/






























































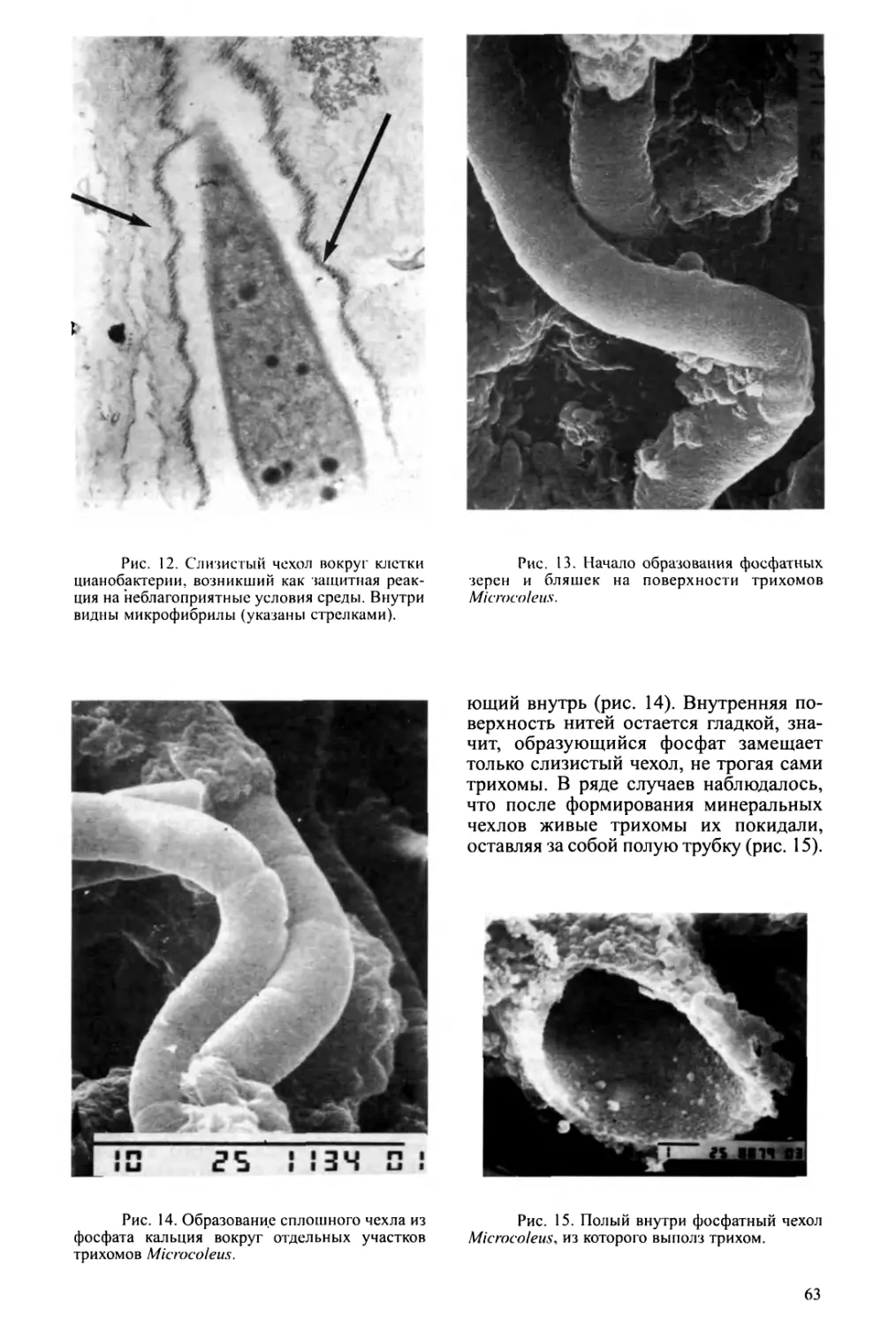

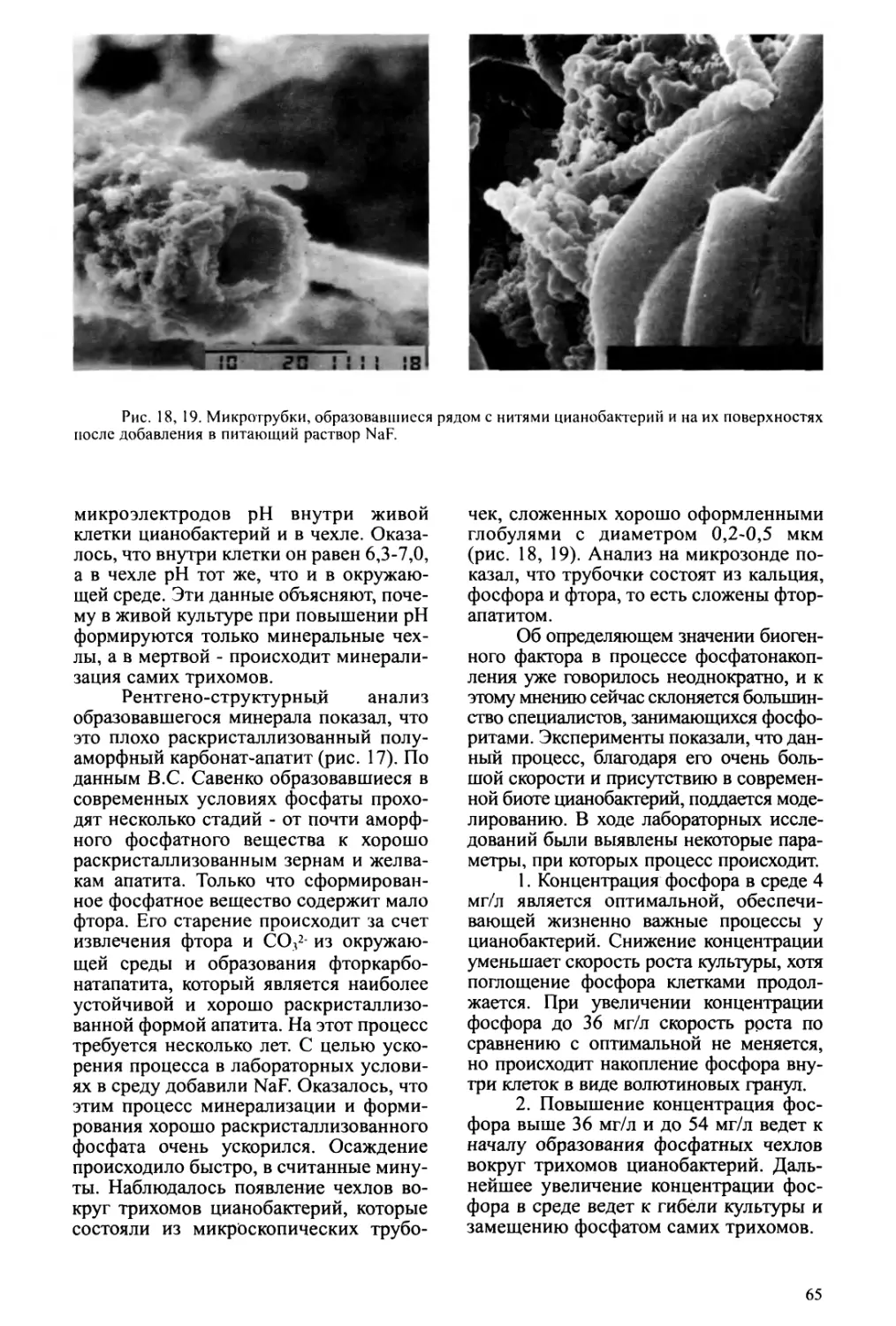













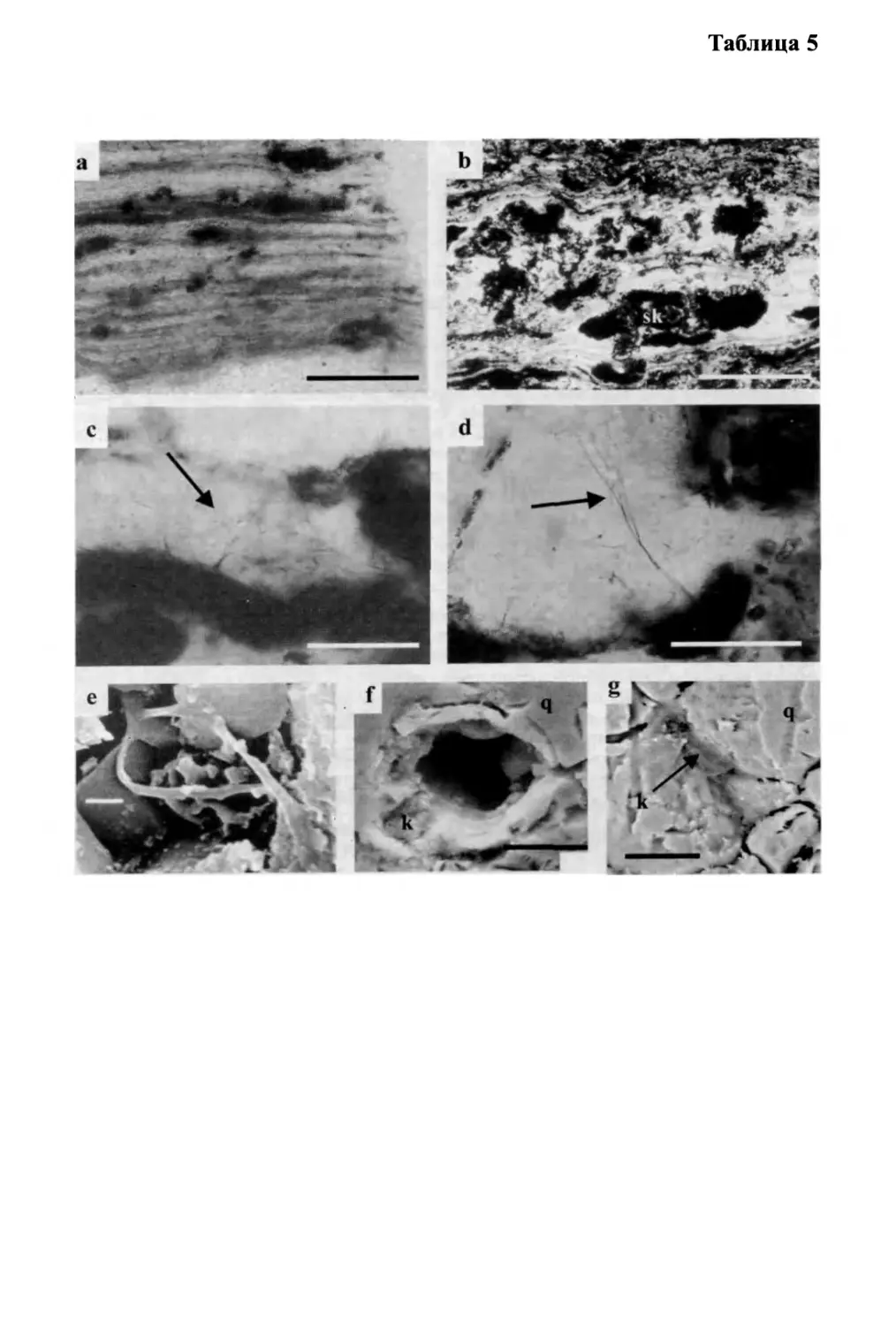





























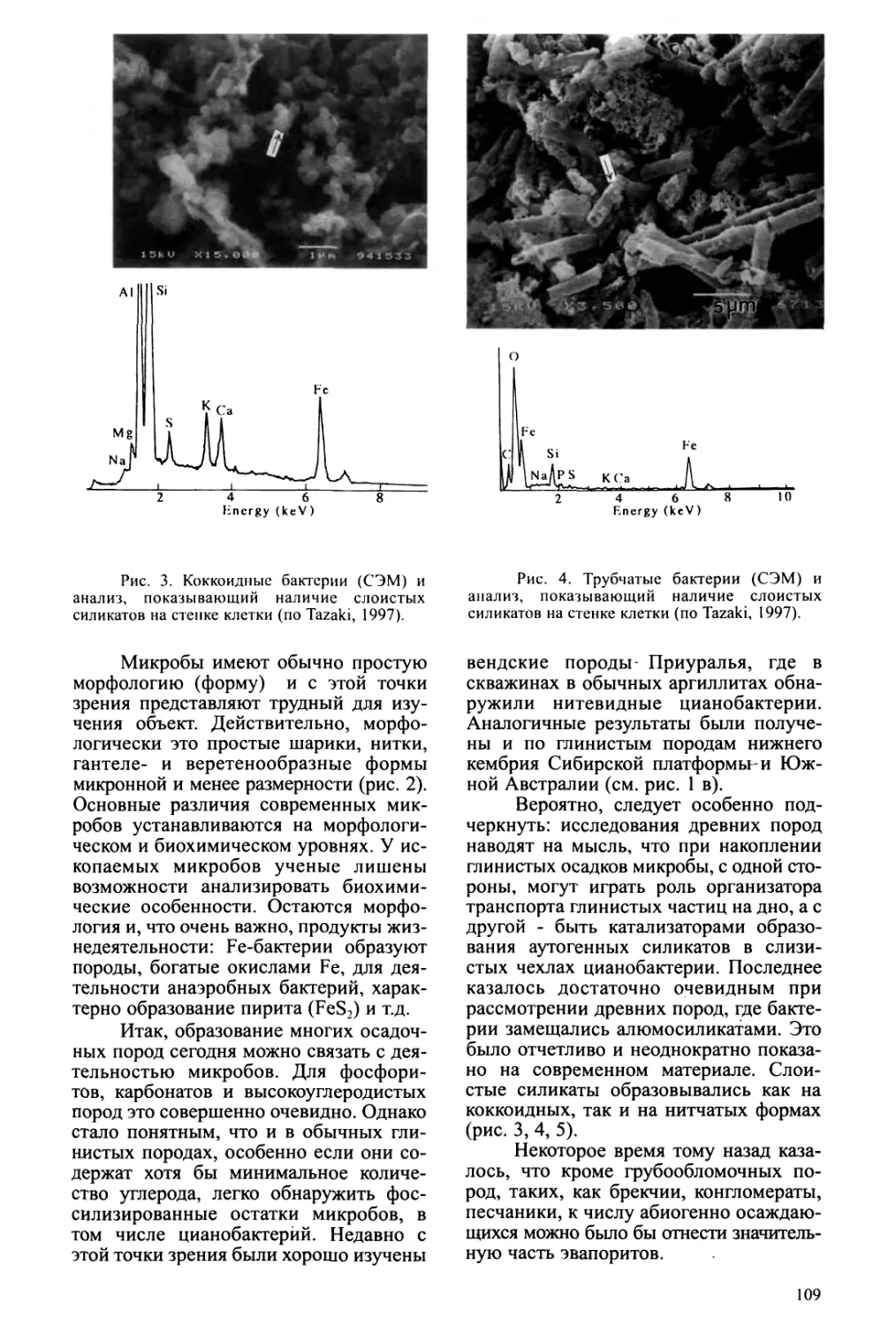



















































































Text
РОССИЙСКАЯ АКАДЕМ ИЯ НАУК ИАЛЕОИТОЛОГИЧЕСКИ Й ИНСТИТУТ МОСКОВСКИЙ ГОСУДАРСТВЕ ЫНЫЙ УН ИВЕРСИТЕТ КАФЕДРА ПАЛ ЕОНТОЛОГИИ РЕДАКТОР А.Ю. РОЗАНОВ С.С. Абызов М. Велш Ф. Вестал Е.А. Воробьева А. Р. Гептнер Л. М. Герасименко Д.А. Гилич инский E.À. Жегалло Г.А. Заварзин Д. Г. Звягинцев В.К. Орлеанский М.Е. Раабен А.IO. Розанов В.Н. Сергеев В.С. Соина Г.Т. Ушатинская Р. Хувер Э.Л. Школьник МОСКВА 2002 ПИН РАН
УДК 552.6 ISBN 5-201-15405-0 Бактериальная палеонтология. Российская академия наук, Палеонтологический институт; Московский государственный университет, Кафедра палеонтологии. Под ред. А.Ю. Розанова. — М.: ПИН РАН, 2002. — 188 с., илл. ISBN 5-201-15405-0. Пособие по бактериальной палеонтологии составлено из нового курса лекций, прочитанных в 1999-2000 гг. на кафедре палеонтологии МГУ. Книга написана для студентов и магистрантов высших учебных заведений — биологов, геологов, палеонтологов. � Коллектив авторов, 2002 � ПИН РАН, 2002 � МГУ, Кафе~да палеонтологии, 2002 ISBN 5-201-15405-0 Утверждено к печати Ученым советом Палеонтологического института РАН
СОДЕРЖАНИЕ ПРЕДИСЛОВИЕ (А.Ю. Розанов) ... 4 ВВЕДЕНИЕ (А.Ю. Розанов) ... 5 СОВРЕМЕННЫЕ БАКТЕРИИ И БАКТЕРИАЛЬНЫЕ СООБЩЕСТВА ПРОКАРИОТНАЯ КЛЕТКА КАК СИСТЕМА (Г.А. Заварзин) ... 6 ЦИАНОБАКТЕРИИ, ЦИАНО-БАКТЕРИАЛЬНЫЕ СООБЩЕСТВА, МАТЫ, БИОПЛЕНКИ (Л.М. Герасименко, Г.Т. Ушатинская) ... 36 ЦИАНО-БАКТЕРИАЛЬНЫЕ МАТЫ - АНАЛОГИ ИСКОПАЕМЫХ СТРОМАТОЛИТОВ (ЛАБОРАТОРНОЕ МОДЕЛИРОВАНИЕ) (В.К. Орлеанский) ... 47 СТРОМАТОЛИТЫ (М.Е. Раабен) ... 52 ЭКСПЕРИМЕНТЫ ПО ФОССИЛИЗАЦИИ ФОСФАТИЗАЦИЯ (Л.М. Герасименко, Г.Т. Ушатинская) ... 59 ПРОЦЕСС ОКРЕМНЕНИЯ (Г.Т. Ушатинская) ... 66 ИСКОПАЕМЫЕ БАКТЕРИИ И БАКТЕРИАЛЬНЫЕ БИОПЛЕНКИ (Ф. Весталл, М. Велш) ... 68 НЕКОТОРЫЕ ИСКОПАЕМЫЕ МИКРОБНЫЕ СООБЩЕСТВА РАННЕАРХЕЙСКИЕ ИСКОПАЕМЫЕ БАКТЕРИИ (Ф. Весталл, М. Велш) ... 84 ОКРЕМНЕННЫЕ МИКРОФОССИЛИИ (В.Н. Сергеев) ... 91 ФОСФОРИТЫ (Е.А. Жегалло, А.Ю. Розанов, Г.Т. Ушатинская) .;. 97 5OKCHTbt (Е.А. Жегалло, А.Ю. Розанов, 3.Л. Школьник) ... 102 ИСКОПАЕМЫЕ БАКТЕРИИ, ОСАДКООБРАЗОВАНИЕ И РУДОГЕНЕЗ (А.Ю. Розанов) ... 107 ЦИАНО-БАКТЕРИАЛЬНЫЕ МАТЫ - ИСТОЧНИК МАТЕРИНСКОГО ВЕЩЕСТВА НЕФТИ (А.Ю. Розанов) ... 114 ШУНГИТЫ - МОДЕЛЬНЫЙ ОБЪЕКТ ДЛЯ ИЗУЧЕНИЯ ГЛОБУЛИЗАЦИИ (А.Ю. Розанов„Г.Т. Ушатинская„Е.А. Жегалло) ... 116 МИНЕРАЛИЗОВАННЫЕ СВИДЕТЕЛИ СУЩЕСТВОВАНИЯ ГИДРОТЕРМАЛЬНОЙ ЭНДОБИОСФЕРЫ (А.Р. Гептнер) ... 123 БАКТЕРИАЛЬНАЯ ПАЛЕОНТОЛОГИЯ И АСТРОБИОЛОГИЯ ИССЛЕДОВАНИЕ ВНЕЗЕМНОГО ВЕЩЕСТВА (А.Ю. Розанов, Р. Хувер, Е.А. Жегалло) ... 136 ЖИЗНЕСПОСОБНЫЕ ЭКОСИСТЕМ Ы КРИОЛ ИТОСФЕРЫ (Е.А. Воробьева, В.С. Соина, Д.Г. Звягинцев, Д.А. Гиличинский) ... 155 ДЛИТЕЛЬНЫЙ АНАБИОЗ МИКРООРГАНИЗМОВ В ЛЕДНИКЕ ЦЕНТРАЛЬНОЙ АНТАРКТИДЫ (С.С. Абызов) ... 169 РАЗВИТИЕ ЖИЗНИ НА ЗЕМЛЕ (Г.А. Заварзин) ... 179 ЛИТЕРАТУРА ... 183 СЛОВАРЬ ТЕРМИНОВ .... 186
ПРЕДИСЛОВИЕ А.IO; Розанов Бактериальная палеонтология стремительно ворвалась в науку последние несколько лет и сегодня весь мир одержим исследованиями иско- паемых микроорганизмов. Бактериальная палеонтология рождалась в муках и долгое время не получала должного признания и разви- тия. Мне представляется, что это происходило лишь потому, что техни- ческие возможности исследователеи были ограничены, а самые прозорли- вые догадки не "возбуждали" научную общественность, поскольку она хотела "видеть и потрогать" ископаемые бак- терии. В этой обстановке могучий авто- ритет некоторых ученых, скептически оценивавших возможную деятельность микробов в формировании пород и, особенно, осадочных полезных иско- паемых, сыграл роковую роль и задер- жал развитие бактериальной палеон- тологии на несколько десятилетии. Тем, кто заинтересуется историей становления бактериальной палеонто- логии, очень рекомендую посмотреть работы первой половины ХХ века и прочесть статью А.Г. Вологдина, опубликованную a ! 947 г. Удивительно, что против призна- ния серьезной роли микроорганизмов в формировании пород и осадочных по- лезных ископаемых выступили единым фронтом идейные противники по ос- новным проблемам седиментологии. Настоящее пособие составлено из коротких очерков на темы тех лекций, которые были прочитаны в 1999-2000 гг. моими коллегами и мною с дополнением разделов, написанных ГА. Заварзиным (см. "Содержание" ) и Ф. Весталл и М. Велш (F. Westall, М. Walsh). Я счел необходимым хотя бы в таком виде дать возможность студентам и магистрантам знакомиться с ре- зультатами изучения ископаемых бактерий. Мне представляется, что наравне с другими разделами пале- онтологии, бактериальная палеонтоло- гия должна постепенно стать пред- метом специальной подготовки студен- тов. Особенно важно и интересно, что бактериальная палеонтология в значи- тельной мере зародилась в России. Составители разделов указаны в "Содержании". Понимая, что этот первый опыт создания проекта учебного пособия по бактериальной палеонтологии может оказаться не самым удачным, авторы примут с благодарностью все замечания, даже самые строгие. Авторы отдают себе отчет в том, что всего через несколько лет многое из изложенного здесь устареет, поскольку бактериальная палеонтология и смежные дисциплины развиваются очень стремительно. Но необходимость даже такого пособия для нас совер- шенно очевидна. Я благодарен первым слушателям этого курса — студентам и магистрантам Д. Барановой, И. Зюгановой, С. Рож- нову, Ю. Шуваловой и др., искренняя заинтересованность которых подтолк- нула меня на создание этого пособия, а также слушателям второй волны, сде- лавшим ценные замечания к рукописи: В. Зейфас, И. Марданян, А. Канюкину, Е. Кирилишиной и др.
ВВЕДЕНИЕ Задачей бактериальной палеонто- логии является изучение ископаемых микробов. Традиционная палеонтология изучает остатки ископаемых организмов, систематизируя их по морфологическим признакам с учетом филогенетических взаимоотношений. Основными "пользо- вателями" палеонтологии являются эво- люционная тео ия, история органичес- кого мира и иостратиграфия. В по- следнее время палеонтология дала не- оценимые данные для изучения эволю- ции биосферы. Значителен вклад пале- онтологии и в палеогеографию. Бактериальная палеонтология ограничена в своих исходных данных очень простой морфологией объектов и спецификой систематизации бактери- ального материала (прежде всего совре- менного). Поэтому для систематизации ископаемых объектов мор ология важ- на, но недостаточна. Осо ое значение приобретают продукты жизнедеятель- ности бактерий, выраженные в литоло- гических и геохимических особенно- стях пород. Важнейшее значение имеет изотопия углерода и строение различ- ных биомаркеров. Резко различаются и основные сферы применения "традиционной" и бактериальной палеонтологии. Бактери- альная палеонтология необходима преж- де всего седиментологии и, соответст- венно, при изучении генезиса осадочных полезных ископаемых, включая нефть и газ. Важна роль бактериальной палеон- тологии в палеогеографии, при изуче- нии эпиконтинентальных бассейнов. Огромное значение имеют бакте- риально-палеонтологические данные при обсуждении проблем эволюции биосферы, особенно в докембрии и ран- нем палеозое. И, наконец, бактериальная палеонтология - один из ключевых ас- пектов изучения астроматериалов. Значе- ние же бактериальной палеонтологии для стратиграфии пока не очень ясно. Для более четкого понимания, что такое бактериальная палеонтология, необходимо остановиться на понятии "геомикробиология". Основное поле деятельности гео- микробиолога - взаимоотношение пород и минералов с бактериями (точнее, микроорганизмами). И, естественно, главным объектом являются современ- ные о ганизмы. еомикробиологические исследо- вания совершенно необходимы не толь- ко для задач изучения современного воздействия микроорганизмов на геоло- гическую среду, но и как актуалистичес- кий материал для интерпретации иско- паемых объектов. Геомикробиологические исследо- вания особенно успешно показали: 1) Что микроо ганизмы активно воздействуют на все ез исключения ми- нералы, образуя на их поверхности био- пленки. Именно микроорганизмы явля- ются неотъемлемой частью процессов выве ивания в широком смысле. ) Что многие микроорганизмы накапливают некоторые ионы металлов. 3) Что микроорганизмы формиру- ют целый ряд минералов, которые явля- ются основой многих осадочных пород, включая доломит, слоистые силикаты и т.д., не говоря уже о карбонате кальция и фосфатах. Бактериальная палеонтология и геомикробиология имеют некоторое об- щее "поле деятельности". Несомненно, часть работ, проводимых в рамках гео- микробиологических исследований, яв- ляется по существу бакте иально-пале- онтологическими и нао орот. Но это совершенно нормально для погранич- ных наук, так же как нормальны подоб- ные взаимоотношения между палеон- тологиеи и неоонтологиеи Нужно иметь в виду чрезвычай- ную молодость бактериальной палеон- тологии. Таким образом, если цель бак- териальной палеонтологии, сформули- рованная в общем виде, достаточно ясна, то трудности бактериально-пале- онтологических исследовании еще со- вершенно не обрисованы. Однако ясно одно, образование сту- дентов-палеонтологов, седиментологов и "полезников" должно быть дополнено хотя бы общими знаниями об ископае- мьп~ микроорганизмах. Напомним, что еще несколько лет назад не было и речи о том, что бактерии хорошо сохраняются в ископаемом - состоянии и их можно специально и обстоятельно изучать.
СОВРЕМЕННЫЕ БАКТЕРИИИ БАКТЕРИАЛЬНЫЕ СООБЩЕСТВА ПРОКАРИОТНАЯ КЛЕТКА КАК СЙС ТЕМА РОСТ И РАЗМНОЖЕНИЕ БАКТЕРИЙ Под понятием роста подразумева- ется увеличение биомассы независимо от того, происходит ли при этом увели- чение численности организмов или нет. Типичным примером роста служит рост мицелия гриба, происходящего из одной единственной споры. Для одноклеточ- ных организмов рост и размножение приблизительно совпадают и означают увеличение численности. Поэтому в бактериологии обычно пользуются сло- вом рост, хотя это и неточно. Размно- жение бактерий происходит путем удвоения. Прокариотная клетка может рас- сматриваться как система, сложенная четырьмя подсистемами: (1) геномом- хромосомой с аппаратом репликации; (2) аппаратом синтеза белка (рибосо- мой); (3) цитоплазмой, включающей сеть метаболических путей с обслужи- вающими их ферментами; (4) мембра- ной с энергодающим аппаратом синтеза АТФ и транспортными системами, осу- ществляющими взаимодействие клетки с внеклеточной средой (рис. 1). 1. Хромосома содержит генетичес- кую информацию для собственной ре- пликации и информацию синтеза белка РНК-полимеразным блоком. Геном бак- терии постоянен для вида и может быть оценен анализом ДНК. Важным итогом изучения генетической информации бактерий в 1990-х гг. было установление последовахельностеи основании, соот- ветствующих отдельным генам. 2. Рибосома является РНК-содер- жащим аппаратом синтеза белка из ами- нокислот-предшественников, поступаю- щих из цитоплазмы, и расходует энер- гию, поступающую от энергодающего мембранного аппарата. 3. Цитоплазма представляет собою (1) "котел" метаболических превраще- ний поступающих извне веществ с обра- зованием энергетических субстратов для АТФ-продуцирующего блока (ката- болизм) и соединений-предшественни- ков для синтеза комлонентов клетки (анаболизм), и (2) собственно, место действия ферментов. Взаимодействие , между подсистемами клетки ведет к то- му, что ни одна из них не способна к самостоятельному существованию. 4. Энергодающая мембрана осуще- ствляет электрохимическое преобразо- вание энергии во внутреннюю энергию мембраны и синтез АТФ. В мембране расположен механизм трансмембранно- го переноса веществ: субстратов внутрь клетки и продуктов наружу. Для более подробного ознаком- ления с работой бактериальной клетки и ее подсистем следует обратиться к учеб- ным пособиям по общей микробиоло- гии, Шлегель Г. "Общая микробиоло- гия" (1987), Готтшалк Г. "Метаболизм бактерий" (1982), Скулачев В.П. "Биоэ- нергетика. Мембранные преобразовате- ли энергии" (1989), Спирин А.С. "Моле- кулярная биология. Структура рибосо- мы и синтез белка" (1986). Особенно рекомендуются доступные обзорные статьи А.С. Спирина, В.П. Скулачева, А.Д. Виноградова, О.Н. Кулаевой и ряда других авторов в Соросовском общеоб- разователномм журнале (1998-2000).
Рис. (. Схема строения прокариотической клетки (бактериальная клетка в продольном разре- зе). Гли - гранулы гилкогена; Ж - жгутик; Кпс — капсула; КСт - клеточная стенка; Ли - липидные ка- пельки; ПГМ - поли+гидроксимасляная кислота; Пи — пили; Плз - плазматическая мембрана; ПФ- гранулы полифосфата; Ри - рибосомы и полисомы; Я — ядро (нуклеоид); S - включения серы; ЦПл- цито плазма. Клеточный цикл Клеточный цикл размножения бак- терий осуществляется путем бинарного деления. При этом все компоненты клет- ки распределяются между сестрински- ми клетками поровну. Исключение со- ставляет хромосома, которая находится одна в каждой клетке. Репликация хро- мосомы начинается с образования двух вилок, на которых идет синтез навстре- чу друг другу комплементарных нитей. Скорость процесса составляет тысячу пар оснований в секунду и для хромосо- мы длиной в 1 мм занимает примерно 40 мин. При более быстром размножении на хромосоме возникает несколько ви- лок. Образование перегородки между клетками и их разъединение начинается после завершения расхождения хромо- сом и требует еще около 20 мин. Вариации в процессе разъединения приводят к образованию разного вида агрегатов клеток в виде цепочек, нитей, трихомов, мицелия, объединений кокковидных клеток. КЛАССИФИКАЦИЯ Современная система бактерий построена на основе последовательно- сти нуклеотидов в гене 16S рибосомной РНК. Из последовательности нуклеоти- дов выведено дерево филогении бакте- рий или прокариот, важнейшей особен- ностью которого является разделение живых организмов на три крупные ветви собственно бактерий (Eubacteria), архебактерий или архей (Archaeobac- teria) и эукариот (Eukarya). Порядок ветвления в классификационном дереве рибосомального гена принимается за порядок происхождения групп организ- мов. В настоящее время имеются последовательности всех культивируе- мых бактерий и многих некультиви- руемых, часто представляющих массо- вые формы, как, например, планктонные археи открытого океана. Эта молекуляр- ная систематика сменила существовав- шую ранее и основанную на чистой жи вой культуре микроорганизмов в признанной коллекции культур, где типовые штаммы видов бактерий сохра- няются в неразмножающемся состоянии либо лиофильно высушенные, либо в жидком азоте. Прямые определения выживаемости микробов в вечной мерз- лоте показывают, что они могут сохра- няться в таком виде десятки тысяч лет, а отдельные находки имеют возраст до миллиона лет. Систематика, основанная на моле- кулярных характеристиках гена рибосо- мальной РНК, очень мало может дать для понимания функциональной роли организма и, тем более, его морфологии. Корреляция признаков, имеющих фено- типическое значение, с положением на генеалогическом дереве рибосомально- го гена невелика, и устанавливается для немногочисленных и однородных групп
как цианобактерии. Однако и для них нет корреляции филогенетического по- ложения с морфологией. Геологи вынуждены в большей степени ориентироваться на практичес- кую классификацию, выработанную за прошедшее столетие и применяемую для понимания функциональной роли бактерий в природе и прежде всего для биогеохимических циклов. Палеонтоло- ги нуждаются в еще более ранней клас- сификации, основанной на морфологии бактерий, которая крайне однообразна, а в ископаемом материале дает представ- ление лишь об общей морфологии кле- ток и их агрегатов. Функциональное разнообразиемикроорганизмов Описание групп бактерий по их функционально значимым характери- стикам было очень удобно для практи- ческих работников. Названия родов бактерий соответствовали их функции- например, Cytophaga - пожиратель кле- точных стенок (целлюлозы). В основу было положено представление о роде бактерий как морфо-физиологическом единстве. Далее роды объединялись в более крупные группировки, например, азотфиксаторов, способных ассимили- ровать азот атмосферы. Важным таксо- номическим признаком, с которым кор- релируют другие свойства бактерий, является способность или, наоборот, не- способность их клеточной стенки окра- шиваться при использовании метода, предложенного в 1884 г. Грамом. Грам- положительные клетки после опреде- ленной обработки красителем имеют темно-фиолетовый, а грамотрицатель- ные обесцвечиваются. Природоведчес- кая микробиология основывается преж- де всего на комплексе таких функцио- нальных характеристик микроорганиз- мов. Поэтому были созданы руководст- ва, основанные на этом принципе и но- сящие практический характер, с не имеющими таксономического значения условными группировками, и книги, посвященные, например, анаэробным микроорганизмам, хемоавтотрофам, пропионовым бактериям, фотосинте- зирующим бактериям, термофилам, представляющие давнюю и очень удобную традицию в микробиологии. До появления филогенетической си- стемы бактерий (см. ниже) была сделана попытка систематизировать этот практический подход. ОСНОВНЫЕ ГРУППЫ БАКТЕРИЙ(ПО РУКОВОДСТВУ БЕРДЖИ,9-Е ИЗДАНИЕ) Ознакомление с биоразнообразием бактерий означает тяжелое упражнение для памяти: множество усваивается на память, а система - по логике. Впрочем, кажется, И. Канту принадлежит утверж- дение: "мы знаем то, что помним". Попытка создать морфо-физиологичес- кую классификацию, наиболее удобную для практических работников, нуждаю- щихся в быстрой идентификации ор- ганизмов, была предпринята американс- ким Комитетом Берджи по систематике бактерий. Она привела к объединению организмов в искусственные группы (19-35 в разных изданиях), выделенные в не имеющие таксономического статуса "секции", в тривиальном словоупотреб- лении часто обозначаемые по наиболее характерному роду или названию, выде- ленному подчеркиванием. На русский язык переведены 8-е и 9-е издания "Краткого определителя Берджи", где бактерии были разбиты на группы, или секции, с задачей их определения по фенотипическим признакам. Перечис- ленные группы были созданы не по единой схеме и, например, одну группу составили изогнутые бактерии, а дру- гую — хемолитотрофы. На английском существуют соответствующие 4-хтом- ные "Bergey's Manual of Determinative Bacteriology" с подробными описаниями видов. Сейчас выходит 10-е издание. Их дополняют 2-х и 4-хтоиные издания "The Prokaryotes", где приводятся и ме- тоды выделения, и культивирования, вместе с характеристиками групп. Кроме того, публикуются сводки, посвя- щенные отдельным группам бактерий, составляющие важную часть литерату- ры по общей микробиологии. Для естествоиспытателя необходи- мо знать функциональную групповую классифкацию, так как именно она поз- воляет анализировать концептуальные связи в сложных природных системах. Попытка определить бактерии по мор- фологии совершенно безнадежна. Есть несколько распознаваемых форм клеток и их объединений: 1) трихом как у циа- нобактери, 2) мицелий ветвящийся у актиномицетов и нити у скользящих флексибактерий, 3) одиночные палочки,
вибрионы, спириллы, которые могут быть собраны в цепочки, 4) кокки, 5) почкующиеся, 6) с выростами-просте- ками и стебельками. Чтобы ориентиро- ваться в микробиологической литерату- ре, эти группы и их основных предста- вителей необходимо знать (таблица 1). Спирохеты (группа I) ° К ним относятся грам-отрицательные бактерии со своеобразным строением клетки: протоплазматический цилиндр навер- нут на центральную ось из жгутиков, и в результате клетка имеет вид длинной гибкой подвижной спирали, способной сворачиваться и развертываться. Среди них имеются свободноживущие анаэ- робные брод ил ьщики-диссипотрофы Spirochaeta, аэробные водные Leptospira с характерным крючком на конце, пара- зитические Treponema, Borrelia, Cristi- spira. Свободноживущие Spirochaeta осуществляют функции сахаролитичес- ких диссипотрофов в анаэробном цел- люлозоразлагающем сообществе. Этот род представляет интересный пример адаптации к разным физическим усло- виям: S. thermophila приспособилась к морским гидротермам, а S. caldaria — к континентальным; в метантенках рас- пространена факультативная S. aurantia; в море S. zuelzerae; в высокоминерализо- ванных крайне щелочных водоемах — S. alkalica, S. аЯсаиа, а в низкоминерали- зованных - S. asiatica. Вместе с тем спи- рохеты представляют не только легко распознаваемую морфологическую груп- пу, но и филогенетически определен- ную отдельную ветвь. Под микроско- пом со спирохетами можно спутать не- которые организмы, образующие тугие спирали (их называют ...-spira), но не имеющие аксиальных жгутиков. Изогнутые палочки и спириллы (группа 11). В эту группу попали спи- риллы с полярным жгутиком, морские Oceanospirillum и пресноводные Aqua- spirillum. Это типично водные органо- трофные организмы, приуроченные к условиям пониженного содержания ки- слорода и развивающиеся в зоне окси- клина в водоемах. Морфологически от- носящиеся к спириллам литотрофные организмы рассматриваются в других группах. Бделловибрионы составляют довольно разнообразную группу микро- организмов, наблюдаемую в разных вод- ных местообитаниях. Bdellovibrio пред- ставляет очень мелкии подвижныи ви- брион, который атакует грам-отрица- тельные бактерии, располагается у них в периплазматическом пространстве, не проникая под цитоплазматическую мем- брану, и размножается здесь, лизируя клетку хозяина. Неподвижные изогнутые палоч- ки - "микроциклусы" (группа III) ° Это грамотрицательные аэробные орга- нотрофы, изогнутые в виде кренделя и легко опознаваемые микроскопически в планктоне водоемов по своей морфоло- гии, но отнюдь не обычные обитатели лабораторных культур. К ним относится собственно микроциклус, сейчас полу- чивший название Ancyclobacter, В~асЬ~- arcus, "Renobacter", Tetrarcus — преиму- щественно олиготрофные планктонные организмы. Грамотрицательные аэробные палочки и кокки (группа IV) состав- ляют группу наиболее массовых пред- ставителей лабораторных культур, полу- чивших суммарное название "псевдомо- нады" по форме клеток с полярным жгу- тиком. Они представляют массу копио- трофов-колонизаторов, быстро развива- ющихся на богатых средах. Многие из них способны использовать в качестве единственного источника углерода и энергии разнообразные (до сотни) орга- нические вещества. Эта их особенность приводит к тому, что в накопительных культурах, особенно на плотных средах, они вырастают в первую очередь. Ти- пичным родом служит Pseudomonas, сеичас дополненныи множеством отли- чающихся от него фенотипически и фи- логенетически родов. Близок к нему Al- caligenes - палочка с редкими перитри- хиально расположенными жгутиками. В море типичной псевдомонадой является Alteromonas. Считается, что псевдомо- надам не свойственно брожение. Ис- ключение составляет Zymomonas. Среди псевдомонад выделяются несколько физиологически обособлен- ных групп. Одну из них составляют симбиотические азотфиксаторы ризо- бии с типовым родом Rhizobium. Другую - свободноживущие азотфикса- торы азотобактеры, к которым относят- ся Azotobacter, Azomonas, Beji'erinckia, но не Azospirillum. Азотфиксаторы состав- ляют важнейшую для биогеохимичес- кой машины планеты функциональную группировку микроорганизмов. Другую группу псевдомонад пред- ставляют ацетобактерии, аэробно про- дуцирующие ацетат из этанола и спо- собные развиваться в кислой среде. Типичным представителем их является
° ° ° ° ° ° ° ° ° ° ° ° X Ъ'. Б О CL Ф З Я CL LA1СОС7 CV1СОCD CV V 1 Ж СЭ О О СЧ~ О СЧ LO С) О Щ Б О2а ? ? Ф~» О О ЛОБ а о Ж Ф ? о ? О Ц; Ф ? О с О Ш Ф И Х Я О K Ю О О О О Ф Я 3 О C Я Ш 3 О с Ш CL Ф О G3 ~С О М Я 6l 3 О с Я О М Я Ш 3 О С Я С~ О Ш Я 6) 3 О с Ф CL О й Я М 3 О Ю Ф CL О З Я М 3 О C С K Щ О 6l З О Ф О T Ф С Ф Ф Tl I- Я K Ф Ю О С Ю О I- X Г) .0 1- Ф :У И З С; Х K Ю О 0 О > С„' И :::Г О CL hC K Б О Ф 3' Q 3 О CL О I- Я 1» K :? A О С: lU М О Ф K О О CL 03 О I- Ф Х О Q. K с О Я с Г ~=Г О C О С„' С И CL И с О Ф 0 Q2 О Х О Х 1- 0 Х О K CL Ю K СП |D :У О (D 2 I- Щ :Г О K О О О LA 1 ~ац I О О>с~гО QО ОО щОО сФ1-3ЖCLЯ О Ю 1 I L0 C) О Ф О ? О (91 Ф О 0 K д- С9 С9 Я Я с О О .Е Ж CL О Я С„' с Ф О Z' cf О Ш Ф И О 0 а ш О:Г О Щ о r О Ф Ф О О CL Щ 0 ? L Ц; О Ш ОЯ Щ2CL -)ХО ОCLС9Я K hC Ч. Я CL О )Я O ? 3 Ф 3 K hC Ф K 5С О CL О О а О C О О Ф Ф 0 с r О Щ ц 2 ж Х О ц О О 6l .'Е hC ~Г О Ф:у - Я > 0 CL О :. Ф K Ф 2 Х Ю О CL С9 Ж Х < "' сч С) 1 1 с~Э сО О О Х K -~ 61 Ы О О Cl (т) — K О 0 Я Я CL О Я с S K О О (D '3 m ~Д Ф О Х CL Ф О Ф .0 Х CL > с CL [: Щ 2 X CL |D О Ф Х Ф A Х CL > с CL ~ъ .0 х Ф С„' Ф K CL (D Ж О Ф О K И Ю О С) О CL I- О 1- С„' О 2 Ф Х Ф X Ю О CL .0 CL О О CQ hC CL О :? Ф CL О З О О а О с Ф О Х т О Я О О CQ С~ О 6) .0 ~Х О CL О C Щ .0 Х .Т О С~ Я О О Я ~С О 61 2 ~:К О а О С. Щ 0 X 3 О ~=Г Я О О Я Ц; О tQ З K Х Ф О С С О Ф Б 0 Ч. О а О C' Щ 0 :д 3 О ~Х Я О О Я О О) 2 Ч. О а О с Щ A Х 3 О С~ Я О О Я сЕ О 6l Ф K О Ф 3 K I- О И х Я Б K K Ю О hC О О
род Acetobacter. В высокоминерализо- ванных соленых водах развивается аэробный органотрофный Halomonas (с которым не следует путать экстре- мально галофильные архебактерии Halobacteriaceae). В термальных источ- никах и компостах характерной псевдо- монадой является аэробный органо- трофный Thermus, некоторые виды которого ярко пигментированы. Неподвижные овальные "коккоба- циллы" Paracoccus послужили приме- ром широкой метаболической приспо- собляемости к разным типам питания. Этот организм использует аэробно в качестве единственного источника угле- рода и энергии разнообразные низкомо- лекулярные вещества — сахара и орга- нические кислоты. Кроме того, в анаэ- робных условиях в присутствии нитрата он способен к денитрификации, а в при- сутствии Н, может расти как хемолито- автотроф, окисляя водород и усваивая углекислоту. Цепь переноса электрона у паракокка оказалась сходной с мито- хондриальной и поэтому допускают, что организм, подобный ему, был когда-то предшественником митохондрий. Среди псевдомонад много орга- низмов с подобным паракокку набором способностей, особо нужно отметить водородные бактерии, раньше объеди- ненные в род "Hydrogenomonas", а те- перь распределенные по разным родам, в частности, род Acidovorax — организ- мы, использующие Н, или органические кислоты (см. ниже Водородные бакте- рии). К псевдомонадам следовало бы отнести и тиобациллы, окисляющие соединения серы. К псевдомонадам отнесены и резко отличающиеся от них морфологи- чески и по типу обмена метанотрофы, представляющие самостоятельную группу организмов и рассматриваемые отдельно (см. ниже). Факультативно-аиюробиые грам- отрицательные палочки (группа V) объединяют энтеробактерии с централь- ным представителем Escherichia со1~. Количество работ по выяснению уни- версальных механизмов бактерий, вы- полненных на Е. coli, превратило этот организм в символ бактерий вообще. Энтеробактерии представляют со- бой органотрофные факультативно- аэробные организмы, способные к окис- лению в аэробных условиях и брожению в анаэробных. Большинство их относит- ся к сахаролитическим организмам. (2 Среди энтеробактерий следует выделить азотфиксирующую свободноживущую Klebsi ella. Азотистые вещества разлага- ют гнилостные бактерии, например, Proteus. Другую группу здесь представля- ют вибрионы, изогнутые подвижные грам-отрицательные палочки с типовым родом Vibrio, имеющие преимуществен- но бродильный тип метаболизма. В группу энтеробактерий входят также светящиеся бактерии рода Photobacte- rium. К энтеробактериям относятся мно- гие патогенные организмы, особенно обитатели кишечного тракта, хотя они там и не составляют основную массу бактерий. Бактероиды (группа VI) состав- ляют группу грамотрицательных не- споровых облигатно анаэробных орга- нотрофных бактерий, осуществляющих брожение. Типовым родом среди них является Bacteroides, массовый обита- тель кишечного тракта. Бактероиды представляют группу типичных первич- ных анаэробов, сбраживающих углево- ды в органические кислоты. Сюда от- носятся также Fusobacterium, Succinivi- brio, Selenomonas, изогнутые клетки которого имеют мощный пучок жгути- ков на вогнутой стороне. Бактероиды вместе с цитофагами и флавобакте- риями образуют совершенно самостоя- тельную филогенетическую ветвь. Сульфатредуцирующие и серо- редуцирующие бактерии (группа VII) представляют довольно компактную группировку с типовым родом Desul- fovibrio, открытым М. Бейеринком и долгое время остававшимся единствен- ным описанным представителем суль- фатредуцирующих бактерий (СРБ). Это облигатно анаэробный организм, обла- дающий диссимиляторным путем вос- становления сульфата с образованием Н S. Он использует крайне ограничен- ное число субстратов, в первую очередь лактат, этанол, с неполным окислением их в ацетат. Сульфатредукторы способ- ны к окислению Н, и в этом случае осу- ществляют литогетеротрофный рост. Комбинация субстратов: Н, или форми- ат, лактат, этанол наиболее обычна для большинства СРБ. Анаболизм их осно- ван на использовании ацетата с после- дующим карбоксилированием в трех- углеродное соединение. Ацетат как суб- страт катаболизма использует Desul- /оЬас~ег - прямая палочка с заметно бо- лее медленным ростом, чем Desulfovi-
brio. Открытие сульфатредуцирующих организмов, способных осуществлять полное окисление органических кислот, было сделано Ф. Видделем (F. Widdel), обнаружившим целый ряд морфоло- гически разнообразных сульфатредук- торов (Desulfosercina, Desulfonema и др.). Соответственно сульфатредуциру- ющие бактерии разделяются на под- группу 1 с неполным окислением суб- страта (ацетогенов) и подгруппу 11 с полным окислением. Число родов орга- низмов, относящихся к этой группе, ко- торая составляет самостоятельную фи- логенетическую ветвь, быстро возраста- ет. Например, были открыты экстре- мально галофильный Desulfomicrobium, экстремально алкалофильные гидроге- нотрофные Desul~înat опотЬгiо и Desul- fonatronum. Все перечисленные роды относятся к Proteobacteria. Грамположительные споровые клостридии Desulfotomaculum представ- ляют другую группу сульфатредукторов с далеко не таким жестким типом обме- на. Среди них есть и умеренно термо- фильные виды. Экстремально термофиль- ная сульфатредуцирующая Thermodesul- fobacterium, в массовом количестве развивающаяся в гидротермах, не вхо- дит в те же филогенетические ветви, что перечисленные сульфатредукторы. В еще большей степени это относится к сульфатредуцирующей архебактерии Archaeoglobus. Сульфатредуцирующие бактерии представляют важнейшую группу вторичных анаэробов, осущест- вляющих удаление из анаэробной системы Н. В биогеохимическом отно- шении они входят в группу сульфидо- генов, осуществляющих образование H S и, таким образом, инициирующих серный цикл на современной Земле. В группу сульфидогенов входят также организмы, восстанавливающие молекулярную серу. К ним относятся "гидрогентиобактерии", описанные в 1930-х гг. А. Пельшем в соленых лагу- нах Сиваша и Кара-Богаз-Гола. Типич- ным примером среди сероредукторов считается открытый Н. Пфеннигом (N. Pfennig) ВезиЦитотопаз. Термофильная Desulfurella, окисляющая ацетат, входит в отдельный подкласс филогенети- ческой системы. В отличие от катабо- лической сульфатредукции сероредук- ция представляет довольно распро- страненное свойство среди разных групп анаэробов. К этой же группе при- надлежат и многие из магнетитобра- зующих анаэробов с характерным родом Geobacter. Анаэробные грамотрицатель- ные кокки (группа VIII) осуществля- ют брожение органических веществ и составляют отдельную группу вейло- нелл. Риккетсии и хламидии (группа IX) представляют внутриклеточных па- разитов эукариотных организмов и от- носятся к объектам медицинской мик- робиологии, в противоположность эндо- симбионтам, входящим в отдельную группу. Аноксигенные фототрофные бактерии (группа Х) четко разделяются на несколько групп по физиологии. П п ные се ные бакте ии (сем. Chromatiaceae) характеризуются зависи- мостью от Н S как донора-восстанови- теля и окисляют его на свету после- довательно в серу и сульфат. Им свойст- венно внутриклеточное запасание серы как субстрата окисления, что, наряду с окраской клеток, служит диагностичес- ким признаком группы. Относительно крупные пурпурные серобактерии с самого начала описания их систематики С.Н. Виноградским на основе микроско- пических наблюдений получили четкие морфологически обоснованные назва- ния. Важнейшие роды: Chromatium, Thiocystis, Thiospirillum, Thiocapsa, Thiodictyon, Amoebobacter, Thiopedia. Обитатель щелочных высокоминерали- зованных водоемов Ectothiorhodospira (сем. Ectothiorhodaceae) откладывает серу вне клетки. Несе ные п п ные бакте ии объединяют преимущественно фотоор- ганотрофных или Н-использующих анаэробных бактерий с широкими мета- болическими возможностями, разли- чающимися по морфологии. Наиболее изучены роды Rhodospirillum, Rhodo- pseudomonas, Rhodobacter. Некоторые виды, например, Rhodopseudomonas palustris, осуществляют неравномерное деление или почкование с четким раз- делением на материнскую и дочернюю клетки. Такая же грушевидная форма свойствена нитрифицирующему Nitro- bacter, метанокисляющему Methylosi- nus. В наибольшей степени почкование выражено у гифомикробов, где почка образуется на конце длинного выроста- "гифы" или простеки. Типичную форму гифомикроба имеет Rhodomicrobium. Вместо подвижных дочерних клеток он может образовывать устойчивые к высу- 13
шиванию треугольные экзоспоры. Фото- синтезирующий аппарат располагается в цитоплазматической и/или внутри- клеточных мембранах, различным обра- зом упакованных у разных родов пур- пурных серных и несерных бактерий, что позволяет различать их на ультра- тонких срезах клеток. Зеленые се ные бакте ии пред- ставляют совершенно отличную от пур- пурных бактерий группу строго анаэ- робных облигатно фототрофных орга- низмов, хотя также анаэробно окисляют сероводород на свету. Первым откры- тым представителем этой группы был Chlorobi um. Светособирающие пигмен- ты фотосинтетического аппарата лока- лизуются у этих бактерий в хлоросомах — эллипсовидных телах, прилежащих к мембране. Развиваясь при низкой интен- сивности света, зеленые бактерии уве- личивают улавливающую свет поверх- ность, образуя трехмерную сеть из кле- ток, как у Pelodictyon. Зеленые нитчатые бакте ии пред- ставлены прежде всего термофильным фотоорганотрофным Chloroflexus, пред- ставляющим обособленную группу с не- обычным путем ассимиляции углеки- слоты через оксипропионат и занимаю- щую совершенно самостоятельное фи- логенетическое положение. Будучи фа- культативным фототрофом и факуль- тативным анаэробом, Chloroflexus, по- добно пурпурным несерным бактериям, обладает широкими метаболическими возможностями. Другой изученный представитель этой группы, Oscillochlo- ris trichoides, напротив, является обли- гатным фототрофом и ассимилирует уг- лекислоту через рибулозобисфосфат- ный путь. К этой же группе принадле- жит еще ряд скользящих трихомных фо- тотрофных организмов: Heliothrix, Chlo- 7"ОШИ20. единственные грамположительные фототрофы, спо- собные к образованию настоящих эндо- спор. Это облигатные анаэробы и гете- ротрофы, содержащие в клетках уни- кальный бактериохлорофилл g. Как и большинство аноксигенных фототроф- ных бактерий, гелиобактерии являются активными азотфиксаторами. Они встре- чаются в почвах и содовых озерах. Пока известно немного родов гелиобактерий, из них типичные Heliobacterium и Heliobacillus. В содовых озерах Цент- ральной Азии недавно В.М. Горленко открыл спиральную форму (Heli orestis), образующую обильную розовую пену из туго свернутых пружинок. Э ит обакте ии и другие аэроб- ные бактерии, образующие бактерио- хлорофилл а, являются облигатными ор- ганогетеротрофами, аэробами, лишь огра- ниченно способными использовать энергию света. Фототрофный метаболизм их совершенно не укладывается в раз- работанную схему. Сюда относятся ро- ды Erytrobacter, Roseobacter, Porphyro- bacter, обитающие в морских и пресных водах, в том числе на поверхности водо- рослей и в циано-бактериальных матах. Оксигенные фототрофы (группа XI) или анобакте ии, раньше (а у альгологов и до сих пор) назывались синезеленые во о осли. Считалось, что они вполне сходны функционально с водорослями, отличаясь строением клетки и набором дополнительных пи- гментов. Систематика этой обширной и разнообразной морфологически груп- пы была разработана на основе микро- скопии природного материала по прин- ципам ботанической номенклатуры в 1920-х гг., и компилятивная классифика- ция, созданная Л. Гейтлером (Ь. Geitler) и А.А. Еленкиным, сохранилась как основа до настоящего времени, потому что оказалась удобной для гидробио- логов (см. "Определитель пресноводных водорослей", т.2, ред. М.М. Голлербах) и, особенно, для микропалеонтологов. Попытка ввести правила бактериологи- ческои хаксономии на основе типовых чистых культур оказалась лишь частич- но удачной, потому что многие предста- вители обширной группы цианобакте- рий еще не поддаются культивирова- нию, а те, которые поддаются, нередко теряют в культуре характерные призна- ки, часто основанные на агрегировании клеток. Тем не менее, после исследова- ний Р. Стейниера, Р. Риппки, Ватербари (R. Stanier et а1., 1979) чистая культура цианобактерий — нормальный объект лабораторной работы. По строению клетки цианобак- терии - типичные прокариоты с фото- синтетическими мембранами, собран- ными в пачки ламелл, называемые тилакоидами. В них располагается хло- рофилл а, как у зеленых растений, но отсутствует хлорофилл b. В качестве дополнительных пигментов цианобак- терии имеют фикобилины, представ- ляющие развернутые в цепь порфири- ны, которые собраны в фикобилисомы- тельца в виде кнопок на внутренней 14
стороне фотосинтезирующей мембра- ны. Именно эта энзиматическая система ответственна за выделение О, и все далеко идущие последствия для биоты и всей биосферы. Ассимиляция СО осу- ществляется по классическому рибулоз- обисфосфатному пути с характерным фракционированием Ь~-'С = -25 %o. Запас рибулозобисфосфаткарбоксилазы (рубиско) обнаруживается в клетках в виде гексагональных карбоксисом. Фи- зиология фототрофных организмов в связи с неравномерностью получения световой энергии в течение суток отли- чается значительной ролью внутрикле- точных запасов. Поэтому при описании кинетики роста таких организмов при- ходится опираться не на концентрацию вещества в среде, например, фосфатов, а учитывать и весь фосфор, в том числе и внутриклеточный. Углерод накапливает- ся в виде полиглюкозы, фосфор в виде полифосфатов, азот в виде цианофицина — полимерной цепи аспарагиновой кисло- ты, содержащей боковые цепи аргинина. В темноте идет аэробное дыхание, в анок- сических условиях сбраживание поли- глюкозы, например, по пути молочноки- слого брожения; цианофицин разлага- ется по орнитиновому циклу. Запасание азотистых соединений цианобактерия- ми возможно, потому что многие из них способны к азотфиксации, особенно в отсутствие О; в аэробных условиях азотфиксация возможна в гетероцистах. Филогенетически цианобактерии составляют отдельную ветвь 16S рРНК дерева, в которую входят также хлоро- пласты эукариот. Морфология цианобактерий до- статочно разнообразна, чтобы опозна- вать их микроскопически, и на ней была основана их классификация. Сейчас вводится филогенетическая система, не совпадающая с морфологической. Про- стейшие формы либо одноклеточные, либо объединены в специфические для родов микроколонии-агрегаты наруж- ными слоями оболочки. Для многокле- точных характерной формой является трихом - нить из цилиндрических кле- ток с очень варьирующим отношением высоты к диаметру. Трихом ведет себя как многоклеточный организм, размно- жение его осуществляется распадением на короткие, но многоклеточные гормо- гонии. Трихомы обладают скользящим движением. Цианобактерии в основном пресноводные организмы, но часть их характерна для прибрежной зоны моря, а Trichodesmium вызывает цветение в открытом океане. Цианобактерии разделены бакте- риологами на крупные порядки по мор- фологии, альгологи дают гораздо более детализированную систематику, и ниже приведены лишь наиболее часто упо- минаемые роды, выделяемые условно как пять секций (таблица 2). 1. Chroococcales — одноклеточные или колониальные, но не нитчатые. Ти- пичными представителями служат одно- клеточные Synechococcus, объединен- ные общей капсулой кокки Gloeothece и Entophysales, слизистые агрегаты Mic- rocystis с газовыми вакуолями, почкую- щийся прикрепленный Chaemosiphon в характерных бокалах. 2. Pleurocapsales образуют при размножении мелкие клетки-беоциты, как у Pleurocapsa. 3. Oscillatoriales имеют трихомы без дифференцированных клеток и в большинстве представляют бентосные формы, образующие маты. Типичные роды представлены скользящими голы- ми трихомами Oscillatoria, заключенны- ми в трубчатое влагалище Phormidium, крупными трихомами Lyngbya, упако- ванными в общее влагалище пучками нитчатых трихомов Microcoleus. В план- ктоне щелочных озер распространены спиральные трихомы Spirulina. К морс- ким осцилляториям относится уже упо- мянутый Tri chodesmi um. 4. Nostocales имеют трихомы с гете- роцистами и способны к азотфиксации. К ним относится заключенный в слизи- стый чехол и образующий крупные плотные колонии Nostoc, пресноводная Anabaena в виде нити крупных бус. Сюда же относятся Ri vularia, Gloeotrichia. 5. Stigonematales имеют ветвящие- ся трихомы с дифференцированными клетками. Морфология их наиболее сложна и трихомы дифференцированы по длине. Они легко опознаются мор- фологически, но с трудом поддаются ла- бораторному культивированию. В тер- мальных источниках характерен ветвя- щийся термофильный Masti gocladus (=Fischerella, как называют почвенные и пресноводные формы). Цианобактерии, будучи основны- ми первичными продуцентами в мире прокариот, служат организмами-эдифи- каторами (от edifice - здание), создаю- щими физическую структуру циано-бак- териального сообщества, включающего разнообразный набор взаимодействую- 15
Таблица 2. Морфология клеток цианобактерий (группа XI, по Берджи, 9-е издание) 16 щих между собой прокариот. (Обратите внимание, что цианобактериальное оз- начает сообщество функционально сходных цианобактерий, а циано-бакте- риальное сообщество означает сообще- ство цианобактерий и функционально очень различных бактерий.) Прохлорофиты образуют очень маленькую группу оксигенных прока- риот — из морского симбиотического Prochloron и свободноживущего Pro- chlorothrix, внешне похожего на осцил- латорию, за которую он и был принят сначала. В море распространен Prochlo- rococcus. Большинство прохлорофит имеют ту же комбинацию хлорофиллов, что и зеленые растения: хлорофиллы а + b. Поэтому они вызвали большой интерес как возможные предшественники зеле- ных водорослей и были выделены в таксон высокого ранга. Однако на осно- вании анализа гена 16S рРНК они были отнесены к цианобактериям и интерес к ним упал. Аэробные хемолитотрофы (груп- па XII) четко разделяются на группы по
используемому субстрату: соединениям азота, серы, железа, H„CO. Вместе с ними нередко рассматриваются окис- ляющие СН, метанотрофы, а с послед- ними — метилотрофы. Объединение ме- танотрофов с нитрификаторами не слу- чайно, поскольку организмы имеют много общего в морфологии, обладая развитой системой внутрицитоплазма- тических мембран и сходным первым ферментом подготовительного обмена- метан- (или аммоний- ) монооксигена- зой. Хемолитотрофы относятся к высо- коспециализированным организмам (монотрофам) и находятся вне конку- ренции за донор электрона в экологи- ческих нишах. Большинство из них- строгие автотрофы, использующие СО как единственный источник углерода по рибулозобисфосфатному пути. Как пра- вило, никакие другие организмы не спо- собны к осуществлению реакций хемо- литотрофов. Нитрификаторы разделены на две группы, соответственно этапам окисле- ния: нитрификаторы 1 фазы окисляют аммоний в нитрит, нитрификаторы II фазы — нитрит в нитрат. Обе группы развиваются в поле термодинамической устойчивости окисленных соединений азота. 1 фаза нитрификации протекает соответственно суммарной реакции: 4NH„- + 70 = 4NO - + 4Н+ + 6Н,О, и разделяется на две энзиматические ступени: окисление аммония в гидро- ксиламин (подготовительная) и гидро- ксиламина в нитрит (энергетическая): NH, + О + НАДН = NH ОН + НО + НАД аммоний-монооксигеназа (АМО); NH OH = HNO + НО = HNO + 2Н'+ 2е — гидроксиламин оксидоредуктаза. Промежуточный продукт реакции HNO может спонтанно распадаться с образованием устойчивой закиси азота N O, важнейшего парникового газа. К нитрификаторам 1 фазы относятся: наи- более обычный обитатель нейтральных и слабощелочных почв Nitrosomonas— довольно крупная овальная подвижная благодаря единственному жгутику па- лочка, Nitrosospira, свернутая в настоль- ко тугую спираль, что выглядит как па- лочка, крупный океанский кокк Nitroso- coccus, и разделенный на доли кокк Ni trosolobus. II фаза нитрификации обеспечи- вает окисление нестойкого и реакционо- способного нитрита в устойчивый ни- трат по реакции: 21ЧаЫО, + О, = 2NaNO, - с энзимати- ческой реакцией; NO - + НО = NO,— + 2е- + 2Н+ - нитрит оксидоредуктаза. Нитрит появляется только в очень низкой концентрации как промежуточ- ный продукт в сообществе. Наиболее известным представителем нитрифика- торов II фазы является почкующаяся бактерия Nitrobacter, внешне напоми- нающая виноградные косточки; осталь- ные организмы гораздо менее изучены, это очень тонкая палочка Nitrospina, морской кокк Nitrococcus, тугая спираль Nitrospira — распространенный морской род. Тионовые бакте ии окисляют вос- становленные соединения серы в суль- фат. Основным представителем этой группы являтся род Thiobacillus, морфо- логически представляющий типичных псевдомонад. Для этого рода особенно четко прослеживается специализация видов в зависимости от физико- химических условий среды. По отно- шению к рН тиобациллы разделяются на ацидофильные и нейтрофильные. В ки- слых условиях H S неустойчив, и основ- ной формой служит молекулярная сера, которую окисляет T. thiooxidans и сход- ные с ним организмы, создающие высо- кую кислотность и развивающиеся при рН 2-4. В нейтральных условиях устой- чивы Н,S и тиосульфат, которые окисля- ются Т. hioparus до молочно-белой серы и сульфата. Один из видов тиобацилл, Tdenitrificans, способен к денитрифи- кации. Этому ряду тионовых хемоли- тоавтотрофов соответствует ряд хемоли- тогетеротрофов (T. acidiphilus, T. novel- lus) и органотрофных псевдомонад, спо- собных к окислению соединений серы. Типичным морским представителем группы служит вибрион Thiomicrospira. В гидротермальных источниках при температуре от 75 до 55 'С развивается кренофильный Thermothrix thiopara, изогнутые палочки которого в потоке свиваются в длинные иногда метровые цепи, покрытые снежно-белой серой. В термальных условиях, кроме собственно тионовых бактерий, ту же функцию окисления соединений серы могут вы- полнять архебактерии группы Sulfolobus - Acidianus, развивающиеся в крайне ки- слых условиях при высокой темпера- туре. 17
В группу XII входят железобак- терии, которые были самыми первыми организмами, еще в начале XIX в. послужившими Х.Г. Эренбергу (C}1.G. Ehrenberg) основанием для утвержде- ния, что микробы могут играть геоло- гическую роль, образуя обширные отло- жения охры. Так же, как и в случае с серобактериями, их культивирование крайне трудно, так как только при крайне кислом рН железо устойчиво к окислению воздухом. Железобактерии довольно строго следуют полям устой- чивости соединений железа. Продуктом их деятельности служат аморфные гидроокислы железа и слабокристал- лизованный минерал ферри гидрит, представляющий ключевое соединение в цикле железа. Примером ацидофильных железо- бактерий может служить открытая Г. Маркосяном мелкая спирилла или ви- брион Leptospirillum ferrooxidans, со- ставляющая отдельную филогенетичес- кую линию. Организм окисляет в серно- кислой среде либо Fe2, либо вместе с ацидофильными тионовыми бактериями пирит. Наиболее изученным представи- телем ацидофильных железобактерий служит Thiobacillus ferrooxidans, объе- диняющий свойства Leptospirillum и Т. thi ooxidans. Благодаря объединению свойств железо- и сероокисляющей бак- терии T. ferrooxidans может окислять в пирите и других сульфидах чередую- щиеся в кристаллической решетке слои металла и серы. Окисление сульфидов составляет важнейшую геохимическую функцию, ведя к сернокислотному вы- щелачиванию. Примером собственно железобак- терий служит Gallionella — исключитель- но своеобразный организм, напоминаю- щий боб, с вогнутой стороны которого выделяются нити гидроокислов железа (ферригидрита). Gallion ella окисляет железо в бикарбонатной среде, большей частью в холодной воде, часто в микро- аэрофильных условиях. Gallionella уда- ется культивировать в градиентных ус- ловиях, где она образует пояса роста на стенках пробирки. Gallionella относится к хемоавтотрофам или миксотрофам. В морфологии этого организма много за- гадочного. Нити, сходные с нитями, экс- кретируемыми Gallion ella, образует скользящая нитчатая бактерия Toxothrix. Leptothrix ferruginea представляет цепочку псевдомонад, заключенную в трубчатый чехол из ферригидрита. Этот основной агент образования болотных руд до сих пор не поддается культиви- рованию, несмотря на массовое разви- тие в природе. Морфологически он очень близок к массовой бактерии за- грязненных вод Sphaerotilus. Считается, но не доказано, что окисление железа происходит под действием внеклеточ- ной пероксидазы, как у других культи- вируемых лептотриксов, поверхность которых покрыта отложениями гидро- окислов железа или марганца. Загадочную группу "железобакте- рий" составляют открытые Б.В. Перфи- льевым формы, получившие название Metallogenium. Это тонкие извитые ни- ти, расходящиеся в виде паучка от цент- рального округлого тела и покрытые конической шубой из окислов марганца. Металлогениум составляет массовую форму в отложениях марганца и хорошо прослежен во вторичном диагенети- ческом профиле в осадках карельских озер. Он обнаружен в отложениях с воз- растом 2 млрд. лет тому назад и описан под названием Eoastrion. Металлоге- ниум легко получить в лаборатории на среде с карбонатом марганца в со- вместной культуре, например, с грибом. Американские исследователи считают его продуктом внеклеточной перокси- дазы (то есть неживым образованием), но, как показала Г.А. Дубинина, им лег- ко заразить культуру, ранее не окисляв- шую марганца. Наконец, еще одну группу железо- бактерий составляют "сидерокапсы" разнообразные микроорганизмы, на поверхности клеток которых отклады- ваются гидроокислы железа. Вероятным источником этих окислов служит оста- точный продукт окисления органичес- ких комплексов железа. Отдельную группу составляют ор- ганотрофные магнитотактические бак- терии, образующие внутри клеток кри- сталлы магнетита — магнето сомы — и поэтому чувствительные к магнитному полю Земли. От магнитотактических бактерий следует отличать анаэробные магнетит- образующие анаэробы, для которых образование магнетита служит катабо- лическим процессом. Они были откры- ты Д. Ловли (D. Lovley). Часть из них, как упомянутый Geobacter, близки сульфатредукторам, но многие отно- сятся к совсем другим группам. Среди анаэробных магнетит-образующих бак- терий многие используют водород для 18
восстановления ферригидрита и явля- ются эффективными гидрогенотрофами. Как показала Д. Заварзина, образование магнетита происходит у них вне клетки в процессе диагенеза из аморфного предшественника. Почкующиеся бактерии и про- стекобактерии (группа ХШ). В эту группу попали бактерии с выраженным жизненным циклом, в котором разли- чаются материнская и дочерняя клетки. В типичном случае материнская клетка последовательно образует подвижные дочерние клетки, которые образуют стадию расселения и должны претер- петь созревание, прежде чем приступить к размножению. Поч ю иеся бакте ии в наибо- лее типичном случае представлены ги- омик обами, у которых имеется длин- ный вырост клетки диаметром около 0,2мкм (гифа-простека), на конце кото- рой образуется небольшое вздутие, пре- вращающееся в подвижную клетку со жгутиком. Эта клетка отрывается и уплывает, а на ее месте образуется но- вая. Подвижная клетка прикрепляется, теряет жгутик и образует гифу, на конце которой образуется почка. В некоторых случаях гифы ветвятся. Вместо почки иногда образуются устойчивые к высу- шиванию экзоспоры. К гифомикробам принадлежит метилотрофный Hypho- mi crobium, органотрофный Hyphomonas, разлагающий почвенные гумусовые комплексы Pedomicrobium, упомянутый выше пурпурный несерный Rhodomic- robium, галофильный Dichotomicrobium. К микроорганизмам, у которых почка образуется непосредственно на материнской клетке, относятся физио- логически разнообразные Nitrobacter, Blastobacter, Methylosinus, Rhodopseu- domonas, и такой тип строения часто связан с наличием чашевиднои пачки мембран, которая не может быть раз- делена надвое при делении и остается материнской клетке, в то время как до- черние синтезируют новую. П остекобакте ии представлены грам-отрицательными организмами с нитевидными выростами клетки. В эти выросты входит цитоплазма, и таким образом поверхность клетки и пространство перипласта существенно увеличиваются. Морфология этих орга- низмов сначала была изучена в элек- тронном микроскопе Д.И. Никитиным как "необычных микробов", но потом их научились различать и в световой ми- кроскоп, и сейчас имеется хорошее представление о распространении и дея- тельности этих организмов в почве и водоемах благодаря работам П. Хирша (P. Hirsch), Дж. Сталей (J. Staley), Л. Ва- сильевой, которым удалось получить их культуры. Большинство простекобак- терий относится к олиготрофам, раз- вивающимся при крайне низкой кон- центрации питательных веществ в среде. Типичным представителем их служат каулобактеры — удлиненные изогнутые клетки которых несут одну простеку с прикрепительным диском на конце. Дочерняя клетка со жгутиком образуется делением и освобождается от прикрепленной материнской клетки, плавает, затем прикрепляется и образует простеку, часто объединяясь в виде розетки. Каулобактеры были обнаруже- ны в олиготрофных озерах и в дистиллированной воде лабораторий, где они попадали на сетки для элек- тронной микроскопии. К каулобактерам относятся несколько видов Caulobacter, Asticcacaulis — олиготрофных аэробных органотрофов с широким спектром ис- пользуемых веществ. Prosthecobacter не имеет подвижнои стадии и напоминает две кегли, соединенные донышками. Не- сколько простек имеют виды Prosthe- comicrobium, Ancalomicrobium. Плоская Stella имеет вид шестилучевой звезды. Labrys имеет форму двулезвийного топора. Плаиктомицеты (подгруппа 2 группы 13) - своеобразные водные ор- ганизмы, размножающиеся почковани- ем, тоже рассматриваются как олигот- рофные органотрофы. Они очень трудно поддаются культивированию, но неко- торые из них удалось выделить Х. Шлесснеру (Н. Schlesner), и они были отнесены в самостоятельныи класс в l6S рРНК системе. Наиболее известен из них Planctomyces (= "Gallionella kljas- ~нипяи"), представляющий собой кокки, собранные в розетку на слизистых сте- бельках, и размножающийся образова- нием почек на противоположном сте- бельку конце. Планктомицеты имеют много особенностей, отличающих их от типовых бактерий, но очень плохо изу- чены из-за трудностей культивирова- ния. Одной из принципиальных осо- бенностей планктомицетов служит на- личие у них мембраны вокруг нукле- оида, двойной у Gemmata и одиночной у Pi rellula. Наличие мембраны обусловли- вает их сходство с эукариотами. 19
