатых питательны-
ми веществами, окружается плотной хити-
новой капсулой, а затем воздухоносным
слоем, содержащим обычно особые гем-
мул ьные микросклеры, которые часто рас-
положены на поверхности капсулы пра-
вильными рядами. Обычно капсула снаб-
жена мелким отверстием для выхода ее
200
содержимого наружу. Геммулы представ-
ляют собой покоящуюся стадию и долгое
время могут сохраняться при неблаго-
приятных условиях, вызывающих гибель
самой губки. В живой или отмершей губке
такие геммулы, около 0,3 мм в диаметре,
иногда встречаются в очень большом ко-
личестве. При наступлении благоприят-
ных условий в геммулах начинается диф-
ференциация клеток, которые в виде бес-
форменной массы выходят наружу и за-
тем формируют молодую губку. Геммулы
встречаются также у некоторых морских
губок (Suberites, Cliona, Haliclona, Dysi-
dea и др.), но здесь они проще устроены,
чем у бадяг, и не имеют специальных ске-
летных элементов.
ГУБКИ—КОЛОНИАЛЬНЫЕ ОРГАНИЗМЫ
Сравнительно немногие губки пред-
ставляют собой одиночные организмы.
Таковы, например, известковые губки
различного типа строения, с одним
устьем на вершине, а также стеклян-
ные и некоторые обыкновенные губки
довольно правильной и симметричной фор-
мы тела. Вообще всякая губка, имеющая
одно устье и связанную с ним единую ка-
нальную систему, рассматривается как
одиночный организм. Большая часть гу-
бок, однако, представлена разного рода
колониальными образованиями. Принято
считать, что колонии возникают в резуль-
тате не доведенного до конца беспо-
лого размножения. Можно себе предста-
вить, что на поверхности одиночной
губки путем почкования формируются
маленькие губочки, которые не отделя-
ются от материнского организма. Они про-
должают существовать совместно, обра-
зуя колонию, объединяющую различное
число отдельных особей, или индивидуу-
мов.
Такие колонии действительно встре-
чаются у некоторых известковых (Leuco-
solenia, Sycon и др.) и стеклянных губок
(Rhabdocalyptus, Sympagella и др.), об-
разующих небольшие группы особей, свя-
занных своими основаниями. Но обычно
в колониях у губок отдельные особи в раз-
Рис. 131. Геммулы и сорит пресноводных губок
(по Резвому):
1 — геммула (Spongilla lacustris) с тонким зерни-
стым воздухоносным слоем и малым количеством
микросклер; 2 — то же (Ephydatia blembingia), вид-
ны клеточное содержимое геммулы,хитиновая кап-
сула, воздухоносный слой с рядами лежащих в нем
микросклер; 3 — сорит байкальской губки (Baika-
lospongia bacillifera) в капсуле из плоских клеток, в
средней части сорита имеется единственное ядро, на
периферии заметны остатки дегенерирующих ядер,
питающих клетки; сорит переполнен зоохлореллами.
личной степени сливаются между собой.
Это слияние происходит очень рано и за-
частую настолько полно, что трудно, даже
невозможно бывает отличить особи коло-
нии друг от друга. На поверхности коло-
нии от каждой особи в подобных случаях
201
сохраняется лишь устьевое отверстие. По-
этому условно принято считать у таких
колоний за отдельную особь участок тела
губки с одним устьем. В образовании
подобного рода колоний сказывается боль-
шая простота строения губок, низкий уро-
вень целостности и слабо выраженная ин-
дивидуальность их организма. Не только
отдельные особи, входящие в состав коло-
нии, но часто и сами колонии, имеющие
вид бесформенных образований, отли-
чаются исключительно слабой индивиду-
альностью. Таковы корковые, комкооб-
разные, стебельчатые, кустистые и прочие
губки неправильной и неопределенной
формы тела, отличающиеся большой из-
менчивостью внешнего вида. Особенно
показательны они для кремнероговых и
четырехлучевых губок. Характерно, что
подобные колонии могут образовываться
не только из одной особи губки, но и при
слиянии растущих рядом губок одного
вида. Более того, даже личинки их спо-
собны соединяться вместе и давать начало
колонии.
Иначе обстоит дело, когда губка приоб-
ретает определенную или правильную фор-
му тела. Устья, которые служат для рас-
познавания отдельных особей колонии,
представляют здесь образования, в той
или иной степени подчиненные губке как
оформленному целому. Происходит про-
цесс индивидуализации колонии при пол-
ном растворении в ней отдельных особей.
Это наблюдается у многих четырехлуче-
вых и некоторых кремнероговых губок,
имеющих более или менее правильную
и симметричную форму тела. Таковы,
например, чашевидные, бокаловидные или
воронковидные формы, часто снабженные
ножкой. Устьевые отверстия у них рас-
положены на внутренней поверхности во-
ронки, а поры — снаружи. Эти губки
представляют собой уже образования бо-
лее высокого порядка, чем обычные бес-
форменные колонии. Но процесс индиви-
дуализации колонии может идти дальше.
Края бокала или воронки, вытягиваясь
кверху, срастаются таким образом, что
внутри губки образуется полость (псев-
доатриальная), открывающаяся на верши-
не отверстием, которое теперь выполняет
функцию единого устья. И внешне такая
трубчатая или мешковидная губка напо-
минает многие одиночные стеклянные губ-
ки. Сходный процесс наблюдается у шаро-
образных или близких к ним форм. Устья
здесь могут быть рассеяны по всей поверх-
ности, собраны различным образом в не-
большие группы или даже слиты в одно
отверстие. В последнем случае (как, на-
пример, у некоторых представителей с е-
м е й с т в a Tetillidae, Geodiidae и др.)
от прежней колониальное™ не остается
и следов. С самого начала развития такие
правильные формы растут как единое
целое. Здесь мы имеем пример возникно-
вения вторичноодиночных особей гу-
бок. Такие одиночные губки встречаются,
кроме того, среди подушковидных и дис-
ковидных представителей губок — поли-
мастий (семейство Polymastiidae),
имеющих на поверхности одну устьевую
папиллу, и у ряда других четырехлуче-
вых губок. Очень близки к ним высоко-
индивидуализированные колонии крем-
нероговых губок, имеющих правиль-
ную радиально-симметричную форму тела.
Таковы многие губки морские ершики,
трубчатые, воронковидные и иные формы.
Но индивидуальность подобных губок
весьма несовершенна и малоустойчива.
Нередко уже вторичноодиночные формы
образуют дополнительные устья, прояв-
ляя тем самым свою исходную колониаль-
ную сущность. Хорошим примером подоб-
ного явления может служить губка мор-
ской гриб, на вершине которого имеется
одно устье. Такая губка есть вторично-
одиночный организм.
При определенных условиях, однако,
появляются экземпляры с двумя или
несколькими устьями. То же самое можно
наблюдать у губок из семейства
Tetillidae.
Таким образом, обыкновенные губки
в основном представлены либо бесфор-
менными колониальными образованиями,
либо вторичноодиночными особями и пере-
ходными между ними формами в виде
высокоиндивидуализированных колоний.
Известковые и стеклянные губки содер-
жат некоторое количество одиночных
форм, могут образовывать разного рода
колонии, и в том числе с хорошо обособ-
ленными особями.
202
ЯВЛЕНИЕ РЕГЕНЕРАЦИИ
У ГУБОК
Под регенерацией подра-
зумевается восстановле-
ние организмом утраченных
частей тела. Многие живот-
ные способны к регенера-
ции, и, чем проще устроен
организм, тем с большей си-
лой проявляется эта способ-
ность. Известно, например,
что гидра может быть раз-
резана на множество кусоч-
ков и из каждого кусочка со
временем восстанавливается
новая гидра. Губки обладают
еще большими возможностя-
ми к регенерации. Об этом
свидетельствуют классиче-
ские опыты Г. Уильсона
по восстановлению губок из
отдельных клеток. Если ку-
сочки губок протирать через
мелкоячеистую ткань, то в
результате получится фильт-
рат, содержащий изолиро-
ванные клетки. Эти клетки
сохраняют жизнеспособность
в течение нескольких дней,
проявляя оживленное аме-
боидное движение, т. е. вы-
пуская ложноножки и пере-
двигаясь с их помощью. Помещенные на
дне сосуда, они собираются группами,
образуя бесформенные скопления, кото-
рые через 6—7 дней превращаются в ма-
ленькие губки. Интересно, что при сме-
шении фильтратов, полученных от раз-
ных губок, собираются вместе лишь од-
нородные клетки, формируя губки со-
ответствующего впда.
Несомненно, приведенные опыты в рав-
ной, если не в большей, степени характе-
ризуют также процесс искусственно вы-
званного бесполого размножения губок,
который, как мы уже знаем, очень часто
происходит путем скопления воспроизво-
дящих клеточных масс. И в этом прояв-
ляется особенность регенеративных про-
цессов у губок. Они настолько тесно пере-
плетаются с явлением бесполого размно-
жения, что порой трудно провести между
Рис. 132. Колониальные и вторичноодиночные губки:
1 — колония с хорошо обособленными особями (Sycon cilia turn);
2 — аморфная колония с крайней степенью слияния особей (Му-
cale ochotensis); 3—5 — высокоиндивидуализированные колонии
губок (Geodia phlegraei, Chondrocladia gigantea, Semisuberites arcti-
ca), 6, 7— вторичноодиночные губки (Tentorium semisuberites, Po-
lymastia hemisphaericum).
ними четкие границы, подобно тому как
трудно установить иногда разницу между
обычным ростом и размножением путем
почкования. В особенности это касается
колониальных губок, не имеющих опре-
деленной формы тела.
Поэтому нередко при повреждении губ-
ки процесс, начавшийся как восстано-
вительный, завершается бесполым раз-
множением. Так, наблюдали, как на по-
верхности губки морской каравай (Hali-
chondria panicea) на месте глубокой ра-
ны при восстановлении поврежденных
частей формировались многочисленные
устьевые отверстия и папиллы. Извест-
но также, что при определенных услови-
ях искусственно можно вызвать почкооб-
разование у известковых и пресновод-
ных губок в результате механического
повреждения или ожога.
203
В наиболее чистом виде регенерацион-
ный процесс можно наблюдать на оди-
ночных губках или при повреждении
каких-либо оформленных образований
(устьевых трубок, или папилл, дермаль-
ной мембраны) на теле колониальных
губок. Вообще губки довольно легко
регенерируют поврежденные участки на
поверхности тела. Рана быстро заруб-
цовывается, затягивается мембраной, и
восстанавливается прежняя структура,
так что очень скоро место повреждения
становится незаметным. Неглубокий раз-
рез через губку морской каравай, на-
пример, ликвидируется через 5—7 дней,
а отверстие около 1 кв. мм, проделанное
вблизи устья известковой губки (Leuco-
solenia), зарастает за 7—10 дней. Одна-
ко при более существенных поврежде-
ниях восстановительный процесс нередко
протекает очень медленно. Так, если у
одиночной известковой губки (Sycon) от-
резать верхнюю часть, несущую устье,
то у оставшегося основания губки за
сутки регенерирует дермальная мем-
брана и образуется новое устье; но толь-
ко через 15 дней здесь образуются жгу-
тиковые трубки. При более значительном
и глубоком повреждении тела губки
морской каравай заживление также
протекает довольно медленно и к тому
же нередко восстановление происходит
не полностью. Очевидно, большая реге-
неративная способность губок не может
здесь проявиться в достаточной степени
ввиду того, что целостность, или степень
интеграции, самих губок как многоклеточ-
ных животных еще крайне незначи-
тельна.
При разрезании губки на две части и
доследующем их тесном соединении про-
исходит очень быстрое срастание этих
частей друг с другом. Срастаться могут
также куски, взятые от разных экземп-
ляров одного и того же вида. Харак-
терно, что в некоторых случаях, когда
разрез прошел через устьевую папиллу,
при срастании вместо одной формируют-
ся две маленькие папиллы, т. е. регене-
рация завершается образованием новой
особи колонии. Живая губка может быть
разрезана на много кусков, и каждый
кусок сохраняет жизнедеятельность. На
поврежденной его поверхности восстанав-
ливается дермальная мембрана, образует-
ся устье, и каждый отрезанный кусочек
продолжает в дальнейшем свое сущест-
вование и рост, как целая губка.
На способности губок к регенерации
основан метод искусственного разведе-
ния промысловых туалетных губок. Та-
кую губку разрезают на куски и при-
крепляют их проволочкой к какому-либо
твердому субстрату. Чаще всего для
этого применяют специальные цемент-
ные диски, которые помещают на дне.
Иногда кусочки губки нанизываются на
шнур, натянутый в горизонтальном поло-
жении у самого дна между двумя колья-
ми. Через несколько лет из кусочка
губки вырастает экземпляр, достигаю-
щий промыслового размера. Правда, от-
мечают, что при подобном способе раз-
множения губка растет значительно мед-
леннее, чем при развитии ее из личинки.
ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ ГУБОК
Продолжительность жизни, или воз-
раст, которого достигают губки, колеб-
лется у разных видов от нескольких не-
дель и месяцев до многих лет. Известко-
вые губки обычно живут в среднем до
одного года. Некоторые из них (Sycon
coronatum, Grantia compressa) отмирают
сразу же по достижении половой зрело-
сти, как только сформировавшиеся ли-
чинки нового поколения покидают их
организм. Большинство мелких четырех-
лучевых и кремнероговых губок живет
в пределах 1—2 лет. Более крупные
стеклянные и обыкновенные губки отно-
сятся к долгоживущим организмам. Осо-
бой долговечностью отличаются те из них,
которые достигают величины 0,5 д и
более. Так, экземпляры конской губки
(Hippospongia communis) около 1 м в
диаметре, по мнению специалистов, дости-
гают возраста не менее 50 лет.
В общем губки растут довольно мед-
ленно. Наибольшая скорость роста
у форм с коротким сроком жизни. Не-
которые известковые губки (Sycon cilia-
tum) за 14 дней вырастали до 3,5 см в
высоту, т. е. достигали почти максималь-
204
ной своей величины. Отделившаяся поч-
ка морского апельсина приобретает раз-
меры материнского организма B—3 см в
диаметре) в течение одного месяца. На-
оборот, долгоживущая конская губка
вырастает до 30 см в диаметре за 4—7
лет. Надо полагать, что и другие круп-
ные морские губки имеют примерно та-
кую же скорость роста. Конечно, в каж-
дом отдельном случае скорость роста и
продолжительность жизни губок во мно-
гом зависят от различных факторов
внешней среды, в том числе от обилия
пищи, температурных условий и пр.
Пресноводные губки сравнительно не-
долговечны и живут обычно всего не-
сколько месяцев. Но в некоторых случаях
они способны создавать многолетние об-
разования особого рода. Такие образо-
вания, достигающие значительной вели-
чины и веса более 1 кг, состоят из внут-
ренней массы отмерших частей губки,
покрытых снаружи жизнедеятельным
слоем. Происходит это следующим образом.
Возникшая половым путем личинка губ-
ки прикрепляется к подходящему субст-
рату и вырастает в небольшую коло-
нию. Образовав геммулы, такая губка
отмирает. По прошествии некоторого
времени с наступлением благоприятных
условий из геммул выходят молодые губ-
ки. Они поднимаются на поверхность
отмершей губки и, сливаясь друг с дру-
гом, образуют молодую колонию. Та-
кая восстановленная колония, достиг-
нув определенного возраста, присту-
пает к половому размножению. Позднее
внутри нее формируются новые гемму-
лы, а сама губка отмирает. На следую-
щий год цикл повторяется, и таким об-
разом постепенно могут создаваться
объемистые колонии пресноводных губок.
РАСПРОСТРАНЕНИЕ И ЭКОЛОГИЯ ГУБОК
В настоящее время насчитывают до
5000 видов губок. В наших арктических
и дальневосточных морях известно около
350 видов; примерно такое же количест-
во их обитает в антарктических водах.
Наибольшего же разнообразия губки
достигают в тропических и субтропиче-
ских районах Мирового океана. Иначе
обстоит дело с плотностью и массовостью
поселений губок на дне. Большие скоп-
ления губок одного или нескольких ви-
дов известны во многих местах северного
и южного полушария. Поскольку губки
ведут сидячий, прикрепленный образ жиз-
ни, они нуждаются для своего развития
и роста в твердом субстрате. Поэтому
скопления губок наблюдаются там, где на
дне имеется много каменистого материа-
ла: валунов, камней, гальки и пр. Вдаль-
невосточных морях встречаются обшир-
ные участки дна, где образуются сплош-
ные заросли кремнероговых губок во-
ронковидной, стебельчатой или иной фор-
мы. Рыбакам, промышляющим рыбу у
Кольского полуострова (в Баренцевом
море), хорошо известны места скопле-
ний крупных шаровидных губок геодий.
Попадая в трал иногда в количестве
свыше тонны, такие геодий сплошь за-
бивают сети и ввиду наличия у них
щетинистого покрова с большим тру-
дом извлекаются из трала. Это, не-
сомненно, создает помехи в работе рыба-
ков, не говоря уж о том, что в таких
случаях рыбы в тралах почти не бывает.
Подобные же геодиевые банки имеются
у берегов Норвегии, Исландии, у восточ-
ных берегов Северной Америки и в дру-
гих местах. Богаты и качественно раз-
нообразны массовые поселения губок в
Средиземном и Карибском морях, в райо-
не островов Малайского архипелага и в
ряде других мест, где на дне имеются ка-
менистые грунты. По обширности участ-
ков дна, занятых скоплениями губок,
пожалуй, первое место занимают антар-
ктические воды. Фактически вокруг все-
го антарктического материка на глубине
100—500 м расположен широкий пояс мас-
совых поселений губок. Это вполне по-
нятно, поскольку многочисленные айс-
берги и ледники постоянно выносят в
океан и откладывают на дне большое
количество камней и валунов, что создает
благоприятные условия для развития
здесь фауны губок. Преобладающее зна-
чение в образовании массовых скопле-
ний губок в Антарктике приходится на
долю крупных стеклянных губок из
семейства Rossellidae.
205
Рис. 133. Глубоководные губки:
I, 2 — губки гиалонемы (Hyalostylus dives, Hyalonema elegans); 3—6 — морские ершики (Cladorhiza
longipinna, Axoniderma mirabile, Asbestopluma sp., Cladorhiza rectangularis).
На дне морей губки могут встречаться
на различной глубине, начиная от при-
ливо-отливной зоны вплоть до боль-
ших глубин океана. Значительная часть
губок относится к сравнительно мелко-
водным существам, обитающим главным
образом на глубине до 400—500 м.
С увеличением глубины количество видов
губок уменьшается, а на глубине свыше
1000—1500 м, как правило, губки встре-
чаются довольно редко. В общем извест-
ковые губки в своем распространении
приурочены к мелководью; стеклянные
губки, наоборот, можно считать глубо-
ководными организмами. Обыкновенные
губки — четырехлучевые и кремнерого-
вые — имеют более широкий диапазон
вертикального распространения, встре-
чаясь как на мелководье, так и на
больших глубинах.
Недавними морскими исследованиями
установлено, что некоторые виды губок
живут даже на дне океанических впадпн,
т. е. на глубине свыше 6 тыс. метров. Эти
губки приспособлены к жизни в таких
условиях, где господствует близкая к ну-
лю положительная или отрицательная
температура воды, очень высокое дав-
ление, а дно обычно илистое. Глубо-
ководные губки отличаются вытянутой
формой тела или часто сидят на длинных
ножках, что предохраняет их от заиле-
206
ния. Такими наиболее характерными фор-
мами являются различные морские ер-
шики (Asbestopluma, Cladorhiza, Chond-
rocladia и др.) из числа кремнероговых
губок и стеклянные губки — гиалонемы
(Hyalonema).
Опасность заиления для губок сущест-
вует и на менее значительной глубине.
Здесь губки также имеют ряд приспособ-
лений, помогающих им при неподвижном
образе жизни избегать заиления. Кроме
вытянутой формы тела, этой цели отве-
чает образование на поверхности неко-
торых подушковидных губок полима-
стий особых длинных выростов тела —
п а п и л л, на концах которых помещаются
поры или устья: тем самым жизненно
важные отверстия губок, с помощью ко-
торых осуществляется их связь с внеш-
ней средой, значительно приподняты над
илистым дном. Несомненно также, что
дисковидная форма тела ряда полима-
стий, лежащих свободно на дне, являет-
ся приспособлением, помогающим им
удерживаться на поверхности мягкого
грунта. Нередко эти же губки приобре-
тают вокруг дисковидного или подушко-
видного тела краевой венчик из длинных
игл, что еще больше увеличивает сопро-
тивляемость губки к погружению ее в
илистый грунт.
Большое влияние на распространение
губок оказывают температура и соленость
воды. В целом губки — теплолюбивые
животные, и поэтому в суровых услови-
ях Арктики и Антарктики разнообразие
видов сравнительно невелико. Отрица-
тельная температура воды на дне пре-
пятствует размножению и развитию в
этих районах значительной части гу-
бок. Но некоторые из них довольно хо-
рошо приспособились к жизни в та-
ких крайних температурных условиях
существования. Об этом свидетельствуют
уже упоминавшиеся массовые поселения
крупных стеклянных губок Росса, об-
наруженные в антарктических водах.
Очень ярким примером зависимости рас-
пространения губок от температуры мо-
гут служить некоторые атлантические виды,
которые проникают далеко в Арктику,
следуя за теплым течением Гольфстрим.
Там, где благодаря этому течению на дне
преобладает сравнительно высокая тем-
пература воды, выше 2—3°С, обнаружи-
вают и эти атлантические теплолюбивые
виды; они встречаются даже у северных
берегов Шпицбергена. Известно, что об-
разование рогового вещества у губок на-
ходится в прямой зависимости от темпе-
ратуры воды. Поэтому настоящие рого-
вые губки обильно представлены лишь
в тропических и субтропических водах,
в поясе от экватора на север и на юг до
45° широты. За пределами этих границ
губки бедны спонгином, а настоящие
роговые губки почти не встречаются.
С другой стороны, замечено, что по на-
правлению от экватора на север и на
юг размеры игл губок в общем становят-
ся крупнее, что также связывают с темпе-
ратурными условиями. Температура во-
ды, несомненно, сказывается на цикле
развития пресноводных губок. Образо-
вание геммул у этих губок в северных
странах вызвано исключительно пони-
жением температуры воды в осенне-
зимнее время. Наоборот, в тропиках и
субтропиках неблагоприятными для су-
ществования пресноводных губок счита-
ют наиболее жаркие месяцы, когда обра-
зуются геммуллы и отмирают губки.
Не менее важным фактором, опреде-
ляющим существование губок, является
соленость воды. Морские губки лучше
всего развиваются в воде с нормальной
морской соленостью. Опреснение воды
приводит к их вымиранию, сокращению
численности видов и особей. Так, если в
Баренцевом море обитает около 150 ви-
дов губок, то в несколько опресненном
Белом море количество их определяет-
ся всего 50 видами; в еще более опрес-
ненном Черном море — около 30 видов
губок, а в Каспийском море — всего не-
сколько близких разновидностей одного
вида; в Балтийском море, в большей его
части, морские губки совершенно не
представлены. Пресноводные губки в
свою очередь не выносят морской воды
и лишь заходят в эстуарии рек (впадаю-
щих, например, в Финский залив, Ка-
спийское и Черное моря).
Исключительное значение для губок
имеет движение воды. С водой приносят-
ся к губке различный пищевой материал
207
и вещества, необходимые для ее роста и
нормального функционирования. В то
же время, что особенно важно, с током
воды удаляются выделяемые губкой про-
дукты жизнедеятельности, которые ока-
зываются очень вредными в первую оче-
редь для самих губок. Поэтому лучше
всего губки развиваются в местах, где
имеется достаточно интенсивная смена
окружающей воды. Многие губки очень
хорошо приспособлены к жизни при
сильных течениях. Они плотно прирас-
тают основанием к твердому субстрату,
имея вид невысоких подушек или корок
либо вытянутых в длину пальцевидных
стебельчатых образований, очень эла-
стичных и гибких. Такая форма тела
и эластичность позволяют губке противо-
стоять напору движущейся воды. Даже
в прибрежной зоне на скалистых бере-
гах в условиях сильного прибоя успеш-
но развивается фауна подушковидных
или корковых губок. Правда, сильный
шторм все-таки может приносить зна-
чительные опустошения в зарослях гу-
бок. На морском берегу, в особенности
в местах открытых, часто можно на-
ходить губок, выброшенных во время
шторма. Известны случаи, как, напри-
мер, у берегов Бискайского залива, когда
после очень сильного шторма губка
морской каравай была выброшена на
берег в таком большом количестве, что
местные жители вывозили разлагающую-
ся массу губок тачками, используя ее
как удобрение на своих полях. На по-
бережье тихоокеанских островов после
сильных ураганов, или тайфунов, нахо-
дят иногда в виде выбросов даже экземп-
ляры стеклянных губок, которые обыч-
но обитают на глубине свыше 100 м.
Форма тела губок зависит от преобла-
дающих течений, характера субстрата,
глубины и других причин. На большой
глубине, где колебания температуры не-
значительны и течение воды очень слабо,
форма тела губок оказывается более по-
стоянной и более симметричной и пра-
вильной. В самом деле, роговые губки,
обычно не встречающиеся на глубине
свыше 200 м, редко имеют постоянную
или правильную форму; наоборот, глу-
боководные стеклянные губки, как пра-
вило, вполне определенны по внешнему
виду. То же следует сказать про другие,
кремнероговые губки: глубоководные
виды имеют форму тела значительно
более правильную и постоянную, чем
виды, обитающие на мелководье.
Примитивность губок проявляется в
чрезвычайной изменчивости формы их
тела даже у представителей одного и
того же вида. В особенности это касает-
ся губок колониальных. Наблюдения за
некоторыми кремнероговыми губками по-
казали, что одна и та же губка, имею-
щая в спокойной воде залива или бухты
вид вееровидной или складчатой пласти-
Таблица 6. Губки.
Известковые губки:
i — Clathrina primordialis;
2 — Clathrina blanca;
3 — Leucosolenia botryoides;
4 — Leuconia aepera;
5, 6— Clathrina primordialis;
7 — Grantia mirabilis.
Стеклянные,
или шестилучевые, губк!
8 — Pheronema giganteunr,
9 — Farrea haeckelii;
10 — корзинка Венеры (Euplectella
aspergillum);
11 — Monorhaphis chuni (часть);
12 — губка Росса (Rossella);
13 — губка гиалонема (Hyalonema
sieboldi).
ны или пучка сплюснутых ветвей, осно-
вания которых срастаются в широкие
пластины, в бурных водах открытого
берега представлена формами пальча-
тыми, с узкими цилиндрическими ветвя-
ми. Внешняя форма может варьировать
и в соответствии с местом, на котором
растет губка. Морской каравай, широ-
ко распространенная губка мелково-
дья, на ровном месте имеет тело в виде
плоской корки или подушечки, от ко-
торой поднимается кверху ряд низких
трубчатых выростов, несущих на кон-
цах устья; обрастая водоросли, эта губ-
ка имеет вид неправильных лопастных
комков или даже разветвленных веточек.
При росте в узком пространстве между
большими камнями трубчатые выросты
значительно увеличиваются, пока не зай-
мут практически все тело губки, которая
представлена теперь рядом длинных вер-
тикальных трубок, открывающихся на
свободном конце широким устьем. Не
менее интересны наблюдения за изме-
нением формы тела у некоторых пред-
ставителей четырехлучевых губок, кото-
рые обычно растут на каком-нибудь твер-
дом субстрате. Таковы многие губки —
полимастии. Когда личинки их посе-
ляются на большом камне, из них выра-
стают губки подушковидной формы, осно-
ванием своим плотно обрастающие суб-
страт. Если же вблизи плавающей личин-
ки не оказывается такого подходящего
твердого субстрата и личинка оседает
на мелкую гальку или частицу гравия,
то при росте губки края ее поднимаются,
и она приобретает чашевидную форму.
О большой пластичности губок свиде-
тельствуют интересные опыты по изме-
нению ориентации тела губок в простран-
стве. Если губку морской каравай от-
делить от субстрата и содержать в аква-
риуме в таком положении, чтобы устья
ее были обращены вниз, то со временем
произойдет полная перестройка орга-
низма губки. На бывшей базальной сто-
роне ее, обращенной теперь в сторону
поверхности воды, формируются новые
устьевые трубки, а старые устья пере-
стают функционировать и зарастают.
Губки, обитающие в приливо-отлив-
ной зоне моря, хорошо приспособлены
к непродолжительному пребыванию на
воздухе, когда во время отлива они вы-
ступают из воды. Устья и поры таких
губок закрываются, что предохраняет
их от излишней потери влаги и высыха-
ния. После прилива губка открывает
свои отверстия и продолжает нормаль-
но функционировать. При более дли-
тельном нахождении на воздухе неко-
торые губки, как известно, образуют
редукционные тела, которые, попав
в воду, снова развиваются в маленькие
губочки.
Таблица 7. Губки.
Четырехлучевые губки:
1 — Polymastia corticata;
2 — Tetilla cranium;
3 — геодия (Geodia macandrewii);
4 — Disyringa dissimilis,
5 — чаша Нептуна (Poterion neptuni).
Кремнероговые губки:
6 — байкальская губка (Lubomirskia
baikalensis);
7 — морской каравай (Halichondria
panicea);
8 — Semisuberites arctica;
9 — роговая губка (Hircinia
variabilis).
14 Жизнь животных, т. 1
Рис. 134. Сожительство губок с другими животными:
1 — стеклянная губка Euplectella oweni и живущие в ее полости самец и самка рачков Spongicola
venusta; 2 —стеклянная губка гиалонема (Hyalonema indicum), на корневом пучке которой поселились
коралловые полипы (Palythoa); 3 — губка Dysidea fragilis, растущая на панцире живого краба Pisa
gibbsi; 4 — губка Halichondria sitiens на раковине двустворчатого моллюска; 5 — губка миксилла
(Myxilla parasitica), обросшая раковину моллюска из рода Chlamis; 6— роговая губка Cryptospongia
enigmafica, растущая на трубке многощетинкового червя Potamilla symbiotica.
Замечательна способность пресновод-
ных губок к перенесению неблагоприят-
ных условий существования. Их гемму-
лы очень хорошо сохраняются даже в те-
чение нескольких лет. При полном высы-
хании водоема, например, они могут са-
мым различным способом, в том числе с
помощью ветра или прикрепившись к
ногам птиц, переноситься в другие места.
И если такие геммулы попадают в воду,
они дают начало новому поселению губок.
При одном из сильнейших извержений
вулкана Кракатау в результате оседа-
ния в окрестностях его большого коли-
чества пепла вся пресноводная фауна
губок соседних водоемов оказалась погиб-
шей. Тем не менее через некоторое время
пресноводные водоемы вновь были засе-
лены губками, очевидно, благодаря со-
хранившимся здесь геммулам.
Цвет губок, их окраска находятся в
прямой зависимости от света. Как пра-
вило, губки, живущие на глубине менее
100 м и, следовательно, в той или иной
мере находящиеся под воздействием све-
та, более ярко окрашены. Глубоковод-
ные губки обычно лишены пигмента.
Враги губок. У губок сравни-
тельно немного врагов. Очень хорошей
защитой им от нападения хищников слу-
жит минеральный скелет, состоящий из
большого количества игл. Поэтому жи-
210
вотных, питающихся губками, очень ма-
ло. В желудках некоторых рыб и тю-
леней находят остатки губок, в особен-
ности таких, как пробковая губка, мор-
ской каравай, мясистая губка и др. Но
вряд ли в пищевом рационе этих живот-
ных губки играют сколько-нибудь суще-
ственную роль. Значительно больше
мелких животных, которые паразити-
руют на губках или внутри их. Это не-
которые голожаберные моллюски, го-
лотурии, ракообразные. К их числу
присоединяются также личинки ручей-
ников и других насекомых, личинки
водяных клещей, встречающиеся на пре-
сноводных губках. Эти животные в той
или иной степени используют тело губок
в пищу. Правда, существенного вреда
они губкам, как правило, не приносят
и обычно не вызывают их гибели. Вы-
еденные и поврежденные участки тела
быстро зарубцовываются или восстанав-
ливаются.
Симбиоз губок с другими орга-
низмами. Очень многие животные, встре-
чающиеся на поверхности и внутри
губок, вступают с ними в безобидные
или обоюдовыгодные отношения. Это
комменсалы (квартиранты, или
нахлебники) и симбионты (сожите-
ли). Первые из них используют тело губок
как убежище и защиту от врагов. Та-
ковы многие ракообразные, некоторые
многощетинковые черви, змеехвостки
и др. Нередко в тканях губок отклады-
вают яйца другие животные, как, напри-
мер, мелкие головоногие моллюски, мно-
гощетинковые черви, некоторые рачки,
рыбы и др. Известным примером сим-
биоза может служить сожительство рака-
отшельника с пробковой губкой (Suberi-
tes domuncula). Губка поселяется на не-
большой пустой раковине брюхоногого
моллюска. В эту же раковину залезает
молодой рак-отшельник, пользующийся
ею для защиты своей мягкой задней
части тела. Постепенно вся раковина
обрастает губкой, остается свободным
лишь вход в жилище рака-отшельника.
Растет губка, растет и рак-отшельник.
В результате внутри губки образуется
спиральная полость, в которой живет
рак-отшельник, выставив наружу перед-
нюю часть тела. При необходимости он
может полностью скрываться внутри губ-
ки. Такой симбиоз выгоден обоим жи-
вотным: рак-отшельник имеет надежное
убежище, а губка получает возможность
передвижения.
Биологические взаимоотношения меж-
ду ракообразными и губками отличают-
ся особенным разнообразием. Наблю-
дается около 500 различных случаев
более или менее тесных связей, которые
рассматриваются как явления коммен-
сализма, симбиоза, паразитизма и отно-
шения смешанного характера. Многие
мелкие веслоногие и равноногие рачки,
бокоплавы и близкие к ним морские ко-
зочки квартируют или паразитируют в
губках. Они встречаются как внутри —
в каналах и полостях губок, так и на
поверхности — в многочисленных ямках
и углублениях, питаясь отмершими ча-
стями тела губки. Иногда такие рачки
встречаются в очень большом количестве
и насчитываются десятками и сотнями
экземпляров в одной губке.
Различные десятиногие раки кре-
ветки используют губок в основном как
убежище от врагов. При этом рачки вы-
ходят из полости губок лишь ночью и
редко покидают их поверхность. Инте-
ресно, что обитающие в губках рач-
ки по сравнению со свободно живущи-
ми близкими родичами откладывают зна-
чительно меньше яиц. Но зато яйца их
гораздо большего размера, что сказы-
вается на сокращении личиночного пе-
риода рачков, а стало быть, и времени,
которое они проводят вне защиты губок.
При небольшом количестве яиц такое ус-
коренное развитие рачков, несомненно,
важное приспособление, отражающееся
на сохранности потомства. Часто кре-
ветки и бокоплавы встречаются в по-
лости губок парами. Очень любопытные
отношения сложились между десятино-
гими рачками спонгикола (Spongicola
venusta) и стеклянной губкой, известной
под названием корзинка Венеры (Eup-
lectella oweni). В обширной полости поч-
ти каждой губки живет пара таких рач-
ков, самец и самка. Они забираются
туда в стадии личинки и во взрослом
состоянии уже не могут выйти на сво-
14*
211
боду через решетчатый остов губки. Так
эти рачки всю жизнь и проводят вместе,
заключенные в полости губки. В связи с
этим в Японии, у берегов которой
эта губка обитает, издавна существует
обычай дарить при свадебных церемо-
ниях экземпляры губок с находящейся
внутри парой рачков как символ вечной
и нерушимой любви и супружеской вер-
ности.
Довольно забавно используют непри-
влекательность губок для хищников не-
которые крабы. Они таскают на своей
спине куски губок, поддерживая их зад-
ней парой конечностей. Такие крабы
(Dromia) отрезают клешнями соответ-
ствующие величине своего тела куски
губок и общипывают эти куски так, что
они помещаются у них на спине. Со
временем мягкая и эластичная губка,
прижатая к телу краба, приобретает в
этом месте форму панциря. Губка служит
крабу своего рода щитом, предохраняю-
щим его от нападения врагов. Вполне
вероятно, что здесь имеет значение также
неприятный отпугивающий запах губок.
При появлении крупного хищника краб
иногда сбрасывает со спины губку, от-
влекая внимание врага, а сам быстро
скрывается. Избегнув опасности, краб
отыскивает другую подходящую губку
и приспосабливает ее таким же образом
на свою спину. Иногда он использует
пробковую губку с сидящим в ней раком-
отшельником. Часто на панцире некото-
рых крабов находят плотно приросших к
ним губок. В данном случае крабы также
активно действуют, откусывая клешнями
кусочки губки и пристраивая их на своей
спине до тех пор, пока они не прирастут
к панцирю.
Гораздо реже на губках поселяются
сидячие, прикрепленные животные. К их
числу, например, относятся различные
усоногие раки (Cirripedia), в частности
морские желуди, или балянусы (Balanus).
Эти рачки имеют наружный скелет из
известковых пластинок, образующих
своеобразный домик; верхние пластин-
ки домика могут расходиться, и в поя-
вившееся отверстие рачок высовывает
наружу свои передние конечности, на-
ходящиеся в непрерывном движении.
Балянусы чаще всего встречаются на
комкообразных губках, которые в про-
цессе естественного роста развивают во-
круг них образования, подобные галлам
растений. Отдельные губки могут нести
свыше сотни таких галлов. Балянусы
достаточно долго сопротивляются пол-
ному обрастанию их губкой, двигая ко-
нечностями, снабженными зубчиками.
Так или иначе до полного обрастания
губкой балянусы успевают достичь по-
ловозрелости и дать потомство. То по-
стоянство, с которым некоторые виды
балянусов поселяются на губках, за-
ставляет предполагать, что обитание на
губках дает им ряд выгод. Очевидно,
имеют значение такие моменты, как за-
щита от врагов, токи воды, создаваемые
губками, слабая пищевая конкуренция
с ними. Конечно, никакой пользы эти
балянусы губкам не приносят. Чрезмерное
засилье балянусов вызывает иногда не-
способность губок к половому размно-
жению и даже их гибель. Другие пред-
ставители усоногих раков, так называе-
мые морские уточки (Lepas), регулярно
встречаются на длинном корневом пучке
игл некоторых стеклянных губок — гиа-
лонем, который раки используют как
очень подходящий субстрат, возвышаю-
щийся над сильно заиленным дном. На
этом же корневом пучке игл могут посе-
ляться колониальные кораллы (Palythoa),
образующие вокруг него плотный чехол
со многими сидящими на нем небольши-
ми полипами. Исключительно постоянно
поселяется на некоторых губках (Axi-
nella) корковая актиния (Parazoanthus
axinellae). Личинки ее обладают изби-
рательной способностью к субстрату,
оседая именно на тех губках, поверхность
которых обычно используется ими в ка-
честве места поселения. Губка играет
здесь пассивную роль и, очевидно, ни-
какой выгоды от такого квартиранта не
имеет.
В очень тесный контакт вступают губ-
ки также с некоторыми гидроидными
полипами, что приводит к особым обра-
зованиям на теле губок, меняющим их
облик. Так, на некоторых стеклянных
губках (Walteria) в углублениях поверх-
ности их тела сидят отдельные полипы,
212
общий ствол которых распространен
внутри тела губки. Вокруг каждого из
этих полипов губка образует защитный
спонгиновый чехол в виде трубки.
Некоторые животные в качестве защи-
ты используют даже отдельные иглы гу-
бок, которые после их гибели в массе
откладываются на дне водоема. Эти жи-
вотные строят свои домики частично или
полностью из игл губок (фораминиферы,
черви и пр.). Интересна голотурия
псевдостихопус (Pseudostichopus trachus),
у которой наружный скелет целиком об-
разован из длинных игл губок, покры-
вающих тело голотурии густым ворси-
стым войлоком, что служит ей хорошей
защитой от врагов.
Иногда губки сами селятся на живом
субстрате. Известно, что пресноводными
губками часто обрастают стебли трост-
ника, кувшинок, осоки; губки встре-
чаются на моховых подушках, обнажен-
ных корнях деревьев и т. п. Некоторы-
ми морскими губками обрастают корни
крупных водорослей ламинарий, в ре-
зультате чего здесь создаются исключи-
тельно благоприятные условия для раз-
вития характерных сообществ донных
животных (червей, моллюсков, мелких
ракообразных, голотурий и др.)- Губки
могут расти также на животных. Дву-
створчатые моллюски, например, в осо-
бенности сравнительно крупные, очень
часто являются субстратом, на котором
охотно поселяются различные губки. В
некоторых случаях губки обрастают в ви-
де невысоких подушек верхние створки
вполне определенных видов моллюсков,
вступая с ними, очевидно, в какие-
то симбиотические отношения. Кроме
упомянутого выше случая, когда крабы
сами культивируют на спине кусочки
губок, наблюдаются примеры самостоя-
тельного поселения губок на их панцире.
На спине некоторых крабов иногда вы-
растает такая большая колония губки,
что она почти совершенно скрывает под
собой краба. Чрезвычайно характерен
постоянный симбиоз роговой губки
криптоспонгии (Cryptospongia enigmati-
са) и многощетинкового червя (Potamilla
symbiotica). Эта губка обитает на зна-
чительной глубине, имеет очень мягкое
лепешковидное тело. Она не смогла бы
существовать на илистом грунте, если
бы не прикреплялась к трубке червя,
которая высоко торчит над дном, что
спасает губку от заиления. Вместо труб-
ки червя эта губка иногда использует
веточки гидроидных полипов.
Нередко губки поселяются на поверх-
ности других губок. Большей частью
такие поселения носят случайный ха-
рактер. Плавающая личинка губки за
неимением подходящего твердого суб-
страта оседает на поверхность другой
губки. Это может привести к образованию
сложных сообществ губок. Так, у бере-
гов Японии был поднят со дна моря
большой экземпляр стеклянной губки
(Chonelasma calyx), на котором росло 13
других стеклянных губок, 4 известко-
вых, несколько корковых и подушковид-
ных четырехлучевых и кремнероговых
губок; здесь же было обнаружено свыше
70 мелких плеченогих, двустворчатых и
брюхоногих моллюсков, некоторое ко-
личество офиур и мшанок.
Губки, в особенности пресноводные,
часто находятся в симбиотических отно-
шениях с различными водорослями. Очень
характерны мелкие одноклеточные водо-
росли — зоохлореллы, встречающиеся в
большом количестве в протоплазме кле-
ток губок. Именно эти водоросли обус-
ловливают зеленую окраску большинст-
ва пресноводных губок. Зоохлореллы
поступают в организм губки вместе с то-
ком воды и захватываются клетками по-
добно другим пищевым частицам. Но в
отличие от последних водоросли не под-
вергаются немедленному перевариванию,
а продолжают жить в клетках губки.
Губка использует кислород, выделяе-
мый водорослями, и избыток веществ,
ассимилируемых ими на свету. Водорос-
ли же находят здесь защиту, питательные
соли и углекислоту, выделяемую губкой
при дыхании. Но в конце концов эти
водоросли постепенно перевариваются в
протоплазме клеток губок. С другой
стороны, нитчатые водоросли, живущие
в мезоглее некоторых пресноводных гу-
бок, иногда разрастаются до такой сте-
пени, что закупоривают каналы и вы-
зывают гибель губки.
213
СВЕРЛЯЩИЕ ГУБКИ
Очень интересный образ жизни ведут
четырех лучевые губки клионы (Cliona).
Их называют еще сверлящими губками,
так как они обладают способностью про-
делывать ходы в твердом известковом
субстрате. Довольно часто таким субст-
ратом являются раковины моллюсков,
массивный скелет кораллов, известковые
водоросли или имеющиеся на дне морей
какие-либо известковые породы. Личин-
ка клионы прикрепляется к подобному
субстрату, и одновременно с превра-
щением ее в маленькую плоскую губку
начинается процесс «сверления» изве-
сткового материала. Вначале образует-
ся небольшое углубление, куда захо-
дит тело губки. Затем по мере роста
клионы в толще субстрата, в основном
вблизи его поверхности, проделывают-
ся ходы, или каналы, сообщающиеся
между собой. Мягкое и нежное тело губ-
ки, обычно желтоватого или коричне-
ватого цвета, заполняет эти каналы, а
изнутри их в сторону поверхности про-
сверливаются округлые отверстия, при
помощи которых губка сообщается с
внешней средой. Из этих отверстий слег-
ка выступают небольшие сосочковидные
выросты тела губки, папиллы, несущие
на концах устья или поры.
О поражении известкового субстрата
сверлящей губкой можно судить по на-
личию на его поверхности округлых от-
верстий, имеющих в среднем около 1 мм
в диаметре.
Благодаря деятельности сверлящих
губок известковый субстрат в значитель-
ной степени разрушается, а разросшаяся
губка уже не помещается в проделанных
ею каналах. Тогда клиона начинает
расти как обычная губка, образуя поду-
шковидные или комкообразные наросты,
иногда достигающие величины 30 см и
более. В некоторых случаях губка мо-
жет выходить наружу еще до разрушения
ею известкового субстрата. Клионы раз-
множаются не только половым путем.
По отмирании губки в каналах извест-
кового субстрата можно обнаружить ша-
ровидные красноватого цвета геммулы.
При благоприятных условиях из них
выходят маленькие губки, также обла-
дающие способностью к сверлению извест-
кового субстрата.
Скелет клион состоит из мелких изо-
лированных кремниевых игл микроскопи-
ческого размера. Раньше даже предпола-
гали,что процесс сверления осуществляет-
ся благодаря механическому действию
этих иголочек. Однако подобная мысль
была отвергнута, когда обнаружили, что
уже личинка клионы, еще не имеющая
скелетных образований, оказывает свер-
лящее воздействие на известковый суб-
страт. При более тщательном наблюде-
нии за деятельностью клион было уста-
новлено, что процесс сверления ими из-
весткового материала заключается в не-
прерывном «выгрызании» очень мелких
чешуек извести. Эти чешуйки скапли-
ваются у оснований устьевых отверстий
и затем выводятся наружу. Но каким
образом происходит это отделение мел-
ких чешуек от известкового субстрата,
окончательно не установлено. Правда,
наблюдалось, что амебоидные клетки ме-
зоглеи, приближаясь к поверхности губ-
ки, направляют в сторону известкового
субстрата нитевидные ложноножки. Поэ-
тому в настоящее время полагают, что
механизм сверления у клион состоит в
одновременном воздействии на субстрат
углекислоты, выделяемой отдельными по-
верхностными клетками губок, и сокра-
тимых усилий этих клеток.
Сверлящие губки являются опасны-
ми вредителями устричных банок. По-
селяясь на раковинах устриц и делая в
них ходы, клионы вызывают так назы-
ваемую пряничную болезнь устриц, ко-
торая приводит их в конце концов к ги-
бели. Тем самым сверлящие губки при-
носят определенный вред человеку, про-
изводя огромные опустошения устрич-
ных плантаций. Для уничтожения этих
вредителей устриц применяются различ-
ные способы борьбы, основанные на зна-
нии их биологии. Известно, что клионы,
являясь морскими животными, не вы-
носят пресной воды. Поэтому поражен-
ных пряничной болезнью устриц на не-
которое время погружают в пресную
воду. В результате такого купания свер-
лящие губки обычно погибают. Другой
214
способ, ограничивающий развитие пря-
ничной болезни устриц, заключается в
создании вокруг их поселения защитного
вала из пустых раковинок моллюсков.
На эти раковины сверлящие губки на-
падают в первую очередь, оставляя не-
тронутой большую часть живых устриц.
Один из новейших способов предохране-
ния устричных плантаций от нападения
сверлящих губок состоит в подборе для
разведения устриц подходящего неизвест-
кового субстрата. Дело в том, что при
создании искусственных устричных ба-
нок обычно в качестве субстрата, на ко-
тором поселяются личинки устриц, ис-
пользуют пустые раковины устриц или
других моллюсков. Но этот субстрат
обильно заселяют сверлящие губки, и в
дальнейшем он служит источником за-
ражения молодых устриц, обычно тесно
растущих друг подле друга. Поэтому
вместо известкового субстрата применяют
обломки черепицы и кирпичей, что дает
хорошие результаты. С этой же целью
используют раковины, покрытые темным
лаком, что препятствует заражению их
сверлящими губками и в то же время
является более привлекательным для
личинок устриц.
Сверлящие губки постоянно встречают-
ся на коралловых рифах, причем они
поселяются лишь на отмерших частях
кораллов. Здесь благодаря деятельности
клион и других сверлящих организмов
происходит разрыхление известкового ма-
териала. В силу этого большие куски
отмерших кораллов легко отламываются
от верхних участков рифа, падают вниз,
постепенно заполняя впадины и углуб-
ления на дне и способствуя тем самым
дальнейшему росту коралловых рифов и
расширению занятой ими площади.
Поселяясь также на прибрежных изве-
стковых породах, сверлящие губки, про-
делывая в них ходы, участвуют в разру-
шении берегов. О роли губок в этом про-
цессе могут свидетельствовать известко-
вые берега Далмации, где почва повсюду
усеяна множеством обломков скал, изъе-
денных сверлящими губками. Известко-
вый субстрат в результате деятельности
этих губок настолько разрыхляется, что
обломки скал можно раздавить рукой.
Рис. 135. Раковины устрицы, пораженные свер-
лящей губкой:
1 — на поверхности раковины видны многочислен-
ные отверстия, просверленные губкой; 2 — часть верх-
него слоя раковины удалена, чтобы показать ходы и
каналы, проделанные в ней сверлящей губкой.
Клионы обитают на сравнительно неболь-
шой глубине. Изредка они встречаются
глубже 100 м. Наиболее богаты ими мел-
ководные участки теплых морей. У нас
сверлящие губки хорошо известны в
Черном море, обнаружены они также в
дальневосточных морях и в юго-западной
части Баренцева моря.
ГУБКИ КАК БИОФИЛЬТРАТОРЫ
Живые губки непрерывно процежи-
вают через свое тело окружающую
их воду. При этом в канальной си-
стеме губок создается довольно силь-
215
ный напор воды. Специальные наблюде-
ния над одной из известковых губок
(Leuconia aspera) показали, что вода у нее
вытекает из устья со скоростью 7—8,5 см
в секунду. У пресноводных губок из
устьевых отверстий бьет хорошо заметная
струя воды на расстояние 15—20 см.
Суточное количество воды, которое про-
пускается через губку, зависит в первую
очередь от величины и сорта губки. Упо-
мянутая выше известковая губка, около
7 см в высоту, процеживает за сутки
22,5 л воды; несколько меньшего разме-
ра пробковая губка— 12 л, а большая
колония морской кремнероговой губки
(Callyspongia sororia), имеющая около
20 устьевых трубок, пропускает через
свое тело за сутки 1575 л воды. Подсчи-
тано, что суточная пропускная способ-
ность губок в пересчете на вес их в су-
хом виде составляет примерно 5 л на 1 г
губки. По другим обобщенным дан-
ным, различные губки объемом около
10 куб. см процеживают за сутки от 100
до 2000 л воды.
Как известно, мелкие органические и
неорганические частицы, взвешенные в
толще воды, попадая через поры в ор-
ганизм губки, задерживаются в нем.
Органические частицы усваиваются губ-
кой, а неорганические вновь поступают
наружу, постепенно оседая на дно. Губ-
ка извлекает из проходящей через ее тело
воды также различные растворенные в
ней вещества, в том числе содержащие
кремний и кальций, потребляемые в зна-
чительном количестве для построения
скелета. В этом заключается роль губок
как биологических фильтраторов, жиз-
недеятельность которых отражается на
плотности и составе окружающей воды.
Ряд опытов свидетельствует о большой
фильтрационной способности губок. На-
блюдалось, например, что экземпляр пре-
сноводной губки (Spongilla lacustris,
Ephydatia mulleri), величиной с палец, за
сутки очищает три литра воды, замут-
ненные двумя кубическими сантиметра-
ми молока. В другом случае отмечается,
что примерно такой же величины прес-
новодная губка делает прозрачными за
трое суток три литра воды, наполовину
смешанные с выжимкой из другой губки.
Поскольку многие губки достигают
значительного размера и образуют к
тому же массовые поселения на дне во-
доемов, роль их в процессах биофильтра-
ции придонной воды должна быть приз-
нана весьма существенной. Особенно прак-
тически важное значение имеют губки в
процессах самоочищения пресных водое-
мов от гниющих органических остатков,
бактерий и планктонных организмов. От-
мечено, что в реках, протекающих через
большие города и индустриальные по-
селки, на некотором расстоянии от на-
селенного пункта вода быстро очищается
от многих загрязняющих ее веществ. В
этом важная роль принадлежит губкам,
которые здесь пышно развиваются, соз-
давая своего рода фильтрационный
барьер.
ЯДОВИТЫЕ СВОЙСТВА ГУБОК
Большинство губок имеет очень рез-
кий и неприятный запах, с которым, как
предполагают, связано выделение в ок-
ружающую их воду каких-то ядовитых
веществ. Еще Э. Геккель наблюдал,
что мелкие одноклеточные организмы,
приближаясь к известковым губкам, по-
гибают и после этого увлекаются током
воды через поры внутрь губки. Большой
интерес представляют опыты по воздей-
ствию на бактерии разного рода вытя-
жек, приготовленных из губок. Оказа-
лось, что даже простая водная вытяж-
ка из морских губок препятствует росту
некоторых бактерий и обладает, следо-
вательно, антибиотическими свойствами.
Испытывались различные вытяжки из
губок и на крупных животных. Было
установлено, что вещество пресноводных
губок, несомненно, обладает ядовитыми
свойствами. Так, введение вытяжки из
губок в брюшину некоторых теплокров-
ных животных (белых мышей, морских
свинок, кроликов и др.) вызывает ряд
болезненных симптомов, в том числе
прогрессивное удушье, частичное раст-
ворение эритроцитов крови. Это ядови-
тое вещество губок оказывается довольно
стойким и не теряет своей силы даже при
кипячении в течение одной минуты. О
216
силе яда можно судить по
следующим данным. Для то-
го чтобы умертвить белую
мышь, достаточно ввести
внутрь ее брюшины 0,5 куб.
см вытяжки из пресновод-
ной губки, причем смерть
наступает через 3—6 часов.
Такое же действие оказыва-
ет вытяжка на морских
свинок, большинство кото-
рых погибает в течение 1 —
2 дней после введения им 1
куб. см вытяжки. С другой
стороны, холоднокровные жи-
вотные (лягушки и рыбы,
например) нечувствительны
к инъекции этой вытяжки,
а введение в пищеваритель-
ный тракт животных вы-
тяжки из пресноводных губок не оказы-
вает на них сколько-нибудь заметного
воздействия.
В дополнение к сказанному следует
отметить случаи кожных заболеваний
человека, вызываемых губками. В водах
Карибского моря имеется несколько ви-
дов губок (Fibula notilangere и др.),
простое прикосновение к которым вызы-
вает сильнейший зуд, покраснение и даже
опухоль кожного покрова. Применение в
качестве примочки разбавленной уксус-
ной кислоты заглушает боль, и через не-
сколько дней последствия такого кон-
такта с губкой обычно проходят. Несом-
ненно, здесь имеет место воздействие
какого-то ядовитого вещества губки на
кожу человека. Но такие губки являются
исключением. Обычно же губки безвред-
ны для человека. Их можно брать голы-
ми руками, и, если иногда особенно ко-
лючие губки при этом вонзаются своими
иглами в тело, никаких последствий это
не вызывает. Однако в особых условиях
иголочки губок могут служить источни-
ком больших неприятностей, вызывая
сильнейшее раздражение кожи. Известно,
что по отмирании губок их иглы оседают
на дно водоема, в большом количестве
насыщая грунт. Длительное пребывание
человека на таком грунте обычно при-
водит к появлению жжения и сильного
зуда кожи, вызванного механическим воз-
..<-.V;;.4?~::-.>,
Рис. 136. Туалетная губка зимокка (Spongia zimocca).
действием на нее многочисленных иголо-
чек. Пострадавший невольно расчесы-
вает эти места, что еще больше усили-
вает болезненные ощущения. При этом
нередко в ранки заносится инфекция.
Подобные кожные заболевания особенно
распространены среди рыбаков и в не-
которых районах, богатых губками, счи-
таются даже профессиональными заболе-
ваниями. Точно такие же последствия
имеют место при производстве разного
рода земляных работ на дне высохших
прудов, где до этого обильно была пред-
ставлена фауна губок.
Природа специфического запаха губок,
а также определенная ядовитость веще-
ства их тела до сих пор мало изучены.
Предполагают, что это обусловлено на-
личием в теле губок и в отдельных его
клетках значительного количества про-
дуктов обмена, которые сами по себе
являются сильным ядом.
ПРОМЫСЛОВЫЕ ГУБКИ
Объектом промысла служат очень не-
многие губки. К их числу относятся не-
которые кремнероговые губки, употреб-
ляемые для туалета, медицинских и тех-
нических целей, а также стеклянные
губки, используемые в качестве укра-
шений.
217
Туалетные губки. Под таким названи-
ем известны губки (Spongia officinalis,
Hippospongia communis и др.), скелет
которых состоит из густой сети роговых
волокон. Этот скелет, имеющий характер
эластичного войлока, освобожденный от
остальных частей тела губки, находит
довольно разнообразное применение. Туа-
летные губки имеют вид ноздреватых ком-
ков, округлых толстых лопастей или ков-
риг, достигающих 20 см величины и зна-
чительно более. Цвет их желтый, ко-
ричневый, серый или иногда черный.
Имеется много сортов туалетных губок.
Наиболее ценные из них — левантий-
ская и долматинская губки — отличают-
ся особой мягкостью и нежностью скеле-
та. Чем меньше в роговых волокнах губки
содержится посторонних минеральных
включений (песчинок, обломков игл, ра-
ковин простейших и пр.), тем дороже
ценится такая губка. Самой же дешевой
туалетной губкой считается конская губ-
ка, имеющая среди них наиболее жест-
кий скелет.
С глубокой древности, по крайней ме-
ре с античных времен, туалетная губка
используется человеком для мытья тела.
Позднее, в средние века, она стала при-
меняться также в медицине в виде там-
понов для остановки кровотечения и при
лечении заболеваний зобной железы. В
последнем случае препарат жженой губ-
ки прописывался как внутреннее сред-
ство. Действие такого препарата вполне
объяснимо, поскольку в составе рогового
вещества губки содержится иод. В то
время еще не знали причин зобных за-
болеваний и лечебные свойства жженой
губки были установлены исключительно
опытным путем.
В настоящее время область применения
туалетной губки значительно расшири-
лась. Так, она используется для мытья
и нанесения красок в полиграфическом
и других производствах, применяется
как тонкий шлифовочный и полировоч-
ный материал в ювелирном деле, в зер-
кальном и оптическом производствах.
Из этих губок делаются также разнооб-
разные фильтры: в Италии, например,
ими пользуются для фильтрации эссен-
ций и масел. С помощью губок произво-
дят по особому способу воронение ста-
ли. И во многих других случаях находят
применение такие свойства туалетной
губки, как эластичность, мягкость, гиг-
роскопичность, пористость и пр. Промы-
сел туалетных губок широко развит в
Средиземном и Красном морях, в Мекси-
канском заливе и Карибском море, а так-
же в Индийском океане, на Филиппинах
и у берегов Австралии. В ряде мест, как,
например, в Адриатическом море, у бе-
регов Флориды и Японии, известны ис-
кусственно созданные плантации туа-
летных губок. Для этого используют
большую способность губок к регенера-
ции и бесполому вегетативному размно-
жению. Как уже упоминалось, при ис-
кусственном разведении туалетные губки
разрезают на куски, которые культиви-
руют в подходящих местах на дне.Через
несколько лет из отдельных кусков вы-
растают губки, достигающие промысло-
вого размера. Губки собирают пловцы-
ныряльщики, водолазы, а также их вы-
лавливают при помощи острог, трезуб-
цев или драг с борта судна или лодки.
Ежегодный мировой промысел туалетных
губок до второй мировой войны состав-
лял в среднем около 300 т (сухой вес).
Эта цифра довольно значительная, по-
скольку вес сухих губок ничтожен: одна
туалетная губка подушковидной формы
до 20 см величины весит всего около 40 г.
Пресноводные губки — бадяги. В от-
личие от туалетных губок бадяги (Spon-
gilla, Ephydatia и др.) имеют скелет,
состоящий из массы мелких кремниевых
иголочек, склеенных спонгином. Внеш-
ний вид бадяг очень разнообразен: это
чаще всего колонии корковой, комкооб-
разной или кустистой формы. Окраска
их варьирует от серой и бурой до зеле-
новатой. Бадяги встречаются на неболь-
шой глубине вблизи берегов в озерах,
реках, прудах, ручьях и т. д., поселяясь
на камнях и скалах, на корягах, ство-
лах и ветвях затонувших деревьев и на
других предметах.
Сушеную и растертую в порошок бадя-
гу применяют во многих странах как
средство народной медицины при ревма-
тизме, ушибах и в ряде других слу-
чаев. Как гомеопатическое лекарство на-
218
Рис. 137.
1 — украшение, изготовленное из корневых пучков
игл губки гиалонемы (Hyalonema sieboldi); 2 — скелет
губки корзинка Венеры (Euplectella aspergillum).
стойку из бадяги употребляют против
невралгии, скрофулеза и других заболе-
ваний. Раньше сбор бадяг в России был
широко распространен. Они даже слу-
жили предметом вывоза за границу, в
особенности в Германию. В Южной Аме-
рике некоторые пресноводные губки из-
давна применяют в гончарном деле. Для
этого порошок из бадяг, содержащий
большое количество иголочек, смешивает-
ся с гончарной глиной. Изготовленные
из такой смеси сосуды отличаются боль-
шой прочностью.
На территории СССР в различных во-
доемах обитает около 20 видов пресновод-
ных губок. Кроме бадяг, известны еще
особые байкальские губки подушковидной
(Baikalospongia) или кустистой формы
(Lubomirskia) высотой до 1 м. Байкаль-
ские губки содержат более жесткий скелет
и используются местным населением для
чистки и полировки металлических изде-
лий.
Стеклянные губки. Некоторые стек-
лянные губки имеют очень красивый и
изящный скелет. Очищенный от органи-
ческого вещества скелет таких губок
используют как украшение и сувениры.
Особенно красива уже упоминавшаяся
губка корзинка Венеры (Euplectella). Ее
скелет имеет вид нежного ажурного ци-
линдра настолько замысловатого и тон-
кого строения, что он кажется сделанным
искусной рукой человека. Рассказывают,
что первый экземпляр этой губки, при-
везенный в Европу, был куплен за басно-
словную сумму в 600 марок. И до настоя-
щего времени корзинка Венеры считается
очень ценным украшением.
Другая стеклянная губка, гиалонема
(Hyalonema), имеет округлое тело, сидя-
щее на конце стержня из очень длинных
толстых игл. Скелет этой губки исполь-
зуют целиком или из его отдельных ча-
стей склеивают причудливые искусствен-
ные украшения. Промысел обеих этих
губок сосредоточен главным образом у бе-
регов Японии и Филиппинских островов.
Добыча стеклянных губок сопряжена с
большими трудностями, так как они
живут на значительной глубине и имеют
очень ломкий скелет.
ГЕОЛОГИЧЕСКОЕ ПРОШЛОЕ ГУБОК
Губки — очень древние животные. Уже
в кембрийском море существовала до-
вольно богатая их фауна, представлен-
ная почти всеми классами и отрядами
губок, известными и в настоящее время.
Лишь очень немногие ископаемые губки
принадлежат к вымершим отрядам восъ-
милучевых (Octactinellida) и разнолуче-
вых губок (Heteractinellida); окаменев-
шие остатки их находят в палеозой-
ских отложениях. Периодом наиболь-
шего расцвета губок считают юрское и
219
меловое время. Тогда же, очевидно,
появились первые пресноводные губки.
Интересно, что некоторые роды и даже
виды губок, которые обнаруживают в
меловых отложениях, дожили до настоя-
щего времени.
Лучше всего в ископаемом состоянии
сохраняются губки, у которых иглы сра-
стаются друг с другом, образуя прочный
скелетный каркас. Остатки этих губок в
виде окаменелостей находят во многих
отложениях разного геологического воз-
раста. Губки со скелетом из изолирован-
ных игл встречаются в ископаемом со-
стоянии гораздо реже. Но зато хорошо в
осадочных породах сохраняются отдель-
ные кремниевые иглы губок. В настоящее
время изучению ископаемых игл губок
уделяют большое внимание. По отдель-
ным сохранившимся иглам можно полу-
чить представление о семействе, а иногда
роде и даже виде губок, которые оби-
тали в местах их захоронения. Находки
ископаемых губок, а также их игл имеют
важное значение для палеонтологии, так
как дают возможность восстанавливать
фауну губок древних морей и условия
существования в них. Подобно другим
животным, ископаемые губки используют
в геологии как показательные формы при
определении относительного возраста оса-
дочных пород.
Кишечнополостные
Coelenterata
Как было указано во введении, многоклеточные могут
быть разделены на две неравные группы. К одной из них
(Parazoa) относится тип Губки, к другой — все остальные
животные, получившие общее название Настоящие много-
клеточные (Eumetazoa). В надраздел настоящих многоклеточ-
ных включают разделы: Радиальные (Radiata) и Двусто-
роннесимметричные (Bilateria). Раздел радиальных вклю-
чает типы: Кишечнополостные (Coelenterata) и Гребневики
(Ctenophora).
ТИП КИШЕЧНОПОЛОСТНЫЕ
(COELENTERATA)
ОБЩАЯ ХАРАКТЕРИСТИКА
Кишечнополостные — самые низко ор-
ганизованные из числа настоящих мно-
гоклеточных животных. Тело кишечно-
полостных состоит из двух слоев кле-
ток — эктодермы и энтодермы, между
которыми имеется более или менее раз-
витый неклеточный слой, называемый
мезоглеей. Свое название эти жи-
вотные получили в связи с тем, что у
них имеется всего одна полость, называ-
емая кишечной или г астраль-
ной полостью. Все кишечнопо-
лостные — водные, за исключением не-
многих видов, морские организмы. Их
тело построено по типу радиально-осе-
вой симметрии.
Несмотря на простоту организации,
внешне кишечнополостные очень разнооб-
разны. Это зависит от двух причин.
Первая причина заключается в способ-
ности кишечнополостных образовывать
колонии. Как правило, отдельная особь
колонии очень невелика, и потому в
первую очередь внимание невольно обра-
щается на всю колонию в целом. Одни
колонии имеют вид кустиков или малень-
ких деревьев. Другие похожи на ершик,
третьи напоминают фантастические пти-
чьи перья. Кроме таких нежных гибких
колоний, есть колонии массивные с мощ-
ным известковым скелетом. Они имеют
то форму шара, то кубка, то гриба, то
колючей елочки. Наконец, встречаются
нежные плавающие колонии.
Вторая причина разнообразия внеш-
него вида кишечнополостных зависит от
того, что у этого типа животных отдель-
ная особь имеет форму либо полипа, ли-
бо медузы. Тело полипа обычно цилинд-
рическое, на верхнем его конце распо-
ложен рот, окруженный щупальцами.
Полипы — малоподвижные или даже
прикрепленные животные, часто они об-
разуют колонии. Медузы — одиночные
плавающие, подвижные организмы. Их
тело имеет форму зонтика со щупальца-
ми по краям. Плавают медузы ртом
вниз.
Наличие у кишечнополостных двух
форм, двух состояний — полипоидного и
медузоидного, способность этих живот-
ных образовывать колонии разнообраз-
ной формы, а также присущая им яркая
окраска делают кишечнополостных очень
многообразной по формам группой жи-
вотных, несмотря на то что их внутрен-
нее строение имеет единый, общий план
и устроены они довольно просто.
В настоящее время известно около 9 тыс.
видов, относящихся к этому типу. Самые
маленькие из них (например, полипы на
колониях гидроидов) едва достигают 1 мм,
наиболее крупные, такие, как медуза
цианея (Cyanea arctica), имеют зонтик до
2 м в диаметре, а щупальца этой медузы
растягиваются на 30 м.
223
Рис. 138. Внешний вид колоний кишечнополостных.
1 — гидроид евдендриум (Eudendrium); 2 — гидроид туярия (Thuiaria); з —морское перо пеннатула
(Pennatula); 4 — коралл акропора (Асгорога); 5 — сифонофора физофора (Physophora).
Кишечнополостные — самые древние
из настоящих многоклеточных животных.
За время длительной истории развития
этого типа его представители сумели очень
хорошо приспособиться к самым разно-
образным условиям обитания. Они засе-
лили буквально весь океан от его поверх-
ности до предельных глубин, их можно
обнаружить и в полярных областях и в
тропиках. Поселяются кишечнополост-
ные на самых разнообразных грунтах,
некоторые из них способны выносить
значительное изменение солености мор-
ской воды, а отдельные виды проникли
даже в пресные воды. Почти повсеместно
они играют очень важную роль в образо-
вании сообществ морских животных и ра-
стений — биоценозов моря. Познакомим-
ся же поближе с этими интересными су-
ществами.
Для того чтобы читатель легче мог
разобраться в систематическом положе-
нии отдельных видов кишечнополостных
и получил более полное представление о
составе этой группы животных, ниже
приводится краткая система типа.
Тип Кишечнополостные (Coelenterata)
Класс Гидроидные (Hydrozoa)
Отдельная особь имеет форму либо полипа,
либо медузы. Кишечная полость полипов ли-
шена радиальных перегородок. Половые железы
развиваются в эктодерме. Около 2800 видов
обитает в море, но имеется несколько пресновод-
ных форм.
Подкласс Гидроиды (Hydroidea)
Колонии донные, приросшие. У некоторых не-
колониальных видов полипы способны плавать
у поверхности воды. В пределах каждого вида
все особи медузоидного строения одинаковы.
Отряд Лептолиды (Leptolida)
Имеются особи и полипоидного и медузоидного
происхождения. Преимущественно морские,
очень редко пресноводные организмы.
Отряд Гидрокораллы (Hydrocorallia)
Ствол и ветви колонии известковые), часто ок-
рашены в красивый желтоватый, розовый или
красный цвет. Медузоидные особи недоразвиты
и погружены в глубь скелета. Исключительно
морские организмы.
Отряд Хондрофоры (Chondrophora)
Колония состоит из плавающего полипа и
прикрепленных к нему медузоидных особей.
Исключительно морские животные. Ранее при-
числялись к подклассу сифонофор.
Отряд Трахилиды (Trachylida)
Исключительно морские гидроидные, имею-
щие форму медузы, полипов нет.
224
Отряд Гидры (Hydrida)
Одиночные пресноводные полипы,
медуз не образуют.
Подкласс Сифонофоры (Si-
phonophora)
Плавающие колонии, в состав ко-
торых входят полипоидные и меду-
зоидные особи разнообразного стро-
ения. Обитают исключительно в
море.
Класс Сцифоидные (Scyphozoa)
Отдельная особь имеет вид либо
маленького полипа, либо крупной
медузы, или же животное несет при-
знаки обоих поколений. Кишечная
полость полипов имеет 4 неполные
радиальные перегородки. Половые
железы развиваются в энтодерме ме-
дуз. Около 200 видов. Исключитель-
но морские организмы.
Отряд Корономедузы (Coronata)
Преимущественно глубоководные
медузы, зонтик которых разделен пе-
ретяжкой на центральный диск и ко-
рону. Полип образует вокруг себя
защитную хитиноидную трубочку.
Отряд Дискомедузы (Discome-
dusae)
Зонтик медуз сплошной, имеются
радиальные каналы. Полипы лише-
ны защитной трубочки.
Отряд Кубомедузы (Cubome-
dusae)
Зонтик медузы сплошной, но ли-
шен радиальных каналов, функцию
которых выполняют далеко выдаю-
щиеся карманы желудка. Полип без
защитной трубочки.
Отряд Ставромедузы (Stau-
romedusae)
Своеобразные донные организмы,
сочетающие в своем строении при-
знаки медузы и полипа.
Класс Коралловые полипы
(Anthozoa)
Отдельная особь имеет форму по-
липа, медуз не образуется. Поло-
вые железы развиваются в энтодер-
ме. Кишечная полость поделена на
камеры радиальными перегородка-
ми. Около 6000 видов. Исключитель-
но морские организмы.
Подкласс Восьмилучевые ко-
раллы (Octocorallia)
Формы колониальные, как пра-
вило, приросшие к грунту. Полип
имеет 8 щупалец, на боковых сторо-
нах которых расположены выросты—
п и н н у л ы.
Отряд Солнечные кораллы (Не-
lioporida)
Скелет сплошной, массивный.
Рис. 139. Полипы
и медузы.
Отряд Альционарии (Alcyo-
naria)
Мягкие кораллы, скелет в виде
известковых игл.
Отряд Роговые кораллы (Gor-
gonaria)
Скелет в виде известковых игл,
обычно имеется также осевой скелет
из рогоподобного или обызвествлен-
ного органического вещества, про-
ходящий через ствол и ветви ко-
лонии.
Отряд Морские перья (Реппа-
tularia)
Своеобразная колония, состоящая
из крупного полипа, на боковых вы-
ростах которого развиваются вторич-
ные полипы. Основание колонии
внедрено в грунт. Некоторые виды
способны передвигаться.
Подкласс Шестилучевые ко-
раллы (Hexacorallia)
Формы колониальные и одиноч-
ные. Щупальца без боковых вырос-
тов, их число обычно равно или
кратно шести.
Отряд Актинии (Actiniaria)
Одиночные, свободноподвижные
бесскелетные полипы, живущие на
поверхности морского дна (имеется
небольшое число роющих видов).
Отряд Мадрепоровые кораллы
(Madreporaria)
Преимущественно колониальные,
реже одиночные (но неподвижные)
кораллы с мощным наружным из-
вестковым скелетом.
Отряд Корковые кораллы (Zo-
antharia)
Одиночные или колониальные
формы, прирастающие к грунту.
Колонии стелющиеся.
Отряд Антипатарии (Anti-
patharia)
Колониальные, разветвленные при-
растающие кораллы с осевым ске-
летом из рогоподобного вещества.
Отряд Цериантарии (Cerian-
tharia)
Одиночные, бесскелетные поли-
пы, живущие в илистом грунте. Стро-
ят себе трубки из ила, скрепляя
его слизистыми выделениями. По-
лип способен передвигаться внутри
трубки.
Сверху вниз: гидроидный полип
гидрактиния (Hydractinia), коралловый
полип актиния (Actinia), красный ко-
ралл (Corallium), гидромедуза битотиара
(Bythotiara), сцифоидная медуза хризао-
ра (Chrysaora).
W 15 Жизнь животных, т. 1
225
ИСТОРИЯ ИЗУЧЕНИЯ
КИШЕЧНОПОЛОСТНЫХ
Интерес к изучению фауны морских
беспозвоночных возник еще в глубокой
древности.
Первые сведения о кишечнополостных
животных мы находим в трудах Аристо-
теля. Правда, в его системе они не
представляют собой единой группы жи-
вотного царства. Отдельные кишечнопо-
лостные описывались Аристотелем в раз-
ных разделах системы. Основная же
часть кишечнополостных объединялась в
группу Cnidae, или стрекающих. Ари-
стотель подробно описывает их неподвиж-
ность, отмечает наличие в теле стрекаю-
щих только одного отверстия — рта, име-
ющего центральное положение и окру-
женного кольцом подвижных щупалец.
Древнегреческому ученому уже было из-
вестно, что в щупальцах находятся осо-
бые стрекающие органы, служащие для
защиты от врагов и для схватывания
добычи. Аристотелю принадлежит и очень
интересное утверждение о том, что ки-
шечнополостные и вместе с ними другие
просто устроенные беспозвоночные, ве-
дущие прикрепленный образ жизни (губ-
ки, асцидии, мшанки), представляют со-
бой организмы, в которых неожиданно
смешивается природа животных и расте-
ний. Эта двойственность природы кишеч-
нополостных, по его мнению, выражалась
в том, что они своей способностью к
питанию животной пищей и к восприятию
внешних раздражений напоминают жи-
вотных, а общим планом строения, про-
стотой организации и прикрепленным об-
разом жизни — растения. Вслед за Ари-
стотелем и другой древний философ и
натуралист Плиний также усматривал
двойственную природу кишечнополост-
ных.
Восемнадцатый и девятнадцатый века
были временем накопления новых сведе-
ний о строении и образе жизни кишечно-
полостных. В 1723 г. интересное открытие
было сделано французским судовым вра-
чом Пейсоннелем. Он установил, что
рифообразующие кораллы — колониаль-
ные кишечнополостные животные, обла-
дают массивным известковым скелетом.
А ведь до его открытия и мореплаватели и
ученые считали, что коралловые полип-
няки принадлежат к неорганическому
миру и представляют собой особый извест-
ковый минерал. Пейсоннелю удалось на-
блюдать, как полипы рифообразующих
кораллов захватывали своими щупальца-
ми проплывавших мимо мелких рачков
и затем поедали их.
Но самой яркой страницей в истории
изучения кишечнополостных явились ис-
следования натуралиста Трамбле, швей-
царца по происхождению, увлекавшего-
ся, как и многие другие в то время,
наблюдением при помощи микроскопа
поведения различных мелких живот-
ных — инфузорий и «насекомых»1.
Случайно он обнаружил прикрепивших-
ся к водорослям пресноводных гидр —
мелких одиночных гидроидных полипов,
которых принял сначала за водные расте-
ния. Трамбле ничего не знал тогда о
гидрах. Да это и не удивительно, так как
вообще почти ничего не было известно о
них и гидры не имели даже зоологического
названия. Трамбле назвал гидру «поли-
пом с руками в виде рогов». Результатом
длительных наблюдений над гидрами яви-
лась книга «Мемуары о пресноводном
полипе», вышедшая в 1744 г. Трамбле с
удивительной полнотой и точностью опи-
сал строение гидры, ее размножение,
питание, способы передвижения и т. д.
Но главным достижением натуралиста
следует считать открытые эксперимен-
тальным исследованием способности гид-
ры к регенерации, т. е. к самовосстанов-
лению. Регенерация у других организ-
мов была известна и раньше, но в пред-
шествовавших экспериментах удавалось
наблюдать восстановление лишь отдель-
ных утраченных частей животного, на-
пример лапок или усиков у ракообраз-
ных. У гидры же Трамбле наблюдал
нечто большее — восстановление целого
организма из отдельных его частей. Эта
способность рассматривалась Трамбле и
его современниками как особого рода
размножение животных путем рассечения
1 Так назывались в то время не только настоя-
щие насекомые, но и многие другие мелкие жи-
вотные — коловратки, черви и т. д.
226
его на части. Работы Трамбле представ-
ляли собой большой интерес. С одной
стороны, его наблюдения окончательно
доказали животную природу гидры и
вообще остальных зоофитов. С другой
стороны, эти исследования привели к от-
рицанию мнения об основном различии
между животными и растениями, до того
времени господствовавшего и заключав-
шегося в том, что растения могут раз-
множаться черенками и отводками, а
животные — нет.
В середине прошлого века зоолог Л е й-
к а р т создает самостоятельный тип
животных — кишечнополостных, выде-
ляя его из типа Zoophita.
СРАВНИТЕЛЬНОАНАТОМИЧЕСКИЙ
ОБЗОР КИШЕЧНОПОЛОСТНЫХ
Строение животных, относящихся к
каждому из классов кишечнополостных,
отличается своими особенностями. В свя-
зи с этим удобнее рассмотреть строение
представителей каждого класса в отдель-
ности.
Гидроидные (Hydrozoa). Тело гидроид-
ного полипа цилиндрической или яйце-
видной формы, в своей нижней части
снабжено обычно ножкой. При помощи
ножки одиночные полипы прикрепляют-
ся к грунту, водным растениям, ракови-
нам моллюсков, т. е. вообще к каким-
либо подводным предметам. У колониаль-
ных форм ножка соединяет полипа с ко-
лонией. На верхнем конце тела полипа
помещается ротовое отверстие, окружен-
ное щупальцами. Щупальца могут рас-
полагаться правильными венчиками или
без строгого порядка. Число щупалец
различно у разных видов. У гигантского
одиночного гидроидного полипа бранхио-
цериантуса (Branchiocerianthus impera-
tor), достигающего метра в высоту, на-
считывается до 380 щупалец, а у малень-
кого монобрахиума (Monobrachium para-
siticum) — всего одно щупальце. Чаще
же каждый полип снабжен 10—30 щу-
пальцами, которые могут быть или про-
стыми, или снабженными на конце була-
вовидным утолщением. У одного вида
(Cladocoryne) щупальца даже ветвятся.
Существуют также полипы вовсе лишен-
ные щупалец. К ним относится пресно-
водная микрогидра (Microhydra).
Щупальцами полипы захватывают пи-
щу, а такие одиночные полипы, как
пресноводная гидра, пользуются ими при
передвижении.
Внутреннее строение гидроидного по-
липа очень простое. Он имеет вид дву-
слойного мешка. Снаружи полип покрыт
слоем клеток эктодермы, а его кишечная
полость выстлана энтодермой.
Эктодерма образована особыми эпите-
лиально-мускульными клетками. Тело
такой клетки имеет вид многогранной
призмочки, клетки расположены тесно,
как пчелиные соты. Призматическая
часть выполняет покровную функцию.
На своем внутреннем конце эпителиаль-
ные клетки имеют длинный вырост, тя-
нущийся вверх и вниз вдоль тела поли-
па. Внутри отростка проходит тонкое
мускульное волокно. Такие же сократи-
тельные отростки отходят и от клеток эн-
тодермы, но они расположены перпен-
дикулярно к отросткам эктодермальных
Рис. 140. Схема продольного разреза гидроид-
ного полипа:
1 — ротовое отверстие, 2 — щупальце; 3 — гастраль-
ная полость; 4 — эктодерма; 5 — энтодерма, 6 —
мезоглея.
15»
227
клеток. Совокупность волокон многих
эктодермальных и энтодермальных кле-
ток дает возможность полипу и его щу-
пальцам вытягиваться и сокращаться.
Между эпителиально-мускульными
клетками располагаются особые стре-
кательные клетки, они сложно
устроены и имеют для всех кишечнополо-
стных очень важное значение как орудие
нападения и защиты. Описание строения
и действия этих клеток будет дано ниже,
в специальном разделе о стрекательных
клетках и действии их яда. В эктодерме
есть и особые нервные клетки,
снабженные длинными тонкими отрост-
ками, образующими в своей совокупно-
сти сетевидное нервное сплетение, не-
сколько более плотное на концах щупа-
лец и вокруг рта. У полипов, способных
размножаться половым путем (например,
у гидры), в эктодерме развиваются также
половые клетки. Обычно они
скапливаются в нижней или средней
части тела и получают название поло-
вых желез или гонад. Нако-
нец, здесь же в эпителии имеются резерв-
ные клетки (они называются промежуточ-
ными или интерстициальными), из кото-
рых развиваются эпителиально-мускуль-
ные, стрекательные, нервные и половые
клетки гидроидных.
Эктодерма многих видов полипов вы-
деляет снаружи тонкую оболочку, со-
стоящую из хитиноподобного вещества.
Эта оболочка выполняет у гидроидных
полипов роль наружного скеле-
т а, служащего опорой и защитой.
У некоторых видов оболочка вокруг
полипа образует род чашечки, в которую
полипы могут втягиваться в случае опас-
ности (рис. 158, 3).
Энтодерма состоит из железистых
и эпителиально-мускуль-
ных клеток. Железистые клетки
вырабатывают пищеварительный сок
(протеолитический фермент), способству-
ющий усвоению белков. Мелкие живот-
ные, попавшие в гастральную полость
полипа, под влиянием пищеварительных
соков перевариваются, распадаются на
части. Клетки энтодермы имеют по 2—5
тонких жгутиков, которые все время из-
виваются и перемешивают содержимое
гастральной полости. Частицы пищи,
оказавшиеся при этом вблизи стенок
гастральной полости, захватываются псев-
доподиями, образующимися на поверхности
эпителиально-мускульных клеток. Даль-
нейшее пищеварение происходит уже
внутри клетки, подобно тому как это
происходит у одноклеточных животных.
Гастральные полости отдельных полипов
колонии сообщаются друг с другом, об-
разуя единую пищеварительную полость
колонии.
В энтодерме имеются и нервные клет-
ки, но их меньше, чем в эктодерме; стре-
кательные же клетки в энтодерме вообще
отсутствуют. Наличие нервных клеток в
энтодерме свойственно кишечнополост-
ным, отчасти — иглокожим, но несвой-
ственно всем другим животным.
Между слоями эктодермы и энтодермы
у гидроидных полипов расположен тон-
кий слой неклеточного вещества — ме-
зоглеи.
228
Гораздо сложнее устроены гид-
роидные медузы. Внешне гидро-
медуза имеет вид прозрачного
диска, зонтика или колокола.
Встречаются и причудливые фор-
мы медуз с кольцевыми перетяж-
ками посередине тела или меду-
зы почти шарообразной формы.
От внутреннего центра зонти-
ка свешивается ротовой хоботок
со ртом на конце. Края рта могут
быть гладкими или снабженными
четырьмя более или менее бах-
ромчатыми ротовыми лопастями.
У некоторых гидромедуз по кра-
ям рта сидят маленькие булаво-
видные ротовые щупальца.
Рот ведет в желудок, зани-
мающий всю полость ротового
хоботка, от желудка к периферии
зонтика отходят 4 (изредка боль-
ше) радиальных к а н а -
л а. На краю зонтика они впа-
дают в кольцевой канал. Совокупность
желудка и каналов носит название г а-
строваскулярной системы. По
краю зонтика гидромедуз располагаются
щупальца и органы чувств.
Щупальца служат для осязания и лов-
ли добычи, они густо усажены стрекатель-
ными клетками.Часть щупалец может быть
видоизменена в особые чувствительные
органы, называемые в зависимости от их
формы колбочками или усика-
ми. У одной группы гидромедуз (тра-
хилид) щупальца видоизменяются в ор-
ганы равновесия. Такое щупальце силь-
но укорочено и сидит как бы на тонком
стебельке. На его конце помещается
известковое зернышко — статолит.
Снаружи щупальце окружено длинны-
ми чувствительными волосками (рис. 143,
А). Когда тело медузы наклоняется,
щупальце под действием силы тяжести
остается висеть отвесно и при этом ка-
сается чувствительных волосков, кото-
рые передают раздражение через нерв-
ную систему к эпителиально-мускуль-
ным клеткам, что вызывает сокращение
их мускульных волокон. Существует пред-
положение, что органы равновесия (они
называются статоцистами) служат глав-
ным образом для того, чтобы мускулату-
Рис. 142. Схема продольного разреза
гидроидной медузы:
А — в плоскости радиального канала; Б — между радиальны-
ми каналами. 1 — ротовое отверстие; 2 — желудок; 3 — рото-
вые щупальца; 4 — радиальный канал, 5 — парус, 6 — краевое
щупальце, 7 — двигательное нервное кольцо; * — глазок,
9 — чувствительное нервное кольцо, 10 — половая железа,
11 — эктодерма, 12 — энтодерма; 13 — мезоглея; 14 — коль-
цевой канал.
ра работала ритмично. Движение медузы
осуществляется благодаря сокращению
мускульных волокон на краю зонтика.
Выталкивая воду из полости зонтика,
медуза получает реактивный толчок и
движется верхней стороной зонтика впе-
ред. Усиление реактивной способности
достигается благодаря наличию на внут-
ренней стороне зонтика кольцевидного
выроста, называемого парусом, ко-
торый сужает выход из полости зонтика.
Каждое сокращение кольцевых мускуль-
ных волокон вызывает колебания стато-
цистов, которые раздражают при этом
клетки нервной системы и вызывают
новое сокращение. У медуз с вырезан-
ными статоцистами регулярность сокра-
щений зонтика резко нарушается, а их
частота уменьшается. У гидромедуз из
группы лептолид статоцисты отсутству-
ют или устроены в виде пузырька, внут-
ри которого находится несколько стато-
литов, а стенки покрыты чувствительны-
ми клетками. Статоцисты лептолид не
имеют ничего общего с щупальцами, но
несут ту же функцию, что и статоцисты
трахилид.
Некоторые гидромедузы обладают све-
точувствительными органами — глаз -
к а м и, которые всегда расположены на
229
Рис. 143. Органы чувств гидромедуз:
А — статоцпст медузы кунина (Gunina); (по О. и Р.
Гертвигам); Б — глазок медузы Корине (Corine) (по
Линко). 1 — статолит; 2 — чувствительные волоски;
3 — светочувствительные клетки; 4 — пигментные
клетки; 5 — хрусталик.
основании щупалец и хорошо заметны
благодаря темной окраске. Глазок состо-
ит из двух родов клеток — свето-
чувствительных и пигме-
нтных, т. е. несущих в себе красящее
вещество. Благодаря наличию пигмент-
ных клеток свет падает на чувствительные
клетки только с одной стороны. Световое
раздражение чувствительные клетки пе-
редают нервной системе. Наиболее про-
стые глазки имеют вид пятен, более
сложные устроены в виде ямок. В наи-
более сложно устроенных глазках по-
лость ямки заполнена прозрачным веще-
ством, выполняющим роль хрусталика
(рис. 143, Б).
В связи со свободноподвижным образом
жизни гидромедуз их нервная система
развита несравненно выше, чем у гидро-
полипов. Хотя сплетение имеет также вид
сети, но на краю зонтика нервные клет-
ки скапливаются очень густо и образуют
два кольца. Одно из них (наружное)
чувствительное, другое (внутреннее)
двигательное.
Чувствительное кольцо проходит вбли-
зи статоцистов, глазков и оснований щу-
палец и воспринимает полученные от
них раздражения. Двигательное кольцо
лежит в основании паруса, где сосредото-
Рис. 144. Схема строения сцифи-
стомы:
1 — щупальце; 2 — чашечка; з — от-
верстие в септе, 4 — септа; 5 — мус-
кульный тяж; б— воронка. Эктодер-
ма на рисунке заштрихована, энто-
дерма зачернена.
чено большое количество кольцевых му-
скульных волоконец, которые иннер-
вируются от двигательного нервного
кольца.
Медузы раздельнополы, их половые
железы располагаются либо в эктодерме
ротового хоботка, либо в эктодерме зон-
тика под радиальными каналами. Здесь
они ближе всего к питательным вещест-
вам, необходимым для развития половых
продуктов. Строение клеток эктодермы
и энтодермы медуз такое же, как и у по-
липов, и потому не требует дополнитель-
ного описания, зато мезоглея у медуз
развита несравненно сильнее. Она богата
водой и имеет студенистый характер,
благодаря чему гидромедузы очень про-
зрачны, многих, даже довольно крупных,
медуз трудно увидеть в воде. Особенно
сильно развита мезоглея в зонтике.
Общая характеристика, данная выше,
относится только к одному подклас-
с у гидроидов (Hydroidea), но в
класс гидроидных (Hydrozoa) входит
также весьма своеобразный подкласс
сифонофор (Siphonophora). Сифонофоры
живут только в море. Они похожи на
колонии гидроидных полипов, целиком,
всей колонией перешедших к пелагиче-
скому существованию.
230
Колониям сифонофор свойственно в
наибольшей степени явление полимор-
физма, равное которому можно найти толь-
ко среди муравьев и термитов. В состав
колонии сифонофор входят особи, имею-
щие особое строение и выполняющие
различные функции. Одни из них вы-
полняют функции движения, другие —
питания, третьи—выделения, четвертые—
размножения, пятые — защиты. Сифо-
нофоры — это одни из самых удивитель-
ных и самых красивых созданий морской
стихии.
Сцифоидные (Scyphozoa). Полипы, от-
носящиеся к этому классу, очень мелкие,
всего несколько миллиметров высотой.
Как правило, они не образуют колоний
и в большинстве случаев лишены скелет-
ной оболочки. Тело сцифоидного полипа
(он также называется сцифистомой) под-
разделяется на чашечку и ножку. В
верхней части тела, по краю чашечки,
сцифистома снабжена венчиком щупалец,
между которыми на ротовом конце поме-
щается четырехугольное ротовое отвер-
стие. Гастральная полость сцифистомы
разделена четырьмя неполными
внутренними перегородками
(септы, т е н и о л и) на цент-
ральную часть и 4 перифери-
ческие камеры. В верхней час-
ти тениоли несут по небольшо-
му отверстию, через которое
соседние камеры сообщаются
между собой. Внутрь каждой
тениоли со стороны рото-
вого конуса вдается кониче-
ская полость — воронка.
Со дна воронки вниз тянется
Рис. 145. Схема продольного раз-
реза сцифомедузы:
А — разрез между радиальными канала-
ми; Б — разрез в плоскости радиального
канала. I — зонтик, 2 — гастральные
нити, з — радиальный канал, 4 — коль-
цевой канал; 5 — глазное пятно; в —
статолит ропалия; 7 — глазок; 8 — нерв-
ный ганглий; 9 — глотка; 10 — ротовая
лопасть; и —ротовое отверстие; 12 —
субгенитальная ямка, 13 — щупальце,
14 — краевая лопасть; 15 — половая
железа; 16 — желудок. Эктодерма за-
штрихована, энтодерма отмечена час-
тым пунктиром, мезоглея — редким пун-
ктиром.
мускульный тяж. Строение эктодермы и
энтодермы сцифистомы в общих чер-
тах напоминает таковое гидроидных по-
липов.
Внешне сцифоидные медузы имеют зна-
чительное сходство с гидроидными, но
значительно превышают их по размерам.
Форма колокола, его расцветка и бах-
ромчатые края, ротовые лопасти и мно-
гочисленные щупальца сцифоидных медуз
делают их самыми красивыми организ-
мами, обитающими в море. Тело сцифо-
медузы имеет форму диска, зонтика или
колокола. В центре нижней стороны рас-
положено ротовое отверстие, окруженное
4 ротовыми лопастями. В промежутках
между ротовыми лопастями у многих
сцифомедуз имеются впячивания, так
называемые субгенитальные
ямки. Они расположены под поло-
выми железами и служат, по-видимому,
для того, чтобы облегчить доступ кисло-
рода к развивающимся половым про-
дуктам.
Край зонтика сцифомедузы имеет ро-
зетковидную форму, так как расчленен
231
на 8 или 16 краевых лопастей, к их ниж-
ней стороне прикрепляются щупальца, а
между ними находятся ропалии —
маленькие, видоизмененные щупальца, не-
сущие органы чувств — глазки и статоцис-
ты. Число щупалец у разных групп сци-
фомедуз различно и колеблется от 4 до
нескольких сотен. У корнеротых медуз
щупальца отсутствуют.
Гастральная полость сцифоидных ме-
дуз устроена довольно сложно. Рот ведет
в короткую трубковидную глотку, кото-
рая открывается в желудок, имеющий
4 боковых кармана. Внутри карманов
стенки желудка образуют многочислен-
ные длинные энтодермальные выросты,
так называемые гастральные нити. Они
выделяют в кишечную полость пищева-
рительные соки. От желудка к периферии
зонтика отходят радиальные каналы,
число которых обычно равно или крат-
но 8.
У ряда видов радиальные каналы
ветвятся или даже образуют сеть. На
краю зонтика радиальные каналы впа-
Рис. 146. Строение ропалия ушастой
медузы (Aurelia) на продольном разре-
зе (по Шевякову):
1 — глазок, 2 — статолиты, 3 — глазное
пятно; 4 — чувствительные волоски; 5 —
участок краевой лопасти.
дают в кольцевой, но иногда последний
отсутствует.
Нервная система сцифоме-
дуз представляет собой сплетение нерв-
ных клеток. По краю колокола идет
нервное кольцо, а вблизи ропалиев кон-
центрируются нервные элементы, обра-
зующие здесь рыхлые скопления, сходные
с ганглиями. Р о п а л и й представляет
собой маленькое недоразвитое видоиз-
мененное щупальце, несущее в своей
концевой части группу известковых те-
лец — статолитов. Прикосновение
кончика ропалия к чувствительным клет-
кам краевой лопасти вызывает раздраже-
ние, передающееся через ближайшее
скопление нервных клеток мускулатуре
зонтика, вызывая ее сокращение. Таким
образом регулируется темп пульсации
зонтика. У экземпляров с удаленными
ропалиями регулярность сокращений на-
рушается. Ропалий, также как гидроста-
тический орган, дает возможность медузе
ориентировать тело ротовым отверстием
вниз.
У многих сцифоидных медуз на ропалии
располагаются также светочувствитель-
ные органы — глазные пятна и
более сложно устроенные глазки, напо-
минающие по строению описанные выше
глазки гидромедуз.
Сцифоидные медузы раздельнополы, их
половые железы развиваются в энтодер-
ме и располагаются в карманах желудка.
Половые продукты выводятся наружу
через ротовое отверстие.
Мезоглея сцифоидных медуз, так же
как и у гидромедуз, развита очень
сильно.
Коралловые полипы (Anthozoa). Тело
кораллового полипа обычно имеет цилинд-
рическую форму и не подразделяется на
туловище и ножку. У колониальных форм
нижний конец тела полипа прикреплен
к колонии, а у одиночных полипов снаб-
жен прикрепительной подошвой. Щупаль-
ца у коралловых полипов расположены
в один или несколько тесно сближенных
венчиков. Различают две большие груп-
пы коралловых полипов: восъмилучевые
(Octocorallia) и шестилучевые (Нехасо-
rallia). У первых всегда 8 щупалец, и они
снабжены по краям маленькими выроста-
232
ми — пиннулами, у вторых число
щупалец обычно бывает довольно боль-
шим и, как правило, кратно шести. Щу-
пальца шестилучевых кораллов гладкие,
без пиннул.
Верхняя часть полипа, между щупаль-
цами, называется ротовым диском.
В его середине расположено щелевидное
ротовое отверстие. Рот ведет в глотку,
выстланную эктодермой. Один из нраев
ротовой щели и нисходящей от него глотки
получили название сифоноглифа.
Эктодерма сифоноглифа покрыта эпите-
лиальными клетками с очень крупными
ресничками, которые находятся в беспре-
рывном движении и гонят воду внутрь
кишечной полости полипа. Кишечная по-
лость кораллового полипа разделена про-
дольными энтодермальными перегород-
ками (септами) на камеры. В верхней
части тела полипа перегородки прира-
стают одним краем к стенке тела, а дру-
гим к глотке. В нижней части полипа,
ниже глотки, септы прикрепляются лишь
к стенке тела, вследствие чего централь-
ная часть гастральной полости — желу-
Рис. 147. Схема строения кораллового полипа:
1 — ротовое отверстие, 2 — глотка, з — септа; 4 —
мезентериальная нить, 5 — отверстие в септе; 6 —
щупальце. Эктодерма заштрихована, энтодерма от-
мечена пунктиром.
док — остается неразделенной. Число
септ соответствует числу щупалец. По
каждой септе, вдоль одной из ее сторон,
проходит мускульный валик. Свободные
края септ утолщены и получили назва-
ния мезентериальных ни-
тей. Две из этих нитей, расположенных
на паре соседних септ, противостоящих
сифоноглифу, покрыты особыми клетка-
ми, несущими длинные реснички. Реснич-
ки находятся в постоянном движении и
гонят воду из гастральной полости на-
ружу. Совместная работа ресничного
эпителия этих двух мезентериальных ни-
тей и сифоноглифа обеспечивает постоян-
ную смену воды в гастральной полости.
Благодаря им в кишечную полость все
время поступает свежая, богатая кисло-
родом вода. Виды, питающиеся мельчай-
шими планктонными организмами, полу-
чают при этом также и пищу. Остальные
мезентериальные нити играют важную
роль в пищеварении, так как образова-
ны железистыми энтодермальными клет-
ками, выделяющими пищеварительные
соки.
Половые железы развиваются в эн-
тодерме септ. У коралловых полипов
очень большую роль играет скелет.
Восьмилучевые кораллы имеют скелет,
состоящий из отдельных известковых
иголочек — с п и к у л, находящихся в
мезоглее. Иногда спикулы соединяются
между собой, сливаясь или же объеди-
няясь органическим рогоподобным веще-
ством.
Среди шестилучевых кораллов имеют-
ся бесскелетные формы, такие, как ак-
тинии. Чаще, однако, скелет у них
имеется, причем он может быть либо
внутренним — в виде стержня из
рогоподобного вещества, либо наруж-
ным — известковым.
Особенно большой сложности достигает
скелет у представителей группы мадре-
поровых. Он выделяется эктодермой поли-
пов и на первых порах имеет вид пластин-
ки или низкой чашечки, в которой сидит
сам полип. Далее скелет начинает разра-
статься, на нем возникают радиальные
ребра, соответствующие септам полипа.
Вскоре полип оказывается как бы наса-
женным на скелетную основу, которая
233
глубоко вдается снизу в его тело, хотя и
отграничена всюду эктодермой. Скелет
мадрепоровых кораллов получает очень
сильное развитие: мягкие ткани покры-
вают его в виде тонкой пленки.
Скелет кишечнополостных играет роль
опорной системы, а вместе со стрекатель-
ным аппаратом представляет мощную
защиту от врагов, что способствовало
их существованию в течение длительных
геологических периодов.
Для коралловых полипов характерно
только полипоидное состояние, медуз они
не образуют.
СТРЕКАТЕЛЬНЫЕ КЛЕТКИ
КИШЕЧНОПОЛОСТНЫХ
И ДЕЙСТВИЕ ИХ ЯДА
Характернейшей особенностью всех
кишечнополостных является наличие у
них особых стрекательных, или
крапивных, клеток. Эти клетки
развиваются из недифференцированных
промежуточных клеток эктодермы. Каж-
дая стрекательная клетка содержит хи-
тиноидную капсулу овальной или
продолговатой формы. Стенка наружного
конца капсулы впячивается внутрь ее
и имеет вид тонкой спирально закручен-
ной трубочки, называемой стрека-
тельной нитью. Полость капсулы
заполнена ядовитой жидкостью.
На внешней поверхности стрекатель-
ной клетки имеется чувствительный воло-
сок — книдоциль. Прикосновение к
волоску вызывает моментальное раздра-
жение клетки, проявляющееся в стреми-
тельном, подобном выстрелу выворачи-
вании стрекательной нити. В зависимо-
сти от деталей строения и способа воздей-
ствия на врага или жертву различают
несколько типов стрекательных капсул.
Часть стрекательных капсул имеет длин-
ную, снабженную шипами нить. При
«выстреле» такой капсулы нить вонзает-
ся в тело жертвы, а через ее полость из-
ливается содержимое капсулы, вызываю-
щее общее или местное отравление.
У других капсул нить короткая, лишенная
шипов. Такие нити только опутывают
жертву. Наконец, имеются липкие нити,
которые удерживают добычу приклеива-
нием. Они же могут служить и для вре-
менного прикрепления самих носителей
стрекательных клеток при передвиже-
нии.
Стрекательные клетки располагаются
по всей поверхности тела кишечнополост-
ных, но их наибольшие скопления на-
блюдаются на щупальцах и вокруг ро-
тового отверстия, т. е. в местах, где они
нужнее всего. После «выстрела» стрека-
тельная клетка погибает и на ее месте
или поблизости от него развивается но-
вая стрекательная клетка.
Весьма интересно, что стрекательные
капсулы могут действовать и после гибе-
ли животного. Так, прикосновение к
мертвым, выброшенным на берег меду-
зам может вызвать довольно сильные по-
краснения кожи, сопровождающиеся ощу-
щением жжения.
При поедании кишечнополостных жи-
вотных некоторыми турбелляриями и
заднежаберными моллюсками часть
стрекательных клеток переходит в тело
хищника. Здесь стрекательные клетки
располагаются также в наружных покро-
вах и могут нормально функциониро-
вать.
Стрекательные клетки, особенно содер-
жащие ядовитую жидкость в капсулах,
являются грозным оружием, которое
используется кишечнополостными и
для защиты и для нападения. Несом-
ненно, что процветание этих низко ор-
ганизованных животных в значительной
мере объясняется наличием у них стре-
кательных клеток. Мелкие животные,
соприкасаясь с кишечнополостными, при-
клеиваются, опутываются и пронзаются
стрекательными нитями, а в их тело по-
падает яд, вызывающий паралич и
смерть. После этого жертва переносится
щупальцами к ротовому отверстию и про-
глатывается. Даже довольно крупные
животные часто оказываются поражен-
ными ядом стрекательных капсул, вызы-
вающим к тому же жгучую боль. Сила
воздействия яда разных видов кишечно-
полостных на человека неодинакова: не-
которые из них совершенно безвредны,
другие представляют собой серьезную
опасность.
234
После прикосновения к нашим черно-
морским актиниям и к медузам цианея,
обитающим в арктических и дальнево-
сточных морях, ощущается легкое жже-
ние, особенно там, где кожа нежнее,
например между пальцами рук. Несколь-
ко сильнее «жжется» корнерот — обыч-
ная для Черного моря беловатая медуза
с фиолетовым краем зонтика. Тем не менее
эти неприятные ощущения, похожие на
легкий ожог крапивой, быстро проходят,
не оставляя следа. Однако в море имеет-
ся много гораздо более опасных кишечно-
полостных, прикосновение к которым
может привести к тяжелому заболеванию.
Так, многие актинии, в том числе оби-
тающие в Японском море, вызывают не
только «ожоги», но и длительное опуха-
ние руки неосторожно дотронувшегося
до них человека.
Однако наиболее неприятные послед-
ствия заключаются не в «ожогах» и опу-
холях, а в общем отравлении организма
ядом стрекательных клеток. У моряков,
плавающих в тропических морях, уже
издавна пользуется дурной славой кра-
сивая сифонофора физалия. Фпза-
лия снабжена большим, до 20 см длиной,
плавательным пузырем, поднимающимся
над поверхностью воды, от которого
свешиваются вниз длиннейшие (до 30 м\)
ловчие щупальца, снабженные много-
численными стрекательными клетками.
Физалия очень ярко окрашена. Пузырь
отливает голубым, фиолетовым и пурпур-
ным цветом, все свешивающиеся вниз
придатки ультрамариновые. За яркую
окраску физалия получила также назва-
ние португальского военного кораблика:
в средние века португальцы любили пе-
стро окрашивать свои военные суда.
Плавающая на поверхности моря физа-
лия вследствие яркой окраски и значи-
тельной величины видна издалека, и
пловцы всегда остерегаются соприкосно-
вения с ней, так как можно получить
очень сильный «ожог», вызывающий силь-
нейшую боль. Пораженный физалией
человек, даже если он хорошо плавает,
с трудом держится на воде. Вскоре после
этого может наступить и общее тяжелое
заболевание с повышением температуры,
длящееся несколько дней. Физалии рас-
Рнс. 148. Стрекательные клетки:
А — в покоящемся состоянии; Б — с выстреленной
стрекательной нитью 1 — ядро, 2 — капсула; з —
чувствительный волосок, 4 — стрекательная нить;
5 — шипы.
пространены исключительно в тропиче-
ских морях, и очень редко отдельные их
экземпляры заносятся в более теплые
участки умеренной зоны.
В последние годы в ряде медицинских
и биологических журналов стали по-
являться сообщения о серьезных отрав-
лениях, вызываемых крупными сцифоид-
ными медузами хиродропус (Chirodropus)
из отряда кубомедуз.
Это также обитатель тропических мо-
рей. Высота колокола медузы достигает
10—12 см, на краю зонтика сидят 4 щу-
пальца, каждое из которых разветвлено
и имеет 9—12 концов. Медуза полупроз-
рачна и плохо видна в воде. Конечно,
ядовитые свойства этой медузы прояви-
лись не в последние годы, но прежде на
нее просто не обращали внимания в свя-
зи с общей плохой изученностью кишеч-
нополостных тропических морей.
235
Впервые на ядовитость этой медузы
обратили внимание в годы второй миро-
вой войны в Австралии, куда было
эвакуировано значительное число евро-
пейцев. Стало известно несколько слу-
чаев загадочной гибели людей во время
купания, причем на теле погибших не
находили следов зубов акул или других
явных признаков повреждения. Долгое
время тайна оставалась неразгаданной,
но потом удалось установить, что причи-
ной гибели является медуза. Получивший
«ожог» человек, едва успев крикнуть от
пронизывающей боли, захлебывался и
тонул. Было отмечено, что среди постра-
давших или погибших были преимущест-
венно вновь прибывшие люди. Местные
жители, особенно из числа коренного
населения Австралии, могли купаться
безбоязненно. По-видимому, у них имел-
ся иммунитет к яду этой медузы.
В фауне наших дальневосточных морей
также имеется одна медуза, вызывающая
серьезные заболевания при соприкосно-
вении с ней. Местные жители называют
эту медузу «крестовичком» за крестооб-
разное расположение четырех темных
радиальных каналов, вдоль которых тя-
нутся четыре также темноокрашенные
половые железы. Зонтик медузы про-
зрачный, слабого желтовато-зеленого цве-
та. Размеры медузы невелики: зонтик
отдельных экземпляров достигает 25 мм
в диаметре, но обычно они значительно
меньше, всего 15—18 мм. На краю зон-
тика крестовичка (научное название —
Gonionemus vertens) имеется до 80 щу-
палец, способных сильно вытягиваться
и сокращаться. Щупальца густо усажены
стрекательными клетками, которые рас-
полагаются поясками. Посередине длины
щупальца имеется небольшая присоска,
при помощи которой медуза прикрепляет-
ся к различным подводным предметам.
Крестовички обитают в Японском море
и вблизи Курильских островов. Они
обычно держатся на мелководье. Их
излюбленные места — заросли морской
травы зостеры. Здесь они плавают и по-
висают на травинках, прикрепившись
своими присосками.Иногда они попадают-
ся и в чистой воде, но обычно невдалеке
от зарослей зостеры. Во время дождей,
когда морская вода у берегов значитель-
но опресняется, медузы погибают. В дож-
дливые годы их почти нет, зато к концу
засушливого лета крестовички появляют-
ся массами.
Хотя крестовички могут свободно пла-
вать, обычно они предпочитают подсте-
регать добычу, прикрепившись к какому-
либо предмету. Поэтому, когда одно из
щупалец крестовичка случайно коснется
тела купающегося человека, медуза уст-
ремляется в этом направлении и пытает-
ся прикрепиться при помощи присосок и
стрекательных капсул. В этот момент
купальщик ощущает сильный «ожог»,
через несколько минут кожа на месте
прикосновения щупальца краснеет, по-
крывается волдырями. Почувствовав
«ожог», нужно немедленно выходить из
воды. Уже через 10—30 минут наступает
общая слабость, появляются боли в поя-
снице, затрудняется дыхание, немеют
руки и ноги. Хорошо, если берег близко,
иначе можно и утонуть. Пораженного
нужно удобно положить и немедленно
вызвать врача. Для лечения применяют
подкожные инъекции адреналина и эфед-
рина; в наиболее тяжелых случаях при-
меняется искусственное дыхание. Бо-
лезнь длится 4—5 суток, но и после этого
срока люди, пораженные маленькой ме-
дузой, долго еще не могут окончательно
поправиться.
Особенно опасны повторные ожоги.
Установлено, что яд крестовичка не толь-
ко не вырабатывает иммунитета, а, на-
против, делает организм повышенно чув-
ствительным даже к небольшим дозам
того же яда. Это явление известно в ме-
дицине под названием а н а ф и л о к с и и.
Уберечься от крестовичка довольно
трудно. В местах, где обычно купается
много людей, для борьбы с крестовичком
выкашивают зостеру, огораживают ку-
пальни мелкоячеистой сеткой, вылавли-
вают крестовичков специальными сетя-
ми.
Интересно отметить, что такими ядови-
тыми свойствами обладают крестовички,
обитающие только в бассейне Тихого
океана. Очень близкая форма, относя-
щаяся к тому же виду, но к другому под-
виду, живущая на американском и ев-
236
ропейском побережьях Атлантического
океана, совершенно безвредна.
Наряду с животными, которые легко
поражаются стрекательными клетками,
имеются такие, на которых яд кишечнопо-
лостных совершенно не действует. Обыч-
но эти животные поселяются в сообщест-
ве с кишечнополостными, которые слу-
жат для них надежной защитой, иногда
же подобное сожительство взаимно вы-
годно для обоих.
ЖИЗНЕННЫЙ ЦИКЛ
КИШЕЧНОПОЛОСТНЫХ,
ЧЕРЕДОВАНИЕ ПОКОЛЕНИЙ
Из современных нам кишечнополостных
животных наиболее просто завершается
жизненный цикл коралловых полипов.
Оплодотворенное яйцо начинает дро-
биться. Сначала оно делится надвое, затем
каждая из образовавшихся клеточек
в свою очередь также разделяется, и так
далее. В результате образуется большое
количество мелких клеток, расположен-
ных в один слой и имеющих вид малень-
кого полого шарика. Вслед за этим часть
клеток начинает погружаться внутрь,
в результате чего получается двуслойный
зародыш. Из его внутреннего слоя впо-
следствии формируется энтодерма, а из
наружного — эктодерма будущего полипа.
Эктодерма покрыта многочисленными мел-
кими ресничками, при помощи которых
зародыш получает способность плавать;
с этого момента он превращается в личин-
ку, называемую планулой. Планула не спо-
собна питаться и размножаться. Она неко-
торое время плавает в толще воды, затем
садится на дно, прикрепляясь к нему перед-
ним концом. Вскоре после этого на зад-
нем (теперь уже верхнем) конце планулы
прорывается ротовое отверстие и образует-
ся венчик щупалец. Так возникает первый
полип. У колониальных форм этот полип
вскоре выпочковывает на себе других
полипов, те в свою очередь следующих
и т. д. Возникает колония. По достиже-
нии колонией определенной степени раз-
вития составляющие ее полипы начинают
размножаться также и половым путем,
образуя яйца. На этом цикл замыкается.
Иначе протекает жизненный цикл
у гидроидных и сцифоидных. Дробление
яйца и развитие планулы происходит
у них так же, как у коралловых полипов,
аналогичным способом возникает и первый
полип — основатель колонии, а потом и
вся колония (у сцифоидных колонии
обычно не образуется). Однако полипы
гидроидных и сцифоидных совершенно
лишены способности к половому размно-
жению, зато они выпочковывают или
отделяют не только подобных себе поли-
пов, но совершенно не похожих на них
медуз. Медузы и являются половыми
особями. Образование медуз у гидроид-
ных и сцифоидных происходит по-раз-
ному. У гидрополипов медузы возникают
из почек. Вначале такая почка внешне
похожа на обычную почку, из которой
развивается полип, но затем появляются
отличия. Медузоидная почка сильно уве-
личивается в размере, становится полу-
прозрачной, внутри ее появляются 4 ра-
диально расположенных канала и отходя-
щий от места их скрещивания ротовой
хоботок. Теперь молодая медузка внешне
напоминает маленький колокольчик или
зонтик. Вскоре она отрывается от коло-
нии полипов и уплывает. Медузы само-
стоятельно питаются и увеличиваются
в размерах, затем у них возникают поло-
вые железы. Яйца и сперма выбрасыва-
Рис. 149. Схема жизненного цикла гидроидных
(Согупе):
1 — полипы, випочковывающие медуз; 2 — свобод-
ноплавающая медуза; з — яйцо, 4 — планула.
237
ются прямо в морскую воду, где происхо-
дит оплодотворение яиц. Из яиц возникает
личинка — планула. На рисунке 149 пока-
зан типичный жизненный цикл на при-
мере гидроида из рода Корине (Согупе
sarsii).
У сцифоидных, как правило, полипы
(их называют сцифистомами) не образуют
колоний: возникшие в результате почко-
вания полипчики отделяются от материн-
ской сцифистомы и ведут самостоятель-
ный образ жизни. Вскоре каждый такой
полип претерпевает серьезные изменения:
его щупальца укорачиваются, а на теле
появляется ряд кольцевидных перетяжек,
которые становятся все более и более
глубокими, как это показано на рисунке
150. Затем начиная с верхнего конца от
полипа отделяются дисковидные медузы
(на этой стадии их называют эфирами),
которые переворачиваются ротовой сто-
роной вниз и уплывают. Эфиры питаются,
Рис. 150. Схема жизненного цикла сцифоидных
(Chrysaora):
J — сцифистома; 2 — сцифистома, выпочковывающая
молодых сцифистом; з — сцифистома в стадии строби-
лы; 4 — эфира; 5 — медува; 6 — яйцо; 7 — планула.
растут и превращаются в крупных поло-
возрелых сцифоидных медуз, размножаю-
щихся исключительно половым путем.
Таким образом, на примере гидроидных
и сцифоидных мы имеем возможность по-
знакомиться с весьма своеобразным явле-
нием, называемым чередованием
поколений. Действительно, в жиз-
ненном цикле этих двух групп кишечно-
полостных наблюдается правильная смена
донного, сидячего, часто колониального
полипоидного поколения, размножающе-
гося вегетативно, с поколением свободно-
плавающим, одиночным, медузоидным, раз-
множающимся половым путем. Разные
поколения ведут различный образ жизни
и устроены неодинаково, поэтому до от-
крытия явления чередования поколений
их считали самостоятельными группами
животных.
Чередование поколений имеет очень
большое значение для жизни гидроидных
и сцифоидных: прикрепленный образ
жизни полипов препятствует их расселе-
нию, а медузы, покинув колонию, могут
далеко уноситься морскими течениями
и разбрасывать яйца на очень большом
пространстве, заселяя все новые и новые
области. Чередование различно устроен-
ных поколений довольно широко встре-
чается в животном мире. Кроме кишечно-
полостных, оно обнаружено у некоторых
плоских, круглых и кольчатых червей,
у ряда ракообразных и даже у низших
хордовых.
Именно у представителей хордовых —
салъп — известный зоолог и поэт Ш а м и с-
с о в 1819 г. открыл чередование поколе-
ний. Это открытие повлекло за собой по-
иски новых случаев чередования поколе-
ний. В 1841 г. М. С а р с описал чередо-
вание поколений у двух сцифоидных ме-
дуз— аурелии (Aurelia) и цианеи (Суапеа),
а годом позже Стенструп установил,
что гидроидные также развиваются с чере-
дованием поколений.
Исследования Шамиссо, Сарса и Стен-
струпа явились значительным событием
для зоологов прошлого века, благодаря
им пришлось пересмотреть многие старые
представления о биологии и систематике.
Описанный выше способ чередования
поколений у кишечнополостных назы-
238
вают метагенезом, а животных
с этим типом жизненного цикла— мета-
генетическими. Для метагенеза
характерно правильное чередование поло-
вого поколения с поколением, размно-
жающимся вегетативно, путем деления
или почкования.
Не следует думать, что жизненный цикл
у всех видов гидроидных и сцифоидных
идет строго по той схеме, с которой мы
уже ознакомились. В большинстве слу-
чаев от такого типичного проявления мета-
генеза имеются более или менее заметные
отклонения, зависящие от целого ряда
причин.
Главной причиной, вызывающей на-
рушение правильного метагенеза, следует
считать различие условий среды, в кото-
рых обитают полипы и медузы. В самом
деле, придонный, неподвижный образ
жизни полипов неизбежно вызывает у них
появление целого ряда приспособлений,
отвечающих условиям их существования.
Так как полипы неподвижно сидят на дне,
они не нуждаются в особых органах
чувств, позволяющих им на расстоянии
узнавать о приближении врага или добычи:
ни приблизиться к добыче, ни уйти от
врагов они все равно не могут. Поэтому
полипы сохранили одно только чувство
осязания, позволяющее им ощупью ло-
вить добычу, прикоснувшуюся к расстав-
ленным щупальцам. Другой способ защи-
ты полипов заключается в образовании
наружного скелета. Чем прочнее и тяжелее
скелет, тем надежнее он защищает поли-
пов и лучше удерживает всю колонию на
морском дне. Указанные выше особенности
полипов характерны не для одних только
кишечнополостных — они вырабатывают-
ся у многих прикрепленных водных
организмов. Общим свойством большин-
ства сидячих животных является также
образование колоний. Очень характерно
оно и для полипов кишечнополостных и
для сифонофор. С образованием колоний
часто связано разделение функций ее чле-
нами. Особенно ярко это проявляется
у гидроидных полипов. Только у очень
немногих видов все полипы устроены
одинаково и каждый из них может отпоч-
ковывать медуз. Гораздо чаще одни
полипы, снабженные щупальцами и ртом,
ловят для колонии добычу, но не способны
размножаться, тогда как другие лишены
щупалец и рта, зато приспособлены исклю-
чительно к почкованию медуз. Так дело
обстоит у широко распространенного гид-
роида обелии (Obelia geniculata). У таких
видов правильность метагенеза нару-
шается, так как часть полипов (кормящие
особи) не дает полового потомства.
Чем сложнее становится колония, тем
большее значение приобретает она в жиз-
ненном цикле. Роль медуз при этом умень-
шается. Большая колония способна про-
кормить не только полипов, но и медуз.
Поэтому медузы долгое время не отры-
ваются от колонии и покидают ее только
тогда, когда их половые продукты созре-
вают. Наконец, имеется много таких
видов гидроидных, колония которых на-
столько усложнена, что медузы вообще
прекращают отрываться и становятся
такими же прикрепленными организмами,
как полипы. При этом характерные при-
знаки медузы — зонтик, глазки, ка-
налы гастроваскулярной системы — по-
степенно утрачиваются и медуза ста-
новится похожей на набитый яйцами ме-
шочек. У пресноводных гидр медузы вовсе
не образуются.
Но вернемся к типичному метагенезу
и проследим за свободноплавающими меду-
зами. Их образ жизни резко отличается
от того, который ведут полипы. Медузы—
подвижные животные, и потому у них
вырабатываются приспособления для дви-
жения и ориентировки в пространстве.
В первую очередь это касается развития
зонтика и мускулатуры, а также органов
чувств. У отдельных видов эти приспособ-
ления развиты настолько хорошо, что
медузы могут далеко уплывать от тех мест,
где они начали свою самостоятельную
жизнь, отделившись от полипа. Часто
они попадают в такие места, где нет под-
ходящих условий для развития полипо-
идного поколения, например в открытое
море, над большими глубинами. Планулы
таких медуз, не найдя подходящего места
для прикрепления, погибают. Но так обсто-
ит дело далеко не со всеми видами гидро-
идных и сцифоидных медуз. Часть из них
приспособилась к жизни именно в поверх-
ностных слоях воды в открытом море, но
239
при этом изменился и их жизненный цикл.
У гидромедузы бугенвиллии (Bougain-
villia platygaster) яйца не выбрасываются
в воду, а развиваются в половой железе
самки, здесь же происходит и образование
маленьких полипчиков, которые выпоч-
ковывают новых медуз. Случай этот не
единственный, развитие полипов прямо
на половой железе медузы характерно
также и для одного вида кампанулярии
(Campanularia maccrady), и для некото-
рых других медуз. У этих видов поли-
поидное поколение хотя и имеется, но оно
перестало быть донным и не образует
больших колоний. У некоторых видов
подавление полипоидной стадии пошло
еще дальше и полипы вовсе исчезли из
жизненного цикла. Из планулы таких
медуз возникают не полипы, а такие же
медузы. Так протекает жизненный цикл
большой группы гидроидных медуз—тра-
хилид, а из сцифоидных у видов рода
пелагия (Pelagia), живущих в открытом
море.
Как видно из приведенных примеров,
метагенетический жизненный цикл весьма
пластичен. Пока он является выгодным
для процветания вида, он сохраняется,
но как только условия меняются и одно
из поколений получает некоторое пре-
имущество, другое поколение подавляется
и может совсем исчезнуть.
ОБРАЗОВАНИЕ И СТРОЕНИЕ КОЛОНИЙ
КИШЕЧНОПОЛОСТНЫХ
У многих беспозвоночных животных
существует особый тип размножения, при
котором на поверхности тела материн-
ского организма образуется бугорок в виде
разросшейся группы клеток. Этот буго-
рок увеличивается в размерах и изменяет
свою форму до тех пор, пока не станет
уменьшенной копией материнского орга-
низма. Этот тип размножения называется
почкованием. Почки могут обра-
зовываться как на внутренней, так и на
внешней поверхности тела. Внешнее
почкование гораздо шире распространено
в природе и служит весьма обычным спо-
собом размножения у кишечнополостных.
У одиночных полипов почка, достигшая
определенной стадии развития, отшнуро-
вывается и переходит к самостоятельному
существованию. Именно так размножается
обычная пресноводная гидра. На мешко-
видно-цилиндрическом теле гидры обра-
зуется вырост, в образовании которого
принимает участие как эктодермальный,
так и энтодермальный слой. Вырост, уве-
личиваясь в размерах, принимает цилинд-
рическую форму, на его верхнем конце
образуется венчик щупалец и проры-
вается ротовое отверстие. Таким образом,
на теле гидры образуется дочерняя гидра,
Таблица 8. Ядовитые кишечнополостные:
1 — «португальский кораблик» (Physalia);
2 — корнерот (Rhizostoma);
3 — хиродропус (Chirodropus);
4 — крестовичок (Gonionemus).
уже способная к самостоятельному образу
жизни, но обладающая несколько мень-
шими размерами. Часто, однако, на теле
такой дочерней гидры, еще не успевшей
отделиться от материнской, в свою оче-
редь образуется новая почка. Процесс
почкования может пойти еще дальше,
и тогда возникает уже довольно слож-
ный комплекс из нескольких соединенных
между собой гидр с общей для всех них
гастральной полостью.
Описанный комплекс гидр обладает
большой целостностью и физиологической
самостоятельностью. С другой стороны,
это сложное целое заведомо делимо, ибо
каждая пз молодых гидр представляет
собой потенциально самостоятельный орга-
низм. И действительно, когда наступает
какой-то момент, отдельные полипы
начинают один за другим отшнуровы-
ваться и переходить к самостоятельному
образу жизни. Рассмотренный комплекс
называют временной колонией.
Гораздо чаще, однако, у кишечнопо-
лостных наблюдается образование на-
стоящих, или постоянных, колоний.
Колонии кишечнополостных удивительно
разнообразны. Среди них могут быть сте-
лющиеся и древовидные, перистые и в фор-
ме ершика, изящные и тонкие колонии
Таблица 9. Коралловые полипы на дне Средизем-
ного моря:
1 — лошадиные актинии (Actinia equina),
2 — сидячая сцифоидная медуза (Lucernaria
campanula),
3 — благородный коралл (Corallium rubrum);
4 — актиния анемон (Anemonia sulcata),
5 — актиния метридиум (Metridium senile);
6 — актинии церактис (Ceractis aurantiaca),
7 — актиния кладактис (Cladactis costata);
8 — коралл органчик (Tubipora);
9 — цериантария (Cerianthus membranaceus);
10 — коралл морская рука (Alcyonium),
11 — морское перо (Pennatula phosphorea);
12 — рак-отшельник пагурус (Pagurus arrosor)
с актиниями каллиактис (Calliactis pa-
rasitica);
13 — рак-отшельник (Eupagurus prideaxi) с ак-
тинией адамсия (Adamsia palliata).
и колонии с массивным известковым ске-
летом. Разнообразна и окраска колоний.
Форма колоний зависит прежде всего
от характера скелета и степени интегра-
ции колонии. Все полипоидные колонии
(за исключением морских перьев) прочно
прикрепляются к грунту или к каким-
нибудь твердым предметам, лежащим на
грунте. Прикрепляются колонии при
помощи расширенного основания. У гид-
роидов таким основанием служит перепле-
тение ползучих нитей, илп г и д-
р о р и з ы. Оно образуется сразу после
оседания личинки гидроида на грунт или
иной субстрат. От гидроризы отпочко-
вываются первые полипы колонии. У ко-
ралловых полипов образование специаль-
ных нитей основания не наблюдается.
Колонии их прикрепляются при помощи
мясистой мягкой подошвы или при помощи
расширенной известковой или роговой
базальной пластинки. Среди глубоковод-
ных шипастых кораллов (Antipatharia)
имеется широко распространенный в океа-
не вид Bathypathes patula, представ-
ленный двумя экологическими формами.
У одной из них основание в виде базаль-
ной пластинки, основание другой крючко-
образно загнуто. По этому признаку обе
формы раньше рассматривались даже как
10 16 Жизиь животных, т. 1
два самостоятельных вида. Образование
той или иной формы основания этих корал-
лов определяется тем, на какой субстрат
осела их личинка. Большую изменчивость
формы основания колоний можно найти
и среди других кишечнополостных. Напри-
мер, мягкий коралл Eunephthya, обычно
поселяющийся на камнях и скалах мелко-
водных горизонтов северных морей, имеет
мясистую широкую подошву. У того же
коралла, развивающегося на жидких
илистых грунтах, основание разрастается
в несколько пузырчатых полостей, захва-
тывающих порции грунта. Эти полости
похожи на заполненные песком мешочки
и служат колонии тем балластом, который
помогает ей сохранять вертикальное поло-
жение (рис. 151).
Прикрепительная пластинка не обра-
зуется только у представителей отряда
морских перьев. У этих животных на ниж-
нем конце стебля образуется мускулистое
расширение, при помощи которого коло-
ния внедряется в грунт. Морские перья —
единственные колониальные коралловые
Рис. 151. Коралл Eunephtia, поселившийся на
илистом грунте.
полипы, могущие самостоятельно пере-
двигаться с одного места на другое. Дви-
жение по грунту осуществляется волно-
образными сокращениями расширенного
основания колонии.
Наиболее просто устроены стелющиеся
и коркообразные колонии. В таких коло-
ниях отдельные полипы отходят непосред-
ственно от сетчатой, нитевидной гидроризы
(у гидроидов) или от мембрановидной
мягкой пластинки, покрывающей суб-
страт (у мягких восьмилучевых корал-
лов или зоантарий). В этих случаях ни
нити гидроризы, ни мембрановидная пла-
стинка не принадлежат какому-нибудь
одному полипу, а представляют собой
общее тело всей колонии — ц е н о с а р к.
Наиболее яркими примерами таких коло-
ний являются гидроиды Hydractinia или
Perigonimus и мягкие восьмилучевые
кораллы Clavularia. Гораздо чаще,
однако, кишечнополостные образуют дре-
вовидные колонии. Тогда от базальной
пластинки или гидроризы отходит несущий
ветви ствол, состоящий из одной или не-
скольких соединенных между собой тру-
бок.
Тип ветвления, а следовательно, и об-
щая форма колонии зависят от места рас-
положения почек или зон роста. У некото-
рых гидроидов, чаще всего у тех, полипы
которых не защищены чашечкой, и у мор-
ских перьев центральный ствол колонии
образуется путем увеличения в длину
ножки самого первого после оседания
личинки полипа. Он остается всегда на
вершине колонии. Непосредственно под
ним локализуется зона роста и почкова-
ния новых, вторичных, полипов. В свою
очередь на ножках вторичных полипов,,
непосредственно под ними также имеется
зона роста и почкования. За счет этих
зон образуются ветви колоний и покры-
вающие их полипы (рис. 152). Ветвление
таких моноподиальных, как их называют,
колоний может быть в простейших слу-
чаях беспорядочным, тогда ветви и полипы
отходят от ствола во все стороны.
Так ветвятся колонии многих гидро-
идов и наиболее просто организованные
колонии морских перьев. У более слож-
ных колоний морских перьев вторичные
полипы отходят от ствола в одной или
242
Рис. 152. Схема роста и почкования колоний кишечнополостных (на примере гидроидных полипов):
% — стелющаяся колония; 2 — моноподиальная колония с полипопдными зонами роста; з — симподиальная
колония; 4 — моноподиальная колония с ценосаркальными зонами роста и почкования. Черным обозначены зоны
роста, пунктиром — места почкования.
нескольких плоскостях и на равном друг
от друга расстоянии или от так называе-
мых «листьев», т. е. боковых выростов,
также расположенных правильными ря-
дами. Такое правильное расположение вто-
ричных полипов и «листьев» колоний
обеспечивается тем, что почкование про-
исходит только в одной плоскости.
Некоторые более высокоорганизованные
гидроиды и коралловые полипы обладают
другим типом ветвления колоний. Зона
почкования у них также расположена
ниже чашечки полипа, но ножка послед-
него имеет ограниченный рост. Поэтому,
после того как первый полип сформиро-
вался, под его основанием начинает обра-
зовываться ножка следующего полипа,
растущего вверх и в сторону. Иногда
в зоне роста одновременно возникает
несколько ножек, дающих начало не-
скольким веточкам. При таком типе вет-
вления, называемом симподиаль-
н ы м, наиболее старые полипы остаются
у основания колоний и их ветвей. Для
симподиальных колоний характерен также
постоянный перелом продольной оси
ствола и ветвей в местах отхождения новых
полипов (рис. 152). Так растут и ветвятся
многие гидроиды, полипы которых защи-
щены чашечками, и некоторые коралловые
полипы. Но высшим типом ветвления
колоний гидроидов и коралловых поли-
пов является такой, при котором зона
роста и почкования находится не ниже
первичного полипа, а на вершине выроста
гидроризы или базальной мембраны.
В этом случае функция роста и почкова-
ния переходит к общему телу коло-
нии, или ценосарку. Основной
ствол растет вверх, постепенно отпочко-
вывая от себя полипов или образуя новые
ценосаркальные выросты, боковые ветви.
Для колоний гидроидов с ценосаркальным
ростом очень характерно то, что отдельные
полипы лишены ножек и в большей или
меньшей степени погружены в толщу
ветвей колонии. Ценосаркальный рост
колоний мы встречаем, кроме гидроидов,
у большинства коралловых полипов.
У более высокоорганизованных коло-
ний ветви и полипы, как правило, упоря-
дочены в своем расположении, у некоторых
гидроидов ветви расположены группами,
в пределах которых они расходятся ра-
диально во все стороны и лежат в одной
плоскости, перпендикулярной стволу.
Такие колонии называются мутовча-
тыми. Весьма обычен тип ветвления,
при котором боковые веточки отходят
от противоположных сторон ствола. При
этом основания ветвей могут находиться
точно друг против друга, или ветви одного
ряда могут быть смещены вверх от осно-
ваний ветвей противоположного ряда.
У гидроидов в таком случае образуются
изящные перистые колонии, а у некоторых
горгониевых кораллов и антипатарий,
обладающих более крепким известковым
или роговым скелетом, вееровидные тг
пластинчатые колонии. При этом сосед-
16*
243
пие веточки часто сливаются между собой,
и тогда эти обычно очень ярко окрашен-
ные вееровидные колонии принимают
форму решетчатой пластинки. Иногда
такие пластинки волнообразно изги-
баются, напоминая складки занавеса.
Встречаются колонии, изогнутые пластин-
ки которых образуют еще побочные пла-
стинки. Среди восьмилучевых кораллорых
полипов очень любопытные колонии
образует так называемый коралл орган-
чик (Tubipora). Скелет органчика со-
стоит из длинных, поднимающихся от
базальноп пластинки известковых тру-
бок, образованных из слившихся извест-
ковых скелетных телец. Эти трубки, напо-
минающие трубки органа, кроме того,
соединены между собой на разных уровнях
поперечными пластинками. И трубки
и соединяющие их пластинки окрашены
в красивый малиново-красный цвет.
При сильном развитии известкового
скелета отдельные ветки сливаются друг
с другом, и вся колония тогда приобре-
тает форму массового монолита. Такие
колонии образуются среди группы гидро-
кораллов, у которых обычный хитиноид-
ный скелет пропитан известью и значи-
тельно утолщен, и у шестилучевых рифо-
образующих кораллов. Мощные коралло-
вые полипняки образуют рифы и даже це-
лые коралловые острова. Недаром же
всего 200 лет назад считалось, что корал-
ловые рифы принадлежат к неорганиче-
ской природе, представляя собой особые
минеральные конкреции! У наиболее мас-
сивных мадрепоровых кораллов скелетные
чашечки полипов, сближаясь, сливаются
между собой, и тогда на поверхности по-
липняка возникают причудливо изви-
вающиеся линии с красивой поперечной
исчерченностью.
Строение полипов и характер располо-
жения их на ветвях колоний тесно свя-
заны с формой самой колонии, а также
с питанием. Все кишечнополостные — пло-
тоядные животные. Они питаются планк-
тонными организмами, мелкими беспо-
звоночными, ведущими придонный образ
жизни, взвешенными в воде остатками
животных и даже мелкими рыбками и их
личинками. Большинство колониальных
кишечнополостных питается мелкими
планктонными животными. У наиболее
примитивных колоний (в виде стелющей-
ся на грунте пластинки) полипы приобре-
тают вытянутую цилиндрическую форму
или высоко поднимаются на тонкой ножке.
Такое приподнятое над пластинкой поло-
жение полипов облегчает им ловлю добычи.
С другой стороны, длинная ножка или
вытянутая форма полипа делает их более
уязвимыми для тех хищников, которые
ими могут питаться. Поэтому эволюция
колониальных форм шла в направлении
приобретения древовидных стволов и одно-
временно сокращения длины ножки. Тем
не менее еще у многих более просто орга-
низованных гидроидов с поднимающи-
мися вверх стволами сохраняются
тонкие длинные ножки полипов. Многие
стелющиеся колонии находят более удоб-
ный способ для успешного облова при-
донных вод, поселяясь на стволах дру-
гих, более высокоорганизованных коло-
ний кишечнополостных. При этом необ-
ходимость образования ножек или соб-
ственных прямостоящих стволов у таких
кишечнополостных отпадает, а отдаление
от грунта осуществляется благодаря ис-
пользованию стволов и ветвей других
колоний. Колони и-э п и ф и т ы встре-
чаются среди гидроидов и зоантарий.
У наиболее высокоорганизованных гидро-
идов, образующих крепкие древовидные
или простые колонии, ножки полипов
исчезают полностью, а чашечки полипов
или прирастают одной стороной к стволу,
или же полностью погружаются в толщу
ветви или ствола. Полипы восьмилучевых
кораллов или способны втягиваться в тело
колонии, или защищены специальными
скелетными образованиями.
Успешность облова водного простран-
ства полипами зависит не только от того,
насколько высоко они приподняты над
грунтом, но и от общей формы колонии.
Представим себе густо и беспорядочно
разросшийся куст гидроида и сравним
положение полипов, сидящих на концах
ветвей и в самой их чаще. Естественно, что
те полипы, которые расположены на кон-
цах ветвей, находятся в более выгодных
условиях и скорее могут поймать добычу,
чем те, которые расположены в чаще
куста. Поэтому дальнейшее упорядоче-
244
ние ветвей, о котором мы уже говорили,
способствует лучшему питанию колоний.
Но одного этого тоже недостаточно. Упо-
рядочение ветвей коррелятивно связано
с упорядочением положения на них отдель-
ных полипов. Если полипы будут сидеть
слишком тесно, они будут мешать друг
другу при ловле добычи. Гораздо успеш-
нее должен проходить лов добычи при
расположении полипов в продольных
рядах и в шахматном порядке. И действи-
тельно, большинство колоний кишечно-
полостных имеет двухрядное положение
полипов. У многих гидроидов, кроме того,
часто встречается такое положение поли-
пов, при котором отверстия их чашечек
попеременно направлены в разные сто-
роны. Путем удвоения рядов могли воз-
никнуть колонии с четырьмя и более ря-
дами полипов. В наиболее высокоорга-
низованных колониях наблюдается много-
рядное положение полипов. В таких рядах
полипы располагаются в шахматном
порядке. Однако известны случаи, когда
расположение полипов в один ряд дает
колонии значительно большие преимуще-
ства, чем многорядное. В частности, та-
кое положение полипов удалось обнару-
жить на глубоководных колониях антипа-
тарии (Bathypathes lyra). Строение ее
будет описано в соответствующем разделе.
Среди колоний коралловых полипов мы
находим и такие, полипы которых распо-
ложены в полном беспорядке. Однако
в этом случае наблюдается отчетливая
разноэтажность венчиков щупалец, обес-
печенная разной длиной ловчих полипов.
При этом ни один из полипов, даже пол-
ностью расправивший свои щупальца,
не мешает своему соседу — в одном и том
же этаже полипы сидят на достаточном
расстоянии друг от друга.
Итак, исследование строения колоний
гидроидных и коралловых полипов пока-
зывает, что в природе отчетливо выражено
стремление к упорядочению ветвей и по-
липов и увеличению их числа. Для неко-
торых групп кишечнополостных обнару-
жена определенная связь между числом
особей в колонии и размерами отдель-
ных полипов. Как правило, чем больше
особей в колонии, тем меньше их раз-
меры.
Колонии одного и того же вида, обита-
ющие в полярных районах, обладают
меньшим числом ветвей и полипов, чем
колонии, живущие в теплых водах. Раз-
мер полипов у первых заметно крупнее,
чем у вторых. Особенно отчетливо про-
текает увеличение размеров у видов, завое-
вывающих большие глубины океана. Ис-
пользуя многие роды морских перьев,
имеющих своих представителей как в мел-
ководной, так и глубоководных зонах
океанов, можно построить яркие морфо-
логические ряды, иллюстрирующие этот
процесс. Так, например, у преимущест-
венно мелководного вида из рода
Kophobelemnon — К. stelliberum, высота
колоний которого достигает 300—350 мм,
на стволе можно насчитать несколько
десятков полипов. В расправленном со-
стоянии они достигают длины 6—10 мм.
У близкого, но более глубоководного
вида К. polyflorum, обитающего на глу-
бине 1000—3500 м, колонии достигают
140 мм в высоту, количество же полипов
при этом сокращается в десятки раз. Чаще
всего на стволе этих кодоний образуется
всего лишь три полипа, длина которых в
расправленном состоянии достигает 22 мм.
Наконец, экспедицией на судне «Витязь»
во многих местах Тихого океана и в Бе-
ринговом море ка глубинах от 2843 до
4070 м были найдены многочисленные
колонии дотоле неизвестного вида К. bif-
lorum, на стволе которого имелось только
два очень крупных полипа. Высота этих
колоний достигала 150 мм, а длина поли-
пов в расправленном состоянии — 33 мм.
Два года назад, во время плавания «Витя-
зя» в Индийском океане, удалось под-
нять с глубины 3500 и 4800 м две очень
интересные колонии умбеллул, также ока-
завшиеся представителями нового вида.
На вершине ствола этих умбеллул имелся
всего лишь один полип, но огромных
размеров. В расправленном виде длина
его достигает 125 мм\ У близких видов
умбеллул — U. thomsoni и U. dutissima,
обитающих на глубине 1300—5600 и
1000—3000 м, имеется соответственно 4—7
и 3—14 полипов гораздо более мелких
размеров. Таким образом, новый вид
умбеллул, названный U. monocephalus
(одноглавая), представляет собой крайний
245
пример этого ряда. Процесс со-
кращения количества полипов
при одновременном возрастании
их размеров прослеживается и
на представителях одного и то-
го же вида, обитающего на
разных глубинах океана. Раз-
личие в строении колоний,
числе и размерах полипов от-
дельных форм, обитающих на
мелководье и на больших глу-
бинах океана, бывает настолько
велико, что такие сильно отли-
чающиеся друг от друга фор-
мы рассматривались многими
учеными как самостоятельные
виды.
До сих пор говорилось о
колониях с точки зрения их об-
щего плана строения. Были рас-
смотрены способы укрепления
колоний и их ветвление, поря-
док расположения. Теперь
нужно остановиться на еще од-
ном очень важном свойстве ко-
лоний кишечнополостных. У
этих своеобразных животных
полипы далеко не всегда бы-
вают одного и того же строения.
Очень часто отдельные поли-
пы имеют различную форму и
играют различную роль в жиз-
ни колонии. Колонии, на ко-
торых образуются разные по
строению и функциям полипы,
называют полиморфны -
м и.
Чаще всего полиморфизм на-
блюдается среди гидроидов. В
силу весьма характерного для
этих кишечнополостных чередо-
вания поколений, или метаге-
неза, на колониях, кроме питаю-
щих полипов (о которых говори-
ли выше), отпочковываются и
особи медузоидного поколения.
Полипы, которые приспособлены для от-
почковывания медуз, значительно отлича-
ются от обычных питающих колонию по-
липов. Они лишены щупалец и ротового
отверстия. Но полиморфизм может идти
гораздо дальше. Тогда на ценосарке об-
разуются чрезвычайно сильно видоизме-
Рис. 153. Строение верх-
ней части колонии сифо-
нофоры (схематизиро-
ванно):
1 — пневматофор; 2 — не-
ктофоры; 3 — пальпаку-
ла; 4 — пальпон; 5 — ар-
канчик; 6 — гастрозоид;
7 — ветвь аркаичика со
стрекательной батареей;
8 — кроющие пластинки,
9 — ствол колонии; 10 —
места прикрепления крою-
щих пластинок; 11 — гоно-
форы.
КОЛОНИИ.
ненные полипы, несущие защит-
ную функцию. Они имеют фор-
му простых или разветвленных
нитей, также без рта и щу-
палец. Но у них особое раз-
витие приобретают стрекатель-
ные капсулы, с помощью кото-
рых полипы могут поражать
врага или добычу. Такие по-
липы, называемые дактило-
зоидами, располагаются в
определенном порядке вокруг
питающих полипов и полипов,
производящих медуз.
Полиморфизм свойствен и
многим восьмилучевым корал-
лам. Но здесь он имеет ограни-
ченный размах, поскольку функ-
ция полового размножения пе-
реходит к обычным кормящим
полипам, или аутозоидам, аме-
дузоидное поколение вообще
отсутствует. В отношении ко-
ралловых полипов правильнее
говорить не о их полиморфизме,
а о диморфизме, так как на
большинстве их колоний обра-
зуются полипы лишь двух ти-
пов: упомянутые выше аутозои-
ды и очень мелкие полипы, час-
то совсем лишенные щупалец,
носящие название с и ф о н о -
з о и д о в. У этих зооидов пе-
регородки в кишечной полости,
столь характерные для корал-
ловых полипов, значительно
редуцированы, зато сифоно-
глиф развит очень сильно. Сифо-
нозоиды не могут добывать пи-
щу и переваривать ее. Функ-
ция сифонозоидов заключает-
ся в том, что они при помощи
биения ресничек сифоноглифов
создают мощный ток воды в
системе каналов, связывающих
между собой отдельные части
С этим током воды по колонии
разносятся питательные вещества и необ-
ходимый для дыхания кислород. Таким об-
разом, мы видим, что у восьмилучевых корал-
лов полиморфизм полипов выражен довольно
слабо. У колониальных шестилучевых ко-
раллов полиморфизм и вовсе не наблюдается.
246
Высшим типом полиморфных коло-
ний являются не гидроиды и коралловые
полипы, а очень своеобразная группа
свободноплавающих кишечнополостных —
сифонофоры. Эти колониальные
животные, которые чрезвычайно разно-
образны по своему строению, окраске
и внешнему виду, состоят из полого ствола,
на котором выпочковываются многочис-
ленные полипоидные и медузоидные особи,
выполняющие различные функции и отли-
чающиеся друг от друга своей формой
и строением.
В самой верхней части колонии выпоч-
ковывается медузоидная особь — пнев-
матофор, или плавательный
пузырь. Он может быть очень малень-
ким (не более 1—20 мм), но иногда до-
стигает 20—30 см. У одних сифонофор
пневматофор замкнут, у других он имеет
пору. Форма пневматофора варьирует от
почти шаровидной, овальной или груше-
видной до асимметричной. Иногда на
поверхности пневматофора образуются раз-
личные выросты, или гребни. Пневмато-
фор служит для большинства колоний сифо-
нофор гидростатическим аппаратом. Жи-
вотное может регулировать количество
газа, содержащегося в пневматофоре, вы-
пуская его через пору, снабженную замы-
кательной мышцей, либо вновь выраба-
тывая особыми железистыми клетками.
Увеличивая или уменьшая объем пневма-
тофора, сифонофора способна подни-
маться к водной поверхности либо опу-
скаться в глубь океана. Для некоторых
сифонофор, обитающих на поверхности
моря, пневматофор служит парусом.
Сразу же под пневматофором у боль-
шинства сифонофор выпочковываются дру-
гие медузоидные особи — нектофоры, или
плавательные колокола (в количестве от
10 до 400 в каждой колонии). У некоторых
видов пневматофор отсутствует, и тогда
нектофоры располагаются в самой верх-
ней части колонии. Нектофоры, последо-
вательно сокращаясь и расширяясь, пере-
двигают колонию в горизонтальном направ-
лении, но могут, так же как пневматофор,
способствовать перемещению сифонофоры
вверх или вниз.
В нижней половине ствола размещаются
остальные особи колонии. Среди них мож-
но различить питающие полипы, или га-
строзоиды, у основания каждого из кото-
рых имеется лишь одно длинное щупальце,
называемое арканчиком. Арканчик, как
правило, ветвится, и на его ветвях нахо-
дятся скопления стрекательных клеток.
Если ветви отсутствуют, то стрекательные
клетки располагаются в виде бляшек
на самом арканчике.
Кроме гастрозоидов, у многих сифоно-
фор имеются полипы другого сорта —
пальпоны и цистозоиды. Они
устроены проще гастрозоидов и иногда
даже лишены рта. Их щупальца не вет-
вятся, но, так же как арканчики, снаб-
жены стрекательными клетками. Предпо-
лагается, что пальпоны и цистозоиды
выполняют чувствительные и выдели-
тельные функции.
У большинства колоний сифонофор
имеются медузоидные особи, играющие
защитную роль. Это кроющие пластинки.
Они очень сильно изменены и совсем не
похожи на медуз.
Наконец, постоянной составной частью
колонии являются особи, несущие функ-
цию полового размножения. Они могут
иметь вид хорошо развитых медузок с ра-
дикальными каналами и ротовым хобот-
ком, в стенках которого формируются
половые продукты. У некоторых сифоно-
фор такие медузоиды даже могут отры-
ваться и свободно плавать, пока половые
продукты в них не созреют. Но гораздо
чаще зонтик медузоидов редуцируется
и даже срастается с ротовым хоботком.
Очень важно отметить, что у наиболее
сложно устроенных сифонофор часть из
перечисленных особей располагается на
стволе небольшими группами, называе-
мыми кормидиями. В каждый
кормидий обязательно входят полипы,
выполняющие функции питания, и поло-
вые медузоиды. Кормидий могут от-
деляться от основной колонии и вести
самостоятельную жизнь. В этом случае
кормидий называют эвдоксиями или эр-
сеями.
Прежде отдельные кормидий считались
даже самостоятельными видами сифоно-
фор. Теперь, после того как рассмотрено
строение различных колоний кишечно-
полостных, интересно вновь вернуться
247
к вопросу о подчинении отдельных частей
индивидуальности всей колонии, который
был уже затронут в начале этого раздела.
В процессе постепенного усложнения
строения колоний отдельные полипы все
больше и больше теряют свое индиви-
дуальное значение и лишь выполняют ту
роль, которая диктуется всей колонией
в целом. Особенно отчетливо такая подчи-
ненность отдельных полипов проявляется
в полиморфных колониях. Полипы в таких
колониях уже и потенциально не способны
нести самостоятельный образ жизни. В по-
лиморфных колониях функции ее членов
строго распределены между собой и вместе
с тем тесно связаны. Постепенная инте-
грация колоний наблюдается во всех
основных группах кишечнополостных.
В классе гидроидных наиболее совершен-
ными в этом отношении колониями явля-
ются сифонофоры. По сути дела, их поли-
морфная колония превратилась в настоя-
щий самостоятельный организм, способ-
ный к активному направленному пере-
движению. Среди коралловых полипов
высшей степенью подчинения отдельных
членов всей колонии в целом обладают
морские перья, также превращающиеся
в самостоятельный прекрасно слаженный
организм. И для морских перьев чрезвы-
чайно характерна способность к само-
стоятельному, активному передвижению
по грунту, способность укрепляться в нем.
К сифонофорам и морским перьям уже
вполне приложимо понятие особи, ибо
эти колонии представляют собой единое
морфо-физиологическое целое, в котором
индивидуальность отдельных частей пол-
ностью отсутствует.
Как же осуществляется общая регуля-
ция деятельности колоний у кишечно-
полостных, этих наиболее просто устроен-
ных животных? Этот вопрос еще ждет
своего решения, так как в настоящее
время знания в области анатомии и физио-
логии кишечнополостных еще недостаточ-
ны для этого. Но многие эксперименты
и наблюдения говорят о том, что общий
регуляторный механизм в колониях суще-
ствует и оказывается тем сложнее, чем
больше степень интеграции колонии.
В простейших случаях, например в коло-
ниях гидроидов, у которых нервная си-
стема находится на очень низкой стадии
развития, регуляторная деятельность едва
выражена. Это объясняется, видимо, тем,
что отдельные полипы колоний связаны
между собой ценосаркальными
каналами. Во всяком случае, о нали-
чии общеколониальной нервной системы
у гидроидов никаких сведений до сих
пор нет, раздражение одного из полипов
колонии не передается остальным. В наи-
более просто устроенных колониях корал-
ловых полипов общеколониальная нервная
система также отсутствует и передача
раздражений также осуществляется толь-
ко по ценосаркальным каналам. Но бла-
годаря особенностям проводящей системы
передача раздражения осуществляется
на более высоком, чем у гидроидов, уров-
не. Например, раздражение одного из поли-
пов колонии Alcyonium уже передается
соседним полипам. Правда, волна сокра-
щения полипов при этом распространяется
удивительно медленно и, как правило,
захватывает только веточки, расположен-
ные в непосредственной близости от того
полипа, к которому было приложено раз-
дражение. Но по мере усложнения строе-
ния колоний и их интеграции услож-
няется и нервная система кишечнополост-
ных. У полиморфных колоний высшего
типа уже образуется общеколониальная
нервная система, связывающая самые
отдаленные друг от друга участки коло-
нии. Нервная система, пронизывающая
все тело колонии, наблюдается, как и
следовало ожидать, у сифонофор среди
гидроидных полипов и у морских перьев
среди коралловых. У этих кишечнополост-
ных животных раздражение одного по-
липа вызывает резкую реакцию всей
колонии. Как правило, эта реакция выра-
жается в сокращении колонии. У сифоно-
фор быстро втягиваются щупальца, сокра-
щается пневматофор или плавательные
колокола. Организм начинает погру-
жаться или меняет направление своего
движения, стремясь уйти от опасности.
Раздражение одного из многочисленных
полипов морского пера в течение несколь-
ких секунд распространяется по всей коло-
нии. Несколькими сильными сократи-
тельными движениями базального рас-
ширения колония втягивается глубоко
248
в грунт. Сокращается и вся верхняя часть
колонии, остающаяся над грунтом. Поли-
пы быстро исчезают в толще ствола, а по
всей колонии при этом пробегают волны
фосфоресцирующего света.
СОЖИТЕЛЬСТВО
КИШЕЧНОПОЛОСТНЫХ
С ДРУГИМИ ОРГАНИЗМАМИ
Среди кишечнополостных довольно мно-
го примеров сожительства (си-
нойкии1 и симбиоза), тогда как
число паразитов очень незначительно.
Объяснить возникновение различных
форм сожительства не столь сложно. У та-
ких организмов, как кишечнополостные,
ведущих прикрепленное существование
и имеющих ветвистую древовидную форму,
легко возникают обрастания одних орга-
низмов другими. В море всегда можно
найти высоко поднимающуюся над грун-
том крепкую древовидную колонию, ветви
которой покрыты разнообразными коло-
ниями гидроидов, альционарий, мшанок
или одиночными организмами — акти-
ниями, асцидиями, брахиоподами. Ки-
шечнополостные, как колониальные, так
и одиночные, часто обрастают шерохова-
тую грубую поверхность асцидий и их
длинных ножек, прикрепляются к поверх-
ности и к отдельным длинным спикулам
губок, покрывают слоевища водорослей
и листья морских трав и т. д. Но большин-
ство из них предпочитает какой-нибудь
один определенный вид, который они и
используют в качестве субстрата. Так,
например, колонии гидроида Filellum
serpens можно встретить на стволах почти
всех других гидроидов, но обычно эта
стелющаяся форма развивается на не-
скольких видах морских елочек — серту-
лярий. Другой гидроид — Obelia fle-
xuosa — обычно живет на слоевищах бу-
рой водоросли Fucus, хотя колонии обе-
лии можно легко встретить и на других
водорослях, и на листьях морской травы
Zostera. Видимо, таким путем совершается
1 Синойкия — сосуществование, от
греческих слов syn — вместе, oikos — дом.
в природе переход от безвыборочного
обрастания к постоянным формам сосу-
ществования (синойкии).
Гидроид Perigonimus — очень харак-
терный сожитель некоторых двустворча-
тых моллюсков. Обычно этот гидроид
поселяется на том краю раковины мол-
люска, который прикрывает его сифоны.
Моллюски, служащие хозяином для Peri-
gonimus, по типу своего питания отно-
сятся к организмам-фильтраторам, пита-
ющимся мельчайшими органическими ча-
стичками, взвешенными в морской воде-
Для того чтобы обеспечить себя пищейг
моллюск должен непрерывно прогонять
через свою мантийную полость воду, по-
ступающую через вводный и выходящую
через выводной сифон. Создаваемые при
этом токи воды, очевидно, облегчают гид-
роиду улавливание пищи. Другой стелю-
щийся гидроид — Hydractinia echinata—
является сожителем брюхоногих моллю-
сков, чаще всего хищного моллюска
Natica. Гидроид Ptilocedium repens, оби-
тающий в Тасмановом море, приспосо-
бился к существованию на морском пере
Sarcophyllum. Это очень любопытный слу-
чай комменсализма, ибо морские перья
крайне редко обрастают.
Синойкия имеется и среди мелких
восьмилучевых кораллов — альционарий.
Например, обитающий в Адриатическом
море Parerythropodium coralloides обитает
исключительно на ветвях живых колоний
горгонарии Eunicella verrucosa. В Ла-Ман-
ше эта же горгонария опять-таки служит
хозяином, но теперь уже другому ком-
менсалу — актинии Amphianthus dohrnii.
Среди самих же горгонарии настоящей
синойкии, видимо, нет, и они могут быть
лишь хозяевами для других морских орга-
низмов. Так, например, два других вида
горгонарии—Primnoa resedaeformis и Рага-
gordia arborea, широко распространен-
ные в умеренной зоне северного полушария,
в Норвежском море почти всегда служат
хозяевами колоний зоантарии Epizoanthus
norvegicus. Желтая Primnoa является,
кроме того, хозяином желтоокрашенной
формы актинии Ptichodactys patula.
Особая форма этой актинии, имеющая
синюю окраску, поселяется на синей же
горгонарии — Paramuricea placomus.
249
Очень много синойкий среди зоантарий
колониальных шестилучевых кораллов,
никогда не образующих собственного ске-
лета, но внешняя поверхность которых
бывает часто инкрустирована всевозмож-
ными посторонними частицами как орга-
нического (раковинами фораминифер и
диатомовых водорослей, спикулами губок
и коралловых полипов), так и неоргани-
ческого происхождения (песчинками, мел-
кими обломками известковых пород и т. д.).
Palythoa, один из видов этих кишечно-
полостных, является сожителем обитаю-
щей в японских водах горгонарип Coral-
lium inutile. Коралл-хозяин служит для
Palythoa не только субстратом для при-
крепления и отдаления от грунта, но, что
гораздо любопытнее, и поставщиком мате-
риала для строительства внешнего ске-
лета: Palythoa инкрустирует поверхность
своей колонии известковыми склеритами
горгонарии. Другой вид зоантарий, рас-
пространенный в Адриатическом море,—
Parazoanthus axinellae — сожитель двух
видов губок из р о д a Axinella *, хотя,
как исключение, и поселяющийся на неко-
торых других губках.
Синойкия широко распространена и
среди актиний. Выше сообщалось уже о
том, что желтоокрашенная и синяя формы
актинии Птиходактис являются соответ-
ственно сожителями горгонарии Primnoa
и Paramuricea. Примеров подобного рода
можно привести очень много. Поэтому
остановимся на наиболее интересных. При
глубоководных тралениях, проведенных
с советского исследовательского судна
«Витязь» в северо-западной части Тихого
океана на глубине 3—4 тыс. ж, очень
часто попадались лопатоногие моллюски
Scaphopoda, чьи конические саблевидно
изогнутые раковины удивительно напо-
минают по форме моржовый клык. Почти
на всех раковинах собранных моллю-
сков были найдены бесцветные тонкостен-
ные актинии с небольшим количеством
щупалец. Актинии эти поселяются на
уровне нижней трети раковины, т. е. чуть
выше той части раковины, которая посто-
янно погружена в грунт. Можно предпо-
1 См. цветную таблицу «Поселение губок на
дне Средиземного моря».
ложить, что при движении моллюска
по грунту мельчайшие организмы, живу-
щие в верхнем слое грунта, всплывают
потревоженные и становятся жертвой
актинии. Сожительство этой актинии с
лопатоногим моллюском настолько обычно,
что случайные находки раковин, свобод-
ных от актиний, каждый раз вызывали
удивление. На другом субстрате эти
животные, по-видимому, никогда не по
селяются.
Известны и еще более интересные слу-
чаи синойкий среди глубоководных акти-
ний. В начале XX в. в результате глубо-
ководных тралений в Атлантическом океа-
не, проведенных Монакским океаногра-
фическим институтом, было поймано два
экземпляра голотурии Pseudostichopus
villosus. Нетрудно представить себе удив-
ление ученых, когда они на нижней по-
верхности этих голотурий обнаружили
укоренившихся там двух небольших плот-
ных актиний). Однако сожительство акти-
нии, названной Sicyopus commensalis, ви-
димо, не случайно. На обоих экземплярах
голотурий актинии поселились в одном и
том же месте — спереди и немного сбоку от
окруженного короткими щупальцами ро-
тового отверстия. Эти уплощенные голоту-
рпи непрерывно ползают на поверхности
грунта, собирая щупальцами частички ор-
ганического детрита. Sicyopus commensalis
при этом оказывается в положении аван-
гарда, первой осваивающей новые «охот-
ничьи угодья». Любопытно, что под сня-
той с тела голотурии актинией была обна-
ружена небольшая ямка, в которую акти-
ния была как бы запрессована. В связи
с этим интересно отметить наблюдения,
проведенные над рядом других актиний,
которые показали, что их подошва выде-
ляет особый секрет, легко растворяющий
тот органический по происхождению суб-
страт, на котором они поселяются. Так,
например, актиния Cereus pedunculatus,
укрепившаяся на раковине моллюска,
постепенно растворяет ее непосредственно
под своей подошвой и через некоторое
время проваливается внутрь раковины
через образовавшееся в ней отверстие.
Возможно, что ямка в теле голотурии,
в которой укрепляется актиния, имеет
такое же происхождение. Сожитель в этом
250
случае не очень-то щадит своего хозяина.
Но еще более безжалостно обращаются
со своим хозяином актинии, поселяю-
щиеся на глубоководном морском пере
Kophobelemnon biflorum. Эти морские
перья были впервые и в большом количе-
стве найдены экспедиционным судном
«Витязь». По имеющимся данным, они
обитают вдоль побережья северной части
Тихого океана на глубине 3—4 тыс. де,
предпочитая заиленные склоны глу-
боководных впадин. На каждом из соб-
ранных экземпляров К. biflorum были
обнаружены одна или несколько актиний,
близких к Sicyopus. Так же как и в случае
сожительства с голотурией, эти актинии
разрушают своей подошвой мягкие ткани
морского пера, все глубже и глубже про-
никая в его тело. Наконец, их подошва
достигает известкового осевого стержня
колонии и плотно его обволакивает. Ком-
менсализм подобного типа оказывается
губительным для морского пера. Все его
мягкие ткани отмирают и отваливаются.
И тогда комменсал-победитель оказы-
вается сидящим на гладком осевом стерж-
не отмершей колонии на высоте 7—10 см
над уровнем грунта. В просмотренной
нами коллекции К. biflorum, собранной
экспедицией «Витязя», можно найти все
стадии описанного процесса.Имеются вели-
колепные экземпляры пера, на котором
молодые актинии только что поселились,
есть и колонии, уже приходящие в упадок.
На двух экземплярах К. biflorum от мяг-
ких его тканей сохранилась лишь самая
верхняя часть с двумя крупными аутозо-
идами, а нижняя полностью разрушена
актиниями, ставшими уже очень круп-
ными.
Наконец, в коллекции есть и не-
сколько стержней отмерших колоний,
плотно охваченных подошвой бывших со-
жителей.
Не только взрослые актинии могут
быть сожителями других животных. Ли-
чинки обитающих в северных морях Ed-
wardsia, Peachia, Haloclava и некоторых
других родов проникают под колокол
медуз и при помощи своего рта прикреп-
ляются к ротовому конусу, гонадам или
внутренней поверхности колокола. При-
крепившись к медузе, личинки плавают
вместе с ней до момента, когда наступает
время превращения их во взрослые стадии.
Если личинка по какой-либо причине теряет
своего хозяина, она способна самостоя-
тельно плавать на поверхности воды, пока
вновь не проникнет под колокол проплы-
вающей мимо медузы. Среди личинок акти-
ний, кроме сожителей медуз, существуют
и сожители гребневиков. Так, например,
некоторые личинки в Северном море часто
поселяются на гребневике Bolinopsis infun-
dibulum. Личинки одной австралийской
актинии, однако, не ограничиваются
жизнью под колоколом медузы, а про-
никают даже в радиальные каналы. Соб-
ственно говоря, такой образ жизни скорее
напоминает паразитизм, чем синойкию.