Молекулярными методами установлено, что планктомицеты представляют мас- совые формы в донных осадках и почве вокруг нуклеоида. Нитчатые бактерии с чехлом (группа XIV) представляют массовые водные формы. Типичным предста- вителем их служит "водный гриб" Sphaeroti lus, образующий белесые, а чаще серые обрастания в загрязненных водах. Он представляет цепь псевдо- монад, заключенных в общий трубчатый чехол и благодаря этому удерживаю- щихся в потоке. Аналогичные орга- низмы, чехлы которых инкрустируются окислами металлов, относятся к Lepto- thrix. В аэротенках вспухание активного ила обусловливает нитчатый Haliscame- nobacter. Наконец, в текучих водах и колодцах встречается Crenothrix с рас- ширяющимся к концу чехлом, где про- исходит образование мелких клеток дроблением, как у хамесифоновых циа- нобактерий. Скользящие бактерии (группы XV, XVI) разделяются на большие группы одноклеточных цитофаг и три- хомных бактерий. Причиной объедине- ния их послужил способ движения: при контакте с твердой фазой они могут скользить по неи, как это своиственно и цианобактериям. Жгутиков у них нет. ~ито аги разлагают полимеры, такие как целлюлоза или хитин. Коло- нии их на агаризованных средах рас- ползаются и часто окрашены в желтый цвет, который при обработке 20% КОН становится пурпурным, указывая на присутствие характерного каротинода флексирубина. Цитофаги относятся к важнейшим аэробным целлюлозоразла- гающим организмам. На фильтроваль- ной бумаге они образуют желтоватые колонии с разложившейся бумагой в середине. За гидролиз целлюлозы ответ- ственны связанные с мембраной эндо- глюконазы и периплазматические экзо- глюконазы. Поэтому клетки должны осу- ществлять тесный физический контакт с волокнами полимера. Клетки цитофаг обычно длинные гибкие с приострен- ными концами. К цитофагам относятся собственно Cytophaga и Sporocytophaga, образующая круглые микроцисты. Ци- тофаги могут разлагать многие полиме- ры растительного происхождения, а не- которые и хитин. К цитофагам относят также длин- ные нитевидные пресноводные Flexi- bacter и морские Microscilla. Многие из этих организмов имеют активные гид- ролитические ферменты, позволяющие им лизировать структурные белки вплоть до кератина. Некоторые скользя- щие бактерии, например, Lysobacter, способны лизировать клетки бактерий, образуя на газоне расползающиеся нега- тивные пятна, подобные тем, которые оставляют за собой амебы. Нитчатые серобактерии были од- ними из первых обнаружены микроско- пистами как массовые крупные водные организмы с включениями серы и си- стематизация их, наряду с наблюдени- ями за физиологией в проточной ми- кроскопической культуре, послужили для С.Н. Виноградского прелюдией к открытию хемосинтеза. Прошедшие 100 лет добавили мало знания из-за труд- ностей культивирования. Морфологи- чески серобактерии, бесцветные нит- чатые бактерии, цианобактерии состав- ляют паралелльные ряды форм. Из серо- бактерий следует отметить Beggiatoa со скользящими трихомами как у осцил- ляторий, сидячий прикрепленный Thio- thrix с органотрофным аналогом морс- ким Leucothrix. У побережья Чили мощ- ные маты образует крупная серобак- терия Thioploca, аналогичная по плану строения цианобактерии Mi crocoleus, накапливающая внутри клетки нитраты как запас окислителя. Из одноклеточных серобактерий следует отметить очень крупный Macromonas. Все эти организ- мы опознаются по включениям в клетки серы, которую они способны окислять. Обладая трихомным строением, эти организмы наряду с гетеротрофными флексибактериями образуют параллель- ный ряд форм с цианобактериями. Миксобактерии (группа XVI) привлекли к себе внимание способно- стью образовывать "плодовые тела", состоящие из экскретируемых ими по- лисахаридов и входящих в них устой- чивых миксоцист (или, иначе, микро- цист). Миксобактерии обладают не только индивидуальной способностью к дифференциации от вегетативной клет- ки до покоящихся микроцист, но и кол- лективной дифференциацией, приводя- щей к образованию из колонии оформ- ленного плодового тела, как у эукариот- ных миксомицетов. Вегетативные клет- ки миксобактерий представлены оди- ночными скользящими палочками с за- остренными или тупыми концами, раз- множающимися делением перетяжкой и образующими "шварм" — координиро- 20
ванно движущуюся колонию. Передо- вые клетки выделяют слизь, и последу- ющие движутся за ними по слизистому следу. Опознаются колонии миксобак- терий по ярко окрашенным плодовым телам, составляющим спорангии, иног- да приподнятые над поверхностью на стебельках и имеющие размеры до 1мм. Миксобактерии - аэробные орга- нотрофы с выраженной гидролитичес- кой активностью. Особенностью миксо- бактерий является их способность кол- лективно лизировать твердые частицы с помощью гидролитических ферментов. Часть из них обладает протеолитичес- кой способностью и лизирует клетки бактерий, попадающие под шварм. По- этому миксобактерий относят к хищным бактериям. Однако некоторые миксобак- терии способны лизировать также цел- люлозу. Часто они развиваются на на- возе. В чистой культуре поддерживать удается только часть родов, и в природе их идентифицируют по спорангиям. Основные роды их следующие: не обра- зуют спорангиев миксобактерии с вере- теновидными клетками рода Myxococ- cus, у которого наиболее подробно изу- чена дифференциация в круглые микро- цисты; Archangium с палочковидными микроцистами. Cystobacter образует спорангии без стебелька, Mellitangium и Stigmatella — на стебельке. Клетки с пря- мыми концами имеет целлюлозолити- ческий Polyangium, образующий округ- лые спорангии. Сложные плодовые тела имеют Nannocystis и Chondro'>ces. Грамположительные кокки (группа XVII). Микрококки составляют обширную группу органотрофных бак- терий с хорошо известными родами аэробных кокков Micrococcus, Staphylo- coccus, Streptococcus, издавна различае- мыми по форме агрегатов клеток и зна- комыми всем патогенными представите- лями. К анаэробным коккам, осуществ- ляющим разные брожения, относятся молочнокислые стрептококки, крупная анаэробная Sarcina ventriculi — обитатель желудка при крайне низком рН, обитате- ли рубца разнообразные Ruminococcus, сбраживающие азотистые соединения Peptococcus. Совершенно самостоятель- ную филогенетическую ветвь образуют крайне радиоустойчивые дейнококки (род Deinococcus). Образующие эндоспоры грам- положительные организмы (группа XVIII). Бациллы и клостридии отно- сили к двум родам: аэробным Bacillus и анаэробным Clostridium, обычно пред- ставленным довольно крупными палоч- ками с перитрихиальным жгутикова- нием. Благодаря наличию термоустой- чивых спор они относительно легко вы- деляются в лабораторные культуры после прогревания 5 мин при с 80 'С. Это массовые формы, споры которых составляют резерв готовых к размноже- нию зародышей. По своим физиологи- ческим функциям виды этих морфоло- гических родов чрезвычайно разнооб- разны. Спорообразующие организмы представляют преимущественно поч- венные формы, поскольку эндоспоры являются приспособлением к высуши- ванию. В большинстве бациллы пред- ставлены органотрофами, использую- щими широкий круг органических ве- ществ. Клостридии более специализи- рованы и осуществляют разнообразные брожения, в типичном случае — масля- нокислое, а некоторые — гомоацетатное. Они послужили моделью для изучения разных брожений — пуринов, аминокис- лот. Многие из них обладают мощными гидролазами и сбраживают полимерные углеводы, например, целлюлозу как Clo- stridium thermocellum. Первым из сво- бодноживущих азотфиксаторов был от- крыт C pasteurianum, но вообще это свойство присуще многим анаэробным клостридиям. Специализированную груп- пу среди анаэробных спорообразую- щих грамположительных организмов составляют сульфатредуцирующие De- sulfotomaculum, рассмотренные выше. Неспорообразующие грам-поло- жительные палочки правильной формы (группа Х1Х) имеют основным представителем род Lactobacillus и по нему обычно называются лактобацил- лами. Они осуществляют молочнокис- лое брожение и относятся к аэротоле- рантным анаэробам, развивающимся, например, в растительном компосте. Сюда же относится ряд менее известных родов, часто отличающихся по форме, как, например, образующий многокле- точные трихомы Caryophanon. Коринеформы (группа ХХ). Это грамположительные палочки непра- вильной формы, иногда склонные к вет- влению. Наиболее многочисленный и разнообразный род Corynebacterium представлен аэробными органотрофами. Очень разнообразны артробактеры Arth- robacter, аналогичные по своим функ- циям псевдомонадам и широко распро- страненные в холодных почвах тундры. 21
К коринеформам принадлежит и целлю- лозолитический Cellulomonas. Анаэроб- ные организмы в этой группе представ- лены органотрофными неспоровыми бактериями: маслянокислыми Eubacte- rium, Butyrivibrio, гомоацетатными Ace- tobacterium, молочнокислыми Bi fidobac- terium, пропионовокислыми Propioniba- cterium, термофильными Thermoanaero- bacter, осуществляющими разнообраз- ные брожения с выделением Н, и обра- зованием различных жирных кислот. Микобактерии (группа XXI). Группа включает кислотоустойчивые (по окраске) органотрофные палочки Mycobacterium с характерным составом липидов оболочки. Нокардиеформы (группа ХХП). Эта группа организмов близка олиго- споровым актиномицетам и включает склонные к образованию ветвящихся форм и слабо развитого мицелия грам- положительные организмы, опознавае- мые по "сухим", не покрытым слизью колониям, выдерживающим прямой контакт с воздухом. Типовым родом группы служит Nocardia, которую мож- но рассматривать как типичного пред- ставителя "автохтонной" микрофлоры, способной использовать органические вещества "черного пара", то есть устой- чивые вещества, остающиеся в конце процесса деструкции. Органотрофные Rhodococcus, несмотря на свое назва- ние, не имеют ничего общего с пурпур- ными несерными бактериями, представ- ляя аэробных относительно медленно растущих органотрофов, известных сво- ей способностью эффективно исполь- зовать углеводороды нефти. Корине- бактерии, нокардиеформы, стрептоми- цеты составляют ряд морфологически усложняющихся организмов, приспо- собленных к существованию в засуш- ливых почвенных условиях и приобрет- ших способность к образованию воз- душного мицелия и конидий как стадии расселения. В 9-м издании Берджи остальные актиномицеты попали в группы 23-28. Геодерматофилы и франкии (группа XXIII) рассматриваются как не образующие мицелия актиномицеты. Представители рода Geodermatophilus широко распространены в почве и раз- виваются в виде микроколоний, сложен- ных паренхиматозной массой клеток, часто окрашенной в черный цвет. Это аэробные органотрофы, скорее всего олигокарбофильные. На поверхности микроколоний некоторые клетки почко- ванием образуют подвижные грушевид- ные клетки, которые служат стадиями расселения. Вследствие своего сходства с патогенным Dermatophilus этот орга- низм не стал объектом детальных иссле- дований дифференциации у прокариот. Frankia представляет азотфикси- рующий симбионт ольхи, образующий на ее корнях клубеньки подобно ризо- биям. У нее имеется типичный воздуш- ный мицелий со спорангием, разделен- ным на много спор. Этот эффективный азотфиксатор с трудом поддается куль- тивированию. Актинопланы (группа XXIV) или водные актиномицеты имеют раз- витый мицелий, как субстратный, так и воздушный. Споры образуются в меш- ковидном спорангии и в нем пережи- вают высушивание. Однако при увлаж- нении споры выходят наружу в виде подвижных жгутиковых клеток. Акти- нопланы часто развиваются на пыльце растений, плавающей на поверхности воды в нейстоне, и способны разлагать воскоподобные соединения кутикулы. Сюда относятся роды Actinoplanes, Dac- tylosporangi um. Стрептомицеты (группа XXV). Именно для них прочно укоренилось тривиальное название актиноми еты, составляющее резкое противоречие с бактериологической номенклатурой, поскольку родовое название Actinomyces было изначально присвоено анаэроб- ным патогенным организмам. Актино- мицеты характеризуются развитым ми- целием из ветвящихся гиф и имеют общее строение, совершенно аналогич- ное эукариотным грибам, но с гораздо более тонкими прокариотными гифами, содержащими много нуклеоидов и не всегда разделенными на отдельные клетки. Это прежде всего почвенные организмы. Актиномицеты приспособ- лены к развитию в относительно за- сушливых условиях на твердом суб- страте и, соответственно, обладают вы- раженной гидролитической активно- стью по отношению к разным поли- мерам, но особенно выражена хитино- литическая активность с развитием актиномицетов по отмершему грибному мицелию. Мицелий актиномицетов делится на субстратный и предназ- наченный для размножения воздушный. Гифы воздушного мицелия имеют пре- дохраняющую от высушивания оболоч- ку и на них образуются конидиеспоры
(термины конидии и споры тоже упо- требляются). Конидиеспоры одновре- менно служат и переживающей высу- шивание стадией и стадией размноже- ния, отличаясь в этом от эндоспор бак- терий, и стадией расселения, распрост- раняемой воздушными потоками. Акти- номицеты относительно медленно ра- стущие организмы. Все они органотроф- ные аэробы и в значительной части гидролитики, использующие очень ши- рокий круг разнообразных органичес- ких веществ. Выделяют их на средах с крахмалом и/или глицерином при вы- соком отношении С/N. Соответственно форме споронос- цев различают роды Streptomyces с более 500 видами, Streptoverticillum, Kineospo- ra, Sporichthya. Актиномицеты явились объектом крайне интенсивных исследо- ваний в поисках образуемых ими анти- биотиков, которые, предположительно, в природе обеспечивают их конкуренто- способность. В биотехнологии они очень важны, поскольку примерно половина антибиотиков образуется этой группой. Мадуромицеты или олигоспоро- вые актиномицеты (группа XXVI) имеют такое же мицелиальное строение, как и другие актиномицеты, но в от- личие от них образуют небольшое число кон иди й. Роды различаются прежде всего по форме конидиеносцев. Это Actinomadura, Microbispora, Microtet- raspora, Planornonospora, Spirillospora, Streptosporangium, названия которых говорят сами за себя. Термоактиномицеты (группа XXVII и ХХУШ) выделены по их способности к умеренной термофилии. Микоплазмы (группа XXX). К ним относят мелкие организмы, лишен- ные клеточной стенки и ограниченные только мембраной. По~тому форма кле- ток у них менее определенная, чем у других бактерий. Обычно это очень мел- кие организмы до 0,2 мкм в диаметре. На ультратонких срезах они опознаются по отсутствию клеточной стенки. 3то кокковидные элементы, иногда с ните- видными выростами, как у Mycoplasma mycoides. Паразит растений Spiroplasma получила название из-за спиральной формы. Большинство .микоплазм разви- вается в межклеточном пространстве тканей животных, растений, колоний бактерий и является паразитами, хотя Acholeplasma laidlawii - распространен- ный сапрофит, а Thermoplasma встреча- ется в гидротермах и само азо еваю- щихея кучах угля. Термоллазма может служить хорошим примером паралле- лизма в морфологии, потому что эта "микоплазма" на самом деле относится к архебактериям. Культивируют мико- плазмы на сложных средах с включе- нием сыворотки, на плотных средах они образуют колонии в виде "яичницы- глазуньи". Сначала микоплазмы рассма- тривали как наиболее примитивных бактерий с М. mycoides, обладатель- ницеи самого маленького генома, и от- носили к порядку Mollicutes. По фило- генетическои системе они относятся к ветви клостридий. В микоплазмопо- добное состояние переходят грамполо- жительные бактерии с подавленным синтезом клеточной стенки, образую- щие сферопласты. А хебакте ии Морфологически архебактерии, кратко обозначаемые как археи, не име- ют резких отличий от эубактерий. При большом навыке можно отличить их клетки по некоторой угловатости, но эти отличия не выходят за пределы, напри- мер, отличий азотобактера от псевдомо- над. Цитологические отличия их клеток с типично прокариотным строением также не существенны. Физиологически многие архебактерии относятся к орга- низмам, развивающимся в экстремаль- ных условиях, экстремофилам, и осуще- ствляют такие уникальные процессы как метаногенез, особый тип усвоения све- товой энергии с участием родопсина, использование серных соединений. Ос- новное различие архе- от эубактерий заключается в аппарате синтеза белка. Архебактерии отличаются от эубак- терий по ряду хемотаксономических признаков. Их мембраны содержат не эфиры глицерина и жирных кислот, а этерифицированы изопреноидами, что дает возможность их идентификации по биомаркерам в органическом углероде осадочных пород. В эубактериях, как и у эукариотах, изопреноиды содержатся в боковых цепях хинонов, хлорофиллах, образуются при синтезе холестерина и каучука, но не входят в мембраны. В клеточной стенке архебактерий цепи полисахаридов и глюкозамина соедине- ны пептидами с иным набором амино- кислот, чем у бактерий с муреином, и поэтому получили название псевдому- реина. У многих архебактерий клеточ- ная стенка сложена белковыми 23
глобулами, образующими так называе- мый S-слой, хорошо различимый на электронных фотографиях как упорядо- ченная поверхность, напоминающая вы- шивку бисером. Архебактерии не чувст- вительны к ряду антибиотиков, которые являются ингибиторами клеточной стенки и синтеза PHK у эубактерий. Архебактерии входят в три фено- типические группы: 1) метанобразую- щих организмов, 2) экстремальных га- лофилов, 3) экстремальных термофилов. В 9-м издании Определителя бактерий Берджи архебактерии входят в группы 31-35. Метаногены (группа XXXI) осуществляют уникальную в живом мире реакцию синтеза метана и играют важнейшую роль в биосферных меха- низмах. Это высокоспециализирован- ные анаэробные организмы, для кото- рых реакция метаногенеза служит един- ственным источником энергии. Окисли- телем в реакциях метаногенеза является СО. Соответственно донорам электрона метаногены разделяются на три группы: 1) водородных (гидрогенотрофных) ме- таногенов; 2) ацетокластических; 3) ме- тилотрофных, окисляющих метилиро- ванные соединения с группой О, N, S; а также политрофную метаносарцину, способную использовать все три пути. Метаногены относятся к строгим ана- эробам и быстро отмирают при контакте с воздухом, т.к. не образуют таких вос- становителей как Н или H S. Морфоло- гически метаногены очень разнообраз- ны: среди них есть палочки Methano- bacterium с клеточной стенкой из псев- домуреина, кокки Methanococcus с бел- ковой клеточной стенкой, плоские угло- ватые формы Methanohalobium, псевдо- паренхиматозные агрегаты Methano- sarcina с гетерополисахаридом, цепи палочек в трубчатых чехлах Мейапо- thrix (Methanosaeta) и спириллы Metha- nospirillum с белковой клеточной стен- кой. В таксономии метаногенов упо- требляется обозначение Methano-, в от- личие от аэробных метанокисляющих метанотрофов Methylo-; словосочетание "метановые бактерии", хотя и должно относиться только к метанобразующим, может быть неправильно понято. Обычным местообитанием мета- ногенов являются донные отложения во- доемов, пищеварительный тракт расти- тельноядных животных„в особенности рубец жвачных, гидротермы с Н. За ис- ключением последнего случая метано- гены зависят от анаэробного микроб- ного сообщества, продуцирующего те немногие субстраты, которые они могут использовать„ в особенности ацетат. Особый случай составляет симбиоз ме- таногенов, в том числе внутриклеточ- ный, с анаэробными протистами, для ко- торых они составляют терминальный сток Н. В анаэробном микробном сооб- ществе они замыкают трофическую цепь, потребляя ацетат и Н,. Метано- гены чрезвычайно важны для биогеохи- мии, потому что служат главным источ- ником метана на Земле. Широкое применение анализа по 16S рРНК привело к очень дробной си- стематике метаногенов на уровне круп- ных таксонов с созданием порядков ги- дрогенотрофных Methanobacteriales, Methanococcales, морских Methanomicro- bi ales, ацетокластических и метилотроф- ных Methanosarcinales. Внутриклеточ- ный путь образования метана оказался совершено своеобразным и проходит в виде ступенчатого восстановления СО с участием специфических ферментов, переносчиков С-1 группы, переносчиков электрона. Основным переносчиком слу- жит производное флавина фактор F„„, получивший свое название по максиму- му поглощения. В окисленном состоя- нии он обладает сильной зеленой флуо- ресценцией и по этому признаку метан- образующие бактерии, кроме ацетокла- стичесхих, пегхо опознаются в люми- несцентном микроскопе. Однако некото- рые другие организмы, богатые флави- нами, дают аналогичное свечение, поэто- му автофлуоресценцию следует считать сильным указанием на присутствие ме- таногенов, но не доказательством. Метаногены не нуждаются в экзо- генном органическом веществе и отно- сятся к автотрофам. Образование био- массы идет в общем по обычным для анаэробов путям через синтез ацетата с участием СО-дегидрогеназы, аналогич- но процессам у гомоацетатных бакте- рий. Центральный метаболизм не имеет существенных отличий. В восстанови- тельной зоне, где развиваются метано- гены, для азотфиксации нет таких пре- пятствий, как в аэробной зоне, и она распространена у метаногенов. Метаногены развиваются при очень низком ЕН, в термодинамическом поле устойчивости метана. Поле рН для них совпадает с областью бикарбонат иона и, соответственно, они являются нейтрофилами. Исключение составляет
метилотрофный алкалофил Methanosal- sus, развивающийся в содовых водое- мах. Ацидофильные метаногены пока не идентифицированы. По отношению к температуре большинство метаногенов является мезофилами с нижней границей около 15 'С. Поиски психрофилов или психро- активных форм затруднены из-за крайне медленного роста, но такие организмы существуют. Так, психрофильный мета- ноген Methanococcoides burtonii имеет минимальную температуру -2,5 'С при максимальной 28 'С. Среди метаногенов много термофилов, в том числе и экст- ремальных, развивающихся в гидротер- мах при температуре до 98 'С как Metha nopyrus, 85 'С как Methanothermus, Me- thanocaldococcus, 70 'С как Methanobac- terium thermoautotrophicum, послужив шая важнеишеи моделью для исследо- вания метаногенов. По отношению к минерализации среды метаногены де- лятся на группы пресноводных, к ко- торым относятся большинство метано- бактерий и метаносарцин, морских ме- таномикробов, галофильных метило- трофных метаногенов, в том числе экст- ремальных как Methanohalobium. Экстремальные галофилы (груп- па XXXIII) представлены небольшой группой архебактерий (галобакте ий), развивающихся в рассолах при содер- жании NaC1 не менее 10% (или 1,8М) и вплоть до насыщения, когда они доми- нируют. При понижении концентрации солей они лизируются. К ним относятся палочковидные Halobacteri um, кокко- видные Halococcus, плеоморфные, ино- гда в виде "кусочков битого стекла" HaIoareaIa, HaIoferax. Группа натроно- бактерий Чае опоЬаеteritam и IVatrotto- coccus развивается при рН10 и насыще- нии раствора содой. Галобактерии раз- виваются в лагунах или соляных чеках, где испарение приводит к концентрации солей и сопутствующему этому накоп- лению органических веществ, позволя- ющему отнести эти организмы к копио- трофам. Галобактерии окрашены в виш- невый цвет каротиноидами и придают рассолу яркую окраску, по которой они были впервые обнаружены на соленой рыбе. Галобактерии вынуждены поддер- живать высокое осмотическое давление в клетке и осуществляют это посредст- вом создания в клетке вь~сокои концен- трации KCl вплоть до 5 М, в то время как Na выкачивается из клетки. Все фер- менты организма должны работать при такой концентрации солей. Галобакте- рии представляют аэробных органотро- фов, использующих широкий круг ор- ганических веществ: аминокислот, угле- водов, органических кислот. Они обла- дают несколько измененными биохи- мическими путями использования саха- ров и полным циклом трикарбоновых кислот. Некоторые способны развивать- ся анаэробно, сбраживая аргинин как Halobacterium salinarium. Halobacterium не составляет большинства среди оби- тателей гиперсоленых водоемов и архей, но виды этого рода могут быть селек- тивно выделены в анаэробных условиях на среде с аргинином. Особое внимание привлек своеоб- разный тип фотосинтеза, осуществляе- мый галобактериями с участием бакте- риородопсина, комплекса белка с пиг- ментом ретиналем, сходным со свето- чувствительным пигментом глаза. Под воздействием света бактериородопсин претерпевает изменения, позволяющие ему функционировать как протонная помпа и вести к энергизации мембраны, осуществлению транспорта и образова- нию АТФ. Этот тип фотосинтеза связан с высокими интенсивностями света, которые и наблюдаются в испаритель- ных прудах и лагунах. Фотосинтез у гало- бактерий выполняет роль дополнитель- ного источника энергии при органоге- теротрофном типе обмена. Развиваясь в очень своеобразных условиях лагун и образования отложений эвапоритов, галобактерии представляют типичный пример экст емо илов, организмов, растущих в условиях непригодных для большинства других живых существ. Экстремальные термофилы (груп- па XXXV), Cvenarchaeota, представ- ляют собой обитателей гидротерм и наиболее тесно связаны с эндогенными процессами в геосфере. Первоначаль- нои моделью для исследования этих организмов для Т. Брока (Т. Вгос1) по- служил термальный источник Октопус Спринг в Иеллоустонском парке в США, а в России нами была изучена кальдера Узок на Камчатке, где развиваются термофильные циано-бактериальные маты. Затем В. Циллиг (W. Zillig) и К. Штеттер (К. Stetter) исследовали, каза- лось бы, безжизненные кипящие грязе- 'вые котлы в Исландии и обнаружили в них массу кокковидных бактерий. Это послужило стартовой точкой для интен- сивных исследований К. Штеттера, опи- 25
савшего множество гипертермофилов, развивающихся в диапазоне 80-110 'С и относящихся к организмам с наиболее глубокой точкой ветвления по 16S рРНК филогении. Особое значение гипертер- мофилы приобрели с изучением подзем- ной биосферы, открытой при бурении глубоких скважин на нефть и газ, как организмы, развивающиеся в подзем- ных водах на большой глубине вплоть до 3 км, где геотермический градиент приводит к высоким температурам вплоть до 100 'С. Организмы, развиваю- щиеся при температуре выше 90 'С, Штеттер предложил называть гипертер- мофилами и обозначать префиксом Руго-. Экстремально термофильные ар- хеи составляют самостоятельную ветвь на филогенетическом дереве архей и получили название Crenarchaeota в свя- зи с тем, что растут в потоке гидротер- мальной воды и газа. Действительно, для многих из них важно удаление продуктов обмена. Большинство (но не все, например упомянутые выше мета- ногены) гипертермофилов связаны с обменом серы и относится к сульфи- догенам. Многие способны использо- вать Н и растут литотрофно, другие от- носятся к органотрофам, использующим небольшие полимеры как пелтон или крахмал. Среди кренархеот следует выде- лить группу аэробных ацидофильных организмов, развивающихся на богатых серой вулканических площадках, полу- чивших название сольфатар. Эти аци- дофильные термофилы были открыты Т. Броком и названы Sulfolobus. Впослед- ствии к этой группе присоединили еще несколько родов. Сульфолобусы имеют форму неправильных кокков и растут при рН1-5, предпочтительно при рН2-3 и температуре от 60 до 90 'С. Обмен сульфолобусов очень разнообразен. Они могут расти органотрофно, окисляя органические вещества в СО; литоавто- трофно, восстанавливая серу в Н,S водо- родом; окислять серу в серную кислоту подобно тиобациллам; окислять сульфи- ды железа в окисное железо и серную кислоту. Сходный с Sulfolobus другой род ацидофильных термофильных архе- бактерий, Acidianus, может расти и анаэ- робно, обладая способностью к сероре- дукции и восстанавливая S< в H~S. Анаэробный нейтрофильный орга- нотрофный Оеяи1/йюсоссиь принадле- жит к сероредуцирующим органотрофам. Такими же свойствами обладают Ther- mococcus, палочка Thermoproteus, длин- ная нить Thermo filum, гипертермофил Pyrococcus fuïosès. Органотрофные экс- тремальные термофилы используют преимущественно разнообразные протеи- ны и по типовому роду Thermoproteus образуют порядок Thermoproteales. Все эти организмы, различающиеся по опти- мальной температуре роста, восстанав- ливают серу и используют ее как сток для водорода при анаэробном полном окислении органических веществ. Sta- phylotermus, в виде цепочки кокков ра- стущий на поверхности "черных ку- рильщиков", сбраживает органические вещества до ацетата и изовалерата. Сульфатвосстанавливающая термофильная архебактерия Archaeo- globus (группа XXXIl) с оптимумом около 80 'С восстанавливает сульфаты в H S подобно термофильной бактерии Thermodesulfobacteri um, используя Н, лактат, пируват, глюкозу, смеси органи- ческих веществ. Наибольший интерес вызвала находка того, что Archaeoglobus содержит компоненты, до сих пор обна- руженные только у метаногенов. Pyrodictium имеет температурный оптимум при 105 'С и выделен из морс- ких гидротерм. Он представлен округ- лыми дисковидными клетками, соеди- ненными длинными нитями, которые состоят из белковых трубок, прикреп- ляющих организм к твердому субстрату. Это строгий гидрогенотрофный анаэ- роб, восстанавливающий серу и расту- щий литотрофно, но стимулируемый ор- ганическими веществами. Морфологи- чески его напоминает органотрофный Therm odiscus. Своеобразный организм представ- ляет выделенная Броком Thermoplasma (группа XXXlV) из куч саморазогре- вающегося угля лишенная клеточной стенки архебактерия, подобная мико- плазмам. Ferriplasma, аналогичная по обмену 1.ерйокргс11ит ferrooxidans, не- давно открыта Г.И. Каравайко. Если для гидрогенотрофных ги- пертермофильных организмов источни- ком энергии может служить эндогенный водород, то откуда они могут получать сложные органические вещества? Воз- можным ответом на вопрос служит ги- потеза "бойлера", предложенная Г.А. За- варзиным, по которой кренархеоты по- лучают органические вещества с кон- векционным потоком воды, проходящим через их местообитание, от организмов, 26
которые свариваются при этом, отнюдь не напоминая "первичный бульон". Молекулярные методы идентифи- кации обнаружили характерные для Crenarchaeota клоны рРНК и показали, что организмы, филогенетически отно- сящиеся к архебактериям, многочислен- ны в морском планктоне, где они со- ставляют доминирующую форму бакте- риопланктона олиготрофного океана морских и пресноводных осадках, а также в числе симбионтов Metazoa. Все эти организмы пока относятся к некуль- тивируемым. Газотрофы Сборная группа газотрофов, окис- ляющих Н, СО, СН, и метилированные соединения, относится к литотрофным организмам; сюда же входят и водород- ные бактерии, принадлежащие разным таксономическим группам. Водородные бактерии в 9-м изда- нии Берджи отдельной группы не обра- зуют. Энергетически обмен этих орга- низмов основан на обратимой реакции гидрогеназы Н = 2Н + 2е, которая лег- ко сочетается с электронтранспортной цепью. Основным продуктом обмена является вода. Субстрат роста, водород, служит характерным продуктом обмена первичных анаэробов. Для литоавто- трофного роста необходима еще реак- ция автотрофной ассимиляции СО, ко- торая осуществляется через рибулозо- бисфосфатный цикл. Поэтому гидроге- нотрофная автотрофия широко распро- странена среди бактерий, получивших название "гидрогеномонады". На самом деле среди них много очень разнообраз- ных по положению организмов. Почти все они органотрофы, использующие преимущественно органические кисло- ты, и, что характерно, формиат, но не метанол. Основной моделью водород- ных бактерий послужил Аlcaligenes eutrophus, обмен которого был изучен Г. Шлегелем (Н. Schlegel) и его сотрудни- ками в Гёттингене. В числе водородных бактерий следует отметить псевдомо- нады Acidovorax ( Hydrogenomonas facilis"), Hydrogenophaga, Pseudomonas, anthomonas, Paracoccus. Среди других форм Aquaspirillum, Ancyclobacter, Fla- vobacterium, азотфиксирующие организ- мы Rhizobium, Derxia, обязательно обла- дающие гидрогеназой. К грам-положи- тельным водородным бактериям отно- сятся Arthrobacter, Bacillus, Nocardia. Штамм Alcaligenes eutrophus Z-l был ис- пользован Т. Валовой в Красноярске для производства белка на водороде в сопря- женной реакции с электролизом воды. Особую группу составили экстре- мально термофильные роды Hydroge- nobacter, Calderobacterium, Aquifex. Они развиваются при температуре более 70 'С, относятся к очень глубокой филогене- тической ветви бактерий и обладают необычным путем ассимиляции СО,. Выделенный из термофильного циано- бактериального мата на Камчатке штамм Calderobacterium был исполь- зован для производства биомассы на во- дороде при 70 'С, когда исключен рост условно патогенных форм. Штамм ока- зался очень технологичным, с высоким содержанием цитохромома с, сходного с бычьим. В этом разделе мы не будем об- суждать обширную группу анаэробных гидрогенотрофов, включающую гомо- ацетатные бактерии, водородные мета- ногены, сульфатредукторы, магнетит- образующие бактерии. Карбоксидобактери и осущест- вляют суммарную реакцию 2СО + О = 2СО. В большинстве это аэробные водород- ные бактерии, способные к использова- нию СО под действием специфического металл содержащего фермента. Типич- ными представителями этой медленно растущей группы являются Carboxy- domonas, Zavarzinia и некоторые другие роды. От аэробных карбоксидобактерий следует отличать недавно открытую у нас В. Светличным группу анаэробных карбоксидотрофов, использующих в качестве окислителя воду по реакции СО+Н,О=СО,+Н, с типичным представителем Carboxy- dothermus hydrogenoformans, термо- фильным организмом, использующим в качестве субстрата только СО. Это мед- ленно растущие организмы. Метанотрофы осуществляют ре- акцию: СН,+О=СО,+Н,О, которая, несмотря на свою кажущуюся простоту, требует высокой энергии акти- вации и поэтому доступна лишь специ- фической группе организмов. Открытые в 1906 г. Н. Зёнгеном (N. Sohngen), они долгое время не были объектом интенсивных исследовании и лишь с 27
ар)ебактер грамположительные П~вгеноты грамотрицательные — протеобактерии укариоты эубактери ианобактерии Схема филогенетического древа бактерий палочки с сериковым путем анаболизма, склонные к микроаэрофилии. Для Me- thylosinus известна способность образо- вывать экзоспоры. До последнего вре- мени не были известны метанотрофы способные развиваться в экстремальных условиях. Только сейчас С. Дедыш най- дены ацидофильные формы, растущие при рН 4-5 и психрофильные организмы с оптимумами вплоть до 2 'С (Василье- ва, Омельченко). Метилотрофы разделяются на облигатных, таких как Methylobacteri um, Hyphomicrobium, и факультативных, к которым принадлежат представители различных групп бактерий и другие ми- кроорганизмы, вплоть до дрожжей Candida. В настоящее время приведенная выше фенотипическая классификация заменяется филогенетической, которая была разработана К. Вёзе (К. Woese) в середине 1970-х гг. В ее основе лежит сопоставление всех организмов по одно- му гену малой рибосомальной РНК, 16S для прокариот и 18S для эукариот, что дало возможность построить единую классификацию всего живого мира, а не переходить к разным наборам призна- ков, как это неизбежно при сравнении, например, насекомых и грибов. Сопо- ставление картин, полученных по 5S-, 16S-, 23S- рРНК, показало их сходство, и, следовательно, все РНК малой части- цы рибосомы, отвечающие за трансля- цию, эволюционировали согласовано. Анализ по 16S рРНК предоставил воз- можность создать единую классифика- ционную систему, которую можно отра- зить на графе сходства последователь- ностей оснований в гене. Последова- тельность оснований устанавливается аналитически с большой точностью и 1960-х гг. привлекли интерес после описания Р. Виттенбари (R. Whittenbury) их разнообразия, с последующим изуче- нием биохимических путей метилотро- фии и использованием для производства белка одноклеточных на природном газе, а также для контроля за эмиссией парникового газа метана. Ключевым ферментом обмена служит медьсодер- жащая метанмонооксигеназа (ММО), акисляющая СН, в метанол по реакции СН, + О+ ХН = СН,ОН + Н,О+ Х, где Х — клеточный донор электрона. От образования метанола пути обмена оказываются сходными для метило- трофов — организмов, использующих метилированные соединения для обме- на. Метанол окисляется метанолдеги- дрогеназой в формальдегид, который используется в анаболизме для постро- ения углеродных скелетов по двум разным путям — рибулозомонофосфат- ному или сериновому. Метанотрофы хорошо распозна- ются под микроскопом, представляя довольно крупные клетки, темные под фазовым контрастом из-за пачек вну- триклеточных мембран. Метанотрофы- строго специализированные организмы, использующие только метан и кислород, факультативные формы среди них неиз- вестны. Сродство к метану очень высо- ко, метанотрофы способны извлекать его из атмосферы при концентрации 1700 частей на миллиард (р.р.b.). Из- вестные метанотрофы представлены нейтрофиль ными и мезофильными организмами. Они разделяются на две группы: Группа 1 Methylobacter, Methylo- monas, Methylococcus - аэробные па- лочки и кокки с рибулозомонофос- фатным путем анаболизма, образую- щие цисты. Группа II Methylocystis, Мейу1о- sinus - почкующиеся или изогнутые ФИЛОГЕНЕТИЧЕСКАЯСИСТЕМАТИКА
Таблица 3. Сходство и различия основных групп бактерий rc aeo acteria и acteria ar ota изнар Метаногены экстре мальн ые термофилы. гало илы Типичные организмы Протисты, грибы, растения, животные 35 групп Типичные размеры 0,5-4 мкм ,5-4 мкм >5 и»5 мкм Ядро с многими сложными хромосомами Есть Кольцевая хромосома Кольцевая хромосома Геном Гистоны Есть Нет Обратная гираза Клеточная с генка Есть Нет Нет Протеин, псевдо муреин Муреин, липопол исахарид Эфиры жирных кислот и глицерина Различная Этери ицированные изоиреноиды иры жирных кислот, стеролы Мембрана Обычны для компартментализации клетки 80S + 70S рибосомы органелл, как у бактерий Внутрицитоплазма- тические мембраны Нет Обычно нет, или белковые Рибосомы 70S 70S Синтез белка ингибируется: Хлорам ениколом Нет Нет иклогекси мидом Нет Нет РН-поли мераза Сложная Сложная Простая 29 расхождений между лабораториями ма- ло. Сопоставление последовательно- стеи отчасти зависит от взятого для ана- лиза набора организмов, и здесь могут получаться разные деревья. По последовательности нуклеоти- дов в малых рибосомальных РНК было выделено три группы организмов "Bacteria", "Archaea", "Еисагуа" как три равноправных линии в графе отноше- нии сходства в последовательностях рРНК. Такое деление стало общеприня- той догмой классификации, под кото- рую подгоняются остальные характери- стики. Для утверждения филогенети- ческой систематики потребовалась прежде всего корреляция с другими при- знаками, например со строением клеточ- ной стенки, составом мембраны, далее признание митохондрий и хлоропластов эукариот потомками прокар иоты ых эндосимбионтов в соответствии с гипо- тезой симбиогенеза Л. Маргулис (L. Mal- gulis). Свободноживущего организма, соответствующего "Еисагуа", но не име- ющего симбиогенетических органелл, пока нет, хотя поиск его среди анаэроб- ных протистов велся интенсивно. Поэтому предшественник(и) цитоплаз- мы и ядра эукариот представляют гипо- тетическую реконструкцию; восходя- щую к "прогеноту" - предку всех живых организмов, давшему три ветви живого мира, или, как их называли, три "домена" (см. схему). Эту номенклатуру предлагается с 1999 г. заменить на уза- коненную и признать существование таксона высшей категории Imperium (мн. число Imperia), а не на "надцарст- ва". Множество живых существ распре- леляется между тремя империями: Eukaryota (не Еисагуа); Archaeobacteria (не Archaea); Eubacteria (не Bacteria). Соответствующие русские термины:
эукариоты, архебактерии, эубактерии. Применение этой номенклатуры, поми- мо соответствия правилам Кодекса, имеет то преимущество, что прокариоты имеют общий определяющий термин "-bacteria" в отличие от эукариот и, таким образом, закрепляет тривиальное поня- тие бактерий как организмов, противо- поставленных эукариотам. Из списка сходств и различий Eubacteria, Archaeo- bacteria, Eukaryota, представленных в таблице 3, видно, что они носят специ- фический характер, относящийся к де- талям ("коррелирующим признакам"), а не к свойствам целого организма. Если различие между прокарио- тами и эукариотами очевидно как раз- личие между простыми (элементар- ными) и сложными (комбинированны- ми) организмами, то различие между архебактериями и эубактериями далеко не столь ясно. Эубактерии и архебак- терии имеют очевидное сходство в стро- ении клетки, клеточном цикле, 60% сходства в последовательности рРНК, различие в метаболизме не больше, чем между разными группами эубактерий при большом сходстве в анаболизме. Сопоставление ряда общих ферментов архебактерий и эубактерий, часто вы- полненные на отдельных представите- лях без уверенности в приложимости вывода ко всему исследуемому множе- ству, показывает разные комбинации сходств и различий. Существенны раз- личия в аппарате синтеза белка. Посту- лирование различного происхождения эубактерий и архебактерий вызывает серию вопросов о причинах сходства, на которые трудно ответить без большой серии допущений, лежащих вне филогенетического подхода. Ответ требует анализа полных последователь- ностей ДНК архебактерий и эубактерий и сопоставления не только генов синтеза белка, но и других. Материал для такого сопоставления быстро накапливается и трудности заключаются в его анализе и выборе приоритетов. Следующим вопросом было сопо- ставление этого классификационного дерева с эволюцией. Постулировалось, что мутации, ведущие к замене нуклео- тидов, происходят случайно и число замен находится в прямой зависимости от времени; наибольшее число замен должно накопиться у самых древних ор- ганизмов. Приняв это допущение, мож- но реконструировать на основе класси- фикационного дерева рРНК гена фило- генетическое дерево организмов. Ре- зультат такой реконструкции в общем совпал с картиной эволюции, установ- ленной на основе палеонтологии и срав- нительного подхода: многоклеточные растения, животные, грибы заняли вер- шину дерева. Упорядочение множества прокариот было субъективным, и те со- впадения в группировании, которые об- наружились, например, у грамотрица- тельных и грамположительных бакте- рий, послужили основанием для приня- тия Н0В0Н систематики как естествен- ной. Следующим вопросом является со- отношение филогении и эволюции. Оба понятия подразумевают последователь- ность во времени, но филогения отно- сится к генеалогии организма, или груп- пы организмов, а эволюция может под- разумевать и более широкие понятия, например, эволюцию осадочного процес- са, который сохраняет свою историю в виде последовательных слоев. Палеон- тологические данные, когда их удается твердо установить, носят силу факта, а филогения — реконструкции. Все эти со- ображения становятся особенно важны, когда рассматриваются наиболее простые организмы — прокариоты. Молчаливо до- пускается, что древние организмы сохра- няются без существенных изменений. Различия во внутренней сфере организма могут иметь слабое выра- жение в его направленных к среде оби- тания функциональных или фенотипи- ческих характеристиках. Для экоси- стемы не важно, какие внутренние изменения происходят в клетках и ап- парате синтеза белка, коль скоро он функционирует, но очень важны хими- ческие функции организма. При анализе противопоставляются два подхода: фило генетических последовательных рядов и комбинаторный для сопостав- ления фенотипических функций. В одни и те же филогенетические ветви попадают крайне разнообразные по физиологии организмы, например, анаэробные фотоавтотрофы пурпурные бактерии и энтеробактерии, цитофаги и бактероиды. При переходе от крупных ветвей грамположительных и грам- отрицательных бактерий к низшим иерархическим уровням таких несовпа- дений становится все больше. Из при- веденных групп бактерий подавляющее большинство таксонов входит в груп- пировки цианобактерий, протеобакте- рий, грамположительных организмов. Эти три ветви хорошо различимы друг 30
от друга. Самостоятельное положение ветвей спирохет, зеленых фототрофов также согласуется с традиционным представлением о единстве и своеобра- зии этих групп. Зеленые бактерии обыч- но функционируют в сходной эконише с серными пурпурными, но сильно разли- чаются по строению фотосинтетичес- кого аппарата. Однако объединение ци- тофаг, традиционно считавшихся близ- кими аэробным миксобактериям, и облигатно анаэробных бактериоидов не имеет функционального обоснования. О. Кандлер, К. Вёзе, и Штеттер (О. Kandler, К. Woese, K. Stetter) нашли, что наиболее уклоняющиеся по последова- тельностям рРНК ("древние") группы принадлежат экстремальным термофи- лам и на этом основании предположили происхождение прокариот из гидротер- мальных мест обитания. С позиций природоведческой мик- робиологии эубактерии и архебактерии рассматриваются вместе как функцио- нальные группы прокариот. Среди архе- бактерий пока выделены крупные таксо- ны "Euryarchaeota" и "Crenarchaeota" по признаку повсеместного присутствия (Eury-) и термофильных, развивающих- ся в потоке (Cren-) в газогидротермах, но эти названия не узаконены. Специа- лизация архебактерий позволяет рассма- тривать отдельно их функциональные группы метаногенов, галофилов, экстре- мальных термофилов. Роды экстремаль- ных термофилов вследствие своего уни- кального положения на 16S рРНК де- реве получают высокие таксономичес- кие ранги. Наибольшую трудность в корреля- ции филогенетической и функциональ- ной систематики представляет наиболее многочисленный и разнообразный комплекс цианобактерий, протеобакте- рий, грамположительных организмов. Было сделано предположение Г.А. За- варзиным, Э. Штакебрандтом (Е. Яа- kebrandt) и Р. Мюрреем (R. Murray), что этот комплекс отражает свое происхож- дение из циано-бактериального сообще- ства как стволовой эволюционной груп- пировки микроорганизмов. Для грам- отрицательных организмов вместо обо- значения "пурпурные бактерии и их род- ственники" был предложен класс Pro- teobacteria. Протеобактерии, ветви кото- рых оцениваются в ранге подклассов, пока обозначаемых греческими буква- ми, попытались разделить на экологи- ческие группировки. Альфа-подкласс включает группу несерных пурпурных бактерий, почкующихся и простекобак- терий, ризобии и агробактерии; среди органотрофов этого подкласса относи- тельно много олиготрофных организмов и связанных с высшими растениями. В бета-подкласс входят "газотрофы", на- пример, метанотрофы и нитрификато- ры, составляющие естественную группу по особенностям строения клетки с мощным мембранным аппаратом и об- меном, основанным на окислении газов анаэробного происхождения. Гамма- подкласс включает органотрофных ко- пиотрофов: факультативных анаэробов- бродильщиков энтеробактерий и вибри- онов, типичных аэробных окислителей псевдомонад, а также бесцветных серо- бактерий, серных пурпурных бактерий. Дельта-подкласс объединяет крайне раз- личных функционально сульфат-реду- цирующих бактерий, бделловибрионов, миксобактерий, занимающих противо- положные позиции в трофической си- стеме. Корреляция филогенетических групп протеобактерий с экологическим положением в системе носит характер лишь общей тенденции с многочис- ленными отклонениями. Классификация прокариот по гену малой субъединицы рибосомальной РНК имеет преимущество идентифика- ции родов прокариот с помощью слож- ных технически молекулярно-биологи- ческих методов и последующей ком- пьютерной обработки результатов опре- деления последовательностей нуклео- тидов. Быстрая идентификация прово- дится с помощью генных проб. Не- удобством классификации по 16S рРНК оказывается появление случаев, когда фенотипически сходные организмы ока- зываются принадлежащими разным ветвям и возникает дробление родов по единственному критерию с возрастаю- щей неустойчивостью систематики на родовом уровне. Из положения организ- ма на филогенетическом дереве можно сделать лишь ограниченные заключения о его функции. В качестве общего пред- ставления можно полагать, что прокари- оты имеют некие сердцевинные компо- ненты, такие как репликацию генома и синтез белка, и периферические, опре- деляющие их функции, то есть значи- мые для занятия функциональной (эко- логической) ниши и выживания в тече- ние эволюции. Итак, микробиолог имеет дело с множеством родов прокариот и разными 31
Таблица 4. Распределение родов Proteobacteria по типам питания и подклассам ганотро ы итотро ы ототро ы ь а-подкласс atro aeter о opseu omonas, odonuсгоЬшт, О OS Dl ИШ Um um, О ЯС1СГ, izobium, cker, псе а, аи ra coccus, osthecomicrobi um, anthobacter, Stella, ymomonas ета-подкласс tothrix, с rorno aeter, ethy1omonas, ei sseria, Methylo romobacteri um, ct ococcus, С' о itrcosci л, амма-подкласс i atoa, i othrix, iomicros ira, erichia, cineto aeter, tothior о 1ra, on iocapsa, ios ~гЫит, тОПаЗ, mona. ~оп сПа, еи omonas,ceanos irillum, so aeter, ucot rix nt omonas, льта-подкласс ces, Bde ои по, Soranóèm, ococcus, Знак « * » означает, что разные виды этого рода попадают в разные ветви. 32 ceto aeter, astobacter, Ас~ zomicrobi um, ro bacterium, omonas, edomicrobi um, uaspirillum"", eij erinckia, Вгл ncalomicrobium, teromon as, a ebsiella, Azo otobacter, i trobacter, Halo crratia, Proteus, по, Moraxclia otobactcri um, ondromy stobacter, obacter, atella ca enes, aeter, SiiuonsieIla, mam onas, osinus, itrosococcus, i trosolobus, i trosospira, iobacr us" esulfo vibrio, sulfobacter, sulfobulbus, esulfococcus, esvlfonema, esu uromonas ое о aeter, amprocystis, iodiction, Th Chromatium
способами их идентификации. Задача сводится к поиску рода, т.е. соответст- вующего ему слова, а не 1500-значного обозначения нуклеотидной последова- тельности. Роды обозначаются назва- ниями и трудность заключается в не- устойчивости номенклатуры с множе- ством синонимов, затрудняющей ис- пользование старой литературы и све- дений из непрофессиональных изданий. Удобнее всего для установления соот- ветствия алфавитные списки названий с указанием синонимов, которые перио- дически выпускает Немецкая коллекция культур (DSMZ). Они служат хорошим примером удобства однозначного упо- рядочения множества объектов (в дан- ном случае родов прокариот) по алфа- виту, как это делается в библиотеках, где предметные каталоги не всегда дают нужную классификацию. В нашем случае важно рассмо- треть прежде всего главных первичных продуцентов (1) цианобактерий; затем (2) протеобактерий, разделив их на группы органотрофов, фототрофов, литотрофов; (3) грамположительных организмов, разделив их на группы одноклеточных бактерий и мицели- альных актиномицетов. Распределение важнейших родов Pro teobacteria по подклассам и типам питания представ- лено ниже (таблица 4). Грамположительные организмы, таксономия которых находится в стадии формирования, разделились на ветви с "низким Г+Ц" и "высоким Г+Ц". В ре- зультате виды старого рода Clostridium, под которым понимали строго анаэробные спорообразующие палочки, распредели- лись по множеству новых родов, обоз- начаемых как "клостридиальная ветвь с высоким Г+Ц" или "клостридиальная ветвь с низким Г+Ц". К последней ветви примыкают галофильные грамотрица- тельные анаэробы порядка Haloanae- robiales, часть которых спорообразую- щая (олигоспоровые). Такая же таксо- номическая судьба поститла множество родов грамположительных облигатных анаэробов (большая часть которых по- лучила новые названия и их переклас- сификация создает ложное впечатление увеличения знаний) и поэтому сущест- вующая их систематика носит времен- ный характер. Более устойчива система аэробных грамположительных организ- мов, близких к актиномицетам. Коротко следует перечислить ма- лые ветви филогенетической системы. Ближе всего к корню дерева собственно бактерий располагаются экстремально терм офильные водородные бактерии родов Hydrogenobacter (Calderobacteri- ит, Aquifex), Thermocrinus, растущие при температуре )70 'С за счет окисле- ния Н хемолитоавтотрофно с необыч- ным путем ассимиляции углекислоты и очень богатые цитохромом. Следующую группу составляют экстремально термофильные органо- трофные Thermotoga, грамотрицатель- ные организмы, имеющие на концах клеток характерные вздутия наружной мембраны, создающие пузыри перипла- ста. Термотоги относительно легко вы- деляются из разных мест обитания, как морских, так и пресноводных, а также из подземной гидросферы. Нитчатые фотогетеротрофные зеленые хлорофлек- сусы обычно обнаруживаются в тер- мальных источниках при более высокой температуре, чем термофильные циано- бактерии. Дейнококки в общем напоми- нают аэробных органотрофных микро- кокков, но отличаются необычайной радиоустойчивостью. Цитофаги и бактероиды состав- ляют многочисленные и важные группы и обсуждаются по отдельности, не имея общих фенотипических свойств. Цито- фаги служат важнейшими аэробными деструкторами целлюлозы и благодаря их способности к скольжению долго рассматривались как не имеющие пло- довых тел миксобактерии. Бактероиды- пептолитические анаэробы, составляю- щие основную часть бактериального населения кишечника. Заканчивая обзор филогенетических ветвей прокариот следует заметить, что он дает очень мало опорных пунктов для логическои систематизации сведении, а тем более руководящих принципов в по- строении большой системы. Филогене- тическая система, основанная на одном рибосомальном гене, не более чем одна из технически удобных и разработан- ных систем упорядочения множества организмов с целью их идентификации. МИКРОБНОЕ СООБЩЕСТВОКАКЦЕЛОСТНОСТЬ Микробное сообщество представ- ляет совокупность взаимодействующих между собой организмов, а не мно- жество видов, находящихся в одном месте, и представленных аналогом фло- ристического перечисления, например, 33
идентифицированных клонов ДНК. Способы взаимодействия микроорга- низмов между собой различны. Для всех организмов первоочеред- ное значение имеет питание. Взаимо- действующие микроорганизмы связаны между собой либо конкуренцией за об- щие субстраты, либо кооперацией в их использовании. Слово "субстрат" в ми- кробиологии многозначно и может обо- значать как твердый носитель, напри- мер, поверхность растения, так и сумму используемых веществ, но чаще всего употребляется как "субстрат питания", т.е. источник питания, индивидуальное химическое вещество, реактант в хими- ческом смысле. В микробном сообще- стве в связи со специализацией функций кооперация имеет гораздо большее зна- чение, чем, например, в сообществе ра- стений, принадлежащих к одной физио- логической группе и конкурирующих за свет, воду, биогены. Конкуренция у бактерий проявляется в пределах одной функциональной группы, например при колонизации общего субстрата, скажем, молока, молочнокислыми бактериями. Кооперация нагляднее всего проявляет- ся в последовательном разложении сложного субстрата как мортмасса ра- стении, где каждыи компонент исполь- зуется своим специалистом, а продукты его обмена в свою очередь использу- ются другими специалистами. Химичес- кие взаимодеиствия настолько домини- руют в микробном сообществе, что его можно представить как гомогенную си- стему, бактериальную взвесь или сус- пензию. Именно эта система охотнее всего исследуется микробиологами по соображениям удобства и дает наиболее глубокое представление о природе ми- кробного сообщества. Однако в природе организмы развиваются на поверхности твердых субстратов, образуя обраста- ния, которые называют биопленками. Типичным примером их служит циано- бактериальный мат. Именно обрастания становятся объектом фоссилизации и распознаются палеонтологами. СИСТЕМА БИОГЕОХИМИЧЕСКИХЦИКЛОВ Связь между биогеохимическими циклами в биосферной системе осуще- ствляется серией частных реакций в этих циклах. Традиционная общая ми- кробиология оказалась подготовленной к решению такой задачи, разработав в течение столетия систему "физиологи- ческих групп" и "морфо-физиологи- ческую" систематику бактерий, преиму- щественно на уровне родов и, иногда, семейств. По своим принципам эта си- стема совпадает до известнои степени с системой "жизненных форм" ботаников и зоологов. Система биогеохимических цик- лов (рис.1) определяется ведущим цик- лом органического углерода C„„,. и со- пряженными с ним в эквимолекулярном отношении 1:1:1 циклами углекислоты СО и кислорода О,. Эквимолярное соотношение между этими циклами сле- дует из хорошо известного уравнения, описывающего фотосинтез и дыхание: СО, + НО = [СНО] + СО. Цикл органического углерода обуслов- лен (1) первичной продукцией за счет использования внешнеи солнечнои энергии, прежде всего, фотоавтотроф- ными оксигенными организмами (циа- нобактериями, водорослями, растения- ми) и в небольшой степени хемоавто- трофами — за счет поступления эндоген- ного водорода в газогидротермах, и (2) деструкцией, осуществляемой органо- трофными организмами, аэробными и анаэробными. Деструкционная ветвь цикла органического углерода наиболее сложна и поэтому её следует рассмо- треть подробнее отдельно, ознакомив- шись с общими принципами органи- зации участия сообществ микробов в циклах. Конечным продуктом деструк- ции служит углекислота, замыкающая цикл органического углерода и сопря- гающая его с циклом неорганического углерода и циклом кислорода. Цикл органического уптерода до- полняется циклом азота, в который азот органических соединений входит в соотношении С:И=6:1, а также происхо- дят превращения неорганических форм азота. Цикл азота с его этапами — азот- фиксацией, аммонификацией, нитрифи- кацией, денитрификацией - целиком определяется деятельностью бактерий. Цикл фосфора стехиометрически связан с циклом органического углерода в отношении С:Р=100:1 (в реакциях анаболизма). В цикле фосфора, как уже отмечалось, в отличие от других био- генных макроэлементов отсутствует стадия воздушной миграции, обеспечи- вающая равномерное распределение по всему земному шару с воздушными потоками.