Для того чтобы выйти затем на свободу,
этим личинкам приходится разрушать
часть колокола. Да и питание этих сожи-
телей скорее всего происходит за счет
хозяина.
Высшим типом сожительства двух орга-
низмов, несомненно, является симбиоз,
приносящий обоим партнерам взаимную
выгоду. Симбиоз значительно реже встре-
чается в природе, чем синойкия, а тем
более паразитизм. Справедливо это и для
кишечнополостных животных. Если сожи-
тельство можно найти почти во всех си-
стематических группах кишечнополост-
ных, среди обитателей любых морей и
любых глубин, то этого уже нельзя ска-
зать об организмах, ведущих симбио-
тический образ жизни.
Так же как и синойкия, симбиоз возник
из простого обрастания одних организмов
другими. В случае обоюдной выгодности
такого сожительства в дальнейшем мог
выработаться инстинкт или определен-
ные приспособления в строении, облег-
чающие встречу организмов-симбионтов.
Наконец, симбиоз становится уже необ-
ходимостью для таких организмов: суще-
ствование каждого из них настолько тесно
зависит от существования другого, что
жизнь их в отдельности оказывается уже
затруднительной или невозможной.
Сказанное можно проиллюстрировать
рядом примеров. Так, гидроид Hydra-
ctinia echinata, как уже было упомянуто
выше, обычно обрастает раковины хищных
брюхоногих моллюсков, являясь их ком-
251
менсалом. Но часто Н. echinata обрастает
и раковины, служащие домиком ракам-
отшельникам. Разрастающаяся со вре-
менем гидрориза гидроида постепенно
превращается в сплошную пластинку,
причем край ее при этом перерастает устье
раковины и нависает над телом рака-
отшельника. Такое сожительство стано-
вится выгодным как для гидроида, так
и для рака-отшельника. Постоянное пере-
движение рака по дну водоема в поисках
пищи способствует лучшему газообмену
гидроида, а взмученные остатки пищи,
пожираемой раком, легко улавливаются
и поедаются полипами гидр актиний.
Вместе с тем поросль гидроида маскирует
раковину рака-отшельника, что для него,
безусловно, полезно. Кроме того, раку
не приходится часто менять свой домик,
ибо по мере роста рака разрастается и пла-
стинка гидроризы, служащая раку-от-
шельнику достаточно надежной защитой.
Легко заметить, что описанное сейчас
сожительство представляет собой про-
стейший случай симбиоза. Здесь нет
активной заинтересованной стороны: гид-
роид случайно поселяется на раковине-
домике, занятой раком-отшельником. По-
следний же не ищет для себя раковины,
уже обросшей колонией гидрактинии.
Каждый из партнеров, и рак, и гидроид,
может существовать вполне самостоя-
тельно.
Классические примеры симбиоза двух
организмов дает сожительство раков-от-
шельников и актиний. Но и в этих слу-
чаях можно различить три ступени по-
степенного осложнения отношений меж-
ду партнерами. В простейшем варианте
рак-отшельник еще не в состоянии снять
поселившуюся на каком-нибудь субстрате
актинию и перенести на свой домик. Он
лишь бросает свой старый домик, если
случайно найдет подходящую пустую
раковину с уже поселившейся на ней
актинией. Так ведет себя, например, рак-
отшельник Eupagurus excavatus, если он
случайно натолкнется на подходящую для
него пустую раковину, на которой посе-
лилась актиния Calliactis parasitica. Совер-
шенно так же поступает рак, найдя пустую
раковину с сидящей на ней Hormathia
corenata или даже Actinia equina, хотя
обе эти актинии ведут вполне самостоя-
тельный образ жизни и не вступают
в симбиоз с другими животными.
Совместная жизнь актинии и рака-от-
шельника дает несомненные преимуще-
ства каждому из партнеров. Актинии
теперь не нужно находиться в постоян-
ном и напряженном ожидании поимки
пищи. Она без труда улавливает щупаль-
цами взмученные остатки пищи рака-
отшельника, в поисках которой он непре-
рывно движется по дну водоема. Рак же
находится теперь под надежной защитой
стрекательных капсул актинии — гроз-
ного оружия для большинства врагов
раков-отшельников.
Более высокую ступень в развитии сим-
биотических взаимоотношений показывает
сожительство той же Calliactis parasitica
с другим раком-отшельником — Pagurus
arrosor *. Рак-отшельник не должен
искать для себя пустую раковину с уже
сидящей на ней актинией. Он способен
снять ее с любого субстрата, перенести
на свой домик и укрепить на нем. Когда
раку-отшельнику старый домик оказы-
вается мал и он вынужден сменить его
на новый, он снимает актинию со старого
и переносит на свой новый домик. Проис-
ходит это следующим образом. Своими
ходильными ногами первой и третьей пар
рак-отшельник начинает поглаживать и
похлопывать актинию. Казалось бы, акти-
ния, которая в ответ на любое раздраже-
ние отвечает сокращением своего тела
и выбрасыванием стрекательных нитей,
должна была бы поступить аналогичным
образом, когда рак начинает ее похлопы-
вать. В данном же случае актиния ведет
себя не совсем так. При первом прикосно-
вении рака она, правда, начинает сжи-
маться, но затем вновь распускается и
стрекательных нитей не выбрасывает.
Даже полностью сжавшаяся актиния
начинает распускаться, если ее начинает
поглаживать рак-отшельник. После того
как актиния полностью распустилась, ее
будущий спутник начинает поглаживать
подошву актинии. Этим поглаживанием
рак добивается сокращения подошвы и
отделения ее от субстрата. Тогда раку
1 См. цветную таблицу 9.
252
остается только подставить свою раковину
поближе к подошве актинии и перекатить
на нее актинию.
Впрочем, имеются указания, что после
того, как произошло отделение подошвы,
актиния сама наклоняется в сторону рако-
вины отшельника и охватывает ее своими
щупальцами. Затем она вновь перевора-
чивается и прикрепляется к раковине
уже подошвой. В тех случаях, когда
рак-отшельник находит не вполне плот-
но укрепившуюся актинию, ему даже
не приходится ее похлопывать и погла-
живать — описанный процесс пересадки
актинии протекает без его помощи.
Актиния выделяет своей подошвой
слизистую перепонку, которая способ-
ствует более надежному укреплению акти-
нии на домике рака-отшельника. Однако
эта тонкая мембрана не может перерасти
за край раковины и тем самым увеличи-
вать ее объем. Отшельнику приходится
через некоторое время менять свой ста-
рый домик, ставший ему тесным, на но-
вый, более просторный, и вновь пересажи-
вать на него свою спутницу. Если на рако-
вине остается достаточно места, предпри-
имчивый рак может обзавестись еще одной
актинией. Описаны случаи, когда Pagu-
rus arrosor носил на своей раковине до 8
акт л ний.
Лучшим примером высшего типа симби-
оза актинии и рака-отшельника служит
сожительство Adamsia palliata с Eupa-
gurus prideaxi. Адамсия, распространен-
ная в Средиземном море и в умеренных
водах восточной части Атлантического
океана, ведет самостоятельное существо-
вание только в самом начале своей жизни.
Молодые адамсии имеют обычное для акти-
ний цилиндрическое тело с довольно широ-
кой подошвой, при помощи которой они
прикрепляются к камням или пустым
раковинам моллюсков. Очень редко они
успевают дорасти до 1 см в высоту: как
правило, еще до этого их захватывают
раки-отшельники. До сих пор еще не
вполне установлено, способна ли эта
актиния и во взрослом состоянии вести
самостоятельный образ жизни, если она
не вступает по какой-нибудь случайности
в симбиоз с отшельником. Однако пред-
ставить себе адамсию-одиночку очень
трудно, зная повадки отшельника
Е. prideaxi. Среди своих сородичей он
самый подвижный и драчливый. Поэтому
он и предпочитает самые мелкие раковины
в качестве своего домика. Раковины хищ-
ных брюхоногих моллюсков Nassa и Na-
tica, которыми он обычно пользуется,
едва прикрывают его мягкое брюшко.
У взрослого, крупного рака такой до-
мик защищает только самый задний отдел
брюшка. Что же касается головогруди
рака, то она никогда не прикрывается
раковиной. Защитить себя от врагов и
хищников Eupagurus может только одним
способом — найти и вступить в симбиоз
с адамсией. Молодой рак находится в по-
стоянных поисках адамсии. В поиске
актинии ему помогает не столько зрение,
сколько осязание и способность улавли-
вать определенные химические раздра-
жения, исходящие от актинии. В аква-
риуме этот рак-отшельник обнаруживает
молодую адамсию и в том случае, если
она прикрыта колпачком из ткани. Как
только он обнаруживает одиночную моло-
дую актинию, он стремительно бросается
к ней. В это время его, кажется, ничто
уже не может остановить. Подойдя к адам-
сии, Eupagurus крепко схватывает ее
своими клешнями и не отпускает в тече-
ние 10 минут. Казалось бы, что такое обра-
щение должно заставить адамсию сжаться.
На самом же деле этого не происходит.
Напротив, подошва молодой адамсии на-
чинает отделяться от субстрата. После это-
го рак, не разжимая клешней, переносит
ее на свой домик.
Когда молодой рак-отшельник, еще не
успевший обзавестись спутницей, встре-
чается со своим более удачливым сороди-
чем, он моментально вступает с ним в бой
за обладание адамсией. Если обороняю-
щийся оказывается слабее, он бросает
раковину и пускается в бегство, оставляя
агрессору и свой дом и свою актинию.
Рак-отшельник Е. prideaxi укрепляет
адамсию на раковине так, чтобы ее рото-
вой диск был направлен вперед, а все
ее тело находилось сзади и чуть ниже
ротового аппарата самого рака. При та-
ком положении актиния без труда может
улавливать и поедать остатки пищи, пожи-
раемой раком. Вместе с тем такое поло-
253
жение актинии заставляет ее со временем
изменить форму тела. Дело в том, что
рост ее не может идти равномерно во
все стороны. Распространению подошвы
по раковине дальше вниз и назад пре-
пятствует постоянное трение раковины о
грунт. Не может актиния расти и вперед—
этому мешают находящиеся в движении
ноги рака. Актиния разрастается вбок
и вверх, вдоль края устья раковины. Тело
ее поэтому приобретает полулунную фор-
му. Если рак-отшельник не обзавелся
второй актинией, то боковые лопасти
подошвы, поднимаясь все выше и выше
по раковине, могут сомкнуться, окольцо-
вывая устье раковины. Если на раковине
находятся две актинии, то края их подошв
также смыкаются друг с другом на верх-
ней и нижней поверхностях раковины.
У адамспи есть и другое свойство, крайне
полезное для рака-отшельника: подошва
адамсии, так же как и подошва калиак-
тис, выделяет плотную слизистую пла-
стинку, которая, однако, у адамсии бы-
стро твердеет и принимает роговую струк-
туру. Эта пластинка по мере роста акти-
нии перерастает край устья раковины
и нависает над телом рака. Объем домика
тем самым все время увеличивается, и
быстрорастущему раку не приходится
менять своего домика. Крупные взрослые
адамсии достигают в ширину 5—7 см.
Домик же рака-отшельника имеет гораз-
до более скромные размеры. Получается,
что адамсия в конце концов сидит не
столько на поверхности первоначального
убежища рака, сколько на роговом кольце
вокруг его тела, которое сама и построила.
Рис. 154. Крабик, носящий в клешнях актиний
бунодеопсис (Bunodeopsis, по Дардену).
Такой домик очень удобен и для рака-
отшельника, ибо он несоизмеримо легче
толстостенной раковины моллюска и до-
статочно прочен. В то же время он эла-
стичен и нисколько не затрудняет движе-
ний рака.
После укрепления адамсии на раковине
меняется не только форма ее тела, но и ее
окраска. Молодые адамсии окрашены в
яркий розово-красный цвет. Но у взрослых
адамсий, вступивших в симбиоз с пагуру-
сом, окраска тела меняется и становится
желто-розовой, различных оттенков, но
такой же, как и окраска головогруди и
верхних члеников ходильных ног рака.
Наиболее ярко окрашены спинные лопа-
сти подошвы актинии; ближе к бокам они
светлеют. Ротовой диск и щупальца, неви-
димые сверху, почти совсем лишены окра-
ски (см. цветную таблицу 9).
Взаимная выгодность симбиоза Adam-
sia palliata и Eupagurus prideaxi очевидна.
Рак-отшельник, почти все тело которого
оказывается не прикрытым раковиной,
давно стал бы жертвой какого-либо хищ-
ника, если бы он не был надежно защищен
актинией. Даже такие свирепые хищники,
как головоногие моллюски, не смеют напа-
дать на отшельника, боясь страшного
оружия адамсии — ее стрекательных ни-
тей. Адамсия в свою очередь не забо-
тится о пище — это прямая обязанность
ее хозяина. Постоянное передвижение
рака по грунту также весьма выгодно
для актинии, так как это улучшает усло-
вия газообмена. Не обязательно, однако,
укрепление актинии на раковине или
панцирях ракообразных. В природе выра-
батываются и другие формы симбиоза
между актиниями и раками. Актиния
может быть использована тогда не только
как оружие защиты, но и как орудие для
добывания пищи. Один из средиземномор-
ских крабов известен тем, что он постоян-
но держит в своих клешнях двух актиний,
которые служат ему одновременно и защи-
той от врагов и орудием добывания пищи.
Своими щупальцами, вооруженными стре-
кательными капсулами, актиния схватыва-
ет проплывающую добычу. Но ее хозяин-
краб беззастенчиво отбирает часть добычи
для себя. Единственное преимущество
актинии, ведущей такой образ жизни,
254
заключается в том, что она все время пере-
мещается с одного места на другое. Сим-
биоз в данном случае носит несколько
однобокий характер.
Во всех до сих пор описанных сообще-
ствах активной стороной были рако-
образные и пассивной — актинии. Но бы-
вает и наоборот. Так, например, актиния
Autholoba reticulata, обитающая на запад-
ном побережье Южной Америки, сама стре-
мится вступить в симбиоз с ракообразны-
ми, предпочитая другим краба Hepatus»
Эта актиния, как и другие ее сородичи,
вначале поселяется на камнях, прикреп-
ляясь к ним своей подошвой. Но затем
она переворачивается ротовым диском
вниз и держится за субстрат своими щу-
пальцами. Подошва ее отрывается от суб-
страта. В таком необычном для актинии
положении она ожидает приближения
своего будущего спутника. Через некото-
рое время терпение ее может быть награж-
дено, и тогда ее подошва крепко схваты-
вает лапку проползающего мимо краба.
Затем актиния переползает на его пан-
цирь.
Существует и не менее интересный сим-
биоз актинии и рыб. На мелководье
западного побережья Австралии обитают
самые крупные в мире актинии Stoichac-
tis, ротовой диск которых достигает
в диаметре 1,5 м. На поверхности диска
несколькими концентрическими рядами
располагаются бесчисленные щупальца
этих актиний. Среди щупалец или в не-
посредственной близости от них держится
парочка очень ярко окрашенных рыбок—
амфиприонов. По наблюдениям ряда авто-
ров, невозможно найти ни одной акти-
нии, возле которой бы не держались эти
рыбки, так же как и рыбок вдали от акти-
ний. Даже в поисках добычи рыбки
никогда не удаляются от актиний более
чем на метр. При малейшей опасности
они прячутся в лесу длинных щупалец
актиний. Если в аквариум с актинией
и амфиприонами посадить хищных рыб,
то они могут жить вместе очень долго,
но стоит убрать актинию, как амфипрно-
ны становятся жертвой хищных рыб. Амфи-
прионы не умеют ни быстро плавать, ни
находить защиту в водорослях, а их очень
яркая окраска и красивый рисунок из
Рис. 155. Личинки трахилнды кунина
(Cunina), паразитирующие на гидро-
идной медузе битотиара (Bythotiara).
белых и черных полос на красном или
золотистом фоне слишком заметны для
врага.
В естественных условиях эта же яркая
окраска идет на пользу рыбкам, привле-
кая к ним добычу. Так как рыбки хва-
тают добычу в непосредственной близи
от щупалец и ротового отверстия актинии,
здесь же поедают ее и, естественно, те-
ряют ее остатки, они тем самым снабжают
пищей и актинию. Кроме того, держась
среди щупалец, рыбки постоянным бие-
нием своих плавников создают токи воды,
улучшающие условия газообмена акти-
нии.
Кроме этих рыбок, среди щупалец акти-
ний часто держится несколько видов кре-
255
веток и небольшой краб, подражающий
в окраске рыбкам.
О симбиозе кишечнополостных с одно-
клеточными водорослями зооксантел-
лами будет сказано в главе о коралло-
вых постройках.
Среди кишечнополостных имеется не-
большое число настоящих паразитов.У гид-
роидных медуз купана (Cunina) и куноктан-
та (Cunoctantha) образовавшаяся из яйца
п л а н у л а оседает не на дно, а на край
зонтика некоторых видов других гидроид-
ных медуз из отряда лептолид. Здесь
планула превращается в двущупальце-
вую личинку, которая пробирается к ро-
товому хоботку хозяина. Вскоре у ли-
чинки появляется вторая пара щупалец
и ротовой хоботок. Придерживаясь щу-
пальцами за нижнюю сторону зонтика,
личинка запускает свой ротовой хоботок
в желудок медузы-хозяина и питается за
ее счет. Личинка быстро растет и, размно-
жаясь почкованием, образует подобных
себе паразитических четырехщупальце-
вых личинок. Вскоре внутри зонтика
медузы-хозяина оказывается целая груп-
па паразитов. После этого все личинки
начинают превращаться в медуз. У них
развивается зонтик, укорачивается рото-
вой хоботок; они покидают своего хозяина
и ведут свободноплавающий образ жизни.
Еще более яркий пример паразитизма
дает своеобразное кишечнополостное жи-
Таблица 10. Распространение рпфообразующпх
кораллов в Мировом океане:
1 — поциллопора (Pocillopora);
2 — тридакофиллия (Tridacophyllia),
3 — павона (Pavona);
4 — порит (Porites),
5 — акропора (Асгорога),
6 — грибовидный коралл (Fun^ia),
7 — саркофитум (Sarcophytum),
8 — фавия (Favia).
вотное из гидроидных — полиподиум
(Polypodium hydriforme).
Полиподпум — единственный предста-
витель кишечнополостных, паразитирую-
щий в теле позвоночного животного, он
поселяется в икре некоторых осетровых
рыб.
Впервые полиподиум был обнаружен
академиком Ф. В. Овсянниковым
в 1871 г. в икре волжской стерляди.
Овсянников обратил внимание на то,
что, кроме нормальных икринок, попа-
даются также больные, несколько более
крупные и светлее окрашенные. Внутри
такой икринки сидит полиподиум. Самая
ранняя из известных стадий развития
паразита — это маленькая двуслойная
личинка, имеющая червеобразную форму.
Личинки поселяются в развивающейся
икринке. По мере роста икринки и накоп-
ления в ней желтка развивается и личин-
ка; она вытягивается в длину, и на ней
образуются многочисленные выросты—
почки. Вскоре в каждой почке появ-
ляется по 12 щупалец, обращенных внутрь.
На этой стадии паразит обладает весьма
своеобразным строением — он как бы вы-
вернут наизнанку. Его пищеваритель-
ный слой — энтодерма — расположен сна-
ружи и непосредственно соприкасается
с желтком икринки, служащим полипо-
диуму пищей. Покровный же слой — экто-
дерма — обращен внутрь.
Перед наступлением периода нереста
рыбы-хозяина слои клеток у паразита
принимают нормальное положение. По-
чки полиподиума при этом вворачи-
ваются, захватывая внутрь гастральной
полости остатки желтка из икринки. Когда
рыба выметывает икру в реку, вместе со
здоровыми икринками в воду попадают
и зараженные. В воде паразит выходит
из икринки, разорвав ее оболочку, и
вскоре после этого распадается на части
по числу почек — из каждой развивается
двенадцатищупальцевый полип около
1 мм высотой. Из одной зараженной
икринки выходит 60—90 полипов.
Первое время полипы лишены рта и
питаются за счет запасов желтка, но на
4—5-й день самостоятельной жизни
у них появляется рот и полипы при
помощи щупалец, усаженных стрекатель-
ными клетками, начинают ловить мало-
щетинковых червей, коловраток и дру-
гих маленьких пресноводных животных.
Четыре коротких щупальца полипа слу-
жат для прикрепления ко дну и для пере-
движения, более длинными щупальцами
полип ловит добычу.
В течение всего лета полипы размножа-
ются делением. Перед делением число щу-
палец у полипа удваивается, а затем на
его теле появляется продольная пере-
тяжка, и вскоре он разделяется надвое.
К осени в теле полипа развиваются
половые железы. Полиподиумы раздель-
нополы, но изредка попадаются герма-
фродитные особи. Весь дальнейший цикл
развития полиподиума пока еще неясен.
Известно только, что мужские полипы
способны оседать на молоди осетровых
рыб и на их тело прикреплять свои го-
нады. Способы оплодотворения и пути
проникновения паразита в икринку по-
ловозрелой рыбы пока совершенно непо-
нятны.
Полиподиум приносит значительный
ущерб, так как зараженная им дорого-
стоящая икра осетровых рыб теряет свою
товарную ценность. Кроме того, пара-
зит значительно снижает продуктивность
осетровых, вследствие того что зара-
женные икринки погибают, не дав потом-
ства.
Поселяется полиподиум в икре стерляди,
осетра, севрюги и аральского шипа. Этот
паразит был обнаружен в рыбах из Волги
и ее притоков, Дона, Днепра, Днестра,
Дуная, Кубани, Северной Двины, Сыр-
дарьи и Амура.
Рыбы, зараженные полиподиумом, лече-
нию не поддаются, но имеются широкие
Таблица 11. Жизнь на коралловом рифе.
Кораллы:
1, 2, 3, 7 — разные виды акропора (Асгорога);
4 — поциллопора (РосШорога),
5 — монтипора (Montipora);
6 — лобофиллия (Lobophyllia);
8 — Coenopsammia.
Морские 8везды:
9 — протореастер (Protoreacter);
10 — акантастер (Acanthaster).
Морские ежи:
11 — диадема (Diadema setosum);
12 — гетероцентротус (Heterocentrotus mammilatus).
Моллюски:
13 — пятнистая ципрея (Сургаеа tigris);
14 — арабская ципрея (Сургаеа arabica);
15 — нильский трохус (Trochus niloticus);
16 — тридакна (Tridacna crocea).
Рыбы:
17 — возничий (Heniochus macrolepidotus);
18 — дая (Daya);
19 — помаценорус (Pomacenorus);
20 — рыба-бабочка (Haetodon).
17 Жизнь животных, т. 1
возможности по проведению профилакти-
ческих мероприятий с целью снизить зара-
женность рыб в данном бассейне и пре-
пятствовать проникновению паразита в
другие реки. Для этого необходимо тща-
тельно просматривать икру при ее заготов-
ке для засола и на рыбоводных станциях.
Ни в коем случае не следует выбрасывать
зараженную икру в воду, где из нее разо-
вьются полипы, которые будут заражать
Рис. 156. Паразит икры осетровых рыб поли-
подиум (Polypodium):
1 — паразит в икринке; 2 — столон; 3 — полип из
реки (по Райковой).
здоровых рыб. При акклиматизации осет-
ровых в новых бассейнах нельзя ввозить
в них взрослых рыб, так как они могут
быть заражены полиподиумом. Лучше
всего для целей акклиматизации использо-
вать тщательно просмотренную здоровую
икру осетровых.
ЗНАЧЕНИЕ КИШЕЧНОПОЛОСТНЫХ
В ПАЛЕОНТОЛОГИИ
Среди наиболее древних кишечнополост-
ных преобладали организмы, лишенные
хорошо развитого скелета. Известны на-
ходки нескольких сцифоидных медуз
из кембрийских и меловых отложений.
Только остатки вымерших рифообразу-
ющих кораллов, обладавших массивным
известковым скелетом, часто встречаются
в самых различных геологических отло-
жениях. Описано более 5000 видов иско-
паемых кораллов. Находили их в виде
отдельных отпечатков, в виде крупных
ископаемых рифов. Наиболее древними
кораллами были археоциатиды из нижних
кембрийских и гелиотиды из силурий-
ских отложений.
Начиная с силурийского периода ста-
новится чрезвычайно распространенной
группа очень своеобразных четырехлуче-
вых кораллов (Tetracorallia). Это были
одиночные или колониальные кораллы
конической формы, с расширенной верх-
ней чашечкой. На ее поверхности отчет-
ливо различимы четыре первичные пере-
городки. Перегородки (или септы) делили
чашечку на четыре сектора, каждый из
которых в свою очередь делился вторич-
ными перегородками.
Настоящие мадрепоровые рифообразу-
ющие кораллы появились только в мело-
вом периоде, а современные нам семейства
этих кораллов — только в четвертичном
периоде.
Несмотря на многочисленные и разно-
образные находки ископаемых кораллов,
группа кишечнополостных в целом состав-
ляла весьма небольшую часть накоплен-
ного палеонтологического материала и не
могла поэтому играть сколько-нибудь
существенной роли в палеонтологических
реконструкциях. Однако недавние иссле-
258
дования, проведенные в долине Эдиакара,
в Южной Австралии, заставили ученых
изменить такое мнение о значении кишеч-
нополостных в палеонтологической лето-
писи.
Еще совсем недавно никто не знал,
какими были предки тех беспозвоночных
животных, которые достигли значительного
развития уже в начале кембрийского
периода.
И вот при раскопках докембрийских
пород в долине Эдиакара удалось нако-
нец собрать более 6000 образцов ископае-
мых беспозвоночных, населявших моря
в протерозойскую эру. Среди собранных
образцов были отпечатки медуз, принад-
лежавших к шести различным родам, мяг-
ких восьмилучевых кораллов и корал-
ловых полипов, напоминавших современ-
ные морские перья, и множество других
животных. Среди многочисленных отпе-
чатков кораллов не было ни одного, ко-
торый свидетельствовал бы о наличии из-
весткового массивного скелета. Ткани древ-
нейших коралловых полипов поддержива-
лись лишь простыми известковыми тель-
цами — спикулами.
Все эти находки еще требуют присталь-
ного изучения. Однако уже сейчас можно
сказать, что предположение о том, что
докембрийские животные были лишены
скелета или не прикрывались панцирем
и потому и оставались до сих пор неиз-
вестными, по-видимому, оказалось спра-
ведливым. Кроме того, изучение собран-
ных остатков позволяет предположить,
что отсутствие раковин или внутреннего
скелета у докембрийских животных объяс-
няется не какими-нибудь особыми усло-
виями обитания в то удаленное от нас на
сотни миллионов лет время и не каким-
нибудь резким отличием образа жизни
этих животных. Отсутствие скелета, ви-
димо, должно объясняться тем, что у
древнейших беспозвоночных животных
еще не выработался механизм кальциевого
обмена, в результате которого организмы
оказались в последующем способны к обра-
зованию массового известкового скелета,
панцирей и раковин.
Теперь благодаря трудам зоологов рус-
ской школы — И.И.Мечникова, В. Н.
Беклемишева и А. А. Захватки-
Рис. 157. Жизненный цикл полиподиума
(по Райковой).
н а—стали понятны самые ранние периоды
в эволюции кишечнополостных. Можно
считать установленным, что предками
современных нам кишечнополостных были
примитивные многоклеточные животные,
напоминавшие планул современных ви-
дов.
К сожалению, среди палеонтологиче-
ских находок нет остатков этих крошеч-
ных, лишенных скелета предков много-
клеточных, и поэтому представить себе
картину ранней эволюции кишечнополо-
стных можно, лишь проводя аналогии
и детально изучая индивидуальное раз-
витие.
Мы вправе предположить, что следующим
за планулой этапом эволюции кишечно-
полостных был полип.
Для многих сидячих организмов очень
характерна способность к вегетативному раз-
множению и образованию колоний. Эта
особенность позволяет им лучше бороться
за существование: полипы сообща ловят и
усваивают добычу, сообща защищаются ог
врагов.
На ранних ступенях развития кишечно-
полостных произошло разделение их на
две крупные ветви. Появились полипы,,
гастральная полость которых была разде-
лена перегородками на камеры. Это спо-
собствовало более интенсивному пищева-
рению. Впоследствии от этих полипов
17*
259
произошли коралловые полипы и сцифо-
идные. Те же полипчики, у которых пере-
городок в кишечной полости не было, далп
гидроидных. Однако прикрепленный образ
жизни имел и свои недостатки: потомство
сидячих животных всегда поселялось ря-
дом с родителями. Это создавало перена-
селенность и препятствовало животным
завоевывать новые жизненные простран-
ства.
Появление подвижных половых особей
медуз было очень существенным шагом
на пути эволюции кишечнополостных, эта
их особенность наряду с развитием стре-
кательного аппарата является одной из
главнейших причин процветания типа.
Мы уже убедились, что кишечнополост-
ные сумели приспособиться к самым раз-
личным условиям жизни в воде. Их нахо-
дят среди бурных волн прибоя на берегу
моря и в спокойных водах океанских глу-
бин. Они поселяются в холодных морях
Арктики и Антарктики и под палящими
лучами тропического солнца. Кишечно-
Бродя по берегу моря, мы часто видим
гряды выброшенных волнами зеленоватых,
бурых или коричневых спутанных комков
жестких нитей. Очень мало кто знает,
что значительная часть этой «морской
травы» имеет не растительное, а живот-
ное происхождение. Всякий, кто бывал
на море, конечно, видел, что все камни,
сваи и другие подводные предметы обра-
стают какими-то нежными кустиками,
извивающимися в волнах. Если собрать
такие кустики и посмотреть их под микро-
скопом, то наряду с настоящими водоро-
слями можно увидеть и нечто совсем
особенное. Вот перед нами коричневая
членистая веточка с розовыми комочками
на концах. Вначале розовые комочки
неподвижны, но стоит им несколько минут
постоять спокойно, и они начинают шеве-
литься, вытягиваться в длину, приобре-
полостные живут на дне, в толще воды и
плавают на ее поверхности. Некоторые
из них перешли к паразитизму, и их жиз-
ненной средой стал организм животного-
хозяина . Кишечнополостные проникают
даже в пресные воды, и только воздушная
среда, наземное существование оказались
для них совершенно недоступными.
Эта очень древняя группа организмов,
несомненно, является в то же время одной
из наиболее процветающих. Конечно,
нельзя ожидать, что в дальнейшем из
кишечнополостных возникнут какие-то
высокоорганизованные животные, но их
способность приспосабливаться к разно-
образным условиям жизни позволит им
и впредь успешно конкурировать с гораздо
более высокоорганизованными обитате-
лями моря.
После данной выше общей характери-
стики типа кишечнополостных мы оста-
новимся подробнее на основных классах
и отрядах этого обширного типа обитате-
лей моря.
тая форму маленького кувшинчика с вен-
чиком щупалец на верхнем конце тела.
Это полипы гидроида эудендриум (Euden-
drium), живущего в наших северных мо-
рях, в Черном море и в морях на Даль-
нем Востоке. Рядом другая, также чле-
нистая, но более светлая веточка. Полипы
на ней также розовые, но по форме напо-
минают веретено. Щупальца сидят на теле
полипа без всякого порядка, и каждое
снабжено на конце маленькой головкой —
скоплением стрекательных клеток. Дви-
жения полипов медлительны, они то
сгибают свое тело, то медленно покачи-
ваются из стороны в сторону, но чаще
сидят неподвижно, широко расставив
щупальца — подстерегают добычу. На
некоторых полипах можно заметить почки
или молодых развивающихся медуз. Под-
росшие медузки энергично сжимают и раз-
КЛАСС ГИДРОИДНЫЕ (HYDROZOA)
(Растения или животные?)
260
жимают свой зонтик, тонкая нить, связы-
вающая медузу с полипом, при этом обры-
вается, и медузка толчками уплывает
прочь. Это полипы корине (Согупе) и их
медузы. Они также обитают и в арктиче-
ских и в умеренных морях.
А вот еще кустик, полипы на нем сидят
внутри прозрачных колокольчиков. Внеш-
не они очень похожи на полипов эуден-
дриум, но ведут себя совершенно иначе.
Стоит слегка дотронуться до полипа кон-
цом иголки, как он стремительно втяги-
вается в глубь своей защитной оболочки—
колокольчика. На этом же кустике можно
найти и медузок: они так же, как полипы,
скрыты внутри прозрачной защитной
оболочки. Медузы плотно сидят на тон-
ком бесщупальцевом полипе. Это колония
гидроида обелии (Obelia).
Теперь, когда мы можем отличать гид-
роидов от водорослей, следует обратить
внимание на перовидную колонию аглао-
фении (Aglaophenia). У этого вида, очень
обычного у нас на Черном море, кормя-
щие полипы сидят на веточке в один ряд.
Каждый заключен в чашечку — г и д-
р о т е к у и окружен тремя защитными
полипами.
Свободноплавающих медуз у аглаофе-
нии не образуется, а недоразвитые особи
медузоидного поколения спрятаны
внутрь очень сложного образования—
корзиночки (видоизмененной вето-
чки колонии).
Колонии гидроидов поселяются чаще
всего на небольших глубинах — от лито-
рали до 200—250 м и предпочитают каме-
нистый грунт или же прикрепляются
к различным деревянным и металличе-
ским предметам. Нередко они очень густо
разрастаются на подводных частях судов,
покрывая их мохнатой «шубкой». В этих
случаях гидроиды приносят значительный
вред судоходству, так как такая «шуба»
резко снижает скорость судна. Известно
немало случаев, когда гидроиды, посе-
ляясь внутри труб морского водопровода,
почти совсем закрывали их просвет и
препятствовали подаче воды. Бороться
с гидроидами довольно трудно, так как
эти животные неприхотливы и вполне хоро-
шо развиваются, казалось бы, в неблаго-
приятных условиях. Кроме того, они отли-
чаются быстрым ростом — за месяц выра-
стают кустики 5—7 см высотой. Чтобы
очистить от них днище корабля, прихо-
дится ставить его в сухой док. Здесь ко-
рабль очищают от наросших гидроидов,
полихет, мшанок, морских желудей и
других животных-обрастателей.
В последнее время стали применять
специальные ядовитые краски—покрытые
ими подводные части корабля подверже-
ны обрастаниям в значительно меньшей
степени.
Гидроиды, поселяющиеся в литораль-
ной зоне, совершенно не боятся прибоя.
У многих из них полипчики защищены
от ударов скелетной чашечкой — т е к о й;
на колониях, растущих в самой прибой-
ной зоне, теки всегда значительно толще,
чем у тех же видов, живущих поглубже,
где прибойные волны не ощущаются
(рис. 159).
У других гидроидов из прибойной зоны
колонии имеют длинные, очень гибкие
ствол и ветви, или же они поделены на
членики. Такие колонии извиваются
вместе с волнами и потому не ломаются
и не рвутся.
На больших глубинах живут особые
гидроиды, не похожие на литоральные
виды. Здесь преобладают колонии в форме
елочки или пера, многие похожи на дерев-
ца, а есть виды, напоминающие ершик.
Они достигают высоты 15—20 см и по-
крывают морское дно густым лесом. В за-
рослях гидроидов живут черви, моллюс-
ки, ракообразные, иглокожие. Многие
из них, например рачки морские козо-
чки, находят среди гидроидов убежище,
другие, как, например, морские «пауки»
(многоколенчатые), не только прячутся
в их зарослях, но и питаются гидрополи-
пами.
Если поводить вокруг поселений гидрои-
дов мелкоячеистым сачком или, что еще
лучше, использовать для этого специаль-
ную, так называемую планктонную, сеть,
то среди массы маленьких рачков и личи-
нок различных других беспозвоночных
животных попадутся гидроидные медузы.
Большинство видов гидромедуз — не очень
крупные животные, редко они дости-
гают более 10 см в диаметре зонтика,
обычно же размеры гидромедузы 2—3 см,
261
I — еувдендриум
Рис. 158. Так выглядят под микроскопом колонии гидроидов:
(Eudendrium); 2 — Корине (Согупе); 3— обелия (Obelia); 4 — аглаофения (Aglaophenia).
а часто всего 1 — 2 мм. Гидроидные меду-
зы очень прозрачны. Даже пойманных и
помещенных в стеклянную посуду медузок
сразу и не заметишь: видны лишь белова-
тые ниточки каналов и ротовой хоботок.
Только внимательно приглядевшись,
можно заметить контуры зонтика.
Рассматривая колонию гидроида корине
(Согупе), мы уже видели только что отпо-
чковавшихся маленьких медузок этого
вида. У вполне сформированной медузы
колокольчатый зонтик 1—8 см высо-
той, четыре щупальца и длинный, черве-
образный ротовой хоботок. Резкими со-
кращениями зонтика медуза быстро пере-
двигается в горизонтальной плоскости
или поднимается вверх. Вниз она медлен-
но опускается под влиянием тяжести,
застыв в воде с распущенными щупаль-
цами. Морские планктонные рачки, со-
ставляющие главную пищу медузы, по-
стоянно совершают вертикальные пере-
мещения: днем погружаются в глубины,
а к ночи поднимаются к поверхности.
Они опускаются в более глубокие, спо-
койные слои воды также и во время вол-
нения. Медузы постоянно двигаются
вслед за ними, преследовать свою добычу
им помогают два чувства — осязание и
зрение. В спокойной воде зонтик медузы
все время ритмично сокращается, под-
нимая животное к поверхности. Как только
медуза начинает ощущать вызванное вол-
нами движение воды, ее зонтик перестает
сокращаться и она медленно погружается
в глубину. Свет она различает при помо-
щи глазков, находящихся в основании
щупалец. Слишком яркий свет действует
на нее подобно волнению — зонтик
перестает сокращаться и животное погру-
жается в более темную глубину. Эти про-
стые рефлексы помогают медузе пресле-
довать добычу и спасаться от гибельного
для нее волнения.
Как уже было сказано выше, медуза
корине питается планктонными организ-
мами, преимущественно веслоногими ра-
чками. Глаза медузы не настолько совер-
шенны, чтобы она могла видеть свою добы-
чу, ловит она ее вслепую. Ее щупальца
могут очень значительно растягиваться,
превосходя высоту зонтика в десятки раз.
Вся поверхность щупальца усеяна много-
численными стрекательными клетками.
Как только к щупальцу прикоснется ра-
чок или какое-нибудь другое маленькое
планктонное животное, оно сразу же
поражается стрекательными клетками.
262
Щупальце при этом быстро сокращается
и подтягивает добычу ко рту. Длинный
хоботок вытягивается в направлении добы-
чи. Если попался более крупный рачок,
медуза оплетает его не одним, а двумя,
тремя или всеми четырьмя щупальцами.
Совсем иначе ловят свою добычу медузы
с плоским зонтиком и многочисленными
щупальцами, например тиаропсис (Tiaro-
psis) — гидромедуза размером с двухко-
пеечную монету, очень обычная в наших
северных морях. По краям ее зонтика нахо-
дится до 300 тонких щупалец. У покоя-
щейся медузы щупальца широко расстав-
лены и охватывают значительное простран-
ство. При сокращении зонтика медуза
как бы сметает ими рачков, подгоняя их
к середине нижней стороны зонтика (см.
рис. 160). Рот у тиаропсис широкий, снаб-
женный четырьмя большими бахромча-
тыми лопастями, которыми медуза захва-
тывает подогнанных рачков.
Несмотря на незначительную величину,
гидроидные медузы очень прожорливы.
Они поедают массу рачков и потому счи-
таются вредными животными — конку-
рентами планктоноядных рыб. Обильная
пища необходима медузам для развития
половых продуктов. Плавая, они разбра-
сывают в море огромное количество яиц,
которые впоследствии дают начало поли-
поидному поколению гидроидов.
Выше мы назвали кишечнополостных
типичными обитателями моря. Это дей-
ствительно так для 9000 видов, относя-
щихся к этому типу, но около полутора-
двух десятков видов кишечнополостных
живет в пресных водах п в морях уже не
встречается. Видимо, их предки очень
давно переселились в пресные воды.
Очень характерно, что все эти формы
как пресноводных, так и солоноватовод-
ных бассейнов относятся только к классу
гидроидных и даже только к одному его
подклассу— гидроидей (Hydroidea).
Среди всех других кишечнополостных
никакой склонности к воде пониженной
солености не наблюдается.
К самым типичным обитателям пресных
вод всего земного шара, часто обра-
зующим очень плотные популяции, отно-
сится несколько видов гидр, составляю-
щих отряд гидровых (Hydrida).
Р
Рис. 159. Литоральные (наверху) и сублитораль-
ные формы гидроидов:
I — морщинистая сертулярелла (Sertularella rugosa);
2 — кампанулярия (Campanularia).
Рис. 160. Гидроидные медузы:
I — корине (Согупе); 2 — тиаропсис (Tiaropsis).
263
ПРЕСНОВОДНАЯ ГИДРА
В каждой группе животно-
го царства имеются излюблен-
ные зоологами представители,
которых они используют в ка-
честве основных объектов при
описании развития и строения
животных и над которыми
ставят многочисленные опыты
по физиологии. В типе кишеч-
нополостных таким классиче-
ским объектом служит гидра.
Это и понятно. Гидр легко най-
ти в природе и сравнительно
просто содержать в лаборатор-
ных условиях. Они быстро раз-
множаются, и потому в корот-
кий срок можно получить мас-
совый материал. Гидра —типич-
ный представитель кишечнопо-
лостных животных, стоящих
у основания эволюционного
древа многоклеточных. Поэто-
му ее используют при выясне-
нии всех вопросов, касающих-
ся изучения анатомии, реф-
лексов и поведения низших
многоклеточных. Это в свою
очередь помогает понять проис-
хождение более высокоорга-
низованных животных и эволю-
цию их физиологических процессов. Кро-
ме того, гидра служит прекрасным объек-
том при разработке таких общебиологи-
ческих проблем, как регенерация, бес-
полое размножение, пищеварение, осевой
физиологический градиент и многое дру-
гое. Все это делает ее незаменимым живот-
ным как для учебного процесса — от
средней школы до старших курсов универ-
ситета, так и в научной лаборатории, где
решаются проблемы современной биоло-
гии и медицины в разных их отраслях.
Первым человеком, который увидел гид-
ру, был изобретатель микроскопа и круп-
нейший натуралист XVII—XVIII вв.
Антон Левенгук.
Разглядывая водные растения, Левен-
гук увидел среди других мелких орга-
низмов странное животное с многочислен-
ными «рогами». Он наблюдал также рост
почек на его теле, образование у них щу-
Рис. 161. Первое изобра-
жение гидры и кончика
ее щупальца (по Левен-
гуку).
Рис. 162. Схема передви-
жения окрашенных кле-
ток в теле гидры.
палец и отделение молодого жи-
вотного от материнского орга-
низма. Левенгук изобразил
гидру с двумя почками, а так-
же нарисовал кончик ее щу-
пальца со стрекательными кап-
сулами, каким он видел его под
своим микроскопом.
Однако находка Левенгука
почти не привлекла внимания
его современников. Лишь через
40 лет гидрой заинтересовались
в связи с необычайным откры-
тием молодого учителя Т р а м б-
ле (ТгатЫеу). Занимаясь в
свободное время изучением ма-
лоизвестных тогда водных жи-
вотных, Трамбле обнаружил су-
щество, похожее и на живот-
ное и на растение. Чтобы ус-
тановить его природу, Трамбле
разрезал это существо пополам.
Регенеративные способности
низших животных тогда были
еще почти неизвестны и счита-
лось, что восстанавливать ут-
раченные части могут только
растения. К удивлению Трамб-
ле, из каждой половинки вырос-
ла целая гидра, обе они шеве-
лились, хватали добычу, значит,
это было не растение. Возмож-
ность превращения куска тела гидры в
целое животное была воспринята как значи-
тельное открытие в науке о жизни, и Трамб-
ле занялся глубоким и серьезным изучени-
ем гидры. В 1744 г. он опубликовал книгу
«Мемуары к истории одного рода пресно-
водных полипов с руками в виде рогов».
В книге было очень подробно изложено
строение гидры, ее поведение (движения,
ловля добычи), размножение почкованием,
некоторые моменты физиологии. Для про-
верки своих предположений Трамбле проде-
лал с гидрой ряд опытов, положив начало
новой науке—экспериментальной зоологии.
Несмотря на несовершенство тогдаш-
ней оптики и слабое развитие зоологии,
книга Трамбле написана на таком высо-
ком научном уровне, что не потеряла свое-
го значения до настоящего времени, а
рисунки из этой книги можно найти во
многих учебниках по зоологии.
264
Сейчас научная литература о гидре
нсчисляется многими сотнями статей и
книг, но тем не менее гидра и по сей день
занимает умы исследователей. Маленькое
примитивное животное служит для них
пробным камнем, на котором решаются мно-
гие вопросы современной науки о жизни.
Бели собрать в прибрежной части озера
или реки водные растения и поместить
их в аквариум с чистой водой, то вскоре
на них можно увидеть гидр. Вначале они
почти незаметны. Потревоженные живот-
ные сильно сжимаются, их щупальца со-
кращаются. Но по истечении некоторого
времени тело гидры начинает вытягивать-
ся, ее щупальца удлиняются. Теперь
гидру можно как следует разглядеть.
Форма ее тела трубковидная, на перед-
нем конце находится ротовое отверстие,
окруженное венчиком из 5—12 щупалец.
Сразу под щупальцами у гидр большин-
ства видов имеется небольшое сужение—
шейка, отделяющая «голову» от туловища.
Задний конец гидры сужен в более или
менее длинную ножку, или стебелек,
с подошвой на конце (у некоторых видов
ножка не выражена). Посередине подо-
швы находится отверстие, так называе-
мая аборальная пора. Гастральная по-
лость гидры сплошная, перегородок в ней
нет, щупальца полые, похожие на пальцы
перчатки.
Стенка тела гидры, как и у всех кишеч-
нополостных, состоит из двух слоев кле-
ток, их тонкое строение уже было описано
выше, и потому здесь мы остановимся
только на одной особенности клеток тела
гидры, которая полностью изучена пока
лишь на этом объекте и не обнаружена
у других кишечнополостных.
Структура эктодермы (и энтодермы)
в разных частях тела гидры неравнозначна.
Так, на головном конце клетки эктодер-
мы мельче, чем на туловище, здесь меньше
стрекательных и промежуточных клеток,
но резкой границы между покровами «го-
ловы» и туловища провести нельзя, так
как изменение эктодермы от туловища
к «голове» происходит очень постепенно.
Эктодерма подошвы гидры состоит из круп-
ных железистых клеток, в месте перехода
подошвы в стебелек железистый харак-
тер покровных клеток постепенно утра-
чивается. То же самое можно сказать
и о клетках энтодермы, Пищеварительные
процессы происходят в средней части
тела гидры, здесь ее энтодерма имеет
большое количество пищеварительных
железистых клеток, а эпителиально-му-
скульные клетки энтодермы срединной
части туловища образуют многочислен-
ные псевдоподии. В головном отделе гаст-
ральной полости, в стебельке и в щупаль-
цах переваривания пищи не происходит.
В этих отделах тела эктодерма имеет вид
выстилающего эпителия, почти лишен-
ного пищеварительных железистых кле-
ток. Опять-таки резкой границы между
клетками пищеварительного отдела гаст-
ральной полости, с одной стороны, и та-
кими клетками «головы», стебелька и щу-
палец, с другой стороны, провести нельзя.
Несмотря на различие в строении кле-
точных слоев в разных частях тела гидры,
все ее клетки не находятся на строго опре-
деленных постоянных местах, а непре-
рывно передвигаются, причем их движе-
ние строго закономерно.
Используя высокую способность гидры
к заживлению ран, можно проделать та-
кой интересный опыт. Берут двух гидр
одинаковой величины и одну из них ок-
рашивают какой-нибудь прижизненной
краской, т. е. таким красящим веществом,
которое проникает в ткани гидры, не уби-
вая ее. Обычно для этого применяют сла-
бый водный раствор нильблаусульфата,
окрашивающий ткани гидры в синий цвет.
После этого гидры подвергаются опера-
ции: каждую из них разрезают на три
части в поперечном направлении. Затем
к срединной части «синей» гидры прира-
щивают головной и нижний концы неокра-
шенного экземпляра. Срезы быстро сра-
стаются друг с другом, и мы получаем экс-
периментальную гидру с синим пояском
посередине тела. Вскоре после операции
можно наблюдать, как синий поясок
распространяется в двух направлениях—
к головному концу и стебельку. При этом
по телу гидры передвигается не краска,
а именно сами клетки. Слои эктодермы
и энтодермы как бы «текут» из середины
тела к его концам, при этом постепенно
меняется характер составляющих их кле-
ток (см. рис. 162).
265
у±*Щ$*=~
Рис. 163. Гидра
В срединной части тела гидры клетки
размножаются наиболее интенсивно, и
отсюда они передвигаются в двух проти-
воположных направлениях. Таким обра-
зом, состав клеток постоянно обновляется,
хотя внешне животное остается почти
неизменным. Эта особенность гидры имеет
очень большое значение при решении во-
просов о ее регенеративных способностях
и для оценки данных о длительности
жизни.
Гидра—типичное пресноводное животное,
лишь в очень редких случаях гидр нахо-
дили в слабо осолоненных водоемах, на-
пример в Финском заливе Балтийского
моря, и в некоторых солоноватоводных
озерах, если содержание солей в них не
превышало 0,5%. Гидры живут в озерах,
реках, ручьях, прудах и даже в канавах,
если вода в них достаточно чистая и со-
держит большое количество растворен-
ного кислорода. Держатся гидры обычно
вблизи берегов, в неглубоких местах, так
как они светолюбивы. При содержании
гидр в аквариуме они всегда перебираются
на его освещенную сторону.
Гидры — малоподвижные животные,
большую часть времени они сидят на
одном месте, прикрепившись подошвой
к веточке водного растения, камню и т. д.
Излюбленная поза гидры в спокойном
состоянии — висеть вниз «головой», спу-
стив несколько расставленные щупальца.
Прикрепляется гидра к субстрату бла-
годаря клейким выделениям железистых
клеток эктодермы подошвы, а также исполь-
зуя подошву в качестве присоски. Дер-
жится гидра очень прочно, зачастую ее
«шагает».
легче разорвать, чем отделить от субстрата.
Если долго наблюдать за сидящей гидрой,
то можно увидеть, что ее тело все время
медленно раскачивается, описывая перед-
ним концом круг. Гидра может произволь-
но очень быстро оставлять место, на кото-
ром она сидит. При этом, по-видимому,
она раскрывает аборальную пору, находя-
щуюся в середине подошвы, и присасы-
вающее действие прекращается. Иногда
можно наблюдать, как гидра «шагает».
Вначале она пригибает тело к субстрату
и укрепляется на нем при помощи щупа-
лец, затем подтягивает задний конец и
укрепляется им на новом месте. После
первого «шага» делает второй и т. д.,
пока не остановится на новом месте.
Таким образом гидра передвигается
относительно быстро, но существует и
другой, гораздо более медленный, способ
передвижения — скольжение на подошве.
Усилием мускулатуры подошвы гидра еле
заметно передвигается с места на место.
Нужно очень много времени, чтобы заме-
тить перемещение животного. Гидры могут
некоторое время плавать в толще воды.
Открепившись от субстрата и широко рас-
ставив щупальца, гидра очень медленно
падает на дно, она способна образовать на
подошве маленький пузырек газа, который
увлекает животное вверх. Тем не менее
гидры редко прибегают к этим способам
передвижения.
Гидра—прожорливый хищник, она пи-
тается инфузориями, планктонными ра-
чками, малощетинковыми червями, напа-
дает также на мальков рыб. Гидры под-
стерегают свою добычу, подвесившись на
266
какой-нибудь сучок или стебель водного
растения, и, широко расставив щупальца,
постоянно делают круговые поисковые
движения. Как только одно из щупалец
гидры коснется жертвы, к ней устремляют-
ся остальные щупальца и парализуют
животное стрекательными клетками. Те-
перь от медлительности гидры не остается
и следа, она действует быстро и «решитель-
но». Добыча подтягивается щупальцами
ко рту и быстро заглатывается. Мелких
животных гидра глотает целиком. Если
жертва несколько крупнее самой гидры,
она также может ее заглотить. При этом
рот хищницы широко раскрывается, а
стенки тела сильно растягиваются. Если
добыча не помещается в гастральную по-
лость целиком, гидра заглатывает лишь
один ее конец, по мере переваривания
проталкивая жертву все глубже и глубже.
Сытая гидра несколько съеживается, и ее
щупальца сокращаются.
В гастральноп полости, где пищевари-
тельные процессы только начинаются,
реакция среды слабощелочная, а в пище-
варительных вакуолях энтодермы, где
пищеварение заканчивается, — слабокис-
лая. Гидра может усваивать жиры,
белки и животные углеводы
(гликоген). Крахмал и цел-
люлоза, имеющие растительное про-
исхождение, гидрой не усваиваются.
Непереваренные остатки пищи выбрасы-
ваются через рот.
Гидры размножаются двумя способами:
вегетативным и половым.
Вегетативное размножение у гидр носит
характер почкования. Почки возникают
в нижней части туловищного отдела тела
гидры над стебельком, последующие
почки находятся несколько выше преды-
дущих, иногда они сидят на противопо-
ложных сторонах тела гидры, иногда рас-
полагаются по спирали (порядок возникно-
вения и расположения почек зависит от
вида гидры). Одновременно на теле гидры
развивается 1—3, редко большее количе-
ство почек, однако наблюдали гидр
с 8 и более почками.
На первых стадиях почка возникает
как едва заметный конический бугорок,
затем она вытягивается, принимая более
или менее цилиндрическую форму. На
наружном конце почки появляются зачат-
ки щупалец, вначале они имеют вид корот-
ких тупых выростов, но постепенно вытяги-
ваются, и на них развиваются стрека-
тельные клетки. Наконец, нижняя часть
тела почки утончается в ножку, а между
щупальцами прорывается ротовое отвер-
стие. Молодая гидра некоторое время
еще остается соединенной с материнским
организмом, иногда на ней даже заклады-
ваются почки следующего поколения.
Отделение выпочковывающихся гидр про-
исходит в той же последовательности,
в какой возникают почки. Молодая гидра
размером несколько меньше материнской
п имеет неполное число щупалец. Недо-
стающие щупальца появляются позднее.
После обильного почкования материн-
ская гидра истощается и в течение неко-
торого времени почек на ней не возникает.
Некоторые исследователи наблюдали
также деление гидр, но этот способ раз-
множения, по-видимому, должен быть от-
несен к разряду ненормальных (патологи-
ческих) процессов. Деление у гидры воз-
никает после повреждения ее тела и может
быть объяснено высокой регенеративной
способностью этого животного.
Рис. 164. Почкование гидры.
267
При обильном питании весь теплый
период года гидры размножаются почко-
ванием, к половому размножению они
приступают с наступлением осени. Боль-
шинство видов гидр раздельнополы, но
есть и гермафродиты, т. е. такие, у кото-
рых на одной особи развиваются и мужские
и женские половые клетки.
Гонады образуются в эктодерме и
имеют вид небольших бугорков, конусов
или округлых тел. Порядок появления
и характер расположения гонад такие же,
как и почек. В каждой женской гонаде
образуется по одному яйцу.
В развивающихся гонадах скапливается
большое количество промежуточных, не-
дифференцированных клеток, из которых
образуются как будущие половые клетки,
так и «питательные» клетки, за счет кото-
рых увеличивается будущее яйцо. На пер-
вых стадиях развития яйца промежуточ-
ные клетки приобретают характер по-
движных амебоидов. Вскоре одна из них
начинает поглощать другие и значительно
увеличивается в размерах, достигая 1,5 мм
в поперечнике. После этого крупный аме-
боид подбирает свои псевдоподии и его
очертания округляются. Вслед за тем
происходят два деления созревания, при
которых клетка делится на две неравные
части, причем на наружной стороне яйца
Рис. 165. Половозрелые гидры:
слева — с семенниками, справа — с яйцами,
остаются два маленьких так называемых
редукционных тельца — клеточки, отде-
лившиеся от яйца в результате деления.
При первом делении созревания число
хромосом яйца сокращается вдвое. Созрев-
шее яйцо выходит наружу из гонады
через разрыв в ее стенке, но остается соеди-
ненным с телом гидры при помощи тон-
кой протоплазматической ножки.
К этому времени в семенниках
других гидр развиваются спермин,
которые покидают гонаду и плавают
в воде, один из них проникает в яйцо,
после чего сразу же начинается дроб-
ление.
В то время когда клетки развиваю-
щегося зародыша делятся, снаружи он
одевается двумя оболочками, внешняя
из которых имеет довольно толстые хитино-
идные стенки и часто бывает покрыта шипи-
ками. В таком состоянии зародыш под
защитой двойной оболочки—эмбриотеки—
перезимовывает. (Взрослые гидры с на-
ступлением холодов погибают.) К весне
внутри эмбриотеки уже имеется почти
сформированная маленькая гидра, кото-
рая покидает свою зимнюю оболочку
через разрыв ее стенки.
В настоящее время известно около
десятка видов гидр, населяющих пресные
воды материков и многих островов. Раз-
личные виды гидр отлича-
ются друг от друга очень не-
значительно. Один из видов
характеризуется яркой зеле-
ной окраской, которая обус-
ловлена наличием в теле этих
животных симбиотических
водорослей — зоохлорелл.
Среди наших гидр наиболее
известны стебельчатая, или
бурая, гидра (Hydra oligactis)
и бесстебельчатая, или-обык-
новенная, гидра (Hydra vul-
garis).
Естественных врагов у
гидры мало, так как она
хорошо защищена стрека-
тельными клетками. Все же
гидр поедают некоторые рес-
ничные черви и брюхоногие
моллюски — прудовики. Име-
ются и специфичные пара-
268
зиты гидр. Из простейших упомянем гид-
рамебу (Hydramoeba hydroxena) — эк-
топаразитическую амебу, которая по-
селяется на теле гидры и питается клет-
ками ее эктодермы. К наружным пара-
зитам гидры надо отнести также и
один вид инфузорий— триходин, или
гидровую «вошь» (Trichodina pediculus).
Наконец, на гидрах паразитируют малень-
кие ветвистоусые рачки анхистропусы
(Anchistropus). Эти рачки прикрепляются
к переднему концу гидры и ее щупальцам
при помощи специальных крючков на
первой паре ног. Зараженные этими рачка-
ми гидры вскоре погибают. Стрекатель-
ные клетки гидры, по-видимому, не дей-
ствуют на ее паразитов. Более того, отме-
чено, что прикосновение паразита к стре-
кательной клетке не вызывает ее «выстре-
ливания».
Как же гидра ведет себя в окружающей
ее среде, как она воспринимает раздраже-
ния и отвечает на них?
Как и большинство других кишечно-
полостных, гидра отвечает на всякое не-
благоприятное раздражение сокращением
тела. Если сосуд, в котором сидят гидры,
слегка тряхнуть, то одни из животных
сократятся сразу же, на других такой тол-
чок не подействует вовсе, часть гидр
только слегка подожмет свои щупальца.
Значит, степень реакции на раздраже-
ние у гидр очень индивидуальна. Гидра
совершенно лишена способности «запо-
минать»: можно часами ее колоть тонкой
булавкой, но после каждого сокращения
она снова вытягивается в том же направ-
лении. Если же уколы будут очень ча-
стыми, то гидра перестает на них реаги-
ровать.
Хотя у гидр нет специальных органов
для восприятия света, они совершенно
определенно реагируют на свет. К свето-
вым лучам наиболее чувствителен перед-
ний конец гидры, тогда как ее стебелек
световых лучей почти не воспринимает.
Если затенить зеленую гидру целиком, то
она через 15—30 секунд сократится,
если же затенять обезглавленную гидру
или притенить только стебелек целой
гидры, то она сократится лишь через 6—12
минут. Гидры способны различать направ-
ление потока света и двигаются в сторону
его источника. Скорость передвижения
гидр по направлению к источнику света
очень невелика. В одном из опытов 50 зеле-
ных и такое же количество бурых гидр были
помещены в сосуд на расстоянии 20 см от
стеклянной стенки, через которую падал
свет. Первыми двинулись к свету зеленые
гидры; через 4 часа 8 из них достигли
светлой стенки аквариума, через 5 часов
здесь их было уже 21, а через 6 часов—44.
К этому сроку туда же пришло 7 первых
бурых гидр. Вообще оказалось, что бурые
гидры шли на свет хуже, только через
10 часов у светлой стенки собралось 39
бурых гидр. Остальные подопытные живот-
ные к этому времени все еще находились
в пути.
Способность гидр двигаться в сторону
источника света или просто перемещаться
в более светлые участки бассейна очень
важна для этих животных. Гидры пита-
ются преимущественно планктонными
рачками — циклопами и дафниями, а эти
рачки всегда держатся в светлых и хорошо
прогретых солнцем местах. Таким обра-
зом, идя навстречу свету, гидры прибли-
жаются к своей добыче.
Для исследователя, изучающего реак-
ции низших организмов на свет, гидры
открывают самое широкое поле деятель-
ности. Можно ставить опыты по выявле-
нию того, насколько животные чувстви-
тельны к слабым или, напротив, очень
сильным источникам света. Оказалось,
что на слишком слабый свет гидры вовсе
не реагируют. Очень сильный свет застав-
ляет гидру уходить в затененные места
и может даже убить животное. Ставились
опыты по выявлению того, насколько чув-
ствительна гидра к изменению силы света,
как она ведет себя между двумя источни-
ками света, различает ли отдельные части
спектра. В одном из опытов стенка аква-
риума была окрашена во все цвета спек-
тра, при этом зеленые гидры собрались
в области сине-фиолетовых, а бурые в об-
ласти сине-зеленых лучей. Значит, гидры
различают цвет,- и разные их виды обла-
дают разным «вкусом» к нему.
Гидры (кроме зеленой) не нуждаются
в свете для нормальной жизнедеятельности.
Если их хорошо кормить, они прекрасно
живут и в темноте. Зеленая гидра, в теле
269
которой живут симбиотические водоросли
зоохлореллы, даже при обилии пищи
в темноте чувствует себя плохо и сильно
сокращается.
На гидрах можно производить опыты
по воздействию на организм различного
рода вредных излучений. Так, выясни-
лось, что бурые гидры погибают уже после
минутного освещения их ультрафиолето-
выми лучами. Зеленая гидра оказалась
к этим лучам более стойкой — она поги-
бает только на 5—6-й минуте облуче-
ния.
Очень интересны опыты по воздействию
на гидр лучей Рентгена. Небольшие дози-
ровки рентгеновских лучей вызывают
у гидр усиление почкования. Облученные
гидры по сравнению с необлученными
дают примерно в 2,5 раза больше потом-
ков за один и тот же срок. Увеличение
дозы облучения вызывает подавление
размножения; если же гидры получают
слишком большую дозу лучей Рентгена,
то они вскоре после этого погибают. Важно
отметить, что слабые дозы облучения
повышают у гидр регенеративные способ-
ности.
При воздействии на гидр радиоактив-
ного излучения был получен совершенно
необычный результат. Общеизвестно, что
животные никак не ощущают радио-
активных лучей и потому, попав в их
зону, могут получить смертельную дозу
и погибнуть. Зеленая гидра, реагируя
на излучения радия, стремится уйти от
его источника.
Из приведенных выше примеров видно,
что такие опыты с гидрами, как изучение
влияния на них различных факторов
внешней среды, не пустая забава, не нау-
ка ради науки, а серьезное и очень
важное дело, результаты которого могут
дать весьма существенные практические
выводы.
Конечно, проводилось изучение влияния
на гидру температуры, концентрации угле-
кислого газа, кислорода, а также целого
ряда ядов, лекарственных препаратов
и т. д.
Гидра оказалась очень удобным объек-
том для проведения целого ряда экспери-
ментальных исследований по изучению
явления регенерации у животных.
Как уже неоднократно упоминалось,
гидра легко восстанавливает
утраченные части тела. Жи-
вотное, разрезанное пополам, вскоре вос-
станавливает недостающие части. Но ста-
новится непонятно: почему на перед-
нем конце отрезка всегда вырастает «го-
лова» со щупальцами, а на заднем стебе-
лек? Какие законы управляют процес-
сами восстановления? Вполне вероятно
предположенпе, что некоторые из этих
законов могут быть общими и для гидры,
и для более высокоорганизованных жи-
вотных. Узнав их, можно сделать важные
выводы, приложимые даже к медицине.
Делать операции на гидрах очень про-
сто, для этого не нужно ни анестезирую-
щих средств, ни сложных хирургических
инструментов. Все оборудование «опера-
ционной» состоит из иглы, вделанной
ушком в деревянную ручку, острого глаз-
ного скальпеля, маленьких ножниц и тон-
ких стеклянных трубочек. Первые опыты
по выяснению регенеративных способно-
стей гидры были проведены более 200 лет
назад Трамбле. Этот кропотливый иссле-
дователь наблюдал, как из продольных
и поперечных половинок гидр возникают
целые животные. Затем он стал делать про-
дольные надрезы и увидел, что из лоскут-
ков в нижней части полипа образуются
стебельки, а из лоскутков в его верхней
части — «головы». Многократно оперируя
одного из подопытных полипов, Трамбле
получил семиглавого полипа. Отрезав ему
все семь «голов», Трамбле стал ждать
результатов и вскоре увидел, что на месте
каждой отрезанной «головы» появилась
новая. Семиглавый полип, у которого
вновь вырастают отрубленные «головы»,
был как две капли воды похож на мифи-
ческое существо — лернейскую гидру,
сраженную великим героем древней Гре-
ции Гераклом. С тех пор за пресноводным
полипом и сохранилось название—гидра.
Попутно Трамбле установил, что гидра
восстанавливается не только из полови-
нок, но и из совсем маленьких кусочков
тела. Теперь установлено, что даже из
V2oo части тела гидры может развиться
целый полип. Однако позднее выяснилось,
что регенеративная способность таких ма-
леньких кусочков из разных частей тела
270
гидры неодинакова. Участок подошвы или
стебелька восстанавливается в целую гид-
ру значительно медленнее, чем участок
из средней части тела. Однако этот факт
долго оставался необъясненным.
Внутренние силы, регулирующие и на-
правляющие процессы нормальной реге-
нерации, были вскрыты много позднее
знаменитым американским физиологом
Чайлдом (СМ. Child). Чайлд установил,
что у целого ряда низших животных в теле
имеется ярко выраженная физиологическая
полярность. Так, под действием ядови-
тых веществ клетки на теле животного
погибают и разрушаются во вполне опре-
деленной последовательности, а именно
от переднего конца к заднему (у гидры от
«головы» к «подошве»). Стало быть, клет-
ки, находящиеся в различных частях
тела, физиологически неравнозначны. Раз-
личие между ними заключается и во мно-
гих других проявлениях их физиологии,
в том числе и в воздействии на развиваю-
щиеся молодые клетки на месте травмы.
Постепенное изменение физиологиче-
ской активности клеток от одного полюса
к другому (вдоль оси тела) получило на-
звание осевого физиологиче-
ского градиента.
Теперь становится понятно, почему
кусочки, вырезанные из подошвы гидры,
очень медленно восстанавливают гипос-
том и щупальца — образующие их клетки
физиологически очень далеки от клеток,
образующих «голову». Осевой градиент
играет очень большую роль при регене-
рации, но на этот процесс оказывают за-
метное влияние также и другие факторы.
При регенерации очень большое значение
имеет наличие на регенерирующей части
развивающейся почки или искусственно
подсаженного участка ткани из другого
отдела тела животного, особенно из его
передней части. Обладая высокой физиоло-
гической активностью, развивающаяся
почка или клетки «головы» определенным
образом воздействуют на рост регенериру-
ющих клеток и подчиняют их развитие
своему влиянию. Такие группы клеток
или органы, которые вносят свои коррек-
тивы в действие осевого градиента, полу-
чили название организаторов.
Выяснение этих особенностей регенера-
ции помогло понять много неясных вопро-
сов в развитии животного организма.
В крупнейшем центре физиологии —
в созданном академиком Павловым
институте в Колтушах стоит памятник
собаке. Большая часть законов, изложен-
ных в учении Павлова, была открыта при
постановке опытов на собаках. Возмож-
но, такой же памятник заслуживает и ма-
ленький пресноводный полип.
ПРЕСНОВОДНАЯ МЕДУЗА
В 1880 г. в бассейне с тропическими ра-
стениями лондонского ботанического об-
щества вдруг появились медузы. Сразу два
зоолога Ланкестер (Lankester) и круп-
ный знаток кишечнополостных О л м е н (А1 ] -
man) сообщили об этой находке на страни-
цах журнала «Нечур» («Природа»). Медуз-
ки были очень маленькие, самые крупные
из них едва достигали 2 см в диаметре зон-
тика, однако их появление взволновало
тогдашних зоологов: до этого и не предпо-
лагали, что могут существовать пресно-
водные медузы. Медуз считали типичными
обитателями моря. Незадолго перед этим
в бассейн было посажено великолепное
южноамериканское водное растение
виктория-регия, поэтому высказывались
предположения, что медузы были завезе-
ны в Лондон вместе с посадочным мате-
риалом из Амазонки. Через некоторое
время медузы исчезли из бассейна так же
Рис. 166. Пресноводная медуза краспедакуста
(Craspedacusta).
271
таинственно, как и появились. Их обна-
ружили снова лишь через пять лет тоже
в Лондоне, но уже в другом бассейне с тем
же тропическим растением. В 1901 г.
эти медузы появились в Лионе (Франция),
также в оранжерейном бассейне с викто-
рией-регией. Затем их стали находить в
Мюнхене, Вашингтоне, Петербурге, Мо-
скве. Медуз обнаруживали то в бассейнах
ботанических садов, то в аквариумах
с тропическими рыбками. К удивлению
любителей-аквариумистов, у них неожи-
данно появились новые питомцы. Крохот-
ные медузки (часто всего 1—2 мм в диа-
метре зонтика) вдруг оказывались в боль-
шом количестве в аквариуме, в котором
накануне не было ни одной. Несколько
дней можно было наблюдать, как медуз-
ки толчками передвигаются в воде и охот-
но поедают маленьких рачков. Но в один
прекрасный день, заглянув в свой аква-
риум, хозяин находил в нем только рыб,
никаких медуз там не было.
К этому времени пресноводная ме-
дуза была подробно описана в специаль-
ной зоологической литературе. Оказа-
лось, что она принадлежит к классу
гидроидных. Назвали ее краспедакустой
(Craspedacusta). Самые маленькие медузки
имеют полусферический зонтик, 4 радиаль-
ных канала и 8 щупалец. По мере роста
медузы форма ее зонтика становится все
более плоской, а число щупалец увеличи-
вается.
Половозрелые медузы достигают 2 см
в диаметре и несут по краю зонтика ши-
рокий парус и около 400 тонких щупа-
лец, усаженных стрекательными клетка-
ми. Ротовой хоботок четырехгранный,
с крестообразным ротовым отверстием,
края рта слабоскладчатые. В месте отхож-
дения от ротового хоботка радиальных ка-
налов развиваются 4 гонады. Медузы
очень прозрачны, их мезоглея бесцветна,
а щупальца, радиальные каналы, ротовой
хоботок и гонады имеют беловатую или
кремовую окраску.
Эта медузка загадала зоологам сложную
загадку. Если согласиться с мнением о
том, что она попадает в оранжереи вместе
с растениями из тропиков, то как она вы-
живает при перевозке? Виктория-регия
транспортировалась с берегов Амазонки
в виде семян или корневищ. Нежные ме-
дузы, случайно захваченные вместе с кор-
невищами, несомненно, должны погиб-
нуть во время длительного пути через
океан. Но если даже предположить, что
медуза, несмотря на подсыхание, может
выжить, то как она попадает в маленькие
аквариумы любителей экзотических ры-
бок?
Вскоре медуз стали находить и в при-
родных водоемах. Первый раз ее поймали
Таблица 12. Животные, населяющие коралловый риф.
Моллюски:
1 — арабская ципрея (Сургаеа arabica);
2 — пятнистая ципрея (Сургаеа tigris),
3 — монетария-монета (Monetaria moneta);
4 — монетария-колечко (Monetaria annulus);
5 — монетария змеиная голова (Monetaria
caputserpentis);
6 — текстильный конус (Conus textile);
7 — пятнистый конус (Conus hebraeus);
8 — нильский трохуо (Trochus niloticus);
9 — мраморный турбо (Turbo marmoratus);
10 — элизия (Elysia);
11 — хромодорис (Chromodoris).
Ракообразные:
12 — краб Atergates;
13 — рак-отшельник Dardanus,
14 — рак-отшельник Calcynus,
15 — рак-отшельник Pagurus в раковине
Conus betuloidas.
Иглокожие:
16 — морской еж Echinometra matthaei;
17 — морской еж Tripneustes gratille.
в реке Янцзы в Китае, потом в Германии,
затем в США. Однако и в естественных
и в искусственных водоемах находки бы-
ли очень редкими и всегда неожиданными:
так, однажды медуз обнаружили в храни-
лищах вашингтонского водопровода.
Наблюдениями над медузой удалось
установить, что она отпочковывается от
крошечных бесщупальцевых полипчиков,
названных микрогидрами (Microhydra).
Эти полипы были найдены еще в 1884 г.
в тех же бассейнах в Лондоне, где ловили
и медуз, но тогда никто не предполагал
о связи между этими двумя столь непохо-
жими существами. Полипы микрогидры
видны простым глазом как белые точки
на фоне зеленых листьев водных растений,
на которых они обычно поселяются. Их
высота обычно не превышает 0,5—1 мм,
форма тела напоминает кеглю: туловище
в виде бутылки, а на короткой шейке сидит
шаровидная «головка» с ртом посереди-
не. Головка густо усажена стрекатель-
ными клетками, щупалец нет. Полипы
иногда образуют примитивные колонии
из 2—7 особей. Микрогидра размножается
почкованием и образует подобных себе
бесщупальцевых полипов. Время от вре-
мени от одной из сторон тела полипа отде-
ляется группа клеток, имеющая форму
маленького червячка. Такие группы кле-
ток называются фрустулами. Фру-
стула способна, извиваясь, ползать по
дну и забираться на водные растения,
здесь она превращается в молодую микро-
гидру.
Однажды удалось наблюдать, как на те-
ле микрогидры из почки стала развиваться
медуза; когда она отделилась от полипа
и начала плавать, то в ней легко было уз-
нать молодую краспедакусту. Удалось
также проследить за развитием яиц крас-
педакусты. Вначале из яйца образуется
червеобразная личинка, лишенная рес-
ничек и очень похожая на фрустулу мик-
рогидры. После некоторого периода пол-
зания по субстрату личинка прикрепля-
ется к нему и превращается в бесщупаль-
цевого полипа. Так было установлено,
что медуза краспедакуста и полип микро-
гидра принадлежат к одному виду кишеч-
нополостных, но к разным его поколе-
ниям.
Рис. 167. Жизненный цикл краспедакусты.
Проведенные опыты показали, что на
смену поколений у этого вида гидроид-
ных чрезвычайно большое влияние ока-
зывают условия среды. Выпочковывание
медуз на полипах происходит только при
температуре воды не ниже 26—33°С, а вы-
почковывание полипов и отделение фру-
стул — при температуре 12— 20°С. После
этого стало ясно, что существование вида
может длительное время поддерживаться
за счет размножения полипов. На малень-
ких неподвижных микрогидр ни аквари-
умисты, ни ботаники в оранжереях не об-
ращают внимания, так как они почти
и не видны простым глазом, очень трудно
найти их и в природе. Полипы могут долго
жить в аквариуме, а при повышении тем-
пературы у всех полипов появляются ме-
ду зоидные почки и они отделяют медуз.
Медузы краспедакусты подвижны и мо-
гут быть замечены в воде невооруженным
глазом. Теперь становится понятным, по-
чему их почти всегда находили в бассей-
V 18 Жизнь животных, т. i
273
нах с тропическими растениями и рыб-
ками: эти бассейны искусственно подогре-
вались. Неясно только одно: всегда ли
медузы жили в Европе или были привезе-
ны туда? (Полипы, возможно, способны
перенести некоторое высыхание и дли-
тельный путь в неблагоприятных усло-
виях.) И где же родина микрогидры—крас-
педакусты?
Ответить на этот вопрос довольно труд-
но. С момента первого обнаружения ме-
дуз в Лондоне было описано свыше 100
случаев их нахождения в самых различ-
ных частях мира. Вот краткое описание
распространения вида. В СССР местом
их обитания является Любовское водохра-
нилище вблизи Тулы, река Дон, озеро
Караязы около Тбилиси (на высоте почти
2000 ж над уровнем моря), река Кура, ис-
кусственные водоемы в Старой Бухаре.
Кроме того, медузы и полипы неоднократ-
но появлялись в аквариумах у любите-
лей-рыбоводов и в университетах Москвы
и Ленинграда. За пределами нашей стра-
Рис. 168. Кордилофора
(Cordylophora).
ны этот вид был обнаружен почти во всех
странах Европы, в Индии, Китае и Япо-
нии, в Австралии, Северной и Южной
Америке. Указать теперь, где его родина
и куда он был завезен, невозможно.
Совсем недавно этот вид кишечнополост-
ных снова заставил зоологов задуматься.
Теперь, когда распространение, образ
жизни, строение полипов и медуз, каза-
лось, были хорошо изучены, вдруг от-
крылось, что из яиц краспедакусты могут
развиваться полипы двух родов — описан-
ные выше бесщупальцевые и снабженные
щупальцами. Оба рода полипов образуют
фрустулы. Щупальценосные полипы при
помощи почкования образуют и подобных
себе и бесщупальцевых полипов, медуз
они выпочковывать не могут. Бесщупаль-
цевые полипы образуют подобных себе поли-
пов и медуз, но не способны выпочковы-
вать полипов, снабженных щупальцами.
Из фрустул образуются обе формы поли-
пов. Щупал ьценосные полипы пока были
обнаружены только два раза: в 1960 г.
в Венгрии и в 1964 г. в аквариуме Ленин-
градского университета. Условия, вызы-
вающие их появление, пока неясны. В ре-
ках Индии и великих озерах Африки оби-
тает еще два вида пресноводных медузу
близких родственников краспедакусты. Хо-
рошо известна медуза из африканского озе-
ра Танганьика, называемая лимнокнидой
(Limnocnida tanganjice).
ПРОИСХОЖДЕНИЕ ПРЕСНОВОДНЫХ
КИШЕЧНОПОЛОСТНЫХ
Как уже говорилось выше, число чисто
пресноводных кишечнополостных огра-
ничивается тремя видами медуз (и их
полипов), десятком видов гидр и па-
разитом икры осетровых рыб полиподиу-
мом, свободная стадия которого живет
в реках. Кроме этого, имеются также не-
которые виды гидроидных, способные вы-
держивать значительное опреснение и да-
же проникающие в пресные воды, но здесь
они не могут размножаться, и потому их
нельзя считать полноценными пресновод-
ными животными.
Из числа таких гидроидов в первую оче-
редь нужно сказать о кордилофоре.
274
Кордилофора образует небольшие неж-
ные колонии в виде кустиков высотой до
10 см. Полипы сидят на концах ветвей
и имеют веретеновидную форму. У каж-
дого полипа 12—15 щупалец, сидящих без
строгого порядка в срединной части тела.
Свободноплавающих медуз у кордилофо-
ры нет, особи медузоидного поколения
прикреплены к колонии.
Этот вид был впервые обнаружен акаде-
миком Российской академии П. С. Дал-
ласом в 1771 г. в северной части Кас-
пийского моря, потому-то кордилофора
и называется каспийской (Cordylophora
caspia). Однако ее распространение вовсе
не ограничивается этим бассейном, она
обитает в Балтийском, Черном и Азовском
морях, а также встречается вдоль всего
атлантического побережья Европы и в
устьях всех крупных рек Азии, Америки
и Австралии. Этот вид поселяется только
в сильно опресненных участках моря
и живет на небольшой глубине, обычно не
глубже 20 м.
В названии, данном Палласом кордило-
форе,—каспийская—имеется и свой смысл.
Дело в том, что родина кордилофоры —
Каспийское море. Только в середине про-
шлого столетия кордилофора по Волге
и Мариинской системе проникла в Балтий-
ское море, где в силу его малой солено-
сти @,8%) нашла свою вторую родину.
Кордилофора— о р г а н и з м-о б р а с т а-
т е л ь; она селится на всех твердых
подводных предметах, как неподвижных,
так и подвижных. Дальнейшую помощь
в расселении ей оказали бесчисленные ко-
рабли, стекающиеся со всех сторон в Бал-
тийское море. Возвращаясь домой, они уво-
зили из Балтийского моря на своем днище
незваного гостя, «нарушителя границ».
По руслам рек кордилофора проникает
иногда довольно высоко, но здесь она не
получает полного развития, ее колонии
по мере удаления от моря становятся ма-
ленькими, на них уже не возникают особи
медузоидного поколения.
Попробуем представить себе, какие же
у кишечнополостных есть возможности
попасть в пресноводный водоем. Таких
возможностей три. Одна из них—это пере-
ход к паразитизму. Возможно, что так по-
пал в наши реки полиподиум. Предки со-
временных осетровых рыб были настоящими
морскими животными, в море они и полу-
чили этого паразита. Затем, когда осетро-
вые стали проходными рыбами (такими,
которые часть жизни проводят в море, а
часть в пресноводных водоемах), полипо-
диум вместе с хозяином попал в реку.
Известно, что паразитический образ жизни
в значительной мере изменяет физиологию
паразита, вынуждая его приспосабливать-
ся к новым условиям жизни. Вместе с осет-
ровыми рыбами к жизни в пресной воде
приспособились и их паразиты.
А как же попадали в пресные водоемы
свободноживущие кишечнополостные? Не
могут ли они использовать для этого устья
рек, впадающих в море? Конечно, могут,
но при этом им придется преодолеть два
препятствия. Одно из них — это пониже-
ние солености. В реки могут попасть
только виды, способные выдерживать
очень значительное опреснение.
Среди типичных морских обитателей
есть такие, на которых даже самое незна-
чительное понижение процентного со-
держания соли в морской воде действует
губительно. К ним относятся почти все
коралловые полипы, сцифоидные медузы
и большинство гидроидных. Но часть
гидроидных все же может существовать
и при некотором опреснении. Из кишеч-
нополостных, упомянутых в этой книге,
к эвригалинным относится Корине. Этот
вид может жить как в воде с нормальной
океанической соленостью, так и в опрес-
ненных морях, например в Белом и Черном.
Из числа эвригалинных видов и вышли
те, потомки которых активно пробирались
в пресноводные водоемы. Процесс завоева-
ния рек и озер шел постепенно. Вначале
выделилась группа солоноватоводных гид-
роидов, которые уже не могли вернуться
в океан, так как не выносили высокой со-
лености его вод. Затем уже солоновато-
водные вплотную приблизились к устьям
рек. Далеко не все из них смогли пре-
одолеть этот «барьер», большинство так
и осталось в речном устье. В настоящее
время по этому пути идет кордилофора.
Попав в реку, морские животные встре-
тили на своем пути и другой «барьер» —
течение. При активном проникновении
морских или солоноватоводных кишечно-
18*
275
полостных в пресные воды им неизбежно
приходилось преодолевать встречный по-
ток воды, сносивший обратно в море
планктонных медуз и неспособных к само-
стоятельному движению прикреплен-
ных полипов или их колонии. Продвиже-
ние таких прикрепительных полипов на-
встречу течению было затруднено.
В отдаленные геологические эпохи кар-
та Земли была иной, чем мы видим ее
сейчас. Во многих местах современную
сушу покрывало море. Когда море ушло,
остались замкнутые соленые бассейны,
а в них сохранились морские животные.
Некоторые из этих бассейнов постепенно
опреснялись, причем животные либо по-
гибали, либо приспосабливались к новым
условиям. Замкнутое теперь Каспийское
море, которое по сути дела является ог-
Рис. 169. Два вида меризий (Moerisia):
2 — черноморская; 2 — ее полип; з — Палласова; 4, 5 — полипы
этой медузы.
ромным солоноватоводным озером, раньше
было соединено с океаном, и в нем сохра-
нилось много животных морского проис-
хождения. Среди них интересное кишечно-
полостное — Палласова меризия (Moerisia
pallasi). У этого вида гидроидных две
формы полипов: одни живут колонией на
дне, другие ведут планктонный образ
жизни. Плавающие полипы образуют ко-
лонии из двух особей, соединенных друг
с другом своими ножками. Время от вре-
мени колония разрывается пополам, и на
месте разрыва у каждого полипа обра-
зуются новый венчик, щупальце и рот.
Кроме того, полипы размножаются и поч-
кованием, отделяя от себя маленьких сво-
бодноплавающих медуз. Один близкий
вид меризий обитает в Черном и Азовском
морях, другой—в соленых озерах Северо-
Восточной Африки.
Совершенно ясно, что все три
вида меризий произошли от од-
ного общего предка, который
когда-то обитал в древнем Сар-
матском море. Когда Сармат-
ское море ушло, на его месте
остался ряд водоемов, в том
числе замкнутые Каспийское мо-
ре и озера Египта. В них раз-
вились самостоятельные виды
меризий.
Если представить себе, что
опреснение водоема идет еще
дальше, то можно понять, как
могут возникнуть пресноводные
медузы. Их способ завоевания
пресноводных бассейнов — дли-
тельное приспособление к уве-
личивающемуся опреснению.
Двигаться при этом им никуда
не нужно, они проделывают путь
из моря в пресную воду не в
пространстве, а во времени.
В 1910 г. на Атлантическом
побережье Северной Америки
было поймано несколько ма-
леньких гидромедуз. Оказалось,
что они принадлежат к ранее
неизвестному виду. Сам по се-
бе этот факт особого значения
не имеет. И теперь ежегодно
описывают несколько новых ви-
дов кишечнополостных — в мо-
276
ре еще много неизученного. Интересно дру-
гое. Эта медуза— ее назвали блакфордией
(Blackfordia) — через 15 лет была пойма-
на в Черном море. Ни в Средиземном море,
фауна которого известна очень хорошо, ни
на европейском побережье Атлантическо-
го океана этот вид не живет. Как же ока-
залась американская блакфордия в Чер-
ном море? Второй случай произошел со-
всем недавно. Один из видов гидроидов,
обитающих в Кильском канале,— буген-
виллия — неожиданно был обнаружен
опять-таки в Черном море. И блак-
фордия и упомянутый балтийский гидроид
(Bougainvillia megas) — солоноватовод-
ные виды; чтобы попасть из одного бас-
сейна с пониженной соленостью в другой,
они должны, подобно кордилофоре, преодо-
леть препятствие — море с его высокой
соленостью.
До постройки канала между Волгой и
Доном в Каспийском море было всего два
вида кишечнополостных — каспийская
меризия и кордилофора. Когда канал был
готов и по нему началось судоходство, из
Азово-Черноморского бассейна в Каспий-
ское море перебрались еще три вида. Уже
через год после ввода канала в строй пере-
селилась в Каспийское море блакфордия,
еще через год черноморская меризия,
а вслед за ней и тот балтийский гидроид
(Bougainvillia megas), который незадолго
перед тем попал в Черное море из Киль-
ской бухты. Конечно, так путешествуют
не одни только кишечнополостные, но и
моллюски, и ракообразные, и черви, и дру-
гие солоноватоводные организмы.
«ПАРУСНЫЙ ФЛОТ»
КИШЕЧНОПОЛОСТНЫХ
Класс гидроидных делится на два
подкласса — гидроидов и сифонофор.
К описанию этих удивительных пелагиче-
ских колониальных кишечнополостных мы
и переходим.
Целый мир живых существ обитает на
грани двух стихий — воды и воздуха. На
плавающих водорослях, обломках древе-
сины, кусках пемзы и других предметах
можно обнаружить разнообразных при-
росших или крепко уцепившихся живот-
ных. Не следует думать, что они попали
сюда случайно — «терпят бедствие». На-
против, многие из них теснейшим обра-
зом связаны и с водной и с воздушной сре-
дой и в иных усяовиях они существовать
не могут. Кроме таких «пассивных пас-
сажиров», здесь же можно видеть живот-
ных, активно плавающих у самой поверх-
ности, снабженных различно устроенными
органами — поплавками, или животных,
которые удерживаются, используя плен-
ку поверхностного натяжения воды. Весь
этот комплекс организмов (плейстон) осо-
бенно богат в субтропиках и тропиках,
где не ощущается губительного действия
низких температур.
Выше, когда речь шла о действии стре-
кательных клеток, уже был упомянут
«португальский военный кораблик» —
крупная сифонофора физалия (Physalia,
см. цветную таблицу 8).
Как и все сифонофоры, физалия пред-
ставляет собой колонию, в состав которой
входят как полипоидные, так и
медузоидные особи. Над поверх-
ностью воды возвышается воздушный
пузырь, или пневматофор,— видо-
измененная медузоидная особь колонии.
У крупных экземпляров пневматофор до-
стигает 30 см, он обычно имеет ярко-
голубой или красноватый цвет. Воздушный
пузырь плавает по поверхности моря как
туго надутый резиновый шар. Наполняю-
щий его газ близок по составу к воздуху*
но отличается повышенным содержанием
азота и углекислого газа и уменьшенным
количеством кислорода. Этот газ выраба-
тывается специальными газовыми желе-
зами, находящимися внутри пузыря.
Стенки пневматофора выдерживают до-
вольно сильное давление газа, так как
образованы двумя слоями эктодермы, дву-
мя слоями энтодермы и двумя слоями ме-
зоглеи. Кроме того, эктодерма выделяет
тонкую хитиноидную оболочку, благода-
ря которой прочность пневматофора также
значительно увеличивается, хотя его стен-
ки остаются очень тонкими. Верхняя часть
пневматофора имеет вырост в виде гребня.
Гребень расположен на пневматофоре не-
сколько по диагонали и имеет слегка вы-
гнутую S-образную форму. Все остальные
277
особи колонии расположены на нижней
стороне пневматофора и погружены в
воду.
Питающие полипы, или гастрозои-
д ы, сидят в один ряд. Они имеют более
или менее бутылковидную форму и обра-
щены ротовым отверстием вниз. Каждый
питающий полип снабжен одним длинным
щупальцем — арканчиком. По всей длине
арканчик густо покрыт стрекательными
клетками. Рядом с каждым питающим по-
липом на нижней стороне пузыря прикреп-
ляется основание гонодендра —
особи полипоидного поколения. На гоно-
дендре и его боковых выростах распо-
ложены грозди редуцированных медузо-
идных особей — гонофоров, в которых
развиваются половые продукты. Здесь же
сидят и защитные бесщупальцевые поли-
пы — п а л ь п о н ы. На каждом гоноденд-
ре имеется одна медузоидная особь, назы-
ваемая нектофором или плава-
тельным колоколом. Половые клет-
ки у нектофора не образуются, а его зонтик
достигает значительной величины и спосо-
бен сокращаться, как у свободноплавающих
медуз. Перед наступлением половой зре-
лости гонофоров гонодендры отрываются
от колонии и плавают у поверхности моря,
причем нектофор выполняет локомотор-
ные функции.
Направление Направление
ветра ветра
Рис. 170. Так под влиянием ветра «правые» и
елевые» «португальские кораблики» расплываются
в разные стороны.
Благодаря косому расположению греб-
ня на плавательном пузыре физалия асим-
метрична, причем известны две формы
физалий — «правая» и «левая», которые
являются как бы зеркальным отображени-
ем друг друга. Было замечено, что все
физалий, обитающие в одном участке мо-
ря, имеют одинаковое строение, т. е. все
они либо «правые», либо «левые». В связи
с этим высказывалось предположение, что
существует два вида или две географиче-
ские расы физалий.
Однако, когда стали изучать развитие
этих сифонофор, обнаружилось, что
среди потомства одной физалий всегда
имеется равное число и «правых» и «ле-
вых». Значит, никаких особых рас у фи-
залий нет. Но как возникают скопления
«левых» и «правых» сифонофор и почему
эти две формы не встречаются вместе?
Ответ на этот вопрос был получен после
детального изучения строения воздушного
пузыря физалий. Оказалось, что форма
и расположение гребня на его вершине
имеют для физалий очень большое значе-
ние. Как было сказано выше, гребень
физалий слегка изогнут в виде буквы S.
Фпзалия передвигается по поверхности
моря благодаря тому, что ветер ударяет
в ее воздушный пузырь. Если бы не было
гребня, сифонофора постоянно двигалась
бы по прямой и ее в конце концов выбро-
сило бы на берег. Но наличие гребня вно-
сит существенные изменения в парусную
оснастку «португальского кораблика». Ко-
со поставленный и изогнутый гребень за-
ставляет животное плыть под острым уг-
лом к ветру и время от времени делать
поворот вокруг своей оси против ветра.
Если наблюдать за физалией, плаваю-
щей вблизи берега, в сторону которого
дует ветер, то можно увидеть, как она то
приближается к берегу, то, неожиданно
повернувшись к наблюдателю другим
боком, медленно отплывает от него. Так
маневрируют целые армады «португаль-
ских корабликов», напоминая действия
парусного флота периода средневековых
войн. При движении «правые» и «левые»
«португальские кораблики» ведут себя по-
разному. Под влиянием ветра, дующего в
одном направлении, они расходятся в раз-
ные стороны — «правые» налево, а «ле-
278
вые» направо. Вот поэтому-то и возникают
скопления одинаковых форм физалий.
К плейстонным организмам от-
носятся также весьма своеобразные ки-
шечнополостные — порпита (Porpita) и
еелелла (Velella), которую также называют
парусником.
Долгое время этих животных относили
к сифонофорам, причем отдельные при-
датки их считали специализированными
особями колонии. Теперь все больше зоо-
логов склоняется к мнению о том, что пор-
пита и парусник представляют собой не
колонию, а крупного одиночного плаваю-
щего полипа, и относит их к отряду
хондрофор (Chondrophora) из к л а с с а
гидроидных. Тело у них уплощенное;
у пюрпиты оно имеет форму круга, у па-
русника — овала. Верхняя сторона дис-
ка покрыта хитиноидной оболочкой, под
которой помещается сложно устроенный
воздушный колокол — пневмато-
ф о р. Он состоит из центральной
камеры, большого количества окру-
жающих его кольцевых камер
и отходящих от них ко всем частям тела
тонких трубочек — трахей, служа-
щих для дыхания. На нижней стороне
диска расположены органы полипа. В цен-
тре находится ротовой конус, а
по периферии располагаются многочис-
ленные щупальца. Между ротовым
конусом и щупальцами имеются особые
выросты тела — гонодендры, на ко-
торых выпочковываются особи медузоид-
ного поколения. Верхняя сторона диска
прибрежной порпиты гладкая; у велеллы,
живущей в открытом океане, на ней на-
ходится высокий вырост треугольной
формы — парус. Парус велеллы имеет
такое же значение, что и гребень на воз-
душном пузыре физалий. Он расположен
на овальном теле парусника асиммет-
рично и слегка S-образно изогнут. Парус
позволяет животному двигаться не по
прямой, а маневрировать, хотя, конечно,
не произвольно, а более или менее слу-
чайно.
В субтропических частях океана, где
температура не падает ниже 15°С, парус-
ники встречаются в очень больших коли-
чествах. Местами эти крупные кишечно-
полостные (они достигают 12 еж по длин-
ной оси диска) собираются в огромные
стаи протяженностью в несколько десят-
ков миль, причем на каждый квадратный
метр поверхности океана приходится по
паруснику. Вместе с крупными парусни-
ками плавают и молодые, величина кото-
рых измеряется миллиметрами.
Ветер, ударяя в парус, гонит стаю ве-
лелл по морю, и они могут проходить
многие сотни миль.
Обитая в открытом океане, парусники
не боятся воды: они не могут утонуть,
так как имеют очень совершенный пнев-
матофор, состоящий из большого числа
независимых камер. Если волна все же
опрокинет велеллу, то при помощи движе-
ний краев диска она принимает нормаль-
ное положение и снова подставляет парус
ветру. Кроме парусников, здесь же мож-
но встретить и много других животных,
которые, однако, вначале почти незаметны.
Общеизвестно, что открытое море тро-
пиков имеет интенсивный синий цвет.
В связи с этим парусники и большинство
животных, которые обитают вместе с ни-
ми, также окрашены в голубые или синие
тона — это служит им хорошей защитой.
Парусники и обитающие среди них
другие животные создают в открытом
море особый тесно связанный мирок —
плейстонный биоценоз, который по воле
течения и ветра все время плывет по по-
верхности океана.
Велелла, как и все кишечнополостные,—
хищник; она питается планктоном, в со-
став ее пищи входят рачки, личинки раз-
личных беспозвоночных, мальки рыб. Все
же остальные животные, которые входят
в состав плавающего биоценоза, либо
питаются парусниками, либо используют
их в качестве постоянного или временного
субстрата для прикрепления. Таким обра-
зом, весь биоценоз существует за счет
планктона, но непосредственно исполь-
зуют планктон только одни парусники.
На верхней стороне диска велеллы, как
на палубе корабля, путешествуют малень-
кие голубые крабики планес (Planes).
Здесь они находят защиту от врагов, а
также получают пищу. Голодный крабик
быстро перебирается на нижнюю сторону
диска парусника и отнимает у него за-
хваченных планктонных рачков. Наевшись,
279
крабик опять взбирается на верхнюю сто-
рону диска и устраивается под парусом,
тесно прижавшись к нему. Крабики ни-
когда не пожирают свой корабль, чего
нельзя сказать о многих других плейстон-
ных животных.
На нижней стороне парусника часто
можно найти хищного брюхоногого мол-
люска янтину (Janthina). Янтины объ-
едают мягкие ткани до тех пор, пока от
парусника не останется один хитиноид-
ный скелет. Потеряв опору, янтина не
тонет, так как хорошо приспособлена
к жизни у поверхности воды. Как только
поедаемый парусник начинает тонуть, ян-
тина выпускает обильную слизь, обра-
зующую пузырьки, наполненные возду-
хом. Слизь эта очень быстро затвердевает,
и получается хороший поплавок, на ко-
тором моллюск может самостоятельно пла-
вать, передвигаясь от одного парусника
к другому. Подплыв к новой жертве,
янтина покидает ненужный ей теперь по-
плавок и быстро переползает на велеллу.
Брошенный поплавок янтины вскоре за-
селяется гидроидами, мшанками, мор-
скими уточками и другими прикреплен-
ными животными, а также маленькими
крабикамщ иногда они поселяются и на
раковине самого моллюска.
Вместе с янтиной на парусниках поселя-
ется и другой хищный моллюск — голо-
жаберник эолис (Aeolis).
Иногда рядом с парусником можно за-
метить сопровождающих его голожабер-
ных моллюсков глаукусов (Glaucus). Тело
этого безраковинного моллюска вытяну-
тое, рыбообразное, по бокам имеется три
пары разветвленных щупальцевидных вы-
ростов, при помощи которых моллюск
прикрепляется к поверхностной пленке
воды. Плавает он темно-синей брюшной
стороной кверху, его спинная сторона
серебристо-белого цвета. Это делает плы-
вущего глаукуса незаметным и с воздуха
и из воды. Проголодавшийся глаукус, под-
гребая щупальцевидными выростами, под-
плывает к паруснику и, придерживаясь
за него, вырывает и поедает большие
куски края диска.
Объеденные моллюсками, парусники
гибнут, но от них остается хитиноидный
скелет, в котором еще сохраняется система
воздушных камер. Такие мертвые парус-
ники плавают некоторое время на поверх-
ности, и на них поселяются личинки усо-
ногих рачков — морских уточек (Lepas
fasciculatus). По мере роста новых посе-
ленцев скелет парусника погружается все
глубже, а на ножке, при помощи которой
морская уточка прикрепляется к суб-
страту, развивается дополнительный ша-
ровидный поплавок, увеличивающий пла-
вучесть рачка.
Все свободноживущие усоногие раки —
прикрепленные животные, исключение со-
ставляет только указанный выше вид
морской уточки. Когда ее шаровидный
поплавок достигает значительной величи-
ны, он отделяется от парусника, и после
этого морская уточка может самостоятель-
но держаться на поверхности воды и даже
плавать, размахивая ножками. У осталь-
ных усоногих рачков взмахи ножек под-
гоняют к рачку пищу — мелкие планктон-
ные организмы, но этот вид морской
уточни, в отличие от всех своих сороди-
чей, ведет хищнический образ жизни.
Подплыв к паруснику, морская уточка
захватывает край его диска ножками и,
передвигаясь вдоль края, быстро выедает
значительную часть велеллы.
Кроме описанных здесь животных,
в биоценоз велеллы входят также неко-
торые креветки, ресничные черви, клопы-
водомерки и ряд других животных, в том
числе один вид летучей рыбы прогнихтис
(Prognichthys agae), которая откладывает
на парусниках икру. Клопы-водомерки
галобатесы живут в тесном контакте с ве-
леллой и порпитой, используя их и как
«пирог» и как «плот».
Носящийся в открытом океане мирок
велеллы очень ограничен, но все его оби-
татели тесно связаны друг с другом. Инте-
ресно отметить, что большинство видов,
составляющих этот биоценоз, относится
к таким группам животных, которые обыч-
но ведут донный образ жизни. Исходя
из этого, можно с уверенностью сказать,
что плейстонные животные происходят
от донных (а не от планктонных) организ-
мов, которые потеряли связь с дном и ста-
ли прикрепляться к различным плаваю-
щим предметам или использовать в каче-
стве опоры поверхностную пленку воды.
280
КЛАСС СЦИФОИДНЫЕ (SCYPHOZOA)
(«Хрустальное мясо»)
Когда в общежитии говорят о медузах,
то обычно имеют в виду не маленьких
гидромедуз, а крупных медуз, относящих-
ся к классу сцифоидных. Именно эти
медузы хорошо видны и с борта корабля
и во время купания; некоторые из них
больно жгутся. Часто их можно видеть
после шторма уже мертвыми на берегу
моря. Одним из самых обычных и наи-
более широко распространенных видов
сцифоидных медуз следует считать уша-
стую медузу, или аурелию (Aurelia aurita),
которая обитает почти во всех умеренных
и тропических морях обоих полушарий
и заходит также и в арктические области.
Из морей, омывающих нашу страну, она
живет в Балтийском, Белом, Баренцевом,
Черном, Азовском, Японском и Беринго-
вом, причем часто встречается в массовых
количествах.
Уплощенный зонтик ушастой медузы
достигает иногда 40 см в диаметре. Ее
легко узнать по розоватому или слегка
фиолетовому цвету и четырем темным под-
ковкам в средней части зонтика — поло-
вым железам. По краям рта медузы рас-
положены 4 длинные ротовые лопасти,
напоминающие по форме ослиные уши; от
них медуза и получила свое название.
Летом в тихую спокойную погоду во
время отлива или прилива можно видеть
большое количество этих красивых медуз,
медленно переносимых течением. Их тела
спокойно колышутся в воде. Ушастая
медуза — плохой пловец, благодаря со-
кращениям зонтика она может лишь мед-
ленно подниматься к поверхности, а затем,
неподвижно застыв, погружаться в глу-
бину.
На краю зонтика аурелии имеется 8
ропалиев, несущих глазки и
статоцисты. Эти органы чувств по-
зволяют медузе держаться на известном
расстоянии от поверхности моря, где ее
нежное тело быстро разорвут волны. Пи-
щу ушастая медуза захватывает так же,
как маленький тиаропсис. По краям ее
вонтика сидят сотни длинных и очень
тонких щупалец, которые «сметают» в рот
медузе мелких планктонных животных.
Заглоченная пища вначале попадает в
глотку, а потом в желудок. Отсюда берут
начало 8 прямых радиальных каналов
и такое же количество ветвящихся. Если
при помощи пипетки ввести в желудок
медузе раствор туши, то можно просле-
дить, как жгутиковый эпителий энтодер-
мы гонит по каналам гастральной системы
пищевые частицы. Вначале тушь прони-
кает в неветвящиеся каналы, затем она
входит в кольцевой канал и по ветвящимся
каналам возвращается обратно в желудок.
Отсюда непереваренные частицы пищи
через ротовое отверстие выбрасываются
наружу.
Половые железы аурелии, имеющие фор-
му четырех незамкнутых или полных ко-
лец, расположены в карманах желудка.
Когда яйца в них созреют, стенка поло-
вой железы разрывается и яйца через рот
выбрасываются наружу. В отличие от
большинства сцифомедуз, аурелия прояв-
ляет своеобразную заботу о потом-
стве. Ротовые лопасти этой медузы не-
сут по своей внутренней стороне глубокий
продольный желоб, начинающийся от ро-
тового отверстия и проходящий до самого
конца лопасти. По обеим сторонам желоба
расположены многочисленные маленькие
отверстия, которые ведут в небольшие
полости—к арманчики. У плаваю-
щей медузы ее ротовые лопасти опущены
вниз, так что выходящие из ротового
отверстия яйца неизбежно попадают в же-
лоба и, продвигаясь вдоль них, задержи-
ваются в карманчиках. Здесь происхо-
дит оплодотворение и развитие яиц. Из
карманчиков наружу выходят вполне
сформированные планулы. Если поме-
стить крупную самку аурелии в аква-
риум, то уже через несколько минут в воде
можно заметить массу светлых точек.
Это планулы, покинувшие карманчи-
ки и плавающие при помощи ресничек.
Молодые планулы обнаруживают стрем-
ление двигаться в направлении к источ-
281
Рис. 171. Сцифоидные медузы цианея (Суапеа) и аурелия (Aurelia).
нику света, вскоре они скапливаются
в верхней части освещенной стороны ак-
вариума. Вероятно, это свойство помогает
им выбраться из затемненных карманчи-
ков на волю и держаться вблизи поверх-
ности, не уходя в глубину.
Вскоре у планул появляется тенденция
к погружению на дно, но обязательно
в светлых местах. Здесь они продолжают
оживленно плавать. Период свободнопод-
вижной жизни планулы длится от 2 до 7
суток, после чего они оседают на дно и
прикрепляются передним концом к ка-
кому-нибудь твердому предмету.
Через двое-трое суток осевшая планула
превращается в маленького полипа —
сцифистому, имеющего 4 щупаль-
ца. Вскоре между первыми щупальцами
появляется 4 новых, а затем еще 8 щупа-
лец. Сцифистомы активно питаются, захва-
тывая инфузорий и рачков. Наблюдается
также каннибализм — поедание
282
сцифистомами планул того же вида. Сци-
фистомы могут размножаться поч-
кованием, образуя подобных себе полип-
чиков. Сцифистома зимует, а следующей
весной, с наступлением потепления, в ней
происходят серьезные изменения. Щупаль-
ца сцифистомы укорачиваются, а на теле
появляются кольцевидные перетяжки.
Вскоре от верхнего конца сцифистомы отде-
ляется первая эфира — маленькая совер-
шенно прозрачная звездообразная ли-
чинка медузы. К середине лета из эфир
развивается новое поколение ушастых ме-
ДУз.
Очень сходный образ жизни ведут и дру-
гие крупные сцифоидные медузы, напри-
мер цианея (Суапеа). Эти гиганты среди
кишечнополостных (известны экземпляры
до 2 м в диаметре зонтика) обитают толь-
ко в холодных водах, и потому их нет
в Черном море. Внешне цианея очень кра-
сива. Зонтик в центре обычно желтова-
тый, к краям темно-красный. Ротовые ло-
пасти имеют вид широких малиново-крас-
ных занавесок, щупальца окрашены
в светло-розовый цвет. Особенно яркой
окраской отличаются молодые медузы.
Всем, кто бывал на Черном и Азовском
морях, хорошо знакома большая сцифо-
идная медуза ризостома, или корнерот
(Rhizostoma pulmo, см. таблицу 8). Зон-
тик этой медузы полусферической или
конической формы с закругленной вер-
шиной. Крупные экземпляры ризостомы
с трудом помещаются в ведро. Цвет меду-
зы беловатый, но по краю зонтика прохо-
дит очень яркая голубая или фиолетовая
каемка.
Щупалец у этой медузы нет, зато ее ро-
товые лопасти разветвляются надвое, при-
чем их боковые стороны образуют много-
численные складки и срастаются между
собой. Концы ротовых лопастей складок
не несут и заканчиваются восемью кор-
невидными выростами, от которых медуза
и получила свое название. Рот у взро-
слых корнеротов зарастает, и его роль
выполняют многочисленные мелкие от-
верстия в складках ротовых лопастей.
Здесь же, в ротовых лопастях,
происходит и пищеварение. В верхней
части ротовых лопастей корнерота имеют-
ся дополнительные складки, так называе-
мые эполеты, усиливающие пищева-
рительную функцию. Питаются корнероты
самыми мелкими планктонными организ-
мами, засасывая их вместе с водой в гаст-
ральную полость.
Корнероты — довольно хорошие плов-
цы. Обтекаемая форма тела и сильная мус-
кулатура зонтика позволяют им продви-
гаться вперед быстрыми частыми толч-
ками. Интересно отметить, что, в отличие
от большинства медуз, корнерот может
изменять свое движение в любом направ-
лении, в том числе и вниз.
Купальщики не очень радуются встрече
с корнеротом: прикоснувшись к нему,
можно получить довольно сильный болез-
ненный «ожог». Держатся корнероты обыч-
но на небольшой глубине вблизи берегов,
часто в большом количестве встречаются
в причерноморских лиманах.
В тропических морях обитает несколь-
ко других видов корнеротых медуз, неко-
торые из них употребляются в пищу1
в Японии и Китае. Одна из этих медуз так
и называется съедобная ропилема (Rhopi-
lema esculenta). Внешне ропилема очень
похожа на нашего черноморского корне-
рота, но отличается от него наличием
очень большого числа концевидных выро-
стов и желтоватой или красноватой
окраской ротовых лопастей и их придат-
ков (см. рис. 172).
Ропилема живет в теплых прибрежных
водах, скапливаясь массами вблизи усть-
ев рек. Замечено, что наиболее интенсив-
но эти медузы растут после наступления
сезона летних ливневых тропических дож-
дей. В период дождей реки выносят в море
большое количество органических веществ,
способствующих развитию планктона,
которым и питаются медузы.
Ропилемы малоподвижны. Их переме-
щения зависят главным образом от морских
течений и ветров. Иногда под влиянием
течения и ветра скопления медуз обра-
зуют пояса по 2,5—3 км длиной. В неко-
торых местах побережья Южного Китая
летом море становится белым от скопив-
шихся там ропилем, которые колышутся
у самой поверхности.
1 В пищу употребляют также ушастых медуз —
аурелий.
283
Ловят медуз сачками или спе-
циальными орудиями лова, имею-
щими вид большого мешка из мел-
коячеистой сети, надетого на об-
руч. Во время прилива или отли-
ва мешок надувается течением и в
него попадают медузы, которые не
могут выбраться вследствие своей
малоподвижности.
У добытых медуз отделяют ро-
товые лопасти и промывают зонтик
до полного удаления внутренних
органов и слизи. Таким образом,
в дальнейшую обработку посту-
пает, по сути дела, только мезог-
лея зонтика. По образному выра-
жению китайцев, мясо у медуз
«хрустальное». Солят медуз поваренной
солью в смеси с квасцами. Соленых медуз
прибавляют к различным салатам, а также
едят в вареном и жареном виде, приправ-
ляя перцем, корицей и мускатным орехом.
Конечно, медуза — малопитательный про-
дукт, но все же в соленых ропилемах
содержится некоторое количество белков,
жиров и углеводов, а также витаминов
Вх, В2 и никотиновой кислоты.
Ушастая медуза, съедобная ропилема и
некоторые близкие к ней виды сцифомедуз,
по всей вероятности, единственные кишеч-
нополостные, которые употребляются в
пищу людьми. В Японии и Китае имеется
даже специальный промысел этих медуз, и
там ежегодно добывают тысячи тонн «хру-
стального мяса».
СИДЯЧИЕ МЕДУЗЫ
Среди большого количества разнооб-
разных сцифоидных медуз несколько особ-
няком стоит группа ставромедуз, предста-
вители которой не способны плавать и ве-
дут донный образ жизни. Форма этих
животных довольно своеобразна и внешне
несколько напоминает воронку. Тело под-
разделяется на чашечку и нож-
ку. В срединной части чашечки находит-
ся ротовое отверстие, окружен-
ное четырьмя складчатыми лопастями.
Края чашечки вытянуты в 8 так называе-
мых рук. На конце каждой «руки» имеет-
ся пучок маленьких головчатых
щупалец. У некоторых ста-
вромедуз между руками на
краю зонтика помещается по осо-
бому видоизмененному головчато-
му щупальцу. Это р о п а л и о и-
ды — органы, служащие для вре-
менного прикрепления медуз при
движении. У некоторых видов в
основании ропалиоида помещается
небольшое пигментное пятно, ко-
торое, возможно, несет светочувст-
вительную функцию.
При помощи ножки ставромеду-
зы прикрепляются к различным
подводным предметам. Мелковод-
ные формы обычно поселяются
на водорослях. Тело животного полу-
прозрачное, мезоглея развита очень силь-
но. Интересно отметить, что цвет живот-
ных может быть светло-желтым, оранже-
вым или бурым, причем окраска тканей
ставромедуз всегда соответствует цвету
того вида водорослей, на которых данный
экземпляр поселился. Высказывалось пред-
положение, что ставромедузы каким-то
образом извлекают из водорослей красящее
вещество, но механизм этого процесса по-
ка еще совершенно неясен.
Известно около 30 видов этих сцифоид-
ных. Наиболее обычны представители р о-
д о в люцернария (Lucernaria) и хали-
клистус (Haliclystus).
Люцернарии могут быть довольно круп-
ными — 12—15 смв высоту. Они живут в
северной части Атлантического океана и в
наших северных морях. Один вид люцер-
нарии, изредка встречающийся в Черном
море, а также большинство других став-
ромедуз, в том числе и упомянутый хали-
клистус, имеют всего 1—3 см в высоту
вместе с ножкой. Интересен восъмирук
(Octomanus), ропалиоиды которого пре-
вратились в хватательные органы, напоми-
нающие человеческую руку в рукавице.
Ставромедузы большую часть времени
сидят на одном месте, плотно прикрепив-
шись к субстрату присоской, находящейся
на конце мускулистой ножки. Они спо-
собны медленно передвигаться. Чтобы пе-
рейти на новое место, ставромедуза дугооб-
разно изгибает ножку и прикрепляется к
водоросли при помощи ропалиоидов. За-
Рис. 172. Съедоб-
ная ропилема
(Rhopilema escu-
lenta).
284
тем она передвигает ножку в направлении
движения и прикрепляется на новом ме-
сте. Один «шаг» сделан, за ним следует
второй. Движения ставромедузы отличают-
ся медлительностью. Как и все кишечно-
полостные, ставромедузы — типичные хи-
щники. Они подстерегают свою добычу,
свесившись с водных растений и широко
раскинув «руки» с многочисленными щу-
пальцами. Своим поведением во время
передвижения и охоты они очень напоми-
нают пресноводных полипов — гидр.
Ставромедузы отличаются от остальных
сцифоидных не только строением и обра-
зом жизни, но и своеобразным жизненным
циклом. Из яиц ставромедузы развивают-
ся не планулы, а лишенные ресничек чер-
веобразные личинки. Они некоторое вре-
мя ползают по дну, а затем собираются
небольшими группами по 3—20 особей, и
каждая из них прикрепляется к субстра-
ту передним концом. После этого личинки
при помощи стрекательных клеток начи-
нают ловить добычу. Действуя сообща,
они могут поймать и убить животных, ко-
торые значительно крупнее их самих.
После некоторого периода питания и
роста личинки выпочковывают по 4 подоб-
ных себе ползающих личинки и превра-
щаются в маленьких полипчиков. Вначале
такой полип снабжен четырьмя головча-
тыми щупальцами, затем появляются еще
4 таких же щупальца. В дальнейшем на
верхней части полипа вырастают 8 «рук».
Таким образом, взрослое животное раз-
вивается из личинки постепенно. Чередо-
вания полипоидного и меду-
зоидного поколений в жиз-
ненном цикле ставромедуз нет.
Как же следует трактовать строение
ставромедузы, что это — медуза или по-
лип? Известный зоолог прошлого века
Эрнест Г е к к е л ь считал их медузами.
Он предполагал, что они приобрели нож-
ку и утратили зонтик в связи с переходом
к донному образу жизни. Геккель даже
описал несколько медуз, строение кото-
рых было как бы промежуточным между
сидячими и свободноплавающими сцифо-
идными. Однако после Геккеля никто та-
ких медуз не видел, да и, судя по его ри-
сункам, эти «промежуточные» медузы ско-
рее всего были просто поврежденными эк-
земплярами корономедуз. Тем не менее
в рассуждениях сторонников медузной
природы ставромедуз довольно много спра-
ведливого. Ставромедузы имеют половые
железы, гастральные нити в желудке и
складчатые ротовые лопасти. Все эти приз-
наки характерны для сцифоидных медуз
и отсутствуют у сцифистом. С другой сто-
роны, в строении ставромедуз явственно
проступают черты организации сцифисто-
мы. Сходна их форма тела. Гастральная
полость тех и других разделена на 4 боко-
вые камеры и центральный желудок. Внут-
ри септ они имеют впячивания —ворон-
к и, которые заканчиваются мускуль-
ными тяжами, идущими вдоль
ножки.
Рис. 173. Сидячая сцифомедуза — восьмирук
(Octomanus).
285
Несомненно, что в строении ставромедуз
имеется какая-то двойственность: по од-
ним признакам это медузы, по другим —
сцифистомы.
Но возможно и третье решение вопроса.
Попробуем представить себе ставроме-
дузу как сцифоидного полипа, который
начал отделять от себя медузу, как это
характерно для полипов других групп
сцифоидных. Медуза только начала фор-
мироваться на верхнем конце полипа, но
не отделилась. У нее возникли складча-
тые ротовые лопасти, появились гастраль-
ные нити, образовались половые железы, но
отделения от сцифистомы не произошло.
Если это предположение правильно, тогда
становится понятным и жизненный цикл
ставромедуз, в котором нет смены двух
различно устроенных поколений.
Таким образом, ставромедуза может
рассматриваться как очень необычный
организм, соединяющий в себе признаки
и медузы и полипа.
ЖИВЫЕ ФОНАРИ
Каждый раз, когда из глубин океана
поднимают планктонную сеть —специаль-
ный прибор для ловли планктонных орга-
низмов, в ней оказываются разнообразные
медузы, живущие на многокилометровой
глубине. Их форма довольно разнообраз-
на: есть медузы в форме тарелочки и в ви-
де шлема, попадаются полушаровидные и
конические; у одних медуз многочислен-
ные тонкие щупальца сидят в несколько
рядов по краю зонтика, у других щупалец
мало, они имеют вид толстых бичей и
прикрепляются к спинной стороне зон-
тика. Здесь есть представители как гид-
роидных (преимущественно из отряда
трахилид), так и сцифоидных (относящих-
ся котряду корономедуз). На тоновой
таблице 14 изображены некоторые наибо-
лее характерные глубоководные медузы.
У трахилидных медуз кроссота (Crosso-
ta) и пантахогон (Pantachogon) на краю
зонтика много тонких длинных щупалец.
Зонтик этих медуз тонкостенный, но му-
скулистый. Они плавают короткими быст-
рыми толчками. Все остальные глубоко-
водные медузы, изображенные на этой же
таблице, плавают очень медленно. Их зон-
тик имеет толстую, хрящевидную мезог-
лею, которая затрудняет пульсирующие
движения, столь характерные для других
медуз. Маленькая глубоководная гидроме-
дуза меатор (Meator) вовсе утратила ти-
пичную медузоидную форму. Она имеет
вид прозрачного шарика с темной сердце-
виной.
Эти медузы живут на глубине от 1 до
6 км, во мраке и холоде. Здесь совершен-
но нет растений, и потому все глубоковод-
ные обитатели либо ведут хищнический
образ жизни, либо им приходится доволь-
ствоваться мертвыми, часто даже разла-
гающимися организмами, которые опу-
скаются на дно из верхних, богатых
жизнью слоев воды. На глубинах океана
всегда ощущается острый недостаток в
пище, и потому все обитатели абиссали
постоянно заняты ее поисками. Ограничен-
ные пищевые возможности— одна из глав-
нейших причин бедности глубоководного
населения. Если подсчитать биомассу
планктонных организмов океана на раз-
личных его глубинах, то окажется, что
99,9 % приходится на глубину менее 1000 ж,
а вся толща воды с глубины 1000 м до
дна содержит лишь 0,1% биомассы планк-
тона. Совершенно очевидно, что глубоко-
водные животные должны иметь какие-то
специальные приспособления, помогаю-
щие им отыскивать пищу. Чем лучше эти
приспособления обеспечивают питание, тем
быстрее размножается глубоководный
хищник. Поимка большеротой абиссаль-
ной рыбы или полупрозрачного глубоко-
водного головоногого моллюска — это
важное событие для исследователей. Мно-
гие виды глубоководных планктонных чер-
вей немертин известны всего лишь по
1—2 экземплярам, а абиссальные медузы
попадаются почти в каждой пробе, подня-
той с глубины. Что же позволило им так
размножиться, занять одно из первых по
численности мест среди глубоководных
обитателей? На первый взгляд это мало-
понятно, особенно если учесть медлитель-
ность их движений и общую примитив-
ность организации. Глубоководные ме-
дузы не преследуют свою добычу, а под-
манивают ее. Питаются они главным об-
286
разом рачками, но при случае поедают
любых других абиссальных животных,
привлекая их к себе ярким светом. Все
глубоководные медузы имеют краснова-
тую или коричневатую окраску. Иногда
красящее вещество расположено только
вдоль радиальных каналов, иногда оно
покрывает всю нижнюю сторону зонтика.
Наличие красно-коричневого пигмента
связано со способностью излучать свет.
В этот же цвет окрашены и многие другие
светящиеся морские организмы или те
части их тела, которые излучают свет.
Находящееся здесь жироподобное веще-
ство люциферин под воздействием
фермента люциферазы медленно
окисляется, излучая яркий свет. Подобно
тому как на свет фонаря слетаются ноч-
ные бабочки, на свет медуз собираются
рачки, а вслед за ними и другие глубоко-
водные животные, питающиеся рачками.
Все они вскоре оказываются в непосред-
ственной близости от длинных подвижных
щупалец и становятся добычей светящей-
ся медузы. Таким образом глубоковод-
ные медузы собирают улов из большого
пространства окружающей их воды.
Следует отметить очень высокий коэф-
фициент полезного действия, достигаемый
ОТРЯД ГОРГОНОВЫЕ КОРАЛЛЫ
(GORGONARIA)
Горгонарий, или роговые кораллы, внеш-
не очень похожи на колонии гидроидных
полипов. Их скелет состоит из образую-
щихся в толще мезоглеального слоя из-
вестковых спикул в виде покрытых бу-
горками палочек и чешуек.
Но для горгонарий более характерен
плотный осевой скелет, про-
низывающий ствол и ветви колоний. В его
в реакции окисления люциферина — он
равен 50%. При любых других реакциях,
дающих свет, на его долю приходятся
лишь десятые части процента, тогда как
основное количество энергии выделяется
в виде тепла.
Способностью светиться обладают так-
же некоторые медузы, живущие у поверх-
ности моря, среди них маленькая гидро-
медуза раткеа (Rathkea), довольно круп-
ная (до 10—15 см в диаметре зонтика)
экворея (Aequorea) и сцифоидная медуза
пелагия ночесветка (Pelagia noctiluca). Ча-
сто эти медузы появляются в очень боль-
ших количествах, и тогда ночью волны
кажутся пламенеющими, а на лопастях
весел возникают огненные шары — так
ярко светятся прилипшие к ним медузы.
Из донных кишечнополостных светятся
некоторые гидроиды и многие морские
перья. Однако свечение этих животных,
по-видимому, не связано с их питанием,
так как они вспыхивают ярким светом
только при механическом раздражении.
Возможно, способность неожиданно из-
лучать сильный свет является защитной
реакцией и служит для отпугивания жи-
вотных, случайно наткнувшихся в темно-
те на морское перо.
образовании в простейшем случае участ-
вуют те же известковые спи-
кул ы, соединенные между собой проме-
жуточным веществом. У большинства же
горгонарий осевой скелет слагается из
концентрических роговых пластинок, об-
разующихся в мезоглеальном слое. Эти
роговые пластинки обычно пропитаны из-
вестью. У многих горгонарий спикулы, из
которых состоит скелет отдельных поли-
пов, скапливаются вокруг ротового диска,
образуя венчик торчащих наружу защит-
ных игл и чешуек.
КЛАСС КОРАЛЛОВЫЕ ПОЛИПЫ (ANTHOZOA)
ПОДКЛАСС ВОСЬМИЛУЧЕВЫЕ КОРАЛЛЫ (OCTOCORALL1A)
287
Колонии горгонарий прочно прикрепля-
ются к твердому субстрату при помощи
округлого расширения основания. Лишь
немногие виды способны развиваться на
мягких песчаных или илистых грунтах.
Тогда в базальной части колонии обра-
зуются корневые выросты. На мелководье,
основном месте обитания горгонарий, они
особенно многочисленны и разнообразны
лишь там, где находят необходимый для
своего укрепления скалистый грунт. На
соседних участках, где дно песчаное или
илистое, можно встретить единичные, к
тому же обычно чахлые и недоразвитые
колонии горгонарий. Однако у глубоко-
водных представителей корневидные вы-
росты основания колоний — более обыч-
ное образование. У некоторых глубоко-
водных видов, приспособившихся к обита-
нию на илистых грунтах, нижняя часть
ствола колонии разрастается в ширину и
принимает характер лопатовидной пла-
стинки, свободно втыкающейся в грунт.
Горгонарий распространены всесветно
от полярных районов до экватора. Но
особенно многочисленны и разнообразны
они в водах тропической зоны. К обита-
нию в холодных водах полярных районов
приспособились очень немногие виды.
Так, в водах Арктической области встре-
чено только 5 видов горгонарий, а в Ан-
Таблица 13. Парусники (Velella) в Тихом океане:
1 — парусник с сидящим на нем крабиком
планес (Planes) и пожирающими мягкие
ткани парусника морской уточкой (Le-
pas fasciculatus) и моллюском глауку-
сом (Glaucus);
2— парусник с сидящим на его нижней
стороне моллюском янтиной (Janthina);
3 — парусник с икрой летучей рыбы;
4 — скелет парусника с молодью морской
уточки;
5 — плывущая янтина, прикрепившаяся к
собственному поплавку;
6 — крабик планес на поплавке, брошен-
ном янтиной;
7 — клоп-водомерка (Halobates).
тарктике их не более 30. Общее же число
видов роговых кораллов достигает 1200.
На мелководье обитает около 70% всех
известных видов горгонарий. Только 16%
их опускается на глубину более 1000 м.
В абиссаль Мирового океана проникло
всего лишь 11 видов горгонарий. Подав-
ляющее большинство роговых кораллов
может жить только в воде с нормальной
морской соленостью, и лишь несколько
видов способно переносить незначитель-
ное опреснение. Поэтому в таких морях,
как Черное или Балтийское, горгонарий
не встречаются.
Способ питанияи состав пищи
горгонарий остаются до сих пор неизвест-
ными. В гастральных полостях почти ни-
когда не удается найти остатки прогло-
ченной пищи. Возможно, что пищей гор-
гонариям служат мельчайшие частицы
органического вещества животного про-
исхождения, взвешенные в морской воде.
В таком случае остатки мелких рако-
образных, которые бывали изредка обна-
ружены в гастральных полостях, очевид-
но, явились случайным приловом для
полипов-фильтраторов.
Колонии роговых кораллов отличают-
ся удивительным разнообразием. Среди
них имеются простые неветвящиеся и
сложные ветвящиеся колонии. Последние
могут быть древовидными, перистыми и
вееровидными. Пожалуй, наиболее краси-
вы именно вееровидные колонии горгона-
рий. Таковы обитающие в теплых водах
различные виды Gorgonia или Rhipidi-
gorgia. Для них характерно ветвление
только в одной плоскости. Многочислен-
ные веточки спаиваются между собой, и
вся колония приобретает вид перфориро-
ванной округлой пластинки. Эти изящные
горгонарии к тому же чрезвычайно ярко
окрашены в желтый, красный и даже фио-
летовый цвет. Веера Rhipidigorgia дости-
гают иногда весьма больших размеров.
Известны колонии, достигающие двух
метров в высоту и полутора в ширину.
Вместе с другими столь же красивы-
ми и ярко окрашенными горгонариями
они образуют целые подводные леса, как
бы окрашенные в осенние краски — жел-
тые, оранжевые, красные и коричневые.
Не менее изящны и перистые колонии.
В наших дальневосточных морях на глу-
бине от 100 до 500 м часто встречается
Plumarella longispina, образующая коло-
нии до полуметра высотой. От основного
ствола ее отходят длинные боковые ветви,
от которых в свою очередь отходят боко-
вые короткие веточки. Вся колония вет-
вится в одной плоскости. Цвет колонии
нежно-розовый, иногда с желтым оттен-
ком.
В европейских водах очень обычна гор-
гонария Eunicella verrucosa, обитающая
в Средиземном море и у берегов Испа-
нии, Португалии и Франции на глубине
до 5 м. Ветвление ее не столь правиль-
но, но тем не менее все ветви этой гор-
гонарии также лежат в одной плоскости.
Кусты Eunicella, достигающей нескольких
десятков сантиметров, обычно окрашены
в красный цвет, но встречаются желтые и
коричневые колонии. Eunicella — излюб-
ленный объект для наблюдений в аква-
риумных условиях. Благодаря особой жиз-
ненной энергии этого вида колонии ее уда-
лось содержать в проточной, насыщенной
кислородом воде в течение нескольких меся-
цев. В отличие от мелководных альциона-
рий, в жизнедеятельности которых наблю-
дается правильный суточный ритм, полипы
Eunicella распускаются и сокращаются
независимо друг от друга. Поэтому на
живой колонии часть полипов находится
почти все время в активном состоянии.
При слабом механическом воздействии
на какой-нибудь один полип только он и
отвечает на это воздействие медленным
сокращением. Лишь сильное раздражение
может привести к сокращению и соседних
полипов.
Колониям, описанным выше, свойствен
осевой скелет, состоящий из концентри-
Рис. 174. Колония горгонарии лептогоргия
(Leptogorgia).
^19 жизнь животных, т. i
289
ческих роговых пластинок, равномерно
пропитанных известью. Однако у неко-
торых роговых кораллов осевой скелет со-
стоит из чередующихся чисто роговых и
сильно пропитанных известью участков.
Такой скелет, в частности, образуется у
горгонарии Isidella elongata, или белого
коралла, весьма обычного в Средиземном
море и в Бискайском заливе на глубине
220—1000 м. Колония Isidella состоит из
длинного, до 1 м, тонкого ствола, от ко-
торого отходят вверх столь же длинные
и тонкие ветви. В целом изиделла имеет
вид изящного и высокого деревца. По-
липы расположены на ветвях и стволе в
два ряда. Они крупные и не способны втя-
гиваться внутрь мягких тканей колонии.
Коралл чисто белого цвета, и только на
концах ветвей, где мягкие ткани почти
прозрачны, сквозь них просвечивают тем-
ные, роговые участки осевого скелета.
Полипы окрашены в белый цвет, но сквозь
их стенки просвечивает ярко-красная глот-
ка. Вокруг ротового отверстия имеется
желто-красное кольцо.
Как у многих глубоководных видов, у
Isidella elongata базальная пластинка не-
правильной формы и часто образует кор-
невидные выросты, облегчающие укреп-
ление коралла на мягких илистых и пес-
чаных грунтах.
Среди горгонарии существуют и виды,
у которых роговое вещество не образует-
ся совсем. У таких кораллов осевой ске-
лет слагается лишь из крепко спаянных
друг с другом известковых спикул. Такой
скелет присущ кораллам из р о д a Coral-
lium. К этому роду, в частности, принад-
лежит так называемый красный, или бла-
городный, коралл — Corallium rubrum (см.
цветную таблицу 11), хорошо известный
по различным украшениям, которые из
него изготовляют. Благодаря концентра-
ции окиси железа известковые спикулы
благородного коралла окрашены в различ-
ные оттенки красного цвета — от почти
белого до почти черного. Красный цвет
спикул придает окраску не только осе-
вому стержню колонии, но и покрываю-
щим его тканям. На красном фоне ветвей
колонии красиво выделяются чисто белые
полипы. Это аутозоиды, или кор-
мящие полипы. Кроме них, как
и у некоторых других альционарий и
горгонарии, имеются и сифонозоиды. Си-
фонозоиды обеспечивают циркуляцию во-
ды в теле колонии. Любопытно отметить,
что у других горгонарии, имеющих си-
фонозоиды, половые клетки образуются
именно в них, как, например, у Anthomas-
tus. Что касается С. rubrum, то половые
клетки образуются у него только в ауто-
зоидах. Колонии благородного коралла
обычно раздельнополы, но бывают и гер-
мафродитные.
С. rubrum встречается в Средиземном
море и в Атлантическом океане у берегов
Канарских островов, на глубине от 30 до
500 м. Однако чаще всего колонии благо-
родного коралла развиваются на глубине
50—200 м. Приспособление для добычи
кораллов состоит из двух сбитых крест-
накрест балок, к которым привязывают-
ся куски крупноячеистых сетей. При
протягивании такой снасти по дну ко-
раллы запутываются ветвями в ячее сетей
и отламываются от субстрата.
С добытых кораллов счищают мягкие
ткани и верхний слой осевого скелета, а
его наиболее плотную сердцевину исполь-
зуют для изготовления различных укра-
шений.
Красивые поделки изготовляют не толь-
ко из благородного коралла. На прибреж-
ном мелководье в Индийском океане и в
Красном море прибыльным делом считают
промысел черного коралла Euplexaura
antipathes, из коричневато-черных осевых
стержней которого вытачивают четки, мунд-
штуки, различные украшения и амулеты.
ОТРЯД МЯГКИЕ КОРАЛЛЫ
(ALCYONARIA)
Слово «коралл» в нашем представлении
всегда связано с чем-то твердым, хрупким.
Таковы в действительности многие корал-
лы, но имеются и мягкие кораллы —
альционарий,
К альционариям относятся наибЪлее
просто организованные колониальные
коралловые полипы, в колониях которых
никогда не образуется центральный, или
осевой, скелет. Многочисленные склериты
290
или беспорядочно разбросаны в толще ме-
зоглеального слоя, или образуют корко-
вый слой непосредственно под эктодер-
мальным слоем клеток. В более редких
случаях склериты, сливаясь между собой,
образуют вокруг полипов изящные трубоч-
ки. Дольчатые, стелющиеся, грибовидные
и шаровидные колонии альционарий проч-
но прикрепляются к субстрату мясистым
основанием. Но некоторые альционарий
способны укрепляться и на мягких или-
стых грунтах. Различно устроенные вы-
росты основания служат тогда колониям
хорошими якорями.
Альционарий — наиболее многочислен-
ный отряд восьмилучевых кораллов. Они
широко распространены в морях и океа-
нах, встречаясь от полярных районов до
экватора и от литорали до больших глу-
бин. Однако в Мировом океане альцио-
нарий распределены весьма неравномер-
но. Более 80% всех известных в настоящее
время видов обитает на мелководье тро-
пических морей Малайского архипелага
и вдоль северных берегов Австралии.
В холодные воды полярных районов север-
ного и южного полушарий проникает всего
несколько видов альционарий. Не более 2%
видов встречается на глубине свыше 1000 м,
а в абиссаль Мирового океана (на глуби-
ну более 3 тыс. м) проникло всего лишь
шесть видов альционарий.
В отряде Alcyonaria различают до
40 родов и 1200 видов мягких кораллов,
неравномерно распределяющихся между
тремя подотрядами — Stolonifera,
Telestacea и Alcyonacea.
Наименее многочисленны первые два
подотряда, объединяющие самых прими-
тивных альционарий. Их стелющиеся ко-
лонии состоят или из мембрановидной
пластинки, покрывающей субстрат, и от-
ходящих от нее отдельных полипов (Clavu-
laria, Anthelia), или из корневидных
нитей, от которых поднимаются вверх
прямые, обычно неветвящиеся стволы с
несколькими боковыми полипами (Teles-
to). Интересно отметить, что многие виды
именно этих наиболее примитивных и
архаичных альционарий проникли в боль-
шие глубины океана и обитают в нижней
батиали B—3 тыс. м) и абиссали (свыше
3 тыс. м). Из шести видов альционарий,
встречающихся на глубине более 3 тыс. му
пять видов относятся к подотрядам Sto-
lonifera и Telestacea. Это свидетельствует
в пользу высказываемого в настоящее
время предположения, что большие глу-
бины океана населены фауной, носящей
черты примитивности и большой древно-
сти.
Колонии Alcyonacea, более высокоор-
ганизованных альцпонарий, чрезвычайно
разнообразны в отношении своей формы,
однако всем им присуще значительное
развитие, мезоглеального слоя,
а также трубчатая форма вытянутых га-
стральных полостей полипов. У Alcyoni-
um digitatum, например, гастральные по-
лости первичных полипов тянутся от по-
верхности колонии до ее основания.
И только более молодые вторичные полипы
обладают более короткими трубками, кон-
цы которых связаны поперечными цено-
саркальными каналами с длинными труб-
ками гастральных полостей первичных по-
липов.
Дольчатые колонии A. digitatum отда-
ленно напоминают кисть человеческой ру-
ки (см. цветную таблицу 9), и не только
формой, но и бледной желтовато-оранже-
вой или желтовато-коричневой окраской.
Не удивительно поэтому, что жители по-
бережий различных европейских стран
независимо друг от друга назвали эту
альционарию «рукой» или «пальцами мерт-
веца». Даже первое научное ее название —
Manus marina — обозначает в переводе
«морская рука». A. digitatum — наиболее
известный вид альционарий в европейских
водах. Колонии ее встречаются в Среди-
земном море, вдоль берегов Испании,
Португалии и Франции, в Английском
канале, в Северном и Норвежском морях.
Изредка они попадаются в проливах Ска-
геррак и Каттегат. Но опресненных вод
Балтийского моря эти альционарий уже
не переносят. A. digitatum — преимуще-
ственно мелководный вид, обычно населяю-
щий мелководные банки, где местные ры-
баки ведут траловый промысел рыбы. Ко-
лонии альциониума поэтому оказываются
самым обычным компонентом тралового
прилова. Однако A. digitatum нередко
проникает в нижнюю сублитораль и даже
в батиаль. В Бискайском заливе, напри-
19*
291
мер, колонии A. digitatum были обнару-
жены на глубине 718 м.
Колонии A. digitatum в состоянии покоя
малозаметны — они сливаются с краска-
ми подводного ландшафта. Но через неко-
торое время невзрачные комья сжавшего-
ся коралла начинают как бы набухать
и расправляться, а короткие и толстые
«пальцы» колонии вытягиваются. При этом
над поверхностью расправившейся коло-
нии поднимаются и распускаются очень
красивые «цветы» — полипы, достигающие
3 мм в длину. Их нежные, почти прозрач-
ные стенки окрашены в легкий желтый
цвет; перистые щупальца белые. Но через
определенное время все полипы охваты-
ваются волной сокращения. Первыми на-
чинают сжиматься щупальца. Они уко-
рачиваются и сильно утолщаются, и тог-
да полип при взгляде сверху напоминает
восьмиконечную звезду. Затем сжимается
и весь полип. Под конец он полностью
погружается в тело колонии, оставляя на
ее поверхности маленькое кратерообраз-
ное углубление. Через некоторое время
распустившаяся красивая колония вновь
приобретает вид бесформенного комка.
Набухание и сокращение колоний про-
исходит регулярно два раза в день — ут-
ром и после полудня. Такой ежедневный
ритм, видимо, связан с ежедневными при-
ливо-отливными колебаниями воды, а не
со сменой дня и ночи, как у некоторых
других мелководных кишечнополостных.
В условиях аквариума правильный ритм
набухания и сокращения колоний быстро
теряется. При искусственном обновлении
воды один раз в сутки набухание и рас-
правление полипов происходит также один
раз. Колонии альционарий нуждаются в
притоке свежей воды, насыщенной кисло-
родом, необходимым для дыхания. В ак-
тивном состоянии через колонию непре-
рывно прогоняется вода, снабжающая ко-
лонию кислородом. Одновременно поли-
пы находятся в напряженном ожидании
пищи. Экспериментально показано, чт.о в
воде с недостатком кислорода колонии
остаются в расправленном состоянии це-
лые сутки.
Правильный суточный режим набухания
и сжатия наблюдается и у другого среди-
земноморского вида—A. palmatum. В ста-
дии сокращения колония этой альцио-
нарий напоминает своими очертаниями
молодой кактус опунцию, но она окрашена
не в зеленый, а в мясо-красный или ко-
ричневый цвет. Распустившаяся колония
A. palmatum очень красива. Она стано-
вится древовидной, стенки ее почти про-
зрачны. Меняется и окраска — основание
сохраняет красный цвет, но ветви ее
становятся нежно-розовыми или желтыми.
Над поверхностью ветвей возвышаются
белые изящные полипы.
Питание Alcyonium сравнительно хоро-
шо изучено. В активном состоянии рас-
пустившиеся полипы находятся в ожидании
пищи. Их жертвой становятся проплываю-
щие мимо мелкие ракообразные и их ли-
чинки — науплиусы. Стрекательные нити
парализуют, а вытянутые тонкие щупаль-
ца крепко схватывают добычу. Затем щу-
пальца несколько сокращаются, подтяги-
ваясь к ротовому диску. Концы их изги-
баются и проталкивают схваченную пищу
в ротовое отверстие. Через глотку пище-
вой комок продвигается уже благодаря
волнообразным перистальтическим сокра-
щениям ее стенок. Комок, попавший в
гастральную полость, обволакивается ме-
зентериальными нитями, расщепляется на
части и наконец переваривается. Любопыт-
но, что переваривание пищи осуществляет-
ся только брюшными и боковыми гаст-
ральными перегородками. Две спинные
перегородки никогда не участвуют в про-
цессе питания.
Красивые, чаще всего древовидные ко-
лонии образуют альционарий из семей-
ства Nephthidae. Это семейство наибо-
лее богато видами. Они довольно широко
распространены в морях и океанах, оби-
тая преимущественно на мелководье тро-
пического пояса. В наших северных и
дальневосточных морях, а также в север-
ных частях Атлантического и Тихого океа-
нов встречается три вида нефтид из р о д а
Eunephthia, с трудом отличающиеся друг
от друга. Колонии их, обычно ярко окра-
шенные в желтый, красный или корич-
невый цвет, состоят из массивного основа-
ния, от которого поднимается вверх тол-
стый неправильно сидящий ствол. На ко-
ротких ветвях колонии группами распо-
лагаются многочисленные, очень густо си-
292
дящие полипы. В состоянии
покоя они целиком втягива-
ются внутрь ветвей, в ак-
тивном состоянии они рас-
правляются и становятся
почти прозрачными.
Эти альционарии обычно
поселяются на выступах скал
или на камнях, образуя
иногда густые заросли. В
подходящих гидрологических
условиях Eunephthia охотно
живут на песке и даже на
илистых грунтах, но тогда
форма основания колонии
претерпевает значительные
изменения, уже описанные
раньше.
Чрезвычайно своеобразным
строением обладают колонии
альционарии из рода Anthomastus.
Некоторые виды антомастуса образуют
обратнокеглевидные колонии, но чаще
всего они удивительно напоминают гриб.
Колония тогда состоит из морщинистой
высокой ножки и округлой или плоской
шляпки. Ножка не имеет полипов, они
образуются только на верхней поверхно-
сти шляпки. Таким необычным способом
и решается задача отделения полипов от
грунта (рис. 175).
Исследуя строение колоний Anthomas-
tus, мы впервые сталкиваемся с явлением
диморфизма полипов у восьмилучевых
кораллов. Различаются крупные кормя-
щие полипы — аутозоиды и мелкие
недоразвитые полипы — сифонозои-
д ы. Аутозоиды, количество которых, как
правило, не превышает 15—30, обладают
обычным для Octocorallia строением, и их
ротовой диск окружен венчиком из восьми
длинных перистых щупалец. Размеры ау-
тозоидов у разных видов очень изменчивы.
У вида A. grandiflorus, встречающегося
в северной части Атлантического океана,
длина распустившегося кормящего поли-
па достигает 36 мм, более мелкими разме-
рами — до 15 мм в длину — обладают
полипы A. rilovi, обитающего у Куриль-
ских островов в Тихом океане.
Аутозоиды покрывают поверхность
шляпки неравномерно и располагаются
преимущественно на ее краю.
Рис. 175. Грибовидная коло-
ния мягких коралловых поли
нов антомастус (Anthomastus)
Оставшееся пространство
занимают сифонозоиды, имею-
щие вид небольших бугор-
ков. Мелкие сифонозоиды,
диаметр которых не более
1 мм, сидят очень густо,
число их превышает сот-
ни. Эти полипы лишены щу-
палец и приспособлены к со-
зданию тока воды внутри
колоний. Любопытно, что у
Anthomastus сифонозоиды
несут еще одну функцию —
на их радиальных перего-
родках образуются половые
клетки. Напомним, что обыч-
но они развиваются в ауто-
зоидах. Сифонозоиды погру-
жены в тело колонии и не
способны выпячиваться.
Ярко окрашенные в красный цвет с раз-
личными оттенками колонии Anthomastus
встречаются во многих морях и океанах
земного шара и обитают как на мелково-
дье, так и на сравнительно большой глу-
бине.
Однако это довольно редкие животные
и попадаются они не часто.
Склериты, образующие мезоглеальный
скелет альционарии, как уже отмечалось
выше, способны сливаться между собой,
образуя вокруг отдельных полипов изящ-
ные трубочки. Таким своеобразным ске-
летом обладают кораллы-органчики из
семейства Tubiporidae. На песчаных пля-
жах атоллов и коралловых островов мож-
но найти выброшенные прибоем куски
кораллов, окрашенных в яркий малино-
вый цвет. Такие куски, лишенные мягких
тканей, состоят из многочисленных тру-
бочек, поднимающихся от общего основа-
ния и связанных на равном расстоянии
друг от друга несколькими поперечными
пластинками. Эти пластинки получаются
также путем слияния спикул в переклади-
нах ценосарка. Внутри пластинок имеется
сеть боковых каналов, соединяющих га-
стральные полости отдельных полипов.
От поперечных пластинок отходят трубоч-
ки вторичных полипов. Длинные трубоч-
ки первичных полипов не совсем парал-
лельны друг другу и слегка расходятся
в верхней части колонии. Однако про-
293
межутки между трубочками здесь не уве-
личиваются благодаря тому, что от каждой
очередной поперечной пластинки подни-
маются все новые и новые трубочки. В це-
лом структура колонии напоминает орган,
что и отражено в названии этого коралла
(см. цветную таблицу 9).
Органчики довольно широко распрост-
ранены на рифах тропической зоны. Со-
временных видов Tubipora немного, но в
прошедшие геологические периоды это
была обычная группа кораллов. Вместе
с мадрепоровыми шестилучевыми корал-
лами Tubiporidae относятся к кораллам-
рифообразователям. Во время прилива
полипы органчика высовываются из
окружающих их трубочек и расправляют
свои щупальца. Тогда колония окраши-
вается в зеленый цвет — в цвет самих
полипов. При опасности полипы быстро
втягиваются внутрь трубочек и коралл
вновь приобретает яркую малиновую
окраску.
Так же как и у многих других кишечно-
полостных, у Alcyonaria возникают оп-
ределенные приспособления к существо-
ванию в различных условиях. У форм,
обитающих на мелководье, в полосе при-
боя, скелет, как правило, бывает очень
слабо развит. Гибкость колонии спасает
ее от ударов набегающих волн. Но иногда
скелет альционарий, обитающих в при-
бойной зоне, бывает и весьма массивным,
что также предохраняет колонию от
волн.
Размеры колоний очень изменчивы. Фор-
мы, обитающие на мелководье, образуют
более крупные колонии, чем те, которые
развиваются на больших глубинах. Пред-
ставители одного и того же вида, живущие
в холодных и теплых водах, заметно от-
личаются друг от друга. У первых наблю-
дается увеличение размеров полипов, хотя
число их оказывается меньшим, чем у
вторых.
В условиях повышенной загрязненно-
сти количество полипов колонии резко
возрастает.
Практическое значение альционарий
ничтожно. Высушенные и перемолотые
колонии Alcyonium, добытые во время
тралового промысла, используют в каче-
стве удобрений.
ОТРЯД МОРСКИЕ ПЕРЬЯ
(PENNATULARIA)
Из всех кишечнополостных самые фан-
тастические по форме колонии образуют
морские перья, относящиеся к восьмилу-
чевым кораллам (рис. 176 и табл. 9).
В отличие от других колониальных ки-
шечнополостных морские перья никогда
не прикрепляются к какому-нибудь суб-
страту. Их нижний конец, имеющий обыч-
но мускулистое расширение, свободно вты-
кается в мягкий грунт, выстилающий дно.
Хотя морские перья ведут неподвижный
образ жизни, они могут в случае необхо-
димости передвигаться по грунту и вновь
укрепляться на новом месте.
Морские перья образуют неразветвлен-
ные, часто довольно крупные и ярко ок-
рашенные колонии. Они состоят из мя-
систого ствола, представляющего собой
разросшееся и сильно видоизмененное те-
ло первичного полипа. Остальные полипы
вырастают на стенках ствола или на его
выростах. Для всех морских перьев ха-
рактерен диморфизм вторичных полипов.
Имеются крупные кормящие полипы —
аутозоиды — и мелкие недоразвитые поли-
пы — сифонозоиды. Каждая колония мор-
ского пера делится на два отдела:
верхний, или полипоидный, и
нижний — стебель. На полипоидной
части колонии образуются аутозоиды и си-
фонозоиды. Стебель свободен от полипов,
хотя у некоторых видов и здесь развивают-
ся мелкие сифонозоиды.
Для морских перьев характерна высо-
коорганизованная система кана-
лов, связывающая различные отделы ко-
лонии. Она состоит из центральной и пе-
риферической систем. Центральная систе-
ма представлена четырьмя основными про-
дольными каналами, пронизывающими всю
колонию. Два из них являются продол-
жением радиальных карманов гастральной
полости первичного полипа. Два других
канала — вторичные образования. Пери-
ферическая система состоит из сложной
густой сети каналов, связывающих между
собой гастральные полости отдельных по-
липов. Каналы периферической системы
открываются в основные продольные ка-
налы колонии.
294
Скелет морских перьев слагается
из многочисленных известковых спикул в
виде палочек и игл, обычно имеющих
три продольных ребрышка. Кроме спикул,
образующих, как правило, корковый слой
тела колонии, имеется и осевой скелет,
образующийся, так же как и у роговых
кораллов, из концентрических роговых
пластинок, пропитанных известью.
Морским перьям свойственна хорошо
развитая мышечная система.
В ней различают мощную кольцевую му-
скулатуру, отражающую продольные ка-
налы, слой продольных мышечных тяжей
и слой косых мускулов, прикрепляющих-
ся одним концом к стержню осевого ске-
лета. Косые мускулы обеспечивают про-
извольное изгибание колонии.
Нервная система морских
перьев достигает высокого для кишечно-
полостных совершенства. Основная черта
ее прогресса заключается в образовании
общеколониальной нервной системы, свя-
зывающей между собой все отделы коло-
нии. Однако специальные органы чувств
отсутствуют и у морских перьев.
Питание пеннатулярий изучено
еще недостаточно. В гастральных полостях
аутозоидов Sclerobelemnon были обнару-
жены мелкие многощетинковые черви из
семейства Syllidae. Имеются ука-
зания на то, что пищей для Protoptilum
служат крылоногие моллюски, а для Vir-
gularia — личинки различных двуствор-
чатых моллюсков. Возможно, что морские
перья питаются и мелкими взвешенными
частицами органического вещества живот-
ного.
Морские перья раздельнополы.
Как правило, женские колонии оказывают-
ся гораздо более многочисленными, чем
мужские. Половые клетки образуются,
в отличие от Alcyonaria и Gorgonaria,
обладающих диморфизмом полипов, ис-
Рис. 176. Морские перья:
1 — веретиллум (Veretillum); 2 — виргулярия (Virgularia); 3 — умбеллула
4 — кофобелемнон (Kophobelemnon).
(Umbellula);
ключительно в аутозоидах. Сифонозоиды
морских перьев обеспечивают лишь цир-
куляцию воды и, следовательно, газооб-
мен колонии. В большинстве случаев
оплодотворение зрелых яиц пеннатулярии
происходит в наружной среде. Зрелые
яйца попадают сначала в гастраль-
ную полость, а затем антиперистальтиче-
скими сокращениями глотки выбрасывают-
ся в воду. Яйца у многих видов пеннату-
лярии выбрасываются ранним утром, не-
зависимо от температуры воды. Нерест
колоний происходит одновременно, при
этом мужские колонии выбрасывают поло-
вые продукты в таком количестве, что вода
вокруг них становится молочно-белой.
У северных видов пеннатулярии нерест мо-
жет продолжаться весь летний период.
Лишь немногие виды пеннатулярии мож-
но считать живородящими. У та-
ких видов оплодотворение происходит в
гастральной полости, а в воду выпу-
скаются личинки на стадии планулы.
Как правило, почкование приводит у
морских перьев, так же как и у других
колониальных полипов, лишь к росту
колоний, ибо новообразованные полипы
не теряют связи с материнским организ-
мом. Только у одного вида — Pennatula
phosphorea — известно размноже-
ние делением, при котором сразу
образуется новая колония. Происходит
это следующим образом. Верхняя часть
молодой колонии, на которой имеется
несколько полипов и куда не доходит осе-
вой стержень, начинает отшнуровываться
и в конце концов полностью отделяется
от материнской колонии. У отделившейся
части образуется вначале стебель, при
помощи которого новообразованная коло-
ния укрепляется в грунте, а затем и осе-
вой стержень.
Морские перья образуют сравнительно
небольшую группу кишечнополостных. Об-
щее количество их видов не достигает 300,
они распространены всесветно. Однако в
приантарктических водах, за исключени-
ем трех видов умбеллул, морские перья не
встречаются. Большинство видов обитает
в тропическом и умеренном поясах север-
ного полушария. В северных частях Ат-
лантического и Тихого океанов число
видов пеннатулярии резко снижается, а
в холодных водах полярного бассейна
встречается только три вида — Umbellu-
la encrinus, Virgularia cladiscus и V. gla-
cialis. На мелководье обитает 42% всех
известных видов пеннатулярии. Однако
распределение подавляющего большинства
мелководных видов, относящихся всего
лишь к нескольким родам, ограничено
теплыми водами тропической зоны. В ниж-
ней сублиторали и в батиали обитает
более половины всех видов пеннатулярии.
На глубину более 1000 м проникает 32
вида, а на глубину более 3000 м — 16,
или 7% всех известных пеннатулярии.
Некоторые глубоководные пеннатулярии
представляют собой весьма характерный
элемент абиссальной донной фауны. За-
воевать большие глубины позволили им
необычайная экологическая пластичность,
а также то, что морские перья приспособи-
лись к обитанию на мягких илистых грун-
тах, которые, естественно, и преобладают
на больших глубинах.
Несколько видов пеннатулярии спо-
собны переносить значительное опресне-
ние воды. Один из таких видов — Virgu-
laria mirabilis, широко распространенный
в Северной Атлантике и Средиземном мо-
ре, смог проникнуть поэтому даже в Чер-
ное море. Другой вид — Renilla patula —
был найден в устье реки Миссисипи.
Рассмотрим несколько наиболее харак-
терных представителей отряда. В Атлан-
тическом океане, вдоль берегов Европы и
Африки от Бискайского залива до Анго-
лы, а также в Средиземном море на глу-
бине 20—40 м часто встречается Veretil-
lum cynomurium. Этот вид принадлежит
к наиболее просто организованным мор-
ским перьям. Колония веретиллума, до-
стигающая 40 см в длину, имеет цилинд-
рическую форму. На поверхности поли-
поидной ее части, составляющей три чет-
верти общей длины колонии, беспорядоч-
но разбросаны десятки крупных, до 7,5 мм,
аутозоидов с длинными щупальцами. Сифо-
нозоиды еще более многочисленны, но они
расположены в продольных рядах. Осе-
вой скелет редуцирован до гладкой па-
лочки длиной в 10—15 мм, а у некоторых
экземпляров вообще отсутствует. Значи-
тельно редуцированы испикулы. Они очень
мелки и имеют овальную форму.
296
Веретиллум — излюбленный объект для
наблюдения в аквариуме. В состоянии
покоя колонии этого морского пера вы-
глядят довольно невзрачно: они имеют
вид коротких желто-коричневых палочек,
косо торчащих в грунте. Однако ночью
веретиллум преображается. Колония вса-
сывает воду, расправляется и превращает-
ся в почти прозрачный желто-оранжевый
высокий цилиндр, на стенках которого рас-
пускаются прозрачные, совершенно белые
полипы. Через некоторое время распу-
стившаяся колония начинает фосфорес-
цировать. При незначительном раздра-
жении колонии свечение усиливается,
и по всей колонии пробегают волны
света. К утру свечение прекращается,
полипы втягиваются, а сама колония
сжимается и приобретает свой обычный
вид сморщенной палочки.
Наличие общеколониальной нервной
системы обеспечивает быструю и согласо-
ванную реакцию всего организма на ме-
ханические или электрохимические раз-
дражения, приложенные к различным от-
делам колонии. Достаточно 20—30 се-
кунд после нанесения раздражения, чтобы
вся колония веретиллума оказалась в
сокращенном состоянии.
К числу примитивных морских перьев
относятся виды из семейства Kopho-
belemnonidae, положение аутозоидов ко-
торых не совсем упорядоченно. Однако их
колониям уже присуща двусторонняя сим-
метрия — признак некоторого усовершен-
ствования колоний. Все представители
семейства имеют уже хорошо развитый
осевой скелет, а известковые спикулы
многих видов приобретают обычную для
остальных пеннатулярий форму длинных
палочек с тремя продольными ребрышка-
ми. Многие виды рода Kophobelemnon
приспособились к обитанию на больших
глубинах океанов. По мере завоевания
глубин происходило и значительное видо-
изменение колоний кофобелемнон. Если
наиболее мелководные представители рода
обладают несколькими десятками мелких
полипов, рядами расположенных на по-
липоидной части колонии, то у наиболее
глубоководных видов — К. pauciflorum
и К. biflorum, обитающих в абиссали
Индийского и Тихого океанов, образуется
соответственно только три и два крупных
аутозоида (рис. 176). Уменьшение числа
аутозоидов при переходе к глубоководно-
му образу жизни — одно из проявлений
приспособительной олигомеризации, о ко-
торой говорилось выше, в разделе, касаю-
щемся общего строения колоний кишечно-
полостных.
Более высокой организации достигли
колонии умбеллул — чрезвычайно свое-
образных морских перьев. Колония ум-
беллулы состоит из необычайно длинного
и тонкого стебля, на вершине которого
помещается чашевидный рахис (так на-
зывается у морских перьев полипоидный
отдел колонии). Его верхняя поверхность
несет несколько очень крупных, не втя-
гивающихся аутозоидов с длинными щу-
пальцами. Многочисленные сифонозоиды
почти сплошь покрывают нижнюю по-
верхность рахиса и пространство между
аутозоидами. Любопытно, что они имеют-
ся на стволе колонии и даже на мясистом
его расширении, при помощи которого
колония укрепляется в грунте. В целом
колония умбеллулы напоминает цветок
амариллиса. Большинство видов умбел-
лул — обитатели холодноводных районов
и больших глубин океана. Только эти
морские перья населяют антарктические
воды южного полушария (Umbellula еп-
crinus, U. magniflora и U. thomsoni).
U. encrinus, кроме того, широко рас-
пространены в Северном Ледовитом океа-
не (рис. 180). Рекордная глубина обита-
ния восьмилучевых кораллов — 6120 м —
также принадлежит умбеллуле U. thom-
soni. Представители рода Umbellula, так
же как и Kophobelemnon, дают яркие
примеры приспособительной олигоме-
ризации, связанной с переходом
к глубоководному образу жизни. Так,
на рахисе нового вида U. monocepha-
lus, найденного недавно в Индийском оке-
ане на глубине свыше 3000 м, имеется
всего лишь один аутозоид необычайно
крупного размера.
Наиболее мелкие виды умбеллул дости-
гают 40—60 см, а более крупные, такие,
как U. encrinus или U. magniflora,— даже
200 см в длину. Большой длиной стебля
A79 см) обладает и U. monocephalus.
Размеры аутозоидов умбеллул, как пра-
297
вило, не превышают 2—3 см
в длину. Единственный тринад-
цатисантиметровый аутозоидИ.
monocephalus кажется исполи-
ном по сравнению с обычными
аутозоидами умбеллул.
Однако высшим типом орга-
низации обладают те морские
перья, у которых аутозоиды
отходят не непосредственно от
боковой поверхности главной
оси, а от особых ее выростов,
называемых листьями. Именно
такие колонии по внешнему
виду наиболее сравнимы с пе-
ром птицы. Это сходство и при-
вело к возникновению названия
рассматриваемой группы ки-
шечнополостных .
К морским перьям, аутозои-
ды которых сидят на боковых
листьях колонии, относится
очень изящный вид Scitaliopsis
djiboutiensis, обитающий в Индийском
океане у берегов Сомали. Наблюдения
над тем, как вытащенное из грунта мор-
ское перо вновь укрепляется в нем, про-
изводились именно над этим мелковод-
ным видом. Положенная на грунт, коло-
ния некоторое время оставалась совер-
шенно неподвижной. Затем нижний ко-
нец «стебля» начал медленно изгибаться
подобно геотропическому изгибу корня
растения. При этом по нему стала распро-
страняться волна мышечных сокращений.
То он вытягивался и становился очень
тонким, то расширялся и сокращался в
длину. При этом нижний конец «стебля»
колонпп постепенно начал вклиниваться
в грунт.
По мере того как стебель все глубже и
глубже проникал в грунт, колония все
больше и больше приподнималась над ним,
пока не приняла свое обычное вертикаль-
ное положение.
Чарлз Дарвин во время своего путеше-
ствия на корабле «Бигль» наблюдал пове-
дение морских перьев Stylatula darvinii,
образующих плотные поселения на мел-
ководье залива Байя Бланка. Он писал,
что там во время отлива можно видеть
сотни этих «зоофитов», неподвижно тор-
чащих над поверхностью грунта. Но до-
Рис. 177. Морское перо
пеннатула (Pennatula
phosphorea):
1 — мелководная форма;
2—глубоководная форма (по
Паксу и Юнгерсену).
статочно только прикоснуться
к такой имеющей несколько
дюймов в длину колонии, как
она с силой начинает вбура-
вливаться в ил и быстро в нем
исчезает.
Очень красивые колонии обра-
зует морское перо Pennatula
phosphorea. Этот вид чрезвычай-
но широко географически рас-
пространен. В Атлантическом
океане он встречается от Грен-
ландии до субантарктических
вод, в Тихом океане — от Бе-
рингова моря до берегов Япо-
нии. Колонии P. phosphorea бы-
ли найдены и в Индийском
океане. Это морское перо об-
ладает широким диапазоном
вертикального распределения,
обитая как на мелководье, так
и в абиссальных глубинах океа-
на. Мелководные экземпляры Р.
phosphorea достигают 30 см в длину и со-
стоят из мясистой главной оси, от которой
в полипоидной части отходят на близком
расстоянии друг от друга многочисленные
узкие треугольные «листья». По краям
последних сидят мелкие аутозоиды, коли-
чество которых определяется длиной ли-
ста. На самых длинных листьях насчи-
тывается до 25 полипов. Каждый аутозоид
защищен чашечкой из длинных палочко-
видных спикул, заканчивающейся не-
сколькими острыми зубцами. Глубоковод-
ные экземпляры не столь красивы. Они
гораздо мельче, листья их более редки.
На верхнем крае листа, скорее имеющего
вид тонкой веточки, располагается всего
3—4 аутозоида (рис. 177).
Колонии P. phosphorea обычно окраше-
ны в ярко-красный цвет, хотя встречают-
ся экземпляры с более светлой окраской—
оранжево-красные, желтые и почти белые.
Своей окраской колонии P. phosphorea
обязаны спикулам, ибо только они окра-
шены в красный или желтый цвет. Мяг-
кие же ткани совершенно бесцветны. Бе-
лые полипы распустившейся колонии кра-
сиво контрастируют с яркой окраской
листьев.
В пассивном состоянии пеннатула поч-
ти плашмя лежит на грунте, но в актив-
298
ном состоянии она начинает быстро впи-
тывать воду и принимает обычное для
морских перьев вертикальное положение.