Цикл неорганического углерода смыкается через углекислоту воздуха и её растворенные формы в гидросфере с циклом органического углерода. В ли- тосфере неорганический углерод пред- ставлен в основном карбонатными поро- дами, прежде всего карбонатом кальция. Цикл кальция определяется преж- де всего биологически опосредованны- ми реакциями растворения (вьпцелачи- вания) и осаждения карбонатов, а также биоминералогией образования скелета некоторыми протистами и макроорга- низмами. Цикл кальция сопряжен также с циклом фосфора и образованием и растворением фосфатов кальция. Первичное поступление кальция и магния в биологические циклы свя- зано с циклом кремния и выщелачива- нием силикатных изверженных пород, вообще говоря, идущим под воздейст- вием углекислоты, но ускоряемым при- мерно в 100 раз под воздействием микроорганизмов и продуктов их об- мена в биологически опосредованных процессах. Выщелачивание обусловли- вает поступление в водную фазу микро- элементов. С циклом органического углерода сопрягается цикл серы в катализируемых только бактериями реакциях сульфат- и сероредукции (сульфидогенеза), окис- ления соединений серы аноксигенными фототрофными и аэробными хемотроф- ными организмами. В биогеохимичес- ком цикле серы участвуют следующие формы соединений серы, создающие значительные резервуары: 1) сульфаты, преимущественно сульфаты моря; 2) сульфиды в виде растворенного H S и нерастворимых сульфидов металлов, ча- стично эндогенного (вулканического) и в основном экзогенного (биогенного) происхождения; 3) сера, в значительной части эндогенного происхождения. Раз- нообразные промежуточные соединения неполного OKHGJIOHHsl серы, как тиосуль- фат или SO, ïîÿâëÿþòñÿ в транзитных формах и незначительной концентра- ции, не образуя резервуары. В цикле серы бактерии осуществляют окисление сероводорода и сульфидов либо при фотосинтезе, либо за счет внешних доноров электрона. Цикл железа сопрягается с циклом кислорода деятельностью аэро быых железобактерий, окисляющих закисное железо в гидрат окиси железа, и с циклом органического углерода деятель- ностью железоредуцирующих бактерий, образующих восстановленное железо и магнетит. Цикл железа связан с циклом серы через образование сульфидов же- леза и их окисление бактериями. Биогеохимическая машина плане- ты представляется системой взаимосвя- занных циклов элементов. Эти циклы действуют как в планетарном масштабе, так и в конкретных ландшафтах-экоси- стемах.
ЦИАНОБАКТЕРИИ,ЦИАНО-БАКТЕРИАЛЬНЫЕ СООБЩЕСТВА,МАТЫ, БИО ПЛЁНКИ 36 Цианобактерии, или синезеленые водоросли, стоят особняком среди царства эубактерий. С эубактериями их объединяют: прокариотное строение клетки, состав клеточнои стенки, наличие газовых вакуолей, наличие 70$- рибосом, так же как в хлоропластах и митохондриях (вместо 80S), чувстви- тельность клетки к пенициллину и стрептомицину, запасное вещество оксимасляная кислота, фиксация азота, рост и выживаемость при высоких тем- пературах. С эвкариотами сближают: оксигенный фотосинтез, присутствие многоклеточных дифференцированных форм, отсутствие жгутиковой стадии, наличие хлорофилла "а", строение кара- тиноидов, наличие специфических ли- пидов в мембранах. Цианобактерии представляют со- бой целый мир (1500 видов) разнооб- разных одноклеточных, колониальных, нитчатых форм (см. таблица 2, с.16), размеры клеток у которых колеблются от 2 до 50 микрон. Это грамотрица- тельные организмы с 4-хслойной кле- точной стенкой; слои стенки обознача- ются символами L-1, L-2, L-3, L-4. Слой L-1 богат ферментами, участвующими в полимеризации предшественников кле- точной стенки. Слой L-2 — муреиновый мешок, обеспечивающий ригидность клеточной стенки. Слой L-3 состоит из фибрилл, участвующих в процессе дви- жения. Слой 1.-4 представляет собой двухслойную мембрану. Покровы. Помимо клеточной стенки у ряда цианобактерий присутст- вуют различные покровы, часто слизи- стые. На наружной мембране клеточной стенки может образовываться S-слой, состоящий из белковых глобул гекса- гональной структуры (состав - глико- протеин). Выделяемая клетками слизь носит название гликокаликс (кислые полисахариды), имеет фибриллярное строение и выполняет защитную роль. Чехол (или влагалище), окружающий трихомы многих нитчатых цианобакте- рий, может быть гомогенным или фи- бриллярным, однослойным или слои- стым. Чехол выполняет защитную роль, но у Плеврокапсовых он также играет роль клеточной оболочки. На поверх- ности клеток цианобактерий могут развиваться различные образования: пили, фибрии, иглы. Пили и фибрии представляют собой полые белковые трубочки, проходящие через клеточную стенку и прикрепляющиеся к цитоплаз- матической мембране. Иглы, или спины, описаны для водных планктонных форм и предназначены для увеличения поверхности клетки. Это цилиндры 100 х 1000 нм. Таким образом, поверхност- ные структуры выполняют различные функции и играют экологическую роль. Включения. Клетки цианобакте- рий содержат различные включения. Газовые вакуоли присутствуют на от- дельных стадиях жизненного цикла по- стоянно или возникают в определенных условиях. Они состоят из правильных рядов газовых пузырьков, напоминаю- щих пчелиные соты, и окружены мем- браной. При избытке углекислоты и де- фиците азота в качестве запасного ве- щества образуются гранулы гликана, который расходуется при дефиците углерода и в темноте. Цианофициновые гранулы имеют овальную или непра- вильную форму, в некоторых случаях
они могут достигать размера до 1 мкм в диаметре. Этот запасной продукт слу- жит источником азота. Полифосфаты в виде гранул волютина имеют жизненно важное значение, так как являются источником фосфора для клетки при де- фиците его в пресных водах. Карбо- ксисомы - полиэдральные тела — являют- ся депо ключевого фермента фотосинте- за — рибулозобифосфат-карбоксилазы. Фотосинтетический аппарат образован сложнои системои плоских дисков тилакоидов, в мембранах которых со- держатся хлорофилл и каратиноиды. Фикобилиновые пигменты, свойствен- ные только цианобактериям, локализо- ваны в особых структурах — фикобили- сомах и представляют собой высокоор- ганизованные белковые комплексы, при- крепленные к наружной поверхности тилакоидной мембраны. Репродуктивные клетки. Для большинства цианобактерий в основе размножения лежит процесс деления клеток надвое. В отличие от бактерий и эвкариот оно начинается с втягивания плазмалеммы и слоя L-2. У некоторых видов размножение происходит путем почкования или множественного деле- ния, то есть деления в нескольких пло- скостях. Такой процесс деления не со- провождается ростом клеток, а приво- дит к образованию маленьких сферичес- ких дочерних клеток - эндоспор, или, согласно современной терминологии, баеоцит. Бинарное деление и почкова- ние характерно для представителей 1 секции цианобактерий, в то время как множественное деление — для 2 секции (см. таблицу 2, стр.16). Обе секции, как указывалось выше, объединяют одно- клеточные цианобактерии, в то время как остальные состоят из нитчатых форм. 3-я секция включает нитчатые (трихомные) формы, растущие, благо- даря интеркалярному делению клеток, лишь в одной плоскости. Для этой секции характерно образование гормо- гониев — фрагментов, на которые распадается трихом. Они состоят из двух-трех или большего числа клеток. Нить распадается на такие куски в результате гибели нескольких средних клеток трихома, и гормогонии освобож- даются, выходя из влагалища наружу, и движутся вперед скольжением. Из гормогониев вырастают новые нити. Многие виды (секция 4 и 5) образуют неподвижные покоящиеся клетки аки- неты. Эти клетки крупнее вегета- тивных, толстостенны, заполнены бел- ком и запасными веществами. Особые клетки — гетероиисты — выделяются своим сильным светопреломлением. Обычно это очень большие клетки с утолщенной 3-хслойной оболочкой цел- люлозной природы. По бокам они име- ют полярные шапочки — светопрелом- ляющие гранулы. По содержанию фер- ментов и метаболизму гетероцисты от- личаются от вегетативных клеток. Са- мое главное их предназначение — фикса- ция молекулярного азота. Кроме того, они являются репродуктивными клетка- ми, т.к. прорастают и дают нормальную нить. В 4-й секции акинеты - укрупнен- ные клетки измененнои пигментации, располагаются чаще всего около гетеро- цист. Некоторые виды могут образовы- вать гормоспоры - короткие ряды грану- лированных клеток. Экология. Цианобактерии разде- ляются на водные формы и почвенные, растущие на поверхности почвы, в поч- ве или в каменистых породах. Среди водных форм различаются планктонные и бентосные. Планктон может бытытрес- новодным и морским. Продуктивность пресноводных цианобактерий падает с глубиной в 6 раз при погружении на 1 м. Из морского планктона продуктивную роль играет только пикопланктон (одно- клеточные цианобактерии с диаметром клетки меньше 1 мкм). Распределение бентосного планктона по глубине зави- сит от содержания О, H S и рН. Самое интенсивное развитие цианобактерий наблюдается в литоральной зоне. Особое внимание обращает на себя гидрофитон горных речек, богатый СаСОЗ и являющийся туфообразовате- лем, и эндофитон, развивающийся внут- ри каменистого субстрата, в раковинах и старых костях (сверлящие водоросли). Последние извлекают для нужд фото- синтеза СО> и освобождают Са. Так, например, цианобактерия Scytonem a образует кустики, инкрустированные СаСО>, причем на субстрате,.не содер- жащем карбонатных солей. В этом слу- чае водоросли способны выбирать из среды Са, из воздуха СО~, накапливать на своих чехлах СаСО>, образуя мине- ральные футляры. Таким образом, в природе цианобактерии могут играть как разрушительную, так и созидатель- ную роль. Последнее время большее внима- ние стали уделять изучению не отдель- йых цианобактерий, а циано-бактери- 37
альных сообществ, куда входят и циано- бактерии, и бактерии, в связи с морфо- логическим сходством этих сообществ с их древними аналогами, обнаружи- ваемыми во многих древних породах. Сообщество синезеленых водорос- лей обычно включает разнообразные бактериальные организмы, которые, на- ряду с цианобактериями, определяют протекающие в сообществе геохими- ческие процессы. Такое сообщество на- зывается циано-бактериальным, в кото- ром циано- относится к синезеленой, а бактериальный — к бактериальной ком- поненте. По текстуре циано-бактери- альное сообщество представляет собою плотную слоистую ткань, сложенную множеством переплетенных нитей три- хомных организмов, погруженных в слизь, в которую заключены и другие бактерии. Это образование названо ци- ано-бактериальныл~ матом. В истории развития науки циано-бактериальные маты имели различные определения: слизь, растущая на поверхности, сое- диняющая частички и разрушающая породы (XVI в.); микробный или водорослевый ковер (XIX в.); стромато- литы (т.с. литифицированный микроб- ный ковер) от латинского слова "stromat- actis", или искусство делать ковры (начало ХХ в.); водорослевый мат, водорослевая кожа, водорослевый ковер (середина ХХ в.); бактериальный фильм или биопленка (середина ХХ в.); бак- териальный или микробный мат (конец ХХ в.). При интенсивном изучении циано- бактериальных матов в последние десятилетия ХХ в. термин "мат" зани- мал умы ученых на многих симпозиу- мах, прежде чем стало ясно, что циано- бактериальные маты - парагистологи- ческая кожа Земли — были и есть осно- вой жизни Земли, включая образование пород, их разрушение, изменение кли- мата и т.д. Цианобактерии в сообщест- вах с другими бактериями определили жизнь на Земле и динамику всех хими- ческих и физических процессов на нашей планете более чем 3 млрд. лет назад. И СеГОДНЯ ОНИ ЯВЛЯЮТСЯ ОСНОВНЫМИ КОН- тролирующими факторами циклов та- ких элементов, как Р, С, О, H2, N и S в глобальных процессах. Структура мата. Циано-бактери- альные маты являются бентосными по- пуляциями микроорганизмов с домини- рующими фототрофными бактериями, среди которых цианобактерии являются основными продуцентами органичес- кого вещества и отвечают за структуру мата (рис.1). Среди них доминирующи- ми формами являются, как правило, осциллаториевые цианобактерии, пред- ставляющие собой нитчатые подвижные организмы. В термофильных матах это Phormidium laminosum, в галофильных и алкалофильных - Microcoleus chthono- plastes. Фототрофные бактерии (зеленые Clzloroflexus aurantiacus - в термо- фильных, пурпурные — Thiocapsa, Ecto- thiorhodospira — в галофильных и алка- лофильных) осуществляют аноксиген- ный фотосинтез в экстремальных для роста цианобактерий условиях (рис. 2). Их вклад в первичную продукцию ор- ганического вещества в мате не превы- шает 20%. В условиях, благоприятных для роста цианобактерий, они осущест- вляют органотрофный рост, окисляя продукты жизнедеятельности цианобак- терий, или окисляют сероводород и, таким образом, участвуют в цикле серы. Занимая среднюю часть мата, эти мик- роорганизмы, с одной стороны, задер- живают фотосинтетический кислород, поступающий от цианобактерий, с дру- гой, защищают последних от сероводо- рода, который продуцируется сульфат- редукторами в нижних слоях мата. Сульфидогенез и метаногенез находятся в конкурентной зависимости, преобла- дание того или другого процесса опре- деляется содержанием сульфатов. По морфологии циано-бактери- альные маты представляют собою плот- ные структурные образования с ясно вы- раженными слоями, окраска которых определяется или цветом пигментов микроорганизмов, или образуемыми между ними минералами. Верхняя зона мата, занятая цианобакгериями, имеет зеленую окраску; зона развития фото- трофных бактерий имеет желтый (при развитии Chloroflexus aurantiacus — в термофильных матах) или вишневый цвет (при развитии пурпурных бактерий в галофильных и алкалофильных ма- тах). Черная зона — анаэробная зона раз- вития сульфидогенов - является мощ- ным генератором сероводорода, кото- рый, соединяясь с железом, образует гидротроилит, окрашивающий зону в черный цвет. Для нижних слоев зоны анаэробной деструкции характерна сложная цепь реакций, осуществляемых различными функциональными группа- ми организмов. Гидролиз полимеров водорослей и последующее сбражива- 38
Рис. 1. Термофильный мат. А, Б - плавающие термофильные маты теплых болот (кальдера Узон, Камчатка). Образование минерализованных "вулканчиков" - современных строматолитов (А). В- поперечный рарез мата (а - зона фотосинтеза с развитием цианобактерий, 0 — прослои карбоната кальция, в - минеральные слои с бляшками опала). Г - поперечный срез современного строматолита- "вулканчика". Д - верхний слой мата с цианобактериями (Phonaidium laminosum). 39
АФ~Ф ECV 8VВЬ8 V8 ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° е ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° а ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° IV Рис. 2. Схема строения галофильного мата. - цианобактерии — слон кареоната кальинл - слой развития пурпурных бак-терий с отложением гипса — сульфатредуцирующие бактерии 1 - зона оксигенного фотосинтеза с развитием цианобактери й П- зона аноксигенного фотосинтеза с развитием пурпурных бактерий Ш — зона захоронения цианобактерий с просло-ями карбоната кальция IV — аноксигенная зона развития сульфатре-дуцирующих бактерий ние продуктов гидролиза завершается полным окислением продуктов броже- ния в терминальных реакциях метано- генеза, сульфидогенеза и сероредукции с полной минерализацией органичес- кого вещества до СН4, СО, Í~. В гало- фильных сообществах эти газы проду- цируются самим сообществом, в термо- фильных они выносятся из гидротерм. Но в обоих случаях эти газы не попа- дают в атмосферу, они удаляются самим же сообществом. Таким образом, соз- дание физико-химических градиентов внутри мата обусловлено жизнедея- тельностью микрофлоры сообщества и ведет к образованию вертикальной зо- нальности мата. Во всех сообществах, независимо от места обитания, можно выделить три зоны: аэробную - с развитием цианобак- терий, строго анаэробную — с развитием сульфидогенов и метаногенов, и проме- жуточную - с развитием факультатив- ных аэробов. Чем же объясняется сходство столь различных в экологическом плане сообществ? По всей видимости, струк- тура сообществ задается, прежде всего, сходством функций населяющих его организмов. Организация трофических цепей в сообществе, в общем, одна и та же. Оксигенный фотосинтез прижат к верхнему слою толщиной в 1-2 мм, и здесь же происходит почти полное по- глощение света. В деструкционной вет- ви важное значение имеет серный цикл. Продуктивность матов. В усло- виях, благоприятных для развития циа- нобактерий, т.е. при солености не выше 16%, щелочности не выше 8%, темпе- ратуре не выше 45 'С, циано-бактери- альные сообщества являются высоко- продуктивной системой, первичная про- дукция которых коррелирует с содержа- нием хлорофилла. По количеству хло- рофилла циано-бактериальные сообще- ства не уступают, а иногда превосходят плотность хлорофилла в листовой пла- стинке деревьев и трав. Общая продук- тивность мата зависит от соотношения продуктивных и деструкционных про- цессов. В благоприятных для циано- бактерий условиях продукционные про- цессы преобладают, в неблагоприятных деструкцив выше продукции. Геохимические барьеры. Разви- тие определенных групп микроорганиз- мов в мате создает различные геохими- ческие барьеры: окислительно-восста- новительный, щелочной, сероводород- ный. Содержание кислорода, наивысшее в верхних слоях мата, благодаря фото- синтезу цианобактерий, внутри мата в пределах нескольких миллиметров па- дает до О, в то время как содержание се- роводорода возрастает. Содержание ки- слорода зависит от температуры и вре- мени суток. Оно наименьшее или равно 0 в промежутке от 20 до 8 ч и наиболь- шее в 12 ч. Значение ЕН в различных матах колеблется в пределах от +200 до -200 mV на глубине 10 мм. Активный фотосинтез цианобактерий ведет к на- рушению карбонатно-бикарбонатного равновесия и созданию щелочного ба- рьера. Значение рН в фотозоне воз- растает до 9,6 а ниже 1-миллиметровой зоны падает до 7. Образование геохими- ческих барьеров в мате ведет к осажде- нию ряда минералов. 40
Минералообразование. Все циа- нобактериальные сообщества образуют явно выраженную слоистую структуру с характерным чередованием зон разви- тия определенных групп микроорганиз- мов со слоями минералов. Минералооб- разованию именно в тонких прослоях, приуроченному к определенной группе микроорганизмов, способствует крайне низкая проницаемость матов. Низкая проницаемость предполагает молеку- лярную диффузию в мате и сводит к ми- нимуму эффект разбавления. В резуль- тате резкие изменения рН и ЕН, связан- ные с жизнедеятельностью микроорга- низмов, приводят к изменению равно- весия раствора и вызывают садку тех соединений, которые не способны нахо- диться в растворе в этих условиях. Несмотря на различия галофиль- ных и термофильных матов, между ни- ми наблюдатся не только структурное и функциональное сходство, но и сходное минералообразование: в обоих случаях осаждаются карбонат кальция и суль- фид железа. Осаждение карбонатов может быть обусловлено усиленным притоком минерализованных вод из глубины и резким изменением рН в зоне фотосин- теза в связи с удалением СО. на мелко- водье, где обмен воды невелик и быст- рое использование растворенного би- карбоната приводит к нарушению рав- новесия. Наилучшие условия для этого создаются под слоем активно растущих цианобактерий в связи с появлением ло- кальных зон с высоким рН и подтоком раствора, несущего Са++ и СО снизу. Таких прослоев может быть несколько, что зависит от периодов активного ро- ста цианобактерий. Прослои карбоната кальция, обра- зованные в фотозоне, при захоронении претерпевают ряд превращений, связан- ных с жизнедеятельностью присутст- вующих здесь микроорганизмов. С од- ной стороны, здесь возможна мобилиза- ция Са при образовании органических кислот первичными анаэробами. С дру- гой стороны, разложение органических кислот вторичными анаэробами, мета- ногенами и сульфидогенами может при- вести к новому отложению СОСО>. Сульфатредукция под матом созда- ет сероводородный барьер на пути миг- рации минерализованных вод и обус- лавливает отложение сульфидов железа гидротроилита или фрам 6 о идно го пирита. Ряд минералов в матах осажда- ется хемогенным путем. В галофильных матах это галит и гипс, в термофильных - мелкодисперсный кремнезем. Указанные минералы могут лити- фицировать цианобактерии или другие микроорганизмы, которые могут сохра- няться в таком виде неопределенно долгое время. Иногда литификация про- исходит за счет более редких минералов, например, сульфидов мышьяка в тер- мофильных матах. Обнаруженные в оча- ге разгрузки современной гидротер- мальной системы на Камчатке мине- ральные слоистые образования (рис. 1) ("вулканчики") сопоставимы со слои- стостью ископаемых пластовых и жел- ваковых строматолитов, широко рас- пространенных в докембрии. Это дает основание считать наблюдаемые по- стройки современными аналогами иско- паемых строматолитов. Циано-бактериальный мат фильтр газов. В естественных местах обитания под матом или между слоями часто образуются пузыри газа, иногда достигающие внушительных размеров (до 50 см в диаметре). Известно, что га- зовая смесь гидротерм, выходя на поверх- ность, состоит на 70-90% из СО2. Состав газов в пузыре существенно отличается как по составу газов, так и по их соотно- шению и от гидротерм и от воздуха. Та- ким образом, можно считать, что циано- бактериальный мат является модифика- тором газов. В лаборатории было показано, что как сообщество в целом, так и чистые культуры цианобактерий, помимо обыч- ной способности поглощать СО. и выде- лять О., оказываются способными по- глощать Н2Я с отложением серы, погло- щать Н в оксигидрогенной реакции. Если цианобактерии поместить в смесь газов, имитирующих газовые выделения гидротерм, или смеси, соответствующие рецептуре, предлагаемой для первичной атмосферы, то цианобактерии изменяют ее состав таким образом, что содержа- ние 02 становится около 20%, а восста- новители, кроме СН4, удаляются. При изучении роста цианобакте- рий в смеси различных газов стало совер- шенно ясно, что все они предпочитают микроаэрофильные условия, и в присут- ствии восстановителей (Н~, H~S) или в атмосфере азота и метана их скорость роста выше, чем в атмосфере кислорода. Пышное развитие цианобактери- альных биоценозов на выходах термаль- ных вод в современной обстановке гово- 41
рит о том, что и в геологическом прош- лом в кальдерах могли наблюдаться подобные ситуации. При этом из пред- ставленных даннътх видно, что кисло- родная атмосфера не только не является необходимым условием для их сущест- вования, но и тормозит развитие циано- бактерий Таким образом, цианобактерии яв- ляются главными продуцентами органи- ческого вещества всего сообщества, по содержанию хлорофилла они не усту- пают высшим растениям, способны к большим скоростям роста и фиксации СО. в восстановительных условиях. Раз- ложение органики в мате идет по анаэ- робному пути без расхода кислорода с конечным результатом — образованием метана или сероводорода. Изучаемые сообщества способны к производству кислорода в значительных количествах за счет фотосинтеза, а значит, цианобак- терии могли создать кислородную атмо- сферу в далеком прошлом. Остатки ископаемых цианобакте- рий известны на Земле из очень древних отложений. Например, они присутству- ют в кремнистых породах Австралии, возраст которых около 3500 млн. лет, на Урале. Оказалось, что кремнезем спосо- бен фиксировать структуру бактериаль- ной клетки, и, таким образом, мы можем наблюдать картину уникальной сохран- ности морфологии микроорганизмов. Выяснилось, что биоценозы цианобак- терий и бактерий обладали удивитель- ной резистентностью на протяжении всей истории нашей планеты. Такие долгожители привлекают пристальное внимание исследователеи, так как помогают понять причины и характер крупнейших преобразований, которые претерпели земная атмосфера, литосфе- ра и гидросфера. Существование циано- бактерий в докембрии подтверждается и широким развитием строматолитов. Природа строматолитов была выяснена еще в начале нашего века. Они образуются в результате жизнедея- тельности цианобактерий и бактерий. Детальное изучение строматолитов началось с середины бО-х гг. с развитием электронно-микроскопической техники. Развитие и господство древних строма- толитов приурочено к определенному геологическому времени. Появившиеся высокоорганизованные конкуренты эвкариоты — вытеснили их с морских просторов в экологические ниши с экстремальными условиями, в которых сами развиваться не могли - в места с активной вулканической деятельностью, литоральные части морей, пересыхаю- щие водоемы с высокои соленостью или щелочно стью. Лабораторное моделирование. Проводя аналогию между древними строматолитами и современными циано-бактериальными сообществами, необходимо учитывать, что геохимичес- кие условия на Земле в эпоху развития строматолитов были существенно от- личными от нынешних. Поэтому ока- зывается невозможным опираться толь- ко на современные аналоги, развиваю- щиеся в естественных условиях, но приходится прибегать дополнительно к моделированию жизнедеятельности со- обществ в контролируемых лаборатор- ных условиях. С этой целью в лаборато- рии были созданы модельные установ- ки конвенционного и лоткового типа. Исследование мата, полученного в ла- боратории, показало, что по общей структуре и видовому составу он соот- ветствует природным и представляет единое сообщество, куда, наряду с циа- нобактериями, входят и другие микро- организмы. Мат имеет слоистую струк- туру, где слои с цианобактериями пере- межались минеральными прослоями (рис. 3). Биопленки стали предметом на- учного изучения в конце XIX в. Однако систематические их исследования на- чались только в 80-х гг. нашего столе- тия в различных областях, включая ми- кробиологию, инженерию, медицину. В ископаемом состоянии биопленки не описывались. Биопленки — ~то совокупность ми- кробных клеток, других мельчайших частиц, воды и в большинстве случаев внеклеточного полимерного органичес- кого вещества (EPS), которое удержи- вает клетки вместе и на поверхности. Клетки и мельчайшие обломки могут быть представлены частично разло- жившимися цианобактериями, разнооб- разными бактериальными клетками и их микроколониями, нередко включают мелкие глинистые частицы и продукты эрозии субстрата. EPS также имеет мик- робиальное происхождение и представ- ляет собою открытую пористую струк- туру (подобную губке) с пустыми про- странствами, заполненными водой (рис. 4). Субстрат может быть пористый и беспоровый. Если поры достаточного размера (микрон и более), то микроорга- 42
Рис. 3. Галофильный мат. А — общий вид мата в лагуне Сиваша с газовыми пузырями; Б — живая часть мата; В - вертикальный срез мата (а - зона фотосинтеза с развитием цианобактерий, б - прослои карбоната кальция, в - зона аноксигенного фотосинтеза с развитием пурпурных бактерий, г- захороненные слои цианобактерий, д - зона сульфатредукции); Г — верхний слой природного мата; Д- вертикальный срез лабораторного мата. 43
направление движения воды РОУ вода РОУ NO2 NO-3 Рис. 4. Схема современной естественной биопленки: ~Б — цианобактерии: Д — диатомовые водоросли; РОУ — растворенный органичекий углерол; ОК — ослизненные клетки; МК — микроколонии (по Costerton et al., 1985). низмы могут проникать и в них. Физи- ческие, химические и биологические свойства биопленок зависят от окружа- ющей среды, в которой биопленка нахо- дится. Факторы могут быть биотичес- кие и абиогенные. Самым важным фак- тором являются сами микроорганизмы, их присутствие и активность, взаимное влияние популяций, образующих сооб- щество. Биопленки играют важную роль в природе и технологии. В современных условиях биопленки очень широко рас- пространены. Они возникают, когда ми- кроорганизмы колонизируют поверх- ность и образуют моно- или мультислои клеток. Появление биопленок - резуль- тат транспортных и ростовых процессов. Биопленки могут быть найдены почти на любой твердой поверхности, грани- чащеи с водои, независимо от состава и строения субстрата. Они аккумулируют- ся на дне морей, рек, озер, развиваются на поверхностях отдельных песчинок, галек, на обломках раковин, на скелет- ных остатках животных, обволакивают взвешенные в воде обломочные частицы (рис. 5). Нередко биопленки появляются и на временно покрываемых водой твердых субстратах, после высыхания Рис. 5. Биопленка из скопления дрожже- вых клеток на стенке метанотенка (резервуара очистного сооружения сточных вод). 44
Рис. 6. Слизь, органические нити и гликокаликс у соврсменных микроорганизмов: а - слизь и тонкие органические тяжи у современного Microcoleus; б - минерализованные нити слизи среди пурпурных бактерий (совр.). Рис. 7. Гликокаликс, образованный дрсвними микроорганизмами: а - минерализованный глико- каликс в бокситах Соколовского месторождения; нижний мел, Урал; б - минерализованный гликокаликс в континентальных фосфоритах; верхний мел, район г.Благовещенска. Рис. 8. Минерализованные микроорганизмы, органические пленки и тяжи внутри раковин кембрийских брахиопод: и - тонкие минерализованные органические пленки между слоями раковины; б - минерализованные тела микроорганизмов и нити между слоями раковины. 45
они могут превращаться в тонкую п ную пленку. Субстрат для биопленок может быть не только твердым, но и жидким, как, например, в масляно- водных эмульсиях, и водно-газообраз- ным. Биопленками, только структуриро- ванными, являются и циано-бакте и- альные маты, в которых, кроме собст- венно цианобактерий, присутствуют другие бактериальные организмы. округ клеток бактерий и циано- бакте ерий или тканей животных нередко на людается появление слизи (гликока- ликса). Выделение слизи, с одной сторо- ны, является естественным процессом жизнедеятельности бактерий, необходи- мым, например, для их движения. С д гой стороны, слизь несет защитную ф нк- цию. Например, у цианобактерий слизь появляется вокруг клеток и нитей при подсыхании или при появлении в окр- жающей среде высоких концентраций каких-либо элементов. Ослизнение, наблю- дающееся при разложении животных и растительных тканей, происходит благо- даря развитию гетеротрофных бактерий. Гл икокаликс циано-бактериальных матов, так же как слизистый матрикс биопленок, состоит из кислых полисахаридов и име- ет фибриллярное строение (рис. 6 а,б). Изучение с помощью СКАН'а об- разцов древних пород, в образовании ко- торых принимали участие микроорга- низмы, таких как фосфориты, бокситы, шунгиты, черные глинистые сланцы и пр. показывает, что биопленки были ши- роко распространены и в далеком п п о- ом. Нередко, будучи минерализован- ными, они сохраняются в ископаемом состоянии. В ископаемом состоян можн Т ЯНИИ жно наблюдать и фоссилизирован- ные участки гликокаликса (рис. 7 а,б). когда, благодаря посмертной мине- рализации, во многих случаях происхо- дящей почти мгновенно, могут сох а- няться разные стадии развития биопле- нок и слизи, то есть на ископаемом ма- териале можно восстанавливать процесс их появления, преобразования и мине- рализации. Бывает, что минерализации подвергаются участки частично аз жи вшихся органических тканей вн ло- ск елетных остатков у ископаемых жи- ри вотных вместе с принимавшими участие в их разложении бактериальными телами (рис. 8 а,б). 46
циАно-БАктЕриАльны~ мАт ы АНАЛОГИ ИСКОПАЕМЫХ СТРОМАТОЛИТОВ (ЛАБОРАТОРНОЕ МОДЕЛИРОВАНИЕ) 47 Изучение водорослей и цианобак- терий в полевых условиях всегда сопро- вождалось стремлением исследователей культивировать эти объекты в лабора- торных условиях, позволяющих более полно и целенаправленно решать те или иные задачи. Когда палеонтологи пришли к за- ключению, что строматолитовые обра- зования - это продукт жизнедеятельно- сти синезеленых водорослей (циано- бактерий), то неоднократно были по- пытки использовать эти организмы для получения в лаборатории живых анало- гов ископаемых строматолитов. Однако эти попытки не дали ре- зультатов до тех пор, пока микробиоло- ги не связали свои исследования с изу- чением такого явления природы, как образование цианобактериями слоистых матов. Маты стали считать потенциаль- ными строматолитообразователями, пред- полагая, что в определенных условиях они литифицируются и сохраняются как строматолиты, хотя в современных ус- ловиях это практически не происходит. Необходимо было сделать вывод, что в ранней истории планеты Земля, когда эти структуры были доминирующими формами жизни, экологические условия отличались от современных. В частно- сти, необходимо было допустить, что водная среда обитания была более мине- рализована, а это приводило к массовой и постоянной разгрузке в виде выпаде- ния минеральных осадков (карбонатов, фосфатов и т.д.). Такая концепция легла в основу ме- тодики, разработанной коллективом ис- следователей — геологов и микробиологов — под руководством акад. ГА. Заварзина (Институт микробиологии РАН). Пос- тавленная задача заключалась в пол- чении живых растущих биомоделей Ъ морфологически схожих с ископаемы- ми строматолитами. Строматолиты име- ют ярко выраженную особенность, заклю- чающуюся в том, что эти образования сформированы постоянной и чередую- щейся парной слоистостью, различаю- щейся по цвету (белесой и более тем- ной). Такое парное чередование слоев трактуется как чередование органоген- ного и минерального слоя. В этой связи целью выращивания лабораторной био- модели было получение такой чередую- щеися последовательности, состоящеи из органогенных и минеральных прослоев. О ганогенный слой представлен ростом лабораторных культур цианобак- терий. В экспериментах использовались культуры, состоящие из представителей различных видов осциллаториевых циа- нобактерий. Эти организмы имеют ряд интересных биологических особенно- стей, которые являются существенными для формирования биомоделей — анало- гов. Так осциллатории — подвижные ни- ти, двигающиеся со скоростью до 100 мкм в мин. Их движение весьма специфич- ное — маятникообразное, которое так и называется осциллаторным. При этом кончик нити, раскачиваясь маятнико- образно, как бы ощупывает впереди се- бя пространство, что позволяет им про- ползать между минеральными частич- ками при выпадении осадка. Важно при этом то, что нити двигаются целена- правленно к свету, то есть практически 11~ 11 солнцу, на поверхность выпавшего осадка. Оказавшись на поверхности ми- нерального слоя, цианобактерии растут,
размножаются и заселяют эту поверх- ность, образуя на ней налет в виде плен- ки-ткани. Эта пленка, состоящая из пе- реплетенных нитей, обладает опреде- ленной гидромеханической устойчи- востью, что позволяет ей удерживать ле- жащий под ней минеральный слой. При создании биомоделей обнару- жилась еще одна особенность циано- бактерий — матообразователей, а именно, популяция выросших нитей не равно- значна. Имеются более активные по- движные нити и нити менее подвижные (более старые и более ослизненные). Менее подвижные нити остаются засы- панными выпадающим минеральным осадком и остаются под ним, указывая на бывший рост в этом месте цианобак- терий. Более активные нити и их части, называемыми гормогониями, ползут наверх, где до нового цикла выпадения MHH.ðàëüíîão осадка должны дать но- вое поколение, новый рост, новую плен- ку из цианобактерий — практически по- пуляцию, способную к выживанию. Мине альный слой. В природных осадочных толщах карбонатные осадки и карбонатные строматолиты встреча- ются наиболее часто. Поэтому карбонат- ный материал активно использовался нами в лабораторных экспериментах. Выпадение карбоната кальция из насы- щенных растворов достигается многими способами: испарением раствора, уве- личением рН среды, добавлением избы- точного количества хлорида кальция и т.д. Опыт работы показал, что для уско- ренного получения карбонатного про- слоя можно на поверхность растущих ци- анобактерий нанести кристаллы СаСО~, имитируя тем самым природную хемо- садку в лабораторном сосуде. В про- цессе экспериментов получали и другие минеральные прослои. А именно: слои фосфата кальция, карбоната магния, сульфата кальция, железистых соедине- ний. Использовались глинистые минера- лы, такие как монтмориллонит, каоли- нит, речной песок и даже такие техно- генные вещества, как окись алюминия, окись титана. Выяснилось, что циано- бактерии работают по единой схеме, описанной выше. Они проникают меж- ду частицами минерального осадка или раздвигают аморфные взвеси глин. При повторении циклов они снова и снова заселяют свободную территорию. Эксперименты показали, что вы- падение минерального осадка должно иметь пульсирующий характер, так как затем необходим период для роста и развития цианобактерий. Показано, что цианобактерии способны оплетать лю- бые минеральные осадки и взвеси, лишь бы вещества и условия, при которых происходит их образование, не угнетали рост и развитие цианобактерий. Есте- ственно, мощность выпадающего мине- рального слоя должна быть соразмерна с расстоянием, через которое могут проникнуть нити цианобактерий, чтобы успеть размножиться до следующего "залпа". Мета ы мо ели ования. При обычной лабораторной скорости роста цианобактериальных пленок и форми- рования корочек карбоната, выпадаю- щего в осадок из раствора, один визу- ально различимый слой толщиной в до- ли миллиметра образуется за несколько дней, иногда за неделю. Получение по- стройки в несколько сантиметров высо- той заняло бы годы. Этот процесс мож- но ускорить, если стимулировать актив- ность пленок и дополнительно вносить порошкообразный карбонат. В цикле экспериментов "строите- лями" были цианобактерии — то сооб- щество, которое формирует маты в тер- мальных источниках вулкана Узон (Кам- чатка), где преобладают два вида осцил- латориевых цианобактерий — Osci llatoria terebriformis (Ag.) Elenk. emend. и Phor- midium angustissimum W. et G.S. West. Чтобы воспроизвести присущую узонс- ким матам тонкую ритмическую сло- истость из "темных" (бактериальных) и "светлых" (минеральных) пленок, ин- тенсивный рост микроорганизмов чере- довали с ускоренным осаждением ча- стиц карбоната кальция. Интенсивное образование пленок достигалось внесе- нием на образующийся бактериальный слой дополнительной биомассы той же культуры, выращенной отдельно, в дру- гих сосудах. После того, как из перво- начальнои пленки и дополнително вне- сенной биомассы цианобактерий возни- кла единая, сплетенная нитями отно- сительно прочная ткань, в жидкую среду вносили заранее рассчитанное количе- ство поршкообразного карбоната каль- ция. Оседая на поверхность растущей бактериальной пленки, он имитирует выпадение карбонатного осадка при образовании строматолитовой построй- ки в естественных условиях. Средняя скорость формирования пары слоев по такому интенсивному методу составляет от нескольких часов до суток, а моделей
Рис. 1. Пластовые строматолиты: а — палеонтологический образец (фото Н,П. Голованова); б - лабораторная биомодель, вертикальные срезы. 49 построек разных форм — от нескольких недель до трех месяцев. В первых опытах были получены нестолбчатые формы - многослойный го- ризонтально-слоистый мат, похожий на безрельефные пластовые строматолиты (рис. 1), и желваковая постройка с при- знаками, характерными для докембрийс- ких купольно-желваковых форм. Приме- чательно, что последнюю форму удалось получить из первоначально образовав- шегося бугорка водорослевой пленки, по- крытой слоем минерального осадка, при верхнем искусственном освещении. Ока- завшись засьтанными минеральным осад- ком, подвижные цианобактерии тяну- лись к источнику света, проникали между зернами кальцита и разрастались на его слое, образуя пленку из переплетенных нитей. Поочередное внесение бактери- .альной массы и карбоната кальция, мно- гократно повторенное, приводило к обра- зованию многослойной куполовидной cTp)'êòÿ)û с выпуклыми слоями. Их из- гиб сначала воспроизводил форму исход- ного буторка, но по мере роста купола уси- ливался за счет кислорода, выделяемого цианобактериями на свету в процессе фо- тоинтезаа, твк как газ накапливался в плен- ке в виде пузыриков и приподнимал "ткань". Экспериментальные куполовид- ные структуры достигали з-7 см в диаме- тре и 2-3 см в высоту, общими очертания- ми и формой арочных слоев походили на купольно-желваковые строматолиты Paniscollenia и Nucleella (рис. 2). Чтобы вырастить столбчатые вет- вящиеся строматолиты с разными типами ветвления, понадобились некоторые до- полнительные ухищрения. Для получе- ния пассивно ветвящихся построек, у ко- торых нижние вертикальные почти ци- линдрические колонки разветвляются на более узкие, минеральный осадок распре- деляли на поверхности бактериального слоя неравномерно. На одних участках плен- ку покрывали тонким слоем кальцита, на других — толстым. Цианобактерии легко и быстро прорастали сквозь тонкий слой и формировали на его поверхности новую органическую пленку. Это были как бы островки роста будущих колонок. В тех местах, где осадок лежал более толстым слоем, рост цианобактериальной пленки сильно замедлялся или совершенно пре- кращался. Повторив неравномерное вне- сение кальцита много раз, удалось в ко- нечном счете получить серию слоев в форме колонок или столбиков. Получен- ные таким способом постройки имеют
большое сходство с ископаемыми пассив- но ветвящимися строматолитами над- группы куссиеллид (рис. 3). Для получения аналогов ветвя- щихся ископаемых строматолитов, в ко- торых колонки расширяются снизу вверх или "разбегаются" в стороны, по- требовалось дополнительно контроли- ровать площадь нарастающих органи- ческих слоев. Если нужно было "заста- вить" цианобактерии расти вбок, то на край пленки вносили дополнительную био- массу, а оставшуюся часть засыпали слоем минерального порошка. В резуль- тате постепенно возникала колонка, ко- торая сильно отходила от центральной части. Образовавшиеся искусственные стро- MaToJIHTbI весьма походили на ископае- мые активно ветвящиеся формы над- группы тунгуссид (рис. 4). Были получе- ны также онколиты (рис. 5). Размеры ла- бораторных биомоделей определяются пожеланиями экспериментатора. Экспериментальные живые моде- ли довольно точно копируют принятые за образцы постройки древних стромато- литов. Поэтому есть основания полагать, что лабораторный процесс формиро- вания и природный механизм морфоге- неза сходны. Рис. 3. Пассивно ветвящиеся стромато- литы: а — палеонтологический образец (фото И.Н. Крылова ); б - вертикальный срез лабо- раторной биомодели; вертикальные срезы. 50 Рис. 2. Желваковые строматолиты: а — палеонтологический образец — вертикальный срез (фото В.А. Комара ); б - лабораторная биомодель; в — вертикальный срез биомодели.
Рис. 4. "Активно" ветвящиеся строматолиты: и - палеонтологический образец — вертикальный срез (фото М.Е. Раабен ); o — вертикальный срез лабораторной биомодели в карбонатном осадке: темные слои сверху и снизу — агаровый фиксатор; а — то же крупным планом (отпрепарировано). Рис. 5. Онколиты: а — палеонтологический образец - срез (фото Р.К. Пауль ); б - срез лабора- торной биомодели. 51 Таким образом, изучая и исполь- зуя биологические свойства современных цианобактерий, можно не только понять механизм образования ископаемых стро- матолитов, но и в какой-то мере предста- вить экологические условия, существо- вавшие в те далекие времена. В самое последнее время появилось уже значительное количество данных, показывающих возможность наблюдения бактерий, как живых, так и субфоссиль- ных в современных и антропогенных строматолитах.