Поникшие листья ее при этом расправля-
ются. Как и у других мелководных форм,
у P. phosphorea существует правильное
чередование в течение суток активного и
пассивного состояния. В активном состоя-
нии колонии пеннатулы находятся в ноч-
ное время.
Подобно веретиллумам и многим другим
пеннатуляриям, колонии пеннатулы спо-
собны к свечению. Свет излучается осо-
бой слизью, выделяемой при раздраже-
нии железистыми эктодермальными клет-
ками. У некоторых видов светятся все
отделы колонии, у других — только по-
липоидная часть, у третьих — только
полипы. У веретиллум, в частности,
способна излучать свет поверхность ко-
лонии, у пеннатулы фосфоресцируют толь-
ко аутозоиды и сифонозоиды. Если не-
которые виды излучают свет все время,
как, видимо, многие глубоководные мор-
ские перья, то другие светятся или только
в ночное время, или в условиях полной
темноты.
ОТРЯД АКТИНИИ, ИЛИ
МОРСКИЕ ЦВЕТЫ (ACTINIARIA)
Если колонии гидроидов и горгонарий
похожи на причудливые кусты и деревья,
то крупные коралловые полипы актинии
(Actiniaria) напоминают фантастические
цветы. На многих языках они и называют-
ся морскими анемонами (см. цветную таб-
лицу 9).
К отряду актиний относятся одиночные,
лишь изредка колониальные животные,
ведущие подвижный образ жизни. Толь-
ко немногие глубоководные виды непод-
вижно прикреплены к субстрату. Актинии
имеют цилиндрическую форму тела с
уплощенным верхним (ротовой диск)
и нижним концом (подошва). Но у
некоторых актиний, в основном у тех,
В дневное время такие животные начи-
нают светиться в ответ на какое-нибудь
раздражение — механическое, электриче-
ское или химическое. Слабое раздражение
вызывает вспышку света лишь в том месте,
к которому был приложен раздражитель.
Но если оно достаточно сильно, то по всей
колонии начинают пробегать концентриче-
ские «волны» световых потоков. Световая
«волна» может распространяться по коло-
нии со скоростью 5 см в секунду. Отсечен-
ные от колонии части тела также начинают
светиться в ответ на их раздражение.
Любопытно, что даже высушенная слизь
морских перьев не теряет способности к
люминесценции. Чтобы вызвать свечение,
достаточно слегка смочить высушенную
слизь водой. Экспериментально доказано,
что для вызывания и поддерживания со-
стояния люминесценции необходим постоян-
ный приток воды, насыщенной в достаточ-
ной степени кислородом. У большинства
видов пеннатулярий излучается синий или
фиолетовый поток света, но у некоторых
видов он может быть желтоватым и даже
зеленым.
которые ведут роющий образ жизни, по-
дошва может и не образовываться.
Число гастральных перегородок у боль-
шинства актиний по меньшей мере равно
шести парам или кратно шести. Образова-
ние новых пар перегородок происходит
почти всегда в промежуточных гастраль-
ных камерах. Однако от такого рас-
положения перегородок встречаются от-
клонения, при которых число перегородок
оказывается равным восьми или кратным
восьми или десяти. Обычно такие откло-
нения особенно характерны для наиболее
примитивных актиний. Известно, что в
процессе индивидуального развития все
актинии проходят стадию четырехлучевой
симметрии, что, возможно, указывает на
родство актиний с восьмилучевыми ко-
ралловыми полипами. Наибольшее сход-
ПОДКЛАСС ШЕСТИЛУЧЕВЫЕ КОРАЛЛЫ (HEXACORALLIA)
299
ство с современными восьмилучевыми ко-
раллами у актиний из р о д a Edwardsia.
Эти актинии ведут роющий образ жизни,
обитая в илистых песчаных грунтах при-
брежного мелководья. Их тело, на поверх-
ности которого различимы восемь про-
дольных валиков, имеет вытянутую чер-
веобразную форму. Углубления между
ними соответствуют восьми гастральным
перегородкам. Кроме восьми полных пе-
регородок, у старых экземпляров Edward-
sia в верхней части тела образуются еще
четыре, но уже неполные перегородки.
Валики продольных мышечных тяжей ле-
жат у этих актиний, так же как и у вось-
милучевых кораллов, на вентральных сто-
ронах перегородок. Восемь полных и
восемь неполных перегородок образуются
и у другой архаичной актинии Gonactinia.
Наиболее хорошо известный европейский
вид гонактинии G. prolifera имеет вид ма-
ленького, 2—3 мм в длину и 1—2 мм
в ширину, прозрачного столбика с нежно-
розовой или красной окраской. Ротовой
диск актинии окружен шестнадцатью неж-
ными щупальцами, расположенными в
два ряда. Глотка ее столь коротка, что
при открытом рте легко различимы в ее
гастральной полости восемь основных ра-
диальных перегородок. Гонактинии при-
крепляются своей подошвой к субстрату,
чаще всего к раковинам моллюсков, а ино-
гда даже к стволам гидроидных полипов.
Число перегородок, кратное десяти, на-
блюдается у представителей семей-
ства Myniadidae, весьма своеобразных
актиний, перешедших к свободноплаваю-
щему образу жизни. Их поддерживает в
воде особая воздушная камера, анало-
гичная пневматофору сифонофор, назы-
ваемая пневмоцистой. Она обра-
зуется в результате сильного впячивания
подошвы. Края подошвы при этом сбли-
жаются и смыкаются над центром углуб-
ления диска. Актиния поэтому плавает
у поверхности воды ртом вниз. Как мно-
гие другие плавающие кишечнополост-
ные, Myniadidae окрашены в голубой
цвет. У остальных актиний число пере-
городок, как уже говорилось, равно шести
парам или кратно шести.
Свободные края гастральных перегоро-
док имеют мезентериальные нити, богатые
железистыми и стрекательными клетками.
У некоторых актиний образуются еще и
особые нити — аконции, на которых
стрекательные капсулы особенно много-
численны. Для защиты от нападения эти
нити выбрасываются актиниями через рот
или через особые отверстия в стенках тела
или щупалец. Ротовой диск актиний окру-
жен щупальцами. В зависимости от чис-
ла щупалец они располагаются в один или
два и даже более концентрических рядов.
В каждом круге щупальца бывают одного
размера и формы, но щупальца, лежащие
в разных кругах, часто довольно сильно
отличаются друг от друга. Как правило,
щупальца соответствуют пространствам
между гастральными перегородками.Обыч-
но щупальца имеют простую коническую
форму, но иногда наблюдаются и значи-
тельные отклонения от нее. У некоторых
видов на концах щупалец образуются
вздутия благодаря тому, что там разви-
ваются многочисленные батареи стрека-
тельных капсул. У некоторых тропиче-
ских мелководных актиний образуются
ветвящиеся или перистые щупальца. На
их концах образуются одна или две пары,
служащие дополнительным средством для
быстрого опорожнения полости тела.
Ротовое отверстие высших актиний
имеет овальную или щелевидную форму.
Глотка сильно сжата с боков и имеет два
сифоноглифа. Только у описанных прими-
тивных видов имеется лишь один слабо-
развитый сифоноглиф или он вообще от-
сутствует. Биением ресничек сифоноглифа
создаются два тока воды: один — направ-
ленный внутрь гастральной полости и при-
носящий кислород (у некоторых актиний—
и частицы пищи), и другой — движущий-
ся в обратном направлении и выносящий
углекислый газ и продукты выделения.
Мышечная система актиний
достигает высокого для кишечнополостных
животных развития. Эктодермальная си-
стема складывается из продольных воло-
кон, лежащих в щупальцах, и радиальных
волокон вокруг ротового отверстия. Эн-
тодермальная система состоит из кольце-
вой мускулатуры щупалец, ротового ди-
ска, глотки, стенок тела и ножного диска.
На гастральных перегородках лежат про-
дольные мышечные валики.
300
Нервная система актиний
складывается из имеющихся во всех от-
делах тела эктодермальной сети нервных
клеток и слаборазвитой энтодермальной
сети, охватывающей только гастральные
перегородки. Особенно много нервных
клеток сконцентрировано у оснований щу-
палец и на ротовом диске. Однако это не
приводит к образованию околоротового
нервного кольца, так как нервные клетки
расположены здесь очень рыхло. Другое скоп-
ление нервных клеток имеется около по-
дошвы. Интересно отметить, что разные
отделы тела оказываются особенно чув-
ствительными к определенным раздраже-
ниям. Подошва, например, чувствитель-
на к механическим раздражениям и не
воспринимает химических. Ротовой диск,
наоборот, весьма чувствителен к химиче-
ским раздражениям и почти не реагирует
на механические. Пожалуй, лишь стенки
тела и щупалец реагируют на механиче-
ские, химические и электрические раздра-
жения, но щупальца оказываются гораздо
чувствительнее к ним, чем стенки тела.
Обычная реакция актинии на раздра-
жение заключается в сокращении тела.
Ротовой диск и щупальца при этом втяги-
ваются, и над ними смыкаются стенки
тела, сжатые особым мышечным кольцом.
Актинии, ведущие роющий образ жизни,
как описанная выше Edwardsia, при этом
быстро закапываются в грунт. При дли-
тельном воздействии раздражителя акти-
нии стремятся уползти как можно даль-
ше от него.
У актиний не образуется скелета, хотя
эктодерма некоторых видов выделяет хи-
тиноидную кутикулу, покрывающую бо-
ковую поверхность тела и подошву. По-
жалуй, только у глубоководных актиний
из семейства Galatheanthemidae, ве-
дущих неподвижный, прикрепленный об-
раз жизни, крепкий кутикулярный чехол,
в который заключено длинное червеобраз-
ное тело актинии, принимает характер
защитного скелета, аналогичного эктодер-
мальному скелету большинства гидроид-
ных полипов. Темно-коричневые защитные
чехлы галатеантемид поднимаются на
высоту от 2—3 до 150 мм. Над их устьем
диаметром около 1 см выступает верхняя
часть тела актинии с венчиком из много-
численных тонких щупалец. Галатеан-
темиды — один из самых глубоководных
кишечнополостных. Они впервые были
обнаружены несколько лет назад, когда
начался период планомерного исследова-
ния максимальных глубин океана. Эти
актинии чаще всего обитают на дне и
склонах глубоководных океанических впа-
дин — Курило-Камчатской, Филиппин-
ской, Японской и других — на глубине
6—10 тыс. м. Образ жизни их еще совер-
шенно не изучен.
Тело актиний подчас бывает очень креп-
ким, хотя они и лишены скелета. Дело в
том, что мезоглея актиний обычно дости-
гает значительного развития и часто при-
обретает плотность хряща благодаря появ-
лению в ней плотного волокнистого соеди-
нительного вещества.
Морские анемоны размножаются как
бесполым, так иполовым путем.
Однако бесполое размножение играет у
них значительно меньшую роль. Случаи
почкования у Actiniaria вообще очень ред-
ки. Чаще наблюдается деление одной особи
на 2 и даже на 3—6 неравноценных частей.
Поперечное деление отмечено только у
примитивных актиний Gonactinia. У G.
prolifera, например, оно протекает следую-
щим образом: на определенной высоте из
стенок тела сначала вырастает венчик щу-
палец, затем верхняя часть отшнуровы-
вается и отделяется от нижней. У верхней
части восстанавливается подошва, а у
нижней образуются ротовой диск и глот-
ка, а также второй круг щупалец. Второе
деление гонактиний иногда начинается до
того, как закончилось первое.
Чаще у актиний встречается про-
дольное деление. При этом
сначала разделяется на две ротовая щель,
а затем такому же делению подвергается
весь ротовой диск, а потом уже расчле-
няется и тело актинии. Продольное деле-
ние оказывается очень длительным про-
цессом. С момента его начала и до полного
отделения новообразованных актиний мо-
жет пройти несколько месяцев. Изредка
отмечается продольное деление актиний,
протекающее в обратном направлении —
от подошвы к ротовому диску. В этих
случаях деление идет очень быстро и за-
вершается через 2—3 часа (рис. 178).
301
Кроме описанных способов бесполого
размножения, у актиний выработался еще
один весьма своеобразный способ — так
называемая лацерация, при кото-
ром сразу образуется несколько малень-
ких особей. При лацерации от подошвы
взрослой актинии отделяется небольшой ее
участок, содержащий в себе остатки гаст-
ральных перегородок. Этот участок затем
дает начало новым актиниям (рис. 178).
Несмотря на то что деление при помо-
щи лацерации известно с 1744 г., сложный
процесс, приводящий к образованию мо-
лодых актиний, до сих пор еще не изу-
чен.
Способность актиний крегенер а -
ц и и очень велика, хотя и не может
сравниться с таковой у пресноводных гидр.
Основной способ размножения актиний—
половой процесс. Половые клет-
ки актиний имеют энтодермальное про-
исхождение и вызревают в мезоглеальном
слое гастральных перегородок. Акти-
нии, как правило, раздельнополы, хотя
и встречаются случаи гермафродитизма.
В этих случаях мужские половые клетки
образуются прежде, чем женские (так на-
зываемый протандрический гермафро-
дитизм). Оплодотворение может быть как
наружным, так и внутренним. В послед-
нем случае молодые актинии достигают в
гастральной полости материнского орга-
низма на стадии планулы или стадии фор-
мирования щупалец и гастральных пере-
городок.
Многим актиниям свойственна защи-
та потомства. У арктических и
антарктических видов, например, моло-
дые актинии заканчивают свое развитие
или под прикрытием специальных выро-
стов на внешней поверхности тела мате-
ринского организма (Epiactis marsu pialis),
или прикрепившись к нижней части тела
и к расширению ножного диска (Epiactis
prolifera). Раньше уже сообщалось, что
личинки некоторых актиний (например,
Peachia и Edwardsia) ведут почти парази-
тический образ жизни, проникая под
колокол и даже в радиальные каналы
медуз и гребневиков.
Размножение актиний, обитаю-
щих в холодных водах северных и южных
широт, начинается обычно весной и за-
канчивается к лету. Наоборот, в тропиче-
ских водах актинии начинают размножать-
ся в разгаре лета. Плавающие личинки-
планулы держатся в планктоне 7—8 дней
и за это время разносятся течениями на
значительные расстояния.
Морские анемоны населяют почти все
моря земного шара, но, как и остальные
коралловые полипы, они особенно много-
численны и разнообразны в теплых водах.
По направлению к холодным приполяр-
ным районам количество видов актиний
быстро сокращается. По своему образу
жизни актинии могут быть разбиты на
донных и пелагических. Myniadidae —
исключительно пелагическая группа. Дон-
ные актинии обладают весьма широким
Рис. 178. Бесполое размножение актиний:
слева — продольное деление (по Деляжу и Еруару); справа — лацерарпя: вверху — постепенное отшнуровы-
вание части подошвы актинии; в н и 8 у — превращение отшнуровавшихся кусков в молодую актинию
(по Коршельту и Гайдеру).
диапазоном вертикального распределения,
встречаясь от прибойной полосы до мак-
симальных глубин океана. Но подавляю-
щее большинство видов актиний приспо-
собилось к обитанию на небольших глу-
бинах прибрежного мелководья. Это ти-
пичные компоненты скалистой фауны, об-
разующие плотные поселения, к тому
же часто представленные одним видом.
Распространение мелководных актиний
в значительной степени зависит от темпе-
ратуры морской воды и ее солености. В хо-
лодных приполярных областях распро-
странение актиний более или менее кру-
гополярно. Некоторые холодноводные ак-
тинии встречаются как в Арктике, так и в
Антарктике, т. е. образуют так называе-
мые биполярные ареалы. В тропической
зоне имеются циркумтропиче-
с к и е виды, но они встречаются много
реже, чем циркумполярные. Объ-
ясняется это тем, что тропические мелко-
водные участки обычно отделены друг от
друга обширными пространствами океана
с его большими глубинами. Типичным
циркумтропическим распространением об-
ладают крупные актинии Stoichactis.
Некоторые виды актиний, впрочем, мало-
чувствительны к изменениям температуры
воды. Такие актинии обычно более широ-
ко распространены. Actinia equina, обыч-
ный вид в наших северных морях, встре-
чается, например, в Атлантическом океа-
не вплоть до Гвинейского залива. Обшир-
ные ареалы, как правило, и у абиссаль-
ных видов актиний. Узкие локализован-
ные ареалы, однако, характерны для ульт-
раабиссальных, обитающих на глубинах
свыше 6000 ж, видов актиний. Отдельные
виды из рода Galatheanthemum, например,
обитают, видимо, в определенных глубоко-
водных впадинах Тихого океана.
Хотя актинии — типичные морские жи-
вотные, многие из них переносят значи-
тельное опреснение воды. Несколько видов
актиний встречается в Кильской бухте и в
Остзее, четыре вида проникли в Черное
море. В Азовском и Балтийском же морях
актинии уже не встречаются. Любопытно,
что даже в реликтовом озере Могильном
на острове Кильдин обнаружена живущая
там измельчавшая форма весьма обычного
в северных морях Metridium dianthus.
Роющие актинии, такие, как Edwardsia
или Haloclava, закапываются более или
менее отвесно в ил или заиленный песок
и в активном состоянии лишь высовывают
из норки верхний конец своего тела с вен-
чиком немногочисленных щупалец. Они
предпочитают не покидать своей норки,
но в случае необходимости могут перепол-
зать на новое место при помощи волнооб-
разных сокращений червеобразного те-
ла. Найдя подходящий грунт, актиния
прекращает движение и быстро наполняет
свою гастральную полость водой. Затем
она выпускает часть воды и плотно за-
крывает рот. Этим она избегает в процессе
закапывания случайной потери оставшейся
в гастральной полости воды. При закапы-
вании задний конец тела изгибается вниз,
в сторону грунта, а по телу начинают про-
бегать ритмичные волны сокращений коль-
цевых мышц. При этом оставшаяся в по-
лости вода все время перекачивается из
переднего отдела в задний и наоборот. При
помощи перистальтических сокращений
тело актинии все глубже pi глубже про-
талкивается в грунт. Примерно после часа
напряженной работы животное полностью
исчезает в своей новой норке.
Большинство актиний имеет подошву
и ведет сидячий образ жизни. Но при
необходимости и они могут медленно про-
двигаться по субстрату. Обычно поступа-
тельное движение актиний осуществляется
при помощи мясистой подошвы. Часть ее
отделяется тогда от субстрата, выдвигает-
ся вперед, в направлении движения, и
там вновь закрепляется. После этого от-
деляется от субстрата и подтягивается
другая часть подошвы. В частности, так
передвигается Actinia equina — широко
распространенный и весьма обычный в
наших северных морях вид. В аквариуме
наблюдалось перемещение A. equina со
стенок аквариума на находившиеся рядом
камни. Отделившийся от стеклянной стен-
ки край подошвы сильно вытягивался и
наклонялся в сторону камней. Затем ак-
тиния повисала щупальцами вниз между
стенкой аквариума и камнем, к которому
уже прикрепился край подошвы. Через
некоторое время отделялся и подтяги-
вался к камню и другой ее край. На ро-
товом диске этой актинии располагают-
303
ся в 6 рядов 192 щупальца. Эти ак-
тинии, ярко окрашенные в красный или
зеленый цвет, очень красивы, особенно в
распустившемся виде с венцом нежно ок-
рашенных, слегка прозрачных щупалец.
В северных морях преобладающая окрас-
ка этих актиний зеленая, а в южных —
красная. A. equina благодаря своей уди-
вительной невзыскательности — один из
излюбленных объектов для наблюдений в
аквариумных условиях. Любопытно, что
живые актинии можно даже пересылать по
почте, во влажном состоянии или оберну-
тыми влажными водорослями.
Актинии других видов передвигаются
по грунту иным способом. Так, например,
Aiptasia carnea полностью отделяет от
субстрата свою подошву и падает на бок.
В таком лежачем на грунте положении эта
актиния начинает передвигаться задним
концом вперед с помощью перистальти-
ческих ритмичных сокращений тела со-
вершенно так же, как передвигаются рою-
щие актинии. Для своих путешествий А.
carnea всегда выбирает ночное время.
Мелкие актинии, так же как Gonactinia
prolifera, могут даже плавать, ритмично
закидывая назад свои щупальца.
Большинство мелководных актиний из-
бегает дневного света и переползает с
Таблица 14. Глубоководные медузы:
1 — Botrynema brucei;
2 — Atolla wyvillei;
3 — Crossota brunnea;
4 — Pantachogon haeckeli;
5 — Periphylla hyacinthina;
6 — Halicreas minimum;
7 — Meator rubator;
8 — Aeginura grimaldi.
освещенных солнечными лучами мест в
затененные расщелины скал. Если поме-
щенную в аквариум актинию внезапно
осветить ярким светом, она быстро сокра-
щается. Большинство мелководных акти-
ний поэтому днем находится в пассивном
состоянии. Они расправляют свои щу-
пальца ночью или в сумерки. Однако ли-
торальные виды актиний относятся к све-
ту или безразлично, или даже стремятся к
нему, переползая на освещенные места
или поворачивая к свету свой ротовой
диск. В пассивном состоянии они находят-
ся ночью.
Литоральные виды, которые относятся
к свету безразлично, вырабатывают иной
суточный ритм жизнедеятельности, свя-
занный с приливо-отливными измене-
ниями уровня воды. A. equina, напри-
мер, распускает свои щупальца с прили-
вом и сокращается в отлив. Суточный
ритм этой актинии оказывается настоль-
ко стойким, что после помещения ее в
аквариум сохраняется еще в течение не-
скольких дней. Сытые актинии могут дли-
тельное время оставаться в сокращенном
состоянии. Напротив, голод и понижен-
ная температура воды заставляют акти-
ний более суток находиться в активном
состоянии.
Питание актиний изучено сравни-
тельно хорошо. У одних актиний основ-
ную роль в питании играют хватательные
движения щупалец, у других — мерцате-
льное движение реснитчатых клеток, раз-
бросанных в эктодерме. Первые питаются
различными мелкими живыми организма-
ми, вторые — органическими частицами,
взвешенными в морской воде. Существуют
два основных типа движения ресничек.
У примитивных актиний, например у
Gonactinia, реснитчатые клетки которых
равномерно покрывают все тело, попадаю-
щие на тело органические частицы обвола-
киваются слизью и перегоняются бие-
нием ресничек снизу вверх, в сторону ро-
тового диска, а затем и в рот. Биение
ресничек идет в том же направлении и на
щупальцах. В том случае, если пищевой
комок попадает на щупальце, то и здесь
он перегоняется в сторону верхнего кон-
ца его. Щупальце наклоняется в сторону
рта, и пища подхватывается потоком, на-
правленным уже в сторону глотки. Ча-
стицы, непригодные для пищи, захваты-
ваются потоком, создаваемым ресничками
щупалец, и, так же как частицы пищи,
передвигаются к верхнему концу щупаль-
ца. Однако это щупальце наклоняется
уже не ко рту, а в обратную сторону.
С конца щупальца эти частицы смываются
течением воды.
У более высокоразвитых актиний рес-
нички образуются только на ротовом ди-
ске и щупальцах. В частности, такой рес-
нитчатый аппарат мы находим у Metridium
dianthus, или морской гвоздики, одной из
самых красивых актиний, встречающихся
в наших водах (цветная таблица 9). На
длинном столбчатом теле ее отдельными
группами располагаются многочисленные,
свыше тысячи, нитевидные щупальца. Окра-
ска М. dianthus чрезвычайно разнооб-
разна — от чисто-белого до темно-красно-
го цвета. Движение ресничек на щупаль-
цах и ротовом диске этих актиний направ-
лено всегда к верхушке щупалец. Все ча-
стицы, попавшие на ротовой диск или
щупальца, поэтому перемещаются в том
же направлении. Щупальце, после того
как пищевой комок достигает его верши-
ны, изгибается в сторону рта. Затем комок
подхватывается ресничками, выстилаю-
щими глотку, и передвигается внутрь га-
стральной полости. Частицы, непригод-
ные в пищу, также передвигаются к верх-
ним концам щупалец, откуда смываются
водой или сбрасываются.
Актинии, схватывающие пищу щупаль-
цами, питаются различными живыми ор-
ганизмами, а также кусками мяса, остав-
шимися после трапезы какого-нибудь дру-
гого хищника. Проводившиеся многочис-
ленные опыты дают хорошее представле-
ние о механизме схватывания жертвы и
транспортировке ее в гастральную полость.
Обычно голодные актинии сидят совершен-
но спокойно, с широко расставленными
щупальцами. Но достаточно самых сла-
бых изменений, происходящих в воде,
чтобы щупальца стали производить коле-
бательные «ищущие» движения. Когда ак-
тиния чует пищу, к ней вытягивается не
только часть или все щупальца, но часто
в сторону пищи наклоняется и все тело ак-
тинии. Поймав жертву, щупальца акти-
нии сокращаются и изгибаются в сторону
рта. Очень любопытно отметить, что под-
тягивание щупалец ко рту протекает часто
рефлекторно, даже независимо от того,
схвачена ли жертва или нет. Если захва-
тывается крупная добыча, например ма-
ленькая рыбка, то к ней направляются
все щупальца хищника, и все они прини-
мают участие в транспортировке жертвы
к ротовому отверстию. Мелкая добыча
вводится в глотку при помощи тока воды,
вызванного биением реснитчатых клеток
в эктодерме глотки, более крупная — при
помощи перистальтических сокращений
глоточной трубки. У актиний, обладающих
короткими щупальцами, глотка слегка
выворачивается наружу и подтягивается
к пище, которую удерживают над ротовым
диском неспособные пригнуться к рото-
вому отверстию щупальца. Так питается,
в частности, толстоногая актиния— Urti-
cina crassicornis, встречающаяся от Среди-
земного моря до Северного и Норвежского
морей. Многочисленные (до 160) короткие
и толстые щупальца этой актинии окру-
жают ее низкое и толстое туловище.
Окраска U. crassicornis чрезвычайно раз-
нообразна, и вряд ли можно сразу встре-
тить два одинаково окрашенных экземпля-
ра этой актинии.
20 Жизнь животных, т. 1
305
U. crassicornis весьма примечательна
еще и в том отношении, что способ ее
размножения зависит от климатических
условий: в более теплых водах эта акти-
ния выметывает яйца, а в холодных во-
дах (например, у берегов Шпицбергена)
она становится живородящей.
Некоторые актинии сразу чувствуют
различия между пищей и непригодными
для питания частицами и никогда не
схватывают их. Другие, особенно в состоя-
нии голода, схватывают любые предметы—
камни, пустые раковины, фильтровальную
бумагу и т. д. После насыщения столь не-
разборчивые до этого актинии уже не
вводят в свою глотку объекты, непригод-
ные в пищу. Если пропитать мясным
экстрактом фильтровальную бумагу, то в
первое время актиния охотно схватывает
ее. Но с течением времени актиния пере-
стает быть слишком доверчивой. На обман
она сможет попасться лишь через некото-
рый промежуток времени, почувствовав
голод.
При неоднократном повторении такого
эксперимента актиния совсем перестает
реагировать на пропитанную мясным
экстрактом бумагу.
Виды актиний, питающихся органиче-
скими частицами, взвешенными в морской
воде, обладают слаборазвитым стрекатель-
ным аппаратом щупалец. У этих актиний
обычно образуются длинные а к о н ц и и,
отлично защищающие их от нападения.
Напротив, у хищных видов актиний стре-
кательные батареи щупалец становятся
весьма многочисленными. Залп выброшен-
ных стрекательных нитей не только уби-
вает мелкие организмы, но часто вызывает
сильные ожоги и у более крупных живот-
ных, и даже у человека. Часто сильно
обжигаются актиниями ловцы туалетных
губок. После ожога кожа рук начинает
краснеть, зуд и жжение в поврежденном
месте сопровождаются головной болью
и ознобом. Через некоторое время боль-
ные места кожи отмирают и образуются
глубокие язвы.
Очень многие виды актиний являются
комменсалами других животных или всту-
пают с ними в мирный симбиоз. Эти отно-
шения актиний к другим животным под-
робно рассматривались ранее.
ОТРЯД ЦЕРИАНТАРИИ
(CERIANTHARIA)
Известным внешним сходством с акти-
ниями обладают представители отряда
цериантарий.
Цериантарии — малочисленная и срав-
нительно плохо изученная группа шести-
лучевых кораллов. К ним относятся ис-
ключительно одиночные бесскелетные ки-
шечнополостные, ведущие закапывающий-
ся образ жизни. Длинное цилиндрическое
тело цериантарий мускулистое и совер-
шенно гладкое. На переднем его конце
имеется расширенный ротовой диск, зад-
ний конец заострен и снабжен порой, не
играющей, однако, роли порошицы. Ро-
товой диск окружен двумя группами щу-
палец, между которыми остается неболь-
шое свободное пространство. В каждой
группе щупальца могут располагаться в
одном или четырех концентрических кру-
гах. Размеры щупалец в пределах каждой
группы одинаковые, но могут быть весьма
различными в разных группах. Чаще всего
более длинные щупальца образуются в
наружной группе. В целом внешний вид
цериантарий почти не отличается от внеш-
него вида многих актиний, ведущих рою-
щий образ жизни (рис. 179). По внутрен-
нему же строению цериантарии резко от-
личаются от актиний.
На поперечном разрезе животного на
уровне глотки отчетливо видны много-
численные и всегда полные гастральные
перегородки, расположенные на равном
расстоянии друг от друга. Однако радиаль-
ная симметрия нарушается сдавленной с
боков глоткой с одним сифоноглифом, ле-
жащим не на брюшной, как у остальных
коралловых полипов, а на спинной сторо-
не. Валики продольной мускулатуры пере-
городок выражены слабо, тем не менее
видно, что они лежат на их вентральной
стороне. Все новые перегородки всегда
образуются на вентральной стороне тела.
Свободные края перегородок, кроме на-
правляющих, заканчиваются мезентериаль-
ной нитью, снабженной железистыми клет-
ками и стрекательными капсулами.
Мышечный слой цериантарий
сильно развит и лежит сразу под эктодер-
306
мой. Мезоглеальный слой представлен тон-
кой гомогенной пластинкой.
Цериантарии не имеют скелета. Обык-
новенно они сидят более или менее отвесно
в грунте, в трубках, образованных из
затвердевающей слизи, выделяемой много-
численными железистыми клетками экто-
дермального слоя. Внутренняя поверх-
ность трубки совершенно гладкая и до-
вольно скользкая, наружная же поверх-
ность становится грубой и шероховатой
на ощупь из-за обилия различных по-
сторонних включений — песчинок, облом-
ков раковин моллюсков, игл губок и ра-
ковинок фораминифер. Трубки церианта-
рии несколько возвышаются над грунтом.
В активном состоянии животное высовы-
вает из трубки свой верхний конец. Щу-
пальца наружной группы обычно расправ-
ляются и лежат в одной плоскости. Более
короткие щупальца внутренней группы
поднимаются вверх. При малейшем раз-
дражении животное быстро втягивается
внутрь своего домика. Длина трубок у
разных видов весьма изменчива — от не-
скольких до 30—40 см. Но известны
трубки, достигавшие 1 м в длину. Цериан-
тарии не способны втягивать щупальца
внутрь своего тела. Прячась в трубку,
животное складывает щупальца над своим
ротовым диском.
Все цериантарии — гермафроди-
т ы, однако мужские и женские половые
клетки образуются у них в разное время.
Как правило, этим животным свойствен
протандрический гермафродитизм. Как
протекает размножение цериантарии, до
сих пор неизвестно. Изучены только пе-
лагические личиночные стадии, называе-
мые церинулой. Они обладают вы-
тянутым телом и плавают ртом вверх,
с расправленными длинными и короткими
щупальцами. В отличие от планул осталь-
ных коралловых полипов, быстро оседаю-
щих на дно, плавающие личинки цериан-
тарии проводят в планктоне длительный
период жизни. Так, например, церинулы
Arachnactis albida в большом количестве
появляются весной в поверхностном планк-
тоне Северного моря. В начале лета в них
образуется 4 больших и 2 маленьких щу-
пальца и 4 пары гастральных перегородок
(стадия церинулы оказывается тем самым
Рис. 179. Цериантария цериантус
(Gerianthus solitaris) в трубке.
уже пройденной). Осенью личинки имеют
уже до 30 щупалец и 2 гастральные пере-
городки. Длина тела личинок достигает
15 мм. С борта кораблей эти личинки вы-
глядят как маленькие белые звездочки.
По толщине тела и значительному разви-
тию наружного мускульного слоя они
почти не отличаются от взрослых живот-
ных. В начале зимы личинки мигрируют
на глубину 800—1000 м и здесь проходят
свою последнюю личиночную стадию. Ин-
тересно, что в это время у личинок уже
начинают созревать половые продукты.
К концу зимы личинки опускаются на дно,
причем длинные щупальца, служившие им
плавательным аппаратом, видимо, отбра-
сываются.
Бесполое размножение у
цериантарии наблюдается редко, но зато
эти животные обладают исключительной
способностью к регенерации.
Цериантарии обитают во всех морях
с нормальной соленостью. В таких морях,
как Черное и Балтийское, эти животные
20*
307
не встречаются. Большинство из 50 из-
вестных в настоящее время видов цериан-
тарий обитает в теплых водах тропическо-
го пояса и в субтропиках. Лишь немногие
виды проникают в холодные воды припо-
лярных районов. Диапазон вертикального
распределения цериантарий довольно ши-
рок.
Образ жизни цериантарий изучен ма-
ло. Многие планктонные личинки их пи-
таются мелкими рачками — копеподами.
Взрослые же животные используют в пи-
щу органические частицы, взвешенные в
придонном слое воды.
Для нормальной жизнедеятельности це-
риантариям необходимо принять более
или менее отвесное положение, при кото-
ром ротовой конец должен быть наверху.
Если поместить пойманного цериантуса
в узкую стеклянную пробирку ротовым
концом вниз, то уже через несколько ми-
нут животное начинает изгибаться, под-
нимая вверх ротовой конец. Примерно
через час цериантус принимает нормаль-
ное положение. Еще более показателен
эксперимент, когда животное кладут на
горизонтальную металлическую сетку с
мелкой ячеей. Через несколько минут
нижний конец тела начинает изгибаться
вниз и протискиваться через ближайшую
ячею. Скоро животное принимает обыч-
ное отвесное положение. Если тогда по-
вернуть сетку на другую сторону, то жи-
вотное вновь направляет вниз задний ко-
нец. Можно еще раз повернуть сетку на
180°, и еще раз задний конец тела будет из-
гибаться вниз и пробиваться сквозь ее ячею.
Длинное тело цериантуса тогда примет
форму буквы S. Однако если оставить жи-
вотное в таком положении, то уже через
6 часов оно выпутывается из сети и нахо-
дит себе более подходящее место в аква-
риуме.
Вообще в аквариумных условиях при
хорошем уходе цериантусы могут жить
годами. Известны случаи, когда отдельные
животные жили в аквариуме до 40 лет.
Живые, только что пойманные цериантусы
выглядят очень эффектно. Они ярко окра-
шены в различные цвета: в фиолетовый
цвет с оттенками от розоватого до почти
черного, в красивый изумрудно-зеленый
цвет с металлическим отливом, в красный
и коричневый цвета. Щупальца их окра-
шены в тот же цвет, что и тело, но на них
еще бывают чередующиеся более светлые
и более темные кольца, что особенно от-
четливо заметно на щупальцах внешней
группы.
ОТРЯД ЗОАНТАРИИ (ZOANTHARIA)
Зоантарии представляют собой очень
небольшую группу кишечнополостных,
иногда одиночных, иногда колониаль-
ных, лишенных скелета и напоминающих
внешним своим видом мелких актиний.
У колониальных зоантарии отдельные
по липы объединяются, как у альционарий,
или мембрановидной, стелющейся по суб-
страту пластинкой, или при помощи сети
коротких отростков.
Расположение гастральных перегородок
отличается от их расположения у всех
современных коралловых полипов. Од-
нако внутреннее строение зоантарии от-
даленно напоминает строение вымерших
четырехлучевых кораллов. Все полные
перегородки расположены парами. Между
полными перегородками образуются корот-
кие неполные перегородки. Имеются две
пары направляющих перегородок, из ко-
торых дорзальная состоит из неполных
сект, достигающих глотки. В каждой па-
ре гастральных перегородок мышечные
валики продольной мускулатуры обраще-
ны друг к другу, за исключением направ-
ляющих, где они лежат на противолежа-
щих сторонах.
Зоантарии не имеют собственного ске-
лета. Эктодермальные клетки поверхности
тела способны выделять лишь довольно
толстую кутикулу. Любопытно, что эти жи-
вотные способны укреплять кутикулу при
помощи всевозможных мелких чужеродных
предметов — песчинок, игл губок, ра-
ковин фораминифер и т. д. Почти все эти
частицы, прорвав эктодермальный слой
клеток, проникают и в мезоглеальный
слой, достигающий у зоантарии значи-
тельного развития. Посторонние частицы
отлагаются в мезоглеальном слое иногда
в столь большом количестве, что вполне
заменяют опорный скелет. Обычно разные
308
Рис. 180. Зоантария политоа (Polythoa).
зоантарии прикрепляют к наружной стороне
тела какие-нибудь определенные чужерод-
ные частицы: Epizoanthus macintoschi,
например, предпочитают раковины фора-
минифер, a Parazoanthus anquicomus —
только мелкие песчинки. Мягкие ткани
зоантарии бесцветны, и окраску колониям
придают лишь эти многочисленные чуже-
родные включения. Не удивительно по-
этому, что в окраске зоантарии преобла-
дают золотистые, желтые, коричневые и
серые цвета.
Мышечная система зоантарии
развита очень слабо, особенно в базаль-
ной части полипов. О нервной системе до
сих пор почти ничего не известно.
Эти животные чаще раздельнополы.
В таких случаях и колонии слагаются толь-
ко из мужских или только из женских осо-
бей. Но имеются и гермафродитные особи
зоантарии. Развитие зоантарии также еще
полностью не прослежено, известны лишь
их своеобразные плавающие личинки.
Зоантарии распространены всесветно,
однако это сравнительно редкие животные
и попадаются они нечасто. Из 300 извест-
ных в настоящее время видов большинство
обитает в тропической зоне. Только отдель-
ные представители зоантарии проникли
далеко на север и юг, встречаясь в холод-
ных водах Арктики и Антарктики. Среди
зоантарии имеются обитатели прибрежно-
го мелководья и больших глубин океана.
Однако в абиссаль океана на глубину
более 3 тыс. м они не смогли проникнуть.
Зоантарии ведут исключительно донный
образ жизни. Одиночные формы живут на
песчаном или заиленном грунте, погружая
в него задний конец своего тела. Примером
такой зоантарии может быть Isozoanthus
giganteus — наиболее крупный вид, до-
стигающий 20 см в длину. Колониальные
зоантарии селятся на твердом субстрате,
причем большинство их предпочитает об-
растать колониальные постройки других
беспозвоночных. Колонии Epizoanthus,
Parazoanthus, Zoanthus и Isozoanthus обыч-
но можно встретить на губках, гидроид-
ных полипах, мшанках, горгонариях,
рифообразующих кораллах, различных
раковинах и трубках червей. Многие зоан-
тарии становятся комменсалами опреде-
ленных видов животных. Часто встречают-
ся и случаи симбиоза зоантарии с ра-
ками-отшельниками и другими животны-
ми. В тканях мелководных зоантарии
часто поселяются зеленые симбиотические
водоросли Zooxanthellae.
ОТРЯД АНТИПАТАРИЙ
(ANTIPATARIA)
Имеется еще одна группа кишечнополо-
стных, внешне напоминающих растения,—
антипатарий.
Это изящные перистые, вееровидные,
древовидные колонии и колонии, напоми-
нающие ершики. Подобно роговым корал-
лам, они имеют осевой роговой скелет, но,
в отличие от роговых кораллов, полностью
лишенный извести.
По главному стволу скелета и по его
боковым отросткам тянутся каналы, а
поверхность его сплошь покрыта кониче-
скими шипами, иногда достигающими зна-
чительной высоты. Шипы могут отсутство-
вать у основания колонии или на концах
молодых ветвей. Все ветви колонии рав-
номерно покрыты тонким, лишенным ок-
раски ценосарком, в котором образуются
мелкие невзрачные, небольшого размера,
удивительно некрасивые и всегда бесцвет-
ные полипы. Вокруг их ротового отверстия
образуется всего лишь шесть коротких
толстых щупалец. Только у представи-
телей одного рода — Dendribranchia — об-
разуется восемь к тому же перистых щупа-
лец. У некоторых антипатарий полипы
309
Рис. 181. Колония глубоководных антипатарий
батипатес (Bathypates) похожа на рыболовную
вершу.
как бы расстилаются вдоль осевых стерж-
ней колонии, и тогда они подразделяются
на три отдела: в нижнем и верхнем обычно
развиваются половые продукты, в середи-
не среднего отдела образуется ротовой
конус с ротовым отверстием на его верши-
не. Каждому отделу соответствует одна
пара коротких щупалец. Гастральных пе-
регородок в теле полипа может быть 6,
10 или 12. Чаще бывает 10 перегородок.
Шесть перегородок первичные, остальные
представляют собой вторичное образова-
ние. Глотка овальной формы. Все перего-
родки полные, т. е. достигают глотки.
Полости щупалец являются продолжением
боковых камер полости тела, образован-
ных первичными перегородками.
Тонкая цененхима, а иногда и тело поли-
пов часто пронизывается почти насквозь
острыми шипами скелета.
Мускулатура у антипатарий поч-
ти совсем не развита. Ничего не известно
о строении нервной системы. Полипы
антипатарий раздельнополы, но на одной
и той же колонии могут развиваться как
женские, так и мужские особи.
Питание антипатарий совершенно
не изучено. В их гастральной полости не
удавалось найти следов схваченной и пе-
реваренной пищи. Однако эктодермаль-
ный слой тела полипов имеет многочислен-
ные реснитчатые клетки, и это позволяет
предполагать, что антипатарий питаются
органическими частицами, взвешенными
в морской воде и переносимыми в глотку
биением ресничек. Биология антипатарий
почти совсем неизвестна.
Из 120 видов антипатарий около 100 ви-
дов обитает на небольших глубинах тропи-
ческого и субтропического поясов океана.
Очень немногие виды смогли проникнуть
в умеренные воды и на большие глубины
океана. Глубже 2 тыс. м встречается 5 ви-
дов антипатарий и глубже 3 тыс. м —
только три вида. Среди них — Bathypa-
thes patula и В. lyra. Первый образует
красивые перистые колонии, прикрепля-
ющиеся к субстрату базальной пластин-
кой или свободно торчащие в грунте.
Колонии второго вида удивительно напо-
минают ловчую сеть, сходство с которой
увеличивается тем, что полипы, сидящие
на боковых ветвях, ориентированы свои-
ми ртами и щупальцами внутрь «ловчей
сети» (рис. 181). Максимальная глубина
обитания В. patula — 8120 м. Глубоко-
водные виды антипатарий распространены
во впадинах всего Мирового океана.
ОТРЯД МАДРЕПОРОВЫЕ КОРАЛЛЫ
(MADREPORARIA)
Мадрепоровые кораллы — самая об-
ширная группа кишечнополостных. Она
насчитывает свыше 2500 видов, обита-
ющих исключительно в море. Среди них
имеются как одиночные, так и колони-
альные формы, но последних подавля-
ющее большинство. Самая характерная
черта кораллов этого отряда—наличие мощ-
ного известкового скелета, образуемого
эктодермой. Скелет мадрешэровых ко-
раллов сплошной, т. е. он не состоит
из отдельных элементов — спикул или
игл. Подошва полипа образует скелетную
пластинку, от которой поднимаются
вверх (внутри септ) радиальные скелет-
ные перегородки. Толща скелета пос-
тоянно наращивается, и у сформирован-
ного полипа живые мягкие ткани (по срав-
нению с толстым скелетом) выглядят как
полупрозрачная пленочка. У колониаль-
ных форм скелетные чашечки отдельных
полипов сливаются в единый скелет
колонии. Полипы одиночных видов мадре-
шэровых кораллов могут достигать до-
вольно большой величины. Так, диаметр
подошвы некоторых видов грибовидных ко-
310
раллов Fungia у наиболее крупных осо-
бей бывает до 25 см. Напротив, поли-
пы колониальных форм обычно очень
маленькие, как правило, всего 1—5 мм
в диаметре. Форма колоний, образуемых
мадрепоровыми кораллами, может быть
очень разнообразной: одни виды образу-
ют кустики, другие — плоские решетчатые
плиты, третьи — похожи на цветную ка-
пусту, имеются и шаровидные колонии.
Во многом форма колонии этих корал-
лов зависит главным образом от силы
прибоя, течений, заиленности и темпе-
ратуры воды.
Многие колониальные мадрепоровые
кораллы проявляют отчетливую тенден-
цию к объединению колонии в единый
организм путем слияния полипов. Рас-
смотреть это явление можно на примере
некоторых шаровидных колоний. Так,
колония фавии (Favia) имеет вид полу-
шария с ячеистой поверхностью. Каж-
дая ячейка — это чашечка полипа. По-
лип сидит внутри ее и похож на малень-
кую актинию — в его центре имеется
рот, окруженный венчиком щупалец. У
некоторых видов цейлорий (Coeloria)
в чашечке сидит уже не один, а два-
три полипа. Такая чашечка имеет не
круглую, а вытянутую форму, рты по-
липов расположены в один ряд и окру-
жены все вместе одним венчиком щупа-
лец. У этого коралла уже трудно уста-
новить, какое щупальце принадлежит
какому полипу. У лобофиллий (Lobop-
hyllia) в ряд сидят уже десятки полипов,
причем границы между отдельными по-
липами утрачиваются еще больше, так
как у некоторых соседних особей рты
сливаются в общую щель. Наконец, у
кораллов из рода меандрин (Meandri-
via) все полипы сливаются воедино. Ко-
лония имеет вид полушария, по поверх-
ности которого проходят причудливо
извитые борозды, что придает кораллу
вид мозга (он и называется мозговиком).
По центру борозды проходит сплошная
щель, образовавшаяся из слившихся
ртов полипов, а по сторонам этого еди-
ного рта колонии сидят рядами щупаль-
ца. У мозговиков вообще невозможно
установить границы полипов — вся ко-
лония превратилась в единый организм.
Мадрепоровые кораллы обитают во
всех зонах Мирового океана и на различ-
ных глубинах; отдельные виды встре-
чаются в полярных и умеренных по-
ясах. Но наибольшего разнообразия эти
кораллы достигают только в тропиках,
где они составляют основу знаменитых
коралловых рифов.
КОРАЛЛОВЫЕ ПОСТРОЙКИ
Коралловыми рифами назы-
вают массовые поселения (заросли) мор-
ских организмов, обладающих твердыми
известковыми скелетами. Основу такого
рифа составляют мадрепоровые кораллы,
но далеко не всегда они являются его
единственными создателями. Наряду с
мадрепоровыми здесь могут быть и мягкие
кораллы — альционарии, и гидрокораллы,
и мшанки, и даже известковые водоросли.
Кроме того, существует большое количе-
ство прикрепленных полихет, моллюсков
и усоногих раков, также обладающих
мощным известковым скелетом и участвую-
щих в построении рифа. Однако главен-
ствующая роль все же остается за мад-
репоровыми кораллами.
Они являются не только самыми много-
численными, но, что гораздо более важно,
непременными компонентами кораллового
рифа, тогда как представители любой из
других перечисленных выше групп жи-
вотных могут отсутствовать или встре-
чаться в небольшом числе экземпляров.
В связи с этим процветание жизни на
коралловом рифе в первую очередь зави-
сит от процветания самих мадрепоровых
кораллов.
Отдельные виды мадрепоровых корал-
лов встречаются в различных частях Ми-
рового океана и на различных глубинах,
но далеко не везде кораллы образуют
обширные поселения, которые получили
название рифов. Для роста и процветания
кораллового рифа необходимо наличие
целого ряда благоприятных условий. Все
рифообразующие кораллы нуждаются в
морской воде с нормальной океанической
соленостью, т. е. с содержанием в ней
около 3,5% солей. Кораллы могут вынести
311
некоторое повышение солености (так, они
хорошо развиваются в водах Красного
моря, соленость которого достигает 3,8 —
4%), но всякое уменьшение количества
солей в морской воде действует на мадре-
поровые кораллы губительно. В связи с
этим они не поселяются вблизи устьев
крупных рек, где вода распреснена. Во
время сильных дождей, принимающих
характер стихийного бедствия, когда реч-
ной сток усиливается и соленость в при-
брежных частях моря падает, наблюдает-
ся массовая гибель мадрепоровых корал-
лов. Особенно боятся кораллы прямого по-
падания на них дождевой воды, что иногда
случается, если дождь идет во время от-
лива. Известно много случаев, когда обиль-
ные дожди вызывали гибель мадрепоро-
вых кораллов, а вместе с ними и всего
населения рифа. Последний раз массовая
гибель мадрепоровых кораллов в резуль-
тате сильнейших ливней наблюдалась в
1956 г. на некоторых участках восточного
побережья Австралии. Гибель кораллов
влечет за собой серьезные последствия для
различных обитателей моря. Разлагающие-
ся мягкие ткани кораллов отравляют воду
и вызывают смерть тех морских живот-
ных, которые уцелели при распреснении.
Даже после того как море похоронит их
трупы и начисто вымоет скелеты кораллов,
еще долго такие участки остаются безжиз-
ненными. Большинство животных, насе-
ляющих коралловый риф, либо питается
кораллами, либо находит в их зарослях
свое убежище, либо получает и то и дру-
гое. Поэтому, пока заново не разрастут-
ся кораллы, жизнь здесь замирает.
Другим не менее важным условием для
жизни рифообразующих кораллов яв-
ляется высокая и постоянная температура
морской воды. Установлено, что корал-
ловые рифы встречаются лишь там, где
температура воды никогда не падает ни-
же 20,5° С. В связи с этим распространение
коралловых рифов ограничено тропически-
ми частями Тихого, Индийского и Атлан-
тического океанов. На карте распростра-
нения коралловых рифов (см. цветную
таблицу 10) почти полностью совпадают
границы зоны, где встречаются коралловые
рифы, с изотермой 20° С. Обращает на
себя внимание то обстоятельство, что в
Атлантическом океане зона рифов у бере-
гов Америки значительно больше, чем на
африканском побережье. То же самое на-
блюдается и в Тихом океане. В его запад-
ной части зона рифов тянется от юга Япо-
нии до Австралии и включает всю север-
ную половину ее береговой линии, тогда
как на тихоокеанском побережье Америки
эта зона расположена в относительно ко-
роткой полосе между тропиком Рака и эк-
ватором и совершенно не заходит в южное
полушарие. Это обстоятельство не случай-
но, оно объясняется особенностями океа-
нических течений. Вдоль экватора с во-
стока на запад круглый год движутся во-
ды поверхностного экваториального те-
чения. Они сильно нагреваются под лу-
чами тропического солнца. Конечно, тро-
пическое течение не имеет вида цельного
кольца: оно разбивается на три дуги ма-
териками Африки, Америки и островами,
расположенными между Азией и Австра-
лией. Дойдя до суши, дуга течения раз-
бивается на два рукава, один из которых
несет теплые воды на север, а другой на
юг. Это происходит и в Тихом, и в Индий-
ском, и в Атлантическом океанах. Теплые
течения обогревают восточные берега всех
материков, в то же время их западное
побережье охлаждается. Это происходит
вследствие подхода к экватору с севера и
с юга вдоль западных берегов более холод-
ных струй, восполняющих убыль воды,
уносимой экваториальным течением. Раз-
ница между поселениями кораллов на запад-
ном и восточном побережье Индийского
океана выражена меньше, так как в этом
океане нет притока холодных вод с севера.
Температура воды и недостаток освеще-
ния ограничивают распространение ко-
раллов также и по вертикали. В связи с
этим рифообразующие кораллы не захо-
дят на глубину более 50 м.
Из этого правила известно пока только
одно исключение: холодноводный север-
ный мадрепоровый коралл лофохелия (Lo-
phohelia prolifera Pall.) образует в Нор-
вежском море значительные заросли на
глубине около 200 м. Конечно, этот вид
мадрепоровых кораллов никогда не встре-
чается в тропиках, так как приспособлен
к жизни в совершенно других условиях.
Отличается своеобразием и состав жи-
312
вотных, населяющих заросли лофохелии,
но имеется и ряд общих черт, сближающих
их с сообществами животных тропических
коралловых рифов.
Вертикальное распределение многих ма-
дрепоровых кораллов ограничивается так-
же и условиями освещенности. В теле
большинства таких кораллов поселяются
микроскопические одноклеточные жгути-
ковые из отряда перидиней (Peridinea),
так называемые зооксантеллы, окрашиваю-
щие живые ткани кораллов в зеленоватый
или желтоватый цвет. До последнего вре-
мени предполагали, что кораллы могут
получать дополнительное питание, пере-
варивая эти водоросли. Какое преимуще-
Рис. 182. Холодноводный мадрепоровый
коралл лофохелия (Lophohelisa).
ство получают водоросли, поселяющиеся
в теле кораллов, оставалось совершенно
неясным. Известно, что такие же одно-
клеточные водоросли поселяются также в
теле некоторых других тропических мор-
ских животных, в частности они всегда
имеются в тканях двустворчатых моллю-
сков тридакна (Tridacna), обитающих
среди зарослей кораллов. Большинство
кораллов образует с набивающими их
ткани зооксантеллами очень сложное фи-
зиологическое целое, загадка которого еще
не разрешена полностью.
Недавно шотландский зоолог Й о н г
(С. М. Yonge, 1958) дал более полное объяс-
нение причин симбиоза между ко-
раллами и зооксантеллами. Несомненно,
что зооксантеллы оказывают существен-
ное общее стимулирующее действие на
метаболизм своих хозяев-кораллов и для
некоторых из них имеют очень большое
значение в процессе питания. Шестилуче-
вые кораллы, которые, как и все кишечно-
полостные, являются хищниками, совер-
шенно не способны усваивать пищу расти-
тельного происхождения. При любых не-
благоприятных условиях (голодании, не-
достатке кислорода или понижении темпе-
ратуры внешней среды) в их тканях про-
исходят серьезные болезненные изменения,
при этом зооксантеллы гибнут и изгоняют-
ся из тела коралла. Вначале они попадают
в кишечную полость, а затем через ротовое
отверстие выводятся наружу. Однако даже
в случае заболевания, вызванного про-
должительным голоданием, зооксантеллы
совершенно не усваиваются кораллами,
хотя и попадают в кишечную полость, в
которой происходит пищеварение. Если
поместить коралл в темноту, то зооксантел-
лы также гибнут, хотя некоторые кораллы
при этом продолжают нормально разви-
ваться.
Установлено также, что алъциониевые
кораллы (Alcyonaria), обильно заселенные
зооксантеллами, утрачивают свободное
питание, не реагируют на животную пищу,
некоторые части их пищеварительной
полости испытывают редукцию, и в тем-
ноте они погибают. Видимо, зооксантеллы
служат для них источником питания.
На основании этих наблюдений Й о н г
делает следующий вывод. Зооксантеллы
313
потому поселяются в теле кораллов (и
некоторых других тропических морских
животных), что они получают здесь уг-
лекислоту (как продукт дыхания корал-
лов), которую используют в процессе
фотосинтеза углеводов. Еще более важ-
ной следует считать возможность исполь-
зования зооксантеллами других продук-
тов обмена, образующихся в результате
жизнедеятельности кораллов, в первую
очередь производных азота и фосфора.
Эти вещества зооксантеллы используют
при синтезе белков — протеинов. Таким
образом, зооксантеллы как бы заменяют
кораллам органы выделения, аккумули-
руя в себе продукты распада. В свою
очередь кораллы благодаря наличию в их
теле зооксантелл получают в дневные часы
дополнительный кислород, образующийся
в результате фотосинтеза. Й о н г считает,
что отдельные колонии кораллов вполне
могут существовать и без зооксантелл, но
наличие этих симбионтов совершенно не-
обходимо для образования мощных за-
рослей — рифов, так как избыток кисло-
рода и интенсивное удаление продуктов
обмена из тела кораллов дают им преиму-
щества для быстрого роста и развития
мощного скелета.
Зооксантеллы могут развиваться толь-
ко при достаточном освещении. Поэтому
мадрепоровые кораллы даже при бла-
гоприятной температуре не опускаются
на такую глубину, куда проникает мало
света; они редко поселяются также в за-
тененных местах — нишах и щелях.
Однако виды рифообразующих мадрепо-
ровых кораллов, в теле которых нет зоо-
ксантелл, напротив, предпочитают укрытые,
затененные места рифа, таков киноварно-
красный коралл Dendrophyllia.
В последнее время в связи с возможно-
стью применения новых методов изучения
процессов, происходящих в организме,
выдвинуто новое объяснение роли зоо-
ксантелл для кораллов.
Г о р о (Goreau, 1963) с помощью радио-
активного изотопа углерода исследовал
биохимическую сторону механизма отло-
жения извести кораллами. Им была уста-
новлена прямая зависимость между фото-
синтезом зооксантелл и скоростью отложе-
ния извести. Оказалось, что на свету от-
ложение извести идет в 9 раз быстрее,
чем в темноте. У э н р а й т (Wainwright,
1963) добавляет, что зооксантеллы выра-
батывают хитиноподобное вещество, ко-
торое служит основой для отлагающейся
извести скелета. Горо считает, что корал-
лы, поселяющиеся в затененных местах
и на некоторой глубине, куда попадает
меньше света, потому обладают более
нежным скелетом, что фотосинтез здесь
замедлен.
Следующим, важнейшим условием, спо-
собствующим нормальной жизнедеятель-
ности рифообразующих кораллов, являет-
ся чистота и прозрачность морской воды.
Мадрепоровые кораллы предпочитают про-
зрачную воду не только потому, что она
лучше пропускает солнечные лучи, не-
сущие тепло и свет, но и потому, что они
не выносят попадания на свое тело частиц
ила. Особенно это касается видов, спо-
собных образовывать сплошные густые
заросли. Для них заиление не менее гу-
бительно, чем понижение температуры
(ниже 20°) или распреснение морской во-
ды. Тем не менее в ряде случаев и в заи-
ленных местах (в бухтах, лагунах) встре-
чаются значительные поселения мадрепо-
ровых кораллов. Однако они никогда не
образуют здесь густых зарослей: обычно
в таких случаях колонии кораллов от-
стоят друг от друга на известном расстоя-
нии, их видовой состав значительно отли-
чается от видового состава кораллов,
обитающих в незамутненной воде.
Если колонию мадрепоровых кораллов
поместить в сосуд с чистой теплой мор-
ской водой, то она все равно через корот-
кий срок неизбежно погибнет вследствие
недостатка кислорода. Дело в том, что с
повышением температуры растворимость
газов в воде понижается; кораллы же
очень быстро поглощают из воды раство-
ренный кислород и, если воду не сменить,
погибают. Вот почему в природе наиболь-
шего развития кораллы достигают в при-
бойных участках моря, где обогащение
воды кислородом облегчено, а сами ко-
раллы постоянно омываются все новыми
порциями воды.
Наконец, для роста кораллов, как и
всех других животных, требуется пища.
Мадрепоровые кораллы питаются мик-
314
роскопическими животными, населяющи-
ми морской планктон: простейшими и раз-
личными мелкими ракообразными. Но-
вые порции пищи также приносятся к
рифу морем.
Казалось бы, немного можно найти мест,
где все эти условия были бы в наличии
одновременно. И тем не менее это не так.
Значительные пространства тропической
части Мирового океана оказываются впол-
не благоприятными для процветания
мадрепоровых кораллов. Общая площадь
всех коралловых сооружений превышает
27 млн. кв. км. Площадь одних только ост-
ровов и обнажающихся при отливе рифов
равна приблизительно 8 млн. кв. км, т. е.
она несколько более площади Австралии,
равной 7,7 млн. кв. км. Уже одно это
говорит о большом значении коралловых
поселений, требующих их всестороннего
изучения.
Коралловые рифы представляют собой
как бы насыщенные жизнью оазисы среди
малопродуктивных вод тропического океа-
на. Вычисления показали, что они дают
1500—3500 г первичной продукции угле-
рода на 1 кв. м в год. Это в 100 раз боль-
ше, чем в окружающем океане, и в 4—8
раз больше, чем в морях умеренной зоны
(не считая ее самой прибрежной части).
Все многообразие построек, создавае-
мых кораллами и другими рифообразую-
щими организмами, можно разделить на
несколько основных типов. Различают
береговые рифы, расположен-
ные непосредственно по берегам островов
или материков, барьерные рифы,
отстоящие от берега на некотором рас-
стоянии, и атоллы — кольцеобразные
коралловые острова.
Наконец, имеются также значительные
коралловые банки, которые
возникают на отмелях, и густые коралловые
поселения в лагунах и бухтах. Процесс
формирования всех этих коралловых по-
строек интересовал геологов и зоологов
уже очень давно, особенно непонятным
казалось происхождение кольцевидных
островов—а т о л л о в. Было предложено
несколько теорий, объясняющих образо-
вание этих островов, причем многие из
них довольно наивны. Так, до середины
прошлого века господствовало предполо-
жение, что атоллы являются коралловыми
обрастаниями кратеров подводных вул-
канов.
Первая убедительная теория происхож-
дения коралловых построек различных
типов была выдвинута крупнейшим на-
туралистом прошлого века Чарлзом
Дарвином. В своей книге «Строение
и распространение коралловых рифов»,
вышедшей в 1842 г., Дарвин дал не только
подробнейшее описание различных корал-
ловых построек, но и показал, как один
тип коралловых поселений по мере разви-
тия переходит в другой.
Дарвин собрал огромный материал,
касающийся особенностей жизнедеятельно-
сти организмов, образующих коралло-
вый риф, их отношения к условиям внеш-
ней среды, интенсивности роста и распро-
странения в Мировом океане. Часть сведе-
ний он получил от капитанов кораблей,
плававших в тропических морях, и от
ученых, занимающихся изучением корал-
лов. Ценнейшие наблюдения провел сам
во время кругосветного путешествия на
«Б игле».
По мнению Дарвина, первой стадией
образования коралловых островов являет-
ся окаймляющий береговой риф. Кораллы
в этом случае используют берега островов
в качестве опоры, или, как говорят специа-
листы, субстрата. Если условия благо-
приятствуют развитию кораллов, а ост-
ров не испытывает подъема или опуска-
ния, риф так и остается окаймляющим
береговым рифом. В тех случаях, когда
дно моря в результате процессов, про-
исходящих в земной коре, начинает под-
ниматься и остров как бы выходит из воды,
окаймляющий риф разрастается вдоль
его новой береговой линии. Участки ри-
фа, которые оказываются поднятыми над
уровнем моря, погибают, а с морской сто-
роны риф нарастает и увеличивается. Но
общая картина при этом не меняется.
Совсем иначе обстоит дело в тех слу-
чаях, когда морское дно опускается и ост-
ров погружается в воду. Как уже указы-
валось выше, рифообразующие организ-
мы требуют для своего развития много пи-
щи и чистой морской воды, богатой кисло-
родом. Благодаря этому нарастание рифа
всегда идет по его периферии, омываемой
315
морем. Вследствие этого между растущим
внешним краем берегового рифа и погру-
жающимся островом вскоре образуется
затопленное водой пространство, на ко-
тором кораллы растут менее интенсивно.
Так возникает барьерный риф. Чем доль-
ше длится этот процесс, тем дальше барьер
отступает от острова. Наконец, может
наступить такой момент, когда остров окон-
чательно погружается в море, а барьер-
ный риф превращается в атолл — кольце-
вой остров с заключенной внутри его
лагуной.
Современные коралловые постройки да-
ют возможность проследить все стадии
постепенного превращения берегового ри-
фа в атолл. По мере дальнейшего опуска-
ния морского дна кольцо атолла посте-
пенно расширяется, причем с его наруж-
ной стороны сразу начинается крутой
свал. Внутренняя лагуна обычно бывает
мелководной, так как она все время за-
растает менее прихотливыми, но и медленно
растущими видами кораллов. Скорость
роста различных кораллов неодинакова.
Массивные виды, как, например, мозго-
вики, нарастают в год на 1—2 см; ветви-
стые формы, если ничто не препятствует
их развитию, дают за это же время ветки
длиной 20—30 см. Вероятно, внутренние
возможности роста кораллов более зна-
чительны, чем это допускается внешними
условиями. Так, рост рифа вверх резко
ограничивается уровнем моря; но как бы
быстро ни опускалось морское дно, корал-
лы всегда успевают разрастись до той
верхней границы, при которой они могут
существовать. Часть современных корал-
ловых островов возвышается со дна моря
на очень значительную высоту. Попытки
бурить, с тем чтобы определить, как глу-
боко идет известковая порода, показали,
что даже на глубине 50—70 м коралловые
острова сложены из известняка.
Позднее некоторые исследователи вы-
двинули новые теории происхождения ко-
ралловых островов. Дело в том, что тео-
рия Дарвина базируется на одном непре-
менном условии — опускании морского
дна. Между тем противники этой теории
утверждают, что отношения между под-
нятиями и опусканиями морского дна в
действительности несравненно сложнее,
чем это полагал Дарвин.
В связи с этим в 1880 г. Джон Мэр-
рей (Murray) выдвинул новое предполо-
жение о происхождении атоллов. Мэррей
полагал, что над подводными возвышен-
ностями (которые могут возникнуть бла-
годаря вулканической деятельности) жизнь
гораздо богаче. Здесь на отмелях поселяют-
ся различные донные морские организмы,
а в воде над ними развивается более бога-
тый планктон. Отмирая, все эти животные
оставляют много скелетных остатков, бла-
годаря чему напластование осадков на
такой возвышенности идет значительно
быстрее, чем на более глубоких участках
морского дна. Когда вершина такого под-
водного бугра приблизится к поверхно-
сти моря на 50 лс, на ней начинают посе-
ляться мадрепоровые кораллы. Они ра-
стут вверх и образуют коралловую банку.
Последняя нарастает по периферии, а ее
внутренние части разрушаются, благода-
ря чему и образуется кольцо атолла. Хо-
тя теория Мэррея на некоторый срок полу-
чила признание, но она не объясняла проис-
хождения барьерных рифов и была забыта.
Позднейшие исследователи опять вер-
нулись к теории Дарвина, но пытались
внести в нее некоторые дополнения. Очень
остроумное решение вопроса предложил
американский ученый Д ели (Dely, 1915).
1 2 3
Рис. 183. Так постепенно из берегового рифа A) образуется барьерный риф B), а из него — атолл C).
316
Он совершенно правильно рассудил, что
для теории Дарвина нет особой разницы,
опускается ли морское дно, или повышает-
ся общий уровень воды в океане. Предло-
женная Дели «теория ледникового конт-
роля» основывается на том, что в послед-
нюю ледниковую эпоху значительные мас-
сы воды вышли из кругооборота и оста-
лись на континентах в виде льда. В это
время наблюдалось общее похолодание на
всем земном шаре и условий для произ-
растания рифообразующих кораллов не
было. С наступлением потепления в тро-
пических частях моря появились и тепло-
любивые кораллы, которые селились на
отмелях и вокруг островов. Потепление
вызвало таяние ледников, что повлекло
за собой повышение уровня Мирового океа-
на примерно на 70 м. Именно на эту глу-
бину залегают достоверные остатки скеле-
тов мадрепоровых кораллов.
Теория Дели, поддержанная известным
исследователем коралловых рифов Д е-
висом (Davis, 1928), в значительной
мере укрепляет теорию Дарвина, хотя и
вносит в нее существенные изменения.
Однако и «ледниковый контроль» не дает
исчерпывающего объяснения происхож-
дению коралловых островов. В последнее
время появилось много новых данных,
касающихся геоморфологии, океаногра-
фии, палеонтологии и биологии современ-
ных рифообразующих животных. Все они
используются для уточнения дарвинов-
ской теории, но в основе наших современ-
ных представлений о происхождении и
развитии коралловых сооружений остает-
ся гениальное творение Чарлза Дарвина.
Совсем недавно теория Ч. Дарвина о
происхождении и развитии коралловых
островов получила новое подтверждение.
Известный английский исследователь ки-
шечнополостных Вильям Рисе (Riss,
1962) сообщил, что проведенные сейсми-
ческие исследования четырех атоллов пол-
ностью подтвердили все основные положе-
ния дарвиновской теории.
Внешне все коралловые острова очень
сходны между собой. При подходе к
такому острову уже издали становятся
видны ряды кокосовых пальм, белая по-
лоса прибрежного пляжа и буруны на
краю рифа.
Коралловые острова обычно очень не-
значительно поднимаются над уровнем мо-
ря, их растительность довольно однооб-
разна: кроме кокосовых пальм, здесь
растут высокие кусты пандануса. Листья
этого растения широкие и длинные, снаб-
женные по краям многочисленными очень
острыми зазубринами-шипами. На кустах
висят плоды, похожие по цвету, величине
и форме на ананас. Ближе к берегу можно
видеть высокую жесткую траву и мясистые
опунции, похожие на некоторые виды как-
тусов. Вся эта растительность удовлет-
воряется скудной почвой и может обхо-
диться минимальным количеством пресной
воды, которая выпадает во время редких
дождей.
Пляж резко отграничен от полосы
зеленых растений, он состоит почти исклю-
чительно из кораллового песка — пере-
молотых волнами скелетов
мадрепоровых кораллов, но
здесь же имеются и раковины
морских простейших фора-
минифер, и обломки раковин
моллюсков. Часто попадаются
целые, обмытые морем рако-
вины, многие из них имеют
очень красивую форму, ярко
Рис. 184. Схема разреза берегового кораллового рифа. Пунктиром показан уровень
воды в прилив
• Живые БШ коралловый песок
^ m шо Отмершие кораллы
ЭЬ кораллыШЛ почва
317
окрашены, блестят перламутром. На
поверхности песка бегают жуки-скаку-
ны, в самом песке роют норы крабы
оципода (Ocypoda). Это очень осторож-
ные животные, укрывающиеся в нору при
малейшей опасности. Если такого краба
застать вдали от норы, он стремительно
несется на высоко поднятых ногах по пля-
жу и с разбегу смело бросается в набегаю-
щую волну. Остановившись на мелково-
дье, оципода высовывает из воды глаза,
сидящие на длинных стебельках, как бы
выполняющих роль перископа.
Ширина пляжа обычно не превышает
10—20 м, его верхняя часть только во
время самого сильного прилива обдается
брызгами морской воды, тогда как ниж-
няя часть пляжа заливается приливной
волной очень регулярно. В приливо-
отливной части пляжа поселяются раз-
личные роющие двустворчатые моллюски:
донаксы (Donax) и атактодея (Atactodea),
здесь живут также ползающие брюхоно-
гие моллюски клипеоморус (Clypeomorus),
планаксис (Planaxis), мелкие раки-отшель-
ники и некоторые виды полихет.
Ниже расположен отмерший участок
рифа, состоящий из кораллового известня-
ка, покрытого многочисленными мелкими
и крупными обломками кораллов. Здесь
в щелях и под кустами мертвых полипня-
ков поселяются змеехвостки макрофиот-
риксы (Macrophiotrix), гигантские поли-
хеты эунице (Eunice aphroditoides), дости-
гающие более 1 д в длину. В толще из-
вестняка обитают животные, сверлящие
и разрушающие скелет мертвых корал-
лов. Среди них наиболее активны неболь-
шие двустворчатые моллюски литофага
(Lithophaga) и гастрохена (Gastrochaena),
а также маленькие морские ежи эхино-
метра (Echinometra matthaei). Отмерший
участок рифа покрыт различными водорос-
лями, которые служат пищей некоторым
его обитателям. Гигантские полихеты эуни-
це далеко высовываются из своих убежищ
и пожирают водоросли. При малейшей
опасности они быстро скрываются в щели.
Если продвигаться далее в сторону
моря, можно увидеть первые колонии
живых мадрепоровых кораллов. Ближе
всего к берегу селятся буро-зеленые мас-
сивные шаровидные или уплощенные ко-
лонии поритес (Porites), фавитес (Favites)
и орбицелла (Orbicella). Они относятся
к разным семействам, но имеют одну
общую особенность: полипы этих видов
могут глубоко втягиваться внутрь мас-
сивных колоний и потому способны пере-
носить непродолжительное обсыхание (этот
участок рифа во время отлива на некото-
рый период остается не покрытым водой).
Отдельные виды живущих здесь корал-
лов не выносят обсыхания, в связи с чем
у них выработалась своеобразная форма
колонии, позволяющая запасать воду на
время отлива. Внешне такая колония
больше всего похожа на чашу или бокал.
Когда уровень воды при отливе понижает-
ся, в полости чаши остается вода. Все по-
липы таких колоний обращены внутрь и
потому постоянно находятся в водной
среде. Это приспособление выработалось
у некоторых видов турбинарий (Turbi-
naria) и подобаций (Podobacia). Колонии
этих же видов при поселении в более глу-
боких местах, где они не подвергаются
обсыханию, имеют плоскую или даже не-
сколько выпуклую форму. В этой зоне
рифа обитают многочисленные брюхоно-
гие моллюски — ципрея (Сургаеа) и ко-
нус (Conus), много змеехвосток, голотурий,
раков-отшельников и крабов. Иногда по-
падаются большие брюхоногие моллю-
ски птероцера (Pterocera), несущие на
раковине длинные изогнутые шипы.
На краю рифа, где даже во время само-
го низкого отлива бьются волны прибоя,
кораллы образуют сплошные многоярус-
ные заросли. Волны наката достигают
здесь огромной силы, они обрушиваются
на риф и разбиваются о него в белую пену.
Действие постоянно движущихся масс во-
ды откладывает на все коралловое поселе-
ние края рифа глубокий отпечаток. Кроме
волн, здесь все прочно и неподвижно.
Обломки кораллов, раковины мертвых
моллюсков прочно заклинены в щелях:
все подвижные предметы неминуемо уно-
сятся волнами. Они либо выбрасываются
на более высокие участки рифа, либо
смываются в море. Колонии кораллов здесь
плотно срастаются с основой рифа и меж-
ду собой. Одни из них имеют вид гигант-
ских шаров, другие комковатые, есть ко-
лонии плоские, как лепешки. Для этой
318
Рис. 185. Литоральная (наверху) и субли-
торальная формы мадрепорового коралла тур-
бина рии (Turbinaria).
части рифа наиболее типичны кораллы-
мозговики (Coeloria и Meandra), шаровид-
ные гладкие пориты (Porites), орбицеллы
(Orbicella), колючие галаксеи (Galaxea),
симфиллия (Symphyllia), лобофиллия (Lo-
bophyllia). Ветвистые формы имеют ко-
роткие толстые ветви. Все они не боятся
разрушительного действия волн, так как
обладают толстым и очень прочным из-
вестковым скелетом. Другие виды корал-
лов, например некоторые представители
рода акропор (Асгорога), приспособились
к жизни в прибойной полосе иначе. Они
имеют вид огромных округлых плит и по-
хожи на гигантские грибы-трутовики. Эти
кораллы растут в несколько ярусов по
наружному склону рифа. Диаметр таких
плит достигает 2—5 м. Вся толща плиты
пронизана многочисленными отверстия-
ми и напоминает собой решето. Набегаю-
щие и уходящие волны свободно проходят
сквозь отверстия, не разрушая колонии.
Все колонии кораллов, поселяющиеся на
краю рифа, окрашены в яркие, сочные
цвета. Благодаря наличию в тканях по-
липов симбиотических водорослей зоо-
ксантелл большинство видов кораллов жел-
товатой или зеленой окраски. Цвет, осо-
бенно зеленый, очень сочный, похожий
на цвет молодой весенней травы. Края
ситовидных плит акропор, как более мо-
лодые участки колонии, окрашены светлее.
Между бурыми и зелеными кораллами
яркими пятнами выделяются карминно-
красные, иногда вишнево-красные коло-
нии поциллопор (РосШорога). Здесь же
поселяются коричневые с голубым или
сиреневым налетом Montipora foliosa,
колония которых по форме напоминает
цветную капусту. В темных щелях сидят
маленькие колонии коралла Dendrophyl-
lia киноварно-красного цвета.
Если опуститься в воду по внешнему краю
рифа на глубину 3—4 м, взору представит-
ся совершенно необыкновенный подвод-
ный лес. На этой глубине сила прибой-
ных волн несколько ослабевает, поэтому
здесь поселяются более нежные ветвящие-
ся колонии, причем преобладают предста-
вители многообразного рода акропор (Асго-
рога). Здесь есть колонии, похожие по
форме на елочки, густые колючие кусты,
грибы-трутовики, имеются и различные
кораллы, причудливая форма которых
трудно поддается описанию. Преобла-
дают яркие краски: лимонно-желтая, изу-
мрудно-зеленая , светло-коричневая, ма-
линовая. Очень часто кончики ветвей
несут голубой налет мелких молодых по-
липов. В ряде случаев к мадрепоровым
кораллам прибавляются мягкие рифооб-
разующие кораллы алъционарии (Alcyo-
naria). Чаще других встречаются здесь
виды рода саркофитум (Sarcophytum).
Колонии саркофитум образуют обширные
скользкие ковры площадью в несколько
квадратных метров, сплошь покрытые со-
сковидными выростами. Когда идешь по
такому ковру, ноги скользят и погружают-
ся в податливую колонию. Цвет мягких
кораллов обычно серый, мутно-зеленый
или серо-фиолетовый. Некоторые виды,
разрастаясь, ниже уровня отлива приоб-
ретают причудливую форму граммофон-
ных труб с волнистыми краями, гигант-
ских воронок, ваз или чаш. В прибой-
ной части рифа поселяются также неко-
торые гидрокораллы, среди которых глав-
ное место занимает миллепора (Millepora),
образующая густые сплетения кремово-
319
желтых ветвей. Здесь же попадаются и
ярко-голубые солнечные кораллы (Helio-
рога).
Очень богато и разнообразно население
прибойной части рифа. В густых зарослях
кораллов находят себе убежище и пищу
многочисленные черви, моллюски, рако-
образные, иглокожие, рыбы и многие
другие животные. Часть из них всю жизнь
прячется внутри колонии, иногда они про-
сверливают ее скелет, в ряде случаев ко-
лония обрастает такое животное со всех
сторон и оно оказывается навсегда заму-
рованным в толщу коралла, получая
пищу через небольшое отверстие. Другие
укрываются в заросли только при опасно-
сти, третьи постоянно ползают по поверх-
ности колоний или держатся вблизи от
них. Приспособления животных к жизни
на рифе также многообразны. Многие
имеют толстый скелет или раковину, за-
щищающую их от ударов волн. Такие
организмы прочно держатся за риф при
помощи крючковатых конечностей, при-
сосок, острых выростов, крепких шипов
или же просто прирастают к кораллам.
Если эти животные подвижны, то они
всегда медлительны. Их тело имеет обте-
каемую форму, и вода не смывает их с
рифа. Из крупных прикрепленных обита-
телей рифа нужно отметить двустворча-
тых моллюсков тридакна (Tridacna). Раз-
ные виды этого рода укрепляются на ри-
фе различно. Некоторые из них прираста-
ют к рифу при помощи мощного пучка
биссуса — особых волокон, выделяемых
специальной биссусовой железой. Рако-
вина таких видов имеет очень толстые
стенки и обычно достигает значительной
величины. Живущая на рифах Индийско-
го океана и Красного моря гигантская
тридакна (Tridacna gigas) имеет ракови-
ну до полутора метров длиной при весе
свыше 250 кг.В спокойном состоянии створ-
ки раковины приоткрыты и наружу вы-
ступает ярко окрашенный край мантии.
Тридакна непрерывно пропускает через
свои сифоны воду и улавливает пищу, ко-
торой ей служат планктонные организмы.
Потревоженная тридакна быстро закры-
вает створки раковины. Ее мускул-замы-
катель обладает очень большой силой, и
случайное попадание руки или ноги ны-
ряльщика в щель между створками грозит
ему увечьем или гибелью. Маленькая,
всего в несколько сантиметров длиной,
тридакна кроцея (Tridacna crocea) живет
в толще кораллового известняка и сооб-
щается с внешней средой через узкую
щель, из которой наружу выступают толь-
ко края раковины и складки мантии.
Очень любопытен брюхоногий моллюск вер-
метус (Vermetus). Его раковина имеет
вид длинной извитой трубки, приросшей
к поверхности кораллов. Иногда колония
обрастает вокруг всю раковину и наружу
выступает только край устья. Верметус
улавливает добычу, выделяя вокруг себя
большое количество слизи. Время от вре-
мени моллюск поедает эту слизь вместе с
прилипшими к ней мелкими организмами.
Подвижные моллюски, живущие на
прибойной части рифа, имеют обычно
крупные размеры и толстую раковину.
Они прочно присасываются к поверхности
рифа, а их тело даже при движении пол-
ностью покрыто раковиной, из-под кото-
рой выдаются лишь кончики щупалец.
Если прибойная волна все же оторвет
такого моллюска от рифа, он или глубо-
ко втягивает мягкие ткани внутрь (так
ведет себя пятнистый трохус — Trochus
maculatus), или же закрывает вход в
устье толстой сферической известковой
крышечкой до 200 г весом {мраморный
турбо — Turbo marmoratus). В самых при-
бойных местах рифа живет очень интерес-
ный морской еж гетероцентротус (Hete-
rocentrotus mammilatus). Его тело имеет
овальную форму, позволяющую ежу за-
бираться в довольно узкие щели и не-
большие полости рифа. Часть игл на спи-
не животного сильно утолщена и служит
для удерживания тела в расселинах.
Основания игл снабжены мощной мус-
кулатурой, позволяющей ежу прочно упи-
раться в стенки своего убежища. Здесь
же между ветвями и вдоль щелей медлен-
Таблица 15. Кораллы в хозяйстве человека:
1 — постройка из кораллового известняка; 2 — печь для обжига кораллов; 3 — участок набережной, облицован—у
ной кирпичами из кораллов; 4 — кладка стены здания из кораллового известняка; 5 — в коралловой лавке.
но ползают округлые коротконогие крабы
калаппа (Calappa) и атергатес (Aterga-
tes). Имеется много других крабов, мол-
люсков, иглокожих, обладающих толсты-
ми защитными скелетными образованиями.
Другой способ защиты от разрушитель-
ного действия волн заключается в том,
что тело приобретает предельную гиб-
кость, оно изгибается и бьется вместе с
волной, благодаря чему сохраняется его
целость. Таковы поселяющиеся на прибой-
ных рифах иглокожие животные морские
лилии. Маленькое тело их прикреплено
к рифу при помощи цирр — корневидных
выростов нижней стороны тела, а длин-
ные «руки» этого животного, имеющие чле-
нистое строение, свободно извиваются
вместе с движущейся водой.
Все обитатели кораллового рифа ярко
и пестро окрашены; это надежно скры-
вает их на разноцветном фоне кораллов.
Особой яркости достигают коралловые
рыбки. Эти быстрые рыбки поодиночке и
стайками «бродят» между кораллами, от-
щипывая полипов, которыми питаются.
Некоторые из них обладают мощными
челюстями, позволяющими им отгрызать
и измельчать куски коралла. Если под
водой разбить на мелкие куски колонию
ветвистого коралла, то вскоре вокруг
обломков собирается много рыбок, от
крошечных до крупных в 20—30 см дли-
ной. Все они набрасываются на обломки
коралла и быстро растаскивают их в
Таблица 16. Гребневики:
1 — берое (Вегое cucumis),
2 — болинопсис (Bolinopsis in-
fundibulum),
3 — Венерин пояс (Cestus ve-
neris);
4 — евплокамис (Euplocamis
cucumis).
W 21 Жизнь животных, т. 1
разные стороны. Большинство коралловых
рыбок очень пугливы, они постоянно жи-
вут среди неподвижных кораллов и вся-
кий шевелящийся предмет заставляет их
искать убежище в зарослях.
Окраска и форма коралловых рыбок
чрезвычайно разнообразны: здесь есть и
красные, и зеленые, и голубые, и желтые, и
черные. Почти все они имеют яркие пятна
или полосы. В коралловых лесах укры-
ваются хищные мурены, иногда достигаю-
щие 3 ж в длину. Змеевидное тело мурены
скрыто в щели или под нависающим ко-
раллом, наружу выставлена маленькая
голова с приоткрытым ртом, усаженным
острыми, как бритва, зубами. Заметив
добычу, мурена покидает засаду и стреми-
тельно нападает на свою жертву. В глу-
бине пасти мурены имеются ядовитые
зубы; попадание яда в кровь ускоряет
гибель животного.
Совсем иначе выглядят коралловые по-
селения в бухтах и лагунах, защищенных
от действия прибоя. В таких местах ко-
раллы не образуют сплошного монолита,
здесь они растут отдельными деревьями,
кустами или лежат на дне, не прикрепив-
шись к нему. Отсутствие волн способствует
развитию нежных ветвистых колоний. Ин-
тересно отметить, что одни и те же виды
при поселении на прибойной части рифа и
в лагуне образуют колонии различной
формы. Широко распространенная на всех
коралловых островах поциллопора оле-
ньи рога (Pocillopora damicornis) в прибой-
ной зоне образует приземистые колонии с
короткими толстыми веточками, а при по-
селении в лагунах этот вид образует ред-
коветвистые колонии с тонкими длинными
ветвями. Изменяется и цвет кораллов. Ла-
гунные формы всегда окрашены менее
интенсивно, здесь преобладают желтова-
тые, бурые и грязно-зеленые тона колоний.
Виды, которые на прибойном рифе имеют
интенсивный красный цвет, в лагунах ок-
рашены в розовато-желтый.
Многие виды кораллов, обитающие на
прибойном рифе, никогда не встречаются
в лагунах; зато здесь имеются свои лагун-
ные виды, которые могут жить только в
спокойной воде. Очень своеобразны полу-
шаровидные колонии эуфиллий (Euphyl-
lia), иногда достигающие 1 л в диаметре.
Каждая такая колония состоит из тесно
расположенных ветвей — секторов, от-
ходящих от центра полушария. Основания
ветвей старых колоний разрушаются, по-
сле чего ветви теряют связь между собой.
Большинство ветвистых колоний акропо-
ры (Асгорога), стилофоры (Stylophora) и
других кораллов имеет длинные корне-
видные выросты, которые внедряются в ил,
благодаря чему колония удерживается на
месте. Веточки этих видов тонкие, чрез-
вычайно ломкие. Даже слабые волны обла-
мывают кончики таких ветвей, однако
поврежденные участки колонии быстро
зарастают. В лагунах поселяются ячеи-
стые колонии павона (Pavona), а также
одиночные (не образующие колоний) гри-
бовидные фунгии (Fungia). Молодые фун-
гии снабжены ножкой, на которой растет
тело коралла. Оно похоже на шляпку
сыроежки, обращенную пластинками
вверх. Когда тело фунгии достигает в
диаметре 2—3 см, ножка обламывается
и коралл падает на дно, где продолжает
расти. На ножке тем временем развивает-
ся новый коралл. Некоторые экземпляры
достигают 10 см в диаметре, а вытяну-
тая в длину колючая фунгия (Fungia echi-
nata) вырастает до 30—35 см. Цвет фун-
гии обычно охристый или нежно-желтый,
некоторые виды имеют розовый край.
Население лагунного рифа отличается
своеобразием, здесь обитает много живот-
ных со слаборазвитым скелетом, с длин-
ными ломкими придатками, а также мяг-
кие, бесскелетные организмы. Встречает-
ся много плавающих и вообще быстро
двигающихся животных. Живущие в ла-
гунах крабы проворно плавают, их зад-
ние конечности уплощены и служат ве-
слами. Плавают также и некоторые мол-
люски. Небольшой двустворчатый мол-
люск лима (Lima) передвигается, быстро
хлопая створками, широкая мантия и
длинные щупальца придают плавающей
лиме внешнее сходство с медузой. Дру-
гой двустворчатый моллюск — гребешок
(Chlammis), набрав воды в мантийную
полость, выбрасывает ее резким толчком,
при этом сам моллюск силой обратного
толчка делает большой скачок. Многие
моллюски, например молоточек (Malleus)
и кренатула (Crenatula), подвешиваются
на ветвистые колонии при помощи биссуса.
На дне лагун попадаются жемчужницы
(Pinctada), в илистом грунте живет боль-
шое количество различных закапывающих-
ся двустворок. Среди них наиболее ин-
тересна пинна (Pinna), достигающая иног-
да 40 см. Раковина пинны, имеющая фор-
му наконечника копья, погружена в ил
на всю свою длину, причем нижний конец
ее при помощи нитей биссуса прикреплен
к обломку коралла. Наружу выступает
лишь самый верхний край раковины.
Брюхоногих моллюсков в лагунах обыч-
но мало, однако нельзя не отметить по-
падающихся здесь различных ципрей (Сур-
гаеа), имеющих красивую глянцевую рако-
вину. В ячейках, образованных колония-
ми кораллов павона (Pavona), укрывают-
ся маленькие осьминоги.
В лагунах живет много разнообразных
иглокожих — голотурий, морских ежей и
звезд. Лагунные морские ежи имеют тон-
кий, ломкий панцирь. Встречаются так-
же мягкие ежи, например токсопустес
(Toxopneustes). Этот еж, достигающий
величины кокосового ореха, сильно ядо-
вит. «Укус» щипчиков (педициллярий),
во множестве сидящих на теле ежа, вы-
зывает сильнейшую боль и продолжитель-
ное сердцебиение.
Ядовиты и ежи диадемы (Diadema seto-
sum). Величина тела такого ежа достигает
размеров антоновского яблока, со всех
сторон он покрыт почти полуметровыми
322
тонкими иглами. Эти черные ежи сотнями
и тысячами лежат на прогалинах между
кораллами. Случайный укол их иглой
также вызывает сильную боль, зависящую
от попадания в ранку ядовитой слизи.
В бухтах массами живут безвредные ко-
роткоиглые морские ежи трипнеустесы
(Tripneustes), окраска которых подверже-
на значительной индивидуальной измен-
чивости. Встречаются и белые экземпля-
ры, и особи с красным или оранжевым
звездообразным рисунком. Лагунные
морские звезды обычно достигают величи-
ны в размахе лучей 20—30 см. Они мед-
ленно ползают между кораллами, поедая
моллюсков и других обитателей лагуны.
Наиболее характерны для лагунного ри-
фа неуклюжие кульциты (Culcita), похо-
жие по форме, величине и цвету на круг-
лый хлеб; очень обычны серо-зеленые с
розовыми шипами звезды ореастеры (Огеа-
ster) и коричневые, также шиповатые,
протореастеры (Protoreaster).
В лагунах живут бурые очень крупные
актинии Stoichactis kentii, диаметр рото-
вого диска которых достигает полуметра.
Вблизи каждой актинии постоянно дер-
жится пара маленьких креветок — самец
и самка. В случае опасности креветки пря-
чутся между щупальцами актинии.
И на прибойных рифах, и в лагунах,
кроме животных, обитающих в зарослях
кораллов и поселяющихся между их ко-
лониями, живет большое число организ-
мов, еще более тесно связанных с мадрепо-
ровыми кораллами. К ним относятся пре-
имущественно мелкие животные: черви, иг-
локожие, моллюски и главным образом
ракообразные. Все они обладают крючка-
ми, большими клешнями, выростами тела
и другими приспособлениями, позволяющи-
ми им прочно удерживаться на ветвях
кораллов. Окраска таких животных обыч-
но бывает покровительственной. Так, все
крабики и рачки, обитающие в красных
колониях поциллопоры, сами окрашены
в красноватый цвет. На бурых и корич-
невых кораллах поселяются темноокра-
шенные виды ракообразных.
В ячейках полипов некоторых видов
мадрепоровых кораллов поселяются ли-
чинки усоногих рачков из рода пиргома
(Pyrgoma). Полип вскоре погибает, а
ячейка по мере развития рачка увеличи-
вается в размерах, сохраняя основные
пропорции. Такие уродливые ячейки по-
хожи на галлы, вызываемые у растений
насекомыми и клещами.
В толще полипняков поселяются раз-
рушители кораллов — двустворчатые мол-
люски. Морские финики (Lithophaga) име-
ют почти цилиндрическую форму. Оны
проделывают в полипняке трубчатые хо-
ды, растворяя известь выделениями спе-
циальной железы. Раковина самого мор-
ского финика, также состоящая из изве-
сти, защищена от действия кислоты слоем
органического вещества — конхиали-
н а. Другие двустворчатые моллюски —
фоласы (Pholas) протачивают коралл своей
раковиной, поверхность которой покрыта
многочисленными крепкими зубцами и
похожа на напильник.
ПРАКТИЧЕСКОЕ ИСПОЛЬЗОВАНИЕ
КОРАЛЛОВ
Как уже отмечалось, живые ткани мад-
репоровых кораллов составляют незначи-
тельную часть веса этих животных, тогда
как главная масса приходится на долю
мощного известкового скелета. Скелет от-
лагается эктодермой кораллов, образую-
щая его известь имеет особую некристал-
лическую форму (а р а г о н и т).
Для населения тропических стран ко-
ралловые рифы имеют чрезвычайно важ-
ное значение. Все обширное пространст-
во, занятое коралловыми рифами, яв-
ляется огромной природной фабрикой из-
вести. Изо дня в день, из года в год кро-
шечные полипы мадрепоровых кораллов,
а также другие морские животные извле-
кают известь из морской воды и отклады-
вают ее в своем теле в виде скелета. Вы-
сокая температура воды в поверхностных
слоях тропических морей способствует*
образованию массивного известкового ске-
лета. Встречаются массивные колонии мад-
репорового коралла Coeloria до 3—6 м в
диаметре, а некоторые виды рода Pon-
tes образуют колонии с диаметром 8—9 м.
В тропической зоне моря известковый
скелет отмерших организмов не подвер-
гается разрушению. Напротив, в холодных
21*
323
водах морские организмы, как правило,
лишены массивного известкового скелета,
который целиком или частично заменяет-
ся скелетом кремниевым или хитиноидным.
Известь скелета отмерших организмов в
холодной морской воде быстро разрушает-
ся. Известно, что мадрепоровые кораллы
являются древними животными, и на
местах их поселения в течение многих
тысячелетий из скелетов отмерших коло-
ний образовались значительные отложе-
ния извести, которые и в настоящее время
продолжают увеличиваться. Благодаря то-
му что рифообразующие мадрепоровые
кораллы поселяются у поверхности моря,
т. е. по берегам островов, либо сами об-
разуют острова, эта известь легко доступ-
на для разработки, а запасы ее практи-
чески неограниченны, хотя и распола-
гаются пятнами на местах, где были или
имеются коралловые рифы.
Скелет мадрепоровых кораллов имеет
довольно широкое хозяйственное приме-
нение. Во многих приморских тропиче-
ских странах его используют как хороший
строительный материал для постройки
домов, молов, набережных, мощения улиц
и т. д. Очень велика ценность мадрепоро-
вых кораллов как сырья для получения
извести. Кроме того, скелет мадрепоро-
вых кораллов употребляют иногда для
заполнения водопроводных фильтров, для
полировки и шлифовки деревянных и
металлических изделий, а также для из-
готовления лекарственных препаратов. Не-
которые мадрепоровые кораллы вследст-
вие красивой формы их колонии употреб-
ляются для устройства искусственных скал
и гротов в садах и парках и в аквариу-
мах, а также используются в качестве
комнатных украшений.
Способы добычи кораллов в значитель-
ной мере зависят от места их обитания.
В лагунах и бухтах, где дно обычно бывает
заилено, колонии кораллов имеют форму
кустов с редкими ломкими ветвями. Они
не прикрепляются ко дну, а держатся
на нем при помощи корневидных выростов,
либо свободно лежат на поверхности ила.
В мелководных лагунах, поросших ко-
раллами, промысел их ведется с лодок.
Кораллы собирают со дна при помощи
трезубца на длинном древке. Вовремя
прилива добытые кораллы подвозят воз-
можно ближе к берегу и сваливают там в
воду, а затем, когда наступает отлив,
транспортируют на телегах к месту даль-
нейшей переработки.
Гораздо сложнее добывать кораллы из
прибойных рифов, где они образуют мас-
сивные колонии, или на мертвых участ-
ках рифа, где из скелетов отмерших ко-
раллов образовался известняк. Здесь ма-
териал добывают откалыванием кусков
при помощи железных клиньев, ломов,
молотов и других инструментов.
Массивные колонии мадрепоровых ко-
раллов употребляют для постройки раз-
личных сооружений: изгородей, дамб, на-
бережных, пристаней и т. д. Обработка
строительного материала весьма неслож-
ная: края шаровидных колоний обкалы-
вают, придавая им форму куба или бру-
са. Величина брусьев строго не определе-
на, чаще она колеблется между такими
размерами: 20—25—30 см и 25—30—35 см.
Такие грубо обработанные брусья корал-
ла укладывают прямой либо косой клад-
кой. С наружной, обычно необработан-
ной поверхности стены хорошо различима
структура колоний, часто позволяющая
судить о родовой и даже видовой принад-
лежности коралла. В большинстве слу-
чаев отдельные брусья в кладке ничем не
связаны и стена удерживается вследствие
плотного прилегания их шероховатых по-
верхностей. Однако для придания построй-
ке большей прочности глыбы коралла
иногда скреплены раствором коралловой
извести, о производстве и употреблении
которой будет сказано несколько ниже.
Строительные брусья делают из мас-
сивных колоний кораллов, относящихся
к родам Favia, Galaxea, Symphyllia,
Coeloria, Porites, Goniastrea и другим.
Свежие и недавно отмершие кораллы—
вполне подходящий материал для по-
стройки различных сооружений на бере-
гу моря (дамбы, пирсы, волноломы, на-
бережные): они не разрушаются под хи-
мическим воздействием морской воды. Од-
нако эти кораллы никогда не употребля-
ют для постройки зданий. Причина этого
заключается в значительной пористости
материала, подверженного разрушению на
воздухе и в результате действия пресной
324
дождевой воды. Для наземных построек
успешно применяют известняк, образо-
вавшийся из отмерших кораллов. Для
такого типа строений коралловый извест-
няк колется на сравнительно небольшие
узкие плитки (в среднем 8X 15 X 20 см),
которые укладывают друг на друга подоб-
но кирпичам и связывают известковым
раствором. Плитки, употребляемые для
строительства зданий, почти лишены сле-
дов структуры колонии. Стена, сложен-
ная из таких плиток, снаружи имеет прият-
ную для глаза поверхность и не требует
штукатурных и других отделочных раГот.
Изнутри же стены штукатурят.
Скелет всех мадрепоровых кораллов с
успехом можно употреблять для обжига
на известь. Сырьем служат как перечис-
ленные выше массивные, так и ветвистые
(Асгорога, Pocillopora, Stylophora), ячеи-
стые (Pavona) колонии и даже одиночные
формы (Fungia), а также обломки любых
кораллов. Вероятно, мадрепоровые ко-
раллы обжигались на известь с древнейших
времен, причем во многих местах сохра-
нились очень примитивные методы обжига.
Химический состав кораллового извест-
няка очень благоприятен для обжига его
на известь. В высушенных мадрепоровых
кораллах углекислый кальций состав-
ляет 90—97%, соединения фосфора и
кремния содержатся в очень небольшом
количестве @,3—2,6%). Последнее очень
важно, так как примесь солей кремние-
вой кислоты снижает качество извести.
Известно несколько типов печей для
обжига кораллов. В наиболее примитив-
ном случае печь складывают из самого
обжигаемого материала, т. е. из ничем не
связанных между собой и не обработанных
колоний мадрепоровых кораллов. Стены
печи укладывают в форме широкого коль-
ца. Сверху печь открыта. Обычно диаметр
такой печи около 10 м, высота стен 2 м.
Стены печи для предохранения их от раз-
валивания подпирают снаружи деревян-
ными кольями. Вследствие неплотного
прилегания кусков коралла, образующих
стену печи, между ними остаются боль-
шие отверстия, через которые при обжиге
поступает воздух. Если поддув оказывает-
ся избыточным (при сильном ветре), стены
печи прикрываются плотным плетнем и
циновками из листьев кокосовой пальмы.
Печь загружают дровами и засыпают не-
сколькими тоннами разбитого коралла
(один кусок обычно не превышает по весу
1 —1,5 кг). Разжигают печь сверху через
отверстие в засыпанных кораллах. Обжиг
производится в течение четырех-пяти су-
ток. Затем около трех суток печь остывает.
После остывания ее разгружают, для чего
разбирают часть стены и выносят известь
через образовавшийся проход. Затем обож-
женный материал просеивают на грохоте.
Такие примитивные печи устанавливают
поблизости от места добычи коралла и
после окончания разработок разрушают.
Более широко используют маленькие по-
стоянные печи. Такая печь имеет форму
полого, открытого сверху цилиндра или
усеченного конуса, обращенного вверх
основанием. Почь ставят на кирпичном
фундаменте, она имеет снизу поддувало.
Под печи снабжен колосниками. Стены
складывают из ничем не связанного кирпи-
ча и для предохранения от рассыпания
оплетают снаружи гибкими ветвями, при-
дающими печи сходство с гигантской кор-
зиной около 2 м в диаметре. На дно печи
укладывают слой топлива, затем слой
мелко битого коралла, далее снова топ-
ливо и т. д. Всего кладется 3—4 слоя
кораллов. Процесс обжига длится около
15 часов. Обожженные кораллы извле-
кают из печи через широкое отверстие в ее
стене.
Гораздо более продуктивны печи не-
прерывного действия. Такая печь имеет
вид высокой башни и построена из кирпи-
ча. Топливо и коралловый известняк за-
гружают через окно в верхней части пе-
чи. В качестве топлива используют камен-
ный уголь. Выем обожженной извести про-
изводят через окна в нижней части печи.
Интенсивность горения регулируют искус-
ственным поддувом от механической воз-
духодувки. Для усиления тяги на верху
печи установлена труба. По мере выема
извести слои горящего угля и обжигае-
мых кораллов под влиянием своего веса
опускаются из верхней части печи в сред-
нюю, где и происходит обжиг. В нижней
части печи обожженная масса остывает.
Отсюда она и извлекается, а через верхнее
окно в печь добавляют новые порции топ-
325
лива и сырья. Печь работает непрерывно.
При использовании в качестве топлива
каменного угля обожженная известь по-
лучается более твердой, чем при топке пе-
чи дровами. Поэтому ее приходится раз-
малывать на шаровой мельнице.
Готовую известь используют в строи-
тельном деле для приготовления связываю-
щих растворов и штукатурных работ.
Качество ее очень высокое. Готовый про-
дукт имеет вид чисто-белого жирного на
ощупь порошка, состоящего почти из
одной окиси кальция.
Как видно из сказанного, мадрепоро-
вые кораллы используют главным обра-
зом для получения строительных мате-
риалов, однако этим применение скелета
кораллов не ограничивается.
Благодаря пористому строению ске-
лет мадрепоров оказался очень удобным
материалом для заполнения водопровод-
ных фильтров. Теперь в большинстве
стран, расположенных у берегов тропи-
ческих морей, фильтры заполняют не
коксом, как это делали раньше, а хоро-
шо промытыми кусками кораллов.
Известь, из которой построен скелет
мадрепоровых, очень легко измельчить
в тонкий порошок, который с успехом
применяют как полировочный материал.
Куском коралла, как напильником, мож-
но обрабатывать деревянные изделия.
На островах Самоа из пластинок ко-
раллового известняка изготовляют блес-
ны для ловли рыбы.
Как уже было сказано, в тропиках
имеется немало островов, возникших из
отмерших коралловых рифов. На них
вообще нет и не может быть природного
камня, а потому здесь кораллы упот-
ребляются как тяжелые предметы. При
их помощи раздавливают плоды для по-
лучения сока, перетирают семена. Куски
коралла употребляют как балласт в лод-
ках и как груз в различных механичес-
ких приспособлениях.
Издавна не только в тропиках, но и в
Европе мадрепоровым кораллам при-
писывали магические свойства, из них
делали амулеты, будто бы предохраняв-
шие от «дурного глаза» и колдовства.
В старину в Европе скелет мадрепо-
ровых кораллов под названием «корал-
лиум альбум» служил для приготовления
некоторых лечебных препаратов. В неко-
торых странах Юго-Восточной Азии и сей-
час толченый скелет этих кораллов можно
найти в любой аптеке.
Довольно большое значение имеет
употребление мадрепоровых кораллов в
качестве украшений и сувениров. Для
этой цели используют преимущественно
ветвистые колонии (Acropora, Pocillopora),
похожие на причудливое растение три-
дакофиллию (Tridacophyllia), а также на
одиночные грибовидные кораллы (Fungia).
Ловцы собирают особенно красивые ко-
лонии, выбирая их среди зарослей. Ото-
бранную колонию осторожно отделяют от
рифа и затем удаляют из нее мягкие тка-
ни. Известно несколько способов отделе-
ния мягких тканей от скелета. Если на
судне имеется водяная помпа, то проще
всего разложить собранные кораллы на
палубе и поливать их из шланга струей
воды под большим напором. При этом
нежные ткани легко смываются и остает-
ся белоснежный известковый скелет, ко-
торый затем высушивается на тропическом
солнце. Более простой и, вероятно, са-
мый древний способ отбеливания корал-
лов состоит в том, что собранные колонии
привязывают одну за другой на длинную
веревку и тянут ее за парусной лодкой.
При этом мягкие ткани также смываются
струями воды. Нужно только следить,
чтобы кораллы не опускались на дно, где
они неминуемо поломаются. Когда лодка
пройдет несколько миль, веревку с отбе-
ленными кораллами вытаскивают на борт.
Наконец, мягкие ткани могут быть уда-
лены продолжительным вымыванием ко-
раллов в пресной воде. Выбеленные и про-
сушенные скелеты мадрепоровых корал-
лов очень твердые, хотя и хрупкие. Они
звенят, как хороший фарфор, да и внеш-
не кажутся сделанными из чистейшего
неглазированного фарфора.
В качестве украшений и сувениров ко-
раллы наравне с раковинами тропических
моллюсков охотно покупаются и местным
населением и приезжими. В Китае и Вьет-
наме из мадрепоровых кораллов делают де-
коративные горки и помещают их в садах
и парках. Иногда перед продажей белые
скелеты окрашивают яркими красками.
Гребневики
Ctenophora
ТИП ГРЕБНЕВИКИ (CTENOPHORA)
Тело гребневика студенистое, обычно
очень прозрачное; часто имеются щупаль-
ца. Чисто внешнее сходство гребневиков
с медузами долгое время служило причи-
ной того, что зоологи считали их медуза-
ми или животными, очень близко род-
ственными медузам.
Первые упоминания о гребневиках мож-
но найти у древнегреческого ученого Ари-
стотеля, жившего в V—IV вв. до нашей
эры. Однако в те времена зоологические
познания были еще очень примитивны,
и Аристотель смешивал гребневиков с го-
лотуриями, относящимися к совсем друго-
му т и п у животных — иглокожим. Толь-
ко в 1671 г. Мартене (Martens) в
своей книге о животных, населяющих
море вблизи Шпицбергена, привел первое
описание и рисунок гребневика. Лин-
ней, создатель первой научной системы
органического мира, уделил гребневикам
очень небольшое внимание, упомянув в
своей «Системе природы» один вид гребне-
виков. В 1829 г. вышла в свет первая в
мире большая работа, посвященная меду-
зам. Ее автор, немецкий зоолог Э ш-
ш о л ь ц (Eschscholtz), описал в ней и
несколько видов известных ему гребневи-
ков. Он считал их особым классом медуз,
который назвал гребневиками (Ctenopho-
га1). Это название сохранилось за ними
1 Ctenos (греч.) — гребешок.
и в настоящее время. Позднее появилось
несколько больших исследований, посвя-
щенных изучению гребневиков. Из них не-
обходимо отметить книгу Карла X у н а
(Chun, 1880), в которой детально описана
анатомия средиземноморских гребневиков,
а также работы А.О.Ковалевского,
И. И. Мечникова и А. Корот-
н е в а.
Подробные сведения о гребневиках при-
ведены в немецком руководстве по зоо-
логии. Автор раздела о гребневиках —
проф. К р у м б а х (Krumbach, 1925).
Тем не менее и сейчас эта группа живот-
ных остается еще слабо изученной; осо-
бенно мало было известно об образе жиз-
ни и поведении гребневиков. Только в
самое последнее время благодаря трудам
советского зоолога проф. М. М. К а м-
шилова многие важные и очень инте-
ресные особенности биологии гребневиков
получили объяснение.
Гребневики исключительно морские,
преимущественно свободноплавающие ор-
ганизмы. Обычно тело гребневика округ-
лой или мешковидной формы. На одном
конце помещается рот. По поверхности
тела гребневика в меридиональном на-
правлении проходит восемь рядов греб-
ных пластинок. Каждая пластинка расщеп-
лена по наружному краю и имеет вид
гребня, за что гребневики и получили
свое название. Каждая гребная пластинка
состоит из ряда слившихся между собой
329
Рис. 186.
(Cydi
крупных ресничек эпителия.
Пластинки бьют по воде и, дей-
ствуя наподобие весел, медлен-
но передвигают животное ро-
товым концом вперед.
Большинство видов гребне-
виков имеет щупальца. Hlyna-
лец всегда два, и располагают-
ся они по бокам тела, причем
могут втягиваться в специаль-
ные щупальцевые карманы.
Щупальца способны далеко
вытягиваться и обычно превы-
шают длину животного во мно-
го раз. От одной из сторон щу-
пальца отходят нитевидные от-
ветвления. Вся поверхность
щупальца и его ответвлений
покрыта многочисленными клей-
кими клетками. Отдельная клет-
ка имеет вид полушария, уса-
женного липкими сосочками.
Под микроскопом такая клетка
снаружи напоминает по строе-
нию ягоду малины. От основа-
ния полушария внутрь щупаль-
ца отходит спирально закру-
ченная нить, прикрепленная своим другим
концом к продольному пучку мышц, про-
ходящих и вдоль щупальца, и вдоль
его ветвей (рис. 187).
Мелкие морские организмы при сопри-
косновении с щупальцами оказываются
приклеенными к ним липкими полушария-
ми. Если добыча пытается вырваться,
то липкие клетки отделяются от поверх-
ности щупальца, но остаются соединен-
ными с ним посредством эластичной спи-
ральной нити. Жертва прочно удержи-
вается многочисленными клейкими клет-
ками, нити которых служат своеобразны-
ми пружинками, смягчающими рывки пой-
манного животного.Подобным образом при
помощи эластичного удилища рыбак удер-
живает на тонкой леске крупную рыбу.
Запутавшуюся в щупальцах добычу греб-
невик захватывает краями рта и поедает.
Рот ведет в трубковидную глотку, ко-
торая в свою очередь открывается в же-
лудок. От желудка отходят каналы га-
строваскулярной системы. Один канал
продолжается в сторону, противополож-
ную рту, и здесь разделяется на четыре
Цидиппе
рре).
коротких канальца, два из ко-
торых заканчиваются слепо, а
два других открываются нару-
жу порами. В стороны от же-
лудка отходят два канала, ко-
торые вскоре дважды разветв-
ляются, благодаря чему к кра-
ям тела проходит уже восемь
каналов. Каждый из них впада-
ет в один из восьми меридио-
нальных каналов, проходящих
под рядами гребных пластинок.
Меридиональные каналы тянут-
ся от ротового конца тела до
противоположного и слепо за-
канчиваются на обоих концах.
По ним питательные вещества
поступают к месту их наиболее
интенсивного расходования —
к гребным пластинкам.
Нервная система
гребневиков очень примитивна,
она состоит из подкожного
нервного сплетения, имеющего
вид сети, и небольшого, но до-
вольно густого скопления ган-
глиозных клеток на абораль-
ной (противоположной ротовому отвер-
стию) стороне тела. Здесь же находит-
ся сложно устроенный, так называемый
аборальный, орган, который, по-видимо-
му, играет роль органа равновесия и ре-
гулирует интенсивность работы отдель-
ных рядов гребных пластинок.
Основу аборального органа составляют
четыре крестообразно расположенные эла-
стичные дужки, образовавшиеся из длин-
ных, слившихся между собой ресничек.
На дужках подвешен статолит—
шарик, состоящий из известковых зерен.
Под статолитом стенка тела образует
впячивание, причем эктодерма здесь утол-
щена в виде подушечки. Снаружи стато-
лит прикрыт коническим шатром из слип-
шихся длинных ресничек. Основания ду-
жек, удерживающих статолит, при помо-
щи особых бороздок, покрытых мерцатель-
ным эпителием, сообщаются с рядами
гребных пластинок. Висящий на пружи-
нящих дужках статолит беспрерывно ко-
леблется, его движения через дужки и
мерцательные бороздки передаются рядам
гребных пластинок, вызывая в них волну
330
биения, которая идет в направлении от
аборального органа к ротовому концу тела.
Если удалить аборальный орган, движе-
ния гребных пластинок нарушаются.
Недавно (Камшилов, 1964) было
установлено, что гребневики способны от-
личать свет от темноты, хотя у них еще не
были обнаружены специальные светочув-
ствительные органы.
На свету тело гребневика берое (Вегое)
имеет желтовато-розовый цвет. Окраска
обусловлена наличием в покровах живот-
ного маленьких клеток, наполненных кра-
сящим веществом. Эти клетки способны
растягиваться и сжиматься. Если поме-
стить берое в темноту, то уже через 12 —
15 минут красящие клетки (хроматофоры)
начинают сокращаться. Через час живот-
ное становится молочно-белым. На свету
цвет быстро восстанавливается и уже че-
рез 6—7 минут гребневик приобретает
первоначальную желтовато-розовую ок-
раску.
У гребневиков очень сильно развита мезо-
глея, придающая их телу значительную
прозрачность. Обычно гребневики бес-
цветны, только немногие из них, как упо-
мянутый выше берое, имеют желтовато-
розовую окраску, лишь ряды движущих-
ся гребных пластинок переливают всеми
цветами радуги.
Гребневпки размножаются исключитель-
но половым путем, причем одно животное
образует и яйца и спермин. Половые
железы имеют вид длинных колбасовид-
ных тел, располагающихся вдоль мери-
диональных каналов. С одной стороны
канала лежит семенник, с другой — яич-
ник. Таким образом, ктенофора имеет 8
женских и 8 мужских половых желез, не
связанных друг с другом. Когда половые
клетки созревают, стенка, отделяющая
половую железу от меридионального ка-
нала, прорывается, после чего половые
продукты через просвет канала и ротовое
отверстие попадают в морскую воду, где
и происходит их развитие.
В процессе дробления яйца образуются
клетки неодинаковой величины. На одном
полюсе возникает большое число быстро
делящихся мелких клеток, которые впо-
следствии образуют эктодерму, на другом
полюсе клетки крупнее и делятся более
Рис. 187. Строение липкой клетки
гребневика:
I — клейкие сосочки; 2 — спиральная
4 5
Рис. 188. Схема строения гребне-
вика:
1 — ротовое отверстие; 2 — глотка; 3 —
желудок; 4 — аборальный канал гастро-
васкулярной системы; 5 — аборальный
орган; 6 —меридиональный канал гастро-
васкулярной системы; 7 — карман щупа-
льца; 8 — щупальце, 9 — ряд гребных
пластинок.
331
медленно. Из них образуется энтодерма
будущего животного. Ротовое отверстие и
гастральная полость возникают вследст-
вие впячивания слоя крупных клеток
внутрь. Впячивается также и часть мел-
ких клеток, из которых формируется
впоследствии глотка. Очень важно отме-
тить, что часть крупных клеток из глу-
бины гастральной полости зародыша по-
гружается в пространство между эктодер-
мой и энтодермой. Здесь из них впослед-
Рис. 189. Строение аборального органа гребневика
(по Хуну):
А — в профиль, Б — сверху. 1 — статолит; 2 — пружинящие
дужки; 3 — мерцательные бороздки; 4 — эктодермальная
подушечка; 5 — шатер из ресничек.
ствии формируется мускулатура. Эти клет-
ки представляют собой зачаток третьего
зародышевого слоя — мезодермы.
В настоящее время известно около 80
видов гребневиков. Самые маленькие из
них, например черноморская плевробрахия
(Pleurobrachia pileus), едва достигают 5 мм
в длину (не считая щупалец, которые мо-
гут вытягиваться очень значительно). Наи-
более крупные виды, такие, как венерик
пояс (Cestus veneris), бывают до 1,5 д
длиной.
Большинство гребневиков живет в тол-
ще воды. Из-за нежности их тканей они
предпочитают держаться в спокойной во-
де, во время волнения опускаются в глу-
бину. Отдельные виды встречаются на
глубине 3 км. Многие гребневики в ноч-
ное время излучают яркий голубоватый
свет. Существует предположение, что све-
тятся не сами гребневики, а живущие в их
кишечнике бактерии.
Все гребневики (за одним исключением) —
хищники, причем до недавнего времени
считалось, что они питаются одними толь-
ко планктонными рачками и являются
конкурентами сельди, сардины и других
планктоноядных рыб. Поэтому всех без
исключения гребневиков относили
к вредным животным, и очень час-
то отсутствие в отдельных участ-
ках моря планктоноядных рыб
объясняли тем, что гребневики
выели всю их пищу. Это каза-
лось тем более правдоподобным,
что гребневики очень быстро раз-
множаются и встречаются в мас-
совых количествах. Такие круп-
ные гребневики, как болинопсис
(Bolinopsis infundibulim) и берое
(Beroe cucumis), 12 — 15 см дли-
ной, за один раз выметывают от
1500 до 3 тыс. яиц. При этом мо-
лодь быстро растет, достигая за
месяц длины 8—10 см, и в свою
очередь приступает к размноже-
нию.
Изучение жизни гребневиков
показало, что каждый из них име-
ет свои особые приспособления
для ловли добычи и что разные
виды гребневиков питаются раз-
личными животными.
332
Гребневики, снабженные щупальцами,
ловят свою добычу — мелких планктон-
ных рачков — при помощи клейких кле-
ток. Болинопсис, у которого нет щупалец,
имеет по краям рта две большие лопасти.
Размахивая ими в воде, болинопсис заго-
няет в рот мелкие планктонные организ-
мы. В желудке этого вида были обнаруже-
ны личинки кольчатых червей, различные
планктонные рачки, личинки моллюсков
и другие мелкие планктонные организмы.
Эти гребневики, несомненно, являются
конкурентами планктоноядных рыб. Дол-
гое время по аналогии берое также счи-
тался вредным животным, выедающим
планктон и нападающим даже на мальков
рыб.
Однако такое мнение не было подкреп-
лено фактическими данными. Изучая био-
логию гребневиков, проф. М. М. К а м-
шилов обратил внимание на одно очень
важное обстоятельство в строении берое.
Этот гребневик малоподвижен и лишен
ловчих щупалец и лопастей. Как же он
ловит быстрых и вертких рачков, а тем
более рыбок? Изучение содержимого же-
лудка берое не дало никаких результа-
тов — остатков рачков обнаружено не бы-
ло. Тогда стали предлагать берое, поме-
щенным в аквариум, самую разную пищу.
Но снова неудача — гребневики отказы-
вались от предложенных им рачков и дру-
гих планктонных организмов. Более того,
оказалось, что, если искусственно ввести
рачка в желудок берое, он вскоре выбрасы-
вает его через рот, причем рачок продол-
жает спокойно плавать — пищеваритель-
ные соки берое на него не действуют.
В одном из опытов в сосуд, где содер-
жался берое, был помещен гребневик
болинопсис. Тогда поведение берое резко
изменилось, он не спеша подплыл к бо-
линопсису, широко раскрыл рот и про-
глотил гребневика, величина которого была
почти такой же, как величина самого
хищника. Оказалось, что берое питаются
исключительно другими гребневиками.
Таким образом, они не только не являют-
ся конкурентами планктоноядных рыб, но,
наоборот, их следует считать полезными
животными, так как они интенсивно ис-
требляют действительно вредных болино-
псисов.
Рис. 190. Особые формы гребневиков:
1 — паразитические гребневики гастродес (Gas-
trodes parasitica) в теле сальпы (по Комаи); 2 —
ползающий гребневик целоплана (Coeloplana)
(по Комаи), 3 — сидячий гребневик тьяльфиел-
ла (Tjalfiella) (по Мортенсену).
Выяснилось и другое не менее важное
обстоятельство: берое служат пищей та-
ким промысловым рыбам, как пикша
и треска; поедают их и другие рыбы,
едят гребневиков крупные сцифоидные
медузы.
Таким образом, раскрывается одна из
сторон сложнейших взаимоотношений
между морскими животными. Когда к
берегам подходит большое количество тре-
ски, она выедает почти всех берое. Пер-
вым следствием этого является массовое
размножение болинопсисов; они в свою
очередь поедают планктонных рачков и
таким образом лишают пищи сельдь.
Как уже отмечалось выше, один из
гребневиков— гастродес (Gastrodes parasi-
tica) не является хищником. Он ведет
паразитический образ жизни, поселяясь в
333
планктонном морском животном салъпе,
относящемся к оболочниковым, стоящим
близко к хордовым. В сальпу проникает
личинка, которая уже внутри хозяина
превращается во взрослого гребневика.
Форма тела гребневиков в значительной
степени зависит от их образа жизни и
поэтому очень разнообразна. Наиболее
типичной является овальная, яйцевидная
или сигарообразная форма тела, характер-
ная для всех гребневиков, снабженных
щупальцами, и для личинок многих видов,
которые во взрослом состоянии щупалец
лишены. Выше уже говорилось о гребневи-
ке болинопсисе, который не имеет щупалец,
но снабжен двумя большими ротовыми
лопастями. Не имеет щупалец и взрослый
гребневик венерин пояс (Cestus veneris),
обитающий в Средиземном море. Тело
этого животного имеет лентовидную форму
и может достигать полутора метров дли-
ны. Венерин пояс плавает при помощи
гребных пластинок и вследствие змее-
видных движений тела, расположенного
всегда в горизонтальной плоскости. Он про-
зрачен и переливает всеми цветами раду-
ги, благодаря чему ему и дано такое
поэтическое название.
Весьма своеобразны платиктениды—
маленькие уплощенные гребневики, спо-
собные не только плавать в толще воды, но
и ползать по дну. К этой группе относят-
ся описанная А. О. Ковалевским
целоплана Мечникова (Coeloplana metsch-
nicowi) и описанная Коротневым
ктеноплана Ковалевского (Ctenoplana ko-
walewskii), а также еще около двух десят-
ков видов. В строении этих гребневиков
можно усмотреть очень много черт строе-
ния, общих с некоторыми морскими пло-
скими червями.Высказывалось даже пред-
положение, что плоские черви произошли
от ползающих гребневиков. Однако в по-
следнее время благодаря исследованиям
проф. В. Н. Беклемишева стало ясно, что
сходство между ползающими гребневиками
и плоскими червями чисто внешнее, за-
висящее от одинакового образа жизни этих
животных, а также от сходства механизма
их эмбрионального развития.
Один арктический вид гребневиков —
тьялъфиелла (Tjalfiella) является сидячим
донным животным. Молодь этого вида ве-
дет планктонный образ жизни, но затем
садится на дно ртом вниз, причем середина
рта зарастает, а его края в виде двух тру-
бок загибаются кверху. К бесщупальце-
вым гребневикам относятся также упо-
мянутый выше берое и другие близкие к
нему виды. Тело бесщупальцевых гребне-
виков имеет вид мешка. Оно снабжено
огромным ротовым отверстием, служащим
для заглатывания крупной добычи— греб-
невиков других видов.
Тип Гребневики включает единствен-
ный класс с шестью отрядами. Предста-
вители первых пяти отрядов (п о д-
класс щупальцевых — Tentaculifera) —
цидипповые (Cydippea), лопастеносные (Lo-
bata), цестиды (Cestidea), платиктениды
(Platictenidea) и тьялъфеллиды (Tjalfiel-
lidea) — в течение всей жизни или на
ранних стадиях развития имеют щупаль-
ца. К подклассу бесщупальцевых
(Atentaculata) относится всего один о т-
р я д морских огурцов (Beroidea), включаю-
щий гребневиков, лишенных щупалец на
всех стадиях их развития.
Низшие черви
Plathelminthes
Acanthocephala
Nemathelminthes
Nemertini
Как указывалось во «Введении», все многоклеточные жи-
вотные, стоящие по своему развитию выше кишечнополостных,
объединяются в раздел двустороннесимметричных (Bilateria),
или трехслойных (Triplastica), животных. Ткани и органы их
тела развиваются из всех трех зародышевых листков—эктодер-
мы, энтодермы и мезодермы. У этих животных вырабатыва-
ется быстрый способ передвижения и в связи с этим — дву-
сторонняя симметрия тела с передним и задним концом, со
спинной, брюшной, правой и левой сторонами тела.
В процессе эволюции Bilateria сформировались две ветви
двустороннесимметричных животных — первичноротые и вто-
ричноротые.
* * *
Относящиеся к первичноротым (Protostomia) И типов жи-
вотных характеризуются общим признаком — на месте личиноч-
ного рта формируется дефинитивное ротовое отверстие взрос-
лой формы, а анальное отверстие, если оно имеется, проры-
вается на противоположном (анимальном) полюсе зародыша.
К первичноротым относятся все «черви», моллюски, члени-
стоногие и некоторые другие группы.
Термин «черви» до недавнего времени занимал видное
место в системе беспозвоночных. Под этим термином объединяли
в один общий тип червей (Vermes) не только плоских, круглых
и кольчатых червей, но и ряд примыкающих к ним групп.
В настоящее время черви потеряли свое классификационное
значение. Однако само понятие не утратило обиходного
зоологического смысла.
Под «червями» подразумевают организмы, имеющие удли-
ненную форму тела, лишенные конечностей и ясно оформлен-
ной головы, снабженные сильной мускулатурой в виде кожно-
мускульного мешка, обеспечивающей типичное для червей
движение изгибанием тела. Самые низкоорганизованные чер-
ви — низшие турбеллярии — по многим особенностям строе-
ния или физиологии близки к кишечнополостным, а высшие —
кольчатые черви — столь же близки к членистоногим.
Червей подразделяют на низших и высших. Выс-
шие черви имеют сегментированное строение, вторичную
полость тела — целом — и кровеносную систему. Низших
червей иначе называют сколецидами (Scolecida) *. К ним от-
носят 13 классов: турбеллярии, трематоды, темноцефалы, удо-
неллиды, моногенетические сосальщики, нематоды, волосатики,
ресничнобрюхие, гирокотилиды, цестоды, колючеголовые, ко-
ловратки и киноринхи.
1 Scolex (греч.) — червь.
ТИП ПЛОСКИЕ ЧЕРВИ
(PLATHELMINTHES)
ОБЩАЯ ХАРАКТЕРИСТИКА
Если поблизости от вас, читатель,
имеется пруд со стоячей водой, заросшей
тростником и покрытой широкими ли-
стьями водяных лилий, попробуйте найти
здесь маленького плоского червя — пла-
нарию. Иной раз достаточно перевернуть
несколько лежащих на заиленном дне
камней, чтобы найти на нижней стороне
одного из них зеленоватую или зеленова-
то-коричневую планарию (Euplanaria go-
nocephala).
Выставив вперед похожие на уши бо-
ковые лопасти, планария плавно сколь-
зит по камню, прижавшись брюшком
к его поверхности. Движения эти, уди-
вительно плавные и как бы скользящие,
обусловлены тем, что кожный покров
нежного тельца планарии (в длину она
достигает едва 2,5 мм) покрыт ресничками.
Они-то и сообщают телу планарии этот
плавный ход.
Будьте осторожны при попытке снять
планарию с камня. Ваши пальцы —
грубый инструмент для этого. Осто-
рожно отделив планарию от камня с
помощью пинцета, столкните ее в под-
ставленную банку с водой, взятой обя-
зательно из того же пруда. Первое же
знакомство с этим маленьким и нежным
существом позволяет констатировать не-
которые признаки, присущие всем пло-
ским червям, всем платодам.
Прежде всего обращает на себя внимание
действительно плоское тело. В нем можно
легко различить передний и задний концы,
брюшную и спинную стороны, ну и, ко-
нечно, правый и левый бока.
Разумеется, организация планарии от-
носительно простая.
Ротовое отверстие у нее расположено на
брюшной стороне тела. Заднепроходного
отверстия нет совсем, и плотные отбросы
пищи выбрасываются через ротовое отвер-
стие. Нет и органов дыхания и крово-
обращения.
Таковы все плоские черви.Тело у них
обычно значительно сплюснутое в спинно-
брюшном направлении, цельное или раз-
деленное на членики. Кожно-мускульный
мешок плоских червей состоит из кожного
покрова (эпителия) и многослойной му-
скулатуры. Все внутренние органы по-
гружены в паренхиму — нежную
рыхлую ткань, наполняющую весь кожно-
мускульный мешок. У плоских червей
имеется центральная нервная
система в виде продольных нервных
стволов; развиты органы чувств, в
том числе органы химического чувства и
фоторецепторы, а также специальные ор-
ганы равновесия. Органы пищеварения
начинаются ротовым отверстием, располо-
женным на брюшной стороне тела. Имеет-
ся глотка и кишка в виде одинарной или
раздвоенной трубки; заднепроходного от-
верстия нет. Развита выделительная систе-
ма, имеющая форму каналов, по которым
жидкие продукты выделяются наружу
^0 22 ЖИЗНЬ ЖИВОТНЫХ, Т. 1
337
через особое выделительное отверстие,
расположенное обычно на заднем конце
тела.
Плоские черви — обоеполые живот-
ные, или гермафродиты. Кровеносной и
дыхательной систем у этих животных нет.
Свыше 5 тыс. видов морских, пресновод-
ных и паразитических плоских червей
известно современной науке. Многие из
паразитических плоских червей опасны
для жизни человека и животных. Зна-
комство с этим типом животных уже по
одной этой причине представляет большой
интерес.
Каждый человек должен иметь пред-
ставление об этих животных хотя бы в
самых основных чертах. Эти полезные зна-
ния пригодятся людям и практически —
необходимо знать, как защитить себя от
опасных паразитических плоских червей.
Плоские черви, или платоды, очень
многообразная группа животных. Плана-
рия, с которой мы начали наше знаком-
ство с этими животными,— свободножи-
вущая форма. Как правило, все родичи
этой планарии — хищники, опасные, разу-
меется, для соразмерных с ними живых
существ. Хищность во многих группах
животных, в том числе и в пределах типа
плоских червей, развивается в другую
форму взаимоотношений — паразитизм,
когда одни животные поселяются на теле
или во внутренних органах других, пи-
таясь и развиваясь за их счет. Многие
плоские черви—типичные наружные (экто-
паразиты) и внутренние (эндопаразиты)
паразиты.
Хищность, эктопаразитизм и эндопа-
разитизм плоских червей привели к фор-
мированию нескольких классов этих жи-
вотных. Наличие в этом мире
паразитов класса свобод-
ноживущих турбеллярий — жи-
вое свидетельство происхожде-
ния паразитических плоских
червей — моногеноидей, дигеней
и ленточных червей — от свобод-
ноживших предков.
К типу плоских червей от-
носится 6 классов: Реснич-
ные черви, или Турбеллярий
(Turbellaria), Моногеноидей (Мо-
nogenoidea), Сосальщики, или
Трематоды (Trematoda), Темно-
цефалы (Temnocephala), Гироко-
тилиды (Gyrocotyloidea) и Лен-
точные черви, или Цестоды (Ces-
toidea).
В организации всех предста-
вителей этих классов в силу
их родства имеется ряд общих
признаков, которые были под-
робно рассмотрены на предыду-
щей странице.
Рис. 191. Планарии:
1 — Mesostoma ehrenbergii на листе, 2 —
она же, схватившая дафнию. Оба экземп-
ляра держатся на водной поверхностной
пленке, подвесившись к ней с помощью
слизевой нити, з — Polycelis cornuta,
4 — Dendrocoelum lacteum* 5 — Eupla-
naria gonocephala; 6 — Polycelis nigra.
338
КЛАСС РЕСНИЧНЫЕ ЧЕРВИ, ИЛИ ТУРБЕЛЛЯРИЙ
(TURBELLARIA)
Следует прежде всего подчеркнуть, что
именно организация этих маленьких жи-
вотных, иногда, впрочем, и довольно круп-
ных, должна рассматриваться как типич-
ная. Это представление далеко не слу-
чайно. Оно определяется тем фактом, что
турбеллярий — свободноживущие орга-
низмы. Поэтому именно они обладают ти-
пичными для плоских червей признаками.
Все остальные классы плоских червей —
паразиты, а паразитизм ведет к специа-
лизованным преобразованиям организа-
ции, а нередко и к ее упрощению. Озна-
комление с организацией турбеллярий
важно для понимания их природы.
Можно начать их описание с плана орга-
низации, и прежде всего перечислить их
морфологические признаки. Это, несом-
ненно, важно. Однако не менее важно
понимать причины, вызвавшие к жизни
ту или иную организацию живых существ.
Итак, первый и главнейший признак рес-
ничных червей — это тот, что они сво-
бодноживущие организмы. Вот что их
определяет! А если это так, то для них долж-
ны быть типичны признаки, присущие сво-
бодным организмам: это прежде всего
органы свободного передвижения в про-
странстве, способность определять в нем
положение собственного тела и, нако-
нец, способность вовремя заметить
других животных — и врагов, и до-
бычу — и вовремя среагировать на
тех и на других. Для ресничных
червей прежде всего характерны сле-
дующие признаки. Тело их покрыто
ресничками, согласованное движе-
ние которых обеспечивает плавное
передвижение в пространстве. У рес-
ничных червей имеется особый орган
равновесия — статоцист — в
виде пузырька с плотным свободным
ядрышком внутри, подобный тем, ка-
кие имеются у кишечнополостных;
этот орган позволяет червю ориен-
Г/9 тироваться в пространстве. Перевер-
ните ресничного червя, и он тотчас
повернется брюшной стороной вниз.
Информацию о своем положении в
пространстве ресничный плоский
червь получил от статоциста. Рес-
ничным червям присущи развитые ор-
ганы обоняния (химические органы)
Рис. 192. Половая система планарий:
А — половая система молочной планарии: 1 —
семенные пузырьки; 2 — семяпровод, 3 — сово-
купительный орган; 4 — железистый орган;
5 — совокупительная бурса; 6 — яичник; 7 —
желточники. Б — схема половой системы пла-
нарии: 1 — семенники; 2 — семяпровод, 3 — се-
менной пузырь; 4 — совокупительный орган;
5 — совокупительная сумка; б— непарный про-
ток; 7 — яичники; 8 — яйцеводы; 9 — желточ-
ники.
22*
339
и органы восприятия света (фо-
торецепторы). Химические ор-
ганы представлены обонятель-
ными ямками по бокам головы,
тогда как фоторецепторы пред-
ставлены глазками, расположен-
ными на переднем крае тела.
Наконец, отметим еще один
признак свободного существо-
вания ресничных червей, а
именно покровы их тела окра-
шены в различные цвета — зе-
леный, желтый, розовый, свет-
ло- и темно-коричневый, почти
черный, красный, фиолетовый,
синевато-серый и др. Таковы
главные признаки ресничных
червей, определяемые их сво-
бодной жизнью. На таблице
17 показаны некоторые пред-
ставители ресничных червей.
Кто скажет, что они некраси-
вы, по крайней мере по ок-
раске?
В остальном ресничным чер-
вям свойственны признаки пло-
ских червей: кожно-мускуль-
ный мешок, состоящий из кож-
ных покровов и сложной систе-
мы продольных, кольцевых, ко-
сых и спинно-брюшных мышц;
паренхима, заполняющая тело;
продольные нервные стволы;
брюшное ротовое отверстие; сле-
пая кишка — прямая или раз-
ветвленная на два ствола; вы-
делительные органы в виде вет-
вящихся трубочек с мерцатель-
ными колбочками на концах и
экскреторным наружным отвер-
стием на заднем конце тела; мощно раз-
витая половая система, в которой комби-
нируются всегда мужские и женские поло-
вые органы. Все это как бы вправлено в
общий «архитектонический план» двубоко-
вой симметрии (рис. 192).
Ресничные черви — хищники. Они на-
падают на мелких животных, например
мелких рачков, и высасывают их, или даже
рвут нежное тельце рачков на части, или
тлотают целиком. Живут они в различных
средах. Известны морские ресничные черви,
пресноводные и, наконец, почвенные.
Рис. 193. Нервная и
пищеварительная систе-
мы молочной планарии
(Dendrocoelum lacteum):
I — нервные стволы; 2 —
ротовое отверстие; 3— глот-
ка; 4 — ветви кишечника.
ОТРЯД БЕСКИШЕЧНЫЕ
РЕСНИЧНЫЕ ЧЕРВИ (ACOELA)
У берегов Бретани (Фран-
ция) начинается отлив. Отсту-
пает море. Обнажается морское
дно. И на обнаженном дне по-
являются зеленые пятна. Они
как живые. Постепенно пятна
становятся темнее и изменяют
свою форму. И действительно,
это скопления живых существ,
бесчисленных маленьких тур-
беллярий Convoluta roskoffen-
sis, относящихся к р о д у кон-
волют (Convoluta). Зеленая
окраска животных вызвана тем,
что в их паренхиме живут зе-
леные водоросли — зоохлорел-
лы, принадлежащие к жгути-
ковым простейшим. Как только
начнется прилив, конволюты
зароются в песок, уходя от
напора воды.
Этот ритм чередующегося за-
рывания в песок и появления
на его поверхности конволют
можно наблюдать и в аква-
риуме.
Как у всех бескишечных тур-
беллярий, у этого червя нет ки-
шечника. Вместо него в парен-
химе развита нежная плазма-
тическая ткань с многочислен-
ными ядрами в ней, но без
клеточных границ. Ткань, в
которой сливаются в общий
комплекс клеточные элементы,
лишенные клеточных границ,
называют синцитием
Мо-
лодые конволюты способны питаться; они
захватывают пищу при помощи ресничек,
которыми усажено ротовое отверстие. Пи-
щевые комочки попадают в синцитий, и
здесь происходит процесс внутриклеточ-
ного пищеварения, столь характерный для
всех турбеллярий, в том числе и имеющих
кишечник. Однако взрослые конволюты
питаются иначе.Они питаются веществами,
1 Синцитий — от греческих слов syn — вме-
сте и cytos — клетка.
340
ассимилируемыми водорослями, которые
живут в их теле. Водоросли (зоохлореллы)
проникают в организм конволют тогда,
когда они еще только развиваются из яйца.
Взрослые конволюты откладывают яйца
в кокон. Эти коконы выделяют вещества,
химически воздействующие на зоохло-
релл и привлекающие их. Можно сказать,
что яйца конволют буквально инфициро-
ваны (заражены) зоохлореллами. Но это
заражение — источник жизни конволют.
Из развивающихся яиц формируется мо-
лодая конволюта, и она оказывается но-
сительницей зоохлорелл, которые раз-
множаются в ее теле и как бы входят в его
состав, обеспечивая процессы ассимиляции
органических веществ и энергии солнеч-
ного света. Так зоохлореллы снабжают
конволюту питанием. С другой стороны,
зоохлореллы зависят от конволют и не
могут жить без них. И конволюты ведут
себя так, что существование зоохлорелл
обеспечено. Зоохлореллы зеленого цвета.
Они носители хлоропластов и
нуждаются в солнечном свете.
Конволюты в течение всего дня
неподвижно сидят на месте,
облитые яркими лучами солн-
ца — источником жизни и энер-
гии зоохлорелл, но вместе с
тем и источником питания са-
мих конволют. Конволюты по-
лезны для зоохлорелл, а эти
последние жизненно необходи-
мы для конволют. Перед нами
типичный пример симбио-
за — обоюдополезного сожи-
тельства свободных организмов.
Как уже говорилось, у бес-
кишечных турбеллярий нет ки-
шечника. На брюшной стороне
тела конволют расположена во-
ронка, ведущая в ротовое от-
верстие, а оно в свою очередь
ведет в синцитий. Кпереди от
ротового отверстия расположен
маленький пузырек, просвечи-
вающий сквозь покровы тела.
Это статоцист. Известный
специалист по турбелляриям
Б р е с л а у так разъясняет
функции статоциста: если кон-
волют, содержащихся в сосу-
Рис. 194. Планария
Convoluta convoluta:
1 — глазки; 2 — ротовое
отверстие.
де, оставить в покое, все они сооерутся
на поверхности воды. Однако даже лег-
кое сотрясение заставляет их опускать-
ся на дно. Этп реакции обусловлены
наличием статоциста. «Реакции,— пишет
Бреслау,— исчезают, если животное обез-
главлено или если соленая морская вода,
в которой находится червь, быстро заме-
няется пресной. В последнем случае про-
исходит повреждение тонкой структуры
статоциста». Итак, статоцист, т. е. орган
равновесия, определяет и упоминавшуюся
выше реакцию конволют на приливы и от-
ливы. На рисунке 194 показан другой вид
конволюты — Convoluta convoluta, в теле
которой видны зоохлореллы, а также
округлые яйца. В отличие от ранее назван-
ного вида, эта конволюта питается не
только за счет зоохлорелл, но и через
ротовое отверстие.
В заключение отметим, что бескишечные
турбеллярий, или ацелы, различно трак-
туются специалистами. В системе их обыч-
но ставят на первое место как
наиболее примитивных турбел-
лярий. Однако некоторые спе-
циалисты считают, что ацелы
скорее всего упрощенные фор-
мы. Известный специалист по
турбелляриям Отто Штейн-
бок A958) считает, что бески-
шечные турбеллярий образуют
примитивную ветвь, от которой
берут свое начало высшие тур-
беллярий. К ним мы и обра-
тимся.
ОТРЯД ПРЯМОКИШЕЧНЫЕ
ТУРБЕЛЛЯРИЙ
(RHABDOCOELA)
И этот отряд представлен мел-
кими животными. Однако, в от-
личие от ацел, у прямокишеч-
ных турбеллярий имеется ки-
шечник в виде прямой трубки,
за что отряд и получил свое на-
звание. Ротовое отверстие, рас-
положенное всегда на брюш-
ной стороне, ведет в мускули-
стую глотку. Ротовое отверс-
341
тие лежит то посередине брюшной сто-
роны тела, то ближе к переднему или зад-
нему концу его.
У прямокишечных турбеллярий имеют-
ся фоторецепторы, т. е. органы,
воспринимающие свет и обычно называе-
мые глазами. И в самом деле, эти органы
пигментированы и несут светопре-
ломляющую линзу — своего ро-
да хрусталик. Глаза могут, однако, и
отсутствовать. Около 400 видов этого от-
ряда живет в пресной воде, в море и ча-
стью в почве.
Чтобы читатель составил себе некоторое
представление о формах этого отряда,
мы ограничимся характеристикой неко-
торых видов.
Род Mesostomum — один из наиболее
богатых видами. У представителей этого
рода обычно плоское тело. Рот помещается
примерно в середине брюшной стороны.
Ротовая полость снабжена сильным со-
сущим аппаратом, с помощью которого эти
маленькие хищники хватают своих жертв
и высасывают их.
Один из красивейших видов этого
рода — Mesostoma ehrenbergii —достигает
в длину 1 см. Назван так в честь
знаменитого зоолога, опубликовавшего
в первой половине XIX в. (Эренберг,
1795—1876) ценные исследования, посвя-
щенные микроскопическим животным. Вид
этот можно обнаружить весной на за-
топленных половодьем лугах, а также в
прудах с илистым грунтом и в водоемах,
заросших тростником и ситником. Живот-
ное прозрачно, как стекло, и кажется, что
оно такое же хрупкое!
Наблюдая за животным, можно заме-
тить, что в воде оно совершает медлен-
ные и, видимо, свободные колебания, ос-
таваясь при этом подвешенным на тон-
кой и невидимой нити, образовавшейся из
его собственных слизистых выделений
(рис. 191).
Достаточно, однако, потревожить по-
кой животного, как тотчас турбеллярия
начинает вздрагивать и изгибаться с та-
ким же проворством, как какая-нибудь
пиявка.
Мезостома — хищник. В высшей сте-
пени интересно наблюдать, как она ата-
кует водяную блоху — маленького пресно-
водного рачка. Мезостома хватает его
примерно так же, как мы ловим рукой
муху. При этом мезостома стремительно
загибает задний конец тела до его сопри-
косновения с передним концом, в то время
как боковые края тела прижимаются друг
к другу. Рачок оказывается в живой ло-
вушке. Он беспомощно бьется в ней неко-
торое время, затем утихает, а мезостома
высасывает его содержимое. Маленькая
трагедия заканчивается, и хищник вытя-
гивается как бы в знак полного удовлет-
ворения.
Некоторые мезостомы (других видов)
способны действовать, как пауки, со-
здавая слизистую паутину, в которую по-
падают их жертвы.
В средней части Европы водится другой
интересный представитель того же р о-
д а — Mesostomum tetragonum. Эта мезо-
стома достигает 7—10 мм в длину и оби-
тает в маленьких пересыхающих или чи-
стых, но заросших водной раститель-
ностью прудах. Она коричневого цвета,
с небольшим красноватым отливом. По
бокам тела развиты лопасти, волнообраз-
но изгибающиеся от переднего к заднему
концу тела. С помощью этих лопастей
и ресничек, покрывающих кожу, мезосто-
ма плавает в воде.
Одной из интересных особенностей био-
логии прямокишечных турбеллярий надо
считать их способность жить в пересыхаю-
щих мелких водоемах. Видимо, они при-
способлены к перенесению сухости. Неко-
торые виды были констатированы в мел-
ких водоемах и даже маленьких лужах,
на недели высыхавших жарким летом.
Яйца одной из таких мезостом, извлечен-
ные из засохшего ила и положенные затем
в воду, через несколько дней стали раз-
виваться.
Отметим, что яйца мезостом бывают
двух типов — с толстой и тонкой скор-
лупой. Последняя характерна для лет-
них яиц. По-видимому, у мезостом на-
мечается закономерная смена форм яиц:
летние яйца развиваются в результате
самооплодотворения мезостомы, а зим-
ние — только в результате перекрестного
оплодотворения двух особей. Яйца мезо-
стом имеют обычно форму дисков с цент-
ральным углублением. Зимние яйца вы-
342
свобождаются из тела матери только после
ее смерти и могут переносить зимнюю сту-
жу и летнюю сушь. Это приспособление
имеет значение для выживания вида.
Из зимних яиц рождаются зимние особи,
встречающиеся ранней весной и летом.
Особи зимней генерации летом отклады-
вают яйца с тонкой скорлупкой, а в даль-
нейшем снова с плотной. Из летних яиц
развиваются летние животные, дающие
два сорта яиц — тонкоскорлуповые и
толстоскорлуповые; таким образом, за
зимними формами следуют летние, проду-
цирующие эти два сорта яиц. Когда
наступают холода, из тонкоскорлупо-
вых яиц рождаются осенние формы,
развивающие зимние яйца. Из них и
рождаются упомянутые выше зимние
формы.
Все, что было сказано выше, станет нам
понятнее, если мы хотя бы сжато позна-
комимся с морфологической организацией
мезостом. Как у всех турбеллярий, тело
их покрыто ресничками — органом движе-
ния. Как уже было сказано выше, кишеч-
ник их представлен прямой трубкой (рис.
195). Плотные отбросы питания выбрасы-
ваются через рот, жидкие — через спе-
циальную выделительную систему— про-
тонефридии, представленные ко-
нечными клетками с «ресничным пламе-
нем» и отходящими от них выделитель-
ными каналами (рис. 195).
Наиболее сложную организацию имеет
половая система. Все мезостомы, да и пря-
мокишечные турбеллярий вообще, как пра-
вило, гермафродиты, т. е. обое-
полые формы.
На рисунке 195 показана половая си-
стема Mesostoma ehrenbergii. He вникая
в подробности ее описания, укажем
лишь на сложность организации половой
системы и на совмещение в ней органов
мужской и женской природы. На рисунке
показан яичник, связанный с семеприем-
ником. По бокам этой системы лежат
семенники. От семенников идут два
семяпровода, по которым сперматозоиды
проникают в мужской совокупительный
орган, имеющий у мезостомы форму опро-
кинутой реторты. Через «горлышко» этой
«реторты» сперма попадает в так называе-
мый общий проток, из которого
она поступает в упомянутый семеприем-
ник, где семя сохраняется. Яйца также
проникают в семеприемник и затем, уже
оплодотворенные, перемещаются через об-
щий проток в половую клоаку. В дальней-
шем яйца, достигшие клоаки, покры-
ваются здесь желтком из желточников и
скорлупой. У мезостом происходит пере-
крестное оплодотворение, т. е. две особи
взаимно оплодотворяют друг друга. Такое
оплодотворение повышает жизнеспособ-
ность потомства и обеспечивает его боль-
шее, наследственно обусловленное много-
образие. Все это показывает, что половые
функции у прямокишечных турбелля-
рий достигают высокого уровня разви-
тия.
Следует отметить, что прямокишечным
турбелляриям присуще не только половое
размножение, но также и бесполое, до-
стигаемое посредством деления особей.
Вот как описывает Бреслау этот процесс
у Microstomum lineare: «У микростомы
деление начинается образованием попе-
речной перегородки, развивающейся меж-
ду кожно-мускульным мешком и мышеч-
ным слоем кишечника, и притом примерно
посередине тела. Последнее оказывается,
таким образом, разделенным на две части,
Рис. 195. Mesostoma ehrenbergii:
А — общий вид; Б — половая система. 1 — канала
экскреторной системы; 2 — кишечник; з — ротовое
отверстие; 4 — глотка; 5 — желточник; 6 — желточ-
ный проток; 7 — проток матки; 8 — матка; 9 — яи-
чник; ю — семяприемник; 11 — общий проток; 12 —
совокупительная сумка; 13 — половое отверстие; 14 —
совокупительный орган; IS — семяпроводы.
343
соответствующие двум будущим дочерним
особям. В дальнейшем ходе процесса де-
ления на месте возникшей перегородки
тело перешнуровывается, перетяжка уг-
лубляется, так что в конце концов обе
половины червя оказываются связанными
только не разделившимся еще кишечни-
ком. Наконец и этот последний разде-
ляется, и оба зооида (таково общее назва-
ние образующихся подобным способом
дочерних индивидуумов) оказываются сво-
бодными. Но прежде чем это произойдет,
кроме описанного первого деления, под-
готовляются и дальнейшие. Обе дочерние
особи, оставаясь еще связанными друг
с другом, сами начинают делиться надвое,
и из первоначально целостного живот-
ного получается цепочка из четырех зоо-
идов, у которых, в свою очередь, разви-
ваются поперечные перегородки, указы-
вающие на наступление третьей и дальней-
ших стадий деления». Впрочем, обычно
в естественных условиях наблюдаются
цепочки из 6—8 зооидов, так как более
длинные цепочки легко разрываются.
«Развитие поперечных перегородок, пре-
вращающих животное в цепочку зооидов,—
продолжает Бреслау,— само по себе
не обеспечивает, конечно, жизнеспособ-
ности дочерних индивидуумов. Процесс
деления необходимо дополняется новооб-
разованиями, возникающими в результате
регенерации и ведущими к тому, что отдель-
ные участки делящегося червя дости-
гают организации самостоятельных жи-
вотных». Бреслау нашел, что обе формы
размножения — половое и бесполое — че-
редуются: после неопределенного числа
поколений, возникших в течение весны
посредством деления, осенью происходит
половая дифференцировка; с наступлением
половозрелости бесполое размножение
прекращается.
Кроме упомянутых двух родов, к пря-
мокишечным турбелляриям относится
и ряд других. Среди них упомянем об
интересных представителях рода Daly-
ellia, для которых характерны симбиоти-
ческие отношения с зоохлореллами. Их
присутствием в теле видов данного рода
обусловлена зеленая окраска у Dalyellia
viridis, особенно типичная для почвы
луговых низин и для заболоченной воды.
ОТРЯД ТРЕХВЕТВИСТОКИШЕЧНЫЕ
ТУРБЕЛЛЯРИИ (TRICLADIDA)
Роды и виды этого отряда обладают,
несомненно, более высокой организацией.
Прежде всего формы этого отряда круп-
нее. Так, широко распространенная молоч-
ная планария, названная так за цвет
тела, достигает 15—26 мм в длину. Име-
ются и более крупные формы.
Расположенное на брюшной стороне те-
ла ротовое отверстие ведет у этих форм
в полость, в которой лежит выдвижная
глотка, в состоянии покоя далеко оття-
нутая назад. Когда животное захватывает
пищу, глотка наподобие хобота выдви-
гается наружу. Она снабжена не только
собственной мускулатурой, но и собствен-
ной иннервацией. Если глотку оторвать,
она продолжает двигаться и извиваться,
как червь. Из глотки пища поступает в ки-
шечник, состоящий из трех ветвей. Одна
из них направлена вперед, а две — назад,
справа и слева. Все они многократно де-
лятся, и последние боковые разветвления
заканчиваются слепо.
Форма тела трикладид обычно удли-
ненная, даже листовидная, иногда тело
расширенное в средней*части или круглое,
у некоторых форм лентовидное. По-
кровы и ткани прозрачны, но вместе с тем
для трикладид достаточно характерны раз-
личные окраски, то более светлые, то более
темные. Большинство форм имеет глазки,
расположенные на спинной стороне перед-
него конца тела. Такие глазки имеют прес-
новодные формы и многие морские пред-
ставители отряда. Однако у ряда форм
имеются многочисленные мелкие глазки,
распределенные по периферии переднего
конца тела параллельно внешнему кон-
туру его (рис. 191, 6). Иногда глаз-
ков нет. Глазки и органы химического
чувства, также расположенные на голов-
ном конце в виде боковых ресничных ямок,
иннервируются от нервных ядер «голов-
ного мозга», состоящего из скоплений
нервных клеток. «Мозг» — парное обра-
зование. От него назад — справа и слева—
идут нервные стволы, более мощные
на брюшной стороне тела и несколько
менее развитые на спинной. Из сказан-
ного видно, что у турбеллярий развива-
344
ется центральная нервная система, стро-
го ориентированная и отличающаяся пе-
ремещением наиболее ответственных орга-
нов чувств на передний конец тела, ориен-
тирующий животное в пространстве. Боль-
шинство трикладид размножается поло-
вым путем и обладает сложной половой
системой (рис. 195), включающей женские
и мужские генитальные органы.
Наука знает свыше 500 видов этого от-
ряда, заселяющих дно морей и пресных
вод и известных также в почве. Около
100 видов трикладид живет в пресной
воде. Остановимся лишь на немногих пред-
ставителях этой группы турбеллярий.
Среди трикладид имеются настоящие
гиганты. Байкальский представитель это-
го о т р я д а Polycotylus достигает в дли-
ну 30 см.
Значительно уступает по размерам бай-
кальскому поликотилусу другая трикла-
дида — молочная планария (Dendrocoelum
lacteum), которая достигает всего 15—26
мм в длину при поперечнике примерно
в 6 мм. Головной конец тупо срезан, зад-
ний округлен. Тотчас позади передне-
го края тела расположены черные глаза.
Под основанием переднего края развита
присасывательная бороздка. Как и дру-
гие виды планарий, молочная планария
прячется под камнями, среди листьев
тростника или на нижней стороне листьев
водяной лилии. Этот вид особенно хорош
для изучения строения кишечника. В па-
дающем свете он кажется почти черным,
в проходящем — несколько светлее.
Чаще темно-коричневая Euplanaria go-
nocephala имеет треугольный головной ко-
нец тела, по бокам которого расположены
несколько выступающие углы, известные
под названием ушей. Другая плана-
рия — Euplanaria polychroa —- характе-
ризуется индивидуальной изменчивостью
окраски: наблюдаются особи коричневого
цвета или темно-коричневые вплоть
до черного, иногда зеленые темных оттен-
ков, известны пестрые формы — черные
со светлыми пятнами. В связи с этим точ-
ное определение вида достигается лишь
изучением его анатомических признаков.
Голова горной планарий (Crenobia alpina)
украшена головными отростками, похо-
жими на щупальца. Эти виды известны
для Европы. От упомянутых видов рога-
тая многоглазка (Polycelis cornuta) и чер-
ная многоглазка (Polycelis nigra) отличают-
ся тем, что у них по переднему краю го-
ловы видны серии глазков.
Планарий — хищники. Они питаются
мелкими рачками, н&падают даже на ули-
ток, на личинок некоторых насекомых,
хотя не отказываются и от разлагающихся
остатков других животных. У планарий
сильно развито химическое чувство (обо-
няние). Почуяв добычу, планария направ-
ляется к ней, выдвигает свою глотку и
сильными сосательными движениями рвет
тело жертвы. Однако планарий выдержи-
вают длительную голодовку и при этом
«худеют», уменьшаясь в размерах, но не
теряя типичных для данного вида пропор-
ций тела.
Яйца планарий заключены в плотную
скорлупку: иногда они лежат в капсулах,
сидящих на тонкой ножке, или в коконахt
которые откладываются в защищенных
местах. В каждой такой кладке содер-
жится несколько десятков яйцевых и сот-
ни желточных клеток, обеспечивающих
питание развивающихся эмбрионов. Из
яиц вылупляются белесые молодые фор-
мы, еще лишенные пигментов.
Замечательной чертой биологии плана-
рий надо считать оригинальную реакцию
на наступление неблагоприятных усло-
вий — повышение температуры воды, не-
достаток кислорода и т. п. В подобных
случаях планарий способны распадаться
на куски, регенерирующие при наступле-
нии благоприятных условий в целых жи-
вотных с полной и типичной организа-
цией. Этот процесс известен под назва-
нием самокалечения, или авто-
томии. Многие формы даже в обычных
условиях способны делиться на части, что
можно рассматривать как особую форму
размножения.
В этой связи необходимо указать на по-
разительную способность планарий, да
и турбеллярий вообще, к регенерации—ре-
конструктивному восстановлению утра-
ченных частей тела. Опыты показали, что
у планарий даже 1/279 часть тела сохра-
няет способность к восстановлению пол-
ной организации со всеми присущими
планариям органами. Едва ли можно
345
сомневаться в том, что эта особенность
имеет важное защитное значение, обеспе-
чивающее сохранение жизни. Здесь уме-
стно познакомить читателя и с другими
защитными приспособлениями у тур-
беллярий. К ним принадлежат прежде
всего кожные железы, позволяющие пла-
ларии плотно прикрепляться к поверх-
ности, по которой она ползет. Какое же
защитное значение имеют особые кожные
образования турбеллярий, известные
под названием р а б д и т? Эти палоч-
кообразные образования лежат в кож-
ном эпителии и могут выбрасываться на-
ружу, расплываясь здесь в слизь, покры-
вающую поверхность тела турбеллярий.
Эта слизь также имеет защитное значение.
Так, рабдиты выбрасываются животным из
его кожи при поранениях; в этих случаях
расплывающиеся в слизь рабдиты закры-
вают ранки, а способность к регенерации
быстро обеспечивает заживление повреж-
денного участка тела. Интересно, что у не-
которых турбеллярий (Mesostoma ehren-
bergii) обнаружены стрекательные клетки
в коже, совершенно сходные с такими же
клетками кишечнополостных. Еще инте-
реснее, что эти клетки, по-видимому,
действительно принадлежат кишечнопо-
лостным, захваченным и съеденным тур-
белляриями. Известно, что планарии на-
падают на пресноводных гидр и охотно
поедают их.
Турбеллярий живут не только в прес-
ных водах. Известен целый ряд видов
трехветвистокишечных турбеллярий (три-
кладид), которые приспособлены к жизни
на почве, обычно под камнями, во влаж-
ных местах. Таков, например, Rhynchode-
mus terrestris. Этот червь имеет цилиндри-
ческое тело и достигает 16 мм в длину.
Спинная сторона его окрашена в темно-
серый цвет, тогда как брюшко окрашено
в белый цвет. На переднем конце тела вид-
ны два черных глаза. Другой вид этого
рода — Rh. bilineatus — попадается в
цветочных горшках. Если земля в них
недостаточно влажная на поверхности,
то животное уползает в глубину. Как
только земля увлажняется, ринходемус
вновь появляется на поверхности, ощу-
пывая свой путь головным концом тела.
Крупные экземпляры этого вида дости-
гают 12 мм в длину. Спина у этого вида
красно-коричневая с мраморным рисун-
ком на этом фоне, причем вдоль спины вид-
ны продольные линии, окрашенные в
красно-коричневый цвет. Примерно в се-
редине тела, на спинной стороне, распо-
ложено темное пятно. Оно соответствует
расположению глотки.
Описанные турбеллярий принадлежат
к группе почвенных трехветвистокишеч-
ных турбеллярий (Tricladida terricola).
Большая часть видов этой экологической
группы приурочена к тропическим и суб-
тропическим странам, где они обитают во
влажной почве. Путешествия замечатель-
ного биолога XIX в. Чарлза Дарвина
познакомили нас с богатейшей фауной
почвенных планарии влажных лесов Юж-
ной Америки. Среди экзотических плана-
рии встречаются настоящие гиганты мира
ресничных червей, достигающие 60 мм
в длину. Следует отметить, что у таких
крупных форм, да и более мелких, но все
же превосходящих скромных турбелля-
рий европейских стран, реснички теряют
значение органов движения. Чем круп-
нее турбеллярия, тем большее значение
в ее движениях приобретают мышцы кож-
но-мускульного мешка. Их волнообраз-
ные сокращения позволяют крупным тур-
белляриям скользить по поверхности суб-
страта примерно так же, как это делают
улитки.
Большинство упомянутых выше почвен-
ных форм размножается половым путем.
Известно около 400 видов почвенных три-
к л ад ид. На острове Цейлон живет инте-
ресный представитель этой группы из
рода бипалий (Bipalium). Этот реснич-
ный червь выделяет слизь из кожи, при-
чем солидная капля ее вытягивается под
тяжестью турбеллярий в нить, и реснич-
ный червь оказывается подвешенным во
влажном воздухе цейлонских лесов.
Один из видов бипалий — Bipalium ne-
wence — хороший объект для изучения
механизма движений с помощью ресни-
чек. Названная бипалия завезена чуть
ли не во все страны света и встречается
в оранжереях с экзотической флорой.
Здесь можно наблюдать, как бипалий
легко ползают по наклонным плоскостям
и даже по вертикальной поверхности.
346
Движения обеспечиваются, с
одной стороны, змеевидными
изгибаниями тела, а с другой—
волнообразными сокращениями
мышц брюшной стороны тела и
работой ресничек, покрываю-
щих его. При этом выясняется,
что животное не может исполь-
зовать для движения реснич-
ки, если брюшная сторона тела
(«подошва») не выделяет слизи.
Таким образом, работа ресни-
чек обеспечивается наличием
выделяемой из кожи слизи. По-
этому позади червя всегда ви-
ден слизистый след. Интересно,
что если животное намеревает-
ся спуститься вниз, то слизи-
стые выделения кожного по-
крова собираются, как уже ука-
зывалось, в комок: червь опус-
кается отвесно вниз, а комок
превращается в нить, удлиняю-
щуюся по мере этого движения.
Естественно, что для такого
«путешествия» слизи нужно мно-
го. И если турбеллярий застав-
ляли несколько раз проделать
такой путь, движение задер-
живалось из-за недостатка сли-
зи. Расход ее был слишком ве-
лик! Таково значение слизис-
тых выделений кожных покро-
вов турбеллярий в движениях
этих животных.
Бипалии, как и другие тур-
беллярий,— хищники, способ-
ные хватать и поедать малень-
ких дождевых червей.
ОТРЯД МНОГОВЕТВИСТО-
КИШЕЧНЫЕ ТУРБЕЛЛЯРИЙ
<POLYCLADIDA)
В этот отряд входит примерно
300 видов. Все они живут в мо-
ре. Это довольно крупные рес-
ничные черви, достигающие 16
см в длину. Они имеют широ-
кое листовидное тело, обычно
окрашенное в великолепные яр-
Рис. 196. Мюллеров-
ская личинка:
J — глотка, 2 — лопа-
сти
Рис. 197. Морская
планария Thysanozo-
on brochii.
кие цвета. Наиболее типичный
признак организации этих жи-
вотных — оригинальное строе-
ние кишечника. Пищеваритель-
ная система начинается мощ-
ной глоткой, которая перехо-
дит в кишечник, имеющий фор-
му трубки с отходящими от нее
по радиусам разветвленными
стволами. Разумеется, и у этих
форм ануса нет, однако у
некоторых форм он присут-
ствует. Личинка поликлад по-
крыта ресничками и способна
плавать в воде. Это так назы-
ваемая мюллеровская
личинка, снабженная мус-
кулистой глоткой и восемью
лопастями. В дальнейшем она
превращается во взрослую по-
ликладу. Таким образом, для
поликлад характерно развитие
с превращением. К этой группе
принадлежит ряд семейств,
представленных многими рода-
ми и видами. У одних поли-
клад развита брюшная присос-
ка, у других ее нет. К фор-
мам с брюшной присоской при-
надлежит, например, живущая
в Северном и Средиземном мо-
рях Planocera folium, несущая
два затылочных щупальца. К
этой же группе поликлад отно-
сится в европейских морях и
известная для фауны СССР Lep-
toplana tremellaris, способная
плавать. Во время отлива она
прячется в песке или под кам-
нями, вновь появляясь с на-
ступлением прилива. Одним из
наиболее интересных предста-
вителей поликлад с брюшной
присоской можно считать Thy-
sanozoon brochii, или мохнатую
планарию. Русское название об-
условлено тем, что на спин-
ной стороне этого животного
(рис. 197) вся поверхность ко-
жи покрыта своеобразными кож-
ными сосочками. На головном
конце тела развиты две похо-
жие на уши складки, имеющие,
347
по-видимому, значение органов осязания.
Брюшная сторона тела белая. Интересно,
что в спинные сосочки проникают разветв-
ления кишечника.
Некоторое представление о многообра-
зии и красоте окрасок поликлад читатель
получит, рассмотрев цветную таблицу 17.
* * *
Фауна нашей страны богата представи-
телями класса турбеллярий, и при-
том всех четырех охарактеризованных вы-
ше отрядов. Особенно поражает фауна
турбеллярий озера Байкал, которая вклю-
чает 13 родов и 90 видов, в огромном боль-
шинстве относящихся к отряду трех-
ветвистокишечных (Tricladida). В этой
группе очень резко выражен эндемизм *
фауны Байкала. Все 13 родов и 90 видов —
эндемики Байкала, т. е. нигде, кроме Бай-
кала, не встречаются. Столь же резкий
эндемизм свойствен байкальским губкам,
олигохетам, брюхоногим моллюскам, неко-
торым группам рыб и ряду других обитате-
лей Байкала. Эндемизм байкальской фау-
ны —свидетельство большой ее древности и
своеобразия этого самого глубокого озе-
ра в мире, его температурного режима
и химических свойств воды.
1 Эндемизм — от греческого endemos, что
значит местный, постоянно живу-
щий.
Название класса — трематоды (Тге-
matoda) — заимствовано из греческого
языка. Оно означает «имеющие присоски».
Название подчеркивает один из характер-
ных признаков представителей данного
класса червей, а именно наличие двух при-
сосок: ротовой, которая находится на пе-
реднем конце тела, и брюшной, располо-
женной обычно в средней части тела на
брюшной стороне. По-русски этот класс
животных называют сосальщиками.
ПРОИСХОЖДЕНИЕ ТУРБЕЛЛЯРИЙ
У турбеллярий сохраняются некоторые
элементы сходства с гребневиками. В ча-
стности, у бескишечных турбеллярий на
одной из стадий формообразования моло-
дого животного обнаруживаются четыре
зачатка «мозга». Четыре корешка брюш-
ных и спинных нервов наблюдаются
у взрослых форм. Таким образом, у бески-
шечных турбеллярий обнаруживаются
элементы лучевой симметрии в организа-
ции нервной системы. Известный совет-
ский специалист по турбелляриям и круп-
ный зоолог В. Н. Беклемишев, ана-
лизируя организацию бескишечных тур-
беллярий, приходит к выводу, что «сим-
метрия турбеллярий легко выводится из
симметрии гребневиков». В. Н. Бекле-
мишев, следуя в этом отношении за
Графом, приходит к выводу, что и
гребневики и турбеллярий, и в част-
ности бескишечные ацелы, произошли от
планулоподобных предков, т. е. форм,
сходных с личинкой (планулой) кишеч-
нополостных животных (см. стр. 237).
Планула кишечнополостных — двуслой-
ный организм, тело которого состоит из
наружной эктодермы и внутренней энто-
дермы, образующей стенку примитивного
кишечника. Тело планулы покрыто рес-
ничками. Примерно так рисуется Графу
и предок турбеллярий, отличающийся от
планулоподобной формы развитием поло-
вых органов.
Все представители класса трематод ве-
дут паразитический образ жизни. Во взро-
слой стадии они поселяются в теле позво-
ночных животных и человека, в личиноч-
ной — преимущественно в беспозвоноч-
ных, главным образом в моллюсках.
Трематоды паразитируют у всех позво-
ночных животных — рыб, амфибий, реп-
тилий, птиц и млекопитающих — и при-
способились к обитанию почти во всех ор-
ганах и тканях их тела. Каждый вид чер-
КЛАСС ТРЕМАТОДЫ (TREMATODA)
348
вя паразитирует обычно у одного или не-
большого числа видов животных и доволь-
но строго придерживается определенной
локализации (места обитания) в орга-
низме своего хозяина.
Большая часть трематод обитает в пи-
щеварительном тракте животных и в свя-
занных с ним железах — печени и подже-
лудочной железе. Некоторые группы при-
способились к жизни в органах дыхания
(в тканях легких, в трахее, в воздушных
мешках), в органах выделения (в почках
и мочеточниках), в кровеносных сосу-
дах. Имеются формы, которые поселяются
в глазах, в носовой полости, в подкожной
клетчатке. Некоторые виды могут прони-
кать даже в мозг. Ни одна группа парази-
тических червей не расселилась так широ-
ко по организму животного, как тремато-
ды, и, пожалуй, ни у одной другой группы
паразитических червей нет таких много-
численных и сложных приспособлений
к существованию в столь разнообразных
условиях. Приспособления эти проявля-
ются и в строении тела паразитов, и в их
своеобразных циклах развития.
Важнейшим приспособлением к парази-
тическому образу жизни у трематод,
как и у других гельминтов, является их
колоссальная плодовитость. Мало того,
что эти черви производят огромное коли-
чество яиц, они обладают еще способно-
стью размножаться в личиночной стадии,
при этом каждая личинка, развивающая-
ся из яйца, превращается во множество
новых зародышей. В результате все по-
томство взрослого червя исчисляется аст-
рономическими числами. Огромная пло-
довитость трематод обеспечивает им боль-
шие возможности расселения по своим
хозяевам.
С паразитическим образом жизни трема-
тод связано и то, что все они, за очень не-
многими исключениями, гермафродиты,
т. е. обоеполые организмы.
Среди трематод имеется немало весьма
опасных паразитов человека и хозяйст-
венно полезных животных. Поселяясь
в организме этих хозяев, они могут вы-
зывать серьезные заболевания, резко ска-
зывающиеся на здоровье человека и со-
стоянии животных и приводящие в неко-
торых случаях даже к смерти. Часто та-
кими болезнями бывают поражены сель-
скохозяйственные животные, особенно ов-
цы, крупный рогатый скот и птицы. Эти
болезни приносят огромный ущерб живот-
новодству.
Общее название болезней, вызываемых
трематодами,— трематодозы. Каж-
дый трематодоз, кроме того, имеет свое
название, оно производится от родового
названия (на латинском языке) трематоды,
которая является возбудителем данного
заболевания. Подробнее об этом мы рас-
скажем ниже, когда будем говорить об
отдельных видах трематод. Здесь же отме-
тим, что в медицине и ветеринарии изу-
чению трематод, как и других гельминтов,
и вызываемых ими заболеваний придается
очень большое значение.
Строение. Тело трематод обычно плос-
кое, листовидное. Окраска тела невзрач-
ная, грязно-белая, сероватая или розова-
тая, часто с темными пятнами.
Типичная листовидная форма тела мо-
жет меняться в зависимости от места оби-
тания паразитов. У трематод, живущих
в кровеносных сосудах и в протоках же-
лез, тело имеет удлиненную и более
округлую в поперечном сечении форму;
у трематод, обитающих в тканях органов
и заключенных в капсулы, тело почти ша-
рообразной формы. Листовидная форма
тела свойственна паразитам, обитающим
главным образом в различных отделах
пищеварительного тракта. Эти трематоды
плотно прилегают широкой брюшной по-
верхностью тела к внутренним стенкам
кишечника или других отделов тракта
и таким образом легко удерживаются.
Подавляющее большинство трематод —
мелкие паразиты. Длина тела их измеряется
в пределах от миллиметра до сантиметра
и редко более. Однако, как и у каждой
группы животных, здесь есть свои кар-
лики и свои великаны. Самые мелкие тре-
матоды измеряются несколькими десяты-
ми долей миллиметра, например некото-
рые представители семейства Microphalli-
dae. Самая длинная трематода (Nematobot-
hrium filarina из семейства Didymosoomi-
dae) паразитирует в ротовой полости аку-
лы (Scyllium aquila) в крупных цистах,
имеющих размер кулака. Длина этой
трематоды достигает 1,5 м. Большие тре-
349
матоды обитают в печени крупного рога-
того скота, овец и оленей в Северной Аме-
рике (Fasciolopsis magna —- до 10 см дли-
ной при ширине 2—3 см) и в печени ки-
тов (Lecithodesmus goliath — до 9 см дли-
ной и I см шириной). Все большие трема-
тоды паразитируют у самых крупных
животных, хотя строгой зависимости меж-
ду размерами трематод и размерами хо-
зяев нет.
Выше мы указывали, что в зависимости
от места обитания в организме хозяина
трематоды имеют различную форму тела,
что является приспособлением к сущест-
вованию в определенных условиях. Все
другие особенности внешнего строения
трематод носят также приспособительный
характер. Они служат главным образом
для фиксации паразита в теле животного,
для перемещений по его органам или для
проникновения в ткани. Рассмотрим важ-
нейшие из них.
Как отмечалось, характерным для тре-
матод является наличие двух присосок.
С их помощью паразит прикрепляется во
внутренних органах хозяина и передви-
гается. Присоски особенно сильно развиты
у тех форм, которые обитают в просвете
пищеварительного тракта, здесь как раз
и необходима прочная фиксация паразита,
иначе он может быть выброшен перисталь-
тическими движениями кишечника и то-
ком пищевых масс. И наоборот, у парази-
тов, обитающих в замкнутых полостях
органов, в воздушных мешках птиц, в тка-
нях и капсулах, присоски очень слабо раз-
виты, а иногда и полностью редуциро-
ваны. Фиксации паразита в теле хозяина
способствуют также острые шипики на
теле, которые имеются у многих видов
трематод и особенно развиты у кишечных
форм.
Рассмотрим кратко внутреннее строение
трематод. Тело трематод покрыто плотной
кутикулой. Под ней располагается
субкутикулярный слой, слагающийся из
редко разбросанных вытянутых клеток
и мышц. Кутикула с субкутикулярным
слоем и мышцами образует вместе так
называемый к о ж н о-м ускульный
мешок. В нем и помещаются внутрен-
ние органы, промежутки между которыми
заполнены паренхимой — особой тканью,
представляющей собой рыхлую массу сое-
динительнотканых клеток.
У трематод имеются следующие сис-
темы внутренних органов: нерв-
ная, пищеварительная, выделительная и по-
ловая. В связи с паразитическим образом
жизни внутренние органы трематод по-
строены сравнительно просто, за исклю-
чением половой системы, которая полу-
чила у них очень сильное развитие.
Нервная система трематод со-
стоит из нервных узлов, расположенных
в передней части тела возле глотки, и от-
ходящих от них нервных стволов, направ-
ляющихся в различные участки тела.
Органы пищеварения на-
чинаются ротовым отверстием, которое
открывается обычно на переднем конце
тела, на дне присоски (ротовой). У неко-
торых трематод ротовое отверстие откры-
вается на брюшной стороне тела. За ро-
товым отверстием следует глотка — мы-
шечный орган округлой или овальной
формы, за ней удлиненный канал пище-
вода, который чаще всего подразделяется
на два ствола кишечника. Стволы направ-
ляются назад и заканчиваются слепо. Зад-
непроходного отверстия у большинства
трематод нет, и пищевые остатки у них
удаляются через рот.
Органы выделения представ-
лены обычно двумя каналами, тянущимися
вдоль тела. Эти каналы сливаются в не-
парный выделительный пузырь, который
открывается на заднем конце выделитель-
ной порой.
В каждый канал впадает множество
мелких ветвящихся боковых канальцев,
несущих на концах так называемые пла-
менные клетки, которые, собст-
венно, и выполняют функцию выделения.
Половая система в связи
с сильным ее развитием занимает большую
часть тела. Как указывалось, трематоды,
за редкими исключениями,— обоеполые
организмы. Мужской половой аппарат
состоит чаще всего из двух семенников.
От каждого отходят семявыносящие про-
токи, которые сливаются в общий семя-
провод. Конечная часть семяпровода обра-
зует циррус, или совокупительный орган,
заключенный обычно в особый мышечный
мешок — половую бурсу. В бурсу за-
350
ключена также и расширенная часть се-
мяпровода — внутренний семенной пузы-
рек и окружающие его здесь многочис-
ленные простатические железы. Другое
расширение семяпровода — наружный се-
менной пузырек — находится вне бурсы.
Мужское половое отверстие открывается
наружу на брюшной поверхности пара-
зита, обычно впереди брюшной при-
соски.
Женский половой аппарат значительно
сложнее. Он состоит из яичника, яйцево-
да, оотипа, матки, желточников, скорлу-
повой железы и семеприемника. В яич-
нике образуются яйцеклетки, которые по
яйцеводу поступают в оотип. Сюда же по
протоку скорлуповой железы поступает
вещество, участвующее в образовании
скорлупы яйца, а по протокам желточ-
ника — питательный материал для яиц.
Часто с оотипом соединяется пузырько-
видный семеприемник (он имеется не у
всех трематод), в нем находится сперма,
полученная от другой особи. В оотипе
происходит оплодотворение яйцеклеток и
формирование яиц, которые затем посту-
пают в матку. Трубчатая, более или ме-
нее длинная матка заканчивается половым
отверстием, расположенным чаще всего
впереди брюшной присоски. Яйца трема-
тод обычно очень мелкие, и увидеть их
можно только с помощью микроскопа.
Скорлупа яйца состоит из нескольких
слоев, она может быть бесцветная или
окрашенная. На одном конце яйца имеется
крышечка.
Развитие. Для развития трематод ха-
рактерны две особенности: смена хозяев
и чередование поколений. Эти особенности
несвойственны только небольшой группе
трематод, отличающихся более простой
организацией.
Явление смены хозяев заключается
в том, что паразит в период своего биоло-
гического цикла живет и развивается в ор-
ганизме не у одного животного, а после-
довательно у двух, иные — у трех или
даже у четырех животных, различных по
своему систематическому положению и по
экологии. Животных, у которых парази-
тирует взрослая форма трематоды, назы-
вают окончательными хозяевами; обычно
это позвоночные — рыбы, земноводные,
пресмыкающиеся, птицы и млекопитаю-
щие. Животных, в теле которых разви-
ваются личиночные стадии паразита, на-
зывают промежуточными хозяевами. У не-
которых видов трематод промежуточных
хозяев может быть два, в таком случае
их называют: первый промежуточный хо-
зяин и второй промежуточный хозяин.
Есть и другие наименования: первый на-
зывается промежуточным, второй — до-
полнительным. Промежуточными хозяе-
вами для трематод всегда являются мол-
люски, дополнительными могут быть мол-
люски, щетинконогие черви, ракообраз-
ные, насекомые, рыбы, земноводные и ре-
же другие животные. Примеров, когда
развитие паразита идет со сменой четырех
хозяев, немного. В этих циклах, помимо
окончательного и промежуточного, или
дополнительного, хозяев, участвуют так
называемые резервуарные хозяева. Они
вклиниваются между промежуточным, или
дополнительным, хозяином и окончатель-
ным. Резервуарный хозяин, в отличие
от всех других, не является обязательным
звеном в жизненном цикле паразита, раз-
витие может быть завершено и без его уча-
стия.
Другая особенность развития трема-
тод — чередование поколений — состоит
в том, что у этих организмов размно-
жаются не только взрослые формы, но
и личинки. Взрослые формы откладывают
оплодотворенные яйца, в них развиваются
личинки. Эти личинки дают начало дру-
гому поколению личинок — партеногене-
тическому (образовавшемуся путем девст-
венного размножения). Последние выра-
стают во взрослые формы.
Размножение, наблюдаемое у личинок
трематод, во много раз увеличивает пло-
довитость паразита, что повышает воз-
можность заражения хозяев.
В процессе развития трематоды прохо-
дят ряд последовательных стадий. Откла-
дываемые трематодой зрелые яйца вместе
с выделениями животного, у которого
паразитирует червь (с экскрементами, мок-
ротой, мочой), попадают во внешнюю сре-
ду, т. е. в воду, на землю. Здесь в яйце
развивается зародыш — мирацидий.
Тело его покрыто ресничками, с помощью
которых мирацидий легко передвигается.
351
У одних видов трематод мирацидий выхо-
дит из оболочки яйца, у других остается
в яйце. Дальнейшее развитие зародыша
может происходить только в организме
промежуточного хозяина — моллюска.
Если зародыш не попадает в моллюска,
то он погибает. В теле промежуточного
хозяина мирацидий превращается в с п о-
роцисту — бесформенную, мешковид-
ную, неподвижную личинку. В спороци-
сте из особых клеток, находящихся на
внутренней поверхности стенок ее тела,
образуются личинки следующей стадии —
р е д и и. Они имеют удлиненную фор-
му, подвижны, лежат в полости тела спо-
роцисты. В редиях в свою очередь обра-
зуются другие личинки — так называе-
мые ц е р к а р и и. Церкарии характери-
зуются наличием хвоста и рядом особен-
ностей, свойственных уже половозрелым
трематодам: у них имеются присоски, ор-
ганы пищеварения, выделительная систе-
ма и зачатки половой. У большинства
церкариев имеются особые железистые
клетки, называемые железами про-
никновения. Они выделяют веще-
ство, способное растворять животные тка-
ни. На переднем конце некоторых церка-
риев помещается твердый и острый стилет.
Эти приспособления облегчают внедрение
паразита в тело хозяина. Церкарии поки-
дают моллюска и плавают в воде до встре-
чи с окончательным хозяином, если пара-
зит развивается со сменой двух хозяев,
или с дополнительным, если для завер-
шения цикла необходима смена трех
хозяев.
Не у всех видов трематод развитие в те-
ле промежуточного хозяина происходит
так, как описано. У отдельных видов
паразитов из мирацидия развивается не
спороциста, а сразу редия. Иногда в спо-
роцистах развиваются не редии, а цер-
карии. Размножение может протекать
и таким образом, что в спороцистах раз-
виваются не сразу редии, а сначала дочер-
ние спороцисты, а уже в них редии.
В редиях также могут развиваться не
церкарии, а дочерние редии. Церкарии
в этом случае развиваются в дочерних ре-
диях. Указанные изменения в ходе раз-
вития обусловлены как особенностями
самих паразитов, так и внешними усло-
виями, главным образом температурой.
Выйдя из тела промежуточного хозяи-
на — моллюска, церкарии недолго оста-
Таблица 17. Различные виды турбеллярий:
1 — Lungia aurantiaca;
2 — Haploplana ornata;
3 — Bipalium strubelli;
4 — Rhynchodemus rubrocinctus;
5 — Pseudoceros maximus;
6 — Bipalium simrothi.
ется во внешней среде. У одних видов тре-
матод он оседает на водных растениях,
теряет хвост и покрывается оболочкой
(инцистируется), превращаясь при этом
в метацеркария. С пищей или
водой метацеркарий попадает в организм
окончательного хозяина. Иногда церка-
рий проникает в тело окончательного хо-
зяина активно, через кожу. У трематод,
которым свойственна смена трех хозяев,
церкарий должен попасть к дополнитель-
ному хозяину, здесь он также превраща-
ется в метацеркария. К окончательному
хозяину в этом случае личинка паразита
обычно попадает вместе с дополнительным
хозяином, которого он поедает. В орга-
низме окончательного хозяина личинка
пробирается в тот орган, где паразити-
руют трематоды данного вида, и разви-
вается там в половозрелую форму. Как
видно из сказанного, циклы развития тре-
матод сложны, для завершения цикла
необходимо благоприятное сочетание це-
лого ряда моментов. Прежде всего нужно,
чтобы яйцо паразита попало в воду или
влажное место, иначе оно погибнет. Необ-
ходимо, чтобы мирацидий встретился
с моллюском — промежуточным хозяи-
ном (причем каждому виду трематод соот-
Таблица 18. Различные виды немертин:
1 — Nectonemertes mirabilis;
2 — Eunemertes echinoderma
marion;
3 — Tubulanus;
4 — Malacobdella grossa;
5 — Lineus geniculates;
6 — Tetrastemma rusticum.
ветствуют только определенные виды мол-
люсков). Выходящие из тела моллюска
церкарий должны попасть в промежуточ-
ного хозяина или в окончательного. Срок
жизни церкариев невелик, и поэтому ре-
шить эту задачу не так просто. Понятно,
что при столь «трудной жизни» немногие
личинки становятся взрослыми тремато-
дами. Большая часть их гибнет на том
или ином этапе жизненного пути. Однако
в природе, если нет специального вмеша-
тельства человека, не происходит замет-
ного уменьшения численности паразита.
Больше того, не всегда применение разра-
ботанных мер борьбы с ними приводит
к достаточно эффективным результатам.
Объясняется такое противоречие просто:
паразит, как указывалось, обладает огром-
ной плодовитостью, усиленной во много
раз размножением его в личиночной ста-
дии. В результате в природе рассеивается
колоссальное количество зародышей и,
как бы много их ни гибло, какая-то часть
достигает взрослой стадии и снова дает
начало многочисленному потомству.
Класс трематод делится на три
подкласса, представители которых четко
различаются по строению, экологии и
биологии.
W 23 Жизнь животных, т. 1
ПОДКЛАСС АСПИДОГАСТРАТА (ASPIDOGASTRATA)
Трематоды этого подкласса лишены
ротовой и брюшной присосок. На брюш-
ной поверхности их тела ткань разраста-
ется в виде диска со складками и ямками
на поверхности. Этим диском паразит
присасывается к тканям хозяина. Указан-
ная особенность отражена в названии
подкласса, которое на русский язык
переводится как складчатобрюхие. Во
взрослой стадии аспидогастрати парази-
тируют у беспозвоночных — в двустворча-
тых моллюсках. Известно несколько ви-
дов, паразитирующих в кишечнике рыб.
Развитие протекает без смены хозяев
и без чередования поколений.
В подклассе только один отряд и одно
семейство с небольшим количеством видов.
Типичным представителем является аспи-
догастер конхиколя.
Аспидогастер конхиколя (Aspidogaster
conchicola) впервые обнаружен и описан
в 1827 г. известным русским ученым
К. М. Б э р о м. Трематода обитает в око-
лосердечной сумке пластинчатожаберных
моллюсков (в беззубках и перловицах).
Распространена в умеренных широтах Ев-
ропы, Азии и Северной Америки.
Тело трематоды достигает 3 мм длины
при ширине до 1 мм. Оно имеет форму на-
ковальни. В передней части тела поме-
щаются внутренние органы, а в задней,
на брюшной стороне, расположен огром-
ный присасывательный диск — наиболее
характерный признак этой трематоды.
Развитие аспидогастер а протекает в ор-
ганизме одного хозяина. Паразит откла-
дывает очень крупные яйца @,128—
0,134x0,048—0,050 мм), в которых на-
ходится подвижная личинка. Покинув
Как показывает название п о д к л а с-
с а (гастеростомата в переводе на
русский значит брюхоротые), ротовое от-
верстие у этих трематод открывается на
брюшной поверхности. Кишечник меш-
ковидный, не раздваивается. Взрослые
Рис. 198. Аспидогастер цефал (Висе-
(Aspidogaster conchico- phalus polymor-
la). phus).
оболочки яйца, личинка здесь же, в орга-
низме моллюска, не испытывая каких-либо
сложных превращений, начинает разви-
ваться во взрослую форму. Некоторые ли-
чинки покидают моллюска и током воды
переносятся к другому хозяину. В нем
они заканчивают свое развитие. Молодые
формы, которые не покинули своего пер-
воначального хозяина, могут развиваться
в нем до половозрелой стадии.
трематоды паразитируют в пищеваритель-
ном тракте холоднокровных животных —
у рыб и изредка у амфибий. Развитие
протекает со сменой трех хозяев — про-
межуточного, дополнительного и оконча-
тельного — и с чередованием поколений.
ПОДКЛАСС ГАСТЕРОСТОМАТА (GASTEROSTOMATA)
354
В подклассе один отряд и несколько
семейств, видовой состав сравнительно
небольшой.
Наиболее распространенный и харак-
терный представитель подкласса — буце-
фал изменчивый.
Буцефал изменчивый (Bucephalus poly-
morphs) — паразит кишечника многих
хищных пресноводных рыб. Это сравни-
тельно мелкая трематода (длина тела
до 2,2 мм, ширина до 0,4 мм) с сильно раз-
витым фиксаторным аппаратом на перед-
нем конце тела. Аппарат представлен
мощной присоской, окруженной семью
мышечными пальцевидными отростками,
которые могут вытягиваться и втягивать-
ся. Ротовое отверстие находится на брюш-
ной стороне в средней части тела. Кишеч-
ник мешковидный. Половые органы рас-
положены в задней половине тела. Поло-
вые отверстия — близ заднего конца.
Развитие буцефала протекает со сме-
ной трех хозяев и чередованием поколе-
ний. Яйца паразита, выделенные из орга-
низма зараженных хищных рыб (судак,
налим, щука и др.)» должны быть загло-
чены промежуточным хозяином. Эту роль
выполняют пластинчатожаберные моллю-
ски (беззубка и перловица). Из моллюска
выходят церкарии. Они попадают к кар-
повым рыбам и превращаются в метацер-
кариев, которые инцистируются в жабрах
или под кожей. Когда рыбы (дополни-
тельные хозяева), зараженные личинками
буцефала, будут проглочены другими,
хищными рыбами (окончательными хозяе-
вами), личинка в их кишечнике вырастет
во взрослую форму паразита.
Рис. 200. Фасциола печеночная
(Fasciola hepatica).
ПОДКЛАСС ПРОЗОСТОМАТА (PROSOSTOMATA)
В переводе на русский язык название
этого подкласса означает передне-
ротые. В названии отражена характерная
особенность строения его представителей:
ротовое отверстие трематод открывается
на переднем конце тела на дне ротовой
присоски. Ветвящийся кишечник состоит
из двух стволов. Окончательными хозяе-
вами трематод могут быть все позвоночные
животные. Паразиты локализуются почти
во всех их органах. Для развития харак-
терно чередование поколений и разнооб-
разие жизненных циклов — возможна сме-
на двух, трех и четырех хозяев.
Подкласс охватывает подавляющее
большинство известных видов трематод,
он представлен большим количеством
отрядов, семейств и родов. Среди пред ста-
23*
355
вителеи подкласса много опас-
ных паразитов животных и че-
ловека.
Расскажем о нескольких се-
кзйствах данного подкласса и
отдельных его видах, которые
имеют важное значение в ме-
дицине и ветеринарии и ко-
торые представляют большой
интерес в биологическом отно-
шении.
Семейство Fasciolidae.
Фасциолиды — паразиты печени,
реже кишечника, домашних и
диких травоядных животных,
а также человека. Они широ-
ко распространены во всех
странах земного шара и при-
чиняют огромный ущерб жи-
вотноводству.
Самым известным видом яв-
ляется фасциола печеночная (Fasciola he-
patica), несколько реже встречается фас-
циола гигантская (Fasciola gigantica). В на-
шей стране последняя распространена пре-
имущественно в южных зонах (Закав-
казье, среднеазиатские республики, юг
Казахстана, Нижнее Поволжье и др.),
тогда как фасциола печеночная встре-
чается повсеместно. Оба вида легко
отличаются друг от друга по внешнему
виду: фасциола печеночная меньше по
размерам (длина тела до 30 мм, ширина
8—13 мм), имеет листовидную форму
тела, постепенно суживающуюся к зад-
нему концу; фасциола гигантская боль-
ших размеров (длина тела до 75 мм, ши-
рина до 12 мм), с удлиненным телом, бо-
ковые края которого почти параллельны.
Близки к названным видам и также
очень патогенны (вредоносны) упоминав-
шаяся уже трематода фасциолоидес (Fas-
ciolopsis magna) — паразит печени круп-
ного рогатого скота, овец и оленей, рас-
пространенный в Северной Америке, и
фасциола пейс (FascioJopsis buski) — пара-
зит кишечника свинец (зарегистрирован
и у человека), распространенный в стра-
нах Юго-Восточной Азии.
Остановимся подробнее на фасциолр
печеночной.
Фасциола печеночная (Fasciola hepatica)
обитает в половозрелой стадии в желчных
Рис. 201. Фасцнолы
печеночная и гигант-
ская в натуральную
величину.
кодах печени у овец, коз, круп-
ного рогатого скота, буйволов,
верблюдов, свиней, лошадей и
у некоторых грызунов (зайцев,
кроликов, морских свинок, нут-
рий и др.). Известны случаи
паразитирования фасциолы у
человека. Особенно сильно по-
ражены этой трематодой овцы
и крупный рогатый скот.
Заболевание, вызываемое рас-
сматриваемым паразитом, носит
название фасциолез.
Р. С. Ш у л ь ц A959) сле-
дующим образом описывает со-
стояние животных, страдающих
фасциолезом: «Вспышки забо-
левания большей частью быва-
ют зимой, поздней осенью или
весной в зависимости от клима-
та, условий местности и хозяй-
ственной обстановки. Животные начинают
заметно худеть и слабеть. Шерсть стано-
вится сухой, у овец она легко выпадает,
особенно на боковых стенках груди. По-
немногу перечисленные признаки болез-
ни могут охватить большую часть пого-
ловья. При дальнейшем развитии болезни
появляется отечность в подчелюстной об-
ласти, на груди и на животе. Появляется
расстройство пищеварения, наблюдаются
поносы, иногда запоры. Область печени
болезненна. Аппетит пониженный. При
возрастающем истощении и водянке мно-
гие погибают, если своевременно не при-
нять меры».
В связи с огромным ущербом, прино-
симым фасциолезом животноводству, осо-
бенно овцеводству, много внимания уде-
ляется разработке и осуществлению мер
борьбы с этой болезнью. Мероприятия
по борьбе с фасциолезом и предохране-
ние животных от заражения паразитом
основаны на данных детального изучения
биологического цикла возбудителя, т. е.
фасциолы. Поэтому, прежде чем говорить
о том, как борются ветеринарные и зоотех-
нические специалисты с фасциолезом в жи-
вотноводческих хозяйствах, рассмотрим,
как протекает развитие паразита.
Зрелые фасциолы откладывают яйца
в желчные ходы печени. С током желчи
яйца попадают в кишечник и выбрасы-
356
ваются затем с экскрементами во внеш-
нюю среду. Яйцо будет развиваться толь-
ко в том случае, если оно попадет в прес-
ную воду: в пруд, лужу или небольшой
ручеек. Через 8—25 дней, в зависимости
от температуры внешней среды, в яйце
формируется подвижный зародыш — м и-
р а ц и д и й. Он покидает затем скорлу-
пу яйца и плавает в воде, отыскивая про-
межуточного хозяина —¦ моллюска, пру-
довика малого (Galba truncatulla). Уста-
новлено, что и некоторые другие прудо-
вики могут служить для фасцйолы про-
межуточными хозяевами (прудовик тин-
ный, овальный, странствующий и др.),
однако основным является прудовик ма-
лый. Встретив моллюска, мирацидий внед-
ряется в его тело и проникает
в печень прудовика. Здесь он
сбрасывает свой ресничный по-
кров и превращается в споро-
цисту. В спороцисте развивает-
ся новое поколение личинок —
редий, в которых в свою оче-
редь могут развиваться дочер-
ние редии. В редиях формиру-
ются церкарий, они покидают
тело моллюска и выходят в во-
ду. Поплавав в воде, церкарий
прикрепляются к подводному
растению или опускаются на
дно водоема и, отбросив хвост,
обволакиваются плотной защит-
ной оболочкой, предохраняю-
щей их от быстрого высыхания
и гибели. Личинка этой стадии
носит общее название мета-
церкарий, однако у фас-
цйолы и некоторых других
трематод она имеет еще свое
название — адолескарий.
Весь процесс развития фасцйо-
лы в малом прудовике—от внед-
рения мирацидия до выхода
церкария — длится при благо-
приятных условиях 29—30 дней,
в теле других моллюсков он
затягивается до 80 дней.
Адолескарий попадает в ор-
ганизм окончательного хозяина
(овцы, коровы и др.) с травой
или с водой. Заражение может
происходить и через сено с за-
болоченных лугов, если оно дается в корм
ранее чем через шесть месяцев после ска-
шивания. Человек может заразиться фас-
циолезом при употреблении для питья во-
ды из мелких водоемов, где обитает ма-
лый прудовик, или когда он берет в рот
траву, сорванную на болотистых местах.
Попав в окончательного хозяина, адоле-
скарий освобождаются от своей оболочки
и внедряются в толщу кишечной стенки.
Затем они попадают в кровеносное русло
и током крови заносятся в сосуды печени,
откуда пробираются в ткань этого органа
и потом в желчные ходы. Созревание фас-
цйолы и выделение яиц во внешнюю сре-
ду наступает на 67-й день или несколько
позже с момента заражения.
Рис. 202. Стадии развития фасцйолы печеночной.
I — мирацидий: 1 — кишечник; 2 — протонефридий; 3 — вывод-
ной канал протонефридия; 4 — зародышевые клетки; II — III — спо-
роцисты, в которых развиваются редии; IV — редия, в которой по-
вторно развиваются редии; V — редия, в которой развиваются церка-
рий; VI — церкарий: J — передняя, или ротовая, присоска; 2 —
брюшная присоска; 3 — кишечник; 4 — железы; 5 — хвост; VII —
адолескарий: i, 2> 3 — см. VI\ 5 — выделительная система.
357
Рис. 203. Справа — дикроцелий лан-
цетовидный (Dicrocoeliumlanceatum); сле-
ва — описторх кошачий (Opistorchis feli-
neus); в середине — мирацидий Opi-
storchis felineus.
Из описания жизненного цикла фасцио-
лы видно, что паразит может развиваться
только там, где водятся подходящие мол-
люски — прудовики. Малый прудовик,
наиболее частый промежуточный хозяин
фасциолы, обитает в мелких водоемах,
на низких заболоченных пастбищах.
В глубоких водоемах, в быстро текущих
ручьях и в воде, сильно загрязненной
гниющими веществами, прудовик жить
не может. Где нет прудовика, там и не
может распространяться фасциолез. В
дождливые годы фасциолез встречается
чаще, особенно если два дождливых года
следуют один за другим.
Из сказанного понятно, что основные
мероприятия по предохранению живот-
ных от заражения фасциолами должны
быть направлены на борьбу с моллюс-
ками — промежуточными хозяевами па-
разита. Такая работа проводится в двух
направлениях. Во-первых, осуществляют
мероприятия по ликвидации заболочен-
ности территории, где выпасают скот:
проводят соответствующие мелиоратив-
ные работы, что создает неблагоприятные
условия для жизни моллюсков; во-вторых,
непосредственно уничтожают моллюсков
химическими средствами. Наилучшим пре-
паратом для этой цели оказался медный
купорос.
В хозяйствах, где проведение указан-
ных мероприятий представляет трудности
и где фасциолез сильно распространен,
рекомендуют не пускать скот на заболочен-
ные территории, используя их под сено-
кос, и периодически менять выпасные уча-
стки. Смену выпасов проводят с учетом
сроков развития личиночных стадий фас-
циолы и регулируют таким образом, что-
бы животные пускались на новый участок
только после того, как там произошла
естественная гибель паразита. И наконец,
важным мероприятием в борьбе с фасцио-
лезом является уничтожение самих пара-
зитов в организме животных. Осуще-
ствляется это ветеринарными врачами пу-
тем применения специальных лечебных
средств.
Борьбе с фасциолезом в нашей страде
уделяют очень большое внимание. Работа
эта проводится в больших масштабах по
специально разработанному государствен-
ному плану.
Семейство Дикроцелииды (Dicrocoel-
lidae) объединяет много видов трематод,
паразитирующих в протоках печени и в
поджелудочной железе млекопитающих и
птиц.
Характерным представителем семейства
является дикроцелий ланцетовидный, или,
как его иначе называют, двуустка ланце-
товидная.
Дикроцелий ланцетовидный (Dicrocoelli-
um lanceatum) достигает в длину не более
10 мм при максимальной ширине 2,5 мм,
имеет плоское, овальное в очертании тело.
Как и фасциола, паразитирует преиму-
358
щественно у травоядных домашних и ди-
ких животных. Причем круг хозяев зна-
чительно шире, чем у фасциолы. Очень
редко может встречаться и у человека.
Распространен почти во всех странах
земного шара.
Дикроцелии не вызывают у животных
такого тяжелого заболевания, как фас-
циолы. Однако известны случаи, когда
у овец и у некоторых грызунов (у зайцев,
например) скоплялось по нескольку де-
сятков тысяч паразитов. Понятно, что
животные при таком заражении гибнут,
если их не лечат.
Долгое время ученые не могли полно-
стью расшифровать биологический цикл
дикроцелия ланцетовидного. Предполага-
лось, что развитие его идет со сменой
двух хозяев (моллюск — промежуточный
хозяин и позвоночное животное — окон-
чательный хозяин), однако опыты не под-
тверждали этого. И только совсем недавно
удалось установить, что развитие дикро-
целия происходит при участии промежу-
точного и дополнительного хозяев, причем
дополнительным хозяином оказался му-
равей.
Полный жизненный цикл этой тре-
матоды, таким образом, представляется
в следующем виде. Яйцо паразита, выде-
ленное из организма окончательного хо-
зяина, заглатывается наземным моллюс-
ком. В теле его формируются церкарии.
Они концентрируются в легочной полости
моллюска, инцистируются здесь и склеи-
ваются в небольшие слизистые комочки.
Эти комочки через дыхательные отвер-
стия моллюска выбрасываются наружу,
где их поедают муравьи. В теле муравьев
церкарии превращаются в метацеркариев.
Зараженных муравьев вместе с травой
заглатывают окончательные хозяева — ов-
цы, коровы и другие животные, в орга-
низме которых метацеркарии вырастают
во взрослых трематод.
Как и при фасциолезе, борьба с д и к-
роцелиозом (заболевание, вызывае-
мое дикроцелием) направлена главным
образом на предохранение домашних жи-
вотных от контакта с промежуточными
хозяевами с одновременным уничтожением
паразитов в организме больных животных
лечебными средствами.
Эвритрема (Euritrema pancreaticum)—
еще один представитель семейства дикро-
целиид, о котором следует коротко упомя-
нуть. Трематода обитает в протоках под-
желудочной железы крупного рогатого
скота и овец, вызывает патологические
изменения в железе и иногда является при-
чиной тяжелых заболеваний. В нашей
стране особенно часто встречается в сред-
неазиатских республиках и на Дальнем
Востоке. Промежуточными хозяевами тре-
матоды являются наземные моллюски.
Однако полностью жизненный цикл еще
не расшифрован. Видимо, он протекает
таким же путем, как и у дикроцелия.
Представители семейства Опи-
сторхиды (Opisthorchidae) — паразиты
млекопитающих и птиц, которые пита-
ются рыбой. Некоторые виды паразити-
руют у человека и вызывают опасные за-
болевания. Они обитают в протоках пе-
чени и в желчном пузыре. Названы так
по характерной особенности их внутрен-
него строения, а именно по расположению
семенников в задней части тела (описто —
сзади, орхис — семенник). Этим
они отличаются от других трематод, пара-
зитирующих в печени, в частности от дик-
роцелиид, на которых внешне похожи
(у дикроцелиид семенники находятся в пе-
редней части тела).
Рассмотрим подробнее двух представи-
телей этого семейства.
Описторх сибирский, или, как его иначе
называют, описторх кошачий (Opistorchis
felineus), очень часто встречается в неко-
торых районах Западной Сибири, особен-
но в бассейне реки Оби и ее притоков, за-
регистрирован также в Поволжье, на
Украине и в Прибалтике. Помимо нашей
страны, известен в некоторых странах
Западной Европы. Очаги распростране-
ния описторха приурочены к берегам рек
или озер, что связано с особенностями раз-
вития паразита — дополнительным хозяи-
ном его являются рыбы.
Взрослая трематода паразитирует в
желчных протоках печени у кошек, со-
бак, лисиц, песцов и соболей. Может
паразитировать и у человека.
Описторхи — паразиты небольших раз-
меров, около 1 см в длину, хвостовой ко-
нец их расширен, передний, несущий
359
ротовую присоску, сужен. На небольшом
расстоянии от ротовой присоски располо-
жена брюшная. Тело описторха полупро-
зрачное, поэтому хорошо видны кишечные
трубки и темная матка, наполненная ог-
ромным количеством очень мелких ко-
ричневых яиц.
Жизненный цикл описторха протекает
следующим образом. Яйца, выделенные
паразитами, попадают с желчью в кишеч-
ник и оттуда с испражнениями выделя-
ются наружу. В яйце заключен уже сфор-
мированный мирацидий, который вылуп-
ляется из скорлупы яйца лишь в кишеч-
нике промежуточного хозяина — пресно-
водного моллюска битинии (Bithinia liachi).
Мирацидий проникает через кишечную
стенку в полость тела моллюска, где пре-
вращается в спороцисту. Примерно через
месяц из спороцисты образуются редии,
в которых начинают развиваться церка-
рии. Они мигрируют в пищеварительную
железу моллюска и дозревают там. Через
два месяца церкарии покидают организм
моллюска и плавают близ дна водоема, где
нападают на дополнительного хозяина —
различных видов рыб, преимущественно кар-
повых (язь, лещ, плотва, елец, жерех, карп,
красноперка), проникают через их кож-
ные покровы в мышечную и соединитель-
ную ткани и инцистируются (покрываются
оболочкой), превращаясь в метацеркариев.
Заражение описторхозом плотоядных
животных происходит при поедании ими сы-
рой рыбы, содержащей метацеркариев пара-
зита. Таким же образом заражается и человек.
Проглоченные вместе с рыбой, метацеркарии
освобождаются от цист и проникают в желч-
ные протоки печени, где в течение трех-четы-
рех недель завершается их развитие. Раз-
витие описторха от яйца до половозрелой
стадии продолжается 4—4,5 месяца. В ор-
ганизме кошки паразит может жить свыше
трех лет.
Люди, страдающие описторхозом, жалу-
ются на боли в подложечной области
или в правом подреберье. Наблюдается
также тошнота, нередко рвота, отрыжка,
обильное слюнотечение. Постепенно раз-
вивается слабость, раздражительность, го-
ловные боли. Печень часто увеличивается
и становится болезненной при прощупы-
вании. Иногда бывает желтуха.
Большое значение в распространении
описторха среди людей имеют профессио-
нальные и бытовые навыки. Нередко лю-
ди, связанные с рыбным промыслом, едят
сырую или непрожаренную рыбу. Это
ведет к заражению их описторхозом. В ря-
де районов Сибири широко распростра-
нен обычай употреблять в пищу мало-
сольную рыбу и так называемую «строга-
нину» (мелко изрубленная свежемороже-
ная рыба), что также небезопасно в от-
ношении заражения паразитом.
Меры предохранения от заражения опи-
сторхисами очень просты: нужно употреб-
лять в пищу только тщательно проварен-
ную и прожаренную рыбу. Особенно строго
следует держаться этого правила в местах,
неблагополучных по описторхозу. В борьбе
с описторхозом большое значение имеет
охрана рек и озер от загрязнения испраж-
нениями человека и животных. Нужно
принимать все меры, чтобы нечистоты не
попадали в воду. Это предохранит рыб от
заражения личинками описторха.
Кланорх китайский (Clanorchis sinen-
sis) по строению очень сходен с опистор-
хом, но несколько больше его и четко
отличается конфигурацией семенников (у
кланорха они ветвистые, у описторха —
цельнокрайние). Отличаются эти виды по
географическому распространению. Опи-
сторх сибирский, как указывалось, осо-
бенно широко распространен по Оби и во-
сточнее Енисея не встречается. Кланорх
китайский обычен на Дальнем Востоке
нашей страны, особенно в бассейне реки
Амура, часто встречается также в Японии,
Китае, Корее и других восточных странах.
Эта трематода, как и описторх, парази-
тирует в печени плотоядных млекопитаю-
щих и у человека и также при интенсив-
ном заражении вызывает тяжелое заболе-
вание (кланорхоз). Биологический цикл
кланорха протекает по той же схеме, что
и цикл описторха, однако в связи с иным
географическим распространением пара-
зита роль промежуточных и дополни-
тельных хозяев для него выполняют дру-
гие виды моллюсков и рыб.
Семейство Шистозоматиды (Schi-
stosomatidae) — своеобразная группа тре-
матод, многими чертами отличающихся от
других представителей класса.
360
Самым, пожалуй, характерным отличи-
тельным признаком следует считать их
раздельнополость. У трематод это крайне
редкое явление. Кстати, отсюда и назва-
ние этих трематод: шисто — значит раз-
двоенный, зома — тело. Впрочем,
при раздельности полов самец и самка почти
постоянно находятся вместе. Самец удер-
живает самку в особом продольном желоб-
ке на его теле (гинекофорный канал, что
в переводе значит канал, «несущий сам-
ку»). Желобок образован боковыми края-
ми плоского тела самца, загнутыми на
брюшную сторону. У некоторых видов
шистозоматид этот желобок тянется вдоль
всего тела, у других — занимает только
часть его. Поверхность желобка обычно
покрыта шипиками, которые крупнее на
боковых краях. Это способствует удержа-
нию самки. Самка обычно несколько длин-
нее самца, имеет нитевидное тело более
или менее равной толщины на всем протя-
жении.
Другая особенность шистозоматид за-
ключается в том, что они, в отличие от
других трематод, не продуцируют боль-
шого количества яиц. Обычно в матке на-
ходится только одно зрелое яйцо. Оно
очень крупное и у некоторых видов имеет
необычную форму: у одних похоже на
колбу, у других — на челнок. Незави-
симо от формы яйца всех шистозоматид
имеют острый шип на одном из полюсов
или несколько сбоку. Шип облегчает про-
никновение яйца сквозь ткани хозяина
при выходе его во внешнюю среду.
Третья отличительная черта шистозо-
матид — они паразитируют в крови.
(Очень немногие из трематод являются
обитателями кровеносной системы, по-
давляющее большинство их имеет иную
локализацию.)
Шистозоматиды паразитируют у чело-
века, у многих домашних и диких млеко-
питающих и у птиц. Большой интерес
из-за своей исключительной вредоносно-
сти представляют формы шистозоматид,
которые паразитируют у человека. Дело
в том, что шистозомозы (заболе-
вание, вызываемое шистозомами) челове-
ка имеют чрезвычайно широкое распрост-
ранение в тропических и некоторых суб-
тропических странах Азии, Африки и Юж-
ной Америки, являясь в отдельных райо-
нах этих стран настоящим бедствием. Не-
безынтересно отметить, что люди страдают
здесь от шистозомозов с очень древних
времен: обызвествленные яйца шистозом
найдены в египетских мумиях, которые
были захоронены за 1300 лет до н. э.
В странах умеренного климата люди
не страдают шистозомозами. Возбудите-
лей этого заболевания нет ни в одном
районе нашей страны.
Зараженность людей шистозомами в от-
дельных странах чрезвычайно велика.
В Африке, например, в некоторых мест-
ностях шистозомами поражено 80—90%
населения. Считают, что в Египте более
60% людей заражено шистозомами.
В Бразилии насчитывается 3 млн. шисто-
зоматозных больных (при 45-миллионном
населении в стране). Всего в странах,
где встречается этот паразит, насчитывают
114 млн. человек, больных шистозомо-
зами. И поэтому вполне понятно, что воп-
росам изучения шистозомозов и изыска-
ния мер борьбы с ними в этих странах
уделяют очень много внимания. Этим за-
няты многие специалисты медицинских
и биологических институтов, с этой целью
созданы специальные научные учрежде-
ния, расходуются большие средства на
мероприятия, связанные с предохране-
нием людей от заражения паразитом.
И тем не менее шистозомозы остаются
пока страшным бичом для людей в стра-
нах, где возбудитель этих заболеваний
находит благоприятные условия для свое-
го развития.
У человека паразитируют три широ-
ко распространенных вида шистозом:
шистозома японская (Schistosoma japo-
nica), шистозома Максона (Schistosoma
mansoni) и шистозома кровяная (Schisto-
soma haematobium). Шистозома японская
встречается в странах Азии — главным
образом в Японии, Китае и на Филиппи-
нах. В Южной Америке распространена
шистозома Мансона. А в странах Африки
могут встречаться два вида: шисто-
зома Мансона и шистозома кровяная.
Шистозома кровяная (Schistosoma hae-
matobium) была найдена в крови человека
и описана более ста лет назад A852) не-
мецким ученым Бильхарцем, ра-
361
ботавшим в Египте. К настоя-
щему времени усилиями многих
исследователей в деталях выяс-
нена биология этой трематоды
и изучены вызываемые ею пато-
логические явления в организ-
ме хозяина.
Шистозомы кровяные кон-
центрируются преимуществен-
но в венозных сосудах брюш-
ной полости и мочевого пузыря.
С их паразитированием связа-
но расстройство мочевыдели-
тельной и половой систем. При
заболевании в мочевом пузыре,
почках и мочеточниках наблю-
даются воспалительные процес-
сы, образуются камни мочевого
пузыря, могут возникать язвы,
больной выделяет мочу с кро-
вью. Поэтому заболевание, вы-
зываемое этим видом шистозо-
мы, называют шистозомоз
половой.
Взрослые формы паразитов
крупные. Тело самца нежное
длиной до 1,5 см, с желобком
всем его протяжении, покрыто бугорка-
ми и шипиками. На переднем конце тела
близко друг от друга находятся ротовая
и брюшная присоски. Пищеварительный
канал, начинающийся пищеводом, вскоре
делится на две кишечные ветви, которые
затем снова сливаются, продолжаясь не-
парной извитой трубкой, заканчивающей-
ся слепо у заднего конца тела. Семенни-
ков — 4—5 (большое число семенников —
особенность всех шистозоматид). Самка
несколько длиннее (до 2 см), но примерно
в четыре раза тоньше самца. На теле у нее
нет шипов. Тело заполнено в основном
органами половой системы: сзади нахо-
дятся многочисленные желточники, кпе-
реди от них удлиненный яичник и затем
петли матки, в которой заключены оди-
ночные зрелые яйца овальной формы
с шипиком на полюсе. Яйца очень круп-
ные — 0,120—0,160 мм в длину и 0,040—
0,060 мм в ширину.
В период размножения шистозомы за-
бираются в мелкие венозные сосуды, нахо-
дящиеся близ просвета толстого отдела
кишечника и мочевого пузыря. Здесь сам-
Рис. 204. Шистозома
кровяная (Schistosoma
haematobium).
моче-
довольно
светлое,
почти на
ка откладывает яйца. Под влия-
нием сократительных движений
стенок кровеносных сосудов ки-
шечника и пузыря яйца попа-
дают в просвет пищеваритель-
ного тракта или протоков мо-
чеполовой системы и вместе с
испражнениями и мочой выде-
ляются наружу. При попада-
нии в воду скорлупа яйца раз-
рывается и из него выходит ми-
рацидий, который разыскивает
моллюска—промежуточного хо-
зяина — и внедряется в его те-
ло. Мирацидий может жить
лишь в течение суток или не-
много больше. Роль промежу-
точных хозяев выполняет око-
ло десяти видов пресноводных
моллюсков, относящихся к трем
родам (Bullinus, Physopsis,
Planorbis).
Развитие шистозомы кровяной
(как и всех других шистозоматид) проте-
кает с участием одного промежуточного
хозяина. В теле моллюска мирацидий раз-
вивается в спороцисты (материнскую и до-
черние), внутри которых формируются
церкарии. После созревания церкарий
разрывает оболочку спороцисты и выхо-
дит из тела моллюска в воду. Развитие
шистозомы в промежуточном хозяине про-
должается около одного месяца. Отличи-
тельная особенность церкариев данного
вида, как и других шистозоматид,— раз-
двоенный на конце хвост.
В организм человека церкарий прони-
кает, вбуравливаясь чаще в кожу ног
или рук. Происходит это обычно в то вре-
мя, когда люди ведут работы на орошае-
мых землях, например на рисовых полях.
С этим связано широкое распространение
шистозомозов именно в тех странах, где
орошаемое земледелие является ведущей
отраслью хозяйства. Этим же объясняется
тот факт, что страдает от шистозомозов
в основном бедное население, постоянно
связанное по роду своих занятий с ороси-
тельными каналами, болотами, зарос-
шими речками, рисовыми полями и т. д.,
т.е. с местами, являющимися самыми благо-
приятными для развития моллюсков —
промежуточных хозяев шистозом.
362
Проникнув через кожу в тело человека,
церкарий теряет хвост, попадает в крове-
носное русло и по мере созревания про-
двигается к месту своего постоянного оби-
тания в организме хозяина. Половозре-
лым паразит становится не менее как
через месяц после заражения. Взрослая
шистозома может жить в организме чело-
века очень долго — годами и даже десят-
ками лет. Это установлено наблюдениями
за людьми, заразившимися в жарких
странах и затем жившими долгое время
в местностях, где шистозомоза нет. Есть
сведения о случаях, когда люди выделяли
яйца шистозомы кровяной с живыми
мирацидиями в течение 28 и 29 лет.
Борьба с заболеваниями, вызываемыми
шистозомами, основывается на проведе-
нии следующих мероприятий: ликвидации
очагов выплода моллюсков, осуществля-
емой путем проведения мелиоративных
работ, обезвреживании экскрементов и мо-
чи больных людей, воздействии на церка-
риев химическими веществами, лечении
лиц, являющихся носителями паразитов,
и, наконец, на предохранении людей от за-
ражения паразитом. Для этого исполь-
зуют специальную одежду, пропитанную
химическими веществами, отпугивающими
церкариев. Следует сказать, что в связи
с широким распространением шистозомо-
зов и особенностями биологии паразита
успешная борьба с шистозомозами — дело
крайне трудное и сложное, Нельзя за-
кончить рассказ о шистозоматидах, не
отметив еще одного явления, связанного
с этими трематодами. Речь идет о так
называемом шистозомном, или
церкариозном, дерматите
(дерматит — заболевание кожи), который
в народе называется «зуд купальщиков»,
«зуд пловцов», «водный зуд» и т. д. Забо-
левание это наблюдается у лиц, купаю-
щихся в стоячих, заросших растительно-
стью прудах или заводях рек: у рыбаков,
охотников и всех других, кто так или
иначе соприкасается с такими водоемами.
Проявляется оно в том, что, находясь
в воде, люди чувствуют зуд на коже,
иногда наблюдается сыпь, покраснение
кожи.
Долго не была известна причина, вызы-
вающая указанную реакцию. Оказалось,
что повинны в этом церкарий шистозома-
тид, паразитирующих у водоплавающих
птиц — уток, нырков, чаек — и у неко-
торых млекопитающих. Дело в том, что
почти все водоплавающие птицы обычно
сильно заражены шистозоматидами, и в
водоемах, где они постоянно держатся,
в летний период всегда плавает масса
церкариев этих трематод. Они-то и внед-
ряются в кожу человека, как только он
погружается в воду. Раздражение кожи
наступает как от механического действия
личинок (вбуравливание), так и от выде-
ляемого при этом церкарием особого ве-
щества, растворяющего ткань.
Внедрившись в кожу, церкарий поги-
бают, поскольку человек не является
для них подходящим хозяином, и зуд
кожи обычно скоро проходит. Только
в редких случаях могут быть более или
менее тяжелые осложнения, требующие
лечения.
Шистозомный дерматит распространен
в США. Там он хорошо изучен. В нашей
стране он отмечался на Украине, в дельте
Волги, в некоторых районах Казахстана.
Несомненно, что шистозомный дерматит
распространен значительно шире. Прак-
тически каждый водоем в средних и юж-
ных районах нашей страны, где нет силь-
ного тока воды и имеется скопление водо-
плавающих птиц, таит в себе опасность
шистозомного дерматита.
Семейство Брахилемиды (Brachy-
laemidae). Представители его паразитиру-
ют главным образом в кишечнике птиц.
Лейкохлоридий парадоксальный (Leuco-
chloridium paradoxum) — весьма интерес-
ный в биологическом отношении предста-
витель этого семейства. В половозрелой
форме обитает в задней кишке и клоаке
ряда воробьиных птиц. Характерным при-
знаком для внешнего вида трематоды яв-
ляются огромные ротовая и брюшная при-
соски. При маленьких размерах паразита
(длина — до 2 мм, ширина — до 1 мм)
такие присоски являются вполне надеж-
ным органом фиксации даже в таком «опас-
ном» месте, как клоака. Однако самое ин-
тересное у этой трематоды — ее биология „
Яйца паразита с экскрементами птицы
попадают на траву или листья деревьев.
Промежуточный хозяин лейкохлоридия —
363
сухопутный моллюск янтарка (Succinea).
Питаясь растениями, янтарка заглаты-
вает и яйца трематоды. В теле моллюска
из яйца выходит мирацидий, который пре-
вращается в спороцисту. Далее развитие
идет укороченным путем: в спороцисте
сразу формируются бесхвостые личинки,
внешне очень похожие на взрослую фор-
му. Они не покидают спороцисты. Споро-
циста же разрастается, образуя большое
количество отростков, распространяю-
щихся по телу янтарки. Отдельные из
них проникают в щупальца улитки, силь-
но раздуваются и приобретают яркую
окраску, хорошо заметную сквозь тонкую
кожицу щупальца янтарки. На концах
спороцисты появляются темные пятна,
а на теле — темные круги. При такой
окраске спороциста, а вместе с ней и щупаль-
це моллюска становятся очень похожими
на гусеницу насекомого. Сходство еще
больше усиливается благодаря периоди-
ческим сократительным движениям споро-
цисты. Мелкие воробьиные птицы прини-
мают щупальце моллюска с ярко окра-
шенной и двигающейся спороцистой за
гусеницу насекомого и склевывают его,
ток спороцисты, также «наряженный»
под личинку насекомого. И снова вместо
желанной гусеницы птица примет порцию
зародышей паразита. Такова довольна
необычная биология этой трематоды.
Несомненно, что именно отмеченное ори-
гинальное приспособление паразита к
проникновению в окончательного хозяина
дало повод назвать его paradoxum, что
значит «необыкновенный», «удивитель-
ный». В родовом же названии трематоды
Leucochloridium — бело-зеленый, пест-
рый — отражено свойство спороцисты па-
разита расцвечиваться разными тонами.
Семейство Диплостоматиды (Dip-
lostomatidae). Взрослые формы живут
в кишечнике птиц и млекопитающих,
преимущественно тех, которые питаются
рыбой. Рыба участвует в биологическом
цикле многих диплостоматид как допол-
нительный хозяин. Паразитирующие у нее
метацеркарии нередко вызывают опасные
заболевания, особенно среди молоди рыб,
причиняя этим большой ущерб рыбному
хозяйству. О двух видах таких трематод
мы и расскажем детальнее. Третий вид
этого семейства, с которым мы также ре-
шили познакомить читателя, интересен
усложненным биологическим циклом —
развитие его может протекать со сменой
четырех хозяев.
Диплостома спатацеум (Diplostomum
spathaceum) в половозрелой стадии пара-
зитирует в кишечнике чаек и реже у не-
которых других рыбоядных птиц.
Тело вытянутое, длиной до 0,5 см,
в средней части перетянуто (это особен-
ность всех диплостоматид), передняя часть
плоская, лопатообразная, задняя — утол-
1 — спороциста; 2 — янтарка, промежуточный хозяин
паразита; в одном из щупалец ее находится спороцис-
та лейкохлоридия.
364
щенная. По бокам ротовой при-
соски находятся две маленькие
присосочкл и небольшие ушко-
видные выступы. Возле брюшной
присоски расположен особый же-
лезистый фиксаторный ор-
ган— орган Брандеса,
характерный и для других дип-
лостоматид. Органы половой сис-
темы расположены в задней части
тела.
Развитие паразита протекает
следующим образом. Яйца трема-
тоды с испражнениями чаек по-
падают в воду. Здесь из яйца
выходит мирацидий. Поплавав в
воде, он проникает в большого
прудовика (Limnea stagnalis). В
организме промежуточного хозяи-
на мирацидий превращается в
спороцисту, в которой образуются
редии, а внутри них формируют-
ся вилохвостые церкарий. Церка-
рий покидают прудовика и отыс-
кивают рыб. В организм рыбы
церкарий может проникать двоя-
ким способом: или вместе с водой
в пищеварительный тракт, или
через жабры и кожу. В том и другом слу-
чае церкарий попадает в кровь. Током
крови он заносится в глаза и проникает
в хрусталик глаза, превращаясь здесь в ме-
тацеркария. В отличие от других трематод,
метацеркарий диплостомы не инкапсули-
руется, в течение всей своей жизни он
подвижен. Находясь в хрусталике, личин-
ки затрудняют проникновение света во
внутреннюю камеру глаза. При длитель-
ном паразитировании метацеркариев в гла-
зу начинает откладываться известь, хрус-
талик постепенно мутнеет и приобретает
молочную окраску. Глаз при этом увели-
чивается в размере и выпячивается из
орбиты. Рыба слепнет. При небольшом
количестве паразитов в глазу слепота
может не наступить. Это заболевание рыб
называется паразитарной ката-
рактой. Поражаются ею главным об-
разом карповые рыбы в самом молодом воз-
расте. Метацеркарий в глазу рыбы живет
не менее 8 месяцев. Чаще всего больные
катарактой рыбы слепнут на оба глаза.
Слепая рыба не может нормально пита-
ться, слабеет, подплывает к по-
верхности воды, ложится на бок
и легко становится жертвой чаек
и других рыбоядных птиц.
В кишечнике чайки, проглотив-
шей зараженную личинками дип-
лостомы рыбу, вырастают взрос-
лые трематоды. Созревание па-
разитов идет очень быстро, к
4—5-му дню паразиты становятся
половозрелыми.
Биологический цикл диплосто-
мы представляет собой яркий
пример приспособления паразита
к проникновению в окончательно-
го хозяина. Используя пищевые
связи между хозяевами, параз:тт
в данном случае «активно содей-
ствует» осуществлению этих свя-
зей, «гарантируя» тем самым про-
дление своего рода.
Постодиплостома кутикульная
(Postodiplostomum cuticula)BO мно-
гом сходна с диплостомой. Основ-
ное отличие паразита состоит в
том, что постодиплостома не име-
ет на переднем конце по бокам
ротовой присоски двух дополни-
тельных присосочек и ушковидных вы-
ростов, характерных для диплостомы. По
этому признаку рассматриваемая тремато-
да причислена к другому роду. Иные так-
же и окончательные хозяева у этой трема-
тоды: взрослые паразиты живут в кишеч-
нике цаплевых птиц. Так как цапли
встречаются преимущественно в южных
районах, то и основные места распростра-
нения постодиплостомы также связаны с
южными водоемами.
Развитие постодиплостомы в общих чер-
тах протекает таким же образом, как
и развитие диплостомы. Роль первого
промежуточного хозяина для нее выпол-
няют также прудовики (Lymnea), допол-
нительным хозяином являются рыбы, в ос-
новном карповые. Различия касаются глав-
ным образом сроков развития паразита
на отдельных стадиях в промежуточном,
дополнительном и окончательном хозяе-
вах и, что особенно существенно, иного
характера повреждений, вызванных ли-
чинкой трематоды у дополнительных хо-
зяев, т. е. у рыб. Метацеркарий постодип-
Рис. 206. Дипло-
стома (Diplosto-
mum spathaceum).
365
лостомы локализуются не в глазах рыбы,
как у диплостомы, а в коже. Заражению
подвергаются, как правило, мальки рыб.
Церкарий внедряется в кожу малька, те-
ряет хвост и превращается в метацерка-
рия. Метацеркарий быстро покрывается
оболочкой (инцистируется), и вокруг него
(это очень характерно) откладывается чер-
ный пигмент. На теле рыбы появляется
черное пятнышко, хорошо заметное нево-
оруженным глазом. При сильном зараже-
нии таких пятен появляется много. При-
чем отдельные из них, когда личинки на-
ходятся рядом, сливаются, образуя боль-
шое черное пятно. Все тело больной рыбы
выглядит как бы обрызганным чернилами.
Отсюда названия болезни — черно-
пятнистая, чернильная. Бо-
лезнь очень опасна для мальков, так как
вызывает их гибель. В организме цапель,
которые заражаются паразитом, поедая
ослабленных болезнью рыб, трематода
очень быстро—на вторые-третьи сутки—до-
стигает половой зрелости и начинает откла-
дывать яйца. Подсчитано, что масса тела
паразита за указанный срок увеличива-
ется в 27—34 тыс. раз. Такие невероятные
«рекорды» роста под силу только парази-
тическому организму. Срок жизни посто-
диплостомы в организме окончательного
хозяина непродолжителен — всего 10—12
суток.
Очаги чернопятнистой болезни рыб свя-
заны с местами поселения цапель,
которые, как известно, живут колониями.
В нашей стране очаги этой болезни отме-
чены в устьях Дуная, Днепра, Волги,
Аму-Дарьи и в некоторых других местах.
Алярия алата (Alaria alata), в отличие
от двух описанных выше диплостоматид,
в половозрелой стадии паразитирует
у млекопитающих. Трематода поселяется
в кишечнике плотоядных животных —
собак, волков, лисиц, енотовидных собак
и др. Особенно часто встречается она
у лисиц. Распространена на Евразиат-
ском материке почти повсеместно.
Характерным признаком внешнего
строения трематоды являются довольно
крупные крыловидные выросты на перед-
нем конце, послужившие, видимо, пово-
дом для названия паразита (алярия —
значит крылатая). Наиболее же при-
мечательное у алярии — это ее биологи-
ческий цикл.
Находясь в кишечнике, половозрелый
червь откладывает яйца. Хотя оконча-
тельными хозяевами алярии являются
вполне сухопутные животные, яйца пара-
зита должны попасть в воду, так как про-
межуточные хозяева его — водные мол-
люски катушки (Planorbis). Яйца, не по-
павшие в воду, гибнут. Развитие в мол-
люсках идет обычным для диплостоматид
путем. Церкарий начинают выделяться
из моллюска через месяц или два (в зави-
симости от температуры внешней среды)
с момента заражения их мирацидием.
Дополнительные хозяева алярии — лягуш-
ки и другие амфибии в их личиноч-
ной и во взрослой стадии. Церкарий
внедряется обычно в тело головастика
и развивается в нем в метацеркария.
Когда головастики превращаются в лягу-
шек, личинки паразита остаются в их
теле. Таким образом осуществляется вы-
ход личинок паразита из воды на сушу.
Когда лягушка, зараженная личинками
алярии, будет съедена окончательным хо-
зяином, лисицей например, в теле послед-
него развивается взрослая алярия. Но
возможен и другой путь заражения окон-
чательного хозяина паразитом. Заражен-
ная лягушка может быть съедена другим
животным, не являющимся окончатель-
ным хозяином алярии, например змеей,
ежом, хищной птицей и т. д. Попав в ор-
ганизм этих животных, личинки алярии
не погибают там. Освободившись после
переваривания из тела лягушки, личинки
внедряются сквозь стенки кишечника но-
вого хозяина и, проникнув в ткани раз-
личных органов его (главным образом
в подкожную клетчатку), оседают здесь,
оставаясь долгое время жизнеспособны-
ми. Как только такое животное будет
съедено хищником — окончательным хо-
зяином алярии (собакой, волком, лисицей
и т. д.), личинка в его организме разви-
вается во взрослого червя. И цикл раз-
вития паразита таким образом замы-
кается.
Указанные животные (змея, еж, хищ-
ная птица и др.) не являются обязатель-
ными в цикле паразита, в них не проис-
ходит развития трематоды, они лишь на-
366
копляют в себе личинок паразита, сохра-
няют и способствуют передаче их оконча-
тельным хозяевам. Как уже указывалось,
такие животные носят название резервуар-
ных хозяев.
Участие в жизненном цикле алярии ре-
зервуарных хозяев — одна из интересных
особенностей развития этого паразита.
Другая особенность биологического ци-
кла алярии — ее миграция в теле оконча-
тельного хозяина. Оказывается, попав
в кишечник плотоядного животного, когда
он съедает дополнительного или резер-
ву арного хозяина, личинка, прежде чем
стать взрослой особью, совершает долгое
и сложное «путешествие» по организму
своего хозяина. Сквозь стенки пищевари-
тельного тракта она проникает в брюш-
ную полость животного, затем в грудную
и здесь внедряется в легкие. Претерпев
в тканях легких определенное развитие,
молодая алярия переходит в просвет брон-
хов, по дыхательной трубке достигает
ротовой полости и оттуда снова попадает
в кишечник. И уже здесь на 30—32-е сут-
ки с момента заражения становится по-
ловозрелой.
Общая продолжительность биологиче-
ского цикла алярии составляет три или
немного более месяцев.
КЛАСС ТЕМНОЦЕФАЛЫ (TEMNOCEPHALA)
40° с. ш. Кроме того, темноцефалы обнару-
жены также в холодных подземных водах
многих пещер Балканского полуострова.
Небольшая своеобразная группа прес-
новодных плоских червей, генетически
связанных с турбелляриями,— темноце-
фалы живут на поверхности тела
пресноводных высших ракообраз-
ных (амфипод, изопод и десятино-
гих), реже на черепахах и в редких
случаях в мантийной полости
двустворчатых моллюсков, посе-
ляясь главным образом на жабрах
и ротовых органах.
Размеры тела темноцефалов ма-
лы @,8—14,0 мм). На переднем
конце тела расположены щупаль-
ца, на заднем — присоска. Тело
темноцефалов полностью или час-
тично покрыто ресничками.
Темноцефалы — гермафродиты.
Все они обладают хорошо развитой
нервной и протонефридиальной
(выделительной) системами (рис.
207).
В большинстве случаев темно-
цефалы не причиняют вреда сво-
им хозяевам, но часто становятся
комменсалами и в редких слу-
чаях паразитами.
Темноцефалы — обитатели тро-
пической и субтропической зон,
исключая Африку, и ограничены
в своем распространении на север Рис. 207. Представители класса темноцефалов (Temnocephala).
367
КЛАСС УДОНЕЛЛИДЫ (UDONELLOIDEA)
Удонеллиды — своеобразные парази-
тические черви, стоящие в системе плос-
ких червей близко к турбелляриям, темно-
цефалам и моногеноидеям. Тело удонеллид
более или менее цилиндрической формы,
размером в несколько миллиметров (рис.
208). На переднем конце тела располо-
жены два железистых органа, а на заднем
одна большая круглая присоска, лишен-
ная крючков. Удонеллиды, как и прочие
плоские черви,— гермафродиты. Крупные
овальные яйца их прикрепляются к телу
хозяина длинными ножками.
Интересна биология удонеллид. Они
являются вторичными паразитами. Их
хозяева — веслоногие ракообразные, па-
разитирующие в свою очередь на различ-
ных морских рыбах. Личинки удонеллид
лишены мерцательного покрова.
Некоторые исследователи считают удо-
неллид только отрядом класса моногене-
тических сосальщиков.
КЛАСС МОНОГЕНОИДЕИ (MONOGENOIDEA)
В системе типа плоских червей моно-
геноидеи только в недавнее время стали
рассматриваться как класс. Ранее они
составляли подкласс в классе трематод
наряду с другим подклассом — ди-
генеями.
Указанные изменения внесены в систе-
му плоских червей в результате деталь-
ного изучения биологии моногеноидей,
проведенного в последнее время совет-
ским ученым Б. Е. Б ы х о в с к и м.
Он установил, что по характеру своего
индивидуального развития моногеноидей
стоят ближе к другим группам плоских
червей — к гирокотилидам и цестодам,
чем к дигенетическим трематодам. Об-
щим признаком, сближающим гирокотилид,
цестод и моногеноидей, оказалось строение
Таблица 19.
Различные донные и пела-
гические полихеты:
1 — томоптерис (Tomopteris renata);
2 — спирографис (Spirograms
spallanzanii);
3 — протула (Protula protula);
4 — серпула (Serpula vermicularis);
5 — нереис (Nereis pelagica);
6 — морская мышь (Aphrodite
aculeata);
7 — эвное (Eunoe nodosa),
8 — нереис (Nereis virens),
9 — пескожил (Arenicola grubei);
10 — автолитус (Autolytus pictus);
11 — амфитрита (Amphitrita Jonstoni);
12 — онуфис (Onuphis conchilega).
личинок этих групп животных, а именно на-
личие на ранних стадиях развития у всех их
своеобразного хвостового отростка, снаб-
женного зародышевыми крючьями. Ли-
чинки дигенетических трематод подоб-
ного отростка с зародышевыми крючьями
не имеют.
От дигенетических трематод моногеноидеи
отличаются также и рядом других сущест-
венных признаков. По-иному, прежде всего,
протекает жизненный цикл моногенетичес-
ких сосальщиков. В отличие от дигеней, их
развитие не связано со сменой хозяев и со
сменой поколений. Значительные отличия
имеются в строении взрослых форм предста-
вителей этих двух групп сосальщиков. Вы-
делительная система, например, у моноге-
ноидеи открывается наружу отверстиями,
расположенными в передней части тела па-
разита, у дигеней же выделительное отверстие
находится в задней части тела. Это важное
отличие. Отмеченные особенности и послу-
жили основанием для выделения моногено-
идеи из класса трематод.
С другой стороны, хотя и было установ-
лено их родство с гирокотилидами и цесто-
дами, они не могли быть объединены ни с
одним из указанных классов. Этому пре-
пятствовало своеобразие их строения и био-
логии. В результате был основан новый
класс.
Таблица 20. Трохофорные личинки нолихет:
1
2
3
4
5
6
7
8
9
10
11
12
—
—
—
—
—
—
—
—
Harmothoe imbricata;
Anaitides citrina;
Nereis virens;
Nephthys
Disoma;
Polydora
ciliata;
ciliata;
Chaetopteridae;
Magelona
Capitella
Arenicola
papillicornis;
capitata;
marina;
Pectinaria koreni;
Spirorbis
spirorbis.
• 24 Жизнь животных, т. 1
Название класса — моногеноидеи — в
переводе с латинского языка означает
«имеющие одно поколение». Названием
подчеркивалось отличие их от дигенети-
ческих трематод, которым свойственна
смена поколений.
По-русски данный класс называют
Сосальщики-многоустки.
Моногеноидеи — сравнительно неболь-
шой класс плоских червей, он включает
около 1000 видов, паразитирующих у мор-
ских и пресноводных рыб и очень редко
у амфибий, рептилий и водных мле-
копитающих.
В отличие от других классов паразити-
ческих плоских червей—трематод и цестод,
поселяющихся во внутренних органах
своих хозяев, моногеноидеи — наружные
паразиты (эктопаразиты). Основное место
их обитания — жабры и кожа рыб.
Моногеноидеи приносят большой эко-
номический ущерб рыбному хозяйству,
так как многие представители их являют-
ся возбудителями очень серьезных забо-
леваний рыб. Особенно ощутимы потери
от этих паразитов в прудовых хозяйствах.
Строение. Почти все моногеноидеи име-
ют уплощенное, вытянутое в длину тело,
расширенное в задней части.
Размеры моногеноидеи в большинстве
случаев незначительны — длина тела ко-
леблется в пределах 0,03—20 мм,
в редких случаях несколько
больше. Паразиты морских рыб
обычно крупнее, чем паразиты
пресноводных.
Передний конец червя с дву-
мя — четырьмя лопастями.
Иногда лопасти несут ямки
или присоски.
Задний конец червя более
или менее обособлен от тела и
превращен в прикрепительный
диск, который снабжен раз-
личного рода хитиноидными
образованиями — крючьями и
соединительными пластинками.
На нем же могут находиться
особые клапаны, ямки и выро-
сты, способствующие более проч-
ной фиксации паразита. Иног-
да весь диск преобразован в
мощную присоску. Строение
прикрепительных органов мо-
ногеноидей отличается большим
разнообразием. Различие в
строении этих органов широко
используется при систематиче-
ском определении представите-
лей класса.
Снаружи тело моногеноидей
покрыто кутикулой. Под ней
находятся субкутикулярный и
мышечные слои, составляющие
кожно-мускульный мешок.
Внутренняя часть тела запол-
нена паренхимной тканью.
В паренхиме располагаются
органы пищеварительной, выделительной,
нервной и половой систем.
Пищеварительный канал на-
чинается ротовым отверстием, расположен-
ным на переднем конце тела. Отверстие
ведет в околоротовую воронку, переходя-
щую в мускулистую глотку, за ней сле-
дует пищевод (он имеется не у всех форм)
и далее кишечная трубка, которая закан-
чивается слепо.
Кишечник состоит из одного или двух
стволов. Они могут быть гладкие или
с боковыми ответвлениями.
Выделительная система
представлена сетью каналов. Главные бо-
ковые каналы открываются наружу в пе-
редней части тела двумя выде-
лительными порами.
Нервная система
состоит из парных головных:
ганглиев, от которых вперед,
и назад отходят 3—4 пары нерв-
ных стволов. У некоторых ви-
дов во взрослой и личиночной
стадии, чаще только в личи-
ночной, имеются особые ор-
ганы чувств — «глазные пят-
на», находящиеся на головном
конце паразита.
Половая система
построена по гермафродитному
типу. Мужская система состоит
из одного, двух или многих се-
менников. Отходящие от них
семявыносящие канальцы объ-
единяются затем в общий се-
мяпровод, оканчивающийся ко-
пулятивным органом. Яичник
всегда один. Отходящий от не-
го короткий яйцевод принимает
протоки желточников и влага-
лища. Яйцевод расширяется в
оотип, куда впадает проток
скорлуповой железы (тельце
Мелиса). От оотипа берет на-
чало сравнительно короткая
трубка матки. Наружу матка
открывается общим с мужской
половой системой гермафродит-
ным отверстием, расположен-
ным в передней части тела.
Трубка влагалища открывается
самостоятельным отверстием.
Яйца моногеноидей очень разнообраз-
ны по форме, они могут быть шаровидные,
овальные, пирамидальные и иной конфи-
гурации. На одном полюсе яйца обычно
имеется крышечка с отростками, на дру-
гом — ножки с филаментами. Некоторые
виды моногеноидей выделяют не яйца, а
личинку.
Развитие моногеноидей протекает без
смены хозяев и чередования поколений.
Однако вышедшая из яйца личинка не
походит на взрослую форму и в процессе
своего развития претерпевает довольна
сложные изменения. Она имеет сигарооб-
разную форму, покрыта полностью или
частично ресничками, обладает головными:
Рис. 208. Представи-
тель класса удонел-
лид — Udonella cali-
gorum:
1 — прикрепительные
ямки; 2 — кишечник;
3 — выделительная си-
стема; 4 — яичник; 5 —
семенник; 6 —присоска.
370
железами, у большинства видов два — че-
тыре глазка, глотка, мешковидный ки-
шечник и слаборазвитая нервная и выде-
лительная системы. Свободно плавает в
воде.
Попав к своему хозяину, личинка при-
крепляется к телу его в месте своего оби-
тания, сбрасывает ресничный покров и,
переходя к паразитическому образу жиз-
ни, начинает расти, превращаясь посте-
пенно во взрослую форму.
Моногеноидеи отличаются резко выра-
женной специфичностью в отношении сво-
их хозяев: обычно каждый вид паразита
приурочен к определенному виду хозя-
ина; случаи, когда представители одного
вида поселяются в нескольких хозяевах,
редки.
Классификация. Класс моногеноидеи
подразделяется на два подкласса
(Polyonchoinea и Oligonchoinea). Каждый
подкласс объединяет по три отряда. Под-
разделение на систематические единицы
Рис. 209. Дакти- Рис. 210.
логирус (Dactylo- Гиродактилюс
girus vastator). (Gyrodactylus).
основывается главным образом на морфо-
логических особенностях представителей
класса, преимущественно на строении эле-
ментов прикрепительного аппарата.
Ниже мы расскажем о нескольких наи-
более обычных видах данного класса.
Дактилогирус вастатор (Dactylogyrus
vastator) паразитирует на жабрах, редко
на коже карпов, отличается большой вре-
доносностью, в связи с чем представляет
очень серьезную опасность для прудовых
хозяйств.
Тело паразита имеет длину от 0,5 до
1 мм, ширину 0,15—0,38 мм. На переднем
конце его — четыре головные лопасти.
Взрослые формы с четырьмя глазками.При-
крепительный диск снабжен двумя цент-
ральными большими крючьями и четыр-
надцатью мелкими краевыми крючочками.
Рот не окружен присоской.
Размножается дактилогирус путем от-
кладки яиц. В матке одновременно нахо-
дится всего лишь одно яйцо, однако пло-
довитость паразита довольно высокая.
Подсчитано, что за шесть часов он откла-
дывает от 14 до 29 яиц.
Из яйца выходит личинка удлиненной
формы с пятью группами ресничек и че-
тырьмя глазками. Ее зачаточный прикре-
пительный диск снабжен четырнадцатью
краевыми крючочками. Большие крючья
отсутствуют.
Поплавав некоторое время в воде, ли-
чинка прикрепляется с помощью своих
крючков к рыбе и сбрасывает реснички.
Вначале она поселяется на коже карпа,
но через некоторое время переползает на
его жабры. Здесь личинка формируется
во взрослого червя.
Дактилогирус вастатор паразитирует
преимущественно у молодняка карпов.
Наибольшей восприимчивостью отлича-
ются мальки длиной от 2 до 5 см. У более
крупных карпов паразит встречается в
меньшем количестве и поражает меньший
процент рыб.
Внешними признаками зараженности
рыб дактилогирусами являются неравно-
мерное окрашивание жабр и обильное вы-
деление слизи. Зараженная рыба делается
беспокойной, устремляется к притоку во-
ды, поднимается к поверхности, заглаты-
вает воздух.
24*
371
При дальнейшем развитии
болезни, если рыба заражена
большим количеством пара-
зитов, наблюдается омерт-
вление пораженных участков
жабр с последующим раз-
рушением их.
Заболевание, вызываемое
дактилогирусами, достигает
наибольшей интенсивности в
прудовых хозяйствах обыч-
но в июле — августе. Особен-
но сильно болезнь развивает-
ся при густой посадке рыб
в пруду, при плохом пита-
нии и неблагоприятных для
рыбы условиях окружающей
среды.
Для лечения больных рыб
в прудовых хозяйствах при-
меняют ванны с растворами
химических веществ, губи-
тельно действующих на пара-
зитов и существенно не от-
ражающихся на состоянии
их хозяев. Чаще других при-
меняют растворы поваренной
соли и медного купороса.
Гиродактилюс элеганс (Gy-
rodactylus elegans) парази-
тирует на коже и плавни-
ках карпов и некоторых дру-
гих близкородственных им
рыб. Это один из наиболее
опасных паразитов в прудо-
вых хозяйствах.
Длина взрослой особи гиродактилюса
часто колеблется от 0,5 до 1 мм при ширине
0,12—0,15 мм. Передний конец с дву-
мя лопастями, способными сильно сокра-
щаться. На вершинах лопастей открывают-
ся отверстия головных железок. Глазные
пятна отсутствуют. Прикрепительный диск,
расположенный на заднем конце, воору-
жен двумя большими центральными крю-
чьями и шестнадцатью мелкими краевыми.
Между большими крючьями имеются две
пластинки. Ротовое отверстие помещается
на брюшной стороне в передней трети
тела. Кишечник подразделяется на два
ствола.
Гиродактилюс принадлежит к группе
живородящих моногеноидей.
Рис. 211. Многоуст (Polystoma
integerrimum).
Формирующийся внутри
тела паразита зародыш от-
личается крупными размера-
ми. Зародыш имеет два боль-
ших крючка. Особенно ин-
тересно, что внутри его по-
мещается другой зародыш
несколько меньших размеров
и также с двумя крючками,
а в нем в свою очередь на-
ходится третий зародыш ги-
родактилюса.
Долгое время не могли
разгадать, каким образом
осуществляется подобное раз-
витие зародышей. Тайна про-
цесса была раскрыта только
в последнее время. Происхо-
дит это следующим обра-
зом. Оплодотворенное яйцо
делится пополам. Из одной
половинки образуется заро-
дыш. Он обхватывает собой
другую половинку яйца. Эта
половинка в свою очередь
снова делится пополам. Одна
из образовавшихся частей
дает начало новому зароды-
шу, который также обхваты-
вает собой оставшуюся часть
яйца. Эта четвертая часть
яйца опять-таки превращает-
ся в нового зародыша. Таким
образом возникают три заро-
дыша, как бы вложенных
друг в друга.
Выделяемый гиродактилюсом зародыш
представляет собой вполне развитую ли-
чинку. Долго существовать как сво-
бодноживущий организм он не может.
Встретившись с хозяином, личинка при-
крепляется к поверхности тела и к плав-
никам рыбы, переходит к паразитиче-
скому образу жизни и начинает раз-
виваться во взрослого червя.
У рыб, сильно зараженных гиродак-
тилюсами, тело покрывается голубовато-
серым налетом, состоящим из обильно
выделяющейся слизи, которой и питаются
паразиты. При дальнейшем развитии бо-
лезни участки кожи могут подвергаться
разрушению и омертвлению. В некоторых
случаях на поверхности кожи появляются
372
небольшие изъязвления. Плавники, на ко-
торых паразитируют гиродактилюсы, мо-
гут разрушаться настолько сильно, что
от них остаются только свободно торчащие
отдельные лучи.
Заражаются гиродактилюсами глав-
ным образом мальки и годовики рыб.
Для лечения больных рыб применяют
ванны с раствором веществ, которые уби-
вают паразитов и от которых не гибнут
рыбы. Обычно для этого используют пова-
ренную соль. Как меру предохранения от
заноса паразита в пруды, где выращива-
ется молодь, применяют обязательное ис-
пользование солевых ванн для произво-
дителей карпов перед посадкой их на не-
рест.
Многоуст (Polystoma integerrimum) па-
разитирует на жабрах головастиков и в
мочевом пузыре лягушек, главным об-
разом травяной лягушки. Отличается
своеобразной биологией, являющейся яр-
ким примером зависимости развития па-
разита от жизненного цикла хозяина.
Взрослый многоуст достигает в длину
6—8 мм при максимальной ширине в 2—
4 мм. Прикрепительный диск его снабжен
шестью присосками, внешне напоминаю-
щими рты (отсюда название «многоуст»).
На диске находятся также два больших
крючка. Ротовое отверстие расположено
на переднем конце. Оно окружено неболь-
шой присоской.
Биологический цикл многоуста проте-
кает следующим образом. Достигнув в мо-
чевом пузыре лягушки половозрелого со-
стояния, паразит начинает выделять яй-
ца. Наиболее интенсивно этот процесс про-
текает весной, когда лягушки, покинув
места зимовок, переходят к активному
образу жизни и приступают к размноже-
нию. Из яиц паразита, попавших в воду,
вылупляется личинка, снабженная пуч-
ками ресничек, с помощью которых она
свободно плавает в воде.
Личинки отыскивают развившихся
к этому времени из икры лягушек голова-
стиков и прикрепляются к их жабрам.
Если головастик оказался молодым, то
личинка паразита при благоприятных
внешних условиях успевает вырасти на
жабрах своего хозяина во взрослую фор-
му, до того как произойдет превращение
головастика в лягушонка. Взрослый пара-
зит откладывает яйца, которые сразу по-
падают в воду и дают личинок. Эта форма
многоуста называется жаберной, она мень-
ше той, которая развивается в мочевом
пузыре, внутренние органы ее развиты
слабее.
Личинки паразита, которые попадают
на жабры более зрелых головастиков, не
успевают вырасти во взрослого многоуста
до того, как жабры начинают атрофиро-
ваться в связи с превращением голова-
стика в лягушонка. Эти личинки после
зарастания жаберных отверстий попадают
в переднюю часть кишечника головастика,
спускаются затем по кишке до клоаки и из
клоаки забираются в мочевой пузырь. Фор-
мирование взрослого паразита в мочевом
пузыре протекает длительное время:
оно завершается только на третий год,
т. е. к тому времени, когда хозяин пара-
зита — лягушка — тоже достигает поло-
возрелое™ и впервые начинает отклады-
вать икру.
Биология многоуста характеризуется не
только сложностью и своей довольно тон-
кой адаптацией к особенностям жизни
373
хозяина, но и другой чертой — переходом
за время своего индивидуального разви-
тия от наружного паразитизма (обитание
на жабрах) к внутреннему паразитизму
(поселение в мочевом пузыре). В этом от-
ношении многоуст отличается от других
моногеноидей, являющихся типичными
эктопаразитами.
Спайник парадоксальный (Diplozoon ра-
radoxum) — это, как показывает название
животного, несколько необычный предста-
витель класса. Своеобразие его заключа-
ется в том, что паразиты живут попарно.
Соединяются они в молодом возрасте.
При этом брюшная присоска одной особи
охватывает специальный спинной бугорок
другой, и наоборот. В местах соединения
животные полностью срастаются, остава-
ясь в таком состоянии всю жизнь. В месте
срастания мужской проток одного червя
соединяется с женским протоком другого:
этим обеспечивается перекрестное оплодо-
творение. Если молодой спайник не
встретится с другой особью, он погибает.
Спайник парадоксальный паразитирует
на жабрах леща. Хозяевами его могут
быть и некоторые другие виды карповых
рыб. Однако в последнее время высказано
мнение, что спайники, паразитирующие
не на лещах, принадлежат к другим ви-
дам данного рода.
По внешнему виду спайник представ-
ляет собой крестообразную фигуру. Тело
каждого червя из сросшейся пары доволь-
но четко подразделяется на две части: пе-
реднюю—листообразную и заднюю —
значительно меньшую, цилиндрическую
по форме. В передней части располагаются
Мелкие, до нескольких сантиметров,
листовидные паразитические гермафро-
дитные черви. Обнаружены в кишечнике
глубоководных химеровых рыб. На перед-
нем конце тела гирокотилид располо-
жена присоска, на заднем — складчатый
прикрепительный диск в виде розетки.
На спине обычна поперечная складча-
тость. Яйца выводятся наружу, и в них
желточники и основная часть кишечника,
в задней находятся органы половой си-
стемы и прикрепительный аппарат, состо-
ящий из восьми ущемляющих клапанов и
срединных крючочков. Длина особи спай-
ника 3,8 — 6,7 мм, ширина 0,7—1,7 мм.
Размножается спайник путем откладки
яиц, из которых в воде выходят личинки.
Прикрепившись к жабрам леща, личин-
ки затем соединяются попарно и выраста-
ют во взрослых паразитов.
развиваются личинки, покрытые реснич-
ками и активно плавающие в воде. Задняя
часть личинки вооружена 10 крючьями.
Класс гирокотилид включает всего
3 рода с четырьмя видами. По своей орга-
низации они занимают как бы промежу-
точное положение между моногенетиче-
скими сосальщиками и цестодами и про-
являют некоторое родство с амфилинами.
КЛАСС ГИРОКОТИЛИДЫ (GYROCOTILOIDEA)
374
КЛАСС ЦЕСТОДЫ (CESTOIDEA)
Цестоды — слово греческого происхож-
дения. Корень этого слова на русский
язык переводится как «пояс», «лента».
Отсюда русское название класса —
Ленточные черви. И действительно, форма
тела животных, объединяемых данным
классом, в типичных случаях очень на-
поминает ленту.
Все цестоды ведут паразитический об-
раз жизни. В половозрелой стадии они
обитают в теле рыб, амфибий, рептилий,
птиц и млекопитающих. Личинки же их
поселяются в различных беспозвоночных,
главным образом членистоногих. Есть и
такие виды — это наиболее опасные пара-
зиты, личинки которых, как и взрослые
особи, обитают в организме позвоночных
животных.
В теле окончательных хозяев — у по-
звоночных животных и человека — поло-
возрелые цестоды поселяются обычно в
просвете кишечника, прикрепляясь перед-
ним концом к его внутренней стенке.
Известно лишь несколько видов, приспо-
собившихся к обитанию в желудке, в
клоаке (у птиц) и в протоках печени.
В этом отношении цестоды отличаются от
родственного им класса трематод, которые,
как отмечалось выше, приспособились
к обитанию почти во всех органах своих
хозяев.
Давние предки цестод, как и всех дру-
гих классов плоских червей, принадле-
жали к свободноживущим организмам.
Считают, что ими были животные, близ-
кие к современным турбелляриям.
В процессе эволюции цестоды очень
резко, пожалуй, более, чем другие группы
паразитических червей, уклонились от
своих прародителей. Происшедшие изме-
нения касаются как их внешнего облика,
строения, так и особенностей развития.
Ниже мы будем подробнее говорить об
организации и развитии цестод.
Одна из самых характерных черт цестод
как высокоспециализированной группы
паразитических организмов —полная утра-
та ими органов пищеварения. Питание
их осуществляется посредством всасыва-
ния питательных веществ поверхностными
тканями тела. Далее, как и другие пара-
зиты, цестоды обладают чрезвычайно вы-
сокой плодовитостью. С этим связано
сильное развитие у цестод половой систе-
мы и их гермафродитизм. И наконец, еще
одна особенность, возникшая, несомненно,
в процессе приспособления ленточных
червей к паразитизму,— это сложность
их жизненных циклов. Сложность эта
проявляется в том, что развитие цестод
протекает со сменой хозяев, а ряд видов,
кроме того, обладает способностью к раз-
множению в личиночной стадии.
Немалое число видов цестод паразити-
рует у человека и хозяйственно полез-
ных животных. У человека паразитирует
32 вида цестод, у крупного рогатого ско-
та, например, может паразитировать свы-
ше 30 видов цестод, несколько больше у
овец и коз. Поселяясь в организме челове-
ка или животного, ленточные черви во
взрослой или личиночной стадии вызы-
вают заболевание. Общее название этих
заболеваний —ц е с т о д о з ы. Отдельные
заболевания называются по родовому наи-
менованию возбудителя.
Многие цестодозы протекают очень тя-
жело, резко отражаются на состоянии ор-
ганизма и иногда могут кончаться смертью
хозяина. Поэтому борьбе с цестодозами
человека и животных, особенно домашних,
уделяется большое внимание. Особенно
широко проводится эта борьба в нашей
стране, где она носит характер плановых
общегосударственных мероприятий.
Строение. Длинное лентовидное тело
цестод подразделяется на три отдела: го-
ловку, или сколекс, шейку и стробилу,
состоящую в типичных случаях из мно-
гих члеников (проглоттид).
Головка несет на себе органы прикреп-
ления. Это могут быть продольные ще-
ли — ботрии, присоски и хи-
тинизированные крючья. Ботрии
характерны для низших цестод. Их чаще
всего две, располагаются ботрии на брюш-
ной и спинной стороне головки. Цестоды,
имеющие ботрии, как правило, лишены
375
крючьев. Присосками — полуша-
ровидными полыми мышечными органа-
ми — вооружены головки более высокоор-
ганизованных ленточных червей. При-
сосок обычно четыре. Головка, обладаю-
щая присосками, имеет еще хоботок,
который у многих цестод вооружен крю-
чьями. У некоторых видов ленточных чер-
вей стенки присосок покрыты мелкими
шипиками.
Строение головки, особенно органов
прикрепления, отличается большим раз-
нообразием, поэтому указанные органы
часто используются при систематическом
определении цестод.
С помощью вооруженной головки це-
стоды прикрепляются к внутренней стен-
ке кишечника хозяина.
Шейка — короткий, более тонкий,
несегментированный участок тела, сое-
диняющий головку с начинающимися по-
зади нее члениками.На заднем конце шейки
происходит образование новых члеников.
Самые молодые членики расположены око-
ло шейки, самые старые — на заднем кон-
це паразита.
Количество члеников, составляющих те-
ло цестоды, колеблется в очень широких
пределах. Есть цестоды, которые состоят
из одного членика, таких видов очень ма-
ло; у большинства же число члеников
исчисляется десятками и сотнями, у от-
дельных видов оно может доходить до не-
скольких тысяч. В соответствии с этим
и общая длина тела цестод очень различ-
на. Она колеблется от нескольких деся-
тых долей миллиметра (например, Davai-
nea proglottina из кишечника домашней
курицы достигает длины всего лишь 0,5 мм)
до двух десятков метров (некоторые
лентецы).
Снаружи тело цестод покрыто кутику-
лой. Под ней расположены слои мышц,
круговых и продольных,— веретенообраз-
ные субкутикулярные клетки. Все эти
ткани образуют к о ж н о-м у с к у л ь-
ный мешок. Помимо указанных мы-
шечных волокон, цестоды имеют еще ле-
жащий глубже них слой поперечных мы-
шечных волокон и тянущиеся в спинно-
брюшном направлении мышечные пучки.
Внутренняя часть тела ленточных чер-
вей заполнена паренхимой, в которой ле-
жат органы нервной, выделительной и по-
ловой систем. Пищеварительные органы,
свойственные предкам цестод, они, как
отмечалось, утратили. Питательные веще-
ства ленточные черви всасывают всей по-
верхностью тела.
Нервная система у цестод
развита слабо. Она состоит из централь-
ного узла, расположенного в головке,
и отходящих от него продольных ство-
лов. Два из них, наиболее развитые, про-
ходят вдоль всего тела в боковых частях
члеников кнаружи от каналов выдели-
тельной системы. Продольные каналы со-
единяются между собой поперечными тя-
жами. Органы чувств у цестод отсутст-
вуют.
Выделительная еистема
ленточных червей представлена множест-
вом мелких, пронизывающих паренхиму
канальцев, которые впадают в четыре
главных продольных канала.
Каждый из мелких канальцев заканчи-
вается особыми пульсирующими клетка-
ми. Эти клетки нагнетают в канальцы
образующиеся в теле червя вредные ве-
щества.
Продольные выделительные каналы по-
парно проходят по бокам тела рядом со
стволами нервной системы. Один из каж-
дой пары каналов — брюшной, широкий,
другой — спинной, более узкий. В голов-
ке червя брюшной и спинной каналы сое-
диняются друг с другом. Кроме того, ка-
налы правой и левой стороны соединяются
между собой в каждом членике, в задней
его части, еще поперечной перемычкой.
В последнем членике цестоды (который
первый отделился от шейки) все четыре
канала соединяются в один общий, кото-
рый открывается наружу на конечном
крае членика. После того как этот членик
отпадает, каждый канал открывается на-
ружу своим отверстием.
Половая система цестод, за
единичными исключениями, гермафро-
дитная, т. е. каждый червь содержит в се-
бе как мужской половой аппарат, так и
женский. Органы половой системы распо-
ложены в каждом членике, и с органами
данной системы соседнего членика они не
связаны. Как правило, каждый членик
имеет одинарный комплект половых орга-
376
нов, но немало и таких видов цестод, чле-
ники которых содержат удвоенный поло-
вой аппарат.
В развитии мужских и женских органов
наблюдается определенная последователь-
ность. Обычно вначале в члениках разви-
ваются органы мужской половой системы,
затем в них же образуются органы жен-
ской системы. После оплодотворения муж-
ская система постепенно исчезает, а по
мере образования яиц начинает интенсив-
но развиваться матка. Конечные зрелые
членики цестоды почти полностью запол-
нены маткой, которая содержит большое
число яиц.
Мужской половой аппарат состоит из
семенников, вырабатывающих мужские по-
ловые клетки — сперматозоиды, и вывод-
ных протоков.
Семенников может быть самое различное
количество — от одного до нескольких
сотен.
От каждого семенника отходят семенные
канальцы, сливающиеся затем в один об-
щий канал — семяпровод. У одних видов
семяпровод представляет собой более или
менее прямую трубку, у других — он
сильно извитой. Семяпровод оканчива-
ется мужским половым отверстием, откры-
вающимся обычно в половую кло-
аку рядом с женским половым отвер-
стием.
Конечная часть семяпровода, превра-
щенная в совокупительный орган — ц и р-
р у с, часто бывает покрыта шипиками
или щетинками. Перед циррусом трубка
семяпровода образует одно или два рас-
ширения, в которых скопляются спермато-
зоиды. Эти расширения носят название
семенных пузырьков. Циррус и один из
семенных пузырьков — внутренний се-
мейной пузырек — заключены в особую
мышечную сумку, называемую половой
бурсой.
Женский половой аппарат имеет более
сложное строение. Основной частью его
является яичник, расположенный обычно
в задней части членика и имеющий чаще
всего двулопастную форму. Яичник выра-
батывает женские половые клетки — яй-
цеклетки.
От яичника отходит короткий яйцевод,
который в свою очередь связан с трубкой
влагалища. Последняя начинается жен-
ским половым отверстием, открывающим-
ся, как и мужское отверстие, в половую
клоаку. На своем пути трубка влагалища
образует расширение — семеприемник.
В нем хранятся сперматозоиды, получен-
ные при совокуплении.
Из яичника в яйцевод поступают яйце-
клетки, из семеприемника — сперматозои-
ды, при слиянии этих половых клеток про-
исходит процесс оплодотворения. Оплодо-
творенные яйцеклетки продвигаются да-
лее по яйцеводу в следующий отдел жен-
ского полового аппарата — в оотип, пред-
ставляющий собой небольшое расшире-
ние половой трубки. Сюда же впадает
желточный проток и проток скорлуповой
железы (тельце Мелиса).
Желточный проток образуется от слия-
ния мелких желточных канальцев, беру-
щих начало от желточников. Желточники
либо разбросаны по паренхиме членика,
либо сконцентрированы в более или менее-
компактные образования, находящиеся у
одних видов по бокам членика, у других —
у задней его стенки возле яичника. По
желточным протокам в оотип поступают
питательные вещества, необходимые для
формирования яиц.
Скорлуповая железа вырабатывает про-
дукт, который поступает в оотип и участ-
вует в образовании скорлупы яиц. Та-
ким образом, в оотипе завершается пол-
ное формирование яиц — они получают
здесь питательные вещества (желточные
клетки) и покрываются оболочками.
От оотипа берет начало матка. В нее
и поступают сформированные яйца. Фор-
ма и строение матки у цестод весьма раз-
нообразны. У низших цестод, у лентецов
например, она представляет собой изви-
той канал, идущий от оотипа к наружно-
му отверстию, расположенному обычно
на брюшной поверхности членика. Это
так называемая открытая матка. Парази-
ты, обладающие такой маткой, выделяют
яйца по мере их формирования. У более
высокоорганизованных червей — цеп-
ней — матка не имеет наружного отвер-
стия, она представляет собой замкнутое
мешковидное образование. Такая матка
заполняет собой весь зрелый членик, ко-
торый, по существу, превращается в ме-
377
шок, набитый яйцами. Выделение яиц
происходит при разрыве стенок членика.
Процесс этот совершается чаще всего во
внешней среде, куда зрелые, отделившиеся
от стробилы членики выходят с испражне-
ниями хозяина.
Развитие. Все цестоды развиваются со
сменой хозяев, причем у одних групп
в биологическом цикле происходит смена
двух хозяев (промежуточный и оконча-
тельный), у других — трех (промежуточ-
ный, дополнительный и окончательный).
У некоторых форм в жизненном цикле мо-
гут принимать участие резервуарные хо-
зяева. Однако явление это среди цестод
встречается сравнительно редко.
Классификация. Класс цестод делится
на два подкласса.
Первый подкласс — цестодообраз-
ные (Cestodaria) — включает небольшое
число видов, которые отличаются следую-
щими важнейшими признаками: 1) тело
их не поделено на членики и содержит
один комплект половых органов; 2) раз-
вивающаяся в яйце личинка этих цестод,
так называемая ликофора, вооружена де-
сятью зародышевыми крючьями.
Наиболее известным представителем
данного подкласса является амфи-
лина (Amphilina foliacea) — паразит по-
лости тела осетровых рыб.
Распространена амфилина у рыб в ре-
ках Каспийского бассейна и в реках
Сибири.
В связи с необычной для цестод локали-
зацией этого гельминта (полость тела хо-
зяина), а также отсутствием в его цикле
развития плероцеркоида высказано пред-
положение, что взрослая амфилина пред-
ставляет собой плероцеркоид, приобрет-
ший способность размножаться половым
путем. Настоящая же половозрелая фор-
ма амфилины обитала в кишечнике ка-
ких-то древних вымерших животных, ви-
димо рептилий, питавшихся рыбами. Воз-
можно, это были ихтиозавры.
Предки современных осетровых рыб,
широко представленные в тот период, бы-
ли дополнительными хозяевами парази-
та. А ракообразные — промежуточные хо-
зяева современных амфилин — выполня-
ли эту роль в цикле паразита, по-види-
мому, и в древние времена.
Второй подкласс — настоящие це-
стоды (Cestoda) — объединяет все осталь-
ные виды данного класса; их довольно
большое количество, и они представлены
разнообразными формами. Для всех их
характерно расчлененное тело (за немно-
гими исключениями), содержащее боль-
шое количество комплектов половых ор-
ганов, и наличие шести зародышевых
крючьев у развивающейся в яйце личин-
ки (так называемой онкосферы).
Подкласс подразделяется на несколько
отрядов. Наибольшее число видов, в том
числе обычных паразитов человека и до-
машних животных, представлено в двух
отрядах: отряде лентецов (Pseudo-
phyllidea) и отряде цепней (Cyclo-
phyllidea).
ОТРЯД ЛЕНТЕЦЫ
<PSEUDOPHYLLIDEA)
Лентецы хотя и паразитируют у пред-
ставителей всех классов позвоночных, од-
нако в большинстве своем связаны с ры-
бами. Для одних видов этих червей рыбы
являются окончательными хозяевами, дру-
гие избрали их дополнительными хозяе-
вами. Роль промежуточных хозяев вы-
полняют ракообразные.
Для лентецов характерно строение го-
ловного конца. Головка этих паразитов
снабжена двумя продольными щелями
или двумя присасывательными ямками.
Крючья на сколексе обычно отсутствуют.
Другая особенность строения ленте-
цов — это открытая матка. От-
верстие матки расположено обычно на
брюшной стороне членика. Через него вы-
деляются яйца по мере их созревания.
Каждый членик, кроме отверстия матки,
имеет еще два — мужское половое отвер-
стие и женское, ведущее в вагину.
Познакомимся с представителями трех
семейств этого отряда, довольно различ-
ных по своему строению и биологии.
Семейство Гвоздичники (Сагуо-
phyllaeidae) — мелкие, примитивные фор-
мы лентецов с нерасчлененным телом
и одним комплектом половых органов,
паразиты кишечника рыб.
378
Кариофиллеус лятицепс, или гвоздич-
ник широкоголовый (Caryophyllaeus lati-
ceps) — наиболее распространенный пред-
ставитель семейства, очень часто встре-
чается у карповых рыб.
Длина тела цестоды 2—4 мм. Головной
конец веерообразно расширен, ботрии из-
менены в своеобразный прикрепительный
аппарат. В передней половине тела рас-
положены многочисленные семенники и
фолликулы желточников. Яичник и матка
с многочисленными яйцами находятся в
последней трети тела. Половые отверстия
открываются в задней части паразита.
У молодых гвоздичников сперва разви-
ваются семенники, а затем органы жен-
ской половой системы.
Развитие этого вида и других гвоздич-
ников протекает при смене двух хозяев —
окончательного и промежуточного.
Промежуточными хозяевами являются
малощетинковые черви — тубифексы.
В их теле из заглоченного яйца паразита
развивается личинка — процеркоид.
В кишечнике рыбы, когда она поедает
зараженных личинками малощетинко-
вых червей, развивается взрослый гвоз-
дичник.
Обычно рыбы бывают заражены боль-
шим количеством гвоздичников — до не-
скольких сотен и даже тысяч экземпля-
ров. Особенно опасен этот паразит для
молоди.
Семейство Ремнецы (Ligulidae)—
довольно крупные паразиты с ремневид-
ным сильномускулистым телом. Во взрос-
лой стадии обитают в кишечнике рыбояд-
ных птиц. Развитие проходит при участии
промежуточных (ракообразные) и допол-
нительных (рыбы) хозяев.
В теле рыб развивается очень крупная
личинка плероцеркоид, по раз-
меру почти не уступающая взрослому
червю. Большая часть зараженных ли-
чинками ремнецов рыб гибнет.
Лигула, или ремнец обыкновенный (Ligula
intestinalis),— один из наиболее широко
распространенных представителей семей-
ства. Окончательными хозяевами его яв-
ляются преимущественно чайки, но он
встречается и у других рыбоядных птиц.
Взрослая форма паразита может дости-
гать 1 м в длину, хотя обычно размеры
лигулы значительно меньше, максималь-
ная ширина ее — 1,2 см. Тело имеет толь-
ко внутреннюю расчлененность, не прояв-
ляющуюся снаружи. На суженном перед-
нем конце червя помещаются две ботрии.
Вдоль всего червя на поверхности тянется
желобок. Внутри тела метамерно распо-
Рис. 214. Гвоздичник
(Caryophyllaeus laticeps):
1 — личинка из тела туби-
фекса; 2 — взрослый пара-
зит из кишечника карпа.
Рис. 215. Ремнец (Ligula intestinalis):
1 — личинка-плероцеркоид, извлеченная из полости
тела рыбы; 2 — рыба, зараженная личинкой ремнеца;
3 — личинки ремнеца, вышедшие из тела рыбы через
разрыв.
379
ложено огромное количество — от 700 до
2000 — комплектов половых органов.
Носители взрослых лигул — чайки и
другие птицы — рассеивают над водое-
мами огромное число яиц паразита. В во-
де из яйца выходит личинка — кора-
ц и д и й. Он покрыт ресничками, с по-
мощью которых активно передвигается.
Корацидия заглатывают веслоногие ра-
чки — промежуточные хозяева лигулы.
В теле рачка корацидий превращается
в процеркоида — личинку удлиненной
формы, лишенную ресничек.
Вместе с рачком процеркоид должен
попасть в рыбу — дополнительного хозяи-
на. Роль эту для данного вида лигулы
выполняют различные виды карповых
рыб. Личинки других видов лигул пара-
зитируют у других групп рыб. Лигулы,
как и все ремнецы, обладают довольно
строгой специфичностью в отношении
рыб — своих дополнительных хозяев. Из
кишечника рыбы процеркоид проникает
в полость и вырастает здесь в личинку
следующей стадии развития — в плеро-
церкоида. Плероцеркоид в теле рыбы до-
стигает, как указывалось, очень крупных
размеров, почти равных половозрелой
форме паразита. При этом в плероцеркои-
де формируются почти все органы, в част-
ности высокой степени зрелости дости-
гает половая система. Недаром для разви-
тия плероцеркоида в организме рыбы
необходим сравнительно долгий срок —
до 14 месяцев.
По мере роста плероцеркоида лигулы
в теле рыбы брюшко ее вздувается, пара-
зит давит на внутренние органы рыб, вы-
зывает иногда их атрофию.
Больная рыба теряет равновесие, дер-
жится у поверхности воды и становится
легкой добычей птиц. Иногда лигула
разрывает стенку тела рыбы и выпадает
в воду,
Л и г у л о з (так называется это забо-
левание) причиняет большой ущерб рыб-
ному хозяйству.
В кишечнике птицы, проглотившей за-
раженную плероцеркоидом рыбу, разви-
тие паразита до половозрелой стадии про-
исходит очень быстро — в течение 35—60
часов. Лигула начинает интенсивно выде-
лять яйца и через 2—4 дня гибнет.
Семейство Дифиллоботрииды (Di-
phyllobothriidae) объединяет очень круп-
ных цестод (некоторые достигают длины
20 м) с полной и четко выраженной члени-
стостью тела, паразитирующих в полово-
зрелой стадии у человека и хищных мле-
копитающих, редко у птиц. Развитие их
протекает при участии промежуточного
и дополнительного хозяев. Роль промежу-
точного хозяина выполняют ракообраз-
ные, дополнительного — чаще всего рыбы,
иногда земноводные и пресмыкающиеся,
редко млекопитающие,
Лентец широкий (Diphyllobothrium la-
tum) — один из обычных представителен
дифиллоботриид. Взрослый червь пара-
зитирует у человека и у многих хищных
млекопитающих, особенно у собак, ли-
сиц, песцов, волков и кошек. Широко рас-
пространен в северных районах земного
шара. В Советском Союзе встречается
в бассейнах нижнего течения рек Лены,
Енисея, Оби, Печоры, в реках Карелии,
на Кольском полуострове и в некоторых
прибалтийских районах.
Половозрелый лентец может достигать
15 м в длину. Из гельминтов, паразити-
рующих у человека, это самый крупный
вид. Длинное тело лентеца состоит из
множества члеников, количество их исчис-
ляется несколькими тысячами. Зрелые
членики почти квадратной формы, наи-
большие из них имеют ширину 1,5 см.
Передний конец паразита суженный, го-
ловка несет две ботрии, В центре зрелых
члеников расположена розетковидная
матка.
В каждом членике формируется одинар-
ный комплект половых органов. Вначале
в молодых члениках развивается мужская
половая система, затем в них же образуют-
ся еще органы женской половой системы,
зрелые членики содержат только матку,
наполненную яйцами.
Матка расположена в центре членика.
Она заметна невооруженным глазом, име-
ет характерную розетковидную форму,
благодаря чему лентец легко отличается
от других крупных цестод, паразитирую-
щих у человека,— бычьего и свиного
цепней. У этих паразитов матка имеет
вид продольного стержня с боковыми от-
ветвлениями.
380
Рис. 216. Цестоды, паразитирующие во взрослой
стадии в кишечнике человека:
1 — цепень свиной (Taenia solium); 2 — цепень бычий
<Taeniarhynchus saginatus); 3 — лентец широкий (Di-
phyllobothrium latum). Верхний ряд — головки
цестод, нижний —зрелые членики.
Яйца, откладываемые лентецом (матка
лентеца имеет отверстие, у цепней матка
замкнутая), овальной формы, с крышечкой
на одном полюсе. Во внешнюю среду яйца
попадают вместе с испражнениями хо-
зяина.
Развитие лентеца широкого протекает
следующим образом. Яйцо попадает в во-
ду, Здесь в нем формируется личинка —
корацидий, который вскоре через открыв-
шуюся крышечку покидает оболочки яй-
ца. Корацидий с помощью ресничек ак-
тивно плавает в воде. Дальнейшее развитие
паразита протекает в теле рачков-цикло-
пов — промежуточных хозяев лентеца.
Заглоченный циклопом корацидий из ки-
шечника рачка проникает в полость и
превращается здесь в процеркоида.
Зараженные циклопы вместе с другим
кормом поедаются рыбами — дополни-
тельными хозяевами лентеца широкого.
Чаще других эту роль выполняют щука,
ерш, окунь, налим, лосось, форель, хариус,
сиг. Процеркоиды паразита, попав в желу-
док рыбы, проникают в полость ее тела,
в мышцы и другие ткани, превращаясь
в личинку следующей стадии — в плеро-
церкоида. Личинка этой стадии вытянутой
формы, длиной от 0,5 до 1 см, молочно-
белого цвета, на головке ее уже
видны присасывательные щели. Извлечен-
ная из рыбы, живая личинка постоянно то
сокращается, то вытягивается.
Заражение окончательных хозяев ленте-
цом широким (человека, хищных млеко-
питающих) происходит в том случае, если
они поедают сырую или недоваренную
рыбу, в теле которой находятся живые
плероцеркоиды. В кишечнике окончатель-
ного хозяина личинки паразита прикреп-
ляются головкой к стенке и начинают быст-
ро расти. Через 5—6 недель лентец
становится взрослым и приступает к от-
кладке яиц.
Заболевание, вызываемое лентецом ши-
роким,— дифиллоботриоз у чело-
Рис. 217. Отдельные стадии развития лентеца
широкого (Diphyllobotrium latum):
1 — яйцо; 2 — сформировавшийся корацидий в яй-
це; 3 — корацидий, вышедший из яйца; 4 — цик-
лоп с процеркоидоы; 5 — процеркоид; 6 — плеро-
церкоид.
381
века обычно протекает довольно тяжело.
У больных появляется тошнота, боли
в животе, нарушается аппетит, ненормаль-
но работает кишечник; нередко больные
жалуются на быструю утомляемость, сла-
бость, головные боли, раздражительность.
В более тяжелых случаях у больных раз-
вивается сильное малокровие.
При своевременном лечении, т. е. уда-
лении паразита, все эти явления более
или менее быстро исчезают.
Заражаются дифиллоботриозом чаще
всего рыбаки и рабочие рыбоконсервных
заводов, которые нередко едят совсем
сырую рыбу. Часто это заболевание встре-
чается у населения тех районов, где рас-
пространен обычай употреблять в пищу
свежемороженую или слегка поджарен-
ную на вертеле рыбу. Плероцеркоиды
лентеца у такой рыбы часто остаются жи-
выми.
Борьба с дифиллоботриозом ведется
прежде всего путем выявления и лечения
больных и затем путем охраны пресновод-
ных водоемов от загрязнения нечистотами,
с которыми туда попадают и яйца пара-
зита.
ОТРЯД ЦЕПНИ (CyCLOPHYLLIDEA)
К о т р я д у цепней относится большая
часть всех цестод. Это наиболее совершен-
ная и специализированная группа ленточ-
ных червей.
Цепни паразитируют преимущественно
у теплокровных животных — у птиц и
млекопитающих. Несколько видов пара-
зитирует у человека.
Развитие представителей этого отряда
протекает при смене двух хозяев: проме-
жуточного и окончательного. Роль проме-
жуточного хозяина для одних видов вы-
полняют беспозвоночные, главным обра-
зом ракообразные и насекомые, для дру-
гих — позвоночные животные. Имеются
цестоды, промежуточным хозяином кото-
рых может быть человек. В таком случае
личинки паразитов являются причиной
очень серьезных заболеваний.
Для строения цепней характерно нали-
чие на головке четырех присосок и хо-
ботка, часто вооруженного одинарной или
двойной короной крючьев. Своеобразное
строение имеет матка паразита: она не
имеет наружного отверстия, замкнутая,
в типичных случаях мешковидной формы.
Каждый членик цепней, таким образом,
имеет только два, а не три, как это свой-
ственно лентецам, половых отверстия;
в одно открывается мужская половая
трубка, другое — отверстие вагины. Скап-
ливающиеся в матке яйца выходят наружу
при разрыве стенок членика.
Более подробно познакомимся с пред-
ставителями трех семейств цепней — тех,
которые имеют большое значение в меди-
цине и ветеринарии.
Семейство Тенииды (Taeniidae)—
наиболее важная в практическом отно-
шении группа цепней. Среди них ряд
весьма опасных паразитов человека и жи-
вотных, в том числе домашних и промыс-
ловых. У человека тенииды вызывают,
пожалуй, самые тяжелые заболевания,
какие вообще могут причинять паразити-
ческие черви. Заболевания эти часто со-
провождаются сильными страданиями и не-
редко заканчиваются смертью. Причиной
болезней могут быть как взрослые пара-
зиты, так и их личиночные стадии.
Роль промежуточных хозяев в биологи-
ческом цикле тениид всегда выполняют
позвоночные животные или человек.
Бычий, или невооруженный, цепень
(Taeniarhynchus saginatus) в половозрелом
состоянии паразитирует у человека, про-
межуточным хозяином служит крупный
рогатый скот (отсюда название «бычий»).
Паразит распространен почти во всех стра-
нах земного шара.
Взрослая цестода достигает длины 4—
10 м; это один из самых длинных ленточ-
ных червей. Ширина конечных члеников—
5—7 мм. Головка, как и у всех цепней,
несет четыре присоски, однако она не
имеет хорошо развитого хоботка и лише-
на крючьев (отсюда название «невоору-
женный»). Стробила состоит более
чем из тысячи члеников. Характерно
строение матки, находящейся в зрелых
члениках: она имеет продольный ствол,
от которого отходят 17—30 боковых слабо-
ветвящихся отростков. Матка одного чле-
ника вмещает в себе до 175 тыс. яиц.
382
Жизнь бычьего цепня в кишечнике че-
ловека подробно изучена советскими уче-
ными Ж. К. Ш т р о м о м и Ф. Ф. Т а-
л ы з и н ы м. Эти ученые проглотили
зрелые личинки цепня, заразили себя
этим паразитом и в течение нескольких лет
вели тщательные наблюдения за состоя-
нием своего организма и жизнью развив-
шихся в них цепней.
Находясь в кишечнике человека, цепень
периодически отделяет от тела зрелые
членики. У члеников при этом открыва-
ется конец центрального канала, и яйца
могут выходить через образовавшееся от-
верстие наружу. Большая часть члеников
выделяется наружу с испражнениями, но
некоторая часть может выходить активно,
так как оторвавшийся членик обладает
способностью к медленному передвиже-
нию. Самостоятельное выхождение на-
блюдается обычно ночью во время сна
больного, но иногда это может происхо-
дить и днем.
Покинувшие кишечник членики могут
ползать по телу и по белью. За сутки
больной выделяет до 28 члеников, если
учесть активно вышедшие и с испражне-
ниями. В этих члениках из организма
человека каждые сутки выделяется око-
ло 5 млн. яиц цепня.
В кишечнике человека бычий цепень мо-
жет жить более десяти лет.
Крупный рогатый скот — промежуточ-
ный хозяин бычьего цепня — заражается
паразитом, заглатывая яйца его вместе
с загрязненной травой. В зрелом яйце
содержится шестикрючная личинка — о н-
косфера. В кишечнике животного
онкосфера освобождается от оболочек и
проникает в кровеносные сосуды кишеч-
ника. С кровью она попадает в различные
органы и в тканях их превращается в
следующую стадию личинки — в ц и с т и-
ц е р к, которая представляет собой пу-
зырек размером приблизительно с горо-
шину, внутри которого находится с к о-
л е к с цестоды, ввернутый в шейку
наподобие пальца перчатки.
Чаще всего цистицерки, иначе они на-
зываются финны, локализуются в му-
скулах. Однако они могут находиться
под кожей, в глазах, в различных желе-
зах, в мозгу и других органах.
Человек заражается бычьим цепнем,
потребляя в пищу сырую или плохо про-
варенную говядину, пораженную цисти-
церками.
У людей, зараженных бычьим цепнем,
наблюдается тошнота, рвота, боли в жи-
воте, неустойчивый стул. Больные люди
страдают повышенной раздражитель-
ностью , бессонницей, головокружением,
иногда припадками, схожими с эпилепти-
ческими. Известны случаи закупорки
кишечника клубками паразита и проник-
новения их тел в желчный пузырь и под-
желудочную железу. Особенно тяжело-
переносят заболевание дети.
Поражение цистицерками бычьего цепня
крупного рогатого скота проходит обычна
незаметно. Но при сильном заражении,
когда наряду с мышцами поражается мозг
и сердце животных, болезнь проявляется
резко и животные, особенно молодняк,
иногда погибают.
Борьба с тениаринхозом
(заболевание, вызываемое взрослой фор-
мой бычьего цепня) проводится путем выяв-
ления и лечения больных людей, а также
проведения общих санитарно-бытовых
мероприятий. Большое значение в борьбе
с этим заболеванием имеет выявление
и выбраковка зараженных цистицерками
туш убиваемого на бойне крупного рога-
того скота. Эти же меры способствуют
предохранению животных от заражения
личиночными формами паразита.
Свиной, или вооруженный, цепень (Tae-
nia solium) отличается от бычьего мень-
шими размерами тела (длина его 2—3 м)
и строением сколекса. У этого вида цепня
сколекс имеет хорошо развитый хоботок,
который вооружен двумя рядами крючьев
(отсюда название цепня— «вооруженный»).
Имеются отличия и в строении половой
системы: двулопастной яичник снабжен
маленькими добавочными дольками, матка
с меньшим числом боковых отростков —
их всего 7—12. Оторвавшиеся от тела
зрелые членики свиного цепня не способны
к самостоятельным передвижениям.
Во взрослой стадии свиной цепень пара-
зитирует в кишечнике человека. Промежу-
точными хозяевами его являются домаш-
ние и дикие свиньи (отсюда название—
«свиной»).
383
Свиньи заражаются личинка-
ми цепня при поедании корма,
в который попадают яйца пара-
зита. Особенно часто заражают-
ся свиньи в неблагоустроен-
ных хозяйствах, где они не-
редко питаются нечистотами,
содержащими иногда не только
отдельные яйца, но и целые
членики цестоды.
Личинки свиного цепня, на-
зываемые также цистицер-
к а м и или финнами, ус-
троены по такому же типу, как
и у бычьего, локализуются они
главным образом в мышцах
свиней. Помимо мышц, личин-
ки могут поражать и другие
органы.
Человек заражается свиным
цепнем при употреблении в пи-
щу необезвреженного свиного
мяса, содержащего цистицерки
паразита. При заболевании про-
являются симптомы, сходные с
теми, какие описаны для боль-
ных, зараженных бычьим цеп-
нем. Однако для человека свиной цепень
представляет большую опасность, чем бы-
чий. Дело в том, что для свиного цепня че-
ловек может быть не только окончатель-
ным, но и промежуточным хозяином. Попав
тем или иным путем в организм человека,
яйцо свиного цепня способно развиваться
здесь в цистицерка, вызывая заболевание,
называемое цистицеркозом. Характер
заболевания цистицеркозом человека
зависит оттого, в каком органе и в каком
количестве находятся личинки пара-
зита. Если они локализуются в под-
кожной клетчатке, в мышечной соедини-
тельной ткани или в мышцах, тяжелого
заболевания может не быть. Но цисти-
церки могут проникать в мозг или в глаза.
Заболевание в этих случаях сопровождает-
ся сильными головными болями, рвотой,
припадками, подобными эпилептическим,
и другими нервными явлениями, веду-
щими нередко к смертельному исходу. Пора-
Рис. 218.
(Echinococcus
1 — паразиты
ную величину;
лый паразит;
4 — крючок
жение глаз вызывает частичную
или полную потерю зрения.
Удаление цистицерков из моз-
га или глаз возможно только
хирургическим путем.
Борьба с тениидозом
человека и цистицеркозом сви-
ней проводится таким же обра-
зом, как и с заболеваниями,
вызываемыми взрослой и личи-
ночной формами бычьего цепня.
О них мы говорили выше.
Эхинококк (Echinococcus gra-
nulosus), в отличие от бычьего
и свиного цепней,— мелкая цес-
тода. Взрослый червь длиной
всего 2,7—5,4 мм, тело его со-
стоит из 3—4 члеников. Ско-
лекс имеет четыре присоски и
хоботок, вооруженный двумя
рядами крючьев, общим числом
36—40. Матка с яйцами нахо-
дится только в заднем членике.
Взрослые эхинококки пара-
зитируют в кишечнике собак,
волков и других плотоядных
животных. Личинки паразита
обитают в различных органах главным
образом травоядных животных: у овец,
коз, крупного рогатого скота, свиней,
лошадей, верблюдов, оленей, грызунов
и др. Личиночной формой может зара-
жаться и человек, для которого эхинококк
представляет очень большую опасность.
Жизненный цикл паразита протекает
следующим образом. Паразитирующие
в кишечнике собаки взрослые эхинококки
(обычно в числе тысяч экземпляров) отде-
ляют членики, заполненные яйцами.
С испражнениями собаки членики выде-
ляются во внешнюю среду. С загрязнен-
ной пищей — травой, сеном или водой —
яйца попадают в пищеварительный тракт
названных выше промежуточных хозяев.
Особенно легко и часто это происходит
на пастбище, где вместе со скотом бегают
сопровождающие пастухов собаки. Зара-
жению травоядных животных способ-
ствует то, что членики паразита способны
Эхинококк
granulosus):
в натураль-
2 — взрос-
3 — яйцо;
хоботка.
Таблица 21. Свиной солитер (Taenia solium).
Вверху, слева направо, головки свиного и бычьего солитеров.
384
передвигаться. Они могут выходить из
испражнений собаки и расползаться по
почве, по подстилке, могут даже подни-
маться на траву.
К человеку яйца паразита попадают
обычно непосредственно от собаки. Уха-
живая за своим другом, человек ласкает
его, гладит и при этом загрязняет руки
яйцами эхинококка, прилипшими к шер-
сти собаки. Яйца попадают затем в пищу
и с ней в кишечник человека. Особенно
легко могут заражаться дети, которые
часто не только ласкают, но и целуют
собак.
Из проглоченного яйца в пищеваритель-
ном тракте промежуточного хозяина —
травоядного животного или человека —
выходит личинка — онкосфера, кото-
рая проникает в кровеносные сосуды
и с кровью заносится в различные органы,
преимущественно в печень и легкие. Из
онкосферы здесь вырастают эхинококко-
вые пузыри.
Эхинококковый пузырь, или, как его
короче называют, эхинококк-личинка,
по своему строению отличается от описан-
ного цистицерка (личинки бычьего или
свиного цепня). Эхинококк — это плот-
ный шар, наполненный бесцветной жид-
костью, внутри него имеются пузыри мень-
шего размера, так называемые дочерние
пузыри, которые в свою очередь могут
содержать внучатные пузыри. Все они
также заполнены жидкостью. На внутрен-
ней поверхности как материнского, так
и других пузырей вырастают выводковые
капсулы, в которых находятся зародыше-
вые сколексы. Множество зародышевых
сколексов скопляется и вне выводных
капсул, в жидкости, заполняющей пузыри.
Из каждого зародышевого сколекса в
организме окончательного хозяина выра-
стает взрослая цестода.
Эхинококковые пузыри имеют размер
обычно от горошины до арбуза, но в от-
дельных случаях они могут достигать зна-
чительно больших размеров. Ветеринар-
ные врачи находили в печени крупного
рогатого скота эхинококковые пузыри,
которые весили свыше 60 кг; в таком пузы-
ре содержалось около 43 л жидкости.
Личинка эхинококка растет медленно.
Через 5 месяцев она вырастает всего лишь
до 1 еле в диаметре, а предельных разме-
ров может достичь только через 2—3 десят-
ка лет.
Жизненный цикл эхинококка замыкает-
ся, когда эхинококковые пузыри с живыми
головками поедают окончательные хозяе-
ва — собаки и другие плотоядные. Слу-
чается это, когда владельцы собак кормят
их внутренностями и бракованными орга-
нами убиваемых эхинококкозных живот-
ных или когда собаки и дикие хищные
животные поедают туши павшего домаш-
него скота и диких травоядных.
В кишечнике окончательных хозяев
цестоды становятся половозрелыми к кон-
цу третьего месяца после заражения.
Заражение человека эхинококковым
пузырем представляет большую опасность.
Чаще всего личинка локализуется в пече-
ни и легких. Разрастающийся пузырь
разрушает ткань пораженного органа и да-
вит на окружающие ткани. Возможны
разрывы пузыря, при которых содержимое
изливается в полость тела,в легкие, в желч-
ный пузырь и т. д. В полости тела при раз-
рыве пузыря из его содержимого иногда
развиваются новые эхинококковые пузы-
ри, усугубляющие заболевание. Разрывы
пузыря ведут часто к смерти хозяина.
Эхинококкоз животных, вы-
званный личиночной стадией, протекает
обычно хронически и бессимптомно. Одна-
ко при очень сильном заражении поражен-
ное животное резко худеет и может по-
гибнуть.
Алъвеококк (Alveococcus multilocularis)
по строению взрослых цестод очень бли-
зок к эхинококку. Более четко различают-
ся личиночные формы этих видов. У аль-
веококка личинка имеет вид грозди пу-
зырьков, общий размер которой достигает
грецкого ореха. Отдельные пузырьки,
величиной с горошину, содержат зароды-
шевые сколексы.
Окончательными хозяевами альвеокок-
ка являются главным образом лисицы,
хотя он может паразитировать у собак,
волков и других хищных животных. Ли-
чиночная форма поселяется преимущест-
венно у грызунов. Ею может заражаться
и человек.
В некоторых районах Сибири альвео-
кокк у людей встречается чаще, чем
Щ0 25 ЖИЗНЬ ЖИВОТНЫХ, Т. 1
385
эхинококк. У человека личинка альвео-
кокка поражает главным образом печень
и легкие.
Заболевание, вызванное альвеококком,
у человека протекает обычно более зло-
качественно, чем при заражении эхино-
кокком. Личинка альвеококка имеет
свойство неудержимо разрастаться, про-
никая при этом из одного органа в дру-
гой. В этом отношении заболевание напо-
минает рак.
Избавление от личиночных форм как
эхинококка, так и альвеококка возможно
только путем хирургической операции.
Чтобы уменьшить опасность заражения
этими паразитами, нужно регулярно обсле-
довать собак и при обнаружении у них
взрослых паразитов лечить их. Следует
строго следить, чтобы собаки не могли
съесть мясо и внутренние органы живот-
ных, зараженных эхинококковыми пузы-
рями.
Рис. 219. Мозг овцы, пораженный мозго-
виком (Multiceps multiceps). Ценур
расположен в передней части левого по-
лушария мозга A).
Мулыпицепс, или мозговик овечий (Multi-
ceps multiceps). Взрослая цестода живет
в кишечнике собак, волков, лисиц и некото-
рых других плотоядных семейства псовых.
Паразит достигает 40—100 см длины
и 5 см ширины. Стробила состоит из
200—250 члеников. Головка с присо-
сками и хоботком, вооруженным 22—
23 крючьями. В матке зрелых члеников от
центрального ствола отходит в каждую
сторону по 9—26 отростков.
Личиночная стадия мозговика — она
называется ценур — локализуется в
головном, иногда в спинном мозгу овец.
Находили ценура и у некоторых других
парнокопытных животных, а также у чело-
века.
Ценур представляет собой наполнен-
ный жидкостью пузырь размером с голу-
биное яйцо, иногда больше. На внутрен-
ней стороне его находятся небольшие бело-
ватые утолщения. Это головки паразита,
которых в одном пузыре может быть иног-
да до нескольких сотен.
Овца заражается ценурами, когда в же-
лудок ее с травой или водой попадают
яйца мозговика. Яйца на пастбище рас-
сеивают собаки, зараженные взрослой
формой паразита. Из яйца в пищевари-
тельном тракте овцы выходит о н к о-
сфера, которая проникает в кровенос-
ные сосуды и с кровью достигает голов-
ного мозга, превращаясь здесь в ценура.
Поедая голову овцы, пораженной личинка-
ми паразита, собака заражается мульти-
цепсами.
Мозговик распространен в тех районах,
где разводятся овцы и где не ведется до-
статочная борьба с бродячими собаками.
В этих же районах могут заражаться цену-
ром и люди. Наибольшей опасности зара-
жения подвержены работники овцевод-
ческих хозяйств, особенно те, которые
близко соприкасаются с собаками.
У человека ценуры могут локализо-
ваться в головном или спинном мозгу,
в глазах, под кожей, в мышцах и в меж-
мышечной соединительной ткани. При
поражении мозга заболевание длится
несколько лет и сопровождается острыми
болезненными явлениями. Лечат людей
путем удаления ценура хирургическим
путем.
386
У овец ценуры локализуются главным
образом в мозгу. Паразит давит на неж-
пую мозговую ткань и вызывает ее отми-
рание. Если пузырь находится на поверх-
ности мозга, он давит на кости черепа, они
истончаются, иногда даже продырявли-
ваются. Заражается ценуром преимуще-
ственно молодняк в возрасте до года.
Внешне болезнь проявляется по-раз-
ному в зависимости от места нахождения
личинки в мозгу, от количества пузырей
и состояния овцы.
При поражении лобных долей мозга
овца стремительно бежит или подолгу
стоит, опершись опущенной головой в ка-
кой-либо предмет. Если ценур локали-
зуется в височно-теменной доле, то овца
совершает круговые движения. Указан-
ный симптом особенно характерен, и поэ-
тому в народе болезнь получила назва-
ние «вертячка». При локализации в ви-
сочно-теменной области мозга у животных
часто нарушается зрение на стороне, про-
тивоположной пораженной половине.
В том случае, если ценур находится в за-
тылочной доле, овца с высоко поднятой
или запрокинутой на спину головой пятит-
ся назад или падает. Поражение мозже-
чка вызывает нарушение равновесия, ко-
ординации движений и частичный паралич
мускулатуры.
Больные овцы отказываются от корма,
худеют и обычно погибают. Случаи само-
стоятельного выздоровления их исклю-
чительно редки.
Лекарственное лечение овец, заражен-
ных ценурами, успеха не имеет и по-
этому не применяется. В последние годы
в Советском Союзе разработан и широко
применяется метод хирургического лече-
ния, заключающийся в оперативном уда-
лении ценуров. В результате такого лече-
ния около 80% больных овец возвращает-
ся колхозам здоровыми.
Для предохранения овец от заражения
ценурами нужно не допускать, чтобы соба-
ки могли поедать головы больных живот-
ных. Собак, обслуживающих овцеводче-
ские хозяйства, необходимо периодически
обследовать на зараженность их взрос-
лыми формами паразита и лечить.
Семейство Аноплоцефалиди (Апор-
locephalidae) включает большое количе-
25*
ство цепней, хозяевами которых являются
главным образом млекопитающие. Мно-
гие из них паразитируют у жвачных жи-
вотных, вызывая тяжелые заболевания.
По своему строению аноплоцефалпды
характеризуются следующими призна-
ками. Сколекс лишен хоботка и крючьев,
стробила обычно большая. Ширина ее
члеников в несколько раз превышает
длину. Семенников много, яичник лопаст-
ной, зрелая матка мешковидная с много-
численными выпячиваниями. Яйца содер-
жат так называемый грушевидный
аппарат, который образован внутрен-
ними оболочками яйца. Аппарат заклю-
чает в себе онкосферу.
Цикл развития аноплоцефалид проте-
кает со сменой двух хозяев. Промежуточ-
ными хозяевами являются мелкие почвен-
ные, или панцирные, клещики.
Мониезия экспанза (Moniezia expansa)—
наиболее распространенный представи-
тель аноплоцефалид. Взрослая цестода
паразитирует в кишечнике овец, коз, круп-
ного рогатого скота и некоторых диких
жвачных.
Паразит достигает 1—5 м длины при:
максимальной ширине до 16 мм. Для
члеников характерно наличие близ их
передних краев розетковидных желез, так
называемых «межпроглоттидных желез».
В каждом членике двойной комплект
женских и мужских половых органов,
располагающихся двумя группами возле
каждого бокового края членика. Соответ-
ственно этому на каждой боковой стороне
членика имеется половая клоака, куда
открываются женские и мужские половые
протоки.
Яйца паразита или целые зрелые чле-
ники с фекалиями овец рассеиваются по
пастбищу. Почвенные клещики — проме-
жуточные хозяева мониезии — заглаты-
вают яйца. В теле клещиков из яиц раз-
виваются цистицерки. Процесс этот длит-
ся довольно долго—2—4 месяца.
К овцам, телятам и другим окончатель-
ным хозяевам зараженные клещики попа-
дают вместе с травой. В кишечнике ука-
занных хозяев через 4—6 месяцев из
цистицерков вырастает взрослая мониезия,
способная отделять членики со зрелыми
яйцами.
387
Мониезиоз (заболевание, вызы-
ваемое мониезиями) наносит огромный
ущерб животноводческим хозяйствам.
Иногда гибель молодняка скота в отдель-
ных хозяйствах может достигать 80%.
Тяжесть заболевания зависит от того,
каким количеством паразитов заражено
животное. Известны случаи, когда у телят
или молодняка овец мониезии почти пол-
ностью на большом протяжении занимали
просвет тонкого кишечника. Естествен-
но, что при таком заражении животные
крайне истощаются и, если не приме-
няется лечение, гибнут.
Борьба с мониезиозом проводится глав-
ным образом путем так называемой пре-
имагинальной дегельминтизации. Меро-
приятие это заключается в том, что живот-
ным (в хозяйствах, где наблюдается ука-
занное заболевание) дают лекарственные
препараты в то время, когда паразиты не
достигли еще половой зрелости и не нача-
ли выделять во внешнюю среду яйца.
В течение пастбищного сезона животные
получают лекарства дважды.
Семейство Гименолепидиды (Ну-
menolepididae) — одна из наиболее совер-
шенных групп цепней, объединяющая
очень большое количество видов, парази-
тирующих у птиц, млекопитающих, а
также у человека.
Для строения гименолепидид
характерны следующие особенности: ма-
лое число семенников (обычно три, иногда
даже два или один); небольшое число крю-
чьев на хоботке сколекса (обычно 8—10),
которые расположены в один ряд; в муж-
ской половой системе — наличие круп-
ного наружного семенного пузырька и
для ряда видов хитинового стилета, про-
низывающего циррус; в женской половой
системе — крупный яичник, компакт-
ный желточник и хорошо развитый семе-
приемник, занимающий иногда значитель-
ную долю членика.
Развитие гименолепидид проте-
кает со сменой двух хозяев. Промежуточ-
ными хозяевами в большинстве случаев
являются ракообразные или насекомые.
Карликовый цепень (Hymenolepis nana —
один из немногих видов гименолепидид,
паразитирующих у человека. Отличается
от других представителей семейства тем,
что человек является для него одновре-
менно и окончательным и промежуточным
хозяином.
Из яиц, выделяемых паразитом в ки-
шечник человека, выходят личинки —
онкосферы, которые проникают
в ворсинки. Здесь из них развиваются
другие личинки — цистицерко-
и д ы. Через некоторое время цистицер-
коиды выходят в кишечник и достигают
половозрелой стадии. Часть яиц выде-
ляется с фекалиями наружу, и если тем
или иным путем они попадают затем в пи-
щеварительный тракт человека, то тоже
развиваются в нем во взрослого червя.
Карликовым цепнем заражаются чаще
дети, чем взрослые. Это связано с более
слабым развитием у детей гигиенических
навыков и меньшей сопротивляемостью
детского организма к заражению.
* * *
Большое число видов гименолепидид
паразитирует у птиц, в том числе и у до-
машних. Особенно большую опасность
представляют эти гельминты для водо-
плавающих птиц — гусей и уток. У гусей
чаще других паразитирует гименолепис
ланцетовидный (Drepanidotaenia lanceo-
lata). Развитие паразита происходит
при участии рачков циклопов (промежу-
точные хозяева).Паразит нередко вызывает
массовый падеж молодняка гусей.
У уток очень часто паразитируют гиме-
нолепидиды родов Dicranoraenia,
Diorchis, Aploparaxis. Промежуточными
хозяевами представителей двух первых
родов являются рачки, последнего —
кольчатые черви.
ТИП НБМАТЕЛЬМИНТЫ
(NEMATHELMINTHES)'
На предшествующих страницах чита-
тель имел возможность ознакомиться с
различными группами платод. Он мог убе-
диться в том, что в пределах этого типа
свободный образ жизни представлен лишь
одним классом турбеллярий, тогда как
три других класса платод включают слож-
ный и многообразный мир паразитов жи-
вотных и человека, нередко весьма опас-
ных для их здоровья и существования.
Тип немателъминтов также содержит
в своем составе большое число форм, опас-
ных для жизни и здоровья животных
и человека, т. е. большое число паразитов.
Но вместе с тем нельзя не отметить, что
в пределах типа немателъминтов размах
многообразия форм жизни становится еще
шире. Среди немателъминтов много форм,
заселивших в качестве свободноживущих
организмов море и пресные водоемы, почву
всех материков света и вместе с тем овла-
девших органами и тканями не только всех
животных и человека, но равным образом
всех известных нам растений — от грибов
и водорослей до высших растений вклю-
чительно. Можно смело сказать, что нет
таких биотопов на нашей планете, в кото-
рых не было бы представителей этого
типа, и нет таких животных и растений,
1 Немателъминты — от греческих слов пета —
нить и helmins — червь.
в органах которых нельзя было бы найти
его представителей. Какова же организа-
ция этих животных?
Прежде чем ответить на этот вопрос,
мы должны ознакомиться с основными
признаками этого типа животных.
Пожалуй, самым интересным моментом
характеристики основных признаков не-
мателъминтов можно считать их происхож-
дение от турбеллярий. Эта точка зрения
является наиболее обоснованной. Однако
первое же знакомство с организацией не-
мателъминтов приводит наблюдателя к кон-
статации у них таких признаков, которые
отсутствуют у турбеллярий. Во-первых,
все тело немателъминтов одето в плотный
кутикулярный покров. Во-вторых, мор-
фологическое исследование этих животных
тотчас обнаруживает, что у них нет раз-
витого кожно-мускульного мешка в том
виде, который наблюдается у турбеллярий
и вообще у платод- Если мышечный слой
имеется, то он всегда состоит только из
продольных мышц, образованных удлинен-
ными клетками. Но у многих представи-
телей этого типа животных нет и системы
мышечного подкожного слоя; развиты
только разобщенные, отдельные мышечные
тяжи, а не сплошной мышечный слой. Та-
ким образом, либо кожно-мускульный
мешок «обеднен» и ограничен только про-
дольной мускулатурой, либо она оказы-
вается представленной только отдельными
389
тяжами продольных мышц, к которым
присоединяются, и притом не всегда, се-
рийно расположенные кольцевые мышцы
(рис. 220). Даже в тех случаях, когда
мышцы образуют сплошной слой, он все
же прерывается вдоль спинной, двух боко-
вых и брюшной сторон, образуя, таким
образом, четыре разобщенных мышечных
сектора. Следовательно, для нематель-
минтов характерна рудиментация и даже
частичная редукция кожно-мускульного
мешка их платодных предков. Вместе с тем
кожно-мускульный мешок этих живот-
ных приобретает более мощно развитый
кутикулярный покров. Таким образом,
кожно-мускульный мешок состоит из ку-
тикулы, слоя живых эпителиальных кле-
ток, получившего наименование гипо-
дермы, и частично или полностью де-
зинтегрированного мышечного
слоя.
Другая особенность организации нема-
тельминтов состоит в том, что у этих
животных появляется на первый взгляд
совершенно новый признак: почти пол-
ностью утрачивается паренхима и между
Рис. 220. Поперечный разрез через тело
аскариды:
J — полость тела, 2 — кожно-мускульный мешок;
3, 4 — спинной и брюшной нервный ствол; 5 —
кишечник; 6 — матка; 7 — яичники.
стенкой тела (дезинтегрированным кожно-
мускульным мешком) и внутренними
органами появились щели, заполненные
жидкостью. Эти щели образуют так назы-
ваемую первичную полость
тела, которая никогда не имеет соб-
ственных стенок, так как их роль выпол-
няется стенкой тела, а не каким-либо
специальным слоем клеток, ограничиваю-
щих эту полость.
Третьим важным новым признаком надо
считать четкую дифференцировку пищева-
рительной системы на три отдела: перед-
нюю, среднюю и заднюю кишку, откры-
вающуюся наружу заднепроходным отвер-
стием (анусом), лежащим иногда на
спине, но чаще на брюшной стороне тела.
Наличие ануса вызвало образование еще
одной части тела — хвостового отдела,
под которым понимается участок тела,
лежащий позади ануса.
Четвертый важный признак — переме-
щение ротового отверстия на передний
конец тела, и притом либо несколько
наклонно к брюшной стороне тела, либо
строго на конце его. Отметим также, что
для немательминтов очень характерно
заметное увеличение длины тела и умень-
шение его диаметров, почему очень часто
возникает веретеновидная или даже цилин-
дрическая форма его.
В заключение несколько слов об орга-
нах, которых у немательминтов никогда
не бывает. Нет у них дыхательной и кро-
веносной систем. Дыхание осуществляется
всей поверхностью тела через кожные по-
кровы, а также через полость кишечника.
Внутренние же органы «омываются» жид-
костью, заполняющей первичную полость
тела, участвующей в сложных процессах
обмена и переработки его продуктов
в связи с функциями выделительных орга-
нов.
В состав типа первичнополостных вхо-
дит несколько классов, из которых рас-
смотрим следующие: класс Брюхорес-
ничные, или Гастротрихи (Gastrotricha),
класс Киноринхи (Kinorhyncha),
класс Нематоды (Nematoda), класс
Коловратки (Rotatoria). Обзор этих клас-
сов позволит составить более четкое и бо-
лее конкретное представление о нематель-
минтах.
390
КЛАСС БРЮХОРЕСНИЧНЫЕ, ИЛИ ГАСТРОТРИХИ
(GASTROTRICHA)
Эти крохотные многоклеточные жи-
вотные были обнаружены еще в XVIII в.
Несовершенство микроскопов и ограничен-
ность знаний о микроскопических живот-
ных привели к тому, что гастротрихи рас-
сматривались как инфузории, на которых
они похожи по величине. Наука должна
быть благодарна ученым (Отто Б ю ч л и,
И.И.Мечников, 3 е л и н к а и др.),
раскрывшим природу гастротрих. Зели-
нк а еще в 1889 г. сообщил ряд ценных
сведений, раскрывших главные особен-
ности организации гастротрих. Большое
значение в изучении их имели работы
Ш у л ь ц е (Schultze). Однако наиболь-
шие заслуги в полном анализе органи-
зации гастротрих, их систематики, эко-
логии и биологии принадлежат ныне ра-
ботающему исследователю Р е м а н е
(Remane), ценные работы которого сыг-
рали решающую роль в прогрессе зна-
ний об этих животных.
* * *
Гастротрихи — весьма небольшая
группа животных, их известно около 160
видов. Самые крупные из них достигают
всего 1,5 мм, все другие — около 0,5 мм
и меньше. Тело гастротрих покрыто кути-
кулой. Форма тела многообразная: иногда
она напоминает микроскопические буты-
лочки или приближается к цилиндру; у не-
которых гастротрих задняя часть тела
вытянута в длинный хвост; у многих форм
на заднем конце тела двувершинный виль-
чатый хвостовой конец. Головной конец
тела несет щетинки и осязательные органы.
По бокам головы, слева и справа, распо-
ложены снабженные ресничками обоня-
тельные ямки, напоминающие подобные
же органы у турбеллярий (см. выше).
Ротовое отверстие находится на перед-
нем крае головного конца тела, смещаясь,
однако, несколько на брюшную сторону.
Оно ведет в длинную глотку (фаринкс),
которую часто называют пищеводом. Хотя
это действительно глотка, а не пищевод,
оставим последний термин, так как в дан-
ном случае он очень удобен. Ротовое отвер-
стие обычно открывается в расширенную
часть глотки, функционирующую как
ротовая полость (см. рис. 221). Ротовая
полость переходит в лежащую позади нее
длинную часть глотки, которая функцио-
нально напоминает пищевод, откуда и воз-
никло это наименование. Будем только
помнить, что пищевод гастротрих гомо-
логичен (т. е. равнозначен по происхож-
дению) глотке. У разных видов этот пище-
вод многообразен по форме. У одних
видов он может быть цилиндрическим,
у других — снабжен сферическим расши-
рением — бульбусом, или луковицей.
Стенки пищевода и бульбуса (иногда их
несколько) снабжены сильной радиальной
мускулатурой. Просвет пищевода всегда
трехгранный. У части видов гастротрих
просвет пищевода сообщается с наружной
средой (морской или пресной водой, в ко-
торой эти виды живут) с помощью боковых
пищеводных пор. Ни у одной
другой группы животных таких пищевод-
ных пор нет.
За пищеводом следует средняя кишка,
стенка которой состоит из одного слоя
клеток: на внутренней стороне этих кле-
ток, обращенной в полость кишки, у мно-
гих гастротрих имеются реснички, рас-
пределяющие пищевые массы в полости
кишки, так как они работают и переме-
щают пищевые частицы. Пищеваре-
ние у гастротрих внутриклеточ-
ное. На брюшной стороне тела развито
ресничное поле, покрытое тонкими рес-
ничками. Эти реснички, так же как у рес-
ничных червей, согласованно работают
и обеспечивают довольно быстрое пере-
движение гастротрих по песчинкам и
другому субстрату морского или речного
дна. Гастротрихи могут плавать. Нередко
они совершают быстрые скачки, очень
для них характерные. Реснички служат
391
Рис. 221. Организация гастротрих.
А — пресноводная гастротриха Chaetonotus maximus;
Б — анатомия пресноводной гастротрихи: J — рото-
вое отверстие, 2 — пищевод; 3 — средняя кишка; 4 —
протонефридии; 5 — яйцо. В — морская гастротриха
Macrodasys buddenbrocki: l — обонятельные ямки;
2 — пищевод; 3 — пищеводные поры; 4 — средняя
кишка; 5 — яичник; 6 — яйца; 7 — семеприемник;
8 — анальное отверстие; 9 — мужской совокупитель-
ный орган; 10 — семенник; и — семяпровод; 12 —
присасывательные ножки.
также механизмом, который вовлекает
пищу гастротрих в ротовое отверстие.
Питаются они различными мелкими орга-
низмами, в том числе и мелкими инфузо-
риями. Ресничные поля создают ток воды,
направленный спереди назад. «Этим пу-
тем,—пишет П. Щульце (Schultze),—
частицы пищи достигают головы. Распо-
ложенные внизу рта серповидные, на-
клоненные вперед реснички проявляют
в это время своеобразную деятельность:
короткими ударами они направляют
частицы пищи к ротовому отверстию, при-
чем не исключено предположение, что они
выполняют роль чувствующих волосков
с определенной избирательной функ-
цией».
Отбросы пищи выбрасываются наружу
через анус. Жидкие продукты обмена
веществ выделяются слаборазвитой выде-
лительной системой. Она принципиально
построена так же, как у турбеллярий,
т. е. состоит из клеток с мерцательным
пучком жгутов, работа которых обеспечи-
вает отток жидких продуктов обмена по
выделительным каналам. У гастротрих
таких каналов всегда два — справа и сле-
ва. Выделительные клетки с мерцатель-
ным аппаратом немногочисленны, обычно
их всего две, по одной на каждый ка-
нал.
Гастротрихи— гермафродиты, т. е. обое-
полые животные. Поэтому у каждой особи
имеются мужские и женские половые
органы (рис. 221). Организация половой
системы гастротрих в известной мере на-
поминает строение ее у ресничных червей.
На рисунке 221 видно, что в организме
гастротрих формируются яйца, из которых
развиваются молодые животные.
Хотя гастротрихи и движутся при по-
мощи ресничек, у них все же имеется
система телесных (соматических) мышц.
Однако, в отличие от кожно-мускульного
мешка ресничных червей, у гастротрих
мышцы не образуют сплошного слоя и
лежат обособленными лентами. Это озна-
чает, что мышечные элементы кожно-мус-
кульного мешка гастротрих подверглись
регрессивному развитию. Едва ли можно
сомневаться, что главная причина этого
заключается в том, что гастротрихи очень
маленькие животные и их передвижения
полностью обеспечиваются работой рес-
ничек. Это, видимо, и было тем фактором,
который вызвал редукцию мышечной
системы и ее частичное рудиментирова-
ние. На вероятность этого предположения
указывает тот факт, что у гастротрих,
достигающих более крупных размеров,
до 1,4 мм в длину, реснички рудименти-
руются, так как не могут преодолеть со-
противления длинного тела. Возникает
новый, весьма интересный способ движе-
ния (не за счет работы ресничек и, ко-
нечно, мышц, поскольку они уже рудимен-
тированы) с помощью особых «хвататель-
ных трубочек» (см. рис. 221). Эти трубо-
чки снабжены прикрепительными дисками,
с помощью которых гастротрихи как бы
«присасываются» к донным предметам
и передвигаются, «шагая» с помощью
трубочек по донному субстрату морско-
го или пресноводного бассейна.
392
Когда смотришь на гастротриху в мик-
роскоп под слабым увеличением, прежде
всего замечаешь ее движения, то плавные,
то переходящие в стремительные и неожи-
данные броски. Кроме того, сразу же
можно увидеть некоторые особенности
внешней организации гастротрих. На
крохотном тельце, внешне похожем на
облик брюхоресничной инфузории, видны
длинные щетинки, а кутикула при пово-
ротах зеркала микроскопа порой мер-
цает в лучах света. Это объясняется тем,
что у многих форм кутикула имеет слож-
ное строение и состоит из отдельных «че-
шуек» различной формы. В пресных водах
гастротрихи ползают по поверхности
подводных предметов или по листьям
водных растений. Однако они способны и
плавать. На морском дне гастротрихи
образуют одну из самых интересных
групп микроскопического мира, поражая
многообразием своих форм.
Этот маленький класс немателъмин-
тов представляет значительный интерес
для каждого специалиста, изучающего
историю животного мира. Выше вскользь
уже отмечались некоторые элементы сход-
ства гастротрих с турбелляриями. Сюда
надо отнести наличие ресничного покрова
тела, развитие обонятельных ямок, при-
сутствие у гастротрих типичных протоне-
фридиев, известное сходство половой
системы этих групп. Отметим еще, что
центральная нервная система гастротрих
напоминает центральную нервную систему
турбеллярий. Как у этих животных,
у гастротрих развит «мозг», состоящий
из скопления нервных клеток; и как
у видов класса турбеллярий, у гастрот-
рих имеются два продольных нервных
ствола.
Современная наука не без основания
принимает взгляд, согласно которому
гастротрихи произошли от прямо-
кишечных турбеллярий. Но если среди
последних встречаются не только мелкие
представители этого класса, но и относи-
тельно крупные, то эволюция гастротрих
сопровождалась уменьшением размеров
тела. Поэтому главным органом движения
у них остались реснички, покрывающие
поля брюшной стороны тела. Мышцы
кожно-мускульного мешка, вероятно,
именно поэтому утратили то значение, ко-
торое они имеют у турбеллярий, и одни из
них подверглись полной редукции, а дру-
гие — известной рудиментации. В резуль-
тате мышечный слой кожно-мускульного
мешка резко деградировал, и у гастротрих,
вместо мышечного слоя турбеллярий, со-
хранились лишь изолированные мышечные
тяжи. Эта утрата сплошного мышечного
слоя оказалась необратимой для класса
немательминтов.
Рис. 222. Пресноводные гастротрихи:
J — Ichthydium forcipatum; 2 — Lepi-
doderma squamatum, 3 — Chaetonotuo
chuni; 4 — Dasydytes ornatus.
393
КЛАСС НЕМАТОДЫ, ИЛИ КРУГЛЫЕ ЧЕРВИ
(NEMATODA)
Если бы тип немательминтов был огра-
ничен только рассмотренными выше клас-
сами животных, то едва ли была бы по-
строена та общая характеристика этого
типа, которая возникла в современной
науке. Кроме того, его значение и в эко-
номике природы, и в хозяйственной дея-
тельности человека было бы настолько
ограниченным, что изучение нематель-
минтов не достигло бы того прогресса,
который наблюдается в нашей современ-
ности. Глубокий интерес к типу нематель-
минтов не столько определяется характери-
стиками гастротрих, киноринх и даже
коловраток, сколько становится следст-
вием изучения центральной группы, глав-
ного и наиболее крупного класса нема-
тельминтов — класса нематод (Nematoda).
Естественно, что этот класс нематель-
минтов будет в центре и нашего внимания.
Биологический прогресс. Известный
•советский ученый академик А. Н. Се-
ве р цов A866—1936), исследуя эво-
люцию позвоночных, подверг глубокому
изучению проблему основных направле-
ний филогенеза животных. Он установил,
в частности, что для многих естествен-
ных групп животных типично состояние
эволюционного развития, характеризу-
ющееся рядом ведущих признаков, опреде-
ляющих жизненный успех этих групп.
А. Н. Северцов обозначил это как специ-
фическое проявление биологичес-
кого прогресса.
Биологический прогресс, указывает
А. Н. Северцов, характеризуется сле-
дующими признаками: 1) численным
увеличением особей данной систематиче-
ской (таксономической) группы; 2) про-
грессирующим расселением, т. е. захва-
том новых сред обитания; 3) возрастающим
многообразием форм (подвидов, видов,
родов и т. п.).
Класс нематод характеризуется
именно этими общими биологическими
лризнаками — признаками биологического
прогресса. Нематоды овладели всеми
лзвестными науке местообитаниями.
Мы не знаем таких сред обитания, таких
биотопов, в которых не было бы нематод.
Дно морей и океанов от северного до
южного полюса (об этом можно говорить
с полной уверенностью) заселено огром-
ным количеством видов и особей нематод.
Свободноживущие нематоды известны ре-
шительно всюду, во всех точках морского
дна, которые подверглись специальным
исследованиям. Завоевав дно всех морей
и океанов, нематоды проникли, видимо
позднее, в солоноватые водоемы. Поэтому
очень большое число видов нематод оби-
тает на дне солоноватоводных бассейнов,
в том числе в лиманах — этих преддве-
риях рек, впадающих в моря. Известен
ряд фактов, свидетельствующих о том,
что в древней истории класса нематод
наступил такой важный этап развития,
когда они стали проникать в пресные
водоемы, и в конце концов многие группы
свободноживущих нематод заселили реки.
В дальнейшем нематоды совершили еще
один важный шаг исторического развития—
они проникли в почвенные воды и стали
компонентами почвенной фауны — комп-
лекса почвенных биоценозов. Отметим,
что на этом экологическое развитие части
групп и остановилось. Для некоторых
групп нематод оказался открытым путь
в сапробиотические очаги. Органические
останки растений и животных подверга-
ются гниению, обусловленному жизне-
деятельностью сменяющих друг друга
групп гнилостных бактерий, под влиянием
которых в почве образуются очаги гнилост-
ного распада. В этих очагах соответствую-
щие группы сапрофитных бактерий после-
довательно разлагают органический мате-
риал на более простые составные части.
В частности, белки расщепляются на
более простые, растворимые в воде компо-
ненты; полисахариды трансформируются
в растворимые ди- и моносахариды; рас-
падаются клетчатка, жиры, пектиновые
оболочки клеток растительной ткани
и т. д. Поэтому сапробиотические очаги
оказались наиболее доступными источ-
394
тайками питания нематод. Так
сформировалась богатая видами
группа сапробиотических нематод.
Именно эта группа стала источ-
ником развития других групп не-
матод, перешедших к паразитиче-
скому существованию за счет ор-
ганизмов животных и растений.
В конечном счете формируются
два больших потока родов и ви-
дов нематод, приспособившихся,
с одной стороны, к паразитирова-
бию в органах человека и живот-
ных, а с другой—в органах рас-
тений.
Из сказанного выше можно ви-
деть, что все биотопы нашей пла-
неты, все среды жизни, вероятно,
почти все многоклеточные живот-
ные и все растения оказались
средой обитания нематод. Это яв-
ление демонстрирует нам один из
наиболее ярких примеров биоло-
гического прогресса. Общее чис-
ло видов нематод в настоящее
время еще не выяснено наукой.
Известный американский специа-
лист по нематодам Н. А. К о б б
(N. A. Cobb) полагал, что общее
число видов нематод — свободных
и паразитических — приближает-
ся к одному миллиону.
Сказанное выше оправдывает
интерес, который проявляют мно-
гие зоологи к этой группе живот-
ных. Ввиду этого знакомство с
их организацией и биологией по-
лезно и читателю книги.
Строение. Как мы видели, дру-
гие классы ыемательминтов пред-
ставлены очень мелкими форма-
ми: большая часть из них меньше
1 мм, реже встречаются более
крупные формы. Среди нематод
имеются очень мелкие формы. Одна из
них — Trichoderma minutum — достигает
всего 80 мк в длину. Такие мелкие формы
известны в морях и в почве, где встреча-
ются нематоды длиной 200—300 мк. Однако
наряду с этими микроскопическими фор-
мами известны настоящие гиганты из мира
нематод. Самки лошадиной аскариды до-
стигают 37 см в длину. Свайник-великан
Рис. 223. Нематоды.
А — организация свободноживущей морской нематоды: 1 — пище-
вод; 2 — задний бульбус; 3 — средняя кишка; 4 — семенник;
5 — спикулы; 6 — хвостовые железы, 7 — шейная железа; 8 — про-
ток шейной железы. Б — Steineria mirabilis.
(Dioctophyme renale) длиной в 1 м,
a Placentonema gigantissima — паразит
кашалота, этого огромного китообраз-
ного, по величине тела, без преувеличения,
напоминает удава, самки его до 8 с лиш-
ним метров в длину! Таким образом, у не-
матод мы наблюдаем очень значительные
колебания длины тела — от 80 мк до
8 м. Эти колебания длины тела — одно из
395
свидетельств многообразия тех сред жиз-
ни, которыми овладели нематоды.
Каков же диаметр тела этих животных?
Известный русский зоолог, автор много-
численных специальных работ и учебников
и вместе с тем талантливый переводчик
проф. Н. А. Холодковский
называл нематод «струнцами». Теперь этот
термин получил более узкое значение,
но он все же хорош тем, что дает образное
представление об общей форме тела нема-
тод. Действительно, они длиннотелые и
тонкие, как струна (рис. 223). При этом
в поперечном сечении их вытянутое тон-
кое тело, как правило, образует правиль-
ный круг (рис. 220).
Мелкие формы часто уподобляются тон-
кой нити. Да и самое слово «нематода»
произошло от греческого слова nemas,
что означает нить.
Форме тела нематод соответствует и
главный, типичный, способ перемещения
нематод в пространстве: они движутся
наподобие микроскопических или видимых
и невооруженным глазом змей. Лежа
всегда на боку, нематоды изгибаются
в спинно-брюшной плоскости и переме-
щаются по дну водоемов, в тесных вод-
ных пленках почвы, в кишечнике и дру-
гих органах человека и животных, меж
клетками корней, стеблей, листьев и дру-
гих частей растений.
Все тело нематод покрыто гибкой, эла-
стичной и прочной кутикулой. Эта кути-
кула — производное лежащего под ней
тонкого слоя кожного эпителия, назы-
ваемого у нематод гиподермой.