СТРОМАТОЛИТЫ 52 Строматолиты тесно связаны с альго-бактериальными сообществами. Они являются одной из категорий био- литов - органа-седиментационных структур, возникающих в результате взаимодействия микроорганизмов и процессов осадконакопления. Стромато- литовые структуры образованы сериями тонких слоев, нарастающих друг на. друга, основанием серии прикреплены к подстилающим породам. Тонкая сло- истость отличает их от тромболитов, с которыми их нередко смешивали, связь с субстратом - от онколитов, - еще одной категории биолитовых структур. Совре- менные строматолиты представляют собой слабо литифицированные альго- бактериальные, в основном циано-бак- териальные маты. Они распространены в немногих районах, формируются на мелководье, в приливно-отливкой зоне морских побережий, в лагунах, озерах и в озерцах при вулканических источ- никах и гейзерах. Верное представление о биолито- вой природе сгроматолитов возникло в начале ХХ в., но получило распростра- нение лишь в конце 30-х гг., после работ В. Блэка по современным строматолитам Багамской банки, а стало господствую- щим в 60-х гг. в результате широких исследований таких форм в Шарк-Бее (Австралия) и капитальных работ В.П. Маслова по древним строматолитам, которые несравненно разнообразнее со- временных. Это особенно относится к протерозойским постройкам. Парадок- сально, но ссылаясь на такое отличие, от- дельные авторы приписывают абиоген- ное происхождение большинству или всем докембрийским строматолитам. Диаметрально противоположное представление о строматолитах как о на- стоящих водорослях тоже можно встре- тить в литературе последних десяти- летий, хотя и очень редко. Но если сто- ронники их биолитовой природы со- ставляют абсолютное большинство, то вопрос об относительной роли биоген- ного и абиогенного факторов в строма- толитообразовании пока остается край- не спорным. Облик строматолитов Слоистые, преимущественно кар- бонатные постройки строматолитов часто хорошо видны на поверхности вмещающих пород благодаря особен- ностям текстуры слоев, их формы, окраски и особенностям выветривания. Их можно встретить в породах любого возраста, но они особенно характерны для протерозоя. Строматолиты залегают чаще всего в известняках, доломитах, окремненных карбонатных породах, из- вестны крупнейшие залежи фосфори- тов, образованные в основном стромато- литовыми фосфатсодержащими по- стройками; в силикокластических поро- дах они очень редки. Обычно стромато- литы слагают более или менее крупные пластообразные или выпуклые тела, ко- торые могут представлять собой либо одиночные постройки, либо скопления многочисленных построек того или ино- го облика. Строматолитовые постройки мно- гообразны, но укладываются, в целом, в несколько основных модификаций, хо- рошо различающихся по очертаниям и размерности, или иначе — морфотипов. Ими являются пластовые строматолиты - пластинчатые и плоско-линзовидные постройки большой протяженности; желваковые — куполовидные и субсфе- рические постройки, сильно варьиру- ющей, неустойчивой величины; столб- чатые неветвящиеся строматолиты с цилиндрическими, столбообразными постройками, ветвящиеся столбчатые строматолиты - кустообразные и древо-
видные постройки, образованные рядом колонок-строматоидов; микрострома- титы - их постройки состоят из мель- чайших, большей частью колонковых строматоидов миллиметровой размер- ности. Каждая из этих основных моди- фикаций представлена множеством бо- лее мелких. Здесь надо заметить, что термин "морфотип" является неформальным и не вошел как ранговая единица в клас- сификационные схемы, за исключением своеобразной и ныне неупотреби- тельной схемы В.П. Маслова 1960 года. Как единица неформальная он исполь- зован в серии работ М.А. Семихатова и М.Е. Раабен по динамике разнообразия протерозойских строматолитов. Классификация строматолитов К началу 2000 г. описано около 1S00 таксонов различного ранга. Из них более 90%, в том числе около 300 родовых и 1000 — видовых выделены в рамках единой, традиционной системы; она наследует от раннего периода ис- следования строматолитов порядок, обычный для палеонтологических опи- саний: иерархическое построение, лати- низированную бинарную номенклатуру низших таксонов, обязательность голо- типов, правила приоритета. В своем со- временном виде она независима от си- стем водорослей или бактерий и рассма- тривается как самостоятельная система с условными (формальными) таксонами. В описательных работах, русских и иностранных, формальные виды стро- матолитов чаще всего названы "форма- ми", роды — "группами", таксоны ранга семейств — "надгруппами". Из терминов, отвечающих более высоким ранговым категориям, часто встречаются "тип", "надтип", "подтип", то как категории соподчиненные, то в качестве однои и той же высшей ранговой единицы. В целом, номенклатура категорий высо- кого ранга не упорядочена, а названия самих высших таксонов обычно не лати- низированы, что вызывает разнобой при их переводах. Правомерность применения "лин- неевской" номенклатуры к строматоли- там встретила ряд возражений; было сделано несколько попыток ввести ту или иную альтернативную классифи- кацию, однако ни одна из них не при- меняется при исследованиях древних строматолитов, так как в отличие от традиционной системы, не обеспечивает возможности использования стромато- литов в стратиграфии. В качестве независимой и фор- мальной, традиционная система стро- матолитов развивается уже на протя- жении 40 лет. За это время, благодаря работам многочисленных исследова- телеи, количество таксонов, видовых и родовых, выросло в ней более чем в 10 раз. Как прямое следствие такого разрастания количества родов, разви- валась и усложнялась классификация надродовых единиц; впервые они были введены в рассматриваемую искусствен- ную систему в 1960 г. И.К. Королюк. В ее самой ранней схеме выделены над- родовые таксоны двух соподчиненных рангов, названные типами и подтипами. Типы: столбчатые, пластовые, желва- ковые строматолиты — широко исполь- зованы в описательных работах 60-х- начала 80-х годов как единицы наи- высшего ранга, и уже в тот же период времени наметился ряд надродовых таксонов меньших рангов: они учтены так или иначе в классификационных схемах, опубликованных И.Н. Крыло- вым, В.А. Комаром, М.Е. Раабен и другими авторами на протяжении этих лет. Эти схемы отнюдь не являются конкурирующими классификациями строматолитов, как считали иногда. Они отражают последовательные этапы развития единой традиционной систе- мы. То же относится к позднейшим схемам, опубликованным К.Н: Конюш- ковым, Ю. Лианем с соавторами, М.Е. Раабен и А.К. Сина во второй половине 80-х гг.. В этих наиболее развернутых, более или менее формализованных схе- мах различаются высшие таксоны чет- ырех рангов, младший из которых оди- наково обозначен как "семейство"; но- менклатура прочих все еще не унифици- рована. Все надродовые таксоны, выде- ленные в рамках традиционной систе- мы, установлены только по морфо- логическим признакам. В формализо- ванных схемах в категории двух высших рангов входят рассмотренные выше морфотипы и их сочетания. Таксоны двух следующих рангов установлены на основе морфологических признаков, об- щих для нескольких или многих родов. Род (группа) занимает централь- ное место в рассматриваемой системе. Вопросы диагностики рода обсужда- лись, пожалуй, наиболее интенсивно, и 53
Рис. 1. Объемные рекоиструкции построек столбчатых ветвящихся стромдтолитов. , в результате он понимается сейчас довольно однозначно. Родовые таксоны выделяются и идентифицируются по совокупности морфологических призна- ков, и лишь в диагнозы редких групп входит микроструктура слоев. Морфо- логические признаки варьируют в соответствии с морфотипом: для вет- вящихся столбчатых форм в комплекс входят тип ветвления построек и различные параметры колонок-строма- тоидов, для неветвящихся столбчатых и
для нестолбчатых — различные характе- ристики рельефа слоев, образующих постройки, для микростроматолитов- степень обособленности их миниатюр- ных строматоидов, их ориентировка и размерность. Изучение морфологических родов требует лабораторных исследований, с распиловкой штуфов, изготовлением при- щлифовок и больших палеонтологи- ческих шлифов (табл. I) и т.д. Для ветвя- щихся строматолитов часто необходимы ориентированные серийные срезы для получения методом графического препари- рования, введенным И.Н. Крыловым, объемных реконструкций (рис. 1). Формальные виды (формы) уста- навливаются внутри родов, и сейчас главным, а чаще — единственным кри- терием служит микроструктура слоев, наблюдаемая в шлифах. Микрострук- туры строматолитовых слоев довольно разнообразны (табл. П). Наиболее ха- рактерны и узнаваемы структуры с крапчатым, штриховатым, волокни- стым, канальцевым рисунками; в таких слоях нередко различимы очень мелкие структурные элементы, образованные криптомикритом и имеющие форму тончайших пленок, нитевидную или сферическую форму. Они различимы, однако, лишь при особенно хорошей сохранности, перекристаллизация кар- боната слоев слишком часто не только стирает такие элементы микрострук- туры, но и изменяет, порой до неузна- ваемости, даже ее общий рисунок. Все это составляет одно из главных затруд- нений в диагностики видов. Для опре- деления видового таксона у строма- толитов совершенно необходимо знать принадлежность к тому или иному роду (группе), так как однотипные, близкие микроструктуры слоев могут наблю- даться в постройках различной родовой принадлежности, и даже у разных над- родовых группировок. Распространено мнение, что микроструктура слоев отражает особен- ности микроорганизмов стромато- литообразователей; вероятно, это верно, но лишь при условии хорошей со- хранности. Микроструктурные эле- менты карбонатных слоев, даже когда они с большей вероятностью могут считаться реликтами альго-бактери- альных структур, редко дают возмож- ность оценить систематическии состав строматолитообразователей. Определи- мые микрофоссилии хорошо изучены в окремненных строматолитовых слоях и представлены в основном цианобак- териями. Соотношения между составом строматолитообразующих биот и мор- фологией построек пока неясны, так как хорошо изучены пока лишь в слоях пластовых строматолитов, и не хватает сравнительного материала по формам различной морфологии. Изменение строматолитовогокомплекса во времени Строматолиты проходят через всю геологическую летопись, появившись на самых ранних стадиях развития биосферы. Их древнейшие постройки, куполовидные и пластообразные, из- вестны в Австралии и Южной Африке в раннеархейских породах с изотопным возрастом около 3,5 млрд. лет. Однако только в протерозое они достигли не- обычайного расцвета, и уже на конец докембрия приходится их глубокий упадок. Фанерозой же, в целом, беден строматолитами. Их протерозойский расцвет на- ступил лишь после глобального оледе- нения, приходящегося на начало палео- протерозоя. Богатейший протерозойс- кий строматолитовый комплекс испы- тывал на протяжении протерозоя зна- чительные изменения. Не только общее систематическое разнообразие на "ро- довом" и "видовом" уровнях, но и соот- ношение строматолитов различных морфотипов значительно и резко варьи- ровали в различные его интервалы (рис. 2). На фоне общей тенденции к возра- станию вырисовывается отчетливый максимум разнообразия в середине па- леопротерозоя и два таких максимума— в рифее. Главный из них и самый зна- чительныи во всеи геологическои лето- писи падает на поздний рифей и явно связан с огромным увеличением количе- ства "родов" и "видов" столбчатых вет- вящихся строматолитов. Распространен- ность строматолитов, отражающая их обилие, варьирует на протяжении проте- розоя в тех же основных направлениях, что и разнообразие, хотя и не совсем синхронно. Помимо различий в общем разнообразии и преобладании тех или иных морфотипов, комплекс обнару- живает на разных интервалах протеро- зоя изменения систематического состава на уровнях от "видового" до "надродо- вого". Практика биостратиграфических исследований показала их примени- «SS
80 60 50 40 30 20 l0 240 220 200 180 160 140 120 100 80 60 40 20 "ЯХ) 2 50 l6Co + 5() i~5o * ZO ИХХ~ ~ 5О 650 + 20 млн. лет Нижний протерозой Верхний протерозой Рис. 2. Динамика глобального разнообразия нротерозойских строматолитов на уровне формаль- ных видов (форм) и формальных родов (групп) и изменения структуры их возрастных ассоциаций. 1- 5 - выраженные в процентах доли видов и родов, принадлежащих следующим надродовым категориям строматолитов: 1 - пластовым, 2 - желваковым, 3 - столбчатым нсветвящимся, 4 — столбчатым ветвя- щимся, 5 - микростроматитам. 56 мость для корреляции хроностратигра- фических подразделений протерозоя, так, например, они послужили одним из главных инструментов в комплексе ме- тодов, позволивших наметить ту отно- сительно дробную стратиграфическую Количество формальных родов ( рупп) Кол ичество формальных идов орм) канву, которая дана для рифея на гра- фике (см. рис. 2). Возможности страти- графического использования стромато- литов имеют ряд ограничений, но имен- но они обусловили интенсивные иссле- дования их докембрийского комплекса.
т~влицА 1 Вид строматолитов в палеонтологических шлифах. 1, 2, 3 - колонки ветвящихся столбчатых строматолитов; 4 - постройки микростроматитов, увеличено в 1О раз; 5 - небольшой фрагмент пластовой постройки. 57
ТАБЛИЦА II Микроструктуры строматолитовых слоев в шлифах (слева - х4, справа -х2).
ЭКСПЕРИМЕНТЫ ПО ФОССИЛИЗАЦИИ ФОСФАТИЗАЦИЯ Рис. 2. Archaeooides sp. - планктонный организм неясного систематического положения из фосфоритов Хубсугульского бассейна. Рис. 1. Tasmanites sp. - акритарха из фос- форитов Хубсугульского бассейна. 59 Время образования крупнейших фосфоритовых месторождений (Хубсу- гульского в Монголии, Каратауского в Казахстане, Южно-Китайской группы месторождений) приходится на том- мотский век раннего кембрия и совпа- дает со временем массового появления первых скелетных организмов на Земле. Поэтому неудивительно, что в Лабора- тории древнейших организмов Палеон- тологического института начали зани- маться исследованием древних фосфо- ритов на примере Хубсугулького фос- форитового месторождения в надежде найти в них органические остатки. Ока- залось, что остатки организмов в дан- ных фосфоритах очень редки и пред- ставлены единичными планктонными формами Tasmanites и Archaeooi des (рис. 1, 2). Но очень скоро изучение хуб- сугульских фосфоритов с помощью ска- нирующего электронного микроскопа позволило обнаружить, что они почти нацело сложены фоссилизированными цианобактериальными матами, состоя- щими из трихомов цианобактерий и их обломков и из комплекса других мик- роорганизмов, прежде всего пурпурных бактерий (рис. 3). Более того, оказалось, что иногда сохранность цианобактерий совершенно удивительна. С помощью С ЭМ в фас форитах можно увидеть отдельные трихомы цианобактерий и их группы, бывает, что сохраняются даже перегородки между клетками в трихо- мах. По морфологии и по размерам эти ископаемые цианобактерии не отли- чаются от тех, которые обитают в совре- менных водоемах (рис. 4). В результате возникли предположения, что, во-пер- вых, данные фосфориты образовались вследствие фосфатизации древних циа- нобактериальных матов и, во-вторых, скорость их фосфатизации была очень большой. К этому времени уже были из- вестны работы К. Мюллера и его коллег, описавших прекрасно сохранившиеся личинки артропод из верхнего кембрия Швеции. Личинки при ' жизни были органическими, а фосфатизировались сразу после смерти, вероятно, во время оседания на дно (Ми11ег, 1983). Велико- лепная сохранность даже тончайших элементов их морфологии говорит о том, что процесс фосфатизации был очень быстрым (рис. 5). И уже были по- лучены данные по фосфатизации мяг- ких тканей у некоторых кембрийских
Рис. 5. Фрагмент фосфатизированной личинки артроподы Waldoria sp. (из работы Muller & Wallossek, 1983). 60 Рис. 3. Фосфорит, состоящий из скопления фосфатизированных цианобактерий. фосфатных брахиопод, когда в ракови- нах этих животных в СЭМ наблюда- лись клетки внешнего эпителия мантии иногда даже с мелкими ворсинками на поверхности клеток - микровиллями или фосфатизированные бактериальные тела, принимавшие участие в посмерт- ном разложении органического вещест- ва (рис. 6, 7). И тоже был сделан вывод, что процесс фосфатизации происходил сразу после смерти и был очень кратко- временным, так как иначе тончайшие прижизненные структуры (около 1 ми- Рис. 4. Современный цианобакгериаль- ный мат. крона) не смогли бы сохраниться из-за тления. Все эти данные натолкнули на мысль попробовать поставить в лабора- тории эксперименты по фосфатизации органических структур. Это было осу- ществлено Палеонтологическим инсти- тутом вместе с Институтом микробио- логии в лаборатории академика Г.А. За- варзина. Надо отметить, что большая экспе- риментальная работа по выяснению усло- вий, необходимых для синтеза фосфат- ных минералов в водной среде в 80-е го-
Рис. 6. Фосфатизированная клетка внешнего эпителия на внутренней поверхности раковины брахиоподы Ыппагыоша sp. Верхняя часть среднего кембрия Сибирской платформы. Рис. 7. Фосфатизированные бактери- альные тела на сколе раковины брахиоподы Etbotreta sp. Средний кембрий Алтае-Саянской складчатой области. ды, была проделана французскими иссле- дователями Ж. Лука и Л. Прево. Они уста- новили, что осаждение апатита и в морс- кой и в пресной воде происходит за счет органических фосфатсодержащих соеди- нений в присутствии бактерий. Бактерии разлагают органику на сахара, дающие необходимую энергию, азотные соедине- ния, участвующие в утилизации магния из морской воды, и фосфорную кислоту, которая, используя присутствующий в во- де кальций, трансформируется в апатит. Таким образом, для осаждения фосфата необходимы органическое вещество и микробиальная деятельность. Задачей наших эксперименталь- ных исследований было определение условий, при которых может проис- ходить фосфатизация цианобактерий, и изучение сопровождающих этот про- цесс морфологических изменений в клетках. В качестве тест-объекта была выбрана нитчатая цианобактерия Micro- coleus chthonoplastes, которая домини- рует в галофильных матах в водоемах аридной зоны, например, в лагуне Сива- ша, в Крыму. Это типичный бентосный организм, массовая форма в гиперсо- леных водоемах аридной зоны. Экспериментально было устано- влено, что М.chthonoplastes способна ра- сти в широком диапазоне концентраций неорганического фосфора. Оптималь- ной, обеспечивающей жизненно-важ- ные процессы является концентрация фосфора 4 мг/л. Снижение концентра- ции ведет к уменьшению скорости роста культуры, а при увеличении концентра- ции биомасса цианобактерий не прира- стает, но происходит накопление вну- триклеточного фосфора..Предельное ко- личество фосфора клетка набирает при концентрации его в среде от 18 до 36 мг/л. Причем активное поступление фосфора из среды, так же, как и накопление его в клетке (рис. 8), происходит в первые три часа после начала эксперимента и заканчивается к концу первых суток. Фосфор накапливается внутри клеток в виде волютиновых зерен (рис. 9, 10) и представлен кислоторастворимыми по- лифосфатами. Последние являются наи- более легко мобилизуемой фракцией и могут быстро переходить обратно в раствор после гибели культуры. Было измерено количество высвобождающе- гося фосфора при разложнии цианобак- терий. Оказалось, что при анаэробном разложении некромассы М. chthonoplas- tes выход фосфора в раствор доводит его концентрацию до 30 мг/л. А при аэроб- ном разложении кроме растворенного фосфора (тех же 30 мг/л) наблюдается накопление нерастворимого фосфатного осадка, что вместе с растворенным фос- фором составляет около 50 мг/л. То есть при разложении биомассы цианобакте- рий происходит выход фосфора в среду Ч мкг~ Р г MHH 14 10 2 315 4560мии3 6 12 18 24 24 ч Рис. 8. График скорости накопления 32Ð в клетках Mi'crocoleus chthonoplasles. 61
Рис. 9. 7рихомы современной циано- бактерии Microcoleus chthonoplastes, не содер- жащие волютиновых зерен. Рис. 10. Трихомы современной циано- бактерии Mi;crocoleus chthonoplastes, наполнен- ные волютиновыми зернами. в количествах, достаточных для начала мин ерализации. В экспериментах были использо- ваны концентрации фосфора, которые рекомендуются для лабораторного куль- тивирования цианобактерий, начиная с Р, мкг/л 50 40 30 20 10 360180 мин 1 Рис. 11. Динамика поглощения фосфора из питательного раствора культурой Microcolcus chth0n0plastes при разных концентрациях. 62 18 мг/л. Данная концентрация обеспечи- вает оптимальный рост цианобактерий в лабораторных условиях. Что же происходит при повыше- нии в питающем растворе концентрации фосфора от 18 до 36 мг/л? Как уже гово- рилось, сначала клетки очень интенсив- но и с большой скоростью начинают за- пасать фосфор в виде волютиновых зе- рен (рис.10,11), одновременно на по- верхности у них появляется слизистый чехол, толщина которого иногда вдвое превышает толщину самой нити Micro- coleus. Ослизнение у цианобактерий, как и у многих других организмов, явля- ется ответом на неблагоприятные усло- вия среды, то есть слизистый чехол но- сит защитную функцию. Он состоит из аморфной массы, в которой заключены микрофибриллы из целлюлозы (рис. 12). При дальнейшем повышении концентра- ции фосфора выше 36 мг/л на поверхно- сти клеток в слизистом чехле возныкани отдельные пюбули (рис. 13). Затем эти глобули oR единяются сперва в бляшки, потом в муфты, а затем в сплошной ми- неральный чехол, покрывающий нити цианобактерий снаружи, но не проника-
63 Рис. 12. Слизиспый чехол вокруг клетки цианобактерии, возникший как защитная реак- ция на неблагоприятные условия среды. Внутри видны микрофибрилы (указаны стрелками}. Рис. 14. Образование сплошного чехла из фосфата кальция вокруг отдельных участков трихомов Мк ocoleus. Рис. 13. Начало образования фосфатных зерен и бляшек на поверхности трихомов Мк'юсоlеи5. ющий внутрь (рис. 14). Внутренняя по- верхность нитей остается гладкой, зна- чит, образующийся фосфат замещает только слизистый чехол, не трогая сами трихомы. В ряде случаев наблюдалось, что после формирования минеральных чехлов живые трихомы их покидали, оставляя за собой полую трубку (рис. 15). Рис. 15. Полый внутри фосфатный чехол М/сюсо/еия, из которого выполз трнхом.
минеральных чехлов вокруг одних и тех же особей может происходить неодно- кратно. При увеличении концентрации выше 36 мг/л толщина чехлов возра- стает, уже выползшие трихомы гибнут. Фосфат кальция замещает сами трихо- мы, а затем и образовавшуюся детрит- ную массу, и осаждается, возможно, хи- мическим путем в пустых пространст- вах внутри нее (рис. 16). Наступает момент, когда вся масса представляет собою минеральный агрегат, состоящий из мелкозернистого фосфата кальция, в котором хорошо видны отдельные по- лые трубки, образовавшиеся вокруг цианобактерий, или целиком минерали- зованные трихомы. Фото этой массы очень напоминает фосфорит из Хубсу- гульского фосфоритового бассейна. Выпадение минерального осадка и образование фосфатных чехлов сильно зависят от величины рН в растворе. В природных условиях для осаждения аморфного фосфата кальция требуется рН, равный 7,7. В экспериментах исход- ный рН среды был равен 8,3, но при ро- сте культуры цианобактерий в резуль- тате их фотосинтеза рН повышался до 8,6-9,0. Вероятно, с этим в значительной степени связано наблюдающееся интен- сивное осаждение фосфата кальция на нитях цианобактерий и в полых прост- ранствах между ними. Немецкие ученые (Merz et а1., 1993) промерили с помощью Рис. 16. Фосфатные чехлы и замещен- ные фосфатом трихомы Мкюсо!еиз. Они двигались в направлении света и в свежий раствор. Выход трихомов из ми- нерализованных чехлов был показан в лаборатории с помощью простого опы- та. Одна половина чашки Петри с мине- рализованной культурой была закрыта черной бумагой, а вторая, свободная от нитей, освещалась лампой. В течение 20-30 минут в темной части остались лишь минерализованные трубки, а в освещеннои скопились живые зеленые трихомы Microcoleus без чехлов. Из этого можно сделать вывод, что благо- даря движению трихомов, образование 2,79 30 Co K 40 50 60 2,785 35' Со К„ 40 Рис. 17. Рентгеновские дифрактограммы порошка осадка, образовавшегося на чехлах цианобактерии.
Рис. 18, 19. Микротрубки, образовавшиеся рядом с нитями цианобактерий и на их поверхностях после добавления в питающий раствор NaF. 65 микро электродов рН внутри живой клетки цианобактерий и в чехле. Оказа- лось, что внутри клетки он равен 6,3-7,0, а в чехле рН тот же, что и в окружаю- щей среде. Эти данные объясняют, поче- му в живой культуре при повышении рН формируются только минеральные чех- лы, а в мертвой — происходит минерали- зация самих трихомов. Рентгена-структурный анализ образовавшегося минерала показал, что это плохо раскристаллизованный полу- аморфный карбонат-апатит (рис. 17). По данным В.С. Савенко образовавшиеся в современных условиях фосфаты прохо- дят несколько стадий - от почти аморф- ного фосфатного вещества к хорошо раскристаллизованным зернам и желва- кам апатита. Только что сформирован- ное фосфатное вещество содержит мало фтора. Его старение происходит за счет извлечения фтора и СО,'- из окружаю- щей среды и образования фторкарбо- натапатита, который является наиболее устойчивой и хорошо раскристаллизо- ванной формой апатита. На этот процесс требуется несколько лет. С целью уско- рения процесса в лабораторных услови- ях в среду добавили NaF. Оказалось, что этим процесс минерализации и форми- рования хорошо раскристаллизованного фосфата очень ускорился. Осаждение происходило быстро, в считанные мину- ты. Наблюдалось появление чехлов во- круг трихомов цианобактерий, которые состояли из микроскопических трубо- чек, сложенных хорошо оформленными глобулями с диаметром 0,2-0,5 мкм (рис. 18, 19). Анализ на микрозонде по- казал, что трубочки состоят из кальция, фосфора и фтора, то есть сложены фтор- апатитом. Об определяющем значении биоген- ного фактора в процессе фосфатонакоп- ления уже говорилось неоднократно, и к этому мнению сейчас склоняется большин- ство специалистов, занимающихся фосфо- ритами. Эксперименты показали, что дан- ный процесс, благодаря его очень боль- шой скорости и присутствию в современ- ной биоте цианобактерий, поддается моде- лированию. В ходе лабораторных иссле- дований были выявлены некоторые пара- метры, при которых процесс происходит. 1. Концентрация фосфора в среде 4 мг/л является оптимальной, обеспечи- вающей жизненно важные процессы у цианобактерий. Снижение концентрации уменьшает скорость роста культуры, хотя поглощение фосфора клетками продол- жается. При увеличении концентрации фосфора до 36 мг/л скорость роста по сравнению с оптимальной не меняется, но происходит накопление фосфора вну- три клеток в виде волютиновых гранул. 2. Повышение концентрация фос- фора выше 36 Mr/ë и до 54 мг/л ведет к началу образования фосфатных чехлов вокруг трихомов цианобактерий. Даль- нейшее увеличение концентрации фос- фора в среде ведет к гибели культуры и замещению фосфатом самих трихомов.
ПРОЦЕСС ОКРЕМНЕНИЯ Процесс окремнения микроорга- низмов был изучен на современных естественных объектах из термальных источников кальдеры Узон (Кроноцкий заповедник) на Камчатке. Основными продуцентами Узонских циано-бакте- риальных сообществ являются циано- бактерии Phormidi um, Lyngbya, Masti- gocladus, Oscillatoria, одноклеточные Synechococcus и зеленые бактерии Chlo- roflexus. Они растут вблизи источников с температурой около 50 'С и ниже и образуют массу слоистых слизистых пленок. Местами эти пленки частично или полностью минерализуются с обра- зованием карбонатов и опала. Окрем- нелые пленки распространены на участках, которые испытывали хотя бы временное осушение. Это объясняется тем, что количество кремнезема, ра- створенного в источниках Узона„дости- гает 360 Mr/ë. Такая концентрация меньше предела насыщения кремнезема и свободного его выпадения не проис- ходит. Когда же отдельные участки ока- зываются выше уровня воды и подсу- шиваются, концентрация кремнезема в растворе растет и на поверхности циа- нобактериального мата появляется тон- кая белесая опаловая корочка (Орле- анский и др., 1982). Образцы окремне- лых пленок (все рисунки в этом разделе взяты из статьи Крылова и Тихомиро- вой, 1988) были собраны В.К. Орлеанс- ким и изучены на сканирующем элек- тронном микроскопе И.Н. Крыловым и Н.С. Тихомировой (1988), которые рас- шифровали, каким образом происходи- ло окремнение цианобактерий (рис. 1). Растворы, содержащие кремне- зем, проникают внутрь стенки и пере- Рис. 1. Схема окремнсния цианобакте- рии: з - нить живой водоросли; б - проникнове- ние крсмнеземсодержащих растворов в стенки и перегородки клеток цианобактерии, начальная стадия выпадения опала в виде OTKpblThtx ост- ровков; в — срастание островков с образование сплошной корочки вокруг клетки. г — хемоген- ное осаждение глобулей опала как снаружи, так и внутри клстки; д - дальнейшее cpacIBHHc гло- булсй; е - полное замещение нити цианобакте- рии крсмнсземом.
Рис. 2. Начальная стадия выпадения опа- ла, в клеточной оболочке образуются островки из сросшихся сферических гранул. Рис. 3. Полное замещение кремнеземом стенок и перегородок цианобактерий, на вну- тренней поверхности клеточных полостей вид- ны сферические гранулы. городок клеток цианобактерий (рис. 1 а,б, рис. 2) и на ранней стадии опал на- чинает выпадать в виде островков сфе- рических гранул диаметром 0,1 — 0,3 мкм. Затем островки сливаются, образуя очень тонкие опаловые корочки (рис. 1в, рис. 3). На следующей стадии опал полностью за- мещает клеточные стенки (рис. 1 г). Поз- же происходит хемогенное осаждение гло- булей опала, нарастающих как снаружи так и внутри нитей цианобактерий и в кон- це концов полностью их замещающих (рис. 1 д,е, рис. 4). В результате образуется сплошной кремнезем, в шлифах из ко- торого можно видеть разнообразные ми- крофоссилии. Скорость процесса окрем- нения на начальных стадиях, как и в слу- чаях с фосфатизацией, весьма высокая- первые дни или даже первые часы после гибели цианобактерий. На это указывает полностью сохранившиеся морфологи- ческие особенности окремнелых остатков цианобактерий. Продолжительность сле- дующих стадий, вплоть до образования сплошного кремнезема зависит, вероятно, от количества кремнезема и скорости его хемогенного осаждения в пустотах поро- ды. Этот процесс скорее всего происходил в позднем диагенезе и эжп.енезе и мог сцсватывать тысячи и даже миллионы лет. Рис. 4. Сплошное заполнение пусто~ кремнеземом с образованием плотной крем- нистой породы. 67
ИСКОПАЕМЫЕ БАКТЕРИИИ БАКТЕРИАЛЬНЫЕ БИОПЛЕНКИ Ископаемые микрофоссилии, био- пленки и микробные маты, в которых они находятся, морфологические сход- ны с современными бактериями, био- пленками и матами. Естественно, что для распознавания и интерпретации ископаемых необходимо некоторое зна- ние современного мира микробов. Современные методы микробио- логических исследовании только в незначительной степени используют особенности морфологии для опреде- ления микроорганизмов, обычно приме- няются или биохимические измерения метаболизма колоний организмов или клонирование части их репродуктивно- го механизма, такого как 16s РНК. Одна- ко эти характеристики не могут сохра- няться в породах в течение геологичес- кого времени. По этой причине для идентификации потенциальных иско- паемых бактерий наряду с биохимичес- кими показателями микробного биоге- незиса используются морфологические характеристики. Биохимическими пока- зателями потенциального биогенезиса являются соотношение изотопов опре- деленных элементов, например, С, Fe, и S, характерное для фракционирования микробов, и химические биомаркеры распавшихся производных первоначаль- ного органического углерода, из кото- рого образуются организмы (кероген). Фоссилизируемые морфологические характеристики бактерий показаны на таблице 1 и включают: 1) Размер - размер большинства бактерий составляет 1-2 цт, хотя в определенных экстремальных условиях они могут быть меньше, 0,1 рт, а неко- торые виды Beggiatoa могут также достигать 500 рт (таблица 1,а). 2) Форма — большинство бактерий представляют собой простые сферы (коккоиды), палочки, нити и изогнутые (вибрионы) и спиральные тела (таблица 1,а). Бактерии могут иметь также аморфную форму. 3) Текстура внешней стенки клет- ки — внешняя стенка бактерии может иметь гладкую, неровную или грануляр- ную текстуру (таблица 1,b). 4) Чехлы - некоторые бактерии образуют толстый чехол из внекле- точных полимерных веществ (ВПВ) (например, цианобактерии). 5) Деление - бактерии размножа- ются делением клеток, что отражается в особенностях морфологии (таблица 1,с). Другие фоссилизируемые характе- ристики бактерий связаны с тем, что последние являются колониальными организмами: 1) Колонии — бактерии образуют колонии, состоящие из нескольких форм до миллионов форм (таблица 1,а, е). 2) Консорциумы - колонии бакте- рий обычно состоят из более чем одного вида организмов, живущих в консорциу- ме (таблица 1,а). 3) ВПВ и биопленка — колонии бак- терий образуют большое количество в неклеточных полимерных веществ (таблица 1,g), которые служат для защи- ты колонии, а также исгочником мест- ных и привнесенных питательных ве- ществ (таблица 1,i,k). Эти полимерные пленки могут варьировать от "простых" слоев бактерий и ВПВ (биопленок) до очень сложных структур, которые также
включают осажденные аутигенные ми- нералы и аллохтонные захваченные ча- стицы (полимеры являются "липкими"). В последнем случае они известны как бактериальные или микробные маты. Полимерные пленки могут быть очень толстыми (таблица 1,i,k) или могут со- стоять из смесей частиц осадка, покры- тых полимерным веществом (таблица l,f,h), и бактерий (таблица l,е,f,g). Фоссилизация бактерий, биопле- нок и ВПВ (таблица 2) происходит, когда ионы растворенного минерала хелатизируются до функциональных групп, таких как группы карбоксила, гидроксила или фосфорила, или в органическое вещество. Эти группы находятся в определенных макромоле- кулярных компонентах оболочек клеток бактерий (таблица 2,а,Ь), в частности в пептидогликане. Они чаще встречаются в частично распавшихся бактериях, чем в неизмененных микроорганизмах, что объясняется высвобождением функцио- нальных групп, вызванным деградацией макромолекул. Они также обычны в полимерах, обогащенных полисахари- дами, которые являются важными ком- понентами ВПВ. За хелатизацией ионов минерала до этих групп следует деги- дратация и полимеризация минерала. Таким образом, органическая структура действует как шаблон для образования ядер минералов в растворе. Бактерии и бактериальные биопленки, сохраняю- щиеся в кремнеземе, фосфорите, карбо- нате, сидерите, гематите и пирите, ука- зываются при описании породы (Folk, 1993; Knoll, 1985; МсКау et al., 1998). Эксперименты по фоссилизации (окремнению) бактерий (Westall et al., 1995; см. также разделы "Процесс окремнения" и "Окремненные микро- фоссилии") показывают, что микроор- ганизмы сохраняются в следующих формах: (а) как пустая корка (таблица 2,с); (б) как корка, наполненная мине- ральным осадком (таблица 2Д или содержащая комок разрушенной фосси- лизированной цитоплазмы (таблица 2,d); (в) как ядро (например, заме- щенное минералом) (таблица 2,d); (г) как пустой отпечаток в мине- ральном матриксе (в этом случае ми- нерал выпадает вокруг организма, кото- рый впоследствии совершенно разруша- ется, или минерал, замещающий орга- низм, позже растворяется под воздей- ствием пост-диагенетических процес- сов) (таблица 6Д. Бактерии также могут сохраняться как углеродистые отпечатки в мелкозер- нистых осадках, таких как сланцы (на- пример, мезопротерозойские сланцы Билляхской серии на севере Сибири (Sergeev et al., 1995). Эксперименты по окремнению микроорганизмов обнаружили интерес- ную разницу между фоссилизацией грамположительных и грамотрицатель- ных бактерий: вокруг грамположитель- ных бактерий образуется толстая креп- кая корка (таблица 2,k), а грамотри- цательные бактерии замещаются только тонкой слабой коркой (таблица 2,1). Причина различного поведения двух типов бактерий связана с разницей в составе и структуре оболочек их клеток: грамположительные бактерии имеют толстую оболочку клетки, содержащую много пептидогликана (и поэтому много свободных функциональных групп). Наоборот, грамотрицательные бактерии имеют тонкии слои пептидогликана, зажатый между двумя другими слоями (и таким образом, меньше свободных функциональных групп). Что случается с органическим ве- ществом, когда бактерия или ВПВ фоссилизируется? Степень сохранности органического углерода в фоссилизи- рованных микроорганизмах и биоплен- ках представляет собой непрерывную шкалу от хорошей сохранности органи- ческого вещества на одном конце до небольшой или никакой сохранности на другом конце. Термин "перминерали- зация" используют в тех случаях, когда значительное количество органическо- го вещества сохраняется в минераль- ном матриксе окремненных организмов (например, раннеархейские микрофос- силии на таблице 5,с-g). В других случа- ях органический углерод распадается и исчезает, как показывают эксперименты по окремнению (таблица 2,g,h) и неко- торые раннеархейские микрофоссилии (например, таблица 7,а-d). ВПВ биопленок и микробных матов само по себе является важной характеристикой микроорганизмов, так как оно может сохраняться, когда проду- цирующие микроорганизмы исчезли. Это происходит благодаря тому, что ВПВ состоит преимущественно из полисахаридов, которые содержат мно- гие функциональные группы, доступ- ные для хелатизации. Поэтому ВПВ 69
биопленок и микробных матов является важным показателем микробной актив- ности даже при отсутствии ископаемых бактерий. Биопленки и микробные маты могут сформировать крепкие макроско- пические фоссилии, такие как строма- толиты (которые могут быть пластовы- ми и желваковыми или куполовидными по форме) (таблица 3,а-f; см. раздел "Строматолиты"). Они также могут быть представлены более тонкими слоями на поверхности осадка (таблица З,g-k), которые включают частицы осадка (таблица 7, е-g) Объяснение к таблице 1 Характеристика бактерий и биопленок на фотографиях, сделанных с помощью сканирующего (SEM) и трансмиссионного (TEM) микроскопов. 70 а) Разнообразные бактерии (консорциум) на диатомовом субстрате. Видно разнообразие морфологических форм: (1) палочки, (2) сферы — коккоиды, (3) цепочки коккоидов, (4) спиральные тела. Шкала 1 рт. Ь) Текстура поверхности внешней стенки бактериальной клетки: клеточная стенка этой бактерии имеет неровную частично морщинистую текстуру. Шкала 1 рт. с) Деление коккоидной клетки (показано стрелками), в результате образуется цепочка клеток, окруженная клеточной стенкой. Шкала 1 рт. d) Палочковидная бактерия (Ь), покрытая тонкой пленкой внеклеточного полимерного вещества (ВПВ) и прикрепленная к субстрату тонкими полимерными нитями. Шкала 1 рт. е) ТЕМ фотография маленькой колонии коккоидных бактерий (Ь), покрытой птинистым минералом (с). Шкала 1 рт. f) Коккоидные бактерии, покрытые пленкой из ВПВ в глинистом осадке (с). Шкала 1 р.т. g) Бактерия овальной формы (Ь), прикрепленная полимерными нитями к глинистому минералу в пленке из ВПВ (с). Шкала 1 рт. h) ТЕМ фотография участка глинистого минерала (с), покрытого пленкой из внеклеточного полимерного вещества (cps). Шкала 1 рт. 1) Тонкая микробиальная биопленка, сформированная диатомовыми водорослями (d), бактериями (не видны при данном низком увеличении) и ВПВ. Шкала 1 рт. k) Участок ВПВ микробиальной биопленки, изображенной на предыдущей фотографии (i). Шкала 1 рт.
Таблица 1
Обьяснение к таблице 2 Экспериментальная фоссилизация (окремнение) бактерий, фотографии сделаны с помощью SEM и ТЕМ микроскопов. а) Начало хелатизации минеральных ионов на внешней стенке бактериальной клетки (стрелка). Шкала 0,1 цт. Ь) Появление минерального ядра (mn) на внешней оболочке (се) бактерии. Участок предыдущей фотографии (а). Обратите внимание на двухслойную внешнюю клеточную оболочку у грамотрицательной бактерии. Аморфный материал в центре- бактериальная цитоплазма (с). Цитоплазматическая мембрана отделена от остальной клеточной оболочки (стрелка). Шкала 0,2 рт. с) Пустая корка. Шкала 0,2 рт. d) Внутренний слепок бактерии (т). Округлые полости внутри слепка могут быть газо- или жидкость содержащими включениями в цитоплазме при жизни. Шкала 0,2 pm. e) Кремнезем в виде мелких шариков заместил клеточную стенку этой бакте- рии, сформировав корку (с). После смерти организма внутренняя цитоплазма сжалась и также была замещена кремнеземом (сс). Органическое вещество не сохранилось. Шкала 0,2 р.т. f) Бактерия покрыта коркой (с) и замещена кремнеземом, образовавшим внутренний слепок (т). Шкала 0,2 цт. g) Тонкая корка из кремнезема (с) вокруг бактерии. Первичное органическое вещество (or) сильно деградировано, большая его часть исчезла. Шкала 0,2 цт. Ь) Начало фоссилизации частично деградированной спиральной бактерии. Оставшаяся цитоплазма (с) сжалась и окружена кристаллитами кремнезема (s), на остальной части клетки видна пятнистость. Шкала 0,2 п,т. i) Окремненная палочковидная бактерия (SEM фотография). Бугорчатая поверхность связана с неравномерным ростом опалиновых кремневых сфер, инкрустирующих организм. Шкала 0,2 цт. k) Толстая корка (стрелка), заместившая оболочку клетки грамположительной бактерии. Шкала 1 pm. !) Тонкая корка, заместившая оболочку клетки грамотрицательной бактерии. Шкала 1 p,т. m) Окремненная бактерия (b), внеклеточное полимерное вещество (cps) и гриб (f) (SEM фотография). Гриб сломан, это показывает, что он был замещен палой коркой. Шкала 1 рт. 72
Таблица 2
Объяснение к таблице 3 Строматолиты, современные и древние. а) Современные куполовидные строматолиты (s) в заливе Шарк, Западная Авст- ралия. Вершина (коричневая) состоит из живущих водорослей и бактерий. Черная, нижняя часть — отмершие организмы. Шкала — 30 см. (Фото Д.Марчесини). b) Между куполовидным и пластовыми (ts) строматолитами (пластообразные микробиальные биопленки) Шкала 20 см. (Фото Д.Марчесини). с) Карбонатно-кремнистые строматолиты возрастом 3,45 млрд. лет в попереч- ном разрезе из группы Варравуна, Пилбара, Западная Австралия. Шкала - молоток справа. (Фото Д.Марчесини). d) Те же строматолиты, что и на фото (с) в плане. Шкала - карандаш в центре. (Фото Д.Марчесини). е) Кремнистые строматолиты (s) возрастом 3,45 млрд. лет в поперечном сечении из группы Варравуна, Пилбара, Западная Австралия (см. также рис. 1). Строматоли- товые слои переслаиваются со слоями розовых гидротермальных кремней (с), проис- ходящих из жилы, располагавшейся непосредственно справа от строматолита. Этот строматолит был, очевидно, еще живым и мягким, когда был перекрыт жидким пото- ком, обогащенным кремнеземом. В результате строматолит был уплощен и деформи- рован в направлении, совпадающим с движением потока из жилы (стрелка). Это ясно показывает биогенное происхождение строматолита на макроскопическом уровне. Шкала 10 см. (Фото Д.Марчесини). f) Кремнистый микростроматолит возрастом 3,45 млрд. лет в поперечном сечении из группы Онфервахт, зеленокаменный пояс Барбертон, Южная Африка (см. также рис. 2). Строматолит состоит из чередования серо-черных и белых слоев. Серо-черные слои представляют собою богатые керогеном бактериальные маты слоистых или волнисто- слоистых строматолитов (s), в то время как белые слои являются химически осажден- ными кремнями (с). Стрелка указывает на отдельные вертикальные выросты. Шкала 1 см. g) Тонкослоистые, мелководные, вулканокластические или биокластические (фрагменты микробиальных матов) осадки возрастом 3,45 млрд. лет из группы Онфервахт, зеленокаменный пояс Барбертон, Южная Африка. Тонкие микробиальные маты (пластовые строматолиты) залегают на поверхностях таких же слоев (см. также таблицу б,а,Ь). Для масштаба справа имеется молоток. (Фото Д.Марчесини). h) Тонкослоистые мелководные вулканокластические осадки возрастом 3,45 млрд. лет из группы Варравуна, Пилбара, Западная Австралия. Стрелка указывает на тонко морщинистые поверхности на кровле некоторых слоев вулканического пепла, кото- рые представляют собою микробиальные маты (см. также таблицу 7,i,k). Светлые слои обогащены пеплом, темные слои - гидротермальный кремнезем. Для масштаба показана лупа. (Фото Д.Марчесини). i) Участок предыдущей фотографии (h) с обогащенными пеплом светлыми слоями и с различимыми волнистыми поверхностями между ними (стрелки). Шкала 1 см. k) Деталь фотографии (i), на которой видна волнистая поверхность. Изучение с помощью SEM показывает, что эта поверхность состоит из бактериального мата (см. 74
Таблица 1
Объяснение к таблице 4 Микростроматолиты из группы Онфервахт возрастом 3,45 млрд. лет из зеленокаменного пояса Барбертон, Южная Африка. а) Столбчатый строматолит, состоящий из темно-пятнистой сердцевины (пятна органического углерода), покрытой тонкими слоями гидротермального кремня. Химически осажденные слои кремня в свою очередь перекрываются слоями микро- биальных матов, богатых органическим углеродом. Шкала 1 мм. b) Микростроматолит, напоминающий по форме цветную капусту. Шкала 2 мм. с) Листовидная обогащенная органическим углеродом сердцевина микростро- матолита. Шкала 200 рт. (Фото Р.Л.Фолк). d) Очень тонкие нити углеродистого материала в микростроматолите, которые могут представлять собою остатки нитчатых бактерий. Шкала 100 рт. (Фото Р.Л.Фолк). е) Темный обогащенный органическим углеродом слой микростроматолита при большом увеличении. Видна гребневидная текстура. Стрелка показывает направле- ния роста организмов, сформировавших маты вверх и наружу (возможно, это были аноксигенные, фотосинтезирующие, нитчатые бактерии). Шкала 200 рт. f) Деталь от (е). При большом увеличении участок с гребневидной текстурой становится диффузным (расплывчатым). Шкала 50 рт. g) SEM фотография (в лучах отраженных электронов) подтравленного тонкого среза того же образца. Темные обогащенные углеродом слои керогена (k) указывают на первоначально микробиальный состав; светлые, обогащенные кварцем слои (q) являются прослоями химически осажденного гидротермального кремня. Пары HF при осторожном травлении удаляют слои гидротермального кремнезема, делая видимыми участки, обогащенные керогеном. Шкала 100 рт. h) SEM фотография (в лучах вторичных электронов), показывающая, что тонко зернистое, обогащенное керогеном вещество вкраплено в кварцевый матрикс. Шкала 0,5 рт.
Таблица 4 77
Обьяснение к таблице 5 Микробиальные биопленки и нитчатые бактерии из группы Онфе вахт, зеленокаменный пояс Барбертон, Южная Африка. a) лиф, на котором видны толстые пачки очень тонких протяженных, вол- нистых, огатых органическим углеродом (керогеном) пластин, представляющих собою микробиальные маты. Шкала 500 рт. ) ф, на котором видны неправильнои формы пятна осадочного керогена \.У (s ), заключенные внутри карбонатных пластин. Бесцветный материал межд слоями — кремень. Шкала 100 рт. с) лиф с тонкими (1,5-2?м ширины) нитевидными перминерализованными актериями (то есть пропитанными кремнеземом) (стрелка), растущими вертикально от нижележащей керогеновой пластины (фоссилизированного мик обиального мата). ерминерализованные бактерии лучше видны в кремневых слоях, заключенных между слоями микробиального мата. Шкала 50 рт. ) Другая перминерализованная нитчатая бактерия длиною более 100 1.т. Шкала 50 рт. e) фотография перминерализованной нитчатой бактерии из тех же об аз- цов. Шкала 1 рт. f) фотография шлифа из того же образца, подтравленного HF. На ней видна одна актериальная нить в поперечном сечении, представляющая собою по т б- ку с гранулярной, пропитанной кремнеземом керогеновой (k) стенкой, заключенная в кварце (q). Шкала 2 рт. ) SEM g) М фотография того же подтравленного шлифа, на котором виден участок керогеновой нити (k) в продольном сечении (стрелка), окруженной ква цем ( ). Шкала 5 рт.
Таблица 5
Объяснение к таблице 6 фотографии ископаемых микробиальных матов, биопленок и бакте ий из группы Онфервахт, зеленокаменный пояс Барбертон, Южная Аф ика. ) оверхность напластования толстого перминерализованного растрес гося мик о иал ро иального мата (формирующего пластовые строматолиты). Шкала 20 т. р стрескавше- ) Деталь фотографии (а), показывающая разнородность мата: слева гладкая поверхность внеклеточного полимерного вещества — ВПВ (е s), сп ава- (cps), справа - гранулярная р сть. рисутствие кристаллов галита (h) в ассоциации с другими минера- лами такими как кальцит (не показан) и трещиноватая поверхность мата скорее всего указывают на условия приливно-отливной зоны. Отметьте также овальные ст и (стрелка): они могут являться сжавшимися пузырьками газа, выделяемого при жизни микробами, живущими внутри мата. Шкала 10 т. с) часток другого толстого микробиального мата, показывающий лип ю текстуру и короткий сегмент одного из нитчатых организмов (стрелка), которые принимали участие в строении мата. Шкала 2 рт. Цепь из трех сфер, упакованных в обычную морщинистую оболочку. Очевид- но, ыло сла ое сдавливание обычной клеточной стенки до ее окремнения в р зашло сжатие оболочки между каждой клеткой (белые стрелки) "3 б ия, в езуль- ~о мы ква ф р рца над микробнои цепью представляют собою с-оси ква ца, п оявив- ~Ф елки . у чатые шиеся после травления в парах HF этого шлифа. Вверху справа видна другая цепь, которая разрезана при изготовлении шлифа (это демонстрирует, что данные мик а- р рт факт), на ней видно, что микрофоссилии представляют Ъ со ою заполненные сферы и являются слепками. Маленькая сфе я сфе а, прикрепленная к цепи двоиной нитью (черная стрелка), возможно, является оставшейся цитоплазмой от бактерии, распавшейся до фоссилизации. Шкала 1 рт. e) кремненные палочковидные бактерии (стрелки), заключенные в частично гладком, частично гранулярном ВПВ (cps). Биогенное происхождение этих структу подтверждается их округлой морфологией, гибкостью (белые ст елки), клеточным делением (cd) и ассоциацией с ВПВ. Шкала 5 рт. f) Полый слепок (т) в форме цепи овальных структур, кото ые могли остаться от овальных бактерий. Шкала 1 рт.
Таблица 6
Объяснение к таблице 7 Окремненные бактерии и внеклеточное полимерное вещество (ВПВ) из формации Варравуна, Пилбара, Австралия. а) Скрученные нитчатые бактерии. Шкала 2 рт. b) Та же фотография (а) в отраженных электронах, на которой видно, что струк- тура содержит очень мало органического углерода: микрофоссилии имеют почти тот же оттенок серого цвета, что и окружающий кварц. Они должны были бы более темными, если бы содержали много углерода (сравнить с таблицей 7,h). Шкала 2 рт. с) Участок фотографии (а), показывающий морщинистую текстуру (стрелка) на поверхности фоссилизированной бактерии. Эта текстура явилась результатом частич- ной деградации и сжатия организма после смерти, но до фоссилизации. Шкала 1 рт. d) Окремненные коккоидные бактерии (стрелки), многие из которых собраны в цепи. Они тоже имеют морщинистую текстуру внешних стенок. Шкала 1 рт. е) Скрученные нитчатые бактерии. Обратите внимание на окремненное ВПВ покрытие частиц (cps) в правой части фотографии. Шкала 2 рт. f) Окремненное ВПВ, покрывающее частицы осадка. Шкала 1 рт. g) Участок фотографии (f); ВПВ (cps). Шкала 0,5 рт. h) Осажденные частицы обломков, обогащенных органическим углеродом (теперь кероген — k) в кварцевом матриксе (q). Шкала 5 рт.