Гиподерма — живая эпителиальная ткань,
которая выделяет на своей поверхности
кутикулу. Кутикула у нематод может
быть гладкой или кольчатой, причем
кольца построены совершенно правильног
все они у каждого вида определенного
размера и часто несут различные уплот-
нения — склероции, имеющие фор-
му правильно расположенных точек, ли-
ний (палочек), пластин и т. д. Гиподерма
очень тонкая. Но по бокам тела, а также
вдоль спины и брюха она утолщена, осо-
бенно по бокам, где образованы правый
и левый гиподермальные валики, извест-
ные под названием «хорд» или полей
(не имеющих, конечно, никакого отноше-
ния к хорде хордовых животных). Внутри
боковых «хорд» у части нематод лежат
правый и левый выделительные каналы.
Кутикула и гиподерма составляют пери-
ферию кожно-мускульного мешка тела
нематод. Под гиподермой расположена
продольная мускулатура. Однако мышечный
слой не сплошной. Он тянется вдоль тела
в виде четырех мышечных тяжей — двух
спинно-боковых и двух брюшно-боковых,
отделенных друг от друга четырьмя упо-
Рис. 224. Головы морских нематод:
I — тангорецепторы; 2 — ротовая полость; з — онхи; 4 — фоторецепторы (глазки),
на себя внимание сильно развитые рецепторы у свободных нематод.
Обращают
396
мянутыми «хордами» (рис. 223). Мы-
шечные клетки удлинены и всегда рас-
положены в одном направлении,что
очень характерно для так называе-
мых поляризованных клеточных ком-
понентов ткани. В этих случаях длин-
ные и перпендикулярные им оси кле-
ток одинаково ориентированы в пре-
делах всего тела. Поэтому все клетки
мышц работают согласованно, син-
хронно, что естественно повышает
их кинетическую энергию. Недаром
тонкие нематоды легко проникают в
узкие пространства между нитями
водорослей, между гифами мицелия
грибов, между частицами почвы,
облеченными капиллярной пленкой
воды, в поры тела животных, в усть-
ица листьев, межклетники корневых,
стеблевых и других тканей растений
и т. п.
Головной конец тела нематод снаб-
жен головной капсулой, опираю-
щейся на внутренний опорный ске-
лет из плотной кутикулы. Голов-
ная капсула состоит из двух основ-
ных частей — головных бугров и
подвижных губ. Но у многих форм
губы и головные бугры сливаются
в общую головную капсулу. На ней
расположены органы осязания —
тангорецепторы, имеющие
форму либо щетинок (рис. 224), ли-
бо сосочков, т. е. п а п и л л. На
переднем конце головной капсулы, стро-
го посередине и лишь изредка сместив-
шись несколько на брюшную сторону,
лежит ротовое отверстие, окруженное
губами. На головной капсуле, или сза-
ди от нее, или на боковых губах лежат
боковые обонятельные ямки, известные
у нематод под названием боковых
органов или амфид. От амфид
отходят обонятельные нервы (рис. 225).
У некоторых свободноживущих нематод
развиты также глаза, снабженные у ря-
да форм линзой и глазным пигменти-
рованным бокалом — зеленым, оранже-
вым, фиолетовым, красным, черным
(рис. 224). Иногда вдоль всего тела тор-
чат щетинки (рис. 223).
Тело нематод очень ясно дифференци-
ровано на три участка. Передний участок
Рис. 225. Представители мор-
ских нематод.
Обращают на себя внимание силь-
но развитые тангорецепторы A)
и органы химического чувства —
круглые B), петлеобразные C),
спиральные D) и карманообраз-
ные E) амфиды.
Рис. 22G. Организа-
ция дитиленхов:
1 — стилет; 2 — пище-
вод; 3 — средний буль-
бус; 4 — железистый
бульбус; 5 — нервное
кольцо; 6 — нервные
ганглиозные клетки.
несет органы чувств, о которых сказано
выше, и ему соответствует передний отре-
зок кишечника — передняя кишка. Вто-
рой отрезок тела соответствует средней
кишке и включает, кроме нее, половые
трубки. Третий — образует хвост, огра-
ниченный на брюшной стороне тела задне-
проходной щелью (анусом). Конец хвоста
имеет у разных видов различную форму
(рис. 223).
Центральная нервная си-
стема состоит из нервных стволов,
расположенных вдоль тела, и кольцевых
комиссур, связывающих продольные
стволы в единую систему. У нематод наи-
более типична кольцевая нерв-
ная комиссур а, охватывающая
пищевод. Она обравует «нервное кольцо»
нематод, которое не несет нервных ядер
397
Рис. 227. Строение нематод. Стилет и копье.
А — эктопаразитическая корневая нематода Hoplola-
imus tylenchiformis: J — головная капсула; 2 — сти-
лет; 3 — головки стилета; 4 — мышцы-протракторы;
5 — начало пищевода; в — место впадения спинной
железы пищевода. Б — Dorylaimus striatus: I —
копье, 2 — амфида.
и состоит из неврофибрилл. Однако впе-*
реди от него и позади него, к пищеводу,
расположена сложная система ганглиоз-
ных клеток (рис. 226). Именно эта система
ганглиозных клеток в своей совокупности
образует нечто подобное «мозгу» турбел-
лярий и гастротрих. В этом «мозгу»
можно различить несколько ганглиев
(рис. 226). От этих ганглиев вперед, к
тангорецепторам и амфидам головы, отхо-
дят нервные тяжи. Кроме того, нервные
ядра заложены в самой ткани пищевода;
нервные ядра регулируют движение му-
скулатуры пищевода, работу зубов, ко-
пий и стилетов, которыми вооружены
многие нематоды, и выделение экскретов
пищеводных желез.
Пищеварительная систе-
м а нематод сложнее, чем у форм пред-
шествующих классов. Передняя кишка
делится на ротовую полость,
или стому, и пищевод. Хотя и
стома и пищевод, строго говоря, представ-
ляют собой не что иное, как глотку, тем
не менее в системе знаний о нематодах,
или нематологии, утвердилась указанная
номенклатура: стома, или ротовая полость,
и пищевод. Для этого имеются солидные
основания. Стома — это часть глотки,
которая функционирует как ротовая по-
лость и часто вооружена различными особо
дифференцированными придатками, за-
служивающими наименования органов.
Пищевод — это часть глотки, способная
к перистальтическим движениям, протал-
кивающим комок пищи в среднюю кишку
(рис. 226). Поэтому нематологи (специа-
листы по нематодам) не называют стому
и пищевод глоткой (фаринксом). Пусть
правы сравнительные морфологи, что
и стома и пищевод нематод — это фаринкс,
глотка. Однако функционально это стома
и пищевод. В стоме различают непо-
движные придатки, или о н х и„
и подвижные зубы; у некоторых
нематод имеются особо дифференцирован-
ные «челюсти», у других — острый с о-
сущий стилет и, наконец,
копье (рис. 227).
Средняя кишка такая же, как
у гастротрих. Стенка ее состоит из одного
слоя клеток. Задняя часть кишки пере-
ходит в прямую кишку, открывающуюся
наружу уже упоминавшимся заднепроход-
ным отверстием. Пищеварение у нематод
своеобразное. В пищеводе лежат спе-
циальные железы, которые выделяют
экскреты, содержащие ферменты. Эти
ферменты либо поступают с пищей в сред-
нюю кишку, где пища перевариваетсяг
либо выделяются наружу, и тогда воз-
никает своеобразный процесс перевари-
вания пищи в наружной среде, в капле
ферментов нематоды, после чего быстро
переваренная пища попадает в просвет сто-
мы и пищевода и усваивается в кишке.
Выделительная система
нематод бывает двух типов. У одних форм
она состоит всего лишь из одной шейной
железистой клетки, проток которой откры-
вается наружу брюшной порой. У других,
кроме этой шейной железы, имеются боко-
вые выделительные каналы. Содержимое
их выделяется наружу через брюшную
выделительную пору (рис. 228). Мы не
будем вдаваться в подробности процессов
выделения. Отметим, что продукты обмена
проникают в полостную жидкость. Здесь
они с помощью особых клеточных систем
обезвреживаются, диффундируют в шей-
ную железу и выделяются наружу.
398
Все нематоды, как правило, раздельно-
полые животные. У самцов развиты се-
менники, семяпроводы и семяизверга-
тельный канал. Семенников может быть
два или один. Кроме того, у самцов имеют-
ся специальные совокупительные органы—
спикулы и руле к, направляю-
щий их движения. Женские половые ор-
ганы состоят из яичников, яйце проводов
и матки. Женское половое отверстие рас-
положено на брюшной стороне тела. Сам-
цы вводят спикулы в женское половое
отверстие и оплодотворяют самок. Сперма
нематод не имеет подвижных жгутиков.
Ни в одном органе нематод вообще нет
подвижных клеточных органелл, и в част-
ности ресничек. Нет, как выше сказано,
Рис. 228. Нематода Aphelenchoides compos-
ticola:
I — стилет; 2 — пищевод; 3 — средний бульбус; 4 —
железы пищевода; 5 — нервное кольцо; 6 — выдели-
тельное отверстие; 7 — средняя кишка; 8 — задняя
кишка; 9 — заднепроходное отверстие; 10 — яичник;
II — семе приемник со сперматозоидами; 12 — перед-
няя матка; 13 — задняя матка; 14 — женское половое
отверстие.
и хвостов у спермы. Она перемещается
амебоидными движениями. В половых
путях самок формируются яйца. Они опло-
дотворяются спермой самцов в женских
половых путях, и в частности в особых
семеприемниках (рис. 228). Затем оплодо-
творенные яйца выделяются наружу через
женское половое отверстие или же раз-
виваются внутри половых трубок. В этом
случае из женского полового отверстия
выходят наружу личинки (живорожде-
ние). Яйца нематод заключены в яйцевые
оболочки, предохраняющие их от физиче-
ских повреждений и химических воздей-
ствий среды. Личинки четыре раза ли-
няют, последовательно, после каждой
линьки переходят в следующую стадиьо
Рис. 229. Онтогенетическое
развитие и формообразова-
ние у Rabditis anomala:
I — личинка первого возраста;
II — личинка второго возра-
ста; III — личинка третьего
возраста; IV —личинка четвер-
того возраста; V — взрослая
самка; J — половая система.
399
развития, превращаясь в личинок вто-
рого, третьего и четвертого возрастов.
Из личинки четвертого возраста разви-
ваются молодые формы — самец или
самка (рис. 229). Очень часто личинки не
похожи на взрослые формы. В таких слу-
чаях нематологи говорят о развитии с пре-
вращением.
Таковы самые общие представления
об организации нематод. Во многом их
организация сходна с организацией га-
стротрих, киноринх и коловраток. Од-
нако нематоды существенно отличаются
от любых представителей этих групп
следующими важными признаками: фор-
мой тела; способом движения; отсутствием
протонефридий; отсутствием в любой
системе органов ресничного эпителия и
подвижных жгутов в каких бы то ни было
клетках, в том числе половых; четкой поло-
вой дифференцировкой (самцы и самки),
^несвойственной по крайней мере гастро-
трихам; большим числом видов и их групп
тз. ярко выраженными признаками биоло-
гического прогресса. Как и у других
групп немательминтов, у нематод нет
ни органов дыхания, ни кровообращения.
Систематика нематод. Это, пожалуй,
один из наиболее сложных вопросов.
Большинство аденофорей, как сказано
выше, живет в морях и океанах. Часть из
них обитает в пресных водах и в почве.
Некоторые группы представлены парази-
тами растений и животных.
Большое число видов различных родов
и семейств аденофорей ведет свободное
существование на дне водоемов, т. е.
в бентосе. Это типичные нематоды, с наи-
более полной и примитивной организацией.
Среди них множество очень мелких форм,
длина тела которых равна долям милли-
метра. Упомянутая выше Trichoderma
minutum принадлежит к этому подклас-
Нематоды — это непрерывно расширяю-
щийся поток вновь открываемых видов
и родов. Приведем пример этого. В 1949 г.
числилось 200 видов нематод из отряда
тиленхид, среди которых много паразитов
растений. Американский исследователь
этих нематод Торн (Thome, 1949) ука-
зывал, что это число, вероятно, состав-
ляет 5% числа видов названного отряда,
в действительности живущих в почве
и в растениях. К 1962 г. число их достигло
800 видов, т. е. с 1949 по 1961 г. было вновь
открыто 600 видов тиленхид.
Большинством ученых класс нема-
тод делится на два подкласса — под-
класс Аденофорей (Adenophorea)
и подкласс Сецерненты (Secernentea).
Главная масса аденофорей — жители
бентоса морей и океанов; многие (хотя
и меньшинство из них) обитают в почве
и в пресных водах. Известны среди них
сравнительно немногие группы, приспо-
собленные к паразитизму в органах расте-
ний и животных. Сецерненты, напротив,
в большинстве приспособлены к паразити-
ческому существованию в тканях и в орга-
нах растений и животных.
Остановимся на некоторых важнейших
группах этих подклассов.
су и обитает в море. Имеются среди аде-
нофорей и относительно крупные формы.
Их длина измеряется не миллиметрами,
а сантиметрами, нередко десятками сан-
тиметров. Но таких все же меньше. Основ-
ная масса аденофорей представлена более
мелкими формами, достигающими длины
J. — О ЛьЛь.
Какими же признаками характеризует-
ся подкласс аденофорей, его многочислен-
ные свободноживущие представители? Пер-
вое — прогрессивным развитием органов
чувств. На рисунке 230 показаны голов-
ные концы нескольких форм, у которых
ПОДКЛАСС АДЕНОФОРЕИ (ADENOPHOREA)
Таблица 22. Немертина Lineus longissimus. -*
хорошо заметно прежде всего
мощное развитие органов осяза-
ния, которые у морских адено-
форей обычно имеют форму
длинных щетинок. Каждая ще-
тинка состоит из кутикуляр-
ного чехла, по оси которого
располагается нерв (танго-
рецепторы). Таких танго-
рецепторов может быть четыре,
шесть, десять, двенадцать. Очень
часто по одному тангорецептору
расположено по бокам голов-
ной капсулы — один справа,
другой слева; кроме того, по
паре рецепторов — по субвент-
ральным и субдорсальным сек-
торам головы (рис. 230). Всего,
следовательно, шесть рецепто-
ров. Однако это число у дру-
гих форм либо возрастает до
десяти (если и по бокам голо-
вы развито по два рецептора),
либо падает до четырех. Раз-
витие тангорецепторов в виде
длинных щетинок очень харак-
терно именно для свободных
аденофорей. Это прямое следст-
вие и приспособления к сво-
бодному существованию, в ус-
ловиях которого подвижная
морская нематода сталкивается
со многими другими донными
беспозвоночными, в том числе
и с хищными нематодами.
Однако длинные щетинковид-
ные рецепторы развиты только
на головных буграх, тогда как на губах
развит внутренний круг рецепторов, всег-
да имеющих форму сосочков или очень
коротких щетинок. Это круг губных ре-
цепторов. Можно предполагать, что го-
ловные рецепторы ориентируют нематоду
во внешней среде, губные — в источни-
ках питания.
Второе, что обращает на себя внимание,
это наличие у свободных аденофорей хоро-
шо развитых обонятельных органов —
а м ф и д. На рисунке 225 видно, что они
могут быть различных типов. Нематоды
воспринимают этими органами химиче-
ские вещества окружающих предметов
и либо уходят, либо приближаются
Рис. 230.
зания
к ним. У ряда морских форм
имеются также глаза, состоя-
щие из глазного бокала, ча-
сто окрашенного в различные
цвета, и хрусталика. Предметов
эти глаза, разумеется, не ви-
дят, но реагируют на свет.
Впрочем, глаза скорее исклю-
чение, чем правило.
Стома у аденофорей очень
многообразна по форме и функ-
циям. Часто она мала, просто
устроена: в виде призмы, труб-
ки или бокала, воронки и т. п.
У многих форм она вооружена
неподвижными острыми придат-
ками (о н х и). У других форм
острые придатки подвижны.
Это «зубы» и «челюсти». Пище-
вод простой. Стенка кишки
большей частью состоит из од-
ного слоя крупных многоуголь-
ных клеток. Прямая кишка от-
крывается на брюшной сто-
роне поперечной анальной
щелью. Характерным призна-
ком аденофорей надо считать
развитие шейной желе-
з ы, рассматриваемой как ос-
морегуляторно - экскреторный
аппарат. Чаще этот орган,
представленный одной клеткой,
называют просто выдели-
тельной железой. Она
имеет проток, лежащий обычно
на брюшной стороне тела, под
пищеводом. Проток открывается
в брюшную выделительную пору (рис. 223).
Сама железа бывает различных типов: труб-
чатая, грушевидная, мешковидная. Кроме
этой железы, получившей также название
шейной, аденофореям присущи и дру-
гие кожные железы. Для них характер-
но серийное расположение прибоковых
(паралатеральных) желез, лежащих ря-
дами (нередко парными) вдоль всего тела.
Эти железы имеют короткие протоки, кото-
рыми они открываются в боковых полях
тела наружу. Это характерный признак
аденофорей. Наконец, большинству из
них, кроме почвенных и некоторых дру-
гих групп, в том числе паразитических,
присущи лежащие в тканях хвоста три
Органы ося
аденофорей.
ф 26 Жизнь животных, т. i
401
хвостовые железы, открывающиеся нару-
жу тремя каналами, которые впадают
в хвостовую выделительную пору на кон-
це хвоста (рис. 231). Таким образом, у
аденофорей, в особенности свободных
морских форм, которые должны рассмат-
риваться как типичные нематоды, имеется
очень большое число кожных желез, или,
как говорят нематологи, им присущ гро-
моздкий железистый аппарат. Несомнен-
но, что все эти железы экскретируют,
т. е. выделяют свои экскреты наружу.
Некоторые специалисты не без основания
рассматривают эти железы как часть
выделительной системы.
Половые органы либо парные (в том
числе и мужские половые железы, или
семенники), либо одинарные. Следует
отметить, что кутикула аденофорей легко
проницаема для растворенных в воде со-
лей. Делятся аденофорей на два отряда:
хромадорид (Chromadorida) и эноплид
(Enoplida).
ОТРЯД ХРОМАДОРИДЫ
(CHROMADORIDA)
Обычно мелкие формы. Этот признак
считается их характерной особенностью.
Многообразие видов хромадорид трудно
исчерпаемо. У большей части хромадорид
на голове расположены более или менее
длинные щетинки (рис. 223). Губные орга-
ны в виде папилл. В стоме видны «зубы»
(подвижные органы) и онхи (неподвижное
вооружение). Пищевод обычно с бульбу-
сом на заднем конце. Бульбус чаще имеет
внутреннюю полость, не снабженную ка-
ким-либо внутренним вооружением. В тка-
ни пищевода лежат три пищеварительные
железы. Яичников один или два; чаще
один, но бывает и два семенника. У сам-
цов вдоль брюшной стороны тела, перед
анусом, нередко развиты типичные для
хромадорид органы, с помощью которых
самцы ориентируются при совокуплении
в положении женского полового отвер-
стия. Спикулы обычно изогнуты и на-
правляются особым опорным органом—
р у л ь к о м (рис. 231). В хвосте распо-
ложены три хвостовые железы, а на конце
хвоста — характерная хвостовая выдели-
тельная трубочка (рис. 231). Содержимое
желез вытекает в морскую или пресную
воду и тотчас застывает в паутинную
нить. В этих случаях хромадорида торчит
на дне вертикально, прикрепившись
к какой-либо песчинке хвостом и помахи-
вая из стороны в сторону своим стройным
телом. Хромадориды часто имеют глаза—
черные, красные, рубиновые, оранжевые,
фиолетовые и других цветов. Кутикула
хромадорид построена очень сложно.
Обычно она состоит из правильно повто-
ряющихся колец определенной ширины.
Кольца инкрустированы уплотнениями,
или склероциями, различной формы, мер-
цающими в лучах микроскопа, как драго-
ценные камни.
Питаются эти нематоды морскими одно-
клеточными водорослями, а многие зуба-
стые виды их — хищники.
Часть хромадорид в далеком прошлом
нашей планеты проникла в пресные воды
и на сушу. Среди почвенных хромадорид
надо отметить семейство плектид
(Plectidae). Это интересная группа мел-
ких хромадорид, нашедших себе в почве
богатый источник питания — очаги загни-
вающего органического материала. В этих
очагах под влиянием сапробиотических
бактерий и сапрофитных грибов происходят
сложные процессы распада нерастворимых
органических веществ, входящих в состав
растительных трупов (остатки корней,
лежалые стебли, мертвые листья и т. п.).
Нерастворимые органические вещества под
влиянием ферментов бактерий и грибов
расщепляются и превращаются в раствори-
мые (в воде) продукты распада белков
и в растворимые углеводы (моно- и диса-
хариды) и другие более простые вещества.
Сапробиотическая среда, кроме того,
всегда богата водой, в которой растворе-
ны названные продукты. Сапробиос всегда
содержит также плотные комочки остав-
шихся «обломков» растительных тканей.
Наконец, в этой среде всегда много сап-
робиотических бактерий, спор сапрофит-
ных грибов и т. п. В этой среде плектиды
и нашли источники питания. Поэтому
и организация этих сапробиотических
хромадорид несет черты приспособлен-
ности к сапробиотическому существова-
402
/" -* Рис. 231. Хвосто-
У ^г^ вой конец тела
||stf> 3 морской нематоды
||> Hypodontolaimus
ST biitschlii.
f 1 Показан хвост самца:
' l — спикулы; 2 —
рулек, «з — дополни-
тельные органы, 4 —
мышцы спикул; 5 —
хвостовые железы;
6 — выводная трубка
хвостовых желез; 7—
-4 строение кутикулы.
6
нию. Не вдаваясь в детали, отметим, что
особенно типично эти черты приспособлен-
ности выражены в организации ротовой
полости и пищевода плектид. Ротовая по-
лость (с т о м а) имеет форму гладкой во-
ронки или даже цилиндра, в полости
которого пищевой комочек без задержки
проскальзывает в пищевод. Сильная мус-
кулатура пищевода обеспечивает быстрые
глотательные движения. На заднем конце
пищевода есть особое утолщение в форме
луковицы, или бульбуса, снабженного
внутренним «дробильным» аппаратом, раз-
дробляющим пищевой комок. Обычно плек-
тиды глотают сапробиотический детрит
и все, что в нем содержится: жидкость с
растворенными в ней продуктами распада
названных выше питательных веществ, и
в том числе бактерий, и т. п.
Из сказанного видно, что плектиды
приспособлены к использованию сапробио-
тического субстрата — источника их пи-
тания. В этом субстрате почвенные хро-
мадориды нашли доступный источник пи-
тания, и именно в этом причина их при-
способленности к жизни в сапробиотиче-
скои среде. Однако жизнь в этой среде—
не единственный источник существова-
ния плектид. Об этих маленьких немато-
дах нельзя сказать, что они узко приспо-
соблены только к сапробиотическои среде.
Они сделали только первый шаг к освое-
нию сапробиотическои среды, но не стали
ее непременными обитателями. Их много
в почве, вокруг корней. Они могут даже
проникать внутрь корневой ткани, исполь-
зуя в ней отдельные очажки гнилостного
распада живой растительной ткани. Плек-
тиды как бы «на распутье». Они утратили
биологические черты своих предков —
морских хромадорид, стали почвенными
формами, связали свою жизнь с сапробио-
тическими очагами, но не вошли в сапро-
биотическую среду как непременные ее
обитатели. Плектиды как бы «временные
гости» этой среды. Поэтому сплошь и ря-
дом их не находят в сапробиосе, и они
гораздо чаще встречаются в средах, где
сапробиотический процесс находится на
начальных и ограниченно выраженных
стадиях своего развития. Дальше мы уви-
дим, что полное завоевание сапробиотиче-
скои среды осуществила другая группа
нематод, с которой нам еще предстоит
ниже ознакомиться.
ОТРЯД ЭНОПЛИДЫ (ENOPLIDA)
Нематоды этого отряда существенно от-
личаются от хромадорид, и многие из
них интересуют нас в связи с их народно-
хозяйственным значением. В отличие от
хромадорид кутикула у эноплид гладкая,
не кольчатая. Голова, как у хромадорид,
несет длинные осязательные головные
щетинки и губные сосочки. Боковые
органы, или а м ф и д ы, т. е. органы
химического чувства (обонятельные),
обычно в виде поперечных карманов, по
одному справа и слева. Пищевод снабжен
системой пищеварительных желез. В хво-
сте развиты три терминальные хвостовые
железы, однако трубочки на конце хвоста
нет. Многие эноплиды утрачивают голов-
ные щетинки, замененные у них сосочками
(папиллами). Это особенно типично
для почвенных эноплид и паразитических
представителей этого отряда. Появление
среди представителей этого отряда парази-
тических форм — одно из типичных отли-
чий его от хромадорид.
В морях и океанах эноплид очень много.
Большинство из них мелкие формы, дру-
гие достигают значительной величины,
измеряемой даже сантиметрами. Морские
эноплиды — хранители типичной орга-
низации форм этого отряда (рис. 224).
Многие среди них —хищники, вооружен-
26*
403
ные зубовидными придатками, лежащими
в ротовой полости и известными под на-
званием онхов. Другие вооружены по-
движными зубами, действующими напо-
добие мощных челюстей (рис. 224). Поч-
венные, пресноводные и некоторые дру-
гие формы вооружены мощным копьем,
способным далеко выдвигаться наружу.
Копье обладает значительной (конечно,
в масштабах микромира) пробивной си-
лой и служит или как орудие хищника,
или как сосущий орган (рис. 227).
Морские эноплиды, так же как и поч-
венные и пресноводные представители этого
отряда, безгранично многообразны. Кроме
того, среди почвенных эноплид наряду
с хищниками известны паразиты живот-
ных и растений. Поэтому мы остановимся
на почвенных и паразитических группах
эноплид.
Эноплиды — хищники. Среди эноплид
очень много видов, представленных про-
жорливыми хищниками. Это своего рода
Рис. 232. Мононх, «однозуб» (Mononchus papil-
latus):
1 — папиллы; 2 — ротовая полость с большим онхом;
3 —пищевод; 4 — нервное кольцо; 5 — кишечник с
крупными клетками; 6 — яичник; 7 — яйцо, 8 — жен-
ское половое отверстие.
«львы» и «тигры» нематодного мира, ви-
димые лишь с помощью микроскопа.
Рассмотрим одного из хищных предста-
вителей эноплид, принадлежащего к р о д у
мононхов (Mononchus, рис. 232). Родовое
название это в переводе на русский язык
означает «однозуб». Мононх, о котором
пойдет речь, относится к виду Mononchus
papillatus. Самки этого вида достигают
в длину 1,8 мм, самцы несколько меньше.
На голове мононха развиты пирамидаль-
ные папиллы, в каждой из которых лежит
нерв. Это чувствительные тангорецепторы.
По бокам головы — карманообразные
амфиды (химические орга-
н ы). Губы также снабжены нервными
сосочками. На переднем конце тела лежит
ротовое отверстие. Оно ведет в обширную
ротовую полость, в которой торчит на-
правленный острием в глубь стомы боль-
шой и острый онх. Довольно длинный
пищевод имеет мощную мускулатуру, хотя
и лишен бульбуса. Просвет пищевода
выстлан плотной кутикулой. Плотное
тело отличается большой силой. Средняя
кишка довольно емкая; ее стенка состоит
из полигональных клеток, сравнительно
крупных. У самок два яичника. Очень
крупные яйца лежат по одному в задней
и передней матке. Плотные яичники рас-
положены спереди и сзади от женского
полового отверстия. Самцы редки. Живет
Mononchus papillatus в почве. Это про-
жорливый хищник. В его кишке нередко
можно найти остатки плотных частей
тела различных нематод, коловраток и
т. д. Было также обнаружено, что этот
мононх способен поедать личинок некото-
рых опасных нематод, паразитирующих
на ценных сельскохозяйственных расте-
ниях. Ввиду этого нематологи заинтересо-
вались, каково же значение названного
мононха в ограничении численности нема-
тод, повреждающих растения и вызываю-
щих у них специфические нематодные
болезни. Начались опыты. Чтобы выяс-
нить вопрос, Штайнер и Хейнли
поместили мононхов в специальную куль-
туру. В культуре этим ученым удалось
содержать мононхов до 18 недель. Ока-
залось, что мононх очень прожорлив. Так,
наблюдение за одним из этих «тигров» мик-
ромира показало, что в течение одного дня
404
мононх съел 83 личинки опасной галловой
нематоды (см. ниже). Другой экземпляр
мононха был подвергнут наблюдению в те-
чение 12 недель, и выяснилось, что за этот
промежуток времени он съел 1332 экзем-
пляра различных нематод!
Как же он это делает? Как ловит он
свою добычу? Надо сказать, что добыча
никогда не «подозревает», как грозен
мононх. Представим себе исследователя,
сидящего за микроскопом и наблюдаю-
щего за мононхом. Мононх плавает в воде
под покровным стеклом. Солидно и мед-
ленно изгибает он свое тело. Вокруг него,
размахивая головой и хвостом, плавают
другие нематоды. Многие из них сталки-
ваются с хищником, прикасаются к нему
и даже толкают его. Никаких результатов!
Мононх их «не замечает». И это действи-
тельно так. Но вот один из неудачников,
конечно совершенно случайно, поступил
иначе. Проплывая мимо мононха, он при-
коснулся своим телом к его ротовым сосоч-
кам. Следует мгновенная информация!
И вслед за ней в какие-то доли секунды
внезапно вздрогнула мускулатура пище-
вода мононха, и, словно мощным водоворо-
том, жертва оказалась стремительно втя-
нутой в вооруженную стому хищника.
Вслед затем жертва — стройная нематода—
уже проскользнула сквозь просвет пище-
вода и попала в кишку мононха. Здесь
она переваривается, и через некоторое
время от нее остаются буквально лишь
«рожки да ножки» — в кишке хищника
можно распознать спикулы проглоченного
самца, стенки ротовой полости жертвы и
другие остатки.
Хищничество мононхов так активно, что
ученые стали думать о возможности исполь-
зования их с целью ограничения числен-
ности паразитических нематод растений.
Мононхи глотают свою добычу. Другие
хищные эноплиды вооружены выдвиж-
ным копьем, напоминающим кривой
острый кинжал. Они способны пронзать
тело своих жертв и высасывать их соки.
Эноплиды —- паразиты растений. Среди
форм отряда эноплид, кроме хищников,
рассмотренных выше, известны и парази-
ты растений. Эти эноплиды вооружены
длинным копьем. Фактически это
копье не что иное, как удлиненный зуб,
способный далеко выдвигаться из ротовой
полости наружу. Нематода пронзает копь-
ем оболочки растительных клеток и выса-
сывает растительные соки. Сама нематода
остается при этом в почве, и только голов-
ной конец ее тела бывает погружен
в ткань нежного питающего корешка ра-
стения. Как правило, эти нематоды
живут в более глубоких слоях почвы —
ниже и глубже пахотного слоя, достига-
ющего обычно 25 см. Многие из этих нема-
тод принадлежат к семейству Longidori-
dae. Один из представителей этого семей-
ства — Xiphinema americanum. Эта па-
разитическая нематода достигает 3 мм
в длину. Основание ее очень длинного
копья утолщено, как у всех представи-
телей этого рода, а пищевод на заднем
конце заметно расширен. Копье имеет
внутренний канал, а вершина его косо
срезана, как кончик иглы медицинского
шприца. Длинное и тонкое тело несколько
напоминает тело змеи. Xiphinema ame-
ricanum, высасывая соки из корневых
клеток различных растений, вызывает
гнилостный распад растительной ткани
и нематодную болезнь, выраженную в рез-
ком угнетении растения.
Как было сказано, эта нематода пора-
жает корни растений, оставаясь при этом
в почве и не погружаясь в растительную
ткань целиком, т. е. принадлежит к эко-
логической группе наружных паразитов
растений (эктопаразитов).
К эктопаразитической группе нематод
принадлежат и оригинальные представи-
тели семейства триходорид (Tri-
chodoridae). У форм этого семейства
копье в средней части как бы расщеплено
на три части. Очень характерны органы
химического чувства — а м ф и д ы: они
у этих форм двуполостные, причем в зад-
ней полости расположены «обонятельные
нервы», лежащие в ней пучком. Пищевод
в задней части резко расширен. В сильно
развитых половых трубках формируются
крупные яйца. Триходорусы, как и кси-
финемы, вонзают свое длинное копье
в ткани корней, вызывая в корешках
растений патогенные процессы, но, кроме
того, они вносят своим длинным копьем
в ткани растений вирусы, вызывая опас-
ные вирусные заболевания.
405
Эноплиды — паразиты беспозвоночных
животных. Среди эноплид известны, одна-
ко, не только паразиты растений, но так-
же и группы, представленные паразитами
животных.
Большое и богатое видами и родами
семейство мермитид (Mermitidae)
представлено паразитами ракообразных,
водных и наземных насекомых и некото-
рых других групп беспозвоночных. Взрос-
лые мермитиды, достигающие в длину
50 см, ведут свободное существование, па-
разитизм же присущ личинкам этих нема-
тод. У мермитид очень толстая много-
слойная кутикула, обычно округленная
голова, несущая сосочковидные танго-
рецепторы и заостренный либо кониче-
ский, либо тупоокругленный хвост.
По бокам головы видны большие или средней
величины амфиды. Иногда они редуци-
рованы. Посередине головного конца, как
у всех нематод, лежит ротовое отверстие,
ведущее в пищевод, который большей
частью непосредственно переходит в длин-
ный кишечник. Тщетно стали бы мы искать
у мермитид кишечник, подобный тому,
который мы видим у других нематод.
Паразитическое существование привело
к развитию у этих нематод совершенно
своеобразной организации средней кишки.
У мермитид нет кишечной полости. Это
в особенности заметно в тех случаях,
когда пищевод почти достигает хвоста.
Весь он состоит из кутикулярной трубки,
стенки которой склеротизированы (уплот-
нены), причем пищеводная трубка окру-
жена рядами плазматических клеток
с крупными ядрами. Это и есть кишечник,
фактически не имеющий собственной по-
лости. Клетки заполнены запасными пита-
тельными веществами. Заднепроходное от-
верстие ясно развито только у самцов
и служит отверстием, через которое на-
ружу выдвигаются спикулы. Яичники
длинные, вершины их светлые, в осталь-
ной части темные вследствие накопления
в половых клетках — будущих яйцах —
значительных количеств желтка. Развив-
шиеся яйца попадают в яйцеводы и оттуда
в мускулистую матку. Яйца выделяются
наружу — в воду или в землю — в коли-
честве нескольких тысяч или сотен. В выс-
шей степени интересно поведение bq вре-
мя яйцекладки у Mermis. Увлажнение
земли стимулирует выход самок наружу,
на поверхность почвы. Мермитида взби-
рается, подобно змее, на растение, какую-
нибудь травинку, совершая колебатель-
ные движения головным концом. При этом
червь обращен головным концом к свету.
Эта реакция объясняется наличием у мер-
митид глазков (род Mermis и др.). Резуль-
татом освещения лучами солнца является
рефлекторное выделение яиц. У Mermis
яйца покрыты оболочкой, несущей отро-
стки. Эти отростки прилегают к скорлупке
яйца, пока оно лежит в матке. При выде-
лении яиц наружу они мгновенно рас-
прямляются и служат тогда приспособле-
нием для прикрепления яйца к субстрату.
У других мермитид выделение яиц и про-
никновение их в хозяев мермитид, в ко-
торых они паразитируют в личиночном
состоянии, достигается другими средст-
вами. Насекомые проглатывают яйца Mer-
mis вместе с растительной пищей. Попав
в насекомое-хозяина, личинка, вышедшая
из яйца, обычно внедряется в полость
тела своего хозяина, где питается эндо-
смотически через кутикулярные покровы.
В дальнейшем личинки покидают тело
насекомого и переходят к свободной жиз-
ни в почве или в воде (в зависимости от
вида и рода мермитиды). К моменту дости-
жения независимой жизни у них разви-
ваются половые органы. Это обычно про-
исходит после внедрения молодой мерми-
тиды в почву.
«Выход личинок из тела насекомого-
хозяина,— пишет известный советский
нематолог И. Н. Ф и л и п ь е в, — яв-
ляется роковым для насекомого, оно гиб-
нет от полного истощения, вызванного
предыдущей жизнью червя, или от зара-
жения гнилостными бактериями». Пора-
жение насекомых мермитидами, как пра-
вило, влечет за собой их гибель или каст-
рацию, т. е. поражение их половых орга-
нов и потерю способности размножаться.
Поэтому мермитиды принадлежат к числу
полезных нематод, ограничивающих чис-
ленность вредных насекомых.
Эноплиды — паразиты позвоночных.
Подавляющее большинство эноплид сво-
бодны, часть из них, как мы видели,—
паразиты растений (корней). Познакоми-
406
лись мы и с немногими видами группы,
представленной паразитами насекомых
и других беспозвоночных. Теперь рассмот-
рим эноплид, паразитирующих в орга-
нах позвоночных.
Остановимся прежде всего на одном из
представителей интересного отряда ди-
октофимат (Dioctophymata). Мы имеем
в виду очень крупную нематоду — насто-
ящего гиганта по сравнению с свободными
формами — свайника-великана (Diocto-
phyme renale). Эта нематода — паразит
лочек и полости тела млекопитающих.
Развитие этой нематоды протекает со сме-
ной двух хозяев — олигохеты и млеко-
питающего — собаки или других хищных
(Carnivora). Из яйца, попавшего в благо-
приятные условия (к которым, в частно-
сти, относится достаточная аэрация), раз-
вивается личинка первого возраста. Если
яйцо будет проглочено олигохетой Lum-
briculus variegatus, то из него высвобожда-
ется личинка, которая попадает в полость
кишечника этого червя и затем в полость
его тела. Отсюда личинка проникает в
брюшной кровеносный сосуд олигохеты,
где она растет и дважды линяет, стано-
вясь, следовательно, личинкой третьей
стадии. Такая личинка уже опасна для
млекопитающего, например собаки, т. е.
она становится инвазионной, способной
развиваться в органах млекопитающего
во взрослого свайника-великана. Но для
этого она должна попасть в кишечник со-
баки.
Собака может проглотить олигохету,
пораженную личинкой свайника-велика-
на, например, с водой. В этом случае
личинка свайника-великана внедряется
в мышечный слой желудка, вызывая обра-
зование гематомы вследствие ограничен-
ного кровоизлияния в месте локализации
паразита. Через две недели личинка внед-
ряется в печень, перемещаясь в ней и на-
рушая целостность тканей этого органа.
Наконец, личинка линяет в последний
раз C-я линька) и, уже в виде личинки
четвертого возраста, проникает в полость
тела млекопитающего. Отсюда она попа-
дает в почку, чаще — в правую, предвари-
тельно завершив последнюю, четвертую,
линьку и превратившись во взрослого
паразита.
Рис. 233. Свайник-великан (Dioctophyme renale).
Паразит вызывает у собаки общее исто-
щение. Находясь в почечной лоханке,
он может вызвать кровотечение. Моча
в этих случаях становится кровавой. Вме-
сте с мочой во внешнюю среду выделя-
ются многочисленные яйца — источник
новых заражений (инвазий).
В случае этой инвазии в почечной ло-
ханке собаки при вскрытии обнаружи-
вают огромную нематоду.
Представители другого подотря-
да — трихоцефалат (Trichocephalata) —
опасные паразиты млекопитающих и чело-
века. Наиболее известны формы, относя-
щиеся к семейству трихоцефалид
(Trichocephalidae) и роду Trichoce-
phalus. Род содержит ряд видов, парази-
тирующих в толстой и слепой кишке многих
животных — у косули, оленя, джей-
рана, лани, овец, верблюдов, у круп-
ного рогатого скота и других млекопитаю-
щих. Паразитирует один вид и у чело-
века, в толстой кишке. Этот вид известен
под названием Trichocephalus trichiuris,
а в медицинской практике его обычно на-
зывают власоглавом. Это наименование
связано с тем, что передний конец тела
власоглава так тонок, что похож на волос.
Задний же конец его резко утолщен.
Длина самца достигает 30—40 мм, а сам-
ки — 35—50 мм (самки у всех нематод
крупнее). Эта нематода белесого цвета,
407
иногда с красноватым оттенком. Тонкий
участок тела содержит лишь переднюю
кишку, т. е. стому и пищевод. Пищевод
вооружен 1—2 рядами крупных желе-
зистых клеток, но мускулатура его раз-
вита слабо. В задней, более толстой части
тела расположены кишечник и половые
органы самца или самки. У самок только
один яичник, у самцов — лишь одна спи-
кула. Самки продуцируют очень типич-
ные яйца власоглава, нахождение которых
в стуле человека безошибочно указывает
на зараженность этой нематодой (рис. 234).
Власоглав паразитирует не в полости
толстой кишки человека, но, как принято
говорить у гельминтологов, «прошивает»
слизистую оболочку ее, внедряясь в нее
головным концом и питаясь за счет крови
человека. Это характерная черта биоло-
гии власоглава. Таким образом, власо-
глав — типичный паразит, живущий за
Рис. 234. Trichocephalus trichiurus.
счет соков организма, в органах которого
он живет. Способность «прошивать» сли-
зистую толстой кишки объясняется нали-
чием упомянутых выше пищеводных же-
лез, обладающих лизирующей функцией,
т. е. функцией химического воздействия
на ткани хозяина с помощью ферментов.
Ферменты — это сложные белковые сое-
динения различных типов, способные рас-
щеплять животные или растительные бел-
ки, переводить нерастворимые сахара (по-
лисахариды) в растворимые (моно- и диса-
хариды и др.). Железы (пищеводные) не-
матод богаты ферментами, и именно они
обеспечивают способность власоглава
«растворять» животную ткань и исполь-
зовать ее как источник питания. «Проши-
вая» ткань, власоглав неизбежно вступает
в контакт с кровью хозяина, обеспечиваю-
щей не только питание паразита, но и ча-
стичное использование им кислорода, свя-
занного с гемоглобином красных кровя-
ных телец крови человека.
Как же заражается человек власогла-
вом? Или, как принято говорить в науке
о паразитических червях, или гельминтах
(отсюда и название науки — гельминтоло-
гия), как достигается инвазия кишечника
человека власоглавом? Оказывается, чело-
век заражается им при потреблении воды,
в которой могут находиться яйца власо-
глава. Это случается, если человеческие
экскременты попадают в воду. В воде,
особенно в теплой, яйца развиваются при-
мерно в течение одного или полутора меся-
цев. Внутри оболочки яиц после истече-
ния этого срока становятся видными ли-
чинки, способные жить в яйцевых оболоч-
ках в течение нескольких месяцев. После
того как человек сделает несколько глот-
ков воды, содержащей яйца паразита, из
яиц в кишечнике человека выходят моло-
дые власоглавы (личинки), развивающие-
ся во взрослые формы по достижении
своей нормальной локализации, т. е. в
толстой кишке. При этом они «прошива-
ют» слизистую оболочку кишки.
Изгнание власоглава — нелегкая задача
именно потому, что он «прошивает» сли-
зистую и прочно удерживается на месте.
Больным назначают специальное лечение;
успех считается достигнутым, если в стуле
прекратилось выделение яиц этого пара-
408
зита. Власоглав может быть при-
чиной расстройств пищеварения,
анемии, нервных явлений.
Много опаснее другой пред-
ставитель семейства Tricho-
cephalidae — трихинелла (Trichi-
nella trichinella), часто называе-
мая «трихиной». Эта нематода до-
стигла очень высокой специали-
зации. Очень простой и примитив-
ный цикл развития власоглава
может быть выражен формулой:
«орган хозяина — вода — орган
хозяина». Трихинелла полностью
утратила следование этой схеме;
ни на одной стадии своего разви-
тия она не выходит во внешнюю
среду (почву, воду и т. п.).
Человек заражается трихинеллой, как
правило, в результате употребления в
пищу недостаточно прожаренного и про-
варенного мяса свиньи, пораженной этим
паразитом. Но что означает это понятие—
«достаточно» или «недостаточно» прова-
ренное или прожаренное мясо? Если ва-
рится или жарится большой кусок сви-
ного мяса или коптится окорок, то «кули-
нарно» он может быть «готов», а гельмин-
толог скажет нам, что такой «готовый»
окорок или большой кусок мяса, если он
содержит трихинелл, становится опасней-
шим источником заболевания человека —
заболевания, известного под названием
трихинеллоза.
Трихинелла сидит в мышечной ткани
свиньи (и в «кулинарно» готовом окороке!)
в известковых капсулах (рис. 235). Если
человек съест кусок мяса с инкапсулиро-
ванными трихинеллами, то в желудке его
мышечные волокна свиного мяса распа-
даются под влиянием желудочного сока;
вместе с тем под влиянием содержащейся
в нем соляной кислоты растворяется и
известковая капсула. Освободившиеся из
капсулы трихинеллы внедряются в ткань
тонкого кишечника и быстро достигают
половозрелого состояния. Уже через
48 часов после внедрения в ткань кишеч-
ника самцы оплодотворяют самок, в поло-
вых органах которых развиваются яйца
и молодые трихинеллы. Потомство одной
самки обычно достигает двух тысяч. Моло-
дые трихинеллы выходят из полового от-
верстия самки и попадают непо-
средственно в ткани кишечникау
где находились и самки. Микро-
скопические враги — молодые три-
хинеллы —попадают теперь в лим-
фатические сосуды стенки тонкого
кишечника человека и затем в его
кровеносное русло. Период, когда
массовые количества личинок три-
хинелл проникают в кишечную
стенку человека, далеко не безраз-
личен для него. В этот период,
часто протекающий очень бурног
у человека поднимается темпера-
тура, развиваются отеки лица и
особенно век, изменяется состав
крови и т. п. Если трихинелл
очень много, этот период может
иметь смертельный исход. Допустим, этого
не случилось — человек выжил. Однако
болезнь далеко не закончилась. Личинки
трихинеллы через кровяное русло зано-
сятся, как правило, в наиболее интенсив-
но работающие мышцы и здесь оседают.
Этот период проникновения в мышцы
также может сопровождаться болезнен-
ными явлениями — болями в мышцах,
иногда временными параличами различ-
ных групп мышц и т. п. Затем болезнь
затухает. Но человек на многие годы остает-
ся носителем трихинелл. В мышцах
вокруг паразитов развивается известко-
вая капсула. Эти замурованные трихи-
неллы в мышцах человека естественно
обречены на гибель.
В мышечных тканях свиней, заражаю-
щихся трихинеллой при поедании крыс,
возникает такой же процесс развития три-
хинелл, как в мышечной ткани человека,
но конечная «судьба» трихинелл в мыш-
цах свиньи другая. Свиное мясо гото-
вит и ест человек. Поэтому свинья —
главный источник заражения человека
трихинеллозом.
Как же быть? Как уберечься от зара-
жения трихинеллозом?
Крысы заражаются, поедая трихинел-
лозное свиное мясо; свиньи заболевают
трихинеллозом, поедая крыс. Поэтому
борьба с крысами, их непременное унич-
тожение в свиноводческих хозяйствах —
одно из важных противотрихинеллозных
мероприятий. Человек охраняется от три-
Рис. 235. Трихи-
нелла в мышечной
ткани свиньи.
409
хинеллоза законом, согласно которому
ни одна свиная туша не допускается в
продажу, пока не подвергнется исследо-
ванию на трихинелл. На рынках и в мага-
зинах продают окорока и свиное мясо,
проверенные на трихинеллоз. Поэтому
и покупка свинины должна проводиться
только через государственную торговую
сеть или рынки, контролируемые ветери-
нарно-медицинским пунктом. Всякая
свинья, идущая на убой, должна обяза-
тельно пройти ветеринарно-медицинскии
осмотр.
Не случайно знакомство с подклас-
сом сецернентов мы начинаем во вто-
рую очередь. Аденофореи — свободные
формы моря — типичные нематоды. Под-
класс сецернентов включает множество
групп нематод, на организации которых
лежит печать специализации.
В пределах отряда хромадорид,
проникших в пресные воды и в почву,
развивается семейство плектид
(Plectidae), представители которого про-
никли в сапробиотические очаги, где илек-
гиды нашли готовый «полуфабрикат» —
доступный источник их питания
(см. стр. 402). Когда возникло семей-
ств о плектид, неизвестно, так как фак-
тически нематоды «не оставили» каких-
либо палеонтологических документов. Но
во всяком случае, они должны были воз-
никнуть тогда, когда уже существовали
растительные организмы и когда в почве
началось накопление органического мате-
риала. Фактически это очень древний про-
цесс. Как указывает выдающийся совет-
ский ученый акад. В. И. Вернад-
ский A926), «чем более мы изучаем
химические явления биосферы, тем более
мы убеждаемся, что в ней нет случаев,
где они были бы независимы от жизни...
И так,— продолжает акад. Вернадский,—
длилось в течение всей геологической
истории». Нам приблизительно известно,
когда на Земле появились грибы, и в част-
ности низшие, гифомицеты. Вероятно,
они появились еще в каменноугольном
Для зоолога трихинелла остается, кро-
ме того, интересным биологическим объек-
том — примером далеко зашедшей спе-
циализации в направлении развития па-
разитизма. Трихинелла, попав в человека,
проходит в его теле все этапы своего раз-
вития, и для их прохождения — от ли-
чинки до взрослого состояния и даже
от яйца (от самок, развившихся в стенке
кишечника) до взрослого состояния — она
может не выходить во внешнюю среду.
Это свидетельствует о далеко зашедшей
приспособленности к паразитизму.
периоде. Бактерии появились много рань-
ше. Но сапробиотические очаги, возни-
кающие в результате загнивания трупов
растений, появились в почве уже тогда,
когда развилась пышная наземная флора
каменноугольного периода. Весьма веро-
ятно, что именно к этому времени, ввиду
сказанного выше, начинается становле-
ние семейства плектид в его современном
облике, т. е. в виде форм, приспособлен-
ных к питанию за счет продуктов распада
белков и растворимых Сахаров, содержа-
щихся в сапробиотических очагах. В во-
доемах процесс мог возникнуть и раньше.
Мы здесь напомнили о семействе
Plectidae потому, что современная наука
установила наличие черт сходства между
Plectidae, с одной стороны, и формами
семейства Rhabditidae — с другой. Инте-
рес этого вопроса связан с тем, что с е-
м е й с т в о Rhabditidae принадлежит не
к подклассу Adenophorea, с кото-
рым выше мы ознакомили читателя, а
к другому подклассу нематод —
Secernentea. Таким образом, оказывается,
что эти два семейства — Plectidae
и Rhabditidae — связывают узами род-
ства оба названных подкласса нематод,
а это восполняет наши представления об
эволюции класса нематод.
Но семейство Rhabditidae глу-
боко интересно и независимо от вопросов
эволюции развития класса нематод.
Сапробиос. Что такое сапробиотическая
среда? Прежде всего это сложное биоло-
ПОДКЛАСС СЕЦЕРНЕНТЫ (SECERNENTEA)
410
гическое явление, сложный биологический
процесс, связанный с биологическими цик-
лами азота, углерода и минеральных эле-
ментов. Циклы названных элементов обус-
ловлены деятельностью организмов, с од-
ной стороны, синтезирующих сложные
органические соединения, характерные
для живых форм (белки, углеводы, жиры,
пектин, клетчатку и т. д.), и, с другой
стороны, возвращающих продукты рас-
пада этих веществ в почву в результате
гнилостного распада тканей растений и
животных. Синтетические процессы всегда
связаны с процессами разрушения, рас-
пада, и обе эти, диалектически связанные,
стороны жизненных процессов присущи
прежде всего почве как биологическому
явлению. Распад органического материа-
ла, возникающий при разложении трупов
животных и растений, и составляет основу
характеристики сапробиотической среды.
В почве сапробиотическая среда возни-
кает в форме отдельных сапробиотических
очагов. В конце вегетации культурных
растений нередко можно наблюдать, такие
очаги после сбора урожая сельскохозяй-
ственных культур. Так, на свекольном
поле, особенно после дождей, можно най-
ти разлагающиеся листья свеклы, причем
особенно ясно следы сапробиотического
распада видны на нижней стороне листьев,
обращенной к почве. Если взять каплю
распадающегося материала (от листа) и
рассмотреть ее под микроскопом, можно
легко убедиться в том, что в этой капле
видны различные одноклеточные организ-
мы, в том числе бактерии, низшие грибы,
простейшие и многочисленные энергично
двигающиеся нематоды. Все эти организ-
мы всегда весьма специфичны, т. е. обра-
зуют группы, встречающиеся только в
данной гнилостной среде. Поэтому их
обычно называют сапрофитными бакте-
риями, сапрофитными грибами, сапрозой-
ными нематодами и т. д. Все эти организ-
мы играют определенную роль в сложных
процессах распада органического мате-
риала и составляют действующие факторы
сапробиотической среды. Их деятельность
обеспечивает возвращение в почву эле-
ментов, за счет которых синтезируются
живые ткани животных и растений. На
языке биохимика этот сложный процесс
созидания и разрушения выражается в
циклах главных элементов, участвующих
в синтезе и распаде органических соеди-
нений, присущих живым организмам и
определяющих само понятие о жизни.
Таким образом, сапробиотическая среда
есть процесс распада органического мате-
риала, причем в этом распаде принимают
участие и даже неизбежно обеспечивают
его специфические, приспособленные к
жизни в сапробиотической среде организ-
мы, в первую очередь бактерии и грибы.
Эти организмы выделяют специфические
ферменты, обеспечивающие распад орга-
нических соединений — белков, углево-
дов, клетчатки и др.— на более элемен-
тарные соединения.
Характерный признак сапробиотиче-
ской среды — ее динамичность. Она всегда
существует в ее изменениях и превраще-
ниях, конечным звеном которых является
минерализация органического материа-
ла — возвращение бывших живых тел в
почву. Сапробиотические процессы всегда
происходят в почве, и в ней живут орга-
низмы, в том числе и нематоды, которые
так или иначе участвуют и в этих слож-
ных процессах распада органического ма-
териала. Эти сапробиотические нема-
тоды — корень всех сецернентов, всего
этого подкласса.
Все основные группы сапробиотических
нематод принадлежат к отряду Rhab-
ditida. В составе этого отряда очень много
различных групп — от сапробиотических
и до специализированных паразитов. Сре-
ди представителей подкласса сецернен-
тов очень много паразитов животных,
человека и растений—гораздо больше, чем
в пределах подкласса аденофорей.
Причина этого заключается в том,
что в пределах подкласса аденофорей
нет ни одного семейства, которое было бы
так тесно связано с сапробиотической сре-
дой, как семейство рабдитид. Правда, чи-
татель познакомился с семейством плек-
тид, принадлежащим к аденофореям. Но
плектиды — источник развития рабдитид,
а не каких-либо аденофорей.
Первый, наиболее характерный признак
представителей семейства рабди-
тид (Rhabditidae) — небольшие размеры
форм. Длина их тела около 1 мм, а часто
411
и меньше. Вторым внешним признаком
этих сапробиотических нематод назовем
свойственную им веретеновидную форму
тела: к голове и к хвосту тело рабдитид
обычно сужено, в среднем отделе заметно
расширено. Голова, как правило, несет
осязательные сосочки (п а п и л л ы), а
не щетинки, как у плектид. А м ф и д ы,
или боковые органы, всегда
расположены на передней плоскости голо-
вы, т. е. на губах. Ротовая полость имеет
форму удлиненного цилиндра (рис. 229).
В глубинном отделе этого цилиндра видны
«глоточные бугры». В суженной части ро-
товой полости, лежащей между вдающи-
мися в нее упомянутыми выше «глоточ-
ными буграми», у большинства предста-
вителей семейства рабдитид расположены
мелкие зубчики, названные онхами.
Цилиндрическая ротовая полость раб-
дитид гладкостенная и достаточно широ-
кая, чтобы легко пропустить комочек сап-
робиотического субстрата, который обыч-
но глотают рабдитиды. Пройдя сквозь
ротовую полость, комочек пищи попадает
в просвет пищевода. Стенки его муску-
листы и несут обычно два утолщения,
известные под названием бульбусов.
Один из них средний, а другой —
задний. Задний бульбус обладает уси-
ленной мускулатурой и особым внутрен-
ним дробильным аппаратом, который
раздробляет проглоченный комок пищи
(рис. 229). За пищеводом следует средняя
кишка, переходящая в заднюю кишку.
Последняя открывается наружу заднепро-
ходным отверстием, лежащим, как у всех
нематод, на брюшной стороне тела, в
основании хвоста.
Половые органы самок хорошо развиты.
Оба яичника (бывает и один) содержат
большое число зародышевых половых кле-
ток — овогониев, за счет которых
формируются яйца (рис. 236). Самцы име-
ют один семенник, удлиненный семяпро-
вод, за которым следует семяизвергатель-
ный канал, впадающий в заднюю кишку.
В ней же лежат выдвижные совокупитель-
ные органы самца — спикулы. Самым за-
мечательным органом самца, несомненно,
являются хвостовые бурсальные
крылья. Одно из них лежит справа,
а другое слева, начинаясь несколько впе-
реди от хвоста и часто до-
стигая его кончика.
Таковы основные осо-
бенности организации раб-
дитид, которые повторяют-
ся в различных не слиш-
ком резких вариантах у
более чем 240 видов этого
семейства. Мы не случай-
но остановились на описа-
нии организации рабдитид.
Задача, которую мы долж-
ны теперь решить, состоит
в том, чтобы попытаться
объяснить, почему сфор-
мировались описанные вы-
ше признаки.
Оказывается, что все пе-
речисленные признаки лег-
ко и естественно объясня-
ются тем, что они полно-
стью отвечают основным
особенностям сапробиоти-
ческой среды. К тем при-
знакам этой среды, кото-
рые выше были описаны,
надо присоединить еще
один: сапробиотическая
среда, как правило, пред-
ставлена не крупными пло-
щадями почвы, но значи-
тельно чаще отдельными
сапробиотическими очага-
ми. Каждый кусочек ор-
ганического материала,
попавший на землю, каж-
дый лист, обрывок стебля и т. п. ста-
новится причиной развития маленького
сапробиотического очага. В почве могут
быть, и фактически имеются, большие ко-
личества таких отдельных, взаимно изоли-
рованных сапробиотических очагов, и в
каждом из них под влиянием бактерий
и грибов в первую очередь развивается
сапробиотический процесс. Здесь же, в
этом крохотном подчас очажке, в почве
покоится, быть может, несколько крохот-
ных личинок рабдитид, покрытых плотной
шкуркой. Они ждут возникновения сапро-
биотического очага. Когда гнилостные,
сапрофитные бактерии начинают свою раз-
рушительную работу и когда начинается
формирование и развитие сапробиотиче-
Рис. 236. По-
ловые трубки
Rhabditis aber-
rans:
l — яичник; 2 —
яйца.
412
ского очага в почве, этот процесс ста-
новится стимулом к развитию личинок
рабдитид. Быстро развиваясь, рабдитиды
превращаются в самок и самцов, и на-
чинается быстротечная, стремительная
жизнь.
С биологической точки зрения, если вос-
пользоваться спортивной терминологией,
можно сказать, что рабдитиды настоящие
спринтеры, но не в смысле быстроты дви-
жения, а в смысле необычайной быстроты
развития.
Известны виды рабдитид, весь цикл
индивидуального развития которых — от
яйца до взрослой плодущей самки — про-
текает всего лишь за несколько часов.
Сроки, равные примерно 12 или 24 ча-
сам,— обычное явление. Если маленький
сапробиотический очажок просущество-
вал, скажем, 20 дней, за этот срок в нем
перебывает до 10—15 видов рабдитид, ко-
торые примерно через каждые 2—3 дня
сменяют друг друга. Эта быстрая смена
видов просто поразительна. Немецкая ис-
следовательница Р а й т е р A928) на-
блюдала картину смены видов в одном
из своих опытов. На третий день в экспе-
риментальном сапробиотическом субстра-
те содержались тысячи особей Rhabditis
inermis, от нескольких дюжин до сотен
Rhabditis elongata и еще три других вида.
Через два дня первый из названных видов
угас, три других размножились в тыся-
чах экземпляров. Еще через три дня кар-
тина снова изменилась и в массовых коли-
чествах появился один из видов, ранее
малочисленный. Таким образом, восьми
дней оказалось достаточно для развития
в небольшом экспериментально организо-
ванном «микрокосмосе» огромных коли-
честв рабдитид, исчисляемых многими ты-
сячами, возникших из ограниченного числа
личинок, имевшихся в комочке почвы,
использованной для опыта. Наблюдая за
этими тысячами рабдитид под покровным
стеклом, можно заметить, что большин-
ство этих маленьких животных находится
в состоянии почти неутомимого движения.
При более сильном увеличении микро-
скопа видно, что все они очень часто рабо-
тают средним бульбусом своего пищевода
и «дробильным» аппаратом второго, кар-
диального, бульбуса и заглатывают массу
пищи, состоящей из мелких органических
частиц сапробиоса — сапробиотического
детрита, бактерий и спор грибов. При
сильном увеличении мы обнаружим, что
на хвосте рабдитид оседают целые коло-
нии бактерий сапробиотической среды.
Кишечник часто заполнен, и нередко мож-
но видеть в нем темные массы пищи. Одна-
ко эта пища содержит много воды; кон-
центрация растворенных в ней продуктов
расщепления белков, Сахаров и других
веществ невелика, и рабдитиды должны
энергично глотать пищу, чтобы обеспечить
не только свое питание, но и «спринтер-
ское» развитие яиц. А ведь каждая самка,
едва достигающая 1 мм в длину, дает
за свою жизнь, измеряемую днями, до
250—300 яиц, из которых за 1—2 дня
развиваются новые самки и самцы. Пища
малопитательна, ее нужно много, и поэто-
му рабдитиды должны непрерывно дви-
гаться, чтобы всегда иметь эту пищу около
себя. Вот почему рабдитиды почти непре-
рывно поглощают пищу и столь же непре-
рывно двигаются. При малом увеличении
микроскопа поле наблюдения буквально
мерцает — все оно в динамике, в движении
от множества движущихся маленьких
змеевидных животных! Здесь возникает
стремительный оборот питания, усвоения
пищи, выделения отбросов, стремительный
рост зародышевых половых клеток,
«спринтерские» скорости развития, смерти
и новых рождений. Самка рабдитид нахо-
дится в непрерывном движении. Даже
когда самец осеменяет ее, она все-таки
хватает пищу и, извиваясь и суетясь, дви-
жется вперед или в стороны под покров-
ным стеклом в свете лучей микроскопа
(или в темноте). Вот почему самцу и нуж-
ны бурсальные крылья. Он прилаживает
их плотно к телу самки и выполняет свою
жизненную функцию все в той же симфо-
нии движения!
Как видим, вся организация рабдитид
отвечает условиям сапробиотической сре-
ды. Эта среда быстротечна, так как в ней
под влиянием различных групп сапробио-
тических бактерий идет распад органи-
ческого материала и каждый день меняет-
ся химическая характеристика сапро-
биоса. Этим объясняется столь быстрое
развитие рабдитид. И именно поэтому они
413
столь малых размеров — малые размеры
упрощают проблему быстрого развития.
Рабдитидам нужно все время питаться,
так как пища непитательна; надо нако-
пить огромное поколение, обеспечиваю-
щее будущее, т. е. жизнь личинок, кото-
рые останутся в почве в ожидании воз-
никновения нового сапробиотического
очага, когда вновь возобновится эта фее-
рическая смена поколений. Вей органи-
зация рабдитид подчинена этому требо-
ванию сапробиотической среды.
Кинооператор мог бы сделать изумитель-
ную картину, рисующую эту неисчерпае-
мую энергию жизни рабдитид и их орга-
низации, во всех деталях приспособлен-
ную к стремительной феерии реальной
жизни!
Таковы рабдитиды. Подчиняясь ритму
распада органического материала, они
участвуют в великом процессе возвраще-
ния почве азота, углерода, серы и других
химических элементов. Под «руководст-
вом» великих химиков природы — бак-
терий сапробиотической среды и с по-
мощью низших грибов — обладателей
мощных ферментов сапробиотические раб-
дитиды участвуют в процессах конечной
минерализации органического материала,
возвращая его почве и становясь тем са-
мым одним из источников новых бесчис-
ленных поколений живых форм. Так по-
гибшие организмы, через распад, стано-
вятся ступенью к новой жизни.
Какая красота и какой оптимизм жиз-
ни в этом творческом круговороте ве-
ществ !
В настоящее время можно с известной
уверенностью предполагать, что рабдитиды
произошли от древних плектид. Основ-
ным фактором развития рабдитид была
сапробиотическая среда, в которой в ре-
зультате влияния естественного отбо-
ра, открытого Чарлзом Дарвином
(см. Ч.Дарвин, Происхождение видов
или К. А. Тимирязев, Чарлз Дар-
вин и его учение), рабдитиды и при-
обрели свои особенности организации,
столь полно отвечающие условиям сапро-
биоса.
Рабдитиды сыграли важную роль в эво-
люции группы нематод, с которой человек
вынужден был серьезно считаться.
ОТРЯД РАБДИТИДЫ (RHABDITIDA)
Рабдитиды, приспособившиеся к жизни
в сапробиотической среде и ставшие участ-
никами круговорота органического мате-
риала под главенствующим воздействием
сапрофитных бактерий и грибов, стали
источником эволюции всего отряда Rhab-
ditida и, более того, всего подкласса
сецернентов (Secernentea). Приспособлен-
ность форм семейства рабдитид
к сапробиотической среде привела уже
самих представителей семейства рабдитид
к приобретению физиологических особен-
ностей, облегчивших переход к существо-
ванию в качестве поселенцев в органах
животных, и прежде всего в их кишечни-
ке. Мысль акад. К. И. Скрябина
A946), что проникновение рабдитид в ки-
шечник животных явилось первым шагом
к развитию зоопаразитических сецернен-
тов, абсолютно достоверна. Известен сле-
дующий интересный научный эпизод. Пче-
лы садились на пологий берег пруда и
лакали воду. С водой они проглотили
живых рабдитид. Рабдитиды не погибли
в кишечнике пчел. Напротив, они жили
в нем и даже вызвали явления катараль-
ного воспаления у пчел. Этот случай и
рабдитиды, в нем участвовавшие, зареги-
стрированы во Всесоюзном научно-иссле-
довательском институте гельминтологии
имени К. И. Скрябина. Рабдитиды повели
себя в этом случае как «зачинающиеся»
паразиты животных. Известны и описаны
виды рабдитид, которые были обнаружены
у человека и оказались способными суще-
ствовать в его кишечнике, и в частности
в толстой кишке. Некоторые рабдитиды
были найдены в человеческих испражне-
ниях. Пусть эти формы еще не «настоя-
щие» паразиты, но, видимо, пребывать
некоторое время в толстой кишке чело-
века они в состоянии, и это, конечно,
объясняется тем, что в толстой кишке
идут процессы, до некоторой степени сход-
ные с сапробиотическими.
В пределах отряда рабдитид,
с которым мы сейчас знакомимся, в част-
ности, на примере семейства Rhab-
ditidae, имеются группы, сходные с пред-
ставителями этого семейства.
414
Рис. 23
loides
Семейство Strongyloidi-
dae представлено нематодами, при-
надлежащими либо к гермафро-
дитам, либо к видам, самки ко-
торых способны к девственному,
или партеногенетическому, раз-
множению. В пределах этого се-
мейства известен вид, в индивиду-
альном развитии которого сочета-
ются два поколения — одно сво-
бодноживущее, другое паразитиче-
ское. Цикл развития включает оба
поколения. Начнем с одного из
них — паразитического.
У человека в слизистой оболоч-
ке двенадцатиперстной и тонкой
кишок могут встречаться в каче-
стве паразитов партеногенетиче-
ские самки. Это так называемая
кишечная, или стронгилоидная, форма
нематоды, известная под названием Stron-
gyloides stercoralis. Она характеризует-
ся мелкими размерами, длинным тон-
ким пищеводом, а также коническим хво-
стом. Женское половое отверстие смещено
назад, ближе к анусу. В двух яичниках
формируются яйца, развившиеся парте-
ногенетически, т. е. без участия самцов.
Эти яйца выделяются наружу, и из них
могут развиться личинки двух типов.
Часть этих личинок, оставаясь в почве,
линяет и затем превращается в «филярие-
видных» личинок (с очень тонким пище-
водом). Эти личинки с питьевой водой
или через кожу проникают в организм
человека и развиваются вновь в герма-
фродитных самок, ведущих паразитиче-
ское существование. Другие личинки, с
рабдитоидным пищеводом, развиваются по
типу гетерогонии, т. е. дают самок и сам-
цов, пребывающих в почве; после опло-
дотворения самцами самок рождаются ли-
чинки, попадающие и в этом случае через
рот или через кожу в организм человека.
Они проникают в конечном счете в сли-
зистую двенадцатиперстной и тонкой ки-
шок. Таким образом, у этой нематоды
имеются два поколения — паразитическое
и свободное. Наличие у части личинок
рабдитоидного пищевода свидетельствует
о филогенетической близости Strongyloi-
didae к рабдитидам. Наличие двух поко-
лений — паразитического и свободного —
7. Strongy-
stercoralis.
также указывает на близость к
свободноживущим рабдитидам.
Strongyloididae — один из началь-
ных шагов развития паразитизма
в пределах подкласса се-
цернентов.
Семейство Oxyuridae —
более поздний и более специали-
зованный этап развития парази-
тизма. В качестве примера рас-
смотрим биологию широко изве-
стного паразита толстой и слепой
кишок человека — Enterobius ver-
micularis. Эта нематода известна
под названием острицы; действи-
тельно, ее тело заострено к хво-
сту. Однако на суженном перед-
нем конце тела имеется кутику-
ляргюе вздутие. Ротовое отверстие
ведет в узкую стому, переходящую в пи-
щевод, снабженный бульбусом. Имеются
два яичника у самок и один семенник
у самцов. Хвост самок резко заострен.
Острица паразитирует своеобразно. Эта
нематода вызывает раздражение и вос-
паление слизистой оболочки толстой
кишки; ей приписывают также опреде-
ленную роль в развитии воспаления
червеобразного отростка слепой кишки
(аппендицит). В кишечнике человека этот
паразит все же не вызывает серьез-
ных поражений (за исключением, разу-
меется, аппендицита). Питается острица
содержимым толстой кишки, и, видимо,
в ее питании большую роль играет бакте-
риальная флора толстой кишки. По край-
ней мере известно, что если человек, инва-
зированный (зараженный) острицей, бу-
дет принимать два-три дня подряд по две
таблетки биомицина (утром и вечером),
то острицы гибнут. Механизм воздейст-
вия, по-видимому, сводится к тому, что
биомицин способствует уничтожению бак-
териальной флоры толстой кишки, а это,
вероятно, лишает источников питания и
остриц. Этот эксперимент указывает на
значение бактериальной флоры в пита-
нии остриц. Такой же эффект можно полу-
чить, если уничтожить биомицином бак-
терий в гнилостной культуре рабдитид.
Таким образом, острицы — это, конечно,
очень примитивная стадия развития пара-
зитизма, физиологически близкая раб-
415
дитидам. Главная неприятность,
испытываемая человеком от при-
сутствия в его кишечнике остриц,
не столько в расстройствах кишеч-
ного тракта, сколько в некоторых
особенностях биологии остриц. По
вечерам самки выходят через анус
человека наружу и вызывают зуд
в области промежности. У детей
этот зуд влечет за собой расчесы.
Под ногтями остаются яйца ост-
риц, которые выходят из ануса
человека и откладывают на его ко-
же яйца. Яйца паразита обяза-
тельно должны пробыть некоторое
время на воздухе, так как кисло-
род воздуха — непременное усло-
вие, стимулирующее их развитие.
С пальцев яйца легко могут по-
пасть в рот ребенка. Отсюда яй-
ца с личинками проникают в ки-
шечник и, достигнув толстой киш-
ки, снова становятся взрослыми
острицами. В таких случаях зара-
жение может продолжаться до-
вольно долго, и не только у детей,
но и у взрослых. Острицами мож-
но заразиться и при контакте с
человеком, пораженным этой не-
матодой, если не соблюдать пра-
вил гигиены. Яйца остриц могут
переносить на пищу человека му-
хи, тараканы-пруссаки и т. д. Мож-
но даже проглотить их с пылью,
если на полу имеются яйца этой
нематоды. При обнаружении зара-
жения необходимо обратиться к врачу.
Так как острицы выползают из ануса
вечером, когда человек собирается или
уже лег спать, то во избежание повтор-
ного самозаражения необходимо на ночь
надевать трусы, чтобы предотвратить по-
падание яиц острицы на простыню. А ут-
ром трусы надо прогладить горячим утю-
гом, чтобы убить яйца. Эти меры необхо-
димо применять как до лечения, так и
во время лечения и до тех пор, пока ана-
лиз испражнений на наличие яиц не по-
кажет, что их больше нет.
Человеческая аскарида (Ascaris lumbri-
coides). Много неприятнее и серьезнее
другая болезнь человека, а именно свя-
занная с паразитированием в его кишеч-
нике человеческой аскариды. Толь-
ко что рассмотренная острица до-
стигает 1 еж в длину (самки). Ас-
карида— это нематода, достигаю-
щая очень крупных размеров:
самка достигает 20—40 см в дли-
ну, а самец—15—25 см при ши-
рине соответственно 6 и 3 мм. Это
уже гигант, особенно по сравне-
нию с крохотными рабдитидами.
Ротовое отверстие аскариды окру-
жено тремя «губами». В конце пер-
вой трети плотного тела виден
«поясок».На брюшной стороне это-
го «пояска» имеется женское по-
ловое отверстие (у самок). Поясок
представляет собой легкую пере-
тяжку тела и служит опорой для
самца во время осеменения сам-
ки. Тело от головы к середине рас-
ширяется и снова сужается к хво-
сту. Хвост очень короткий, причем
у самцов он загнут на брюшную
сторону. Кутикула, покрывающая
тело аскариды, плотная. Мышцы
сильные, но движения медленные.
На вскрытой самке виден дубин-
ковидный короткий пищевод, не-
сколько уплощенная средняя киш-
ка и короткая задняя кишка, от-
крывающаяся наружу анусом. Но
самое характерное в организации
аскариды-самки — половые орга-
ны. Женское половое отверстие,
расположенное на брюшной сто-
роне «пояска», ведет в трубку вла-
галища (вагины), которое соединено с
двумя матками — правой и левой. Мат-
ки постепенно сужаются и переходят в
яйцеводы, а они в свою очередь — в
длинные и тонкие яичники. Половые труб-
ки аскариды достигают значительной дли-
ны, превосходя в несколько раз длину
тела. У свободноживущих нематод яич-
ники никогда не достигают такой длины
и всегда короче общей длины тела или
немногим превышают ее. Известный не-
мецкий зоолог Л е й к а р т (Leuckart,
1876) писал по поводу длины половых
трубок аскариды следующее: «У одной
самки 280 мм длиной каждая из по-
ловых трубок достигала 1500 мм, а обе
вместе были в 11 раз больше общей дли-
Рис. 238. Остри-
ца (Enterobius ver-
micularis).
416
ны тела». Это мощное развитие
полового аппарата—приспособ-
ление к паразитическому суще-
ствованию, которое связано с
колоссальной плодовитостью че-
ловеческой аскариды. Каждая
самка в течение суток выделя-
ет примерно 200тыс. яиц. Био-
логический смысл этой «астро-
номической» плодовитости в
том, что на пути развития —
от внешней среды в организм
человека — встречается множе-
ство факторов, приводящих зна-
чительную часть потомства к
гибели, и только очень боль-
шая плодовитость может «га-
рантировать» продолжение жиз-
ни вида.
Аскарида паразитирует в тон-
кой кишке человека. Однако
потомство не обеспечивается
существованием самки в тонкой
кишке хозяина. Самцы опло-
дотворяют яйца, но не обеспе-
чивают их развитие, как это
имеет место у свободноживу-
щих животных. Для того чтобы
яйцо вступило в начальные ста-
дии развития, оно обязательно
должно попасть во внешнюю
среду, так как нуждается в кис-
лороде. Находясь во внешней
среде, яйца примерно за 9—13
дней развиваются до стадии ли-
чинки второго возраста.
В яйцевых оболочках личин-
ка претерпевает первую линьку. Затем
развитие прекращается и не возобновля-
ется, если яйцо с личинкой второго воз-
раста не попадет в организм человека.
Яйцо должно быть проглочено. Заражение
происходит примерно так же, как и при
заражении острицами, — с пищей, через
грязные руки, в результате грызения ног-
тей; на пищу яйца могут быть занесены
мухами, или тараканами, или в результа-
те небрежного, негигиеничного обращения
с продуктами питания. Яйцо аскариды,
попавшее в рот человека и содержащее
личинку второго возраста, получает новый
стимул для развития. В тонкой кишке
человека личинки освобождаются из скор-
лупки яйца и с этого момента
начинают свой сложный путь
дальнейшего развития. Прежде
всего они проникают в слизи-
стую оболочку тонкой кишки,
а затем — в кровеносные со-
суды.
Вскользь отметим, что в по-
следнее время появились в на-
учной литературе некоторые со-
мнения в том, принадлежит ли
аскарида и ее родичи к сецер-
нентам и не следует ли всех
аскаридат (подотряд As-
caridata) перенести в отряд
аденофорей. Свободные адено-
фореи—кислородолюбивые фор-
мы. Как мы уже отмечали, для
зоопаразитических аденофорей
в значительной степени типич-
на склонность «прошивать» тка-
ни, вступая тем самым в кон-
такт с кровеносными сосудами
и, следовательно, с кислородом
крови. Быть может, путь ас-
каридных личинок по тканям
человека, также связанный с
кровяным руслом нашего орга-
низма, может рассматриваться
как следствие исторических свя-
зей со свободными аденофоре-
ями? Сейчас этот вопрос еще
не решен окончательно.
По кровяному руслу личин-
ки аскарид проникают в печень,
из печени — в сердце, со-
ответственно кругообороту кро-
ви. Из правого желудочка сердца личинки
проникают в легкие. В легких личинки
вскоре оседают в капиллярах, нередко
обусловливая их разрывы и кровотече-
ния. В дальнейшем путь личинок изме-
няется. Из капилляров легочной ткани
личинки проникают в бронхи, из них —
в трахею, а из трахеи — в глотку. Теперь
открыт путь в пищевод и оттуда в кишеч-
ник (через желудок). Это конечный этап
длинного пути. В тонком кишечнике ли-
чинки завершают свое развитие и превра-
щаются во взрослых самок и самцов.
Начинается новый цикл развития.
К сожалению, аскарида — один из
обычнейших паразитов человека. Зара-
Рис. 239. Аскарида
человеческая (Ascaris
lumbricoides): сле-
ва— самка, спра-
ва — самец, вни-
зу— яйцо.
ЩР 27 Жизнь животных, т. 1
417
женность этой нематодой может дости-
гать внушительных размеров. Выделение
аскарид из кишечника вместе с испражне-
ниями — неприятное, иногда пугающее
открытие. Но дело далеко не только в том
естественном чувстве отвращения, или
страха, или беспокойства, которое испы-
тывает человек, и далеко не редко — дети.
Во многих случаях болезнь, которую вы-
зывают аскариды, особенно если их в тон-
кой кишке оказалось много, приобретает
тяжелый и даже опасный для здоровья
и жизни человека характер. Аскариды
обладают очень характерной чертой пове-
дения — они стремятся проникнуть в уз-
кие щели и ходы. Известны случаи про-
никновения аскарид в желчные протоки
печени. Иногда аскариды проникают в
желудок, а затем в результате рвотных
движений поднимаются в глотку, а отсюда
в лобные пазухи и в полость носа. Изве-
стны неоднократные случаи выхождения
живых или мертвых аскарид через нос.
«Сюда,— говорит доктор Браун,— отно-
сится случай, сообщенный Альбрехтом,
согласно которому аскарида была уда-
лена из носа десятилетней девочки...
Из носоглотки,— продолжает Браун,— ас-
кариды могут проникать в евстахиеву
трубу и оттуда — в среднее ухо». Гораз-
до опаснее случаи проникновения аска-
рид в гортань и трахею, так как в этих
случаях возможно даже смертельное
удушье. Аскариды могут проникать в мо-
чевые органы. Описаны случаи прободе-
ния стенок тонкой кишки с последующим
проникновением в полость тела, что вле-
чет за собой перитонит, т. е. воспаление
брюшины.
Из сказанного видно, что не следует
допускать эту нематоду в пищевари-
тельные органы человека. А уж если ас-
кариды проникли в кишечник человека,
приходится немедленно принимать меры
к их удалению, т. е. проводить специаль-
ную дегельминтизацию — уда-
ление или изгнание этого паразита из
тонкого кишечника человека.
Еще лучше — заботиться о том, чтобы
аскариды не могли попасть в наш кишеч-
ник. Важнейшая предупредительная ме-
ра — содержание в чистоте рук и тела,
а также носильного и постельного белья,
если человек уже заражен, чтобы избе-
жать повторного самозаражения боль-
ного аскаридозом. Большое значение име-
ют предупредительные меры, принимае-
мые в отношении возможности заражения
через продукты.
Известны случаи нахождения живых
яиц аскариды на поверхности пирожных
и других пищевых товаров с влажной
поверхностью. Строго говоря, вовсе не
плохо прогревать хлеб перед едой на
огне газовой горелки. Не следует есть не-
мытые овощи, например морковь, салат,
поднятые с земли огурцы и т. п., так
как яйца аскарид могут быть занесены
в почву и на ее поверхность.
С целью исключения заболевания гель-
минтозами, в том числе аскаридозом, необ-
ходимо вести борьбу со скверной привыч-
кой некоторых людей грызть ногти.
Аскаридоз всегда вероятен там, где лето
длительное и теплое, в периоды, когда в
почве много влаги.
Опасно употребление овощей с ин-
дивидуальных огородных участков, так
как все еще бытует привычка удобрять
почву экскрементами человека. В этих
случаях яйца аскарид заражают почву
и растения. Использование немытых ово-
щей с таких индивидуальных участков
всегда чревато последствиями. Нужно во-
обще отвыкать от распространенной при-
вычки есть на улице, в метро, в вагоне
трамвая и т. п. Нет никакой гарантии
против возможности заражения в этих
случаях аскаридой, да и другими немато-
дами и прочими гельминтами, в том числе
и смертельно опасными, вроде эхинококка
или альвеококка.
Таким образом, гигиенический режим
жизни, сознательное отношение к вопро-
сам культурного поведения — главный
враг и аскариды, и других паразитических
червей.
Представители семейства аска-
рид поражают не только человека. Близ-
кая к человеческой свиная аскарида пора-
жает свиней, а крупная лошадиная аска-
рида (Parascaris equorum) — обычный
паразит домашней лошади. Все эти формы
относятся к семейству Ascaridae.
Близкое к нему семейство Ascari-
diidae с типичным родом Ascaridia содер-
418
жит довольно большое число видов нема-
тод, поражающих кишечник птиц, пре-
имущественно сухопутных. В народном
хозяйстве значительную опасность пред-
ставляет Ascaridia galli — паразит кур.
Этот паразит особенно опасен для молод-
няка.
Известный советский специалист по
аскаридам А. А. Мозговой указы-
вает, что «аскаридоз... протекает неред-
ко в виде эпизоотии с большой смерт-
ностью». Аскаридоз кур снижает яйце-
носкость; цыплята если не умирают, то
отстают в росте. В борьбе с аскаридозом
кур большое значение имеет обеспечение
птиц витаминами, повышающими сопро-
тивляемость их организма инвазии. По-
мимо этого, необходимы особые меры пре-
дотвращения этой опасной болезни. В хо-
зяйствах содержат кур в чистоте: помет
подвергают обеззараживанию термическим
воздействием; кормят птицу из специаль-
ных кормушек; поилки должны быть чи-
стыми; выгульные дворики подвергают
ежегодному перепахиванию; весь инвен-
тарь курятников — поилки, кормушки,
да и само помещение — тщательно обезза-
раживают; помещение обрабатывают 3—
5-процентным раствором карболовой кис-
лоты и т. п. Все это необходимо, чтобы
победить болезнь и предотвратить ее рас-
пространение, грозящее большими поте-
рями поголовья кур.
ПОДОТРЯД СТРОНГИЛЯТЫ (STRONGILATA)
В толстых кишках лошади живет нема-
тода из семейства стронгйлид
(Strongylidae) — Strongylus vulgaris. Это
довольно крупные нематоды желтого цве-
та, самки которых достигают в длину 20—
21 мм (самцы — 14—16 мм). Передняя
часть стомы несет «корону» лепестков —
наружную и внутреннюю. Стома боль-
шая, бокаловидная (рис. 240). У самцов
имеется характерная трехлопастная бурса.
Самки этой крупной нематоды выделяют
яйца, которые попадают в наружную сре-
ду вместе с экскрементами лошадей (а так-
же осла и зебры). Из яиц уже в течение
первых суток выходят личинки первого
возраста. Они растут очень быстро:
уже на вторые сутки претерпевают пер-
вую линьку, становясь личинками вто-
рого возраста. Через неделю следует вто-
рая линька, во время которой личинка
достигает третьего возраста. Эти личинки,
не освобождаясь от своей шкурки, ста-
новятся очень устойчивыми к внешним
условиям и в то же время инвазионными,
т. е. заразными для животных. Лошади,
ослы и зебры, проглатывая таких личи-
нок с водой, заражаются паразитом. У ло-
шадей личинки этой стронгилиды парази-
тируют в поджелудочной железе, что вле-
чет за собой тяжелый катар, последствия
которого могут вызвать гибель живот-
ного. Лучшее средство уберечь лошадь
от этого заболевания — чистота пойла
и корма, а также своевременная дегель-
минтизация, т. е. изгнание паразита спе-
циалистом врачом-ветеринаром.
К стронгилятам принадлежит также
очень опасный паразит человека — кри-
воголовка двенадцатиперстная (Ancylosto-
ma duodenale). Нематода достигает 8—
18 мм длины и живет в тонкой кишке
человека. Название — кривоголовка —
объясняется тем, что ротовое отверстие
у этого паразита несколько сдвинуто на
брюшную сторону тела (рис. 241). Боль-
шая ротовая полость вооружена острыми
онхами (неподвижными зубами), поражаю-
щими слизистую оболочку тонкой кишки
человека. Кровоточащие ранки — одно
из последствий паразитирования криво-
головки в кишечнике человека. Криво-
головка широко распространена в ряде
стран. В СССР она известна на Кавказе.
Взрослые самки, паразитируя в тонкой
кишке человека, выделяют яйца, выпа-
дающие наружу вместе с испражнениями
человека. Личинки имеют рабдитоидный
пищевод. В почве они ведут сначала
образ жизни сапробионтов подобно раб-
дитидам. Затем прекращают сапробиоти-
ческое питание и превращаются в «строн-
гилоидную» личинку, которая после линь-
ки переходит в третью стадию развития.
Такие личинки, находясь в почве, стано-
вятся инвазионными, т. е. способными
проникать в организм человека. Самым
серьезным моментом биологии этих личи-
нок надо считать способность проникать
27*
419
Рис. 240. Delofondia vulgaris.
в кровяное русло человека через кожу
(перкутанный путь инвазии). Личинки
кривоголовки попадают в венозную систе-
му человека и затем пассивно заносятся
в правое предсердие и правый желудочек
сердца. В дальнейшем через легочную
артерию личинки попадают в легкие чело-
века. В этот период возможно крово-
харканье. Из легких нематоды попадают
в глотку и оттуда в кишечник человека,
в частности в двенадцатиперстную киш-
ку, где кривоголовка достигает взрос-
лого, половозрелого, состояния и начи-
нает выделять яйца.
Понятно, что пребывание паразита в
организме человека не остается без послед-
ствий, «У анкилостоматиков,— пишет
акад. Е. Н. Павловский,— отмечается
боль под ложечкой, усиливающаяся при
давлении. Эти болезненные ощущения ос-
лабевают при поедании таких веществ,
как земля или мел; у анкилостоматиков
(т. е. больных анкилостомозом.—А. П.) на-
блюдается часто извращение вкуса, и они
инстинктивно ощущают потребность
поедания земли (геофагия), что можно
поставить в связь с умеряющим дейст-
вием земли на боли под ложечкой». «Бы-
вает,— продолжает автор,— рвота водя-
нистыми, слизистыми или желчными мас-
сами. Понос наблюдается чаще запоров.
В стуле может быть примесь крови. Быва-
ет прерывистая лихорадка с поднятием
температуры до 38—39° С». В дальнейшем
могут появиться явления анемии, слабо-
сти, головокружения, сердцебиение, повы-
420
Рис. 241. Головной конец
кривоголовки двенадцати-
перстной (Ancylostoma duo-
denale). В ротовой полости
видны крупные зубы.
шенный пульс, одышка. Не исключен и
смертельный исход.
Следует иметь в виду, что заражение
возможно, например, при контакте голой
кожи с землей. Отсюда связь анкилосто-
моза (как называется болезнь) с профес-
сией землекопов, земледельцев, горнора-
бочих шахт и т. д.
Тяжесть заболевания диктует необхо-
димость борьбы с этим опасным пара-
зитом и специального лечения в случа-
ях заболеваний. Широкая дегельминти-
зация населения районов нахождения
анкилостом — важнейшее звено в борьбе
с этим паразитом.
Семейство Нитчатки (Filariata).
К этому семейству относится нематода,
которую у нас в стране удалось уничто-
жить полностью. Это один из наиболее
ярких примеров возможности полного или
частичного уничтожения паразитических
нематод на основе точного знания их био-
логии и циклов развития. Мы имеем в виду
паразита, достигающего длины 120 см
при ширине тела 1,0—1,7 мм. Такова
ришта, или Dracunculus medinensis. Это
один из опаснейших паразитов человека.
Взрослая самка паразитирует в лимфати-
ческих сосудах, лимфатических железах
и в подкожной клетчатке преимуществен-
но ног человека. Болезнь начинается с
зуда в определенных местах кожных по-
кровов, причем в этих местах ощущается
затвердение. Позднее развивается язва,
из глубины которой, через разрыв
и изъязвление кожи, может высовываться
петля тела ришты. Через язву нематода
выделяет массы личинок. Позднее в зави-
симости от локализации ришты могут
появляться боли в суставах. Возможны
и вторичные инфекции, нарывы и т. п.
Как же заражается человек риштой? Если
яйца ришты попадут в воду, из них выхо-
дят личинки. Впрочем, это же происходит
и во влажной почве. В воде личинки
живут недолго, около трех дней. В тече-
ние этого времени они проникают в орга-
низм мелкого веслоногого рачка —
циклопа. Известно несколько видов цикло-
пов, и все они (Cyclops coronatus, С. quad-
ricornis, С. oitonoides, С. vicinus, С. ulja-
nini) могут быть промежуточными хозяе-
вами личинки ришты. Циклопы проглаты-
вают этих личинок и заражаются. Из
желудка циклопа личинки проникают в
полость тела рачков. Здесь они линяют,
продолжают свое развитие в течение 1 —
1,5 месяца и, оставаясь в организме рач-
ков, оформляются в типичных микрофи-
лярий. Если человек вместе с питьевой
водой проглотит такого зараженного рач-
ка, паразиты проникнут в конечном счете
в подкожную клетчатку, и тогда возникнет
болезнь, которая по родовому наименова-
нию паразита названа дракункуле-
з о м.
Ришта очень широко распространена.
Она известна в Иране, в Индии, по бере-
гам Красного моря, на побережьях Нила,
в Азии и в Африке. Человек давно знает
болезнь, вызываемую риштой. Знамени-
тый анатом Гален A31—201 гг. до н. э.)
уже указывал на болезнь, вызываемую
этим паразитом. Ришта была известна
арабским писателям и древним евреям
с берегов Красного моря, называвшим
длинную паразитическую нематоду «ог-
ненной змеей».
Единственным средством лечения в ста-
рые времена было извлечение ришты пу-
тем наматывания длинного червя на па-
лочку. Метод этот был известен очень
давно.
В СССР дракункулез ликвидирован в
результате применения плановых меро-
приятий. Они состояли в том, что преж-
де всего подвергли дегельминтизации всех
людей, больных дракункулезом. Покон-
чив со взрослой риштой у людей, взя-
лись за циклопов. Для этого водоемы
(х а у з ы) подверглись последователь-
ному осушению. Солнечные лучи уничто-
жили всех циклопов. Затем водоемы (ха-
узы) были отремонтированы, щели меж-
ду отдельными плитами хаузов были за-
литы цементом, а в городе (Бухаре)
построили водопровод. Людям было стро-
жайше запрещено пить сырую воду из во-
доемов без фильтра. Для этого было ре-
комендовано пить воду из чайников, носик
которых был накрыт плотным платком,
чтобы уцелевшие в водоемах циклопы
не могли быть проглочены человеком.
Итак, были предприняты меры предуп-
реждения возможных заражений и меры
лечения больных людей. Единый комплекс
мероприятий, примененный Советской
властью на основе данных науки, позво-
лил ликвидировать дракункулез в пре-
делах нашей страны. Это был большой
успех советской науки о паразитических
червях — гельминтах, известной под об-
щим названием гельминтологии.
Этот пример показывает, насколько важ-
но точное изучение развития гельминтов
(паразитических червей). На точном науч-
ном изучении гельминтов и их развития
базируется учение о девастации
гельминтов — их искоренении из жизни
и быта советских людей.
Это заветная мечта крупнейшего совет-
ского гельминтолога акад. К. И. Скря-
бина.
Диктиокаулез. Овца кашляет
сухим, отрывистым кашлем. Позднее ка-
шель усиливается, становится трудным
и тяжелым. Изо рта и ноздрей овцы время
от времени выделяется зеленовато-серая
слизь. В конечном итоге животное гиб-
нет. Причина этой смерти — нематода,
известная в науке под названием Dictyo-
caulus filaria. Белое тело этой нематоды
достигает 30—80 мм в длину у самцов
и 50—112 мм у самок при толщине тела,
равной 0,35—0,59 мм. У самцов имеется
бурса.
Нематоды паразитируют в бронхах
и в трахее овцы нередко в огромных мас-
сах, вызывая поражения слизистых обо-
лочек и описанные выше внешние прояв-
ления болезни, именуемой д и к т и о-
каулезом.
421
В бронхах и частично в трахее взрослые
паразиты откладывают множество яиц.
Яйца откашливаются овцой в ротовую
полость. Во внешнюю среду яйца посту-
пают лишь частично; здесь из них выхо-
дят личинки. Большая же часть яиц про-
никает в кишечник овцы. Здесь из яиц
также выходят личинки. Они растут, ста-
новятся очень подвижными и, попав из
кишечника животного на пастбище, при-
обретают значение инвазионных личинок,
т. е. способных к паразитическому суще-
ствованию в организме овцы. Инвазион-
ные личинки — главная опасность. Они
и служат источником заражения овцы дик-
тиокаулезом. Как только такие личинки
вместе с подножным кормом попадут в
ротовую полость овцы, начинается даль-
нейшее развитие паразита. В кишечнике
овцы личинки линяют, сбрасывают шкур-
ку, остающуюся после второй линьки
(первая была в яйце), и затем проникают
в лимфатические сосуды овцы. По руслу
этих сосудов личинки проникают в серд-
це, а оттуда в легкие овцы. Разрывая
мелкие кровеносные сосуды легких, ли-
чинки попадают в бронхи. Исход зара-
жения зависит от степени инвазирован-
ности овцы. Если она велика, смерть
животного — обычный исход болезни.
Диктиокаулез — бич овцеводства, и
поэтому с этой болезнью овец ведется
упорная борьба, в системе которой, по-
мимо специального лечения овец, большое
значение имеет и режим выпасов овец.
Учитывая, что личинки диктиокаулюса
становятся инвазионными через 6—7 дней,
овец держат на данном пастбище не более
пяти дней, а затем перегоняют на новое
пастбище, на котором они пасутся тоже
не более пяти дней, переходя в дальней-
шем на новые участки. В результате таких
плановых перемещений овец происходит
следующее: личинки диктиокаулюса раз-
виваются в кишечнике овцы, а в почве,
попав туда вместе с экскрементами живот-
ных, достигают инвазионной стадии, но
хозяина, т. е. овец, не находят (они уже
перемещены на чистое пастбище). «Поле
битвы» — пастбище — вовремя покида-
ется овцами, а оставшиеся на нем личин-
ки диктиокаулюса гибнут. Понятно, что
это остроумное мероприятие возможно
только в условиях крупных хозяйств на-
шей Родины с их обширными пастбищны-
ми угодьями.
Диктиокаулезом болеют и другие жи-
вотные, в том числе крупный рогатый
скот, но действующим началом в этом
случае является другой вид.
ОТРЯД ТИЛЕНХИДЫ (TYLENCHIDAE)
Мир сецернентов, паразитирующих в
органах человека и животных, многообра-
зен и включает огромное число видов
нематод. Мы не имеем здесь возможности
вести дальнейшее изложение сведений об
этих гельминтах. Остается задержаться
на некоторых представителях сецернен-
тов, приспособившихся к паразитирова-
нию в органах растений. Все эти нема-
тоды, общим числом около 1000 видов,
принадлежат к отряду тиленхид (Ту-
lenchidae).
Стеблевые нематоды принадлежат к с е-
м е й с т в у Tylenchidae ироду Dity-
lenchus. Остановимся на описании стеб-
левой нематоды, поражающей чеснок и
лук. Выбор наш определяется тем боль-
шим ущербом, который эта нематода при-
чиняет урожаям лука в хозяйствах нашей
страны. В хранилищах потери могут до-
стигать 40—60% хранимых луковиц; пос-
ледние поражаются луковой стеблевой
нематодой и гибнут в результате гнилост-
ного распада.
Автору этих строк пришлось наблюдать
большие потери чеснока, вызванные этой
стеблевой нематодой. Они достигали 60%
урожая чеснока осенней посадки и почти
100% весенней. Вместе с тем дитилен-
х о з лука широко распространен и об-
наруживает тенденцию к дальнейшему
росту.
Стеблевая нематода, о которой здесь
идет речь, называется Ditylenchus dip-
saci. Вид этот относится к числу полити-
пических, т. е. видов с очень широкой
экологической приспособляемостью, или
валентностью. Это выражено, в частности,
в том, что Ditylenchus dipsaci дает боль-
шое число экологических рас, приспособ-
ленных к паразитированию на многих
422
растениях. Так, луковая раса способна
паразитировать не только в тканях лука
и чеснока, но равным образом на фасоли,
гречихе, шпинате, кормовой свекле, гор-
чице, сельдерее и других растениях.
Как же питается дитиленх? Прежде
всего для него, как и для всех других
представителей отряда тиленхид, харак-
терно превращение ротовой полости в тон-
кое острое оружие — стилет. У Ditylen-
chus dipsaci стилет достигает 11 мм в дли-
ну. Передний конец стилета косо срезан
и тем самым заострен, как игла медицин-
ского шприца. На заднем конце стилет
несет три «головки». К этим «головкам»
прикрепляются три пучка мышц, про-
тивоположные концы которых связаны с
тремя головными базальными перегород-
ками. Эти базальные перегородки обра-
зуют основание стилета головной капсулы,
составляющей передний конец тела дити-
ленха. Три пучка мышц стилета образуют
три протрактора, которые обла-
дают большой двигательной силой и спо-
собны стремительно выбрасывать стилет
сквозь узкое ротовое отверстие наружу.
Дитиленх буквально «стреляет» своим сти-
летом, пробивая им узкое отверстие в неж-
ной и тонкой оболочке. Стилет служит,
однако, не только как «перфорирующий»
орган, но и как орган, через внутренний
канал которого дитиленх высасывает рас-
творенный клеточный сок. Весь этот точ-
ный и подвижный механизм имеет при
этом микроскопические размеры. Не бу-
дет ошибкой, если мы скажем, что перед
нами тончайшая «ювелирная» работа при-
роды! Это и есть оружие дитиленха и лю-
бого другого представителя нематод из
отряда тиленхид (рис. 242). Само назва-
ние этого отряда—Tylenchida—составлено
английским нематологом Бастианом
в 1865 г. путем комбинирования двух гре-
ческих слов: tylos — мозоль («голов-
ки» стилета) и enchos — игла. Просвет
стилета переходит в просвет пищевода,
имеющий вначале, т. е. по соседству со
стилетом, такой же диаметр, как и про-
свет стилета.
Пищевод делится на три отдела: кор-
пус, истмус (перешеек) и
кардиальный бульбус. Кор-
пус в свою очередь делится на перед-
ний отдел, или прокорпус, и задний —
метакорпус, расширенный у большин-
ства тиленхид, в том числе и у дитилен-
ха лука, в средний бульбус.
Чтобы понять смысл перфорирующей
(пронзающей) работы стилета, остановим-
ся на функциях этого бульбуса. Метакор-
пальный бульбус снабжен мышцами и спо-
собен к сократительным сосущим движе-
ниям, что, собственно, и превращает сти-
лет не только в колющий, но и сосущий
орган. Поэтому стилет тиленхид имеет
значение колюще-сосущего органа и в гру-
бой форме может быть сравнен с колюще-
сосущими аппаратами некоторых насеко-
мых (тли, комары, клопы и т. п.).
Однако стилет может выполнять свою
функцию только тогда и в том случае,
если его колющий кончик погружен в
жидкость, и притом в жидкость с невысо-
кой вязкостью. В противном случае его
работа будет затруднена или станет не-
возможной. Значит, мало воткнуть сти-
лет в такую вязкую массу, как плазма
растительной клетки. Надо эту плазму
растворить. Для этого необходимо иметь
аппарат, способный эту функцию выпол-
нить. А это означает, что необходимо
превратить клеточный сок растительной
клетки в продукты, растворимые в воде.
Эта функция выполняется тремя железа-
ми, лежащими в кардиальном бульбусе
(рис. 241). Ведущее значение имеет спин-
ная железа, которая у дитиленха имеет
длинный выводной проток, впадающий в
просвет пищевода позади стилета. Экскрет
этой железы течет по выводному протоку,
вливается в полость стилета и впрыски-
вается в плазму растительной клетки.
Действие этого экскрета спинной железы
почти мгновенно: он растворяет или изме-
няет элементы плазмы растительной клет-
ки и все содержимое ее, и в частности
вещества белкового происхождения и по-
лисахариды (помимо других веществ, о
которых здесь мы не будем говорить).
Белки расщепляются на более простые,
растворимые вещества, полисахариды
трансформируются в моно- и дисахариды,
также растворимые в воде. В этом, в сущ-
ности, основа процесса усвоения таких
сложных веществ, как белки и полисаха-
риды. Доведение их до более простых
423
и растворимых в воде веществ превращает
эти более простые элементы в вещества,
которые животный организм, в том числе
и организмы наших микроскопических
фитопаразитических нематод, в состоянии
усвоить. Это относится и к другим веще-
ствам органического происхождения, ко-
торыми так богат растительный мир, —
жирам, клетчатке, пектиновым оболоч-
кам, «склеивающим» растительные клетки
в целостную ткань и образующим как бы
канву, на которой вышивается самой при-
родой рисунок живой растительной тка-
ни. Дитиленх и другие тиленхиды в раз-
Рис. 242. Луково-чесночный дитиленх (Dity-
lenchus dipsaci):
1 — стилет; 2 — пищевод, 3 — средний бульбус, 4 —
железы пищевода; 5 — нервное кольцо; 6 — экскре-
торный проток; 7 — средняя кишка; 8 — анальное
отверстие; 9 — яичник; ю — яйцо; 11 — преуте-
ральная железа; 12 — передняя и 13 —задняя матка.
ной форме и в разной степени разрушают
эти сложные органические системы, дово-
дят их до более простых и усвояемых, и
прежде всего до растворимых тел.
Что же лежит в основе этой замечатель-
ной способности наших маленьких «чер-
вячков»? Наука дает на этот вопрос ясный
общий ответ: за это превращение сложных
компонентов живых растительных клеток
и тканей в растворимые и усвояемые ве-
щества ответственны ферменты, выделяе-
мые тиленхидами из трех пищеводных
желез, включенных в задний отдел пище-
вода или связанных с ним. Железы выде-
ляют жидкий экскрет. Спинная железа
имеет, по-видимому, ведущее значение.
Ее экскрет, содержащий ферменты, при-
текает либо к основанию стилета, либо
в просвет переднего сектора метакорпаль-
ного бульбуса (рис. 242). Отсюда фермен-
тированный экскрет проникает в полость
стилета и затем в растительную клетку.
Почти мгновенно ближайший участок
плазмы клетки, обработанный каплей фер-
ментативного экскрета, растворяется и
сосущей силой метакорпального бульбуса
вовлекается в просвет пищевода. Впрочем,
стилет и сам «сосет», так как выполняет
роль микрокапилляра с огромной сосу-
щей силой. Следовательно, фактически
тиленхидам присуще внеклеточное пище-
варение. Всасывание готовой, растворен-
ной и переваренной еще вне нематоды
капли питательного вещества происходит
в средней кишке. Так питаются эти фито-
паразитические нематоды.
Луковый дитиленх, или луковая раса
(Ditylenchus dipsaci), обладает способ-
ностью питаться несколько иначе. Эта
нематода не вонзает стилет в клетки луко-
вицы. Она только нарушает стилетом це-
лостность ткани и действует своим фер-
ментом на пектиновые оболочки расти-
тельных клеток. Оболочки растворяются,
и ткань луковицы подвергается так назы-
ваемой мацерации, т. е. распа-
дается на теперь уже взаимно не
связанные клетки. Их содержимое диф-
фундирует наружу, в жижицу, состоя-
щую из воды с растворенными в ней
питательными веществами, извлеченными
диффузией из клеток. Этим раствором и
питается нематода.
424
Мацерация позволяет нематодам все
глубже проникать в ткань луковицы и
постепенно разрушать ее, питаясь продук-
тами биохимического распада питательно-
го материала.
Однако нематода не только питается, но
и начинает производить яйца. Из яиц
здесь же, в этой ферментированной среде,
рождаются личинки. Еще в яйце личинки
первого возраста линяют. Выйдя из яйце-
вых оболочек, они извиваются в среде
мацерированнои луковой ткани вместе со
своими взрослыми родичами. После линь-
ки в яйцевых оболочках они становятся
личинками второго возраста. Затем они
снова линяют и переходят в третий воз-
раст. В дальнейшем происходит третья
линька и личинки достигают четвертого
возраста. После четвертой линьки нема-
тоды превращаются во взрослые формы.
Все это развитие протекает приблизитель-
но 10—12 дней. Оно протекает, следова-
тельно, относительно быстро, хотя и не-
сравненно медленнее, чем у рабдитид.
Развитие достаточно быстро для того,
чтобы из 250 яиц, которые дает одна
самка за свою жизнь, развилось относи-
тельно большое потомство. Все это потом-
ство — ив этом характерная черта дити-
ленха — не покидает луковицы, в кото-
рой началось развитие. Напротив, в ее
тканях развиваются ряды последователь-
ных поколений, и так как продолжитель-
ность жизни дитиленха велика (не менее
года), то в ткани луковицы за время ее
нахождения в почве, а затем в хранилище
накапливается множество нематод. Так,
в одном зубке чеснока однажды было
подсчитано 7186 экземпляров дитиленха!
Если разрезать луковицу лука или зу-
бок чеснока на мелкие кусочки и погру-
зить их в воронку с водой, в которой они
будут лежать на металлической сетке, то
даже простым глазом, а еще лучше под
двукратной лупой можно увидеть карти-
ну, поражающую впервые видящего ее.
Из кусочков луковицы или чеснока спу-
стя несколько минут начинают выделять-
ся, медленно погружаясь в горлышко
воронки, тысячи дитиленхов. Извиваясь,
они уходят в резиновую трубку, наса-
женную на горло воронки. Оттуда, раз-
двинув лапки зажима, можно выделить
эту взвесь нематод в подставленную про-
бирку. Просмотр препарата этих нематод
под покровным стеклом даст нам возмож-
ность ознакомиться с организацией дити-
ленха (рис. 242). Мы видим все типичные
черты этого микроскопического живот-
ного, самки которого едва превышают
1 мм в длину: стилет, протракторы его,
пищеводный бульбус, мощный пакет же-
лез в задней части пищевода, половые ор-
ганы, спикулы и буреальные крылья у
самца в области хвоста. У самок в области
яйцевода (рис. 242) виден особый орган—
преутеральная железа, со-
стоящая из четырех рядов округлых кле-
ток. Предполагают, что эта железа выде-
ляет экскрет, стимулирующий развитие
одной из яйцевых оболочек. По наличию
этой железы легко узнать дитиленха.
К стеблевым нематодам, или дитилен-
хам, принадлежит также дитиленх карто-
феля, очень сходный с ранее описанным.
Дитиленх картофеля — один из опасней-
ших паразитов клубней картофеля, нано-
сящий большие потери хозяйству.
Галловые нематоды. Среди форм ти-
ленхид известны еще более опасные нема-
тоды, принадлежащие к роду галловых
нематод — Meloidogyne. Самки этих не-
матод имеют вздутое тело, самцы более
стройные. Суженный и несколько вытяну-
тый передний конец тела самок несет не-
большую головную капсулу, вооружен-
ную стилетом. Во вздутой задней части
тела лежит огромный кишечник и две
длинные половые трубки, производящие
большое число яиц,— свыше тысячи за
двухмесячное существование самки. Ли-
чинки галловых нематод по выходе из
яйцевых оболочек попадают в почву и про-
никают в корни многих видов растений.
Обычно личинки внедряются в корешок
вблизи от чехлика. Они продвигаются
дальше и затем оседают на месте, повер-
нувшись головным концом к сосудистому
пучку корня. С этого момента подвиж-
ность личинок прекращается. Они ста-
новятся сидяче-прикрепленными, или се-
дентарными, животными. Начинается раз-
витие, проходящее через серию четырех
личиночных и одну взрослую стадию.
По мере развития диаметр тела личинки
увеличивается и она превращается во
425
вздутую взрослую самку или в самца. Сам-
цы появляются обычно редко и преиму-
щественно при ухудшении условий суще-
ствования.
Пока идет развитие, под влиянием выде-
лений (ферментов) нематод происходят
глубокие изменения физиологического со-
стояния клеточных элементов корневой
ткани, и притом тех, которые примыкают
к головному концу развивающейся личин-
ки. Клетки увеличиваются в размерах
и теряют способность делиться, несмотря
на то что ядро таких клеток неоднократно
делится. Поэтому возникают многоядер-
ные крупные клетки, получившие назва-
ние гигантских. Параллельно идет
прогрессивное разрастание клеток, окру-
жающих нематоду, и развитие корневой
опухоли, имеющей округлую форму и из-
вестную под названием галла, откуда и
название «галловые нематоды». Такие гал-
лы, как бисер, усеивают корешки, пора-
женные галловой нематодой. Здесь необ-
ходимо рассказать о другой стороне
галлообразования. Дело в том, что галл
образуется по мере развития самки, заклю-
ченной в его ткань. Галл -- как бы дот
галловой нематоды, ее «опорный пункт», ее
защита, среда, в которой она живет, раз-
вивается и дает потомство. В жизни сам-
ки, достигшей половозрелого состояния,
наступает самый ответственный момент —
подготовка к выделению яиц. К этому
моменту из анальных желез ее начинает
выделяться фиброзная масса, которая пор-
циями накапливается на заднем конце те-
ла самки. Фиброзная масса увеличивается
в результате ритмичных выделений все
новых ее порций. Приблизительно каж-
дые 10 секунд выделяется новая капля
фиброзных выделений. Когда вся область
женского полового отверстия покроется
фиброзной массой, в нее начинают выде-
ляться яйца. Таким образом образуется
как бы сумка с яйцами, или о о т е к а.
Когда из яиц выходят личинки, их судь-
ба может быть различной. Часть личинок,
ближайших к внешней поверхности галла,
уходит в почву и заражает новые кореш-
ки. Другие остаются в галле, оседают в
нем или вблизи него. Поэтому галл посте-
пенно увеличивается. Возникает с л о ж-
Рис. 243. Галловая нематода:
А — три самки галловой нематоды A) в
корневой ткани огурца; 2 — гигантская
клетка; 3 — ядра гигантских клеток. Б —
кончик корня огуречного растения. Личин-
ки галловой нематоды проникают в корне-
вую ткань.
н ы й галл, или с и н г а л л. Сингал-
лы могут достигать очень крупных раз-
меров, иногда до 2—3 см в диаметре.
Из таких галлов в воронку с водой можно
выделить множество личинок.
Судьба галлов всегда одна и та же.
Сперва они белые, с опалесцирующей
поверхностью. Позднее в них начинаются
процессы некроза — реакции расте-
ния на пребывание паразитов в корневой
ткани. Тогда галл постепенно буреет.
Некротические процессы в нем привле-
кают почвенных гнилостных бактерий.
Вслед за бактериями и процессами гние-
ния, вызванными ими, в загнивающий
галл проникают уже известные нам сапро-
биотические рабдитиды. В итоге разру-
шается и галл и участки пораженной кор-
невой системы. Растение либо угнетено
и не дает плодов, либо гибнет полностью.
В тепличных хозяйствах в результате
размножения галловых нематод разви-
вается массовый галловый нема-
то д о з, следствием которого во многих
случаях становится уничтожение до 40—
60% урожая основной культуры этих
хозяйств (огуречной). Таковы галловые
нематоды, поражающие сотни различных
видов растений, в том числе тепличных,
огородных, бахчевых, плодово-ягодных
и технических.
Борьба с этими нематодами весьма труд-
на и дорого стоит. До настоящего времени
еще не найдено надежных средств борьбы,
которые полностью гарантировали бы ист-
ребление галловых нематод, хотя и име-
ются успехи в применении различных
противонематодных препаратов, извест-
ных под общим наименованием н е м а-
т о ц и д о в. Они позволяют снизить
потери и дать хозяйству некоторую пере-
дышку, хотя приходится вновь и вновь
подвергать почву специальной химиче-
ской обработке, повторяя эту дорогостоя-
щую «химизацию» неблагополучных поч-
венных площадей неоднократно, т. е. в
течение нескольких вегетационных перио-
дов.
Большой интерес представляет ориги-
нальная по своей организации группа
почвенных нематод, принадлежащих к се-
мейству Cephalobidae. Их блестящая
кутикула состоит из крупных колец, не-
Рис. 244. Представители почвенных Ce-
phalobidae:
А — Acrobeles complexus; Б — голова Асго-
beles ciliatus; В — голова Acrobeles ctenoce-
phalus, Г — голова Acrobeles ornatus. Видны
хорошо развитые проболы.
редко украшенных блестящими точечными
инкрустациями. Однако особое внимание
привлекает к себе организация головы:
она вооружена причудливо разветвляю-
щимися придатками, которые получили
название п р о б о л. Проболы образуют
сложные аппараты, с помощью которых
нематоды рвут разлагающиеся отходы
корней, служащие источником питания
Cephalobidae. Многие из этих нематод
способны проникать внутрь корней. По-
этому некоторые ученые рассматривают
эту группу нематод как животных, став-
ших на путь овладения растительной
тканью с помощью своего оригинального
вооружения.
427
Кроме названных выше фитопарази-
тических нематод, известны многие дру-
гие виды, наносящие сельскому хозяйству
очень большой ущерб. Изучением этих
нематод (тиленхид) занимаются фито-
гельминтологи — представители
молодой и ныне быстро развивающейся
науки — фитогельминтологии,
или фитонематологии, как эту
науку называют во многих странах.
ЗАКЛЮЧЕНИЕ
Трудно ответить на вопрос, где нет
нематод. Ну конечно, их нет в горячих
источниках. Однако в водоемах с темпера-
турой около 40°С они известны. Немато-
ды живут во всех средах, где возможна
жизнь. Не будет преувеличением, если
мы скажем, что биосфера в целом — это
среда жизни нематод. В почве по числу
особей нематоды — господствующая груп-
па многоклеточных почвенных организмов.
Дно морей и океанов «усеяно» нематода-
Коловратками называют своеобразную
группу в основном пресноводных микро-
скопических животных, выделенных в до-
бавочный класс типа Nemathelminthes.
Коловратки — самые мелкие из много-
клеточных животных (от 0,04 до 2 мм).
По своему систематическому положению
они стоят среди низших сколецид (низ-
ших червей — турбеллярий, нематод, га-
стротрих). Вместе с тем они поражают
наблюдателя своей красотой, изумитель-
ным разнообразием и тонкостью органи-
зации и биологии. Поражает исключи-
тельное разнообразие их движений
(плавание, ползание, «шагание», непо-
движное существование) и связанных с ними
строения покровов тела и «домиков» (у
ми. Нет таких многоклеточных животных,
в органах которых нельзя было бы найти
нематод. Нематоды паразитируют в пара-
зитах. Их можно обнаружить в члениках
цестод. Несомненно, что можно было бы
удивляться растениям, в которых не ока-
залось бы ни одной нематоды. Известно,
что нематоды и их яйца ветром переносятся
по воздуху. Они распространены повсеме-
стно. Ни одна другая группа животных
не может «похвастаться» такой изумитель-
ной распространенностью. Роль нематод
в природе, несомненно, очень велика.
Может ли быть иначе, если вся почва про-
низана ими, если в каждом растении ве-
роятна встреча с ними, если они стали
паразитами почти всех животных, если
они нападают на наши растения, если они
проникли всюду? Эта распространенность
нематод — самая интересная черта их ис-
тории, их жизни.
Изучение секретов необычайного успеха
нематод — этих большей частью мелких
животных — одна из важных проблем
современной зоологии и всего народного
хозяйства.
сидячих форм), а также ресничного аппа-
рата. У коловраток широко распростра-
нена колониальность.
Многообразие форм движения связано у
коловраток с чрезвычайной специализа-
цией ресничного вооружения и развити-
ем поперечнополосатой мускулатуры, со-
стоящей из отдельных тяжей. Тем более
удивительна свойственная коловраткам ма-
локлеточность при постоянстве клеточно-
го состава. Эти особенности соответствуют
столь же исключительной по многообра-
зию реактивности коловраток на внешние
раздражения. Не менее характерно для
коловраток разнообразие приспособлений
к смене климатических условий, выражаю-
щееся в явлениях сезонного цикломорфо-
КЛАСС КОЛОВРАТКИ (ROTATORIA)
(ДОБАВОЧНЫЙ КЛАСС NEMATHELMINTHES)
428
за, в многообразии половых циклов и в
способности переходить в покоящееся со-
стояние.
Всеми этими особенностями коловрат-
ки резко выделяются не только среди низ-
ших беспозвоночных, но и вообще среди
всех животных.
Наиболее типичными образованиями, по
которым сразу можно отличить коло-
враток от других животных, являются к о-
ловращательный аппарат
и жевательная глотка, так
называемый «мастаке». Коловращатель-
ный аппарат представляет собой совокуп-
ность различно расположенных на перед-
нем конце тела ресничек, которые у не-
которых видов располагаются по краям
дисковидных выростов головы и своим
биением, напоминающим мерцание спиц
быстро вращающегося колеса, вызывают
коловращение, за что они получили свое
название. Коловраток называют также
«ротатории», от латинских слов rota —
колесо и fего—носить (rotiphera — колесо-
несущие). Этот ресничный аппарат слу-
жит для передвижения и захватывания
пищи.
Мастаке — расширенная передняя
часть пищеварительной трубки (глотки) со
специальной мускулатурой, с железами и
нервами. Внутри мастакса находится же-
вательный аппарат из твердых частей,
называемых челюстями.
В процессе эволюции у коловраток выра-
ботались самые разнообразные приспособ-
ления к различным условиям существова-
ния. Такая пластичность организации со-
действовала тому, что коловратки населяют
не только разнообразные пресные, солоно-
ватоводные и соленые бассейны как с очень
высокой, так и с очень низкой температу-
рой воды, но многие из них приспособились
к существованию в местах, где вода бы-
вает или в ничтожном количестве, или
только временно: в прибрежном песке
водоемов, во мху, в лесной подстилке, в
дуплах деревьев, в почве. Основная масса
видов коловраток населяет пресные водое-
мы, в морях коловраток мало, но все же
есть живущие только в море. Помимо
видов своеобразного отряда Seisoni-
dea, есть исключительно морские предста-
вители родов: Synchaeta, Trichocerca, Lin-
dia, Proales, Keratella и др. Значительно
больше видов встречается в опресненных
частях морей; это те формы, которые по-
падают в море из рек и хорошо приспо-
сабливаются к повышенной солености,
причем некоторые виды в новых условиях
даже интенсивнее развиваются, чем в прес-
ной воде (Asplanchna brightwellii, Filinia
longiseta, F. brachiata). Немногие коло-
вратки ведут паразитический образ жизни.
Так, в червях паразитируют Drilophaga
(рис. 245, 12), Albertia и Balatro; в отло-
женных яйцах прудовиков встречается
Proales gigantaea, в солнечниках — Proales
latrunculus; есть паразиты водорослей,
например: в Vaucheria живет Proales wer-
necki, в колониях Volvox — Proales pa-
rasita. У позвоночных животных и у че-
ловека коловратки не паразитируют. Не-
которые виды коловраток сожительствуют
с другими организмами. Так, на дафниях
живут Proales daphnicola (рис. 245, 2),
питаясь водорослями, обрастающими пан-
цирь рачка; Brachionus rubens (рис. 245,
3) часто и надолго прикрепляется ногой
тоже к панцирю дафний, где, не затрачи-
вая энергии на передвижение, в спокойном
состоянии отфильтровывает пищевые ча-
стицы, все время меняя при помощи рачка
место облова.
Подавляющее большинство видов коло-
враток космополиты, они встречаются на
всем земном шаре, но есть и свойственные
только Америке, например Brachionus ges-
sneri (рис. 245, 5), Br. dolabratus (рис. 245,
6), Br. havanaensis и др., или только Ста-
рому Свету, как Brachionus diversicornis
(рис. 245, 4), Br. forficula; в Байкале жи-
вут виды, встречающиеся только в этом
озере, например Notholca baicalensis, N.
olehonensis (рис. 245, 7), Synchaeta pachi-
poda (рис. 245, 8) и др.; в Антарктиде
найдены коловратки, нигде больше не
встреченные. Некоторые обычные виды
коловраток живут в самых разнообразных
водоемах, они нетребовательны к условиям
существования — была бы вода и пища,
им неважно, соленая или пресная вода,
проточная или стоячая, теплая или холод-
ная, они почти при всех условиях хорошо
себя чувствуют и размножаются. Такие
организмы называются убиквистами,
т. е. встречающимися везде, повсюду. Ши-
429
Рис. 245. Коловратки:
1 — Seison nebaliae Grube; 2 — Proales daphnicola, 3 — Brachionus rubens; 4 — очертание панциря Br. di-
versicornis; 5 — очертание панциря Br. gessneri, 6 — то же у Br. dolabratus; 7 — Notholca olclionensis, 8 —
Synchaeta pachipoda; 9 — Trichocerca cylindrica, ю — Macrochaetus collinsi; 11 — Filinia longiseta, 12 —
Drilophaga delagei, 13 — Philodina brevipes; 14 — Rotaria neptunia; 15 — Conochiloides coenodasis (амфотер-
ная самка с тремя амиктическими и пятью миктическими яйцами; внизу — только что вышедший самец);
J6—Monommata appendiculata, 17 — Lindia brotzkayae.
рокое распространение многих видов свя-
зано с легкостью их переноса на большие
расстояния в сухом виде: у пиявковидных
коловраток — взрослых самок, а у дру-
гих — покоящихся яиц. Но немало ко-
ловраток, которые предъявляют строгие
требования к условиям существования, к
качеству пищи, к химическому составу
воды и ее физическим показателям, и да-
леко не в каждом водоеме бывают нужные
им условия. Если попавшая в данный во-
доем коловратка не находит в нем необ-
ходимых ей условий, она просто погибает.
Такие виды встречаются не часто и отно-
сятся к «редким». Таким образом, распро-
странение коловраток по земному шару в
большинстве случаев проблема не столь-
ко зоогеографическая, сколько экологи-
ческая, так же как распределение их по
водоемам разных типов и их отдельным
местам (прибрежные заросли, дно, толща
воды).
Естественно, что коловраток стали изу-
чать со времени изобретения микроскопа,
т. е. с конца XVII в. Естествоиспытатели
XVIII и начала XIX в. объединяли коло-
враток с простейшими, и только в 1817 г.
Кювье впервые выделил коловраток в
самостоятельную группу животных, объе-
диненных им в р а з д е л Radiata. Наи-
более полное современное руководство по
коловраткам написано Р е м а н е и опубли-
ковано в 1929 г., а в 1957 г. вышел подроб-
нейший определитель коловраток Ф о й г-
т а. В настоящее время известно около
1500 видов коловраток, но ежегодно опи-
сываются все новые виды, и можно с уве-
ренностью сказать, что всего на земном
шаре имеется не меньше 2 тыс. видов.
Класс коловраток делится на три
отряда. Отряд Seisonidea содержит
всего несколько видов, живущих на жаб-
рах морского рачка Nebalia (рис. 245, 1).
Эти виды характеризуются тем, что раз-
множаются всегда с оплодотворением.
Самцы существуют постоянно наряду с сам-
ками. Они имеют два семенника. Все осталь-
ные органы такие же, как у самки. У са-
мок два яичника, желточник отсутствует.
У обоих полов коловращательный аппарат
редуцирован. Отряд Bdelloidea, или
пиявковидные коловратки, назван так по-
тому, что они передвигаются, как пиявки.
Их называют также Digononta, так как у
них два яичника и два желточника. Сам-
цов у этих видов нет. Размножаются исклю-
чительно партеногенетически. У многих
видов выработалось замечательное при-
способление к сохранению жизни при
исчезновении воды. Они могут высыхать,
впадая в анабиотическое состояние, т. е.
в состояние скрытой жизни. У некоторых
видов такая способность настолько резко
закрепилась в процессе эволюции, что
коловратки не могут уже существовать
в водоемах без периодического их высыха-
ния.
И наконец, отряд Monogononta,
т.е. однояичниковые. У самок имеются один
яичник и один желточник. Самцы появ-
ляются спорадически, у многих видов да-
же не каждый год. Самцы сильно редуци-
рованы, пищеварительная система у них
отсутствует; у некоторых видов отсутст-
вует и выделительная система; имеют
один большой семенник. У большинства
видов самцы в несколько раз меньше са-
мок, живут один-два дня, в то время как
жизнь самки в среднем продолжается две-
три недели. Без самцов самки размножают-
ся партеногенетически. С появлением сам-
цов происходит двуполое размножение и у
оплодотворенных самок формируются по-
коящиеся яйца, которые способны пере-
носить неблагоприятные условия и из
которых не сразу развивается коло-
вратка.
У некоторых видов Monogononta ясно
выражена гетерогония, т. е. чередование
однополого размножения (партеногетиче-
ского) с двуполым, после чего все самки
погибают, а сохраняются только покоя-
щиеся яйца, из которых обычно через не-
сколько месяцев, преимущественно зим-
них, выходят самки, начинающие новый
цикл размножения. Но у многих видов
двуполое размножение идет параллельно с
однополым (смешанное размножение), и су-
ществование вида в активной форме с об-
разованием покоящихся яиц не прекра-
щается. Есть виды, занимающие как бы
промежуточное положение в отношении
размножения: в одном водоеме или в одном
году они размножаются и партеногенети-
чески и с оплодотворением, а в другом во-
доеме или в другом году однополое раз-
431
множение полностью сменяется двуполым,
или, наконец, размножение происходит
только партеногенетическим путем в те-
чение всего года.
Однояичниковые коловратки наиболее раз-
нообразны и многочисленны. Многие их
представители заслуживают особого вни-
мания, так как в прудах, озерах, реках
они часто значительно преобладают над
всеми животными планктона, составляя
до 99% от общего их числа, развиваясь
временами в огромных количествах. В пе-
риод массового развития в 1 куб. см воды
может быть несколько сот экземпляров,
и конечно, при таком количестве коло-
вратки не могут не играть существенной
роли в переработке органических веществ
водоема. Они очищают воду, уничтожая
массы бактерий, водорослей и детрита
(разложившиеся остатки отмирающих жи-
вотных и растений), которые им служат
пищей. В то же время коловратки сами
служат пищей другим организмам, и осо-
бенно только что вылупившимся из икры
личинкам многих видов рыб. При искус-
ственном рыборазведении вопрос о живых
кормах специально для личинок рыб, когда
наблюдается наибольший процент гибели
молоди, стоит очень остро, и с этой точки
зрения коловратки — одни из первых кан-
Таблица 23. Коловратки:
1 — Testudinella patina;
2 — Polyarthra dolichoptera;
3 — Trichotria pocillum;
4 — Synchaeta pectinata;
5 — Trichocerca rattus;
6 — Asplanchna priodonta;
7 — Kellicottia longispina;
8 — Collotheca heptabrachiata;
9 — Microcodon clavus;
10 — Conochilus unicornis;
11 — Notommata copeus;
12 — Stephanoceros fimbriatus;
13 — Keratella quadrata.
дидатов на детальное, всестороннее изу-
чение, особенно изучение их биологии,
что до сих пор еще не сделано с такой пол-
нотой, которая позволила бы в практиче-
ских целях управлять их жизнью.
Познакомимся же поподробнее с этими
крошечными существами. Прежде всего,
каковы размеры их тела? Оказывается,
среди коловраток есть и «большие», хоро-
шо заметные простым глазом виды, до-
стигающие 2 мм. Некоторые из них об-
разуют колонии, которые часто состоят
из нескольких сотен экземпляров. Одна
колония достигает в диаметре примерно
5 мм. Но есть и такие коловратки, кото-
рые среди всех многоклеточных живот-
ных являются самыми маленькими, имея
в длину около 40—50 мк (Ascomorpha
minima). Это крайние величины. Подавляю-
щее же большинство видов коловраток
размером от 80 до 400 мк.
Большинство коловраток бесцветны, но
у многих видов отдельные части тела и
полостная жидкость бывают окрашены.
Так, Gastropus stylifer изумительно ярко
окрашен в фиолетовые, синие и оранже-
вые цвета со многими оттенками; гиподер-
ма и полостная жидкость этой коловратки
розового цвета. У многих видов пищевари-
тельный канал часто бывает окрашен в
зеленый, бурый, желтый цвет в зависимо-
сти от заглоченной пищи и стадии ее пере-
варивания. Часто запасы питательных
веществ сохраняются в виде красных и жел-
тых жировых капель. Некоторые пиявко-
видные коловратки окрашены в красный,
желто-зеленый и коричневый цвета.
Как уже отмечалось, форма тела коло-
враток очень разнообразна. Среди плаваю-
щих встречаются виды, имеющие форму
мешка (табл. 23,6), или почти круглой пла-
стинки (табл. 23 J), или волчка (табл. 23,9 и
4); часто встречаются коловратки, «закован-
ные» в панцирь, словно в средневековые
латы, причудливой формы, с шипами, ко-
лючками, выступами (табл. 23,7 и 3 и др.).
Есть такие виды коловраток, которые
имеют придатки, позволяющие им делать
прыжки. Эти придатки могут быть или
широкие, напоминающие руки (рис. 250,
2), или довольно узкие, зазубренные,
словно пильчатые ножи (табл. 23,2),
или тонкие, как хлысты (рис. 245, 11).
У многих видов коловраток вокруг тела
образуются футляры, чехлики, домики,
которые служат им как защитные образо-
вания. Прикрепленные коловратки не мо-
гут активно избежать опасности и спасают-
ся от врагов, целиком втягиваясь в домик
и закупориваясь свернутым коловраща-
тельным аппаратом, словно пробкой. Чрез-
вычайно разнообразны сооружаемые ко-
ловратками постройки. У одних имеется
совершенно прозрачный, почти невиди-
мый футляр из студенистого вещества,
выделенного кожей (рис. 245,9), у других
этот футляр хорошо заметен. Бывают до-
мики плотные, бутылкообразные (рис. 246);
у одних видов к липким наружным стен-
кам футляра приклеиваются посторонние
частицы (рис. 246,1), у других сооружают-
ся трубки из мягких частей ила (рис.
246, 3), из плотных шариков или колец
(рис. 246, 4 и 6). Домики и футляры за-
щищают не только коловратку, но также
и откладываемые ею яички. Вполне сформи-
рованная личинка, развившись в домике,
покидает его и после недолгого плавания
посредством выделений железок ноги при-
крепляется к какому-либо предмету и
начинает сооружать вокруг себя собствен-
ное «помещение». Прослежено, как строит
свой домик-трубу молодая самка флоску-
лярии (рис. 246, 4). Молодые флоскуля-
рии часто прикрепляются к материнскому
домику, образуя целые гроздья трубок.
Стенки трубки сооружаются из одинако-
вой величины шариков, состоящих из
экскрементов коловратки, смешанных с
посторонними частицами. Для изготовле-
ния таких шариков флоскулярия имеет
так называемый «пилюльный» орган, ко-
торый представляет собой углубление,
выстланное ресничками и лежащее под
лопастью коловращательного аппарата;
на дне углубления открывается проток
железы, выделяющей секрет, склеиваю-
щий материал для шариков. Этот материал
под действием токов воды, вызванных
мерцанием ресничек, все время крутясь
на одном месте, уплотняется и округляется.
Затем коловратка, изгибая свой коловра-
щательный аппарат, приклеивает готовый
шарик в точно определенное место среди
уже прикрепленных ранее шариков, и в
конце концов .тело коловратки окружается
изящной и прочной трубкой. Глядя на
такие домики, поражаешься сложности
поведения крохотного существа, бессозна-
тельно сооружающего такие точные кон-
струкции! Здесь невольно вспоминаются
постройки перепончатокрылых насекомых.
Вспомним хотя бы постройку пчелой сот —
изумительно точных шестигранных воско-
вых ячеек! И вот оказывается, что у зна-
чительно ниже организованных существ,
какими являются коловратки, уже имеют-
ся такие способности. Многие из пиявко-
видных коловраток могут покидать свои
домики и сооружать новые, чего не
могут делать прикрепленные виды.
Разнообразны и чрезвычайно красивы
нежные коловращательные аппараты при-
крепленных коловраток: то в виде верши
с неподвижными щетинками (табл. 23,12),
сидящими на длинных пальцевидных вы-
ростах или на лопастях с булавовидными
окончаниями (табл. 23,8), то в виде
сердцевидных пластинок или колесиков
(рис. 246,6), одного или двух (рис. 246,2),
или реснички располагаются на четырех-
лопастных выростах (рис. 246, 4). Как
увлекательно следить за работой этих
аппаратов у живых коловраток! Вот перед
нами флоскулярия. Медленно и плавно
поворачиваются ее лопасти, по краям
^ 28 Жизнь животных, т. i
433
Рис. 246. Домики коловраток:
t — Habrotrocha visa; 2 — Ptygura tihanyensis, 3 — Gephalodella forficula;
5 — Habrotrocha angusticollis, 6
Limnias melicerta.
4 — Floscularia ringens;
которых, как спицы в колесе, мерцают
реснички, загоняя в ротовую воронку
пищевые частицы; вдруг внезапное сжатие
лопастей, резкое сокращение ноги мол-
ниеносно втягивает тело в домик. Что
случилось? Оказывается, какая-то инфу-
зория задела коловратку, на что последова-
ла такая защитная реакция. Но вскоре
из трубки появляется медленно раскры-
вающийся коловращательный аппарат и
вновь начинается «коловращение». Не-
даром немцы называют прикрепленных ко-
ловраток «цветочными» (Blumenradchen).
Действительно, они напоминают цветок, а
в момент развертывания коловращатель-
ного аппарата — распускающийся бутон.
У коловраток с мягкой кожей наблюдается
большое разнообразие формы тела. Это
объясняется изменчивостью по физиологи-
ческим причинам: наполнен или опорож-
нен желудок, имеются ли в наличии или
отсутствуют развивающиеся яйца в яич-
нике и каково их количество, плавает или
ползает коловратка, соскабливает ли пи-
щу и т. п. У видов с панцирем часто очень
изменчивы шипы, особенно их длина. В
разное время года и в разных водоемах
один и тот же вид имеет разную форму
панциря, например Keratella quadrata,
Brachionus calyciflorus встречаются с длин-
ными задними шипами, с короткими и
совсем без шипов. Червеобразная форма
тела ползающих коловраток может счи-
таться наиболее примитивной. Когда ко-
ловратка не сжата, обычно ясно видно,
что ее тело делится на четыре отдела —
голову, шею, туловище и ногу. Такое рас-
членение тела коловратки то в более
четком, то в менее четком виде можно при-
нять как типичное для всего класса. Это
расчленение осуществляется наружными
перетяжками кожи, создавая впечатление
сегментации тела, но эта сегментация не
имеет ничего общего с истинной сегмента-
цией высших червей и членистоногих,
которая обусловлена наличием внутрен-
них перегородок. У коловраток таковых
не имеется. Ложная кольчатость коло-
враток вызывается утончением кожи в оп-
ределенных местах, которое позволяет
телу быть подвижным и при помощи спе-
циальных мышечных тяжей втягивать один
сегмент в другой. В силу этого голова и
шея, а у многих и нога могут втягиваться
в туловище. Коловратки, имеющие пан-
цирь, таким способом прекрасно защищают-
ся от своих врагов подобно черепахам;
не имеющие панциря втягиванием пе-
реднего и заднего конца тела превращают
себя в неподвижный компактный комочек,
значительно менее уязвимый, чем когда
коловратка бывает в вытянутом состоянии.
На голове у коловраток находится самый
замечательный для них орган — коловра-
щательный аппарат, затем органы чувств,
начало пищеварительной и выделительной
систем и у многих своеобразный ретро-
церебральный орган. Коло-
вращательный аппарат очень разнообразен
не только у прикрепленных коловраток,
что уже было отмечено. Наиболее простое
устройство его мы находим у Notommati-
dae, Dicranophoridae и Lindiidae, где
одинаковой длины реснички образуют во-
круг ротового отверстия ресничное
(буккальное) поле и отходящий
от него кольцевой ресничный (цир-
кумапикальный) пояс, окайм-
ляющий теменное (апикальное)
поле, лишенное ресничек. По верхнему
краю ресничного пояса над ротовым от-
верстием расположен ряд массивных рес-
ничек, образующих надротовое кольцо,
так называемый т р о х у с. Такого же
типа ресничное кольцо имеется и на ниж-
нем крае пояса под ротовым отверстием,
называемое цингулюм. Изменение в
расположении ресничек и их количества
в этих кольцах и в конфигурации выро-
стов головы, где они располагаются, в
основном и создает большое разнообразие
типов коловращательного аппарата (рис.
247), а в связи с этим и большое разно-
образие в характере движения коловраток
и в способе захватывания пищи.
По способу движения коловраток можно
сгруппировать следующим образом.
К первой группе относят толь-
ко плавающие формы (Asplanchna, Syn-
chaeta, Keratella и др.). Эти типично
планктонные животные всю жизнь прово-
дят в толще воды, находясь в непрерыв-
ном движении и не имея ни минуты покоя.
Характер движения у большинства планк-
тонных коловраток очень своеобразный:
они плывут, вращаясь вокруг собственной
28*
435
Рис. 247. Схема основных типов коло-
вращательных аппаратов:
1 — Notommata (вид снизу и сбоку); 2 —
Bdelloidea; 3 — Asplanchna; 4 — Conochilus;
5 — Hexarthra; 6 — Euchlanis; 7 — Epiphanes.
Крупные точки обозначают реснички трохуса,
мелкие — цингулюма; an — апикальное поле;
р — рот, бу -— буккальное поле; ца — циркум-
апикальный ресничный пояс.
оси, вперед по винтовой линии. Только
немногие виды передвигаются без враща-
тельного движения — одни вперед голо-
вой, например Asplanchna (табл. 23,6),
другие вперед ногой, например Conochi-
loides (рис. 245, 15). При сравнении с
другими микроскопическими животными,
которые передвигаются тоже при помощи
ресничек (инфузории, турбеллярии), мед-
леннее всего оказывается движение ко-
ловраток. Эта медлительность позволяет
коловратке во время движения питаться,
захватывать ресничками пищевые частицы,
взвешенные в воде. Чрезвычайно интерес-
но, что виды, плавающие очень быстро
(Brachionus bennini), для отыскания пищи
выпускают длинную нить из затвердеваю-
щей в воде слизи ножных желез, которая,
зацепившись за что-либо, останавливает
коловратку, как на якоре. Во время такой
стоянки коловратка отфильтровывает пи-
щевые частицы, что трудно делать при
быстром движении, особенно при неболь-
шом количестве пищи в данном водоеме.
Через 1—2 минуты коловратка резким дви-
жением ноги отрывает нить и быстро
уносится на другое место, выделяя новую
нить, и т. д. Есть данные, обнаруживаю-
щие связь между длиной нижних шипов у
Brachionus calyciflorus и количеством пищи
в окружающей воде. При небольшой кон-
центрации кормовой взвеси шипы длин-
нее, и наоборот. По-видимому, длинные
шипы являются как бы тормозами, кото-
рые замедляют движение коловратки и
облегчают отфильтровывание пищевых
частиц. Таким образом, эти данные дают
возможность рассматривать явление ц и к-
ломорфоза (периодическое изменение
формы тела, в данном случае измене-
ние задних шипов панциря) как приспо-
собление для лучшего улавливания пищи.
Многие из плавающих коловраток спо-
собны делать прыжки, которые не яв-
ляются нормальным способом движения,
а представляют собой защитное приспо-
собление. Внезапные довольно длинные
прыжки, резко меняющие направление
движения, значительно содействуют спа-
сению от врагов, охотящихся за коло-
вратками,— хищных рачков, личинок ко-
маров и других мелких хищников.
Вторую группу, наиболее мно-
гочисленную, образуют коловратки, кото-
рые и ползают, и плавают, и временно
прикрепляются к какому-либо предмету.
Сюда относятся виды, живущие в берего-
вой зоне различных водоемов от больших
глубин до прибрежных песчаных отмелей.
В этой группе можно встретить самые раз-
нообразные формы движения. Есть много
видов, которые большей частью плавают
и только временно прикрепляются ногой
к предметам (многие Brachionidae, Testudi-
nella, Euchlanis, Lepadella и др.). Это
типичные формы береговых зарослей. У
донных форм преобладает ползающее дви-
жение. У Notommatidae — путем глав-
ным образом скольжения при помощи
ресничек коловращательного аппарата,
расположенных вокруг рта; у Bdelloidea
ползающее движение осуществляется но-
гой и хоботком, находящимся на голове.
Это ползание происходит при втянутом
коловращательном аппарате следующим
образом: хоботок своей конечной чашеч-
кой с ресничками прикрепляется к какому-
либо предмету, затем конец вытянутой
436
ноги подтягивается к хоботку и клейкими
выделениями желез фиксируется, после
чего хоботок отрывается, тело вытягивает-
ся на всю длину и хоботок снова прикреп-
ляется, а нога вновь подтягивается и т.д.
Так осуществляется «шагающее» движение,
как у пиявки или гусениц пяденицы.
Виды, живущие во влажном песке, мху и
в почве, передвигаются ползанием и толь-
ко редко, при попадании в места, где
много воды, плавают; возможно, некото-
рые вообще лишены способности плавать,
например Lindia (рис. 245, 17). Есть пе-
счаные коловратки, имеющие на голове
длинные щетинки, которые помогают ла-
зить им между песчинками. И наконец,
многие коловратки перешли к сидячему
образу жизни, они плавают только не-
сколько часов по выходе из яйца, а затем
прикрепляются обычно к веточке какого-
либо водяного растения. Шея имеется у
многих коловраток, особенно хорошо она
выражена у морских Seisonidae (рис. 245,
1), у Lindia (рис. 245,77) и у пиявковид-
ных коловраток (рис. 246, 1).
Ногой у коловраток называется та
часть тела, которая находится ниже аналь-
ного отверстия. Ее строение также очень
разнообразно и в значительной степени
определяет образ жизни коловратки. Нога
оканчивается или пальцами, на конце
которых открываются протоки специаль-
ных ножных желез, выделяющих клейкий
секрет, или пластинкой с отверстиями для
протоков этих желез, или, наконец, пуч-
ком ресничек. Значительно вытянутую
и сильно кольчатую ногу имеют пиявко-
видные коловратки. У многих из них при
сокращении ноги ее членики вдвигаются
один в другой, как колена подзорной тру-
бы. Особенно резко это сходство выраже-
но у Rotaria neptunia (рис. 245, 14), у
которой членики ноги очень длинные. У
Notommatidae нога тоже членистая, но
значительно короче, так как имеет мень-
шее количество члеников. Notommatidae
также имеют на конце ноги пальцы и клей-
кие железы, что обеспечивает им ползаю-
щее движение или скольжение, причем в
последнем случае нога служит рулем. У
многих плавающих видов (Asplanchna,
Keratella, Filinia, Pompholyx и др.) нога
совсем отсутствует, а у тех, у которых
она имеется, выполняет функцию руля,
например широкие пальцы Euchlanis, Се-
phalodella. У некоторых из плавающих
нога или ее пальцы снабжены сильными
тяжами поперечнополосатой мускулатуры,
позволяющими животным делать резкие
прыжки (Monommata, рис. 245,16, Scandi-
um, Eudactylota), как это проделывают
виды, имеющие специальные прыгательные
придатки. У прикрепленных коловраток
нога длинная, может сильно сокращаться
и втягивать все тело в домик, в котором
они живут; пальцы не развиты. Только что
вылупившаяся из яйца коловратка при-
крепленных видов на конце ноги имеет
пучок ресничек, а в самой ноге массивные
клейкие железы, при помощи которых
она прикрепляется к субстрату. Многие
виды из этой группы коловраток обра-
зуют колонии, одни прикрепленные, дру-
гие плавающие. Способность образовывать
колонии появилась только с переходом к
неподвижному существованию. В образо-
вании колонии большую роль играет нога
коловратки. У Lacinularia flosculosa при-
крепленная колония формируется из сво-
бодноплавающих молодых особей, выходя-
щих из материнской колонии, где они вы-
лупляются из яиц, отложенных в студени-
стую массу. Эти молодые особи держатся
вместе, образуя скопление, которое нахо-
дится все время в движении и меняет
свою форму, то сжимаясь в виде клубка,
то растекаясь в виде ленты. Вскоре отдель-
ные коловратки начинают сцепляться рес-
ничками конца ноги и склеиваться секре-
том ножных желез, образуя пачки, состоя-
щие из небольшого количества особей.
Такие пачки начинают прикрепляться к
субстрату одна возле другой и, выделяя
большое количество студенистого веще-
ства, образуют новую прикрепленную ко-
лонию. В период массового размножения
колонии ляцинулярии состоят более чем
из 2500 особей. Ножные железы, массив-
ные у молодых, у взрослых редуцируются,
так как они уже выполнили свое назначе-
ние. Несколько иначе образуются пла-
вающие колонии у Conochilus unicornis.
Молодые коловратки, вышедшие из яиц,
находящихся также в студенистой массе
колонии, не покидают материнской коло-
нии, а остаются в ней, увеличивая число
437
особей примерно до 25 экземпляров, скон-
центрированных в одном центре. Вновь
появляющиеся коловратки, склеиваясь но-
гами, образуют внутри этой же колонии
новые центры, распадающиеся в дальней-
шем на новые самостоятельные колонии;
происходит как бы почкование их. Необы-
чайно оригинально формируется колония
у свободноплавающей коловратки Lacinu-
laria ismailoviensisI. Ее колония яйцевид-
ной формы с центральной осью, вокруг
которой расположено большое количество
(несколько сот) самок. Здесь резко вы-
ражены два полюса — один, где сосредо-
точены взрослые, обычно уже половозре-
лые особи с отложенными ими яйцами, и
второй — на противоположном конце, где
находятся только молодые особи, причем
в точке, расположенной по прямой линии
от конца оси к периферии, находится са-
мая молодая особь, с еще не окончатель-
но развитым коловращательным аппаратом
и еще не достигшая размеров взрослой.
Между этими полюсами располагаются сам-
ки промежуточных стадий развития и
роста. Такое закономерное распределение
коловраток по возрасту обусловлено свое-
образной миграцией самки, только что
вылупившейся из яйца, лежащего у
ноги матери. Она пробирается через сту-
день колонии на его поверхность, затем
пиявковидным движением переползает по
студню к противоположному полюсу ко-
лонии и точно против оси прикрепления
проникает ногой между другими колов-
ратками к центру колонии, где и прикреп-
ляется. По-видимому, проникновение мо-
лодой коловратки в определенном месте к
центру колонии объясняется тем, что
только в этом месте студенистая масса
колонии доступна для проникновения че-
рез нее, так как находящаяся здесь
молодь еще не успела выделить всего
секрета своих ножных желез, а выделен-
ный не успел еще уплотниться. Обычно
такие колонии медленно двигаются, плав-
но катясь как шарики. Но часто встречают-
ся колонии с дисгармоничным движением.
1 Видовое название этой коловратки происхо-
дит от названия района Измайлово, где впер-
вые этот вид был найден и описан М. Ю. Пог-
генполлем.
Они передвигаются не как единое целое,
без неуловимых движений отдельных осо-
бей, как это имеет место у нормальных.
Особи такой колонии обнаруживают поры-
вистые беспрестанные сокращения всего
тела, направленные к преодолению со-
противления движению колонии, которое
вызывают отмирающие и иногда уже умер-
шие коловратки на полюсе «стариков».
Колония L. ismailoviensis образуется толь-
ко однажды основательницей, вышедшей
из покоящегося яйца, за счет своих до-
черних и внучатных особей, прикрепляю-
щихся ногами у ноги основательницы. В
дальнейшем происходит лишь постепен-
ное обновление этой колонии — старые ко-
ловратки на одном конце отмирают, а на
другом колония пополняется молодыми.
Тело коловраток покрыто кожей, состоя-
щей из двух слоев: гиподермы, пред-
ставленной живыми клетками, сильно вы-
тянутыми и слившимися друг с другом,
и, снаружи от них, кутикулы, об-
разованной гиподермой. Кутикула может
быть тонкой, мягкой и прозрачной у
беспанцирных коловраток, но у многих
видов она утолщается и твердеет, образуя
панцирь. Он состоит или из пластинок,
или из колец, или из спинного и брюш-
ного щитков, которые соединены по бокам
более тонкой кутикулой. Передний и зад-
ний отделы тела остаются мягкими.
У некоторых видов и членики ноги по-
крыты панцирем, а на голове имеется
плотный щит (табл. 23,3). Прыгательные
придатки на теле некоторых коловраток
кутикулярного происхождения. Кутикула
также образует на панцирях всевозмож-
ные шипы, выросты, бугорки, ямки, ха-
рактерные для каждого вида коловраток.
Число клеток, из которых состоит все
тело коловраток, как у круглых червей,
небольшое и постоянное. Так, у Epipha-
nes senta тело состоит из 959 клеток, у
Asplanchna — из 900. Постоянство клеточ-
ных элементов обусловлено тем, что после
эмбрионального развития клетки уже не
делятся. Из яйца выходит коловратка зна-
чительно меньших размеров, чем взрос-
лая; в течение одного-двух дней происхо-
дит ее рост, во время которого клетки не
размножаются, а только растягиваются и
во многих тканях сливаются, образуя син-
438
цитий. Под кожей находится первичная
полость тела, которая заполнена жидко-
стью, выполняющей функцию кровенос-
ной и дыхательной систем, отсутствующих
у коловраток. В полости тела находятся
внутренние органы. Пищеварительная си-
стема начинается ротовым отверстием, на-
ходящимся в середине ресничного поля
коловращательного аппарата, часто в во-
ронкообразном углублении, выстланном
ресничками. Ротовое отверстие ведет в
глоточную трубку и в ее расширенную
часть — мастаке. В мастаксе находят-
ся челюсти, состоящие из плотных кути-
кулярных образований — наковал ь-
н и (инкус) и двух молоточков (мал-
леус, рис. 248). Наковальня состоит из
непарной подпорки (фулькрум) и
двух ветвей (рамус), молоточек сла-
гается из рукоятки (манубриум) и
крючка (ункус). Несмотря на единую
схему строения, у разных видов отдель-
ные части челюстей очень различны и
служат хорошим признаком для опреде-
ления вида (рис. 248, 2—7).
Глотка продолжается в пищевод, впа-
дающий в желудок. Желудок представ-
ляет собой у большинства коловраток
объемистый мешок, который занимает
большую часть полости тела. В него
впадают протоки парных пищеваритель-
ных желез, обычно лежащих по бокам
переднего края желудка. Желудок, су-
жаясь, переходит в кишку, выстланную
длинными ресничками, в то время как они
в желудке короткие. Кишка оканчивается
анальным отверстием, лежащим на спин-
ной стороне тела. Несколько выше него
в кишку впадают протоки выделительной
системы и яйцевод, поэтому эта часть
кишки называется клоакой. Некото-
рые коловратки не имеют кишки и аналь-
ного отверстия (Asplanchna, Ascomorpha),
и вся система заканчивается желудком;
непереваренные остатки пищи выбрасы-
ваются через ротовое отверстие.
Как уже отмечалось, коловратки пи-
таются водорослями, бактериями и детри-
том, кроме того, есть хищники, питающие-
ся животными — простейшими и коло-
вратками, которые значительно меньше
хищника, но хищники могут питаться и во-
дорослями. Подавляющее большинство ко-
Рис. 248. Основные типы челюстного аппарата
коловраток:
1 — общая схема аппарата; у — ункус (крючок); м —
манубриум (рукоятка); ункус и манубриум вместе об-
разуют маллеус (молоточек); р — рамус (ветвь) и
ф —фулькрум (подпорка), образующие инкус (нако-
вальня); 2 — форципатный тип у Encentrum; з —
маллеатный у Dapidia, 4 — виргатный у Notom-
mata copeus; 5 — раматный у Bdelloidea; 6 —инку-
датный у Asplanchna; 7 — унцинатный у Collotheca.
ловраток полифаги, т. е. мало огра-
ниченные в своем выборе пищи, но есть
и монофаги, т. е. такие, которые пита-
ются только одним родом пищи, например
детритом или только определенным видом
водоросли. По способу захватывания пищи
коловраток можно распределить по сле-
дующим группам: 1 — коловратки, у ко-
торых захватывание пищи происходит пу-
тем привлечения ее в ротовую воронку
токами воды, вызванными мерцанием рес-
ничек коловращательного аппарата; 2 —
свободноплавающие хищники, которые,
помимо водорослей, пригнанных реснич-
ками, активно хватают живые организмы
своими челюстями, высовывая их наружу,
или, проколов жертву, высасывают ее со-
ки; 3 — коловратки, использующие свой
своеобразный коловращательный аппарат
439
Рис. 249. Схема строения коловрат-
ки из отряда Monogononta.
Вид со спинной стороны (справа) и сбоку
(слева): 1 —проток ретроцеребрального
органа; 2 — проток субцеребральной же-
лезы; 3 — наружное теменное щупаль-
це; 4 — спиннобоковая чувствительная
иембранелла коловращательного аппара-
та; 5 — боковой орган чувств с боковым
глазком и ресничной ямкой; 6 — утолще-
ние гиподермы под коловращательным
аппаратом; 7 — корешок главного спин-
ного нерва; 8 — внутреннее теменное щу-
пальце; 9 — лобный, или теменной, гла-
вок; 10 — подмозговая железа; 11 — цир-
кумапикальный ресничный пояс; 12 —
мускул-сжиматель переднего конца тела;
13 — ретроцеребральный мешок; 14 —
мозг; 15 — спинное щупальце; 16 — моз-
говой глазок; 17 — спинной главный
нерв; 18 — передний спинной мускул-
втягиватель; 19 —безъядерный кольцевой
мускул; 20 — железистая часть прото-
нефридия; 21 — задний спинной мус-
кул-втягиватель; 22—капиллярная часть
протонефридия; 23 — яйцо; 24 — боко-
вое (латеральное) щупальце; 25 — пла-
менковые клетки протонефридия, 26 —
анальное отверстие; 27 — ножная клей-
кая железа; 28 — ножное щупальце;
29 — диафрагма пальцев; 30 — резерву-
ар ножной железы; 31 — палец; 32 —
анастомоз Гексли; 33 — спинная слюнная
железа; 34 — мастаке; 35 — передняя
часть пищевода без ресничек; 36—брюш-
ная слюнная железа; 37 — задняя часть
пищевода с ресничками; 38 — желудоч-
ная (пищеварительная) железа; 39— же-
лудок; 40 — желточник; 41 — яйцевод;
42 — кишка; 43 — мочевой пузырь, 44 —
клоака; 45 — отверстие мочевого пузыря
в клоаку; 46 — боковой мускул-втяги-
ватель; 47 — главный брюшной нерв;
48 — ротовое отверстие; 49 — утолщение
гиподермы под буккальным (ротовым)
полем; 50 — боковая брюшная чувстви-
тельная мембранелла коловращательного
аппарата; 51 — ганглий мастакса; 52 —
ножной ганглий; 53 — мастаке, 54 —
яичник; 55 — брюшной мускул-втягива-
тель; 56 — малый мускул ноги; 57 —под-
порка (фулькрум); 58 — спинные органы
чувств глотки; 59 — гиподерма; 60 —по-
лость тела.
440
с длинными неподвижными ресничками в
качестве ловчего аппарата наподобие
верши.
У коловраток отсутствует кожно-мус-
кульный мешок и мышцы образуют тяжи
(рис. 249). Основные из них: кольце-
вые, сжимающие тело; продольные, уко-
рачивающие и сгибающие его; мышцы
коловращательного аппарата и ноги, втя-
гивающие и развертывающие их. Движе-
ние ресничек коловращательного аппара-
та осуществляется не мышцами, а самой
клеткой, на которой находятся реснички.
Кроме мускулов тела, имеются мускулы
органов — в мастаксе, в пищеварительной
трубке, в мочевом пузыре. Обычно у ко-
ловраток мышцы гладкие, но встречаются
и поперечнополосатые, особенно в придат-
ках тела, служащих для прыжков, и в
пальцах прыгательных ног (рис. 245,ii
и 250,2).
Нервная система коловраток
(рис. 249) состоит из мозга, лежащего над
глоткой в головном отделе (мозговой ган-
глий), ганглиев мастакса и ноги, мелких
ганглиев, разбросанных по многим местам
тела, и нервов, отходящих от мозга, свя-
зывающих ганглии и идущих к органам
чувств и мускульным тяжам, Органы чувств
представлены глазами и щупальцами.
Глаза — красные или черные пигментные
пятна. У некоторых видов перед пятном
имеется линза (светопреломляющее тель-
це). У большинства коловраток один «за-
тылочный» глаз, лежащий вблизи мозга;
у многих по два глаза по бокам головы;
есть «лобные» глаза; у некоторых пияв-
ковидных коловраток глаза находятся на
хоботке; часть видов коловраток совсем
лишена глаз, а у прикрепленных форм
глаза имеют только молодые особи.
Щупальца представлены особыми клетка-
ми с чувствительными щетинками, рес-
ничками или сосочками. Они многочислен-
ны в коловращательном аппарате; на
спинной стороне находится одно дорзаль-
ное щупальце, у некоторых видов очень
длинное; по бокам тела лежат латераль-
ные щупальца; есть щупальца в полости
глотки и ножные щупальца. Щупальца
выполняют функции органа осязания,
статического чувства и восприятия хими-
ческих раздражений.
Над мозгом у многих видов коловраток,
но далеко не у всех, имеется орган неизвест-
ного значения, называемый ретро-
церебральным, т. е. органом «на-
против мозга». Он состоит из довольно
большого мешка, часто заполненного чер-
ными крупными зернами, и лежащих по
бокам его двух субцеребральных желез.
У некоторых видов имеются выводные
протоки этого органа, открывающиеся на
тыльной стороне головы. Этот загадочный
орган по происхождению относится к кож-
ным железам.
Выделительная система
представлена типичными протонефридия-
ми (рис. 249). В полости тела по бокам про-
ходят каналы, начинающиеся в голове и
открывающиеся в пульсирующий моче-
вой пузырь, представляющий собой у
некоторых видов расширенную клоаку,
у других — расширенные концы каналов.
Содержимое пузыря выделяется в клоаку.
У некоторых видов в голове боковые ка-
налы соединяются посредством а н а -
стомозаГексли. В боковые каналы,
которые представляют железистую часть
протонефридиев,впадают замкнутые капил-
лярные трубки, дающие веточки, оканчи-
вающиеся терминальными клетками с рес-
ничками внутри их просвета. Таких клеток
с «мерцательным пламенем» у каждого
вида свое количество, может быть от 3 до
50 с каждой стороны. При сильном увели-
чении микроскопа у прозрачных коло-
враток хорошо видно мерцание ресничек
терминальных клеток. Эти клетки втяги-
вают в себя из полостной жидкости вред-
ные продукты обмена и выводят их через
боковые каналы наружу. Помимо этого,
выделительная система выполняет функ-
цию регулятора осмотического давления
и функцию дыхания, пропуская воду, по-
падающую в полость тела через пищева-
рительную систему.
Половая система самки, от-
носящейся к отряду Monogononta,
состоит из яичника, желточника и яйце-
вода, открывающегося в клоаку (рис. 249).
При рассматривании коловратки под ми-
кроскопом сразу виден очень большой
желточник с крупными ядрами, лежащий
рядом с желудком. У разных видов он
имеет различную форму — овальную, круг-
441
Рис. 250. Keratella quadrata A) и Hexarthra
raira B):
1 — отложенные амиктические яйца связаны меж-
ду собой, последнее прикреплено к самке; 2 —
отложенные амиктические яйца прикреплены к ве-
точкам раздвоенного стебелька.
лую, подковообразную, лентовидную и
даже лапчатую. Его назначение — снаб-
жать питательными веществами яйце-
клетки. Яичник представляет собой скоп-
ление мельчайших ооцитов, имеющих в
диаметре 1—5 мк, лежащих на желточни-
ке или даже погруженных в него. По мере
роста ооциты передвигаются к яйцеводу
и на этом пути получают питательные ве-
щества из желточника через временное
отверстие или своеобразную трубку в
оболочке яйца. Желточник вместе с яич-
ником одеты общей оболочкой, продол-
жение которой образует трубку — яйце-
вод, впадающий в клоаку. Большинство
видов коловраток откладывает яйца, толь-
ко немногие — живородящие; у них раз-
витие яйца происходит в теле матери. У
многих планктонных коловраток отложен-
ные яйца остаются прикрепленными к
основанию ноги матери при помощи сте-
белька, образованного слизистым выде-
лением ножных желез, как это хорошо
видно на рисунке 250. Обычно каждое
яйцо имеет свой стебелек (рис. 252), но
иногда у некоторых видов стебельки свя-
зывают несколько яиц и самки таскают
их за собой как цепочку (рис. 250, 1).
Бывают случаи, что один стебелек делит-
ся на две веточки и на каждой подвешено
по яйцу (рис. 250, 2). В таком прикреплен-
ном состоянии яйца развиваются во время
плавания матери. После выхода из яйца
молодой особи пустая скорлупка еще дол-
го путешествует с коловраткой. Другие
планктонные виды выпускают яйца прямо
в воду, оболочка этих яиц покрыта длин-
ными тонкими щетинками, что позволяет
им парить в воде; при беглом взгляде
такие яйца очень напоминают солнеч-
ников. Некоторые виды коловраток при-
клеивают яйца к растениям или к панцирю
других коловраток или рачков. Покоя-
щиеся яйца у большинства видов тоже
откладываются, но есть виды, у которых
они остаются в полости тела матери и ос-
вобождаются только после ее смерти,
когда труп уничтожится инфузориями и
бактериями. Половая система самца (рис.
251) состоит из большого семенника —
половой железы, вырабатывающей муж-
ские половые клетки — сперматозо-
иды, дополнительных (предстательных)
железок, выводного протока и копулятив-
ного органа. У большинства коловраток
самцы рождаются уже с вполне развиты-
ми сперматозоидами. Сперматозоиды от-
носительно крупные, состоят из головки
и вибрирующего хвостика; их немного,
наибольшее число (около 300) известно у
Asplanchna. У многих видов семенник под-
вешен на широкой ленте, которая являет-
ся рудиментом пищеварительного канала.
У самцов некоторых видов планктонных
коловраток на спинной стороне, примерно
в середине тела, лежит крупная жировая
капля, которая препятствует вращатель-
ному движению самца вокруг оси своего
тела. Самцы плавают прямо и по сравне-
нию с самками очень быстро. Самцы жи-
вут не больше двух суток, ведь они ниче-
го не едят, расходуя за свою короткую
жизнь только те питательные вещества,
которые им достались от матери. Наиболее
активными они бывают в первые часы
после вылупления. Процесс копуляции,
т. е. передачи сперматозоидов самке, у
некоторых видов происходит через клоаку,
у других — через любое место тела самки:
442
в последнем случае у самцов над копуля-
тивным органом имеется присасывательная
чашечка, которой самец прикрепляется
к самке. Очень вероятно, что имеющиеся
в нижнем отделе семенника стилетовид-
ные клетки, которые рассматриваются как
второй вид сперматозоидов, служат для
прокалывания кожи самки во время ко-
пуляции. При таком способе осеменения
сперматозоиды попадают в полость тела
самки и активно проникают в яичник,
отбрасывая хвост, остающийся в полостной
жидкости. Оплодотворение происходит в
яичнике.