Таблица 7
НЕКОТОРЫЕ ИСКОПАЕМЫЕ МИКРОБНЫЕ СООБЩЕСТВА РАННЕАРХЕЙСКИЕ ИСКОПАЕМЫЕ БАКТЕРИИ МЕТОДЫ ИЗУЧЕНИЯ РАНК ЕАРХЕЙСКИХ МИКРОФОССИЛИЙ Раннеархейские ископаемые бак- терии, найденные в отложениях с возра- стом 3,5-3,3 млрд. лет серии Онфервахт зеленокаменного пояса Барбертон Юж- ной Африки и серии Варравуна кратона Пилбара Австралии, являются самым ранним свидетельством жизни на Земле (рис. 1). Эти организмы появились через один миллиард лет после формирования Земли, и у нас пока нет более ранних сведении, не считая данных по изотопам углерода из высоко метаморфизованных пород с возрастом 3750 млн. лет из Аки- лия/Исуа зеленокаменных поясов Запад- ной Гренландии. Все первичные формы с различимой морфологией в этих отло- жениях были разрушены сильными ме- таморфическими процессами, которыми они подвергались. Раннеархейские микрофоссилии Южной Африки и Австралии морфологи- чески неотличимы от современных бакте- рий. Следует напомнить, что с точки зре- ния эволюции бактерии являются относи- тельно сложными организмами и пред- ставляют высокий уровень эволюции. У нас нет свидетельств перехода от не-жиз- ни к жизни или от "первых" клеток к бак- териям. В связи с этим самые ранние ис- копаемые имеют огромное значение. Породы, содержащие микрофосси- лии, необходимо анализировать в кон- тексте широкого изучения среды, поско- льку микроорганизмы занимали особую микронишу в специфических условиях. Поэтому бактериально-палеонтологи- ческие исследования требуют общей геологическои подготовки для понима- ния условий седиментации. Как замечено выше, микробные биопленки и маты способны создавать макроскопические видимые структуры, такие как строматолиты (таблица 3,с4). (Строматолито-подобные структуры мо- гут также появиться в результате роста некоторых минералов). Мелкие неров- ности на поверхности осадка также мо- гут указывать на присутствие микроб- ной пленки или мата (таблица З,g-k). Изучение шлифов, потенциально содер- жащих микрофоссилии, дает фоновую информацию о происхождении и диаге- нетической истории породы. Бактерио- логические структуры, такие как био- пленки или маты и ископаемые бакте- рии, могут также изучаться в шлифах (но не всегда). Толстые, хорошо разви- тые микробные маты могут просматри- ваться в шлифах как связки темных обо- гащенных углеродом пластов, которые могут быть вздутыми или извилистыми (таблицы 4 и 5,а,b). Если микробные ма- ты очень тонкие и состоят из переплета- ющихся бактерий, ВПВ и частиц осадка, они могут не просматриваться в шлифах (особенно если присутствует мало керо- гена). Микрофоссилии большего разме- ра, такие как нитчатые бактерии, могут наблюдаться в шлифах, если они состо- ят из материала, контраспгрующего с окру- жающим веществом, например, углерод в кремнеземе (таблица 5,с-d). Это не касается того случая, когда фоссилии со-
Рис. I. (а) Местоположение зеленокаменного пояса Барбертон в Южной Африкс и (Ь) и кратона Пилбара на северо-западе Австралии. держат малое количество контрасти- рующего материала (например, они замещены кремнеземом и заключены в кремнезем). Еще труднее точно распо- знавать в шлифах мелкие микрофос- силии (< 2 pm) из-за ограниченности разрешения светового микроскопа. Изучение с помощью микроанали- тической техники (светового микроско- па и сканирующего электронного ми- кроскопа (SEM), наряду с анализом рас- сеяния энергии элементов, показывает, что раннеархейские микрофоссилии и биопленки окремнены и заключены в кремнеземный матрикс.(Walsh, 1992). С течением времени в процессе диагенеза первоначально гидратированный крем- незем, который заместил микроорганиз- мы, становится дегидрированным и отвердевшим, образуя кремень (и даже кварц в случае диагенетической пере- кристаллизации). Эти породы крайне твердые, и необходимы специальные методы распознавания в них микрофос- силий. Для детального исследования раннеархейских кремней с микрофос- силиями требуется осторожное травле- ние породы в парах HF для обнаружения
структур с микрофоссилиями. При ис- пользовании этого метода необходимо проявлять большую осторожность в целях избежания артефактов. Особен- ности, наблюдаемые в шлифах, могут быть в дальнейшем изучены под SEM (если они находятся на поверхности шлифа) после тщательного травления шлифа (таблица 5,е-g). Изучение трав- леных кусочков породы и шлифов по- зволит распознавать микроструктуры, заключенные в породе, и их взаимо- отношения с породой и другими ча- стицами (таблицы 6,7). При изучении кусочков пород следует избегать по- верхностей со старыми трещинами, в которых могут содержаться более моло- дые эндолитические микроорганизмы. Во время травления HF воздействует преимущественно на матрикс породы, оставляя нетронутым кремнезем-орга- нический комплекс микрофоссилий и фоссилизированных биопленок и матов. Это явление объясняется двумя причи- нами: 1) остатки молекул органического вещества в кремнеземе, который прони- кает в микроорганизмы или замещает их, являются контаминантами, и 2) орга- нический шаблон, на котором образу- ются ядра кремнезема, изменяет кри- сталлическую структуру. РАННЕАРХЕЙСКИЕ БАКТЕРИИИ БИОПЛЕНКИ Геологические условия иокружающая среда Самые ранние бактерии из зелено- каменных поясов Барбертон в Южной Африке и Пилбара Австралии (рис. 1) создавали микробные маты на поверх- ности мелководного осадка (Walsh, 1992). На сегодня мы не знаем, сущест- вовали ли они в других условиях, напри- мер глубоководных или подземных. Это происходит частично из-за недостаточ- ной изученности древних микрофосси- лйй и частично из-за уничтожения древ- них пород, вызванных позднейшей тек- тонической активностью. Современные детальные седиментологические и мик- ропалеонтологические исследования об- наружили тесную связь микробных матов с гидротермальной деятельно- стью в мелководных условиях. Геоло- гические условия этих сред обитания сильно отличаются от существовавших на Земле в фанерозое или даже в протерозое. Маты, сформированные на вершинах конусов экструзивов, интру- зивов и вулканокластических осадков, находятся на поверхности большей ча- стью затопленных протоконтинентов. Надводных континентальных масс и континентальных платформ, продуктов плеиттектоническои деятельности, в раннем архее еще не существовало. В атмосфере, которая состояла большей частью из С02 и, возможно, некоторого количества СН4, было мало кислорода (в действительности для ранних этапов жизни кислород был ядом) (Kastling, 1993). Эти "парниковые" газы, вероятно, были необходимы для предохранения Земли от вымерзания, т.к. от Солнца тогда поступало почти на 30% меньше света и, следовательно, тепла. Микрофоссилии и биопленки Раннеархейские кремни из серии Барбертон Южной Африки и серии Пил- бара Австралии содержат макроскопи- ческие свидетельства биологической ак- тивности в форме строматолитов, сохра- нившихся большей частью в кремнеземе (таблица З,е,f) и также в смеси кремне- зем/барит и кремнезем/карбонат (табли- ца 3,с,d) (Walter, 1983; Walsh, 1992; Hof- mann et аl., 1999). Однако Д.Лоув и Дж. Гротцингер и Д.Ратман указывают, что со строматолитами могут быть смешаны некоторые минералогические структуры и поэтому для определения стромато- литов всякий раз необходимо свидетель- ство их биогенного происхождения. Это может быть проиллюстрировано двумя примерами из серии Варравуна района Пилбара Южной Австралии и из серии Овервоч зеленокаменного пояса Барбер- тон в Южной Африке. (1) На фотографии таблицы З,е, сделанной в поле (серия Варравуна, Австралии), видно изображение слои- стой куполообразной структуры, лежа- щей на толще окремненных прослоев и под пластом аморфного кремнезема. Внутри этой куполообразной структуры находятся тонкие выпуклые прослои, которые собраны в горизонтальные
contort layerâ Рис.2. Искривление тонких слоев вн и полоо Стромат толитовые слои ~ ~ переслаиваются с гидротермальными к емнями. Нап а 'S) утр куполообразного строматолита (см. таблицу 3 e). Ъ соседней гидротермальной жилы альными кремнями. аправление течения от направления изгиба слоев. ы показано стпелкой с п нкти ом. р у ром. Стрелки слева указывают на 87 складки и изогнутые на одной стороне купола (см. изображение на рис. 2). Ис- кривление внутренних прослоев купо- ла очевидно связано с пластическои деформацией еще мягких прослоев, что свидетельствует о том, что купол был образован скорее мягкими пластичными слоями, чем вертикальным ростом кристаллов. Оказывается, что причиной деформации был вес выпавшего аморф- ного кремнезема перекрывающего слоя. Подобная пластическая деформация ха- рактерна для микробных матов в связи с их преимущественно полимерным со- ставом. Более того, купола, сформиро- ванные мягкими пластичными прослоя- ми, указывают, что биогенное, стромато- литовое происхождение связано с ро- стом вверх микробных матов (Krumbein, 1983). Причиной искривления прослоев на одной стороне строматолитового купола может быть его положение в нижнем течении гидротермального по- тока, расположенного непосредственно направо от купола (т.е. на противопо- ложной стороне строматолита). (2) На таблице З,f изображены желваковые микростроматолитовые структуры из зеленокаменного пояса Бар берто н, для которых характерны чередующиеся зубчатые черные и белые слои (см. также рис. 3). При детальном изучении шлифов и анализе в элек- тронном сканирующем микроскопе и EDS, было обнаружено, что черные слои состоят из керогена, а белые — из чистого кремнезема (таблица 4,g,h). Кероген имеет форму ветки или пятнистых ша- риков в центре желвака и слоев гребен- чатой текстуры вокруг него (табли- ца 4,с). Гребенчатая текстура керогено- вых слоев ориентирована вертикально по отношению к базальному слою, ука- зывая на рост вверх и в стороны (таблица 4,е-g). Очень редко бактерии, создающие желваково-купольные струк- туры, сохраняются в. форме нитей (таблица 4,d). На биогенное происхож- дение указывает связь керогена, пред- ставляющего собой остаточный органи- ческий углерод, с ядрами желваков и создающего гребенчатую текстуру сло- ев. Пятнистые или в форме листа ядра возможно представляют собой колонии микробов на поверхности, которая позднее будет покрыта слоем(ями)
„иц 3, f Можно видеть зоб аженяого на таблице к емней м ми осгроматолита, изо р оями гидротермальнь "р рс.З. Прорисовка слоев м~кр литовых слоев (s) со слоям в деталях видна на ых ке огеном строматоли оматолитовых слоях в ев. Он п р слаивание богаты ,cs) в пластовых стр пластовых стро оматолитовых сло б ло цветы. ребы~ж~~ ру Ь . Тонкие кварцевые жилы (q ) v 4 К. Верткьный строматолит таблице,е, . рного цвета в виде вь содержит материал р ке огена че пересе кают строматоли новые слои. 88 ж енного кремнезем . а. В химически осажд кремне- е ь обогащенные свою очередь, окрываются слои впоследствии пок пленкой. Первоначальное микробной пленкои. ядрами и взаимо отнош ение между я огащенными керо геном, и слоями, обог щ о тонком кремнистыми слоями говорит остом микроб- аимодействии между ростом ных желваков и окру- силизации микро ных ж В настоящее время аннеархейских бактерий бными ~~ю- связано с плоскими микро ны ст омаголиты но пленками ст В польными и или желваковыми ст азвитые микробные маты со- литами. азви ительно алые ое количест- из-за ольшо как 3 замечено выше, из- ьных групп в мате- личе у ства нкциональны полисахаридами, ле обогащенном по ют ионы минерала они легко хелатизируют м об азом фоссилизируются. в виде темных тон~х вол 5 b) Hx темная м эле онным сканирующим необы- пом можно наблюд ать такои же о сложныи состав, текс р чаино оистой поверхно- е из ет современные матов со слои стью, какой характериз ет
микробные биопленки и маты (таблица б,а-с). Образцы на таблице 6 представ- ляют собой толстые растрескавшиеся пленки с частично гладкои, частично гранулированной до тягучей текстурой поверхности. В пленках заключены минералы, такие как кристаллы галита, кальцита и турмалина. Определенные своиства этои пленки имеют важное значение для понимания условий осад- конакопления и обитания микробов, которые сформировали пленку. Присут- ствие комплекса эвапоритовых мине- ралов (включая не изображенные здесь галитовые воронки, возникающие ис- ключительно в эвапоритовых условиях), наряду с трещинами мата, указывают на то, что эта частная пленка подвергалась воздействию атмосферы. В свою оче- редь, это предполагает, что осадок, на поверхности которого она сформиро- валась, накапливался на мелководье, возможно даже в приливно-отливной зоне. Кроме особенностей текстуры, со- става и структуры, сравнимых с совре- менными микробными матами, пленка также обладает другими признаками, указывающими на ее вероятное биоген- ное происхождение: овальные структу- ры с поднятыми краями напоминают газовые пузырьки (таблица б,b), которые возникают внутри слизистых слоев мата как следствие бактериального метабо- лизма. Реже в матах заключены иско- паемые бактерии (таблица б,с,е). Микробные маты часто содержат большое количество частиц осадка (как показано на современных образ- цах на таблице 1,f,g,i), создавая таким образом запутанную сеть из микро- организмов, ВПВ и осадочных частиц с полимерной оболочкой (Westal1, Rinc~, 1994). Подобные маты также были обнаружены в раннеархейских кремнях серии Варравуна района Пил- бара Австралии (таблица 7). Очевидно, что микробные фоссилии (таблица 7,а-е) и частицы детрита с полимерной оболочкой (последние в основном вулканического происхождения, таб- лица 7,f,g) взаимодействовали на начальных этапах. Частицы углеро- дистого вещества (таблицы 5,b; 7,h) также нередки в этих древних отло- жениях. В раннеархейских микрофоссили- ях можно выделить несколько морфоло- гических типов. Остатки микроорганиз- мов, которые, возможно', были основны- ми строителями матов, состоят из длин- ных тонких нитей 1-2 рт шириной и 100 рт длиной. Они представляют со- бой как перминерализованные ископае- мые (со значительным количеством остаточного органического углерода или керогена, таблица 5,с-g), так и фосси- лии, замещенные кремнеземом с неболь- шим количеством остаточного керогена (таблица 7,а-с,е). Перминирализованые фоссилии представляют собой стенки, состоящие из смеси керогена и крем- незема наннометровой микрогрануляр- ной размерности. Многие из замещен- ных кремнеземом микрофоссилий име- ют продольно ребристые внешние стен- ки (таблица 7,с), свидетельствующие, что микроорганизмы уже сморщились и частично распались перед фоссилизаци- ей. Такие фоссилии идентифицируются как нитчатые бактерии на основе их (а) тонкой вытянутой и закрученной мор- фологии, (б) ассоциации с органическим углеродом и/или текстурой внешних стенок (как, например, сморщенная тек- стура микрофоссилий на таблице 7,с, указывающая на распад органического материала), (в) ассоциации с ВПВ и биопленками/матами и (г) с другими микрофоссилиями. Выше отмечалось, что микроб- ные маты в основном состоят из консорциума различных видов микро- организмов. Так, другие морфотипы, связанные с матами, включают палоч- кообразные бактерии 1 рт шириной и до 3,8 рт длиной (таблица б,с) и коккоидальными или овальными фор- мами размером О,б-1 рт (таблицы б,d-f, 7,d). Так же как и нити, эти структуры идентифицируются как ис- копаемые бактерии на основе их сход- ства с современными бактериями по (а) размеру и форме, (б) свидетельству клеточного деления (таблицы б,с,d, 7,d), (в) сморщенной текстуре наруж- ной стенки клетки (таблицы б,d, 7,d), (r) связи с биопленками и ВПВ, (д) связи с другими ископаемыми бакте- риями и (е) присутствию углерода в перминерализованных структурах. Что за бактерии представлены раннеархейскими микрофоссилиями Барбертона и Пилбара? Существование сводовых/желваковых и пластовых строматолитов в условиях мелководья и приливно-отливкой зоны позволяет сде- лать вывод о том, что микробные маты формировались организмами, для кото- рых свет был важным источником энергии, такими как фотосинтезирую- 89
щие организмы. Коккоидальные, овальные и палочкообразные бактерии, связанные с первичными нитчатыми строителями матов, возможно, являются гетеротрофными или хемолитическими микроорганизмами. Однако нет ни одно- го свидетельства присутствия ископае- мых цианобактерий, предполагаемого предыдущей гипотезой о раннеар- хейских микрофоссилиях (например, Schopf 1993). (Цианобактерии харак- теризуются высокоразвитым метабо- лизмом фотосинтеза, при котором выделяется О, в качестве побочного продукта). Выше было отмечено, что ранне- архейская атмосфера, возможно, со- стояла из СО~ с небольшой примесью парникового газа, например, СН4 (Kasting, 1993). Содержание О>, воз- можно, составляло не более 1N от се- годняшнего уровня, соответственно тогда не было озонового слоя для за- щиты микроорганизмов от вредного ультрафиолетового излучения. Однако это излучение не играло такую важ- ную роль в эволюции жизни, как дру- гие факторы, например, воздействие воды. Толстые биопленки и микроб- ные маты, в которых жили микроорга- низмы, за.цитили бы их как от ультра- фиолетовой радиации, так и от деги- драции (как в современных экстре- мальных условиях).
ОКРЕМНЕ ННЫЕ МИКРОФОСС ИЛИИ Методика исследований 91 Остатки докембрийских микро- организмов сохраняются в основном либо как муммифицированные остат- ки, либо как псевдоморфозы. Методика сбора и последующего лабораторного изучения окремненных микрофосси- лий достаточно проста, хотя и требует определенных навыков при отборе кремней в поле и последующего поис- ка самих микроостатков. Наиболее перспективными являются конкреции диагенетических кремней иссиня-чер- ной окраски со стеклянным блеском; необходимые размеры отобранных об- разцов могут не превышать спичечно- го коробка, что необходимо для изго- товления нескольких шлифов. При ла- бораторной обработке окремненные микрофоссилии изучаются в стандарт- ных петрографических шлифах в про- ходящем свете. Растворять кремни для выделения остатков микроорганизмов в целом нецелесообразно, если этого не требуют специальные исследования, или диаметр микроостатков превыша- ет 100 мкм. При растворении в плави- ковой кислоте удается выделить остат- ки микроорганизмов из породы, однако их содержание резко снижается по сравнению с концентрацией в шлифе, а сохранившиеся прижизненные струк- туры дезинтегрируются. Биологическая интерпретация Встреченные в составе докемб- рийских окремненных микробиот мик- рофоссилии представляют собой остат- ки прокариотных микроорганизмов, в основном цианобактерий, а также низ- ших эукариот - протист. Большинство остатков до кембрийских микроорга- низмов имеют аналоги среди 5-6 раз- личных групп современных морфологи- чески сходных, но биологически отлич- ных организмов, или не имеют анало- гов совсем, что делает прямое сравнение современных и ископаемых форм весьма сложным. Кроме того, на морфо- логию фоссилизированных остатков на- ложились постмортальные преобразо- вания, иногда меняющие внешний об- лик микроорганизмов до неузнаваемо- сти. Например, долгое время за ядра внутри микрофоссилий принимались темные шаровидные образования, пред- ставляющие собой сколлапсировавший после гибели клетки протопласт. Поэто- му в основу биологической интепрета- ции древнейших микрофоссилий был положен комплексный подход, кроме морфологии анализирующий структуру ископаемого микробного сообщества, характер распределения в нем различ- ных микроостатков, паЛеофациальное положение встреченной ископаемой по- пуляции,и современные биологические данные о соотношениях различных групп микромира. Эколого — а алистический по- ход, Большинство окремненных докемб- рийских микрофоссилий представляют собой остатки цианобактерий. Этот вывод был получен в основном путем сравнения окремненных микробиот и современных цианобактериальных со- обществ, обитавших в пределах лито- рали и сублиторали аридных бассейнов. Как показывают данные седиментоло- гического анализа, большинство до- кембрийских микробных сообществ обитало в сходных эколого-фациальных обстановках, что подтверждается нали- чием трещин усыхания, знаков ряби, глиптоморфоз по гипсу и галиту, и при- родой самих доломитовых илов, харак- терных для аридных условий. При этом структура и состав ископаемых и совре- менных циано-бактериальных сообществ оказались очень близкими, включая так- сономическии состав как основных ма- тообразующих микроорганизмов, так и обитавших в матах микроорганизмов- симбионтов, причем это сходство наблю- дается на родовом или даже видовом уровне. Более точное сопоставление ци-
анобактерий и протист с современными мик- роорганизмами достигается путем срав- нительно-морфологического анализа. С авнительно-мо алогический анализ. Благодаря точной палеоэколо- гической локализации циано-бактери- альных сообществ, демонстрирующих идентичность структуры и сходство ос- новных таксонов, сравнительно-морфо- логический подход вполне приемлем с учетом постмортальных преобразова- ний. На первоначальном этапе исследо- ваний игнорирование влияния постмор- тального изменения на морфологию по- гибших микроорганизмов привело к серии ошибок. Однако для наиболее простых форм микроорганизмов метод морфологического анализа малопродук- тивен: здесь начинают работать законы бактериального микромира, в основу ко- торого положены различия в типе мета- болизма его многочисленных групп. Более приемлемым сравнительно-мор- фологический метод является для остат- ков дифференцированных микроорга- низмов, как цианобактерий, так в боль- шей степени эукариотных водорослей. Это позволило легко установить таксо- йы морфологически характерных гормо- гониевых цианобактерии, а также выя- вить представителей протист. В послед- нем случае сложность связана с тем, что ввиду древности обнаруженных эука- риотных остатков они в значительной степени относятся к полностью вымер- шим группам ранних ядерных орга- низмов. Более успешно сравнительно- морфологический подход проявился при анализе в целом более разнообразных органостенных биот, в составе которых удалось выявить представителей ос- новных отделов водорослей и грибов. Стратиграфическое распространение А хе и анний п оте азой. В архее указывается несколько проблема- тичных остатков микрофоссилий. Толь- ко начиная с протерозоя остатки окрем- ненных микроорганизмов становятся многочисленны, разнообразны и имеют хорошую сохранность. В нижнем проте- розое присутствуют почти все основные морфотипы современных цианобакте- рий и остатки достаточно крупных фос- силий - вероятно, ранних протист. По- скольку в этих же отложениях обнаруже- ны стерины-биомаркеры, характерные для клеточных мембран зеленых водо- рослей, то считается, что эукариоты поя- вились, вероятно, даже ранее 1,7-1,9 млрд. лет назад. Видимо, эукариотные орга- низмы были уже достаточно широко распространены в нижнем протерозое, что согласуется с имеющимися доказа- тельствами существования в это время кислородной атмосферы. Раннепротерозойские окремнен- ные микробиоты подразделяются на бел- черский и ганфлинтский типы. В микро- биотах белчерского типа, развитого в мелководных обстановках карбонатных платформ, представлены морфологичес- ки простые микроорганизмы, имеющие аналоги среди современных цианобак- терий на родовом или даже на видовом уровнях. Наиболее яркой чертой микро- биот этого типа является преобладание в них энтофизалесовых цианобактерий ро- да Eoentophysalis при низком разнообра- зии прочих хроококковых, а также гормо- гониевых цианобактерий. Микробиоты ганфлинтского типа представлены преи- мущественно остатками микроорганиз- мов сложной морфологии (зонтичной, гантелевидной, звездчатой и др.), кото- рые имеют своих аналогов среди раз- личных групп прокариот или эукариот, хотя большинство проблематичных остат- ков протист из нижнего протерозоя приу- рочено к сланцевым толщам, ще они встре- чаются как отпечатки на поверхности слоев. Микробиоты ганфлинтского типа характерны только для определенного отрезка раннего протерозоя (2.0-1.8 млрд. лет); узкое стратиграфическое распрост- ранение ассоциаций этих микроорга- низмов обусловлено тем, что они ассоци- ируют с железорудными формациями, практически исчезающими из геологичес- кой летописи в конце раннего протерозоя. Нижний-с е ний и ей мезоп о- те азой . В нижнем и среднем рифее, как и в нижнем протерозое, широко рас- пространены энтофизалесовые циано- бактерии (род Eoentophysalis); большим распространением пользуются синехо- коккусовые цианобактерии (род Eosyne- chococcus), представляющие собой, по- видимому, генетически гетерогенную группу одноклеточных фотосинтези у- ющих прокариот. Среди других мор о- типов следует отметить остатки однокле- очных хроококковых цианобактерий (род Gloeodiniopsis), сходных с предста- вителями современных родов Gloeocap- sa или Chroococcus, è полые чехлы Lyn- gbya- или Phormidium-подобных циано- фит, формально выделяемых в род Sipho- nophycus, встречающийся, пожалуй, во всех микробиотах рифейского возраста. Интересной формой, широко распрост- раненной в нижнем и среднем рифее при единичных находках в более молодых от- ложениях, является род Circumvaginalis полые чехлы с характерными черными кольцами, представляющими собой пу- стые чехлы Scytonema-подобных форм. 92
Рис. l. Основные морфотипы фоссилизированных в кремнях циапобактерий: 1, 3 — остатки хро- ококковых цианооактерий рода Gloeodilni opsiis с темными включениями внутри — сколлапсировавшей цитоплазмой; 2 — глеокапсовидные колонии хроококковых или энтофизалесовых цианобактерий; 4 — эл- липсовидные акинеты рода Aichaeoellipsoides; 5 - палмеллоидные колонии энтофизалесовых циано- баггерий рода Eoentoph>sèIIü; 6 — многорядные колонии плеурокапсовых цианобактерий рода Ра1аео- pleuiocapsa; 7, 8 — трихомы осциллаториевых цианобакгерий рода О~сИа(опор~~~; 9 — спирально- цилиндрические нити рода Obruchevella; 1О — фоссилизироваиные чехлы гормогониевых цианобактерий с остатками клеток трихома внутри; 11 - полые чехлы гормогониевых цианобактерий рода ~~р ЬопорЬч~~'пк. 93
Рис. 2. Некоторые докембрийские окремненные цианобактерии (фиг. 1-5). протисты и водоросли (фиг. 6-9): ! — полые чехлы из вложенных друг в друга сегментов Circumvagittalis elongatus; 2 - грибовидные колонии Eoeiitoph»salis heleherensis; 3 — короткий трихом Fili'constri'c tosus cephalon; 4 - эллипсовидные акинеты Archaeoellipsoides grandis (микрофоссилии, показанные на фигурах 1-4, про- иаходят из раннерифейской котуйканской свиты Анабарского поднятия); 5 — стебельковая цианобак- терия Polybessurus bipartitus из средперифсйской авзянской свиты Южного Урала; 6 - акантоморфная (с шипами) акритарха ТгасЬуЬуппсЬоьрЬаии раг~а: 7 — толстостенный гладкий фитопланктонный ми- кроорганизм Leiosphaeridiaj acutica; 8 - толстостенная двуслойная циста эукариотного фитопланктона Pterospermopsimorpha? sp. (микрофоссилии, показанные на фигурах 6-8, происходят из позднери- фейской шорихинской свиты Туруханского поднятия); 9 - остатки бангиевых красных водорослей из позднесреднерифейской или позднерифейской формации Хантинг Арктической Канады. Масштабная линейка на всех фотографиях равняется 20 мкм. 94
Рис. 3. Распространение различных микроорганизмов в докембрии. 1-6 — морфологически сложные прокариотные микроорганизмы из микробиот ганфлинтского типа: 1 — Еолр/гаеги 2 - Kalcabe- Aia, 3 — АгсИаеогез!Ь, 4 — Хенойпх, 5 — Erochobrachim, 6 - Еоал~ ion', 7. 8 — мелкие (< 10 мкм) одиночные сферические микро оссилии и нитчатые микроорганизмы диамет ом < 10 мкм; '9 - одноклеточные хроококковые циано актериии типа Gloeocapsa (род Gloeodiniopsis; 10 — одноклеточные эллипсовид- ные хроококковые цианобактерии типа Synechococcus (род ЕоюулесАососсию); 11 — остатки нитей ос- циллаториевых цианобактерий (рода Si~honophycus, Oscillatoriopsis или Palaeolyngbya); 12 - остатки энтофизалссовых цианобактерии (род Foentophysalis); 13 - остатки акинет цианобактерий (род Archa- eoelitpsoides): 14 - остатки крупных сфе ических микрофоссилии типа Chuaria; 15 — остатки крупных нитчатых микро оссилий нечеткой мор ологии; 16 — остатки многорядных нитей стигонемоподобных цианобактерии; 7 - остатки плеурокапсоподобных цианобакгерий 1 ~ - ветвящиеся нити с перегород- ками; 19 - остатки стебельковых цианобактерий рода Ро1уЬеыи~ия; 20 - акантоморфные акритархи рода ТгисйуЬлйпсйозрЬс~ега; 21 — микрофоссилии вазовидной формы рода MelanocvriIltutn: 22 — остатки вет- вящихся нитей вошериевых водорослей (рода Proterocladus и lophyton); 23 - спирально-цилиндри- ческие микрофоссили рода Obruchevella; 24 — крупные акантоморфные акритархи из ассоциации пср- тататакского типа 25 - мелкие акантоморфные акритархи рода М~сгЫ.кМЙит; 6 - акантоморфные акритархи родов kiagia и Baltisphaeridium: 27 - остатки бангиевых водорослей. 95
Во многих ассоциациях нижне- среднерифейского возраста доминируют эллипсоидальные микрофоссилии рода Archaeoellipsoides, являющиеся аки- нетами (спорами) анабеновых циано- бактерий; сходные эллипсовидные мик- роостатки известны и в отложениях нижнего протерозоя, а также верхнего рифея и палеозоя. Анабеновые и их спо- ры многочисленны в современных прес- новодных и опресненных морских мел- ководных водоемах, но ассоциации, в которых доминируют подобные эллип- совидные микрофоссилии, обнаружены лишь в нижнем-среднем рифее. Не оста- навливаясь на всех возможных причи- нах господства Archaeoellipsoides в от- ложениях среднего-нижнего рифея, сто- ит отметить, что обилие их несомненно связано с какими-то физико-химичес- кими особенностями нижне-среднери- фейских бассейнов - возможно, с суще- ствованием в это время обширных лито- рально-сублиторальных мелководных областей, что вело к массовому споро- образованию у колонизирующих эти ни- ши цианофит. В этих же нижне-средне-. рифейских толщах многочисленны ко- роткие трихомы, возможно, являющиеся начальными стадиями прорастания (гер- минации) цианобактериальных спор. Ве хний и ей неоп оте азой и "неоп оте озойская еволю ия". Если об отличиях в составе среднерифейских и нижнерифейских микробиот говорить сейчас не приходится, то граница сред- него-верхнего рифея является одним из наиболее значительных биостратигра- фических рубежей докембрия. Главное событие, приуроченное к этому рубежу и имевшее огромное значение как для развития биосферы, так и для биостра- тиграфии докембрия, и получившее название "неопротерозойской револю- ции" - это взрывообразная диверсифи- кация эукариотных одноклеточных ми- кроорганизмов и, как следствие, появ- ление в отложениях верхнего рифея морфологически значительно более сложно построенных форм. Эта вспыш- ка разнообразия согласуется с данными молекулярной биологии, предполагаю- щей существование "большого взрыва" в эволюционном развитии эукариот, ко- торый произошел около миллиарда лет назад. Только начиная с отложений позд- него рифея или позднего среднего рифея появляются формы с шипами и выро- стами, в частности, род Trachyhystri- chosphaera. В позднем рифее появляется и ряд других сложнопостроенных форм, настоящие ветвящиеся талломы (рода Ulophyton и Proterocladus), сравнимых с современными зелеными вошериевыми водорослями, а также красные бангие- вые водоросли. В отложениях только верхней части верхнего рифея не древ- нее 850 млн. лет обнаружены своеобраз- ные шиповатые вазовидные микрофос- силии рода Melanocyrillium. К сожалению, стратиграфическое использование микрофоссилий и созда- ние эволюционных моделей развития древних микроорганизмов осложняется особенностями их фациального распре- деления в бассейнах. Наряду с разнооб- разными и мор алогически дифферен- цированными ормами преимущест- венно из открыто-морских фаций, в верхнем рифее присутствуют и мелко- водные ассоциации морфологически простых прокариотных микроорганизмов, напоминающие более древние микро- биоты, в том числе и белчерского типа. Вен ский пе ио . В конце позд- него рифея происходит резкое сокраще- ние или вымирание ряда морфологичес- ки сложных эукариотных фитопланк- тонных микроорганизмов с шипами и выростами (род Trachyhystrichosphaera и другие); изменений в составе про- кариотных организмов практически не отмечается. В нижнем венде следует отметить появление и широкое распро- странение так называемой ассоциации пертататакского типа, в которой доми- нируют крупные акритархи с шипами и выростами, встреченные в Австралии, в Китае, в Индии и ряде других местона- хождений. В верхнем венде ассоциации пертататакского типа исчезают и боль- шинство акантоморфных акритарх пред- ставлено мелкими формами рода Mic- rhystridium со слабо развитыми шипи- ками. Однако на представления об ассо- циациях МФ всрхнего венда могла нал- житься палеогеографическая специфика бассейна его типового разреза на Во- сточно-Европейской плат орме, где, ве- роятно, преобладали не лагоприятные фации. Следует заметить, что шипова- тые формы, появившиеся в верхнем ри- фее и распространившиеся в нижнем венде, начинают доминировать только с кембрия и настоящая экспансия проис- ходит примерно с атдабанского яруса, когда представители родов Skiagia, Bal- tisphaeridium, Micrhystridium и прочих становятся доминирующими среди как органостенных, так и окремненных мик- робиот, отражая переход от марфологи- чески простых, преимущественно прока- риотных микроорганизмов, составляю- щих докембрийскую биосферу, к биосфе- 'ре фанерозойского типа, в которой доми- нировали эукариотные организмы. 96
ФОСФОРИТЫ Одним из наиболее интересных объектов, в которых хорошо сохра- няются ископаемые микробные сооб- щества, являются фосфориты. Форми- рование осадочных фосфоритов чаще всего происходило в ходе замещения биогенных остатков фосфатом кальция. Особенно крупномасштабной была роль цианобактериальных сообществ в обра- зовании древних фосфоритов. Так, в начале кембрия происхо- дило накопление крупнейших фосфори- товых месторождений Монголии (Хубсугульское), Казахстана (Каратаус- кое), Южного Китая. Одним из главных модельных объектов изучения иско- паемых бактериальных сообществ является Хубсугульский фосфорито- носный бассейн. В современной гео- логической структуре он представляет собою синклинорий, выполненный комплексом верхнерифейско-кембрийс- ких отложений (рис. 1, 2). В средней части комплекса выделяется хэсэнская свита томмотского возраста, которая сложена слоистыми и массивными известняками и доломитами, заклю- чающими линзы и пласты фосфоритов. Мощность свиты от 350 до 600 м, фос- форитовые пласты суммарно состав- ляют до 10-15% ее объема. Они состоят из чередования темных, почти черных тонких (доли мм или первые мм) слоев собственно фосфорита и светлых слоев известняка или доломита, иногда с прослоями черных кремней. При изу- чении фосфоритов с помощью элек- тронного сканирующего микроскопа было обнаружено, что они сложены мел- кими желвачками размером десятки или сотни микрон, которые являются скоп- лениями фосфатизированных обломков цианобактериальных матов, микроооли- тов, микростроматолитов и псевдомор- фоз по пурпурным и иным бактериям (рис. 3). Как правило наблюдается соче- тание нитчатых форм с разным диамет- ром нитей и микроорганизмов округло- го, веретеновидного и гантелевидного габитуса (рис. 4). Анализ геологичес- ких, палеонтологических и палеогеогра- фических данных позволил предполо- жить, что в начале раннего кембрия район Хубсугульского бассейна пред- ставлял собою очень мелководный бас- сейн типа залива или пролива со слабой гидродинамикой и был заселен циано- бактериальными матами, которые состо- яли из сообществ нитчатых цианобак- терий и пурпурных бактерий. Периоди- чески при возникновении подходящих условий, происходила фосфатизация матов. Скорость процесса была очень велика, что определяло прекрасную со- хранность микроорганизмов в образую- щихся фосфоритах. Остатки фосфатизированных циа- нобактериальных матов и бактерио- морфных тел в большом количестве присутствуют также в Каратауских и Южно-Китайских фосфоритах (рис. 5). Среди древних фосфоритов неред- ко встречаются мелкие фрагменты или целые прослои, сложенные фосфатизиро- ванными строматолитами и онколитами, которые также являются продуктами жизнедеятельности цианобактериаль- ных матов. Строматолиты были особен- но широко распространены в докемб- рии. С начала фанерозоя их роль в биоте стала уменьшиться и в настоящее время строматолитовые постройки распрост-
бо ~Я ° ° ° гФ ~ / ' ° l ° ° ~l ° ° ./И' [ I ° е Ф~ Сщ( ( ) Ю,Ьз"/Г г ° )50 )5$ ° ° ° ° ° ° Рб5 ° ° F40 'f, an.° ° ! °° °М/ l �Ф ~ /): ° ) ° . ° ° Ф Ф ° '~3 1ц, 1 >ей! L ° ° Э чС,ez lг,3 « °° ° L ° ° б. ° ° ° ° . юг ° / // 1 j70 р )Я ~,,~~~,с~~~ ° ~ с„~ 1~lр)гг )/: J~ saf. ° r ~~~5) .' . 5 5 /' юл Ъ,Ф п о илем и легендои- ac~pro дна с геологичес~~~~ ПРОФ p„~ ~ [ еОЛОгическ» <~P~'~ 1 К 1 (. (/1 1 ° ) l г R3at 1 l 1 1 (( ~э ) г(( L L
УСЛОВНЫЕ ОБОЗНАЧЕНИЯ Диориты, гранодиориты Доломиты с линзами кремней 6 Рт1 Сиениты и граносиениты ь Известняковые брекчии Гран иты биотитовыв бион ит-роговообманковыв Известня ки глинистые, углеродистые пиритоносмые с праслоями карбонатно-глинистых сланцев е1 (oÎ f~l . >I ° ' / D Туфогравели ~ ы /1 'I 1. z l. 'i 1- Главный фосфоритоносный горизонт Туфопесчаники Горизонт черных кремней Алевроли~ы )1инзы окремненных пород с жалезомарганцевым оруднением Сланцы Г,! ~!/ ~]1б ~ ) г г г Разрывные нарушения Туфы средне-основного состава Элементы залегания слои стости Конгломераты, гравелить~ полимиктовые Карбонвтные конгломераты, граеелить, конглобрекчии Археоциаты Трилобиты Известняки On Он колиты Доломиты Песчани ки 99 ранены лишь в экстремальных усло- виях. Фосфатизированные строматоли- ты известны в синииских и нижне- кембрийских отложениях Южного Ки- тая, в протерозойских и нижнекемб- рийских отложениях Индии, имеются меловые фосфатизированные стромато- литы среди фосфоритов Израиля. Онко- литы - маленькие от нескольких милли- метров до 1-2 см в диаметре округлые тела с концентрической слоистостью. Они образуются в неспокойной обста- новке в пределах мелководных баров и отмелей. Известны месторождения, где основная масса фосфоритов представ- лена сгруженными фосфатными онко- литами (палеоцен, Алжир, м-ние Дже- бел-Онк). Часто встречаются фосфати- зированные онколиты в фосфоритах Хубсугульского бассейна, в формации Фосфория (США) (рис. б,7). Цианобактериальные маты играли большую роль и при накоплении более о о д Известняки брекчиевидные молодых фосфоритов. Например, в ран- ней перми на западе Северной Америки существовал огромный фосфоритонос- ный бассейн Фосфория. Хотя фосфо- риты этого региона образовались глав- ным образом за счет фосфатизации крупных губковых поселений, но в них часто встречаются фрагменты фосфа- тизированных цианобактериаьных ма- тов. Вероятно, обстановки широкого распространения губковых поселений и цианобактериальных матов были близки и переходили друг в друга. Мел-палеоген-неогеновые фос- фориты, богатые месторождения ко- торых известны в Северной Африки, на Ближнем Востоке, в Северной и Южной Америке, на Сахалине и в других местах также заключают зна- чительное число обломков, сложенных фосфатизированными цианобактери- альными матами и скоплениями бак- терий (рис.8).
Stratigraphic column C О Е О U ° Щ Ф И С2. О Оезсп родоп Deposits л Яиаг~вггагу Q1 о 150 1ОМЮГ о 100 Sheets ог apicai basl~ пуавуавв аав ~ва в siltstones аг~Ь з~~ез green~ey. ~ the Ьазе- и а Ф ав'апвувппв васса 500 Е О е12 Uh Lrrestones grey, dms grey to black wlh sparse олсойез авЬ tNobltes. At the base-паыв ва 500 Dolores grey, гпизЬе. UIT�stones grey. dark -~~Г4у, Ьгеса1а46се 200 Umestones grey, d3Fk grey, rrl|ssive WNh lenses of ЬЮВ-Ягеу сЬюпз. 400 б1 er ОоЬггйез НдН-дгеу and grey, зотеМтвз Ьгессщ49(э. (nterbeds eNh archaeocyath ax( trNobite Нггвз3опез. 260 Umestones дгеу, диас-уау елй rterbeds of shales азЬ phosph~e е~е~з Ьа~геу аЬ lenses of зНЮЯеЬ rocks wNh Ьггоютнща- ~1 в%\а абп. ~ aa в а 350 О Dohrrites дгеу, Эю ргайе4 wNh йим4Ве interbeds of dark yey chelts. 200 Slllclhed sandstone s and gravelstones, дгеуЫз4м'оев cherts. 350- 600 О Е Е О О 61 hS — [ — i С: Щ '6 С > 300- 400 О О Чоо йа аг 1500 Рис. 2. Стратиграфическая колонка верхнедокембрийких - нижнекембрийских отложений Хубсугульского бассейн. 100 l 10 Е О C 63 ° ~ C 63 Lh 63 U Ф~ < Oolorraes grey, bedded, rrasstve cream соЬогеЬ. ееаЫу зйсОеб, with lenses of phesphoNes. 1и1айж3з of дгеу мк] Ими-уеу а пв.о а вав а вп апа bearing bedded and йаззЬе, grey and dark-аву.ув ау ув»апвууезва в пс and еаИ '4оЮгтйс (2-5). At the Ьазв-~пва~ вава п~ па» with phchlng ай u9ureous~orrNc breccia and conglobrecc!a. Dolordtes grey. bedded and гтаззЬе irregularly sNicihed еЮ) sparse strometolites. Urnestones grey to dark~ey, rrasslve. Lenses Ю sRstones and shales with pebbles авапваау пв . в а са ВВВвааапа с 34юбз(0043. ваа вваааввваав gravelstones polymlctlc, green, ягеуЬп-Ьгоеп, grey, in places рцгрЫЙЬГЬэбз ог dolordtes у в вававу.~а ~в а lenses (1-2mj of grey Эзе-уаве4 limsstones wilh oncoites.
101 Рис. 7. Фосфатный онколит на срезе, США, формация Фосфория. Рис. 3. Фрагмент хубсугульского фосфо- рита, ны котором видно, ч и) в значи гсльнои мс[)с он состоит из обломков фосфатизированного цианооактсри;пьного мл а. Рис. 5. Ьактсриомор палые фосфатизиро- ванные тела в Каратаусьих < н>сфоритах. Рис. 8. Скопление фосфатизированных бактериальных тел в плиоценовых фосфоритах о. Сахалин. Рис. 4. Фрагмент фосфатизированного цианобак~е1~~~лльиого мата из хубсугульских фосфоритов. Рис. 6. Ф агмснз фосфатного онколита из хубсу~ ульских ()oc([)opuтов.
БОКС ИТЫ 102 Бокситы, являющиеся сырьем для получения алюминия, распространены в отложениях различного возраста — от до- кембрия до современных, но наиболь- шие концентрации их характерны для мезо-кайнозоя. Большинство исследова- телей выделяют латеритные бокситы, образование которых связано с корой выветривания в условиях жаркого кли- мата, и бокситы переотложенные, содер- жащие переотложенные продукты коры выветривания в озерных, болотных или прибрежно-морских обстановках. Еще в 40-50-х гг. высказывались (Л.С. Берг, А.Г. Вологдин) представления об уча- стии в формировании бокситов биологи- ческих образований - озерно-болотной растительности и микробов. Но подав- ляющее большинство исследователей рассматривает латеритный процесс как чисто химическии. Применение для изучения бокси- тов сканирующего электронного микро- скопа (СЭМ) вначале не дало сущест- венно новых результатов, но уже с конца 80-х гг. было установлено (А.Д. Слукин и др.) присутствие в них микробных остатков. В самое последнее время в ПИН РАН было начато систематическое изучение бокситов в СЭМ, которое по- казало, что бокситы многих месторож- дений мира (Индия, Бразилия, Гвинея, Урал, Сибирь, США) содержат в огром- ных количествах бактериальные формы, йреимущественно коккоидные. Особенно подробно были изучены образцы бокситов Соколовского место- рождения и арканзаских бокситов райо- на пос. Бокситы (США) (рис. 1). Соколовское месторождение нахо- дится на восточном склоне Среднего Урала в 120 км к юго-востоку от Екате- ринбурга. Нижнемеловые бокситы этого ме- сторождения образовались в прибреж- ной зоне болотно-озерного бассейна. Бок- ситоносная толща залегает на рэтских сланцах и девонских порфиритах, облом- mr которых встречаются в продтчсгяаньо~ слоях. Собственно бокситовые залежи подстилаются пестрыми и бокситовыми птинами, а перекрываются нижнемело- выми континентальными и верхнемело- выми морскими отложениями. Преобладающие пизолитовые (бо- бовые) разности слагают в общем пла- стообразные залежи, но иногда заполня- ют значительные карманы в подсти- лающих породах. Утверждается, что свежие пизолиты образованы магнети- том и гематитом, но обычно замещены гидратами алюминия, в меньшей сте- пени железа, причем последние более распространены в заполняющей массе. Пизолиты часто округлые, сферические, но частично и остроугольные, вытяну- тые, неправильные. Редко встречаются обломки каолиновых пород, замещен- ных в разной степени гидратами глино- зема. В порах, кавернах на стенках при- сутствуют щетки бемита, выделения ге- тита, сидерита, лептохлорита. Относительно генезиса отработан- ного Соколовского месторождения су- ществуют различные точки зрения. Боль- шая часть исследователей рассматрива- ет бокситы как переотложенные образо- вания латеритной коры выветривания на рэтских сланцах и девонских порфири- тах, их туфах. Ранее представлялось, что образование бокситов идет чисто хими- ческим путем, при непосредственном
с) ,. vv„чоч Ъ Ъ ч ° „ф ° ( Ф о о 'л @ I~)ф С (.) ' ° 'I ') ) (, , 1 ).).1.) ).).1 )., ! l..l.').) )',', .I (.I .): ' I' . ' ' ).)'у' ;1' ') I . .I ) е!. I ' I ). 1. - 'I' ° 4).1. l ., ) I,g. !)) I) Ю( I (1) I 1 1")с( 'ЯР' "( )O I ! ".l. i ' 1:!.'С l:t':) e' > ('.I:):) ). ° ) (.I.1','(lk) I I '). ° . ( l ). ° )) II p)X (..' ).I ),I hO (..( ° ('I .',11.' t: Ю)) I ° I ) ) >а) > I ) ), , .I Ю> ° ~3 4аа в в с~ с ~Ъ ° > с rl Ю С ! r / с \ r1 а ) .,)!.II ) ). . '! '. l; I ) ~, 1' .I (: ..' ~) . '); 1' (':i 1'(а:е,':); t,'.1 t. l'I'(.!.1 ° ) ° (. ! . r> ° ( I ° I,1, )Фа I. (.') ..М С ° 1r ° I..:!' ° ° ю' Се \ Ф О 1 ° с > .,~~у ( в ~Ъ ° Ф ° а ° r 1 с с ° 1 ° 4 - ° ,а r!r % ° С 4> ив сс Щ юъв ° Ъв ю в ° в ° в в в Sr% r 1 с в p ° p ' с 1 р в >p ° 4P с с p p ° всма ф ! с а >1 СЪ- .-О ф,4 '~~, х ; Ъ)сгФ > Ъ ° 4 ° Ъ 1СС в. р рС с~ о 4 х Фй ci М О Е 65 Ж ° Ъ ъ '.' ° 1 У Iа / ° ъ ъа ю в I4 а р в ° 1 с Вс с в >>в'- 1 в Ъ с с>с '~са с авю 1,са 1С pir / ю ° с с с в ~с с \' в ~ ° '. T С с A в в Ф с ,в с Ф I ° 4 р I 4 (>) а3 О~ Фф D ° Яс I'I 1'1' ).1 ° I'I )(.1('I° I ..> с в в в Ъ 1 A с ю ° 1 I'! '1 ).>.( ! ! ° ! I. I 1 Вс'вс' аваавс1 в'4са,Ъ1'а °с' 4 ав 'I С >Ъ- Ф О 1 ° вс % -' Ъ Ъ 1, Ъ в I р O D 4 ° а С ва сс Ъ 1 в 1 Ъ Э6 g) 11 I ! 1 1 1 1 1 1 1 С л юв Х Э , 1!. ° ° ' ° с ю ' ° ° 1° !1 с У ° t 1 (' ( 1 I ))] 11 ()! (1 > I ( °- В1 l I I I I I I I I I >- И ) К :Э V) 3 < О О 'е е> ') l ~! ' ')> С( t Ъ,|4 ' S-.- P (q I 1 I I ъс rI в в с в с ю Ъ е 1 I+ ф Ж 3 Л >) 1 с 1r а 1 С с 1 ,1 С с Ъ I Ф Ъ ° С ° l 1 1 ° С ю \ ° а С> 111 I ( I lI 1) I I 1 I l( Il 1( tI I I I I ф И О Z < 3С < С° с Ч 4 Ю О О Л O Л 103 ° С Ч чо о о о о ф~~ о о о o Q ч~~ еч о 1 С4 4 4 g 4 4Я Я ф ф чо о е о Ч О й 2 й о -'4 О и~ v О И 65 О 65 Ж 65 М CL < 65 [м Ж О М О 1О 6~ X М й:[ М О CL О Р ) О Р О О М ."Г' Ц -2- О CL [: О X hC о М :T Ж (-4 О й,' О
Bauute - 2, Ural, Sekolovsk 0 0.00 т.оо Ь.оо 2.00 i.0O 4.00 а.оо Э.ео в.оо keV Saux ate-2 Ьх2М2 4Мх, Ьиаяоой сгувМИ Anal' JNTrlarl Риса Зб - спектр количественного анализа кристаллов гиббита или бемита. 104 Рис. 2. Фоссилизированные палочковид- ные и кокков~~дные бактерии. отложении алюмогелеи на дне конти- нентальных бассейнов. В бокситах Соколовского место- рождения довольно часто присутствуют бобовины с концентрической перифе- рией. Часто концентры перемежаются с участками, сложенными коккоидными формами. Чаще бобовины состоят из скоп- Рис. За — кристаллы гиббита или бемита среди палочковидных фоссилизированных бактерий. лений коккоидных форм диаметром до 5-10 мкм. Отдельные формы достигают 20 мкм в поперечнике. Иногда наблю- дается срастание небольших коккоидов в крупные сферические глобули, воз- можно, колонии, диаметром до 40-50 мкм. Реже боксит сложен скоплениями столб- чктьт червеобразных и коккоилных форм.
1642 1381 Э21 4В0 0 0.00 7.00 10.00 8. 00 9. 00 2.00 1.00 3. 00 4.00 S.00 1сеЧ В.00 Bauxite-2 bx2h34 8kx, Surface around the small Ьа! Ь Analyst: Jomen 105 Рис. 4. а - общий вид глобулы (шарика) пиролюзита; б - фрагмент 4а, ячеистая поверхность является оссилизированным гликокаликсом, содержащим Мп, а мелкие кокки иреи ественно содержат е; в - спектры количественных анализов гликокаликса (два больших пика Ее и n) и мелких кокков (один большои пик Fe).
Рис. 5. а - общий вид онкоидного строения боксита из Арканзасского месторождения; o - фраг- мент боксита с фоссилизированными гифами грибов. 10б В боксите систематически встре- чаются пустотки размером от десятков микрон до первых миллиметров, в ко- торых развиты существенно желе- зистые, преимущественно палочковид- ные коккоидные формы (рис. 2), кото- рые иногда покрыты гликокаликсом (?). Обнаружены также хорошо выраженные кристаллы гиббсита [Al(OH),] или бемита [A10(OH)] (рис. 3a,б). Иногда присутствуют, располагаясь послойно линзочками, шарики (50-70 мкм в диаметре) пиролюзита, представляю- щие собой массу, возможно, глико- каликса, содержащего марганец и мел- кие кокки (0,5-1 мкм в диаметре), содержащие преимущественно железо (рис. 4а-в). В 60-х гг. Г.А. Заварзин показал, что окислителем Мп может быть симбиоз грибов и бактерий рода Metallogeni'um. В этой связи не исклю- чено, что то, что мы здесь принимаем за гликокаликс, является грибом, тем более, что весь Мп содержится именно в этои части. Бокситы арканзасского региона являются несомненным продуктом выветривания сиенитов ого массива (рис. 5a,б). Детальное изучение об- разцов в СЭМ в ПИНе показало ана- логичную картину, как и в бокситах Соколовского месторождения Урала. Таким образом, в случае с бок- ситами Соколовского и арканзаского ме- сторождений отчетливо можно распоз- нать псевдоморфозы по коккоидным железобактериям и специфическим бак- териям, производящим огромное ко- личество гликокаликса в виде шаров, или, возможно, симбиоз бактерий с грибами. С последними связаны по- вышенное содержание марганца и формирование пиролюзита. Не вызы- вает особых сомнений очевидная роль железо-марганцевых (?) бактерий или симбиозов. Однако осталось не очень понятно, с чем связан алюминии. Поскольку в пустотах были обнару- жены хорошо выраженные кристаллы, а масса железобактерий содержит алюминий, вероятно, нельзя исключить существование на каком-то этапе "алюмогеля", пронизывающего весь осадок, включая бактерии, и имевшего химическую или бактериально-хими- ческую природу.
ИСКОПАЕМЫЕ БАКТЕРИИ,ОСАДКООБРАЗОВАНИЕ И РУДОГЕНЕЗ 107 О том, что микроорганизмы, и прежде всего археи и бактерии, играют огромную роль в геологических процес- сах, ученые догадывались давно. Не вы- зывало сомнений огромное значение микроорганизмов в процессах выветри- вания горных пород, в формировании ме- сторождений полезных ископаемых, раз- рушении нефтей, переотложении некото- рых пород и полезных ископаемых и т.д. В конце прошлого века Н.И. Ан- друсов (1897 г.) в своих лекциях обра- щал внимание на накопление S и Fe бла- годаря серо- и железобактериям. Однако в процессах седиментации терригенных и даже карбонатных пород, которые со- ставляют подавляющее большинство всех осадочных пород, роль микроор- ганизмов считалась весьма скромной или лишь теоретически допустимой. Совсем недавно, всего два-три десятка лет тому назад, мысль о том, что мик- робы могут сохраняться в ископаемом состоянии, особенно в массовых коли- чествах, могла бы показаться абсурдной. Когда в 1943 г. член-корреспон- дент АН СССР А.Г. Вологдин описал округлые тельца как железобактерии из железистых кварцитов Курской маг- нитной аномалии (КМА), это вызвало огромный скепсис. В 60-е гг. сначала американские, а затем российские и австралийские ученые открыли и опи- сали случаи сохранения микроор- ганизмов в кремнях древних докемб- рийских пород. Тогда считалось, что окремнение бактерий — явление доста- точно уникальное. Среди окремненных микроорганизмов часть была признана цианобактериями (в то время они на- зывались синезелеными водорослями). Серьезный прорыв произошел, когда более или менее одновременно стали изучать, но уже с помощью элек- тронного микроскопа древние фосфо- риты и высокоуглеродистые породы (черные сланцы, битуминозные кар- бонатные породы). В результате выяс- нилось, что эти породы буквально на- пичканы окаменевшими микроорга- низмами (рис. 1). Основой этих ассо- циаций микроорганизмов были циа- нобактерии и пурпурные бактерии, со- ставляющие основную массу циано- бактериальных матов. Особенно иссле- дователей потрясла удивительная со- хранность цианобактерий в древних фосфоритах, которые всегда считались хемогенными, то есть осажденными химическим путем. Сегодня можно видеть, что в боль- шинстве научных журналов публику- ются статьи о роли бактерий в фор- мировании тех или иных пород. Боль- шое внимание уделяется так называе- мым карбонатным микритам (очень мелкозернистым породам). Большин- ство исследователеи сходятся во мне- нии, что образование микритов, осо- бенно распространенных в биогермных и рифовых ассоциациях, является ре- зультатом деятельности бактерий. Ко- нечно, следует помнить, что подавляю- щее количество карбонатов всегда обра- зовывалось за счет деятельности орга- низмов, накопления их самих (напри- мер, писчий мел) или за счет скопления обломков разных организмов с карбо- натным скелетом (органогенно-обло- мочные известняки). Изучение современных микроор- ганизмов — широко разветвленная
Рис. 2. Ископаемые бактерии: а — нитча- тые (Монголия, нижний кембрий) и б - гантеле- видные (Казахстан, нижний кембрий). Рис. l. Фоссилизированные цианобакте- рии в дрсвних иородах: а - слаицы среди протс- розойски х джес пилитов Курской магн итной аномалии (фото С.И. Жмура); о — глинистые сланцы венда (евролейская часть России); в- глинистый известняк (кембрий, Австралия). область исследований, которыми зани- маются не только отдельные ученые, но и институты. Не касаясь гграктического значения этих исследован~ для геннои инженерии, использования для получе- ния биопродуктов, для очистки пром- стоков, а также вредного воздействия микроорганизмов на человека, на ка- менные и металлические конструкции, загрязнение водоемов, несомненный 108 интерес представляет возможность участия микроорганизмов в формирова- нии тех или иных соединений, минера- лов, пород. Так, например, пурпурные, некоторые зеленые бактерии и циано- бактерии накапливают в клетках поли- фосфат в виде так называемых волю- тиновых гранул. Некоторые пурпурные серобактерии накапливают в клетках се- ру. Однако, вероятно, гораздо чаще бак- терии не являются элементоспецифи- ческими, а способствуют накоплению тех или иных элементов или соединении как катализаторы или фильтры. Крайне интересными были иссле- дования микробных сообществ в ано- мальных условиях, вероятно похожих на условия на Земле в архейское и проте- розойское время. Это, с одной стороны, мелководные бассейны типа Сиваша, а с другой - условия вокруг термальных источников. Справедливо предполага- лось, что такие микробные сообщества из-за эволюционного консерватизма многих микроорганизмов и сходства условий могут более всего напоминать древние биоты Земли.
4 6 Еncr gy (1се Ч ) 10 4 6 Епсгду (keV) 109 Рис. 3. Коккоидные бактерии (C3M) и анализ, показы вающи й наличие слоистых силикатов на стенке клетки (по Tazaki, 1997). Микробы имеют обычно простую морфологию (форму) и с этой точки зрения представляют трудный для изу- чения объект. Действительно, морфо- логически это простые шарики, нитки, гантеле- и веретенообразные формы микронной и менее размерности (рис. 2). Основные различия современных мик- робов устанавливаются на морфологи- ческом и биохимическом уровнях. У ис- копаемых микробов ученые лишены возможности анализировать биохими- ческие особенности. Остаются морфо- логия и, что очень важно, продукты жиз- недеятельности: Fe-бактерии образуют породы, богатые окислами Fe, для дея- тельности анаэробных бактерий, харак- терно образование пирита (FeS ) и т.д. Итак, образование многих осадоч- ных пород сегодня можно связать с дея- тельностью микробов. Для фосфори- тов, карбонатов и высокоуглеродистых пород это совершенно очевидно. Однако стало понятным, что и в обычных гли- нистых породах, особенно если они со- держат хотя бы минимальное количе- ство углерода, легко обнаружить фос- силизированные остатки микробов, в том числе цианобактерий. Недавно с этой точки зрения были хорошо изучены Рис. 4. Трубчатые бактерии (C3M) и анализ, показывающий наличие слоистых силикатов на стенке клетки (по Такай, 1997). вендские породы- Приуралья, где в скважинах в обычных аргиллитах обна- ружили нитевидные цианобактерии. Аналогичные результаты были получе- ны и по глинистым породам нижнего кембрия Сибирской платформы-и Юж- ной Австралии (см. рис. 1 в). Вероятно, следует особенно под- черкнуть: исследования древних пород наводят на мысль, что при накоплении глинистых осадков микробы, с одной сто- роны, могут играть роль организатора транспорта глинистых частиц на дно, а с другой — быть катализаторами образо- вания аутогенных силикатов в слизи- стых чехлах цианобактерии. Последнее казалось достаточно очевидным при рассмотрении древних пород, где бакте- рии замещались алюмосиликатами. Это было отчетливо и неоднократно показа- но на современном материале. Слои- стые силикаты образовывались как на коккоидных, так и на нитчатых формах (рис. 3, 4, 5). Некоторое время тому назад каза- лось, что кроме грубообломочных по- род, таких, как брекчии, конгломераты, песчаники, к числу абиогенно осаждаю- щихся можно было бы отнести значитель- ную часть эвапоритов.
Рис. 5. Срез трубчатой бак-герии. Черное - слоистые силикаты, покрывающие стенку клеток (ТЭМ) (по Tazaki, 1997). 110 Во всяком случае считается, что осаждение доломита (CaMg(CO,) ) и тем более солей не имеет отношения к деятельности микроорганизмов. Однако известно, что галобактерии растут при очень высоких содержаниях NaCI и оп- тимальных температурах роста 30-50 'С. Отмечен рос г галобактерии на по- верхности рапы (насыщенного соляно- го раствора), которая становится крас- ной. Галобактерии образуют в большом количестве каротиноиды. В результате поглощения света этими пигментами повышается температура и ускоряются испарение воды и кристаллизация соли. Можно предположить, что обычные для отложений солей розовые и красные прослои могут быть связаны с перио- дическим увеличением активности га- лобактерий. Самые последние результаты швей- царских и французских исследователей показали возможность садки доломита анаэробными бактериями. Морфоло- тически получаются гантелевидные и звездообразные псевдоморфозы. Особенным является вопрос о зна- чении микробных сообществ в форми- ровании месторождений полезных иско- паемых. Бактериальные сообщества мо- гут выступать в этих случаях в самых разных ролях. Во-первых, органическое веще- ство цианобактериальных матов и дру- гих микробов может служить источни- ком органического вещества нефтема- теринских пород, то есть пород, орга- ническое вещество которых после пере- работки в течение геологического вре- мени преобразуется в нефть и газ. Было показано, что для доке мбрийских и раннепалсозойских нефтематеринских толщ характерно присутствие в них ископаемых цианобактериальных матов. Таким образом, решается проблема орган и чес кого вещества, необходимого для образования нефти в древних толщах. В этой связи существенно меня- ются и перспективы нахождения неф- тяных месторождений в докембрийских породах, которые ранее считались мало- вероятными. Более того, долгое время находки нефти в древних толщах счи- тались показателем более молодого, чем докембрийский, возраста пород. Во-вторых, цианобактериальный мат может служить своеобразным филь- тром, осаждая на себе определенные компоненты. Так, сейчас известно, что ураганные содержания более 10'Ы ред-
коземельных элементов (P33) и Nb Томторского месторождения в Сибири, обязаны своим происхождением циа- нобактериальным матам. В этом случае накопление РЗЭ происходило в каком-то бассейне типа мелкого озера, где циано- бактериальные маты постепенно увели- чивали концентрации РЗЭ из вод, посту- пающих с окружающей суши. В-третьих, эта работа элементо- специфических микробов, накапливаю- щих, например, Fe. Mn, S. Выше были упомянуты железистые кварциты Курс- кой магнитной аномалии. Вероятно, все джеспилиты (железистые кварциты) в мире имеют отношение к активности железобактерий. Интересно, что макси- мум образования джеспилитов прихо- дится на всех континентах более или ме- нее на одно время в начале протерозоя (1,9-2 млрд. лет тому назад). Следует помнить, что многие годы некоторые авторы настаивали на гидро- термальном происхождении Ее в джеспи- литах. Это нисколько не противоречит сказанному выше. Бактериальный спо- соб накопления Ее и вулканический источник поступления Fe в воды Миро- вого океана вполне могут быть совме- щены в единую концепцию. Не так дав- но в рамках Международной програм- мы геологической корреляции обсуж- дался специальный проект по биогенно- му накоплению Ее и Мп. Результаты этих исследовании не оставляют сомне- ний в огромной роли бактерий в накоп- лении Ее и Мп. Огромную работу проделывают Fe- бактерии и другие микробы в образова- нии латеритов (кор выветривания), поэто- му образование некоторых месторожде- нии осадочных полезных ископаемых, связанных с латеритами, таких, как бок- ситы (алюминиевое сырье), вполне веро- ятно. Предстоят новые открытия в иой области (подробнее о бокситах см. выше). В-четвертых, образование тех или иных пород или полезных ископаемых неэлементноспецифическими организма- ми. Например, цианобактерии в состоя- нии накапливать не только растворимый Р в волютиновых гранулах внутри клетки. При соответствующих условиях Р волю- тиновых гранул может преобразовы- ваться в нерастворимые соединения (апа- титы различных модификаций). А неко- торые бактерии способны, например, образовывать внутри клеток магнетит. В-пятых, сейчас известно по на- ходкам бактерий, что с их участием Рис. 6. Палеогеографичекие схемы ран- него кембрия (а), раннего ордовика (б) и позднего девона (в). Показаны эпиконти- нентальные моря на современных KQHTHHQHTBx (зач pHcHo). 111
Рис. 7. Палеогеография Сибирской платформы в раннем кембрии. А — традиционная трактовка (по В.Е. Савицкому, В.А. Асташкину). 1 - отложения лагунного типа; 2 — рифогенные отложения; 3 — доманиковые отло- жения шельфа открытого моря; 4 - граница Сибирской платформы; 5 - граница рифовой зоны; 6 - типовые разрезы и их номера.; Б — после обнаружения ЦБМ (по А.Ю. Розанову и Г.А. Заварзину). 112
могут образовываться соединения ура- на, меди, а также золотые псевдомор- фозы. Изучение метаболизма (обмена веществ) бактерий и механизмов, влия- ющих яа осаждение тех или иных эле- ментов, еще только начинается. Оставим в стороне все вопросы, связанные с участием микробов в обра- зовании всевозможных газов, посту- пающих в атмосферу и затем имеющих естественное отношение к образованию некоторых пород, или газов, посту- пающих в воды бассейнов и создающих, например, "аноксические" условия. Естественен, однако, вопрос: почему, зная все это, седиментологи (специа- листы, занимающиеся изучением зако- номерности осадкообразования) мало уделяли внимания роли микроорга- низмов? Здесь две основные причины. Первая — это то, что только в последнее время после внедрения сканирующего электронного микроскопа и выяснения практически мгновенной скорости фос- силизации бактерий стали доступны их непосредственные наблюдения. И таким образом ученые осознали их повсе- местное присутствие в прошлом. А вто- рая причина состоит в том, что актуа- листические модели седимен'гации не могут в полной мере объяснить законо- мерности седиментации прошлого. Дело в том, что сегодняшнее обилие суши с резкими контрастными глуби- нами океанов и высотой гор (высокое стояние материков) является достаточно уникальным в истории Земли. Гораздо чаще материки в значительной мере были покрыты морями, аналогов кото- рых по своим параметрам практически сегодня нет. Эти бассейны называются эпиконтинентальными. Точно так же до определенного времени, вероятно, не было столь глубоких огромных океа- нов. Особенно не схожая с современ- ностью картина была в палеозое (кемб- рий, ордовик, силур, девон, карбон и пермь). На палеогеографических кар- тах можно хорошо видеть, сколь об- ширные участки современных конти- нентов были заняты морями (рис. 6). Сравните современную картину с па- леогеографическими картами, напри- мер, начала раннего кембрия (рис. 6а), начала раннего ордовика (рис. 6б) и конца девона (рис. 6в). Чем же отличались древние эпикон- тинентальные бассейны? Прежде всего это были огромные по территории, иск- лючительно мелководные (десятки. ино- гда первые сотни метров глубины) бас- сейны, у которых значительная часть дна находилась в пределах фотической зоны. Часто на этих акваториях возникали мно- гочисленные и короткоживущие мигри- рующие острова. Специфика древних бассейнов способствовала широкому раз- витию микроорганизмов и особенно циа- нобактериальных матов, которые могли иногда функционировать на тысячах квадратных километров (например, Си- бирская платформа в докембрии). Поэто- му в этих бассейнах роль бактерий и вообще микроорганимов была огромна не только в формировании трофических связей, но и в формировании, а затем и преобразовании осадков. Интересны возможные послед- ствия для интерпретации палеогео- графической ситуации в результате обнаружения в породах микробных со- обществ, особенно если это цианобак- териальные бентосные маты. Так, например, в синской свите ботомского яруса и ее аналогах на Сибирской платформе были обнаружены ЦБМ, что заставило совершенно иначе пред- ставить палеобиогеографию того вре- мени (сравни рис. 7А и Б). Вместо глубоководного открытого в "океан" бассейна мы вынуждены рисовать мел- ководный полузамкнутый бассейн с аноксической приданной ситуацией. Это нашло подтверждение и в анализе геологической ситуации по территории восточнее Верхоянского складчатого пояса, где были обнаружены разрезы, типичные для "переходной" зоны Си- бирской платформы (мелководные с ар- хеоциато-водорослевыми биогермами). Перед седиментологией стоит сложная, но интересная задача разра- ботки принципиально отличных от современных моделей седиментации в древних эпиконтинентальных бассей- нах. Совершенно очевидно при этом, что огромная роль биогенного, и осо- бенно бактериального, фактора должна быть учтена, поскольку за редким ис- ключением все осадочные породы в той или иной степени образовывались с участием микробов. 113
ЦИАНО-БАКТЕРИАЛЬНЫЕ МАТЫИСТОЧНИК МАТЕРИНСКОГОВЕЩЕСТВА НЕФТИ 114 До последнего времени подавляю- щее большинство исследователей до- статчно единодушно считало, что источ- ником органического вещества (ОВ) нефтематеринских пород является план- ктон и в значительно меньшеи степени- бентос. Однако обнаружение докембрийс- кой нефти поставило под сомнение эти убеждения. Исследователи стали счи- тать, что источником углеводородов (УВ) может быть любое органическое вещество. В случае с докембрием и нижним палеозоем необходимо было вычислить такую "систему", которая была бы и ши- роко распространена, и достаточно про- дуктивна. В этом смысле наиболее интерес- ными являются бактериальные маты (БМ) и прежде всего — бентосные циано- бактериальные маты ((ЦБМ). Именно в докембрии они были широко распрост- ранены и появились уже как минимум 3,5 млрд. лет тому назад. Сегодня ЦБМ развиваются глав- ным образом в узкой зоне прибрежно- морских условий (литораль, сублито- раль, себхи, лагуны) и в континен- тальных озерах. Однако современная географическая ситуация достаточно аномальна — сегодня нет обширных эпи- континентальных бассейнов, столь ха- рактерных для многих прошлых эпох, когда ЦБМ могли занимать простран- ство в миллионы квадратных кило- метров. Жизнедеятельность ЦБМ чрезвы- чайно благоприятна для перехода в погребенное состояние ОВ. Продук- тивность может составлять 2,5-4,5 до 8-10 С >/м'- в сутки. Таким образом, за 1 млн. лет такая система на площади 1 км2 способна была продуцировать миллиа ды т Co> . О бактериальных матов обога- щено липидными (битуминозными) компонентами. Кроме того, ОВ бактери- альных матов в повышенных концен- трациях содержит кислород, азот, серу, фосфор. Это связано с тем, что среди микроорганизмов ЦБМ присутствует большое количество сульфатредукто- ров, фиксаторов азота и фосфора, кото- рые способны накапливать полифос- фаты. Обогащенность бактериального ОВ порфиринами предопределена высо- ким содержанием хлорофилла в мате, плотность которого в пересчете на пло- щадь может превышать таковую в ли- стовой пластине в несколько раз (Гера- сименко, Заварзин, 1993). Гетероэле- менты О, N, S разрыхляют структуру керогена, находясь в мостиковых связях, они обусловливают деструкцию его при менее жестких РТ-условиях (P — дав- ление, Т — температура) и тем самым могут способствовать раннему (прото- катагенетическому) нефтеобразованию (Баженова, 1990). Древность бактериальных матов, широкая география областей их развития в эпиконтинентальных бассейнах про- шлого, способность к созданию и накоп-
Рис.1. Схема соотношения участия бентоса и планктона в формировании УВ в геологической истории Земли. 1 — планктон, 2 — бентос. 115 лению преобразованного ОВ, при мини- мальном рассеивании его в простран- стве, делают эти бентосные системы наиболее приспособленными для форми- рования исходного ОВ нефти в течение всей геологической истории их суп1ест- вования. Особенно велика была их роль в протерозое и раннем палеозое (рис.1). Изложенное подкрепляется элек- тронно-микроскопическим изучением древних высокоуглеродных пород, в которых всегда обнаруживаются фосси- лизированные коккоидные и нитчатые микробы.
шунгиты - модЕльныЙ оБъЕкт ДЛЯ ИЗУЧЕНИЯ ГЛОБУЛИЗАЦИИ 116 Шунгит — это твердый некри- сталлический углерод со структурой, неспособной к графитизации. Внешне он напоминает антрацит. Название происходит от пос. Шуньга (Карелия). Специфический состав, использование шунгитов в качестве разнообразного сырья и, наконец, обнаружение в них фуллеренов, определило огромный к ним интерес и стимулировало их детальное изучение. Применительно к интересам бактериальной палеонтоло- гии и изучению биоморфных структур внеземного вещества, шунгиты инте- ресны выявленным в них гпобупярным строением. Генезис шунгитов тракту- ется по-разному. Некоторые считают шунгиты эндогенными, многие считают шунгиты биогенными, а ряд исследова- телей считает шунгиты полигенными. Самые масштабные местонахо- ждения (месторождения) шунгитов отмечены в Карелии, однако в незна- чительных количествах шунгиты изве- стны на Урале, в Якутии и Казахстане. Карельские шунгиты приурочены к нижнепротерозойским породам (рис. 1) и, как правило, распространены на двух уровнях — людиковийском и калевий- ском, хотя встречаются и выше вплоть до широко известных кварцитов шок- шинской свиты. В последнем случае они изучены значительно слабее. По представлениям многих иссле- дователей шунгитовое вещество, преоб- разованное из первоначально сапро- пелиевого вещества, накапливалось на мелководье в застойно-морских усло- виях. Этим выводам соответствуют геологические данные о характере зале- гания (рис. 2) шунгитных пород, а также многочисленные данные исследования органического вещества (ОВ) шунгитов, обстоятельно проанализированные в монографии "Органическое вещество шунгитоносных пород Карелии", Петро- заводск, 1994. Из разобранных в этой работе особо отметим данные по 5C'-', сравнительную характеристику ОВ разного генезиса (табл. 1) и сравнительные данные по средним содержаниям элементов высокоуглеро- дистых шунгитовых пород (табл. П). В этой же работе проведен полный обзор палеонтологических данных по шунгитоносным породам. Из них осо- бенно следует выделить нахождение достоверных строматолитов и онколи- тов, цианобактериальных образований и акритарх. Цианобактерии и акритархи обнаружены непосредственно в углеро- дистых разностях. С.И. Жмуром с соавторами ранее были также предприняты попытки изучения в СЭМ шунгитоносных пород этого региона, в результате чего сложи- лось впечатление об участии в формиро- вании ОВ шунгитов цианобактериаль- ного коккоидного мата (ЦБМ). В связи с представлениями об участии коккоид- ного ЦБМ в формировании шунгитов необходимо коснуться проблемы глобу- лярной модели строения шунгитов. Н.П. Юшкин обобщил известные сведения и на основании исследования спектров комбинационного рассеяния шунгитов показал, что на молекулярном уровне шунгит представляет собой "слабоупорядоченную постройку из правильных или деформированных гек- сагонапьных графитовых молекул с включениями фуллереновых блоков или отдельных фуллеренов, а может быть, и других углеродистых молекул". Как показали В.В. Ковалевский, Е.А. Голубев и Н.П. Юшкин, целый ряд разновидностей шунгитов имеет глобу- лярную надмолекупярную структуру. При этом следует особо обратить
Таблица 1. Сопоставление некоторых характеристик ОВ разного генезиса Таблица 2. Среднее содержание элементов и доверительные интервалы для доманикоидов и шунгитовых пород нгитовые по оды ) лемент' ++ +! + Ъ ,51+0,11 + ++ 10 + +! 122+ +111 ,1+,1 7, +, ! + ++ ет данных +! +! 1 1+91 132+1 11+ 1 +11 18+6, 'C„,„, К, Na, Fe — в '4, Au - в мг/т, остальные элементы - в г/т. -' Коэффициент концентрации по отношению к кларкам осадочных пород 117 ленекские горючие сланцы (от, до 1, ) 6,5+2,1 + 1,1 1,2 гиллиты женовскои свиты (30) +! ст данных 1,1 +, l,35+0, 2,3+1, + + ,3+ 1,! ++
внимание на то, что размеры глобул, которыми этими авторами характери- зуется "надмолекулярная структура" шунгита в пределах первых десятков нм (рис. 3). Н.П. Юшкин получил досто- верную реальную картину TRM глобу- лярной структуры шунгитов (рис. 4). И в этом случае размер глобул не превышает десятков нм, а их агрегаты 200-300 нм. Н.П. Юшкин говорит о двух уровнях глобулярных агрегатов. Полученные после термической обработки шунгитов (рис. 5) глобулы и полиэдры также не превышают 100 нм. Вместе с тем структуры, напоминающие глобулы циа- нобактерии, имеют размерность мик- ронную или даже десятки микрон в случае объединения нескольких "кле- ток" в одну глобулу или колонию. Дополнительные исследования дали возможность получить картину глобулярной структуры шунгитов (рис. 6-9), в которой можно наблюдать и "надмолекулярный уровень" глобул Н.П.Юшкина и глобулы цианобакте- риальной размерности, и практически непрерывный размерностный ряд. Таким образом остается непонят- ным вопрос о природе разных уровней глобулизации шунгита. Можно сегодня с равной вероятностью считать: 1) что многоуровневая глобулизация — это отражение процесса самоорганизации шунгитового вещества или 2) что мы наблюдаем фоссилизированный мат или биопленки, где разного размера глобули соответствуют разным типам или разным стадиям развития клеток или других образований внутри бактери- ального сообщества мата и 3) то, что наномстрический уровень (или "надмо- лекулярный уровень" Юшкина) — это огражение абиогенного процесса гло- булизации, а микронный уровень — это псевдоморфозы по бактериям и фосси- лизированным биопленкам (рис. 14). K2i ERs ~ñÐ Я ~Др КЬ EvHv10 [ч ч~~ ~rz кЫНФ Ийю 'K$ ая~д 256/9 Рис. 1. Сводная стратиграфическая колонка образований нижнего протерозоя Онежской мульды. 1 - полевошпат-кварцевые песчаники и конгломераты (кварцевые и полимиктовые), 2- полевошпат-кварцевые алевролиты, 3 — кварциты, 4 - туфы и кремнистые сланцы, 5 - гравелиты, 6- хлорит-плагиоклаз-кварцевые сланцы, 7 - шунгитсодержащие плагиоклаз-хлоритовые сланцы, 8- туфопесчаники и туфоалевролиты, 9 — туфоконгломераты и туфобрекчии, 10 - шаровые лавы основного состава, 11 - лавы основного состава, 12 - лавы ультраосновного состава, 13 - шунгитсодержащие и шунгитистые кварц-биотитовые сланцы и туфопелиты, 14 — доломиты (а) и шунгитсодержащие доломиты (б), 15 - лидиты, 16 - шунгитовые прады калиевого (а) и натриевого (б) ряда, 17 - туфы литокластические (а) и витрокластическис (б), 18 - слоистые доломиты и кварц-биотит-карбонатные сланцы, 19 - доломиты с органичсскими остатками, 20 - доломиты и сланцы, 21 - красноцветные доломиты, 22 — вывстрелые граниты. 118
Рис. 2. Разрез Кондопожской структуры. 1 — витрокластические туфы (верхняя попсвита), 2- сероцветная толща кулмукских сланцев (пачка В), 3 - бесшунгитовые аналоги нигозеритов (пачка Б), 4 - шунгитсодержащие сланцы (пачка Б), 5 - пирокластотефроидные песчаники (пачка А), 6- туфоалевролиты, 7 - переслаивание сланцев и туфоалевролитов, 8 - полимиктовые конгломераты, 9- пикритовые и пироксеновые пофириты, 10 - миндалекаменные диабазы, 11 - скважины. Ранее вместе с Д. Маккеем, Ф. Ве- сталл и Р. Хувером мы исследовали глобулярную структуру нижнекембрийс- ких фосфоритов Монголии. И в случае с фосфоритами наблюдались глобули нан- ометровой размерности, образующие псевдоморфозы по "пурпурным (?) бак- териям", которые в свою очередь со- ставляют стенки реально опознаваемых цианобактерий (рис. 10-13). Друтая группа образований, на которую следует обратить внимание при изучении шунгитов, напоминает псев- доморфозы по конидиям низших грибов (рис. 15-16). Это вытянутые веретено- образные тела длиной в несколько мк. Достоверность такого заключения, ко- нечно, очень низкая, но подобная интер- претация была подсказана массовыми находками М.Ю. Беловой и А.М. Ахме- довым, вероятно, грибов в углеродистых порЬдах печенгской серии того же региона. Находки остатков предположи- тельно низших грибов в нижнепро- терозойских породах указывались и ранее Д. Дерби, Е.С. Баргхурном, С.А. Тайлером, Б.В. Тимофеевым и др. В этой связи представляется возможным, во-первых, переинтерпретировать неко- торые коккоидные маты в грибные сообщества и, во-вторых, предполо- жить, что спецификой раннего про- терозоя является вспышка развития низших грибов, а ОВ шунгитов является производной от ОВ не только разно- образных бактерий, но и низших грибов. ЯоЬи1аг роге G but Рис.3. Электронно-микроскопический снимок шунгитового углерода на просвет (а) и соответствующая ему мЬдель глобулярного строения (Ь). 119
Рис. 4. Глобулярная субструкгур» шунппд (рас гровьш туннслы~ый микроскон). Рис. 5. Элекгронно-микроскопичсскис снимки на нросвст глобулярного шунгитового углсрода нослс тсриической обработки. 120
Рис. 10. Полуаморфный фосфат кальция, слагаюший стенку трубки цианобактерии снаружи. Рис. 6. Рис. 8. Рис. 6-9. Глобулярные структуры шунгитов в ESEM. Рис. 11. То же, увеличено. Рис. 7. Рис. 9.
МИНЕРАЛИЗОВАННЫЕ СВИДЕТЕЛИСУЩЕСТВОВАНИЯ ГИДРОТЕРМАЛЬНОЙЭНДОБИОСФЕРЫ Гндротермальные зоны вторичнойминерализации 123 В настоящее время установлено большое разнообразие состава минера- лов, формирующихся при активном уча- стии бактерий. Исследование литификации бакте- рий в природной обстановке и в усло- виях лабораторного эксперимента пока- зали, что формирование минерального вещества происходит очень быстро. Наиболее определенно скорость бакте- риальной минерализации была выявле- на при формировании фосфатов и кар- бонатов. Лабораторные эксперименты показали, что минерализация фосфатом бактериальных нитей происходит в теч- ение нескольких часов (см. главу "Фосфатизация). Осаждение карбонатов кальция в морской воде при участии бактерий происходит также очень быстро. В последнее время появились данные, указывающие на быструю минерализацию и замещение бактери- альных нитчатых структур слоистыми силикатами в современных горячих источниках на дне океана и в пресно- водной обстановке. Бактериальные сообщества хорошо изучены в гидротермальных водах на поверхности Земли в пределах фотичес- кой зоны. Значительно меньше сведений имеется о существовании и формах жиз- ни в глубоких горизонтах пород, промы- ваемых нагретыми подземными водами. В мощных наземных толщах мио- ценовых платобазальтов Исландии, по- гружавшихся ниже уровня подземных вод и промывавшихся нагретыми вода- ми, выявлены минерализованные ми- кроорганизмы и их колонии, свидетель- ствующие о существовавшей глубоко в толще вулканогенных пород жизни (эндобиосферы) (Гептнер и др., 1997). Исландия — очень хороший объект для изучения возникновения и существо- вания жизни в гидротермальных систе- мах глубоких подземных горизонтов. Остров сложен практически целиком вулканогенными породами. В основном это базальты, не более 10% составляют кислые породы. Роль осадочных пород, содержащих какие-либо биогенные остатки, ничтожно мала. Поэтому с большой долей вероятности можно го- ворить о том, что появление биоты в гидротермальных системах Исландии вряд ли может быть связано непосред- ственно с существованием здесь осадоч- ной оболочки литосферы. О характере гидротермальных сис гем, оставивших минеральные метки в миоценовых платобазальтах, мы мо- жем судить по минеральным парагене- зам и по аналогии с современными гидротермами. В прошлом, как и в настоящее время, крупнейшие водоносные си- стемы были связаны с трещинными роя- ми, трассирующимися на вулканические постройки центрального типа. Тепловой nouveau,Has? nocaegl,HHx ~an~sics~ ocvoa~o~ причиной возникновения гидротермаль- ных систем. По составу и распределению в вертикальном разрезе вторичные мине- ралы в вулканических толщах Исландии принципиально не отлчаются от зо- нальности, установленной в других районах современной вулканической
Рис. 1. Результаты лабораторного эксперимента. Продукты преобразования базальтов и базальтового (сидеромеланового) стскла, сформировавшихся в контейнере, закрытом от света в тече- ние 9 месяцев при температуре = +75' С' с участием бактериальных термофилов. СЭМ. а - нитчатые и шарообразные бактериальные структуры; б — крупная, минерализованная нитчатая бактериальная структура; в, г - нанобактерии на поверхности листочков слоистых силикатов; д - микропюбулярная структура частички слоистых силикатов - начальная стадия преобразования силикатного коллоида. l24 активности. По минералам-индикаторам выделяются следующие зоны: (1) зона смектитов-цеолитов; (2) зона смешан- нослойных смектит-хлоритов; (3) зона хлорит-эпидота; (4) зона хлорит-акти- нолита, характеризующие разные тем- пературные интервалы. Помимо смек- титов, характерным глинистым мине- ралом верхней зоны является селадонит. Наличие этого минерала в породе благо- даря типичной яркой голубовато-зеле- ной окраске легко распознается в поле и может быть закартировано. Особенно важно отметить, что в состав селадони- та входит значительное количество ка- лия, важного элемента живой природы. Смектит-селадонитовые минералы в толще платобазальтов образуют протя- женные зоны от нескольких десятков до 100 м мощности, располагающиеся согласно с напластованием лав. Позднее образовавшиеся гидротермальные ми- нералы - цеолиты и кремнистые мине- ралы - образуют тоже стратиформно
располагающиеся зоны, которые секут моноклинально залегающую сейчас толщу платобазальтов и согласно распо- ложенную с ней зону смектит-селадо- нита. Это указывает на то, что формиро- вание комплекса смектит-селадонит- овых минералов происходило на началь- ной стадии погружения базальтовых толщ ниже уровня подземных вод, вско- ре после накопления лав. Время форми- рования смектит-селадонитовой мине- рализации таким образом можно счи- тать близким к миоценовому возрасту платобазальтов. Смектит-селадонитовая минерализация отделена от более позд- него этапа минерализации значитель- ным периодом времени. Это означает, что формирование комплекса смектит- селадонитов и более поздних мине- ралов происходило в разных физико- химических условиях. Современные данные позволяют считать, что темпе- ратура образования смектитов и селадо- нита была ниже 100 'С. Биоморфные структурыв слоистых силикатах Методы изучения Диагностика биоморфных или биоморфноподобных микроструктур в слоистых силикатах проводилась при сравнении и сопоставлении с уже изве- стными минерализованными в разных условиях бактериальными формами жизни. Это современные примеры фос- силизации в естественных условиях и проводящиеся в лаборатории. Подав- ляющее большинство имеющихся дан- ных относится к микробиоте, жившей на поверхности Земли. Хорошо извест- ны примеры минерализации микроор- ганизмов железистыми и марганцевыми минералами, карбонатами, фосфатами, кремнистыми минералами. Недавно бы- ли обнаружены микроорганизмы, мине- рализованные глинистыми минералами разного состава в зоне разгрузки тер- мальных вод на дне океана и на поверх- ности Земли. Интересные результаты, под- тверждающие активную роль бактерий при формировании глинистых мине- ралов, получены при исследовании воз- действия бактерий на базальты в искус- ственной гидротермальной системе. Эти исследования проводились А.Р. Гептне- ром совместно с исландскими гидрохи- миками и биологами (Hrefna Krist- mannsdottir, Jakob К. Kristjansson, Viggo Th. Marteinsson) в 1997-98 гг. Образцы базальтов и базальтового (сидеромела- нового) стекла, погруженные в спе- циально подготовленные растворы с жи- выми бактериальными сообществами, аналогичные обитающим в современ- ных термальных системах Исландии, выдерживались в течение 9 месяцев при температуре 75' С. Полученные резуль- таты свидетельствуют о большой роли бактерий при разрушении базальтового материала и формировании слоистых силикатов (рис. 1). Ниже рассматриваются примеры формирования биоморфных микро- и наноструктур, сложенных слоистыми силикатами смекти г-селадонитового состава, формировавшиеся на началь- ной стадии изменения лав, погрузив- шихся ниже уровня подземных вод. Необходимо заметить, что селадониты- характерные минералы гидротермаль- ных систем — являются "ближайшими родственниками" глауконитов. Участие микроорганизмов в формировании смектитов и глауконитов в зоне диа- генеза современных осадков сейчас можно считать доказанным. Все исследованные образцы были отобраны из разрезов платобазальтов, измененных -низкотемпературными (менее 100 'С) пресными подземными водами. Очень важно подчеркнуть, что в современных низкотемпературных гид- ротермальных системах подобного типа формирования глинистых минералов не происходит. Общая минерализация в них не превышает 250-300 мг/л и из них вообще ничего не осаждается. В Ислан- дии они нередко используются непо- средственно в системах горячего водо- снабжения и отопления в городах и фер- мах, и отложения солеи в системе водо- снабжения не наблюдается. Метаколлоидный характер минерализации биоморфных структур Наблюдения в поле и исследо- вания в лаборатории выявили ряд характерных микроструктур слоистых силикатов селадонитового и смектит- селадонитового состава, располагаю- щихся по стенкам и в свободном про- странстве трещин, газовых пустот и разнообразных полостей в лавах. Важно подчеркнуть, что одинаковые по форме, составу и размерам структуры были 125
СМ сел ~ — 0,8 см — ~в» нит ,5ММ— Ш Шб ° ° ° ° селадонит ° ° ° 1 ° ° ~1 l 1 ° ~ е~ I е: Ф ° ° ° е ° ° ° кремнистые минералы е ° ° ° ° ° ° е ° ° ° е ° с ° ° ~~ ° ° е ° у ° ° ° ° ° ° ° ° ° ° ° ° ~ ° ° 1 ° ° ° ° ° ° ° ° ° ~ ° е ~ ° в ° ° ° ° ° ° е ° ° ° ° ° ° ° ° ° ° ~ ' ° е ° ' ° ° ~ ~ ° ~ ° ° ° ° ° ° ° ° ° 1.'.' ~ 1: 1 ° ° ° - смектиты=)é1 1,~ см ° ~ ю ~ ~faр ° °° ° ъ° ~в ° ° °, ° ° ° ° ° ° ° ° J% е ~ ~~ ~ ~~Q ~~В° ° ° ° ° ° ° ° ° °° ° ° ° Рис. 2. Хемогеш1ыс и био . 1 Оиох~.Mol L.11ныi. гицы в!opllillloll мицсра~!изд11ии в !103!ос! ях ~1авовых llo- токов, зарисовки со шлифов. 1. — хемогспнос. с1рати<ю мпос ип > иформпос иполнепис газовой полости глинистыми длйми. - оиох ' ' ' ' ', ' ' . ' ' ' ) смс1!'1'с. - мс'!'аколло!1дн ыс злами. - оиохсмигснllыс ссладо11и'1'овь!с ! Ilп'и в смек1'и10в()м Il," ' ' 111 лалонитовыс полусферы иа cTcllKèõ га швой полости: 111а - об1пий виз, lll6- ния. IV - биохемогенцыс сел» ) .) ° ' ° ' -,, - ~- . а — о 'дони'!ов1.1с !!И1! !!1!с с!руктчр1,! в г;!зоной !1Олос1и: Ша — о " . !"б - деталь стпоения. V - соотношс ~ - б р - . - ( шсние биохсмогс1и1ь1х ссладоцитовых !И111а1 hlx структур стратиформных смекгитов и крсмнист» Ix мин ) Ч — Г р и хсмогенных i-[ «ЛОВ: cl - 0611 СИИ ВИД, О - ДСТс!ЛЬ CTPOL.'IIÈß. ном (коллоидном) осаждении матери- ала, последующей его раскристаллиза- ции и превращении в слоистые силикаты селадонитового и смектит-селадонито- вого состава (рис. 2) (Чухров, 1955). Подтверждением этого служат дан- ные, полученные о составе и структуре слоистых силикатов, формирующихся сейчас у горячего подводного источника 126 встречены в разных районах. Метакол- лоидный характер этих структур (микро- глобулярные формы, ветвящиеся и изги- бающиеся нити, сферы и полусферы на стенках пустот и т.д.) и взаимоотноше- ние их с более поздними, запечатываю- щими минералами и вмещающими по- родами определенно свидетельствуют о первично микроскопически-аморф- 'L11ll)IIIII(I1ЪЪ\1 оссилизированная ! ° ] ' ] нить И: ю.' ! J у i ° f Крустификационная f. ° 1ка l ° ° $ ° ° 1 ° el
Рис. 3. Биохемогенные структуры в отло- жениях современного подводного горячего источника, нитчатые и кокоидные структуры, сложеиныс сапонитом. СЭМ. Описание биоморфныхмикроструктур Рис. 4. Фоссилизированные нанобактерии (некоторые указаны стрелками) в смектит- селадонитовом матриксе, заполняющем газовую полость в базальтовом лавовом пласте. СЗМ. а— нано-бактерии на поверхности и гранях глинистых чешуек; о - плотное скопление нанобактерий. на дне Эйьяфьорда, на севере Исландии. Два образца, отобранные исландскими учеными в мае и июне 1998 года в раз- ных местах подводной термальной по- стройки, минералогически исследова- лись в Геологическом институте РАН. Они оказались идентичными по хими- ческому составу, но один представлен по данным рентгеновского анализа сапонитом (магнезиальный слоистый силикат), а другой, судя по рентгено- грамме, видимо, сложен рентгена-аморф- ным предшественником этого минерала, с еще не сложившейся трехмерной струк- турой. Биоморфный характер структур, минерализованных сапонитом и его предшественником, не оставляет сомне- ний в активной роли микробиоты в процессе формирования постройки подводного горячего источника (рис. 3). Многие из изученных микрост- руктур слоистых силикатов современ- ной подводной термальной постройки и из гидротермально измененных миоце- новых базальтов похожи и могут срав- ниваться по размерам с бактериями и их колониями„минерализованными в раз- ных условиях. К такому выводу можно придти, если сравнивать с минерали- зованными бактериальными структу- рами, изученными из современных по- строек океанских курильщиков, из дру- гих поверхностных обстановок или фос- силизировавшихся в лабораторных условиях. Необходимо подчеркнуть, что на- блюдаемые сейчас размеры биоморф- ных структур не всегда точно соответ- ствуют прижизненным формам. Процес- сы фоссилизации часто значительно изменяют их размеры. В настоящее время выявлены и исследованы следующие морфологи- ческие типы биоморфной минерали- зации в подземных гидротермальных условиях: палочки, шары, нити, пленки, полусферы. Эти морфоло- гические структуры встречаются от- дельно или образуют разнообразные ассоциации в виде неправильных по- ристых масс в газовых полостях и открытых трещинах наземных лавовых ПОТОКОВ.
Рис. 5. Фоссилизированные шарообразные бактсриальные структуры. СЭМ: и - кокоидные нанобактерии (отмечены стрелками) в микрозернистой, пористой глинистой массе, заполняющей крупную полость в лавовом потоке (обратить внимание на вытянутьи поры с характерной круглой и овальной формой на поперечном срезе); б — плотная шарообразная структура, сложенная чешуйками смектита, в окружающем пространстве — вытянутыс игольчатые кристаллы селадонита. 128 Важно отметить, что при анализе пространственной ориентировки рас- сматриваемых образований не удалось обнаружить какой-либо закономерности или особенностей их расположения, связанных, например, с действием силы тяжести. Это позволяет считать, что рассматриваемые морфологические ти- пы слоистых силикатов не могли воз- никнуть в результате чисто химических мембранных процессов и расслоения жидкостей разной плотности. Палочкообразные структуры это слегка вытянутые с закругленными контурами образования, располагаю- щиеся на гранях и реже на плоскостях глинистых чешуек. Иногда их скопление достаточно велико, и они заполняют промежутки между глинистыми чешуй- ками. Размер изолированных тел такого типа не превышает 0,7 микрона (рис. 4). Эти структуры по форме и размеру сход- ны с фассилизированны ми нан
Рис. б. Шарообразные структуры смсктит-ссладонитового состава s строении нитей. а - общий вид нитей, сложенных глинистыми минералами и запечатанных кремнистыми минералами. Внутрсннис части нитсй сложены кокоидными струк- тууами, а с поверхности покрыты пленкой глинистого вещества зонального стросния. Оптичсский микеоскон, шлиф. о - нитс- ооразные структуры в крупной газовой полости лавового потока, фото образца породы, диаметр газовои полости около 0,5 см. в - нитеобразная структура, сложенная смектитовыми шарообразными структурами. покрыта с поверхности слоем метаколлоид- ных крсмнистых минералов; обратить внимание на круглые отвсрстия (часть помсчсна стрелками) в'кремнистой оторочкс. включавшие шарообразные смсктитовые образования. Скол образца, '3M. г - кокоидная форма (сложенная смектитами) из колломорфной кремнистой оторочки нитеобразной структуры, деталь рис.бв. СЗМ. д - смскт товыс кокоидныс структуры, меж- ду которыми располагаются крупныс вытянутые кристаллы ссладонита, деталь строения центральной части нитсобразной структуры, показанной на рис.бв. 'СЭМ. е - смсктитовые кокоидныс образования, располагающиеся на повсрхности нитчатой структуры с отчетливым секционным строением, стрелкой показана ссладонитовая оторочка, благодаря которой поперечный размер нитчатой структуры значительно увеличен. СЭМ. ж - смсктитовые кокоидныс (шарообразные) теда разного размера, располагающиеся в плотной глинистой массс. СЭМ. 129
Рис. 7. Некоторые особенности строения структур нитей, сложенных глинистыми минералами. и - нить с полым центральным каналом. б - поперечный срез, демонстрирующий зональное строение нити, на поверхности которой видны кокоидные образования. в - нить, состоящая из плотно соеди- ненных пористых шарообразных структур, обратить внимание на круглые и овальные поры в этих структурах; цементируется нить кремнистыми метаколлоидными минералами. СЭМ. г - ветвящиеся нити, состоящие из плотно прижатых кокоидных образований, оптический микроскоп, шлиф. 130 Поперечный размер минерали- зованных нитей (10-40 микрон) зависит от толщины крустификационной глини- стой или кремнистой каймы. Мини- мальный размер без такой каймы, на- пример, вытянутых круглых полостей (рис. 7а), колеблется от 4-7 до микрон. Максимальная длина нитей зависит от размера полости, в котором она фор- мировалась. Важно подчеркнуть, что в нитях, наблюдавшихся в разных райо- нах, иногда можно видеть отчетливо выраженное секционное строение (рис. бе, 7в). Характерно сочетание ветвящихся нитей и скоплений глобулярных тел, встречающееся в глинистых массах, заполняющих в лавах большие по размеру пустоты (рис. 9). Пленки - это сложные образова- ния заполняющие частично или пол- ностью самые крупные полости в ба- зальтах. В шлифах на срезе видно, что это серия параллельных зон, каждая из которых имеет ровное, плавно изги- бающееся основание, сменяющееся си- стемой субпараллельных нитей, ориен- тированных перпендикулярно к осно- ванию пленки. При изучении на ска- нирующем электронном микроскопе этой части пленки обнаружено, что она состоит из агрегатов круглых и непра- вильной формы тел с диаметром от 6 до 20 микрон. При большом увеличении видно, что пленка состоит из сильно по- ристой массы, в которой можно разли- чать отдельные крупные шаровые структуры и цепочки мелких глобуляр- ных тел, образующие фигуры, подобные кружевам. В отдельной полости плен- ки образуют параллельно ориентиро- ванную систему, но в соседних поло- стях их пространственная ориентиров- ка не совпадает. Полусферы — это структуры 1-2 мм диаметром с тонкой концентрической
Рис. 8. Ветвящиеся и изгибающиеся нити, сложенные смектитами. Оптический микроскоп, фотография со шлифов. а -нити, в структуре которых при небольшом увеличении видна неоднород- ность внутреннего строения. б - ветвящиеся нити, внутреннее строение которых показано на рис.7 а,б. Пористость - характерная чертарассмотренных тинов биоморфныхструктур 131 слоистостью и четкой внешней грани- цей. Располагаются они хаотически, не- посредственно на вмещающих породах, слагающих стенки разнообразных по- лостей в базальтах, а также были встре- чены внутри этих полостей в общей глинистой массе. В месте скопления по- лусфер они налегают друг на друга (рис.2 Ш). Характерная черта разнообразных скоплений рассмотренных типов био- морфных структур, а также глинистой массы, залегающей между ними - силь- ная пористость. Важно обратить внима-
Рис.9. Ассоциация нитевидных и шарообразных (глобулярных) структур в смектит-селадонито- вом заполнении крупной полости в лавовом потоке, оптический микроскоп, фотография со шлифа. микроскопе, помимо прочих особенно- стей их строения, обнаружены много- численные круглые и овальные отвер- стия и вытянутые каналы, по размеру и форме чрезвычайно напоминающие те, которые были встречены в скоплениях слоистых силикатов с разнообразными биоморфными структурами в миоцено- вых базальтах. В некоторых образцах оксалатных конкриментов в порах были обнаружены фоссилизированные чехлы микроорганизмов (рис. 11а, 6). Что подтверждает биогеннуюприроду морфоструктур врассмотренныхслоистых силиматах? О природе рассматриваемой по- ристости можно судить не только срав- нивая их с миоценовыми фоссилизиро- ванными биоморфными структурами. Сходный характер пористости был об- наружен при изучении строения конкри- ментов, т.е. минеральных тел, образо- вавшихся в теле человека. При иссле- довании оксалатных и фосфатных обра- зований на сканирующем электронном 132 ние на форму и размер пор, характер ми- нерализации стенок пор. Удивительно, но во многих изученных образцах, отобранных из разных местонахож- дений, характер пористости сходен по размерам и форме. Вытянутые планко- образные кристаллы селадонита плавно изгибаются, обрамляя паровые отвер- стия. Характер этого взаимоотношения таков, что невольно напрашивается предположение, что кристаллы глини- стых минералов формировались вокруг каких-то тел, теперь исчезнувших. Фор- ма этих тел в поперечном разрезе была круглая или овальная, а в продольном они были сильно вытянуты, ветвились и прихотливо изгибались. По форме, ха- рактеру ветвления и размеру они впол- не соответствуют рассмотренным выше минерализованным нитям. Диаметр та- ких полостей, оставаясь близким для одного образца, варьирует в разных образцах от 3-5 до 14-18 микрон (рис. 10 а-в). Суммируя вышесказанное, отме- тим, что помимо общего морфологи- ческого сходства рассмотренных струк- тур с современными фоссилизиро- ванными бактериями и их колониями следует обратить внимание на сле- дующее. (1) Рассмотренные микро- структуры обнаружены в разных райо- нах Исландии, в горизонтах разного возраста, но везде были встречены уди- вительно похожие по форме и размерам "сообщества" структур. Это явление трудно объяснить простым совпадением при чисто химическом способе их образования. (2) В основном для био- морфноподобных образований харак-
Рис. 10. Характер пористости смектит-селадонитовых скоплений в полостях лавовых потоков. СЭМ. а - общий вид пористого участка; б — хорошо раскристаллизованное глинистое вещество между порами (видны игольчатые кристаллы), стенки пор облекаются тонкой пленкой, в которой кристаллы располагаются согласно с поверхностьк~ пор. g - деталь строения пленки, облекающей стенки пор, хорошо видиы илаикообразш и: кристаллы ссладоиита, "облекающие" иолость иоры. 133 терны одинаковые или узкие пределы колебаний размеров. Диаметр нитей, без учета крустификационной каймы, равен 5-7 микрон. Для шарообразных струк- тур существуют два наиболее часто встречающихся размера диаметра - 20- 30 микрон и 50-70 микрон. В последнем случае шаровые структуры имеют, как правило, концентрическое строение, а их большой размер, возможно, связан с интенсивной гидротермальной минера- лизацией. (3) Прямым подтверждением фоссилизации органики является на- блюдавшийся секционный характер ни- тей и наличие в их центре круглых или овальных каналов. (4) Биоморфные структуры состоят из разных глини- стых минералов, в составе которых ка- лия намного больше (например, в села- донитах до 7-8'4~) по сравнению с фоно- вым содержанием этого элемента (0,2- 0,4/o) во вмещающих базальтах. Из- вестно, что этот элемент играет важней- шую роль в метаболизме живых орга- низмов и мог аккумулироваться в процессе жизнедеятельности микроор- ганизмов в слоистых силикатах. (5) Косвенным подтверждением биохемо- генного генезиса рассматриваемых объектов может быть также тот факт, что гидротермальные слоистые сили- каты с биоморф ными микрострук- турами оказались обогащенными угле- водородами по сравнению с другими вторичными минералами и свежими вулканитами (Гептнер и др., 1999).
Рис. 11. Характер иористости в оксалатных конкриментах. С'ЭМ. а - фоссилизированные остатки палочкообразных микроорганизмов, торчащис из обломк» конкримента; б - пористость, оставленная палочкообразными микроорганизмами 8 обломке конкримснта. 134 Обогащение гидротермальных минера- лов углеводородами и появление здесь же биоморфных структур, сложенных слоистыми силикатами, скорее всего не случайно. Логично допустить, что при абиогенном, вулканогенном синтезе углеводородов и последующей их транс- портировке в верхние слои литосферы гидротермальными растворами созда- вались особенно благоприятные усло- вия для существования хемолитотроф- ных сообществ микроорганизмов. (6). Наконец, данные лабораторного экспе- римента показывают, что за 9 месяцев при температуре около 75 'С базаль- товое стекло оказалось в значительнои степени разрушено бактериями, и поя- вились структуры, морфологически сходные с биоморфными образованиями из миоценовых платобазальтов. В со- ставе этих структур обнаружены ми- нералы, сходные, судя по составу и ми- кроструктуре, с природными слоистыми силикатами. В контрольных образцах без бактерий стекло также подверглось изменению, но значительно более слабо- му, и нет никаких признаков ново- образованных слоистых силикатов.
Заключение 1. Глубоко в литосфере в гидро- термальных системах микроорганизмы оказывают существенное влияние на процессы минералообразования. На ос- новании полученых данных можно го- ворить о биохемогенном извлечении из слабо минерализованных вод химичес- ких элементов, необходимых для фор- мирования слоистых силикатов селадо- нит-смектитового состава. Каково происхождение микроор- ганизмов подземных гидротермальных систем? Можно предложить два воз- можных варианта. 2. Согласно первому предпола- гается, что бактерии в глубокие го- ризонты гидротермальных систем могли проникнуть с поверхности Земли. Хоро- шо известны термофилы, обитающие в горячих термальных озерах или обнару- женные у выхода горячих вод на дне океана. Однако трудно представить, насколько успешно распространение жизни вглубь литосферы могло проис- ходить по восходящей гидротермальной системе. Более вероятным кажется пред- положение о проникновении микро- организмов с поверхности с холодными инфильтрационными водами. Остается неясным, могут ли микроорганизмы- обитатели холодных поверхностных вод адаптироваться к высоким температу- рам, высоким значениям РН, отсутст- вию кислорода и органического углеро- да, и превратиться в хемолитотрофных термофилов, не нуждающихся в солнеч- ной энергии. 3. Можно рассматривать сущест- вование хемолитотрофных термофиль- ных микроорганизмов в термальных системах платобазальтов как постоян- ных обитателей подземных микро- биологических сообществ, унаследо- ванных от тех, которые возникли и адаптировались для жизни глубоко под землей еще тогда, когда на ее поверх- ности условия для существования были неподходящими. Вероятно, не послед- нюю роль для обеспечения жизнедея- тельности микроорганизмов рассма- триваемой обстановки всегда, как и сейчас, могли играть углеводороды, об- разующиеся при высокотемператур- ных вулканических процессах и транс- портирующиеся к поверхности Земли с газами и подземными водами. Может быть, рассмотренные биоморфные структуры являются ископаемыми сви- детелями одной из древнейших экоси- стем, существующих на нашей планете. Не нуждаясь в солнечном свете, они могли возникнуть и существовать на Земле даже до появления атмосферы и первых фотоавтотрофов. 135
БАКТЕРИАЛЬНАЯ ПАЛЕОНТОЛОГИЯИ АСТРОБИОЛОГИЯ ИССЛЕДОВАНИЕ ВНЕЗЕМНОГО ВЕЩЕСТВА ИСТОРИЯ ИЗУЧЕНИЯБИОМОРФНЫХ СТРУКТУРМЕТЕОРИТОВ В 1806 году Л. Дж. Тенард обна- ружил необыкновенно высокое содер- 136 Одним из значительных стимулов развития бактериальной палеонтологии явились развивающиеся исследования космического (метеоритного) материала. Обнаружение российскими и американс- кими учеными в метеоритах биоморф- ных структур, сходных с различного рода бактериями и, возможно, низшими гри- бами, вернуло человечество к необходи- мости переосмысления идей панспермии. Следует при этом отметить, что обнаруженные в метеоритах псевдо- морфозы по микроорганизмам не имеют отношения к проблеме происхождения жизни на Земле, поскольку в метеоритах Наиболее интригующие резуль- таты бактериальной палеонтологии получены при изучении внеземного вещества. Последние несколько лет, после публикаций (МсКау et al., 1996, 1997; Жмур и др., 1997; Hoover, 1997; Герасименко и др., 1999 и др.) вновь зазвучал вопрос о присутствии в метео- ритах ископаемых микроорганизмов. мы находим фоссилизированные их ОСтатки. Возможности заноса микроорга- низмов на Землю упирались всегда в про- блему их сохранения и транспорта в кос- мическом пространстве. С этой точки зре- ния важнейшим элементом стали работы по д,пительному анабиозу микроорганиз- мов в условиях антарктических льдов и вечной мерзлоты. Поэтому в этом разделе описаны и проведенные работы по длительному со- хранению микроорганизмов при низких температурах и, соответственно, резуль- таты исследования самих метеоритов. жание углерода (2,5%) в каменном ме- теорите, который упал 15 марта 1806 г. в Алаисе (Франция). Метеорит из Алаиса, первый известный углистый хондрит, подвергся интенсивному изучению. В 1834 г. шведский химик Дж. Берцелиус изучил образец Алаиса. Он был удив- лен, когда обнаружил воду в этом образ- це, и почти уверился, что это не образец метеорита, поскольку он был убежден, что вода в метеоритах не сохраняется. Когда он узнал, что Алаис действитель- но содержит воду, он продолжил изуче- ние и установил, что этот метеорит со- стоит главным образом из глинистопо- добных минералов. Берцелиус также от-
метил наличие органического вещества в Алаисе, но аналитические методы XIX столетия не позволили ему выяснить, какие именно органические вещества присутствовали в метеорите. Однако он отметил сходство углистого вещества Алаиса с земным биологическим мате- риалом. Метеорит Оргэй после падения в 1864 г. был немедленно подвергнут тщательному химическому и минера- логическому анализу Клоэцом и Пи- саки. Было установлено, что он состоит из мягкого, черного горючего материала, с солями аммония, гумусом, магне- титом, кремниевой кислотой. Силикат- ные минералы скорее всего представ- лены серпентином. Было обнаружено, что минералы метеорита Оргэй напоми- нают глины и минералы со структурой, напоминающей пирокластические осад- ки, похожие на земные пепловые пот- оки, которые сформированы вулкани- ческим пеплом при осаждении в воду. Однако состав метеорита Оргэй совер- шенно отличен, поскольку вулканичес- кий пепел обычно содержит меньше магния и больше кремния, алюминия и окислов кальция, чем в метеоритах. Содержание углерода в обломках метеорита Оргэй было таким высоким, что поначалу это рассматривалось как следствие загрязнения земным вещест- вом. Соединения углерода в углистых хондритах представлены главным обра- зом полимероподобными материалами, органическими соединениями (сходны- ми с торфом или лигнитом), которые не растворяются обычными растворителя- ми. Соединения углерода также обн- аружены в форме алифатических и ароматических углеводородов, расеми- ческих и хиральных аминокислот, пу- риновых и пиримидиновых оснований, жирных кислот и липидов, порфири- нов, а также представлены широким спектром сложной органики, которая не вписывается в рамки земной биохимии. Надь (Nagy, 1975) подробно рассмотрел углистые метеориты, геохимию нефти и комплексные полимеры, керогены и уг- леводороды, содержащиеся в углистых хондритах. Ходжсон и Бейкер обнару- жили порфириновые пигменты в ме- теорите Оргэй, которые спектрально схожи с земными осадочными порода- ми. Они обнаружили зоны абсорбции, похожие на ванадиловые порфирины (встречающиеся в угле, нефти и слан- цах). Нефтяники рассматривают ванади- ловые порфирины как важные показа- тели биогенного происхождения нефти. Они также пришли к выводу, что инфракрасные спектры напоминают органику в древних земных осадочных породах (например, посидониевые слан- цы). Эти близко связанные пигменты присутствуют практически во всех зем- ных организмах и должны были бы ука- зывать на современное загрязнение зем- ным веществом. Хлорины медленно преобразуются в порфирины и также отсутствуют в древних земных осадках. Открытие порфиринов в метеорите Ор- гэй свидетельствует о "весьма вероят- ном участии живого материала в фор- мировании метеоритов". Более того, отсутствие хлоринов свидетельствует о том, что метеоритные порфирины воз- никают либо в результате пребиоти- ческого синтеза, либо в результате древ- ней микробной жизни, и не могут быть результатом современного загрязнения земным веществом. В 1961 г. Б. Надь и его коллеги со- общили, что масспектральный анализ показал, что органика в метеорите Ор- гэй имеет ту же организацию молеку- лярных фрагментов (включая фрагмен- ты алифатических и ароматических сое- динений углерода), которые найдены в ископаемом веществе на Земле и в неф- ти. Клаус и Надь провели микробиоло- гическое изучение углистых хондритов Оргэй и Ивуна. Это исследование выя- вило большое число (-1600/мг) очень маленьких (-10 мкм в диаметре) тел, ко- торые напоминали ископаемые водорос- ли, пыльцу и гистрихосферы. Эти тела различной морфологии были обнаруже- ны во многих углистых хондритах, но они отсутствовали в неуглистых метео- ритах Холтрук и Брудерхайм. Изучая метеориты, эти исследователи нашли и исключили распознаваемые земные за- грязнения в каждом образце, но при этом пришли к выводу, что в метеоритах присутствует свой комплекс микрофос- силий. После решительных возражений противников этих взглядов Клаус и Надь решили назвать эти тела (которые, как они считали, являлись микрофоссилия- ми метеоритов) "организованными эле- ментами" до тех пор, пока их точная природа не будет установлена. Андерс и Фитч (Anders, Fitch, 1962) также обнаружили эти структу- ры, но в меньшем количестве и только простой морфологии в их образце ме- теорита. Они посчитали сложные струк- 137
туры артефактами или загрязнением. Андерс (Anders, 1962a) выступил про- тив биогенной природы материала, заявив, что ископаемые в метеоритах "не имеют аналогов" и что биогенную теорию следует отвергнуть. Фитч и Андерс были самыми активными кри- тиками того, что "организованные эле- менты" свидетельствуют о присутствии микрофоссилий в метеоритах. Они пред- положили, что эти тела не были биоло- гическими объектами и посчитали все другие биологически-подобные тела зем- ными загрязнениями, такими как "му- зейная пыль" или "пыльца". Клаус и Надь вполне отдавали себе отчет в про- блеме загрязнения и они очень детально рассмотрели ее в своей первой статье. Детальное изучение биологичес- ких материалов в почве и осадочных породах в окрестностях Оргэй позво- лило оценить возможное действие зара- жения земным веществом. Они также обнаружили идентичные микрострук- туры ("организованные элементы") в метеоритах Танк п Алаис. Это показало, что источником данных структур не бы- ло локальное земное загрязнение. Клаус и Надь использовали стандартные мик- ропалеонтологические и палеонтоло- гические методы и провели биологи- ческое окрашивание этих микрострук- тур, чтобы различить органические и минералогические частицы. Характер окрашивания и ультрафиолетовые спект- ры микроструктур, обнаруженных в метеоритах, привели их к выводу, что "организованные элементы" имеют био- логическое происхождение. Таш (Tasch) тщательно изучил явления загрязнения и отметил, что хотя в углистых хонд- ритах были обнаружены распознавае- мые земные загрязнения, они не могли объединить большинство инситных и очевидно биогенных микроструктур. Более того, эти формы не смогли опре- делить высококвалифицированные мик- ропалеонтологи, которые исследовали препараты и проводили свои собствен- ные исследования углистых хондритов. Возможно, наиболее важное неза- висимое исследование природы и "ор- ганизованных элементов" Клауса и Надя было проведено известными палиноло- гами М. Россигнол-Стрик и 3. Барг- хурном, ведущими специалистами по древним микрофоссилиям. Используя стандартные палинологические методы, Россигнол-Стрик и Баргхурн обрабаты- вали внутренние, чистые образцы из метеорита Оргэй. Они обнаружили мно- жество полых сфер и ооидов, мембраны, "воронковидные" и "грибовидные" структуры и т.п. (Rossignol-Strick, Barg- hoom, 1971). Многие из этих тел из ме- теорита Оргэй, как было установлено, являются близкими по размеру, форме и внешнему облику к микрофоссилиям (Huroniospora sp.) из ганфлинтских кремней. Эти сферы имеют тот же цвет, плотность и состав, как и включающии коричневый органический осадок. Бо- лее того, они имеют хорошо очерченные стенки (похожие на клеточные стенки) с гладкими внутренними поверхностями и неровными внешними поверхностями из-за присутствия частиц породы на внешней стороне сфер. Некоторые из этих стенок были обломаны и создают впечатление эластичности. Эти органи- ческие тела негативно реагируют на палинологический краситель сафранин и поэтому не являются пыльцевыми зернами. Их также нельзя спутать со спорами грибов или волокнами тканей. Эти кислота-устойчивые микрострук- туры имели близкий размер и черты с "организованными элементами", изу- ченными Клаусом и Надем. Баргхурн и Россигнол-Стрик сообщили, что "оби- лие органических объектов в метеорите Оргэй может быть свидетельством био- генного происхождения". Полые сферы метеорита Оргэй также похожи морфо- логически на кислотоустойчивые полые сферы размером 6-20 мкм, которые наблюдаются в осадочных породах, подстилающих серию Фиг Три (серия Онфервахт (Onverwacht) возрастом более 3.2 млрд. лет). Гистограммы выявили, что распределение по размеру полых микроструктур в метеорите Оргэй такое же, как в популяциях микро- фоссилий из железистых кварцитов Ган- флита в Онтарио (Канада) возрастом два миллиарда лет. Многие из "организо- ванных элементов" похожи по размеру, форме и внешнему облику на разнооб- разные докембрийские микрофоссилии (например, Eosphaera tvleri, Melasmato- sphaera sp., Huroniospora sp. Они отме- тили, что "на Земле фоссилии обна- ружены только в осадочных породах" и поэтому считали, что биогенная приро- да должна быть связана с окружающей средой, а поскольку метеоритная порода не была осадочной, то данные формы имели абиогенное происхождение. В то время микробные сообщества гидротер-
мальных источников, глубинных базаль- товых и гранитных пород и вулкани- ческих туфов были неизвестны, и пале- онтологи не ожидали встретить ископае- мые в неосадочных породах. Другие исследователи загадочные микроструктуры в углистых метеоритах относили к микрофоссилиям биологи- ческого происхождения, которые при- надлежат данным метеоритам. Бриггс и Китто также нашли сложные однокле- точные тела, напоминающие микроорга- низмы, в метеорите Макая. Надь и др., использовав микроанализатор, пока- зали, что некоторые "организованные элементы" были насыщены лимонитом, содержащим немного никеля, а другие- силикатами. Используя НС1 и HF, чтобы растворить минералы, они обнаружили, что большинство оставшихся углистых тел не содержали элементов с атомным номером выше магния. Это они расце- нили как доказательство фоссилизации, которое ясно исключает земное загряз- нение, -как источник этих тел. Росс (Ross, 1963) также встретил грибовид- ные "полые трубки с рваными мемб- ранами в самой легкой части материала метеорита Оргэй, который он выделил с помощью суспензий кадмия. Чтобы свести к минимуму возможные за- грязнения, в его анализе применялись стерильные инструменты, которыми вскрывался нетронутый метеорит Оргэй из Британского музея. Тан и ВанЛадингхам (Tan, VanLa- dingham, 1967) получили фотографии с электронного микроскопа многих ки- слотоустойчивых тел, которые они рас- сматривали как микрофоссилии, кото- рые входили в состав метеоритов Оргэй, Ивуна и Тонк. Использовавшиеся силь- ные окислители - азотная, соляная и плавиковая кислоты уничтожили бы карбонатные и кремниевые микрофос- силии, которые содержатся в большин- стве земных сообществ (диатомовые, силикофлагеллаты, фораминиферы и кокколиты). Они обнаружили многие морфологические формы, включая "гри- бообразные" структуры с темными под электронным микроскопом "ножками" и более прозрачные, мембрановидные шляпки со складками. Некоторые из этих форм они посчитали очень схожи- ми по размеру, форме и особенностям изображений под электронным микро- скопом с углистымии наночастицами, которые Пфлюг и Хайнц (Phlug, Heinz, 1997) обнаружили в барите из формации Фиг Три возрастом 3.3 млрд. лет. Они также посчитали нити с одним закруг- ленным и другим заостренным конца- ми, которые содержали цепь непрозрач- ных образований (вероятно, кристаллов) вдоль длиннои оси, очень сходными со структурами, которые теперь описыва- ются из магнетотактических бактерий. Палик (Palik, 1962) обнаружила и изо- бразила сложные структуры, похожие на нитчатые водоросли, в механически разрушенных (не обработанных кисло- той" образцах меторита Оргэй. Некото- рые нити были загнуты, их ширина ме- нялась от 3 до 16 мкм, а длина от 40 до 170 мкм. Она описала необыкновенную нить N 2 как "часть нити шириной 4 мкм и длиной 55 мкм. Существование внеземных "жиз- неподобных" форм было достаточно широко принято до середины 60-х годов, и некоторые палеонтологи начали изучать проблемы систематического положения внеземных микрофоссилии (Claus, Nagy, 1962; Van Landingham, 1963). Однако последовала интенсивная критика, и противоречия, связанные с "организованными элементами", были такими сильными, что к середине 70-х годов большинство исследователей пре- кратили какое-либо изучение возмож- ных микрофоссилий в метеоритах. В 1966 г. Х.С. Урей (Urey, 1966) (лауреат Нобелевской премии по химии 1934 г.) сделал обзор доказательств биологических материалов в метео- ритах. Он отметил, что в метеоритах встречаются органические субстанции, которые близко напоминают таковые из древних земных пород, что органика, встреченная в углистых хондритах, не напоминает ту, которая присутствует в современных загрязнениях. Урей отме- чал: "...некоторые вещества в метео- ритах, если бы их нашли в земных объектах, несомненно бы считались биогенными". Он также заметил: "... те из нас, к.го в течение нескольких лет работали с метеоритами, были уверены, что там не могло быть остатков никаких живых организмов. Если бы метеориты имели состав осадочных пород Земли, это не вызвало бы большого удивления". МЕТЕОРИТЫ На Землю постоянно поступает огромное количество космического ма- териала. Он поступает в виде косми- ческой пыли и, значительно реже, в виде 139
Таблица 1. Химические составы хондритов (по Р.Т. Додду, 1986, с сокращениями) Обыкновенные (О) Энствтитовые (Е) глистые( ) ласс L(LL) Н О Группа % 15. 12. 15. 18. 10. .31 14. 15.01 10. Л .61 140 достаточно крупных обломков. Оценки количества внеземного вещества, по-, ступающего на Землю, весьма противо- речивы. Но наиболее принятые — это 100-1000 т в сутки. По существующим представлениям метеориты имеют астероидальное или ко- метное происхождение. Все они по со- ставу разделяются на (1) каменные, (2) железокаменные и (3) железные в зави- симости от содержания силикатных мине- ралов и никелистого железа. Подавляю- щее большинство обнаруженных метео- ритов — каменные, среди которых также преобладают так называемые хондриты. Каждый из трех упоывиутьо~ выше типов метеоритов подразделяется на классы. Среди железных метеоритов уста- новлено два класса: палласиты и мезо- сидериты. Среди каменных метеоритов разли- чают хондриты и ахондриты. Хондриты получили свое название от хондр — спе- цифических сфероидальных образований. Среди хондритов выделяются три класса [энстатитовые (Е), обыкновенные (0) и углистые (С)] и ряд групп, различаю- щихся по химическому составу (табл. 1). Для наших целей наиболее инте- ресными оказываются углистые хонд- риты, среди которых выделено четыре группы (I, M, О, Ч). Обозначения 1, М, О, V соответствуют первым буквам ти- пичных метеоритов (Ivuna, Mighea, Omans и Vigarano). Таким образом, на- пример, сочетание СО означает угли- стый хондрит типа Omans. Названия ме- теоритов даются по местности. Cl хондриты довольно редки и мелки, и материалы по ним немного- численны. В отличие от всех остальных хондритов, CI хондриты не имеют на- стоящих хондр. Основной материал сло- жен из мельчайших зерен монтморилло- нита (глинистый минерал); септохлори- та (водный силикат, общая формула Fe,(Si„0„)(0H),) и магнетита. Иногда присутствуют в незначительных количе- ствах и другие минералы. СМ хондриты распространены значительно шире. Известные находки достаточно крупные. Хондриты СМ груп-
пы содержат ярко выраженные хандры и непереплавленные агрегаты, погружен- ные в матрицу. Отдельные минералы представлены оливином, пироксенами, хромитом, Са-А l силикатными стеклами и некоторыми другими минералами (кальцит, гипс, магнетит). В матрице наиболее распространены септохлорит и в меньшей степени монтмориллонит. Углерод СМ хондритов находится в виде сложных углеводородов, реже кальцита. СО хондриты так же, как и СМ хондриты, содержат хандры, непере- плавленные агрегаты и одиночные кри- сталлы и их обломки. Наибольшие различия СМ и СО в содержании хондр (значительно больше у СО). Хандры мо- гут состоять: из гранулярного оливина, погруженного в чистое стекло, низко- кальциевых пироксеновых волокон в стекле, зонарного оливина и железисто- хромистой шпинели в стекловатой ос- новной массе. Агрегаты неправильной амебовидной формы, как правило, сло- жены оливином с незначительной при- месью пироксена, шпинели и Na-Al-си- ликатов. В матрице были отмечены гли- нистоподобные водные силикаты и оли- вин. Углистая составляющая в виде сложных углеводородов. CV хондриты — последняя группа, отличается от СО хондритов более всего по структуре. Несколько слов о возрасте хондри- тов. Более или менее достоверные опре- деления рубидиево-стронциевым мето- дом были сделаны для групп обыкно- венных и энстатитовых (Н, L, LL и Е) хондритов, а также для всех классов свинцово-свинцовым методом. Полу- ченные значения находятся в интервале 4.39-4.59 млрд. лет тому назад. СОВРЕМЕННЫЕ ИССЛЕДОВАНИЯМЕТЕОРИТОВ В последние годы в углеродистых метеоритах Мурчисон, Ефремов ка, Алленде, Оргей и др. были выявлены литифицированные остатки коккоидных и нитчатых форм по морфологии и раз- мерам аналогичные современным бакте- риальным формам и их древним ана- логам (Жмур и др., 1997; Hoover, 1997; Hoover et аl, 1998; Герасименко и др., 1999; Розанов, 2000). Остановимся на двух опублико- ванных примерах из метеорита Мур- чисон. Р.Хувером была обнаружена (рис. 1) длинная (до 300мк) тонкая (3 мк) нить, тесно связанная с породой, у которой местами наблюдаются пережимы. Создается впечатление клеточного стро- ения. Вместе с нитью наблюдаются то- роидальные формы, которые напомина- ют гормогонии. Другая форма, пред- ставленная несколькими экземплярами (рис. 2), более всего напоминает Micro- coleus, поскольку состоит из многочис- ленных параллельных трубочек, объеди- ненных одним чехлом. Хотя из Мурчисона и Ефремовки были описаны многочисленные кокко- идные формы, именно они вызывали более всего сомнений. Рис. l. Рис. 2.