Жизнь коловраток, особенно относя-
щихся к отряду Monogononta, очень
сложна, своеобразна, во многом еще не
изучена. Она все больше привлекает вни-
мание исследователей, так как коловратки
имеют значение, как уже отмечалось вы-
ше, в рыбном хозяйстве как живой корм
для молоди, только что вышедшей из
икринок. Дело в том, что у промысловых
рыб, икринки которых содержат мало
желтка (сельдевые, карповые и др.), вы-
клюнувшиеся личинки очень скоро рас-
ходуют питательные вещества своего ма-
ленького желточного мешка и бывают вы-
нуждены переходить на активное питание.
Ну, а что может схватить своим малень-
ким ртом рыбешка, имеющая в длину все-
го несколько миллиметров? Мельчайшие
водоросли и бактерии она может схваты-
вать, но они ей недостаточны для пита-
ния, ей нужны животные корма. Мелкие
рачки — циклопы и дафнии — ей недо-
ступны по величине, личинки цикло-
пов — науплиусы — по величине под-
ходящи, но по быстроте движения не могут
быть схвачены медленно двигающимися
рыбками. Только одноклеточные (глав-
ным образом инфузории и жгутиковые)
и коловратки полностью удовлетворяли
бы запросы рыбок и по величине, и по
характеру своего медленного и плавного
движения. Но дело осложняется тем, что
эти «живые корма» далеко не всегда бывают
в достаточном количестве в водоеме, где
находятся личинки рыб. Для пышного
размножения простейших обычно такие
водоемы непригодны, и если они и встре-
чаются, то в таких ничтожных количест-
вах, что «прокормить» личинок не в со-
стоянии. С коловратками трудность дру-
гая, она целиком зависит от их биологии.
Оказывается, коловраток в водоеме не
всегда бывает много. Обычно они появ-
ляются внезапно, быстро достигают мас-
сового развития и так же внезапно могут
исчезнуть или остаться в ничтожном коли-
честве. И вот дальнейшая судьба рыбок
зависит от времени их выклева из икри-
нок: если он совпадет с наличием в во-
доеме массового развития коловраток, то
личинки рыбок будут процветать за их
счет и обеспечат прекрасный урожай, ес-
ли же выклев произойдет, когда в водоеме
не будет коловраток, масса рыбок погиб-
нет от голода и ждать хорошего урожая
нельзя. Естественно возникает вопрос:
чем же объясняется такое непостоянство
«кормовой базы» для личинок рыб? Чем
Рис. 251. Строение самцов коловраток.
А — Keratella quadrata (длина тела около 70 мк):
1 — мозг, 2 — глазок; 3 — жировая капля; 4 — се-
менник; 5 — рудимент панциря, 6 — копулятивный
орган; 7 — железы копулятивного органа. Б — No-
tommata copeus (вид с брюшной стороны, длина те-
ла около 300 мк): 1 — ресничное поле; 2 — ретро-
церебральный орган; 3 — пламенковые клетки про-
тонефридия, 5 — семенник; 6 — боковое щупальце;
7 — предстательная железа; 8 — канал протонефри-
дия; 9 — слизистая оболочка вокруг тела; ю — нож-
ная железа; 11 — палец; 12 — рудимент пищевари-
тельного канала. В — вид сбоку: 1 — спинное щу-
пальце; 2 — копулятивный орган.
443
Рис. 252. Brachionus calyciflorus:
1 — амиктическая самка с партеногенетическими яйцами на самку;
2 — миктическая самка с партеногенетическими яйцами на самца, 3 —
миктическая самка с оплодотворенными покоящимися яйцами на самку.
объяснить появление и исчезновение ко-
ловраток? Чтобы ответить на эти вопросы,
нужно проследить весь жизненный путь
какой-либо коловратки и выявить основ-
ные моменты ее биологии, от которых за-
висит интересующее нас явление. Для это-
го самым подходящим местом будет ста-
рый искусственный пруд, в котором есть
рыба; еще лучше — пруд, куда весной
высаживают мальков карпа, с тем чтобы
к осени выловить товарную рыбу, и где
в течение лета карпа подкармливают жмы-
хом, пищевыми отходами и другими кор-
мами. Все это содействует пышному про-
цветанию некоторых видов коловраток.
Если периодически, хотя бы два раза в
неделю, с начала освобождения пруда от
льда зачерпывать стеклянной банкой во-
ду и просматривать ее на свет, то в один
прекрасный день можно заметить, что вода
стала мутной, серого цвета, словно кто-то
взбаламутил со дна ил. При тщательном
рассматривании можно уловить, что муть
состоит из мельчайших подвижных точе-
чек. Под микроскопом становится очевид-
но, что это коловратки. Аквариумисты
хорошо знают ценность для выкармлива-
ния личинок рыб прудовой воды с мутью,
которую они называют водой с «пылью» или
водой с «живой пылью». В руководствах
по аквариумному рыбоводству упоминает-
ся такая пыль, но не указывается, из чего
она состоит. Такие внезапные «вспышки»
могут дать в прудах целый ряд
планктонных коловраток, чаще
всего представители родов Bra-
chionus, Keratella, Filinia, Po-
lyartra и др. В начале массово-
го развития почти каждая сам-
ка имеет при себе по несколь-
ку довольно крупных яиц с
тонкой оболочкой (рис. 252, 1)у
через которую видны эмбрионы
на разных стадиях развития.
Из таких яиц всегда выходяг
только самки. Эти яйца разви-
ваются без оплодотворения,т.е.
партеногенетически. При на-
личии благоприятных условий
окружающей среды коловратки
очень быстро размножаются,.
несмотря на то что каждая
самка откладывает за всю свою
короткую жизнь всего около десятка яиц.
Быстрота размножения обусловлена ко-
роткими сроками созревания самки — уже
на второй день после рождения она при-
ступает к откладке яиц — и коротким
сроком развития яйца — всего около двух
дней проходит с момента откладки яйца
до выхода из него коловратки. Это приво-
дит к тому, что вода вскоре после появле-
ния в пруду коловраток буквально кишит
ими. Но уже следующая порция воды, взя-
тая из пруда, может дать нам иную карти-
ну размножения коловраток. Мы можем
заметить, что стали появляться самки, у
которых имеются яйца значительно мень-
ших размеров, чем в прошлой пробе, но
в большем количестве (рис. 252, 2). Об-
наружив таких самок, нужно каждый
день зачерпывать воду, чтобы не упустить
дальнейших изменений, которые теперь
будут происходить очень быстро. Ежеднев-
но просматривая состав микроскопическо-
го населения нашего пруда, мы заметимг
что самок с мелкими яичками с каждым
днем становится все больше и больше. Из
таких мелких яиц партеногенетическим
путем развиваются самцы. Они осеме-
няют самок, у которых после оплодотворе-
ния развиваются покоящиеся яйца. После
того как начали встречаться самки с та-
кими яйцами, количество коловраток в
пруду стало уменьшаться. Может слу-
читься так, что спустя несколько дней
444
Рис. 253. Покоящиеся яйца коловраток:
1,2 — Filinia longiseta; 3 — Filinia passa, 4 — Filinia terrainalls; 5 — Brachionus
calyciflorus (оранжевое); 6 — от той же самки — серое; 7 — Hexarthra mira, 8 — Tri-
chocerca cylitidrica; 9, 10, 11 — Polyarthra dolichoptera; 12 — Keratella testudo; 13 —
Asplanchna priodonta, 14 — Asplanchna girodi, 15 — Lacinularia ismailoviensis.
все самки будут с оплодотворенными яйца-
ми, а еще через неделю взятая вода может
оказаться совсем прозрачной, так как ко-
ловраток там уже не будет. Самки отми-
рают, образовав всего по нескольку штук
покоящихся яиц, и вид в данном во-
доеме прекращает свое существование в
активной форме. Таким образом, видно, ка-
кое большое значение в жизни коловраток
имеет их своеобразное размножение с обра-
зованием трех типов яиц. Первый тип —
партеногенетически развивающиеся яйца,
дающие самок, которые обусловливают
увеличение количества коловраток в дан-
ном водоеме. Второй тип — яйца, разви-
вающиеся тоже партеногенетически, но
дающие самцов, необходимых для образо-
вания третьего типа яиц — оплодотворен-
ных, играющих, как мы увидим, очень
существенную роль в жизни коловраток.
Оплодотворенные яйца крупнее партеноге-
нетических и, что самое замечательное,
они имеют сложную систему оболочек,
прекрасно защищающих эмбрион от все-
возможных неблагоприятных воздействий.
Покоящиеся яйца одеты двумя внутрен-
ними — одной тонкой, другой пергамент-
ной — оболочками, а снаружи покрыты
твердой непрозрачной скорлупой с очень
разнообразными скульптурными образо-
ваниями (рис. 253). Скорлупа появляется
в конце формирования яйца, так как она
образуется эктодермальными клетками уже
развившегося к этому времени эмбриона.
У некоторых видов поверх скорлупы имеет-
ся еще четвертая тонкая оболочка (рис.
253, 15), которая, правда, через несколько
дней исчезает. Для вылупления из яйца
готовой коловратки на скорлупе имеется
полоска, по которой яйцо трескается под
давлением растущей коловратки. У од-
них видов полоска окружает один конец
яйца, при разрыве образуя крышечку (рис.
253, 5, 6), у других она расположена по
бокам яйца, окружая один полюс и не
доходя до другого (рис. 253, 1, 2, 4, 7).
У таких яиц при вылуплении коловратки
скорлупка распадается на две створки.
После появления скорлупы дальнейшее
развитие эмбриона останавливается, на-
ступает стадия покоя, которая у многих
яиц продолжается длительный период вре-
мени (есть данные о сохранении жизнеспо-
собности яиц Polyarthra, лежащих в иле
озера в течение 30 лет). В стадии покоя
яйца могут переносить и промерзание и
высыхание, что для сохранения вида имеет
огромное значение, так как сами колов-
ратки, относящиеся к отряду Monogo-
nonta, не могут переживать такие резкие
изменения окружающей среды. Обычно
давать оплодотворенные покоящиеся яйца
могут только те самки, которые партеноге-
нетически дают самцов; их называют
миктическими самками, а их яй-
ца — миктическими (от слова
м и к с т о — смешивание, т. е. подразу-
мевается слияние мужской и женской по-
ловых клеток). Те же самки, которые пар-
теногенетически дают самок, не могут
быть оплодотворены и называются а м и к-
тическими самками, т. е. неспособ-
ными к слиянию половых клеток, а их
яйца — амиктическими. Цитоло-
гические исследования яйцевых клеток
показали, что у миктических самок во
время созревания половина хромосом вы-
брасывается и развивающиеся партеноге-
нетически яйца дают самцов с половин-
ным числом хромосом в каждом ядре, с чем
связана карликовая форма самца и редук-
ция многих органов. В случае если такое
яйцо будет оплодотворено, то нормальное
число хромосом восстановится и образуется
покоящееся яйцо, из которого разовьется
самка. Яйцеклетки амиктических самок
не испытывают процесса редукции хро-
мосом, которые полностью остаются в
ядре яйца, и оно не способно к оплодотво-
рению. Такое физиологическое разграни-
чение самок было установлено в 1897 г.
Эрлангером и Лаутерборном
на Asplanchna priodonta. Затем в последую-
щие годы это было обнаружено еще на
пяти видах коловраток и на основании это-
го был сделан вывод о том, что все самки
Monogononta делятся на амиктических и
миктических. Но в 1958 г. у одного вида
колониальной коловратки (Sinantherina
socialis), а в 1960 г. у другого (Conochiloi-
des coenobasis) А. С. Богослов-
ским было обнаружено, что, кроме
амиктических и миктических самок, имеют-
ся такие, которые могут давать и микти-
ческие и амиктические яйца, т. е. была
установлена третья физиологическая ка-
446
тегория самок, названных амфотер-
н ы м и (рис. 245, 15). Наличие амфотер-
ных самок позволяет нам представить себе
возникновение миктических и амиктиче-
ских самок.
По-видимому, амфотерные самки более
древние и под влиянием каких-то воздей-
ствий внешней среды в процессе эволю-
ции некоторые самки стали образовывать
только амиктические яйца, а другие
только миктические.
Часто массовое размножение и макси-
мальное количество коловраток предше-
ствует появлению самцов и формированию
покоящихся яиц, после чего появление
новых самок прекращается, что приводит
к исчезновению коловраток в данном водое-
ме. Таким образом, уменьшение числен-
ности коловраток связано с увеличением
числа миктических самок, которые обра-
зуют покоящиеся яйца, переводя вид в
неактивное состояние. В практических це-
лях важно, чтобы в водоеме преобладали
амиктические самки, так как они обеспе-
чивают увеличение количества коловра-
ток. Предотвратить появление самцов мы
сможем только тогда, когда будем знать
точно, что заставляет яйцеклетку выбра-
сывать половину хромосом. Возможно,
что причиной появления миктических
самок является воздействие на самку про-
дуктов их выделений, которые при пере-
населении водоема коловратками накап-
ливаются в большом количестве. Есть дан-
ные (Gilbert, 1963), показывающие, что в
уплотненных культурах Brachionus caly-
ciflorus миктические самки появляются в
большем количестве, чем в разреженных.
Но точно механизм этого воздействия
не установлен и у всех ли видов коло-
враток это наблюдается, неизвестно. Прак-
тика ждет от зоологов решения этой проб-
лемы. Наряду с этим для практики важна
другая, тоже не решенная, проблема:
вскрыть причину внезапного появления
коловраток в водоеме. Эта проблема, по-
жалуй, даже актуальнее первой, так как
решение ее даст нам оружие для получе-
ния коловраток в нужное время, т. е.
за несколько дней до выклева молоди
рыб, питающейся коловратками.
Решение этой проблемы осложняется не-
знанием причин развития покоящихся яиц.
А ведь как раз развитие их и обусловли-
вает появление коловраток в водоеме.
Есть много данных, говорящих о том,
что сроки продолжительности покоя яиц
очень различны даже для одного вида ко-
ловраток. Наблюдения над развитием по-
коящихся яиц Filinia longiseta показали,
что яйца, находясь в одних и тех же
условиях, развиваются небольшими пор-
циями в течение ряда лет, т. е., иными сло-
вами, очевидно, что стимулируют развитие
(выход из стадии покоя) не окружаю-
щие яйцо условия, а какие-то внутрен-
ние свойства самого яйца. При тщатель-
ном обследовании яиц нескольких видов
коловраток оказалось, что яйца одного
вида, отложенные одновременно в одном
водоеме, не одинаковы; они разнокачест-
венны как физиологически, так и морфоло-
гически. Так, одна и та же самка Brachio-
nus calyciflorus откладывает первое оп-
лодотворенное яйцо (рис. 253, 5) с резко
выраженными складками на скорлупке, с
большим пространством между скорлупой
и эмбрионом, и при рассматривании в за-
темненном поле зрения микроскопа вы-
является его оранжевая окраска. После-
дующие яйца серые, с почти гладкой
поверхностью скорлупы, с более крупным
эмбрионом, поэтому пространство между
ним и скорлупой значительно меньше
(рис. 253, 6). Покоящиеся яйца Polyar-
thra dolichoptera обычно серого цвета,
имеют на скорлупе характерные палочко-
видные выросты, своеобразные штифтики,
связанные с внутренней оболочкой (рис.
253, 9). Но иногда встречаются у этого же
вида коловратки яйца красного цвета, со
штифтиками, сидящими на наружной обо-
лочке (рис. 253, 10). У некоторых яиц
эти штифтики не развиваются до конца,
и получается яйцо третьей формы (рис.
253, 11). Такое недоразвитие скульптур-
ных образований на скорлупе наблюдает-
ся и у других видов коловраток, что также
создает ди- или даже полиморфизм покоя-
щихся яиц. Каждое покоящееся яйцо
Filinia longiseta имеет свой рисунок на
скорлупе, что вызывается разнообразием
формы, объема и расположения скульп-
турных образований (рис. 253, 1, 2).
Очень вероятно, что морфологические приз-
наки с ясно выраженной индивидуальной.
447
изменчивостью небезразличны для даль-
нейшего развития зародыша. Возможно,
разная толщина скорлупы в определенных
ее местах изменяет газообмен эмбриона,
а может быть, и другие физиологические
процессы, влияющие на темп развития.
Вероятно, на развитие влияет и количе-
ство питательных веществ, переданных
самкой яйцу. Наблюдения за развитием
яиц Brachionus calyciflorus, отличающих-
ся друг от друга, показали, что оно идет
неодинаково — серые оказались более
стойкими, и резкие изменения среды
(промораживание, высыхание, насыщение
воды кислородом) не стимулируют их раз-
вития. В то же время из некоторых оран-
жевых яиц, даже без резких воздействий
на них, развиваются коловратки уже через
несколько дней после их формирования.
Изменчивость яиц в пределах одного вида,
вероятнее всего, зависит от биохимических
процессов, совершающихся в яичнике сам-
ки, которые изменяются с изменением ее
пищи. И этой изменчивостью можно объяс-
нить разные сроки развития яиц у одного
и того же вида в одном и том же водоеме,
что в свою очередь объясняет совершенно
неожиданное появление в водоеме коло-
враток в различное время года. Но дале-
ко не всегда коловратки, вышедшие из
покоящихся яиц, остаются в водоеме и
успешно размножаются. Это бывает ред-
ко, обычно же вышедшие из некоторых
покоящихся яиц самки быстро умирают
или размножаются очень медленно, и
число их поэтому невелико. Процветание,
гибель или прозябание коловраток зави-
сит от того, насколько окружающая среда
в момент их выхода будет для них подхо-
дяща. Многие виды, как уже отмечалось,
очень требовательны по отношению к
условиям существования. Нужные усло-
вия в водоемах для некоторых видов бы-
вают не так уж часто, иногда даже не
каждый год. Разные сроки развития
покоящихся яиц позволяют виду не
упустить наступление нужных для него
условий существования. Таким образом,
спорадичность появления коловраток объяс-
няется тем, что покоящиеся яйца развива-
ются порциями со значительными интер-
валами между ними, зависящими от
индивидуальных особенностей яиц, а пыш-
ное процветание их зависит непосредственно
от условий окружающей среды. И еще
очень существенную роль в жизни коло-
враток играют покоящиеся яйца. Они поз-
воляют виду широко расселяться по водо-
емам, как местным, так и других стран.
Как бы в компенсацию за то, что взрослые
коловратки Monogononta не могут оста-
ваться без воды (на что способны пиявко-
видные коловратки), их покоящиеся яйца
прекрасно переносят высыхание. При-
крепившись вместе с комочком ила, часто
микроскопической величины, к какому-
либо животному, которое может активно
менять водоемы, как, например, водные
насекомые, водоплавающие птицы, амфи-
бии, некоторые млекопитающие, покоя-
щиеся яйца переселяются с ними из водое-
ма в водоем часто на огромные расстояния,
например с птицами во время их осеннего
и весеннего перелета.
От каких животных и когда возникли
коловратки, точно не установлено. Остат-
ки их не могли сохраниться в земле, и
поэтому палеонтологический метод дока-
зательства происхождения данных су-
ществ отпадает. Эмбриология имеет мало
данных для установления родственных
связей коловраток с другими организма-
ми, тем более что эмбриональное развитие
их изучено только для нескольких видов
и только на партеногенетическом яйце.
Развитие оплодотворенного яйца вовсе не
изучено, а оно, очевидно, является первич-
ным. Остается сравнительноанатомиче-
ский метод. Наличие ресничного поля на
брюшной стороне у примитивных коловра-
ток и отсутствие кожно-мышечного мешка
сближает их с гастротрихами, постоян-
ство клеточных элементов и билатераль-
ный тип дробления — с нематодами. Нали-
чие протонефридиев, кольцевых мышц и
брюшное положение рта сближает коло-
враток с турбелляриями. Таким образом,
можно предположить, что много миллио-
нов лет назад от каких-то предков реснич-
ных червей отделились организмы, дав-
шие гастротрих, затем отдельная ветвь
пошла к нематодам и где-то между ними
возникла такая форма, которая дала ко-
ловраток, по своей организации стоявших
близко к современным нотомматидам.
Очень возможно, что это произошло в мор-
448
скои песчаной литорали, где и сейчас су-
ществуют примитивные нотомматиды,
очень близкие по своему строению к мор-
ским прямокишечным турбелляриям. Это
не исключает возможности предполагать
В дословном переводе на русский язык
название класса означает нечто близкое к
«подвижнохоботным». Собственно хобота у
киноринх нет, но головной конец этих жи-
вотных действительно способен выдвигать-
ся вперед. Когда киноринха ползет по
морскому дну, а точнее по мелким донным
предметам, она не может двигаться впе-
ред так, как это делают гастротрихи.
Вместо ловких, быстрых, иногда как бы
скачущих микроскопических животных мы
видим перед собой странных существ, дви-
гающихся вперед довольно несовершен-
ным способом. Вытянув вперед головной
конец, вооруженный венцом плотных ще-
тинок, киноринха использует голову как
своего рода якорь, к которому вслед за
выдвижением ее вперед подтягивается все
тело.
Механизм движения состоит в том,
что жидкость, заполняющая первичную
полость этих животных, нагнетается сокра-
щением задней части туловища в его пе-
редний (головной) конец. Голова с помо-
щью головных щетинок закрепляется на
достигнутом участке поверхности субст-
рата, а вслед за тем к ней подтягивает-
ся туловище животного. Так, «шаг за
шагом» киноринха перемещается по суб-
страту.
Естественно, что описанный способ дви-
жения требует совершенно особой орга-
низации мускулатуры. И действительно,
кроме продольных мышц, животное обла-
дает специальными спинно-брюшными па-
рами мышц. При этом все тело явственно
1 Киноринхи—от греческих слов kinos — дви-
жение, rhynchos — х о б о т.
возникновение коловраток в морской лп-
торали, а не в пресной воде, как это обыч-
но считается на основании того, что в
пресных водах обитает наибольшее коли-
чество видов.
расчленено на «кольца», и каждому тако-
му кольцу тела соответствует пара упо-
мянутых спинно-брюшных мышц. Кольча-
тость возникла как необходимое следствие
описанного способа движения. Но самым
замечательным следствием кольчатости яви-
лась, как мы видим, закономерная повтор-
ность спинно-брюшных мышц. Этот факт
позволяет — пусть с известной осторож-
ностью — говорить о метамерной органи-
зации спинно-брюшных мышечных тяжей.
И ей, как показывает наш рисунок 254,
отвечает наружная метамерия члеников
тела.
Мало того, как у гастротрих, у ки-
норинх имеется скопление нервных кле-
ток, охватывающее, однако, глотку пол-
ным кольцом, в то время как у гастро-
трих нервное кольцо неполное. От этого
«мозга» отходит брюшной нервный ствол,
который в каждом «кольце» тела несет
утолщение, состоящее из нервных клеток,
связанных продольно расположенным спин-
ным нервом. Таким образом, и в органи-
зации центральной нервной системы на-
блюдаются признаки повторности эле-
ментов ее, в частности скоплений нервных
клеток.
Имеются у киноринх и органы пери-
ферической нервной системы.
Это осязательные щетинки, группы кото-
рых также повторяются в каждом коль-
це тела. Кроме того, к органам чувств
этих животных можно отнести примитив-
ные «глазки».
Ротовое отверстие лежит на переднем
конце тела. На заднем конце тела, строго
на его заостренном конце, расположено
анальное отверстие. Пищеварительный ап-
парат, в общем, очень похож на такой же
КЛАСС КИНОРИНХИ (KINORHYNCHA)
40 29 Жизнь животных, т. 1
449
Рис. 254. Киноринхи.
I — внутреннее строение: 1 — ротовое отвер-
стие, 2 — глотка; 3 — нервное кольцо; 4 — ки-
шечник; 5 — анальное отверстие; 6 — половое
отверстие, 7 — половая железа. II — Semno-
dera armigera с втянутым (вверху) и расправ-
ленным (внизу) передним концом.
аппарат у гастротрих. И у киноринх мы
видим мышечную глотку, которая перехо-
дит в среднюю кишку. Однако у киноринх
в ней нет ресничек, так как пища распре-
деляется по пищеварительным клеткам
сократительными движениями тела.
Как и у гастротрих, у киноринх имеет-
ся одна пара протонефридиальных кана-
лов с одним мерцательным пламенем
внутри каждого из них.
В отличие от гастротрих киноринхи —
раздельнополые организмы. У самок
имеется яичник, у самцов — семенник.
Через половые протоки у самок выде-
ляются яйца, а у самцов — сперма; из
оплодотворенных яиц развиваются личин-
ки, претерпевающие превращение и до-
стигающие организации взрослых форм,
величина которых не превышает 1 мм в
длину.
Питаются киноринхи одноклеточными
морскими водорослями и органическим
осадком, покрывающим донные предметы.
Продолжительность жизни киноринх при-
мерно около одного года.
* * *
Некоторые исследователи допускают,
что киноринхи произошли от уп-
рощенных форм кольчатых червей (ан-
нелид). Эта мысль едва ли может считать-
ся правильной. Несомненна принадлеж-
ность киноринх к первичнополостным жи-
вотным. Известный советский морфолог
В.Н.Беклемишев A964) справедли-
во утверждает, что «сходство с нематода-
ми и гастротрихами настолько велико,
что заставляет нас признать единство
основного плана строения киноринх с
остальными немательминтами и, следова-
тельно, с турбелляриями». Остается лишь
не совсем ясным, от каких именно тур-
беллярий возник класс киноринх. Если
часть авторов полагают, что источник
развития киноринх и гастротрих — пря-
мокишечные турбеллярии, как это счи-
тает и В. Н. Беклемишев, то другие (на-
пример, Steinbeck, 1958) думают, что
источником развития гастротрих
и киноринх были бескишечные турбелля-
рии. Предоставим будущему решать этот
вопрос. Однако несомненно, что гастро-
трихи возникли от прямокишечных
турбеллярий, что же касается кино-
ринх, то, возможно, прав Штепнбок,
считающий, что эта группа, несомненно
специализированная, возникла от гастро-
трих. В морях их немного, чуть более
100 видов.
КЛАСС ВОЛОСАТИКОВЫЕ (NEMATOMORPHA)
В заключение обзора первичнополост-
ных форм и групп, т. е. немательминтов,
коротко остановимся на волосатиковых.
Сюда принадлежат организмы, отличаю-
щиеся длинным и очень тонким телом,
оправдывающим наименование этой груп-
пы животных. Они достигают в длину от
0,5 до 1 м, тогда как в диаметре их тело
составляет всего около 3 мм. Создается
впечатление, будто в водоеме, запутавшись
среди стеблей и листьев какого-нибудь
водяного растения, лежит длинный кон-
ский волос (рис. 255). Окраска тела во-
лосатиков варьирует от светлого до тем-
но-коричневого, переходящего в черный.
Под плотной кутикулой расположен слой
гиподермы, представленной однослойным
эпителием. В отличие от нематод, гиподер-
ма волосатиков не несет боковых валиков.
Но как у нематод, мышцы образуют слой
продольных мышечных тяжей, ограничи-
вающих первичную полость тела — прото-
цель; по-видимому, эта полость тела пред-
ставляет собой щели в мезенхиме, так как
у взрослых животных эта полость тела в
значительной степени заполнена мезен-
химными клетками. Нервная система пред-
ставлена глоточным нервным кольцом и
брюшным нервным стволом. В гиподерме
обнаружены сенсорные (чувствующие) клет-
ки, рассеянные в гиподерме. Несомнен-
но, что периферическая нервная система
функционально весьма примитивна.
Кишечник у малоподвижных волосати-
ковых почти полностью редуцирован.
Более развит пищевод. Средняя кишка
узкая и не функционирует. Однако анус
открыт и служит у самцов половым про-
током, как у нематод.
Волосатиковые — раздельнополые не-
мательминты. Половые трубки парные.
Яичники самок несут многочисленные бо-
ковые выпячивания (дивертикулы).
Волосатиковые во взрослом состоянии
свободны. Они живут в пресных водах,
особенно в канавах, в которых можно на-
блюдать значительные скопления этих
животных, нередко образующих клубки
из переплетенных взрослых особей. Имен-
но в пресных водоемах происходит и
оплодотворение самок самцами, которые
выделяют вблизи или непосредственно
над женским половым отверстием капли,
содержащие сперму.
Установлено, что яйца самок в большом
количестве погружены в студнеобразные
тяжи и связаны ими. В результате раз-
вития из яйца формируется крохотная
личинка, длина которой едва дости-
гает 0,1—0,4 мм в длину. У таких личи-
нок головной конец вооружен венчиком
крючьев и тремя стилетами, расположен-
ными на выдвижном хоботке. Такая орга-
низация напоминает акантора скребней,
тем более что подобно им эти личинки
волосатиковых проникают сквозь места
Рис. 255. Волосатик:
1 — Gordius aquaticus, 2 — головной ко-
нец; з — вильчато раздвоенный задний ко-
нец тела волосатика; 4 — личинка воло-
сатика в яйцевой оболочке и без нее.
29*
451
с тонкими хитинистыми покровами в тело
насекомых и локализуются в их полости
тела. Поражаются ими также многонож-
ки. В дальнейшем личинки утрачивают
свое вооружение и превращаются в блед-
ноокрашенных волосатиков, которые бы-
стро растут и выходят из тела насекомых
наружу, где их покровы приобретают тем-
ное окрашивание. Растут волосатиковые в
течение нескольких месяцев. Момент вы-
хода их из тела насекомых может окончить-
ся для последних трагически: насекомые
часто гибнут. Интересно, что даже назем-
ные насекомые, к моменту выхода воло-
сатиков, держатся вблизи водоемов, и
таким образом взрослые волосатики попа-
дают обычно в свою родную стихию — в
воду. В полости тела насекомых волоса-
тики питаются не через кишечник, а
В большинстве случаев паразитическое
существование многих нематод относитель-
но мало влияет на их общую морфологи-
ческую организацию, так как она близка
к организации свободноживущих форм
этого обширного класса животных. Скреб-
ни в этом отношении существенно отли-
чаются от нематод. В самом деле, среди
скребней неизвестны свободные формы: все
они паразиты рыб, птиц и млекопитающих.
Паразитизм наложил на скребней глубо-
кую печать.
Тело скребней в поперечном сечении
круглое и может быть сравнено с цилинд-
ром. Передний конец тела несет сильно
развитый втяжной хобот, вооруженный
загнутыми назад острыми крючьями. Разме-
ры тела значительно варьируют: пример-
но от 1,5 мм у мелких форм до 40 мм
с лишним в длину у наиболее круп-
ных.
Втяжной хобот выдвигается наружу или
втягивается в корпус тела с помощью осо-
бой мускулатуры (рис. 256, 2). Строение
хобота значительно изменяется у различ-
ных видов и родов скребней и имеет боль-
осмотически — через кожные покровы.
К наиболее обычным представителям во-
лосатиковых принадлежат виды родов
Gordius и Parachordodes. На рисунке 255
показан самец Gordius aquaticus. Всего
известно около 100 видов этих животных.
Их филогенетические отношения к дру-
гим немательминтам недостаточно изуче-
ны. В отличие от нематод волосатиковые
полностью лишены экскреторных аппара-
тов, у них нет типичных для нематод
боковых валиков кутикулы, а также ли-
теральных боковых валиков. Все это за-
ставляет считать, что волосатиковые об-
разуют отдельный класс немательминтов и
что их эволюционное (филогенетическое)
развитие претерпело особый путь, незави-
симый от путей развития основного класса
немательминтов — нематод.
шое значение в систематике этой группы
паразитических животных. У некоторых
видов хобот удлинен, у других очень то-
нок, иногда он вздут в виде шарообраз-
ного образования или имеет веретенооб-
разную форму. Крючья, которые хобот
несет, погружены своим основанием в
кожные покровы, а их заостренные концы,
как выше сказано, обращены назад и
вонзаются в слизистую оболочку кишеч-
ника животных, в тонкой кишке которых
скребни поселяются. Крючья расположены
в определенном порядке и числе на по-
верхности хобота, и притом так, что каж-
дый крючок может быть использован для
фиксации в слизистой оболочке кишечника
хозяина.
Хоботок переходит в короткую шейку,
обособленную от тела. Тело скребня не-
сет нередко явственную, но только наруж-
ную кольчатость. Изредка у самцов на
поверхности тела развиты иглы, распо-
ложенные на переднем или заднем конце
тела. Кожа покрыта тонкой кутикулой,
под которой, подобно тому, как это мы
видели у нематод, развита гиподерма, про-
КЛАСС СКРЕБНИ (ACANTHOCEPHALA)
452
низанная волокнами и системой каналов.
В основании хобота от гиподермы отходят
особые выросты — лемниски; в эти
лемниски проникают упоминавшиеся ги-
подермальные каналы. Под гиподермой
залегает мускулатура, состоящая из на-
ружных, кольцевых, и внутренних, про-
дольных, мышечных тяжей. Считают, что
гиподермальные каналы служат провод-
никами питательных веществ. Лемниски,
видимо, также несут эту функцию, так
как и их ткань пронизана гиподермаль-
ными каналами. Пищеварительная система
у скребней отсутствует. Центральная нерв-
ная система примитивна: она состоит из
нервного узла, лежащего у основания хо-
бота, и боковых нервных стволов, тянущих-
ся к заднему концу тела. Хобот также ин-
нервирован. От боковых нервных стволов
отходят боковые ветви, идущие в различ-
ные участки тела.
Скребни раздельнополы. У самцов два
семенника выделяют семя в соответствую-
щие семяпроводы, а через них — в специаль-
ный семенной пузырек (рис. 256, 8).
Он расположен в задней части тела. Ко-
нец семенного пузырька вытянут в особый
отросток, который выполняет функцию
совокупительного органа, выдвигающего-
ся наружу.
Половая система самок значительно
сложнее. Оба яичника лежат внутри осо-
бого образования — лигамента. Он
представлен мышцей, идущей от основа-
ния хоботного влагалища к заднему кон-
цу тела самки. Выпавшие из лигамента
яйца попадают в полость тела в виде
комьев яиц, которые плавают в полостной
жидкости. В этой жидкости находятся
как оплодотворенные, так и неоплодотво-
ренные яйца. Выведение оплодотворенных
яиц наружу производится при помощи
своеобразного «дифференциатора», как
можно назвать соответствующий орган.
Это оригинальный аппарат, известный под
названием колокола. Этот орган спо-
собен производить глотательные движения,
в результате которых яйца попадают в
его внутреннюю полость. В стенке колоко-
ла имеются три отверстия. Два из них
узкие, и они ведут в матку. Третье, более
крупное, ведет в полость тела. Колокол
заглатывает и узкие, тонкие, т. е.
оплодотворенные, яйца, и комья неоплодот-
воренных яиц. Комья не могут проникнуть
в узкие отверстия колокола и выталки-
ваются через более широкое отверстие
обратно в полость тела. Узкие оплодотво-
ренные яйца проходят сквозь отверстия
стенок колокола, ведущие в яйцеводы, и
оттуда проходят в матку, далее — во вла-
галище и наконец — наружу.
Заключенный в яйцевую скорлупку эм-
брион для своего последующего развития
должен попасть в тело промежуточных
хозяев. Обычно эмбрионы проглатываются
некоторыми насекомыми. В их кишечнике
эмбрионы освобождаются от яйцевых обо-
лочек, пробуравливают стенку кишечника
промежуточного хозяина и попадают в
полость его тела. Здесь они вырастают в
маленьких личинок, достигая нормальной
величины уже в кишечнике позвоночного,
проглотившего это насекомое.
Из изложенного выше видно, что раз-
витие скребней протекает с превраще-
нием: сначала формируется личинка, от-
даленно напоминающая цистицерка цестод,
а затем эта личинка превращается во
взрослого скребня. Этот процесс сопро-
вождается сменой хозяев: личинки жи-
Рис. 256. Скребни.
А — Macrocanthorhynchus hirudinaceus. Б — Echi-
norynchus (схема): 1 — хобот; 2 — ретрактор хобота;
3 — лемниски; 4 — лигамент (связка), 5 — семен-
ник; б — семяпроводы; 7 — железы; 8 — семенной
пузырь; 9 — совокупительный орган В — хобот
(увел ), видны загнутые назад крючья. Г — личин-
ка скребня.
453
вут в промежуточных хозяевах — насеко-
мых, а взрослые скребни паразитируют в
органах названных выше позвоночных.
Личинка получила особое наименование—
акантор (acanthor), т. е. «носитель
колючек». Отсюда произошло и наимено-
вание всей группы, так как слово «акан-
тоцефалы» в переводе означает <колючего-
ловые».
Следует указать, что обрисованный вы-
ше путь развития: яйцо (наружная среда) —
акантор (насекомое)—взрослый скребень
(рыбы, птицы, млекопитающие) в ряде
случаев искажается. Так, акантор может
попасть не в кишечник рыбы, птицы или
млекопитающего, а в кишку лягушки или
ящерицы, т. е. животных, которые пи-
таются насекомыми. В кишечнике лягуш-
ки или ящерицы акантор инцистируется
и не развивается дальше. В сущности,
это один из примеров резервуарного пара-
зитизма, так как в организме ящерицы
или лягушки акантор не превращается
во взрослого скребня, а только пребы-
вает в организме этих животных и может
достичь конечного (дефинитивного) раз-
вития только в том случае, если лягушка
или ящерица окажутся жертвой какого-
либо млекопитающего или птицы.
Ознакомившись с организацией скреб-
ней, читатель, быть может, спросит:
каково же происхождение скребней? Ведь
они мало похожи на других червей. С ка-
кой группой можно сравнить этих жи-
вотных? Вопрос этот и в настоящее время
нельзя считать полностью решенным. И
дело совсем не в женском половом аппара-
те с его «глотательным» колоколом. По-
добные, хотя и не идентичные «яйцегло-
тающие» приспособления имеются у пла-
тод (плоских червей). Это признак взрос-
лых форм, а взрослые формы — плохие
ч<советчики» в решении вопросов филоге-
нии, т. е. истории происхождения и
исторического развития живых форм, осо-
бенно животных. Главное значение в реше-
нии проблемы происхождения любой
группы животных имеет история индиви-
дуального развития животных, т. е.
закономерности их онтогенеза. Если с этой
стороны взглянуть на характеристику
скребней, с их в высшей степени ориги-
нальной организацией, то тайна их про-
исхождения несколько приоткрывается.
Изучение закономерностей дробления яиц
скребней показало, что для них характер-
но спиральное дробление, принявшее фор-
му билатерального. Билатеральное дроб-
ление свойственно и яйцам нематод, у
которых оно также является особым вари-
антом спирального дробления яиц. Этот
факт заставляет исследователей искать
предков скребней среди групп, от которых
произошли нематоды. В некоторых от-
ношениях скребни близки к нематодам,
являясь первичнополостными формами. Од-
нако нет основания считать, что скребни
произошли от нематод. Скорее всего пра-
вильна точка зрения, согласно которой
скребни произошли от группы, общей и
для нематод с гастротрихами, и для скреб-
ней. Такой узловой предковой группой
при настоящем состоянии знании можно
считать ресничных червей, т. е. турбел-
лярий, для которых очень типично и спи-
ральное дробление яиц. В этих аспектах
ряд признаков скребней (наличие пер-
вичной полости тела, архитектоника цент-
ральной нервной системы, особенности
организации кожно-мускульного мешка)
сближает их с нематодами.
Скребни — особая, специализированная
ветвь первичнополостных форм (нематель-
минтов), глубоко измененная паразитизмом.
Рассмотрим скребня-великана (Масгосап-
thorhynchus hirudinaceus) как представи-
теля группы скребней. Самцы его дости-
гают 70—150 мм, а самки—300—650 мм.
Хобот несет 36 крючьев. Промежуточны-
ми хозяевами этого скребня служат май-
ский хрущ и бронзовка (Cetonia aurata),
а окончательными собака, а также домаш-
ние и дикие свиньи. Вполне достоверных
случаев паразитирования скребня-вели-
кана в человеке неизвестно. «Оно возмож-
но,— пишет акад. Е. Н. Павловский,—
лишь при условии проглатывания личи-
нок пластинчатоусых жуков, что едва ли
кому-либо придет в голову». В кишечнике
рыб часто можно наблюдать Echinorhyn-
chus proteus, а у утиных птиц — Echino-
rhynchus polymorphic.
454
КЛАСС ПРИАПУЛИДЫ (PRIAPULIDA)
К классу приапулид относится всего
два рода морских раздельнополых
свободноживущих червей: Приапулюс
(Priapulus) с пятью видами и Галикрип-
тус (Halicryptus) с одним видом. Приапу-
лиды широко распространены в умерен-
ных зонах Мирового океана и на больших
глубинах в экваториальной зоне, что до
недавнего времени давало основание счи-
тать их биполярными формами.
Приапулиды имеют вальковатое тело
длиной до 10—15 см и ветвистую метель-
чатую жабру на заднем конце тела. Гали-
криптус совсем лишен придатков. Орга-
ны кровообращения, а у галикриптуса и
органы дыхания, отсутствуют; дыхание
осуществляется всей поверхностью тела.
Кожно-мускульный ме-
шок у приапулид состоит из двух слоев
мышц — наружного, кольцевого, и внут-
реннего, продольного,— и окружает об-
ширную свободную полость тела, зани-
мающую больше половины общего объема
тела. Нервная система приапулид весь-
ма примитивна. В передней части тела
имеется окологлоточное нервное кольцо,
от которого отходит брюшной нервный
тяж и подкожное нервное сплетение.
Органы чувств отсутствуют.
Кишечник у приапу-
лид прямой и сквозной, ро-
товое и анальное отверстия
расположены на концах
тела.
Характерная особенность
приапулид — их образ жиз-
ни и связанные с ним черты
организации, которые выра-
батывались у них, видимо, в
течение долгих геологиче-
ских периодов. Приапулиды
живут в плотных песчани-
стых грунтах. В связи с этим
у них выработалась особая
форма передвижения в них,
несколько напоминающая
движение дождевых червей
в почве, но в гораздо более
ярко выраженной форме.
Как приапулюсы, так и галикриптусы
имеют окружающий глотку сильный, во-
оруженный направленными назад крючья-
ми и шипами вывертывающийся хобот.
Раздув тело и заклинившись таким обра-
зом в ходе, червь с силой выбрасывает
вперед выворачивающийся при этом хобот
и пробивает ход. Затем, раздув хобот и
заклинив его в грунте, червь при помощи
продольной мускулатуры подтягивает те-
ло, чтобы в следующий момент опять вы-
бросить хобот вперед. Подтягиванию тела
вперед помогают специальные мышцы рет-
ракторы, прикрепленные одним концом к
переднему отделу кишечника, а другим —
к стенке тела в средней его части. Следует
при этом отметить, что в результате опи-
санного способа движения у приапулид
выработались элементы радиальной сим-
метрии в строении покровов тела. Ряды
крючков и шипиков, покрывающих тело,
расположены радиально одинаковыми про-
дольными рядами. Количество этих рядов
кратно 5, и эта пятилучевая симметрия
передней части тела выражена очень ясно
(рис. 257). Это давало основание некото-
рым зоологам ошибочно искать у приапу-
лид родственные связи с иглокожими.
Рис. 257. Приапулидовые черви:
приапулюс (Priapulus), 2 — галикриптус (Halicryptus); 3 — перед-
ний конец галикриптуса (вид сбоку); 4 — то же, вид сверху; 5 — схе-
ма расположения крючьев на переднем конце галикриптуса.
1 —
455
Очевидно, эта пятилучевая симметрия —
самобытный признак и выработался са-
мостоятельно и независимо от иглоко-
жих.
Объяснение способа движения приапу-
лид в плотном грунте было бы неполным,
если бы мы не обратили внимания на
значение в этом процессе обширной поло-
сти тела и заполняющей ее полостной жид-
кости.
Сжатие кольцевой мускулатуры в зад-
ней части тела направляет вперед ток
полостной жидкости и обусловливает вы-
Рис. 258. Галикриптус:
I — в передней части тела вскрытого галикрип-
туса видны глотка и четыре ретрактора, в задней
части тела — мочеполовые органы; 2 — личинка
галикриптуса; 3 — пучок протонефридиальных
мерцательных клеток на мочеполовых органах.
брасывание хобота. Такой способ движе-
ния может быть назван гидравличе-
ски м, о чем говорилось во введении. Из-
за плотности грунта движение приапулид
очень медленное, они проходят не более
1 м в час. Вместе с тем такое движение
требует затраты большой энергии. Сила,
с которой приапулюс продавливает ход в
грунте, в несколько десятков раз превы-
шает вес его собственного тела. Полость
хвостовой железы у приапулюсов сооб-
щается с общей полостью тела, и перекач-
ка полостной жидкости способствует и об-
мену жидкости в жабре.
В задний отдел кишечника у приапулид
открываются выделительные железы (рис.
258), представляющие собой органы типа
протонефридиев.
Все перечисленные признаки дают ос-
нование относить приапулид к низшим
червям, и имеется только один признак,
который как бы противоречит этому —
наличие у них эпителиального слоя, вы-
стилающего изнутри полость тела, т. е.
так называемого целотелия. Наличие
его как будто дает основание считать
внутреннюю полость вторичной по-
лостью тела, или ц е л о м о м,
который свойствен высшим червям и при-
мыкающим к ним группам. Личинка приа-
пулид (рис. 258) не имеет ничего общего
с трохофорой, и процесс дробления не
имеет особенностей спирального. Все это
укрепило положение приапулид среди низ-
ших червей, хотя ани и представляют очень
резко обособленную группу. Отпечатки
близкого к приапулидам организма обна-
ружены в среднекембрийских отложе-
ниях.
Морфологическое своеобразие и малое
число видов приапулид дают основа-
ние считать их очень древней группой
червей.
До недавнего времени географическое
распространение приапулид считалось би-
полярным (см. введение), но глубоковод-
ные исследования показали, что приапу-
лиды в экваториальном поясе обитают на
больших глубинах и, таким образом, не
являются биполярными.
Priapulus candatus — одна из массовых
форм фауны литорали северных морей.
Приапулиды в основном — хищники.
456
КЛАСС КАМПТОЗОА, ИЛИ СГИБАЮЩИЕСЯ
(KAMPTOZOA)
Небольшая группа, объединяющая око-
ло 60 видов в основном морских, донных,
преимущественно колониальных живот-
ных, образующих небольшие тонкие и неж-
ные колонии, состоящие из основной части
в виде паутинки, покрывающей субстрат
(камни, ракушки и др.). От этой основы
поднимаются вверх стебельки высотой в
2—3 мм, имеющие тонкую органическую
эластичную оболочку. На конце стебель-
ков находятся чашечки, в которых поме-
щаются все внутренние органы — ки-
шечник, выделительная система и половые
железы. Вторичной полости тела (целома)
у них нет и промежутки между органами
заполнены особой рыхлой тканью —
паренхимой. Чашечки сверху плоские и
по окружности несут венец коротких про-
стых щупалец.
Изогнутый кишечник начинается рото-
вым отверстием, находящимся на верхней
поверхности чашечки, на одном ее конце.
Пищевод идет вниз к основанию чашечки
и переходит в довольно объемистый же-
лудок, который лежит поперек нее. На
противоположном конце от желудка от-
кодит задняя кишка, которая идет вверх
и открывается анальным отверстием внут-
ри кольца щупалец.
Выделительными органа-
м и являются протонефридпи.
Кровеносная и дыхательная системы от-
сутствуют. Обмен газов происходит через
щупальца. Нервная система довольно
Рис. 259. Камптозоа, или сгибающиеся (Kamptozoa).
Pedicelina: 1 — часть колонии; 2 — продольный разрез через чашечку; 3 — личинка; 4 — Urnatella
gracilis — чашечка с частью стебелька. Внутри виден пищевой комок; 5,6 — Arthropodaria kowalp-
vski E — выпрямившаяся, 6 — сгибающаяся).
457
примитивная и представлена одним узлом,
лежащим неглубоко в паренхиме между
ротовым и анальным отверстиями, ближе к
первому. Органами чувств являются щу-
пальца.
Камптозоа бывают как раздельно-
полые, так и гермафродитные.
Половые железы в виде небольших ме-
шочков лежат в паренхиме. Яйцеклетка,
прежде чем выйти наружу, дозревает в
особом углублении, выводковой камере,
на верхней поверхности чашечки. Разви-
тие с метаморфозом: имеется свободнопла-
вающая довольно сложная личинка, ко-
торая некоторое время активно плавает в
толще воды при помощи ресничек, затем
опускается на дно и превращается в пер-
вичную особь, которая после бесполого
размножения (почкованием) образует ко-
лонию.
Положение камптозоа в системе животно-
го мира было неясным. Долгое время их
соединяли вместе с мшанками в единую
группу, выделяя внутри этой группы в виде
подкласса под названием внутрипороши-
цевые (Endoprocta) на основании положе-
ния анального отверстия внутри кольца
щупалец, в отличие от мшанок, у которых
оно располагается вне такового. Когда бы-
ло установлено, что у них нет настоящей
полости тела и имеются протонефридии,
такое соединение, естественно, отпало.
В настоящее время «сгибающихся» рас-
сматривают как особую группу, близкую
к низшим червям, причем, в отличие от
свободнодвигающихся брюхоресничных и
особенно коловраток, они занимают особое
положение, как перешедшие к сидячему
образу жизни.
Название «сгибающиеся» основано на
интересном свойстве этих животных: когда
они испытывают какое-либо раздражение,
стебелек при помощи продольных мышц
сгибается и животное выводится из сферы
раздражения. Когда раздражение прохо-
дит, стебелек благодаря эластичной оболоч-
ке вновь выпрямляется и поднимает ча-
шечку над субстратом, где ее щупальца
могут опять нормально фильтровать окру-
жающую воду. Сгибающиеся имеют еще
одну интересную биологическую особен-
ность — большую способность к регенера-
ции: когда чашечка по каким-либо причи-
нам отмирает, стебелек может восстано-
вить ее и все внутренние органы.
Распространены камптозоа широко, они
встречаются во всех морях, но благодаря
своим мелким размерам и миниатюрности
колоний часто, по-видимому, не замеча-
ются исследователями.
Для того чтобы обнаружить камптозоа,
необходимо тщательно просматривать суб-
страт, на котором они могут образовы-
вать свои колонии (мелкие камни, водорос-
ли и особенно мертвые ракушки, а также
другие подводные предметы). Соленость
и температура воды их, по-видимому, не
лимитируют, так как они встречены в це-
лом ряде опресненных мест Черного моря,
например в глубине Севастопольской бух-
ты около устья Черной речки. В последнее
время камптозоа обнаружены в Каспий-
ском море. В этом случае они, конечно,
мигранты, попавшие пассивно через Волго-
Донской канал, и являются в Каспии
представителями азово-черноморской, а
возможно, и средиземноморской фауны.
Такие длительные «путешествия» мож-
но объяснить интересной биологической
особенностью камптозоа. Попав в новые,
неблагоприятные для них условия суще-
ствования, они способны сбрасывать ча-
шечки, а затем, приспособившись к новым
условиям, регенерировать их и все внут-
ренние органы (Н. В. Насонов, 1926).
Что касается пресных вод, то в них
камптозоа встречаются редко. До конца
прошлого века они были известны только
в Северной Америке (Urnatella gracilis),
затем были пассивно, по-видимому при
помощи судов, перенесены в Европу, спер-
ва в Бельгию, а затем быстро расселились
по водным путям Центральной Европы
и уже в 1960 г. были обнаружены в реке
Тиссе. В наших пределах они обнаружены
пока только в низовьях Днестра.
ТИП НЕМЕРТИНЫ (NEMERTINI)
Краткая история открытия немертин.
Выдающийся советский зоолог Н. А. Ли-
ванов A955) открывает свой анализ
организации этого типа животных с ука-
зания, что немертины составляют этап
исторического развития живых форм, не-
посредственно следующий за развитием
типа плоских червей. Однако надо под-
черкнуть, что немертины — существенно
новый этап эволюции жизни. Правда,
в течение долгого времени основные осо-
бенности немертин оставались неизвест-
ными. Поэтому зоологи XVIII в., обна-
ружившие немертин и впервые попытав-
шиеся их изучить, ошибочно оценивали
положение этой группы животных в си-
стеме. Еще в 1758 г. Барлейз описал
одну из немертин, дал ее изображение
и назвал ее... «длинным морским червем»
(Sea Long Worm). В XVIII в. сведения об
организации немертин постепенно расши-
рялись. Однако в «Системе природы»
Карла Линнея о них ничего не гово-
рилось, и только в 13-м издании этого
знаменитого труда, выпущенного в свет
в обработке Г м е л и н а, появляются
сведения об этой группе животных. Гме-
лин назвал немертин червями и указал
на сходство их с планариями. В XIX в.
1 Название это взято из греческой мифологии:
Nemertes—одна из нереид — дочерей морского
царя Нерея.
значительно пополнились сведения о гео-
графическом распространении немертин
и данные об их организации. Многие
крупные зоологи этого века привнесли
в науку новые сведения об этих ориги-
нальных и интересных животных, обитаю-
щих преимущественно в морях всего све-
та, и лишь частично — в пресных водах.
Только в XIX в. было предложено и само
название — «немертины». Впервые знаме-
нитый французский зоолог Жорж К ю-
в ь е A815) дал это название одной из
немертин, известной ныне как Lineus
longissimus. Кювье назвал этого длин-
ного «червя» — Nemertes, а в 1831 г. дру-
гой знаменитый ученый — Эренберг
обозначил так всю группу и дал ей назва-
ние, сохранившееся поныне, — Nemertini.
Долгое время оставалось неясным поло-
жение немертин в зоологической системе.
О. Ф. Мюллер в 1776 г. причислил
немертин к р о д у аскарид (круглые чер-
ви). Кювье был первым ученым, который
выделил немертин из группы ресничных
червей. Однако еще в течение длительного
периода времени положение немертин в зо-
ологической системе оставалось неясным.
Некоторые ученые склонялись к сближе-
нию немертин с кольчатыми червями —
аннелидами. Однако после тщательного
изучения организации немертин стало яс-
ным, что они образуют обособленную
группу животных, все же родственных
459
ресничным червям — турбел-
ляриям. Главная заслуга в
выяснении характеристики
немертин принадлежит немец-
кому зоологу Отто Бюр-
геру.
Основные особенности ор-
ганизации немертин. Немер-
тины, несомненно, облада-
ют оригинальной организа-
цией, и она долгое время
препятствовала установле-
нию их положения в зооло-
гической системе. У немертин
очень длинное тело, хотя из-
вестны и формы небольших
размеров, и похожие на пия-
вок. Однако в большинстве
случаев это очень тонкие и
длинные формы. Обычно тело
их круглое в поперечном се-
чении. Наименьшие формы
достигают всего 3—10 мм, од-
нако английский зоолог Мак
И н т о ш нередко обнаружи-
вал у берегов Великобрита-
нии немертину Lineus longis-
simus, достигающую 5—8 м
в длину, при ширине тела,
равной 2,5 — 9 мм. Однако
большинство видов достигает
всего 2—20 см в длину при
ширине в 1—5 мм. Эти соот-
ношения очень характерны
для немертин — все они тон-
кие, шнуровидные. Более
толстые формы, напоминаю-
щие пиявок, — исключение.
Однако главнейшие отличи-
тельные признаки организа-
ции немертин не в размерах и
не в пропорциях тела. Еще важнее некото-
рые особенности их организации. Тело не-
мертин покрыто эпителием, очень богатым
железами и несущим, как у турбеллярий,
ресничный покров. Под эпителием раз-
вита многослойная мускулатура. Как у
плоских червей, у немертин нет по-
лости тела и пространство между
внутренними органами и стенками тела
заполнено паренхимой. Нервная
система—в виде головного узла («мозга»)
и двух продольных нервных стволов.
В этих признаках немертины,
как видим, обладают значи-
тельным сходством с плоски-
ми червями. Однако у них
имеются органы, ставящие не-
мертин на более высокую сту~
пень развития. Во-первых,
кишечник всегда снабжен зад-
непроходным отверстием. Во-
вторых, хотя у немертин нет
специальных орга-
нов дыхания, зато раз-
вита кровеносная система, ко-
торой у плоских червей нет.
Нужно далее отметить, что
кровеносная сис-
тема немертин замкнутая,
т. е. что кровь обращается
только по кровеносным со-
судам. Это, несомненно, при-
знак более высокой органи-
зации. Характернейший ор-
ган немертин — это хобот.
Он лежит в особом влагали-
ще над кишечником. Наружу
он высовывается или через
особое отверстие, или через
рот. При этом хобот ввернут
в хоботную полость, или
ринхоцелом. Сокра-
щением специальной муску-
латуры хобот выдвигается на-
ружу как вывернутый па-
лец перчатки. Особые мыш-
цы ретракторы (втя-
гиватели) могут вер-
нуть хобот в исходное поло-
жение. Хобот — это грозное
оружие немертин. Немерти-
ны — хищники. У некоторых
из них хобот снабжен специ-
альной ядовитой железой и острым стиле-
том, пронзающим жертву. Крупные немер-
тины нападают даже на мелких рыб, но
главной их пищей служат кольчатые черви
(аннелиды). Хобот, однако, только орудие
убийства. Питание осуществляется обыч-
ным путем. У немертин имеется ротовое
отверстие, расположенное на переднем
конце брюшной стороны тела. Оно ведет
в глотку, переходящую в длинную кишку,
над передней частью которой и лежит хо-
бот. Кишка имеет боковые карманы и пере-
Рис. 260. Организация не-
мертины:
1 — надглоточная и под-
глоточная нервные комиссуры;
2 — хобот; 3 — церебральные
органы; 4 — спинной крове-
носный сосуд; 5 — боковой
кровеносный сосуд; 6 — кар-
маны кишечника; 7 — яичники.
460
ходит в заднюю кишку, открывающуюся,
как уже выше сказано, наружу анальным
отверстием.
Кровеносная система со-
стоит из двух боковых продольных ство-
лов и одного спинного, проходящего
вдоль средней кишки. Кровь у немертин
бесцветная или красная, и течет она от
заднего конца тела к переднему. Два ко-
ротких сосуда с мерцательными клет-
ками выполняют функции выделительного
аппарата. Головные нервные узлы до-
стигают значительного развития и обра-
зуют «мозг» немертин. От этого «мозга»
берут свое начало два продольно-боко-
вых нервных ствола. Следует отметить,
что головные нервные узлы, образующие
«мозг» немертин, связаны нервными тя-
жами (комиссурами), образующими коль-
цо, сквозь которое проходит уже извест-
ный нам хобот.
В отличие от плоских червей немерти-
ны — раздельнополые живот-
ные, т. е. у них известны самцы и самки,
хотя внешне они неразличимы. Половые
железы лежат по бокам кишечной трубки
и правильно повторяются, располагаясь
в промежутках между карманами средней
кишки. Половые отверстия желез лежат
на спинной стороне тела. Зрелые яйца
выделяются наружу и здесь оплодотво-
ряются спермой самцов. У многих немер-
тин, прежде чем это случится, яйца со-
храняются в кольцевых коконах на теле
самки.
У многих видов немертин развитие со-
провождается формированием особой ли-
чинки. Несведущий человек никогда не
скажет, что эта личинка, так называемый
пилидий,— будущая немертина. Она
не похожа на взрослую немертину, поэто-
му и следует считать, что развитие немер-
тин может происходить с превращением.
Пилидий похож на каску или шлем, по-
чему и назван шлемовидной ли-
чинкой. Само название происходит,
строго говоря, от другого корня — От
греческого слова pylidion, что значит
«войлочная шляпа».
Мелкие реснички покрывают все тело
пилидия, а края «шляпы», или «шлема»,
несут более крупные реснички. Над «шле-
мом» развит длинный пучок теменных
жгутов, тогда как по бокам свешиваются
две широкие лопасти. Между ними рас-
положено ротовое отверстие, которое ведет
в кишечник, не имеющий у личинки задне-
проходного отверстия. В результате очень
сложных перестроек внутренней организа-
ции пилидия образуется молодая немер-
тина, высвобождающаяся из покровов пи-
лидия.
Многие немертины имеют поразительно
красивую окраску. «Немертины,— пишет
Отто Бюргер,— частью великолепно окра-
шенные черви». Нередко они окрашены
в желтые, коричневые, красные тона.
Иногда немертины окрашены в чистый
белый или темно-зеленый цвет. Краски
отличаются блеском, животные словно
полированные. Кроме того, основной кра-
сочный фон оживляется характерным ри-
сунком в виде сетчатой или мраморной
фактуры. Нередко голова несет особую
окраску или рисунок. Эти красивые ри-
сунки зависят от наличия красочных
секретов кожных желез и пигментов. Сле-
дует отметить, что очень часто окраска
немертин имеет значение покровительст-
венной, т. е. защитной.
Немертины, как и турбеллярии, спо-
собны к регенерации.
Образ жизни немертин. Море — типич-
ная стация немертин. Большинство ви-
дов приурочено к береговой зоне, где они
забираются в галечник, в щели скал,
в отверстия кораллового известняка или
в гущу морских водорослей. Некоторые
немертины зарываются в песок, другие
живут в трубках, построенных за счет
выделений кожных желез. Немногие виды
приспособились к жизни на земле, а пред-
ставители одного рода проникли в прес-
ную воду. Как уже было сказано, немер-
тины—хищники. Однако некоторые из них
ведут существование «нахлебников», или
комменсалистов, пользуясь пищей дру-
гих животных. Впрочем, они не причи-
няют этим животным какого-либо вреда.
Однако некоторые немертины стали пара-
зитами. В настоящее время известно
примерно около 1000 видов этих живот-
ных.
Избранные представители. Великолеп-
ный по окраске Tubulanus superbus при-
надлежит ксемейству Tubulanidae.
461
Представители этого семейства характе-
ризуются отсутствием спинного крове-
носного сосуда и тем, что их хобот лишен
стилета. Животное достигает 75 см в дли-
ну, тогда как ширина его тела равна
всего 5 мм. Животное окрашено в корич-
нево-красный цвет. На этом фоне резко
выделяются достигающие заднего конца
тела белые продольные полосы, пересе-
ченные многочисленными поперечными по-
лосками того же цвета, создающими изящ-
ный рисунок. Этот вид распространен
в Атлантическом океане, у берегов Шот-
ландии, Англии и Франции, а также
в Средиземном море. Обычно эта изящная
немертина живет на песчаном дне, в при-
брежной зоне, хотя и попадается на глу-
бинах, достигающих 40—50 м.
Наиболее крупные среди немертин фор-
мы наблюдались у берегов Англии. Таковы
представители семейства Lineidae.
Они близки к предшествующей форме,
так как тоже обладают невооруженным
хоботом. Lineus longissimus, достигаю-
щий, как выше указывалось, 10—15 м
в длину,— гигант среди немертин. Изве-
стны экземпляры, имеющие почти 30 м
в длину! Животные иногда встречаются
целыми скоплениями. Во время прили-
ва они покидают свои убежища и ловят
добычу.
Другой вид — Cerebratulus margi-
natus — много меньше предшествующего.
На заднем конце его широкого и сильного
тела развит небольшой придаток, напоми-
нающий хвост. Этот вид обычен не только
на севере Атлантического океана, но и в
Северном море. Особи этого вида хорошо
плавают, напоминая по своим движениям
угря.
К невооруженным немертинам отно-
сятся также представители с е м е й с т-
в a Pelagonemertidae. Они хорошо пла-
вают, чем объясняется и название этого
семейства. На больших глубинах Индий-
ского океана, к юго-востоку от Австралии,
была найдена немертина Pelagonemertes
moseleyi, достигающая в длину 4 см при
ширине тела 2 см. У этого животного со-
вершенно прозрачное тело. Сквозь наруж-
ные ткани немертины отчетливо просвечи-
вает темный, окрашенный в каштановый
цвет кишечник. Сзади тело немертины су-
живается и перетяжками делится на пять
отрезков. Передний отдел по длине равен
всем остальным и расширен в виде кры-
ловидных придатков, позволяющих жи-
вотному «парить» в воде.
К своеобразным формам принадлежат
виды рода Malacobdella. Один из видов
этого рода — Malacobdella grossa — жи-
вет в мантийной полости моллюсков и рас-
пространен в Северном море, в северной
части Атлантики и в Средиземном море.
Немертина питается мелкими организ-
мами, которые попадают вместе с водой,
омывающей жабры моллюсков, в мантий-
ную полость. Настоящим паразитом этот
вид не может быть назван, а скорее заслу-
живает наименования нахлебника.
В Кильской гавани чуть ли не 80°/0 более
крупных Cyprina islandica содержат в сво-
ей мантийной полости одну или несколько
этих своеобразных немертин.
Из наземных немертин назовем здесь
представителей рода Geonemertes. Фор-
мы этого рода относятся ксемейству
Prosorhochmidae. Некоторые из этих форм
достигают 7 см в длину. Все они обладают
сильным хоботом, высовывающимся через
ротовое отверстие наружу. Большинство
этих немертин имеет четыре глаза.
К этому семейству принадлежит, в част-
ности, Geonemertes agricola. Бюргер пишет
по поводу этой немертины следующее:
«Этот вид в большом числе особей встре-
чается в различных частях Бермудских
островов, и притом, поскольку это пока-
зывают наблюдения, только вдоль берего-
вой полосы, переходящей в мангровые бо-
лота. Летом этих наземных немертин
можно найти только во влажной почве,
вблизи от линии воды, тогда как весной
они встречаются немного выше, на скло-
нах холмов. Вполне возможно, что летом
в местах, становящихся сухими, они ис-
пользуют ходы, проделанные дождевыми
червями, и уходят в более глубокие слои
почвы. Впрочем, их излюбленное место-
обитание находится в полосе, граничащей
с наивысшими пределами прилива. Здесь,
на границе с густыми мангровыми заро-
слями, где почва состоит из черного ила,
переходящего выше к берегу в темную
или красную глину, они прячутся под
камнями, кусками дерева и другими пред-
462
метами, выброшенными морем». Бюргер
указывает, что, по его мнению, почвенные
немертины происходят не от пресновод-
ных, а от морских прибрежных форм.
Описываемая немертина ползает наподо-
бие пиявок, пользуясь своим ротовым
диском как присоской.
Представители семейства Prosto-
matidae живут в море и в различных мед-
ленно текущих или стоячих водоемах
с пресной водой. Хотя они не могут быть
названы часто встречающимися формами,
все же они и не редкость. Это очень ма-
ленькие формы, всего 1 —1,5 мм в длину,
изредка до 3,5 мм. Они стройные и пло-
ские. У них имеются четыре глаза. Кишеч-
ные карманы у этих форм не ветвятся.
Бюргер указывает, что эти маленькие не-
мертины распространены, вероятно, все-
светно. Хобот у них вооружен стилетом.
Пресноводные формы этого семейства при-
надлежат к роду Stichostema. Сюда
относятся относительно крупные формы,
достигающие 3—30 мм. Они отличаются
необычайно длинной хоботной полостью,
простирающейся назад до последней трети
тела, но не достигающей ануса; эти формы
широко распространены в Европе, в том
числе в Англии, Германии, Австрии и дру-
гих странах, по пресным водоемам. Рас-
пространены они также в Азии, Африке
и Северной Америке.
Кровеносная система, пред-
ставленная замкнутыми кровеносными со-
судами, не позволяет рассматривать не-
мертин как примитивные формы, несмот-
ря на некоторые черты близости их к тур-
белляриям. С последними немертин сбли-
жает наличие ресничного эпителия, что,
несомненно, должно рассматриваться как
черта примитивной организации. Об этом
же говорит отсутствие полости тела и на-
личие паренхимы, в которую погружены
внутренние органы; о примитивности не-
мертин свидетельствуют также протоне-
фридии, т. е. выделительные органы,
представленные трубками с конечными про-
тонефридиальными клетками. Но наряду
с этими примитивными чертами организа-
ции мы видим ряд прогрессивных призна-
ков — развитие вышеупомянутой крове-
носной системы, перемещение ротового
отверстия, в отличие от плоских червей,
на передний конец тела, значительное
развитие центральной нервной системы
и в особенности «мозга» немертин, повы-
шение общей энергии жизнедеятельности
и подвижности, что связано с развитием
хищничества,и другие признаки прогресса.
Все это наряду с оригинальной струк-
турой хобота заставляет зоологов рассмат-
ривать немертин как особый тип живот-
ного царства, образующий одну из эво-
люционных ветвей животного мира.
Высшие черви
Annelides
Как указывалось во введении, термин «черви» потерял свое
классификационное значение, хотя еще недавно они рассмат-
ривались как отдельный тип. Сейчас они подразделяются
на 4 самостоятельных типа. Однако все же названием «черви»,
как мы уже указывали, широко пользуются. Так же часто
червей делят на низших (Scolecida) и высших (Annelides).
К первым относят типы плоских и круглых червей, немер-
тин и ряд добавочных групп, а ко вторым — один тип —
кольчатых (Annelides).
Основные характерные особенности высших червей — это
сегментация тела, наличие кровеносной системы и целома
(вторичная полость тела), спиральный тип дробления
и личинка (если она имеется) типа трохофоры.
В качестве добавочного к типу кольчатых червей относят
класс сипункулид (Sipunculoidea).
ТИП КОЛЬЧАТЫЕ ЧЕРВИ (ANNELIDA)
ОБЩАЯ ХАРАКТЕРИСТИКА
К кольчатым червям принадлежат пер-
вичные кольчецы, многощетинковые и мало-
щетинковые черви, пиявки и эхиуриды.
В типе кольчатых червей насчитывают
около 8 тыс. видов. Это наиболее высо-
коорганизованные представители группы
червей. Размеры кольчецов колеблются
от долей миллиметра до 2,5 м. Преимуще-
ственно это свободноживущие формы. Тело
кольчецов подразделено на три части:
голова, туловище, состоящее из колец,
и анальная лопасть. Такого четкого деле-
ния тела на отделы нет у стоящих ниже
по своей организации животных.
Голова кольчецов снабжена различными
органами чувств. У многих кольчецов
хорошо развиты глаза. Некоторые имеют
особо острое зрение, и их хрусталик спо-
собен к аккомодации. Правда, глаза могут
быть расположены не только на голове,
но и на щупальцах, на теле и на хвосте.
У кольчецов развиты и вкусовые ощуще-
ния. На голове и щупальцах у многих
из них есть особые обонятельные клетки
и ресничные ямки, которые воспринимают
различные запахи и действие многих хими-
ческих раздражителей. Хорошо развиты
у кольчецов органы слуха, устроенные
по типу локаторов. Недавно у морских
кольчецов эхиурид открыты органы слу-
ха, очень похожие на органы боковой
линии у рыб. С помощью этих органов
животное тонко различает малейшие шо-
рохи и звуки, которые в воде слышны
гораздо лучше, чем в воздухе.
Тело кольчецов состоит из колец, или
сегментов. Число колец может достигать
нескольких сотен. Другие кольчецы состо-
ят всего из нескольких сегментов. Каждый
сегмент до некоторой степени представ-
ляет самостоятельную единицу целого ор-
ганизма. Каждый сегмент включает части
жизненно важных систем органов.
Очень характерны для кольчецов осо-
бые органы движения. Они располагаются
по бокам каждого сегмента и называются
параподиями. Слово «параподии»
означает «похожие на ноги». Параподии —
это лопастевидные выросты тела, из кото-
рых наружу торчат пучки щетинок. У не-
которых пелагических полихет длина па-
раподии равна диаметру тела. Парапо-
дии развиты не у всех кольчецов. Есть
они у первичных кольчецов и многоще-
тинковых червей. У малощетинковых ос-
таются только щетинки. Примитивная пи-
явка, акантобделла имеет щетинки. Ос-
тальные пиявки обходятся в движении
без параподии и щетинок. У эхиурид
параподии нет, а щетинки есть только
на заднем конце тела.
Параподии, узлы нервной системы, ор-
ганы выделения, половые железы и, у не-
30*
467
которых полихет, парные карманы кишеч-
ника планомерно повторяются в каждом
сегменте. Эта внутренняя сегментация сов-
падает с наружной кольчатостью. Много-
кратное повторение сегментов тела на-
звано греческим словом «метамерия». М е-
т а м е р и я возникла в процессе эволю-
ции в связи с удлинением тела предков
кольчецов. Удлинение тела вызвало необ-
ходимость многократного повторения
сначала органов движения с их муску-
латурой и нервной системой, а затем
и внутренних органов.
Чрезвычайно характерна для кольче-
цов сегментированная вторичная полость
тела, или целом. Эта полость находится
между кишечником и стенкой тела. По-
лость тела выстлана сплошным слоем эпи-
телиальных клеток, или целотелием.
Эти клетки образуют слой, покрывающий
кишечник, мышцы и все другие внутрен-
ние органы. Полость тела разделена на
сегменты поперечными перегородками —
диссепиментами. По средней
линии тела проходит продольная перего-
родка — мезентерий, которая де-
лит каждый отсек полости на правую
и левую части.
Полость тела заполнена жидко-
стью, которая по своему химическо-
му составу очень близка к морской
воде. Жидкость, заполняющая по-
лость тела, находится в непрерыв-
ном движении. Полость тела и по-
лостная жидкость выполняют важ-
ные функции. Полостная жидкость
(как и всякая жидкость вообще) не
сжимается и поэтому служит хоро-
Рис. 261. Анатомия кольчецов.
А — вскрытый Nereis: 1 — голова (простомиум);
2 — головные тактильные усики, 3 — глаза,
4 — органы химического восприятия (пальпы);
5 — ротовой сегмент (пернстомиум), 6 — усики
ротового сегмента; 7 — параподни с пучками
щетинок; 8 — сегменты тела, 9 — стенка те-
ла, 10 — глотка; 11 — желудок, 12 — боковые
выросты кишечника, 13 — средняя кишка (на
рисунке срезана, чтобы показать брюшную нерв-
ную цепочку), 14 — брюшная нервная цепоч-
ка, 15 — брюшной кровеносный сосуд, 16 —
спинной кровеносный сосуд, 17 — кольцевые
кровеносные сосуды; 18— органы выделения (ме-
танефридии); 19 — перегородки вторичной по-
лости тела, 20 — вторичная полость тела (це-
лом). Б —_глаз пелагической полихеты Alcio-
ра: 1 — линза; 2 — клетка, производящая стек-
ловидное тело, 3 — стекловидное тело, 4 — слой
палочек ретины; 5 — чувствительный слой рети-
ны; 6 — оптический нерв. В —поперечный раз-
рез через жаберную нить и сложный глаз Вгап-
chiomma: 1 — хрящевой стержень жаберной ни-
ти, 2 — кровеносный сосуд, 3 — кроющий эпи-
телий сложного глаза, 4 — отдельные линзы
глазков; 5 — отдельные ретинальные клетки
сложного глаза. Г — поперечный разрез голов-
ного мозга Sthenelais (Aphroditidae): l — паль-
парный нерв, 2 — окологлоточный коннектив;
3 — грибовидные тела (глобули). Д — попереч-
ный разрез сегмента тела Nereis: 1 — спинная
ветвь параподии, 2 — спинной усик; 3 — пучок
спинных щетинок; 4 — брюшная ветвь парапо-
дии; 5 — брюшной усик; 6 — пучок брюшных
щетинок; 7 — апикулярная щетинка; 8 — спин-
ная лента мышц, 9 — брюшная лента мышц;
10 — кишечник; 11 — спинной кровеносный
сосуд; 12 — брюшной кровеносный сосуд;
13 — брюшная нервная цепочка.
468
шим «гидравлическим скелетом». Движе-
нием полостной жидкости могут перено-
ситься внутри тела кольчецов различные
питательные продукты, выделения желез
внутренней секреции, а также кислород
и углекислый газ, участвующие в про-
цессе дыхания.
Внутренние перегородки защищают ор-
ганизм при тяжелых ранениях и разрывах
стенки тела. Например, дождевой червь,
разрезанный пополам, не погибает. Пере-
городки не позволяют полостной жидкости
вытекать из тела. Внутренние перегород-
ки кольчецов, таким образом, предохра-
няют их от гибели. Морские корабли
и подводные лодки тоже имеют внутрен-
ние герметические перегородки. Если борт
пробит, то вода, хлынувшая в пробоину,
заливает только один поврежденный от-
сек. Остальные отсеки, не залитые водой,
сохраняют плавучесть поврежденного ко-
рабля. Так и у кольчецов нарушение од-
ного сегмента их тела не влечет за собой
гибели всего животного. Но не все коль-
чатые черви имеют хорошо развитые пере-
городки в полости тела. Например, у эхиу-
рид полость тела не имеет перегородок.
Прокол стенки тела эхиуриды может при-
вести ее к гибели. Кроме дыхательной
и защитной роли, вторичная полость вы-
полняет роль вместилища для половых
продуктов, которые вызревают там, преж-
де чем выводятся наружу.
Кольчецы, за немногими исключениями,
имеют кровеносную систему.
Однако сердца у них нет. Стенки крупных
сосудов сами сокращаются и проталки-
вают кровь через тончайшие капилляры. У
пиявок функции кровеносной системы и вто-
ричной полости настолько совпадают, что
эти две системы совмещаются в единую
сеть лакун, по которым течет кровь. У од-
них кольчецов кровь бесцветная, у дру-
гих окрашена в зеленый цвет пигментом,
который называется хлоркруорин. Часто
кольчецы имеют красную кровь, по со-
ставу сходную с кровью позвоночных.
Красная кровь содержит железо, которое
входит в состав пигмента гемоглобина.
Некоторые кольчецы, зарываясь в грунт,
испытывают острый дефицит кислорода.
Поэтому у них кровь приспособлена к то-
му, чтобы особенно интенсивно связывать
кислород. Например, у полихеты Magelo-
na papillicornis развит пигмент гемэ-
р и т р и н, который содержит железа
в пять раз больше, чем гемоглобин.
У кольчецов по сравнению с низшими
беспозвоночными обмен веществ и дыха-
ние протекают гораздо интенсивнее. У не-
которых многощетинковых кольчецов раз-
виваются специальные органы дыхания —
жабры. В жабрах разветвляется сеть
кровеносных сосудов, и сквозь их стенку
кислород проникает в кровь, а затем раз-
носится по всему телу. Жабры могут быть
расположены на голове, на параподиях
и на хвосте.
Сквозной кишечник кольчецов состоит
из нескольких отделов. Каждый отдел
кишечника выполняет свою особую функ-
цию. Рот ведет в глотку. У некоторых
кольчецов в глотке располагаются силь-
ные роговые челюсти и зубчики, помогаю-
щие крепче схватывать живую добычу.
У многих хищных кольчецов глотка слу-
жит мощным орудием нападения и защи-
ты. За глоткой следует пищевод. Этот
отдел часто снабжен мышечной стенкой.
Перистальтические движения мышц мед-
ленно проталкивают пищу в следующие
отделы. В стенке пищевода располага-
ются железы, фермент которых служит для
первичной переработки пищи. За пище-
водом следует средняя кишка. В отдельных
случаях бывают развиты зоб и желудок.
Стенка средней кишки образована эпите-
лием, очень богатым железистыми клет-
ками, которые вырабатывают пищевари-
тельный фермент. Другие клетки средней
кишки всасывают переваренную пищу.
У одних кольчецов средняя кишка в виде
прямой трубки, у других она изогнута
петлями, третьи имеют с боков кишеч-
ника метамерные выросты. Задняя кишка
заканчивается анальным отверстием.
Специальные органы—м етанефри-
д и п—служат для выделения жидких про-
дуктов обмена веществ. Часто они служат
для выведения наружу половых клеток —
сперматозоидов и яйцеклеток. Метанефри-
дии начинаются воронкой в полости тела;
от воронки идет извитой канал, который
в следующем сегменте открывается нару-
жу. В каждом сегменте располагаются
два метанефридия.
469
Размножаются кольчецы бес-
полым и половым путем. У водных коль-
чецов часто встречается бесполое размно-
жение. При этом их длинное тело распа-
дается на несколько частей. Через неко-
торое время каждая часть восстанавли-
вает голову и хвост. Иногда голова с гла-
зами, щупальцами и мозгом образуется
в середине тела червя еще до того, как
он разделится на части. В этом случае
отделившиеся части уже имеют голову
со всеми необходимыми органами чувств.
Полихеты и олигохеты сравнительно хо-
рошо восстанавливают утраченные части
тела. Такой способностью не обладают
пиявки и эхиуриды. Эти кольчецы утра-
тили сегментированную полость тела. От-
части поэтому, видимо, способность к бес-
полому размножению и восстановлению
утраченных частей у них отсутствует.
Оплодотворение яиц у морских кольче-
цов происходит чаще всего вне тела мате-
ринского организма. В этом случае самцы
и самки одновременно выбрасывают поло-
вые клетки в воду, где и происходит опло-
дотворение.
У морских многощетинковых кольче-
цов и эхиурид дробление оплодотворен-
ных яиц приводит к развитию личинки,
которая нисколько не похожа на взро-
слых животных и носит название трохо-
форы. Трохофора короткое время
живет в поверхностных слоях воды, а за-
тем оседает на дно и постепенно превра-
щается во взрослый организм.
Пресноводные и наземные кольчецы ча-
ще всего гермафродиты и имеют прямое
развитие. Свободной личинки у пресно-
водных и наземных кольчецов нет. Это
связано с тем, что пресная вода имеет
солевой состав совсем иного свойства, чем
морская. Для развития жизни морская
вода более благоприятна. Пресная же во-
да содержит даже некоторые ядовитые
ионы (например, магния) и для развития
организмов менее пригодна. Поэтому раз-
витие пресноводных животных почти все-
гда происходит под прикрытием специ-
альных малопроницаемых оболочек. Еще
более плотные оболочки — скорлупки —
образуются у яиц наземных кольчецов.
Плотные оболочки предохраняют здесь
яйцеклетки от механических повреждений
и от высыхания под палящими лучами
солнца.
Практическое значение кольчатых чер-
вей все более возрастает в связи с разви-
тием интенсивности биологических иссле-
дований.
У нас в СССР впервые в истории
мировой науки осуществлена акклимати-
зация некоторых беспозвоночных для уси-
ления кормовой базы моря. Например,
полихета Nereis, акклиматизированная
в Каспийском море, стала важнейшим пи-
щевым объектом осетра и других рыб.
Дождевые черви не только служат на-
живкой при рыбной ловле и пищей для
птиц. Они приносят огромную пользу
человеку, разрыхляя почву, делая ее
более пористой. Это благоприятствует сво-
бодному проникновению воздуха и воды
к корням растений и увеличивает урожаи
сельскохозяйственных культур. Роясь
в земле, черви проглатывают кусочки
почвы, измельчают их и выбрасывают на
поверхность хорошо перемешанными с ор-
ганическим веществом. Выносимое на по-
верхность червями количество грунта по-
разительно велико. Если бы распределить
по всей поверхности суши почву, перепа-
хиваемую дождевыми червями за каждые
10 лет, то получился бы слой плодород-
ной земли толщиной в 5 см.
Пиявки применяются в медицинской
практике при гипертонических заболева-
ниях и угрозе кровоизлияния. Они впу-
скают в кровь вещество гирудин, которое
препятствует свертыванию крови и спо-
собствует расширению кровеносных сосу-
дов.
Тип кольчецов включает несколько
классов. Наиболее примитивными явля-
ются морские первичные кольчецы — ар-
хианнелиды. Многощетинковые кольчецы
и эхиуриды — обитатели моря. Малоще-
тинковые кольчецы и пиявки — в основ-
ном обитатели пресных вод и почвы.
470
КЛАСС ПЕРВИЧНЫЕ КОЛЬЧЕЦЫ
(ARCHIANNELIDA)
Архианнелиды — наиболее примитив-
ный класс кольчатых червей. Кольча-
тость их тела имеет первичный и прими-
тивный характер. Первые признаки коль-
чатости возникают в поверхностных слоях
тела. Процесс возникновения кольчато-
сти исторически, видимо, начинается с ор-
ганов движения. На удлиненном теле пер-
вичных кольчецов возникает несколько
ресничных колец, с помощью которых эти
организмы плавают в воде или скользят
в грунте.
Архианнелиды — чаще всего мелкие
кольчецы. Некоторые из них очень малы
и не превышают 2—3 мм. Число сегментов,
или колец их тела, не превышает 7.
Другие состоят из большего чис-
ла сегментов и достигают боль-
шей длины— до 2—3 см. Архи-
аннелиды живут преимущест-
венно в прибрежной части мо-
ря. Одни из них скользят по
поверхности грунта или водо-
рослей с помощью ресничек.
Другие движутся в промежут-
ках между частицами песка. В
этом случае у них могут быть
одновременно pi ресничные коль-
ца, и параподии со щетинками.
Щетинки образуются из реснич-
ных клеток как приспособление
к жизни в грунте. Класс архи-
аннелид состоит из нескольких
семейств — Dinophilidae, Proto-
drilidae, Polygordiidae, Saccocir-
ridae, Nerillidae.
Наиболее примитивны среди
первичных кольчецов предста-
вители семейства дино-
филид. Динофилиды очень малы
и чаще всего окрашены в оран-
жевые или красноватые тона.
Например, Dinophilus maris albi
из Белого моря размером все-
го около 1 мм. Динофилиды
живут среди нитчатых водорос-
лей. Они питаются илом, бога-
Рис.
ди-
262. Строение
нофилуса:
1 — головной мозг, 2 —
глаз, 3 — боковые нервные
стволы; 4 — комиссуры;
5 — рот, в — глотка, 7 —
кишечник, 8 — анальное
отверстие, 9 — ресничные
кольца, 10 — органы вы-
деления; 11 — гонады; 12 —
половое отверстие.
тым органическим веществом, а также
одноклеточными водорослями, инфузо-
риями и другими мельчайшими организ-
мами.
Тело динофилид опоясано несколькими
венчиками ресничек (рис. 262). Два вен-
чика находятся на голове и еще пять рас-
полагаются на туловище. Нервная систе-
ма состоит из головного мозга и двух ши-
роко расставленных брюшных нервных
стволов. Соответственно пяти ресничным
кольцам на каждом из нервных стволов
имеются скопления нервных клеток —
ганглии. В течение лета самки
откладывают несколько десятков яиц, из
которых выходят молодые динофилиды.
Другое семейство — про-
тодрилид представлено мелки-
ми кольчецами длиной от 1 до
20 мм. Тело их состоит из не-
скольких десятков сегментов,
опоясанных ресничными коль-
цами. У некоторых видов наря-
ду с ресничными кольцами раз-
виты и щетинки. На голове па-
ра длинных усиков. Протодри-
лиды живут в песке в прибреж-
ной зоне моря. С помощью рес-
ничной бороздки на брюшной
стороне тела они пробираются
в свободных промежутках меж-
ду песчинками и головными уси-
ками вылавливают ил и мелких
животных, которыми питаются.
Глаз у них нет. Зато имеются
специальные сейсмические орга-
ны — с т а т о ц и с т ы, кото-
рые отлично улавливают малей-
шие шорохи. У Protodrilus ста-
тоцисты расположены у основа-
ния головных щупалец. Воспри-
нимая шорохи, эти кольчецы
могут реагировать на присутст-
вие поблизости жертвы или
врага. Своей мускулистой глот-
кой они раздавливают добы-
чу.
471
Взрослые протодрилиды откладывают
между частицами песка маленькие коко-
ны с несколькими яйцеклетками. Оплодо-
творенные яйцеклетки дробятся и посте-
пенно превращаются в личинок — т р о-
х о ф о р.
Сначала трохофоры имеют только один
ресничный пояс. Тело их набито эмбрио-
нальным желтком. Позднее появляется
второй ресничный пояс, глаза и ротовое
отверстие. Когда через некоторое время
сформируется сквозной кишечник, тро-
хофоры покидают кокон и уплывают в мо-
ре. Они имеют размер около 70 мк и не вид-
ны невооруженным глазом. Около рото-
вого отверстия у них формируются огром-
ные губы, которые могут выпячиваться
далеко вперед и схватывать добычу. Они
могут также складываться или совсем
Рис. 263. Личинки протодрилуса:
А — трохофора, втянувшая ловчий аппарат, Б — тро-
хофора с вытянутым аппаратом для отлавливания до-
бычи, В — метатрохофора с брюшной стороны, Г —
она же слева с вытянутым ловчим аппаратом: 1 — те-
менной хохолок; 2 — глаз, 3 —рот; 4 — верхняя гу-
ба, 5 — хобот, б — кишечник; 7 — анальное отвер-
стие; 8 — органы выделения; 9 — ресничные кольца.
втягиваться внутрь (рис. 263). Личинки
питаются очень мелкими одноклеточными
водорослями и быстро растут. Тело личи-
нок заметно вытягивается, и на нем появ-
ляются сначала третье, а затем четвертое
ресничные кольца. В это время личинки
напоминают взрослых динофилид с непол-
ным числом ресничных колец. Личинки
длиной около 0,5 мм оседают на дно, про-
никают в промежутки между частицами
песка и постепенно превращаются во
взрослых червячков. Число сегментов по-
степенно увеличивается.
Рассмотренный здесь цикл индивидуаль-
ного развития одного из первичных коль-
чецов показывает возможный путь исто-
рического развития всей группы кольча-
тых червей. Известно, что в индивидуаль-
ном развитии отражается в какой-то мере
путь эволюционного развития той же
группы животных. Эта закономерность
носит название биогенетического закона.
Представители семейства поли-
гордиусов ведут роющий образ жизни
в песчаных грунтах. Обычно они живут
в море на глубине от 10 до 20 м. Длина
полигордиусов около 5 см.
Сильно вытянутое цилиндрическое тело
состоит из многочисленных одинаковых
сегментов. На голове развиты щупальца
и две обонятельные ямки. С помощью
этих органов животное воспринимает раз-
личные механические и химические раз-
дражения. Глаза отсутствуют.
Если динофилиды и протодрилиды дви-
жутся главным образом с помощью рес-
ничек и щетинок, то полигордиусы пере-
двигаются в грунте только с помощью
мышц. В стенке их тела хорошо развита
продольная мускулатура, а кольцевая
отсутствует. Нервная система со-
стоит из головного и брюшного мозга.
Головной мозг устроен очень сложно.
Питаются полигордиусы органиче-
скими частицами животного и раститель-
ного происхождения. Не брезгают они
и мелкими животными. Специальной ды-
хательной системы у полигордиусов нет.
Зато хорошо развиты кровеносные сосу-
ды, по которым течет красная кровь.
Личиночное развитие полигордиусов
протекает в толще воды. Трохофора описа-
на в каждом учебнике зоологии.
472
Представители семейства сакко-
циррусов также обитают в морских пес-
чаных грунтах. В прибрежной зоне Чер-
ного моря на каждый квадратный метр
песчаного дна приходится в среднем не-
сколько сотен тысяч этих кольчецов.
Саккоциррусы имеют тело до 8 см дли-
ной, с большим количеством сегментов.
Сегменты с каждой стороны несут по
одному пучку простых щетинок. Разви-
тие этих кольчецов проходит с превраще-
нием. Трохофора сначала превращается
в личинку с небольшим числом сегментов,
Полихеты названы так потому, что па-
раподии у них включают пучки много-
численных щетинок.
По-гречески poly означает много,
chaeta — щетинка. Полихеты — ти-
пичные морские животные и очень редко
встречаются в пресной воде.
Всего насчитывается более 5 тыс. видов
полихет. Основная масса их населяет
соленые моря и океаны. Сравнительно
немногие приспособились к жизни в соло-
новатых водоемах, к которым относятся,
например, Азовское и Каспийское моря.
Встречаются полихеты в солоноватой воде
среди мангровых зарослей на побережьях
Индии и Малайских островов. Еще мень-
ше полихет в пресных водах. Такова,
например, Manayunkia, обитающая в озе-
рах Таймыра, реках Северной Америки,
в озере Байкал. В пресной воде пещер
Герцеговины обнаружена полихета Meri-
fugia из семейства Serpulidae. Это
единственный случай приспособления по-
лихеты к пещерному существованию.
В исключительных случаях полихеты при-
спосабливаются к наземному существова-
нию. Известен один вид полихеты Lycas-
topsis catarractarum из семейства
Nereidae, которая живет в тропиках, за
многие километры от морского берега, сре-
ди пальм на плантациях бананов и какао.
количество которых постепенно увеличи-
вается.
К семейству нериллид относятся
архианнелиды не более 2 мм длиной,
с небольшим числом сегментов. Сегменты
снабжены пучками щетинок. Один из
представителей этого семейства найден
в пещерах Швейцарии. Называется он
Troglochaetus beranecki, что в переводе
означает «пещерный щетинконос бера-
нека». Это исключительный случай при-
способления архианнелиды к обитанию
в пресных грунтовых водах пещер.
Она питается гниющими листьями и слад-
кими плодами.
Как правило, полихеты живут свободно.
Только в некоторых случаях они связы-
вают место своего обитания с другими
животными и делят с ними пищу. Таких
партнеров называют сотрапезниками или
комменсалами. Полихеты-комменсалы жи-
вут внутри губок, где находят дом и пи-
щу, в раковинах раков-отшельников или
на морских звездах.
В исключительных случаях полихеты
паразитируют на рыбах. Например,
Ichthyotomus sanguinarius, который ртом
прикрепляется к плавникам морских
угрей, путешествует с ними через Атлан-
тический океан и сосет их кровь.
Морские полихеты встречаются во всех
частях и на всех глубинах Мирового океа-
на. Они обитают на дне, в толще грунта
и в толще воды. Полихеты особенно
обильны в прибрежной зоне моря и немно-
гочисленны на больших глубинах. На
больших глубинах обитают особые глу-
боководные (абиссальные) виды.
Наиболее богата и разнообразна фауна
полихет в прибрежной зоне тропических
морей. Но и в северных морях их довольно
много. Иногда они встречаются большими
скоплениями. Например, в Баренцевом
море плотность поселения полихеты Mai-
КЛАСС МНОГОЩЕТИНКОВЫЕ КОЛЬЧЕЦЫ
(POLYCHAETA)
473
dane sarsi достигает 90 тыс. эк-
земпляров на каждом квадрат-
ном метре морского дна. На
больших площадях морского
дна полихеты образуют живые
ковры удивительных узоров, пе-
реливающиеся всеми цветами
радуги.
Многие полихеты живут сре-
ди морских водорослей. Дру-
гие прячутся в укромных угол-
ках среди зарослей коралловых
рифов. Немалое количество по-
лихет зарывается в песок или
в мягкий ил. Очень часто поли-
хеты строят себе роговые, пес-
чаные или известковые труб-
ки. Некоторые способны жить
в очень трудных условиях. По-
лихета Capitella capitata, на-
пример, обитает в самых гряз-
ных гаванях, где нефть и ма-
зут плавают на поверхности во-
ды, покрывают все дно, все под-
водные предметы. В таких га-
ванях гибнут от нефти рыбы,
тюлени и даже птицы. Только у
берегов Англии погибает от
нефти ежегодно четверть мил-
лиона водоплавающих птиц.
Трудно представить себе, как
Capitella может существовать в таких
неестественных условиях. Другая группа
полихет приспособлена к обитанию в
толще воды. Таких животных называют
пелагическими.
Весь свой жизненный цикл они совер-
шают вдали от берегов и переносятся на
большие расстояния морскими течениями.
Пелагических полихет, например, много
в теплом течении Гольфстрим.
Движутся полихеты двумя способами.
Они или волнообразно изгибаются, или
попеременно то сокращают, то вытяги-
вают свое тело. Полихеты имеют по бокам
каждого сегмента придатки — параподии,
которые служат для передвижения. Пара-
подии несут пучки многочисленных ще-
тинок. Щетинки образуются внутри рес-
ничных клеток еще у личинок полихет.
Параподии со щетинками приспособлены
для движения в толще воды, на поверх-
ности грунта, в толще грунта и внутри
Рис. 2G4. Типы щетинок
полихет:
Верхний ряд — ще-
тинки свободноживущих по-
лихет, средний ряд —
щетинки полихет, рою-
щих грунт; нижний
ряд — щетинки полихет,
обитающих в трубках.
трубок. В зависимости от места
обитания животного щетинки
имеют соответствующую форму
(рис. 264). Полихеты, обитаю-
щие в толще воды или времен-
но поднимающиеся со дна на
поверхность, имеют на конце
щетинки подвижный членик.
Этот членик поворачивается в
суставе рукоятки так, что впе-
ред движется ребром, а назад
широкой плоскостью. Точно так
же работают веслами гребцы на
академических спортивных лод-
ках-скифах. Только у полихет
гребные движения щетинок-ве-
сел сочетаются с изгибанием
всего тела.
Полихеты, обитающие на по-
верхности грунта, имеют два
вида щетинок. Одни острые,
как шипы у розы, легко вка-
лывающиеся в стебли и листья
водорослей. Другие щетинки
торчат вверх и зазубрены, как
гарпуны. Они представляют на-
дежную защиту и часто ядовиты.
У кольчецов, роющих грунт,
щетинки похожи на острые
клинки, вилы или копья.
Наконец, у обитателей песча-
ных и известковых трубок щетинки похожи
на птичьи клювы, рыболовные крючки,
гребни для расчесывания волос или хок-
кейные клюшки.
Форма щетинок настолько характерна,
что опытный специалист может только
по одним щетинкам определить вид мор-
ского червя. Это приходится делать тогда,
когда по полупереваренным остаткам в ки-
шечниках рыб специалисту необходимо
определить, какими именно червями пи-
тается рыба, и ответить на вопрос, хватит
ли ей пищи в том или другом участке
моря.
В движении плавающие и ползающие
полихеты волнообразно изгибают свое те-
ло. Голова и передний конец тела при
этом остаются вытянутыми прямо вперед.
Параподии участвуют в движении как
весла или плавники. При ударе назад
пучки щетинок высовываются, что значи-
тельно увеличивает их рабочую поверх-
474
ность, а при возвращении вперед щетинки
втягиваются. Спасаясь от преследования
или догоняя свою добычу, полихеты раз-
вивают такую скорость, что могут сопер-
ничать с некоторыми рыбами. В грунте
полихеты тоже движутся очень быстро.
В этом случае они с силой перекачивают
из заднего конца тела в передний свою
полостную жидкость, а щетинками упи-
раются в грунт. Гидравлический способ
движения животных в грунте очень эко-
номичен и вполне может быть перенесен
в технику.
Питаются полихеты по-разному. Од-
ни из них — стремительные хищники.
К таким относится большинство пелаги-
ческих полихет, а также некоторые из
донных. И те и другие имеют длинную,
выбрасывающуюся вперед глотку. Глотка
снабжена хватательными придатками, ост-
рыми шипами или крепкими челюстями.
Пелагические полихеты настигают свою
добычу и хватают ее придатками глотки,
похожими на пальцы руки. Vanadis, To-
mopteris и другие пелагические хищные
полихеты нередко настигают мальков рыб
и съедают их. Особенно часто гибнут в
пасти полихет личинки и мальки сельди.
Донные хищники имеют огромную мус-
кулистую глотку, вооруженную двумя
парами сильных челюстей. Например,
морская мышь (Aphrodite aculeata) питается
мелкими рачками, моллюсками, кольча-
тыми червями и гидроидами, которых
хватает челюстями. Другие полихеты пи-
таются в основном растениями. Нереиды,
например, также имеют глотку с сильны-
ми зазубренными челюстями. Однако эти
челюсти служат порой для того, чтобы
отгрызать кусочки водорослей. Nereis pela-
gica питается стеблями и листьями мор-
ской водоросли Laminaria. В морях эта
водоросль встречается очень часто. На
Дальнем Востоке многие жители употреб-
ляют морскую капусту в пищу. Она отно-
сится к лекарственным растениям и упот-
ребляется при склерозе.
У полихет много врагов. В море их пое-
дают ракообразные, рыбы, иглокожие и
кишечнополостные. С воздуха на них охо-
тятся чайки и другие морские птицы.
Поэтому в процессе эволюции полихеты
приспособились к защите.
Наиболее надежный способ защиты от
крупных врагов — зарывание в грунт.
Способность к зарыванию сопровождается
развитием ряда специальных приспособ-
лений. Например, развиты железы, выде-
ляющие клейкий и слизистый секрет.
Слизь предохраняет тело от ранений.
Клейкий секрет позволяет укреплять стен-
ки норы или склеивать песчинки и куски
растений, из которых полихеты строят
трубку-домик.
Роющий образ жизни ведет обычный на
мелководье в наших морях пескожил.
Пескожил (Arenicola) живет в изогнутых
норах. Оба конца норы подходят к поверх-
ности грунта. Своей глоткой пескожил
захватывает и проглатывает песок вместе
с органической пищей. Песок проходит
сквозь кишечник и выбрасывается поза-
ди. Поэтому около рта песок оплывает,
и на поверхности грунта образуется во-
ронка. В воронку попадают гниющие во-
доросли, которыми питается пескожил.
Таким образом, зарываясь в грунт, пес-
кожил обеспечивает себе защиту от врагов
и непрерывный приток пищи. Однако
рыбы приспособились подстерегать песко-
жила в тот момент, когда он выставляет
задний конец из норки, чтобы выбросить
очередную порцию песка, прошедшего че-
рез кишечник. Треска, навага и другие
рыбы хватают в этот момент пескожила за
хвост в надежде вытащить его из норы. Но
он упирается щетинками в песчаные стен-
ки норы, хвост обрывается и остается
в пасти рыбы, а пескожил ускользает
в нору (рис. 265). Через некоторое время
у спасшегося пескожила отрастает новый
хвост.
Так возникшая в процессе эволюции
цепь взаимных приспособлений позволяет
пескожилу остаться живым, а рыбе —
сытой.
У полихет, обитающих открыто на по-
верхности грунта, развиты специальные
щетинки, зазубренные, как гарпуны. Они
легко входят в тело врага и обламываются.
Такие щетинки-гарпуны часто бывают ядо-
виты; они хорошо защищают животное.
Особенно характерны гарпунообразные ще-
тинки для полихет семейства Amphi-
nomidae, представители которого распро-
странены в тропических морях.
475
Рис. 265. Пескожил жертвует хвостом, спасая себе жизнь.
Некоторые полихеты больно кусаются
своими челюстями. В странах, где развито
морское и спортивное рыболовство, обыч-
ной наживкой рыбакам служат полихеты.
Их добывают в больших количествах заго-
товители, чтобы продать рыбакам на рын-
ке. Специалисты-заготовители во время
отлива ходят по колено в иле на морском
берегу и руками нащупывают крупных
червей — глицерид и других полихет.
Если неосторожно схватить глицериду,
она может больно укусить. К концу ра-
бочего дня руки ловцов сильно опухают
от укусов, а пальцы перестают сги-
баться.
Особым видом защиты у полихет явля-
ется свечение, широко развитое у мор-
ских животных. Некоторые светящиеся
в темноте полихеты вспыхивают особенно
ярко, когда их потревожат. Яркая вспыш-
ка света отпугивает врагов. Но враги
бывают разные. Некоторых, напротив,
привлекает яркое свечение жертвы. На-
пример, крабы охотно поедают некоторых
светящихся афродитид. А рыбы извлекают
из трубок светящихся там Chaetopterus va-
riopedatus (см. табл. 24). Свечение поли-
хет и других животных привлекает внима-
ние электриков и энергетиков. Живое све-
чение в 4—5 раз эффективнее люмине-
476
сцентных электроламп и стоит вне всякой
конкуренции с ними по всем показателям.
Человек должен научиться у природы
создавать столь экономные и яркие све-
тильники.
Покровительственная окраска тоже за-
щищает полихет. Пятнистая расцветка
спинных чешуек афродитид делает их
незаметными среди камней и водных расте-
ний. У некоторых афродитид спинные че-
шуи покрыты мелкими водорослями, сре-
ди которых шныряют микроскопические
ракушковые рачки и другие животные.
Такой «лес» на спине служит афродитидам
надежной маскировкой и позволяет не-
заметно приблизиться к жертве.
Как установил недавно Я. Д. К и р-
шенблат, у многих животных, а также
и у полихет существует взаимная сигна-
лизация. Они могут «разговаривать» меж-
ду собой и другими животными на своем,
особом, только им понятном языке. Чело-
век еще не владеет таким языком живот-
ных, но догадывается о нем и даже разби-
рает смысл «разговора». Этот «разговор»
происходит с помощью особых веществ
телергонов, которые вырабатыва-
ются кожными железами и выделяются
в воду. Одни из этих удивительных веществ
возбуждают и привлекают особей про-
тивоположного пола. Благодаря таким
веществам самцы быстро находят самок.
Другие вещества имеют неприятный запах
и отпугивают врагов. Иные же возбуж-
дают тревогу и рефлекс бегства. Появле-
ние таких веществ в воде равносильно
сигналу опасности. Тончайшие оттенки
сигналов полихеты улавливают особыми
чувствительными органами различного
строения. Удаление этих органов у Nereis
приводило к тому, что полихета не могла
находить пищу и становилась беспомощ-
ной жертвой врагов.
Размножаются полихеты бесполым и по-
ловым путем. Иногда оба эти способа
размножения правильно следуют один за
другим.
Бесполое размножение
у полихет сопровождается делением тела
червя на части. Отделившаяся часть тела
восстанавливает недостающие голову или
хвост. Бесполое размножение у полихет
обычно связано с явлением эпитокии.
Эпитокия — это резкое изменение внеш-
ней формы и внутреннего строения тела
полихеты во время созревания половых
продуктов. Морфология червя, обитаю-
щего на дне, меняется так, что у него
появляются приспособления к жизни
в толще воды: широкие плавательные ло-
пасти на параподиях; старые щетинки
выпадают и заменяются новыми, плава-
тельными; появляются большие глаза
и длинные чувствующие щупальца на го-
лове. При этом внешний вид меняется до
неузнаваемости. Систематики прошлого
века, введенные в заблуждение этим
маскарадом животных, дали им отдель-
ные видовые названия. Только гораздо
позже стало известно, что это не самостоя-
тельные виды животных, а всего-навсего
их эпитокные формы.
У Platynereis dumerilii донная форма
становится половозрелой, не изменяя сво-
ей организации. После размножения та-
кие формы не погибают. У них наступает
некоторый перерыв половой деятельности,
после которого они превращаются в эпи-
Рис. 266. Жизненный цикл Platyne-
reis dumerilii:
1 — исходная донная форма; 2 — полово-
зрелая атокная форма; 3 — подросшая аток-
ная форма; 4 — малая гетеронереидная фор-
ма; 5 — большая атокная форма; в — боль-
шая гетеронереидная форма; 7 — личинка.
477
Рис. 267. Охота на палоло.
токные формы и всплывают на поверхность
моря. Здесь они снова размножаются.
Цикл размножения может быть осложнен
наличием двух эпитокных форм — боль-
шой и малой (рис. 266). В исключитель-
ных условиях у Platynereis dumerilii
может развиваться гермафродитизм. Тог-
да одна и та же особь может попеременно
быть то самцом, то самкой. Эпитокные
формы полихет появляются на поверхно-
сти моря в определенное время года. Мас-
совое появление размножающихся эпи-
токных полихет обычно бывает связано
с определенными фазами луны. Напри-
мер, тихоокеанский палоло (Eunice viri-
dis), который живет на коралловых ри-
фах у островов Фиджи и Самоа, подни-
мается к поверхности в октябре или нояб-
ре в день новолуния. Палоло появляются
в таких количествах, что вода становится
почти непрозрачной. Передний конец па-
лоло остается на дне, а наиболее крупная
эпитокная задняя часть тела, длиной око-
ло 40 см, набитая зрелыми половыми клет-
ками, всплывает на поверхность. Мест-
ное население островов Фиджи с нетерпе-
нием ждет времени размножения пало-
ло. Сотни лодок устремляются в это время
в море. Начиненные «икрой» палоло счи-
таются лакомством в сушеном и жареном
виде (рис. 267). Атлантический палоло
(Eunice fucata) живет у Антильских ост-
ровов и массами поднимается на поверх-
ность также в новолуние, но в июле. Бело-
морский эпитокный Nereis virens длиной
до 60 см. Эта нереида также поднимается
на поверхность для размножения в июле
478
в новолуние. Игра нереид при лунном
свете представляет захватывающее зре-
лище, от которого часами невозможно
оторвать взгляда. Палоло и нереиды выме-
тывают половые продукты в воду, а сами
погибают. Гибель происходит потому, что
половые продукты могут выходить наружу
только через разрывы тела. Каждый сег-
мент рвется на боковой стороне. Из этих
тяжелых ран вытекают яйцеклетки и спер-
матозоиды. В воде происходит оплодо-
творение. Оставшиеся шкурки, обреченные
на гибель, становятся легкой добычей
многих рыб. Чайки и морские утки в это
время также питаются эпитокными поли-
хетами.
Бесполое размножение у Syllidae при-
водит к образованию особых почек, кото-
рые превращаются в новые особи, отры-
ваются от материнского организма и ведут
самостоятельный образ жизни. У Myri-
anida на заднем конце тела образуется
длинная цепочка таких почек. Силлиды
Autolytus имеют бесполое размножение,
которое сопровождается эпитокией особо-
го рода. У взрослой Autolytus, обитаю-
щей на дне среди гидроидов и губок, в се-
редине тела формируется новая голова
с глазами и головными щупальцами. Ког-
да формирование новой головы закончено,
эта часть тела отрывается и всплывает на
поверхность. Всплывшие особи ярко окра-
шены в желтый, красный или фиоле-
товый цвет и обладают половым димор-
физмом. Самцы отличаются от самок раз-
мером, окраской и длиной усов. Самка
носит на брюшной стороне сумку с дете-
нышами и старательно прикрывает их
своим телом. Заботясь о потомстве, роди-
тели не питаются, потому что у них нет
кишечника. Они живут, используя запасы
энергии своих мышц, которых хватает на
две или три недели. За это время потомст-
во успевает подрасти и покинуть мате-
ринскую сумку, а родители после этого
погибают.
Основной способ размножения у поли-
хет — половое размножение.
Оплодотворенные яйцеклетки проходят
свое развитие свободно в толще воды или
в специальных капсулах. Только в исклю-
чительных случаях у полихет встречается
живорождение. Тогда яйца развиваются
в полости тела (Syllis, Vivipara, Dodeca-
ceria).
Полихеты откладывают разное количе-
ство яиц. Пескожил Arenicola marina
в Белом море выбрасывает в воду до мил-
лиона яиц, тогда как средняя плотность
взрослых пескожилов составляет всего
несколько десятков особей на квадратный
метр морского дна. Из соотношения этих
цифр ясно, какое огромное количество
яиц и молодых животных погибает, не
достигнув половой зрелости. Плодови-
тость нереид несколько ниже. Например,
черноморские Platynereis dumerilii выбра-
сывают в воду до 37 тыс. яиц. Полихеты,
которые проявляют заботу о потомстве,
откладывают несколько сотен или всего
несколько десятков яиц. Например, спио-
ниды откладывают в специальную вывод-
ковую капсулу несколько десятков опло-
дотворенных яиц, которые развиваются
с разной скоростью. Личинки, опережаю-
щие в развитии своих меньших собратьев,
Рис. 268. Роторная подводная лодка
и трохофора.
479
начинают пожирать их и растут еще бы-
стрее. Поэтому в живых остается несколь-
ко самых крупных и прожорливых особей.
Наиболее часто забота о потомстве наблю-
дается у Syllidae. Например, Sphaerosyl-
lis прикрепляет яйцеклетки к себе на спи-
ну и носит их продолжительное время,
пока не появится молодое поколение.
Некоторые нереиды откладывают яйце-
клетки в свои норки. После того как яйца
отложены, самка покидает норку и в ней
поселяется самец, который охраняет яйца.
Волнообразными движениями своего тела
он обеспечивает потомство постоянным
притоком свежей воды.
Так или иначе развивающиеся яйце-
клетки претерпевают сложный путь пре-
образований, который называют метамор-
фозом. Полихеты в своем развитии про-
ходят весьма характерную для них личи-
ночную стадию трохофоры (табл. 20).
Трохофоры полихет имеют специальные
приспособления, которые сохраняются
только на время их личиночной жизни,
например пояски ресничек, с помощью
которых личинка плавает. У трохофоры
может быть один, два или несколько рес-
ничных поясов. Реснички работают так,
что вдоль всего ресничного пояса пробе-
гает волна, увлекающая трохофору
вперед. Движение трохофор чрезвычайно
маневренно. Недавно в США предложена
новая модель подводной лодки, по прин-
ципу действия двигателя напоминающая
трохофору (рис. 268). В корпусе лодки
установлены два так называемых роторных
двигателя, расположенных на носу и кор-
ме. Диаметр роторов равен диаметру кор-
пуса лодки. Роторы могут вращаться в од-
ном и в противоположных направлениях,
а их лопасти могут поворачиваться. Из-
меняя положение лопастей, можно заста-
вить лодку двигаться боком, вверх и вниз
при сохранении горизонтального или вер-
тикального положения. Этот подводный
корабль гораздо маневреннее обычных
подводных лодок и обходится без верти-
кальных и горизонтальных рулей.
Моделированием машин и устройств,
принципы которых взяты из природы,
занимается специальная наука — био-
ника.
Некоторые трохофоры малоподвижны
и лишены обычных ресничных поясков.
Скорость их движения столь мала, что
они не могут ускользнуть от многочислен-
ных врагов — медуз, мальков рыб и про-
чих хищников. У них вместо ресничек
развиваются пучки очень длинных вееро-
образно расставленных щетинок. Щетинки
создают большую площадь опоры и по-
стоянно поддерживают личинку в верх-
Таблица 24. Светящиеся морские кольчецы:
1 — краб, поедающий светящуюся
Aphroditidae B);
3 — угорь, извлекающий из трубки Chaetopte-
rus D);
4 — светящийся Chaetopterus в норке;
5 — светящийся Pterosyllis.
них горизонтах моря. Одновременно пучки
длинных, остро зазубренных щетинок хо-
рошо защищают личинку от врагов. Та-
кие личинки похожи на пушинки одуван-
чика, несущиеся по ветру на большие рас-
стояния. Например, личинка митра-
р и я, названная так за сходство с шап-
кой-митрой, может несколько месяцев на-
ходиться в толще воды и переноситься
морскими течениями на огромные расстоя-
ния. Некоторые митрарии без всякого тру-
да переплывают Атлантический океан от
американских берегов к берегам Европы.
Питаются личинки очень мелкими одно-
клеточными водорослями, пользуясь раз-
личными ловчими приспособлениями. Од-
ни движутся с достаточной скоростью,
проходя значительные пространства и
встречая на пути необходимое количество
пищи. Другие имеют небольшую скорость
движения и поэтому реже могут встре-
титься со своей жертвой. Но у них раз-
виты длинные ловчие щупальца, которые
изгибаются в разные стороны и тщатель-
но ощупывают пространство. Таковы, на-
пример, личинки Amphinomidae. За длин-
ный нос их называют рострариями.
(Рострария — значит носатая.)
У них, как и у митрарии, развиты пучки
длинных щетинок, помогающих им по-
движно парить в воде. Щупальца их снаб-
жены продольными ресничными борозд-
ками. Реснички создают ток, направлен-
ный прямо в рот. Если щупальце ощутит
поблизости одноклеточную водоросль или
другой объект питания, реснички начи-
нают усиленно работать, и водоросль по
борозде неизбежно попадает в рот. Если
водорослей в окружающем пространстве
много, то личинки без труда набивают
свой желудок пищей.
Многие личинки — хищники. Они пи-
таются мелкими рачками, а при случае
глотают и своих зазевавшихся собратьев.
Рот таких личинок очень велик и может
сильно растягиваться. Хищная личинка
с таким ртом не смущается большим раз-
мером своей жертвы и буквально силой на-
тягивает себя на лакомую пищу. Так по-
ступает личинка Chaetopterus. За огром-
ный рот ее называют стомарией,
что в переводе с латинского языка озна-
чает ротастая (см. табл. 20).
Питаясь, трохофора растет и в своем
развитии проходит ряд стадий. В про-
цессе превращения личинки происходит
замена прежних личиночных органов но-
выми, свойственными взрослому живот-
ному.
После стадии трохофоры следует новая
стадия — метатрохофоры. (Слово «мета»
означает «следующий за чем-либо».) На
этой стадии появляются первые сегменты
тела. Число их ограниченно и совершенно
определенно для каждого семейства. У ме-
татрохофор семейства Aphroditidae закла-
дывается чаще всего 7 сегментов. Для ли-
чинок семейства Nereidae характер-
ны 4 сегмента. У Capitellidae одновремен-
но закладывается 13 личиночных сег-
ментов.
В процессе индивидуального развития
могут повторяться отдельные детали про-
цесса исторического развития. Этот закон
биологии дает основание думать, что пред-
ки полихет сначала имели сравнительно
небольшое число сегментов. Вспомним,
что многие первичные кольчецы также
имеют небольшое число сегментов.
Следующая стадия развития носит на-
звание нектохета. (В переводе сло-
во «нектохета» означает «плавающий с по-
мощью щетинок».) Личинка продолжает
жить в толще воды. Она уже имеет при-
знаки взрослого животного, хотя частич-
но сохраняет и личиночные признаки.
Например, движение уже только частично
осуществляется с помощью ресничных по-
ясков. В это время начинают вступать
в строй параподии. Питаются нектохеты
по-прежнему мелкими растениями и жи-
вотными. Активное питание личинок по-
зволяет им накопить некоторое количе-
ство жира в стенке кишечника. Жир лег-
че воды и, как поплавок, поддерживает
личинку в верхних слоях моря.
Постепенно у личинки формируются но-
вые органы, характерные для взрослого
животного, например: глотка с челюстями,
новые пучки щетинок и вторичная полость
тела. Пока происходит формирование но-
вой глотки, личинка временно не может
питаться, так как зачаток глотки не мо-
жет функционировать. В этот период
расходуется запас жира. Жир, играю-
щий роль поплавка, исчезает и больше
31 Жизнь животных, т. 1
481
не удерживает личинку в толще воды.
Тогда она опускается на дно.
Некоторые личинки, опускаясь на дно,
могут подыскивать себе подходящий суб-
страт. Они охотно оседают на те участки
дна, где живут взрослые особи того же
вида. С меньшей поспешностью личинки
оседают на участки дна, на которых та-
кие особи отсутствуют и которые вообще
населены какими-нибудь растениями
и животными. Эксперимент показал, что
личинки неделями и месяцами не оседают
на дно, искусственно лишенное жизни.
Несомненно, что личинки обладают спе-
циальными приспособлениями, которые
позволяют им воспринимать тончайшие
запахи. Ориентируясь на эти запахи, ли-
чинки распознают на расстоянии пригод-
ность тех или иных участков морского
дна для жизни (см. стр. 477 — о телер-
гонах).
После оседания наступает период, в те-
чение которого образуются остальные сег-
менты. Однако на всю жизнь сохраняется
разница между первыми, личиночными
сегментами, возникшими еще в толще
воды, и всеми остальными сегментами,
возникающими позже на дне. Передние
личиночные сегменты никогда не несут
половых желез.
Продолжительность жизни у полихет
невелика. Обычно они живут 2—3 года.
Многие полихеты после размножения по-
гибают, так как вывод половых продук-
тов у них связан со значительными раз-
рывами стенок тела.
На всех стадиях развития полихеты
в больших количествах поедаются раз-
личными рыбами. Калорийность полихет
весьма высока. Например, 1 г сухого
вещества Nereis diversicolor содержит 5578
калорий, Nereis virens — 4494 калории,
Arenicola marina — 2552 калории.
Полихеты не имеют скелета и могут
усваиваться в кишечнике других живот-
ных почти полностью. Рыбы добывают
полихет сравнительно легко. Все это де-
лает полихет излюбленной пищей морских
рыб. Конечно, наибольшую роль в пита-
нии рыб играют массовые виды полихет.
В период размножения полихет и появле-
ния их на поверхности моря многие рыбы
переходят почти исключительно на пита-
ние одними только эпитокными полихета-
ми. В этот период ими питаются треска,
навага, камбала, нерка, корюшка и даже
сельдь, которая обычно употребляет в пи-
щу только мелкий планктон.
В целях усиления кормовой базы для
осетровых рыб Каспийского моря в СССР
впервые в мире была проведена акклима-
тизация азовской нереиды— Nereis diver-
sicolor. Специальным транспортом в хо-
лодильниках было переброшено несколько
десятков тысяч нереид. В Каспийском
море вселенцы отлично прижились, раз-
множились и распространились на огром-
ные пространства морского дна. Теперь
общая масса нереид в Каспийском море
составляет миллионы тонн в живом весе.
Ценнейшие каспийские породы осетро-
вых рыб, и в первую очередь осетры, пере-
шли на питание нереидами и другими все-
ленными в Каспийское море кормовыми
беспозвоночными животными. Таким об-
разом, значение полихет как кормовых
объектов рыб очень велико. В Японии
нереида Nereis japonica служит земле-
удобрительным туком.
Лишь в исключительных случаях поли-
хеты приносят вред. Например, некоторые
нереиды, хорошо переносящие опресне-
ние, могут через устья рек проникать
из моря на рисовые поля. Здесь они под-
грызают молодые ростки риса и портят
всходы. Такие случаи известны на рисо-
вых плантациях в Японии и Китае.
Вред иного характера приносят серпу-
лиды. Эти мелкие, но очень многочислен-
ные полихеты имеют крепкие известковые
домики, которые прикрепляются к под-
водным предметам. Когда личинки серпу-
лид массами оседают на днищах судов,
они вместе с другими животными и расте-
ниями-обрастателями образуют шерша-
вый живой слой, который значительно
снижает скорость движения морских ко-
раблей.
В ископаемом состоянии морские коль-
чецы встречаются не так часто, как мол-
люски, иглокожие и другие беспозвоноч-
ные с хорошо развитым скелетом. Прав-
да, известковые трубки полихет серпулид,
прикрепленные на кораллах, раковинах
брахиопод и моллюсков, встречаются в ис-
копаемом состоянии постоянно. Иногда
482
известковые трубки серпулид об-
разуют значительные скопления.
Например, в меловых отложениях
Германии известковые породы сер-
пулиды залегают слоем мощностью
до 50 м. Эти породы сплошь со-
стоят из трубок морских серпу-
лид.
От червей сохраняются не толь-
ко их трубки. В ископаемом со-
стоянии изредка находят челюсти
(сколекодонты), щетинки, а чаще
следы ползания, ходы и норы коль-
чецов.
В 1947 г. в Южной Австралии
были найдены отпечатки мягких
частей морских кольчецов, а так-
же медуз и других беспозвоноч-
ных. Они пролежали в земле не
менее 600 млн. лет. Это самые
древние ископаемые остатки жи-
вотных, известные палеонтологам.
Совершенно исключительные
условия нужны были для того,
чтобы силы природы так долго не кос-
нулись этих отпечатков. Нужны были
также определенные стечения обстоя-
тельств, чтобы отпечатки эти оказа-
лись в поле зрения специалистов. Со-
хранность отпечатков оказалась отлич-
ной. В некоторых отпечатках удалось
узнать животных, близких к тем, которые
живут в морях и теперь. Другие отпечат-
ки оказались столь странного вида, что
в них не удалось обнаружить сходства
с какими-либо современными животными.
Видимо, они должны быть отнесены к но-
вым, неизвестным еще типам животных.
Наиболее интересной находкой среди
найденных в Австралии были мелкие чер-
ви величиной около 4,5 см. Они были
названы Spriggino floundersi (рис. 269).
Это отпечатки кольчецов с узким гиб-
ким телом. Тело состоит из сегментов.
Каждый сегмент несет параподии, которые
оканчиваются игловидными шипами. Го-
лова имеет форму подковы. От острых кон-
цов подковы отходят назад длинные тон-
кие нити. Отпечатки чрезвычайно похожи
на современных пелагических полихет
рода Tomopteris (табл. 19 и 25).
Кроме пелагических кольчецов, в древ-
нейших австралийских пластах найдены
отпечатки многих других пела-
гических животных. Это позволя-
ет думать, что древнейшая морская
фауна состояла из животных, на-
селяющих толщу воды и лишен-
ных скелетных образований.
В современной фауне известно
около 50 семейств полихет. Мы
рассмотрим наиболее интересные,
массовые и практически важные
группы.
Семейство Phyllodocidae.
Представители этого семейства в
больших количествах встречаются
в морях. Название семейства за-
имствовано из мифологии. Фил-
лодока была морской нимфой,
дочерью Нерея и Дориды. Мор-
ские нимфы, по преданию, по-
стоянно сопровождали морского
царя Нептуна. В самом деле, эти
полихеты очень характерны во всех
морях мира. Они имеют тонкое
длинное тело, число сегментов
которого иногда достигает 700, Хорошо
обособленная голова несет 4 или 5 го-
ловных усиков. Глотка у полихет очень
длинная, похожая на хобот п может
выбрасываться далеко вперед. Параподии
обычно одноветвистые с одним пучком
сложных щетинок. Окрашены филлодо-
циды обычно в яркие цвета зеленого или
оранжевого тона. Кровь бесцветна. Ды-
хательный пигмент — гемоглобин — за-
ключей в нервной системе.
Филлодоциди откладывают слизистые
шарообразные кладки, в которых первое
время развиваются личинки. Затем они
прорывают слизистую оболочку и в огром-
ных количествах выходят в толщу воды.
В каждом кубическом метре морской во-
ды в это время находится несколько десят-
ков тысяч личинок только одного этого
семейства. Личинки очень подвижны
и прожорливы. Они поедают мелких рач-
ков, которые часто больших размеров, чем
сами личинки. Личинки с трудом натяги-
вают себя на рачка и на некоторое время
принимают его очертания, Трохофора
становится метатрохофорой, когда появ-
ляется личиночная сегментация (см. табл.
20). Число первых сегментов обычно не
меньше семи. Остальные сегменты появ-
Рис. 269. Отпеча-
ток древнего пе-
лагического червя
Spriggino flounder-
si, найденный в
докембрийских от-
ложениях Австра
лии.
31*
483
ляются только после оседания личинки
на дно.
Некоторые представители филлодоцид
на всех стадиях развития живут в толще
воды. Иногда они имеют небольшое число
сегментов и сохраняют в течение всей
жизни личиночные признаки. Таковы, на-
пример, Maupasia и Pedinosoma.
Семейство Alciopidae. Исключи-
тельно пелагические полихеты. Встреча-
ются в поверхностных слоях моря вдали
от берегов. Голова с пятью усиками.
Огромные глаза имеют хрусталик, спо-
собный к аккомодации. Такими глазами
хищные алъциопы могут отличать свою
жертву от врага. Длинная глотка выбра-
сывается далеко вперед. Хватательные
придатки на конце глотки способны цепко
схватывать жертву. Параподии одновет-
вистые. Личиночное развитие альциопы
проходят в толще воды.
Семейство Tomopteridae. Исклю-
чительно пелагические полихеты. Живут
в открытых частях морей и океанов. Они
свободно переносятся морскими течения-
ми на огромные пространства. Латинское
название семейства означает, что они,
как птицы на крыльях, свободно носятся
в океане. Тело томоптер прозрачно, как
стекло. Это скрывает их в воде от много-
численных врагов и позволяет им неза-
метно подкрадываться к малькам рыб и
другим животным, которыми они пита-
ются. На подковообразной голове развиты
нитевидные усы, длина которых превос-
ходит длину тела. Две длинные щетинки
значительно увеличивают площадь опоры
животного и не дают ему опускаться на
дно. Вместо обычных пучков щетинок
на боках сегментов развиты широкие ло-
пасти, похожие на весла. Весь цикл раз-
вития протекает в толще воды.
Некоторые представители пелагических
полихет из других семейств показаны на
таблице 25.
Найденные в древнейших породах Авст-
ралии отпечатки полихет напоминают со-
временные формы томоптер (рис. 269 и
и табл. 19). Удивительно, что приспособ-
ления к пелагическому образу жизни у то-
моптер развиты так, что они почти не из-
менились в течение сотен миллионов лет.
Приспособления некоторых древних форм
к определенной среде обитания обеспечи-
вали им выживание.
Семейство Aphroditidae. Широко
распространенные донные морские поли-
хеты. Название семейства заимствовано из
мифологии. Афродита — богиня красоты,
покровительница моряков, возникла из мор-
ской пены. Полихеты этого семейства дей-
ствительно очень красивы. Щетинки у не-
которых из них ярко переливают всеми цве-
тами радуги (см. табл. 19). Тело их плос-
кое и относительно короткое. Голова снаб-
жена тремя — пятью щупальцами. Креп-
кая, мускулистая глотка с силой выбра-
сывается вперед. На конце глотки располо-
жены 4 челюсти, приспособленные для за-
хвата жертвы. Питаются афродитиды мол-
люсками и другими животными. На спине
у них двумя рядами располагаются круг-
лые чешуи, которые защищают и маски-
руют животное. На этих чешуях посе-
ляются мелкие водоросли, а на них—
масса мелких животных. Такой естествен-
ный «лес» позволяет хищным афродитидам
незаметно приближаться к своим жертвам
и маскироваться от врагов. Некоторые
из афродитид в темноте излучают свет
и таким образом пытаются отпугивать вра-
гов (см. табл. 24). Иные же в момент не-
минуемой опасности сбрасывают спинные
чешуйки, а сами убегают. Через некоторое
время чешуйки вырастают снова.
Многие представители этого семейства
живут совместно с другими животными.
Некоторые афродитиды живут в домиках
других полихет, например в трубках Chae-
topterus. Иные живут внутри тела гу-
бок, где хорошо защищены и всегда
обеспечены пищей. Здесь они могут ла-
комиться добычей, которую собирают для
себя губки, или даже откусить кусочек
самой губки, от чего гостеприимная хо-
зяйка не страдает. Другие афродитиды
живут между многочисленными ножками
морских звезд. Морская звезда прикрыва-
ет гостью своим телом и предоставляет
ей остатки от своего стола. Отдельные
виды афродитид ведут пелагический об-
раз жизни, например Nectochaeta gri-
maldii. Афродитиды используют спинные
чешуи в виде инкубатора для выведения
потомства. Они откладывают под чешуей
яйца и носят их там первое время. Из яиц
484
выводятся трохофоры, которые вылезают
из-под чешуи и уплывают (см. табл. 20).
Ранней весной они в огромном количест-
ве появляются в морях умеренной зоны
и служат излюбленной пищей для рыб и их
личинок.
Семейство Syllidae. Сравнитель-
но мелкие и ярко окрашенные полихеты.
Они обитают на дне среди густых зарослей
губок, гидроидов, мшанок и других жи-
вотных. Некоторые виды силлид питаются
гидроидами, которых высасывают спе-
циально приспособленным хоботом. Хо-
рошо обособленная голова снабжена гла-
зами и чувствительными придатками.
Способы размножения у силлид весьма
разнообразны. У некоторых форм встре-
чается бесполое размножение. Например,
у Syllis ramosa, паразитирующей внутри
губок, почки новых особей образуются
на всем теле. Из одних почек образуются
силлиды обычного типа, похожие на ма-
теринскую особь. Другие почки дают
начало половым особям. Из каждой поч-
ки образуется сначала только задний ко-
нец. В результате возникает очень сложно
разветвленная колония с одной головой
и множеством задних концов, в которые
входят ветви кишечника. Такая колония
живет внутри тела губки и питается ее
тканями. Паразитический способ пита-
ния позволяет этой силлиде одним своим
ртом прокормить многочисленных детей,
пока еще не имеющих собственной головы.
У Trypanosyllis почкование сосредоточено
близ заднего конца материнской особи.
Здесь образуется целый пучок половых
особей разного возраста. Старшие и наи-
более зрелые постепенно отшнуровыва-
ются и уплывают. Материнская особь не
имеет пола и всегда остается на дне.
У Autolytus и Myrianida на заднем конце
возникают цепочки молодых особей. Пе-
редняя материнская особь здесь также не
имеет определенного пола. Зато молодые
особи становятся половыми. Отделив-
шись, они уплывают в верхние слои
воды для свершения брачного танца.
Самцы имеют большие разветвленные
усы, огромные черные глаза и длин-
ные плавательные щетинки. Они стре-
мительно кружатся около более скромных
самок (рис. 270). На брюшке самки нахо-
Рис. 270. Брачный танец Autolytus (Syllidae):
наверху — самка с сумкой, наполненной яйцами;
внизу — кружащийся около нее самец.
дится большая выводковая сумка, в кото-
рой помещается несколько сотен яйцекле-
ток. Своим телом самка защищает потом-
ство, пока оно находится в сумке. Чаще
всего силлиды проходят обычные стадии
трохофоры и метатрохофоры под прикры-
тием материнского тела внутри выводковой
сумки. Когда молодые силлиды оконча-
тельно сформируются для полноценной
жизни на дне, они прорывают стенку
сумки, выходят из нее и опускаются на
дно. К этому времени силы материнской
особи совершенно истощаются и она по-
гибает.
Семейство Nereidae. Название это-
го семейства также заимствовано из гре-
ческой мифологии. От брака бога Нерея
с дочерью Океана Доридой родились 50 не-
реид — веселых морских нимф. С игра-
ми и танцами они повсюду сопровождали
властелина моря Нептуна, который путе-
шествовал по океану на дельфине.
Обширное семейство нереид пред-
ставлено во всех морях мира многочислен-
ными мелкими и крупными формами. Тело
нереид чаще всего окрашено в зеленые
тона и отливает всеми цветами радуги.
Нереиды живут в норах, которые роют
в иле и песке. К тому времени, когда
у нереид начинают созревать половые про-
485
дукты, внешний вид этих полихет резко
изменяется. Обе лопасти параподий вес-
лообразно расширяются. Обычные щетин-
ки заменяются плавательными с широ-
кими и плоскими конечными члениками.
Сегменты тела тоже становятся шире. Та-
кие особи приобретают способность пла-
вать и переходят к планктонному образу
жизни. Для размножения многие нереиды
массами поднимаются со дна на поверх-
ность моря, где совершают свой брачный
танец. Самки, змеевидно изгибаясь, плы-
вут у самой поверхности, а самцы кружат-
ся вокруг них. В это время они стано-
вятся легкой добычей рыб и птиц, кото-
рые собираются большими стаями, чтобы
полакомиться икряными нереидами.
Из оплодотворенных яиц развиваются
трохофоры, тело которых перепоясано
четырьмя ресничными кольцами. Трохо-
форы самостоятельно добывают пищу и бы-
стро совершают свой метаморфоз (см.
табл. 20). Через две-три недели после опло-
дотворения новое поколение нереид осе-
дает на дно моря.
Семейство Eunicidae. Длинные,
красивые полихеты, обитающие на дне.
Некоторые из них живут в трубках, кото-
рые они сами делают из песчинок, оскол-
ков раковинок моллюсков и другого строи-
тельного материала. В связи с необходи-
мостью более интенсивного дыхания в
норках у них на параподиях образуются
перистые жабры. Глотка эвницид снабжена
крепкими челюстями. Верхняя челюсть
состоит из нескольких мелко зазубрен-
ных пластинок. Роговые или хитиновые
пластинки челюстей очень крепки и слу-
жат для откусывания и измельчения ку-
сочков растений, а иногда и животных.
Челюстные пластинки у каждого вида
эвницид имеют свою специфичную форму.
Опытные специалисты могут только по
одним челюстям определить вид эвници-
ды. Челюсти эвницид — сколеко-
д о н т ы — сохраняются в ископаемом со-
стоянии в течение многих миллионов лет.
Хитин — очень прочное вещество. Это
вещество не поддается действию времени.
Изучение челюстей ископаемых эвницид,
пролежавших в земле более 350 млн. лет,
показало, что с того времени эвнициды
мало изменились. Челюсти ископаемых
и современных эвницид имеют очень боль-
шое сходство. Этот факт позволяет нам
представить себе, как медленно в отдель-
ных случаях совершаются морфологиче-
ские изменения в процессе эволюции. Ос-
новная масса современных эвницид оби-
тает на дне моря в норках или трубках.
В исключительных случаях эвнициды пе-
реходят к паразитическому образу жизни.
Некоторые из них паразитируют в поло-
сти тела уже знакомых нам силлид, а так-
же спионид, теребеллид и эхиурид. Все
эти паразитические эвнициды, конечно,
очень мелкие формы, размер которых не
превышает 1 см. Организация их в связи
с паразитическим образом жизни значи-
тельно упрощается. О способах размноже-
ния эвницид мы уже говорили. Знамени-
тый червь пололо, на которого охотятся
жители тихоокеанских островов Фиджи
и Самоа, принадлежит к этому семейству.
Семейство Spionidae. Спиониды —
мелкие полихеты, преимущественно оби-
тающие на дне. На голове у спионид нахо-
дятся глаза и два очень длинных и под-
вижных усика. Этими усиками спиониды
ощупывают вокруг себя пространство и от-
лавливают годные в пищу частицы. На го-
лове и на нескольких следующих сегмен-
тах тела находится спинной орган чувств.
Этот чувствительный орган воспринимает
информацию от веществ — телергонов (см.
стр. 477). Спинной орган чувств имеет вид
округлых или вытянутых углублений, на
дне которых специальные клетки с нерв-
ными окончаниями воспринимают тончай-
шие оттенки запахов, распространенных
в воде. Реагируя на раздражения, спио-
ниды или выходят из своих норок, или еще
глубже забираются в них. Некоторые спио-
ниды сверлят раковины моллюсков. Ли-
чинка спионид в этом случае находит себе
подходящего моллюска, оседает на его
раковину и просверливает в ней ход.
Спрятавшись в просверленном ходе, моло-
дая спионида выставляет наружу щупаль-
ца и начинает сотрапезничать со своим
хозяином. Если подходящего места для
оседания личинка не находит, она надол-
го может задержаться в толще воды. Изве-
стны случаи, когда спиониды так и оста-
ются жить в толще воды. Например, ли-
чинка Prionospio может достигать полово-
486
зрелого состояния и размножаться, не опу-
скаясь на дно. Личинки спионид удержи-
ваются во взвешенном состоянии с помо-
щью длинных волосовидных щетинок и
и многочисленных ресничных поясов
(см. табл. 20). Личинки прожорливы
и часто нападают друг на друга, пое-
дая своих братьев и близких родствен-
ников. Плотность поселений спионид, осо-
бенно в прибрежной зоне, бывает огром-
ной и может достигать нескольких тысяч
особей на 1 м2.
Семейство Arenicolidae. Пескожи-
лы — крупные черви, вся морфология ко-
торых приспособлена к жизни в песке.
Название Arenicola происходит от слова
«арена», что значит «песок». Площади ам-
фитеатра, а иногда и цирковые арены по-
сыпают песком. В русском языке слово
«арена» употребляют только для обозна-
чения места цирковых представлений. Мы
уже рассказали, что пескожилы делают
в песке ходы и живут в них, заглатывая
и пропуская через свой кишечник грунт.
На плоских песчаных отмелях морского
дна обычно встречаются обширные про-
странства, занятые поселениями песко-
жилов. На таком берегу во время отлива
видны сотни тысяч ловчих воронок и ко-
нических возвышений песка, выброшен-
ного пескожилами. Среднего размера взро-
слый пескожил выбрасывает около 40 г
грунта в сутки. На каждом квадратном
метре морского песчаного берега обычно
можно встретить в среднем 40 пескожи-
лов. Значит, они в сутки выбросят более
1,5 кг песчаного грунта. Таким обра-
зом, только на 1 га морского побережья
пескожилы пропускают через свои кишеч-
ники 16 т грунта в сутки. Так пескожилы
переворачивают и обновляют слой песка,
в котором живут. Пескожил считается
лучшей наживкой у рыбаков. Треска,
навага и другие морские рыбы охотно клю-
ют на пескожила. Ловля трески и наваги
на удочку очень увлекательна.
Несмотря на то что пескожилов в огром-
ных количествах истребляют, плотность
их поселений остается ежегодно почти
одинаковой из-за их огромной плодовито-
сти. В полости тела самки развивается до
миллиона яиц. Трохофоры плавают около
прибрежных водорослей, а затем опус-
каются на дно и зарываются в песок (см.
табл. 19). Личинки охотнее оседают на
площадях, уже занятых взрослыми песко-
жилами, которые привлекают их своим
запахом. Таким образом, наиболее удоб-
ные площади оказываются заселенными
гораздо более плотно.
Семейство Chaetopteridae. Xemon-
териды живут на дне в плотных роговых
трубках. Вся их организация глубоко
приспособлена к жизни в закрытом поме-
щении. Только на стадии трохофоры они
ведут свободный образ жизни в толще
воды (см. табл. 20).
Личинка хетоптеруса прожорлива и бы-
стро совершает свой метаморфоз. В толще
воды она выделяет вокруг себя тонкую
прозрачную трубку и вместе с ней оседает
на дно, превращаясь со временем во взро-
слого кольчеца. Некоторые хетоптериды
достигают 20 см в длину, а трубки их —
0,5 м и более. Трубки могут быть прямыми
или изогнутыми дугой так, что оба откры-
тых конца подходят к поверхности грун-
та. Входное отверстие трубки по мере ро-
ста червя оказывается уже, чем ее сред-
няя часть. Хетоптерус как бы замуровы-
вает себя в трубке и уже никогда не может
вылезти из нее. Дыхание и питание
хетоптеруса затруднены, так как осу-
ществляются через узкое входное отвер-
стие в его жилище. Чтобы постоянно осве-
жать воду внутри трубки, хетоптерусы по-
махивают своими параподиями, похожими
на крылья. Отсюда и происходит название
«хетоптерус», которое означает «щетино-
крыл». Но этих помахивающих движений все
же,видимо,недостаточно для полного ритма
дыхания. Усиление дыхания достигается
симбиозом хетоптерусов с одноклеточными
зелеными водорослями. Водоросли, поселив-
шиеся в трубке, выделяют на свету кисло-
род, которым дышит хетоптерус. Углеки-
слоту и продукты выделения кольчеца
водоросли употребляют в пищу. Так
хетоптерус решает проблему биологиче-
ской совместимости.
В будущих длительных космических по-
летах человек также предполагает исполь-
зовать одноклеточные водоросли для
непрерывного получения кислорода и по-
глощения углекислоты внутри тесных по-
мещений космических кораблей. В косми-
487
ческом пространстве солнце светит всег-
да. Там нет ночи. Поэтому водоросли на
свету круглосуточно будут производить
кислород и поглощать углекислоту, на-
копившуюся в ракете. Все знают, что
в космосе солнечный свет льется безудерж-
но. Но откуда же свет внутри темных тру-
бок хетоптерид, зарытых в грунт? Ради
кислорода, который им доставляют водо-
росли, хетоптерусы освещают их своим
собственным фосфорическим светом. Па-
раподии, особенно хвостовой части тела,
снабжены специальными клетками, кото-
рые выделяют на поверхность тела интен-
сивно светящееся вещество. Оно-то и осве-
щает водоросли в течение круглых суток.
Этот свет, струящийся из узких отверстий
трубок, привлекает внимание рыб, и хето-
птерусы становятся их жертвами (см.
табл. 24).
Семейство Sabellidae. Сабелли-
ды — одни из красивейших полихет. Это
обитатели трубок, передний конец кото-
рых приподнят над грунтом. Из открытого
отверстия трубки сабеллиды выставляют
головной конец с крупным ярким вен-
чиком щупалец, похожим на веер. Иногда
этот венчик свернут широкой воронкой.
Тело сабеллид спрятано в трубку, а щу-
пальца торчат наружу. Они выполняют
роль органов дыхания и одновременно
захватывают пищу. Обитателям трубок
бывает нелегко добыть себе пищу.
Покинуть трубку — значит немедленно
стать жертвой прожорливой рыбы. Си-
деть на месте тоже безрадостно: еду никто
не принесет. Сабеллиды выходят из по-
ложения, пользуясь «дождем» трупов, ко-
торый постоянно идет в крупных водое-
мах. «Дождь» трупов образуется в связи
с кратковременностью жизни большин-
ства мелких организмов, обитающих в тол-
ще воды. Отмирая, эти животные и расте-
ния медленно опускаются на дно. Полу-
чается непрерывный поток, состоящий из
отмерших одноклеточных водорослей, про-
стейших, остатков медуз, ракообразных
и других животных. Отдельные капли это-
го «дождя» падают на щупальца сабел-
лид и других подобных обитателей моря,
ожидающих своего блага с «неба». Са-
беллиды во все глаза следят за происхо-
дящим вокруг, наблюдая за хищниками,
готовыми откусить их щупальца. На каж-
дом щупальце развито по нескольку глаз,
а общее число щупалец доходит до 50 и
и более. Вполне понятно, что сотнями
глаз можно издали заметить врага и во-
время спрятаться в трубку.
Одна из сабеллид— Manayunkia — удив-
ляет своим необычным распространением
в наших морях. Она обитает в устьях се-
верных рек Америки и Евразии, а также
в предустьевых пространствах рек Азов-
ского и Каспийского морей. Но самое уди-
вительное то, что Manayunkia найдена
в озере Байкал. Такое распространение
сабеллиды доказывает, что все простран-
ство между Северным Ледовитым океаном,
Каспийским морем и озером Байкал в про-
шлые геологические эпохи было занято
огромным морем. Исчезая, это море разъ-
единило, своих обитателей тысячами кило-
метров, занятых теперь дремучими сибир-
скими лесами. Так маленькая сабеллида
помогает познать геологическую историю.
Семейство Serpulidae. Серпулиды
постоянно живут в известковых трубках.
Вынутые из трубок, они быстро поги-
бают. Крепкие, иногда массивные трубки
серпулид служат надежной защитой от
врагов. Они обычно прикреплены к рако-
винам моллюсков, брахиопод, часто встре-
чаются среди коралловых рифов. Многие
виды обрастают днища морских кораблей
и образуют весьма крупные сростки тру-
бок. Трубки могут быть расположены
в плоскости субстрата или приподняты
вертикально вверх, могут быть завернуты
в красивую спираль или змеевидно изогну-
ты в самых изящных позах. Отсюда проис-
ходит название всего семейства (serpo —
«змеевидно ползать»). Серпулиды окра-
шены в розовые, фиолетовые и красные
тона. Голова их очень мала, а вместо усов
на ней располагаются изумительной кра-
соты веерообразные жаберные лучи.
У спирографис основания лучей располо-
жены по спирали, отчего весь пучок жабр
напоминает штопор (см. табл. 19). Жабры
служат одновременно для нескольких це-
лей. С их помощью животное дышит и ло-
вит частицы, годные в пищу. Жабры
расправлены веером, и серпулиды напря-
женно ждут, когда сверху на них упадет
что-либо съедобное. Они питаются тру-
488
нами, так же как сабеллиды и другие
животные с подобным способом питания.
Жабры пестро окрашены и снабжены мно-
гочисленными глазами. Глаза серпулид
очень сложно устроены и обладают острым
зрением. Увидев тень приближающегося
хищника, они молниеносно передают сиг-
нал тревоги. Серпулида мгновенно втя-
гивается в трубку и ждет, когда хищник
снимет осаду.
Глаза некоторых серпулид напоминают
совершенные глаза ракообразных, насе-
комых и других членистоногих. Удивитель-
ны, например, многочисленные сложные
глаза на жабрах серпулиды бранхиоммы.
Шаровидный сложный глаз бранхиом-
мы состоит из многих сотен отдельных
глазков, оптические оси которых радиаль-
но направлены в разные стороны. Каждый
сложный глаз просматривает пространст-
во под углом почти 360°, т. е. почти пол-
ную сферу (рис. 270). Если учесть, что
таких сложных глаз у бранхиоммы не-
сколько десятков, то можно представить
себе, как зорко она охраняет свою непри-
ПОДКЛАСС MYZOSTOMIDA
Особое место среди многощетинковых
кольчецов занимают мизостомиды, кото-
рые паразитируют на морских лилиях,
морских звездах и офиурах. Паразитиче-
ский образ жизни всегда накладывает
резкий отпечаток на организацию живот-
ных.
Тело мизостомид уплощается и уко-
рачивается, принимая форму диска. Коль-
чатость, столь обычная для кольчецов,
утрачивается. Голова сливается с телом
и не несет сложных органов чувств. Одна-
ко принадлежность к кольчецам выдают
многие признаки. Развитие у мизостомид
протекает с трохофорой. У взрослых ми-
зостомид развиты 5 пар параподий со ще-
тинками. Есть также и остатки полости
тела.
Мизостомиды сравнительно невелики.
Они сидят на теле иглокожих и сосут
соки своих хозяев.
косновенность. Сложные, или фасеточ-
ные, глаза позволяют точно определять
направление и скорость движения объек-
тов наблюдения. По принципу строения
и действия такие глаза очень близки к не-
которым типам обзорных станций, контро-
лирующих движение спутников и ракет
разного радиуса действия. Однако миниа-
тюрность и экономичность конструкции
сложных глаз животных могут соперни-
чать с последними моделями, применяе-
мыми в технике.
У некоторых серпулид один из лучей
жаберного аппарата превращен в кры-
шечку, которая плотно закрывает вход-
ное отверстие трубки, после того как сер-
пулида спрячется в ней. Иногда специ-
альная полость внутри такой крышечки
служит выводковой камерой. В этом инку-
баторе серпулиды бережно выращивают
свое подрастающее поколение. Трохофоры
серпулид называются л и м б а р и я м и
(см. табл. 19) потому, что тело их укра-
шено воротником, похожим на модное
жабо (limbus — «воротник»).
Итак, многощетинковые кольчецы пред-
ставляют собой очень разнообразную груп-
пу животных, которая играет свою немало-
важную роль в общих биологических про-
цессах, протекающих в океане. Полихеты
составляют пищу многих полезных рыб,
входят в состав группы обрастателей мор-
ских судов, участвуют в образовании мор-
ских осадочных пород — биолитов.
Человек, насколько это возможно, исполь-
зует в своих интересах наряду с другими
и эту группу животных. Акклиматизируя
полихет в водоемах, где ведется рыбный
промысел, человек изменяет и подчи-
няет себе природу морей. Однако много
других, еще не использованных воз-
можностей таит в себе природа полихет.
Мы рассказали о новой науке — бионике,
которая со временем найдет среди поли-
хет и других животных многие объекты,
достойные быть образцом для моделиро-
489
вания. Упомянутая роторная подводная
лодка или попытки получить яркий хо-
лодный и очень экономный источник све-
та — это только первые шаги, которые
приоткрывают для фантазии ученых неис-
числимые тайны природы.
Вместе с тем изучение полихет, как
и других животных, позволяет нако-
пить огромное количество фактов. С по-
мощью этих фактов человек разгадывает
историю климата Земли прошлых геологи-
Большинство олигохет —обитатели поч-
вы: более крупные из них хорошо всем
известны под названием земляных или дож-
девых червей. Есть, однако, и пресновод-
ные и даже морские олигохеты. Из общего
числа (около 3 тыс.) описанных до сих пор
видов примерно 400 — обитатели рек,
озер, болот и других пресных водоемов,
несколько десятков видов встречается в со-
лоноватых водоемах и прибрежной зоне
морей, все остальные — обитатели почвы
(но некоторые из них, встречающиеся
преимущественно по берегам водоемов,
отчасти в воде, ведут земноводный образ
жизни). В пределах СССР пока зарегист-
рировано немногим более 300 видов; циф-
ра эта, однако, далеко не отражает дейст-
вительного положения вещей, так как
некоторые семейства, и в частности одно
из наиболее богатых видами — энхитреи-
ды, остаются у нас еще почти не изучен-
ными.
Среди олигохет есть и карлики и ги-
ганты. Длина тела самых мелких пресно-
водных измеряется долями миллиметра,
так что это в полном смысле слова микро-
скопические животные; а у наиболее круп-
ных тропических земляных червей она
превышает 2,5 м. Но это исключение.
Большинство же водных олигохет имеет
размеры от нескольких миллиметров до
10—15 см, а почвенных — от полусанти-
метра до 30—40 см.
ческих эпох, убеждается в поразительной
пластичности организмов в процессе при-
способления их к условиям обитания. Уче-
ные стремятся разгадать сложные родст-
венные отношения животных и найти сре-
ди них формы, которые могли бы служить
прообразом древних предков.
Развитие эволюционной теории имеет
огромное значение в борьбе против ре-
лигиозных утверждений о неизменяе-
мости мира.
Важнейшей особенностью организации
олигохет, как и полихет, является мета-
мерия, т. е. правильная повторяемость
органов вдоль оси тела животного. Внешне
это выражается в том, что все червеобраз-
ное тело поделено перетяжками на от-
дельные участки — кольца (рис. 271),
называемые сегментами или сомитами
(«члениками»). Число сегментов тела у раз-
ных видов олигохет различно, от 5 —6 до
500—600; в отличие от полихет они никог-
да не имеют парных выростов — парапо-
дий, но несут по 4 пучка щетинок.
На переднем конце тела находится го-
ловная, или предротовая, ло-
пасть; у некоторых водных видов она
бывает вытянута в более или менее длин-
ный щупальцевидный придаток («хоботок»),
у немногих — почти исчезла, у большин-
ства же представляет небольшой округ-
ленный выступ впереди рта. Далее сле-
дует первый, или ротовой, сегмент, ни-
когда не несущий щетинок; на его нижней
передней стороне находится рот. Все ос-
тальные сегменты, начиная со второго,
как правило, снабжены четырьмя пучками
щетинок каждый: двумя спинными (спра-
ва и слева) и двумя брюшными. У неко-
торых видов спинные пучки начинаются не
вместе с брюшными, со второго сегмента,
а несколько отступя от переднего конца
тела — четвертого, пятого, шестого и даже
двадцатого сегмента. Как исключение,
КЛАСС ОЛИГОХЕТЫ, ИЛИ МАЛОЩЕТИНКОВЫЕ
КОЛЬЧЕЦЫ (OLIGOCHAETA)
490
имеются виды лишь с брюшными щетин-
ками и даже вовсе без них.
В каждом пучке часто бывает по две
более или менее одинаковых щетинки
(например, у большинства дождевых чер-
вей). У тропических червей рода Phere-
tima число щетинок на каждом сегменте
значительно больше — несколько десят-
ков, а у некоторых видов до 150 — и рас-
положены они не пучками, а в один ряд
вокруг всего сегмента. У водных олиго-
хет, как правило, в каждом пучке по
нескольку щетинок, от 3—4 до 10—15,
причем эти щетинки бывают различной
формы: игловидные, крючковидные с про-
стым или двузубчатым концом, веерные,
простые и перистые волосовидные и др.
(рис. 272).
Стенка тела олигохет состоит из пяти
основных слоев: тонкой неклеточной ку-
тикулы, выделяемой кожей, кожного эпи-
телия, состоящего из одного слоя клеток,
двух слоев — мускулатуры наружного
кольцевого и внутреннего продольного и,
наконец, внутреннего слоя целомическо-
го эпителия, ограничивающего вторичную
полость тела, или целом, в которой рас-
полагаются внутренние органы. Эта по-
лость, заполненная более или менее
прозрачной жидкостью, содержащей взве-
шенные в ней клетки, не тянется непре-
рывно вдоль всего тела, а разделена тон-
кими мускульными перегородками — дис-
сепиментами — на отдельные отсеки, соот-
ветствующие наружной сегментации. Та-
ким образом, в каждом сегменте находит-
ся свой обособленный участок полости
тела, а в нем — пара своих выделитель-
ных органов — нефридиев, один справа,
другой слева (рис. 273). Имеется замкну-
тая кровеносная система: продольные со-
суды идут вдоль всего тела, связываю-
щие их кольцевые сосуды имеются в каж-
дом сегменте. Пищеварительный канал
проходит вдоль всего тела от ротового
отверстия до анального; обычно он под-
разделяется на отделы: ротовая полость,
глотка, пищевод, иногда зоб, один или
Рис. 273. Поперечный разрез дождевого червя:
1 — брюшной кровеносный сосуд; 2 — брюшная нерв-
ная цепочка; з — полость тела; 4 — полость кишеч-
ника; 5 — нефридий; 6 — спинной кровеносный сосуд.
Рис. 271. Различные олигохеты.
Рис. 274. Продольный разрез передней части
дождевого червя:
1 — глотка; 2 — пищевод; 3 — зоб; 4 — мускульный
желудок; 5 — средняя кишка.
Рис. 275. Схема строения половой системы Lum-
bricus:
1 — передний семеприемник; 2 — передний семенник;
S — задняя воронка семяпровода; 4 — яичник; 5 —
яйцевод; 6 — семенные пузырьки (мешки).
Рис. 277. Коконы олигохет.
несколько желудков, средняя кишка (са-
мая длинная часть), задняя кишка (рис.
274).
В каждом сегменте, в брюшной его ча-
сти, находится двойной нервный узел
(состоящий из сближенных и почти слив-
шихся правого и левого) с отходящими
от него нервами. Узлы всех сегментов
соединяются продольными нервными тя-
жами — коннективами — в еди-
ную брюшную нервную цепочку. В голов-
ной лопасти или несколько позади нее,
над глоткой, находится головной мозг
(надглоточный ганглий), соединенный с
первым узлом брюшной цепочки — под-
глоточным— двумя тяжами, охва-
тывающими глотку,— окологлоточ-
ными коннективами. Глаз у
большинства олигохет нет, они имеются
только у части видов одного семей-
ства водных олигохет — наидид; од-
нако, как правило, олигохеты реагируют
на освещение, что связано с наличием в
коже, особенно на головном конце, особых
светочувствительных клеток -фото-
рецепторов. Дыхание у подавляю-
щего большинства кожное, но у неко-
торых пресноводных олигохет имеются
жабры.
Все олигохеты гермафродиты, т. е. у
каждого взрослого червя имеются одно-
временно и мужская и женская половые
системы, строение которых довольно слож-
но (рис. 275). Оплодотворению предшест-
вует спаривание, во время которого спер-
матозоиды каждого из двух червей пере-
ходят в семеприемники другого (рис. 276).
После этого черви расходятся. Затем у
каждого из них на пояске, представляю-
щем железистое утолщение кожи несколь-
ких определенных сегментов (у разных
олигохет различных, но всегда ближе к
головному концу), выделяется кокон, пер-
воначально имеющий форму бочонка (или
муфты), открытого с обоих концов. Об-
разовавшийся кокон сокращениями му-
скулатуры тела сдвигается постепенно
вперед и наконец сбрасывается «через го-
лову», после чего концы его смыкаются и
он принимает форму лимона (рис. 277).
Но прежде чем кокон будет сброшен, по-
ка он медленно скользит от пояска к голов-
ному концу, в него откладываются яйца
492
(когда он продвигается над отверстиями
яйцеводов), а затем из семеприемников
поступают сперматозоиды, полученные при
спаривании от другого червя. Таким обра-
зом, оплодотворение яиц происходит не в
теле червя и не во внешней среде, а в
коконе, внутри которого происходит и
дальнейшее развитие. Из кокона выходят
маленькие червячки, в общем уже похо-
жие на взрослых (рис. 278). У некоторых
олигохет описано самооплодотворение, а
другие способны к партеногенезу, т. е.
девственному, без оплодотворения, раз-
витию отложенных яиц.
Кроме полового, у олигохет наблюдает-
ся и бесполое, вегетативное, размножение,
но оно свойственно представителям толь-
ко некоторых семейств водных червей. Бес-
полое размножение может происходить
либо путем архитомии, либо путем
паратомии. При архитомии тело чер-
вя самопроизвольно (иногда в результате
внешнего раздражения) распадается на
два или большее количество кусков —
фрагментов, после чего у каждого куска
развиваются недостающие части и он
постепенно превращается в целого червя.
При паратомии (рис. 279) на теле червя,
на некотором определенном расстоянии от
головного конца намечается перетяжка,
подразделяющая его на две части; после
этого задняя часть, не отделяясь сначала
от передней, образует свой головной конец,
а передняя — свой задний. Теперь два
образовавшихся таким образом червя мо-
гут разделиться окончательно, но часто,
прежде чем это произойдет, каждый из
них опять начинает делиться. Таким об-
разом возникают «цепочки», состоящие из
четырех-пяти, а иногда и большего числа
особей.
Деление класса олигохет на отряды
еще окончательно не установилось; мы
рассмотрим последовательно важнейшие
семейства.
Семейство Эолосомы (Aeolosomati-
dae) включает около полутора десятков
видов самых мелких пресноводных оли-
гохет, длина тела которых редко превы-
шает 2— Змм, а у некоторых даже мень-
ше 1 мм.Поэтому, хотя они встречаются до-
вольно часто в болотах, канавах и при-
брежных зарослях прудов, озер и рек,
обнаружить их не так легко, а хорошо
рассмотреть можно только под микроско-
пом. Зато если среди водорослей и про-
стейших в капле воды удастся заметить
одно из этих изящных животных, напри-
мер гемприхову эолосому (Aeolosoma hem-
prichi), одну из самых обычных у нас,
понаблюдать за ним стоит. Маленький
прозрачный червячок сразу узнается по
характерным оранжевым кожным желез-
кам, которые разбросаны по всей поверх-
ности тела в виде округлых пятнышек
(рис. 280). У других видов эолосом желез-
ки окрашены иначе — в желтоватый, зе-
леный или голубоватый цвета, а могут быть
и бесцветными. Число сегментов их тела
невелико — от 7 до 20, сегментация выра-
Рис. 279. Бесполое размножение (паратомия) у Nais.
Рис. 278. Вылупление молодого Tubifex из кокона.
493
Рис. 280. Aeolosoma hemprichi.
жена не резко. На каждом сегменте тела
имеется по четыре пучка тонких волосо-
видных щетинок, а на нижней стороне
головной лопасти и около рта — много-
численные реснички, благодаря которым
эти черви могут не только ползать, но и
плавать; для них характерно скользящее
движение. Пищей им служат микроско-
пические одноклеточные водоросли и бак-
терии, мельчайшие частицы разлагающих-
ся органических веществ, поэтому особен-
но благоприятные условия для размноже-
ния большинство из них находит в уме-
ренно загрязненных водоемах, в местах
скопления гниющих растений у берегов,
в болотах и т. п. Нередко эолосомы раз-
множаются в большом количестве в бан-
ках, где разводят в лаборатории инфузо-
рий и других простейших, а также иногда
в аквариумах с рыбами, для которых они,
конечно, никакой опасности не представ-
ляют. Половое размножение у эолосом
наблюдается редко, обычно они размно-
жаются вегетативно, путем паратомии (см.
выше), причем образуются цепочки из
двух—пяти особей.
Семейство Наидиды, или Водя-
ные змейки (Naididae), гораздо богаче ви-
дами, которых насчитывается свыше сот-
ни. Все они — водные животные, при-
чем большинство является обитателями
пресных водоемов и лишь немногие встре-
чаются в солоноватых водоемах и в при-
брежной зоне морей. Хотя среди наидид
имеются и очень мелкие, длиной всего в
1 — 2 мм, большинство из них крупнее,
10—20 мм, и хорошо заметны простым гла-
зом, а самые крупные достигают длины
35—50 мм. Сегментация выражена хоро-
шо, число сегментов у большинства в
пределах от 10—15 до 30—40, но есть ви-
ды, имеющие до 100 сегментов. По форме
тела, внешнему виду, окраске наидиды
очень разнообразны. Головная лопасть у
большинства округлая или удлиненная, но
у некоторых вытянута в более или менее
длинный щупальцевидный придаток («хо-
боток»), а у других укорочена и почти сов-
сем не развита. Наидиды — единственное
семейство олигохет, у большинства пред-
ставителей которого имеются глаза. Ще-
тинки, как правило, расположены на
каждом сегменте четырьмя пучками B спин-
ных и 2 брюшных), причем форма щетинок
очень разнообразна: в брюшных пучках
чаще всего имеются вильчатые крючковид-
ные, а в спинных — игловидные и более
длинные волосовидные; но есть виды, ли-
шенные волосовидных щетинок, а есть и
такие, у которых вообще отсутствуют
спинные пучки, так что щетинки имеются
только на брюшной стороне. Спинные
щетинки у большинства, в том числе у
видов рода Nais, начинаются с шестого
сегмента, но у пристин (род Pristina) —
со второго, а у некоторых других — с
третьего, четвертого, пятого или позади
шестого.
Некоторые виды наидид имеют жабры,
расположенные у одних на заднем конце,
а у других вдоль большей части тела, по
бокам.
У наидид тоже имеются две формы раз-
множения: половое, обычно приуроченное
к определенным сезонам, и вегетативное,
путем паратомии, которое преобладает;
поэтому чаще встречаются не одиночные
черви, а «цепочки», состоящие из двух —
пяти особей.
494
Наидиды встречаются во всех странах
мира; в СССР обнаружено свыше 50 видов.
Род щетинкобрюхов (Chaetogaster)
включает несколько имеющих только брю-
шные щетинки видов, из которых самый
крупный щетинкобрюх прозрачный (Chae-
togaster diaphanus). Длина его цепочек
достигает иногда 25 мм. Это довольно не-
уклюжий, сравнительно толстый червяк,
совершенно прозрачный, бледного розо-
вато-желтого цвета. Он медленно ползает
по водным растениям или по поверхно-
сти дна в поисках добычи. Как и дру-
гие виды этого рода, он хищник; пи-
тается крупными простейшими, коловрат-
ками, мелкими рачками и другими не-
большими животными, которых заглаты-
вает целиком. Головная лопасть у него
не развита, передний конец выглядит как
бы обрубленным и большой рот направлен
прямо вперед. Рот ведет в объемистую
мускулистую глотку, способную частично
выпячиваться наружу; с ее помощью и
захватывается добыча. Глаз у щетинко-
брюхов нет. Другой вид этого рода —
щетинкобрюх улитковый (Chaetogaster lim-
naei, рис. 281) нередко встречается на ра-
ковинах, на поверхности тела и в дыха-
тельной полости различных пресновод-
ных улиток — прудовиков, физ, катушек
и др.; иногда, однако, он попадается и на
водных растениях. Настоящим паразитом
моллюсков он не является, скорее это со-
житель, нахлебник (комменсал); его пищу
составляют различные инфузории, колов-
ратки и т. п., а если моллюск заражен
личинками паразитических червей — со-
сальщиков, то он охотно поедает выхо-
дящих из тела моллюска этих личи-
нок — церкарий. Известны случаи, когда
на одном моллюске находили до тысячи
щетинкобрюхов.
К числу наиболее обычных в наших во-
доемах принадлежат многие виды рода
наид (Nais). Редко случается, чтобы их не
удалось обнаружить при рассматривании
в банке воды, зачерпнутой вместе с вод-
ными растениями у берега пруда, озера,
речки или в болоте. Большинство видов
этого рода — нежные беловатые прозрач-
ные червячки длиной около 10—15 мм,
с торчащими в стороны пучками тонких
щетинок и парой темных глаз на округло-
удлиненной головной лопасти (рис. 281).
Они оживленно ползают, поочередно вы-
тягивая и сокращая свое тело, нередко
обвиваясь вокруг нитчатых водорослей;
многие виды этого рода хорошо плавают.
Наиды питаются растительной пищей и
мельчайшими частицами разлагающихся
остатков других организмов; в их ки-
шечнике постоянно можно видеть одно-
клеточные зеленые и диатомовые водорос-
ли, а иногда и некоторых простейших
животных. По-видимому, немалую роль
в их питании играют и бактерии. В аква-
риумах многие наиды хорошо живут и
энергично размножаются при подкормке
дрожжевой взвесью. Отдельные виды этого
рода различаются с трудом, поэтому мы
не приводим здесь их описания; отметим
только, что есть среди них виды, встре-
чающиеся преимущественно в реках (на-
пример, Nais behningi), есть связанные
преимущественно с холодными ручьями
и родниками (Nais elingius), большинство
же встречается в прибрежной зоне как
различных стоячих водоемов, так и рек
(Nais variabilis, N. pseudoobtusa и др.).
По характерной уплощенной головной
лопасти, вытянутой в длинный щупальце-
видный придаток — «хоботок», и довольно
длинным и толстым волосовидным щетин-
кам, торчащим в стороны под прямым
углом к туловищу, легко узнать стилярию
(Stylaria lacustris, рис. 281). Она не-
сколько крупнее наид, до 20 мм длиной,
и обладает хорошо развитыми глазами.
Передний конец ее прозрачного тела имеет
обычно желтоватую окраску. Стилярия
часто встречается среди прибрежных за-
рослей озер и рек; она хорошо плавает и
иногда попадается в планктоне. Очень
своеобразна небольшая наида Ripistes
parasita, несущая на шестом — восьмом
сегментах три пары густых пучков очень
длинных волосовидных щетинок (рис. 281).
Кстати сказать, ее видовое название мо-
жет ввести в заблуждение: она вовсе не
паразит, а свое название получила слу-
чайно, потому что впервые была найдена
в колонии мшанок. Ее головная лопасть,
снабженная глазами, вытянута в корот-
кий «хобот», а передняя часть тела имеет
обычно коричневато-желтую окраску. Во
второй половине лета в некоторых озерах,
495
особенно торфяных, этот червь встречает-
ся иногда массами. Ripistes parasita строит
на стеблях тростника, камыша и т. п.
небольшие трубочки из слизи и частиц
ила, внутри которых и живет, высунув
наружу только передний конец тела с
пучками длинных щетинок. Пражский
профессор Кори, занимавшийся наблю-
дениями над этой наидой, говорит, что она
создает своими длинными щетинками дви-
жение воды вдоль тела в трубке, необхо-
димое для дыхания, и одновременно как
бы отцеживает из этой воды мелкие пи-
тательные частицы, которые затем «сли-
зывает» с этих щетинок.
Как уже упоминалось, у большинства
олигохет дыхание кожное, но у некото-
рых имеются специальные органы дыха-
ния — жабры. Среди семейства наидид
обладателями жабр являются виды широ-
ко распространенных родов Dero и Aulo-
phorus (рис. 281), а также сравнительно
редкий Branchiodrilus hortensis, найден-
ный в СССР только в бассейне реки Амура
и в озере Ханка (рис. 281).
Dero и Aulophorus — обитатели дна во-
доемов. Передним концом они зарывают-
ся в ил (которым питаются), а задний
выставляют над поверхностью дна. Хво-
стовой конец у них расширен в виде ди-
ска, на котором сидят парные листовид-
ные жабры. Эти жаберные листочки густо
Таблица 25. Пелагические полихеты:
1 — Alciopidae,
2 — Tomopteris,
3 — Lopadorhynchus,
4 — Sagittella,
5 — Nectochaeta,
6 — Typhloscolecs.
покрыты ресничками, благодаря работе
которых свежая вода все время омывает
жабры.
Виды родов Dero и Aulophorus име-
ют красноватую окраску, зависящую от
наличия в их крови гемоглобина, что в
свою очередь связано с необходимостью
лучше использовать кислород, которого
в придонных слоях воды может быть очень
мало. Глаз эти донные виды не имеют.
Некоторые из них хорошо плавают.
Некоторые мелкие наидиды, особен-
но виды рода Prist ina, лишенные глаз
(рис. 281), нередко в большом количестве
размножаются в аквариумах с рыбами.
У всех пристин спинные щетинки начинают-
ся со второго сегмента, как и брюшные; у
некоторых головная лопасть вытянута в
небольшой хоботок. Никакой опасности
для рыб они не представляют, скорее
даже наоборот: питаясь иловыми частица-
ми, бактериями, одноклеточными водорос-
лями, они являются хорошими санитара-
ми, а кроме того, ими самими могут пи-
таться мальки рыб.
Семейство Трубочники, или Туби-
фициды (Tubificidae), включает около 130
видов олигохет, подавляющее большин-
ство которых обитает на дне пресных
водоемов и лишь немногие встречаются в
прибрежной зоне морей. В СССР найдено
около 70 видов этого семейства. Трубочни-
Рис. 281. Представители семейства Naididae:
I — Stylaria lacustris; 2 — Chaetogaster Hmnaei, 3 — Nais pseudobtusa; 4 — Ripister parasita;
5 — Aulophorus, 6 — Branchiodrilus, 7 — Pristina longiseta.
ки крупнее наидид; их тонкое нитевидное
тело (рис. 282) достигает у некоторых
видов длины 100—180 мм, большинство
же имеет длину 30—60 мм, а самые мел-
кие — всего 5—10 мм. Все они лишены
глаз. Тело состоит из довольно большого
количества сегментов, обычно 60—100,
у некоторых — свыше 200. Щетинки рас-
положены четырьмя пучками на каждом
сегменте,причем в брюшных пучках имеют-
ся только простые, крючковидные или
вильчатые, а в спинных у многих видов
еще и волосовидные. Трубочники — ти-
пичные донные животные, особенно мно-
гочисленные на илистых грунтах; неред-
ко они образуют массовые скопления >
32 Жизнь животных, т. 1
497
выглядящие как красноватые «по-
душки» на дне и состоящие из ты-
сяч особей. Некоторые виды вы-
держивают очень сильное загряз-
нение и могут жить при мини-
мальном количестве растворенно-
го в воде кислорода, перенося в
течение многих суток даже пол-
ное его отсутствие. Все трубочники
имеют красную или розоватую
окраску, обусловленную наличи-
ем в их крови гемоглобина; имен-
но наличие гемоглобина, а также
свойственная им способность про-
изводить особые дыхательные дви-
жения, вызывающие приток к их
телу свежей воды (рис. 283), и
позволили им заселить дно или-
стых водоемов. Следует помнить,
что илы благодаря происходя-
щим в них процессам разложе-
ния органических остатков по-
глощают кислород из придонных
слоев воды, снижая здесь его ко-
личество до минимальных величин.
Передним концом трубочники
обычно погружены в ил, а зад-
ний выставляют над поверхностью
дна, производя им колебательные
(дыхательные) движения (рис. 283).
Именно в коже заднего конца
тела особенно густо разветвлены
кровеносные сосуды, так что
хвост червя является в известном
смысле органом дыхания. Чем
меньше в окружающей воде ки-
слорода, тем больше высовывают-
ся хвосты червей из ила и тем
энергичнее они производят дыха-
тельные движения. Наоборот, при
высоком содержании кислорода
черви почти полностью погружа-
ются в ил. Вокруг высовываю-
щейся из грунта части тела
многие виды трубочников обра- *
вуют из склеенных слизью ило- :Щ
вых частиц короткую гибкую труб- \Щ
ку — отсюда и их название. По- Щ
ТреВОЖеННЫе ТрубоЧНИКИ МГНО- !^/,.v.V:,;?..v-:»..;-.v..v;..-
венно скрываются в иле. 'л.-лл;? > -
Трубочники — типичные грун- „ 000 т ,
FJ тт rj Рис. 283. Трубоч-
тоеды. Погружаясь передним кон- ники (Tubifex) в
цом в грунт дна водоема на глу- грунте.
Рис. 282. Tubifex.
бину до 50—100 мм, они все вре-
мя заглатывают ил с песком, про-
пуская через кишечник за сутки
количество материала, во много раз
превышающее вес их собственного
тела. При этом, естественно, про-
исходит минерализация пропускае-
мого через кишечник грунта, так
как входящие в его состав орга-
нические вещества частично ис-
пользуются червями для питания.
Характерно, что, чем беднее грунт
органическим веществом, тем боль-
ше его приходится заглатывать и
пропускать через свой кишечник
червям.
Если учесть, что в сильно за-
грязняемых водоемах, например
реках и прудах в черте городов,
количество трубочников может до-
стигать более ста тысяч на 1 м2
дна, легко представить себе мас-
штабы их деятельности и ее зна-
чение в процессах биологическо-
го самоочищения этих водоемов.
Хотя таких высоких цифр коли-
чество трубочников достигает
только в сильно загрязненных,
богатых органическими вещест-
вами местах, их все же достаточ-
но и на дне более или менее чи-
стых рек, озер и водохранилищ:
сотни и тысячи на каждый квад-
ратный метр. Это значит, что они
играют немалую роль в питании
рыб, тем более что тело их, бога-
тое ценными питательными веще-
ствами, очень легко переваривает-
ся в кишечнике рыб. Трубочники
являются очень удобным и хоро-
шим кормом и для аквариумных
рыб.
Размножение у них только по-
ловое. После спаривания черви
откладывают коконы, содержащие
по нескольку яиц каждый. Если
рассматривать трубочников нево-
оруженным глазом, то различные
их виды трудно отличить друг от
друга; только изучив под микро-
скопом особенности строения спин-
ных и брюшных щетинок, можно
в них разобраться, а для точного
498
определения вида часто и этого бывает
недостаточно, приходится изучать строе-
ние половой системы. Поэтому мы не
приводим здесь описания различных ви-
дов (для этого существуют определители,
например, О. В. Ч е к а н о в с к а я, «Вод-
ные малощетинковые черви фауны Совет-
ского Союза»), укажем только, что наиболее
часто встречающимися являются виды р о-
да Tubifex, имеющие в спинных пучках
волосовидные и крючковидные, или веер-
ные, щетинки, например самый обычный
из них и особенно стойкий к загрязне-
нию трубочник обыкновенный (Tubifex
tubifex).
Виды рода Limnodrilus лишены воло-
совидных щетинок; из них чаще других,
нередко вместе с Т. tubifex, встречается
лимнодрил Гоффмейстера (Limnodrilus
hoffmeisteri), у которого все щетинки, и
спинные и брюшные, имеют по 2 загнутых
одинаковых зубца на конце. Назовем еще
самого крупного из наших лимнодрилов —
невского (L. newaensis). Он впервые был
найден в реке Неве, но оказался широко
распространенным в реках, водохранили-
щах и некоторых озерах Ев-
ропейской части СССР.Это до-
вольно толстый, до 2 мм, червь,
длиной до 90 мм * обычно тем-
ной серовато-красной окраски;
щетинки у него с короткими
зубцами, причем верхний ко-
роче нижнего.
В прибрежной полосе морей,
в частности Белого и Баренце-
ва, при солености до 20°/00,
обитает не встречающийся в
пресных водоемах Clitellio are-
narius, как и лимнодрилы, ли-
шенный волосовидных щетинок.
Есть среди трубочников и об-
ладающие наружными жабра-
ми. Это довольно редкий вид—
бранхиура (Branchiura sower-
byi, рис. 284). Водится бран-
хиура в Индии, Индонезии, Ки-
тае и Японии, была найдена
во Франции, а у нас обнару-
жена в бассейне реки Амура и
в Закавказье. Интересно, что gJ^J^
этот вид много раз находили в ura sowe.
тепловодных бассейнах ботани- rbyi.
ческих садов, в которых выращивают зна-
менитую викторию (Victoria regia). Впер-
вые он был найден именно в таком бассей-
не в Лондоне и описан; позже он был най-
ден в бассейне оранжереи ботанического
сада Московского университета.
Семейство Энхитреиды (Enchy-
traeidae) содержит как водных, так и су-
хопутных (почвенных) червей, причем по-
следних больше, а общее количество ви-
дов этого семейства достигает почти 400.
Энхитреиды — небольшие или мелкие, ма-
лопрозрачные черви, в большинстве слу-
чаев беловатые, реже желтоватые или ро-
зоватые. Длина их от 2—3 мм до 40—45 мму
большинство видов от 10 до 25 мм. Сег-
ментация выражена хорошо, число сег-
ментов, как правило, в пределах от 25
до 50, у некоторых больше, до 70—80.
Глаз нет.
Щетинки расположены четырьмя пуч-
ками на каждом сегменте, начиная со вто-
рого; все щетинки более или менее оди-
наковые, игловидные или палочковидные,
с простым, не вильчатым (за одним ис-
ключением) кончиком. Размножение толь-
ко половое; у некоторых на-
блюдается своеобразная форма
бесполого размножения, при
котором червь распадается на
две или большее число частей,
восстанавливающих затем пу-
тем регенерации недостающие
головные или хвостовые отделы
(так называемая архитомия).
При благоприятных условиях
некоторые виды почвенных и
литоральных энхитреид размно-
жаются в огромном количестве,
до десятков тысяч на 1 м2 (а
в очень богатых перегноем поч-
вах даже до 150—200 тыс.), и
играют очень существенную
роль в разложении органиче-
ских остатков и почвообразо-
вании. Благодаря тому что их
легко разводить в ящиках или
других сосудах с землей (раз-
множаются они очень быстро),
некоторые виды, главным об-
разом белый энхитрей (Enchy-
traeus albidus), стали использо-
ваться как хороший корм
32*
499
для рыб, притом не только аквариумных,
но и в более крупных масштабах, на рыбо-
водных заводах. Поскольку эти энхитреи-
ды разводились аквариумистами часто в
цветочных горшках с землей, они получи-
ли название «горшечных червей». На мор-
ских побережьях, в частности на Белом
море, под выброшенными волнами гнию-
щими водорослями некоторые энхитреиды,
особенно Enchytraeus albidus и Lumbri-
cillus lineatus, образуют большие скопле-
ния, в которых черви буквально кишат.
Распространены энхитреиды очень ши-
роко, преимущественно в северном полу-
шарии, в умеренной и субарктической зо-
нах; на север они проникают дальше дру-
гих олигохет и, в частности, известны со
Шпицбергена, Новой Земли, Новосибир-
ских островов и Гренландии. Встречаются
и высоко в горах, до высоты 3—4 км над
уровнем моря, найдены и в пещерах.
Известно, что некоторые виды этого
семейства выдерживают продолжительное
воздействие температуры ниже нуля. Так,
ряд ученых наблюдал их оживание при
оттаивании мерзлого грунта или кусков
льда, в который они вмерзли. Американ-
ский зоолог М у р рассказывает, что он
наблюдал один из видов этих червей
{Mesenchytraeus solifugus) на снегах гор-
ных ледников Аляски; в начале лета, ког-
да снег был плотным, они вяло ползали
по его поверхности вечером и ночью, но в
середине дня, когда солнце светило ярко,
они зарывались вглубь; позже, когда снег
стал подтаивать и пропитался водой,
они были уже более активны и нередко
их можно было наблюдать в образовавших-
ся на поверхности лужицах. Подобные
же наблюдения над этим и другими вида-
ми были сделаны и на других ледниках.
Интересно, что многие виды энхитреид,
живущие постоянно на снегах, имеют,
как и встречающиеся там же мелкие на-
секомые, черную или темную красно-
коричневую окраску; возможно, что это
имеет значение для поглощения тепловых
лучей и как защита от ультрафиолетового
излучения. Пищей им могут служить
одноклеточные красные водоросли, часто
в несметном количестве размножающиеся
на горных снегах, а также, возможно,
пыльца хвойных.
Семейство Гаплотаксиды (Нар-
lotaxidae) включает всего около 20 видов
водных олигохет. Из этого небольшого
семейства мы упомянем один широко-
распространенный вид — Haplotaxis gor-
dioides. Это очень длинный, до 300 мм,
и тонкий червь, тело которого состоит
из 200—480 сегментов (рис. 285); каждый
сегмент несет всего по 4 или даже толь-
ко по 2 (брюшные) довольно крупные
простые щетинки. Встречается этот червь,
однако, нечасто и только местами; дер-
жится преимущественно в водоемах с хо-
лодной водой — колодцах, источниках,
ручьях, реках и совершенно отсутствует
в сильно прогреваемых прудах и т. п.;
в больших озерах отмечен до глубины
свыше 300 м. Иногда попадается и не в
воде, а в глинистом грунте по берегам
ручьев и других водоемов. Чаще его на-
ходят в весенние месяцы. Есть указание,
что, попав в аквариум, эти черви могут
повреждать корни растений, например
валлиснерии. В СССР Haplotaxis gordioi-
des найден в озере Байкал, где водится и
другой представитель этого рода —
Haplotaxis ascaridoides (нигде, кроме Бай-
кала, не найденный), в Телецком озере на
Алтае (где, кроме него, водится другой
представитель этого семейства — Pelo-
drilus ignatovi), а также в озерах Камчат-
ки, реках и озерах Кавказа, Средней Азии.
Семейство Люмбрикулиды (Lumbri-
culidae) включает свыше 60 видов. Они
имеют только крючковидные щетинки, од-
но- или двухзубчатые, по 4 пары на каж-
дом сегменте, начиная со второго. Головная
лопасть у большинства округленная, у
некоторых вытянута в небольшой хоботок,
глаз нет. Окраска обычно красноватая
или серовато-красная, иногда с фиоле-
товым оттенком, иногда, преимущественно
на переднем конце, с зеленоватой или чер-
новатой пигментацией. Тело нередко полу-
прозрачное, его поверхность у некоторых
видов при сильном освещении отливает
радужными тонами (иризирует). Люм-
брикулиды — обитатели пресных вод.
Большинство из них имеет средние раз-
меры от 20—30 мм до 80—100 мм при
толщине 1—2 мм, но наиболее крупные
виды достигают длины свыше 180 мм и
толщины до 4,5 мм. Число сегментов у
500
Рис. 285. Haplotaxis gordioides.
большинства видов около 80—150, но у
некоторых больше 200.
Самым обычным представителем семей-
ства является Lumbriculus variegatus,
встречающийся почти во всех водоемах
и очень широко распространенный. Осо-
бенно часто он попадается в небольших
заболоченных водоемах, лужах, канавах.
Его длина — 40—90 мм, окраска темная,
красноватая. Легче всего узнать люмбри-
кулюса по характерным движениям: буду-
чи потревожен, он начинает быстро и
резко извиваться и уплывает в сторону.
Особенно ярко эта реакция бывает вы-
ражена в скоплениях червей: такой клу-
бок, если его потревожить, буквально
«взрывается», и все черви бросаются в
разные стороны. Однако сколько-нибудь
долго люмбрикулюсы плыть не могут и
вскоре снова опускаются на дно. Другой
особенностью этого вида является то, что
он почти утратил способность к половому
размножению; найти половозрелых червей
почти никому не удавалось. Зато он приоб-
рел способность очень быстро размножать-
ся бесполым путем, распадаясь на отдель-
ные куски (размножение фрагментацией,
или архитомией), которые затем восстанав-
ливают головные и хвостовые концы путем
регенерации. Люмбрикулюса почти невоз-
можно схватить пинцетом не повредив: он
сейчас же распадается на 2 или несколь-
ко кусков. Способность к регенерации у
этого вида очень велика, и червь мо-
жет восстановиться даже из небольшого
куска.
Другой широко распространенный, но
более редкий вид этого семейства— Rhyn-
chelmis limosella — значительно крупнее:
80—150 мм длиной, 2—3 мм толщиной.
Встречается преимущественно в холод-
ных или сильно затененных водоемах,
обладает «хоботком».
Оба вида послужили объектами многих
специальных исследований по изучению
закономерностей процесса регенерации.
Но самой, может быть, замечательной
особенностью этого семейства является
то, что больше половины всех его видов
(около 40) водится только в озере Байкал
и нигде больше на земном шаре не встре-
чается! Люмбрикулиды заселяют Байкал
от прибрежной полосы до глубин свыше
1 км, причем каждой зоне свойственны
свои виды. Они очень разнообразны и по
размерам и по внешнему виду, некоторые
обладают характерной окраской и силь-
но иризируют. Это обстоятельство, т. е.
наличие в Байкале большого количества
эндемичных (только в нем встречающихся)
видов люмбрикулид и их своеобразие,
говорит о большой древности Байкала
именно как пресноводного озера.
Семейство рачьих пиявок (Вгап-
chiobdellidae) — небольшое и очень свое-
образное семейство, включающее пример-
но 30 довольно мелких видов. Действи-
тельно, все виды этого семейства, распро-
страненного как в Европе, так и в Азии,
Северной и Южной Америке, паразити-
руют на жабрах и на поверхности тела
различных видов пресноводных раков.
Паразитический образ жизни наложил
на их внешний вид и всю их организацию
столь сильный отпечаток, что многие зоо-
логи причисляли их раньше к классу
пиявок. Однако, несмотря на полную
утрату щетинок, это все же настоящие
олигохеты, и притом по своей организации
стоящие ближе всего к предыдущему се-
мейству люмбрикулид. Тело рачьих пия-
вок, не превышающее 12 мм в длину, от-
501
носительно толстое и поделено на два
главных отдела: несегментированный го-
ловной, образовавшийся слиянием четы-
рех первых сегментов, и туловищный, со-
стоящий из одиннадцати сегментов, имею-
щих обычно вторичную кольчатость. На
заднем конце тела находится довольно
сильная мускулистая присоска. В рото-
вой полости имеются две твердые роговые
челюсти (верхняя и нижняя), у большин-
ства видов снабженные зубчиками. У не-
которых видов на переднем конце разви-
ты особые выросты.
В молодом возрасте рачьи пиявки пи-
таются водорослями и детритом, следо-
вательно, не вредят своему «хозяину», на
котором они сидят, но, выросши, они на-
чинают питаться кровью рака, т. е. ста-
новятся настоящими паразитами.
Семейство Настоящие дождевые
черви, или Люмбрициды (Lumbricidae).
Всем хорошо известны земляные, или дож-
девые, черви. Иногда в сырую погоду,
особенно после дождей (отсюда и назва-
ние), они в большом количестве появляют-
ся на поверхности, ползают по дорожкам
садов и парков, лежат на дне лужиц дож-
девой, а весной талой воды. Не следует,
однако, думать, что название «дождевые
черви» обозначает какое-то одно опреде-
ленное семейство олигохет; оно применяет-
ся ко всем более или менее крупным пред-
ставителям пяти различных семейств этого
класса, обитающим в почве, т. е. является
сборным. На первый взгляд, они действи-
тельно до некоторой степени сходны меж-
ду собой; зоологи различают эти семей-
ства главным образом по особенностям
строения внутренних органов, в частности
органов размножения, по расположению
пояска, щетинок и по другим признакам.
В учебниках зоологии в качестве приме-
ра для класса олигохет приводится «обык-
новенный дождевой червь» — Lumbri-
cus terrestris. Этот довольно крупный
червь, до 30 см длиной и до 1 см тол-
щиной, принадлежит к наиболее полно
изученному семейству настоящих
дождевых червей, или люмбрицид (Lumbri-
cidae), включающему около 200 видов,
самые мелкие из которых имеют всего
2—3 см в длину и около 1 мм в толщину,
а самые крупные достигают полуметровой
длины при толщине до 1,5—2 см.Размноже-
ние у них только половое.
Упомянутый нами Lumbricus terrestris,
носящий местами народные названия
«большой выползок» или «большой красный
червь», легко отличается от других дожде-
вых червей своими крупными размерами,
уплощенным и расширенным («лопатообраз-
ным») хвостовым концом и окраской: пе-
редняя треть его тела со спинной стороны
довольно темного, тусклого красно-фио-
летового цвета, а остальное тело светлее,
грязно-розоватого оттенка, с более тем-
ной полосой, тянущейся вдоль середины
спины почти до самого конца тела. На
солнце передний конец этого червя отли-
вает радужными тонами (иризирует),
что связано с особенностями строения
кутикулы. Щетинки у него расположены
на сегментах четырьмя парами, а поясок —
с 31-го или 32-го по 37-й сегмент.
Этот вид распространен по всей Евро-
пе, от Атлантического побережья до Ура-
ла; в Сибири, Казахстане, Средней Азии,
на Кавказе не найден, хотя не исключено,
что может быть местами обнаружен, как
завезенный, в городских садах и пар-
ках; известен он и в Северной Америке.
В земле он роет глубокие ходы («норы»)
уходящие от поверхности на 2 м и больше.
В теплую и сырую погоду, по ночам1
эти черви вылезают из своих норок, но не
совсем, а цепляясь хвостом за отверстие,
так что в случае опасности могут быстро
скрыться. Вытягиваясь, они обшаривают
окружающее пространство, захватывают
своим ртом (при этом слегка выворачи-
вается наружу, а затем втягивается обрат-
но мускулистая глотка) сырые опавшие
листья, полусгнившие травинки и т. п. и
затаскивают их в свои норы. Но черви
заглатывают и землю, которая поэтому
всегда имеется в их кишечнике. Органи-
ческие вещества почвы, перегной, исполь-
зуются в качестве пищи, песчинки способ-
1 Дождевые черви вообще очень чувствительны
к свету. При внезапном освещении дневным
светом, например при переворачивании камней,
под которыми они скопились, или ночью при
свете фонаря они быстро скрываются в норки.
Однако на очень слабый свет они реагируют поло-
жительно. К красному свету практически нечув-
ствительны.
502
Рис. 286. Зарывающийся в землю дождевой
червь.
ствуют измельчению и перетиранию про-
глоченных кусочков листьев, наконец, ча-
стично почва заглатывается при прорыва-
нии ходов, хотя главная работа при
этом проделывается мускулатурой стенки
тела червя. Когда червь зарывается в зем-
лю с поверхности (рис. 286) или когда
прокладывает новый ход в толще почвы,
он действует передним концом как кли-
ном, попеременно то сужая и вытягивая
его, то вздувая и сокращая, и таким об-
разом расталкивает частицы почвы в сто-
роны. Большую роль при этом играют
щетинки, цепляющиеся за стенки хода и
не дающие возможности телу червя со-
скальзывать назад при его сокращении. По
поверхности земли, не имея опоры со
всех сторон, черви передвигаются сравни-
тельно медленно, но внутри своих ходов —
довольно быстро: если перевернуть ка-
мень или другой предмет, под которым
скопились черви, и попытаться схватить
их, они почти мгновенно скрываются в
норки.
Переработанную ими, прошедшую через
кишечник землю черви выбрасывают на
поверхность над отверстиями ходов в виде
характерных кучек — выбросов, или
копролитов. Появление таких ку-
чек на оголенной земле весной свидетель-
ствует о начале деятельности червей.
Подсчитано, что за сутки каждый червь
пропускает через свой кишечник коли-
чество земли, примерно равное весу его
тела (а средний вес Lumbricus terrestris —
4—5 г). Зимуют эти черви в нижней части
своих ходов, ниже глубины промерзания
почвы; при высыхании почвы во время
летней засухи они также спускаются в
глубокие ее горизонты. Продолжитель-
ность жизни этого вида точно неизвестна,
но, по некоторым наблюдениям, не мень-
ше 5—6 лет.
К роду Lumbricus принадлежит 10
видов. Самым обычным у нас является
малый красный червь, или малый вы-
ползок (Lumbricus rubellus). Он пример-
но вдвое короче и тоньше предыдущего,
но окрашен ярче, вишнево-красного цвета.
Задний конец его тела тоже уплощен и
расширен, а поясок у него обычно распо-
ложен на сегментах с 27-го по 32-й. В
средней полосе это, пожалуй, второй по
встречаемости вид. Таких глубоких ходов,
как большой выползок, он не делает;
обычно держится в самых поверхностных
горизонтах почвы, а в лесах — в сырова-
той подстилке, среди полугнилых листьев.
Но самым распространенным и часто
встречающимся у нас видом является
серая аллолобофора (Allolobophora caligino-
sa), называемая иногда пашенным червем,
так как на пашнях и полях попадается
чаще других; найти ее можно также и в
лесах, на огородах и на побережьях водое-
мов. Это средних размеров, до 15—16 см
длиной, сероватый червь, совершенно ли-
шенный пурпурной пигментации, с пояс-
ком, расположенным обычно на сегментах
с 27-го по 34-й. Задний конец тела не
уплощен. Как у других видов этого рода
(а их насчитывается около 50), щетинки
на каждом сегменте сближены попарно.
В отличие от выползков аллолобофора дер-
жится обычно в толще почвы, на глубине
5—15 см, где и питается главным образом
почвенным перегноем. На поверхность ее
могут выгнать только сильные дожди.
При благоприятных условиях размножает-
ся в огромном количестве: с площади в
1 м2 удается иногда выкопать до 400—500
экземпляров. Во время летней засухи
серая аллолобофора уходит на глубину
40—60 см, свертывается там клубком, иног-
да буквально завязываясь узлом, выде-
ляет слизь и с ее помощью строит вокруг
себя из почвенных частиц довольно проч-
ную защитную капсулу, впадая затем
в неактивное состояние, причем про-
цессы развития приостанавливаются. Та-
кое состояние, диапауза, может продол-
503
жаться больше двух месяцев. Благодаря
этой способности аллолобофоры более ус-
тойчивы к засухе, чем выползки, и могут
заселять более сухие, открытые места.
К этому роду принадлежит и один из
самых крупных наших червей — алтай-
ская гигантская аллолобофора (АН о lobо-
phora magnifica), носящая у местных жи-
телей названия «щура» и «салазана». Дли-
на заспиртованных червей этого вида до-
стигает 25—30 см при толщине около 1,5 см;
живые черви могут растянуться до 40 см
и больше. Их ходы, до 2 см диаметром,
проникают в почву на глубину 4—5 м.
Местами вся почва бывает пронизана та-
кими ходами, а над отверстиями обитае-
мых норок образуются кучки выбросов,
достигающие 8 см в высоту и 10 си* в попе-
речнике. С сельскохозяйственной точки
зрения воздействие на почву этих червей
(как и других видов) положительно, но
строители смотрят на это дело часто иначе,
так как сильное рыхление червями земли
около фундаментов доставляет им лишние
хлопоты. Пока этот вид известен только
из районов Северо-Западного Алтая.
Род Дендробена (Dendrobaena) ха-
рактеризуется широко расставленными (не
сближенными попарно) щетинками. На-
звание рода указывает на обитание его
представителей в лесах; описанный уже
давно вид этого рода — небольшой, очень
подвижный, темный, красно-фиолетовый
червь восьмигранная дендробена (Dendro-
baena octaedra)—настолько характерен для
лесной зоны, что нередко его называют
«таежным червем». Держится он главным
образом под сырыми опавшими листьями,
в лесной подстилке, и распространен
широко — от Атлантического океана до
Тихого, встречаясь на далеком Севере,
в Заполярье и в горах Крыма и Кавказа.
Однако далеко не все виды этого рода,
а их насчитывается свыше 40, являются
обитателями лесов. Мы упомянем еще
только один из них — мариупольскую ден-
дробену (Dendrobaena mariupolienis), рас-
пространенную в Приазовье, Крыму и
на Кавказе. Это очень крупный червь,
до 35 см длиной, довольно толстый и
сильный, темной, черновато-бурой окраски;
поясок у него захватывает с 25-го по 32-й
сегмент. По наблюдениям Г. Н. Вы-
соцкого, мариупольская дендробена
довольно многочисленна в почвах целин-
ных степей. Замечательно, что ходы этого
червя удалось проследить на глубине
8 м; часто по ним проникают в нижние го-
ризонты почвы корни растений (рис. 289).
Одним из наиболее ярко окрашенных
червей является навозный (Eisenia foetida),
обладающий характерным неприятным за-
пахом, что отражено и в его названии —
foetida, что значит «дурно пахнущая»,
«вонючая». Это сравнительно небольшой
червь, длиной 6—10 см, с попарно сбли-
женными щетинками. Цвет его темно-
красный или красно-коричневый, но ок-
раска не сплошная, а кольчатая, так как
бороздки, разделяющие сегменты, светлые,
что особенно хорошо видно, когда червь
сильно растянется. Навозный червь —
очень широко распространенный вид; на
севере и в средней полосе он встречается
почти исключительно недалеко от жилья
человека, в скоплениях навоза, гнилой
соломы, в парниках и т. п. (нередко мас-
сами, до 1000 и более на 1 м2). Но уже в ле-
состепной полосе, а также в Крыму и на
Кавказе он становится довольно обычным
и в лесах, далеких от жилья, где его мож-
но найти в старых, трухлявых пнях, дуп-
лах, скоплениях прелых листьев и по бе-
регам речек и ручьев. При раздражении
навозный червь выделяет довольно силь-
но пахнущую желтую жидкость.
К этому же роду относится и норденшелъ-
дов червь, или эйзения Норденшельда (Eise-
nia nordenskioldi), названная так в честь
известного шведского путешественника,
исследователя полярных стран Норден-
шельда, собравшего большие зоологи-
ческие коллекции на севере Сибири. Этот
вид распространен по всей Сибири, встре-
чается на Алтае и в Маньчжурии, а также
в восточной части Европейской территории
СССР, доходя до Крыма. По окраске и
размерам он очень изменчив; типична для
него окраска, похожая на таковую у
Lumbricus rubellus: довольно темная,
вишнево-красная, причем задний конец
несколько светлее; характерны два свет-
лых пятна по бокам (на 9—11-м сегмен-
тах). Встречаются иногда и очень слабо
окрашенные, почти совсем светлые черви.
Местами, особенно в Сибири, норденшель-
504
дов червь достигает 25—30 см длины; в
Европейской части СССР он обычно мель-
че, всего 6—12 см. Интересно, что этот
вид довольно многочислен в тундрах, в
районе вечной мерзлоты; недавно он был
найден даже на Ляховских островах, на
вечных льдах. Последний вид семейства
люмбрицид, который мы упомянем, — че-
тырехгранная эйзениелла (Eiseniella tet-
raedra) — один из самых мелких, длиной
всего 2,5—6 см. Ее окраска серовато-
коричневая, с желтоватым или оливко-
вым оттенком; поясок с 22—23-го по 26 —
27-й сегмент; задний конец тела четырех-
гранный. Это земноводный вид, обитаю-
щий по берегам водоемов; иногда его мож-
но найти и в воде у берега.
В умеренной и холодной зонах север-
ного полушария, в частности на террито-
рии СССР, распространены почти исклю-
чительно только «настоящие» дождевые
черви, т. е. представители семейства
Lumbricidae, с которым мы только что
познакомились. В странах южного полу-
шария, а также в тропической и отчасти
субтропической зоне северного полуша-
рия дождевые черви представлены четырь-
мя другими семействами, включающими в
общей сложности около 1800 видов, лишь
немногие из которых проникают в южные
районы северного умеренного пояса. С дру-
гой стороны, некоторые виды люмбри-
см ю
Рис. 287. Гигантские дождевые черви Megascoli-
des australis. (В руке, для сравнения, обычный
в Подмосковье Lumbricus terrestris.)
цид, будучи завезены случайно челове-
ком в страны южного полушария, нашли
в ряде случаев подходящие для себя ус-
ловия и размножились в большом коли-
честве, успешно конкурируя с местными
видами других олигохет (например, в Ав-
стралии и Новой Зеландии).
Семейство Глоссосколециды (Glos-
soscolecidae). Большинство из 200 с лиш-
ним видов этого семейства распространено
в Южной Америке, некоторые встречаются
в Центральной и даже Северной Америке,
а также Африке, Юго-Восточной Азии и Ав-
стралии. В Европу проникли лишь не-
многие: несколько видов рода Ногто-
gaster известны в Италии, Сицилии и Сар-
динии, а один вид этого семейства —
Criodrilus lacuum — распространен доволь-
но широко в странах Южной и Средней
Европы, встречается и в Европейской ча-
сти СССР. По внешнему виду этот доволь-
но крупный A2—32 см длиной, 3—5 мм тол-
щиной), коричневато-серой окраски, ино-
гда с зеленоватым или красноватым оттен-
ком, червь похож на обычных дождевых
червей, но по образу жизни он является
водным животным и держится в иле среди
корней водных растений в прибрежной
части рек и озер, а также в слабосоленых
водоемах, например в Азовском море.
Найден также в водоемах Закавказья и в
Амуре. Очень своеобразны откладываемые
им коконы.
Хотя среди видов этого семейства есть
и другие, ведущие водный образ жизни,
большинство глоссосколецид, как и наши
люмбрициды, обитает в почве. Среди них
есть несколько видов, достигающих длины
около 1 м, а южноамериканский Rhinodri-
lus fafner даже 2 м с лишним длиной и поч-
ти 2,5 см толщиной!
Семейство Мегасколециды (Mega-
scolecidae) — самое богатое видами семей-
ство олигохет, распространенное в тропи-
ках и южном полушарии. Среди его пред-
ставителей есть довольно мелкие, всего
2—3 см длиной, есть и очень крупные,
свыше 1 м; к этому семейству принадлежит
и самый длинный земляной червь — ав-
стралийский Megascolides australis, дости-
гающий длины 2,5 м ( а по некоторым дан-
ным, даже до 3 м). Издали такого червя
можно принять за крупную змею (рис. 287).
505
Крупные виды мегасколецид
перерабатывают большое коли-
чество земли; их выбросы (рис.
288), имеющие иногда вид баше-
нок, бывают до 20—25 см высо-
той. У одного из африканских
видов этого семейства (Dicho-
gaster jaculatrix) глинистые вы-
бросы, торчащие местами в боль-
шом количестве среди опавших
листьев, напоминают своим ви-
дом миниатюрные красноватые
печные трубы; кстати, местные
жители делают из этих выбро-
сов трубки для курения, пред-
варительно обжигая их.
Большинство мегасколецид
по размерам и окраске слабо
отличается от европейских люм-
брицид; однако среди них встре-
чаются виды довольно ярко ок-
рашенные, иногда с кольчатым
и даже мраморным рисунком.
Преобладающие тона — красно-
коричневые, пурпурные, сероватые; реже
встречаются желтые, оливково-зеленые,
синеватые, почти черные и даже перла-
мутровые. Кроме обычного для большин-
ства земляных червей расположения ще-
тинок — по 4 пары на каждом сегменте, у
многих видов мегасколецид наблюдается
пное — в виде сплошного ряда или цепоч-
ки, опоясывающей примерно посредине
каждый сегмент, причем количество ще-
тинок в такой цепочке, т. е. на каждом
сегменте, нередко бывает равно 60—70 , а
в отдельных случаях доходит до 150; иног-
да эти ряды прерваны на спинной и брюш-
ной сторонах.
Почти все мегасколециды — обитатели
почвы; их обилие и интенсивность деятель-
ности, как и у европейских дождевых чер-
вей, зависит от влажности почвы, темпе-
ратуры, обилия органических веществ и
т. д.; при неблагоприятных условиях они
уходят в глубокие слои и их деятельность
замедляется или вовсе прекращается, на-
пример в периоды засухи. Было, однако,
замечено, что один вид — Pheretima pos-
thuma — встречается и в довольно сухих
местах, причем оказывается активным на
поверхности даже в засушливые периоды.
Это объясняется тем, что у него вырабо-
Рис. 288. Башенко-
образные выбросы
Pheretima.
тался особый физиологический
механизм для сбережения воды
в теле, подобный тому, который
наблюдается у насекомых: жид-
кие продукты обмена веществ,
экскреты, не выделяются поч-
ти совсем во внешнюю среду,
а попадают в кишечник, где во-
да из них подвергается обрат-
ному всасыванию клетками
стенки кишечной трубки. Таким
образом, этот вид представляет
редкий среди олигохет пример
высокосовершенного приспособ-
ления к наземному образу жиз-
ни.
Многие мегасколециды часто
попадаются в лежащих на зем-
ле гниющих стволах деревьев,
под корой, а некоторые заби-
раются довольно высоко, до
7—8 м, на живые деревья, где
держатся в углублениях коры,
среди эпифитных папоротни-
ков и даже (в сырых сильно затененных
джунглях) на листьях; ряд видов в связи
с этим получил соответственные названия,
например: древесный перионикс (Perionyx
arboricola), пальмовый дихогастер (Dicho-
gaster bolaui palmicola).
Пресноводных и литоральных (живущих
в прибрежной полосе морей) видов среди
представителей этого семейства мало; не-
которые виды, живущие обычно во влаж-
ных почвах, встречаются иногда и в во-
доемах, например маленький A,5—3 см
длиной) Ocnerodrilus occidentalis, недав-
но найденный и у нас в Средней Азии, на
рисовых полях и в некоторых водоемах.
Несколько видов этого семейства было
найдено в Южной Европе, а один из них,
широко распространенный в Америке, Ин-
дии, Африке, Microsco lex phosphoreus,
встречается и в Средней Европе. Этот
червь, имеющий очень небольшие размеры
A—3,5 см), замечателен своей способно-
стью светиться в темноте. Обычно свече-
ние начинается, если червя потревожить,
например даже при небольшом сотрясении
почвы. Цвет свечения желтовато-зеленый.
Светится слизь, выделяемая червем; но у
погибающих червей свечение возникает и
внутри тела. Большое количество этих
506
червей наблюдалось в одной из заброшен-
ных угольных шахт под Краковом на
глубине 230 м. Зрелище было очень эф-
фектное: в темноте двигались сотни све-
тящихся точек. Способность светиться на-
блюдается и у некоторых других видов. У
нас в Закавказье, в Батумском ботаниче-
ском саду, встречается много червей рода
Pheretima, большинство видов которого
(а их свыше 300) водится в Юго-Восточной
Азии. Это очень подвижные, похожие на
маленьких змеек черви, в сырую погоду
быстро ползающие по поверхности земли.
Найдены они и в Рионской низменности
(длина их 10—15 см).
Семейство Монилигастриды (Moni-
ligastridae). Представители его распро-
странены в Индии, Индонезии, Индокитае,
Китае и Японии, встречаются у нас на
Дальнем Востоке, в южных районах При-
морья. Эти, средних размеров, обычно
темноокрашенные и часто отливающие ра-
дужными тонами, земляные черви попа-
даются иногда вместе с люмбрицидами
преимущественно в сырых местах, по бе-
регам рек и других водоемов.
Первым ученым, обратившим внимание
на обилие дождевых червей в почвах, за-
нявшимся изучением образа их жизни и
пришедшим к выводу об очень большой
их роли в процессах почвообразования,
был Ч. Дарвин. Впервые он сообщил
о своих наблюдениях над их деятельно-
стью в небольшой статье, опубликованной
еще в 1837 г., а подробно описал резуль-
таты своих многолетних исследований над
ними в отдельной книге, вышедшей за
год до его смерти, в 1881 г.
«Дождевые черви в истории образова-
ния земной коры играли гораздо более
важную роль,— пишет Дарвин,— нежели
это может казатвся большинству с пер-
вого взгляда. Почти во всех влажных ме-
стностях они необыкновенно многочислен-
ны и сравнительно с их величиной обла-
дают весьма значительной мускульной си-
лой. Во многих местностях Англии на
площадь в акр г ежегодно выбрасывается
более 10 тонн A0 516 килограммов) сухой
земли, прошедшей через их тело, так что
Акр равен 0,405 га. (Прим. автора.)
весь поверхностный слой растительной зем-
ли в течение нескольких лет проходит через
их тело. Вследствие спадения стенок ста-
рых норок червей растительный слой на-
ходится хотя и в медленном, но постоян-
ном движении, причем составные частицы
перетираются друг о друга. В силу этих
явлений свежие поверхностные слои по-
стоянно подвергаются действию углекисло-
ты, содержащейся в почве, и гумусовых ки-
слот, роль которых в разрушении горных
пород является еще более значительной.
Образование гумусовых кислот обуслов-
ливается, вероятно, перевариванием полу-
разложившихся листьев, заглатываемых
дождевыми червями. Таким образом, ча-
стицы земли, составляющие верхний рас-
тительный слой, находятся при таких ус-
ловиях, которые вполне благоприятствуют
их разрушению и распадению.
... Черви наилучшим образом приготов-
ляют почву для произрастания растений с
мочковатыми корнями и для всякого рода
семенных растений. Они периодически
подвергают растительную землю действию
воздуха, просеивают ее так, что в ней не
остается ни одного камешка крупнее тех,
которые они могут заглотить. Они рав-
номерно перемешивают все, подобно садов-
нику, готовящему мелкозернистую землю
для своих избранных растений. В таком
виде она становится одинаково хорошо
приспособленной как к сохранению влаж-
ности и поглощению растворимых веществ,
так равно и для процессов нитрификации...
Листья, стаскиваемые в норки червей на
пищу, после того как они разорваны на
мельчайшие кусочки, отчасти переварен-
ные, отчасти смоченные жидкими выделе-
ниями кишечного канала и мочевыми выде-
лениями, перемешиваются с большими ко-
личествами земли. Эта земля образует тог-
да тот темный, плодородный гумус, кото-
рый почти сплошь покрывает поверхность
страны хорошо обособленным слоем...»
Дарвин показал, что черви, поселившие-
ся под отдельными камнями, перекапывают
под ними землю и, пропустив ее через свой
кишечник, выбрасывают вокруг, причем
прорытые ими ходы с течением времени
спадаются; в результате сами камни по-
степенно как бы тонут, погружаясь всо
глубже в землю. Таким же образом про-
507
Рис. 289. Корни деревьев, растущие
по ходам дождевых червей.
исходит, по Дарвину, в течение столетий
погребение остатков древних построек (по-
чему и приходится производить раскопки
для их обнаружения), хотя, конечно, в
этом процессе играют свою немалую роль
также вода и ветер, образующие наносы.
Заключает свою замечательную книгу о
дождевых червях Дарвин следующими
словами: «Нельзя не удивиться, когда
подумаешь о том, что весь растительный
слой уже прошел через тело дождевых чер-
вей и через несколько лет снова пройдет
через них. Плуг принадлежит к числу
древнейших и имеющих наибольшее зна-
чение изобретений человека; но еще за-
долго до его изобретения почва правильно
обрабатывалась червями и всегда будет
обрабатываться ими. Весьма сомнительно,
чтобы нашлись еще другие животные, ко-
торые в истории земной коры заняли бы
столь видное место...»
Книга Дарвина дала толчок новым ис-
следованиям, число которых продолжает
увеличиваться. И если вначале многие ду-
мали, что он переоценил до некоторой сте-
пени роль червей, то теперь становится
ясно, что в действительности во многих
случаях значение их еще больше. Нужно
только внести одну существенную поправ-
ку: в процессах почвообразования, кроме
дождевых червей, участвуют многие дру-
гие беспозвоночные животные, главным
образом некоторые насекомые и их личин-
ки. И хотя в отдельных случаях, особенно
там, где червей мало или вовсе нет, другие
животные играют заметную роль, в общем
все же первое место в процессах почвооб-
разования среди животных принадлежит
дождевым червям.
О том, что мнение Дарвина о масштабах
деятельности дождевых червей не было
преувеличенным, можно судить по следу-
ющим цифрам. Дарвин исходил из того,
что количество червей на 1 га было равно
60—133 тыс. Позднейшие работы показа-
ли, что хотя в некоторых местах числен-
ность их может быть иногда даже несколь-
ко меньшей, но, как правило, она значи-
тельно выше, порядка 500 тыс.—2 млн. на
1 га, а в отдельных случаях достигает
5—7 млн. и даже 20 млн. Соответственно
живой вес (биомасса) будет измеряться
для слабо заселенных ими территорий ве-
личинами порядка от нескольких десятков
килограммов до 100—120 кг на 1 га, а
для тех мест, где их много,— до 2—3 т
на 1 га. Если мы вспомним, что за сутки
каждый червь пропускает через кишечник
количество земли, равное весу его тела
(а некоторые виды даже больше), то пой-
мем, что на каждом гектаре черви пере-
рабатывают ежесуточно от нескольких
десятков килограммов земли до 2—3 га, а
в среднем около четверти тонны. Расчеты,
сделанные для различных мест умеренно-
го и тропического поясов, показывают,
что количество выбрасываемой червями
земли таково, что оно образует за один
год слой от 1 до 7 мм толщиной, в среднем
около 3 мм.
Влияние деятельности дождевых червей
на особенности почвы многообразно. Преж-
де всего, в результате перекапывания ими
земли и прокладывания ходов повышает-
ся скважность почвы, она делается более
рыхлой, ее объем увеличивается (на 15 —
30%). Вследствие этого облегчается доступ
в глубокие слои почвы воды и атмосферно-
го воздуха, необходимых как для корней
травянистых растений и деревьев, так и
для деятельности полезных микроорга-
низмов. На глубину около 1 м почва обыч-
508
но бывает вся пронизана ходами червей,
а у ряда видов их ходы идут гораздо глуб-
же — до 4—8 м. Общая длина ходов под
1 м2 поверхности земли нередко превышает
1 км, а в отдельных случаях достигает
8 км. По ходам червей легче проникают в
глубину корни растений, особенно в более
плотных почвах (рис. 289). Далее, черви
постоянно перемешивают поверхностные
слои, занося полуразложившиеся листья
и другие органические остатки с поверх-
ности в глубь почвы и вынося землю из
нижних горизонтов на поверхность, а так-
же откладывая экскременты внутри своих
ходов. Расчеты показывают, что верхние
слои почвы, по крайней мере на глубину
0,5—1 ле, полностью перерабатываются
червями, проходя через их кишечник, при-
мерно за 100—200 лет.
Под влиянием червей изменяются и хи-
мические особенности почвы. В перера-
ботанной ими, смешанной с кишечной
слизью и выброшенной в виде копролитов
земле повышается содержание кальция,
магния, аммиака, нитратов, фосфорной
кислоты. Многие соединения переводятся
в другую, более доступную для использо-
вания растениями форму. Благодаря вы-
работке червями в особых известковых
железках пищевода сростков кристалли-
ков углекислого кальция — кальци-
та — происходит нейтрализация вредных
почвенных кислот и выбрасываемая чер-
Пиявки ... Это слово обычно вызывает
неприятное чувство: воображение рисует
длинных, темных червей, живущих в бо-
лотистых водоемах, нападающих на че-
ловека и сосущих его кровь. Многие люди
знают только одну пиявку — медицин-
скую, применяемую при лечении некото-
рых заболеваний, нередко очень тяжелых.
Между тем на земном шаре насчитывается
около 400 видов пиявок, их строение раз-
нообразно, они обитают не только в бо-
лотистых водоемах, но и в реках, озерах,
вями земля имеет щелочную реакцию.
Наконец, копролиты червей, высыхая,
постепенно распадаются на небольшие
микропористые комочки, долго не размы-
ваемые водой. Благодаря этому перера-
ботанная червями земля приобретает
стойкую мелкокомковатую структуру.
Кроме того, в копролитах повышается
количество полезных почвенных микро-
организмов. Все эти процессы, происходя-
щие в почве под влиянием дождевых чер-
вей, заметно изменяют и улучшают ее
состав, структуру и свойства, повышают
ее плодородие.
Однако, кроме положительной, дожде-
вые черви могут в определенных условиях
играть и отрицательную роль. Как прави-
ло, живых здоровых растений они не
трогают, но могут иногда втягивать в
норки концы листьев травянистых расте-
ний и в какой-то мере повреждать всходы.
Приносимый ими таким образом вред не
может иметь существенного значения. Го-
раздо важнее то, что дождевые черви яв-
ляются промежуточными хозяевами для
нескольких видов паразитических червей,
вызывающих заболевания сельскохозяй-
ственных животных. Так, ими распро-
страняется довольно тяжелое заболевание
свиней, от которого особенно страдает
молодняк,— метастронгилез, или
легочно-глистная болезнь, а также ряд
глистных заболеваний кур и других птиц.
горных ручьях и даже в морях и океанах.
Пиявки-кровососы действительно состав-
ляют большинство этого класса типа коль-
чецов, но живут они за счет разных жи-
вотных (представителей всех классов по-
звоночных, мягкотелых, ракообразных, вод-
ных насекомых, червей и др.), а не только
млекопитающих и человека. И совсем уж
мало кому известно, что многие виды пия-
вок не могут сосать кровь, а принадлежат
к числу хищников, которые целиком или
по частям заглатывают мелких животных.
КЛАСС ПИЯВЩ1 (HIRUDINEA)
509
Рис. 290. Глаза пиявок:
1 — птичьей; 2 — улитковой; 3 — рыбьей;
4 — челюстной; 5 — глоточной.
Правда, хищные формы произошли от
кровососущих и сохранили основные
признаки своих предков, но по характеру
питания они коренным образом отличают-
ся от них. Что касается пиявок, сосущих
кровь млекопитающих и человека, то в
тропиках их изрядное количество, но в
нашей стране таких видов всего два-
три (из 50 пресноводных видов) и распро-
странены они преимущественно в южных
районах.
Принадлежность пиявок к т и пу коль-
чатых червей не вызывает никаких сомне-
ний. Тело их сегментировано, централь-
ная нервная система состоит из головных
узлов, окологлоточных тяжей и брюшной
цепочки; в кожно-муокульном мешке есть
те же основные элементы, что и у других
кольчецов, перенос веществ осуществляет-
ся при помощи кровеносной системы, орга-
ны выделения — метанефридии, кишечник
сквозной, заканчивающийся анальным
отверстием, мускулистый, обильно снаб-
женный кровеносными сосудами, и т. д. В
то же время для пиявок характерны мно-
гие особенности, которые позволяют лег-
ко отличить их от других групп типа.
В связи с необходимостью прикреплять-
ся к телу других животных для сосания
крови у пиявок развились две присоски
(передняя, окружающая рот, и задняя),
тело стало более или менее уплощенным.
Щетинок, за одним исключением, нет, так
как ползают эти черви при помощи при-
сосок. Число сегментов, или сомитов, в
отличие от других кольчецов, постоянное
и у всех видов, за исключением одного,
равно 33, из которых последние семь обра-
зуют заднюю присоску. Сравнительно не-
большое количество сегментов, вероятно,
уменьшало гибкость тела, и у пиявок воз-
никла очень характерная для них вторич-
ная кольчатость тела: сомиты подразде-
ляются на определенное для каждой
группы видов число колец. У медицинской
и других челюстных пиявок в сомите пять
колец, у улитковой и подавляющего боль-
шинства видов семейства плоских
пиявок — три и т. д. Вторичная кольча-
тость затрагивает только наружные по-
кровы и на внутренние органы не распро-
страняется. Центральным кольцом счи-
тается то, которое несет узел брюшной
нервной цепочки.
Среди пиявок почти нет настоящих па-
разитов, т. е. таких видов, которые по-
стоянно обитают на теле хозяина или внут-
ри его, что всегда влечет за собой упро-
щение организации паразита. Наоборот,
большинство их ведет активный образ
жизни в поисках своих жертв, сопротив-
ление которых они должны преодолеть.
Поэтому у пиявок хорошо развита нерв-
ная система, и у всех червей, живущих на
свету, имеются глаза, число и расположе-
ние которых характерны для групп видов
или даже для отдельных видов. Тело очень
мускулисто, и в этом отношении пиявки
превосходят всех червей. Так, например,
у дождевых червей мускулатура состав-
ляет 29,7% объема всего тела, а у пиявок-
до 65,5%. Многие виды хорошо плавают.
Промежутки между органами заполнены
соединительной тканью, и полость тела
(вторичная), характерная для всех коль-
чецов, у пиявок отсутствует. Тело их
очень плотное, с трудом поддающееся
разрыву. Кровеносная система, прекрасно
развитая, претерпела существенные из-
менения. Изменились и метанефридии,
лишившиеся в связи с отсутствием поло-
сти тела воронок.
Существенным изменениям подвергся пи-
щеварительный аппарат. Все пиявки, за
исключением одного древнего вида, делят-
ся на два о т ряда: хоботных и челюстных
(бесхоботных), У первых в передней части
пищеварительной трубки развился му-
510
скулистый хобот, у вторых — челюсти (их
обычно три), усаженные зубчиками. При
помощи хобота или челюстей кровососущие
виды повреждают кожу или слизистые
оболочки своих жертв. У хищных челю-
стных пиявок, целиком заглатывающих
свою добычу, челюсти уменьшаются или
даже совсем исчезают. За глоткой, слу-
жащей для сосания крови, и коротким пи-
щеводом идет желудок, объем которого у
кровососущих видов сильно увеличен бла-
годаря парным боковым отросткам. У
хищных видов отростки желудка пол-
ностью или частично исчезают. Усвоение
пищи совершается в идущей за желудком
кишке, которая у многих пиявок тоже
имеет отростки. Кал удаляется через зад-
нюю кишку и анальное отверстие, лежащее
на спинной стороне, у задней присоски.
Хорошо известно, что после укусов
пиявок раны долго кровоточат. Это объяс-
няется тем, что в раны из слюнных желез,
открывающихся в ротовую полость пиявок,
попадает особое белковое вещество ги-
рудин (от греческого слова «гирудо»—
пиявка), которое препятствует сверты-
ванию крови. Если бы гирудин не выде-
лялся, то быстро образовались бы кровя-
ные сгустки (тромбы) и сосание крови ста-
новилось бы невозможным. Благодаря
гирудину и другим веществам, выделяе-
мым слюнными железами, кровь месяцами
сохраняется в желудке пиявок в жидком
состоянии, не подвергаясь гниению.
Все пиявки гермафродиты (у взрослых
червей развиты и мужские и женские поло-
вые органы) и размножаются только поло-
вым путем. На брюшной стороне этих чер-
вей, выше средней части тела, довольно
хорошо заметны два половых отверстия:
переднее, более крупное — мужское, зад-
нее — женское. Расстояние между этими
отверстиями, измеряемое количеством ко-
лец, у различных видов неодинаковое и
имеет значение для определения видов
пиявок. Оплодотворение, т. е. слияние
живчиков и яйцеклеток, происходит внут-
ри организма. Семя передается при по-
ловом акте двумя способами. У одних
видов (в том числе у медицинской) оно
вводится в женское половое отверстие при
помощи совокупительного органа, имею-
щего вид тонкой нити, а у остальных жив-
чики выделяются в особых мешочках
(сперматофорах), которые прикрепляются к
коже«другой пиявки в разных местах. Жив-
чики через ранки, образующиеся на коже
после прикрепления сперматофоров, про-
никают внутрь тела, отыскивают зрелые
яйцеклетки и оплодотворяют их.
Оплодотворенные яйца выделяются в
коконах, строение которых будет описано
дальше. У половозрелых пиявок, как и у
малощетинковых червей, в области поло-
вых органов на коже образуется «поясок»,
часто хорошо заметный. Из выделений
желез пояска образуются стенки коконов.
По окончании развития, которое обычно
длится несколько недель, из коконов вы-
ходят маленькие черви, в основном похо-
жие на взрослых.
Рис. 291. Медицинская пиявка (Hirudo medi-
cinalis).
А — схема пищеварительного аппарата: I — перед-
няя присоска; 2 — глотка, 3 — пищевод, 4—7 — раз-
ные части желудка; 8 — переход желудка в кишку;
9 — железистые придатки кишки; ю — кишка; 11 —
задний отросток желудка; 12 — начало задней киш-
ки; 13 — задняя кишка, 14 — анальное отверстие;
15 — задняя присоска. Б — передняя часть пищева-
рительного канала: 1, 2, 3 — челюсти, 4 — глотка;
5—пищевод. В — отдельная челюсть (сильно уве-
личена).
511
Пиявки распространены во всех частях
света. Каждая зоогеографическая область
характеризуется своим составом видов
этих червей. Видов, обитающих в двух
или более областях, очень мало. Морские
пиявки разделяются на ряд групп, из
которых каждая характерна для опреде-
ленного района Мирового океана и при-
легающих к нему морей. В Черном море
В сороковых годах прошлого столетия
выдающийся русский натуралист
А. Ф. Миддендорфво время путеше-
ствия по Енисею собрал с пеледи,или сырка
(Coregonus peled — из сигов, семей-
ство лососевых рыб), странных червей,
разъедавших мягкие части спинного плав-
ника. Уже поверхностное изучение этих
червей, проведенное известным зоологом
Э. Г р у б е, показало, что в их организа-
ции удивительным образом сочетаются
признаки малощетинковых червей и пия-
вок. На переднем конце их тела имеются
щетинки, которые выполняют роль перед-
ней присоски (отсутствующей у этих чер-
вей) и вместе с несовершенной задней
присоской служат для прикрепления к те-
лу хозяина. Грубе отнес описанных им
червей к новому р о д,у Acanthobdella,
что значит «вооруженная пияпка», а ви-
довое название дал по имени рыб, с ко-
Рис. 292. Щетинковая пиявка (Acanthobdella):
1 — целая пиявка; 2 — передний конец тела с ще-
тинками.
пиявок нет совсем, так как его соленость
(в два раза меньше солености океана) недо-
статочна для настоящих морских пиявок
и слишком велика для видов, обитающих
в пресных и солоноватых водах.
Пиявки имеют практическое значение,
как полезное, так и вредное, о чем вкрат-
це будет рассказано при описании отдель-
ных видов.
торых они были сняты. Впоследствии
акантобделла, или щетинковая пиявка,
была обстоятельно изучена известным рус-
ским зоологом Н. А. Ливановым,
который выяснил, что во внутреннем строе-
нии ее тоже наблюдается сочетание при-
знаков малощетинковых червей и пиявок,
но признаки последних преобладают, и
акантобделла была отнесена к особой,
низшей группе класса пиявок. Сейчас ще-
тинковых пиявок выделяют в особый под-
класс древних пиявок, существование
которых является блестящим подтвержде-
нием эволюционной теории.
Длина акантобделлы обычно не превы-
шает 30 мм, а толщина — 3 мм. Цвет тела
серый со слабым мясо-красным оттенком.
Часто на теле заметны темные поперечные
полоски. Эта пиявка распространена на
крайнем севере Европы и Азии, от Нор-
вегии до Колымы; самое южное ее место-
нахождение — Онежское озеро. Парази-
тирует щетинковая пиявка на разных лосо-
севых рыбах, а также на хариусах и пи-
тается не только кровью, но и размягчен-
ными наружными тканями хозяев и, сле-
довательно, еще не является специализиро-
ванным кровососом. На каждой рыбе обыч-
но обитает несколько акантобделл, которые
причиняют существенный вред своему хо-
зяину. Они появляются на рыбах поздней
весной или в начале лета, когда вес их
равен всего 5—10 мг\ и в конце осени или
в начале зимы достигают половой зрело-
сти и веса больше 200 мг. Затем акантобдел-
лы покидают хозяев и, по-видимому, раз-
множаются и развиваются на мелких ме-
ПОДКЛАСС ДРЕВНИЕ ПИЯВКИ (ARCHIHIRUDINEA)
512
стах, заросших подводной растительностью.
Но никто еще не ловил акантобделл в
свободном состоянии и не наблюдал, как
происходит размножение и развитие этих
В этот подкласс входят все виды пия-
вок, кроме щетинковой. Передняя при-
соска у них всегда хорошо развита, ще-
тинок нет; все особенности, отличающие
пиявок от малощетинковых червей, хоро-
шо выражены. Подкласс делится на два
отряда: отряд хоботных и отряд
челюстных, или бесхоботных.
ОТРЯД ХОБОТНЫЕ ПИЯВКИ
(RHYNCHOBDELLAE)
Название отряда говорит о главной осо-
бенности входящих в его состав видов:
все они имеют хобот. Заметить у живой
пиявки хобот очень трудно, у убитых чер-
вей он иногда торчит изо рта. Отряд в
свою очередь подразделяется на два резко
отличающихся между собой семейства:
семейство плоских пиявок и с е м е й-
с т в о рыбьих пиявок.
Семейство Плоские пиявки, или
Глоссифониды (Glossiphonidae). Русское
название этого семейства не совсем удач-
но, так как уплощенное тело вообще ха-
рактерно для пиявок. Правда, у глоссифо-
нид (или, как их раньше называли, клеп-
син) уплощенность выражена особенно
сильно, но и здесь бывают исключения.
Правильнее было бы назвать этих пия-
вок широкими или листообразными, по-
тому что тело у них относительно широкое,
суживающееся к концам. Размеры глос-
сифонид, как правило, незначительны (от
нескольких миллиметров до нескольких
сантиметров). Не плавают. Сосут кровь
(а некоторые виды и разжиженные ткани)
разных животных — беспозвоночных и
позвоночных.
Все виды глоссифонид проявляют забо-
ту о потомстве. Пиявки прикрывают своим
пиявок, хотя их удавалось содержать неко-
торое время в аквариумах. Можно только
утверждать, что жизненный цикл этих ин-
тересных червей продолжается около года.
телом отложенные ими тонкостенные бес-
форменные коконы, в которых заключе-
но множество яиц. Молодые пиявки после
вылупления прикрепляются к брюху ма-
тери и передвигаются вместе с ней. В
случае опасности пиявка-мать перестает
двигаться, защищая детей своим телом.
Подрастая, молодь переходит к самостоя-
тельной жизни, сначала временно, потом
совсем.
Первые глоссифониды, как и древние
пиявки, вероятно, сосали кровь рыб. В
наших водоемах встречается довольно по-
движная Hemiclepsis marginata, которую
иногда находят на разных рыбах. Ее легко
отличить от других глоссифонид по силь-
но расширенному переднему концу тела,
что помогает ей удерживаться на быстро
передвигающихся хозяевах. Длина ее до
30 мм, цвет тела зеленоватый с примесью
коричневого. Она также охотно сосет
кровь земноводных.
Особенно многочисленна гемиклепсис
в бассейне Амура. В связи с этим следует
отметить, что она обитает и в Южной
Азии.
Родственными предыдущему виду яв-
ляются птичьи пиявки, которые выделены
в особый род Protoclepsis (в иностранной
литературе этот род обычно носит назва-
ние Theromyzon). Они достигают при пол-
ном развитии значительной для глосси-
фонид величины: длина до 50 мм, ширина
до 10—15 мм] однако чаще попадаются
более мелкие экземпляры. Окраска тела
различная: от зеленовато-прозрачной до
почти черной. Их можно распознать по
числу (четыре пары) и расположению глаз
и удивительно мягкому телу. Они легко
проникают в ротовую полость и дыхатель-
ные органы птиц, где сосут кровь из сли-
зистых оболочек; могут находить себе
пристанище и в конъюнктивном мешке
ПОДКЛАСС НАСТОЯЩИЕ ПИЯВКИ (EUHIRUDINEA)
33 Жизнь животных, т. i
513
глаз. В некоторых стоячих водоемах пти-
чьи пиявки иногда размножаются в ог-
ромном количестве, что может привести
к массовой гибели уток и гусей. Домаш-
нюю водоплавающую птицу из стоячих
водоемов, где сильно размножилась про-
токлепсис, нужно перевести в проточные
водоемы, в которых такие явления не на-
блюдаются. Для изгнания паразитов из
организмов в ноздри их хозяев вводят
10-процентный раствор поваренной соли,
люголевский раствор, чистый скипидар
или его водные растворы и другие веще-
ства.
В пределах нашей страны встречаются
два вида Protoclepsis: широко распростра-
ненная обыкновенная птичья пиявка (Р.
tessulata) (она известна также в Северной
и Южной Америке) и пятнистая птичья
пиявка (P. maculosa), встречающаяся в
северной половине Европы и в Северной
Азии. Опасна главным образом первая.
Интересны особенности жизни птичьих
пиявок, изученные главным образом у
второй из них. Первый раз она сосет кровь
птиц через 1 —1,5 месяца после выхода из
кокона, второй раз — через 20—30 дней
после первого питания и третий раз —
через 1,5—2 месяца после второго. Через
4—6 месяцев после третьего питания про-
токлепсис становится половозрелой и раз-
множается. После откладки коконов пияв-
ки могут прожить еще некоторое вре-
мя, но крови больше не сосут. Только те
из них, которые по какой-то причине не
отложили коконов, могут питаться в чет-
вертый раз.
Обыкновенная птичья пиявка отклады-
вает три — пять коконов. Общее число
отложенных яиц колеблется в широких
пределах: от 65 до 611. Молодь может жить
на теле материнского организма до двух-
трех месяцев.
Возможно, что в родстве с птичьими
пиявками находится черепашья пиявка
(Haementeria costata). Ее довольно легко
узнать по прерывистой (пунктирной) уз-
кой светлой полоске и ряду мелких сосоч-
ков, идущих посередине спины (остальные
ряды сосочков проходят снаружи от сре-
динного ряда). Обычная длина взрослых
червей около 20 мм, но встречаются и
более крупные экземпляры. Окраска от
зеленоватой до почти коричневой. Глаз
одна пара. Паразитирует главным образом
на болотной черепахе (Emys orbicularis),
Рис. 293. Черепашья пиявка (Haementeria
costata):
I — со спинной стороны; 2 — с брюшной стороны
(видны прикрепленные к брюху зародыши пиявок).
Рис. 294. Улитковая
пиявка (Glossiphonia
complanata) со
спинной стороны.
514
но довольно часто попадается и в свобод-
ном состоянии. В период размножения
всегда оставляет своего хозяина. Иначе
во время нередких выходов черепахи на
сушу молодые пиявки, кожные покровы
которых очень нежны, быстро погибали
бы. Черепашьи пиявки могут в некото-
рых случаях сосать кровь птиц, млекопи-
тающих и человека. Выдающийся русский
зоолог А. О. Ковалевский, посвя-
тивший этому виду прекрасную моногра-
фию, рассказывает, как пиявки сосали у
него кровь между пальцами ног, когда
он ходил в воде около берега, собирая
материал для своей работы. Черепашья
пиявка в нашей стране часто встречается
только на юге (Украина, Кавказ и т. д.).
Других видов этого рода у нас нет. Меж-
ду тем в Африке (южнее Сахары), Юж-
ной Азии и Южной Америке, а также
на юге Северной Америки видов Haemen-
teria много, сосут они кровь главным
образом водных пресмыкающихся и зем-
новодных, которые, как известно, много-
численны в теплых и жарких странах.
Некоторые виды гементерий из Централь-
ной и Южной Америки имеют большие
размеры, чем наша черепашья пиявка,
и используются там для медицинских
целей. Эти пиявки сосут также и кровь
птиц.
Южного происхождения и глоссифони-
ды, принадлежащие к роду Batracobdella
(«батрахос» по-гречески — лягушка,
«бделла» — пиявка). Они сосут кровь
земноводных. В Крыму водится в большом
количестве маленькая (обычная длина око-
ло 8 мм) зеленовато-бурая пиявка с парой
крупных глаз — В. algira. Почти всю
жизнь она проводит на больших лягуш-
ках и покидает их только тогда, когда
наступает период размножения (ср. с
черепашьей пиявкой). Она впервые была
найдена в Алжире, чем объясняется ее
видовое название, и распространена на
севере Африки, в Передней Азии, а в
Европе — на Пиренейском полуострове и
юге Балканского. У нас, кроме Крымского
полуострова, она нигде не найдена. Пред-
полагают, что когда-то Крым был соеди-
нен с Малой Азией. Возможно, что
именно тогда эта интересная пиявка про-
никла в Крым. Другой вид того же рода—
четырехглазая Batracobdella paludosa —
тоже тяготеет к югу Европы, но заходит
довольно далеко на север (Англия, Поль-
ша и др.) и никогда не встречается вместе
с первым видом. Главный источник пита-
ния этой пиявки — лягушки и другие
земноводные, но она сосет и кровь катушек
(из брюхоногих моллюсков), у которых,
как и у позвоночных, кровь красного
цвета, т. е. содержит гемоглобин.
Самая известная представительница
глоссифонид в наших пресных водоемах,
по родовому названию которой названо
и все семейство,— улитковая пиявка (Glos-
siphonia complanata). Тело ее, длина ко-
торого редко превосходит 15—20 мм, от-
носительно очень широко. Окраска зеле-
новато-коричневая, очень изменчивая,
иногда весьма пестрая. На спинной сторо-
не три пары продольных рядов сосочков,
из которых срединные развиты лучше
других. Взрослая пиявка на редкость ле-
нива и много времени неподвижно лежит,
присосавшись к подводным предметам и к
широколистным растениям *. Благодаря
своей окраске и неподвижности, покрытая
нередко частицами ила, она почти или
совсем незаметна. Ее главные жертвы —
преимущественно легочные брюхоногие,
мягкотелые (прудовики и др.), которые
нередко после ее нападения погибают в
результате потери крови и других соков
или закупорки дыхательного отверстия.
Овладение пиявкой своими жертвами об-
легчается медлительностью названных мол-
люсков.
Возможно, что глоссифониды, живу-
щие за счет мягкотелых, возникли по-
зднее представителей того же семейства,
паразитирующих на быстро передвигаю-
щихся позвоночных животных.
Улитковая пиявка живет около двух
лет. Размножается дважды: в конце пер-
вого и второго года жизни. Откладывает
до 120 яиц, по 20 в каждом коконе. После
второй кладки яиц пиявки обычно поги-
бают, лишь немногие из них могут достиг-
нуть трехлетнего возраста. G. complanata
обитает и в Северной Америке.
1 Если улитковых пиявок отделить от суб-
страта, то они, как и некоторые другие глосси-
фониды, свертываются наподобие ежей.
33*
515
Еще чаще встречается в на-
ших водоемах, в особенности
в стоячих, маленькая (обыч-
ная длина — 5—6 мм) се-
ровато-белая глоссифонида—
Helobdella stagnalis. Ее от-
личительный признак — лин-
зообразная пластиночка жел-
того или коричневого цвета,
расположенная на спине ме-
жду 12-м и 13-м кольцами.
Поэтому по-русски ее следо-
вало бы назвать пластинко-
носной пиявкой. Глаз одна па-
ра, довольно крупных. В от-
личие от улитковой пиявки
Н. stagnalis очень подвиж-
на, что облегчает ей нападе-
ние на личинок водных на-
секомых, рачков, малощетин-
ковых червей, других пиявок
и мелких беспозвоночных,
которых она нередко выса-
сывает целиком. В то же вре-
мя благодаря своей подвиж-
ности она значительно чаще
улитковой пиявки попадаете
желудки рыб. Живет пластинконосная пи-
явка всего год. Вылупившиеся из коконов
весной пиявочки быстро растут и в июле —
августе уже могут размножаться; затем,
следующей весной, они еще раз откладыва-
ют яйца и погибают. В одной кладке от 7 до
37 яиц, которые находятся в двух коконах.
Н. stagnalis — одна из самых распростра-
ненных пиявок: кроме северной полови-
ны Азии, Европы и севера Африки, она
живет в Северной и Южной Америке, где
насчитывается не менее двух десятков
видов того же рода, в то время как из
других частей света известно по 1 — 3 вида.
Возможно, что столь обычная у нас пияв-
ка южноамериканского происхождения.
В заключение обзора плоских пиявок
следует вкратце остановиться на байкаль-
ских видах этого семейства, которых всего
три: Baicaloclepsis grubei, В. echinulata,
Paratorix baicalensis.
Фауна Байкала удивительна и привле-
кает внимание зоологов уже в течение
ста лет. Подавляющее большинство жи-
вотных, обитающих в этом самом глубоком
и древнем озере на земном шаре, встречает-
Рис. 295. Пиявка торикс
(Torix cotylifer) с брюшной
стороны.
ся только здесь и резко от-
личается от животных тех же
групп, населяющих водоемы
Сибири. Многие байкальские
виды принадлежат к особым
родам и даже семействам. Пе-
речисленные пиявки тоже
относятся к родам, которые
не имеют представителей вне
Байкала. Особенно интерес-
на Baicaloclepsis echinulata.
Ее видовое название («еже-
видная») вполне заслуженно:
вся спина покрыта сосочка-
ми, и эта серовато-белая пи-
явка (длина ее не более 15 мм)
имеет мохнатый вид. Более
крупная (длина до 40 мм) —
Baicaloclepsis grubei желто-
ватого цвета с нежно-розо-
вым оттенком, имеет на спин-
ной стороне шесть рядов
больших сосочков. У пер-
вой пиявки совсем нет глаз,
а у второй они слабо развиты.
Недоразвитие или отсутствие
глаз и беловатая окраска
тела обеих пиявок, возможно, объясняет-
ся тем, что они обитают на довольно
большой глубине, где света очень мало.
Третья— байкальская глоссифонида (Para-
torix baicalensis) — живет, вероятно, в
условиях лучшего освещения, так как
окраска ее коричневатая и глаза развиты.
О питании, размножении и развитии этих
интересных пиявок никаких достоверных
данных нет. Все они имеют маленькие зад-
ние присоски и, очевидно, сосут кровь ма-
лоподвижных животных, каких именно —
неизвестно. Как почти все байкальские
животные, они могут жить только в хо-
лодной воде, хорошо насыщенной кисло-
родом.
Описанные виды чужды современной
фауне пиявок. Известно лишь несколько
видов, которые находятся с ними в род-
стве. Из них назовем паразита мягкокожей
черепахи (Amyda sinensis) — Torix cotyli-
fer — маленькую беловатую пиявку, ко-
торая вместе со своим хозяином встречает-
ся у нас в озере Ханка, а за пределами
СССР — в Китае, и сильно похожую на
нее Oligobdella orientalis (Южные Курилы,
516
^ввввв«»»«ааавда»
Рис. 296. Обыкновенные рыбьи пиявки (Piscicola
geometra), паразитирующие на карпе.
Япония) — паразита лягушек и, возмож-
но, других земноводных. У обеих пиявок,
паразитирующих на очень подвижных,
сильных животных, огромные задние при-
соски. В Корее, Китае, Японии, США
и Бразилии найдено еще пять видов,
близких к предыдущим. Эта своеобразная
группа пиявок, по-видимому, много мил-
лионов лет назад была широко распро-
странена в Восточной Азии и проникла
оттуда в Америку, а потом в изменивших-
ся условиях большей частью вымерла.
Таким образом, байкальские глоссифони-
ды весьма древнего происхождения.
Семейство Рыбьи пиявки, или Их-
тиобделлиды (Ichthyobdellidae, или Pisci-
colidae). Подавляющее большинство видов
этого семейства принадлежит к паразитам
различных рыб, и лишь некоторые виды
имеют других хозяев: ракообразных, мно-
гоколенчатых (пантопод), мягкотелых п че-
репах. Только в этом семействе есть пияв-
ки, обитающие в морях и океанах, причем
морских видов рыбьих пиявок раза в три
больше, чем пресноводных и солоновато-
водных. Интересно, что почти все пресно-
водные ихтиобделлиды распространены в
северном полушарии. Поэтому в водоемах
СССР численность обеих групп рыбьих
пиявок приблизительно равна. Известно
всего лишь несколько видов рассматривае-
Рис. 297. Трахелобделла китай-
ская (Trachelobdella sinensis).
мого семейства, живущих как в морской,
так и в пресной воде.
Отличить рыбьих пиявок от других очень
легко: их передняя присоска резко отгра-
ничена от остальной части тела и часто
имеет форму диска, чашечки и т. д. Оче-
видно, сильное развитие передней присос-
ки объясняется необходимостью крепко
держаться на таких быстро плавающих
животных, как рыбы. На передней при-
соске у большинства видов имеются две
пары глаз, а на задней присоске часто
расположены глазоподобные точки. У мор-
ских видов еще одна пара глаз может на-
ходиться на одном из передних колец за
присоской, а по бокам некоторых сомитов
нередко развиты сегментальные глазки.
Количество колец в сомите колеблется от
трех (что характерно, как мы видели, для
большинства глоссифонид, от которых
произошли ихтиобделлиды) до четырнад-
цати. У ряда видов по бокам тела распо-
ложены дыхательные пузырьки разных
размеров: от почти незаметных до очень
больших. Пузырьки ритмически сокра-
щаются и усиливают газообмен. Они име-
ются у всех пресноводных видов и лишь у
некоторых морских. Это объясняется тем,
что условия дыхания в пресной воде менее
благоприятны, чем в морской. Оплодотво-
ренные яйца откладываются в маленьких
твердых коконах, которые, как правило,
прикрепляются к растениям или к раз-
личным предметам, а у морских видов,
паразитирующих на раках и многоколен-
чатых, к панцирям хозяев. Некоторые
виды плавают.
В наших пресных водоемах широко рас-
пространена обычная рыбья пиявка (Pi-
se icola geometra), или писцикола. Длина
ее до 20—50 мм, а ширина— 2,5 мм.
Окраска очень изменчивая, чаще всего
серовато-зеленая, иногда желтоватая. Пи-
сцикола сосет кровь самых разных кости-
стых рыб. Живет она только в водах, до-
статочно насыщенных кислородом. В обыч-
ных прудах писцикола встречается очень
редко, но иногда может размножаться в
огромном количестве в проточных прудах.
Неоднократно описывались случаи, когда
на одной рыбе находили десятки и даже
сотни этих пиявок, что вело к массовой
гибели рыб. Паразиты не только истощают
хозяев, но часто заражают их опасными
болезнями. Поэтому рыб перед посадкой
в пруды необходимо тщательно осматри-
вать и в случае обнаружения пиявок на
полчаса погружать в ванны с 2,5-процент-
ным раствором поваренной соли или 0,005-
Рис. 298. Морские
пиявки:
1 — карцинобделла (Са-
rcinobdella cyclostoma)—
паразит камчатского
краба; 2 — левинсения
(Levinsenia rectongula-
ta) — паразит трески.
Рис. 299. Пре-
сноводная пи-
явка озобран-
хус (Ozobran-
chus jantsea-
nus). По бокам
тела — жабры.
процентным раствором двухлористои меди.
Указанные растворы не опасны для рыб,
но гибельно действуют на пиявок, кото-
рые оставляют своих хозяев и вскоре уми-
рают. Размножается писцикола преиму-
щественно весной. Ее овальные коконы
(длина их 1—1,5 мм, ширина около 0,5 мм)
красновато-коричневого цвета.
В Каспийском море часто попадается
маленькая Piscicola caspica, сильно от-
личающаяся по наружному и внутренне-
му строению от обычной рыбьей пиявки.
Средняя длина ее 8 мм, ширина 1 мм,
цвет тела зеленый со слабым розовым
оттенком или реже розовый, серый, свет-
ло-коричневый. Паразитирует она на раз-
ных осетровых и костистых рыбах. К
каспийской писциколе очень близка писци-
кола Фадеева (Piscicola fadejewi) — паразит
разных костистых рыб, и в особенности
леща. Она распространена, как и целый
ряд других каспийских животных (глав-
ным образом раков), в реках Азово-Чер-
номорского бассейна. В последнее время
эта пиявка сильно размножилась в неко-
торых водохранилищах и, несомненно,
причиняет существенный вред.
В противоположность писциколам виды
рода Cystobranchus обычно паразитируют
на одном каком-нибудь хозяине. Назовем
здесь прежде всего крупную (длина до
75 мм) сомовую пиявку (Cystobranchus
fasicatus), отличающуюся огромной зад-
ней присоской. Особенно много находят
ее в дельте Волги на сомах в ямах, где эти
рыбы скопляются во время зимовки. Мень-
шие размеры (длина до 20 мм), более ко-
роткое тело и недоразвитые глаза имеет
налимовая пиявка (Cystobranchus mammil-
latus), которая прикрепляется к жабрам
хозяина. Распространена она на севере.
От предыдущих видов резко отличается
очень крупная (длина до 44 мм, ширина
около 10 мм) трахелобделла китайская
(Trachelobdella sinensis). Родовое название
этой и близких к ней пиявок объясняется
тем, что тело их состоит из более узкой
шеи (по-гречески «трахелос» — шея) и
широкого туловища. Паразитирует она
на жабрах сазанов и иногда карасей, т.е.
обитает в местах, где мало света. Поэтому
ее окраска желтовато-белая, обычная для
внутренних паразитов. Дыхательные пу-
518
зырьки огромны. У нас она распростра-
нена только в бассейне Амура и особенно
многочисленна в озере Ханка, а за преде-
лами СССР — в Китае и Монголии. Ее
нередко находят на рыбах в большом ко-
личестве, особенно зимой и весной. По-
нятно, что такие паразиты могут причи-
нить своим хозяевам серьезный вред. По-
этому при акклиматизации дальневосточ-
ных сазанов и карасей в других местах
Советского Союза следует опасаться зано-
са трахелобделл. Между тем известен слу-
чай, когда эта огромная пиявка была об-
наружена на сазанах, завезенных в Лит-
ву с Дальнего Востока.
В Средней Азии (бассейн Аральского
моря и река Или) на рыбах маринке и
усаче паразитирует более мелкая (длина
не более 20 мм) форма — трахелобделла
туркестанская (Trachelobdella turkesta-
nica).
Совсем мала байкальская трахелобдел-
ла (Trachelobdella torquata), обычная
длина которой 4—6 мм. Ее хозяева —
мелкие рачки-бокоплавы и бычки, т. е.
самые многочисленные группы байкаль-
ских животных.
Байкальская трахелобделла очень мно-
гочисленна в прибрежной зоне Байкала.
Это единственная байкальская пиявка,
которая встречается и за пределами
озера, в вытекающей из него Ангаре, но
только в самой верхней части реки,
где вода еще холодна и очень насы-
щена кислородом. В этой части Ангары
обычные пиявки не встречаются. Вообще
виды обеих групп (обыкновенной и бай-
кальской) вместе не живут. Следует от-
метить, что байкальские животные очень
чувствительны к изменению условий жиз-
ни (ухудшение кислородного режима,
сток различных химических веществ, заи-
ление грунтов и т. д.). Так, например,
после сооружения плотины Иркутской
гидроэлектростанции байкальская трахе-
лобделла почти не встречается в Иркут-
ском водохранилище, образовавшемся в
верхней части Ангары, где эта пиявка бы-
ла многочисленна. В названном водохра-
нилище вода сравнительно чиста, но дно
стало заиляться и резко уменьшилась
скорость течения. Это показывает, как
опасны различные изменения режима озе-
ра Байкал, животный мир которого уни-
кален и должен быть сохранен во что бы
то ни стало.
Известна еще одна байкальская ихтиоб-
делла — Codonobdella truncata, отли-
чающаяся очень маленькой задней при-
соской. Все тело ее покрыто, в отличие от
других пресноводных видов того же се-
мейства, мелкими сосочками. Обитает она
чаще всего на большой глубине (вплоть
до 1100 ле, где она встречается ча-
сто) и быстро погибает, когда попадает
на поверхность. Паразитирует она, по-
видимому, на бокоплавах.
Из видов рассматриваемого семейства,
обитающих в наших северных и дальне-
восточных морях, упомянем лишь о не-
которых.
На креветках (Sclerocrangon boreas)
и, возможно, на бычках находят не-
большую пиявку (длина 10—20 мм) —
Platybdella fabricii, встречающуюся от
Гренландского до Японского моря. Коко-
ны она прикрепляет обычно к панцирям
раков.
На знаменитых гигантских камчатских
«крабах» (Paralithodes camtschatica), ко-
торые на самом деле находятся в родстве
с раками-отшельниками, а на настоящих
крабов похожи лишь внешне, паразити-
рует крупная пиявка Carcinobdella cyclos-
toma. Она может достигать в длину 125 мм
при ширине 0,7 мм; откладывает коконы
на панцирях хозяев. Этот вид распростра-
нен от Берингова моря до Японского и
вдоль Тихоокеанского побережья Север-
ной Америки. На одном из видов трески
(Gadus macrocephalus) тоже в северной
части Тихого океана встречается доволь-
но большая (длина до 43 мм) и относитель-
но широкая Levinsenia rectangulata с
очень хорошо развитой задней присоской.
В арктических морях от Гренландии до
Аляски распространена Johanssonia arcti-
са, которая «облюбовала» хозяев из двух
весьма далеких друг от друга групп жи-
вотных: многоколенчатых и скатов. Толь-
ко на скате Raja radiata в северной части
Атлантического океана и на западе Ледо-
витого океана паразитирует Oxytonosto-
ma typica, средняя длина которой около
17 мм, имеющая, как и предыдущий вид,
чашевидную присоску. На разных видах
519
скатов встречается очень большая, дли-
ной до 200 мм, стройная пиявка, вся по-
крытая сосочками,— Pontobdella murica-
ta. Она распространена от Средиземного
моря, где она многочисленна, до Гренлан-
дии; в водах СССР не встречается.
Нельзя не упомянуть пиявок тропиче-
ского рода Ozobranchus, которые интерес-
ны в разных отношениях. Эти пиявки, те-
ло которых разделено на шею и туловище,
имеют на передних туловищных сомитах
ветвистые жабры. Дыхательных пузырь-
ков у них нет. Одни виды рода живут в
морях, другие — в пресных водах Юж-
ной и Юго-Восточной Азии. Паразити-
руют они на различных видах черепах.
Большой интерес представляет Ozobran-
chus jantseanus, обитающий в реке Янцзы-
цзян, в Китае. Он способен подвергаться
значительному высыханию, превращаясь
в сморщенный комочек, а потом опять
оживать в воде. Очевидно, такое приспо-
собление позволяет ему хорошо перено-
сить путешествия вместе с его хозяевами
на суше. Жабры имеются и у видов мор-
ского рода Branchelion, но здесь они
иначе устроены и образуются около дыха-
тельных пузырьков.
В последнее время ряд видов ихтиоб-
деллид найден также в антарктических
водах. Вообще же представители этого
семейства обитают во всех океанах.
ОТРЯД ЧЕЛЮСТНЫЕ,
ИЛИ БЕСХОБОТНЫЕ, ПИЯВКИ
(GNATHOBDELLEA,
ARHYNCHOBDELLEA)
У видов этого отряда нет хобота, а раз-
виты челюсти, которые у хищных форм
менее развиты или совсем рудиментарны.
Семейство Челюстные пиявки (Gna-
thobdellidae). Крупные (длина более 100 мм)
или средней величины (длина более 30 —
50 мм) черви. Глаз обычно пять пар, рас-
положенных в виде дуги. В ротовой поло-
сти три челюсти. Сомит пятиколечный.
Имеется совокупительный орган. Коконы
с яйцами откладываются во влажном грун-
те в прибрежной полосе. Большинство ви-
дов — кровососы, живущие за счет раз-
ных позвоночных, меньшинство—хищники,
заглатывающие свою добычу.
Самая известная представительница
этого семейства — медицинская пиявка
(Hirudo medicinalis), которую уже в глу-
бокой древности использовали для лече-
ния людей. Средняя длина ее около 120 мм
(при ширине около 10 мм), но она может
достигать и значительно больших разме-
ров B50—300 мм), а в лаборатории извест-
ного советского специалиста по этим чер-
вям Г. Г. Щеголева путем усиленно-
го кормления была выращена за полтора
года гигантская пиявка в длину 440 мм\
Таблица 26. Пиявки:
1—пиявка, выползшая на берег для откладки яиц;
2 — коконы медицинской пиявки; слева —целый,
в центре — в продольном разрезе, справ а—
в поперечном разрезе.
3 — рыбья пиявка (Piscicola geometra);
4 — пиявка гелобделла (Helobdella stagnalis, уве-
личена).
5 — медицинская пиявка (Hirudo medicinalis, вид
снизу и сверху);
6 — улитковая пиявка (Glossiphonia complanata);
7 — конская пиявка (Limnatis nilotica, вид сни-
8у и сверху);
8 — ложноконская пиявка (Haemopis sanguisuga,
вид сниэу и сверху).
Это показывает, что не всегда можно су-
дить о возрасте пиявки по ее величине.
Между тем раньше считали, что медицин-
ские пиявки, достигшие максимальных
размеров, имеют возраст около 20 лет.
Сейчас это предположение нуждается в
проверке. Для медицинских целей обыч-
но употребляют сравнительно небольших
червей, длиной в несколько сантиметров.
Окраска Hirudo medicinalis очень измен-
чива, и описано множество цветовых форм
ее. Основной фон спинной стороны может
быть коричневым (разных оттенков), ры-
жеватым, оливково-черным, оливково-зе-
леноватым и т. д. Но как бы ни была ве-
лика изменчивость окраски, медицинскую
пиявку всегда можно узнать по двум про-
дольным узорчатым узким полоскам на
спине, которые заметны даже у очень
темных экземпляров. Боковые края (со
спинной и брюшной стороны) желтовато-
оранжевые. Брюхо обычно очень пестрое,
но может быть и однотонным. Поверхность
тела покрыта очень мелкими сосочками.
Тело довольно плотное. Задняя присоска
велика, ее диаметр превышает половину
наибольшей ширины тела. Пиявки про-
кусывают кожу тремя челюстями, усажен-
ными по краям острыми зубчиками (до
100 на каждой челюсти). Анальное отвер-
стие маленькое.
Медицинские пиявки обычно живут в
небольших мелких водоемах. Могут пере-
носить пересыхание, если грунт остается
достаточно влажным. Сосут кровь пред-
ставителей всех классов позвоночных, но
главными источниками их питания являют-
ся лягушки и млекопитающие (чаще всего
крупный рогатый скот, приходящий
на водопой). Опыты в лаборатории показа-
ли, что при кормлении на лягушках пияв-
ки достигают состояния, допускающего их
использование, через 17—20 месяцев, а
при кормлении на кроликах или сначала
на лягушках, а потом на кроликах —
через 8—10 месяцев. Пиявки могут до-
стичь половой зрелости и при питании за
счет холоднокровных животных, но тогда
развитие их протекает очень долго, они
откладывают всего один кокон (вместо
трех—восьми) и с меньшим количеством
яиц. Наиболее эффективно, по-видимому,
комбинированное кормление, т. е. на ля-
гушках и млекопитающих, что и происхо-
дит в природе. Таким образом, эволюция
медицинской и ряда других челюстных
пиявок совершалась в тесной связи с мле-
копитающими.
Медицинские пиявки — очень подвиж-
ные черви, особенно в голодном состоянии.
Они,как и большинство челюстных пиявок,
хорошо плавают, совершая волнообразные
движения. Содержать их нужно в хорошо
закрытой (марлей, сеткой и т. д.) посуде,
ибо они выползают из воды. Если кислоро-
да в воде недостаточно, то они, как и мно-
гие бесхоботные пиявки, укрепившись зад-
ней присоской, совершают дыхательные
движения, похожие на плавательные. Эти
пиявки хорошо реагируют на разные раз-
дражения. Так, если в воде производить
шум палкой, опусканием фанерного листа,
просто хождением, то они быстро при-
плывают к источнику шума. Если в сосуд с
пиявками бросить два одинаковых пред-
мета, из которых один был в руках чело-
века, а другой — нет, то около первого
скопляется больше червей, чем около вто-
рого. На некоторые же запахи (например,
одеколона) они реагируют отрицательно.
Они предпочитают теплую поверхность
холодной. Понятно, что чувствительность
к разным раздражениям помогает этим
кровососам отыскивать свои жертвы.
В природе Hirudo medicinalis достигает
половой зрелости, по-видимому, только
на третьем году жизни и откладывает ко-
коны один раз в год, летом. В лаборато-
рии половозрелых пиявок при благоприят-
ных условиях содержания и кормления
можно вырастить за 12—18 месяцев и,
содержа их при температуре 18—22° зи-
мой и 24—27° летом, заставить размно-
жаться в любое время и откладывать
коконы каждые 6—8 месяцев. В естест-
венной обстановке пиявки откладывают
коконы несколько выше уровня воды в
прибрежной полосе, для чего им нужно
преодолеть большое сопротивление грун-
та. Известен случай, когда коконы были
найдены в ста метрах от водоема. Коконы
очень похожи на коконы шелковичных
червей, их стенка состоит из сплетенных
волокон, выделенных железами пояска,
средняя длина кокона 20 мм, ширина
16 мм, цвет рыжевато-серый. В одном ко-
521
коне в среднем 15—20 яиц, продолжитель-
ность развития — около месяца. Вылу-
пившиеся пиявочки называются «нитчат-
ками», длина их всего 7—8 мм, челюсти у
них еще очень слабы, и прокусить кожу
млекопитающих они не в состоянии, но
вскоре могут прокусывать кожу земновод-
ных и сосать их кровь.
Медицинская пиявка южного происхож-
дения. В нашей стране она распростране-
на главным образом в Молдавии, на Украи-
не, Кавказе и в Средней Азии (правда, ее
нет в Туркмении). В северной половине
Европейской территории СССР, почти на
всем протяжении Западной Сибири, во
всей Восточной Сибири и на Дальнем Во-
стоке она отсутствует.
В течение многих столетий Hirudo medi-
cinalis применялась при лечении самых
разнообразных заболеваний, она была од-
ним из излюбленных средств народной ме-
дицины и врачей. Еще в середине прошло-
го столетия из Восточной Европы в запад-
ные страны, где запасы пиявок были исто-
щены, ввозились сотни миллионов этих
червей. Например, во Францию ввезли в
1850 г. около 100 млн. штук. Экспорт
пиявок из России считался выгоднейшей
статьей дохода. Во второй половине XIX в.
по мере развития научной медицины при-
менение пиявок врачами стало быстро со-
кращаться и ими почти перестали пользо-
ваться, хотя в народной медицине их про-
должали употреблять. Однако в 20-х го-
дах текущего столетия лечение пиявками
начало возрождаться. Различные исследо-
вания медиков и физиологов показали, что
выделяемые пиявками гирудин и, возмож-
но, другие вещества оказывают благотвор-
ное влияние при некоторых заболеваниях,
особенно при тромбофлебитах, гипертонии
и т. д. Конечно, сейчас не рассматривают
пиявок как панацею от всех недугов,
как это полагали когда-то, но в ряде слу-
чаев их применение целесообразно. Спрос
на пиявок в последнее время опять возрос,
и часто аптеки не могут его удовлетво-
рить. В связи с этим московские специа-
листы разработали методы быстрого выра-
щивания в лабораторных условиях пиявок,
как выловленных в природе, так и полу-
ченных из коконов в лаборатории. Широ-
кое применение методов искусственного
выращивания пиявок не исключает пла-
номерного вылова их в естественных водое-
мах, но при этом необходимо оградить
этих полезных червей от полного истреб-
ления, обеспечив их размножение.
Медицинские пиявки иногда причиняют
и вред. В некоторых мелких водоемах на
юге люди могут подвергнуться массовому
нападению этих кровососов. В таких слу-
чаях нужно немедленно выйти из воды и
снять червей, при этом лучше их не отры-
вать, а посыпать солью или смазать спир-
том, йодом и др., после чего они сами
отпадут. При небрежном содержании пия-
вок, употребляемых для лечения, они мо-
гут заползти в рот и другие отверстия
человека и даже присосаться к глазу.
Серьезную опасность для людей и до-
машних млекопитающих представляет дру-
гая челюстная пиявка — лимнатис (Lim-
natis nilotica). По-русски ее называют
конской или нильской, египетской, хотя
она сосет кровь разных млекопитающих
и живет не только в Египте, а во всех среди-
земноморских странах, в Абиссинии, Кон-
го, Танганьике, у нас — в Закавказье
и Средней Азии. Во взрослом состоянии
она достигает почти такой же величины,
как и Hirudo medicinalis. Спина ее зеле-
новато-коричневая, брюхо, в отличие от
других пиявок, окрашено темнее спинной
стороны и отливает синим или лиловым.
По бокам тела желтые полосы. Задняя
присоска велика, ее диаметр заметно пре-
восходит половину наибольшей ширины
тела. Челюсти малы и слабы и прокусить
кожу не могут. Поэтому лимнатис может
сосать кровь только из слизистых оболо-
чек, чем и объясняется ее серьезная
вредоносность. Она обитает в источниках,
в разных других небольших водоемах и
попадает в ротовую полость, а затем в
глотку, носоглотку, гортань млекопитаю-
щих и человека, когда они пьют воду
прямо из водоемов. В некоторых случаях
пиявка может закупорить гортань и вы-
звать удушение. Во время купания лю-
дей в водоемах, где водится описываемая
пиявка, она может проникнуть в моче-
вые и женские половые органы, в конъюн-
ктивный мешок глаз. Присутствие лимна-
тис вызывает кровохарканье и кровотече-
ние, нередко обильные. По некоторым дан-
522
ным, в Средней Азии иногда до 30% ско-
та, пригоняемого на бойни, заражено ею.
Аналогичные данные приводятся для Бол-
гарии, переднеазиатских стран и т. д.
Проникновение лимнатис в организмы
людей и животных облегчается тем, что
поверхность ее тела очень гладкая и вы-
деляет колоссальное количество слизи,
а благодаря мощной задней присоске она
прочно удерживается в тех или иных ор-
ганах. Известен случай, когда эта пиявка
находилась в человеке 3 месяца 20 дней.
Конская пиявка может также сосать и
лягушек. Размножение и развитие ее
во многом сходно с теми же процессами
Hirudo medicinalis.
Челюстные кровососущие пиявки мно-
гочисленны в жарких странах. Так, в
Африке (южнее Сахары) описано 9 видов
рода Hirudo и 14 видов рода Lim-
natis.
Среди видов описываемого семейства
насчитывается довольно большое количе-
ство некровососущих, хищных пиявок. В
водоемах СССР широко распространен
только один такой вид — большая ложно-
конская пиявка (Haemopis sanguisuga).
Ложноконской ее назвали, чтобы отличить
от лимнатис, на которую она похожа внеш-
не, а большой — в отличие от некоторых
глоточных пиявок, называемых малыми
ложноконскими пиявками (см. дальше). По
величине она не уступает медицинской
пиявке, а нередко ее превосходит. Спина
взрослой Н. sanguisuga черная с коричне-
ватым оттенком, на ее поверхности могут
быть разбросаны темные пятна, а у моло-
дых экземпляров основной фон спины
более светлый, чем у взрослых, и на нем
нередко виден правильный узор. Брюхо
серое или зеленовато-серое, боковые жел-
тые ленты часто отсутствуют. Задняя при-
соска невелика (меньше половины наи-
большей ширины тела). Челюсти у нее
развиты значительно хуже, чем у Н. medi-
cinalis. Анальное отверстие большое, так
как через него выделяются крупные куски
непереваренной пищи. Большая ложно-
конская пиявка— сильный и прожорли-
вый хищник, поедающий червей (в том
числе и пиявок), мягкотелых, личинок
водных насекомых и других водных беспоз-
воночных, а также мелких позвоночных
(например, головастиков), которых она
может одолеть. Иногда можно наблюдать,
как два хищника с двух противоположных
концов пытаются проглотить длинного
червя. Если они не справляются с добы-
чей, то отрывают от нее куски.
Большая ложноконская пиявка обитает
преимущественно в мелких водоемах, не-
редко — в лужах, которые иногда пере-
сыхают, но дно их остается влажным;
попадается она и в прибрежной зоне озер
и рек. Описываемый вид распространен
на всей территории нашей страны (вплоть
до Коми АССР и севера Сибири), но в
холодных местностях встречается редко
и обитает в более крупных водоемах, ибо
мелкие водоемы в условиях сурового кли-
мата промерзают до дна. Особенно много-
численна Н. sanguisuga на юге СССР (Мол-
давия, Украина, Кавказ и др.). Размноже-
ние и развитие ее сходны с теми же про-
цессами медицинской пиявки. Выхухоль,
а возможно, и другие водные позвоночные
охотно поедают эту большую пиявку.
В бассейне Амура, особенно на юге При-
морского края, часто попадается в неболь-
ших водоемах хищная челюстная пиявка
Whitmania leavis, достигающая иногда
еще большей величины, чем Н. sanguisu-
ga. Она отличается от последней сильно
суженным передним концом тела, рыжева-
то-коричневой узорчатой окраской, весь-
ма изменяющейся с возрастом. Этот вид и
другие виды того же рода распространены
в Китае, Японии и в Южной Азии. В свя-
зи с этим следует заметить, что в бассейне
Амура известно несколько видов пиявок,
происходящих из Юго-Восточной Азии и
отсутствующих на остальной территории
СССР. Значительное число видов хищных
челюстных пиявок обитает в Северной и
Южной Америке. В большем количестве,
чем в Европе и Северной Азии, они встре-
чаются в Африке, Южной и Юго-Восточ-
ной Азии. Некоторые из этих видов во
влажных тропиках ведут полуназемный
образ жизни.
Наземные челюстные кровососущие пияв-
ки. Эти пиявки особенно многочисленны
в Южной и Юго-Восточной Азии, а также
живут на юге Японии, в Австралии, на
острове Мадагаскар, в Южной и Централь-
ной Америке. Наиболее известны виды
523
рода Haemadipsa. Они обитают там,
где влажность воздуха настолько велика,
что высыхание им не угрожает. Величи-
на их по сравнению с другими челюстны-
ми пиявками незначительная (средняя
длина 30—40 мм). Они находят себе при-
ют на кустах, деревьях, в траве, где, бу-
дучи голодными, сидят в выжидательной
позе, прикрепившись задней присоской.
Обладая острым обонянием, они легко
отыскивают свои жертвы — млекопитаю-
щих и людей. Это на редкость неприятные
кровососы, забирающиеся в разные щели
одежды. Недавно изобретены специальные
отпугивающие пиявок химические вещест-
ва (репелленты), которыми пропитывают
одежду. Даже после нескольких стирок
такая одежда охраняет людей от напа-
дения пиявок.
Наземные челюстные кровососущие пияв-
ки, вероятно далекие «родственники» тро-
пических видов, найдены в горах Австрии
и Югославии. Их два вида, принадлежа-
щих к роду Xerobdella («ксерос» по-гре-
чески — сухой). Полагают, что они сосут
кровь водящихся в горах саламандр 1. Воз-
можно, что в прошедшие геологические
эпохи, когда климат в Европе был более
теплый и влажный, эти пиявки были рас-
пространены широко, а после ухудшения
климатических условий сохранились толь-
ко в горных местностях, защищенные
благодаря изоляции от борьбы за суще-
ствование с другими пиявками или конку-
рентами из разных групп животных.
Семейство Глоточные пиявки (Нег-
pobdellidae). Все виды этого семейства —
хищники и крови не сосут. Они, несом-
ненно, произошли от челюстных пиявок,
о чем свидетельствуют рудименты челю-
стей у многих из них. Желудок без отрост-
ков. Весьма характерно расположение
глаз: четыре расположены на краю, а по
два — по бокам переднего конца тела. В со-
мите пять или больше колец. Оплодотво-
рение совершается при помощи спермато-
форов. Коконы чаще всего овальные, ко-
ричневого цвета, прикрепляются к под-
водным растениям, камням и т. д. Плава-
ют и совершают дыхательные движения
1 Некоторые авторы считают, что ксеробдел-
лы питаются мелкими беспозвоночными.
(см. «Медицинская пиявка»). В водоемах
СССР живет большое количество видов
этого семейства (около 28% от общего
числа видов наших пресноводных и соло-
новатоводных пиявок), и почти везде по
количеству экземпляров они стоят на
первом месте. Однако их нет в Байкале.
Также нужно отметить, что лишь три-
четыре вида рассматриваемого семейства
широко распространены на территории
нашей страны, а остальные встречаются
только в некоторых местах на юге. По-
давляющее большинство глоточных пия-
вок, обитающих на территории СССР,
принадлежит к р о д у Herpobdella (дру-
гие названия рода: Erpobdella и Nephe-
lis). Все виды этого рода называются малы-
ми ложноконскими пиявками.
У нас наиболее известна глоточная пияв-
ка — герпобделла обыкновенная (Herpob-
della octoculata). Она самая распростра-
ненная и многочисленная пиявка Европы
и Северной Азии. Длина ее редко превос-
ходит 40—50 мм. Типичная форма имеет
коричневую или серовато-коричневую
спину, покрытую поперечными рядами
желтых пятнышек. Однако темный пиг-
мент может в той или иной степени исче-
зать, и пятнышки остаются видными толь-
ко на части спины или совсем отсутствуют,
в результате чего спинная поверхность
становится одноцветной, сероватой, на
которой кое-где сохраняются темные пят-
524
Sara
+4*
на. Жизненный цикл Н.
octoculata сходен с цик-
лом Glossiphonia comp-
lanata (см. выше), т. е.
эта пиявка живет около
двух лет. Ее коричнева-
то-желтые коконы часто
1 попадаются на водных
растениях и различных
подводных предметах.
Число яиц в одном ко-
коне может достигать 24, в среднем
оно равно 11 — 12. Питается эта пиявка
мелкими червями, личинками насекомых
и особенно комаров хирономид. Сама
она подвергается нападению разных
хищных беспозвоночных; находят ее и в
желудках рыб. Карликовая (длина поло-
возрелых особей в среднем 16—17 мм),
почти одноцветная форма описываемого ви-
да глоточной пиявки обитает в озере Севан.
Это озеро, несмотря на свое высокогорное
расположение, очень богато пиявками,
представленными, однако, тремя самыми
обычными видами: кроме Herpobdella octo-
culata, там встречаются описанные раньше
Glossiphonia complanata и Helobdella
stagnalis, которые тоже отличаются мел-
кими размерами и светлой окраской.
Другой вид того же рода — герпобделла
линейная (Herpobdella lineata), названная
так потому, что у нее посередине спины
проходят две продольные черные узкие
полоски, часто и в большом количестве
встречается на юге и обитает в лужах,
нередко совсем пересыхающих. Родствен-
ны ей две своеобразные пиявки: герпоб-
делла пещерная (Herpobdella absoloni) и
археобделла каспийская (Archaeobdella
esmonti).
Первая пиявка, обитающая в пещерах
Балканского полуострова и недавно най-
денная в пещерах Грузии, как и большин-
ство животных, живущих в полной темноте,
Рис. 302. Герпобделла обыкновен-
ная (Herpobdella octoculata):
1 — внешний вид; 2 — изменчивость ок-
раски спинной стороны; з — кокон.
беловатого цвета и лишена глаз. Те же при-
знаки характеризуют и вторую пиявку, обыч-
ную обитательницу ила в Каспийском море,
т. е тоже живущую в темноте или почти без
света. Наиболее же характерный признак
археобделлы, отличающий ее от всех пия-
вок, на основании которого она выделена
в особый род,— почти полная редукция
задней присоски. Она передвигается в тол-
ще ила, изгибая свое червеобразное тело,
и задняя присоска стала излишней.
Коконы археобделлы похожи на коконы
герпобделл.
Археобделла встречается и в устьях рек
Азово-Черноморского и Каспийского бас-
сейнов.
Среди глоточных пиявок есть и крупные
черви, которые по величине не уступают
большим челюстным пиявкам и даже пре-
восходят их. Все они южного происхож-
дения.
В плавнях Днестра, недалеко от Одес-
сы многочисленна огромная (длина до
250 мм\) трохета (Trocheta subviridis),
которая распространена по всей Южной
Европе и на севере Африки. Цвет ее тела
Рис. 303. Археобделла каспийская
(Archaeobdella esmonti).
525
коричневый или серый. Это сильная му-
скулистая пиявка, способная в поисках
дождевых червей, которых она пожирает,
глубоко зарываться в сырую землю при-
брежной полосы. Ее же поедают некото-
рые птицы и, вероятно, другие позвоноч-
ные животные. Так, в зобу одной каравай-
ки было найдено много крупных трохет.
Эти черви используются также в качестве
прекрасной наживки для рыб. Коконы
трохет похожи на коконы герпобделл, но,
понятно, крупнее их. Интересно отметить,
что другой вид того же рода, Trocheta
bykowskii, приспособлен к жизни в гор-
ных потоках (например, в Карпатах) и
небольших текучих водоемах (в Англии,
К классу эхиуровых червей относит-
ся не менее 100 видов свободноживущих
типично морских раздельнополых червей,
стоящих близко к морским кольчецам-
полихетам, хотя по многим признакам
они резко от них отличаются.
Ряд других признаков дает полное
основание сближать их с кольчецами и
включать в т и п аннелид в качестве
четвертого класса.
Размеры тела эхиурид колеблются от
нескольких миллиметров до 20—30 см.
Геологически это очень древняя группа
роющихся в грунте червей, получивших
в связи с таким образом жизни ряд ха-
рактерных особенностей.
Мешковидно-вальковатое тело эхиуриды
(рис. 304) спрятано в грунте. На переднем
конце его, за редким исключением, поме-
щается хобот, способный сильно вытяги-
ваться и сжиматься. В нормальном поло-
жении хобот поднимается над грунтом и у
некоторых форм имеет в длину свыше 1 м.
Других придатков тела у эхиурид, кроме
щетинок, нет. У отверстия нефридиев у
части эхиурид находятся две так называе-
мые половые щетинки, а у некоторых эхиу-
Голландии и других западноевропейских
странах). Недавно она указана для Аф-
ганистана и, вероятно, встречается в Кры-
му и на Кавказе. К двум предыдущим
видам близка более примитивная Fadeje-
wobdella quinqueannulata, найденная толь-
ко в некоторых местах Украины и на се-
веро-западе Кавказа. Она обитает в лужах
и подобно трохетам переносит пересыха-
ние водоемов, если грунт последних ос-
тается достаточно влажным. Значитель-
ное число видов крупных глоточных пия-
вок, ведущих полуназемный образ жизни,
известно в Японии и Юго-Восточной Азии,
где климат отличается большой влаж-
ностью.
рид вокруг анального отверстия распола-
гается кольцо щетинок в один или два
ряда.
У взрослых форм совершенно от-
сутствуют сегментация, параподии и па-
раподиальные щетинки. Кожа тонкая и
прозрачная, особенно у форм, живущих
на глубинах; у мелководных видов —
иногда толстая и покрытая папиллами.
Кишечник у эхиурид сквозной, часто
образующий петли (рис. 304); ротовое и
анальное отверстия расположены на про-
тивоположных концах тела, ротовое — у
основания хобота. Хобот обычно имеет
желобчатый характер и покрыт мерцанием.
Передний конец хобота иногда расши-
ряется в большую лопасть, у некоторых
видов цельную, у других — разделенную
на два лепестка. Внутри тела эхиурид
помещается обширная полость. Сам
червь, особенно взрослый, почти непо-
движен, хобот же его обладает большой
подвижностью. Внутри его и вдоль тянут-
ся три канала, сообщающихся с общей
полостью тела. Сокращение мышц кожно-
мускульного мешка создает внутренний
тургор: под некоторым давлением полост-
КЛАСС ЭХИУРИДЫ (ECHIUROIDEA)
526
ная жидкость нагнетается в каналы и по-
лости хобота, и он приобретает твердость
(гидравлический скелет). При ослаблении
внутреннего давления хобот спадается и
избегает таким образом опасности быть
съеденным другими животными. По пе-
реднему краю хобота иногда располагают-
ся особые органы чувств, возможно свое-
образные «радары», предупреждающие об
опасности. В случае утери хобота он до-
вольно быстро регенерирует вновь. По-
лость тела обширная и несегментирован-
ная. Единственный след сегментированно-
сти предков у взрослых форм — это нали-
чие у некоторых представителей эхиурид
2—3—4 пар нефридиев. Нефридии эхиу-
рид очень сходны с нефридиями сипунку-
лид, но в еще большей степени теряют
железистые функции и служат для выве-
дения наружу зрелых половых продуктов.
Гроздевидные скопления половых продук-
тов развиваются внутри полости тела, и
яйца по достижении половозрелости за-
хватываются нефростомом, а иногда пол-
ностью набивают нефридий.
Наружное отверстие нефридиев
(нефропор) снабжено двумя
крепкими половыми щетин-
ками, помогающими при
оплодотворении (рис. 304,5).
Для части эхиурид (сем. Ве-
nelliidae) известен резко вы-
раженный половой димор-
физм с развитием самцового
паразитизма. Микроскопи-
ческие самцы паразитируют
в особых отделах нефридиев
самок, где и осуществляют
оплодотворение яиц. Они
уже настоящие паразиты,
лишенные пищеварительной
системы и ряда других ор-
ганов. Экспериментально бы-
ло показано, что из большей
части яиц, развивающихся
на хоботе самки, выходят
самцы, которые спускаются
по хоботу к телу самки и
проникают затем через по-
ловое отверстие, располага-
ющееся вблизи хобота на
брюшной стороне тела, внутрь
нефридиев.
Из яиц, развивающихся вне контакта
с самками, выходят главным образом
самки.
У эхиурид хорошо развита крове-
носная система. В самый задний
отдел кишки открываются две анальные
железы, выполняющие выделительные
функции. Они состоят из одного или не-
скольких иногда ветвистых канальцев,
обычно покрытых большим количеством
железистых папилл с воронками на кон-
цах.
Эхиуриды — типичные детритоеды, их
кишечник обычно бывает набит большим
количеством детрита.
Очень типично для эхиурид спиральное
дробление и личиночные формы, не остав-
ляющие сомнений в близости эхиурид к
полихетам. Это весьма типичные трохофо-
ры с хорошо выраженной зачаточной сег-
ментацией, которая в дальнейшем исчезает.
Значительное своеобразие строения, не
сходного с какой-либо другой группой
кольчатых червей, малое число видов эхиу-
рид, приуроченность большого количества
видов к абиссальным глубинам океана
могут быть истолкованы как
доказательства большой их
древности.
В северных морях обитает
всего 3—4 вида эхиурид.
5 Главное разнообразие их при-
урочено к тропическим водам.
Около трех десятков видов ха-
рактерно для больших глу-
бин океана, где иногда они
являются характерной груп-
пой населения.
Рис. 304. Представители класса
эхиурид.
А — Echiurus echiurus: l — хобот;
2 — придаток хобота; 3 — брюшные
щетинки; 4 — петли кишечника,
просвечивающие сквозь стенку тела;
5 — половые отверстия; 6 — брюш-
ная медиальная линия; 7 — аналь-
ное отверстие; 8 — анальные щетин-
ки; Б — Bonellia viridis; В — Tha-
lassema gigas.
527
КЛАСС СИПУНКУЛИДЫ (SIPUNCULOIDEA)
(ДОБАВОЧНЫЙ КЛАСС К ВЫСШИМ ЧЕРВЯМ)
Сипункулиды — сравнительно неболь-
шая группа (около 300 видов) типично
морских червей. По ряду признаков их
можно отнести к высшим червям как до-
полнение к типу аннелид.
Сипункулиды в процессе эволюционно-
го развития далеко отошли от основного
ствола кольчатых червей. Тело сипунку-
лид гладкое, лишенное совсем каких-ни-
будь придатков, в том числе параподий и
щетинок. Оно вытянуто и снабжено хобо-
том, который может выбрасываться дале-
ко вперед, удлиняя ход в грунте. На кон-
це хобота находится ротовое отверстие,
окруженное венцом щупалец. Шупальца
служат для сортировки частиц, годных в
пищу. Анальное отверстие находится в
передней трети тела. Кишечник значи-
тельно длиннее тела, и его нисходящая и
восходящая части закручены винтом одна
вокруг другой. Тело сипункулид погруже-
но в грунт или в пористую массу известко-
вых водорослей или кораллов, а хобот вы-
ставлен наружу. При таком положении
тела анальное отверстие оказывается над
поверхностью грунта и фекалии выделяют-
ся в окружающую воду.
1 t
Рис. 305. Представители класса сипункулид:
Слева направо: Golfingia vulgare; Aspidosiphon
steenstrupii; Dendrostomum pyroides. 1 — венчик щу-
палец вокруг ротового отверстия; 2 — хоботок.
В передней части тела помещаются один,
два или несколько пар нефридиев, имею-
щих железистую стенку и сложенный
вдвое канал. Нефростомами неф-
ридии открываются в полость тела, а
нефропорами — наружу.
Нервная система сипункулид
развита слабо, что соответствует их ма-
лоподвижному и скрытому существованию.
От окологлоточного нервного кольца по
брюшной стороне тела отходит нервный
тяж. Органы чувств отсутствуют.
Полость тела поделена на три
части. Однако перегородки выражены сла-
бо. Полостная жидкость перегоняется му-
скулатурой, и сипункулиды движутся гид-
равлическим способом.
Сипункулиды раздельнополы.
Половые продукты формируются в поло-
сти тела и по созревании выводятся нару-
жу через нефридии. Оплодотворение яиц
происходит в воде.
Спиральное дробление яиц и наличие
личинок типа трохофоры сближает сипун-
кулид с полихетами. У некоторых сипун-
кулид на стадии поздней трохофоры раз-
виваются щетинки, которые потом исче-
зают. Таким образом обнаруживается
связь этих животных с кольчатыми червя-
ми, особенно на ранних стадиях развития.
Но по мере развития признаки сходства
все более утрачиваются. Поэтому в послед-
нее время все большее число исследова-
телей склонно выделять сипункулид в
самостоятельную группу как добавление
к высшим червям. В щупальцевом аппа-
рате и в передней части тела сипункулид
имеется особая срютема каналов, несколь-
ко напоминающая амбулакральную сис-
тему иглокожих. Иногда их считали род-
ственными мшанкам и форонидам. Но и до
настоящего времени систематическое по-
ложение сипункулид остается не ясным.
Сипункулиды обитают во всех морях и
океанах (вплоть до больших глубин),
образуя значительную и характерную часть
донного населения.
528
Форониды
Phoronoidea
Мшанки
Bryozoa
Плеченогие
Brachiopoda
*
Описываемые ниже три группы беспозвоночных — мшанки
(Bryozoa), плеченогие (Brachiopoda) и форониды (Phoronoidedi)—
занимают среди первичноротых (см. введение) особое поло-
жение. Состоя в несомненном родстве с высшими червями, они
характеризуются малосегментностью (Oligomera), которая пред-
ставляет собой один из основных признаков в стволе вторич-
норотых. Положение всех трех групп в системе животных не
может считаться твердо установленным. Это нашло свое
отражение и в очень большом количестве названий, которые
им давались. Их называли червеобразными (Vermoidea), сбли-
жая с червями, моллюскообразными (Molluscoidea), сближая с
моллюсками, шупалъцевыми (Tentaculata), переднепорошицевыми
(Prosopygii). Чаще их объединяли в один тип, однако тоже без
достаточно веских к тому оснований. Поэтому мы считаем
возможным дать их описание как самостоятельных групп.
ТИП ФОРОНИДЫ (PHORONOIDEA)
Форониды — очень небольшая, охва-
тывающая всего несколько видов группа
морских донных одиночных животных.
Живут форониды в трубочках из орга-
нического вещества, обычно инкрустиро-
ванных песчинками, образуя скопления —
дерновинки из переплетающихся, но не
соединенных особей (рис. 306, 1). Из тру-
бочки высовывается только передний ко-
нец животного, несущий сложный, двой-
ной, спиральный щупальцевый аппарат
(рис. 306, 2).
Полость тела вторичная. Ки-
шечник начинается ротовым отверстием
внутри спирали щупальцевого аппарата
(рис. 306, 2), проходит вдоль всего тела
до заднего конца, поворачивает обратно
и открывается анальным отверстием вне
трубочки (рис. 306, 3). Питаются форо-
ниды различными микроорганизмами и
частичками детрита, отфильтрованными
щупальцевым аппаратом из окружающей
воды.
У форонид имеется замкнутая кро-
веносная система, состоящая
из спинного и брюшного сосудов. Спин-
ной сосуд сокращается, и кровь течет от
заднего конца к переднему. В заднем
конце сосуды переходят один в другой
(рис. 306, 3).
Выделительными органа-
м и является пара метанефридиев, откры-
вающихся воронками в полость тела. На-
ружу продукты выделения выводятся
через отверстия в переднем конце тела
(рис. 306, 4), они же служат для выве-
дения половых продуктов, образующихся
в заднем конце тела. Форониды — герма-
фродиты.
Нервная система очень при-
митивная, но, по-видимому, эта прими-
тивность первичная, а не связанная с не-
подвижным образом жизни форонид. Име-
ются подкожные нервные сплетения диф-
фузного типа и только в переднем конце
тела этих животных, между ротовым и
анальным отверстиями, образуется более
плотное нервное сплетение (нервный
«ганглий», рис. 306, 2). Первичная прими-
тивность нервной системы была, возмож-
но, одной из причин, почему форониды не
эволюционировали прогрессивно и не дали
высших форм, а остались маленькой, но
самостоятельной эволюционной веточкой
между первичноротыми и вторичноротыми.
Развитие происходит с резко вы-
раженным метаморфозом: имеется своеоб-
разная личинка — актинотроха
(рис. 306, 2), которая сначала свободно
плавает в планктоне, а затем после слож-
ного превращения из нее развивается взрос-
лое животное. Надо отметить, что развитие
форонид из личинки до взрослого животно-
го было исследовано великим русским
биологом И. И. Мечниковым, кото-
рый, как известно, в начале своей науч-
ной деятельности был зоологом-эмбрио-
логом.
34*
531
Рис. 306. Различные виды форонид:
J — группа особей Phoronis sp., образующих харак-
терные для всего типа дерновинки; видны выпячен-
ные из трубочек передние концы с щупальцевым ап-
паратом (Черное море); 2 — личинка форонид акти-
нотроха (Черное море); «3 —Phoronis sp. (крупный
экземпляр из Охотского моря); 4 — передний конец
Phoronis sp., виден спирально закрученный щупаль-
цевый аппарат и эпистом. Ниже свисает в полость те-
ла метанефридий.
Форониды распространены понемногу
во всех морях. Излюбленным субстратом,
на котором они образуют свои дерновинки,
являются мертвые раковины моллюсков
(«ракуша»). Соленость, по-видимому, не
лимитирует распространение форэнид, так
как у нас в Черном море, даже в его
опресненных частях, они встречаются.
Более отрицательно форониды относятся
к низкой температуре, так как в северных
морях они не найдены.
В период размножения актинотрохи
могут быть массовой формой планктона,
как это бывает, например, у нас в Черном
море. Взрослые формы благодаря их ма-
лой заметности и небольшой величине
(черноморские формы достигают вместе с
трубочкой 1 —1,5 см) встречаются редко.
Более крупные формы форонид распрост-
ранены у нас в дальневосточных морях
(рис. 306).
Положение форонид в системе животно-
го мира до сих пор неясно. Некоторые
исследователи сближают их, а некоторые
даже соединяют с мшанками, причем в
связи с наличием у них эпистома — с по-
крыторотыми. В таком сближении есть
некоторые основания, но все же форони-
ды имеют ряд специфических черт в строе-
нии, как прогрессивных, так и примитив-
ных, на основании которых соединять их
с мшанками вряд ли возможно. Прогрес-
сивной чертой является наличие у них
замкнутой кровеносной системы с опреде-
ленным движением крови. Примитивная
черта организации — очень примитивная
нервная система, не образующая настоя-
щего нервного ганглия. Кроме того, личин-
ки форонид и мшанок резко различаются.
В последнее время было высказано пред-
положение о связи форонид с погонофора-
ми. Группа эта относится к низшим хор-
довым. Однако форониды резко отличают-
ся от погонофор наличием кишечника,
которого, как известно, у погонофор нет,
и переваривание и всасывание пищи про-
исходит у них щупальцами.
Форониды, по-видимому, очень древняя
группа, правда, в связи с отсутствием
у них скелетных элементов они плохо
сохраняются в ископаемом состоянии,
но есть данные, позволяющие считать,
что они существовали даже в древнем па-
леозое.
Практического значения
форониды не имеют, но возможно, что
крупные формы, не имеющие скелета и
довольно мясистые, могут служить пи-
щей некоторым донным рыбам (хотя кон-
кретных данных по этому поводу нет).
ТИП МШАНКИ (BRYOZOA)
Мшанки — своеобразная группа. В ос-
новном это неподвижные сидячие живот-
ные. Однако среди них встречаются и от-
дельные подвижные формы. Например,
пресноводная Cristatella mucedo, червеоб-
разные колонии которой имеют широ-
кую мускулистую подошву; при ее по-
мощи они медленно ползают по подвод-
ным предметам, например по стеблям
водных растений (рис. 307, 14).
Скорость движения Cristatella — около
1 —15 мм в день. Могут двигаться и мо-
лодые колонии Lophopus, Lophopodella и
Pectinatella.
В громадном большинстве мшанки —
колониальные животные (рис. 308), внеш-
не похожие даже на растительные орга-
низмы (отсюда и название — мшанки, т. е.
похожие на мох).
Положение мшанок в системе животного
мира было долгое время неясным. Старые
авторы (Линней и др.), основываясь на
чисто внешнем сходстве, относили их к
кишечнополостным; причем имеющих мяг-
кий скелет (рис. 307) — к гидроидам, а
имеющих известковый (рис. 307) — к ко-
раллам.
Более детальное изучение мшанок пока-
зало, что они построены значительно
сложнее, чем полипы (рис. 308). Тогда их
стали — без достаточных оснований — сое-
динять вместе с некоторыми другими груп-
пами в один тип под различными названи-
ями: червеобразные, моллюскообразные,
щупальцевые. Такое соединение искус-
ственно, и в настоящее время как зоологи,
так и палеонтологи от него отказались.
Мшанки рассматриваются теперь как осо-
бый, древний тип животного мира, по
эволюционному развитию стоящий между
первичноротыми и вторичноротьгми. Нали-
чие у них вторичной полости (рис. 308)
и некоторых других признаков позволяет
ставить их на уровне с кольчатыми,
высшими, червями.
В связи с сидячим образом жизни у них
действительно выработались некоторые
признаки, сходные с полипами, что и при-
вело ранее к ошибочному пониманию их.
Мшанки обитают как в морях, так и в
пресных водах, хотя в морях их значи-
тельно больше.
Колонии морских мшанок по форме очень
разнообразны (рис. 307): с одной стороны,
имеются мягкие с роговидным скелетом —
кустистые (рис. 307), листовидные (рис.
307) или мясистые без сколько-нибудь оп-
ределенной формы (рис. 307); с другой
стороны, имеются виды с твердым (извест-
ковым) скелетом, иногда сложные по фор-
ме (рис. 307).
В некоторых случаях колонии в виде
тонких корочек покрывают подводные
предметы, например камни (рис. 307),
раковины моллюсков и пр. Все эти коло-
нии очень изменчивы и зависят от условий,
в которых развиваются и обитают: от
глубины, субстрата, движения воды и
прочих факторов внешней среды (эколо-
гическая изменчивость).
%Г 35 жизнь животных, т. 1
533
Рис. 307. Мшанки:
1 — Flustra foliacea (северные и дальневосточные моря, очень обычен в Белом море); 2 — Alcyonidium disci-
forme (северные и дальневосточные моря), з — Paiella sp., 4 — Fredericella sultana (пресные воды), 5 — Meni-
реа zelandica; 6 — Retepora cellulosa (северные и дальневосточные моря), 7 — Defrancia lucernaria (северные
моря); 8 — Eucrata loricata (северные и дальневосточные моря); 9 — Australella indica (другая форма колонии
этого вида — на рис. 15); 10 — Hornera lichenoides (северные и дальневосточные моря), 11 — Membranipora
ер. (все моря); 12 — Pectinatella sp.; 13 — AJcyonidiura gelatinosum (северные и дальневосточные моря);
14 — Cristatella mucedo (пресные воды); 15 — Australella indica (пресные воды Южно-Уссурийского края).
Колонии ряда морских мшанок замеча-
тельны тем, что составляющие их особи
не одинаковы, а различны как по строе-
нию, так и по функции, т. е. в таких ко-
лониях имеется специализация и «разделе-
ние труда» между особями. Такие колонии
получили название полиморфных или мно-
гоформенных, в отличие от мономорфных
или одноформенных, у которых все особи
одинаковы.
У пресноводных мшанок имеются толь-
ко мономорфные колонии.
У полиморфных колоний различаются
следующие группы особей.
Первая группа — обычные особи,
т. е. имеющие лофофор, венец щупалец,
нормально устроенный кишечник и обра-
зующие половые продукты. Таких особей во
всех колониях всегда больше остальных —
они улавливают пищевые частицы, пере-
Рис. 308-309.
1 — отдельная особь голоротой мшанки
(продольный разрез) с выпяченным ло-
фофором, несущим венец щупалец, рас-
положенных по кругу. Над ротовым от-
верстием эпистома нет; 2 — отдельная
особь покрыторотой мшанки (продоль-
ный разрез). Виден выпяченный лофофор
с подковообразным венцом щупалец. Над
ротовым отверстием виден эпистом. Чер-
ная точка между ротовым и анальным
отверстием — нервный ганглий.
Рис. 310. Особенности строения мор-
ских мшанок:
А — авикулярия морской мшанки с откры-
той «нижней челюстью». Черным показа-
ны мышцы, захлопывающие «челюсть». Б —
вибракулярий морской мшанки. Виден жгут
B) с двигающими его мышцами. С л е в а—
чувствующий шпенек (J).
варивают, усваивают их и питают всю ко-
лонию. В связи с этим они получили на-
звание питающих или обычных особей. Ча-
сто около или над ними образуются особые
особи, так называемые оеции, т. е.
особи, служащие выводковыми камерами,
в которых развиваются яйцеклетки.
Вторая группа резко изменен-
ных особей, которые несут функцию за-
щиты колоний от различных «непроше-
ных гостей» — мелких червей, рачков и
других мелких хищников. Среди этих за-
щитных особей различаются а в и к у -
л я р и и, имеющие вид «птичьих голов»
(рис. 310, А). Они не имеют щупалец и
поэтому не могут сами питаться, а получа-
ют питание от обычных особей. У них
сильно развивается особый придаток в виде
«нижней челюсти», который при сокраще-
нии особых мышц может захлопываться.
При помощи такого приспособления авику-
лярии захватывают «непрошеных гостей»
и, таким образом, освобождают от них
колонию. Второй формой защитных особей
(реже встречающейся) являются виб-
ракулярий (рис. 310, Б). У них
развивается особый длинный подвижный
придаток, который может при помощи
особых мышц вибрировать и отгонять
«непрошеных гостей», не давая им даже
забраться на колонию.
У авикулярий и вибракулярий имеются
особые чувствующие образования, кото-
рые связаны с нервной системой и сигна-
лизируют о наличии врага. У некоторых
форм имеются особые особи, при помощи
которых колонии прикрепляются к суб-
страту.
Кроме этих защитных особей, «активно»
защищающих колонии, у многих форм
имеются образования пассивной защиты —
различные выросты наружной стенки —
шипы, колючки и пр. У некоторых форм
они покрывают всю колонию, делая ее
колючей и этим отпугивая врагов. У дру-
гих шипы развиваются вокруг отверстий
ячеек, не давая врагам забраться в них.
Примером первого случая может быть
описанный нашим известным исследова-
телем мшанок Г. А. К л ю г е из сибир-
ских морей вид Uschakovia gorbunovi.
Колонии его покрыты длинными ши-
повидно-колосистыми выростами (рис. 311),
делающими колонпп этого вида недо-
ступными для врагов. Встречается этот
вид на глубине до 700 м при температурах
от —0,9° до —1,4°С и рассматривается
как высокоарктический. Вид этот интере-
сен и в другом отношении: в нижней части
его колоний имеются особи (рис. 311),
лишенные щупальцевого аппарата и не
образующие половых продуктов; ячейки
их почти всегда наполнены белой зерни-
стой массой. Эти своеобразные ячейки
представляют собой видоизменения особей
колонии, «склады» запасного питатель-
ного вещества для поддержания роста
этих сложных колоний.
Примером второго случая, когда шипы
располагаются главным образом вокруг
отверстий ячеек, могут служить: во-пер-
вых, Flustrella hispida — вид очень обыч-
ный в приливо-отливной зоне мурманско-
го побережья Белого моря и других бо-
реальных и арктических морей, где они
образуют довольно толстые коричневые
колонии, главным образом на фукоидах;
во-вторых, черноморская мшанка Disco-
рога, образующая красноватые тонкие из-
вестковые корочки на водорослях, рако-
винах моллюсков и другом субстрате на
глубине 20—80 м в Севастопольской бухте,
по Южному берегу Крыма и в северо-за-
падной части Черного моря.
536
Колонии пресноводных мшанок менее
разнообразны. Они или в виде разветвлен-
ных тонких лежащих трубочек распола-
гаются на подводных предметах и подвод-
ной растительности, например на нижней
поверхности листьев кувшинок и кубышек
(рис. 307), или образуют массивные ко-
лонии на подводных предметах — камнях,
затонувших бревнах, растениях (рис. 307),
а иногда и на животных — моллюсках
(анадонта, живородка), изредка даже на
раках.
Колонии мшанок состоят из большого
числа отдельных очень мелких особей.
Так, например, кусочек колонии Flustra
foliacea (рис. 307) весом в 1 г содержит
1330 особей. Каждая особь помещается в
отдельной ячейке, имеющей довольно об-
ширную полость (рис. 308). Из ячейки
может выпячиваться передняя часть осо-
би, так называемый лофофор с входя-
щей в него полостью тела.
На лофофоре помещается ротовое отвер-
стие, окруженное венцом щупалец, кото-
рые располагаются кругом или подково-
образно. Рот может быть открытым
(голоротые мшанки) или прикрываться
особым выростом, так называемым эписто-
м о м (покрыторотые мшанки, рис. 308).
Ротовое отверстие ведет в глотку, перехо-
дящую в пищевод, идущий вниз, т. е. в
глубину ячейки, и переходящий в до-
вольно объемистый желудок; от последнего
отходит задняя кишка, идущая вверх и
открывающаяся анальным отверстием на
лофофоре, вне венца щупалец (рис. 308).
Питаются мшанки при помощи
щупалец, покрытых ресничками. Послед-
ние находятся в непрерывном движении,
создавая ток воды, идущий по направле-
нию ко рту, а затем выходящий между щу-
Рпс. 312. Спинобласт Lophopodella carteri.
На вершинах видны хитиновые выросты с крючочка-
ми (экземпляр из водоема авандельты Волги).
537
Рис. 313. Личинки различных морских мшанок:
1 — Cyphonautes, 2 — Microporella ciliata; 3 — Hippotoa hyali-
na, 4 — Porella concinea; 5 — Bowerbankia pystolosa (вид
спереди), б — Crisia eburnea; 7 — Flustrella hispida.
пальцами. При этом из воды отфильтро-
вываются различные микроорганизмы и
органическая взвесь (детрит). Все это
подгоняется ресничками к ротовому от-
верстию, заглатывается, поступает в же-
лудок и там переваривается. Таким обра-
зом, по характеру питания мшанки — ти-
пичные фильтраторы и в этом отношении
приносят некоторую пользу, очищая во-
доемы, но вместе с тем приносят более
значительный вред, обрастая и засоряя
различные гидротехнические и водопро-
водные сооружения. Особенно это касает-
ся массивных пресноводных форм мша-
нок (рис. 307).
Дыхательный аппарат,
кровеносная и выделитель-
ная системы у мшанок отсутст-
вуют. Обмен газов происходит через щу-
пальца. Кровью служит полостная жид-
кость, а выделительные функции выпол-
няют особые клетки, находящиеся в ней.
Нервная система в свя-
зи с сидячим образом жизни сильно
упрощена и состоит у каждой особи
только из одного нервного узла,
расположенного между ротовым и
анальным отверстиями (рис. 308, 309).
От него отходят нервы к щупальцам
и ко всем органам особи. Единой
нервной системы для всей колонии
нет. Органами чувств являются щу-
пальца.
Размножаются мшанки как
половым, так и бесполым путем. Темп
почкования у некоторых мшанок
очень высокий, особенно в теплых
морях. Имеются сведения, что у Га-
вайских островов мшанки образуют
колонии до 2 м высоты всего за
несколько часов. В Одесском порту
затонувшее судно покрылось сплош-
ной корой из мшанок за 3—4 ме-
сяца.
Половые продукты образуются в
полости тела ячеек из клеток цело-
мического эпителия, причем большин-
ство мшанок гермафродиты. У неко-
торых морских мшанок имеются осо-
бо измененные особи, которые слу-
жат выводковыми камерами, так на-
зываемые о е ц и и, в которых про-
исходит развитие яйцеклетки до ли-
чинки. Если оеций нет, то личинка разви-
вается в полости тела материнской особи.
Сформированная личинка выходит нару-
жу и некоторое время ведет планктонный
образ жизни; таким образом, развитие
мшанок происходит с метаморфозом.
Личинки мшанок сильно отличаются
от личинок других групп (кольчатых чер-
вей, брахиопод и др.) как по строению, так
и по их превращению в первичную особь.
Это еще раз доказывает, что мшанки —
своеобразная и самостоятельная группа
(тип) животного мира.
Личинки мшанок очень разнообразны
(рис. 313), все они имеют более или менее
развитый ресничный аппарат, при помо-
щи которого могут активно плавать в
планктоне. Затем они садятся на субстрат
и превращаются в первичную особь, кото-
рая путем почкования образует колонии.
Наиболее известная личинка — ц и ф о-
н а у т е с, треугольной формы, имеет
538
тонкую прозрачную двустворчатую рако-
вину (рис. 313, 1).
Личинки пресноводных (покрыторо-
тых) мшанок имеют значительно более
простое строение, чем личинки морских (го-
лоротых). Они представляют собой микро-
скопические овальные тела без каких-либо
выростов. Личинки образуются в каждой
ячейке из мезодермического эпителия. По-
чти в каждой ячейке, вырастая, они зани-
мают всю полость ее, вытесняя питающую
особь. Таким образом, у покрыторотых
все особи являются своего рода оециями,
но, в отличие от таковых у голоротых,
они представляют собой временное образо-
вание. Когда личинка вылупляется, все
приходит в норму и питающая особь вос-
станавливается.
Обычно личинки живут очень недолго,
всего несколько часов, и благодаря своим
мелким размерам и простоте строения они
часто не замечаются исследователями. Ха-
рактерной особенностью личинок покры-
торотых является то, что в них заложена
не одна первичная особь, а две. Когда
личинки садятся на субстрат (под Москвой
это происходит обычно в конце лета), обе
особи начинают развиваться, причем одна
вправо, другая влево, и образуют ха-
рактерные двухлопастные колонии. Эти
колонии настолько своеобразны по форме,
что ранее выделялись как особые виды.
Такие формы в некоторые годы буквально
осыпают нижнюю поверхность листьев
кувшинок и кубышек.
Затем колонии разрастаются, образуют
флотобласты, и весной из них полу-
чаются типичные колонии. Так обра-
зуются многолетние, иногда очень круп-
ные колонии, достигающие, например,
у Plumatella fungosa (рис. 307) размеров
более 1 м в высоту и 25—30 см в попереч-
нике.
Бесполое размножение происходит пу-
тем почкования: каждая особь отпочковы-
вает (рис. 309) новые; таким образом уве-
личивается число особей в колонии и она
растет; колонии некоторых мшанок до-
стигают в высоту нескольких десятков сан-
тиметров.
Особый случай бесполого размножения
имеется у пресноводных мшанок. Он за-
ключается в том, что внутри особи (рис.
Рис. 314. Развитие мшанки.
309) образуются особые внутренние почки,
так называемые статобласты, по-
крытые плотной оболочкой. Статобласты
возникают тогда, когда в водоеме создают-
ся неблагоприятные для жизни колоний
условия (у нас — осенью, в тропических
странах — при наступлении засушливого
периода).
Статобласты благодаря своей прочной
оболочке переживают неблагоприятные
условия, и весной из них выходит молодая
особь, из которой путем почкования об-
разуется новая колония. Статобласты бы-
вают различные. Пайптобласты,
имеющие простую овальную или бобовид-
ную форму, покрыты плотной оболочкой
и свободно лежат в трубочках колоний.
Когда колонии распадаются, пайптобласты
выпадают наружу и из них образуются
новые колонии на том же месте. Таким
образом сохраняется вид при наступивших
неблагоприятных условиях.
Следующая форма — флотобла-
с т ы, или плавающие статоб-
539
л а с т ы. У них вокруг капсулы образует-
ся особое хитиновое ячеистое кольцо, в
ячейках которого скопляются пузырьки
воздуха. При помощи этого «гидростати-
ческого» аппарата флотобласты могут
всплывать и некоторое время пассивно
плавать в толще воды и разноситься тече-
ниями в новые места, где из них образуют-
ся новые колонии. Таким образом флото-
бласты не только обеспечивают сохранение
вида, но и его расселение.
Наиболее сложно устроенные статобла-
сты — спинобласты, у которых на
капсуле или на плавающем кольце об-
разуются хитиновые крючочки, при помо-
щи которых они могут прицепляться к
подвижным подводным предметам или к
другим животным, например к перьям и
ногам водоплавающих птиц, и при их
помощи разноситься на далекие расстоя-
ния. Так, например, летом 1962 г. много-
численные спинобласты индо-африкан-
ской мшанки Lophopodella carteri (рис. 312)
были неожиданно обнаружены в водоемах
авандельты Волги, куда они были, види-
мо, занесены перелетными птицами. По-
скольку в водоемах авандельты Волги,
особенно летом, температурные условия
близки к таковым в субтропических водое-
мах, возможно, что из этих занесенных
спннобластов разовьются новые колонии и
этот вид мшанок приживется в водоемах
дельты Волги.
Как уже говорилось, большинство мша-
нок обитает в морях, в пресных водах их
немного — в основном покрыторотые (рис.
308). Морские мшанки распространены
очень широко по всем морям и океанам, на-
чиная от приливо-отливной зоны (Flustrel-
la hispida) до больших глубин, почти до
8 тыс. му например Bugula sp. Наибольшее
число видов встречается на глубинах не
свыше 200—300 м. Мшанки обитают в воде
при самой различной температуре (от
—2° до 29°С). Литоральные виды переносят
и значительно более низкую температуру,
правда, впадая в состояние анабиоза.
Всего в настоящее время насчитывается
около 4 тыс. современных видов мшанок
и почти 15 тыс. ископаемых.
Из наших морей мшанки особенно мно-
гочисленны в дальневосточных и северных,
так, например, в Белом море их насчиты-
вается 132 вида. Причем там особенно надо
отметить вид Flustra foliacea (рис. 307),
который в некоторых районах моря, на-
пример в Великой Салме, образует, по
наблюдениям Беломорской биостанции
МГУ, мощные заросли. Этот арктическо-
бореальный вид широко распространен в
морях, омывающих Западную Европу, но
не встречается на мурманском побережье,
хотя и отмечен в Чешской губе Баренцева
моря и в Карском море.
В наших южных морях в связи с их
малосоленостью фауна мшанок беднее: в
Черном море их известно около 30 видов,
в Азовском — 7 видов, а в Каспийском —
всего 6, причем один из них, а именно
Membranipora crustulenta (рис. 307), про-
ник туда в последние годы вместе с другими
азово-черноморскими организмами, по-
видимому, через Волго-Донской канал.
Особая фауна мшанок имеется в соло-
новатых водоемах, например в Каспийском
и Аральском морях. Их распространение
связано с геологическим прошлым некото-
рых водоемов, например с океаном «Тэ-
тис» и его производных. Подобное распро-
странение имеют и некоторые солоновато-
водные кишечнополостные.
В пресноводных и солоноватых водоемах
нашей страны известно 20 видов мшанок:
четыре из класса голоротых, остальные —
покрыторотые, главным образом предста-
вители рода Plumatella. Встречаются
они в самых различных водоемах: в боль-
ших озерах — Байкал, Онежское, Севан;
в маленьких прудах, в больших реках
и маленьких речушках1.
Пресноводные мшанки распространены
очень широко, хотя они более обильны и
разнообразны в водоемах тропических и
субтропических стран (Индия, Индонезия);
почти не встречаются в водоемах Севера,
известно только несколько случаев нахож-
дения их в водоемах севернее полярного
круга, например в водоемах Гренландии,
Исландии, на Кольском полуострове, око-
1 В озере Севан на прибрежных камнях очень
обычна особая форма широко распространенной
Plumatella fungosa, образующая довольно
плотные корки, состоящие из толстых, уплотнен-
ных, с толстой коричневой кутикулой трубочек.
Флотобласты ее по строению и форме такие же,
как у типичной формы.
540
ло 66° с. ш. Статобласты мшанок найдены
на Шпицбергене и Новой Земле. Наи-
более южное нахождение пресноводных
мшанок — водоемы Огненной Земли.
Некоторые виды, например Plumatella
emarginata, Cristatella mucedo, имеют
очень широкое распространение, другие
виды, наоборот, имеют очень узкие ареалы,
связанные с водоемами определенных рай-
онов. Например, Stephanella hina встречает-
ся только в водоемах Японии, Lophopusel-
1а — только в водоемах Южной Африки.
Географическое распространение мша-
нок иногда сильно расширяется. Наличие
у них статобластов, особенно спинобла-
стов, позволяет им преодолевать большие
пространства с помощью судов, плавающе-
го дерева, рыб и других животных. На-
пример, Lophopodella carteri считалась до
последнего времени индо-африканским
видом, затем была обнаружена в Индоне-
зии, Австралии, Китае, Японии, а в по-
следние годы ее спинобласты были зане-
сены в Северную Америку и в дельту
Волги.
В географическом распространении прес-
новодных мшанок необходимо отметить
интересную черту — параллельное разви-
тие одинаковых форм в водоемах с при-
мерно одинаковыми абиотическими и
биотическими условиями существования.
Например, в алтайском озере Телецкое и
балканском Охрида встречается один и
тот же подвид обычной Fredericella sultana
subsp. lepnevae.
Особенно интересны мшанки водоемов
южной части нашего Дальнего Востока,
где наряду с обычными широко рас-
пространенными формами встречаются
формы южного характера, например Aust-
ralella indica, с крупными массивными,
студенистыми, полупрозрачными коло-
ниями, в которых просвечивают цепочки
отдельных особей (рис. 307).
Интересна также фауна озера Байкал,
где встречается вид из рода Hislopia
(из голоротых мшанок), тоже распростра-
ненного в субтропических странах. В Бай-
кале этот вид рассматривается как реликт
прошлых, более теплых, геологических
периодов. У этого вида особенно хорошо
выражена экологическая изменчивость
(рис. 315).
Рис. 315. Байкальская мшанка и ее экологиче-
ская изменчивость:
1 — прибрежная форма с плоскими ячейками, обра-
зующая тонкие корочки на прибрежных камнях, дре-
весных обломках и других прибрежных предметах;
2 — форма, обитающая на подводной растительности,
с яйцевидными ячейками, образующая довольно ком-
пактные колонии; 3 — форма, обитающая на подвод-
ной растительности, но глубже, чем предыдущая, и
образующая ветвистые колонии с удлиненными ячей-
ками; 4 — форма, обитающая на илистых грунтах,
образующая тонкие нежные деревца в несколько сан-
тиметров высотой и с удлиненными ячейками.
Мшанки как пресноводные, так и мор-
ские вместе с другими сидячими организ-
мами входят в состав обрастаний судов,
портовых и гидротехнических сооружений,
водоводов, принося этим довольно сущест-
венный вред. Особенно в этом отношении
следует отметить пресноводные массивы
формы Plumatella fungosa. Они не только
нарушают нормальную работу упомянутых
сооружений, но после гибели мшанок ча-
стицы засоряют водопроводную сеть.
Борьба с ними ведется путем уничтоже-
ния колоний, счисткой их; но при этом на-
до иметь в виду, что эта счистка должна
быть очень тщательной, чтобы не остались
статобласты (особенно прикрепленные к
субстрату), так как в противном случае
541
из них вновь разовьются колонии. В основ-
ном же для защиты от обрастания мшанка-
ми, как и другими организмами, применя-
ют предупреждающие меры — покрытие
поверхности противообрастающими крас-
ками, ультразвуковая защита и др.
Довольно сильное обрастание мшанками
встречается в Каспийском море. Предста-
вители родов Victorella и Bowerbankia
вместе с некоторыми другими формами
образуют густые дерновинки, покрываю-
щие портовые сооружения в загрязненных
водах Бакинского и Красноводского пор-
тов, a Membranipora плотной известковой
корочкой обрастает гидротехнические со-
оружения и суда в чистых водах Среднего
и Южного Каспия.
Пищевое значение мшанок очень неболь-
шое. О с б о р н A921) приводит некоторые
данные о том, что мшанки могут служить
пищей для некоторых птиц и рыб. Особен-
но, конечно, это относится к формам, не
имеющим скелета. Некоторые мягкие фор-
мы вроде Alcyonidium используются жите-
лями северо-восточных районов (Чукотка)
на корм домашним животным (собакам).
Геологическая история мшанок. Наибо-
лее древние остатки ископаемых мшанок,
принадлежащих к классу голоротых, из-
вестны с начала ордовика. В отложениях
этой системы найдены представители четы-
рех отрядов, в том числе двух в настоящее
время вымерших. До настоящего времени
неизвестны достоверные остатки кембрий-
ских мшанок. Однако можно предполо-
жить, что они существовали уже с начала
этого периода. Развитие их в разных на-
правлениях в течение кембрия и привело
к формированию отрядов.
В современных морях и пресных водах
известны представители двух подклас-
сов — голоротые и покрыторотые. В со-
став первого входят отряды Ctenostomata,
Cheilostomata, Cyclostomata. Остальные, а
именно Cryptostomata, Tipostomata, вымер-
ли, как уже указывалось, в мезозое.
В некоторых местах ископаемые мшанки
образовывали довольно мощные рифы
наподобие современных коралловых. Так,
например, отложения Керченского полу-
острова в большинстве состоят из мшан-
ковых рифов.
ТИП ПЛБЧБНОГИЕ (BRACHIOPODA)
Это группа исключительно морских оди-
ночных донных животных, имеющих дву-
створчатую раковину и ведущих, прикреп-
ленный образ жизни.
Положение брахиопод в системе живот-
ного мира было долгое время неясным и
спорным. На основании имеющейся у
брахиопод раковины их относили ранее к
моллюскам, выделяя среди последних в
особый класс. Тогда-то по аналогии с дру-
гими классами моллюсков (брюхоногие,
головоногие, лопатоногие и т. п.) рассмат-
риваемая группа и получила свое название,
которое в переводе на русский язык зна-
чит «руконогие», но обычно заменяется
термином «плеченогие». Это название,
морфологически мало обоснованное, до
сих пор остается в употреблении, но мно-
гие авторы предпочитают называть их ла-
тинским словом в русской транскрипции—
брахиоподами.
Дальнейшее изучение внутреннего строе-
ния брахиопод показало, что они резко
отличаются от всех моллюсков. В течение
долгого периода брахиопод вместе с не-
которыми другими группами объединяли
в тип червеобразных, или щупальцевых.
В настоящее время на основании глав-
ным образом эмбриологических данных
большинство зоологов и палеонтологов
рассматривает брахиопод как особый древ-
ний тип двустороннесимметричных цело-
мических животных, занимающих промежу-
точное положение между первичноротыми
(червями, членистоногими, моллюсками)
и вторичноротыми (иглокожими, полу-
кордовыми, хордовыми).
Филогенетические взаимоотношения
брахиопод с другими группами до сих пор
недостаточно выяснены.
О брахиоподах принято говорить, что у
них «все в прошлом». Действительно, в
морях прошлых геологических периодов
брахиоподы были разнообразнее и много-
численнее. Современная их фауна, насчи-
тывающая около 280 видов, представляет
собой лишь самую вершину огромного
дерева их родословной, мощная крона
которого, образованная более чем 10 тыс.
ископаемых видов, скрыта в глубине геоло-
гической истории (рис. 320). Из 39 над-
семейств, известных в типе брахиопод, в
современных морях обитает только 7.
Самая крупная современная форма Magel-
lania venosa (до 8,4 см в длину) не может
соперничать по величине с палеозойским
Gigantoproductus (до 40 см в длину).
Однако брахиоподы остаются и сейчас
широко распространенной группой, встре-
чаясь в морях с нормальной соленостью от
тропических до полярных областей и от
приливо-отливной зоны побережий до ше-
стикилометровых глубин океана. В неко-
торых районах при благоприятных усло-
виях существования брахиоподы образуют
большие скопления. В юго-западной части
Баренцева моря, например к северу от
Нордкапа, Macandrevia cranium встречает-
543
Рис. 316. Представители плеченогих в естест-
венной обстановке. Видны стебельки, при по-
мощи которых животные прикрепляются к суб-
страту.
ся в количестве более 100 экземпляров на
1 кв. м дна, являясь преобладающей
формой в донном сообществе. У берегов
Южной Калифорнии Glottidia albida по-
селяется с плотностью более 200 экземпля-
ров на 1 кв. м.
Створки раковины брахиопод, в отличие
от двустворчатых моллюсков, закрывают
тело животного не с боков, а со спинной
и брюшной сторон (рис. 316). По способу
сочленения створок, а также по ряду
других признаков среди плеченогих разли-
чают два класса: Беззамковые (Inar-
ticulata), у которых створки соединены
только мышцами, и Замковые (Articulata),
у которых выросты на заднем краю брюш-
ной створки входят в соответствующие
углубления на спинной створке, образуя
«замок» — прочное сочленение, разнять
которое можно только сломав раковину.
В связи с этой особенностью раковины
ископаемых замковых брахиопод находят
обычно целыми, состоящими из обеих
створок.
Открывание и закрывание створок ра-
ковины обеспечивается работой сложной
системы мышц.
Прикрепляются плеченогие чаще всего
при помощи «ноги» — особого стебелька,
который выходит наружу или просто меж-
ду створками (семейство Lingulidae),
или через отверстие в брюшной створке.
При этом у беззамковых плеченогих се-
мейства Discinidae (рис. 317) спин-
ная створка оказывается сверху, а у зам-
ковых — снизу, так как в месте выхода
ноги брюшная створка у них вытянута в
так называемый клюв, загибающийся на
спинную сторону, и раковина оказывается
в перевернутом положении (рис. 318).
Подошва ноги прочно сочленяется с жест-
ким субстратом, врастая в его шерохова-
тости либо всей поверхностью (Discinisca),
либо ризоидоподобными отростками (Те-
rebratulina). Глубоководная форма Chli-
donophora chuni имеет особенно сильно
разветвленную ногу, способную укрепить-
ся среди мелких частиц фораминиферовых
и глобигериновых илов.
Наибольшего развития достигает нога у
представителей семейства Linguli-
dae беззамковых брахиопод, представленно-
го в современной фауне двумя родам и—
Glottidia и Lingula. Эти формы, в отличие
от других брахиопод, ведут роющий образ
жизни (рис. 318). В приливо-отливной зоне
или на небольших глубинах на песчаных
и илистых грунтах они обитают в верти-
кальных норах, выстланных слизью. Мощ-
ная червеобразная нога укреплена в ниж-
нем отделе норы, плоская раковина как
бы закрывает щелевидный вход в нору. В
случае опасности (сигналом может слу-
жить даже промелькнувшая тень) мускули-
стая нога сокращается и животное быстро
втягивается в нору. При обвале животное
восстанавливает нору роющими движе-
ниями створок и ноги.
Некоторые брахиоподы (семейство
Craniidae из беззамковых и семей-
ство Thecideidae из замковых), приспо-
собленные к обитанию исключительно на
жестких грунтах, вообще не имеют ноги и
прирастают к камням брюшной створкой.
Мягкие части тела животного занимают
только одну заднюю треть полости ракови-
ны, близкую к стебельку (рис. 317). Ос-
544
тальное пространство в передней части
раковины, выстланное тонкой двухслойной
складкой — мантией, а потому имену-
емое мантийной полостью, за-
нимает фильтровальный аппарат — л о -
ф о ф о р. Мантия обеспечивает питание и
рост раковины, а также, по-видимому,
служит органом дыхания. Вдоль края ра-
ковины мантия вооружена хитиновыми
щетинками, защищающими фильтроваль-
ный аппарат от засорения крупными ча-
стицами.
Лофофор состоит из неподвижных сим-
метричных выростов, получивших назва-
ние рук. Вдоль рук расположен ряд щу-
палец, покрытых ресничками, которые
колебательными движениями прогоняют
воду между щупальцами. Руки поддержи-
ваются особыми скелетными выростами
спинной створки, так называемым руч-
ным аппаратом, иногда очень слож-
но устроенным (рис. 317). Строению лофо-
фора и ручного аппарата придается боль-
Рис. 317. Продольный разрез плеченогого.
Внизу виден стебелек B), при помощи которого
животное прикрепляется к грунту. В центре
виден спирально завитой жаберный аппарат B).
шое значение в систематике брахиопод.
У основания рук, в середине стенки,
отделяющей мантийную полость от поло-
сти тела, помещается рот, который ведет
в короткий пищевод, переходящий в сред-
нюю кишку. В расширенную переднюю
часть средней кишки впадают с боков
протоки двух пищеварительных желез.
У замковых брахиопод средняя кишка
заканчивается слепо, у беззамковых имеет-
ся задняя кишка, которая открывается
анальным отверстием в мантийную по-
лость.
Полость тела, в которой разме-
щены мышечная, пищеварительная, кро-
веносная, выделительная и половая систе-
мы органов, представляет собой замкнутое,
наполненное жидкостью пространство —
целом, или вторичную полость тела.
Каналы целома заходят в лофофор и обра-
зуют сложно разветвленную сеть в ман-
тии. Целомические каналы сопровожда-
ются каналами кровеносной системы,
Рис. 318. Плеченогое Lingular
слева — втянувшееся в норку, справа —
высунувшееся. Стрелками показано движение
воды во внутреннюю полость и из нее.
545
которые от пульсирующего пузырька (сердца),
расположенного ближе к спинной стороне,
идут ко всем внутренним органам, где и
теряются в межклеточных пространствах.
Функцию выделения выполняют
одна или две пары метанефридиев —
трубкообразных органов, открывающихся
одним концом в целом, а другим — в ман-
тийную полость.
Половые железы располага-
ются в целомических полостях спинной
и брюшной складок мантии. Они имеют
гроздевидную или решетковидную форму
и иногда видны сквозь тонкую раковину.
Зрелые половые клетки попадают в це-
лом и выводятся оттуда наружу метане-
фридиями. Развитие сложное, со свободно
плавающей личинкой (рис. 317).
Нервная система упрощена
в связи с сидячим образом жизни. Она
состоит из окологлоточного кольца с отхо-
дящими от него стволами в мантию и лофо-
фор. На переднем краю мантии у некото-
рых видов имеются пигментированные
участки, реагирующие на свет. Краевые
реснички мантии чувствительны к при-
косновению . Свободноплавающие личин-
ки брахиопод имеют глазки и статоцисты,
которые утрачиваются в процессе разви-
тия в связи с переходом к сидячему образу
жизни. В этом отношении развитие брахио-
под можно рассматривать как регрессив-
ный метаморфоз. Необходимо отметить,
что развитие брахиопод одним из первых
изучил и правильно объяснил русский
эмбриолог А. О. Ковалевский.
У взрослых особей брахиопод в спокой-
ном состоянии створки раковины при-
открыты. Щупальца лофофора, сомкнув-
Рис. 319. Ископаемое плеченогое Spirifer. Ра-
ковина взломана так, что видны известковые ко-
нические спирали скелета рук.
шпсь друг с другом, образуют фильтрующую
стенку, разделяющую всю мантийную
полость на две камеры, в одну из кото-
рых вода поступает, из другой выхо-
дит профильтрованная. Щель между
створками при этом разделяется краями
фильтрующей стенки на участки, соответ-
ствующие входному и выходному отвер-
стиям. Отфильтрованные частицы попа-
дают в желобок, идущий вдоль лофофора
у основания щупалец, и прогоняются по
нему в рот. В случае неблагоприятных
обстоятельств (внезапное увеличение мут-
ности воды, попадание на лофофор круп-
ных частиц, механическое раздражение)
процесс фильтрации прекращается и
створки быстро захлопываются, выбра-
сывая наружу воду с нежелательными
частицами. Пищу брахиопод составляют
мелкие планктонные водоросли и живот-
ные, а также, по-видимому, взвешенные
в воде частицы детрита. Небольшое тело,
скрытое в прочной раковине,— незавид-
ная добыча для хищника, поэтому сами
брахиоиоды редко становятся жертвой
обитателей моря. Известен лишь один
вид морской звезды (Coscinasterias cala-
maria), который питается преимуществен-
но брахиоподами. Нахождения брахио-
под в желудках рыб рассматривают обычно
как случайные.