1597 Fe К„ 2228 1278 SiK 1671 639 [ 1114 557-1ОК„1 РеК 1 I 111 319 CaK„, AIK О'" Се„А Рис. 3. Рис. 4. 1361 32Ь9 -а (I 1089 2607 К„ 817 1 955 1304 272 652 ( К„С uIVI„ .l' 7.00 8.00 9.00 10.00 Рис. 5. Рис. 6. 142 Важный результат, как нам кажет- ся, был получен в результате изучения метеорита Мигея. Образец утлистого хондрита Ми- гея был получен для изучения в комитете по метеоритам РАН и исследован на ESEM в MSFC-NASA (Хантсвилл, Ала- бама). Образец был исследован обычным методом. Под микроскопом просматри- вался свежий скол, обработанный азоти- стым бором (BN). Рабочие увеличения со- ставляли 2-20 тысяч. Химический состав образца можно видеть на спектрах (рис. 3-б). Обычны Mg, Si, Fe, Ni. Однако обращает на себя внимание большее, чем обычно, в углистых хондритах содержа- ние S. Вероятно, S входит в состав трои- лита. Однако есть подозрение, что при- сутствует и чистая S. В метеорите Mighei обнаружены трубчатые и мешковидные образования, а также шарообразные, шарообразные с ое» 0.00 1.00 2.00 8.00 4.00 5.00 8.00 Т 00 8,00 8.00 10 00 Mig hei keV mighei general overall 60х Analyst' влетал 0.00 1.00 2.00 3.00 4.00 5.00 Mig hei mighei1„122kx filament head Апа!увй]епмап выростами (?). Все они имеют микрон- ную размерность. Первый из трубчатых объектов (рис. 7) имеет длину — 20 мк и диаметр — 3 мк. Трубка слоистая. Толщина слойков - доли мк. Создается впечатление, что трубка либо расколота, либо состоит из более тонких трубок меньшего диаметра (< 1 мк). Трубчатую или нитевидную форму диаметром чуть менее 1 мк имеет объект, изображенный на рис. 8. Наблю- даемые пережимы иммитируют клеточ- ное строение. Химический состав ин- тересен необычно высоким содержа- нием Ее, минералогическое объяснение которому дать трудно (рис. 4). Сходный по диаметру фрагмент нити, имеющий общую крючкообразную форму, был также обнаружен в другой части об- разца. Длина фрагмента 8-9 мк. Инте- ресно в этом случае высокое содержание серы (рис. 9-11). Близкий по диаметру 1 SiK„ 1 РеК~, мдк,,1, 6 к„сик„, cIK„~ jlIIIK с" ЯЯ„,15 1Ф ' са 1.00 2.00 3.00 4 ОО 6.00 6 00 2.00 6 00 0.00 10.00 Mighei mighei1093500x, bnghter Ьр of large filament Analyst: Jerman 0.00 1.00 2.00 3.00 4,00 5.00 6.00 7.00 8.00 9.00 10.00 Migh8i mighei1 033kx. triangular head Ала!уэй Jerrnan
Рис. К Рис. 7. 4574 В161 3659 6529 2744 <897 1830 3264 1632 О О. Nlighel mighei114 12kx, filarnent Analyst: Jerman 4.00 keV Рис. 1Î. Рис. 9. 143 фрагмент изображен также на рис. 12. Его химический состав близок к среднему составу матрицы метеорита (рис. 13). Были встречены также обрыв- ки, возможно, филаментов гораздо большего диаметра (6-7 мк) (рис. 14). И в этом случае химический состав прин- ципиально не отличается от матрицы метеорита (рис. 5). Следует заметить, что описанные выше структуры не могут быть од- нозначно сравнены с какими-либо кон- кретными микроорганизмами. Яаже в первом случае, когда есть подозрение на находку пучка нитей. Однако, судя по морфологии, веро- ятность того, что мы имеем дело с псевдоморфозами по бактериям, весьма велика. Повьппенные содержания серы могут, возможно, дать некоторые осно- вания для отнесения некоторых из ука- занных образований к фоссилизирован- ным серобактериям. Обнаружены также мешков идные формы (рис. 15, 16). В первом случае это довольно крупная форма длинною не менее 20-25 мк при диаметре (?) 8-9 мк в самом широком месте. Постепенно ширина уменьшается, образуя не совсем правильной формы конус. Тонкий конец, вероятно, округлый. Химический состав (рис. 6) отличается небольшим содер- жанием Fe. Основу составляет Mg и Si. Во втором случае (рис. 16) обнаружен- ные формы напоминают округлый меш- ковидный (?) сосуд с искривленным гор- 0.00 1.00 2.00 3.00 5.00 6.00 7.00 mghei МеЧ rnighei11412kx, lighter matrix next to filament Analyst Jerman
2842 22)4 1705 1137 568 0 0.00 1.00 2.00 3.0С 4.00 5.0С 6.00 7.0 Mighei keV ° mighei1 14 12kx, dark matrix to left of filament Analyst. )еггап Рис. 11. 2391 1913 1435 478 Рис. 13. Рис. 16. Рис. 15. 144 0.00 1.00 2.00 3.00 4.00 5 00 6.00 7.00 8.00 9.00 10.00 MIghei mighei130 10kx, rod Analyst. Jårmап
Рис. 17. Рис. 18. 647 518 SK„ 388 259 СЖ„ СаК„ Ге 129 Рис. 19. Рис. 20. 145 лом. Диаметр мешка 11-12 мк, диаметр горла около 2,5 мк. Поскольку такой эк- земпляр был найден в единственном чи- сле, очень трудно быть уверенным в его истинной форме. Однако, если морфо- логия воспринимается нами правильно, вероятно, следует искать аналогов этой формы среди экстермофилов типа Cryptoc0ccus. Две группы совершенно иного морфологического типа объектов, ближе всего напоминающие часто описывае- мые в последнее время нанобактерии, представляет, вероятно, наибольший интерес. Первая группа — это сферические образования "клеток" размером 200nm— 1 mk, обнаруживаемые группами. Часто 0 '! 0.00 1.00 2.00 3.00 4.00 5.00 6.00 7.00 8.00 9.СО 10.00 +lgh+i rnighei1 05 6kx, open matrix between globules Analyst: Jåïíап они образуют скопления по несколько штук или "колонии" из десятков эк- земпляров. В некоторых случаях наблю- даются диплококки или делящиеся "клетки" (рис. 17, 18). Химический со- став достаточно обычный (рис. 19) для матрикса метеоритов. Однако надо иметь в виду, что этот результат может скорее относиться к поверхности, на которой расположены бактериоморфы. Вторая группа чрезвычайно специ- фичных образований (рис. 20-24), пред- ставляет собой войлокоподобные по структуре пластины, в которых наблю- даются скопления или единичные гло- булы. Толщина ниточек войлока не бо- лее 50 nm, диаметр глобул 200-400 nm. Нитки войлока образуют запутанную
ткань, компоновка ниточек очень рых- лая. Иногда создается впечатление, что глобулы больше всего локализуются в линии (хотя это впечатление может быть обманчивым) и возможно, прикрепле- ны к концам ниточек. Вероятно, возмож- ны две версии интерпретации этой био- морфной структуры: 1) глобулы — это на- нобактерии, а войлочные пластинки- биопленки, создаваемые этими бакте- риями; 2) и нитки войлока и глобулы яв- ляются самостоятельными разновидно- стями нанобактерий, образующих не- кую ассоциацию. Очень важно отме- тить, что воилочные пластины имеют резкие ограничения по кристаллогра- фическим граням (рис. 21-23), что со- вершенно исключает возможность трак- товать эти образования как засорения. Однако это может быть воспринято как химические нанноструктуры кристаллов. Анализы показывают обычную для данного метеорита картину (рис.24). Таким образом можем констати- ровать, что в метеорите Mighei удалось, вероятно, обнаружить весьма специфи- ческий комплекс биоморфных структур, который отличается от ряда других ме- теоритов (Мурчисон, Ефремовка, Ал- ленде) меньшими размерами объектов и присутствием достаточно вероятных на- нобактерий. При всей трудности интер- претации этих материалов, складывает- ся впечатление, что обнаруженные ас- социации, скорее всего, характеризова- ли какие-то экстремальные ситуации, в которых они развивались. Три последующих примера метео- ритов Murray, A11ende и Nagoya отличает прежде всего высокое в них содержание S, причем в ряде случаев есть реальные подозрения на присутствие серы не только в троилите, но и в чистом виде. Метеорит Murray имеет общий хи- мический состав обычного метеорита с оливиновой или серпентиновой осно- вой, однако необычно высокое содер- жание серы (рис. 25), вероятно, входя- щей в троилит. В метеорите обнаружены нитеобразные формы, иногда с пережи- мами, и шарообразные. Нитевидные формы (рис. 26-30) имеют диаметр око- ло 2-3 мк, расположены пучками или субпараллельно. Иногда наблюдается субпараллельное скопление таких обра- зований внутри округлых тел диаметром 100-150 мк (рис. 27). Во всех случаях присутствуют Mg, Si, Fe и повышенное содержание серы (спектры 31, 32). Шарообразные тела диаметром более 20 мк имеют чистый Ее состав, однако более мелкие, имея в составе большое количество Fe, вместе с тем содержат нормальный набор элементов Mg, Si, Fe, но и Аи, Ni. Cu. У этих последних (рис. 33, 34) наблюда- ются интересные отростки. К метеориту Алленде мы еще вер- немся, когда будем обсуждать трудности бактериальной палеонтологии при изу- чении метеоритов. Здесь лишь скажем, что в нем встречены мешковидные фор- мы диаметром около 10 мк (рис. 35), со- стоящие из чистой S и, возможно, явля- ющиеся ископаемыми серобактериями. Гораздо больше и более странных данных получено по метеориту Nagoya. Здесь были обнаружены многочис- ленные формы колбасообразной (со- сисковидной) формы, соединяющиеся часто с облаковидными или мешко- видными образованиями. В некоторых случаях картина почти тождественная той, которую мы наблюдаем в округлых телах метеорита Миггау (рис. 36-38). Именно внутри такого поля распо- лагаются описанные выше образования (рис.39-41). Все они содержат значи- тельные количества S и Са. Пока трудно предположить - что это, хотя прежде всего напрашивается предположение о каком-то типе серобактерий. Химичес- кий состав одной из форм показан на рис. 42. Последний пример, который за- служивает особого внимания, это метео- рит Оргей в котором Палик обнаружила структуры, подобные доменам магнети- та у бактерий магнитотактиков . Значительное морфологическое единство земных микробных организ- мов как современных, так и древних, с псевдоморфозами по микроорганизмам из углеродистых метеоритов, дает осно- вание полагать принципиальное един- ство микробиологического мира Земли и космических объектов. При этом, ко- нечно, не исключено обнаружение и таких форм, которые не будут иметь земных аналогов. Набор встреченных в метеоритах микрофоссилий показывает, что это были не отдельные микроорганизмы, а сооб- щества, часть из которых была подобна цианобактериальному мату. Эти сообще- ства, вероятнее всего, принимали уча- стие в формировании углеродистого вещества исследованных метеоритов.
Рис. 22. Рис. 21. 1510 1256 942 628 314 7. 0 00 6.keV Рис. 24. Рис. 23. 3226 2581 1936 1290 3.00 Рис. 26. Рис. 25. 147 0 0.00 1.00 2.00 Murray General Overall 50х Ana1yst Jerman ОС 0.00 1.00 2 00 3.00 Mighei mighei1 24 20kx, small ball Analyst Jermsn
Рис. 27. Рис. 28. Рис. 29. Рис. 30. 1906 FeK„ 1525 921 МЦК„ 1174- 681 762 460 S KÄ 381 230 Рис. 31. Рис. 32. 148 0 0.00 1.00 2.00 3.00 4.00 5.00 6.00 7.00 8.00 9.00 10.00 Murray keV еытау1 022500х, fuzzy filament surface to right of smooth surface Analyst Jerman ><а М 0.00 1.00 2.00 3.00 4.00 5.00 6,00 7. Murray murray1 03 kx, filament Analyst Jårrriал
253 127 Рис. 34. Рис. 33. Рис. 36. Рис. 35. S ГеК„ 1186 СаК„ 889 1482 1 | iK„ MgK„ NaK„A аKOI l=- x СгК„ 0.00 1.00 2.00 Э.ОО 4.00 5.00 6 00 7.00 8.00 9.00 10.0 Nogoya nogoya1 08 1Кх, radiation stalk Analyst. Jerman 0 0.00 1.00 2.00 3,00 4.00 5.00 6.00 7. Murray гпнггзу1 0610Мх, Fe rich ball Analyst; Jerman
Рис. 40. Рис. 39. 2212: S K„1 СаК„ 132 / SiK„ 442 0.00 100 200 300 Nogoya поцоуэ1 04 4kx, rod Analyst. Jårn~an Рис. 42. Рис. 41. 150 ОСНОВНЫЕ ТРУДНОСТИ БАКТЕРИАЛЪНО- ПАЛЕОНТОЛОГИЧЕСКОГО ИЗУЧЕНИЯ МЕТЕОРИТОВ Наиболее распространенные воз- ражения против интерпретации "би- морфных" структур в метеоритах как псевдоморфоз по микроорганизмам мо- гут быть разбиты на 4 группы. 1) Утверждение, что все "био- морфные структуры" являются засоре- нием, которое возникает после падения метеоритов на Землю. 2) Утверждение, что нанометро- вый уровень объектов, продемонст- рированный при изучении марсианского аК„~ ГеК„ СГК. д реКВ д.у.~ 4 00 500 600 y CC В CG- 900 10СС keV метеорита ALH84001, невозможен для земных живых объектов. 3) Соображение, что мы очень плохо знаем морфологию абиогенных материалов на нанометровом уровне и поэтому можем столкнуться со струк- турами, неотличимыми от тех, которые принимаются нами за псевдоморфозы по микробам и низшим грибам. 4) Утверждение о том, что заведо- мо абиогенные полимерные кристаллы, например, керита, не отличимы от фос- силизированных микробных, а точнее даже цианобактериальных форм. Эта группа возражений относится прежде всего к проблемам морфоло- гического изучения "биогенных струк-
1'cK 110.00 62.00 49.76 88.00 37.32 24.88 44.03 ! 2.44 22.02 1 2 3 4 5 6 7 8 9 10 0 1 2 3 4 5 6 7 8 9 !О 075.00 SiK 83.00 Ьб.00 49.70 45.24 33.13 30.16 15.08 ! 2 ! 2 3 4 6 7 Я 9 !О 5 keV 3 4 5 6 7 8 9 10 keV Рис. 43. Графики химических анализов метеорита Ефремовка, полученных на микроана- лизаторе Link: а - матрикс, б, в - филаменты и нити; г - засорение. 151 тур" и является сферой "бактериальной палеонгологии". Остановимся несколько подробнее на этих возражениях. 1) То, что пребывание метеоритов на Земле, в том числе и в музеях, сопря- жено с их освоением микроорганизмами — несомненно. Несомненно и то, что при приготовлении препаратов создать исклю- чительно стерильную среду очень труд- но. Однако следует помнить, что мы пы- таемся изучать не современные нефос- силизированные остатки, а те, которые уже являются псевдоморфозами, образо- ванными теми или иными минералами. Следовательно, если состав иссле- дуемого объекта не отличается от соста- ва самого метеорита, то нет оснований говорить о засорении. Если же отличает- ся, то подозрение о наличии засорения становится очень весомым. Вопрос лишь в точности химических исследовании, например, на микроанализаторах. О мно- гих проблемах выявления засорений гово- рилось выше (см. историческую справку). При исследовании метеорита Мур- чисон на сканирующем микроскопе CamScan c Link были проанализированы б объектов, принятых за псевдоморфозы по цианобактериям. Сравнение с ре- зультатами анализа Мурчисона (Додд, 1986), показало, что все проанализи- рованные объекты по составу не отли- чаются от матрикса метеорита. Были также проанализированы на сканирующем микроскопе с микроана- лизатором некоторые объекты из метео- рита Ефремовка (рис. 43). В тех из них, которые являются псевдоморфозами, не было обнаружено ничего, выходящего за рамки обычного состава углистого хондрита. В тех же случаях, когда было подозрение на засорение, были обна- ружены очень высокие содержания, например, Cl и К. Показательный ре- зультат был получен при исследовании метеорита Алленде (Allende). Один из объектов при исследовании SEM на протяжении короткого времени (не- сколько минут) менял форму (рис. 44). Проделанный анализ выявил высокие содержания О и Са, но самое главное- присутствие значительного количества К и V (рис. 45). Нужно при этом иметь в виду, что диаметр точки, в которой производится определение, соизмерим с
3416 2 <33 2005 1366 683 10 8 9 4 5 6 keV Рис. 46. Распределение O„C и N в образце с актиномицетами. 152 Рис. 44. Метеорит Алленде, в цент- ральной части расположено вертикальное тем- ное пятно — засорен ие. Рис. 45. Графики химичсских анализов метеорита Аллснде; всрхпяя линиия — засоре- нис, нижняя — матрикс метеорита.
Рис. 48. Рис. 47. Рис. 50. Рис. 49. Рис. 47-50: современные актиномицеты, обрастающие образец метеорита Мурчисон из Филд музея, Чикаго. 153 диаметром кокков или нитей. Поэтому элементный фон породы будет присут- ствовать почти всегда. Важно для опре- деления засорений присутствие необыч- ных для метеоритов элементов. Очень наглядно могут выглядеть карты рас- пределения элементов, когда N, О, С следуют распределению современных организмов (рис. 46). Другой аспект проблемы засоре- ний состоит в том, что при поисках "биоморфных структур" изначально мы ищем такие, которые тесно связаны с породой и не лежат на поверхности ис- следуемого образца. Еще раз повторимся: засорения есть и очень часты. Нередки случаи про- растания метеоритов актиномицетами или грибами по микротрещинам. Так в образце Мурчисона из Филд Музея (Чи- каго) было обнаружено, что вся поверх- ность образца заражена современными актиномицетами (рис. 47-50). Но, во- первых, это распознаваемо ггриемами, описанными выше, и, во-вторых, это не может иметь отношения к цианобак-
териям. Последние — фотосинтетики, и внедряться внутрь метеорита по трещи- нам было бы для них "алогичным". 2) Утверждение о невозможности существования нанометровых микроор- ганизмов является очевидным недора- зумением. Существует даже специаль- ное понятие — нанобактерии. Можно привести множество работ с описанием нанобактерии в различных условиях существования и с описанием их мета- болизма (Uwins et аl., 1998; Posfai et аl., 1998; Kajander et аl., 1998; Vainshtein et аl., 1998; Hjerklund et аl., 1998; Qiftgioglu et аl., 1998а, b). Более того, существуют работы, показывающие принципиальную неотличимость совре- менных нанобактерий от объектов из АЬН84001 (Folk, Lynch,1998). 3) Совершенно иной вопрос о мор- фологии абиогенных кристаллов нано- метровой размерности. Думается, что нас могут подстерегать здесь неожидан- ности и, несомненно, требуется обоб- щение уже опубликованного материала, которое, вероятно, еще не делалось. Очевидно, что иногда среди син- тезированных материалов и, в частно- сти, гётита и гематита наблюдаются выделения, весьма напоминающие бак- териальные (Zachara et аl., 1998). 4) Вопрос о морфологии полимер- ных кристаллов действительно очень сложен, и на их природу существует три разные точки зрения. Уточним, что речь идет прежде всего о керитах из грани- тов, точнее камерных пегматитов (Во- лынь, Украина), переописанных в по- следнее время Н.П. Юшкиным (Yushkin, 1996, 1998). Состав керита C„, Н,,„„O»S(N). Юшкин приводит следующие пара- метры формирования кристаллизации минералов (включая керит): тем- пература — 730-120'С, давление — 110-20 МПа. Традиционно при таких пара- метрах речь идет исключительно об аби- огенном происхождении таких кристал- лов, в том числе и керита. Однако Юш- кин на основании прежде всего при- сутствия многих (18) аминокислот и формы кристаллов выдвинул концеп- цию "предбиологических организмов". Сходство войлокоподобной струк- туры керитов со структурой современ- ных и древних нитчатых цианобак- териальных матов наводит на мысль, что исследования керита и вообще гра- фитов необходимо продолжить, тем бо- лее, что нельзя исключить, что образо- вание керита в упомянутом случае про- исходило позднее формирования мине- ралов на последней стадии гидротер- мального процесса. Следует отнестись с большим вниманием к публикации В.М.Горленко и др. (Gorlenko et аl., 1999), в которой показаны многие клеточ- ные атрибуты именно в нитях керита. К числу трудностей бактериально -палеонтологического изучения внезем- ного вещества необходимо отнести и неясность возраста возникновения псев- доморфоз по микроорганизмам. Если иметь в виду, что фосси- лизация, как говорилось выше, про- исходит почти мгновенно, то фоссили- зированные (твердые) остатки в метео- ритах могут быть и очень молодыми. Однако фоссилизированные на Земле остатки микроорганизмов должны так же резко отличаться по химизму от состава метеорита. Кроме возражений "бактериально- палеонтологических", существуют воз- ражения по поводу значения изотопных данных, данных о температурах форми- рования ряда минералов, значения при- сутствия различных "биомаркеров", в том числе керогена, пуринов, изопреноидов, циклоалламенов, различных аминокис- лот и т.п., а также специфической хираль- ности аминокислот метеоритов. На этих вопросах мы не будем подробно останав- ливаться и отошлем читателя к работам Hodgson, Baker, 1964; Nagy, 1975. 154
ЖИЗНЕСПОСОБНЫЕ ЭКОСИСТЕМЫ КРИОЛИТОСФЕРЫ 155 Обычно при обсуждении присут- ствия в метеоритах бактерий спорящие забывают о том, что в работах послед- него времени обсуждается присутствие только ископаемых фоссилизированных микроорганизмов. К проблеме появле- ния жизни на Земле, к проблеме ее пе- реноса из космического пространства это не имеет прямого отношения. К проблемам переноса микроор- ганизмов в космическом пространстве имеет отношение проблема длительно- сти пребывания микроорганизмов в анабиозе, в чем речь пойдет ниже. До недавнего времени полагали, что микробная жизнь быстро иссякает по мере удаления от поверхности вглубь земной коры. Представления о биосфере ограничивались оценкой наземной и почвенной биомассы с учетом распро- странения организмов в приземные слои атмосферы. Сведения об обнаружении жизнеспособных микроорганизмов в подпочвенных слоях, осадочных поро- дах, залегающих на достаточно больших глубинах, игнорировались, т.к. методы бурения и отбора образцов не выдер- живали критики с позиции микробиоло- гического анализа. Другие данные, по- лученные микробиологами, несомненно представляли интерес, но были редки, касались неглубоких подпочвенных сло- ев или не содержали строгого подтверж- дения асептики отб6ра образцов. Ряд сенсационных сообщений об извлече- нии жизнеспособных клеток из древних пород привлекли внимание в связи с широко обсуждаемыми проблемами воз- никновения и сохранения жизни. В их числе следует упомянуть сообщения об обнаружении прокариот в морских осадках на глубине до 750 м, литоав- тотрофных сообществ в базальтах, тер- мофильных микроорганизмов в песча- никах на глубине 2000 м, микробных сообществ в гранитных породах на глу- бине до 1240 м, выделении жизнеспо- собных споровых бактерий из тела пче- лы, заключенной в древнем янтаре (25-40 млн. лет), а также микроорга- низмов из пород возрастом 100 млн. и даже 650 млн. лет. Обзорные обобщения убеждают в повсеместном распространении жизне- способных микроорганизмов в недрах Земли и доминирующей роли темпера- турного фактора, ограничивающего экс- пансию биологических систем посред- ством механизма тепловой деструкции клеток. Но при этом следует учитывать, что устойчивость клеток к повышенным температурам в природных местообита- ниях существенно превосходит показа- тели, которые получают в лаборатории при исследовании чистых культур мик- роорганизмов. Микроорганизмы живут и активно размножаются, образуя спе- цифические сообщества в горячих источниках в условиях повышенного давления при температуре растворов, превышающей 100 'С. Например, выде- лены архебактерии, живущие в глубо- ководных термальных экосистемах при температуре 113 'С. Некоторые авторы предполагают возможность сохранения жизнеспособных клеток в глубоких сло- ях при значительно более высоких температурах: 120 'С, 150 'С. Таким образом, представления о биосфере Земли меняются, хотя нижняя граница распространения жизни осознается еще весьма условно.
ГЕТЕРОФАЗНАЯ СРЕДА ОБИТАНИЯ Подземный макрокосм, заселен- ный микроорганизмами, есть совокуп- ность множества сред обитания, пред- ставляющих собой разнообразные ком- бинации гетерофазных систем, варьиру- ющих по физико-химическим, грануло- метрическим, минералогическим и дру- гим свойствам. Градиенты температур увеличиваются в приповерхностном и прогреваемом глубинном слоях, тогда как мощный срединный слой характе- ризуется стабильными низкими положи- тельными температурами. Именно ста- бильность температурного фактора и от- сутствие прямого или косвенного (в виде растительного опада) притока сол- нечной энергии является главным отли- чием подземных биотопов от поверхно- стных. Почвенные микросреды пред- ставляют собой структурные элементы, непрерывно меняющиеся в пространст- ве-времени в силу взаимосвязанных внешних и внутренних (метаболизм) причин. В условиях глубокого грунта следует ожидать формирования относи- тельно устойчивых в течение длитель- ного времени микробных комплексов. Заселение земных глубин мик- роорганизмами теоретически возможно двумя путями: 1. В процессе формирования оса- дочных толщ, т.е. погребения поверх- ностных слоев, заселенных микроор- ганизмами. В этом случае формируются "сообщества выживших", когда микро- организмы поставлены перед необхо- димостью адаптироваться или неопре- деленно долгое время переживать не- благоприятные условия. 2. Как результат возникновения или активизации эволюции в литосфере в условиях постоянной температуры, давления, уровня радиации, наличия хи- мических и органических источников энергии живых самореплицирующихся систем, не зависимых от энергии солнца и фотосинтеза. Очевидно, в этом случае речь могла бы идти о более активных со- обществах или популяциях микроор- ганизмов, вполне приспособленных к жизни в глубоких горизонтах, когда ос- новными факторами, лимитирующими выживание, являются температура и на- личие доступной воды. Сообщения об обнаружении кле- ток в породах возрастом более 100 млн. лет и даже 650 млн. лет инициируют дискуссии о продолжительности жизни на клеточном уровне. Очевидно, что возраст пород не позволяет однозначно датировать возраст микроорганизмов. Необходима оценка физиологического состояния сообществ (анабиоз или мета- болизм и смена генераций), при этом для исследуемых объектов должно быть доказано отсутствие притока фильтруе- мых растворов, содержащих микроор- ганизмы из более молодых толщ. Труд- ность исследовании заключается еще и в том, что прямой in situ анализ глубоко залегающих пород невозможен. Обычно исследуют образцы ненарушенного кер- на, в которых сразу по извлечении начи- наются необратимые физико-химичес- кие процессы, отражающиеся на физио- логическом состоянии или даже жизне- способности микроорганизмов. Среди земных природных сред обитания живых организмов вечная мерзлота представляет собой совокуп- ность уникальных биотопов, где воз- действие отрицательных температур в течение геологического времени, веро- ятно, максимально стабилизирует физи- ко-химические условия и препятствует миграционным потокам. Отдельные сообщения о присут- ствии в мерзлоте микроорганизмов дол- гое время не принимались во внимание. Лишь начавшиеся в 80-х гг. система- тические биологические исследования мерзлых осадков в восточном секторе Арктики убедительно продемонстриро- вали их заселенность жизнеспособными клетками. ВЕЧНОМЕРЗЛЫЕ ОСАДОЧНЫЕПОРОДЫ АРКТИКИИ АНТАРКТИДЫ — ПРИРОДНЫЕМЕСТООБИТАНИЯМИКРООРГАНИЗМОВ Обьекты исследования Мерзлота занимает почти 40% площади суши Земли. Основные терри- тории расположены в Северном полу- шарии и Антарктиде. Результатом про- цессов эволюции криолитозоны явилось формирование разнообразных по гене- зису ледовых комплексов, несущих в себе признаки смены геологических эпох. Объектами систематических мик- робиологических исследований мерзло- ты служили образцы позднекайнозойс- ких отложений Ямала, Аляски, Канадс- кой Арктики (долина р.Маккензи), Се- веро-восточной Сибири (Колымская 156
низменность) и Антарктиды (р-н Сухие Долины). Последние два района явля- ются полигонами многолетних исследо- вании, что позволяет сопоставить стати- стические данные микробиологичес- кого и биохимического анализов одно- возрастных осадков, формировавшихся в условиях существенно различных от- рицательных температур. Немаловажно что район Колымской низменности, как и Антарктида, удален от индустриаль- ных центров, не загрязнен химическими и тепловыми стоками. Зарождение криосферы в Арктике обусловлено изоляцией Северного По- лярного бассейна в середине-конце мио- цена. Развитие арктической криосферы происходило под воздействием транс- грессивно-регрессивных циклов Аркти- ческого океана — главного регулятора палеоклиматических изменений в Се- верном полушарии. При этом законо- мерно менялись площадные соотноше- ния, мощность и температурный режим всех составных частей криосферы Арк- тики (атмосферной, гидросферной, гля- циосферной и литосферной). Вечная мерзлота в районе Северо-востока Евра- зии имеет сплошное распространение, достигает 400 м в глубину, представляя собой наиболее древние в Арктике мерзлые толщи позднеплиоценового возраста (свыше 3 млн. лет), не протаи- вавшие в плейстоцене. Промерзание осадочных толщ шло в основном синге- нетично с накоплением осадков. В плей- стоценовую эпоху, в связи с прогрес- сирующим похолоданием, объем более поздних оледенений увеличивался. В настоящее время среднегодовые темпе- ратуры составляют -7-12 'С. Геотермический коэффициент в верхних 100 м равен О, среднее его зна- чение в мерзлой толще — 1,5 'С/100 м. Содержание органического углерода в этих осадках не зависит от глубины залегания и возраста пород и составляет 0.5-7%. Основная масса запасов органи- ческого углерода имеет растительное происхождение. Предполагается, что формирова- ние криосферы в Антарктиде началось 35-40 млн лет назад, на границе эоцена и олигоцена. Начало оледенения материка одни исследователи датируют тем же временем, другие — серединой миоцена, 14 млн. лет назад. Свободная от ледника мерзлота занимает не более 2% терри- тории, локализована на восточных участ- ках антарктического щита. Исследован- ные осадочные толщи относятся к числу наиболее экстремальных местообита- ний. Район отбора образцов Ross Desert (Dry Valleys) характеризуется наиболее низкими температурами пород, аридно- стью климата, значительным содержа- нием солей в поверхностном слое. Про- анализированы образцы, датируемые возрастом до 2 млн лет (по иной дати- ровке — до 15 млн лет). Глубина отбора образцов до 17 м. Содержание органи- ческого вещества в образцах не превы- шало 0,5%, растительные остатки обна- руживались лишь при микроскопичес- ком набпюдении (фитопиты, пыпьца). Особого внимания заслуживают данные по содержанию водного льда антарктических образцов. Традицион- ное мнение о "сухой" мерзлоте Антарк- тиды, предусматривающее иссушение пород вследствие низкотемпературной сублимации льда, не получило под- тверждения. Лишь в отдельных горизон- тах льдистость была менее 10%. В сред- нем содержание льда в образцах было на уровне 34-50%. Доступность воды длямикроорганизмов Вода в доступной форме - важней- шее условие сохранения жизнеспособ- ности клеток. Концентрация незамерз- шей воды в мерзлом грунте зависит от гранулометрического состава и является функцией температуры. Резкое сниже- ние содержания незамерзшей воды в мерзлоте происходит лишь в узком диа- пазоне отрицательных температур 0-(-3 'С). При последующем снижении температуры процесс льдообразования тормозится, а содержание незамерзшей воды в различных породах составляет 7-2% и менее (Антарктические осадки). Доступность этой воды клеткам харак- теризуется степенью ее активности — ве- личиной, определяемой силой взаимо- действия воды с гидрофильными по- верхностями (минеральные частицы, лед), макромолекулами или ионами в ра- створе, т.е. всей совокупностью физико- химических свойств среды. В общем случае условия мерзлоты как среды оби- тания микроорганизмов следует рассма- тривать как условия физиологической сухости. При максимальных значениях активности воды, по-видимому, еще осуществляется обмен с окружающей средой, тогда как при минимальной до- ступности воды клетки вынуждены ми- 157
2 3 4 5 6 7 8 9 io ii i2 i3 i4 i5 i6 i7 i8 i9 ванию мерзлых кернов, отобранных в территориально удаленных друг от друга полярных районах Северного и Южного полушарий, убеждает в том, что показатели численности и биомассы не зависят существенным образом от генезиса осадков и их возраста (рис. 1). По обилию бактериальных клеток веч- ная мерзлота не отличается от фильтруе- мых осадочных толщ и минеральных горизонтов современных почв (табл. 1). Распределение клеток по глубине носит неравномерный характер, однако содер- жание бактериальной биомассы в оса- дочной толще не убывает вплоть до ис- следованной максимальной глубины (300 м). Наблюдение периодических пи- ков численности клеток по профилю, возможно, связано с процессами частич- ного протаивания в период формиро- вания мерзлоты, когда на границе мерз- лого слоя количество микроорганизмов увеличивалось как в результате передви- жения влаги, так и вследствие акгивиза- ции и размножения клеток. Количество бактериальных кле- ток, способных к размножению на пи- тательных средах (от 10 до 10' КОЕ*/г абс.сух.веса), значительно уступает нимизировать взаимодействие со средой и перейти на эндогенный метаболизм. При обсуждении обеспеченности клеток доступной влагой в мерзлых по- родах следует учитывать, что толщина пленок незамерзшей воды является функцией не только отрицательной тем- пературы, но и давления. Повышенное давление понижает температуру плавле- ния льда, образующаяся вода пополняет пленочную воду вплоть до возможного образования перемычек и возникнове- ния условий для локальной фильтраци- оннои консолидации. Микроорганизмы в мерзлотеП ока иоты Вечномерзлые осадки полярных областей Земли содержат большое ко- личество жизнеспособных микроор- ганизмов. Общее число бактерий в образцах различного литологического и химического состава, возраста и гене- зиса, по результатам эпифлуорес- центного анализа, колеблется от 10' до 10'" кл./r абс.сух.веса. Обширный статистический материал по исследо- 158 Рис. 1. Биомасса бактерий в мерзлых осадочных породах Арктики и Антарктиды (прямой учет методом эпифлуоресцентной микроскопии). 1- современная тундровая почва; мерзлые осадочные породы Арктики (Сев.-Вост. Сибирь): 2, 3 - 5-10 тыс. лет; 4, 5 - 15-50 тыс. лет; б, 7 - до 150 тыс. лет; 8, 9 - 200-600 тыс. лет; 10-12 - О,б-1,8 млн. лет; 13, 14 — 3,0 млн. лет. мерзлые осадочные породы Антарктиды: 15 - поверхностный слой 0,1 м; 1б, 17 - 0,4-0,5 м; 18 - 50 тыс. лет; 19 — 170 тыс. лет
Таблица 1. Численность бактерий в природных местообитаниях Число бактерийЧисло культивируемыхклеток (KOE~/г)1 '- 10' С да обитаиия О щее кол-во (кл./г) Почвы 10в 1010 садочные и другие породы:до 10мдо 1240 м 10' - 10'< l0'- 10' 10' - 10'<1 2 ) 8 Мерзлые осадки:Арктика (300 м)Антарктида (20 м)Мерзлые погребенные почвыОзерные донные осадки:верхние слоим 10' - 10' 1О7 1 Ps 102 108 1О 1О4 10' - 10" 104- 10' 10" - 10'10' 4 1У 102 10з ентральная часть морей и океанов:водаверхние слои донных осадковглубокие слои донных осадков(500 м)Грунтовые воды(до 2000 м)Антарктический лед(до 3 бОО м) 10' - 10'кл/мл10' О - 10 кл/мл10'-104 10' 10' - 10' кл/мл 105- 106 кл/мл ед кл - 1 3 кл/мл * КОЕ — колониеобразующие единицы 159 общим показателям обилия, составляя 0,1-10% в образцах арктической мерз- лоты и лишь 0,001-0,01% в мерзлых осадках Антарктиды. В наиболее древ- них осадках арктического возрастного ряда (2-3 млн. лет) число покусов с высоким содержанием культивируемых клеток снижается. В мерзлоте Антарк- тиды количество колониеобразующих клеток невелико (10'-10' KOE/г). Осо- бенностью антарктических бактериаль- ных сообществ является и пролонги- рованный период их оживления. В усло- виях возрастающей "физиологической сухости" возрастает число некультиви- руемых клеток, наблюдаемых лишь пря- мыми микроскопическими методами. Феномен заметного различия по- казателей численности клеток, учиты- ваемых микроскопическими методами, и количества гетеротрофных клеток, способных к росту на питательных сре- дах, давно известен. Высказывались предположения, что в глубоких слоях должна преобладать анаэробная хемоли- тотрофная микрофлора ввиду удален- ности от поверхности, обеспечивающей приток кислорода и органического ве- щества. Такая микрофлора действи- тельно обнаруживает себя при культи- вировании мерзлых осадков на специ- фических питательных средах, а также проявлением потенциальной биогеохи- мическои активности после оттаивания: восстановлению сульфатов, продукции метана, денитрификации и др. В от- дельных случаях из образцов аркти- ческои зоны выделялось значительное количество анаэробов. Тем не менее, стоит подчеркнуть повсеместное при- сутствие культивируемых гетеротроф- ных азробных бактерий в количестве, не уступающем или превышающем чис- ленность выявляемых анаэробов. Это согласуется с мнением о повышении чувствительности вегетативных расту- щих форм микроорганизмов к глубоко- му охлаждению в условиях аноксиби- оза. Следует также учитывать, что низ- кие температуры способствуют повы- шению оксигенированности растворов. Переход вегетативных клеток в некультивируемое состояние иницииру-
Рис. 2. Бактерии с грамотрицательным типом клеточной стенки, сконцентрированные из мерз- лых пород олерской свиты (2 млн. лет, 54 м). Клетки расположены внутри гелевого матрикса, окружен- ного органо-минеральными частицами среды, Т х 40 000. 160 ется изменением параметров среды, при этом отмечается уменьшение размеров клеток, утолщение клеточных стенок. Электронно-микроскопические исследования (ТЭМ) показали, что боль- шинство бактериальных клеток (около 80'4) в образцах мерзлых осадков сразу после оттаивания представлены ультра- микроформами (0,1-0,4 мк в диаметре), которые защищены от внешнего воздей- ствия капсулярными слоями и органо- минеральными частицами (рис. 2). По- лисахаридные капсулы способны удер- живать большое количество воды и, очевидно, служат надежным резерву- аром незамерзшей воды для клеток. Внутри клеток видимых нарушений клеточных структур не обнаруживает- ся. Среди бактерий наблюдаются формы, не образующие специализированных покоящихся клеток, с разными типами клеточных стенок. Нередко можно наблю- дать конгломераты клеток, погруженные в единый матрикс. В таких микропопу- ляциях принципиально возможно разви- тие синхронизованных процессов акти- визации и даже пролиферация одних клеток за счет гибели других. Предпола- гается, что высокая жизнеспособность бактерий в условиях длительной консер- вации при отрицательных температу- рах обеспечивается дифференциацией клеточных структур, изменением соот- ношения насыщенных и ненасыщен- ных жирных кислот в мембранах кле- ток, накоплением внутриклеточного пу- ла веществ с асмо- и криопротекторны- ми свойствами. Эти вещества стабили- зируют мембраны и внутриклеточные структуры, предотвращают уменьшение объема клеток до критического состоя- ния как следствия удаления воды, преду- преждают замерзание цитоплазмы. Исследование образцов мерзлоты сразу после оттаивания методами скани- рующей электронной микроскопии (СЭМ) позволяет наблюдать биоформы в естественной минеральной среде. Сложность гетерофазной среды порож- дает дискуссии о дифференциации био- логических и минеральных объектов, нередко весьма близких по морфологии.
161 Рис. 3. Редко встречающийся в естествен- ных мерзлых осадках мицелий гриба (Антарк- тида, глубина 4 5 м; 50 тыс. лет), х500. Наиболее крупные формы, размером 10- 30 мк и более (крупные клетки дрожжей, мицелий и споры грибов, макроводоро- сли, пыльца растений, фитолиты) обычно не вызывают сомнений в своем биологи- ческом происхождении, так как имеют вполне различимые характерные особен- ности морфологии (рис. 3). Более мелкие объекты микронного уровня нередко "вписаны" в среду, тем не менее в боль- шинстве случаев тоже достаточно хорошо визуально дифференцируемы, как по мор- фологии, так и по характеру распределе- ния в среде. Затруднения возникают в слу- чае сорбции клеток вследствие присутст- вия на поверхности минеральных частиц или образования минеральных чехлов. Бактерии неравномерно распреде- лены в грунте, встречаются единичные клетки или их скопления — микроколо- нии. Осадочная порода представляет со- бой сложную комбинацию минеральных частиц, в разной степени благоприятных для колонизации. Поры и каверны - удоб- ные экологические микрониши. Неред- ко бактерии прикрепляются к поверх- ности более крупных биоформ (рис. 4). При анализе образцов в сканиру- ющем электронном микроскопе наибо- лее сложна визуальная интерпретация объектов субмикронного уровня. Сейчас уже не вызывает сомнения широкое рас- пространение в природных местообита- ниях таких малых микроорганизмов, установлена зависимость размеров кле- ток от воздействия среды. Тем не менее, дискуссия о возможности существова- ния микроорганизмов размером 0,2-0,1 мк и меньше и пределов допустимых раз- Рис. 4. Бактерии на поверхности более крупной эукариотной клетки в образце мерзлых осадков Антарктиды с глубины 4.5 м (50 000 лет), х5000. меров клетки продолжается. Возникаю- щие противоречия, на наш взгляд, во многом обусловлены автоматической экстраполяцией "лабораторного" знания на природные обьекты, тогда как особен- ности существования микроорганизмов в естественной среде пока слабо изучены. Электронно-микроскопическое ска- нирование оттаявших осадков выявляет присутствие так называемого биогеля (биопленки), описанного исследовате- лями поверхностей минералов. По-ви- димому, можно выделить несколько мор- фологических вариаций биогеля, в ряде случаев его трудно дифференцировать от глинистых частиц. Наибольший инте- рес представляют морфоформы биогенно- го вида с размером субъединиц 50-200 нм, дискретно распределенные по поверх- ности минералов или скученные в об- разования типа микроколоний (рис. 5а). В отдельных случаях обильно "обсеме- ненные" участки соседствуют с абсо- лютно чистыми поверхностями. Неред- ко наблюдаются "зоны отсутствия ро- ста" вокруг более крупных морфоформ, что напоминает антагонистические от- ношения среди микроорганизмов и по- зволяет предполагать биогенное проис- хождение обьектов. Наконец, инкубиро- вание с питательными субстратами при- водит к заметному увеличению коли- чества биогеля вплоть до образования сплошной биопленки. Все это убеждает в том, что биогель представляет собой популяции (или совокупность популя- ций - сообщества) бактерий, нечеткость морфологии которых обусловлена нали- чием вокруг клеток и конгломератов
Рис. 5а. "Биогель" (биопленка) в антарктических осадках (глубина l 7 м, 170 тыс. лет), х30 000. 162 Рис. 5в. После прогрева при 600 'С (3 часа) видны фрагменты "биогеля" с элемента- ми мицелиального ветвления и более крупные бактериальные клетки, х 10 000. Рис. 5б. Развитие "биогеля" при инкуби- ровании образца с питательным субстратом, х 5 000.
Рис. бб. Грам-отрицательная бактерия из мерзлых осадков Арктики (4 млн. лет), ТЭМ, х80 000. 163 полисахаридных гелевых чехлов (кап- сул). При высоких разрешениях видно, что фрагменты биогеля, как правило, представляют собой псевдомицелиальные образования или изогнутые цепочки глобул длиной около 0,25 мк с размером субъединиц 0,04-0,05 мк. Подобная мор- фология весьма напоминает морфологию бактерий порядка Actinomycetales. На- блюдаемые биоформы чрезвычайно тер- мостабильны: после прогревания образ- цов антарктической мерзлоты в течение 3 часов при 180 'С, 400 'С и даже 600 'С сохраняются четко различимые много- численные фрагменты биогеля и более крупные (0,5 мк) одиночные и сдвоен- ные клетки типа микрококков (рис. 5б, в). Бактериальное биоразнообразие в мерзлых осадках исследовано пока лишь на родовом уровне для культи- вируемых гетеротрофных аэробных со- обществ. Описан широкий спектр грам- положительных и грамотрицательных (рис. 6 а, б) родов бактерий (около 50 ро- дов), среди которых актиномицетная ли- ния (за исключением истинных мицели- альных .актиномицетов) занимает веду- щее положение. Обращает на себя вни- мание разнообразие неспорообразую- Рис. ба. Грам-положительная бактерия из мерзлых осадков Арктики (4 млн лет). ТЭМ, х80 000. щих бактерий, хотя нередко выделяют- ся и спороносные формы, которые при- нято считать наиболее устойчивыми к воздействию неблагоприятных факторов. Среди изолированных штаммов бактерий - представители разнообраз- ных физиологических групп, потенци- ально способные к осуществлению це-
Рис. 7б. Бактерия рода Nitrobacter, тонкое строение, ТЭМ, х60000. 164 Рис. 7а. Нитрифицирующая бактерия рода Ni~trohailсг из мерзлоты олсрской свиты (2 мли. лет), целые клетки, ТЭМ, х 10 000. лого спектра биохимических процессов: нитрификаторы (рис. 7а, б) и денитри- фикаторы, азотфиксирующие бактерии, целлюлозоразрушители, фототрофы, сульфатредукторы, газообразующие клостридии и др. В числе прокариот в мерзлых осадках присутствуют цианобактерии и относительно редко - мицелиальные актиномицеты (p. Streptomyces). Особого внимания заслуживает факт обнаружения в мерзлоте, в том чи- сле наиболее древней мерзлоте Арктики и в Антарктиде, значительного числа эукариотных клеток: спор грибов-мик- ро-мицетов, дрожжей, водорослей. Глу- бокие осадки принято считать бакте- риальными биотопами. Однако эпифлу- оресцентный микроскопический анализ (краситель — калькофлюор белый) сви- детельствует, что грибные споры в количестве 10-' — 10' кл/г абс.сух.веса присутствуют во всех проанализиро- ванных образцах мерзлых осадков Арк- тики и Антарктиды. Эти показатели
уступают показателям обилия бактери- альных клеток, однако, вследствие раз- ницы в размерах клеток, содержание грибной биомассы в осадках (1-60 мкг/г абс.сух.веса) на порядок превышает со- держание биомассы бактерий. Числен- ность спор не снижалась с глубиной в более древних осадках. Подобно распре- делению бактериальных клеток в мерз- лых толщах, на обилии спор микроми- цетов не отразились заметным образом различия в генезисе накопления осадков или их возраст. Грибной мицелий, кото- рый постоянно встречается в поверхност- ном почвенном слое, в мерзлоте практи- чески не обнаруживается. Это убеждает в неактивном состоянии грибов при температурах (-7) - (-27) 'С. Морфологи- ческое разнообразие спор весьма вели- ко, жизнеспособность подтверждается прорастанием спор при инкубировании образцов в стерильных влажных каме- рах или при активизации субстратом с последующим эпифлуоресцентным или СЭМ-анализом проб грунта (рис. 8 а-в). Температурные параметры роста штаммов микромицетов (рис. 9) харак- теризуют их, подобно бактериям, как психротрофные организмы, растущие в широком интервале температур. При этом одни штаммы лучше вегетируют при низких температурах (+4 'С), дру- гие — при более высоких (+20 'С). Наряду с микромицетами в мерз- лых осадках, в том числе наиболее древ- них (олерская и томусярская свиты, 3 млн. лет) обнаруживаются дрожжи (рис. 10) и жизнеспособные зеленые во- доросли. Рис. 8. Активизированный in situ мице- лий грибов в антарктических осадках (4,5 м, l70 тыс. лет; сусло I:3): а - 3 суток инкубации, х 500; 6, в - 2 месяца инкубации, х 20 000. 165
Рис. 9. Колонии грибов, выделенных из олерских осадков (2 млн. л~т). 166 ПОТЕНЦИАЛЬНАЯБИОЛОГИЧЕСКАЯ АКТИВНОСТЬ Структура микробных сообществ мерзлоты определяется тем, какова сте- пень покоя или активности микроорга- низмов при отрицательных температу- рах, стабильно поддерживаемых в мерз- лых толщах Арктики и Антарктиды в течение геологически длительного вре- мени. В случае консервации клеток в анабиотическом состоянии сообщества мерзлоты следует рассматривать как "сообщества выживших" древних ми- кроорганизмов, возраст которых соот- ветствует возрасту мерзлоты. Теорети- чески удержание жизнеспособного со- стояния клеткой при наличии в ней де- структивных процессов без репарации и обновления (медленная смерть) воз- можно в течение продолжительного вре- мени, превышающего время пребыва- ния исследованных осадков в мерзлом состоянии. Выдвигались предположе- ния о пос'гепенном.накоплении в мик- роорганизмах дефектов, вызванных по- током естественной радиации. Однако установлено, что устойчивость к облу- чению микроорганизмов в условиях естественной гетерофазной среды дале- ко превосходит дозу, которую они наби- рают за несколько миллионов лет. При наличии же в клетках гипометаболизма, обеспечивающего медленные репараци- онные процессы в геологическом мас- штабе времени, жизнеспособность кле- ток, очевидно, может поддерживаться неопределенно долго при минимальных энергетических ресурсах. Более активная модель сообщества должна учитывать популяционные меж- клеточные взаимодействия. Гомеостаз в этом случае может поддерживаться не торможением пролиферации микроор- ганизмов, а сбалансированным меха- низмом регуляции процессов гибели и размножения клеток в популяции (условно говоря, гибель клетки-альтру- иста — удвоение клетки-эгоиста). Такая гомеостазная система может минималь- но нуждаться в энергетической под- питке. При постоянстве обилия клеток такая система будет развиваться в про- странстве-времени, приобретая новые
качества вследствие процессов физио- логической адаптации к условиям среды. Метаболическая активность в мерзлоте была проанализирована по включению меченого по углероду суб- страта в липиды микробной биомассы. Установлено, что процесс утилизации субстрата происходил при температу- рах -5 'С и -10 'С, при -20 'С отмечены лишь следы активности. Во всех случаях процесс останавливался в тече- ние года. Возможно, процесс утилиза- ции происходил по мере выравнивания гомеостатического состояния системы после оттаивания с последующим пере- ходом микроорганизмов к глубокому анабиозу. С другой стороны, наличие метаболической активности не обяза- тельно обеспечивает активность репро- дуктивную. Тем не менее, эти данные подтверждают сообщения о возможно- сти метаболически активного состояния клеток при отрицательных температу- рах по крайней мере до -10-12 'С. Таким образом, можно предполо- жить, что бактериальные сообщества в арктической мерзлоте содержат динамич- ную метаболически активную компонен- ту и, возможно, являют собой не закон- сервированный продукт времен плиоце- на, а современные сообщества микроор- ганизмов, адаптированных к существова- нию при отрицательных температурах. Широко известным способом адаптации микрооргаизмов к низким температурам является психрофилия (рост при пониженных температурах и неспособность размножаться при тем- пературе более 20-21 'С). Психрофиль- ные микроорганизмы распространены в холодных морях, океанах и других водных средах обитания. Для почв и грунтов психрофилия оказалась явлени- ем нетипичным. Адаптация к условиям среды здесь проявляется как стабилиза- ция клеток в широком диапазоне изменения воздействующего фактора. Причем стабилизация, как правило, носит множественный характер, то есть микроорганизмы приобретают высокую устойчивость к изменению целого ряда факторов: температуры, рН, концентра- ции солей и др. Основа стабилизации- иммобилизованное состояние клеток и универсальность механизмов ответа на стресс. Микробные сообщества мерзло- ты проявляют себя как психротрофные- способные к росту в широком диапазоне Рис. IO. дрожжевая клетка, выделенная из мерзлых осадков маастаской свиты (Восточ- ная Сибирь), 600 тыс. лет, х 20 000. температур, от отрицательных (-1-8 'С) до 65 'С. Некоторые гидролитические ферменты, синтезируемые микроорга- низмами, выделенными из мерзлоты, также проявляют высокую термоста- бильность. Сообщества мерзлоты обнаружи- вают широкий спектр потенциальной биогеохимической активности. Сразу после оттаивания активизируются про- цессы синтеза и репарации в клетках, быстро нарастает дыхательная, фермен- тативная и репродуктивная активность. Интенсивность биохимических процес- сов в пересчете на единицу биомассы очень велика, заметно выше, чем в та- лых почвах. Спектр утилизируемых после оттаивания источников углерода, оцененный методом мультисубстрат- ного тестирования, существенно отли- чает сообщества мерзлоты от микроб- ных сообществ современных почв, при- чем отмечены различия и по возрастной шкале осадков. Это также служит аргу- ментом в пользу постепенных адаптив- ных изменений микробных сообществ по мере их пребывания в мерзлоте. 167
БИОМАРКЕРЫ Несмотря на то, что ряд косвенных данных свидетельствует о биохимичес- ких процессах в мерзлоте, вопрос о на- личии и скорости метаболических про- цессов в клетках при отрицательных температурах ill Бйи остается открытым. Интерпретация получаемых данных до- статочно сложна. В гетерофазной среде некоторые низкотемпературные биохи- мические процессы могут протекать благодаря, например, активности иммо- билизованных внеклеточных ферментов при наличии анабиотически неактивных микроорганизмов. Иммобилизован кое состояние макромолекул, присутствие криопротекторов (растворимых органи- ческих веществ: углеводов, аминокис- лот, многоатомных спиртов и пр.) пред- отвращают низкотемпературную дена- турацию ферментов. Более того, возмож- на активизация биохимических процес- сов при структурных превращениях льда. Мерзлые осадки с первых момен- тов таяния обнаруживают нарастающую активность гидролитических и окисли- тельно-восстановительных ферментов. Ряд ферментов проявляет высокую тер- мостабильность. Другая группа биомаркеров, по- стоянно обнаруживаемая в мерзлоте- это пигменты. В частности, хлорофилл присутствует практически повсеместно. В числе биомаркеров принято рас- сматривать и биогенные гюы: двуокись уг- лерода, метан, окислы азота, сероводород и др. Мерзлота является мощным резер- вуаром биогенных газов. Установлено, что газы в мерзлоте могут находиться в виде твердой фазы, образуя скопления газо- гидратов. О взаимодействии микроорга- низмов с газогидратами ничего не изве- стно. На примере метана показано. что распределение концентраций газов по профилю арктической толщи мерзлоты носит неравномерный и дискретный характер: зоны накопления сменяются слоями, в которых метан отсутствует. В метаносодержащих слоях выявлены бактерии-метаногены. Наличие биомаркеров - фермен- тов, растворимых метаболитов, пиг- ментов, биогазов и др. - основа для бы- строго восстановления функциониро- вания экосистем при таянии мерзлоты. С другой стороны, не исключено, что продолжительность жизни биоактивных макромолекул ггри низких температурах превышает возможности клетки. ЗАКЛЮЧЕНИЕ Мерзлота, занимающая значи- тельную часть территории земной суши, заселена вполне полноценными и жизнеспособными сообществами микроорганизмов, почти не уступаю- щими по обилию поверхностным поч- венным слоям. Гетерофазная среда, стабильные температуры и высокая стабильность физико-химической сре- ды создают оптимальные условия для длительного поддержания гомеостаза. Это обеспечивает возможности для реализации механизмов устойчивого поддержания жизни в клетках. Физи- ческие параметры мерзлоты, по край- ней мере арктической зоны, не про- тиворечат возможности физиологи- чески активного состояния прокариот: гипометаболизма или даже репродук- тивных процессов, регулируемых по- пуляционными взаимосвязями. Для эукариот мерзлота, очевидно, надежно обеспечивает глубокий покой клеток с сохранением генетического потенци- ала и возможности реверсии. Следо- вательно, в пределах геологически продолжительного времени в мерзлых осадках могут формироваться экоси- стемы, обладающие специфическими особенностями. Признаки таких спе- цифических экосистем можно усма- тривать в динамике процессов акти- визации клеток при оттаивании, спектре их физиологической актив- ности, структурных особенностях ци- томорфологии и др. Жизнеспособ- ность микроорганизмов в мерзлоте в условиях низких температур обеспечи- вается структурными перестройками, свойственными клеткам при переходе в состояние переживания, которое может характеризоваться разным уровнем снижения метаболической активности вплоть до глубокого покоя. Естественные пол идисперсные гетерофазные системы обеспечивают чрезвычайно высокую стабильность клеток и биологически активных моле- кул как результат процессов иммоби- лизации. Отрицательные температуры являются дополнительным фактором. способствующим поддержанию ста- бильности биологических систем.
ДЛИТЕЛЬНЫЙ АНАБИОЗ МИКРООРГАНИЗМОВ В ЛЕДНИКЕ ЦЕНТРАЛЬНОЙ АНТАРКТИДЫ 169 Возможность длительного сохра- нения жизнеспособности микроорга- низмов в состоянии анабиоза при низ- ких температурах в природе долгое вре- мя вызывала горячие споры. Возмож- ность "экспорта" и "импорта" жизни в межпланетном пространстве стало пред- метом прямых экспериментальных ис- следований. Опираясь на результаты многочисленных опытов по изучению влияния космической среды на микро- организмы, ряд крупных ученых 60-х годов (Опарин, Rush, Firsoff, Шкловс- кий, Имшенецкий) пришли к выводу, что низкие температуры, близкие к абсолютному нулю, отсутствие влаги, отсутствие кислорода, ультравысокий вакуум порядка 10 ' мм рт. ст., раз- личные виды радиации, существующие в космическом пространстве, не могут служить препятствием для преодоления микроорганизмами огромных расстоя- ний космоса в состоянии анабиоза. Фактор времени, по некоторым теоретическим соображениям, также не является губительным. Некоторые ис- следователи считают, что нет принци- пиальной разницы между организмами, просуществовавшими в состоянии ана- биоза 200 или 2000 и более лет, осо- бенно если они находились при низких температурах. Однако, несмотря на ряд имевшихся по этому поводу публи- каций, "не было ни одной работы, ко- торая доказывала бы с помощью бе- зупречных методов возможность ожив- ления водорослей или бактерий, нахо- дившихся сотни тысяч и миллион JToT в состоянии анабиоза в каменной соли, горных породах или ископаемых жи- вотных Толщу ледников Антарктиды и Гренландии можно рассматривать как удачный природный материал для ре- шения проблемы сверхдлительного ана- биоза. Удаленность от районов интен- сивного антропогенного воздействия, стабильность температурного фактора в режиме отрицательных значений и надежная защищенность сформировав- шихся в геологическом прошлом го- ризонтов толщи ледника от последу- ющих влияний окружающей среды де- лают ледниковый щит Антарктиды иде- альным объектом для эксперименталь- ного решения проблемы существования в природе сверхдлительного анабиоза. Несмотря на активные микробио- логические исследования, проводив- шиеся в Антарктиде в последние деся- тилетия, толща ледника оставалась со- вершенно не изученной микробиоло- гами. Первостепенной задачей являлась разработка метода асептического отбора проб из толщи ледника Центральной Ан- тарктиды для микробиологических ис- следований. Объектом исследований служила толща ледника Центральной Антарктиды в районе российской стан- ции "Восток", находящейся на полюсе холода нашей планеты, с координатами 78'27' южной широты и 106'52' восточ- Н0Н ДОЛГОТЫ. Средняя температура августа со- ставляет -68 'С, абсолютный максимум температур зафиксирован в 1974 г. -13 'С, средняя же температура значи- тельно ниже. Таяние поверхности снега и льда здесь совершенно исключено. Толщина ледникового покрова превы- шает 3750 м. Скорость аккумуляции осадков за последние 5000 лет состав-
Рис. 1. Общий вид микробиологической буровой установки (МБУ) на. станции "Восток". МЕТОДИКА АСЕПТИЧЕСКОГООТБОРА ПРОБ 170 ляет около 2,4 см льда в год. Темпера- тура льда обычно -50 -60 'С. Посте- пенно повышаясь по мере углубления, температура ледника у ледникового ложа достигает примерно -2 -3 'С. По данным гляциологов характер первичной структуры ледниковых отло- жений и морфологические особенности ледника в его внутриматериковой части исключают возможяос'уь элиан(.'тичос- кого, т.е. вторичного, проникновения микрофлоры в толщу ледникового по- крова. Следы ранее существовавших трещин или каких-либо других отступ- лений от классического для районов Центральной Антарктиды пути разви- тия ледниковой толщи в исследуемой зоне не обнаруживаются. Был создан комплекс технических средств колонкового бурения-плавления скважин во льду и отбора проб из полу- ченного ледяного керна, а также разра- ботана методика асептического отбора проб для последующих микробиологи- ческих анализов. Передвижная микробиологическая буровая установка представляет собой смонтированное на металлических са- нях помещение размером 9х3,7х2,5 м, состоящее из 3-х отсеков - буровой ла- боратории (I), микробиологической ла- боратории (Ц) и тамбура (Ш). Общий вид установки и схема расположения от- секов представлена на рис. 1 и 2. Отсек для проведения микробио- логических работ оснащен всем необ- ходимым оборудованием (термостат, сушильный шкаф, насос и УФ лампы для стерилизации воздуха и наружных поверхностей оборудования и инст- румента). Для отбора проб из отрезков ледя- ного керна смонтированы специальные боксы с перчатками, обеспечивающие надежную защиту от загрязнения проб посторонней микрофлорой, что доказы- валось контрольными экспериментами. В боксах создавалось избыточное дав- ление подачей стерильного воздуха для обеспечения надежного заслона против проникновения туда посторонней мик- рофлоры в момент работы. Буровая лаборатория имеет утеп- ленную вышку высотой 7 м над устьем скважины, лебедку с кабель-тросом, бу- ровой снаряд, допускающий нагрев до 200 'С с целью его стерилизации, пульт управления буровым снарядом и на-
4 1 3 2 Рис. 2. План внутренних помещений МБУ-1: I - буровой отсек; II микробиологический отсек; III - тамбур; IV. - мойка. 1 — устьс скважины; 2 - лебедка; 3 — пульт управления буровым снарядом; 4- опоры для вышки; 5 - стол; 6 - бокс с установкой для асептического отбора проб из ледникового керна: 7 - бокс для анализа отобранных проб; 8 - рабочий стол; ) — тсрмостат; 10 - автоклав; 11 - бак для воды. стольную установку для асептического отбора проб из отрезков ледяного керна. Асептический отбор проб осуще- ствляется путем выплавления из цент- ральной части ледяного керна. При этом наружный, не растаявший слой льда служит надежным экраном, изолирую- щим отбираемые из центральной части керна пробы от окружающей среды и препятствующим попаданию в пробу посторонней микрофлоры. Непосредственно перед отбором пробы на образце ледяного керна полу- чают не загрязненный окружающей микрофлорой участок поверхности пу- тем откалывания торцевой части керна. Этот метод обладает тем преимущест- вом, что исключает контакт получаемой поверхности с инструментом. С этой це- лью в установке предусмотрено специ- альное кольцевое скалывающее устрой- ство. После откалывания торец керна быстро опускается на стерильную по- верхность освобожденного от крышки нагревателя и при включении послед- него начинается отбор пробы. Темпе- ратура, измеренная термопарой на кон- такте лед-нагреватель (на поверхности вогнутого конуса) и, следовательно, от- бираемой пробы воды, колеблется от б' до 13 'С в зависимости от первоначаль- ной температуры льда. Многократно проведенные испы- тания показали, что тест-культура мик- роорганизмов, нанесенная на поверх- ность ледяных кернов, не может прони- кать в его внутренние слои, и разра- ботанная методика гарантирует стериль- ный процесс отбора проб из централь- ной части керна. Второй способ прямого микробио- логического контроля осуществлялся в процессе отбора проб из ледяного керна. Он заключается в том, что первая проба высевается на богатую питательную среду, например, мясопептонный агар (МПБ), сразу после опускания свеже- сколотого торца керна на стерильную поверхность головки нагревателя и нач- ала вытаивания центральной части керна. Отсутствие роста микроорганиз- мов в первой контрольной пробе при об- наружении микроорганизмов в одной или нескольких последующих пробах, отобранных из замкнутой системы, яв- ляется показателем надежности разра- ботанного метода (схема отбора, рис.3). Выплавляемую из центральной части керна талую воду собирали в под- готовленные литровые колбы с перетя- нутым горлышком, которые стерильно присоединяли к установке посредством резиновой трубки перед началом отбора пробы. Колбы содержали концентри-
дста такюрва а о а о б о Рис. 3. Схема отбора проб из центральной части ледяного керна. А - Г - варианты проб, а-отбор пробы, б - запаянная колба, 1-5 - исследуемые участки отрезка керна; (МПБ — мясопептонный бульон). 172 рованные питательные среды, приго- товленные с таким расчетом, чтобы по- сле добавления пробы воды среда раз- бавлялась до наружной концентрации. Таким образом, отбираемая проба слу- жила одновременно и анализируемым материалом, и разбавителем питатель- ной среды. Поступление отбираемой пробы воды в приемную колбу прекра- щали отключением нагревателя. После этого колбы, не отсоединяя от прибора, в месте перетяжки запаивали при помо- щи газовой горелки. Затем к установке асептически присоединяли следующую колбу, и процесс отбора проб возобнов- лялся. Это позволяло проводить высевы из одного и того же отрезка керна на раз- личные жидкие питательные среды, а также отбирать пробы для последую- щих исследований с применением дру- гих ме годик. Третий способ контроля надежно- сти отбираемых проб связан с петрост- руктурными исследованиями ледяных шлифов, изготовленных из остатков ле- дяного керна после отбора из него проб для микробиологических анализов. Эти остатки представляли собой полые ци- линдры с толщиной стенки около 20 мм и дном толщиной около 50 им. Дно этих "стаканов" и послужило объектом де- тальных макро- и микроскопических петроструктурных исследований. Результаты изучения этих шлифов показали, что в ледяном керне, во время бурения не происходят ни относи- тельные подаижки отдельных частеи керна, ни проникновение вглубь керна талой воды, образующейся при бурении скважины. Возможные трещины на поверхности керна, глубина которых не превышала 5-8 мм от поверхности кер- на, мгновенно залечивались водои на поверхности керна, имевшего темпера- туру -55'С, что также приводит к мгно- венной изоляции центральных частей керна. Следов ранее существовавших трещин или каких-либо отклонений от классического для районов центральной Антарктиды пути развития ледниковой толщи в изученных образцах керна не обнаружено.
Таблица 1. Результаты микробиологических анализов проб из ледяного керна и водосборного бака на присутствие тест-культуры Serratia marcescens Справедливость этих выводов пол- ностью подтвердилась при дополнитель- ном прямом контроле с использованием тест-культуры в момент бурения с отбо- ром керна. Для этого культуру Serratia marcescens вносили на забой скважины непосредственно в момент бурения. После окончания процесса буре- ния и подъема снаряда керн извлекали из керноприемника и подвергали мик- робиологическому анализу. Одновре- менно проводили микробиологический анализ воды, которая в процессе буре- ~сия откачивается в специальныи водо- сборный бак бурового снаряда. После фильтрации проб через мембранные фильтры последние накладывали на МПА в чашках Петри и помещали в термастат при температуре 26-28 'С. Ре- зультаты опыта оценивали по появле- нию колоний Serratia marcescens на мембранных фильтрах после инкубации в термостате (табл.1). Из таблицы видно, что в воде, образующейся в процессе бурения вокруг ледяного керна и собранной в водосборный бак, содержание клеток тест-культуры в 1 мл даже после значи- тельного разбавления первоначальной суспензии водой, образующейся в про- цессе бурения ледника, было довольно высоким. В пробах же воды объемом 1000 мл, отобранных из центрального стержня керна, ни в одном случае при- сутствие тест-культуры бактерий не обнаружено. Таким образом, прямые микро- биологические контрольные исследо- вания на забое скважины показали, что возможность проникновения посто- ронних организмов из талой воды в керн в процессе бурения полностью исключена и достоверность материа- лов, полученных из центрального стержня ледяного керна, не вызывает сомнении. 173
Таблица 2. Количество жизнеспособных микроорганизмов в толще ледника центральной Антарктиды по мере углубления в древнейшие слои КОЛИЧЕСТВЕННАЯ ОЦЕНКАСОДЕРЖАНИЯ ЖИЗНЕСПОСОБНЫХМИКРООРГАНИЗМОВ В ТОЛЩЕАНТАРКТИЧЕСКОГО ЛЕДНИКА Анализ полученных данных по распределению жизнеспособных микро- организмов в различных слоях толщи лед- ника показал, что их распространение име- ет неравномерный и случайный харак- тер. По мере уптубления в ледник, пример- Обозначения: - неспорообразующие бактерии 100 80 )) - спорообразующие ф бактерии (- и О CL - дрожжи - мицелиальные грибы у О О l - снижение количества жизнеспособных микроорганизмов Глубина в метрах Рис. 4. Распределение различных систематических групп жизнеспособных микроорганизмов (и их процентное содержание от общего количества) по мере углубления в ледник Антарктиды. 174 О Б о ~ О ~ Z с щ с ~- ,CL „-о !О > r S о 2 е~ х ~ Б ф Я Z Б Ф e e о ~~ х < .й Z >- %в 1 С г С9 О3 CL~ С О но со 114 метров (около 3000 лет), веро- ятность обнаружения жизнеспособных микроорганизмов в пробах снижается. Анализы керна из сверхглубокой скважины, проведенные до глубины 2405 м, подтвердили первоначальный ВЫВОД. Все оценки возраста исследуемых слоев ледника до птубины 320 м сделаны на основании п~яциологических данных по среднегодовому накоплению снега и 100 200 300 600 1500 2400 (3 000 (7 000 (12 000 (35 000 (107 000 (окопа 200 000 лет) лет) пет) пет) пет) пет)
Таблица 3. Количество микроорганизмов в толще ледника по данным эпи- флуоресцентной микроскопии и метода проращивания на питательных средах Количество клеток в 1 ДОЗ Рост на Количество клеток в 1 мл x10' Рост на Глубина, м питательныхС ДЯХзацрсаа плотная Глубина, м питательных с дах)KjlllXRR llJ1o'nlaa 1,00 2002 1502 l,40 0,76 2025 1,10 1525 l,40 2055 1,80 1548 0,43 2074 1,10 1575 0,47 2100 1.30 1599 0,79 2125 0,34 1625 О 0,72 2149 0,65 1650 2176 0,38 1675 0,83 2200 1,00 1699 0,46 2224 1,20 1726 1,50 2250 0,76 1749 2,20 2275 1,20 1775 1,30 2300 1,50 1798 0,79 2326 0,58 1824 0,86 2349 0,54 1850 0,83 2375 1,40 1875 1,40 2395 1,30 1899 6,80 2402 3,90 1925 0,86 2405 1,90 1949 1,40 1975 При мечанис. "+" — рост микроорганизмов на жидкой срсдс, "0" отсутствие роста. На плоп ой среде указано количество выросших колоний. 175 льда в районе станции "Восток". Возраст слоев ледника ниже 320 м оценивался по данным совместных советско-французс- ких птящиологичес~сих исследоваюы. Данные по изменению содержания жизнеспособных микроорганизмов по мере углубления в древнейшие слои ледника приведены в табл. 2. Из таблицы видно, что по мере углубления в ледник с отбором проб че- рез определенные интервалы наблюда- ется снижение количества жизнеспо- собных микроорганизмов, обнаруживае- мых в обследованных отрезках керна. Таким образом, по мере углубле- ния в древнейшие слои ледника вероят- ность обнаружения жизнеспособных микроорганизмов на питательных сре- дах снижается. Однако с помощью эпи- флуоресцентной и сканирующей элект- ронной микроскопии в пробах талой во- ды, полученной из тех же горизонтов ледника, были обнаружены микроор- ганизмы, принадлежащие к различным таксономическим группам и отличаю- щиеся большим морфологическим раз- нообразием. Общее количество микро- организмов в пересчете на 1 см-' пробы
176 Рис. 5. Различные дрожжи, встречаю- щиеся в толще ледника (по данным М.Н. П о глазовой. льда составляет 10-'-10' клеток. Среди выделенных из толщи ледника микроор- ганизмов обнаружены как прокариоты (неспорообразующие и спорообразую- щие бактерии и актиномицеты), так и эукариоты - мииелиииьиые грибы и дрожжи. В относительно молодых горизон- тах ледника наблюдается более широ- кое разнообразие микроорганизмов. В более древних горизонтах до 2405 м и глубже жизнеспособная микрофлора встречается значительно реже и преиму- щественно спорообразующие бактерии. Ыа рис. 4 схематически показано рас- пределение представителей различных систематических групп жизнеспособ- ных микроорганизмов по глубинам. При дальнейших исследованиях глубоких горизонтов ледника с помо- щью оптической и электронной микро- скопии было обнаружено значительно большее количество микроорганизмов, чем при высевах на питательные среды (табл. 3). Из 39 посевов на питательные среды только в 6 случаях есть рост. Под- счет общего количества клеток показал присутствие клеток во всех пробах. Удалось установить, что микро- флора древних горизонтов ледника в интервале глубин от 1500 до 3344 м со- стоит почти из тех же групп микроор- ганизмов, что и современная микрофло- ра. Обнаруженные микроорганизмы Рис. 6. Грибы из толщи ледника (по дан н ы м М.Н. Иоглазо вой. Рис. 7. Бактериальная клетка, лежащая на осколке панциря диатомовой водоросли (по данным И.Н. Мицкевич).
Таблица 4. Потребление радиоактивного субстрата (14C-гидролизата белка) в пробах талой воды, полученной из образцов ледяного керна на станции Восток (по данным И.Н. Мицкевич) * Измерения не производили. 177 прннщщежат к различньпч TRKcoHQMH- ческим группам и характеризуются зна- чителыым морфологическим разнооб- разием. По всей толще исследуемого слоя встречались бактерии различной формы и ~измеров. Среди них были кокки, ди- плококки, палочки разной длины и тол- щины, прямые и изогнутые, а также клетки овальной формы. Нередко обна- руживались актиномицеты. Среди зука- риотных организмов встречались раз- личные виды дрожжей, как почкую- щихся, так и делящихся грие. 5). В некоторых пробах встречались обрывки мицелия различных грибов, гифы которых были частично или пол- ностью лизированы (рис. 6а). В отличие от мицелия обнаруженные нами кони-дии грибов сохранились достаточно хо-роша (рис. бб,в). В некоторых горизон-тах была найдена пыльца растений. С помощью люминесцентной и сканирую-щей электронной микроскопии было об- наружено большое разнообразие одно- клеточных водорослей. Были найдены диатомовые водо- росли, количество которых было осо- бенно велико в слое ледника на глубине 2035 м. Из горизонта 2375 м, возраст ко- торого составляет примерно 180 тыс. лет, были найдены остатки диатомовых водо- рослей и бактериальные клетки (рис. 7). Что же касается общего коли-чества микробных клеток в 1 мл талой воды вз различных гориюнтов, оно, как мы вндгпг из Ta6JI. 3, колебалось в IIp0-делах 10'-10-' клеток на 1 мл. Эти ре-зультаты показывают, что значительная часть микроорганизмов, находящихся в толще ледника, либо уже погибли, либо не pacryr на применяемых нами пита-тельных средах. Дополнительные анализы с при- менением меченого по С14 гидролизата белка в качестве источника питания показали, по в результате инкуб;щии m 4 до 40 часов наблюдается использо-вание субстрата, что свидетельствует о присутствии жизнеспособных микро- организмов в горизонтах ледника, воз- раст которых превышает 240 тыс. лет (табл. 4). Очевидно, совершенствуя ме- тоды, возможно добиться получения культур микроорганизмов, сохранивших ж)изнеспособность. Сопоставление данных ггрямого счета по распределению микробов в рез- ных горизонтах с результатами гляцио- логических исследований по распреде- лению микрочаспац папи, попавщей из атмосферы в эти же горизонты, показы- вает, что колебания тех и других имеют аналогичный характер грие. 8б,в). В свою очередь, колебания в концентра-ции микрочастиц пыли зависят от кли-
hgc. Ie' уъ 150 173 I I0 409 (а) о Р 9М -2 -480 ° ° :11 t ° Ф в ° е ° Т tOOOOOO (а) ИМтоо~рвйвв ° ° % ° ° i1ЪФ 1 ° '1 ° 1 ° ° е1 1 t ф1 ° ' ' ' 1 а ° е ь ° ° ~ ~° ° hlmmptgric miaoparticks 4 Ъ ° ° Ф ;11 Ф 1 ° ° ° 11 в1 ° !50 Е v Ъ 50 (а) 1500 Рис. 8. Количество клеток микроорганизмов и пылсвых частиц в толще ледника Центральной Антарктиды в различные климатические периоды (см. объяснение в тексте). ЗАКЛЮЧЕНИЕ 178 матических условий, существовавших в те же периоды, что является очевидным при сравнении данных, представленных на рис. За, в. В эти периоды, т.е. 240-110 тыс. лет назад (8а) происходило форми- рование исследуемой нами части ледни- ка. Анализ данных микробиологических и гляциологических исследовании пока- зал, что наибольшая концентрация пы- ли, а также микробов обнаруживается в периоды глобального похолодания. Так, на рис. Зб,в видно, что максимумы, со- ответствующие наибольшему количест- ву пыли, а также сохранившихся клеток микроорганизмов, соответствуют гори- зонтам, расположенным на глубине при- мерно 2000-2150 м и 2300-2400 м. Следует отметить, что в пробах с этих же горизонтов с помощью скани- рующего электронного микроскопа бы- ло обнаружено наиболее высокое содер- жание остатков диатомовых водорослей, а также пыльцы высших растений. В периоды максимального похо- лодания, обеспечившие наибольшую со- хранность пылевых частиц и биологи- ческих объектов, по данным гляциоло- гов, происходило увеличение аридных территорий, соответствующих высохшим участкам шельфа Мирового океана, уси- ление ветровой деятельности и меридио- нального перемещения воздушных масс. Эти условия, по всей вероятнсти, спо- собствовали интенсивному поступле- нию биологических и минеральных ча- стиц в атмосферу, их дальнейшему пе- реносу и стоку на поверхность ледни- кового покрова, тем более, что размеры большинства микробных клеток (0,4-1,5 мкм) и микроводорослей (15-30 мкм) примерно соответствуют размерам мел- ких пылевых частиц. Результаты микробиологических и гляциологических исследовании толщи ледника Центральной Антарктиды пока- зывают, что этот объект является уни- кальной моделью для решения различ- ных проблем экзобиологии. Не только следы прошлых климатических ката- клизмов на Земле запечатлены в древ- них горизонтах ледника. Там прекрасно сохранились представители различных таксономических групп микроорганиз- мов. Часть микроорганизмов сохранила свою жизнеспособность в течение мно- гих десятков и сотен тысяч лет. Велика вероятность того, что лед- никовые образования, часто встречаю- щиеся во Вселенной, могут служить но- сителями жизнеспособных микробов, надежно сохраняющихся при длитель- ном путешествии в межпланетном про- странстве.
РАЗВИТИЕ ЖИЗНИ НА ЗЕМЛЕ п1СО+ Н,О = [СНО]+ О,) иm ([CH0 + О = СО + Н 0}, 179 Биосфера современного типа с растительным покровом на суше и эвкариотным фитопланктоном в море с основанными на этих первичных про- дуцентах пищевыми пирамидами жи- вотных сложилась в фанерозое. Расти- тельный покров распространился с конца силура в девоне около 300 млн. лет назад. Появление эвкариот дати- руется сейчас 2,2-2,5 млрд. лет (грибы?). Начало распространения эвкариотного фитопланктона датируется по микро- фоссилиям (акритархам), сопоставляе- мым с празинофитами, не менее 1 млрд. лет назад в течение неопротерозойской революции. Несколько позже обнаружи- ваются многоклеточные красные водо- росли. Предшествующий период, зани- мающий 4/5 истории биосферы, отно- сится к её становлению при господстве прокариот (рис. 1). Жизнь существует не сама по себе, а в составе биосферы, где она взаи- модействует с внешними факторами: космическими в виде излучения Солнца и геосферными. Взаимодействие обус- ловлено прежде всего биогеохимичес- ким и процессам и. Обычно становлен ие биосферы пытаются излагать с возник- новения жизни на Земле. Такой подход неудобен тем, что он начинает изложе- ние с наименее известного времени и посвящен разного рода гипотезам, ни одна из которых не подтверждена фак- тическим палеонтологическим мате- риалом. Большинство гипотез исходно ошибочно, потому что подменяет жизнь, существующую только в виде дискрет- ных целостных организмов, анализом происхождения их- частей, например нуклеиновых кислот. Наиболее простым является прокариотный организм-клет- ка. Живые предшественники её неизве- стны. Поэтому историю биосферы нуж- но начинать со свидетельств появления на Земле первых клеток прокариот. Пря- мые свидетельства их существования относятся к 3,5 млрд. лет назад (Варра- вуна, Австралия), косвенные — к 3,8 млрд. (Исуа, Гренландия) и совпадают с появ- лением осадочных пород и началом гео- логической осадочной летописи. Бакте- риоморфные микрофоссилии в метеори- тах имеют возраст более 4,6 млрд. лет, но их интерпретация не общепринята и они не относятся к земным объектам. Становление биосферы целесообразно анализировать сверху вниз от извест- ного состояния ее в фанерозое. Биосферные механизмы нагляднее всего описываются, исходя из системы сопряженных биогеохимических цик- лов, центральную роль в которых играет цикл органического углерода. Этот цикл включает продукционную ветвь, обус- ловленную автотрофной ассимиляцией СО главным образом при оксигенном фотосинтезе, и деструкционную ветвь при дыхании по реакциям: причем п='тп, поскольку противополож- ное невозможно. Из неравенства n>m следует биогеохимическая сукцессия, обусловленная накоплением неразло- женного С pr в осадочных породах в виде керотена. Разность n — т соответствует избыточному О„остающемуся в атмо- сфере, который может расходоваться на окисление эндогенных восстановителеи
млрд. лет 2, Рис. 1. Главнейшие события в эволюции биосферы докембрия (по Розанов, 1992 с допол- нениями). 180 пз геосферных источввп<ов и приводит к переходу биосферы от нейтрального или восстановленного состояния к окислен- ному. Переход этот осуществлялся от 2,5 до 1 млрд. лет назад и привел к возникновению О -содержащей атмо- сферы, служащей необходимым пред- варительным условием для появления аэробных эвкариот (см. рис.). В переход- ный период происходило образование резервуаров окисленньm форм Ее в виде отложений железистых кварцитов и S в виде сульфатов океана, представляющих основные резервуары кислорода, соот- ветствующие рассеянному С р- осадоч- ных отложений; в современной атмо- сфере по балансу с Сор. остается около 5% О. С [CHO] биомассы связаны сте- хиометрическими соотношениями дру- пзе биогенные элементьс за счет синтеза запасных веществ и клетоп~ых стенок происходит увеличение отношений в пользу С. Обычно для биомассы прини- мают состав [СН О]„„,[1ЧН,] б[Н,РО,], определяющий соотношение циклов биогенных элементов в продукционных процессах. Цикл Сорг связан с циклом С1 кчзр~ через лабильные формы: СО, атмо-сферы и находящимся в равновесии с ним бикарбонатом океана. Источником CO считается дегазация Земли. Сток СО, осуществлялся при образовании от- ложений карбонатов Са и Mg, посту- павших за счет yrJIeKHcJIQTHoro вьпце- лачивания изверясенных пород. Карбо- натная система обусловливает близкое к нейтральному рН состояние поверхно- спьи оболочек Земли. В истории Земли можно отметить этапы последователь- ного доминирования пропеосов атлоясе- ния окислов железа, доломитов, извест- няков. Все эчапы сопряженных биогеохи- мических циклов катализируются про-
кариотами и только прокариотами. Клю- чевую реакцию оксигенного фотосин- теза осуществляют первичные проду- центы — цианобактерии. При фотоасси- миляции происходит преимущественное включение в биомассу изотопа С'-' и ор- ганическое вещество древнейших оса- дочных пород несет изотопное отноше- ние автотрофной ассимиляции. Циано- бактерии способны так же к фиксации атмосферного азота и зависят в первую очередь от света, развиваясь только на дневной поверхности, и доступного фосфора. Деструкцию мортмассы циа- нобактерий осуществляют бактерии-де- структоры, образующие сложную тро- фическую сеть специализированных ор- ганизмов, которые используют (1) био- полимеры в сахаролитическом, протео- литическом, липолитическом путях, и, (2) продукты первичной деградации, как например летучие жирные кислоты (ЛЖК), для полного разложения до СО и НО, замыкая цикл С,~. Для ранних этапов эволюции с бескислородной ат- мосферой особое значение имеют анаэ- робные организмы, причем первично анаэробный обмен свойственен только прокариотам, включая окисление за счет неорганических акцепторов электрона, таких как СО и соединения серы. Аэробы не обязательно связаны с появлением О в атмосфере, поскольку они должны были появиться одновре- менно с оксигенными цианобактерия- ми, участвуя в детоксикации кислорода, задерживавшегося в виде пузырьков в фотосинтезирующем сообществе. Наблюдать эти процессы можно в так называемых реликтовых сооб- ществах бактерий, развивающихся в экстремальных условиях, где отсутст- вуют эвкариоты. Реликтовые сообще- ства бактерий служат современными аналогами прокариотной биосферы про- терозоя. Термофильные цианобактерии обильно развиваются в низкотемпера- турных газогидротермах от 65 до 45 'С, преобразуя вулканические выделения в газовую смесь, близкую по составу к воздуху. Этот процесс аналогичен пре- образованию продуктов дегазации Зем- ли в воздух кислородной атмосферы и его легко воспроизвести в лаборатории. Другим примерам циано-бактери- ального сообщества могут служить дон- ные сообщества соленых морских ла- гун, образующие слоистые маты с по- следователъным расположением сверху вниз функциональных группировок ор- ганизмов от первичных продуцентов к организмам заключительных этапов де- струкции. Такие маты обусловили тек- стуру преимущественно карбонатных слоистых биогенных пород — стромато- литов — прослеживающихся на протяже- нии геологической истории Земли от архея и резко выклинивающихся в конце протерозоя. Наряду с одиночными на- ходками микрофосслий строматолиты относятся к наиболее ранним палеонто- логическим свидетельствам жизни на Земле, демонстрируя, что жизнь возник- ла и поддерживалась в виде сообществ. Циано-бактериальные сообщества матов содержат все необходимые компоненты, чтобы катализировать полную систему биогеохимических циклов, как она опи- сана выше. Аналогичные сообщества обнаружены в высокоминерализован- ных содовых озерах, которые представ- ляют континентальные экосистемы, и, таким образом могут рассматриваться как аналоги ранних наземных экоси- стем. Замкнутость биогеохимических циклов служит основным критерием возможности существования экосисте- мы а течение геологически длительного времени. Бактерии и только бактерии спо- собны катализировать циклы азота, же- леза и серы. Для серы возможен вариант альтернативный циано-бактериальному сообществу с участием аноксигенных фототрофных серных бак герий как первичных продуцентов. В таком анаэ- робном цикле можно предполагать эн- догенный источник H S, сток продуктов в бассейны и регенерацию сульфида сульфатвосстанавливающими бактерия- ми, судя ло изотопным соотношениям начавшуюся менее 2 млрд. лет назад. Однако доказательств работы такого цикла пока нет, даже для Черного моря, представляющего аналог древнего "сульфидного океана". В большинстве современных экосистем, за исключени- ем пресных наземных с метаногенезом, серный цикл с сульфат-редукторами и пурпурными фототрофами замыкает де- струкцию, например в содовых озерах, соленых лагунах, морском мелководье. Для термофилъных сообществ характер- но отсутствие пурпурных фототрофных серных бактерий. Сульфаты образуются путем аэробного окисления Н S бакте- риями. 181
Особую группу реликтовых экоси- стем могут составлять представители подземной биосферы, использующие эн- догенные вещества геотермального про- исхождения, прежде всего Н, как источ- ник энергии для хемосинтеза. Большин- ство этих организмов относится к гипер- термофилам, способным развиваться вплоть до температуры несколько выше температуры кипения с установленным верхним пределом 110 'С. Продуктами их обмена являются СН, и H S. Как пример сообщества, основанного на хе- мосинтезе, приводят глубоководные "оазисы" жизни на выходах термальных вод с "курильщиками". Этот пример не- корректен для начала биосферы, по- скольку используется резервуар раство- ренного О океана, образованного фото- синтезом. Поскольку гипертермофилы, среди которых большинство составляют археи, неспособны к фотосинтезу, то это сообщество нельзя считать исходным для ствола биологической эволюции. Таким стволом служит циано-бак- териальное сообщество, где цианобак- терии составляют продукционную, а бактерии — деструкционную ветви. Из этого сообщества хорошо прослежива- ется как биогеохимическая, так и фило- генетическая эволюция Земли и биоти- ческого разнообразия. Более того, раз- нообразие прокариот и созданная ими экосистема служит необходимым пред- варительным условием появления эвкариотпротист и последующего эво- люционного дерева живых существ. Начиная с эвкариот живые организмы могут существовать только в условиях биосферы, созданной и поддерживае- мой прокариотами. В этом смысле эво- люция происходит аддитивно. Основ- ными этапами биологической эволюции служат прокариотная биосфера с начала геологической летописи, распростране- ние эвкариот-протист начиная с неопро- терозойской революции около 1 млрд. лет назад (хотя они отмечаются и ранее 2 млрд. лет), распространение много- клеточных нетканевых организмов (Pa- razoa, губки) ранее 0,6 млрд. лет в венде и позднем рифее и появление бесскелетных тканевых многоклеточных, начало ске- летной фауны в кембрии, распростране- ние наземной растительности в девоне (см. рис.1). В пределах прокариотн ой био- сферы, насколько можно судить по ископаемым микрофоссилиям циано- бактерий, не было существенных изме- нений: появившись ранее 2,5 млрд. лет назад цианобактерии в своих крупных таксонах не изменились до настоящего времени. Эволюция прокариот и про- тист реконструируется на основе фило- генетического анализа одного гена 16S (18S) рибосомальной РНК. Реконструк- ция устанавливает последовательность изменения гена, но не время. 182
ЛИТЕ PATУPA 183 Баженова О.К. Геология нефти и газа. 1990. № 7. С.2-5. Валентон И. Бокситы. М. 197. 215 с. Вологдин А.Г. Геологическая деятельность ми- кроорганизмов. Известия AH СССР. Сер. Геол. 1947. № 3. С.19-36. Гептнер A.Р., Алексеева Т.А., Пиковский Ю.И. Полициклические ароматические углево- дороды в вулканических породах и гидро- термальиых мииералах Исландии. 1999. Литология и полезн. ископаемые. № 6. Р.619-631. Гептнср А.Р., Петрова В.В., Соколова А.Л., Горькова Н.В. Биохсмогенное форми- рование слоистых силикатов при гидро- термальном изменении базальтов, Ислан- дия // Ли.гология и полезн. ископаемые. № 3. 1997. С.249-259. Герасименко Л.М., Гончарова И.В., Жегалло Е.А, Заварзин Г.A., Зайцева Л.В., Орле- анский В 1~., Розанов А.Ю., Тихомирова Н.С., Ушатинская Г.Т. Процесс минерали- зации (фосфатизации) нитчатых циано- бактерий // Литология и полезн. иско- паемые. 1996. № 2. С.208-214. Герасименко Л.М., Жегалло Е.А., Жмур С.H., Розанов A.Þ., Хувер Р. Бактериальная палеонтология и исследования углистых хондритов // Палеонтол. журн. 1999. № 4. С.103-125. Герасименко Л.М., Заварзин Г.А. Реликтовые цианобактериальные сообщества // Про- блемы доантропогенной эволюции био- сферы. М.: Наука. 1993. С.222-254. Гиличинский Д.А, Воробьева Е.А., Соина В.С. и др. Микробиология вечной мерзлоть!. 1996. Материалы 1 конф. геокриологов России. Кн.1. С.174-185. Готтшалк Г. Метаболизм бактерий. 1982. Громов Б.В. Поведение бактерий // Соросовский образовательный журн. 1997. № 9. С.28-32. Громов 6.В. Удивительный мир архей // Соросовс- кий образовательный журн. 1997. № 4. С.23-26. Громов Б.В. Цианобактсрии в биосфере // Горо- совский образовательный журн. 1996. № 9. С.33-39. Додд Р.Т. Метеориты. Пстрология и гсохимия. М.: Мир. 1986. 384 с. Жмур С.И., Горленко В.М., Розанов А.Ю., Же- галло Е.А., Лобзова Р.В. Цианобактери- альная бентосная система — продуцент углеродистого вещества щунгнтов нижнего иротерозоя Карелии // Литология и полези. ископ. 1993. № 2. С.122-124. Жмур С.И., Розанов А.Ю., Горленко В.М. Лити- фицированные остатки микроорганизмов в углистых хондритах // Геохимия. 1993. № 1. С.66-68. Заварзин Г.А. Бактерии и состав атмосферы. М.: Наука. 1984. 199 с. Заварзин Г.A. Развитие микробиальнь!х сооб- ществ в истории Земли 0 Ироблемы доан- тропогенной эволюции биосферы. М.: Наука. 1993. С.212-222. Звягинцев Д.Г. Микроорганизмы в вечной мерзлоте. Успехи микробиологии. 1992. Т.25. С.3-27. Звягинцев Д.Г. Почва и микроорганизмы. Изд. Моск. ун-та. 1987. 256 с. Звягинцев Д.Г., Гиличинский Д.А., Благодатс- кий С.А., Воробьева Е.А. и др. Длитель- ность сохранения микроорганизмов в постоянно мерзлых осадочных породах и погребенных почвах // Микробиология. 1985. Т.54. № 1. 155 с. Крылов И.Н., Гихомирова Н.С. К образованию кремнистых микрофоссилий // Палеон- тол. журн. 1988. № 3. С.3-9. Кузнецов С.И. Геохимическая активность микрофлоры озер. Л.: Наука. 1970. 175 с. Кулаев H.Ñ. Ироисхождение эукариотических клеток // Соросовский образовательный журн. 1998. № 5. С.17-22. Определитель пресноводных водорослей. Под ред. М.М.Голлербах, .гом 2. М.: Советская наука. 1953. 652 с. Органическое вещество шунгитоносных пород Карелии (генезис, эволюция, методы изу- чения) // М.М. Филиппов, А.И. Голубев, П.В. Медведев и др. Петрозаводск: Ка- рельский научный центр PAH. 1994. 208 с. Орлеанский В.К., Герасименко В.К. Лабора- торное моделирование термофильного цианобактериального сообщества // Ми- кробиология. 1982. Т.51. № 4. С.538-542. Орлеанский В.К., Раабеи М.Е.. Строматоличъ~ живые буквы каменной летописи Земли // Природа. 1998. № 11. С.68-85. Розанов A.Þ. История становления скелетных фаун // Соросовский образовательный журн. 1996. № 12. С.62-68. Розанов A.Þ. Современная палеонтология // Соросовский образовательный журн. ]999. № 1. С.47-55. Розанов А.Ю. Цианобактерии и, возможно, низшие грибы в метеоритах ll Соросовский образо- вательный я<урн. 1996. № 11. С.б1-65. Розанов A.Þ. Бактериально-палеонтологичес- кий подход к изучению метеоритов // Вестник PAH. 2000. Т.70, № 3. С.214-233. Розанов А.Ю., Заварзин Г.А. Бактериальная палеонтология // Вестник PAH. 1997. Т.67. № 3. С.241-245. Сергеев В.Н. Окремненные микрофоссилии докембрия и кембрия Урала и С~дней Азии. М.: Наука. 1992. 134 с. Сергеев В.Н., Нолл Э.Х., Заварзин Г.A. Первые три миллиарда лет жизни: от прокариот к эукариотам // Природа. 1996. № 6. С.54-67.
Скулачев В.П. Биоэнергетика. Мембранныс преобразователи энергии. 1989. Скулачев В.П. Элсктродвигатель бактерий /i Соросовский образовательный журн. 1998. № 9. С.2-7. Спирин A.Ñ. Молекулярная биология. Структура рибосомы и синтез белка. 1986. Тимофеев Б.В. Микрофитофоссилии раннего докембрия. Л.: 1982. 128 с. Тимофеев Б.В. Микрофоссилии печентской се- рии. В кн.: Палеонтолоп~я докембрия и ран- него кембрия. Л.: Наука. 1979. С.119-120. Чухров Ф.В. Коллоиды в земной коре. Изд. AH СССР. М. 1953. С.671. Шлегель Г. Общая микробиология. 1987. Юшкин Н.П. Глобулярная надмолекулярная структура шунгита: данные растровой туннельной микроскопии // Доклалы АН. 1994. Т.337. ¹ 6. С.800-803. Янкаускас Т.В., Герман Т.Н., Михайлова H.С., Сергеев В.Н. и лр. Микрофоссилии докембрия СССР. Л.: Наука. 1989. 190 с. Abyzov, S.S. Microorganisms in Antarctic ice. Io: Antarctic Microbiology. Ed. ЕЛ. Friedmann, Wiley-Liss. 1993. Р.265-297. Anders Е. Meteoritic hydrocarbons and extraterres- trial life// Ann. N.Ó. Acad. Sci. Lett. 1962. Ч.93. Р.651-657. Anders Е., Fitch F. Search or organized clemcnts in carbonaceous chondrites // Science. 1962. Ч.138. Р.1392-1399. Bardossy G., Аleva G.J. Lateritic Bauxitcs. Akademia Kiado. 1990. 624 р. Barghoorn Е.S., Tyler S.A. Microorganisms from the Gunflint cherts // Science. 1965. Ч.147. № 3658. Р.563-577. Bj8rklund М., giftr;ioglu N., Kajander Е.О. Extraordinary survival of nanobacteria under extreme conditions // Instruments, Methods, and Missions for Astrobiology..Proc. SPIE. 1998. Ч.3441. Р.123-139. Bramlette М.N. Geology of Arkansas Bauxite region // Arkansas geological survey. In1ог- mation circular 8. Little Rock. 1936. Qiftqioglu N., Qiftr;ioglu Ч., Vali Н. Sedimentary rocks in our mouth: dental pulp stones made by nanobacteria // Instruments, Methods, and Missions for Astrobiology. Proc. SР1Е, 1998Ь. Ч.3441. P.130-136. Qiftpioglu N., Bjarklund М.. Kajander Е.О. Stone formation and calcification by nano- bacteria in the human body. Instruments, Methods, and Missions for Astrobiology // Proc. SPIE. 1998. Ч.3441. Рр.105-111. Claus G., Nagy В. A microbiological examination of some carbonaceous chondrites // Nature. 1961. Ч.192. Р.594-596. Claus G., Nagy В. Consideration of Extraterrestrial Таха // Taxon. 1962. V. I I. P.160-161. Colwell, F., Fredrickson, J.K., Chandler, D., Del- viche, М., Onstott, Т.С, et al. Microorga- nisms from deep, high temperature, cretace- ous sandstones. ISSM'96, 15-21 Sept.,1996. Davos-Switzerland. Abstracts. Costerton J.W., Marrie Т.J., Cheng К3. Phenomena of bacterial adhesion. In: Bacterial Adhe- sion: Mechanisms and Phisiological Signi- ficance. Savage D.Ñ. & Fletcher М, (eds.). 1985. New-York, Plenum. Pp.3-43. Ehrlich H.!.. Geomicrobiology. New York: Marcel Dekker. 1996. 719 р. Folk КЛ . $ЕМ imaging of bacteria and nannobac- teria in carbonate sediments and rocks // J. Sediment. Petrol. 1993. Ч.63. Рр.990-999. Folk R.L., Lynch Н . The possible role of nan- nobacteria (Dwarf bacteria) in clay-mineral diagenesis and the importance of careful sample prepartion in high-magnification SEM study // Journal of Sedimentary Research. 1997. Ч.67. № 3. Р.583-589. Folk R.L., Lynch Е1. Carbonaceous objects resem- bling nanobacteria in the Allende meteorite. instruments, Methods, and Missions for Astrobiology // Proc. SPIE. 1998. Ч.3441. Рр.112-122. Gold, Т. The deep hot biosphere. Proc. Nat. Acad. Sci. USA 89. 1992. Р.6045-6049. Gorlenko Ч.М., Zhmur S.I., Osipov G.A., Suzina N.E., Dmitriev ЧЧ. Microbial nature of fibrous kerite of Чо1уп. Instruments, Methods, and Missions for Astrobiology // Proc. SPIE. 1999. Ч.3755. Рр.83-95. Hodgson С.W., Baker В.L. Evidence of porphyrins in the Orgueil meteorite // Nature. 1964. Ч.202. Р.125-131, Hofmann Н.J., Grey К., Hickman, A.H. Thorpe R.l. Origin of 3.45 Ga coniform stromatolites in the Warrawoona Group, Western Australia // Geol. Soc. A mer. Bull. 1999. Ч.111. Pp.1256-1262. Hoover R.B. Meteorites, microfossils, and Exo- biology. Instruments, Methods, and Missions for Extraterrestrial Microorganisms // Proc. SPIE. 1997. Ч.3111. P.115-136. Hoover R., 1<ozanov А.Yu., Zhniur $.1., Gorlcnko V.М. Further cvidcnce of Microfossils in 'arhonaccous Chondrites. Instruments, Me- hods, and Missions for Astrobiology // Proc. SPIE. 1998. Ч.3441. Рр.203-213. Kajander Е.О., Bjог)с1ип4 М., ~~йроа1и N. Mineralization by nanobacteria // Instru- ments, Methods, and Missions for Astrobio- logy. Proc. SPIE. 1998. Ч.3441. Р.86-94 Kasting 3.F. Earth' s early atmosphere // Science. 1993. V.259. Рр.920-926. Knoll А.H. Exceptional preservation of photosyn- thetic organisms in silicified carbonates and pcats I/ Phil. Trans. R. Soc. London, Scr. В311. 1985. Рр.11-22. Krumbein W.Е. Stromatolites - the challenge of а term iri space and time // Precambrian Res. 1983. Ч20. Рр.493-531. Marteinsson V., Kristjansson J., Kristmannsdottir Н., Geptner А. Discovery and Description of Giant Submarine Smectite Cones on the Seaf1o0r in Eyjafjordur, N-Iceland and а Novel Thermal Microbial Habitat. 2000 (in press). Manuscript submitted to Applied and Environ>riental Microbiology. 184
McKay D.S., Gibson Е.К., Thomas-Keprta 5г.К.L., Vali Н., Romanek S., Clemett S.J., Chillier Х.D.F., Maedling С.R., Zare КМ Search for past life on Mars: Possible relic biogenic activity in Martian meteorite ALN 84001 // Science. 1996. Ч.273. Р,924-930, МсКау D.S., Gibbson E.К., Thomas-Kerpta К., Romanek С.S. Evidence for possible past life in а Martian meteorite: а current assessment. Instruments, Methods, and Missions for Ast- robiology // Proc. SPIE. 1997. V.3111. Рр.2-6. Merz М., Schlue ЖК., Zankl Н. PH-measurements in the sheath of calcifying filamentous cyanobacteria // Bull. De ГInstitut oceano- graphique. Monaco. 1993. P.45. Muller К. Crustacea with preserved soft part &om the Upper Cambrian of Sweden//Lethaia. 1983. V.16. № 2. Р,93-109. Muller К.J., Wallosek D. A remarcable arthropod fauna from the Upper Cambrian "Orsten" of Sweden // Trans. of the Royal Soc. of Edin- burgh. 1985. Ч.76. Рр.161-172. Nagy В. Carbonaceous Meteorites. N. Y.: Elsevier Sci. Publ. Со., 1975. P.1-747. Palik P. Further 1ifeforms in the Orgueil meteorite // Nature. 1962. Ч.194. P.1065. Pflug Н.D., Heinz В. Analysis of Fossil-Organic Nanostructures, Terrestrial and Extraterrestrial // Instruments, Methods, and Missions for the Investigation of Extraterrestrial Micro- orgnisms. Proc. SPIE. 1997. V.3111. P.86-97. Posfai М., Buseck P.R., Bazylinski D., Frankel R. Iron sulfides from magnetotactic bacteria: Structure, composition, and phase transitions // American Mineralogist. 1998. Ч.83. № 11- 12. P.1469-1481. Ross R. Panel Discussion: The identity of the 'organized elements' // Ann. МХ. Acad. Sci. 1963. V.108. Рр.608-609. RossignoI-Strcik М., Barghoorn E.S. Extraterres- trial abiogenic organization of organic mat- ter: The hollow spheres of the OrgueiI mete- orite // Space Life Sci. 1971. Ч.З. P.89-107. Rozanov A.Yu. The Cambrian Radiation of SheIIy fossils. TREE. 1992. Ч.7. № 3. Рр.84-87. Schopf J.W. Microfossils of the Early Archean Apex Chert: new evidence of the antiquity of life // Science. 1993. Ч.260. Рр.640-646. Schopf J.W., Klein С. (eds.) The Proterozoic bio- sphaere: а multidiscipIinary study. Camb- ridge: Cambridge Univ. Press. 1992. 1348 р. Sergeev Ч.N., KnoII, A.Н., Grotzinger J.P. Paleo- bioIogy of the Mesoproterozoic Billiakh Group, Anabar UpliA, Northeastern Siberia // Journal of Paleontology. 1995. Ч.69. Memoir 39. 37 р. The Shorter Bergey's Manual of Determinative Bacteriology, 9-th Edition. 1994. Baltimora, Maryland. 787 р. Stanier R.W., Sistrom W.R., Hansen Т.А. et а1. Proposal to place the nomenclature of the Cyanobacteria (blue-green algae) under the rules of the International Code of Nomen- clature of Bacteria // Int. J. Syst. Bact. 1978. Ч.28. Рр.335-336. Stevens, Т.О. & McKinley, J.P. Lithoautotrophic microbial ecosystems in deep basalt aquifer // Science. 1995. V.270. P.450-454. Tan W.Ñ., VanLandingham ЯЛ . Electron micro- scopy of biological-like structures in the Orgueil carbonaceous meteorite // J. Roy. Atr. Soc. 1967. Ч.12. P.237. Tazaki К. Biomineralization of layer silicates and hydrated Fe/Mn oxides in microbial mats: an electron microscopical study // Clays and Clay minerals. 1997. V.45. № 2. P.203-212. Urey Н.С. Biological Material in Meteorites: а Review // Science. 1966. V.151. P.157-166. Uwins P.J.R., Webb R.I., Taylor A.P. Novel nanoor- ganisms from Australian sandstones // American Mineralogist. 1998. V.83. № 11- 12. P.1541-1563. Vainshtein М., Kudryashova Е., Suzina N., Ariskina Е., Voronkov V. Formation of bacterial nano- cells // Proc. SPIE. 1998. V.3441. P.95-104. VanLandingham. The naming of estraterrestrial taxa // Taxon. 1963. Ч.12. P.282. Vorobyova Е., Soina V., Gorlenko М., Minkovskaya N. The deep cold biosphere: facts and hypoth- esis // FEMS Microbiol. Rev. 1997. V.20. P.277-290. Walsh М.М. Microfossils and possible microfossils from the Early Archean Onver-wacht Group, Barberton Mountain Land, South Africa // Precambrian Res. 1992. V.54. Рр.271-293. Walte М.R. Archaean stromatolites: evidence of the еаг�s oldest benthos. In: Earth' s Earliest Biosphere, edited by J.W. Schopf, Princeton Univ. Press, New Jersey. 1983. Рр.187-213. Westall F. and Rince Y. Biofilms, microbial mats and microbe particle interactions: electron microscope observations from diatomaceous sediments // Sedimentology. 1994. V.41. Рр.147-162. Westa11 F., Boni. L. and Guerzoni М.Е. The experi- mental silicification of microorganisms // Palaeontol. 1995. Ч.38. Рр.495-528. Yushkin N.Р. Natural poIymer crystals of hydrocar- bons as models of prebiological organisms О J. Crystal Growth. 1996. Ч.167. Р.237-247. Yushkin N.P. Nydrocarbon crystals as protoorgan- isms and biological systems predessors // Proc. SPIE. 1998. Ч.3441. P.234-246. Zachara J.М., Frederickson J.Ê., Li Shu-Mei, Kennedy D.W., Smith S.С., Gassman P.Ь. BacteriaI reduction of crystaIline FE' oxides n singIe phase suspension and subsuface materials // American Mineralogist. 1998. Ч.83. № 11-12. P.1426-1443. Zavarzin G.A. Metalloginium symbioticum // Zeitschrift fur A1lg. Mikrobiologie. 1964. V.4. № 5. S.390-395. Zhegallo Е.А., Rozanov АХи., Ushatinskaya G.Т., et а1. Atlas of Microorganisms from Ansient Phosphorites of Khusugul (Mongolia). Hunts- ville, Alabama. USA. NASA. 2000. 167 р. 185
СЛОВАРЬ ТЕРМИНОВ 186 Актинолит — моноклинный амфибол, промежуточный член изоморфного ряда тремолит-ферроактинолит. АТФ — аденозинтрифосфат. Ацетат — анион уксусной кислоты (СН,СОО-) или соль этой кислоты. Форма, в которой уксусная кислота присутствует в живых клетках. Бактериохлорофиллы- тетрапиррольные Mg- содержащие пигменты пурпурных и зеленых бактерий, обуславливающие их способность к фотосинтезу. Локализованы во внутриклеточных мембранах. Имеются бактериохло- рофиллы а, b, с, d, е, g, каждый с характерным спектром поглощения. Волютиновые гранулы- цитоплазматические включения в виде гранул полифосфата у микроорганизмов. В.г. — внутренний резерв фосфатов, за счет которого клетка может при недостатке фосфора в среде осуществлять еще несколько делении. Гликокаликс — внеклеточная поли- сахаридная слизь, однородная или с включением целлюлозных фибрилл. Грамотрицательные бактерии— прокариоты, клетки которых не окрашиваются по методу Грама. Они характеризуются т.н. грамотри- цательным типом строения клеточ- ных стенок: присутствием наруж- ной мембраны и зоны периплазмы, некоторыми особенностями состава и строения мембранного аппарата, устойчивостью к ряду антибиотиков. Грамположительные бактерии— прокариоты, клетки которых окрашиваются по методу Грама. Они характеризуются т.н. грамполо- жительным типом строения клеточ- ных стенок. Для них характерны чувствительность к некоторым антибиотикам, не действующих на грамотрицательные бактерии, некоторые особенности состава и строения мембранного аппарата. Изопреноиды — природные соединения из группы липидов, образующиеся в живых организмах. Среди изопренои- дов много физиологически активных веществ: антибиотики, витамины А, D, Е, К, гормоны, пигменты. Каротиноиды — желтые, оранжевые или красные пигменты, синтезируе- мые главным образом бактериями, грибами и высшими растениями; полиненасыщенные углеводороды. Катагенез — процесс в земной коре, протекающий в условиях низкой температуры и давления. Отвечает тои стадии в жизни осадочнои породы, которая наступает после диагенеза, но предшествует метаморфизму. Кероген — органическое вещество горючих сланцев, а также сингенетичное рассеянное органическое вещество любого генетического типа. Лактат — анион молочной кислоты или СОЛЬ ЭТОИ КИСЛОТЫ. Липиды — жироподобные вещества, входящие в состав всех живых клеток и играющие важную роль в жизненных процессах. Перминерализация — не полностью прошедший процесс минерализа- ции, когда значительное количество органического вещества сохраня- ется в минеральном матриксе окремненных организмов. Полисахариды — крахмал, гликоген, запасные утлеводы микроорганизмов. У некоторых микробов при помощи цветной реакции с раствором Люголя можно идентифицировать крахмал (синяя окраска) или гликоген (коричневая окраска).
Полифосфаты — многие бактерии и зеленые водоросли способны накап- ливать фосфорную кислоту в виде анул полифосфата. Такие гранулы ыли впервые описаны у Spirillum volutans, поэтому их называют волютиновыми гранулами. О О О II II II I I I ОН ОН ОН Порфирины — пигменты растений и животных, в основе молекулы которых лежит структура из четырех пиррольных колец — порфин. Протокатагенез — начальный этап катагенеза, отвечающ~ буроугольной стадии углефикации углеродистого органического вещества. Родопсин — сложный белок, комплекс гликопротеида и липидов. В клетках некоторых бактерий родопсин участвует в фототрофном питании, преобразуя энергию света в энергию макроэргических соединении. Сапонит — магнезиальный слоистый силикат. Селадонит-глауконит — полигенная группа гетерогенных слоистых силикатов, представлена рядом соединений, переходных от смектитов до почти чистои гидрослюды. Селадонит обычно гидротермальный. Смектиты — сложная группа слоистых силикатов (монтмориллонит, бейделлит, нонтронит, сапонит), общим свойством которых является ярко выраженная способность к внутрикристаллическому набуха- нию. Встречаются в самых разнооб- разных геологических обстановках. СО-дегидрогеназа — ферменты катализирующие реакции отщепления водорода от одного субстрата и переносящие его на другие. Фикобилин — пигменты красных водорослей и цианобактерий; по химической природе — белки из группы хромопротеидов. Формиат — анион муравьиной кислоты (HCOO ) или соль этой кислоты. Хелаты — внутрикомплексные соли, в которых центральный атом или ион связаны одновременно с двумя или более атомами органической молекулы. Образование хелатного комплекса называется хелатированием или хелатизацией. Хитин — материал наружного скелета членистоногих, брахиопод и неко- торых других животных. Он являет- ся также важнеишим компонентом клеточных стенок больших групп грибов, базидиомицетов, аскомицетов, зигомицетов. Хлориты — водные метаалюмосилика- ты Mg и Fe, слоистой структуры. Хлорофиллы — зеленые пигменты растений, с помощью которых они улавливают энергию солнечного света и осуществляют фотосинтез. Основу молекулы хлорофилла со- ставляет Mg-порфириновый комп- лекс. Существ ет несколько типов хлорофиллов а, Ь, с, d), отличаю- щихся системой сопряженных связей и спектрами поглощения. Целлюлоза — главный компонент клеточных стенок у высших растений и водорослей. сн~он он Цеолиты — водные алюмосиликаты щелочеи и щелочных земель. Цикл трикарбоновых кислот — общий заключительныи этап окислительного распада продуктов обмена углеводов, жиров и белков (или аминокислот) до СО и Н,О. Эпидот — Са(Без+, Al)A1[O/ОН//SiO,/Si O,] — минерал низкой и средней степени метаморфизма. Этанол — этиловый спирт. 187
УЧЕБНОЕ ПОСОБИЕ ВАКТЕРИАЛЬНАЯ ПАЛЕОНТОЛОГИЯ С.С. Абызов М. Велш Ф. Весталл Е.А. Воробьева А.Р. Гептнер Л.М. Герасименко Д.А. Гиличинский Е.А. Жегалло Г.А. Заварзин Д.Г. Звягинцев В.К. Орлеанский М.Е. Раабен А.Ю. Розанов В.Н. Сергеев В.С. Саина Г.Т. Ушатинская Р. Хувер Э.Л. Школьник Ответственный редактор: А.Ю. Розанов Художник: И.С. Сергеенкова Компьютерная верстка: М.К. Емельянова Изд.лиц. ИД № 00783 от 20 января 2000 г. Подписано в печать 28.06.2001 г. Формат 60х88/8. Гарнитура «Таймс». Печать офсетная. Бумага мелованная. Усл.печ.л.22. Уч.-изд.л.10. 'Гираж 600 экз. Заказ № Палеонтологический институт РАН 117868 Москва, Профсоюзная ул., д.123 Отпечатано в типографии 188