Биология брахиопод до сих пор изу-
чена явно недостаточно. О продолжитель-
ности их жизни, например, имеются лишь
отрывочные сведения. О североамерикан-
ском виде Glottidia pyramidata известно,
что его особи живут только один год. Про-
должительность жизни новозеландского
вида Terebratella incospicua определяют
в 4 года. Для индопацифического вида
Lingula unguis хорошо известны семи-
летние особи, возраст же крупных экзем-
пляров, достигающих 5 см в длину, пред-
положительно определяют в 12 лет.
Столь же отрывочны сведения и о сезоне
размножения брахиопод. Известно, что
чаще всего он приурочен к летним меся-
цам и в ряде случаев значительно растянут
во времени. Для некоторых форм указы-
вают круглогодичное размножение. У лин-
гулид периоды вымета, по-видимому,
связаны со временем суток и фазами
луны.
546
Рис. 320. Геологическая ис-
тория плеченогих. Видно боль-
шое разнообразие групп в
палеозое и постепенное умень-
шение их к современной эпохе.
В подавляющем большинстве брохио-
поды раздельнополы. Исключение составля-
ют три средиземноморских вида Argyrothe-
са. Полового диморфизма у брахиопод
не наблюдается, т. е. самки по внешнему
виду неотличимы от самцов. Оплодо-
творение яиц нроисходит либо вне рако-
вины, либо в мантийной полости самки.
У антарктического вида Liothyrella antar-
ctica развитие зародышей протекает
в складках лофофора, а у средиземномор-
ских видов рода Lacarella — в спе-
циальной сумке под лофофором, откуда
выходят уже сформировавшиеся личинки.
Жизнь личинки продолжается от не-
скольких часов до 10—12 суток, после
чего личинка оседает и развивается во
взрослую особь. Личинки замковых бра-
хиопод очень редко встречаются в планк-
тоне, по-видимому, из-за кратковремен-
ности своего существования. Личинки
беззамковых семейства Lingulidae
исемейства Discinidae встречаются
в планктоне чаще. Они живут более про-
должительное время, имеют личиночные
пищеварительные органы и, по-видимому,
могут преодолевать значительные рас-
стояния.
Короткий период жизни свободнопла-
вающей личинки представляет единствен-
ную возможность для расселения брахио-
под. Тем, что возможность эта невелика,
зоологи и объясняют образование больших
скоплений плеченогих одного вида. В ме-
стах, благоприятных для обитания, ли-
чинки оседают тут же, рядом с роди-
телями. Десятки особей облепляют каж-
дый камешек, молодые экземпляры по
нескольку штук прикрепляются к рако-
винам старых, образуя целые грозди.
У берегов Южной Калифорнии в одной
такой грозди насчитывали до 30 экземпля-
ров (Laquens californicus).
Ограниченная способность брахиопод
к расселению проявляется также в чрез-
вычайно высокой степени обособленности
брахиоподных фаун в отдельных районах
Мирового океана. Видов со всесветным
распространением известно очень мало.
Наибольшее видовое разнообразие бра-
хиопод в современных морях наблюда-
ется у берегов Японии, Австралии и Новой
Зеландии, в Северной Атлантике и у бере-
гов Антарктиды. В наших северных
и дальневосточных морях известно около
15 видов, в южных морях с пониженной
соленостью брахиоподы отсутствуют.
Большинство современных видов бра-
хиопод обитает на материковом шельфе
и в верхней части склона, в районах
с быстрым течением и не слишком мутной
водой. Со времени глубоководной экспе-
547
диции на «Челленджере» стали известны
плеченогие с больших океанических глу-
бин. Сейчас наука насчитывает 29 видов,
опускающихся на глубину более 2 тыс. м,
но лишь 2 из них — Pelagodiscus atlanti-
cus и Abyssothyris wivillei достигают
наибольших глубин океанического ложа
порядка 5,5—6,0 тыс. м.
Наиболее древние остатки ископаемых
брахиопод известны из верхних горизон-
тов протерозоя, где встречены хитиново-
фосфатные раковинки беззамковых. По-
видимому, брахиоподы существовали
длительное время еще в докембрии, так
как в отложениях нижнего кембрия они
представлены уже почти всеми отрядами
беззамковых. Основное развитие беззам-
ковых происходило в раннем палеозое.
Известные, однако, с нижнего кембрия
замковые брахиоподы, родоначальниками
которых были представители отряда
Orthida, как наиболее прогрессивная
группа, приобретали все больший вес,
и уже в ордовике они сыграли ведущую
роль. Формирование всех основных групп
брахиопод закончилось в силуре, и в даль-
нейшем происходили лишь специализация
и формообразование внутри отрядов. Наи-
больший расцвет брахиопод относится
к палеозою, когда они занимали господ-
ствующее положение среди морской дон-
ной фауны. С начала мезозойской эры
свое место в донных сообществах брахио-
поды уступили двустворчатым моллю-
скам. Из 8 палеозойских отрядов замко-
вых брахиопод в кайнозое известно лишь
2: Rhynchonellida и Terebratulida. Послед-
ний из них в настоящее время насчитывает
семейств больше, чем в какую-либо из
прошлых геологических эпох, т. е. нахо-
дится в расцвете. Из беззамковых плече-
ногих до наших дней дошли 3 специали-
зированные группы, представленные с е-
мействами Lingulidae, Craniidae
и Discinidae, не претерпевшие за всю
долгую историю своего существования
особенно больших изменений.
Тормозящими факторами в эволюцион-
ном развитии довольно сложно организо-
ванной группы брахиопод явились, по-
видимому, прикрепленный образ жизни
взрослых особей, а также наличие у них
раковины, которая, надежно защищая от
врагов, долгое время была залогом про-
цветания группы. Но в конце концов эта
раковина превратилась в свою противо-
положность, так как ограничивает воз-
можность развития размещенного в ней
фильтровального аппарата, который по
характеру своего устройства не может
соперничать в эффективности с хорошо
развитым аппаратом других фильтраторов.
Брахиоподы представляют большой на-
учный интерес при изучении закономер-
ностей эволюционного процесса в царстве
животных как пример значительной по
объему, систематически обособленной,
высокоспециализированной группы, про-
шедшей долгую историю развития. Иско-
паемые виды имеют большое практическое
значение: встречаясь с самых древних
времен и в больших количествах, они
служат геологам руководящими вехами
при определении возраста того или иного
слоя земли, что необходимо при разведке
полезных ископаемых. Современные формы
не имеют столь большого практического
применения. Известно только, что муску-
листая нога лингулы употребляется в пищу
в Японии и в Квинсленде.
УКАЗАТЕЛЬ ТЕРМИНОВ
(Звездочкой отмечены страницы, на которых помещены рисунки, жирным шрифтом —
цветные или тоновые таблицы с изображением данного животного.)
Абиссаль 46*, 47, 291, 309
Абиссальные кишечнополостные
286, 288, 291, 296, 298,
303
— радиолярии 92
— эхиуриды 527
Абиссопелагиаль 46*, 47
Аборальная пора 265, 266
Аборальный канал 331*
— орган 330, 331, 331*, 332*
— полюс 27
Авикулярии 536, 536*
Автотомия (самокалечение) 345,
527
Агамогония 25
Агглютинированная раковина 81
Аденозинтрифосфат (АТФ) 60
Адолескарий 357, 357*
Адоральные мембранеллы 145
Акантор 451, 454
Акклиматизация 39, 41, 42, 482,
489, 519
Аккомодация 467, 484
Аконции 300, 306
Аксоподии 93, 94, 94*
Аксостиль 106, 106*, 107*, 112,
113*
Актинотроха 531, 532, 532*
Амебиаз 78
Амебоид 130, 131*, 133, 133*,
268
Амебоидное (псевдоподиальное)
движение 67, 203
Амебоциты 184, 184*, 186, 187,
196, 199
Аминокислоты 59
Амитоз 138
Амфибластула 198, 198*
Амфидиски 187*, 192
Амфиды 397, 397*, 398, 398*,
401, 403, 404, 405, 406
Анабиоз 38, 540
Анальная лопасть 467
— щель 401
Анальное отверстие кольчатых
червей 471*, 510, 511*, 526,
527*, 528
— — мшанок 535*, 537
— — нематод 396*, 397
Анальное отверстие турбеллярий
345, 347
Анастомоз Геке ли 440*, 441
Анафаза 61, 61*, 62
Анаэробиоз 37
Ангиосомит 29
Анимальное питание 101, 103,
104
Анимальный полюс 336
Анкилостомоз 420
Анафилоксия 236
Антапикальный вырост 101, 101к
Анус (анальное отверстие) 390,
392, 392*, 397, 451
Апикальный вырост 101, 101*
Апикальная игла 90
Апикальное (теменное) поле 435,
436*
Аппарат Гольджи 56, 57*
Арагонит 323
Ареал 39, 40
Арканчик 246*, 247, 278
Арктический солоноватоводный
реликт 39
Ароморфоз 176, 177
Археоциты 184, 196, 199, 200,
200*
Архитомия 493, 499, 501
Аскаридоз 418, 419
Аскон 183, 185*
Асконоидный тип строения 186,
189, 190, 198, 199
Атокная форма 477*
Атолл 315, 316, 316*, 317
Атриальная синцитиальная пере-
мычка 191*
— полость 183, 185, 198
АТФ (аденозинтрифосфат) 60
Аутозоиды 246, 251, 290, 293,
294, 296, 297, 298
Ауторепродукция 58, 62
Аутотрофные организмы 43
Аутотрофный тип обмена (пита-
ния) 69, 95, 103, 104, 116
Ахроматиновое веретено 61
Байкальские пиявки 519
— турбеллярий 345, 348
Бактериальная флора 415
Балантидиоз 170
Батиаль (материковый склоы)
46*, 47, 291, 296
Батипелагиаль 46*, 47
Бенталь 46*
Бентос 48, 49
Бесщупальцевые полипы 274
Бинарная номенклатура 10
Биогенетический закон 22, 472
Биогеоценоз 42
Биолиты 489
Биологические индикаторы 96
Биосфера 40, 42, 43
Биотический фактор 45
Биотоп 40, 42, 45, 46, 51, 394
Биофильтраторы 215, 216
Биоценоз 42
Биоценология 45
Биполярность 53, 456
Биполярные ареалы 303
Биссус 320, 322
Биссусная железа 320
Бластула 198
Блефаропласт 67, 105
Боковые органы 403, 412, 440*
Ботрии 375, 379, 380
Бродяжки 91, 149, 149*, 150,
152, 153*, 158, 160*, 161
Буккальное (ресничное) поле 435,
436*, 440*
Бульбус (луковица) 391, 395*,
397*, 399*, 402, 403, 412, 413,
415, 423, 424, 424*
Бурса (выводковая сумка) 419, 421
Бурсальные крылья 412, 413
В
Б
Базальные зерна 105, 106*, 107
Байкальские губки 219
— мшанки 541, 541*
Вагина (влагалище) 378, 382, 417
Вакуоли пищеварительные 26, 75,
76, 76*, 102*, 134*, 136,136*,
146*, 187
— сократительные 69, 76, 76*,
93, 95*, 97, 97*, 98*, 103,
103*, 104*, 105, 105*, 111,
134*, 137*, 141*, 145, 146*,
148*, 161*, 162*, 163*, 164,
170, 173, 173*
^р36 Жизнь животных, т. 1
549
«Вертячка» 387
Вибракулярии 536, 536*
Вирусы 53
Влагалище (вагина) 370, 377,
378, 417, 453, 460
Воронка кишечнополостных
230*, 231, 285
— околоротовая 370
Воротничковые клетки (хоаноци-
ты) 184
Вторичноодиночные формы губок
202, 203*
Выбросы червей 503, 506
Выводковая камера 489, 536
— капсула 479
— сумка 485
Выделительная железа 401
— пора 350
— система брахиопод 545
— — мшанок 538
— — немате л ьминтов 392, 398,
402, 431, 435, 439, 441, 457
— — плоских червей 337, 343,
350, 357*, 367, 370, 370*, 376
Выделительный пузырь 350
Вымершие животные 219, 308,
Г
Гаксастры 187*, 192, 193
Галлы 426, 427
— сложные (сингаллы) 426, 427
Галосфера 17
Гальванотаксис 135
Гаметогенез 131
Гаметогония 121*
Гаметоциты 125, 127
Гаметы 70, 86, 97, 100, 111, 119,
119*, 120, 121, 125, 176
Гамогония 25, 119*
Гамонт 119*
Ганглии кольчатых червей 471,
492
— круглых червей 398, 440*, 441
— мшанок 535*
— плоских червей 370
— сцифоидных 231*
— фор они д 531
Ганглиозные клетки 330, 397*, 398
Гастральная мембрана 191*
Гастральная полость 223, 227*,
228, 231, 232, 233, 257, 259,
265, 288, 291, 293, 296, 300,
305, 310, 332
Гастральные нити 231*, 232, 286
— перегородки 299, 300, 302,
307, 309, 310
Гастроваскулярная система 229,
239 330
Гастрозоиды 246*, 247, 278
Гельминтозы 418
Гельминтология 421
Гельминты 349, 380, 408, 421, 422
Гематохром 97
Геммулы 38, 199, 200, 201, 201*,
205, 207, 210, 214
Гемоглобин 124, 408, 469, 483,
496, 498, 515
Гемэритрин 469
Генеративные клетки 181
Гермафродитизм 23
— губок 196
— кишечнополостных 290, 302,
307, 309
— кольчатых червей 470, 478,
492, 511
— круглых червей 392, 415
— мшанок 538
— плоских червей 338, 343, 349,
367, 368, 376
— протандрический 302, 307
— фор они д 531
Гермафродитное отверстие 370
Гетерогония 181, 431
Гетеронереидная форма 477*
Гетеротрофные организмы 43
Гетеротрофный тип обмена (пита-
ния) 69, 95, 103, 116
Гигантские клетки 426, 426*
Гидравлический скелет 469, 527
— способ движения 28, 456, 475,
528
Гидроризы 241, 242, 252
Гидростатический аппарат 540
Гидросфера 42
Гидротека 261
Гинекофорный канал 361
Гипергалинные организмы 37
Гиподерма 390, 396, 432, 438,
440*, 451, 452
Гиподермальныс каналы 452, 453
Гиподермальный валик 396
Гипонейстон 48, 49, 50*
Гипостом 271
Гирудин 470, 511
Глаза кольчатых червей 467, 468*,
470, 471, 472
— круглых червей 402, 441,
462, 463
— олигохет 492, 494, 495, 496
— пиявок 510, 510*, 515, 516,
519, 520, 524
— полихет 479, 484, 485, 488,
489
— турбеллярий 345
— фасеточные 489
Глазки кишечнополостных 229,
229*, 230, 231*, 232, 232*,
239, 281
— кольчатых червей 468*, 517
— круглых червей 406, 440*,
443*, 450
— плеченогих 546
— плоских червей 341*, 344,
345, 371
Глазное пятно 231*, 232, 232*
Глазной бокал 397, 401
Глазок (стигма) 97, 98*, 99, 103,
103*
Гликоген 57, 118
Глобигериновые илы 544
Глобулп (грибовидные тела) 468*
Глотка гребневиков 330, 331*
332
— жевательная (мастаке) 429
— кишечнополостных 233, 233*,
300, 305, 310
Глотка кольчатых червей 468*,
469, 471*, 475, 481, 483, 484,
491, 492*, 511*
— круглых червей 391, 398, 439,
450*, 450, 455, 456*, 460
— плоских червей 337, 340*,
341, 343*, 344, 347, 347*, 350,
370
Глубоководная зона 46*
Глубоководные актинии 301, 303
— альционарии 291
— антипатарии 310
— брахиоподы 544, 547
— горгонарии 288
— губки 191, 206, 208, 210
— кишечнополостные 241, 245,
260, 261, 286
— медузы 287, 304*
— полихеты 473
— радиолярии 88, 92
— эхиуриды 526, 527
Голова кольчатых червей 468*.
470, 474, 479, 483, 484, 485
Головка (сколекс) 375, 376, 378,
381*, 382, 384, 386
Головная капсула 397, 398*,
401, 425
Головной мозг кольчатых червей
468*, 470, 471, 471*, 472, 492
— — немертин 460, 461, 463
— — турбеллярий 344, 348
Головные лопасти 371, 492, 493,
495, 501
— рецепторы 401
Гонады (половые железы) 228,
268, 272, 471*
Гонодендр 278, 279
Гонофоры 246*, 278
Гранулы 56
Гребные пластинки гребневиков
329, 330, 331, 331*, 334
Грибовидные тела (глобули) 468*
Грунтоеды 34, 498
Губные рецепторы 401
Губы 397, 401, 404, 412, 416, 472
д
Дактилозоиды 246
Двустороннесимметричные живот-
ные 297
Девастация гельминтов 421
Дезоксирибонуклеиновая кислота
(ДНК) 58, 58*, 59, 62, 138
Дейтомерит 117, 117*, 118
Деление кишечнополостных 267,
296, 301, 302, 302*
— множественное (шизогония)
70, 120
— поперечное 139, 139*, 301
Деление продольное 301, 302,
302*
— простейших 70, 76, 81*, 85,
111*, 120, 138, 138*, 140*, 144
— турбеллярий 343
— ядра 77, 81*, 85, 140*
Депигментация 48
550
Дермальная мембрана 185, 191*,
204
Детритоеды 527
Диапауза 503
Дикроцелиоз 359
Диктиокаулез 422
Диморфизм 246, 293, 294
— половой 479, 527, 547
Диссепименты (поперечные пере-
городки сегментов) 29, 468,
468*, 491
Дитиленхоз 422
Дифиллоботриоз 381, 382
ДНК (дезоксирибонуклеиновая
кислота) 58, 58*, 59, 62, 138
«Дождь» трупов 488
Домики коловраток 428, 433,
434*, 435
Донная фауна 52
Донные брахиоподы 543
— гребневики 334
— кишечнополостные 225, 280,
284, 303, 309
— олигохеты 496, 497
— полихеты 475, 484
— простейшие 81
— сипункулиды 528
— форониды 531
— эхиуриды 526
Дополнительный носитель 37
Дракункулез 421
Древние животные 92, 219, 224,
258, 260, 483*
Дробильный аппарат нематод 412,
413
Дробление яйца 268, 470
— билатеральное 454
— спирального типа 454, 466,
527, 528
•ЯР?
smm>
Жаберные нити 468*
Жабры наружные 499
— олигохет 492, 496, 499
Жгутики простейших 67, 68*,
95, 95*, 97*, 98*, 101*,
102*, 103*, 104*, 105*, 106*,
111*, 115*, 122*
— губок 196, 198
Жгутиковое движение 24, 67
Жгутиковые камеры 185, 191*
— трубки 185, 191, 193, 196, 198
Железа ножная 440*, 443*
— преутеральная 424*, 425
— скорлуповая 351, 370, 377
— субцеребральная 440*, 441
— хвостовая 395*, 402, 403,
403*
— шейная 395*, 398, 401
— ядовитая 460
Железистые клетки кишечнопо-
лостных 228, 265, 266, 299
Железистый орган 339*, 368
Железка предстательная 442,
443*
Железы анальные 527
— клейкие 437, 440*
Железы ножные 437
— пищеварительные 545
— проникновения 352
— слюнные 511
Желточники коловраток 431,
440*, 441, 442
Желточники моногеноидей 370,
374
— трематод 351, 362
— турбеллярий 339*, 343, 343*
— цестод 377, 379, 388
Желточный проток 343, 343 *,
377
Желудок гребневиков 330, 331*
— кишечнополостных 229, 229*,
231*, 232, 285
— кольчатых червей 457, 468*,
469, 492*, 511, 511*, 524
— круглых червей 439, 440*,
441*, 457
— мшанок 537
Живорождение 479
Живородящие 296
Жировая капля 442, 443*
Л
Забота о потомстве кишечнопо-
лостных 281, 302
— — червей 479, 480, 484, 485,
513
Закапывающиеся (зарывающиеся)
кишечнополостные 303
Замок брахиопод 544
Зародыш 154, 372, 514*
Зародышевые пласты (листки) 182,
183, 198
Зигота 71, 86, 97, 100, 119, 119*,
120, 122, 125, 127*
Зоб 469, 491, 492*
Зообентос 49
Зоопланктон 48
Зооиды 344
II
Иглы губок 183, 184, 186*, 187*,
188*, 190, 194*, 195, 219*
— кишечнополостных 225
Идиоадаптация 177
Изгибательное движение 28, 28*
Индивидуализация колонии 202
Инкус 439, 439*
Инволютная раковина 83
Интерстициальная фауна 51
Интерстициальные (промежуточ-
ные) клетки 228
Инцистирование 79, 80*, 118,
145, 159, 170
Ихтиофтириазит 160*
Ископаемые брахиоподы 543,
546*, 547*
— губки 219
— кишечнополостные 258, 259
Ископаемые мшанки 540, 542, 548
— полихеты 482
— простейшие 72, 87, 92, 96
К
Кала-азар (висцеральный лейшма-
ниоз) 110, 110*
Калимма 88
Каннибализм 282
Капилляры 469
Кариокинез (митоз) 60, 61, 61*,
77
Кариолимфа (ядерный сок) 58, 59,
62, 70
Кефализация 31
Кинетозома 68, 70*
Кинетонуклеус 109
Кинетопласт 107, 108, 109, 115
Кишечник кольчатых червей 468*,
471*, 472, 510, 511*, 526,
527*, 528
— моногеноидей 370, 370*, 372
— мшанок 535
— немательминтов 390, 390*,
397, 404*, 408, 449*, 451,
455, 457
— немертин 460*
— трематод 350, 357, 357*
— турбеллярий 340*, 341, 343*г
344, 345, 347
— форонид 532
— цестод 374
Кишечные боковые выросты 468*,
469, 511
— карманы 468
Кишка 337, 340
— задняя 398, 399*, 412, 457,
511*, 537, 545
— средняя 392*, 395*, 398, 399*,
404, 406, 412, 417, 450, 461,
468*, 469, 545
Кланорхоз 360
Клейкие (липкие) клетки 330,
331*, 333
— сосочки 331*
Клетка 53, 55*, 57*, 61*, 65,
73, 74, 176
Клеточная мембрана 55
Клоака коловраток 439, 440*,
441
— плоских червей 363, 373
Книдоциль 234
Кожа эхиурид 526
Кожное дыхание 492, 496
Кожно-мускульный мешок 24, 28
— — немательминтов 390, 390*,
392, 393, 441, 454, 455
пиявок 510
— — плоских червей 337, 340,
343, 346, 350, 370, 376
— — эхиурид 526
Кожные железы 493
Коконы олигохет 492
— пиявок 505, 511, 514, 516, 518,
519, 520, 521, 525*, 526
Кокцидиоз 123
Колбочки кишечнополостных 229
Колленциты 184, 184*
Коловращательный аппарат 429,
431, 433, 435, 436, 436*, 438,
439, 440*, 441
36*
551
Колокол скребней 453
Колониальность 21, 116, 150,
201, 202, 428, 533
Колонии временные 241
Колонии губок 182, 190, 201,
202
— двулопастпые 539
— кишечнополостных 223, 224*,
239, 241, 243, 243*, 248, 202*,
276, 287, 293*. 309, 318
— коловраток 437
— мономорфные 535
— моноподиальные 242, 243*
— мутовчатые 243
— мшанок 533, 535, 536, 537,
541, 541*
— полиморфные 246, 248, 535
— полихет 485
Колонии постоянные 241
— простейших 98, 99, 99*, 100,
100*, 149, 150, 150*, 151
— сгибающихся 457
— симподиальные 243, 243*
— стелющиеся 243*
— — эпифиты 244
Кольца (сегменты) 449, 467, 490
Кольцевой канал кишечнополост-
ных 229, 229*, 231*, 281
Кольчатость 449, 468, 471, 489
— вторичная 502, 510
— ложная 435, 452
Комменсализм 33, 34, 37, 211, 249
Комменсалы губки 211
— кишечнополостные 251, 306,
309
— кольчатые черви 473, 495
— немертины 461
— плоские черви 367
Конвергенция (конвергентное
сходство) 173
Коннективы (нервные тяжи) 466,
492
— окологлоточные 468*, 492,
510
Консументы 43
Конули 194
Конхиолин 322
Конъюгация 25, 111, 139, 140,
140*, 152
Копролиты 503, 506, 509
Копулятивный орган 442, 443*
Копуляция 25, 111, 115, 119,
119*, 127*
«Копье» нематод 398, 398*, 404,
405
Коралловые рифы 310, 312, 313,
314
— — , происхождение 314, 315,
316
— — , обитатели 215, 310, 318,
320
Корацидий 380, 381, 381*
Корзиночка гидроидных 261
Кормидии 247
Корневая нить жгутика 67
Космополитизм 38
Космополиты 275, 429
Краевая лопасть сцифомедуз
231*, 232*
Крапивные (стрекательные) клет-
ки 234
Крахмальные зерна 57
Кристы 56
Критидиальная стадия 108
Кровеносная система брахиопод
545
— — замкнутая 531, 532
— — кольчецов 469
— — мшанок 538
— — форонид 531
— — червей 460, 463, 466, 469,
491, 510, 527
Кроющие пластинки кишечно-
полостных 246*
Крючья волосатиковых 451, 452
— плоских червей 370, 371, 372,
375, 382, 384*, 386
— приапулид 455, 455*
Кутикула 15
Кутикула кишечнополостных 301,
308
— немательминтов 391, 392, 396,
402, 406, 416, 427, 436, 451
Личинка нематод 399*, 400, 406,
407, 409, 415, 419, 421, 425.
426, 426*
— немертин 461
— плеченогих 545, 546, 547
— полихет 369, 477, 479, 480.
481, 483, 487
— приапулид 456, 456*
— сгибающихся 457, 458*
— скребней 453, 453*
— трематод 351, 354, 364, 366
— форонид 531, 532, 532*
— цестод 375, 378, 379, 379*,
380, 381, 383, 384
— эхиурид 527
Ложные ножки (ложноножки) гу-
бок 186, 187, 203, 214
— — простейших 66, 75, 77
Лофофор мшанок 535, 535*, 537
Люминесценция (свечение) 299
Люцифераза 287
Люциферин 287
Л
Латентное состояние 38
Лацерация 302, 302*
Легкие 15
Легочные мешки 15
Лейкон 183, 185, 185*
Лейконоидный тип строения губок
185, 189, 190, 191, 193, 198
Лейшманиоз висцеральный (кала-
азар) НО, 111
— кожный (пендинская язва)
109, 109*, НО
Лемниски скребней 453, 453*
Лигамент скребней 453, 453*
Лигулоз 380
Лизосомы 56, 57*
Ликофора 378
Лимбарии 489
Линза 468*
Линька 114, 115, 399, 407, 415,
417, 419, 421, 425
Литораль (осушная зона) 46, 46*,
291
Литоральная фауна 46, 52
Литоральные брахиоподы 543,
544
— губки 209
— кишечнополостные 261, 263*,
304, 318
— мшанки 540
— олигохеты 499, 500, 506
Литосфера 42
Личинка волосатиковых 451, 451 *
— губок 196, 198, 198*, 199,
202, 204, 205, 214
— киноринх 450
— кишечнополостных 237, 255*,
256, 273, 285, 296, 302, 307
— коловраток 433
— кольчатых червей 466, 470, 472
— моногеноидей 370, 373, 374
— мшанок 538, 538*, 539
II
Макрогамета 71*
Макрогаметоциты 125, 127*
Макрогаметы 121, 121*, 123*»
125, 127*
Макрозоиды 150*, 151, 151*
Макромеры 196
Макронуклеус 70, 71*, 134*,
138, 139*, 140, 140*, 142,
143*, 145, 146*, 149, 163*,
170*, 173, 175*, 177
Макросклеры 187*, 188*, 189,
190*, 191*, 192, 195
Макросферическое поколение 85,
86
Маллеус 439, 439*
Малярия 37, 41, 74, 124,
127
Мантийная полость брахиопод
545
Мантия брахиопод 545, 546
— моллюсков 320
Манубриум 439, 439*
Мастаке (жевательная глотка)
429, 439, 440*, 441
Матка моногеноидей 370, 371
— нематод 390*, 399, 399*, 417,
424*
— скребней 453
— трематод 351, 361, 362
— турбеллярий 343*
— цестод 378
Медузоидная форма (поколение)
223, 224, 225, 238, 247, 261,
277 285
Медузы 223, 224, 225*, 238, 238*,
239, 260, 273, 275
Межпроглоттидные железы 387
Мезентериальные нити 233, 233*,
300, 307
Мезентерий 468
Мезенхима 451
Мвзенхимные клетки 451
552
Мезоглеальный скелет 293
— слой 302, 307, 309
Мезоглея губок 183, 185, 186,
187, 191, 196, 198, 213, 214
— гребневиков 331
— кишечнополостных 223, 227*,
228, 229*, 230, 231*, 272,
277
Мезодерма 11, 332, 336
Мезодермический эпителий 539
Меланин 124
Мелководные губки 206, 208,
209
— кишечнополостные 245, 289,
296, 298, 300, 309
— эхиуриды 526
Мембрана базальная 243
Мембранеллы 68, 141, 141*, 142,
143*, 144*, 145, 146*, 167
Мембраны 141, 161*
Меридиональные каналы 330,
331 331*
Мерозоит 120, 121, 121*, 122*,
124, 125, 127*
Мерцательное пламя 450
Мерцательные бороздки 330, 332*
— клетки 456*, 461
— колбочки 340
Метагенез (чередование поколе-
ний) 239, 246
Метаболизм 313
Метамерия (сегментация) 29, 450,
468
— гетерономная 31*
— гомономная 30, 31*
— истинная 29, 31*
— наружная 554
Метамерное строение 29, 449
Метамеры (сомиты) 29
Метаморфоз у губок 196, 198,
199
— мшанок 538
— полихет 480, 486, 487
— регрессивный 546
— сгибающихся 457
— форонид 531
Метанефридии 468*, 469, 510,
531, 532*, 546
Метастронгилез 509
Метатрохофора 481, 483, 485
Метафаза 61, 61*
Метацеркарий 353, 355, 357, 359,
360, 365, 366
Метациклические формы 108
Миграции вертикальные 33, 33*,
48
— генеративно-алиментативные
32
— горизонтальные 32, 33*
Микрогаметы 121, 121*, 122*,
125, 127*
Микрогаметоциты 121, 122*, 125,
127*
Микрозоиды 151, 151*
Микромеры 196
Микронуклеус 70, 71*, 134*,
138, 139*, 140, 140*, 142,
146*, 149, 153, 163*, 170, 173,
175*, 177
Микропиле 121
Микросклеры 187*, 188*, 189,
190*, 191, 192, 193, 194, 194*,
195, 200, 201*
Микросферическое поколение 85,
86
Микрофилярии 37, 421
Миксоспоридиоз 132
Минеральный скелет 67
Мионемы 68
Миосомит 29
Миоциты 186
Мирацидий 351, 352, 353, 357,
357*, 358*, 360, 362, 364,
365, 366
Митоз (кариокинез) 60, 61, 61*
Митохондрии 56, 57*, 59
Митрария 481
Многокамерная раковина 83
Множественное деление (шизого-
ния) 120
Мониезиоз 388
Монофаги 34, 439
Монофилия 17
Морские брахиоподы 543
— губки 183
— кишечнополостные 223, 263
Морские мшанки 535, 536*, 538*,
540, 541
— пиявки 517, 518*
— простейшие 72, 88, 93, 96,
100, 134
— сипункулиды 528
— форониды 537
— эхиуриды 526
Моториум 69
Мочевой пузырь 440*, 441
Мускул втягиватель 440*
Мускул замыкатель ноги 440*
Мускулатура (мышцы, мускулы)
65
— кишечнополостных 295, 301,
303, 309
— червей 440*, 449, 451, 452,
455, 456, 457, 468*
Мускулы протракторы 398*, 423,
425
— ретракторы 455, 460
Мутуализм 34
Мюллеровская личинка 347, 347*
Н
Нагана 108
Наземная фауна 53
Наземные животные 523
Невросомит 29
Неврофибриллы 398
Нейстон 48, 49
Неклеточные (доклеточные) фор-
мы жизни 53
Некроз 427
— флоэмы 115
Нектон 48
Нектофоры (плавательные коло-
кола) 246*, 247, 278
Нектохета 481
Нематодоз галловый 427
Нервная система брахиопод 546
— — гребневиков 330, 340*,
350
— — кишечнополостных 229*,
230, 248, 295, 301
— — мшанок 538
Нервная система немательминтов
393, 397, 440*, 441*, 451, 455,
460
— — плоских червей 340*,
345, 348, 350, 370, 376
— — форонид 531, 532
— ткань 65
— цепочка брюшная 468*, 491*,
492, 510
Нервное кольцо (кольцевая ко-
миссура) 397, 399, 404*, 449*,
450, 455
двигательное 229*, 230
— — окологлоточное 451, 528,
546
— — чувствительное 229*, 230
Нервные клетки 228
— комиссуры 460*, 461, 471*
— — головные 460, 461, 510
— стволы брахиопод 546
— — кольчатых червей 471*
— — немательминтов 397, 451,
453
— узлы 467, 492
— — немертин 460, 461
Нервных клеток скопление 450,
471, 531
Нефропор 527, 528
Нефридии 491, 491*, 526, 527,
528
Нефростом 527, 528
Нижняя челюсть мшанок 536,
536*
Нога брахиопод 544, 548
Норы брахиопод 544, 545*
Нуклеиновые кислоты 54, 56
Нуклеолы 70
Нуклеотиды 58, 59
Нуклеопротеиды 58, 59
О
Оболочка клетки 66
Обонятельные клетки" 467
— ямки 340, 392*, 393, 397, 472
Обрастатели 261, 275, 488, 538, 541
Одиночные 201, 223, 225, 227,
259, 299, 306, 308, 309
Оеции 536, 538, 539
Ожоги кишечнополостных 235,
236, 283
Олигогалинный 37
Олигомеризация 31, 297
Олигомикстный биоценоз 44
Олинтус (рагон) 198
Онкосфера 378, 383, 385, 386,
387 388
Онхи'396*, 398, 401, 404, 404*,
412, 419
Оокинета 125, 127*
553
Оотека 426
Оотип 351, 370, 377
Ооцисты 119, 119*, 120, 121*,
122, 123, 125, 127*, 177
Ооциты 442
Описторхоз 360
Оплодотворение 25
Оплодотворение кишечнополост-
ных 238, 296
— перекрестное 342, 374
— простейших 119, 121, 122,
238, 296
— червей 377, 470, 479, 492, 511,
524, 527
Оральный полюс 27
Орган Брандеса 365
Организаторы 271
Органоиды 56, 176
Органы обоняния 339, 403
— осязания (химического чувст-
ва) 401, 401*
— чувств кишечнополостных
230*
— — червей 345, 440*, 455,
457, 467
Осевой физиологический гради-
ент 271
Оскулум (устье) 183
Осмотическое (сапрофитное) пи-
тание 103, 104, 107, 120, 174
Осушная зона (литораль) 44
Осязательные сосочки (папиллы)
412
П
Пайптобласты 539
Пальпакула 246*
Пальпоны 246*, 247, 278
Пальпы 468*
«Пальцы» коловраток 440*, 443*
Панцирь губок 194
— коловраток 430*, 433, 435,
438, 443*
Папиллы губок 194, 203, 204,
207, 214
— нематод 397, 402, 403, 404,
404*
Парабазальное тело 104, 104 *,
107*
Парагастральная полость 183
Паразитарная катаракта 365
Паразитические амебы 77
— гребневики 333
— жгутиконосцы 105, 107, 107*,
109*, 110, 115
— инфузории 134, 159, 164, 177
— кишечнополостные 255, 256,
275
— книдоспоридии 129, 132
— олигохеты 501, 502
— пиявки 512, 513, 515, 517,
518
— споровики 116, 117, 120, 124
— черви 338, 348, 354, 368, 382,
408, 418, 421, 484, 495, 509
Паразитизм 34, 35, 36, 37
Паразитизм внутренний 374
— губок 181, 211
— кишечнополостных 251, 256,
275
— наружный 374
— простейших 116, 133, 164, 177
— самцовый 527
Паразитизм червей 338, 339,
375, 400, 406, 410, 415, 452, 454,
527
Паразитический образ жизни 128,
371, 375, 415, 429, 486, 501
Паразиты внутренние (эндопара-
зиты) 338
Паразиты наружные (эктопара-
зиты) 338
Паратомия 493, 493*, 494
Пароксизм 37
Параподии 24, 467, 468*, 471,
474, 481, 484, 489, 526
Паренхима 337, 340, 350, 370,
377, 390, 457, 460, 463
Паренхимула 196, 198, 198*
Партеногенез 25, 493
Партеногенетическое поколение
351
— размножение 36, 181, 199,
415, 431, 444, 446
Парус кишечнополостных 229,
229*, 230, 272, 279
Педицеллярии 321
Педосфера 43
Пелагиаль 46*, 47
Пелагическая фауна 52
Пелагические кишечнополостные
303
Пелагические полихеты 474, 483,
484, 496
Пелагический образ жизни 231,
484
Пелликула 66, 95, 103, 103*,
112, 136, 137
Пендинская язва (леишманиоз)
109, 109*, НО
Перистом 136, 143, 144*, 146*,
170
Перистомальная губа 149
Перистомальное поле 142, 149
Перистомиум 468*
Пещерное существование 473
Пигмент 183, 210, 461
Пигментные клетки 230, 230*
«Пилюльный» орган коловраток
433
Пилидий 461
Пинакоциты 184, 184*, 185
Пиннулы 225, 232
Пищеварение внутриклеточное 93,
187, 340, 391
Пищеварительная система бра-
хиопод 547
— — кольчатых червей 491,
510, 527
— — немате л ьминтов 390, 391,
398, 431, 435, 439, 450, 453
— плоских червей 337, 340*,
347, 350, 370, 375
Пищевод брахиопод 545
— мшанок 537
Пищевод червей 370, 391, 395*,
397*, 399*, 404*, 412, 424*, 439,
440, 451, 469, 492*, 511,
511*
Пищеводные поры 391, 392*
Плавательные колокола (некто-
форы) 246*, 247
Плавательные лопасти 477
Плавательный пузырь (пневма-
тофор) 246*, 247
Плазматический воротничок 182
Пламенковые клетки 440*, 443*
Пламенные клетки 350
Планктон 48, 283, 532
Планктонные 86, 87*, 100, 154,
283, 435
Планула 237, 237*, 238, 238*,
256, 259, 281, 302, 348
Плейстон 48, 277
Плейстонные животные 279, 280
Плейстонный биоценоз 280
Плероцеркоид 378, 379, 379*, 380,
381*, 382
Пневматофор (плавательный пу-
зырь) 246*, 247, 277, 279
Пневмоциста 300
Подвижные зубы 398, 402, 404
Подошва кишечнополостных 233,
266, 299, 302, 303
— мшанок 533
— турбеллярий 347
Полимеризация 30
Полимерия гомономная 31
Полимерные структуры 30
Полимикстный биоценоз 44
Полиморфизм 21, 151, 231, 246
Полиплоидия 177
Полипоидная особь 277
— форма 223, 224, 225
Полипоидное поколение 285
Полипы 223, 224, 225*, 239, 246,
273, 275, 276, 276*
Политипические виды 422
Полифаги 34, 439
Полифилия 17
Половая клоака 343, 377
Половая система брахиопод 545,
546
— — немательминов 392 399*,
441, 453, 492*, 527
— — плоских червей 339, 339*,
343*, 350, 362, 370, 374, 375,
376, 380, 383
Половое отверстие 343, 379, 382,
409, 449, 471*, 527*
— — женское 377, 399*, 402,
404*, 415, 426, 451, 511
мужское 377, 378, 511
Половые железы (гонады) греб-
невиков 331
— — кишечнополостных 224,
229*, 231*, 233, 286, 286*
— — червей 449, 457, 461,467,
482
— — клетки 228, 290, 293, 333,
412, 442, 469, 478
— протоки 450
Полостная жидкость 468, 469,
491, 526, 528, 538
554
Полость тела вторичная (целом)
336, 456, 468*, 481, 491, 510,
526, 545
— — несегментированная 527
— — сегментированная 468,
470, 491
— — первичная (протоцель)
390, 390*, 439, 449, 451, 454
Популяция 44
Пороциты 184, 184*
Почвенные нематоды 400, 401
— олигохеты 490, 499, 506
— простейшие 75, 134
— турбеллярии 340, 346
Почвообразование 499, 507, 508
Почкование внешнее 240
— внутреннее 154, 199
— губок 199, 201, 203
— кишечнополостных 240, 243*,
264, 267*, 273, 301
— мшанок 538, 539
— простейших 25, 153, 173
— червей 438, 457, 485
«Поясок» 416, 417, 492, 511, 521
Правило рекапитуляции 22
Пред ротовая лопасть (головная)
490
Пресноводная фауна 51
Пресноводные губки 205, 207,
210, 218, 220
— инфузории 134, 154
— кишечнополостные 224, 227,
263, 268, 271, 274, 287
— мшанки 535, 537, 540, 541
— олигохеты 506
— пиявки 510, 518*, 519
— простейшие 79, 81, 93, 96,
100
— турбеллярии 340, 344
Прибойные (обитающие в зоне
прибоя) 260, 294, 314, 320
Прибрежные кишечнополостные
283, 309
— простейшие 81
— черви 473, 506, 519, 526
Прикрепительный аппарат 172*,
173, 371, 374, 379
— диск 370, 371, 372, 373, 392
Прикрепительные ямки 370, 370*
Прикрепленные формы 148, 183,
205, 239
Присасывательная ямка 378
Присасывательные ножки 392*
Присасывательный диск 354
Присоска брюшная 347, 348, 351,
355, 357*, 360, 365, 374
— ротовая 348, 350, 357*, 360,
362, 365, 373, 510
Присоски 36
— кишечнополостных 236, 266
— кольчатых червей 510, 511*,
512, 516, 519, 521
— плоских червей 350, 352, 355,
365, 367, 370*, 376
— простейших 107, 161, 173
Проболы 427, 427*
Проглотиды 375
Промысел губок 217, 218
— кишечнополостных 290, 323
Промысловые губки 204, 217,
290
Простатические железы 351
Простомиум 468*
Протозойная фауна 167
Протомерит, 117, 117*, 118
Протонеф радиальная система
367
Протонефридиальные каналы 450
Протонефридий 357, 392*, 393,
400, 440*, 441, 443*, 448,
456, 457, 463
Протоплазма 38, 54, 77
Протоплазматическое тело фора-
минифер 83, 84, 85
Протопласт 97, 98
Прототрофные организмы 43
Протоцель (первичная полость
тела) 451
Профаза 60, 61*
Процеркоид 379, 380, 381, 381*
«Пряничная» болезнь устриц 214
Псаммофильная фауна 51, 156,
157
Псевдоатриальная полость 202
Псевдометамерия 29, 29*
Псевдоподиальное (амебоидное)
движение 23, 67
Псевдоподии (ложноножки) 23, 67
— губок 186
— кишечнополостных 228, 268
— простейших 67, 75, 78*, 80*,
81*, 89*, 95, 105*, 127*,
129*, 130*
Пульсирующие вакуоли 26
— клетки 376
Пульсирующий пузырек плече-
ногих 546
Рабдитоидный пищевод 415, 419
Рабдиты 346
Рабды 189, 190
Рагон (олинтус) 198
Радиальные выросты кишечно-
полостных 229, 229*, 231*,
232, 251, 272, 281, 300
— перегородки 224, 225
Радиоляриты 92
Разведение губок 204
Развитие коловраток 438, 444
— кольчатых червей 470, 472, 493
— мшанок 539
— нематод 413, 417, 421
— плеченогих 546
— плоских червей 351, 356, 357,
357*, 359, 364, 370, 372, 378,
381, 388
— форонид 531
Размножение губок 196
-— кишечнополостных 273, 240,
267, 302
— мшанок 538
— простейших 80, 84, 91, 118,
120, 131, 139, 153
— червей 446, 470, 477, 485, 498,
502, 514
Размножение бесполое 25
— — губок 196, 199, 200*, 203
¦— — жгутиконосцев 97, 99*
115
— — инфузорий 138, 138*.
145, 150, 173
— — кишечнополостных 301,
302, 302*, 308
— — миксоспоридий 131
— — мшанок 538, 539
— — олигохет 493, 493*, 499,
501
— — полихет 479, 485
— — саркодовых 70, 81*, 85,
94
— — сгибающихся 457
— — споровиков 118, 120,
127*
— — турбеллярии 343
Размножение вегетативное 259,
267, 493, 494
Размножение половое 25
— — губок 196
— — кишечнополостных 267,
301, 302
— — мшанок 538
— — олигохет 494, 499, 501,
502
— — полихет 479
—- — турбеллярии 343
Размножение смешанное 431
Раковина брахиопод 543, 544,
545, 548
— простейших 79, 80, 80*,
81*, 82, 83, 83*, 87
— хитиновофосфатная 548
Рамус 439, 439*
Рахис 297
Реактивное движение 28
Регенерация (самовосстановле-
ние) губок 202, 203, 204,
218
— кишечнополостных 226, 270,
271, 302, 308
— немертин 461
— олигохет 499, 501
— сгибающихся 457
— турбеллярии 344, 345
— эхиурид 526
Редии 352, 357, 357*, 360, 365
Редукционные тельца 199, 209,
268
Редукция 22, 23
Редуценты 43
Реликты 39, 276, 541
Репелленты 524
Репликация 58
Реснички 23, 68
— брюхоресничных 391, 393
— инфузорий 134*, 135, 143*,
149, 170, 172, 176
— кишечнополостных 233, 305
— коловраток 429,433, 435, 437,
446*
— кольчатых червей 472, 480,
494, 496
— мшанок 537, 538
— немертин 461
— плеченогих 545
555
Реснички плоских червей 337,
342,346,349, 367, 370, 380
— простейших 134*, 135, 143*,
149, 170, 172
Ресничное движение 24, 28, 67
«Ресничное пламя» 343
Ресничное (буккальное) поле 435,
443*, 448
Ресничные клетки 305, 310, 471
Ресничные кольца 471, 471*, 472
— ямки 440*, 467
Ресничный аппарат 428, 538
— пояс 472, 480, 481, 487
Ретина 468*
Ретинальные клетки 468*
Ретроцеребральный орган 435,
440*, 441, 443*
— мешок 440*
Рибонуклеиновая кислота
(РНК) 56, 58, 59, 62
Рибосомы 56, 58, 59
Ризоиподобные отростки 544
Ризоподии 84, 84*
Ринхоцелом 460
Рифообразующие кораллы 258,
294, 310, 311, 316, 319
Рифы барьерные 314, 315, 315*,
316
— береговые 315
— коралловые 37, 311, 313, 322,
488
— мшанковые 542
РНК (рибонуклеиновая кисло-
та) 58, 59, 62
Ропалий 231*, 232, 232*, 281
Ропалиоиды 284
Рострария 481
Рот личиночный 336
— мшанок 537
— плеченогих 545
— червей 469, 471*
Ротовая полость (стома) 396*,
398, 403, 404*, 491
Ротовое отверстие брюхореснич-
ных 391, 392*
— — гребневиков 331*, 332,
334
— — киноринх 449*
— — кишечнополостных 229,
229*, 231*, 233*, 267, 284,
300, 310
— — коловраток 439, 440*
— — моногеноидей 370, 372,
390
— — мшанок 535*
— — нематод 397, 406, 415, 419
— — немертин 460, 461, 463
— — приапулид 455
— — трематод 350, 354, 355
— — турбеллярий 340, 340*,
341*, 343*, 344
— — червей 336, 337
Ротовой диск 233, 287, 300, 304,
306
— конус 279, 310
Ротовые лопасти 229, 231*, 281,
283, 286
Роющий образ жизни брахиопод
544
Роющий образ жизни кишечнопо-
лостных 299, 300, 301
Руки брахиопод 545
Рулек 399, 402, 403*
Ручной аппарат (скелет рук) 545
С
Самки амиктические 444*, 446
— амфотерные 430*, 447
— миктические 444*, 446
Самовосстановление (регенера-
ция) 226
Самокалечение (автотомия) 345,
527
Самооплодотворение 342, 493
Самоочищение водоемов 216, 498
Сапробионты 419
Сапробиос 402, 403, 410, 413
Сапробиотическая среда 402, 403,
410, 411, 412, 413
Сапробиотические бактерии 402
— очаги 394, 403, 410, 412
— хромадориды 402, 411
Сапрозойные нематоды 411
Сапрофитные бактерии 411
— грибы 402, 411
Сапрофитный способ питания
103, 104, 107
Саркода 73
Сверлящие губки 195, 214, 215,
215*
— полихеты 486
Сверхпаразитизм 36
Светочувствительность 502
Светочувствительные клетки 230,
230*
Светящиеся гребневики 332
— кишечнополостные 287, 297,
299
— олигохеты 506
— полихеты 476
Свечение (люминесценция) 103,
287, 297, 299, 332, 476, 506
Свободноживущие 22
— гребневики 329
— кишечнополостные 230, 239,
275, 278, 285
— простейшие 72, 93, 104
— турбеллярий 338, 339
Сегментация (метамерия) 29,
435, 468, 493, 499, 526
— гетерономная 31
— гомономная 30, 31
— внутренняя 468
— личиночная 483, 527
Сегменты (кольца) личиночные
481, 482
— червей 467, 468*, 471, 481,
490, 496, 500, 510
Секрет слизистый 475
Семенники брюхоресничных 392*
— гребневиков 331
— киноринх 450
— кишечнополостных 268
— коловраток 431, 442, 443*
— нематод 395*, 399, 402
— скребней 453, 453*
Семенники трематод 350, 359, 360.
362
— турбеллярий 339, 343
— цестод 377, 379, 387, 388
Семенные пузырьки 351, 377,
388, 453, 453*, 492*
Семявыносящие протоки 350, 377
Семе приемник 343, 343*, 351,
377, 388, 392*, 399*, 492, 492*
Семяпровод 339*, 343, 343*, 350,
377, 392*, 399, 412, 453, 453*,
492*
Септы 230*, 231, 233, 233*, 259
Сердце брахиопод 546
Серпулиты 483
Сигмы 188*, 190*
Сидячий образ жизни 303, 334,
546
Сизигий 118, 119, 119*
Сикон 183, 185, 185*
Сиконоидный тип строения 185,
186, 189, 190, 198
Силикобласты 193
Симбиоз 33, 34, 35, 37, 88, 113,
142, 211, 213, 249, 251, 252,
253, 306, 309, 313, 341, 487
Симбионты 35, 211, 251, 314
Симметрии полюс 27
Симметрия 25
— гетерополярная 26
— гомополярная 26
— двухбоковая 26, 27, 340
— двухсторонняя И, 26, 26*
297, 336
— пятилучевая 455, 456
— радиальная 66, 190, 194, 306,
455
— радиально-лучевая 25, 26,
26*, 223*
— радиально-осевая 11, 25, 26,
26*
— четырехлучевая 300
Сингаллы (галлы сложные) 427
Синкарион 140, 140*
Синойкия 249, 250, 251
Синцитий 185, 191, 192, 340, 341,
438
Сифоноглифы 27, 233, 246, 300,
306
Сифонозоиды 246, 290, 293, 294,
296, 297
Скелет внутренний 29, 188, 233
— губок 189
— известковый 188, 189, 223, 233,
310, 311, 323
— кишечнополостных 233, 234,
287, 295
— кремниевый 90, 94, 96, 188,
189, 192, 218
— мшанок 533
— наружный 29, 228, 233, 239,
250
— осевой 225, 289, 290, 295, 296,
309
— простейших 89, 90, 94, 95
— роговой 188, 189, 218, 309
Скелет рук брахиопод 545, 546*
Скелетные лучи 172*, 174, 175*
— пластинки 81*, 163*, 164
556
Склериты 250, 291, 293
Склеробласты 38, 184, 190, 193
Склероции 396, 402
Сколекодонты 483, 486
Сколекс (головка) 375, 378, 383,
385, 387, 388
Случная болезнь 108
Смена хозяев 351, 352, 354, 355,
359, 364, 375, 378, 379, 382,
387, 388, 407, 453
Совокупительная бурса 339*,
350, 377
— сумка 339*, 343*
Совокупительные спикулы 395,
399, 402, 403*, 406, 408, 412
Совокупительный орган (циррус)
339*, 343, 343*, 350, 377, 392*,
399, 453, 453*, 511, 520
Сожительство 249, 250, 251, 341
Солоноватоводная фауна 49, 51*
Солоноватоводные животные 263,
275, 540
Сомиты (метамеры) 29, 490, 510,
517, 520, 524
Сонная болезнь 107, 108
Сориты 199, 200, 201*
Сосущий аппарат турбеллярий 342
Сперма 450, 451
Сперматозоиды гребневиков 331
— губок 196
— кишечнополостных 268
— червей 343, 377, 399*, 442,
469, 479, 493
Сперматофоры 511, 524
Спикулы губок 189
— кишечнополостных 233, 259,
287, 290, 293, 298
— простейших 67, 90, 173
Спиральная нить 330, 331*
Спонгин 189, 193, 195, 198*, 207,
218
Спонгиобласты 195
Споробласты 121*, 122
Спорогония 119, 119*, 120, 121*,
122 123 124 131
Спорозоиты 119', 119*, 120, 121*,
122, 124, 125, 127*
Спороцисты 352, 357, 357*, 360,
362, 364, 364*
Споруляция 122
Споры 38, 121*, 122, 129, 130,
130*, 131, 131*, 133*
— зимующие 38
Спячка 38
Стадия покоя 38, 40
Статобласты 539, 541
Статолит 229, 230*, 231*, 232,
232*
Статоцисты кишечнополостных
229, 230, 230*, 281
— кольчатых червей 471
— плеченогих 546
— турбеллярий 339, 341
Стационарное ядро 139, 140*
Стебель (стебелек) плеченогих
544, 545*
Стеблевые нематоды 422
Стекловидное тело 468*
Стенобатные радиолярии 92
Стеногалинные организмы 37
Стенооксибионтные организмы 37
Стенотермные организмы 37
Стеррастры 194*
Стигма (глазок) 95*, 97, 98*,
99 103*
Стилет 352, 397*, 398*, 399*,
423, 424*, 451, 460
Стилетовидные клетки 443
Столон 258 *
Стома (ротовая полость) 396*,
401, 402, 405, 408, 415, 419
Стомария 481
Стрекательная батарея 246*
Стрекательная капсула кишеч-
нополостных 234, 236, 246,
252, 264, 300, 307
— — простейших 129, 130*,
131*, 133*
Стрекательные клетки кишечно-
полостных 228, 234, 235*, 247,
260, 265, 272, 278
— — турбеллярий 346
— нити кишечнополостных 234,
235*, 252, 254, 292, 306
— — простейших 131, 131*,
133 133*
Стробила 375, 378, 382, 386, 387
Стронгилоидная личинка 419
— форма нематод 415
Субатриальные синцитиальные
перемычки 191*
Субдермальные синцитиальные
перемычки
Субгенитальные ямки 231, 231*
Субкутикулярные клетки 376
Субкутикулярный слой 350, 370
Сублитораль 46*, 47, 291, 296
Сублиторальная фауна 52
Супралитораль 46, 46*, 291, 296
Сурра 107
Сцифистома (сцифоидный полип)
230*, 231, 238, 238*, 282,
283, 285
Сцифоидный полип (сцифистома)
231, 286
Съедобные кишечнополостные
284, 284*
Т
Тангорецепторы 396*, 397, 397*,
398, 401, 404, 406
Таксис 69, 135
Тегма 31
Тегматизация 31
Тека 261
Телергоны 477, 482, 486
Телефаза 61*, 62
Тельце Мелиса (скорлуповая
железа) 370, 377
Тениаринхоз 383
Тенидоз 384
Тениоли 231
Тепловодные формы 91, 294, 306
Тепловодный реликт 39
Терминальные клетки 441
Тигмотаксис 156
Томиты 158, 158*
Томонт 158
Трансмиссивное заболевание 108
Трахеи 15, 279, 349
Трематодоз 349
Трипанозомоз 108
Трихинеллоз 409, 410
Трихоцисты 134*, 136, 136*
Тропические губки 205, 207
— иглокожие 370
— кишечнополостные 235, 281,
283, 300, 308, 322
— простейшие 88
— турбеллярий 346
Трофонт 158*
Трохофора кольчатых червей
369, 466, 472, 479, 480, 527.
528
Трохус 435, 436*
Трубки (трубочки) мшанок 537.
539*
— полихет 474, 475, 485, 487,
488
— форонид 531, 532, 532*
У
Убиквисты 429
Ультраабиссаль 46*, 47
Ультраабиссальная зона 46*, 47
Ультраабиссальные кишечнопо-
лостные 303
Ундулирующая мембрана 28*,
105, 106, 106*, 107, 109, 148*
Ункус 439, 439*
Усики брюшные 468*
— головные 468*, 471, 483, 484,
486
— ротового сегмента 468*
— тактильные 468*
Устричные банки 214, 215
Устье (оскулум) губок 183, 185,
194, 198, 201, 204, 207, 20Ь,
214
— простейших 81
О»
Фасциолез 356, 357, 358, 359
Феодиум 91
Фибриллы 66, 68, 68*, 70*
Фильтраторы 34
— брахиоподы 546, 547
— губки 215, 216
— кишечнополостные 288
— мшанки 538
Фильтровальный аппарат плече-
ногих 545, 547
Фиксаторный аппарат (орган
Брандеса) 355, 365
Финка 37
Финны (цистицерки) 383
Фитобентос 49
Фитопланктон 48
557
Флотобласты 539, 540
Фораминиферовые илы 87, 544
Фосфорилирование 60
Фоторецепторы 337, 340, 342,
396, 492
Фотосинтез 57, 103, 314
Фототропизм 97
Фрустулы 273, 274
Фулькрум 439, 439*, 440*
X
Хаузы 421
Хвостовая выделительная пора
401
Хелы 190*
Хемотаксис 135, 135*
Хитин 486
Хищные гребневики 332
— инфузории 147, 158, 164, 166*
— кишечнополостные 266, 279,
285, 305, 306, 313
— полихеты 481, 484
— пиявки 509, 511, 520, 523,
524
Хлоропласты 57
Хлоркруорин 469
Хлорофилл 18, 57, 60, 97, 100,
104, 115
Хоаноциты (воротничковые клет-
ки) И, 182, 184, 184*, 185,
190, 192, 196, 198
Хобот (хоботок) коловраток 436
— кольчатых червей 485, 490,
495, 501, 511
— немертин 460, 462, 463
— приапулид 455, 456
— сипункулид 528, 528*
— скребней 452, 453*, 454
— цестод 376, 382, 383
— эхиурид 526, 527*
Хозяин 35, 36, 77, 78, 79
— дополнительный 351, 354,
359, 365, 378
— окончательный 351, 354, 359,
365, 378, 381, 384, 388
— промежуточный 37, 351, 357,
362, 378, 388, 421, 453, 509
— резервуарный 351, 367, 378
Хозяйственное значение губок
218, 219
— — кишечнополостных 322,
323
— — червей 349
Холодноводная фауна 40
Холодноводные губки 207
— кишечнополостные 288, 297,
303, 306, 312, 313*, 322
— простейшие 88
Хроматин (хромосомы) 58, 59,
70, 138
Хроматофоры 95, 97*, 100, 103,
331
Хромосомы (хроматин) 58, 58*,
60, 62, 70, 138, 177, 446, 447
Хрусталик 230, 230*, 342, 401,
467, 484
ц
Целлюлоза 97
Целобластула 196, 198*
Целом (вторичная полость тела)
336
— кольчатых червей 468, 468*
— плеченогих 545
— приапулид 456
— сгибающихся 457
— эхиурид 526
Цел омические каналы 545
Цел отел ий 456, 468
Цененхима 310
Ценосарк 242, 243, 293, 310
Ценосаркальные каналы 248, 291
Центральная камера 279
— капсула 88, 89*
Центросома 55 *, 56, 57*, 62
Ценур 386, 386*, 387
Церебральные органы 460*
Церинула 307
Церкарии 352, 355, 357, 357*,
359, 362, 365, 495
Церкариозный (шистозомный)
дерматит 363
Цестодозы 375
Циклическая раковина 83, 84
Цикломорфоз 428, 436
Цингулюм 435, 436*
Циркумапикальный ресничный
пояс 435, 436*, 440*
Циркумполярные виды 303
Циркумтропические виды 303
Цирры 23, 68
— простейших 68,141, 146, 162,
167, 171
Циррус (совокупительный орган)
350, 377
Цистицерки (финны) 383, 384,
385, 387
Цистицеркоз 384
Цистицеркоиды 388
Цистозоиды 247
Цисты 38, 80*, 118, 119*, 133*,
144*, 158*, 160*, 165, 170*, 360
— зимующие 38
Цитоплазма 54, 55*, 56, 57*
— динофлагеллят 100
— инфузорий 138,150, 154, 173
— книдоспоридий 129
— простейших 66, 69, 70
— споровиков 119, 121
Цифонаутес 538
Ч
Чашечка 457, 458*
Челюсти 429, 439, 469, 475, 484,
511*, 520
Челюстной аппарат коловраток
виргатный 439*
— — инкудатный 439*
— — маллеатный 439*
— — раматный 439*
— — унцинатный 439*
— — форципатный 439*
Чередование поколений кишечно-
полостных 237*, 238, 285
— — моногеноидей 369
— — нематод 415
— — оболочников 238
— — простейших 86, 120
— — трематод 351, 354, 355
— — турбеллярий 344
Чернильная (чернопятнистая)
болезнь рыб 366
Членики червей 375, 380, 381*,
386, 428, 450, 490
Чувствительные волоски 229-
230*, 232*
Ш
Шейка цестод 375, 376
Шизогония (множественное де-
ление) 25, 36,37,120, 121, 122*,
124, 125, 127*, 131
Шизонт 120, 121*, 122*, 124,
125, 127*, 133*
Шистозомный (церкариозный)
дерматит 363
Шистозомоз 361, 363
— мочеполовой 362
Шпильки 190*
щ
Щетинки анальные 527*
— брюшные 468*, 491, 527
— гарпуны полихет 475
— немате л ьминтов 391, 397, 401,
433 449
— олигохет 490, 491*, 496, 497,
503, 506
— параподиальные 526
— пиявок 510, 512, 512*
— полихет 467, 471, 474*, 480,
489
— половые 526, 527
Щупальца гребневиков 329, 330,
331*, 334
— кишечнополостных 227, 227*,
229*, 230*, 231*, 233*, 300, 305
— кольчатых червей 467, 470*
472, 479, 484
— мшанок 537
— немательминтов 441, 457
— плеченогих 546
Щупальцевый аппарат 531, 532*,
535, 535*, 536
Щупальценосные полипы 274
Э
Эвдоксии (эрсеи) 247
Эволютная раковина 83
Эврибатные организмы 91
Эвригалинная фауна 49
Эвригалинные организмы 37
Эвриоксибионтные организмы 37
558
Эвритермные организмы 37
Эвфотическая зона 47
Эдафон 51
Экваториальная пластинка 61
Экзоциста 38
Эктодерма 11
— гребневиков 331, 332
— губок 182, 198
— кишечнополостных 223, 227*,
229*, 230*, 231*, 233*, 237,
256, 265, 277, 301, 310, 322
Эктодермальная подушечка 330,
332*
Эктопаразиты (паразиты наруж-
ные) 160, 269, 338, 369, 374, 405
Эктоплазма жгутиконосцев 105,
105*, 111
— инфузорий 136, 143*, 162,
173
— книдоспоридий 129
— простейших 66
— саркодовых 76, 76*, 78, 78*,
88, 93, 93*
Эмбриональная камера 85, 86
Эмбрионы коловраток 446, 447,
448
— скребней 453
Эмбриотека 268
Эндемизм байкальской фауны 348
Эндемичные олигохеты 501
Эндемик 40
Эдопаразитизм 164, 338
Эндопаразиты (паразиты внут-
ренние) 338
Эндоплазма жгутиконосцев 102,
105, 105*, 111
— инфузорий 136, 143*, 162,
173
— книдоспоридий 129, 130
— саркодовых 76, 76*, 78, 78*,
93, 93*
Эндоплазматическая сеть 56, 57*
Эндоциста 38
Энтодерма И, 336
— гребневиков 332
Энтодерма губок И, 181, 182,198,
198 *
Энтодерма кишечнополостных 223,
227, 227*, 228, 229*, 230, 231*,
233*, 237, 277
Эпимерит 117*, 118, 118*
Эпипелагиаль (поверхностная зо-
на) 46*, 47
Эпистом 532, 532*, 535*, 537
Эпителиально-мышечные клетки
24, 227, 228, 228*, 265
Эпителиальные клетки 233, 645
Эпителий 78, 181, 184, 185, 192,
337, 460
— мерцательный 28, 330
— однослойный 451
Эпитокия 477
Эпитокные формы 477, 478
Эполеты 283
Эритроциты 79, 79*, 112, 124,
125, 127, 127*, 170, 216
Эрсеи (эвдоксии) 247
Эфира 238, 238*
Эхинококк 385
Эхинококковый пузырь 385, 386
Эхинококкоз 385
Я
Яд губок 217
— кишечнополостных 234, 236
Ядерная оболочка 57, 62, 70
Ядерный сок (кариолимфа) 58,
59, 62, 70, 118
Ядовитые губки 216
— кишечнополостные 236
— морские ежи 321
Ядро жгутиконосцев 95*, 98*,
101*, 104*, 107*, 108*, 111,
112, ИЗ*, 115
Ядро клеточное 26, 54, 55*, 57*,
58, 60, 61*, 70, 71*
— книдоспоридий 129, 131
— корненожек 76*, 77, 78*,
79*, 80, 80*, 81*
— мигрирующее 139, 140*
— солнечников 93, 93*, 94, 95*
— споровиков 117, 117*, 119,
121, 122*, 125, 127*
Ядро стационарное 139
— фораминифер 84, 85
Ядрышко 55*, 57*, 58, 62, 70,
118, 121
Яичники гребневиков 331
— моногеноидей 370, 370*
— немательминтов 390*, 392*,
399*, 402, 404*, 408, 412*,
417, 424*, 431, 440*, 451, 453
— немертин 460*
— трематод 351, 362
— турбеллярий 339, 343, 343*
— цестод 379, 383, 387, 388
Яйца амиктические 430*, 442*,
446
— гребневиков 331
— зимующие 38
—- кишечнополостных 237, 237*,
238*, 268*, 281, 296
— кольчатых червей 470, 479,
484, 492, 511, 514, 527
— миктические 430*, 446
— моногеноидей 370, 371, 373,
374
— немательминтов 392*, 399,
404*, 412*, 416*, 424*, 429,
440*, 442*, 450, 453
— немертин 461
— партеногенетические 444*
— покоящиеся 431, 438, 442,
444*, 445*, 446, 447
— трематод 351, 354, 355, 359,
360, 365, 366
— турбеллярий 341, 342
— удонеллид 368
— цестод 377, 380, 381*, 383,
384 *, 386
Яйцеводы кольчатых червей 492*
— круглых червей 406, 417, 439,
440, 441, 453
— плоских червей 339*, 350,
370, 377
Яйцеклетка кольчатых червей
469, 472, 479, 480, 485, 511
— мшанок 536
— сгибающихся 457
Якорьки 190*, 195
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
А
Автолитус 368
Аглаофения 261, 262*
Адамсия 241, 253
Аденофореи 400, 401*, 410, 411,
417
Акантарии 90, 92
Акантастер 257
Акантобделла (пиявка щетинко-
вая) 467, 512
Акантоцефалы (скребни) 454
Акропора 224*, 256, 257, 319,
322
Актинии (морские анемоны) 225,
233, 235, 249, 250, 251, 252,
299, 300, 302*
Актиния 34, 241
— корковая 212
— лошадиная 241
— толсторогая 306
Аллантозома 169
Аллолобофора серая 503
— алтайская гигантская 504
Альвеококк 385, 418
Альционарии (мягкие кораллы)
225, 249, 289, 290, 308, 310,
313, 319
Альционария (альциониум —
рука мертвеца, или пальцы
мертвеца) 291
Альциониум (альционария) 291
Альционы 484
Алярия 366, 367
— алата 366
Амеба дизентерийная 77, 78, 78 *,
79, 79*, 80*, 170
— протей 75, 76, 76*, 77
Амебы 66, 69, 76*, 78*, 136, 176
— голые 75, 80
Аммониты 19
Аммоноидеи 16
Амфибии 14, 16, 17, 51, 108, 348,
354, 366, 369, 375
Амфиболиды 617
Амфилина 374, 378
Амфиподы 367
Амфиприоны 255
Амфитрита 368
Аннелиды (черви кольчатые) И,
12, 21, 29, 459, 526, 528
Аноплодиний 164
Аноплофридий 172
Аноплоцефалиды 387
Анофелес 112, 125, 127
Антипатарии (кораллы шипастые)
225, 241, 243, 309
Антозоа 30
Антомастус 293, 293*
Анхистропусы 269
Аплекса 618, 618*
Аппендикулярии 14
Археобделла каспийская 525,
525*
Археоциатиды 258
Архианнелиды (кольчецы первич-
ные) 470, 471
Арцелла 80
Аскарида 390*, 416, 418, 459
— лошадиная 395, 418
— свиная 418
— человеческая 416, 416*, 417
Аскаридаты 417
Аскариды 417
Асцидии 249
Аспидогастер 354, 354*
— конхиколя 354
Аспидогастраты (складчатобрю-
хие) 354
Астоматы 171, 173, 174
Асцидии 14, 49, 226, 249
Атактодея 318
Атергатес 321
Аурелия (медуза ушастая) 238,
281, 282*
Афродитиды 476, 486
Ацелы (Черви ресничные бески-
шечные) 341
Б
Бадяги (губки бадяги) 51
Байкалия 582, 583*
Бактерии 78, 79, 164, 216, 410,
411, 412, 427, 432, 494
Балантидий 169
— коли 169, 170
Балянусы (морские желуди) 212
Батипатес 310*
Беззубка (аподонта) 354, 355
Белемниты 19
Белемноидеи 16
Берое 321, 331, 332, 333, 334
Бесчерепные 14
Бипалий 346
Битиния 360
Битотиара 225*, 255*
Блакфордия 276, 277
Бокоплавы 51, 211, 519
Болинопсис 328, 332, 333, 334
Бонеллиды 527
Бороздчатобрюхие 14
Бранхиомма 489
Бранхиоцериантус 227
Бранхиура 499
Брахилемиды 363
Брахиоподы (плеченогие) 16,
249, 543, 544, 546
— беззамковые 544, 545, 547,
548
— замковые 544, 545, 547, 548
Брюхоногие 14, 16
Брюхоротые (гастеростомата)
354
Бугенвиллия 240, 277
Бунодеопсис 254*
Буцефал 354*
— изменчивый 355*
В
Велелла (парусник) 278, 279, 280
Венерин пояс 321, 332, 334
Веретиллум 295*, 296, 297
Верметус 320
Виктория-регия 271, 499
Виргулярия 295*
Власоглав 407, 408
Внутрипорошицевые 457
Водоросли 16, 174, 429, 432
— диатомовые 48, 156, 174,
250
— жгутиковые 48
— нитчатые 213, 471
— одноклеточные 48, 73, 84,
88, 156, 471, 487, 494
— перидиниевые (динофлагел-
ляты, жгутиковые панцирные,
перидинеи) 35, 48, 88, 100,
116, 156, 313
— сине-зеленые 48
Водяная блоха 342
Водяной ослик 51
Волосатик 451*
Волосатики 13, 336, 451
Волосатиковые 451, 452
Вольвокс 26, 98, 99, 100*, 116
Восьмирук 284, 285*
Вошь гидровая 269
Вторичнополостные (целомиче-
ские) 543, 551, 554
560
Вторичноротые 11, 12, 15, 18,
24, 29, 31, 330, 530, 531, 533,
543
Выдра морская (калан) 19
Выползок большой 503
— малый (червь малый красный)
503
Г
Галаксея 319
Галатеантемиды 301
Галикриптус 455, 455*, 456*
Гамбузия 41, 41*, 42, 127
Гаплотаксиды 500
Гаптофрия 173
Гастеростомата (брюхоротые)
354
Тасродес 333, 333*
Гастротрихи (черви брюхорес-
ничные) 390—392, 392*, 393*,
398, 400, 428, 448-450, 454
Гастрохена 318
Гвоздичник 379*
Гвоздичники 378, 379
Гелобделла 520
Гементерия (пиявка черепашья)
515
Гелиотиды 258
Гемадипса цейлонская 524*
Геодия 192, 194, 194*, 199, 205,
209
Герпобделла линейная 525
— обыкновенная 524, 525*
— пещерная 524, 525
Гетероцентротус 257, 320
Гиалонема 193, 206*, 207, 208,
210, 212, 219, 219*
Гигантостраки 19
Гидра 203, 227, 240, 264, 264*,
266*, 267*
— бесстебельчатая (обыкновен-
ная) 268
— зеленая 269, 270
— стебельчатая (бурая) 268
Гидры 51, 225, 263, 285
Гидрактиния 225*, 252
Гидрамеба 269
Гидровые 263
Гид розой 13, 30
Гидроидеи 263
Гидроидные 224, 230, 237, 260,
272, 277, 279, 286
Гидроидные медузы (гидроме-
дузы) 229, 229*, 231, 261,
276
Гидроидные полипы (гидрополи-
пы) 73, 213, 227*, 237, 239,
309
Гидроиды 224, 230, 242, 244, 249,
252, 262*, 277, 280, 533
Гидромедузы (гидроидные меду-
зы) 229, 230, 230*, 231, 232,
261, 276
Гидрополипы (гидроидные поли-
пы) 230, 239
Гименолепидиды 388
Гименолепис ланцетовидный 388
Гпродактилюс 371, 372
— элеганс 372
Гирокотилиды 336, 338, 368, 374,
374*
Глаукус 280, 288
Глицериды 476
Глобигерина 86
Глоссифонида байкальская 516
Глоссифониды (пиявки плоские —
клепсины) 513, 515
Глоссосколециды 505
Головоногие 7, 14
Головохордовые 12
Голожаберные 211
Голотурии (морские кубышки) 14,
211, 250, 251, 318, 322
Гонактинии 300, 301
Гониум 98
Горгонарии (кораллы роговые)
243, 249, 287, 309
Горилла 171
Гребешок 322
Гребневики (ктенофоры) 12, 13,
222, 251, 327, 329, 331*, 333*,
348
Гребневики бесщупальцевые 334
— лопастеносные 334
— цидипповые 334
— щупальцевые 334
Грегарина 117*
Грегарины 23, 117
— настоящие 118
Грибы 412
Грибы низшие 410, 411, 414
Грызуны 110
Губка байкальская 196,199,209,219
— гиалонема 193
— далматинская 201*, 218
— конская 204
— корзинка Венеры 193, 208,
211, 219, 219*
— кубок Нептуна 195, 209
— левантийская 218
— монорафис 193
— морской ершик 206*, 207
— морской апельсин 192, 194,
199, 204
— морской гриб 194, 202
— морской каравай 196, 203,
204, 208, 209, 209, 211
— Росса 193, 207, 208
— туалетная 218
— туалетная Зимокка 217*
Губки 11, 12, 13, 15, 34, 38, 49,
179, 182, 184*, 203, 203*, 206*,
210*, 219, 222, 226, 250, 309,
473, 484, 485
Губки бадяги 196, 218, 219
— восьмилучевые 219
— известковые 189, 190, 196,
198, 199, 201, 206, 216
— кремнероговые 195, 196, 197,
199, 202, 204, 206, 208, 217
— миксиллы 196, 210
— мясистые 193, 211
— обыкновенные 189, 193, 204
— полимастии 194, 202, 207, 209
— пробковые 192, 194, 211
— разнолучевые 219
Губки роговые 196, 207, 208
— сверлящие (клионы) 195, 214,
215, 215*
— стеклянные (шестплучевые)
189, 191, 193, 196, 199, 201,
204, 206, 208, 217, 219
— факеллии 196
— четырехлучевые 193, 196, 199,
202, 204, 206, 209
— шестилучевые (стеклянные)
189, 208
д
Дактилогирус 371, 371*
— вастатор 371
Дафнии 429
Дая 257
Двуслойные 11, 18
Двустворчатые (безголовые, плас-
тинчатожаберные, топороногие)
14, 16
Двустороннесимметричные (трех-
слойные) 12, 222, 336
Двуустка ланцетовидная (дикро-
целий ланцетовидный) 358, 358*
Дендробена восьмигранная (та-
ежный червь) 504
— мариупольская 504
Дендрофиллия 318
Диадема 257, 322
Дигенеи (трематоды дигенетиче-
ские) 338, 368
Дидиний 147, 147*
Дикроцелииды 358
Дикроцелий 359
— ланцетовидный (двуустка лан-
цетовидная) 358, 358 *, 359
Диктиокаулюс 422
Динофилиды 471
Динофилус 471*
Динофлагелляты (перидинеи,
жгутиковые панцирные, периди-
ниевые водоросли) 35, 48, 88,
100, 116, 156, 312
Диоктофимата 407
Диплостома 365, 365*
— спатацеум 364
Диплостоматиды 364, 365, 366
Дискомедузы 225
Дисконанты 224
Дитиленх 423, 424, 425
— картофеля 425
— луковый (луковая раса) 424,
424*
Дитиленхи 397*
Диффлюгия 80, 81
Дихогастер пальмовый 506
Дициемиды 13
Додо (дронт) 19, 40
Донаксы 317
Дронт (додо) 19
Е
Евдендриум (эудендриум) 224*
Евплокамис 321
561
till/
Жвачные 161
Жгутиковые 13, 17, 20, 25, 26
— панцирные (динофлагелляты,
перидинеи — перидиниевые во-
доросли—перидинеи) 35, 48, 88,
100, 116, 156, 312
Жгутиконосцы 23, 66, 67, 68, 69,
71, 74, 94, 95, 136, 176, 182
Жгутиконосцы животные (зоо-
мастигины) 96, 104
— растительные (фитомастиги-
ны) 96, 97
Жемчужницы морские настоящие
(крыловидки) 322
Жук-вертячка 117
— колорадский 41
— майский 42
Жуки-скакуны 318
3
Заяц-беляк 9
— европейский 9
Земноводные 39, 173, 351, 380,
515, 522
— панцирные (стегоцефалы) 19
Змеехвостка 192, 211, 318
Змеехвостки (офиуры) 14
Змеи 25
Зоантарии242, 244, 250, 308, 309
Зооксантеллы 35, 88, 256, 313,
318
Зоомастигины (жгутиконосцы
животные) 104
Зоотамний 151
Зоофиты (радиаты) 11, 182, 227
Зоохлореллы 35, 88, 201*, 213,
268, 340, 341, 344
Зостера 39, 236
Зубр 40
И
Иглокожие 10, 12, 16, 19, 21,
31, 33, 117, 172, 174, 261, 329,
455
Изиделла (коралл белый) 290
Изоподы 367
Инфузории 13, 17, 23, 26, 38, 51,
66, 68, 69, 70, 73, 74, 81, 94,
134, 436
— апостоматы 158
— брюхоресничные 146, 174
— кругоресничные 67*, 148,
149 *, 161
— малоресничные 174
— равноресничные 141, 164, 167,
174
— разноресничные 143
— сосущие (суктории) 153, 169,
169 *
Инфузория бурсария 143
— спирофрия 158
Инфузория трубач 141, 141*
— — голубой 142
— туфелька 134, 134*, 136*
Ихтиозавр 378
Ихтиофтириус 159
Ихтиобделла байкальская 519
Ихтиобделлиды (пиявки рыбьи)
517, 520
К
Калан (выдра морская) 19
Калаппа 321
Калиактис 241, 254
Кампанулярия 240, 263*
Камптозоа (сгибающиеся) 13, 457,
458*
Карповые рыбы 51, 131, 360, 365,
374, 380
Карпоидеи 15
Карцинобделла 518*
Катушки 366, 495, 515
Кашалот 395
Киноринхи 13, 336, 390, 400, 449,
449*, 450
Кистеперые рыбы 17, 18
Китообразные 395
Киты 20
Кишечнодышащие 14
Кишечнополостные 12, 15, 21, 24,
25, 26, 48, 49, 182, 183, 221,
223, 224, 224*, 249
Кладактис 241
Кланорх китайский 360
Клепсины (глоссифониды, пияв-
ки плоские) 513
Клещи 51
— водяные 211
Клещики почвенные (панцирные)
387
Клиона 192
Клионы (губки сверлящие) 214,
215, 215*
Клипеоморус 318
Клопы водомерки 280, 288
Книдоспоридии 13, 74, 129, 177
Кокцидии 72, 74, 120, 128, 177
Коленчатоногие (пауки морские)
14
Коловратки (ротатории) 13, 17,
23, 26, 30, 38, 51, 73, 94, 257,
336, 390, 400, 428, 430*, 432,
434*, 448, 457
— морские 49, 437
— однояичниковые 431, 432
— пиявковидные 431, 433, 437,
448
Кольчецы 468*, 470, 480
— малощетинковые (олигохеты—
черви—мал ощетинковые) 470,
490
— многощетинковые (полихеты—
черви многощетинковые) 13, 30,
470, 473, 526
— первичые (архианнелиды) 467,
470, 471, 481
Колюшка трехиглая 132, 133*
Колючеголовые 12, 13, 35, 336
Конволюта 340
Конус 318
— пятнистый 272
— текстильный 272
Коралл белый (изиделла) 290
— благородный (красный) 241,
290
— грибовидный 256
— красный (благородный) 241
— морская рука 241
— органчик 241, 244, 293
— черный 290
Кораллы 13, 320, 533
— восьмилучевые 225, 232, 242,
259, 287
— корковые 225
— мадрепоровые 53, 225, 234,
259, 310, 312, 314, 316, 317,
318*, 323
— мягкие (альционарии) 290
— роговые (горгонарии) 225, 287
— солнечные 225, 320
—- четырехлучевые 259, 308
— шестилучевые 225, 232, 244,
299
Кордилофора 274, 274*, 277
— каспийская 275
Корине 238, 261, 262, 262*
Корненожки 23, 38, 51, 75, 79,
81*
— раковинные 79, 80, 80*, 81,81*
Корнерот (ризостома) 240, 283
Корономедузы 225, 286
Корюшка 132
Корюшковые 49
Кофобелемнон 295*, 297
Краб китайский мохнатоногий 41
Крабы 49, 212, 318, 480*
Краспедакуста 271*, 272, 273,
273*
Креветки 211, 280, 323, 519
Кренатула 322
Крестовички 236, 240
Кривоголовка двенадцатиперст-
ная 419, 420*
Криптоспонгия 213
Кроссота 286
Кролик 42
Круглоротые 14, 15
Крыложаберные 14
Ксифинема 405
Ктеноплана Ковалевского 334
Ктенофоры (гребневики) 24, 27,
30, 48
Кубомедузы 225, 235
Кульциты 323
Кунина 255*, 256
Куноктата 256
Л
Ламинария 213
Латимерия 9, 19
Левинсения 518*
Легочные (пульмонаты) 51
Лейкохлоридий 363, 364
— парадоксальный 363, 364*
Лейшмании 66, 96, 109
562
Лентец широкий 36, 380, 381,
381*
Лентецы 378, 380
Лептогоргия 289*
Лептолиды 224, 229, 256
Лигула (ремнец обыкновенный)
379, 380
Лима 322
Лимнатис (пиявка египетская,
конская, нильская) 522
Лимнодрил Гоффмейстера 499
— невский 499
Лингулиды 546
Литофага (камнеточец) 318
Лобофиллия 257, 311, 319
Лопатоногие 14
Лососевые 49, 512
Лососи 33, 131
Лофохелия 312, 313*
Лошадиные 167
Лошадь Пржевальского 40
Луковая раса (дитиленх луковый)
424, 424*
Лучевики (радиолярии) 75
Люмбрикулиды 501
Люмбрикулюс 501
Люмбрициды (черви дождевые на-
стоящие) 502, 505, 507
Люцернария 284
Лямблии 66, 96, 106, 107*, 112
Лягушки 39, 217
Ляцинулярия 437
М
Макрофиотриксы 318
Малосегментные (олигомерные)
11, 12
Малярийный паразит 36
Меатор 286
Мегасколециды 505
Медуза ушастая (аурелия) 232,
281
Медузы 24, 26, 27, 48
— гидроидные 229, 229*, 231,
261, 276
— корнеротые 232
— сцифоидные (сцифомедузы)
231, 231*, 232, 258, 333
Мезозой 179, 181
Мезостома 342
Менилелла 173
Меризия Палласова 276, 276*, 277
— черноморская 276*, 277
Мермитиды 406
Меростомовые 14
Метридиум 241
Мечехвост 18
Мечехвосты 15, 19
Мешкогрудые 35
Мидия 41
Мизостомиды 489
Микрогидра 227, 273
Микроспоридии 129, 132
Микростома 343
Миксоспоридии 129, 132*, 133*
Миллепора 319
Мил иол иды 83
Млекопитающие 14, 16, 17, 51,
108, 123, 124, 348, 351, 355,
359, 366, 369, 375, 380, 387, 407,
452, 454, 515
Многоглазка рогатая 345
— черная 345
Многоклеточные И, 12, 65, 259
— настоящие 12, 222, 223
Многоножки 14, 17, 25, 452
Многосегментные (полимерные)
И, 12
Многоуст 372 *, 373, 374
Мозговик овечий (мультицепс)
386, 386 *
Мозговики 316, 318
Моллюски (мягкотелые) 11, 12,
15, 17, 21, 30, 33, 46, 49, 53, 120,
170, 261, 317, 351, 352, 353,
354, 358, 462, 530, 543
— брюхоногие (улитки) 14, 16,
23, 36, 37, 51, 213, 251, 310
— головоногие 28, 211, 254
— двустворчатые (пластинчато-
жаберные) 28, 34, 354
— лопатоногие 250
— пластинчатожаберные (дву-
створчатые) 355
Молоточек 322
Монетария змеиная голова 272
— колечко 272
— монета 272
Мониезия 387
— экспанза 387
Монилигастриды 507
Монобрахиум 227
Моногенеи 338
Моногеноидеи (моногенетические
сосальщики) 338, 368, 369
Мононх (однозуб) 404, 404*
Монтипора 257, 318
Морская выдра (калан) 19, 40
— гвоздика 305
— мышь 368, 475
Морские анемоны (актинии) 299,
301, 303
— ежи 14, 174, 272, 322
— желуди 46, 261
— звезды 473, 484
— козочки 211, 261
— лилии 14, 321
— огурцы (гребневики) 334
— пауки (многоколенчатые) 14
— перья (пеннатулярии) 225,
241, 242, 248, 287, 294, 295,
295* 296
— уточки 212, 280, 288
— финики (камнеточцы) 323
Морское перо (пеннатула) 224*,
241, 248, 251, 298, 298*
Морской апельсин 192, 194, 199
— таракан 39
Москиты 109
Мультицепс (мозговик овечий)
386
Муравьи 21, 34, 359
Мурена 321
Муха цеце 107, 108, 109*
Мухи 416
Мшанка байкальская 541, 541*
Мшанки 11, 12, 13, 15, 16, 21,
25, 29, 30, 38, 49, 73, 182, 213,
226, 249, 261, 280, 309, 310,
457, 495, 529, 530, 532, 533,
534*, 538
— голоротые 535*, 537, 539, 541,
542
— покрыторотые 532, 535*, 537,
539, 540, 542
Мягкотелые (моллюски) И, 17
Н
Наидиды (водяные змейки) 492,
494
Наиды 495
Насекомые 14, 16, 17, 21, 25, 31,
36, 39, 382, 388, 406, 452, 454
Нассула 148
Наутилиды 18
Наутилоидеи 16, 19
Нематоды (черви круглые) 13,
22, 30, 36, 37, 38, 51, 336, 390,
394, 395, 395*, 396*, 397*,
398*, 400, 410, 411, 428, 448,
450, 451, 452, 454
— галловые 405, 425, 426, 426*
Немательминты 389, 393, 450, 451
Нематоморфы 11
Немертины 11,12,13, 15, 24, 29,
29*, 30, 286,336, 353,459
Непарнокопытные 162
Нереиды 475, 482, 485
Нереис 368
Нериллиды 473
Несегментированные 12
Нефтиды 292
Нитчатки 420
Нотомматиды 448
Ночесветка 101, 102*
Нуммулиты 66
О
Обелия 239, 249, 261, 262*
Оболочники (оболочниковые —
туникаты) 10
Оболочниковые (оболочники —
туникаты) 12, 21, 25, 48, 117
Однозуб (мононх) 404, 404*
Одноклеточные 11, 12, 18, 182
Однокрыгаечковые 13, 19
Озобранхус 518*
Олигомерные (малосегментные)
И, 12
Олигохеты (черви малощетинко-
вые) 13, 17, 49, 51, 348, 407,
470, 490, 491, 491*
Ондатра 41, 41*
Онихофоры 14
Онуфис 368
Опалина 111
Опалиниды НО
Описторх 359
— сибирский (кошачийK58*, 359
Описторхиды 359
Ореастер 322
563
Ортонектиды 13
Орбицелла 318, 319
Осетр 34
Осетровые 49, 132, 256, 257, 274,
275, 378, 482, 518
Остракоды (ракообразные ракуш-
ковые) 16
Острица 415, 416, 416*
Осьминоги 321
Оципода 318
Офиуры (змеехвостки) 213
Офриосколекс 164
Офриосколециды 162, 167
П
Павона 256, 322
Пагурус 241, 254
Палитоа 309*
Палоло 478, 478*, 486
— атлантический 478
— тихоокеанский 478
Пандорина 26
Пантахогон 286
Панцирные 14
Парусник (велелла) 278, 279, 280,
288
Парусница 49
Паукообразные 14, 17, 25, 31
Пелагия 240
— ночесветка 287
Пеннатула (морское перо) 224*,
298, 298*
Пеннатулярии (морские перья)
295, 296
Первичнополостные 18, 390, 454
Первичноротые 11, 12, 15, 18,
31, 336, 530, 531, 533, 543
Первичнотрахейные 11, 12
Переднепорошицевые 15, 530
Переднеротые (прозостомата) 355
Перидинеи (динофлагелляты,
жгутиковые панцирные, водо-
росли перидиниевые) 35, 48,
88, 100, 116, 156, 313
Перидиний 100
ТТерионикс древесный 506
Перловица 354, 355
Пескожил 368, 475, 476*, 479
Пескожилы 487
Пикша 333
Пинна 322
Пироплазмиды 66
ТТиргома 323
Писцикола каспийская 518
Писцикола обычная рыбья(пияв-
ка рыбья) 517*, 518, 520
— Фадеева 518
Пиявка конская (нильская, или
египетская—лимнатис) 520, 522
— ложноконская большая 520,
523, 524*
— — малая 523, 524*
— налимовая 518
— медицинская 509, 510, 511*,
520, 520, 521
— пластинконосная 516
— птичья обыкновенная 510*, 514
Пиявка птичья пятнистая 514
— рыбья (писцикола) 517*, 518,
520
— сомовая 518
— улитковая 510, 510*, 514*,
515, 520
— черепашья 514, 514*
— щетинковая (акантобделла)
512, 512*
Пиявки 13, 24, 49, 51, 467, 470,
509, 510*
— бесхоботные (челюстные) 510,
510*, 513, 520, 524
— глоточные 510*, 524
— древние 512
— настоящие 513
— плоские (глоссифониды, клеп-
сины) 510, 513
— птичьи 513
— рыбьи (ихтиобделлиды) 510*,
513, 517, 517*
— хоботные 510, 513
— челюстные (бесхоботные)
510, 510*, 513, 520, 524
Плазмодий малярийный 125
Планаксис 318
Планарии (черви плоские, пла-
тодыJ7,338,338*,459
Планария 337, 339*
— горная 345
— молочная 339*, 340*, 344
— мохнатая 347, 347 *
Планес 279, 288
Пластинчатожаберные (безголо-
вые, двустворчатые, топороно-
гие) 14
Платиктениды 344
Платоды (планарии, плоские чер-
ви) 337, 338, 389, 454
Плевроврахия черноморская 332
Плектиды 402, 403, 410, 411, 412,
414
Плеченогие (брахиоподы) 11, 12,
13, 15, 16, 29, 30, 49, 213, 529,
530, 543, 544*, 545*
Погонофоры 12, 14, 19, 532
Подобация 318
Позвоночные 11, 21, 25, 29, 31,
105, 124, 382, 406, 429
Полигордиус 472
Поликлады (турбеллярии много-
ветвистокишечные) 347
Поликотилус байкальский 345
Полимерные (многосегментные)
И, 12
Полипластрон 164
Полиподиум 256, 257, 258, 258*,
274, 275
Полипы гидроидные (гидрополи-
пы) 73, 213, 227*, 237, 239, 309
— коралловые (кораллы) 225,
232, 237, 250, 287
— сцифоидные (сцифистомы) 231
Полихеты (черви многощетинко-
вые) 13, 15, 31, 49, 261, 310,
318, 368, 396, 467, 470, 474,
479, 526
Полухордовые 12
Помаценорус 257
Порит 256
Поритес 317, 318
Пориты 319
Порпита 278, 279, 280
Постодиплостома кутикульная
365
Поциллопора 256, 257, 319, 323
— оленьи рога 321
Пресмыкающиеся 14, 39, 108, 351,
380, 515
Приапулиды 13, 30, 455, 455*,
456
Приапулюс 455, 455*, 456
Пристины 494
Прогнихтис 280
Прозостомата (переднеротые) 355
Простейшие 12, 21, 23, 24, 35, 65,
71, 72, 73,74,176,182,411,431
Протей 51
Протодрилиды 471, 472
Протодрилус 472*
Протоклепсис (пиявка птичья)
514
Протомонады 104
Протореастер 257, 323
Протула 368
Прудовик большой 365
— малый 357, 358
— овальный 357
— странствующий 357
— тинный 357
Прудовики 268, 358, 365, 425,
495, 515
Псевдостихопус 213
Птеродактили 19, 20
Птероцера 318
Птицы 14, 16, 17, 20, 108, 123,
124, 348, 351, 359, 363, 375, 388,
452, 454, 513, 515
Птиходактис 250
Йчела 132, 133
— медоносная 132
Пятиустки 14
Р
Рабдитиды 411, 412, 414, 427
Радиальные 12, 222
Радиолярии (лучевики) 16, 25, 26,
35, 48, 66, 68, 72, 72*, 74, 88,
154
Радиаты (зоофиты) 11
Радиофрия 173, 174
Ракообразные 14, 17, 20, 25, 33,
46, 48, 49, 51, 156, 211, 261,
367, 378, 380, 382, 388, 406
— веслоногие 211, 262, 380
— ветвистоусые 38
— высшие 31
— десятиногие 367
— листоногие 38
— равноногие 211
— ракушковые (остракоды) 51
Рак-отшельник 34, 192, 211, 241,
252, 272, 309, 318, 473
Ракообразные 36, 378
Рапана 41
Раткеа 287
564
Ремнец обыкновенный (лигула)
379, 379*
Ремнецы 379
Рептилии 16, 17, 25, 348, 375, 378
Ресничнобрюхие 13
Ризомастигины 105
Ризостома (корнерот) 283
Ринходемус 346
Ритропанопеус 41
Ришта 420, 421
Ропилема съедобная 283, 284*
Ротатории (коловратки) 429
Ручейники 211
Рыба-бабочка 257
— возничий 257
Рыбы 14, 16, 33, 36, 39, 108,
217, 348, 351, 354, 355, 359,
369, 372, 380, 381, 452, 454,
473, 519
— костистые 518
— лоцманы 34
— прилипалы 34
— удильщики 527
— химеровые 374, 374*
С
Сабеллиды 488
Саккоциррусы 473
Салака 42
Сальпы 14, 333, 333*
Сардины 53, 55*
Саркодовые 13, 17, 23, 25, 26, 67,
69, 74, 75, 176
Саркофитум 256, 319
Свайник-великан 395, 407, 407*
Свинья 409, 410
Сгибающиеся (камптозоа) 457,
458*
Севрюга 34
Сельди 49
Серпула 368
Серпулиды 482, 488
Сертулярелла 263*
Сертулярии 249
Сецерненты 400, 410, 411, 414,
415, 422
Сиговые 49
Силлиды 479, 485, 486
Симфиллия 319
Синдесмия 34
Сипункулиды 336, 466, 527, 528,
528*
Сифонофоры 24, 48, 49, 224, 225,
230, 231, 239, 246*, 247, 277
Скворец европейский 41
Складчатобрюхие (аспидогастра-
ты) 354
Сколециды (черви низшие) 11, 29,
30, 336
Скребень-великан 454
Скребни (акантоцефалы) 452, 453*
Слепни 108
Снеток 132
Солитер бычий (цепень бычий)
384
— свиной (цепень свиной) 37, 384
Солнечники 66, 75, 93, 429
Сосальщики (трематоды) 13, 348,
495
Сосальщики моногенетические
(моногеноидеи) 336
Спайник 373*
— парадоксальный 374
Спиониды 479, 486
Спирографис 368, 488
Спонгикола 211
Споровики 13, 25, 35, 71, 74, 116,
177
— кровяные (гемоспоридии) 74,
124, 127*, 128
Ставромедузы 225, 284, 285
Стегоцефалы (земноводные пан-
цирные) 19
Стеллерова корова 19, 40
Стилонихия 146
Стилофора 322
Стилярия 495
Стробилидий 174
Стронгилиды 419
Стронгиляты 419
Сувойка 148
Судак 49, 131
Суктории (сосущие инфузории)
153, 169
Сцифистома (сцифоидный полип)
230*, 231
Сцифозоа 30
Сцифоидные 13, 225, 231, 237,
281, 286
— медузы (сцифомедузы) 231,
231*, 232,258,333
— полипы (сцифистомы) 231
Сцифомедузы (медузы сцифоид-
ные) 231, 231*, 232, 285*
Т
Тенииды 382
Темноцефалы 13, 336, 338, 367,
367*
Теребеллиды 486
Термиты 21, 111, 112, 113, 114
Тиаропсис 263, 263*, 281
Тиленхиды 400, 422, 423, 424,
425
Тинтинниды 154
Тихоходки 14, 38
Токсопнеустес 322
Томоптерис 368
Томоптеры 484
Топороногие (безголовые — дву-
створчатые—пластинчатожабер-
ные) 14
Торикс 516*
Трахеаты 17
Трахелобделла байкальская 517*,
519
— китайская 517*, 518
— туркестанская 517*, 519
Трахилиды 224, 229, 240, 286
Трематоды (сосальщики) 35, 36,
336, 348, 355, 375
— диагенетические (дигенеи)
368, 369
Треска 333
Трехслойные (двустороннесим-
метричные) 11, 18, 181, 336
Тридакна 257, 312, 320
— гигантская 320
— кроцея 320
Тридакофиллия 326
Трикладиды (турбеллярии трех-
ветвистокишечные) 344
Трилобиты 14, 16, 19, 20
Трипаносомиды 109, 115
Трипаносомы 23, 28*, 96, 107
Трипнеустес 322
Трихина (трихинелла) 36, 409
Трихинелла (трихина) 409, 409*,
410
Триходины 160, 161 *, 269
Триходориды 405
Триходорус 405
Трихомонады 105, 106, 112
Трихонимфа 113
Трихоцефалиды 407
Троглодителла 171
Трохета 525, 526
Трохус нильский 257, 272
— пятнистый 320
Трубочник обыкновенный 499
Трубочники (тубифициды) 496,497
Тубифициды (трубочники) 499
Тубифекс 379
Тунцы 33
Турбеллярии (черви ресничные)
13, 17, 23, 24, 30, 36, 51, 336,
338, 339, 348, 352, 375, 389,
391, 393, 428, 436, 448, 450,
454, 460, 463
— бескишечные (ацелы) 341, 348,
450
— многоветвистокишечные (по-
ликлады) 347
— прямокишечные 341, 393, 449,
450
— трехветвистокишечные (три-
клады) 344, 346, 348
Турбинарии 318, 319*
Турбо мраморный 272, 320
Туярия 224*
Тьяльфиелла 333, 334
Тьяльфеллиды 334
У
Угри 33, 473
Удонеллиды 336, 368
Униониды 51
Умбеллула 245, 295*, 296, 297
Урцеларииды 160
Усоногие 23, 34, 212, 310
Устрица 41
Устрицы 214, 215,215*
Ф
Фавитес 318
Фавия 256, 311
Фасциола гигантская 356, 356 °
— печеночная 355*, 356, 356*.
357*
^^ 37 Жизнь животных, т. 1
565
Фасциолиды 356
Фасциолоидес 356
Фасциолопсис 356
Физалия (португальский военный
кораблик) 240, 277, 278*
Физа 495
Филлодоциды 483
Филоксера 41
Фитомастигины (жгутиконосцы
растительные) 96, 97
Флоскулярия 433
Фоласы 323
Фораминиферы (корненожки мор-
ские) 16, 25, 29, 29*, 48, 49,
66, 67, 70, 71, 72, 81, 82*, 83,
83*, 84*, 176, 213, 250, 317
Форель 131, 133*
Форониды И, 12, 13, 15, 529, 530,
531, 532*
Фукоиды 536
Фунгия 322
— колючая 322
X
Халиклистус 284
Хелицеровые 15, 17, 19
Хетоптериды 487
Хетоптерус 487
Хиродропус 235, 240
Хламидомонада 97, 97*
Хламидомонады 97, 99
Хоанофлагеллаты 182
Хондрокладия гигантская 196
Хондрофоры 224, 279
Хордовые низшие 532
Хризаора 225*
Хромадориды 402, 403, 410
Хромодорис 272
ц
Цейлория 311
Цел омические (вторичной олост-
ные) 543, 543*
Целоплана 333
— Мечникова 334
Цепень бычий (невооруженный —
солитер бычий) 381*, 382, 383,
384, 384, 385
— карликовый 388
— свиной (вооруженный — со-
литер свиной) 381*, 382, 383,
384, 384, 385
Цепни 378, 382
Церактис 241
Церациум 101
Цериантарии 225, 306
Цериантария 241, 307*
Цериантус 307*, 308
Цестиды 334
Цестодообразные 378
Цестоды (черви ленточные) 13, 30,
35, 36, 336, 368, 369, 375, 378,
381, 381*
— настоящие 378
Цианея 223, 238, 282, 283
Цидиппе 330*
Циклопы 36, 381, 381*, 388, 421
Ципрея (улитка фарфоровая)
318, 322
— арабская 257, 272
— пятнистая 257, 272
Ч
Человекообразные обезьяны 171
Червеобразные 530, 533, 543
Червь австралийский 505
— дождевой обыкновенный 172,
469, 490, 491*, 492*, 502*,
508
— малый красный (выползок ма-
лый) 503
— навозный 504
— норденшельдов (эйзения Нор-
деншельда) 504
— таежный (дендробена восьми-
гранная) 504
Черви 24, 33, 35, 46, 48, 213, 257,
261, 336, 543
— брюхоресничные (гастротри-
хи) 390, 391, 457
— высшие 465, 466
— дождевые, или земляные насто-
ящие (люмбрициды), 502, 507,
510
— круглые (нематоды) 10, 12,
73, 156, 336, 394, 438, 459,
466
— кольчатые (аннелиды) 10, 10,
И, 12, 15,25, 29, 117, 120, 156,
172, 336, 338, 450, 459, 466,
467, 470, 510, 528
— ленточные (цестоды) 13, 30,
173, 338, 375
— малощетинковые (кольчецы
малощетинковые — олигохеты)
13, 17, 172, 379, 467
— многощетинковые (кольчецы
многощетинковые — полихеты)
15, 31, 49, 211, 261, 295, 310,
317, 368, 369, 467, 470, 473,
496, 526
— низшие (сколециды) И, 15, 29,
30, 335, 457, 466
— плоские (планарии—платоды)
10, И, 12, 334, 336, 337, 454,
466
— ресничнобрюхие 336
— ресничные (турбеллярии) 73,
94, 172, 173, 268, 280, 338, 339,
459, 460
— ресничные бескишечные (аце-
лы) 340
Черепаха болотная 515
— мягкокожая 516
Членистоногие И, 12, 15, 21,
22, 25, 29, 31, 35, 120, 378,
543
Ш
Шелкопряд тутовый 132, 133
Шизогрегарины 118
Шимпанзе 171
Шистозома кровяная 361. 362»
362*
— Мансони 361
— японская 361
Шистозоматиды 360, 361, 362
ШистозомыЗб!, 362
щ
Щетинкобрюх прозрачный 495
— улитковый 495
Щетинконос пещерный беранека
473
Щетинкочелюстные (морские
стрелки — хетогнаты) 12, 14
Щитень 19
Щука 131
Щупальцевые 15, 530, 533, 543
Э
Эвгленовые 103
Эвглена 103
Эвдорина 26, 98
Эвнициды 486
Эвное 368
Эвритрема 359
Эйзения Норденшельда (червь
норденшельдов) 504
Эйзениелла четырехгранная 505
Эймерия 120, 122
Экворея 287
Элизия 272
Эноплиды 402, 403, 405
Энтодиниоморфы 161, 162, 167,
171, 174
Энтодиний 162, 164
Энторипидиум 174
Энхитреиды 490, 499
Энхитрей белый 499
Эолис 280
Эолосомы 493
Эолосома гемприхова 493
Эпидиний 164
Эуглифа 80
Эудендриум (евдендриумJ60,262*
Эудиплодиний 164
Эунице 318
Эуфиллия 322
Эхинококк 37, 384, 384*, 418
Эхинометра 318
Эхиуриды (эхиуровые) 467, 469,
470, 526, 527, 527*
Эхиуровые (эхиуриды) 13, 526
Я
Янтарка 364, 364*
Янтина 280,288
УКАЗАТЕЛЬ ЛАТ
А
Abyssothyris wivillei 548
Acantharia 80, 90, 91, 92
Acanthaster 257
Acanthella 192
Acanthobdella 512, 512*
Acanthocephala 12, 13, 18, 335,
452
Acanthodesmia prismatium 80
Acanthometra tetracopa 80
Acoela 340
Acrania 14
Acrobeles ciliatus 427*
— complexus 427*
— complexus 427*
— ctenocephalus 427*
— ornatus 427*
Acropora 224*, 256, 257, 319,
322, 325, 326
Actinia 225*
— equina 241, 252, 303,304
Actiniaria 225, 299, 301
Actinophrys sol 94
Actinosphaerium 94
— eichhorni 93, 93*
Adamsia palliata 241, 253, 254
Adenophorea 400
Aedes 128
Aeginura grimaldi 304
Aeolis 280
Aelosoma hemprichi 493, 494*
Aelosomatidae 493
Aequorea 287
Aggregata eberthi 71*
Aglaophenia 261, 262*
Agptasia carnea 304
Alaria alata 366
Albertia 429
Alciopa 468*
Alciopidae 484, 496
Alcyonacea 291
Alcyonaria 225, 290, 291, 294,
295, 313, 319
Alcyonidium 542
— disciforme 534*
— gelatinosum 534*
Alcyonium 241, 248, 292, 294
— digitatum 291, 292
— pal ma turn 292
Allantosoma 169
— cucumis 170*
— intestinalis 170*
Allolobophora caliginosa 503
НСКИХ НАЗВАНИЙ
Allolobophora magnifica 504
Alveococcus multilocularis 385
Amera (Scolecida) 11, 12, 15, 29
Ammodiscus 83
— incertus 82*
Amoeba limax 78*
— polypodia 78*
— proteus 75, 76*, 78*
— radiosa 78*
— sphaeronuclei 71*
— terricola 77*
— verrucosa 78*
Amoebina 26, 75
Amphianthus dohrnii 249
Amphibia 14
Amphidiscophora 189, 192, 193
Amphilina foliacea 378
Amphinomidae 475
Amphitrita jonstoni 368
Amyda sinensis 516
Anaitides citrina 369
Anchistropus 269
Ancylostoma duodenale 419, 420
Andreninae 8
Anemonia sulcata 241
Annelides 12, 13, 18, 465, 466,
467
Anopheles 36, 124, 127
— maculipennis 112, 128*
— superpictus 112
Anoplocephalidae 387
Anoplodinium 164
— bubalidis 165*
— costatum 166*
— denticulatum 162
— gracile 165*
Anoplophrya 172
— teleuscolicis 173*
Anoxycalyx ijimae 200*
Anthelia 291
Anthomastus 290, 293, 293*
— grandiflorus 293
— rilovi 293
Anthozoa 13, 225, 232, 287
Antipatharia 225, 241, 309
Aphelenchoides composticola 399*
Aphrodite aculeata 368, 475
Aphroditidae 468*, 480, 481, 484
Aploparaxis 388
Apostomata 158
Appendicularia (Copelata) 14
Apus cancrioformis 18
Arachnactis albina 307
Arachnocorys circumtexta 80, 96
Arachnida 14
Arcella 79*, 80
Archaeobdella esmonti 525, 525*
Archiacina 84
— verworni 83*
Archiannelida 471
Archihirudinea 512
Arenicola 475, 487
— grubei 368
— marina 369, 479, 482
Arenicolidae 487
Argyrotheca 547
Arnynchobdellea (Gnathobdellea)
520
Artemia 44
— salina 37, 38
Arthropoda 8, 12, 14, 18
Arthropodaria kowalevski 457*
Articulata 544, 547*
Asbestopluma 196, 207
— sp. 206*
Ascaridae 418
Ascaridata 417
Ascaridia 418
— galli 419
Ascaridiidae 418
Ascaris lumbricoides 416, 416*
Ascidiacea 14
Ascomorpha 439
— mini ma 432
Ascothoracida 35
Aspidogaster conchicola 354, 354*
Aspidogastrata 354
Aspidosiphon steenstrupn 528*
Asplanchna 435, 436, 436*, 437—
439, 439*, 442
— brightwellii 429
— girodi 445*
— priodonta 432, 445*, 446
Asterias panopla 37
Asteroidea 14
Astomata 171, 172*, 174
Astrorhiza 82
— limicola 82*
Atactodea 318
Atentaculata 334
Atergates 272, 321
Atolla wyvillei 304
Atrypida 547*
Aulacantha scolymantha 71*
Auloceras arborescens 80
Aulopiiorus 496, 497*
Aurelia 232*, 238, 282*
— aurUa 281
Australella indica 534*, 541
Autholoba reticulata 255
37*
567
Autolytus 479, 485, 495*
— pictus 368
Aves 14
Avicula (Pteria) 19
Axinella 192, 212, 250
Axinellidae 196
Axinoderma mirabile 206*
В
Baicaloclepsis echinulata 516
— grubei 516
Baikalospongia 219
— bacillifera 201*
Balantidium coli 169, 170*
Balanus 212
Balatro 429
Bathypathes 310*
— lyra 245, 310
— patula 241, 310
Batracobdella 515
— algira 515
— paludosa 515
Bdelloidea 431, 436, 436*, 439*
Beloides firnus 118*
Beroe 331
— cucumis 321, 332
Beroidea 334
Bilateria (Triplastica) 11, 12, 222,
336
Bipalium 346
— newense 346
— simrothi 352
— strubelli 352
Bivalvia (Lamellibranchiata-Pele-
cyroda-Acephala) 14
Blackfordia 276
Blaps mortisaga 119*
Blepharoprosthium pireum 168*
Bod о 104
— edax 104*
Bolinopsis infundibulum 251, 321,
332
Bombinae 8
Bonellia viridis 527*
Bonelliidae 527
Botrynema brucei 304
Bougainvillia megas 277
— platygaster 240
Bowerbankia 542
— pystolosa 538*
Brachionidae 436
Brachionus 444
— bennini 436
— calyciflorus 435, 436, 444*,
445*, 448
— diversicornis 429, 430*
— dolabratus 429, 430*
— forficula 429
— gessneri 429, 430*
— havanaensis 429
— rybens 429, 430*
Brachiopoda 12, 18, 529, 530, 543
Brachylaemidae 363
Branchelion 520
Branchiobdellidae 501
Branchiocerianthus imperator 227
Branchiodrilus 497*
Branchiodrilus hortensis 496
Branchiomma 468*
Branchiura sowerbyi 499, 499*
Bryozoa 12, 13, 18, 529, 530, 533
Bucephalus polymorphus 354*,
355
Bullinus 362
Bunodeopsis 254*
Bursaria 66
— truncatella 143, 143*, 144
Bythotiara 225*, 255
Bythotrephes 37
С
Calappa 321
Calcarea (Calcispongia) 189
Calcispongia (Calcarea) 189
Calcynus 272
Galliactis parasitica 241, 252
Callyspongia sororia 216
Calocyclus monumentum 96
Calonympha grassi 113*
Calyx 192
Campanella umbellaria 149*
Campanularia 263*
— maccrady 240
Camptonema nutans 94
Capitella capitata 369
Capitellidae 481
Carchesium 150
— polypinum 149*
Carcinobdella cyclostoma 518*, 519
Carnivora 407
Carpoidea 15
Carteria 35
Caryophyllaeidae 378
Caryophyllaeus laticeps 379, 379*
Centrophorella fasciolata 157*
— grandis 157*
Cephalobidae 427, 427*
Cephalochordata 12,14, 18
Cephalodella 437
— forficula 434*
Cephalopoda 14
Ceractis aurantiaca 241
Ceratium 101
— hirudinella 101, 101*
— fuscus 71*
— palmatum 102*
— voltur 102*
Ceratomyxa appendiculata 130*
— truncata 131*
Cerebratulus marginatus 462
Cereus pedunculatus 250
Ceriantharia 225, 306
Cerianthus membranaceus 241
— solitaris 307*
Cestidea 334
Cestoda 378
Cestodaria 378
Cestoidea 13, 338, 375
Cestus veneris 321, 332, 334
Cetonia aurata 454
Chaetogaster 495
— diaphanus 495
— limnaei 495, 497*
Chaetognatha 12, 14, 18
Chaetonotus chuni 393*
— maximus 392*
Chaetopteridae 369, 487
Chaetopterus 480, 484
— variopedatus 476
Cheilostomata 542
Chirodropus 235, 240
Chlamydomonas 97
— angulosa 97*
Chlamys 210*, 322
Chlidonophora chuni 544
Chloromyxum leidigi 130*
Choanoflagellata 11
Chondrilla 192
Chondrocladia 196, 207
— gigantea 196, 203*
Chondrophora 224, 279
Chonelasma calyx 213
Chordaria 12
Chromadorida 402
Chromodoris 272
Chrysaora 225*, 238
Ciliata (infusoria) 13, 134
Cirripedia 212
Cladactis costata 241
Cladocoryne 227
Cladorhiza 196, 207
— longipinna 206*
— rectangularis 206*
Clanorchis sinensis 360
Clathrina 190
— blanca 208
— reticulum 198*
— primordialis 208
Clathrulina 94
— elegans 94, 94*
Clavularia 242, 291
Cliona 192, 196, 201, 214
Clionidae 195
Clitellio arenarius 499
Clypeomorus 318
Cnidae 226
Cnidosporidia 13, 74, 129
Coccidia 120
Cocconema sulsi 132
Codonobdella truncata 519
Coelenterata 12, 13, 18, 221
224
Coeloplana 333*
— metschnikowi 27, 334
Coeloria 311, 319, 323, 324
Coenopsammia 257
Collotheca 439*
— neptabrachiata 432
Cometoides crinitus 118*
Condylostoma 154
— remanei 157*
Conochiloides 436
— coenobasis 430*, 446
Conochilus 26, 436*
— unicornis 432, 437
Conus 318
— betuloidas 272
— hebraeus 272
— textilis 272
Convoluta 35, 340
— convoluta 341, 341 *
— roscoffensis 340
Corallium 225*, 290
568
Corallium inutile 250
— rubrum 241, 290
Cordylophora 274*
— caspia 40, 275
Coregonus peled 512
Cornacuspongida 189, 193,
195
Cornuspira 82
— involvens 82*
Coronata 225
Coronidium acacia 96
Coronympha octonaria 113*
Corycella armata 117, 117*
Coryne 230*, 237*, 261, 262,262*,
263*
— sarsii 238
Coscinasterias calamaria 546
Crania 18
Craniidae 544, 548
Craspedacusta 271*, 272
Crenatula 322
Crinoidea 14
Criodrilus lacuum 505
Crisia eburnea 538*
Cristatella 533
— mucedo 533, 534*,
541
Crossota 286
— brunnea 304
Crustacea 14
Cryptocercus 114
Cryptospongia enigmatica 210*,
213
Cryptostomata 542
Ctenophora 12, 13, 222, 327,
329
Ctenoplana kowalewskii 27,
334
Ctenostomata 542
Cubomedusae 225
Culcita 323
Cunina 230*, 255*, 256
Cunoctantha 256
Cyanea 238, 282*, 283
— arctica 223
Cyclophyllidea 378, 382
Cycloposthium 170*
— bipalmatum 168*
Cyclops coronatus 421
— oithonoides 421
— quadricornis 421
— uljanini 421
— vicinus 421
Cyclostomata 14, 542
Cydippe 330*
Cydippea 334
Cyphonautes 538*
Cypraea 318, 322
— annulus (Monet aria annulus)
272
— arabica 257, 272
— caputserpentis 272
— moneta (Monetaria moneta)
272
— tigris 257, 272
Cyprina islandica 462
Cystobranchus 518
— fasciatus 518
— mamillatus 518
D
Dactylogyrus vastator 371, 371*
Dalyellia 344
— viridis 343
Dapidia 439*
Dardanus 272
Dasydytes ornatus 393*
Davainea proglottina 376
Daya 257
Defrancia lucernaria 534*
Delofondia vulgaris 420*
Demospongia 189, 193
Dendrobaena 504
— mariupoliensis 504
— octaedra 504
Dendrobrancbia 310
Dendrocoelum lacteum 338*, 340*,
345
Dendrogaster 23
Dendrophyllia 314, 319
Dendrosoma radians 152*
Dendrostomum pyroides 528*
Dero 496
Deuterostomia 11, 12
Diadema setosum 257, 322
Dichogaster bolaui palmicola 506
— jaculatrix 506
Dicranophoridae 435
Dicranotaenia 388
Dicrocoellidae 358
Dicrocoellium lanceatum 358, 358*
Dictyocaulus filaria 421
Dicyemida 13, 181
Didinium 66
— nasutum 147, 147*
Didymosoomidae 349
Difflugia 79*, 80
Digononta 431
Dinoflagellata (Peridinea) 88, 100,
101
Dinophillidae 471
Dinophilus maris albi 471
Dioctophymata 407
Dioctophyme renale 395, 407,
407*
Diorchis 388
Diphyllobothriidae 380
Diphyllobothrium latum 380, 381*
Diplocercus 90
— fuscus 80
Diplostomatidae 364
Diplostomum spathaceum 364,
365*
Diplozoon paradoxum 373*, 374
Discinidae 544, 547, 548
Discinisca 544
Discomedusae 225
Disconantha 224
Discopora 536
Discorbis vesicularis 82*
Disoma 369
Disyringa dissimilis 209
Ditylenchus 422
— dipsaci 422, 423, 424*
Dividualia 11
Dodecaceria 479
Donax 318
Dorcadospyris dinocerae 96
Dorylaimus striatus 398*
Dracunculus medinensis 420
Drepanidotaenia lanceolata 388
Drilophaga 429
— delagei 430*
Dromia 212
Dunaliella 37, 44
Dysidea 201
— fragilis 210*
— palescens 197*
Ё
Echinococcus granulosus 384,
384*
Echinodermata 12, 14, 18
Echinoidea 14
Echinomera hispida 118*
Echinometra matthaei 272, 318
Echinorynchus 453*
— polymorphus 454
— proteus 454
Echiuroidea 13, 526
Echiurus echiurus 527*
Edwardsia 251, 300—303
Eimeria 120, 123*
— coecicola 123*
— cyprini 123
— intestinalis 123*
— irresidua 123*
— magna 120, 121*, 122, 122*,
123*
— media 123*
— perforans 123*
— piriformis 123*
— praecox 123
— tenella 123
— smithi 123
— zurni 123
Eisenia foetida 504
— nordenskioldi 504
Eiseniella tetraedra 505
Elphidium 85
— crispa 85, 85*
— strigilata 83*
Elysia 272
Emys orbicularis 515
Enantiozoa 11
Encentrum 439*
Enchytraeidae 499
Enchytraeus albidus 499, 500
Endoprocta 457
Enoplida 402, 403
Entamoeba histolytica 77, 78*,
79*
Enterobius vermicularis 415, 416*
Enteropneusta 14, 18
Enteroxenos 23
Entodiniomorpha 161, 167, 168*,
171, 174
Entodinium 162, 166*
— simplex 163*, 166*
— vorax 166*
Entodiscus indomitus 175*
Entorhipidium 174
— tenue 175*
569
Entorhipidium triangularis 175*
Ephelota gemmipara 152*, 153*
Ephydatia 218
— blembingia 201*
— mulleri 216
Epiactis marsupialis 302
— prolifera 302
Epidinium 164
— ecaudatum 71*, 163*
Epiphanes 436*
— senta 438
Epistylis 150
Epizoanthus 309
— macintoschi 309
— norvegicus 249
Eriocheir sinensis 41
Euchitonia virchovi 80
Euchlanis 436, 436*. 437
Eucrata loricata 534*
Eudactylota 437
Eudendrium 224*, 260, 262*
Eudiplodinium 164
— neglectum 163*
Eudorina 21
— elegans 98, 99*
Euglandina rosea 631*
Euglena 103
— spirogyra 103*
— viridis 95*
Euglenoidea 103
Euglypha 79*, 80
— alveolata 81*
Eugregarinida 118
Eunirudinea 513
Eumetazoa 11, 12, 183, 222
Eunemertes echinoderma marion
353
Eunephthia 242, 242*, 292, 293
Eunice aphroditoides 318
— fucata 478
— viridis 478
Eunicella 289
— verrucosa 249, 289
Eunicidae 486
Eunoe nodosa 368
Eupagurus 253
— exavatus 252
— prideaxi 241, 523, 254
Euphyllia 322
Euplanaria gonocephala 337, 338*,
345
— polychroa 345
Euplectella 219
— aspergillum 191*, 208, 219*
— oweni 210*. 211
Euplectellidae 193
Euplexaura antipathes 290
Euplocamis cucumis 321
Euretidae 193
Euritrema pancreaticum 359
F
Fadejewobdella quinqueannulata
526
Farrea haeckelii 208
Fasciola gigantica 356
— hepatfca 355*, 356
Fasciolidae 356
Fasciolopsis buski 356
— magna 350
Favia 256, 311, 324
Favites 318
Felis leo 10
Fibula notilangere 217
Filariata 420
Filellum serpens 249
Filinia 437, 444
— brachiata 429
— longiseta 429, 430*, 445*, 447
Filinia passa 445*
— terminal's 445*
Fissurella graeca 572, 576
Fissurellidae 572
Flagellata (Mastigophora) 13, 95
Floscularia ringens 434*
Flustra foliacea 534*, 540
Flustrella hispida 536, 538*, 540
Foraminifera 81
Fredericella sultana 534*
— — subsp. lepnevae 541
Fucus 249
Fungia 256, 311, 322, 325
— echinata 322
G
Gad us macrocephalus 519
Galatheanthemidae 301
Galatheanthemum 303
Galaxea 319, 324
Galba truncatula 357
Gambusia affinis 127 128*
Gasterostomta 354
Gastrochaena 318
Gastrodes parasitica 333,
Gastropoda 14
Gastropus stylifer 432
Gastrotricha 13, 390, 391
Gemellaria loricata 534*
Geniorhynchus monnieri 118*
Geodia 192
— barretti 194*, 199
— macandrewii 209
— phlegraei 203*
Geodiidae 202
Geonemertes 462
— agricola 462
Gigantoproductus 543
Girocotyloidea 13, 374
Glaucus 280, 288
Globigerina 86
— bulloides 87*
— sp. 82*
Glossina 108
— morsitans 37
— palpalis 107, 109*
Glossiphonia complanata ?
515, 520, 525
Glossiphonidae 513
Glossoscolecidae 505
Glottidia 544
— albida 544
— pyramidata 546
Glugea 132
— anomala 133*
570
Glugea hertwigi 132
Gnathobdellea (Arhynchobdellea)
520
Gnathobdellidae 520
Gnathostomata 14
Golfingia vulgare 528*
Gonactinia 300, 301, 305
— prolifera 300, 301
Goniastrea 324
Gonionemus 240
— vertens 236
Gonium pectorale 98, 98*
Gordiacea(Nemathomorpha) 13
Gordius 452
— aquaticus 451*, 452
Gorgonaria 225, 287, 295
Gorgonia 289
Gorgonocephalus arcticus 37
Grantia compressa 204
— mirabilis 208
Gregarina longa 118*
Gregarinida 177
Grenobia alpina 345
Gyrinus natator 117
Gyrocotiloidea 338
Gyiodactylus 371*
— elegans 372
H
Habrotrocha angusticollis 434*
— visa 434*
Haemadipsa 524
— ceylonica 524*
Haementeria costata 514, 514*
Haemopis sanguisuga 520, 523,
524*
3* Haemosporidia 124
Haetodon 257
Halichondria 184*, 192
— panicea 203, 209
— sitiens 210*
Halichondriidae 196
Haliclona 192, 201
— gracilis 197*
— schmidti 197*
Haliclystus 284
Halicreas minimum 304
Halycryptus 455, 455*
Halisarca 193, 199
Haloclava 251, 303
Haplophragmoides canadensis
82*
Haploplana ornata 352
Haplotaxidae 500
Haplotaxis ascaridoides 500
— gordioides 500, 500*
Haptophrya 172
Harmothoe imbricata 369
, Helicoprorodon multinucleatum
157*
Heliopora 320
Helioporida 225
Heliozoa 74, 93
Helobdella stagnalis 516, 520, 525
Hemichordata 12, 14
Hemiclepsis marginata 513
Heniochus macrolepidotus 257
Henneguya 131
— acerina 131*
— lobosa 132*
Hepatus 255
Herpobdella (Eipobdella, Nephelis)
524
— absoloni 525
—— linpata ^S25
— octoculata 520, 520*, 524, 525,
525*
Herpobdellidae 524
Heteractinellidae 219
Heterocentrotus mammilatus 257,
320
Heterocoela 189, 190, 198
Heterotricha 143
Hexacorallia 225, 232, 299
Hexactinellidae (Hyalospongia)
189
Hexancistra quadricuspis 96
Hexarthra 436*
— mira 442*, 445*
Hexasterophora 189, 192, 193
Hippospongia communis 204,
218
Hippotoa hyalina 538*
Hircinia variabilis 209
Hirudinea 13, 509
Hirudo 523
— medicinalis 511*, 520, 520, 521,
522, 523
Hislopia 541
Holocephala 374
Holotricha 141, 168*, 174
Holothuria tubulosa 35
Holothurioidea 14
Homo sapiens 17
Homocoela 189, 190, 196, 198
Hoplolaimus tylenchiformis 398*
Hormathia corenata 252
Hornera lichenoides 534*
Hyalonema 207, 219
— elegans 206*
— indicum 210
— sieboldi 208, 219*
Hyalonematidae 193
Hyalospongia (Hexactinellidae)
189, 191
Hyalostylus dives 206*
Hydra oligactis 268
Hydra vulgaris 263
Hydractinia 225*, 242
— echinata 249, 251, 252
Hydra moeba hydroxena 269
Hydrida 225, 263
Hydrocorallia 224
Hydroidea 224, 230, 263
Hydrozoa 13, 224, 227, 230,
260
Hymedesmia 192
Hymedesmidae 196
Hymenolepididae 388
Hymonolepis nana 388
Hymenoptera 8
Hyperammina 82
— elongata 82*
Hyper mastigina 112
Hypodontolaimus butschlii 403 *
Hypotricha 146, 174
I
Ichthydium forcipatum 393*
Ichthyobdellidae (Piscicolidae) 517
Ichthyophthirius multifiliis 159,
160*
Ichthyotomus sanguinarius 473
Idia 158
Inarticulata 544, 547
Infusoria (Ciliata) 13, 74, 134
Insecta 14
Isidella 290
— elongata 290
Isospora aksaica 123
Isozoanthus 309
— giganteus 309
¦I
Janthina 280, 288
Johansonia arctica 519
К
Kamptozoa 13, 457, 457*
Kellicottia longispina 432
Keratella 429, 435, 437, 444
— quadrata 432, 435, 442*, 443*
— testudo 445*
Kinorhyncha 13, 390, 449, 450*
Kophobelemnon 245, 295*, 297
— biflorum 245, 251, 297
— pauciflorum 297
— polyflorum 245
— stelliberum 245
Kophobelemnonidae 297
L
Lacarella 547
Lacinularia flosculosa 437
— ismailoviensis 438, 445*
Lagena 83
— plurigera 82*
Lamblia intestinalis 106, 107*
Lamellibranchiata (Acephala —
Bivalvia — Pelecypoda) 14
Laminaria 475
Laquens californicus 547
Leander 42
Lecithodesmus goliath 350
Leda (Nuculana) 19
Leishmania 66, 109
— donovani 110, 110*
— tropica 109, 109*, 110
Lepadella 436
Lepas 212
— fasciculatus 280, 288
Lepidoderma squamatum 397*
Leptogorgia 289*
Leptolida 224
Leptomonas 110, 115
— davidi 115*
Leptotheca agilis 130*
Lepus europaeus 9
571
Lepus timidus 9
Leucochloridium 364
— paradoxum 363, 364*
Leuconia 190
— aspera 208, 216
Leucosolenia 190, 201, 204
— botryoides 208
Levinsenia rectongulata 518*, 519
Ligula intestinalis 379, 379*
Ligulidae 379
Liliomorpha 154
— viridis 154*
Lima 19, 322
Limnea stagnalis 365
Limnatis 523
— nilotica 520, 522
Limnias melicerta 434*
Limnocnida tanganjice 274
Limnodrilus 499
— hoffmeisteri 499
— newaensis 499
Limuius 18, 19, 20
Lindia 429, 437
— brotzkayae 430*
Lindiidae 435
Lineidae 462
Lineus geniculatus 353
— longissimus 400, 459, 460, 462
Lingula 18, 544, 545*
— unguis 546
Lingulidae 544, 547, 548
Liothyrella antarctica 547
Lithophaga 318, 323
Lithoptera 90
— mulleri 80
Lobata 334
Lobophyllia 257, 311, 319
Longidoridae 405
Lopadorhynchus 496
Lophohelia 313*
— prolifera 312
Lophopodella 533
— carteri 537*, 540, 541
Lophopus 533
Lophopussella 541
Loricata 14
Loxodes 155
Lubomirskia baikalensis 209
Lubomirskiidae 196
Lucernaria 284
— campanula 241
Lucernariida 27
Lumbricidae 502, 505
Lumbricillus lineatus 500
Lumbriculidae 500
Lumbriculus variegatus 407, 501
Lumbricus 492*, 503, 504
— rubellus 503, 504
— terrestris 502, 503
Lungia aurantiaca 352
Lycastopsis catarractarum 473
M
Macandrevia cranium 543
Macrocanthorhynchus hirundina-
ceus 453 *, 454
Macrochaetus collinsi 430*
Macrodasys buddenbrocki 392*
Macrophiotrix 318
Madreporaria 225, 310
Magellania venosa 543
Magelona papillicornis 369, 469
Malakia 7
Malakostraca 7
Malacobdella 462
— grossa 353, 462
Maldane sarsi 473
Malleus 322
Mammalia 14
Manayunkia 473, 488
Manus marina 291
Marcierella enigma 41
Mastigamoeba aspersa 105, 105*
Mastigophora (Flagellata) 13, 74,
95
Maupasia 484
Meandra 319
Meator 286
— rubator 304
Magascolecidae 505
Magascolides australis 505, 505*
Mellifera 8
Meloidogyne 425
Membranipora 542
— crustu]enta 540
— sp. 534*
Menipea zelandica 534*
Merifugia 473
Mermis 406
Mermitidae 406
Mesenchytraeus solifugus 500
Mesidothea entomon 39
— sajDini 37
Mesnilella 172
— maritui 173*
Mesostomum 342
— ehrenbergii 338*, 342, 343,
343*, 346
— tetragonum 342
Mesozoa 11, 12, 13, 179, 181, 181*
Metazoa 12, 65, 183
Metridium dianthus 303, 305
— senile 241
Microcodon clavus 432
Microhydra 227, 273
Microphallidae 349
Microporella ciliata 538*
Microscolex phosphoreus 506
Microsporidia 129, 132
Microstomum lineare 343
Miliolidae 83
Millepora 319
Modiolus 19
Moerisia 276*
— pallasi 276, 276*
Mollusca 12, 13, 18
Molluscoidea 530
Monetaria annulus (Cypraea annu-
lus) 272
— caputserpentis (Cypraea caput-
serpentis) 272
— moneta (Cypraea moneta) 272
Moniezia expansa 387
Moniligastridae 507
Monobrachium 27
Monobrachium parasiticum 227
Monocystidae 118
Monogenoidea 13, 338, 368
Monogononta 431, 440*, 441, 443,
446, 448
Monommata 437
— appendiculata 430*
Mononchus 404
— papillatus 404, 404*
Monoplacophora 9, 13, 19
Monorhaphis chuni 193, 208
Montipora 257
— foliosa 319
Mucophria pelagica 154, 155*
Multiceps multiceps 386, 386*
Mycale ochotensis 203*
Myniadidae 300, 303
Myrianida 479, 485
Myriapoda 14
Mysis relicta 39
Mytilaster lineatus (Brachyodontes
lineatus) 42
Myxillidae 195
Myxilla parasitica 210*
Myxobolus 131
— cyprini 131, 131*
— sp. 131
Myxosoma cerebralis 131, 132, 133*
Myxosporidia 129
Myzostomida 489
Naididae 494
Nais 493*, 494, 495
— behningi 495
— elingius 495
— pseudoobtusa 495
— variabilis 495
Nassa 253
Nasselaria 80, 90, 92
Nassula 148, 148*
Natica 249, 253
Nautiloidea 19
Nautilus 20
Nebalia 431
Nectochaeta 496
— grimaldii 484
Nectonemertes mirabilis 353
Nemathelminthes 12, 13, 18, 335,
389, 428
Nemathomorpha (Gordiacea) 13
Nematobothrium filarina 349
Nematoda 13, 390, 394
Nemertes 459
Nemertini 12, 13, 18, 335, 459
Neopilina 9, 18, 19
Nephthidae 292
Nephthys ciliata 369
Nereidae 473, 481, 485
Nereis 468*, 470, 477
— diversicolor 42, 482
— japonica 482
— pelagica 368, 475
— virens 368, 369, 478, 482
Nerillidae 471
Noctiluca miliaris 101, 102*
Nodosaria 83
Nodosaria hispida 82*
Nonion umbilicatulus 82*
Nose ma 133 *
— apis 132
— bombycis 132, 133, 133*
Notholca baicalensis 429
— olchonensis 429, 430*
Notommata 436 *
— copeus 432, 439*, 443*
Notommatidae 435, 436, 437
Nucula 19
О
Obelia 261, 262*
— flexuosa 249
— geniculata 239
Ocnerodrilus occidentalis 506
Octactinellidae 219
Octocorallia 225, 232, 287, 293
Octomanus 284, 285
Ocypoda 318
Oligobdella orientalis 516
Oligochaeta 13, 172, 490
Oligomera 11, 12, 15, 29, 530
Oligonchoinea 371
Oligotricha 174
Onuphis conchilega 368
Onychophora (Protracheata) 12,
14, 15
Opalina 111
— ranarum 111, 111*
Opalinidae 110
Opercularia 150
Ophiopleura borealis 37
Ophiuroidea 14
Ophryoscolecidae 162, 163*, 165*,
166*
Ophryoscolex 164
— caudatus 162*
Opisthotrichum janus 165*
Opistorchidae 359
Opistorchis felineus 358*, 359
Orbicella 318, 319
Orbitolites 84
— complanatus 84*
Oreaster 323
Ornithocercus 101
— magnificus 102*
Orthida 547*, 548
Orthonectida 181
Orthonectidae 13
Oscarella 193, 196
Ostracodinium 165*
Oxytonostoma typica 519
Oxyuridae 415
Ozobranchus 520
— jantseanus 518*, 520
P
Pagurus 272
— arrosor 241, 252, 253
Palythoa 210*, 212, 250, 309*
Pandorina 21
Pantachogon 286
— haeckeli 304
572
Pantopoda 14
Parachordodes 452
Paracineta patula 153*
Paragordia arborea 249
Paralithoides camtschatica 519
Paramecium 134
— aurelia 71*
— caudatum 134, 134*, 138
Paramuricea 250
— placomus 249
Parascaris equorum 418
Paratorix baicalensis 516
Parazoa 12, 183, 222
Parazoanthus 309
— anguicomus 309
— axinellae 192, 212, 250
Parella sp. 534*
Parenteroxenos 23, 36
Parerythropodium corralloides
249
Pavona 256, 322, 325
Peachia 251, 302
Pectinaria koreni 369
Pectinatella 533
— sp. 534*
Pedicelina 458*
Pedinosoma 484
Pelagia 240
— noctiluca 287
Pelagodiscus atlanticus 548
Pelagonemertes moseleyi 462
Pelagonemertidae 462
Pelecypoda (Lamellibranchiata —
Bivalvia — Acephala) 14
Pelodrilus ignatovi 501
Pelomyxa binucleata 78*
Pennatula 224*
— phosphorea 241, 296, 298, 298*
299
Pennatularia 225, 294
Pentamerida 547*
Pentastomida 14
Peranema 104
Perforatella 534
Peridinea (Dinoflagellata) 88, 100,
313
Peridinium tabulatum 100, 101*
Perigonimus 242, 249
Perionyx arboricola 506
Periphylla hyacinthina 304
Peritricha 148, 155
Perypanicium amphocorona 96
Pheodaria 80, 90, 91, 92
Pheretima 491, 507
— posthuma 506
Pheronema giganteum 208
Philodina brevipes 430*
Phlebotomus pappatasii 110
Pholas 323
Phoronoidea 12, 13, 18, 529, 530,
531 532*
Phyllodocidae 483
Physalia 49, 240, 277
Physophora 224*
Physopsis 362
Phytomastigina 96
Pileocephalus heeri 118*
Pinctada 322
Pinna 322
Pisa gibbsi 210*
Piscos 14
Piscicola caspica 518
— fadejewi 518
— geometra 517*, 518, 520
Piscicolidae (Ichthyobdellidae)
517
Placentonema gigantissima 395
Plakina 196
Planaria alpina 37
Planaxis 318
Planes 279, 288
Planocera folium 347
Planorbis 362, 366
Plasmodium 36, 124, 125, 128
— falciparum 112, 124, 125
— gallinaceum 128
— malariae 112, 124, 125
— ovale 124, 125
— vivax 112, 124, 125, 127*
Plathelminthes 12, 13, 18, 335, 337
Platybdella fabricii 519
Platyctenidae 27
Platyctenidea 334
Platynereis dumerilii 477, 477*,
478, 479
Plectidae 402, 410
Pleurobrachia pileus 332
Plumarella longispina 289
Plumatella 540
— emarginata 541
— fungosa 540, 541
Pocillopora 256, 257, 319, 325, 326
— damicornis 322
Podobacia 318
Pogonophora 12, 14, 18
Polyarthra 444, 446
— dolichoptera 432, 445*, 447
Polycelis cornuta 338*, 345
— nigra 338*, 345
Polychaeta 13, 473
Polycladida 347
Polycotilus 345
Polycystidae 118
Polydora ciliata 369
Polygordiidae 471
Polylophus philippiensis 200*
Polymastia 199
— corticana 209
— hemisphaericum 203*
— kurilensis 200*
— mammillaris 200*
Polymastigina 112
Polymastiidae 194, 202
Polymera 12, 29
Polyonchoinea 371
Polyplastron 164
— multivesiculatum 163*
Polypodium 258 *
— hydriforme 256
Polystoma integerrimum 372*, 373
Pomacenorus 257
Pompholyx 437
Pontaster tenuispinus 37
Pontobdella muricata 520
Popillia japonica 42
Porella concinea 538*
Porifera (Spongia) 12, 13, 18,
182, 183, 189
Porites 256, 318, 319, 323, 324
Porpita 278
Portlandia arctica 37
Postodiplostomum cuticula 365
Potamilla symbiotica 210*, 213
Poterion neptuni 195, 209
Priapulida 13, 455
Priapuloidea 19
Priapulus 20, 455, 455*
— caudatus 456
Primnoa 249, 250
— resedaeformis 249
Prionospio 486
Pristina 494, 496
— longiseta 497*
Proales 429
— gigantaea 429
— daphnicola 429, 430*
— latrunculus 429
— parasita 429
— wernecki 429
Productida 547*
Prognichthys agae 280
Pronax plumosa 198*
Prosopygii 15, 530
Prosorhochmidae 462
Prosostomata 355
Prospora gigantea 66
Prostomatidae 463
Protoclepsis 513, 514
— maculosa 514
— tessulata 514
Protodrilidae 471
Protodrilus 471
Protomonadina 104
Protoptilum 295
Protoreaster 257, 323
Protostomia 11, 12, 336
Protozoa 12, 13, 18, 65, 71, 73,
176
Protracheata (Onychophora) 12, 14
Protula protula 368
Psammonix 66
Pselaphinidae 34
Pseudoceros maximus 352
Pseudophyllidea 378
Pseudoprorodon arenicola 157*
Pseudostichopus tranchus 213
— villosus 250
Pseudotrichonympha 68*
Pterobranchia 14
Pterocera 318
Pterosyllis 480
Ptilocedium repens 249
Ptychodactys patula 249
Ptygyra tihanyensis 434*
Pyrgoma 323
Pyxudium ventriosa 149*
a
Quinqueloculina seminulum 82*
R
Rabditis anomala 399*
Radiata 10, 11, 12, 222, 431
573
Badiolaria 75, 88
Radiophrya 173
— hoplites 172*
Raja radiata 519
Rapana besoar 41
Raspailia 192
Rathkea 287
Bemanella caudata 157*
Renilla patula 296
Reptilia 14
Retepora cellulosa 534*
Rhabdammina linearis 82*
Rhabditida 411, 414
Rhabditidae 410, 411
Rhabdites aberrans 412*
— elongata 413
— inermis 413
Rhabdocalyptus 201
Rhabdocoela 341
Rheopax nodulosus 82*
Rhinodrilus fafner 505
Rhitropanopeus harrisi 41
Rhizomastigina 105
Rhizopoda 75
Rhizostoma 240
— pulmo 283
Rhopilema esculenta 283, 284*
Rhynchelmis limosella 501
Rhynchobdellea 513
Rhynchodemus bilineatus 346
— rubrocinctus 352
— terrestris 346
Rhynchonellida 547*, 548
Rhynchonympha tarda 113*
Ripistes parasita 495, 496, 497*
Rossella 208
Rosselidae 193, 205
Rotaria neptunia 430*, 437
Rotatoria 13, 30, 390, 428
s
Sabellidae 488
Saccamina sphaerica 82*
Saccocirridae 471
Sacculina 23, 36
Sagittella 496
Sarcodina 13, 74, 75
Sarcophyllum 249
Sarcophytum 256, 319
Scaphopoda 14, 250
Scaridium 437
Schistosoma haematobium 361,
362*
— japonica 361
— mansoni 361
Schistosomatidae 360
Schizogregarinida 118
Sclerobelemnon 295
Sclerocrangon boreas 519
— ferox 37
Scolecida (Amera) 11, 336, 466
Scyllium aquila 349
Scyphozoa 13, 225, 231, 281
Scytaliopsis djiboutiensis 298
Secernentea 400, 410, 414
Seison nebaliae 430*
Seisonidae 437
Seisonidea 429, 431
Semisuberites arctica 203*, 209
Semnodera armigera 449*
Serpula vermicularis 368
Serpulidae 473, 488
Sertularia rugosa 263*
Sicyopus 251
— commensalis 250
Silicoflagellidae 96
Sinantherina socialis 446
Sinuolinea dimorpha 131*
Siphonochalina 192
Siphonophora 225, 230
Sipunculoidea 13, 466, 528
Solenogastres 14
Sphaerospora divergens 131*
Sphaerosyllis 480
Spionidae 486
Spirifer 546*
Spiriferida 547*
Spirodinium equi 168*
Spirograms spallanzanii 368
Spiroloculina depressa 82*
Spirophrya subparasitica 158,
158*
Spirorbis spirorbis 369
Spirostomum 66
Spirotrichonympha flagellata 113*
Spongia (Porifera) 12, 13, 179, 182
Spongia officinalis 197*, 218
— zimocca 217*
Spongicola venusta 210*, 211
Spongiidae 196
Spongilla 184*, 218
— lacustris 201*, 216
Spongillidae 51, 196
Sporozoa 13, 22, 74, 116
Spriggino floundersi 483, 483*
Spumellaria 80, 90, 91, 92
Staphylinidae 34
Stauromedusae 225
Steineria mirabilis 395*
Stellettidae 195
Stentor 141
— coeruleus 141*, 142
— polymorphus 141, 142
Stepnanella nina 541
Stephanoceros fimbriatus 432
Stephanopogon 139
Sthenelais 468*
Stichostema 463
Stoichactis 255, 303
— kentii 323
Stolonifera 291
Strobilidium 174
— rapulum 175*
Strongilidium arenicolus 157*
Strongylata 419
Strongylidae 419
Strongylus vulgaris 419
Strongyloides stercoralis 415, 415*
Strongyloididae 415
Strongylus vulgaris 419
Strophomenida 547*
Sturnus vulgaris 41
Stylaria lacustris 495, 497*
Stylatula darvinii 298
Stylocephalus longicolHs 118*,
191*
574
Stylocordyla 195
Stylonichia mytilus 146, 146*
Stylophora 322, 325
Suberites 201
— domuncula 192, 211
Suberitidae 194
Succinea 364
Suctoria 153, 155, 169, 169*
Sycia inipitata 118*
Sycon 190, 201, 204
— ciliatum 203*
— coronatum 204
— raphanus 198*
Syllidae 295, 479, 480, 485, 485*
Syllis ramosa 485
— vivipara 479
Sympagella 201
Symphyllia 319, 324
Synchaeta 429, 435
— pachipoda 429, 430*
— pectinata 432
T
Taenia solium 381*, 383, 384
Taeniarhynchus saginatus 381*.
382
Taeniidae 382
Tardigrada 14, 38
Telestacea 291
Telesto 291
Temnocephala 13, 338, 367, 367*
Temnocephalidae 367
Tentaculata 15
Tentaculifera 334
Tentorium semisuberites 194,
203*
Teraster umberis 35
Teratonympha mirabilis 113*
Terebratella incospicua 546
Terebratulida 547*, 548
Terebratulina 544
Testacea 80
Testudinella 436
— patina 432
Tethya 196, 199
— aurantium 192, 194, 199,
200*
Tetilla 199
— cranium 209
Tetillidae 194, 202
Tetracorallia 259
Tetrastemma rusticum 353
Tetraxonida 189, 193
Textularia 83
— sagittula 82*
Thalassema gigas 527*
Thalassicola nucleata 89*
Thaliacea 14
Thecideidae 544
Theromyzon 513
Thuiaria 224*
Thysanozoon brochii 347, 347*
Tiaropsis 263, 263*
Tintinnidae 154
Tintinnopsis beroidea 155*
Tipostomata 542
Tjalfiella 333*, 334
Tjalfiellidea 334
Tokophrya quadripartita 152*
— cyclopum 153*
Tomopteridae 484
Tomopteris 475, 483, 496
— renata 368
Torix cotylifer 516, 516*
Toxopneustes 322
Trachelius 154
Trachelobdella sinensis
518
—- torquata 517*, 519
— turkestanica 519
Trachelonema grassei 157*
Trachylida 224
Trematoda 13, 338, 348
Triadinium galea 168*
Trichinella trichinella 409
Trichocephalata 407
Trichocephalidae 407, 409
Trichocephalus 407
— trichiuris 407, 408*
Trichocerca 429
— cylindrica 430*, 445*
— rattus 432
Trichoderma minutum 395,
Trichodina 67*, 160
— domerguei 161*
— pediculus 161*, 269
Trichodoridae 405
Trichomonas 105, 106*
— hominis 105, 106*
Trichonympha 113, 114*
— turkestanica 113*
Trichotria pocillum 432
Tricladiba 344, 348
— terricola 346
Tridacna 35, 313, 320
— crocea 257, 320
— gigas (derasa) 320
Tridacophyllia 256, 326
Trilobita 14
Triplastica (Bilateria) 336
Tripneustes 323
— gratille 272
Trocheta bykowskii 526
— subviridis 525
Trochus maculatus 320
— niloticus 257, 272
Troglochaetus beranecki 473
Troglodytella 170
— gorillae 171*
Tryblidium 18
Trypanosoma 107
— brucei 71*, 108
— equiperdum 108
*, — evansi 108
— gambiense 37, 107, 108, 108*
Trypanosyllis 485
Tubifex 493*, 498*, 499
— tubifex 499
Tubificidae 496
Tubipora 241, 244, 294
Tubiporidae 293, 294
Tubulanidae 461
Tubulanus 353
— superbus 461
Tunica ta (Urochorda) 12, 14, 18
Turbellaria 13, 338, 339
Turbinaria 318, 319*
Turbo marmoratus 272, 320
Tuscarilla nationalis 80
Э0 Tylenchida 422, 423
Tylenchidae 422
Typhloscolex 496
U
Udonella caligorum 370*
Udonelloidea 368
Umbellula 295*, 297
— dutissima 245
— encrinus 296, 297
— magniflora 297
— monocephalus 245, 297, 298
— thomsoni 245, 297
Urcelariidae 160
Urceolus 104
Urnatella gracilis 458, 458*
Urocentrum turbo 70
Urochorda (Tunicata) 14
Uroleptus rattulus 157*
Urticina crassicornis 306
Uschakovia gorbunovi 536, 537*
V
Vanadis 475
Vaucheria 429
Velella 49, 278, 288
Veretillum 295*
— cynomurium 296
Vermes 336
Vermetus 320
Vermoidea 530
Vertebra ta 12, 14, 18
Victorella 542
Victoria regia 499
Virgularia 295, 295*
— cladiscus 296
— glacialis 296
— mirabilis 296
Volvox 98, 116, 429
— aureus 98, 99*
— globator 98
Vorticella 148, 148*
— nebulifera 148
— microstoma 149*
W
Walteria 212
Whitmania laevis 523
X
Xerobdella 524
Xiphinema americanum 405
Xiphosura 14, 15
Z
Zoantharia 225, 308
Zoanthus 309
Zoo mastigina 96, 104
Zoophyta 8, 227
Zoothamnium arbuscula 150*, 151,
151 *
Zooxantellae 309
Zostera 249
ИМЕННОЙ УКАЗАТЕЛЬ
А
Аристотель 7, 10, 226, 329
Б
Барлейз 459
Бастиан 423
Бауер 159
Беклемишев В. Н. 259, 334,
450
Бильхарц 361
Блэнвилль И
Богословский А. С. 446
Браун А. 418
Бреслау 341, 343, 344
Бродский Л. Л. 71
Быховский Б. Е. 368
Бэр К. М. 22, 354
Бюргер О. 460, 461, 463
Бючли О. 74, 391
В
Вернадский В. И. 42, 410
Виноградов А. П. 20
Вудруф 74
Высоцкий Г. Н. 504
Г
Гален 421
Геккель Э. 22, 89, 216, 285
Гертвиг 74
Гмелин 10, 459
Гольдфус 73
Горо 314
Грассе 105
Грасси 74
Граф 348
Грубе Э. 512
д
Данилевский 74
Дарвин Ч. 9, 22, 298, 315, 316,
317, 346, 414, 507
Девис 317
Дели 316
Деляж И. 183
Дженнингс 74
Джильберт 447
Догель В. А. 74, 164, 176
Дюжа рдэн 73
г
Захваткин А. А. 259
Зелинка 391
Зибольд 74
' Зюсс Э. 42
И
Иванов А. В. 19, 23
й
Йонг 313, 314
К
Калабухов Н. И. 39
Калкинс 74
Камшилов М. М. 329, 331,
Кёлликер 74
Киршенблат Я. Д. 447
Кларк Д. 182
Кливленд 114
Клюге Г. А. 536
Кооб Н. А. 395
Ковалевский А. О. 27, 329,
515, 546
Кори 496
Коротнев А. 27, 329, 334
Крумбах 329
Кювье Ж. 10, 17, 18, 431,
459
Л
Ламарк Ж.-Б. 10
Ланкестер 271
Латышев Н. 110
Лаутерборн 446
Левенгук А. 73, 264
Лейкарт 227, 416
Ливанов Н. А. 459, 512
Линней К. 7, 9, 10, 73, 329,
533
М
Мак Интош 460
Малышги 10
Марков 74
Мартене Э. 329
Марциновский 74
Меккель 22
Метальников 74
Мечников И. И. И, 182, 259, 329,
391, 531
Миддендорф А. Ф. 512
Мозговой А. А. 419
Мопа 74
Мур 500
Мэррей Дж. 316
Мюллер О. Ф. 73, 459
Н
Насонов А. В. 458
Норденшельд А. 504
О
Овсянников Ф. В. 256
333 Олмен 271
Осборн 542
П
Павлов 271
334 Павловский Е. Н. 108, 420, 454
Паллас П. С. 9, 275
Пейсоннель 226
Плиний 8, 226
Поггенполь М. Ю. 438
Полканов А. А. 20
Р
Райтер 413
Рей Дж. 10
Ремане 391, 431
С
Саре М. 238
459, Сваммердам Я. 10
Северцов А. Н. 394
576
Скрябин К. И. 414, 421
Стеенструп 238
Стрелков А. 167
Сукачев В. Н. 42
Т
Талызин Ф. Ф. 383
Торн 400
Трамбле 226, 264, 270
У
Уильсон Г. 203
Уоллес А. 40
Уэнрайт 314
Ф
Филипченко 74
Филипьев И. Н. 406
Фойгт 431
X
Хангейт 113
Хейнли 404
Хейсин Е. М. 123
Холодковский Н. А. 396
Хун К. 329
Ч
Чайлд 271
Чокановский О. В. 499
Ш
Шамиссо А. 238
Шаудин 74
Шванн 74
Шевяков В. Т. 74, 92, 118
Шлейден 74
Шмидт П. Ю. 39
Шмидт О. 182
Шмидт О. Ю. 20
Штайнер 404
Штейнбок 341, 450
Штром Ж. К. 383
Щульц Р. С. 356
Шульце П. 391, 392
Шульце Ф. 182
щ
Щеголев Г. Г. 520
Э
Элтон 42
Эпштейн 74
Эренберг 73, 342, 459
Эрлангер 446
Эшшольц 329
Я
Якимов 74
СОДЕРЖАНИЕ
Предисловие 5
Введение. Л. А. Зенкевич ... 7
Системы животного мира .... 8
Филогения и геологическое прош-
лое животного мира 15
Некоторые этапы эволюции живот-
ных 21
Некоторые биологические особен-
ности животных 32
Биосфера и биоценоз 42
Фаунистические области океана
и суши 52
Клетка. Ю. И. Полянский ... 54
ТИП ПРОСТЕЙШИЕ (PROTOZOA).
Ю. И. Полянский 65
КЛАСС САРКОДОВЫЕ (SARCO-
DINA) 74
Подкласс Корненожки (Rhizopoda) 75
Отряд Фораминиферы (Foramini-
fera) 81
Подкласс Лучевики, или Радиоля-
рии (Radiolaria) 88
Подкласс Солнечники (Heliozoa) 93
КЛАСС ЖГУТИКОНОСЦЫ (MAS-
TIGOPHORA, или FLAGELLA-
ТА) 95
Подкласс Растительные жгутико-
носцы, или Фитомастигины (Pbyto-
mastigina) 97
Отряд Панцирные жгутиконосцы,
или Перидинеи (Dinoflagellata,
или Peridinea) 100
Отряд Эвгленовые (Euglenoidea) 103
Подкласс Животные жгутиконос-
цы, или Зоомастигины (Zoom as-
tigina) 104
КЛАСС СПОРОВИКИ (SPORO-
ZOA) 116
Отряд Грегарины (Gregarinida) 117
Отряд Кокцидии (Coccidia) . . .120
Отряд Кровяные споровики
(Haemosporidia) 124
КЛАСС КНИДОСПОРИДИИ (CNI-
DOSPORIDIA) 129
Отряд Миксоспоридии (Myxospo-
ridia) —
Отряд Микроспоридии (Microspo-
ridia) 132
КЛАСС ИНФУЗОРИИ (INFUSO-
RIA, или CILIATA) 134
ТИП МЕЗОЗОЙ (MESOZOA).
Л. А. Зенкевич 181
ТИП ГУБКИ (PORIFERA, или SPO-
NGIA). В. М. Колтун 182
КЛАСС ИЗВЕСТКОВЫЕ ГУБКИ
(CALC1SPONG1A) 189
Отряд Homocoela 190
Отряд Heterocoela —
КЛАСС СТЕКЛЯННЫЕ ГУБКИ
(HYALOSPONGIA) 191
Отряд Hexasterophora 193
Отряд Amphidiscophora —
КЛАСС ОБЫКНОВЕННЫЕ ГУБ-
КИ (DEMOSPONG1A) . . . . —
Отряд Четырехлучевые губки
(Tetraxonida) —
Отряд Кремнероговые губки
(Cornacuspongida) 195
ТИП КИШЕЧНОПОЛОСТНЫЕ
(COELENTERATA). Д.В. Наумов,
Ф. А, Пастернак 223
КЛАСС ГИДРОИДНЫЕ (HYDRO-
ZOA) 260
КЛАСС СЦИФОИДНЫЕ (SCYPHO-
ZOA) 281
КЛАСС КОРАЛЛОВЫЕ ПОЛИПЫ
(ANTHOZOA) 287
Подкласс Восьмилучевые кораллы
(Octocorallia) —
Отряд Горгоновые кораллы (Gor-
gonaria) —
Отряд Мягкие кораллы (Alcyo-
naria) 290
Отряд Морские перья (Реппа-
tularia) 294
Подкласс Шестилучевые кораллы
(Hexacorallia) 299
Отряд Актинии, или Морские цветы
(Actiniaria) —
Отряд Цериантарии (Ceriantharia) 306
Отряд Зоантарии (Zoantharia) . . 308
Отряд Антипатарии (Antipataria) 309
Отряд Мадрепоровые кораллы (Ма-
dreporaria) 310
ТИП ГРЕБНЕВИКИ (CTENOPHO-
RA). Д. В. Наумов 329
НИЗШИЕ ЧЕРВИ 335
ТИП ПЛОСКИЕ ЧЕРВИ (PLATHE-
LMINTHES) 337
КЛАСС РЕСНИЧНЫЕ ЧЕРВИ,
или ТУРБЕЛЛЯРИИ (TURBEL-
LARIA). А. А. Парамонов .... 339
Отряд Бескишечные ресничные
черви (Acoela) 340
Отряд Прямокишечные турбелля-
рии (Rhabdocoela) 341
Отряд Трехветвистокишечные тур-
беллярии (Tricladida) 344
Отряд Многоветвистокишечные тур-
беллярии (Policladida) .... 347
КЛАСС ТРЕМАТОДЫ (TREMATO-
DA). К. М. Рыжиков 348
Подкласс Аспидогастрата (Aspidoga-
strata) 354
Подкласс Гастеростомата (Gastero-
stomata) —
Подкласс Прозостомата (Prososto-
mata) 355
КЛАСС ТЕМНОЦЕФАЛЫ (ТЕМ-
NOCEPHALA). Л. А. Зенкевич . . 367
КЛАСС УДОНЕЛЛИДЫ (UDONE-
LLOIDEA). Л. А. Зенкевич ... 368
КЛАСС МОНОГЕНОИДЕИ (MONO-
GENOIDEA). К. М. Рыжиков . . . —
КЛАСС ГИРОКОТИЛИДЫ (GYRO-
COTYLOIDEA). Л. А. Зенкевич 374
КЛАСС ЦЕСТОДЫ (CESTOIDEA).
К. М. Рыжиков, П. Г. Ошмарин 375
Отряд Лентецы (Pseudophyllidea) 378
Отряд Цепни (Cyclophyllidea) . . 382
ТИП НЁМАТЁЛЬМИНТЫ (NEMA-
THELMINTHES) 389
КЛАСС БРЮХОРЕСНИЧНЫЕ,
или ГАСТРОТРИХИ (GASTROT-
RICHA). А. А. Парамонов .... 391
КЛАСС НЕМАТОДЫ, или
КРУГЛЫЕ ЧЕРВИ (NEMATODA).
А. А. Парамонов 394
Подкласс Аденофореи (Adenopho-
геа) 400
Отряд Хромадориды (Chromado-
rida) 402
Отряд Эноплиды (Enoplida) . . . 403
Подкласс Сецерненты (Secern en tea) 410
Отряд Рабдитиды (Rhabditida) . . 414
Подотряд Стронгиляты (Strongi-
lata) 419
Отряд Тиленхиды (Tylenchidae) . . 422
КЛАСС КОЛОВРАТКИ (ROTATO-
RIA). А. С. Богословский 428
КЛАСС КИНОРИНХИ (KINOR-
HYNCHA). А. А. Парамонов ... 449
КЛАСС ВОЛОСАТИКОВЫЕ (NE-
MATOMORPHA). А. А. Парамонов 451
КЛАСС СКРЕБНИ (ACANTHOCEPH-
ALA). А. А, Парамонов 452
КЛАСС ПРИАПУЛИДЫ (PR1APU-
L1DA). Л.А.Зенкевич 455
КЛАСС КАМПТОЗОА, или СГИ-
БАЮЩИЕСЯ (К AMPTOZOA).
Г. Г. Абрикосов . . • 457
ТИП НЁМЁРТИНЫ (NEMERTINI).
A. А. Парамонов 459
ВЫСШИЕ ЧЕРВИ 465
ТИП КОЛЬЧАТЫЕ ЧЕРВИ (AN-
NEUDES) 467
КЛАСС ПЕРВИЧНЫЕ КОЛЬЧЕ-
ЦЫ (ARCH 1ANNEL IDA).
B. А. Свешников 471
КЛАСС МНОГОЩЕТИНКОВЫЕ
КОЛЬЧЕЦЫ (РОЬУСНАЕТА).
В. А. Свешников 473
Подкласс Myzostomida 489
КЛАСС ОЛИГОХЕТЫ, или МА-
ЛОЩЕТИНКОВЫЕ КОЛЬЧЕЦЫ
(OL1GOCHAETA). И. И. Малевич 490
КЛАСС ПИЯВКИ (H1RUD1NEA)
Е. И. Лукин 509
Подкласс Древние пиявки (Archihi-
rudinea) 512
Подкласс Настоящие пиявки (Euhi-
rudinea) ... , 513
Отряд Хоботные пиявки (Rhyn-
chobdellae) —
Отряд Челюстные, или Бесхобот-
ные, пиявки (Gnathobdellea, или
Arhynchobdellea) 520
КЛАСС ЭХИУРИДЫ (ECH1UR01-
DEA). Л. А. Зенкевич 526
КЛАСС СИПУНКУЛИДЫ (S1PUN-
CUL01DEA). Л. А. Зенкевич 528
ТИП ФОРОНИДЫ (PHORONOI-
DEA). Г. Г. Абрикосов 531
ТИП МШАНКИ (BRYOZOA).
Г. Г. Абрикосов 533
ТИП IIЛЕЧЕНО! НЕ (BRACHIO-
PODA). Г. Г. Абрикосов 543
Указатель терминов 549
Указатель русских названий .... 560
Указатель латинских названий . . . 567
Именной указатель 576
жизнь животных 1
В шести томах ТОМ
Редактор Е. Н. ХУНЦКАРИЯ
Редактор карт О. В. ПАЩЕНКО
Переплет художника М. В. СЕРЕГИНА
Суперобложка художников:
В. И. ПРЕОБРАЖЕНСКОЙ и М. В. СЕРЕГИНА
Цветные и тоновые иллюстрации художников:
Н. Н. КОНДАКОВА, А. В. КОНДРАТЬЕВА,
Н. Г. КОРОБОВОЙ, Т. Н. НАРБЕКОВОЙ,
В. И. ПРЕОБРАЖЕНСКОЙ, О. Ф. ХЛУДОВОЙ.
Макет и художественное редактирование
М. Л. ФРАМА.
Технический редактор Н. Н. МАХОВА.
Корректоры: Н. А. МЯСНИКОВА и
В. С, АНТОНОВА
Составители указателя:
Э. С. КОРМОВСКАЯ, Т. Л. МУРОМЦЕВА
•
Сдано в набор 3/XII 1965 г. Подписано к печати
17/XI-1967 г. 84XI08Vie- Янонис № 1.
Печ. листов 60,9 C6,25)+вкл. 4,2 B,5). Учебн.-изд. л.
56,37 + вкл. 4,97. А 14523. Тираж 300 тыс. экз.
Издательство «Просвещение» Комитета по печати
при Совете Министров РСФСР
Москва, 3-й проезд Марьиной рощи, 41
Заказ № 830
Московская типография №2 Главполиграфпрома Коми-
тета по печати при Совете Министров СССР. Москва,
проспект Мира, 105
Цена 3 р. 60 к.
U
59
Ж 71